Заболевания центральной нервной системы, такие как болезнь Альцгеймера и болезнь Паркинсона, обычно приводят к необратимому повреждению структур нервной ткани, что часто сопровождается серьезными когнитивными или моторными нарушениями. Помимо ограниченного потенциала эндогенной регенерации в центральной нервной системе, методы лечения таких расстройств в значительной степени являются симптоматическими и лишь временно улучшают качество жизни пациентов [Song CG, Zhang YZ, Wu HN, et al. 2018]. Поэтому необходим поиск новых способов лечения заболеваний центральной нервной системы. В 2006 году Яманака продемонстрировал, что соматические клетки можно репрограммировать в индуцированные плюрипотентные стволовые клетки. Это позволило к настоящему времени получать различные типы клеток, в том числе нейроны и глию, от взрослых доноров [Takahashi, Kazutoshi, and Shinya Yamanaka. 2006].
Терапия стволовыми клетками является многообещающей возможностью для лечения неврологических расстройств, обоснованность такого подхода была подтверждена в последние годы во множестве исследований на животных [Zakrzewski W,  Szymonowicz М, Rybak Z. 2019] Механизмы, лежащие в основе положительных эффектов, многообразны и включают в себя: непосредственную замену погибших клеток, восстановление поврежденной ткани путем слияния с эндогенными клетками, паракринный и иммуномодулирующий эффект. В частности, паракринный эффект может быть обусловлен секрецией факторов роста (FGF, EGF, VEGF), нейротрофинов (BDNF, GDNF, NGF, NT3-5, CNTF), экзосом. [Kwak KA, Lee SP, Yang JY, Park YS. 2018] Для практического использования паракринного эффекта стволовых клеток возможно применение кондиционированных сред стволовых клеток (КС). Существуют данные, подтверждающие эффективность КС в моделях in vivo при неврологических заболеваниях, таких как ишемический инсульт - при введении КС нейральных стволовых клеток (НСК) крысам с моделированным ишемическим инсультом показатели оценки неврологического статуса были выше, чем в контрольной группе, объем инфаркта снижался по сравнению с контролем, так же были исследованы изменение уровня апоптотической активности методом TUNEL, который также показал снижение клеточной гибели. В модели спинальной травмы изучалось действие КС НСК на жизнеспособность нейронов и регенерацию аксонов кортикоспинального тракта. Применение КС приводило к экстенсивному росту аксонов в шейном отделе и более чем утроило образование синаптических контактов между коллатералями кортикоспинального тракта и проприоспинальными интернейронами. КС также уменьшала экспрессию каспазы 3 и снижала гибель нейронов через 6 недель после травмы [Peng Liang, Jiaren Liu et al. 2014].
Szymonowicz М, Rybak Z. 2019] Механизмы, лежащие в основе положительных эффектов, многообразны и включают в себя: непосредственную замену погибших клеток, восстановление поврежденной ткани путем слияния с эндогенными клетками, паракринный и иммуномодулирующий эффект. В частности, паракринный эффект может быть обусловлен секрецией факторов роста (FGF, EGF, VEGF), нейротрофинов (BDNF, GDNF, NGF, NT3-5, CNTF), экзосом. [Kwak KA, Lee SP, Yang JY, Park YS. 2018] Для практического использования паракринного эффекта стволовых клеток возможно применение кондиционированных сред стволовых клеток (КС). Существуют данные, подтверждающие эффективность КС в моделях in vivo при неврологических заболеваниях, таких как ишемический инсульт - при введении КС нейральных стволовых клеток (НСК) крысам с моделированным ишемическим инсультом показатели оценки неврологического статуса были выше, чем в контрольной группе, объем инфаркта снижался по сравнению с контролем, так же были исследованы изменение уровня апоптотической активности методом TUNEL, который также показал снижение клеточной гибели. В модели спинальной травмы изучалось действие КС НСК на жизнеспособность нейронов и регенерацию аксонов кортикоспинального тракта. Применение КС приводило к экстенсивному росту аксонов в шейном отделе и более чем утроило образование синаптических контактов между коллатералями кортикоспинального тракта и проприоспинальными интернейронами. КС также уменьшала экспрессию каспазы 3 и снижала гибель нейронов через 6 недель после травмы [Peng Liang, Jiaren Liu et al. 2014].
Также проведенные клинические испытания пептидного препарата Церебролизин, содержащего аминокислоты и полипептиды молекулярной массой до 10 кДа, свидетельствуют о его эффективности при сосудистых деменциях [Chen N, et al. 2013]. Более того, в последнее время появились данные о нейропротекторных свойствах белков теплового шока и возможной их секреции из клеток в окружающую среду. Предположительно, секреция белков теплового шока стволовыми и прогениторными клетками при воздействии повышенной температуры должна усилить нейропротекторное действие.
В настоящее время ИПСК рассматриваются как самый перспективный источник нейральных предшественников для регенеративной медицины, позволяющий получать клеточные популяции различных типов на разных, заранее запланированных этапах эмбрионального развития. И при этом проблемы дефицита донорского материала и этические вопросы, возникающие при применении эмбрионального материала и культур эмбриональных стволовых клеток, полностью отсутствуют. Получение ИПСК уже стало рутинной задачей, хотя экспериментальные исследования проводятся только последние 10 лет. В результате экспериментов по ретровирусной трансдукции генами различных транскрипционных факторов мышиных фибробластов были идентифицированы 4 гена - Oct-4, Sox2, с-Мус и Klf4, которые переводили их в плюрипотентное состояние. Клетки, полученные таким образом, назвали индуцированными плюрипотентными стволовыми клетками (ИПСК) [Takahashi K., Yamanaka S. 2006]. Затем ИПСК были успешно получены из фибробластов человека с использованием того же набора транскрипционных факторов [Takahashi K. et al., 2007]. Другой научной группой были получены ИПСК человека с использованием иного набора транскрипционных факторов: Oct4, Sox2, Nanog и Lin28 [Yu J. et al., 2007]. Дальнейшие исследования продемонстрировали возможность получения ИПСК как от различных млекопитающих: крыс [Li W. et al., 2009], макак-резусов [Liu Н. et al., 2008], свиней [Kwon D.-J. et al., 2013]; так и из различных типов соматических клеток человека: кератиноцитов [Aasen Т. et al., 2008], нейрональных клеток [Eminli S. et al., 2008.], клеток печени и клеток желудка [Aoi, 2008], терминально дифференцированных лимфоцитов, а также из эндотелия [Lagarkova М.А. et al., 2010]. Эти исследования продемонстрировали универсальность процесса репрограммирования соматических клеток до плюрипотентного состояния с помощью введения в клетки генов транскрипционных факторов. Целый ряд подходов был разработан для того чтобы доставлять репрограммирующие факторы в соматические клетки. Были также продемонстрированы и другие методы репрограммирования соматических клеток до плюрипотентного состояния без интеграции экзогенной ДНК в геном клетки-хозяина: с использованием piggyBac транспозона [Woltjen K. et al., 2009], синтетической РНК, аденовирусных векторов [Zhou W., Freed C.R., 2009], вируса Сендай [Fusaki N. et al., 2009], эписомных векторов [Yu J. et al., 2009], белков [Kim D. et al., 2009]. Тесты на способность к дифференцировке считают ключевыми в доказательстве плюрипотентности клеток. Для ИПСК человека в качестве основных тестов используются in vitro дифференцировки, формирование тератом при введении иммунодефицитным мышам и способность к образованию эмбриоидных телец. Плюрипотентные стволовые клетки (ПСК) обладают естественным свойством дифференцироваться во все типы клеток взрослого организма, но важнейшей экспериментальной задачей является осуществление дифференцировки плюрипотентных клеток in vitro в нужном направлении с использованием специальных условий культивирования, биологически активных химических соединений и белковых факторов. Эффективный метод дифференцировки ИПСК в нейральном направлении основан на блокировании BMP и TGF-β сигнальных путей, такими молекулами как Noggin, дорсоморфин, SB431542 и LDN193189 на ранних стадиях дифференцировки [Fingar D.С, et al., 2004]. Такой подход при длительном культивировании позволил получить до 90% клеток, которые экспрессировали маркеры нейроэктодермы РАХ6 и PSA-NCAM (молекула адгезии нервных клеток). Для дальнейшей дифференцировки в нейрональные предшественники необходима активация Shh сигнального пути, а также добавление в среду для культивирования фактора роста фибробластов FGF2. Полученные таким образом предшественники экспрессируют маркер незрелых нейронов β-тубулин III. Рутинный способ получения ИПСК, а также существование известных методов дифференцировки ИПСК в нейральном направлении делают применение нейрональных предшественников наиболее перспективными кандидатами при терапии ИИ. Возможность эффективной экспансии ИПСК позволит решить проблему дефицита донорского материала и снять ряд этических вопрос. На сегодняшний день большое количество научных групп ведут разработки в области нейральной дифференцировке ИПСК. Основными лидерами по данному направлению являются институт генетических болезней и геномных наук, Токио, Япония (Department of Genetic Diseases and Genomic Science, The Jikei University School of Medicine, Tokyo, Japan); Центр исследования и применения ИПСК, Университет Киото, Япония (Centre for iPS Cell Research and Application, Kyoto University, Japan); Центр клеточной биологии, Китай (Department of Cell Biology, Xuanwu Hospital of Capital Medical University, Beijing, China). Данные лаборатории проводят передовые исследования по дифференцировке ИПСК, рационализируют и создают международно признанные протоколы дифференцировки ИПСК, более того проводятся экспериментальные разработки по введению дифференцированных нейральных ИПСК в органы-мишени. В этой связи стоит отметить Лабораторию Института Биоинженерии и нанотехнологии, Сингапур (Institute of Bioengineering and Nanotechnology, 31 Biopolis Way, The Nanos, Singapore 138669, Singapore), осуществляющую инновационные разработки в области дифференцировки ИПСК, и методов целенаправленной доставки в органы-мишени дифференцированных клеток, с использованием 3D микрофиброзных капсул, которые увеличивают эффективность доставки. Европейские лаборатории и лаборатории США, Канады занимают лидирующее положение по общему количеству проведенных исследовательских работ в области нейральной дифференцировки ИПСК. При изучении литературы часто встречаются современные разработки лаборатории Центра Генетики Университета шт. Висконсин совместно с институтом биомедицинских исследований, Кэмбридж (Waisman Center, Genome Center, Department of Anatomy, and Department of Neurology, School of Medicine and Public Health, University of Wisconsin, Madison. Whitehead Institute for Biomedical Research, Cambridge). В данных работах можно отметить передовые исследования по поиску новых регуляторных механизмов в области нейральной дифференцировки ИПСК.
Однако активность клеток зависит от условий окружающей среды и некоторые воздействия, в частности тепловой шок, гипоксия, наличие экзогенных факторов, введение новых генов или увеличение их копийности путем трансдукции или трансфекции, может влиять на синтетическую активность клеток, тем самым увеличивая или уменьшая их секреторную функцию [Mazzio ЕА, Soliman KF. 2012]. Гипоксия - пониженное содержание кислорода в организме или отдельных органах и тканях. На молекулярном уровне гипоксия вызывает высоко скоординированные клеточные реакции, направленные на обеспечение жизнеспособности клеток. Семенза и его коллеги обнаружили фактор, индуцируемый гипоксией, HIF-1, при изучении регуляции экспрессии гена эритропоэтина (ЕРО) во время гипоксического стресса [Ramakrishnan S, Anand V, Roy S. 2014]. HIF представляют собой семейство гетеродимерных белков, состоящих из альфа-субъединицы и бета-субъединицы. Альфа-субъединица экспрессируется во время гипоксии, тогда как бета-субъединица конститутивна. Бета-субъединица называется ядерным транслокатором арильных углеводородных рецепторов (ARNT). Последующие исследования показали, что HIF-1 связывается с консенсусной последовательностью NCGTG, называемой элементом ответа на гипоксию (HRE). HRE-сайты присутствуют в промоторе сотен генов, которые функционально связаны с гипоксической адаптацией. Например, HIF индуцирует экспрессию GLUT-1, переносчика глюкозы, который увеличивает внутриклеточные уровни гликолитического субстрата. В то же время многие ферменты необходимы для повышенного расщепления сахара, гексокиназы-1, глюкозо-6-фосфат-изомеразы, фосфофруктокиназы, PFK1, фермента, ограничивающего скорость гликолиза, фосфоглицерат-мутазы-1 (PGAM1), пируваткиназы типа М- 2 (PKM2), лактатдегидрогеназа (LDHA) увеличивается с помощью HIF-1-опосредованной трансактивации. [Kihira, Yoshitaka, et al. 2015]. В дополнение к поддержанию энергетического гомеостаза, HIF также непосредственно ответственен за увеличение производства ЕРО, фактора роста эндотелия сосудов (VEGF), необходимого для ангиогенеза. Три основных системы лиганд-рецептор активируются при гипоксическом стрессе, VEGF-VEGFR, Angiopoietins-Tie-1 / Tie-2 и Delta-Notch. Скоординированный выход этих сигнальных систем определяет ангиогенез, кровоток, перфузию тканей, экстравазацию воспалительных клеток, ремоделирование и восстановление тканей. [Pham, Isabelle, et al. 2002]. Гипоксия усиливает синтез и секрецию BDNF посредством механизмов мобилизации внутриклеточного кальция через инозитол 1,4,5-трифосфат (IP3) - и рианодин-чувствительные каналы [Hartman W, Helan М, Smelter D, et al. 2015]. Влияние гипоксии на секрецию других нейротрофинов и изменения состава секретируемых экзосом и цитокинов является активно исследуемым вопросом. Создание условий кислородного голодания возможно двумя основными путями - непосредственным изменением газового состава окружающий среды (смесь газов с содержанием кислорода от 1 до 2%.) с помощью герметичной камеры инкубатора или использования химического агента (наиболее часто используемым является хлоридом кобальта - CoCl2). Газовая камера и химические модели используются взаимозаменяемо. [Calvo-Anguiano G, Lugo-Trampe JJ, Camacho A, et al. 2018].
Основная причина, ограничивающая эффективность и безопасность всех существующих нейропептидных препаратов, полученных из головного мозга млекопитающих - это их ксеногенное происхождение. При выделении нейропептидов из тканей головного мозга животных для снижения их иммуногенности (антигенных свойств) прибегают к кислотному или ферментативному гидролизу белков и высокомолекулярных полипептидов, тем самым уменьшая количество нейротрофических факторов роста белковой и полипептидной природы [Хабриева. У., 2005]. Пептидный препарат, полученный таким способом, содержит денатурированные продукты кислотного гидролиза белков и полипептидов с нарушенной четвертичной и третичной структурой, что снижает его специфичность и биологическую активность, повышает его иммуногенность, вызывая аллергические реакции при применении.
Для решения данной проблемы нами предложено получить белково-пептидную композицию с выраженными ноотропными свойствами из секретома нейрональных прогениторных клеток в условиях гипоксии, полученных методом направленной дифференцировки индуцированных плюрипотентных стволовых клеток человека. Данная композиция характеризуется тем, что в ее состав входят белки и полипептиды с молекулярной массой от 5 кДа до 250 кДа, мозговой нейротрофический фактор (BDNF) не менее 130 пг/мл и глиального нейротрофического фактора (GDNF) не менее 30 пг/мл, фактор роста эндотелия сосудов (VEGF) не менее 90 пг/мл, суммарная масса белка не менее 1 мг/мл препарата, при этом клеточная культура нейрональных прогениторных клеток, из которой получают композицию, демонстрирует экспрессию нейронального β-тубулина III, молекулы адгезии нервных клеток (PSA-NCAM) и состоит из 95±5% TUBB3+ - клеток.
Наиболее важным направлением современной медицины и биологической науки является выделение из клеточных культур биологически активных веществ, прежде всего молекул, запускающих каскад процессов регенерации и паракринной индукции репаративных процессов. Применение охарактеризованного и стандартизованного комплекса биологически активных веществ может обладать той же эффективностью, но гораздо большей безопасностью по сравнению с живыми клеточными культурами. Возможность эффективной экспансии ИПСК и их дифференцировки в разных направлениях, в том числе нейрональном, решают проблему дефицита донорского материала и снимают ряд этических вопросов.
Изобретение осуществляется следующим образом:
Пример 1. Получение белково-пептидного комплекса.
Забор биоптата кожи (размером от 0,5 до 1 см2) производят за ушной раковиной или с внутренней стороны предплечья. После отсечения с помощью микрохирургического лезвия, биоптат помещают в стерильный флакон, содержащий транспортную среду (F12 с амикацином, 0,5 г/л). Флакон с содержимым маркируют следующими данными: ФИО донора, дата и время забора. Транспортировку производят при температуре 2-6°С.
Полученный материал в стерильных условиях переносят в чашки Петри и трижды промывают раствором Хенкса с цефазолином (1 г/л), затем дезагрегируют путем инкубации в растворе Версена с добавлением 0,25% трипсина. В течение 1-1,5 часов при 37°С. После инкубации суспензию интенсивно перемешивают и центрифугируют в течение 10 мин при 1400 об/мин. Осадок разводят культуральной средой (DMEM/F12 1:1 с добавлением 10% сыворотки, 500 мг/л амикацина), перемешивают и переносят в культуральную посуду. Культуральные флаконы помещают в CO2-инкубатор (37°С, 5% CO2). Через 2-3 дня осуществляют замену культуральной среды на свежую. Наращивание и пересевы осуществляют раствором Версена с добавлением 0,25% трипсина и переносят в новые чашки Петри. Коэффициент рассева выбирают в зависимости от индивидуальных пролиферативных характеристик фибробластов. Культивируют не более четырех пассажей. Перед программированием необходимо провести контроль стабильности кариотипа и способность фибробластов синтезировать коллаген I типа (с помощью иммунофлуоресцентного окрашивания).
При необходимости длительного хранения полученные фибробласты подвергают криоконсервации. В боксе на ледяной бане (при 4°С) готовят криозащитный раствор, состоящий из 10% ДМСО (диметилсульфоксид) и 90% аутогенной сыворотки. Клеточную культуру дезагрегируют раствором Версена с добавлением 0,25% трипсина, подсчитывают количество клеток с помощью счетчика и осаждают клетки центрифугированием (1400 об/мин, 10 мин). К осадку добавляют свежеприготовленный криозащитный раствор из расчета 1 мл на 5 млн. клеток, перемешивают и помещают полученную суспензию в криопробирки. Маркируют, указывая ФИО донора, номер пассажа, количество клеток, дату криозамораживания, данные ответственного исполнителя.
Образцы замораживают до - 80°С с помощью программного замораживателя, а затем помещают в сосуды Дьюара с жидким азотом и хранят при - 196°С.
Для разморозки криопробирки извлекают из сосуда Дьюара и размораживают при температуре 37°С на водяной бане. Клеточную суспензию из криопробирки переносят в центрифужную пробирку с культуральной средой, наполненной до десятикратного объема и центрифугируют при 1400 об/мин в течение 10 мин. Клеточный осадок переносят в культуральную посуду с полной ростовой средой.
Перед процедурой репрограммирования культуру фибробластов переводят на среду, не содержащую компоненты животного происхождения. Для этого, согласно протоколу фибробласты 1-2 пассажа промывают фосфатно-солевым буфером и открепляют раствором Трипсин/Версен 1:1 в течение 3-5 мин. Суспензию клеток центрифугируют 5 мин при 1100 об/мин, после чего отбирают супернатант, а к осадку добавляют среду DMEM с содержанием 10% эмбриональной телячьей сыворотки, 2 mM L-глутамина, пенициллина-стрептомицина 100 мг/л и рассаживают в количестве 1500-2000 клеток/см2 на чашки Петри диаметром 60 мм, покрытые коллагеном Coating Matrix Kit (Corning, США). Через 24 часа прикрепившиеся клетки промывают дважды фосфатно-солевым буфером и производили замену среды, состоящую из DMEM/F12, HEPES, натрия бикарбоната 4,8 г/мл, 10% заменителя сыворотки Knockout SR, фактора роста фибробластов человека bFGF 10 мкг/мл, эпидермального фактора роста человека EGF 1 мг/мл, 2 мкМ гидрокортизона. Для адаптации клеток к новой среде культивирование проводят в течение 1-2 пассажей.
Для репрограммирования фибробластов используют коммерческий набор для репрограммирования клеток человека «CTS CytoTune-iPS 2.1 Sendai Reprogramming Kit» (Invitrogen, США) на основе вируса Сендай. За день до репрограммирования фибробласты высевают в количестве 200 тысяч на лунку 6-луночного планшета, предварительно покрыв коллагеном, и культивировали в 2,5 мл среды без экзогенных компонентов. На следующий день добавляют вектора, несущие гены Oct4 и Sox2 (KOS), L-Myc (hL-Myc), Klf-4 (hKlf4) в количестве вирусных частиц на клетку (MOI) - 5, 5 и 3 соответственно. Через сутки производят замену среды для фибробластов без экзогенных компонентов, далее смена среды происходит ежедневно в течение 6 дней. На 7-ой день после репрограммирования клетки промывают дважды фосфатно-солевым буфером, открепляют их реагентом TrypLE Select и центрифугируют при 1100 об/мин в течение 5 мин. Полученный супернатант отбирают, а клеточный осадок пересаживают в количестве 100 тысяч на чашку Петри диаметром 60 мм, предварительно покрыв витронектином в концентрации 10 мкг/мл. Через 24 часа после посева производят замену среды на Essential 8 Medium для культивирования плюрипотентных клеток человека. Смену среды осуществляют каждый день.
Отбор образовавшихся колоний проводят по результатам прижизненного иммуноцитохимического окрашивания антителами к поверхностному маркеру плюрипотентных клеток - протеогликану TRA-1-60 с помощью коммерческого набора «TRA-1-60 Alexa Fluor 488 Conjugate Kit» (ThermoFisher Scientific, США). Колонии ИПСК, в которых визуализируется высокая экспрессия протеогликана TRA-1-60 отбирают механически наконечником, предварительно обработав раствором Версена, далее сажают на 6-ти луночные планшеты, покрытые витронектином, и культивировали в среде Essential 8 Medium. Дальнейшие пересевы проводят только с помощью раствора Версена и с добавлением в среду ROCK-ингибитора Y27632 в концентрации 5 мкМ в течение суток.
При культивировании ИПСК ведется визуальный контроль за морфологическими характеристиками культур. ИПСК растут плотными колониями с ровными краями. Клетки мелкие, с крупными ядрышками. Культивирование осуществляют в чашках Петри диаметром 35 мм, используя культуральную среду Essential 8™ (Е8).
Клетки рассаживают по достижении плотности 50-70% монослоя. Из чашек удаляют культуральную среду и промывают 2 мл DPBS. Для дезагрегации добавляют 1 мл раствора Версена и инкубируют 5-8 мин при комнатной температуре. Затем удаляют раствор Версена, добавляют 2 мл среды, взбалтывают и переносят клеточную суспензию на чашки, предварительно покрытые витронектином, и добавляют среды до 2 мл. Замену среды на свежую проводят каждый день.
Стандартизацию ИПСК проводят по экспрессии генов, специфичных для эмбриональных стволовых клеток ОСТ4, SOX2, NANOG, SSEA4, TRA-1-81, TRA-1-60 иммуноцихимическим методом и ПЦР с обратной транскрипцией и.
ИПСК подвергают криоконсервации. Готовят на льду криозащитный раствор из расчета на 1 чашку: 900 мкл среды Е8, 100 мкл DMSO и 1 мкл ингибитора Rho-киназы (сток 5 мМ). Из каждой чашки удаляют среду и промывают 2 мл DPBS. Добавляют 1 мл Версена и инкубируют 5-8 мин при комнатной температуре. Затем удаляют раствор Версена и добавляют 1 мл криозащитного раствора. Клетки снимают с поверхности чашки, перемешивают и переносят в криопробирку объемом 1,8 мл (1 мл на 1 пробирку). Образцы замораживают до - 80°С с помощью программного замораживателя, а затем помещают в сосуды Дьюара с жидким азотом и хранят при - 196°С.
Криопробирки с клеточным материалом извлекают из сосуда Дьюара и размораживают при температуре 37°С на водяной бане.
Клеточную суспензию из криопробирки (1 мл) по каплям переносят в центрифужную пробирку, доливают 9 мл теплой культуральной среды DMEM/F12 до десятикратного объема и центрифугируют при 800 об/мин в течение 5 мин.
Отмытые от криоконсерванта клетки переносят на чашки Петри, предварительно покрытые витронектином, с культуральной средой Е8, содержащей ингибитор Rho-киназы. При переносе сохряняют целостность колоний. Адаптация клеток после криоконсервации продолжается от 1 до 3 недель.
Для дифференцировки в нейрональном направлении ИПСК обрабатывают раствором Версена, центрифугировали при 800 об/мин в течение 5 мин. Супернатант удаляют, а клеточный осадок переносят в новые чашки Петри диаметром 60 мм, покрытые поли-1-орнитином 20 мкг/мл (Sigma-Aldrich, США) и ламинином 2,5 мкг/мл (Gibco, США). По достижению культуры 80% конфлюэнтности среду Essential 8 Medium меняют на DMEM/F12 с 1% N2, 2 мМ глутамина, пенициллин-стрептомицина 100 мг/л, 10 мкМ SB431542, 2 мкМ дорсоморфина. Среду меняют через день. Через 10-14 дней в культуре начинают появляться розеткоподобные структуры. После, полученную культуру пересевают на чашки Петри, покрытые поли-1-орнитином 20 мкг/мл и ламинином 2,5 мкг/мл, в плотности 250-400 тысяч клеток/см2 и культивировали в среде DMEM/F12 (Панэко, РФ), 2% добавки В27 (Gibco, США), 2 мМ глутамина (Панэко, РФ), пенициллин-стрептомицина 100 мг/л (Панэко, РФ), 10 нг/мл FGF-2 (ProSpec, Великобритания), 1 мкМ пурморфамина (Stemcell Technologies, США) в течение 14 суток, производя замену среды каждые 2 суток.
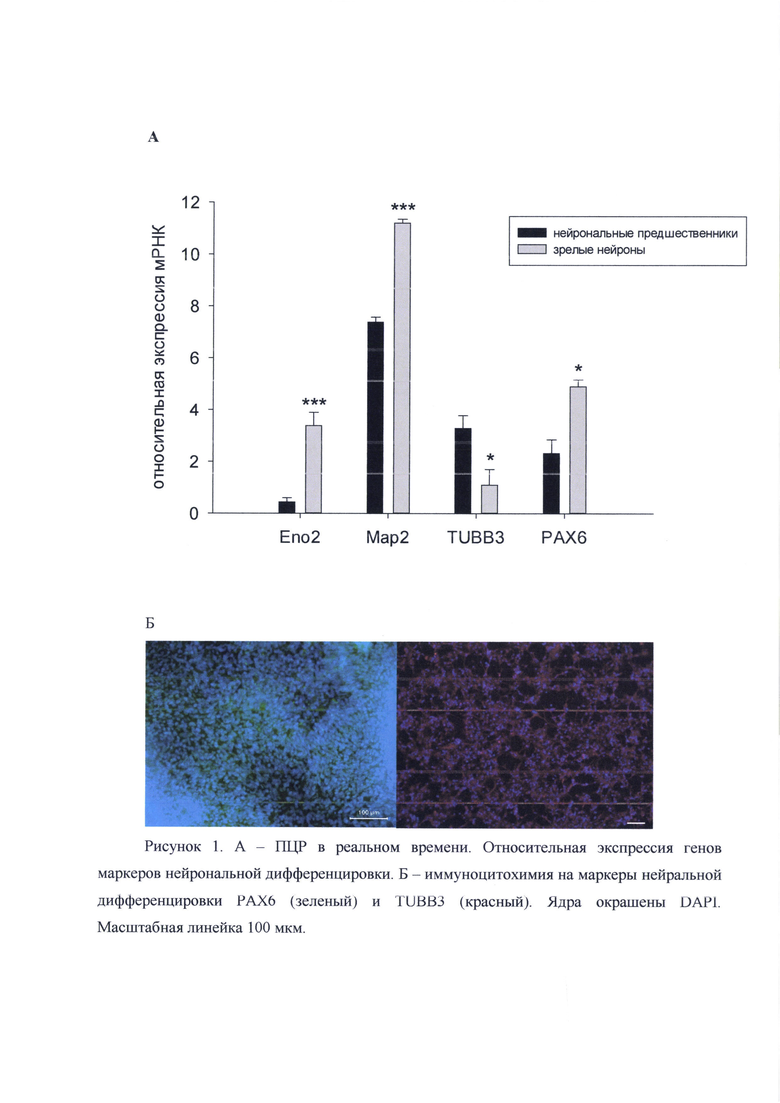
Стандартизацию нейральных клеток проводили по экспрессии генов РАХ6, FOXP2, NCAM1, ENO2, Nestin, TUBB3, SOX2.
Для моделирования гипоксии нейрональные предшественники культивируют в течение 3 часов в газовой камере с содержанием кислорода 2%, после чего производят замену среды на DMEM/F12 и культивируют в течение 6 часов. После чего кондиционированную среду собирают и центрифугируют при 300 об/мин в течение 5 мин. Супернатант отбирают и концентрируют в 24 раза с помощью 3 кДа мембран Amicon Ultra (Sigma-Aldrich, США). Полученный концентрат подвергают стерилизующей фильтрации. Композиция включает в себя белки и пептиды молекулярной массой в пределах от 3 до 250 кДа.
Пример 2. Характеристика нейрональных прогениторных клеток.
Для подтверждения фенотипа, полученных культур нейрональных прогениторных клеток проводили иммуноцитохимическое окрашивание на нейральные маркеры. Клетки фиксировали 4% раствором формальдегида (Merck KGaA, Германия) 10 мин при комнатной температуре, затем промывали фосфатно-солевым буфером, инкубировали в фосфатно-солевом буфере с 0,25% Triton Х-100 (Sigma-Aldrich, США) и 1% бычьим сывороточным альбумином (AppliChem, Германия) 30 мин, а затем обрабатывали первичными антителами ночь при +4°С. Для нейральных стволовых клеток использовались антитела к белкам Pax6, Tubulin-IIIβ, Nestin и SOX2. Затем клетки промывали фосфатно-солевым буфером и инкубировали с вторичными антителами, меченными флуорохромами Alexa Fluor 488 anti-rabbit, Alexa Fluor 555 anti-mouse (Invitrogen, США) в темноте 60 мин. Ядра красили раствором DAPI (4,6-диамино-2-фенилиндол дигидрохлорид) 1 мкг/мл (Sigma-Aldrich, США) в фосфатно-солевом буфере. Для анализа использовали люминесцентный инвертированный микроскоп Axio Observer.D1 с камерой AxioCam HRc (Carl Zeiss, Германия).
Количественную оценку проводят с помощью проточной цитометрии. Культуру снимали с помощью раствора Версена (Панэко, РФ) в случае НП или 0,05% раствора трипсина (Панэко, РФ) в случае ГП, центрифугируют 1600 об/мин в течение 5 мин, супернатант отбирают, а осадок промывают раствором Хенкса. После осаждения клетки 10 мин фиксируют 4%-ым раствором параформальдегида, затем клетки промывают фосфатно-солевым раствором. Пермеабилизацию проводят 70% метанолом на льду в течение 10 мин, после чего клетки промывают дважды в фосфатно-солевом растворе и осаждали центрифугированием 1800 об/мин 5 мин. Далее клетки окрашивают первичными антителами против β3-тубулина (разведение 1:500, Abcam), инкубация проходила при +4°С в течение 12 часов. Затем промывали фосфатно-солевым буфером, центрифугируют 1800 об/мин 5 мин и инкубируют с вторичными антителами, меченными флюорохромом Alexa Fluor 555 anti-mouse (разведение 1:600, Invitrogen, США) в темноте 60 мин. Полученную суспензию анализируют, используя проточный цитофлюориметр CyFlow ML, а количество TUBB3+ - клеток считали с помощью программного обеспечения FloMax. По данным проточной цитометрии культуры нейрональных прогениторных клеток должна состоять не менее 99,5±2,45% TUBB3+ - клеток.
Для анализа экспрессии нейрональных генов маркеров проводили ПЦР. мРНК выделяли с помощью коммерческого набора «RNeasy Mini Kit» согласно инструкции производителя. Далее проводили синтез кДНК набором «RevertAid™ First Strand cDNA Synthesis Kit» по протоколу, приложенному в инструкции. Для ПЦР в реальном времени использовали готовую реакционную смесь с интеркалирующим красителем Sybr Green, амплификацию ставили в термоциклере BioRad iQ cycler. Схема реакции: первичная денатурация 95°С, 5 мин.; денатурация 95°С, 20 сек.; отжиг праймеров 55-63°С, 20 сек.; элонгация 72°С, 20 секунд (40 циклов). Уровень мРНК анализируемых генов выравнивали по отношению к усредненным данным амплификации двух генов домашнего хозяйства GAPDH (глицеральдегид-3-фосфат дегидрогеназа) и ActB (β-актин). Относительное количество мРНК рассчитывали с помощью ΔС (Т) метода.
В результате дифференцировки нейрональные прогениторные клетки демонстрировали экспрессию маркерных генов PAX6, TUBB3, МАР2, ENO2, Nestin (рисунок 1) как на уровне транскрипции, так и трансляции. Анализ показал, что до 95% полученных клеток экспрессировали нейрональный маркер TUBB3 (рис. 2).
Пример 3. Характеристика ноотропной композиции
Композицию характеризовали по количеству в 1 мл общего белка и таких факторов, как мозговой нейротрофический фактор (Brain-Derived Neurotrophic Factor, BDNF), глиальный нейротрофический фактор (GDNF), фактор роста эндотелия сосудов (VEGF). Для этого использовали твердофазный иммуноферментный анализ. 1 мл суспензии центрифугировали, полученные в результате центрифугирования супернатанты апплицировали в лунки планшета с предварительно адсорбированными первичными антителами к BDNF, GDNF, VEGF, далее анализ проводили согласно инструкции производителя. Содержание факторов выражали в пг/мл композиции. Всего было исследовано 10 образцов композиции, получены результаты по содержанию BDNF не менее 130 пг/мл, GDNF не менее 30 пг/мл, VEGF не менее 90 пг/мл. Суммарная масса белка не менее 1 мг/мл препарата. Общий белок в 1 мл суспензии определяли методом Бредфорда.
Пример 4. Оценка эффективности использования композиции на in vitro модели глутаматной токсичности на клеточной линии РС-12.
Для изучения нейротрофического и нейропротекторного действия ноотропных композиций на основе среды НП, полученных в стандартных условиях (1) и подвергнутых гипоксии (2) была, выбрана линия феохромоцитомы крысы РС-12, широко используемая в качестве in vitro модели в нейробиологии. Клетки линии РС-12 (АТСС, США) культивировали в среде DMEM/F12 с добавлением 10% фетальной бычьей сыворотки (ФБС). За 12 часов до эксперимента клетки пересевали в 48-луночные планшеты, покрытые раствором полли-DL-лизина в количестве 20 тысяч клеток на лунку и затем инкубировали в среде Opti-MEM, содержащей 0,5% ФБС. Далее добавляли БПК 1 и БПК 2 (5% от объема лунки, раствор с концентрацией общего белка 0.5 мг/мл), и инкубировали в течение 6 дней, меняя среду каждые 2 дня. Для оценки нейритогенеза рассчитывали среднее значение длины нейритов с использованием программы ImageJ.
В работе использовали модель глутаматной токсичности, воспроизводящий условия повышенного содержания активных форм кислорода. Клетки феохромоцитомы РС-12 культивировали в 96 - луночных планшетах. Для изучения протекторного действия ноотропных композиции (1, 2), производили их добавление в среду культивирование (5% от объема лунки, раствор с концентрацией общего белка 0.5 мг/мл) за 24 часа до моделирования глутаматной токсичности (добавляли глутамат до концентрации 30 мМ).
Для определения жизнеспособности клеточной линии SHSY-5Y спустя 24 часа сокультивирования добавляли МТТ (3-[4,5диметилтиазол-2-ил]-2,5-дифенил-тетразолийбромид, ПанЭко, РФ) в концентрации 0,5 мг/мл и инкубировали в темноте при 37°С в течение 2 часов с последующим растворением кристаллов формазана ДМСО и измерением оптической плотности при 590 нм с помощью планшетного ридера. Все МТТ анализы были выполнены в трех повторах.
В результате инкубация с ноотропной композицией, полученной в стандартных условиях(1) увеличивает число живых клеток РС-12 на 9,58±3,2% по сравнению с контролем. Добавление ноотропной композиции, полученной с применением моделирования гипоксии (2) число живых клеток РС-12 на 16,58±3,2%, при этом оказывая более выраженное действие на стимулирование роста нейритов, средняя длина нейритов больше в 1,47 раза больше в сравнении с группы 1.
Список использованных источников
1. Song CG, Zhang YZ, Wu HN, et al. Stem cells: a promising candidate to treat neurological disorders. Neural Regen Res. 2018; 13(7):1294-1304. doi: 10.4103/1673-5374.235085.
2. Takahashi, Kazutoshi, and Shinya Yamanaka. "Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors." cell 126.4 (2006): 663-676.
3. Zakrzewski W, Szymonowicz M, Rybak Z. Stem cells: past, present, and future. Stem Cell Res Ther. 2019; 10(1):68. Published 2019 Feb 26. doi: 10.1186/s 13287-019-1165-5.
4. Kwak KA, Lee SP, Yang JY, Park YS. Current Perspectives regarding Stem Cell-Based Therapy for Alzheimer's Disease. Stem Cells Int. 2018; 2018:6392986. Published 2018 Mar 1. doi:10.1155/2018/6392986.
5. Neural Stem Cell-Conditioned Medium Protects Neurons and Promotes Propriospinal Neurons Relay Neural Circuit Reconnection After Spinal Cord Injury. Peng Liang, Jiaren Liu, Jinsheng Xiong, Qing Liu, Jiaxin Zhao, Hongsheng Liang, Liwei Zhao, and Haitao Tang. Cell Transplantation, Vol.23, Supplement 1, pp. S45-S56, 2014.
6. Cerebrolysin for vascular dementia. Chen N, Yang M, Guo J, Zhou M, Zhu C, He L. Cochrane Database Syst Rev. 2013 Jan 31.
7. Takahashi K., Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors // cell. - 2006. - T. 126. - №. 4. - C. 663-676.
8. Takahashi K. et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors // cell. - 2007. - T. 131. - №. 5. - C. 861-872.
9. Yu J. et al. Induced pluripotent stem cell lines derived from human somatic cells // science. - 2007. - T. 318. - №5858. - C. 1917-1920.
10. Liu H. et al. Generation of induced pluripotent stem cells from adult rhesus monkey fibroblasts // Cell stem cell. - 2008. - T. 3. - №6. - C. 587-590.
11. West F.D. et al. Porcine induced pluripotent stem cells produce chimeric offspring // Stem cells and development. - 2010. - T. 19. - №8. - C. 1211-1220.
12. Aasen T. et al. Efficient and rapid generation of induced pluripotent stem cells from human keratinocytes // Nature biotechnology. - 2008. - T. 26. - №11. - C. 1276.
13. Eminli S. et al. Reprogramming of neural progenitor cells into induced pluripotent stem cells in the absence of exogenous Sox2 expression // Stem cells. - 2008. - T. 26. - №10. - C. 2467-2474.
14. Shi Y. et al. Induction of pluripotent stem cells from mouse embryonic fibroblasts by Oct4 and Klf4 with small-molecule compounds // Cell stem cell. - 2008. - T. 3. - №5. - C. 568-574.
15. Lagarkova M.A. et al. Induction of pluripotency in human endothelial cells resets epigenetic profile on genome scale // Cell cycle. - 2010. - T. 9. - №5. - C. 937-946.
16. Woltjen K. et al. piggyBac transposition reprograms fibroblasts to induced pluripotent stem cells // Nature. - 2009. - T. 458. - №7239. - C. 766.
17. Zhou W., Freed C.R. Adenoviral gene delivery can reprogram human fibroblasts to induced pluripotent stem cells // Stem cells. - 2009. - T. 27. - №11. - C. 2667-2674.
18. Fusaki N. et al. Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai virus, an RNA virus that does not integrate into the host genome // Proceedings of the Japan Academy, Series B. - 2009. - T. 85. - №8. - C. 348-362.
19. Yu J. et al. Human induced pluripotent stem cells free of vector and transgene sequences // Science. - 2009. - T. 324. - №5928. - C. 797-801.
20. Zhou H. et al. Generation of induced pluripotent stem cells using recombinant proteins // Cell stem cell. - 2009. - T. 4. - №. 5. - C. 381-384.
21. Mazzio EA, Soliman KF. Basic concepts of epigenetics: impact of environmental signals on gene expression. Epigenetics. 2012;7(2): 119-130. doi:10.4161/epi.7.2.18764.
22. Ramakrishnan S, Anand V, Roy S. Vascular endothelial growth factor signaling in hypoxia and inflammation. J Neuroimmune Pharmacol. 2014;9(2):142-160. doi:10.1007/s11481-014-9531-7.
23. Kihira, Yoshitaka, et al. "Hypoxia decreases glucagon-like peptide-1 secretion from the GLUTag cell line." Biological and Pharmaceutical Bulletin 38.4 (2015): 514-521.
24. Pham, Isabelle, et al. "Hypoxia upregulates VEGF expression in alveolar epithelial cells in vitro and in vivo." American Journal of Physiology-Lung Cellular and Molecular Physiology 283.5 (2002): L1133-L1142.
25. Hartman W, Helan M, Smelter D, et al. Role of Hypoxia-Induced Brain Derived Neurotrophic Factor in Human Pulmonary Artery Smooth Muscle. PLoS One. 2015; 10(7):e0129489. Published 2015 Jul 20. doi: 10.1371/journal.pone.0129489.
26. Calvo-Anguiano G, Lugo-Trampe JJ, Camacho A, et al. Comparison of specific expression profile in two in vitro hypoxia models. Exp Ther Med. 2018;15(6):4777-4784. doi:10.3892/etm.2018.6048.
27. Хабриев P.У. и др. Руководство по экспериментальному (доклиническому) изучению новых фармакологических веществ. - Открытое акционерное общество Издательство Медицина, 2005.
28. Song CG, Zhang YZ, Wu HN, et al. Stem cells: a promising candidate to treat neurological disorders. Neural Regen Res. 2018; 13(7): 1294-1304. doi:10.4103/1673-5374.235085.
29. Takahashi, Kazutoshi, and Shinya Yamanaka. "Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors." cell 126.4 (2006): 663-676.
30. Zakrzewski W, Szymonowicz M, Rybak Z. Stem cells: past, present, and future. Stem Cell Res Ther. 2019;10(1):68. Published 2019 Feb 26. doi: 10.1186/s13287-019-1165-5.
31. Kwak KA, Lee SP, Yang JY, Park YS. Current Perspectives regarding Stem Cell-Based Therapy for Alzheimer's Disease. Stem Cells Int. 2018; 2018:6392986. Published 2018 Mar 1. doi:10.1155/2018/6392986.
32. Neural Stem Cell-Conditioned Medium Protects Neurons and Promotes Propriospinal Neurons Relay Neural Circuit Reconnection After Spinal Cord Injury. Peng Liang, Jiaren Liu, Jinsheng Xiong, Qing Liu, Jiaxin Zhao, Hongsheng Liang, Liwei Zhao, and Haitao Tang. Cell Transplantation, Vol. 23, Supplement 1, pp. S45-S56, 2014.
33. Cerebrolysin for vascular dementia. Chen N, Yang M, Guo J, Zhou M, Zhu C, He L. Cochrane Database Syst Rev. 2013 Jan 31.
34. Takahashi K., Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors //cell. - 2006. - T. 126. - №4. - C. 663-676.
35. Takahashi K. et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors // cell. - 2007. - T. 131. - №5. - C. 861-872.
36. Yu J. et al. Induced pluripotent stem cell lines derived from human somatic cells // science. - 2007. - T. 318. - №5858. - C. 1917-1920.
37. Liu H. et al. Generation of induced pluripotent stem cells from adult rhesus monkey fibroblasts // Cell stem cell. - 2008. - T. 3. - №6. - C. 587-590.
38. West F.D. et al. Porcine induced pluripotent stem cells produce chimeric offspring // Stem cells and development. - 2010. - T. 19. - №8. - C. 1211-1220.
39. Aasen T. et al. Efficient and rapid generation of induced pluripotent stem cells from human keratinocytes // Nature biotechnology. - 2008. - T. 26. - №11. - C. 1276.
40. Eminli S. et al. Reprogramming of neural progenitor cells into induced pluripotent stem cells in the absence of exogenous Sox2 expression // Stem cells. - 2008. - T. 26. - №10. - C. 2467-2474.
41. Shi Y. et al. Induction of pluripotent stem cells from mouse embryonic fibroblasts by Oct4 and Klf4 with small-molecule compounds //Cell stem cell. - 2008. - T. 3. - №5. - C. 568-574.
42. Lagarkova M.A. et al. Induction of pluripotency in human endothelial cells resets epigenetic profile on genome scale // Cell cycle. - 2010. - T. 9. - №5. - C. 937-946.
43. Woltjen K. et al. piggyBac transposition reprograms fibroblasts to induced pluripotent stem cells // Nature. - 2009. - T. 458. - №7239. - C. 766.
44. Zhou W., Freed C.R. Adenoviral gene delivery can reprogram human fibroblasts to induced pluripotent stem cells // Stem cells. - 2009. - T. 27. - №11. - C. 2667-2674.
45. Fusaki N. et al. Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai virus, an RNA virus that does not integrate into the host genome // Proceedings of the Japan Academy, Series B. - 2009. - T. 85. - №8. - C. 348-362.
46. Yu J. et al. Human induced pluripotent stem cells free of vector and transgene sequences // Science. - 2009. - T. 324. - №5928. - C. 797-801.
47. Zhou H. et al. Generation of induced pluripotent stem cells using recombinant proteins // Cell stem cell. - 2009. - T. 4. - №5. - C. 381-384.
48. Mazzio EA, Soliman KF. Basic concepts of epigenetics: impact of environmental signals on gene expression. Epigenetics. 2012; 7(2): 119-130. doi: 10.416l/epi.7.2.18764.
49. Ramakrishnan S, Anand V, Roy S. Vascular endothelial growth factor signaling in hypoxia and inflammation. J Neuroimmune Pharmacol. 2014; 9(2): 142-160. doi: 10.1007/s11481-014-9531 -7.
50. Kihira, Yoshitaka, et al. "Hypoxia decreases glucagon-like peptide-1 secretion from the GLUTag cell line." Biological and Pharmaceutical Bulletin 38.4 (2015): 514-521.
51. Pham, Isabelle, et al. "Hypoxia upregulates VEGF expression in alveolar epithelial cells in vitro and in vivo." American Journal of Physiology-Lung Cellular and Molecular Physiology 283.5 (2002): L1133-L1142.
52. Hartman W, Helan M, Smelter D, et al. Role of Hypoxia-Induced Brain Derived Neurotrophic Factor in Human Pulmonary Artery Smooth Muscle. PLoS One. 2015; 10(7):e0129489. Published 2015 Jul 20. doi: 10.1371/jouraal.pone.0129489.
53. Calvo-Anguiano G, Lugo-Trampe JJ, Camacho A, et al. Comparison of specific expression profile in two in vitro hypoxia models. Exp Ther Med. 2018; 15(6):4777-4784. doi:10.3892/etm.2018.6048.
54. Хабриев P.У. и др. Руководство по экспериментальному (доклиническому) изучению новых фармакологических веществ. - Открытое акционерное общество Издательство Медицина, 2005.
| название | год | авторы | номер документа |
|---|---|---|---|
| Ноотропная композиция на основе полипептидных комплексов, выделенных из нейрональных прогениторных клеток в условиях теплового шока, и способ ее получения | 2019 |
|
RU2752906C2 |
| Ноотропная композиция на основе полипептидных комплексов, выделенных из глиальных прогениторных клеток в условиях теплового шока, и способ её получения | 2019 |
|
RU2732599C1 |
| УЛУЧШЕННЫЙ СПОСОБ ДИФФЕРЕНЦИРОВКИ ИНДУЦИРОВАННЫХ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА В ДОФАМИНЭРГИЧЕСКИЕ НЕЙРОНЫ | 2023 |
|
RU2823290C1 |
| Способ получения глиальных производных индуцированных плюрипотентных стволовых клеток с повышенной экспрессией NGF для терапии нейродегенеративных заболеваний | 2023 |
|
RU2829363C1 |
| УЛУЧШЕННЫЙ СПОСОБ ПОЛУЧЕНИЯ МОЗГОВЫХ ОРГАНОИДОВ | 2022 |
|
RU2784072C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pAd-SM, КОДИРУЮЩАЯ БЕЛКИ SOX2 И C-MYC ЧЕЛОВЕКА, ЯВЛЯЮЩАЯСЯ ОСНОВОЙ ДЛЯ ПОЛУЧЕНИЯ ВИРУЛЕНТНЫХ АДЕНОВИРУСОВ, ПРЕДНАЗНАЧЕННЫХ ДЛЯ ПОЛУЧЕНИЯ ИНДУЦИРОВАННЫХ ПЛЮРИПОТЕНТНЫХ КЛЕТОК ЧЕЛОВЕКА | 2012 |
|
RU2495127C2 |
| СПОСОБ ПОЛУЧЕНИЯ ХРЯЩЕПОДОБНЫХ СТРУКТУР ИЗ СТВОЛОВЫХ КЛЕТОК С ИНДУЦИРОВАННОЙ ПЛЮРИПОТЕНТНОСТЬЮ И СТРУКТУРЫ, ПОЛУЧЕННЫЕ УКАЗАННЫМ СПОСОБОМ | 2023 |
|
RU2814248C1 |
| СПОСОБ ПОЛУЧЕНИЯ ПАЦИЕНТ (ДОНОР)-СПЕЦИФИЧЕСКИХ ФИБРОБЛАСТОПОДОБНЫХ КЛЕТОК ИЗ ИНДУЦИРОВАННЫХ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА | 2014 |
|
RU2568059C1 |
| КОНВЕРСИЯ СОМАТИЧЕСКИХ КЛЕТОК В ИНДУЦИРОВАННЫЕ РЕПРОГРАМИРОВАННЫЕ НЕЙРОНАЛЬНЫЕ СТВОЛОВЫЕ КЛЕТКИ (ИРНСК) | 2011 |
|
RU2562111C2 |
| СПОСОБ ОЦЕНКИ МОРФОФУНКЦИОНАЛЬНОГО СОСТОЯНИЯ ДИФЕРЕНЦИРОВАННЫХ В ДОФАМИНЕРГИЧЕСКИЕ НЕЙРОНЫ ИНДУЦИРОВАННЫХ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК БОЛЬНЫХ ПАРКИНСОНИЗМОМ | 2012 |
|
RU2501853C1 |

Изобретение относится к области биохимии, в частности к способу получения ноотропной композиции на основе полипептидных комплексов, выделенных из нейрональных прогениторных клеток в условиях гипоксии, а также к ноотропной композиции на основе полипептидных комплексов, выделенных из нейрональных прогениторных клеток в условиях гипоксии. Указанная композиция характеризуется тем, что в ее состав входят белки и полипептиды с молекулярной массой от 5 кДа до 250 кДа, мозговой нейротрофический фактор (BDNF) не менее 130 пг/мл и глиального нейротрофического фактора (GDNF) не менее 30 пг/мл, фактор роста эндотелия сосудов (VEGF) не менее 90 пг/мл, суммарная масса белка не менее 1 мг/мл препарата. При этом клеточная культура нейрональных прогениторных клеток, из которой получают композицию, демонстрирует экспрессию нейронального β - тубулина III, молекулы адгезии нервных клеток (PSA-NCAM) и состоит из 95±5% TUBB3+ - клеток. Изобретение позволяет эффективно получать белково-пептидную композицию, в которую включены не только небольшие пептиды, но и полноразмерные белковые нейрональные ростовые и дифференцировочные факторы, эффективность действия которых заведомо выше, чем у коротких пептидов, а иммуногенность отсутствует. 2 н.п. ф-лы, 4 пр., 2 ил.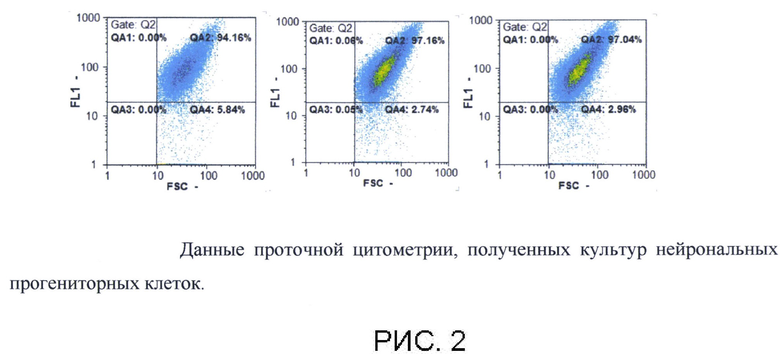
1. Способ получения ноотропной композиции на основе полипептидных комплексов, выделенных из нейрональных прогениторных клеток в условиях гипоксии, включающий: забор биоптата кожи, получение культуры фибробластов, криоконсервацию культуры фибробластов, размораживание культуры фибробластов, адаптирование культуры фибробластов к xeno free условиям культивирования, репрограммирование фибробластов для получения индуцированных плюрипотентных стволовых клеток (ИПСК), культивирование ИПСК, криоконсервацию ИПСК, размораживание ИПСК, дифференцировку ИПСК в глиальные прогениторные клетки, культивирование нейрональных прогениторных клеток в условиях гипоксии, сбор кондиционированной среды и получение белково-пептидного комплекса,
где для моделирования гипоксии нейрональные прогениторные клетки культивируют в течение 3 ч в газовой камере с содержанием кислорода 2%, после чего производят замену среды на DMEM/F12 и культивируют в течение 6 ч, после чего кондиционированную среду собирают и центрифугируют при 300 об/мин в течение 5 мин, супернатант отбирают и концентрируют в 24 раза с помощью 3 кДа мембран Amicon Ultra, полученный концентрат подвергают стерилизующей фильтрации и получают ноотропную композицию.
2. Ноотропная композиция на основе полипептидных комплексов, выделенных из нейрональных прогениторных клеток в условиях гипоксии, характеризующаяся тем, что в ее состав входят белки и полипептиды с молекулярной массой от 5 кДа до 250 кДа, мозговой нейротрофический фактор (BDNF) не менее 130 пг/мл и глиального нейротрофического фактора (GDNF) не менее 30 пг/мл, фактор роста эндотелия сосудов (VEGF) не менее 90 пг/мл, суммарная масса белка не менее 1 мг/мл препарата, при этом клеточная культура нейрональных прогениторных клеток, из которой получают композицию, демонстрирует экспрессию нейронального β - тубулина III, молекулы адгезии нервных клеток (PSA-NCAM) и состоит из 95±5% TUBB3+ - клеток.
| Способ получения ноотропной композиции на основе полипептидных комплексов, выделенных из нейронов и глиальных клеток, полученных методом направленной дифференцировки индуцированных плюрипотентных стволовых клеток человека | 2018 |
|
RU2690498C1 |
| Ноотропная композиция на основе полипептидных комплексов, выделенных из нейронов и глиальных клеток, полученных методом направленной дифференцировки индуцированных плюрипотентных стволовых клеток человека | 2018 |
|
RU2690846C1 |
| WO2001088104 A2, 16.05.2001 | |||
| US6455277 B1, 24.09.2002. | |||

