Настоящее изобретение относится к новому белку, в частности мутантным ферментам люциферазам, которые проявляют отличительные свойства по сравнению с соответствующим ферментом дикого типа, к ДНК, кодирующей данные белки, к применению данного фермента в аналитических тестах и к наборам, включающим их.
Люцифераза светляков катализирует окисление люциферина в присутствии АТФ, Мg2+ и молекулярного кислорода с испусканием света. Данная реакция имеет квантовый выход примерно 0,88. Свойство испускания света привело к ее применению в самых разнообразных люминометрических аналитических тестах, в которых определяют уровни АТФ. Примеры таких тестов включают таковые, которые основаны на описанных в ЕР-В-680515 и WO 96/02665, и многие другие, которые обычно используют в лабораториях.
Люциферазу получают непосредственно из насекомых, в частности жуков, таких как светляки или личинки светляков. Определенные виды, из которых получали люциферазы, включают японских светляков GENJI или KEIKE, Luciola cruciata и Luciola lateralis, восточно-европейского светляка Luciola mingrelica, северо-американского светляка Photinus pyralis и личинок светляка Lampyrus noctiluca.
Однако поскольку многие из генов, кодирующих данные ферменты, были клонированы и секвенированы, их можно также получить с использованием технологии рекомбинантных ДНК. Последовательности рекомбинантной ДНК, кодирующие ферменты, используют для трансформации микроорганизмов, таких как Е.coli, которые затем экспрессируют желаемый ферментный продукт.
Цвет испускаемого данными ферментами света при использовании в тестах в лаборатории совершенно одинаковый. Было бы полезно, если бы длина волны менялась, либо для того, чтобы легче улавливать специфическим детектором, либо для применения в системах, где требуются многочисленные репортеры, например для мониторинга различных событий, происходящих в одном и том же образце. Одним аспектом отличительных репортерных молекул является применение молекул люциферазы, которые испускают свет при различных длинах волн. Это можно достичь с использованием репортерных молекул, включающих люциферазы, полученные из различных видов светляков или личинок светляков. Однако альтернативной стратегией является получение мутантных люцифераз с использованием технологии рекомбинантных ДНК для того, чтобы получить различие в длине волны сигнала. Примеры таких мутантов предоставлены в WO 95/18853.
Кроме того, стабильность при нагревании люцифераз дикого и рекомбинантного типа такова, что они очень быстро теряют активность при воздействии температуры выше примерно 30°С, особенно выше 35°С. Данная нестабильность вызывает проблемы, когда фермент используется или хранится при высокой окружающей температуре, или если анализ проводится в условиях реакции при высокой температуре, например, для того, чтобы повысить скорость реакции.
Известны из ЕР-А-524448 и WO/95/25798 мутантные люциферазы, обладающие повышенной термостабильностью. В первой из них описывается мутантная люцифераза, имеющая мутацию в положении 217 люциферазы японского светляка, в частности, заменой остатка треонина на остаток изолейцина. В последней описываются мутантные люциферазы, обладающие 60% гомологией по отношению к люциферазе из Photinus pyralis, Luciola mingrelica, Luciola cruciata или Luciola lateralis, но в которых аминокислотный остаток, соответствующий остатку 354 Photinus pyralis или 356 вида Luciola, мутирован таким образом, что он является отличным от глутамата и, в частности, отличным от глутамата, аспартата, пролина или глицина.
В совместно рассматриваемой заявке на выдачу патента Великобритании №9823468.5 и на ее основе международной патентной заявки дополнительно описаны такие мутанты. В данном случае описаны белки, которые обладают люциферазной активностью и по меньшей мере 60% гомологией по отношению к люциферазе дикого типа, такой как фермент из Photinus pyralis, Luciola mingrelica, Luciola cruciata или Luciola lateralis, но которые включают мутации в различных положениях в белке, в том числе среди прочего:
(a) аминокислотный остаток, соответствующий остатку 214 в люциферазе Photinus pyralis и остатку 216 люциферазы Luciola mingrelica, Luciola cruciata или Luciola lateralis; или
(b) аминокислотный остаток, соответствующий остатку 232 в люциферазе Photinus pyralis и остатку 234 люциферазы Luciola mingrelica, Luciola cruciata или Luciola lateralis; или
(с) аминокислотный остаток, соответствующий остатку 295 люциферазы Photinus pyralis и остатку 297 люциферазы Luciola mingrelica, Luciola cruciata или Luciola lateralis.
Заявители установили, что с использованием мутирования (или введения) аминокислоты в различное положение в белке люциферазы можно достичь существенных сдвигов в длине волны испускаемого света и/или повышенной термостабильности фермента. Кроме того, может быть усилен протонный поток испускаемого света, делая фермент более подходящим для анализов in vivo, где устраняется свойственная личинке светляка кинетика фермента, или анализов in vitro, где отсутствуют СоА или другие вещества, “индуцирующие свойственную личинке светляка кинетику фермента”.
Настоящее изобретение относится к рекомбинантному белку, обладающему люциферазной активностью и по меньшей мере 60% гомологии по отношению к люциферазе дикого типа, где в последовательности фермента аминокислотный остаток, соответствующий остатку 357 в люциферазе Photinus pyralis, мутирован по сравнению с соответствующей люциферазой дикого типа так, что люцифераза способна испускать свет при другой длине волны по сравнению с соответствующей люциферазой дикого типа и/или обладает повышенной термостабильностью по сравнению с соответствующей люциферазой дикого типа.
Последовательности люциферазы дикого типа, которые могут составить основу рекомбинантных форм по изобретению, включают Photuris pyralis, Luciola mingrelica, Luciola cruciata или Luciola lateralis, Hotaria parvula, Pyrophorus plagiophthalamus, Lampyris noctiluca, Pyrocoelia miyako, Photuris pennsylvanica или Phrixothrix (“железнодорожные” светляки - смотри Biochem. 38 (1999) 8271-8279).
Биолюминесцентные ферменты из видов, которые могут использовать субстрат D-люциферин (4,5-дигидро-2-[6-гидрокси-2-бензо-тиазолил]-4-тиазолкарбоновая кислоту), для испускания света, могут составить основу мутантных ферментов по изобретению.
Определенные последовательности люцифераз дикого типа, которые могут составить основу рекомбинантных форм по изобретению, включают Photuris pyralis, Luciola mingrelica, Luciola cruciata или Luciola lateralis, Hotaria parvula, Pyrophorus plagiophthalamus, Lampyris noctiluca, Pyrocoelia miyako и Photuris pennsylvanica.
В частности, люциферазы являются ферментами, получаемыми из фермента Photuris pyralis, Luciola mingrelica, Luciola cruciata или Luciola lateralis. В ферментах Luciola mingrelica, Luciola cruciata или Luciola lateralis соответствующий аминокислотный остаток находится в положении 359 в последовательности.
Показано, что последовательности всех различных люцифераз являются высоко консервативными, обладающими высокой степенью гомологии между ними. Это означает, что соответствующие области последовательностей ферментов легко определяются при исследовании последовательностей для установления наиболее сходных областей, хотя, если необходимо, можно использовать коммерчески доступное программное обеспечение (например, “Bestfit” от Генетической компьютерной группы Университета шт. Висконсин; смотри Devereux et al. (1984) Nucleic Acid Research 12: 387-395) для того, чтобы определить соответствующие области или определенные аминокислоты в различных последовательностях. Альтернативно или дополнительно можно определить соответствующие аминокислоты со ссылкой на L.Ye et al., Biochim. Biophys Acta 1339 (1997) 39-52, где приведены последовательности ферментов с нумерацией, которая также используется и в настоящей заявке.
В отношении возможной замены аминокислотного остатка, соответствующего остатку 357 в люциферазе Photuris pyralis, то большинство последовательностей дикого типа имеют кислый остаток (аспарагиновую кислоту или глутаминовую кислоту) в данном положении. Исключением являются некоторые формы люциферазы Photuris pennsylvanica, в которых соответствующий остаток (356) является неполярным остатком, валином, или некоторые формы люциферазы Phrixothrix, где соответствующее положение является V354 в PvGR или в PhRE, где это лейцин L354. Таким образом, в целом аминокислота, использованная в качестве заменяющей аминокислоты в данном положении, является другой, чем аспарагиновая кислота, глутаминовая кислота, валин или лейцин.
Следовательно, в большинстве случаев остаток кислой аминокислоты заменяется на некислый остаток, включая основные аминокислоты, такие как лизин или аргинин, неполярные аминокислоты, такие как лейцин, валин или изолейцин, незаряженные полярные аминокислоты, такие как тирозин, аспарагин, глутамин, фенилаланин, серин, триптофан или треонин. В частности, его можно заменить незаряженной полярной аминокислотой, такой как тирозин, аспарагин, серин или треонин. Особенно предпочтительными аминокислотными остатками для замены в данном положении являются тирозин, фенилаланин или триптофан и наиболее предпочтительно тирозин. Говоря в общем, ароматические остатки в данном положении приводят к наибольшим сдвигам и также могут способствовать возникновению термостабильности.
Там, где последовательности дикого типа включают некислые аминокислотные остатки в данном положении, их соответствующим образом мутируют в другие некислые остатки.
Было установлено, что при мутировании фермента таким образом, сдвигается длина волны испускаемого люциферазой света, в некоторых случаях до 50 нм к красной области спектра. Так, мутантная люцифераза Photinus pyralis D357Y испускает свет при длине волны около 612 нм по сравнению с ферментом дикого типа, который испускает свет при длине волны 562 нм.
Сдвиг длины волны на 50 нм представляет существенную потенциальную возможность для применения в анализах, поскольку сдвиг такой величины может быть легко определен спектрально. Различные окрашенные люциферазы можно использовать в качестве репортерных молекул в исследованиях по экспрессии генов, делая возможным прослеживать более чем за одним геном, например, как описано в WO 95/18853. Множественное аналитическое исследование можно также проводить с использованием люциферазы в качестве меток.
Тот факт, что свет в данном случае окрашен в интенсивный красный цвет, особенно полезен в аналитической методологии. Красный мутант может быть полезен при анализе раствора на АТФ, который содержит пигменты или другие соединения, которые могут абсорбировать более короткие длины волн света. Например, окрашенный в красный цвет раствор не будет абсорбировать красный свет. Примеры окрашенных в красный цвет растворов, которые часто являются объектом такого анализа, включают образцы крови или раствор культуральной среды с эукариотическими клетками, который может содержать окрашенный в красный цвет индикатор рН.
При использовании смеси колориметрических агентов, таких как люциферазы, может быть полезной способность образовывать интенсивный красный сигнал, особенно, где другой агент в образце образует зеленый сигнал. Фотоумножитель, используемый в спектральном анализе с фотокатодом, можно установить для детектирования либо одного, либо обоих пиков, образующихся в одной образце. Другими словами, возможно различать фотонный поток от красного или зеленого эмиттера в одном и том же образце.
Кроме того, было установлено, что на сдвиг длины волны может оказывать влияние присутствие кофактора кофермента А(СоА). В результате данного свойства возникает возможность того, что данный фермент можно использовать в тесте на кофактор.
Как описано ниже, было исследовано влияние кофактора кофермента А на спектр испускаемого света в условиях in vitro. По мере увеличения концентрации кофермента А изменяется спектральное распределение, и при наиболее высоких концентрациях СоА в спектре преобладают длины волн в области 590-630 нм с выраженным пиком при 610 нм.
Таким образом, в соответствии с дополнительным аспектом изобретения обеспечивается аналитический тест для определения присутствия в образце СоА, где данный тест включает добавление к образцу, в котором, как полагается, находится СоА, люциферазы, как описано выше, вместе с другими реагентами, которые необходимы для проведения реакции люцифераза/люциферин, определение длины волны испускаемого пробой света и соотнесение этого с присутствием или отсутствием СоА.
Такой аналитический тест может быть полезен при определении состояния роста или активности клеток, например микроорганизмов или эукариотических клеток.
Например, концентрация СоА в клетках Е. coli является относительно высокой и в значительной мере варьирует в зависимости от метаболического статуса. Мутантные ферменты по изобретению можно использовать для мониторинга метаболического статуса организма, в частности концентрации СоА in vivo, поскольку длина волны испускания варьирует в зависимости от концентрации СоА. Такие аналитические тесты могут быть особенно полезными в ситуациях, где СоА является важным первичным метаболитом при продукции антибиотиков (например, у стрептомицетов).
Концентрации СоА в клетках также являются важным индикатором биосинтеза жирных кислот и варьируют в зависимости от потребности клетки в том или ином компоненте. При ряде метаболических нарушений, таких как карциногенез и сахарный диабет, имеют место нарушения метаболитов жирных кислот и, соответственно, необычные уровни СоА. Тесты по изобретению можно использовать при диагностике таких состояний. Например, можно определить концентрации СоА в клеточном образце, таком как образец крови, от пациента, при определении длины волны света, испускаемого люциферазой по изобретению, использованной в тесте. Данный результат можно сравнить с таковым, полученным с пробой здоровых клеток, для определения того, изменяется ли длина волны, и таким образом, что СоА присутствует в измененной концентрации. Это может указывать на наличие болезненного состояния у пациента. Клетки перед анализом соответствующим образом лизируют с использованием известного литического агента.
Полагается, что аминокислотный остаток в положении 357 критически связан с сайтом связывания кофермента А. Когда поверхность фермента люциферазы была смоделирована (с использованием программного обеспечения для моделирования белков SYBL, Tripos Ltd.) до разрешения 1 ангстрем (Å), было отмечено наличие полярного “кармана”. Оказалось, что данный “карман” образован остатками Н310, Е354 и D357 и составляет 8-10 Å. При обзоре молекулы сверху данный “карман” является частью более крупного “кармана”, образованного остатками Н310, Е354, D357 и 1232. Оказалось, что остатки Н310 и Е354 образуют мостик через углубление, что создает вид двух меньших “карманов” (смотри фигуру 8).
Не ограничиваясь конкретной теорией, представляется возможным, что мостиковые остатки могут быть достаточно гибкими, чтобы высвобождаться, когда фермент находится в растворе для обеспечения более крупного кармана (≈12 Å глубиной и ≈8 Å шириной), который позволяет связывать СоА. Это совпадает с энергетическими расчетами.
Когда клетки Е. coli, экспрессирующие мутанты Photuris-люциферин-4-монооксигеназы по изобретению, росли при смене различных источников углерода, определяли спектр испускаемого света в условиях in vivo. Переключение от богатой среды (LB) до определенной минимальной среды с ацетатом или глюкозой в качестве единственного источника углерода приводило к сдвигу к более длинным длинам волн испускаемого света и снижению вклада более коротких длин волн. Это может обеспечить еще дополнительно средство контролирования длины волны испускаемого света для аналитических целей.
Было установлено, что мутация положения 357 в белке приводит к повышенной термостабильности.
Белки могут включать дополнительные мутации в последовательности, при условии, что люциферазная активность белка в значительной мере не нарушается. Мутации стабильно усиливают свойства фермента или определенным образом лучше приспосабливают его для предназначенной цели. Это может означать, что они приводят к повышенной термостабильности и/или сдвигам цветовых свойств и/или Km для АТФ ферментов. Примеры мутаций, которые приводят к цветовым сдвигам описаны в WO 95/18853. Мутации, которые влияют на значения Km, описаны, например, в WO 96/22376 и заявке на международный патент № PCT/GB 98/01026.
В основном было установлено, что воздействие мутаций является аддитивным в плане изменений в свойствах.
Мутантные люциферазы по изобретению могут включать другие специфические мутации, которые повышают термостабильность по сравнению с люциферазой дикого типа, например мутации, описанные в WO 00/24878. В частности, по меньшей мере один:
(a) аминокислотный остаток, соответствующий аминокислоте 354 люциферазы Photuris pyralis (356 в люциферазе Luciola) мутирован;
(b) аминокислотный остаток, соответствующий положению 215 в люциферазе Photuris pyralis или (217 в люциферазе Luciola), является другой гидрофобной аминокислотой; или
(c) аминокислотный остаток, соответствующий остатку 214 в люциферазе Photuris pyralis или остатку 216 люциферазы Luciola mingrelica, Luciola cruciata или Luciola lateralis;
(d) аминокислотный остаток, соответствующий остатку 232 в люциферазе Photuris pyralis или остатку 234 люциферазы Luciola mingrelica, Luciola cruciata или Luciola lateralis;
(e) аминокислотный остаток, соответствующий остатку 295 в люциферазе Photuris pyralis или остатку 297 люциферазы Luciola mingrelica, Luciola cruciata или Luciola lateralis;
(f) аминокислотный остаток, соответствующий аминокислоте 14 люциферазы Photinus pyralis или остатку 16 Luciola mingrelica или 17 Luciola cruciata или Luciola lateralis;
(g) аминокислотный остаток, соответствующий аминокислоте 35 люциферазы Photinus pyralis, или остатку 37 Luciola mingrelica, или остатку 38 Luciola cruciata или Luciola lateralis;
(h) аминокислотный остаток, соответствующий аминокислотному остатку 105 люциферазы Photinus pyralis или остатку 106 гена Luciola mingrelica, 107 Luciola cruciata или 108 Luciola lateralis;
(i) аминокислотный остаток, соответствующий аминокислотному остатку 234 люциферазы Photinus pyralis или остатку 236 Luciola mingrelica, Luciola cruciata или Luciola lateralis;
(j) аминокислотный остаток, соответствующий аминокислотному остатку 420 люциферазы Photinus pyralis или остатку 422 Luciola mingrelica, Luciola cruciata или Luciola lateralis;
(k) аминокислотный остаток, соответствующий аминокислотному остатку 310 люциферазы Photinus pyralis или остатку 312 Luciola mingrelica, Luciola cruciata или Luciola lateralis;
является отличным от аминокислоты, которая имеется в соответствующей последовательности дикого типа и где фермент люциферазы обладает повышенной термостабильностью по сравнению с ферментом, имеющим в данном положении аминокислоту соответствующей люциферазы дикого типа.
Таким образом, предпочтительными примерами белков по изобретению являются мутированные люциферазы дикого типа, где более чем одна аминокислота, например до 100 аминокислотных остатков, предпочтительно не более чем 40 аминокислот и более предпочтительно до 30 аминокислот, являются другими по отношению к аминокислоте в соответствующем положении в соответствующем ферменте дикого типа.
Таким образом, в предпочтительном воплощении белок по изобретению включает люциферазу Photinus pyralis, где в дополнении к мутации в положении 357, как описано выше, по меньшей мере один:
(a) аминокислотный остаток, соответствующий аминокислоте 354 люциферазы Photinus pyralis, является отличным от глутамата;
(b) аминокислотный остаток, соответствующий положению 215 в люциферазе Photinus pyralis, является гидрофобной аминокислотой, отличной от аланина;
(c) аминокислотный остаток, соответствующий остатку 214 в люциферазе Photinus pyralis, является отличным от треонина;
(d) аминокислотный остаток, соответствующий остатку 232 в люциферазе Photinus pyralis, является отличным от изолейцина;
(e) аминокислотный остаток, соответствующий остатку 295 в люциферазе Photinus pyralis, является отличным от фенилаланина;
(f) аминокислотный остаток, соответствующий аминокислоте 14 люциферазы Photinus pyralis, является отличным от фенилаланина;
(g) аминокислотный остаток, соответствующий аминокислоте 35 люциферазы Photinus pyralis, является отличным от лейцина;
(h) аминокислотный остаток, соответствующий аминокислотному остатку 105 люциферазы Photinus pyralis, является отличным от аланина;
(i) аминокислотный остаток, соответствующий аминокислотному остатку 234 люциферазы Photinus pyralis, является отличным от аспарагиновой кислоты;
(j) аминокислотный остаток, соответствующий аминокислотному остатку 420 люциферазы Photinus pyralis, является отличным от серина;
(k) аминокислотный остаток, соответствующий аминокислотному остатку 310 люциферазы Photinus pyralis, является отличным от гистидина.
Альтернативно белок по изобретению включает белковую люциферазную последовательность фермента Luciola mingrelica, Luciola cruciata или Luciola lateralis, и где в дополнении к мутации в положении 359, как описано выше, по меньшей мере один:
(a) аминокислотный остаток, соответствующий аминокислоте 356 люциферазы Photinus pyralis, является отличным от глутамата;
(b) аминокислотный остаток, соответствующий положению 215 в люциферазе Photinus pyralis, является гидрофобной аминокислотой, отличной от аланина или треонина;
(c) аминокислотный остаток, соответствующий остатку 216 люциферазы Luciola mingrelica, Luciola cruciata или Luciola lateralis, является отличным от глицина (для Luciola mingrelica основные последовательности) или аспарагина (Luciola cruciata или Luciola lateralis) основные последовательности;
(d) аминокислотный остаток, соответствующий остатку 234 люциферазы Luciola mingrelica, Luciola cruciata или Luciola lateralis, является отличным от серина;
(e) аминокислотный остаток, соответствующий остатку 297 люциферазы Luciola mingrelica, Luciola cruciata или Luciola lateralis, является отличным от лейцина;
(f) аминокислотный остаток, соответствующий аминокислоте 16 Luciola mingrelica, или аминокислоте 17 Luciola cruciata или Luciola lateralis, является отличным от фенилаланина;
(g) аминокислотный остаток, соответствующий остатку 37 Luciola mingrelica или 38 в Luciola cruciata или Luciola lateralis, является отличным от лизина;
(h) аминокислотный остаток, соответствующий аминокислотному остатку 106 Luciola mingrelica, или аминокислотный остаток Luciola cruciata или 108 Luciola lateralis, является отличным от глицина;
(i) аминокислотный остаток, соответствующий аминокислотному остатку 236 Luciola mingrelica, Luciola cruciata или Luciola lateralis, является отличным от глицина;
(j) аминокислотный остаток, соответствующий остатку 422 Luciola mingreiica, Luciola cruciata или Luciola lateralis, является отличным от треонина;
(k) аминокислотный остаток, соответствующий аминокислотному остатку 312 Luciola mingrelica, Luciola cruciata или Luciola lateralis, является отличным от треонина (для Luciola mingrelica основные последовательности) или валина (для Luciola cruciata или Luciola lateralis) основные последовательности.
Конкретные в каждом случае замещенные аминокислоты, приводящие к появлению повышенной термостабильности, можно определить обычными способами, как показано ниже. В каждом случае различные замены могут приводить к повышенной термостабильности. Замену можно проводить направленным на сайт мутагенезом ДНК, кодирующей нативный или подходящий мутантный белки, что, очевидно, понятно специалисту в данной области. Изобретение в данном случае связано с определением положений, которые связаны с термостабильностью.
Однако в основном может быть желательным планировать замену аминокислотой других свойств вместо аминокислоты дикого типа. Так, гидрофильные аминокислотные остатки можно в некоторых случаях предпочтительно заменять на гидрофобные аминокислотные остатки и наоборот. Аналогично кислые аминокислотные остатки можно заменять на основные остатки.
Например, белок может включать белок, обладающий люциферазной активностью и по меньшей мере 60% гомологией по отношению к люциферазе из Photinus pyralis, Luciola mingrelica, Luciola cruciata или Luciola lateralis, где в последовательности фермента по меньшей мере один:
(а) аминокислотный остаток, соответствующий остатку 214 в люциферазе Photinus pyralis и остатку 216 люциферазы Luciola mingrelica, Luciola cruciata или Luciola lateralis, мутирован и является отличным от треонина в случае люциферазы Photinus pyralis; или
(b) аминокислотный остаток, соответствующий остатку 232 в люциферазе Photinus pyralis и остатку 234 люциферазы Luciola mingrelica, Luciola cruciata или Luciola lateralis мутирован и является отличным от изолейцина в случае люциферазы Photinus pyralis; или
(c) аминокислотный остаток, соответствующий остатку 295 в люциферазе Photinus pyralis и остатку 297 люциферазы Luciola mingrelica, Luciola cruciata или Luciola lateralis, мутирован и является, например, отличным от фенилаланина в случае люциферазы Photinus pyralis;
и фермент люциферазы обладает повышенной термостабильностью по сравнению с люциферазой дикого типа.
Показано, что последовательности всех различных люцифераз являются высококонсервативными, обладающими значительной гомологией между ними. Это означает, что соответствующие области последовательностей ферментов легко определяются при исследовании последовательностей для установления наиболее сходных областей, хотя, если необходимо, можно использовать коммерчески доступное программное обеспечение (например, “Bestfit” от Генетической компьютерной группы Университета шт. Висконсин; смотри Devereux et al. (1984) Nucleic Acid Research 12: 387-395) для того, чтобы определить соответствующие области или определенные аминокислоты в различных последовательностях.
Альтернативно или дополнительно можно определить соответствующие аминокислоты при обращении к L. Ye et al., Biochim. Biophys Acta 1339 (1997), 39-52.
В отношении возможной замены аминокислотного остатка, соответствующего остатку 214 в люциферазе Photinus pyralis, то полярную аминокислоту треонин соответственно заменяют на неполярную аминокислоту, такую как аланин, глицин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан или цистеин. Особенно предпочтительной заменой остатка треонина, соответствующего остатку 214 в Photinus pyralis, является аланин. Более предпочтительной заменой является цистеин. Однако другие полярные остатки, такие как аспарагин в данном положении, могут также усиливать термостабильность соответствующего фермента, имеющего треонин в данном положении. Другие аминокислоты, имеющиеся в данном положении в люциферазах дикого типа, включают глицин (Luciola mingrelica, Hotaria parvula), аспарагин (Pyrophorus plagiophthalamus, GR, YG, YE и OR, Luciola cruciata, Luciola lateralis, Lampyris noctiluca, Pyrocelia miyako, Photuris pennsylvanica LY, KW, J19) и серин (Phrixothix). Их можно преимущественно заменить на неполярные или другие неполярные боковые цепи, такие как аланин и цистеин.
В отношении возможной замены аминокислотного остатка, соответствующего остатку 232 в люциферазе Photinus pyralis, то неполярную аминокислоту изолейцин соответственно заменяют на другую неполярную аминокислоту, такую как аланин, глицин, валин, лейцин, пролин, фенилаланин, метионин, триптофан или цистеин. Другие аминокислоты, имеющиеся в данном положении в последовательностях дикого типа, включают серин и аспарагин. Соответственно, данные полярные остатки заменяют на неполярные остатки, такие, как указаны выше. Особенно предпочтительной заменой остатка, соответствующего остатку 232 в люциферазе Photinus pyralis и остатку люциферазы 234 Luciola mingrelica, Luciola cruciata или Luciola lateralis, в группе является аланин.
Замены аминокислотного остатка, соответствующего остатку 295 в люциферазе Photinus pyralis и остатку 297 люциферазы Luciola mingrelica, Luciola cruciata или Luciola lateralis, также могут оказывать влияние на термостабильность белка. (Это относится к положению 292 в люциферазе Phrixothix). В основном аминокислота в данном положении является неполярной аминокислотой фенилаланином или лейцином. Их соответственно заменяют на другие неполярные аминокислоты. Например, у Photinus pyralis неполярную аминокислоту фенилаланин соответственно заменяют на другую неполярную аминокислоту, такую как аланин, лейцин, глицин, валин, изолейцин, пролин, метионин, триптофан или цистеин. Особенно предпочтительной заменой остатка фенилаланина, соответствующего остатку 214 в люциферазе Photinus pyralis, является лейцин.
Также возможна мутация аминокислотного остатка, соответствующего аминокислоте 14 люциферазы Photinus pyralis или аминокислоте 16 или 17 в люциферазе Luciola (13 в люциферазе Phrixothix). Данный аминокислотный остаток (который обычно является фенилаланином, но также может быть лейцином, серином, аргинином или в некоторых случаях тирозином) соответственно заменяют на другую аминокислоту, в частности на другую неполярную аминокислоту, такую как аланин, валин, лейцин, изолейцин, пролин, метионин или триптофан, предпочтительно аланин.
Также может быть эффективной мутация аминокислотного остатка, соответствующего аминокислоте 35 люциферазы Photinus pyralis или аминокислотному остатку 37 в люциферазе Luciola mingrelica (38 в других видах Luciola spp.). Данная аминокислота варьирует среди ферментов дикого типа, которые могут включать в данном положении лейцин (Photinus pyralis), но также лизин, гистидин, глицин, аланин, глутамин или аспарагиновую кислоту. Соответственно, аминокислотный остаток в данном положении заменяют на неполярный аминокислотный остаток или другую неполярную аминокислоту, такую как аланин, валин, фенилаланин, изолейцин, пролин, метионин или триптофан. Предпочтительной аминокислотой в данном положении является аланин, что отличается от фермента дикого типа.
Мутации по аминокислоте, соответствующей положению 14 последовательности Photinus pyralis, и/или мутация по аминокислотному остатку, соответствующему аминокислоте 35 люциферазы Photinus pyralis, предпочтительно являются не единственными мутациями в ферменте. Соответственно, они сопровождаются другими мутациями, определенными выше, в частности таковыми в положениях, соответствующих положениям 214, 395 или 232 люциферазы Photinus pyralis.
Замены аминокислотного остатка, соответствующего остатку 105 в люциферазе Photinus pyralis и остатку 106 люциферазы Luciola mingrelica, или остатку 107 Luciola cruciata или 108 Luciola lateralis (102 Phrixothix), также могут оказывать влияние на термостабильность белка. В основном аминокислота в данном положении является неполярной аминокислотой аланином, или глицином, или серином у Phrixothix. Их соответственно заменяют на другие неполярные аминокислоты. Например, у Photinus pyralis неполярную аминокислоту аланин соответственно заменяют на другую неполярную аминокислоту, такую как фенилаланин, лейцин, глицин, валин, изолейцин, пролин, метионин или триптофан. Особенно предпочтительной заменой остатка аланина, соответствующего остатку 105 в люциферазе Photinus pyralis, является валин.
Замены аминокислотного остатка, соответствующего остатку 234 в люциферазе Photinus pyralis и остатку 236 люциферазы Luciola mingrelica, Luciola cruciata или Luciola lateralis (231 Phrixothix), также могут оказывать влияние на термостабильность белка. В основном аминокислота в данном положении является аспарагиновой кислотой или глицином и в некоторых случаях глутамином или треонином. Их соответственно заменяют на неполярные или другие неполярные аминокислоты, какие подходят. Например, у Photinus pyralis аминокислотный остаток, являющийся аспарагиновой кислотой, соответственно заменяют на неполярную аминокислоту, такую как аланин, лейцин, глицин, валин, изолейцин, пролин, метионин или триптофан. Особенно предпочтительной заменой остатка фенилаланина, соответствующего остатку 234 в люциферазе Photinus pyralis, является глицин. Там, где в данном положении находится неполярный аминокислотный остаток, такой как глицин (например, в люциферазе Luciola), его можно заменить на другую неполярную аминокислоту.
Замены аминокислотного остатка, соответствующего остатку 420 в люциферазе Photinus pyralis и остатку 422 люциферазы Luciola mingrelica, Luciola cruciata или Luciola lateralis (417 в зеленых Phrixothix и 418 в красных Phrixothix), также могут оказывать влияние на термостабильность белка. В основном аминокислота в данном положении является незаряженной полярной аминокислотой серином, или треонином, или глицином. Их соответственно заменяют на другие незаряженные полярные аминокислоты. Например, в Photinus pyralis серин можно заменить на аспарагин, глутамин, треонин или тирозин и, особенно, треонин.
Замены аминокислотного остатка, соответствующего остатку 310 в люциферазе Photinus pyralis и остатку 312 люциферазы Luciola mingrelica, Luciola cruciata или Luciola lateralis, также могут оказывать влияние на термостабильность белка. Аминокислотный остаток в данном положении варьирует среди известных люциферазных белков, будучи гистидином в люциферазе Photinus pyralis, Pyrocoelia miyako, Lampyris noctiluca и некоторых формах Photuris pennsylvanica, треонином - в люциферазе Luciola mingrelica, Hotaria parvula и Phrixothix (где он является аминокислотой 307), валином - в люциферазе Luciola cruciata и Luciola lateralis, и аспарагином - в люциферазе некоторых Pyrophorus plagiophthalamus. Таким образом, в основном аминокислота в данном положении является гидрофильной аминокислотой, которая может быть замещена на другой аминокислотный остаток, который повышает термостабильность фермента. Особенно предпочтительной заменой остатка гистидина, соответствующего остатку 310 в люциферазе Photinus pyralis, является аргинин.
Другие мутации также могут присутствовать в ферменте. Например, в предпочтительном воплощении белок также имеет аминокислоту в положении, соответствующем аминокислоте 354 люциферазы Photinus pyralis (356 в люциферазе Luciola), замененную с глутамата, в частности, на аминокислоту, отличную от глицина, пролина или аспарагиновой кислоты. Соответственно, аминокислотой в данном положении является триптофан, валин, лейцин, изолейцин и аспарагин, но наиболее предпочтительно лизин или аргинин. Данная мутация описана в WO 95/25798. Было установлено, что гидрофобные остатки в данном положении усиливают сдвиг длины волны фермента. Кроме того, присутствие крупной гидрофобной (V или I), полярной (N) или положительной заряженной (К или R) аминокислоты в положении 354 повышает термостабильность.
В альтернативном предпочтительном воплощении белок также обладает аминокислотой в положении, соответствующем аминокислоте 217 в люциферазе Luciola (215 в Photinus pyralis), замещенной на гидрофобную аминокислоту, в частности на изолейцин, лейцин или валин, как описано в ЕР-А-052448.
Белки по изобретению включают как люциферазы дикого типа, так и рекомбинантные. Они обладают 60% гомологией по отношению к последовательностям дикого типа, как таковые у фермента Photinus pyralis, Luciola mingrelica, Luciola cruciata или Luciola lateralis, в том смысле, что по меньшей мере 60% аминокислот, находящихся в ферментах дикого типа, находятся в белках по изобретению. Такие белки могут обладать более высокой степенью гомологии, в частности по меньшей мере 70%, более предпочтительно по меньшей мере 80% и наиболее предпочтительно по меньшей мере 90% по отношению к ферментам дикого типа, перечисленным выше. Подобные белки данного типа включают аллельные варианты, белки от других видов насекомых, а также ферменты, полученные рекомбинантными методами. Их можно легко идентифицировать, так как они кодируются нуклеиновыми кислотами, которые гибридизируют с последовательностями, которые кодируют ферменты дикого типа в условиях гибридизации с определенной ионной силой. Такие условия, очевидно, понятны специалистам в данной области, и их примеры приводятся у Sambrook et al. (1989) Molecular Cloning, Cold Spring Harbor Laboratory Press). В общих чертах, условия с низкой ионной силой можно определить, как 3×SCC примерно при от комнатной температуры до примерно 65°С, и условия с высокой ионной силой, как 0,1×SSC при примерно 65°С. SSC является названием буфера из 0,15 М NaCl, 0,015 М цитрата натрия. 3×SSC является в три раза сильнее, чем SSC, и так далее.
В частности, гомологию определенной последовательности по отношению к последовательностям по изобретению можно оценить с использованием метода множественного сравнительного анализа первичной структуры, описанного Lipman and Pearson (Lipman D.J. & Pearson W.R. (1985) Rapid and Sensitive Protein Similarity Searches, Science, vol.227, pp.1435-1441). Следует высчитать “оптимизированную” процентную схему по следующим параметрам для алгоритма Липмана-Пирсона: ktup = 1, пробел = 4 и длина пробела = 12. Последовательности, для которых требуется оценить гомологию, следует использовать в качестве “испытуемой последовательности”, что означает, что основную последовательность для сравнения, такую как последовательность Photinus pyralis, или любую из других последовательностей, о которых сообщали Ye et al. выше, следует вводить в алгоритм первой.
Конкретными примерами белков по изобретению являются последовательности люциферазы дикого типа с одной или более мутацией, как указано выше.
Изобретение дополнительно обеспечивает нуклеиновые кислоты, которые кодируют люциферазы, как описано выше. Соответственно, нуклеиновые кислоты основаны на последовательностях дикого типа, которые хорошо известны в данной области. Подходящий метод мутации для проведения желаемой мутации в аминокислотной последовательности легко определить, основываясь на сведениях о генетическом коде.
В предпочтительном воплощении изобретения нуклеиновая кислота является синтетическим геном. Соответственно, синтетический ген конструируют с удалением кодонов, редко обнаруживаемых в генах с высоким уровнем экспрессии из обычных экспрессирующих хозяев, таких как Е.coli, и в то же время избегая введения кодонов, редко обнаруживаемых в генах, кодирующих люциферазы светляков. Данный подход обеспечивает то, что новый ген имеет применение кодона, которое является оптимальным как для Е. coli, так и экспрессирующих систем насекомых.
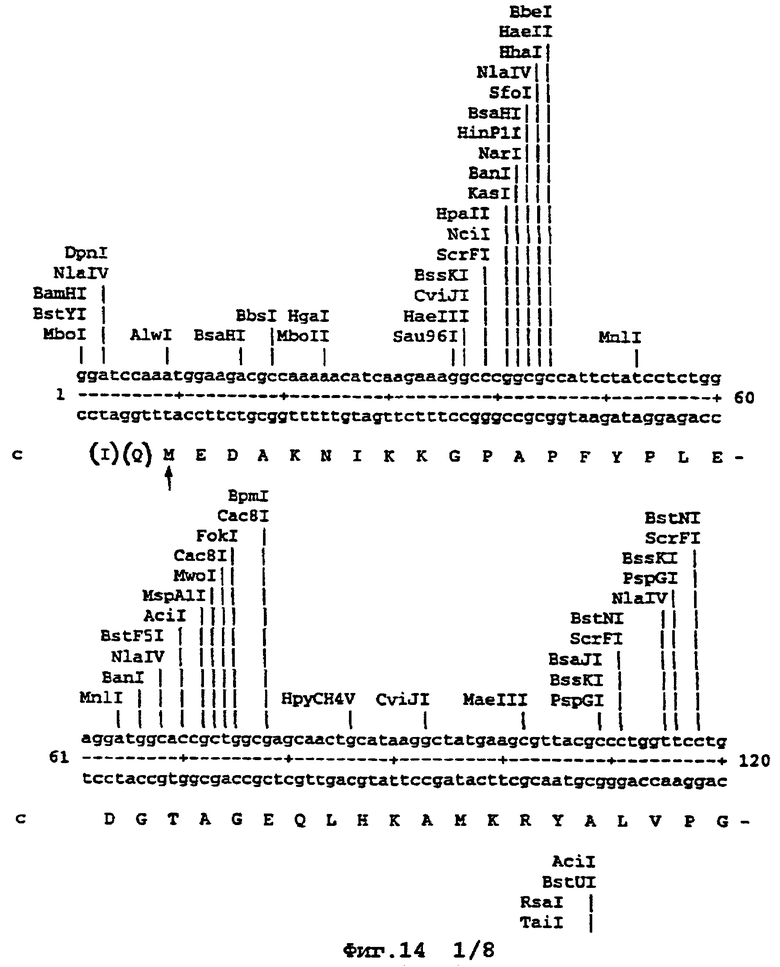
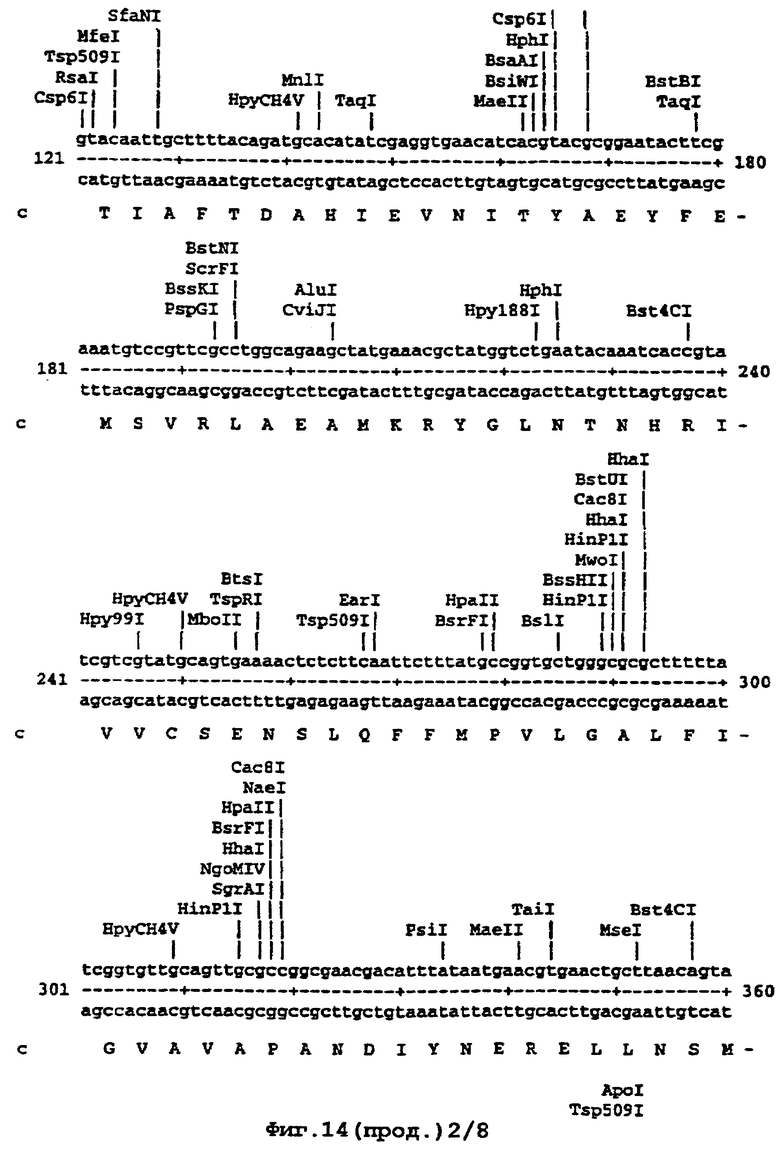
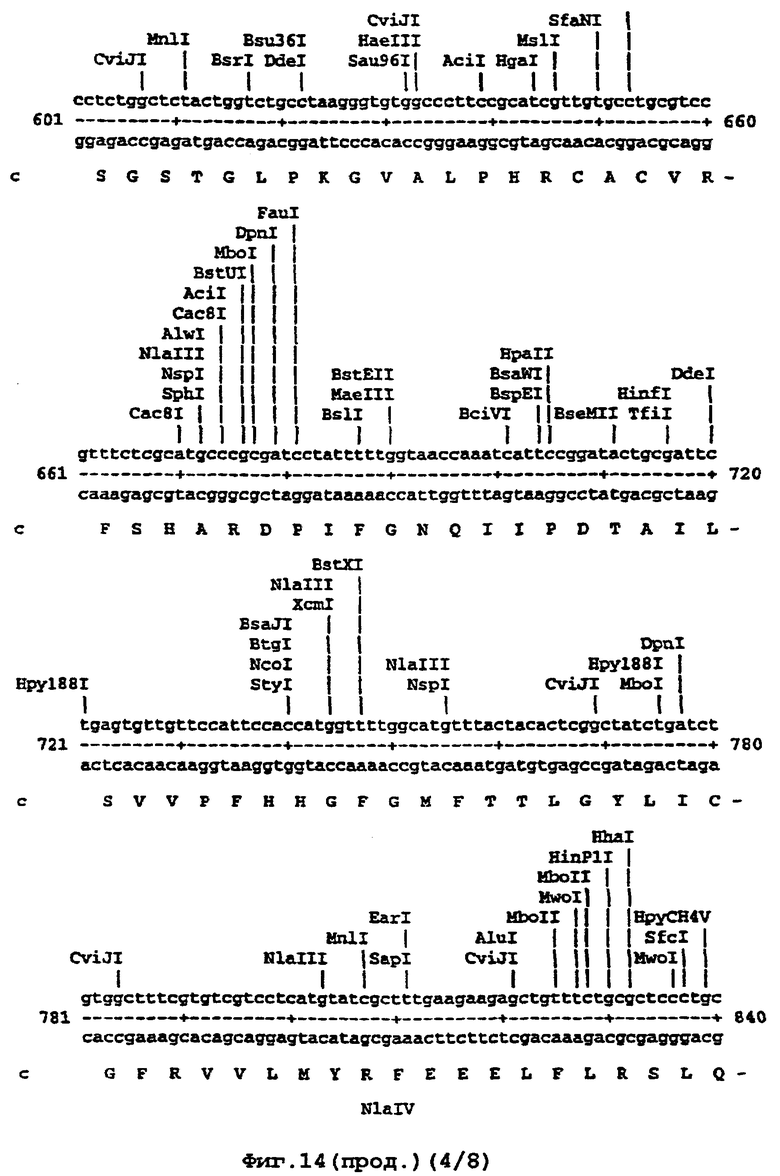
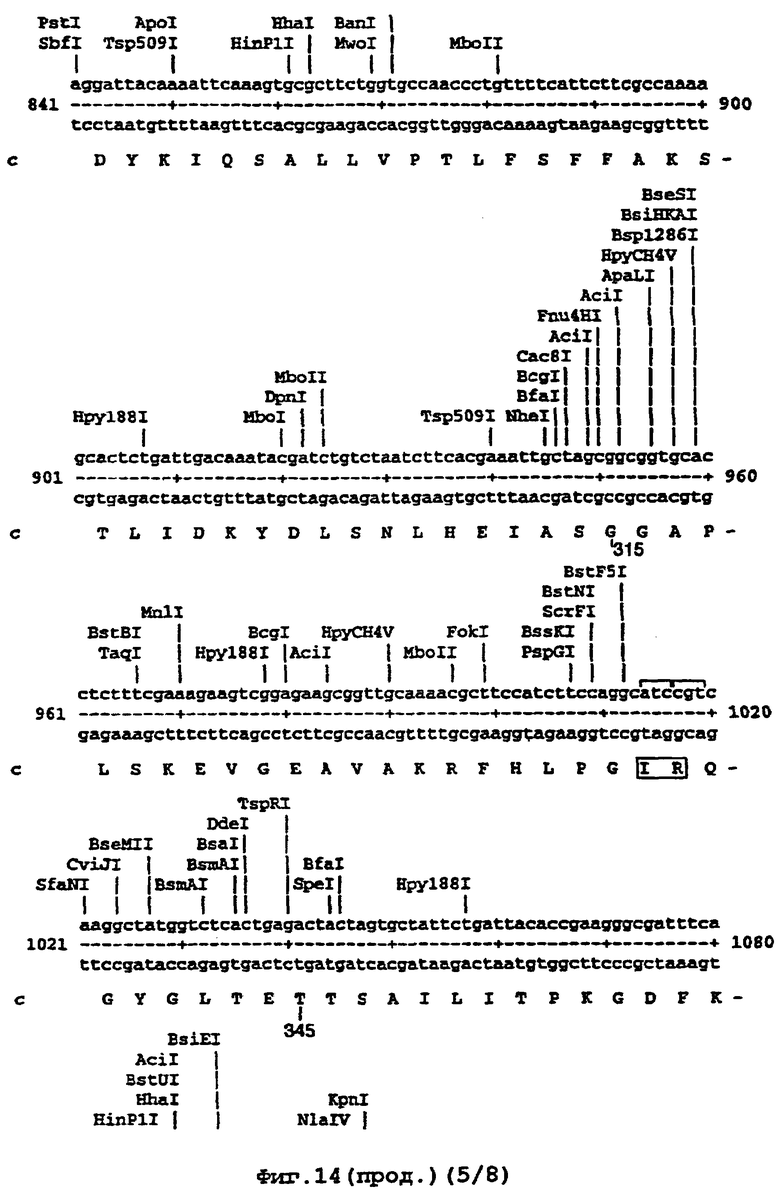
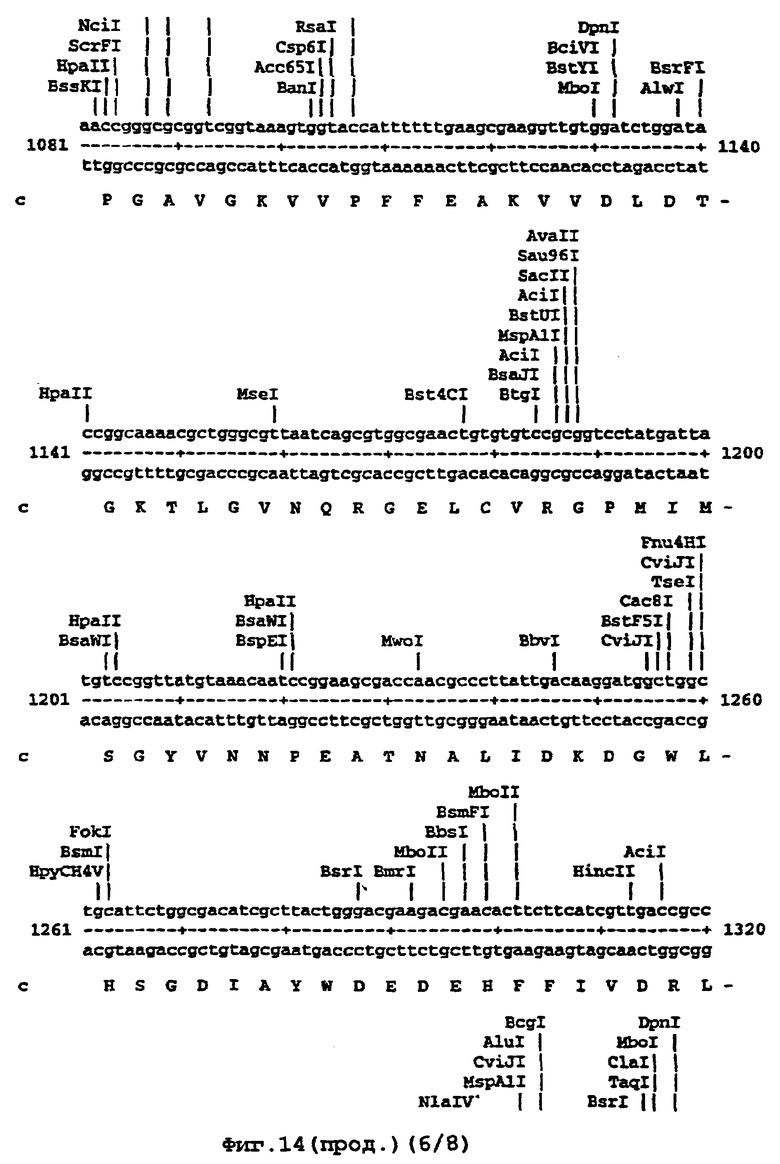
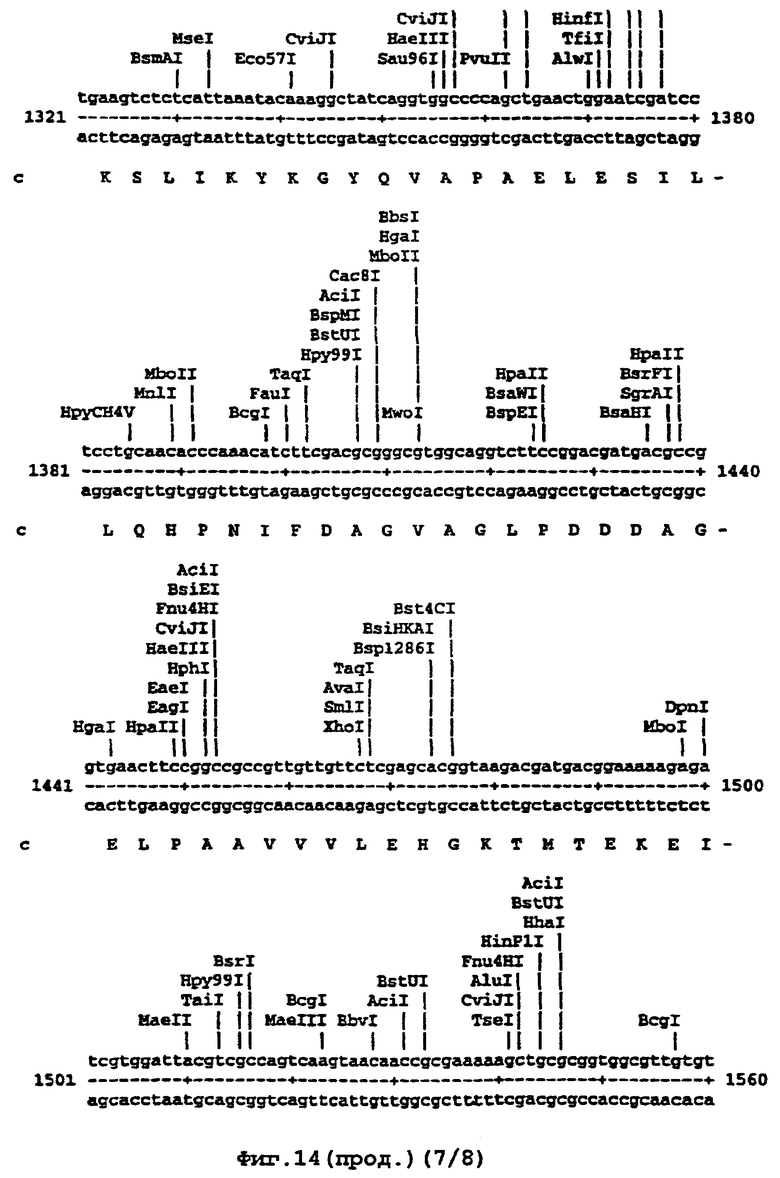
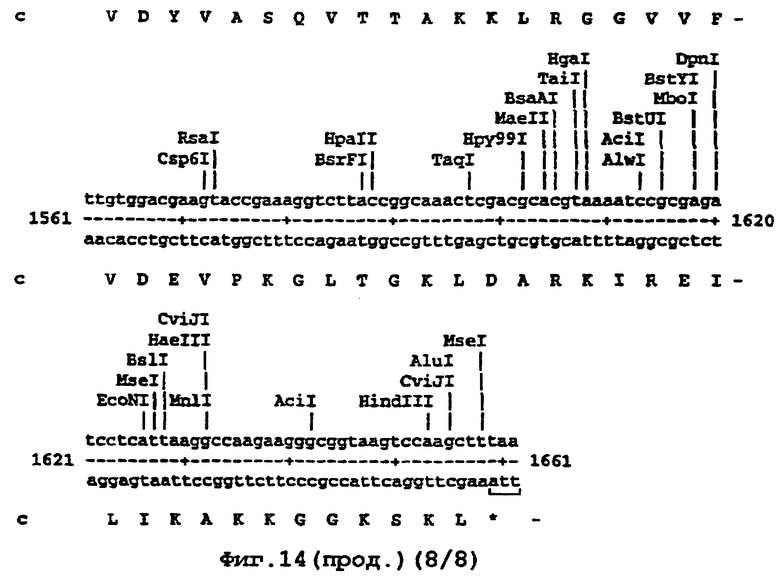
Например, там, где это возможно, кодоны аминокислот аrg, leu, ile, glу и pro замещали на CGT или CGC (аrg), CTG, СТТ или СТС (leu), АТС или АТТ (ile), GGT или GGC (gly) и CCG ССА или ССТ (pro), элиминируя таким образом редкие кодоны. В случае синтетического гена, показанного ниже (SEQ ID No 1) и на фигуре 14, это приводило в целом к 139 молчащим мутациям, создающим 62 новых нередких кодона (11% в целом). Первые 8 нуклеотидов, представленных на фигуре 14, образуют часть рибосомного сайта связывания и таким образом не кодируют. Кодирующая последовательность начинается с остатка метионина, указанного стрелкой. Данная кодирующая последовательность и очень близкие последовательности, например последовательности, которые имеют по меньшей мере 90% гомологию или предпочтительно 95% гомологию, образуют предпочтительный аспект изобретения.
Другим полезным свойством, которое можно использовать при получении синтетической упорядоченной структуры, является включение новых уникальных сайтов рестрикции. Данные сайты делают мутагенез, в частности комбинаторный мутагенез кассеты, гена проще и более эффективным. В частности, может быть желательным встроить уникальные сайты рестрикции внутри кДНК, кодирующей субдомен В в ферменте. Кроме того, может быть преимущественным создание уникального сайта рестрикции на противоположном 3’-конце гена для создания простых слитых конструкций и/или удаления пероксисомных целевых последовательностей.
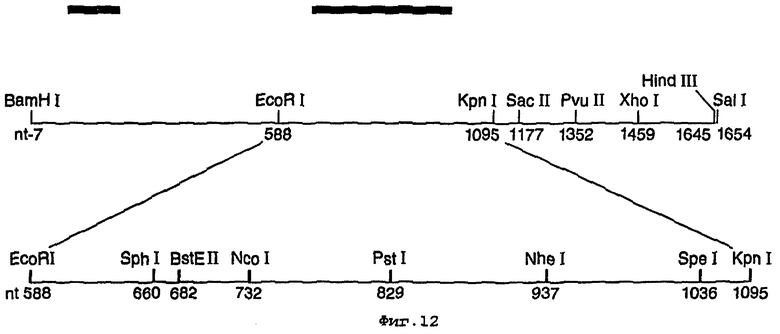
В примере, представленном ниже, было сконструировано девять новых уникальных сайтов рестрикции, в основном в центральной трети гена, и был получен уникальный сайт Hind III на противоположном 3’-конце гена для создания простых С-концевых слитых конструкций (фигура 12).
Наконец, применение синтетического гена предоставляет возможность для введения мутаций для повышения термостабильности генного продукта или иначе модификации свойств продукта, как желательно. В примере, представленном ниже, например, было получено три не молчащих мутации для введения в полипептид термостабилизирующих аминокислотных заменов Т214С, Е354К и D357F.
Нуклеиновые кислоты по изобретению соответственно включают в экспрессирующий вектор, такой как плазмида, под контролем контролирующих элементов, таких как промоторы, усилители, терминаторы и т.д. Затем данные векторы можно использовать для трансформации клетки-хозяина, например прокариотической или эукариотической клетки, такой как растительная или животная клетка, и, в частности, прокариотической клетки, такой как Е.coli, для того чтобы клетка экспрессировала желаемый фермент люциферазу. Культивирование трансформированных таким образом клеток с использованием условий, которые хорошо известны в данной области, будет приводить к продукции фермента люциферазы, который затем можно выделить из культуральной среды. Там, где клетки являются растительными или животными клетками, из указанных клеток можно получить растения или животных. Затем белок можно экстрагировать из растений или в случае трансгенных животных белки можно выделить из молока. Векторы, трансформированные клетки, трансгенные растения и животные и способы получения фермента при культивировании данных клеток образуют дополнительные аспекты изобретения.
Мутантную люциферазу Photinus pyralis D357Y получали случайным мутагенезом, как описано ниже. Было установлено, что единичная точечная мутация D357Y приводит к большому цветовому сдвигу длины волны испускаемого света и также повышенной термостабильности по сравнению с люциферазой дикого типа. В дополнительных исследованиях было установлено, что ряд замен в данном положении приводит к хорошей термостабильности и/или большим цветовым сдвигам.
Конкретные примеры мутантных ферментов Photinus pyralis, которые попадают в объем изобретения, включают следующие:
D357Y
D357F
D357W
D357K
D357N
D357I
E354I/D357Y
E354V/D357Y
E354C/D357Y
E354R/D357Y
E354S/D357Y
E354N/D357Y
E354K/D357M
E354R/D357L
E354W/D357W
E354H/D357W
E354R/D357F
E354K/D357F
E354S/D357F
E354M/D357F
E354A/D357R
E354A/D357F
E354T/D357Y
E354A/D357N
I351M/E354R/D357V
E354S/D357V
E354R/D357W
E354R/D357M
E354R/D357S
E354N/D357S
или равноценные варианты любого из них, когда они происходят из люцифераз других видов.
Мутации для получения вышеуказанных мутантов вводили в ген люциферазы в плазмиде рЕТ23 направленным на сайт мутагенезом, (ПЦР) или комбинаторным мутагенезом кассеты. Олигонуклеотиды, добавленные в ПЦР для того, чтобы провести соответствующие мутации, представлены ниже.
Ранее сообщалось, что воздействие точечных мутаций в положениях 354 и 215 является аддитивным. Данное изобретение обеспечивает возможность сочетания трех или более таких мутаций для обеспечения высокой термостабильности в мутантном ферменте, который имеет большой цветовой сдвиг.
Люциферазные белки по изобретению будут преимущественно использоваться в любом биолюминесцентном тесте, в котором применяется реакция люцифераза/люциферин в качестве сигнальных средств. По литературе известны многие такие аналитические тесты. Следовательно, белки можно включить в наборы, приготовленные для постановки таких тестов, необязательно с люциферином и любыми другими реагентами, необходимыми для постановки определенного теста.
Сейчас изобретение будет подробно описано с помощью примера при обращении к сопровождающим схематическим фигурам, где:
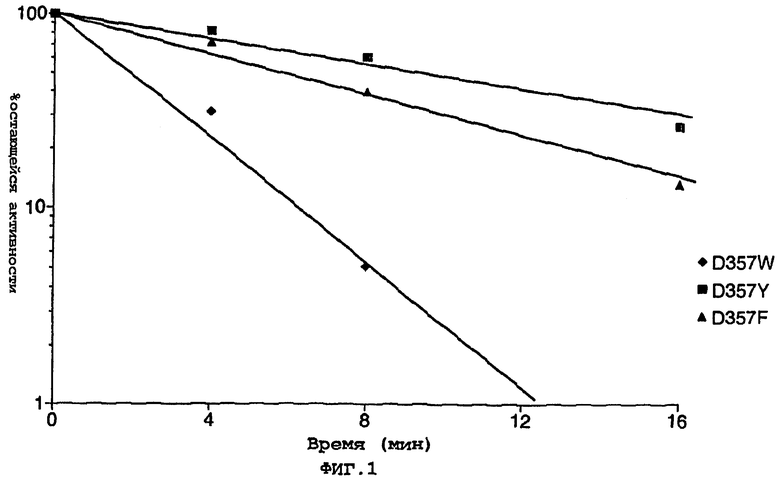
На фигуре 1 представлен log-график, показывающий % остающейся активности по отношению ко времени, при инкубации при 45°С нескольких мутантных ферментов по изобретению;
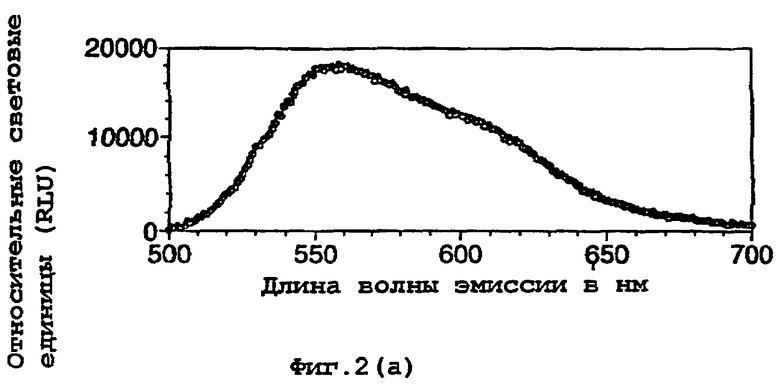
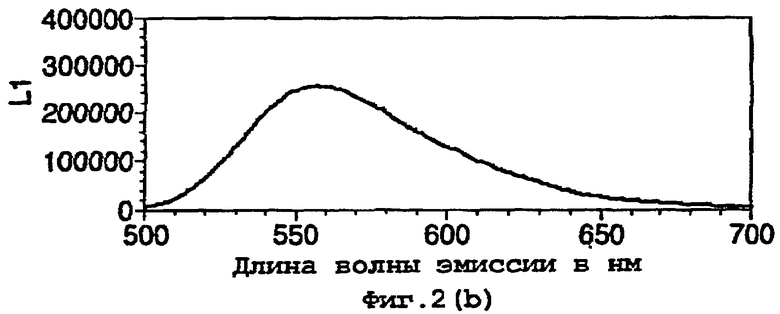
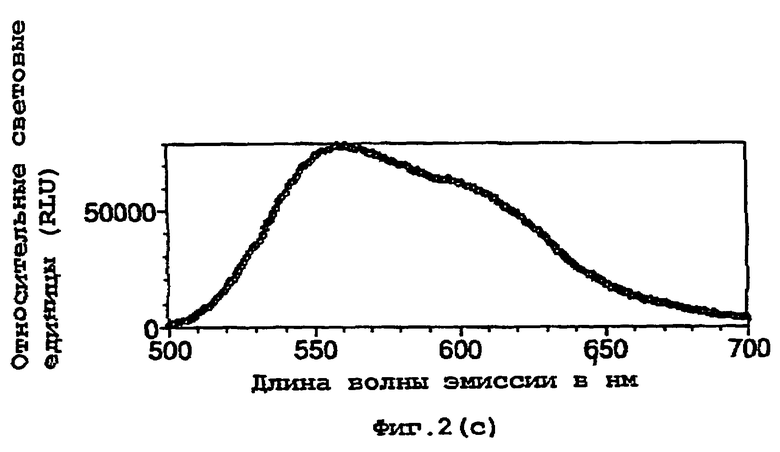
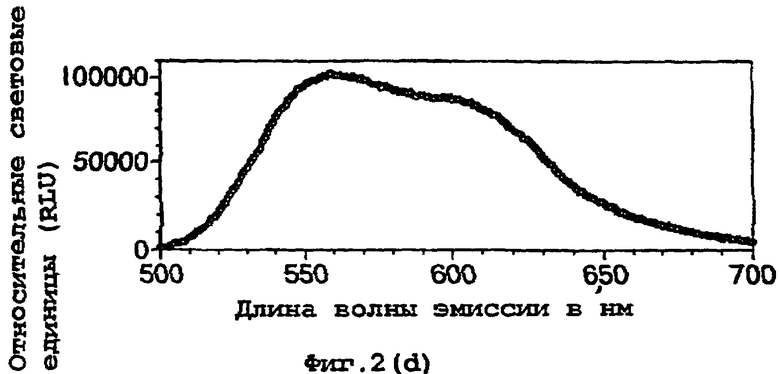
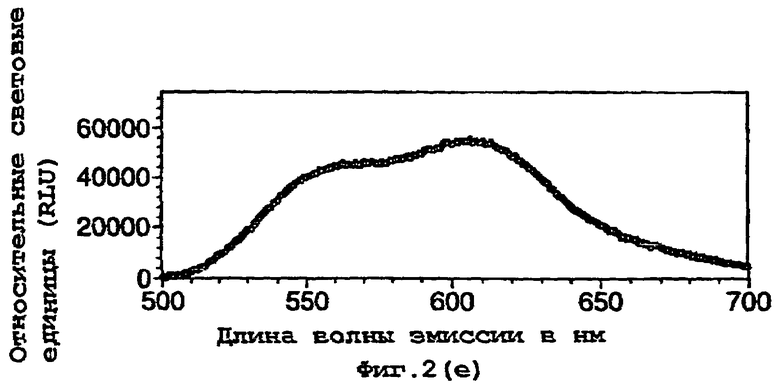
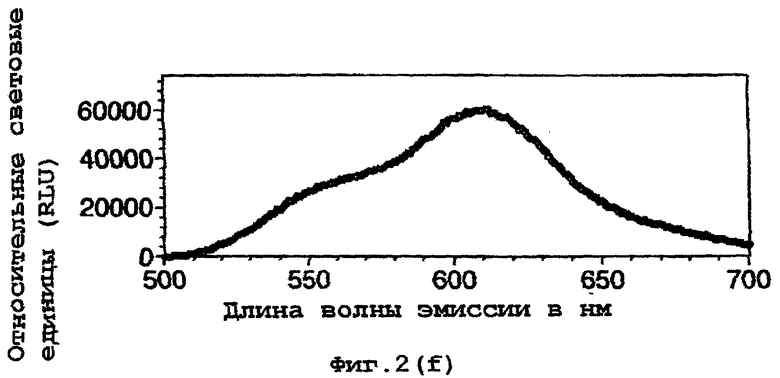
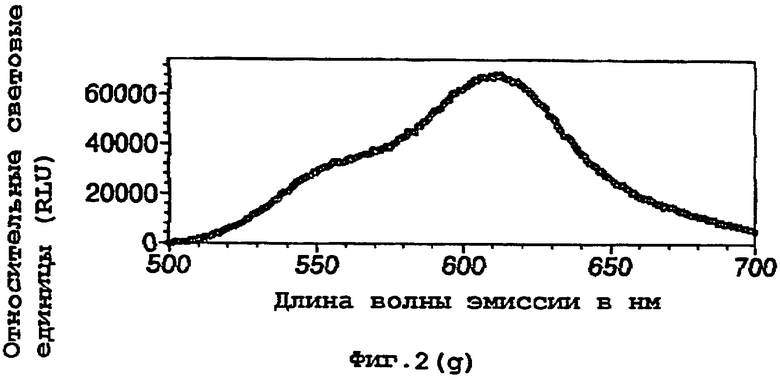
На фигуре 2 представлены спектральные пики, полученные при культивировании клеток Е.coli, экспрессирующих люциферазы, в цитратном буфере с D-люциферином, где использованный фермент представляет (а) рекомбинантную люциферазу Photinus pyralis дикого типа, (b) мутант D357K, (с) мутант D357N, (d) мутант D357W, (е) мутант D357I, (f) мутант D357F, (g) мутант D357Y и (h) двойной мутант E354I/D357Y;
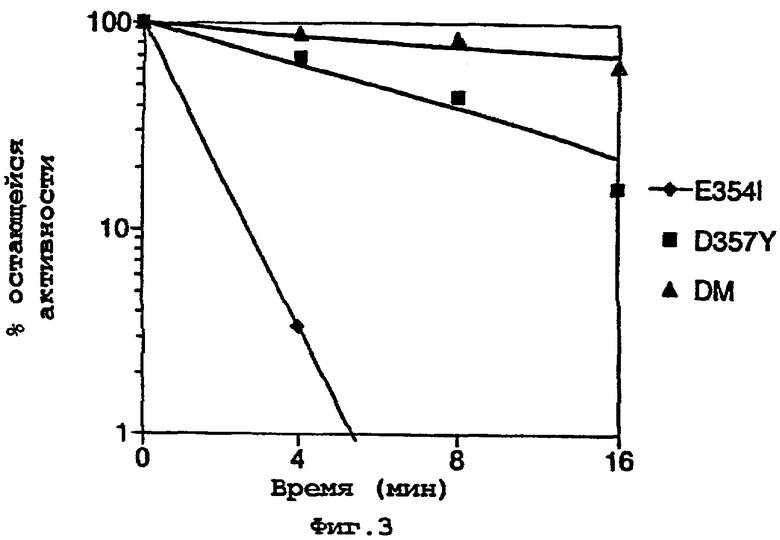
На фигуре 3 представлен график, показывающий % остающейся активности по отношению ко времени, для трех мутантных ферментов, E354I, D357Y и двойного мутанта (DM) E354I/D357Y;
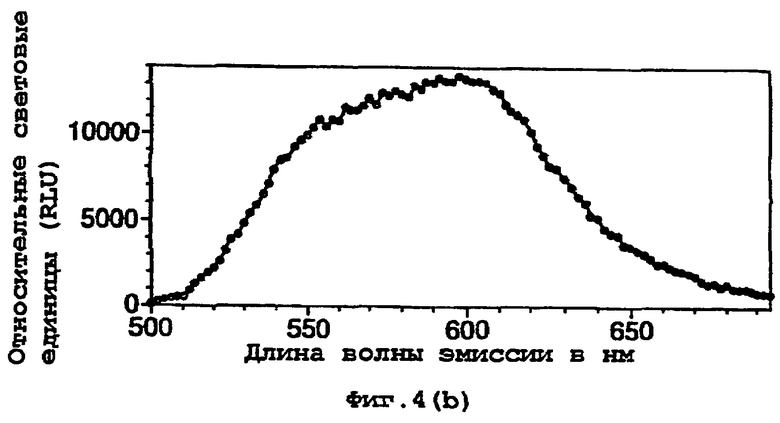
На фигуре 4 показаны спектры испускания (а) рекомбинантного фермента дикого типа и (b) двойного мутанта (DM) E354I/D357Y;
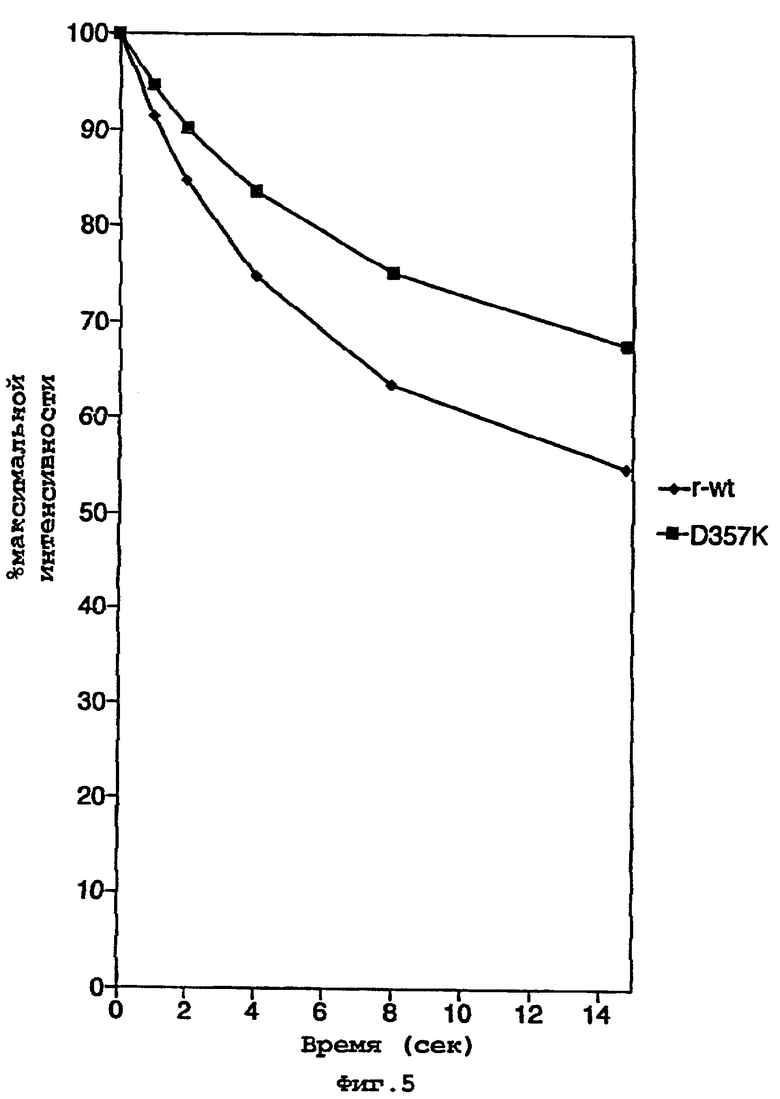
На фигуре 5 представлен график, показывающий скорость затухания фотонных эмиссий рекомбинантного фермента дикого типа (◆) r-wt и мутантного фермента D357K  ;
;
На фигуре 6 представлена диаграмма молекулярного моделирования, показывающая потенциальный “карман” связывания СоА в люциферазе;
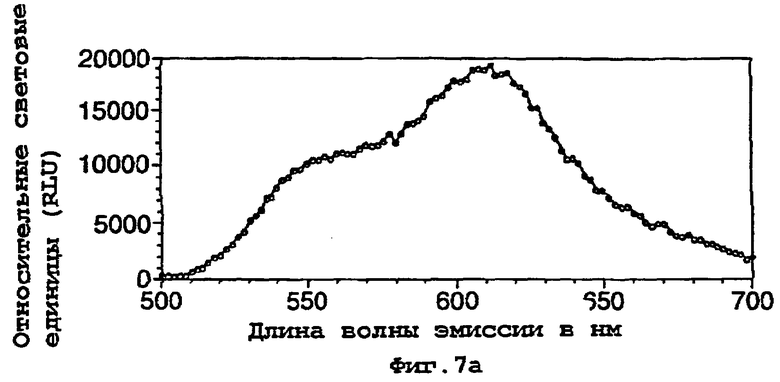
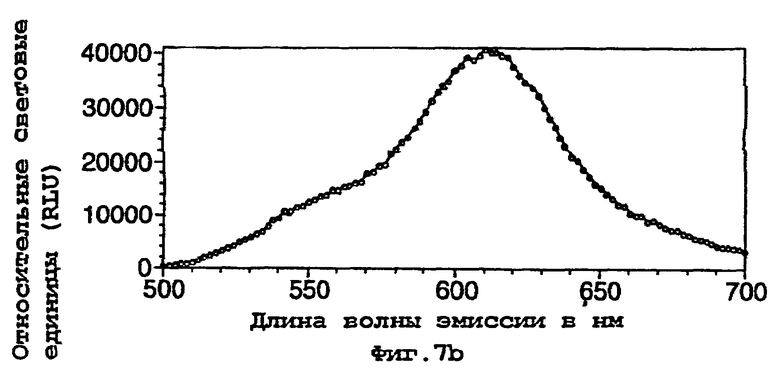
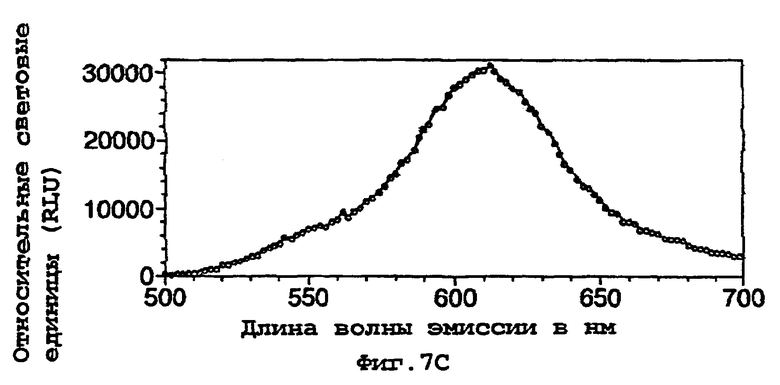
На фигуре 7 представлены спектры биолюминесценции in vivo, испускаемые клетками Е.coli, экспрессирующими мутантную люциферазу Р.pyralis D357Y (а) рост на LB; (b) рост на минимальной среде и ацетате натрия; (с) рост на минимальной среде и глюкозе;
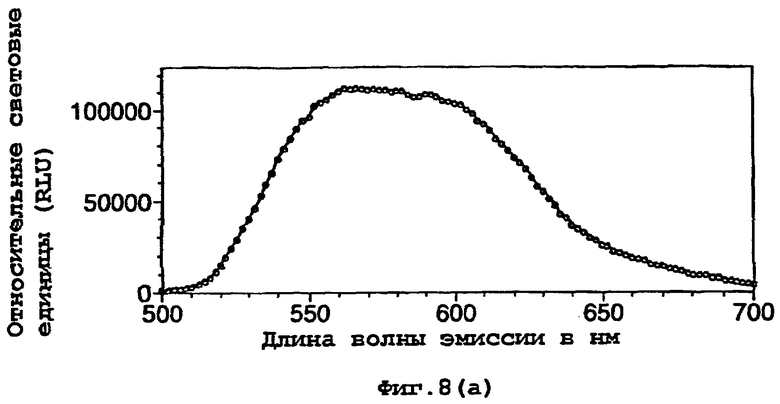
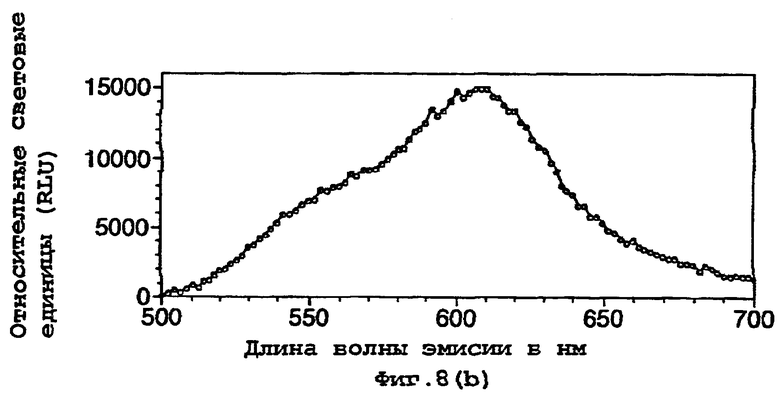
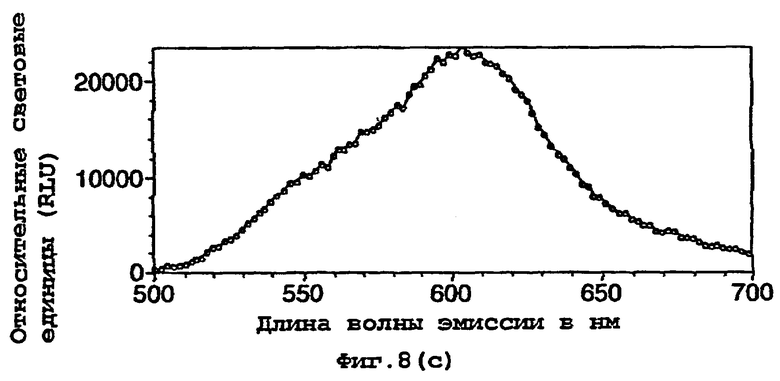
На фигуре 8 представлены спектры биолюминесценции in vivo, испускаемые клетками Е.coli, экспрессирующими мутантную люциферазу Р.pyralis E354K/D357M (а) рост на LB, (b) рост на минимальной среде и ацетате натрия; (с) рост на минимальной среде и глюкозе;
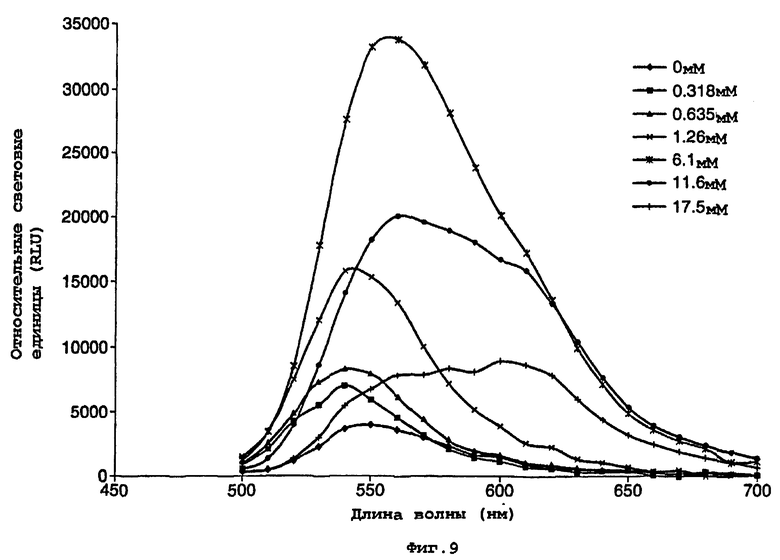
На фигуре 9 представлен график, показывающий воздействие СоА на спектральное распределение света, испускаемого мутантной люциферазой Р.pyralis D357Y;
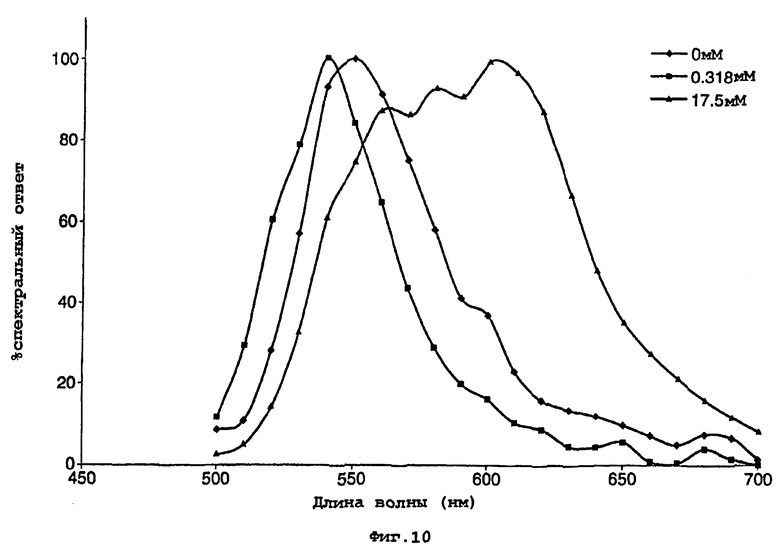
На фигуре 10 представлен график, показывающий нормализованные данные по воздействию СоА на спектральное распределение света, испускаемого мутантной люциферазой Р.pyralis D357Y;
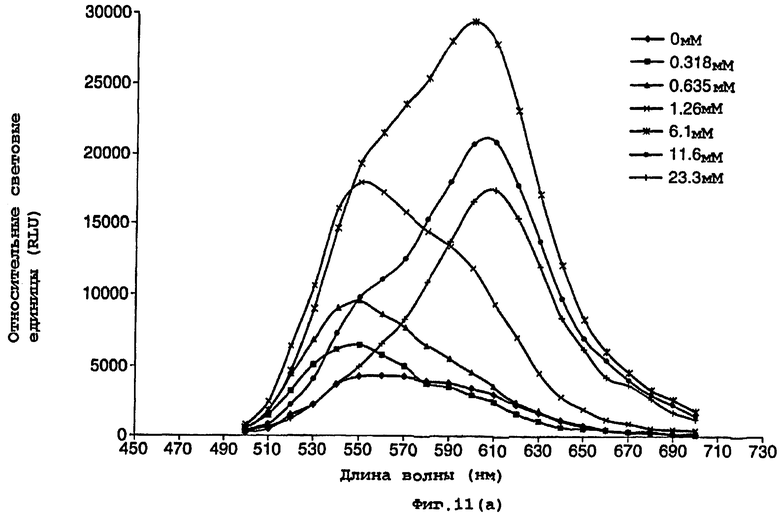
На фигуре 11 представлен график, показывающий воздействие СоА на спектральное распределение света, испускаемого мутантной люциферазой Р.pyralis E354I/D357Y (фигура 11а) и нормализованные данные (фигура 11b);
На фигуре 12 представлены модификации сайтов рестрикции, использованные при конструировании синтетического гена люциферазы;
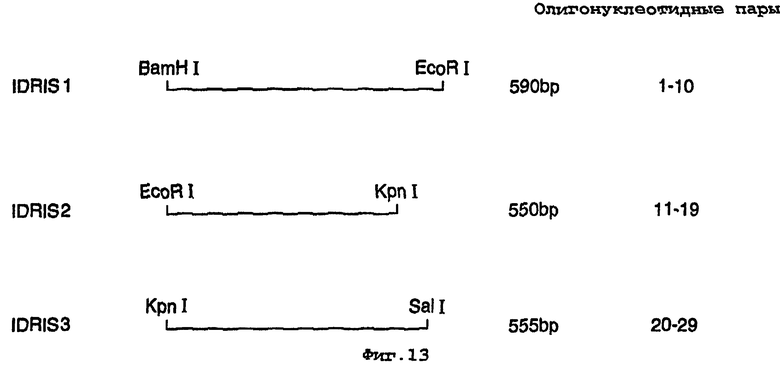
На фигуре 13 представлены конструкции, использованные в синтезе гена люциферазы;
На фигуре 14 представлена последовательность кДНК (SEQ ID No 1) синтетического гена люциферазы (включая нуклеотиды 1-8, которые образуют часть рибосомного сайта связывания, но не являются кодирующими) и кодированная аминокислотная последовательность, которая начинается с остатка метионина, указанного стрелкой (SEQ ID No 2); и
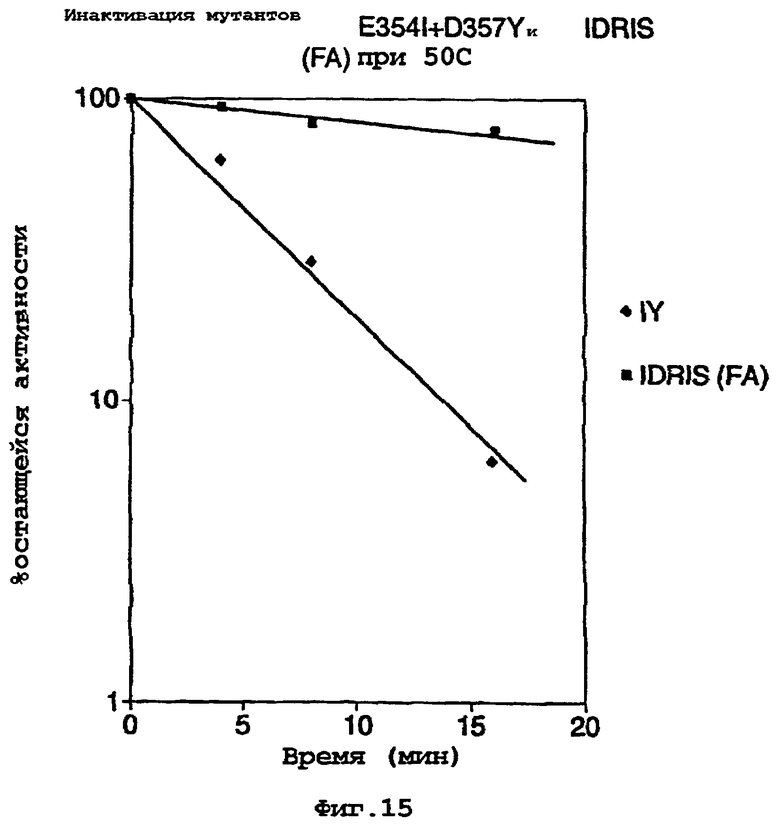
На фигуре 15 представлена термостабильность мутантов, включая мутанты, кодированные синтетическим геном, при 50°С.
Пример 1
Идентификация и характеристика мутантной люциферазы
Приготовили две библиотеки Photuris-люцифирин-4-монооксигеназы (Photinus pyralis), полученных ПЦР [M.Fromant et al., Anal. Biochem. (1995) 224, 347-353]. Одну библиотеку, включающую продукты ПЦР полной длины гена luc, клонировали в экспрессирующую систему рЕТ23а Т7 (Novagen Inc., Madison, WI, США). Вторую библиотеку, включающую продукты ПЦР короткого участка гена luc, охватывающего аминокислоты 199-353, клонировали в вектор pBSK(+) (Stratagene, La Jolla, CA, США).
Библиотеку рЕТ23а экспрессировали в штамме Е. coli BL21(DE3), (Е.coli В F dcm ompT hsdS(rB-mB-) galλ (DE3)).
Библиотеку pBSK(+) экспрессировали в клетках Е. coli НВ101 (supE44 ara14 galK2 lacY1 ∀(gpt-proA)62 prsL20 (Strr) xyl-5 mtl-1 recA13 ∀(mrcC-mrr) HsdS- (r-m-). Обе рЕТ23а и pBSK(+) имеют ген β-лактамазы и придают клеткам Е. coli, включающим плазмиду, резистентность к ампициллину.
Штамм Е. coli трансформировали полученной библиотекой электропорацией с использованием генератора импульсов BIORAD для Е. coli и выращивали в течение ночи при 37°С на агаре LB, содержащем ампициллин в концентрации 50 мкг/мл. Клетки переносили на нейлоновые мембраны (Osmonics, Minnetonka, Minnesota, США) и опрыскивали раствором люциферина (500 мкМ D-люциферин, соль калия в 100 мМ натриевым цитратном буфере, рН 5,0). Колонии оценивали с использованием документации и аналитической системы Alphalmager™ 1200 (Flowgen, Lichfield, Staffordshire, Великобритания). Это давало биолюминесценцию, испускаемую в течение определенного периода времени, с получением изображения света, испускаемого колониями. Яркость люминесценции принимали в качестве показателя термостабильности люциферазы.
Затем колонии тестировали на термостабильность. Колонии отбирали на основе яркости испускаемого света и выделяли для дальнейшей характеристики. В некоторых тестах скрининга колонии Е. coli инкубировали при 42°С в течение 2 ч перед скринингом для того, чтобы можно было отобрать термостабильные мутанты. Колонии, отобранные в первичном скрининге, переносили на нейлоновые мембраны и также культивировали в течение ночи в среде LB, содержащей ампициллин. “Пэтчи” опрыскивали раствором люциферина и оценивали в Alphalmager™. Данный вторичный скрининг помогал положительно идентифицировать клоны для анализа активности люциферазы in vitro. Клоны Е.coli, экспрессирующие возможные термостабильные ферменты, анализировали in vitro на активность люциферазы и термостабильность.
В тестах in vitro активность люциферазы определяли при комнатной температуре с использованием аналитической системы для определения активности люциферазы Promega (Promega Corporation, Madison, WI, США).
Люциферазную реакцию начинали с добавления 10 мкл сырого клеточного экстракта к 100 мкл аналитической смеси для определения активности люциферазы Promega (1 в 2 разведении). Полученную биолюминесценцию определяли с использованием люминометра Biotrace M3.
Сырые клеточные экстракты готовили, как описано в техническом бюллетене Promega no. 101. Аликвоты ночных культур Е.coli лизировали реагентом для лизиса клеточных культур (25 мМ Трис-фосфата, рН 7,8, 1 мМ дитиотреитола (DTT), 2 мМ 1,2-диаминоциклогексам-N,N,N’,N’-тетрауксусной кислоты, 10% глицерина, 1% тритона Х-100, 1,25 мг/мл куриного лизоцима) в течение 10 мин при комнатной температуре. Затем сырые лизаты хранили на льду перед анализом.
Свойства ферментов дополнительно тестировали в опытах по зависимой от времени инактивации. Пробирки Эппендорфа, содержащие 50 мкл аликвоты сырого клеточного экстракта, инкубировали на водяной бане при данной температуре. В определенные временные точки пробирки вынимали и охлаждали на льду перед анализом. Остающуюся активность люциферазы выражали в виде процента от первоначальной активности.
Строили log-графики процента остающейся активности по отношению ко времени инкубации и использовали для расчета значений t1/2. T1/2 представляет время, необходимое для того, чтобы фермент потерял 50% его первоначальной активности, после инкубации при данной температуре. Значения t1/2 (время для уменьшения активности до 50% от первоначальной активности) определяли в сырых экстрактах при 37°С по log-графикам % остающейся активности по отношению ко времени (не показано).
Плазмидная ДНК из клонов Е.coli, экспрессирующих в основном термостабильную люциферазу, как определено выше, секвенировали для того, чтобы определить мутации, ответственные за термостабильность фермента.
Плазмидную ДНК готовили с использованием набора QIAGEN QIAprep Spin Miniprep (QIAGEN Ltd., Crawley, W. Sussex, Великобритания), следуя протоколу для использования микроцентрифуги (QIAprep Miniprep Handbook 04/98).
Секвенирование ДНК во всех случаях проводила Babraham Technix, Cambridge, Великобритания с использованием секвенатора ДНК ABI PRISM™ 377 и реакционного набора для циклического секвенирования с терминатором ABI PRISM™ BigDye™ (Perkin Elmer Applied Biosystems), который основан на дидезокситерминационном методе [F. Sanger et al., Proc. Natl. Acad. Sci. USA 74, (1977) 5463-5467].
В результате данной работы идентифицировали новый мутант D357Y.
Кристаллическая структура люциферазы [Е.Conti et al., Structure, 4 (1996) 287-298] показала, что положение 357 находится на поверхности белка и близко к положению 354, которое может оказывать влияние как на термостабильность, так и на спектральные свойства. Это указывает на то, что данная область может быть важной в плане термостабильности фермента.
D357Y является особенно термостабильным мутантом, будучи наиболее термостабильной люциферазой с единичной аминокислотной заменой.
Пример 2
Сайт-направленный мутагенез для получения других мутантов по положению 357
Для того чтобы оценить различные мутации в положении 357 проводили сайт-направленный мутагенез с использованием набора для направленного на сайт мутагенеза Stratagene QuikChange™ (Stratagene, La Jolla, CA, США). Во всех опытах с направленным на сайт мутагенезом использовали плазмиду pPW601a J54 (PJW, MoD Report, 3/96). Все продукты реакций мутагенеза трансформировали в штамм Е.coli XLl-Blue [e14-(mcrA-)  (mcrCB-hsdSMR-mrr)171 endA1 supE44 thi-1 gyrA96 relA1 lac recB recJ sbcC umuC::Tn5 (Kanr) uvrC [F’ proAB laclqZ
(mcrCB-hsdSMR-mrr)171 endA1 supE44 thi-1 gyrA96 relA1 lac recB recJ sbcC umuC::Tn5 (Kanr) uvrC [F’ proAB laclqZ M15 Tn 10 (Tetr) Amy Camr]]. Олигонуклеотидные праймеры синтезировала Sigma-Genosys Ltd., Cambridge, Великобритания, и их конструировали с использованием смысловой допинговой системы [A.R.Arkin et al., Biotechnology, (1992)10, 297-300, W., Huang et al., Anal. Biochem. 218, 454-457] и использовали для конструкции дегенеративных олигонуклеотидных праймеров с получением групп возможных мутаций в большей степени, чем использовали отдельные праймеры для каждой аминокислотной замены.
M15 Tn 10 (Tetr) Amy Camr]]. Олигонуклеотидные праймеры синтезировала Sigma-Genosys Ltd., Cambridge, Великобритания, и их конструировали с использованием смысловой допинговой системы [A.R.Arkin et al., Biotechnology, (1992)10, 297-300, W., Huang et al., Anal. Biochem. 218, 454-457] и использовали для конструкции дегенеративных олигонуклеотидных праймеров с получением групп возможных мутаций в большей степени, чем использовали отдельные праймеры для каждой аминокислотной замены.
Таким образом получали библиотеки мутантов люциферазы с замещенными аминокислотами.
Использовали следующие олигонуклеотиды (и их комплементарные партнеры):
Библиотеки мутантов подвергали скринингу, как описано ранее, на термостабильность. Число колоний для скрининга вычисляли с использованием уравнения [S.Climie et al., J. Biol. Chem. 265 (1990) 18776-18779]
N=[ln(1-P)]/[Ln((n-1)/n)],
где N представляет число колоний для скрининга, n представляет число возможных кодонов в целевом положении, и Р представляет вероятность того, что каждый кодон в смеси отобран для скрининга по меньшей мере один раз. Уравнение основано на значении Р=0,95. Полученные в результате направленного на сайт мутагенеза мутанты анализировали на люциферазную активность и характеризовали в исследованиях по зависимой от времени термоинактивации.
Мутанты, идентифицированные таким путем, как желаемые, культивировали в 400 мл среды LB, содержащей ампициллин до А260≈0,5. Экспрессию люциферазы затем индуцировали добавлением изопропил-β-тиогалактозида (IPTG) до конечной концентрации 1 мМ. Затем клетки инкубировали при 30°С при встряхивании в течение 3 ч перед осаждением центрифугированием. Полученный при центрифугировании клеточный осадок ресуспендировали в 10 мл реагента для экстракции белка B-PER™ (Pierce Chemical Company, Rochford, США), 1 мМ DTT с получением сырого экстракта, следуя протоколу B-PER™ для максимальной экстракции бактериального белка. Восстановленную смесь ингибитора протеазы, 500 мкл (продукт №Р8465, Sigma, Saint Louis, Missouri, США) добавляли к раствору B-PER™ для ингибирования эндогенных протеаз. Затем клеточный лизат центрифугировали при 30000 g в течение 30 мин.
Супернатант сырого экстракта подвергали фракционированию с сульфатом аммония. Фракция, которая осаждалась между 30% и 55% насыщения, включала люциферазную активность. Данный материал ресуспендировали в 0,5 мл Трис-HCl, рН 8,0, 1 мМ DDT и использовали для термоинактивации и спектральных исследований.
Вводили замещения D357L, Т, V, W, R, I, S, К, N и F. Данные мутанты были охарактеризованы в исследованиях сырых экстрактов in vitro на термоинактивацию.
Частично очищенные экстракты разбавляли, 1 в 11, в буфере для термоинактивации: 50 мМ калиевый фосфатный буфер, рН 7,8, содержащий 10% насыщенного сульфата аммония, 1 мМ дитиотреитола и 0,2% BSA.
110 мкл аликвоты белкового раствора инкубировали при 4°С или 45°С в течение определенных периодов времени и охлаждали на льду перед анализом. Люциферазную активность затем определяли, как описано в примере 1, с использованием реагента для теста определения активности люциферазы Promega (1 в 2 разведении).
Результаты представлены в таблицах 2 и 3 на фигуре 1. Значения t1/2 определяли в сырых экстрактах при 40°С (таблица 2) и 45°С (таблица 3).
Все проведенные замены повышали термостабильность по сравнению с рекомбинантным диким типом.
Пример 3
Изменения длины волны испускаемого света
Также наблюдали, что замены аминокислот в положении 357 оказывают воздействие на спектр испускаемого ферментом света in vivo. Аликвоту (250 мкл) клеточных культур Е.coli, как описано в примере 2, культивировали в течение ночи при 37°С, центрифугировали в микроцентрифуге и супернатант удаляли. Клетки, экспрессирующие различные мутантные люциферазы, инкубировали в цитратном буфере (рН 5,0), содержащем 150 мкл D-люциферина, и анализировали свет, испускаемый в результате реакции, in vivo при определении спектров испускания с использованием спектрофлуориметра для микропланшетов SPECTRAmax® (Molecular Devices Corp. Калифорния, США). Для мутантов D357Y, F и I, наблюдали значительные изменения спектрального пика, а также распределения длин волн (фигура 2(а)-(g)). Эти результаты обобщены в таблице 4 ниже.
Кроме того, оценивали глазом в темной комнате люминесценцию мутантов in vivo. Мутанты D357 показывали различные цвета их спектров люминесценции. В частности, D357Y, F и I, показывали значительные сдвиги в сторону более длинных волн испускаемого света.
Оказалось, что в некоторых случаях (например, D357F) изменение цвета светового испускания имело место не только за счет сдвига в λmах, но также за счет различия вклада в спектр со стороны различных длин волн видимого света.
Рекомбинантный фермент дикого типа (r-wt) использовали для сравнения λmах световой испускания с некоторыми из мутантов 357 in vivo. D357Y, F и I, показывали значительные сдвиги в их максимальной длине волны.
Пример 4
Свойства фермента в присутствии или отсутствии СоА
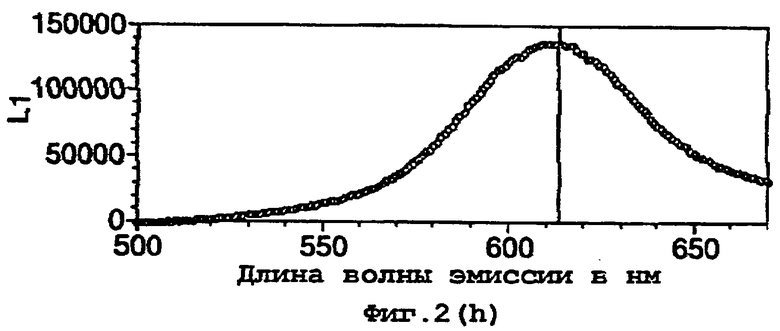
D357Y частично очищали преципитацией с сульфатом аммония, как описано в примере 1. Данный частично очищенный фермент D357Y (5 мкл) смешивали с 150 мкл реагента для теста определения активности люциферазы Promega. Другую аликвоту смешивали с эквивалентным буфером для анализа, в котором отсутствовал СоА (25 мМ Трис-трицин, рН 7,8, 5,0 мм MgSO4, 0,1 мМ ЭДТА, 2 мМ DTT, 470 мкм D-люциферина, 530 мМ АТФ). Снимали спектры испускания двух реакций, и они представлены на фигурах 9 и 10.
Спектры показывают выраженное различие в биолюминесцентной испускания в отсутствии и присутствии СоА со значительным сдвигом в λmax. Воздействие СоА на кинетику люциферазной реакции также можно видеть по различию в величинах RLU. (RLU - относительные световые единицы).
Данное различие в испускании дало основание для возможности использования фермента в тесте для обнаружения присутствия СоА.
Пример 5
Получение и свойства двойного мутанта
С использованием направленного на сайт мутагенеза, как описано в примере 2, конструировали двойной мутант E354I+D357Y для того, чтобы изучить любые кумулятивные эффекты на термостабильность и цвет светового испускания.
Частично очищенный двойной мутант E354I+D357Y разбавляли 1 к 11 в буфере для термоинактивации: 50 мМ калиевый фосфатный буфер, рН 7,8, содержащий 10% насыщенного сульфата аммония, 1 мМ дитиотреитола и 0,2% BSA.
110 мкл аликвоты белкового раствора инкубировали при 45°С в течение определенных периодов времени и охлаждали на льду перед анализом. Затем определяли активность люциферазы, как описано ранее, с использованием реагента для определения активности люциферазы Promega (1 в 2 разведении).
Двойной мутант показывал заметное повышение термостабильности по сравнению с одиночными мутантами E354I и D357Y по отдельности (смотри фигуру 3). Исследования с термоинактивацией частично очищенного двойного мутанта подтвердили повышенную термостабильность мутанта со значением t1/2, равным 7,7 мин при инактивации при 45°С.
Было отмечено, что двойной мутант показывает значительно более интенсивный красный цвет люминесценции, чем отдельные мутанты E354I и D357Y, показывая аддитивность цвета люминесценции.
Также были сняты спектры испускания рекомбинантного дикого типа и сырого экстракта двойного мутанта E354I+D357Y с использованием буфера для анализа, описанного в примере 3.
Спектры испускания, снятые in vivo, показывали λmax при 611 нм. Однако спектр имел значительно больший вклад люминесценции из красной области длин волн, приводя к ее более интенсивному красному виду при визуальной оценке глазом. Спектры испускания сырых экстрактов показывали определенное изменение спектральной формы и сдвиг длин волн, равный 44 нм, по сравнению с rWT (смотри фигуру 4).
Спектр испускания двойного мутанта in vivo показывал как заострение ширины полосы максимальной длины волны испускаемого света (613 нм), так и уменьшение вклада со стороны длин волн света, находящихся в области 540-560 нм.
Выраженное воздействие данных мутаций указывает на важность данной области фермента для цвета биолюминесцентного света.
Пример 6
Усиленный фотонный поток
Наблюдали, что биолюминесценция клеток Е.coli, экспрессирующих мутант D357K, является очень яркой по сравнению с другими мутантами по данному положению. Кинетику свечения данного фермента анализировали с использованием люминометра, с помощью которого можно определить интенсивность фотонного испускания в течение времени. Аликвоты не содержащих клеток экстрактов Е.coli, содержащих рекомбинантный фермент дикого типа или мутант D357, добавляли к смеси для теста определения активности люциферазы, которая не включала каких-либо реагентов, способствующих свойственной личинке светляка кинетике фермента, например кофермент А. Наблюдали, что скорость затухания фотонного испускания, определенная в течение времени (15 с) для обоих ферментов, была значительно ниже для мутанта D357K (фигура 4). Другими словами, мутантный фермент имеет кинетику реакции, которая ингибируется в меньшей степени в течение по меньшей мере первых 15 с реакции по сравнению с рекомбинантным ферментом дикого типа.
Пример 7
Комбинаторный мутагенез кассеты в положениях Е354 и D357
Стадия 1
Конструирование плазмиды pPW601aJ54 для мутагенеза кассеты
Два новых уникальных сайта рестрикции встраивали в ген luc в плазмиде pPW601a/J54 с использованием двух пар синтетических олигонуклеотидов (смотри ниже). В целом с помощью шести молчащих мутаций ввели в ген сайты рестрикции Spel и Kpnl, 63 пары оснований по отдельности. Плазмиду, включающую данные новые сайты, назвали pPW601aJ54SpeI/KpnI. Наличие и пространственная близость данных сайтов рестрикции сделали возможным применять комбинаторный мутагенез кассеты для исследования воздействия случайных замен аминокислот в положениях 354 и 357 в первичной последовательности Рhоturis-люциферин-4-монооксигеназы.
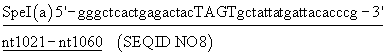
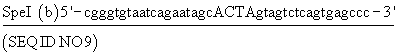
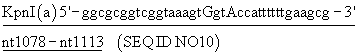
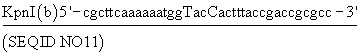
Нуклеотиды, отмеченные в конструкции, образуют сайт узнавания эндонуклеозы и в верхнем случае положение точечных мутаций, необходимых для создания сайта.
Стадия 2
Конструирование кассеты и конструкция библиотек
Синтезировали пару синтетических олигонуклеотидов, которые при отжиге давали двухцепочечную кассету, которую можно было непосредственно лигировать в плазмиду pPW601aJ54SpeI/KpnI, расщепленную по новым сайтам рестрикции. Кассету построили для введения всех возможных комбинаций 20 имеющихся в естественных условиях аминокислот, в положения 354 и 357 в первичной последовательности.
Петлевая библиотека 2А
5’-ctagtgctattctgattacacccNNG/CggggatNNG/Caaaccgggcgcggtcggtaaagtggta-3’
(SEQ ID NO 12)
Петлевая библиотека 2В
5’-cactttaccgaccgcgcccggtttG/CNNatccccG/CNNgggtgtaatcagaatagca-3’
(SEQ ID NO 13)
2 мкг каждого из петлевых библиотечных олигонуклеотидов смешивали в буфере, содержащем 50 мМ Трис-HCl, рН 7,4, 25 мМ NaCl, и нагревали до 100°С в течение 3 мин. Данный раствор медленно охлаждали в нагревательном блоке до <50°С для отжига комплементарных последовательностей. Подвергнутые отжигу олигонуклеотиды затем лигировали в плазмиду pPW601aJ54SpeI/KpnI, которую расщепляли Spel и Kpnl. Затем аликвоты реакционной смеси при лигировании использовали для трансформации клеток Е.coli HB101 электропорацией. После электропорации трансформированные клетки высевали на чашки с агаром LB, содержащим 50 мкг/мл ампициллина, и культивировали в течение ночи при 37°С. На следующий день произвольно отбирали с чашек 869 колоний и использовали для посева в 1 мл LB, содержащей ампициллин, в 96-луночных планшетах (Beckman). Планшеты накрывали и клетки культивировали в течение ночи при 37°С при встряхивании.
Стадия 3
Скрининг произвольно выбранных клонов в условиях in vivo
На следующее утро 50 мкл аликвоты ночных культур в стационарной фазе роста переносили в два прозрачных пластмассовых с круглым дном 96-луночных планшета для микротитрования (Dynex). Один планшет покрывали и инкубировали в нагревательном блоке в течение 8 мин (температура поверхности блока 45°С), в то время как другой содержали при 37°С. Затем определяли и записывали активность люциферазы в клетках из обоих планшетов в условиях in vivo при комнатной температуре при добавлении в лунки 50 мкл 100 мМ натриевого цитратного буфера, рН 5,0, содержащего 0,5 мМ D-люциферина и затем при переносе планшета в систему захвата с видеокамерой (Alpha Imager). Свет, испускаемый нагретыми и контрольными культурами, интегрировали в течение 1 или 2 мин и изображение записывали на термическую бумажную пленку.
Для второго этапа скрининга отбирали семьдесят девять культур, показывающих наиболее высокую биолюминесцентность, что определяли по яркости записанного на пленку изображения. На этот раз культуры инкубировали в течение 16 мин в нагревательном блоке перед постановкой анализа. Из 55 клонов, отобранных по результатам скрининга на термостабильность in vivo, отобрали 25 для проведения спектрального анализа in vivo. Данные клоны культивировали в течение ночи в среде LB при 37°С и на следующее утро 200 мкл ночных культур центрифугировали, и клеточные осадки Е.coli ресуспендировали в 150 мкл 100 мМ натриевого нитратного буфера, рН 5,0, содержащего 0,5 мМ D-люциферина. Затем ресуспендированные клетки помещали в белый пластмассовый планшет для микротитрования и анализировали в условиях in vivo спектр биолюминесцентной испускания, испускаемой каждой из мутантных люцифераз с использованием флуориметра для 96-луночных планшетов Molecular Devices Spectramax. Результаты обобщены в таблице 1 ниже.
Стадия 4
Идентификация мутантов
Плазмидную ДНК готовили из 25 клонов, отобранных при скрининге in vivo, и секвенировали с использованием специфических праймеров для секвенирования. Идентифицировали мутации, приводящие к аминокислотным заменам в положениях 354 и 357 в первичной последовательности. Один мутант также включал дополнительную мутацию, полученную в результате замены аминокислоты в положении 1351 (таблица 5).
где rWT означает рекомбинант дикого типа.
Ряд мутантных люцифераз отбирали на основе результатов анализа в условиях in vivo на термостабильность. Большинство данных люцифераз также показывает значительные изменения в спектре испускаемого света in vivo во многих случаях с большим вкладом со стороны более длинных длин волн света (>580 нм). Ряд спектров также показывал существенное сужение ширины полосы вокруг единственного пика при 610-614 нм.
Замены Е354 и D357 соответственно гидрофобной и ароматической аминокислотой, например E354V, D357Y, приводят к самому большому изменению в спектре in vivo, который показывает один пик с узкой шириной полосы, около 612 нм.
Пример 8
Скрининг в условиях in vitro на термостабильность
Готовили лизисом не содержащие клеток экстракты отобранных клонов и определяли термостабильность люциферазы из каждого экстракта в опыте по термической инактивации. 50 мкл каждого экстракта помещали в пробирку Эппендорфа и инкубировали на водяной бане, нагретой до 45°С, в течение 4, 9 и 16 мин. В соответствующей временной точке отбирали аликвоту и определяли остающуюся активность люциферазы. В таблице 6 показан процент остающейся активности по отношению ко времени для всех мутантных ферментов, а также рекомбинантных ферментов дикого типа.
где “nd” означает не делали.
Данные показывают, что наиболее термостабильными люциферазами были те, которые имели ароматическую аминокислоту в положении 357 (Y, F или W) и крупную гидрофобную (V или I), полярную (N) или положительно заряженную (К и R) аминокислоту в положении 354.
Пример 9
Воздействие условий культивирования на спектр испускаемого света in vivo
Оценивали воздействие различных источников углерода на спектр света, испускаемого клетками Е.coli BL21(DE3), экспрессирующими мутантные люциферазы D357Y или Е354К+D357M (7 выше).
50 мл культуры клеток инкубировали до фазы логарифмического роста на среде LB и затем собирали центрифугированием. Клеточный осадок ресуспендировали в 1 мл стерильной дистиллированной воды и затем 100 мкл аликвоту данной суспензии использовали для посева в 5 мл свежей LB, минимальной среды М9+2 мМ ацетата натрия или минимальной среды + 2 мМ глюкозы в стерильной пробирке емкостью 25 мл. Культурам давали расти при 37°С при встряхивании и через 90 мин (D357Y) или 120 мин (фермент 7) отбирали 200 мкл аликвоту клеток, центрифугировали и ресуспендировали в 150 мкл 100 мМ натриевого цитратного буфера, рН 5,0, содержащего 0,5 мМ D-люциферина. Затем ресуспендированные клетки переносили в планшет для микротитрования и анализировали спектр биолюминесцентной испускания, испускаемой каждой из мутантных люцифераз с использованием флуориметра для 96-луночных планшетов Molecular Devices Spectramax. Результаты представлены на фигурах 7 и 8.
Результаты показывают, что переключение от богатой среды (LB) (фигуры 7а, 8а) к определенной минимальной среде либо с ацетатом (фигуры 7b, 8b), либо с глюкозой (фигуры 7с, 8с) в качестве единственного источника углерода приводило к сдвигам к более длинным длинам волн испускаемого света и снижению вклада со стороны более коротких длин волн.
Пример 10
Очистка и спектральная характеристика рекомбинантной дикого типа и мутантной люцифераз
Рекомбинантный дикого типа фермент Photinus pyralis и мутантные люциферазы D357Y и E354I+D357Y очищали до гомогенности для того, чтобы анализировать воздействие кофактора кофермента А на спектр биолюминесцентной реакции. Все три люциферазы выделяли в виде слитых конструкций с модулем связывания углеводов из 143 аминокислот из анаэробного гриба Piromyces equii. Было показано, что данный СВМ селективно связывается с набухшей под действием кислоты целлюлозой и растворимыми углеводами галактоманнаном и глюкоманнаном, образуя основу для простой в одну стадию схемы аффинной очистки.
Люциферазы, слитые с СМВ, связывали с целлюлозой в сырых, не содержащих клеток экстрактах, промывали и затем селективно элюировали с использованием растворимых полисахаридов. Слитые белки, очищенные таким образом, использовали в тестах для определения длин волн испускаемого света в реакционных смесях, содержащих различные количества кофермента А. Фермент (5 мкл) добавляли к 100 мкл реагента для анализа, 25 мМ трис-трицин, рН 7,8, 5,0 мМ MgSO4, 0,1 мМ ЭДТА, 530 мкМ АТФ и 470 мкМ D-люциферина, содержащего различные количества кофермента А. На фигурах 9-11 показано воздействие возрастающих концентраций кофермента А на спектр света, испускаемого очищенными люциферазами D357 и E354I + D357Y.
При анализе in vivo спектра биолюминесцентного света, испускаемого клетками Е. coli, экспрессирующими Photuris-люциферин-4-монооксигеназу, слитую с С-концом грибкового СВМ, не было показано каких-либо существенных различий по сравнению с клетками, экспрессирующими нативную люциферазу. Аналогичным образом при анализе in vitro спектр биолюминесцентного света, испускаемого коммерческим источником очищенной рекомбинантной люциферазы (Promega), был идентичен спектру света, испускаемого слитым белком.
Наблюдаемые различия, следовательно, связаны с концентрациями СоА. По мере возрастания концентрации СоА менялось спектральное распределение, и при самых высоких концентрациях СоА в спектре преобладали длины волн в области 590-630 нм с выраженным пиком при 610 нм. Спектральный сдвиг наиболее выражен для двойного мутанта, где имеется значительное сужение ширины полосы около единственного пика при длине волны 610 нм (фигура 11).
Пример 12
Получение синтетической люциферазы Photinus pyralis, мутированной таким образом, что она имеет 214C/354K/357F
Был сконструирован и собран синтетический ген luc из пары олигонуклеотидов с использованием стратегии синтеза, описанной выше. Последовательность гена была сконструирована для создания люциферазы с аминокислотами 214С, 354К и 357F.
Синтезировали двадцать девять пар перекрывающихся синтетических олигонуклеотидов с участием Sigma-Genosyc Ltd, очищали PAGE и лигировали в три упорядоченные структуры размером примерно 550 п.н. (IDRIS 1, 2 & 3, фигура 3). Каждую упорядоченную структуру затем по отдельности лигировали в вектор рВSК(+) и полученные конструкции использовали для трансформации клеток Е.coli XL1-Blue. Плазмидную ДНК готовили из клонов, включающих собранные вставки, и секвенировали для подтверждения привязанности ОРС. Присутствие n-1 олигонуклеотидов (побочные продукты олигосинтеза) в упорядоченных структурах осложняли процесс построения. Секвенирование ДНК выявило единственную правильную упорядоченную структуру IDRIS 2, и использовали ПЦР для поправки упорядоченной структуры IDRIS 3, которая включала делецию из одной пары оснований на 5’-конце конструкции. Упорядоченная структура полной ОРС достигалась лигированием смеси плазмид, включающих IDRIS 1 с IDRIS 2 и 3.
Затем лигированную ДНК использовали для трансформации клеток Е.coli XL1-Blue и отбирали клоны, экспрессирующие активный фермент с использованием анализа in vivo. Отбирали несколько клонов и секвенировали для подтверждения присутствия и привязанности синтетического гена luc, имеющего последовательность, представленную на фигуре 14. Полная ОРС была названа IDRIS (FA).
Синтетический ген встраивали в вектор pBSK(+) между сайтами ВаmН I и Sal I в полилинкер. В данном положении ген не находится в рамке считывания с альфапептидом и имеет значительную удаленность от промотора lac. Однако продуцировалось достаточное количество люциферазы для предварительной характеристики фермента. Готовили сырые не содержащие клеток экстракты Е.coli XL1-Blue, экспрессирующие IDRIS (FA), из ночных культур с использованием метода лизиса Promega.
Затем тестировали термостабильность фермента в экстракте при 50°С в течение 20 мин и сравнивали с термостабильным мутантом E354I+D357Y. Оптимизированный с новым кодоном тройной мутант был значительно более термостабильным по сравнению с мутантом E354I+D357Y (фигура 15).
| название | год | авторы | номер документа |
|---|---|---|---|
| МУТАНТНАЯ ЛЮЦИФЕРАЗА (ВАРИАНТЫ), ДНК, КОДИРУЮЩАЯ УКАЗАННУЮ ЛЮЦИФЕРАЗУ, И ВЕКТОР ДЛЯ ЭКСПРЕССИИ УКАЗАННОГО БЕЛКА | 1996 |
|
RU2210594C2 |
| ЛЮЦИФЕРАЗЫ | 1995 |
|
RU2192467C2 |
| РЕАГЕНТ ДЛЯ ОПРЕДЕЛЕНИЯ АДЕНОЗИН-5'-ТРИФОСФАТА | 2009 |
|
RU2420594C2 |
| НОВЫЕ ЛЮЦИФЕРАЗЫ И СПОСОБЫ ИХ ИСПОЛЬЗОВАНИЯ | 2017 |
|
RU2674894C2 |
| РЕАГЕНТ ДЛЯ ОПРЕДЕЛЕНИЯ АДЕНОЗИН-5'-ТРИФОСФАТА | 2004 |
|
RU2268944C2 |
| УКОРОЧЕННАЯ МУТАНТНАЯ ЛЮЦИФЕРАЗА ИЗ Metridia longa ДЛЯ ПРИМЕНЕНИЯ В КАЧЕСТВЕ БИОЛЮМИНЕСЦЕНТНОГО РЕПОРТЕРА В ЖИВЫХ КЛЕТКАХ | 2012 |
|
RU2495929C1 |
| МУТАНТНАЯ ОКСИДАЗА D-АМИНОКИСЛОТ | 2009 |
|
RU2412998C1 |
| МУТАНТНЫЙ ФОТОПРОТЕИН (ВАРИАНТЫ) ДЛЯ ОПРЕДЕЛЕНИЯ ВНУТРИКЛЕТОЧНОГО КАЛЬЦИЯ ОДНОВРЕМЕННО В РАЗНЫХ ОРГАНЕЛЛАХ КЛЕТКИ | 2008 |
|
RU2402566C2 |
| МУТАНТНЫЕ ОКСИДАЗЫ D-АМИНОКИСЛОТ | 2007 |
|
RU2362806C2 |
| ТЕРМОСТАБИЛЬНЫЕ МУТАНТЫ ФЕРМЕНТОВ БИОСИНТЕЗА КРАХМАЛА | 2002 |
|
RU2303633C2 |












Изобретение относится к области генной и белковой инженерии и может быть использовано в биолюминесцентных анализах различного назначения. Получены новые мутантные формы люциферазы, характеризующиеся повышенной по сравнению с соответствующими ферментами дикого типа термостабильностью и в ряде случаев изменением длины волны испускаемого света. Все предложенные мутеины отличаются тем, что природный аминокислотный остаток в положении, соответствующем положению 357 в последовательности люциферазы Photinus pyralis, заменен другим, предпочтительно остатком незаряженной полярной аминокислоты, в частности тирозина. Приобретаемые мутантными люциферазами по изобретению новые свойства позволяют эффективно использовать их в качестве репортерного агента в различных аналитических системах. 6 с. и 17 з.п. ф-лы, 15 ил., 6 табл.
(a) замену аминокислотного остатка, соответствующего аминокислоте 354 люциферазы Photinus pyralis (356 в люциферазе Luciola);
(b) замену аминокислотного остатка, соответствующего положению 215 в люциферазе Photinus pyralis (217 в люциферазе Luciola), другой гидрофобной аминокислотой или
(c) замену аминокислотного остатка, соответствующего остатку 214 в люциферазе Photinus pyralis или остатку 216 люциферазы Luciola mingrelica, Luciola cruciata или Luciola lateralis;
(d) замену аминокислотного остатка, соответствующего остатку 232 в люциферазе Photinus pyralis или остатку 234 люциферазы Luciola mingrelica, Luciola cruciata или Luciola lateralis;
(e) замену аминокислотного остатка, соответствующего остатку 295 в люциферазе Photinus pyralis или остатку 297 люциферазы Luciola mingrelica, Luciola cruciata или Luciola lateralis;
(f) замену аминокислотного остатка, соответствующего аминокислоте 14 люциферазы Photinus pyralis или остатку 16 люциферазы Luciola mingrelica или остатку 17 люциферазы Luciola cruciata или Luciola lateralis;
g) замену аминокислотного остатка, соответствующего аминокислоте 35 люциферазы Photinus pyralis или остатку 37 люциферазы Luciola mingrelica или остатку 38 люциферазы Luciola cruciata или Luciola lateralis;
(h) замену аминокислотного остатка, соответствующего аминокислотному остатку 105 люциферазы Photinus pyralis или остатку 106 люциферазы Luciola mingrelica, остатку 107 люциферазы Luciola cruciata или остатку 108 Luciola lateralis;
(i) замену аминокислотного остатка, соответствующего аминокислотному остатку 234 люциферазы Photinus pyralis или остатку 236 люциферазы Luciola mingrelica, Luciola cruciata или Luciola lateralis;
(j) замену аминокислотного остатка, соответствующего аминокислотному остатку 420 люциферазы Photinus pyralis или остатку 422 люциферазы Luciola mingrelica, Luciola cruciata или Luciola lateralis;
(k) замену аминокислотного остатка, соответствующего аминокислотному остатку 310 люциферазы Photinus pyralis или остатку 312 люциферазы Luciola mingrelica, Luciola cruciata или Luciola lateralis.
| WO 9914336 А, 25.03.1999 | |||
| WHITE P.J | |||
| ET AL., Biochemical journal, v.319, р.343-350, 1996 | |||
| WO 9525798 A, 28.09.1995. |

