Область техники, к которой относится настоящее изобретение
Не относящиеся к человеку животные, которые генетически сконструированы в локусе (или в трансгене) вариабельной области (V) тяжелой цепи иммуноглобулина для получения антител из ограниченного ряда вариабельных сегментов (VH) (или одного сегмента VH) тяжелой цепи иммуноглобулинов и/или их вариантов. Не относящиеся к человеку животные, которые содержат вариабельный домен тяжелой цепи человека, полученный из единственного вариабельного генного сегмента тяжелой цепи иммуноглобулина, например, генного сегмента VH1-69 иммуноглобулина человека или генного сегмента VH1-2 человека. Способы получения последовательностей антител у не относящихся к человеку животных, которые применимы для связывания патогенов, включая в себя патогены человека.
Предшествующий уровень техники настоящего изобретения
Не относящиеся к человеку животные, например мыши, были генетически модифицированы, чтобы быть пригодными инструментами в способах получения последовательностей антител для использования в терапевтических средствах для людей на основе антител. Мышей с гуманизированными локусами вариабельной области (например, VH, DH и JH гены, и VL и JL гены) используют для создания родственных вариабельных доменов тяжелой и легкой цепи для использования в терапевтических средствах на основе антител. Доступны другие мыши, которые продуцируют полностью человеческие антитела с родственными тяжелыми и легкими цепями.
Терапевтические средства на основе антител для лечения человека разрабатывают на основе требуемых характеристик в отношении определенных заранее выбранных антигенов. Гуманизированных мышей иммунизируют предварительно выбранными антигенами, и иммунизированных мышей используют для получения популяций антител, из которых идентифицируют высокоаффинные родственные тяжелые и легкие вариабельные домены с требуемыми характеристиками связывания. Некоторые гуманизированные мыши, такие как те, у которых гуманизированы только вариабельные области в эндогенных локусах мыши, продуцируют популяции В-клеток, которые схожи по характеру и количеству с популяциями В-клеток мыши дикого типа. В результате чрезвычайно большая и разнородная популяция В-клеток доступна у тех мышей, у которых подвергают скринингу антитела, отражая большое количество различных реаранжировок иммуноглобулина для определения тяжелых и легких вариабельных доменов с наиболее необходимыми характеристиками.
Но не все антигены вызывают иммунный ответ, который проявляет очень большое количество реаранжировок из широкого выбора вариабельных сегментов (V). Другими словами, гуморальный иммунный ответ человека на определенные антигены практически ограничен. Ограничение отражается в клональной селекции В-клеток, которые экспрессируют только определенные сегменты V, связывающие определенный антиген с достаточно высокой аффинностью и специфичностью. Некоторые такие антигены клинически значимы, т.е. ряд из них представляет собой хорошо известные патогены человека. Возникает предположение, что сегмент V, экспрессированный при иммунном ответе человека, представляет собой сегмент V, который в сочетании с человеческим сегментом D и J более вероятно продуцирует применимое высоаффинное антитело, чем случайно выбранный сегмент V, который не наблюдался в ответе антитела человека на этот антиген.
Предполагается, что естественный отбор в течение тысячелетий выбрал наиболее эффективную основу или основание, из которого разрабатывается наиболее эффективное оружие для нейтрализации человеческих патогенов - клонально выбранный сегмент V. В настоящей области техники существует потребность в увеличении и улучшении антител, которые связываются и/или нейтрализуют такие антигены, как рассмотренные выше патогены. Существует потребность в более быстром получении применимых последовательностей из определенных сегментов V, включающих полиморфные и/или соматически мутированные определенные V сегменты, и в более быстром получении применимых популяций В-клеток, содержащих реаранжировки V сегментов с различными сегментами D и J, в том числе соматически мутированными их модификациями, и в частности, реаранжировки с уникальными и применимыми CDR3. Существует потребность в биологических системах, например, не относящихся к человеку животных (таких как, например, мыши, крысы, кролики и т.д.), которые могут продуцировать терапевтически применимые последовательности вариабельной области антитела из предварительно выбранных сегментов V в увеличенном количестве и разнообразии, чем, например, может быть получено у существующих модифицированных животных. Существует потребность в конструкциях биологических систем, содержащих коммитированную гуморальную иммунную систему для клонального выбора вариабельных последовательностей антител, полученных из ограниченных, заранее выбранных сегментов V, включая без ограничения родственные вариабельные области тяжелой и легкой цепи человека, применимые в производстве терапевтических средств на основе антител человека против определенных антигенов, включая определенные патогены человека.
В настоящей области техники существует потребность в терапевтических антителах, которые способны нейтрализовать вирусные антигены, например, ВИЧ и ВГС, включая антигенспецифические антитела, содержащие тяжелые цепи, полученные из одного вариабельного сегмента человека, и в системе, которая продуцирует разнообразный источник антител, из которых выбирают последовательности терапевтических антител. Существует также потребность в дополнительных способах и не относящихся к человеку животных для получения применимых антител, включающих антитела, которые содержат репертуар тяжелых цепей, полученных из одного VH сегмента человека, и содержащие разнообразный набор последовательностей CDR, и включая такие тяжелые цепи, которые экспрессируют с родственными вариабельными областями легкой цепи человека. Требуются способы для отбора CDR для связывающих белков на основе иммуноглобулинов, которые предусматривают повышенное разнообразие связывающих белков, из которых производят выбор, и повышенное разнообразие вариабельных доменов иммуноглобулинов, включая композиции и способы для создания соматически мутированных и клонально отобранных вариабельных доменов иммуноглобулинов, для использования, например, в получении терапевтических средств для людей.
Краткое раскрытие изобретения
Предусматриваются генетически модифицированные локусы иммуноглобулинов, которые содержат некоторое ограниченное количество различных генных сегментов вариабельной области тяжелой цепи (т.е. генов V, генов VH, генных сегментов VH или генных сегментов V), например, не более одного, двух или трех различных генов V; или не более одного представителя семейства генных сегментов V присутствующего, например, в единственной копии или во множественных копиях и/или содержащий один или несколько полиморфизмов.
Предусматриваются локусы, которые способны к реаранжировке и образованию гена, кодирующего вариабельную область тяжелой цепи, полученного из рестриктированного репертуара генов VH, например, представляющих собой один генный сегмент VH или выбранных из множества полиморфных вариантов одного генного сегмента VH. Предусматриваются модифицированные локусы иммуноглобулинов, которые включают в себя локусы, содержащие последовательности иммуноглобулинов человека, например, сегмент V человека, функционально связанный с константной 4 последовательностью человеческого (или химеры человеческого/не относящегося к человеку) или не относящегося к человеку иммуноглобулина (и функционально связанный, например, с сегментом D и/или J). Предусматриваются модифицированные локусы, которые содержат множественные копии одного генного сегмента VH, включая при этом одну или несколько копий, содержащих полиморфный вариант. Предусматриваются модифицированные локусы, которые включают множественные копии одного сегмента VH, функционально связанного с одним или несколькими сегментами D и одним или несколькими сегментами J, функционально связанными с константной последовательностью не относящегося к человеку иммуноглобулина, например, последовательностью мыши или крысы. Также предусматриваются не относящиеся к человеку животные, содержащие такие гуманизированные локусы.
Предусматриваются не относящиеся к человеку животные, которые содержат редуцированную вариабельность генного сегмента тяжелой цепи иммуноглобулина (т.е. ограниченное количество вариабельных генных сегментов тяжелой цепи или ограниченный репертуар вариабельных генов тяжелой цепи), причем редуцированная вариабельность генного сегмента тяжелой цепи иммуноглобулина характеризуется наличием не более одного или не более двух вариабельных генных сегментов тяжелой цепи, и при этом присутствующие вариабельные гены тяжелой цепи функционально связаны с человеческой или не относящейся к человеку последовательностью константной области.
Предусматриваются не относящиеся к человеку животные, которые содержат редуцированную вариабельность генного сегмента тяжелой цепи иммуноглобулина (например, один генный сегмент VH или ограниченное число генных сегментов VH, которые представляют собой полиморфные варианты одного генного сегмента VH), причем редуцированная вариабельность генного сегмента тяжелой цепи иммуноглобулина характеризуется наличием единственного генного сегмента VH или множества генных сегментов VH, которые представляют собой полиморфные формы единственного генного сегмента VH (например, генные сегменты VH, ассоциированные с высоким числом копий и/или полиморфизмом у людей), и при этом присутствующие вариабельные гены тяжелой цепи функционально связаны с человеческой или не относящейся к человеку последовательностью константной области. Согласно различным вариантам осуществления присутствующие вариабельные гены тяжелой цепи функционально связаны с одним или несколькими генными сегментами D и/или одним или несколькими генными сегментами J в зародышевой линии не относящегося к человеку животного.
Предусматриваются не относящиеся к человеку животные, которые содержат вариабельный локус тяжелой цепи иммуноглобулина (например, на трансгене или в качестве вставки или замещения в эндогенном вариабельном локусе тяжелой цепи не относящегося к человеку животного), который содержит единственный сегмент VH, функционально связанный с генным сегментом D и/или J. Согласно различным вариантам осуществления единственный сегмент VH функционально связан с одним или несколькими генными сегментами D и/или с одним или несколькими генными сегментами J в эндогенном вариабельном локусе тяжелой цепи иммуноглобулина не относящегося к человеку животного.
Предусматриваются не относящиеся к человеку животные, которые модифицированы в локусах вариабельной области тяжелой цепи их иммуноглобулина для удаления всех или по существу всех (например, всех функциональных сегментов или практически всех функциональных сегментов) эндогенных сегментов VH иммуноглобулина и которые содержат сегмент VH1-69 человека (или сегмент VH1-2 человека), функционально связанный с сегментом D и J или сегментом J в эндогенном вариабельном локусе тяжелой цепи иммуноглобулина не относящегося к человеку животного.
Также предусматриваются не относящиеся к человеку животные, которые модифицированы в локусах вариабельной области тяжелой цепи их иммуноглобулина для приведения эндогенных локусов вариабельной области в состояние неспособности к реаранжировке с образованием функциональной тяжелой цепи, содержащей эндогенные генные сегменты вариабельной области; при этом не относящиеся к человеку животные содержат единственный вариабельный генный сегмент человека (VH1-2 человека или генный сегмент VH1-69 человека), функционально связанный с сегментом D и J или сегментом J в эндогенном вариабельном локусе тяжелой цепи иммуноглобулина не относящегося к человеку животного.
Предусматриваются не относящиеся к человеку животные, которые содержат ограниченное количество (например, не более одного или не более двух) генных сегментов тяжелой цепи, функционально связанных с последовательностью человеческой или не относящейся к человеку константной области. Согласно одному варианту осуществления не более одного или не более двух генных сегментов тяжелой цепи, связанных с последовательностью константной области, находятся на трансгене, например, находятся в положении, отличном от эндогенного локуса тяжелой цепи.
Предусматриваются способы получения последовательностей иммуноглобулина человека от не относящихся к человеку животных. Согласно различным вариантам осуществления последовательности иммуноглобулина человека получают из репертуара последовательностей V иммуноглобулина, которые состоят преимущественно из единственного человеческого сегмента V, например, VH1-69 или VH1-2, и одного или нескольких сегментов D и J или одного или нескольких сегментов J. Предусматриваются способы получения последовательностей иммуноглобулина человека от не относящихся к человеку животных, тканей и клеток, причем последовательности человеческого иммуноглобулина связывают патоген.
Предусматриваются способы получения мышей, характеризующихся ограниченным локусом тяжелой цепи иммуноглобулина, причем ограничение касается числа генных сегментов VH иммуноглобулина. Согласно различным аспектам ограничение относится к одному или не более двум, или к единственному представителю семейства генов VH (например, к одному или нескольким аллелям VH, вариантам или их полиморфным вариантам). Согласно различным аспектам локус тяжелой цепи дополнительно содержит один или несколько генных сегментов DH и один или несколько генных сегментов jh. Согласно различным аспектам генные сегменты VH, DH и JH представляют собой генные сегменты человека. Согласно различным аспектам генные сегменты VH, DH и JH функционально связаны с не относящейся к человеку константной областью (например, IgM и/или IgG). Согласно различным аспектам константная область представляет собой константную область мыши или крысы.
Согласно одному аспекту предусмотрен способ получения мыши, содержащей ограниченный локус тяжелой цепи иммуноглобулина, предусматривающий введение описанной в настоящем документе конструкции нуклеиновой кислоты в эмбриональную стволовую (ES) клетку мыши и выделение или идентификацию ES клетки мыши, содержащей конструкцию нуклеиновой кислоты.
Согласно одному варианту осуществления конструкция нуклеиновой кислоты содержит единственный генный сегмент VH человека, один или несколько генных сегментов DH человека, а также один или несколько генных сегментов JH человека. Согласно одному варианту осуществления конструкция нуклеиновой кислоты содержит один или несколько сайтов сайт-специфической рекомбинации (например, сайт LoxP или Frt).
Согласно одному аспекту предусматривается мышь, полученная с использованием описанного в настоящем документе нацеливающего вектора, нуклеотидной последовательности или клетки. Согласно различным вариантам осуществления нацеливающий вектор, нуклеотидная последовательность или клетка содержит последовательность ДНК, которая содержит единственный генный сегмент VH человека (или его полиморфные варианты), один или несколько генных сегментов DH человека, и один или несколько генных сегментов JH человека, функционально связанные с не относящимся к человеку константным геном.
Согласно одному аспекту предусматривается способ получения мыши, содержащей ограниченный локус тяжелой цепи иммуноглобулина, включающий замещение локуса тяжелой цепи иммуноглобулина мыши человеческой геномной последовательностью, содержащей единственный генный сегмент VH человека (или его полиморфные варианты), один или несколько генных сегментов DH человека, и один или несколько генных сегментов JH человека, причем генные сегменты VH, DH и JH человека, способны к реаранжировке с формированием химерной тяжелой цепи, содержащей вариабельную область человека, функционально связанную с не относящейся к человеку константной областью. Согласно одному варианту осуществления не относящаяся к человеку константная область представляет собой константную область мыши или крысы.
Согласно различным аспектам не относящиеся к человеку животные представляют собой грызунов. Согласно различным аспектам грызуны представляют собой мышей и/или крыс.
Согласно одному аспекту предусматривается модифицированный локус тяжелой цепи иммуноглобулина, который содержит репертуар сегментов V тяжелой цепи, ограниченный по отношению к идентичности V сегмента, и который содержит один или несколько D сегментов и один или несколько сегментов J или один или несколько сегментов J. Согласно одному варианту осуществления сегмент V тяжелой цепи представляет собой сегмент человека. Согласно одному варианту осуществления один или несколько сегментов D представляют собой сегменты D человека. Согласно одному варианту осуществления один или несколько сегментов J представляют собой сегменты J человека. Согласно одному варианту осуществления один или несколько сегментов D и один или несколько сегментов J представляет собой сегменты D и J человека.
Согласно одному варианту осуществления модифицированный локус представляет собой не относящийся к человеку локус. Согласно одному варианту осуществления не относящийся к человеку локус модифицируют по меньшей мере с одной последовательностью иммуноглобулина человека.
Согласно одному варианту осуществления ограничение касается одного представителя семейства сегментов V. Согласно одному варианту осуществления один представитель семейства сегментов V присутствует в двух или более копиях. Согласно одному варианту осуществления один представитель семейства сегментов V присутствует в виде двух или более вариантов (например, две или более полиморфные формы представителя семейства сегментов V). Согласно одному варианту осуществления один сегмент V представляет собой представителя семейства сегментов V человека. Согласно одному варианту осуществления один представитель семейства сегментов V присутствует в нескольких вариантах, как это наблюдается в человеческой популяции, по отношению к этому варианту. Согласно одному варианту осуществления представителя семейства сегментов V выбирают из таблицы 1. Согласно одному варианту осуществления представитель семейства сегментов V присутствует в нескольких вариантах, как показано для каждого сегмента V, в нескольких аллелях от одной аллели до нескольких аллелей, показанных в правой колонке таблицы 1.
Согласно одному варианту осуществления ограничение относится к генному сегменту VH1-69 человека. Согласно одному варианту осуществления генный сегмент VH1-69 человека присутствует в двух или более копиях. Согласно одному варианту осуществления генный сегмент VH1-69 человека присутствует в двух или более вариантах (например, две или более полиморфные формы гена VH1-69 человека). Согласно одному варианту осуществления генный сегмент VH1-69 человека присутствует в нескольких вариантах, как это наблюдается в человеческой популяции, по отношению к генному сегменту VH1-69 человека. Согласно одному варианту осуществления генный сегмент VH1-69 человека выбирают из таблицы 2. Согласно одному варианту осуществления генный сегмент VH1-69 человека присутствует в нескольких вариантах, как показано для каждого генного сегмента VH1-69, в нескольких аллелях от одной аллели до нескольких аллелей, показанных в таблице 2.
Согласно одному варианту осуществления ограничение относится к генному сегменту VH1-2 человека. Согласно одному варианту осуществления генный сегмент VH1-2 человека присутствует в двух или более копиях. Согласно одному варианту осуществления генный сегмент VH1-2 человека присутствует в двух или более вариантов (например, две или более полиморфные формы гена VH1-2 человека). Согласно одному варианту осуществления генный сегмент VH1-2 человека присутствует в нескольких вариантах, как это наблюдается в человеческой популяции, по отношению к генному сегменту VH1-2 человека. Согласно одному варианту осуществления генный сегмент VH1-2 человека выбирают из таблицы 3. Согласно одному варианту осуществления генный сегмент VH1-2 человека присутствует в нескольких вариантах, как показано для каждого генного сегмента VH1-2, в нескольких аллелях от одной аллели до нескольких аллелей, показанных в таблице 3.
Согласно одному аспекту предусматривается локус тяжелой цепи иммуноглобулина, который содержит единственный функциональный сегмент V человека. Согласно одному варианту осуществления единственный функциональный сегмент V человека выбирают из сегментов VH1-2, VH1-3, VH1-8, VH1-18, VH1-24, VH1-45, VH1-46, VH1-58, VH1-69, VH2-5, VH2-26, VH2-70, VH3-7, VH3-9, VH3-11, VH3-13, VH3-15, VH3-16, VH3-20, VH3-21, VH3-23, VH3-30, VH3-30-3, VH3-30-5, VH3-33, VH3-35, VH3-38, VH3-43, VH3-48, VH3-49, VH3-53, VH3-64, VH3-66, VH3-72, VH3-73, VH3-74, VH4-4, VH4-28, VH4-30-1, VH4-30-2, VH4-30-4, VH4-31, VH4-34, VH4-39, VH4-59, VH4-61, VH5-51, VH6-1, VH7-4-1 и VH7-81. Согласно одному варианту осуществления единственный функциональный сегмент V человека представляет собой сегмент VH1-69; согласно конкретному варианту осуществления единственный функциональный сегмент V человека присутствует в 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или 13 полиморфных формах, обнаруженных в человеческой популяции. Согласно одному варианту осуществления единственный функциональный сегмент V человека представляет собой сегмент VH1-2; согласно конкретному варианту осуществления один функциональный сегмент V человека присутствует в 1, 2, 3, 4 или 5 полиморфных формах, обнаруженных в человеческой популяции.
Согласно одному варианту осуществления локус тяжелой цепи иммуноглобулина представляет собой модифицированный локус не относящегося к человеку животного. Согласно одному варианту осуществления модифицированный не относящийся к человеку локус тяжелой цепи иммуноглобулина присутствует у не относящегося к человеку животного в положении в геноме, в котором находится соответствующий немодифицированный не относящийся к человеку локус у не относящегося к человеку животного дикого типа. Согласно одному варианту осуществления модифицированный не относящийся к человеку локус тяжелой цепи иммуноглобулина присутствует на трансгене у не относящегося к человеку животного.
Согласно одному варианту осуществления один функциональный генный сегмент V человека представляет собой генный сегмент VH1-69. Согласно одному варианту осуществления генный сегмент VH1-69 содержит SEQ ID NO: 34. Согласно одному варианту осуществления генный сегмент VH1-69 происходит из SEQ ID NO: 34. Согласно одному варианту осуществления генный сегмент VH1-69 по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 98% идентичен SEQ Ю NO: 34.
Согласно одному варианту осуществления один функциональный генный сегмент V человека кодируется нуклеотидной последовательностью SEQ ID NO: 34.
Согласно одному варианту осуществления один функциональный генный сегмент V человека представляет собой генный сегмент VH1-2. Согласно одному варианту осуществления генный сегмент VH1-2 содержит SEQ ID NO: 60. Согласно одному варианту осуществления генный сегмент VH1-2 происходит из SEQ ID NO: 60. Согласно одному варианту осуществления генный сегмент VH1-2 по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 98% идентичен SEQ ID NO: 60.
Согласно одному варианту осуществления один функциональный генный сегмент V человека кодируется нуклеотидной последовательностью SEQ ID NO: 60.
Согласно одному варианту осуществления один функциональный генный сегмент V человека функционально связан с одним или несколькими сегментами D и одним или несколькими сегментами J, или одним или несколькими сегментами J. Согласно одному варианту осуществления сегмент V и один или несколько сегментов D и/или J функционально связаны с последовательностью константной области тяжелой цепи иммуноглобулина. Согласно одному варианту осуществления последовательность константной области тяжелой цепи иммуноглобулина выбирают из последовательности CH1, шарнира, CH2, CH3 и их комбинации. Согласно одному варианту осуществления каждый из CH1, шарнира, CH2, CH3 или их комбинации представляет собой не относящиеся к человеку эндогенные константные последовательности. Согласно одному варианту осуществления по меньшей мере один из CH1, шарнира, CH2, CH3 или их комбинации представляет собой последовательность человека. Согласно конкретному варианту осуществления CH1 и/или шарнир представляют собой последовательности человека.
Согласно одному аспекту предусматривается модифицированный эндогенный не относящийся к человеку локус тяжелой цепи иммуноглобулина, содержащий замещение всех функциональных генных сегментов V одним генным сегментом V человека (или один генный сегмент V человека присутствует во множественных полиморфных формах или числе копий), причем локус тяжелой цепи не относящегося к человеку иммуноглобулина не способен к реаранжировке с образованием вариабельного гена тяжелой цепи, который происходит от генного сегмента V, отличного от единственного генного сегмента V человека (или одного из полиморфных форм или копий).
Согласно одному варианту осуществления единственный генный сегмент V человека представляет собой VH1-69. Согласно одному варианту осуществления единственный генный сегмент V человека представляет собой VH1-2.
Согласно одному варианту осуществления локус содержит по меньшей мере один генный сегмент DH человека или не относящийся к человеку, и один генный сегмент JH человека или не относящийся к человеку. Согласно конкретному варианту осуществления локус содержит генный сегмент DH человека и генный сегмент JH человека. Согласно конкретному варианту осуществления локус содержит генный сегмент JH человека. Согласно другому конкретному варианту осуществления локус содержит генный сегмент VH1-69 человека (присутствует в виде единственной копии или множественных копий различных полиморфных вариантов), все функциональные генные сегменты DH человека, и все функциональные генные сегменты JH человека. Согласно другому конкретному варианту осуществления локус содержит генный сегмент VH1-2 человека (присутствует в виде единственной копии или множественных копий различных полиморфных форм), все функциональные генные сегменты DH человека и все функциональные генные сегменты JH человека. Согласно одному варианту осуществления генные сегменты V, D и J человека (или генные сегменты V и J) функционально связаны с геном константной области в эндогенном локусе тяжелой цепи мыши. Согласно конкретному варианту осуществления локус тяжелой цепи мыши содержит репертуар последовательностей константной области иммуноглобулина мыши дикого типа.
Согласно одному аспекту предусматривается генетически модифицированное не относящееся к человеку животное, причем только функциональный генный сегмент V тяжелой цепи иммуноглобулина не относящегося к человеку животного выбирают из генных сегментов VH1-2, VH1-3, VH1-8, VH1-18, VH1-24, VH1-45, VH1-46, VH1-58, VH1-69, VH2-5, VH2-26, VH2-70, VH3-7, VH3-9, VH3-11, VH3-13, VH3-15, VH3-16, VH3-20, VH3-21, VH3-23, VH3-ЗО, VH3-ЗО-З, VH3-30-5, VH3-33, VH3-35, VH3-38, VH3-43, VH3-48, VH3-49, VH3-53, VH3-64, VH3-66, VH3-72, VH3-73, VH3-74, VH4-4, VH4-28, VH4-30-1, VH4-30-2, VH4-30-4, VH4-31, VH4-34, VH4-39, VH4-59, VH4-61, VH5-51, VH6-1, VH7-4-1 и VH7-81 человека. Согласно одному варианту осуществления генный сегмент V тяжелой цепи представляет собой генный сегмент VH1-69 человека. Согласно одному варианту осуществления генный сегмент V тяжелой цепи представляет собой генный сегмент VH1-2 человека.
Согласно одному аспекту предусматривается генетически модифицированное не относящееся к человеку животное, причем не относящееся к человеку животное содержит один функциональный генный сегмент VH человека (присутствует в виде единственной копии или множественных копий различных полиморфных форм), и причем не относящееся к человеку животное по существу не способно к образованию реаранжированного гена вариабельного домена тяжелой цепи иммуноглобулина, который не содержит единственного функционального генного сегмента VH человека (или одну из полиморфных форм или копий).
Согласно одному аспекту предусматривается генетически модифицированное не относящееся к человеку животное, причем только вариабельная область тяжелой цепи иммуноглобулина, экспрессируемая у не относящегося к человеку животного, происходит от одного сегмента человека, выбранного из генных сегментов VH1-2, VH1-3, VH1-8, VH1-18, VH1-24, VH1-45, VH1-46, VH1-58, VH1-69, VH2-5, VH2-26, VH2-70, VH3-7, VH3-9, VH3-11, VH3-13, VH3-15, VH3-16, VH3-20, VH3-21, VH3-23, VH3-30, VH3-30-3, VH3-30-5, VH3-33, VH3-35, VH3-38, VH3-43, VH3-48, VH3-49, VH3-53, VH3-64, VH3-66, VH3-72, VH3-73, VH3-74, VH4-4, VH4-28, VH4-30-1, VH4-30-2, VH4-30-4, VH4-31, VH4-34, VH4-39, VH4-59, VH4-б1, VH5-51, VH6-1, VH7-4-1 и VH7-81 человека. Согласно одному варианту осуществления сегмент человека представляет собой сегмент VH1-69. Согласно одному варианту осуществления сегмент человека представляет собой сегмент VH1-2. Согласно одному варианту осуществления единственный вариабельный участок тяжелой цепи иммуноглобулина, экспрессируемый мышью, происходит от единственного представителя семейства сегментов V, и согласно одному варианту осуществления только вариабельную область тяжелой цепи иммуноглобулина получают из полиморфного варианта единственного представителя семейства сегментов V.
Согласно одному аспекту предусматривается не относящееся к человеку животное, содержащее ограниченный репертуар генных сегментов V тяжелой цепи иммуноглобулина, причем не относящееся к человеку животное дополнительно содержит один или несколько вариабельных сегментов (Vκ) легкой цепи κ иммуноглобулина человека. Согласно одному варианту осуществления один или несколько сегментов Vκ функционально связаны с одним или несколькими сегментами J человека. Согласно конкретному варианту осуществления сегменты J представляют собой сегменты J человека. Согласно другому конкретному варианту осуществления не относящееся к человеку животное не экспрессирует легкую цепь λ иммуноглобулина. Согласно другому конкретному варианту осуществления не относящееся к человеку животное не содержит функциональный человеческий или функциональный эндогенный вариабельный локус легкой цепи λ иммуноглобулина.
Согласно одному варианту осуществления не относящееся к человеку животное представляет собой грызуна. Согласно одному варианту осуществления грызун представляет собой мышь.
Согласно одному варианту осуществления не относящееся к человеку животное содержит замещение в эндогенном локусе Vκ не относящегося к человеку иммуноглобулина всех или по существу всех функциональных эндогенных сегментов Vκ одним или несколькими функциональными сегментами Vκ человека. Согласно дополнительному конкретному варианту осуществления замещение представляет собой замещение всеми или по существу всеми функциональными сегментами Vκ иммуноглобулина человека.
Согласно одному варианту осуществления не относящееся к человеку животное содержит замещение в эндогенном локусе Vκ не относящегося к человеку иммуноглобулина всех или по существу всех функциональных эндогенных генных сегментов Vκ генными сегментами Vκ человека, выбранными из Vκ4-1, Vκ5-2, Vκ7-3, Vκ2-4, Vκ1-5, Vκ1-6, Vκ3-7, Vκ1-8, Vκ1-9, Vκ2-10, Vκ3-11, Vκ1-12, Vκ1-13, Vκ2-14, Vκ3-15, VK-1-16, VK1-17, VK2-18, VK2-19, Vκ3-20, Vκ6-21, Vκ1-22, Vκ1-23, Vκ2-24, Vκ3-25, Vκ2-26, Vκ1-27, Vκ2-28, W2-29, Vκ2-30, Vκ3-31, Vκ1-32, Vκ1-33, Vκ3-34, Vκ1-35, Vκ2-36, Vκ1-37, Vκ2-38, Vκ1-39, Vκ2-40 и их комбинации.
Согласно одному варианту осуществления не относящееся к человеку животное содержит замещение в эндогенном локусе Jκ не относящегося к человеку иммуноглобулина всех или по существу всех функциональных эндогенных сегментов Jκ не относящегося к человеку иммуноглобулина одним или несколькими функциональными сегментами Jκ иммуноглобулина человека. Согласно дополнительному конкретному варианту осуществления замещение представляет собой замещение всеми или по существу всеми функциональными сегментами Jκ иммуноглобулина человека.
Согласно одному варианту осуществления не относящееся к человеку животное содержит замещение в эндогенном локусе Jκ не относящегося к человеку иммуноглобулина всех или по существу всех функциональных эндогенных генных сегментов Jκ не относящегося к человеку иммуноглобулина генными сегментами Jκ человека, выбранными из Jκ1, Jκ2, Jκ3, Jκ4, Jκ5 и их комбинации.
Согласно конкретному варианту осуществления не относящееся к человеку животное содержит локус вариабельной области тяжелой цепи иммуноглобулина, который содержит репертуар сегментов V, состоящих преимущественно из единственного сегмента V и/или его полиморфных вариантов. Согласно конкретному варианту осуществления единственный сегмент V тяжелой цепи иммуноглобулина представляет собой VH1-69 сегмент человека, и не относящееся к человеку животное дополнительно содержит замещение всех не относящихся к человеку функциональных сегментов DH всеми функциональными сегментами DH человека, и дополнительно содержит замещение всех не относящихся к человеку функциональных сегментов JH всеми функциональными сегментами JH человека, и причем локус вариабельной области тяжелой цепи иммуноглобулина функционально связан с генной последовательностью константной области человека или не относящейся к человеку. Согласно конкретному варианту осуществления генная последовательность константной области представляет собой эндогенную генную последовательность не относящейся к человеку константной области. Согласно конкретному варианту осуществления не относящееся к человеку животное реаранжирует сегменты в локусе тяжелой цепи не относящегося к человеку иммуноглобулина с образованием гена, кодирующего вариабельную область тяжелой цепи, содержащую последовательность VH1-69 человека, последовательность DH человека, последовательность JH человека и последовательность константной области мыши.
Согласно конкретному варианту осуществления не относящееся к человеку животное содержит локус вариабельной области тяжелой цепи иммуноглобулина, который содержит репертуар сегментов V, состоящих преимущественно из единственного сегмента V и/или его полиморфных вариантов. Согласно конкретному варианту осуществления единственный сегмент V тяжелой цепи иммуноглобулина представляет собой сегмент VH1-2 человека, и не относящееся к человеку животное дополнительно содержит замещение всех не относящихся к человеку функциональных сегментов DH всеми функциональными сегментами DH человека, и дополнительно содержит замещение всех не относящихся к человеку функциональных сегментов JH всеми функциональными сегментами JH человека, и причем локус вариабельной области тяжелой цепи иммуноглобулин функционально связан с генной последовательностью константной области человека или не относящейся к человеку. Согласно конкретному варианту осуществления генная последовательность константной области представляет собой эндогенную генную последовательность не относящейся к человеку константной области. Согласно конкретному варианту осуществления не относящееся к человеку животное реаранжирует сегменты в локусе тяжелой цепи не относящегося к человеку иммуноглобулина с образованием гена, кодирующего вариабельную область тяжелой цепи, содержащую последовательность VH1-2 человека, последовательность DH человека, последовательность JH человека и последовательность константной области мыши.
Согласно одному варианту осуществления предусматривается В-клетка, которая содержит реаранжированный ген. Согласно конкретному варианту осуществления В-клетка представляет собой В-клетку описанной в настоящем документе мыши которая была иммунизирована представляющим интерес антигеном, и В-клетка кодирует антитело, которое специфически связывается с представляющим интерес антигеном. Согласно одному варианту осуществления представляющий интерес антиген представляет собой патоген. Согласно конкретному варианту осуществления патоген выбирают из вируса гриппа, вируса гепатита (например, вируса гепатита В или гепатита С) и вируса иммунодефицита человека. Согласно конкретному варианту осуществления В-клетка кодирует соматически мутированное высокоаффинное (например, приблизительно 10-9 KD или ниже) антитело, содержащее вариабельную область легкой цепи человека (например, вариабельную область легкой цепи κ человека), которое специфически связывается с представляющим интерес антигеном.
Согласно одному аспекту предусматривается не относящееся к человеку животное, содержащее ограниченный репертуар сегментов V тяжелой цепи иммуноглобулина, причем не относящееся к человеку животное содержит один или несколько вариабельных сегментов (Vλ) легкой цепи λ человека. Согласно одному варианту осуществления один или несколько сегментов Vλ человека функционально связаны с одним или несколькими сегментами J человека. Согласно конкретному варианту осуществления сегменты J представляют собой сегменты Jλ, человека. Согласно другому конкретному варианту осуществления не относящееся к человеку животное не экспрессируют легкую цепь κ. Согласно другому конкретному варианту осуществления не относящееся к человеку животное не содержит функциональный вариабельный локус легкой цепи κ человека или не относящегося к человеку.
Согласно одному варианту осуществления не относящееся к человеку животное содержит замещение всех или по существу всех функциональных сегментов Vλ, не относящегося к человеку иммуноглобулина. Согласно дополнительному конкретному варианту осуществления замещение осуществляется всеми или по существу всеми функциональными сегментами Vλ иммуноглобулина человека.
Согласно одному варианту осуществления не относящееся к человеку животное содержит замещение всех или по существу всех функциональных не относящихся к человеку сегментов Vλ фрагментом кластера А локуса легкой цепи λ человека. Согласно конкретному варианту осуществления фрагмент кластера А локуса легкой цепи λ, человека содержит сегменты от Vλ3-27 до Vλ3-1 гена Vλ, человека.
Согласно одному варианту осуществления не относящееся к человеку животное содержит замещение всех или по существу всех функциональных не относящихся к человеку сегментов Vλ фрагментом кластера В локуса легкой цепи λ, человека. Согласно конкретному варианту осуществления фрагмент кластера В локуса легкой цепи λ человека содержит сегменты от V7.5-52 до Vλ1-40 гена Vλ человека.
Согласно одному варианту осуществления не относящееся к человеку животное содержит замещение всех или по существу всех функциональных не относящихся к человеку сегментов Vλ, фрагментом кластера А и фрагментом кластера В локуса легкой цепи λ человека, причем в результате замещения содержит сегменты от Vλ5-52 до Vλ3-1 гена Vλ, человека.
Согласно одному варианту осуществления не относящееся к человеку животное содержит замещение всех или по существу всех функциональных не относящихся к человеку сегментов Vλ по меньшей мере 12 генными сегментами Vλ, человека, по меньшей мере 28 генными сегментами VA, человека или по меньшей мере 40 генными сегментами Vλ человека.
Согласно одному варианту осуществления не относящееся к человеку животное содержит замещение всех или по существу всех функциональных генных сегментов Jλ не относящегося к человеку иммуноглобулина. Согласно дополнительному конкретному варианту осуществления замещение осуществляется всеми или по существу всеми функциональными генными сегментами Jλ иммуноглобулина человека. Согласно различным вариантам осуществления функциональные генные сегменты Jλ человека включают в себя Jλ1, Jλ2, Jλ3 и Jλ7.
Согласно конкретному варианту осуществления не относящееся к человеку животное содержит локус вариабельной области (VH) тяжелой цепи иммуноглобулина, который содержит только единственный сегмент VH, причем единственный сегмент VH представляет собой сегмент VH1-69 человека или сегмент VH1-2 человека, и дополнительно содержит замещение всех функциональных не относящихся к человеку сегментов DH всеми функциональными сегментами DH человека, и дополнительно содержит замещение всех функциональных не относящихся к человеку сегментов JH всеми функциональными сегментами JH человека, и причем локус VH области функционально связан с генной последовательностью константной области человека или не относящейся к человеку. Согласно конкретному варианту осуществления генная последовательность константной области представляет собой генную последовательность не относящейся к человеку константной области, например, эндогенную генную последовательность не относящейся к человеку константной области. Согласно конкретному варианту осуществления не относящееся к человеку животное реаранжирует сегменты в локусе тяжелой цепи не относящегося к человеку иммуноглобулина с образованием гена, кодирующего вариабельную область тяжелой цепи иммуноглобулина, содержащую последовательность VH1-69 человека (или последовательность VH1-2 человека), последовательность DH человека, последовательность JH человека и эндогенную последовательность не относящейся к человеку константной области.
Согласно одному варианту осуществления предусматривается В-клетка, которая содержит реаранжированный ген. Согласно конкретному варианту осуществления В-клетка представляет собой клетку описанного в настоящем документе не относящегося к человеку животного, которое иммунизировали представляющим интерес антигеном, и В- клетка кодирует антитело, которое специфически связывается с представляющим интерес антигеном. Согласно одному варианту осуществления антиген представляет собой белок человека, выбранный из лиганда, рецептора клеточной поверхности и внутриклеточного белка. Согласно одному варианту осуществления представляющий интерес антиген представляет собой патоген. Согласно конкретному варианту осуществления патоген выбирают из вируса гриппа, вирус гепатита (например, вируса гепатита В или гепатита С) и вируса иммунодефицита человека. Согласно конкретному варианту осуществления В-клетка кодирует соматически мутированное высокоаффинное (например, приблизительно 10-9 KD или ниже) антитело, содержащее вариабельную область легкой цепи человека (например, вариабельную область легкой цепи λ человека), которое специфически связывается с представляющим интерес антигеном.
Согласно одному аспекту предусматривается не относящееся к человеку животное, содержащее ограниченный репертуар сегментов VH иммуноглобулина, причем не относящееся к человеку животное содержит сегмент VH1-69 человека (или сегмент VH1-2 человека) на трансгене, причем сегмент VH1-69 человека функционально связан на трансгене с сегментом DH человека или не относящимся к человеку и/или с сегментом J человека или не относящимся к человеку, и трансген дополнительно содержит ген константной области человека или не относящейся к человеку или химерную константную область человека/не относящуюся к человеку (например, CH1, шарнир, CH2, CH3 или их комбинацию, где по меньшей мере одна последовательность представляет собой не относящуюся к человеку последовательность, например, выбранную из шарнира, CH2 и CH3 и/или шарнира). Согласно одному варианту осуществления не относящееся к человеку животное представляет собой мышь или крысу, и ген не относящегося к человеку D, J и/или константной области представляет собой ген мыши или крысы или химерный ген человека/мыши или крысы.
Согласно одному варианту осуществления не относящееся к человеку животное содержит трансген, который содержит локус вариабельной области легкой цепи иммуноглобулина, который содержит один или несколько генных сегментов Vλ и генных сегментов Jλ иммуноглобулина человека или один или несколько генных сегментов Vκ и генных сегментов Jκ иммуноглобулина человека и ген константной области легкой цепи κ или λ иммуноглобулина человека, в результате чего трансген реаранжирует у не относящегося к человеку животного для образования реаранжированного гена легкой цепи κ или λ иммуноглобулина. Согласно различным вариантам осуществления генные сегменты Vκ и Jκ человека представляют собой описанные в настоящем документе сегменты. Согласно различным вариантам осуществления генные сегменты Vλ и Jλ человека представляют собой описанные в настоящем документе сегменты.
Согласно конкретному варианту осуществления не относящееся к человеку животное содержит трансген, содержащий вариабельный локус тяжелой цепи иммуноглобулина, который содержат единственный сегмент V, представляющий собой сегмент VH1-69 человека (или сегмент VH1-2 человека), один или несколько сегментов D человека, один или несколько сегментов J человека и константный ген человека, функционально связанный с вариабельным локусом тяжелой цепи, в результате чего мышь экспрессирует с трансгена полное антитело человека, полученное из сегмента VH1-69 (или сегмента VH1-2). Согласно одному варианту осуществления не относящееся к человеку животное не содержит функциональный эндогенный локус вариабельной области тяжелой цепи иммуноглобулина. Согласно конкретному варианту осуществления не относящееся к человеку животное содержит нефункциональный эндогенный локус вариабельной области тяжелой цепи иммуноглобулина, который содержит делецию эндогенного не относящегося к человеку сегмента DH и/или эндогенного не относящегося к человеку сегмента JH, в результате чего не относящееся к человеку животное не способно к реаранжировке локуса вариабельной области тяжелой цепи иммуноглобулина с образованием реаранжированного гена не относящегося к человеку антитела. Согласно конкретному варианту осуществления не относящееся к человеку животное содержит делецию последовательности переключения (switch sequence), функционально связанную с эндогенной константной областью тяжелой цепи мыши. Согласно конкретному варианту осуществления последовательность переключения представляет собой не относящуюся к человеку (например, мыши) последовательность переключения µ. Согласно другому варианту осуществления не относящееся к человеку животное дополнительно не содержит функциональный эндогенный вариабельный локус легкой цепи, выбранный из локуса к иммуноглобулина и локуса λ иммуноглобулина. Согласно конкретному варианту осуществления не относящееся к человеку животное содержит делецию последовательности Jκ и/или Jλ, так что не относящееся к человеку животное не способно к реаранжировке эндогенной вариабельной области легкой цепи κ не относящегося к человеку иммуноглобулина и/или эндогенной вариабельной области легкой цепи λ не относящегося к человеку иммуноглобулина с образованием реаранжированного эндогенного гена легкой цепи κ не относящегося к человеку иммуноглобулина и/или легкой цепи λ не относящегося к человеку иммуноглобулина.
Согласно одному варианту осуществления не относящееся к человеку животное содержит делецию эндогенной последовательности легкой цепи κ не относящегося к человеку иммуноглобулина, что приводит к функциональному нокауту эндогенной легкой цепи κ не относящегося к человеку иммуноглобулина. Согласно одному варианту осуществления не относящееся к человеку животное содержит делецию эндогенной последовательности легкой цепи λ не относящегося к человеку иммуноглобулина, что приводит к функциональному нокауту эндогенной легкой цепи κ не относящегося к человеку иммуноглобулина.
Согласно одному варианту осуществления не относящееся к человеку животное содержит функционально выключенный эндогенный локус вариабельного гена тяжелой цепи иммуноглобулина, и содержит ограниченный репертуар вариабельных генных сегментов тяжелой цепи человека (например, не более одного или не более двух). Согласно одному варианту осуществления функциональное выключение содержит модификацию эндогенного локуса вариабельного гена не относящейся к человеку тяжелой цепи, выбранную из делеции, вставки, инверсии и их комбинации.
Согласно одному аспекту предусматривается грызун, который содержит репертуар VH иммуноглобулина, полученный из не более одного сегмента VH человека или одной или нескольких его полиморф, из сегмента D, выбранного из репертуара одного или нескольких сегментов D, и из сегмента J, полученного из репертуара одного или нескольких сегментов J. Согласно одному варианту осуществления грызун реаранжирует сегмент VH человека, сегмент D человека и сегмент J человека и образует реаранжированную последовательность тяжелой цепи человека, которая функционально связана с последовательностью константной области человека или грызуна. Согласно одному варианту осуществления последовательность константной области человека и/или грызуна выбирают из CH1, шарнира, CH2, CH3 и их комбинации. Согласно одному варианту осуществления грызун экспрессирует легкую цепь иммуноглобулина, которая содержит вариабельную область человека, причем легкая цепь состоит в родстве с доменом тяжелой цепи человека, полученным из реаранжированной последовательности тяжелой цепи человека. Согласно одному варианту осуществления грызун не экспрессирует полипептидную последовательность, выбранную из вариабельного домена не относящейся к человеку тяжелой цепи, вариабельного домена не относящейся к человеку легкой цепи и их комбинации.
Согласно одному варианту осуществления сегмент VH человека присутствует в 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18 или 19 или более полиморфных вариантах, причем каждый полиморфный вариант функционально связан с сегментом D и/или J, так что каждый полиморфный вариант способен к реаранжировке и образованию реаранжированного вариабельного домена тяжелой цепи с любым из одного или нескольких сегментов D и любым из одного или нескольких сегментов J. Согласно одному варианту осуществления грызуна представляет собой мышь или крысу. Согласно одному варианту осуществления репертуар сегментов D содержит два или более сегмента D. Согласно одному варианту осуществления репертуар сегментов J содержит два или более сегмента J. Согласно одному варианту осуществления сегменты D и/или J представляют собой сегменты человека.
Согласно одному аспекту предусматривается конструкция нуклеиновой кислоты, который содержит последовательность, кодирующую один сегмент VH иммуноглобулина человека и/или его полиморфные варианты и одну или несколько последовательностей DH, и одну или несколько последовательностей J, причем конструкция содержит по меньшей мере одно гомологичное плечо, гомологичное вариабельному локусу тяжелой цепи не относящегося к человеку иммуноглобулина, или сайт рестрикции рекомбиназы (например, сайт lox). Согласно одному варианту осуществления сегмент V представляет собой сегмент VH1-69 или сегмент VH1-2.
Согласно одному аспекту предусматривается конструкция нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты, кодирующую единственный сегмент V тяжелой цепи иммуноглобулина человека, причем единственный сегмент VH представляет собой сегмент VH1-69 (или VH1-2). Согласно одному варианту осуществления конструкция содержит сайт-специфический сайт рестрикции рекомбиназы. Согласно одному варианту осуществления конструкция содержит первое мышиное гомологичное плечо выше по ходу транскрипции от сегмента VH1-69 (или VH1-2), и второе мышиное гомологичное плечо ниже по ходу транскрипции от сегмента VH1-69 (или VH1-2), и причем первое мышиное гомологичное плечо гомологично области хромосомы мыши, непосредственно выше по ходу транскрипции от вариабельной области тяжелой цепи иммуноглобулина мыши, но не включая функциональный вариабельный сегмент тяжелой цепи иммуноглобулина мыши. Согласно одному варианту осуществления конструкция содержит SEQ ID NO: 3. Согласно одному варианту осуществления конструкция содержит SEQ ID NO: 70.
Согласно одному аспекту ограниченный единственный сегмент VH находится в не относящемся к человеку животном или ограниченный сегмент VH находится в локусе тяжелой цепи не относящегося к человеку иммуноглобулина (например, in situ или в трансгене), и не относящееся к человеку животное или локус тяжелой цепи не относящегося к человеку иммуноглобулина выбирают из локуса или животного: мыши, крысы, кролика, свиньи, крупного рогатого скота (например, коровы, быка, буйвола), оленя, овцы, козы, курицы, кошки, собаки, хорька, примата (например, мармазетки, макака-резуса). Согласно конкретному варианту осуществления не относящееся к человеку животное или локус представляет собой локус мыши или крысы.
Согласно одному аспекту предусматривается клетка или ткань, причем клетка или ткань происходит из описанного в настоящем документе не относящегося к человеку животного и содержит ограниченный репертуар сегментов VH. Согласно одному варианту осуществления репертуар сегментов VH ограничивают до единственного представителя семейства сегментов VH и/или их полиморфных вариантов. Согласно конкретному варианту осуществления единственный сегмент VH представляет собой сегмент VH1-69 человека или сегмент VH1-2 человеческого. Согласно одному варианту осуществления клетку или ткань получают из селезенки, лимфатического узла или костного мозга не относящегося к человеку животного.
Согласно одному варианту осуществления клетка представляет собой ES клетку. Согласно одному варианту осуществления клетка представляет собой В-клетку. Согласно одному варианту осуществления клетка представляет собой зародышевую клетку.
Согласно одному варианту осуществления ткань выбирают из соединительной, мышечной, нервной и эпителиальной ткани. Согласно конкретному варианту осуществления ткань представляет собой репродуктивную ткань.
Согласно одному варианту осуществления клетку и/или ткань, полученные из описанной в настоящем документе мыши, изолируют для использования в одном или нескольких анализах ex vivo. Согласно различным вариантам осуществления один или несколько анализов ех vivo включают измерения физических, тепловых, электрических, механических или оптических свойств, хирургическую процедуру, измерения взаимодействий различных типов тканей, развитие техник визуализации или их комбинацию.
Согласно одному варианту осуществления не относящееся к человеку животное представляет собой мышь.
Согласно одному аспекту предусматривается не относящийся к человеку эмбрион, содержащий ограниченные описанные в настоящем документе сегменты VH тяжелой цепи. Согласно одному варианту осуществления эмбрион содержит донорскую ES клетку, которая содержит ограниченный сегмент VH, и эмбриональные клетки-хозяева.
Согласно одному варианту осуществления не относящееся к человеку животное представляет собой мышь.
Согласно одному аспекту предусматривается не относящаяся к человеку клетка, содержащая хромосому или ее фрагмент описанного в настоящем документе не относящегося к человеку животного. Согласно одному варианту осуществления не относящаяся к человеку клетка содержит ядро описанного в настоящем документе не относящегося к человеку животного. Согласно одному варианту осуществления не относящаяся к человеку клетка содержит хромосому или ее фрагмент в результате ядерной передачи.
Согласно одному аспекту предусматривается ядре, полученное из описанного в настоящем документе не относящегося к человеку животного. Согласно одному варианту осуществления ядро представляет собой ядро из диплоидной клетки, которая представляет собой не В-клетку.
Согласно одному аспекту предусматривается плюрипотентная, индуцированная плюрипотентная или тотипотентная клетка, полученная из описанного в настоящем документе не относящегося к человеку животного. Согласно конкретному варианту осуществления клетка представляет собой эмбриональную стволовую (ES) клетку мыши.
Согласно одному аспекту предусматривается не относящаяся к человеку клетка индуцированная плюрипотентная клетка, содержащая 01раниченпый репертуар сегментов VH. Согласно одному варианту осуществления индуцированную плюрипотентную клетку получают от описанного в настоящем документе не относящегося к человеку животного.
Согласно одному аспекту предусматривается гибридома или квадрома, полученная из клетки описанного в настоящем документе не относящегося к человеку животного. Согласно одному варианту осуществления не относящееся к человеку животное представляет собой мышь или крысу.
Согласно одному аспекту предусматривается лимфоцит из описанного в настоящем документе не относящегося к человеку животного. Согласно одному варианту осуществления лимфоцит представляет собой В-клетку.
Согласно одному аспекту предусматриваются клетки мыши и мышиные эмбрионы, включая в себя без ограничения ES клетки, плюрипотентные клетки и индуцированные плюрипотентные клетки, которые содержат описанные в настоящем документе генетические модификации. Предусматриваются клетки, которые представляют собой XX и XY клетки. Также предусматриваются клетки, которые содержат ядро, содержащее описанную в настоящем документе модификацию, например, модификация, введенная в клетку путем пронуклеарной инъекции.
Согласно одному аспекту предусматривается последовательность вариабельного домена антитела, полученная у описанного в настоящем документе не относящегося к человеку животного.
Согласно одному аспекту предусматривается терапевтическое средство для людей, содержащее вариабельный домен антитела, содержащий последовательность, полученную от описанного в настоящем документе не относящегося к человеку животного.
Согласно одному аспекту предусматривается способ получения последовательности вариабельной области антитела от не относящегося к человеку животного, причем последовательность вариабельной области антитела происходит из сегмента VH1-69 или сегмента VH1-2 человека, причем способ предусматривает (а) иммунизацию не относящегося к человеку животного представляющим интерес антигеном, причем не относящееся к человеку животное содержит замещение в эндогенном локусе тяжелой цепи иммуноглобулина всех или по существу всех не относящихся к человеку вариабельных сегментов единственным вариабельным сегментом человека, причем единственный вариабельный сегмент человека представляет собой сегмент VH1-69 или сегмент VH1-2, и причем не относящееся к человеку животное по существу не способно к образованию последовательности вариабельной области тяжелой цепи иммуноглобулина, который не происходит от VH1-69 сегмента или VH1-2 сегмента человека; (b) предоставление не относящемуся к человеку животному возможности развивать иммунный ответ в отношении представляющего интерес антигена; и (с) идентификацию или выделение последовательности вариабельной области тяжелой цепи иммуноглобулина не относящегося к человеку животного, причем антитело связывается с представляющим интерес антигеном.
Согласно одному варианту осуществления единственный вариабельный сегмент человека представляет собой сегмент VH1-69.
Согласно одному варианту осуществления последовательность вариабельной области антитела происходит из SEQ ID NO: 34. Согласно одному варианту осуществления последовательность вариабельной области антитела идентична SEQ ID NO: 34 по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 98%. Согласно одному варианту осуществления последовательность вариабельной области антитела содержит SEQ ID NO: 34.
Согласно одному варианту осуществления единственный вариабельный сегмент человека представляет собой сегмент VH1-2.
Согласно одному варианту осуществления последовательность вариабельной области антитела происходит из SEQ ID NO: 60. Согласно одному варианту осуществления последовательность вариабельной области антитела идентична SEQ ID NO: 60 по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 98%. Согласно одному варианту осуществления последовательность вариабельной области антитела содержит SEQ ID NO: 60.
Согласно одному варианту осуществления иммунный ответ на антиген характеризуется титром антител, который приблизительно в 6×104-5×105 раз больше, чем двухкратный фон, как определено в анализе ELISA. Согласно конкретному варианту осуществления титр антител приблизительно в 1×105-2×105 раз больше, чем двухкратный фон, как определено в анализе ELISA. Согласно конкретному варианту осуществления титр антител приблизительно в 1,5×105 раз больше, чем двухкратный фон, как определено в анализе ELISA. Согласно одному варианту осуществления антиген представляет собой рецептор клеточной поверхности человека.
Согласно одному аспекту предусматривается способ для получения репертуара вариабельных областей антитела человека у не относящегося к человеку животного, причем вариабельные области тяжелой цепи человека репертуара происходят из того же представителя семейства генов VH и одного из множества сегментов DH, и одного из множества сегментов JH, причем репертуар характеризуется наличием последовательностей FR1 (каркасная область 1), CDR1, FR2, CDR2 и FR3 тяжелой цепи иммуноглобулина из единственного представителя семейства гена VH. Согласно одному варианту осуществления репертуар дополнительно характеризуется наличием множества различных последовательностей CDR3+FR4.
Согласно одному варианту осуществления единственное семейство генов VH выбирают из семейства VH 1, 2, 3, 4, 5, 6 и 7. Согласно конкретному варианту осуществления единственное семейство генов VH представляет собой семейство VH1. Согласно одному варианту осуществления единственное семейство генов VH выбирают из VH1-2, VH1-69, VH2-26, VH2-70 и VH3-23. Согласно конкретному варианту осуществления единственный представитель семейства генов VH представляет собой VH1-69. Согласно конкретному варианту осуществления единственный представитель семейства генов VH представляет собой VH1-2.
Согласно одному варианту осуществления репертуар содержит последовательности FR1, CDR1, FR2, CDR2 и FR3 тяжелой цепи, полученные из сегмента VH1-69. Согласно конкретному варианту осуществления репертуар содержит последовательности FR1, CDR1, FR2, CDR2 и FR3 тяжелой цепи, полученные из SEQ ID NO: 35. Согласно конкретному варианту осуществления репертуар содержит последовательности FR1, CDR1, FR2, CDR2 и FR3 тяжелой цепи из SEQ ID NO: 35.
Согласно одному варианту осуществления репертуар содержит последовательности FR1, CDR1, FR2, CDR2 и FR3 тяжелой цепи, полученные из сегмента VH1-2. Согласно конкретному варианту осуществления репертуар содержит последовательности FR1, CDR1, FR2, CDR2 и FR3 тяжелой цепи, полученные из SEQ ID NO: 61. Согласно конкретному варианту осуществления репертуар содержит последовательности FR1, CDR1, FR2, CDR2 и FR3 тяжелой цепи из SEQ ID NO: 61.
Согласно одному аспекту предусматривается биологическая (т.е. in vivo) система для получения множества различных последовательностей CDR3 человека, отражающая множество реаранжировок единственного генного сегмента VH человека с множеством сегментов D и J человека, причем система продуцирует вариабельные домены тяжелой цепи человека, характеризующиеся наличием последовательностей FR1-CDR1-FR2-CDR2-FR3 человека, которые идентичны, но для соматических гипермутаций, причем вариабельные домены тяжелой цепи характеризуются тем, что они соматически гипермутированы и получены из единственного генного сегмента VH человека и множества сегментов D и J человека; причем система содержит генетически модифицированное описанное в настоящем документе не относящееся к человеку животное (например, грызуна, например, мышь или крысу).
Согласно одному варианту осуществления единственный генный сегмент VH человека выбирают из VH1-2, VH1-69, VH2-26, VH2-70 и VH3-23. Согласно одному варианту осуществления единственный генный сегмент VH человека представляет собой VH1-69. Согласно одному варианту осуществления единственный генный сегмент VH человека представляет собой VH1-2. Согласно одному варианту осуществления единственный генный сегмент VH человека идентифицируют в таблице 1. Согласно одному варианту осуществления единственный генный сегмент VH человека идентифицируют в таблице 2. Согласно одному варианту осуществления единственный генный сегмент VH человека идентифицируют в таблице 3.
Согласно одному аспекту предусматривается способ in vivo для получения множества последовательностей CDR тяжелых цепей, полученных из реаранжировок единственного генного сегмента VH человека с множеством сегментов D и J человека, причем способ производит вариабельные домены тяжелой цепи человека, характеризующиеся наличием последовательностей FR1-CDR1-FR2-CDR2-FR3 человека, которые идентичны, но для соматических гипермутаций, причем вариабельные домены тяжелой цепи характеризуются тем, что они соматически гипермутированы и получены из единственного генного сегмента VH человека и множества сегментов D и J человека; причем система содержит генетически модифицированное описанное в настоящем документе не относящееся к человеку животное (например, грызуна, например, мышь или крысу).
Согласно одному варианту осуществления способ предусматривает воздействие на описанное в настоящем документе не относящееся к человеку животное представляющим интерес антигеном, предоставление не относящемуся к человеку животному возможности развивать иммунный ответ на антиген, причем иммунный ответ производит множество последовательностей CDR тяжелой цепи, полученные из реаранжировок единственного генного сегмента VH человека с одним из сегментов D человека и одним из сегментов J человека, и идентификацию набора CDR тяжелой цепи, которые связываются с антигеном. Согласно одному варианту осуществления способ предусматривает выделение из животного последовательности нуклеиновой кислоты, которая кодирует домен VH человека, который содержит CDR тяжелой цепи.
Согласно одному варианту осуществления последовательности CDR тяжелой цепи происходят из реаранжировки генного сегмента VH1-69 человека. Согласно одному варианту осуществления последовательности CDR тяжелой цепи происходят из реаранжировки генного сегмента VH1-2 человека.
Согласно одному аспекту предусматривается способ получения множества различных последовательностей CDR3 и FR4 у не относящегося к человеку животного, предусматривающий воздействие на не относящееся к человеку животное, которое содержит локус вариабельного гена тяжелой цепи иммуноглобулина с репертуаром сегментов VH, ограниченных до единственного представителя семейства сегментов VH, представляющего интерес антигена, предоставление не относящемуся к человеку животному возможности развивать иммунный ответ на антиген, причем иммунный ответ производит репертуар В-клеток, чьи вариабельные домены тяжелой цепи получены из единственного представителя семейства сегментов VH и содержат множество различных последовательностей CDR3 и FR4.
Согласно одному варианту осуществления единственный представитель семейства сегментов VH является человеческим. Согласно одному варианту осуществления не относящееся к человеку животное выбирают из мыши, крысы и кролика. Согласно одному варианту осуществления представляющий интерес антиген выбирают из лиганда, рецептора, внутриклеточного белка и секретируемого белка. Согласно одному варианту осуществления представляющий интерес антиген представляет собой описанный в настоящем документе патоген человека.
Согласно одному варианту осуществления единственного представителя семейства генов VH человека выбирают из VH1-2, VH1-69, VH2-26, VH2-70 и VH3-23. Согласно одному варианту осуществления единственный представитель семейства генов VH человека представляет собой VH1-69. Согласно одному варианту осуществления единственный представитель семейства генов VH человека представляет собой VH1-2. Согласно одному варианту осуществления единственный представитель семейства генов VH человека идентифицируют в таблице 1. Согласно одному варианту осуществления единственный представитель семейства генов VH человека идентифицируют в таблице 2. Согласно одному варианту осуществления единственный представитель семейства генов VH человека идентифицируют в таблице 3.
Согласно одному аспекту предусматривается нуклеотидная последовательность, кодирующая вариабельную область иммуноглобулина, полученную от описанного в настоящем документе не относящегося к человеку животного.
Согласно одному аспекту предусматривается аминокислотная последовательность вариабельной области тяжелой цепи или легкой цепи иммуноглобулина, полученного от описанного в настоящем документе не относящегося к человеку животного.
Согласно одному аспекту предусматривается нуклеотидная последовательность вариабельной области тяжелой цепи или легкой цепи иммуноглобулина, кодирующая вариабельную область антитела, полученного от описанного в настоящем документе не относящегося к человеку животного.
Согласно одному аспекту предусматривается антитело или его антигенсвязывающий фрагмент (например. Fab, F(ab)2, scFv), полученный от описанного в настоящем документе не относящегося к человеку животного.
Согласно одному аспекту предусматривается мышь, содержащая ограниченный локус тяжелой цепи иммуноглобулина, характеризующийся наличием единственного генного сегмента VH человека, одного или нескольких генных сегментов DH человека и одного или нескольких генных сегментов JH человека, причем единственный генный сегмент VH человека находится в эндогенном локусе мыши и генный сегмент VH функционально связан с одним или несколькими генными сегментами DH человека, одним или несколькими генными сегментами JH человека и эндогенным константным геном тяжелой цепи иммуноглобулина.
Согласно одному варианту осуществления мышь дополнительно содержит гуманизированный локус легкой цепи иммуноглобулина, содержащий один или несколько генных сегментов VL человека и один или несколько генных сегментов JL человека, причем генные сегменты VL человека и генные сегменты JL человека функционально связаны с не относящимся к человеку геном константной области легкой цепи иммуноглобулина. Согласно конкретному варианту осуществления генные сегменты VL и JL человека находятся в эндогенном локусе легкой цепи мыши, и причем не относящийся к человеку ген константной области легкой цепи иммуноглобулина представляет собой ген мыши.
Согласно одному варианту осуществления гуманизированный локус легкой цепи иммуноглобулина находится на трансгене, и ген константной области выбирают из мыши, крысы и человека.
Согласно одному варианту осуществления генные сегменты VL и JL человека представляют собой генные сегменты Vκ и Jκ. Согласно одному варианту осуществления генные сегменты VL и JL человека представляют собой генные сегменты Vλ и Jλ.
Согласно одному аспекту предусматривается не относящееся к человеку животное, причем не относящееся к человеку животное содержит репертуар В-клеток, который экспрессирует вариабельные домены тяжелой цепи иммуноглобулина, полученные из единственного представителя семейства сегментов V. Согласно одному варианту осуществления по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 95% репертуара В-клеток вариабельного домена тяжелой цепи не относящегося к человеку иммуноглобулина, экспрессированного в репертуаре В-клеток, происходит от того же представителя семейства сегментов V. Согласно конкретному варианту осуществления процент составляет по меньшей мере 90%. Согласно одному варианту осуществления репертуар В-клеток состоит преимущественно из периферических В-клеток (крови). Согласно одному варианту осуществления репертуар В-клеток состоит преимущественно из В-клеток селезенки. Согласно одному варианту осуществления репертуар В-клеток состоит преимущественно из В-клеток костного мозга. Согласно одному варианту осуществления репертуар В-клеток состоит преимущественно из периферических В-клеток, В-клеток селезенки и В-клеток костного мозга.
Согласно одному аспекту предусматривается генетически модифицированное не относящееся к человеку животное, причем более 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80% или более 90% В-клеток не относящегося к человеку животного, которые экспрессируют вариабельный домен тяжелый цепи иммуноглобулина, экспрессируют вариабельный домен тяжелый цепи иммуноглобулина, полученный из единственного представителя семейства генных сегментов VH. Согласно одному варианту осуществления по меньшей мере 75% В-клеток не относящегося к человеку животного, которые экспрессируют вариабельный домен тяжелый цепи иммуноглобулина, экспрессируют вариабельный домен тяжелый цепи иммуноглобулина, полученный из единственного представителя семейства генных сегментов VH. Согласно конкретному варианту осуществления процент составляет по меньшей мере 90%. Согласно одному варианту осуществления все В-клетки, которые экспрессируют домен тяжелой цепи, который происходит из единственного представителя семейства гена VH.
Согласно одному аспекту предусматривается генетически модифицированная мышь, которая продуцирует антигенспецифическую популяцию В-клеток в ответ на иммунизацию представляющим интерес антигеном, причем по меньшей мере 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80% или более 90% указанной антигенспецифической популяции В-клеток экспрессирует тяжелые цепи иммуноглобулина, все из которых получены из того же генного сегмента VH. Согласно одному варианту осуществления по меньшей мере 75% антигенспецифической популяции В-клеток экспрессирует тяжелые цепи иммуноглобулина, полученные из того же генного сегмента VH. Согласно одному варианту осуществления все антигенспецифические В-клетки экспрессируют тяжелые цепи иммуноглобулина, полученные из того же генного сегмента VH.
Согласно одному аспекту предусматривается не относящееся к человеку животное, содержащее ограниченный репертуар генных сегментов VH, причем ограничение относится к генному сегменту VH1-69 человека или генному сегменту VH1-69, который по меньшей мере на приблизительно 75,5%, 76,5%, 86,7%, 87,8%, 94,9%, 96,9%, 98% или 99% идентичен генному сегменту VH1-69*01. Согласно конкретному варианту осуществления ограниченный репертуар выбирают из одного или нескольких вариантов VH1-69 фигуры 15.
Согласно одному аспекту предусматривается не относящееся к человеку животное, содержащее ограниченный репертуар генных сегментов VH, причем ограничение относится к генному сегменту VH1-2 человека или генному сегменту VH1-2, который по меньшей мере на приблизительно 94,9%, 95,9%, 96,9%, 98% или 99% идентичен генному сегменту VH1-2. Согласно конкретному варианту осуществления ограниченный репертуар выбирают из одного или нескольких вариантов VH1-2 фигуры 18.
Согласно одному варианту осуществления не относящееся к человеку животное представляет собой мышь.
Согласно одному варианту осуществления мышь проявляет иммунофенотип, характеризующийся более высоким соотношением зрелых В-клеток к незрелым В-клеткам, по сравнению с мышью дикого типа. Согласно конкретному варианту осуществления соотношение вычисляют у В-клеток, собранных из селезенки. Согласно одному варианту осуществления мышь проявляет популяцию зрелых В-клеток размером приблизительно 1×107. Согласно одному варианту осуществления мышь проявляет популяцию незрелых В-клеток размером приблизительно 0,5×107. Согласно одному варианту осуществления мышь проявляет соотношение зрелых В-клеток к незрелым В-клеткам в селезенке мыши, превышающее приблизительно в 1,5-2 раза проявляемое мышью дикого типа.
Согласно одному варианту осуществления соотношение вычисляют у В-клеток, собранных из костного мозга. Согласно конкретному варианту осуществления мышь проявляет популяцию зрелых В-клеток размером приблизительно 3×105. Согласно одному варианту осуществления мышь проявляет популяцию незрелых В-клеток размером приблизительно 7×105. Согласно одному варианту осуществления мышь проявляет соотношение зрелых В-клеток к незрелым В-клеткам в селезенке мыши, превышающее приблизительно в 3 раза или приблизительно в 3,3 раза проявляемое мышью дикого типа.
Согласно одному варианту осуществления мышь проявляет иммунофенотип, характеризующийся более высоким числом пре-В-клеток в костном мозге, по сравнению с мышью дикого типа. Согласно конкретному варианту осуществления мышь проявляет популяцию пре-В-клеток в костном мозге мыши, которая превышает от приблизительно 2,5 раз до приблизительно 3 раз проявляемую в костном мозге мыши дикого типа. Согласно конкретному варианту осуществления мышь проявляет популяцию пре-В-клеток в костном мозге мыши, которая превышает приблизительно в 2,75 раза проявляемую в костном мозге мыши дикого типа.
Согласно одному варианту осуществления мышь проявляет иммунофенотип, характеризующийся выбранным из группы, состоящей из популяции CD19+ В-клеток селезенки, составляющей приблизительно 80% В-клеток дикого типа, популяции CD3+ Т-клеток селезенки, составляющей приблизительно столько же, как и у мыши дикого типа, и их комбинации.
Согласно одному варианту осуществления мышь содержит популяцию лимфоцитов, чье количество CD19+ В-клеток в селезенке в % приблизительно такое же, как у мыши дикого типа. Согласно одному варианту осуществления количество CD 19+ B-клеток в селезенке мыши составляет по меньшей мере 50% от числа CD19+ В-клеток в селезенке мыши дикого типа.
Согласно одному варианту осуществления не относящееся к человеку животное содержит по меньшей мере от приблизительно 75% до приблизительно 80% CD19+ B-клеток в костном мозге, по сравнению с мышью дикого типа.
Согласно одному варианту осуществления общее количество CD19+ костных клеток в бедренной кости мыши составляет не менее, чем приблизительно 30%, 40%, 50%, 60% или 75% от общего числа CD19+ клеток костного мозга у мыши дикого типа.
Согласно одному варианту осуществления мышь экспрессирует IgD и IgM приблизительно на том же уровне, который наблюдался у мыши дикого типа.
Согласно одному аспекту предусматривается мышь, содержащая ограниченный репертуар сегментов VH человека, дополнительно содержащая гуманизированный локус вариабельного сегмента легкой цепи иммуноглобулина, причем соотношение легких цепей λ к κ, экспрессируемое у мыши, приблизительно такое же, как и у мыши дикого типа.
Согласно одному аспекту предусматривается мышь, содержащая ограниченный локус тяжелой цепи иммуноглобулина, характеризующийся наличием единственного генного сегмента VH, одного или нескольких генных сегментов DH и одного или нескольких генных сегментов JH, причем единственный генный сегмент VH представляет собой полиморфный генный сегмент VH.
Согласно одному варианту осуществления полиморфный генный сегмент VH представляет собой генный сегмент VH человека, который связан с большим числом копий в популяциях человека. Согласно одному варианту осуществления генный сегмент VH человека выбирают из VH1-2, VH1-69, VH2-26, VH2-70, VH3-23 или их полиморфного варианта. Согласно конкретному варианту осуществления генный сегмент VH человека представляет собой генный сегмент VH1-69. Согласно другому конкретному варианту осуществления генный сегмент VH человека представляет собой генный сегмент VH1-2.
Согласно одному варианту осуществления единственный генный сегмент VH функционально связан с геном константной области иммуноглобулина человека, мыши или химеры человека/мыши. Согласно конкретному варианту осуществления ген константной области иммуноглобулина представляет собой ген константной области мыши. Согласно одному варианту осуществления константный ген иммуноглобулина содержит последовательность человека, выбранную из CH1 человека, шарнира человека, CH2 человека, CH3 человека и их комбинации Согласно одному варианту осуществления константный ген мыши находится в эндогенном локусе тяжелой цепи иммуноглобулина.
Согласно одному варианту осуществления мышь дополнительно содержит генный сегмент VL иммуноглобулина человека, функционально связанный с генным сегментом J и константным геном легкой цепи. Согласно конкретному варианту осуществления генный сегмент VL и/или генный сегмент J выбирают из генного сегмента к человека и генного сегмента λ человека. Согласно одному варианту осуществления генные сегменты VL и/или J представляют собой генные сегменты к человека.
Согласно различным вариантам осуществления мышь содержит делецию всех или по существу всех эндогенных генных сегментов VH.
Согласно различным вариантам осуществления не относящееся к человеку животное содержит инактивированный эндогенный локус вариабельного гена тяжелой цепи. Согласно различным вариантам осуществления инактивированный эндогенный локус вариабельного гена тяжелой цепи не функционально связан с эндогенным геном константной области тяжелой цепи.
Согласно одному аспекту предусматривается мышь, причем мышь характеризуется экспрессией иммуноглобулина сыворотки, причем более 80% иммуноглобулина сыворотки содержит вариабельный домен тяжелой цепи человека и родственный вариабельный домен легкой цепи человека, причем вариабельный домен тяжелой цепи человека происходит из репертуара генных сегментов VH, состоящего преимущественно из единственного генного сегмента VH человека и/или его полиморфных вариантов.
Согласно одному варианту осуществления единственный генный сегмент VH человека представляет собой генный сегмент VH1-69 человека и/или его полиморфные варианты. Согласно одному варианту осуществления единственный генный сегмент VH человека представляет собой генный сегмент VH1-2 человека и/или его полиморфные варианты.
Согласно одному аспекту предусматривается мышь, содержащая в своей зародышевой линии, замещение в эндогенном локусе тяжелой цепи иммуноглобулина всех или по существу всех эндогенных генных сегментов VH единственным генным сегментом VH человека и/или его полиморфными вариантами. Согласно одному варианту осуществления единственный генный сегмент VH человека представляет собой генный сегмент VH1-69 человека и/или его полиморфные варианты. Согласно одному варианту осуществления единственный генный сегмент VH человека представляет собой генный сегмент VH1-2 человека и/или его полиморфные варианты.
Согласно одному варианту осуществления мышь дополнительно содержит замещение в эндогенном локусе легкой цепи им всех или по существу всех эндогенных генных сегментов VL одним или несколькими генными сегментами VL человека. Согласно конкретному варианту осуществления мышь дополнительно содержит один или несколько генных сегментов JL человека, функционально связанные с генными сегментами VL человека.
Согласно одному аспекту предусматривается использование описанной в настоящем документе мыши для получения нуклеотидной последовательности вариабельной области иммуноглобулина. Согласно одному варианту осуществления последовательность содержит реаранжированный генный сегмент VH1-69. Согласно одному варианту осуществления последовательность содержит реаранжированный генный сегмент VH 1-2.
Согласно одному варианту осуществления нуклеотидная последовательность вариабельной области иммуноглобулина по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 98% идентична генному сегменту VH1-69 человека. Согласно конкретному варианту осуществления нуклеотидная последовательность вариабельной области иммуноглобулина по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 98% идентична SEQ ID NO: 34. Согласно различным вариантам осуществления генный сегмент VH1-69 человека идентифицируют из таблицы 2.
Согласно одному варианту осуществления нуклеотидная последовательность вариабельной области иммуноглобулина кодирует аминокислотную последовательность, которая по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 98% идентична SEQ ID NO: 35.
Согласно одному варианту осуществления нуклеотидная последовательность вариабельной области иммуноглобулина по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 98% идентична генному сегменту VH1-2 человека. Согласно конкретному варианту осуществления нуклеотидная последовательность вариабельной области иммуноглобулина по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 98% идентична SEQ ID NO: 60 Согласно различным вариантам осуществления генный сегмент VH1-2 человека идентифицируют из таблицы 3.
Согласно одному варианту осуществления нуклеотидная последовательность вариабельной области иммуноглобулина кодирует аминокислотную последовательность, которая по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 98% идентична SEQ ID NO: 61.
Согласно одному аспекту предусматривается использование описанной в настоящем документе мыши для получения полностью человеческого Fab или полностью человеческого F(ab)2. Согласно одному варианту осуществления полностью человеческий Fab или полностью человеческий F(ab)2 содержит вариабельную область тяжелой цепи, которая содержит реаранжированный генный сегмент VH1-69 человека. Согласно одному варианту осуществления полностью человеческий Fab или полностью человеческий F(ab)2 содержит вариабельную область тяжелой цепи, которая содержит реаранжированный генный сегмент VH1-2 человека.
Согласно одному аспекту предусматривается использование описанной в настоящем документе мыши для получения иммортализованной клеточной линии.
Согласно одному аспекту предусматривается использование описанной в настоящем документе мыши для получения гибридомы или квадромы.
Согласно одному аспекту предусматривается использование описанной в настоящем документе мыши для получения библиотеки фагов, содержащей вариабельные области тяжелой цепи человека и вариабельные области легкой цепи человека.
Согласно одному варианту осуществления вариабельные области тяжелой цепи человека получают из генного сегмента VH1-69 человека, который содержит последовательность, выбранную из SEQ ID NO: 34, SEQ ID NO: 36, SEQ ID NO: 38, SEQ ID NO: 40, SEQ ID NO: 42, SEQ ID NO: 44, SEQ ID NO: 46, SEQ ID NO: 48, SEQ ID NO: 50, SEQ ID NO: 52, SEQ ID NO: 54, SEQ ID NO: 56 и SEQ ID NO: 58.
Согласно одному варианту осуществления вариабельные области тяжелой цепи человека получают из генного сегмента VH1-69 человека, который содержит последовательность, выбранную из SEQ ID NO: 35, SEQ ID NO: 37, SEQ ID NO: 39, SEQ ID NO: 41, SEQ ID NO: 43, SEQ ID NO: 45, SEQ ID NO: 47, SEQ ID NO: 49, SEQ ID NO: 51, SEQ ID NO: 53, SEQ ID NO: 55, SEQ ID NO: 57 и SEQ ID NO: 59.
Согласно одному варианту осуществления все вариабельные области тяжелой цепи человека получают из генного сегмента VH1-2 человека, который содержит последовательность, выбранную из SEQ ID NO: 60, SEQ ID NO: 62, SEQ ID NO: 64, SEQ ID NO: 66 и SEQ ID NO: 68.
Согласно одному варианту осуществления вариабельные области тяжелой цепи человека получают из генного сегмента VH1-2 человека, который содержит последовательность, выбранную из SEQ ID NO: 61, SEQ Ю NO: 63, SEQ ID NO: 65, SEQ ID NO: 67 и SEQ ID NO: 69.
Согласно одному аспекту предусматривается применение описанной в настоящем документе мыши для продукции последовательности вариабельной области для получения антитела человека, предусматривающее (а) иммунизацию описанной в настоящем документе мыши представляющим интерес антигеном, (b) выделение лимфоцита из иммунизированной мыши (а), (с) воздействие на лимфоцит одним или несколькими мечеными антителами, (d) идентификацию лимфоцита, который способен к связыванию с представляющим интерес антигеном и (е) амплификацию одной или нескольких последовательностей вариабельной области нуклеиновой кислоты из лимфоцита, таким образом получая последовательность вариабельной области.
Согласно одному варианту осуществления лимфоцит получают или выделяют из селезенки мыши. Согласно одному варианту осуществления лимфоцит получают или выделяют из лимфатического узла мыши. Согласно одному варианту осуществления лимфоцит получают или выделяют из костного мозга мыши. Согласно одному варианту осуществления лимфоцит получают или выделяют из крови мыши.
Согласно одному варианту осуществления меченое антитело представляет собой конъюгированное с флуорофором антитело. Согласно одному варианту осуществления одно или несколько конъюгированных с флуорофором антител выбирают из IgM, IgG и/или их комбинации.
Согласно одному варианту осуществления лимфоцит представляет собой В-клетку.
Согласно одному варианту осуществления одна или несколько последовательностей вариабельной области нуклеиновой кислоты содержит последовательность вариабельной области тяжелой цепи. Согласно одному варианту осуществления одна или несколько последовательностей вариабельной области нуклеиновой кислоты содержит последовательность вариабельной области легкой цепи. Согласно конкретному варианту осуществления последовательность вариабельной области легкой цепи представляет собой последовательность вариабельной области легкой цепи κ иммуноглобулина. Согласно одному варианту осуществления одна или несколько последовательностей вариабельной области нуклеиновой кислоты содержит последовательность вариабельной области тяжелой цепи и легкой цепи κ.
Согласно одному варианту осуществления предусматривается применение описанной в настоящем документе мыши для продукции последовательности вариабельной области тяжелой цепи и легкой цепи κ для получения антитела человека, предусматривающее (а) иммунизацию описанной в настоящем документе мыши представляющим интерес антигеном, (b) выделение селезенки из иммунизированной мыши (а), (с) воздействие на В-лимфоциты из селезенки одним или несколькими мечеными антителами, (d) идентификацию В-лимфоцита (с), который способен к связыванию с представляющим интерес антигеном и (е) амплификацию нуклеотидной последовательности вариабельной области тяжелой цепи и нуклеотидной последовательности вариабельной области легкой цепи κ из В-лимфоцита, таким образом получая последовательности вариабельной области тяжелой цепи и легкой цепи κ.
Согласно одному варианту осуществления предусматривается применение описанной в настоящем документе мыши для продукции последовательности вариабельной области тяжелой цепи и легкой цепи κ для получения антитела человека, предусматривающее (а) иммунизацию описанной в настоящем документе мыши представляющим интерес антигеном, (b) выделение одного или нескольких лимфатических узлов из иммунизированной мыши (а), (с) воздействие на В-лимфоциты из одного или нескольких лимфатических узлов одним или несколькими мечеными антителами, (d) идентификацию В-лимфоцита (с), который способен к связыванию с представляющим интерес антигеном и (е) амплификацию нуклеотидной последовательности вариабельной области тяжелой цепи и нуклеотидной последовательности вариабельной области легкой цепи κ из В-лимфоцита, таким образом получая последовательности вариабельной области тяжелой цепи и легкой цепи κ.
Согласно одному варианту осуществления предусматривается применение описанной в настоящем документе мыши для продукции последовательности вариабельной области тяжелой цепи и легкой цепи κ для получения антитела человека, предусматривающее (а) иммунизацию описанной в настоящем документе мыши представляющим интерес антигеном, (b) выделение костного мозга из иммунизированной мыши (а), (с) воздействие на В-лимфоциты из костного мозга одним или несколькими мечеными антителами, (d) идентификацию В-лимфоцита (с), который способен к связыванию с представляющим интерес антигеном и (е) амплификацию нуклеотидной последовательности вариабельной области тяжелой цепи и нуклеотидной последовательности вариабельной области легкой цепи κ из В-лимфоцита, таким образом получая последовательности вариабельной области тяжелой цепи и легкой цепи κ. Согласно различным вариантам осуществления одно или несколько меченых антител выбирают из IgM, IgG и/или их комбинации.
Согласно различным вариантам осуществления представляющий интерес антиген представляет собой патоген, который поражает людей, включающий, например, вирусный антиген. Иллюстративные вирусные патогены включают в себя, например, главным образом патогены из семейств Adenoviridae, bacteria Picomaviridae, Herpesviridae, Hepadnaviridae, Flaviviridae, Retroviridae, Orthomyxoviridae, Paramyxoviridae, Papovaviridae, Polyomavirus, Rhabdoviridae и Togaviridae. Длина таких иллюстративных вирусов обычно находится в пределах 20-300 нанометров. Согласно различным вариантам осуществления представляющий интерес антиген представляет собой вирусный антиген, выбранный из вируса гепатита (например, вируса гепатита С, вируса гепатита В и т.п.), вируса иммунодефицита человека (ВИЧ) или вируса гриппа (например, H1N1).
Согласно различным вариантам осуществления предусматривается использование описанной в настоящем документе мыши для продукции последовательности вариабельной области тяжелой цепи и легкой цепи κ для получения антитела человека, дополнительно предусматривающее слияние амплифицированных последовательности вариабельной области тяжелой и легкой цепи с последовательностями константной области тяжелой и легкой цепи человека, экспрессию слитых последовательностей тяжелой и легкой цепи в клетке и восстановление экспрессированных последовательностей тяжелой и легкой цепи, таким образом получая антитело человека.
Согласно различным вариантам осуществления константные области тяжелой цепи человека выбирают из IgM, IgD, IgA, IgE и IgG. Согласно различным конкретным вариантам осуществления IgG выбирают из IgGI, IgG2, IgG3 и IgG4. Согласно различным вариантам осуществления константная область тяжелой цепи человека содержит CH1, шарнир, CH2, СнЗ, Сн4 или их комбинацию. Согласно различным вариантам осуществления константная область легкой цепи представляет собой константную область к иммуноглобулина. Согласно различным вариантам осуществления клетку выбирают из клетки HeLa, клетки DU145, клетки Lncap, клетки MCF-7, клетки MDA-MB-438, клетки РС3, клетки T47D, клетки ТНР-1, клеткик U87, клетки SHSY5Y (нейробластомы человека), клетки Saos-2, клетки Vero, клетки СНО, клетки GH3, клетки PC 12, клетки сетчатки человек (например, клетки PER.C6™) и клетки МС3Т3. Согласно конкретному варианту осуществления клетка представляет собой клетку СНО.
Согласно одному аспекту предусматривается способ для получения обратного химерного антитела грызуна/человека, специфичного против представляющего интерес антигена, предусматривающий стадии иммунизации антигеном описанной в настоящем документе мыши, выделения по меньшей мере одной клетки от мыши, продуцирующей обратное химерное антитело мыши/человека, специфичное к антигену, культивирования по меньшей мере одной клетки, продуцирующей обратное химерное антитело мыши/человека, специфичное к антигену, а также получение указанного антитела.
Согласно одному варианту осуществления обратное химерное антитело мыши/человека содержит вариабельный домен тяжелой цепи человека, слитый с константным геном тяжелой цепи мыши или крысы, и вариабельный домен легкой цепи человека, слитый с константным геном легкой цепи мыши или крысы, или человека. Согласно конкретному варианту осуществления вариабельный домен тяжелой цепи человека содержит реаранжированный генный сегмент VH1-б9 человека или генный сегмент VH1-2 человека.
Согласно одному варианту осуществления культивирование по меньшей мере одной клетки, продуцирующей обратное химерное антитело грызуна/человека, специфичное к антигену, проводят по меньшей мере на одной клетке гибридомы, полученной по меньшей мере из одной клетки, выделенной из мыши.
Согласно одному варианту осуществления представляющий интерес антиген представляет собой патоген, который поражает описанных в настоящем документе людей.
Согласно одному аспекту предусматривается способ для получения полностью человеческого антитела, специфичного в отношении представляющего интерес антигена, предусматривающий стадии иммунизации антигеном описанной в настоящем документе мыши, выделения по меньшей мере одной клетки от мыши, продуцирующей обратное химерное человеческое/относящееся к грызуну антитело, специфичное к антигену, получения по меньшей мере одной клетки, продуцирующей полностью человеческое антитело, полученное от обратного химерного антитела грызун/человек, специфичного к антигену, и культивирования по меньшей мере одной клетки, продуцирующей полностью человеческое антитело, а также получение указанного полностью человеческого антитела.
Согласно различным вариантам осуществления по меньшей мере одна клетка, выделенная из мыши, продуцирующая обратное химерное антитело грызун/человек, специфичное к антигену, представляет собой спленоцит или В-клетку.
Согласно различным вариантам осуществления антитело представляет собой моноклональное антитело.
Согласно различным вариантам осуществления антитело содержит вариабельную область тяжелой цепи, которая содержит реаранжированный генный сегмент Vn1-69 человека или генный сегмент VH1-2 человека.
Согласно различным вариантам осуществления иммунизацию представляющим интерес антигеном проводят белком, ДНК, комбинацией ДНК и белка или клетками, экспрессирующими антиген. Согласно одному варианту осуществления представляющий интерес антиген представляет собой патоген, который поражает описанных в настоящем документе людей.
Согласно одному аспекту предусматривается использование описанной в настоящем документе мыши для получения последовательности нуклеиновой кислоты, кодирующей вариабельную область иммуноглобулина или ее фрагмент. Согласно одному варианту осуществления используют последовательность нуклеиновой кислоты для получения антитела человека или его антигенсвязывающего фрагмента. Согласно одному варианту осуществления используют мышь для получения антигенсвязывающего белка, выбранного из антитела, мультиспецифического антитела (например, биспецифического антитела), ScFv, биспецифического ScFv, диатела, триатела, тетратела, V-NAR, VHH, VL., F(ab), F(ab)2, DVD (т.е. антигенсвязывающий белок двойного вариабельного домена), SVD (т.е. антигенсвязывающий белок единственного вариабельного домена) или биспецифические активаторы Т-клеток (BiTE).
Согласно одному аспекту предусматривается способ получения антигенсвязывающего белка человека, предусматривающий воздействие на генетически модифицированное описанное в настоящем документе не относящееся к человеку животное представляющим интерес антигеном, предоставление генетически модифицированному не относящемуся к человеку животному возможности усилить иммунный ответ на антиген, получение из генетически модифицированного не относящегося к человеку животного последовательности нуклеиновой кислоты вариабельного домена тяжелой цепи, кодирующей вариабельный домен тяжелой цепи человека, который специфически связывается с представляющим интерес антигеном, клонирование последовательности нуклеиновой кислоты вариабельного домена тяжелой цепи до последовательности константной области человека, и экспрессирование в клетке млекопитающего антитела, содержащего последовательность вариабельного домена тяжелой цепи человека и последовательность константной области человека. Согласно одному варианту осуществления клетка млекопитающего представляет собой клетку СНО. Согласно одному варианту осуществления генетически модифицированное не относящееся к человеку животное содержит репертуар генных сегментов VH человека, который содержит преимущественно единственный генный сегмента VH человека, необязательно присутствующий в двух или более его полиморфных вариантах, функционально связанный с одним или несколькими сегментами D и/или J человека. Согласно одному варианту осуществления репертуар генных сегментов VH человека находится в эндогенном локусе не относящегося к человеку сегмента VH. Согласно одному варианту осуществления репертуар генных сегментов VH человека находится в локусе, который не представляет собой эндогенный локус сегмента VH. Согласно одному варианту осуществления генный сегмент VH человека реаранжирует с сегментом D человека и сегментом J человека с образованием реаранжированного гена VDJ человека, функционально связанного с последовательностью константной области, причем последовательность константной области выбирают из последовательности человека и последовательности грызуна (например, последовательности мыши или крысы, или хомяка). Согласно одному варианту осуществления последовательность константной области содержит последовательность, выбранную из CH1, шарнира, CH2, CH3 и их комбинации; согласно конкретному варианту осуществления последовательность константной области содержит CH1, шарнир, CH2 и CH3. Согласно одному варианту осуществления вариабельный домен человека и константная последовательность экспрессируют в клетке млекопитающего с родственным вариабельным доменом легкой цепи человека, полученным от той же мыши (например, последовательностью, полученной из той же В-клетки, что и последовательность вариабельного домена человека); согласно одному варианту осуществления последовательность, кодирующая вариабельный домен легкой цепи человека, полученную от мыши, затем сливают с последовательностью, кодирующей константную последовательность легкой цепи человека, и последовательность легкой цепи и последовательность тяжелой цепи экспрессируют в клетке млекопитающих.
Согласно одному варианту осуществления представляющий интерес антиген представляет собой патоген, который поражает описанных в настоящем документе людей.
Согласно одному аспекту предусматривается способ для получения вариабельного домена тяжелой цепи антитела, которое связывается с представляющим интерес антигеном, предусматривающий экспрессию в единственной клетке (а) первой последовательности VH иммунизированного описанного в настоящем документе не относящегося к человеку животного, причем первая последовательность VH слита с генной последовательностью Сн; и (b) генной последовательности VL иммунизированного описанного в настоящем документе не относящегося к человеку животного, причем генная последовательность VL слита с генной последовательностью CL человека; поддержание клетки в условиях, достаточных для экспрессии антитела; и выделение вариабельного домена тяжелой цепи антитела. Согласно одному варианту осуществления генная последовательность VL представляет собой родственную последовательность первой последовательности VH.
Согласно одному варианту осуществления клетка содержит вторую генную последовательность VH иммунизированного описанного в настоящем документе не относящегося к человеку животного, причем вторая генная последовательность VH слита с генной последовательностью Сн, причем первая генная последовательность VH кодирует домен VH, который специфически связывается с первым эпитопом, и вторая генная последовательность VH кодирует домен VH, который специфически связывается со вторым эпитопом, причем первый эпитоп и второй эпитоп не идентичны.
Согласно одному варианту осуществления все последовательности константной области представляют собой последовательности константной области человека. Согласно одному варианту осуществления представляющий интерес антиген представляет собой патоген, который поражает описанных в настоящем документе людей.
Согласно одному аспекту предусматривается способ получения биспецифического антитела человека, предусматривающий получение биспецифического антитела с использованием генных последовательностей вариабельной области человека из В-клеток описанного в настоящем документе не относящегося к человеку животного.
Согласно одному варианту осуществления способ предусматривает (а) идентификацию клонально выбранного лимфоцита не относящегося к человеку животного, причем не относящееся к человеку животное подвергли воздействию представляющим интерес антигеном и позволили развить иммунный ответ на представляющий интерес антиген, и причем лимфоцит экспрессирует антитело, которое специфически связывается с представляющим интерес антигеном, (b) получение из лимфоцита или антитела нуклеотидной последовательности, которая кодирует вариабельную область тяжелой цепи человека, которая специфически связывается с представляющим интерес антигеном, и (с) использование нуклеотидной последовательности, кодирующей вариабельную область тяжелой цепи человека, которая специфически связывается с представляющим интерес антигеном, в создании биспецифического антитела. Согласно конкретному варианту осуществления вариабельная область тяжелой цепи человека содержит реаранжированный генный сегмент VH1-2 или VH1-69.
Согласно одному варианту осуществления стадии от (а) до (с) выполняют в первый раз для первого представляющего интерес антигена для получения первой последовательности вариабельной области тяжелой цепи человека, и стадии от (а) до (с) выполняют второй раз для второго представляющего интерес антигена для получения второй последовательности вариабельной области тяжелой цепи человека, и причем первую последовательность вариабельной области тяжелой цепи человека экспрессируют слитой с первой константной областью тяжелой цепи человека с образованием первой тяжелой цепи человека, вторую последовательность вариабельной области тяжелой цепи человека экспрессируют слитой с константной областью тяжелой цепи человека с образованием второй тяжелой цепи человека, причем первая и вторая тяжелые цепи человека экспрессируют в присутствии единственной легкой цепи человека, экспрессированной из реаранжированного генного сегмента Vκl-39 или Vκ3-20 человека. Согласно конкретному варианту осуществления единственная легкая цепь человека содержит последовательность зародышевой линии.
Согласно одному варианту осуществления способ предусматривает (а) клонирование вариабельных областей тяжелой цепи из В-клеток описанного в настоящем документе не относящегося к человеку животного, на которое воздействовали первым представляющим интерес антигеном, и того же животного или другого не относящегося к человеку животного, которое является генетически таким же и на которое воздействовали вторым представляющим интерес антигеном; и (b) экспрессирование в клетке вариабельных областей тяжелой цепи (а) с той же константной областью тяжелой цепи и той же легкой цепью для получения биспецифического антитела.
Согласно одному аспекту предусматривается использование описанного в настоящем документе не относящегося к человеку животного для получения последовательности нуклеиновой кислоты, которая кодирует вариабельный домен тяжелой цепи человека. Согласно одному варианту осуществления вариабельный домен тяжелой цепи содержит реаранжированный генный сегмент VH человека, выбранный из VH1-2 и VH1-69.
Согласно одному аспекту предусматривается использование описанного в настоящем документе не относящегося к человеку животного для получения клетки, которая кодирует вариабельный домен тяжелой цепи человека. Согласно одному варианту осуществления вариабельный домен тяжелой цепи человека содержит реаранжированный генный сегмент VH человека, выбранный из VH1-2 и VH1-69.
Согласно одному аспекту предусматривается использование описанного в настоящем документе не относящегося к человеку животного для получения вариабельного домена антитела человека. Согласно одному варианту осуществления вариабельный домен содержит реаранжированный генный сегмент VH человека, выбранный из VH1-2 и VH1-69.
Согласно одному аспекту предусматривается использование описанного в настоящем документе не относящегося к человеку животного для получения антитела человека, предусматривающее получение антитела с использованием генной последовательности вариабельной области человека из В-клеток описанного в настоящем документе не относящегося к человеку животного. Согласно одному варианту осуществления антитело человека представляет собой биспецифическое антитело человека. Согласно конкретному варианту осуществления биспецифическое антитело содержит один вариабельный домен тяжелой цепи, полученный из реаранжированного генного сегмента VH1-2 или VH1-69 человека. Согласно одному варианту осуществления генные последовательности вариабельной области человека содержат реаранжированный генный сегмент VH1-2 или VH1-69 человека.
Согласно одному аспекту предусматривается использования описанного в настоящем документе не относящегося к человеку животного для выбора вариабельного домена тяжелой цепи иммуноглобулина человека. Согласно одному варианту осуществления вариабельный домен тяжелой цепи содержит реаранжированный генный сегмент VH человека, выбранный из VH1-2 или VH1-69.
Согласно одному аспекту предусматривается использование описанной в настоящем документе мыши для производства лекарственного средства (например, антигенсвязывающего белка) или для продукции последовательности, кодирующей вариабельную последовательность лекарственного средства (например, антигенсвязывающего белка), для лечения заболевания или нарушения человека. Согласно одному варианту осуществления вариабельная последовательность лекарственного средства содержит полиморфный генный сегмент VH человека. Согласно одному варианту осуществления вариабельная последовательность лекарственного средства содержит генный сегмент VH1-69 человека. Согласно одному варианту осуществления вариабельная последовательность лекарственного средства содержит генный сегмент VH1-2 человека.
Согласно одному аспекту предусматривается конструкция нуклеиновой кислоты, кодирующий вариабельный домен иммуноглобулина, полученный у описанной в настоящем документе мыши. Согласно одному варианту осуществления вариабельный домен представляет собой вариабельный домен тяжелой цепи. Согласно конкретному варианту осуществления вариабельный домен тяжелой цепи содержит реаранжированный генный сегмент VH человека, выбранный из VH1-2, VH1-69, VH2-26, VH2-70 или VH3-23. Согласно другому конкретному варианту осуществления вариабельный домен тяжелой цепи содержит реаранжированный генный сегмент VH1-2 человека. Согласно другому конкретному варианту осуществления вариабельный домен тяжелой цепи содержит реаранжированный генный сегмент VH1-69 человека.
Согласно одному варианту осуществления вариабельный домен представляет собой вариабельный домен легкой цепи. Согласно конкретному варианту осуществления вариабельный домен представляет собой вариабельный домен легкой цепи κ, который родственен вариабельному домену тяжелой цепи человека, который содержит реаранжированный генный сегмент VH1-69 человека. Согласно конкретному варианту осуществления вариабельный домен представляет собой вариабельный домен легкой цепи κ, который родственен вариабельному домену тяжелой цепи человека, который содержит реаранжированный генный сегмент VH1-2 человека.
Согласно одному аспекту предусматривается использование описанной в настоящем документе мыши для получения конструкции нуклеиновой кислоты, кодирующего вариабельный домен иммуноглобулина человека. Согласно одному варианту осуществления вариабельный домен представляет собой вариабельный домен легкой цепи. Согласно одному варианту осуществления вариабельный домен представляет собой вариабельный домен легкой цепи κ, который содержит реаранжированный генный сегмент Vκ человека, выбранный из Vκ4-l, Vκ5-2, Vκ7-3, Vκ2-4, Vκ1-5, Vκ1-6, Vκ3-7, Vκ1-8, Vκ1-9, Vκ2-10, Vκ3-11, Vκ1-12, Vκ1-13, Vκ2-14, Vκ3-15, Vκ1-16, Vκ1-17, Vκ2-18, Vκ2-19, Vκ3-20, Vκ6-21, Vκ1-22, Vκ1-23, Vκ2-24, Vκ3-25, Vκ2-26, Vκ1-27, Vκ2-28, Vκ2-29, Vκ2-30, Vκ3-31, Vκ1-32, Vκ1-33, Vκ3-34, Vκ1-35, Vκ2-36, Vκ1-37. Vκ2-38, Vκ1-39 и Vκ2-40.
Согласно одному варианту осуществления вариабельный домен представляет собой вариабельный домен тяжелой цепи. Согласно конкретному варианту осуществления вариабельный домен тяжелой цепи содержит реаранжированный генный сегмент VH, выбранный из VH1-2, VH1-69, VH2-26, VH2-70 или VH3-23. Согласно конкретному варианту осуществления вариабельный домен тяжелой цепи содержит реаранжированный генный сегмент VH1-69 человека. Согласно конкретному варианту осуществления вариабельный домен тяжелой цепи содержит реаранжированный генный сегмент VH1-2 человека.
Согласно одному аспекту предусматривается использование описанной в настоящем документе мыши для получения вариабельного домена иммуноглобулина человека. Согласно одному варианту осуществления вариабельный домен представляет собой вариабельный домен легкой цепи. Согласно одному варианту осуществления вариабельный домен представляет собой вариабельный домен легкой цепи κ, который содержит реаранжированный генный сегмент Vκ человека, выбранный из Vκ4-1, Vκ5-2, Vκ7-3, Vκ2-4, Vκ1-5, Vκ1-6, Vκ3-7, Vκ1-8, Vκ1-9, Vκ2-10, Vκ3-11, Vκ1-12, Vκ1-13, Vκ2-14, Vκ3-15, Vκ1-16, Vκ1-17, Vκ2-18, Vκ2-19, Vκ3-20, Vκ6-21, Vκ1-22, Vκ1-23, Vκ2-24, Vκ3-25, Vκ2-26, Vκ1-27, Vκ2-28, Vκ2-29, Vκ2-30, Vκ3-31, Vκ1-32, Vκ1-33, Vκ3-34, Vκ1-35, Vκ2-36, Vκ1-37, Vκ2-38, Vκ1-39 и Vκ2-40.
Согласно одному варианту осуществления вариабельный домен представляет собой вариабельный домен тяжелой цепи. Согласно конкретному варианту осуществления вариабельный домен тяжелой цепи содержит реаранжированный генный сегмент VH, выбранный из VH1-2, VH1-69, VH2-26, VH2-70 или VH3-23. Согласно конкретному варианту осуществления вариабельный домен тяжелой цепи содержит реаранжированный генный сегмент VH1-69 человека. Согласно конкретному варианту осуществления вариабельный домен тяжелой цепи содержит реаранжированный генный сегмент VH1-2 человека.
Согласно одному аспекту предусматривается применение описанного в настоящем документе не относящегося к человеку животного для получения последовательности нуклеиновой кислоты, кодирующей вариабельный домен тяжелой цепи человека. Согласно одному варианту осуществления вариабельный домен тяжелой цепи человека характеризуется наличием последовательностей FR1-CDR1-FR2-CDR2-FR3 человека, полученных из полиморфного генного сегмента VH человека. Согласно конкретному варианту осуществления генный сегмент VH человека выбирают из генного сегмента VH1-2, VH1-69, VH2-26, VH2-70 или VH3-23 человека. Согласно одному варианту осуществления генный сегмент VH человека представляет собой генный сегмент VH1-69 человека. Согласно одному варианту осуществления генный сегмент VH человека представляет собой генный сегмент VH1-2 человека.
Согласно одному аспекту предусматривается способ получения последовательности нуклеиновой кислоты, кодирующей домен VH человека, способ, предусматривающий иммунизацию описанного в настоящем документе не относящегося к человеку животного представляющим интерес антигеном, позволяя не относящемуся к человеку животному развивать иммунный ответ на представляющий интерес антиген, и получение из него последовательности нуклеиновой кислоты, кодирующей домен VH человека, который связывается с представляющим интерес антигеном. Согласно одному варианту осуществления способ дополнительно предусматривает получение последовательности нуклеиновой кислоты, кодирующей домен VL человека, который родственен домену VH человека, предусматривающий выделение В-клетки, кодирующей домен VH человека и домен VL человека, и получение из него последовательности вариабельных доменов тяжелой и легкой цепи. Согласно различным вариантам осуществления домен VH человека получают от реаранжированного генного сегмента VH1-69 или VH1-2 человека. Согласно различным вариантам осуществления домен VL человека выбирают из домена Vκ или VX, человека.
Согласно одному аспекту предусматривается применение описанного в настоящем документе не относящегося к человеку животного для получения терапевтического средства для людей, предусматривающее иммунизацию не относящегося к человеку животного представляющим интерес антигеном, предоставление не относящемуся к человеку животному возможности развивать иммунный ответ, и получение из животного последовательности нуклеиновой кислоты, кодирующей вариабельный домен иммуноглобулина, который связывается с представляющим интерес антигеном, а также применение вариабельного домена иммуноглобулина в терапевтическом средстве для людей. Согласно одному варианту осуществления вариабельный домен представляет собой вариабельный домен тяжелой цепи. Согласно конкретному варианту осуществления вариабельный домен тяжелой цепи получают от реаранжированного генного сегмента VH1-69 или VH1-2 человека. Согласно одному варианту осуществления вариабельный домен представляет собой вариабельный домен легкой цепи. Согласно конкретному варианту осуществления вариабельный домен легкой цепи получают из реаранжированного генного сегмента Vκ или VA, человека.
Согласно одному аспекту предусматривается способ получения терапевтического средства для людей, предусматривающий иммунизацию описанного в настоящем документе не относящегося к человеку животного представляющим интерес антигеном, предоставление не относящемуся к человеку животному возможности развивать иммунный ответ, и получение из животного последовательности нуклеиновой кислоты, кодирующей вариабельный домен иммуноглобулина, который связывается с представляющим интерес антигеном, а также применение вариабельного домена иммуноглобулина в терапевтическом средстве для людей. Согласно одному варианту осуществления вариабельный домен представляет собой вариабельный домен тяжелой цепи. Согласно конкретному варианту осуществления вариабельный домен тяжелой цепи получают от реаранжированного генного сегмента VH1-69 или VH1-2 человека. Согласно одному варианту осуществления вариабельный домен представляет собой вариабельный домен легкой цепи. Согласно конкретному варианту осуществления вариабельный домен легкой цепи получают из реаранжированного генного сегмента Vκ или Vλ человека.
Согласно одному аспекту предусматривается способ получения антигенсвязывающего белка человека, предусматривающий иммунизацию описанного в настоящем документе не относящегося к человеку животного представляющим интерес антигеном, позволяя не относящемуся к человеку животному развивать иммунный ответ, получение из мыши последовательности нуклеиновой кислоты, кодирующей вариабельный домен иммуноглобулина, который специфически связывается с представляющим интерес антигеном, клонирование последовательности нуклеиновой кислоты в вектор, подходящий для экспрессии нуклеиновой кислоты, причем последовательность нуклеиновой кислоты клонируют в рамке считывания с последовательностью нуклеиновой кислоты, кодирующей константную область иммуноглобулина человека или ее функциональный фрагмент, и внедрение вектора в клетку млекопитающего, и поддержание клетки в условиях, подходящих для экспрессии антигенсвязывающего белка, который содержит вариабельный домен иммуноглобулина и константную область иммуноглобулина или его функциональный фрагмент. Согласно одному варианту осуществления антигенсвязывающий белок представляет собой антитело человека. Согласно конкретному варианту осуществления антитело содержит вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, полученные от описанной в настоящем документе мыши. Согласно конкретному варианту осуществления антитело содержит вариабельный домен тяжелой цепи, полученный от описанной в настоящем документе мыши. Согласно различным вариантам осуществления вариабельный домен тяжелой цепи происходит от реаранжированного генного сегмента VH1-69 или VH1-2 человека.
Согласно одному аспекту предусматривается последовательность нуклеиновой кислоты, кодирующая антигенсвязывающий домен человека, полученный от описанного в настоящем документе не относящегося к человеку животного. Согласно одному варианту осуществления последовательность нуклеиновой кислоты кодирует домен VH иммуноглобулина человека. Согласно одному варианту осуществления последовательность нуклеиновой кислоты кодирует домен VH иммуноглобулина человека и родственный домен VL человека. Согласно различным вариантам осуществления домен VH человека происходит от реаранжированного генного сегмента VH1-69 или VH1-2 человека.
Согласно одному аспекту предусматривается способ получения антитела человека, предусматривающий иммунизацию описанного в настоящем документе не относящегося к человеку животного представляющим интерес антигеном, позволяя не относящемуся к человеку животному развивать иммунный ответ, сбор лимфоцита (например, В-клетки) из иммунизированного животного, слияние лимфоцита с клеткой миеломы с образованием клетки гибридомы, получение из клетки гибридомы последовательности нуклеиновой кислоты, которая кодирует домен VH человека и домен VL человека, клонирование последовательности нуклеиновой кислоты в рамке считывания (т.е. в функциональной связи) с последовательностью константной области человека для создания тяжелой цепи иммуноглобулина и легкой цепи иммуноглобулина, и экспрессию тяжелой и легкой цепей в клетке, способной экспрессировать полностью человеческое антитело. Согласно одному варианту осуществления клетка представляет собой клетку СНО. Согласно различным вариантам осуществления VH домен человека происходит от реаранжированного генного сегмента VH1-69 или VH1-2 человека.
Согласно одному аспекту предусматривается способ получения антитела человека, предусматривающий иммунизацию описанного в настоящем документе не относящегося к человеку животного представляющим интерес антигеном, позволяя не относящемуся к человеку животному развивать иммунный ответ, сбор лимфоцита (например, В-клетки) из иммунизированного животного, получение из лимфоцита последовательности нуклеиновой кислоты, которая кодирует домен VH человека и домен VL человека, клонирование последовательности нуклеиновой кислоты в рамке считывания (т.е. в функциональной связи) с последовательностью константной области человека для создания тяжелой цепи иммуноглобулина и легкой цепи иммуноглобулина, и экспрессия тяжелой и легкой цепей в клетке, способной экспрессировать полностью человеческое антитело. Согласно одному варианту осуществления клетка представляет собой клетку СНО. Согласно различным вариантам осуществления VH домен человека происходит от реаранжированного генного сегмента VH1-69 или VH1-2 человека.
Согласно различным аспектам представляющий интерес антиген представляет собой патоген, который поражает описанных в настоящем документе людей. Согласно различным аспектам представляющий интерес антиген представляет собой вирус, который способен инфицировать человека. Иллюстративные антигены, которые могут быть использованы в описанных в настоящем документе способах и применениях включают микробы или микроорганизмы, такие как вирус, бактерия, прион или грибок или любой другой патоген, вызывающий заболевание у людей. Специалисту в настоящей области техники после прочтения раскрытия будут понятны те патогены человека, которые будут применимы для описанных в настоящем документе способов и применений. Различные аспекты и варианты осуществлений могут быть использованы совместно, если специально не указано иное или контекст ясно запрещает совместное использование.
Краткое описание графических материалов
На фиг.1 показано общее изображение, без соблюдения масштаба, серии стадий нацеливания и молекулярного конструирования, используемые для создания нацеливающего вектора для конструкции модифицированного локуса тяжелой цепи, содержащего единственный генный сегмент VH1-69 человека, двадцать семь генных сегментов DH человека и шесть генных сегментов JH человека в эндогенном локусе тяжелой цепи иммуноглобулина.
На фиг.2 показано общее изображение, без соблюдения масштаба, серии стадий нацеливания и молекулярного конструирования, используемые для создания нацеливающего вектора для конструкции модифицированного локуса тяжелой цепи, содержащего единственный генный сегмент VH1-2 человека, двадцать семь генных сегментов DH человека и шесть генных сегментов JH человека в эндогенном локусе тяжелой цепи иммуноглобулина.
На фиг.3 показаны контурные графики спленоцитов, гейтированных по отдельным лимфоцитам и окрашенных на наличие CD 19 (В-клетка) и CD3 (Т-клетка), от мыши дикого типа (WT) и мыши, гомозиготной по единственному генному сегменту VH человека, двадцати семи генным сегментам DH человека и шести генным сегментам JH человека в эндогенном локусе тяжелой цепи иммуноглобулина (1 г VH НО).
На фиг.4А слева показан процент CD19+ В-клеток в селезенках, собранных от мышей дикого типа (WT) и мышей, гомозиготных по единственному генному сегменту VH человека, двадцати семи генным сегментам DH человека и шести генным сегментам JH человека в эндогенном локусе тяжелой цепи иммуноглобулина (1 г VH НО). Справа показано количество CD19+ В-клеток на селезенку как для мышей дикого типа (WT), так и мышей, гомозиготных по единственному генному сегменту VH человека, двадцати семи генным сегментам DH человека и шести генным сегментам JH человека в эндогенном локусе тяжелой цепи иммуноглобулина (1 г VH НО).
На фиг.4 В слева показан процент CD19+ В-клеток в костном мозге, собранном из бедренных костей мышей дикого типа (WT) и мышей, гомозиготных по единственному генному сегменту VH человека, двадцати семи генным сегментам DH человека и шести генным сегментам JH человека в эндогенном локусе тяжелой цепи иммуноглобулина (1 г VH НО). Справа показано количество CD19+ В-клеток на бедренную кость как для мышей дикого типа (WT), так и мышей, гомозиготных по единственному генному сегменту VH человека, двадцати семи генным сегментам DH человека и шести генным сегментам JH человека в эндогенном локусе тяжелой цепи иммуноглобулина (1 г VH НО).
На фиг.5 показаны контурные графики спленоцитов, гейтированных по CD19+ В-клеткам и окрашенных на экспрессию lgλ+ + и lgκ+, от мыши дикого типа (WT) и мыши, гомозиготной по единственному генному сегменту VH человека, двадцати семи генным сегментам DH человека и шести генным сегментам JH человека в эндогенном локусе тяжелой цепи иммуноглобулина (1 г VH НО).
На фиг.6 показаны контурные графики спленоцитов, гейтированных по CD19+ В-клеткам и окрашенных на наличие иммуноглобулина D (IgD) и иммуноглобулина М (IgM), от мыши дикого типа (WT) и мыши, гомозиготной по единственному генному сегменту VH человека, двадцати семи генным сегментам DH человека и шести генным сегментам JH человека в эндогенном локусе тяжелой цепи иммуноглобулина (1 г VH НО).
На фиг.7 показано общее количество промежуточных В-клеток (CD19+IgMhiIgDint), зрелых В-клеток (CD19+IgMIgDhi) и соотношение количества зрелых и незрелых В-клеток в собранных селезенках от мышей дикого типа (WT) и мышей, гомозиготных по единственному генному сегменту VH человека, двадцати семи генным сегментам DH человека и шести генным сегментам JH человека в эндогенном локусе тяжелой цепи иммуноглобулина (1 г VH НО).
На фиг.8 показаны контурные графики клеток костного мозга, гейтированных по синглетам, окрашенным на наличие иммуноглобулина М (IgM) и В220, от мыши дикого типа (WT) и мыши, гомозиготной по единственному генному сегменту VH человека, двадцати семи генным сегментам DH человека и шести генным сегментам JH человека в эндогенном локусе тяжелой цепи иммуноглобулина (1 г VH НО).
На фиг.9 показано общее количество незрелых (B220intIgM+) и зрелых В-клеток (B220hi IgM+) в костном мозге, выделенном из бедренной кости мышей дикого типа (WT) и мышей, гомозиготных по единственному генному сегменту VH человека, двадцати семи генным сегментам DH человека и шести генным сегментам JH человека в эндогенном локусе тяжелой цепи иммуноглобулина (1 г VH НО).
На фиг.10 показаны контурные графики клеток костного мозга, гейтированных по CD19+ В-клеткам и окрашенных на наличие ckit+ и CD43+, от мыши дикого типа (WT) и мыши, гомозиготной по единственному генному сегменту VH человека, двадцати семи генным сегментам DH человека и шести генным сегментам JH человека в эндогенном локусе тяжелой цепи иммуноглобулина (1 г VH НО).
На фиг.11А показан процент CD19+ В-клеток в популяциях про-В-клеток (CD19+CD43+ckit+) и пре-В-клеток (CD19+CD43+ckit+) из костного мозга, собранного из бедренных костей мышей дикого типа (WT) и мышей, гомозиготных по единственному генному сегменту VH человека, двадцати семи генным сегментам DH человека и шести генным сегментам JH человека в эндогенном локусе тяжелой цепи иммуноглобулина (1 г VH НО).
На фиг.11 В показано абсолютное количество клеток на бедренную кость в популяциях про-В-клеток (CD19+CD43+ckit+) и пре-В-клеток (CD19+CD43+ckit+) из костного мозга, собранного из мышей дикого типа (WT) и мышей, гомозиготных по единственному генному сегменту VH человека, двадцати семи генным сегментам DH человека и шести генным сегментам JH человека в эндогенном локусе тяжелой цепи иммуноглобулина (1 г VH НО).
На фиг.12 показана относительная экспрессия мРНК (у-ось) VH1-69, полученных из тяжелых цепей очищенных В-клеток селезенки с помощью количественного ПЦР анализа с использованием зонда, специфичного к генному сегменту VH1-69 человека у мышей, гомозиготных по замещению эндогенных VH, dh, JH тяжелой цепи, а также по замещению эндогенных генных сегментов Vκ и Jκ легкой цепи генными сегментами VH, DH, JH, Vκ и Jκ человека (Нκ), мышей дикого типа (WT), мышей гетерозиготных по единственному генному сегменту VH человека, двадцати семи генным сегментам DH человека и шести генным сегментам JH человека в эндогенном локусе тяжелой цепи иммуноглобулина (1 г VH НЕТ) и мышей, гомозиготных единственному генному сегменту VH человека, двадцати семи генным сегментам DH человека и шести генным сегментам JH человека в эндогенном локусе тяжелой цепи иммуноглобулина (1 г VH НО). Сигналы нормируются по отношению к экспрессии мыши Ск.
На фиг.13 показано выравнивание нуклеотидов второго экзона для каждого из тринадцати представленных аллелей для гена VH1-69 человека. Основания строчными буквами обозначают различия нуклеотидов зародышевой линии среди аллелей. Определяющие комплементарность области (CDR) обозначены прямоугольниками вокруг последовательности. Тире обозначают искусственные пробелы для правильного выравнивания последовательностей. VH1-69*01 (SEQ ID NO: 34); VH1-69*02 (SEQ ID NO: 36); VH1-69*03 (SEQ ID NO: 38); VH1-69*04 (SEQ ID NO: 40); VH1-69*05 (SEQ ID NO: 42); VH1-69*06 (SEQ ID NO: 44); VH1-69*07 (SEQ ID NO: 46); VH1-69*08 (SEQ ID NO: 48); VH1-69*09 (SEQ ID NO: 50); VH1-69*10 (SEQ ID NO: 52); VH1-69*11 (SEQ ID NO: 54); VH1-69*12 (SEQ ID NO: 56); VH1-69*13 (SEQ ID NO: 58).
На фиг.14 показано выравнивание белка в зрелой последовательности вариабельного гена тяжелой цепи для каждого из тринадцати представленных аллелей для гена VH1-69 человека. Аминокислоты строчными буквами обозначают различия зародышевой линии среди аллелей. Определяющие комплементарность области (CDR) обозначены прямоугольниками вокруг последовательности. Тире обозначают искусственные пробелы для правильного выравнивания последовательностей. VH1-69*01 (SEQ ID NO: 35); VH1-69*02 (SEQ ID NO: 37); VH1-69*03 (SEQ ID NO: 39); VH1-69*04 (SEQ ID NO: 41); VH1-69*05 (SEQ ID NO: 43); VH1-69*06 (SEQ ID NO: 45); VH1-69*07 (SEQ ID NO: 47); VH1-69*08 (SEQ ID NO: 49); VH1-69*09 (SEQ ID NO: 51); VH1-69*10 (SEQ ID NO: 53); VH1-69*11 (SEQ ID NO: 55); VH1-69*12 (SEQ ID NO: 57); VH1-69*13 (SEQ ID NO: 59).
На фиг.15 показан процент идентичности/процент сходства матрицы для выровненных белковых последовательностей зрелого вариабельного гена для каждого из тринадцати представленных аллелей для гена VH1-69 человека. Процент идентичности среди аллелей VH1-69 указан над затемненными прямоугольниками, и процент сходства указан под затемненными прямоугольниками. Значения процента идентичности и процента сходства определяли с помощью инструмента выравнивания ClustalW (v 1.83), используя программное обеспечение Mac Vector (Mac Vector, Inc, Северная Каролина).
На фиг.16 показано выравнивание нуклеотидов второго экзона для каждого из пяти представленных аллелей гена для гена VH1-2 человека. Основания строчными буквами обозначают различия нуклеотидов зародышевой линии среди аллелей. Определяющие комплементарность области (CDR) обозначены прямоугольниками вокруг последовательности. Тире обозначают искусственные пробелы для правильного выравнивания последовательностей. VH1-2*01 (SEQ ID NO: 60); VH1-2*02 (SEQ ID NO: 62); VH1-2*03 (SEQ ID NO: 64); VH1-2*04 (SEQ ID NO: 66); VH1-2*05 (SEQ ID NO: 68)
На фиг.17 показано выравнивание белка в зрелой последовательности вариабельного гена тяжелой цепи для каждого из пяти представленных аллелей для гена VH1-2 человека. Аминокислоты строчными буквами обозначают различия зародышевой линии среди аллелей. Определяющие комплементарность области (CDR) обозначены прямоугольниками вокруг последовательности. Тире обозначают искусственные пробелы для правильного выравнивания последовательностей. VH1-2*01 (SEQ ID NO: 61); VH1-2*02 (SEQ ID NO: 63); VH1-2*03 (SEQ ID NO: 65); VH1-2*04 (SEQ ID NO: 67); VH1-2*05 (SEQ ID NO: 69).
На фиг.18 показан процент идентичности/процент сходства матрицы для выровненных белковых последовательностей зрелого вариабельного гена для каждого из пяти представленных аллелей для гена VH1-2 человека. Процент идентичности среди аллелей VH1-2 указан над затемненными прямоугольниками, и процент сходства указан под затемненными прямоугольниками. Значения процента идентичности и процента сходства определяли по инструменту выравнивания ClustalW (v 1.83), используя программное обеспечение Mac Vector (Mac Vector, Inc, Северная Каролина).
На фиг.19 показан титр антител от мышей, гомозиготных по локусам вариабельных генов тяжелой и легкой цепи κ человека (Нκ, n=4), и мышей, гомозиготных по единственному генному сегменту VH1-69 человека, двадцати семи генным сегментам DH человека и шести генным сегментам JH человека в эндогенном локусе тяжелой цепи иммуноглобулина (1 г VHHO; n=10), которые были иммунизированы рецептором клеточной поверхности человека (антиген А).
На фиг.20 показан титр антител от мышей, гомозиготных по локусам вариабельных генов тяжелой и легкой цепи κ человека (Нκ, n=5), и мышей, гомозиготных по единственному генному сегменту VH1-69 человека, двадцати семи генным сегментам DH человека и шести генным сегментам JH человека в эндогенном локусе тяжелой цепи иммуноглобулина (1 г VHHO; n=5), которые были иммунизированы двумя различными вакцинами против гриппа.
На фиг.21 показан процент (у-ось) примированных IgM тяжелых цепей, характеризующихся заданной аминокислотной длиной для области VH CDR3 (х-ось), от мышей, гомозиготных по единственному генному сегменту VH1-69 человека, двадцати семи генным сегментам DH человека и шести генным сегментам JH человека в эндогенном локусе тяжелой цепи иммуноглобулина и гомозиготных по замещению эндогенных вариабельных локусов легкой цепи κ вариабельными локусами легкой цепи κ человека, которые были иммунизированны рецептором клеточной поверхности человека (антиген А).
На фиг.22 показан процент (y-ось) примированных IgG тяжелых цепей, характеризующихся заданной аминокислотной длиной для области VH CDR3 (х-ось), от мышей, гомозиготных по единственному генному сегменту VH1-69 человека, двадцати семи генным сегментам DH человека и шести генным сегментам JH человека в эндогенном локусе тяжелой цепи иммуноглобулина и гомозиготных по замещению эндогенных вариабельных локусов легкой цепи κ вариабельными локусами легкой цепи κ человека, которые были иммунизированны рецептором клеточной поверхности человека (антиген А).
Подробное описание изобретения
Настоящее изобретение не ограничивается конкретными способами и описанными экспериментальными условиями, поскольку такие способы и условия могут варьироваться. Кроме того, следует понимать, что используемая в настоящем документе терминология предназначена только для цели описания конкретных вариантов осуществления и не предусматривается для ограничения, поскольку объем настоящего изобретения определяется формулой изобретения.
Если не указано иное, все используемые в настоящем документе термины и фразы, включают в себя те значения, которые термины и фразы имеют в настоящей области техники, если иное прямо не указано или явно не следует из контекста, в котором используется термин или фраза. Хотя любые способы и материалы, подобные или эквивалентные описанным в настоящем документе, могут быть использованы на практике или при тестировании настоящего изобретения, теперь будут описаны конкретные способы и материалы. Все упомянутые публикации включены в настоящий документ посредством ссылки.
Фраза ″существенный″ или ″по существу″ при использовании для обозначения количества генных сегментов (например, ″по существу все″ генные сегменты V) включает в себя как функциональные, так и не функциональные генные сегменты и включает в себя, согласно различным вариантам осуществления, например, 80% или более, 85% или более, 90% или более, 95% или более 96% или более, 97% или более, 98% или более или 99% или более всех генных сегментов; согласно различным вариантам осуществления ″по существу все″ генные сегменты включают в себя, например, по меньшей мере 95%, 96%, 97%, 98% или 99% функциональных (т.е. не псевдогенов) генных сегментов.
[0001] Термин ″замещение″ включает в себя помещение последовательности ДНК в геном клетки таким образом, чтобы заменить последовательность в геноме гетерологичной последовательностью (например, последовательность человека у мыши) в локусе геномной последовательности. Помещенная таким образом последовательность ДНК может включать в себя один или несколько регуляторных последовательностей, которые представляют собой часть исходной ДНК, используемой для получения помещенной таким образом последовательности (например, промоторы, энхансеры, 5′- или 3′- нетранслируемые области, соответствующие последовательности сигналов рекомбинации и т.п.). Например, согласно различным вариантам осуществления замещение представляет собой замену эндогенной последовательности гетерологичной последовательностью, что приводит к производству генного продукта из таким образом помещенной последовательности ДНК (содержащей гетерологичную последовательность), но не экспрессии эндогенной последовательности; замещение эндогенной геномной последовательности последовательностью ДНК, которая кодирует белок, который характеризуется аналогичной функцией, что и белок, кодируемый эндогенной геномной последовательностью (например, эндогенная геномная последовательность кодирует ген иммуноглобулина или домен, и фрагмент ДНК кодирует один или несколько генов иммуноглобулина человека или доменов). Согласно различным вариантам осуществления эндогенный ген или его фрагмент заменяют соответствующим человеческим геном или его фрагментом. Соответствующий человеческий ген или его фрагмент представляет собой человеческий ген или фрагмент, который представляет собой ортолог, гомолог или по существу идентичный или такой же по структуре и/или функции, как эндогенный ген или его фрагмент, который заменен.
Было выполнено точное замещение in situ шести миллионов пар оснований вариабельных областей мышиных локусов тяжелой цепи иммуноглобулина (VH-DH-JH) ограниченным локусом тяжелой цепи иммуноглобулина человека при сохранении окружающих последовательностей мыши интактными и функциональными в гибридных локусах, включая все гены константной цепи мыши и регионы транскрипционного контроля локуса (фиг.1 и фиг.2). В частности, единственный генный сегмент VH, 27 генных сегментов DH и шесть генных сегментов JH человека были введены посредством химерных основанных на ВАС нацеливающих векторов в ES клетки мыши с использованием технологии генетической инженерии VELOCIGENE® (смотрите, например, патент США №6586251 и Valenzuela, D.M., et al. (2003). High-throughput engineering of the mouse genome coupled with high-resolution expression analysis. Nat Biotechnol 21, 652-659).
He относящиеся к человеку животные с ограниченными генными сегментами VH иммуноглобулина
Предусматриваются не относящиеся к человеку животные, содержащие локусы иммуноглобулина, которые содержат ограниченное количество генов VH и один или несколько генов D, и один или несколько генов J, а также способы их получения и использования. После иммунизации представляющим интерес антигеном не относящиеся к человеку животные генерируют популяции В-клеток с вариабельными областями антител, полученными только от ограниченного, заранее выбранного гена VH или набора генов VH (например, заранее выбранный ген VH и его варианты). Согласно различным вариантам осуществления предусматриваются не относящиеся к человеку животные, которые генерируют популяции В-клеток, которые экспрессируют вариабельные домены человеческих антител, которые представляют собой вариабельные домены тяжелой цепи человека, вместе с родственными вариабельными доменами легкой цепи человека. Согласно различным вариантам осуществления не относящиеся к человеку животные реаранжируют вариабельные генные сегменты тяжелой цепи человека и вариабельные генные сегменты легкой цепи человека из модифицированных эндогенных локусов иммуноглобулина мыши, которые включают замещение или вставку нереаранжированных не относящихся к человеку последовательностей вариабельной области нереаранжированными последовательностей вариабельной области человека.
Ранняя работа по организации, структуре и функции генов иммуноглобулина была проведена частично на мышах с недееспособными эндогенными локусами и сконструированными для получения трансгенных локусов (помещенных в случайном порядке) с частичными генами иммуноглобулина человека, например, частичный репертуар генов тяжелой цепи человека, связанный с константным геном человека, случайным образом вставленным в геном в присутствии или в отсутствие трансгена легкой цепи человека. Хотя этих мышей было несколько меньше, чем оптимально для получения применимых высокоаффинных антител, они способствовали определенным функциональным анализам локусов иммуноглобулина. Некоторые из этих мышей содержали всего лишь два или три, или даже всего лишь единственный вариабельный ген тяжелой цепи.
Сообщалось о мышах, которые экспрессируют полностью человеческие тяжелые цепи иммуноглобулина, полученные из единственного гена VH5-51 человека и 10 генов DH человека, и шести генов JH человека, с константными генами µ и γ1 человека, на случайно вставленном трансгене (и недееспособных эндогенных локусах иммуноглобулина) (Xu and Davis, 2000, Diversity in the CDR3 Region of VH Is Sufficient for Most Antibody Specificities, Immunity 13:37-45). Полностью человеческие тяжелые цепи иммуноглобулина этих мышей в основном экспрессируются с одной из всего двух полностью мышиных легких цепей λ, полученных из эндогенного локуса легкой цепи λ мыши (только Vλ1-Jλ1 или Vλ2-Jλ2), и не могут экспрессировать легкую цепь к (мыши представляют собой Igκ-/-). Эти мыши проявляют строго ненормальную дисфункцию в развитии В-клеток и экспрессии антител. Количество В-клеток, как сообщается, составляет 5-10% от дикого типа, содержание IgM 5-10% от дикого типа и содержание IgGI только 0,1-1% от дикого типа. Наблюдаемый репертуар IgM выявил крайне ограниченную множественность J-сегментов. Полностью человеческие тяжелые цепи отображают в основном одинаковую длину CDR3 по антигенам, одну и ту же частоту использования JH (JH2) в антигенах и начальный остаток Q множественности сегментов J, таким образом отражая определенную нехватку разнообразия CDR3. Почти у всех полностью мышиных легких цепей λ были замены W96L в Jλ1 в качестве начального остатка множественности сегментов J. Как сообщается, мыши не в состоянии продуцировать любые антитела против бактериального полисахарида. Поскольку вариабельные домены человека образуют пару с легкими цепями мыши, применимость вариабельных областей человека сильно ограничена.
Сообщалось о других мышах, у которых есть только единственный ген VH3-23 человека, гены DH и JH человека и гены легкой цепи мыши, но они проявляют ограниченное разнообразие (и, следовательно, ограниченную применимость) отчасти из-за потенциала ошибочного спаривания между доменами VH человека и VL мыши (смотрите, например, Mageed et al., 2001, Rearrangement of the human heavy chain variable region gene V3-23 in transgenic mice generates antibodies reactive with a range of antigens on the basis of VHCDR3 and residues intrinsic to the heavy chain variable region, Clin. Exp.Immunol. 123:1-5). Аналогичным образом, мыши, которые несут два гена VH (3-23 и 6-1) вместе с генами DH и JH человека в трансгене, содержащем константный ген µ человека (Bruggemann et al., 1991, Human antibody production in transgenic mice: expression from 100kb of the human IgH locus, Eur. J. Immmunol. 21:1323-1326), и экспрессируют их в цепи IgM человека с легкими цепями мыши, могут проявлять репертуар, ограниченный ошибочным спариванием (Mackworth-Young et al., 2003, The role of antigen in the selection of the human V3-23 immunoglobulin heavy chain variable region gene, din. Exp.Immunol. 134:420-425).
Также сообщалось о других трансгенных мышах, которые экспрессируют ограниченные VH полностью человеческие тяжелые цепи с трансгена человека, случайным образом вставленном в геном, с ограниченным репертуаром А, человека, экспрессированным с полностью человеческого случайным образом вставленного трансгена (смотрите, например, Taylor et al., 1992, A transgenic mouse that expresses a diversity of human sequence heavy and light chain immunoglobulins, Nucleic Acids Res. 20(23):6287-6295; Wagner et al., 1994, Antibodies generated form human immunoglobulin miniloci in transgenic mice. Nucleic Acids Res. 22(8): 1389-1393). Тем не менее, трансгенные мыши, которые экспрессируют полностью человеческие антитела из случайным образом интегрированных в геном мыши трансгенов, и которые содержат поврежденные эндогенные локусы, как известно, проявляют существенные различия в иммунном ответе, по сравнению с мышами дикого типа, которые влияют на разнообразие вариабельных доменов антитела, получаемого из таких мышей.
Применимые не относящиеся к человеку животные, которые продуцируют разнообразную популяцию В-клеток, которые экспрессируют вариабельные домены антитела человека от ограниченного репертуара генов VH и одного или нескольких генов D и одного или нескольких генов J, будут способны продуцировать, предпочтительно согласно некоторым вариантам осуществления репертуары реаранжированных генов вариабельной области, которые будут достаточно разнообразны. Согласно различным вариантам осуществления разнообразие включает в себя разнообразие множественности сегментов J, соматическую гипермутацию и полиморфное разнообразие в последовательности гена VH (для вариантов осуществления, где гены VH присутствуют в полиморфных формах). Комбинаторное разнообразие происходит в спаривании гена VH с одним из множества родственных вариабельных доменов легкой цепи человека (которые, согласно различным вариантам осуществления, содержат разнообразие множественности J-сегментов и/или соматические гипермутации).
Не относящиеся к человеку животные, содержащие ограниченный репертуар генов VH человека и полный или по существу полный репертуар генов VL человека, будут согласно различным вариантам осуществления продуцировать популяции В-клеток, которые отражают различные источники разнообразия, такие как разнообразие множественности сегментов J (например, соединение VDJ, VJ, присоединения Р, присоединения N), комбинаторное разнообразие (например, родственный ограниченный VH тяжелой цепи человека, легкой цепи человека) и соматические гипермутации. Согласно вариантам осуществления, включающим ограничение репертуара VH до одного гена VH человека, один ген VH человека может присутствовать в двух или более вариантах. Согласно различным вариантам осуществления присутствие двух или более полиморфных форм гена VH будет обогащать разнообразие вариабельных доменов популяции В-клеток.
Изменчивости в последовательностях зародышевой линии генных сегментов (например, генов V) вносят вклад в разнообразие ответа антител у людей. Относительный вклад в разнообразие благодаря различиям последовательности гена V изменяется среди генов V. Степень полиморфизма варьирует в семействе генов и отражается во множестве гаплотипов (участки последовательности с унаследованными полиморфизмами), способных производить дополнительное разнообразие, как это наблюдается в различиях гаплотипов VH между связанными и несвязанными индивидуумами в человеческой популяции (смотрите, например, Souroujon et al., 1989, Polymorphisms in Human H Chain V Region Genes from the VHIII Gene Family, J. Immunol. 143(2):706-711). Основываясь на данных от определенных полиморфных семейств генов VH человека, некоторые предположили, что разнообразие гаплотипов в зародышевой линии вносит основной вклад в неоднородность генов VH в человеческой популяции, что отражается в большом разнообразии различных генов VH зародышевой линии в человеческой популяции (смотрите, Sasso et al., 1990, Prevalence and Polymorphism of Human VH3 Genes, J. Immunol. 145(8): 2751-2757).
Хотя человеческая популяция отображает большое разнообразие гаплотипов по отношению к репертуару генов VH благодаря широко распространенному полиморфизму, определенные полиморфизмы отражаются в преобладающих (т.е. консервативных) аллелях, наблюдаемых в человеческой популяции (Sasso et al., 1990). Полиморфизм VH может быть описан в двух основных формах. Первая представляет собой вариацию, вытекающую из вариации аллелей, связанной с различиями в нуклеотидной последовательности между аллелями одного и того же генного сегмента. Вторая вытекает из многочисленных дубликатов, вставок и/или удалений, которые произошли в локусе тяжелой цепи иммуноглобулина. Это привело к уникальной ситуации, в которой гены VH, полученные путем дублирования из идентичных генов, отличаются от их соответствующих аллелей одной или несколькими нуклеотидными заменами. Это также напрямую влияет на количество копий генов VH в локусе тяжелой цепи.
Полиморфные аллели вариабельных генных сегментов тяжелой цепи иммуноглобулина человека (гены VH) в значительной степени были результатом вставки/удаления генных сегментов и единственных нуклеотидных различий в кодирующих областях, оба из которых характеризуются потенциалом иметь функциональные последствия на молекулу иммуноглобулина. В таблице 1 представлены функциональные гены VH, перечисленные по семейству генов VH человека и количеству идентифицированных аллелей для каждого гена VH в локусе тяжелой цепи иммуноглобулина человека. Есть некоторые данные для предположения, что полиморфные гены VH были вовлечены в чувствительность к таким определенным заболеваниям, как, например, ревматоидный артрит, в то время как в других случаях связь между VH и заболеванием была менее ясна. Эта неоднозначность была приписана числу копий и присутствию различных аллелей в различных популяциях человека. На самом деле, несколько генов VH человека демонстрируют вариации числа копий (например, VH1- 59 2, VH1-69, VH2-26, VH2-70 и VH3-23). Согласно различным вариантам осуществления описанные в настоящем документе гуманизированные мыши с ограниченными репертуарами VH содержат множественные полиморфные варианты отдельного представителя семейства VH (например, два или более полиморфных вариантов VH1-2, VH1-69, VH2-26, VH2-70 или VH3-23, заместив все или по существу все функциональные сегменты VH мыши в эндогенном локусе мыши). Согласно конкретному варианту осуществления два или более полиморфных варианта описанных в настоящем документе мышей находятся в количестве вплоть до и включая в себя число, указанное для соответствующего представителя семейства VH в таблице 1 (например, для VH1-69, 13 вариантов; для VH1-2, пять вариантов; и т.д.).
Обычно наблюдаемые варианты определенных генов VH человека известны в настоящей области техники. Например, один из наиболее сложных полиморфизмов в локусе VH принадлежит гену VH1-69. Ген VH1-69 человека содержит 13 известных аллелей (Sasso et al., 1993, A fetally expressed immunoglobulin VH1 gene belongs to a complex set of alleles. Journal of Clinical Investigation 91: 2358-2367; Sasso et al., 1996, Expression of the immunoglobulin VH gene 51 pi is proportional to its germline gene copy number. Journal of Clinical Investigation 97(9): 2074-2080) и существует по меньшей мере в трех гаплотипах, которые несут дубликаты гена VH1-69, что приводит к множественным копиям гена VH в данном локусе. Эти полиморфные аллели включают в себя различия в определяющих комплементарность областях (CDR), которые могут существенно влиять на антигенную специфичность. В таблице 2 представлены известные аллели для VH1-69 человека и SEQ ID NO для последовательностей ДНК и белка зрелых вариабельных областей тяжелой цепи. В таблице 3 представлены известные аллели для VH1-2 человека и SEQ ID NO для последовательностей ДНК и белка зрелых вариабельных областей тяжелой цепи.
Репрезентативные последовательности геномной ДНК и полномерного белка гена VH1-69 представлены в SEQ ID NO: 1 и SEQ ID NO: 2, соответственно. На фиг.13 и фиг.14 представлены выравнивания тринадцати известных VH1-69 аллелей ДНК и белка, соответственно. Репрезентативные последовательности ДНК и белка гена VH1-2 представлены в SEQ ID NO: 60 и SEQ ID NO: 61, соответственно. На фиг.16 и фиг.17 представлены выравнивания пяти известных VH1-2 аллелей, соответственно. На фиг.16 и фиг.17 представлен процент идентичности/сходства матрицы для выравнивания белковых последовательностей, соответствующих тринадцати известным VH1-69 аллелям и пяти известным VH1-2 аллелям человека, соответственно. Согласно различным вариантам осуществления модифицированный локус настоящего изобретения содержит ген VH, выбранный из таблицы 1, присутствующий в двух или более числе копий, причем число копий включает в себя число аллелей, включительно, представленные в таблице 1. Согласно одному варианту осуществлений модифицированный локус настоящего изобретения содержит ген VH1-69, выбранный из таблицы 2, присутствующий в двух или более числе копий, причем число копий включает в себя число аллелей, включительно, представленные в таблице 1. Согласно одному варианту осуществления модифицированный локус изобретения содержит ген VH1-2, выбранный из таблицы 3, присутствующий в двух или более числе копий, причем число копий включает в себя число аллелей, включительно, представленные в таблице 1.
Хотя широко обсуждается варианты осуществления, использующие ограниченный репертуар VH человека в мыши, также предусматриваются другие не относящиеся к человеку животные, которые экспрессируют ограниченный репертуар VH человека. Такие не относящиеся к человеку животные включают в себя любые из тех, которые могут быть генетически модифицированы, чтобы экспрессировать описанный в настоящем документе ограниченный репертуар VH человека, включая в себя, например, мышь, крысу, кролика, свинью, корову (например, корову, быка, буйволу), оленя, овцу, козу, курицу, кошку, собаку, хорька, примата (например, мармазетку, макака-резуса) и т.п. Например, для тех не относящихся к человеку животных, для которых подходящие генетически модифицируемые ES клетки не легкодоступны, используют и другие способы для получения не относящегося к человеку животного, содержащего генетическую модификацию. Такие способы включают в себя, например, модификацию генома не ES клетки (например, фибробластов или индуцированных плюрипотентных клеток) и использования ядерной передачи для передачи модифицированного генома подходящей клетке, например, яйцеклетке, и постепенное формирование модифицированной клетки (например, модифицированного ооцита) у не относящегося к человеку животного при подходящих условиях с образованием эмбриона. Способы модификации генома не относящегося к человеку животного (например, генома свиньи, коровы, грызуны, курицы, и т.п.) включают в себя, например, использование цинк-пальцевой нуклеазы (ZFN) или эффекторной нуклеазы, подобной активаторам транскрипции (TALEN) для изменения генома для включения ограниченного репертуара VH человека. Таким образом, согласно одному варианту осуществления предусматривается способ для редактирования генома не относящегося к человеку животного для включения в него ограниченного репертуара VH человека, содержащего стадию редактирования генома, используя ZFN или TALEN для включения в себя не более одного или не более двух генных сегментов VH человека (или их полиморфных вариантов), причем не более одного или не более двух генных сегментов VH человека функционально связаны с последовательностью константного гена иммуноглобулина. Согласно одному варианту осуществления последовательность константного гена выбирают из константной последовательности тяжелой цепи человека и не относящейся к человеку константной последовательности тяжелой цепи. Согласно одному варианту осуществления константная последовательность представляет собой не относящуюся к человеку, и не более одного или не более двух генных сегментов VH человека функционально связаны с не относящейся к человеку последовательностью константного гена в эндогенном локусе иммуноглобулина человека.
Согласно одному аспекту не относящееся к человеку животное представляет собой небольшое млекопитающее, например, из надсемейства Dipodoidea или Muroidea. Согласно одному варианту осуществления генетически модифицированное животное представляет собой грызуна. Согласно одному варианту осуществления грызуна выбирают из мыши, крысы и хомяка. Согласно одному варианту осуществления грызуна выбирают из надсемейства мышиных. Согласно одному варианту осуществления генетически модифицированное животное представляет собой животное из семейства, выбранного из Calomyscidae (например, мышевидные хомячки), Cricetidae (например, хомячок, крысы и мыши Нового Света, полевки), Muridae (истинные мыши и крысы, песчанки, колючие мыши, хохлатые крысы), Nesomyidae (рипидомисы, скальные крысы, белохвостые крысы. Малагасийские крысы и мыши), Platacanthomyidae (например, колючие соневидные хомяки) и Spalacidae (например, кроты, бамбуковые крысы и цокоры). Согласно конкретному варианту осуществления генетически модифицированных грызунов выбирают из истинной мыши или крысы (семейство Muridae), песчанки, колючей мыши и косматого хомяка. Согласно одному варианту осуществления генетически модифицированная мышь представляет собой представителя семейства Muridae.
Согласно одному варианту осуществления не относящееся к человеку животное представляет собой грызуна, который представляет собой мышь линии C57BL. Согласно одному варианту осуществления линию C57BL выбирают из C57BL/A, C57BL/An, C57BL/GrFa, C57BL/KaLwN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6N, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr и C57BL/01a. Согласно другому варианту осуществления мышь представляет собой мышь линии 129. Согласно одному варианту осуществления линию 129 выбирают из группы, состоящей из 129Р1, 129Р2, 129Р3, 129Х1, 129S1 (например, 129S1/SV, 12981/SvIm), 129S2, 129S4, 129S5. 129S6, 12989/SvEvH (129/SvEvTac), 129S7, 129S8, 129Т1, 129Т2 (смотрите, например, Festing et al. (1999) Revised nomenclature for strain 129 mice. Mammalian Genome 10:836, смотрите также Auerbach et al. (2000) Establishment and Chimera Analysis of 129/SvEv- and C57BL/6- Derived Mouse Embryonic Stem Cell Lines). Согласно одному варианту осуществления генетически модифицированная мышь представляет собой сочетание вышеуказанной линии 129 и вышеуказанной линии C57BL (например, линии C57BL/6). Согласно другому варианту осуществления, мышь представляет собой сочетание вышеуказанных линий 129 или сочетание вышеуказанных линий C57BL/6. Согласно одному варианту осуществления линия 129 сочетания представляет собой линию 129S6 (129/SvEvTac). Согласно другому варианту осуществления мышь представляет собой сочетание из линии, полученной от 129/SvEv и C57BL/6. Согласно конкретному варианту осуществления мышь представляет собой сочетание из линии, полученной от 129/SvEv nC57BL/6, как описано в Auerbach et al. 2000 BioTechniques 29:1024-1032. Согласно другому варианту осуществления мышь представляет собой сочетание линии BALB, например, линия BALB/C. Согласно другому варианту осуществления мышь представляет собой сочетание линии BALB (например, линия BALB/C) и вышеуказанной линии.
Согласно одному варианту осуществления не относящееся к человеку животное представляет собой крысу. Согласно одному варианту осуществления крысу выбирают из крысы линии Wistar, линии LEA, линии Sprague Dawley, линии Fischer, F344, F6 и Dark Agouti. Согласно одному варианту осуществления линия крысы представляет собой сочетание двух или более линий, выбранных из группы, состоящей из Wistar, Lea, Sprague Dawley, Fischer, F344, F6 и Dark Agouti.


Антиген-зависимое использование гена VH
Антиген-зависимое преимущественное использование генов VH может быть использовано в разработке терапевтических средств для людей, направленных на клинически значимые антигены. Способность продуцировать репертуар вариабельных доменов антител с использованием конкретного гена VH может обеспечить значительное преимущество в поиске высокоаффинных вариабельных доменов антител для использования в терапевтических средствах для людей. Исследования, касающиеся частоты использования гена VH наивной мыши и человека в вариабельных доменах антител показывают, что большинство вариабельных доменов тяжелой цепи не происходят от любого определенного единственного или преимущественно используемого гена VH. С другой стороны, исследования ответа антител на определенные антигены показывают, что в некоторых случаях определенный ответ антитела отображает смещенное использование определенного гена VH в репертуаре В-клеток после иммунизации.
Хотя репертуар VH человека довольно разнообразен, по некоторым оценкам ожидаемая частота использования любого данного гена VH, предполагая случайный выбор генов VH, составляет приблизительно 2% (Brezinschek et al., 1995, Analysis of the Heavy Chain Repertoire of Human Peripheral В Cells Using Single-Cell Polymerase Chain Reaction, J. Immunol. 155:190-202). Но использование VH в периферических В-клетках у людей искажается. Согласно одному исследованию функциональное изобилие генов V соответствует схеме VH3>VH4>VH1>VH2>VH5>VH6 (Davidkova et al., 1997, Selective Usage of VH Genes in Adult Human Lymphocyte Repertoires, Scand. J. Immunol. 45:62-73). Одно из ранних исследований показало, что частота использования семейства VH3 была приблизительно 0,65, в то время как частота использования семейства VH1 была приблизительно 0,15; эти и другие наблюдения показывают, что сложность зародышевой линии репертуара VH человека не точно отражена в компартменте периферической В-клетки у людей, имеющих нормальный репертуар VH зародышевой линии, ситуация, которая близка с наблюдаемой у мыши, т.е. экспрессия гена VH нестохастическая (Zouali and These, 1991, Probing VH Gene-Family Utilization in Human Peripheral В Cells by In Situ Hybridization, J. Immunol. 146(8):2855-2864). Согласно одному сообщению частота использования гена VH у людей, от наибольшей к наименьшей представляет собой VH3>VH4>VH1>VH5>VH2>VH6; реаранжировки в периферических В-клетках, показывают, что частота использования семейства VH3 выше, чем можно ожидать, основываясь на относительном числе генов VH3 зародышевой линии (Brezinschek et al., 1995). Согласно другому сообщению частота использования VH у человека соответствует схеме VH3>VH5>VH2>VH1>VH4>VH6, основанной на анализе периферийных небольших иммунокомпетентных В-клеток, активированных митогеном лаконоса (Davidkova et al., 1997, Selective Usage of VH Genes in Adult Human В Lymphocyte Repertoires, Scand. J. Immunol. 45:62-73). В одном из докладов утверждается, что среди наиболее часто используемых представителей семейства VH3 находятся 3-23, 3-30 и 3-54 (Brezinschek et al., 1995). В семействе VH4 представитель 4-59 и 4-4b обнаруживались относительно более часто (там же), а также 4-39 и 4-34 (Brezinscheck et al., 1997, Analysis of the Human VH Gene Repertoire, J. Clin. Invest. 99(10): 2488-2501). Другие утверждают, что активированный репертуар тяжелой цепи искажен в пользу высокой экспрессии VH5 и более низкой экспрессии VH3 (Van DiJκ-Hard and Lundkvist, 2002, Long-term kinetics of adult human antibody repertoires, Immunology 107: 136-144). Другие исследования утверждают, что к наиболее часто используемым генам VH в репертуаре взрослого человека относится VH4-59, а затем VH3-23 и VH3-48 (Amaout et al., 2001, High-Resolution Description of Antibody Heavy-Chain Repertoires in Humans, PLoS ONE 6(8): 108). Хотя исследования частоты использования основаны на относительно небольшом количестве образцов, таким образом, проявляют высокую дисперсию, взятые вместе исследования показывают, что экспрессия V гена не представляет собой чисто стохастический процесс. В действительности, исследования с определенными антигенами установили, что в некоторых случаях результаты свидетельствуют четко против определенных частот использования и в пользу других.
Со временем стало очевидно, что наблюдаемый репертуар вариабельных доменов тяжелой цепи человека, созданных в ответ на определенные антигены, сильно ограничен. Некоторые антигены связаны почти исключительно с нейтрализующими антителами, содержащими только определенные конкретные гены VH, в том смысле, что эффективные нейтрализующие антитела получают, по сути, только из одного гена vh. Так обстоит дело в отношении ряда клинически важных патогенов человека.
Были обнаружены производные VH1-69 тяжелых цепей в различных репертуарах антигенспецифических антител терапевтического значения. Например, VH1-69 часто наблюдались в транскриптах тяжелых цепей репертуара IgE лимфоцитов периферической крови у детей раннего возраста с атопическим заболеванием (Bando et al., 2004, Characterization of VHε gene expressed in PBL from children with atopic diseases: detection of homologous VH1-69 derived transcripts from three unrelated patients, Immunology Letters 94:99-106). Производные VH1-69 тяжелых цепей с высокой степенью соматической гипермутации также встречаются в В-клетках лимфомы (Perez et al., 2009, Primary cutaneous B-cell lymphoma is associated with somatically hypermutated immunoglobulin variable genes and frequent use of VH1-69 and VH4-59 segments, British Journal of Dermatology 162: 611-618), в то время как некоторые производные VH1-69 тяжелых цепей с существенными последовательностями зародышевой линии (т.е. практически без соматической гипермутации) были обнаружены среди аутоантител у пациентов с заболеваниями крови (Pos et al., 2008, VH1-69 germline encoded antibodies directed towards ADAMTS13 in patients with acquired thrombotic thrombocytopenic purpura, Journal of Thrombosis and Haemostasis 7: 421-428).
Дополнительно, были найдены нейтрализующие антитела против таких вирусных антигенов, как ВИЧ, грипп и гепатит С (HCV), чтобы использовать последовательности, полученные из VH1-69 зародышевой линии и/или соматически мутированные (Miklos et al., 2000, Salivary gland mucosa-associated lymphoid tissue lymphoma immunoglobulin VH genes show frequent use of V1-69 with distinctive CDR3 features. Blood 95(12):3878-3884; Kunert et al., 2004, Characterization of molecular features, antigen-binding, and in vitro properties of IgG and IgM variants of 4E10, an anti-HIV type I neutralizing monoclonal antibody. Aids Research and Human Retroviruses 20(7):755-762; Chan et al., 2001, VH1-69 gene is preferentially used by hepatitis С virus-associated В cell lymphomas and by normal В cells responding to the E2 viral antigen. Blood 97(4): 1023-1026; Carbonari et al., 2005, Hepatitis С virus drives the unconstrained monoclonal expansion of VHl-69-expressing memory В cells in type II cryoglobulinemia: A model of infection-driven lymphomagenesis. Journal of Immunology 174:6532-6539; Wang and Palese, 2009, Universal epitopes of influenza virus hemagglutinins?, Nature Structural & Molecular Biology 16(3): 233-234; Sui et al., 2009, Structural and functional bases for broad-spectrum neutralization of avian and human influenza A viruses. Nature Structural & Molecular Biology 16(3):265-273; Marasca et al., 2001, Immunoglobulin Gene Mutations and Frequent Use of VH1-69 and VH4-34 Segments in Hepatitis С Virus-Positive and Hepatitis С Virus-Negative Nodal Marginal Zone B-Cell Lymphoma. Aw. J. Pathol. 159(1): 253-261).
Искажение частоты использования VH наблюдается также в гуморальном иммунном ответе на гемофильные инфекции типа В (Hib PS) у людей. Исследования показывают, что семейство VHIII (подсемейство VHIIIb, в частности, VH9.1) характеризует исключительно гуморальный ответ человека на Hib PS, с разнообразными генами D и J (Adderson et al., 1991, Restricted Ig H Chain V Gene Usage in the Human Antibody Response to Haemophilus influenzae Type b Capsular Polysaccharide, J. Immunol. 147(5): 1667-1674; Adderson et al., 1993, Restricted Immunoglobulin VH Usage and VDJ Combinations in the Human Response to Haemophilus influenzae Type b Capsular Polysaccharide, J. Clin. Invest. 91: 2734-2743). Гены JH человека также отображают искаженную частоту использования; JH4 и JH6 наблюдаются приблизительно в 38-41% в периферических В-клетках у людей (Brezinschek et al., 1995).
Как сообщалось, частота использование VH у ВИЧ-1-инфицированных людей искажена относительно использования VH3 и в пользу семейств генов VH1 и VH4 (Wisnewski et al., 1996, Human Antibody Variable Region Gene Usage in HIV-1 Infection, J. Acquired Immune Deficiency Syndromes & Human Retroviology 11(1):31-38). Однако анализ кДНК костного мозга от инфицированных пациентов показал значительное использование VH3, не экспрессированных в функциональном репертуаре В-клеток, где Fab, отражающие использование VH3, проявляли эффективную нейтрализацию ВИЧ-1 in vitro (там же). Можно предположить, что гуморальный иммунный ответ на ВИЧ-1 инфекцию, возможно, ослабляется вследствие ограничения VH; описанные в настоящем документе не относящиеся к человеку модифицированные животные (не инфицированные ВИЧ-1), могут быть таким образом полезны для получения доменов нейтрализующих антител, полученных из определенных генов VH, присутствующих у описанных в настоящем документе генетически модифицированных животных, но полученных из разных генов VH, чем наблюдаемые в ограниченном репертуаре инфицированных людей.
Таким образом, способность продуцировать высокоаффинные вариабельные домены антител человека у ограниченных по VH мышей, например, (ограниченный, например, представителем семейства VH3 и его полиморфой (полиморфами)) иммунизированных ВИЧ-1, может предусматривать богатый источник для разработки эффективных терапевтических средств для людей, нейтрализующих ВИЧ-1, путем получения ограниченного (например, ограниченный представителем семейства VH3 и его полиморфы (полиморф)) репертуара такой иммунизированной мыши.
Ограничение ответа человеческого антитела на определенные патогены может снижать вероятность получения вариабельных областей антител от инфицированных людей, которые могут служить трамплинами для разработки высокоаффинных нейтрализующих антител против патогена. Например, иммунный ответ человека на инфекцию ВИЧ-1 клонально ограничен по всем инфекциям ВИЧ-1 и в развитии СПИДа (Muller et al., 1993, B-cell abnormalities in AIDS: stable and clonally restricted antibody response in HIV-1 infection, Scand. J. Immunol. 38:327-334; Wisnewski et al., 1996). Дополнительно, гены VH вообще не присутствуют во всех полиморфных формах у любого определенного индивидуума; определенные индивидуумы в определенных популяциях обладают одним вариантом, в то время как индивидуумы в других популяциях обладают другим вариантом. Таким образом, наличие биологической системы, которая ограничена одним геном VH и его вариантами будет согласно различным вариантам осуществления предусматривать до сих пор неиспользованный источник разнообразия для продукции вариабельных областей антител (например, тяжелые и легкие родственные домены человека) на основе ограниченного гена VH. Таким образом, согласно одному аспекту предусматривается генетически модифицированное не относящееся к человеку животное, которое содержит множество полиморфных вариантов не более одного, или не более двух представителей семейства генных сегментов VH человека. Согласно одному варианту осуществления не более одного, или не более двух генных сегментов VH человека функционально связаны с одним или несколькими генными сегментами DH человека, одним или несколькими генными сегментами JH человека и человеческим или не относящимся к человеку генным сегментом константной области. Согласно одному варианту осуществления константная область находится в эндогенном локусе константного гена не относящегося к человеку иммуноглобулина. Согласно одному варианту осуществления, не относящееся к человеку животное дополнительно содержит последовательность нуклеиновой кислоты, полученную из последовательности VL человека, например, реаранжированный или нереаранжированный генный сегмент VL человека или реаранжированная последовательность VL/JL человека. Согласно одному варианту осуществления последовательность нуклеиновой кислоты, полученная из последовательности VL человека, находится в эндогенном локусе не относящегося к человеку гена VL согласно одному варианту осуществления последовательность нуклеиновой кислоты, полученная из последовательности VL человека, находится на трансгене. Согласно конкретному варианту осуществления не относящееся к человеку животное не в состоянии экспрессировать вариабельный домен легкой цепи иммуноглобулина, который сам содержит эндогенный генный сегмент VL или JL и содержит не более одного или не более двух генов легкой цепи, которые кодируют реаранжированные домены VL человека (например, из не более одного или не более реаранжированных последовательностей VL/JL человека).
Генетически модифицированные мыши, которые экспрессируют вариабельные области тяжелой цепи человека с ограниченным использованием генного сегмента VH, применимы для получения относительно большого репертуара разнообразия множественности сегментов J, комбинаторного разнообразия и соматически мутированных высокоаффинных вариабельных областей тяжелой цепи иммуноглобулина человека от иначе ограниченного репертуара. Согласно одному варианту осуществления ограниченный репертуар относится к заранее определенному ограничению в количестве и/или идентичности генов зародышевой линии, что приводит к невозможности образовать у мыши реаранжированный ген тяжелой цепи, который происходит от любого гена V, кроме предварительно выбранного гена V. Согласно вариантам осуществления, которые используют предварительно выбранный ген V, но не предварительно выбранный ген D и/или ген J, репертуар ограничен по отношению к идентичности гена V, но не гена D и/или J (например, репертуар состоит в основном из не более одного или не более двух генных сегментов VH (и/или их полиморф), а также множества генных сегментов D и множества генных сегментов J)). Идентичность предварительно выбранного гена V (и любого предварительно выбранного гена D и/или J) не ограничивается каким-либо конкретным геном V.
Разработка такой мыши, которая реаранжировала бы единственный ген VH (присутствует в виде единственного сегмента или набора вариантов) с различными генными сегментами D и J человека (например, сегментами DH и JH) предусматривает перестановочную машину in vivo разнообразия множественности сегментов J/комбинаторного разнообразия/соматической гипермутации, которая может быть использована для повторения мутаций в результирующих реаранжированных последовательностях вариабельной области тяжелой цепи (например, в зависимости от обстоятельств может быть V/D/J или V/J). У такой мыши процесс клональной селекции работает для выбора подходящих вариабельных областей, которые связывают представляющий интерес антиген, основанные на единственном заранее выбранном гене VH (или его вариантах). Поскольку компоненты клональной селекции мыши предназначены для селекции, основанной на единственном заранее выбранном генном сегменте VH, фоновый шум (например, большое разнообразие не являющихся антигенсвязывающими доменов VH, полученных из многих генных сегментов зародышевой линии) в значительной степени ликвидирован. С помощью правильного отбора генного сегмента VH относительно большее число клонально отобранных антигенспецифических антител может быть подвергнуто скринингу в течение более короткого периода времени, чем с помощью мыши с большим разнообразием сегментов V.
Предварительный отбор ограниченного репертуара и ограничение мыши по единственному сегменту V предусматривает систему для перестановки множественности сегментов V/D/J со скоростью, которая согласно различным вариантам осуществления выше, чем наблюдаемая у мышей, которые иначе должны рекомбинировать до 40 или более сегментов V с областей D и J. Удаление других сегментов V освобождает локус для образования большего количества комбинаций V/D/J для предварительно выбранного сегмента V, чем иначе наблюдается. Увеличенное число транскриптов, к которым приводит рекомбинация предварительно выбранного V с одним из множества сегментов D и одним из множества сегментов J, будет поддерживать эти транскрипты в системе клональной селекции в виде пре-В клеток, и система клональной селекции, таким образом, предназначена для циклирования В-клеток, которые экспрессируют предварительно выбранную область V. Таким образом, более уникальные реаранжировки области V, полученные от предварительно выбранного сегмента V, могут быть подвергнуты скринингу организмом, чем это было бы возможно в ином случае в заданный промежуток времени.
Согласно различным аспектах описываются мыши, которые повышают разнообразие множественности сегментов рекомбинаций V/D/J для предварительно выбранной области V, потому что все или по существу все рекомбинации в вариабельном локусе тяжелой цепи иммуноглобулина будут в предварительно выбранном сегменте V и сегментах D и J, которые помещены в таких мышах. Таким образом, мыши предусматривают способ получения разнообразия сегментов CDR3 с использованием основания или ограниченного репертуара генов VH.
Согласно одному аспекту предусматривается не относящееся к человеку животное, причем популяция В-клеток не относящегося к человеку животного экспрессирует тяжелые цепи иммуноглобулина, которые получают из не более одного или не более двух генных сегментов VH человека. Согласно одному варианту осуществления каждый из не более одного или не более двух генных сегментов VH человека присутствует в двух или более полиморфных формах. Согласно одному варианту осуществления генный сегмент VH человека присутствует в трех, четырех, пяти, шести, семи, восьми, девяти, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 полиморфных формах. Согласно одному варианту осуществления не относящееся к человеку животное экспрессирует вариабельный домен легкой цепи человека, полученный из генного сегмента VL человека.
Согласно одному аспекту предусматривается способ получения популяции В-клеток у не относящегося к человеку животного, причем популяция В-клеток экспрессирует тяжелые цепи человека, полученные из единственного генного сегмента VH зародышевой линии человека и двух или более генных сегментов D человека и двух или более генных сегментов J человека; способ, содержащий стадию иммунизации описанного в настоящем документе не относящегося к человеку животного представляющим интерес антигеном, и позволение не относящемуся к человеку животному развивать иммунный ответ на представляющий интерес антиген, причем иммунный ответ включает в себя экспрессию тяжелых цепей человека на поверхности В-клеток в популяции В-клеток. Согласно одному варианту осуществления не относящееся к человеку животное представляет собой грызуна (например, мышь или крысу). Согласно одному варианту осуществления генный сегмент VH человека, сегмент DH человека и сегмент JH человека функционально связаны с не относящимся к человеку геном константной области. Согласно одному варианту осуществления не относящееся к человеку животное дополнительно содержит последовательность нуклеиновой кислоты, кодирующую домен VL человека. Согласно одному варианту осуществления последовательность нуклеиновой кислоты, кодирующая домен VL человека, связана с генной последовательностью константной области не относящейся к человеку легкой цепи.
Согласно одному аспекту предусматривается способ получения не относящегося к человеку животного, которое экспрессирует популяцию иммуноглобулинов, характеризующихся наличием тяжелых цепей, которые происходят из множества реаранжировок единственного генного сегмента VH человека (или единственного представителя семейство генов VH человека) и одного из множества генных сегментов DH и одного из множества генных сегментов JH. Согласно одному варианту осуществления генный сегмент VH человека представляет собой генный сегмент VH1-69 человека.
Согласно одному варианту осуществления генный сегмент VH человека представляет собой генный сегмент VH1-2 человека.
Согласно одному аспекту предусматривается способ получения популяции вариабельных доменов тяжелой цепи иммуноглобулина человека, чьи CDR1 и CDR2 происходят от того же генного сегмента VH зародышевой линии, и чьи CDR3 происходят от генного сегмента зародышевой линии и двух или более сегментов D человека, и двух или более сегментов J человека; способ, предусматривающий иммунизацию описанного в настоящем документе не относящегося к человеку животного представляющим интерес антигеном и предоставление не относящемуся к человеку животному развивать иммунный ответ на представляющий интерес антиген, причем иммунный ответ включает в себя экспрессирование вариабельных доменов тяжелой цепи человека в тесной связи с вариабельным доменом легкой цепи. Согласно одному варианту осуществления не относящееся к человеку животное представляет собой грызуна (например, мышь или крысу). Согласно одному варианту осуществления генный сегмент VH человека, сегмент D человека и сегмент J человека функционально связаны с не относящимся к человеку геном константной области. Согласно одному варианту осуществления не относящееся к человеку животное дополнительно содержит последовательность нуклеиновой кислоты, кодирующую домен VL человека. Согласно одному варианту осуществления последовательность нуклеиновой кислоты, кодирующая домен VL человека, связана с генной последовательностью константной области не относящейся к человеку легкой цепи.
Согласно одному аспекту предусматривается генетически модифицированное не относящееся к человеку животное, причем не относящееся к человеку животное не способно экспрессировать не относящийся к человеку домен VH, и причем каждая тяжелая цепь иммуноглобулина популяции тяжелых цепей, экспрессированных у животных, содержит домен VH человека, содержащий CDR1 и CDR2, которые идентичны, но для одного или нескольких соматических гипермутаций, и причем популяция тяжелых цепей содержит множество последовательностей CDR3, полученных из множества реаранжировок с множеством генных сегментов D и J.
Согласно одному аспекту предусматривается биологическая система для получения изменения в идентичности и длине CDR3, содержащая генетически модифицированное описанное в настоящем документе не относящееся к человеку животное, причем не относящееся к человеку животное содержит не более одного или не более двух генных сегментов VH человека и два или более генных сегмента D, и один или несколько генных сегментов J, причем не относящееся к человеку животное дополнительно содержит гуманизированный локус легкой цепи иммуноглобулина. Согласно различным вариантам осуществления не относящееся к человеку животное в ответ на иммунизацию представляющим интерес антигеном производит иммунный ответ, который включает экспрессию популяции тяжелых цепей иммуноглобулина, характеризующихся наличием у каждой CDR1 и CDR2, которые отличаются только соматической гипермутацией, и CDR3, которые отличаются реаранжировкой и соматической гипермутацией. Согласно одному варианту осуществления биологическая система представляет собой мышь, которая генетически модифицирована, как описано в настоящем документе. Согласно одному варианту осуществления генный сегмент VH человека и генный сегмент VL человека находятся в эндогенных локусах мышиного тяжелого и легкого иммуноглобулина, соответственно. Согласно одному варианту осуществления один или несколько генных сегментов VH человека и генный сегмент VL человека находятся на трансгенах (т.е. в локусе, отличном от эндогенного локуса иммуноглобулина).
Примеры
Предусматриваются следующие примеры, чтобы описать специалистам в настоящей области техники, как производить и применять способы и композиции согласно настоящему изобретению, и не предназначены для ограничения объема того, что авторы настоящего изобретения считают своим изобретением. Если не указано иное, температура указана в градусах Цельсия, а давление равно или близко к атмосферному. В вышеизложенных примерах, когда указывается использование наборов и реагентов от различных поставщиков, все процедуры были проведены в соответствии с требованиями производителя.
Пример 1. Конструкция ограниченных локусов тяжелой цепи
Уникально сконструированный локус тяжелой цепи человека, содержащий единственный генный сегмент VH человека, расположенный перед всеми генными сегментами DH и JH человека создали с помощью серии реакций гомологичной рекомбинации в бактериальных клетках (BHR) с использованием ДНК бактериальной искусственной хромосомы (ВАС). Несколько нацеливающих конструкций для создания единственного VH, содержащего локус тяжелой цепи, сконструировали с использованием технологии генетической инженерии VELOCIGENE® (смотрите, например, патент США №6586251 и Valenzuela, D.M., et al. (2003). High-throughput engineering of the mouse genome coupled with high-resolution expression analysis. Nat Biotechnol 21, 652-659).
Конструкция ограниченного локуса тяжелой цепи VH1-69 человека. Вкратце, четыре модификации выполнили с использованием ДНК ВАС человека для создания нацеливающей конструкции, содержащей генный сегмент VH1-69 человека со всеми сегментами DH и JH человека (фиг.1). Согласно первой модификации модифицированная ВАС человека, содержащая множесгченные дистальные (5′) генные сегменты VH человека, включая VH1-69, расположенные выше гигромициновой кассеты выбора, и 5 гомологичное плечо мыши было нацелено со второй спектиномициновой кассетой, которое также содержало модифицированную последовательность сигнала рекомбинации (RSS; BHR 1, фиг 1, вверху слева). Эта модифицированная последовательность сигнала рекомбинации (RSS) вводила две точечные мутации (Т в А и G в А) в 3′ области RSS гена VH1-69 человека, изменяя нонамер RSS на оптимальную консенсусную последовательность. Таким образом, первая модификация (BHR 1) создала геномный фрагмент человека, содержащий генный сегмент VH1-69 человеческий с модифицированной 3′ RSS, уникальный сайт рестрикции AsiSI, расположенный приблизительно в 180 п.о. ниже по ходу транскрипции от RSS и спектиномициновую кассету (фиг.1, посередине слева).
Вторая модификация (BHR 2) включала в себя использование неомициновой (Нео) кассеты, окруженной сайтами FRT, для удаления гигромициновой кассеты и 5′ генных сегментов VH человека выше по ходу транскрипции от генного сегмента VH1-69. Эту модификацию нацеливали 5′ на генный сегмент VH1-69 человека, чтобы оставить интактными около 8,2 т.п.о. промоторной области VH1-69 человека и 5″ гомологичного плеча мыши (фиг.1, внизу слева).
Третья модификация (БГР 3) включала в себя другую спектиномициновую кассету, окруженную уникально сконструированными сайтами 5′ PI- Seel и 3′ AsiSI, нацеленными на геномной фрагмент человека, содержащий первые три функциональных генных сегментов VH человека и все генные сегменты DH и JH человека (фиг.1, посередине справа). Геномный фрагмент человека ранее нацеливали на неомициновую кассету, и он содержал гомологичные плечи 5′ и 3′, содержащие мышиную геномную последовательность 5′ и 3′ эндогенного локуса тяжелой цепи, включая в себя интронный энхансер 3′ и ген IgM. Эта модификация удаляла геномную последовательность 5′ мыши и генные сегменты VH человека, оставляя приблизительно 3,3 т.п.о. межгенной области VH-DH выше по ходу транскрипции от генного сегмента DH1-1 человека, все сегменты DH и JH человека и геномный фрагмент 3′ мыши, содержащий интронный энхансер 3′ и ген IgM (фиг.1, внизу справа).
Четвертую модификацию получали с помощью использования уникальных сайтов PI- SeeI и AsiSI (описанные выше) для лигирования двух модифицированных ВАС от BHR 2 и BHR 3 (фиг.1, внизу в центре), которые производили окончательную нацеливающую конструкцию. Окончательная нацеливающая конструкция для создания модифицированного локуса тяжелой цепи, содержащей единственный генный сегмент VH человека и все генные сегменты DH и JH человека в ES клетках, содержала от 5′ до 3′, 5′ гомологичное плечо, содержащее приблизительно 20 т.п.о. мышиной геномной последовательности выше по ходу транскрипции от эндогенного локуса тяжелой цепи, 5′ сайт Frt, неомициновую кассету, 3′ сайт Frt, приблизительно 8,2 т.п.о. промотера VH1-69 человека, VH1-69 человека с модифицированной 3′ RSS, 27 генных сегментов DH человека, шесть сегментов JH человека, а также 3′ гомологичное плечо, содержащее приблизительно 8 т.п.о. мышиной геномной последовательности ниже по ходу транскрипции от генного сегмента JH мыши, включая 3′интронной энхансер и ген IgM (фиг.1, внизу). Нацеливающий вектор VH1-69 человека (SEQ ID NO: 3) линеаризовали и электропорировали в гетерозиготные ES клетки мыши для удаления эндогенного локуса тяжелой цепи.
Конструкция ограниченного локуса тяжелой цепи VH1-2 человека. С использованием описанных выше стадий, другие полиморфные генные сегменты VH, связанные с константными областями тяжелой цепи мыши, используют для конструкции серии мышей, содержащих ограниченное число сегментов V тяжелой цепи иммуноглобулина (например, 1, 2, 3, 4 или 5), причем сегменты V представляют собой полиморфные варианты представителя семейства генов V. Иллюстративные полиморфные генные сегменты VH происходят от генных сегментов VH человека включая в себя, например, VH1 2, VH2-26, VH2-70 и VH3-23. Такие генные сегменты VH человека получают, например, путем синтез de novo (например, технология Blue Heron, Bothell, WA) с использованием последовательности, доступные из опубликованных баз данных. Таким образом, фрагменты ДНК, кодирующие каждый ген VH, согласно некоторым вариантам осуществления, получают независимо для включения в описанный в настоящем документе нацеливающий вектор. Таким образом, множественные модифицированные локусы тяжелой цепи иммуноглобулина, содержащие ограниченное число генных сегментов VH, конструируют связанными с константными областями тяжелой цепи мыши. Иллюстративная стратегия нацеливания для создания ограниченного гуманизированного локуса тяжелой цепи, содержащего генный сегмент VH1-2 человека, 27 генных сегментов DH человека и шесть генных сегментов JH человека, показана на фиг.2.
Вкратце, модифицированный клон ВАС человека, содержащий три генных сегмента VH человека (VH6-1, VH1-2, VH1-3), 27 генных сегментов DH человека и шесть генных сегментов JH человека (смотрите USSN 13/404075; поданный 24 февраля 2012, который включен в настоящий документ посредством ссылки), используют для создания ограниченного гуманизированного локуса тяжелой цепи, содержащего генный сегмент VH1-2 человека. Этот модифицированный клон ВАС функционально связывает вышеупомянутые генные сегменты тяжелой цепи человека с интронным энхансером мыши и константной областью IgM. Ограниченный локус тяжелой цепи на основе VH1-2 человека получают с помощью двух гомологичных рекомбинаций с использованием описанного выше модифицированного клона ВАС человека.
При первой гомологичной рекомбинации, 205 п.о. генного сегмента VH6-1 человека (от приблизительно 10 п.о. выше по ходу транскрипции (5′) от стартового кодона VH6-1 в экзоне 1 до приблизительно 63 п.о. ниже по ходу транскрипции (3*) от начала экзона 2) в модифицированном клоне ВАС человека удаляют с помощью бактериальной гомологичной рекомбинации с использованием спектиномициновой (aadA) кассеты, окруженной уникальными сайтами рестрикции PI-Scel (фиг.2, BHR 1). Это позволяет впоследствии удалять кассету aadA, не нарушая другие генные сегменты человека в ограниченном локусе тяжелой цепи.
Для второй гомологичной рекомбинации, 5′ конец модифицированного клона ВАС человека, включая в себя весь генный сегмент VH1-3 человека и около 60 п.о. ниже по ходу транскрипции (3′) от генного сегмента, удаляют с помощью гомологичной рекомбинации с использованием гигромициновой кассеты, содержащей окруженные 5′ AsiSI и 3′ AscI сайты рестрикции (фиг.2, BHR 2). Как описано выше, спектиномициновую кассету необязательно удаляют после подтверждения финального нацеливающего вектора, включая удаление двух генных сегментов VH человека, окружающих генный сегмент VH1-2 человека (фиг.2, внизу). Иллюстративный нацеливающий вектор VH1-2 человека представлен в SEQ ID NO: 70.
Использование полиморфных генных сегментов VH в ограниченном локусе тяжелой цепи иммуноглобулина представляет собой новый подход к получению антител, популяции антител и популяции В-клеток, которые экспрессируют антитела, содержащие тяжелые цепи с различными CDR, полученными из единственного генного сегмента VH человека. Использование комплекса соматической гипермутации животного-хозяина вместе с комбинаторной ассоциацией с реаранжированными вариабельными доменами легкой цепи иммуноглобулина человека приводит к конструированию уникальных тяжелых цепей и уникальных пар VH/VL, которые расширяют иммунный репертуар генетически модифицированных животных и повышают их применимость в качестве платформы для создания нейтрализующих антител, специфичных к человеческим патогенам.
Таким образом, использование описанной выше стратегии для включения дополнительных и/или других полиморфных генных сегментов VH в локус тяжелой цепи иммуноглобулина мыши позволяет продуцировать новые репертуары антител для использования в нейтрализации человеческих патогенов, которые могли бы эффективно ускользать от иммунной системы хозяина.
Описанные выше нацеленные ES клетки использовали в качестве доноров ES клеток и вводили в 8-клеточный эмбрион мыши с помощью способа VELOCIMOUSE® (смотрите выше). Мышей, несущих гуманизированный локус тяжелой цепи, содержащий единственный генный сегмент VH человека, все генных сегментов DH и JH человеческий, функционально связанные с генами константной области иммуноглобулина мыши, определяли путем генотипирования с использованием модификации анализа аллелей (Valenzuela et al., смотрите выше), который обнаружил присутствие неомициновой кассеты, генного сегмента VH человека и области в генных сегментах DH и JH человека, а также эндогенные последовательности тяжелой цепи. В таблице 4 представлены праймеры и зонды, используемые в этом анализе для подтверждения мышей, несущих ограниченный локус тяжелой цепи, содержащий единственный генный сегмент Vn1-69 человека, 27 генных сегментов DH человека и шесть генных сегментов JH человека.
Мыши, несущие сконструированный локус тяжелой цепи, который содержит единственный генный сегмент VH человека, могут быть отнесены к линии FLPe делеторных мышей (смотрите, например, Rodriguez, C.I. etal. (2000) High-efficiency deleter mice show that FLPe is an alternative to Cre-loxP. Nature Genetics 25: 139-140) для того, чтобы удалить любую неомициновую кассету Frt′ed, введенную нацеливающим вектором, который не удаляют, например, на стадии ES клеток или в зародыше. Необязательно, неомициновая кассета остается в мышах.
Потомство является генотипированным, и выбирают детеныша, гетерозиготного по гуманизированному локусу тяжелой цепи, содержащей единственный генный сегмент VH человека, все сегменты DH и JH человека, функционально связанные с эндогенными константными генами иммуноглобулина мыши, для характеристики репертуара тяжелой цепи иммуноглобулина.

Пример 2. Характеристика мышей, экспрессирующих тяжелые цепи, полученные из единственного генного сегмента VH человека
Описанных в примере 1 мышей, гомозиготных по единственному генному сегменту VH человека в эндогенном локусе тяжелой цепи, оценивали на экспрессию и развитие В-клеток с помощью проточной цитометрии.
Вкратце, селезенки и костный мозг собирали у мышей дикого типа (n=3 в группе; шесть недель, самки или самцы) и у мышей, гомозиготных по единственному генному сегменту VH человека, всем генным сегментам DH и JH человека, функционально связанным с константными областями тяжелой цепи мыши. Эритроциты из селезенок лизировали с помощью лизирующего буфера АСК (Lonza Walkersville) с последующим отмыванием полной средой RPMI.
Проточная цитометрия. Клетки (1×106) инкубировали с мышиными антителами CD16/CD32 (2,4G2, BD PHARMINGEN™) на льду в течение 10 минут, после чего метили следующими панелями антител в течение 30 минут на льду. Панель костного мозга: мышиные антитела FITC-CD43 (1В11, BioLegend), PE-ckit (2B8, BIOLEGEND®), PeCy7-IgM (II/41, EBIOSCIENCE®), PerCP-Cy5.5-IgD (11-26c.2a, BIOLEGEND®), APC-eFluor 780-B220 (RA3-6B2, EBIOSCIENCE®), APC-CD19 (MB19-1, EBIOSCIENCE®). Панель костного мозга и селезенки: мышиные антитела FITC-Igκ (187.1, BD Biosciences), PE-Igλ, (RML-42, BIOLEGEND®), PeCy7-IgM (II/41, EBIOSCIENCE®), PerCP-Cy5.5-IgD (11-26c.2a, BIOLEGEND®), Pacific Blue-CD3 (17A2, BIOLEGEND®), APC-B220 (RA3-6B2, EBIOSCIENCE®), APC-H7-CD19 (ID3, BD Biosciences). Костный мозг: незрелые В-клетки (B220intIgM+), зрелые В-клетки (B220hiIgM+), про-В-клетки (CD19+ckit+CD43+), пре-В-клетки (CD19+ckit-CD43-), незрелые Igκ+ В-клетки (B220intIgM+Igκ+Igλ-), незрелые Igλ+ В-клетки (B220intIgM+Igκ-Igλ+), зрелые Igκ+ В-клетки (B220hiIgM+Igκ+Igλ-), зрелые Igλ+ В-клетки (B220hiIgM+Igκ-Igλ+). Селезенка: В-клетки (CD19+), зрелые В-клетки (CD19+IgDhiIgMint), переходные/незрелые В-клетки (CD19+IgDintIgMhi). Костный мозг и селезенка: Igκ+ В-клетки (CD19+Igκ+Igλ-), Igλ+ В-клетки (CD19+Igκ-′Igλ+).
После окрашивания клетки отмывали и фиксировали в 2% формальдегиде. Сбор данных проводили на проточном цитометре LSRII и анализировали с помощью программного обеспечения FLOWJO™ (Tree Star, Inc.). Результаты для компартмента селезенки показаны на фиг.3, 4А и 5-7. Результаты для компартмента костного мозга показаны на фиг.4В и 8-11В.
Экспрессия VH человека. Экспрессию генного сегмента VH1-69 человека определяли у мышей, гетерозиготных и гомозиготных по генному сегменту VH1-69 человека, всем генных сегментов DH и JH человека, функционально связанными с константными областями тяжелой цепи мыши, с помощью анализа количественной ПЦР с использованием зондов TAQMAN®.
Вкратце, CD19+-клетки очищали из селезенки групп мышей (n=3 в группе), используя мышиные микрогранулы CD 19 (Miltenyi Biotec) в соответствии с требованиями завода-изготовителя. Общую РНК очищали с использованием мининабора RNEASY™ (Qiagen) и геномную РНК удаляли с использованием обработки, не содержащей РНазу ДНКазой на колонке (Qiagen). Приблизительно 200 нг мРНК обратно транскрибировали в кДНК с использованием набора First Stand cDNA Synthesis (Invitrogen) и затем амплифицировали с использованием универсального мастер-микса для ПЦР TAQMAN™ (Applied Biosystems) с использованием системы обнаружения последовательности ABI 7900 (Applied Biosystems). Использовали уникальные комбинации праймер/зонд для специального определения экспрессии производных VH1-69 тяжелых цепей человека (табл.5). Относительную экспрессию нормализовала к константной области к мыши (mCκ). Результаты показаны на фиг.12.
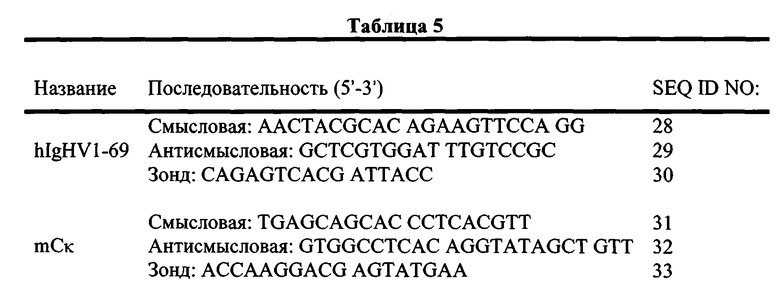
Пример 3. Гуморальный иммунный ответ у мышей, экспрессирующих тяжелые цепи, полученные из единственного генного сегмента VH человека
Гуморальный иммунный ответ определяли у мышей, гомозиготных по локусам вариабельного гена тяжелой и легкой к цепи человека (Нк), и у мышей, гомозиготных по единственному генному сегменту VH человека, всем генным сегментам DH и JH человека, функционально связанным с константными областями тяжелой цепи мыши (1 г VH НО), с помощью сравнительной иммунизации с использованием человеческого рецептора клеточной поверхности (Антигена А).
Иммунизация. Сыворотку собирали у группы мышей до иммунизации описанным выше антигеном. Антиген (2,35 мкг каждого) вводили в начале первичной иммунизации, смешанного с 10 мкг олигонуклеотида CpG (Invivogen) и 25 мкг Adju-phos (Brenntag) в качестве адъювантов. Иммуноген вводили через подушечку лапы (п.л.) в объеме 25 мкл на мышь. Впоследствии мышей повторно иммунизировали через п.л. в количестве 2,3 мкг антигена вместе с 10 мкг CpG и 25 мкг Adju-phos в качестве адъювантов на 3, 6, 11, 13, 17 и 20 день для достижения в общей сложности шести повторных иммунизации. У мышей брали кровь на 15 и 22 дни после четвертой и шестой повторной иммунизации, соответственно, и антисыворотку оценивали на титры антител с антигеном А.
Титры антител определяли в сыворотке иммунизированных мышей с использованием анализа ELISA. Девяносто шести луночные планшеты для микротитрования (Thermo Scientific) покрывали антигеном А (1 мкг/мл) в фосфатном буфере (PBS, Irvine Scientific) в течение ночи при 4°С. На следующий день планшеты отмывали фосфатным буфером, содержащим 0,05% Tween 20 (PBS-T, Sigma-Aldrich) четыре раза, используя машину для мойки планшетов (Molecular Devices). Затем планшеты блокировали с помощью 250 мкл 1% бычьего сывороточного альбумина (BSA, Sigma-Aldrich) в PBS и инкубировали в течение одного часа при комнатной температуре. Затем планшеты отмывали четыре раза в PBS-T. Сыворотки от иммунизированных мышей и предварительно иммунные сыворотки серийко разводили в десять раз в 0,1% BSA PBS-Т, начиная с 1:100, и добавляли к блокированным планшетам в двух экземплярах, и инкубировали в течение одного часа при комнатной температуре. Последние две лунки оставляли пустыми для использования в качестве вторичного антитела контроля. Планшеты снова отмывали четыре раза PBS-T в машине для мойки планшетов. Разведение 1:5000 козьего антитела к мышиному конъюгированному вторичному антителу IgG-Fc-пероксидаза хрена (HRP, Jackson hnmunoresearch) добавляли в планшеты и инкубировали в течение одного часа при комнатной температуре. Планшеты снова отмывали восемь раз PBS-T и развивали с использованием в качестве субстрата ТМВ/Н202. Субстрат инкубировали в течение двадцати минут, и реакцию останавливали с помощью 1 н H2SO4 (VWR). Планшеты считывали на спектрофотометре (Victor, Perkin Elmer) при 450 нм. Титры антител рассчитывали с использованием GRAPHPAD PRISM™ (GraphPad Software, Inc)..
Титр сыворотки рассчитывали как разведение сыворотки в экспериментальном диапазоне титрования при сигнале связывания с антигеном, эквивалентном двухкратному превышению фона. Титр антител для гуморального иммунного ответа против рецептора клеточной поверхности человека (антиген А) приведены на фиг.19.
В аналогичном эксперименте гуморальный иммунный ответ определяли у мышей, гомозиготных по локусам вариабельного гена тяжелой и легкой к цепи человека (Нκ), и у мышей, гомозиготных по единственному генному сегменту VH человека, всем генным сегментам DH и JH человека, функционально связанным с константными областями тяжелой цепи мыши (1 г VH НО)) по сравнительной иммунизации с использованием вакцин против вируса гриппа FLUVIRIN® (Novartis Vaccines) и FLUMIST® (MedImmune LLC).
Вкратце, сыворотку собирали у группы мышей до иммунизации описанным выше антигеном (как описано выше). Мыши (n=5), гомозиготные по единственному генному сегменту VH (VH1-69) человека, всем генным сегментам DH и JH человека, функционально связанным с константными областями тяжелой цепи мыши (1 г VH НО) иммунизировали интраназально (и.н.) FLUMIST® (живой ослабленной вакциной против гриппа) в 1/3 обычной дозы на мышь. Одна нормальная доза FLUMIST® содержит 106,5-7,5 ОЕФ (относительная единица флуоресценции) живой ослабленной вакцины против гриппа. Таким образом, каждую мышь примировали. с помощью 70 мкл FLUMIST® в день 1, после чего повторно иммунизировали на 3, 6, 11, 13, 17, 20 день для достижения в общей сложности шести повторных иммунизации. В этой иммунизации вспомогательные вещества не использовали. У мышей брали кровь на 15 и 22 день после 4 и 6 повторной иммунизации, соответственно, и антисыворотки оценивали на титры антител к FLUMIST® (как описано выше).
Подобным образом, в иммунизациях с FLUVIRIN®, предварительно иммуннизированную сыворотку собирали у мышей до начала иммунизации. Мыши (n=5), гомозиготные по единственному генному сегменту VH (VH1-69) человека, всем генным сегментам DH и JH человека, функционально связанным с константными областями тяжелой цепи мыши (1 г VH НО) иммунизировали FLUVIRIN® (трехвалентной инактивированной вакциной против гриппа) через подушечку лапы (п.л.) в объеме 75 мкг каждого из гемагглютинина/мыши/повторной вакцинации. Мышей примировали на 1 день с последующей повторной вакцинацией через f.p. на 3, 6, 11, 13, 17, 20 день для достижения в общей сложности шести повторных иммунизации. В этой иммунизации вспомогательные вещества не использовали. У мышей брали кровь на 15 и 22 день после 4 и 6 повторной иммунизации, соответственно, и антисыворотки оценивали на титры антител к FLUVIRIN® (как описано выше).
Титр сыворотки рассчитывали как разведение сыворотки в экспериментальном диапазоне титрования при сигнале связывания антигена, эквивалентном двухкратному превышению фона. Титр антител для гуморального иммунного ответа против FLUMIST® и FLUVIRIN® приведены на фиг.20.
Как показано в этом примере, титры антител, полученные у мышей 1 г VH НО, были сопоставимы с полученными у мышей, имеющих множество генных сегментов VH человека (Нκ), как для рецептора клеточной поверхности человека, так и вирусного антигена (например, гриппа). Таким образом, мыши, содержащие локусы тяжелой цепи иммуноглобулина, ограниченные до одного генного сегмента VH, способны развивать устойчивый иммунный ответ на антиген способом, сравнимым с мышами, имеющими локусы тяжелой цепи иммуноглобулина, содержащие множество генных сегментов VH человека (например, 80 VH).
Пример 4. Анализ частоты использования генов антител и длины СОЮ у мышей с ограниченным локусом тяжелой цепи иммуноглобулина
Спленоциты собирают из мышей, гомозиготных по единственному генному сегменту VH человека в эндогенном локусе тяжелой цепи и гомозиготных по замещению эндогенных вариабельных локусов легкой цепи κ вариабельными локусами легкой цепи κ человека, иммунизированными рецептором клеточной поверхности человека (антигена А) анализировали на частоту использования генного сегмента тяжелой и легкой цепей с помощью полимеразной цепной реакции с обратной транскриптазой (ОТ-ПЦР) на мРНК
из В-клеток селезенки.
Вкратце, селезенки собирают и гомогенизируют в 1×PBS (Gibco) с использованием предметных стекол. Клетки осаждали в центрифуге (с ускорением 500g в течение 5 минут), а эритроциты лизировали в лизирующем буфере АСК. (Gibco) в течение 3 минут. Клетки отмывали с помощью 1×PBS и фильтровали с использованием фильтра с ячейкой 0,7 мкм. В-клетки выделяли из клеток селезенки с использованием магнитного позитивной селекции MACS для CD 19 (Miltenyi Biotec). Общую РНК выделяли из осажденных В-клеток с помощью набора RNeasy Plus (Qiagen). ПолиА+ мРНК выделяли из общей РНК с использованием мини-набора прямой мРНК Oligotex ® (Qiagen).
Двухцепочечную кДНК получали из мРНК В-клеток селезенки с помощью 5′-RACE с использованием набора синтеза кДНК SmarTer™ Pico (Clontech) с заменой подаваемой обратной транскриптазы и dNTP с Superscript® II и dNTPs (Invitrogen). Репертуары антител VH и Vκ амплифицировали из кДНК с использованием праймеров, специфичных для константных областей IgM, IgG или IgK и 5′ RACE праймеров SmarTer™ (табл.6). Продукты ПЦР очищали с использованием набора для очистки ПЦР QIAquick ® (Qiagen). Второй раунд ПЦР выполняли с использованием того же праймера 5′ RACE и двухраундового праймера 3′, специфичного для константных областей IgM, IgG или IGκ (табл.7). Продукты второго раунда ПЦР очищали с использованием системы SizeSelect ™ E-Gel® (Invitrogen). Третью ПЦР проводили с праймерами, которые добавляли 454 адаптеров и штрих-кодов. Продукты третьего раунда ПЦР очищали с использованием гранул ХР Agencourt ® AMPure ® (Beckman Coulter). Очищенные продукты ПЦР количественно определяли с помощью кПЦР SYBR® с помощью набора количественной библиотеки КАРА (КАРА Biosystems). Суммарные библиотеки подвергали ПЦР в эмульсии (emPCR) с использованием набора 454 GS Junior Titanium Series Lib-A emPCR (Roche Diagnostics) и двунаправленному секвенированию с использованием инструмента Roche 454 GS Junior в соответствии с требованиями завода-изготовителя.
Биоинформационный анализ. 454 последовательности сортировали на основе идеального совпадения со штрих-кодом образца и обрезали для соответствия определенному качеству. Последовательности аннотировали на основе выравнивания реаранжированных последовательностей иммуноглобулина в базу данных сегментов V(D)J зародышевой линии человека с использованием локальную установки Igblast (NCBI, v2.2.25+). Последовательность помечали как неоднозначную и удаляли из анализа, если были обнаружены множественные лучшие попадания с идентичными сцепленными генами. Набор скриптов Peri разработали для анализа результатов и сохранения данных в базе данных mysq1. Область CDR3 определяла между консервативным кодоном С и мотивом FGXG для легких цепей и мотивом WGXG для тяжелых цепей. Длину CDR3 определяли с использованием только продуктивных антител. Из последовательностей нуклеиновых кислот и предсказанной аминокислотной последовательности антител, частоту использование гена идентифицировали для последовательностей примированных IgM (15650), примированных IgG (18967) и примированных Ig (26804). Результаты показаны в таблице 8, в таблице 9, на фиг.21 и фиг.22.
В таблице 8 представлен процент наблюдаемых генных сегментов DH и JH человека, используемых в последовательностях вариабельных областей производных VH1-69 тяжелой цепи, примированных IgM (15650 последовательностей) и примированных IgG (18967 последовательностей). Генные сегменты DH4-4/DH4-11 и DH5-5/DH5-18 человека представлены в таблице 8 вместе из-за одинаковой идентичности последовательностей между соответствующими парами генных сегментов DH. В таблице 9 представлен процент генных сегментов Vκ и Jκ человека, наблюдаемых среди легких цепей (26804 последовательности), родственных вариабельных областей производных VH1-69 тяжелой цепи. Процентные показатели в таблицах 8 и 9 представляют округленные значения и в некоторых случаях могут не составлять 100%, когда суммируются.
Длина аминокислотной области CDR3 примированных IgM производными VH1-69 тяжелых цепей показана на фиг.21. Длина аминокислотной области CDR3 примированных IgG производных VH1-69 тяжелых цепей показано на фиг.22.
Как показано в таблицах 8 и 9, мыши, согласно настоящему изобретению продуцируют антигенспецифические антитела, содержащие производные VH1-69 тяжелых цепей, которые демонстрируют различные реаранжировки генного сегмента VH1-69 человека с различными сегментами DH человека и сегментами JH человека. Дополнительно, антигенспецифические антитела содержат родственные легкие цепи человека, содержащие домены Vκ человека, получаемые в результате различных реаранжировок генных сегментов Vκ и Jκ человека.
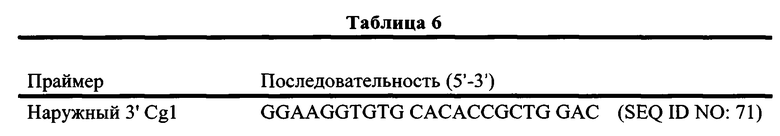

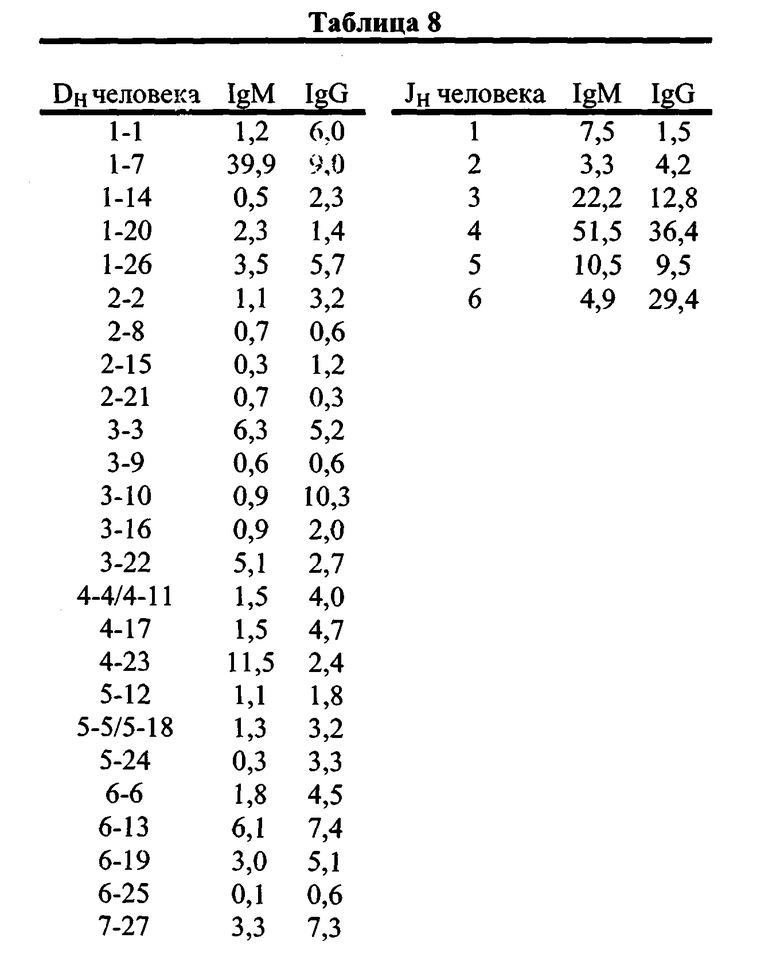

| название | год | авторы | номер документа |
|---|---|---|---|
| ГУМАНИЗИРОВАННЫЕ НЕ ОТНОСЯЩИЕСЯ К ЧЕЛОВЕКУ ЖИВОТНЫЕ С ОГРАНИЧЕННЫМИ ЛОКУСАМИ ТЯЖЕЛОЙ ЦЕПИ ИММУНОГЛОБУЛИНА | 2013 |
|
RU2656155C2 |
| МЫШИ С ОГРАНИЧЕННОЙ ТЯЖЕЛОЙ ЦЕПЬЮ ИММУНОГЛОБУЛИНА | 2012 |
|
RU2743589C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2664181C2 |
| ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА ЖИВОТНЫЕ, ЭКСПРЕССИРУЮЩИЕ ЧУВСТВИТЕЛЬНЫЕ К pН ПОСЛЕДОВАТЕЛЬНОСТИ ИММУНОГЛОБУЛИНОВ | 2013 |
|
RU2664473C2 |
| ЖИВОТНЫЕ, НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ, ИМЕЮЩИЕ СКОНСТРУИРОВАННУЮ ЛЕГКУЮ ЦЕПЬ ЛЯМБДА ИММУНОГЛОБУЛИНА, И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2778410C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2761639C2 |
| МЫШИ ADAM6 | 2012 |
|
RU2582261C2 |
| МЫШЬ С ОБЩЕЙ ЛЕГКОЙ ЦЕПЬЮ | 2013 |
|
RU2683514C2 |
| МЫШИ, ЭКСПРЕССИРУЮЩИЕ ОГРАНИЧЕННЫЙ РЕПЕРТУАР ЛЕГКИХ ЦЕПЕЙ ИММУНОГЛОБУЛИНА | 2014 |
|
RU2689664C2 |
| МЫШИ ADAM6 | 2012 |
|
RU2722373C2 |
















































































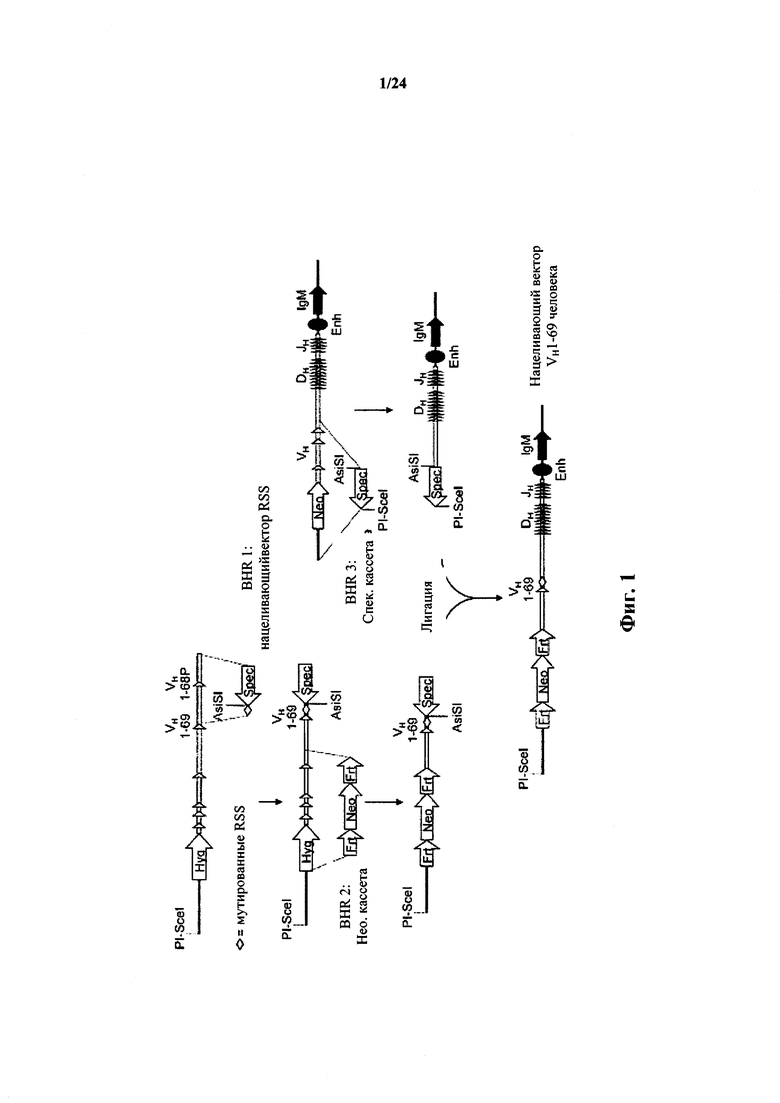
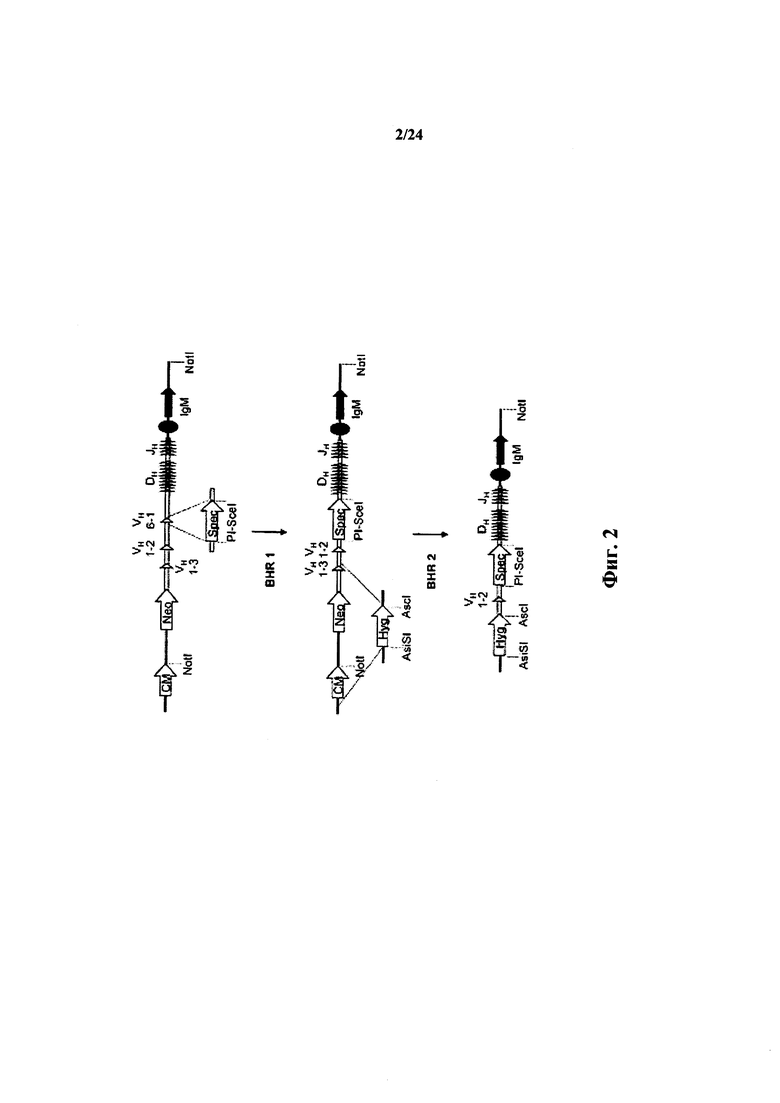
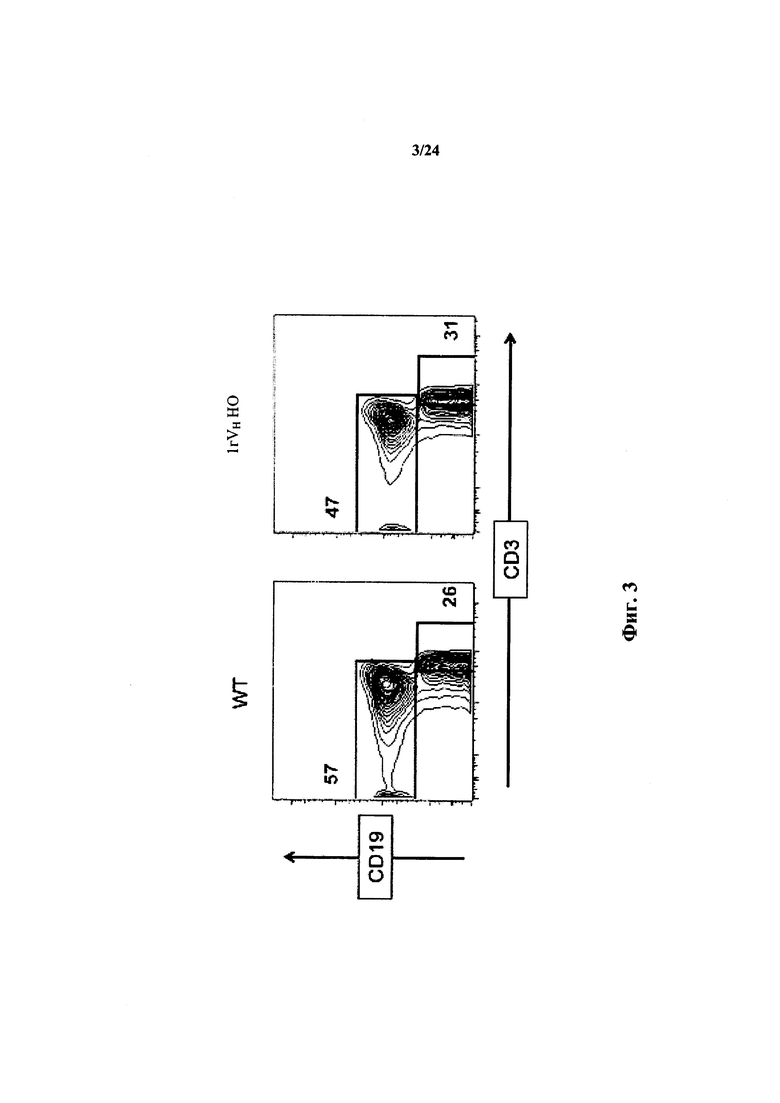
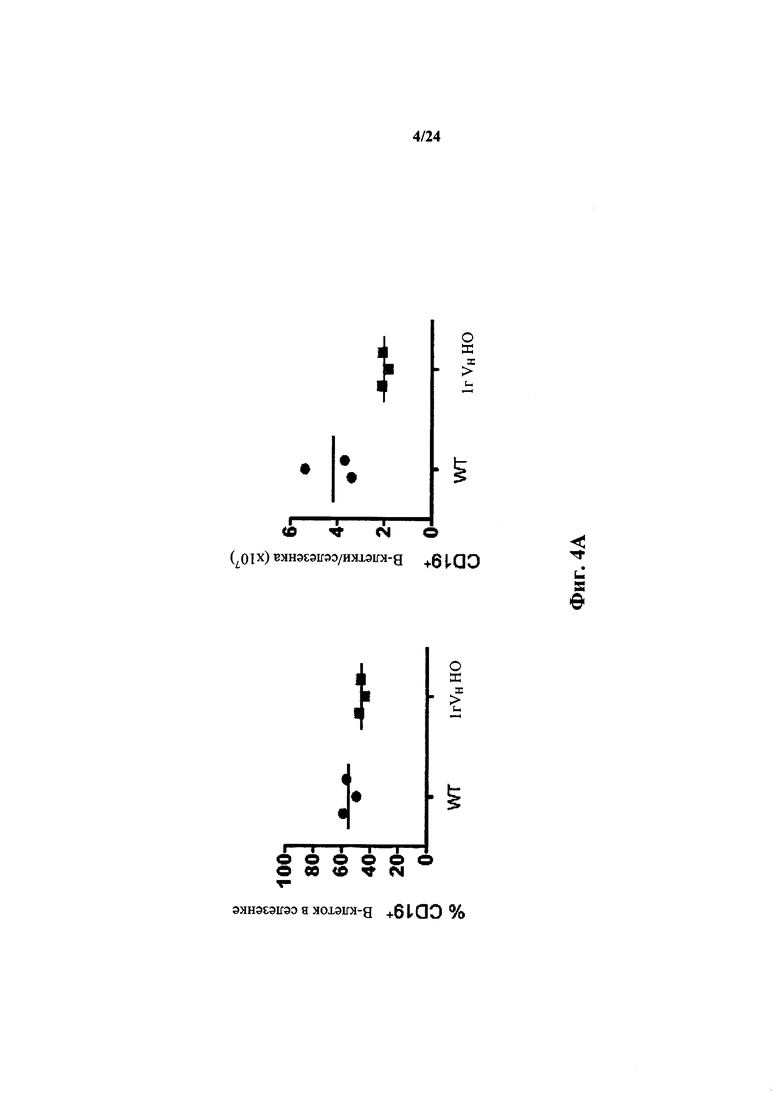
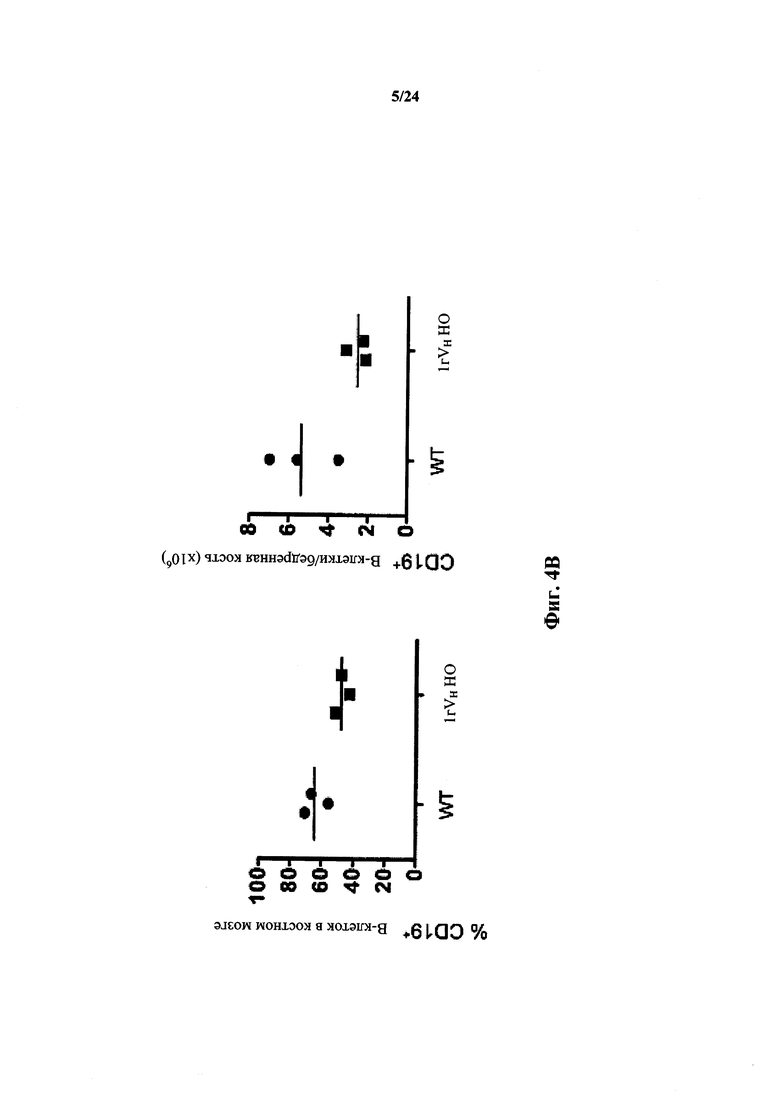
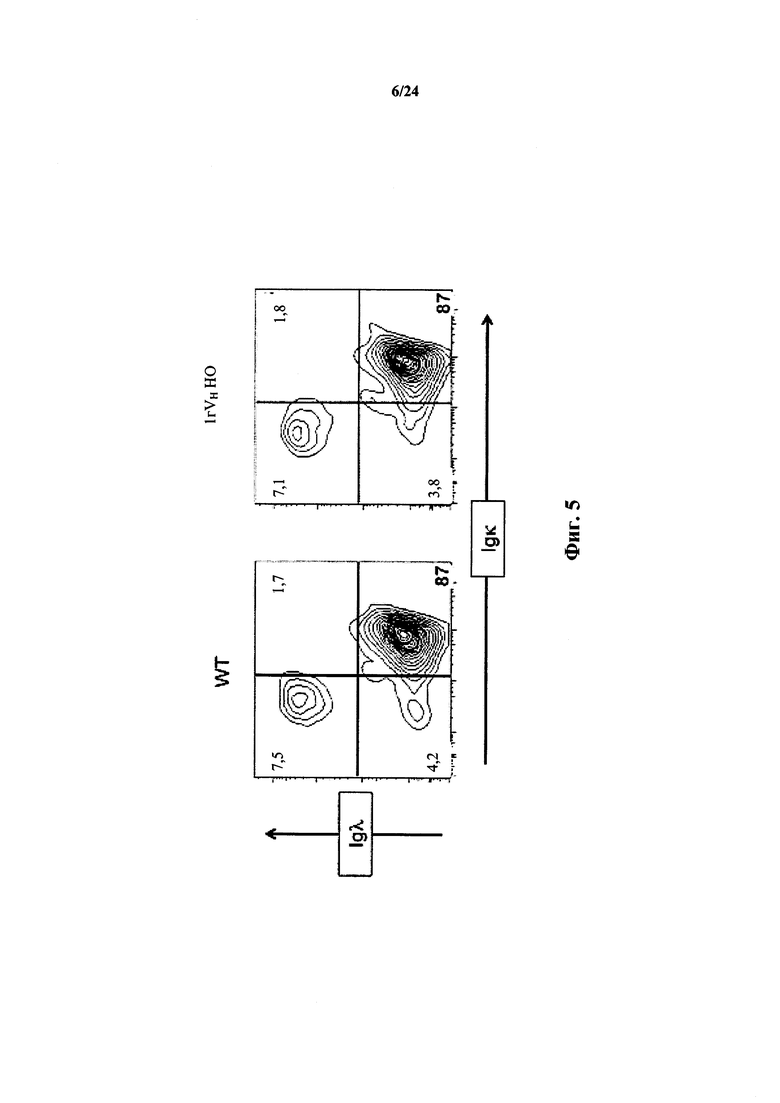
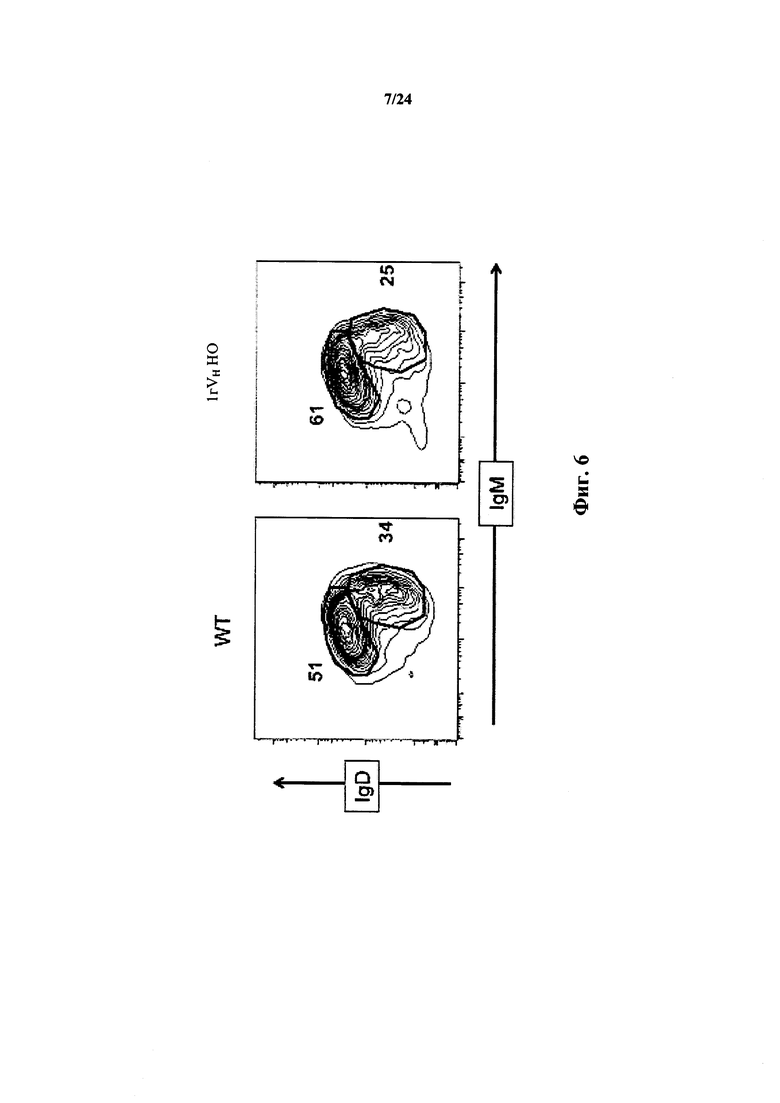
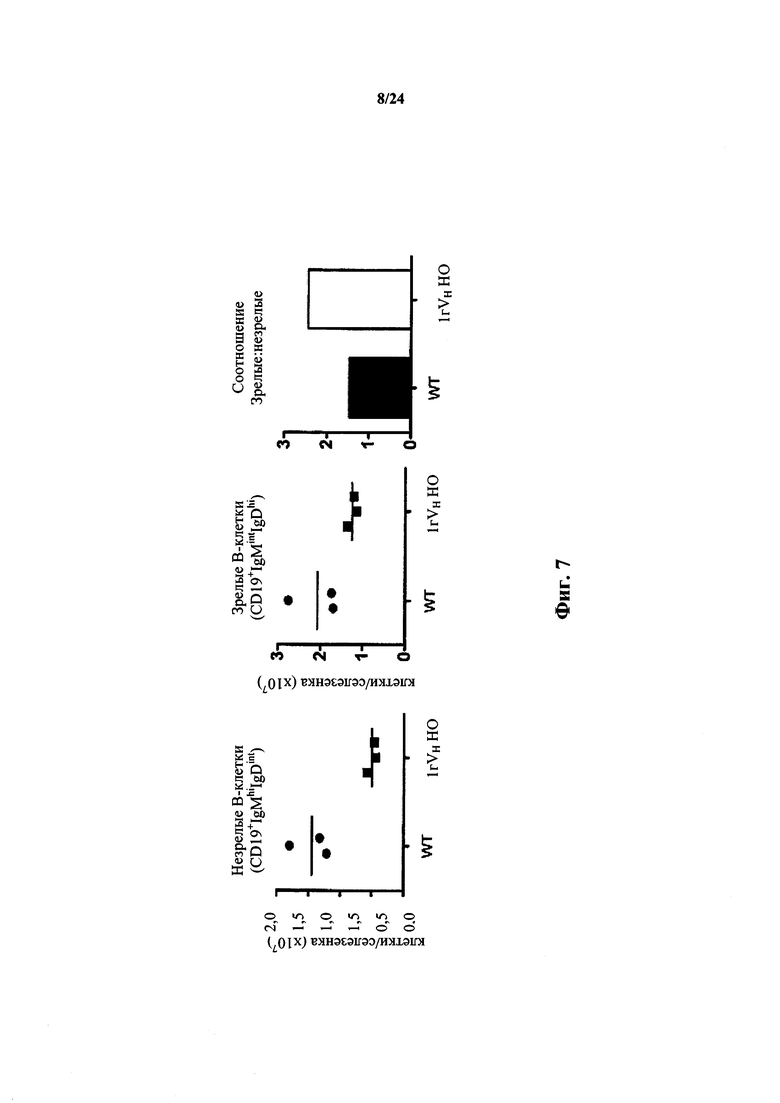
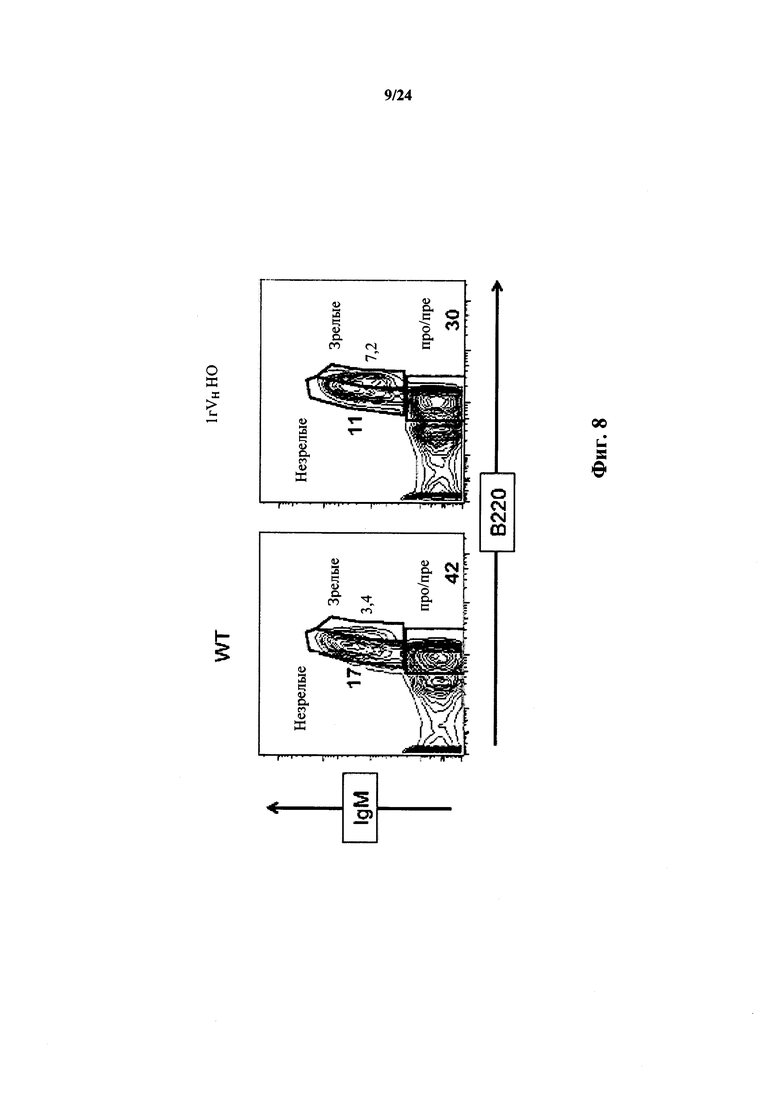
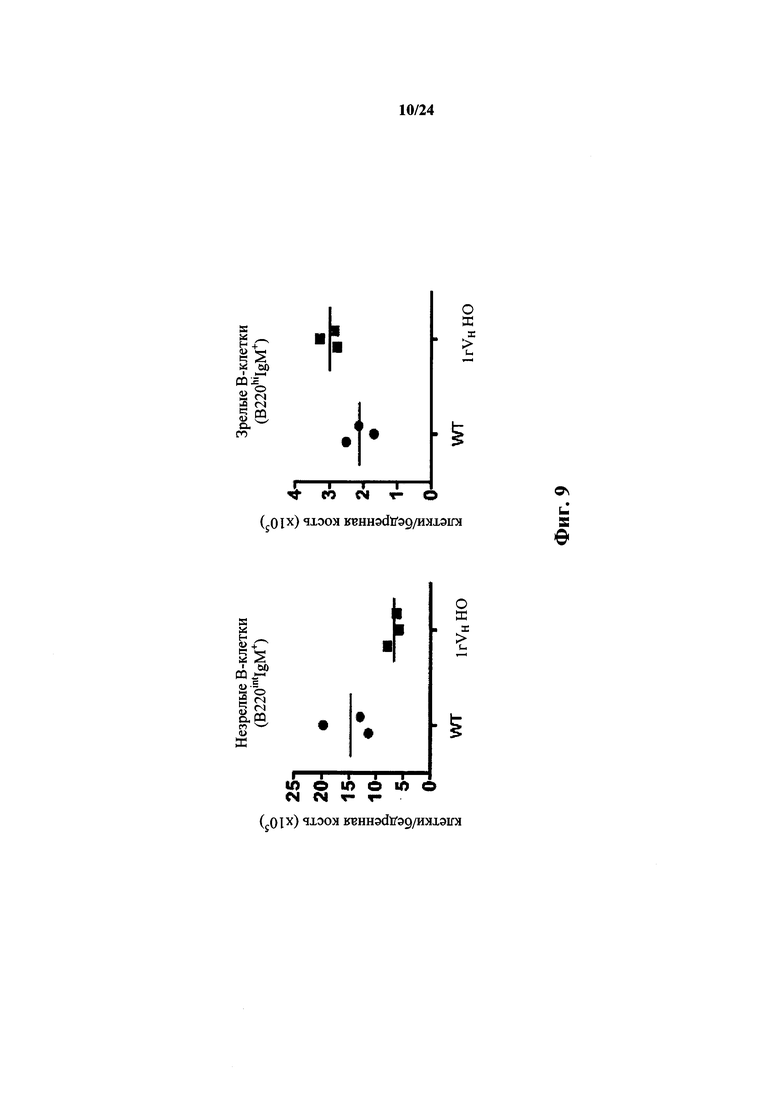
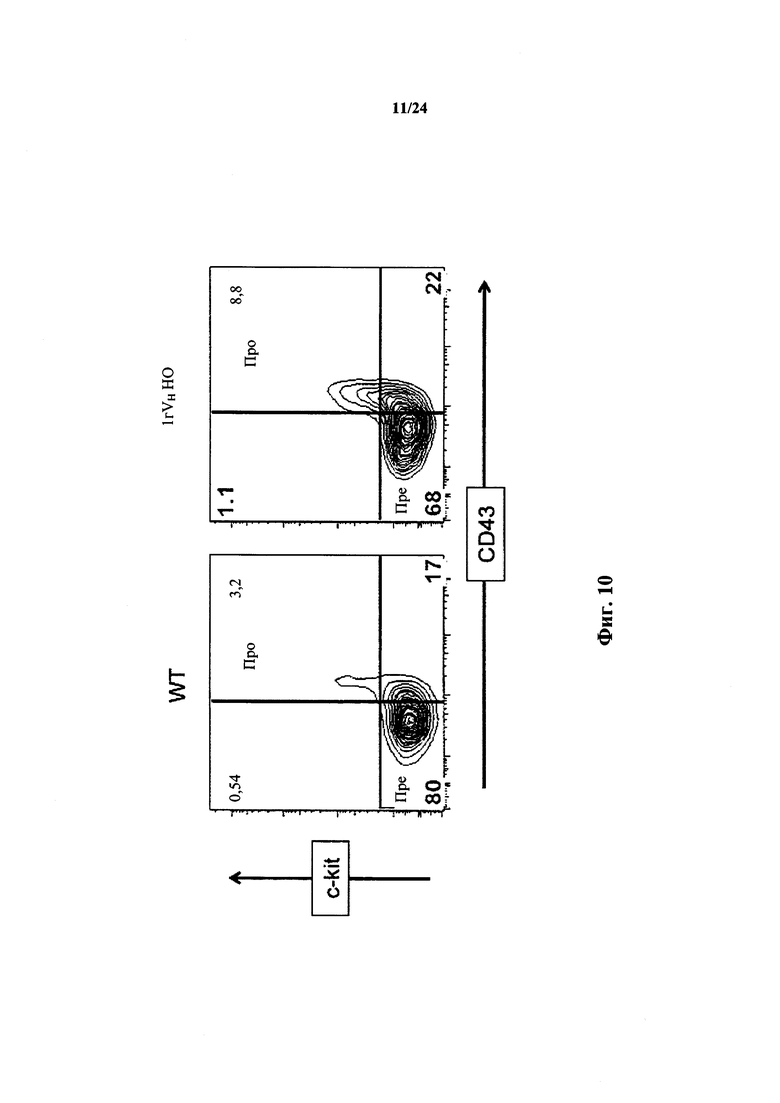
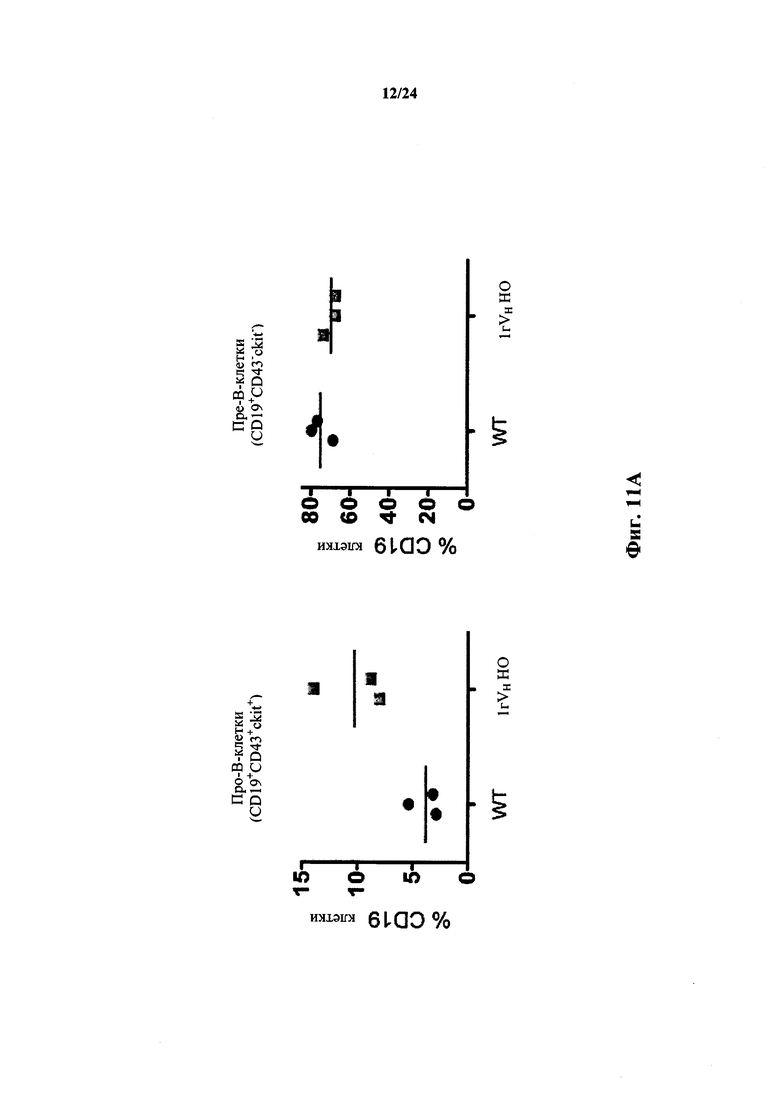
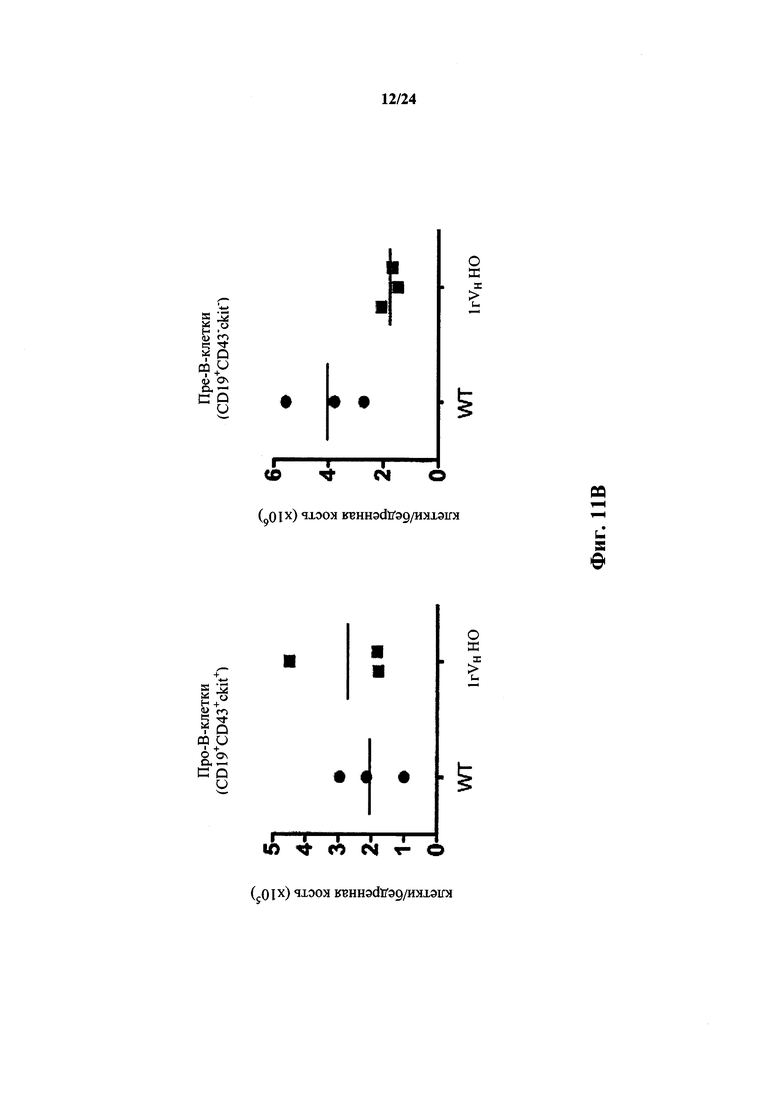
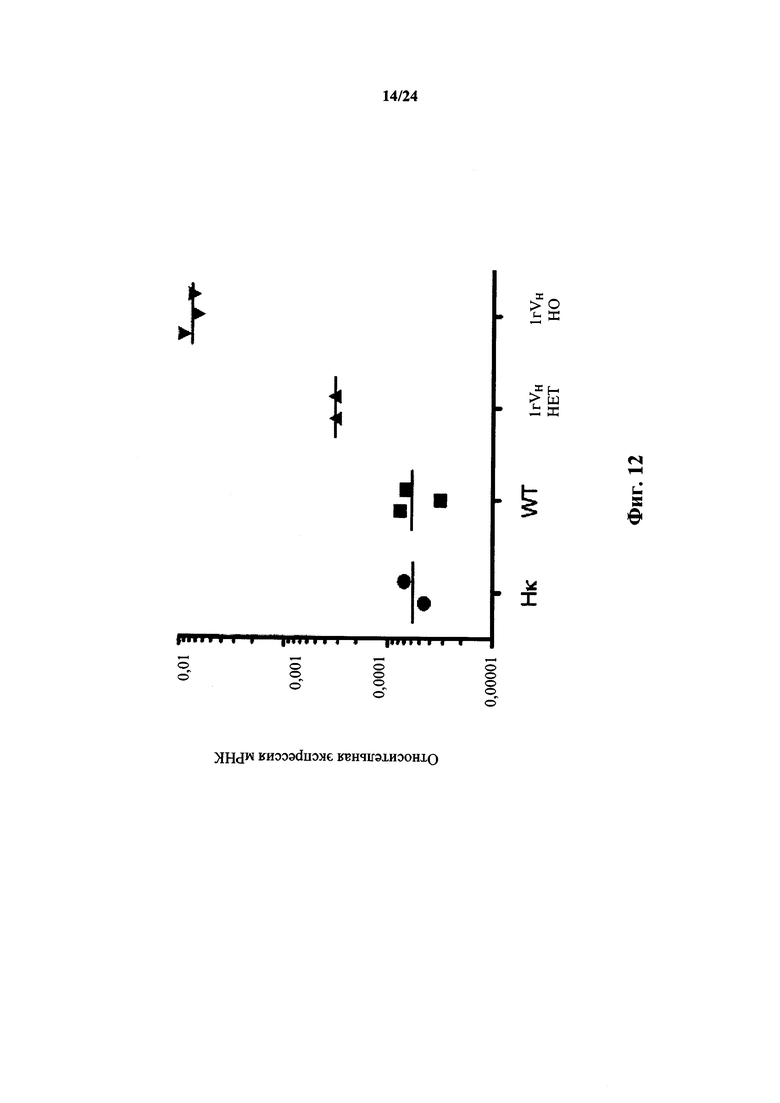

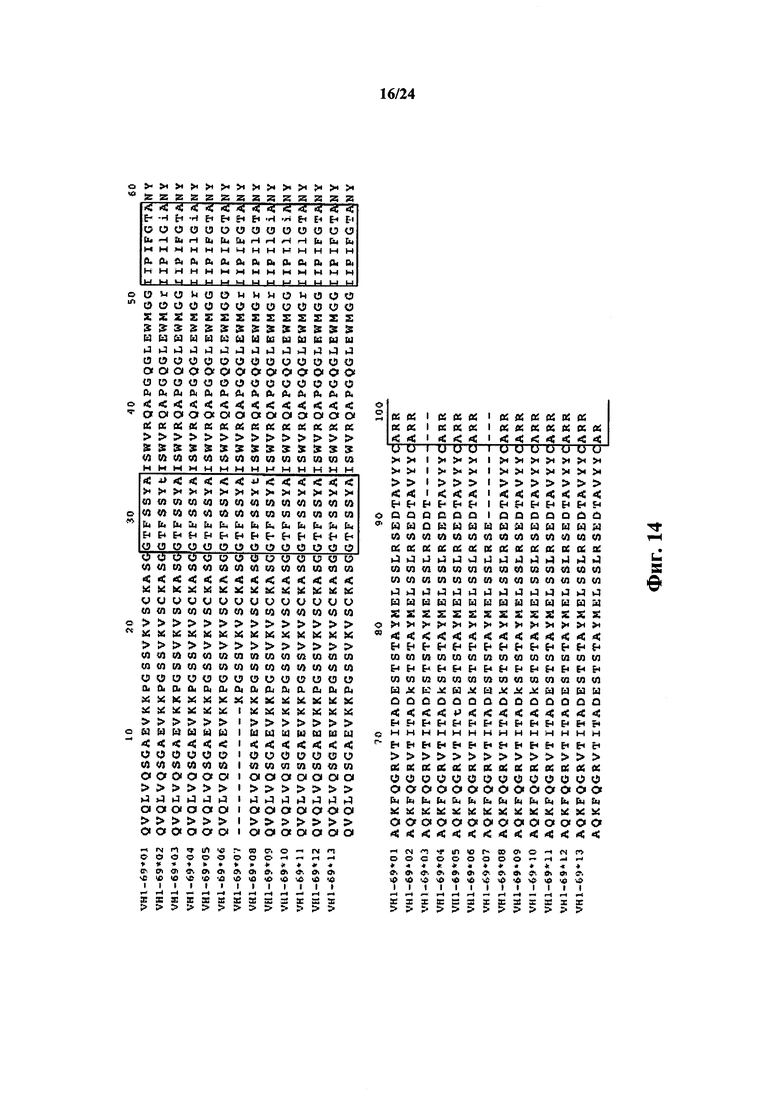
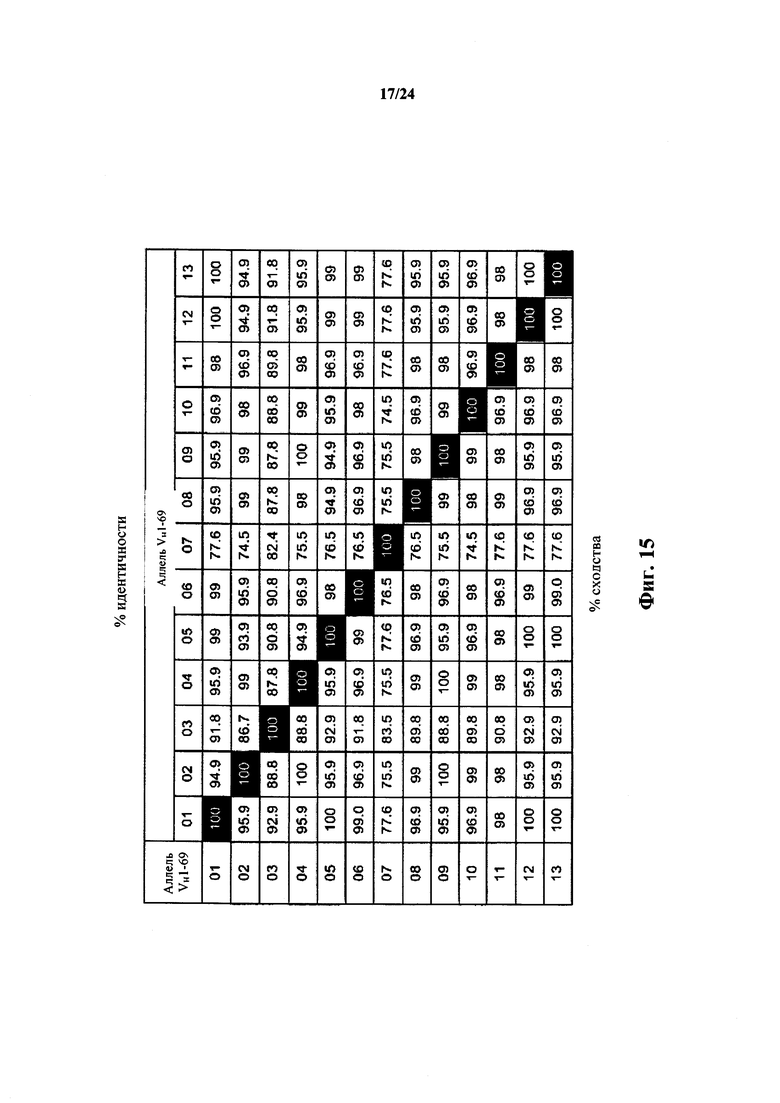
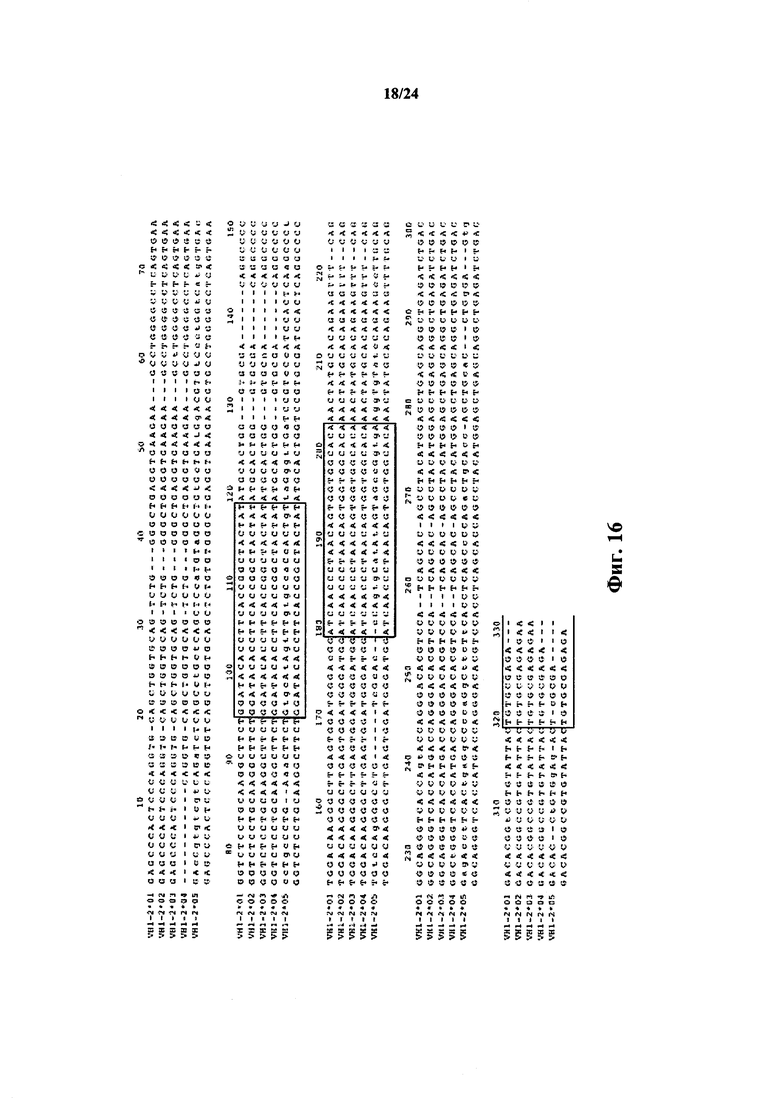
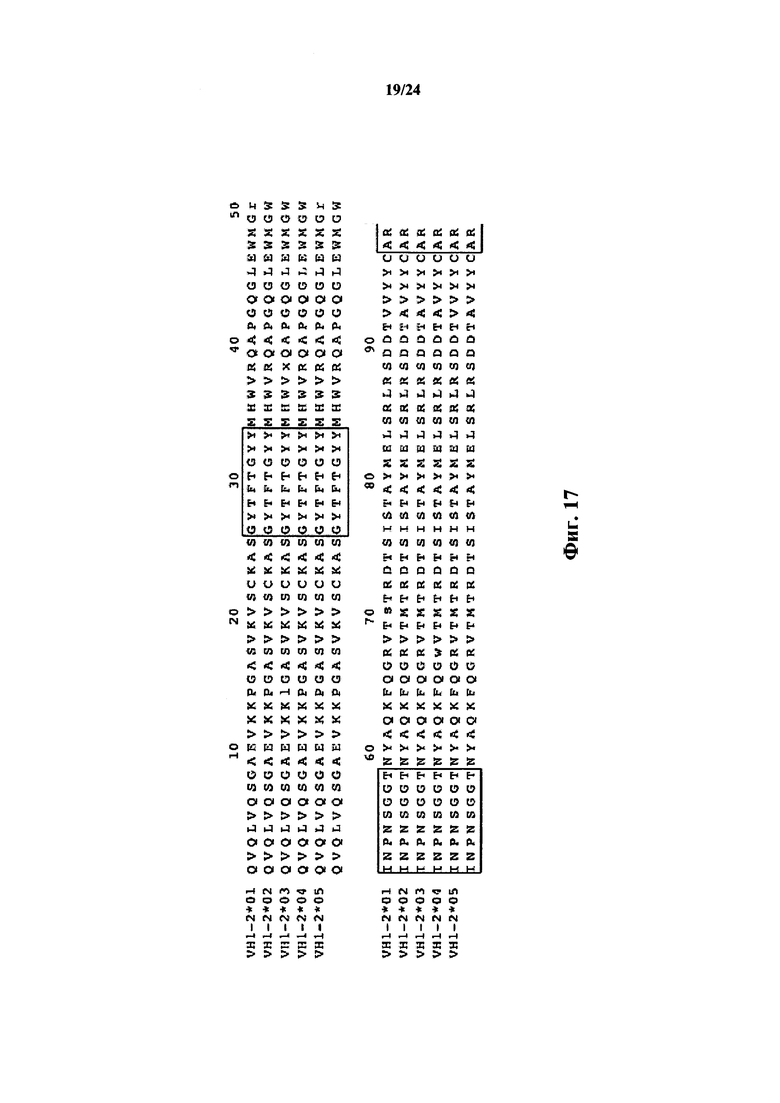
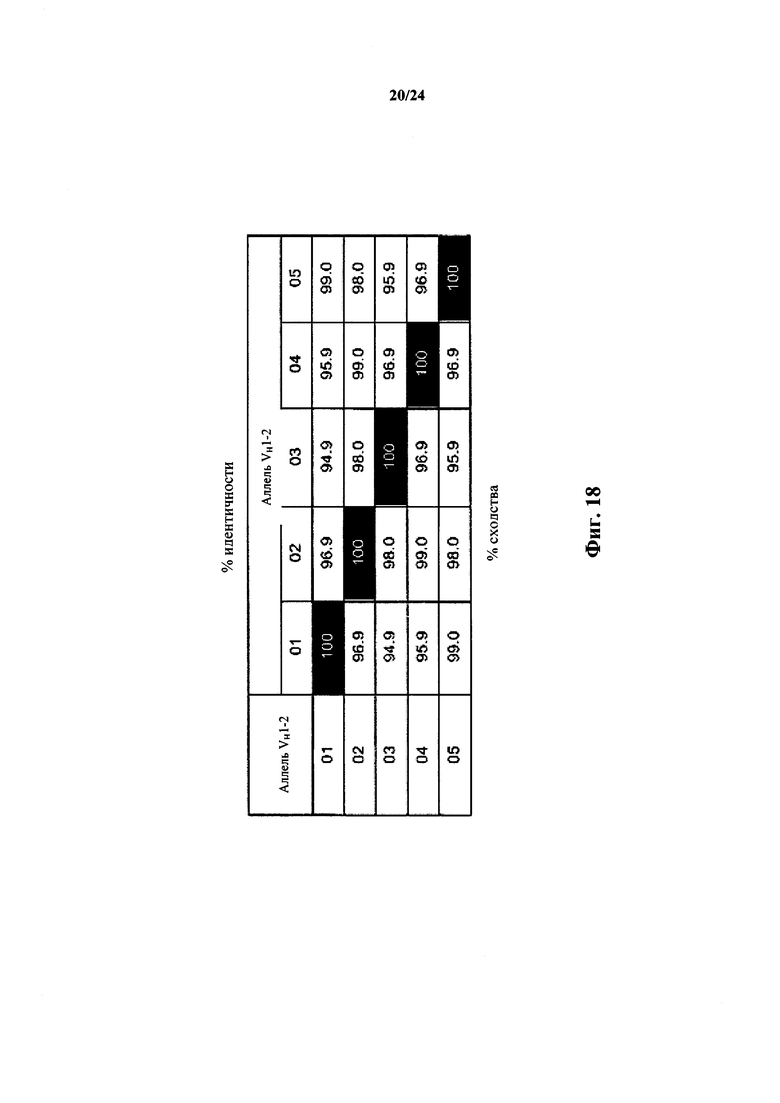
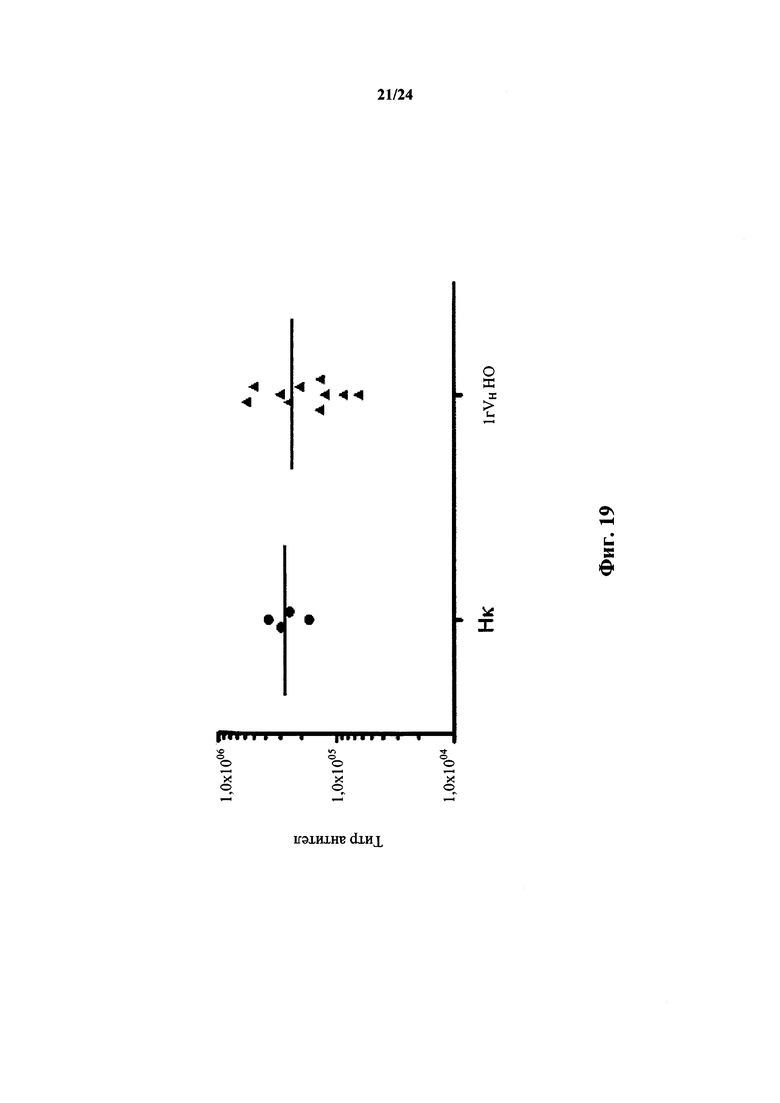
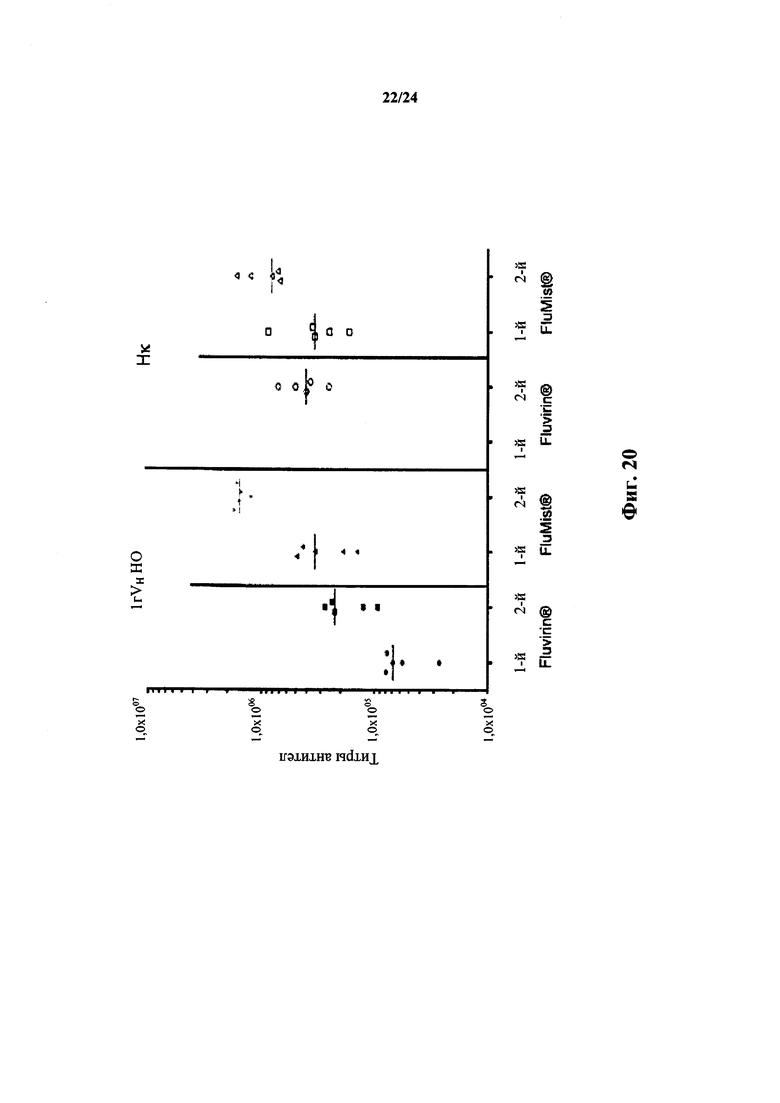
Изобретение относится к области биотехнологии. Описаны мыши, содержащие ограниченный локус тяжелой цепи иммуноглобулина. Локус характеризуется единственным полиморфным генным сегментом VH человека, множеством генных сегментов DH человека и множеством генных сегментов JH. Также описаны способы получения последовательности антител, которые связываются с антигеном, предусматривающие иммунизацию мыши представляющим интерес антигеном. Предложенная группа изобретений может быть использована в медицине. 9 н. и 30 з.п. ф-лы, 24 ил., 9 табл., 4 пр.
1. Мышь для получения вариабельного домена тяжелой цепи человека, содержащая в геноме своей зародышевой линии ограниченный эндогенный локус тяжелой цепи иммуноглобулина, характеризующийся наличием единственного нереаранжированного генного сегмента VH человека, одного или нескольких нереаранжированных генных сегментов DH человека и одного или нескольких нереаранжированных генных сегментов JH человека, функционально связанных с не относящейся к человеку генной последовательностью константной области,
при этом все или по существу все эндогенные генные сегменты VH, DH и JH в ограниченном эндогенном локусе иммуноглобулина приведены в состояние неспособности к реаранжировке, и
при этом мышь дополнительно содержит разнообразный репертуар реаранжированных генов вариабельной области тяжелой цепи иммуноглобулина человека, каждый из которых связан с не относящейся к человеку генной последовательностью константной области и происходит из ограниченного локуса тяжелой цепи иммуноглобулина.
2. Мышь по п. 1, которая гетерозиготна по ограниченному эндогенному локусу тяжелой цепи иммуноглобулина.
3. Мышь по п. 1, у которой единственный нереаранжированный генный сегмент VH человека выбирают из VH1-2, VH1-69, VH2-26, VH2-70 и VH3-23 или их полиморфных вариантов.
4. Мышь по п. 3, у которой единственный нереаранжированный генный сегмент VH человека представляет собой VH1-69 или его полиморфный вариант.
5. Мышь по п. 3, у которой единственный нереаранжированный генный сегмент VH человека представляет собой VH1-2 или его полиморфный вариант.
6. Мышь по п. 1, у которой единственный нереаранжированный генный сегмент VH человека функционально связан с геном константной области иммуноглобулина грызуна.
7. Мышь по любому из пп. 1-6, у которой ген константной области иммуноглобулина представляет собой ген константной области мыши.
8. Мышь по любому из пп. 1-6, дополнительно содержащая один или несколько нереаранжированных генных сегментов VL иммуноглобулина человека, функционально связанных с одним или несколькими нереаранжированными генными сегментами JL человека.
9. Мышь по п. 8, у которой один или несколько нереаранжированных генных сегментов VL человека и/или один или несколько нереаранжированных генных сегментов JL человека выбирают из генных сегментов κ и λ человека.
10. Мышь по п. 8, у которой один или несколько нереаранжированных генных сегментов VL иммуноглобулина человека и один или несколько нереаранжированных генных сегментов JL человека функционально связаны с не относящимся к человеку геном константной области легкой цепи.
11. Мышь по п. 10, у которой не относящийся к человеку ген константной области легкой цепи выбирают из гена константной области κ или λ мыши или крысы.
12. Мышь для получения вариабельного домена тяжелой цепи человека, содержащая в геноме своей зародышевой линии замену в эндогенном локусе тяжелой цепи иммуноглобулина всех или по существу всех эндогенных генных сегментов VH, DH и JH единственным нереаранжированным генным сегментом VH человека, одним или несколькими нереаранжированными генными сегментами DH человека и одним или несколькими нереаранжированными генными сегментами JH человека, которые функционально связанны с не относящейся к человеку генной последовательностью константной области,
при этом мышь дополнительно содержит разнообразный репертуар реаранжированных генов вариабельной области тяжелой цепи иммуноглобулина, каждый из которых связан с не относящейся к человеку генной последовательностью константной области и происходит из единственного нереаранжированного генного сегмента VH человека, одного или нескольких нереаранжированных генных сегментов DH человека и одного или нескольких нереаранжированных генных сегментов JH человека.
13. Мышь по п. 12, дополнительно содержащая замену в эндогенном локусе легкой цепи иммуноглобулина всех или по существу всех эндогенных генных сегментов VL и JL одним или несколькими нереаранжированными генными сегментами VL и одним или несколькими нереаранжированными генными сегментами JL человека.
14. Мышь по п. 12 или 13, у которой единственный генный сегмент VH человека выбран из генного сегмента VH1-69 и VH1-2 человека.
15. Клетка для получения реаранжированной последовательности нуклеиновой кислоты, которая кодирует вариабельный домен тяжелой цепи иммуноглобулина человека, при этом клетка происходит из мыши по любому из пп. 1-6 и 12-13, и при этом клетка содержит один из разнообразного репертуара реаранжированных генов VH иммуноглобулина человека.
16. Ткань для получения реаранжированной последовательности нуклеиновой кислоты, которая кодирует вариабельный домен тяжелой цепи иммуноглобулина человека, при этом ткань происходит из мыши по п. 1 или 12, и при этом ткань содержит клетки, которые экспрессируют один или несколько из разнообразного репертуара реаранжированных генов вариабельной области тяжелой цепи иммуноглобулина человека.
17. Способ получения последовательности нуклеиновой кислоты, которая кодирует вариабельный домен тяжелой цепи иммуноглобулина человека, включающий:
амплификацию нуклеиновой кислоты, содержащей реаранжированную последовательность VH области тяжелой цепи иммуноглобулина человека из лимфоцита мыши по любому из пп. 1-6 и 12-13, или гибридомы, полученной из указанного лимфоцита, с использованием по меньшей мере 3′-праймера, специфичного для константных областей IgM или IgG,
при этом указанный лимфоцит содержит один из разнообразного репертуара реаранжированных генов области VH иммуноглобулина человека.
18. Способ по п. 17, в котором последовательность области VH иммуноглобулина по меньшей мере на 75% идентична SEQ ID NO: 34, SEQ ID NO: 36, SEQ ID NO: 38, SEQ ID NO: 40, SEQ ID NO: 42, SEQ ID NO: 44, SEQ ID NO: 46, SEQ ID NO: 48, SEQ ID NO: 50, SEQ ID NO: 52, SEQ ID NO: 54, SEQ ID NO: 56 или SEQ ID NO: 58.
19. Способ по п. 17, в котором последовательность области VH иммуноглобулина содержит SEQ ID NO: 34, SEQ ID NO: 36, SEQ ID NO: 38, SEQ ID NO: 40, SEQ ID NO: 42, SEQ ID NO: 44, SEQ ID NO: 46, SEQ ID NO: 48, SEQ ID NO: 50, SEQ ID NO: 52, SEQ ID NO: 54, SEQ ID NO: 56, SEQ ID NO: 58 или их полиморфные варианты.
20. Способ по п. 17, в котором последовательность области VH иммуноглобулина кодирует белок, который по меньшей мере на 75% идентичен SEQ ID NO: 35, SEQ ID NO: 37, SEQ ID NO: 39, SEQ ID NO: 41, SEQ ID NO: 43, SEQ ID NO: 45, SEQ ID NO: 47, SEQ ID NO: 49, SEQ ID NO: 51, SEQ ID NO: 53, SEQ ID NO: 55, SEQ ID NO: 57 или SEQ ID NO: 59.
21. Способ по п. 17, в котором последовательность области VH иммуноглобулина по меньшей мере на 95% идентична SEQ ID NO: 60, SEQ ID NO: 62, SEQ ID NO: 64, SEQ ID NO: 66 или SEQ ID NO: 68.
22. Способ по п. 17, в котором последовательность области VH иммуноглобулина содержит SEQ ID NO: 60, SEQ ID NO: 62, SEQ ID NO: 64, SEQ ID NO: 66, SEQ ID NO: 68 или их полиморфный вариант.
23. Способ по п. 17, в котором последовательность области VH иммуноглобулина кодирует белок, который по меньшей мере на 95% идентичен SEQ ID NO: 61, SEQ ID NO: 63, SEQ ID NO: 65, SEQ ID NO: 67 или SEQ ID NO: 69.
24. Применение мыши по любому из пп. 1-6 и 12-13 для получения последовательности нуклеиновой кислоты, кодирующей вариабельный домен тяжелой цепи человека.
25. Применение по п. 24, при котором вариабельный домен тяжелой цепи человека характеризуется наличием последовательностей FR1-CDR1-FR2-CDR2-FR3 человека, происходящих из полиморфного генного сегмента VH человека.
26. Применение по п. 25, при котором генный сегмент VH человека выбирают из генного сегмента VH1-2, VH1-69, VH2-26, VH2-70 или VH3-23 человека.
27. Применение по п. 26, при котором генный сегмент VH человека представляет собой VH1-2.
28. Применение по п. 26, при котором генный сегмент VH человека представляет собой VH1-69.
29. Применение мыши по любому из пп. 1-6 и 12-13 для получения антитела человека, при котором антитело человека содержит вариабельный домен тяжелой цепи, происходящий из реаранжированного генного сегмента VH1-2 человека, генного сегмента VH1-69 человека или их полиморфного варианта.
30. Применение по п. 29, при котором реаранжированный генный сегмент VH1-69 человека по меньшей мере 75% идентичен SEQ ID NO: 34, SEQ ID NO: 36, SEQ ID NO: 38, SEQ ID NO: 40, SEQ ID NO: 42, SEQ ID NO: 44, SEQ ID NO: 46, SEQ ID NO: 48, SEQ ID NO: 50, SEQ ID NO: 52, SEQ ID NO: 54, SEQ ID NO: 56 или SEQ ID NO: 58.
31. Применение по п. 29, при котором реаранжированный генный сегмент VH1-2 человека по меньшей мере на 95% идентичен SEQ ID NO: 60, SEQ ID NO: 62, SEQ ID NO: 64, SEQ ID NO: 66 или SEQ ID NO: 68.
32. Клетка по п. 15, при этом клетка представляет собой лимфоцит или гибридому, полученную из указанного лимфоцита.
33. Способ по п. 17, дополнительно включающий иммунизацию мыши представляющим интерес антигеном перед амплификацией нуклеиновой кислоты.
34. Способ по п. 17 или 33, в котором реаранжированная последовательность гена области VH иммуноглобулина человека содержит по меньшей мере одну соматическую гипермутацию.
35. Способ по п. 17 или 33, в котором лимфоцит специфически связывает представляющий интерес антиген.
36. Нуклеиновая кислота, содержащая последовательность, которая кодирует VH домен иммуноглобулина человека, полученного способом по п. 17 или 33.
37. Тотипотентная клетка мыши, полученная из мыши по любому из пп. 1 или 12 и предназначенная для получения мыши, которая продуцирует вариабельный домен тяжелой цепи (VH) человека, содержащая в своем геноме ограниченный локус тяжелой цепи иммуноглобулина, характеризующийся наличием единственного нереаранжированного генного сегмента VH человека, одного или нескольких нереаранжированных генных сегментов DH человека и одного или нескольких нереаранжированных генных сегментов JH человека, функционально связанных с не относящейся к человеку генной последовательностью константной области тяжелой цепи человека.
38. Тотипотентная клетка мыши по п. 37, в которой не относящаяся к человеку генная последовательность константной области тяжелой цепи представляет собой генную последовательность константной области тяжелой цепи мыши.
39. Тотипотентная клетка мыши по п. 37 или 38, при этом тотипотентная клетка мыши представляет собой эмбриональную стволовую клетку.
| WO 2009097006 A2, 06.08.2009 | |||
| TAYLOR L.D | |||
| et al., "A transgenic mouse that expresses a diversity of human sequence heavy and light chain immunoglobulins", Nucleic Acids Res | |||
| Пуговица для прикрепления ее к материи без пришивки | 1921 |
|
SU1992A1 |
| JANSSENS R | |||
| et al., "Generation of heavy-chain-only antibodies in mice", Proc Natl Acad Sci U S A | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| RIECHMANN L | |||
| et al., | |||

