Область изобретения
Изобретение относится к медицине, в частности к фармакологии, и касается применения нового активатора HIF-1α.
Уровень техники
В связи с тем, что гипоксия является важнейшим патогенетическим фактором широкого спектра патологических состояний, поиск средств, повышающих резистентность организма к дефициту снабжения кислородом, является задачей высокой степени актуальности. Высокая чувствительность мозга к гипоксии проявляется в том, что хотя масса мозга составляет лишь 2% от массы всего организма, мозг потребляет 20% количества кислорода, поглощаемого организмом. Системой, играющей основную роль в реализации гомеостатического ответа мозговой ткани на гипоксическое воздействие, является транскрипционный фактор, индуцируемый гипоксией (Hypoxia Induced Factor, HIF-1α). HIF-1 является гетеродимером, состоящим из двух субъединиц - кислород-чувствительной субъединицы HIF-1α и конститутивно-экспрессирующейся HIF-1β. Гипоксия способствует возрастанию уровня HIF-1α, его димеризации с HIF-1β, мобилизации ко-активаторов (p300/CBP) и связыванию этого комплекса с последовательностью HRE (hypoxia-response element) в регуляторных областях генов-мишеней. В нормоксических условиях гидроксилирование остатков пролина белка HIF-1α пролилгидроксилазами (PHD, prolyl hydroxylase-domain protein; ЕС 1.1411.29) является необходимым условием для связывания компонентом убиквитин протеин лигазы E3 - белком фон Хиппеля-Линдау (VHL). Присутствие кислорода вызывает также гидроксилирование аспарагинового остатка C-терминального домена трансактивации (C-TAD) HIF-1α ферментом аспарагин-гидроксилазой (FIH1, factor inhibiting HIF-1), что блокирует его взаимодействие с ко-активатором транскрипции p300/CBP.
Если при нормальном снабжении тканей кислородом ферменты PHD и FIH инактивируют HIF-1α, в условиях гипоксии активность PHD и FIH снижается, что приводит к уменьшению деградации HIF-1α, его накоплению, и, как следствие - запуску зависимых от HIF-1α генов [Semenza G. // Cell. 2012. V. 148. №3. P. 399-408]. Показано [Zheng K, Fridkin M., Youdim М. // Persp. Med. Chem. 2015. V. 7. P. 1-8], что HIF-1α активирует в общей сложности до 100 генов, в том числе гены, вовлеченные в процессы ангиогенеза за счет активации фактора роста эндотелия сосудов, усиления синтеза эритропоэтина, активации систем транспорта глюкозы через мембраны, цитопротекции нейротрофическими факторами, нормализации клеточного цикла и метаболизма на уровне митохондрий, включая активацию антиоксидантных ферментов - супероксиддисмутазы и каталазы. Таким образом, регулируя экспрессию генов, вовлеченных в энергетический обмен, ангиогенез, эритропоэз, клеточную пролиферацию, HIF определяет реализацию адаптивного ответа на гипоксическое воздействие.
Наряду с этим HIF-1 влияет на состояние многих нейротрансмиттерных систем - он активирует белок, контролирующий ГАМК рецепторы (GABARBP) [Park S.H., Kim B.R., Lee J.H. et al // Cell Signal. 2014. V. 26. №7. P. 1506-7573], повышает активность тирозингидроксилазы [Schnell P.O., Ignacak M.L., Bauer A.L., et al., // J. Neurochem. 2003 V. 85. №2. P. 483-491]. Описаны тесные взаимоотношения HIF-1α с холинорецепторами [Hirota K., Fukuda R., Takabuchi S. Et al. // J. Biol. Chemistry. 2004. V. 279. №40. P. 41521-41528].
Известно, что острый дефицит доставки кислорода и макроэргов при ишемическом инсульте ведет к включению эндогенных протективных механизмов, ключевым из которых является система HIF (Shi, 2009; Xin et al, 2011). У мышей, нокаутных по HIF-1α, перевязка средней мозговой артерии сопровождалась повышением смертности и развитием большего по объему очага поражения, чем у нормальных животных (Baranova et al., 2007).
Хроническая гипоксия играет важную роль в прогрессировании нейродегенеративных заболеваний. Так, уже на ранних фазах болезни Альцгеймера имеет место снижение содержания HIF с сопутствующим замедлением утилизации глюкозы в мозговой ткани (Zhang Z., Yan J., Chang Y., ShiDu Yan S., Shi H. // Curr. Med. Chem. 2011. V. 18. №28. P. 4335-4343). Снижение активности системы HIF с сопутствующим угнетением тирозингидроксилазы и дегенерацией допаминергических нейронов выявлено у пациентов, страдающих болезнью Паркинсона (Borghammer P., Cumming P., Ostergaard K. et al. J. Neurol. Sc. 2012, 313, (1-2), 123-128).
Имеются данные о роли дефицита HIF в бета-клетках поджелудочной железы в патогенезе диабета. Так, сообщается о способности инсулина нарушать образование HIF-1α и о роли дефицита этого фактора в развитии как диабета 1-го типа (абсолютный дефицит инсулина), так и диабета 2-го типа (относительный дефицит инсулина), а также их осложнений [Cheng K., Но K, Stokes R. et al, // J. Clin. Invest. 2010. V. 120. №6. P. 2171-2183]. Нарушение функции систем транспорта глюкозы GLUT1 и GLUT3 через клеточные барьеры, имеющее место при дефиците HIF-1α, способствует развитию инсулиновой резистентности как при болезни Альцгеймера, так и при диабете [Liu Y., Liu F. , Iqbal K., et al, // FEBS Lett. 2008. V. 582. P. 359-364]. Доказано участие HIF-1α в экспрессии инкретинов - важнейших факторов цитопротекции бета-клеток поджелудочной железы [Van de Velde S., Hogan M.F., Montminy M. // PNAS. 2011. V. 108. №41. P. 16876-16882].
Применение веществ, активирующих систему HIF, может быть эффективно при широком спектре патологических состояний, перечисленных выше.
Активаторы системы HIF разделяются на 2 группы: вещества, усиливающие транскрипцию и трансляцию этого транскрипционного фактора, и вещества, ингибирующие его деактивацию.
Описан многофункциональный хелатор железа М30 (5-[N-метил-N-пропаргиламинометил]-8-оксихинолин), который активирует HIF-1α и обладает нейропротективной активностью в экспериментах in vitro и in vivo (Н. Zheng et al., Bioorganic and Medicinal Chemistry, 2005,vol. 13, №. 3, pp. 773-783; Y. Avramovich-Tirosh et al, Curr. Alzheimer Res. 2010. V. 7, №4, p. 300-306).
В патенте US 8524699 описаны замещенные дигидропиразолоны как ингибиторы HIF-пролил-4-гидроксилазы общей формулы:

Эти соединения являются активаторами HIF и могут быть полезными для лечения анемии, хронического заболевания почек, инфаркта миокарда, атеросклероза.
В патенте US 8541455 B2 описаны производные 2-пиридин-2-ил-пиразол-3(2H)-она как активаторы транскрипционного фактора HIF общей формулы:
Показано, что эти вещества усиливают секрецию фактора роста сосудистого эндотелия (VEGF) и эритропоэтина (ЕРО) - двух основных маркеров активации HIF-1α в гепацитах.
Описан N-(2-меркаптопропионил)-глицин как активатор HIF-1α за счет ингибирования HIF-пролилгидроксилазы-2 (S. Yum et al., Biochem. Biophys. Res. Comm. 2014. V. 443, p. 1008-1013). Показано, что это соединение усиливает секрецию фактора роста сосудистого эндотелия (VEGF) и обладает протективным эффектом в условиях экспериментального колита.
Показано (Y. Liu et al., Cell Physiol Biochem 2014; 33: 1036-1046), что известный хелатор железа, дефероксамин, использующийся при заболеваниях, характеризующихся избытком железа, вызывает усиление экспрессии HIF-1α и вовлечен в сигнальный каскад, включающий активацию ERK, что указывает на его инвазивный потенциал.

Обнаружено, что пролин-аргинин-обогащенный пептид PR-39 обладает кардиопротективной активностью за счет стабилизирующих свойств в отношении HIF-1α (E.D. Muinck et al., Antioxid. Redox Signal. 2007; 9(4): 437-445).
В статье X. Ma et al., PLOS One, 2014. V. 9, №4, описаны два пролиновых соединения - N-бензилоксикарбонилпролин (РА2) и бис-(пролил-о-фенилендиамин) (РА1) как ингибиторы пролилгидроксилазы-3, обладающие активирующим влиянием на HIF-1α сигнальный путь, включая усиление синтеза VEGF и ключевых модуляторов гликолиза (пируваткиназных изозимов M1/M2 и альфа-енолазы-1).
В статье J.-J. Min et al., Scientific Reports, 2014. V. 4. P. 1-5 описан D,L-3-н-бутилфталид, который усиливал экспрессию HIF-1α и улучшал обучение и память у крыс в условиях хронической гипоксии. Это соединение также усиливало экспрессию BDNF и фосфорилирование CREB.
Известен ингибитор пролилгидроксилазы GSK360A, производное хинолина формулы:
При пероральном введении GSK360A оказывает кардиопротекторное действие при сердечной недостаточности после инфаркта миокарда и проявляет защитное действие на модели ишемически-реперфузионных повреждений у мышей (S.S. Banerjee et al., Toxicology Mechanisms and Methods, 2012; 22(5): 347-358).
Однако приведенные выше ссылки не открывают и не предполагают, что замещенный дипептид этиловый эфир N-фенилацетил-L-пролилглицина (Дипептид I) оказывает активирующее влияние на транскрипционный фактор HIF-1α.
Сущность изобретения
Задачей изобретения стал поиск способа активации транскрипционного фактора HIF-1α. Эта цель была достигнута с помощью дипептида этилового эфира N-фенилацетил-L-пролилглицина (Патент РФ 2119496).
Описание изобретения
Данное изобретение иллюстрируется следующими примерами.
Пример 1.
Культивирование клеток. Использование клеточных линий, стабильно экспрессирующих люциферазные репортерные конструкции с сайтами связывания определенных ТФ, является одним из наиболее эффективных скрининговых подходов к поиску соединений, влияющих на активность транскрипционных факторов (ТФ), [Phippard D., Manning A.M. (2003) Methods Mol Biol., 225, 19-23]. В работе использовались клетки линии HEK293 (перевиваемые клетки почки эмбриона человека; Российская коллекция клеточных культур, Институт цитологии РАН, Санкт-Петербург). Клетки культивировали при 37°C при 5% CO2 в среде DMEM ("Биолот", Россия), содержащей 10% эмбриональной телячьей сыворотки ("Sigma", США), 2 мМ L-глутамин, 50 мкг/мл гентамицина сульфата, 2.5 мкг/мл амфотерицина В ("ПанЭко", Россия).
Репортерные векторные конструкции. Люциферазные репортерные векторы содержат консенсусные нуклеотидные последовательности для соответствующих ТФ, при этом репортерный ген (ген люциферазы) находится под их контролем. Связывание ТФ с сайтом узнавания приводит к экспрессии гена люциферазы и биолюминесценции, детектируемой при внесении субстрата люциферина. Уровень биолюминесценции прямо пропорционален количеству фермента и связывающей активности того или иного транскрипционного фактора. Принцип in vitro тест-систем для биологического скрининга соединений на предмет их влияния на тот или иной ТФ основан на том, что, если тестируемое соединение стимулирует/ингибирует активность ТФ, то будет наблюдаться увеличение/снижение уровня люминесценции (вследствие изменения ДНК-связывающей активности ТФ). Таким образом, становится возможным отбирать соединения, мишенями для которых являются транскрипционные факторы, и исследовать механизмы действия как новых, так и уже применяющихся лекарств. В качестве специфических респонсивных элементов, узнающих и связывающих транскрипционные факторы (ТФ) CREB, NFAT, NF-кВ, р53, STAT1, VDR, HSF1 и HIF1, и определяющие активацию транскрипции на минимальном промоторе HSVtk (минимальный промотор тимидинкиназы вируса простого герпеса), были использованы следующие последовательности (Таблица 1). Олигонуклеотиды ("Синтол", Россия), содержащие несколько копий последовательностей, соответствующих сайтам связывания отдельных ТФ, были клонированы в плазмидном векторе pTL-Luc ("Panomics", США; несет ген люциферазы Photinus pyralis) по сайтам рестрикции NheI (5′-G↓CTAGC-3′) и BglII (5′-A↓GATCT-3′) ("Fermentas", Литва). Данный репортерный люциферазный вектор содержит минимальный промотор тимидинкиназы вируса простого герпеса, активность которого зависит от вышележащей консенсусной последовательности, способной связывать определенный ТФ. В свою очередь экспрессия гена люциферазы находится под контролем данного модельного промотора. Наличие и ориентацию вставок подтверждали секвенированием согласно протоколу ABI PRISM BigDye™ Terminator Cycle Sequencing Ready Reaction Kit ("Applied Biosystems", США). Выделение и очистку плазмидной ДНК проводили с помощью набора EndoFree Plasmid Maxi Kit ("Qiagen", США) [Салимгареева M.X., Садовников С.В., Фарафонтова Е.И., Зайнуллина Л.Ф., Вахитов В.А., Вахитова Ю.В. Клеточные тест-системы для поиска модуляторов активности транскрипционных факторов. Прикладная биохимия и микробиология, 2014. - № 2. - С. 219-225]. 

Трансфекция. Для проведения транзиентных трансфекций клетки линии HEK293 рассаживали в количестве 45×103 клеток на лунку в 96-луночных планшетах в 100 мкл среды без антибиотика (DMEM, 10% эмбриональной телячьей сыворотки, 2 мМ L-глутамин). Через 24 ч проводили трансфекции данными конструкциями (таблица 1) с помощью реагента Липофектамин 2000 ("Invitrogen", США) согласно протоколу изготовителя. Через 6 ч после трансфекций среду заменяли на содержащую антибиотик (DMEM, 10% эмбриональной телячьей сыворотки, 2 мМ L-глутамин, 50 мкг/мл гентамицин, 2.5 мкг/мл амфотерицин B) и через 18 ч добавляли изучаемые препараты (Дипептид I в концентрации 10 мкМ; Пирацетам в концентрации 1 мМ). Клетки инкубировали в присутствии Дипептида (этиловый эфир N-фенилацетил-L-пролилглицина, I) или Пирацетама еще в течение 24 ч. Детекцию люциферазной активности в клеточных лизатах проводили с помощью набора Dual Luciferase Reporter Assay System (Promega, США) на планшетном анализаторе 2300 EnSpire® Multimode Plate Reader (Perkin Elmer, США). В качестве внутреннего контроля трансфекций применяли ко-трансфекцию с плазмидой pRL-TK (Promega, США), кодирующей ген люциферазы Renilla reniformis. Значения люминесценции для Photinus pyralis нормировали по люминесценции для Renilla reniformis в каждом измерении.
Среднее арифметическое значений, полученных по двум повторам в каждом эксперименте в трех независимых экспериментах, и стандартную ошибку среднего рассчитывали с помощью программы Statistica 6.1 (StatSoft. Inc., США). Сравнение экспериментальных групп проводили с использованием парного t-критерия Стьюдента для зависимых выборок.
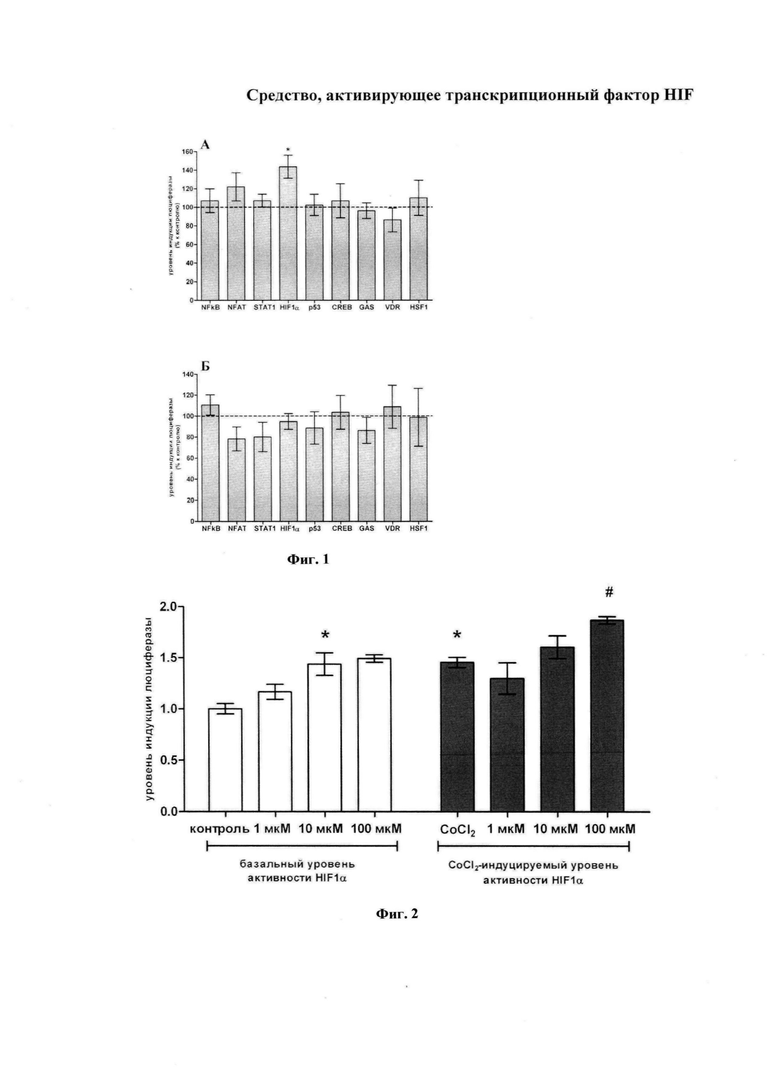
Данные, представленные на фиг. 1A, свидетельствуют о том, что Дипептид I при инкубации в течение 24 часов в концентрации 10 мкМ увеличивал ДНК-связывающую активность HIF-1α на 43% и не влиял на ДНК-связывающую активность факторов CREB, NFAT, NF-кВ, p 53, STAT1, GAS, VDR и HSF1. Как следует из данных, представленных на фиг. 2 (слева - влияние на базальную активность), выраженность HIF-позитивного эффекта Дипептида I зависела от его концентрации.
Эксперименты с препаратом сравнения показали, что Пирацетам ни в эквимолярной (10 мкМ, данные не представлены), ни в более высокой концентрации (1 мМ) не вызывает статистически значимых изменений ДНК-связывающей активности изученных транскрипционных факторов (фиг. 1Б).
ПРИМЕР 2
Экспериментальное моделирование гипоксии осуществляли с использованием CoCl2, который вызывает стабилизацию HIF-1α и является фармакологическим миметиком гипоксии [Piret JP, Mottet D, Raes M, Michiels С. CoCl2, a chemical inducer of hypoxia-inducible factor-1, and hypoxia reduce apoptotic cell death in hepatoma cell line HepG2. Ann N Y Acad Sci. 2002; 973: 443-7]. Механизм действия CoCl2 обусловлен его способностью замещать Fe2+ в активном сайте пролилгидроксилазы, инактивировать, таким образом, фермент и ингибировать гидроксилирование HIF-1α [Epstein АС1, Gleadle JM, McNeill LA, Hewitson KS, O'Rourke J, Mole DR, Mukherji M, Metzen E, Wilson MI, Dhanda A, Tian YM, Masson N, Hamilton DL, Jaakkola P, Barstead R, Hodgkin J, Maxwell PH, Pugh CW, Schofield CJ, Ratcliffe PJ. C. elegans EGL-9 and mammalian homologs define a family of dioxygenases that regulate HIF by prolyl hydroxylation. Cell. 2001; 107(1): 43-54].
Люциферазная конструкция для анализа активности HIF-1 содержит 4 копии консенсусной последовательности 5′ ACGTG 3′, являющейся сайтом связывания для белка HIF-1 (конструкция HIF-1-Luc). Клетки, трансфицированные плазмидным вектором HIF-1-Luc, предварительно инкубировали с Дипептидом I в течение 8 ч (конечные концентрации 1, 10, 100 мкМ; двукратное введение через каждые 4 ч), далее вносили индуктор гипоксии CoCl2 в рабочей концентрации 50 мкМ и продолжали совместную инкубацию Дипептида I и CoCl2 в течение 16 ч, после чего люциферазную активность детектировали, как описано выше. Среднее арифметическое значений, полученных по двум повторам в каждом эксперименте в трех независимых экспериментах, и стандартную ошибку среднего, рассчитывали с помощью программы Statistica 6.1 (StatSoft. Inc., США). Сравнение экспериментальных групп проводили с использованием парного t-критерия Стьюдента для зависимых выборок.
Эксперименты с изучением влияния Дипептида I на ДНК-связывающую активность HIF-1α в присутствии фармакологического миметика гипоксии CoCl2 дали следующие результаты. В полном соответствии с литературными данными, в этом эксперименте на фоне действия CoCl2 отмечается повышение активности HIF-1α. Впервые выявлен зависимый от концентрации эффект Дипептида I на ДНК-связывающую активность HIF-1α (фиг. 2): в концентрации 1 мкМ отмечается лишь недостоверное снижение данного параметра, а в концентрациях 10 и 100 мкМ - Дипептид I вызывал соответствующее увеличение HIF-1-зависимой люциферазной активности.
Таким образом, установлено, что Дипептид I обладает способностью увеличивать как базальную, так и индуцированную фармакологическим миметиком гипоксии активность HIF-1α in vitro.
Совокупность полученных данных свидетельствует о том, что Дипептид I вызывает концентрационно-зависимое увеличение базальной ДНК-связывающей активности HIF-1α. Показано, что в условиях стабилизации HIF-1α с помощью химического индуктора этого транскрипционного фактора CoCl2. Дипептид I вызывает дополнительное нарастание ДНК-связывающей активности HIF-1α. Эффект в отношении HIF-1α специфичен для Дипептида I:, классический ноотропный препарат Пирацетам не влияет на активность данного транскрипционного фактора. Дипептид I увеличивал ДНК-связывающую активность только HIF-1α, тогда как активность других изученных транскрипционных факторов (CREB, NFAT, NF-κВ, p 53, STAT1, GAS, VDR и HSF1) под влиянием Дипептида I не повышалась.
Краткое описание чертежей.
На фиг. 1 изображено влияние Дипептида I а 10 мкМ (А) и препарата сравнения Пирацетам а 1 мМ (Б) на базальную ДНК-связывающую активность транскрипционных факторов NF κВ, NFAT, STAT1, HIF-1, p 53, CREB, GAS, VDR, HSF1 in vitro. Достоверность различий оценивали с помощью парного t-критерия Стьюдента для зависимых выборок (n=3, *p<0.05).
На фиг. 2 изображено влияние Дипептида I на базальную и индуцированную активность HIF-1α. Группа «контроль» - значения базальной активности HIF-1α нестимулированных клетках; группа «CoCl2» - значения активности HIF-1α CoCl2-стимулированных клетках. Данные представлены в виде среднего арифметического ± стандартная ошибка среднего (n=3; *p<0.05 по отношению к группе «контроль»; #p<0.05 по отношению к группе «CoCl2»).
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ прогнозирования плацентарной недостаточности во второй половине беременности у серопозитивных по цитомегаловирусу женщин | 2020 |
|
RU2724395C1 |
| ЛЕКАРСТВЕННОЕ СРЕДСТВО, УСИЛИВАЮЩЕЕ ОКСИГЕНАЦИЮ ТКАНЕЙ ПРИ ДИАБЕТИЧЕСКОЙ СТОПЕ, И СПОСОБ ЕГО ПРИМЕНЕНИЯ | 2017 |
|
RU2717674C1 |
| СПОСОБ ПРОГНОЗИРОВАНИЯ РАЗВИТИЯ ГЕМАТОГЕННЫХ МЕТАСТАЗОВ ПОСЛЕ КОМБИНИРОВАННОГО ЛЕЧЕНИЯ РАКА ПОЧКИ | 2013 |
|
RU2528100C1 |
| Метод определения формирования угрозы выкидыша эмбриона на первом триместре гестации при действии цитомегаловирусной инфекции в период обострения на фактор, индуцируемый гипоксией | 2016 |
|
RU2620565C1 |
| СПОСОБ ПРОГНОЗИРОВАНИЯ ЭФФЕКТИВНОСТИ ТЕРАПИИ ЭВЕРОЛИМУСОМ У БОЛЬНЫХ МЕТАСТАТИЧЕСКИМ РАКОМ ПОЧКИ | 2016 |
|
RU2616533C1 |
| СПОСОБЫ УСИЛЕНИЯ СТАБИЛИЗАЦИИ ИНДУЦИРУЕМОГО ГИПОКСИЕЙ ФАКТОРА-1 АЛЬФА | 2010 |
|
RU2521251C2 |
| СПОСОБ ОЦЕНКИ БИОАКТИВНОСТИ ХИМИЧЕСКИХ СОЕДИНЕНИЙ | 2013 |
|
RU2556823C2 |
| СПОСОБ ПРОГНОЗИРОВАНИЯ РАЗВИТИЯ ФИБРИЛЛЯЦИИ ПРЕДСЕРДИЙ У ПАЦИЕНТОВ С СИНДРОМОМ ОБСТРУКТИВНОГО АПНОЭ СНА | 2022 |
|
RU2793677C1 |
| Ноотропная композиция на основе полипептидных комплексов, выделенных из нейрональных прогениторных клеток в условиях гипоксии, и способ ее получения | 2019 |
|
RU2732600C1 |
| ПРИМЕНЕНИЕ СТАБИЛИЗАТОРОВ HIF-АЛЬФА ДЛЯ УСИЛЕНИЯ ЭРИТРОПОЭЗА | 2004 |
|
RU2414214C2 |
Группа изобретений относится к области фармацевтики и медицины и касается применения этилового эфира N-фенилацетил-L-пролилглицина для активации транскрипционного фактора HIF (фактор индуцируемый гипоксией), а также способа активации HIF с помощью указанного фактора и способа лечения заболеваний, связанных с дефицитом транскрипционного фактора HIF. Группа изобретений обеспечивает повышение резистентности организма к дефициту снабжения кислородом, что является полезным для лечения гипоксических состояний различного генеза. 3 н.п. ф-лы, 1 табл., 2 пр., 2 ил.
1. Применение этилового эфира N-фенилацетил-L-пролилглицина для активации транскрипционного фактора HIF (Hypoxia induced factor, фактор, индуцируемый гипоксией).
2. Способ активации транскрипционного фактора HIF, заключающийся во введении профилактически или терапевтически эффективного количества этилового эфира N-фенилацетил-L-пролилглицина.
3. Способ лечения заболеваний, связанных с дефицитом транскрипционного фактора HIF, заключающийся во введении профилактически или терапевтически эффективного количества этилового эфира N-фенилацетил-глицилпролина, вызывающего активацию транскрипционного фактора HIF.
| НОВИКОВ В.Е | |||
| и др | |||
| Новые направления поиска лекарственных средств с антигипоксической активностью и мишени для их действия | |||
| Ж | |||
| Экспериментальная и клиническая фармакология, 2013,т.76, с.41-45 | |||
| ПРОИЗВОДНЫЕ N-АЦИЛПРОЛИЛДИПЕПТИДОВ | 1993 |
|
RU2119496C1 |
| US20020187977A1, 12.12.2002 | |||
| ЛЕВЧЕНКОВА О.С | |||
| и др | |||
| Индукторы регуляторного фактора адаптации к гипоксии | |||
| Российский медико-биологический вестник им.акад | |||
| И.П.Пвлова, 2014. | |||

