Область техники, к которой относится изобретение
[001] Это изобретение относится к области биологии и химии. В частности, изобретение направлено на биолюминесцентные системы.
Уровень техники
[002] Биолюминесценция - процесс излучения света живыми организмами в ходе биохимической реакции, в которой химическая энергия превращается в световую. Описано несколько типов биолюминесцентных систем.
[003] Например, у ряда морских кишечнополостных описаны системы, включающие белки семейства экворина (Prasher, et al., Biochem. 1987, 26:1326-1332; Tsuji et al., Photochem Photobiol, 1995 62(4):657-661). Семейство экворина (aequorin) включает также обелин (obelin), халистаурин или митрокомин (halistaurin, = mitrocomin), фиалидин или клитин (phiallidin, = clytin) и т.д. Это фотопротеины, содержащие ковалентно связанный с ними люциферин, который в присутствии ионов Са2+ подвергается химическим превращениям с образованием продукта в возбужденном электронном состоянии.
[004] Компоненты биолюминесцентных систем (люциферазы, фотопротеины, люциферины и т.д.) - широко используемые реагенты для множества анализов, включая диагностические системы, системы контроля качества и т.д. Например, белки семейства экворина широко используются для исследований высвобождения и связывания Са2+ в биологических системах, например, во время мышечного сокращения. Использование биолюминесцентных систем в подробностях описано, например, в Cormier, М.L. et al., Photochem. & Photobiol. 49/4, 509-512 (1989), Smith, D.F. et al. in ′′Bioluminescence and Chemiluminescence: Current Status (P. Stanley & L. Krick, eds.), John Wiley and Sons, Chichester, U.K. 1991, 529-532.
[005] Расшифровка новых компонентов биолюминесцентных систем позволяет расширить спектр доступных анализов и приложений для использования.
[006] Биолюминесценция высших грибов хорошо известна. Плодовые тела многих грибов способны продуцировать постоянный свет, хорошо видимый невооруженным глазом. Люминесценция экстрактов биолюминесцентных грибов была впервые продемонстрирована в 1959 году (R.L. Airth and W.D. McElroy, Light emission from extracts of luminous fungi, J. Bacterid., 1959, 77, 249). Свет возникал в ответ на добавление НАДФН к смеси «холодных» и «горячих» экстрактов, приготовленных из мицелия грибов Collybia velutipes и Armillaria mellea.
[007] Как здесь используется, термин «холодный» экстракт относится к экстракту, содержащему ферменты биолюминесцентной системы гриба и свободному от низкомолекулярных компонентов этой системы. Для получения холодного экстракта может быть использован следующий протокол: биомассу грибного мицелия отмыть от среды культивирования и поместить в большое количество дистиллированной воды (1:100-1:200 по массе) на 15-16 часов, при температуре 26°C. После вымачивания биомассу собрать и заморозить при -20°C. Далее биомассу разморозить и несколько раз промыть дистиллированной водой. Мицелий залить 0.01М фосфатным буфером, pH 7.5, в соотношении 1:10 (вес/объем), измельчить с помощью гомогенизатора и обработать ультразвуком (например, на приборе Ultrasonic disintegrator UD-20 (Techpan, Poland) или аналогичном на льду 5 раз по 1 мин с интервалами 1 мин. Полученный гомогенат центрифугировать при 30000 g в течение 20 мин при 4°C.
[008] Как здесь используется, термин «горячий» экстракт относится к экстракту, содержащему низкомолекулярные компоненты биолюминесцентной системы гриба и свободному от активных ферментов этой системы. Для получения горячего экстракта может быть использован следующий протокол: биомассу грибного мицелия отмыть от среды культивирования и поместить в большое количество дистиллированной воды (1:100-1:200 по массе) на 5-6 часов, при температуре 26°C. После вымачивания собрать биомассу, нагреть до кипения, после чего быстро остудить на льду и центрифугировать при 30000 g в течение 20 мин.
[009] Исследования биолюминесценции высших грибов привели к заключению, что в ее основе лежит единый двухстадийный процесс: на первом этапе происходит образование люциферина из прекурсора с помощью НАД(Ф)-Н-зависимого фермента, на втором - окисление люциферина люциферазой с появлением видимого света (R.L. Airth, Characteristics of cell-free fungal bioluminescence, in Light and Life, ed. W.D. McElroy, B. Glass, Johns-Hopkins Press, Baltimore, 1961, pp. 262). Однако до настоящего времени химическая природа компонентов биолюминесцентных систем высших грибов не была определена.
[010] В 1966 г. Кувабара и Вассинк описали выделение люциферина из Omphalia flavida, однако не предоставили данных о его химической структуре (S. Kuwabara and Е.С. Wassink, in Bioluminescence in Progress, ed. F.H. Johnson and E.Y. Haneda, Princeton University Press, Princeton, 1966, p. 233). В 1970 г. Эндо и соавт. выделили флуоресцентный компонент из Pleurotus japonicus с максимумом эмиссии флуоресценции при 530 нм (близко к максимуму эмиссии биолюминесценции гриба), названный иллудин С (illudin S). Однако это вещество не показало биолюминесцентной активности (М. Endo, М. Kajiwara and K. Nakanishi, Chem. Commun., 1970, 309). Позднее, в 1987-1988 гг., Изобе и соавт. выделили из того же источника рибофлавин и ламптерофлавин (lampteroflavin) с максимумом эмиссии флуоресценции при 524 нм, однако эти вещества также не обладали биолюминесцентной активностью (М. Isobe. D. Uyakul and T. Goto, J. Biolumin. Chemilumin., 1987, 1, 181; M. Isobe, D. Uyakul and T. Goto, Tetrahedron Lett., 1988, 44, 1169). Кандидаты на роль люциферина также выделены в качестве люциферинов из Mycena chlorophos (S. Hayashi еу al. 2012. Biophysics Vol. 8, pp. 111-114) и Panellus stipticus (O. Shimomura et al. J Biolumin. Chemilumin. 1993, 8, 201-205; O. Shimomura et al Tetrahedron 1988, 44, 1597-1602, O. Shimomura Bioluminescence: Chemical Principles and Methods. CHAPTER 9 LUMINOUS FUNGI 2006, World Scientific, Singapore), однако их участие в биолюминесценции так и не было обосновано. В 2009 г. Оливейра и Стевани описали выделенные компоненты биолюминесцентных систем нескольких видов грибов: Gerronema viridilucens, Mycena lucentipes и Mycena luxaeterna (A.G. Oliveira and С.V. Stevani, Photochem. Photobiol. Sci., 2009, 8, 1416), указав, что исследование их структур затруднено из-за низкой концентрации и малой стабильности этих веществ. Последние работы этих исследователей на широком спектре грибов показали кросс-реакции «горячих» и «холодных» экстрактов из разных видов, подтвердив единый механизм и сходство биолюминесцентных систем у всех высших грибов (Oliveira et al. Photochemical & Photobiological Sciences 2012, 11 (2): 848-52, Stevani et al. Photochemistry and Photobiology, 2013, 89: 1318-1326).
Сущность изобретения
[011] Заявители показали, что предлюциферином (или прекурсором) люциферина грибов является гиспидин, люциферином - 3-гидроксигиспидин, а ферментом, который превращает предлюциферин в люциферин, является НАД(Ф)Н-зависимая гидроксилаза (далее гиспидин-3-гидроксилаза). Это открытие раскрывает широкие возможности для анализа активности люциферазы грибов в биологических образцах.
[012] Так же Заявители показали, что в качестве предшественника люциферина может также выступать биснориангонин, а люциферина - 3-гидроксибиснориангонин.
[013] Таким образом, предметом изобретения являются методы и реактивы для детекции люциферазы в биологических образцах. Методы и реактивы настоящего изобретения позволяют выявлять люциферазу грибов или ее функциональные аналоги. Подобные системы на основе целентеразина (coelenterazine) разработаны для выявления люцифераз кишечнополостных в биологических образцах, однако они не применимы для анализа люцифераз высших грибов и их функциональных аналогов вследствие иного механизма биолюминесценции.
[014] Настоящее изобретение обеспечивает метод для выявления люциферазы в биологическом образце с помощью 3-гидроксигиспидина или его функционального аналога, включающий
(а) добавление к образцу 3-гидроксигиспидина;
(б) инкубацию получившейся реакционной смеси в условиях, подходящих
для возникновения биолюминесценции;
(в) детекцию биолюминесценции реакционной смеси.
[015] В преимущественных воплощениях 3-гидроксигиспидин или его функциональный аналог (например, 3-гидроксибиснориангонин) добавляют к образцу до конечной концентрации - 0.03-30 мкМ, чаще 1-5 мкМ, например, 2.4 мкМ. В преимущественных воплощениях 3-гидроксигиспидин или его функциональный аналог добавляют к образцу в буферном растворе, имеющем pH в диапазоне от 6.0 до 9.8, чаще от 6.5 до 9.0, например от 7.0 до 8.0. Часто раствор содержит реагенты, увеличивающие растворимость 3-гидроксигиспидина и его функционального аналога в воде, например, детергенты, такие как Triton Х100 или нонилфеноксиполиэтоксиэтанол.
[016] Реакционная смесь может также содержать компоненты, стабилизирующие и защищающие люциферазу от ингибирующего воздействия следовых количеств ионов тяжелых металлов и от деградации протеазами. Например, реакционная смесь может содержать дитиотреитол (ДТТ), бета-меркаптоэтанол и/или ЭДТА.
Также реакционная смесь может содержать ингибиторы протеаз, например, фенилуксусную кислоту или щавелевую кислоту.
[017] Настоящее изобретение также обеспечивает метод для выявления люциферазы в биологическом образце в присутствии гиспидин-3-гидроксилазы с помощью гиспидина или его функционального аналога, включающий
(а) добавление к образцу гиспидина или его функционального аналога и НАД(Ф)Н;
(б) инкубацию получившейся реакционной смеси в условиях, подходящих
для возникновения биолюминесценции;
(в) детекцию биолюминесценции реакционной смеси.
[018] В преимущественных воплощениях гиспидин или его функциональный аналог (например, биснориангонин) добавляют к образцу в конечной концентрации 0.03-30 мкМ, чаще 1-5 мкМ, например, 2.4 мкМ. В преимущественных воплощениях гиспидин или его функциональный аналог добавляют к образцу в буферном растворе, имеющем рН в диапазоне от 6.0 до 9.8, чаще от 6.5 до 9.0, например от 7.2 до 8.0. Часто раствор содержит реагенты, увеличивающие растворимость гиспидина и его функционального аналога в воде, например, детергенты, такие как Triton Х100 или нонилфеноксиполиэтоксиэтанол.
[019] Также в реакционную смесь добавляют НАДФН или НАДН (конечная концентрация не менее 20 мкМ, например, 1-3 мМ), преимущественно в буферном растворе, имеющем pH в диапазоне от 6.0 до 9.8, чаще от 6.5 до 9.0, например от 7.2 до 8.0.
[020] Реакционная смесь может также содержать компоненты, стабилизирующие и защищающие ферменты биолюминесцентной системы от ингибирующего воздействия следовых количеств ионов тяжелых металлов и от деградации протеазами. Например, реакционная смесь может содержать ДТТ, бета-меркаптоэтанол и/или ЭДТА. Также реакционная смесь может содержать ингибиторы протеаз, например, фенилуксусную кислоту или щавелевую кислоту.
[021] Настоящее изобретение также обеспечивает реактивы и наборы реактивов для детекции люциферазы в биологических образцах, включающие 3-гидроксигиспидин (или его функциональный аналог) или гиспидин (или его функциональный аналог) и НАД(Ф)Н.
Краткое описание фигур
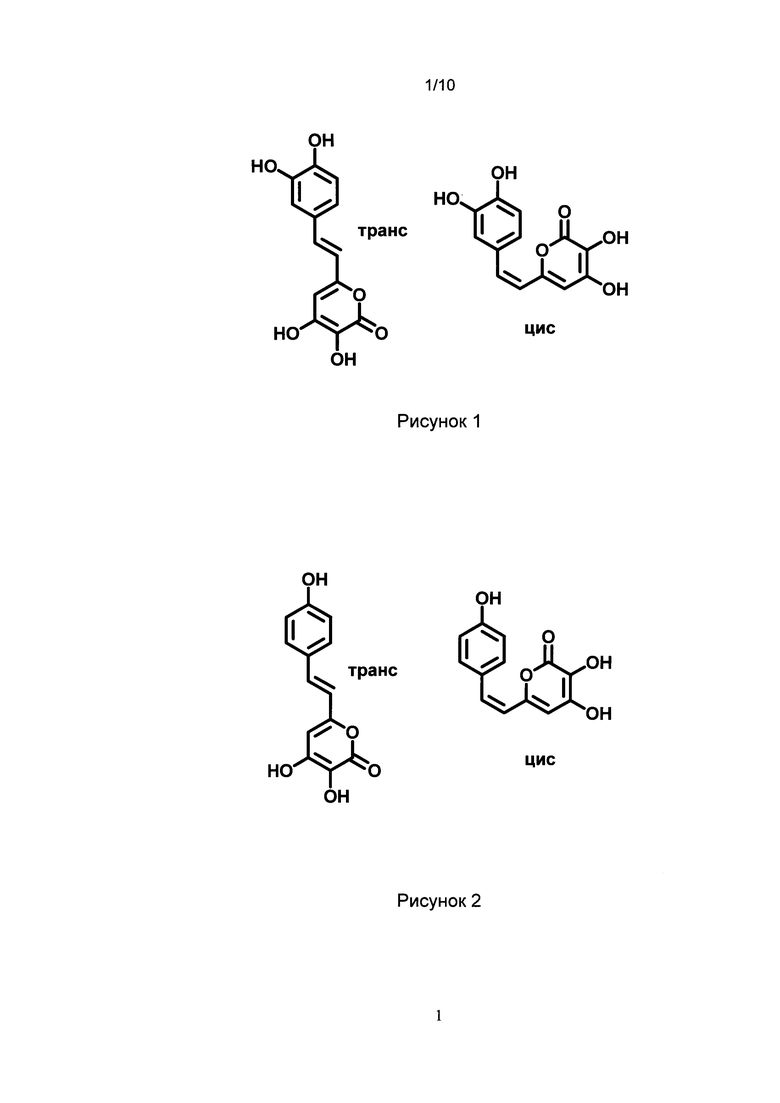
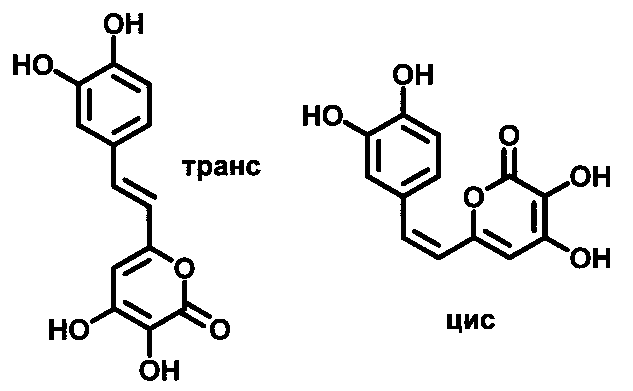
[022] Рис. 1 показывает химическую структуру изомеров по двойной связи 3-гидроксигиспидина.
[023] Рис. 2 показывает химическую структуру изомеров по двойной связи 3-гидроксибиснориангонина.
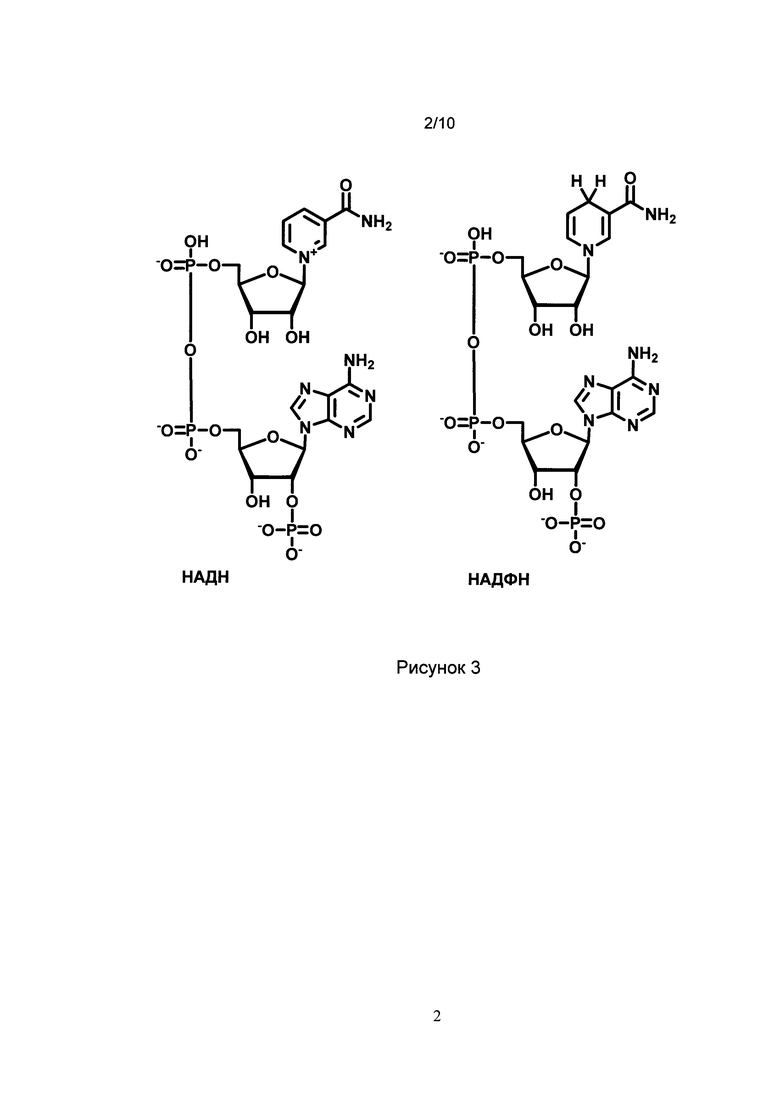
[024] Рис. 3 показывает химическую структуру восстановленных форм никотинамидадениндинуклеотидфосфата (НАДФН) и никотинамидадениндинуклеотида (НАДН).
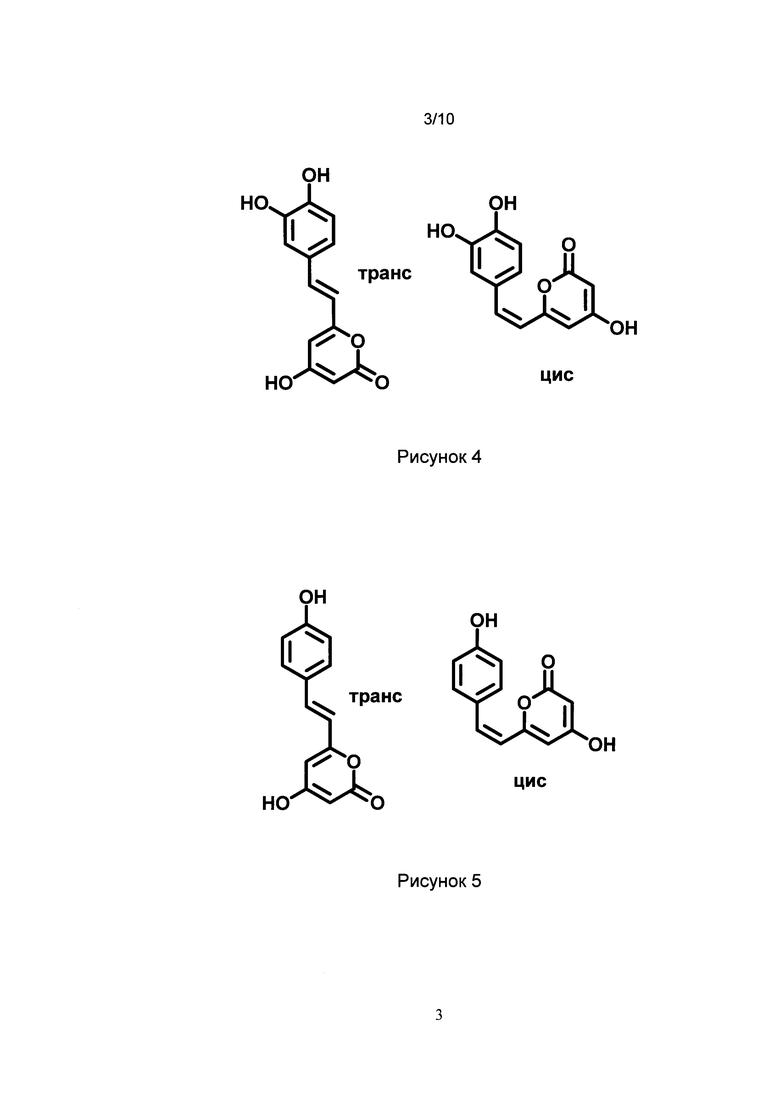
[025] Рис. 4 показывает химическую структуру изомеров по двойной связи гиспидина.
[026] Рис. 5 показывает химическую структуру изомеров по двойной связи биснориангонина.
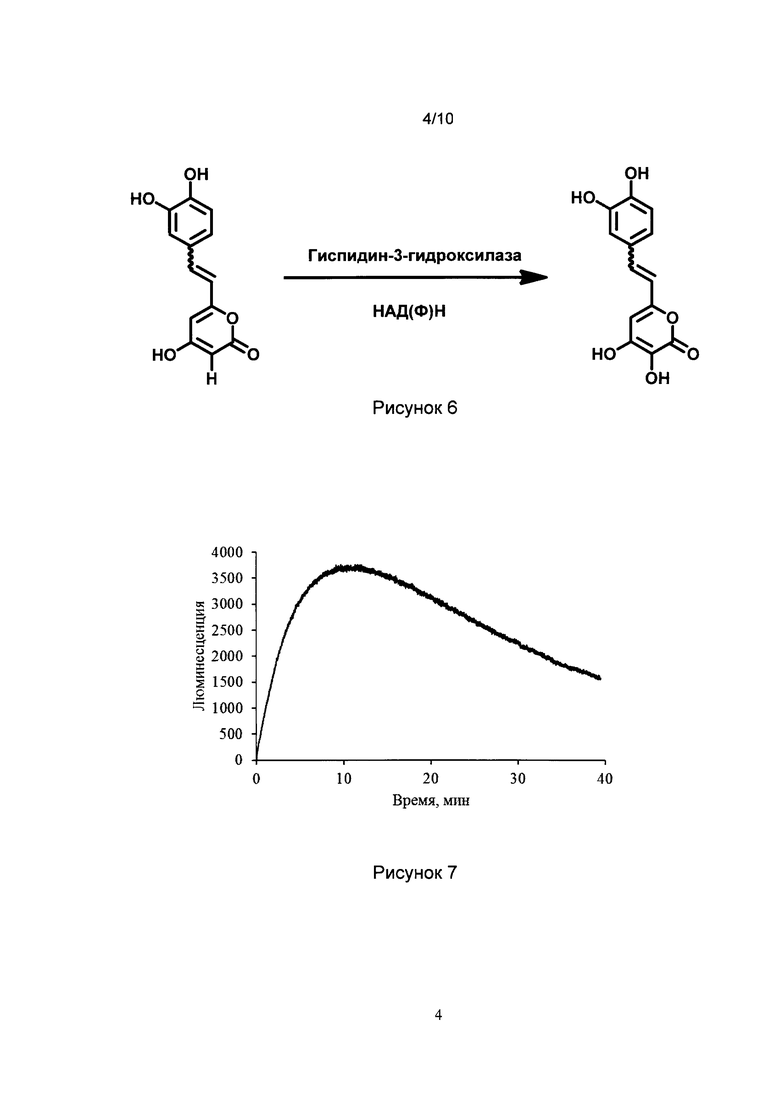
[027] Рис. 6 показывает процесс гидроксилирования гиспидина и его функциональных аналогов по положению 3 пиранонового цикла под действием гиспидин-3-гидроксилазы в присутствии НАД(Ф)Н.
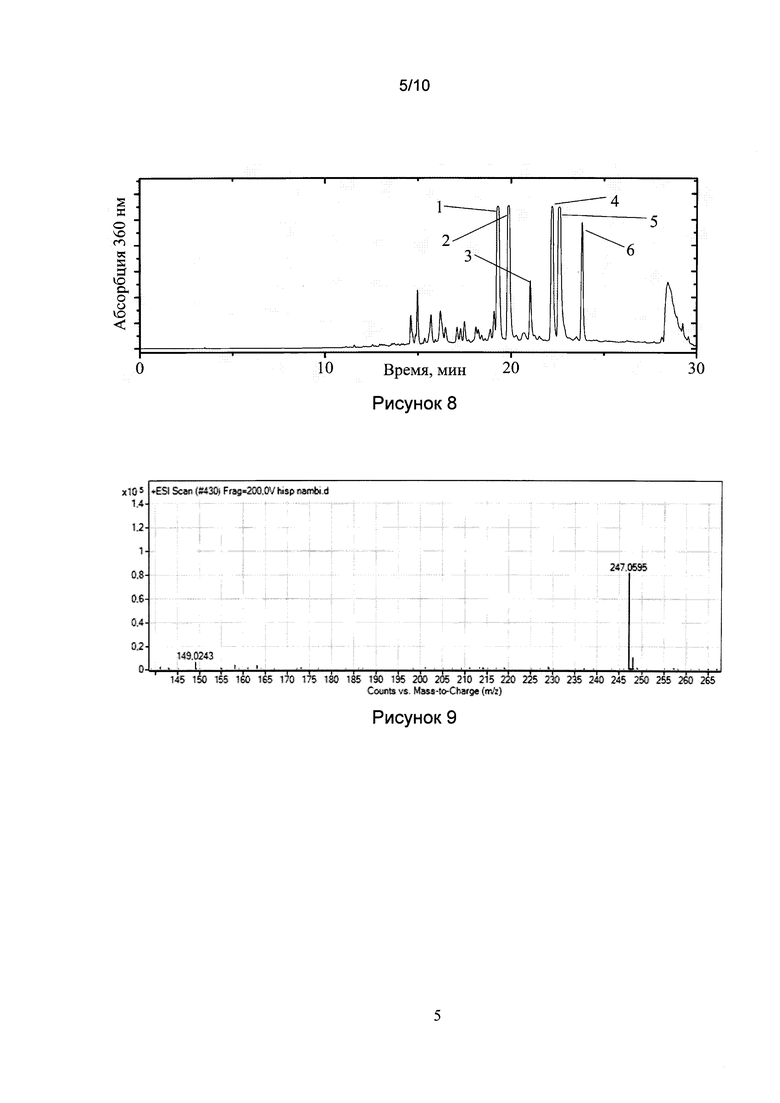
[028] Рис. 7 показывает результат измерения биолюминесценции.
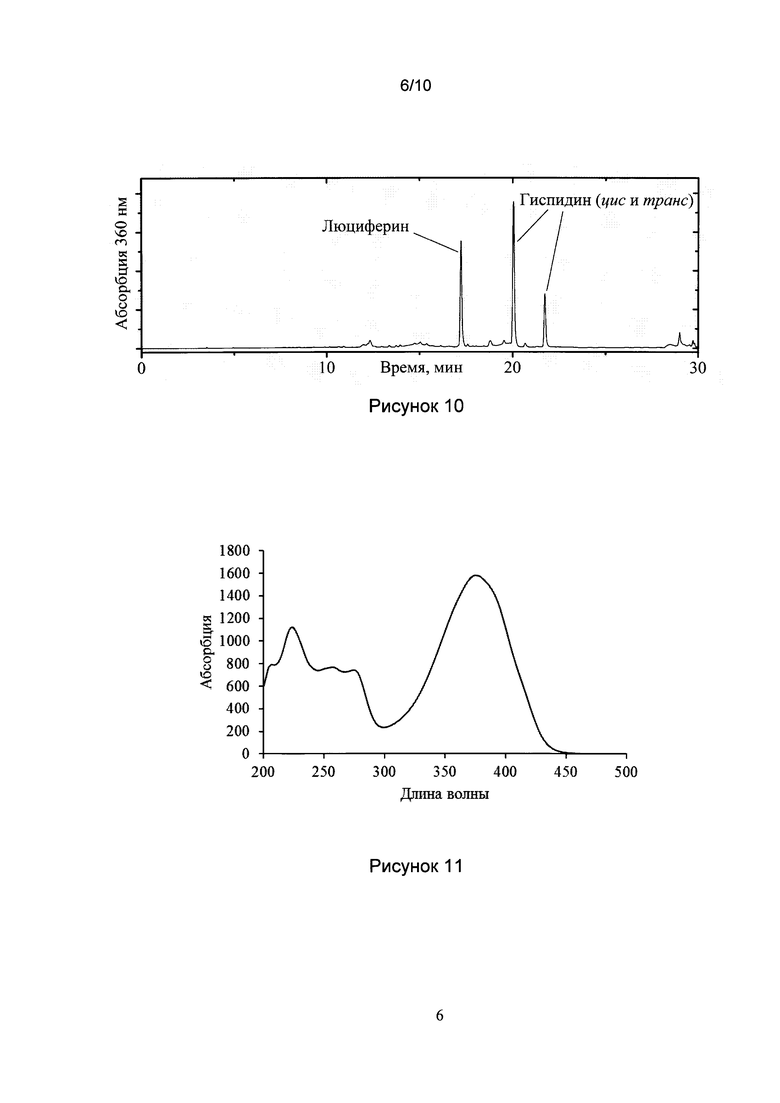
[029] Рис. 8 показывает результат обратнофазовой ВЭЖХ горячего экстракта из плодовых тел Ph. squarrosa.
[030] Рис. 9 показывает HRMS спектр гиспидина, выделенного из N. nambi.
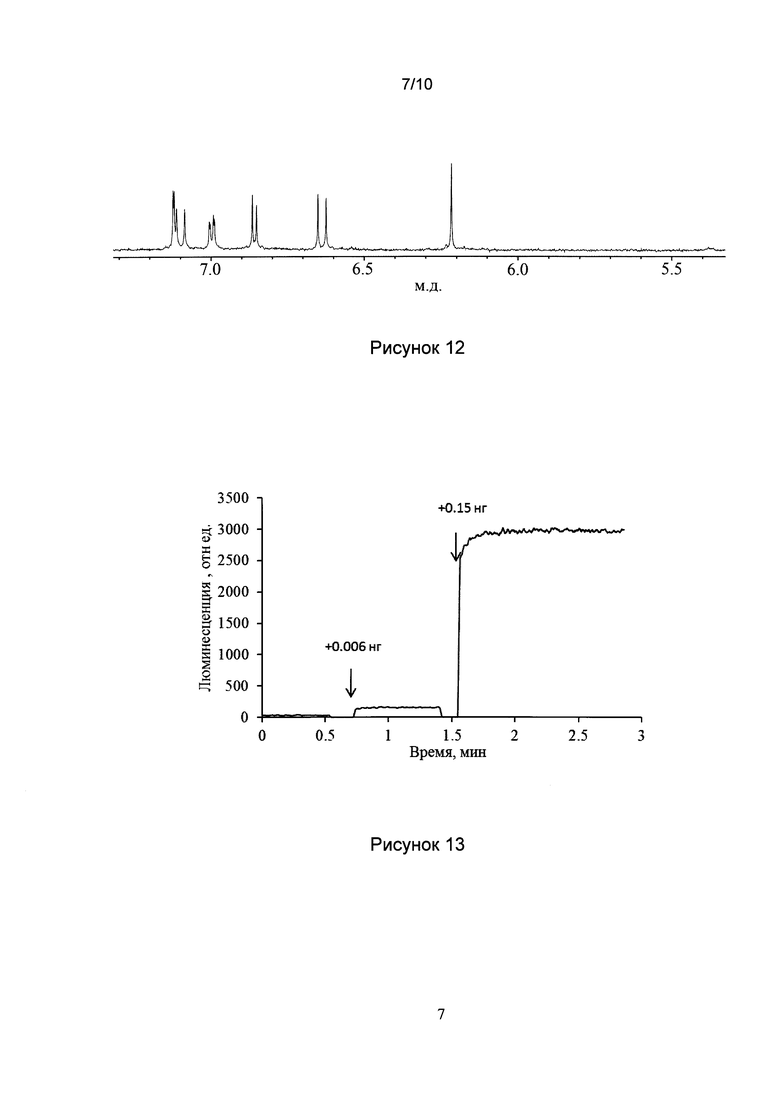
[031] Рис. 10 показывает результаты ВЭЖХ реакционной смеси, проявляющей НАДФН-независимую люминесценцию.
[032] Рис. 11 показывает UV-vis спектр люциферина, выделенного из реакционной смеси, проявляющей НАДФН-независимую люминесценцию.
[033] Рис. 12 показывает 1H ЯМР спектр люциферина, выделенного из реакционной смеси, проявляющей НАДФН-независимую люминесценцию (м.д. - миллионные доли).
[034] Рис. 13 показывает типичный результат детекции люминесценции биологических образцов из грибов при добавлении 3-гидроксигиспидина.
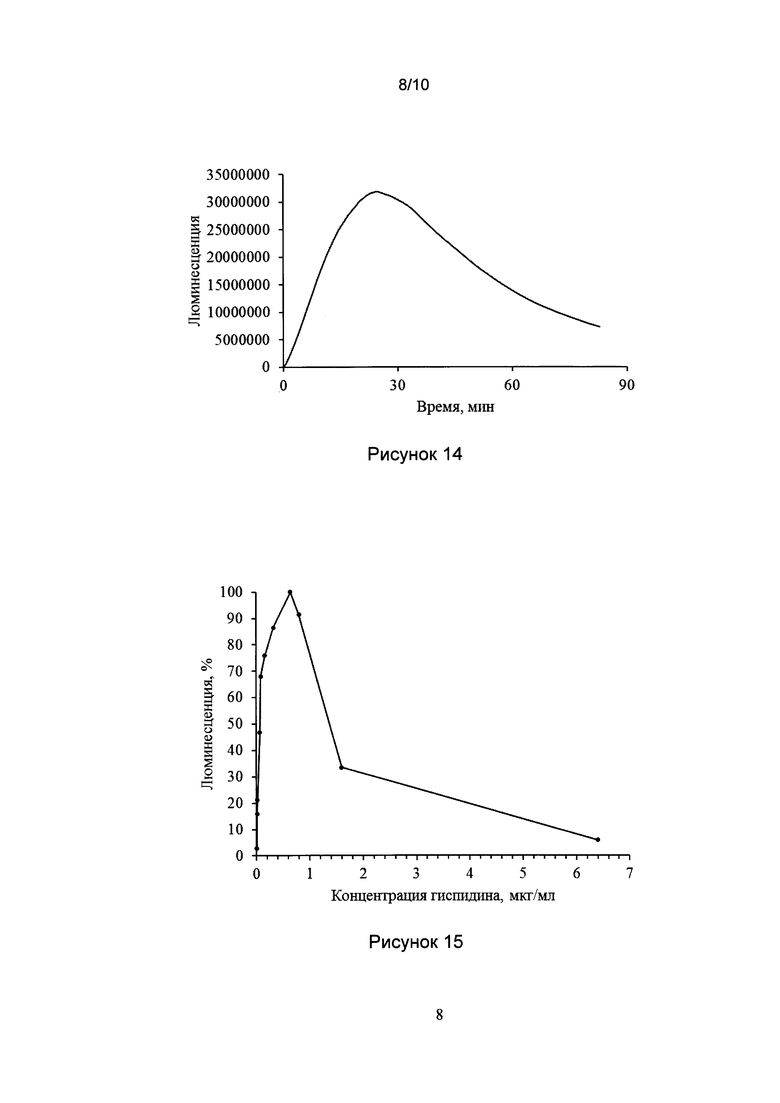
[035] Рис. 14 показывает типичный результат люминесценции биологических образцов из грибов при добавлении гиспидина и НАДФН.
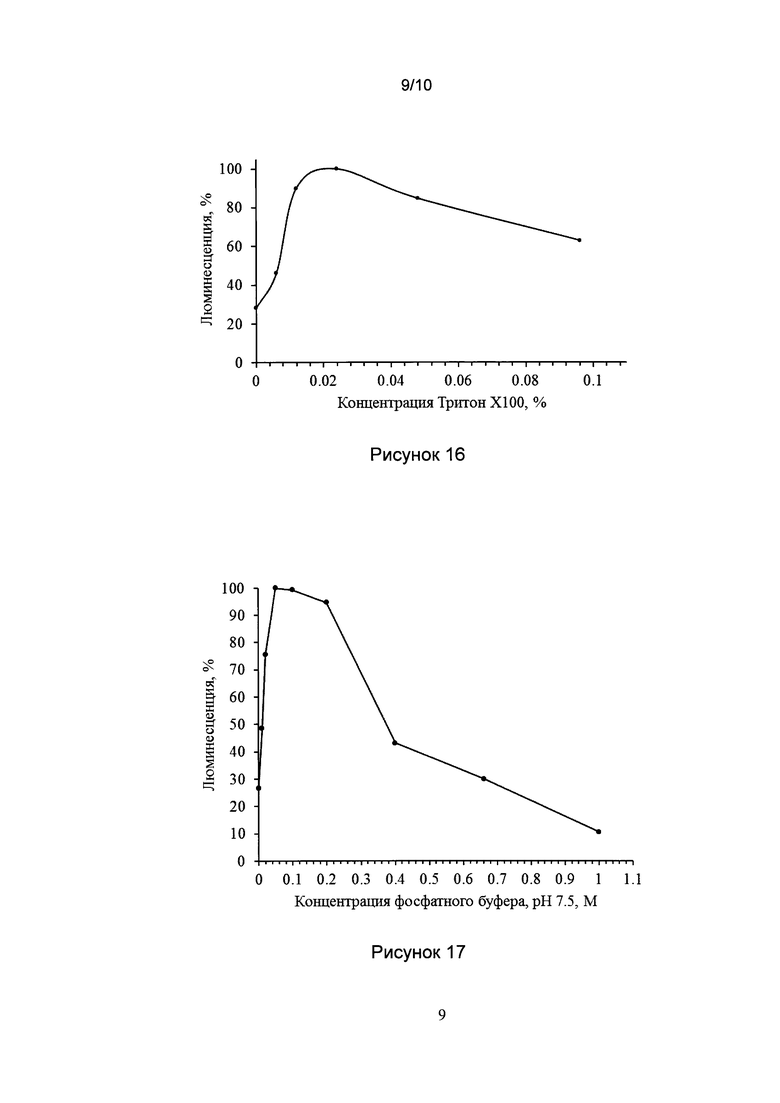
[036] Рис. 15 показывает график зависимости максимальной биолюминесценции биологического образца от концентрации гиспидина в реакционной смеси.
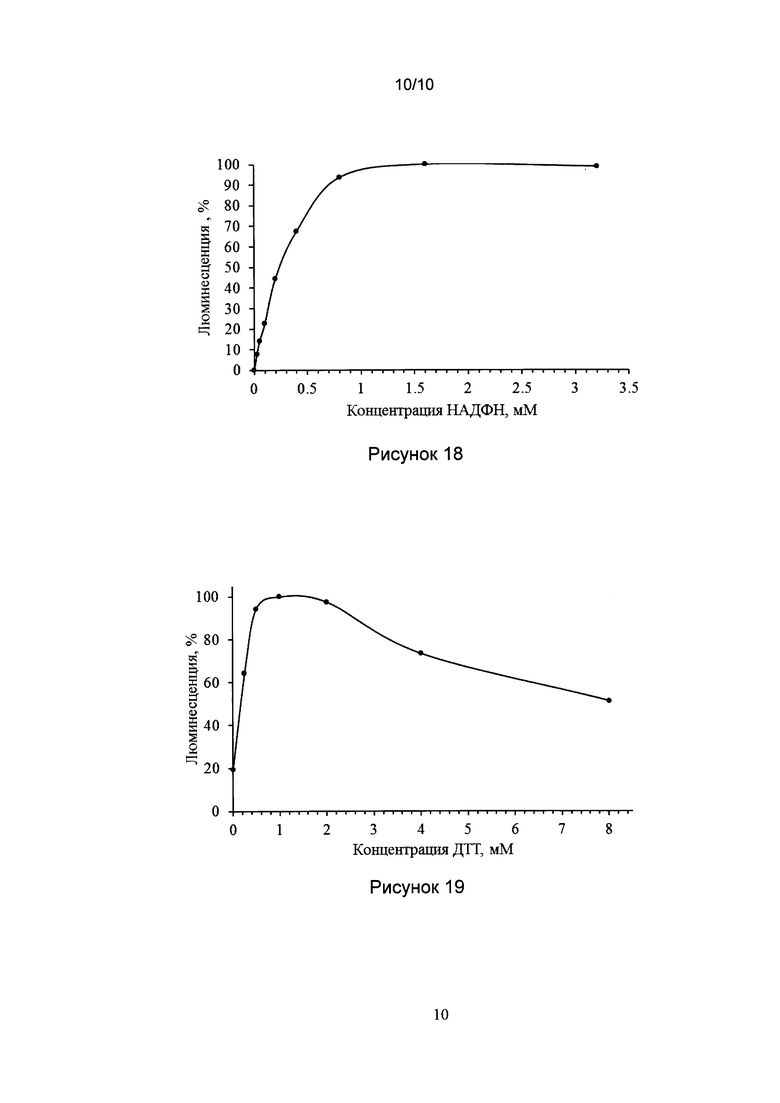
[037] Рис. 16 показывает график зависимости максимальной биолюминесценции биологического образца от концентрации детергента Тритон Х100 в реакционной смеси.
[038] Рис. 17 показывает график зависимости максимальной биолюминесценции биологического образца от концентрации буферного раствора.
[039] Рис. 18 показывает график зависимости максимальной биолюминесценции биологического образца от концентрации НАДФН.
[040] Рис. 19 показывает график зависимости максимальной биолюминесценции биологического образца от концентрации ДТТ в реакционной смеси.
Подробное описание изобретения
[041] Как указано выше, настоящее изобретение направлено на способы детекции активности люциферазы в биологических образцах. [042] В частности, настоящее изобретение обеспечивает метод для выявления люциферазы в биологическом образце с помощью 3-гидроксигиспидина или его функционального аналога, включающий
(а) добавление к образцу 3-гидроксигиспидина или его функционального аналога;
(б) инкубацию получившейся реакционной смеси в условиях, подходящих для возникновения биолюминесценции;
(в) детекцию биолюминесценции реакционной смеси.
[043] Также настоящее изобретение обеспечивает метод для выявления люциферазы и гиспидин-3-гидроксилазы в биологическом образце с помощью гиспидина или его функционального аналога, включающий
(а) добавление к образцу гиспидина или его функционального аналога и НАД(Ф)Н;
(б) инкубацию получившейся реакционной смеси в условиях, подходящих для возникновения биолюминесценции;
(в) детекцию биолюминесценции реакционной смеси.
[044] Также обеспечиваются реактивы и наборы реактивов для реализации методов настоящего изобретения.
[045] Для более полного раскрытия вышеперечисленных характеристик настоящего изобретения ниже предлагается детальное описание изобретения, кратко сформулированного выше, в виде ссылок на воплощения, некоторые из которых проиллюстрированы дополнительными фигурами. При этом следует отметить, что прилагаемые фигуры иллюстрируют лишь типичные воплощения настоящего изобретения и, следовательно, не должны быть восприняты в качестве ограничения объема изобретения, которое может допускать другие, в равной степени эффективные, воплощения.
Биологические образцы
[046] Реализация методов настоящего изобретения обеспечивает возникновение люминесценции реакционной смеси, содержащей биологический образец, если указанный образец содержит люциферазу, использующую в качестве субстрата (то есть люциферина) 3-гидроксигиспидин или его функциональный аналог (например, 3-гидроксибиснориангонин). Такую люциферазу содержат, например, способные к биолюминесценции высшие грибы.
[047] Исследуемый образец может также содержать гиспидин-3-гидроксилазу, способную трансформировать гиспидин или его функциональный аналог в субстрат для люциферазы. Гиспидин-3-гидроксилазу содержат, например, способные к биолюминесценции высшие грибы.
[048] Биологические образцы могут быть получены с помощью различных технологий, известных в биологии, и включают образцы тканей, клеток, экстракты, гомогенаты, белковые смеси различной степени очистки и т.д. Например, биологические образцы могут быть получены из высших грибов.
[049] Биологические образцы могут также содержать выделенные компоненты (люциферазу или люциферазу и гиспидин-3-гидроксилазу) биолюмнесцентных систем высших грибов или их функциональные аналоги.
[050] Как используется в настоящем описании, термин «выделенный» означает, что указанные компоненты находятся в среде, отличной от среды, в которой они находятся в естественных условиях. Например, указанные компоненты могут находиться по существу в очищенной форме. По существу очищенная форма означает, что белки являются, по меньшей мере, приблизительно на 20% чистыми, часто, по меньшей мере, на 30% чистыми, обычно на 50% чистыми, или, по меньшей мере, на 90% чистыми.
[051] Для выделения белков могут быть использованы любые обычные методики очистки белка, описанные, например, в Guide to Protein Purification, (Deuthser ed.) (Academic Press, 1990). Например, из исходного источника может быть приготовлен лизат или холодный экстракт и очищен с использованием ВЭЖХ, эксклюзионной хроматографии, гель-электрофореза, аффинной хроматографии и т.п. Белковые препараты могут быть протестированы на наличие активной люциферазы или комплекса люциферазы и гиспидин-3-гидроксилазы с помощью методов настоящего изобретения.
[052] Биологические образцы могут также экспрессировать рекомбинантные люциферазу или люциферазу и гиспидин-3-гидроксилазу или их функциональные мутанты. Последовательности нуклеиновых кислот для экспрессии указанных белков могут быть получены из природных источников (например, из высших грибов) или синтезированы. В настоящее время известно множество методов для клонирования генов, кодирующих белки, обладающие известной активностью. Частично такие методы описаны в Maniatis, Т., et al. (Molecular Cloning-A Laboratory Manual Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. 1982) и Newman и Campagnoni (Neuromethods, v. 16, 1990, pp 13-48). Например, может быть приготовлена экспрессионная библиотека в подходящих клетках-хозяевах и протестирована на активность люциферазы. Или может быть осуществлено выделение белка из холодного экстракта, определена его частичная аминокислотная последовательность и осуществлено клонирование соответствующей кДНК из образца кДНК из высшего гриба. Последовательности нуклеиновых кислот должны быть встроены в кассету экспрессии. Кассета экспрессии может существовать как внехромосомный элемент или может быть включена в геном клетки в результате введения указанной кассеты экспрессии в клетку. В кассете экспрессии нуклеиновая кислота, кодирующая белок, является функционально связанной с регуляторной последовательностью, которая может включить промоторы, энхансеры, терминаторы, операторы, репрессоры и индукторы. После введения кассеты экспрессии в клетку в ней может образовываться функциональный белок. Системы экспрессии включают, например, бактериальные системы, дрожжевые клетки, насекомых, рыб, земноводных или клетки млекопитающих. Методы изготовления кассет экспрессии или систем для экспрессии желаемого продукта известны специалистам, квалифицированным в данной области. Клеточные линии, которые устойчиво экспрессируют люциферазу или люциферазу вместе с гиспидин-3-гидроксилазой могут быть выбраны способами, известными в данной области (например, ко-трансфекция с селектируемым маркером, таким как dhfr, gpt, неомицин, гигромицин, что делает возможным выявление и выделение трансфецированных клеток, которые содержат ген, включенный в геном). Вышеописанные системы экспрессии могут использоваться в прокариотических или эукариотических хозяевах. Для получения белка могут использоваться клетки-хозяева, такие как Е. coli, В. subtilis, S. cerevisiae, клетки насекомого в комбинации с бакуловирусными векторами, или клетки высшего организма, такого как позвоночные, например, COS 7 клетки, НЕК 293, СНО, ооциты Xenopus и т.д.
[053] Могут быть также экспрессированы функциональные мутанты природных белков. Как используется в настоящем описании, термин «функциональный» по отношению к люциферазе означает, что указанный белок способен использовать 3-гидроксигиспидин или его цис-изомер по двойной связи в качестве люциферина. Как используется в настоящем описании, термин «функциональный» по отношению к гиспидин-3-гидроксилазе означает, что мутантный белок способен в присутствии НАД(Ф)Н и кислорода воздуха превращать гиспидин или биснориангонин в 3-гидроксигиспидин или 3-гидроксибиснориангонин, соответственно.
[054] Как используется в настоящем описании, термин «мутант» относится к белку, в котором одна или несколько добавленных и/или замещенных, и/или делетированных, и/или вставленных аминокислот на N-конце и/или на С-конце, и/или внутри нативных аминокислотных последовательностей белков. Как используется в настоящем описании, термин «мутант» относится к молекуле нуклеиновой кислоты, которая кодирует мутантный белок. Кроме того, термин «мутант» относится к любому более короткому или более длинному варианту соответствующего белка или соответствующей нуклеиновой кислоты.
[055] Модификации, а также добавки или делеции могут быть встроены любым методом, известным в данной области (см., например, Gustin et al., Biotechniques (1992) 14:22; Barany, Gene (1985) 37:11-123; и Colicelli et al., Mol. Gen. Genet. (1985) 199:537-539), Sambrook et al., Molecular Cloning: A Laboratory Manual, (1989), CSH Press, pp. 15.3-15.108), включая ошибочно-направленную ПЦР, перестановку, сайт-направленный мутагенез с использованием олигонуклеотидов, мутагенез с использованием ПЦР на основе спаренных молекул, мутагенез in vivo, кассетный мутагенез, рекурсивный согласованный мутагенез, экспоненциальный согласованный мутагенез, сайт-направленный мутагенез, случайный мутагенез, генную повторную сборку, генный сайт-насыщенный мутагенез (GSSM), повторную сборку при проведении синтеза лигированием (SLR), или их сочетание. Указанные модификации, добавления или делеции могут быть также встроены способом, включающим рекомбинацию, рекурсивную рекомбинацию последовательности, мутагенез ДНК путем фосфотиоатной модификации, мутагенез на основе включения матрицы, содержащей урацил, мутагенез на основе дуплекса, содержащего бреши, репарационный мутагенез с точечными ошибочными спариваниями, мутагенез с использованием штамма-хозяина, дефицитного по репарации, химический мутагенез, радиогенный мутагенез, делеционный мутагенез, мутагенез с использованием ограничения по селекции, мутагенез с использованием ограничения по очистке, искусственный синтез гена, согласованный мутагенез, создание химерного мультимера нуклеиновой кислоты или их сочетание. Ссылка на нуклеотидную последовательность, "кодирующую" полипептид означает, что с нуклеотидной последовательности в ходе трансляции и транскрипции мРНК продуцируется этот полипептид. При этом может быть указана как кодирующая цепь, идентичная мРНК и обычно используемая в списке последовательностей, так и комплементарная цепь, которая используется как матрица при транскрипции. Как очевидно для любого специалиста в данной области техники, термин также включает любые вырожденные нуклеотидные последовательности, кодирующие одинаковую аминокислотную последовательность. Нуклеотидные последовательности, кодирующие полипептид, включают последовательности, содержащие интроны.
Реактивы для выявления активности люциферазы
[056] Методы настоящего изобретения основаны на использовании 3-гидроксигиспидина или его функционального аналога для детекции активности люциферазы в биологических образцах.
[057] Как используется в настоящем описании, термин «3-гидроксигиспидин» относится к соединению (Е)-6-(3,4-дигидроксистририл)-3,4-дигидрокси-2Н-пиран-2-он. Так как вещество склонно к изомеризации в растворе и образует равновесные смеси цис- и транс-изомеров по двойной связи, термин «3-гидроксигиспидин» для нужд настоящего изобретения также относится к его цис-изомеру по двойной связи (Z)-6-(3,4-дигидроксистририл)-3,4-дигидрокси-2Н-пиран-2-он. Химические структуры транс-изомера 3-гидроксигиспидина и его цис-изомера по двойной связи показаны на Рис. 1.
[058] 3-гидроксигиспидин проявляет стабильность при хроматографировании в водно-ацетонитрильном растворе с добавлением муравьиной кислоты (конечная концентрация 0.1%) в течение дня. Лиофильно высушенный субстрат хранится при -20°C без снижения активности не менее 30 дней, чаще не менее 60 дней, обычно не менее года.
[059] 3-гидроксигиспидин хорошо растворим в апротонных растворителях (ДМСО, ацетонитрил, ацетон и др.) и умеренно растворим в воде.
[060] 3-гидроксигиспидин стабилен в широком диапазоне температур, например, он выдерживает кратковременное кипячение в водном растворе.
[061] Также может использоваться функциональный аналог 3-гидроксигиспидина: 3-гидроксибиснориангонин.
[062] Как используется в настоящем описании, термин «3-гидроксибиснориангонин» относится к соединению (Е)-3,4-дигидрокси-6-(4-гидроксистирил)-2Н-пиран-2-он. Так как вещество склонно к изомеризации в растворе и образует равновесные смеси цис- и транс-изомеров по двойной связи, термин «3-гидроксибиснориангонин» для нужд настоящего изобретения также относится к его цис-изомеру по двойной связи (Z)-3,4-дигидрокси-6-(4-гидроксистирил)-2Н-пиран-2-он. Химические структуры транс-изомера 3-гидроксибиснориангонин и его цис-изомера по двойной связи показаны на Рис. 2.
[063] 3-гидроксигиспидин и 3-гидроксибиснориангонин могут быть получены путем энзиматического синтеза из гиспидина или биснориангонина, соответственно, в присутствии гиспидин-3-гидроксилазы, НАДФН (или НАДН) и кислорода воздуха.
[064] Для осуществления синтеза предпочтительными являются следующие условия: рН в пределах от 6.5 до 9.0, концентрация НАД(Ф)Н 1-20 мМ, например, 10 мМ. Реакционная среда может содержать компоненты, стабилизирующие и защищающие гиспидин-3-гидроксилазу от ингибирующего воздействия следовых количеств ионов тяжелых металлов и от деградации протеазами. Например, реакционная смесь может содержать ДТТ, бета-меркаптоэтанол и/или ЭДТА. Также реакционная смесь может содержать ингибиторы протеаз. Также реакционная смесь может содержать детергенты, такие как Тритон Х100 или нонилфеноксиполиэтоксиэтанол. Также реакционная смесь может содержать примеси ДМСО, ацетонитрила или других растворителей. Пример условий для энзиматического синтеза показан в экспериментальной секции ниже.
[065] Полученные вещества могут быть выделены из реакционной смеси с помощью экстракции растворителями: водой, спиртом, ацетонитрилом, ДМСО или их смесями, или растворителями, не смешивающимися с водой: этилацетатом, хлороформом, дихлорметаном и другими, с последующим разделением полученного экстракта жидкостной хроматографией или без такого разделения. Одна из возможных схем выделения 3-гидроксигиспидина описана в деталях в экспериментальной секции ниже.
[066] Альтернативно, 3-гидроксигиспидин и 3-гидроксибиснориангонин могут быть получены путем химического синтеза.
[067] 3-гидроксигиспидин или его функциональный аналог могут быть растворены в воде, буферном растворе, ДМСО, спирте, ацетонитриле или в их смеси. Полученный раствор может быть разведен до рабочей концентрации с помощью воды, буферного раствора или 0.01-1% раствора муравьиной кислоты. Пригодные буферные растворы включают фосфатный буфер, Трис-HCl, HEPES и другие, обычно используемые в диапазоне рН от 6.0 до 9.8, обычно от 6.5 до 9.0, как правило, от 7.0 до 8.0. В преимущественных воплощениях в раствор добавляют реактивы, повышающие растворимость 3-гидроксигиспидина и его функционального аналога в воде, например, детергенты (такие как Тритон Х100 или нонилфеноксиполиэтоксиэтанол).
[068] Термин «рабочая концентрация» используется здесь для обозначения концентрации раствора, который добавляют к биологическому образцу.
[069] В случаях, если биологический образец содержит люциферазу и гиспидин-3-гидроксилазу, для их детекции используется предшественник люциферина, то есть гиспидин или его функциональный аналог вместе с НАД(Ф)Н.
[070] Под действием гиспидин-3-гидроксилазы в присутствии НАД(Ф)Н происходит гидроксилирование гиспидина и его функциональных аналогов по положению 3-пиранонового цикла, как показано на рис. 6. 3-гидроксигиспидин и его функциональные аналоги служат субстратами для люциферазы.
[071] Как используется в настоящем описании, термин «НАД(Ф)Н» означает восстановленную форму никотинамидадениндинуклеотидфосфата (НАДФН) или восстановленную форму никотинамидадениндинуклеотида (НАДН). Формулы никотинамидадениндинуклеотидфосфата и никотинамидадениндинуклеотида представлены на Рис. 3.
[072] Гиспидин (или (Е)-6-(3,4-дигидроксистирил)-4-гидрокси-2Н-пиран-2-он) - хорошо известное соединение, относящиеся к группе стирилпиронов (styrylpyrones), выделенных из ряда грибов и растений (Beckert et al. Phytochemistry, VoL 44, No. 2, pp. 275-283, 1997; In-Kyoung Lee and Bong-Sik Yun The Journal of Antibiotics (2011) 64, 349-359). Так как вещество склонно к изомеризации в растворе и образует равновесные смеси цис- и транс-изомеров по двойной связи, термин «гиспидин» для нужд настоящего изобретения также относится к его цис-изомеру по двойной связи (2)-6-(3,4-дигидроксистирил)-4-гидрокси-2Н-пиран-2-он. Химические структуры транс- и цис изомеров по двойной связи гиспидина показаны на Рис. 4.
[073] Также может использоваться функциональный аналог гиспидина - биснориангонин.
[074] Как здесь используется, термин «биснориангонин» относится к соединению (Е)-4-гидрокси-6-(4-гидроксистирил)-2Н-пиран-2-он. Так как вещество склонно к изомеризации в растворе и образует равновесные смеси цис- и трансизомеров по двойной связи, термин «биснориангонин» для нужд настоящего изобретения также относится к его цис-изомеру по двойной связи (Z)-4-гидрокси-6-(4-гидроксистирил)-2Н-пиран-2-он). Химические структуры транс- и цис изомеров по двойной связи биснориангонина показаны на Рис. 5.
[075] Гиспидин или его функциональный аналог могут быть растворены в воде, буферном растворе, ДМСО, спирте, ацетонитриле, или в их смеси. Полученный раствор может быть разведен до рабочей концентрации с помощью воды, буферного раствора или 0.01-1% раствора муравьиной кислоты. Пригодные буферные растворы включают фосфатный буфер, Трис-HCl, HEPES и другие, обычно используемые в диапазоне pH от 6.0 до 9,8, обычно от 6,5 до 9,0, как правило, от 7,0 до 8,0. В преимущественных воплощениях в раствор добавляют реактивы, повышающие растворимость 3-гидроксигиспидина и его функционального аналога в воде, например, детергенты (такие как Тритон Х100 или нонилфеноксиполиэтоксиэтанол). В преимущественных воплощениях в раствор добавляют реактивы, повышающие растворимость гиспидина и его функционального аналога в воде, например, детергенты (такие как Тритон Х100 или нонилфеноксиполиэтоксиэтанол).
Состав условий для развития биолюминесцентного сигнала
[076] Формирование биолюминесценции зависит от количества и сохранности люциферазы или люциферазы и гиспидин-3-гидроксилазы в биологических образцах.
[077] На формирование сигнала влияет pH реакционной смеси. Формирование биолюминесцентного сигнала происходит в диапазоне pH от 6.0 до 9.8, обычно в диапазоне pH от 6.5 до 9.0, преимущественно, в диапазоне от 7.0 до 8.0. Для обеспечения pH могут быть использованы любые стандартные буферные растворы для данного диапазона pH, включая фосфатный буфер, HEPES, Трис-HCl. В преимущественных воплощениях молярность буферного раствора не превышает 2, например, не превышает 1, чаще находится в диапазоне от 0.05 до 0.4, обычно от 0.1 до 0.2.
[078] Реакционные смеси для нужд настоящего изобретения могут также содержать компоненты, стабилизирующие и защищающие ферменты биолюминесцентной системы от ингибирующего воздействия следовых количеств ионов тяжелых металлов и от деградации протеазами.
[079] Например, реакционная смесь может содержать ДТТ в концентрации не более 20 мМ, чаще в концентрации от 0.1 до 8 мМ, преимущественно в концентрации 0.1-4 мМ.
[080] Реакционная смесь может также содержать бета-меркаптоэтанол и/или ЭДТА в конечной концентрации от 0 до 5 мМ.
[081] Например, реакционная смесь может содержать 0.1-2 мМ ДТТ и 0.1-1 мМ ЭДТА.
[082] Также реакционная смесь может содержать ингибиторы протеаз, например, фенилуксусную кислоту или щавелевую кислоту в стандартно используемых концентрациях.
[083] Для нужд настоящего изобретения 3-гидроксигиспидин или его функциональный аналог добавляют к биологическому образцу до конечной концентрации 0.03-30 мкМ,, чаще 1-5 мкМ.
[084] В некоторых воплощениях к образцу добавляют смесь реагентов, включающих буферный раствор, компоненты, стабилизирующие и защищающие ферменты биолюминесцентной системы от ингибирующего воздействия следовых количеств ионов тяжелых металлов и от деградации протеазами и 3-гидроксигиспидин. В других воплощениях к биологическому образцу сперва добавляют буферный раствор, компоненты, стабилизирующие и защищающие ферменты биолюминесцентной системы от ингибирующего воздействия следовых количеств ионов тяжелых металлов и от деградации протеазами, а затем раствор 3-гидроксигиспидина.
[085] Для нужд настоящего изобретения гиспидин или его функциональный аналог добавляют к биологическому образцу в конечной концентрации 0.03-30 мкМ, чаще 1-5 мкМ, например, 2.4 мкМ.
[086] НАД(Ф)Н добавляют к биологическому образцу вместе, до или после добавления гиспидина или его функционального аналога в конечной концентрации 0.01-10 мМ, чаще в концентрации 0.4-2 мМ, обычно в концентрации 1 мМ.
[087] В некоторых воплощениях к образцу добавляют смесь реагентов, включающих буферный раствор, компоненты, стабилизирующие и защищающие ферменты биолюминесцентной системы от ингибирующего воздействия следовых количеств ионов тяжелых металлов и от деградации протеазами, НАД(Ф)Н и гиспидин или его функциональный аналог.
[088] Например, к образцу (объемом 1-5 мкл) добавляют смесь объемом 100 мкл, содержащую 0.1М фосфатный буфер, pH 7.5, 0.1 мМ ДТТ, 2.4 мкМ гиспидина, 0.01% Тритон Х100, 0.4 мМ НАД(Ф)Н.
[089] В некоторых воплощениях к образцу последовательно добавляют две смеси реагентов, одна из которых включает НАД(Ф)Н, а вторая гиспидин или его функциональный аналог.
[090] Например, сначала к образцу (объемом 1-5 мкл). добавляют смесь объемом 100 мкл, содержащую 0.1М фосфатный буфер, pH 7.5, 0.1 мМ ДТТ, 2.4 мкМ мкг гиспидина (или его функциональный аналог), 0.01% Тритон Х100, а затем добавляют 4 мкл 10 мМ водного раствора НАД(Ф)Н для инициации реакции.
[091] В некоторых воплощениях сначала к образцу (объемом 1-5 мкл) добавляют смесь объемом 100 мкл, содержащую 0.1М фосфатный буфер, pH 7.5, 0.1 мМ ДТТ, 0.4 мМ НАД(Ф)Н, а затем добавляют 4 мкл 64 мкМ водного раствора гиспидина или его функционального аналога в водном растворе, содержащем 0.1% Тритон Х100 и 0.1% муравьиную кислоту.
[092] В зависимости от растворителя, использованного для приготовления раствора гиспидина, 3-гидроксигиспидина или их функциональных аналогов, реакционная смесь может содержать небольшие количества использованных растворителей.
[093] Также реакционная смесь может содержать детергенты, такие как Triton Х100 или нонилфеноксиполиэтоксиэтанол. В преимущественных воплощениях концентрация детергентов в реакционной смеси не превышает 0,2%, чаще не превышает 0,1%, оптимально не превышает 0,06%.
[094] Примеры измерения активности люциферазы в биологических образцах, содержащих также гиспидин-3-гидроксилазу, приведены в экспериментальной секции ниже.
[095] Также реакционная смесь может содержать бычий сывороточный альбумин (БСА) или другие белки в концентрации, не превышающей 2%, чаще не превышающей 1%, оптимально не превышающей 0,5%. БСА используется, когда концентрация биологического образца крайне мала, тогда БСА играет роль стабилизатора белков.
[096] Развитие биолюминесцентного сигнала происходит в широком диапазоне температур - от 0 до 40°C, оптимально при 20-25°C.
[097] Формирование люминесцентного сигнала начинается сразу после инициации реакции при добавлении ключевых реактивов для выявления активности люциферазы, указанных выше.
Интенсивность люминесценции увеличивается со временем после инициации реакции, как правило, в течение 5-30 мин, например, в течение 10 мин. Далее идет экспоненциальный спад, скорость которого определяется активностью ферментов и начальными концентрациями субстратов. При определенных условиях (субстратов много, активность ферментов мала, температура реакции снижена) реакция может наблюдаться в течение 24 и более часов.
Детекция биолюминесценции
[098] Методы настоящего изобретения включают детекцию биолюминесценции, возникающей в биологическом образце, содержащем люциферазу, при появлении в нем люциферина.
[099] Биолюминесценция может быть обнаружена с помощью методов, известных специалистам в данной области, в частности, с помощью визуального скрининга или с использованием люминометра, фотометра, флуориметра, цифровой фотокамеры, с помощью светочувствительной фотопленки. В качестве количественной характеристики может быть использована максимальная интенсивность люминесценции, которая достигается через 5-30 мин после инициации биолюминесцентной реакции или скорость нарастания люминесценции в интервале до 30 мин после инициации биолюминесцентной реакции, например, в течение 5, 10, 20, 30, 60 сек после инициации реакции или дольше.
[0100] В преимущественных воплощениях измеряемая люминесценция представляет собой длительное свечение, скорее, чем световые вспышки. В преимущественных воплощениях интенсивность люминесценции зависит от активности ферментов биолюминесцентной системы, присутствующих в образце, начальных концентраций субстратов и температуры реакционной смеси и обычно колеблется в пределах от 10 кв/сек до 10 млн кв/сек, чаще 100-100000 кв/сек.
[0101] Интенсивность люминесценции зависит от используемых реактивов. Например, максимальная интенсивность люминесценции образца при добавлении биснориангонина, согласно методу настоящего изобретения, примерно в 10 раз ниже, чем при использовании гиспидина. Также максимальная интенсивность люминесценции образца при добавлении НАДН в два-три раза ниже, чем при добавлении НАДФН.
[0102] Реакция длится не менее 30 мин после инициации, чаще 30-60 мин, иногда (в зависимости от условий) часы и даже сутки.
Способы применения
[0103] Методы и реактивы настоящего изобретения могут быть использованы в широком спектре биолюминесцентных анализов in vivo и in vitro.
[0104] В частности, методы и реактивы настоящего изобретения могут быть использованы для выявления активных компонентов биолюминесцентных системы высших грибов в процессе их очистки.
[0105] Также методы и реактивы настоящего изобретения могут быть использованы для выявления функциональных аналогов ферментов биолюминесцентной системы грибов в биологических образцах.
[0106] Также методы и реактивы настоящего изобретения могут быть использованы для выявления активности рекомбинантных люциферазы или люциферазы и гиспидин-3-гидроксилазы в клетках-хозяевах.
[0107] В некоторых воплощениях для осуществления применения должна быть получена нуклеиновая кислота, кодирующая люциферазу. Полученная нуклеиновая кислота должна быть встроена в кассету экспрессии, обеспечивающую временную или постоянную экспрессию этой нуклеиновой кислоты в клетках-хозяевах. Кассета экспрессии может содержать элементы, обеспечивающие адресную доставку конструкции в интересующие клетки или клеточные компартменты, или находится в составе частиц, обеспечивающих адресную доставку. После трансфекции клеток кассетой экспрессии (например в составе экспрессионного вектора) и по истечении времени, необходимого для наработки в клетках продукта экспрессии, может быть осуществлено выявление активности люциферазы внутри клеток или в клеточном лизате.
[0108] В некоторых воплощениях для осуществления применения должны быть получены нуклеиновые кислоты, кодирующие люциферазу и гиспидин-3-гидроксилазу. Полученные нуклеиновая кислота должны быть встроены в кассеты экспрессии, обеспечивающие временную или постоянную экспрессию нуклеиновых кислот в клетках-хозяевах, например под интересующими исследователя промоторами. Кассеты экспрессии могут содержать элементы, обеспечивающие адресную доставку конструкции в интересующие клетки или клеточные компартменты, или находится в составе частиц, обеспечивающих адресную доставку. После трансфекции клеток кассетой экспрессии (например, в составе экспрессионного вектора) и по истечении времени, необходимого для наработки в клетках продукта экспрессии, может быть осуществлено выявление одновременного присутствия люциферазы и гиспидин-3-гидроксилазы внутри клеток или в клеточном лизате.
Наборы
[0109] Также обеспечиваются в соответствии с настоящим изобретением наборы для использования при осуществлении вышеописанных применений.
[0110] В некоторых воплощениях наборы обычно включают 3-гидроксигиспидин и/или его функциональный аналог, предпочтительно с буферным раствором для растворения указанного субстрата и/или его добавления к биологическим образцам. 3-гидроксигиспидин и/или его функциональный аналог может присутствовать в растворенном виде в соответствующей среде для хранения, такой как DMSO, водный или буферный раствор с детергентом, обычно в соответствующей емкости. Альтернативно, 3-гидроксигиспидин и/или его функциональный аналог может присутствовать в наборе в лиофилизованном виде.
[0111] В некоторых воплощениях наборы обычно включают гиспидин и/или его функциональный аналог, предпочтительно с буферным раствором для растворения указанного субстрата и/или его добавления к биологическим образцам. В преимущественных воплощениях наборы так же включают НАД(Ф)Н, предпочтительно с буферным раствором для растворения указанного субстрата и/или его добавления к биологическим образцам.
[0112] В дополнение к описанным выше компонентам, заявленные наборы могут дополнительно включать инструкции для осуществления заявленных способов. Эти инструкции могут присутствовать в заявленных наборах в различных формах (например, в печатном варианте или на электроном носителе в виде текстового и/или графического файла) в количестве одна или более.
[0113] Следующие примеры предлагаются в качестве иллюстративных, но не ограничивающих.
Примеры
Пример 1. Выявление предшественника люциферина в плодовых телах высших грибов
[0114] Для приготовления горячих экстрактов использовали мицелий ряда биолюминесценных грибов: гриб Neonothopanus nambi был собран в лесах на юге Вьетнама, Mycena citricolor был получен из АТСС #12578, Panellus stipticus был собран в Оттаве (Канада), Armillaria borealis был собран вблизи Красноярска (Россия). Мицелий Neonothopanus nambi выращивали на среде, содержащей картофельный бульон в концентрации 200 г/л и сахарозу в концентрации 20 г/л, в течение 5 дней при 24°C при непрерывном перемешивании со скоростью 160-180 оборотов в минуту. Мицелий остальных грибов выращивали на среде, содержащей картофельный бульон в концентрации 200 г/л и глюкозу в концентрации 20 г/л, в сходных условиях: A. borealis в течение 12 дней при 24°C, P. stipticus - 5-7 дней при 24°C и М. citricolor - 5-7 дней при 27°C.
[0115] Также для приготовления горячих экстрактов использовали плодовые тела ряда небиолюминесценных грибов, собранных в лесу в районе Красноярска: Pholiota squarrosa, Thcholoma sp., Phellinus sp., Russula foetens and Clitocybe sp. Для приготовления горячего экстракта в каждом случае 10 г плодового тела или мицелия помещали в 30 мл дистиллированной воды, гомогенизировали и нагревали до кипения в микроволновой печи. Далее экстракты быстро охлаждали на льду и центрифугировали при 20000 g в течение 20 мин при 4°C. Супернатант экстрагировали с помощью 40 мл этилацетата, концентрировали на роторном испарителе и растворяли в 200 мкл 30% ДМСО, содержащего 0,1% муравьиной кислоты.
[0116] Полученные экстракты тестировали в биолюминесцентном тесте, описанном Oliveira и Stevani (Photochem. Photobiol. Sci., 2009, 8, 1416) с помощью холодного экстракта из Neonothopanus nambi.
[0117] Для получения холодного экстракта мицелий был отмыт от среды культивирования, помещен в 200-кратный объем дистиллированной воды на 15 часов при 26°C. Далее мицелий был отделен от воды центрифугированием, гомогенизирован на льду в 10 объемах 0,2 М фосфатного буфера, pH 7,5, и обработан ультразвуком с помощью Ultrasonic disintegrator UD-20 на льду в течение 10 мин. Гомогенат центрифугировали при 30000 g при 4°C в течение 20 мин. Полученный холодный экстракт хранили при - 20°C.
[0118] Для биолюминесцентного теста смешивали в 500 мкл: 25 мкл холодного экстракта, 2 мкл горячего экстракта, 442 мкл 0,2 М фосфатного буфера, pH 7.5, 1 мкл 1М ДТТ и 10 мкл 1% Triton Х-100. Биолюминесцентную реакцию инициировали в кювете люминометра Glomax 20/20 (Promega, США) добавлением 20 мкл 20 мМ НАДФН. Люминесценцию измеряли при 23±1°C. Типичные результаты измерений показаны на рис. 7. Предшественник люциферина был обнаружен в плодовых телах всех небиолюминесцентных грибов, причем, в концентрации существенно большей, чем в мицелии биолюминесцентных грибов.
Пример 2. Определение структуры предшественника люциферина высших грибов
[0119] Горячий экстракт из плодовых тел Ph. squarrosa, полученный как описано в Примере 1, был использован для обратнофазвной ВЭЖХ (HPLC), позволившей идентифицировать шесть различных фракций: 1-6 (рис. 8).
[0120] Все шесть фракций демонстрировали активность в биолюминесцентном тесте в присутствии холодного экстракта из Neonothopanus nambi, как описано в Примере 1. Максимальная люминесценция фракций 1-6 составляла 24000, 80, 6670, 40, 1300 и 1000, соответственно (отношение сигнал/шум люминометра, значения были нормированы на оптическую плотность при 360 нм).
[0121] Кроме того, с помощью повторной хроматографии выделенных фракций, было показано, что пары фракций (1, 3) и (5, 6) представляют собой гиспидин и биснориангонин, образующие равновесные смеси, цис- и транс-изомеров этих веществ по двойной связи (Рис 4, 5).
[0122] Все шесть фракций были подвергнуты повторной хроматографии и использованы для получения спектров ЯМР: 1Н, 2D DQF-COSY, 2D 1Н-13С HSQC и 2D 1Н-13С НМВС. Эти данные, в сочетании со спектрами HRMS (масс-спектры высокого разрешения), позволили определить химические структуры выделенных веществ. Спектры соединений (1, 3) и (5, 6) оказались полностью идентичны и содержали два набора сигналов, соответствующих гиспидину и его цис-изомеру по двойной связи (1, 3) и биснориангонину и его цис-изомеру по двойной связи (5, 6). Соединение 2 было идентифицировано как гомодимер гиспидина (3,3′-бигиспидинил) и соединение 4-гетеродимеру гиспидина и биснориангонина (3-биснориангонил-14-гиспидин).
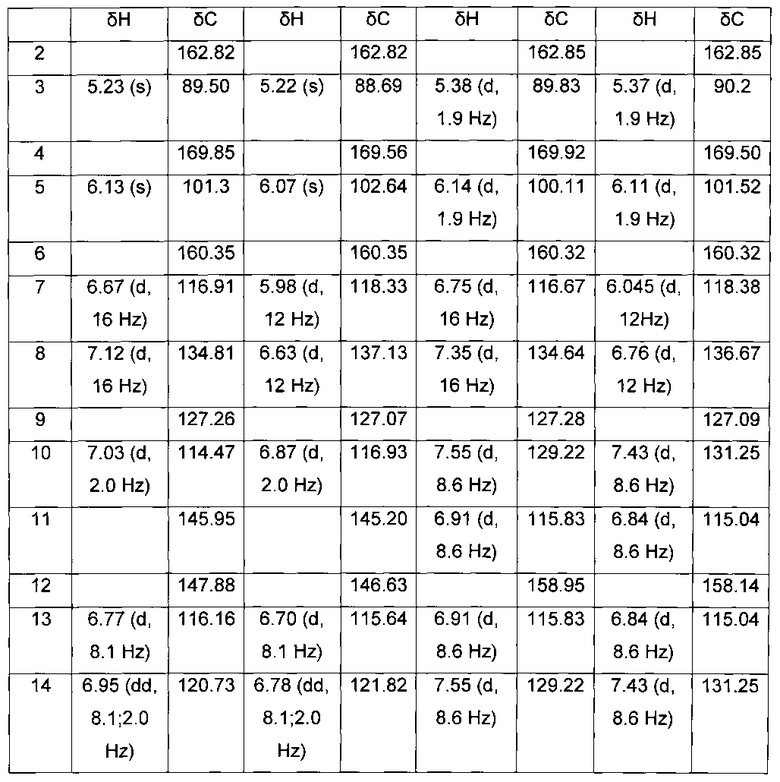
[0123] Структурное соответствие соединений 1, 3, 5 и 6 указанным веществам было подтверждено сравнением их хроматограмм и спектров с хроматограммами и спектрами коммерчески доступного препарата гиспидина (Сигма, США) и синтетического биснориангонина. Так, было обнаружено совпадение хроматографических времен удерживания соединений 1, 3 временами удерживания коммерчески доступного гиспидина и его транс-изомера, и совпадение хроматографических времен удерживания соединений 5, 6 с временами удерживания синтетического биснориангонина и его транс-изомера. Также было обнаружено совпадение химических сдвигов 1Н и 13С в спектрах ЯМР для веществ 1,3 с таковыми коммерчески доступного гиспидина и его трансизомера, а также совпадение химических сдвигов 1Н и 13С в спектрах ЯМР для веществ 5,6 с таковыми синтетического биснориангонина и его транс-изомера. Таблицы химических сдвигов веществ 1, 3, 5, 6 приведены в Табл. 1.

[0124] Чтобы подтвердить роль гиспидина в качестве предшественника люциферина в светящихся грибах, было осуществлено его выделение из светящегося мицелия N. nambi. В ходе предварительных экспериментов было показано, что замачивание мицелия в дистиллированной воде на ночь приводит к резкому увеличению активности горячих и холодных экстрактов (до 250 и 140 раз, соответственно). Приготовление горячего экстракта, полученного из такого вымоченного мицелия N. nambi с последующей ВЭЖХ, привела к выделению двух соединений с временами удерживания и УФ-спектрами, идентичными транс-гиспидину и его цис-изомеру по двойной связи. Кроме того, эти два соединения показали таутомеризацию, специфическую биолюминесцентную активность и спектры HRMS (Рис. 9), идентичные транс-гиспидину и его цис-изомеру, полученным из плодовых тел Ph. squarrosa.
[0125] В целом, из 10 г мицелия было выделено 0,5 мкг смеси таутомеров гиспидина и его цис-изомера.
[0126] Присутствие гиспидина и его цис-изомера было далее подтверждено с помощью хроматографии горячих экстрактов других биолюминесцентных грибов, полученных как описано в Примере 1.
Пример 3. Синтез, очистка и идентификация люциферина высших грибов
[0127] Из холодного экстракта N. nambi, полученного как описано в Примере 2, с помощью хроматографии на Superdex 75 (Sigma-Aldrich, США) получали фракцию ~35 kDa, содержащую частично очищенный препарат НАДФН-зависимого фермента биолюминесцентной системы гриба. Такжее получали микросомальную фракцию (фракцию белковых компонентов с молекулярными массами более 200 кДа) холодного экстракта, способную к люминесценции. Синтетический гиспидин (Sigma-Aldrich, США) инкубировали с фракцией 35 kDa в присутствии НАДФН. Реакционная смесь содержала 5 мл 0,2 М фосфатного буфера (pH 7,5), 200 мкл 0.1М ДТТ, 200 мкл 0,65 мМ водного раствора гиспидина с 0.1% муравьиной кислоты и 0.1% Тритона Х100, 5 мл фракции 35 kDa, 1 мл 10 мМ водного раствора НАДФН.
[0128] Каждые 5 мин аликвоты реакционной смеси объемом 1 мкл добавляли к микросомальной фракции холодного экстракта N. nambi, выявляя НАДФН-независимую люминесценцию люциферазы с собственным субстратом (люциферином). Анализ с помощью ВЭЖХ показал, что после начала инкубации при комнатной температуре в реакционной смеси появляется и накапливается новое соединение, концентрация которого становится максимальной после 35 мин инкубации (Рис. 10).
[0129] Реакцию терминировали закислением до рН 2.0, добавляя 150 мкл концентрированной соляной кислоты, центрифугировали при 30000 g в течение 15 мин и концентрировали с помощью колонок Диапак-С16 (BioChemMak S&T, Россия). Колонки промывали 10 мМ HCl, содержащим 3% MeCN и элюировали 2 мл 75% водного раствора MeCN. Элюат концентрировали до объема 50 мкл с помощью вакуумного центрифугирования. Полученный раствор разводили в 50 мкл ДМСО и использовали для обратнофазной хроматографии на полупрепаративной колонке ZORBAX Eclipse XDB-C18 (9.4 мм ×250 мм, Agilent, США). В качестве растворителя А использовали 0.1% муравьиную кислоту, pH 2.8, и растворителя Б - ацетонитрил. Использовали программу градиента 5-40% растворителя Б в течение 25 мин. Хроматографию проводили при 25°C со скоростью потока 3 мл/мин. Поглощение контролировали при 210, 230, 250, 270, 290, 310, 330 и 360 нм. Пик, появляющийся после 17,2 мин хроматографирования, собирали. В итоге из 32 мкг гиспидина было получено 19 мкг люциферина.
[0130] UV-vis спектр выделенного люциферина был по существу сходен со спектром гиспидина (рис. 11), 1Н ЯМР спектр демонстрировал одинаковый паттерн протонов в люциферине и гиспидине, однако наблюдалось отсутствие сигнала, соответствующего Н-3 гиспидина (рис. 12). HRMS спектр люциферина выявил наличие молекулярного иона с отношением масса/заряд (m/z) 263.0572, соответствующий брутто-формуле  (вычисленное значение m/z 263.0550). Это позволило однозначно идентифицировать выделенный люциферин как 3-гидроксилированный гиспидин, структура которого показана на рис. 1, определить природу НАДФН-зависимой реакции получения люциферина из гиспидина как гидроксилирование, а природу НАДФН-зависимого фермента биолюминесцентной системы гриба как гиспидин-3-гидроксилазы.
(вычисленное значение m/z 263.0550). Это позволило однозначно идентифицировать выделенный люциферин как 3-гидроксилированный гиспидин, структура которого показана на рис. 1, определить природу НАДФН-зависимой реакции получения люциферина из гиспидина как гидроксилирование, а природу НАДФН-зависимого фермента биолюминесцентной системы гриба как гиспидин-3-гидроксилазы.
Пример 4. Использование 3-гидроксигиспидина для выявления люциферазы в биологических образцах
[0131] 3-гидроксигиспидин был получен, как описано в Примере 3. В качестве биологических образцов использовали микросомальные фракции холодных экстрактов из грибов Neonothopanus nambi, Mycena citricolor, Panellus stipticus и Armillaria borealis. Холодные экстракты были получены как описано в Примере 2. Микросомальные фракции были получены как описано в Примере 3.
[0132] 10 мкг 3-гидроксигиспидина было растворено в 1 мл 50% водно-ацетонитрильном растворе, содержащем 0.1% муравьиной кислоты.
[0133] В ходе эксперимента в каждом случае сначала измеряли фоновую люминесценцию микросомальной фракции, а затем в моменты времени 0.7 мин и 1.55 мин добавляли аликвоты раствора 3-гидроксигиспидина, содержащие 0.006 нг и 0.16 нг люциферина, соответственно.
[0134] Во всех случаях была выявлена люминесценция биологических образцов, типичный результат показан на рис. 13.
Пример 5. Использование гиспидина для выявления люциферазы и гиспидин-3 гидроксилазы в биологических образцах
[0135] Коммерчески доступный гиспидин (Sigma-Aldrich, США) был использован для выявления люциферазы и гиспидин-3-гидроксилазы в холодных экстрактах, полученных из мицелиев Neonothopanus nambi, Mycena citricolor, Panellus stipticus, Armillaria borealis как описано в Примере 1.
[0136] Гиспидин растворяли в ДМСО до конечной концентрации 8 мг/мл (32 мМ). Далее полученный концентрированный раствор разводили в 500 раз водой с 0.1% Тритоном Х100 и 0.1% муравьиной кислотой. Концентрация гиспидина в полученном рабочем растворе составляла 64 мкМ.
[0137] Реакционная смесь в кювете биолюминометра Glomax 20/20 (Promega, США) содержала: 100 мкл 0.2 М фосфатного буфера, pH 7.5, содержащего 0.1 мМ ДТТ и 0.1 мМ ЭДТА, 1-5 мкл экстракта грибов, 4 мкл 64 мкМ раствора гиспидина. Люминесцентную реакцию инициировали добавлением 4 мкл 10 мМ НАДФН. До момента добавления НАДФН реакционная смесь не обладала детектируемой люминесценцией.
[0138] Во всех случаях было выявлено появление люминесценции, интенсивность которой достигала максимального значения (Imax) через 5-30 мин после начала реакции. Типичный результат эксперимента показан на рис. 14.
[0139] В другом эксперименте те же образцы были протестированы на присутствие активностей люциферазы и гиспидин-3-гидроксилазы как описано выше, за исключением того, что вместо НАДФН использовали НАДН. В этом случае люминесценция оказалась в 2-3 раза ниже, чем с НАДФН.
[0140] Было исследовано влияние концентрации гиспидина на эффективность люминесценции биологических образцов.
[0141] Из концентрированного раствора гиспидина изготавливали разведения 1/50, 1/500, 1/5000, 1/50000 в водном растворе с 0.1% тритоном Х100 и 0.1% муравьиной кислотой.
[0142] Реакционная смесь в кювете биолюминометра содержала: 100 мкл 0.1М фосфатного буфера, pH 7.5, содержащего 0.1 mM DTT, 4 мкл холодного экстракта N. nambi, по 1-5 мкл разведений гиспидина. Биолюминесцентную реакцию активировали добавлением 4 мкл 10 мМ НАДФН. Биолюминесценцию измеряли при 25°C. Зависимость максимальной биолюминесценции образца от концентрации гиспидина в реакционной смеси показана рис. 15. Оптимальная концентрация гиспидина в реакционной смеси составляет 2.4 мкМ, детектируемая люминесценция наблюдалась при концентрации гиспидина в реакционной смеси 0.002 мкМ и более, а при концентрации гиспидина более 5 мкМ наблюдалось ингибирование люминесцентной реакции, связанное, скорее всего, с субстратным ингибированием.
[0143] Было исследовано влияние концентрация детергента на эффективность люминесценции биологических образцов.
[0144] Реакционная смесь в кювете биолюминометра содержала: 100 мкл 0.1 М фосфатного буфера, pH 7,5 содержащего 0.1 мМ ДТТ, 4 мкл холодного экстракта N. nambi, 4 мкл 64 мкМ раствора гиспидина и различные количества детергента Тритон Х100. Биолюминесцентную реакцию активировали добавлением 4 мкл 10 мМ НАДФН. Зависимость Imax от концентрации детергента показана на рис. 16. Оптимальная концентрация детергента в реакционной смеси была определена как 0.015-0.03%. Максимальная концентрация детергента, при которой наблюдалась биолюминесценция составила 0.5%.
[0145] Было исследовано влияние молярности буферного раствора на эффективность люминесценции биологических образцов.
[0146] Реакционная смесь в кювете биолюминометра содержала: 100 мкл фосфатного буфера различной молярности, pH 7.5, содержащего 0.1 мМ ДТТ, 4 мкл холодного экстракта N. nambi, 4 мкл 64 мкМ раствора гиспидина. Биолюминесцентную реакцию активировали добавлением 4 мкл 10 мМ НАДФН. Биолюминесценцию измеряли при 25°C. Зависимость Imax от молярности буферного раствора показана на рис. 17. Оптимальная в данном эксперименте молярность буферного раствора составила 0.05-0.2 М. Детектируемая биолюминесценция наблюдается при 1 мМ и ниже, в воде без добавления буфера интенсивность люминесценции составляла 25% от максимальной.
[0147] Было исследовано влияние pH буферного раствора на эффективность люминесценции биологических образцов. Реакцию осуществляли, как описано выше, используя 0,1 М Трис-HCl и фосфатный буфер. Согласно полученным данным, люминесценция биологического образца может быть детектирована в диапазоне pH от 6,0 до 9,8, оптимальным является использование буферных растворов с pH от 7,0 до 9,5.
[0148] Было исследовано влияние концентрации НАДФН на эффективность люминесценции биологических образцов.
[0149] Реакционная смесь в кювете биолюминометра содержала: 100 мкл 0.1 М фосфатного буфера, pH 7.5, содержащего 0.1 mM DTT, 4 мкл холодного экстракта N. nambi, 4 мкл 64 мкМ раствора гиспидина. Биолюминесцентную реакцию активировали добавлением различных количеств НАДФН. Зависимость Imax от концентрации НАДФН показана на рис. 18. Минимальная концентрация НАДФН в реакционной смеси, при которой наблюдалась детектируемая биолюминесценция, составила 20 мкМ.
[0150] Было исследовано влияние концентрации ДТТ на эффективность люминесценции биологических образцов.
[0151] Реакционная смесь в кювете биолюминометра содержала 100 мкл 0.1 М фосфатного буфера, pH 7.5, содержащего различные концентрации DTT, 4 мкл холодного экстракта N. nambi, 4 мкл 64 мкМ раствора гиспидина. Биолюминесцентную реакцию активировали добавлением 4 мкл 10 мМ НАДФН. Зависимость Imax от концентрации ДТТ показана на рис. 19. Оптимальная концентрация ДТТ в реакционной смеси 0.5-3 мМ. Максимальная концентрация ДТТ, при которой еще наблюдалась биолюминесценция составила 10 мМ.
Группа изобретений относится к области биохимии. Предложен метод выявления люциферазы в биологических образцах с использованием 3-гидроксигиспидина или 3-гидроксибиснориангонина в качестве люциферина. Предложено соединение для реализации указанного метода, представляющее собой 3-гидроксигиспидин. Кроме того, предложен метод выявления люциферазы в присутствии гиспидин-3-гидроксилазы в биологических образцах с использованием гиспидина или биснориангонина в качестве прекурсора люциферина и НАД(Ф)Н. Также предложен набор реактивов для реализации указанного метода, включающий гиспидин или биснориангонин и НАД(Ф)Н. Группа изобретений позволяет выявлять люциферазу грибов в биологических образцах и в дальнейшем может быть использована в широком спектре биолюминесцентных анализов in vivo и in vitro. 4 н.п. ф-лы, 19 ил., 1 табл., 5 пр.
1. Метод выявления люциферазы в биологических образцах, включающий
(а) добавление к образцу 3-гидроксигиспидина, имеющего структуру:
или 3-гидроксибиснориангонина, имеющего структуру
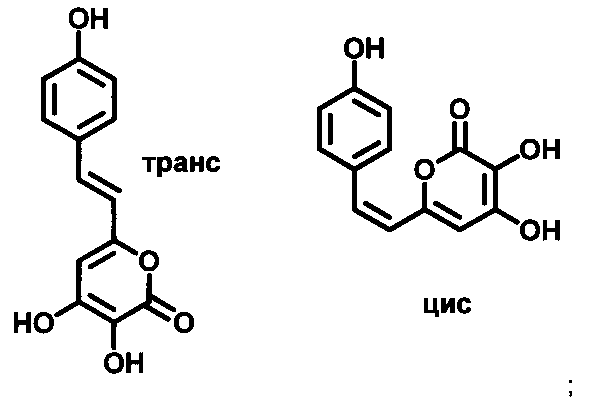
(б) инкубацию получившейся реакционной смеси в условиях, подходящих для возникновения биолюминесценции;
(в) детекцию биолюминесценции реакционной смеси, возникающей в биологическом образце, содержащем люциферазу, при появлении в нем люциферина.
2. Метод выявления люциферазы в присутствии гиспидин-3-гидроксилазы в биологических образцах, включающий
(а) добавление к образцу гиспидина или биснориангонина и НАД(Ф)Н;
(б) инкубацию получившейся реакционной смеси в условиях, подходящих для возникновения биолюминесценции;
(в) детекцию биолюминесценции реакционной смеси, возникающей в биологическом образце, содержащем люциферазу, при появлении в нем люциферина.
3. Соединение для реализации метода по п. 1, представляющее собой 3-гидроксигиспидин, имеющий структуру
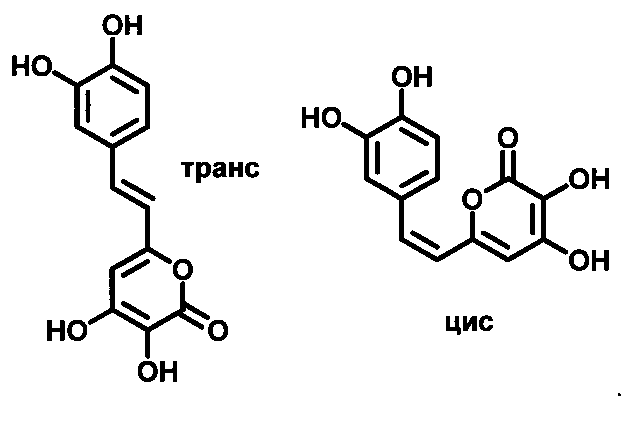
4. Набор реактивов для реализации метода по п. 2, в состав которого входит гиспидин или биснориангонин и НАД(Ф)Н.
| ТЯГЛИК А.Б | |||
| и др., ИССЛЕДОВАНИЕ СВЕЧЕНИЯ ВЫСШЕГО ГРИБА "NEONOTHOPANUS NAMBI", Молодежь и наука: сборник материалов IХ Всероссийской научно-технической конференции студентов, аспирантов и молодых ученых с международным участием, посвященной 385-летию со дня основания г | |||
| Красноярска [Электронный ресурс] | |||
| —? Красноярск: Сибирский федеральный |

