Область техники, к которой относится изобретение
Изобретение относится к средствам для лечения заболевания и, в частности, для лечения посредством активации регуляторных T-клеток CD4+CD25+ с участием рецептора поверхностного рецептора T-клеток CD4. Изобретение относится к способам скрининга для идентификации таких средств, к средствам, способным активировать регуляторные T-клетки CD4+CD25+, и к их применению для лечения заболевания, в частности аутоиммунных заболеваний, а также в способах, осуществляемых in vitro.
Уровень техники
T-клетки относятся к лимфоцитам и ответственны за ряд ключевых функций в иммунной системе. У млекопитающих T-клетки (тимоциты) дифференцируются в тимусе из гематопоэтических клеток-предшественников, образованных в костном мозге. Частью процесса дифференцировки является экспрессия поверхностных рецепторов, главным образом гликопротеидов CD4 и CD8. T-клетки, экспрессирующие CD4, так называемые T-клетки CD4+, связывают комплексы MHC II (Reinerz and Schlossman, Cell 19, 821-827 (1980); Reinerz et al., PNAS USA 77, 1588-1592 (1980)), тогда как T-клетки CD8+ связывают комплексы MHC класса I (Fitch, Microbiol. Rev. 50, 50-69 (1986)). T-клетки высвобождаются в кровь и лимфу.
Позитивные CD4-клетки могут дифференцироваться в субпопуляции T-хелперов (Th1 и Th2), а также в регуляторные T-клетки. Регуляторные T-клетки можно дополнительно разделить на подклассы, при этом наиболее охарактеризованы происходящие из тимуса (nTreg) и индуцированные T-клетки (iTreg).
Хотя существуют другие субпопуляции Treg, такие как, например, Tr1 или Th3, настоящее изобретение относится к CD4-позитивным образуемым в тимусе Treg (nTreg) и индуцированным Treg, при этом оба типа экспрессируют фактор транскрипции Foxp3. В качестве основного отличия Foxp3 стабильно и непрерывно экспрессируется в nTreg, поддерживая необратимый фенотип Treg, тогда как в индуцированных Treg наблюдается индуцируемая или временная экспрессия Foxp3, которая является обратимой.
Treg секретируют иммуномодулирующие цитокины, такие как IL-10, TGF-бета или IL-35, и проявляют супрессорную активность по отношению к эффекторным T-клеткам посредством нескольких механизмов, например, посредством супрессии продукции провоспалительных цитокинов, прямого межклеточного контакта и модулирования состояния активации или функции антигенпрезентирующих клеток (АПК) (Shevach et al. Immunity (2009) 30; 636-645). Основным характерным признаком CD4-позитивных CD25 Treg-клеток является их анергический фенотип, означающий, что они не пролиферируют при стимуляции TCR, что может быть восстановлено при добавлении экзогенного IL-2.
Главная роль Treg состоит в поддержании гомеостаза в отношении иммунных реакций и аутотолерантности. Дисфункция Treg коррелирует с аутоиммунными заболеваниями.
Обычно регуляторные T-клетки могут быть выделены благодаря гликопротеинам поверхностных рецепторов CD4, CD25 и охарактеризованы по внутриклеточному окрашиванию FOXP3. Дополнительным поверхностным белком является CD127 (IL-7 R), который подвергается понижающей регуляции в клетках Treg и может быть использован для дальнейшей очистки Treg. Кроме того, экспрессия CD39 (эндонуклеотидазы) (Borselino et al., Blood (2007) 110, 1225-1232) или GARP (преобладающие повторы гликопротеина A (GARP или LRRC32) (Wang et al., PNAS (2009) 106, 32, 13439-13444).
CD4 человека кодируется хромосомой 12 и относится к надсемейству иммуноглобулинов (Ig). Его природная функция в качестве поверхностного рецептора T-клеток связана с активацией T-клеток при связывании MHC-комплексов класса II. Кроме того, CD4 может связывать белок gp120 ВИЧ-1, белок P4HB/CDI и капсидные белки вируса герпеса человека HHV-7. Также сообщалось о взаимодействии с белками gp120 и Vpu ВИЧ-1. CD4 содержит 458 аминокислот. Пептидная последовательность показана на фигуре 1.
База данных UniProt, вход P01730, выдает доменную структуру CD4, которая показана ниже в таблице 1 и на фигуре 6. Первые 25 аминокислот являются сигнальным пептидом, который отщепляется в биологически активной форме. Положения 26-396 составляют внеклеточный домен, за которым следует трансмембранная область, положения 397-418. Asn296 и Asn325 являются известными сайтами гликозилирования (Konig et al., J. Biol. Chem. 263, 9502-9507 (1988); Carr et al., J. Biol. Chem. 264, 21286-21295 (1989)).
Последняя часть, положения 419-458 относится к цитоплазматическому домену. В этой части находится сайт связывания тирозиновой протеинкиназы LCK (p56lck) (Rudd et al, PNAS USA 85, 5190-5194 (1988); Veillette et al., Cell 55, 301 (1988)), которая является частью пути передачи сигнала, активируемого лигандами, связывающимися с CD4.
Таблица, в которой показана доменная структура CD4 (согласно UniProt P01730)
Внеклеточная часть содержит 4 подобных иммуноглобулину домена. Первый домен, N-концевой домен, содержащий положения 26-125, является Ig-подобным доменом V-типа. На основе гомологии с антителами он имеет три гомологичных области, определяющие комплементарность антигену, CDR1, CDR2 и CDR3 (Ashkenazi et al., PNAS USA 87, 7150-7154 (1990)) (см. фигуру 6). Участки CDR1 и CDR2 вовлечены в связывание молекул MHC класса II (Moebius et al., PNAS USA 89, 12008-120012 (1992)), белка оболочки ВИЧ-1 gp120 (Moebius et al., J. Exp. Med. 176, 507-517 (1992)) и анти-CD4-антител (Lanza et al., PNAS USA 90, 11683-11687 (1993)). Phe68 в CDR2 играет ключевую роль в распознавании и связывании молекул MHC класса II и белка оболочки ВИЧ-1 gp120 (Sharma et al., Biochemistry 44, 16192-16202 (2005)). Все известные лиганды CD4 связываются с N-концевым Ig-подобным доменом V-типа.
Механизм того, как работают регуляторные T-клетки, полностью не выяснен. Treg CD4+CD25+ ингибируют поликлональную и антигенспецифичную активацию T-клеток. Супрессия может быть опосредована, например, зависимым от клеточных контактов механизмом, который регулирует активацию Treg CD4+CD25+ через TCR, но Treg не проявляют пролиферативного ответа при активации TCR или стимуляции митогенными антителами (анергические) (Shevach, Nature Rev. Immunol. 2: 389 (2002). После стимуляции они становятся компетентными в отношении супрессии независимым от антигена образом ответа T-клеток CD4+ и T-клеток CD8+, а также ингибирования активации B-клеток и клональной экспансии.
Способность регуляторных T-клеток CD4+CD25+ оказывать регулирующее влияние на активность иммунной системы натолкнула на мысль о признании их в качестве потенциальной мишени для лечения заболеваний, таких как аутоиммунные заболевания, при которых требуется контролировать иммунную систему.
Аутоиммунитет означает неспособность организма распознавать собственные составляющие части (вплоть до субмолекулярных уровней) как «свое», что приводит к иммунной реакции против его собственных клеток и тканей. Любое заболевание, которое возникает в результате такой аномальной иммунной реакции, называют аутоиммунным заболеванием. Аутоиммунные заболевания включают рассеянный склероз (РС), ревматоидный артрит (РА), псориаз, псориатический артрит, язвенный колит, болезнь Крона, сахарный диабет типа I (T1D), тяжелую псевдопаралитическую миастению (MG), аутоиммунный полигландулярный синдром типа II (APS-II), тиреоидит Хашимото (HT), системную красную волчанку (СКВ), синдром Шегрена и аутоиммунный лимфопролиферативный синдром (ALS).
Аутоиммунное заболевание возникает, когда T-клетки распознают и реагируют на «собственные» молекулы, то есть молекулы, продуцируемые клетками хозяина. Активация «аутореактивных» T-клеток в результате презентации аутоантигенов, процессированными антигенпрезентирующими клетками (АПК), приводит к их клональной экспансии и миграции к специфичным тканям, где они индуцируют воспаление и разрушение ткани.
Супрессия такой функции эффекторных T-клеток с использованием иммуносупрессивных лекарственных средств является основной терапевтической методикой, которую успешно применяли для лечения аутоиммунных заболеваний. Однако такие лекарственные средства индуцируют общую иммунную супрессию вследствие их плохой избирательности, приводя к ингибированию не только опасных функций иммунной системы, но также и полезных функций. Как следствие могут возникать некоторые риски, подобные инфекции, злокачественной опухоли и токсичности лекарственных средств.
В общем, установлено, что T-клетки CD4+ играют основную роль в инициации и поддержании аутоиммунитета. Соответственно, было предложено применение мАт против поверхностных молекул Т-клеток CD4+ и, в частности, анти-CD4-мАт, в качестве иммуносупрессивных средств. Хотя многочисленные клинические исследования подтвердили потенциальный интерес к такому подходу, они также подняли несколько проблем, которые необходимо учитывать, чтобы получать анти-CD4-мАт, более подходящие для применения в обычной клинической практике.
Предложено несколько разных механизмов действия CD4-мАт, включая: (1) антагонизм взаимодействий CD4-MHC II, приводящий к ингибированию активации T-клеток, (2) модулирование рецептора CD4, которое определяют по уменьшению экспрессии CD4 на поверхности клеток, (3) частичная передача сигнала через рецептор CD4 в отсутствие перекрестного связывания T-клеточных рецепторов, что может супрессировать последующую активацию T-клеток и запускает апоптозную гибель T-клеток CD4, (4) Fc-опосредованная зависимая от комплемента цитотоксичность (CDC) или опосредованная антителами клеточная цитотоксичность (ADCC), приводящая к истощению T-клеток CD4, и (5) стимуляцию регуляторных T-клеток.
Несколько анти-CD4-антител, мишенью которых являются T-клетки проходят клинические испытания (Schulze-Koops et al., J Rheumatol. 25(11): 2065-76 (1998); Mason et al., J Rheumatol. 29(2): 220-9 (2002); Choy et al., Rheumatology 39(10): 1139-46 (2000); Herzyk et al., Infect Immun. 69(2): 1032-43 (2001); Kon et al., Eur Respir J. 18(1): 45-52 (2001); Mourad et al., Transplantation 65(5): 632-41 (1998); Skov et al., Arch Dermatol. 139(11): 1433-9 (2003); Jabado et al., J Immunol. 158(1): 94-103 (1997)), главным образом, с целью истощения популяции клеток CD4 с использованием только некоторых CD4-антител, которым приписывают другие механизмы, подобных TRX-1, TNX-355, IDEC-151, OKTcdr4A.
Способ применения средств, направленных на активацию регуляторных T-клеток, для терапии аутоиммунных заболеваний признан чрезвычайно сложным. Активация Treg через TCR с использованием агонистического анти-CD3-антитела OKT-3 (Abramowicz et al., N. Engl. J Med. 1992 Sep 3; 327(10): 736) или через костимулирующую молекулу CD28 с использованием суперагонистического анти-CD28-антитела TGN 1412 приводит к полному истощению популяции регуляторных T-клеток, а также других обычных T-клеток и к системной индукции и высвобождению избыточных количеств провоспалительных цитокинов, включая IFN-γ, TNF-α, IL-1 и IL-2, приводя к клинически выраженному синдрому высвобождения цитокинов (CRS) у человека (Suntharalingam et al., N. Engl. J Med. 2006 Sep 7; 355(10): 1018-28).
Однако недавно в WO2004/083247 были описаны гуманизированные анти-CD4-антитела, которые способы активировать регуляторные T-клетки CD4+CD25+. Антитела, описанные в WO2004/083247, представляют собой гуманизированные варианты антитела mB-F5 мыши, IgG1 мыши против CD4 человека, описанного Racadot et al. (Clin. Exp. Rheum., 10, 365-374 (1992)). Эпитоп mB-F5 сообщен в публикации Racadot et al., как охватывающий Ig-подобные домены C2 типа 1 и типа 2 CD4 человека от аминокислоты 162 до аминокислоты 232, который показан на фигуре 6.
Последующие клинические испытания, описанные в WO2009/112502, WO2009/121690, WO2009/124815 и WO2010/034590, с использованием таких антител, названных BT061 (гуманизированные моноклональные IgG1) привели к успешному лечению пациентов, страдающих от псориаза и ревматоидного артрита, и стали доказательством того, что такими антителами можно лечить аутоиммунные заболевания безопасно и с высокой эффективностью.
Достигнутые многообещающие клинические результаты повысили интерес к получению дополнительных терапевтических средств, обладающих сходными свойствами. Поэтому целью настоящего изобретения являются способы скрининга для идентификации таких средств и получение дополнительных терапевтических средств.
Соответственно, настоящее изобретение относится к способу скрининга молекулы, способной связываться с CD4, включающему:
(a) получение одной или нескольких молекул-кандидатов;
(b) определение того способны ли одна или несколько молекул-кандидатов связываться с одной или несколькими из следующих областей CD4 человека: аминокислотами 148-154, аминокислотами 164-168 и аминокислотами 185-192; и
(c) отбор молекулы, определенной на стадии (b) как способной связываться с CD4.
Авторы настоящего изобретения неожиданно обнаружили, что гуманизированное антитело BT061 связывается с доменом CD4, который ранее не был выявлен в качестве сайта связывания лиганда. Такое открытие особенно неожиданно с учетом того, что было известно в данной области в качестве эпитопа для антитела mB-F5 мыши, из которого получено BT061. Авторы настоящего изобретения также установили остатки BT061, которые вовлечены в связывание молекулы CD4, и неожиданно обнаружили, что не все CDR BT061 вовлечены в связывание CD4.
Идентификация области связывания и подробностей механизма связывания позволила разработать дополнительные способы скрининга и антитела и фрагменты антител, способные активировать регуляторные T-клетки CD4+CD25+.
Соответственно, настоящее изобретение также относится к способу скрининга антитела или фрагмента антитела, способного связываться с CD4, включающему:
(a) получение антитела или фрагмента антитела, содержащего CDR1 и CDR2 легкой цепи BT061 и CDR1 и CDR3 тяжелой цепи BT061, необязательно, с аминокислотными заменами в последовательностях CDR, при условии, что:
(i) CDR1 легкой цепи содержит: Ser32, Gly33 и Tyr 34;
(ii) CDR2 легкой цепи содержит: Leu54 и Ile57;
(iii) CDR1 тяжелой цепи содержит Asp31, Glu31, Thr31, Cys31, Pro31, Met31 или Tyr31; и
(iv) CDR3 тяжелой цепи содержит Tyr103, Phe103 или His103; Arg104; Tyr105; Asp106 и Tip110, Phe110, His110 или Tyr110;
(b) определение того, способно ли антитело или фрагмент антитела связываться с CD4, и
(c) отбор антитела или фрагмента антитела, определяемого на стадии (b) как способного к связыванию с CD4,
при этом антитело или фрагмент антитела не содержит CDR1, CDR2 и CDR3 тяжелой цепи BT061 и CDR1, CDR2 и CDR3 легкой цепи BT061.
Кроме того, настоящее изобретение относится к антителу или фрагменту антитела, способному активировать регуляторные T-клетки CD4+CD25+, включая антитело или фрагмент антитела, способный активировать регуляторные T-клетки CD4+CD25+, содержащий CDR1 и CDR2 легкой цепи BT061 и CDR1 и CDR3 тяжелой цепи BT061 необязательно с аминокислотными заменами в последовательностях CDR, при условии, что:
(i) CDR1 легкой цепи содержит: Ser32, Gly33 и Tyr34;
(ii) CDR2 легкой цепи содержит: Leu54 и Ile57;
(iii) CDR1 тяжелой цепи содержит Asp31, Glu31, Thr31, Cys31, Pro31, Met31 или Tyr31; и
(iv) CDR3 тяжелой цепи содержит Tyr103, Phe103 или His103; Arg104, Tyr105, Asp106; и Trp110, Phe110, His110 или Tyr110,
и при этом антитело или фрагмент антитела не содержит CDR1, CDR2 и CDR3 тяжелой цепи BT061 и CDR1, CDR2 и CDR3 легкой цепи BT061.
Изобретение будет только проиллюстрировано посредством примера со ссылкой на следующие чертежи.
На фигуре 1 показана пептидная последовательность (SEQ ID NO:1) и дисульфидные мостики CD4 человека (UniProt ID P01730).
На фигуре 2A показана пептидная последовательность (SEQ ID NO:2) легкой цепи гуманизированного антитела BT-061. Остатки CDR показаны в прямоугольниках (CDR1: SEQ ID NO:4, CDR2: SEQ ID NO:5 и CDR3: SEQ ID NO:6). Остатки, окруженные пунктирными рамками не представлены кристаллической структурой.
На фигуре 2B показана пептидная последовательность (SEQ ID NO:3) тяжелой цепи гуманизированного антитела BT-061. Остатки CDR показаны в прямоугольниках (CDR 1: SEQ ID NO:7, CDR2: SEQ ID NO:8 и CDR3: SEQ ID NO:9). Остатки, окруженные пунктирными рамками не представлены кристаллической структурой.
На фигуре 3 представлено изображение асимметрической единицы кристаллической структуры CD4-BT061.
На фигуре 4 представлено изображение кристаллической структуры CD4-BT061, совмещенной с кристаллической структурой комплекса CD4 с молекулой MHC класса II (PDB, код 1JL4).
На фигуре 5 представлено изображение кристаллической структуры CD4-BT061, совмещенной с кристаллической структурой комплекса CD4 с белком ВИЧ-1 gp120 (PDB, код 2NY1). Последний, кроме того, связан с антителом 17b.
На фигуре 6 показана пептидная последовательность (SEQ ID NO:1) и доменная структура CD4 человека (Uniprot ID P01730).
На фигуре 7 представлено изображение сайта связывания BT061 на поверхности CD4. Все показанные аминокислоты являются частью Ig-подобного домена С2-типа 1, за которым следует N-концевой Ig-подобный домен V-типа.
На фигуре 8 представлена аминокислотная последовательность легкой цепи BT061 (SEQ ID NO:2). Аминокислоты, вовлеченные в связывание с CD4, указаны круглыми рамками.
На фигуре 9 представлена аминокислотная последовательность тяжелой цепи BT061 (SEQ ID NO:3). Аминокислоты, вовлеченные в связывание с CD4, указаны круглыми рамками.
На фигуре 10 представлено изображение сайта связывания BT061 на поверхности CD4. Аминокислоты, образующие карман связывания для Tyr105 тяжелой цепи BT061, обведены кругом.
На фигуре 11 представлено изображение сайта связывания BT061 на поверхности CD4. Аминокислоты, образующие карман связывания для Arg104-Asp106 тяжелой цепи BT061, обведены кругом.
На фигуре 12 представлено изображение сайта связывания BT061 на поверхности CD4. Аминокислоты, образующие карман связывания для Tyr34 легкой цепи BT061, обведены кругом.
Подробное описание изобретения
Способы скрининга
Настоящее изобретение относится к способам скрининга одной или нескольких молекул, способных связываться с CD4, и предпочтительно, с CD4 человека. Как указано выше, информация, приведенная в настоящей публикации, описывает взаимодействие между CD4 и антителом BT061, которое способно активировать регуляторные T-клетки CD4+CD25+. В частности, BT-061 связывается как с T-хелперными, так и регуляторными T-клетками и избирательно активирует регуляторные T-клетки без активации T-хелперных клеток. Знание структур BT061 и того, как они взаимодействуют с внеклеточной областью CD4 дает возможность конструировать и получать средства со свойствами, сходными со свойствами BT061 в отношении связывания CD4 и избирательной активации регуляторных T-клеток.
В первом аспекте настоящее изобретение относится к способу скрининга молекулы, способно связываться с CD4, включающему: (a) получение одной или нескольких молекул-кандидатов; и (b) определение того, способна ли одна или несколько молекул-кандидатов связываться с одной или несколькими из следующих областей или аминокислот CD4 человека: аминокислотами 148-154, аминокислотами 164-168 и аминокислотами 185, 187, 189, 190 и 192; (c) отбор молекулы, определенной на стадии (b) как способной связываться с CD4. Более конкретно, области CD4 человека представляют собой аминокислоты 148-154, аминокислоты 164-168 и аминокислоты 185-192.
В одном из вариантов осуществления стадии (a)-(c) могут быть осуществлены с использованием компьютерной системы, и взаимодействие между CD4 и одной или несколькими молекулами-кандидатами может быть смоделировано на основе информации, полученной при этом в отношении взаимодействия между BT061 и CD4 человека, т.е. скрининг осуществляют посредством выполняемого с помощью компьютера конструирования молекул. В частности, трехмерную структурную модель CD4, и в частности, его внеклеточной области, создают в компьютерной системе, используя взаимодействие аминокислотной последовательности по меньшей мере для части CD4, и компьютерную программу, известную специалисту в данной области, например, Discovery Studio (Accelrys®) или Benchware 3D Explorer (Tripos). Указанные программы, кроме того, позволяют осуществлять ввод или создание de novo последовательности и/или структурной информации об одной или нескольких молекулах-кандидатах. Затем можно исследовать способность молекулы-кандидата связываться с областями, важными для связывания BT061 (которые обсуждаются ниже).
В частности, способ скрининга молекулы, способной связываться с CD4, может включать стадии (a) и (b): (i) введение в компьютерную систему или программу аминокислотой последовательности CD4, содержащей по меньшей мере аминокислоты 148-154, 164-168 и кислоты 185, 187, 189, 190 и 192; (ii) создание трехмерной модели полипептида или пептида, кодируемого аминокислотной последовательностью; (iii) создание или введение трехмерной структуры одной или нескольких молекул-кандидатов; и (iv) имитация взаимодействия между аминокислотными последовательностями CD4 и молекулы-кандидата, чтобы определить, способна ли молекула-кандидат связываться с CD4 посредством аминокислот 148-154, 164-168 и аминокислот 185, 187, 189, 190 и 192.
Отбор молекул-кандидатов может быть дополнительно ограничен с учетом признаков взаимодействия CD4-BT061, описанных ниже в примере 1. В частности, молекулы-кандидаты могут быть ограничены молекулами, которые связываются с областями CD4 без солевых мостиков, и/или молекулами, которые содержат один или несколько компонентов, которые размещаются в одном или нескольких карманах для связывания на поверхности CD4, включая:
(i) аминокислотные остатки S152, V153, Q154, Q164, G165, T185, V186, L187 и K192 CD4 (которые показаны на фигуре 10);
(ii) аминокислотные остатки S150, S152, G165 и G166 CD4 (которые показаны на фигуре 11); и/или
(iii) аминокислотные остатки S150, P151, S152 и K167 (которые показаны на фигуре 12).
Способ согласно такому осуществляемому с помощью компьютера варианту может дополнительно включать стадию (d), на которой получают молекулу, отобранную на стадии (c). Например, когда отобранная молекула представляет собой пептид, аминокислотную последовательность пептида получают на основе данных компьютера и производят пептид in vitro. Затем активность отобранной молекулы можно оценить in vitro благодаря контакту отобранной молекулы с пептидом или полипептидом, содержащим соответствующие области/аминокислоты CD4, или благодаря контакту отобранной молекулы с клеткой, экспрессирующей CD4. Такие стадии in vitro дополнительно описаны ниже в связи с вариантом осуществления первого аспекта, в случае которого стадии (a)-(c) осуществляют in vitro.
В частности, в качестве альтернативы осуществляемому с помощью компьютера варианту, описанному выше, стадии (a)-(c) способа скрининга могут быть осуществлены in vitro, и стадия (b) может включать осуществление контакта одной или нескольких молекул-кандидатов с соответствующими областями/аминокислотами CD4 и определение того способна ли молекула-кандидат связываться с одной или несколькими такими областями. Пример такого варианта осуществления включает способ скрининга одной или нескольких молекул, способных связываться с CD4, включающий: (a) осуществление контакта одной или нескольких молекул с пептидом, содержащим одну или несколько из следующих областей CD4 человека: аминокислоты 148-154, аминокислоты 164-168 и аминокислоты 185-192; и (b) выявление того связывается ли одна или несколько молекул с одной или несколькими областями пептида,
при этом молекула не содержит CDR1, CDR2 и CDR3 тяжелой цепи BT061 и CDR1, CDR2 и CDR3 легкой цепи BT061.
Способ согласно настоящему изобретению, предпочтительно, включает скрининг библиотеки молекул. В частности, библиотека может представлять собой библиотеку в фаговом дисплее, полученную согласно способам, известным в данной области. Библиотека может представлять собой библиотеку пептидов, отражающую системное сочетание разных аминокислот/пептидов в большом количестве. Обычно библиотеку пептидов синтезируют на твердой фазе, главным образом на смоле, которая может быть изготовлена в виде плоской поверхности или шариков (твердофазный синтез пептидов). Такую библиотеку можно использовать для белок-белковых взаимодействий, обнаружения лекарственных средств, очистки белков и изменения последовательности распознавания антител, чтобы создавать варианты антител с разными аффиностями.
Кроме того, библиотеки кроме фагового дисплея включают дрожжевой дисплей, бактериальный дисплей, мРНК-дисплей, рибосомный и полисомный дисплей. Рибосомный дисплей основан на процессе, результатом которого являются транслированные белки, которые связаны с их мРНК-предшественником, который был использован, в виде комплекса для связывания с иммобилизованным лигандом на стадии отбора. мРНК-дисплей в результате дает транслированные пептиды или белки, которые связаны с их мРНК-предшественником посредством связи с пуромицином.
В случае методики презентирования в виде бактериального дисплея библиотека полипептидов, представляемых на поверхности бактерий, может быть подвергнута скринингу с использованием проточной цитометрии или многократных процедур селекции (биопэннинг).
В способе дрожжевого дисплея (Abbott) представляющий интерес белок презентирован в виде слияния с белком Aga2p на поверхности дрожжей. В природе белок Aga2p используется дрожжами для опосредования межклеточных контактов во время спаривания дрожжевых клеток. Как таковой, дисплей белка через Aga2p отодвигает белок от клеточной поверхности, минимизируя возможные взаимодействия с другими молекулами на клеточной стенке дрожжей. Применение магнитного разделения и проточной цитометрии в сочетании с библиотекой в виде дрожжевого дисплея является высокоэффективным способом выделения высокоаффинных белковых лигандов для почти любого рецептора посредством направленной эволюции.
Полисомный дисплей содержит очень большую библиотеку пептидов, представленных на бактериальных полисомах (Mattheakis et al., 1994). Методику пептидов MULTIPIN® также можно применять для создания библиотек (Tribbick et al., J Immunl. Methods (2002) 267: 27-35).
На стадии контактирования in vitro отобранная молекула или молекула-кандидат может быть подвергнута контакту с пептидом или полипептидом, содержащим одну или несколько из следующих областей или аминокислот («соответствующих областей/аминокислот CD4») CD4 человека: аминокислоты 148-154, аминокислоты 164-168 и аминокислоты 185, 187, 189, 190 и 192, при этом аминокислоты пронумерованы как показано на фигуре 6. Предпочтительно, области CD4 человека представляют собой: аминокислоты 148-154, аминокислоты 164-168 и аминокислоты 185-192. Более предпочтительно, пептид или полипептид содержит все указанные области. Еще более предпочтительно, пептид или полипептид дополнительно содержит по меньшей мере одну из следующих аминокислот CD4 человека: Lys26, Arg156, Arg159, Lys161 и Lys192. Более предпочтительно, пептид или полипептид содержит Ig-подобный домен C2-типа 1 CD4 человека (т.е. аминокислоты 126-205) и необязательно также Ig-подобный домен V-типа. В частности, D1 CD4 используется для стабилизации эпитопа. Соответственно, когда пептид или полипептид необходимо использовать в анализе конкурентного связывания, предпочтительно, использование D1. Однако следует отметить, что одна или две аминокислоты из указанных областей могут быть удалены. Предпочтительно, чтобы пептид имел длину менее 50 аминокислот и, более предпочтительно, имел длину менее 20 аминокислот.
Пептиды могут представлять собой природные пептиды, например пептиды, полученные при ферментативном расщеплении CD4 или непосредственно при экспрессии клеткой-хозяином, или они могут представлять собой синтетические пептиды. Пептиды также могут быть модифицированы, например, пегилированием, фосфорилированием, амидированием, ацетилированием, мечением биотином или флуоресцирующими красителями, такими как ФИТЦ, или мечением изотопами. Дополнительные модификации можно получить с использованием методик, подобных «нанесению множественных антигенных пептидов». С использованием такой методики можно получить высокий титр антител против пептида и синтетические пептидные вакцины. В такой системе используют α- и ε-аминогруппы лизина для образования остова, с которым могут быть связаны множественные пептидные цепи. В зависимости от количества лизиновых ярусов можно синтезировать разное количество пептидных ветвей. Это исключает необходимость в конъюгировании антигена с белком-носителем (Briand et al., J. Immunol. Methods (1992). 156; 2: pp 255-265).
Как показано выше, способы согласно настоящему изобретению, предпочтительно, осуществляют с использованием пептидных последовательностей из CD4 человека. Однако в равной мере они могут быть осуществлены с использованием гомологичных областей белков CD4 других млекопитающих или с использованием других молекул, содержащих Ig-подобный домен C2-типа 1.
Стадию осуществления контакта одной или нескольких молекул, отобранной молекулы или молекулы-кандидата с пептидом и стадию выявлении того, связывается ли одна или несколько молекул с одной или несколькими областями пептида можно осуществить согласно способам, известным в данной области. В частности, в одном варианте осуществления изобретения пептид представляет собой линейный пептид, который нанесен пятнами или фиксирован на мембране. Во время стадии контактирования молекулы, которые способны связываться с пептидной последовательностью CD4, оказываются захваченными.
В альтернативном варианте создают пептиды, которые могут имитировать конформацию эпитопа CD4 человека дикого типа. Это можно осуществить способами конструирования молекул на основе структуры, известными в данной области.
Ниже указаны способы дисплея, которые можно использовать для скрининга:
Для скрининга линейных эпитопов можно использовать методику картирования эпитопов. Аминокислотные последовательности, представляющие части эпитопа мишени (например, 10-15 аминокислот), которые перекрывается одной аминокислотой, наносят пятнами на мембрану (например, целлюлозную). Затем можно осуществлять скрининг в отношении белков или пептидов, распознающих нанесенные в виде пятен аминокислотные последовательности. Можно провести несколько раундов селекции в разных условиях жесткости, чтобы отобрать вещества, связывающиеся с высокой аффинностью.
Для скрининга в отношении прерывистых эпитопов разработаны такие методики, как фаговый дисплей. Современные стандартные библиотеки линейных или циклических пептидов содержат многообразие независимых клонов, примерно 109, при этом подразумевается, что библиотеки, имеющие до семи рандомизированных положений, теоретически могут гарантировать исчерпывающий охват возможного репертуара последовательностей. Системы трансляции in vitro дают библиотеки пептидов с более высоким разнообразием, так как связывание пептида с его мРНК достигается в бесклеточной системе, включающей небольшие частицы комплексов РНК/пептид/рибосома или только мРНК/пептид. Следующие библиотеки включают основанный на полисомах или рибосомах дисплей (Mattheakis et al., PNAS 1994; 91 (19): 9022-6) или методику PROfusion (Roberts and Szostak, PNAS (1997) 94(23): 12297-302). Последняя методика включает ковалентное слияние между мРНК и пептидом или белком, который она кодирует, которое может быть образовано при трансляции in vitro синтетической мРНК, которая несет пуромицин, антибиотик, являющийся акцептором пептидила, на своем 3'-конце.
Также возможен дисплей на основе миниклеток (патент США № 7125679), который содержит препарат пептидов для скрининга, которые экспрессированы на наружной поверхности, содержащей библиотеку олигонуклеотидов. Подобным образом также можно использовать библиотеку случайных пептидов Flitrix (Invitrogen Corp.), в которой используют белок жгутиков бактерий FliC и тиоредоксин.
Кроме того, также как указанные выше способы дисплея, можно использовать масс-спектрометрию или извлечение эпитопов с твердой фазы (SPHERE) (Genzyme) (Lawendowski et al., J. Immunol., (2002) 169: 2414-2421).
В одном из вариантов осуществления выявление связывания включает использованием рентгеновской кристаллографии или ЯМР. В частности, могут быть отобраны молекулы, которые связываются с пептидом без образования солевого мостика, с использованием рентгеновской кристаллографии, которая известна в данной области.
Альтернативно или дополнительно способ согласно первому аспекту может включать осуществление контакта отобранной молекулы или молекулы-кандидата с клеткой, экспрессирующей CD4, и, в частности, регуляторной T-клеткой CD4+CD25+. Это можно осуществлять, в частности, для определения способности отобранной молекулы или молекулы-кандидата модулировать активность и, в частности, активировать регуляторные T-клетки CD4+CD25+ (предпочтительно, избирательно активировать клетки Treg без активации T-хелперных клеток), или для определения способности отобранной молекулы или молекулы-кандидата уменьшать или осуществлять понижающее модулирование экспрессии рецептора CD4, в частности, на специфичных популяциях лимфоцитов в культуре in vitro PBMC (мононуклеарных клеток периферической крови). В таких вариантах осуществления, предпочтительно, отобранная молекула или молекула-кандидат представлена антителами или фрагментами антител, и в частности антителами IgG1-типа, как обсуждается ниже.
Для анализа модулирования Treg, в общем, могут быть выделены с использованием коммерчески доступных наборов для выделения (выделение на магнитных шариках), сортировки в отношении CD25, CD27, CD62L и/или CD127 и дополнительного внутриклеточного окрашивания в отношении FoxP3. Treg являются негативными по CD127, позитивными по CD25 и Foxp3. CD39, связанная с клеточной поверхностью эктонуклеотидаза, также может быть использована для очистки Treg с сильными супрессорными функциями (Mandapathil et al., J Immunol. Methods (2009). 346 (1-2), 55-63). В коммерчески доступных наборах может быть использована негативная селекция CD4+ с последующим позитивным выделением позитивных по CD25 клеток с получением в результате популяции позитивных по CD4 и CD25 клеток. Полученные клетки могут быть дополнительно обработаны. CD25 и фактор транскрипции Foxp3 являются маркерами экспрессии, связанными с супрессорной функцией Treg. Хотя внутриклеточное окрашивание Foxp3 подтверждает регуляторный фенотип, вследствие внутриклеточного окрашивания клетки не жизнеспособны для дальнейшего терапевтического применения. Foxp3 обычно используют в качестве внутриклеточного маркера активированных Treg/функционально активных Treg.
Кроме того, вследствие того факта, что BT061 и молекула, отбираемая при скрининге, связывается с эпитопом, который отличается от эпитопа, связываемого другими коммерчески доступными антителами, можно очистить Treg с использованием коммерчески доступных наборов для выделения (выделение на магнитных шариках), и дополнительно другого CD4-антитела (например, SK-3 OKT4), которое не конкурирует за сайт связывания BT061 на CD4, и затем проанализировать активацию Treg с использованием молекулы-кандидата или отобранной молекулы.
Способность молекулы-кандидата активировать Treg можно анализировать исследуя супрессорную активность Treg после контакта с молекулой-кандидатом посредством совместного культивирования Treg с CD4-позитивными CD25-негативными эффекторными T-клетками. Активированные Treg способны ингибировать пролиферацию эффекторных Т-клеток CD4+CD25-, которые могут быть мечены CFSE (оценка экспансии клеток посредством анализа разведения CFSE). Альтернативно, пролиферацию эффекторных клеток можно определить по включению [3H]-тимидина.
Более конкретно, способность к супрессии можно оценить, например, используя реакцию смешанных лимфоцитов (MLR). Клеточное деление эффекторных T-клеток может быть ингибировано супрессорным действием Treg. Для этого “наивные” аутологичные T-клетки-респондеры CD4+CD25- стимулируют облученными аллогенными стимулирующими PBMC. Treg или обычные T-клетки титруют в культуре, и пролиферацию можно оценить по включению тимидина.
Активацию Treg также можно оценить путем определения продукции циклического АМФ (как описано в WO 2008/092905).
Цитокины, на которые влияют активированные Treg в совместной культуре, также могут быть измерены для определения активности таких клеток по отношению к эффекторным клеткам. Например, Treg проявляют свою супрессорную активность также через поглощение IL-2, что приводит к ингибированию пролиферации эффекторных T-клеток. IL-4 или IFN-гамма также могут быть определены в анализе совместной культуры, и их количество снижается в случае активированных Treg. Кроме того, количество маркеров активации на поверхности эффекторных T-клеток, таких как CD25, снижается, когда Treg-клетки активируются и проявляют супрессорную активность.
Кроме того, определение клеточной гибели (по таким факторам как Bim) в эффекторных клетках (которые также могут быть CD4-позитивными), индуцированной активированными Treg в совместной культуре, представляет собой следующий способ исследования того, являются ли Treg активированными (Pandiyan et al., Nature Immunol. (2007) 8: 1353-1362).
Так как Treg составляют только небольшую часть в крови (2-10%), известно несколько методик их экспансии. Например, стимуляция TCR с использованием анти-CD3 и костимуляция с использованием анти-CD28 и рапамицина могут быть использованы для увеличения количества T-клеток. Рапамицин избирательно стимулирует избирательную жизнеспособность Treg, но не эффекторных T-клеток.
Имеется несколько протоколов поликлональной экспансии, например, после позитивной селекции в отношении CD4/CD25 Treg-клетки могут быть размножены поликлонально in vitro с использованием анти-CD3- и анти-CD28-антитела (для стимуляции) в сочетании с IL-2 и/или IL-15, способным увеличивать количества Treg при сохранении способности к супрессии (Earle et al., Clin. Immunol. (2005) 115: 3-9).
В связи с модулированием экспрессии CD4 следует отметить, что при добавлении анти-CD4-антител экспрессия рецепторов CD4 на поверхности клеток может быть снижена. Указанный признак можно использовать в качестве основы для анализа эффективности, чтобы определить степень связывания CD4. В частности, в таком анализе стадия осуществления контакта может включать (i) инкубацию молекулы-кандидата или отобранной молекулы с мононуклеарными клетками периферической крови (PBMC) из донорской крови человека при 37°C; (ii) окрашивание инкубированных клеток меченым анти-CD4-антителом, которое не конкурирует с BT061 за связывание с CD4; и (iii) количественную регистрацию окрашивания, чтобы определить занятость рецепторов CD4, и следовательно, определение количества молекул CD4, присутствующих на поверхности клеток.
Более конкретно, такой анализ включает выделение PBMC (т.е. лимфоцитов и моноцитов) из донорской крови человека, которые затем инкубируют с разными концентрациями молекулы-кандидата или отобранной молекулы (предпочтительно, антитела или фрагмента антитела) при 37°C. После инкубации в течение 3 часов клетки красят, добавляя меченые флуорохромом антитела, такие как фикоэритрин-анти-CD4, которые связываются с другим эпитопом на CD4, отличным от эпитопа, который связывается BT061. Такие красящие антитела метят CD4-рецепторы на специфичных популяциях лимфоцитов. Так как BT061 и используемое меченое флуорохромом CD4-антитело распознают разные эпитопы на молекуле CD4 и не конкурируют, такой способ позволяет оценить количество определяемых CD4-рецепторов на поверхности клеток, независимо от связывания CD4 молекулой-кандидатом или отобранной молекулой. Измерение осуществляют в проточном цитометре, в котором меченые антителом клетки проходят через лазерный луч и стимулируются. При стимуляции лазерным лучом флуоресценция окрашенных клеток пропорциональна связанному антителу, и испускаемый свет улавливается проточным цитометром и затем происходит оценка с использованием компьютерной программы, известной в данной области, например, компьютерной программы FlowJo (Tree Star, Inc). С использованием компьютерной программы (такой как компьютерная программа Parallel Line Assay, Stegmann Systems) затем можно вычислить относительную активность (эффективность) для образца относительно стандарта, оцениваемого параллельно.
В следующем варианте настоящего аспекта изобретения стадия осуществления контакта включает осуществление контакта пептида или полипептида CD4 (предпочтительно, содержащего D1 и D2 CD4) или клетки, экспрессирующей CD4, и молекулы-кандидата/отобранной молекулы с конкурирующим антителом или фрагментом антитела, содержащим вариабельные домены тяжелой и легкой цепи BT061, чтобы определить, способна ли молекула-кандидат/отобранная молекула блокировать связывание конкурентного антитела или фрагмента антитела с CD4.
Одна или несколько молекул-кандидатов в первом аспекте могут представлять собой пептид или непептидную молекулу. В частности, одна или несколько молекул могут представлять собой мимеотоп, пептидомиметик, малую молекулу, белок распознавания на основе природного или сконструированного липокалина, олигонуклеотид, ми-РНК, DARPin, фибронектин, белок-миметик антител Affibody, ингибитор типа Кунитца, пептидный аптамер, рибозим, токсин, антитело верблюдовых, фрагмент антитела или полученная из антитела молекула.
Мимеотопы представляют собой пептиды, имитирующие белковые, углеводные или липидные эпитопы, и могут быть созданы способом на основе фагового дисплея. В случае селекции антителами, они представляют исключительно B-клеточные эпитопы и лишены антиген/аллерген-специфичных T-клеточных эпитопов. Связанные с носителями или представленные в форме множественных антигенных пептидов мимиотопы достигают иммуногенности и индуцируют специфичные для эпитопа гуморальные ответы при вакцинации.
Пептидомиметик представляет собой небольшую подобную белку цепь, сконструированную для имитации пептида. Обычно они являются результатом модификации существующего пептида, осуществляемой для того, чтобы изменить свойства молекулы. Например, они могут быть результатом модификаций, осуществляемых для изменения стабильности молекулы или биологической активности. Такие модификации включают изменения в пептиде, которые не происходят в природе (такие как измененные остовы и включение неприродных аминокислот).
Примерами пептидомиметиков были пептидомиметики, сконструированные и синтезированные с целью связывания с белками-мишенями, чтобы индуцировать переход злокачественных клеток к форме запрограммированной гибели клеток, называемой апоптозом. По существу они работают, имитируя ключевые взаимодействия, которые активируют апоптозный путь в клетке.
Фолдамер представляет собой молекулу с отдельной цепью или олигомер, который принимает вторичную структуру, стабилизированную нековалентными взаимодействиями (Gellman, Acc. Chem. Res (1998) 31 (4): 173-180; Hill et al., Chem. Rev. (2001) 101 (12): 3893-4012). Они являются искусственными молекулами, которые имитируют способность белков, нуклеиновых кислот и полисахаридов укладываться в отчетливо выраженные конформации, такие как спирали и β-слои. Было показано, что фолдамеры проявляют ряд интересных супрамолекулярных свойств, включая самосборку, распознавание молекул и химию «гость-хозяин». Их исследуют в качестве моделей биологических молекул, и было показано, что они проявляют противомикробную активность. Они также потенциально могут быть широко применимы для разработки новых функциональных материалов.
Малая молекула представляет собой низкомолекулярное органическое соединение, которое по определению не является полимером. Верхний предел молекулярной массы для малой молекулы составляет примерно 800 дальтон, что обеспечивает возможность для быстрой диффузии через клеточные мембраны, так что они могут достигать мест действия внутри клетки. Малая молекула проявляет высокую аффинность к биополимеру, такому как белок, нуклеиновая кислота или полисахарид и, кроме того, изменяет активность или функцию биополимера.
DARPin (сконструированные белки с повторами анкирина) являются искусственными белками, которые также способны распознавать антиген или антигенные структуры. Структурно они получены из анкириновых белков, имеют молекулярную массу примерно 14 кДа (166 аминокислот) и состоят из трех повторяющихся мотивов. Они проявляют сравнимую с антителами аффинность к антигенам. (Stumpp et al., Drug Discov. Today (2008) 13, Nr. 15-16, S. 695-701).
Молекулы Affibody® являются небольшими белковыми молекулами с очень высокой аффинностью, которые могут быть сконструированы для специфичного связывания с большим количеством белков-мишеней.
Термин «верблюдовые» относится к антителам, продуцируемым верблюдовыми и содержащими гомодимер тяжелой цепи, и производным таких молекул (Muyldermans et al., Veterinary Immunology and Immunopathology, 128; 1-3; pp.178-183 (2009)).
Предпочтительно, молекула-кандидат является антителом или фрагментом антитела. Фраза «антитело, фрагмент антитела или полученная из антитела молекула» охватывает моноклональные антитела, поликлональные антитела, полиспецифичные антитела и фрагменты антител. Термин «фрагмент антитела» включает, в частности, фрагменты, содержащие фрагменты Fab, Fab', F(ab)'2, Fv и scFv и диатела или триатела. Предпочтительно, они основаны на гуманизированных антителах или антителах человека. Более предпочтительно, антитело содержит константную область/домен, т.е. Fc-часть. В том случае, когда антитело содержит константную область человека, такая константная область может быть выбрана из константных областей любого класса иммуноглобулинов, включая IgM, IgG, IgD, IgA и IgE, и любого изотипа, включая IgG1, IgG2, IgG3 и IgG4. Предпочтительные константные области выбраны из константных доменов IgG, в частности, IgG1.
Fc-части разных подклассов Ig связываются клеточным FcR, который специфичен для отдельных подклассов. Известны три разных класса FcR, которые связывают изотипы IgG с разными аффинностями, CD16, CD32 и CD64. Разнообразные картины FcγR экспрессируются различными иммунными клетками, такими как моноциты, B-клетки, клетки природные киллеры (NK) и другие. Авторы настоящего изобретения обнаружили, что in vitro способность константной области антитела BT061 связываться с рецепторами Fc (FcR) важна для способности антитела вызывать понижающее модулирование CD4 в T-клетках. В частности, исследования in vitro показывают, что из Fcγ-рецепторов в основном вовлечен рецептор Fcγ1, который главным образом экспрессируется на моноцитах; присутствие моноцитов в культуре PBMC необходимо и достаточно для осуществления понижающего модулирования CD4 в BT061-обработанных T-клетках.
Соответственно, в одном из вариантов осуществления изобретения молекула-кандидат способна связываться с Fc-рецептором, предпочтительно, FcyR1 (т.е. CD64) и, наиболее предпочтительно, содержит Fc-часть антитела IgG1. Кроме того или альтернативно, молекула-кандидат способна связываться с моноцитами через Fc-рецептор.
В данном аспекте настоящего изобретения молекула, подвергаемая скринингу, не является антителом или фрагментом антитела, который содержит CDR1, CDR2 и CDR3 тяжелой цепи BT061 и CDR1, CDR2 и CDR3 легкой цепи BT061. Такие последовательности CDR показаны на фигурах 2A и 2B. В предпочтительном варианте осуществления молекула, подвергаемая скринингу не является молекулой B-F5 мыши, описанного Racadot et al. (Clin. Exp. Rheum., 10, 365-374 (1992)).
В предпочтительном варианте осуществления первого аспекта изобретения, в котором одна или несколько молекул-кандидатов представляют собой антитела или фрагменты антител, антитела или фрагменты антител содержат CDR1 и CDR2 легкой цепи BT061 и CDR1 и CDR3 тяжелой цепи BT061 необязательно с аминокислотными заменами в последовательностях CDR, при условии, что:
(i) CDR1 легкой цепи содержит: Ser32, Gly33 и Tyr 34;
(ii) CDR2 легкой цепи содержит: Leu54 и Ile57;
(iii) CDR1 тяжелой цепи содержит Asp31, Glu31, Thr31, Cys31, Pro31, Met31 или Tyr31; и
(iv) CDR3 тяжелой цепи содержит Tyr103, Phe103 или His103; Arg104; Tyr105; Asp106; и Trp110, Phe110, His110 или Tyr110.
Альтернативно, молекула-кандидат представляет собой антитело или фрагмент антитела, содержащий V-домены, имеющие по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 85%, более предпочтительно, по меньшей мере 90% идентичность последовательности с V-доменами BT061 (т.е. SEQ ID NO:2 и SEQ ID NO:3) и содержащий мотив последовательности SGYSY (SEQ ID NO:10) в CDR1 V-домена легкой цепи, мотив последовательности LASILE (SEQ ID NO:11) в CDR2 V-домена легкой цепи и мотив последовательности SYY/F/HRYD (SEQ ID NO:13) в CDR3 V-домена тяжелой цепи.
В данном способе одна или несколько молекул, подвергаемых скринингу, представляют собой набор молекул-кандидатов, определяемых по их сходству с антителом BT061 и таким остаткам в BT061, которые авторы настоящего изобретения идентифицировали в качестве важных для взаимодействия между антителом BT061 и CD4. В частности, данный способ, предпочтительно, представляет собой способ, посредством которого идентифицируют молекулы, имеющие примерно эквивалентные или улучшенные аффинность/специфичность и активность в отношении активации Treg по сравнению с BT061.
Пептидные последовательности легкой цепи и тяжелой цепи гуманизированного антитела BT061 показаны на фигурах 2A и 2B, соответственно. CDR легкой и тяжелой цепи отмечены на указанных фигурах. В данном случае аминокислотные остатки в таких CDR указаны типом остатка и числом, при этом число указывает положение аминокислоты в вариабельной области легкой или тяжелой цепи антитела BT061, как показано на фигурах 2A и 2B. Тип остатка и число использовали в целях четкой идентификации аминокислотного остатка CDR BT061, в случае указания такого остатка. Однако будет понятно, что число, которым обозначают остаток, не предназначено для ограничения нахождения остатка в данном положении в выбранном для исследования антителе или фрагменте, который подвергают скринингу данным способом. Например, в антителе согласно данному варианту осуществления Ser32 может быть в положении 31 в CDR1 легкой цепи, если несущественный аминокислотный остаток был делетирован из участка 1-30 легкой цепи.
В предпочтительном варианте осуществления аминокислотные замены в последовательности CDR1 и CDR2 легкой цепи BT061 и CDR1 и CDR3 тяжелой цепи BT061 выбраны из замен, указанных в таблице 4 и таблице 5, ниже. Более предпочтительно, антитело или фрагмент антитела содержит легкую цепь, содержащую Tyr53 или Phe53, и тяжелую цепь, содержащую Ser28 (как показано в таблице 7 в Примере 1). Альтернативно или дополнительно антитело или фрагмент антитела содержит легкую цепь, содержащую Asp64, и/или антитело или фрагмент антитела содержит тяжелую цепь, имеющую по меньшей мере один из остатков Asp31 и Glu56 (как показано в таблице 8, в примере 1). Антитело или фрагмент антитела может дополнительно содержать CDR3 легкой цепи BT061 и/или CDR2 тяжелой цепи BT061 необязательно с аминокислотными заменами в последовательностях таких CDR, при этом замены выбраны из замен, указанных в таблице 4 и таблице 5.
Выбранное для исследования антитело или фрагменты антител для использования в способах скрининга могут быть образованы в результате мутации известной последовательности BT061. В частности, когда такие мутации находятся в CDR, они могут быть целенаправленными мутациями, чтобы гарантировать, что сохраняются аминокислоты, перечисленные выше, или получены требуемые замены.
Когда стадию (a) способа скрининга осуществляют in vitro, целенаправленный мутагенез можно использовать для того, чтобы вызвать замену аминокислоты в определенных положениях в CDR. Зная соответствующую последовательность ДНК для аминокислот можно создать библиотеку специфичных мутантов, содержащих ряд требуемых аминокислотных замен. Когда стадии (a)-(c) способа скрининга необходимо осуществить в компьютерной системе, трехмерная структура выбранных для исследований антител или фрагментов антител может быть создана введением аминокислотной последовательности подобно тому, как указано выше в отношении CD4.
Таблица, в которой показаны варианты последовательности CDR легкой цепи BT061
Таблица, в которой показаны варианты последовательности CDR тяжелой цепи BT061
Изостерические варианты представляют собой варианты, которые не оказывают стерического влияния, т.е. они никоим образом не изменяют конформацию антитела.
Во втором аспекте настоящее изобретение относится к способу скрининга молекулы-кандидата, которая представляет собой антитело или фрагмент антитела, которая способна связываться с CD4, включающему: (a) получение антитела или фрагмента антитела, содержащего CDR1 и CDR2 легкой цепи BT061 и CDR1 и CDR3 тяжелой цепи BT061, необязательно с аминокислотными заменами в последовательностях CDR, при условии, что:
(i) CDR1 легкой цепи содержит: Ser32, Gly33 и Tyr 34;
(ii) CDR2 легкой цепи содержит: Leu54 и Ile57;
(iii) CDR1 тяжелой цепи содержит Asp31, Glu31, Thr31, Cys31, Pro31, Met31 или Tyr31; и
(iv) CDR3 тяжелой цепи содержит Tyr103, Phe103 или His103; Arg104; Tyr105; Asp106; и Trp110, Phe110, His110 или Tyr110;
(b) определение того, способно ли антитело или фрагмент антитела связываться с CD4, и (c) отбор антитела или фрагмента антитела, определенного на стадии (b), как способного связываться с CD4, при этом антитело или фрагмент антитела не содержит CDR1, CDR2 и CDR3 тяжелой цепи BT061 и CDR1, CDR2 и CDR3 легкой цепи BT061.
В одном из вариантов осуществления стадии (a)-(c) способа скрининга согласно второму аспекту настоящего изобретения можно осуществить в компьютерной системе подобно тому, как описано выше в отношении первого аспекта изобретения. В частности, может быть создано некоторое количество вариантов BT061 in silico на основе информации, представленной в настоящем описании об остатках, важных для взаимодействия BT061 с его эпитопом CD4, и имитировано взаимодействие таких вариантов с CD4.
В настоящем варианте осуществления способ может дополнительно включать стадию, на которой выбранное для исследования антитело или фрагмент антитела подвергают контакту с клеткой, экспрессирующей CD4.
В варианте, альтернативном варианту, осуществляемому в компьютерной системе, стадии (a)-(c) способа скрининга согласно второму аспекту изобретения можно осуществлять in vitro, и стадия (b) может включать осуществления контакта выбранного для исследования антитела или фрагмента антитела с клеткой, экспрессирующей CD4. Предпочтительно, способность связываться с CD4 определяют на основе способности кандидата активировать регуляторные T-клетки CD4+CD25+.
В частности, данный аспект настоящего изобретение может представлять собой способ скрининга антитела или фрагмента антитела, способного активировать регуляторные T-клетки CD4+CD25+, который включает:
(a) осуществление контакта одного или нескольких антител или фрагментов антител с регуляторными T-клетками CD4+CD25+;
(b) оценку способности антитела или фрагмента антитела активировать указанные регуляторные T-клетки CD4+CD25+; и
(c) идентификацию антитела или фрагмента антитела, которое активирует регуляторные T-клетки CD4+CD25+,
при этом антитело или фрагмент антитела содержит CDR1 и CDR2 легкой цепи BT061 и CDR1 и CDR3 тяжелой цепи BT061 необязательно с аминокислотными заменами в последовательностях CDR, при условии, что:
(i) CDR1 легкой цепи содержит: Ser32, Gly33 и Tyr 34;
(ii) CDR2 легкой цепи содержит: Leu54 и Ile57;
(iii) CDR1 тяжелой цепи содержит Asp31, Glu31, Thr31, Cys31, Pro31, Met31 или Tyr31; и
(iv) CDR3 тяжелой цепи содержит Tyr103, Phe103 или His103; Arg104; Tyr105; Asp106; и Tip110, Phe110, His110 или Tyr 110,
и при этом антитело или фрагмент антитела не содержит CDR1, CDR2 и CDR3 тяжелой цепи BT061 и CDR1, CDR2 и CDR3 легкой цепи BT061.
Следует отметить, что описание вариантов осуществления и предпочтительных признаков, приведенное выше в отношении способа скрининга согласно первому аспекту настоящего изобретения, также применимо к способу скрининга согласно второму аспекту настоящего изобретения.
Оба указанных аспекта изобретения могут дополнительно включать получение выбранной молекулы, и в частности выбранного антитела или фрагмента антитела, в целях их дальнейшего применения или последующего анализа.
Соответственно, изобретение, кроме того, относится к способу получения терапевтической композиции и к терапевтическим композициям, получаемым данным способом. В частности, способ включает:
(a) отбор молекулы, определяемой как способной активировать регуляторные T-клетки CD4+CD25+, с использованием способа согласно способам скрининга, описанным выше; и
(b) получение терапевтической композиции, содержащей молекулу.
В частности, терапевтическая композиция может быть получена посредством объединения молекулы с одним или несколькими фармацевтически приемлемыми носителями или разбавителями.
Антитела и фрагменты антител
Работа, выполненная авторами настоящего изобретения, позволила идентифицировать характерные признаки BT061, которые важны для его взаимодействия и активации регуляторных T-клеток CD4+CD25+. Это привело к идентификации специфичных мутантов/вариантов антитела BT061, которые будут способны активировать регуляторные T-клетки CD4+CD25+.
Соответственно, в третьем аспекте настоящее изобретение относится к антителу или фрагменту антитела, способному активировать регуляторные T-клетки CD4+CD25+, содержащему CDR1 и CDR2 легкой цепи BT061 и CDR1 и CDR3 тяжелой цепи BT061 необязательно с аминокислотными заменами в последовательностях CDR, при условии, что:
(i) CDR1 легкой цепи содержит: Ser32, Gly33 и Tyr 34;
(ii) CDR2 легкой цепи содержит: Leu54 и Ile57;
(iii) CDR1 тяжелой цепи содержит Asp31, Glu31, Thr31, Cys31, Pro31, Met31 или Tyr31; и
(iv) CDR3 тяжелой цепи содержит Tyr103, Phe103 или His103; Arg104; Tyr105; Asp106; и Tip110, Phe110, His110 или Tyr110,
и при этом антитело или фрагмент антитела не содержит CDR1, CDR2 и CDR3 тяжелой цепи BT061 и CDR1, CDR2 и CDR3 легкой цепи BT061.
Альтернативно, третий аспект относится к антителу или фрагменту антитела, способному активировать регуляторные T-клетки CD4+CD25+, содержащие CDR1 и CDR2 легкой цепи BT061 и CDR1 и CDR3 тяжелой цепи BT061 необязательно с аминокислотными заменами в последовательностях CDR, при условии что:
(i) CDR1 легкой цепи содержит: Ser32 или Pro32; Gly33 или Ala33; и Tyr 34;
(ii) CDR2 легкой цепи содержит: Leu54, Ile54 или Thr54; и Ile57, Leu57, Val57 или Thr57;
(iii) CDR1 тяжелой цепи содержит Asp31, Glu31, Thr31, Cys31, Pro31, Met31 или Tyr31; и
(iv) CDR3 тяжелой цепи содержит Tyr103, Phe103 или His103; Arg104; Tyr105; Asp106; и Tip 110, Phe110, His110 или Tyr110,
и при этом антитело или фрагмент антитела не содержит CDR1, CDR2 и CDR3 тяжелой цепи BT061 и CDR1, CDR2 и CDR3 легкой цепи BT061.
Как обсуждалось выше, пептидные последовательности легкой цепи и тяжелой цепи гуманизированного антитела BT061 показаны на фигурах 2A и 2B, соответственно. CDR легкой и тяжелой цепи обозначены на указанных фигурах. Как обсуждалось выше, аминокислотные остатки в таких CDR указаны типом остатка и числом, при этом число указывает положение аминокислоты в вариабельной области легкой или тяжелой цепи антитела BT061, как показано на фигурах 2A и 2B. Тип остатка и число использовали в целях четкой идентификации аминокислотного остатка CDR BT061 в случае указания такого остатка. Однако будет понятно, что число, которым обозначают остаток, не предназначено для ограничения нахождения остатка в данном положении в антителе или фрагменте согласно изобретению. Например, в антителе согласно данному варианту Ser32 может быть в положении 31 в CDR1 легкой цепи, если несущественный аминокислотный остаток был делетирован из участка 1-30 легкой цепи.
В предпочтительном варианте осуществления аминокислотные замены в последовательности CDR1 и CDR2 легкой цепи BT061 и CDR1 и CDR3 тяжелой цепи BT061 выбраны из замен, указанных в таблице 4 и таблице 5 выше. Более предпочтительно, антитело или фрагмент антитела содержит легкую цепь, содержащую Tyr53 или Phe53, и тяжелую цепь, содержащую Ser28. Альтернативно или дополнительно антитело или фрагмент антитела содержит легкую цепь, содержащую Asp64, и/или антитело или фрагмент антитела содержит тяжелую цепь, имеющую по меньшей мере одну из аминокислот, указанных в таблице 8 в Примере 1 в качестве аминокислот, важных для взаимодействия. В частности, тяжелая цепь содержит Asp31 и/или Glu56. Антитело или фрагмент антитела дополнительно может содержать CDR3 легкой цепи BT061 и/или CDR2 тяжелой цепи BT061 необязательно с аминокислотными заменами в последовательностях таких CDR, при этом замены выбраны из замен, указанных в таблице 4 и таблице 5.
В частности, распознавание и связывание эпитопа CD4 антителом BT061 основано на конкретном составе и конформации трех определяющих комплементарность областей CDR1, CDR2 и CDR3. Даже если только CDR1 и CDR2 легкой цепи и CDR1 и CDR3 тяжелой цепи находятся в непосредственном контакте с CD4, все шесть CDR очень плотно упакованы и взаимно поддерживают конформацию друг друга. Как следствие, многие положения в CDR не допускают никакой аминокислотной замены без значительной потери активности и эффективности BT061. Однако в некоторых положениях замены не дестабилизируют структуру.
Примеры таких консервативных замен приведены в таблицах 4 и 5. В указанных таблицах варианты последовательности выбраны так, чтобы сохранить общую сеть взаимодействия. Синтаксия взаимного притяжения, а также взаимного отталкивания сохраняется. Прямые полярные взаимодействия могут быть обратными, например, пара донор-акцептор водородных мостиков, ионные партнеры солевых мостиков или места партнеров диполь-квадрупольных взаимодействий могут меняться.
В следующих вариантах антитело или фрагмент антитела содержит последовательность (SEQ ID NO:14):


где аминокислоты в положениях 24-29, 31, 37, 60, 96-98 и 101, отмеченные «X» выбраны из аминокислот, показанных в соответствующих положениях в таблице 4,
и дополнительно содержит последовательность (SEQ ID NO:15):

где аминокислоты в положениях 31-34, 51-67, 103, 107-110, 111 и 112, отмеченные как «X», выбраны из аминокислот, показанных в соответствующих положениях в таблице 5.
В частности, конкретные мотивы аминокислот в CDR1 и CDR2 легкой цепи и в CDR3 тяжелой цепи BT061 важны для связывания CD4. Соответственно, антитело или фрагмент антитела могут содержать SGYSY (SEQ ID NO:10) из CDR1 легкой цепи BT061 и/или последовательность LASILE (SEQ ID NO:11) из CDR2 легкой цепи BT061 и/или последовательность YYRYD (SEQ ID NO:12) из CDR3 тяжелой цепи BT061.
Кроме того, антитело или фрагмент антитела, способный активировать регуляторные T-клетки CD4+CD25+ может иметь V-домены, которые по меньшей мере на 80% идентичны, более предпочтительно, по меньшей мере на 90% идентичны V-доменам антитела BT061, при этом V-домены содержат:
(i) мотив последовательности SGYSY (SEQ ID NO:10) в CDR1 V-домена легкой цепи;
(ii) мотив последовательности LASILE (SEQ ID NO:11) в CDR2 V-домена легкой цепи; и
(iii) мотив последовательности SYXRYD, где X означает Y, F или H (SEQ ID NO:13) в CDR3 V-домена тяжелой цепи,
при условии, что антитело или фрагмент антитела не содержит V-доменов, которые на 100% идентичны V-доменам антитела BT061.
В конкретных вариантах третьего аспекта настоящего изобретения антитело или фрагмент антитела содержит последовательность CDR легкой цепи BT061 и последовательности CDR тяжелой цепи BT061 с одной аминокислотной заменой, при этом замена представляет собой:
(i) A63G в тяжелой цепи;
(ii) R33K в тяжелой цепи; или
(iii) L98I в легкой цепи;
или антитело или фрагмент антитела содержит последовательности CDR легкой цепи BT061 и последовательности CDR тяжелой цепи BT061 и двойную аминокислотную замену, при этом замены представляют собой:
(i) R33K и A63G в тяжелой цепи; или
(ii) L98I в легкой цепи и R33K в тяжелой цепи.
В указанных конкретных вариантах осуществления изобретения антитело или фрагмент антитела может дополнительно содержать остальные последовательности вариабельных доменов тяжелой и легкой цепей BT061.
Антитела или их фрагменты согласно настоящему изобретению, в частности, могут быть получены посредством мутагенеза полинуклеотидной последовательности, которая, как известно, кодирует вариабельные домены антитела B-F5 мыши и антитела BT061 (которые описаны в WO2004/083247).
Следует отметить, что определения и предпочтительные варианты осуществления антитела и фрагментов антитела, описанные выше в отношении первого и второго аспектов настоящего изобретения также применимы к третьему аспекту изобретения. В частности, антитела и фрагменты, предпочтительно, являются IgG1-антителами и/или, предпочтительно, содержат Fc-часть так, чтобы антитело или фрагмент антитела был способен связываться с Fc-рецептором, предпочтительно, FcγRI (т.е. CD64). Наиболее предпочтительно, антитело или фрагмент антитела содержит Fc-часть IgG1-антитела. Дополнительно или альтернативно антитело или фрагмент антитела способен связываться с моноцитами через Fc-рецептор.
Кроме того, настоящее изобретение относится к выделенному пептиду, содержащему менее 50 аминокислот белка CD4 человека и содержащему одну или несколько из следующих областей CD4 человека: аминокислоты 148-154, аминокислоты 164-168 и аминокислоты 185-192. Предпочтительно, выделенный пептид содержит две из указанных областей, более предпочтительно, три. Дополнительно или альтернативно выделенный пептид содержит мене 30 аминокислот и, наиболее предпочтительно, выделенный пептид содержит менее 20 аминокислот.
Кроме того, настоящее изобретение относится к мимеотопному пептиду выделенного пептида, описанного выше.
Настоящее изобретение также относится к нуклеиновым кислотам, кодирующим антитело или фрагмент антитела, описанного в настоящей публикации. Нуклеиновая кислота может представлять собой РНК или ДНК, но, предпочтительно, ДНК, и наиболее предпочтительно, кодирует V-домен H-цепи или L-цепи антитела или фрагментов. Полинуклеотид может быть слит с полинуклеотидом, кодирующим константную область H- или L-цепи человека, в целях экспрессии полных H- и L-цепей.
В изобретении также используются кассеты экспрессии, в которых полинуклеотид, который описан выше, связан с соответствующими регуляторными последовательностями для обеспечения регуляции его транскрипции и трансляции в выбранной клетке-хозяине. Следующими вариантами осуществления являются рекомбинантные векторы, содержащие полинуклеотид или кассету экспрессии, которая описана выше.
Полинуклеотид, который описан выше, может быть связан в экспрессирующем векторе с соответствующими регуляторными последовательностями, обеспечивающими регуляцию его транскрипции и трансляции в выбранной клетке-хозяине. Такие рекомбинантные конструкции ДНК могут быть получены и введены в клетки-хозяева хорошо известными способами, основанными на рекомбинантной ДНК, и способами генетической инженерии.
Применимыми клетками-хозяевами могут быть прокариотические или эукариотические клетки. Походящими эукариотическими клетками являются растительные клетки, клетки дрожжей, таких как Saccharomyces, клетки насекомых, таких как Drosophila или Spodoptera, и клетки млекопитающих, такие как HeLa, CHO, 3T3, C127, BHK, COS и т.п. Антитела или фрагменты, описанные в настоящей публикации, могут быть получены в результате культивирования клетки-хозяина, содержащей вектор экспрессии, содержащий последовательность нуклеиновой кислоты, кодирующую указанное антитело, в условиях, подходящих для ее экспрессии, и извлечения указанного антитела из культуры клеток-хозяев.
Согласно настоящему изобретению клеткой-хозяином также может быть гибридома, получаемая слиянием клетки, продуцирующей антитело согласно настоящему изобретению с клеткой миеломы.
Антитело или фрагмент антитела имеет медицинское и немедицинское применения, которые описаны далее.
С учетом медицинского применения антитело или фрагмент антитела, описанные в настоящей публикации, могут быть приготовлены в фармацевтической композиции. В частности, фармацевтическая композиция согласно настоящему изобретению содержит антитело или фрагмент антитела и фармацевтически приемлемый носитель или разбавитель.
Более того, антитело или фрагмент антитела могут дополнительно содержать метку. Способы мечения антител хорошо известны в данной области. Соответственно, только в качестве примера антитело может мечено флуоресцирующей меткой, такой как GFP или флуоресцирующий краситель (например, ФИТЦ, высокоэффективный dyLight), радиоактивный изотоп, биотин, HRP и т.д.
Применения антител и эпитопов
Настоящее изобретение, кроме того, относится к способам лечения с применением антитела или фрагмента антитела согласно настоящему изобретению. Так как антитело или фрагмент антитела способны избирательно активировать регуляторные T-клетки CD4+CD25+, это имеет конкретное применение в терапии. Настоящее изобретение относится к способу лечения индивида, страдающего от аутоиммунного заболевания или отторжения трансплантата, или способу профилактики аутоиммунного заболевания или отторжения трансплантата у индивида, включающему введение указанному индивиду антитела или фрагмента антитела согласно настоящему изобретению. Подобным образом, настоящее изобретение также относится к антителу или фрагменту антитела, которые описаны в настоящей публикации, для применения в медицине, и в частности, для применения при лечении аутоиммунного заболевания или отторжения трансплантата. Соответственно, настоящее изобретение относится к применению антитела или фрагмента антитела, которые описаны в настоящей публикации, для производства лекарственного средства для применения при лечении аутоиммунного заболевания или отторжение трансплантата. Подходящими применениями в медицине и способами лечения являются применения и способы, которые описаны для BT061 в WO2009/112502, WO2009/121690, WO2009/124815 и WO2010/034590, описания которых включены в настоящее описание в качестве ссылки.
В предпочтительном варианте осуществления аутоиммунное заболевание выбрано из псориаза, ревматоидного артрита, рассеянного склероза, диабета типа-1, воспалительного заболевания кишечника, болезни Крона, аутоиммунного тиреоидита, аутоиммунной тяжелой псевдопаралитической миастении, системной красной волчанки, язвенного колита, атопического дерматита, миокардита и связанных с трансплантацией заболеваний, таких как реакции «трансплантат против хозяина» или «хозяин против трансплантата», или проблем общей органной толерантности. Лечение псориаза и ревматоидного артрита является особенно предпочтительным.
Настоящее изобретение, кроме того, относится к способу лечения индивида, страдающего от аутоиммунного заболевания или отторжения трансплантата, или профилактики аутоиммунного заболевания или отторжения трансплантата у индивида, включающему извлечение образца, содержащего регуляторные T-клетки CD4+CD25+ из организма индивида, осуществление контакта образца с антителом или фрагментом антитела, которое описано в настоящей публикации, чтобы активировать регуляторные T-клетки CD4+CD25+, и введение активированных клеток индивиду. Такой способ может дополнительно включать стадию in vitro увеличения количества Treg-клеток. Это можно осуществить, используя методику экспансии, описанную в настоящей публикации (Peters et al., 2008).
Подобным образом настоящее изобретение относится к активированным регуляторным T-клеткам CD4+CD25+, которые были активированы in vitro с использованием антитела или фрагмента антитела согласно настоящему изобретению. Такие активированные регуляторные T-клетки могут применяться в медицине, и в частности применяться для лечения аутоиммунного заболевания или отторжения трансплантата. Подобным образом, настоящее изобретение относится к применению регуляторных T-клеток CD4+CD25+, активированных с использованием антитела или фрагмента антитела согласно настоящему изобретению для производства лекарственного средства для применения при лечении аутоиммунного заболевания или отторжения трансплантата.
У пациентов с аутоиммунным заболеванием, таким как ревматоидный артрит, обнаруживают Treg, которые обладают более низкой способностью к супрессии, что может быть следствием влияния провоспалительных цитокинов, таких как TNF-альфа. Настоящее изобретение относится к способу скрининга, выделения и/или активации Treg пациентов, страдающих от аутоиммунного заболевания, которые могут иметь (но не обязательно) неработающую популяцию Treg. Специфичная форма связывания антитела и его фрагментов согласно настоящему изобретению обеспечивает возможность не только для связывания антитела с CD4, но что более важно, для активации Treg. Выделение Treg можно осуществлять, используя BT061 или антитело или фрагмент антитела согласно настоящему изобретению. В случае, когда будет следовать стадия активации, селекция в отношении CD4 должна осуществляться с использованием CD4-антитела, которое не конкурирует с BT061 за связывание с CD4. Альтернативно методика экспансии может быть включена перед стадией активации.
В настоящем изобретении Treg могут быть выделены с использованием BT061 и/или антитела или фрагмента антитела согласно настоящему изобретению и перенесены назад пациенту в ходе «основанной на Treg-клетках иммунотерапии». Альтернативно Treg можно стимулировать непосредственно при введении пациенту либо внутривенно, либо подкожно.
Основанная на Treg иммунотерапия привлекает большой общественный интерес для индукции толерантности при аутоиммунных заболеваниях или трансплантациях. Существует несколько способов получения индуцируемых Treg (Treg) in vitro, таких как использование ретиноевой кислоты, индуцирующей экспрессию FoxP3, или совместное культивирование с полученными из костного мозга ДК в добавление к стимуляции анти-CD3- и анти-CD28-антителами. Например, заявка PCT No. PCT/US2009/054631 относится к способу очистки Treg FoxP3 для лечения аутоиммунных заболеваний («основанная на Treg иммунотерапия») с использованием LAP и CD121b.
Терапевтические применения требуют больших количеств Treg и такие клетки должны сохранять свой регуляторный фенотип. В настоящее время это достигается только посредством селекции природных Treg. Индуцируемые Treg, образованные способами in vitro имеют недостаток, состоящий в том, что их фенотип может возвращаться к фенотипу эффекторных клеток, что является причиной непредсказуемого риска для пациентов.
Однако было показано, что BT061 активирует Treg в реакции смешанной культуры лимфоцитов (WO 2009112502 A1). Такая активация является следствием неожиданного специфичного характера связывания с CD4, описанного в настоящей публикации.
Применения in vitro
Антитело и фрагменты антитела и выделенные пептиды согласно настоящему изобретению также имеют ряд применения in vitro. В частности, антитело или фрагмент антитела, описанные в настоящей публикации, можно применять для активации регуляторных T-клеток CD4+CD25+ in vitro или для идентификации регуляторных T-клеток CD4+CD25+ in vitro.
Более конкретно, антитело или фрагмент антитела согласно настоящему изобретению можно использовать в способе скрининга в отношении присутствия регуляторных T-клеток CD4+CD25+ в образце. Такой способ может включать стадию осуществления контакта меченого антитела или фрагмента антитела с образцом, промывки образца для удаления несвязанного антитела и выявления присутствия метки в образце.
В частности, в таком способе скрининга регуляторные T-клетки CD4+CD25+ являются активированными регуляторными T-клетками CD4+CD25+. Образец, предпочтительно, является образцом крови, взятом у индивида, страдающего от аутоиммунного заболевания.
Настоящее изобретение также относится к набору для выделения регуляторных T-клеток CD4+CD25+, содержащему магнитные шарики, покрытые антителом или фрагментом антитела, описанным в настоящей публикации. Набор может дополнительно содержать второе анти-CD25-антитело и/или анти-CD4+-антитело. Также могут быть включены дополнительные антитела для осуществления дополнительных стадий селекции, например, позитивной селекции в отношении CD39, стадии негативной селекции в отношении CD127, истощения CD19-позитивных клеток. LAP (связанный с латентностью пептид), GARP или CD121b (рецептор II-1 типа 2) можно дополнительно использовать для характеристики фенотипа Treg.
После выделения Treg клетки, выделенные с использованием антитела или фрагмента антитела согласно настоящему изобретению также могут быть подвергнуты криоконсервации (Peters et al., PLoS One (2008) 3; 9: e 3161).
Кроме того, настоящее изобретение относится к способу in vitro для активации регуляторных T-клеток CD4+CD25+, включающему осуществление контакта клеток с антителом или фрагментами антитела, описанными в настоящей публикации. Способы оценки способности к супрессии активированных Treg включают (кроме указанной выше MLR), MLR-анализы, определяющие состояние активации эффекторных T-клеток по высвобождению цитокинов или экспрессии маркеров активации на эффекторных T-клетках (такие как анализ пролиферации и анализ цитокинов, описанные в WO 2009112592 A1, которая включена в настоящее описание в виде ссылки). Указанные способы могут дополнительно включать первую стадию выделения регуляторных T-клеток CD4+CD25+. Если такая стадия выполняется с использованием антитела, то такое антитело не должно быть конкурирующим с CD4-антителом (например, OKT4 или SK3). Это позволяет активировать клетки на основной стадии с использованием антитела или его фрагмента согласно настоящему изобретению, который связывается с другим эпитопом на CD4. В другом сценарии клетки могут быть выделены с использованием других поверхностных маркеров экспрессии Treg, которые отличаются от CD4, таких как CD25 или CD39, или в результате негативной селекции в отношении CD127.
Антитело согласно настоящему изобретению было описано ранее как способное стимулировать Treg, что может быть подтверждено при совместном культивировании с эффекторными T-клетками. Treg способны подавлять пролиферацию CD8-позитивных T-клеток посредством ингибирования продукции IL-2 и IFN-гамма аллореактивными CD8-позитивными T-клетками. Кроме того, было показано (WO 2009112592), что предварительно активированные Treg CD4+CD25+ делают супрессированные клетки CD8+ неспособными экспрессировать CD25 при повторной стимуляции.
Далее изобретение будет описано дополнительно в связи со следующими конкретными вариантами.
Примеры
ПРИМЕР 1. Кристаллическая структура CD4 в комплексе с BT061
Кристаллическую структуру CD4 человека в комплексе с Fab-фрагментом BT061 получали с использованием дифракции рентгеновских лучей.
Способ кристаллизации BT061 (Fab):CD4
Рекомбинантный CD4 человека получали, используя обычные способы: Различные конструкции CD4 клонировали стандартными способами в векторах для гетерологичной экспрессии в клетках насекомых с последующей очисткой с помощью iNTA. Fab-фрагмент BT061 отщепляли от интактного антитела, используя протеазу папаин и очищали на белке A. Затем Fab-фрагмент дополнительно очищали эксклюзионной хроматографией по размеру.
Комплекс CD4-Fab образовывали, смешивая очищенные белки с молярным избытком CD4 и используя дополнительную очистку эксклюзионной хроматографией по размеру.
Кристаллы комплекса CD4:BT061 получали способом сокристаллизации, имея в виду, что очищенный комплекс использовали в испытаниях по кристаллизации, в которых применяли как стандартный скрининг с использованием примерно 1200 разных условий, так и условия кристаллизации, идентифицированные с использованием литературных данных. Изначально полученные условия были оптимизированы с использованием стандартной методики, системного изменения параметров, оказывающих решающее влияние на кристаллизацию, таких как температура, концентрация белка, соотношение в капле и другие. Такие условия также корректировали, системно варьируя pH или концентрации преципитатов.
Кристаллы быстро замораживали и измеряли при температуре 100 K.
Данные рентгеновской дифракции комплекса CD4:BT061 с применением источника излучения SWISS LIGHT SOURCE (SLS, Villigen, Switzerland), используя криогенные условия. Структуру определяли и уточняли до конечного разрешения 2,9 Å.
Кристаллы относятся к пространственной группе P21 с двумя комплексами в ассиметричной единице. Данные обрабатывали, используя программы XDS и XSCALE. Статистическая обработка собранных данных суммирована в таблице 6 ниже.
Статистика сбора и обработки данных
2 Числа в скобках соответствуют элементу разрешения с Rsym=44,3%.
Информацию о фазах, необходимую для определения и анализа структуры, получали при молекулярном замещении. Опубликованные модели CD4 и Fab-фрагмента использовали в качестве модели для поиска.
Последующее построение и уточнение модели осуществляли согласно стандартным протоколам, используя пакет компьютерных программ CCP4 и COOT. Ассиметричная единица (которая показана на фигуре 3) состоит из двух таких комплексов, определенных с разрешением RMS 2,9 Å.
В кристаллической структуре сигнальные пептиды молекулы CD4 отсутствуют, и пептидные цепи содержат только Ig-подобный домен V-типа, Ig-подобный домен C2-типа 1 и часть Ig-подобного домена C2-типа 2. Точные цепи в кристаллической структуре тянутся от аминокислотного остатка 26 до остатка 258, как показано на фигуре 1, на которой аминокислоты, не представленные кристаллической структурой, обозначены внутри пунктирной рамки. Соответственно, молекулы CD4 имеют только первых два дисульфидных мостика, один между Cys41 и Cys109, а другой между Cys155 и Cys184.
Легкие цепи двух единиц BT061 в кристаллической структуре простираются от положения 1 до положения 215 в одной единице, и от положения 1 только до положения 182 в другой единице, как показано на фигуре 2A. Имеется два дисульфидных мостика в легких цепях BT061, Cys23-Cys92 и Cys138-Cys198.
Тяжелые цепи двух единиц BT061 в кристаллической структуре простираются от положения 2 до положения 219 в одной единице, как показано на фигуре 2B, и от положения 2 до положения 220 в другой единице. Также в тяжелых цепях имеются два дисульфидных мостика, один между Cys22 и Cys98, а другой между Cys151 и Cys207.
Анализ кристаллической структуры показывает, что в отличие от других известных лигандов CD4 в связывание BT061 вовлечен только Ig-подобный домен C2-типа 1. Это является совершенно новым способом связывания, документально доказанным на основании кристаллической структуры, приведенной в Приложении.
Как показано на фигуре 3, BT061 не связывается с N-концевым доменом CD4, существует возможность одновременного связывания BT061 и молекулы MHC класса II или белка оболочки ВИЧ-1 gp120 (как показано на фигурах 4 и 5).
Подробный анализ кристаллической структуры выявил взаимодействующие аминокислоты как CD4, так и BT061. Отбор был основан на критерии расстояния. Принимая во внимание разрешение кристаллической структуры, все аминокислоты CD4, имеющие неводородные атомы в сфере радиусом d=4,5 Å вокруг неводородного атома аминокислоты BT061, были отобраны в качестве взаимодействующих партнеров. Радиус d был выбран в виде типичного расстояния невалентного взаимодействия, 3 Å, увеличенного на половину разрешения кристаллической структуры 2,9 Å (d=3 Å+½×2,9 Å=4,45 Å).
Следовательно, кристаллическая структура показывает, что участок связывания на поверхности CD4 состоит из кластера из 7 последовательных аминокислот (Gly148-Gln154), кластера из 6 последовательных аминокислот (Gln164-Thr168), плюс Thr185, Leu187, Asn189, Gln190 и Lys192, как показано на фигуре 7.
В таблице 7 ниже показана матрица взаимодействующих партнеров CD4 и BT061. Существует только небольшое количество чистых взаимодействий 1:1; большинство из перечисленных аминокислот как в CD4, так и в BT061 взаимодействуют более чем с одним партнером. Взаимодействия, главным образом, представлены водородными связями, дополненными контактами Ван-дер-Ваальса и полярными взаимодействиями.
В верхнем ряду таблицы 7 показаны аминокислоты CD4, которые взаимодействуют с аминокислотами BT061. Слева приведены аминокислоты BT061, взаимодействующие с CD4. Каждый «x» соответствует по меньшей мере одному взаимодействию. Так как существует только небольшое количество пар, взаимодействующих 1:1, находят картину, продолжающуюся в рядах и колонках. Ser150 и Pro151 CD4 взаимодействуют как с легкой, так и с тяжелой цепью BT061. Val186, который является важной частью кармана связывания для Tyr105 тяжелой цепи BT061, в данном случае не перечислен, поскольку не имеет прямых взаимодействий с аминокислотами BT061. Все указанные взаимодействия являются либо водородными связями, полярными взаимодействиями, контактами Ван-дер-Ваальса или сочетаниями указанных типов взаимодействий. Идентификация взаимодействующих аминокислот основана на кристаллической структуре, приведенной в Приложении. Критерий расстояния использовали, принимая во внимание все аминокислоты, которые содержат по меньшей мере один неводородный атом, расположенный ближе чем d=4,5 Å к неводородному атому другого молекулярного партнера. Грубо говоря, d соответствует типичному расстоянию для нековалентного взаимодействия 3 Å, увеличенному на половину общего разрешения кристаллической структуры (2,9 Å).
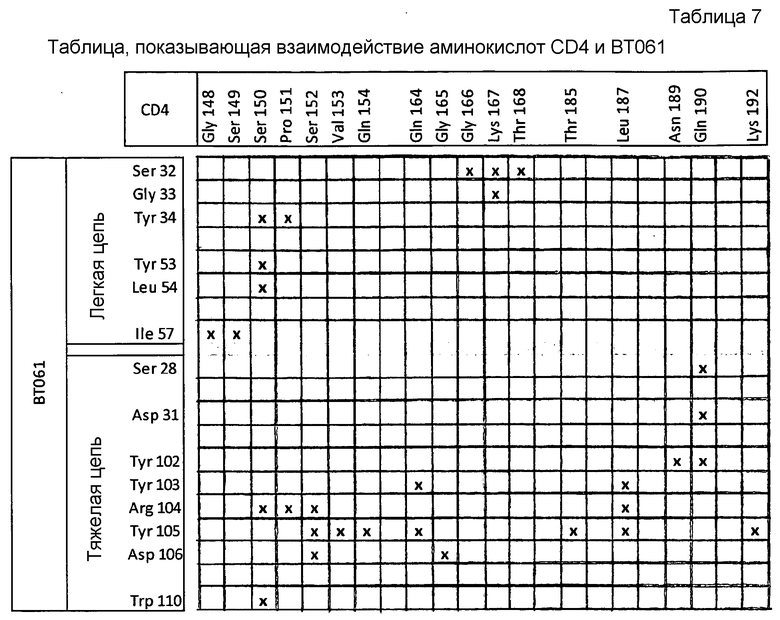
На фигуре 7 показаны аминокислоты CD4 сайта связывания BT061 в виде схематичного представления их относительной ориентации. Кроме аминокислот, непосредственно удовлетворяющих указанному выше критерию, предъявляемому к расстоянию, несколько аминокислот доступны для дополнительных взаимодействий с BT061 при небольших изменениях конформации BT061 и CD4. Такие аминокислоты, Arg156, Gln188, Asn189, Gln190 и Lys192, вероятно вовлечены в механизм сигнальной трансдукции CD4.
BT061 связывается как легкой, так и тяжелой цепью с CD4. Соответствующие аминокислоты легкой цепи показаны на фигуре 8. Аминокислоты тяжелой цепи, вовлеченные в связывание с CD4, показаны на фигуре 9.
Tyr105 тяжелой цепи BT061 играет очень важную роль во взаимодействии с CD4. Его боковая цепь точно подходит для кармана на поверхности CD4 (фигура 10).
Несмотря на то, что солевые мостики являются типичными элементами взаимодействия между антителами, солевых мостиков между CD4 и BT061 не существует в кристаллической структуре. Интересно, что в BT061 обнаружено несколько внутримолекулярных солевых мостиков. Один из них, образованный между Arg104 и Asp106, обеспечивает полярные взаимодействия с карманом на поверхности CD4 (фигура 11). На образование внутримолекулярных солевых мостиков могут влиять вещества с буферной активностью, которые в свою очередь могут индуцировать конформационные изменения BT061 и комплекса с CD4, таким образом внося вклад в сигнальную трансдукцию CD4.
Другим остатком, очень важным для взаимодействия с CD4, является Tyr34 в легкой цепи BT061. Подобно Tyr105 тяжелой цепи Tyr34 укладывается в карман на поверхности CD4 (фигура 12).
Сравнивая кристаллическую структуру комплекса CD4 с BT061 с кристаллическими структурами комплексов CD4 с молекулой MHC класса II и с белком оболочки ВИЧ-1 gp120 можно непосредственно увидеть, что BT061 связывается с совершенно другой частью на поверхности CD4. Что касается других лигандов CD4, то BT061 связывается с противоположной стороны внеклеточной части CD4 (фигуры 4 и 5) и не является помехой для сайтов связывания других лигандов.
Аминокислоты CD4 Lys26, Arg156, Lys161 и Lys192 доступны для дополнительных взаимодействий с заряженными дополнительными аминокислотами BT061, которые указаны в таблице 8, показанной ниже.
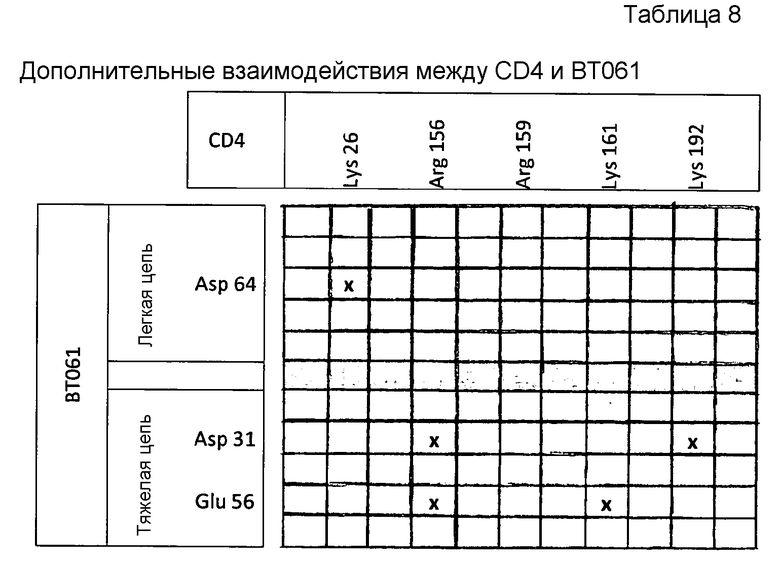
В верхнем ряду таблицы 8 показаны аминокислоты CD4, которые могут образовывать солевые мостики с аминокислотами BT061. Слева приведены аминокислоты BT061, являющиеся возможными партнерами солевых мостиков. Каждый «x» соответствует возможному солевому мостику. Указанные пары не подпадают под критерий расстояния d=4,5 Å, указанный выше. Однако серия небольших конформационных изменений в CD4 и BT061 может привести к образованию указанных солевых мостиков. В случае BT061 сдвиг участков последовательности Ser56-Gly68 легкой цепи, и Ser25-Cys32, а также Ser52-Gly59 тяжелой цепи позволяет образовываться солевым мостикам с Asp31 и Glu56 CD4.
ПРИМЕР 2. Создание и тестирование мутантов BT061
Мутанты вариабельного домена BT061 получали введением специфичных мутаций в нуклеотидные последовательности, кодирующие вариабельные домены BT061, и экспрессией мутантной аминокислотной последовательности и, следовательно, антитела или фрагментов антител с использованием стандартных методик обработки ДНК и получения антител.
Получили следующие варианты BT061:
(1) шесть вариантов, имеющих одну мутацию, каждая в одной из 6 CDR: мутации в CDR1, CDR2 и CDR3 легкой цепи представляют собой мутации G33A, A55G и L98I, соответственно; мутации в CDR1, CDR2 и CDR3 тяжелой цепи представляют собой R33K, A63G и S101T, соответственно;
(2) один вариант, имеющий две мутации в одном вариабельном домене: R33K и A63G в вариабельном домене тяжелой цепи; и
(3) три варианта, имеющие параллельную мутацию и в тяжелой и в легкой цепи (например, сочетания вариантов тяжелой и легкой цепей из части (1): L98I легкой цепи с R33K тяжелой цепи; A55G легкой цепи с R33K тяжелой цепи; и G33A легкой цепи с R33K тяжелой цепи.
Способность вариантов BT061 активировать регуляторные T-клетки CD4+CD25+ определяли в анализе in vitro. В качестве негативного контроля использовали антитело, которое не связывается с CD4. Также создавали вариант BT061, имеющий мутацию в тяжелой цепи CDR3 (Y105W), в качестве негативного контроля; на основании результатов, полученных в примере 1, было сделано предположение, что такая аминокислота является важной для связывания BT061. В качестве позитивных контролей использовали антитело, полученное из исходной последовательности легкой и тяжелой цепи, а также образец BT061, полученного для клинических испытаний.
Анализ in vitro:
Индукцию способности к супрессии регуляторных T-клеток CD4+CD25+ (Treg) здоровых доноров вариантами антитела BT061 оценивали в реакциях смешанных аллогенных культур лимфоцитов (непрямое совместное культивирование).
Регуляторные T-клетки CD4+CD25+ представляли собой свежевыделенные клетки от первого донора (донор A). PBMC получали из EDTA-крови центрифугированием в градиенте плотности. Treg, T-клетки-респондеры (Tresp) и АПК отделяли иммуномагнитным способом от PBMC, как описано ранее (Haas et al., J Immunol. 2007 Jul 15; 179(2): 1322-30). Treg предварительно инкубировали в присутствии или без связанного с планшетом антитела в течение 2 суток. Количество и фенотип Treg определяли, используя шестицветную проточную цитометрию, как описано ранее (Haas et al., 2007). Предварительно инкубированные Treg переносили к T-клеткам-респондерам (второй донор B) в присутствии истощенных по T-клеткам и облученных PBMC (первый донор A). После стимуляции добавляли [3H]-тимидин и через пять дней измеряли пролиферацию T-клеток-респондеров.
Результаты:
Результаты анализа in vitro показаны ниже в таблице 9. Ингибирование в процентах T-клеток-респондеров измеряли три раза для каждого варианта и вычисляли среднее.
Результаты анализа in vitro с использованием вариантов BT061
Как предполагалось, нокаутированный мутант LC-исходная HC Y105W не связывал CD4 и не обладал способностью активировать регуляторные-T клетки. Кроме того, результаты показывают, что отдельные положения в тяжелой цепи могут быть подвергнуты мутациям по меньшей мере в соответствии с заменами, указанными в таблице 5 (варианты с отдельными мутациями HC R33K, A63G и двойные варианты HC R33K и A63G сохраняли активность). Кроме того, как предполагалось на основании работы по определению кристаллической структуры, остатки, окружающие Tyr34 в легкой цепи, которые образуют петлю, по-видимому, также являются ключевыми для связывания.
| название | год | авторы | номер документа |
|---|---|---|---|
| ТЕРАПЕВТИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2002 |
|
RU2328506C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ IL-10 ДЛЯ ЛЕЧЕНИЯ СИСТЕМНОЙ КРАСНОЙ ВОЛЧАНКИ (SLE) | 2010 |
|
RU2587622C2 |
| АНТИТЕЛО ПРОТИВ TNFR2 И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2793165C1 |
| АНТИТЕЛА ПРОТИВ ОХ40 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2562874C1 |
| СПОСОБ ЛЕЧЕНИЯ АУТОИММУННОГО ЗАБОЛЕВАНИЯ (ВАРИАНТЫ) | 2009 |
|
RU2539110C2 |
| ЧЕЛОВЕЧЕСКОЕ ОПУХОЛЕСПЕЦИФИЧЕСКОЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО | 2008 |
|
RU2488593C2 |
| АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ ИНТЕРЛЕЙКИНУ-2 И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2745451C1 |
| ЛЕЧЕНИЕ РАКА С ПОМОЩЬЮ ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА К CD33 | 2015 |
|
RU2747384C2 |
| НОВЫЕ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА К БЕЛКУ ПРОГРАММИРУЕМОЙ СМЕРТИ 1(PD-1) | 2016 |
|
RU2757316C2 |
| СРЕДСТВО ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЯ | 2009 |
|
RU2540013C2 |





























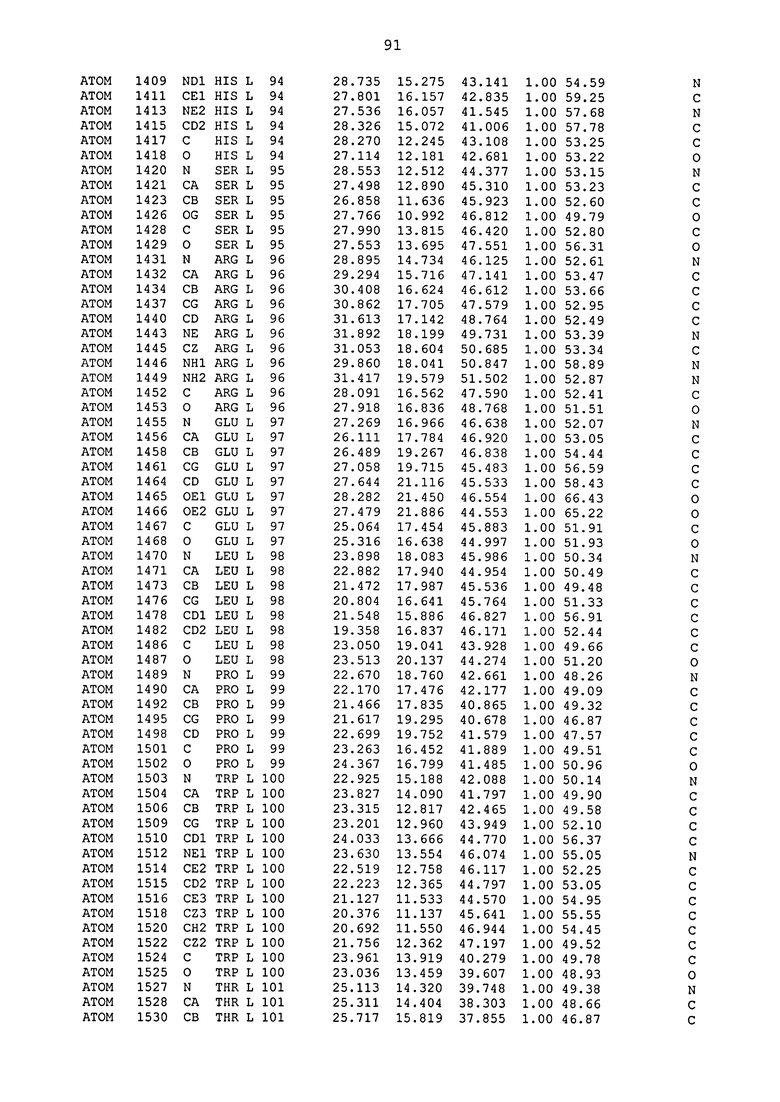
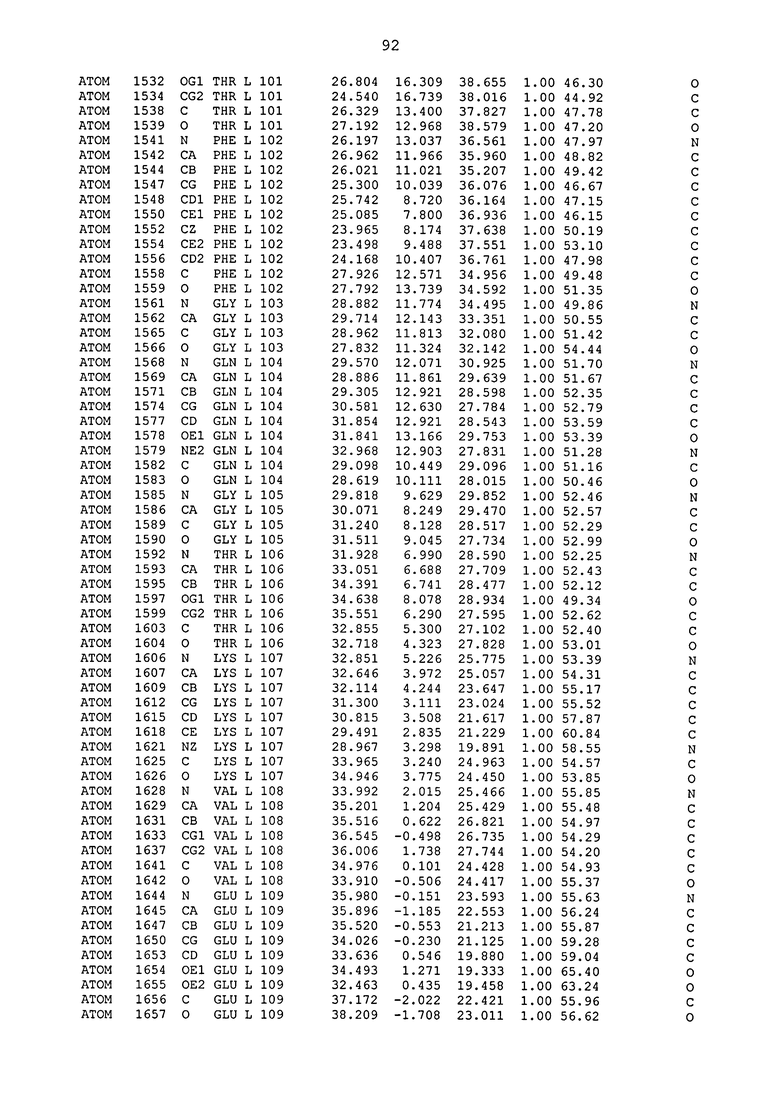
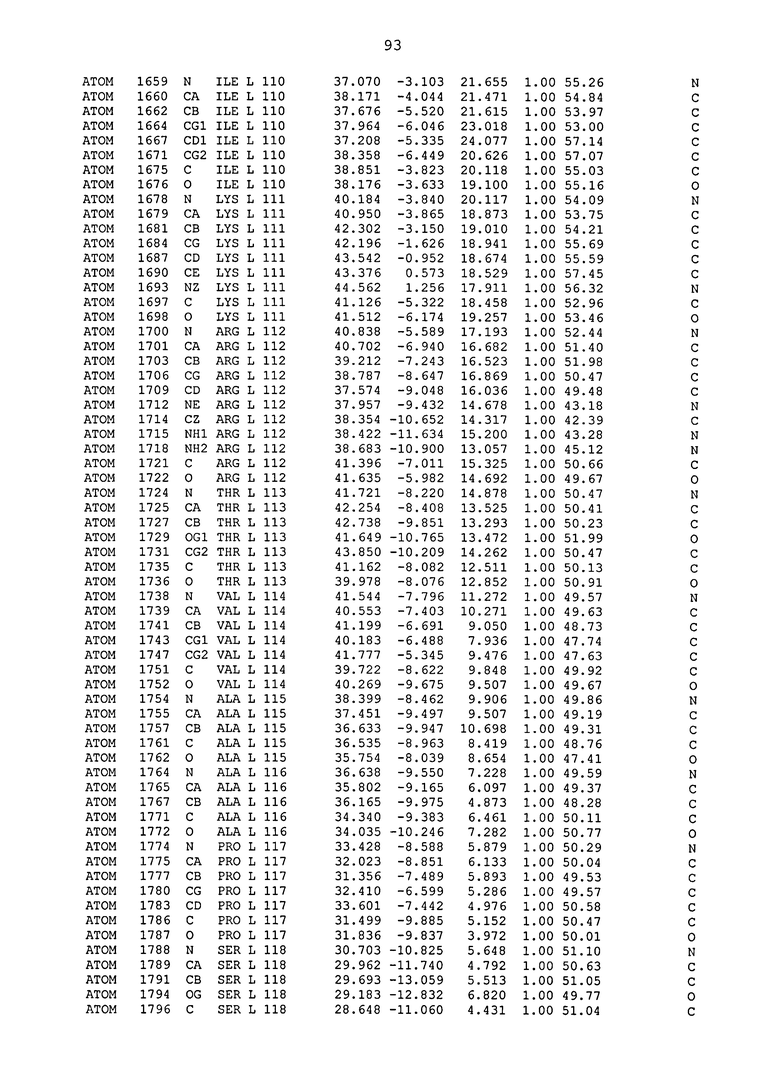

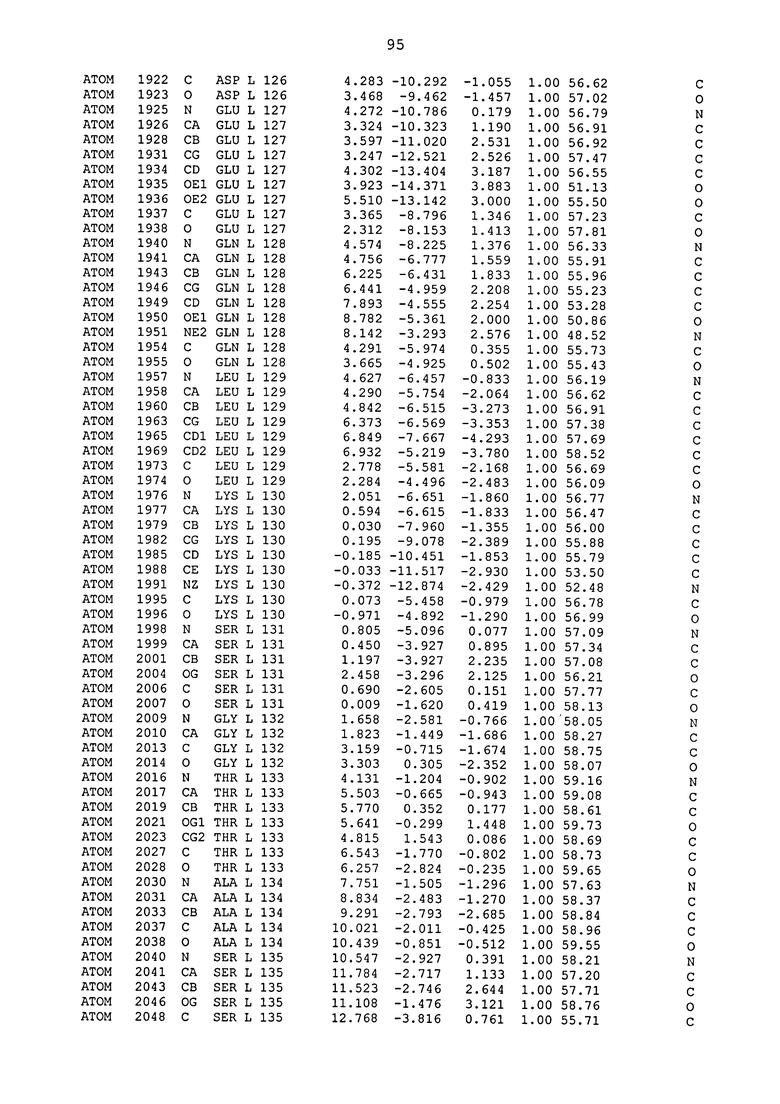







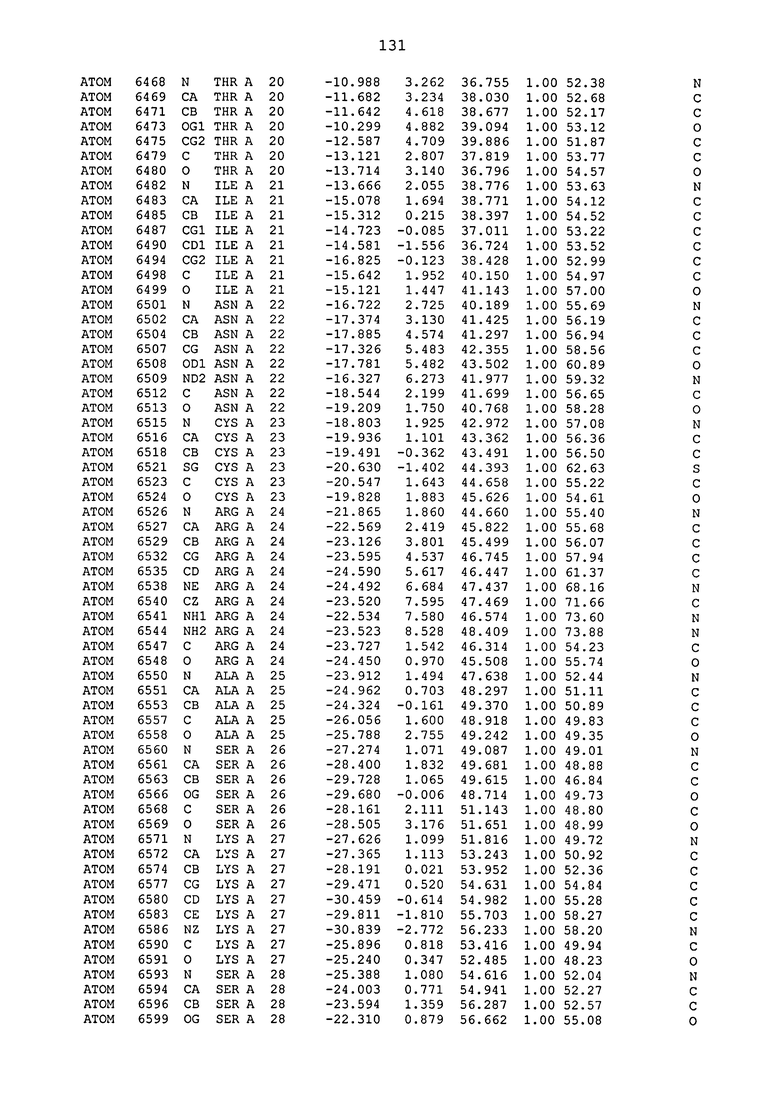
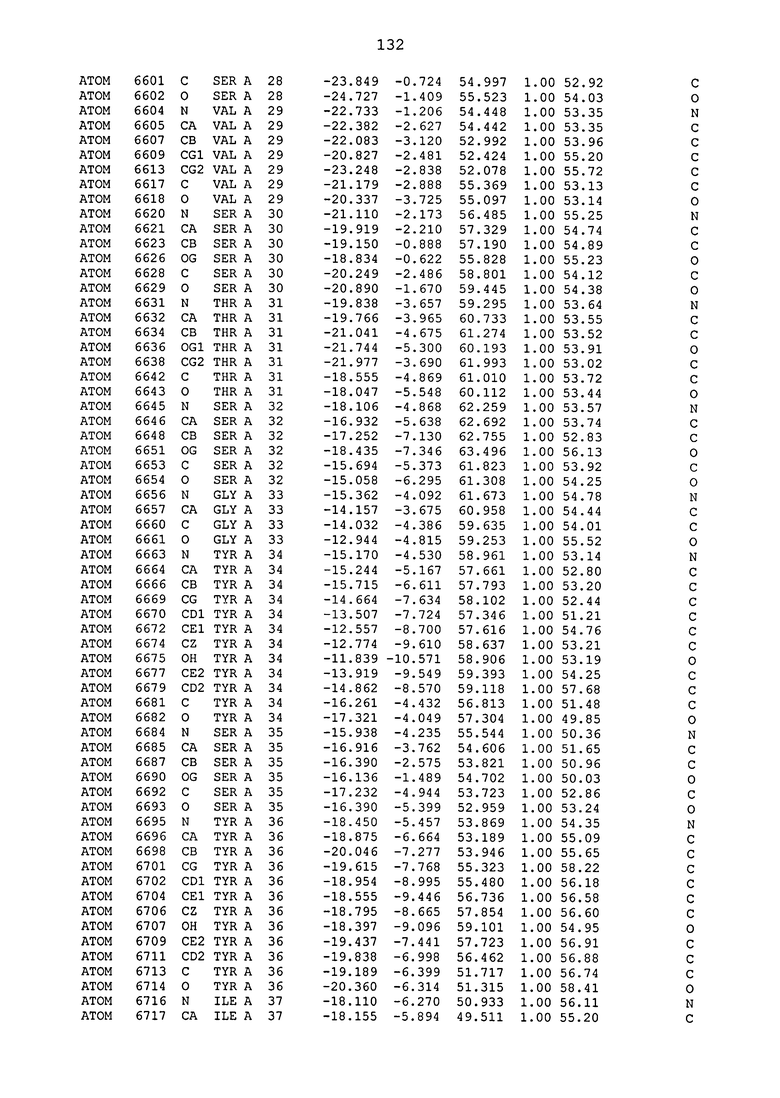

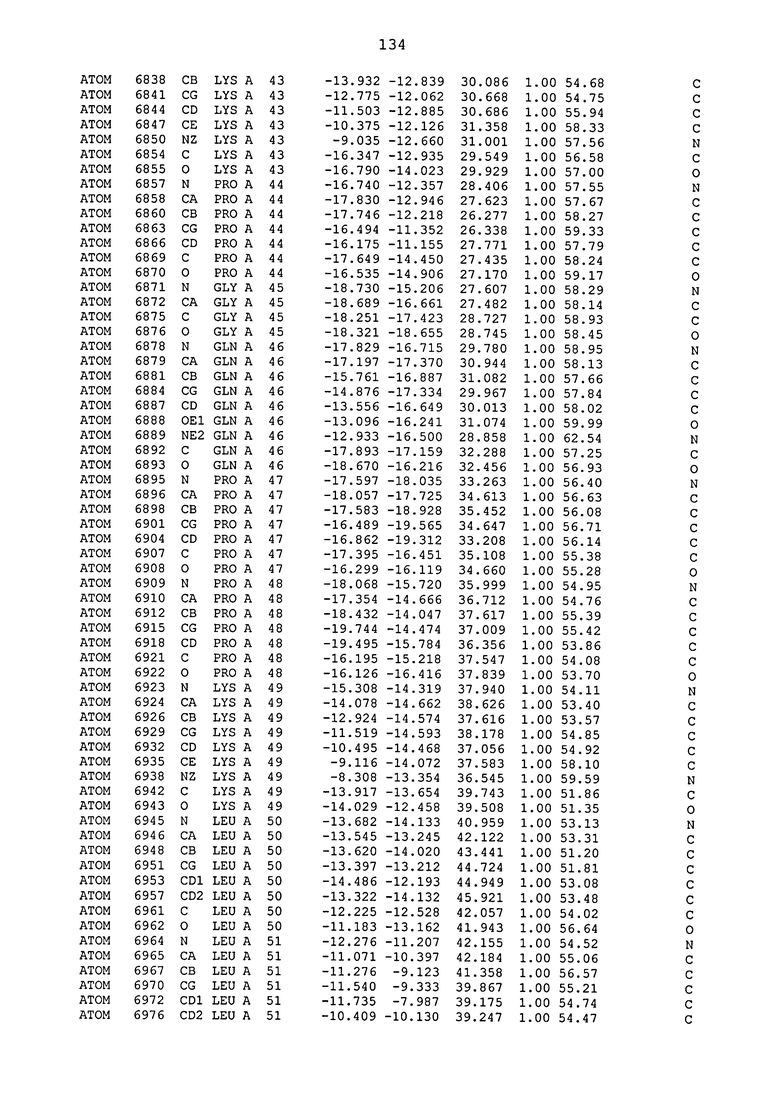
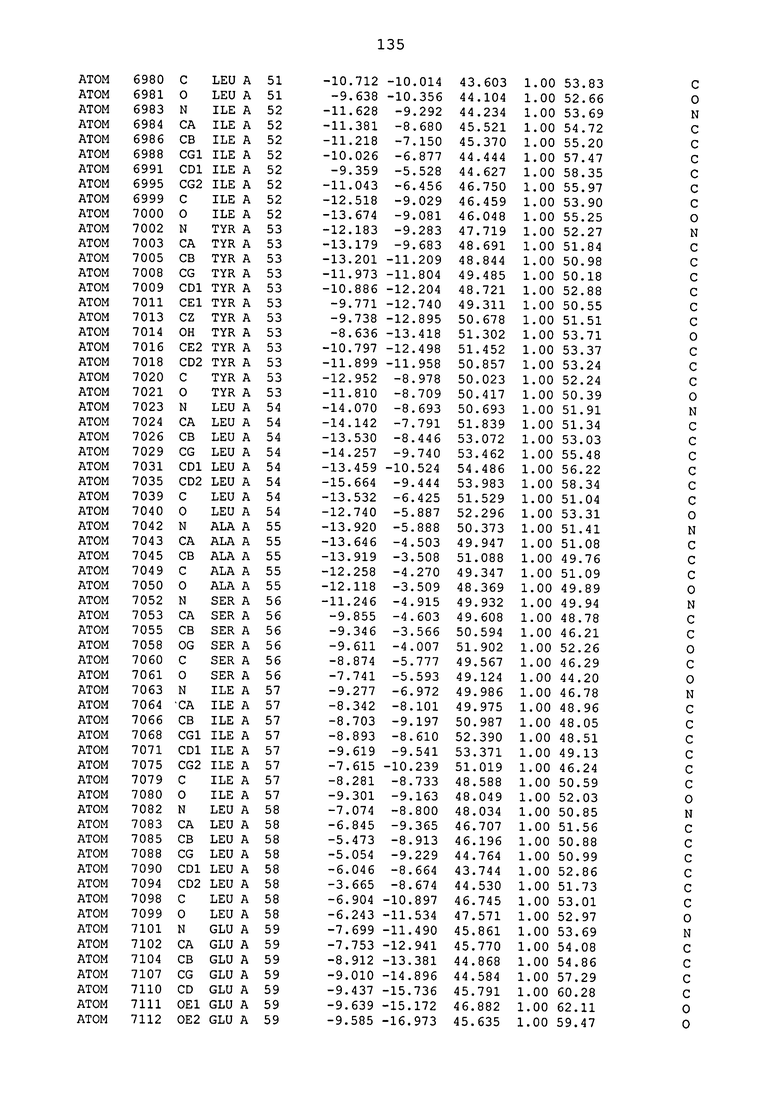












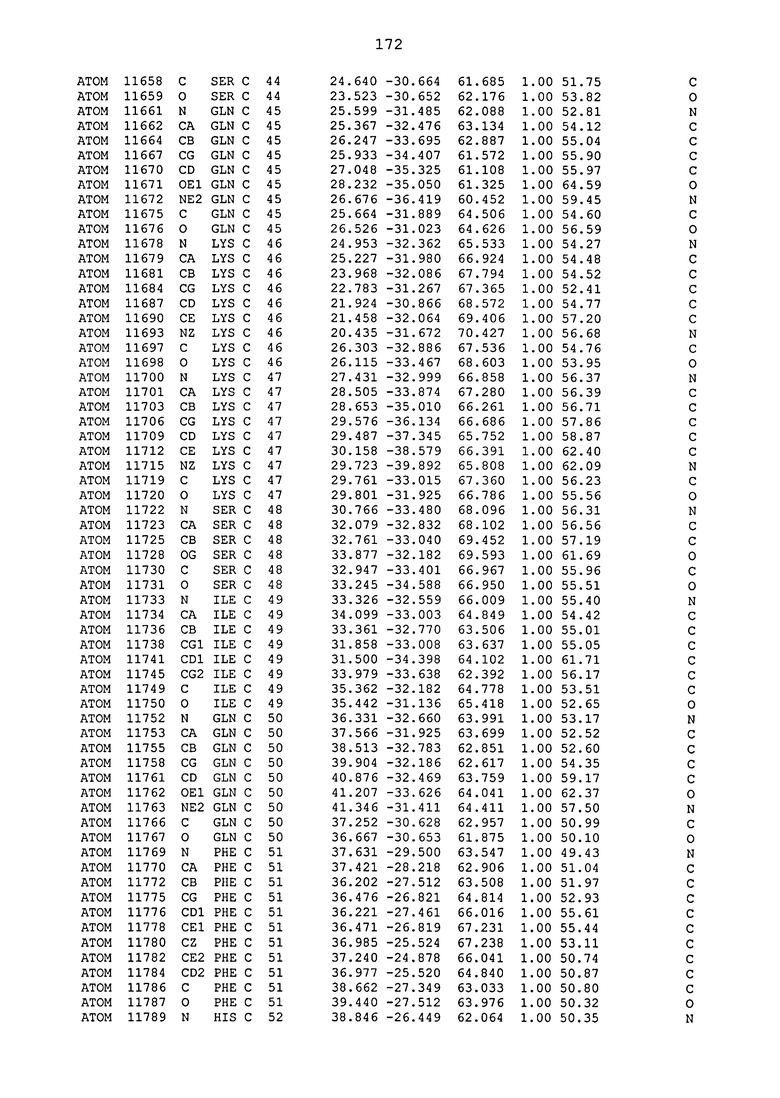
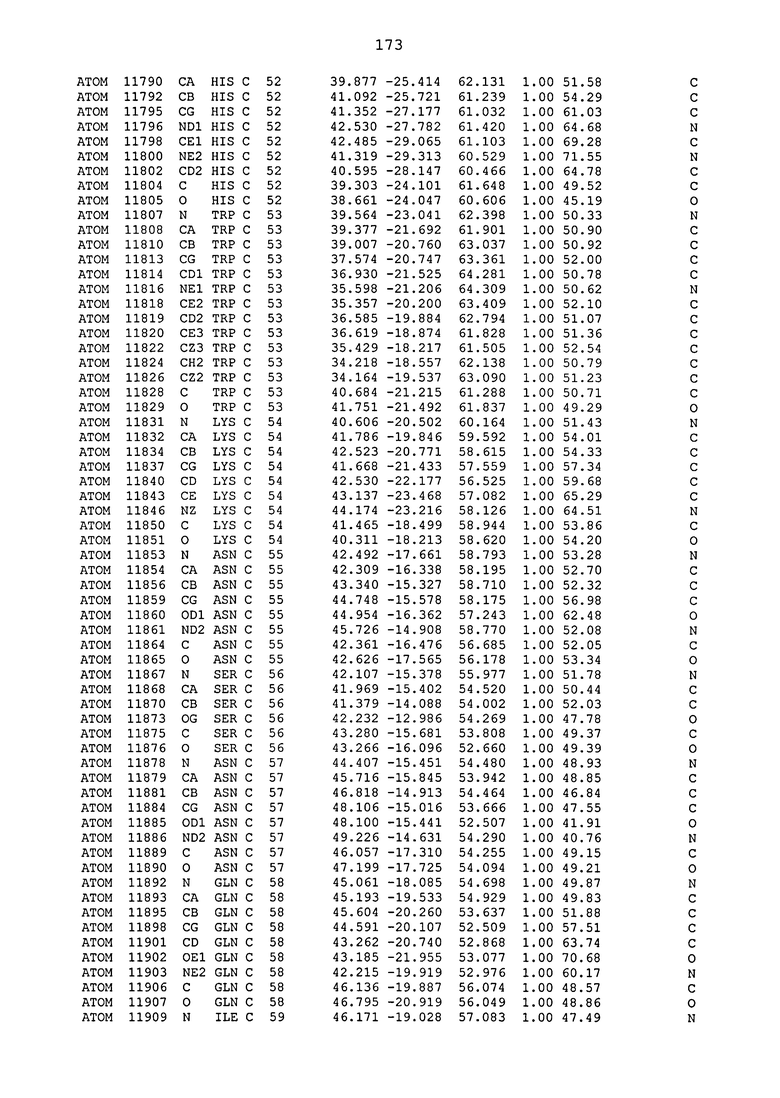
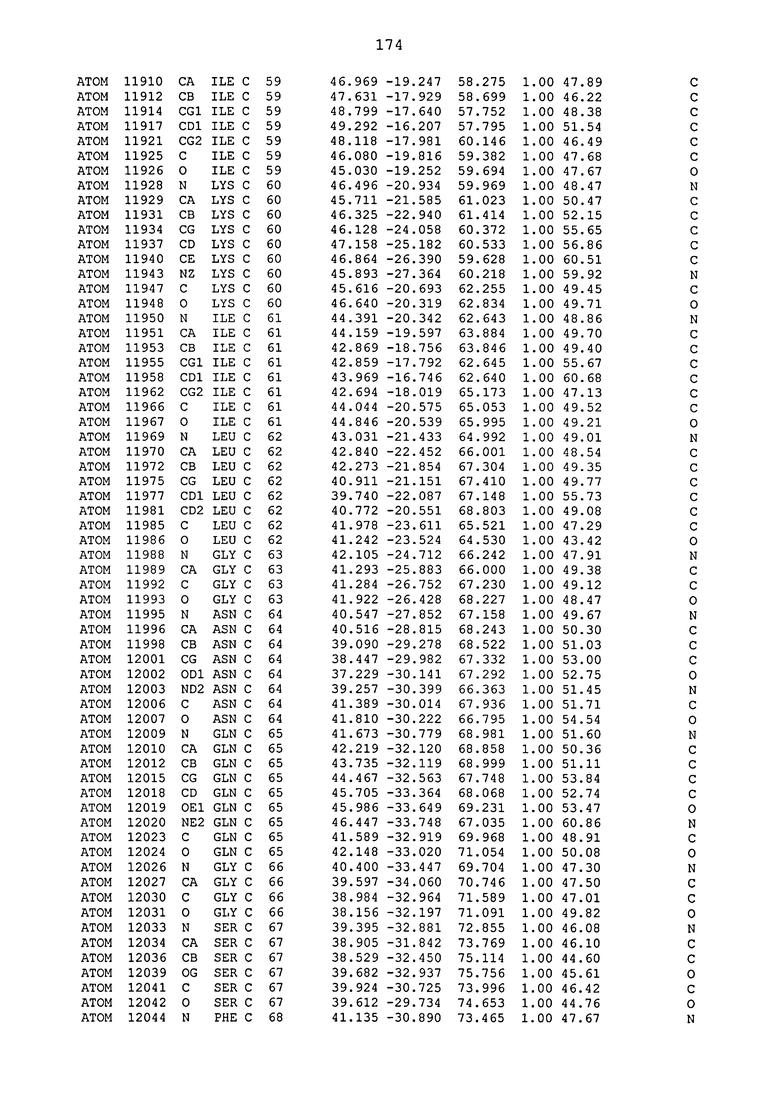

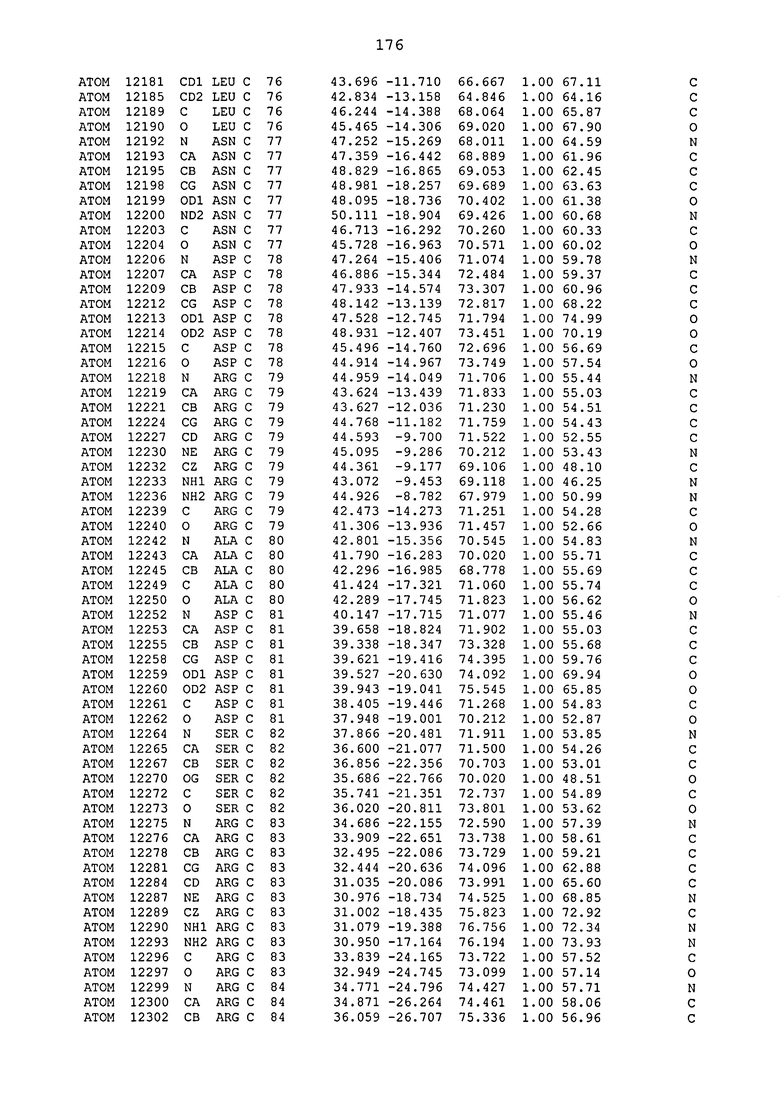

















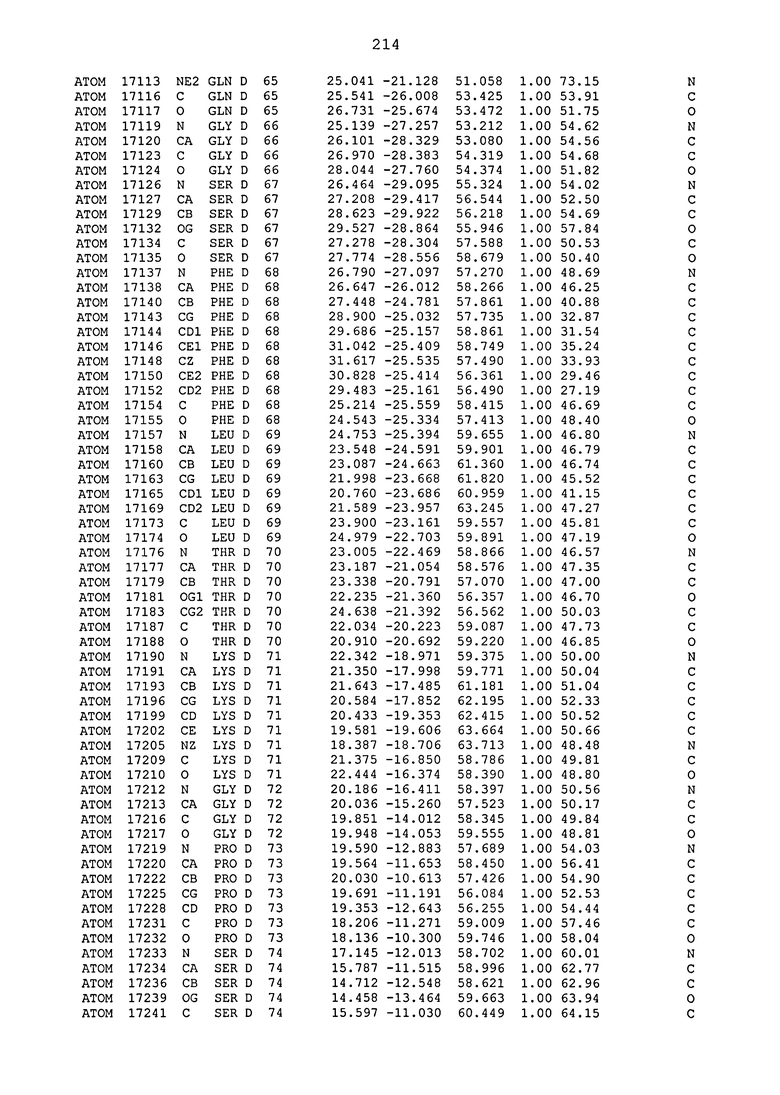
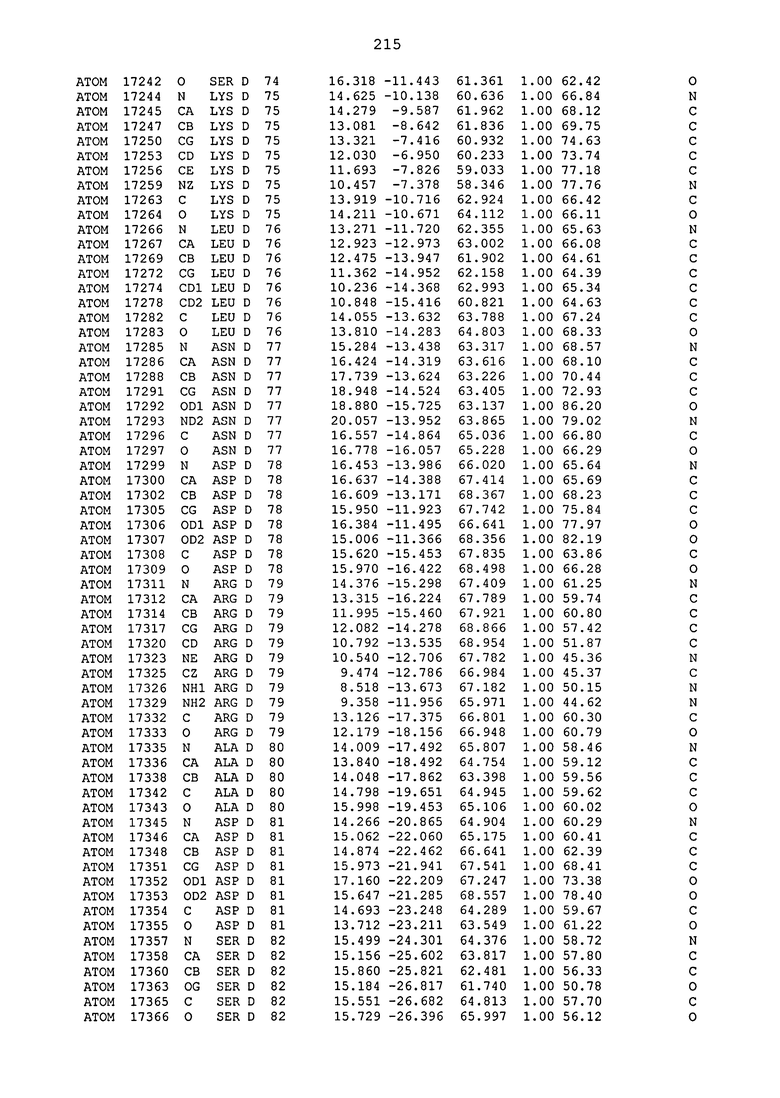
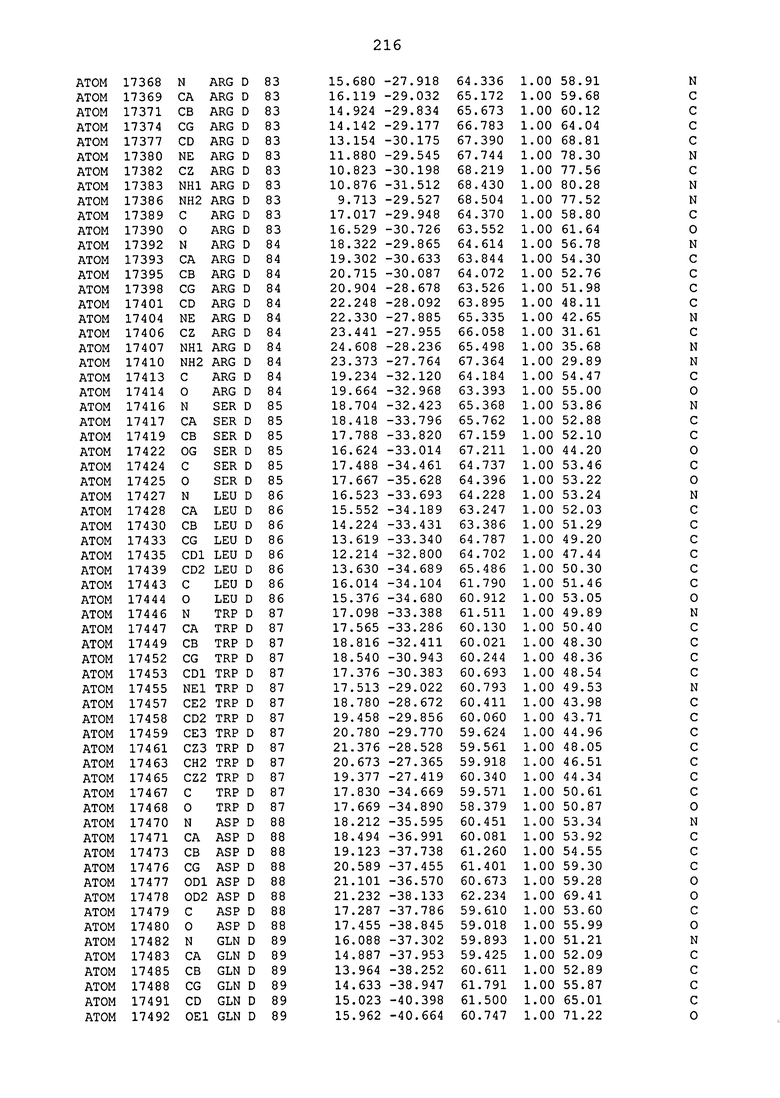
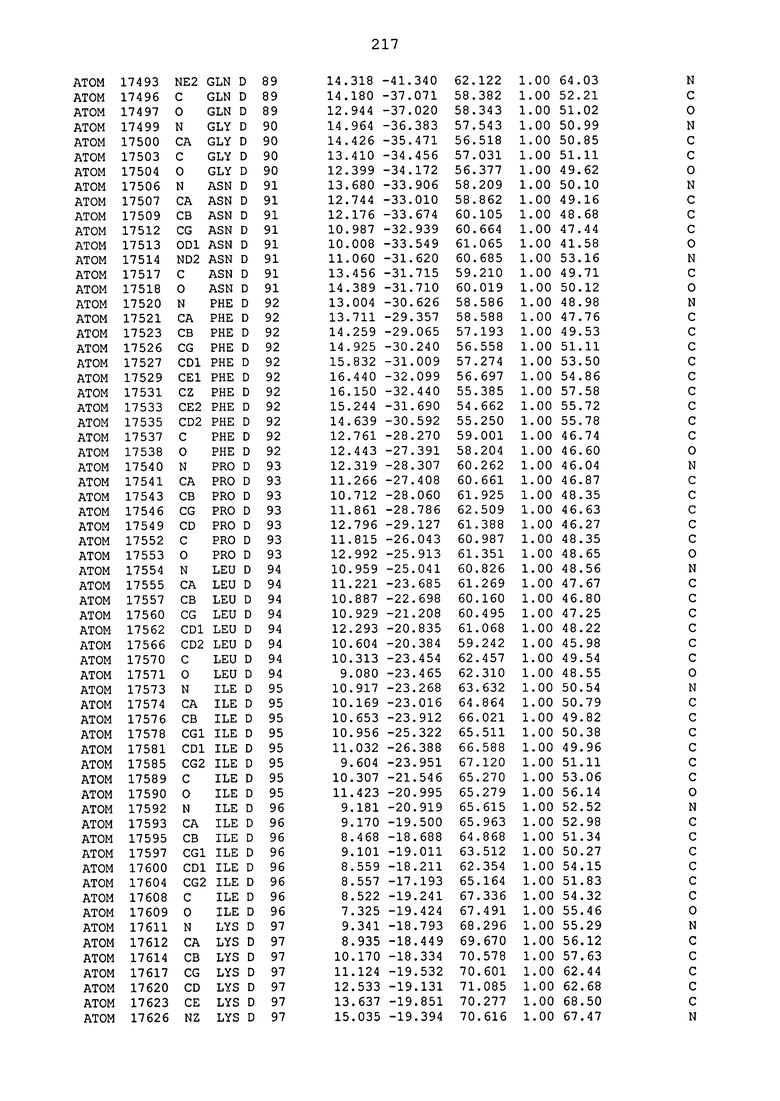








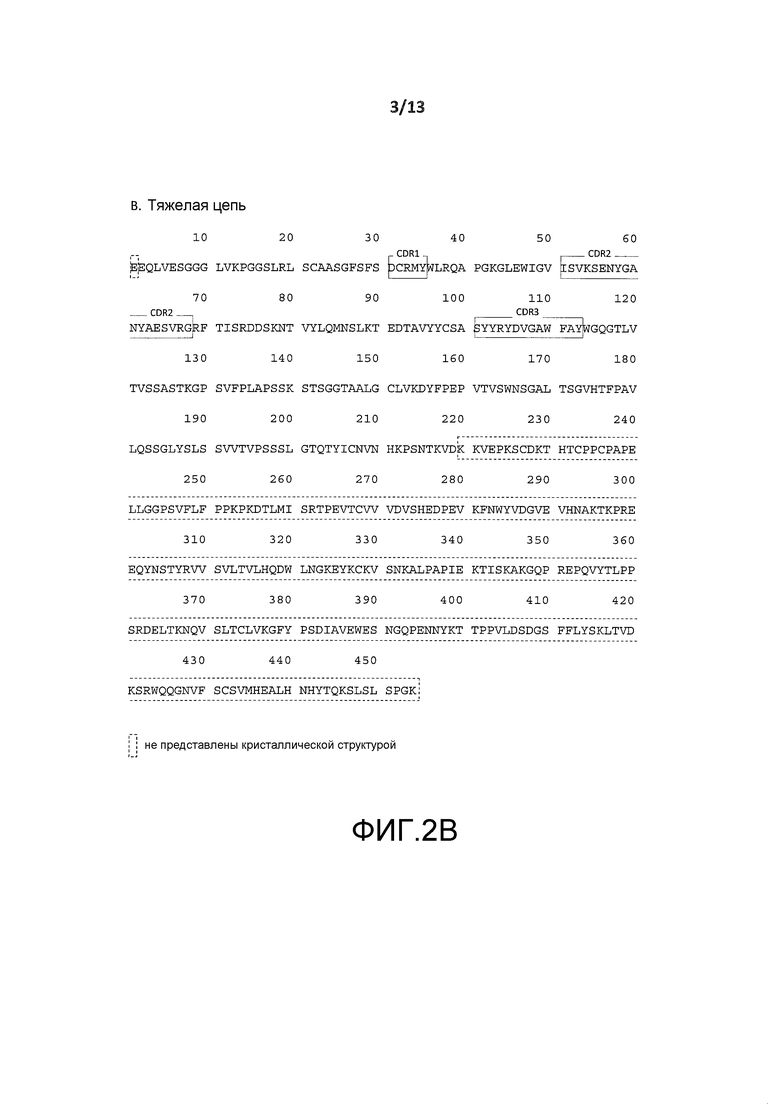
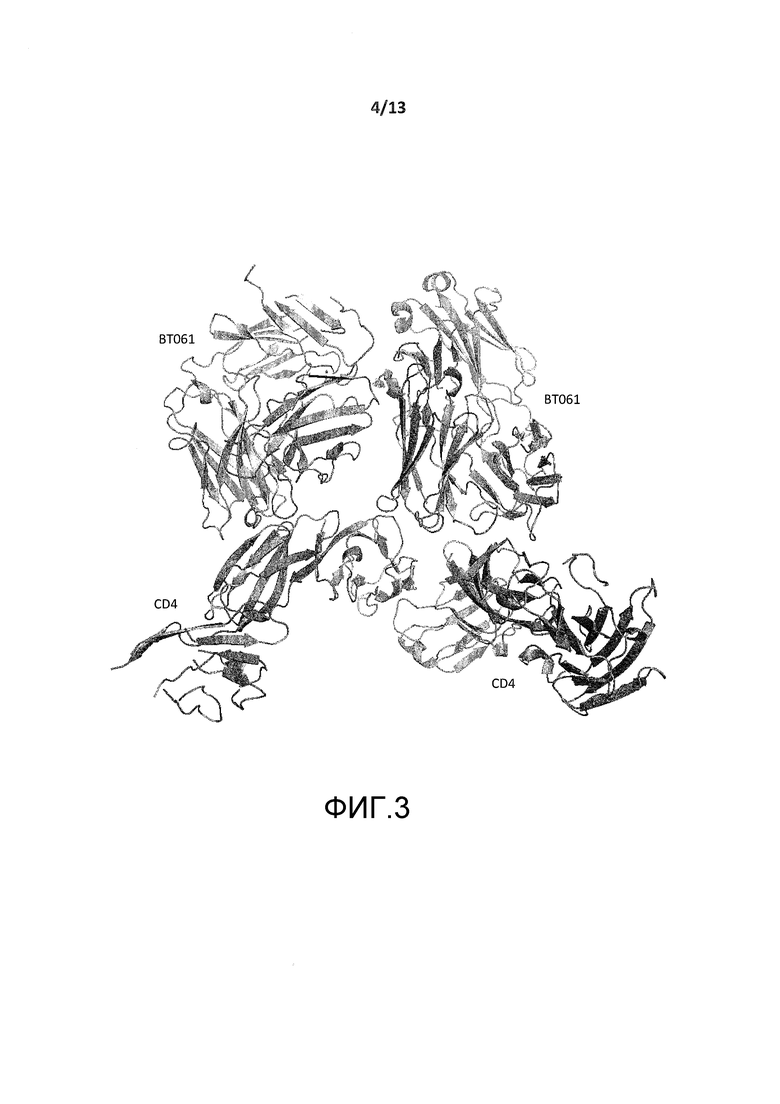
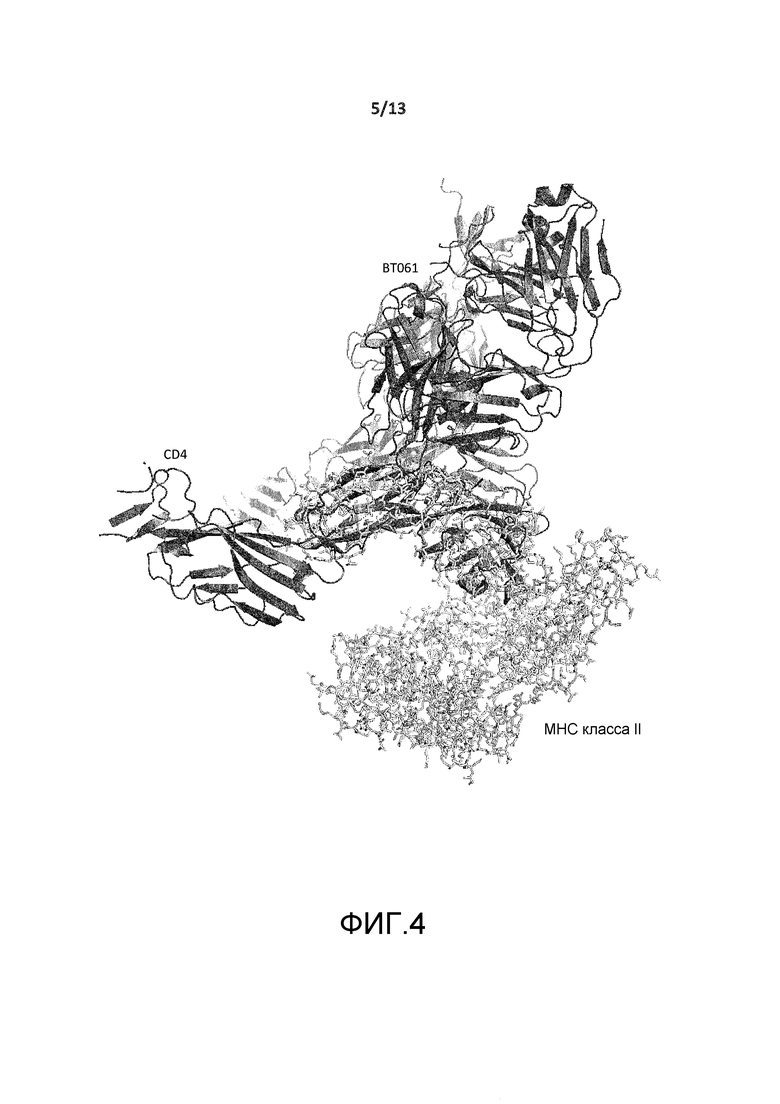
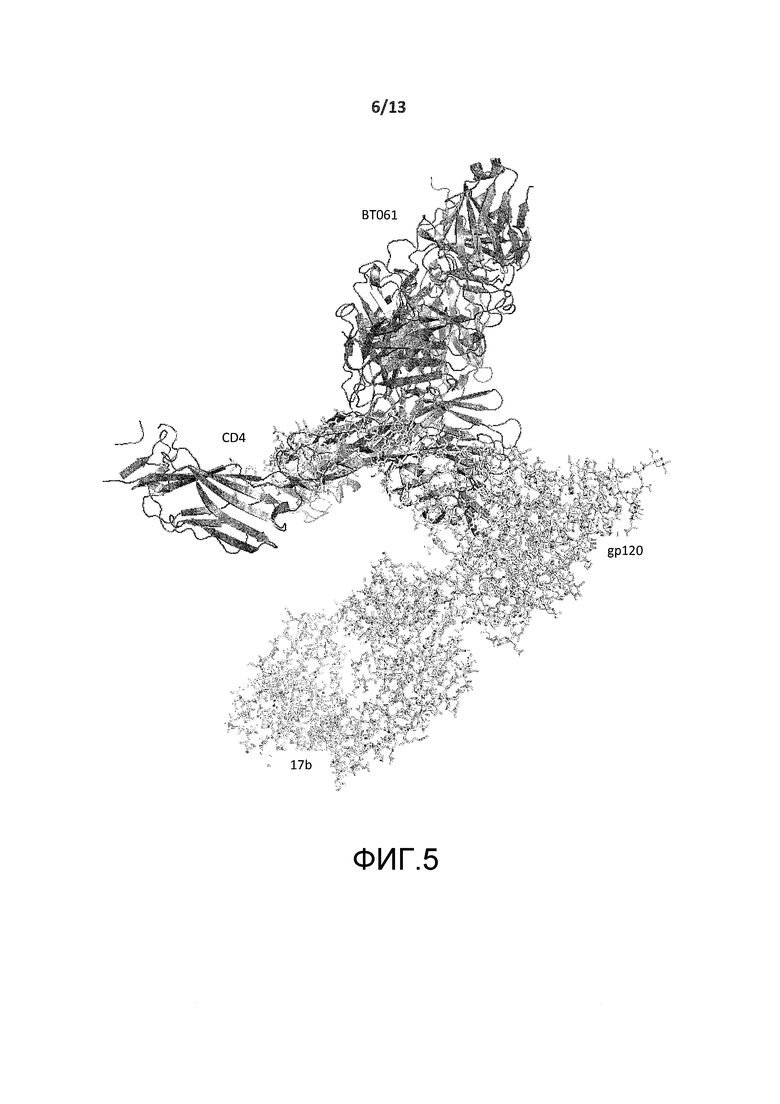

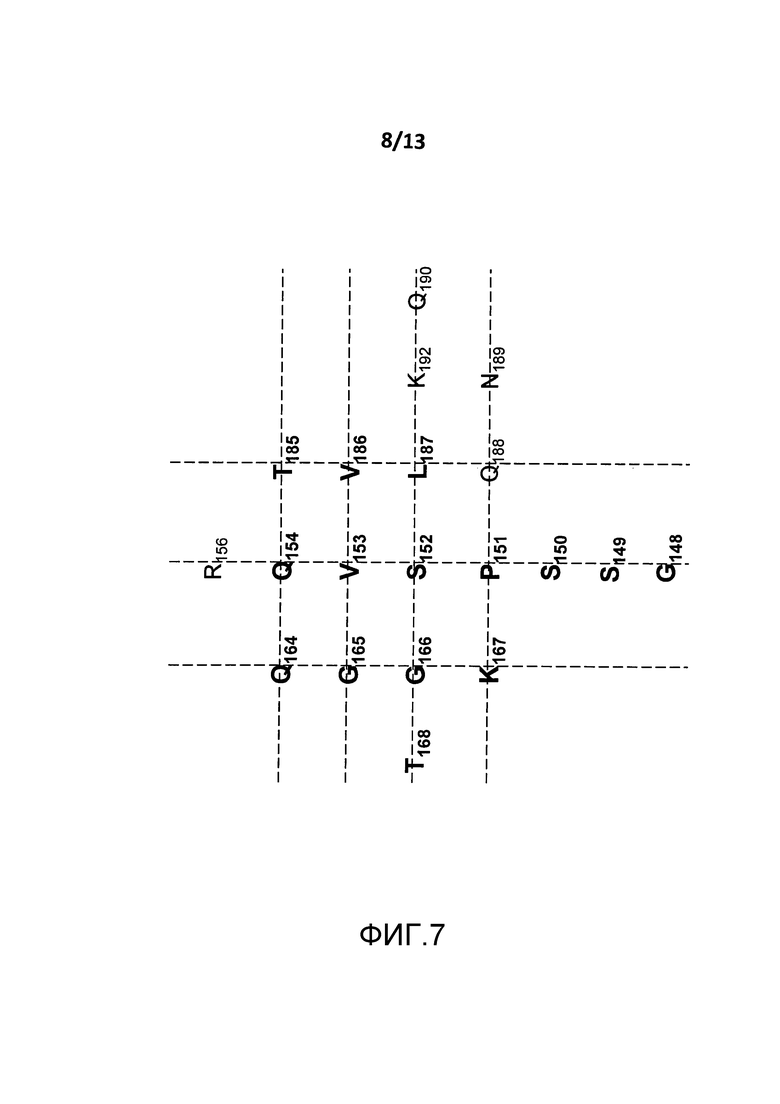
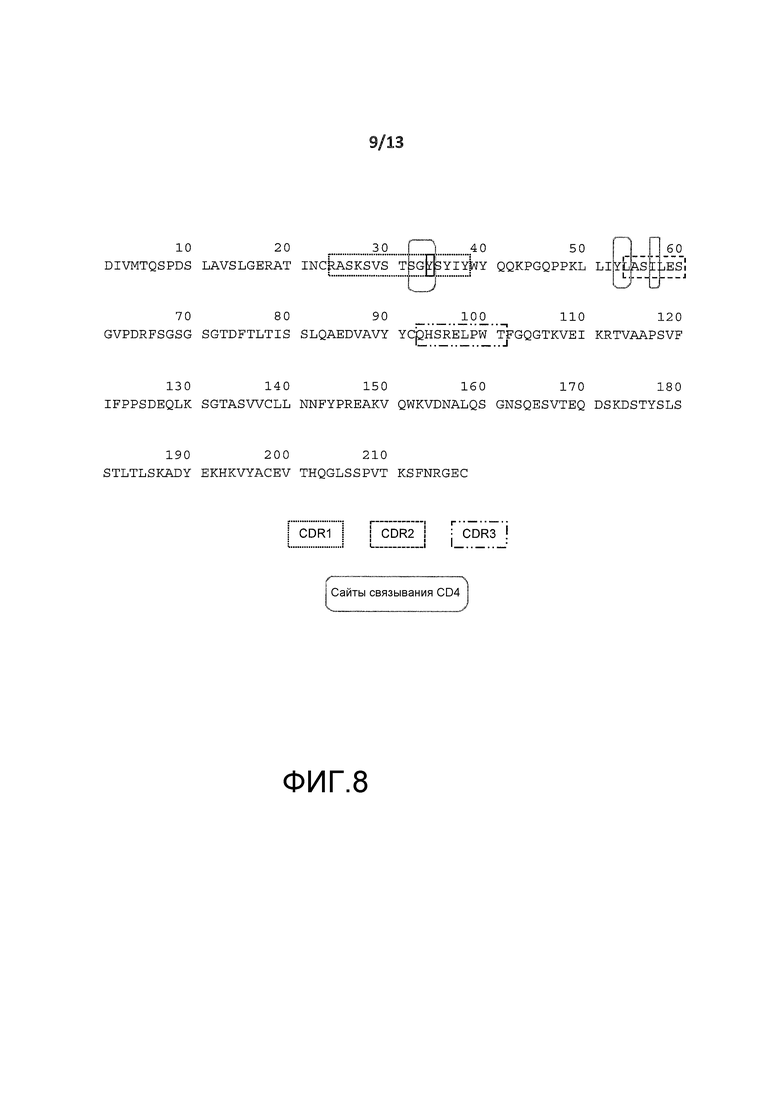
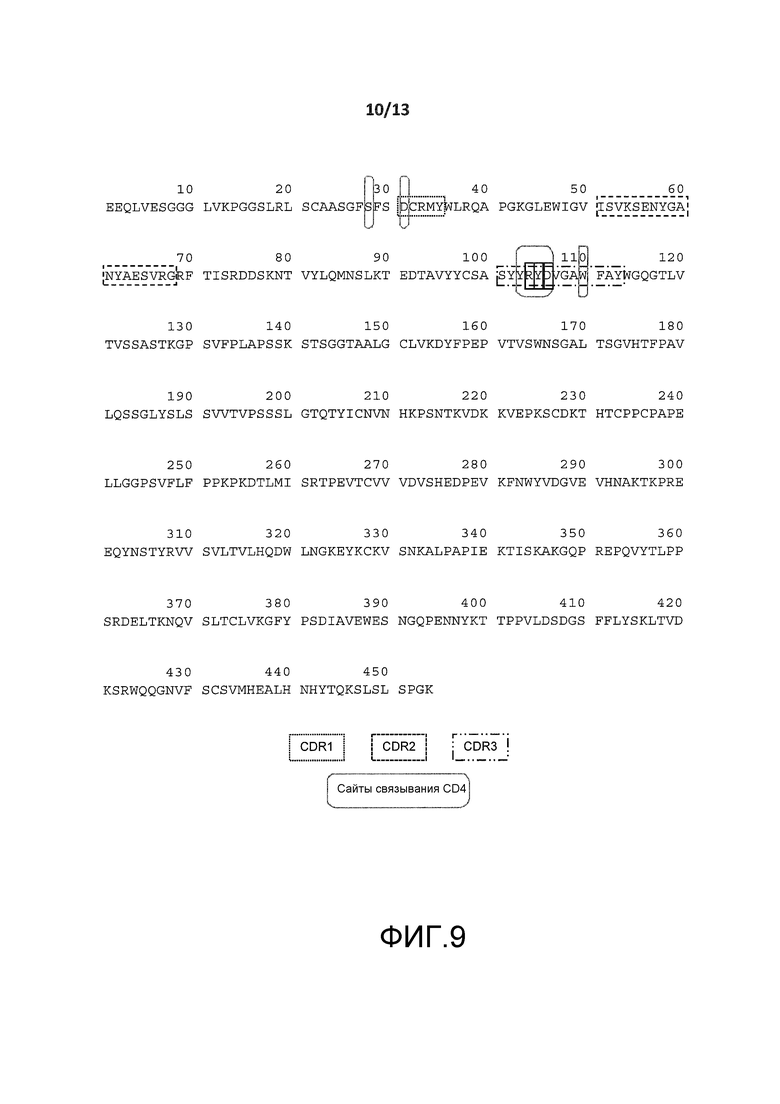
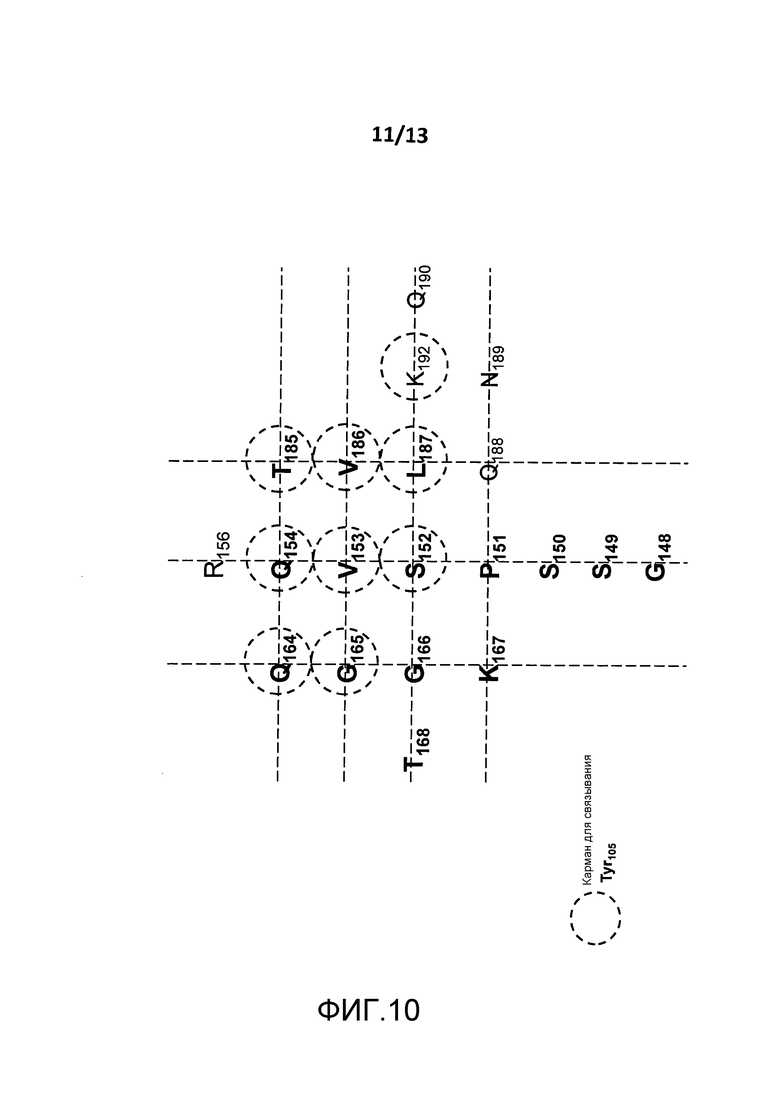
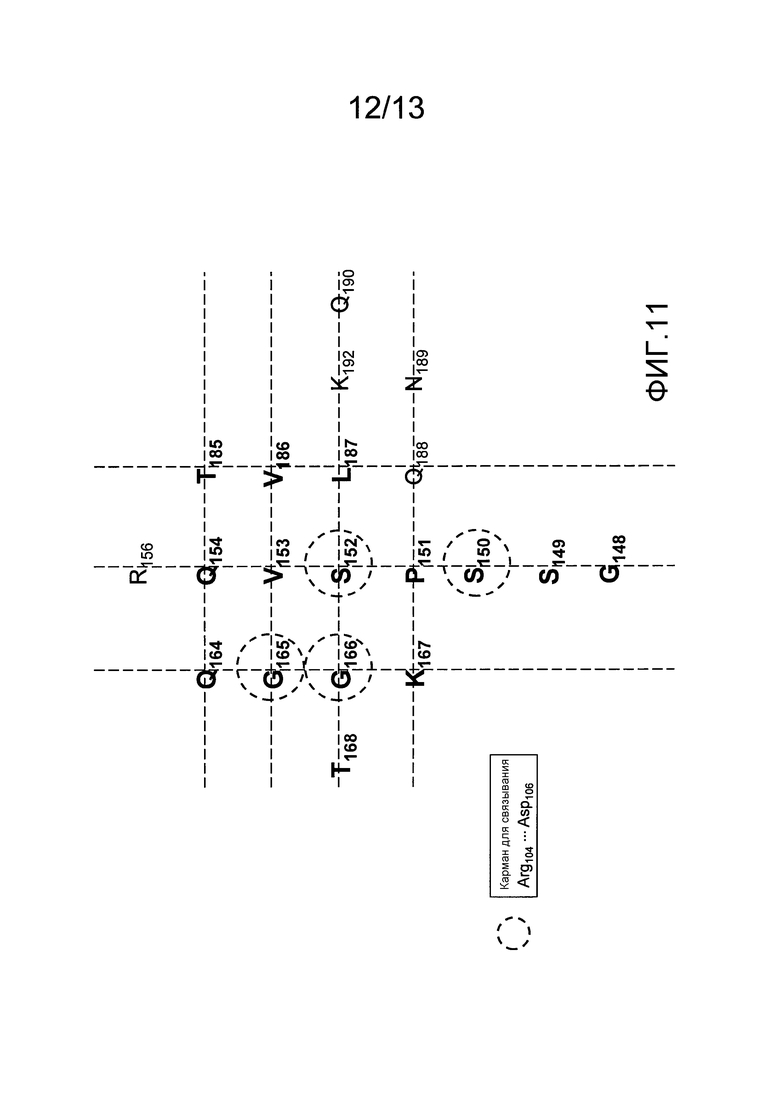
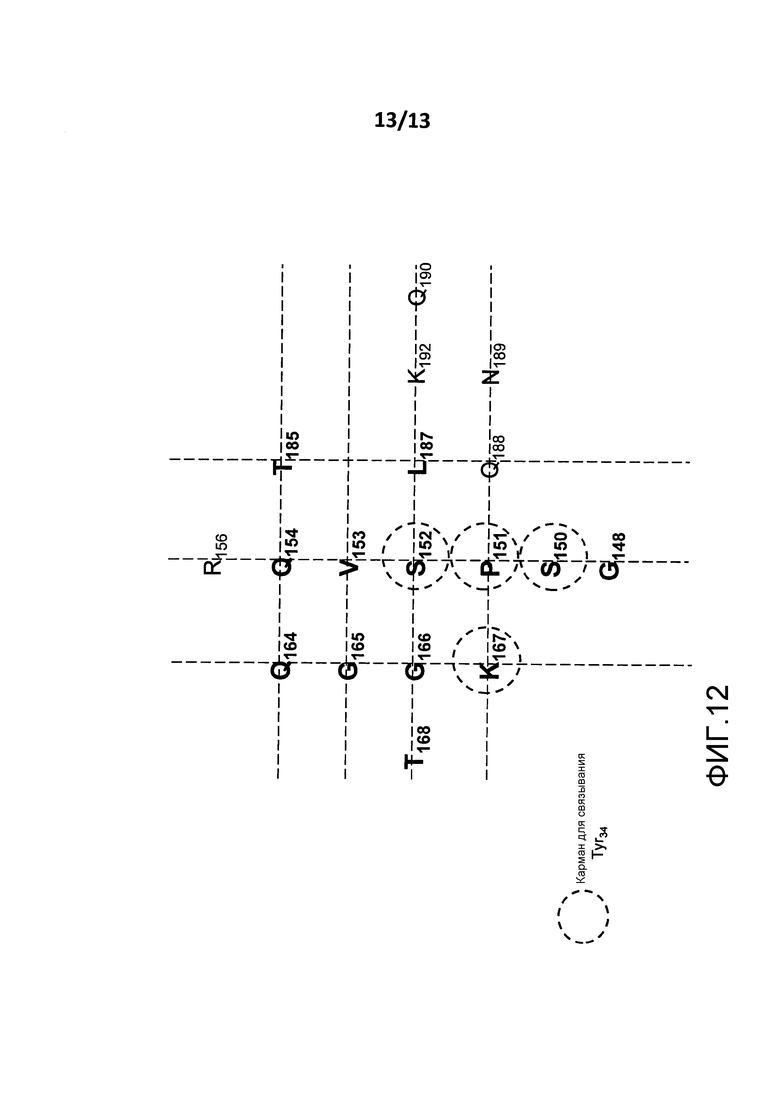
Изобретение относится к области иммунологии. Предложен способ скрининга молекулы, способной связываться с Ig-подобным доменом С2-типа 1 CD4 человека, а именно с аминокислотными остатками в положениях 148-154, 164-168 и 185-192. Также рассмотрены способы получения терапевтической композиции; способ скрининга антитела или его фрагмента; антитела, полученные на основе антитела ВТ061, и их фрагменты; выделенный пептид из CD4 человека; мимотоп; а также нуклеиновая кислота, вектор экспрессии, клетка-хозяин и способ получения антитела по изобретению; фармацевтическая композиция и способы лечения индивида, страдающего от аутоиммунного заболевания или отторжения трансплантата, или профилактики указанного; способ in vitro активации регуляторных Т-клеток; применение антитела или его фрагмента для получения лекарственного средства и для активации регуляторных Т-клеток; применение выделенного пептида или мимотопа для скрининга; способ скрининга в отношении присутствия регуляторных Т-клеток в образце; набор для выделения регуляторных Т-клеток. Данное изобретение относится к новому эпитопу CD4 человека и может найти применение в медицине. 22 н. и 34 з.п. ф-лы, 12 ил., 7 табл., 2 пр.
1. Способ скрининга молекулы, способной связываться с CD4, включающий:
(a) получение одной или нескольких молекул-кандидатов;
(b) определение одной или нескольких молекул-кандидатов, способных связываться со следующими областями CD4 человека с последовательностью SEQ ID NO: 1: аминокислотами 148-154, аминокислотами 164-168 и аминокислотами 185-192; и
(c) отбор молекулы, определенной на стадии (b) как способной связываться с CD4,
где:
(i) стадии (а)-(с) выполняются in vitro; или
(ii) стадии (а)-(с) выполняются в компьютерной системе и способ дополнительно включает:
(d) получение молекулы, выбранной на стадии (с); и
(e) определение in vitro того, что полученная молекула способна связываться с аминокислотами 148-154, аминокислотами 164-168 и аминокислотами 185-192 CD4 человека с последовательностью SEQ ID NO: 1.
2. Способ по п.1, в котором стадия (b) дополнительно включает определение того, связывается ли одна или несколько молекул-кандидатов по меньшей мере с одной из: аминокислоты в положении 26, аминокислоты в положении 156, аминокислоты в положении 159, аминокислоты в положении 161 CD4 человека с последовательностью SEQ ID NO: 1.
3. Способ по п.1 или 2, в котором стадия (b) включает определение того, способна ли одна или несколько молекул-кандидатов связываться с CD4 без солевого мостика.
4. Способ по п.1, в котором одна или несколько молекул-кандидатов представляют собой антитело, полученную из антитела молекулу, пептид, олигонуклеотид, миРНК, мимотоп, пептидомиметик, фолдамер, малую молекулу, белок распознавания на основе природного или сконструированного липокалина, DARPin, фибронектин, белок-миметик антител Affibody, ингибитор типа Кунитца, пептидный аптамер, рибозим или токсин.
5. Способ по п.4, в котором антитело является гуманизированным антителом или антителом верблюдовых.
6. Способ по п.1, в котором одна или несколько молекул-кандидатов представляют собой антитела или фрагменты антител, содержащие CDR1 и CDR2 легкой цепи ВТ061 и CDR1 и CDR3 тяжелой цепи ВТ061 необязательно с аминокислотными заменами в последовательностях CDR, при условии, что:
(i) CDR1 легкой цепи содержит: Ser32, Gly33 и Tyr34;
(ii) CDR2 легкой цепи содержит: Leu54 и Ile57;
(iii) CDR1 тяжелой цепи содержит: Asp31, Glu31, Thr31, Cys31, Pro31, Met31 или Tyr31; и
(iv) CDR3 тяжелой цепи содержит: Tyr103, Phe103 или His103; Arg104; Tyr105; Asp106 и Trp110, Phe110, His110 или Tyr110.
7. Способ по п.6, в котором аминокислотные замены в последовательности CDR1 и CDR2 легкой цепи ВТ061 и CDR1 и CDR3 тяжелой цепи ВТ061 выбраны из замен, указанных в таблице 4 и таблице 5.
8. Способ по п.6, в котором антитело или фрагмент антитела содержит легкую цепь, содержащую Tyr53 или Phe53, и тяжелую цепь, содержащую Ser28.
9. Способ по п.6, в котором антитело или фрагмент антитела содержит легкую цепь, содержащую Asp64, и/или тяжелую цепь, содержащую Glu56.
10. Способ по п.6, в котором антитело или фрагмент антитела дополнительно содержит CDR3 легкой цепи ВТ061 и/или CDR2 тяжелой цепи ВТ061 необязательно с аминокислотными заменами в последовательностях указанных CDR, при этом замены выбраны из замен, указанных в таблице 4 и таблице 5.
11. Способ по п.1, где стадии (а)-(с) выполняются в компьютерной системе и стадия (е) включает осуществление контакта in vitro молекулы, полученной на стадии (d), с клеткой CD4+ или с пептидом или полипептидом, содержащим следующие области CD4 человека с последовательностью SEQ ID NO: 1: аминокислоты 148-154, аминокислоты 164-168 и аминокислоты 185-192.
12. Способ по п.11, в котором стадия (е) включает определение того, что молекула, полученная на стадии (d):
(i) связывается с пептидом, содержащим следующие области CD4 человека с последовательностью SEQ ID NO: 1: аминокислоты 148-154, аминокислоты 164-168 и аминокислоты 185-192;
(ii) активирует регуляторные Т-клетки CD4+CD25+; и/или
(iii) снижает экспрессию рецептора CD4 на CD4-экспрессирующей клетке.
13. Способ по п.1, в котором стадии (а)-(с) проводят in vitro и стадия (b) включает осуществление контакта одной или нескольких молекул-кандидатов с пептидом или полипептидом, содержащим следующие области CD4 человека с последовательностью SEQ ID NO: 1: аминокислоты 148-154, аминокислоты 164-168 и аминокислоты 185-192.
14. Способ по п.12, в котором стадия определения включает осуществление рентгеновской кристаллографии или ЯМР и отбор молекулы, которая связывается с пептидом или полипептидом без солевого мостика.
15. Способ по п.13, дополнительно включающий стадию (d) скрининга молекулы, отобранной на стадии (с), в отношении способности активировать регуляторные Т-клетки CD4+CD25+, и/или стадию (е) скрининга молекулы, отобранной на стадии (с), в отношении способности снижать экспрессию рецептора CD4 на CD4-экспрессирующей клетке.
16. Способ по п.11, в котором пептид или полипептид фиксирован на мембране или эквивалентной подходящей поверхности или связан с магнитным шариком.
17. Способ по п.11 или 15, в котором пептид или полипептид содержит Ig-подобный домен С2-типа 1 CD4 человека или содержит Ig-подобный домен V-типа и Ig-подобный домен С2-типа 1 CD4 человека.
18. Способ по п.11, в котором пептид или полипептид имитирует конформацию эпитопа CD4 дикого типа.
19. Способ по п.13, в котором стадия осуществления контакта включает контактирование пептида и молекулы с конкурирующим антителом или фрагментом антитела, имеющим вариабельные домены тяжелой и легкой цепей ВТ061, чтобы определить, способна ли молекула-кандидат или отобранная молекула блокировать связывание конкурирующего антитела или фрагмента антитела с областями пептида или полипептида.
20. Способ получения терапевтической композиции, включающий:
(a) отбор молекулы, определенной как способной активировать регуляторные Т-клетки CD4+CD25+ с применением способа по п.12 или п.15; и
(b) получение терапевтической композиции, содержащей отобранную молекулу.
21. Способ скрининга антитела или фрагмента антитела, способного связывать CD4, включающий:
(a) получение антитела или фрагмента антитела, содержащего CDR1 и CDR2 легкой цепи ВТ061 и CDR1 и CDR3 тяжелой цепи ВТ061 необязательно с аминокислотными заменами в последовательностях CDR, при условии, что:
(i) CDR1 легкой цепи содержит: Ser32, Gly33 и Tyr 34;
(ii) CDR2 легкой цепи содержит: Leu54 и Ile57;
(iii) CDR1 тяжелой цепи содержит: Asp31, Glu31, Thr31, Cys31, Pro31, Met31 или Tyr31; и
(iv) CDR3 тяжелой цепи содержит: Tyr103, Phe103 или His103; Arg104; Tyr105; Asp106 и Trp110, Phe110, His110 или Tyr110;
(b) определение того, способно ли антитело или фрагмент антитела связываться с аминокислотами 148-154, аминокислотами 164-168 и аминокислотами 185-192 CD4 человека с последовательностью SEQ ID NO: 1, и
(c) отбор антитела или фрагмента антитела, определенного на стадии (b) как способного связываться с CD4,
при этом антитело или фрагмент антитела не содержит все из CDR1, CDR2 и CDR3 тяжелой цепи ВТ061 и CDR1, CDR2 и CDR3 легкой цепи ВТ061,
где:
(i) стадии (а)-(с) выполняются in vitro; или
(ii) стадии (а)-(с) выполняются в компьютерной системе и способ дополнительно включает:
(d) получение молекулы, выбранной на стадии (с); и
(e) определение in vitro того, что полученная молекула способна связываться с аминокислотами 148-154, аминокислотами 164-168 и аминокислотами 185-192 CD4 человека с последовательностью SEQ ID NO: 1.
22. Способ по п.21, в котором аминокислотные замены в последовательностях CDR1 и CDR2 легкой цепи ВТ061 и CDR1 и CDR3 тяжелой цепи ВТ061 выбраны из замен, указанных в таблице 4 и таблице 5.
23. Способ по п.21 или 22, в котором антитело или фрагмент антитела содержит легкую цепь, содержащую Tyr53 или Phe53, и тяжелую цепь, содержащую Ser28.
24. Способ по п.21, в котором антитело или фрагмент антитела содержит легкую цепь, содержащую Asp64, и/или тяжелую цепь, содержащую Glu56.
25. Способ по п.21, в котором антитело или фрагмент антитела дополнительно содержит CDR3 легкой цепи ВТ061 и/или CDR2 тяжелой цепи ВТ061 необязательно с аминокислотными заменами в последовательностях таких CDR, при этом замены выбраны из замен, указанных в таблице 4 и таблице 5.
26. Способ по п.21, где стадии (а)-(с) выполняются в компьютерной системе и стадия (е) включает определение того, что антитело или фрагмент антитела, полученное на стадии (d):
(i) активирует регуляторные Т-клетки CD4+CD25+;
(ii) связывается с одной или несколькими из следующих областей CD4 с последовательностью SEQ ID NO: 1: аминокислотами 148-154; аминокислотами 164-168 и аминокислотами 185-192; и/или
(iii) осуществляет понижающую регуляцию экспрессии CD4 на CD4-экспрессирующей клетке.
27. Способ по п.21, в котором стадии (а)-(с) проводят in vitro.
28. Способ по п.27, в котором стадия (b) включает определение того, что антитело или фрагмент антитела:
(i) активирует регуляторные Т-клетки CD4+CD25+;
(ii) связывается с одной или несколькими из следующих областей CD4 с последовательностью SEQ ID NO: 1: аминокислотами 148-154; аминокислотами 164-168 и аминокислотами 185-192; и/или
(iii) осуществляет понижающую регуляцию экспрессии CD4 на CD4-экспрессирующей клетке.
29. Способ по любому из пп.12, 15, 26 и 28, в котором CD4-экспрессирующими клетками являются РВМС.
30. Способ получения терапевтической композиции, включающий:
(a) отбор антитела или фрагмента антитела, идентифицированного по п.26 или 28 как способного активировать регуляторные Т-клетки CD4+CD25+; и
(b) получение терапевтической композиции, содержащей отобранную молекулу.
31. Антитело или фрагмент антитела, способные активировать регуляторные Т-клетки CD4+CD25+, где антитело или фрагмент антитела содержат вариабельный домен легкой цепи, по меньшей мере на 80% идентичный вариабельному домену легкой цепи антитела ВТ061, и вариабельный домен тяжелой цепи, по меньшей мере на 80% идентичный вариабельному домену тяжелой цепи антитела ВТ061, причем антитело или фрагмент антитела содержат последовательности CDR легкой цепи ВТ061 и последовательности CDR тяжелой цепи ВТ061 с одной аминокислотной заменой, при этом замена представляет собой замену:
(i) A63G в тяжелой цепи;
(ii) R33K в тяжелой цепи; или
(iii) L98I в легкой цепи.
32. Антитело или фрагмент антитела, способные активировать регуляторные Т-клетки CD4+CD25+, где антитело или фрагмент антитела содержат вариабельный домен легкой цепи, по меньшей мере на 80% идентичный вариабельному домену легкой цепи антитела ВТ061, и вариабельный домен тяжелой цепи, по меньшей мере на 80% идентичный вариабельному домену тяжелой цепи антитела ВТ061, причем антитело или фрагмент антитела содержат последовательности CDR легкой цепи ВТ061 и последовательности CDR тяжелой цепи ВТ061 и двойную аминокислотную замену, при этом замены представляют собой замены:
(i) R33K и A63G в тяжелой цепи;
(ii) L98I в легкой цепи и R33K в тяжелой цепи; или
(iii) A55G в легкой цепи и R33K в тяжелой цепи.
33. Антитело или фрагмент антитела по п.31 или 32, дополнительно содержащие остальные последовательности вариабельных доменов ВТ061.
34. Антитело или фрагмент антитела по п.31 или 32, дополнительно содержащие Fc-рецептор, способный связываться с CD64.
35. Антитело по п.31 или 32, где антитело является IgG-антителом.
36. Выделенный пептид для идентификации молекулы, способной связываться с CD4, содержащий фрагмент CD4 человека (белок) из менее 50 аминокислот и включающий следующие области CD4 человека с последовательностью SEQ ID NO: 1: аминокислоты 148-154, аминокислоты 164-168 и аминокислоты 185-192.
37. Мимотоп для идентификации молекулы, способной связываться с CD4, содержащий фрагмент CD4 человека (белок) из менее 50 аминокислот и включающий следующие области CD4 человека с последовательностью SEQ ID NO: 1: аминокислоты 148-154, аминокислоты 164-168 и аминокислоты 185-192.
38. Нуклеиновая кислота, кодирующая антитело или фрагмент антитела по любому из пп.31-35.
39. Вектор экспрессии, содержащий нуклеиновую кислоту по п.38.
40. Клетка-хозяин для продукции антитела или фрагмента антитела по любому из пп.31-35, где клетка-хозяин содержит нуклеиновую кислоту по п.38 или вектор по п.39.
41. Способ получения антитела или фрагмента антитела по любому из пп.31-35, включающий стадию культивирования клетки-хозяина по п.40 в культуральной среде в условиях, обеспечивающих возможность экспрессии антитела или фрагмента антитела, и отделение антитела или фрагмента от культуральной среды.
42. Фармацевтическая композиция для лечения аутоиммунного заболевания и отторжения трансплантата, отличающаяся тем, что содержит антитело или фрагмент антитела по любому из пп.31-35.
43. Антитело или фрагмент антитела по п.31 или 32, дополнительно содержащие метку.
44. Способ in vitro для активации регуляторных Т-клеток CD4+CD25+, включающий осуществление контакта клеток с антителом или фрагментом антитела по любому из пп.31-35 или 43.
45. Способ лечения индивида, страдающего от аутоиммунного заболевания или отторжения трансплантата, или профилактики аутоиммунного заболевания или отторжения трансплантата у индивида, включающий введение указанному индивиду антитела или фрагмента антитела по любому из пп.31-35.
46. Антитело или фрагмент антитела по любому из пп.31-35 для применения в медицине.
47. Антитело или фрагмент антитела для применения в медицине по п.46, при этом применением является лечение аутоиммунного заболевания или отторжения трансплантата.
48. Применение антитела или фрагмента антитела по любому из пп.31-35 для получения лекарственного средства для лечения аутоиммунного заболевания или отторжения трансплантата.
49. Применение антитела или фрагмента антитела по любому из пп.31-35 или 43 для активации регуляторных Т-клеток CD4+CD25+ in vitro.
50. Применение антитела или фрагмента антитела по любому из пп.31-35 или 43 для идентификации регуляторных Т-клеток CD4+CD25+ in vitro.
51. Применение выделенного пептида по п.36 или мимотопа по п.37 для скрининга молекул, способных связываться с CD4 и/или активировать регуляторные Т-клетки CD4+CD25+.
52. Способ скрининга в отношении присутствия регуляторных Т-клеток CD4+CD25+ в образце, включающий стадию осуществления контакта меченого антитела или фрагмента антитела по п.43 с образцом, промывку образца для удаления несвязанного антитела и определение присутствия метки в образце.
53. Способ скрининга по п.52, в котором регуляторные Т-клетки CD4+CD25+ активированы.
54. Способ скрининга по п.52 или 53, в котором образец является биологическим образцом, взятым у индивида, страдающего от аутоиммунного заболевания или отторжения трансплантата.
55. Способ лечения индивида, страдающего от аутоиммунного заболевания или отторжения трансплантата, или профилактики аутоиммунного заболевания или отторжения трансплантата у индивида, включающий извлечение образца, содержащего регуляторные Т-клетки CD4+CD25+, из организма индивида, осуществление контакта образца с антителом или фрагментом антитела по любому из пп.31-35, чтобы активировать регуляторные Т-клетки CD4+CD25+, и введение активированных клеток индивиду.
56. Набор для выделения регуляторных Т-клеток CD4+CD25+, содержащий магнитные шарики, покрытые антителом или фрагментом антитела по любому из пп.31-35 или 43, и второе анти-CD4-антитело или анти-CD25-антитело.
| Колосоуборка | 1923 |
|
SU2009A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| MOORE J | |||
| P., et al | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |

