ПЕРЕКРЕСТНЫЕ ССЫЛКИ НА РОДСТВЕННЫЕ ЗАЯВКИ
Данная заявка истребует приоритет согласно предварительной заявке №61/279279, поданной 16 октября 2009 г., и предварительной заявке №61/322722, поданной 9 апреля 2010 г., и которые включены в настоящую заявку во всей полноте посредством ссылки.
ЗАЯВЛЕНИЕ В ОТНОШЕНИИ ПЕРЕЧНЯ ПОСЛЕДОВАТЕЛЬНОСТЕЙ Перечень последовательностей, связанный с настоящей заявкой, предоставлен в текстовом формате вместо бумажной копии, и включен в настоящую заявку путем отсылки к описанию. Название текстового файла, содержащего перечень последовательностей: 35668_Seq_Final.txt. Размер текстового фала - 109 Кб. Он был создан 15 октября 2010 г., и предоставляется через EFS-Web совместно с подачей спецификаций.
УРОВЕНЬ ТЕХНИКИ
Система комплемента представляет собой механизм раннего реагирования, служащий для инициации и усиления воспалительной реакции в ответ на микробную инфекцию и другие острые поражения (М.K.Liszewski and J.P.Atkinson, 1993, in Fundamental Immunology, Third Edition, edited by W.E. Paul, Raven Press, Ltd., New York). Несмотря на то, что активация комплемента обеспечивает весьма ценную первоочередную защиту от потенциальных патогенов, активность комплемента, которая вызывает защитную воспалительную реакцию, может также представлять потенциальную угрозу для организма хозяина (K.R.Kalli, et al., Springer Semin. Immunopathol. 15:417-431, 1994; B.P.Morgan, Eur. J. Clinical Investig. 24:219-228, 1994). Например, протеолитические продукты С3 и С5 мобилизуют и активируют нейтрофильные лейкоциты. Эти активированные клетки являются неразборчивыми при выработке разрушающих ферментов и могут привести к поражению органов. Кроме того, активация комплемента может привести к накоплению цитолитического комплекса терминальных компонентов комплемента как на микробных клетках-мишенях, так и на соседних клетках организма хозяина, что приводит к лизису клеток организма хозяина.
Считается, что система комплемента принимает участие в патогенезе многочисленных острых и хронических заболеваний, включая: инфаркт миокарда, реваскуляризация вследствие нарушения мозгового кровообращения, респираторный дистресс-синдром у взрослых (ARDS), поражение вследствие реперфузии, септический шок, капиллярное кровотечение вследствие термического ожога, воспаление вследствие посткардиолегочного шунтирования, отторжение трансплантата, ревматоидный артрит, рассеянный склероз, прогрессирующая миастения, и болезнь Альцгеймера. Почти во всех перечисленных случаях комплемент не является непосредственной причиной данных заболеваниях, но представляет собой один из серьезных факторов, вовлеченных в патогенез. Тем не менее, активация комплемента может быть главным патологическим механизмом и представлять собой эффективный показатель для клинического контроля при многих из указанных выше заболеваниях. Растущее признание важности комплемент-опосредованных поражений тканей во множестве болезней подчеркивает необходимость создания эффективных медикаментов для ингибирования комплемента. В настоящее время не существует официально одобренных для использования человеком медикаментов, которые бы имели специфическую нацеленность и ингибировали бы активацию комплемента. В настоящее время широко известно, что система комплемента может быть активирована тремя различными путями: классическим путем активации, пектиновым путем активации и альтернативным путем активации комплемента. Классический путь активации обычно инициируется при связывании антитела с инородной частицей (т.е., антигеном) и, таким образом, требует предварительного воздействия данного антигена для генерации специфического антитела. Поскольку активация классического пути связана с развитием иммунной реакции, классический путь активации является частью приобретенной иммунной системы. В отличие от этого, лектиновый и альтернативный пути не зависят от клонального иммунитета и являются частью врожденной иммунной системы.
Первым шагом в активации классического пути является прикрепление специфической молекулы распознавания, C1q, к антигенно-нагруженным элементам IgG и IgM. Результатом активации системы комплемента является последовательная активация зимогенов сериновой протеазы. C1q связан с проферментами сериновой протеазы C1r и C1s в комплексе, известном как C1u, при связывании C1q с иммунным комплексом, аутопротеолитическое расщепление участка Arg-Ile элемента C1r сопровождается C1r-активацией C1s, которая необходима для возможности расщепления С4 и С2. Расщепление С4 на два фрагмента, обозначенных соответственно С4а и C4b, позволяет фрагментам C4b сформировать ковалентные связи с примыкающим гидроксильными или аминогруппами и последующей генерацией конвертазы С3 (C4b2b) посредством нековалентного взаимодействия с фрагментом C2b активированного элемента С2. Конвертаза С3 (C4b2b) активизирует элемент С3, который направляет генерацию конвертазы С5 (C4b2b3b) и формирование мембраноатакующего комплекса (C5b-9), что может спровоцировать микробный лизис. Активированные формы С3 и С4 (C3b и C4b) ковалентно расположены на чужеродных целевых поверхностях, которые распознаются рецепторами комплемента на множественных фагоцитах.
Независимо, первым шагом в активации системы комплемента при лектиновом пути также является прикрепление специфической молекулы распознавания, за которым следует активация ассоциированных сериновых протеаз. Однако более верным, чем прикрепление к иммунным комплексам при помощи C1q, является то, что молекулы распознавания в лектиновом пути активации представляют собой белки, связывающие углеводороды (маннозосвязывающий лектин (MBL), Н-фиколин, М-фиколин, и L-фиколин) (J.Lu et al., Biochim. Biophys. Acta 1572:387-400, 2002; Holmskov et al., Annu. Rev. Immunol. 21:547-578 (2003); Teh et al., Immunology 101:225-232 (2000)). Икеда (Ikeda et al.) первым продемонстрировал, что подобно C1q, MBL может активировать систему комплемента С4-зависимым путем, прикрепляясь к эритроцитам, покрытым маннанами дрожжей (K.Ikeda et al., J. Biol. Chem. 262:7451-7454, 1987). MBL, член семейства коллектиновых белков, представляет собой кальций-зависимый лектин, который связывает карбогидраты с 3- и 4-гидроксигруппами, ориентированными в экваториальной плоскости пиранозного кольца. Главными лигандами для MBL являются, таким образом, D-манноза и N-ацетил-D-глюкозамин, в то время как для карбогидратов, не соответствующих подобному стерическому требованию, афинность по отношению к MBL не выявлена (Weis, W.I., et al., Nature 360:127-134, 1992). Взаимодействие между MBL и моновалентными сахарами является чрезвычайно слабым, с типичными константами диссоциации порядка 2 ммоль. MBL достигает стойкого специфичного связывания с полисахаридными лигандами в процессе одновременного взаимодействия с множественными остатками моносахаридов (Lee, R.T, et al., Archiv. Biochem. Biophys. 299:129-136, 1992). MBL распознает углеводородные структуры, которые обычно покрывают микроорганизмы, такие как бактерии, дрожжи, паразиты и некоторые вирусы. Однако MBL не распознает D-галактозу и сиаловую кислоту, предпоследний и последний сахара, которые обычно покрывают «зрелый» гликоконъюгатный комплекс гликопротеинов, присутствующих в плазме млекопитающих и находящихся на поверхности их клеток. Данная специфичность связывания может помочь в защите от самопроизвольной активации. Однако MBL не образует высокоаффинных связей со скоплениями высокоразветвленных маннозных предшественников гликанов на N-связанных гликопротеинах и гликолипидах, находящихся в эндоплазмическом ретикулуме и звездчатых невроцитах (Гольджи) клеток млекопитающих (Maynard, Y., et al., J. Biol. Chem. 257:3788-3794, 1982). Следовательно, поврежденные клетки являются потенциальными мишенями для пектинового пути активации посредством MBL-связывания.
Фиколины обладают отличным от MBL типом лектинового домена, известным как фибриноген-подобный домен. Фиколины связывают остатки сахара Са++-независимым путем. У людей идентифицировано три типа фиколинов: L-фиколин, М-фиколин и Н-фиколин. Два сывороточных фиколина, L-фиколин и Н-фиколин имеют общность в специфичности к М-ацетил-D-глюкозамину; однако Н-фиколин также связывается с N-ацетил-D-галактозамином. Специфичность по отношению к различным сахарам у L-фиколина, Н-фиколина и MBL означает, что различные пектины могут быть комплементарны различным, хотя и частично перекрывающимся гликоконъюгатам. Данная концепция поддерживается недавним сообщением о том, что среди известных науке пектинов, принимающих участие в пектиновом пути активации, только L-фиколин специфическим образом связывается с липотейхоевой кислотой, гликоконъюгатом клеточной стенки, обнаруженным во всех Грамм-положительных бактериях (Lynch, N. J., et al., J. Immunol. 172:1198-1202, 2004). Коллектины (т.е., MBL) и фиколины не имеют значительного сходства в аминокислотной последовательности. Однако эти две группы белков имеют схожие организации доменов и, подобно Clq, собираются в олигомерические структуры, которые максимально увеличивают возможность многосайтового связывания. У здоровых людей концентрация MBL в сыворотке сильно варьирует в популяциях, и он генетически контролируется полиморфизмом/мутациями как в промоторе, так и в кодирующей области MBL гена. Так как белок MBL является белком острой фазы, то его дальнейшая экспрессия регулируется во время воспалительного процесса. L-фиколин присутствует в сыворотке в такой же концентрации, как и MBL. Поэтому, роль L-фиколина в пектиновом пути активации по значимости потенциально сравнима с ролью MBL. MBL и фиколины могут также выступать в роли опсонинов, которые требуют взаимодействия между данными белками и рецепторами фагоцитов (Kuhlman, M., et al., J. Exp.Med. 169:1733, 1989; Matsushita, M., et al., J. Biol. Chem. 271:2448-54, 1996). Однако идентичность рецептора (ов) на клетках-фагоцитах не была установлена.
MBL человека при помощи своего коллагеноподобного домена осуществляет высокоаффинное специфичное взаимодействие с уникальными Clr/Cls-подобными сериновыми протеазами, названными MBL-ассоциированными сериновыми протеазами (MASPs). К настоящему времени описано три типа MASPs. Сначала одиночный фермент «MASP» был идентифицирован и охарактеризован как фермент, отвечающий за инициацию классического пути активации комплемента (т.е., расщепление С2 и С4) (Ji, Y.Н., et al., J. Immunol. 150:571-578, 1993). Позже выяснилось, что MASP на самом деле представляет собой комбинацию двух протеаз: MASP-1 и MASP-2 (Thiel, S., et al., Nature 386:506-510, 1997). Однако было продемонстрировано, что комплекс MBL-MASP-2 сам по себе является достаточным для активации комплемента (Vorup-Jensen, Т., et al. J. Immunol. 165:2093-2100, 2000). Кроме того, только MASP-2 расщеплял С2 и С4 с высокой результативностью (Ambrus, G, et al., J. Immunol. 170:1374-1382, 2003). Поэтому, MASP-2 представляет собой протеазу, ответственную за активацию С4 и С2 для генерации конвертазы С3, C4b2b. В этом заключается его значительное отличие от С1 комплекса где скоординированная активность двух специфичных сериновых протеаз (C1r и C1s) приводит к активации системы комплемента. Не так давно была выделена третья новая протеаза, MASP-3 (Dahl, M.R., et al., Immunity 15:127-35, 2001). MASP-1 и MASP-3 представляют собой продукты одного и того же гена, полученные в результате альтернативного сплайсинга. Их биологические функции до сих пор остаются невыясненными.
MASPs имеют одинаковую доменную организацию доменов с белками C1r и C1s, ферментными компонентам комплекса С1 (Sim, R.B., et al., Biochem. Soc. Trans. 28:545, 2000). Данные домены включают в себя N-концевой домен Clr/C1s/морского ежа ФРЭС/костного морфогенного белка (CUB), домен эпидермального фактора роста, второй домен CUB, тандем доменов белков, контролирующих комплемент, и домен сериновой протеазы. Как в протеазах С1, активация MASP-2 протекает при расщеплении связи Arg-Ile, примыкающей к домену сериновой протеазы, это приводит к расщеплению фермента на связанные дисульфидными связями цепочки А и В, последняя из которых состоит из домена сериновой протеазы. Генетически обусловленная неполноценность MASP-2 была описана в недавних исследованиях (Stengaard-Pedersen, K., et al., New Eng. J. Med. 349:554-560, 2003). Мутация одного нуклеотида приводит кАзр-у обмену в домене CUB1, что в свою очередь лишает MASP-2 способности связываться с MBL.
MBL также связывается с неферментным белком, обозначаемым как MBL-связанный белок 19 кДА kDa (МАр19) (Stover, C.M., J. Immunol. 162:3481-90, 1999) или малым MBL-связанным белком (sMAP) (Takahashi, M., et al., Int. Immunol. 11:859-863, 1999). МАр19 формируется при альтернативном сплайсинге продукта гена MASP 2 и содержит два первых домена MASP-2, за которыми следует дополнительная последовательность четырех уникальных аминокислот.Гены MASP I и MASP 2 расположены на хромосомах 3 и 1, соответственно (Schwaeble, W., et al., Immunobiology 205:455-466, 2002).
Ряд данных указывает на то, что существуют различные комплексы MBL-MASPs, a также на то, что значительная часть находящихся в сыворотке MASPs не связана с MBL (Thiel, S., et al., J. Immunol. 165:878-887, 2000). Н- и L-фиколин также связываются с MASP и активируют лектиновый путь также, как MBL (Dahl, M.R., et al., Immunity 15:127-35, 2001; Matsushita, M., et al., J. Immunol. 168:3502-3506, 2002). Оба - лектиновый и классический, - пути активации комплемента формируют общую конвертазу С3 (C4b2b) и оба пути активации сходятся в этом пункте.
Повсеместно считается, что пектиновому пути активации комплемента принадлежит ведущая роль в иммунной защите организма против инфекции. Надежные доказательства вовлеченности MBL в иммунную реакцию были получены в результате наблюдений за субъектами, имеющими пониженный уровень функционального MBL в сыворотке (Kilpatrick, D.C., Biochim. Biophys. Acta 1572:401-413, 2002). Такие субъекты демонстрируют восприимчивость к рецидивирующим бактериальным и грибковым инфекциям (микозу). Данные симптомы обычно проявляются в раннем возрасте, во время определенного промежутка времени, когда титр производных от материнских антител снижается, а собственные антитела еще не вырабатываются в полном объеме. Данный синдром часто является следствием мутаций некоторых сайтов в коллагеновой части MBL, которая препятствует непосредственному образованию олигомеров MBL. Однако, так как MBL способен функционировать в качестве независимого от комплемента опсонина, остается невыясненным, до какого предела повышенная восприимчивость к инфекции обусловлена нарушениями в активации комплемента.
Несмотря на исчерпывающие доказательства участия всех путей активации - классического и альтернативных - в патогенезе неинфекционных человеческих заболеваний, роль пектинового пути активации только начинают оценивать. Недавние исследования показали, что активация пектинового пути может провоцировать активацию комплемента и воспаление, связанное с ишемическим реперфузионным поражением. Коллард (Collard et al.(2000)) сообщает, что культивируемые эндотелиальные клетки, подвергнутые окислительному стрессу, связываются с MBL и демонстрируют отложение С3 в присутствии человеческой сыворотки (Collard, C.D., et al., Am. J. Pathol. 156:1549-1556, 2000). Кроме того, обработка человеческой сыворотки блокирующими анти-MBL моноклональными антителами ингибирует связывание MBL и активацию комплемента. Данные открытия были проверены опытным путем на лабораторных крысах с миокардиальной ишемией-реперфузией, в процессе чего у крыс, подвергшихся лечению с блокирующими крысиный MBL антителами, наблюдали значительно меньше случаев поражения миокарда вследствие закупорки коронарной артерии, чем у крыс, подвергнутых лечению контрольными антителами (Jordan, J.E., et al., Circulation 104:1413-1418, 2001). Молекулярный механизм связывания MBL с эндотелием сосудов после окислительного стресса остается неясным; вследствие недавних исследований было выдвинуто предположение, что активация пектинового пути после окислительного стресса может быть опосредована связыванием MBL с васкулярными эндотелиальными цитокератинами, а не с гликоконьюгатами (Collard, C.D., et al., Am. J. Pathol. 159:1045-1054, 2001). В результате других исследований было выявлено, что роль классического и альтернативных путей активации в развитии патологического процесса ишемии/реперфузионного повреждения и роль пектинового пути в данном заболевании остается спорной (Riedermann, N.C., et al., Am. J. Pathol. 162:363-367, 2003).
В отличие от классического и пектинового путей, не было выявлено инициаторов альтернативного пути активации для осуществления распознавательных функций, выполняемых C1q и пектинами в двух вышеназванных путях. В настоящее время принято считать, что альтернативный путь активации инициируется спонтанно инородными или другими патологическими телами (бактерии, дрожжи, инфицированные вирусами клетки или поврежденные ткани). Существует четыре плазменных белка, напрямую вовлеченных в альтернативный путь активации: С3, факторы В и D, пропердин. Протеолитическая генерация C3b из нативного С3 необходима для функционирования альтернативного пути активации. Учитывая тот факт, что конвертаза альтернативного пути С3 (C3bBb) содержит C3b в качестве основной субъединицы, вопрос происхождения первой C3b посредством альтернативного пути активации представляет собой сложнейшую задачу, что стимулирует проведение соответствующих научных исследований.
Белок С3 принадлежит к семейству белков (совместно с С4 и а-2-макроглобулином), которое имеет редкую посттрансляционную модификацию, известную как тиоэфирное соединение. Тиоэфирная группа представлена глутамином, в котором конечная карбонильная группа связана с сульфгидрильной группой цистеина, который находится через три аминокислоты от глутамина. Данная связь является нестабильной, и электрофильная карбонил-группа глутамина может формировать ковалентную связь с другими молекулами через гидроксильную или аминокислотную группы. Тиоэфирная группа является достаточно устойчивой при секвестировании с гидрофобным участком неповрежденной С3. Однако протеолитическое расщепление С3 на С3а и C3b являет результатом экспозиции высоко реактивной тиоэфирной связи с C3b, и, при помощи подобного механизма, C3b ковалентно присоединяется к молекуле-мишени. Кроме хорошо известной роли в ковалентном прикреплении C3b к молекулярной мишени, тиоэфиру С3 также присваивают главную роль в инициации альтернативного пути активации комплемента. Согласно получившей широкое распространение теории «спящего механизма» (tick-over theory), альтернативный путь активации инициируется генерацией гомогенной конвертазы, iC3Bb, которая формируется из С3 и гидролизованного тиоэфира (iC3; С3(H2O)) и фактора В (Lachmann, PJ., et al., Springer Semin. Immunopathol. 7:143-162, 1984). C3b-подобная iC3 генерируется из исходной С3 путем медленного спонтанного гидролиза внутреннего тиоэфира в белковой молекуле (Pangburn, М.K., et al., J. Exp.Med. 154:856-867, 1981). В продолжение активности конвертазы iC3Bb, молекулы C3b размещаются на поверхности молекулярных мишеней, таким образом, инициируя альтернативный путь активации.
Очень мало известно о природе инициаторов активации альтернативного пути. Считается, что активаторы включают в себя дрожжевые клеточные стенки (зимозаны), множество чистых полисахаридов, кроличьи эритроциты, некоторые иммуноглобулины, вирусы, грибы, бактерии, животные опухолевые клетки, паразиты и поврежденные клетки. Единственной общей чертой у данных активаторов является наличие углевода, однако сложность строения и разнообразие углеводных структур представляют сложность в установлении общих молекулярных детерминант, которые распознаются в процессе активации.
Альтернативный путь активации может также предоставить мощный усиливающий контур для пектинового/классического пути активации конвертазы С3 (C4b2b) после того, как только любая синтезированная C3b начнет участвовать совместно с фактором В в формировании дополнительных конвертаз альтернативного пути С3 (C3bBb). Конвертаза альтернативного пути активации С3 стабилизируется связыванием с пропердином. Пропердин увеличивает период полураспада конвертазы альтернативного пути активации С3 в шести, десять раз. Добавление C3b к конвертазе С3 приводит к формированию С5 конвертазы альтернативного пути.
Считалось, что все три пути активации (т.е., классический, пектиновый и альтернативный) сходятся на С5, которая расщепляется для формирования продуктов с множественными провоцирующими воспаление эффектами. Конвергентный путь активации был отнесен к конечному пути активации комплемента. С5а является самым действенным анафилатоксином, индуцирующим изменения в гладкой мускулатуре и сосудистом тонусе, также как в проницаемости сосудов. Он также представляет собой эффективный хемотаксин и активатор как нейтрофилов, так и моноцитов. С5а-опосредованная клеточная активация может значительным образом усиливать воспалительные реакции путем индуцирования секреции множественных дополнительных воспалительных медиаторов, включая цитокины, гидролитические ферменты, метаболиты арахидоновой кислоты и активные формы кислорода. Расщепление С5 приводит к формированию C5b-9, также известного как мембраноатакующий комплекс (MAC). В настоящее время имеются веские доказательства того, что сублитическое отложение MAC может играть важную роль в воспалительном процессе в дополнение к роли литического пороформирующего комплекса.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Краткое изложение сущности изобретения приводится с целью представления в упрощенной форме выбора концепций, которые в дальнейшем описаны в разделе «Подробное описание». Настоящее краткое изложение не предназначено для идентификации ключевых признаков заявленного предмета изобретения, а также не предназначено для использования в качестве средства для определения объема заявленного предмета изобретения.
В одном своем аспекте настоящее изобретение предлагает способ ингибирования отрицательных эффектов при MASP-2-зависимой активации комплемента в живом организме. Способ включает в себя введение субъекту, который в этом нуждается, дозы агента, ингибирующего MASP-2, достаточной для эффективного подавления MASP-2-зависимой активации комплемента. В данном контексте, словосочетание «MASP-2-зависимая активация комплемента» относится к альтернативному пути активации комплемента, что происходит посредством пектин-зависимой системы MASP-2. В другом своем аспекте настоящее изобретение относится к агенту, ингибирующиму MASP-2, подавляющему активацию комплемента посредством пектин-зависимой системы MASP-2 без существенного ингибирования активации комплемента посредством классической или C1q-зависимой системы, таким образом, что dq-зависимая система остается функциональной.
В некоторых вариантах осуществления данных аспектов изобретения агент, ингибирующий MASP-2, представляет собой анти-MASP-2 антитело или его фрагмент.В следующих вариантах осуществления, анти-MASP-2 антитело обладает редуцированной эффекторной функцией. В некоторых вариантах осуществления, агент, ингибирующий MASP-2, представляет собой пептид, ингибирующий MASP-2, или непептидный ингибитор MASP-2.
В еще одном аспекте настоящее изобретение предлагает композиции для подавления отрицательных эффектов MASP-2-зависимой активации комплемента, содержащей терапевтически эффективную дозу агента, ингибирующего MASP-2, и фармацевтически приемлемый носитель. Также предлагаются способы для производства медикамента для использования с целью ингибирования отрицательных эффектов MASP-2-зависимой активации комплемента в живых субъектах, который в том нуждаются, включая комбинирование терапевтически эффективной дозы агента, ингибирующего MASP-2, и фармацевтического носителя. Также предлагаются способы производства лекарственных средств для использования с целью ингибирования MASP-2-зависимой активации комплемента при лечении каждого из состояний, заболеваний и расстройств, описанных ниже.
Способы, композиции и лекарственные средства по настоящему изобретению, используются для ингибирования отрицательных эффектов MASP-2-зависимой активации комплемента in vivo у объектов исследования, относящихся к млекопитающим, включая людей, страдающих острыми или хроническими патологическими состояниями или поражениями, как описано далее. Такие состояния и поражения включают в себя без ограничений MASP-2-опосредованную активацию комплемента в ассоциированных аутоиммунных расстройствах и/или воспалительных заболеваниях.
В еще одном аспекте настоящего изобретения предлагаются способы ингибирования MASP-2-зависимой активации комплемента у субъекта, страдающего нарушением свертывания крови или предрасположенного к нему, например, нарушением свертывания крови, опосредованным комплементом, или коагулопатией, путем введения терапевтически эффективной дозы агента, ингибирующего MASP-2, и фармацевтического носителя такому субъекту. Состояния, подлежащие лечению по настоящему изобретению, включают, путем примера, не имеющего ограничительного характера, диссеминированное внутрисосудистое свертывание (ДВС), также называемое коагулопатией потребления.
В одном аспекте изобретение предлагаются способы ингибирования MASP-2-зависимой активации комплемента у субъекта, страдающего нарушением свертывания крови или предрасположенного к нему, путем введения субъекту дозы агента ингибирующего MASP-2, достаточной для ингибирования MASP-2-зависимой активации комплемента.
В еще одном аспекте изобретение предлагаются способы ингибирования MASP-2-зависимой активации комплемента для лечения субъекта, страдающего нарушением свертывания крови или предрасположенного к нему, включая введение субъекту дозы агента, ингибирующего MASP-2, достаточной для выборочного ингибирования MASP-2-зависимой активации комплемента без существенного ингибирования C1q-зависимой активации комплемента.
В другом аспекте настоящего изобретения предлагаются способы производства лекарственных средств для использования с целью ингибирования MASP-2-зависимой активации комплемента у живого субъекта, страдающего нарушением свертывания крови, опосредованным комплементом, совмещающих терапевтически эффективную дозу агента, ингибирующего MASP-2, с фармацевтическим носителем.
В другом аспекте настоящего изобретения предлагаются способы лечения, профилактики или снижения тяжести диссеминированного внутрисосудистого свертывания у нуждающегося субъекта, включающие введение субъекту композиции, включающей дозу агента, ингибирующего MASP-2, достаточной для ингибирования MASP-2-зависимой активации комплемента.
ОПИСАНИЕ РИСУНКОВ
Вышеупомянутые аспекты и множество сопутствующих преимуществ данного изобретения смогут быть быстрее оценены и станут более доступны для понимания путем ссылки на нижеследующее подробное описание, рассмотренное совместно с сопроводительными рисунками, как то:
ФИГУРА 1 представляет собой структурную схему, иллюстрирующую новое открытие того, что для активации комплемента посредством альтернативного пути требуется зависимая от пектинового пути MASP-2 активация;
ФИГУРА 2 представляет собой диаграмму, иллюстрирующую геномную структуру MASP-2 человека;
ФИГУРА 3А представляет собой схематичную диаграмму, иллюстрирующую доменную структуру белка MASP-2 человека;
ФИГУРА 3В представляет собой схематичную диаграмму, иллюстрирующую доменную структуру белка М Ар 19 человека;
ФИГУРА 4 представляет собой диаграмму, иллюстрирующую стратегию подавления гена MASP-2 у мыши;
ФИГУРА 5 представляет собой диаграмму, иллюстрирующую минигенную конструкцию MASP-2 человека;
ФИГУРА 6А представляет результаты, демонстрирующие, что MASP-2-недостаточность ведет к потере опосредованной пектиновым путем С4 активации, что обусловлено отсутствием отложения C4b на маннане;
ФИГУРА 6В представляет результаты, демонстрирующие, что MASP-2-недостаточность ведет к потере опосредованной пектиновым путем С4 активации, что обусловлено отсутствием отложения C4b на зимозане;
ФИГУРА 6С представляет результаты, демонстрирующие относительные уровни С4 активации в образцах сыворотки, полученных из MASP-2+/-; MASP-2-/- и линий дикого типа, в результате отложения C4b на маннане и зимозане;
ФИГУРА 7А представляет результаты, демонстрирующие, что MASP-2-недостаточность ведет к потере опосредованной лектиновым путем и альтернативным путем С3 активации в результате отсутствия отложения C3b на маннане;
ФИГУРА 7В представляет результаты, демонстрирующие, что MASP-2-недостаточность ведет к потере опосредованной лектиновым путем и альтернативным путем С3 активации в результате отсутствия отложения C3b на зимозане;
ФИГУРА 8 представляет результаты, демонстрирующие, что добавление рекомбинантного мышиного MASP-2 к MASP-2-/- образцам сыворотки восстанавливает опосредованную лектиновым путем С4 активацию, величина которой находится в зависимости от концентрации белка, в результате отложения C4b на маннане;
ФИГУРА 9 представляет результаты, демонстрирующие, что классический путь активации функционирует у линий MASP-2-/-;
ФИГУРА 10 представляет результаты, демонстрирующие, что MASP-2-зависимая система активации комплемента активируется в фазе ишемии/реперфузии, сопровождаемой репарацией абдоминальной аневризмы аорты;
ФИГУРА 11А представляет результаты, демонстрирующие, что антитело №11 анти-MASP-2 Fab2 ингибирует формирование С3 конвертазы, как описано в Примере 24;
ФИГУРА 11В представляет результаты, демонстрирующие, что антитело №11 анти-MASP-2 Fab2 связывается с нативным MASP-2 крысы, как описано в Примере 24;
ФИГУРА 11С представляет результаты, демонстрирующие, что антитело №41 анти-MASP-2 Fab2 ингибирует расщепление С4, как описано в Примере 24;
ФИГУРА 12 представляет результаты, демонстрирующие, что все протестированные антитела анти-MASP-2 Fab2 подтвердили, что ингибирование формирования конвертазы С3 также явилось фундаментом для ингибирования расщепления С4, как описано в Примере 24;
ФИГУРА 13 представляет собой диаграмму, иллюстрирующую использование рекомбинантных полипептидов, полученных из крысиного MASP-2, для картирования эпитопов анти-MASP-2 антител, блокирующих Fab2, как описано в Примере 25;
ФИГУРА 14 представляет результаты, демонстрирующие связывание антител анти-MASP-2 Fab2 №40 и №60 с полипептидами MASP-2 крысы, как описано в Примере 25;
ФИГУРА 15 представляет результаты, показывающие клиренс азота мочевины крови для «дикого» типа (+/+) и MASP-2(-/-) мыши через 24 и 48 часов после реперфузии в лабораторной модели ишемического реперфузионного повреждения почек, как описано в Примере 26;
ФИГУРА 16А представляет результаты, демонстрирующие масштабы инфаркта для «дикого» типа (+/+) и редуцированные масштабы инфаркта у MASP-2(-/-) мыши после повреждения в лабораторной модели с закупоркой коронарной артерии и реперфузии, как описано в Примере 27;
ФИГУРА 16В представляет результаты, демонстрирующие распределение единичных животных, использованных в лабораторной модели закупорки коронарной артерии и реперфузии, как описано в Примере 27;
ФИГУРА 17А представляет результаты, демонстрирующие исходные данные уровня белков VEGF в RPE-хориоидальном комплексе, выделенном из мышей «дикого» типа (+/+) и мышей MASP-2(-/-), как описано в Примере 28;
ФИГУРА 17В представляет результаты, демонстрирующие уровень белков VEGF в RPE-хориоидальном комплексе на третий день после нанесения поражения лазером на лабораторной модели дегенерации желтого пятна у мышей «дикого» типа (+/+) и MASP-2(-/-), как описано в Примере 28;
ФИГУРА 18 представляет результаты, демонстрирующие средние показатели уровня хориоидальной неоваскуляризации (CNV) на седьмой день после нанесения поражения лазером у мышей «дикого» типа (+/+) и мышей MASP-2(-/-), как описано в Примере 28;
ФИГУРА 19 представляет результаты, демонстрирующие средние показатели клинического артрита у мышей «дикого» типа (+/+) и мышей MASP-2(-/-) после моделирования у них Со12 mAb-индуцированного ревматоидного артрита, как описано в Примере 29;
ФИГУРА 20А представляет собой диаграмму, демонстрирующую направленное разрушение гена sMAP (Map 19), как описано в Примере 30;
ФИГУРА 20В представляет результаты Саузерн-блоттинга геномной ДНК, потомства от спаривания самца sMAP (-/-) химерной мыши и самки мыши линии C57BL/6, как описано в Примере 30;
ФИГУРА 20С представляет результаты генетический анализ методом PCR мышей «дикого» типа (+/+) и мышей sMAP (-/-), как описано в Примере 30;
ФИГУРА 21А представляет результаты Нозерн-блоттинга мРНК MAP и MASP-2 мышей sMAP (-/-), как описано в Примере 30;
ФИГУРА 21В представляет результаты количественного RT-PCR анализа кДНК, кодирующей Н-цепь MASP-2, L- цепь MASP-2 и sMAP, у мышей «дикого» типа (+/+) и мышей sMAP (-/-), как описано в Примере 30;
ФИГУРА 22А представляет иммуноблот sMAP(-/-), а именно МАр19(-/-), демонстрирующий отсутствие MASP-2 и sMAP в сыворотке мыши, как описано в Примере 30;
ФИГУРА 22В представляет результаты, демонстрирующие, что MASP-2 и sMAP были обнаружены в комплексе MBL-MASP-sMAP, как описано в Примере 30;
ФИГУРА 23А представляет результаты, демонстрирующие отложение С4 в покрытых маннаном лунках в сыворотке мышей «дикого» типа (+/+) и sMAP (-/-), как описано в Примере 30;
ФИГУРА 23В представляет результаты, демонстрирующие отложение С3 в покрытых маннаном лунках в сыворотки мышей «дикого» типа (+/+) и sMAP (-/-), как описано в Примере 30;
ФИГУРА 24А представляет результаты, демонстрирующие восстановление комплекса MBL-MASP-sMAP в сыворотке мышей sMAP (-/-), как описано в Примере 30;
ФИГУРА 24B-D представляет результаты, демонстрирующие конкурентное связывание rsMAP и MASP-2I с MBL, как описано в Примере 30;
ФИГУРА 25А-В представляет результаты, демонстрирующие восстановление активности отложения С4 при добавлении к MASP-2, но не rsMAP, как описано в Примере 30;
ФИГУРА 26А-В представляет результаты, демонстрирующие редукцию активности отложения С4 при добавлении rsMAP, как описано в Примере 30; и
ФИГУРА 27А-С представляет результаты, демонстрирующие, что MASP-2 отвечает за С4 активацию С3 по альтернативному пути, как описано в Примере 31.
ФИГУРЫ 28А и 28В представляют кривую зависимости от дозы для ингибирования отложения C4b (ФИГ.28В) и ингибирование активации тромбином после введения антитела MASP-2 Fab2 в нормальную крысиную сыворотку, как описано в Примере 32;
ФИГУРЫ 29А и 29В представляют измеренную агрегацию тромбоцитов (выраженную в области агрегации) у мышей MASP-2(-/-) (ФИГ.29В) по сравнению с агрегацией тромбоцитов у необработанных мышей «дикого» типа, у которых путь активации комплемента ингибирован фактором яда кобры (CVF) кровопускательного средства, и терминального ингибитора пути активации (антагониста C5aR) (ФИГУРА 29А) у локализованной модели реакции Шварцмана диссеминированного внутрисосудистого свертывания, как описано в Примере 33;
ФИГУРЫ 30А-30С иллюстрируют результаты исследования оборота С3 в плазме С4-/- в анализах, характерных для пути активации либо классического, либо пектинового путей;
ФИГУРА 31А графически иллюстрирует среднюю площадь, подверженную риску (AAR), и объемы инфаркта (INF) в процентах от общих объемов миокарда у мышей дикого типа (+/+) и MASP-2(-/-) после проведения окклюзии и реперфузии левой передней нисходящей артерии, как описано в Примере 34;
ФИГУРА 31В графически иллюстрирует отношение между объемом инфаркта (INF), построенного по средней площади, подверженной риску (MR), в процентах от объема миокарда левого желудочка у мышей дикого типа (+/+) и MASP-2(-/-) после проведения закупорки и реперфузии артерии, как описано в Примере 34;
ФИГУРА 31С графически иллюстрирует объем инфаркта (INF) в сердцах мышей дикого типа (+/+) и MASP-2(-/-), залитых буферным раствором, и подготовленных в соответствии с моделью перфузии изолированного сердца мыши по Лангендорфу, в которой общая ишемия и реперфузия проводились в отсутствии сыворотки, как описано в Примере 34;
ФИГУРА 31D графически иллюстрирует отношение между объемом инфаркта (INF) и зоной риска в сердцах мышей дикого типа (+/+) и MASP-2(-/-), залитых буферным раствором, и подготовленных в соответствии с моделью перфузии изолированного сердца мыши по Лангендорфу, как описано в Примере 34;
ФИГУРА 32 графически иллюстрирует уровни азота мочевины крови (BUN), измеренных либо у мышей дикого типа (+/+) (В6) или MASP-2(-/-), реципиентов почек от мышей-доноров дикого типа (+/+), как описано в Примере 35;
ФИГУРА 33 графически иллюстрирует процент выживаемости мышей дикого типа (+/+) и MASP-2(-/-) в зависимости от количества дней после микробной инфекции в модели лигирования и пункции слепой кишки (CLP), как описано в Примере 36;
ФИГУРА 34 графически иллюстрирует количество бактерий, измеренное у мышей дикого типа (+/+) и MASP-2(-/-) после микробной инфекции в модели лигирования и пункции слепой кишки (CLP), как описано в Примере 36;
ФИГУРА 35 представляет собой график Каплана-Мейера, иллюстрирующий процент выживаемости мышей дикого типа (+/+), MASP-2(-/-) и С3 (-/-) через шесть дней после контрольного заражения путем интраназального введения синегнойной палочки (Pseudomonas aeruginosa}, как описано в Примере 37;
ФИГУРА 36 графически иллюстрирует уровень отложения C4b, измеренный в процентах от контрольного, в образцах, отобранных в различные моменты времени после подкожного введения либо 0,3 мг/кг, либо 1,0 мг/кг мышиного анти-MASP-2 моноклонального антитела у мышей дикого типа, как описано в Примере 38;
ФИГУРА 37 графически иллюстрирует уровень отложения C4b, измеренный в процентах от контрольного, в образцах, отобранных в различные моменты времени после внутрибрюшинного введения 0,6 мг/кг мышиного анти-MASP-2 моноклонального антитела у мышей дикого типа, как описано в Примере 38;
ФИГУРА 38 графически иллюстрирует средний объем хориоидальной неоваскуляризации (CNV) на седьмой день после лазерного повреждения у мышей дикого типа (+/+), предварительно обработанных путем внутрибрюшинного введения 0,3 мг/кг или 1,0 мг/кг мышиного aHTH-MASP-2 моноклонального антитела, как описано в Примере 39;
ФИГУРА 39А графически иллюстрирует процент выживаемости мышей MASP-2(-/-) и мышей дикого типа (+/+) после заражения 5×108/100 мкл КОЕ менингококка (N. meningitidis), как описано в Примере 40;
ФИГУРА 39В графически иллюстрирует логарифм КОЕ/мл менингококка (N. meningitidis), полученный в различные моменты времени в образцах крови, отобранных у мышей MASP-2 KO (-/-) и мышей дикого типа (+/+), зараженных 5×108 КОЕ/100 мкл менингококка (N, meningitidis), как описано в Примере 40;
Фигура 40А графически иллюстрирует процент выживаемости мышей MASP-2 KO (-/-) и мышей дикого типа (+/+) после заражения 2×108 КОЕ/100 мкл менингококка (N. meningitidis), как описано в Примере 40;
Фигура 40В графически иллюстрирует логарифм КОЕ/мл менингококка (N. meningitidis), полученный в различные моменты времени в образцах крови, отобранных у мышей дикого типа (+/+), зараженных 2×108 КОЕ/100 мкл менингококка (N. meningitidis),как описано в Примере 40; и
Фигура 40С графически иллюстрирует логарифм КОЕ/мл менингококка (N. meningitidis), полученный в различные моменты времени в образцах крови, отобранных у мышей MASP-2(-/-), зараженных 2×108 КОЕ/100 мкл менингококка (N. meningitidis), как описано в Примере 40.
ОПИСАНИЕ ПЕРЕЧНЯ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO:1 МАр19 кДНК человека
SEQ ID NO:2 человеческий белок МАр19 (с лидерной последовательностью)
SEQ ID NO:3 человеческий белок МАр19 (зрелый)
SEQ ID NO:4 MASP-2 кДНК человека
SEQ ID NO:5 человеческий белок MASP-2 (с лидерной последовательностью)
SEQ ID NO:6 человеческий белок MASP-2 (зрелый)
SEQ ID NO:7 MASP-2 гДНК человека (экзоны 1-6)
АНТИГЕНЫ: (ПО ОТНОШЕНИЮ К ЗРЕЛОМУ БЕЛКУ MASP-2)
SEQ ID NO:8 последовательность CUBI (аминокислоты 1-121)
SEQ ID NO:9 последовательность CUBEGF (аминокислоты 1-166)
SEQ ID NO:10 CUBEGFCUBII (аминокислоты 1-293)
SEQ ID NO:11 участок EOF (аминокислоты 122-166)
SEQ ID NO:12 домен сериновой протеазы (аминокислоты 429-671)
SEQ ID NO:13 неактивный домен серин протеазы (аминокислоты 610-625 с мутацией Ser618 на Ala)
SEQ ID NO:14 TPLGPKWPEPVFGRL (CUBI пептид)
SEQ ID NO:15 TAPPGYRLRLYFTHFDLELSHLCEYDFVKLSSGAKVLATLCGQ (пептид CUBI)
SEQ ID NO:16 TFRSDYSN (связывающий участок MBL, центральная часть)
SEQ ID NO:17 FYSLGSSLDITFRSDYSNEKPFTGF (связывающий участок MBL)
SEQ ID NO:18 IDECQVAPG (пептид EGF)
SEQ ID NO:19 ANMLC AGLESGGKDSCRGDSGGALV (связывающий участок сериновой протеазы, центральная часть)
ДЕТАЛЬНОЕ ОПИСАНИЕ ПЕПТИДОВ-ИНГИБИТОРОВ:
SEQ ID NO:20 полноразмерная кДНК MBL
SEQ ID NO:21 полноразмерный белок MBL
SEQ ID NO:22 OGK-X-GP (консенсус связывания)
SEQ ID NO:23 OGKLG
SEQ ID NO:24 GLR GLQ GPO GKL GPO G
SEQ ID NO:25 GPO GPO GLR GLQ GPO GKL GPO GPO GPO
SEQ ID NO:26 GKDGRDGTKGEKGEPGQGLRGLQGPOGKLGPOG
SEQ ID NO:27 GAOGSOGEKGAOGPQGPOGPOGKMGPKGEOGDO (h-фиколин человека)
SEQ ID NO:28 GCOGLOGAOGDKGEAGTNGKRGERGPOGPOGKAGPOGPNGAOGEO (фиколин р35 человека)
SEQ ID NO:29 LQRALEILPNRVTIKANRPFLVFI (сайт расщепления С4)
ИНГИБИТОРЫ ЭКСПРЕССИИ:
SEQ ID NO:30 кДНК домена CUBI-EGF (нуклеотиды 22-680 в SEQ ID NO:4)
SEQ ID NO:31
5' CGGGCACACCATGAGGCTGCTGACCCTCCTGGGC 3' Нуклеотиды 12-45 в SEQ ID NO:4, включая сайт инициации трансляции MASP-2 (смысловой)
SEQ ID NO:32
5-GACATTACCTTCCGCTCCGACTCCAACGAGAAG3' Нуклеотиды 361-396 в SEQ ID NO:4, кодирующие участок, содержащий сайт связывания MASP-2 MBL (смысловой)
SEQ ID NO:33
5' AGCAGCCCTGAATACCCACGGCCGTATCCCAAA3' Нуклеотиды 610-642 в SEQ ID NO:4, кодирующие участок, содержащий CUBII домен
ПРАЙМЕРЫ КЛОНИРОВАНИЯ:
SEQ ID NO:34 CGGGATCCATGAGGCTGCTGACCCTC (5' ПЦР для CUB)
SEQ ID NO:35 GGAATTCCTAGGCTGCATA (3' ПЦР для CUB)
SEQ ID NO:36 GGAATTCCTACAGGGCGCT (3' ПЦР для CUBIEGF)
SEQ ID NO:37 GGAATTCCTAGTAGTGGAT (3' ПЦР для CUBIEGFCUBII)
SEQ ID NO:38-47 представлены праймерами клонирования для гуманизированных антител
SEQ ID NO:48 представляет собой пептидную связь из 9 аминокислот
ВЕКТОР ЭКСПРЕССИИ:
SEQ ID NO:49 минигенная вставка MASP-2
SEQ ID NO:50 кДНК MASP-2 мыши
SEQ ID NO:51 мышиный белок MASP-2 (с лидерной последовательностью)
SEQ ID NO:52 мышиный белок MASP-2 (зрелый)
SEQ ID NO:53 кДНК MASP-2 крысы
SEQ ID NO:54 крысиный белок MASP-2 (с лидерной последовательностью)
SEQ ID NO:55 крысиный белок MASP-2 (зрелый)
SEQ ID NO:56-59 олигонуклеотиды для сайтнаправленного мутагенеза MASP-2 человека, которые используются для получения MASP-2A человека
SEQ ID NO:60-63 представлена олигонуклеотидами для сайтнаправленного мутагенеза MASP-2 мыши, которые используются для получения MASP-2A мыши
SEQ ID NO:64-65 представлена олигонуклеотидами для сайтнаправленного мутагенеза MASP-2 крысы, которые используются для получения MASP-2A крысы
ПОДРОБНОЕ ОПИСАНИЕ
Настоящее изобретение базируется на неожиданном обнаружении факта, что MASP-2 необходим для инициации альтернативного пути активации комплемента. С использованием модели нокаутной мыши MASP-2-/-, авторами была доказана возможность ингибирования альтернативного пути активации комплемента посредством опосредованного MASP-2 пектинового пути при том, что классический путь активации остается незадействованным, таким образом, устанавливая пектин-зависимую активацию как необходимое условие для альтернативной активации комплемента, не задействуя классический путь. Настоящее изобретение также описывает использование MASP-2 с терапевтической целью для ингибирования клеточных повреждений, связанных с лектин-опосредованным альтернативным путем активации комплемента, не задействуя классический (C1q-зависимого) путь активации компонента иммунной системы.
I. ОПРЕДЕЛЕНИЯ
Если специально не указывается иное значение, то вся используемая терминология в данном описании имеет то же самое значение, которое может быть понято специалистом в области настоящего изобретения. Следующие определения предложены с целью внесения ясности в отношении терминов, которые использованы в описании и пунктах формулы настоящего изобретения.
В настоящем описании, термин «MASP-2-зависимая активация комплемента» относится к альтернативному пути активации комплемента, который осуществляется посредством пектин-зависимой активации MASP-2.
В настоящем описании, термин «альтернативный путь активации» относится к активации комплемента, которая инициируется, например, зимозаном из грибковых и дрожжевых клеточных стенок, липополисахаридом (LPS) из наружной мембраны грамотрицательных бактерий, и кроличьих эритроцитов, а также из множества чистых полисахаридов, кроличьих эритроцитов, вирусов, бактерий, животных опухолевых клеток, паразитов и поврежденных клеток; обычно считалось, что данный вид активации комплемента является результатом спонтанной протеолитической генерации C3b из комплементарного фактора С3.
В настоящем описании, термин «пектиновый путь активации» относится к активации комплемента, которая происходит посредством специфического связывания сывороточных и несывороточных карбогидратсвязывающих белков, включая маннозосвязывающий лектин (MBL) и фиколины.
В настоящем описании, термин «классический путь активации» относится к активации комплемента, которая инициируется прикреплением антитела к инородной частице и требует связывания с молекулой распознавания C1q.
В настоящем описании, термин «агент, ингибирующий MASP-2» относится к любому агенту, который связывается или напрямую взаимодействует с MASP-2 и эффективно ингибирует MASP-2-опосредованную активацию комплемента, включая анти-MASP-2 антитела и их MASP-2-связывающие фрагменты, такие как натуральные и синтетические пептиды, малые молекулы, растворимые MASP-2-рецепторы, ингибиторы экспрессии и выделенные натуральные ингибиторы; он также охватывает пептиды, которые конкурируют с MASP-2 в отношении связывания с другими молекулами распознавания (например, MBL, Н-фиколин, М-фиколин или L-фиколин) в лектиновом пути активации, но не охватывает антитела, которые связываются к другим молекулам распознавания. Агенты, ингибирующие MASP-2, используемые в настоящем изобретении, могут редуцировать MASP-2-опосредованную активацию комплемента более чем на 20%, в некоторых случаях более чем на 50%, в некоторых случаях более чем на 90%. В одном варианте осуществления агент, ингибирующий MASP-2, редуцирует MASP-2-опосредованную активацию комплемента более чем на 90% (т.е., конечная MASP-2-опосредованная активация комплемента составляет только 10% или менее).
В настоящем описании, термин «антитело» охватывает антитела и фрагменты антител, полученных от любого производящего антитела млекопитающего (например, мышь, крыса, кролик и приматы, включая человека); данные антитела специфическим образом связываются с полипептидами MASP-2 или их участками. Типичные антитела включают в себя поликлональные, моноклональные и рекомбинантные антитела; поливалентные антитела (например, бивалентные антитела); гуманизированные антитела; мышиные антитела; химерные, мышино-человеческие, примато-мышиные, примато-человеческие моноклональные антитела; и антиидиотипические антитела, а также любые интактные молекулы или их фрагменты.
В настоящем описании, термин «фрагмент антитела» относится к части, выделенной из или относящейся к полноценному первичному aHTH-MASP-2 антителу, обычно включающей в себя антигенсвязывающий или вариабельный домен. Наглядные примеры фрагментов антител включают в себя фрагменты Fab, Fab', F(ab)2, F(ab')2 и Fv, фрагменты scFv, диммеры, линейные антитела, одноцепочные молекулы антител и мультиспецифические антитела, состоящие из фрагментов антител.
В настоящем описании, термином «одноцепочный Fv» или «scFv» фрагмент антитела обозначает домены VH и VL антитела, в котором данные домены присутствуют в одиночной полипептидной цепи. В основном, полипептид Fv далее включает в себя полипептидный линкер между доменами Vpj и VL, который позволяет scFv сформировать желательную структуру для антигенсвязывающего домена.
В настоящем описании, термин «химерное антитело» обозначает рекомбинантный белок, который содержит вариабельные домены и гипервариабельные участки, выделенные из антител, млекопитающих различных видов, кроме человека (например, грызунов), притом, что остальные фрагменты молекулы антитела извлечены из антител человека.
В настоящем описании, термин «гуманизированное антитело» обозначает химерное антитело, которое включает в себя минимальную последовательность, соответствующую специфичным гипервариабельным участкам, извлеченным из нечеловеческого иммуноглобулина, трансплантированного в каркас человеческого антитела. Гуманизированные антитела представлены типичными рекомбинантными белками, в которых только гипервариабельные участки антител не являются человеческими.
В настоящем описании, термин «маннозосвязывающий лектин» (MBL) является эквивалентом для термина «маннозосвязывающий белок» (МБР).
В настоящем описании, термин «мембраноатакующий комплекс» (MAC) относится к комплексу из пяти конечных компонентов комплемента (С5-С9), который внедряется внутрь и разрушает мембрану. Этот комплекс также обозначается как C5b-9.
В настоящем описании, термин «объект» относится ко всем млекопитающим, включая, помимо прочего, человека, приматов, кроме человека, собак, кошек, лошадей, овец, коз, коров, кроликов, свиней и грызунов.
В настоящем описании, остатки аминокислот имеют следующие обозначения: аланин (Ala;А), аспарагин (Asn;N), аспарагиновая кислота (Asp;D), аргинин (Arg;R), цистеин (Cys;C), глутаминовая кислота (Glu;E), глютамин (Gln;Q), глицин (Gly;G), гистидин (His;H), изолейцин (Ile;I), лейцин (Leu;L), лизин (Lys;K), метионин (Met;M), фенилаланин (Phe;F), пролин (Pro;Р), серин (Ser;S), треонин (Thr;Т), триптофан (Trp;W), тирозин (Tyr;Y) и валин (Val;V).
В широком смысле, встречающиеся в природе аминокислоты могут быть разделены на группы согласно химическим характеристикам боковой цепи соответствующей аминокислоты. Так «гидрофобными» аминокислотами являются: Ile, Leu, Met, Phe, Trp, Tyr, Val, Ala, Cys и Pro, а «гидрофильными» аминокислотами являются: Gly, Asn, Gln, Ser, Thr, Asp, Glu, Lys, Arg и His. Эта разделение аминокислот на группы далее может быть развито следующим образом. Под термином «незаряженная гидрофильная» аминокислота подразумеваются Ser, Thr, Asn или Gln. Под «кислой» аминокислотой подразумеваются Glu или Asp. Под «основной» аминокислотой подразумеваются Lys, Arg или His.
В настоящем описании, термин «консервативное замещение аминокислоты» проиллюстрирован замещением среди аминокислот, разделенных на следующие группы: (1) глицин, аланин, валин, лейцин и изолейцин, (2) фенилаланин, тирозин и триптофан, (3) серин и треонин, (4) аспартат и глютамат, (5) глютамин и аспарагин, (6) лизин, аргинин и гистидин.
Термин «олигонуклеотид», который используется в настоящем описании, относится к олигомеру или полимеру рибонуклеиновой кислоты (РНК) или дезоксирибонуклеиновой кислоты (ДНК) или их миметическим формам. Данный термин также относится к олигонуклеиновым основаниям, образованным связями натуральных нуклеотидов, сахаров и ковалентных интернуклеозидов (главная цепь) так же, как и олигонуклеотидами, имеющие синтетические модификации.
II. АЛЬТЕРНАТИВНЫЙ ПУТЬ АКТИВАЦИИ: НОВОЕ ПОНИМАНИЕ
Впервые альтернативный путь активации комплемента был описан Луисом Пиллмером (Louis Pillemer) и его коллегами в начале 1950-х, данное описание опиралось на исследования, в которых полученный из клеточных стенок дрожжей зимозан использовался в качестве активатора комплемента (Pillemer, L. et al., J. Exp. Med. 103:1-13, 1956; Lepow, I.H., J. Immunol. 125:471-478, 1980). С тех пор зимозан считался каноническим примером специфического активатора для альтернативного пути в сыворотке человека и грызунов (Lachmann, P.J., et al., Springer Semin. Immunopathol. 7: 143-162, 1984; Van Dijk, H., et al., J. Immunol. Methods 85:233-243, 1985; Pangburn, M.K., Methods in Enzymol. 162:639-653, 1988). Наиболее подходящим и широко используемым анализом на активацию альтернативного пути является инкубирование сыворотки с зимозаном, нанесенным на пластиковую оболочку, с последующим определение количества отложения C3b на твердой фазе. Как ожидалось, наблюдается значительное отложение C3b на оболочках, покрытых зимозаном, после инкубации с нормальной сывороткой мыши (Фиг.7В). Однако инкубация сыворотки, полученной от гомозиготной мыши с MASP-2-недостаточностью, с зимозан-покрытыми оболочками провоцирует значительную редукцию отложения C3b, по сравнению с нормальной сывороткой. Далее, при использовании сыворотки от гетерозиготной мыши с дефецитом гена MASP 2 в данном исследовании, наблюдаемые уровни отложения C3b занимают среднее значение между уровнями, полученными при использовании сыворотки от гомозиготной мыши с MASP-2-недостаточностью и от здоровой мыши. Параллельные результаты были получены при использовании оболочек, покрытых мананном, другим полисахаридом, известным в качестве активатора альтернативного пути (Фиг.7А). Учитывая, что нормальная мышь и мышь с MASP-2-недостаточностью имеют общие генетические данные, за исключением гена MASP 2, данные неожиданные результаты демонстрируют, что MASP-2 играют наиважнейшую роль в активации альтернативного пути.
Данные результаты являются неоспоримым доказательством того, что альтернативный путь не является независимым автономным путем активации комплемента, как описано практически во всех ныне существующих медицинских учебниках и недавних обзорных статьях по комплементу. На сегодняшний день существует широко распространенное научное мнение, что альтернативный путь активируется на поверхности определенных объектов-мишеней (микробы, зимозан, кроличьи эритроциты) путем усиления спонтанной активации С3 типа «спящий механизм». Однако отсутствие значительной активации альтернативного пути в сыворотке, полученной от мыши с блокированным геном MASP-2, с двумя хорошо известными «активаторами» альтернативного пути делает несостоятельной данную теорию «спящего механизма» (tick-over theory) функциональной активации, которая описывает важный физиологический механизм для активации комплемента.
Учитывая тот факт, что протеаза MASP-2 выполняет специфичную и четко определенную роль фермента, отвечающего за инициацию пектинового каскада комплемента, данные результаты указывают на активацию пектинового пути при участии зимозана и маннана как первых критических инициаторов для последующей активации альтернативного пути. C4b представляет собой продукт активации, образующийся в пектиновом путем, но без участия альтернативного пути. В соответствии с данной концепцией, инкубация нормальной мышиной сыворотки с лунками, покрытыми зимозаном или маннаном, приводит к отложению C4b в лунках и данное отложение C4b значительно сокращается, когда покрытые лунки подвергаются инкубации с сывороткой мыши с MASP-2-недостаточностью (Фиг.6А, 6В и 6С).
Кроме того, что как считают, альтернативный путь представляет собой независимый путь активации комплемента, он также может являться усиливающим контуром для активации комплемента, изначально спровоцированной посредством классического и пектинового пути (Liszewski, М.K. and J.P.Atkinson, 1993, in Fundamental Immunology, Third Edition, edited by W.E.Paul, Raven Press, Ltd., New York; Schweinie, J.E., et al., J. Clin. Invest. 84:1821-1829, 1989). В данном механизме усиления, опосредованного альтернативным путем, конвертаза С3 (C4b2b), полученная активацией классического или пектинового каскада комплемента, расщепляет С3 на С3а и C3b, и таким образом формирует элемент C3b, который может участвовать в формировании C3bBb, конвертазы С3 альтернативного пути. Подобное объяснение отсутствия активации альтернативного пути в сыворотке с выключенным геном MASP-2 состоит в том, что лектиновый путь необходим для начальной активации комплемента зимозаном, маннаном и прочими предполагаемыми «активаторами» альтернативного пути, в то время, как альтернативный путь играет принципиальную роль в амплификации активации комплемента. Другими словами, альтернативный путь активации скорее всего является не независимым линейным каскадом, а представляет собой предварительный усилительный контур, действующий по принципу положительной обратной связи, и зависящий от лектинового и классического путей активации комплемента.
Несмотря на то, что каскад комплемента активируется посредством трех различных путей активации (классическим, альтернативным и лектиновым путями) как рассматривалось выше, полученные нами результаты указывают на то, что более правильным было бы рассматривать комплемент как две основные системы, которые соответствуют врожденному (лектин) и приобретенному (классический) механизмам иммунозащитной системы комплемента. Лектины (МБР, М-фиколин, Н-фиколин и L-фиколин) представляют собой специфические молекулы распознавания, которые инициируют врожденную систему комплемента и данная система включает в себя лектиновый путь активации и ассоциированный с ней альтернативный путь усиливающего контура. C1q представляет собой специфическую молекулу распознавания, которая активирует приобретенный механизм иммунозащитной системы комплемента и данная система включает в себя классический путь активации и ассоциированный с ним альтернативный путь усиливающего контура. Мы обозначаем эти две основные системы активации комплемента как пектин-зависимая система активации комплемента и C1q-зависимая системе активации комплемента, соответственно.
В дополнение к своей основной роли иммунной защитной реакции, система комплемента активируется при повреждении тканей во многих клинических состояниях. Таким образом, налицо острая необходимость в развитии терапевтически эффективных ингибиторов комплемента для предупреждения данных отрицательных воздействий. Исходя из принятия того факта, что комплемент состоит из двух основных систем активации комплемента, следует вывод о том, что было бы весьма желательно специфическим образом ингибировать только систему активации комплемента, провоцирующую отдельную патологию, без полного отключения иммунозащитных способностей комплемента. Например, при болезненных состояниях, в которых активация комплемента опосредована преимущественно пектин-зависимой системой комплемента, было бы полезно специфичным образом ингибировать только данную систему. Таким образом, C1q-зависимая система активации комплемента осталась бы незатронутой для управления активностью иммунного комплекса, что помогло бы в защитной иммунной реакции против инфекции.
MASP-2 является предпочтительным белковым компонентом для нацеливания в разработке терапевтических агентов для специфического ингибирования лектин-зависимой системы комплемента. Среди всех прочих белковых компонентов пектин-зависимой системы комплемента (MBL, Н-фиколин, М-фиколин, L-фиколин, MASP-2, С2-С9, фактор В, фактор D и пропердин), только MASP-2 является уникальным для пектин-зависимой системы комплемента и необходимым для функционирования системы. Пектины (MBL, Н-фиколин, М-фиколин и L-фиколин) также являются уникальными компонентами в пектин-зависимой системе комплемента. Однако нехватка любого из данных пектиновых компонентов не ингибировала бы активацию системы в достаточной степени по причине избыточности пектинов. Было бы необходимым ингибировать все четыре пектина для гарантированного подавления пектин-зависимой системы активации комплемента. Далее, учитывая факт того, что MBL и фиколины проявляют независимую от комплемента опсоническую активность, ингибирование функции лектина спровоцировало бы снижение благотворного механизма иммунной защитной реакции против инфекции. При этом, данная комплемент-независимая лектиновая опсоническая активность осталась бы незатронутой в случае ингибирования гена MASP-2. Дополнительная ценность MASP-2 в качестве терапевтического гена-мишени при ингибировании пектин-зависимой системы активации комплемента состоит в том, что концентрация MASP-2 в плазме является одной из самых низких по сравнению с другими белками комплемента (~ 500 нг/мл); поэтому, соответственно низкие концентрации высокоафинных ингибиторов MASP-2 могут быть необходимы для полного его 24 подавления (Moller-Kristensen, M, et al., J. Immunol Methods 282:159-167, 2003).
III. РОЛЬ MASP-2 В РАЗЛИЧНЫХ ЗАБОЛЕВАНИЯХ И СОСТОЯНИЯХ И ТЕРАПЕВТИЧЕСКИЕ СПОСОБЫ ИСПОЛЬЗОВАНИЯ АГЕНТОВ, ИНГИБИРУЮЩИХ MASP-2
ИШЕМИЧЕСКОЕ РЕПЕРФУЗИОННОЕ ПОВРЕЖДЕНИЕ
Ишемическое реперфузионное повреждение (I/R) возникает, когда ток крови восстанавливается после длительного периода ишемии. Это является частой причиной заболеваемости и смертности в широком спектре заболеваний. Хирургические пациенты являются весьма восприимчивыми после репарации аневризмы аорты, экстракорпорального кровообращения, они страдают васкулярным реанастомозом вследствие, например, трансплантации органов (например, сердце, легкое, печень, почка) и реимплантации конечностей/пальцев, инсульта, инфаркта миокарда и гемодинамической реанимации с последующим шоком и/или хирургическим вмешательством. Пациенты с атеросклеротическими заболеваниями предрасположены к инфаркту миокарда, инсульту и эмболически индуцированной кишечной ишемии и ишемии нижних конечностей. Пациенты с травмой часто страдают от временной ишемии конечностей. Кроме того, любое повреждение, сопровождающееся значительной кровопотерей приводит к тотальной ишемическому реперфузионному поражению.
Патофизиология ишемического реперфузионному поражения является сложной с как минимум двумя главными факторами, содействующими процессу: активация комплемента и стимуляция нейтрофилов с сопутствующим повреждением, опосредованным кислородным радикалом. Активация комплемента при ишемическом реперфузионном поражении была впервые описана в связи с инфарктом миокарда более 30 лет назад, что повлекло за собой многочисленные исследования вклада системы комплемента в ишемическое реперфузионное поражение тканей (Hill, J.H., et al., J. Exp. Med. 133:885-900, 1971). Накопленные к настоящему времени знания указывают на то, что комплемент выполняет функции центрального посредника при ишемическом реперфузионном поражении. Ингибирование комплемента позволило уменьшить степень ишемического реперфузионного поражения у лабораторных животных, у которых была смоделирована данная патология. В ранних исследованиях снижение количества С3 было достигнуто в результате инъекции фактора из яда кобры, и сообщалось о благотворном эффекте этой инъекции при лечении ишемического реперфузионного поражения сердца и почки (Maroko, P.R., et al., 1978, J. Clin Invest. 61:661-670, 1978; Stein, S.H., et al., Miner Electrolyte Metab. 11:256-61, 1985). Однако растворимая форма рецептора комплемента 1 (sCR1) была первым специфичным ингибитором комплемента, используемым для предотвращения ишемического реперфузионного поражения миокарда (Weisman, H.F, et al., Science 249:146-51, 1990). Применение sCR1 для лечения ишемического реперфузионного поражения миокарда ослабляет инфаркт, связанный с пониженным уровнем отложения комплекса C5b-9 на коронарном эндотелии и пониженной инфильтрацией лейкоцитов после реперфузии.
При экспериментальном ишемическом реперфузионном поражении миокарда ингибитор эстеразы С1 (С1 INH), введенной перед реперфузией, предотвращает отложение C1q и в значительной мере сокращает область некроза сердечной мышцы (Buerke, M., et al., 1995, Circulation 91:393-402, 1995). У животных с генетической недостаточностью С3 менее выражен локальный некроз тканей после скелетно-мышечной либо кишечной ишемии (Weiser, M.R., et al., J. Exp. Med. 183:2343-48, 1996).
Мембраноатакующий комплекс является последним звеном в комплемент-опосредованном повреждении тканей и исследования на животных с С5-недостаточностью показали снижение уровня локального и обширного повреждения в у животных с моделированным ишемическим реперфузионном поражениием (Austen, W.G. Jr., et al., Surgery 126:343-48, 1999). Ингибитор активации комплемента, растворимый Crry (ген Y связанного с рецептором комплемента), проявил себя эффективным против поражения при его введении мышам до и после возникновения кишечной реперфузии (Rehrig, S., et al., J. Immunol. 167:5921-27, 2001). В модели скелетно-мышечной ишемии использование растворимого первого рецептора комплемента (sCR1) также редуцирует мышечное повреждение при введении после начала реперфузии (Kyriakides, С., et al., Am. J. Physiol. Cell Physiol. 281:0244-30, 2001). При использовании свиней в качестве модели ишемического реперфузионного поражения миокарда, животных подвергли лечению моноклональным антителом (MoAb) к анафилатоксину С5а до реперфузии, в результате чего было достигнуто уменьшение степени инфаркта (Amsterdam, E.A., et al., Am. J. Physiol. Heart Circ. Physiol. 268:H448-57, 1995). При лечении крыс с помощью С5 MoAb было достигнуто уменьшение силы инфаркта, нейтрофильной инфильтрации и апоптоза миокарда (Vakeva, A., et al., Circulation 97:2259-67, 1998). Данные результаты экспериментов подчеркивают важность активации комплемента в патогенезе ишемического реперфузионномго поражения.
Остается неясным вопрос, какой из путей активации комплемента (классический, пектиновый или альтернативный) в первую очередь вовлечены в активацию комплемента при ишемическом реперфузионном поражении. Вайзер (Weiser et al.) опытным путем продемонстрировал важнейшую роль пектинового и/или классического пути активации при скелетном ишемическом реперфузионном поражении, показав защищенность мыши с выключенными генами С3 либо С4 от ишемического реперфузионного поражения, которая выражалась в значительном редуцировании васкулярной проницаемости (Weiser, M.R., et al., J. Exp. Med. 183:2343-48, 1996). Между тем, эксперименты по почечной ишемии/реперфузии на мышах с выключенными геном С4 не продемонстрировали значительной протекции тканей, в то время, как мыши с выключенными генами С3, С5 и С6 не пострадали от повреждения, что позволяет выдвинуть предположение об активации комплемента при почечном ишемическом реперфузионном поражении посредством альтернативного пути (Zhou, W., et al., J. Clin. Invest. 105:1363-71, 2000). Используя мыши с недостаточностью D-фактора, Шталь (Stahletal.) недавно подтвердил очевидность важнейшей роли альтернативного пути активации при кишечной ишемии/реперфузии у мыши (Stahl, G., et al., Am. J. Pathol. 162:449-55, 2003). В отличие от него, Уильяме (Williams et al.) выдвинул предположение о доминирующей роли классического пути активации для инициации ишемии/реперфузии в кишечнике мышей, продемонстрировав уменьшение окрашивания органов для С3 и защиту от повреждения у мышей с С4- и IgM-недостаточностью (RagI-/-) (Williams, J.R, et al., J. Appl. Physiol. 86:938-42, 1999).
Лечение крыс при модели ишемического реперфузионного поражения миокарда с использованием моноклональных антител против крысиного маннозосвязывающего лектина (MBL) вызвало уменьшение постишемического реперфузионного поражения (Jordan, J.E., et al., Circulation 104:1413-18, 2001). Антитела MBL также уменьшили отложение комплемента на эндотелиальных клетках in vitro после окислительного стресса, указывая на роль пектинового пути при ишемическом реперфузионном поражении миокарда (Collard, CD., et al., Am. J. Pathol. 156:1549-56, 2000). Так же очевидно, что ишемическое реперфузионное поражение в некоторых органах может быть опосредовано специфической категорией IgM, вышеназванными натуральными антителами и активацией классического пути (Fleming, S.D., et al., J. Immunol. 169:2126-33, 2002; Reid, R.R., et al., J. Immunol. 169:5433-40, 2002).
Некоторые ингибиторы активации комплемента были разработаны как потенциальные терапевтические агенты для предупреждения заболеваемости и смертности от осложнений после ишемического реперфузионного поражения миокардиа. Два из этих ингибиторов, sCRI (TPIO) и гуманизированный анти-С5 scFv (Pexelizumab), прошли Фазу II клинических испытаний. Pexelizumab также прошел Фазу III клинических испытаний. Хотя TP1O хорошо переносился пациентами и оказывал благотворное действие в раннем периоде Фазы I/II клинических испытаний, результаты по Фазе II клинических испытаний в конце февраля 2002 не соответствовали ожидаемым. Однако дополнительный анализ данных, полученным от пациентов мужского пола, относящихся к группе риска, перенесших операции на открытом сердце, продемонстрировал значительное уменьшение заболеваемости и сокращение области, затронутой инфарктом. Введение гуманизированного анти-С5 scFv снизило у всех пациентов уровень смертности, связанной с острым инфарктом миокарда в СОМА и COMPLY Фазе II испытаний, однако конечный результат не соответствовал ожидаемому (Mahaffey, K.W., et al., Circulation 108:1176-83, 2003). Не столь давно были получены результаты по начальному этапу Фазы III клинических испытаний (PREVIO-CABG) анти-С5 scFv по улучшению состояния пациентов после аортокоронарного шунтирования. Хотя ожидаемые результаты не были получены в данном исследовании, в целом оно продемонстрировало снижение заболеваемости и смертности у прооперированных пациентов.
Доктор Уолш (Dr. Walsh) и его коллеги показали, что мыши с отсутствием MBL и, как следствие, лишенные механизма MBL-опосредованного пектинового пути активации комплемента с сохранением полной активности классического пути активации комплемента, защищены от сердечной реперфузии, что обуславливает сохранение сердечной функции (Walsh et al., J. Immunol. 175:541-46, 2005). Мыши с отсутствием C1q, компонента распознавания классического пути активации, но обладающие неповрежденным MBL путем активации комплемента, в значительной степени подвергнуты повреждению. Данные результаты указывают на то, что лектиновый путь активации играет ведущую роль в патогенезе ишемического реперфузионного поражения миокарда.
Активация комплемента известна тем, что она способна играть важную роль при повреждении тканей, опосредованном гастроинтестинальным ишемическим реперфузионным поражением. Используя мышиную модель гастоинтерального ишемического реперфузионного поражения в качестве объекта исследования Харт (Hart) и его коллеги недавно показали, что мыши с генетической MBL-недостаточностью защищены от повреждения кишечника после гастроинтестинального ишемического реперфузионного поражения (Hart et al., J. Immunol. 174:6373-80, 2005). Введение рекомбинантного MBL мышам с MBL-недостаточностью значительно увеличивает повреждение по сравнению с необработанными мышами с MBL-недостаточностью после гастроинтестинального ишемического реперфузионного поражения. В тоже время, мыши с генетической недостаточностью C1q, компонента распознавания классического пути активации, не защищены от повреждения тканей после гастроинтестинального ишемического реперфузионного поражения.
Почечное ишемическое реперфузионное поражение является важной причиной острой почечной недостаточности. Система комплемента, по-видимому, играет важную роль при ишемическом реперфузионном поражении почек. В недавнем исследовании, Де Врие (deVries) и его коллеги показали, что лектиновый путь активируется в ходе эксперимента так же как и клиническое ишемическое реперфузионное поражение почек (de Vries et al., Am. J. Path. 165:1677-88, 2004). Более того, лектиновый путь активации предшествует и локализуется совместно с комлементарным отложением СЗ, С6 и С9 при возникновении ишемического реперфузионного поражения почек. Данные результаты указывают на то, что лектиновый путь активации комплемента вовлечен в ишемическое реперфузионное поражение почек.
Таким образом, один из аспектов настоящего изобретения направлен на лечение ишемического реперфузионного повреждения у субъекта, перенесшего ишемическую реперфузию, при помощи терапевтически эффективной дозой агента, ингибирующего MASP-2, в фармацевтически приемлемом носителе. Агент, ингибирующий MASP-2, может быть введен субъекту посредством внутриартериального, внутривенного, внутричерепного, внутримышечного, подкожного либо другого парентерального способа введения, возможен также оральный прием непептидергических ингибиторов, однако наиболее подходящим является внутриартериальное или внутривенное введение. Введение композиций, ингибирующих MASP-2, по настоящему изобретению соответствующим образом начинается непосредственно после либо как можно скорее после случая ишемической реперфузии. В случаях, когда реперфузия происходит на контролируемом участке (например, вследствие репарации аневризмы аорты, трансплантации органов или реплантации отделенных или поврежденных конечностей или пальцев), агент, ингибирующий MASP-2, может быть введен до и/или во время и/или после реперфузии. Введение может периодически повторяться, если таковое предписано врачом для достижения оптимального терапевтического эффекта.
АТЕРОСКЛЕРОЗ
Вполне очевидно, что активация комплемента вовлечена в атерогенез человека. Некоторое число научных исследований убедительно продемонстрировали, что, несмотря на отсутствие значительной активации комплемента в здоровых артериях, комплемент в значительной степени активируется в атеросклеротических поражениях и особенно сильно в нестабильных и разорванных бляшках. Компоненты конечного пути активации комплемента часто обнаруживаются в атеросклеротических бляшках человека (Niculescu, F, et al., Mol. Immunol. 36:949-55.10-12, 1999; Rus, H.G., et al., Immunol. Lett. 20:305-310, 1989; Torzewski, M., et al., Artehoscler. Thromb. Vase. Biol. 18:369-378, 1998). Было также продемонстрировано отложение С3 и С4 в области артериальных поражений (Hansson, G. К., et al., Acta Pathol. Microbiol. Immunol. Scand. (A) 92:429-35, 1984). Объем отложения C5b-9 был измерен с целью корреляции со степенью тяжести поражения (Vlaicu, R., et al., Atherosclerosis 57:163-77, 1985). Отложение комплемента iC3b, но не C5b-9, было особенно сильным в разорванных и нестабильных бляшках, из чего можно сделать предположение, что активация комплемента может представлять собой фактор при острых коронарных синдромах (Taskinen S., et al., Biochem. J. 367:403-12, 2002). При экспериментальной атероме у кроликов активация комплемента имела место до развития поражения (Seifer, P.S., et al., Lab Invest. 60:747-54, 1989).
При атеросклеротических поражениях комплемент активируется посредством классического и альтернативного путей, но до сих пор существует мало доказательств того, что активация комплемента осуществляется через пектиновый путь. Некоторые компоненты артериальной стенки могут инициировать активацию комплемента. Классический путь активации комплемента может быть активирован связыванием С-реактивного белка (CRP) с ферментативно расщепленным липопротеидом низкой плотности (LDL) (Bhakdi, S., et al., Arterioscler. Thromb. Vase. Biol. 19:2348-54, 1999). Совместимым с данной точкой зрения является открытие того, что конечные белки комплемента солокализуются с CRP во внутренних оболочках сосудов на ранней стадии поражений у человека (Torzewski, J, et al., Arterioscler. Thromb. Vase. Biol. 18:1386-92, 1998). Подобным образом иммуноглобулин М или антитела IgG, специфичные к окисленным LDL, находящимся в области поражения, могут активировать классический путь активации комплемента (Witztum, J. L, Lancet 344:793-95, 1994). Липиды, выделенные из человеческих атеросклеротических поражений, обладают высоким содержанием неэтерифицированного холестерина и способны активизировать альтернативный путь (Seifert Р.S., et al., J. Exp. Med. 172:547-57, 1990). Chlamydia pneumonia, грамотрицательная бактерия, часто связанная с атеросклеротическими поражениями, может также активировать альтернативный путь активации комплемента (Campbell L.A., et al., J. Infect. Dis. 172:585-8, 1995). Другие потенциальные активаторы комплемента, присутствующие в атеросклеротических поражениях, включают в себя кристаллы холестерина и клеточный дебрис, каждый из которых может активировать альтернативный путь (Seifert, P.S., et al., Mot. Immunol. 24:1303-08, 1987).
Побочные продукты активации комплемента известны своими многочисленными биологическими свойствами, которые могут влиять на развитие атеросклеротических поражений. Локальная активация комплемента может индуцировать клеточный лизис и генерировать по меньшей мере клеточный дебрис, обнаруженный в некротическом ядре генерализованных поражений (Niculescu, F. et al., Mol. Immunol. 36:949-55, 10-12, 1999). Сублитическая активация комплемента могла бы стать значимым фактором, способствующим сглаживанию пролиферации мышечных клеток и инфильтрации моноцитов во внутренней оболочке артерий при атерогенезе (Torzewski J., et al., Arterioscler. Thromb. Vase. Biol. 18:673-77, 1996). Стойкая активация комплемента может быть пагубной, так как может инициировать и поддерживать воспаление. Вдобавок к инфильтрации компонентов комплемента из плазмы крови артериальные клетки экспрессируют информационную РНК белков комплемента, а экспрессия различных компонентов комплемента регулируется в атеросклеротических поражениях (Yasojima, K., et al., Arterioscler. Thromb. Vase. Biol. 21:1214-19, 2001).
Было проведено ограниченное число исследований, посвященных изучению влияния недостаточности белков комплемента на атерогенез. Результаты экспериментов на лабораторных животных были противоречивыми. У крысы формирование поражений, подобных атеросклеротическим, причиненных введением токсических доз витамина D, было сокращено у животных с недостаточностью комплемента (Geertinger P., et al., Acta. Pathol. Microbiol. Scand. (A) 78:284-88, 1970). Далее, у кроликов с повышенным уровнем холестерина ингибирование комплемента или при помощи генетической недостаточности С6 (Geertinger, P., et al., Artery 1:177-84, 1977; Schmiedt, W., et al., Arterioscl. Thromb. Vase. Biol. 18:1790-1795, 1998) или антикомплементарным агентом K-76 COONa (Saito, E., et al., J. Drug Dev. 3:147-54, 1990) подавляло развитие атеросклероза, не затрагивая уровни холестерина в сыворотке. С другой стороны, недавнее исследование показало, что С5-недостаточность не уменьшает развитие атеросклеротических поражений у мыши с недостаточностью аполипопротеина E (ApoE) (Patel, S., et al., Biochem. Biophys. Res. Commun. 286:164-70, 2001). Однако другое исследование показало повышенное развитие атеросклеротических поражений у мыши с недостаточностью LDLR (Idir-) с или без С3-недостаточности (Buono, С, et al., Circulation 105:3025-31, 2002). Исследования обнаруживают, что развитие атеромы в поражениях, подобных атеросклеротическим, частично зависит от присутствия интактной системы комплемента.
Таким образом, один аспект настоящего изобретения направлен на лечение либо предупреждение атеросклероза при проведении терапии субъекту, перенесшему либо подверженному атеросклерозу, при помощи терапевтически эффективной дозы агента, ингибирующего MASP-2, в фармацевтически приемлемом носителе. Агент, ингибирующий MASP-2, может быть введен субъекту посредством внутриартериального, внутривенного, интратекального, внутричерепного, внутримышечного, подкожного либо другого парентерального администрирования, возможен также оральный прием непептидергических ингибиторов. Введение состава, ингибирующего MASP-2, может быть начато после установления диагноза атеросклероз у объекта обследования или профилактически у объекта наблюдения, относящегося к группе повышенного риска развития подобного заболевания. Введение может периодически повторяться, если таковое предписано врачом для достижения оптимального терапевтического эффекта.
ПРОЧИЕ СОСУДИСТЫЕ ПОРАЖЕНИЯ И ЗАБОЛЕВАНИЯ
Эндотелий подвергается обширному воздействию со стороны иммунной системы и является особенно чувствительным к присутствующим в плазме крови белкам комплемента. Комплемент-опосредованное васкулярное поражение было продемонстрировано при сравнении патофизиологии нескольких заболеваний сердечно сосудистой системы, включая атеросклероз (Seifert, P. S., et al., Atherosclerosis 73:91-104, 1988), ишемическое реперфузионное поражение (Weisman, H.F, Science 249:146-51, 1990) и инфаркт миокарда (Tada, Т., et al., Virchows Arch 430:327-332, 1997). Будет очевидным предположить, что активация комплемента может распространяться на другие васкулярные заболевания.
Например, имеются доказательства того, что активация комплемента относится к патогенезу многих форм васкулита, включая: болезнь Шенлейн-Геноха, васкулит, обусловленный системной красной волчанкой, васкулит, обусловленный ревматоидным артритом (также известный как злокачественный ревматоидный артрит), иммунокомплексный васкулит, синдром Такаясу. Болезнь Шенлейн-Геноха является формой системного васкулита малых вен с иммунным патогенезом, в котором активация комплемента посредством пектинового пути, управляющего эндотелиальным повреждением, опосредованным C5b-9, является важнейшим механизмом (Kawana, S., et al., Arch. Dermatol. Res. 282:183-7, 1990; Endo, M., et al., Am J. Kidney Dis. 35:401-7, 2000). Системная красная волчанка (SLE) является примером системных аутоиммунных заболеваний, затрагивающих многие органы, включая кожу, почки, суставы, серозные поверхности и центральную нервную систему, и часто является опосредованным тяжелым васкулитом. Анти-эндотелиальные антитела IgG и комплексы IgG, способные к связыванию с эндотелиальными клетками, присутствуют в сыворотке пациентов с активной SLE, а отложения иммунных комплексов IgG и комплемента обнаруживаются в стенках кровяных сосудов у пациентов с васкулитом SLE (Cines, D.В., et al., J. Clin. Invest. 73:611-25, 1984). Ревматоидный артрит, опосредованный васкулитом, также известный как злокачественный ревматоидный артрит (Tomooka, K., Fukuoka Igaku Zasshi 80:456-66, 1989), иммунокомплексный васкулит, васкулит, опосредованный гепатитом А, лейкоцитокластический васкулит и артрит, известный как синдром Такаясу, формируют другую плеоморфную группу заболеваний человека, в которых роль комплемент-зависимой цитотоксичности в отношении эндотелиальных и других клеточных типов также задокументирована (Tripathy, N.K., et al., J. Rheumatol. 28:805-8, 2001).
Также существуют доказательства того, что активация комплемента имеет место при дилатационной кардиомиопатии. Дилатационная кардиомиопатия является синдромом, характеризующимся увеличением сердца и ослаблением систолической сердечной функции. Недавние данные позволили выдвинуть предположение, о том, что постоянное воспаление в сердечной мышце может свидетельствовать о развитии болезни. Известно, что количество конечного мембраноатакующего комплекс комплемента C5b-9 значительно коррелирует с отложением иммуноглобулина и экспрессией миокардиального TNF-альфа. В миокардиальной биопсии от 28 пациентов с дилатационной кардиомиопатией была продемонстрирована миокардиальная аккумуляция C5b-9, что позволило сделать предположение о том, что хроническая иммуноглобулин-опосредованная активация комплемента в миокарде может частично способствовать прогрессированию дилатационной кардиомиопатии (Zwaka, ТР., et al. Am. J. Pathol. 161(2):449-57, 2002).
Таким образом, один аспект настоящего изобретения направлен на лечение сосудистого заболевания, включая сердечно сосудистые заболевания, цереброваскулярные заболевания, периферические (например, скелетно-мышечные) васкулярные заболевания, реноваскулярные заболевания и мезентериальные/энтериальные васкулярные заболевания, путем введения состава, включающего в себя терапевтически эффективную дозу агента, ингибирующего MASP-2, в фармацевтически приемлемом носителе. Заболевания, для которых данное изобретение может быть полезным, представлены следующим перечислением (список заболеваний данным перечислением не ограничивается): васкулит, включая болезнь Шенлейн-Геноха, васкулит, обусловленный системной красной волчанкой, васкулит, обусловленный ревматоидным дискоидным артритом (также известный как злокачественный ревматоидный артрит), иммунокомплексный васкулит и синдром Такаясу; дилатационная кардиомиопатия, диабетическая ангиопатия, синдром Кавасаки (артериит), венозная воздушная эмболия (VGE). Также, учитывая, что активация комплемента происходит в результате люминальной травмы и воспалительной реакции на инородное тело, опосредованной кардиоваскулярными внутривенными процедурами, мы считаем, что составы, ингибирующие MASP-2, настоящего изобретения могут также быть использованы для ингибирования рестеноза вследствие введения стента, ротационной атерэктомии и/или чрескожной транслюминальной коронарной ангиопластики (РТСА), одни либо в комбинации с другими агентами, ингибирующими рестеноз, которые описаны в патенте США No.6,492,332, выданном Демопулосу
Агент, ингибирующий MASP-2, может быть введен субъекту посредством внутриартериального, внутривенного, внутримышечного, интратекального, внутричерепного, подкожного либо другого парентерального администрирования, возможен также оральный прием непептидергических ингибиторов. Введение может периодически повторяться, если таковое предписано медицинским специалистом для достижения оптимального терапевтического эффекта. Для ингибирования рестеноза, композиции, ингибирующие MASP-2, могут быть администрированы до, во время либо после введения стента или проведения процедуры атерэктомии либо ангиопластики. Альтернативно, композиции, ингибирующие MASP-2, могут быть нанесены на стент или инкорпорированы в него.
ЖЕЛУДОЧНО-КИШЕЧНЫЕ ЗАБОЛЕВАНИЯ
Неспецифический язвенный колит и гранулематозная болезнь (болезнь Крона) являются хроническими воспалительными заболеваниями пищеварительного тракта, которые относятся к группе состояний, известных под общим названием воспалительной болезни кишечника (IBD). IBD характеризуется спонтанно возникшим, хроническим, повторяющимся воспалением неизвестного происхождения. Несмотря на обширные исследования в области данного заболевания у людей и подопытных животных, точные механизмы данной патологии остаются неизвестны. Однако считается, что система комплемента активируется у пациентов с IBD и играет важную роль в патогенезе заболевания (Kolios, G, et al., Hepato-Gastroenterology 45:1601-9, 1998; Elmgreen, J, Dan. Meet. Bull. 33:222, 1986).
Было показано, что C3b и другие активированные продукты комплемента обнаруживаются на люминальной поверхности эпителиальных клеток, также как и в кровяных сосудах мышечной, слизистой и подслизистой оболочках у IBD пациентов (Halstensen, Т.S., et al., Immunol. Res. 10:485-92, 1991; Halstensen, T.S., et al„ Gastroenterology 98:1264, 1990). Более того, полиморфно-ядерная клеточная инфильтрация, обычно возникающая в результате генерации С5а, характерным образом обнаруживается в воспаленном кишечнике (Kohl, J., Mol. Immunol. 38:175, 2001). Имеются также сведения о том, что мультифункциональный ингибитор комплемента К-76 способен продуцировать симптоматическое улучшение при неспецифическом язвенном колите в небольшом клиническом исследовании (Kitano, A., et al., Dis. Colon Rectum 35:560, 1992), и в лабораторной модели каррагенин-опосредованного колита у кроликов (Kitano, A., et at., Clin. Exp. Immunol. 94:348-53, 1993).
Было показано, что новый антагонист рецептора С5а человека обладает защитными свойствами в отношении патологических проявлений болезни в модели IBD у крыс (Woodruff, T.M., et al., J. Immunol. 171:5514-20, 2003). Мыши с генетической недостаточностью фактора распада (DAF), мембранного комплемент-регуляторного белка, были использованы в модели IBD с целью продемонстрировать, что DAF-недостаточность приводила к явно повышенному повреждению тканей и повышала уровень провоспалительных цитокинов (Lin, F, et al., J. Immunol. 172:3836-41, 2004). Поэтому, контроль комплемента важен для регулирования гомеостаза кишечника и может представлять собой главный патогенный механизм, вовлеченный в развитие IBD.
Таким образом, настоящее изобретение предлагает способы ингибирования MASP-2-зависимой активации комплемента у субъектов, страдающих воспалительными желудочно-кишечными заболеваниями, включая, помимо прочего, панкреатит, дивертикулит и кишечные расстройства, в том числе гранулематозная болезнь (болезнь Крона), неспецифический язвенный колит и синдром раздраженной толстой кишки, путем введения состава, содержащего терапевтически эффективную дозу агента, ингибирующего MASP-2, в фармацевтически приемлемом носителе пациенту, страдающему от подобного расстройства. Агент, ингибирующий MASP-2, может быть введен субъекту посредством внутриартериального, внутривенного, внутримышечного, подкожного, интратекального, внутричерепного, либо другого парентерального администрирования, возможен также оральный прием непептидергических ингибиторов. Введение может периодически повторяться, если таковое предписано медицинским специалистом для контроля симптомов заболевания, подлежащего лечению.
ЛЕГОЧНЫЕ ЗАБОЛЕВАНИЯ
Комплемент вовлечен в патогенез многих воспалительных легочных заболеваний, включая: синдром острой дыхательной недостаточности (ARDS) (Ware, I, et al., N. Engl. J. Med. 342:1334-49, 2000), трансфузионно-обусловленное поражение легких (TRALI) (Seeger, W., et al., Blood 76:1438-44,1990), острое ишемическое реперфузионное легочное заболевание (Xiao, F., et al., J. Appl. Physiol. 82:1459-65, 1997), хроническое обструктивное заболевание легких (COPD) (Marc, M.M., et al., Am. J. Respir. Cell Mol. Biol. (Epub ahead of print), March 23, 2004), астма (Krug, N, et al., Am. J. Respir. Crit. Care Med. 164:1841-43, 2001), некротический неинфекционный гранулематоз (гранулематоз Вегенера) (Kalluri, R., et al., J. Am. Soc. Nephrol. 8:1795-800, 1997) и заболевание гломерулярной базальной мембраны (болезнь Гудпасчура) (Kondo, С, et al., Clin. Exp. Immunol. 124:323-9, 2001).
Сейчас повсеместно принято мнение, что патофизиология ARDS инициирует нерегулируемый воспалительный каскад, который сначала играет роль нормального иммунного ответа на инфекцию или прочий провоцирующий фактор, и, в конечном счете, провоцирует значительное аутопоражение у реципиента (Stanley, T.R, Emerging Therapeutic Targets 2:1-16, 1998). Практически у всех пациентов с ARDS отчетливо проявляется экстенсивная активация комплемента (повышенные уровни компонентов С3а и С5а комплемента в плазме), и степень активации комплемента была соотнесена с развитием и исходом ARDS (Hammerschmidt, D.F, et al., Lancet 1:947-49, 1980; Solomkin, J.S., et al., J. Surgery 97:668-78, 1985).
Различные экспериментальные и клинические данные свидетельствуют о роли активации комплемента в патофизиологии ARDS. В животных моделях систематическая активация комплемента ведет к острому легочному заболеванием с гистопатологией, схожей с той, что проявляется при ARDS человека (Till, G.O., et al., Am. J. Pathol. 129:44-53, 1987; Ward, P.A, Am. J. Pathol. 149:1081-86, 1996). Ингибирование каскада комплемента общим уменьшением комплемента или специфическим ингибированием С5а обеспечивает защиту от острого легочного заболевания в животных моделях (Mulligan, M.S., et al., J. Clin. Invest. 98:503-512, 1996). У модельных крыс, sCRI обладает защитным эффектом в комплемент- и нейтрофил-опосредованных легочных заболеваниях (Mulligan, M.S., Yeh, et al., J. Immunol. 148:1479-85, 1992). Кроме того, практически все компоненты комплемента могут быть спродуцированы локально в легких эпителиальными клетками легочной альвеолы типа II, альвеолярными макрофагами и легочными фибробластами (Hetland, G., et al., Scand. J. Immunol. 24:603-8, 1986; Rothman, В.I, et al., J. Immunol. 145:592-98, 1990). Таким образом, роль каскада комплемента четко определена и состоит в том, что он значительно способствует легочному воспалению и, как следствие, легочному поражению при ARDS.
Астма представляет собой, прежде всего, воспалительное заболевание. Кардинальные черты аллергической астмы включают в себя гиперчувствительность дыхательных путей к множеству специфических и неспецифических стимуляторов, избыточное выделение дыхательными путями слизи, легочную эозинофилию и повышенную концентрацию сыворотки IgE. Хотя происхождение астмы соотносят со множеством факторов, считается, что данное заболевание возникает главным образом в результате самопроизвольного иммунного ответа на обычные природные антигены у индивидуумов с генетической восприимчивостью. Хорошо известен тот факт, что система комплемента активирована в высокой степени в легких человека, страдающего астмой. (Humbles, A.A., et al., Nature 406:998-01, 2002; van de Graf, E.A., et al., J. Immunol. Methods 147:241-50, 1992). Более того, недавние данные, полученные на животных моделях и человеке, явно свидетельствуют, что активация комплемента является важнейшим механизмом, относящимся к патогенезу заболевания (Кагр, С.L, et al., Nat. Immunol. 1:221-26, 2000; Bautsch, W., et al., J. Immunol. 165:5401-5, 2000; Drouin, S.M., et al., J. Immunol. 169:5926-33, 2002; Walters, D.M, et al., Am. J. Respir. Cell Mol. Biol. 27:413-18, 2002). Роль пектинового пути активации при астме подтверждается исследованиями, в которых используется модель хронической фунгоидной астмы у мышей. У мышей с генетической недостаточностью маннозосвязывающего лектина развивается измененная гиперчувствительность дыхательных путей, по сравнению с нормальными животными в данной астматической модели (Hogaboam, C.M., et al., J. Leukoc. Biol. 75:805-14, 2004).
При астме комплемент может быть активирован посредством нескольких путей, включая:
(a) активацию посредством классического пути в результате формирования комплекса антител к аллергенам;
(b) активацию альтернативного пути на поверхности аллергенов;
(c) активацию пектинового пути через соединение углеводных структур с аллергенами; и
(d) расщепление С3 и С5 протеазами, выделяющимися из клеток воспаления.
Хотя остается еще множество вопросов, нуждающихся в выяснении комплексной роли, которую выполняет комплемент при астме, идентификация путей активации комплемента, вовлеченных в развитие аллергической астмы, может послужить отправной точкой для развития инновационных терапевтических стратегий против данного заболевания, которое становится все более масштабным.
Определенное число исследований с использованием животных моделей продемонстрировало критическую роль С3 и продукта его расщепления, С3а, в развитии аллергического фенотипа. Друин (Drouin) и его коллеги использовали мышей с С3-недостаточностью в модели астмы, вызванной овальбумином (OVA)/Aspergillus fumigatus (Drouin et al., J. Immunol. 167:4141-45, 2001). Они обнаружили, что при контрольном введении аллергена, мыши с С3-недостаточностью проявляют поразительно пониженный уровень AHR и легочной эозинофилии по сравнению с контрольными мышами дикого типа. Далее, у данных мышей с С3-недостаточностью было значительно редуцировано количество клеток, продуцирующих IL-4, и ослаблены Ag-специфичный IgE и IgGI ответы. Тоб (Taube) и его коллеги получили схожие результаты в OVA-модели астмы при блокировании активации комплемента на уровне С3 и С4 с использованием растворимой рекомбинантной формы рецептора комплемента Crry мыши (Taube et al., Am. J. Respir. Crit. Care Med. 168:1333-41, 2003). Умбль (Humbles) и его коллеги удалили у мышей C3aR для изучения роли С3а в эозинофильной функции (Humbles et al., Nature 406:998-1001, 2000). Используя OVA-модель астмы, они наблюдали практически полную защиту от развития AHR под воздействием распыленного метахолина. Друин и его коллеги (2002) использовали мышей с C3aR-недостаточностью в модели астмы OVA/A, fumigatus, и продемонстрировали ослабление аллергического ответа весьма схожего с реакцией животных с С3-недостаточностью, у которых был уменьшен уровень AHR, эозинофильного восстановления, продуцирования цитокинов ТН2 и секреции слизи в легких, а также, ослаблены Ag-специфичные IgE и IgGI ответы (Drouin et al., J. Immunol. 169:5926-33, 2002). Ботч (Bautsch) и его коллеги провели исследования с использованием линии морских свинок с естественной делецией C3aR (Bautsch et al., J. Immunol. 165:5401-05, 2000). Используя OVA-модель аллергической астмы, они отметили значительную защиту от бронхоконстрикции дыхательных путей при взаимодействии с антигеном.
Множество недавних исследований с использованием животных моделей продемонстрировало критическую роль С5 и продукта его расщепления С5а в развитии аллергического фенотипа. Абе (Abe) и его коллеги получили доказательства того, что активация C5aR связана с воспалением дыхательных путей, продуцированием цитокинов и восприимчивостью дыхательных путей (Abe et al., J. Immunol. 167:4651-60, 2001). В их исследованиях ингибирование активации комплемента растворимой формой CRI, футана (один из ингибиторов активации комплемента) или антагонистом синтетического гексапептида С5а блокировало воспалительную реакцию и восприимчивость дыхательных путей к метахолину. В исследованиях с использованием блокирующего анти-С5 моноклонального антитела, Пенг (Peng) и его коллеги обнаружили, что активация С5 относилась в основном как к воспалению дыхательных путей, так и к AHR в OVA-модели астмы (Peng et al., J. Clin. Invest. 115:1590-1600, 2005). Также, Белдер (Baelder) и его коллеги отметили, что блокада C5aR существенно уменьшает AHR в астматической модели A. fumigatus (Baelder et al., J. Immunol. 174:783-89, 2005). Более того, одновременная блокада C3aR и C5aR существенно уменьшает воспаление дыхательных путей, о чем свидетельствует сокращенное количество нейтрофилов и эозинофилов bbal
Хотя все вышеупомянутые исследования подчеркивают важность факторов комплемента С3 и С5 и продуктов их расщепления в патогенезе экспериментальной аллергической астмы, данные исследования не предоставляют информации о вкладе каждого из этих трех путей активации комплемента, так как С3 и С5 присутствуют во всех трех этих путях. Однако недавнее исследование Хогабома (Hogaboam) и его коллег указывает на то, что пектиновый путь активации может играть ведущую роль в патогенезе астмы (Hogaboam et al., J. Leukocytye Biol. 75:805-814, 2004). В этих исследованиях использовали мышей с генетической недостаточностью белка маннозосвязывающего лектина-А (MBL-A), связывающего молекулу углевода, которая выполняет функцию распознавательного компонента для активации пектинового пути активации комплемента. В модели хронической фунгиальной астмы, мыши MBL-A(+/+) и MBL-A(-/-), сенсибилизированные A. fumigatus были обследованы на 4 и 28 дни после взаимодействия с кондидиями A. fumigatus. По результатам обоих обследований AHR у сенсибилизированных мышей MBL-A(-/-) был в значительной мере ослаблен по сравнению с группой сенсибилизированных мышей MBL-A(+/+). Также эти исследователи обнаружили, что на 4 день после сенсибилизации уровни цитокинов ТН2 в легких (IL-4, IL-5 и IL-13) были значительно ниже у мышей MBL-A(-/-), сенсибилизированных A. fumigatus, по сравнению с группой дикого типа. Полученные ими результаты указывают на то, что MBL-A и лектиновый путь активации играют основополагающую роль в развитии и поддержании AHR при хронической грибковой астме.
Результаты, полученные в ходе недавних клинических исследований, в которых продемонстрирована очевидная связь между специфичным MBL-полиморфизмом и развитием астмы, доказывают, что лектиновый путь активации может выполнять важную патологическую роль в данном заболевании (Kaur et al., Clin. Experimental Immunol. 143:414-19, 2006). Концентрация MBL в плазме широко варьируется у индивидуумов, и это в первую очередь связано с полиморфизмом первичной структуры гена MBL. Авторы вышеупомянутой статьи обнаружили, что индивидуумы-носители хотя бы одной разновидности специфичного MBL-полиморфизма, который увеличивает экспрессию MBL от двух до четырех раз, подвергаются пятикратному увеличению риска развития бронхиальной астмы. Была также отмечена возросшая серьезность маркеров заболевания у тех пациентов, страдающих бронхиальной астмой, которые являлись носителями данного MBL-полиморфизма.
Таким образом, один аспект настоящего изобретения предлагает способ лечения легочных заболеваний посредством введения состава, содержащего терапевтически эффективный объем агента, ингибирующего MASP-2, в фармацевтически приемлемом носителе субъекту, страдающему легочными заболеваниями, включающими в себя (список данным перечислением не ограничивается) синдром острой дыхательной недостаточности, трансфузионно-обусловленное поражение легких, острое ишемическое реперфузионное легочное заболевание, хроническое обструктивное заболевание легких, астма, некротический неинфекционный гранулематоз (гранулематоз Вегенера), заболевание гломерулярной базальной мембраны (болезнь Гудпасчура), синдром мекониевой аспирации, синдром облитерирующего бронхиолита, идиопатический легочный фиброз, острое повреждение легких вследствие ожога, некардиогенный отек легких, трансфузионно-обусловленное угнетенное дыхание и эмфизема. Агент, ингибирующий MASP-2, может вводиться субъекту систематически посредством внутриартериального, внутривенного, внутримышечного, при ингаляции, через нос, подкожного либо другого парентерального администрирования, возможен также оральный прием непептидергических ингибиторов. Состав с содержанием агента, ингибирующий MASP-2, может быть смешан с одним и более дополнительными терапевтическими агентами, включая противовоспалительные агенты, антигистаминные, кортикостероидные либо противомикробные агенты. Введение может периодически повторяться, если таковое предписано медицинским специалистом, до полного выздоровления.
ЭКСТРАКОРПОРАЛЬНОЕ КРОВООБРАЩЕНИЕ
Существует множество медицинских процедур, во время которых кровь выводится из системы кровообращения пациента (экстракорпоральная система кровообращения или ЕСС). Такие процедуры включают в себя гемодиализ, плазмаферез, лейкоцитоферез, экстракорпоральный мембранный оксигенатор (ЕСМО), гепарино-индуцированную экстракорпоральную мембранную оксигенацию с преципитацией липопротеинов низкой плотности (HELP) и сердечно-легочное шунтирование (СРВ). При данных процедурах кровь или продукты крови попадают на инородные поверхности, что может изменить нормальную клеточную функцию и гемостаз. В первых исследованиях Крэддок (Craddock et al.) определил активацию комплемента как возможную причину гранулоцитопении при гемодиализе (Craddock, PR., et al., N. Engl. J. Med. 296:769-74, 1977). Результаты многочисленных исследований, начиная с 1977 и по настоящее время, указывают на то, что многие из побочных эффектов, возникающих у пациентов, которых подвергают гемодиализу или СРВ, вызваны активацией системы комплемента (Chenoweth, D.E., Ann. N.Y. Acad. Sci 516:306-313, 1987; Hugli, Т.Е., Complement 3:111-127, 1986; Cheung, A.K, J. Am. Soc. Nephrol. 1:150-161, 1990; Johnson, R.J., Nephrol. Dial. Transplant 9:36-45 1994). Например, потенциал активирования комплемента был продемонстрирован как важный критерий в определении биосовместимости гемодиализаторов в отношении восстановления функции почек, восприимчивости к инфекции, дисфункции легких, заболеваемости и коэффициенту выживаемости пациентов с почечной недостаточностью (Hakim, R.M., Kidney Int. 44:484-4946, 1993).
Ранее считалось, что активация комплемента при мембранном гемодиализе происходит в результате действия механизмов альтернативного пути активации благодаря слабой генерации С4а (Kirklin, J.K., et al., J. Thorac. Cardiovasc. Surg. 86:845-57, 1983; Vallhonrat, H., et al., ASAIO J. 45:113-4, 1999), однако недавние исследования позволяют предположить, что классический путь активации может также быть вовлечен в этот процесс (Wachtfogel, Y.T, et al., Blood 73:468-471, 1989). Однако до сих пор имеет место неправильное понимание факторов инициации и контроля активации комплемента на исскуственных носителях, включая биомедицинские полимеры. Например, мембранный купрофан (Cuprophan membrane), используемый при гемодиализе, был классифицирован как весьма возможный активатор комплемента. Не желая ограничиваться теорией, авторы настоящего изобретения предположили, что это отчасти можно объяснить полисахаридной природой данного соединения. Система MASP-2-зависимой активации комплемента, описанная в настоящем изобретении, предлагает механизм, в котором активация пектинового пути инициирует альтернативный путь активации комплемента.
Пациенты, подвергнутые ЕСС при СРВ, страдают от системной воспалительной реакции, которая частично вызвана контактом крови с искусственными носителями экстракорпоральной цепи, равно как и факторами, не опосредованными поверхностными носителями, как, например, хирургическая травма и ишемическое реперфузионное поражение (Butler, J., et al., Ann. Thorac. Surg. 55:552-9, 1993; Edmunds, L.H., Ann. Thorac. Surg.66 (Suppl):S12-6, 1998;Asimakopoulos, G., Perfusion 14:269-77, 1999). Воспалительная реакция, инициированная СРВ, может также спровоцировать постоперационные осложнения, в основном известные как «постперфузионный синдром». В числе таких послеоперационных случаев называют когнитивные расстройства (Fitch, J., et al., Circulation 100(25):2499-2506, 1999), дыхательную недостаточность, нарушение свертываемости крови, дисфункцию почек и, в самых тяжелых случаях, полиорганную недостаточность (Wan, S., et al., Chest 112:676-692, 1997). Операция коронарного шунтирования с СРВ приводит к обширной активации комплемента, по сравнению с операцией без СРВ, но со сопоставимыми уровнями хирургической травмы (Е. Fosse, 1987). Поэтому, первичной потенциальной причиной данных проблем, опосредованных СРВ, является самопроизвольная активация комплемента при проведении процедуры шунтирования (Chenoweth, K., et al., N. Engl. J. Med. 304:497-503, 1981; P. Haslam, et al., Anaesthesia 25:22-26, 1980; J.K. Kirklin, et al., J. Thorac. Cardiovasc. Surg. 86:845-857, 1983; Moore, F.D., et al., Ann. Surg 208:95-103, 1988; J. Steinberg, et al., J. Thorac. Cardiovasc. Surg 106:1901-1918, 1993). В системах СРВ альтернативный путь активации комплемента играет предоминантную роль, вытекающую из взаимодействия крови с искусственными поверхностями системы СРВ (Kirklin, J. К., et al., J. Thorac. Cardiovasc. Surg., 86:845-57, 1983; Kirklin, J.K., et al., Ann. Thorac. Surg. 41:193-199, 1986; Vallhonrat H., et al., ASAIO J. 45:113-4, 1999). Однако существуют данные о том, что при СРВ происходит активация и классического пути (Wachtfogel, Y.Т., et al., Blood 73:468-471, 1989).
Первичные воспалительные субстанции генерируются после активации системы комплемента, включая анафилатоксины С3а и С5а, опсонин C3b и мембраноатакующий комплекс C5b-9. С3а и С5а являются потенциальными стимуляторами нейтрофилов, моноцитов и тромбоцитов (Haeffner-Cavaillon, N, et al., J. Immunol., 139:794-9, 1987; Fletcher, M.P, et al., Am. J. Physiol. 265:1-11750-61, 1993; Rinder, C.S., et al., J. Clin. Invest. 96:1564-72, 1995; Rinder, C.S., et al., Circulation 100:553-8, 1999). Активация данных клеток приводит к выделению воспалительных цитокинов (IL-I, IL-6, IL-8, TNF альфа), окислительных свободных радикалов и протеаз (Schindler, R., et al., Blood 76:1631-8, 1990; Cruickshank, A.M., et al., Clin Sci (Lond) 79:161-5, 1990; Kawamura, Т., et al., Can. J. Anaesth. 40:1016-21, 1993; Steinberg, J.B., et al., J. Thorac. Cardiovasc. Surg. 106:1008-1, 1993; Finn, A., et al., J. Thorac. Cardiovasc. Surg. 105:234-41, 1993; Ashraf, S.S., et al., J. Cardiothorac. Vase. Anesth. 11:718-22, 1997). Было показано, что С5а обладает способностью регулировать слияние молекул CDUb и CD 18 компонента Mac-1 в полиморфно-ядерных клетках (PMNs) и индуцировать дегрануляцию PMNs для высвобождения воспалительных ферментов. Rinder, С, et al., Cardiovasc Pharmacol. 27 (Suppl 1):S6-12, 1996; Evangelista, V., et al., Blood 93:876-85, 1999; Kinkade, J.M., Jr., et al., Biochem. Biophys. Res. Commun. 114:296-303, 1983; Lamb, N.J, et al., Crit. Care Med. 27:1738-44, 1999; Fujie, К., et al., Eur. J. Pharmacol. 374:117-25, 1999. C5b-9 индуцирует экспрессию адгезивной молекулы Р-селектина (CD62P) в тромбоцитах (Rinder, С.S., et al., J. Thorac. Cardiovasc. Surg. 118:460-6, 1999), в которых оба компонента С5а и C5b-9 индуцируют поверхностную экспрессию Р-селектина на поверхности эндотелиальных клеток (Foreman, K.E., et al., J. Clin. Invest. 94:1147-55, 1994). Данные адгезивные молекулы вовлечены во взаимодействие между лейкоцитами, тромбоцитами и эндотелиальными клетками. Экспрессия адгезивных молекул на активированных эндотелиальных клетках ответственна за секвестрацию активированных лейкоцитов, которая затем приводит к воспалению и повреждению тканей (Evangelista, V., Blood 1999; Foreman, K.E., J. Clin. Invest. 1994; Lentsch, A.B., et al., J. Pathol. 190:343-8, 2000). Именно эти воздействия данных продуктов активации комплемента на нейтрофилы, моноциты, эритроциты и другие циркулирующие клетки управляют различными проблемами, возникающими после СРВ.
Некоторые ингибиторы комплемента изучаются с целью потенциального применения в СРВ. Они включают в себя рекомбинантный растворимый рецептор 1 комплемента (sCRI) (Chai, PJ., et al., Circulation 101:541-6, 2000), гуманизированное одноцепочное анти-С5 антитело (h5G1.1-scFv или Pexelizumab) (Fitch, J.C.K., et al., Circulation 100:3499-506, 1999), рекомбинантный слитый гибрид (САВ-2) кофакторного белка мембраны человека и комплементзависимого стимулятора гемолиза человека (Rinder, CS., et al., Circulation 100:553-8, 1999), 13-остаточный С3-связывающий циклический пептид (Compstatin) (Nilsson, В., et al., Stood'92:1661-7, 1998) и антифактор D MoAb (Fung, M., et al., J. Thoracic Cardiovasc. Surg. 122:113-22, 2001). SCR1 и САВ-2 ингибируют классический и альтернативный пути активации комплемента на этапах активации С3 и С5. Компстатин (Compstatin) ингибирует оба пути активации комплемента на этапе активации С3, в котором h5G1.1-scFv производят данное действие только на этапе активации С5. Антифактор D MoAb ингибирует альтернативный путь на этапе активации С3 и С5. Однако ни один из данных ингибиторов комплемента не ингибирует специфическим образом систему MASP-2-зависимой активации комплемента, описанную в данной заявке.
Были опубликованы результаты фазы III обширного перспективного клинического исследования, имевшего целью выяснить эффективность и безопасность гуманизированного одноцепочного анти-С5 антитела (h5G1.1-scFv, pexelizumab) для уменьшения числа случаев периоперационного инфаркта миокарда и снижения смертности при операциях аортокоронарного шунтирования (CABG) (Verrier, E.D., et al., JAMA 291:2319-27, 2004). В сравнении с плацебо, применение моноклонального антитела пекселизумаба (pexelizumab) не привело к значительному снижению смертности или уменьшению риска развития инфаркта миокарда у 2746 пациентов, которых подвергли операции аортокоронарного шунтирования. Однако имело место статистически значимое уменьшение этих показателей спустя 30 дней после проведения операции у всех 3099 пациентов, перенесших операцию аортокоронарного шунтирования с операцией на клапане или без нее. Учитывая, что моноклональное антитело пекселизумаб ингибирует активацию на этапе С5, оно ингибирует генерацию С5а и sC5b-9, не воздействуя при этом на генерацию двух других потенциальных воспалительных субстанции комплемента, С3а и опсонина C3b, которые также известны способностью принимать участие в воспалительной реакции, инициированной СРВ.
Один аспект настоящего изобретения, таким образом, направлен на предупреждение и лечение воспалительной реакции, инициированной экстракорпоральной экспозицией, путем лечения субъекта, которого подвергают процедуре экстракорпорального кровообращения, составом, включающим в себя терапевтически эффективную дозу агента, ингибирующего MASP-2, в фармацевтически приемлемом носителе, и в число пациентов входят те, кто подвергается гемодиализу, плазмаферезу, лейкоцитоферезу, экстракорпоральныму мембранному оксигенатору (ЕСМО), гепарино-индуцированной экстракорпоральной мембранной оксигенации с преципитацией липопротеинов низкой плотности (HELP) и сердечно-легочному шунтированию (СРВ). Лечение агентом, ингибирующим MASP-2, в соответствии со способами настоящего изобретения рассматривается эффективным для редуцирования или предупреждения когнитивной дисфункции, которая порой возникает из-за проведения процедур СРВ. Агент, ингибирующий MASP-2, может быть введен субъекту перед процедурой, и/или во время процедуры, и/или после процедуры, посредством внутриартериального, внутривенного, внутримышечного, подкожного или другого вида парентерального введения. Альтернативно, агент, ингибирующий MASP-2, может быть введен в кровоток субъекта во время экстракорпорального кровообращения посредством инъекций агента, ингибирующего MASP-2, в трубку или сквозь мембрану, в которой циркулирует кровь, или при контакте крови с поверхностью, покрытой агентом, ингибирующим MASP-2, которая представлена внутренней стенкой трубки, мембраны или другой поверхности оборудования СРВ.
ВОСПАЛИТЕЛЬНЫЕ И НЕВОСПАЛИТЕЛЬНЫЕ АРТРИТЫ И ДРУГИЕ СКЕЛЕТНО-МЫШЕЧНЫЕ ЗАБОЛЕВАНИЯ
Факт активации системы комплемента был установлен в патогенезе широкого спектра ревматологических заболеваний, включающих в себя ревматоидный артрит (Linton, S.M., et al., Molec. Immunol. 36:905-14, 1999), ревматоидный артрит у детей (Mollnes, ТЕ, et al., Arthritis Rheum. 29:1359-64, 1986), остеоартрит (Kemp, PA, et al., J. Clin. Lab. Immunol. 37:147-62, 1992), системную красную волчанку (SLE) (Molina, H., Current Opinion in Rheumatol. 14:492-497, 2002), болезнь Бехчета (Rumfeld, W.R, et al., Br. J. Rheumatol. 25:266-70, 1986) и синдром Шегрена (Sanders, M.E, et al., J. Immunol. 138:2095-9, 1987).
Имеются убедительные доказательства того, что активация комплемента, спровоцированная иммунным комплексом, представляет собой основной патологический механизм, который приводит к поражению тканей при ревматоидном артрите (RA). Многочисленные публикации свидетельствуют о наличии продуктов активации комплемента в плазме у пациентов, страдающих этим заболеванием (Morgan, B.P, et al., Clin. Exp.Immunol, 73:473-478, 1988; Auda, G., et al., Rheumatol. Int. 10:185-189, 1990; Rumfeld, W.R., et al., Br. J. Rheumatol. 25:266-270, 1986). Такие продукты активации, как, например, С3а, С5а, и sC5b-9, также были обнаружены в воспаленных суставах, пораженных ревматизмом, и была установлена положительная корреляция между степенью активации комплемента и тяжестью RA (Makinde, V.A., et al., Ann. Rheum. Dis. 48:302-306, 1989; Brodeur, J.R, et at., Arthritis Rheumatism 34:1531-1537, 1991). При обоих типах ревматоидного артрита, у взрослых и детей, наблюдается повышенное содержание в сыворотке и синовиальной жидкости Bb - продукта активации комплемента при альтернативном пути, по сравнению с C4d (маркер классического пути активации). Этот факт указывает на то, что активация комплемента происходит преимущественно за счет альтернативного пути (EI-Ghobarey.A.F. et al., J. Rheumatology 7:453-460, 1980; Agarwal, A., et al., Rheumatology 39:189-192, 2000). Продукты активации комплемента могут прямым образом поражать ткани (посредством C5b-9) или косвенно вызывать воспаление посредством инициации клеток воспаления анафилатоксинами С3а и С5а.
Для выяснения роли комплемента в патогенезе RA широко использовались животные с моделированным артритом. Истощение комплемента фактором яда кобры у животных моделей предупреждает заболевание артритом (Morgan, К., et al., Arthritis Rheumat. 24:1356-1362, 1981; Van Lent, PL, et al., Am. J. Pathol. 140:1451-1461, 1992). Внутрисуставные инъекции растворимой формы рецептора комплемента 1 (sCR1), который является ингибитором комплемента, подавляло воспаление у крыс с моделированным RA (Goodfellow, R.M., et al., Clin. Exp.Immunol. 110:45-52, 1997). Более того, sCR1 ингибирует развитие и прогрессию коллаген-опосредованного артрита у крыс (Goodfellow, R.M., et al., Clin Exp.Immunol. 119:210-216, 2000). Растворимый CR1 ингибирует классический и альтернативный пути активации комплемента на этапах активации С3 и С5 в обоих путях, альтернативном и классическом, таким образом, ингибируя генерацию С3а, С5а и sC5b-9.
В конце 1970-х было признано, что иммунизация грызунов коллагеном гетерологичного типа II (CII; основной коллагеновый компонент суставного хряща у человека) приводит к развитию аутоиммунного артрита (коллаген-опосредованный артрит или CIA) со значительным сходством с RA человека (Courtenay, J.S., et al., Nature 283:666-68 (1980), Banda et al., J. of Immunol. 171:2109-2115 (2003)). Аутоиммунный ответ у восприимчивых животных включает в себя сложную комбинацию факторов, среди которых специфичные молекулы главного комплекса гистосовместимости (МНС), цитокины и СП-специфичные В- и Т-клеточные ответы (описано Myers, L.K., et al., Life Sciences 61:1861-78, 1997). Наблюдение, что почти 40% инбредных линий мышей полностью лишены компонента С5 комплемента (Cinader, В., et al., J. Exp. Med. 120:897-902, 1964) предоставили дополнительную возможность исследования роли комплемента в модели артрита путем сравнения развития CIA у полноценных и С5-дефицитных линий. Результаты этих исследований указывают на то, что наличие С5 является абсолютной необходимостью для развития CIA (Watson et al., 1987; Wang, Y., et al., J. Immunol. 164:4340-4347, 2000). Дополнительные доказательства важности С5 и комплемента в развитии RA были получены с использованием моноклональных анти-С5 антител (MoAbs). Профилактическое внутрибрюшинное введение анти-С5 MoAbs мышам с моделированным CIA практически полностью предупреждало появление болезни, при том, что лечение в активной фазе артрита проявлялось в значительном улучшении клинической картины и смягчении протекания заболевания на уровне тканей (Wang, Y., et al., Proc. Natl. Acad. Sci. USA 92:8955-59, 1995).
Дополнительные результаты, свидетельствующие о потенциальной роли активации комплемента в патогенезе заболевания, были предоставлены исследованиями с использованием трансгенных мышей с K/BxN Т-клеточным рецептором - недавно полученной животной модели воспалительного артрита (Korganow, A. S., et al., Immunity 10:451-461, 1999). У всех K/BxN животных спонтанно развивается аутоиммунное заболевание с большинством (однако не всеми) клинических, гистологических и иммунологических признаков RA человека. Более того, введение сыворотки от мышей K/B×N, страдающих артритом, здоровым животным в течение дней вызывает у них артрит за счет передачи артритогенных иммуноглобулинов. Для того, чтобы определить какие специфические этапы в активации комплемента необходимы для развития заболевания сыворотку, полученную от мышей K/BxN, страдающих артритом, вводили различным группам мышей с генетической недостаточностью в отношении отдельных продуктов путей активации комплемента (Ji, H., et al., Immunity 16:157-68, 2002). Интерес представляет то, что результаты исследования продемонстрировали, что альтернативный путь активации является критическим, в то время как классический путь активации не играет существенной роли. Кроме того, генерация С5а является критической при защите от развития заболевания у мышей с С5-недостаточностью и мышей с C5aR-недостаточностью. Согласуясь с подобными результатами, предыдущее исследование показало, что генетическое подавление экспрессии рецептора С5а защищает мышей от артрита (Grant, E.R, et al., J, Exp. Med. 196:1461-1471, 2002).
Гуманизированное анти-С5 антитело MoAb (5G1.1), предупреждающее расщепление компонента С5 комплемента человека на провоспалительные компоненты, сейчас находится на стадии разработки компанией «Alexion Pharmaceuticals, Inc.», в Нью-Хейвен, шт. Коннектикут, в качестве потенциального средства для лечения RA.
Две исследовательские группы независимо друг от друга выдвинули предположение о том, что пектиновый путь стимулирует воспаление у пациентов, страдающих RA, при взаимодействии MBL со специфичными гликоформами IgG (Malhotra et al., Nat. Med. 1:237-243, 1995; Cuchacovich et al., J. Rheumatol. 23:44-51, 1996). Ревматоидный артрит связан с заметным повышением уровня гликоформ IgG, у которых на участке Fc молекулы отсутствует галаклактоза, (известные также как гликоформы IgGO) (Rudd et al., Trends Biotechnology 22:524-30, 2004). У пациентов процентное содержание гликоформ IgGO повышается по мере того как прогрессирует заболевание и возвращается к норме при ремиссии. В тестах in vivo, IgGO откладывался на синовиальной ткани, а MBL проявлялся в повышенных уровнях в синовиальной жидкости у индивидуумов с RA. Агалактозил IgG (IgGO), агрегированный на кластерном IgG, ассоциированным с RA, может связывать маннозосвязывающий пектин (MBL) и активировать пектиновый путь активации комплемента. Кроме того, результаты недавнего клинического исследования, целью которого было рассмотрение аллельных вариантов MBL у пациентов с RA, показали, что MBL может исполнять роль усиливающего воспаление фактора в данном заболевании (Garred et al., J. Rheumatol. 27:26-34, 2000). Поэтому, пектиновый путь активации комплемента может исполнять важную роль в патогенезе RA.
Системная красная волчанка (SLE) является аутоиммунным заболеванием неопределенной этиологии, которое заключается в продуцировании аутоантител, генерации циркулирующих иммунных комплексов и эпизодической неконтролируемой активации системы комплемента. Хотя происхождение аутоиммунной реакции при SLE не определено, в настоящее время ученые обладают достаточным объемом информации, указывающей на активацию комплемента как на важный механизм, относящийся к васкулярному поражению приданном заболевании (Abramson, S.B, et al., Hospital Practice 33: 107-122, 1998). При данном заболевании происходит активация обоих путей комплемента, классического и альтернативного, и оба компонента - C4d и Bb - являются чувствительными маркерами прогрессирования заболевания от средней степени тяжести к тяжелой (Manzi, S., et al., Arthrit. Rheumat. 39:1178-1188, 1996). Активация альтернативного пути активации комплемента сопровождается острыми проявлениями болезни при системной красной волчанке во время беременности (Buyon, J.P., et al., Arthritis Rheum. 35:55-61, 1992). Кроме того, при развитии болезни может проявляться пектиновый путь активации комплемента, что подтверждается недавним фактом обнаружения аутоантител к MBL в сыворотке крови пациентов с SLE (Seelen, M.A-, et al., Clin Exp. Immunol. 134:335-343, 2003).
Считается, что опосредованная иммунным комплексом активация комплемента посредством классического пути является одним из механизмов поражения тканей у пациентов при SLE. Однако наследственная недостаточность компонентов комплемента классического пути активации повышает риск заболевания волчанкой и схожими с ней заболеваниями (Pickering, M.C., et al., Adv. Immunol. 76:227-324, 2000). SLE или связанные с ней заболевания обнаруживаются у более чем 80% пациентов с тотальным дефицитом C1q, C1r/C1s, C4 или С3. Этот факт представляет собой несомненный парадокс совмещения пагубного воздействия и защитной функции комплемента при волчанке.
Важная функция классического пути активации комплемента проявляется в инициации устранения иммунных комплексов из системы кровообращения и тканей посредством системы моноядерных фагоцитов (Kohler, P.F, et al., Am. J. Med. 56:406-11, 1974). Кроме того, недавние исследования обнаружили важную роль комплемента в уничтожении и выведении апоптических телец (Mevorarch, D., et al., J. Exp.Med. 188:2313-2320, 1998). Недостаточность функции классического пути активации может являться причиной развития SLE, позволяя болезни развиваться с накоплением иммунных комплексов и апоптических телец в тканях, что провоцирует воспаление и секрецию аутоантигенов, которые в свою очередь стимулируют продуцирование аутоантител и еще большее количество иммунных комплексов и, таким образом, вызывает аутоиммунный ответ (Botto, M., et al., Nat. Genet. 19:56-59, 1998; Botto, M., Arthritis Res. 3:201-10, 2001). Однако данная «тотальная» недостаточность элементов компонентов классического пути активации обнаруживается у одного пациента из 100, страдающих SLE. Следовательно, у подавляющего большинства пациентов с SLE недостаточность компонентов комплемента классического пути активации не относится к этиологии заболевания, и активация комплемента может являться важным механизмом, связанным с патогенезом SLE. Тот факт, что SLE часто развивается у пациентов с редкой формой генетической недостаточности компонентов классического пути активации комплемента в определенный момент их жизни свидетельствует о чрезмерном количестве механизмов, способных провоцировать заболевание.
Результаты, полученные на животных моделях с SLE, подтверждают важную роль активации комплемента в патогенезе этого заболевания. Ингибирование активации С5 с использованием блокирующего анти-С5 антитела MoAb уменьшают риск развития протеинурии и заболевания почек у мышей NZB/NZW FI, являющихся биологической моделью SLE (Wang Y, et al., Proc. Natl. Acad. Sci USA 93:8563-8, 1996). Кроме того, введение анти-С5 антител мышам MoAb с тяжелым комбинированным иммунодефицитным заболеванием, которым имплантировали клетки, выделяющие анти-ДНК антитела, способствует уменьшению протеинурии и улучшению картины ренальной гистологии, что приводит к улучшению показателя выживаемости по сравнению с контрольной группой животных (Ravirajan, C.T, et al., Rheumatology 43:442-7, 2004). Кроме того, было обнаружено, что альтернативный путь активации комплемента также играет важную роль в проявлениях аутоиммунного заболевания при SLE, так как возвратное скрещивания мышей с В-недостаточностью с мышами MRL/lpr, которые являются биологической моделью SLE, позволило выявить, что недостаточность фактора В уменьшает васкулит, гломерулярное заболевание; истощение С3 и уровни IgG3 RF, показатели, которые обычно свойственны данной модели без изменения уровней других аутоантител (Watanabe, H., et al., J. Immunol. 164:786-794, 2000). В данное время проводятся исследования гомонизированного анти-С5 антитела MoAb как потенциального средства для лечения SLE. Данное антитело предупреждает расщепление С5 на С5а и C5b. В Фазе I клинических испытаний не было отмечено побочных эффектов, в настоящее время проводятся многочисленные испытания с участием людей для определения эффективности данного способа при лечении SLE (Strand, V., Lupus 10:216-221, 2001).
Результаты исследований на животных и людях позволяют предположить прямое участие системы комплемента в патогенезе мышечной дистрофии. Исследования биопсических проб страдающих дистрофией людей показали, что С3 и С9 отлагаются как на некротических, так и на ненекротических волокнах дистрофичной мышцы (Cornelio and Dones, Ann. Neurol. 16:694-701, 1984; Spuler and Engel, A.G., Neurology 50:41-46, 1998). Используя методы микрочип-ДНК диагностики, Портер (Porter) и его коллеги обнаружили, что значительное увеличение экспрессии генов во многих связанных с системой комплемента мРНК у мышей с недостаточностью дистрофина (mdx) совпадает с развитием дистрофического заболевания (Porter et al., Hum. Mol. Genet. 17:263-72, 2002).
Мутации в гене человека, кодирующем дисферлин, трансмембранный мышечный белок, были идентифицированы как основные факторы риска развития двух форм мышечно-скелетного заболевания, известных как мышечная дистрофия тазового пояса (LGMD) и дистрофия Миоши (Liu et al., Nat. Genet. 20:31-6, 1998). Было создано несколько мышиных биологических моделей с мутациями дисферлина, где также обнаружили развитие прогрессирующей мышечной дистрофии. У некоторых пациентов, страдающих LGMD, активация каскада комплемента была идентифицирована на поверхности ненекротических мышечных волокон (Spuler and Engel., Neurology 50:41-46, 1998). В недавних исследованиях Уензель (Wenzel) и его коллеги показали отсутствие ингибирующего комплемент фактора CD33/DAF, специфического ингибитора C5b-9 MAC (мембраноатакующего комплекса), в мышечных волокнах с дисферлин дефицитных мышей и человека (Wenzel et al., J. Immunol. 175:6219-25, 2005). Как следствие, не некротические мышечные клетки с дисферлин-недостаточностью являются более чувствительными к клеточному лизису, опосредованному комплементом. Уензель и его коллеги выдвинули предположение, что опосредованный комплементом лизис скелетно-мышечных клеток может представлять собой основной патологический механизм, вовлеченный в развитие LGMD и дистрофии Миоши у пациентов. Конноли (Connolly) и его коллеги исследовали роль комплемента С3 в патогенезе с использованием модели тяжелой врожденной дистрофии dy-/- у мыши с недостаточностью ламинина а2 (Connolly et al., J. Neuroimmunol. 127:80-7, 2002). Они вывели животных с генетической недостаточностью С3 и ламинина а2 и обнаружили, что отсутствие С3 пролонгирует выживание в dy-/- модели мышечной дистрофии. Как следствие, мыши с двумя нокаутированными генами (С3-/-, dy-/-) демонстрировали большую мышечную крепость, чем мыши dy-/-. Данная работа позволяет выдвинуть предположение о том, что система комплемента может быть связана напрямую с патогенезом данной формы врожденной дистрофии.
В одном аспекте настоящее изобретение направлено, таким образом, на предупреждение или лечение воспалительных и невоспалительных поражений кожи ревматического или подагрического происхождения и прочих скелетно-мышечных заболеваний, включая, помимо прочего, остеоартрит, ревматоидный артрит, ревматоидный артрит у детей, подагра, нейрогенная артропатия, псориатический артрит, анкилозирующий спондилоартрит или другие формы спондилоартропатии и фрагментарной артропатии, мускульная дистрофия и системная красная волчанка (SLE) посредством введения композиции, содержащей терапевтически эффективную дозу агента, ингибирующего MASP-2, в фармацевтически приемлемом носителе субъекту, страдающему от подобного перечисленным заболевания. Агент, ингибирующий MASP-2, может вводиться субъекту систематически посредством внутриартериального, внутривенного, внутримышечного, подкожного или другого вида парентерального введения, возможен также оральный прием непептидергических ингибиторов. Альтернативно введение может осуществляться путем локальной передачи, например, внутрисуставными инъекциями. Агент, ингибирующий MASP-2, может вводиться периодически спустя продолжительное время для лечения или контроля хронического заболевания, либо путем однократного или повторяющегося введения в периоде до, во время и/или после тяжелой травмы или поражения, включая хирургические операции на суставах.
ЗАБОЛЕВАНИЯ ПОЧЕК
Было обнаружено вовлечение системы активации комплемента в патогенез широкого спектра заболеваний почек, включая мембранозно-пролиферативный гломерулонефрит (IgA-нефропатия, болезнь Берже) (Endo, M., et al., Clin. Nephrology 55:185-191, 2001), мембранозный гломерулонефрит (Kerjashki, D., Arch B Cell Pathol. 58:253-71, 1990; Brenchley, RE., etal., Kidney Int., 41:933-7, 1992; Salant, D.J., et al., Kidney Int. 35:976-84, 1989), мезангиопролиферативный гломерулонефрит (мезангиокапиллярный гломерулонефрит) (Bartlow, B.G., et al. Kidney Int. 15:294-300, 1979; Meri, S., et al., J. Exp. Med. 175:939-50, 1992), острый постинфекционный гломерулонефрит (постстрептококковый гломерулонефрит), криоглобулинэмический гломерулонефрит (Ohsawa, I, et al., Clin Immunol. 101:59-66, 2001), волчаночный нефрит (Gatenby, P.A, Autoimmunity 11:61-6, 1991), и нефрит как осложнение при болезни Шенлейн-Геноха (Endo, M., et al., Am. J. Kidney Dis. 35:401-407, 2000). Вовлечение комплемента в заболевания почек исследовалось несколько десятилетий, однако и по сей день его точная роль в начальной фазе развития и конечной фазе заболевания почек вызывает горячие споры. В нормальных условиях участие комплемента оказывает благотворное влияние на организм, но самопроизвольная активация и отложение комплемента может привести к поражению тканей.
Имеются доказательства того, что гломерулонефрит, воспаление клубочка, часто инициируется отложением иммунных комплексов в гломерулярных или тубулярных структурах, которое затем инициирует активацию комплемента, воспаление и поражение тканей. Кан (Kahn) и Шинниа (Sinniah) продемонстрировали увеличенное отложение C5b-9 в тубулярных базальных мембранах в биопсической пробе, взятой у пациентов, страдающих различными формами гломерулонефрита (Kahn, T.N, et al., Histopath. 26:351-6, 1995). В исследовании пациентов с IgA нефрологией (Alexopoulos, A., et al., Nephrol. Dial. Transplant 10:1166-1172, 1995), отложение C5b-9 в тубулярных эпителиальных/базальных мембранных структурах связано с уровнем креатинина в плазме. Другое исследование мембранной нефропатии продемонстрировало взаимосвязь между клиническими результатами и уровнями sC5b-9 в моче (Коп, S.P, et al., Kidney Int. 48:1953-58, 1995). Повышенный уровень sC5b-9 коррелировал с неблагоприятным прогнозом. Лето (Lehto) и коллеги провели измерение повышенных уровней регулирующего комплемент фактора CD59, ингибирующего мембраноатакующий комплекс в плазматических мембранах, а также C5b-9 в моче пациентов, страдающих мембранным гломерулонефритом (Lehto, Т., et al., Kidney Int. 47:1403-11, 1995). Гистопатологический анализ образцов биопсии, взятых у тех же пациентов, показал отложение белков С3 и С9 в клубочках, принимая во внимание, что экспрессия CD59 в данных тканях была снижена по сравнению с экспрессией в тканях здоровых почек. Данные различные исследования позволяют предположить, что в результате прогрессирующего комплемент-опосредованного гломерулонефрита белки комплемента выделяются при мочеиспускании, что соответствует степени поражения тканей и прогнозу болезни.
Ингибирование активации комплемента в различных моделях гломерулонефрита на животных также показало важность активации комплемента в этиологии заболевания. В модели мембранозно-пролиферативного гломерулонефрита (MPGN), введение анти-Thy1 антисыворотки крысам с С6-недостаточностью (которые не способны синтезировать C5b-9) привело к уменьшению гломерулярной клеточной пролиферации на 90%, сокращению инфильтрации тромбоцитов и макрофагов на 80%, уменьшению синтеза коллагена IV типа (который является маркером мезангиальной матричной экспансии), и сокращению протеинурии на 50%, по сравнению с животными дикого типа (Brandt, J., et al., Kidney Int. 49:335-343, 1996). Данные результаты указывают на C5b-9 как на основной медиатор при поражении тканей комплементом у крыс в модели антитимоцитарной сыворотки. В другой модели гломерулонефрита введение градуированных доз кроличьей антисыворотки к крысиной гломерулярной базальной мембране вызвало дозозависимый приток полиморфноядерных лейкоцитов (PMN), который был ослаблен предварительным введением фактора яда кобры (для поглощения комплемента) (Scandrett, A.L, et al., Am. J. Physiol. 268:F256-F265, 1995). Крысы, которых подвергали обработке фактором яда кобры, также показали уменьшение гистопатологии, сокращение продолжительной протеинурии и более низкие уровни креатинина при сравнении с контрольными животными. Используя три модели гломерулонефрита (GN) на крысах (антитимоцитарная сыворотка, конкавалин А анти-конкавалин А и пассивный нефрит Хейманна), Коузер (Couser) и его коллеги продемонстрировали потенциальную терапевтическую эффективность подходов по ингибированию комплемента посредством использования рекомбинантного белка sCRI (Couser, W.G., et al., J. Am. Soc. Nephrol. 5:1888-94, 1995). Крысы, которых подвергли обработке sCRI, показали значительное уменьшение притока полиморфноядерных клеток, тромбоцитов и макрофагов, сокращение мезангиолиза и протеинурии по сравнению с контрольными животными. Дальнейшие доказательства важности активации комплемента при гломерулонефрите было получены при использовании анти-С5 антитела MoAb в модели на мышах NZB/W F1. Анти-С5 антитела MoAb ингибируют расщепление С5, блокируя, таким образом, генерацию С5а и C5b-9. Непрерывная терапия с анти-С5 антителами MoAb в течение 6 месяцев имела результатом значительное улучшение течения гломерулонефрита. Гуманизированное моноклональное анти-С5 антитело MoAb (5G1.1), которое предупреждает расщепление компонента С5 комплемента человека на его провоспалительные компоненты, сейчас находится на стадии разработки компанией «Alexion Pharmaceuticals, Inc.», Нью-Хейвен, шт.Коннектикут, в качестве потенциального средства для лечения гломерулонефрита.
Прямые доказательства патологической роли комплемента при повреждении почек были получены при обследовании пациентов с генетической недостаточностью специфических компонентов комплемента. Во множестве публикаций отмечается связь заболевания почек с недостаточностью регулирующего комплемент фактора Н (Ault, B.H., Nephrol. 14:1045-1053, 2000; Levy, M., et al., Kidney Int. 30:949-56, 1986; Pickering, M.C., et al., Nat. Genet. 31:424-8, 2002). Недостаточность фактора Н проявляется в низких уровнях содержания в плазме фактора В и С3 и истощении C5b-9. Атипичный мембранозно-пролиферативный гломерулонефрит (MPGN) как и идиопатический гемолитический уремический синдром (HUS) связаны с недостаточностью фактора Н. Свиньи с недостаточностью фактора Н (Jansen, J.Н., et al., Kidney Int. 53:331-49, 1998), и мыши с нокаутированным фактором Н (Pickering, M.С., 2002) имеют MPGN-подобные симптомы, подтверждая важность фактора Н в регуляции комплемента. Недостаточность других компонентов комплемента приводит к заболеванию почек, которая является вторичным проявлениями при развитии системной красной волчанки (SLE) (Walport, M.J., Davies, et al., Ann. N.Y. Acad. Sci 815:261-81, 1997). Недостаточность C1q, C4 и С2 напрямую способствует развитию SLE посредством включения механизмов, относящихся к неполному выведению иммунных комплексов и апоптотического материала. У многих из пациентов, страдающих SLE, возникает волчаночный нефрит, характеризующийся отложением иммунных комплексом в клубочках.
Дальнейшие доказательства, показывающая связь активации комплемента и заболевания почек, были получены при идентификации у пациентов аутоантител, направленных против компонентов комплемента, некоторые из которых напрямую связаны с заболеванием почек (Trouw, LA, et al., Mol. Immunol. 38: 199-206, 2001). Число данных аутоантител показывает настолько высокую степень корреляции с заболеванием почек, что для обозначения данной активности был введен специальный термин нефритогенный фактор (NeF). В клинических исследованиях у 50% пациентов, положительных в отношении нефритогенных факторов, развивалось заболевание MPGN (Spitzer, R.E., et al., Clin. Immunol. Immunopathol. 64:177-83, 1992). C3NeF представляет собой аутоантитело, направленное против конвертазы С3 альтернативного пути активации (C3bBb), данное аутоантитело стабилизирует эту конвертазу, способствуя, таким образом, активации альтернативного пути (Daha, M.R., et al., J. Immunol. 116:1-7, 1976). Подобным образом, аутоантитело со специфичностью к конвертазе С3 классического пути активации (C4b2a), известная как C4NeF, стабилизирует данную конвертазу и помогает, таким образом, активации классического пути (Daha, M.R. et al., J. Immunol. 125:2051-2054, 1980; Halbwachs, L, et al., J. Clin. Invest. 65:1249-56, 1980). В литературе имеются данные о том, что анти-dq аутоантитела связаны с нефритом у пациентов, страдающих SLE (Hovath, L., et al., Clin. Exp. Rheumatol. 19:667-72, 2001; Siegert, С, et al., J. Rheumatol. 18:230-34, 1991; Siegert, С, et al., Clin. Exp. Rheumatol. 10:19-23, 1992), а повышение титра данных анти-dq аутоантител является предиктором обострения нефрита (Coremans, I.E., et al„ Am. J. Kidney Dis. 26:595-601, 1995). Анализ иммунных наслоений, выделенных из почек пациентов, умерших от SLE, показал аккумуляцию в них данных анти-C1q аутоантител (Mannick, M., et al., Arthritis Rheumatol. 40:1504-11, 1997). Все приведенные факты указывают на патологическую роль данных аутоантител. Однако не у всех пациентов с анти-dq аутоантителами развивается заболевание почек и также некоторые здоровые индивиды обладают низким титром анти-C1q аутоантител (Siegert, С.Е., et al., Clin. Immunol. Immunopathol. 67:204-9, 1993).
В дополнении к альтернативному и классическому путям активации комплемента, пектиновый путь активации комплемента может также играть важную патологическую роль в ренальном заболевании. Повышенные уровни MBL, MBL-ассоциированной сериновой протеазы и продуктов активации комплемента были зафиксированы при помощи иммуногистохимических методов обработки образцов ткани почек, взятых при биопсии у пациентов, страдающих некоторыми заболеваниями почек, включая нефрит при болезни Шенлейн-Геноха (Endo, M., et al., Am. J. Kidney Dis. 35:401-407, 2000), криоглобулемический гломерулонефрит (Ohsawa, I, et al., Clin. Immunol. 101:59-66, 2001) и IgA нейропатию (Endo, M., et al., Clin. Nephrology 55:185-191, 2001). Поэтому, несмотря на тот факт, что связь между комплементом и заболеванием почек известна уже в течение нескольких десятилетий, исследования того, как конкретно комплемент воздействует на данные заболевания почек, еще далеки от своего завершения.
Один аспект настоящего изобретения, таким образом, направлен на лечение заболеваний почек, включающих в себя, помимо прочего, мезангиопролиферативный гломерулонефрит, мембранозный гломерулонефрит, мембранозно-пролиферативный гломерулонефрит (мезангиокаппилярный гломерулонефрит), острый постинфекционный гломерулонефрит (постстрептоккоковый гломерулонефрит), криоглобулинемический гломерулонефрит, волчаночный нефрит, нефрит при болезни Шенлейн-Геноха или IgA нефропатию, путем введения композиции, содержащей терапевтически эффективную дозу агента, ингибирующего MASP-2, в фармацевтически приемлемом носителе субъекту, страдающему подобным расстройством. Агент, ингибирующий MASP-2, может вводиться субъекту систематически посредством внутриартериального, внутривенного, внутримышечного, подкожного или другого вида парентерального введения, возможен также оральный прием непептидергических ингибиторов. Агент, ингибирующий MASP-2, может вводиться периодически спустя продолжительное время для лечения или контроля хронического заболевания, либо путем однократного или повторяющегося введения в периоде до, во время и/или после тяжелой травмы или поражения.
КОЖНЫЕ ЗАБОЛЕВАНИЯ
Псориаз представляет собой хроническое заболевание, поражающее кожу, которым болеют миллионы людей и которое вызывается как генетическими, так экологическими факторами. Средства местного применения, равно как и фототерапия UVB (фототерапия с использованием ультрафиолетовых лучей спектра В) и фототерапия PUVA (фототерапия с использованием псоралена и ультрафиолетовых лучей спектра А), повсеместно считались первоочередным средством лечением от псориаза. Однако при генерализованном или широко распространенном заболевании предписана систематическая терапия, указанная в качестве первичной обработки или, в некоторых случаях, для усиления эффекта фототерапии UVB и PUVA.
Этиология, лежащая в основе многих кожных заболеваний, таких как псориаз, подтверждает роль иммунных и провоспалительных процессов, включая вовлечение системы комплемента. Более того, было установлено, что система комплемента представляет собой важный механизм неспецифической защитной реакции кожи. Его активация приводит к образованию продуктов, которые не только помогают поддерживать нормальную иммунную защиту, но также опосредует воспаление и поражение тканей. Провоспалительные продукты комплемента включают в себя большие фрагменты С3 с опсонической и клеточностимулирующей активностью (C3b и C3bi), анафилатоксины с низкой молекулярной массой (С3а, С4а и С5а) и мембраноатакующие комплексы. Среди них, С5а или продукт его распада С5а des Arg выделяется как наиболее важный медиатор из-за его способности оказывать потенциальный хемотаксический эффект на воспалительные клетки. Внутрикожное введение анафилатоксина С5а вызывает кожные изменения, весьма схожие с изменениями, наблюдаемыми при кожном васкулите с синдромом гиперчувствительности, который возникает вследствие активации комплемента посредством иммунного комплекса. Активация комплемента вовлечена в патогенез воспалительных изменений при аутоиммунном буллезном дерматозе. Активация комплемента посредством внедрения пемфигусного антитела в эпидермис может быть ответственной за развитие характерных воспалительных изменений вследствие эозинофильного межклеточного отека. При буллезном пемфигоиде (ВР), взаимодействие антигена базальной мембранной зоны и антител ВР управляет активацией комплемента, что, по всей видимости, связано с лейкоцитами, выстилающими место дермоэпидермального соединения. Результирующие анафилатоксины не только активируют инфильтрацию лейкоцитов, но и индуцируют дегрануляцию мастоцитов, что облегчает дермоэпидермальное разделение и эозинофильную инфильтрацию. Похожим образом, активация комплемента может играть более точную направленную роль в дермоэпидермальном разделении, отмеченном при приобретенном буллезном эпидермолизе и герпесе беременных.
Доказательства вовлеченности комплемента в развитие псориаза получены в недавних экспериментах, описанных в научной литературе, посвященной патофизиологическим механизмам воспалительных изменений при псориазе и связанных с ним заболеваниях. Растущее число таких доказательств указывает на то, что Т-клеточный иммунитет, играет важную роль в инициации и поддержании псориатических поражений. Было обнаружено, что лимфокины, продуцируемые активированными Т-клетками в псориатических поражениях, оказывают сильное влияние на пролиферацию эпидермиса. Характерная аккумуляция нейтрофилов под роговым слоем кожи может наблюдаться в сильно воспаленных участках псориатических поражений. Нейтрофилы хемотаксически привлекаются и активируются в таких участках посредством синергического действия хемокинов, IL-8 и Gro-альфа, высвобождаемых активированными кератиноцитами, в частности С5а/С5а des-arg, образующимся в альтернативном пути активации комплемента (Terui, Т., Tahoku J. Exp. Med. 190:239-248, 2000; Terui, Т., Exp. Dermatol. 9:1-10, 2000).
Экстракты из псориатических чешуек содержат уникальную фракцию хемотаксических пептидов, которая вероятно принимает участие в индукции циклического трансэпидермального лекоцитарного хемотаксиса. В ходе недавних исследований в данной фракции было идентифицировано присутствие двух несвязанных между собой хемотаксических пептидов, а именно С5а/С5а des Arg и интерлейкина 8 (IL-8) и связанных с ним цитокинов. Для выяснения их относительного вклада в трансэпидермальную лейкоцитарную миграцию, а также для изучения их взаимодействие в псориатических поражениях; были количественно оценены уровни концентрации иммунокреативных С5а/С5а desArg и IL-8 в экстрактах из псориатических чешуек и в экстрактах стерильных гнойничковых дерматозов. Было обнаружено, что уровни концентрации С5а/С5а desArg и IL-8 были значительно выше в экстрактах роговой ткани пораженной кожи, чем в экстрактах невоспаленной кожи при ортокератозе. Данное повышение уровней концентрации С5а/С5а desArg было специфичным для экстрактов из чешуек пораженной кожи. Опираясь на данные результаты, можно предположить, что С5а/С5а desArg генерируется только в воспаленной пораженной коже под влиянием специфических обстоятельств, которые предполагают активацию комплемента. В соответствии с данным предположением было сочтено рациональным использование ингибитора активации комплемента для улучшения состояния при псориатических поражениях.
В то время как широко известно, что классический путь активации системы комплемента инициируется при псориазе, существует очень мало сообщений, свидетельствующих о вовлечении альтернативного пути в воспалительные реакции при псориазе. В соответствии с традиционным взглядом на пути активации комплемента, фрагменты комплемента C4d и ВЬ освобождаются во время активации комплемента по классическому и альтернативному пути, соответственно. Наличие фрагмента C4d или Bb, таким образом, указывает на активацию комплемента, которая протекает посредством классического и/или альтернативного пути. В одном из исследований были измерены уровни C4d и Bb в экстрактах из псориатических чешуек с использованием техники иммуноферментного анализа. Методом иммуноферментного анализа чешуек от данных дерматозов были обнаружены более высокие уровни C4d и Bb, чем в роговом слое невоспаленной кожи (Takematsu, Н., et al., Dermatologica 181:289-292, 1990). Основываясь на данных результатах можно предположить, что альтернативный путь активизируется в добавление к классическому пути активации комплемента в пораженной псориазом коже.
В доказательство очевидности вовлечения комплемента в псориатический и атопический дерматит были измерены уровни нормальных компонентов комплемента и продукты его активации в периферической крови у 35 пациентов с атопическим дерматитов (AD) и у 24 пациентов с псориазом в стадиях от легкой до средней степени тяжести. Уровни инактиваторов С3, С4 и С1 (С1 INA) в сыворотке были определены методом радиальной иммунодиффузии, причем уровни С3а и С5а были измерены методом радиоиммунологического анализа. По сравнению с нормальными неатопическими контрольными уровнями, было обнаружено значительное повышение уровней С3, С4 и С1 INA в случае обоих заболеваний. При AD была обнаружена тенденция к повышению уровней С3а, особенно отметили повышение уровней С3а при псориазе. Результаты указывают на то, что и при AD, и при псориазе система комплемента участвует в воспалительном процессе (Ohkonohchi, К., et al., Dermatologica 179:30-34, 1989).
Активация комплемента при псориатическом поражении кожи также приводит к отложение конечных комплексов комплемента в эпидермисе, что доказано измерением уровней SC5b-9 в плазме и ороговевших тканях у пациентов, страдающих псориазом. Уровни SC5b-9 в псориатической плазме при измерении были значительно выше, чем контрольные уровни, измеренные у пациентов с атопическим дерматитом. Исследования общего белка, выделенного из пораженной кожи, показали, что, несмотря на отсутствие SC5b-9 в невоспаленных ороговевших тканях, были обнаружены высокие уровни SC5b-9 в роговых тканях, пораженных псориазом. При использовании метода иммунофлуоресценции для исследования моноклонального антитела к неоантигену C5b-9 отложение C5b-9 наблюдалось в роговом слое псориатической кожи. В заключение, в пораженной псориазом коже система комплемента активируется и активация комплемента протекает полностью до конечного этапа, генерируя мембраноатакующий комплекс.
Недавно для лечения псориаза стали доступны новые биологические препараты, которые селективным способом нацеливаются на иммунную систему Четыре биологических препарата, которые совсем недавно были либо одобрены Управлением США по контролю качества пищевых продуктов и лекарственных средств (FDA), либо находятся в Фазе III клинических испытаний: алефацепт (Amevive®) и эфализумаб (Raptiva®), которые являются модуляторами Т-клеток; этанерцепт (Enbrel®); растворимый TNF-рецептор, и инфликсимаб (Remicade®) - анти-TNF моноклональное антитело. Раптива (Raptiva) представляет собой модулятор иммунного ответа, в котором целевой механизм действия представляет собой блокаду взаимодействия между LFA-1 на лимфоцитах и ICAM-1 на антиген-представляющих клетках и на клетках эндотелия сосудистой стенки. Связывание CDHa препаратом Раптива приводит к насыщению доступных сайтов связывания CD11a на лимфоцитах и дальнейшее изменение экспрессии CDHa на клеточной поверхности лимфоцитов. Данный механизм действия ингибирует активацию Т-клеток, транспортировку клеток в дермис и эпидермис и реактивацию Т-клеток. Таким образом, многочисленные научные данные доказывают роль комплемента в воспалительных заболеваниях кожи, и современные фармацевтические подходы в лечении таких заболеваний нацелены на иммунную систему или на специфические воспалительные процессы. Однако ни одно исследование не определило MASP-2 своей целью. Базируясь на новом понимании авторами настоящего изобретения роли MASP-2 в активации комплемента, авторы считают MASP-2 эффективной мишенью при лечении псориаза и других кожных заболеваний.
Один аспект настоящее изобретения, таким образом, направлен на лечение псориаза, аутоиммунного буллезного дерматоза, эозинофильного межклеточного отека, буллезного пемфигоида, приобретенного буллезного эпидермолиза, атопического дерматита, герпеса беременных и других заболеваний кожи и лечения термических и химических ожогов, включая вызванное ими капиллярное кровотечение, посредством введения композиции, содержащего терапевтически эффективный объем агента, ингибирующего MASP-2, в фармацевтически приемлемом носителе субъекту, страдающему подобным кожным заболеванием. Агент, ингибирующий MASP-2, может быть введен субъекту локально, посредством нанесения в форме спрея, лосьона, геля, мази, бальзама или орошением раствором, содержащим агент, ингибирующий MASP-2, или систематически посредством внутриартериального, внутривенного, внутримышечного, подкожного или другого вида парентерального введения, возможен также оральный прием непептидергических ингибиторов. Лечение может подразумевать однократное введение или повторяющиеся аппликации или дозировки при остром заболевании или периодические аппликации или дозировки для контроля над хроническим заболеванием.
ТРАНСПЛАНТАЦИЯ
Активация системы комплемента вносит значительный вклад в развитие воспалительной реакции после трансплантации паренхиматозных органов. При аллотрансплантации система комплемента может активироваться посредством ишемии/реперфузии, а также существует возможность активации посредством антител, направленных против трансплантата (Baldwin, W.M, et al., Springer Seminol Immunopathol. 25:181-197, 2003). При ксенотрансплантации органов приматам от неприматов основными активаторами комплемента являются конституциональные антитела. Исследования, выполненные на животных моделях, показали, что использование ингибиторов комплемента значительно пролонгируют срок жизнеспособности трансплантата (см. ниже). Таким образом, определена роль системы комплемента в поражении органов после трансплантации и, поэтому авторы изобретения считают, что ингибиторы комплемента, направленные против MASP-2, могут предупреждать повреждение трансплантата после алло- или ксенотрансплантации.
Врожденные иммунные механизмы, особенно комплемент, играют более значимую роль в воспалении и иммунном ответе на трансплантат, чем предполагалось ранее. Например, альтернативный путь активации комплемента опосредует ишемическое реперфузионное поражение почек, а проксимальные тубулярные клетки могут быть одновременно источником и участком атаки компонентов комплемента в установлении поражения. Комплемент, локально спродуцированный в почках, также влияет на развитие как клеточного, так и опосредованного антителами иммунного ответа на трансплантант.
C4d представляет собой продукт распада С4, фактора активированного комплемента, компонента классического и пектин-зависимого пути активации комплемента. Контрастирование C4d было использовано как практичный маркер гуморального отторжения, как в острой, так и в хронической форме, что привело к возобновлению интереса к изучению процесса формирования антидонорских антител. Связь между C4d и морфологическими признаками острого клеточного отторжения имеет статистически значимую величину C4d обнруживается в 24-43% случаях отторжения Типа I, в 45% случаях отторжения Типа II и 50% случаях отторжения Типа III (Nickeleit, V., et al., J. Am. Soc. Nephrol. 13:242-251, 2002; Nickeleit, V., et al., Nephrol. Dial. Transplant 18:2232-2239, 2003). В данное время в разработке находится несколько видов терапии, ингибирующей комплемент или редуцирующей локальный синтез для достижения улучшения клинического исхода после трансплантации.
Активация каскада комплемента происходит в результате некоторого числа процессов в ходе трансплантации. Существующая в настоящее время терапия является эффективной в ограничении клеточного отторжения, но при этом не полностью решает проблему некоторых барьеров. К ним относятся гуморальное отторжение и хроническая нефропатия или дисфункция аллогенного трансплантата. Хотя полноценный иммунный ответ к трансплантированному органу является результатом определенного числа эффекторных механизмов со стороны рецепиента, комплемент может сыграть ключевую роль в одном из них. В случае трансплантации почек, локальный синтез комплемента проксимальными тубулярными клетками является наиболее важным.
Наличие специфических ингибиторов комплемента может предоставить возможность для улучшения клинического исхода вследствие трансплантации органов. Ингибиторы, которые действуют посредством механизма, блокирующего атаку комплемента, могут быть особенно полезны, так как они поддерживают повышение эффективности и ликвидацию истощения системы комплемента у рецепиента с уже ослабленным иммунитетом.
Комплемент также играет критическую роль в отторжении ксенотрансплантата. Поэтому, эффективные ингибиторы комплемента представляют огромный интерес в качестве потенциальных терапевтических агентов. В трансплантации органов от свиней приматам особенно острое отторжение (HAR) возникало из-за отложения антител и активации комплемента. Многочисленные стратегии и целевые препараты были протестированы для предупреждения особо острого отторжения ксенотрансплантата в комбинации от свиней к приматам. Данные подходы были осуществлены при удалении природных антител, отложения комплемента фактором яда кобры, или предотвращением активации С3 растворимым ингибитором комплемента sCR1. Кроме того, блокатор активации комплемента-2 (САВ-2), рекомбинантный растворимый химерный белок, полученный из фактора человека, усиливающего разложение (DAF) и мембранного кофакторного белка, ингибирует конвертазы С3 и С5 как классического, так и альтернативного пути активации комплемента. САВ-2 уменьшает комплемент-опосредованное поражение тканей в сердце свиньи, через которое ex vivo пропускали кровь человека. Исследование эффективности САВ-2 при гетеротопической трансплантации свиного сердца макаке-резусу не показало иммуносупрессии, продемонстрировав, что срок жизнеспособности трансплантата был значительно пролонгирован у обезьян, получавших CAB-2 (Salerno, C.T, et al., Xenotransplantation 9:125-134, 2002). САВ-2 значительным образом ингибировал активацию комплемента, что подтверждается сильным уменьшением синтеза С3а и SC5b-9. При отторжении трансплантата, отложение iC3b, С4 и С9 в тканях было схожим или незначительно сниженным по сравнению с контролем, а отложение IgG, IgM, C1q и фибрина не изменилось. Таким образом, данный подход к ингибированию комплемента устранил особо острое отторжение в свиных сердцах, трансплантированных макакам-резусам. Данные исследования демонстрируют благотворный эффект от ингибирования комплемента на срок жизнеспособности трансплантата, и авторы изобретения считают, что ингибирование MASP-2 может также быть полезным при ксенотрансплантации.
Другой подход фокусируется на определении того, могут ли моноклональные антитела антикомплемента 5 (С5) предупреждать особо острое отторжение (HAR) в модели трансплантации сердца от крыс предварительно сенсибилизированным мышам и можно ли при помощи данных MoAb, комбинированных с циклоспорином и циклофосфамидом, достичь продолжительного срока жизнеспособности трансплантата. Было обнаружено, что анти-С5 MoAb предупреждает HAR (Wang, H., et al., Transplantation 68:1643-1651, 1999). Таким образом, авторы изобретения считают, что другие мишени в каскаде комплемента, как, например, MASP-2, также могут иметь ценность для предупреждения HAR и острого васкулярного поражения в будущей клинической ксенотрансплантации.
Хотя ключевая роль комплемента в особо остром поражении, наблюдаемом в ксенотрансплантате, уже установлена, проявляется его едва уловимая роль в аллогенетической трансплантации. Связь между комплементом и приобретенной иммунной реакцией известна уже в течение долгого времени, с открытием того, что животные с недостаточностью комплемента обнаруживают субнормальный гуморальный иммунный ответ вследствие антигенной стимуляции. Опсонизация антигена продуктом расщепления комплемента C3d была продемонстрирована как значительно повышающая эффективность презентации антигенов В клеткам, которая происходит посредством вовлечения рецептора второго типа комплемента в этих клетках. Данная работа была применена в модели трансплантации кожи у мышей, в которой мыши с недостаточностью С3- и С4 обладали маркированным дефектом продуцирования алло-антител, благодаря разрушению класса переключения на IgG с высокой аффинностью. Важность данных механизмов в трансплантации почек повышается благодаря значению антидонорских антител и гуморального отторжения.
Предыдущая научная работа уже продемонстрировала регуляцию синтеза С3 проксимальными тубулярными клетками при отторжении аллотрансплантата после трансплантации почек. Роль локального синтезированного комплемента была подвергнута исследованию на модели трансплантации почек у мышей. Трансплантаты от С3-отрицательного донора, пересаженные рецепиентам с недостаточностью С3 продемонстрировали более долгий срок жизнеспособности (более 100 дней), по сравнению с контрольными трансплантатами от С3-положительных доноров, которые отторгались в течение 14 дней. Более того, пролиферативная реакция антидонорских Т-клеток у реципиентов С3-отрицательных трансплантатов была значительно менее интенсивной, чем у контролируемой группы, что указывает на воздействие локально синтезированного С3 на Т-клеточное примирование.
Данные наблюдения позволяют выдвинуть предположение о том, что представление донорского антигена Т-клеткам прежде всего начинается в трансплантате и что локально синтезированный комплемент повышает презентацию антигена посредством опсонизации донорского антигена либо генерированием дополнительных сигналов как клеткам с наличием антигена, так и Т-клеткам. В условиях трансплантации почек тубулярные клетки, продуцирующие комплемент, также демонстрируют отложение комплемента на их клеточной поверхности.
Таким образом, один аспект настоящего изобретения направлен на предупреждение и лечение воспалительной реакции, вызванной трансплантацией ткани или паренхиматозных органов, посредством введения состава с содержанием терапевтически эффективного объема агента, ингибирующего MASP-2, в фармацевтически приемлемом носителе реципиентам, в группу которых включены субъекты, перенесшие аллотрансплантацию или ксенотрансплантацию цельного органа (например, почка, сердце, печень, поджелудочная железа, легкое, роговица, и т.д.) или трансплантата (например, клапан, сухожилие, костный мозг, и т.д.). Агент, ингибирующий MASP-2, может быть введен субъекту посредством внутриартериального, внутривенного, внутримышечного, подкожного или другого вида парентерального введения, возможен также оральный прием непептидергических ингибиторов. Введение может проводиться во время периода обострения, следующего после трансплантации и/или в качестве долгосрочной посттрансплантационной терапии. Дополнительно или вместо посттрансплантационного введения может проводиться лечение субъекта агентом, ингибирующим MASP-2, перед трансплантацией и/или во время процедуры трансплантации, и/или предварительная обработка агентом, ингибирующим MASP-2, органа или ткани, предназначенных для трансплантации. Предварительная обработка органа или ткани может проводиться посредством нанесения раствора, геля или мази с содержанием агента, ингибирующего MASP-2, на поверхность органа или ткани в форме спрея или орошения, либо орган или ткань могут быть помещены в раствор с содержанием ингибитора MASP-2.
ЗАБОЛЕВАНИЯ И НАРУШЕНИЯ ЦЕНТРАЛЬНОЙ И ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
Активация системы комплемента была обнаружена в патогенезе разнообразных заболеваний и повреждений центральной нервной системы (CNS) и периферической нервной системы (PNS), включая, помимо прочего, рассеянный склероз (MS), прогрессирующую миастению (MG), болезнь Гентингтона (HD), боковой амиотрофический склероз (ALS), инфекционный полиневрит (синдром Гийена-Барре), посттравматическую реперфузию, перерожденный диск (остеохондроз), церебральную травму, болезнь Паркинсона (PD) и болезнь Альцгеймера (AD). Первоначальное определение того, что белки комплемента синтезируются в клетках CNS, включая нейроны, астроциты и микроглия, а также открытие того, что анафилатоксины, образованные в CNS после активации комплемента, могут изменять нейрональную функцию, привела к открытию потенциальной роли комплемента в расстройствах CNS (Morgan, В.Р., et al., Immunology Today 17:10: 461-466, 1996). Было показано что, рецепторы С3а и рецепторы С5а обнаруживаются в нейронах и демонстрируют широкую распространенность в определенных участках сенсорного, моторного и лимбического отделов головного мозга (Barum, S.R., Immunologic Research 26:7-13, 2002). Более того, было показано, что анафилатоксины С5а и С3а участвуют в изменении поведенческих процессов принятия пищи и питья у грызунов, и способны запускать индуцированные ионами кальция сигналы в микроглии и нейронах. Данные открытия подтверждают возможность рассмотрения терапевтической полезности для ингибирования активации комплемента в различных воспалительных заболеваниях CNS, включая церебральную травму, демиелинизацию, менингит, посттравматическую реперфузию и болезнь Альцгеймера.
Травма головного мозга или кровотечение представляют собой основную клиническую проблему, а активация комплемента может иметь место и обостряться в результате воспаления и отека. Эффекты ингибирования комплемента были изучены в модели травмы головного мозга у крыс (Kaczorowski et al., J. Cereb. Blood Flow Metab. 15:860-864, 1995). Введение sCR1 непосредственно перед повреждением головного мозга в значительной степени ингибировало инфильтрацию нейтрофилов в поврежденном участке, указывая на важность комплемента для активизации фагоцитов. Подобным образом, активация комплемента у пациентов, переживших церебральное кровотечение, ясно определяется наличием высоких уровней множественных продуктов активации комплемента, как в плазме, так и в спиномозговой жидкости (CSF). Активация комплемента и повышенное содержание комплексов C5b-9 было продемонстрировано в секвестированных тканях поясничных дисков, что позволяет сделать предположение о роли комплемента в ишиалгии, опосредованной междисковым грыжеобразованием (Gronblad, M., et al., Spine 28(2): 114-118, 2003).
Рассеянный склероз (MS) характеризуется прогрессирующей потерей миелин-накапливающих и изолирующих нейтритов в CNS. Хотя изначальная причина не известна, активизация иммунной системы определена весьма ясно (Prineas, J.W., et al., Lab Invest. 38:409-421, 1978; Ryberg, В., J. Neurol. Sci 54:239-261, 1982). Также существуют неоспоримые доказательства того, что комплемент играет ключевую роль в патофизиологии состояний демиелинизации CNS или PNS, включая MS, синдром Гийена-Барре и синдром Миллера-Фишера (Gasque, P., et al., Immunophannacology 49:171-186, 2000; Barnum, S.R. in Bondy S.R. et al. (eds.) Inflammatory events in neurodegeneration, Prominent Press 139-156, 2001). Комплемент способствует разрушению тканей, воспалению, обеднению миелинового слоя и даже ремиелизации аксонов. Несмотря на имеющиеся доказательства вовлеченности комплемента, идентификация терапевтических мишеней комплемента находится на стадии разработки для экспериментального аллергического энцефаломиелита (ЕАЕ) - животной модели рассеянного склероза человека. Исследования показали, что ЕАЕ мыши с недостаточностью С3 или фактора В проявили пониженную демиелинизацию, по сравнению с контрольными ЕАЕ мышами (Barnum, Immunologic Research 26:7-13, 2002). Исследования на ЕАЕ мышах с использованием жидкой формы ингибитора комплемента, обозначенного как «sCrry», и особей С-/- и фактор В-/- показали, что комплемент участвует в развитии и прогрессии модели заболевания на нескольких стадиях. Кроме того, отмеченное уменьшение тяжести ЕАЕ у мышей с фактор В-/- предоставило дополнительное доказательство роли альтернативного пути активации комплемента в ЕАЕ (Natafet al., J. Immunology 165:5867-5873, 2000).
Прогрессирующая миастения (MG) представляет собой заболевание нервно-мышечного соединения с сокращением числа ацетилхолиновых рецепторов и разрушением концевых пластинок двигательного нерва. sCR1 имеет высокую эффективность при лечении животной модели MG, что указывает на роль комплемента в данном заболевании (Piddelesden et al., J. Neuroimmunol. 1997).
Гистологическими признаками болезни Альцгеймера (AD), нейродегенеративного заболевания, являются старческие бляшки и нейрофибриллярные клубки (McGeer et al., Res. Immunol. 143:621-630, 1992). Данные патологические маркеры также относятся к компонентам системы комплемента. Совершенно очевидно, что локальное нервновоспалительное заболевание вызывает гибель нейронов и конгнитивную дисфункцию. Старческие бляшки содержат абнормальный бета-амилоидный пептид (Бета-амилоид, пептид, образующийся из белка-предшественника амилоида. Была показано, что бета-амилоид способен связывать С1 и инициировать активацию комплемента (Rogers et al., Res. Immunol. 143:624-630, 1992). Кроме того, отличительной чертой AD является ассоциация активированых белков классического пути активации комплемента от C1q до C5b-9, которые были обнаружены в старческих бляшках в высокой концентрации (Shen, Y., et al., Brain Research 769:391-395, 1997; Shen, Y., et al., Neurosci. Letters 305(3): 165-168, 2001). Таким образом, бета-амилоид не только инициирует классический путь активации, но и приводит к продолжительному воспалительному заболеванию, которое в свою очередь может привести к гибели нейронных клеток. Более того, тот факт, что активация комплемента при AD прогрессирует к терминальной фазе C5b-9, указывает, что регуляционные механизмы системы комплемента были неспособны остановить процесс активации комплемента.
Некоторые ингибиторы пути активации комплемента были предложены в качестве потенциальных терапевтических средств для лечения AD, включая протеогликан в качестве ингибитора связывания C1Q, нафамстат (Nafamstat) в качестве ингибитора конвертазы СЗ, и блокаторы активации С5 или ингибиторы рецепторов С5а (Shen, Y, et al., Progress in Neurobiology 70:463-472, 2003). Роль MASP-2 как инициирующего звена в активации как врожденного, так и альтернативного пути активации комплемента, предоставляет новые возможности разработки терапевтического подхода и поддерживается ценными данными, позволяющими выдвигать гипотезу о вовлеченности пути активации комплемента при AD.
В пораженных участках головного мозга пациентов, страдающих болезнью Паркинсона (PD) так же, как и при других дегенеративных заболеваниях CNS, выявлено воспаление, характеризующееся глиальной реакцией (особенно, микроглией), а также повышение экспрессии антигенов HLA-DR, цитокинов и компонентов комплемента. Данные наблюдения позволяют выдвинуть предположение о вовлечении механизмов иммунной системы в патогенез нейронного поражения при PD. Хотя клеточные механизмы первичного поражения при PD остаются невыявленными, вероятно, что в этом процессе играют свою роль митохондриальные мутации, окислительный стресс и апоптоз. Боолее того, воспаление, инициированное нейронным поражением в полосатом теле и черной субстанции головного мозга при PD, может осложнить течение болезни. Данные наблюдения позволяют выдвинуть предположение о том, что лечение посредством препаратов, ингибирующих комплемент, может замедлять прогрессию PD (Czlonkowska, A., et al., Med. Sci Monit. 8:165-177, 2002).
Таким образом, один аспект настоящего изобретения направлен на лечение расстройств или поражений периферийной нервной системы (PNS) и/или центральной нервной системы (CNS) при лечении субъекта, страдающего подобным расстройством или поражением, посредством введения ему состава, содержащего терапевтически эффективный объема агента, ингибирующего MASP-2, в фармацевтически приемлемом носителе. Расстройства и поражения CNS и PNS, которые можно лечить в соответствии с настоящим изобретением, включают, помимо прочего, рассеянный склероз (MS), прогрессирующую миастению (MG), болезнь Гентингтона (HD), боковой амиотрофический склероз (ALS), синдром Гийена-Барре, постинсультную реперфузию, перерожденный диск (остеохондроз), церебральную травму, болезнь Паркинсона (PD), болезнь Альцгеймера (AD), синдром Миллера-Фишера, церебральную травму и/или кровоизлияние, демиелинизация и, возможно, менингит.
Для лечения расстройств CNS и церебральной травмы агент, ингибирующий MASP-2, может быть введен субъекту посредством интратекального, внутричерепного, интравентрикулярный, внутриартериального, внутривенного, внутримышечного или подкожного введения, возможен также оральный прием непептидергических ингибиторов. Лечение расстройств PNS и церебральной травмы может проводиться систематическим введением или, альтернативно, локальным введением в участки дисфункции или травмы. Введение композиций, ингибирующих MASP-2, по настоящему изобретению может периодически повторяться, если таковое определено врачом до эффективного улучшения или контроля симптомов.
ЗАБОЛЕВАНИЯ КРОВИ
Сепсис обусловлен генерализованной реакцией пациента на инвазию микроорганизмов. Основной функцией системы комплемента является управление воспалительной реакцией на инвазивные бактерии и прочие патогены. В многочисленных исследованиях было показано, что активация комплемента занимает центральное место в патогенезе сепсиса, что согласуется с физиологической ролью комплемента (Bone, R.C., Annals. Internal. Med. 115:457-469, 1991). Определение клинических проявлений сепсиса находится на стадии развития. Сепсис обычно описывается как системная реакция реципиента в ответ на инфекцию. Однако во многих случаях у пациентов с симптомами сепсиса не обнаруживается очевидных клинических признаков инфекции (например, положительные бактериальные гемокультуры). Данное противоречие впервые было вынесено на обсуждение на Конференции по достижению консенсуса в 1992, на которой было утверждено понятие «синдром систематической воспалительной реакции» (systemic inflammatory response syndrome, SIRS), для которого не требуется выявляемого наличия бактериальной инфекции (Bone, R.C., et al., Crit. Care Med. 20:724-726, 1992). В настоящее время принято основное мнение, что сепсис и SIRS сопровождаются неспособностью регулировать воспалительную реакцию. В данном кратком обзоре мы рассмотрим клиническое определение сепсиса, в которое входит тяжелый сепсис, септический шок и SIRS.
До конца 1980-х гг. доминирующим источником инфекции у пациентов, страдающих от сепсиса, являлись грамотрицательные бактерии. Инъекции липополисахарида (LPS), основного компонента клеточной стенки грамотрицательных бактерий, животным провоцировали секрецию медиаторов воспаления из клеток различных типов и вызывали сиптомы острого инфекционного заболевания (Haeney, M.R., et al., Antimicrobial Chemotherapy 41 (Suppl. A):41-6, 1998). Интерес вызывает тот факт, что спектр ответственных за развитие заболевания микроорганизмов по неизвестным пока причинам сместился от преимущественно грамотрицательных бактерий в конце 1970-х и 1980-х к преимущественно грамположительным бактериям в настоящее время (Martin, G.S., et al., N. Eng. J. Med. 348:1546-54, 2003).
Многочисленные исследования продемонстрировали важность активации комплемента в опосредовании воспаления и способствовании образованию признаков шока, в частности, септического и гемморагического шока. Как грамотрицательные, так и грамоположительные микроорганизмы обычно вызывают септический шок. LPS является потенциальным активатором комплемента, преимущественно действующим посредством альтернативного пути активации, хотя классический путь активации, опосредованный антителами, также сохраняется (Fearon, D.T, et al., N. Engl. J. Med. 292:937-400, 1975). Основными компонентами клеточных стенок грамоположительных бактерий являются пептидогликан и липотейхоевая кислота, и оба компонента являются потенциальными активаторами альтернативного пути активации комплемента, хотя при наличии специфических антител они также могут выполнять роль активаторов классического пути активации комплемента (Joiner, K.A., et al., Ann. Rev. Immunol. 2:461-2, 1984).
Изначально, система комплемента была отнесена к патогенезу сепсиса, так как исследователями было отмечено, что анафилатоксины С3а и С5а опосредуют многочисленные воспалительные ответы, которые также могут возникать при сепсисе. Данные анафилатоксины индуцируют вазодилатацию и повышение микроваскулярной проницаемости, явления, которые играют центральную роль при септическом шоке (Schumacher, W.A., et al., Agents Actions 34:345-349, 1991). Кроме того, анафилатоксины индуцируют бронхоспазм, секрецию гистамина из мастоцитов и аггрегацию тромбоцитов. Более того, они оказывают многочисленные воздействия на гранулоциты, вызывая, например, хемотаксис, аггрегацию, адгезию, высвобождение лизосомальных ферметов, генерацию токсичного супероксид аниона и формирование лейкотриенов (Shin, H.S., et al., Science 162:361-363, 1968; Vogt, W., Complement 3:177-86, 1986). Считается, что данная биологическиая активность принимает участие в развитии осложнений при сепсисе, как, например, шок или синдром острой дыхательной недостаточности (ARDS) (Hammerschmidt, D.E., et al., Lancet 1:947-949, 1980; Slotman, G.T, et al., Surge/y 99:744-50, 1986). Более того, повышенные уровни анафилатоксина С3а связаны с летальным исходом при сепсисе (Hack, С.Е., et al., Am. J. Med. 86:20-26, 1989). В нескольких животных моделях шока некоторые линии с недостаточностью комплемента (наприемр, штамм с С5-недостаточностью) оказались более устойчивыми к эффектам, сопутствующим введению LPS (Hseuh, W., et al., Immunol. 70:309-14, 1990).
Блокирование генерации С5а антителами в начале развития сепсиса у грызунов приводило к значительному повышению уровня выживаемости (Czermak, В. J., et al., Nat. Med. 5:788-792, 1999). Подобные открытия были сделаны при блокировании рецептора С5а (C5aR) антителами или малым молекулярным ингибитором (Huber-Lang, M.S., et al., FASEB J. 16:1567-74, 2002; Riedemann, N.C. et al., J. Clin. Invest, 110:101-8, 2002). Ранние экспериментальные исследования на обезьянах позволили выдвинуть предположение о том, что блокада С5а антителами ослабляла септический шок, вызванный кишечной палочкой, и синдром дыхательной недостаточности у взрослых (Hangen, D.H., et al., J. Surg. Res. 46:195-9, 1989; Stevens, J.H., et al., J. Clin. Invest. 77:1812-16, 1986). У людей, страдающих сепсисом, отмечается повышение уровня С5а, что ассоциируется со значительным снижением уровнями выживаемости вместе с мультиорганной недостаточностью, по сравнению с пациентами с менее тяжелой формой сепсиса и выжившими (Nakae, H., et al., Res. Commun. Спет.Pathol. Pharmacol. 84:189-95, 1994; Nakae, et al., Surg. Today 26:225-29, 1996; Bengtson, A., et al., Arch. Surg. 123:645-649, 1988). Механизмы, при помощи которых С5а оказывает свои вредные воздействия при сепсисе, нуждаются в тщательном детальном исследовании, однако недавно полученные данные позволяют выдвинуть предположение о том, что генерация С5а при сепсисе в значительной степени нарушает врожденные иммунные функции нейтрофилов крови (Huber-Lang, M.S., et al., J. Immunol. 169:3223-31, 2002), их способность вызывать окислительный взрыв и генерировать цитокины (Riedemann, N.C., et al., Immunity 19:193-202, 2003). В дополнении, генерация С5а при сепсисе оказывает прокоагулирующий эффект (Laudes, LJ, et al., Am. J. Pathol. 160:1867-75, 2002). Комплемент-модулирующий белок Cl INH также демонстрирует свою эффективность в животных моделях сепсиса и ARDS (Dickneite, G, Behring Ins. Mitt. 93:299-305, 1993).
Пектиновый путь активации комплемента также может принимать участие в патогенезе сепсиса. Было показано, что MBL может связываться с целым рядом клинически важных микроорганизмов, включая как грамотрицательные, так и грамположительные бактерии, и активировать пектиновый путь (Neth, О., et al., Infect. Immun. (68:688, 2000). Липотейхоевая кислота (LTA) все в большей степени рассматривается как грамположительный аналог LPS. Она является потенциальным иммуностимулятором, который индуцирует высвобождение цитокинов из мононуклеарных фагоцитов и цельной крови (Morath, S. et al., J. Exp.Med. 195:1635, 2002; Morath, S. et al., Infect. Immun. 70:938, 2002). Недавние исследования показали, что L-фиколин специфическим образом связывается с LTA, выделенной из многочисленных видов грамположительных бактерий, включая золотистый стафилококк (Staphylococcus aureus), и активирует пектиновый путь (Lynch, N.J., et al., J. Immunol. 172:1198-02, 2004), Также было показано, что MBL связывается с LTA из разновидности фекального стрептококка (Enterococcus spp), в которой полиглицерофосфатная цепь замещена гликозильными группами, но не с LTA из девяти других видов, включая золотистый стафилококк (Polotsky, V.Y, et al. Infect. Immun. 64:380, 1996).
Таким образом, один аспект настоящего изобретения предоставляет способ лечения сепсиса или заболевания, возникшего в результате сепсиса, посредством введения композиции с содержанием терапевтически эффективного объема агента, ингибирующего MASP-2, в фармацевтически приемлемом носителе субъекту, страдающему от сепсиса или заболевания, возникшего в результате сепсиса, включая, помимо прочего, острый сепсис, септический шок, синдром острой дыхательной недостаточности как сопутствующее заболевание при сепсисе, и синдром системной воспалительной реакции (SIRS). Родственные способы предлагаются для лечения прочих заболеваний крови, включая геморрагический коллапс, гемолитическую анемию, аутоиммунный тромбоцитопенический акроангиотромбоз (ТТР), синдром гемолитической уремии (HUS) или другие деструктивные заболевания костного мозга/крови, посредством введения композиции с содержанием терапевтически эффективного объема агента, ингибирующего MASP-2, в фармацевтически приемлемом носителе субъекту, страдающему подобным заболеванием. Агент, ингибирующий MASP-2, вводится субъекту систематически посредством внутриартериального, внутривенного, внутримышечного, ингаляционного (особенно в случае ARDS), подкожного либо другого вида парентерального введения; потенциально возможен оральный прием непептидергических агентов. Композиция с содержанием агента, ингибирующего MASP-2, может быть комбинирован с одним или более дополнительными терапевтическими агентами для борьбы с осложнениями сепсиса и/или шока. В случае прогрессирующего сепсиса или шока или угнетенного состояния как результата вышеозначенных заболеваний, композиция, ингибирующая MASP-2, может быть введена с использованием форм быстродействующей дозировки, каким является внутривенное или внутриартериальное введение болюсов раствора, содержащего агент, ингибирующий MASP-2. Повторное введение может быть осуществлено по предписанию врача до полного излечения заболевания.
УРОГЕНИТАЛЬНЫЕ ЗАБОЛЕВАНИЯ
Система комплемента вовлечена в некоторые явно выраженные урогенитальные расстройства, включая синдром болезненного мочевого пузыря, гиперактивный мочевой пузырь, хронический абактериальный цистит и интерстициальный цистит (Holm-Bentzen, М., et al., J. Urol. 138:503-507, 1987), бесплодие (Cruz, et al., Biol. Reprod. 54:1217-1228, 1996), беременность (Xu, С., et al., Science 287:498-507, 2000), фетоматеринская толерантность (Xu, С, et al., Science 287:498-507, 2000) и преэклампсия (Haeger, М., Int. J. Gynecol. Obstet. 43:113-127, 1993).
Синдром болезненного мочевого пузыря, гиперактивный мочевой пузырь, хронический абактериальный цистит и интерстициальный цистит являются недостаточно определенными заболеваниями с неизвестной этиологией и патогенезом, и, как следствие, не существует рациональной терапии для данных заболеваний. В настоящее время доминируют патогенетические теории, рассматривающие дефекты в эпителии и/или в слизистой поверхности, покрывающей мочевой пузырь, и теории, рассматривающие иммунологические нарушения (Holm-Bentzen, М., et al., J. Urol. 138:503-507, 1987). Пациенты, страдающие интерстициальным циститом, были протестированы на иммуноглобулин (IgA, G, М), компоненты комплемента (C1q, СЗ, С4) и ингибитор С1-эстеразы. Было обнаружено весьма значительное истощение уровней компонента комплемента С4 в сыворотке (р менее 0,001) и значительное повышение иммуноглобулина G (р менее 0,001). Данное исследование позволяет предположить наличие классического пути активации системы комплемента и выступает в поддержку возможности того, что хронический локальный иммунологический процесс вовлечен в патогенез заболевания (Mattila, J., et al., Eur. Urol. 9:350-352, 1983). Более того, вследствие связывания антител с антигенами в слизистой мочевого пузыря, активация комплемента может быть вовлечена в продуцирование поражения тканей и в хроническое самовозобновляющееся воспаление, типичное при данном заболевании (Helin, H., et al., Clin. Immunol. Immunopathol. 43:88-96, 1987).
В дополнение к роли комплемента в урогенитальных воспалительных заболеваниях, локальная регуляция пути активации комплемента может нарушать репродуктивные функции. Были выделены природные ингибиторы комплемента для протекции клеток реципиента с целью контроля системы комплемента всего организма. Для определения функции регуляторного контроля комплемента в развитии плода были проведены исследования Crry, природного ингибитора комплемента у грызунов, который имеет структуру, подобную ингибиторам комплемента человека, МСР и DAF. Интересным является то, что попытки создать мышь с Crry-/- не увенчались успехом. Вместо этого, было обнаружено, что гомозиготные мыши с Crry-/- погибали в утробе. Эмбрионы Crry-/- доживали до 10 дней после осеменения, затем выжившие быстро отторгались в результате смерти, вызванной остановкой развития. Также в плацентарной ткани эмбрионов Crry-/- была отмечена значительная инвазия клеток воспаления. Между тем, в плаценте эмбрионов Crry+/+ обнаруживается отложение С3. В связи с этим можно предположить, что активация комплемента происходила на плацентарном уровне, при отсутствии регуляции комплемента эмбрион погибал. Подтверждающие исследования выявляли действие мутации Crry при ее введении в генотип с С3-недостаточностью. Данная поддерживающая стратегия оказалась успешной. Рассмотренные вместе приведенные данные показывают, что трансплацентарный комплементарный барьер должен регулироваться. Незначительные изменения в регуляции комплемента в плаценте могут способствовать развитию плацентарной дисфункции и выкидышу (Xu, С, et al., Science 287/498-507, 2000).
Преэклампсия представляет собой обусловленное беременностью гипертензивное расстройство, в котором обнаруживает себя активация системы комплемента, однако степень ее участия остается спорной (Наедег, М., Int. J. Gynecol. Obstet. 43:113-127, 1993). Активация комплемента в большом круге кровообращения тесно связана с развитием заболевания при преэклампсии, но до появления клинических симптомов не выявлено повышения маркерных уровней и, следовательно, компоненты комплемента не могут рассматриваться как прогностические факторы преэклампсии (Наедег, et al., Obstet. Gynecol. 78:46, 1991). Однако повышение активации комплемента в прилегающих к плаценте участках могут подавлять механизмы локального контроля, что приводит к повышению уровней анафилатоксинов и C5b-9 (Haeger, et al., Obstet. Gynecol. 73:551,1989).
Один из предложенных механизмов бесплодия, вызываемого антиспермальными антителами (ASA), как полагают, связан с активацией комплемента в половых путях. Синтез опсонинов C3b и iC3b, которые может усиливать связывание сперматозоидов фагоцитарными клетками при помощи их комплементарных рецепторов и формирование терминального комплекса C5b-9 на поверхности сперматозоидов, что снижает подвижность сперматозоидов, являются потенциальными причинами, вызывающими уменьшение фертильности. Повышенные уровни C5b-9 были также продемонстрированы в фолликулярной жидкости из яичников бесплодной женщины (D'Cruz, OJ., et al., J Immunol. 144:3841-3848, 1990). Прочие исследования показали ослабление миграции сперматозоидов и уменьшение их взаимодействия с яйцеклеткой, что может быть опосредовано комплементом (D'Cruz, O.J., et al., J. Immunol. 146:611-620, 1991; Alexander, N.J., Fertil. Steril. 41:433-439, 1984). Наконец, исследования с sCR1 показали защитный эффект против ASA- и комплемент-опосредованного поражения сперматозоидов человека (D'Cruz, O.J., et al., Biol. Reprod. 54/1217-1228, 1996). Эти данные предоставляют ряд доказательств, свидетельствующих в пользу использования ингибиторов комплемента для лечения урогенитальных заболеваний и расстройств.
Таким образом, один аспект настоящего изобретения предлагает способ ингибирования MASP-2-зависимой активации комплемента у пациента, страдающего от урогенитального нарушения, посредством введения композиции с содержанием терапевтически эффективного объема агента, ингибирующего MASP-2, в фармацевтически приемлемом носителе субъекту, страдающему от подобного нарушения. Считается, что урогенитальные нарушения являются заболеваниями, подлежащими терапевтическому лечению способами и композициями по настоящему изобретению, и в группу данных заболеваний включены (список данным перечислением не ограничивается): синдром болезненного мочевого пузыря, гиперактивный мочевой пузырь, хронический абактериальный цистит и интерстициальный цистит, мужское и женское бесплодие, плацентарная дисфункция/недостаточность и самопроизвольный аборт и преэклампсия. Агент, ингибирующий MASP-2, может вводиться субъекту систематически посредством внутриартериального, внутривенного, внутримышечного, ингаляционного, подкожного или другого вида парентерального введения. Возможен также оральный прием непептидергических агентов. В качестве альтернативы, композиция, ингибирующая MASP-2, может вводиться локально в урогенитальный тракт посредством внутрипузырного орошения или инстилляции с жидким раствором или составам в форме геля. Введение может повторяться, если таковое предписано врачом для контроля состояния или полного излечения.
ДИАБЕТ И СВЯЗАННЫЕ С ДИАБЕТОМ ЗАБОЛЕВАНИЯ
Диабетическая микроангиопатия сетчатки характеризуется повышением проницаемости, лекостазом, микротромбозом и апоптозом капиллярных клеток, и каждое из названных проявлений может быть вызвано или усилено активацией комплемента. Гломерулярные структуры и эндонейронные микрососуды пациентов-диабетиков демонстрируют признаки активации комплемента. Факты снижения доступности и эффективности ингибиторов комплемента при диабете были поддержаны открытиями того, что высокий уровень глюкозы in vitro селективным образом понижает на поверхности эндотелиальных клеток экспрессию CD55 и CD59, двух ингибиторов, которые являются гликозилфосфатидилинозитол-связанными мембранными белками (glycosylphosphatidylinositol (GPI)-anchored membrane proteins), и что CD59 подвергается неферментной гликации, которая препятствует выполнения им функции ингибирования комплемента.
Исследования Занга и его коллег (Zhang et al. (Diabetes 51:3499-3504, 2002)) имели целью изучение активации комплемента как одной из характеристик непролиферативной диабетической ретинопатии человека и его взаимосвязь с изменениями молекул-ингибиторов. Было обнаружено, что отложение C5b-9, конечного продукта активации комплемента, происходит на стенках сосудов сетчатки человеческого глаза у пациента, страдающего диабетом 2-го типа, но не в сосудах у людей того же возраста, но не страдающих диабетом. C1q и С4, уникальные компоненты комплемента классического пути активации, не были обнаружены в сетчатке диабетиков, что указывает на генерирование C5b-9 посредством альтернативного пути активации. У пациентов-диабетиков проявляется значительное уменьшение в сетчатке уровней CD55 и CD59, двух ингибиторов комплемента, связанных с плазматической мембраной при помощи гликозилфосфатидилинозитольного якоря (GPI anchors). Подобная активация комплемента в сосудах сетчатки и выборочное снижение в сетчатке уровней CD55 и CD59 наблюдались у крыс с 10-недельным диабетом, вызванным стрептозотоцином. Таким образом, диабет вызывает дефективную регуляцию ингибиторов комплемента и активацию комплемента, которая предшествует большинству прочих проявлений микроангиопатии сетчатки, опосредованной диабетом.
Жерль и его коллеги (GerI et al. (Investigative Ophthalmology and Visual Science 43:1104-08, 2000)) выявили наличие активированных компонентов комплемента в глазах пациентов, пораженных диабетической ретинопатией. Иммуногистохимические исследования выявили обширное отложение комплексов комплемента C5b-9, в хориокапиллярисе, слое капиллярных сосудов, непосредственно лежащем в основе базальной пластинки и плотно окруженном капиллярами, во всех 50 видах диабетической ретинопатии. Результаты окрашивания C3d положительно коррелировали с результатами окрашивания C5b-9, что указывало на факт наличия активации комплемента in situ. Далее, позитивное окрашивание было установлено для витронектина, который формирует стабильные комплексы с внеклеточным C5b-9. В тоже время, не было обнаружено позитивного окрашивания для С-реактивного белка (CRP), маннозосвязывающего лектина (MBL), C1q или С4, что указывает на отсутствие активации комплемента посредством С4-зависимого пути. Таким образом, наличие C3d, C5b-9 и витронектина указывает на то, что активация комплемента происходит до завершения, возможно посредством альтернативного пути активации в хориокапиллярисе глаз, пораженных диабетической ренопатией. Активация комплемента может быть причинным фактором в патологическом осложнении, что может относиться к поражению ткани глаза и нарушению зрения. Отсюда следует, что использование ингибитора комплемента может быть эффективной терапией для редуцирования или блокирования поражения микрососудов, имеющего место при диабете.
Инсулинозависимый сахарный диабет (IDDM, также называемый диабет Типа I) является аутоиммунным заболеванием, опосредованным наличием антител различного типа (Nicoloff et al., Clin. Dev. Immunol. 11:61-66, 2004). Присутствие данных антител и соответствующих антигенов в кровотоке приводит к формированию циркулирующих иммунных комплексов (CIC), которые известны способностью сохраняться в крови в течение долгого времени. Отложение CIC в малых кровеносных сосудах способно вызывать микроангиопатию с негативными клиническими последствиями. У детей-диабетиков наблюдается корреляция между С1С и развитием микроваскулярных осложнений. Данные открытия позволяют предположить, что повышенные уровни С1С IgG связаны с развитием ранней диабетической нефропатии и что ингибитор пути активации комплемента может быть эффективным при блокировании диабетической нефропатии (Kotnik, et al., Croat. Med. J. 44/707-11, 2003). Кроме того, формирование вторичных белков комплемента и вовлечение альтернативного пути активации комплемента, вероятно, вносят соответствующий вклад в функционирование инсулоцитов при IDDM, а использование ингибитора комплемента рассматривается эффективным для редукции потенциального поражения или ограничения некроза клеток (Caraher, et al., J. Endocrinol. 162:143-53, 1999).
Концентрация циркулирующих MBL значительно повышена у пациентов, страдающих диабетом Типа 1, по сравнению с контрольной группой здоровых индивидов, и данная концентрация MBL положительно коррелирует с уровнем выделения белка с мочой (Hansen et al., J. Clin. Endocrinol. Metab. 88:4857-61, 2003). Недавнее клиническое исследование показало, что частоты генотипов с повышенной и пониженной экспрессией MBL были схожи у пациентов, страдающих диабетом Типа 1, и здоровыми индивидуумами из контрольной группы (Hansen et al., Diabetes 53/1570-76, 2004). Однако риск приобретения нефропатии среди пациентов-диабетиков был значительно выше, если они имели генотип, характеризующийся высоким уровнем экспрессии MBL. Это указывает на то, что высокие уровни MBL и пектиновый путь активации комплемента могут инициировать развитие диабетической нефропатии. Это заключение поддерживается недавним перспективным исследованием, в котором была изучена связь между уровнями MBL и развитием альбуминурии в когорте заново диагностированных пациентов, страдающих диабетом Типа 1 (Hovind et al., Diabetes 54:1523-21, 2005). В ходе данного исследования было обнаружено, что высокие уровни MBL на ранней стадии течения диабета Типа 1 были в значительной степени связаны с последующим развитием устойчивой альбуминурии. Эти результаты указывают, что MBL и пектиновый путь активации могут быть вовлечены в специфический патогенез диабетических васкулярных осложнений в большей степени, чем только провоцировать усиление существующих изменений. В недавнем клиническом исследовании (Hansen et al., Arch. Intern. Med. 166:2007-13, 2006) было измерено базовое значение уровней MBL у хорошо охарактеризованной когорты пациентов, страдающих диабетом Типа 2 и находящихся под наблюдением более 15 лет. Было обнаружено, что даже после устранения известных факторов риска, риск смертности был значительно выше среди пациентов с высокими уровнями MBL в плазме (>1000 мг/л), чем среди пациентов с низкими уровнями MBL (<1000 мг/л).
В еще одном аспекте изобретения предлагаются способы ингибирования MASP-2-зависимой активации комплемента у субъекта, страдающего диабетом без склонности к ожирению (IDDM) или ангиопатией, нейропатией или ретинопатией как осложнениями IDDM или приобретенным диабетом (Тип 2), посредством введения композиции, содержащей терапевтически эффективный объем ингибитора MASP-2 в фармацевтически приемлемом носителе. Агент, ингибирующий MASP-2, может вводиться субъекту систематически посредством внутриартериального, внутривенного, внутримышечного, подкожного либо другого вида парентерального введения; возможен также оральный прием непептидергических агентов. Альтернативно, введение может проводиться локально в участки, проявляющие ангиопатические, нейропатические или ретинопатические симптомы. Агент, ингибирующий MASP-2, может быть введен периодически спустя продолжительное время с целью лечения или контроля хронического заболевания, или однократно либо серийно для лечения в состоянии обострения.
ПЕРИХИМИОТЕРАПЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ И ЛЕЧЕНИЕ ЗЛОКАЧЕСТВЕННЫХ ЗАБОЛЕВАНИЙ
Активация системы комплемента может также быть вовлеченной в патогенез злокачественных заболеваний. В недавних исследованиях с использованием поликлональных и моноклональных антител и техники стрептавидин-биотин-пероксидазы были выявлены неоантигены комплекса комплемента C5b-9, IgG, С3, С4, S-белок/витронектин, фибронектин и макрофаги в 17 образцах рака молочной железы и в 6 образцах доброкачественной опухоли молочной железы. Все образцы тканей с карциномой на каждой стадии TNM показали отложение C5b-9 на мембранах опухолевых клеток, истонченные гранулы в клеточных остатках и диффузию отложений в участках омертвения (Niculescu, F, et al., Am. J. Pathol. 140:1039-1043, 1992).
Кроме того, активация комплемента может являться следствием химиотерапии или радиотерапии, и, таким образом, ингибирование активации комплемента было бы полезным в качестве дополнения к лечению злокачественных заболеваний с целью уменьшения ятрогенного воспаления. В случаях, когда химиотерапия и радиотерапия проводились перед оперативным вмешательством, отложение C5b-9 было более интенсивным и обширным. Отложение C5b-9 отсутствовало во всех образцах доброкачественных новообразований. S-белок/витронектин был обнаружен в виде фибриллярного преципитата в соединительнотканном матриксе и в виде преципитата, рассеянного вокруг опухолевых клеток, менее интенсивного и масштабного, чем фибронектин. Отложения IgG, СЗ, и С4 были обнаружены только в образцах злокачественной ткани. Наличие отложений C5b-9 показательно для активации комплемента и его последующих патогенетических эффектов при раке молочной железы (Niculescu, F, et al., Am. J. Pathol. 740:1039-1043, 1992).
Терапия с импульсным лазером на красителях с перестройкой частоты (577 нм) (PTDL) вызывает коагуляцию гемоглобулина и некроз тканей, что ограничивается в основном кровеносными сосудами. В исследованиях PTDL-облученной здоровой кожи, главными открытиями стали следующие: 1) фрагменты С3, С8, С9 и MAC откладываются на стенках сосудов; 2) данные отложения не являлись результатом денатурации белков, так как они появились только спустя 7 минут после облучения, в отличие от немедленного отложения трансферрина в сайтах коагуляции эритроцитов; 3) было показано, что отложения С3 способны усиливать активацию комплемента посредством альтернативного пути, что указывает на специфическую реакцию, так как сам некроз тканей не вызывает подобной амплификации; и 4) данные реакции предшествуют локальной аккумуляции полиморфноядерных лейкоцитов. Некроз тканей был более ярко выражен в гемангиомах. Не было зафиксировано значительных проявлений активности комплемента в больших ангиоматозных сосудах в центре некроза. Между тем, отложение комплемента в сосудах, расположенных на периферии, было схожим с тем, которое наблюдали в нормальной коже, с одним исключением: С8, С9 и MAC были обнаружены в некоторых кровеносных сосудах сразу после лечения, что согласуется с данными, о сборке MAC, которая происходит напрямую без формирования конвертазы С5. Данные результаты указывают на то, что комплемент активируется в PTDL-опосредованном васкулярном некрозе и может вызывать последующую воспалительную реакцию.
Фотодинамическая терапия (PDT) новообразования вызывает сильную иммунную реакцию пациента и одно из ее проявлений выражается в нейтрофилии. В добавление к фрагментам комплемента (прямым медиаторам), выделенным как следствие PDT-опосредованной активации комплемента, существует как минимум дюжина вторичных медиаторов, все из которых являются результатом активности комплемента. Последние включают в себя цитокины IL-1beta, TNF-alpha, IL-6, IL-10, G-CSF и КС, тромбоксан, простагландины, лейкотриены, гистамин и факторы коагуляции (Cede, I., et al., Cancer Lett. 183:43-51,2002).
Наконец, использование ингибиторов MASP-2-зависимой активации комплемента может быть рассмотрено в связи со стандартным терапевтическим режимом лечения рака. Например, лечение с использованием ритуксимаба, химерного анти-CD20 моноклонального антитела, может быть ассоциировано с побочными эффектами первой дозы от средней до тяжелой степени, особенно у пациентов с большим количеством циркулирующих опухолевых клеток. Недавние исследования во время первого введения ритуксимаба были направлены на замер продуктов активации комплемента (C3b/с и C4b/с) и цитокинов (фактор некроза опухолей альфа (TNF-alpha), интерлейкина 6 (IL-6) и IL-8) в пяти случаях рецидива у пациентов злокачественной лимфомы, не относящейся к группе заболеваний, известных как болезнь Ходжкина (NHL). Введение ритуксимаба вызывала быструю активацию комплемента, предшествующую секреции TNF-alpha, IL-6 и IL-8. Хотя группа испытуемых была небольшой, уровень активации комплемента коррелировал как с количеством циркулирующих клеток В перед инфузией (r=0.85; Р=0.07), так и со степенью серьезности побочных эффектов. Результаты указали на ключевую роль комплемента в патогенезе побочных эффектов при обработке с ритуксимабом. Так как активация комплемента не может быть предупреждена приемом кортикостероидов, это может стать предметом исследования возможной роли ингибиторов комплемента при первичном введении ритуксимаба (van der Kolk, L.E., et al., Br. J. Haematol. 115:807-811, 2001).
В еще одном аспекте настоящего изобретения предлагаются способы ингибирования MASP-2-зависимой активации комплемента у субъекта, которого подвергли химиотерапии и/или радиотерапии, включая, помимо прочего, лечение раковых заболеваний. Данный способ включает введение субъекту композиции, содержащей терапевтически эффективный объем ингибитора MASP-2 в фармацевтически приемлемом носителе перихимиотерапевтически, т.е., до и/или во время и/или после проведения химиотерапии и/или радиотерапии. Например, введение композиции с ингибитором MASP-2 по настоящему изобретению может быть начато перед или одновременно с началом химио- или радиотерапии и продолжено в течение всего курса терапии для уменьшения разрушительных эффектов химио- и/или радиотерапии в здоровых тканях, не являющихся мишенями. Кроме того, композиция, ингибирующая MASP-2, может вводиться после химио- и/или радиотерапии. Абсолютно ясно, что режимы химио- и/или радиотерапии часто требуют повторного лечения и, как следствие, возможно повторяющееся введение композиции, ингибирующей MASP-2, что будет совпадать с химиотерапевтическим и радиотерапевтическим лечением. Также считается, что агенты, ингибирующие MASP-2, могут использоваться в качестве химиотерапевтических агентов, самостоятельно или в комбинации с другими химиотерапевтическими агентами и/или радиотерапевтическими агентами для лечения пациентов, страдающих злокачественными заболеваниями. Введение может проводиться посредством орального приема (для непептидергических), внутривенным, внутримышечным или другим парентеральным способом.
ЭНДОКРИННЫЕ ЗАБОЛЕВАНИЯ
Не так давно была показана связь системы комплемента с несколькими эндокринными заболеваниями и расстройствами, включая тиреоидит Хашимото (Blanchin, S., et al., Exp. Eye Res. 73(6):887-96, 2001), стресс, тревожное расстройство и прочие потенциальные гормональные нарушения, затрагивающие регулируемую секрецию пролактина, рост новообразований или другие факторы, провоцируемые инсулинозависимыми состояниями и адренокортикотропным гормоном гипофиза (Francis, K., et al., FASEB J. 17:2266-2268, 2003; Hansen, Т.K., Endocrinology 144 (12): 5422-9, 2003).
Между эндокринной и иммунной системой существует двустороннее взаимодействие с использованием молекул, таких как гормоны и цитокины. В недавних исследованиях обнаружили новый путь активации, при котором С3а, образованный комплементом цитокин, стимулирует секрецию гормона передней доли гипофиза и активирует гипоталамо-гипофизарно-надпочечниковую систему, что является основным ответом на стрессовую реакцию и на контроль воспаления. Рецепторы С3а выделяются в клетках секреции гормона гипофиза и клетках не секретирующих этот гормон (фолликулостелаты). С3а и C3adesArg (невоспалительные метаболиты) стимулируют гипофизарные клеточные культуры для секреции пролактина, соматотропного гормона и адренокортикотропина. Уровни данных гормонов в сыворотке, вместе с кортикостероном надпочечников, повышаются в зависимости от введенной дозы рекомбинантных С3а и C3adesArg in vivo. Таким образом, можно заключить, что путь активации комплемента регулирует специфичную для тканей и системную воспалительную реакцию посредством взаимодействия с гипофизом (Francis, K., et al., FASEB J. 17:2266-2268, 2003).
Увеличивающееся число исследований на животных и человеке указывает на то, что гормон роста (GH) и инсулиноподобный фактор-1 роста (IGF-1) регулируют иммунную функцию. GH терапия повышает уровень смертности среди тяжелобольных пациентов. Превышение принятого уровня смертности было полностью вызвано септическим шоком или полиорганной недостаточностью, из чего можно сделать предположение о вовлечении GH-опосредованного изменения иммунной и комплементарной функции в этот процесс.Маннозосвязывающий лектин (MBL) представляет собой плазматический белок, который исполняет важнейшую роль во врожденном иммунитете посредством активации каскада комплемента и воспаления вследствие связывания с углеводными структурами. В литературе имеются данные о значительном влиянии гормона роста на уровни MBL, и, следовательно, о его потенциальном влиянии на пектин-зависимую активацию комплемента (Hansen, Т.K., Endocrinology 144 (12):5422-9, 2003).
Тиропероксидаза (ТРО) является одним из основных аутоантигенов, вовлеченных в аутоиммунные заболевания щитовидной железы. ТРО состоит из большого N-концевого миелопероксидаза-подобного модуля, за которым следует ССР - подобный модуль (ССР - контрольный белок комплемента) и модуль, подобный эпидермальному фактору роста. ССР модуль состоит из молекул, вовлеченных в активацию компонента комплемента С4; были проведены исследования по изучению способа, которым С4 связывается с ТРО и инициирует путь активации комплемента в аутоиммунных заболеваниях. ТРО посредством своего модуля ССР напрямую активирует комплемент без всякого опосредования Ig. Более того, у пациентов с тиреоидитом Хашимото тироциты экспрессируют в чрезмерных количествах С4 и все основные компоненты пути активации комплемента. Данные результаты указывают, что из всех прочих механизмов, относящихся к активации комплемента, только ТРО может способствовать массовому клеточному разрушению, которое наблюдается при тиреоидите Хашимото (Blanchin, S, et al.,2001).
Таким образом, аспект настоящего изобретения предлагает способ ингибирования MASP-2-зависимой активации комплемента для лечения эндокринного заболевания посредством введения субъекту, страдающему эндокринным заболеванием, композиции, содержащей терапевтически эффективный объема агента, ингибирующего MASP-2, в фармацевтически приемлемом носителе. Состояния, при которых возникает необходимость проведения подобного лечения в соответствии с настоящим изобретением, включают в себя, помимо прочего, тиреоидит Хашимото, стресс, тревожное расстройство и прочие возможные гормональные осложнения, затрагивающие регулируемую секрецию пролактина, рост новообразований или другие факторы, провоцируемые инсулинозависимыми состояниями и адренокортикотропным гормоном гипофиза. Агент, ингибирующий MAS-2, может вводиться субъекту систематически посредством внутриартериального, внутривенного, внутримышечного, ингаляционного, назального, подкожного либо другого вида парентерального администрирования; возможен также оральный прием непетидергических агентов. Композиция с содержанием агента, ингибирующего MASP-2, может быть комбинирована с одним или более дополнительными терапевтическими агентами. Введение может повторяться, если таковое предписано врачом, до излечения заболевания.
ОФТАЛЬМОЛОГИЧЕСКИЕ ЗАБОЛЕВАНИЯ
Возрастная дегенерация желтого пятна (AMD) представляет собой заболевание, связанное с потерей зрения, которое поражает миллионы взрослых людей. По настоящее время биохимические, клеточные и/или молекулярные механизмы, приводящие к развитию AMD, мало изучены. AMD приводит к прогрессирующему разрушению желтого пятна, что связано с формированием внутриклеточных отложений, известных как друзы, расположенных внутри и вокруг желтого пятна, позади сетчатки и между пигментным эпителием сетчатки (RPE) и сосудистой оболочкой глаза. Недавние исследования открыли, что в составе друз преобладали белки, связанные с воспалением и иммунно-опосредованными процессами. Транскрипты, кодирующие множество данных молекул, были обнаружены в клетках сетчатки, RPE и сосудистой оболочки. Эти данные также демонстрируют, что дендровидные клетки, которые также являются потенциальными антиген-презентующими клетками, находятся в тесной связи с развитием друз, и что активация комплемента является ключевым путем, который действует как в друзах, так и на RPE-хориоидальном интерфейсе (Hageman, G.S., et al., Prog. Retin. Eye Res., 20:705-732, 2001).
Некоторые независимые исследования показали устойчивую связь между AMD и полиморфизмом гена фактора комплемента Н (CFH), где у гомозиготных по рискованному аллелю индивидуумов вероятность AMD повышалась на 7,4 (Klein, RJ. et al., Science, 308:362-364, 2005; Haines et al., Science 308:362-364. 2005; Edwards et al. Science 308:263-264, 2005). Ген CFH был картирован на участке хромосомы 1q31, который, по результатам шести независимых определяемых, оказался сцеплен с AMD (см., например, Schultz D.W. et al., Hum. Mol. Genet. 12:3315, 2003). Известно, что CFH является ключевым регулятором системы комплемента. Было показано, что CFH в клетках и в кровотоке регулирует активность комплемента посредством ингибирования активации расщепления С3 на С3а и C3b и деактивации существующего C3b. Отложение C5b-9 наблюдалось в мембране Бруша (Brusch's membrane), интеркапиллярных перетяжках и внутри друз у пациентов, страдающих AMD (Klein et al.). По результатам экспериментов с использованием методов иммунофлуоресценции было выдвинуто предположение о том, что при AMD, полиморфизм CFH может опосредовать повышение уровня отложения комплемента в хориоидальных капиллярах и хориоидальных сосудах (Klein et al.).
Мембранно-ассоциированный ингибитор комплемента, рецептор комплемента 1, также был обнаружен в друзах, однако он не был выявлен в клетках RPE при использовании иммуногистохимического метода. В тоже время, второй мембрано-ассоциированный ингибитор комплемента, белок мембранного кофактора, присутствует в клетках RPE, ассоциированных с друзами, равно как и малых сферических структурных элементах внутри друз. Данные элементы, в недавнем прошлом еще неидентифицированные, также проявили устойчивую иммунореактивность для протеолитических фрагментов компонента комплемента С3, которые характерным образом откладываются в участках активации комплемента. Было выдвинуто предположение, что данные структуры представляют собой остаточные продукты дегенеративных клеток RPE, которые являются мишенями атаки комплемента (Johnson, L.V., et al., Exp. Eye Res. 73:887-896, 2001).
Идентификация и локализация данных многочисленных регуляторов комплемента, равно как и продуктов активации комплемента (С3а, С5а, C3b, C5b-9), позволили заключить, что хроническая активация комплемента играет важную роль в процессе биогенеза друз и этиологии AMD (Hageman et al., Progress Retinal Eye Res. 20:705-32, 2001). Идентификация продуктов активации С3 и С5 в друзах не дает информации о том, посредством какого пути активируется комплемент: классического, пектинового или альтернативного, так как в соответствии с настоящим исследованием, С3 и С5 присущи всем трем путям активации. Однако были проведены два исследования иммунного маркирование друз с использованием антител, специфичных к C1q, основному компоненту распознавания классического пути активации комплемента (Mullins et al., FASEB J. 14:835-846, 2000; Johnson et al., Exp. Eye Res. 70:441-449, 2000). В результате обоих исследований был сделан вывод о том, что C1q в друзах не обнаружен. Данные отрицательные результаты по C1q позволяют предположить, что активация комплемента в друзах происходит без участия классического пути активации. Кроме того, иммуно-маркирование друз на наличие компонентов иммунного комплекса (легкая цепь IgG, IgM) показало слабую вариабельность количества этих компонентов, на основании чего был сделан вывод о том, что классический путь активации играет незначительную роль в активации комплемента при данном заболевании (Mullins et al., 2000).
Недавно были опубликованы два исследования, в которых изучалась роль комплемента в развитии вызываемой лазером хориоидальной неоваскуляризации (CNV), у мышей, являющихся моделью CNV человека. Используя иммуногистологический метод, Бора и его коллеги (Bora and colleagues (2005)) обнаружили значительное отложение C3b и C5b-9 (MAC), продуктов активации комплемента в неоваскулярных комплексах вследствие проведения лазерной терапии (Bora et al., J. Immunol. 174:491-7, 2005). Важно то, что CNV не развилась у мышей с генетической недостаточностью С3 (мыши С3-/-), основного компонента, который необходим для всех путей активации комплемента. Уровни РНК трех ангиогенных факторов VEGF, TGF-P2, и P-FGF, вовлеченных в развитие CNV, были повышены в ткани глазного яблока у мышей после индуцированной лазером CNV. Истощение комплемента приводило к значительному понижению уровней РНК данных ангиогенных факторов.
Используя метод ИФА, Нозаки и его коллеги (Nozaki et al.) показали, что мощные анафилатоксины С3а и С5а генерируются в ранней стадии индуцируемой лазером CNV (Nozaki et al., Proc. Natl. Acad. ScL U.S.A. 103:2328-33, 2006). Более того, два биоактивных фрагмента С3 и С5 вызывали экспрессию VEGF после их инъекций в стекловидное тело мышам дикого типа. Опираясь на данные результаты, Нозаки и его коллеги также показали, что генетическое удаление рецепторов С3а и С5а уменьшала экспрессию VEGF и формирование CNV после лазерного поражения, и что опосредованная антителами нейтрализация С3а или С5а или фармакологическая блокада их рецепторов также уменьшает CNV. Предыдущие исследования установили, что восстановление лейкоцитов и особенно макрофагов выполняет ключевую роль в индуцированной лазером CNV, (Sakurai et al., Invest. Opthomol. Vis. Sci 44:3578-85, 2003; Espinosa-Heidmann, et al., Invest. Opthomol. Vis. Sci 44:3586-92, 2003). В своей работе 2006 г., Нозаки и его коллеги сообщают, что восстановление лейкоцитов после лазерного поражения значительно редуцировано у мышей C3aR(-/-) и C5aR(-/-).
Таким образом, один аспект настоящего изобретения предлагает способ ингибирования MASP-2-зависимой активации комплемента для лечения возрастной дегенерации желтого пятна или другого опосредованного комплементом офтальмологического заболевания посредством введения композиции с содержанием терапевтически эффективного объема агента, ингибирующего MASP-2, в фармацевтически приемлемом носителе субъекту, страдающему от подобного заболевания или другого офтальмологического заболевания, опосредованного комплементом. Композиция, ингибирующая MASP-2, может быть введена локально в глазное яблоко посредством орошения или аппликации композиции в форме геля, бальзама или капель. В качестве альтернативы, возможно осуществление систематического введения агента, ингибирующего MASP-2, субъекту посредством внутриартериального, внутривенного, внутримышечного, ингаляционного, назального, подкожного либо другого вида парентерального введения. Также возможен оральный прием непетидергических агентов. Композиция с содержанием агента, ингибирующего MASP-2, может быть скомбинировано с одним или более дополнительными терапевтическими агентами, описанными в заявке на патент, опубликованной как публикация патентной заявки США №2004-0072809-А1. Возможно повторяющееся введение, если таковое предписано врачом для контроля или излечения заболевания.
КОАГУЛОПАТИЯ
Представлено доказательство роли системы комплемента в диссеминированном внутрисосудистом свертывании (ДВС), например, ДВС, являющееся осложнением серьезного физического повреждения.
Предыдущие исследования показали, что мыши С4-/- не защищены от реперфузионного повреждения почек (Zhou, W., et al., "Predominant role for C5b-9 in renal ischemia/reperfusion injury," J Clin Invest 105:1363-1371 (2000)). С целью изучения способности мышей С4-/- активировать комплемент через классический или пектиновый пути, обновление С3 в плазме С4-/- было измерено в анализах специфичных для способов активации классического или пектинового путей. Расщепление С3 не наблюдалось при запуске активации через классический путь тогда, как наблюдалась высокоэффективная активация СЗ, зависимая от лектинового пути, в сыворотке с С4-недостаточностью (ФИГУРА 30). Можно увидеть, что отложение C3b на маннаны и зимозаны является сильно нарушенным у мышей MASP-2-/- даже при экспериментальных условиях, что согласно многим ранее опубликованным документам по активации альтернативного пути, должно быть допустимым для всех трех путей. При использовании одинаковой сыворотки в лунках, покрытых иммуноглобулиновыми комплексами вместо маннана или зимозана, отложение C3b и расщепление фактора В наблюдаются в мышиной сыворотке MASP-2+/+ и в сыворотке MASP-2-/-, но не в Clq-обедненной сыворотке. Это указывает на то, что активация альтернативного пути облегчена в сыворотке MASP-2-/- при добавлении начального C3b через классическую активность. На ФИГУРЕ 30С изображается удивительное обнаружение того, что С3 можно эффективно активировать способом зависимым от лектинового пути в плазме с С4-недостаточностью.
Данная «альтернативная активация С4» отменяется ингибированием активации лектинового пути через предварительную инкубацию плазмы с растворимым маннаном или маннозой.
Аберрантная, неиммунная активация системы комплемента представляет потенциальную опасность для человека, а также может играть важную роль в активации гематологического пути, особенно в случаях серьезных повреждений, в которых воспалительный и гематологический пути активированы. При нормальном состоянии здоровья, конверсия С3 составляет <5% от общего С3 белка плазмы. При сильно распространенной инфекции, включающей в себя септицемию и болезнь иммунных комплексов, конверсия С3 восстанавливает сама себя примерно на 30% с уровнями комплемента обычно более низкими, чем нормальные, вследствие повышенного использования и изменений в распределении пула. Незамедлительная активация пути С3 больше 30% обычно предоставляет несомненные клинические данные вазодилятации и потери жидкости в тканях. При конверсии С3 свыше 30%, инициирующие механизмы являются преимущественно неиммунными, а результирующие клинические проявления заболеваний представляют опасность для пациента. Уровни комплемента С5 при нормальном состоянии здоровья и при контролируемом заболевании являются более устойчивыми, чем С3. Существенные сокращения и или конверсия уровней С5 связаны с реакцией пациента на аномальную политравму (например, дорожно-транспортные происшествия) и вероятное развитие синдромов шокового легкого. Таким образом, любое доказательство активации комплемента С3 выше 30% васкулярного пула или любое участие С5, или и то и другое вместе, может рассматриваться, как вероятный предвестник пагубного патологического изменения у пациента.
Свободные анафилатоксины С3 и С5 (С3а и С5а), которые действуют на тучных клетках и базофилах, высвобождают сосудорасширяющие химические вещества. Они устанавливают хематоксические градиенты для направления полиморфно-ядерных клеток (PMN) в центр иммунологических нарушений (положительная реакция), но здесь они отличаются, поскольку С5а оказывает специфический эффект скопления (агрегации) на данные фагоциты, предупреждая их случайное удаление от реакционного участка. При нормальном контроле инфекции С3 активирует С5. Однако при политравме С5 является широко активированным, системно генерирующим анафилатоксины С5а. Данное неконтролируемое действие приводит к скоплению полиморфов в сосудистой системе, затем данные скопления попадают в капилляры легких, которые они закупоривают и порождают локальные эффекты повреждений в результате высвобождения супероксида. Не ограничиваясь теорией, механизм, возможно, является важным в патогенезе респираторного дистресс-синдрома у взрослых (ARDS), несмотря на то, что данная точка зрения недавно подверглась сомнению. Анафилатоксины С3а in vitro можно показать, как сильные агрегаторы тромбоцитов, но их участие in vivo менее четко определено и высвобождение тромбоцитов и плазмина при заживлении раны может только второстепенно задействовать комплемент С3. Возможно, необходим пролонгированный подъем активации С3 для генерации DIC.
В дополнение к клеточным и сосудистым эффектам вышеописанного активированного компонента комплемента, который может объяснить связь между травмой и ДВС, появляющиеся научные открытия идентифицировали прямые молекулярные связи и функциональный обмен между комплементом и системами коагуляции. Подтверждающие данные были получены из исследований на мышах с С3-недостаточностью. Поскольку С3 является общим компонентом для каждого из путей комплемента, у мышей с С3-недостаточностью отсутствует полная функция комплемента. Удивительно, однако, мыши с С3-недостаточностью могут превосходно активировать терминальные компоненты комплемента (Huber-Lang, M., et al., "Generation of C5a in the absence of СЗ: a new complement activation pathway," Nat. Meet 12:682-687 (2006)) Детальные исследования выявили, что С3-независимая активация терминальных компонентов комплемента опосредована тромбином, ферментом, ограничивающим скорость коагуляционного каскада. (Huber et al., 2006) Молекулярные компоненты, опосредующие активацию тромбина после начальной активации комплемента, остались неясными.
Авторы настоящего изобретения объяснили, что представляет собой молекулярный базис для обмена между комплементом и коагуляционными каскадами и идентифицировали MASP-2 в качестве центральной контрольной точки, связывающей две системы. Биохимические исследования в субстратной специфичности MASP-2 идентифицировали протромбин в качестве возможного субстрата в дополнение к широко известным белкам комплемента С2 и С4. MASP-2 специфически расщепляет протромбин на функционально релевантных участках, генерируя тромбин, фермент, ограничивающий скорость коагуляционного каскада (Krarup, A., et al., "Simultaneous Activation of Complement and Coagulation by MBL-Associated Serine Protease 2," PLoS. ONE. 2:e623 (2007)). Тромбин, генерированный MASP-2, способен стимулировать отложение фибрина в определенной восстановленной системе in vitro, демонстрируя функциональную релевантность расщепления MASP-2 (Krarup et al., 2007). Как обсуждается в нижеприведенных примерах в данном документе, изобретатели дополнительно подтвердили физиологическую значимость данного открытия посредством документирования активации тромбина в нормальной сыворотке грызунов после активации пектинового пути, а также продемонстрировали, что данный процесс блокируется нейтрализующими MASP-2 моноклональными антителами.
MASP-2 может представлять центральную точку разветвления в пектиновом пути, способную стимулировать активацию систем комплемента и коагуляции. Поскольку активация лектинового пути представляет собой физиологическую реакцию на многих виды травматических повреждений, авторы настоящего изобретения полагают, что сопутствующее системное воспаление (опосредованное компонентами комплемента) и диссеминированную коагуляцию (опосредованную через коагуляционный путь) можно объяснить способностью MASP-2 активировать оба пути. Данные открытия четко указывают на роль MASP-2 в генерации ДВС и терапевтическую полезность ингибирования MASP-2 при лечении или предупреждении ДВС. MASP-2 может обеспечить молекулярную связь между комплементом и коагуляционной системой, и активация пектинового пути, возникающая при разных травмах, может непосредственно инициировать активацию коагуляционной системы через ось MASP-2-тромбин, обеспечивая механизм связи между травмой и ДВС. В соответствии с аспектом настоящего изобретения ингибирование MASP-2 будет ингибировать активацию пектинового пути и сокращать генерацию анафилатоксинов С3а и С5а. Полагают, что необходим пролонгированный подъем активации С3 для генерации ДВС.
Следовательно, аспект изобретения, таким образом, предлагает способ MASP-2-зависимого ингибирования активации комплемента для лечения диссеминированного внутрисосудистого свертывания или другого комплемент-опосредованного коагуляционного расстройства, посредством введения композиции, содержащей терапевтически эффективную дозу агента, ингибирующего MASP-2 (например, антитело aHTH-MASP-2 или его фрагмент, пептидные ингибиторы или низкомолекулярные ингибиторы), в фармацевтическом носителе субъекту, страдающему заболеванием или прерасположенному к такому состоянию. В некоторых вариантах осуществления агенты, ингибирующие MASP-2, могут блокировать уже активированный MASP-2. Композиция, ингибирующая MASP-2, соответствующим образом вводится субъекту системно, например, внутриартериально, внутривенно, внутримышечно, ингаляционно, назально, подкожно или другим парентеральным введением, или возможно оральное введение для непептидергических ингибиторов. Введение можно повторять, как определяется врачом, пока состояние не разрешится или не будет взято под контроль. Способы данного аспекта настоящего изобретения можно использовать для лечения ДВС, осложняющего сепсис, серьезную травму, включая неврологическую травму (например, острая травма головы, смотрите Kumura, E., et al., Acta Neurochirurgica 85:23-28 (1987), инфекцию (бактериальную, вирусную, грибковую, паразитарную), рак, акушерское осложнение, болезнь печени, тяжелую токсическую реакцию (например, укус змеи, укус насекомого, трансфузионная реакция), шок, тепловой удар, отторжение трансплантатов, аневризму сосудов, печеночную недостаточности, лечение рака посредством химиотерапии или лучевой терапии, ожог, случайное радиационное облучение или другие причины. Смотрите, например, Becker J.U. and Wira C.R. "Disseminated Intravascular Coagulation" emedicine.medscape.com/9/10/2009. Для ДВС, осложняющего травму или другое острое событие, композицию, ингибирующую MASP-2, можно ввести незамедлительно после травматического повреждения или профилактически до, во время, незамедлительно или в течение от одного до семи дней или дольше, например, в течение от 24 часов до 72 часов, после получения травмы или при ситуациях, например, хирургической операции на пациентах, подверженных риску ДВС. В некоторых вариантах осуществления композиция, ингибирующая MASP-2, может соответствующим образом вводиться в быстродействующей лекарственной форме, например, посредством внутривенной или внутриартериальной доставки инъекции раствора, содержащего композицию агента, ингибирующего MASP-2.
IV. АГЕНТЫ, ИНГИБИРУЮЩИЕ MASP-2
В одном аспекте настоящее изобретение предлагает способ ингибирования пагубных эффектов MASP-2-зависимой активации комплемента. Агенты, ингибирующие MASP-2, вводятся в объемах, эффективных для подавления MASP-2-зависимой активации комплемента у живых субъектов. Для практического использования данного аспекта изобретения, представленные в данном описании агенты, ингибирующие MASP-2 включают в себя: молекулы, которые ингибируют биологическую активность MASP-2 (такие как малые молекулярные ингибиторы, анти-MASP-2 антитела или блокирующие пептиды, которые взаимодействуют с MASP-2 или препятствуют белок-белковому взаимодействию), и молекулы, сокращающие экспрессию MASP-2 (такие как молекулы MASP-2 антисмысловой нуклеиновой кислоты, MASP-2 специфические молекулы RNAi и рибозимы MASP-2), таким образом, удерживая MASP-2 от активирования альтернативного пути активации комплемента. Агенты, ингибирующие MASP-2, могут быть использованы самостоятельно в качестве первичной терапии или в комбинации с другими терапевтическими агентами как дополнительная терапия для повышения терапевтической эффективности другого медицинского лечения.
Ингибирование MASP-2-зависимой активации комплемента характеризуется как минимум одним из следующих изменений в компоненте системы комплемента, которые происходят в результате введения агента, ингибирующего MASP-2, в соответствии со способами настоящего изобретения: ингибирование генерации или продуцирования системных продуктов MASP-2-зависимой активации комплемента C4b, С3а, С5а и/или C5b-9 (MAC) (измеренных, например, как описано в Примере 2), снижение альтернативной активации комплемента, определенной в гемолитических исследованиях с использованием несенсибилизированных красных кровяных телец кроликов или морских свинок, снижение расщепления С4 и отложения C4b (измеренная, например, как описано в Примере 2) или снижение расщепления С3 и отложения C3b (измеренная, например, как описано в Примере 2).
В соответствии с настоящим изобретением, агенты, ингибирующие MASP-2, эффективно используются для ингибирования системы MASP-2-зависимой активации комплемента. Агентами, ингибирующими MASP-2, эффективными в практическом применении настоящего аспекта по настоящему изобретению включают: анти-MASP-2 антитела и их фрагменты, пептиды, ингибирующие MASP-2, малые молекулы, MASP-2 растворимые рецепторы и ингибиторы экспрессии. Агенты, ингибирующие MASP-2, могут подавлять систему MASP-2-зависимой активации комплемента посредством блокирования биологической функции MASP-2. Например, ингибитор может эффективно блокировать MASP-2 белок-белковые взаимодействия, интерферировать MASP-2 димеризацию или сборку, блокировать Са2+связывание, интерферировать активный участок MASP-2 сериновой протеазы или редуцировать экспрессию белков MASP-2.
В некоторых вариантах осуществления агенты, ингибирующие MASP-2, выборочным образом ингибируют MASP-2 активацию комплемента, не затрагивая систему C1q-зависимой активации комплемента.
В одном варианте осуществления агент, ингибирующий MASP-2, пригодный для практического использования в способах по настоящему изобретению, представлен ингибитором MASP-2, который специфическим образом связывается с полипептидом, включающим в себя SEQ ID NO:6 с аффинностью как минимум в 10 раз большей, чем у других антигенов в системе комплемента. В другом варианте осуществления агент, ингибирующий MASP-2, специфическим образом связывается с полипептидом, включающим в себя SEQ ID NO:6 со связующей аффинностью как минимум в 100 раз большей, чем у других антигенов в системе комплемента. Связующая аффинность агента, ингибирующего MASP-2, может быть определена при соответствующем анализе связывания.
Полипептид MASP-2 имеет молекулярную структуру, схожую с MASP-I, MASP-3 и C1r и C1s, протеазами системы комплемента С1. Молекула кДНК, представленная в SEQ ID NO:4, кодирует типичный образец MASP-2 (состоящий из аминокислотной последовательности, представленной в SEQ ID NO:5) и является полипептидом MASP-2 человека с лидерной последовательностью (аа 1-15), которая отщепляется после секреции, приводя к образованию зрелой формы MASP-2 человека (SEQ ID NO:6). Как показано на ФИГУРЕ 2, ген MASP 2 человека состоит из двенадцати экзонов. кДНК MASP-2 человека кодируется экзонами В, С, D, F, G, Н, I, J, К и L. Альтернативный сплайсинг приводит к образованию белка, массой 20 кДа, названному MBL-ассоциированным белком 19 («МАр19» или «sMAP») (SEQ ID NO:2), который кодируется нуклеотидной последовательностью SEQ ID NO:1, состоящей из экзонов В, С, D и Е, приведенных на ФИГУРЕ 2. Молекула кДНК, представленная в SEQ ID NO:50, кодирует MASP-2 мыши (состоящий из аминокислотной последовательности, представленной в SEQ ID NO:51) и является полипептидом MASP-2 мыши с лидерной последовательностью, которая отщепляется после секреции, приводя к образованию зрелой формы MASP-2 мыши (SEQ ID NO:52). Молекула кДНК, представленная в SEQ ID NO:53 кодирует MASP-2 крысы (состоит из аминокислотной последовательности, приведенной в SEQ ID NO:54) и предоставляет собой полипептид MASP-2 крысы с лидерной последовательностью, которая отщепляется после секреции, приводя к образованию зрелой формы MASP-2 крысы (SEQ ID NO:55).
Специалисты в данной области техники должны понимать, что последовательности, приведенные в SEQ ID NO:4, SEQ ID NO:50 и SEQ ID NO:53 представляют собой единичные аллельные варианты человеческого, мышиного и крысиного MASP-2, соответственно, и что другие аллельные варианты и альтернативный сплайсинг могут иметь место. Аллельные варианты нуклеотидных последовательностей, представленных в SEQ ID NO:4, SEQ ID NO:50 и SEQ ID NO:53, включающие в себя последовательности с молчащими мутациями и последовательности, в которых мутации вызывают изменения аминокислотных последовательностей, являются областью настоящего изобретения. Аллельные варианты последовательности MASP-2 могут быть клонированы из геномных библиотек или библиотек кДНК, проскринированных при помощи зондов, в соответствии со стандартными процедурам.
Домены белка MASP-2 человека (SEQ ID NO:6) показаны на ФИГУРЕ 3 А и они включают в себя N-терминальный домен C1r/C1s/фактора роста эндотелия сосудов (Vegf) морского ежа/морфогенного белка кости (CUBI) (аминокислоты 1-121 в SEQ ID NO:6), домен, подобный эпидермальному фактору роста, (аминокислоты 122-166), второй CUBI домен (аминокислоты 167-293), тандем доменов контрольных белков комплемента и домен сериновой протеазы. Альтернативный сплайсинг гена MASP2 приводит к образованию МАр19, показанного на ФИГУРЕ 3В. МАр19 является неферментным белком, содержащим N-терминальный CUBI-EGF участок MASP-2 с четырьмя дополнительными аминокислотными остатками (EQSL), относящимися к экзону Е, изображенному на ФИГУРЕ 2.
Было показано, что некоторые белки способны связываться или взаимодействовать с MASP-2 посредством белок-белковых взаимодействий. Например, известно, что MASP-2 способен связываться и формировать Са2+ зависимые комплексы с пектиновыми белками MBL, Н-фиколином и L- фиколином. Было показано, что каждый MASP-2/лектиновый комплекс способен активировать комплемент посредством MASP-2-зависимого расщепления белков С4 и С2 (Ikeda, К., et al., J. Biol. Chem. 262:7451-7454, 1987; Matsushita, M, et al., J. Exp. Med. 176:1497-2284, 2000; Matsushita, M., et al., J. Immunol. 168:3502-3506, 2002). Исследования показали, что CUBI-EGF домены MASP-2 являются существенными для связывания MASP-2 с MBL (Thielens, N.M., et al., J. Immunol. 166:5068, 2001). Также было показано, что CUB1EGFCUBII домены опосредуют димеризацию MASP-2, необходимую для формирования активного комплекса MBL (Wallis, R., et al., J. Biol. Chem. 275:30962-30969, 2000). Отсюда следует, что агенты, ингибирующие MASP-2, определяются своей способностью связываться или взаимодействовать с MASP-2 участками-мишенями, играющими важную роль в процессе MASP-2-зависимой активации комплемента.
АНТИ-MASP-2 АНТИТЕЛА
В некоторых вариантах осуществления данного аспекта настоящего изобретения агент, ингибирующий MASP-2, включает в себя анти-MASP-2 антитело, которое ингибирует систему MASP-2-зависимой активации комплемента. Анти-MASP-2 антитела, пригодные для осуществления данного аспекта настоящего изобретения, включают в себя поликлональные, моноклональные либо рекомбинантные антитела, производные любого спродуцированного млекопитающим антитела, и данные антитела могут быть мультиспецифическими, химерными, гуманизированными, антиидиотипическими антителами и фрагментами антител. Фрагменты антител включают в себя Fab, Fab', F(ab)2, F(ab')2, Fv фрагменты, scFv фрагменты и одноцепочные антитела как описано далее.
Некоторые анти-MASP-2 антитела были описаны в научной литературе, несколько из них перечислены в ТАБЛИЦЕ 1. Эти ранее описанные анти-MASP-2 антитела могут быть проскринированы на способность ингибирования системы MASP-2-зависимой активации комплемента с использованием описанных в данной заявке лабораторных исследований. Например, было показано, что анти-MASP-2 антитела Fab2 крысы блокируют MASP-2-зависимую активацию комплемента, более детальное описание предложено в Примерах 24 и 25 данной заявки. Как только определяется анти-MASP-2 антитело, обладающее признаками ингибитора MASP-2, оно может использоваться для продуцирования антиидиотипических антител и идентификации прочих MASP-2 связующих молекул, как описано ниже.
АНТИ-MASP-2 АНТИТЕЛА С РЕДУЦИРОВАННОЙ ЭФФЕКТОРНОЙ ФУНКЦИЕЙ
В некоторых воплощениях данного аспекта анти-MASP-2 антитела имели редуцированную эффекторную функцию для уменьшения воспаления, которое может возникать при активации классического пути комплемента. Было обнаружено, что способность IgG молекул инициировать классический путь активации комплемента связана с участком Fc молекулы (Duncan, A.R., et al., Nature 332:738-740 1988). Молекулы IgG, у которых участок Fc был удален посредством ферментного расщепления, лишены данной эффекторной функции (см. Harlow, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, New York, 1988). В соответствии с этим, антитела с редуцированной эффекторной функцией могут быть получены в результате недостачи Fc участка молекулы, имеющей генноинженерную Fc последовательность, которая обладает минимальной эффекторной функцией, эта молекула может относится как к изотипу IgG2 так и к изотипу IgG4 человека.
Антитела с редуцированной эффекторной функцией могут быть получены при помощи стандартных молекулярных биологических манипуляций с участком Fc тяжелых цепей IgG, как описано в Примере 9 данного изобретения и также описано в Jolliffe, et al., Int'l Rev. Immunol. 10:241-250, 11993, и Rodrigues, et al., J. Immunol. 151:6954-6961, 1998. Антитела с редуцированной эффекторной функцией также включают в себя изотипы lgG2 и lgG4 человека, которые обладают сниженной способностью активировать комплемент и/или вступать во взаимодействие с Fc рецепторами (Ravetch, J.V., et al., Annu. Rev. Immunol. 9:457-492, 1991; Isaacs, J.D, et al., J. Immunol. 148:3062-3071, 1992; van de Winkel, J.G, et al., Immunol. Today 14:215-221, 1993). Гуманизированные или полноценные человеческие антитела, специфичные к MASP-2 человека и включающие в себя изотипы IgG2 или lgG4, могут быть получены одним из нескольких способов, известных специалисту в данной области техники, как описано у Vaughan, T.J., et al., Nature Biotechnical 16:535-539, 1998.
ПОЛУЧЕНИЕ АНТИ-MASP-2 АНТИТЕЛ
Анти-MASP-2 антитела могут быть получены с использованием полипептидов MASP-2 (например, полноразмерного MASP-2) или антигенных MASP-2 эпитоп-несущих пептидов (например, часть полипептида MASP-2). Иммунногенные пептиды могут состоять всего из пяти аминокислотных остатков. Например, полипептиды MASP-2, содержащие полную аминокислотную последовательность SEQ ID NO:6, могут быть использованы для индукции анти-MASP-2 антител, которые могут быть использованы в способе, заявленном в настоящем изобретении. Известно, что конкретные домены MASP-2 (такие как CUBI и CUBIEGF домены) вовлечены в белок-белковые взаимодействия, равно как и участки, окружающие сайт активной сериновой протеазы, и могут выделяться как рекомбинантные полипептиды, как описано в Примере 5, и использоваться в качестве антигенов. Кроме того, пептиды, содержащие как минимум 6 аминокислот полипептида MASP-2 (SEQ ID NO:6), также используются для получения MASP-2 антител. Дополнительные примеры антигенов, производных MASP-2, обладающих способностью индуцировать MASP-2 антитела, приведены ниже в ТАБЛИЦЕ 2. Пептиды и полипептиды MASP-2, использованные для создания антител, могут быть выделены как натуральные пептиды, или рекомбинантные или синтетические пептиды и каталитически неактивные рекомбинантные полипептиды, такие как MASP-2A, как описано ниже в Примерах 5-7. В некоторых вариантах осуществления данного аспекта настоящего изобретения, анти-MASP-2 антитела получены с использованием трансгенной линии мышей, как описано в Примерах 8 и 9 и далее описано ниже.
Антигены, способные продуцировать анти-MASP-2 антитела, также включают в себя слитые полипептиды, например полипептиды, полученные в результате слияния MASP-2 либо его части с полипептидом иммуноглобулина или с белком, связывающим мальтозу. Полипептидный иммуноген может быть представлен полноразмерной молекулой или ее частью. Если часть полипептида представлена гаптеном, такая часть может быть преимущественно присоединена или связана с носителем макромолекул (таким как гемоцианин фиссуреллы (KLH), бычий сывороточный альбумин (BSA) или столбнячный анатоксин) для иммунизации.
ПОЛИКЛОНАЛЬНЫЕ АНТИТЕЛА
Поликлональные антитела против MASP-2 могут быть приготовлены посредством иммунизации животных полипептидом MASP-2 или его иммуногенным участком при использовании способов, хорошо известных специалисту в данной области техники. См., например, Green, et al., "Production of Polyclonal Antisera", в Immunochemical Protocols (Manson, ed.), страница 105, и, как описано далее в Примере 6. Иммуногенность полипептида MASP-2 может быть повышена при использовании адьюванта, включающего в себя минеральные гели, такие, как гидроксид алюминия или адъювант Фрейнда (полный или неполный), поверхностно активные вещества, такие, как лизолецитин, плюрониловые высокомолекулярные спирты, полианионы, масляные эмульсии, гемоцианин фиссуреллы и динитрофенол. Поликлональные антитела обычно получают у следующих животных: лошади, коровы, собаки, куры, крысы, мыши, кролики, морские свинки, козы или овцы. В качестве альтернативы, анти-MASP-2 антитело, используемое в настоящем изобретении, может также быть получено от человекообразной обезьяны. Общие основы техники получения диагностически и терапевтически полезных антител у бабуинов описываются, например, у Goldenberg et al., Международная публикация патента №WO 91/11465, и у Losman, M. J., et al., Int. J. Cancer 46:310, 1990. Сыворотка с содержанием иммунологически активных антител затем выделяется из крови таких иммунизированных животных с использованием стандартных общеизвестных процедур.
МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА
В некоторых вариантах осуществления агент, ингибирующий MASP-2, представлен анти-MASP-2 моноклональным антителом. Анти-MASP-2 моноклональное антитело является высокоспецифичным эпитопом, направленным против единичного эпитопа MASP-2. Как указано в данном описании, модификатор «моноклональное» указывает на характер антитела, которое было получено от существенно гомогенной популяции антител, и не подразумевает получение антител каким-либо особым способом. Моноклональные антитела могут быть получены с использованием любой техники, которая предусматривает получение молекул антител в культуре стабильной клеточной линий, каковой, например, является гибридома по способу, описанному у Kohler, G., et al., Nature 256:495, 1975, или они могут быть получены при помощи технологии рекомбинантных ДНК (см., например, патент США №4,816,567). Моноклональные антитела могут быть также выделены из библиотеки антител-бактериофагов при использовании технологий, описанных у Clackson, Т., et al., Nature 352:624-628, 1991, и Marks, J.D., et al., J. Mol. Biol. 222:581-597, 1991. Подобные антитела могут принадлежать любому классу иммуноглобулина, включая IgG, IgM, IgE, IgA, IgD и любому их подклассу.
Например, моноклональные антитела могут быть получены путем инъекций композиции, содержащей MASP-2 полипептид или его части, подходящим млекопитающим (например, мыши линии BALB/c). После заранее определенного периода времени, у мыши выделяют спленоциты и ресуспендируют их в культуральной среде. Затем осуществляют слияние спленоцитов с иммортализованной линией клеток для формирования гибридомы. Сформированные гибридомы выращивают в культуре клеток и проверяют на способность к продуцированию моноклональных антител к MASP-2. Дальнейшее описание получения анти-MASP-2 моноклональных антител предоставлено в Примере 7 (см. также Current Protocols in Immunology, Vol.1, John Wiley & Sons, pages 2.5.1-2.6.7,1991.)
Моноклональные антитела человека могут быть получены от трансгенной мыши, созданной при помощи генной инженерии для генерации человеческих антител в ответ на антигенный стимул. В данной технологии элементы локуса тяжелых и легких цепей иммуноглобулина человека вводятся в линии мышей, полученные из стволовых эмбриональных клеточных линий, в которых эндогенные локусы тяжелых и легких цепей иммуноглобулина были направлено разрушены. Трансгенные мыши могут синтезировать человеческие антитела, специфичные к человеческим антигенам, таким как антигены MASP-2, описанные в данном изобретении; также мыши могут использоваться для получения гибридом, секретирующих антитела к MASP-2 человека. Такие гибридомы могут быть получены путем слияния В-клеток таких животных с подходящими клеточными линиями миеломы с использованием обычной технологии Келера-Мильштейна, как описано ниже в Примере 7. Трансгенные мыши с геномом иммуноглобулина человека являются коммерчески доступными (например, от «Abgenix, Inc.», Фремонт, шт.Калифорния, и «Medarex, Inc.», Аннандейл, шт.Нью-Джерси). Способы получения человеческих антител от трансгенной мыши описаны в научной литературе, например Green, L.L, et al., Nature Genet, 7:13, 1994; Lonberg, N., et al., Nature 368:856, 1994; и Taylor, L.D., et al., Int. Immun. 6:579, 1994).
Моноклональные антитела могут быть выделены и очищены от культуры гибридом при помощи многочисленных хорошо известных технологий. Подобные технологии выделения включают в себя аффинную хроматографию посредством белка-А сефарозы, хроматографию по размеру молекул и ионообменную хроматографию (см., например, Coligan стр.2.7.1-2.7.12 и стр.2.9.1-2.9.3; Baines et al., "Purification of Immunoglobulin G (IgG)," в Methods in Molecular Biology, The Humana Press, Inc., Vol.10, p.79-104, 1992).
После получения поликлональные, моноклональные или антитела, произведенные фагами, подвергаются тестированию на специфическое связывание с MASP-2. Множество способов, известных специалисту в данной области техники, может быть использовано для определения антител, специфически связывающихся с MASP-2. В качестве примера таких способов можно назвать стандартные способы вестерн-блоттинга или иммунопреципитации (например, как описано у Ausubel et al.), иммуноэлектрофорез, иммуноферментный анализ (ИФА), дот-блотинг, метод ингибирования или конкурирования (как описано у Harlow и Land, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, 1988). После выявления специфической способности связываться с MASP-2, анти-MASP-2 антитела были протестированы на возможность функционировать в качестве ингибитора MASP-2 в одном из нескольких исследований, как, например, специфичное к пектину расщепление С4 (описано в Примере 2), исследование отложения C3b (описано в Примере 2) или исследование отложения C4b (описано в Примере 2).
Аффинность анти-MASP-2 моноклональных антител может быть легко определена специалистом в данной области техники при помощи стандартных процедур (см., например, Scatchard, A., NY Acad. Sci 51:660-672, 1949). В одном воплощении анти-MASP-2 моноклональные антитела, используемые для способов настоящего изобретения, связываются с MASP-2 со связующей аффинностью <100 нмоль, предпочтительно <10 нмоль и более предпочтительно <2 нмоль.
ХИМЕРНЫЕ/ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА
Моноклональные антитела, используемые в способе настоящего изобретения, включают в себя химерные антитела, в которых участок тяжелой и/или легкой цепи является идентичным или гомологичным соответствующим последовательностям в антителах, полученных из одних видов или принадлежащим к одному классу или подклассу антител, в то время, как остальная часть цепи (цепей) является идентичной или гомологичной соответствующим последовательностям антител, выделенных из других видов или принадлежащих другому классу или подклассу антител, равно как и фрагменты таких антител (патент США №4,816,567, выданные Cabilly, и Morrison, S. L, et al., Proc. Nat'l Acad. Sci USA 81:6851-6855, 1984).
Одна форма химерного антитела, используемого в данном изобретении, представлена гуманизированным моноклональным анти-MASP-2 антителом. Гуманизированные формы нечеловеческих (например, мышиных) антител представлены химерными антителами, которые содержат минимальную последовательность, выделенную из нечеловеческого иммуноглобулина. Гуманизированные моноклональные антитела продуцируются посредством передачи нечеловеческих (например, мышиных) определяющих комплементарность областей (CDR) из тяжелых и легких вариабельных цепей иммуноглобулина мыши в человеческий вариабельный домен. Характерно, что остатки человеческих антител затем замещаются в каркасных областях нечеловеческих прототипов. Более того, Гуманизированные антитела могут содержать остатки, которые не обнаруживаются в антителах рецепиента или в антителах донора. Данные модификации производятся с целью дальнейшей очистки полученных антител. В целом, гуманизированное антитело содержит практически все признаки как минимум одного, а обычно двух вариабельных доменов, в которых все или практически все признаки гипервариабельных петель соответствуют признакам петель нечеловеческого иммуноглобулина и все или практически все признаки каркасных областей Fv соответствуют признакам последовательности иммуноглобулина человека. Факультативно, гуманизированное антитело также содержит как минимум один участок константной области иммуноглобулина (Fc), типичной для человеческого иммуноглобулина. Детальное описание можно найти у Jones, Р.Т, et al., Nature 321:522-525, 1986; Reichmann, L., et al., Nature 332:323-329, 1988; и Presta, Curr. Op.Struct. Biol. 2:593-596, 1992.
Гуманизированные антитела, используемые в настоящем изобретении, включают в себя человеческие моноклональные антитела, содержащие как минимум MASP-2 связывающий CDR3 участок. Кроме того, области Fc могут быть заменены таким образом, чтобы продуцировать IgA или IgM также, как человеческие IgG антитела. Также Гуманизированные антитела будут обладать особой клинической полезностью, так как они будут специфическим образом распознавать MASP-2 человека, но не будут провоцировать иммунную реакцию у человека против самого антитела. Как следствие, они лучше подходят для введения человеку in vivo, особенно, когда введение носит повторяющийся характер или необходимо долгосрочное введение.
Пример получения гуманизированного анти-MASP-2 антитела из мышиного анти-MASP-2 моноклонального антитела приведен в Примере 10. Технологии для получения гуманизированных моноклональных антител также описаны, в следующих работах: Jones, Р.Т, et al., Nature 321:522, 1986; Carter, P., et al., Proc. NatI Acad. Sci USA 89:4285, 1992; Sandhu, J.S., Crit. Rev. Biotech. 12:437, 1992; Singer, I.I., et al., J. Immun. 150:2844, 1993; Sudhir (ed.), Antibody Engineering Protocols, Humana Press, Inc., 1995; Kelley, "Engineering Therapeutic Antibodies," в Protein Engineering: Principles and Practice, Cleland et al. (eds.), John Wiley & Sons, Inc., pages 399-434, 1996; и в патенте США №5,693,762, выданный Queen, 1997. Кроме того, существуют коммерческие организации, которые синтезируют гуманизированные антитела из специфических участков антитела, например, «Protein Design Labs» (Маунтин-Вью, шт.Калифорния).
РЕКОМБИНАНТНЫЕ АНТИТЕЛА
AHTH-MASP-2 антитела могут быть получены при помощи рекомбинантных способов. Например, человеческие антитела могут быть получены с использованием экспрессионной библиотеки человеческого иммуноглобулина (доступные, например, у «Stratagene, Corp.», Ла-Хойа, шт.Калифорния) для получения фрагментов человеческих антител (VH, VL, Fv, Fd, Fab or F(ab')2). Данные фрагмента затем используются для конструирования полноценных человеческих антител с использованием технологий, схожих с технологиями для получения химерных антител.
АНТИИДИОТИПИЧЕСКИЕ АНТИТЕЛА
После идентификации желаемой ингибирующей активности aHTH-MASP-2 антител, данные антитела могут использоваться для получения антиидиотипических антител, которые весьма схожи с частью MASP-2, при использовании технологий, которые хорошо известны науке. См., например, Greenspan, M.S., et al., FASEB J. 7:437, 1993. Например, антитела, которые связываются с MASP-2 и конкурентно ингибируют MASP-2 белковые взаимодействия, необходимые для активации комплемента, могут использоваться для получения антиидиотипических антител, схожих с MBL-связывающим участком белка MASP-2, и, как следствие, связывают и нейтрализуют связывание лигандов MASP-2, как, например, MBL.
ФРАГМЕНТЫ ИММУНОГЛОБУЛИНА
К числу агентов, ингибирующих MASP-2, которые используются в способе данного исследования, относятся не только интактные молекулы иммуноглобулина, но и хорошо известные фрагменты, включая Fab, Fab', F(ab)2, F(ab')2 и Fv фрагменты, scFv фрагменты, димеры, линейные антитела, одноцепочные молекулы антител и мультиспецифические антитела, сформированные из фрагментов антител.
Хорошо известно, что только небольшая область молекулы антитела, паратоп, вовлечена в связывание антитела с его эпитопом (см., например, dark, W.R., The Experimental Foundations of Modem Immunology, Wiley & Sons, Inc., NY, 1986). Участки pFc' и Fc антитела являются эффекторами классического пути активации комплемента, но не вовлекаются в связывание антигенов. Антитело, из которого при помощи ферментного расщепления был извлечен участок pFc', или которое было получено без участка pFc', обозначено как F(ab')2 фрагментом и содержит оба сайта связывания антигенов интактного антитела. Выделенный фрагмент F(ab')2 относится к бивалентному моноклональному фрагменту из-за наличия двух сайтов связывания антигенов. Похожим образом, антитело, из которого при помощи ферментного расщепления был извлечен участок Fc, или которое было получено без участка Fc, обозначено фрагментом Fab, и содержит один из сайтов связывания антигенов интактной молекулы антитела.
Фрагменты антител могут быть получены посредством протеолитического гидролиза, таким как пепсиновый гидролиз или папаиновый гидролиз цельного антитела стандартным способом. Например, фрагменты антитела могут быть получены ферментным расщеплением антител пепсином для получения фрагмента 5S, обозначенного F(ab')2. Данный фрагмент может далее быть расщеплен при использовании агента, восстанавливающего тиол, с целью получения 3,5S Fab' моновалентных фрагментов. Дополнительно, реакция расщепления может быть выполнена при использовании группы, блокирующей сульфгидрильные группы, которые образуются в результате расщепления дисульфидных связей. В качестве альтернативы, ферментное расщепление с использованием пепсина напрямую продуцирует два моновалентных фрагмента Fab и фрагмент Fc. Данные способы описаны, например, в патенте США №4,331,647, выданном Goldenberg; Nisonoff, A., et al., Arch. Biochem. Biophys. 89:230, 1960; Porter, R.R., Biochem. J. 73:119, 1959; Edelman, et al., в Methods in Enzymology 1:422, Academic Press, 1967; и у Coligan на стр.2.8.1-2.8.10 и 2.10.-2.10.4.
В некоторых вариантах осуществления использование фрагментов антител с отсутствующим участком Fc является предпочтительным для подавления активации классического пути комплемента, которая инициируется посредством связывания Fc с рецептором Fcγ. Существует несколько способов продуцирования MoAb с подавлением взаимодействия рецептора Fcγ. Например, участок Fc моноклонального антитела может быть удален химическим способом с использованием частичного гидролиза при помощи протеолитических ферментов (например, гидролиз фицином), вследствие чего происходит генерирование антиген-связующих фрагментов таких антител, как, например, Fab или F(ab)2 фрагменты (Mariani, M., et al., Mol. Immunol. 28:69-71, 1991). Альтернативно, изотип γ4 IgG человека, не связывающий рецепторы Fcγ, может использоваться в конструировании гуманизированного антитела, как описано в данной заявке. Антитела, одноцепочные антитела и антиген-связывающие домены, у которых отсутствует домен Fc также могут быть созданы с использованием описанных в данной заявке рекомбинантных технологий.
ФРАГМЕНТЫ ОДНОЦЕПОЧНЫХ АНТИТЕЛ
Альтернативно, возможно создание молекул связывания с единичной полипептидной цепью, специфичных к MASP-2, в которых тяжелая и легкая цепь участков Fv связаны между собой. Фрагменты Fv могут связываться пептидным линкером для формирования одноцепочного антиген-связывающего белка (scFv). Такие одноцепочные антиген-связывающие белки получаются посредством конструирования структурного гена, включающего в себя последовательности ДНК, кодирующие домены VH и VL, связанные с олигонуклеотидом. Структурный ген встраивают в экспрессионный вектор, который затем вводится в клетку-хозяина, как, например, кишечную палочку (Е. coli). Рекомбинантные клетки синтезируют одиночную полипептидную цепь с пептидным линкером, соединяющим два V домена. Способы для производства scFvs описаны, например, в Whitlow, et al., "Methods: A Companion to Methods in Enzymology" 2:97, 1991; Bird, et al., Science 242:423, 1988; патент США №4,946,778, выданном Ladner; Pack, P., et al., Bio/Technology 11:1271, 1993.
В качестве иллюстрирующего примера, MASP-2 специфичный scFv, может быть получен посредством экспозиции лимфоцитов с MASP-2 полипептидом in vitro и селекцией библиотек, представляющих антитела в фагах или схожих векторах (например, посредством использования иммобилизованных или меченых белков или пептидов MASP-2). Гены, кодирующие полипептиды, обладающие потенциальными доменами связывания полипептидов MASP-2, могут быть получены посредством тщательного скрининга пептидных библиотек в фагах или бактериях, как, например, в кишечной палочке (Е. Coli). Данные рандомизированные библиотеки пептидов могут быть использованы для скрининга пептидов, которые взаимодействуют с MASP-2. Технологии создания и скрининга подобных рандомизированные библиотек пептидов хорошо известны (патент США №.5,223,409, выданный Lardner; патент США №4,946,778, выданный Ladner; патент США №5,403,484, выданный Lardner; патент США №5,571,698, выданный Lardner; и Кау et al., Phage Display ofPeptides and Proteins Academic Press, Inc., 1996) и рандомизированные библиотеки пептидов и наборы для их скрининга являются коммерчески доступными, например он предоставляются «CLONTECH Laboratories, Inc.» (Пало-Альто, шт.Калифорния), «Invitrogen Inc.» (Сан-Диего, шт.Калифорния), «New England Biolabs, Inc.» (Беверли, шт.Массачусетс), и «Pharmacia LKB Biotechnology Inc.» (Пискатавэй, шт.Нью-Джерси).
Другой формой фрагмента анти-MASP-2 антитела, используемого в данном аспекте настоящего изобретения, является пептид, представляющий собой отдельный гипервариабельный участок в молекуле антителаза (CDR), который связывается с эпитопом на антигене MASP-2 и ингибирует MASP-2-зависимую активацию комплемента. CDR пептиды (минимальные единицы распознавания) могут быть получены посредством конструирования генов, кодирующих CDR представляющего интерес антитела. Подобные гены получают, например, с использованием полимеразной цепной реакции для синтеза вариабельного участка из РНК клетки, продуцирующей антитела (см., например, Larrick et al., Methods: A Companion to Methods in Enzymology 2:106, 1991; Courtenay-Luck, "Genetic Manipulation of Monoclonal Antibodies" в Monoclonal Antibodies: Production, Engineering and Clinical Application, Ritter et al. (eds.), page. 166, Cambridge University Press, 1995; и Ward et al., "Genetic Manipulation and Expression of Antibodies" в Monoclonal Antibodies: Principles and Applications, Birch et al. (eds.), page. 137, Wiley-Liss, Inc., 1995). Антитела MASP-2, описанные в данной заявке, вводится субъекту, который в этом нуждается, для ингибирования MASP-2-зависимой активации комплемента. В некоторых воплощениях агент, ингибирующий MASP-2, представлен человеческим или гуманизированным моноклональным анти-MASP-2 антителом высокой аффинности с редуцированной эффекторной функцией.
ПЕПТИДНЫЕ ИНГИБИТОРЫ
В некоторых вариантах осуществления данного аспекта настоящего изобретения, агент, ингибирующий MASP-2, включает в себя выделенные ингибиторы MASP-2 пептидной природы, в том числе выделенные природные пептидные ингибиторы и синтетические пептидные ингибиторы, подавляющие систему MASP-2-зависимой активации комплемента. Как указано в настоящем описании, термин «выделенные пептидные ингибиторы MASP-2» относится к пептидам, которые ингибируют MASP-2 зависимую активацию комплемента посредством связывания с другой распознаваемой молекулой, конкурирующей за связывание с MASP-2 (например, MBL, Н-фиколин, М-фиколин или L-фиколин) в лектиновом пути активации, и/или напрямую взаимодействуют с MASP-2 с целью подавления MASP-2-зависимой активации комплемента. Эти пептиды являются химически чистыми и в основном лишены примеси прочих субстанций, вместе с которыми данные пептиды обнаруживаются в естественных условиях, что определяет рамки их практического использования по назначению.
Были проведены опыты по успешному использованию ингибиторов пептидной природы in vivo с целью подавления каталитического участка и белок-белковых взаимодействий. Например, пептидные ингибиторы молекул адгезии с родственной LFA-1 структурой недавно были одобрены для клинического использования при коагулопатии (Ohman, E.M., et al., European Heart J. 16:50-55, 1995). Короткие линейные пептиды (<30 аминокислот) обладают способностью предупреждать или вмешиваться в интегрин-зависимую адгезию (Murayama, О., et al., J. Biochem. 120:445-51, 1996). Пептиды с большей протяженностью, длина которых колеблется от 25 до 200 аминокислотных остатков, также успешно использовались для блокирования интегрин-зависимой адгезии (Zhang, L, et al., J. Biol. Chem. 271(47):29953-57, 1996). В основном, пептиды с большей протяженностью обладают большей аффинностью и/или более низкими скоростями диссоциации, чем короткие пептиды, вследствие чего, могут стать потенциальными ингибиторами. Циклические пептидные ингибиторы также проявляют эффективность в отношении подавления интегринов in vivo для лечения воспалительных заболеваний человека (Jackson, D.Y., et al., J. Med. Chem. 40:3359-68, 1997). Один из способов производства циклических пептидов предусматривает синтез пептидов, в котором терминальные аминокислоты пептидов представлены цистеинами, что позволяет пептиду существовать в циклической форме за счет дисульфидной связи между терминальными аминокислотами, которая улучшает аффинность и период полувыведения in vivo при лечении гемопоэтических неоплазий (например, патент США №6,649,592, выданный Larson).
СИНТЕТИЧЕСКИЕ ПЕПТИДНЫЕ ИНГИБИТОРЫ MASP-2
Пептиды, ингибирующие MASP-2, используемые в способах данного аспекта настоящего изобретения, представлены аминокислотными последовательностями, которые имитируют участки-мишени, важные для функции MASP-2. Размеры синтетических пептидов-ингибиторов, используемых в способах настоящего изобретения, колеблются от 5 аминокислот до 300 аминокислот. В ТАБЛИЦЕ 3 представлен список типичных пептидных ингибиторов, которые могут быть использованы в данном аспекте настоящего изобретения. Кандидат в ингибиторы MASP-2 может быть протестирован на способность функционировать в качестве агента, ингибирующего MASP-2, при помощи одного из анализов, включая, например, анализ по расщеплению пектин-специфичного С4 (описано в Примере 2), и анализ отложения C3b (описано в Примере 2).
В некоторых вариантах осуществления пептиды, ингибирующие MASP-2, получены из полипептидов MASP-2 и выбраны методом селекции из полноразмерного зрелого белка MASP-2 (SEQ ID NO:6), или из специфического домена белка MASP-2, как, например, домен CUBI (SEQ ID NO:8), домен CUBIEGF (SEQ ID NO:9), домен EGF (SEQ ID NO:11), и домен сериновой протеазы (SEQ ID NO:12). Как описывалось ранее, была показана необходимость участков CUBEGFCUB II для димеризации и связывания с MBL (Thielens et al., supra). В частности, было показано, что пептидная последовательность TFRSDYN (SEQ ID NO:16) в CUBI домене MASP-2 принимает участие в связывании с MBL в исследовании, посвященном идентификации наличия у человека гомозиготной замены Asp105 на Gly105, приводящей к удалению MASP-2 из комплекса MBL (Stengaard-Pedersen, К., et al., New England J. Med. 349:554-560, 2003).
Пептиды, ингибирующие MASP-2, могут также быть получены из МАр19 (SEQ ID NO:3). Как описано в Примере 30, МАр19 (SEQ ID NO:3) (также обозначенный как sMAP), обладает способностью к понижающей регуляции пектинового пути, который активируется посредством комплекса MBL. Iwaki et al., J. Immunol. 177:8626-8632, 2006. He вдаваясь в подробности, можно предположить, что sMAP способен занимать MASP-2/sMAP сайт связывания в MBL и тем самым предотвращать связывания с ним MASP-2. Существуют также данные, что sMAP конкурирует с MASP-2 при образовании связей с фиколином А и ингибирует активацию комплемента при участии комплекса A/MASP-2 фиколина. Endo Y. et al., Immunogenetics 57: 837-844 (2005).
В некоторых воплощениях пептиды, ингибирующие MASP-2, получают из пектиновых белков, которые связываются с MASP-2 и вовлекаются в пектиновый путь активации комплемента. Было идентифицировано несколько различных пектинов, которые вовлечены в данный путь активации, среди них маннозосвязывающий пектин (MBL), L-фиколин, М-фиколин и Н-фиколин. (Ikeda, К., et al., J. Biol. Chem. 262:7451-7454, 1987; Matsushita, M, et al., J. Exp. Med. 176:1497-2284, 2000; Matsushita, M, et al., J. Immunol. 168:3502-3506, 2002). Данные лектины присутствуют в сыворотке как олигомеры гомотримерных субъединиц, каждая из которых обладает N-терминальными коллагеноподобными волокнами с доменами распознавания углеводов. Было показано, что различные лектины способны связываться с MASP-2, a лектиновый/MASP-2 комплекс активирует комплемент посредством расщепления белков С4 и С2. Н-фиколин имеет амино-терминальный участок, состоящий из 24 аминокислот, коллагено-подобный домен с 11 Gly-Xaa-Yaa повторами, шеечный домен из 12 аминокислот и фибриногенноподобный домен из 207 аминокислот (Matsushita, M., et al., J. Immunol. 168:3502-3506, 2002). H-фиколин связывается с GlcNAc и агглютинирует человеческие эритроциты, покрытые LPS, полученными из S. typhimurium, S. minnesota и E. coli. Было показано, что Н-фиколин образует связи с MASP-2 и МАр 19 и активирует лектиновый путь комплемента. Id. L-фиколин/Р35 также связывается GlcNAc и образует связи с MASP-2 и МАр 19 в человеческой сыворотке и данный комплекс проявляет способность к активации пектинового пути комплемента (Matsushita, M., et al., J. Immunol. 164:2281, 2000). Соответственно, пептиды, ингибирующие MASP-2 и используемые в настоящем изобретении, могут включать в себя участок, состоящий из как минимум 5 аминокислот, выбранных из белка MBL (SEQ ID NO:21), белка Н-фиколина (инвентарный номер Genbank NM_173452), белка М- фиколина (инвентарный номер Genbank 000602) и белка L- фиколина (инвентарный номер Genbank NM_015838).
Кроме того, учеными был идентифицирован сайт связывания MASP-2 в MBL с 12 Gly-X-Y триплетами «GKD GRD GTK GEK GEP GQG LRG LQG POG KLG POG NOG PSG SOG PKG QKG DOG KS» (SEQ ID NO:26), которые расположены между шарнирным и шеечным участками в С-терминальной области коллагеноподобного домена МБР (Wallis, R, et al., J. Biol. Chem. 279:14065, 2004). Аминокислотная последовательность сайта связывания MASP-2 является высококонсервативной также в Н-фиколине и L-фиколине человека. Было показано, что сайт связывания имеет консенсусную последовательность, которая является высоко консервативной у всех трех лектиновых белков, и которая состоит из следующих аминокислот: «OGK-X-GP» (SEQ ID NO:22), где буквой «О» обозначен гидроксипролин, а буквой «X» - гидрофобный остаток (Wallis et al., 2004, supra). Соответственно, в некоторых воплощениях пептиды, ингибирующие MASP-2 и используемые в данном аспекте настоящего изобретения, состоят, как минимум, из 6 аминокислот и содержат SEQ ID NO:22. В исследованиях in vitro была показана способность пептидов, производных MBL и содержащих аминокислотную последовательность «GLR GLQ GPO GKL GPO G» (SEQ ID NO:24), связываться с MASP-2 (Wallis, et al., 2004, supra). Для усиления связывания с MASP-2, возможен синтез пептидов с двумя триплетами GPO на каждом из концов («GPO GPO GLR GLQ GPO GKL GPO GGP OGP О» SEQ ID NO:25), что усиливает формирование тройных спиралей в нативном белке MBL (как описано у Wallis, R., et al., J. Biol. Chem. 279:14065, 2004).
Пептиды, ингибирующие MASP-2, также могут быть получены из Н-фиколина человека, содержащего последовательность «GAO GSO GEK GAO GPQ GPO GPO GKM GPK GEO GDO» (SEQ ID NO:27) из консенсусной области MASP-2 связывающего участка в Н-фиколине. Также это могут быть пептиды, полученные из L-фиколина человека, содержащего последовательность «GCO GLO GAO GDK GEA GTN GKR GER GPO GPO GKA GPO GPN GAO GEO» (SEQ ID NO:28), из консенсусной области MASP-2 связывающего участка в L-фиколине.
Пептиды, ингибирующие MASP-2, также могут быть получены из сайта расщепления С4, например, «LQRALEILPNRVTIKANRPFLVFI» (SEQ ID NO:29), который представляет собой сайт расщепления С4, связанный с С-терминальной областью антитромбина III (Glover, G. L, et al., Mol. Immunol. 25:1261 (1988)).
Пептиды, полученные из сайта расщепления С4, равно как и прочие пептиды, ингибирующие сайт MASP-2 сериновой протеазы, могут быть химически модифицированы таким образом, что они необратимым образом конвертируются в ингибиторы протеазы. Например, соответствующие модификации могут включать в себя, помимо прочего, галометил кетон (Br, Cl, I, F) на С-терминальном участке, Asp или Glu, или присоединенный к функциональным боковым цепям; галоацетиловые (или другие α-галоацетиловые) группы на аминогруппах или прочих функциональных боковых цепях; эпоксид или имин-содержащие группы на амино- или карбоксил-терминальном участке или функциональных боковых цепях; или имидат-сложные эфиры в на амино- или карбоксил-терминальных участках или функциональных боковых цепях. Подобные модификации предоставляют преимущество в перманентном ингибировании фермента посредством ковалентного прикрепления пептида. Это может выразиться в уменьшении эффективной дозы и/или снижении частоты администрирования пептидного ингибитора.
В добавление к пептидным ингибиторам, описанным выше, пептиды, ингибирующие MASP-2 и используемые в способе настоящего изобретения, включают в себя пептиды, содержащие MASP-2-связывающий CDR3 участок aHTH-MASP-2 MoAb, полученного вышеописанным способом. Необходимая для синтеза пептидов последовательность участков CDR может быть определена известными современной науке способами. Вариабельный участок тяжелой цепи представлен пептидом, длина которого обычно колеблется от 100 до 150 аминокислот. Вариабельный участок легкой цепи представлен пептидом, длина которого обычно колеблется от 80 до 130 аминокислот. Количество последовательности CDR внутри вариабельных участков тяжелой и легкой цепи колеблется от 3 до 25 аминокислотных последовательностей, и их можно легко определить при помощи стандартных способов, хорошо известных специалисту в данной области техники.
Специалистам в данной области техники известно, что описанные выше существенно гомологичные варианты пептидов, ингибирующих MASP-2, также проявляют активность по ингибированию MASP-2. Как пример подобных вариантов можно называть пептиды с инсерциями, делениями, замещениями и/или дополнительными аминокислотами в карбокси-терминальных или амино-терминальных областях заявленных пептидов и их смесях. Соответственно, данные гомологичные пептиды, обладающие активностью, подавляющей MASP-2, также могут быть использованы в способах данного изобретения. Описанные пептиды также могут включать в себя дуплицированные мотивы и прочие модификации с консервативными заменами. В данной заявке описаны консервативные варианты, которые включают в себя замену одной аминокислоты на другую кислоту с аналогичным зарядом, размером, гидрофильностью и прочими признаками.
Пептиды, ингибирующие MASP-2, могут быть модифицированы для повышения растворимости и/или максимизации положительного или отрицательного заряда для того, чтобы они больше походили на сегмент в интактном белке. Производное может обладать или не обладать точной первичной структурой аминокислотной последовательности пептида, раскрытого в данной заявке, но оно должно сохранять способность к ингибированию MASP-2. Модификации также могут включать в себя замещение аминокислоты одной из широко известных двадцати аминокислот или любой другой аминокислотой, изменение или замещение аминокислотой с дополнительными желаемыми характеристиками, такими как устойчивость к ферментативной деградации, или замещение D-аминокислотой, или замещение другой молекулой или ее составляющей, например молекулой углевода, которая имитирует природную конформацию и функции аминокислоты, аминокислот или пептида; разрушение аминокислоты; инсерцию аминокислоты одной из двадцати широко известных аминокислот или любой другой аминокислотой, производной или замещенной аминокислотой с вспомогательными характеристиками, такими как устойчивость к ферментативной деградации, или D-аминокислотой, или замещением другой молекулой или ее составляющей, например молекулой углеводорода, которая имитирует природную конформацию и функции аминокислоты, аминокислот или пептида; или замещение другой молекулой или ее составляющей, например молекулой углеводорода или мономер нуклеиновой кислоты, которые имитируют природную конформацию, распределение зарядов и функционирование пептида-прототипа. Пептиды также могут быть модифицированы посредством ацетилирования или амидирования.
Синтез производных пептидов-ингибиторов может быть основан на известных техниках пептидного биосинтеза, углеводного биосинтеза и т.п. В качестве отправной точки, технологи могут использовать подходящую компьютерную программу для определения конформации представляющего интерес пептида. После определения конформации пептида технолог может определить рациональную структуру для выявления типа необходимых замещений в одном или более сайтах производного, которое сохраняет базовую конформацию и распределение зарядов пептида-прототипа, но при этом может обладать характеристиками, которые у него отсутствуют или усиливать имеющиеся у пептида-прототипа свойства. Как только происходит идентификация производных молекул-кандидатов, эти молекулы могут быть протестированы для определения того, могут ли они функционировать в качестве агентов, ингибирующих MASP-2, используя анализы, описанные в данном изобретении.
СКРИНИНГ ИНГИБИРУЮЩИХ MASP-2 ПЕПТИДОВ
Для получения и скрининга пептидов, которые имитируют молекулярную структуру ключевых участков связывания MASP-2 и ингибируют MASP-2-зависимую активацию комплемента, применяются технологии молекулярного моделирования и рационального молекулярного дизайна. Молекулярные структуры, используемые для моделирования, включают в себя участки CDR анти-MASP-2 моноклональных антител, а также целевые области, известные своей важностью для функции MASP-2, включая участки, необходимые для димеризации, область, вовлеченную в связывание с MBL, и активный сайт сериновой протеазы, как описано выше. Науке хорошо известны способы для идентификации пептидов, которые связываются с определенными мишенями. Например, молекулярные импринтинг может использоваться для конформационного конструирования макромолекулярных структур, таких как пептиды, связывающиеся с определенными молекулами. См., например, Shea, KJ., «Molecular Imprinting of Synthetic Network Polymers: The De Novo synthesis of Macromolecular Binding and Catalytic Sites» TRIP 2:(5) 1994.
В качестве иллюстрации ниже приведен пример одного из способов получения имитаторов MASP-2-связывающих пептидов. Функциональные мономеры известного MASP-2-связывающего пептида или связывающего участка анти-MASP-2 антитела, которое ингибирует активность MASP-2 (образец) подвергают полимеризации. Затем образец удаляют, после чего следует полимеризация второго класса мономеров в пространстве, оставленном образцом, для создания новой молекулы, которая демонстрирует одно и более необходимых свойств, подобных свойствам образца. Кроме приготовления пептидов подобным способом, прочие MASP-2-связывающие молекулы, представленные агентами, ингибирующими MASP-2, например, полисахариды, нуклеозиды, лекарства, нуклеопротеины, липопротеины, углеводы, гликопротеины, стероиды, липиды и прочие биологически активные материалы, также могут быть приготовлены. Данный способ используется для создания большого многообразия биологических имитаторов, которые являются более стабильными, чем их природные прототипы, так как они приготавливаются посредством полимеризации функциональных мономеров свободными радикалами, образуя в результате сложный состав с неподверженным биохимическому разложению каркасом.
СИНТЕЗ ПЕПТИДОВ
Пептиды, ингибирующие MASP-2, могут быть приготовлены с использованием технологий, хорошо известных современной науке, как, например, техника твердофазного синтеза впервые описанная Merrifield, в J. Amer. Chem. Soc. 85:2149-2154, 1963. Можно применять автоматизированный синтез, используя прибор компании «Applied Biosystems» 431A Peptide Synthesizer (Фостер-Сити, шт.Калифорния) в соответствии с инструкциями от производителя. Прочие технологии описаны, например, у Bodanszky, M., et al., Peptide Synthesis, second edition, John Wiley & Sons, 1976, а также, в других работах, известных специалисту в данной области техники.
Пептиды также могут быть получены с использованием стандартных приемов генной инженерии, известных специалисту в данной области техники. Например, можно синтезировать пептид ферментативным способом, посредством инсерции нуклеиновой кислоты, кодирующей пептид в экспрессионный вектор, осуществления экспрессии ДНК и трансляции ДНК в пептид в присутствии необходимых аминокислот. Пептид затем очищают посредством хроматографии или электрофореза, или при помощи белка-переносчика, который может быть слит с целевым пептидом, а затем отщеплен от него путем встраивания последовательности нуклеиновой кислоты, кодирующей белок-носитель, в экспрессионный вектор, совпадающий по фазе с последовательностью, кодирующей пептид. Слитый белок-пептид может быть выделен при помощи хроматографических, электрофоретических или иммунологических технологий (такие как связывание смолой белка-переносчика через антитела). Пептид может быть расщеплен посредством химических методик или ферментативно, например, при помощи гидролазы.
Пептиды, ингибирующие MASP-2, используемые в данном способе настоящего изобретения, также могут быть получены в рекомбинантных клетках при помощи обычных технологий. Для экспрессии последовательностей, кодирующих пептиды, ингибирующие MASP-2, молекулы нуклеиновой кислоты, кодирующие пептид, должны быть операционно связаны с регуляторными последовательностями, контролирующими уровень транскрипции в экспрессионном векторе, а затем внедрены в клетку-хозяин. В добавление к транскрипционным регулирующим последовательностям, какими являются промоторы и энхансеры (участок ДНК вне гена, усиливающий экспрессию этого гена), векторы экспрессии могут включать в себя трансляционные регулирующие последовательности и маркерный ген, соответственно подходящие для отбора клеток, способных переносить векторы экспрессии.
Молекулы нуклеиновых кислот, кодирующие пептид-ингибитор MASP-2, могут быть синтезированы с использованием «генной машины» по таким протоколам, как, например, фосфорамидный способ. Если необходимо химически синтезировать двухцепочечную молекулу ДНК, например, в случае синтеза гена или его фрагмента, то каждая комплементарная цепь синтезируется отдельно. Получение коротких генов (от 60 до 80 пар оснований) быстро развивается с технической точки зрения и может выполнено посредством синтезирования комплементарных цепей и затем их гибридизации. Для получения более длинных генов, синтетические гены (двунитевые) собираются в модульную форму из одноцепочных фрагментов, которые имеют длину от 20 до 100 нуклеотидов. Обзор темы синтеза полинуклеотидов можно найти, например, у Glick and Pasternak, Molecular Biotechnology, Principles and Applications of Recombinant DNA, ASM Press, 1994; Itakura, K., et al., Annu. Rev. Biochem. 53:323, 1984; и Climie, S., et al., Proc. Nat'l Acad. Sci USA 87:633, 1990.
НИЗКОМОЛЕКУЛЯРНЫЕ ИНГИБИТОРЫ
В некоторых вариантах осуществления данного изобретения агенты, ингибирующие MASP-2, представлены низкомолекулярными ингибиторами, включая природные и синтетические субстанции, обладающие низким молекулярным весом, как, например, пептиды, пептидомиметики и непептидные ингибиторы (включая олигонуклеотиды и органические вещества). Низкомолекулярные ингибиторы MASP-2 могут быть созданы на основе молекулярной структуры вариабельных участков анти-MASP-2 антител.
Низкомолекулярные ингибиторы могут быть смоделированы и созданы на основе кристаллической структуры MASP-2 с использованием программ для компьютерного моделирования лекарственных средств (Kuntz I.D., et al., Science 257:1078, 1992). Была описана кристаллическая структура MASP-2 крысы (Feinberg, H., et al., EMBO J. 22:2348-2359, 2003). Используя способы, описанные у Kuntz et al., координаты кристаллической структуры MASP-2 вводятся в компьютерную программу, например DOCK, которая на выходе выдает список низкомолекулярных структур с ожидаемой способностью связывания с MASP-2. Использование подобных компьютерных программ хорошо известно для специалиста в данной области техники. Например, кристаллическая структура ингибитора протеазы HIV-1 была использована для идентификации уникальных непептидных лигандов, которые являются ингибиторами протеазы HIV-1, путем определения совместимости веществ, из базы данных Cambridge Crystallographic, с сайтом связывания фермента с использованием программы DOCK (Kuntz, LD., et al., J. Mol. Biol. W:269-288, 1982; DesJarlais, R.L, et al., PNAS 87:6644-6648, 1990).
Список низкомолекулярных структур, идентифицированных при помощи компьютерных программ как потенциальные ингибиторы MASP-2, подвергли скринингу в исследовании MASP-2 связывания, как описано в Примере 7. Малые молекулы, проявляющие способность к связыванию с MASP-2, затем были опробованы в функциональном исследовании, как описано в Примере 2, с целью определения их способности ингибировать MASP-2-зависимую активацию комплемента.
РАСТВОРИМЫЕ РЕЦЕПТОРЫ MASP-2
Считается, что подходящие агенты, ингибирующие MASP-2, включают в себя растворимые рецепторы MASP-2, которые могут быть получены при использовании технологий, хорошо известных специалисту в данной области техники.
ИНГИБИТОРЫ ЭКСПРЕССИИ MASP-2
В другом варианте осуществления данного аспекта настоящего изобретения агент, ингибирующий MASP-2, представлен ингибитором экспрессии MASP-2, способным ингибировать MASP-2-зависимую активацию комплемента. В практическом варианте осуществления данного аспекта настоящего изобретения характерные ингибиторы экспрессии MASP-2 включают в себя молекулы антисмысловой нуклеиновой кислоты MASP-2 (такие как антисмысловая мРНК, антисмысловая ДНК или антисмысловые олигонуклеотиды), MASP-2 рибозимы и MASP-2 молекулы РНК-интерференции.
Антисмысловые РНК и ДНК молекулы напрямую блокируют трансляцию мРНК MASP-2 посредством гибридизации с мРНК MASP-2 и предупреждения трансляции белка MASP-2. Молекула антисмысловой нуклеиновой кислоты может быть сконструирована посредством различных способов, и она обладает способностью препятствовать экспрессии MASP-2. Например, молекула антисмысловой нуклеиновой кислоты может быть сконструирована посредством инвертирования кодирующего участка (или его части) кДНК MASP-2 (SEQ ID NO:4) относительно его нормальной ориентации при транскрипции, что позволяет осуществить транскрипцию его комплемента.
Обычно молекула антисмысловой нуклеиновой кислоты является идентичной как минимум части целевого гена или генов. Однако для того, чтобы ингибировать экспрессию целевого гена нуклеиновая кислота не обязательно должна быть абсолютно идентичной его нуклеотидной последовательности. Как правило, чем короче последовательность антисмысловой нуклеиновой кислоты, тем более высокой гомологией она должна обладать. Минимальный процент идентичности обычно составляет не менее 65%, однако чем этот процент выше, тем эффективней осуществляется подавление экспрессии эндогенной последовательности. Процент идентичности, существенно превышающий 80%, является предпочтительным, хотя значения в интервале от 95% до полной идентичности являются самым предпочтительным.
Молекула антисмысловой нуклеиновой кислоты может не иметь той же интронной или экзонной структуры, что и целевой ген, а некодирующие сегменты целевого гена могут быть также эффективны в достижении антисмыслового подавления экспрессии целевого гена, как и кодирующие сегменты. Последовательность ДНК из 8 или более нуклеотидов может быть использована в качестве молекулы антисмысловой нуклеиновой кислоты, хотя предпочтительнее является более длинная последовательность. В настоящем изобретении типичным примером агента, ингибирующего MASP-2, является молекула антисмысловой нуклеиновой кислоты MASP-2, которая как минимум на 90% идентична комплементарной кДНК MASP-2, состоящей из последовательности нуклеиновой кислоты, представленной в SEQ ID NO:4. Последовательность нуклеиновой кислоты, представленной в SEQ ID NO:4, кодирует белок MASP-2, состоящий из последовательности аминокислоты, представленной в SEQ ID NO:5.
Нацеливание антисмысловых олигонуклеотидов на связывание с мРНК MASP-2 представляет собой другой механизм, который может быть использован для понижения уровня синтеза белка MASP-2. Например, синтез полигалактуроназы и ацетилхолинового мускаринового рецептора второго типа ингибируется антисмысловыми олигонуклеотидами, направленными на соответствующие последовательности мРНК (патент США №5,739,119, выданный Cheng, и патент США №5,759,829, выданный Shew/maker). Кроме того, была показана возможность антисмыслового ингибирования для ядерного белка циклина, гена множественной лекарственной устойчивости (MDG1), ICAM-1, Е-селектина, STK-1, стриарного рецептора GABAA и EGF человека (см., например, патент США №5,801,154, выданный Baracchini; патент США №5,789,573, выданный Baker; патент США №5,718,709, выданный Considine; и патент США №5,610,288, выданный Reubenstein).
Была описана система, позволяющая специалисту в данной области техники определить, какие олигонуклеотиды могут быть использованы для целей настоящего изобретения. Эта система предусматривает зондирование подходящих сайтов в целевой мРНК с использованием расщепления РНКазой Н как индикатора доступности последовательностей, внутри транскрипта. Scherr, M., et al., Nucleic Acids Res. 26:5079-5085, 1998; Lloyd, et al., Nucleic Acids Res. 29:3665-3673, 2001. Смесь антисмысловых олигонуклеотидов, являющихся комплементарными по отношению к определенным участкам транскрипта MASP-2, добавляется в клеточные экстракты, экспрессирующие MASP-2, например, гепатоцитам, и подвергаются гибридизации для создания восприимчивых сайтов для РНКазы Н. Данный способ может быть скомбинирован с выбором последовательностей при помощи компьютерных технологий, что определяет оптимальную селекцию последовательностей для антисмысловых композиций, базирующихся на их относительной способности формировать димеры, шпильки или другие вторичные структуры, которые препятствуют специфическому связыванию с целевой мРНК в клетке-хозяине или редуцируют его. Анализ данных вторичных структур и выбор целевых сайтов могут быть выполнены посредством анализа праймера при помощи компьютерной программы OLIGO (Rychlik, I., 1997) и компьютерного алгоритма BLASTN 2.0.5 (Altschul, S.F, et al., Nucl. Acids Res. 25:3389-3402, 1997). Антисмысловые соединения, направленные на целевую последовательность, предпочтительно содержат приблизительно от 8 до 50 нуклеотидов. Антисмысловые олигонуклеотиды, содержащие в среднем от 9 до 35 нуклеотидов, являются особенно предпочтительными. Авторы изобретения рассматривают все олигонуклеотидные композиции в ряду от 9 до 35 нуклеотидов (т.е., состоящие из 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34 или 35 или более в длину) как наиболее предпочтительные для практического употребления в антисмысловых олигонуклеотидных способах настоящего изобретения. Наиболее предпочтительными целевыми участками мРНК MASP-2 являются те, которые расположены в непосредственной близости от инициирующего кодона трансляции AUG, и те последовательности, которые существенно комплементарны участку 5' мРНК, например, между -10 и +10 участками нуклеотидной последовательности гена MASP-2 (SEQ ID NO:4). Примеры ингибиторов экспрессии MASP-2 представлены в ТАБЛИЦЕ 4.
Как отмечено выше, термин «олигонуклеотид», используемый в настоящем изобретении, относится к любому олигомеру или полимеру рибонуклеиновой кислоты (РНК) или дезоксирибонуклеиновой кислоты (ДНК) или их имитаторам. Под данным термином также подразумевают олигонуклетидные основания, состоящие в основном из существующих в природе нуклеотидов, Сахаров и ковалентных межнуклеозидных (структурных) связей, также как и олигонуклеотидов, подвергшихся синтетически сформированным модификациям. Такие модификации позволяют вводить определенные желательные качества, которыми не обладают натуральные олигонуклеотиды, как, например, уменьшение токсических свойств, повышение устойчивости к нуклеазной деградации и повышение уровня поглощения клетками. В наглядных вариантах осуществления антисмысловые соединения данного изобретения отличаются от натуральной ДНК вследствие модификации структуры фосфодиэфирного остова с целью продлить жизненный цикл антисмыслового олигонуклеотида, при которой фосфатные заместители заменены на фосфоротиоатные. Подобным образом, один либо оба конца олигонуклеотида могут быть замещены одним или более акридиновыми производными, которые интеркалируют между соседними парами нуклеотидов в цепи нуклеиновой кислоты.
Другой альтернативой использования антисмысловых элементов является использование «РНК интерференции» (РНКи). Двуцепочные РНК (дсРНК) могут вызывать подавление транскрипции гена у млекопитающих in vivo. Природная функция РНКи и косупрессии проявляется в защите генома от инвазии мобильными генетическими элементами, такими как ретротранспозоны и вирусы, продуцирующие аберрантные РНК или дсДНК в клетке при их активации (см., например, Jensen, J., et al., Nat. Genet. 21:209-12, 1999). Молекула двуцепочной РНК может быть получена в процессе синтеза двух цепей РНК, обладающих способностью формировать молекулы двуцепочной РНК, каждая из которых в длину равна приблизительно от 19 до 25 (например, 19-23 нуклеотидов). Например, молекула дсРНК, используемая в способах настоящего исследования, может включать в себя РНК, относящуюся к последовательности и ее комплементу, приведенной в ТАБЛИЦЕ 4. Предпочтительно, по меньшей мере одна цепь РНК обладает 3' «липким» концом от 1-5 нуклеотидов. Синтезированные цепи РНК скомбинированы в условиях, аналогичных для формирования двухцепочной молекулы. Как минимум, последовательность РНК может включать в себя восьминуклеотидную часть из последовательности SEQ ID NO:4 с общей длиной в 25 нуклеотидов или менее. Конструирование последовательностей малой интерферирующей РНК (миРНК) для заданной цели хорошо известно специалисту в данной области техники. Существуют коммерческие организации, занимающиеся конструированием последовательностей миРНК и гарантирующие нокдаун экспрессии с минимальной эффективностью в 70% («Qiagen», Валенсия, шт.Калифорния).
дсРНК может быть введена в форме фармацевтической композиции, когда введение осуществляется известными способами, при которых нуклеиновая кислота вводится в желаемую клетку-мишень. Общеизвестные способы использования генного трансферта включают в себя фосфат кальция, DEAE-декстран, электропорацию, микроинъекции и вирусные способы. Подобные способы изучены и описаны у Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., 1993.
Рибозимы также могут быть использованы для сокращения объема и/или биологической активности MASP-2, например, такие рибозимы, которые нацеливаются на мРНК MASP-2. Рибозимы представляют собой каталитические молекулы РНК, способные расщеплять молекулы нуклеиновой кислоты с последовательностью, полностью или частично гомологичной последовательности рибозима. Существует возможность конструирования рибозимных трансгенов, кодирующих рибозимы РНК, которые образуют специфические пары с целевой РНК и расщепляют фосфодиэфирные связи в специфическом участке, вследствие чего происходит функциональная инактивация целевой РНК. При проведении подобного расщепления сам рибозим не подвергается видоизменению, сохраняя, таким образом, способность к повторению цикла и расщеплению других молекул. Включение рибозимных последовательностей в антисмысловые РНК обуславливает активность расщепления РНК, вследствие чего повышается активность антисмысловых конструкций.
Используемые в практических вариантах осуществления настоящего изобретения рибозимы обычно включают в себя гибридизующийся участок по меньшей мере из девяти нуклеотидов, который является комплементарным по нуклеотидной последовательности как минимум части целевой мРНК MASP-2, и каталитическую область, которая способна к расщеплению целевой мРНК MASP-2, (основные сведения в ЕРА No.0321201; WO 88/04300; Haseloff, J., et al., Nature 334:585-591, 1988; Fedor, M.J, et al., Proc. Natl. Acad. Sci USA 87:1668-1672, 1990; Cech, T.R., et al., Ann. Rev. Biochem. 55:599-629, 1986).
Рибозимы также могут быть прямым образом нацелены на клетки в форме олигонуклеотидов РНК, инкорпорирующих последовательности РНК или внедряющихся в клетку как вектор экспрессии, кодирующий желаемую рибозимальную РНК. Рибозимы могут использоваться посредством того же способа, который описан для антисмысловых полинуклеотидов.
Антисмысловые РНК и ДНК, рибозимы и молекулы РНКи, используемые в способах настоящего изобретения, могут быть синтезированы посредством любого метода, известного современной науке в данной области техники для синтеза молекул ДНК и РНК. К ним относятся технологии химического синтеза олигодезоксирибонуклеотидов и олигорибонуклеотидов, широко известные в науке, такие как, твердофазный фосфорамидный химический синтез. Альтернативно, молекулы РНК могут быть получены посредством транскрипции in vitro и in vivo последовательностей ДНК, кодирующих молекулу антисмысловой РНК. Подобные последовательности ДНК могут быть инкорпорированы в широкий спектр векторов, которые содержат подходящие промоторы полимеразы РНК, такие как Т7 или SP6 промоторы полимеразы. Альтернативно, конструкции антисмысловой кДНК, синтезирующие антисмысловую РНК конституитивным или индуктивным способом, в зависимости от используемого промотора, могут устойчиво вводиться в клеточные линии.
Различные хорошо известные модификации молекул ДНК могут представлять собой средство для повышения стабильности и периода полувыведения. Пригодные для практического применения модификации включают, помимо прочего, добавление фланкирующих последовательностей рибонуклеотидов или дезоксирибонуклеотидов к 5' и/или 3' концам молекулы или использование фосфоротиоата (phosphorothioate) или 2' O-метила вместо фосфодиэфирных связей с олигодезоксирибонуклеотидным каркасом.
V. ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ И СПОСОБЫ ДОСТАВКИ ДОЗИРОВКА
В другом аспекте настоящее изобретение предлагает композиции для ингибирования побочных эффектов MASP-2-зависимой активации комплемента. Такие композиции включают в себя терапевтически эффективный объем агента, ингибирующего MASP-2, и фармацевтически приемлемый носитель. Агенты, ингибирующие MASP-2, могут вводиться субъекту, который в этом нуждается, в терапевтически эффективных дозах для лечения или улучшения состояний, вызванных MASP-2-зависимой аткивацией комплемента. Под терапевтически эффективными дозами подразумевается объем агента, ингбирующего MASP-2, достаточный для улучшения состояния и уменьшения симптомов заболевания.
Токсичность и терапевтическая эффективность агентов, ингибирующих MASP-2, может быть определена посредством стандартных фармацевтических процедур с использованием экспериментальных животных моделей, таких как мышиная модель MASP-2-/-, экспрессирующая человеческий трансген MASP-2, как описано в Примере 3. Используя подобные животные модели, NOAEL (уровень отсутствия наблюдаемых нежелательных эффектов) и MED (минимальная эффективная доза) могут быть определены стандартными способами. Дозовое соотношение между NOAEL и MED эффектами является терапевтическим соотношением, которое выражается как соотношение NOAEL/MED. Агенты, ингибирующие MASP-2, имеющие значительные терапевтические соотношения или показатели, являются самыми предпочтительными. Данные, полученные в результате изучения клеточных культур и исследований на животных, могут быть использованы для формулирования диапазона доз для человека. Дозирование агента, ингибирующего MASP-2, предпочтительным образом связано с концентрацией агента в кровотоке, которая образована MED с минимальным уровнем или полным отсутствием токсичности. Дозировка может варьироваться внутри данного ряда в зависимости от формы дозирования или используемого типа введения.
Для составления любой композиции терапевтически эффективная доза может быть опробована на животной модели. Например, доза может быть определена на животной модели для достижения такого ряда концентрации в системе циркуляции плазмы, который включает в себя MED. Количественные уровни агента, ингибирующего MASP-2, в плазме также могут быть замерены, например, посредством высокоэффективной жидкостной хроматографии.
Помимо исследований токсичности эффективность дозы также может быть оценена по количеству MASP-2 белка в живом субъекте и аффинности связывания агента, ингибирующего MASP-2. Было показано, что MASP-2 у здоровых людей присутствует в сыворотке в небольших количествах на границе 500 нг/мл, а уровни MASP-2 у определенных субъектов могут быть определены при помощи количественного анализа на MASP-2, как описано в Moller-Kristensen M., et al., J. Immunol. Methods 282:159-167, 2003.
Обычно, дозировка введенных композиций, содержащих агенты, ингибирующие MASP-2, варьируется в зависимости от таких факторов, как возраст субъекта, его вес, рост, пол, основные медицинские состояния и предшествующая медицинская история. В качестве иллюстрации, агенты, ингибирующие MASP-2, такие как aHTH-MASP-2 антитела, могут быть введены в уровнях дозировки от приблизительно 0,010 до 10,0 мг/кг, предпочтительно, 0,010 до 1,0 мг/кг, более предпочтительно 0,010 до 0,1 мг/кг веса тела субъекта. В некоторых вариантах осуществления композиция включает в себя комбинацию aHTH-MASP-2 антител и пептидов, ингибирующих MASP-2.
Терапевтическая эффективность композиций, ингибирующих MASP-2, и способов настоящего изобретения для данного субъекта, а также подходящей дозировки может быть определена в соответствии при помощи исследований комплемента, хорошо известных специалисту в данной области техники. Комплемент генерирует многочисленные специфические продукты. За последние десять лет были разработаны точные и специфичные способы анализа, которые являются коммерчески доступными для большинства из данных продуктов активации, включая малые фрагменты активации С3а, С4а и С5а и большие фрагменты активации iC3b, C4d, Bb и sC5b-9. В большинстве этих способов анализа использовались моноклональные антитела, которые реагируют на новые антигены (неоантигены), расположенные на этих фрагментах, но не на естественных белках, из которых они образовались, что делает данные способы анализа весьма доступными и специфичными. В большинстве случаев используют технологию ИФА, хотя радиоиммунологический анализ до сих пор иногда используется для С3а и С5а. Последний способ используют для измерения уровня непроцессированных фрагментов и их фрагменты 'desArg', которые являются основными формами, обнаруживаемыми в кровяном русле. Непроцессированные фрагменты и C5adesArg быстро выводятся посредством связывания с рецепторами клеточной поверхности и, следовательно, присутствуют в очень низких концентрациях, тогда как C3adesArg не связывается с клетками и аккумулируется в плазме. Измерение С3а представляет собой точный, не зависимый от пути активации комплемента индикатор активации комплемента. Альтернативный путь активации комплемента может быть определен при помощи измерения фрагмента Bb. Детекция растворимых продуктов мембраноатакующего пути активации, sC5b-9, представляет доказательства того, что комплемент активируется полностью. Так как оба, пектиновый и классический, пути активации генерируют одни и те же продукты активации, С4а и C4d, измерение данных двух фрагментов не предоставляет никакой информации о том, какой из данных двух путей активации комплемента сгенерировал продукты активации.
ДОПОЛНИТЕЛЬНЫЕ АГЕНТЫ
Композиции и способы, включающие в себя агенты, ингибирующие MASP-2, могут в качестве дополнения содержать один или более добавочных терапевтических агентов, которые могут повышать активность агента, ингибирующего MASP-2, или выполнять родственные терапевтические функции аддитивным или синергистическим способом. Например, один или более агентов-ингибиторов MASP-2 могут быть администрированы в комбинации с одним или более противовоспалительным и/или анальгетическим агентами. Включение и выбор дополнительного агента(-ов) определяется с целью достижения желаемого терапевтического эффекта. Подходящими противовоспалительным и/или анальгетическим агентами являются: антагонисты серотонинового рецептора; агонисты серотонинового рецептора; антагонисты гистаминового рецептора; антагонисты брадикининового рецептора; ингибиторы калликреина; антагонисты тахикининового рецептора, включая антагонисты подтипа рецепторов нейрокинина1 и нейрокинина2; антагонисты рецептора ген-опосредованного кальцитонинового пептида (CGRP); антагонисты интерлейкинового рецептора; ингибиторы ферментов, активных при синтетическом пути активации для метаболитов арахидоновой кислоты, включая ингибиторы фосфолипазы, включая ингибиторы изоформы PLA2 и ингибиторы изоформы PLCγ, ингибиторы циклооксигеназы (СОХ) (которые могут быть представлены либо СОХ-1, СОХ-2, либо неселективными ингибиторами СОХ-1 и -2), ингибиторы липооксигеназы; антагонисты простаноидных рецепторов, включая антагонисты подтипа рецепторов эйкозаноидов ЕР-1 и ЕР-4 и антагонисты подтипа рецепторов тромбоксана; антагонисты рецептора лейкотриена, включая антагонисты подтипа рецепторов лейкотриена В4 и лейкотриена D4; агонисты опиоидного рецептора, включая агонисты подтипа рецепторов µ-опиоида, δ-опиоида и κ-опиоида; агонисты пуринергического рецептора и антагонисты, включающие в себя антагонисты рецептора P2X и агонисты рецептора P2Y; аденозин трифосфат (АТФ)-чувствительные открыватели калиевого канала; ингибиторы MAP киназы; ингибиторы холиномиметического ацетилхолина; и агонисты альфа-адренергического рецептора (включая альфа-1, альфа-2 и неселективные альфа-1 и 2 агонисты).
При использовании с целью предупреждения или лечения рестеноза, агент, ингибирующий MASP-2, по настоящему изобретению, может быть скомбинирован с одним или более анти-рестенозными агентами для сопутствующего введения. Подходящими анти-рестенозными агентами являются: антитромбоцитарные агенты, включая: ингибиторы тромбина и рецепторные антагонисты, антагонисты рецептора аденозиндифосфата АДФ (ADP) (также известные как антагонисты пуринергического рецептора1, ингибиторы тромбоксана и антагонисты рецептора и антагонисты рецептора гликопротеина оболочки тромбоцитов; ингибиторы молекул клеточной адгезии, включая ингибиторы селектина и интегрина; антихемотаксические агенты; антагонисты интерлейкинового рецептора; и ингибиторы внутриклеточного сигнала, включая: ингибиторы протеинкиназы С (РКС) и протеин-тирозинфосфатазы, модуляторы ингибиторов внутриклеточной протеин-тирозинкиназы, ингибиторы доменов src гомолога2 (SH2), и антагонисты кальциевых каналов.
Агенты, ингибирующие MASP-2, по настоящему изобретению также могут быть введены в комбинации с одним и более другими ингибиторами комплемента. В настоящее время не существует ингибиторов комплемента, одобренных для использования с целью лечения человека, однако некоторые фармакологические агенты продемонстрировали in vivo способность к блокированию комплемента. Многие из данных агентов также токсичны или являются только частичными ингибиторами (Asghar, S.S., Pharmacol. Rev. 36:223-44, 1984), и их использование было ограничено до использования только в качестве инструментов исследования. К76СООН и нафамстат мезилат представляют собой два агента, которые показали определенную эффективность в моделях трансплантации на животных (Miyagawa, S., et al., Transplant Proc. 24:483-484, 1992). Гепарины с низким молекулярным весом также обнаружили эффективность в регулировании активности комплемента (Edens, R.E., et al., Complement Today, pp.96-120, Basel: Karger, 1993). Считается, что данные низкомолекулярные ингибиторы могут быть полезны в качестве агентов для использования в комбинации с агентами, ингибирующими MASP-2, по настоящему изобретению.
Другие существующие в природе ингибиторы комплемента могут быть использованы в комбинации с агентами, ингибирующими MASP-2, по настоящему изобретению. Биологические ингибиторы комплемента включают в себя растворимый фактор комплемента 1 (sCR1). Это полученный натуральным образом ингибитор, который обнаруживается в наружной мембране человеческих клеток. Прочие мембранные ингибиторы включают в себя мембранные ингибиторы DAF, МСР и CD59. Рекомбинантные формы были протестированы на их антикомплементарную активность in vitro и in vivo. sCR1 проявил эффективность при ксенотрансплантации, в которой система комплемента (как альтернативная, так и классическая) является инициатором синдрома гиперактивного отторжения во время перфузии крови в недавно пересаженный орган (Platt J.L, et al., Immunol. Today 11:450-6, 1990; Marino I.R., et al., Transplant Proc. 1071:6, 1990; Johnstone, P.S., et al., Transplantation 54:573-6, 1992). Использование sCR1 защищает и увеличивает время жизнеспособности трансплантированного органа, вовлекая путь активации комплемента в патогенез выживания органа (Leventhal, J. R. et al., Transplantation 55:857-66, 1993; Pruitt, S.K., et al., Transplantation 57:363-70, 1994).
Подходящие дополнительные ингибиторы комплемента для использования в комбинации с композициями по настоящему изобретению также включают в себя, например, MoAbs подобные тем, что были разработаны компанией «Alexion Pharmaceuticals, Inc.», Нью-Хейвен, шт.Коннектикут, и анти-пропердин MoAbs.
Для использования с целью лечения артрита (например, остеоартрит и ревматоидный артрит), агент, ингибирующий MASP-2, по настоящему изобретению, может быть скомбинирован с одним или более хондропротекторными агентами, которые могут включать в себя один или более промоторов хрящевого анаболизма и/или один или более ингибиторов хрящевого катаболизма, и соответственно оба - анаболический агент и катаболический ингибитор - используются для сопутствующего введения. Подходящими анаболическими агентами, обладающими хондропротекторной функцией, являются агонисты рецептора интерлейкина (IL), включая IL-4, IL-10, IL-13, rhIL-4, rhIL-10 и rhIL-13, и химерные IL-4, IL-10, or IL-13; агонисты суперсемейства трансформирующего фактора роста-β, включая TGF-β, TGF-β1, TGF-β2, TGF-β3, структурные морфогенные белки, включая BMP-2, BMP-4, BMP-5, BMP-6, BMP-7 (OP1I) и ОР-2/ВМР-8, факторы дифференциального роста, включая GDF-5, GDF-6 и GDF-7, рекомбинантные TGF-P и BMP, и химерные TGF-p и BMP; инсулиноподобные факторы роста, включая IGF-I; и факторы роста фибробласта, включая bFGF. Подходящими катаболическими хондропротекторными агентами-ингибиторами являются антагонисты рецептора интерлейкина-1 (IL-1) (IL-1ra), включая растворимые человеческие рецепторы IL-1 (shuIL-1R), rshuIL-1R, rhIL-1ra, анти-IL1-антитела, AF11567 и AF12198; антагонисты рецептора фактора некроза опухолей (TNF) (TNF-α), включая растворимые рецепторы, включая STNFR1 и sTNFRII, рекомбинантные растворимые рецепторы TNF, и химерные растворимые рецепторы TNF, включая химерные rhTNFR:Fc, слитые растворимые рецепторы Fc и анти-TNF антитела; ингибиторы циклооксигеназы-2 (специфичный СОХ-2), включая DuP 697, SC-58451, целекоксиб, рофекоксиб, нимезулид, диклофенак, мелоксикам, пироксикам, NS-398, RS-57067, SC-57666, SC-58125, флозулид, этодолак, L-745,337 и DFU-T-614; ингибиторы митоген-активированной протеинкиназы (МАРК), включая ингибиторы ERK1, ERK2, SAPK1, SAPK2a, SAPK2b, SAPK2d, SAPK3, включая SB 203580, SB 203580 iodo, SB202190, SB 242235, SB 220025, RWJ 67657, RWJ 68354, FR 133605, L-167307, PD 98059, PD 169316; ингибиторы ядерного фактора каппа В (NFkB), включая фенилэтиловый эфир кофеиновой кислоты (CAPE), DM-CAPE, SN-50 пептид, гимениальдизин и пиролидон дитиокарбамат; ингибиторы синтеза окисида азота (NOS), включая NG-монометил-L-аргинин, 1400W, дифениленйодиум, S-метил изотиомочевига, S-(аминоэтил) изотиомочевина, L-N6-(1-иминоэтил)лизин, 1,3-PBITU, 2-этил-2-тиопсевдомочевина, аминогуанидин, Nω-нитро-L-аргинин, и Nω-нитро-L-аргинин метилэфир, ингибиторы металлопротеиназ матрицы (MMPs), включая ММР-1, ММР-2, ММР-3, ММР-7, ММР-8, ММР-9, ММР-10, ММР-11, ММР-12, ММР-13, ММР-4 и ММР-15, и включая U-24522, миноциклин, 4-Abz-Gly-Pro-D-Leu-D-Ala-NHOH, Ас-Arg-Cys-Gly-Val-Pro-Asp-NH2, руман TIMP1, руман TIMP2, и фосфорамидон; молекулы клеточной адгезии, включая агонисты и антагонисты интегрина, включая aVp3 MoAb LM 609 и эхистатин; анти-хемотаксические агенты, включая рецепторы F-Met-Leu-Phe, IL-8 рецепторы, МСР-1 рецепторы и MIP1-I/RANTES рецепторы; ингибиторы внутриклеточного сигнала, включая (а) ингибиторы протеинкиназы, включая оба ингибитора протеинкиназы (i) С (РКС) (изозим), включая калфостин С, G-6203 и GF 109203X, и (ii) ингибиторы протеин-тирозинкиназы; (b) модуляторы внутриклеточных протеин-тирозинфосфатаз (PTPases); и (с) ингибиторы доменов src гомолога2 (SH2).
Для некоторых видов применения может быть полезным введение агентов, ингибирующих MASP-2, по настоящему изобретению в комбинации с агентом, ингибирующем спазмы. Например, для применения при урогенитальных заболеваниях, может быть полезным включение как минимум одного агента, ингибирующего спазм гладкой мускулатуры, и/или как минимум одного противовоспалительного агента, а для проведения операций на сосудах может быть полезным включение как минимум одного ингибитора вазоспазма и/или как минимум одного противоспалительного агента и/или как минимум одного антирестенозного агента. Типичными примерами агентов, ингибирующих спазм, являются: антагонисты рецептора подтипа серотонина2; антагонисты рецептора тахикинина; доноры оксида азота; расширитель АТФ-чувствительного калиевого канала; антагонисты кальциевого канала; и антагонисты рецептора эндотелина.
ФАРМАЦЕВТИЧЕСКИЕ НОСИТЕЛИ И СРЕДСТВА ДОСТАВКИ ЛЕКАРСТВА
В основном, составы агентов, ингибирующих MASP-2, по настоящему изобретению, смешанные с любым другим выбранным терапевтическим агентом, содержатся в подходящем фармацевтически приемлемом носителе. Носитель является нетоксичным, биосовместимым и выбран таким образом, чтобы избежать губительного воздействия биологической активности агента, ингибирующего MASP-2 (и любого другого терапевтического агента, включенного в комбинацию). Типичные фармацевтически приемлемые носители для пептидов описаны в патенте США №5,211,657, выданном Yamada. AHTH-MASP-2 антитела и пептиды-ингибиторы, используемые в настоящем изобретении, могут быть сформированы посредством приготовления в твердой, полутвердой, гелеобразной, жидкой и газообразной форме в виде таблеток, капсул, порошка, гранул, мазей, растворов, депозиториев, ингаляций и инъекций, предназначенных для орального, парентерального или хирургического введения. Настоящее изобретение также предусматривает местное введение композиций, посредством покрытия медицинских инструментов и т.п.
Подходящими носителями для парентерального введения посредством инъекции, инфузии или орошения и местного ведения лекарства являются дистиллированная вода, забуференный фосфатом физиологический раствор, раствор Рингера нормальный или с лактатом, раствор Д-глюкозы, раствор Хэнка или пропандиол. Кроме того, стерильные, нелетучие масла могут быть использованы в качестве растворителя или питательной взвеси. Для данной цели может быть использовано любое биосовместимое масло, включая синтетические моно- или диглицериды. Кроме того, жирные кислоты, такие как олеиновая кислота, используются для приготовления инъекций. Носитель и агент могут быть представлены в виде жидких растворов, суспензий, способных или не способных к полимеризации гелей, паст или бальзамов.
Под носителем также может подразумеваться средство (т.е., расширяющее, задерживающее или регулирующее) для доставки агента (ов) или для усиления доставки, накопления, стабилизации или фармакокинетики терапевтических агентов. Подобное средство доставки может быть представлено, например, микрочастицами, микросферами, наносферами или наночастицами, состоящими из белков, липосом, углеводов, синтетических органических веществ, неорганических веществ, полимерных или кополимерных гидрогелей и полимерных мицелл. Подходящие системы доставки на основе гидрогелей и мицелл включают в себя кополимеры РЕО:РНВ:РЕО и кополимерные/циклодекстриновые комплексы, описанные в WO 2004/009664 А2, и РЕО и РЕО/циклодекстриновые комплексы, описанные в публикации заявки на патент США №2002/0019369 А1. Подобные гидрогели могут быть введены путем местных инъекций в область предполагаемого воздействия, или подкожно или внутримышечно для обеспечения замедленного высвобождения агента.
Для внутрисуставной доставки, агент, ингибирующий MASP-2, может быть доставлен посредством вышеописанных жидких или гелевых носителей, предназначенных для инъекций, вышеописанных средств доставки, обеспечивающих замедленное высвобождение, предназначенных для инъекций, или в гиалуроновой кислоте либо ее производных.
Для орального приема непептидергических агентов, агент, ингибирующий MASP-2, может применяться в инертных наполнителях или растворителях, таких как сахароза, кукурузный крахмал или целлюлоза.
Для местного введения агент, ингибирующий MASP-2, может содержаться в составе мази, лосьона, крема, геля, капель, суппозитория, спрея, жидкости или порошка, или в гелевых или микрокапсульных системах доставки лекарственных препаратов при помощи трансдермального пластыря.
Различные назальные и легочные системы доставки лекаства, включая аэрозоли, дозирующие ингаляторы, ингаляторы сухого порошка и распылители, находятся на стадии разработки и могут соответствующим образом быть адаптированы для доставки препарата настоящего изобретения посредством аэрозоля, ингалятора или средства доставки в виде распылителя, соответственно.
Для инратекального (IT) или интрацеребровентрикулярного (ICV) способа доставки препарата могут быть использованы соответствующие стерильные системы доставки препарата, описанные в настоящем изобретении (например, жидкости; гели, суспензии, и т.д.).
Композиции по настоящему изобретению могут также включать в себя биосовместимые формообразующие лекарственные средства, такие как дисперсионные или увлажняющие агенты, суспендирующие агенты, растворители, буфферы, агенты, усиливающие проникновение, эмульгаторы, связующие средства, загустители, ароматизирующие вещества (для орального приема).
ФАРМАЦЕВТИЧЕСКИЕ НОСИТЕЛИ ДЛЯ АНТИТЕЛ И ПЕПТИДОВ
Отдельно для анти-MASP-2 антител и пептидов-ингибиторов, типовые рецептуры приготовления лекарственного средства могут вводиться парентерально посредством дозированных инъекций раствора или суспензии вещества в физиологически приемлемом растворителе с фармацевтическим носителем, который может быть представлен стерильной жидкостью, такой как вода, масла, солевой раствор, глицерин или этанол. Дополнительно, вспомогательные субстанции, такие как увлажняющие или эмульгирующие агенты, поверхностно-активные вещества, рН буферные субстанции и т.п. могут присутствовать в композициях, включающих в себя анти-MASP-2 антитела и пептиды-ингибиторы. Дополнительные компоненты фармацевтических композиций включают в себя петролеум (как-то животного, растительного или синтетического происхождения), например, соевое масло и минеральное масло. Как правило, гликоли, такие как пропиленгликоль и полиэтиленгликоль, являются предпочтительными жидкими носителями для растворов для инъекций.
Анти-MASP-2 антитела и пептиды-ингибиторы также могут вводиться в форме инъекций веществ замедленного высвобождения или при подготовке имплантата, что может быть произведено таким образом, чтобы позволить замедленное или пульсирующей высвобождение активных агентов.
ФАРМАЦЕВТИЧЕСКИ ПРИЕМЛЕМЫЕ НОСИТЕЛИ ДЛЯ ИНГИБИТОРОВ ЭКСПРЕССИИ
Более конкретно, в части ингибиторов экспрессии, используемых в способах данного изобретения, предложенные композиции содержат в себе ингибитор экспрессии, как описано выше, и фармацевтически приемлемый носитель или растворитель. Композиции могут далее содержать коллоидальную дисперсионную систему Фармацевтические композиции, содержащие ингибиторы экспрессии, могут включать в себя, помимо прочего, растворы, эмульсии и содержащие липосомы составы. Данные композиции могут быть получены из различных компонентов, включающих, помимо прочего, заранее пригодовленные жидкости, самоэмульгирующиеся твердые и самоэмульгирующиеся полутвердые носители. Приготовление подобных композиций обычно предусматривает комбинирование ингибиторов экспрессии с одним или несколькими из следующих элементов: буферы, антиоксиданты, низкомолекулярные полипептиды, белки, аминокислоты, углеводы, включая глюкозу, сахарозу или декстрин, хелатообразующий агент, такой как EDTA, глютатион и прочие стабилизаторы и наполнители. Нейтральные соляные буферы или соляные растворы, смешанные с неспецифичным сывороточным альбумином представляют собой примеры соответствующих растворителей.
В некоторых вариантах осуществления композиции могут быть приготовлены и сформированы в виде эмульсий, которые являются типичными гетерогенными системами одного жидкого раствора, диспергированного в другом в форме микрокапель (см., Idson, в Pharmaceutical Dosage Forms, Vol.1, Rieger and Banker (eds.), Marcek Dekker, Inc., N.Y., 1988). Примерами природных эмульгирующих агентов, используемых для формирования эмульсий, являются гуммиарабик, пчелиный воск, ланолин, лецитин и фосфатиды.
В одном варианте осуществления композиции, содержащие нуклеиновые кислоты, могут быть сформированы в виде микроэмульсий. Микроэмульсия, как указано в настоящем описании, представляет собой систему из воды, масла и амфифильного вещества, которая представляет собой одиночный оптически однородный и термодинамически стабильный жидкий состав (см. Rosoff в Pharmaceutical Dosage Forms, Vol.1). Способ данного изобретения может также включать использование липосом для трансферта и доставки антисмысловых олигонуклеотидов в желаемую область.
Фармацевтические композиции и рецептуры ингибиторов экспрессии для локального администрирования могут быть представлены трансдермальными пластырями, мазями, лосьонами, кремами, гелями, каплями, суппозиториями, спреями, жидкостями и порошками. Также могут быть использованы традиционные фармацевтические носители, например, водные, порошковые или маслянные основы и загустители и т.п.
СПОСОБЫ ВВЕДЕНИЯ
Фармацевтические композиции, содержащие агенты, ингибирующие MASP-2, могут быть введены посредством различных путей, в зависимости от того, какой из способов введения - местный или системный - требуется для лечения заболевания. Дополнительно, как описано выше, учитывая специфику процедур экстракорпоральной реперфузии, агенты, ингибирующие MASP-2, могут быть введены посредством интродукции композиций настоящего изобретения в систему циркуляции крови или плазмы. Далее, композиции по настоящему изобретению могут быть доставлены посредством покрытия или инкорпорирования композиций на имплантируемое медицинское оборудование или в него.
СИСТЕМНАЯ ДОСТАВКА ПРЕПАРАТА
В соответствии с настоящим описанием, предполагается, что термины «системная доставка» или «системное введение» включают, помимо прочего, оральные и парентеральные способы, включая внутримышечный (IM), подкожный, внутривенный (IV), внутриартериальный, ингаляционный, подъязычный, буккальный, местный, трансдермальный, назальный, ректальный, вагинальный и прочие способы введения, которые эффективно действуют на дисперсию вводимого агента в одиночную или в многочисленные области предполагаемого терапевтического воздействия. Предпочтительные способы системной доставки настоящих композиций включают внутривенный, внутримышечный, подкожный и ингаляционный. Весьма ценным будет определение точного способа системного введения для отобранных агентов, используемых в отдельных композициях настоящего изобретения, с учетом восприимчивости агента к метаболической трансформации, вызванной назначенным способом введения. Например, самым подходящим способом введения пептидергических агентов является любой, кроме орального.
Антитела и полипептиды, ингибирующие MASP-2, могут быть доставлены субъекту, который в этом нуждается, посредством любого подходящего способа. Способы доставки антител и полипептидов MASP-2 включают в себя оральное, пульмональное, парентеральное введение (например, внутримышечные, интраперитонеальные, внутривенные (IV) или подкожные инъекции), ингаляции (например, в форме тонкоизмельченного порошка), трансдермальное, назальное, вагинальное, ректальное или подкожное введение, и могут быть выполнены в дозировках и формах, соответствующих каждому из способов введения.
Иллюстрирующий пример демонстрирует, что антитела и пептиды, ингибирующие MASP-2, могут быть введены живому субъекту посредством аппликации на телесную оболочку, обладающую способностью абсорбировать полипептиды, например, назальные, гастроинтестинальные и ректальные мембраны. Полипептиды обычно применяются к абсорбирующим мембранам в сочетании с агентом, усиливающим проникание, (см., например, Lee, V.H.L, Crit. Rev. Ther. Drug Carrier Sys. 5:69, 1988; Lee, VH.L, J. Controlled Release 13:213, 1990; Lee, V.H.L, Ed., Peptide and Protein Drug Delivery, Marcel Dekker, New York (1991); DeBoer, A.G, et al., J. Controlled Release, 13:241, 1990). Например, STDHF представляет собой синтетическое производное фусидовой кислоты, стероидное поверхностно-актиное вещество, которое обладает структурой, схожей с солями желчных кислот, и которое использовалось в качестве агента, усиливающего проникание при назальной доставке. (Lee, W.A., Biopharm. 22, Nov./Dec. 1990.)
Антитела и полипептиды, ингибирующие MASP-2, могут участвовать в связывании с другой молекулой, например, с липидом, с целью защиты полипептидов от ферментной деградации. Например, ковалентное прикрепление полимеров, особенно, полиэтиленгликоля (PEG), было использовано для защиты некоторых белков от ферментного гидролиза в организме, и, таким образом, продлило период полувыведения (Fuertges, F, et al., J. Controlled Release 11:139, 1990). Многие системы полимеров были описаны для доставки белков (Bae, Y.H., et al., J. Controlled Release 9:271, 1989; Hori, R., et al., Pharm. Res. 6:813, 1989; Yamakawa, I., et al., J. Pharm. Sci. 79:505, 1990; Yoshihiro, I., et al., J. Controlled Release 10:195, 1989; Asano, M., et al., J. Controlled Release 9:111, 1989; Rosenblatt, J., et al., J. Controlled Release 9:195, 1989; Makino, K., J. Controlled Release 12:235, 1990; Takakura, Y, et al., J. Pharm. Sci. 78:117, 1989; Takakura, Y, et al., J. Pharm. Sci. 78:219, 1989).
Недавно были разработаны липосомы с повышенной стабильностью сыворотки и увеличенным периодом полувыведения (см., например, патент США №5,741,516, выданный Webb). Кроме того, были рассмотрены различные способы приготовления липосом и им подобных элементов в качестве потенциальных носителей лекарственных препаратов (см., например, патент США №5,567,434, выданный Szoka; патент США №5,552,157, выданный Yagi; патент США №5,565,213, выданный Nakamori; патент США №5,738,868, выданный Shinkarenko; и патент США №5,795,587, выданный Gao).
Для трансдермальных аппликаций, антитела и полипептиды, ингибирующие MASP-2, могут быть скомбинированы с прочими подходящими ингредиентами, например, носителями и/или адьювантами. Ограничений касательно природы подобных ингридиентов не существует, за исключением того, что они должны быть фармацевтически приемлемыми для запланированного введения и не могут снижать действие активных ингредиентов композиции. Примерами соответствующих средств являются кремы, мази, гели или суспензии, содержащие или не содержащие очищенный коллаген. Антитела и полипептиды, ингибирующие MASP-2, также могут быть вкраплены в трансдермальные накладки, пластыри и бандажи, предпочтительно в жидкой или полужидкой форме.
Композиции настоящего изобретения могут быть введены систематически на периодической основе в интервалы, определенные для поддержания желаемого уровня терапевтического эффекта. Например, составы могут быть введены посредством подкожных инъекций каждые две-четыре недели или через менее частые интервалы. Схема приема определяется терапевтом с учетом различных факторов, которые могут оказывать влияние на действие комбинации агентов. Данные факторы включают в себя степень прогрессирования наблюдаемого заболевания, возраст пациента, его пол и вес и прочие клинические факторы. Определение доз каждого индивидуального агента варьируется в зависимости от функции агента, ингибирующего MASP-2, включенного в композицию, равно как и от наличия и природы любого средства доставки лекарственного препарата (например, средство доставки препарата с замедленным высвобождением). Кроме того, количественное определение дозы может быть установлено соответственно варьированию частоты введения и фармакокинетического поведения доставляемого(ых) агента(ов).
МЕСТНАЯ ДОСТАВКА ПРЕПАРАТА
Используемый в настоящем описании термин «местное» охватывает аппликацию препарата на или вокруг области предполагаемого местного действия и может подразумевать, например, местную доставку в кожу или другую пораженную ткань, офтальмическую доставку, интратекальное (IT), интрацеребровентрикулярное (ICV), внутрисуставное, внутриполостное, внутричерепное или внутрипузырное введение, наложение или орошение. Местное введение может быть более предпочтительным с целью понижения дозы для того, чтобы избежать системных побочных эффектов, и для более точного контроля времени доставки и концентрации активных агентов в области местной доставки. Местное введение предусматривает известные концентрации в целевой области, безотносительно к индивидуальным отличиям пациентов в плане метаболизма, тока крови, и т.д. Усовершенствованный контроль дозировки также является результатом прямого способа доставки.
Местная доставка агента, ингибирующего MASP-2, может требоваться в контексте хирургических способов лечения болезни или состояния, например, при хирургических процедурах артериального шунтирования, атерэктомии, лазерных процедурах, ультразвуковых процедурах, баллонной ангиопластики и установки стента. Например, ингибитор MASP-2 может быть введен субъекту в связи с процедурой баллонной ангиопластики. Процедура баллонной ангиопластики заключается во введении в артерию катетера со спущенным баллоном. Спущенный баллон располагается в непосредственной близости от атеросклеротической бляшки и наполняется таким образом, что бляшка оказывается прижатой к сосудистой стенке. В результате, поверхность баллона вступает в контакт со слоем васкулярных эндотелиальных клеток на поверхности кровесносных сосудов. Агент, ингибирующий MASP-2, может быть прикреплен к катетеру баллонной ангиопластики таким способом, чтобы сделать возможным выделение агента в область атеросклеротической бляшки. Агент может быть связан с баллонным катетером в соответствии со стандартными процедурами, хорошо известными в данной области техники. Например, агент может быть аккумулирован в отделении баллонного катетера до раскрытия баллона, и в таком случае происходит выделение в местную среду Альтернативно, агент может покрывать поверхность баллона, таким образом, происходит контакт с клетками артериальной стенки при наполнении баллона. Также агент может быть доставлен в перфорированном баллоне катетера, как это описано у Flugelman, M.Y., et al., Circulation 85:1110-1117, 1992. См. также публикацию заявки РСТ WO 95/23161 для типичной процедуры соединения терапевтического белка с баллонным катетером для ангиопластики. Подобным образом, агент, ингибирующий MASP-2, может быть включен в состав геля или полимерного покрытия, нанесенного на поверхность стента, либо может быть инкорпорирован в материал стента таким образом, чтобы стент выделял агент, ингибирующий MASP-2, после помещения в полость сосуда.
Композиции, ингибирующие MASP-2, используются для лечения артритов (поражение кожи ревматического или подагрического происхождения) и других скелетномышечных расстройств и могут быть доставлены местно посредством внутрисуставных инъекций. Подобные композиции могут соответственным образом включать в себя средства доставки с замедленным высвобождением препарата. В качестве дальнейшего примера случаев желательного местного воздействия, композиции, ингибирующие MASP-2, используемые для лечения урогенитальных расстройств, могут вводиться малыми дозами (капельно) внутрь мочевого пузыря или другой урогенитальной структуры.
ПОКРЫТИЕ МЕДИЦИНСКИХ ИНСТРУМЕНТОВ
Агенты, ингибирующие MASP-2, например антитела или пептиды-ингибиторы, могут быть иммобилизированы на (или в) поверхность имплантируемого или присоединяемого медицинского инструмента. Модифицированная поверхность вступит характерным образом в контакт с живой тканью после имплантации в тело животного. Под «имплантируемым или присоединяемым медицинским инструментом» подразумевается любой предмет, имплантированный в или присоединенный к ткани тела животного во время операции (например, стенты и имплантируемые устройства по доставке препарата). Такие имплантируемые или присоединяемые медицинские устройства могут быть изготовлены из нитроцеллюлозы, диазоцеллюлозы, стекла, полистирола, поливинилхлорида, полипропилена, полиэтилена, декстрана, Сефарозы, агара, крахмала, нейлона, нержавеющей стали, титана и биодеградируемых и/или биосовместимых полимеров. Прикрепление белков к устройству может быть осуществлено посредством любой технологии, которая не нарушает биологическую активность связанных белков, например, при помощи связывания одного или обоих N-С-терминальных остатков белка с устройством. Присоединение также может осуществляться в одном или более внутренних сайтах белка. Множественные присоединения (как внутренние, так и с концами белка) также могут быть использованы. Поверхность имплантируемого или присоединяемого медицинского устройства может быть модифицирована для включения функциональных групп (например, карбоксил, амид, амин, эфир, гидроксил, циан, нитрид, сульфаниламид, ацетилин, эпоксид, силанид, ангидрид, сукцинид, азид) для белковой иммоблизации. Связывание химических структур включает, помимо прочего, формирование сложных эфиров, эфиров, амидов, азидо- и сульфаниламидо-производных, цианата и прочих связей с функциональными группами, которые содержаться в антителах и пептидах, ингибирующих MASP-2. Антитела и фрагменты ингибиторов MASP-2 также могут быть присоединены нековалентным образом при помощи добавления к белку последовательности аффинных тагов, например GST (D.B.Smith и K.S.Johnson, Gene 67:31, 1988), полигистидинов (Е.Hochuli et al., J. Chromatog. 411:77, 1987), или биотина. Подобные аффинные таги могут использоваться для обратимого присоединения белка к устройству
Белки также могут присоединяться ковалентным образом к поверхности устройства, например, посредством ковалентной активации поверхности медицинского устройства. Примером для иллюстрации выступает матрицеллюлярный(ые) белок(и), который может присоединяться к устройству при помощи любой из следующих пар реактивных групп (один член пары располагается на поверхности устройства, а другой член пары располагается в матрицеллюлярном(ых) белке(ах)): гидроксил/карбоксильная группа кислоты для образования сложноэфирной связи; гидроксил/ангидрид для образования сложноэфирной связи; гидроксил/изоцианат для образования уретановой связи. Поверхность медицинского устройства, не обладающая необходимыми реактивными группами, может быть обработана способом плазменного высокочастотного разряда (RFGD), который при помощи травления генерирует реактивные группы, позволяющие обеспечить отложение матрицеллюлярного(ых) белка(ов) (например, обработка кислородной плазмой для введения кислород-содержащих групп; обработка пропиламино плазмой для введения амино групп).
Агенты, ингибирующие MASP-2, содержащие молекулы нуклеиновой кислоты, такие как антисмысловые РНКи- или ДНК-кодирующие ингибиторы пептидов, могут быть внедрены в пористые матрицы, присоединенные к устройству. Типичными пористыми матрицами, используемыми в качестве поверхностного слоя, являются матрицы, созданные из коллагена кожи или сухожилий, полученного от разнообразных коммерческих источников (например, компания «Sigma and Collagen Corporation»), или коллагенные матрицы, созданные способом, описанным в патентах США №4,394,370, выданном Jefferies и №4,975,527, выданном Koezuka. Примером коллагенного материала является UltraFiber™, производимый компанией «Norian Corp.» (Маунтин-Вью, шт.Калифорния).
При необходимости, также могут быть использованы некоторые полимерные матрицы, которые включают в себя полиакрилаты сложного эфира и полимеры молочной кислоты; описание может быть найдено в патентах США №4,526,909, выданном Urist и №4,563,489, выданном Urist. Частными примерами используемых полимеров являются ортоэфиры, ангидриды, пропилен-кофумараты или полимеры одного или более мономеров α-гидрокси карбоновой кислоты (например, α-оксиуксусной кислоты (гликолиевой кислоты) и/или α-гидрокси пропионовой кислоты (молочной кислоты).
СХЕМЫ ЛЕЧЕНИЯ
В профилактических целях, фармацевтические композиции вводятся субъекту, подверженному или находящемуся в группе риска заболевания, опосредованного MASP-2-зависимой активацией комплемента в объеме, достаточном для подавления или уменьшения риска развития симптоматики заболевания. В терапевтических целях, фармацевтические композиции вводятся субъекту с подозрением на или уже страдающему заболеванием, опосредованным MASP-2-зависимой активацией комплемента в терапевтически эффективном объеме, достаточном для облегчения или по меньшей мере частичного подавления симптоматики заболевания. Как в профилактических, так и в терапевтических целях, композиции с содержанием агентов, ингибирующих MASP-2, могут вводиться поэтапно до достижения удовлетворительного терапевтического результата. Применение композиций, ингибирующих MASP-2, по настоящему изобретению может быть осуществлено посредством однократного введения композиции или ограниченной серией введений для лечения острого заболевания, например, поражения при реперфузии или при другом травматическом поражении. Альтернативно, композиция может вводиться в периодические интервалы в течение продолжительного времени для лечения хронического заболевания, например, артрита или псориаза.
Способы и композиции по настоящему изобретению могут использоваться с целью подавления воспаления и связанных с ним процессов, которые обычно проявляются в результате проведения диагностических и терапевтических медицинских и хирургических процедур. Для ингибирования подобных процессов, композиция для ингибирования MASP-2 по настоящему изобретению может применяться перипроцедурно. Как употребляется в настоящем описании, словосочетание «перипроцедурно» относится к введению композиций-ингибиторов перидпроцедурно и/или интрапроцедурно и/или постпроцедурно, т.е., до процедуры, до и в течение процедуры, до и после процедуры, до, во время и после процедуры, во время процедуры, во время и после процедуры, или после процедуры. Перипроцедурное применение относится к местному введению композиции в область хирургического или процедурного вмешательства посредством инъекции или продолжительного или интермиттирующего орошения данной области, либо посредством систематического введения. Подходящие способы местной периоперативной доставки растворов, содержащих агенты, ингибирующие MASP-2, описаны в патентах США №6,420,432, выданном Demopulos и №6,645,168, выданном Demopulos. Подходящие способы местной доставки композиций с хондропротекторными свойствами, содержащих агент(ы), ингибирующий (ие) MASP-2, описаны в международной заявке РСТ на патент WO 01/07067 А2. Подходящие способы и композиции целенаправленной системной доставки композиций с хондропротекторными свойствами, содержащих агент(ы), ингибирующий(ие) MASP-2, описаны в международной заявке РСТ на патент WO 03/063799 А2.
VI. ПРИМЕРЫ
Приведенные ниже примеры являются только иллюстрациями наилучшего способа практической реализации данного изобретения, однако не должны быть истолкованы как ограничения для данного изобретения. Все списки цитируемых литературных материалов приведены здесь в качестве ссылки.
ПРИМЕР 1
Данный пример описывает получение линии мышей с недостаточностью MASP-2 (MASP-2-/-), но полноценных по MApl9 (MApl9+/+).
Материалы и методы: Целевой вектор pKO-NTKV 1901 был создан для разрушения трех экзонов, кодирующих С-терминальный конец мышиного MASP-2, включая экзон, который кодирует домен сериновой протеазы, как показано на ФИГУРЕ 4. PKO-NTKV 1901 был использован для трансфекции мышиной ES клеточной линии Е14.1а (SV129 Ola). Были выбраны клоны, устойчивые к неомицину и чувствительные к тимидинкиназе. 600 ES клоны были подвергнуты скринингу и из их числа посредством саузерн-блоттинга были идентифицированы и верифицированы четыре клона, содержащие ожидаемые селективные целевые и рекомбинантные элементы, как показано на ФИГУРЕ 4. При помощи способа переноса эмбриона из данных четырех положительных клонов были получены химеры. Затем химеры подвергли обратному скрещиванию с исходной формой C57/BL6 с целью создания трансгенных особей мужского пола. Затем трансгенные особи мужского пола были скрещены с женскими для генерации F1s с 50% потомства, проявляющего гетерозиготность по нарушенному гену MASP-2. Гетерозиготных мышей подвергли интеркроссированию (скрещивание неродственных особей) для получения гомозиготного потомства с недостаточностью MASP-2, в результате чего были получены гетерозиготные и дикого типа мыши в соотношении 1:2:1, соответственно.
Результаты и фенотип: Полученные в результате гомозиготные мыши с недостаточностью MASP-2-/- проявили жизнеспособность и фертильность и были проверены на отсутствие MASP-2 посредством саузерн-блоттинга для подтверждения правильности целевого трансгеноза, посредством нозерн-блоттинга для подтвержцения отсутствия мРНК MASP-2, и посредством вестерн-блоттинга для подтверждения отсутствия белка MASP-2 (данные не приводятся). Наличие мРНК МАр19 и отсутствие мРНК MASP-2 были далее подтверждены при помощи RT-PCR с разрешением во времени на аппарате LightCycler. Как и ожидалось, мыши MASP-2-/- продолжали экспрессировать МАр19, MASP-1 и мРНК MASP-3 и белок (данные не приводятся). При помощи анализа LightCycler у MASP-2-/- мышей было выявлено присутствие и чрезмерное количество мРНК пропердина, фактора В, фактора D, С4, С2 и СЗ, также было обнаружено, что эти данные идентичны с теми, что обнаруживаются у контрольных однопометных животных дикого типа (данные не приводятся). Плазма гомозиготных MASP-2-/- мышей обладает абсолютной недостаточностью активации комплемента, опосредованной лектиновым путем и альтернативным путем, как описано далее в Примере 2.
Генерация MASP-2-/- линии на чистой основе C57BL6: MASP-2-/- мыши были подвержены обратному скрещиванию с чистой C57BL6 линией в течение девяти поколений прежде, чем использовать MASP-2-/- линию в качестве экспериментальной животной модели.
ПРИМЕР 2
Данный пример демонстрирует, что MASP-2 необходим для активации комплемента посредством альтернативного или пектинового пути.
Материалы и методы:
Исследование специфического расщепления С4 при пектиновом пути активации комплемента: Изучение расщепления С4 было описано Петерсеном (Petersen, et al., J. Immunol. Methods 257:101 (2001)), который измеряет активацию пектинового пути комплемента, опосредованную липотейхоевой кислотой (LTA) из золотистого стафилококка (S. aureus), которая связывает L-фиколин. Исследование, описанное в Примере 11, было адаптировано для измерения уровня активации пектинового пути посредством MBL при помощи покрытия планшета LPS и маннаном или зимозаном перед добавлением сыворотки от MASP-2-/- мышей, как описано далее. Исследование также было модифицировано для исключения возможности расщепления С4 при помощи классического пути. Это было достигнуто посредством использования образца слабого буферного раствора, содержащего 1 моль NaCl, который обеспечивает возможность высокоаффинного связывания компонентов узнавания пектинового пути активации с их лигандами, но предупреждает активацию эндогенного С4, таким образом, исключая участие классического пути в диссоциировании комплекса С1. Вкратце, в модифицированном исследовании образцы сыворотки (отмытые в буфере с большим количестовм соли (1 моль NaCl)) были добавлены в планшеты, покрытые лигандом, после добавления постоянного объема очищенного С4 в буфер с физиологическим уровнем концентрации соли. Связывающие комплексы узнавания, содержащие MASP-2, расщепляют С4, вследствие чего происходит отложение C4b.
Способы анализа:
1) Титрационные микропланшеты Nunc Maxisorb (Maxisorb, Nunc, кат. №442404, «Fisher Scientific») были обработаны раствором маннана (М7504 Sigma) с концентрацией 1 мкг/мл или любым другим лигандом (например, из перечисленных ниже), растворенным в покрывающем буфере (15 ммоль Na2C03, 35 ммоль NaHCO3, pH 9,6).
В исследовании были использованы следующие реагенты:
а. маннан (1 мкг на лунку маннана (М7504 Sigma) в 100 мкл покрывающего буфера);
b. зимозан (1 мкг на лунку зимозан (Sigma) в 100 мкл покрывающего буфера);
с. липотейхоевая кислота LTA (1 мкг на лунку в 100 мкл покрывающего буфера или 2 мкг на лунку в 20 мкл метанола);
d. 1 мкг Н-фиколин специфичного Mab 4H5 в покрывающем буффере
е. PSA из Aerococcus viridans (2 мкг на лунку в 100 мкл покрывающего буфера);
f. 100 мкл на лунку формалин-зафиксированого S. aureus DSM20233 (OD550=0,5) в покрывающем буфере.
2) Планшет подвергли инкубации в течение ночи при 4°С.
3) После инкубации в течение ночи, остаточные участки связывания белков были насыщены при помощи инкубирования планшетов с 0,1% HSA-TBS блокирующим буфером (0,1% (по весу) HSA в 10 ммоль Tris-CL, 140 ммоль NaCl, 1,5 ммоль NaN3, pH 7,4) в течение 1-3 часов, и затем отмыты в палншетах 3Х с TBS/tween/Ca2+ (TBS с 0,05% Tween 20 и 5 ммоль CaCl2, 1 ммоль MgCl2, pH 7,4).
4) Образцы сыворотки для тестирования были разбавлены в MBL-связывающем буфере (1 моль NaCl) и разбавленные образцы были добавлены в планшеты и подвергнуты инкубации в течение ночи при 4°С. Лунки, получающие только буфер, были использованы только для отрицательного контроля.
5) После инкубации в течение ночи при 4°С, планшеты были трижды промыты с TBS/tween/Ca2+. C4 человека (100 мкл на лунку раствора в BBS (4 ммоль барбитала, 145 ммоль NaCl, 2 ммоль CaCl2, 1 ммоль MgCl2, pH 7,4) с концентрацией 1 мкг/мл) затем был добавлен в планшеты и подвергнут инкубации в течение 90 минут при 37°С. Затем планшеты были снова трижды промыты с TBS/tween/Ca2+.
6) Отложение C4b определяли античеловеческими С4 с антителами курицы, конъюгированными с щелочной фосфатазой (разведенными в соотношении 1:1000 в TBS/tween/Ca2+), который был добавлен в планшеты. Планшеты инкубировали в течение 90 минут при комнатной температуре. Затем планшеты снова трижды промыли с TBS/tween/Ca2+.
7) Щелочная фосфатаза определялась посредством добавления 100 мкл раствора субстрата р -нитрофенил фосфата, последующей инкубации при комнатной температуре в течение 20 минут, и определения оптической плотности OD405 в спектрофотометре для прочтения планшетов.
Результаты: ФИГУРЫ 6А-В демонстрируют объем отложения C4b на маннане (ФИГУРА 6А) и зимозане (ФИГУРА 6 В) в растворах сывороток от MASP-2+/+ (скрещивания), MASP-2+/- (замкнутые кольца) и MASP-2-/- (замкнутые треугольники). На ФИГУРЕ 6С показана относительную активность конвертазы С4 на планшетах, покрытых зимозаном (белые столбцы) или маннаном (темные столбцы) от MASP-2-/+ мышей (n=5) и MASP-2-/- мышей (n=4) относительно мышей дикого типа (n=5), основываясь на замере объема отложения C4b, нормализованного в сыворотке дикого типа. Столбцы погрешностей отражают стандартное отклонение. Как показано в ФИГУРАХ 6А-С, плазма от MASP-2-/- мышей совершенно лишена активации комплемента, опосредованной лектиновым путем, в покрытых маннаном и зимозаном планшетах. Данные результаты демонстрируют очевидность того, что MASP-2, а не MASP-1 или MASP-3, является эффекторным компонентом пектинового пути активации комплемента.
Исследование отложения C3b:
1) Nunc Maxisorb титрационные микропланшеты (Maxisorb, Nunc, кат. №442404, «Fisher Scientific») были покрыты маннаном (М7504 Sigma) или любым другим лигандом в количестве 1 мкг на лунку, растворенным в покрывающем буфере (15 ммоль Na2CO3, 35 ммоль NaHCO3, pH 9,6) и инкубированы в течение ночи при 4°С.
2) Остаточные участки связывания белков были обогащены при помощи инкубирования планшета с 0,1% HSA-TBS блокирующим буфером (0,1% (по весу) HSA в 10 ммоль Tris-CL, 140 ммоль NaCl, 1,5 ммоль NaN3, pH 7,4) в течение 1-3 часов.
3) Планшеты были промыты в TBS/tw/Ca++(TBS с 0,05% Tween 20 и 5 ммоль CaCl2) и растворенный BBS был добавлен к образцам сыворотки (4 ммоль барбитала, 145 ммоль NaCl, 2 ммоль CaCI2, 1 ммоль MgCl2, pH 7,4). Лунки, получающие только буфер, были использованы для отрицательного контроля. Контрольные наборы образцов сыворотки, полученные от мышей дикого типа или MASP-2-/- мышей, были освобождены от C1q до использования в исследовании, C1q-истощенная мышиная сыворотка была приготовлена с использованием сопряженных с А-белком микрошариков Dynabeads («Dynal Biotech», Осло, Норвегия), покрытых кроличьими античеловеческими C1q IgG антителами («Dako», Глоструп, Дания), согласно инструкциям производителя.
4) После инкубации в течение ночи при 4°С, и повторного промывания с TBS/tw/Ca++, был обнаружен конвертированный СЗ, связанный с поликлональным античеловеческим-С3с антителом (Dako A 062), растворенным в TBS/tw/Ca++ в соотношении 1:1000). Вторичное антитело представлено козлиным антикроличьим IgG антителом (полноценная молекула), коньюгированным с алкалинфосфатазой (Sigma lmmunochemicalsA-3812), разведенным в соотношении 1:10,000 в TBS/tw/Ca++. Наличие альтернативного пути активации комплемента (АР) определяется при добавлении 100 мкл раствора субстрата (набор таблеток Sigma Fast р-нитрофенил фосфата, Sigma) и инкубации при комнатной температуре. Был проведен мониторинг гидролиза с целью количественного замера абсорбции при 405 нм в пластине микротитратора в спектрофотометре для прочтения планшетов. Для каждого анализа приготовлена калибровочная кривая с использованием последовательных разведении образцов плазмы/сыворотки.
Результаты: На ФИГУРАХ 7А и 7В приведены результаты использования смешанной сыворотки от нескольких мышей. Кресты обозначают сыворотку MASP-2+/+, заштрихованные круги обозначают dq-истощенную сыворотку MASP-2+/+, белые квадраты обозначают сыворотку MASP-2-/- и белые треугольники обозначают C1q-обедненную сыворотку MASP-2-/-. Как показано на ФИГУРАХ 7А-В, сыворотка от MASP-2-/- мышей, протестированная в исследованиях отложения C3b, обнаруживает очень низкие уровни активации С3 на планшетах, покрытых маннаном (ФИГУРА 7А) и зимозаном (ФИГУРА 7 В). Данные результаты являются доказательством того, что MASP-2 необходим для инициации начальной генерации C3b из С3, что активизирует альтернативный путь комплемента. Это неожиданный результат в свете широко распространенных взгляда, что комплементарные факторы С3, В, D и пропердин формируют независимый функциональный альтернативный путь активации комплемента, в котором С3 может подвергаться спонтанному конформационному изменению в «C3b-подобную» форму, которая затем генерирует жидкую фазу конвертазы iC3Bb и откладывает молекулы C3b на активирующих поверхностях, таких как зимозан.
Рекомбинантный MASP-2 восстанавливает активацию С4, опосредованную пектиновым путем комплемента в сыворотке, полученной от MASP-2-/- мышей
Эффект от добавления рекомбинантного белка MASP-2 в образцы сыворотки был рассмотрен в вышеизложенном исследовании специфического расщепления С4 при пектиновом пути с целью установить тот факт, что отсутствие MASP-2 было прямой причиной недостаточности актиации С4, опосредованной лектиновым путем, у MASP-2-/-мышей. Функционально активный мышиный MASP-2 и каталитически неактивный мышиный MASP-2A (в котором активный сайт остатка серина в домене сериновой протеазы был замещен остатком аланина) рекомбинантных белков были получены и очищены, как описано ниже в Примере 5. Смешанная сыворотка от 4 MASP-2-/- мышей была проинкубирована с повышением концентраций белков рекомбинантного мышиного MASP-2 или неактивного рекомбинантного мышиного MASP-2A, и активность конвертазы С4 была оценена, как описано выше.
Результаты: Как показано на ФИГУРЕ 8, добавление функционально активного мышиного рекомбинантного белка MASP-2 (обозначен белыми треугольниками) в сыворотку, полученную от MASP-2-/- мышей, восстанавливало активацию С4, опосредованную пектиновым путем, в зависимости от концентрации белка, при этом каталитически неактивный мышиный белок MASP-2A (обозначен звездочками) не восстанавливал активацию С4. Результаты, показанные на ФИГУРЕ 8, нормализованы по отношению к активации С4, наблюдаемой в смешанной сыворотке от мышей дикого типа (обозначено пунктирной линией).
ПРИМЕР 3
Данный пример описывает получение трансгенной линии мышей, которая имеет мышиный MASP-2-/-, мышиный МАр19+/+ и экспрессирует человеческий трансген MASP-2 (мышиный MASP-2 нокаут (knock-out) и человеческий MASP-2 нокин (knock-in)).
Материалы и методы: Миниген, кодирующий человеческий MASP-2 и названный «"мини hMASP-2» (SEQ ID NO:49), как показано на ФИГУРЕ 5, был сконструирован с включением области промотора человеческого гена MASP-2, в том числе первые три экзона (экзон 1-экзон 3), с последующей последовательностью кДНК, которая представляет кодирующую последовательность со следующими 8 экзонами, и как следствие кодирующую полноразмерный белок MASP-2, под контролем его эндогенного промотора. Конструкция мини hMASP-2 была введена посредством инъекции в оплодотворенные яйцеклетки MASP-2-/- с целью замещения инактивированного мышиного гена MASP-2 на человеческий MASP-2, экспрессируемый трансгенным способом.
ПРИМЕР 4
Данный пример иллюстрирует выделение белка MASP-2 человека в форме профермента из человеческой сыворотки.
Способ выделения MASP-2 человека: Способ выделения MASP-2 из человеческой сыворотки описан у Matsushita et al., J. Immunol. 165:2637-2642, 2000. Вкратце, человеческая сыворотка проходит через колонку сефарозы с дрожжевым маннаном с использованием 10 ммоль буфера имидазола (рН 6,0) с содержанием 0,2 моль NaCl, 20 ммоль CaCI2, 0,2 ммоль NPGB, 20 мкмоль p-APMSF и 2% маннитола. Проферменты MASP-1 и MASP-2 образуют комплексы с MBL и вымываются вышеназванным буфером с содержанием 0,3 моль маннозы. Для отделения проферментов MASP-1 и MASP-2 от MBL, препараты с содержанием данных комплексов применяют анти-MBL-сефарозу и затем MASPs вымываются буфером с имидазолом, содержащим 20 ммоль EDTA и 1 моль NaCl. В конечной стадии, проферменты MASP-1 и MASP-2 разделяются один от другого при прохождении через анти-MASP-1-ce$apo3y в том же буфере, что использовался для анти-МВЬсефарозы. MASP-2 восстанавливается в вытекающих жидкостях, из которых вымыт MASP-1 при помощи 0,1 моль глицинового буфера (рН 2,2).
ПРИМЕР 5
Данный пример иллюстрирует рекомбинантную экспрессию и продуцирование рекомбинантных полноразмерных человеческих, крысиных и мышиных белков MASP-2, полипептидов, производных от MASP-2 и каталитически неактивных мутантных форм MASP-2.
Экспрессия полноразмерного человеческого, крысиного и мышиного MASP-2:
Полноразмерная последовательность кДНК человеческого MASP-2 (SEQ ID NO:4) была также субклонирована в экспрессионный вектор pCI-Neo млекопитающих (Promega), который управляет эукариотической экспрессией под контролем CMV энхансерной/промоторной области (описано у Kaufman RJ. et al., Nucleic Acids Research 19:4485-90, 1991; Kaufman, Methods in Enzymology, 185:531-66 (1991)). Полноразмерная мышиная кДНК (SEQ ID NO:50) и крысиная кДНК MASP-2 (SEQ ID NO:53) были субклонированы в экспрессионный вектор pED. Экспрессионные вектора MASP-2 были затем трансфицированы в адгезивные линии DXB1 клеток яичников китайских хомяков по стандартной процедуре трансфекции с использованием фосфата кальция, описанной у Maniatis et al., 1989. Клетки, трансфицированные такими конструкциями, развивались очень медленно, что объясняется цитотоксичностью кодируемой протеазы.
В другом подходе, конструкция минигена (SEQ ID NO:49), содержащая человеческую кДНК MASP-2, управляемую ее эндогенным промотором, кратковременно трансфицируется в клеки яичников китайских хомяков (СНО). Человеческий белок MASP-2, секретируемый в культуральную среду, выделяют по схеме, описанной ниже.
Экспрессия полноразмерного каталитически неактивного MASP-2:
Обоснование: MASP-2 активируется посредством аутокаталитического расщепления после того, как субкомпоненты распознавания MBL или фиколины (либо L-фиколин, Н-фиколин или М-фиколин) связываются с соответствующими им углеводными молекулами. Аутокаталитическое расщепление, вызванное активацией MASP-2, часто происходит во время процедуры выделения MASP-2 из сыворотки, либо во время очищения после рекомбинантной экспрессии. Для того, чтобы получить более стабильный препарат белка в качестве антигена, каталитически неактивная форма MASP-2, обозначенная как MASP-2A, была создана путем замещения остатка серина, который присутствует в каталитической триаде домена протеазы, на остаток аланина у крыс (SEQ ID NO:55 Ser617 кА1а617); у мышей (SEQ ID NO:52 Ser617 кА1а617); или у человека (SEQ ID NO:3 Ser618 к Ala618).
Для получения каталитически неактивных человеческих и мышиных белков MASP-2A, был осуществлен сайт-направленный мутагенез при помощи олигонуклеотидов, представленных в ТАБЛИЦЕ 5. Олигонуклеотиды по ТАБЛИЦЕ 5 были сконструированы таким образом, чтобы ренатурироваться на участок человеческой и мышиной кДНК, кодирующий ферментативно активный серии, и олигонуклеотид содержит некомплементарные нуклеотиды, с целью замены триплета серина на триплет аланина. Например, PCR олигонуклеотиды SEQ ID NOS:56-59 были использованы в комбинации с человеческой кДНК MASP-2 (SEQ ID NO:4) для амплификации участка между старт-кодоном и ферментативно-активным серином и от серина до стоп-кодона для получения полного открытого считывания из мутировнного MASP-2A, содержащего Ser618/Ala618 мутацию. Продукты PCR были очищены после электрофореза в агарозном геле, выделение окрашенных фрагментов из геля и синтез липких А-концов проводилис использованием стандартной процедуры наращивания. Полученный в результате вышеописанных манипуляций продукт MASP-2A затем был клонирован в вектор pGEM-T easy, и трансформирован в Е. coli.
Каталитически неактивный белок MASP-2A крысы был получен посредством киназинга и ренатурирования SEQ ID NO:64 и SEQ ID NO:65 посредством комбинирования двух данных олигонуклеотидов в равных молярных объемах, нагрева при температуры 100°С в течение 2 минут и медленном охлаждении при комнатной температуре. Полученный в результате ренатурирования фрагмент содержал совместимые с эндонуклеазами рестрикции Pst1 и ХЬа1 концы и был внедрен в область фрагмента Pst1-Xba1 кДНК MASP-2 крысы дикого типа (SEQ ID NO:53) для получения крысиного MASP-2A.
5' GAGGTGACGCAGGAGGGGCATTAGTGTTT 3' (SEQ ID NO:64)
5' CTAGAAACACTAATGCCCCTCCTGCGTCACCTCTGCA3' (SEQ ID NO:65)
Человеческий, мышиный и крысиный MASP-2A были каждый далее субклонированы в экспрессионных векторах млекопитающих pED или pCI-Neo и трансфицированы в линии клеток DXBI яичников китайских хомяков, как описано ниже.
В другом подходе каталитически неактивная форма MASP-2 сконструирована при использовании способа, описанного у Chen et al., J. Biol. Chem., 276(28);25894-25902, 2001. Вкратце, плазмида, содержащая полноцепочную человеческую кДНК MASP-2 (описано у Thiel et al. Nature 386:506, 1997), разрезали рестриктазами Xho1 и EcoR1 и кДНК MASP-2 (описано в данном изобретении как SEQ ID NO:4) клонировали по соответствующим сайтам рестрикции в бакуловирусный трансфертный вектор pFastBad (Life Technologies, NY). Активный сайт MASP-2 сериновой протеазы Ser618 затем видоизменяли в Ala618 путем замещения натурального участка с 610 по 625 аминокислоту двухцепочными олигонуклеотидами, кодирующими область пептида с 610 по 625 аминокислоту (SEQ ID NO:13), с целью создания полноразмерного полипептида MASP-2 с неактивным протеазным доменом.
Конструирование плазмид, содержащих полипептидные области производные от человеческого MASP-2.
Для секретирования различных доменов MASP-2 получают следующие конструкции с использованием сигнального пептида MASP-2 (остатки 1-15 последовательности SEQ ID NO:5).
Конструкция, экспрессирующая домен CUBI человеческого MASP-2 (SEQ ID NO:8), получается при помощи PCR амплификации области, кодирующей остатки 1-121 MASP-2 (SEQ ID NO:6) (соответствует N-терминальному домену CUBI). Конструкция, экспрессирующая домен CUBIEGF человеческого MASP-2 (SEQ ID NO:9), получается при помощи PCR амплификации области, кодирующей остатки 1-166 MASP-2 (SEQ ID NO:6) (соответствует N-терминальному домену CUBIEGF). Конструкция, экспрессирующая домен CUBIEGFCUBII человеческого MASP-2 (SEQ ID NO:10) получается при помощи PCR амплификации области, кодирующей остатки 1-293 MASP-2 (SEQ ID NO:6) (соответствует N-терминальному домену CUBIEGFCUBII). Вышеобозначенные домены амплифицируются посредством PCR с использованием VentR полимеразы и pBS-MASP-2 как матрицы, в соответствии с установленными способами PCR. 5' праймерная последовательность смыслового праймера f5'-CGGGATCCATGAGGCTGCTGACCCTC-3' SEQ ID NO:34) вводит BamHI сайт рестрикции (подчеркнутый) на 5' конце ПЦР продукта. Антисмысловые праймеры для каждого домена MASP-2, показанные ниже в ТАБЛИЦЕ 5, сконструированы для введения стоп-кодона (полужирный шрифт), за которым следует сайт EcoRI (подчеркнутый) на конце каждого ПЦР продукта.
После амплификации, фрагменты ДНК обрабатываются рестриктазами BamHI и EcoRI и клонируются в соответствующие сайты вектора pFastBac1. Полученные в результате конструкции характеризуются рестрикционным картированием и подтверждаются секвенированием дсДНК.
Рекомбинантная эукариотическая экспрессия MASP-2 и получение белков ферментативно неактивного мышиного, крысиного и человеческого MASP-2A.
Вышеописанные экспрессионные конструкции MASP-2 и MASP-2A были трансфецированы в DXB1 клетки с использованием стандартной процедуры кальций-фосфатной трансфекции (Maniatis et al., 1989). MASP-2A получали в бессывороточной питательной среде, чтобы исключить контаминацию препаратов другими сывороточными белками. На каждый второй день производили сбор конфлюэнтных клеток из питательных растворов (в общей сложности процесс произвели четыре раза). Средний уровень рекомбинантного MASP-2A для каждой из трех разновидностей составил приблизительно 1,5 мг/литр культуральной среды.
Очищение белка MASP-2A: MASP-2A (мутантная форма Ser-Ala, описанная выше) был очищен посредством аффинной хроматографии на колонках МВР-А-агарозы. Данная стратегия позволяет проводить быструю очистку без использования внешних тагов. MASP-2A (100-200 мл питательной среды, растворенной в равном объеме загрузочного буфера (50 ммоль Tris-Cl, pH 7,5, с содержанием 150 ммоль NaCl и 25 ммоль СаС12) был загружен в колонку МВР-аффинной-агарозы (4 мл) предварительно уравновешенную 10 мл загрузочного буфера. После процедуры промывания с дополнительными 10 мл загрузочного буфера, белок был элюирован в 1 мл фракций с 50 ммоль Tris-Cl, pH 7,5, содержащих 1,25 моль NaCl и 10 ммоль EDTA. Фракции, содержащие MASP-2A, были идентифицированы при помощи денатурирующего электрофореза в полиакриламидном геле. При необходимости MASP-2А был далее очищен посредством ионообменной хроматографии в колонке MonoQ (MR 5/5). Белок диализировали с 50 ммоль Tris-Cl pH 7,5, содержащим 50 ммоль NaCl, и загружали в колонку, уравновешенную тем же буфером. После промывания, связанный MASP-2А был элюирован при помощи 10 мл 0,05-1 моль градиентного раствора NaCl.
Результаты: Выход белка MASP-2A составил 0,25-0,5 мг на 200 мл питательной среды. Молекулярная масса составила 77,5 кДа, как определено MALDI-MS, что является больше, чем расчетная величина немодифицированного полипептида (73,5 кДа), что обусловлено гликозилированием. Присоединение гликанов к каждому из сайтов N-гликозилирования объясняет наблюдаемое увеличение массы. MASP-2A мигрирует как одиночная окрашенная зона в SDS-полиакриламидном геле, это свидетельствует о том, что в процессе биосинтеза он не подвергается протеолитическому разложению. Средневзвешенная молекулярная масса, определенная посредством равновесного ультрацентрифугирования, согласуется с расчетным значением для гомодимеров гликозилированного полипептида.
ПОЛУЧЕНИЕ РЕКОМБИНАНТНЫХ ПОЛИПЕПТИДОВ MASP-2 ЧЕЛОВЕКА
Другой способ получения рекомбинантных производных полипептидов MASP-2 и MASP2A описан у Thielens, N.M., et al., J. Immunol. 166:5068-5077, 2001. В кратком изложении суть его состоит в следующем: клетки насекомого Spodoptera frugiperda (клетки Ready-Plaque Sf9, полученные от компании «Novagen», Мэдисон, шт.Висконсин) выращиваются и поддерживаются в бессывороточной питательной среде Sf900ll («Life Technologies»), содержащей 50 МЕ/мл пенициллина и 50 мг/мл стрептомицина («Life Technologies»). Клетки насекомого Trichoplusia ni («High Five»), полученные от Jadwiga Chroboczek, Institut de Biologic Structurale, Гренобль, Франция, поддерживаются в питательной среде ТС100 («Life Technologies»), содержащей 10% FCS («Dominique Dutscher», Брумат, Франция), 50 МЕ/мл пеницилина и 50 мг/мл стрептомицина. Рекомбинантные бакуловирусы получают при помощи системы Bac-to-Bac («Life Technologies»). Бакмидная ДНК очищается при помощи системы очистки Qiagen midiprep («Qiagen») и используется для трансфекции клеток насекомых Sf9 при использовании селфектина в питательной среде Sf900 II SFM («Life Technologies»), как описано в протоколе производителя. Рекомбинантные вирусные частицы собирают через 4 дня, титруют методом вирусных бляшек и амплифицируют, как описано в King and Possee, в The Baculovirus Expression System: A Laboratory Guide, Chapman and Hall Ltd., London, pp.111-114, 1992.
Клетки High Five (1.75×107 клеток/175-см2 тканевой культуры в колбе) были инфицированы рекомбинантными вирусами, содержащими полипептиды MASP-2, посредством множественного инфицирования 2 в среде Sf900 II SFM при 28°С в течение 96 часов. Супернатанты собирали при помощи центрифугирования и добавляли диизопропил фосфорофлуоридат до конечной концентрации 1 ммоль.
Полипептиды MASP-2 секретировались клеточной культурой в питательную среду. Культуральные супернатанты подвергли процедуре диализа на фоне 50 ммоль NaCl, 1 ммоль CaCl2, 50 ммоль триэтаноламин гидрохлорида, рН 8,1, и загружали со скоростью 1,5 мл/мин в колонку Q-Sepharose Fast Flow («Amersham Pharmacia Biotech») (2,8×12 см), уравновешенную тем же буфером. Элюирование было проведено путем применения 1,2 литра линейного градиента к 350 ммоль NaCl в этом же буфере. Фракции, содержащие рекомбинантные полипептиды MASP-2, были идентифицированы при помощи вестерн блоттинга, осаждены путем добавления (NH4)2SO4 до 60% (по весу), и оставлены на ночь при 4°С. Осадки были ресуспендированы в 145 ммоль NaCl, 1 ммоль CaCl2, 50 ммоль триетаноламин гидрохлорида, рН 7,4, и добавлены в колонку TSK G3000 SWG (7,5×600 мм) («Tosohaas», Монтгомеривилл, шт.Пенсильвания), уравновешенную тем же буфером. Затем очищенные полипептиды были сконцентрированы до 0,3 мг/мл при помощи ультрафильтрации в микроконцентраторах Microsep (малозначимый молекулярный вес = 10000) («Fiitron», Карлштайн, Германия).
ПРИМЕР 6
Данный пример иллюстрирует способ получения поликлональных антител к полипептидам MASP-2.
Материалы и методы:
Антигены MASP-2: Антисыворотку поликлональных античеловеческих MASP-2 получают посредством иммунизации кроликов следующими выделенными полипептидами: человеческим MASP-2 (SEQ ID NO:6), выделенным из сыворотки, как описано в Примере 4; рекомбинантным человеческим MASP-2 (SEQ ID NO:6), MASP-2A, содержащим домен неактивной протеазы (SEQ ID NO:13), как описано в Примере 4-5; и рекомбинантными CUBI (SEQ ID NO:8), CUBEGFI (SEQ ID NO:9), и CUBEGFCUBII (SEQ ID NO:10), экспрессированными как описано выше в Примере 5.
Поликлональные антитела: Кролики в возрасте шести недель, которых подвергли первичному воздействию вакцины БЦЖ (бацилла Кальметта-Герена), были иммунизированы путем инъекции 100 мкг полипептида MASP-2 в 100 мкг/мл стерильного соляного раствора. Инъекции проводились каждые 4 недели с проведением мониторинга антител при помощи ИФА, как описано в Примере 7. Культуральные супернатанты были собраны с целью очищения антител посредством аффинной хроматографии белком А.
ПРИМЕР 7
Данный пример иллюстрирует способ получения мышиных моноклональных антител против крысиных или человеческих полипептидов MASP-2.
Материалы и методы:
Самцам мышей лини A/J («Harlan», Хьюстон, шт.Техас), в возрасте 8-12 недель, подкожно вводили 100 мкг человеческих или крысиных полипептидов rMASP-2 или rMASP-2A (полученых, как описано в Примере 4 или Примере 5) в полном адъюванте Фрейнда («Difco Laboratories», Детройт, шт.Мичиган) в 200 мкл забуференного фосфатом соляного раствора (PBS) с рН 7,4. С двухнедельными интервалами мыши дважды получили подкожные инъекции 50 мкг человеческого или крысиного полипептида rMASP-2 или rMASP-2A в неполном адъюванте Фрейнда. На четвертой неделе мыши получили инъекцию 50 мкг человеческого или крысиного полипептида rMASP-2 или rMASP-2A в PBS и через 4 дня мыши были использованы для слияния.
Для каждого слияния были приготовлены одиночные клеточные суспензии из селезенки иммунизированной мыши и использованы для слияния с Sp2/0 клетками миеломы. 5×108 клеток Sp2/0 и 5×108 клеток селезенки были слиты в среде, содержащей 50% полиэтиленгликоля (M.W. 1450) («Kodak», Рочестер, шт.Нью-Йорк) и 5% диметилсульфосида («Sigma Chemical Co.», Сент-Луис, шт.Миссури). Затем концентрация клеток селезенки была доведена до 1,5х105 клеток селезенки в 200 мкл суспензии в среде Iscove («Gibco», Гранд-Айленд, шт.Нью-Йорк), дополненной 10% фетальной сыворотки КРС, 100 единиц/мл пеницилина, 100 мкг/мл стрептомицина, 0,1 ммоль гипоксантина, 0,4 мкмоль аминоптерина и 16 мкмоль тимидина. Двести микролитров клеточной суспензии были добавлены в каждую лунку из приблизительно двадцати 96-луночных микропланшетов. После десяти дней проводили скрининга культуральныех супернатантов на предмет их взаимодействия с очищенным фактором MASP-2 при помощи ИФА.
Иммуноферментный анализ (ИФА): Лунки микропланшетов с Immunol 2 («Dynatech Laboratories», Шантильи, шт.Вирджиния) были покрыты посредством добавления 50 мкл очищенного hMASP-2 с концентрацией 50 нг/мл или крысиного rMASP-2 (или rMASP-2A) в течение ночи при комнатной температуре. Низкая концентрация MASP-2 для покрытия создает возможность для отбора высокоаффинных антител. После покрытия, раствор был удален посредством встряхивания планшета, и были добавлены 200 мкл препарата BLOTTO (обезжиренное сухое молоко) в PBS в каждую лунку на один час для блокирования неспецифических сайтов. Час спустя, лунки были отмыты буфером PBST (PBS с содержанием 0,05% Tween 20). Пятьдесят микролитров культуральных супернатантов от каждого слияния были собраны и смешаны с 50 мкл препарата BLOTTO и затем добавлены в индивидуальные лунки микропланшетов. После одного часа инкубации, лунки были промыты PBST. Связанные мышиные антитела затем были детектированы посредством реакции с пероксидазой хрена (HRP) коньюгированной с антимышиным IgG козла (Fc специфичный) («Jackson ImmunoResearch Laboratories», Вест-Гров, шт.Пенсильванния) и разбавлены в соотношении 1:2000 в BLOTTO. Субстратный раствор пероксидазы, содержащий 0,1% 3,3,5,5 тетраметилбензидина («Sigma», Сент-Луис, шт.Миссури) и 0,0003% перекиси водорода («Sigma») был добавлен в лунки и оставлен на 30 минут для проявления окрашивания. Реакция завершилась при добавлении в каждую лунку 50 мкл 2М H2SO4. Оптическая плотность реакционной смеси при 450 нм определяли с помощью прибора BioTek ELISA Reader («BioTek Instruments», Винооски, шт.Вермонт).
Анализ связывания MASP-2:
Культуральные супернатанты, давшие положительный результат в иммуноферментном анализе MASP-2, как было описано выше, могут быть протестированы в анализе связывания для определения связывающей аффинности агентов, ингибирующих MASP-2, с самим MASP-2. Сходный анализ может быть использован для определения того, может ли агент-ингибитор связываться с другими антигенами в системе комплемента.
Лунки полистирольного микропланшета (96-луночные планшеты, связывающие питательные среды, «Corning Costar», Кэмбридж, шт.Массачусетс) были покрыты MASP-2 (20 нг/100 мкл/на лунку, «Advanced Research Technology», Сна-Диего, шт.Калифорния) в забуференном фосфатом соляном растворе (PBS) с рН 7,4 в течение ночи при 4°С. После удаления раствора MASP-2, лунки были блокированы с PBS, содержащим 1% бычьего сывороточного альбумина (BSA; «Sigma Chemical»), в течение 2 часов при комнатной температуре. Лунки без покрытия MASP-2 использовались в качестве контроля. Аликвоты гибридомных супернатантов или очищенных анти-MASP-2 MoAbs, с варьирующей концентрацией в блокирующем растворе, были добавлены в лунки. После двухчасовой инкубации при комнатной температуре лунки обильно промывали PBS. MASP-2-связанное анти-MASP-2 MoAb было детектировано при добавлении коньюгированного с пероксидазой антимышиного IgG козла («Sigma Chemical») в блокирующий раствор, который инкубировали 1 час при комнатной температуре. Планшет снова тщательно промывали PBS, после чего добавляли 100 мкл субстрата 3,3',5,5'-тетраметилбензидина (ТМВ) («Kirkegaard and Perry Laboratories», Гейтерсберг, шт.Мэриленд). Реакцию ТМВ останавливали путем добавления 100 мкл 1 моля фосфорной кислоты, и планшет считывали с помощью прибора SPECTRA MAX 250 («Molecular Devices», Саннивейл, шт.Калифорния) при длине волны 450 нм.
Культуральные супернатанты из положительных лунок затем снова были протестированы на способность к ингибированию активации комплемента в функциональном исследовании, таком, как анализ расщепления С4, как описано в Примере 2. Затем клетки из положительных лунок были клонированы методом серийных разведении. MoAbs были снова протестированы на реагирование с hMASP-2 при помощи ИФА, как описано выше. Отобранные гибридомы выращивались во вращающихся колбах и истощенный культуральный супернатант собирали для очищения антител путем аффинной хроматографии белком А.
ПРИМЕР 8
Данный пример описывает получение нокаутной мыши MASP-2-/-, экспрессирующей человеческий MASP-2 для использования в качестве модели, для скрининга агентов, ингибирующих MASP-2.
Материалы и методы: MASP-2-/- мышь, как описано в Примере 1, и MASP-2-/-мышь, экспрессирующая трансгенную конструкцию MASP-2 человека (нокин MASP-2 человека), как описано в Примере 3, скрестили между собой, и полученное потомство, представленное мышиным MASP-2-/-, мышиным МАр19+, человеческим MASP-2+, было использовано для идентификации агентов, ингибирующих MASP-2 человека.
Подобные животные модели могут быть использованы качестве тестовых субстратов для идентификации эффективности агентов, ингибирующих MASP-2, таких как антитела анти-MASP-2 человека, пептиды и непептиды-ингибиторы MASP-2 и композиции, включающие в себя агенты, ингибирующие MASP-2. Например, животная модель была подвержена воздействию вещества или агента, известного своей способностью инициировать MASP-2-зависимую активацию комплемента; при этом агент, ингибирующий MASP-2, был введен животному в необходимое время и в достаточной концентрации для подавления симптомов заболевания у экспонированного животного.
Кроме того, мыши с генами мышиных MASP-2-/-, МАр19+ и человеческих MASP-2+ могут быть использованы для получения клеточных линий, содержащих один и более клеточных типов, вовлеченных в заболевание, опосредованное MASP-2, что может быть использовано в качестве модели клеточной культуры для данного заболевания. Создание стабильных клеточных линий от трансгенных животных хорошо известно современной науке, см., например, Small, J.A., etal, Mol. Cell Biol., 5:642-48, 1985.
ПРИМЕР 9
В данном примере описывается способ получения человеческих антител против человеческого MASP-2 у нокаутной мыши MASP-2, которые экспрессируют человеческий MASP-2 и человеческие иммуноглобулины.
Материалы и методы:
MASP-2-/- мышь была получена, как описано в Примере 1. Затем была получена мышь, которая экспрессировала человеческий MASP-2, как описано в Примере 3. Гомозиготная MASP-2-/- мышь и MASP-2-/- мышь, экспрессирующая человеческий MASP-2, были скрещены с мышью, полученной из линии стволовых эмбриональных клеток, при помощи методов генной инженерии, у которой имелись целевые разрывы в локусах тяжелой и легкой цепей эндогенного иммуноглобулина, и которая экспрессировала по меньшей мере сегмент локуса человеческого иммуноглобулина. Предпочтительно, сегмент локуса человеческого иммуноглобулина включает в себя неупорядоченные последовательности компонентов тяжелой и легкой цепей. Инактивация генов эндогенного иммуноглобулина и интродукция генов экзогенного иммуноглобулина может быть достигнуты путем целенаправленной гомологической рекомбинации. Трансгенные млекопитающие, полученные в результате данного процесса, способны к функциональной перегруппировке последовательностей компонентов иммуноглобулина и экспрессии спектра антител различных изотипов, кодируемых генами человеческого иммуноглобулина, без экспрессии генов эндогенного иммуноглобулина. Получение и свойства млекопитающих, обладающих подобными качествами, описаны, например, у Thomson, A.D., Nature 148:1547-1553, 1994, и Sloane, B.F, Nature Biotechnology 14:826, 1996. Полученные при помощи генной инженерии мышиные линии, в которых гены антител мыши инактивированы и функционально заменены на гены антител человека, являются коммерчески доступными (например, XenoMouse®, производства «Abgenix», Фремонт, шт.Калифорния). Полученное в результате потомство мышей способно продуцировать MoAb человека на фоне человеческого MASP-2, что является пригодным для использования в терапии человека.
ПРИМЕР 10
Данный пример описывает генерацию и продуцирование гуманизированных мышиных анти-MASP-2 антител и фрагментов антител.
Мышиное анти-MASP-2 моноклональное антитело получают от самца мыши линии A/J, как описано в Примере 7. Затем мышиное антитело подвергают процессу гуманизирования, как описано ниже, для редукции его иммуногенности посредством замены мышиных константных областей их человеческими аналогами для генерации химерного IgG и фрагмента Fab антитела, который может быть использован для ингибирования побочных эффектов MASP-2-зависимой активации комплемента у человеческих индивидуумов в соответствии с настоящим изобретением.
1. Клонирование вариабельных областей генов анти-MASP-2 из клеток мышиной гибридомы. Тотальную РНК выделяют из клеток гибридомы, секретирующих анти-MASP-2 MoAb (полученного, как описано в Примере 7) посредством использования RNAzol в соответствии с протоколом производителя («Biotech», Хьюстон, шт.Техас). Первая цепь кДНК синтезируется на основе тотальной РНК при использовании последовательности олиго dT в качестве праймера. PCR проводили с использованием 3' праймеров, производных от константной С-области иммуноглобулина, и наборов вырожденных праймеров, производных от лидирного пептида, или первой каркасной области мышиных генов VH или VK в качестве 5' праймеров. Зафиксированную PCR осуществляли, как описано у Chen и Platsucas (Chen, P.F, Scand. J. Immunol. 35:539-549, 1992). Для клонирования гена VK, двухцепочная кДНК была получена с использованием праймера Not1-MAK1 (5'-TGCGGCCGCTGTAGGTGCTGTCTTT-3' SEQ ID NO:38). гибридизационные адаптеры AD1 (5'-GGAATTCACTCGTTATTCTCGGA-3' SEQ ID NO:39) и AD2 (5'-TCCGAGAATAACGAGTG-3' SEQ ID NO:40) лигируются к обоим 5' и 3' концам двухцепочной кДНК. Адаптеры на 3' концах удаляются посредством расщепления Not1. Затем продукт расщепления используется как матрица в PCR с олигонуклеотидом AD1 в качестве 5' праймера и МАК2 (5'-CATTGAAAGCTTTGGGGTAGAAGTTGTTC-3' SEQ ID NO:41) в качестве 3' праймера. Фрагменты ДНК, приблизительно 500 пар оснований, клонируют в вектор pUC19. Несколько клонов выбираются для анализа последовательности с целью подтверждения того, что клонированная последовательность охватывает ожидаемую константную область мышиного иммуноглобулина. Олигонуклеотиды Not1-MAK1 и МАК2, получены из области VK и расположены на 182 и 84 паре оснований, соответственно, ниже от первой пары оснований гена С kappa. Были отобраны те клоны, которые содержали полноразмерный VK и лидирный пептид.
Для клонирования гена VH двухцепочная кДНК была получена с использованием праймера Not1 MAG1 (5'-CGCGGCCGCAGCTGCTCAGAGTGTAGA-3' SEQ ID NO:42). Гибридизационные адаптеры AD1 и AD2 лигируются к обоим 5' и 3' концам двухцепочной кДНК. Адаптеры на 3' концах удаляются посредством расщепления Not1. Затем продукт расщепления используется как матрица в PCR с олигонуклеотидом ADI и MAG2 (5'-CGGTAAGCTTCACTGGCTCAGGGAAATA-3' SEQ ID NO:43) в качестве праймеров. Фрагменты ДНК от 500 до 600 пар оснований в длину клонировали в pUC19. Олигонуклеотиды Not1-MAG1 и MAG2 получены из области Су.7.1 мыши, и расположены на 180 и 93 паре оснований, соответственно, ниже от первой пары оснований в гене мышиного Су.7.1. Были отобраны те клоны, которые содержали полноразмерный VH и лидирный пептид.
2. Конструирование экспрессионных векторов для химерного MASP-2 IgG и Fab. Клонированные гены VH и VK, описанные выше, использовали в качестве матрицы в реакции PCR для добавления консенсусной последовательности Козака к 5' концу и донор сплаисированного фрагмента к 3' концу нуклеотидной последовательности. После анализа последовательностей на отсутствие ошибок PCR, гены VH и VK встраивали в кассеты экспрессионных векторов, содержащих человеческий С. γI и С. kappa, соответственно, и получали pSV2neoVH-huCγI и pSV2neoV-huCγ. Очищенные в градиенте CsCl плазмидные ДНК векторов тяжелой и легкой цепи использовали для трансфекции клеток COS посредством электропорации. После 48 часов инкубации, культуральный супернатант тестировали при помощи ИФА анализа для подтверждения наличия приблизительно 200 нг/мл химерного IgG. Из собранных клеток выделяли тотальную РНК. Первая цепь кДНК была синтезирована на матрице тотальной РНК с использованием олиго dT в качестве праймера. Данная кДНК использовали в качестве матрицы в PCR для получения ДНК фрагментов Fd и kappa. Для гена Fd, PCR была проведена с использованием 5'-AAGAAGCTTGCCGCCACCATGGATTGGCTGTGGAACT-3' (SEQ ID NO:44) в качестве 5' праймера и СН1-производного 3' праймера (5'-CGGGATCCTCAAACTTTCTTGTCCACCTTGG-3' SEQ ID NO:45). Было установлено, что последовательность ДНК содержит полноценные VH и домен СН1 человеческого IgG1. После проведения расщепления соответствующими ферментами, фрагменты Fd ДНК были встроены в сайты рестрикции HindIII и BamHI в экспрессионную векторную кассету pSV2dhfr-TUS для получения pSV2dhfrFd. Плазмида pSV2 является коммерчески доступной и состоит из сегментов ДНК, взятых из различных источников: pBR322 ДНК (тонкая линия) содержит сайт начала репликации ДНК pBR322 (pBR ori) и ген лактамаза ампицилин устойчивости (Amp); SV40 ДНК, которая представлена широкими штрихами и маркировкой, содержит сайт начала репликации ДНК SV40 (SV40 ori), ранним промотором (5' к dhfr и неогенам), и сигнал полиаденилирования (3' к dhfr и неогенам). SV40-производный сигнал полиаденилирования (рА) был также помещен на 3' конец гена Fd.
Для гена kappa, PCR проводили с использованием 5'-AAGAAAGCTTGCCGCCACCATGTTCTCACTAGCTCT-3' (SEQ ID NO:46) в качестве 5' праймера и CK-производного 3' праймера (5'-CGGGATCCTTCTCCCTCTAACACTCT-3' SEQ ID NO:47). Было подтверждено, что последовательность ДНК содержит участки полноразмерного VK и человеческого CK. После расщепления соответствующими ферментами рестрикции, ДНК фрагменты kappa встраивали в сайты рестрикции HindIII и BamHI экспрессионной векторной кассеты pSV2neo-TUS для получения pSV2neoK. Экспрессия обоих генов Fd и kappa управляется HCMV-производными энхансером и промотором. Так как ген Fd не включает в себя остаток цистеиновой аминокислоты, принимающего участие в образовании дисульфидной связи между цепями, данный рекомбинантный химерный Fab содержит нековалентным образом связанные тяжелые и легкие цепи. Этот химерный Fab обозначен как cFab. Для получения рекомбинантного Fab с дисульфидной связью между тяжелой и легкой цепями, вышеуказанный ген Fd может быть расширен за счет включения последовательности, кодирующей 9 дополнительных аминокислот (EPKSCDKTH SEQ ID NO:48) от шарнирной области человеческого IgG1. Сегмент ДНК BstEII-BamHI, кодирующий 30 аминокислот на 3' конце гена Fd, может быть замещен сегментами ДНК, кодирующими расширенный Fd, что в результате дает pSV2dhfrFd/9aa.
3. Экспрессия и очистка химерного анти-MASP-2 IgG
Для получения клеточных линий, секретирующих химерный анти-MASP-2 IgG, клетки NSO подвергли процедуре траснфекции очищенной плазмидной ДНК pSV2neoVH-huC.γ1 и pSV2neoV-huC kappa посредством электропорации. Трансфецированные клетки отбирали в присутствии 0,7 мг/мл G418. Клетки выращивали в 250 мл вращающейся колбе в содержащей сыворотку питательной среде. Культуральный супернатант, полученный после центрифугирования 100 мл культуры, было загружен в 10-мл колонку PROSEP-A («Bioprocessing, Inc.», Принстон, шт.Нью-Джерси). Колонка была промыта 10 объемами слоя PBS. Связанное антитело элюировали 50 ммоль цитратным буфером, с рН 3,0. Равный объем 1 моль Hepes, с рН 8,0 был добавлен к фракции, содержащей очищенное антитело, с целью доведения рН до 7,0. Остаточные соли были удалены посредством замены буфера на PBS при помощи процедуры ультрафильтрации с мембраной Millipore (малозначимый молекулярный вес: 3000). Белковая концентрация очищенного антитела была определена методом ВСА (Pierce).
4. Экспрессия и очистка химерного анти-MASP-2 Fab
Для получения клеточных линий, секретирующих химерные анти-MASP-2 Fab, клетки СНО были подвергнуты процедуре трансфекции очищенной плазмидной ДНК pSV2dhfrFd (или pSV2dhfrFd/9aa) и pSV2neokappa посредством электропорации. Трансфецированные клетки отбирали в присутствии G418 и метотрексата. Выбранные клеточные линии были размножены в повышенных концентрациях метотрексата. Клетки были представлены одиночными клетками, субклонированными методом серийных разведении. Высокопродуктивные клеточные линии, полученные от субклонированных одиночных клеток затем выращивались в 100 мл культуры с постоянным перемешиванием на бессывороточной питательной среде.
Химерное анти-MASP-2 Fab очищали при помощи аффинной хромотографии с использованием мышиного анти-идиотипического MoAb к MASP-2 MoAb. Анти-идиотипическое MASP-2 MoAb может быть получено посредством иммунизации мыши мышиным анти-MASP-2 MoAb, коньюгированным с гемоцианином фиссуреллы (KLH), а затем подвергнуто скринингу на специфичное связывание с MoAb, которое может конкурировать с человеческим MASP-2. Для очистки, 100 мл супернатанта из культуры клеток СНО, продуцирующих cFab или cFab/9aa, было загружено в аффинную колонку, спаренную с анти-идиотипическим MASP-2 MoAb. Затем колонка была тщательно промыта PBS до элюции связанного Fab 50 ммоль диэтиламином с рН 11,5. Остаточные соли были удалены посредством замены буфера, как описано выше. Белковая концентрация очищенного Fab была определена методом ВСА (Pierce).
Способность химерных MASP-2 IgG, cFab и cFAb/9aa ингибировать MASP-2-зависимые пути активации комплемента может быть определена при помощи анализа ингибирования, описанного в Примере 2.
ПРИМЕР 11
Данный пример описывает анализ расщепления С4 in vitro, используемый в процедуре функционального скрининга для идентификации способности агентов, ингибирующих MASP-2, к блокированию MASP-2-зависимой активации комплемента при помощи Ьфиколина/Р35, Н-фиколина, М-фиколина или маннана.
Анализ расщепления С4: Анализ расщепления С4 был описан у Petersen, S.V., et al., J. Immunol. Methods 257:107, 2001, где производились измерения уровня активации пектинового пути, вызванного липотейхоевой кислотой (LTA) из S. aureus, которая связывает L-фиколин.
Реагенты: Формалин-зафиксированный препарат S. aureous (DSM20233) готовили следующим образом: бактерии выращивали в течение ночи при 37°С в триптон-соевой кровяной питательной среде, после трижды промывали в PBS, затем фиксировали в течение 1 часа при комнатной температуре в PBS/0,5% формалина, и) трижды отмывали в PBS перед ресуспендированием в покрывающем буффере (15 ммоль Na2CO3, 35 ммоль NaHCO3, pH 9,6.
Анализ: Лунки микропланшета Nunc MaxiSorb («Nalgene Nunc International», Рочестер, шт.Нью-Йорк) были покрыты 100 мкл препарата формалин-зафиксированного S. aureus DSM20233 (00550=0,5) в покрывающем буфере с 1 мкг L-фиколина. После инкубации в течение ночи, лунки были блокированы 0,1% раствором сывороточного альбумина человека (HSA) в TBS (10 ммоль Tris-HCl, 140 ммоль NaCl, pH 7,4), затем были отмыты TBS, содержащим 0,05% Tween 20 и 5 ммоль CaCI2 (промывочный буфер). Образцы человеческой сыворотки были растворены в 20 ммоль Tris-HCl, 1 моль NaCl, 10 ммоль CaCI2, 0,05% Triton X-100, 0,1% HSA, pH 7,4, которые предупреждают активацию эндогенного С4 и разлагают комплекс С1 (состоящий из C1q, C1r и C1s). Агенты, ингибирующие MASP-2, включая aKm-MASP-2 MoAbs и пептиды-ингибиторы, добавляли к образцам сыворотки в различных концентрациях. Разведенные образцы наносили на планшет и инкубировали в течение ночи при 4°С. После 24 часов планшеты тщательно промывали промывочным буфером, затем 0,1 мкг очищенного человеческого С4 (полученного, как описано у Dodds, A.W, Methods Enzymol. 223:46, 1993) в 100 мкл 4 ммоль барбитала, 145 ммоль NaCl, 2 ммоль CaCl2, 1 ммоль MgCl2, pH 7,4 добавляли в каждую лунку. После 1,5 часов инкубации при 37°С, планшеты снова промывали и определяли отложение C4b посредством коньюгированного с щелочной фосфатазой куриного античеловеческого С4 с («Immunsystem», Уппсала, Швеция) и производили замер с использованием колориметрического субстрата р -нитрофенилфосфата.
Анализ С4 на маннане: Вышеописанный анализ был адаптирован для измерения активации пектинового пути посредством MBL при помощи покрытия планшета маннаном и LSP перед добавлением сыворотки, смешанной с различными агентами, ингибирующими MASP-2.
Анализ С4 на Н-фиколине (Hakata Ад): Вышеописанный анализ был адаптирован для измерения активации пектинового пути посредством Н-фиколина при помощи покрытия планшета Н-фиколином и LPS перед добавлением сыворотки, смешанной с различными агентами, ингибирующими MASP-2.
ПРИМЕР 12
Следующий анализ демонстрирует наличие классического пути активации у мышей дикого типа и MASP-2-/- мышей.
Методы: Иммунные комплексы были получены in situ путем покрытия микропланшетов (Maxisorb, Nunc, кат. №442404, «Fisher Scientific») 0,1% раствором сывороточного альбумина человечека в 10 ммоль Tris, 140 ммоль NaCl, pH 7,4 на 1 час при комнатной температуре с последующей инкубацией в течение ночи при 4°С с овечьей антисывороткой на фоне цельной сыворотки («Scottish Antibody Production Unit», Карлюк, Шотландия), разведенной в соотношении 1:1000 с TBS/tween/Ca2+. Образцы сыворотки были получены от мышей дикого типа и MASP-2-/- мышей и добавлены в покрытые планшеты. Были приготовлены контрольные образцы, которые содержали образцы сыворотки мышей дикого типа и MASP-2-/- с истощенным уровнем C1q. Мышиная сыворотка с истощенным уровнем C1q была приготовлена с использованием сопряженного с белком А препарата Dynabeads («Dynal Biotech», Осло, Норвегия), покрытого кроличьими античеловеческими C1q IgG («Dako», Глоструп, Дания), в соответствии с инструкциями производителя. Планшеты инкубировали в течение 90 минут при 37°С. Связанный C3b детектировали с поликлональным античеловеческим С3с антителом (Dako A 062), растворенном в TBS/tw/ Ca++ в соотношении 1:1000. Вторичным антителом является антикроличий IgG козла.
Результаты: На ФИГУРЕ 9 показаны относительные уровни отложения C3b на планшетах, покрытых IgG, в сыворотке дикого типа, сыворотке MASP-2-/-, сыворотке дикого типа с истощенным C1q и сыворотке MASP-2-/- с истощенным C1q. Данные результаты демонстрируют, что классический путь остается незатронутым у мышей линии MASP-2-/-.
ПРИМЕР 13
Следующий анализ был использован для тестирования способности агента, ингибирующего MASP-2, блокировать классический путь активации, посредством изучения действия агента, ингибирующего MASP-2, в условиях, при которых классический путь активируется иммунными комплексами.
Методы: Для тестирования эффективности агента, ингибирующего MASP-2 в условиях активации комплемента, когда классический путь активации инициируется иммунными комплексами, три образца сыворотки по 50 мкл с содержанием 90% NHS были инкубированы при 37°С в присутствии 10 мкг/мл иммунного комплекса (IC) или PBS, и, параллельно, были включены еще три образца (+/-IC), которые содержали 200 нмоль моноклональных анти-пропердин антител во время инкубации при 37°С. После двухчасовой инкубации при 37°С, во все образцы добавили 13 ммоль EDTA с целью прекратить дальнейшую активацию комплемента, после чего все образцы были немедленно охлаждены до 5°С. Затем образцы хранили при температуре -70°С до момента их использования в анализе на продукты активации комплемента (С3а и sC5b-9) с использованием набора ИФА («Quidel», кат. №А015 и А009) в соответствии с инструкциями производителя.
ПРИМЕР 14
Данный пример демонстрирует, что лектин-опосредованная система MASP-2 активации комплемента актируется в фазе ишемии/реперфузии, следующей за репарацией аневризмы брюшной аорты.
Обоснование и планирование эксперимента: Пациенты, перенесшие репарацию аневризмы брюшной аорты (ААА) находятся в группе риска ишемического реперфузионного поражения, которое в основном вызывается активацией комплемента. Мы исследовали роль MASP-2-зависимой активации комплемента посредством пектинового пути при ишемическом реперфузионном поражении у пациентов, перенесших репарацию ААА. Истощение маннозосвязывающего пектина (MBL) в сыворотке было использовано для оценки степени MASP-2-зависимой активации пектинового пути, которая происходит при реперфузии.
Получение образцов сыворотки пациента: В общей сложности, были обследованы 23 пациента, перенесшие элективную инфраренальную реперфузию ААА и 8 контрольных пациентов, перенесших крупное хирургическое вмешательство на органах брюшной полости.
Для пациентов, перенесших репарацию ААА, был произведен забор образцов крови из системного кровотока из лучевой артерии каждого пациента (при помощи внутриартериального катетера) в четыре определенных временных точки во время процедуры: временная точка 1: индукция анестезии; временная точка 2: непосредственно перед проведением аортального зажима; временная точка 3: непосредственно перед удалением аортального зажима; и временная точка 4: во время реперфузии.
Для контрольных пациентов, перенесших крупное хирургическое вмешательство на органах брюшной полости, был произведен забор образцов крови из системного кровотока во время индукции анестезии и два часа после начала процедуры.
Анализ уровней MBL: Образцы плазмы каждого пациента были проанализированы на уровни маннозсвязывающего лектина (MBL) с использованием технологии ИФА.
Результаты: Результаты данного исследования показаны на ФИГУРЕ 10, которая содержит график, демонстрирующий среднее изменение процентного соотношения в уровнях MBL (по оси Y) в каждую из различных временных точек (по оси X). Начальные объемы MBL приняты за 100%, с показанным позже относительным уменьшением. Как показано на ФИГУРЕ 10, пациенты с ААА (n=23) проявили значительное снижение уровней MBL в плазме, которое колебалось приблизительно на уровне 41%-ного понижения в период ишемии/реперфузии, следующей за ААА. В отличие от этих пациентов, в образцах плазмы контрольных пациентов (n=8), перенесших крупное хирургическое вмешательство на органах брюшной полости, было обнаружено достаточно незначительное потребление MBL.
Представленные данные явно указывают на то, что MASP-2-зависимый пектиновый путь активации системы комплемента инициируется в фазе ишемии/реперфузии, следующей за репарацией ААА. Понижение уровней MBL связывают с ишемическим/реперфузионным поражением, так как уровни MBL значительно снижаются при реперфузии зажатого основного кровеносного сосуда в конце операции. В то время как, контрольная сыворотка пациентов, перенесших крупное хирургическое вмешательство на органах брюшной полости без крупного ишемического реперфузионного инсульта, показывает лишь незначительное снижение уровней MBL в плазме. В свете четко установленного вклада активации комплемента при реперфузионном поражении, можно сделать вывод о том, что активация MASP-2-зависимого пути в ишемических эндотелиальных клетках представляет собой основной фактор в патологии ишемического реперфузионного поражения. Следовательно, можно ожидать, что специфическая кратковременная блокада или редукция MASP-2-зависимого пути активации комплемента оказывает значительный благотворный терапевтический эффект для улучшения исхода клинических процедур и самого заболевания, которое связано с кратковременным ишемическим инсультом, например, инфаркт миокарда, интестинальный инфаркт, ожоги, трансплантация и инсульт.
ПРИМЕР 15
Данный пример описывает использование линии MASP-2-/- в качестве животной модели для тестирования агентов, ингибирующих MASP-2, используемых для лечения ревматоидного артрита.
Предпосылки и обоснование: Мышиная модель артрита: трансгенные мыши (tg) с K/B×N Т клеточным рецептором (TCR), являются недавно сконструированной моделью воспалительного артрита (Kouskoff, V., et al., Cell 87:811-822, 1996; Korganow, A.S., et al., Immunity 10:451-461, 1999; Matsumoto, L, et al., Science 286:1732-1735, 1999; Maccioni M. et al., J. Exp. Med. 195(8): 1071-1077, 2002). У мышей K/B×N происходит спонтанное развитие аутоиммунного заболевания, большинство клинических, гистологических и иммунологических признаков которого совпадают с проявлениями ревматоидного артрита у людей (Ji, H., et al., Immunity 16:157-168, 2002). Суставные заболевания мышей отличаются определенной спецификой, однако они инициируются и затем сохраняются в организме посредством аутоиммунной реакции Т-, а затем В-клеток в отношении глюкозо-6-фосфатизомеразы (GPI), повсеместно экспрессируемого антигена. Далее, передача сыворотки (или очищенного анти-GPI Igs) от артритной K/B×N мыши к здоровому животному вызывает артрит спустя несколько дней после заражения. Также было показано, что поликлональные анти-GPI антитела или совокупность анти-GPI моноклональных антител изотипа IgGI индуцирует артрит при проведении инъекций здоровым реципиентам (Maccioni et al., 2002). Мышиная модель является релевантной к человеческому ревматоидному артриту, так как сыворотка, полученная от пациентов с ревматоидным артритом содержит анти-GPI антитела, которые не обнаруживаются у нормальных индивидуумов. Мыши с С5-недостаточностью были протестированы на основе данной системы и проявили способность к блокированию развития артрита (Ji, H., et al., 2002, supra). Также был зафиксирован факт стойкого ингибирования артрита у мышей с нулевым С3, что подразумевает вовлечение альтернативного пути активации, однако, артрит не развивался у мышей с нулевым МВР-А. Однако наличие у мышей МВР-С может компенсировать потерю МВР-А.
На основании приведенного в данной работе утверждения, что MASP-2 исполняет основную роль в инициации как пектинового, так и альтернативного пути, модель артрита K/B×N используется для скрининга агентов, ингибирующие MASP-2, проявляющих эффективность для использования в качестве терапевтических агентов для лечения ревматоидного артрита.
Методы: Образцы сыворотки от K/B×N мышей, страдающих артритом, в возрасте 60 дней, объединили в пул и ввели посредством инъекций (150-200 мкл интраперитонеально) MASP-2-/- реципиентам (получено, как описано в Примере 1); и контрольным однопометным животным с или без агентов, ингибирующих MASP-2 (MoAb, пептиды-ингибиторы и т.п., как описано в данном изобретении) в дни 0 и 2. Группа мышей с нормальным набором генов также подверглась обработке агентом, ингибирующим MASP-2, за два дня до введения им сыворотки. Следующая группа мышей получила инъекцию сыворотки в день 0, с последующим введением агента, ингибирующего MASP-2, в день 6. Клинический показатель оценивался в динамике по времени в баллах: по одному баллу на каждую пораженную артритом лапу, и по половине балла на каждый участок лапы с незначительным воспалением. Также было замерено утолщение голеностопного сустава при помощи циркуля (утолщение определяется как разница по сравнению с измерением в день 0).
ПРИМЕР 16
Данный пример описывает исследование ингибирования комплемент-зависимого поражения тканей ex vivo в животных моделях кроличьих сердец, подвергающихся перфузии человеческой плазмой.
Предпосылки и обоснование: Активация системы комплемента вносит свой вклад в процесс гиперактивного отторжения ксенотрансплантатов. Предыдущие исследования показали, что гиперактивное отторжение может происходить в отсутствие антидонорских антител через активацию альтернативного пути (Johnston, P.S., et al., Transplant Proc. 23:877-879, 1991).
Методы: Для определения способности выделенных анти-MASP-2 ингибирующих агентов, таких как анти-MASP-2 антитела, описанные в Примере 7, ингибировать пути активации комплемента при поражении тканей, анти-MASP-2 MoAbs и фрагменты антител могут быть протестированы с использованием модели ех vivo, в которой изолированные кроличьи сердца подвергаются перфузии разведенной человеческой плазмой. В предыдущих исследованиях данная модель была показана как вызывающая поражение миокарда кролика вследствие активации альтернативного пути комплемента (Gralinski, M.R., et al., Immunopharmacology 34:79-88,1996).
ПРИМЕР 17
Данный пример описывает исследование по измерению активации нейтрофилов, что может быть использовано для определения дозы агента, ингибирующего MASP-2, эффективной для лечения заболевания, вызванного лектин-зависимым путем активации комплемента в соответствии со способами данного изобретения.
Методы: Способ для замерения эластазы нейтрофилов был подробно описан у Gupta-Bansal, R., et al., Molecular Immunol. 37:191-201, 2000. В кратком изложении он выглядит следующим образом: комплекс эластазы и сыворотки α1-антитрипсина был измерен при помощи двухцентрового сэндвич-метода, в котором используются антитела против эластазы и α1-антитрипсина. Полистироловые микропланшеты покрывали раствором антитела античеловеческой эластазы 1:500 («The Binding Site», Бирмингем, Великобритания) в PBS в течение ночи при 4°С. После удаления раствора антител, лунки были блокированы PBS, содержащим 0,4% HAS в течение двух часов, при комнатной температуре. Аликвотные пробы (100 мкл) образцов плазмы, обработанных или необработанных ингибирующим MASP-2 агентом, были нанесены в лунки. После двухчасовой инкубации при комнатной температуре, лунки были тщательно промыты в PBS. Связанный комплекс эластазы-α1-антитрипсина детектировали посредством добавления 1:500 раствора антитела пероксидазы, коньюгированной с антителом к α1-антитрипсином в блокирующем растворе, и последующего инкубирования в течение 1 часа при комнатной температуре. После промывания планшета PBS, в лунки было добавлено 100 мкл субстрата аликвотных проб ТМВ. Реакция ТМВ останавливали посредством добавления 100 мкл фосфорной кислоты, планшеты считывали при длине волны 450 нм при помощи считывающего микропланшеты устройства.
ПРИМЕР 18
Данный пример описывает животную модель для тестирования агентов, ингибирующих MASP-2, применяемых для лечения миокардиальной ишемии/реперфузии.
Методы: Модель миокардиальной ишемии-реперфузии была описана у Vakeva et al. Circulation 97:2259-2267, 1998, и Jordan et al., Circulation 104(12):1413-1418, 2001. Описанная модель может быть модифицирована для использования на MASP-2-/- и MASP-2+/+ мышах следующим образом. Проводили анестезию взрослых особей мышей. Яремную вену и трахею подвергли процедуре катетеризации, и осуществляли вентиляцию 100% кислородом при помощи вентилирующего устройства, настроенного таким образом, чтобы поддерживать выдыхаемый СО2 между 3,5% и 5%. Далее выполняли торакотомию левой доли легкого, а хирургический шов располагали на расстоянии от 3 до 4 мм от места расположения левой венечной артерии. За пять минут до ишемии, животные получали агент, ингибирующий MASP-2, примером которого являются анти-MASP-2 антитела (например, в диапазоне доз от 0,01 до 10 мг/кг). Затем ишемию провоцировали сжатием шва вокруг коронарной артерии и поддерживали в течение 30 минут, за чем следовала четырехчасовая реперфузия. Животные с фиктивной операцией были подготовлены идентичным способом без стягивания шва.
Анализ отложения комплемента С3: После реперфузии образцы для иммуногистохимии были получены из центрального участка левого желудочка сердца, зафиксированного и замороженного при -80°С до начала анализа. Срезы тканей подвергли инкубации с HRP-коньюгированным антикрысиным С3 антителом козла. Срезы тканей анализировали на наличие С3 окрашиванием в присутствии анти-MASP-2 агентов-ингибиторов, и проводили сравнение животных, перенесших симуляцию операции, и MASP-2-/- животных для идентификации агентов, ингибирующих MASP-2, редуцирующих отложение С3 in vivo.
ПРИМЕР 19
Данный пример описывает использование линии MASP-2-/- в качестве животной модели для тестирования агентов, ингибирующих MASP-2, на способность защищать трансплантированную ткань от ишемического реперфузионного поражения.
Предпосылки и обоснование: Известно, что ишемическое реперфузионное поражение происходит в донорских органах при трансплантации. Масштаб тканевого поражения связан с продолжительностью ишемии и опосредован комплементом, что продемонстрировано в многочисленных моделях ишемии и при использовании агентов, ингибирующих комплемент, например, растворимого рецептора типа 1 (CR1) (Weisman et al., Science 249:146-151, 1990; Mulligan et al., J. Immunol. 148:1479-1486, 1992; Pratt et al., Am. J. Path. 163(4): 1457-1465, 2003). Животная модель трансплантации была описана у Pratt et al., Am. J. Path. 163(4): 1457-1465, данная модель может быть модифицирована для использования с MASP-2-/- мышиной моделью и/или в качестве MASP-2+/+ модельной системы, в которой возможен скрининг агентов, ингибирующих MASP-2, на предмет их способности защищать трансплантируемую ткань от ишемического реперфузионного поражения. Промывание донорской почки перфузионной жидкостью перед процедурой трансплантации предоставляет возможность введения агентов, ингибирующих анти-MASP-2, в донорскую почку.
Методы: Проводили анестезию MASP-2-/- и/или MASP-2+/+ мышей. Левая донорская почка была иссечена, аорту лигировали краниально и каудально по отношению к ренальной артерии. Трубчатый катетер («Portex Ltd», Хайт, Великобритания) был внедрен между лигатурами, почку подвергли перфузии 5 мл раствора для почечной перфузии Soltran («Baxter Health Care», Великобритания), содержащего агенты, ингибирующие MASP-2, например, анти-MASP-2 моноклональные антитела (в диапазоне доз от 0,01 мг/кг до 10 мг/кг) на время не менее 5 минут. Затем была проведена трансплантация почки, в течение которой мыши находились под постоянным мониторингом.
Изучение реципиентов трансплантатов: Почечные трансплантаты были собраны в различные временные промежутки и срезы тканей были проанализированы с использованием анти-С3 для определения масштабов отложения С3.
ПРИМЕР 20
Данный пример описывает использование животной модели коллаген-индуцированного артрита (CIA) для тестирования агентов, ингибирующих MASP-2, используемых для лечения ревматоидного артрита.
Предпосылки и обоснование: Коллаген-индуцированный артрит (CIA) представляет собой аутоиммунный полиартрит, индуцируемый в восприимчивых линиях грызунов и приматов после иммунизации коллагеном природного типа II, и признан релевантной моделью человеческого ревматоидного артрита (RA) (см. Courtney et al., Nature 283: 666 (1980); Trenthan et al., J. Exp. Med. 146: 857 (1977)). Оба типа RA и CIA характеризуются воспалением суставов, формированием паннуса и эрозией хряща и кости. Мышиные линии DBA/1LacJ, восприимчивые к CIA, являются выведенными моделями CIA, в которых у мышей развивается артрит со сложными клиническими проявлениями после иммунизации бычьим коллагеном типа II (Wang et al., J. Immunol. 164: 4340-4347 (2000). Линия мышей с С5-недостаточностью была скрещена с DBA/1LacJ, и полученная в результате скрещивания линия обнаружила большую устойчивость к развитию артрита CIA (Wang et al., 2000, supra).
Основываясь на приведенных в данном исследовании наблюдениях, подтверждающих, что MASP-2 играет ведущую роль в инициации как пектинового, так и альтернативного путей активации комплемента; модель артрита CIA может быть использована для проведения скрининга агентов, ингибирующих MASP-2, являющихся эффективными для использования в качестве терапевтических агентов для лечения RA.
Методы: Мышь MASP-2-/- получают способом, описанным в Примере 1. Затем производят скрещивание мыши MASP-2-/- с мышью, представляющей другую линию DBA/1LacJ («The Jackson Laboratory)». F1 и последующее потомство скрещивают между собой для получения гомозиготных MASP-2-/- в линии DBA/1LacJ.
Коллагенная иммунизация проводится по способу, описанному у Wang et al., 2000, supra. В соответствии с ним, мышей DBA/1LacJ дикого типа и мышей MASP-2-/-DBA/1LacJ подвергали иммунизации бычьим коллагеном типа II (BCII) или мышиным коллагеном типа II (MCII) (полученными от «Elastin Products», Оуэнсвилл, шт.Миссури), растворенными в 0,01 моль ацетиловой кислоты в концентрации 4 мг/мл. Каждая мышь получала инъекцию 200 мкг СИ и 100 мкг микобактерий внутрикожно у основания хвоста. Повторная иммунизация производилась через 21 день, каждый день проводилось обследование животных на проявление артрита. Показатели артрита оцениваются с течением времени относительно серьезности проявлений артрита в каждой пораженной конечности.
Скрининг агентов, ингибирующих MASP-2, на мышах дикого типа DBA/1LacJ CIA осуществляли посредством инъекции агентов, ингибирующих MASP-2, таких как анти-MASP-2 моноклональные антитела (в диапазоне доз от 0,01 мг/кг до 10 мг/кг) во время коллагеновой иммунизации, либо системно, либо местно в один или несколько суставов, показатели артрита оценивали в динамике по времени, как описано выше. AHTH-hMASP-2 моноклональные антитела в качестве терапевтических агентов могут быть оценены в мышиных моделях MASP-2-/-, нокине hMASP-+/+ DBA/1LacJ CIA.
ПРИМЕР 21
Данный пример описывает использование животной модели (NZB/W) F1 для тестирования агентов, ингибирующих MASP-2, эффективных для лечения гломерулонефритов, опосредованных иммунным комплексом.
Предпосылки и обоснование: У новозеландских черных мышей, скрещенных с новозеландскими белыми мышами (New Zealand black × New Zealand white) (NZB/W) F1)), спонтанно проявляется аутоиммунный синдром с синдромами, существенными схожими с гломерулонефритом человека, опосредованным иммунными комплексом. Мыши NZB/W F1 всегда погибают от гломерулонефрита в возрасте 12 месяцев. Как обсуждалось ранее, было показано, что активация комплемента играет значительную роль в патогенезе гломерулонефрита, опосредованного иммунным комплексом. В дальнейшем было показано, что введение анти-С5 MoAb мышиной модели NZB/W F1 привело к значительному улучшению течения гломерулонефрита (Wang et al., Proc. Natl. Acad. Sci. 93:8563-8568 (1996)). Основываясь на приведенных в данной заявке наблюдениях, показывающих, что MASP-2 играет существенную роль в инициации как пектинового, так и альтернативного путей активации комплемента, животная модель NZB/W F1 используется для скрининга агентов, ингибирующих MASP-2, проявляющих эффективность в качестве терапевтических агентов для лечения гломерулонефрита.
Методы: Мышь MASP-2-/- получают способом, описанным в Примере 1. Затем мышь MASP-2-/- по-отдельности скрещивают с мышами, полученными от линий NZB и NZW («The Jackson Laboratory»). F1 и последующее потомство затем скрещивают между собой для получения гомозиготной MASP-2-/- на генетических основах как NZB, так и NZW. Для определения роли MASP-2 в патогенезе гломерулонефрита в данной модели было проведено сравнение развития данного заболевания у индивидуумов F1, полученных в результате скрещивания NZB × NZW мышей дикого типа или мышей MASP-2-/-NZB × MASP-2-/-NZW. Образцы мочи собирали через недельные интервалы у мышей MASP-2+/+ и MASP-2-/- F1, уровни белков в моче подвергали мониторингу на наличие анти-ДНК антител (как описано у Wang et al., 1996, supra). Также проводили гистопатологическое исследование почек для контроля объема мезангиального матричного отложения и развития гломерулонефрита.
Животная модель NZB/W F1 также использовалась для скрининга агентов, ингибирующих MASP-2, на эффективность в качестве терапевтических агентов для лечения гломерулонефрита. Мыши дикого типа NZB/W F1 в возрасте 18 недель получали интраперитонеальные инъекции с агентами, ингибирующими анти-MASP-2, например анти-MASP-2 моноклональными антителами (в диапазоне доз от 0,01 мг/кг до 10 мг/кг) раз в неделю или раз в две недели. Вышеупомянутые гистопатологические и биохимические маркеры гломерулонефрита используются для оценки развития заболевания у мышей и идентификации агентов, ингибирующих MASP-2, которые могут быть использованы для лечения данного заболевания.
ПРИМЕР 22
Данный пример описывает использование трубчатого шунта в качестве модели для тестирования агентов, ингибирующих MASP-2, эффективных в предупреждении поражения тканей, полученного в результате экстракорпорального кровообращения (ЕСС), например, при помощи сердечно-легочного шунтирования (СРВ).
Предпосылки и обоснование: Как обсуждалось ранее, пациенты, перенесшие ЕСС во время проведения процедуры с помощью СРВ, страдают систематической воспалительной реакцией, которая частично опосредована контактом крови с искусственными поверхностями аппарата экстракорпорального кровообращения, равно как и не связанными с этим факторами, например, хирургической травмой и ишемическим-реперфузионным поражением (Butler, J., et al., Ann. Thorac. Surg. 55:552-9, 1993; Edmunds, L.H., Ann. Thorac. Surg. 66 (Suppl):S12-6, 1998; Asimakopoulos, G., Perfusion 14:269-77, 1999). Далее было показано, что альтернативный путь активации комплемента играет доминирующую роль в активации комплемента в цепи СРВ, опосредованную взаимодействием крови с искусственными поверхностями в цепи СРВ (см. Kirklin et al., 1983, 1986, упомянутую ранее). Поэтому, основываясь на приведенных в данной заявке наблюдениях, было установлено, что MASP-2 играет существенную роль в инициации как пектинового, так и альтернативного пути активации комплемента, модель трубочного шунтирования была использована для скрининга агентов, ингибирующих MASP-2, проявляющих эффективность при использовании в качестве терапевтических агентов для предупреждения либо лечения воспалительной реакции, вызванной экстракорпоральной экспозицией.
Методы: Модификация вышеописанной модели трубочного шунтирования для циркулирующего экстракорпорального кровообращения используется (см. Gong et al., J. Clinical Immunol. 16(4):222-229 (1996)), как описано у Gupta-Bansal et al., Molecular Immunol. 37:191-201 (2000). Согласно этому описанию, у здорового субъекта производится забор свежей крови в 7 мл вакуумную пробирку (содержащую 7 единиц гепарина на 1 мл цельной крови). Полиэтиленовые трубки, аналогичные тем, что используются при проведении процедур СРВ (например, вн. диам. 2,92 мм; внеш. диам. 3,73 мм, длина: 45 см), наполняют 1 мл крови и закрывают в петле с коротким участком силиконовой трубки. Контрольные трубки, содержащие гепаринизированную кровь с 10 ммоль EDTA, были включены в исследование в качестве контрольного первоисточника. Образцы и контрольные трубки вращали в вертикальном положении в водяной бане в течение 1 часа при 37°С. После инкубации, образцы крови переносили в микроцентрифужные пробирки объемом 1,7 мл, содержащие EDTA, с конечной концентрации 20 ммоль EDTA. Затем производили центрифугирование образцов и отбор плазмы. Агенты, ингибирующие MASP-2, такие как антитела анти-MASP-2, добавляли в гепаринизированную кровь непосредственно перед ротацией. Затем проводили исследование образцов плазмы для замера концентрации С3а и растворимого C5b-9, как описано у Gupta-Bansal et al., 2000, supra.
ПРИМЕР 23
Данный пример описывает использования модели системы лигирования и пункции слепой кишки у грызунов (CLP) для тестирования агентов, ингибирующих MASP-2, эффективных для лечения сепсиса или опосредованного им заболевания, в том числе острый сепсис, септический шок, синдром острой дыхательной недостаточности как сопутствующее заболевание при сепсисе и синдром системной воспалительной реакции.
Предпосылки и обоснование: Как обсуждалось ранее, во многих исследованих была показана ведущая роль активации комплемента в патогенезе сепсиса (см. Bone, R.C., Annals. Internal. Med. 115:457-469, 1991). Модель грызунов CLP является признанной моделью, имитирующей клиническое течение сепсиса у человека, и рассматривается в качестве подходящей суррогатной модели для сепсиса человека (см. Ward, P., Nature Review Immunology Л: 133-142 (2004). Недавнее исследование показало, что лечение животных с CLP анти-С5а антителами выражается в снижении бактериемии и значительном повышении уровня выживаемости (Huber-Lang et al., J. of Immunol. 169: 3223-3231 (2002). Поэтому, основываясь на приведенных в данной заявке наблюдениях, показывающих, что MASP-2 принадлежит доминирующая роль в инициации как пектинового, так и альтернативного пути активации, модель грызунов CLP можно использовать для скрининга агентов, ингибирующих MASP-2, проявляющих эффективность в качестве терапевтических агентов для предупреждения или лечения сепсиса или опосредованного сепсисом заболевания.
Методы: Модель CLP является адаптированной моделью, описанной у Huber-Lang et al., 2004, supra. Животных MASP-2-/- и MASP-2+/+ подвергали анестезии. Делали двухсантиметровый разрез по средней линии брюшной стенки, и слепая кишка была туго лигирована ниже илеоцекального клапана, для избежания кишечной непроходимости. Затем проводили пункцию слепой кишки при помощи иглы калибра 21. После этого разрез брюшной стенки зашивали шелковой нитью и фиксировали при помощи кожных зажимов («Ethicon», Саммервилл, шт.Нью-Джерси). Непосредственно после CLP, животным проводят инъекцию агента, ингибирующего MASP-2, например, aHTH-MASP-2 моноклональных антител (в диапазоне доз от 0,01 мг/кг до 10 мг/кг). AHTH-hMASP-2 моноклональные антитела в качестве терапевтических агентов могут быть оценены в мышиных моделях CLP на MASP-2-/-, hMASP-+/+нокин-мышах. Затем плазму мышей анализируют на предмет уровней анафилатоксинов, производных комплемента, и окислительного взрыва при помощи исследований, описанных у Huber-Lang et al., 2004, supra.
ПРИМЕР 24
Данный пример описывает процесс идентификации высокоаффинных фрагментов антител анти-MASP-2 Fab2, блокирующих активность MASP-2.
Предпосылки и обоснование: MASP-2 представляет собой белковый комплекс со множеством отдельных функциональных доменов, включая: сайт(ы) связывания для MBL и фиколинов, каталитический сайт сериновой протеазы, сайт связывания для протеолитического субстрата С2, сайт связывания для протеолитического субстрата С4, сайт расщепления MASP-2 для самоактивации зимогена MASP-2 и два связывающих сайта Са++. Были идентифицированы фрагменты антител Fab2, которые образуют высокоаффинную связь с MASP-2, идентифицированные фрагменты Fab2 были протестированы в функциональном исследовании для определения их способности к блокированию функциональной активности MASP-2.
Для блокирования функциональной активности MASP-2 антитело или фрагмент антитела Fab2 должны связываться и вступать во взаимодействие со структурным эпитопом на MASP-2, который необходим для функциональной активности MASP-2. Поэтому многие или все высокоаффинные связывающие анти-MASP-2 Fab2 не обладают способностью ингибировать функциональную активность MASP-2, если они не связываются со структурными эпитопами на MASP-2, которые напрямую обуславливает функциональную активность MASP-2.
Функциональное исследование по измерению ингибирования формации конвертазы С3 лектиновым путем было использовано для оценки «блокирующей активности» анти-MASP-2 Fab2. Известно, что первичная физиологическая роль MASP-2 в пектиновом пути активации комплемента состоит в генерировании следующего функционального компонента пектин-зависимой активации комплемента, известной как конвертаза С3 пектинового пути. Конвертаза С3 пектинового пути представляет собой критический ферментный комплекс (C4bC2a), который протеолитическим образом расщепляет С3 на С3а и C3b. MASP-2 не является структурным компонентом конвертазы С3 пектинового пути (C4bC2a); однако функциональная активность MASP-2 необходима для генерирования двух белковых компонентов (C4b, С2а), которые заключают в себе конвертазу С3 пектинового пути. Кроме того, все отдельные виды функциональной активности MASP-2, перечисленные выше, являются необходимыми для генерирования конвертазы С3 лектинового пути посредством MASP-2. По этой причине предпочтительным способом анализа для оценки «блокирующей активности» анти-MASP-2 Fab2 считается функциональное исследование, которое измеряет ингибирование формирования конвертазы С3 лектинового пути.
Получение высокоафинных Fab2: Библиотека фаговых дисплеев последовательностей легких и тяжелых цепей антител человека и технологии автоматизированной селекции антител для идентификации Fab2, которые взаимодействуют с выбранными лигандами, представляющими научный интерес, была использована для создания высокоаффинных Fab2 против крысиного белка MASP-2 (SEQ ID NO:55). Известное количество крысиного MASP-2 (~1 мг, >85% чистоты) белка было использовано для скрининга антител. Для отбора антител с наилучшей аффинностью было проведено три круга амплификации. Приблизительно 250 различных наборов, экспрессирующих фрагменты антител, были использованы для скрининга при помощи ИФА. Высокоаффинные наборы были впоследствии отсеквенированы для определения уникальности различных антител.
Пятьдесят уникальных антител анти-MASP-2 были очищены и 250 мкг каждого очищенного антитела Fab2 было использовано для составления спецификации связывающей аффинности MASP-2 и функционального тестирования пути активации комплемента. Более детальное описание представлено ниже.
Исследования, использованные для оценки ингибирующей (блокирующей) активности анти-MASP-2 Fab2
1. Исследование по измерению ингибирования формирования конвертазы С3 пектинового пути активации:
Предпосылки: Конвертаза С3 лектинового пути активации представляет собой ферментный комплекс (C4bC2a), который протеолитическим образом расщепляет С3 на два потенциальных провоспалительных фрагмента, анафилатоксин С3а и опсонин C3b. Формирование конвертазы С3 является ключевым шагом в лектиновом пути активации в показателях опосредованного воспаления. MASP-2 не является структурным компонентом конвертазы С3 лектинового пути (C4bC2a); следовательно, анти-MASP-2 антитела (или Fab2) не подавляют напрямую активность уже существующей конвертазы СЗ. Однако активность серин протеазы MASP-2 является необходимым фактором для генерации двух белковых компонентов (C4b, С2а), которые включают в себя конвертазу С3 лектинового пути. Поэтому, анти-MASP-2 Fab2, ингибирующий функциональную активность MASP-2 (т.е., блокирующий анти-MASP-2 Fab2) будет ингибировать новую формацию конвертазы С3 лектинового пути. Часть структуры С3 представлена нетипичной и высоко реактивной группой сложного тиоэфира. Что касается расщепления С3 посредством конвертазы СЗ, то группа сложного тиоэфира в C3b может формировать ковалентную связь с гидроксил- или аминогруппами макромолекул, иммобилизованных на дне пластиковых лунок при помощи эфирных или амидных связей, что облегчает детекцию C3b в исследовании ИФА.
Дрожжевой маннан является известным активатором лектинового пути. В нижеприведенном способе измерения формирования конвертазы С3 пластиковые лунки, покрытые маннаном, подвергли инкубации в течение 30 минут при 37°С с разведенной крысиной сывороткой с целью активации лектинового пути. Затем лунки были отмыты и исследованы на C3b, иммобилизированный в лунках, посредством стандартной технологии ИФА. Количество полученного в данном исследовании C3b явился прямым отображением новой формации конвертазы С3 лектинового пути. В данном исследовании отобранные концентрации анти-MASP-2 Fab2 были протестированы на способность к ингибированию формирования конвертазы С3 и последующей генерации C3b.
Методы:
96-луночные планшеты Costar Medium Binding инкубировали в течение ночи при 5°С с маннаном, разведенном в 50 ммоль карбонатного буфера, с уровнем рН 9,5 в соотношении 1 мкг маннана на 50 мкл буфера на лунку После инкубации в течение ночи, каждая лунка была трижды промыта 200 мкл PBS. Затем лунки блокировали 1% раствором бычьего сывороточного альбумина в PBS в 100 мкл на лунку и подвергли инкубации в течение одного часа при комнатной температуре с легким помешиванием. Затем каждая лунка была трижды промыта 200 мкл раствора PBS. Образцы анти-MASP-2 Fab2 разбавили до выбранных концентраций в содержащем Са++ и Mg++ буфере GVB (4,0 ммоль барбитала, 141 ммоль NaCl, 1,0 ммоль MgCl, 2,0 ммоль CaCl2, 0,1% желатин, рН 7,4) при 5°С. После добавления в каждый образец 0,5%-ной крысиной сыворотки при 5°С, 100 мкл были перенесены в каждую лунку. Планшеты были покрыты и инкубированы в течение 30 минут на водяной бане при 37°С с целью инициации активации комплемента. Реакцию остановили посредством перемещения планшетов из водяной бани с температурой 37°С в контейнер, содержащий смесь льда и воды. Каждую лунку промывали пять раз 200 мкл раствора PBS-Tween 20 (0,05% Tween 20 в PBS), затем промывали дважды 200 мкл PBS. Раствор первичных антител 1:10000 (кроличьи античеловеческие С3, DAKO A0062) в объеме 100 мкл на лунку добавляли в PBS, содержащий 2,0 мг/мл бычьего сывороточного альбумина, и подвергли инкубированию в течение 1 часа при комнатной температуре с легким помешиванием. Каждая лунка была промыта 5×200 мкл PBS. Слабый раствор вторичных антител (коньюгат пероксидазы с антикроличими IgG козла, American Qualex A102PU) в объеме 100 мкл на лунку в соотношении 1:10000 добавляли в PBS, содержащий 2,0 мг/мл бычьего сывороточного альбумина, и подвергали инкубированию в течение 1 часа при комнатной температуре в шейкере с легким помешиванием. Каждая лунка была промыта пять раз 200 мкл в растворе PBS. Затем добавляли 100 мкл на лунку субстрата пероксидазы ТМВ («Kirkegaard & Perry Laboratories») и проводили инкубацию при комнатной температуре в течение 10 минут. Пероксидазную реакцию останавливали посредством добавления 1,0 М H3PO4 в объеме 100 мкл на лунку, после чего производили замер OD450.
2. Исследование по измерению ингибирования MASP-2-зависимого расщепления С4
Предпосылки: Активность сериновой протеазы MASP-2 обладает высокой специфичностью, и только два белковых субстрата MASP-2 были идентифицированы -С2 и С4. Расщепление С4 приводит к образованию С4а и C4b. AHTH-MASP-2 Fab2 может связываться со структурными эпитопами на MASP-2, которые напрямую вовлекаются в расщепление С4 (например, MASP-2-связывающий сайт для С4; MASP-2-каталитический сайт сериновой протеазы) и как следствие ингибирует функциональную активность MASP-2 для расщепления С4.
Дрожжевой маннан представляет собой известный активатор лектинового пути активации комплемента. В нижеизложенном способе по измерению активности MASP-2 расщепления С4, пластиковые лунки, покрытые маннаном, подвергли инкубированию в течение 30 минут при температуре 37°С с раствором крысиной сыворотки с целью активации лектинового пути. При использовании первичного антитела в исследовании ИФА возможно только распознавание человеческого С4, разбавленный раствор крысиной сыворотки также был обогащен человеческим С4 (1,0 Тг/мл). Затем лунки были отмыты и изучены на человеческий C4b, иммобилизированный на лунках, с использованием стандартных способов ИФА. Объем полученого в данном исследовании C4b является маркером MASP-2-зависимой активности расщепления С4. Анти-MASP-2 Fab2 в выбранных концентрациях были протестированы в данном исследовании на их способность к ингибированию расщепления С4.
Методы: 96-луночные планшеты Costar Medium Binding подвергли инкубированию в течение ночи при 5°С с маннаном, разведенным в 50 ммоль карбонатного буфера с рН 9,5 в количестве 1,0 Тг/50 Тл на лунку. Каждую лунку трижды промыли 200 Тл PBS. Затем лунки блокировали с 1%-ным бычьим сывороточным альбумином в PBS объемом 100 Тл на лунку и подвергли инкубированию в течение одного часа при комнатной температуре при легком помешивании. Каждую лунку трижды промыли 200 Тл PBS. Образцы анти-MASP-2 Fab2 были разведены в выбранных концентрациях буфера GVB с содержанием Са++ и Mg++ (4,0 ммоль барбитала, 141 ммоль NaCl, 1,0 ммоль MgCl2, 2,0 ммоль CaCl2, 0,1% желатин, рН 7,4) при 5°С. В данные образцы также добавили 1,0 Тг/мл человеческого С4 («Quidel»). Затем в вышеописанные образцы добавили 0,5% крысиную сыворотку при 5°С и 100 Тл были добавлены в каждую лунку. Планшеты были покрыты и инкубированы в течение 30 минут на водяной бане при 37°С для инициации активации комплемента. Реакцию останавливали посредством перемещения планшетов из водяной бани с температурой 37°С в контейнер, содержащий смесь льда и воды. Каждую лунку промывали пять раз 200 Тл PBS-Tween 20 (0,05% Tween 20 в PBS), затем промывали дважды 200 Тл PBS. Раствор коньюгированных с биотином куриных античеловеческих С4 («Immunsystem AB», Уппсала, Швеция) в разведении 1:700 в размере 100 Тл на лунку добавили в PBS, содержащий 2,0 мг/мл бычьего сывороточного альбумина (BSA), и провели инкубацию в течение одного часа при комнатной температуре с легким помешиванием. Затем каждую лунку промыли пять раз 200 Тл PBS. 100 Тл на лунку коньюгированного с пероксидазой стрептавидина с концентрацией 0,1 Тг/мл («Pierce Chemical», №21126) добавили в PBS, содержащий 2,0 мг/мл BSA, и подвергли инкубированию в течение одного часа при комнатной температуре в шейкере со слабым помешиванием. Затем каждую лунку промыли пять раз 200 Тл с PBS. После добавления 100 Тл на лунку пероксидазного субстрата ТМВ («Kirkegaard & Perry Laboratories») была проведена инкубация при комнатной температуре в течение 16 минут.Пероксидазную реакцию остановили посредством добавления 1,0 М H3PO4 в объеме 100 Тл на лунку и затем произвели замер OD450.
3. Анализ связывания антикрысиного MASP-2 Fab2 с «натуральным» крысиным MASP-2
Предпосылки: MASP-2 обычно присутствует в плазме в форме диммерных комплексов MASP-2, которые также включают в себя специфические пектиновые молекулы (маннозосвязывающий белок (MBL) и фиколины). Как следствие, при наличии интереса к исследованию связывания анти-MASP-2 Fab2 с физиологически релевантной формой MASP-2, важность представляет анализ связывания, в котором рассматривается взаимодействие в плазме между Fab2 и «натуральным» MASP-2, а не очищенным рекомбинантным MASP-2. В данном анализе связывания «натуральный» комплекс MASP-2-MBL из 10%-ной крысиной сыворотки был первым иммоболизирован на лунках, покрытых маннаном. Связывающая аффинность различных анти-MASP-2 Fab2 по отношению к иммобилизированному «натуральному» MASP-2 была исследована с использованием стандартной методологии ИФА.
Методы: 96-луночные планшеты Costar Medium Binding инкубировали в течение ночи при 5°С с маннаном, разведенном в 50 ммоль карбонатного буффера, рН 9,5 в количестве 1 Тг/50 Тл на лунку. Каждую лунку трижды промывали 200 Тл PBS. Затем лунки блокировали 0,5%-ным обезжиренным сухим молоком в PBST (PBS с 0,05% Tween 20) в объеме 100 Тл на лунку и инкубировали в течение одного часа при комнатной температуре при легком помешивании. После этого, каждую лунку трижды промыли 200 Тл промывочного буффера TBS/Tween/Ca++ (трис-буфферный солевой раствор, 0,5% Tween 20, содержащий 5,0 ммоль CaCl2, рН 7,4. 10%-ная крысиная сыворотка в связывающем буфере с высоким содержанием солей (High Salt Binding Buffer) (20 ммоль Tris, 1,0 моль NaCl, 10 ммоль CaCl2, 0,05% Triton-Х100, 0,1% (по весу) бычьего сывороточного альбумина с рН 7,4) была приготовлена на льду 100 Тл на лунку добавили и инкубировали в течение ночи при 5°С. Затем лунки были трижды промыты 200 Тл промывочного буффера TBS/Tween/Ca++. Далее лунки были дважды промыты 200 Тл PBS. 100 Тл на лунку выбранных концентраций анти -MASP-2 Fab2, разведенных в буфере, содержащем GVB Са++ и Mg++, (4,0 ммоль барбитала, 141 ммоль NaCl, 1,0 ммоль MgCl, 2,0 ммоль CaCl2, 0,1% желатин, рН 7,4) добавили в лунки и подвергли инкубации в течение одного часа при комнатной температуре с легким помешиванием. Каждую лунку промывали пять раз 200 Тл PBS. Затем добавляли 100 Тл на лунку коньюгированного с HRP анти-Fab2 козла («Biogenesis», кат. №0500-0099), растворенного в соотношении 1:5000 в 2,0 мг/мл бычьего сывороточного альбумина в PBS, и проводили инкубацию в течение часа при комнатной температуре с легким помешиванием. Каждую лунку промывали пять раз 200 Тл PBS. Далее, добавляли 100 Тл на лунку пероксидазного субстрата ТМВ («Kirkegaard & Perry Laboratories»), после чего проводили инкубацию при комнатной температуре в течение 70 минут. Пероксидазную реакцию остановили посредством добавления 1,0 моль H3PO4 в объеме 100 Тл на лунку, затем производили замер OD450.
РЕЗУЛЬТАТЫ:
При проведении скрининга посредством ИФА обнаруживаются приблизительно 250
различных Fab2, которые реагируют с высокой аффинностью с белками крысиного MASP-2. Подобные высоко аффинные Fab2 были упорядочены с целью определения уникальности различных антител. Уникальные анти-MASP-2 антитела в количестве 50 были очищены для дальнейшего исследования. 250 мкг каждого очищенного антитела Fab2 было использовано для описания характерных признаков связывающей аффинности MASP-2 и функционального тестирования пути активации комплемента. Результаты данного исследования представлены в ТАБЛИЦЕ 6.
Как показано выше в ТАБЛИЦЕ 6, из 50 протестированных анти-MASP-2 Fab2, семнадцать Fab2 были идентифицированы как Fab2, блокирующие MASP-2, которые потенциально могут ингибировать формирование конвертазы С3 с IC50 эквивалентным или менее 10 нмоль Fab2 (34%-ный положительный коэффициент эффективности поиска). Восемь из семнадцати идентифицированных Fab2 обладают IC50 в субнаномолярном ряду. Как следствие, все семнадцать Fab2, блокирующих MASP-2, показанные в ТАБЛИЦЕ 6, главным образом, показывают полное ингибирование формирования конвертазы С3 в исследовании конвертазы С3 пектинового пути активации комплемента. ФИГУРА НА представляет собой графические иллюстрации результатов исследования формирования конвертазы С3 для антитела Fab2 №11, которое выбрано в качестве представителя прочих протестированных антител Fab2; результаты данных тестов представлены в ТАБЛИЦЕ 6. Данный факт представляет важность, учитывая теоретическую возможность того, что «блокирующее» Fab2 может только частично ингибировать функцию MASP-2 даже в том случае, когда каждая молекула MASP-2 связывается с Fab2.
Хотя маннан является известным активатором пектинового пути, существует теоретическая возможность того, что наличие анти-маннановых антител в крысиной сыворотке может также активировать классический путь и генерировать C3b посредством конвертазы С3 классического пути. Однако каждое из семнадцати блокирующих анти-MASP-2 Fab2, перечисленных в данном примере, потенциально могут ингибировать генерирование C3b (>95%), таким образом, демонстрируя специфичность данного исследования для конвертазы С3 пектинового пути.
Анализ связывания был также проведен для всех семнадцати блокирующих Fab2 с целью подсчета Kd для каждого. Результаты исследований по связыванию анти-крысиных MASP-2 Fab2 с натуральными крысиными MASP-2 для шести из блокирующих Fab2 также представлены в ТАБЛИЦЕ 6. ФИГУРА 11 В представляет собой графические иллюстрации результатов исследования связывания с антителом Fab2 №11. Подобные исследования по связыванию также были проведены для других Fab2, их результаты представлены в ТАБЛИЦЕ 6. В основном, Kd, полученные для связывания каждого из шести Fab2 с «натуральным» MASP-2, полностью соответствует IC50 для Fab2 в функциональном исследовании конвертазы СЗ. Существуют доказательства того, что MASP-2 подвергается конформационным изменениям от неактивной к активной форме посредством инициации протеазной активности (Feinberg et al., EMBO J 22:2348-59 (2003); Gal et al., J. Biol. Chem. 280:33435-44 (2005)). В нормальной крысиной плазме, используемой в исследовании формирования конвертазы СЗ, MASP-2 проявляются первоначально в форме «неактивной» конформации зимогена. В то время как, при анализе связывания, MASP-2 присутствует как часть комплекса с MBL, связанным с имобилизированным маннаном; поэтому, MASP-2 скорее всего представлен «активной» конформацией (Petersen et al., J. Immunol Methods 257:107-16, 2001). Следовательно, не следует ожидать четкой связи между IC50 и Kd для каждого из семнадцати блокирующих Fab2, протестированных в данных двух функциональных исследованиях, учитывая тот факт, что в каждом исследовании Fab2 связывается с различными конформационными формами MASP-2. Тем не менее, если исключить Fab2 №88, было 6 разумно исключить связь между IC50 и Kd для каждого из прочих шестнадцати Fab2, протестированных в двух исследованиях (см. ТАБЛИЦУ 6).
Некоторые блокирующие Fab2 были оценены на способность к ингибированию MASP-2-зависимого расщепления С4. ФИГУРА 11 С представляет графические иллюстрации результатов исследования расщепления С4, демонстрируя ингибирование с Fab2 №41, с IC50=0,81 нмоль (см. ТАБЛИЦУ 6). Как показано на ФИГУРЕ 12, все из протестированных Fab2 обнаружили способность к ингибированию расщеплению С4 с IC50, подобным тем, что получены в исследовании конвертазы С3 (см. ТАБЛИЦУ 6).
Хотя маннан является известным активатором пектинового пути, существует теоретическая возможность того, что наличие анти-маннановых антител в крысиной сыворотке может также активировать классический путь и генерировать C4b посредством ds-опосредованного расщепления С4. Однако было установлено, что некоторые анти-MASP-2 Fab2 способны ингибировать генерацию C4b (>95%), таким образом, демонстрируя специфичность данного исследования для MASP-2-опосредованного расщепления С4. С4, как и С3, включает в себя необычные и высоко реактивные группы сложного эфира как часть структуры. Вследствие расщепления С4 при помощи MASP-2, как описано в данном исследовании, группа сложного эфира на C4b может формировать ковалентную связь с гидроксильными или аминогруппами на макромолекулах, иммобилизованных на дне пластиковых лунок посредством эфирных или амидных связей, таким образом, облегчая детекцию при помощи ИФА.
Данные исследования явно демонстрируют создание высокоаффинных FAB2 по отношению к белку MASP-2, которые блокирует функциональную активность конвертазы С4 и С3, и как следствие предупреждают активацию пектинового пути.
ПРИМЕР 25
Данный Пример описывает картирование эпитопов нескольких блокирующих антикрысиных антител MASP-2 Fab2, которые были получены способом, описанным в Примере 24.
Методы:
Как показано на ФИГУРЕ 13, следующие белки, все с N-термиальными 6Х His метками, были экспрессированы в клетках СНО с использованием вектора pED4:
крысиный MASP-2A, полноразмерный белок MASP-2, инактивированный посредством замены серина в активном центре на аланин (S613A);
крысиный MASP-2K, полноразмерный белок MASP-2, измененный для редуцирования самоактивации (R424K);
CUBI-II, N-терминальный фрагмент крысиного MASP-2, который включает себя только следующие домены: CUBI, EGF-подобные и CUBII; и
CUBI/EGF-подобные, N-терминальные фрагменты крысиного MASP-2, которые включают в себя только CUBI и EGF-подобный домены.
Данные белки были очищены из культуральных супернатантов посредством никель-аффиной хроматографии, как было описано ранее (Chen et al., J. Biol. Chem. 276:25894-02 (2001)). С-терминальный полипептид (CCPII-SP), содержащий CCPII и домен сериновой протеазы крысиного MASP-2, был экспрессирован в Е. coli как слитый с тиоредоксином белок с использованием pTrxFus («Invitrogen»). Белок был очищен от клеточных лизатов при использовании аффинной смолы Thiobond. Слитый с тиоредоксином белок был экспрессирован в пустой pTrxFus в качестве отрицательного контроля.
Все рекомбинантные белки подвергли диализу в буфере TBS, далее их концентрации определили посредством измерения OD при 280 nm.
ДОТ-БЛОТ АНАЛИЗ:
Серийное разведение пяти рекомбинантных полипептидов MASP-2, описанное выше и показанное на ФИГУРЕ 13 (и полипептид тиоредоксин в качестве отрицательного контроля для полипептида CCPII-сериновой протеазы), было нанесено на нитроцеллюлозную мембрану. Количество нанесенного белка находилось в диапазоне от 100 нг до 6,4 пкг, в пятикратных разведениях. В последующих экспериментах, количество нанесенного белка находилось в диапазоне от 50 нг до 16 пкг, также в пятикратных разведениях. Мембраны были блокированы с 5%-ным снятым сухим молоком в TBS (блокирующий буфер), затем инкубированы с 1,0 мкг/мл анти-MASP-2 Fab2 в блокирующем буфере (содержащем 5,0 нмоль Са2+). Связанные Fab2 были определены при помощи HRP-коньюгированного античеловеческого Fab (AbD/Serotec; разведен в соотношении 1/10000) и набором для детекции ECL («Amersham»). Одна мембрана была инкубирована с поликлональным кроличьим античеловеческим MASP-2 АЬ (описано у Stover et al., J Immunol 163:6848-59 (1999)) в качестве положительного контроля. В данном случае, связанное АЬ было определено при использовании HRP-коньюгированного анти-кроличьиго IgG козла (Dako; разведенных в соотношении 1/2000).
Анализ связывания MASP-2
Планшеты ИФА инкубировали с раствором рекомбинантного MASP-2A или CUBI-II полипептида в карбонатном буфере (уровень рН 9,0) взятом в количестве 1,0 мкг на лунку в течение ночи при 4°С. Лунки были блокированы 1%-ным BSA в TBS, затем последовательные разведения анти-MASP-2 Fab2 были добавлены в TBS, содержащий 5,0 нмоль Са2+. Планшеты инкубировали в течение одного часа при комнатной температуре. После тройного промывания в TBS/tween/Ca2+, добавили HRP-коньюгированное античеловеческое Fab (AbD/Serotec), разведенное в соотношении 1/10000 в TBS/ Са2+, и планшеты снова подвергли инкубированию в течение одного часа при комнатной температуре. Связанное антитело было обнаружено с помощью набора субстрата пероксидазы ТМВ («Biorad»).
РЕЗУЛЬТАТЫ:
Результаты дот-блот анализа, демонстрирующие реактивность Fab2 с различными полипептидами MASP-2, представлены ниже в ТАБЛИЦЕ 7. Численные значения, приведенные в ТАБЛИЦЕ 7, обозначают количество нанесенного белка, необходимого для подачи сигнала приблизительного полумаксимальной интенсивности. Как показано, все полипептиды (исключение составляет только химерный белок тиоредоксина) были распознаны посредством положительного контроля Ab (сыворотка поликлонального античеловеческого MASP-2, полученная от кроликов).
Все антитела Fab2 вступили в реакцию с MASP-2A, как и с MASP-2K (данные не приводятся). Большинство из Fab2 распознают полипептид CCPII-SP, но не N-терминальные фрагменты. Исключения представляют Fab2 №60 и Fab2 №57. Fab2 №60 распознает MASP-2A и фрагмент CUBI-II, но не CUBI/EGF-подобный полипептид или CCPII-SP полипептид, что предполагает его связывание с эпитопом в CUBII, или перекрытие домена CUBII и EGF-подобного домена. Fab2 №57 распознает MASP-2A, но ни один из протестированных фрагментов MASP-2, что может указывать на тот факт, что данное антитело Fab2 распознает эпитоп в CCPI. Fab2 №40 и №49 связывался только с полным MASP-2A. В анализе связывания ИФА, показанном на ФИГУРЕ 14, Fab2 №60 также связывался с полипептидом CUBI-II, хотя при этом уровень очевидной аффинности был незначительно ниже.
Данные результаты демонстрируют идентификацию уникального блокирования Fab2 в отношении многочисленных областей белка MASP-2.
ПРИМЕР 26
Данный пример описывает исследование MASP-2-/- мыши в модели мышиной ренальной ишемии/реперфузии.
Предпосылки и обоснование: Ишемическое реперфузионное (I/R) поражение в почке при температуре тела проявляется в определенном количестве клинических состояний, включая гиповолемический шок, закупорку почечной артерии и процедуры пережатая.
Почечная ишемия-реперфузия (I/R) является важной причиной острой почечной недостаточности, приводящей к смертности в 50% случаев (Levy et al., JAMA 275:1489-94, 1996; Thadhani et al., N. Engl. J. Med. 334:1448-60, 1996). Посттрансплантационная почечная недостаточность представляет собой общее и опасное осложнение после трансплантации почек (Nicholson et al., Kidney Int. 58:2585-91, 2000). В настоящее время не существует доступного эффективного лечения I/R поражения почек, единственной формой поддержания является гемодиализ. Патофизиология I/R поражения почек является весьма сложной. Недавние исследования продемонстрировали, что пектиновый путь активации комплемента может исполнять важную роль в патогенезе I/R поражения почек (deVries et al., Am. J. Path. 165:1677-88, 2004).
Методы:
MASP-2(-/-) мышь была получена, как описано в Примере 1, и подвергнута обратному скрещиванию как минимум в течение 10 поколений с С57 В1/6. Шесть самцов MASP-2(-/-) и шесть мышей дикого типа (+/+) с весом 22-25 г получали интраперитонеальные инъекции Hypnovel (6,64 мг/кг; «Roche products Ltd.», Уэлвин-Гарден-Сити, Великобритания), а затем были подвергнуты анестезии посредством ингалирования изофлюраном («Abbott Laboratories Ltd.», Кент, Великобритания). Изофлюран был выбран потому, что он является мягким ингаляционным анестезирующим средством с минимальной гепатотоксичностью; концентрации были выбраны очень тщательно и животные восстанавливались быстро даже после продолжительной анестезии. Введение Hypnovel проводилось по причине того, что он вызывает состояние нейролептаналгезии у животных, что подразумевает введение меньшей дозы изофлюрана. Животных размещали на теплых подстилках с целью поддержания постоянной температуры тела. Далее, был произведен разрез по средней линии брюшной стенки, и полость тела оставили открытой при помощи пары ранорасширителей. Соединительную ткань очистили выше и ниже почечной вены и артерии обеих правой и левой почек, а почечная питающая ножка была пережата посредством наложения зажимов микроаневризмы на 55 минут.Данный период ишемии первоначально был создан в предыдущем исследовании, проведенном в данной лаборатории (Zhou et al., J. Clin. Invest. 105:1363-71 (2000)). В добавление, стандартный ишемический период в 55 минут был выбран вследствие ишемической титрации, и было обнаружено, что 55 минут провоцировали стойкое поражение, которое также являлось обратимым, с низким уровнем смертности, менее 5%. После окклюзии, 0,4 мл теплого соляного раствора (37°С) поместили в брюшную полость, а затем брюшную полость закрыли на время ишемии. После удаления зажимов микроаневризмы обследование почек проводили до изменения окраски, что означало возобновление кровообращения. Далее в брюшную полость поместили 0,4 мл теплого соляного раствора и на разрез наложили швы, после чего животных немедленно вернули в их клетки. Образцы крови были взяты из хвостов животных спустя 24 часа после удаления зажимов, и спустя 48 часов мышей умертвили и собрали дополнительные образцы крови.
Установление ренального поражения: Почечную функцию оценивали через 24 и 48 часов после реперфузии у шести самцов MASP-2(-/-) и шести мышей дикого типа (+/+). Определение креатинина в крови проводили посредством масс-спектрометрии, которая предоставляет воспроизводимый индекс ренальной функции (чувствительность <1,0 мкмоль/л). ФИГУРА 15 графическим образом иллюстрирует клиренс азота мочевины крови для контрольной группы дикого типа С57 В1/6 и MASP-2(-/-) через 24 часа и 48 часов после реперфузии. Как показано на ФИГУРЕ 15, MASP-2(-/-) мыши проявили значительное сокращение в объеме мочевины крови через 24 и 48 часов, в сравнении с контрольными мышами дикого типа, что указывает на защитное функциональное действие против поражения почек в модели ишемического реперфузионного поражения.
В итоге, повышенный уровень мочевины в крови наблюдался в обоих случаях у мышей дикого типа (+/+) и MASP-2(-/-) мышей спустя 24 и 48 часов после хирургической процедуры ишемического инсульта. Уровень мочевины в крови у неишемических прооперированных животных дикого типа (+/+), установленный по-отдельности, составил 5,8 ммоль/л. В дополнение к данным, представленным на ФИГУРЕ 15, одно MASP-2(-/-) животное продемонстрировало практически полную защиту от ишемического инсульта, с показателями 6,8 и 9,6 ммоль/л через 24 и 48 часов, соответственно. Это животное исключили из группы испытуемых как потенциально аномальное, у которого не могут проявляться признаки ишемического поражения. Как следствие, окончательное исследование, представленное на ФИГУРЕ 15, было проведено на 5 MASP-2(-/-) мышах и 6 мышах дикого типа (+/+), данные продемонстрировали существенное статистическое сокращение мочевины в крови через 24 и 48 часов у MASP-2(-/-) мышей (t-критерий Стьюдента р<0,05). Данные результаты указывают на то, что ингибирование активности MASP-2 должно оказывать защитное или терапевтическое действие против поражения почек вследствие ишемии.
ПРИМЕР 27
Данный пример описывает исследование MASP-2(-/-) мышей в мышиной модели миокардиальной ишемии/реперфузии.
Предпосылки и обоснование:
Маннозосвязывающий лектин (MBL) представляет собой циркулирующую молекулу, которая инициирует активацию комплемента независимо от иммунного комплекса, в ответ на широкое разнообразие углеводных структур. Данные структуры могут быть компонентами инфекционных агентов или переработанных эндогенных углеводных компонентов, в особенности внутри некротических, онкотических или апоптических клеток. Подобные формы некроза клеток имеют место при реперфузии миокарда, когда активация комплемента похожим образом развивает поражение за границей существующего на момент прекращения ишемии вследствие реперфузии. Несмотря на наличие убедительных доказательств того того, что активация комплемента усугубляет миокардиальную реперфузию, механизм подобной активации до конца не выяснен, и существует вероятность того, что ингибирование всех известных путей активации оказывает недопустимые пагубные воздействия. В ходе недавнего исследования было выдвинуто предположение, что активация может вовлекать MBL вместо амплификационной петли классического или альтернативного пути (как определено в настоящем изобретении), учитывая то, что инфаркт был редуцирован у MBL(А/С)-, а не у C1q- нулевых мышей (Walsh M.C. et al., Jour of Immunol. 175:541-546 (2005)). Однако, несмотря на обнадеживающие результаты, у данных мышей все еще сохраняются циркулирующие компоненты, например, Фиколин А, способные к активации комплемента посредством пектинового пути.
Данное исследование было проведено на мышах MASP-2(-/-) по сравнению с мышами котрольной группы дикого типа (+/+) для выяснения того, могут ли мыши MASP-2(-/-) быть менее чувствительными к миокардиальной ишемии и реперфузионному поражению. MASP-2(-/-) мышей подвергли регионарной ишемии, после чего провели сравнение размера инфаркта у этих мышей с размером инфаркта у однопометных животных дикого типа.
Методы: Нижеследующий протокол базируется на процедуре идуцирования ишемического реперфузионного поражения, описанного Marber et al., J. Clin Invest. 95:1446-1456(1995).
MASP-2(-/-) мышь была получена, как описано в Примере 1, затем последовало обратное скрещивание по меньшей мере с 10 поколениями с С57В1/6. Семь мышей MASP-2(-/-) и семь мышей дикого типа (+/+) подвергли анестезии с кетамин/медетомидином (100 мг/кг и 0,2 мг/кг, соответственно) и поместили в положении на спине на подогреваемую подстилку с термостатическим контролем для поддержания ректальной температуры на уровне 37±0,3°С. Мышей интубировали под непосредственным наблюдением и вентилировали воздухом помещения при интенсивности дыхания на уровне 11 О/мин и с дыхательным объемом на уровне 225 мкл/мин (вентилятор - Hugo Sachs Elektronic MiniVent Type 845, Германия).
Шерсть сбрили и произвели переднелатеральный разрез кожи в левой подмышеной впадине в близости от мечевидного отростка. Большая грудная мышца была иссечена, разрезана по ее стернальной границе и перемещена в подмышечную впадину. Малая грудная мышца была разрезана по ее краниальной границе и перемещена в область хвоста. Мышца была позже использована в качестве мышечного лоскута, покрывающего сердце во время окклюзии коронарной артерии. Мышцы из 5-ого межреберного промежутка и париетальная плевра были пенетрированы при помощи микропинцетов по средней границе левого легкого, что таким образом позволяет избежать поражения легких или сердечной мышцы. После пенетрирования плевры микропинцеты были очень осторожно направлены за плевру по направлению к грудине, не затрагивая сердце, а плевра и межреберные мышцы были иссечены при помощи прибора для прижигания с батарейным питанием («Harvard Apparatus», Великобритания). Во избежание кровотечения операции осуществлялись с повышенной осторожностью. Используя ту же технологию, торакотомия была расширена до сердней подмышечной линии. После разреза 4-ого ребра по его стернальной границе, межреберный промежуток расширили настолько, чтобы полностью открыть сердце от основания до апекса. Околосердечная сумка была открыта двумя зажимами малых артерий и была сформирована перикардиальная секция для перемещения сердца вперед. Левая передняя нисходящая артерия (LAD) была экспонирована, затем под LAD наложили шов из моноволокна 8-0 при помощи круглой иглы. Область наложения лигатуры LAD лежит каудально от кончика левого предсердия, приблизительно на 1/4 вдоль линии, проходящей от атриовентрикулярного отростка к апексу левого желудочка.
Все эксперименты были проведены слепым методом, в отсутствие информации о генотипе каждого животного. После завершения инструментальных и хирургических процедур, мышам дали 15 минут на период равновесия. Затем мышей подвергли 30-минутной окклюзии коронарной артерии со временем реперфузии в 120 минут.
Модель окклюзии коронарной артерии и реперфузии
Окклюзия коронарной артерии была создана с использованием системы с подвешенным грузом, как описано выше (Eckle et al., Am J Physiol Heart Circ Physiol 29^H2533-H2540, 2006). Оба конца лигатуры из моноволокна были пропущены сквозь отрезок полиетиленовой трубки РЕ-10 2 мм длиной и присоединены к шву 5-0 при помощи цианоакрилового клея. Затем шов был направлен поверх двух горизонтально приподнятых перемещающихся металических стержней, каждый с грузом в 1 г и присоединенных к обоим концам шва. При поднятии стержней, грузы подвешивались и шов располагался под контролируемым давлением для окклюзии LAD с четко определенным и постоянным давлением. Окклюзия LAD была подтверждена бледностью в области риска, преобразованием цвета зоны перфузии LAD из яркокрасного в фиолетовый, указывающим на прекращение кровоснабжения. Реперфузия была достигнута посредством снижения уровней стержней до положения грузов на область оперативного вмешательства со снижением давления лигатуры. Реперфузия была проверена теми же тремя критериями для подтверждения окклюзии. Мыши были исключены из дальнейшего исследования, если все три критерия не были установлены в начале окклюзии коронарной артерии или в течение 15 минут после репефузии. Во время окклюзии коронарной артерии, температура и влажность сердечной поверхности поддерживалась посредством покрытия сердца лоскутом из малой грудной мышцы и герметизации торакотомии при помощи марли, пропитанной 0,9%-ным соляным раствором.
Измерение масштабов инфаркта миокарда:
Масштаб инфаркта (INF) и область риска (AAR) были определены посредством планиметрии. После внутривенной инъекции 500 ME гепарина, LAD подвергли реокклюзии, а в яремную вену ввели 300 мкл 5% (по весу) препарата Evans Blue («Sigma-Aldrich», Пул, Великобритания) посредством медленного введения для установления границ области риска (AAR). Это способствует проникновению красителя в неишемическую область левого желудочка и оставляет ишемическую AAR неокрашенной. Мышей умерщвляли посредством смещения шейных позвонков после чего немедленно извлекали сердце. Сердце охлаждали на льду и помещали в блок 5%-ной агарозы, а затем разрезали на 8 поперечных срезов толщиной 800 мкм. Все срезы подвергли инкубированию при 37°С в течение 20 минут с 3%-ным раствором 2,3,5-трифенилтетразолум хлорида («Sigma Aldrich», Пул, Великобритания), разведенным в 0,1 моль растворе буфера Na2HPO4/NaH2PO4 с рН 7,4. Затем срезы зафиксировали в 10%-ном формальдегиде на ночь. Срезы были помещены между двумя покровными стеклами, при помощи оптического сканера были сделаны цифровые снимки каждой стороны покровного стекла. Затем цифровые снимки проанализировали с использованием программного обеспечения SigmaScan (SPSS, США). Область пораженной зоны (бледная), область риска (красная) левого желудочка (LV) и область нормальной перфузии LV (голубая) были очерчены в каждой секции посредством идентификации их цветового проявления и цветовых границ. Области были просчитаны на обеих сторонах каждого среза, исследователи вывели среднее значение. Масштаб инфаркта был подсчитан в % от зоны риска у каждого животного.
РЕЗУЛЬТАТЫ: Область зоны инфаркта (бледная), область риска LV (красная) и область нормальной перфузии LV (голубая) были очерчены в каждой секции посредством идентификации их цветового проявления и цветовых границ. Области были просчитаны на обеих сторонах каждого среза, исследователи вывели среднее значение. Масштаб инфаркта был подсчитан в % от зоны риска у каждого животного. ФИГУРА 16А представляет оценку семи мышей дикого типа (+/+) и семи MASP-2(-/-) мышей по определению масштабов инфаркта после проведения процедуры окклюзии коронарной артерии и технической реперфузии, описанных выше. Как показано в ФИГУРЕ 16А, MASP-2(-/-) мыши продемонстрировали статистически устойчивую редукцию (р<0,05) масштабов инфаркта, чем мыши дикого типа (+/+), что указывает на защитное миокардиальное воздействие от поражения в модели ишемического реперфузионного поражения. ФИГУРА 16 В демонстрирует распределение индивидуальных протестированных животных, указывая на явный защитный эффект для мышей MASP-2(-/-).
ПРИМЕР 28
Данный пример описывает результаты, полученные от MASP-2-/- в мышиной модели дегенерации желтого пятна.
Предпосылки и обоснование: Возрастная дегенерация желтого пятна (AMD) представляет собой первостепенную причину слепоты после 55 лет в современном индустриализированном мире. AMD проявляется в двух основных формах: неоваскулярная (влажная) AMD и атрофическая (сухая) AMD. Неоваскулярная (влажная) форма является причиной 90% серьезной потери зрения, опосредованного AMD, хотя только у приблизительно 20% индивидуумов с AMD развивается влажная форма. Клинические признаки AMD включают в себя множественные старческие бляшки, географическую атрофию и хориоидальную неоваскуляризацию (CNV). В декабре 2004, Управление по контролю качества пищевых продуктов и лекарств США (FDA) одобрило препарат Macugen (пегаптаниб), новый класс офтальмических медикаментов для специфическим воздействия и блокирования эффектов фактора роста эндотелия сосудов (VEGF) для лечения влажной (неоваскулярной) формы AMD (Ng et al., Nat Rev. Drug Discov 5:123-32 (2006)). Несмотря на то, что Macugen представляет собой новое терапевтическое средство для подгруппы пациентов с AMD, существует срочная необходимость развития дополнительных способов для лечения комплекса данных заболеваний. Многочисленные независимые исследования указывают на центральную роль активации комплемента в патогенезе AMD. Патогенез хориоидальной неоваскуляризации (CNV), самой серьезной формы AMD, может вовлекать активацию путей комплемента.
Более двадцати пяти лет назад Риан (Ryan) описал модель лазерного поражения CNV на животных (Ryan, S.J., Tr. Am. Opth. Soc. LXXVII: 707-745, 1979). Изначально модель была разработана на макаках-резусах, однако такая же технология была использована для развития подобных моделей CNV на разнообразных лабораторных животных, включая мышей (Tobe et al., Am. J. Pathol. 153:1641-46, 1998). В данной модели использовалась лазерная коагуляция для разрушения мембраны Бруха, результатом данного действия является формирование CNV-подобных мембран. Индуцированная лазером модель охватывает множество важных проявлений человеческого заболевания (для ознакомления с последними исследованиями, см. Ambati et al., Survey Ophthalmology 48:257-293, 2003). Индуцированная лазером мышиная модель хорошо разработана в настоящее время, и широко используется в качестве основы для экспериментов в постоянно растущем количестве научно-исследовательских проектов. Существует общепризнанное мнение, что индуцированная лазером модель обладает достаточной биологической схожестью с CNV человека, поэтому доклинические исследования патогенеза и ингибирование при помощи медецинских препаратов с использованием данной модели являются релевантными по отношению к CNV человека.
Методы:
MASP-2(-/-) мышь была получена как описано в Примере 1, затем последовало обратное скрещивание по меньшей мере с 10 поколениями с С57 В1/6. В настоящей заявке проводится сравнение результатов исследований самцов мышей MASP-2(-/-) и MASP-2(+/+) оцененных по ходу течения индуцированной лазером CNV, ускоренной модели неоваскулярного AMD, с фокусом на масштаб индуцированной лазером CNV при помощи конфокальной лазерной сканирующей микроскопии как средства для измерения тканевого поражения и определения уровней VEGF, потенциального ангиогенного фактора, вовлеченного в CNV, в ретинальном пигментном эпителии (RPE)/ сосудистой оболочке глаза при помощи ИФА после лазерного поражения.
Индукция хориоидальной неоваскуляризации (CNV): Лазерная фотокоагуляция (532 нм, 200 мВт, 100 мс, 75 мкм; «Oculight GL», «Iridex», Маунтин-Вью, шт.Калифорния) была смодулирована в каждом глазу каждого животного на нулевой день посредством установления единичной индивидуальной реакции на группу препаратов. Лазерные пятна были расположены стандартизированным образом вокруг оптического нерва при использовании системы установки щелевой лампы и покровного стекла в качестве контактной линзы. Морфологической конечной точкой лазерного поражения было появление кавитационного пузырька, что считалось признаком корреляции с повреждением мембраны Бруха. Далее следует детальное описание способа и оценка полученных с его помощью результатов.
Флуоресцентная ангиография: Флуоресцентная ангиография была осуществлена при помощи камеры и системы формирования изображений (камера TRC 50 1А; система ImageNet 2.01; «Торсоп», Парамус, шт.Нью-Джерси) через 1 неделю после лазерной фотокоагуляции. Фотографии были сделаны с использованием линз 20-D в контакте с объективом съемочного аппарата с линзой фундус-камеры после интраперитонеальной инъекции 0,1 мл 2,5%-ного флуоресцеин натрия. Специалисты по сетчатой оболочке, не участвующие в лазерной фотокоагуляции или ангиографии, оценивали флуоресцентные ангиограммы в одиночных случаях в латентной манере.
Масштаб хориоидальной неоваскуляризации (CNV): Спустя одну неделю после лазерного поражения, глаза были энуклеированы и зафиксированы 4%-ным параформальдегидом на 30 минут при 4°С. Глазные бокалы были получены посредством удаления внутренних сегментов и затем были промыты трижды в PBS, после дегидратации и регидратации с применением ряда разведении метанола. После двойного блокирования буфером (PBS с содержанием 1%-ного бычьего сывороточного альбумина и 0,5%-ного Triton X-100) в течение 30 минут при комнатной температуре, глазные бокалы подвергли инкубации в течение ночи при 4°С с 0,5%-ным ПТС-изолектином В4 («Vector laboratories», Бурлингейм, шт.Калифорния), разведенным PBS, содержащем 0,2%-ный BSA и 0,1%-ный Triton X-100, который связывает конечные остатки β-D-галактозы на поверхности эндотелиальных клеток и селективным образом маркирует сосудистую сеть мыши. После двойного промывания PBS, содержащем 0„1%-ный Triton X-100, нейросенсорная оболочка глаза была осторожно отслоена и отсоеденена от оптического нерва. Затем были сделаны четыре послабляющих радиальных надреза, и остаточный комплекс RPE-хориоидея-склера был помещен в консервирующую питательную среду (Immu-Mount Vectashield Mounting Medium;
«Vector Laboratories») и закрыты покровным стеклом.
Образцы для микроскопического исследования были изучены при помощи конфокальной лазерной сканирующей микроскопии (TCS SP; «Leica», Гейдельберг, Германия). Сосуды визуализировали посредством возбуждения голубым аргоном с длиной волны 488 нм и поглощающей эмиссии между 515 и 545 нм. Во всех исследованиях изображений был использован 40-кратный масляно-иммерсионный объектив. Горизонтальные оптические секции (с шагом 1 мкм) были получены с поверхности комплекса RPE-хориоидея-склера. Самая глубокая фокальная плоскость в которой может быть определена окружающая хориоидальная васкулярная сеть, связанная с поражением, была рассмотрена в качестве основания поражения. Любой сосуд, находящийся в области-мишени для лазерного воздействия и расположенный на поверхности вышеуказанной области, был оценен как CNV. Изображения каждой секции были сохранены в цифровом виде. Область CNV-связанной флуоресценции были измерена посредством анализа компьютерных изображений с программным обеспечением для электронного микроскопа (TCS SP; «Leica»). Суммирование всей флуоресцентной области в каждой горизонтальной секции было использовано как показатель масштаба CNV. Изображения были выполнены оператором, скрытым от назначения терапевтичекой группы.
Так как возможность каждого лазерного поражения, провоцирующего развитие CNV, попадает под действие группы, к которой он принадлежит (мышь, глаз и лазерное пятно), средние масштабы поражения подвергли сравнению с использованием линейной смешанной модели, поделенной на участки по принципу повторяющихся измерений. Полный фактор участка был представлен генетической группой, к которой принадлежали животные, где фактор расщепленного участка был представлен глазом. Статистическая значимость была определена на уровне 0,05. Ретроспективные сравнения средств были созданы при помощи поправки Бонферрони для множественных сравнений.
Иммуноферментный твердофазный анализ фактора роста эндотелия сосудов (VEGF ELISA). Через три дня после поражения 12 лазерных пятен, комплекс RPE-хориоидея был разрушен ультразвуком в лизисном буфере (20 ммоль имидазол HCI, 10 ммоль KCl, 1 ммоль MgCl2, 10 ммоль EGTA, 1% Triton X-100, 10 ммоль NaF, 1 ммоль молибдат натрия и 1 ммоль EDTA с ингибитором протеазы) на льду в течение 15 минут. Уровни белка VEGF в супернатанте были определены при помощи набора для ИФА («R&D Systems», Миннеаполис, шт.Миннесота), который распознает все сплайсинговые вариации, от 450 до 570 нм (Emax; «Molecular Devices», Саннивейл, шт.Калифорния), и нормализованы по тотальному белку. Дублирующие измерения были выполнены в скрытом режиме оператором, не вовлеченным в фотокоагуляцию, создание изображений или ангиографию. Количество VEGF было представлено как средняя+/- стандартная ошибка по меньшей мере в трех независимых экспериментах с последующим сравнением при использовании U-критерия Манна-Уитни. Нулевая гипотеза была отклонена при Р<0,05.
РЕЗУЛЬТАТЫ:
Оценка уровней VEGF:
ФИГУРА 17А иллюстрирует в графической форме уровни белков VEGF в комплексе RPE-хориоидея, выделенном у мышей дикого типа С57 В16 и MASP-2(-/-) мышей в нулевой день. Как показано на ФИГУРЕ 17А, установление уровней VEGF указывает на сокращение базовых уровней для VEGF у мышей MASP-2(-/-) по сравнению с мышами дикого типа C57b1 контрольной группы. ФИГУРА 17 В иллюстрирует в графической форме уровни белков VEGF, измеренные на третий день после индуцированного лазером поражения. Как представлено на ФИГУРЕ 17 В, уровни VEGF были значительно повышены у мышей дикого типа (+/+) через три дня после индуцированного лазером поражения, что согласуется с опубликованными исследованиями (Nozaki et al., Proc. Natl. Acad. Sci USA 103:2328-33 (2006)). Однако уровни VEGF, наблюдаемые у мышей MASP-2(-/-), были удивительно низкими.
Оценка хориоидальной неоваскуляризации (CNV):
В добавление к редукции уровней VEGF после индуцированной лазером дегенерации желтого пятна, область CNV определяли до и после лазерного поражения. ФИГУРА 18 иллюстрирует в графической форме объем CNV, измеренный у мышей дикого типа C57b1 и MASP-2(-/-) мышей на седьмой день после индуцированного лазером поражения. Как представлено на ФИГУРЕ 18, MASP-2(-/-) мыши показали приблизительно 30%-ное редуцирование в области CNV после индуцированного лазером поражения на седьмой день по сравнению с контрольной группой мышей дикого типа.
Данные открытия указывают на редуцирование в VEGF и CNV, как показывали наблюдения MASP(-/-) мышей по сравнению с контрольными мышами дикого типа (+/+), равно как и на то, что блокирование MASP-2 при помощи ингибитора может оказывать превентивный или терапевтический эффект в лечении дегенерации желтого пятна.
ПРИМЕР 29
Данный пример описывает результаты MASP-2(-/-) в модели ревматоидного артрита, опосредованного мышиными моноклональными антителами.
Предпосылки/обоснование: Наиболее часто используемая модель ревматоидного артрита (RA) на животных представлена коллаген-индуцированным артритом (CIA) (для ознакомления с недавними исследованиями см. Linton and Morgan, Mol. Immunol. 36:905-14, 1999). Коллаген типа II (СИ) представляет собой один из основных компонентов артикулярных матричных белков, а иммунизация натральным СИ в адьюванте вызывает аутоиммунный полиартрит посредством кроссреактивного аутоиммунного ответа на СИ в суставном хряще. Как в случае RA, чувствительность к CIA связана с экспрессией определенных аллельных генов МНС класса II. Некоторые линии мышей, включая линию С57В1/6, обладают устойчивостью к классическому CIA вследствие отсутствия у них соответствующего гаплотипа МНС, и как следствие отсутствие генерирования высоких титров анти-CII антител. Однако было обнаружено, что стойкий артрит может быть индуцирован во всех линиях мышей при помощи внутривенного или внутрибрюшинного введения им смеси из четырех специфических моноклональных антител против коллагена типа II. Подобные артридогенные моноклональные антитела являются коммерчески доступными («Chondrex, Inc.», Редмонд, шт.Вашингтон). Такая пассивная трансфертная модель CIA успешно использовалась в некоторых недавно опубликованных отчетах с использованием линии мышей С57В1/6 (Kagari et al., J. Immunol. 169:1459-66, 2002; Kato et al., J. Rheumatol. 30:241-55, 2003; Banda et al., J. Immunol. 177:1904-12, 2006). Описанное ниже исследование сравнивает чувствительность мышей дикого типа (+/+) и MASP-2(-/-) мышей, где обе линии обладают единой генетической основой С57 В1/6, к развитию артрита с использованием пассивной трансфертной модели CIA.
Методы:
Животные: MASP-2(-/-) мышь получали как описано в Примере 1, затем проводили обратное скрещивание по меньшей мере с 10 поколениями с С57В1/6. Для данного исследования были отобраны четырнадцать самцов и самок мышей дикого типа C57BL/6 в возрасте от семи до восьми недель во время проведения инъекций с антителами и десять самцов и самок мышей MASP-2(-/-) и дикого типа (+/+) С57В1/6 в возрасте от семи до восьми недель во время проведения инъекций с антителами. Двадцать мышей получили инъекции со смесью моноклональных антител для получения 20 устойчивых отвечающих субъектов (две группы из десяти). Животные (десять/на группу) были разделены по пять животных на клетку, и были оставлены для акклиматизации на пять-семь дней до момента начала исследования.
Мышам были проведены внутривенные инъекции со смесью моноклональных антител («Chondrex», Редмонд, шт.Вашингтон) (5 мг) в день 0 и день 1. Тестовый агент был представлен моноклональным антителом + LPS от компании «Chondrex». На второй день, мышам внутрибрюшинно вводили дозу LPS. Мыши были взвешены в день 0, 2, 4, 6, 8, 10, 12 и перед окончанием эксперимента в день 14. На четырнадцатый день мышей анестезировали изофлюраном и окончательно обескровлили для получения сыворотки. После сбора крови, мышей умертвили, затем удалили обе передние конечности и задние конечности с коленями, которые поместили в формалин для последующей обработки.
Группы исследования:
Группа 1 (контрольная): 4 мыши линии C57/BL/6 WT (+/+);
Группа 2 (тестовая): 10 мышей линии C57/BL/6 WT (+/+) (получали смесь mAb плюс LPS); и
Группа 3 (тестовая): 10 мышей линии C57/BL/MASP-2KO/6AJ (-/-) (получали смесь mAb плюс LPS).
Клинические показатели артрита оценивались ежедневно с использованием следующей системы подсчета:
0=норма; 1=1 пораженный сустав задней или передней конечности; 2=2 пораженных сустава задней или передней конечности; 3=3 пораженных сустава задней или передней конечности; 4=среднее (эритема и средняя опухоль, или 4 пораженных сустава пальцев); 5=тяжелое (рассеянная эритема и обширная опухоль всей лапы, неспособность пальцев к движению)
Результаты:
На ФИГУРЕ 19 в форме графика представлены данные ежедневного клинического наблюдения проявлений артрита (в виде среднего балла по группе) в течение двух недель. У контрольной группы, не получавшей лечение с CoL2 MoAb, клинические показатели артрита отсутствовали. MASP (-/-) мыши продемонстрировали более низкие клинические показатели артрита с дня 9 по день 14. Общие клинические показатели артрита с применением анализа площади под кривой (AUC) показали 21%-ную редукцию в группах MASP-2(-/-) против группы мышей дикого типа (+/+). Однако мыши с генетической основой С57В16, описанной ранее, не продемонстрировали сильных суммарных клинических показателей артрита. Благодаря небольшому коэффициенту заболеваемости и размеру группы, в случае положительной тенденции, данные предоставлялись только для тенденции (р=0,1) и не представляли статистической значимости на уровне р<0,05. Дополнительные животные в группах исследования должны обязательно демонстрировать статистическую значимость. Благодаря снижению заболеваемости артритом, пораженные конечности были оценены на тяжесть заболевания. Ни одного клинического показателя проявления артрита выше 3 не было обнаружено ни у одного из представителей MASP-2(-/-) мышей, которые наблюдались в 30% у мышей дикого типа (+/+), что позволяет выдвинуть следующие предположения: (1) тяжесть артрита может быть связана с активацией пути комплемента, и (2) блокирование MASP-2 может оказывать благотворный эффект при артрите.
ПРИМЕР 30
Данный Пример демонстрирует, что малый маннозосвязывающий ассоциированный с пектином белок, (Small Mannose-Binding Lectin- Associated Protein) (Map 19 или sMAP) является ингибитором MASP-2-зависимой активации комплемента.
Предпосылки/обоснование:
Краткое изложение содержания:
Маннозосвязывающий лектин (MBL) и фиколины являются структурными белками распознавания, действующими во врожденном иммунитете и инициирующими активацию пектинового пути комплемента посредством MBL-ассоциированных сериновых протеаз (MASPs). При активации лектинового пути, MASP-2 расщепляет С4 и С2. Малый MBL-ассоциированный белок (sMAP), процессированная форма MASP-2, также ассоциируется с МВификолин-MASP комплексами. Для выяснения роли sMAP, посредством целевого разрушения sMAP-специфичного экзона нами были получены мыши с sMAP-недостаточностью (sMAP-/-). Вследствие генного разрушения, уровень экспрессии MASP-2 у sMAP-/- мышей также был снижен. При замещении комплекса MBL-MASP-sMAP рекомбинантным sMAP (rsMAP) и рекомбинантным MASP-2 (rMASP-2) в дефицитной сыворотке, связывание данных рекомбинант с MBL было конкурирующим, а активность расщепления С4 комплексом MBL-MASP-sMAP восстанавливалась при добавлении rMASP-2, в то время как добавление rsMAP ослабляло активность. Поэтому, MASP-2 является основным элементом для активации С4, а sMAP исполняет роль регулятора в активации пектинового пути.
Введение:
Система комплемента запускает цепную реакцию протеолизиса и сборки белковых комплексов, исполняя ведущую роль в биологической защите как части обеих врожденной и адаптивной иммунных систем. Система комплемента млекопитающих состоит из трех путей активации: классического, альтернативного и пектинового путей активации комплемента (Fujita, Nat. Rev. Immunol. 2: 346-353 (2002); Walport, N Engl J Med 344: 1058-1066 (2001)). Лектиновый путь активации предоставляет первичную линию защиты против инвазивных агентов. Компоненты распознавания патогенов в данном пути, маннозосвязывающий лектин (MBL) и фиколины, связываются с совокупностью углеводов на повехности бактерий, вирусов и паразитов и активируют MBL- ассоциированные сывороточные протеазы (MASPs) для инициации реакции нисходящего каскада. Важность пектинового пути для врожденной иммунной защитной реакции подчеркивается многочисленными клиническими исследованиями, связывающими недостаточность MBL с повышенной чувствительностью к разнообразным инфекционным заболеваниям, особенно в раннем детстве до установления адаптивной иммунной системы (Jack et al., Immunol Rev 180: 86-99 (2001); Neth et al. Infect Immun 68: 688-693 (2000); Summerfield et al., Lancet 345: 886-889 (1995); Super et al., Lancet 2: 1236-1239 (1989)). Однако пектиновый путь активации также спообствует нежелательной активации комплемента, который вовлекается в процесс воспаления и поражения тканей в некоторых патологических состояниях, включая ишемическое перфузионное поражение в сердце и почках (de Vries et al., Am J Pathol 165: 1677-1688 (2004); Fiane et al., Circulation 108: 849-856 (2003); Jordan et al., Circulation 104: 1413-1418 (2001); Walsh et al., J Immunol 175: 541-546 (2005)).
Как упоминалось ранее, лектиновый путь активации вовлекает распознавание углеводов при помощи MBL и фиколинов (Fujita et al., Immunol Rev 198: 185-202 (2004); Holmskov et al., Annu Rev Immunol 21; 547-578 (2003); Matsushita and Fujita, Immunobiology 205: 490-497 (2002) и данные лектины формируют комплексы с MASP-1 (Matsushita and Fujita, J Exp Med 176: 1497-1502 (1992); Sato et al., Int Immunol 6: 665-669 (1994); Takada et al., Biochem Biophys Res Commun 196: 1003-1009 (1993), MASP-2 (Thiel et al., Nature 386: 506-510 (1997), MASP-3 (Dahl et al., Immunity 15: 127-135 (2001), и процессированный белок MASP-2 (малый MBL-ассоциированный белок; sMAP или МАр19) (Stover et al., J Immunol 162: 3481-3490 (1999); Takahashi et al., Int Immunol 11: 8590863 (1999). Члены семейства MASP состоят из шести доменов; два C1r7C1s/Uegf/bone морфогенных белковых (CUB) домена, домен, подобный эпидермальному фактору роста (EGF), два домена комплемент-контролирующего белка (ССР), или домены кратких консенсусных повторов (SCR), и домен сериновой протеазы (Matsushita et al., Curr Opin Immunol 10: 29-35 (1998). MASP-2 и sMAP генерируются посредством альтернативного сплайсинга из одиночного структурного гена, и sMAP состоит из первого домена CUB (CUB1), EGF-подобного домена и дополнительных 4 аминокислот на С-терминальном конце, закодированным sMAP- специфичным экзоном. MASP-1 и MASP-3 также генерируются из одиночного гена посредством альтернативного сплайсинга (Schwaeble et al., Immunobiology 205: 455-466 (2002). Когда MBL и фиколины связываются с углеводами на поверхности микробов, проэнзимная форма MASP расщепляется между вторым ССР и доменом протеазы, результатом чего является активная форма, состоящая из двух полипептидов, известных как тяжелая (Н)- и легкая (L)-цепи, и таким образом, вызывая протеолитическую активность против компонентов комплемента. Накопленные сведения делают очевидным, что MASP-2 расщепляет С4 и С2 (Matsushita et al., J. Immunol 165: 2637-2642 (2000) которые приводят к формированию конвертазы С3 (C4bC2a). Мы предположили, что MASP-1 напрямую расщепляет С3 и впоследствии активирует петлю амплификации (Matsushita and Fujita, Immunobiology 194: 443-448 (1995), но эта функция является противоречивой (Ambrus et al., J. Immunol 170: 1374-1382 (2003). Несмотря на то, что MASP-3 также содержит домен сериновой протеазы в L-цепи и демонстрирует свою протеолитическую активность против синтетического субстрата (Zundel et al., J Immunol 172: 4342-4350 (2004), его физиологические субстраты не идентифицированы. Функция sMAP с отсутствующим доменом сериновой протеазы остается невыясненной.
В настоящем исследовании по выяснению роли sMAP в активации пектинового пути активации комплемента, мы разрушили sMAP-специфический экзон, который кодирует 4 аминокислотных остатка (EQSL) на С-терминальном конце sMAP, и получили sMAP-/- мышей. Здесь мы впервые сообщаем о способности sMAP к понижающей регуляции активации пектинового пути комплемента.
Материалы и методы
Мыши
Был сконструирован целевой вектор, содержащий экзон 1-4 и часть экзона 6 от 129/Sv мышиного MASP-2 гена и неомицин-резистентную генную кассету вместо экзона 5 (ФИГУРА 20А). Ген DT-A был внедрен в 3' конец вектора и три сайта lox p были внедрены для осуществления обусловленного нацеливания для удаления неомицинной кассеты и участка промотора в будущем. Целевой вектор был электропорирован в клетки 129/Sv ES. Целевые клоны ES были внедрены посредством микроинъекций в бластоцисты C57BL/6J, которые были имплантированы в матку суррогатных ICR матерей. Самцы химерных мышей были спарены с самками мышей C57BL/6J для продуцирования гетерозиготных мышей (+/-). Гетерозиготные мыши (+/-) были подвергнуты скринингу при помощи саузерн-блоттинга хвостовой ДНК, обработанной эндонуклеазой рестрикции с BamHI при использовании зонда, как показано на ФИГУРЕ 20А. Саузерн-блоттинг показал две зоны гибридизации размером 6,5 т.п.н. и 11 т.п.н. в ДНК от гетерозиготных мышей (+/-) (ФИГУРА 20 В). Гетерозиготные мыши (+/-) были подвергнуты обратному скрещению с мышами C57BL/6J. Для получения гомозиготных мышей (-/-), Гетерозиготные мыши (+/-) были скрещены между собой. Гомозиготные мыши (-/-) (C57BL/6J генетическое основание) были идентифицированы при помощи PCR-анализа генотипа хвостовой ДНК. PCR-анализ был выполнен с использованием смеси экзон 4-специфичных и неоген-специфичных смысловых праймеров и экзон 6-специфичного антисмыслового праймера. С ДНК от гомозиготных мышей (-/-) получали единичный диск размером 1,8 т.п.н. (ФИГУРА 20С). Во всех экспериментах были использованы мыши в возрасте от 8 до 12 недель, в соответствии с директивой для экспериментальных исследований на животных Медицинского Университета Фукушима (Fukushima Medical University).
Нозерн-блоттингз
Поли(А)+РНК (1 мкг) из печени мышей дикого типа (+/+) и гомозиготных мышей (-/-) была разделена при помощи электрофореза, перенесена на нейлоновую мембрану, и гибридизирована с 32Р-меченным кДНК зондом, специфичным к sMAP, MASP-2 Н-цепи, MASP-2 L-цепи, или неогену. Та же самая мембрана была отмыта и регибридизирована при помощи зонда, специфичного к глицеральдегид-3-фосфат дегидрогеназе (GAPDH).
Количественный RT-PCR анализ
PCR в реальном времени был произведен при помощи системы LightCycler («Roche Diagnostics»). кДНК, синтезированные из 60 нг поли(А)+РНК из печени мышей дикого типа (+/+) и гомозиготных мышей (-/-), использовали в качестве образцов для PCR в реальном времени и фрагменты кДНК из MASP-2 Н- и L-цепей и sMAP были амплифицированы и изучены.
Иммунноблотинг
Электрофорез образца проводили в 10 или 12%-ном SDS-полиакриламидном геле в восстановительных условиях, и белки были перенесены на поливинилиден дифторидные мембраны (PVDF). Белки на мембранах были детектированы при помощи анти-MASP-1 антисыворотки, против L-цепи MASP-1, или при помощи анти-MASP-2/sMAP антисыворотки, против пептида из Н-цепи MASP-2.
Детектирование MASPs и sMAP в комплексе MBL-MASP-sMAP
Мышиная сыворотка (20 мкл) была добавлена к 480 мкл буфера TBS-Ca2+ (20 ммоль Tris-HCl, pH 7,4, 0,15 моль NaCl, и 5 ммоль CaCl2), содержащего 0,1% (по весу) BSA (TBS-Ca2+/BSA), и инкубирована с 40 мкл 50%-ной маннан-агарозы гелевой гидросмеси («Sigma-Afdrich», Сент-Луис, шт.Миссури) в буфере TBS-Ca2+/BSA при 4°С в течение 30 минут. После инкубации каждый гель был отмыт буфером TBS-Ca2+и выборочный буфер для SDS-PAGE добавили в гель. Гель довели до кипения, супернатанты исследовали на SDS-PAGE, после чего проводили иммуноблоттинг на определение MASP-1, MASP-2 и sMAP в комплексе MBL.
Анализ отложения С4
Мышиную сыворотку развели буфером TBS-Ca2+/BSA до 100 мкл. Разведенный образец добавили в лунки микропланшета, покрытые маннаном, и провели инкубацию при комнатной температуре в течение 30 мин. Затем лунки промыли охлажденным промывочным буфером (ТВ8-Са2+ буфер с содержанием 0,05% (по объему) Tween 20). После промывания, в каждую лунку добавляли С4 человека и проводили инкубацию на льду в течение 30 минут. Затем лунки промывали охлажденным промывочным буфером и в каждую лунку добавляли HRP-коньюгированное античеловеческое поликлональное антитело С4 («Biogenesis», Пул, Англия). После инкубации при 37°С в течение 30 минут, лунки промывали промывочным буфером и в каждую лунку добавляли раствор 3,3',5,5'-тетраметилбензидина (ТМВ). После проявления добавляли 1 моль H3PO4 и определяли спектральную поглощающую способность при длине волны 450 нм.
Анализ отложения С3
Мышиную сыворотку разводили буфером BBS (4 ммоль барбитала, 145 ммоль NaCl, 2 ммоль CaCl2, и 1 ммоль MgCl2, рН 7,4), содержащим 0,1% (по весу) Н5А до 100 мкл. Растворенный образец добавляли в лунки микропланшета, покрытые маннаном, и проводили инкубацию при 37°С в течение 1 часа. Затем лунки промыли промывочным буфером. После промывания в каждую лунку добавили HRP-коньюгированное античеловеческое поликлональное антитело С3с («Dako», Глоструп, Дания). После инкубации при комнатной температуре в течение 1 часа лунки промыли промывочным буфером и в каждую лунку добавили раствор ТМВ. Окраска была замерена, как описано выше.
Рекомбинанты
Рекомбинантные мышиные sMAP (rsMAP), rMASP-2, и инактивный мутант (MASP-2i) мышиного MASP-2, чей остаток серина активного сайта в домене сериновой протеазы был замещен остатком аланина, были получены при помощи способов, описанных ранее (Iwaki and Fujita, 2005).
Восстановление комплекса MBL-MASP-sMAP
Сыворотка от гомозиготных мышей (-/-) (20 мкл) и различные объемы MASP-2J и/или rsMAP подвергли инкубации в общем объеме 40 мкл в буфере TBS-Ca2+ на льду в течение ночи. Смесь инкубировали с гелевой гидросмесью маннан-агарозы. Связанные с гелем MASP-2J и rsMAP в комплексе MBL-MASP определяли как описано в «Детектирование MASPs и sMAP в комплексе MBL-MASP-sMAP».
Восстановление активности отложения С4
Сыворотка от гомозиготных мышей (-/-) (0,5 мкл) и различные объемы rMASP-2 и/или rsMAP подвергли инкубации в общем объеме 20 мкл в TBS-Ca2+на льду в течение ночи. Смесь развели 80 мкл буфера TBS-Ca2+/BSA и добавили в лунки, покрытые маннаном. Все последующие процедуры были произведены, как описано в «Исследовании отложения С4».
Результаты:
ФИГУРА 20: Целевое разрушение гена sMAP. (А) Частичные рестриктазные карты гена MASP-2/sMAP, целевой вектор и целевые аллели. sMAP-специфический экзон (экзон 5) был замещен неогенной кассетой. (В) Саузерн-блоттинг геномной ДНК, выделенной у потомства, полученного от скрещивания самца химерной мыши с самкой мыши C57BL/6J. ДНК из хвостов обрабатывали рестриктазой BamHI и гибридизировали с зондом, изображенным на (А). Зона гибридизации размером 11 т.п.н. соответствовует аллелю дикого типа, зона гибридизации размером 6,5 т.п.н. соответствовует целевому аллелю. (С) PCR анализ определения генотипа. Был проведен PCR анализ ДНК из хвостов с использованием смеси экзон 4-специфического и неоген-специфического смысловых праймеров и экзон 6-специфического антисмыслового праймера. Диск размером 2,5 т.п.н. соответствует аллелю дикого типа, а диск размером 1,8 т.п.н. соответствовует целевому аллелю.
ФИГУРА 21: Экспрессия мРНК sMAP и MASP-2 у гомозиготной мыши (-/-). (А) Нозерн-блоттинг. Поли(А)+РНК из печени мыши дикого типа (+/+) и гомозиготной мыши (-/-) была разделена при помощи электрофореза, перенесена на нейлоновую мембрану, и гибридизирована с 32Р-меченным зондом кДНК, спецефичным к sMAP, MASP-2 Н-цепи, MASP-2 L-цепи, или неогену. Зона гибридизации, специфичная для нео (2,2 т.п.н.) наблюдалась у гомозиготных мышей (-/-). (В) Количественный RT-PCR. MASP-2 Н- и L-цепи и sMAP фрагменты ДНК были амплифицированы при помощи PCR в реальном времени на приборе LightCycler («Roche Diagnostics»). В качестве образцов были использованы кДНК, синтезированные из поли(А)+РНК, извлеченные из печени мышей дикого типа (+/+) и гомозиготных мышей (-/-). Представленные данные являются средними для двух экспериментов.
ФИГУРА 22: Отсутствие MASP-2 в сыворотке от гомозиготных мышей (-/-). (А) Имммуноблоттинг MASP-2 и sMAP в мышиной сыворотке. Сыворотку от мышей дикого типа (+/+) или гомозиготных мышей (-/-) (2 мкл) подвергли иммуноблоттингу и диагностировали при помощи анти-MASP-2/sMAP антисыворотки. (В) Детектирование MASPs и sMAP в комплексе MBL-MASP-sMAP. Мышиную сыворотку инкубировали с гелевой гидросмесью маннан-агарозы, затем проводили детектирование sMAP, MASP-I, и MASP-2 в комплексе MBL, связанных с гелем, как описано в «Материалах и методах».
ФИГУРА 23: Снижение расщепления С4 и С3 в сыворотке гомозиготной мыши (-/-). (А) Отложение С4 на покрытых маннаном лунках. Проводили двукратное разведение мышиной сыворотки, которую затем подвергали инкубированию в покрытых маннаном лунках при комнатной температуре в течение 30 минут. После того, как лунки были промыты, в каждую лунку добавляли человеческий С4 и снова инкубировали на льду в течение 30 минут. Объем отложения человеческого С4 на лунках измеряли с использованием HRP-коньюгированного античеловеческого С4 поликлонального антитела. (В) Отложение С3 на покрытых маннаном лунках. Разведенную мышиную сыворотку добавляли в покрытые маннаном лунки, и проводил инкубацию при 37°С в течение 1 часа. Отложение эндогенного С3 на лунках определили при помощи HRP-коньюгированного античеловеческого С3с поликлонального антитела.
FIGURE 24: Конкурирующее связывание sMAP и MASP-2 с MBL. (А) Восстановление комплекса MBL-MASP-sMAP в сыворотке от гомозиготной мыши (-/-). MASP-2J и/или rsMAP (4 мкг) инкубировали с гомозиготной мышиной сывороткой (-/-) (20 мкл). Далее смесь инкубировали с маннан-агарозным гелем, а rsMAP и MASP-2I, связанные во фракции с гелем, были выявлены посредством иммуноблоттинга. (В) Различные количества MASP-2I (0-5 мкг) и постоянное количество rsMAP (5 мкг) проинкубировали с гомозиготной мышиной сывороткой (-/-) (20 мкл) и далее с маннан-агарозным гелем. (С) Постоянное количество MASP-2I (0,5 мкг) и различные количества rsMAP (0-20 мкг) проинкубировали с гомозиготной мышиной сывороткой (-/-) (20 мкл). (D) Различные количества rsMAP (0-20 мкг) проинкубировали с сывороткой от мыши дикого типа (+/+) (20 мкл).
ФИГУРА 25: Восстановление активности отложения С4 посредством добавления rMASP-2. Различные количества rsMAP (0-5 мкг) (А) или rMASP-2 (0-1,5 мкг) (В) проинкубировали с 0,5 мкл сыворотки от гомозиготной мыши (-/-) в общем объеме 20 мкл в буфере TBS-Ca2+ на льду в течение ночи. Затем смесь развели 80 мкл буфера TBS-Ca2+/BSA и добавили в покрытые маннаном лунки, и количество С4, отложившееся на лунках, было измерено.
ФИГУРА 26: Уменьшение активности отложения С4 посредством добавления sMAP. (А) rMASP-2 (1 мкг) и различные количества rsMAP (0-0,5 мкг) проинкубировали с 0,5 мкл сыворотки от гомозиготной мыши (-/-). Смесь добавили к маннан-покрытым лункам, а количество С4, отложившееся на лунках, было измерено. (В) rsMAP (0-0,7 мкг) проинкубировали с сывороткой дикого типа (0,5 мкл), а количество С4, отложившееся на маннан-покрытых лунках, было измерено.
РЕЗУЛЬТАТЫ:
Экспрессия sMAP и MASP -2 у гомозиготных мышей (-/-)
Для установления роли sMAP in vivo, нами была создана мышь, у которой отсутствовал ген sMAP. Был сконструирован целенаправленный вектор для замещения специфического экзона для sMAP (экзон 5) неомицин-резистенетной геной кассетой (Фигура 20А). Положительные клоны ES были введены при помощи инъекций в бластоцисты C57BL/6 и полученные первичные химеры были скрещены с самками C57BL/6J. Саузерн-блоттинг хвостовой ДНК от потомства дикой окраски показал эмбриональную трансмиссию целевых аллелей (Фигура 20 В). Гетерозиготные мыши (+/-) были подвергнуты скринингу посредством саузерн-блоттинга хвостовой ДНК, обработанной BamH I, с использованием зонда, указанного на Фигуре 20А. Саузерн-блопинг показал наличие дисков размером 6,5 т.п.н. и 11 т.п.н. в ДНК от гетерозиготных мышей (+/-) (Фигура 20 В). Провели обратное скрещивание гетерозиготных мышей (+/-) с мышами C57BL/6J. Далее, для получения гомозиготных мышей (-/-), провели скрещивание гетерозиготных мышей (+/-) между собой (интеркросс). Гомозиготные мыши (-/-) (с генетической основой C57BL/6J) были идентифицировы методом PCR, основанным на генотипировании хвостовой ДНК, с образованием одного диска размером 1,8 т.п.н. (Фигура 20С).
Развитие гомозиготных мышей (-/-) протекало в пределах нормы без проявления существенной разницы в весе по сравнению с мышами дикого типа (+/+). Также между данными группами не было отмечено морфологических различий. При нозерн-блоттинге зонд, специфичный к sMAP, обнаружил один диск размером 0,9 т.п.н. у мышей дикого типа (+/+), при отсутствии специфических дисков у гомозиготных мышей (-/-) (Фигура 21А). При использовании зонда, специфичного к MASP-2 Н- или L-цепям, у мышей дикого типа (+/+) были обнаружены несколько специфичных дисков, как было отмечено ранее (Stover et al., 1999), зонд, специфичный к Н-цепи, также выявил диск, специфичный к sMAP. Однако у гомозиготных мышей (-/-) соответствующие диски были весьма слабыми, и было обнаружено несколько дополнительных дисков. Мы также провели количественный анализ RT-PCR для проверки уровней экспрессии мРНК sMAP и MASP-2. У гомозиготных мышей (-/-) экспрессия мРНК sMAP полностью отсутствовала, а экспрессия MASP-2 была значительно снижена: в количественном соотношении это представляет приблизительно 2% от общего числа мышей дикого типа (+/+) в обеих Н- и L-цепях при PCR в реальном времени (Фигура 21 В). Кроме того, мы исследовали экспрессию MASP-2 на уровне белков. Как sMAP, так и MASP-2 не были выявлены в сыворотке гомозиготных мышей (-/-) посредством иммуноблоттинга (Фигура 22А). После инкубации сыворотки гомозиготных мышей (-/-) с маннан-агарозным гелем, оба элемента sMAP и MASP-2 не были обнаружены в связанной с гелем фракции, хотя MASP-1 был обнаружен в комплексе (Фигура 22В).
Активность расщепления С4 и С3 посредством пектинового пути активации комплемента в сыворотке гомозиготных мышей (-/-)
При инкубировании сыворотки гомозиготных мышей (-/-) в покрытых маннаном лунках, количество отложения человеческого С4 на лунках составил 20% от количества в нормальной сыворотке при разведении в пределах от 1/400 до 1/50 (Фигура 23А). Мы также исследовали активность отложения С3 при пектиновом пути в сыворотке гомозиготных мышей (-/-). Мышиную сыворотку добавили в покрытые маннаном лунки и произвели замер количества эндогенного С3, отложившееся на лунках. В дефицитной сыворотке количество С3 было пониженным и составляло 21% от его количества в нормальной сыворотке при разведении в соотношении 1/10 (Фигура 23 В). Восстановление комплекса MBL-MASP-sMAP в сыворотке гомозиготных мышей (-/-)
Когда рекомбинантные мышиные sMAP (rsMAP) или неактивные измененные мышиные MASP-2 (MASP-2i) были добавлены к сыворотке гомозиготных мышей (-/-), обе рекомбинанты обнаружили способность к связыванию с MBL (Фигура 24А, дорожки 3 и 4). Когда rsMAP и MASP-2i были одновременно инкубированы с сывороткой (Фигура 24А, дорожка 5), обе рекомбинанты были обнаружены в комплексе MBL-MASP-sMAP. Однако количество связанного с комплексом sMAP был меньше, чем при инкубации только rsMAP с сывороткой. Затем мы провели дальнейшее исследование конкурирующего связывания sMAP и MASP-2 с MBL. Постоянное количество rsMAP и различные количества MASP-2i были добавлены в дефицитную сыворотку Связывание rsMAP понижалось дозозависимым образом по мере увеличения количества MASP-2i (Фигура 24 В). Обратно пропорционально, объем связанного с MBL элемента MASP-2i уменьшался при добавлении rsMAP (Фигура 24С). Когда rsMAP добавили к сыворотке дикого типа, связывание обоих эндогенного sMAP и MASP-2 с MBL уменьшалось дозозависимым образом (Фигура 24D).
Восстановление активности отложения С4 в сыворотке гомозиготной мыши (-/-)
Нами был выполнен эксперимент по восстановлению отложения С4 на покрытых маннаном лунках с использованием рекомбинант. Когда rsMAP был добавлен к дефицитной сыворотке, количество С4, отложившегося в момент исследования, дозозависимым образом понижалось до базового уровня (Фигура 25А). Когда rMASP-2 добавили к сыворотке, количество С4 восстанавливалось дозозависимым образом до 46% от объема в сыворотке дикого типа и стабилизировалося (ФИГУРА 25 В). Далее, нами было проведено исследование воздействия sMAP на отложение С4. Когда постоянное количество rMASP-2 и различные количества rsMAP добавили к дефицитной сыворотке, количество отложившегося С4 дозозависимым образом понижалось при добавлении rsMAP в (Фигура 26А), добавление rsMAP в сыворотку дикого типа также понижало количество отложившегося С4 (Фигура 26 В), что позволяет выдвинуть предположение о регулирующей роли sMAP в активации пектинового пути комплемента.
Обсуждение
Посредством нацеленного разрушения sMAP- специфического экзона нами были получены мыши sMAP-/-. Уровень экспрессии MASP-2 у данных мышей был также чрезвычайно понижен как на уровне мРНК так и на уровне белков (Фигуры 21 и 22). Нозерн-блоттинг с зондом MASP-2 выявил только дополнительные диски в поли (А)+РНК от sMAP-/- мышей, что позволяет выдвинуть предположение о том, что нормальный сплайсинг гена MASP-2 был изменен посредством нацеливания гена sMAP и как следствие, произошло значительное снижение экспрессионного уровня MASP-2. Как результат, расщепление С4 посредством комплекса MBL-MASP в дефицитной сыворотке было снижено приблизительно на 80% по сравнению с нормальной сывороткой (Фигура 23А). В экспериментах по восстановлению, активность расщепления С4 была восстановлена путем добавления rMASP-2, но не rsMAP (Фигура 25). Редуцирование отложения С4, которое наблюдалось в дефицитной сыворотке, по всей вероятности, вызвано отсутствием MASP-2 в комплексе MBL-MASP (Фигура 22 В). Таким образом, очевидно, что MASP-2 является основным фактором для активации С4 посредством комплекса MBL-MASP. Однако добавление rMASP-2 не полностью восстанавливало активность расщепления, и отложение С4 стабилизировалось. Как сообщалось ранее (Cseh et al., J Immunol 169: 5735-5743 (2002); Iwaki and Fujita, J Endotoxin Res 11: 47-50 (2005), большая часть rMASP-2 была конвертирована в активную форму посредством самооактивации во время процедур по очищению и наблюдалось некоторое снижение его протеазной активности. Учитывая отсутствие значительного влияния активных и неактивных состояний MASP-2 на его связь с MBL (Zundel et al., J Immunol 172: 4342-4350 (2004), существует возможность того, что потерявший свою протеазную активность rMASP-2 связывается с MBL и избирательным способом предупреждает связь в активной форме, таким образом, провоцируя неполное восстановление отложения С4. Активность расщепления С3 пектинового пути также была ослаблена в дефицитной сыворотке (Фигура 23 В). Уменьшение количества отложившегося С3 может быть обусловлено очень низким уровнем активности конвертазы С3, которая состоит из фрагментов C4b и С2а, генерируемых MASP-2.
MASP и sMAP каждый связывались с образованием гомодимеров и формировали комплексы с MBL или L-фиколином посредством N-терминального CUB и EGF-подобного доменов (Chen and Wallis, J Biol Chem 276: 25894-25902 (2001); Cseh et al., J Immunol 169: 5735-5743 (2002); Thielens et al., J Immunol 166: 5068-5077 (2001); Zundel et al., J Immunol 172: 4342-4350 (2004)). Кристаллические структуры sMAP и CUB1-EGF-CUB2 сегмента MASP-2 выявляют их гомодимерную структуру (Feinberg et al., EMBO J 22: 2348-2359 (2003); Gregory et al., J Biol Chem 278: 32157-32164 (2003)). Коллаген-подобный домен MBL вовлечен в связывание с MASPs (Wallis and Cheng, J Immunol 163:4953-4959 (1999); Wallis and Drickamer, J Biol Chem 279: 14065-14073 (1999), а некоторые мутации, введенные в этот домен, снизили связывание MBL с CUB1-EGF-CUB2 сегментами MASP-1 и MASP-2 (Wallis and Dodd, J Biol Chem 275: 30962-30969 (2000)). Связывающие сайты для MASP-2 и для MASP-1/3 перекрывают друг друга, хотя без полной идентичности (Wallis et al., J Biol Chem 279: 14065-13073 (2004)). Хотя sMAP-связывающий сайт MBL еще не был идентифицирован, связывающие сайты для sMAP и MASP-2 являются практически идентичными, так как область CUBI-EGF является одной и той же для sMAP и MASP-2. Таким образом, является вполне обоснованным, что sMAP и MASP-2 конкурируют друг с другом в связывании с MBL в восстановлении комплекса MBL-MASP-sMAP (Фигура 24). Аффинность sMAP для MBL является ниже, чем для MASP-2 (Cseh et al., J Immunol 169: 5735-5743 (2002); Thielens et al., J Immunol 166: 5068-5077 (2001)). Концентрация sMAP в мышиной сыворотке не была определена. Как показано на Фигуре 22А, количество sMAP в сыворотке дикого типа является гораздо более высоким, чем количество MASP-2. Поэтому, sMAP обладает способностью занимать связывающий сайт MASP-2/sMAP и предохранять MASP-2 от связывания с MBL, и, следовательно, активность расщепления С4 комплекса MBL-MASP также снижается. Регулирующий механизм sMAP в лектиновом пути остается вопросом для дальнейшего исследования. До сих пор остается невыясненным, выполняет ли sMAP свою регуляторную функцию до или после активации комплемента. sMAP может предупреждать непроизвольную активацию комплекса MBL-MASP перед микробной инфекцией или подавлять повторную активацию уже инициированного пектинового пути. Существует другой потенциальный регулятор пектинового пути активации комплемента. MASP-3 также является конкурентом для MASP-2 в связывании с MBL и осуществляет пониженную регуляцию активности расщепления С4 и С2 эллемента MASP-2 (Dahl et al., Immunity 15: 127-135 (2001)). Хотя взаимодействие между sMAP и MASP-3 еще не было исследовано, существует возможность того, что они способны совместно подавлять активацию пектинового пути.
В данной работе мы показали, что sMAP и MASP-2 конкурируют друг с другом в связывании MBL, и что sMAP обладает способностью к подавлению пектинового пути, который активизируется при помощи комплекса MBL-MASP. Обоснованным является то, что sMAP также регулирует другой способ инициации пектинового пути, активированного комплексом фиколин-MASP. MASP-2 и sMAP также конкурируют в связывании мышиного фиколина А и осуществляют подавление активности расщепления С4 комплекса фиколин A-MASP (Y Endo et al., в подготовке). Исследование MBL-нулевой мыши было недавно опубликовано (Shi et al., J Exp Med 199: 1379-1390 (2004). MBL-нулевые мыши не проявляют активности расщепления С4 в MBL лектиновом пути и являются чувствительными к инфекции золотистого стафилококка (Staphylococcus aureus). В настоящем исследовании, мыши sMAP-/-, также обладающие недостаточностью MASP-2, продемонстрировали редуцирование в активности расщепления С3 из-за активности расщепления С4 в лектиновом пути. По причине их ослабленной опсонизирующей активности, мыши с sMAP-недостаточностью могут проявлять чувствительность к бактериальным инфекциям. Дальнейшее исследование мышей с sMAP-недостаточностью будет иметь целью прояснение функций пектинового пути в защитном механизме против инфекционных заболеваний.
Другим важным открытием является то, что добавление rsMAP к нормальной сыворотке проявляется в редуцировании активации С4 (Фигура 26 В). Также было показано, что пектиновый путь способен регулировать воспаление и тканевое поражение в некоторых органах (de Vries et al., Am J Pathol 165: 1677-1688 (2004); Fiane et al., Circulation 108: 849-856 (2003); Jordan et al., Circulation 104: 1413-1418 (2001); Walsh et al., J Immunol 175: 541-546 (2005)). У пациентов с MBL-недостаточностью, проходящих лечение от торакальной аневризмы брюшной аорты, не происходило активации комплемента и уровни воспалительных маркеров были снижены вследствие опреативного лечения (Fiane et al., Circulation 108: 849-856 (2003)). Накопленные сведения со всей очевидностью показали возможность MBL играть патофизиологическую роль в состояниях ишемии и реперфузии в разнообразных сосудистых ложах. Далее, специфическая блокада MBL или ингибирование пектинового пути активации комплемента могут представлять собой терапевтически релевантную стратегию для предупреждения поражения, опосредованного ишемией/перфузией. Таким образом, существует возможность того, что sMAP является кандидатом на роль ингибитора, так как действует в качестве аттенюатора активации пектинового пути комплемента.
ПРИМЕР 31
Данный пример показывает, что MASP-2 отвечает за С4-альтернативную активацию С3.
Предпосылки/обоснование: В самых недавних исследованиях было показано, что ингибирование альтернативного пути защищает почку от острого ишемического поражения (Thurman et al., J. Immunol 170: 1517-1523 (2003)). Описанные в настоящей заявке данные подразумевают, что пектиновый путь инициирует активацию альтернативного пути, которая в свою очередь синергетически усиливает активацию комплемента. Мы выдвигаем гипотезу, что временное ингибирование пектинового пути также может затрагивать активацию альтернативного пути, и, таким образом, улучшать долгосрочный исход при трансплантации органов, что выражается в ограничении комплемент-опосредованного поражения трансплантанта и воспаления, и может смягчать нежелательную индукцию адаптивного иммунного ответа на трансплантант и уменьшать риск вторичного отторжения трансплантанта посредством адаптивной иммунной системы. Вышеизложенное поддерживается недавно полученными клиническими данными, демонстрирующими, что частично ослабленный лектиновый путь активации, опосредованный наследственной недостаточностью MBL (представлено приблизительно 30% человеческой популяции), связан с повышением уровня выживаемости почечного аллотрансплантата у людей (Berger, Am J Transplant 5:1361-1366 (2005)).
Вовлечение компонентов комплемента С3 и С4 в ишемическое реперфузионое (I/R) поражение было хорошо изучено в модели временной интестинальной и мышечной ишемии с использованием линий мышей с целевыми генами (Weiser et al., J Exp Med 183: 2342-2348 (1996); Williams et al. J Appl Physiol 86: 938-42, (1999)). Было четко установлено, что С3 играет ярко выраженную роль в ишемическом реперфузионом поражении почек и вторичном отторжении трансплантата (Zhou et al., J Clin Invest 105: 1363-1371 (2000); Pratt et al., Nat Med 8: 582-587 (2002); Farrar, et al., Am J. Pathol 164:133-141 (2004)). Далее был обнаружен неожиданный факт того, что фенотип для С4 недостаточности не наблюдался в опубликованных моделях отторжении почечного трансплантата у мышей (Lin, 2005 In Press). Последующий анализ сыворотки и плазмы от данных мышей с С4-недостаточностью, однако, показал, что данные мыши сохраняют остаточную функциональную активность, показывающую LP-зависимое расщепление С3 и дальнейшую нисходящую активацию комплемента (см. ФИГУРУ 27С).
Существование функционального С4-альтернативной активации (и С2-альтернативной активации) представляет собой феномен, который был ранее описан (но не был полностью охарактеризован) несколькими исследователями (Miller et al., Proc Natl Acad Sci 72: 418-22 (1975); Knutzen Steuer et al., J Immunol 143(7): 2256-61 (1989); Wagner et al., J Immunol 163: 3549-3558 (1999) и относится к С3-обновлению, не зависимому от альтернативного пути, в С4 (и С2)-дефицитной сыворотке.
Методы: Воздействие пектинового и классического путей активации комплемента на отложение СЗ. Мышиная плазма (с EGTA/ Mg2+в качестве антикоагулянта) была разведена и рекальцинирована в 4,0 ммоль барбитала, 145 ммоль NaCl, 2,0 ммоль CaCl2, 1,0 ммоль MgCl, рН 7,4, затем добавлена в микротитрационные планшеты, покрытые маннаном (как показано на ФИГУРЕ 27А и 27С) или зимозаном (как показано в ФИГУРЕ 27 В), и подвергнута инкубации в течение 90 минут при 37°С.Планшеты были трижды отмыты 10 ммоль Tris-Cl, 140 ммоль NaCl, 5,0 ммоль CaCl, 0,05% Tween 20, рН 7,4. Затем отложение C3b было измерено с использованием антимышиного С3с антитела.
Результаты: Результаты, представленные на ФИГУРЕ 27 А-С, получены от трех независимых экспериментов. При использовании той же самой сыворотки в лунках, покрытых иммуноглобулиновыми комплексами вместо маннана или зимозана, отложение C3b и расщепления Фактора В было обнаружено в мышиной сыворотке дикого типа (+/+) и смешанной сыворотке MASP-2(-/-), но не в dq-обедненой сыворотке (данные не представлены). Данное наблюдение указывает на то, что активация альтернативного пути может быть восстановлена в сыворотке MASP-2-/-, когда изначальный C3b предоставляется через активность СР. ФИГУРА 27С изображает неожиданное открытие того, что С3 может быть эффективно активирован способом, зависимым от пектинового пути, в С4 (-/-)-недостаточной плазме. Данная «С4-альтернативная активация» аннулируется ингибированием активации пектинового пути посредством преинкубирования плазмы с жидким маннаном или маннозой.
Было обнаружено, что отложение C3b на маннане и зимозане сильно нарушено у мышей с недостаточностью MASP-2(-/-), даже в условиях эксперимента, что согласно многим ранее опубликованным работам по активации альтернативного пути должно допускаться во всех трех путях активации. Как показано на ФИГУРЕ 27А-С мышиная плазма с недостаточностью MASP-2(-/-) не активирует С4 посредством пектинового пути и не расщепляет С3, ни пектиновым, ни альтернативным путем. Поэтому мы выдвигаем гипотезу, что MASP-2 необходим при С4-альтернативной активации. Дальнейший прогресс в идентификации компонентов, которые могли бы быть вовлечены в С4-альтернативную активацию, опосредованную лектиновым путем, был недавно описан в работе профессора Тейцо Фуджиты (Prof. Teizo Fujita). Плазма мышей с С4-недостаточностью, скрещенных с линией мышей с MASP-1/3-недостаточностью, предложенной Фуджитой, теряет остаточную способность С4-дефицитной плазмы к расщеплению С3 посредством пектинового пути. Эта способность была восстановлена при помощи добавления рекомбинантного MASP-1 к комбинированной С4 и MASP-1/3-дефицитной плазме (Takahashi, Mol Immunol 43: 153 (2006), что позволяет выдвинуть предположение о том, что MASP-1 вовлечен в формирование комплексов, образованных лектиновым путем, которые расщепляют С3 в отсутствие С4 (рекомбинантный MASP-1 не расщепляет С3, но расщепляет С2; Rossi et al., J Biol Chem 276: 40880-7 (2001); Chen et al., J Biol Chem 279: 26058-65 (2004). Мы наблюдали, что MASP-2 требуется для запуска альтернативной активации.
Хотя для обобщения данного экспериментального исследования необходимо больше функциональных и количественных параметров и гистологических данных, его предварительные результаты могут рассматриваться в качестве должной поддержки той гипотезы, что активация комплемента посредством пектинового пути в значительной степени относится к патофизиологии I/R поражения почек, так как MASP-2-/- мыши демонстрируют гораздо более быстрое восстановление ренальной функции.
ПРИМЕР 32
Данный пример демонстрирует, что активация тромбина может возникнуть после активации пектинового пути при физиологических условиях, а также демонстрирует степень участия MASP-2. В нормальной сыворотке крысы активация пектинового пути приводит к активации тромбина (оцененной как отложение тромбина) одновременно с активации комплемента (оцененной как отложениеС4). Как показано на ФИГУРАХ 28А и 28 В, активация тромбина в данной системе ингибируется посредством блокирующего антитела MASP-2 (формат Fab2), показывающего кривую концентрации-эффекта при ингибировании (ФИГУРА 28В), которая проходит параллельно кривой концентрации-эффекта при ингибировании касательно активации комплемента (ФИГУРА 28А). Эти данные показывают, что активация пектинового пути, происходящая при травме, приведет к активации системы комплемента и коагулирующей системы образом полностью зависимым от MASP-2. Отсюда следует вывод, что блокирующие антитела MASP2 могут оказаться эффективными в уменьшении случаев избыточной системной коагуляции, например, диссеминированного внутрисосудистого свертывания, которое является одним из признаков, приводящих к смертности в большинстве случаев травм.
ПРИМЕР 33
В данном Пример приводятся результаты, полученные при использовании модели локальной реакции Шварцмана диссеминированного внутрисосудистого свертывания (ДВС) у мышей с MASP-2-недостаточностью (-/-) и с MASP-2-достаточностью (+/+) с целью оценки роли пектинового пути в ДВС.
Предпосылки/обоснование:
Как описано выше, блокада MASP-2 ингибирует активацию пектинового пути и сокращает генерацию анафилатоксинов С3а и С5а. Анафилатоксины С3а можно показать, как сильные агрегаторы тромбоцитов in vitro, но их участие in vivo менее четко определено и высвобождение тромбоцитов и плазмина при заживлении раны может только второстепенно задействовать комплемент СЗ. В данном примере роль пектинового пути анализировалась на мышах MASP-2(-/-) и дикого типа (+/+) с целью рассмотрения необходимости пролонгированного подъема активации С3 для генерации диссеминированного внутрисосудистого свертывания.
Способы:
Мыши MASP-2(-/-), использованные в данном исследовании, были получены, как описано в Примере 27. В данном эксперименте использовалась модель локальной реакции Шварцмана. Локальная реакция Шварцмана (LSR) представляет собой липополисахарид (LPS) - индуцированную реакцию с хорошо изученными вкладами из клеточных и гуморальных элементов врожденной иммунной системы. Признана зависимость LSR от комплемента (Polak, L, et al., Nature 223:738-739 (1969); Fong J.S. et al., J Exp Med 134:642-655 (1971)). В модели LSR мышей сенсибилизировали в течение 4 часов при помощи ФНО-альфа (500 нг, внутримошоночно), затем мышей анестезировали и подготовили для прижизненной микроскопии мышцы, поднимающей яичко. Сети посткапиллярных венулей (диаметром 15-60 мкм) с хорошим кровотоком (1-4 мм/с) выбрали для наблюдения. Животных обработали флуоресцирующими антителами для избирательного мечения нейтрофилов или тромбоцитов. Сеть сосудов последовательно сканировали и изображения всех сосудов последнего анализа записали в цифровой форме. После регистрации базального состояния микроциркуляции мышам ввели одну внутривенную инъекцию LPS (100 мкг) отдельно или с нижеперечисленными веществами. Ту же сеть сосудов затем сканировали каждые 10 минут в течение 1 часа. Удельная аккумуляция флуорофоров идентифицирована посредством субтракции фоновой флуоресценции и увеличена посредством порогового представления яркости изображения. Величину реакций измерили с записанных изображений. Первичное измерение реакций Шварцмана представляло собой агрегированные данные.
Исследования сравнили мышей с MASP-2-достаточностью (+/+) или мышей дикого типа, подвергшихся воздействию известного кровопускательного вещества комплементарного пути, фактора яда кобры (CVF) или ингибитора терминального пути (антагонист C5aR). Результаты (ФИГ. 29А) демонстрируют, что CVF также как и антагонист C5aR, предупредили появление бляшек в сосудистой сети. Более того, мыши с MASP-2-недостаточностью (-/-) (ФИГ. 29 В) также продемонстрировали полное ингибирование локальной реакции Шварцмана, поддерживающей участие пектинового пути. Данные результаты четко демонстрируют роль MASP-2 в генерации ДВС и поддерживают использование ингибиторов MASP-2 для лечения и предупреждения ДВС.
ПРИМЕР 34
В данном Примере описывается анализ мышей MASP-2(-/-) на мышиной модели ишемии/реперфузии миокарда.
Предпосылки/обоснование:
Для оценки вклада MASP-2 в воспалительное реперфузионное повреждение после ишемического инсульта на коронарной артерии, мышей MASP-2(-/-) и MASP-2(+/+) сравнили на мышиной модели ишемии/реперфузии (MIRP), как описано у Marber et al., J. Clin Invest. 95:1446-1456 (1995), и на модели перфузии изолированного сердца мыши по Лангендорфу.
Способы:
Мыши MASP-2(-/-), использованные в данном исследовании, были получены, как описано в примере 27. Ишемический инсульт на левом желудочке провели на восьми мышах дикого типа (MASP-2(+/+) и одиннадцати мышах MASP-2(-/-), при использовании методов, описанных в Примере 27. Размер инфаркта (INF) и площадь, подверженная риску (AAR), определены посредством планиметрии, как описано в Примере 27.
Модель перфузии изолированного сердца мыши по Лангендорфу: Способ препарирования мышиных сердец по модели перфузии изолированного сердца мыши по Лангендорфу провели, как описано у F.J. Sutherland et al., Pharmacol Res 41: 613 (2000). Смотрите также A.M. Kabir et al., Am J Physio! Heart Circ Physiol 291: H1893 (2006); Y. Nishino et al., Circ Res 103:307 (2008) и I.G. Webb et al., Cardiovasc Res (2010)).
Вкратце, шесть самцов мышей дикого типа (+/+) и девять самцов мышей MASP-2(-/-) анестезировали пентобарбиталом (300 мг/кг) и гепарином (150 единиц) внутрибрюшинно. Сердца быстро изолировали и поместили в ледяной модифицированный буферный раствор Кребса-Хенселейта (КН, 118,5 ммоль/л NaCl, 25,0 ммоль/л NaHCO3, 4,75 ммоль KCl KH2PO4 1,18, MgSO4 1,19, D-глюкоза 11,0 и CaCl2 1,41). Удаленное сердце поместили в аппарат Лангендорфа с водяной рубашкой и катаболически перфузировали при постоянном давлении 80 мм рт.ст. с буферным раствором КН, уравновешенным 95% O2 и 5% CO2. Температура перфузата поддерживалась на уровне 37°С. Баллон, заполненный жидкостью, вставленный в левый желудочек, контролировал сократительную функцию. Баллон постепенно наполняли воздухом, пока конечное диастолическое давление не было установлено в пределах 1 и 7 мм рт.ст. Предсердную стимуляцию осуществили при 580 ударах в минуту при помощи 0,075-мм серебряной проволоки («Advent»). Коронарный кровоток измерили посредством рассчитанного по времени сбора перфузата. Оценка инфаркта in vitro
После начала катаболической перфузии, сердца стабилизировали в течение 30 минут. Для включения в исследование все сердца должны были отвечать нижеследующим критериям: коронарный кровоток в пределах 1,5 и 4,5 мл/мин, частота сердечных сокращений >300 ударов в минуту (без стимуляции), развитое давление левого желудочка >55 мм рт.ст., время от торакотомии до канюлирования аорты <3 минуты, и отсутствие постоянной аритмии в ходе стабилизации. Затем проведена глобальная ишемия и реперфузия при отсутствии сыворотки. Все сердца затем подвергли глобальной ишемии в течение 30 минут посредством зажатия трубки притока аорты с последующей реперфузией в течение 2 часов.
Электрическая стимуляция была остановлена, когда сокращение сердечной мышцы прекратилось во время ишемии и повторно началось при реперфузии в течение 30 минут. После 2 часов реперфузии, сердца перфузировали в течение 1 минуты при помощи 5 мл 1% трифенилтетразолия хлорида (ТТС) в КН, а затем поместили в идентичный раствор при температуре 37°С на 10 минут. Предсердия затем извлекли, провели сухой блоттинг сердец, взвесили и хранили при температуре -20°С в течение до 1 недели.
Затем сердца разморозили, поместили в 2,5% глютаральдегид на 1 минуту и поместили в 5% агарозу. Агарозные блокады сердца затем секционировали от верхушки до основания на 0,7 мм срезы при использовании вибратома («Agar Scientific»). После секционирования срезы поместили на ночь в 10% формальдегид при комнатной температуре до перемещения в забуференный фосфатом физиологический раствор (PBS) еще на один день при температуре 4°С. Срезы затем сжали между пластинами Perspex (на расстоянии 0,57 мм) и получили изображение при использовании сканера (модель Epson G850A). После увеличения провели планиметрию при использовании программного обеспечения для анализа изображений (SigmaScan Pro 5.0, SPSS) общей площади поверхности, и ТТС-отрицательный миокард левого желудочка трансформировали в объем посредством умножения на плотность тканей. В каждом сердце после суммирования индивидуальных срезов, ТТС-отрицательный объем инфаркта выразили как процентное отношение объема левого желудочка или как график левого желудочка.
Результаты:
Размер инфарктной области (тусклый), площадь левого желудочка (LV) с повышенным риском (красная) и зона нормально перфузированного левого желудочка (синяя) выделены в каждом срезе посредством идентификации их цветового признака и цветовых границ. Исследователь подсчитал области с обеих сторон каждого среза и вывел среднее число. Объем инфаркта рассчитан, как % зоны риска (% RZ) для каждого животного.
На ФИГУРЕ 31 А показана оценка восьми мышей дикого типа (+/+) и одиннадцати мышей MASP-2(-/-) для определения их размера инфаркта после проведения окклюзии коронарной артерии и вышеописанного метода реперфузии. На ФИГУРЕ 31А графически изображается средняя область, подверженная риску (AAR, единица измерения области, пораженной ишемией) и объемы инфаркта (INF, единица измерения повреждения миокарда), как процент общего объема миокарда. Как показано на ФИГУРЕ 31А, несмотря на отсутствие отличия в AAR между двумя группами, объемы INF в значительной степени сокращаются у мышей MASP-2(-/-) по сравнению с однопометными мышами дикого типа, указывая, таким образом, на защитный эффект от повреждения миокарда при отсутствии MASP-2 в данной модели MIRP.
На ФИГУРЕ 31 В графически изображается взаимосвязь между графиком INF по отношению к AAR, как % объема миокарда левого желудочка (LV). Как показано на ФИГУРЕ 31 В по каждой AAR, животные MASP-2(-/-) показали высокозначимое сокращение размера их инфаркта по сравнению с однопометными животными дикого типа.
На ФИГУРАХ 31С и 31D показываются результаты инфаркта миокарда сердец мышей дикого типа (+/+) и MASP-2(-/-), перфузируемых в буферном растворе, подготовленных в соответствии с моделью перфузии изолированного сердца мыши по Лангендорфу, в котором глобальную ишемию и реперфузию провели при отсутствии сыворотки. Как показано на ФИГУРАХ 31С и 31D, отличие в результирующем объеме инфаркта (INF) между сердцами мышей MASP-2(-/-) и дикого типа (+/+) не наблюдалось, что указывает на то, что разница в размерах инфарктов, показанная на ФИГУРАХ 31А и 31В, вызвана плазматическими факторами, а не более низкой восприимчивостью ткани миокарда мышей MASP-2(-/-) к ишемическому повреждению.
В своей совокупности данные результаты демонстрируют, что MASP-2-недостаточность в значительной степени сокращает повреждение миокарда при реперфузии ишемии сердца на мышиной модели ишемии миокарда/реперфузии, и поддерживают использование ингибиторов MASP-2 для лечения и предупреждения ишемического реперфузионного повреждения.
ПРИМЕР 35
В данном примере описывается анализ мышей MASP-2 на мышиной модели пересадки почки.
Предпосылки/обоснование:
Роль MASP-2 в функциональном результате пересадки почки была оценена при использовании мышиной модели.
Способы:
Функциональный результат пересадки почки был оценен при использовании изографта одной почки у мышей-реципиентов, у которых удалили одну почку, с шестью реципиентами трансплантата дикого типа (+/+) (В6) и шестью реципиентами трансплантата MASP-2(-/-). Для оценки функции пересаженной почки, оставшаяся нативная почка была извлечена у реципиента через 5 дней после пересадки, и почечная функция была оценена через 24 часа посредством измерения уровней азота мочевины крови (BUN).
Результаты;
На ФИГУРЕ 32 графически изображаются уровни азота мочевины крови (BUN) почки за 6 дней после пересадки почки у реципиентов дикого типа (+/+) и реципиентов MASP-2(-/-). Как показано на ФИГУРЕ 32 сильно повышенные уровни BUN наблюдались у реципиентов трансплантата дикого типа (+/+) (В6) (нормальные уровни BUN у мышей составляют <5 ммоль), указывая на почечную недостаточность. Для сравнения мыши-реципиента изографта MASP-2(-/-) показали существенно более низкие уровни BUN, указывая на улучшенную почечную функцию. Констатировали, что данные результаты были получены при использовании графтов от доноров почек дикого типа (+/+), указывающих на то, что отсутствие только функционального пектинового пути у реципиента трансплантата является достаточным для достижения благоприятного терапевтического эффекта.
В совокупности данные результаты указывают на то, что транзиторное ингибирование пектинового пути через ингибирование MASP-2 предоставляет способ сокращения заболеваемости и отсроченную функцию графта при пересадке почки и что данный подход, вероятно, будет полезен при других условиях трансплантата.
ПРИМЕР 36
В данном примере демонстрируется, что мыши MASP-2(-/-) являются устойчивыми к септическому шоку на мышиной модели полимикробного септического перитонита.
Предпосылки/обоснование:
Для оценки потенциальных эффектов MASP-2(-/-) в инфекции, была оценена модель лигирования и пункции слепой кишки (CLP), модель полимикробного септического перитонита. Данная модель считается наиболее точно имитирующей течение септического перитонита у человека. Модель лигирования и пункции слепой кишки (CLP) представляет собой модель, в которой слепая кишка лигируется и делается пункция иглой, приводя к постоянной утечке бактерий в брюшную полость, которые попадают в кровь через лимфодренаж и затем разносятся ко всем органам брюшной полости, приводя к мультиорганной недостаточности и септическому шоку (Eskandari et al., J Immunol 148(9):2724-2730 (1992)). Модель CLP имитирует течение сепсиса, наблюдаемого у пациентов, и индуцирует раннюю гипервоспалительную реакцию с последующей ярко выраженной гиповоспалительной фазой. Во время данной фазы животные являются высокочувствительными к бактериальным изменениям (Wichterman et al., J. Surg. Res. 29(2); 189-201 (1980)).
Способы:
Смертность полимикробной инфекции при использовании модели лигирования и пункции слепой кишки (CLP) была измерена на мышах дикого типа (+/+) (n=18) и MASP-2 (-/-) (n=16), как описано в Примере 23. Вкратце, мышей с MASP-2-недостаточностью и однопометных мышей дикого типа анестезировали, а слепую кишку вывели и лигировали выше дистального отдела на 30%. После этого на слепой кишке сделали один раз пункцию иглой диаметром 0,4 мм. Слепую кишку затем поместили в брюшную полость и кожу закрыли зажимами. Выживаемость мышей, подверженных CLP, наблюдалась в течение 14 дней после CLP. Перитонеальный диализ был собран у мышей через 16 часов после CLP для измерения бактериальной обсемененности. Серийные разведения перитонеального диализа были приготовлены в PBS и инокулированы в чашках Мюллера-Хинтона с последующей инкубацией при температуре 37°С в анаэробных условиях в течение 24 часов после чего была определена бактериальная обсемененность.
Цитокиновый ответ ФНО-альфа на бактериальную инфекцию также был измерен у мышей дикого типа (+/+) и MASP-2(-/-) через 16 часов после CLP в легких и селезенке через количественную полимеразную цепную реакцию в масштабе реального времени (qRT-PCR). Уровень сыворотки ФНО-альфа через 16 часов после CLP у мышей дикого тпа (+/+) и MASP-2(-/-) также подсчитан посредством иммуноферметного сэндвич-анализа.
Результаты:
На ФИГУРЕ 33 графически изображается процент выживаемости животных, обработанных CLP, как функция дней после процедуры CLP. Как показано на ФИГУРЕ 33 недостаточность пектинового пути у мышей MASP-2(-/-) не повышает смертность мышей после полимикробной инфекции при использовании модели лигирования и пункции слепой кишки по сравнению с мышами дикого типа (+/+). Однако, как показано на ФИГУРЕ 34, мыши MASP-2(-/-) показали в значительной степени более высокую бактериальную обсемененность (примерно 1000-кратное увеличение содержания бактерий) в перитонеальном диализе после CLP при сравнении с однопометными мышами дикого типа (+/+). Данные результаты указывают на то, что мыши с MASP-2-недостаточностью (-/-)являются устойчивыми к септическому шоку. Редуцированный бактериальный клиренс у мышей с MASP-2-недостаточностью в данной модели можно обосновать нарушенным C3b-опосредованным фагоцитозом, как продемонстрировано, что отложение С3 является MASP-2 зависимым.
Было определено, что цитокиновый ответ ФНО-альфа на бактериальную инфекцию не был повышен у мышей MASP-2 (-/-) по сравнению с контрольными мышами дикого типа (+/+) (данные не приведены). Также было определено наличие в существенной степени более высокой концентрации сыворотки ФНО-альфа у мышей дикого типа (+/+) через 16 часов после CLP по сравнению с мышами MASP-2(-/-), где уровни сыворотки ФНО-альфа остались практически неизменными. Данные результаты указывают на то, что интенсивная воспалительная реакция на септическое условие была ослаблена у мышей MASP-2(-/-) и животные выжили при наличии более высокого количества бактерий.
В совокупности данные результаты демонстрируют потенциальные вредные действия пектинового пути активации комплемента в случае септицемии и повышенной смертности пациентов с генерализованным сепсисом. Данные результаты дополнительно демонстрируют, что MASP-2-недостаточность модулирует воспалительную иммунную реакцию и сокращает уровни экспрессии медиаторов воспаления во время сепсиса. Следовательно, полагают, что ингибирование MASP-2(-/-) посредством введения ингибирующих моноклональных антител против MASP-2 будет эффективным в ослаблении воспалительной реакции у субъекта, страдающего септическим шоком.
ПРИМЕР 37
В данном Примере описывается анализ мышей MASP-2(-/-) на мышиной модели интраназальной инфекционности.
Предпосылки/обоснование:
Синегнойная палочка (Pseudomonas aeruginosa) представляет собой грамотрицательный оппортунистический человеческий бактериальный патоген, который возбуждает широкий диапазон инфекций, особенно у людей с нарушенным иммунитетом. Она является основным источником приобретенных нозокомиальных инфекций, особенно внутрибольничной пневмонии. Она также отвечает за существенную заболеваемость и смертность пациентов с кистозным фиброзом поджелудочной железы (CF). Легочная инфекция синегнойной палочкой характеризуется сильным рекрутингом нейтрофилов и значительным воспалением легких, приводящих к обширному повреждению тканей (Palanki M.S. et al., J. Med. Chem 51:1546-1559 (2008)).
В данном Примере было проведено исследование для определения повышения восприимчивости мышей к бактериальным инфекциям при удалении пектинового пути у мышей MASP-2(-/-).
Способы:
Двадцать две мыши дикого типа (+/+), двадцать две мыши MASP-2(-/-) и одиннадцать мышей С3 (-/-) подвергли внутриназальному введению бактериального штамма синегнойной палочки. Мышей наблюдали в течение шести дней после инфицирования и построили графики Каплана-Майера, показывающие процент выживаемости.
Результаты;
ФИГУРА 35 представляет собой график процентного выживания Каплана-Майера мышей дикого типа (+/+), MASP-2(-/-) или С3 (-/-) через шесть дней после инфицирования. Как показано на ФИГУРЕ 35 отличие между мышами MASP-2(-/-) по сравнению с мышами дикого типа (+/+) не наблюдалось. Однако удаление классического пути (C1q) у мышей С3 (-/-) привело к сильной восприимчивости к бактериальной инфекции. Данные результаты демонстрируют, что ингибирование MASP-2 не повышает восприимчивость к бактериальной инфекции, указывая на возможность сокращения нежелательных воспалительных осложнений у травматологических больных посредством ингибирования MASP-2 без ухудшения способности пациента сопротивляться инфекциям при использовании классического пути комплемента.
ПРИМЕР 38
В данном примере описывается фармакодинамический анализ репрезентативных высокоаффинных антител анти-MASP-2 Fab2, которых идентифицировали, как описано в Примере 24.
Предпосылки/обоснование:
Как описано в Примере 24 для идентификации антител с высокой аффинностью, которые блокируют пектиновый путь крыс, использовался крысиный белок MASP-2 для сортировки библиотеки фагового отображения. Библиотеку разработали с целью предоставления высокого иммунологического разнообразия и построили при использовании только человеческих последовательностей иммуноглобулиновых генов. Как показано в Ппримере 24 было идентифицировано примерно 250 индивидуальных клонов фагов, которые связываются с высокой аффинностью с крысиным белком MASP-2 посредством скрининга ИФА. Последовательности данных клонов идентифицировали 50 уникальных фагов, кодирующих антитело MAPS-2. Белок Fab2 экспрессировали из данных клонов, очистили и проанализировали на предмет связывающей способности MASP-2 и функционального ингибирования пектинового пути комплемента.
Как показано в ТАБЛИЦЕ 6 Примера 24, в результате данного анализа было идентифицировано 17 анти-MASP-2 Fab2 с функциональной блокирующей активностью (34% коэффициент совпадений для блокирующих антител). Функциональное ингибирование лектинового пути комплемента посредством Fab2 было заметно на уровне отложения С4, который является прямым показателем расщепление С4 посредством MASP-2. Важно, что ингибирование было в равной мере заметно при оценке активности конвертазы СЗ, демонстрирующее функциональную блокаду лектинового пути комплемента. 17 идентифицированных Fab2, блокирующих MASP-2, как описано в Примере 24, сильно ингибируют образование конвертазы С3 со значениями IC50 равными или меньшими чем 10 нмоль. Восемь из 17 идентифицированных Fab2 имеют значения IC50 в суб-наномолярном диапазоне. Более того, все 17 Fab2, блокирующие MASP-2, показали существенно полное ингибирование образования конвертазы С3 в анализе лектинового пути конвертазы С3, как показано на ФИГУРЕ 11А-С и обобщено в ТАБЛИЦЕ 6 Примера 24. Более того, каждый из 17 Fab2, блокирующих анти-MASP-2, показанных в ТАБЛИЦЕ 6, сильно ингибирует генерацию C3b (П95%), демонстрируя, таким образом, специфичность данного анализа для лектинового пути конвертазы С3.
Варианты крысиных IgG2c и мышиных IgG2a полноразмерных изотипов антител были получены из Fab2 #11. Данный пример описывает характеристику данных изотипов in vivo для фармакодинамических параметров.
Способы:
Как описано в примере 24, использовался крысиный белок MASP-2 для сортировки библиотеки фагового отображения Fab, из которой был идентифицирован Fab2#11. Варианты крысиных IgG2c и мышиных IgG2a полноразмерных изотипов антител были получены из Fab2 #11. Крысиные IgG2c и мышиные IgG2a полноразмерные изотипы антител были характеризированы in vivo для фармакодинамических параметров, как указано ниже.
Исследование in vivo на мышах:
Фармакодинамическое исследование было проведено на мышах с целью изучения эффекта дозирования анти-MASP-2 антитела на активность плазмы лектинового пути in vivo. В данном исследовании отложение С4 было измерено ех vivo в анализе лектинового пути в разные периоды времени после подкожного (sc) и внутрибрюшинного (ip) введения 0,3 мг/кг или 1,0 мг/кг мышиного анти-MASP-2 MoAb (мышиный IgG2a полноразмерный изотип антитела, полученный из Fab2#11),
На ФИГУРЕ 36 графически изображается лектиновый путь специфичного отложения С4, измеренный ех vivo в неразбавленных образцах сыворотки, взятых у мышей (n=3 мыши/группа) в разные периоды времени после подкожного дозирования 0,3 мг/кг или 1,0 мг/кг мышиного анти-MASP-2 MoAb. Образцы сыворотки мышей, собранные перед дозированием антитела, использовались в качестве отрицательных контролей (100% активность) тогда, как сыворотка, дополненная in vitro 100 нмоль того же блокирующего антитела анти-MASP-2, использовалась в качестве положительного контроля (0% активность).
Результаты, показанные на ФИГУРЕ 36, демонстрируют быстрое и полное ингибирование отложения C4b после подкожного введения дозы 1,0 мг/кг мышиного анти-MASP-2 MoAb. Частичное ингибирование отложения С4 наблюдалось после подкожного введения дозы 0,3 мг/кг мышиного анти-MASP-2 MoAb.
Период восстановления пектинового пути продолжался в течение трех недель после одного внутрибрюшинного введения мышиного анти-MASP-2 MoAb дозой 0,6 мг/кг мышам. Как показано на ФИГУРЕ 37 резкое снижение активности пектинового пути возникло после дозирования антитела с последующим полным ингибированием пектинового пути, которое продолжалось примерно 7 дней после внутрибрюшинного введения. Медленное восстановление активности пектинового пути наблюдалось в течение второй и третьей недели с полным восстановлением пектинового пути у мышей через 17 дней после введения анти-MASP-2 MoAb.
Данные результаты демонстрируют, что мышиный анти-MASP-2 Moab, полученный из Fab2 #11, ингибирует пектиновый путь мышей в зависимости от дозы при систематической доставке.
ПРИМЕР 39
В данном примере описывается анализ мышиного анти-MASP-2 Moab, полученного из Fab2 #11 касательно эффективности на мышиной модели возрастной дегенерации желтого пятна.
Предпосылки/обоснование:
Как описано в Примере 24, крысиный белок MASP-2 использовался для сортировки библиотеки фагового отображения Fab, из которой был идентифицирован Fab2#11 в качестве функционально активного антитела. Полноразмерные антитела крысиного IgG2c и мышиного IgG2a изотипов генерировали из Fab2 #11. Полноразмерное антитело анти-MASP-2 мышиного IgG2a изотипа было характеризовано для фармакодинамических параметров, как описано в Примере 38. В данном Примере полноразмерное антитело мышиного анти-MASP-2, полученное из Fab2 #11, анализировалось на мышиной модели возрастной дегенерации желтого пятна (AMD), описанной Bora P.S. et al., J Immunol 174:491-497 (2005).
Способы:
Мышиный IgG2a полноразмерный изотип антитела анти-MASP-2, полученный из Fab2 #11, как описано в Примере 38, исследован на мышиной модели возрастной дегенерации желтого пятна (AMD), как описано в Примере 28 с нижеследующими модификациями.
Введение мышиных анти-MASP-2 MoAbs
Две разные дозы (0,3 мг/кг и 1,0 мг/кг) мышиного анти-МА5Р-2 MoAb вместе с обработкой изотипическим контролем MoAb ввели внутрибрюшинно мышам дикого типа (+/+) (n=8 мышей на группу) за 16 часов до индукции CNV.
Индукция неоваскуляризации хориоидеи (CNV)
Индукция неоваскуляризации хориоидеи (CNV) и измерение объема CNV проведено при использовании лазерной фотокоагуляции, как описано в Примере 28.
Результаты:
На ФИГУРЕ 38 графически изображается область CNV, измеренная через 7 дней после лазерного повреждения у мышей, обработанных изотипическим контролем MoAb или мышинным анти-MASP-2 MoAb (0,3 мг/кг и 1,0 мг/кг). Как показано на ФИГУРЕ 38 у мышей, предварительно обработанных анти-MASP-2 MoAb дозой 1,0 мг/кг, наблюдалось статистически существенное (р<0,01) примерно 50% снижение CNV через семь дней после лечения лазером. Как дополнительно показано на ФИГУРЕ 38 наблюдалось, что доза 0,3 мг/кг анти-MASP-2 MoAb не была эффективной в снижении CNV. Констатировали, что доза 0,3 мг/кг анти-MASP-2 MoAb обладает частичным и транзиторным ингибированием отложения C4b после подкожного введения, как описано в Примере 38 и показано на ФИГУРЕ 36.
Результаты, описанные в данном Примере, демонстрируют, что блокада MASP-2 ингибитором, таким, как анти-MASP-2 MoAb обладает профилактическим и/или терапевтическим эффектом при лечении дегенерации желтого пятна. Констатировали, что данные результаты не расходятся с результатами, наблюдаемыми в исследовании на мышах MASP-2(-/-), описанном в Примере 28, в котором наблюдалось 30% снижение CNV через 7 дней после лазерного лечения у мышей MASP-2(-/-) по сравнению с контрольными мышами дикого типа. Более того, результаты в данном Примере дополнительно демонстрируют, что системная доставка am-n-MASP-2 антитела обеспечивает локальную терапевтическую полезность глазу, подчеркивая тем самым потенциал по системному пути введения с целью лечения пациентов с AMD. В целом данные результаты предоставляют доказательство, поддерживающее использование MASP-2 MoAb при лечении AMD.
ПРИМЕР 40
В данном примере демонстрируется, что мыши с MASP-2-недостаточностью защищены от смертности, индуцированной менингококком (Neissena meningitidis) после инфицирования менингококком (N. meningitidis), и обладают усиленным клиренсом бактериемии по сравнению с контрольными мышами дикого типа.
Обоснование: Менингококк представляет собой гетеротрофную грамотрицательную диплококковую бактерию, известную за свою роль в менингите и других формах менингококковой инфекции такой, как менингококкемия. Менингококк является основной причиной заболеваемости и смертности в детском возрасте. Серьезные осложнения включают в себя септицемию, синдром Уотерхауса-Фридериксена, недостаточность надпочечников и диссеминированную внутрисосудистую свертываемость (ДВС). Смотрите, например, Rintala E. et at., Critical Care Medicine 28(7):2373-2378 (2000). В данном Примере роль пектинового пути анализировалась на мышах MASP-2(-/-) и мышах дикого типа (+/+) с целью рассмотрения восприимчивости мышей с MASP-2-недостаточностью к смертности, вызванной менингококком.
Способы:
MASP-2 нокаутные мыши были получены, как описано в Примере 27. 10-недельных мышей MASP-2 КО (n=10) и мышей С57/В6 дикого типа (n=10) инокулировали посредством внутривенной инъекции дозой 5×108 КОЕ/100 мкл, 2×108 КОЕ/100 мкл или 3×107 КОЕ/100 мкл менингококка серогруппы A Z2491 в 400 мг/кг декстрана железа. Выживаемость мышей после инфицирования наблюдалась в течение 72 часов. Образцы крови брали у мышей с часовым интервалом после инфицирования и анализировали с целью определения уровня сыворотки (логарифм КОЕ/мл) менингококка для подтверждения инфекции и определения скорости клиренса бактерий из сыворотки.
Результаты:
На ФИГУРЕ 39А графически изображается процент выживаемости мышей MASP-2 KO и дикого типа после введения инфекционной дозы менингококка 5×108/100 мкл КОЕ. Как показано на ФИГУРЕ 39А, после инфицирования самой высокой дозой менингококка 5×108/100 мкл КОЕ, 100% мышей MASP-2 KO выжило в течение 72-часового периода после инфицирования. Для сравнения, только 20% мышей дикого типа оставались все еще живыми через 24 часа после инфицирования. Данные результаты демонстрируют, что MASP-2 дефицитные мыши защищены от менингококка, индуцирующего смертность.
На ФИГУРЕ 39 В графически изображается логарифм КОЕ/мл менингококка, восстановленного в разные периоды времени в образцах крови взятых у мышей MASP-2 KO и дикого типа, инфицированных менингококком 5×108 КОЕ/100 мкл. Как показано на ФИГУРЕ 39В, у мышей дикого типа уровень менингококка в крови достиг пика, равного, примерно, 6,5 логарифм КОЕ/мл через 24 часа после инфицирования и упал до нуля через 48 часов после инфицирования. Для сравнения, у мышей MASP-2 КО уровень менингококка достиг пика, равного, примерно, 3,5 логарифм КОЕ/мл через 6 часов после инфицирования и упал до нуля через 36 часов после инфицирования
На ФИГУРЕ 40А графически изображается процент выживаемости мышей MASP-2 КО и дикого типа после инфицирования дозой менингококка 2×108 КОЕ/100 мкл. Как показано на ФИГУРЕ 40А после инфицирования дозой менингококка 2×108 КОЕ/100 мкл, 100% мышей MASP-2 КО выжило в течение 72-часового периода после инфицирования. Для сравнения, только 80% мышей дикого типа оставались все еще живыми через 24 часа после инфицирования. Схожие с результатами, показанными на ФИГУРЕ 39А, данные результаты дополнительно демонстрируют, что мыши с MASP-2-недостаточностью защищены от смертности, индуцированной менингококком.
На ФИГУРЕ 40В графически изображается логарифм КОЕ/мл менингококка, восстановленного в разные периоды времени в образцах крови взятых у мышей дикого типа, инфицированных менингококком 2×108 КОЕ/100 мкл. Как показано на ФИГУРЕ 40 В, уровень менингококка в крови мышей дикого типа, инфицированных 2×108 КОЕ, достиг пика, равного, примерно, 4 логарифм КОЕ/мл через 12 часов после инфицирования, и упал до нуля через 24 часа после инфицирования. ФИГУРА 40С графически изображает логарифм КОЕ/мл менингококка, восстановленного в разные периоды времени в образцах крови взятых у мышей MASP-2 КО, инфицированных менингококком 2×108 КОЕ/100 мкл. Как показано на ФИГУРЕ 40С, уровень менингококка в крови мышей MASP-2 КО, инфицированных 2×108 КОЕ, достиг пикового уровня, равного, примерно, 3,5 логарифм КОЕ/мл через 2 часа после инфицирования, и упал до нуля через 3 часа после инфицирования. Схожие с результатами, показанными на ФИГУРЕ 39 В, данные результаты демонстрируют, что, несмотря на то, что мышей MASP-2 КО инфицировали одинаковой дозой менингококка, как и мышей дикого типа, мыши MASP-2 КО обладают усиленным клиренсом бактериемии по сравнению с мышами дикого типа.
Процент выживаемости мышей MASP-2 КО и дикого типа после инфицирования самой низкой дозой менингококка 3×107 КОЕ/100 мкл составил 100% за 72-часовой период времени (данные не приведены).
Обсуждение
Данные результаты показывают, что мыши с MASP-2-недостаточностью защищены от смертности, индуцированной менингококком, и обладают усиленным клиренсом бактериемии по сравнению с мышами дикого типа. Следовательно, принимая во внимание данные результаты, ожидается, что терапевтическое применение ингибиторов MASP-2 таких, как MASP-2 MoAb, будет эффективным для лечения, предупреждения или ослабления эффектов инфицирования бактериями менингококка (т.е. сепсис и ДВС). Кроме того, данные результаты указывают, что терапевтическое применение ингибиторов MASP-2 таких, как MASP-2 MoAb не предрасполагает субъекта к повышенному риску заражения инфекциями менингококка.
Наряду с тем, что иллюстративные варианты осуществления данного изобретения были проиллюстрированы и описаны, различные изменения могут быть внесены при условии, что они не изменяют сущности и объема изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способы лечения состояний, связанных с masp-2 зависимой активацией комплемента | 2012 |
|
RU2662563C2 |
| СПОСОБЫ ЛЕЧЕНИЯ СОСТОЯНИЙ, СВЯЗАННЫХ С MASP-2 ЗАВИСИМОЙ АКТИВАЦИЕЙ КОМПЛЕМЕНТА | 2012 |
|
RU2743409C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ИНГИБИРОВАНИЯ MASP-1, И/ИЛИ MASP-2, И/ИЛИ MASP-3 ДЛЯ ЛЕЧЕНИЯ ПАРОКСИЗМАЛЬНОЙ НОЧНОЙ ГЕМОГЛОБИНУРИИ | 2013 |
|
RU2655299C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ИНГИБИРОВАНИЯ MASP-1, И/ИЛИ MASP-2, И/ИЛИ MASP-3 ДЛЯ ЛЕЧЕНИЯ РАЗЛИЧНЫХ ЗАБОЛЕВАНИЙ И НАРУШЕНИЙ | 2013 |
|
RU2709351C2 |
| КОМПОЗИЦИИ ДЛЯ ИНГИБИРОВАНИЯ MASP-2-ЗАВИСИМОЙ АКТИВАЦИИ КОМПЛЕМЕНТА | 2012 |
|
RU2636038C2 |
| СПОСОБЫ ЛЕЧЕНИЯ СОСТОЯНИЙ, АССОЦИИРОВАННЫХ С MASP-2-ЗАВИСИМОЙ АКТИВАЦИЕЙ КОМПЛЕМЕНТА | 2014 |
|
RU2718850C2 |
| СПОСОБЫ ЛЕЧЕНИЯ СОСТОЯНИЙ, АССОЦИИРОВАННЫХ С MASP-2-ЗАВИСИМОЙ АКТИВАЦИЕЙ КОМПЛЕМЕНТА | 2014 |
|
RU2830289C2 |
| КОМПОЗИЦИИ ДЛЯ ИНГИБИРОВАНИЯ MASP-2-ЗАВИСИМОЙ АКТИВАЦИИ КОМПЛЕМЕНТА | 2017 |
|
RU2725958C2 |
| Способ определения активности маннан-связывающих лектин-ассоциированных сериновых протеаз в тесте коагуляции фибриногена | 2019 |
|
RU2739113C1 |
| Определение активности классического пути системы комплемента в тесте коагуляции фибриногена | 2019 |
|
RU2707568C1 |




















































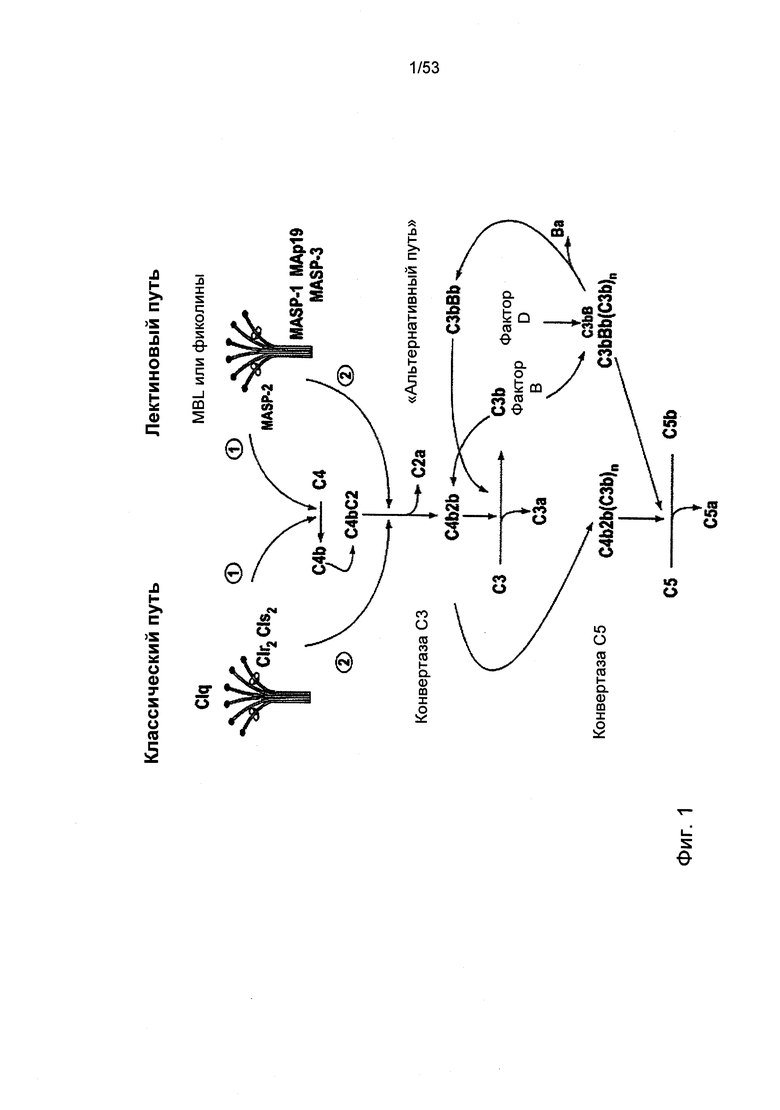
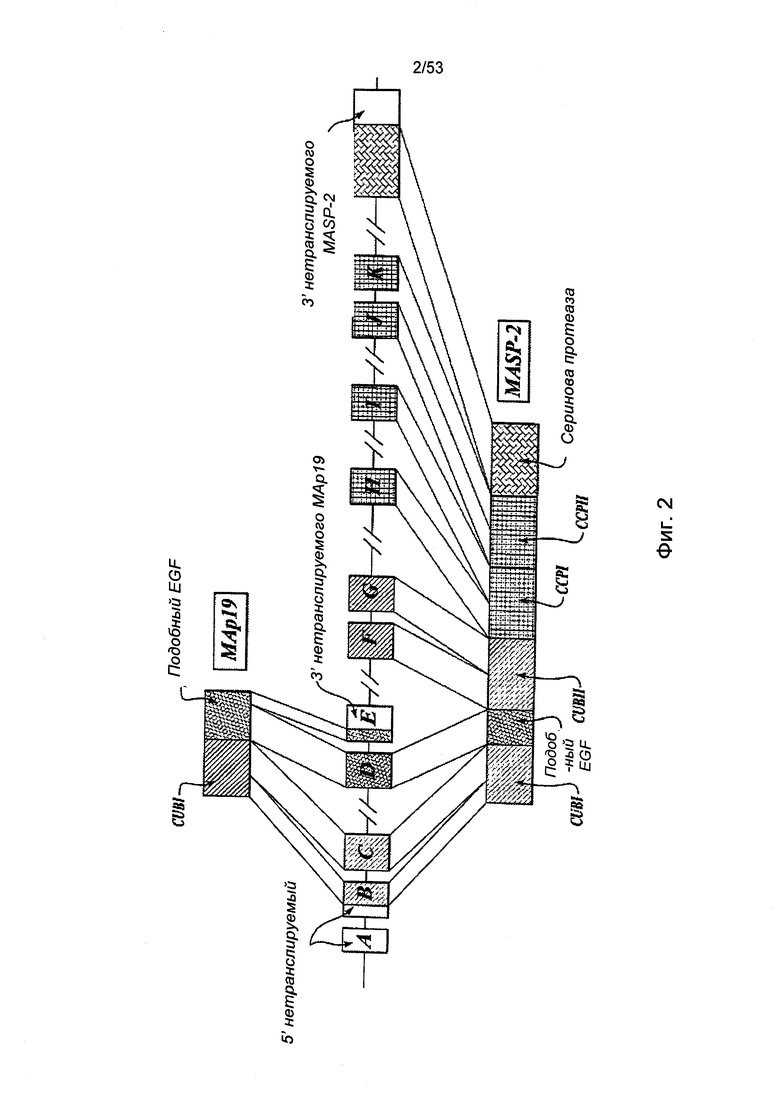
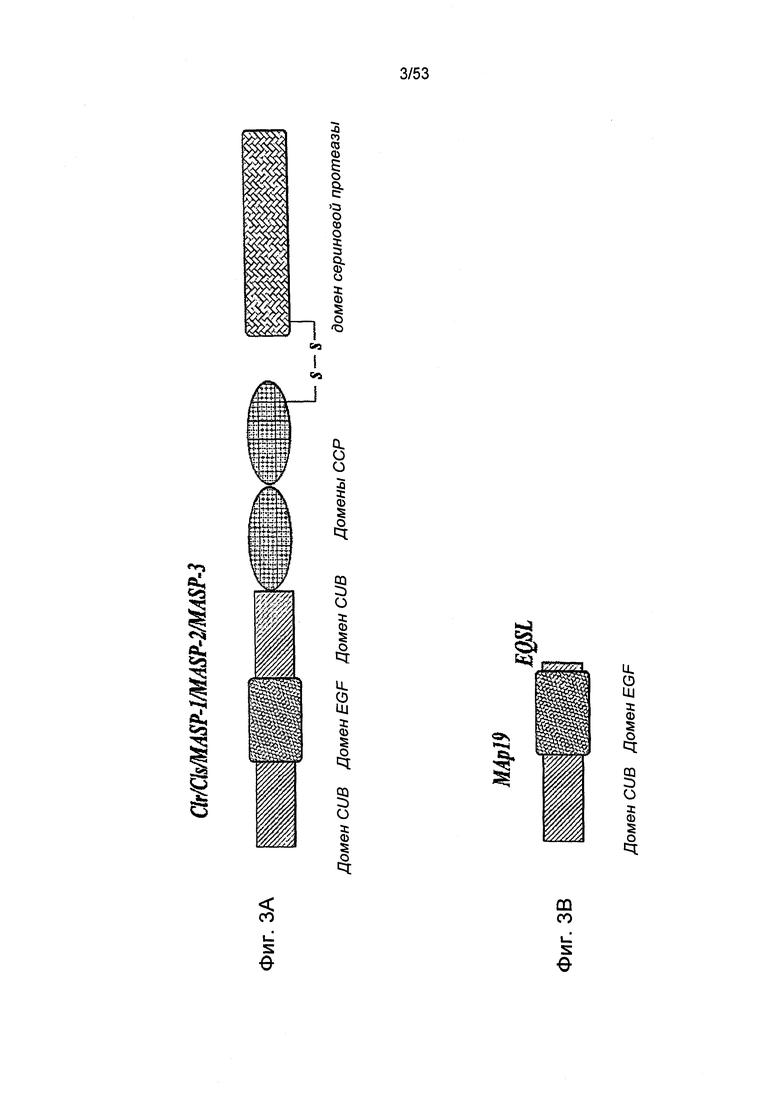
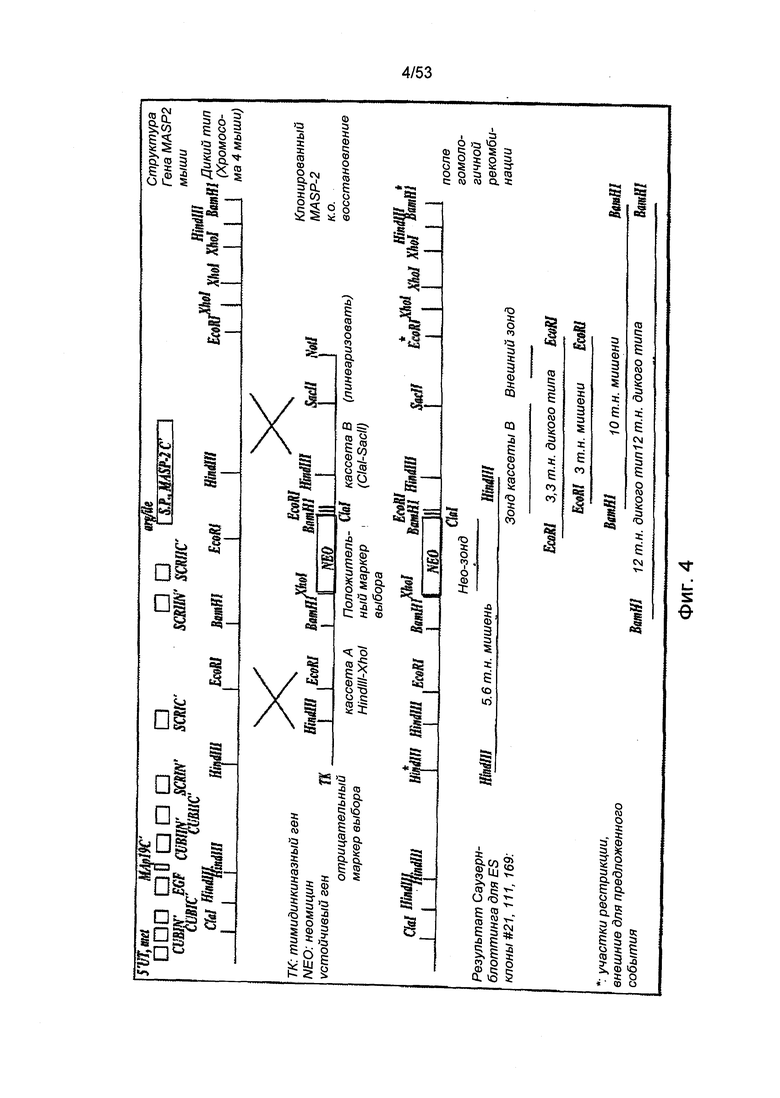
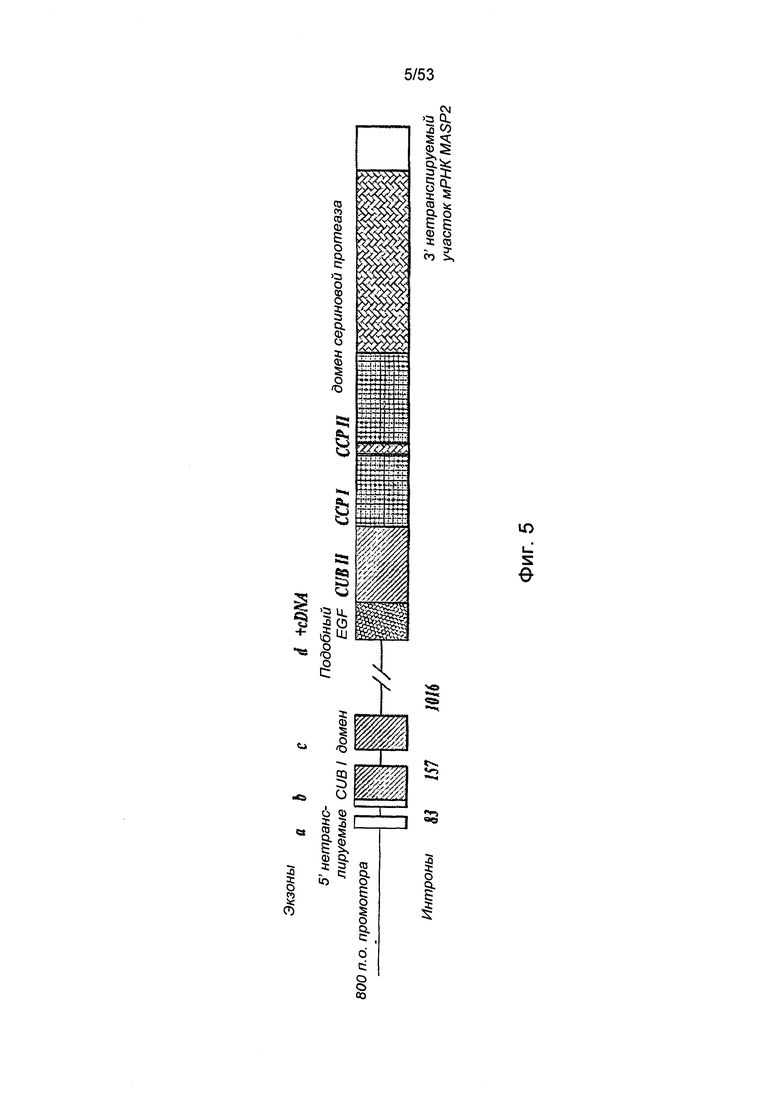
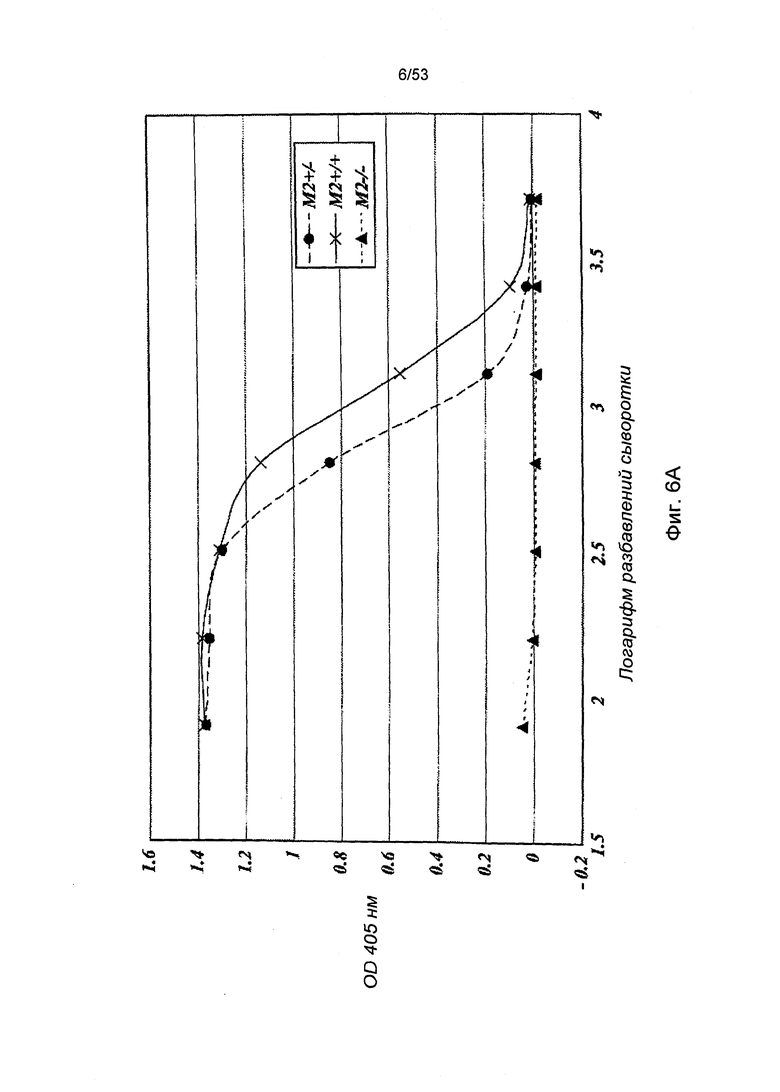
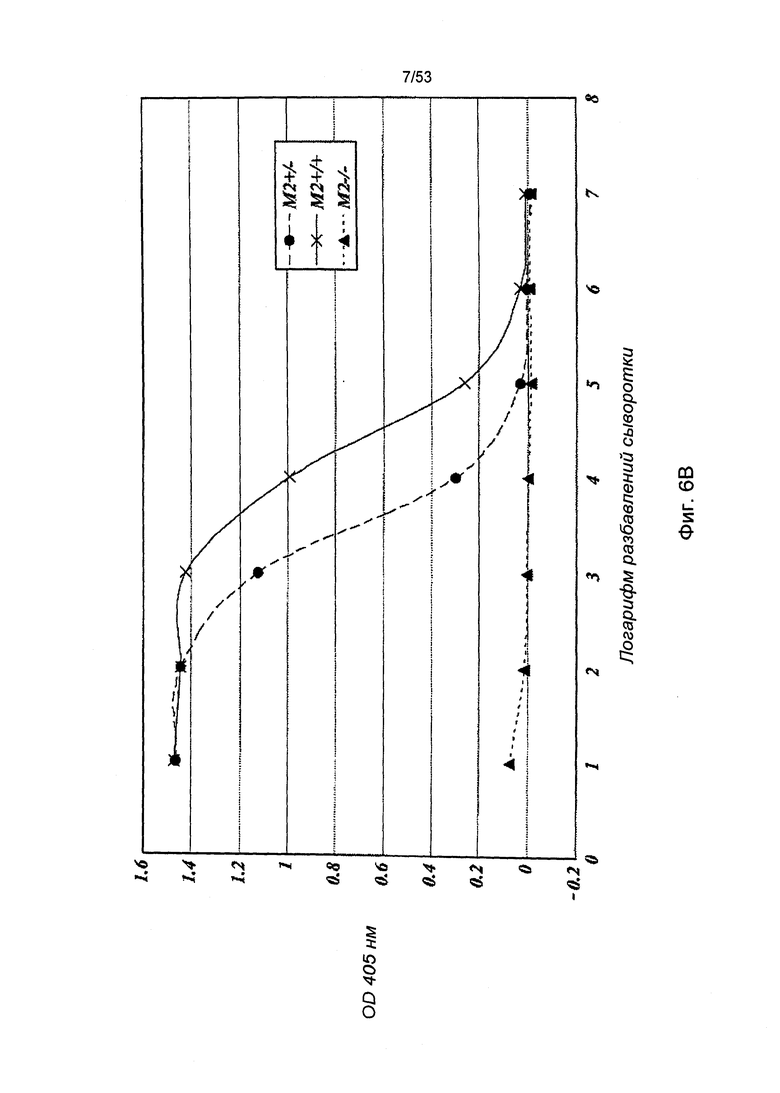
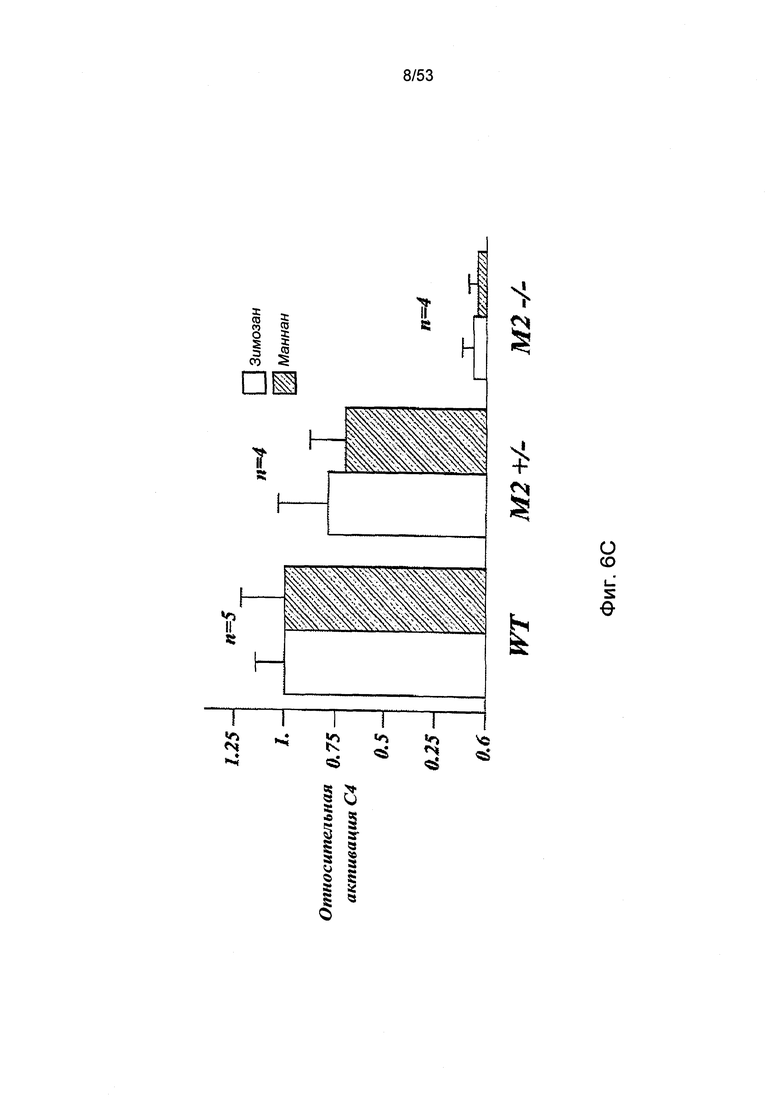


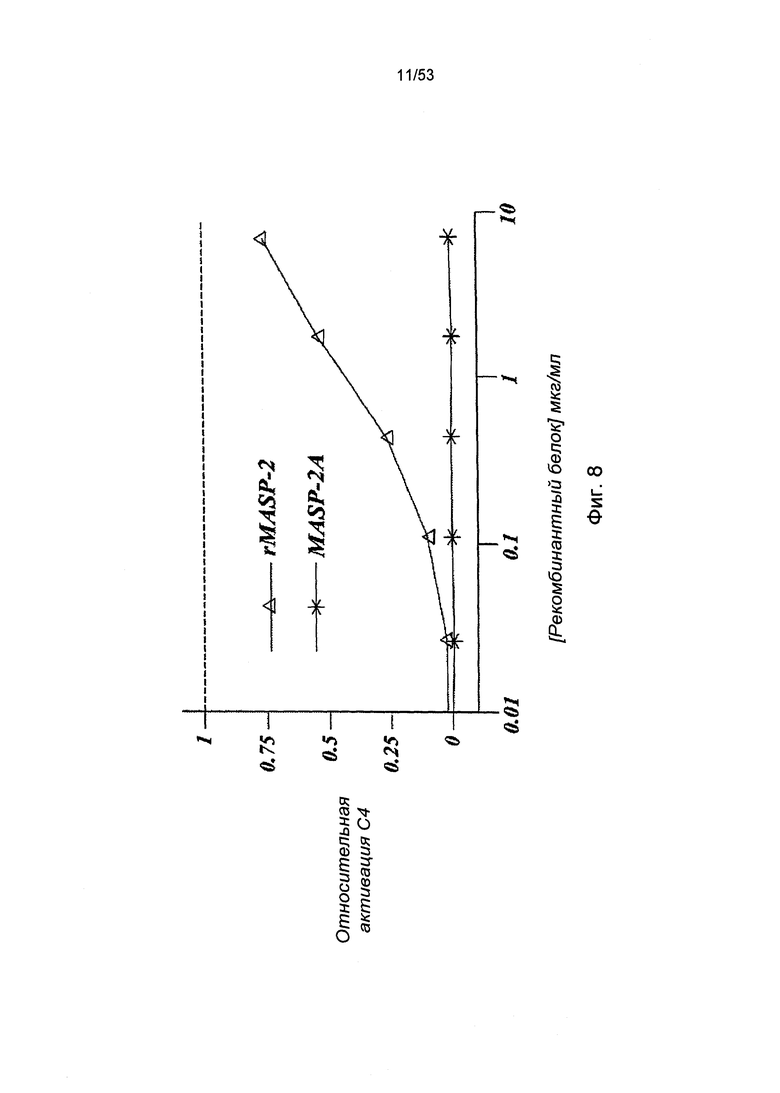

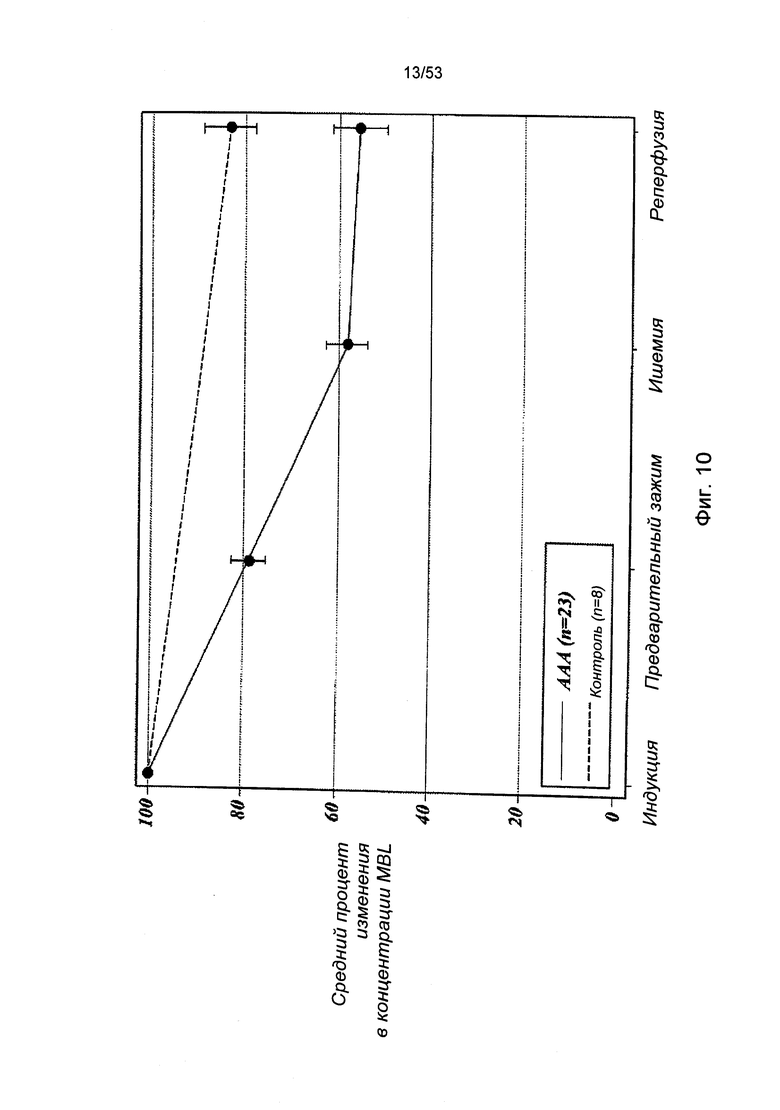
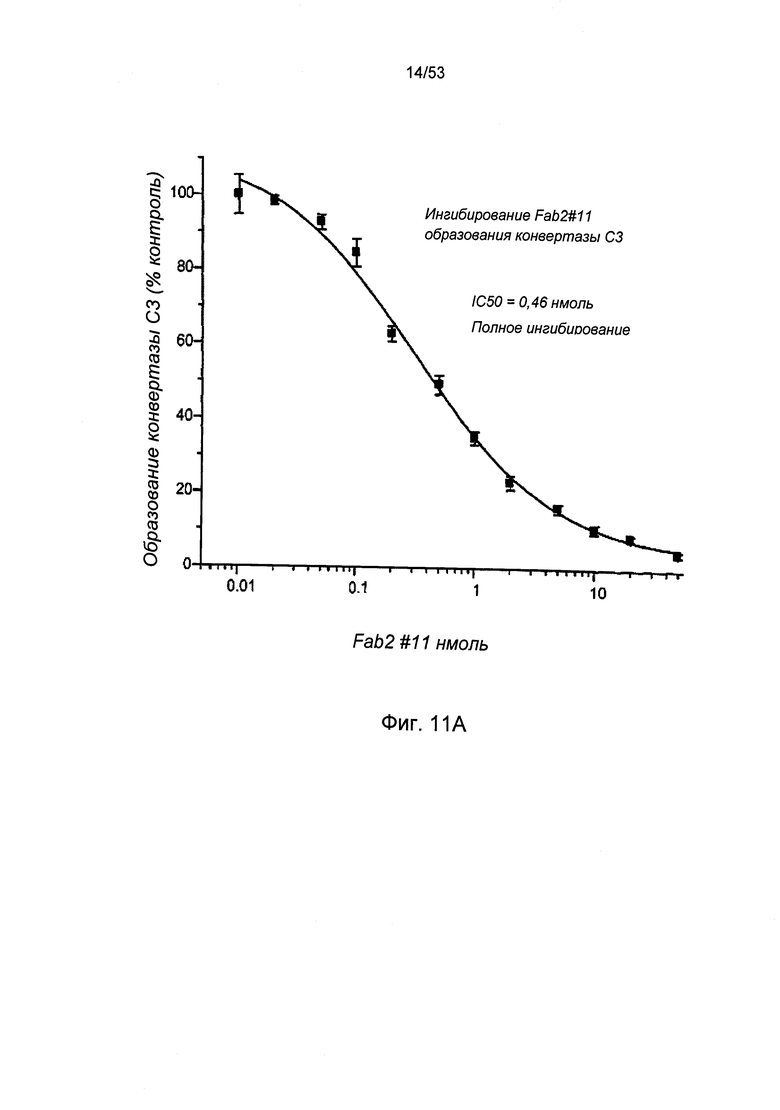
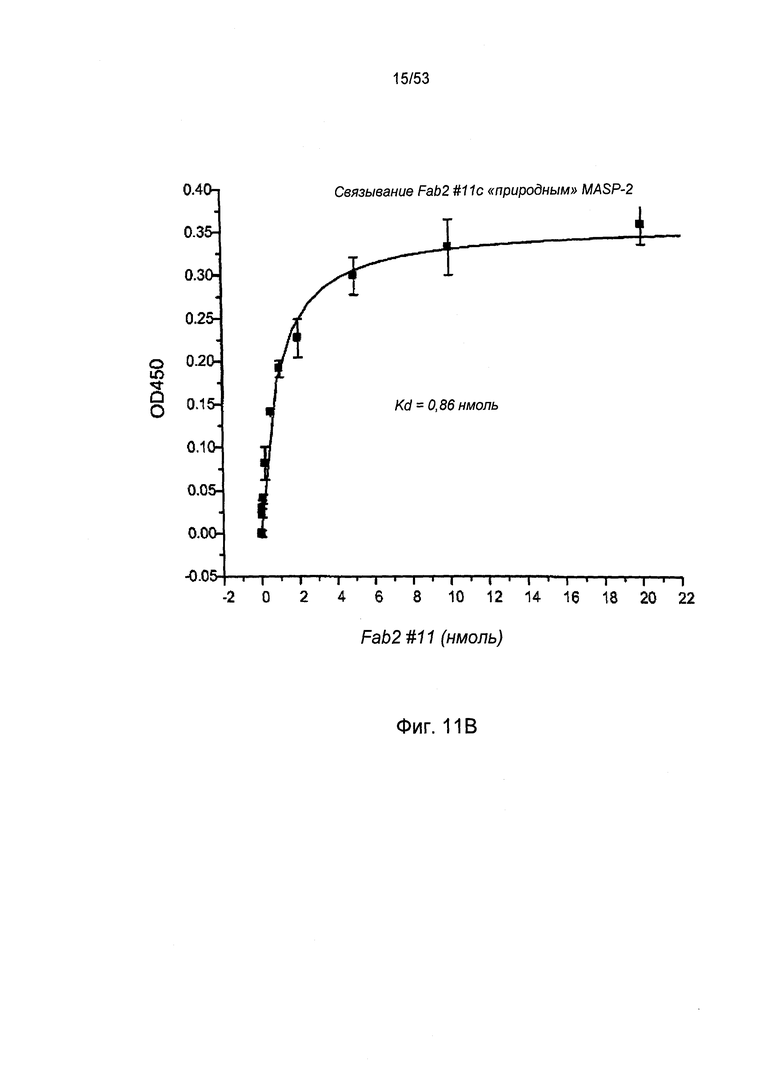
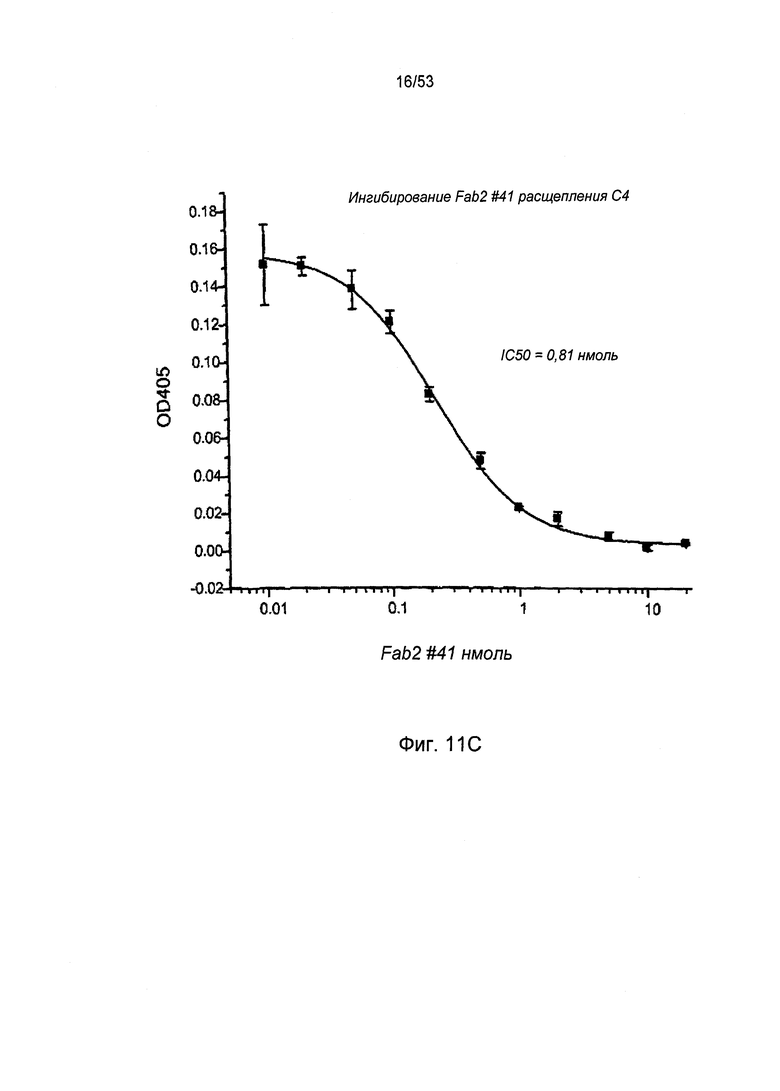
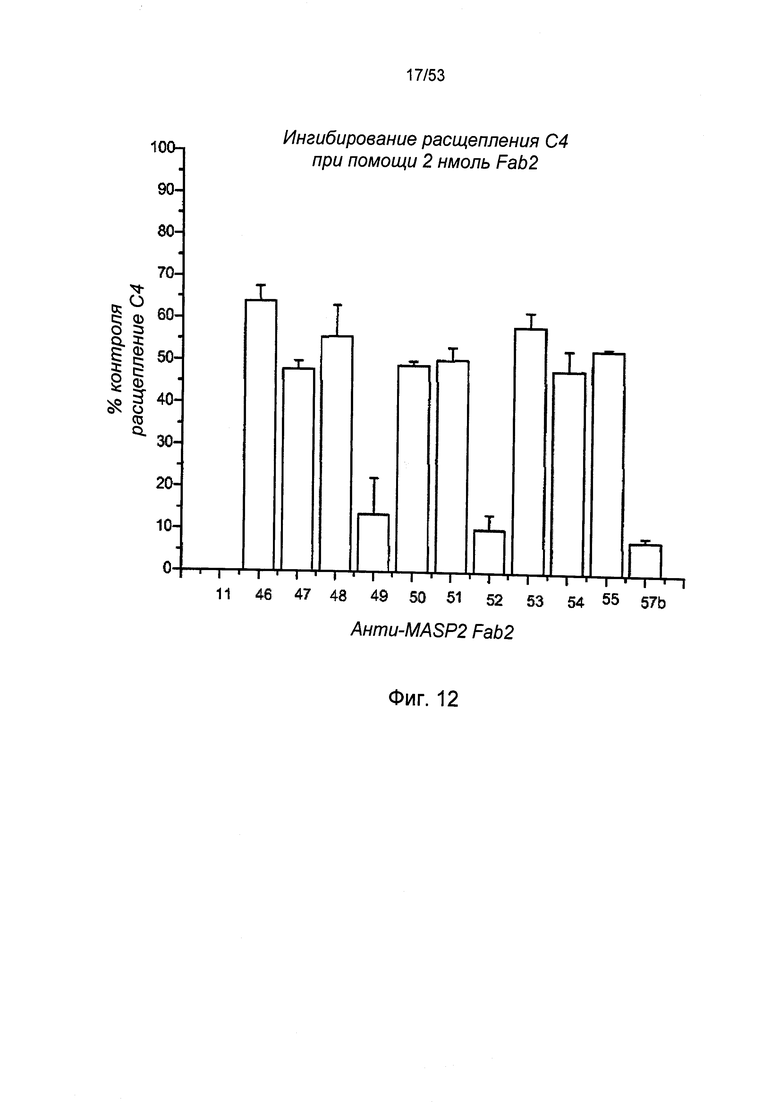
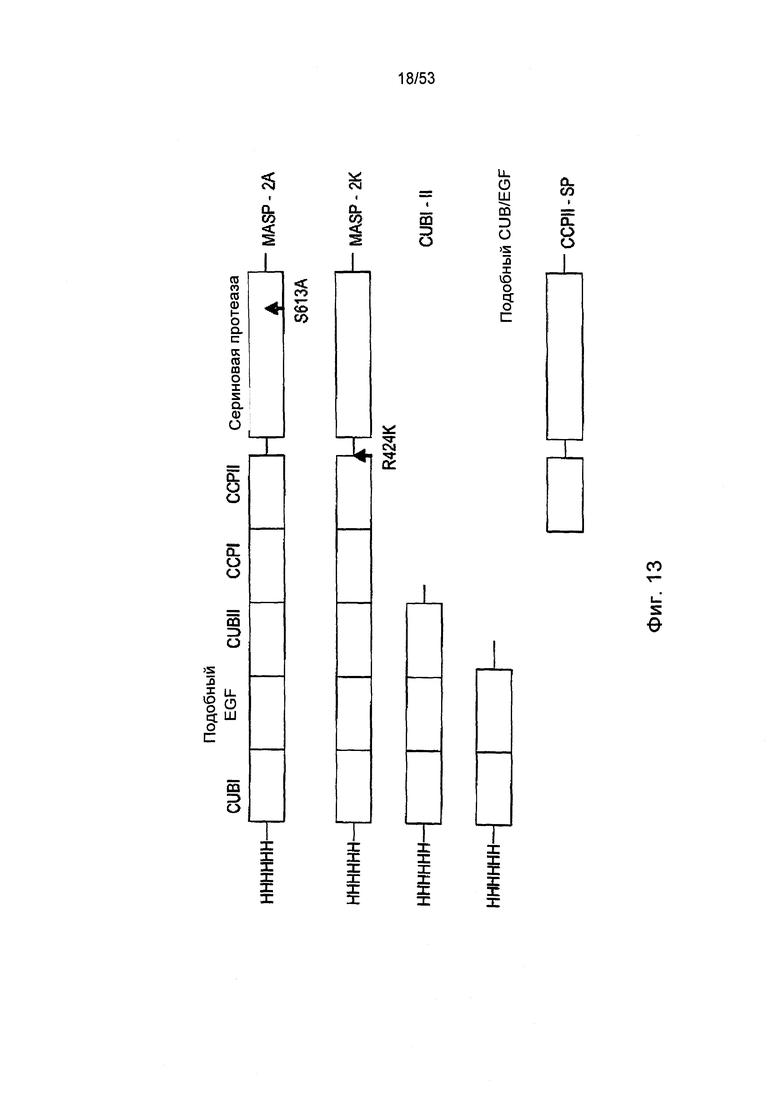
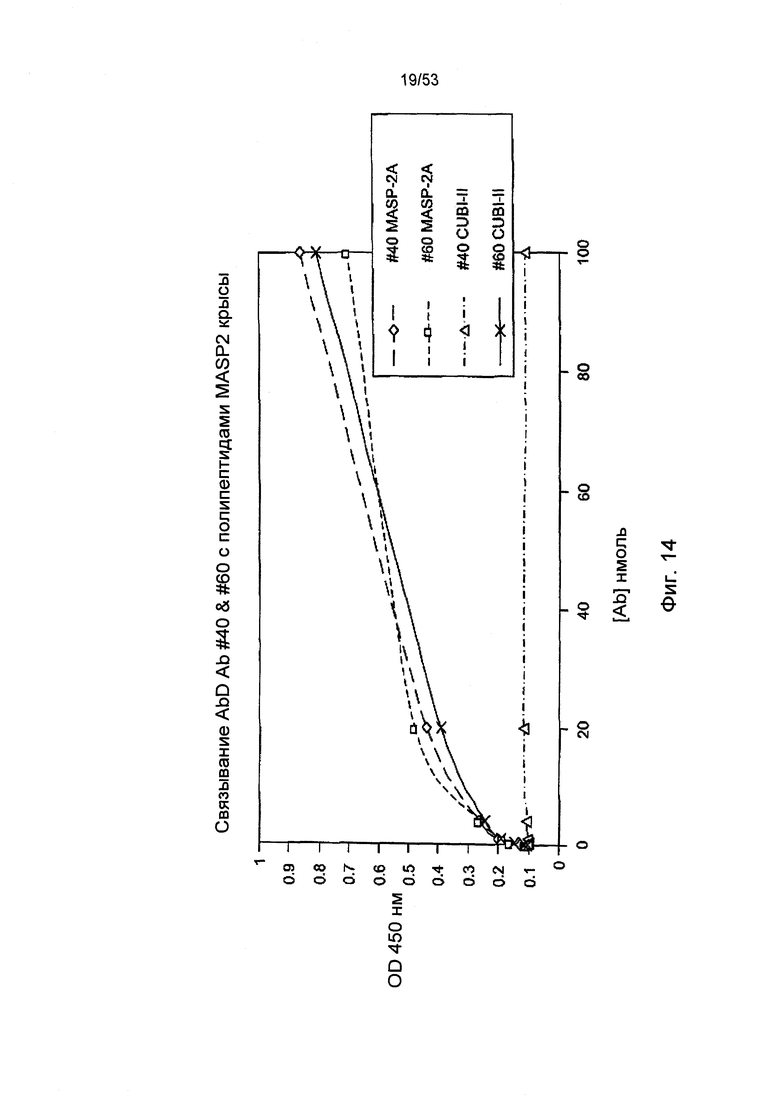
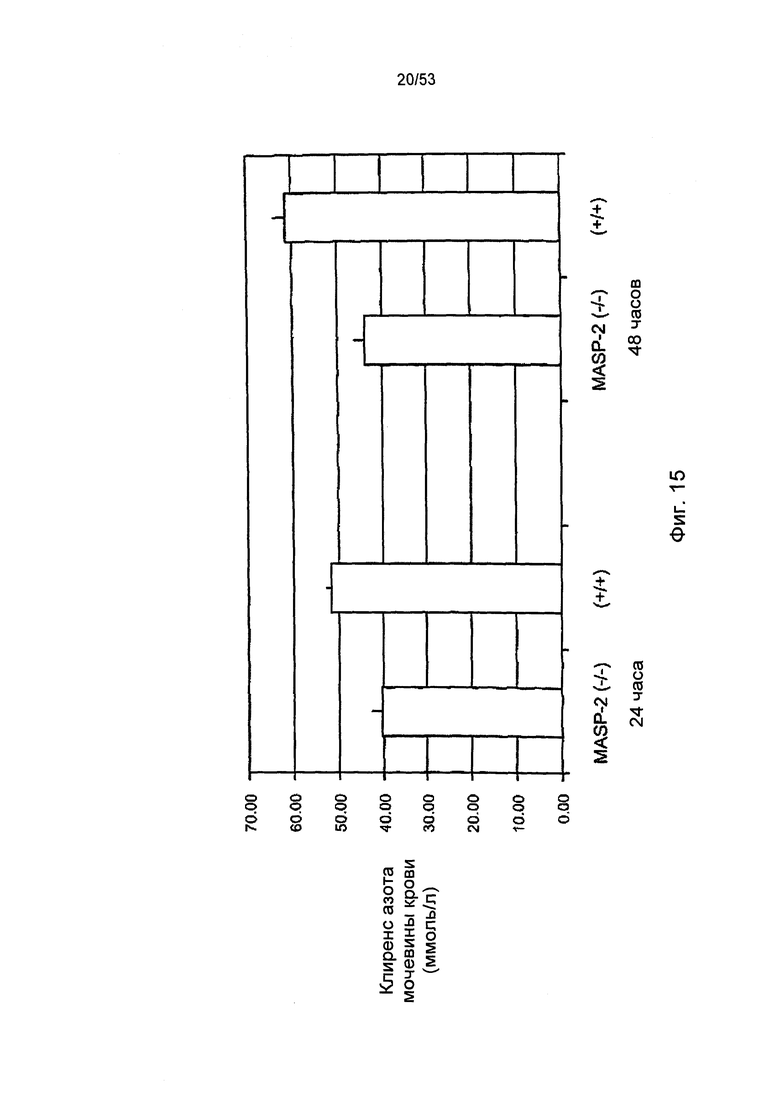
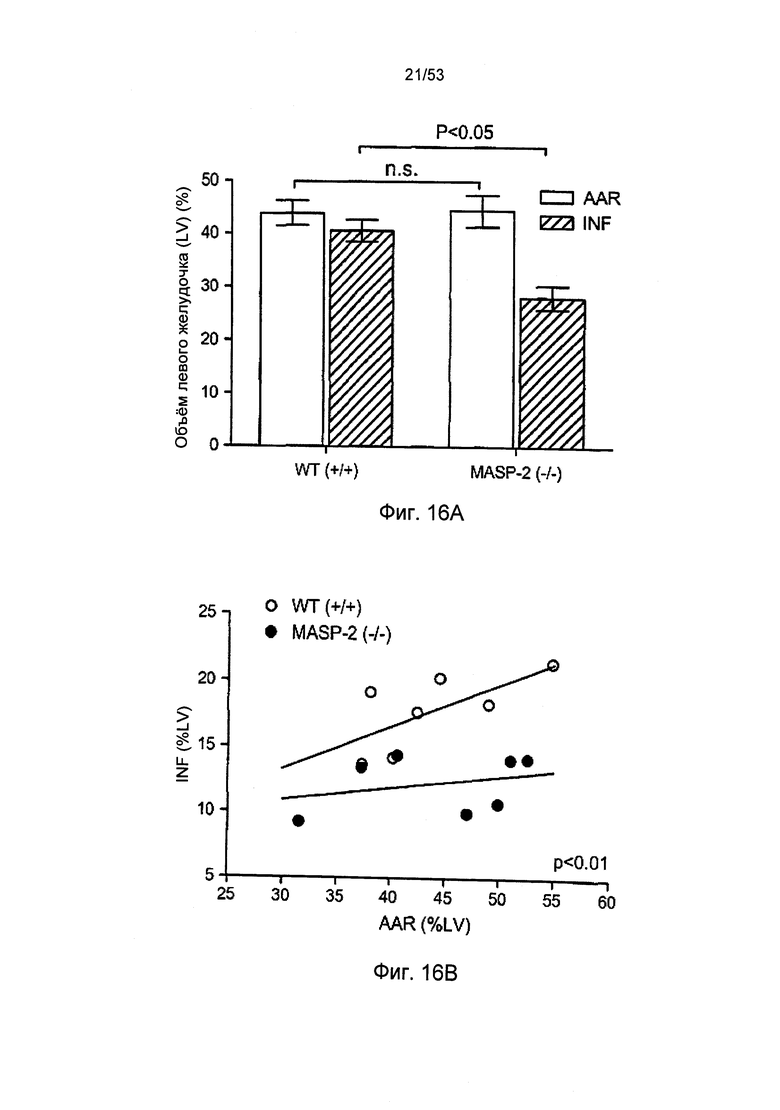
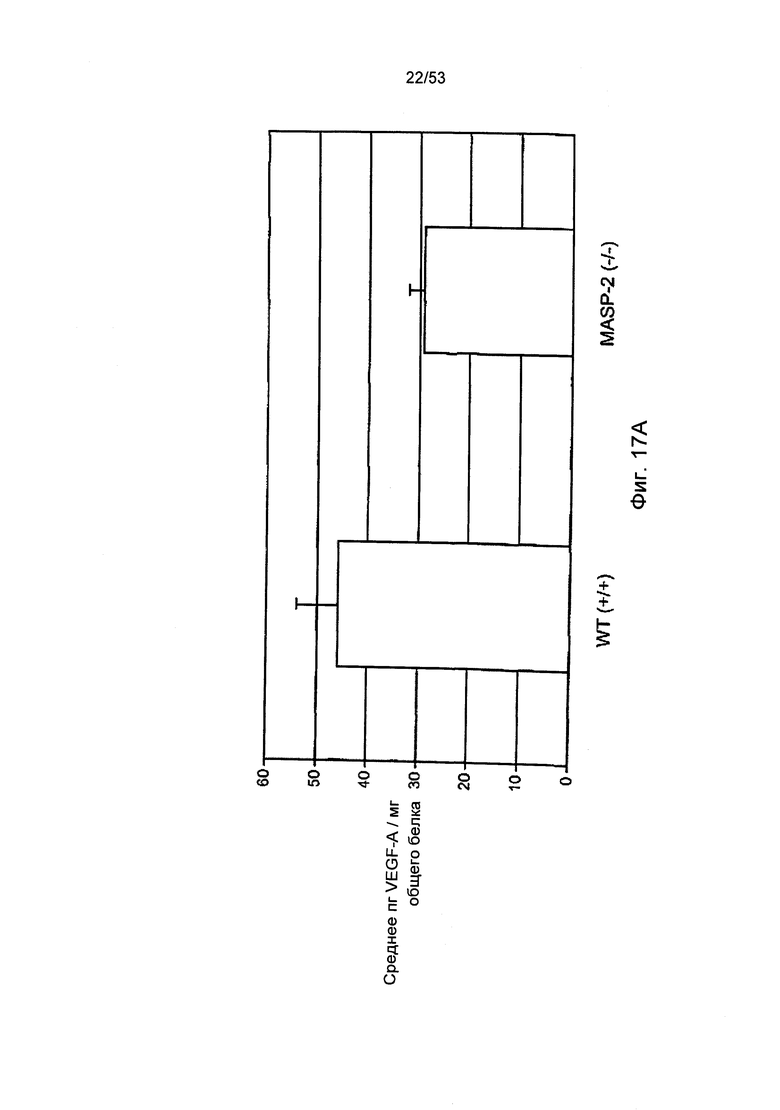
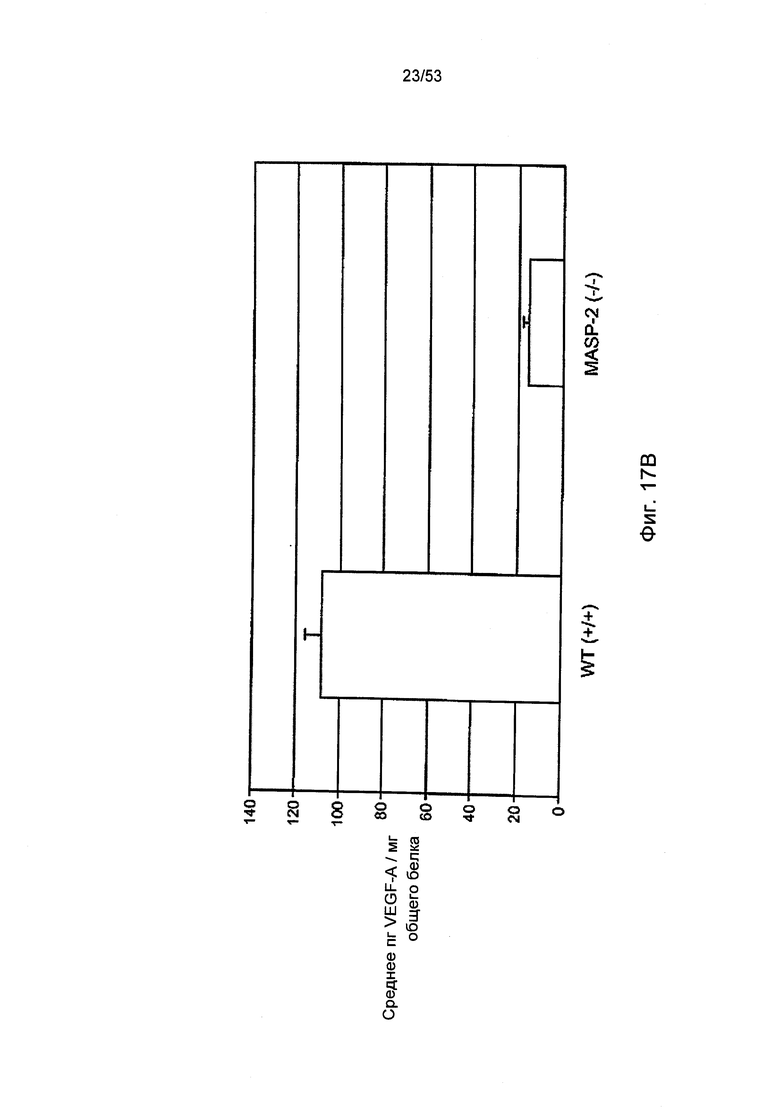
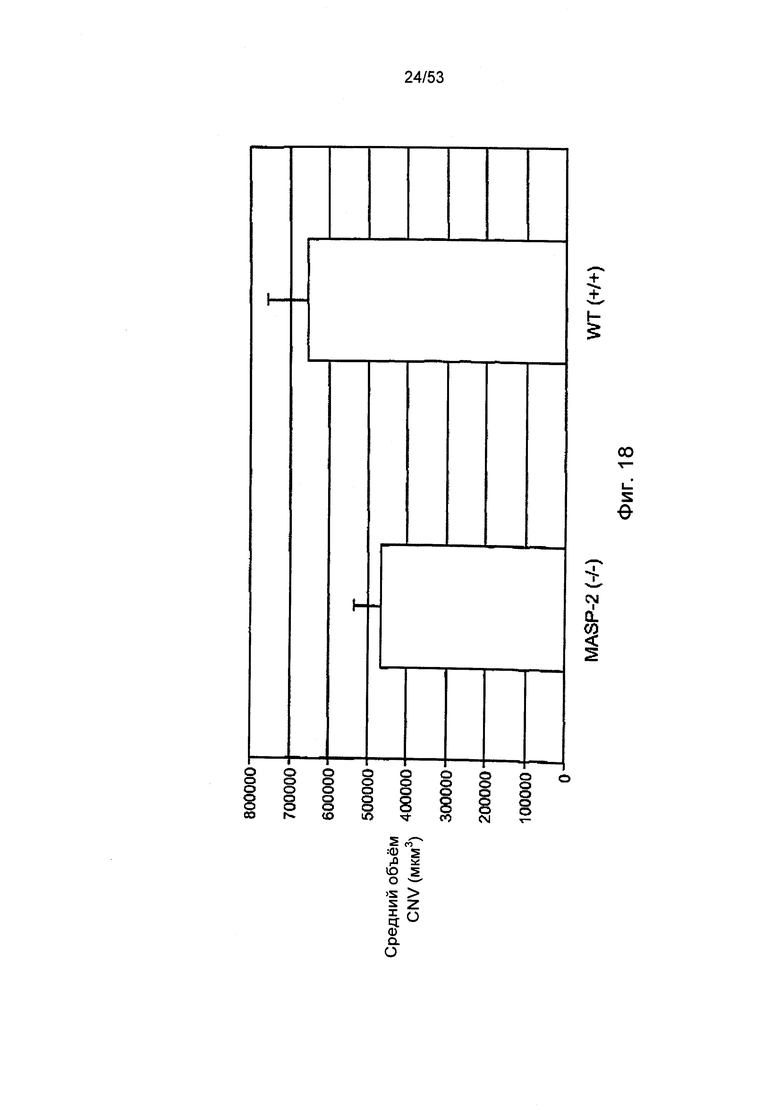
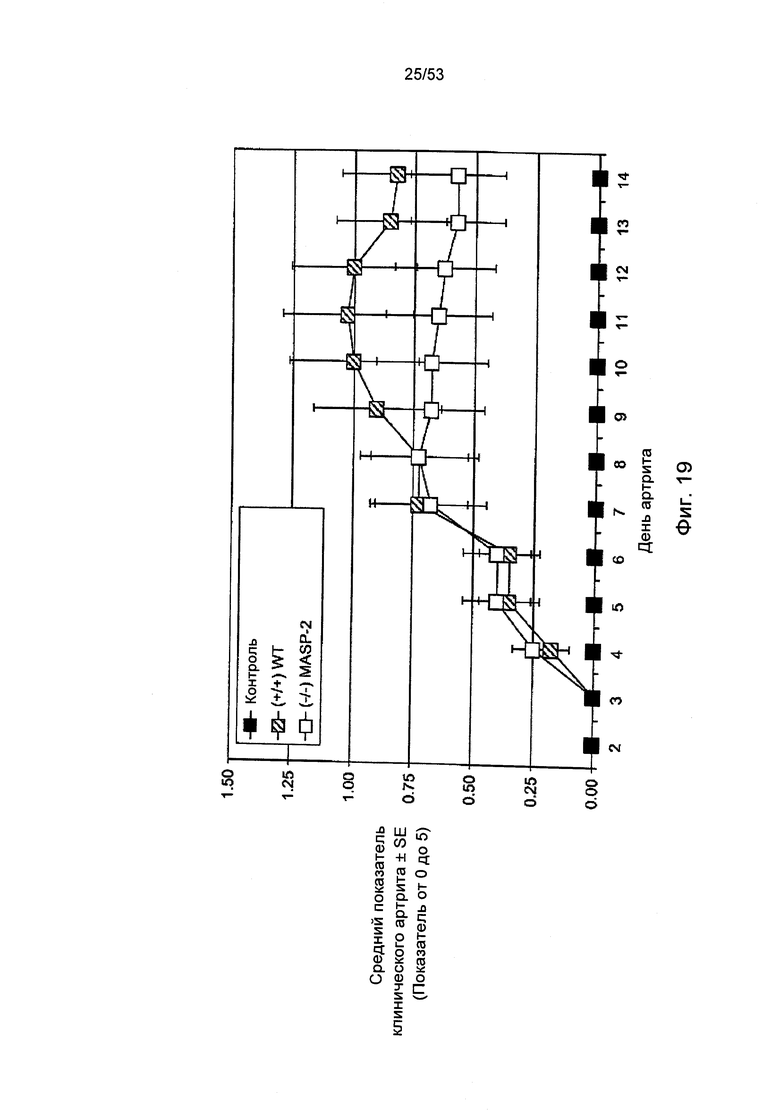
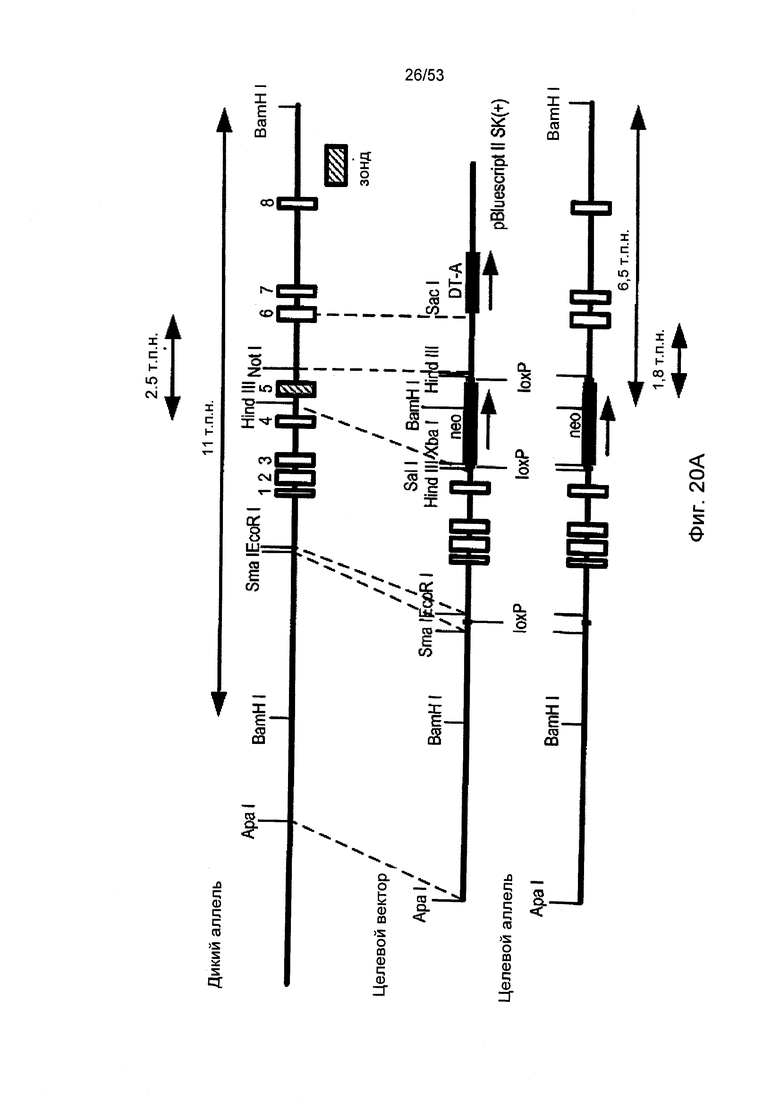
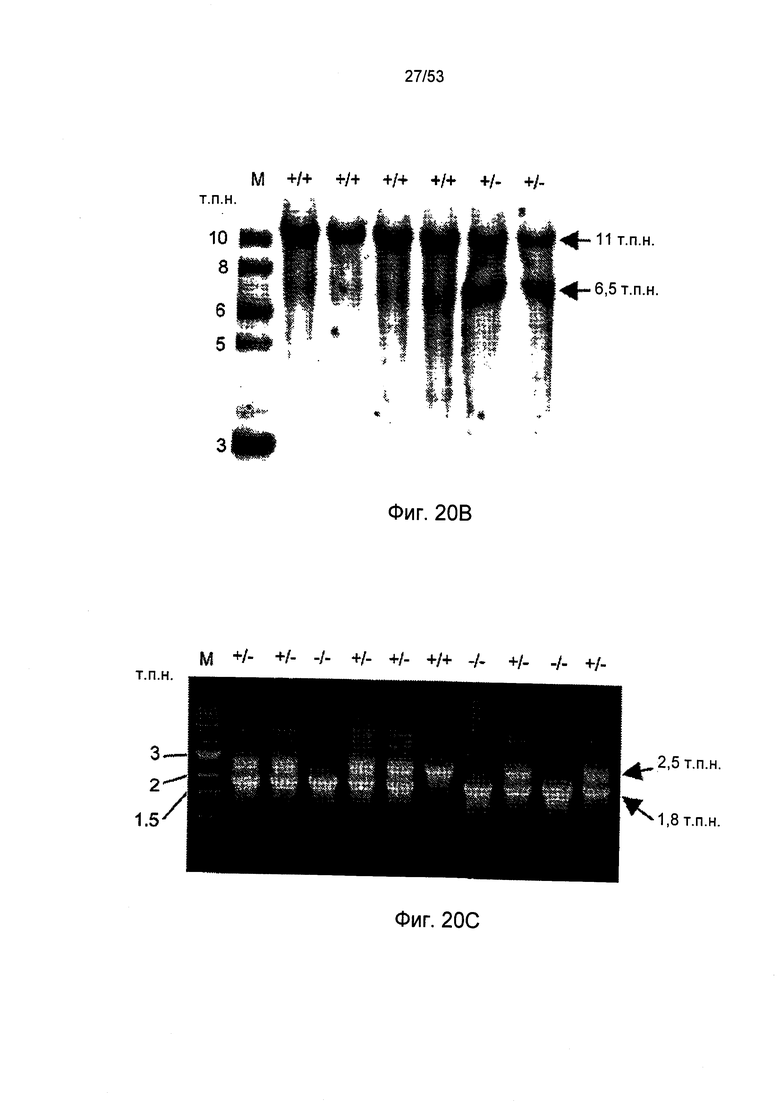
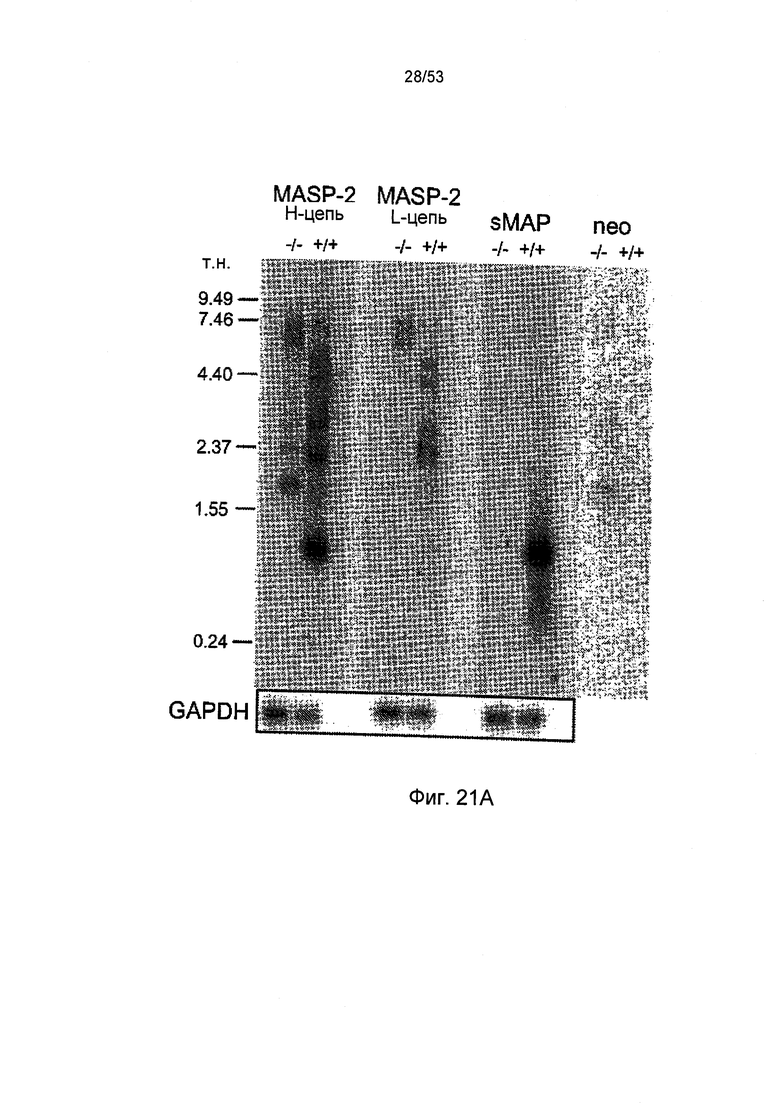
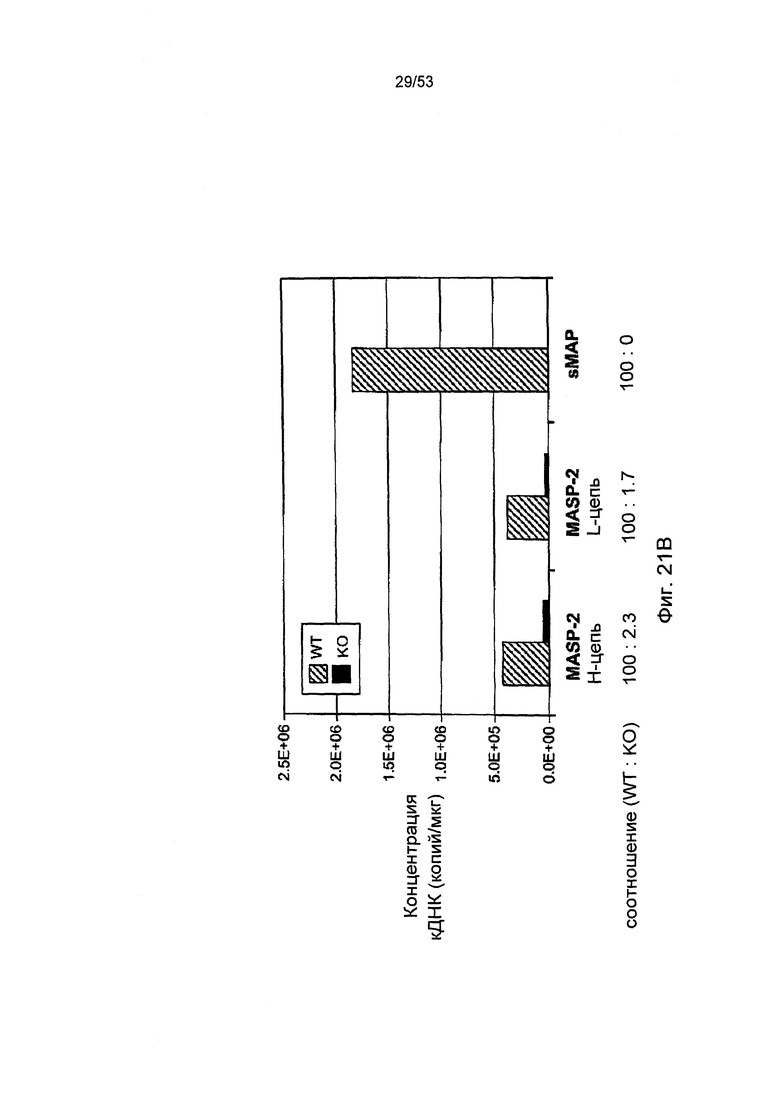
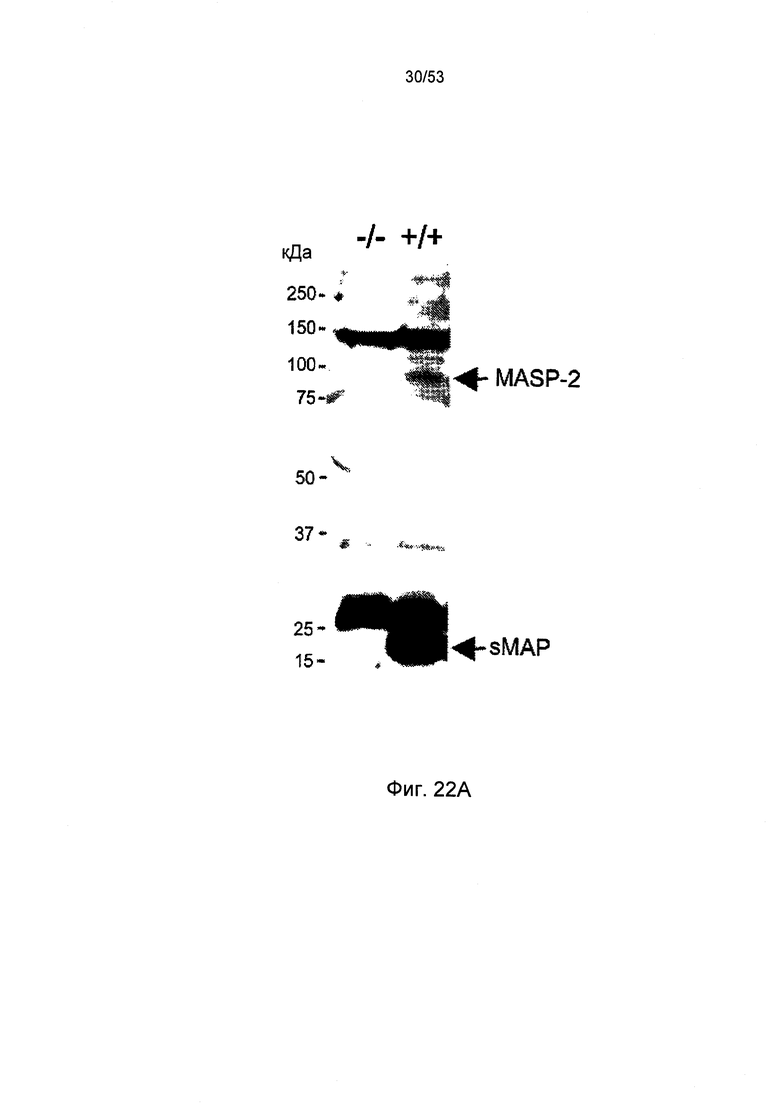
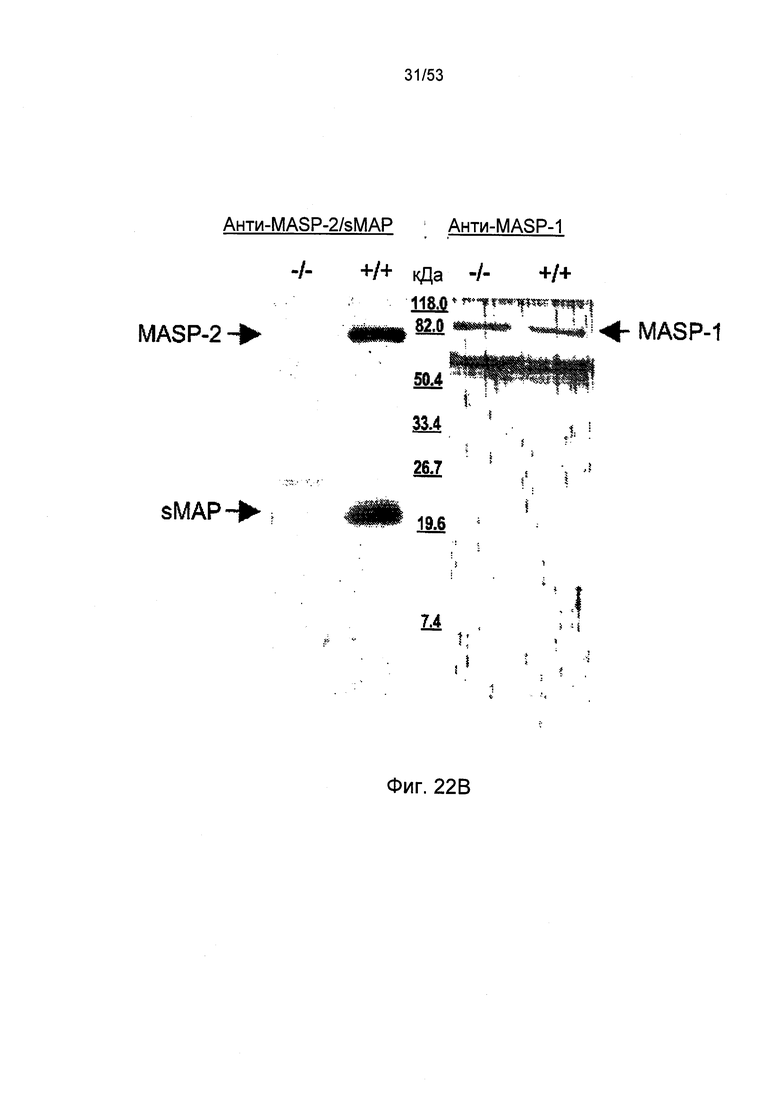
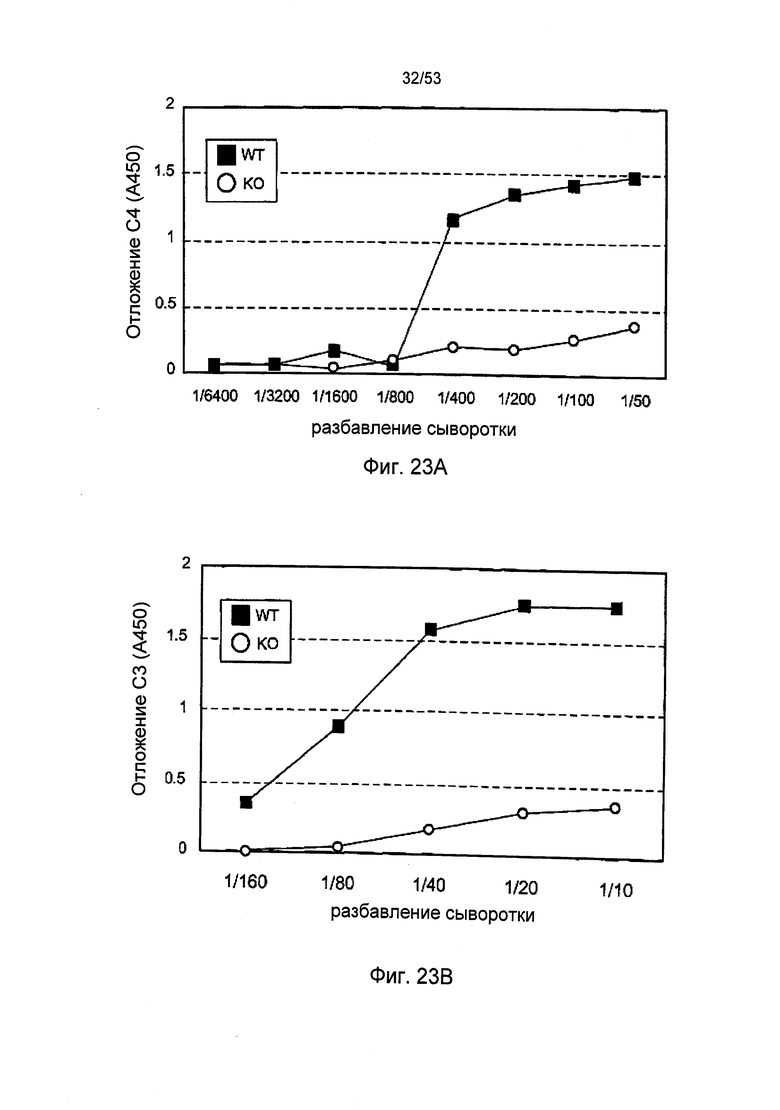
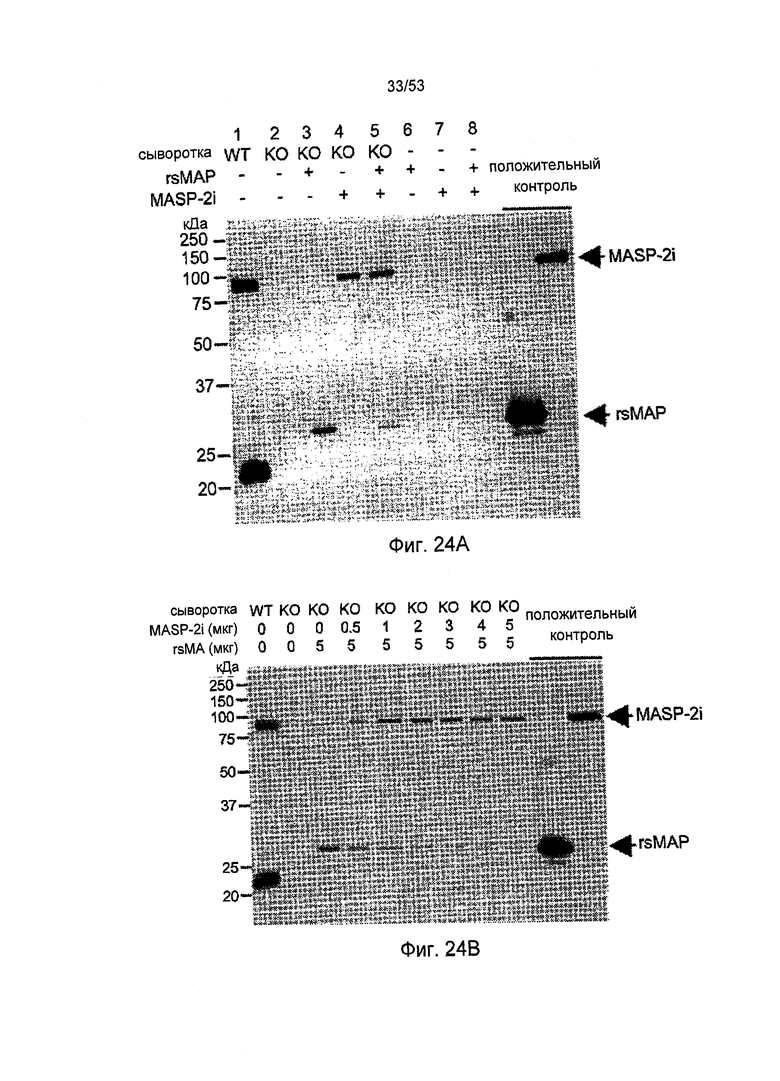
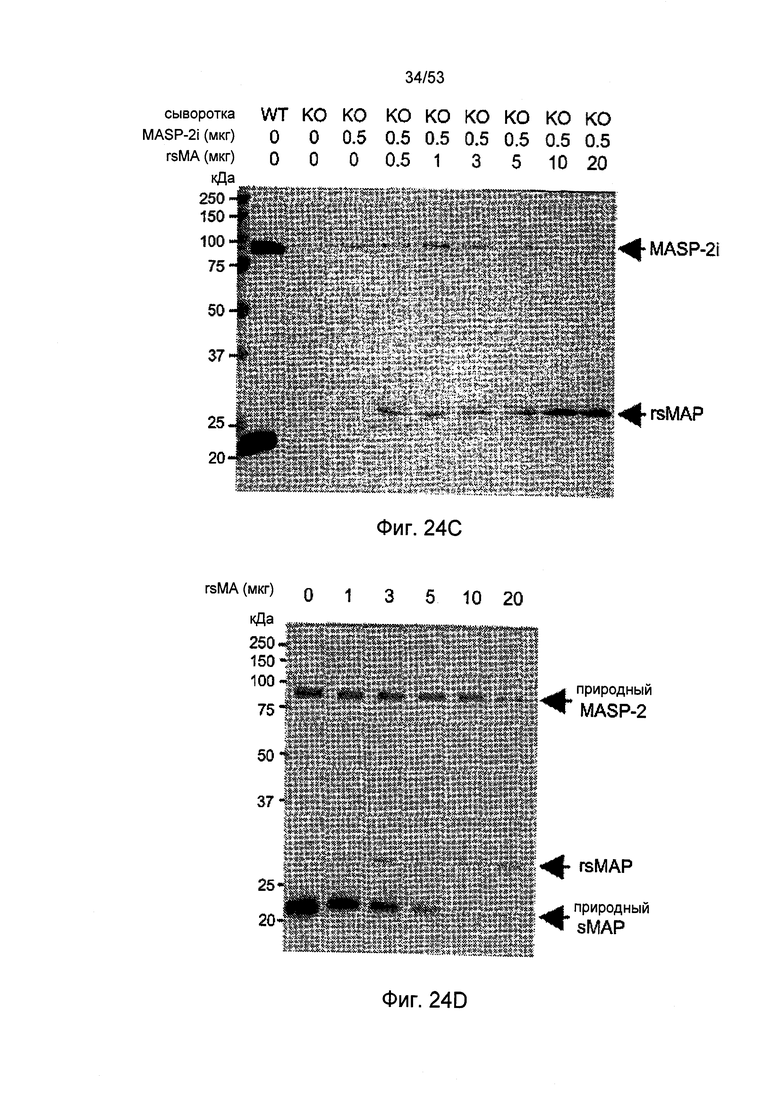
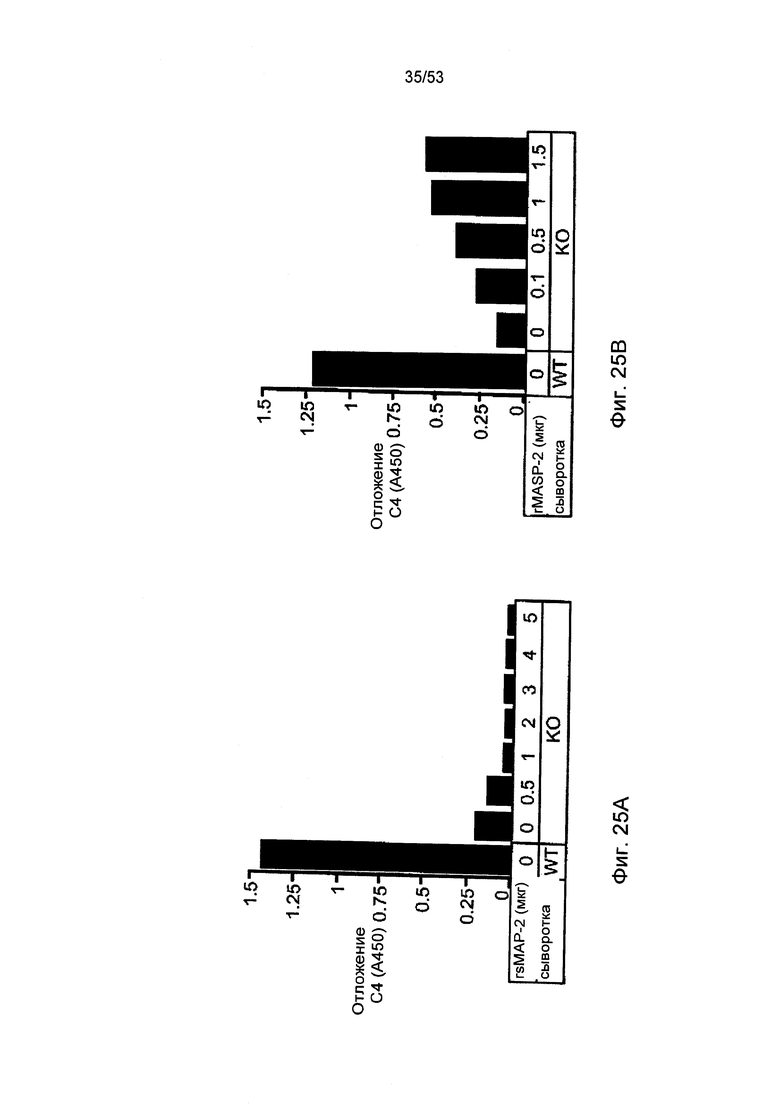
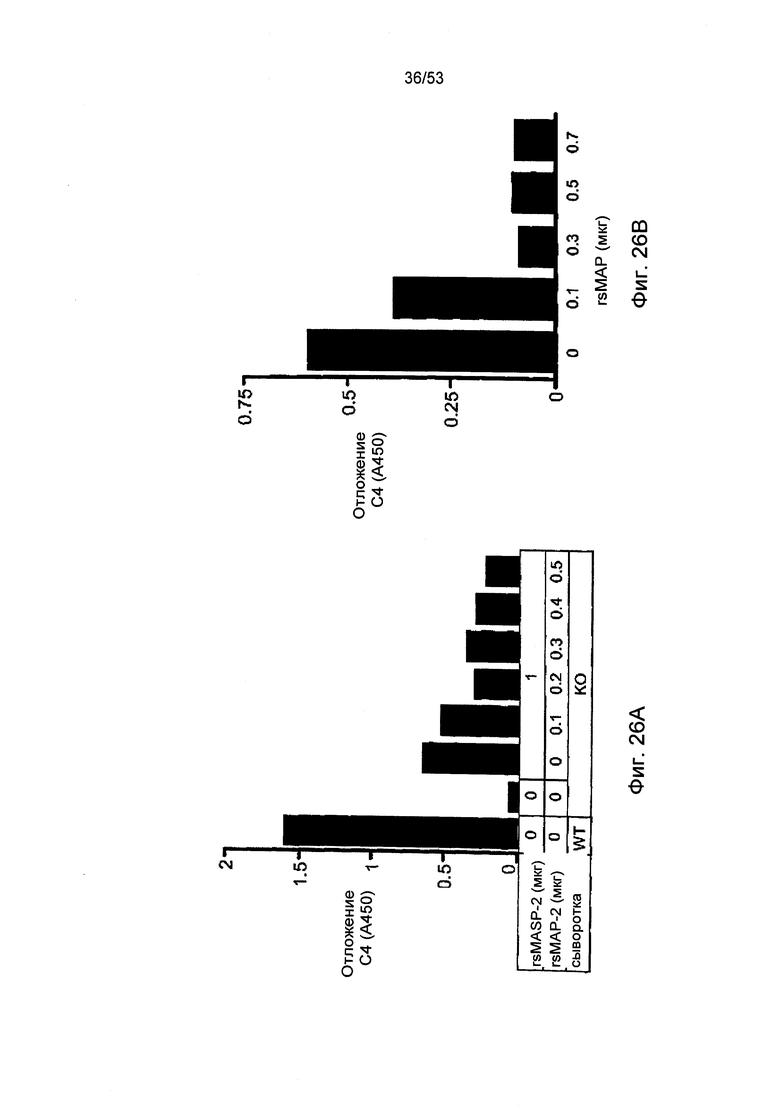
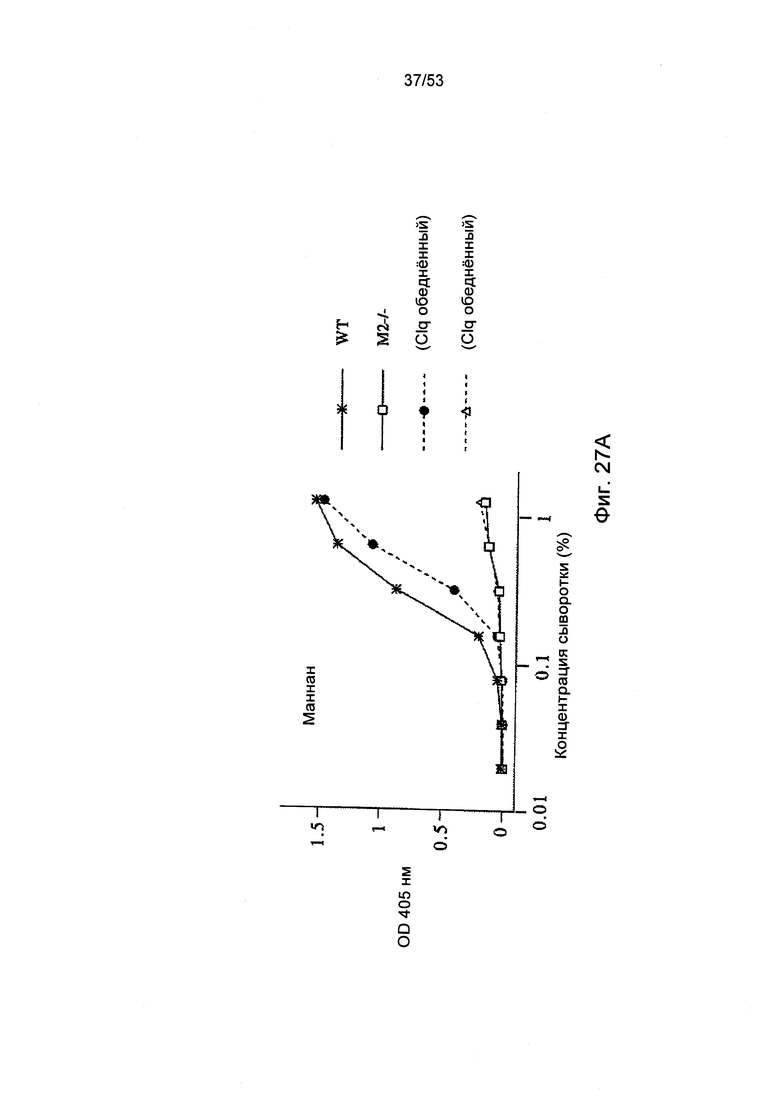
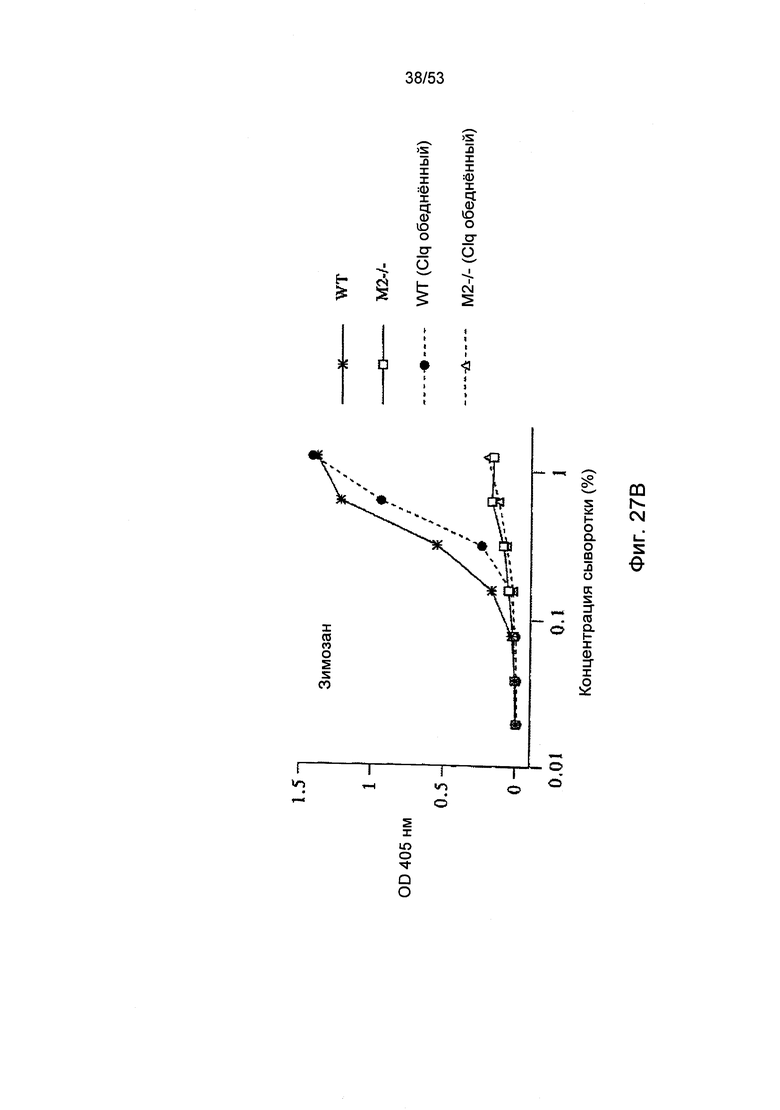
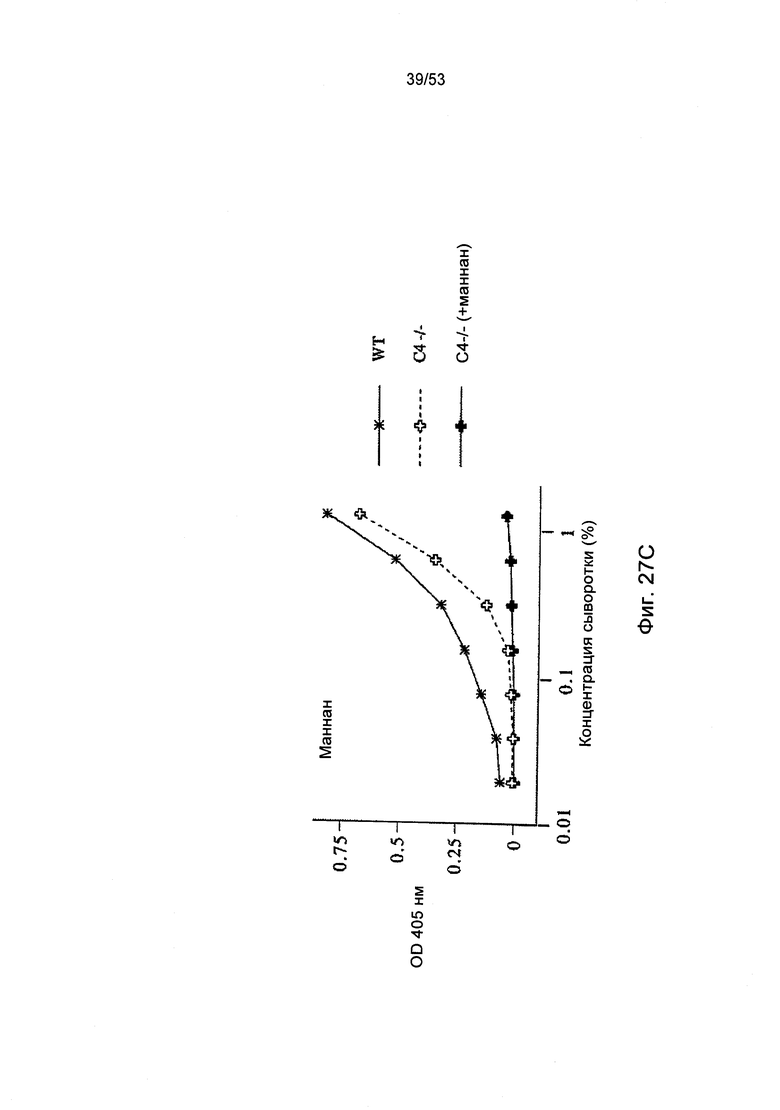
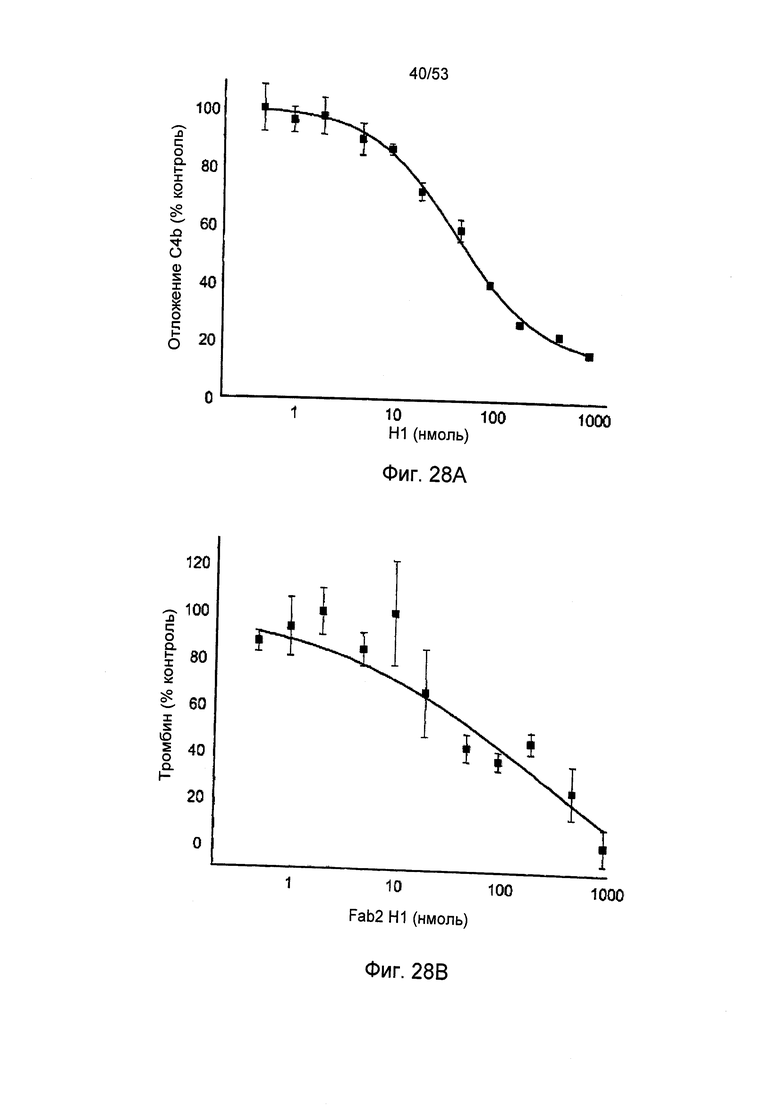
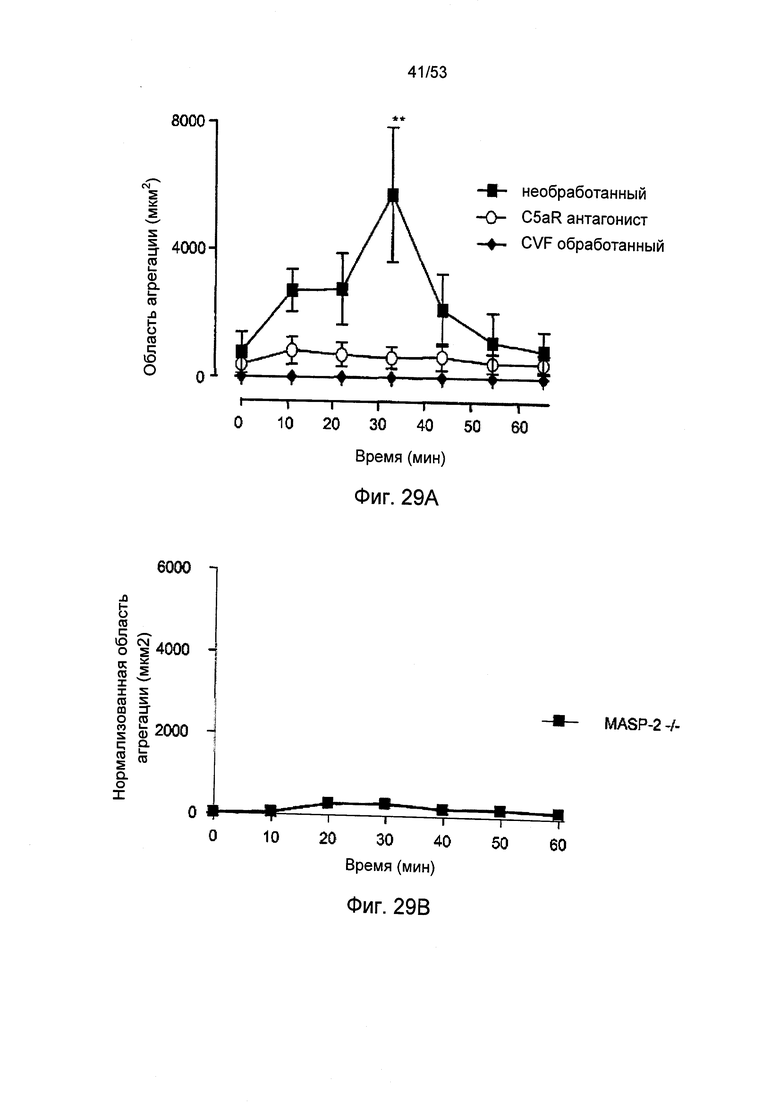
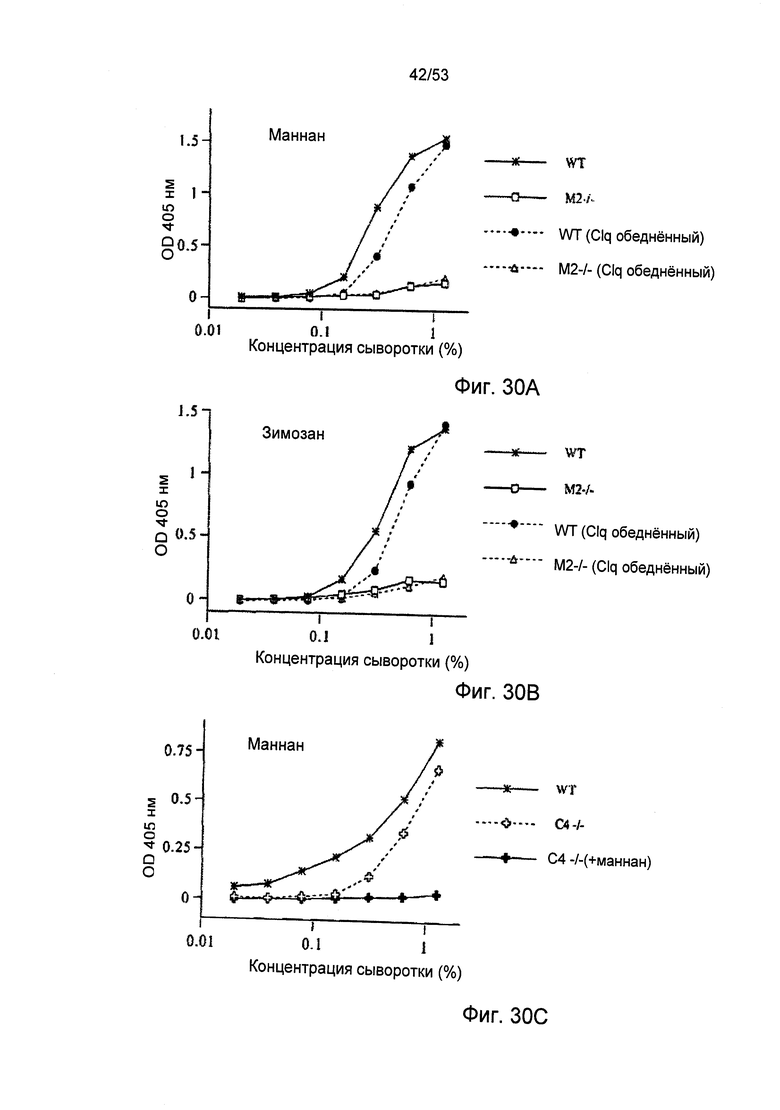
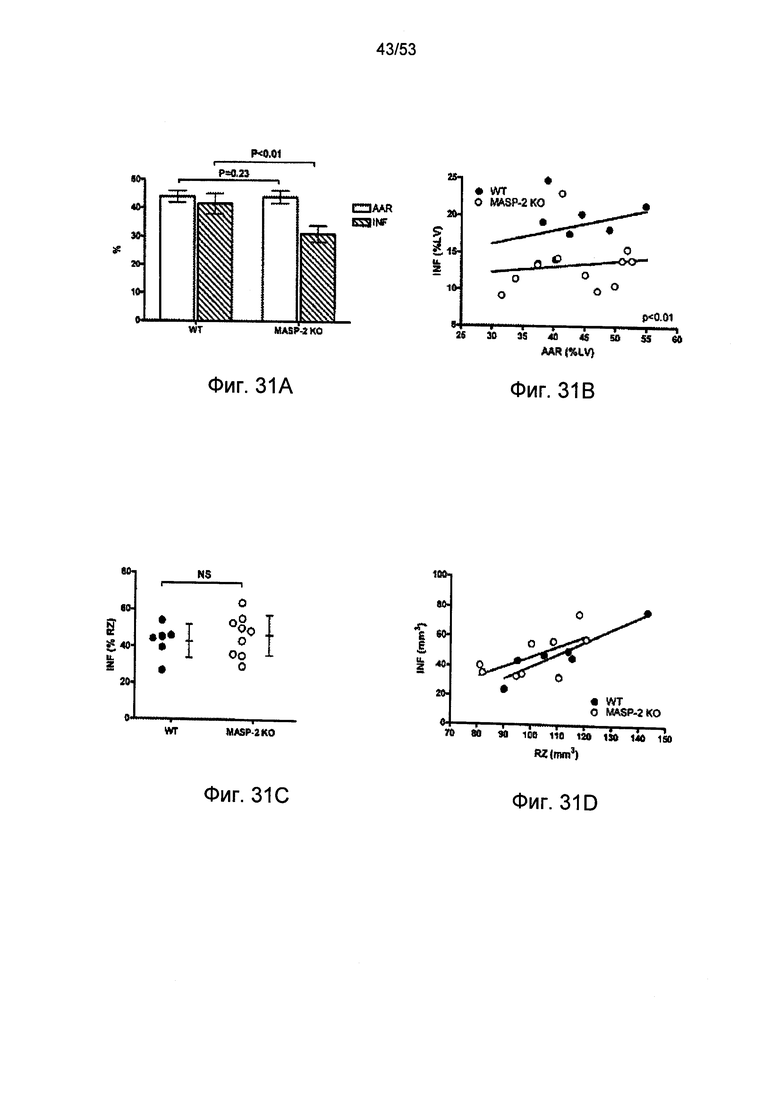
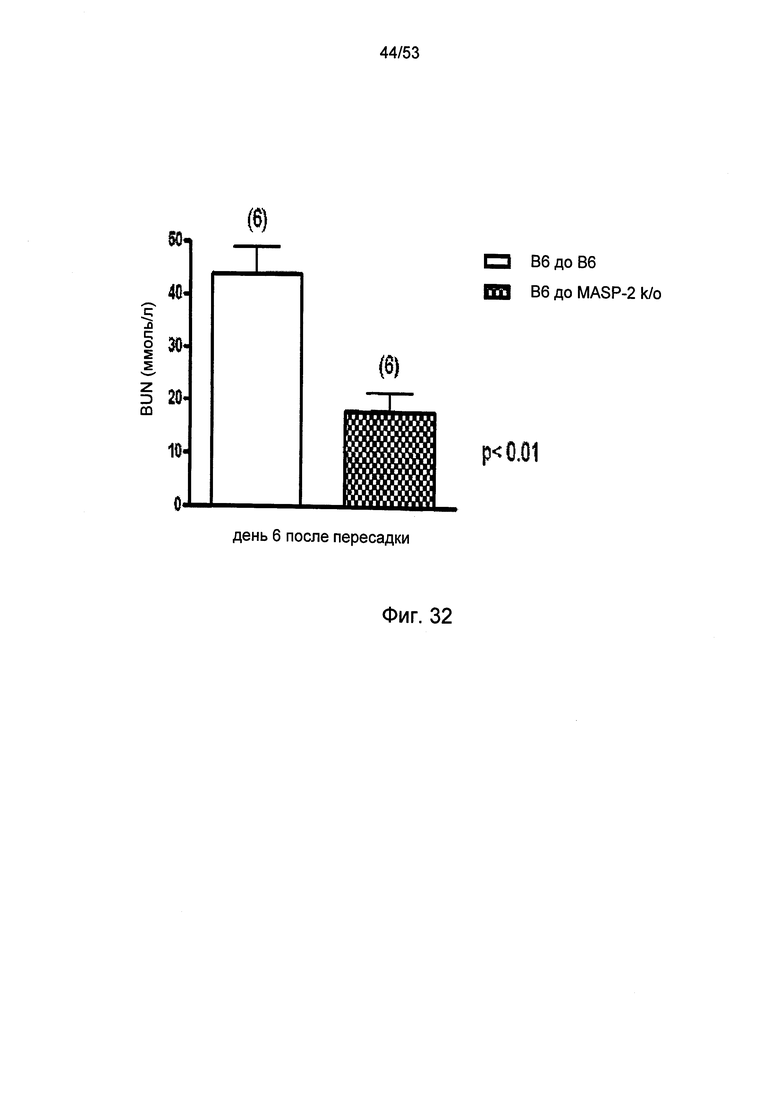
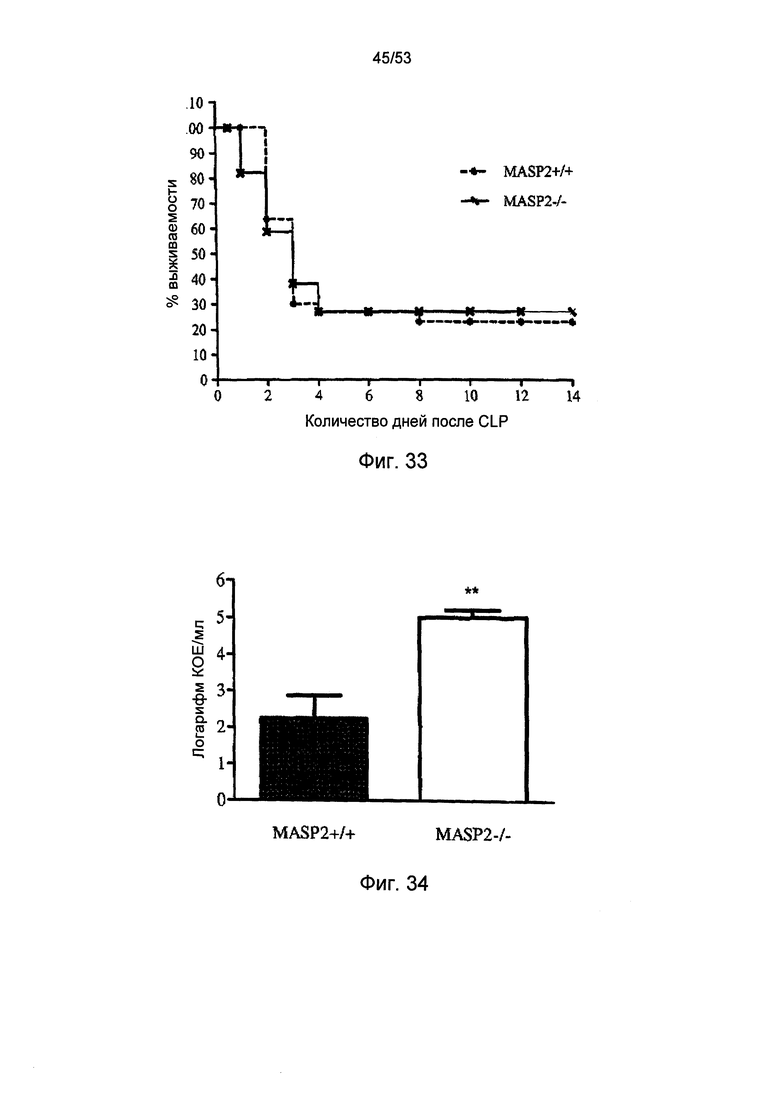
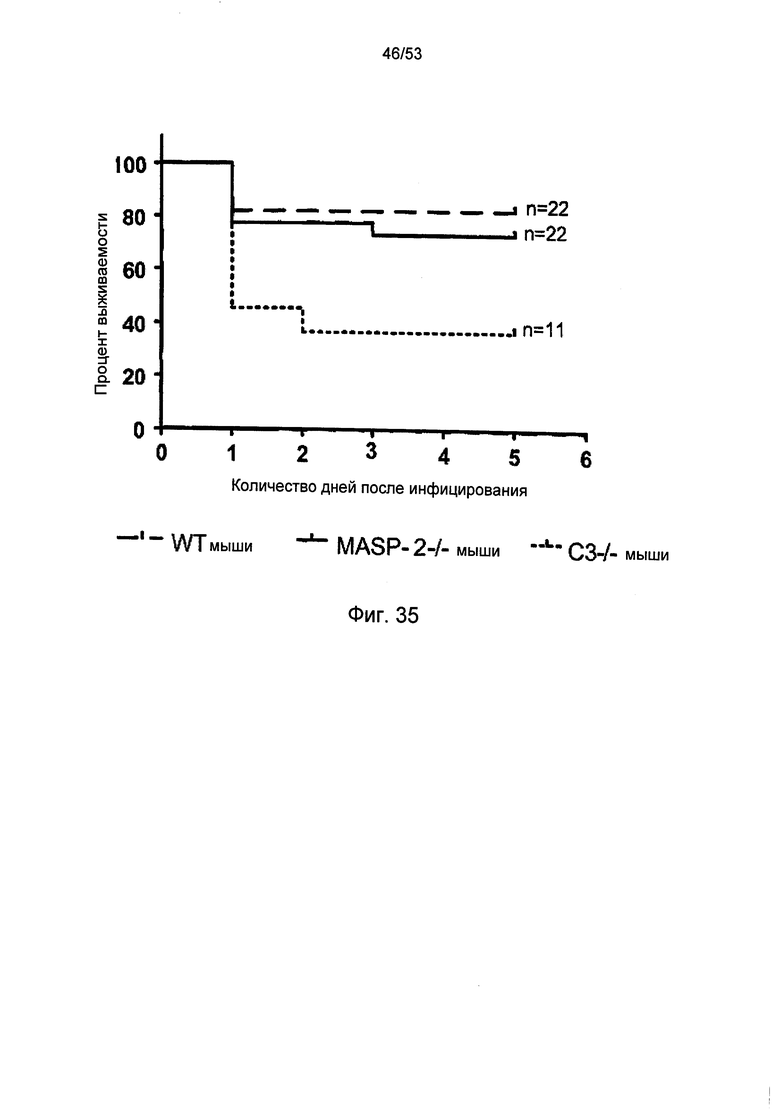
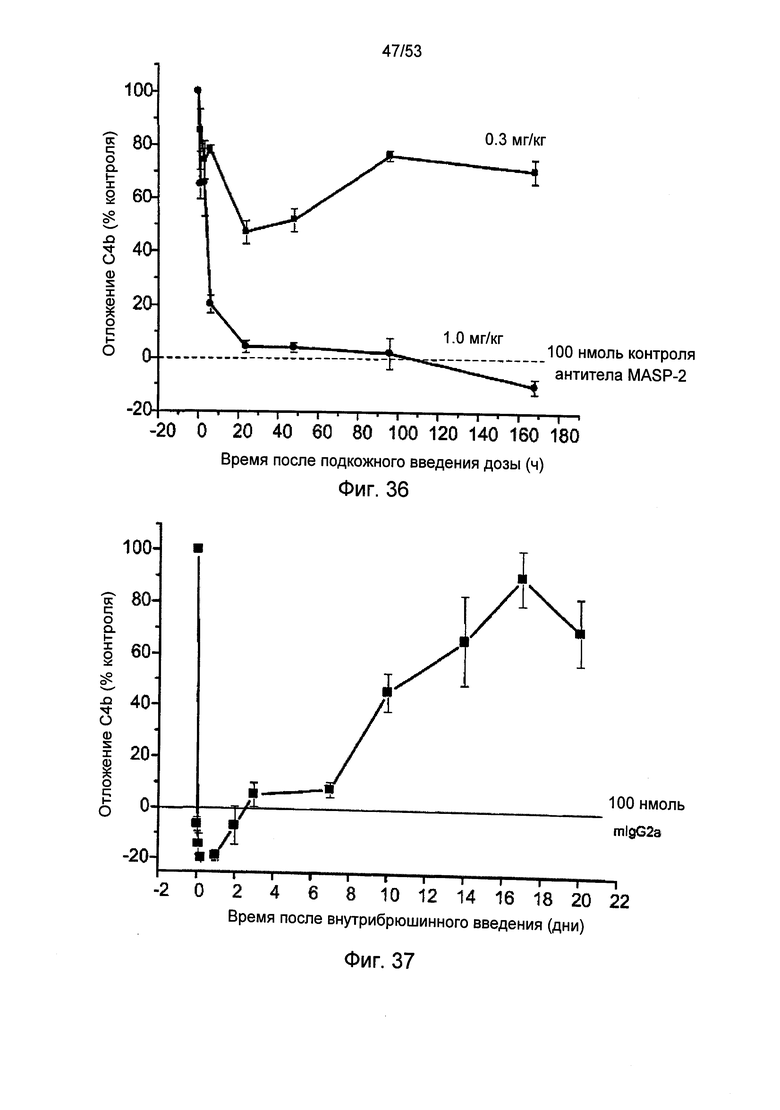

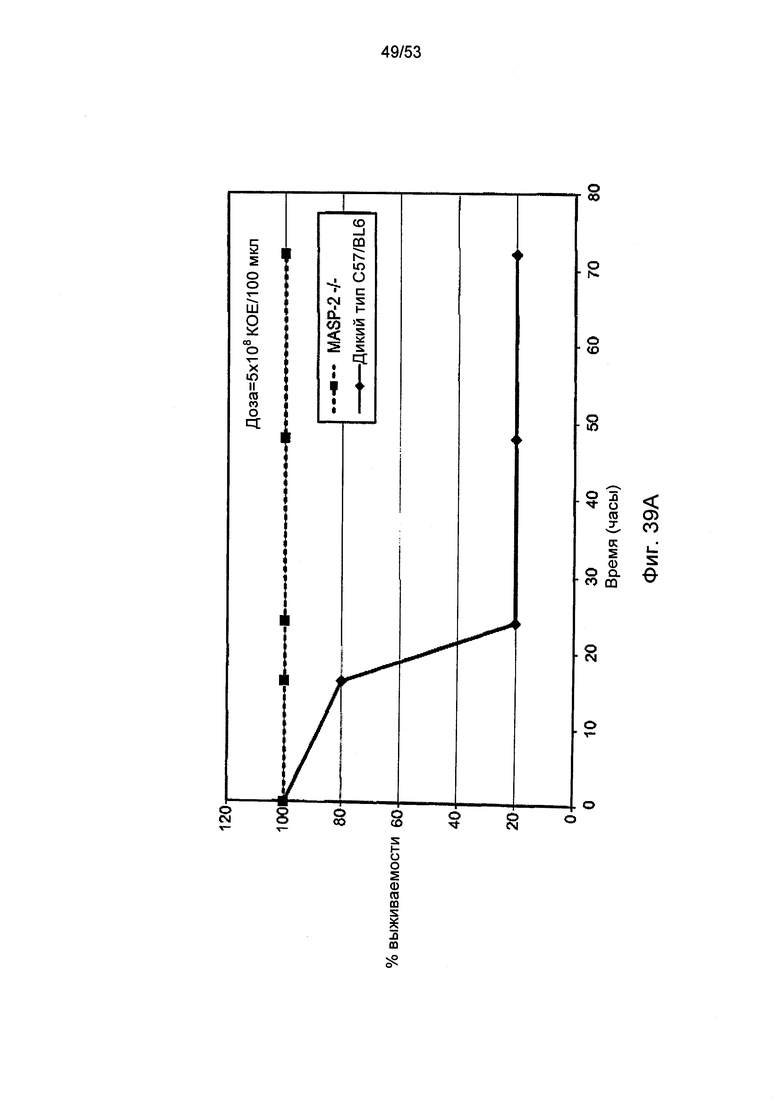
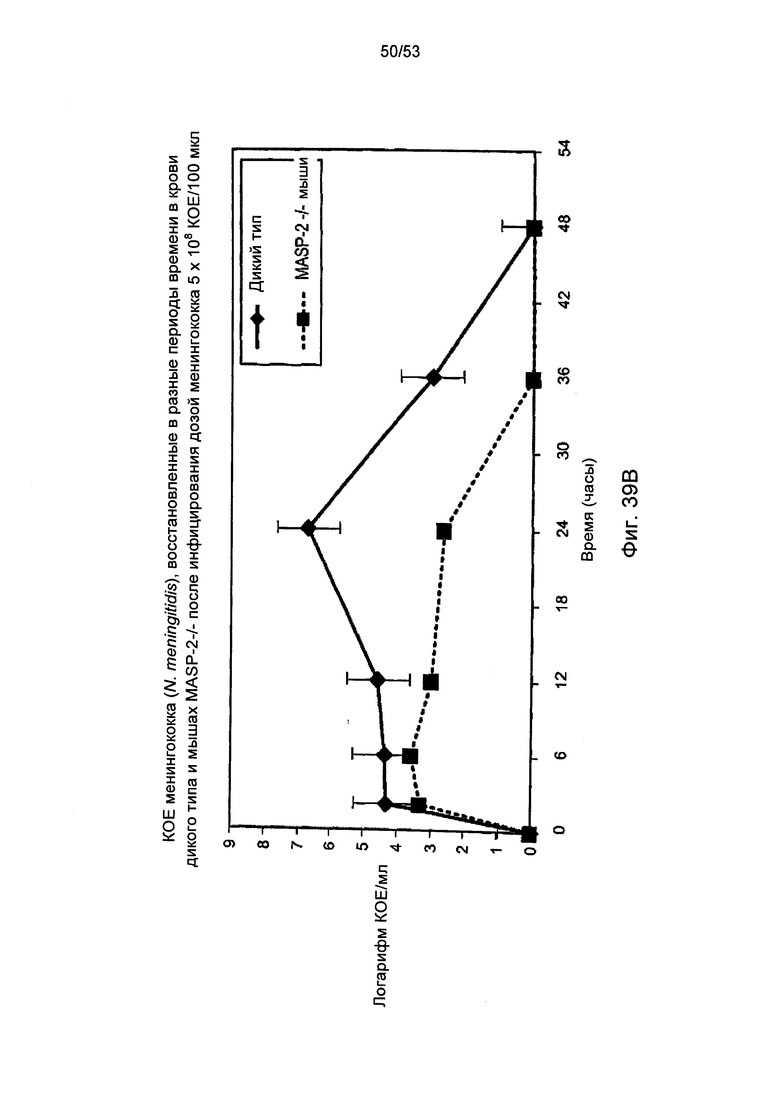
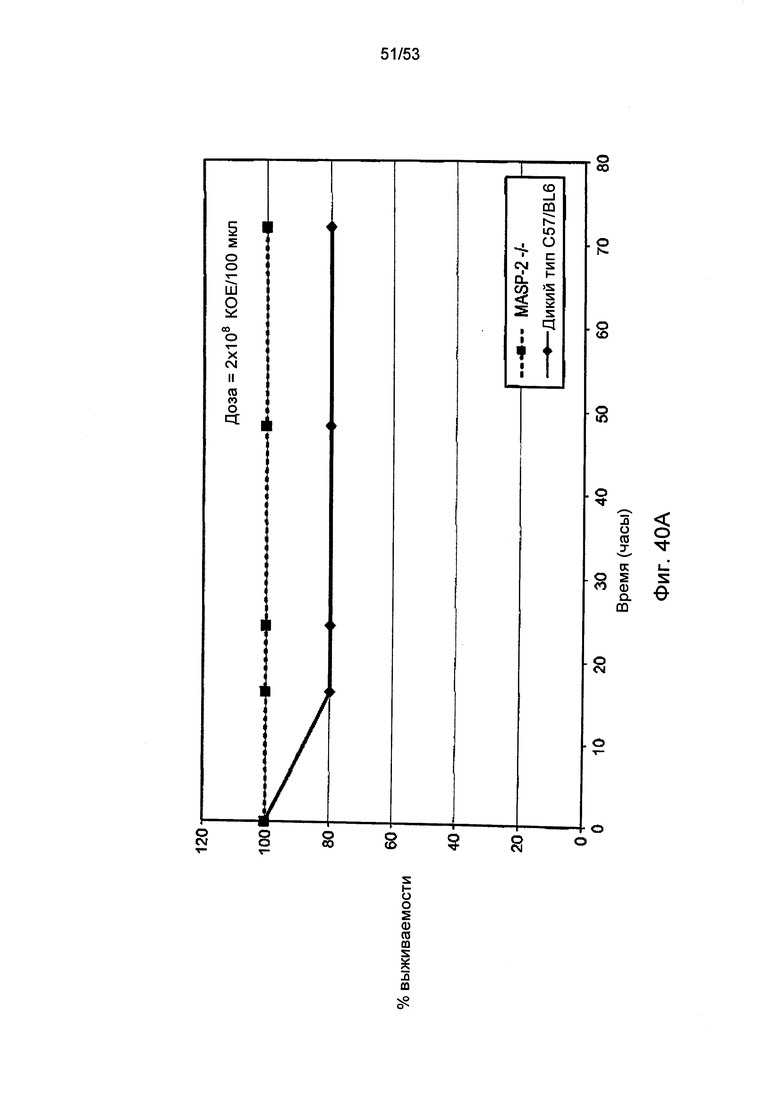
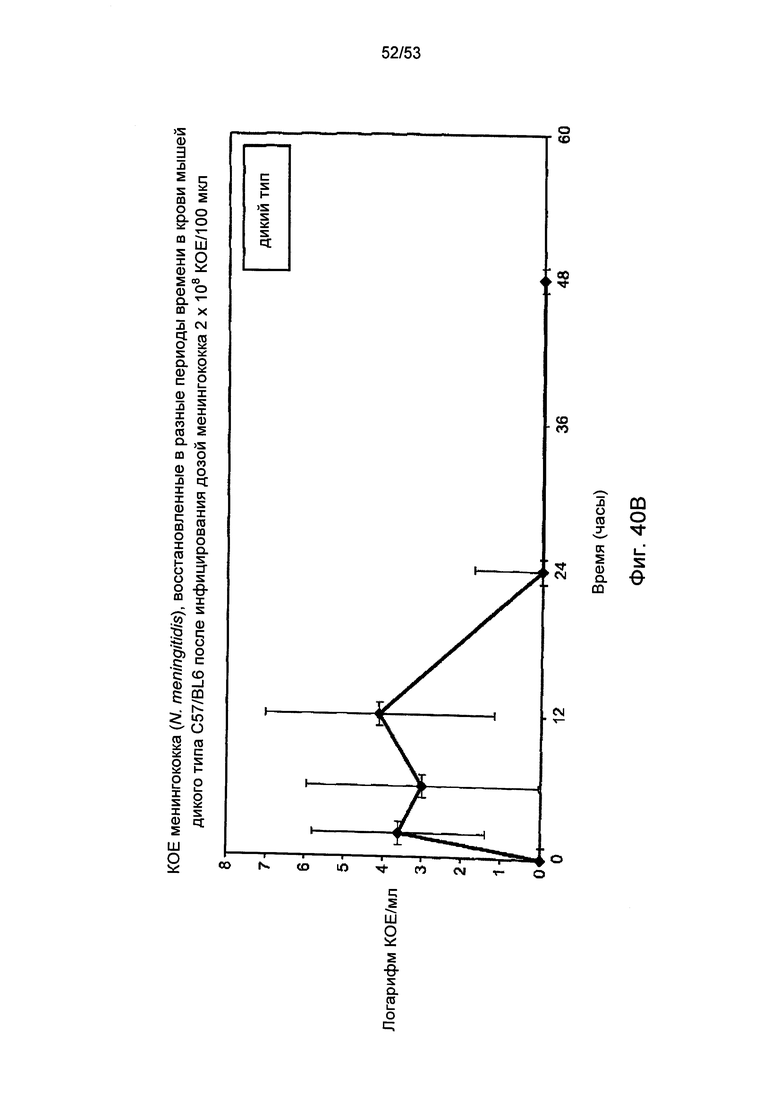

Настоящее изобретение относится к биохимии, в частности к применению композиции, содержащей агент, ингибирующий MASP-2, в количестве, эффективном для ингибирования или предупреждения образования бляшек в сосудистой системе пациента, для приготовления лекарственного средства для лечения указанного пациента, страдающего или подверженного риску развития комплемент-опосредованного нарушения свертывания крови, такого как диссеминированное внутрисосудистое свертывание крови (ДВС). Настоящее изобретение также раскрывает применение указанной композиции в производстве лекарственного средства для лечения или профилактики инфекции менингококка Neisseria meningitidis у пациента. Настоящее изобретение также раскрывает способ лечения комплемент-опосредованного нарушения свертывания крови и способ лечения или профилактики инфекции менингококка Neisseria meningitidis у пациента. Настоящее изобретение раскрывает новые применения агента, ингибирующего MASP-2, и позволяет расширить арсенал средств для лечения ДВС и инфекции Neisseria meningitidis. 5 н. и 34 з.п. ф-лы, 40 ил., 7 табл., 40 пр.
1. Применение композиции, содержащей количество агента, ингибирующего MASP-2, эффективного для ингибирования или предупреждения образования бляшек в сосудистой системе субъекта, страдающего или предрасположенного к риску развития комплемент-опосредованного нарушения свертывания крови, в производстве лекарственного средства для лечения субъекта, страдающего от или подверженного риску развития комплемент-опосредованного нарушения свертывания крови.
2. Применение по п. 1, в котором композиция не повышает восприимчивость у субъекта к инфекции менингококка Neisseria meningitidis.
3. Применение по п. 1 или 2, в котором агент, ингибирующий MASP-2, специфически связывается с полипептидом, содержащим последовательность SEQ ID NO: 6, с аффинностью, по меньшей мере в 10 раз большей, чем при его связывании с другим антигеном в системе комплемента.
4. Применение по п. 3, в котором агент, ингибирующий MASP-2, представляет собой анти-MASP-2 антитело или его фрагмент, который специфически связывается с частью последовательности SEQ ID NO:6.
5. Применение по п. 4, в котором анти-MASP-2 антитело или его фрагмент является моноклональным или рекомбинантным антителом, или является химерным, гуманизированным или человеческим антителом, или в котором антитело имеет пониженную эффекторную функцию.
6. Применение по п. 1, в котором композиция составляется в виде быстродействующей лекарственной формы для системной доставки.
7. Применение по п. 1, в котором комплемент-опосредованное нарушение свертывания крови является осложнением недостаточности регуляторного фактора комплемента, травматического повреждения или инфекции.
8. Применение по п. 7, в котором инфекция представляет собой бактериальную инфекцию.
9. Применение по п. 1, в котором композиция используется в производстве лекарственного средства для лечения, профилактики или снижения тяжести диссеминированного внутрисосудистого свертывания у субъекта, страдающего диссеминированным внутрисосудистым свертыванием или подверженного риску его развития.
10. Применение по п. 9, в котором композиция используется в производстве лекарственного средства для лечения, профилактики или снижения тяжести диссеминированного внутрисосудистого свертывания у субъекта, страдающего диссеминированным внутрисосудистым свертыванием или предрасположенного к риску его развития, являющегося осложнением сепсиса, травмы, инфекции, злокачественного новообразования, акушерского осложнения, болезни печени, тяжелой токсической реакции, шока, теплового удара, отторжения трансплантатов, трансфузионной реакции, аневризмы сосудов, печеночной недостаточности, лечения рака посредством химиотерапии или лучевой терапии, ожога или случайного радиационного облучения.
11. Применение по п. 10, в котором диссеминированное внутрисосудистое свертывание является осложнением бактериальной инфекции.
12. Способ лечения субъекта, страдающего от или предрасположенного к риску развития комплемент-опосредованного нарушения свертывания крови, способ, включающий введение субъекту такого количества агента, ингибирующего MASP-2, которое эффективно для ингибирования или предупреждения образования бляшек в сосудистой системе субъекта, страдающего от комплемент-опосредованного нарушения свертывания крови или предрасположенного к риску его развития.
13. Способ лечения по п. 12, в котором композиция составляется в быстродействующей лекарственной форме для системной доставки.
14. Способ лечения по п. 12 или 13, в котором композиция не повышает восприимчивость к инфекции менингококка Neisseria meningitidis.
15. Способ лечения по п. 12, в котором агент, ингибирующий MASP-2, специфически связывается с полипептидом, содержащим последовательность SEQ ID NO: 6, с аффинностью по меньшей мере в 10 раз большей, чем при его связывании с другим антигеном в системе комплемента.
16. Способ лечения по п. 15, в котором агент, ингибирующий MASP-2, представляет собой анти-MASP-2 антитело или его фрагмент, который специфически связывается с частью последовательности SEQ ID NO: 6.
17. Способ лечения по п. 16, в котором антитело MASP-2 или его фрагмент является моноклональным или рекомбинантным антителом, или является химерным, гуманизированным или человеческим антителом, или в котором антитело имеет пониженную эффекторную функцию.
18. Применение композиции, содержащей количество агента, ингибирующего MASP-2, эффективное для ингибирования или предупреждения образования бляшек в сосудистой системе субъекта, страдающего или предрасположенного к риску развития диссеминированного внутрисосудистого свертывания крови, в производстве лекарственного средства для лечения субъекта, страдающего от диссеминированного внутрисосудистого свертывания крови.
19. Применение по п. 18, в котором композиция не повышает восприимчивость у субъекта к инфекции менингококка Neisseria meningitidis.
20. Применение по п. 18 или 19, в котором агент, ингибирующий MASP-2, специфически связывается с полипептидом, содержащим последовательность SEQ ID NO: 6, с аффинностью по меньшей мере в 10 раз большей, чем при его связывании с другим антигеном в системе комплемента.
21. Применение по п. 20, в котором агент, ингибирующий MASP-2, представляет собой антитело анти-MASP-2 или его фрагмент, который специфически связывается с частью последовательности SEQ ID NO: 6.
22. Применение по п. 21, в котором анти-MASP-2 антитело или его фрагмент является моноклональным или рекомбинантным антителом, или является химерным, гуманизированным или человеческим антителом, или в котором антитело имеет пониженную эффекторную функцию.
23. Применение по п. 18, в котором композиция разрабатывается в виде быстродействующей лекарственной формы для системной доставки.
24. Применение по п. 18, в котором диссеминированное внутрисосудистое свертывание является осложнением сепсиса, травмы, инфекции, злокачественного новообразования, акушерского осложнения, болезни печени, тяжелой токсической реакции, шока, теплового удара, отторжения трансплантатов, трансфузионной реакции, аневризма сосудов, печеночной недостаточности, лечения рака посредством химиотерапии или лучевой терапии, ожога или случайного радиационного облучения.
25. Применение по п. 24, в котором субъект страдает от неврологической травмы.
26. Применение по п. 24, в котором субъект страдает от бактериальной инфекции.
27. Применение композиции, содержащей количество агента, ингибирующего MASP-2, эффективное для лечения или профилактики инфекции менингококка Neisseria meningitidis в производстве лекарственного средства для лечения или профилактики инфекции менингококка Neisseria meningitidis у субъекта, страдающего от или подверженного риску развития инфекции менингококка Neisseria meningitidis.
28. Применение по п. 27, в котором композиция разрабатывается в виде быстродействующей лекарственной формы для системной доставки.
29. Применение по п. 27 или 28, в котором агент, ингибирующий MASP-2, специфически связывается с полипептидом, содержащим последовательность SEQ ID NO: 6, с аффинностью по меньшей мере в 10 раз большей, чем при его связывании с другим антигеном в системе комплемента.
30. Применение по п. 29, в котором агент, ингибирующий MASP-2, представляет собой анти-MASP-2 антитело или его фрагмент, который специфически связывается с частью последовательности SEQ ID NO: 6.
31. Применение по п. 30, в котором анти-MASP-2 антитело или его фрагмент является моноклональным или рекомбинантным антителом, или является химерным, гуманизированным или человеческим антителом, или в котором антитело имеет редуцированную эффекторную функцию.
32. Применение по п. 27, в котором субъект страдает комплемент-опосредованным нарушением свертывания крови или предрасположен к риску его развития.
33. Способ лечения или профилактики инфекции менингококка Neisseria meningitidis, у субъекта, страдающего от инфекции менингококка Neisseria meningitides или предрасположенного к риску заболеть указанной инфекцией, включающий введение субъекту такого количества агента, ингибирующего MASP-2, которое эффективно для лечения или профилактики инфекции менингококка Neisseria meningitides.
34. Способ лечения по п. 33, где композиция разрабатывается в виде быстродействующей лекарственной формы для системной доставки.
35. Способ лечения по п. 33 или 34, в котором агент, ингибирующий MASP-2, специфически связывается с полипептидом, содержащим последовательность SEQ ID NO: 6, с аффинностью по меньшей мере в 10 раз большей, чем при его связывании с другим антигеном в системе комплемента.
36. Способ лечения по п. 35, в котором агент, ингибирующий MASP-2, представляет собой анти-MASP-2 антитело или его фрагмент, который специфически связывается с частью последовательности SEQ ID NO: 6.
37. Способ лечения по п. 36, в котором анти-MASP-2 антитело или его фрагмент является моноклональным или рекомбинантным антителом, или является химерным, гуманизированным или человеческим антителом, или в котором антитело имеет пониженную эффекторную функцию.
38. Применение композиции по п. 18, в котором субъект имеет клиническое подтверждение, что он страдает от или подвержен риску развития диссеминированного внутрисосудистого свертывания крови.
39. Применение композиции по п. 18, в котором клиническое подтверждение включает один или более следующих условий (i) активация комплемента С3 выше 30% васкулярного пула, (ii) понижение или конверсия уровня С5, (iii) вазодилятация и/или, (iv) потеря жидкости в ткани.
| WO 2005123128, 29.12.2005 | |||
| BRUCHIM Y, et al., Disseminated intravascular coagulation, Compend Contin Eud Vet, 2008, Vol.30, No.10, pp.E1-E16 | |||
| WARD JR, Agonists of toll-like receptor (TLR)2 and TLR4 are unable to modulate platelet activation by adenosine diphosphate and platelet activating factor, Thromb Haemost, 2005, Vol.94, No.4, pp.831-838 | |||
| RU 2003104524 A, 20.01.2005. |

