Область техники, к которой относится настоящее изобретение
Предусмотрена генетически модифицированная мышь, которая экспрессирует антитела с общей относящейся к человеку вариабельной/относящейся к мыши константной легкой цепью, ассоциированной с разнообразными относящимися к человеку вариабельными/относящимися к мыши константными тяжелыми цепями. Предусмотрен способ получения биспецифического антитела человека из относящихся к человеку генных последовательностей вариабельной области В-клеток мыши.
Предшествующий уровень техники настоящего изобретения
Антитела, как правило, содержат гомодимерный компонент тяжелой цепи, причем каждый мономер тяжелой цепи ассоциирован с идентичной легкой цепью. Антитела с гетеродимерным компонентом тяжелой цепи (например, биспецифические антитела) необходимы в качестве терапевтических антител. Однако получение биспецифических антител с подходящим компонентом легкой цепи, который может желательным образом ассоциироваться с каждой из тяжелых цепей биспецифического антитела, оказалось затруднительным.
Согласно одному подходу легкую цепь можно выбрать путем исследования статистики использования всех вариабельных доменов легкой цепи, идентифицируя наиболее часто используемую легкую цепь в антителах человека, и создавая пару этой легкой цепи in vitro с двумя тяжелыми цепями отличающейся специфичности.
Согласно другому подходу легкую цепь можно выбрать путем исследования последовательностей легкой цепи в библиотеке фагового дисплея (например, библиотеке фагового дисплея, содержащей последовательности вариабельной области легкой цепи человека, например, библиотеке scFv человека) и выбора наиболее часто используемой вариабельной области легкой цепи из библиотеки. Легкую цепь затем можно исследовать на двух различных представляющих интерес тяжелых цепях.
Согласно другому подходу легкую цепь можно выбрать путем анализа библиотеки фагового дисплея вариабельных последовательностей легкой цепи с использованием вариабельных последовательностей тяжелой цепи обеих представляющих интерес тяжелых цепей в качестве зондов. Легкая цепь, которая ассоциируется с обеими вариабельными последовательностями тяжелой цепи, может быть выбрана в качестве легкой цепи для тяжелых цепей.
Согласно другому подходу кандидатную легкую цепь можно выравнивать с легкими цепями, когнатными тяжелым цепям, и производить в легкой цепи модификации для более близкого совпадения характеристик последовательностей, общих с когнатными легкими цепями обеих тяжелых цепей. Если необходимо минимизировать шансы возникновения иммуногенности, модификации предпочтительно дают в результате последовательности, которые присутствуют в известных последовательностях легкой цепи человека так, что протеолитическая обработка маловероятно создаст Т-клеточный эпитоп на основании параметров и способов, известных в настоящей области техники для оценки вероятности иммуногенности (т.е. in silico, а также мокрые анализы).
Все приведенные выше подходы основаны на способах in vitro, которые включают в себя ряд априорных ограничений, например, идентичность последовательностей, способность ассоциироваться со специфическими предварительно выбранными тяжелыми цепями и т.д. В настоящей области техники существует потребность в композициях и способах, которые не основаны на манипуляции с in vitro условиями, но которые вместо этого используют более биологически целесообразные подходы для получения эпитопсвязывающих белков человека, которые включают в себя общую легкую цепь.
Краткое раскрытие настоящего изобретения
Предусмотрены генетически модифицированные мыши, которые экспрессируют вариабельные домены тяжелой и легкой цепи иммуноглобулина человека, причем мыши содержат ограниченный вариабельный репертуар легкой цепи. Предусмотрена биологическая система для создания вариабельного домена легкой цепи человека, который ассоциируется и экспрессируется с разнообразным репертуаром вариабельных доменов тяжелой цепи человека с созревшей аффинностью. Предусмотрены способы получения связывающих белков, содержащих вариабельные домены иммуноглобулина, предусматривающие иммунизацию мышей, которые содержат ограниченный репертуар легкой цепи иммуноглобулина представляющим интерес антигеном и использование генной последовательности вариабельной области иммуноглобулина мыши в связывающем белке, который специфически связывает представляющий интерес антиген. Способы включают в себя способы получения вариабельных доменов тяжелой цепи иммуноглобулина человека, подходящих для применения в получении мультиспецифических антигенсвязывающих белков.
Предусмотрены генетически сконструированные мыши, которые выбирают подходящие вариабельные домены тяжелой цепи иммуноглобулина человека с созревшей аффинностью, происходящие из репертуара нереаранжированных генных сегментов вариабельной области тяжелой цепи человека, причем вариабельные домены тяжелой цепи человека с созревшей аффинностью ассоциируются и экспрессируются с единственным вариабельным доменом легкой цепи человека, происходящим из одного генного сегмента вариабельной области легкой цепи человека. Также предусмотрены генетически сконструированные мыши, которые представляют выбор двух генных сегментов вариабельной области легкой цепи человека.
Предусмотрены генетически сконструированные мыши, которые экспрессируют ограниченный репертуар вариабельных доменов легкой цепи человека или единственный вариабельный домен легкой цепи человека из ограниченного репертуара генных сегментов вариабельной области легкой цепи человека. Мышей генетически конструируют, чтобы они включали в себя единственный нереаранжированный генный сегмент вариабельной области легкой цепи человека (или два генных сегмента вариабельной области легкой цепи человека), который реаранжируется для образования реаранжированного гена вариабельной области легкой цепи человека (или двух реаранжированных генов вариабельной области легкой цепи), который экспрессирует единственную легкую цепь (или которые экспрессируют любую или обе из двух легких цепей). Реаранжированные вариабельные домены легкой цепи человека способны спариваться с разнообразными выбранными мышами тяжелыми цепями человека с созревшей аффинностью, причем вариабельные области тяжелой цепи специфически связывают различные эпитопы.
Предусмотрены генетически сконструированные мыши, которые экспрессируют ограниченный репертуар вариабельных доменов легкой цепи человека или единственный вариабельный домен легкой цепи человека из ограниченного репертуара последовательностей вариабельной области легкой цепи человека. Мышей генетически конструируют, чтобы они включали в себя единственную последовательность V/J легкой цепи человека (или две последовательности V/J), которая экспрессирует вариабельную область единственной легкой цепи (или которые экспрессируют любую или обе из двух вариабельных областей). Легкая цепь, содержащая вариабельную последовательность, способна спариваться с разнообразными тяжелыми цепями человека с созревшей аффинностью, подвергнутых клональной селекции мышами, причем вариабельные области тяжелой цепи специфически связывают различные эпитопы.
Согласно одному аспекту предусмотрена генетически модифицированная мышь, которая содержит единственный генный сегмент вариабельной области легкой цепи (VL) иммуноглобулина человека, который способен реаранжироваться с генным сегментом J человека (выбранным из одного или множества сегментов JL) и кодировать домен VL человека легкой цепи иммуноглобулина. Согласно другому аспекту мышь содержит не больше двух генных сегментов VL человека, каждый из которых способен реаранжироваться с генным сегментом J человека (выбранным из одного или множества сегментов JL) и кодировать домен VL человека легкой цепи иммуноглобулина.
Согласно одному варианту осуществления единственный генный сегмент VL человека функционально связан с генным сегментом JL человека, выбранным из Jκ1, Jκ2, Jκ3, Jκ4 и Jκ5, причем единственный генный сегмент VL человека способен реаранжироваться для образования последовательности, кодирующей ген вариабельной области легкой цепи, с любым из одного или нескольких генных сегментов JL человека.
Согласно одному варианту осуществления генетически модифицированная мышь содержит локус легкой цепи иммуноглобулина, который не содержит эндогенный генный сегмент VL мыши, который способен реаранжироваться для образования гена легкой цепи иммуноглобулина, причем локус VL содержит единственный генный сегмент VL человека, который способен реаранжироваться, чтобы кодировать область VL гена легкой цепи. Согласно конкретному варианту осуществления генный сегмент VL человека представляет собой генный сегмент Vκ1-39Jκ5 человека или генный сегмент Vκ3-20Jκ1 человека. Согласно одному варианту осуществления генетически модифицированная мышь содержит локус VL, который не содержит эндогенный генный сегмент VL мыши, который способен реаранжироваться для образования гена легкой цепи иммуноглобулина, причем локус VL содержит не больше двух генных сегментов VL человека, которые способны реаранжироваться, чтобы кодировать область VL гена легкой цепи. Согласно конкретному варианту осуществления не больше чем 2 генных сегмента VL человека представляют собой генный сегмент Vκ1-39Jκ5 человека и генный сегмент Vκ3-20Jκ1 человека.
Согласно одному аспекту предусмотрена генетически модифицированная мышь, которая содержит единственную реаранжированную (V/J) вариабельную область легкой цепи (VL) иммуноглобулина человека (т.е. область VL/JL), которая кодирует домен VL человека легкой цепи иммуноглобулина. Согласно другому аспекту мышь содержит не больше двух реаранжированных областей VL человека, которые способны кодировать домен VL человека легкой цепи иммуноглобулина.
Согласно одному варианту осуществления область VL представляет собой реаранжированную последовательность Vκ1-39/J человека или реаранжированную последовательность Vκ3-20/J человека. Согласно одному варианту осуществления сегмент JL человека реаранжированной последовательности VL/JL выбран из Jκ1, Jκ2, Jκ3, Jκ4 и Jκ5. Согласно конкретному варианту осуществления область VL представляет собой последовательность Vκ1-39Jκ5 человека или последовательность Vκ3-20Jκ1 человека. Согласно конкретному варианту осуществления мышь содержит как последовательность Vκ1-39Jκ5 человека, так и последовательность Vκ3-20Jκ1 человека.
Согласно одному варианту осуществления генный сегмент VL человека функционально связан с лидерной последовательностью человека или мыши. Согласно одному варианту осуществления лидерная последовательность представляет собой лидерную последовательность мыши. Согласно конкретному варианту осуществления лидерная последовательность мыши представляет собой лидерную последовательность Vκ3-7 мыши. Согласно конкретному варианту осуществления лидерная последовательность функционально связана с нереаранжированным генным сегментом VL человека. Согласно конкретному варианту осуществления лидерная последовательность функционально связана с реаранжированной последовательностью VL/JL человека.
Согласно одному варианту осуществления генный сегмент VL функционально связан с промоторной последовательностью иммуноглобулина. Согласно одному варианту осуществления промоторная последовательность представляет собой промоторную последовательность человека. Согласно конкретному варианту осуществления промотор иммуноглобулина человека представляет собой промотор Vκ3-15 человека. Согласно конкретному варианту осуществления промотор функционально связан с нереаранжированным генным сегментом VL человека. Согласно конкретному варианту осуществления промотор функционально связан с реаранжированной последовательностью VL/JL человека.
Согласно одному варианту осуществления локус легкой цепи содержит лидерную последовательность, фланкированную 5' (относительно направления транскрипции генного сегмента VL) промотором иммуноглобулина человека и фланкированную 3' генным сегментом VL человека, который реаранжируется с сегментом J человека и кодирует домен VL обратной химерной легкой цепи, содержащей эндогенную константную область легкой цепи мыши (CL). Согласно конкретному варианту осуществления генный сегмент VL находится на локусе Vκ мыши, и CL мыши представляет собой Cκ мыши.
Согласно одному варианту осуществления локус легкой цепи содержит лидерную последовательность, фланкированную 5' (относительно направления транскрипции генного сегмента VL) промотором иммуноглобулина человека и фланкированную 3' реаранжированной областью VL человека (последовательностью VL/JL) и кодирует домен VL обратной химерной легкой цепи, содержащей эндогенную константную область легкой цепи мыши (CL). Согласно конкретному варианту осуществления реаранжированная последовательность VL/JL человека находится на каппа (κ) локусе мыши, и CL мыши представляет собой Cκ мыши.
Согласно одному варианту осуществления локус VL модифицированной мыши представляет собой локус κ легкой цепи, и локус κ легкой цепи содержит κ интронный энхансер мыши, κ 3' энхансер мыши или как интронный энхансер, так и 3' энхансер.
Согласно одному варианту осуществления мышь содержит нефункциональный локус лямбда (λ) легкой цепи иммуноглобулина. Согласно конкретному варианту осуществления локус λ легкой цепи содержит делецию одного или нескольких последовательностей локуса, причем одна или несколько делеций делают локус λ легкой цепи неспособным к реаранжировке для образования гена легкой цепи. Согласно другому варианту осуществления все или по существу все генные сегменты VL локуса λ легкой цепи удалены.
Согласно одному варианту осуществления мышь производит легкую цепь, которая содержит соматически мутированный домен VL, происходящий из генного сегмента VL человека. Согласно одному варианту осуществления легкая цепь содержит соматически мутированный домен VL, происходящий из генного сегмента VL человека, и область Сκ мыши. Согласно одному варианту осуществления мышь не экспрессирует λ легкую цепь.
Согласно одному варианту осуществления генетически модифицированная мышь способна к соматической гипермутации последовательности области VL человека. Согласно конкретному варианту осуществления мышь содержит клетку, которая содержит реаранжированный ген легкой цепи иммуноглобулина, происходящий из генного сегмента VL человека, который способен реаранжироваться и кодировать домен VL, и реаранжированный ген легкой цепи иммуноглобулина содержит соматически мутированный домен VL.
Согласно одному варианту осуществления мышь содержит клетку, которая экспрессирует легкую цепь, содержащую соматически мутированный домен VL человека, связанный с Cκ мыши, причем легкая цепь ассоциируется с тяжелой цепью, содержащей соматически мутированный домен VH, происходящий из генного сегмента VH человека, и причем тяжелая цепь содержит константную область тяжелой цепи (CH) мыши. Согласно конкретному варианту осуществления тяжелая цепь содержит CH1 мыши, шарнир мыши, VH2 мыши и CH3 мыши. Согласно конкретному варианту осуществления тяжелая цепь содержит CH1 человека, шарнир, CH2 мыши и CH3 мыши.
Согласно одному варианту осуществления мышь содержит замещение эндогенных генных сегментов VH мыши одним или несколькими генными сегментами VH человека, причем генные сегменты VH человека функционально связаны с геном области CH мыши так, что мышь реаранжирует генные сегменты VH человека и экспрессирует обратную химерную тяжелую цепь иммуноглобулина, которая содержит домен VH человека и CH мыши. Согласно одному варианту осуществления 90-100% нереаранжированных генных сегментов VH мыши замещают по меньшей мере одним нереаранжированным генным сегментом VH человека. Согласно конкретному варианту осуществления все или по существу все эндогенные генные сегменты VH мыши замещают по меньшей мере одним нереаранжированным генным сегментом VH человека. Согласно одному варианту осуществления замещение осуществляют с помощью по меньшей мере 19, по меньшей мере 39 или по меньшей мере 80 или 81 нереаранжированного генного сегмента VH человека. Согласно одному варианту осуществления замещение осуществляют с помощью по меньшей мере 12 функциональных нереаранжированных генных сегментов VH человека, по меньшей мере 25 функциональных нереаранжированных генных сегментов VH человека или по меньшей мере 43 функциональных нереаранжированных генных сегментов VH человека. Согласно одному варианту осуществления мышь содержит замещение всех сегментов DH и JH мыши по меньшей мере одним нереаранжированным сегментом DH человека и по меньшей мере одним нереаранжированным сегментом JH человека. Согласно одному варианту осуществления по меньшей мере один нереаранжированный человека сегмент DH выбран из 1-1, D1-7, 1-26, 2-8, 2-15, 3-3, 3-10, 3-16, 3-22, 5-5, 5-12, 6-6, 6-13, 7-27 и их комбинации. Согласно одному варианту осуществления по меньшей мере один нереаранжированный сегмент JH человека выбран из 1, 2, 3, 4, 5, 6 и их комбинации. Согласно конкретному варианту осуществления один или несколько генных сегментов VH человека выбран из генного сегмента VH человека 1-2, 1-8, 1-24, 1-69, 2-5, 3-7, 3-9, 3-11, 3-13, 3-15, 3-20, 3-23, 3-30, 3-33, 3-48, 3-53, 4-31, 4-39, 4-59, 5-51,6-1 и их комбинации.
Согласно одному варианту осуществления мышь содержит В-клетку, которая экспрессирует связывающий белок, который специфически связывает представляющий интерес антиген, причем связывающий белок содержит легкую цепь, происходящую из реаранжировки Vκ1-39/Jκ5 человека или реаранжировки Vκ3-20/Jκ1 человека, и причем клетка содержит реаранжированный ген тяжелой цепи иммуноглобулина, происходящий из реаранжировки генных сегментов VH человека, выбранных из генного сегмента 1-69, 2-5, 3-13, 3-23, 3-30, 3-33, 3-53, 4-39, 4-59 и 5-51. Согласно одному варианту осуществления один или несколько генных сегментов VH человека реаранжированы с генным сегментом JH тяжелой цепи человека, выбранным из 1, 2, 3, 4, 5 и 6. Согласно одному варианту осуществления один или несколько генных сегментов VH и JH человека реаранжированы с генным сегментом DH человека, выбранным из 1-1, 1-7, 1-26, 2-8, 2-15, 3-3, 3-10, 3-16, 3-22, 5-5, 5-12, 6-6, 6-13 и 7-27. Согласно конкретному варианту осуществления ген легкой цепи содержит 1, 2, 3, 4 или 5 или больше соматических гипермутаций.
Согласно одному варианту осуществления мышь содержит В-клетку, которая содержит реаранжированную генную последовательность вариабельной области тяжелой цепи иммуноглобулина, содержащую область VH/DH/JH, выбранную из 2-5/6-6/1, 2-5/3-22/1, 3-13/6-6/5, 3-23/2-8/4, 3-23/3-3/4, 3-23/3-10/4, 3-23/6-6/4, 3-23/7-27/4, 3-30/1-1/4, 3-30/1-7/4, 3-30/3-3/3, 3-30/3-3/4, 3-30/3-22/5, 3-30/5-5/2, 3-30/5-12/4, 3-30/6-6/1, 3-30/6-6/3, 3-30/6-6/4, 3-30/6-6/5, 3-30/6-13/4, 3-30/7-27/4, 3-30/7-27/5, 3-30/7-27/6, 3-33/1-7/4, 3-33/2-15/4, 4-39/1-26/3, 4-59/3-16/3, 4-59/3-16/4, 4-59/3-22/3, 5-51/3-16/6, 5-51/5-5/3, 5-51/6-13/5, 3-53/1-1/4, 1-69/6-6/5 и 1-69/6-13/4. Согласно конкретному варианту осуществления В-клетка экспрессирует связывающий белок, содержащий вариабельную область тяжелой цепи иммуноглобулина человека, слитую с константной областью тяжелой цепи мыши, и вариабельную область легкой цепи иммуноглобулина человека, слитую с константной областью легкой цепи мыши.
Согласно одному варианту осуществления реаранжированная область VL человека представляет собой последовательность Vκ1-39Jκ5 человека, и мышь экспрессирует обратную химерную легкую цепь, содержащую (i) домен VL, происходящий из последовательности VL/JL человека и (ii) CL мыши; причем легкая цепь ассоциирована с обратной химерной тяжелой цепью, содержащей (i) CH мыши и (ii) соматически мутированный домен VH человека, происходящий из генного сегмента VH человека, выбранного из генного сегмента VH человека 1-2, 1-8, 1-24, 1-69, 2-5, 3-7, 3-9, 3-11, 3-13, 3-15, 3-20, 3-23, 3-30, 3-33, 3-48, 3-53, 4-31, 4-39, 4-59, 5-51, 6-1 и их комбинации. Согласно одному варианту осуществления мышь экспрессирует легкую цепь, которая является соматически мутированной. Согласно одному варианту осуществления CL представляет собой Сκ мыши. Согласно конкретному варианту осуществления генный сегмент VH человека выбран из генного сегмента 2-5, 3-13, 3-23, 3-30, 4-59, 5-51 и 1-69. Согласно конкретному варианту осуществления соматически мутированный домен VH человека содержит последовательность, происходящую из сегмента DH, выбранного из 1-1, 1-7, 2-8, 3-3, 3-10, 3-16, 3-22, 5-5, 5-12, 6-6, 6-13 и 7-27. Согласно конкретному варианту осуществления соматически мутированный домен VH человека содержит последовательность, происходящую из сегмента JH, выбранного из 1, 2, 3, 4, 5 и 6. Согласно конкретному варианту осуществления соматически мутированный домен VH человека кодируется реаранжированной последовательностью VH/DH/JH человека, выбранной из 2-5/6-6/1, 2-5/3-22/1, 3-13/6-6/5, 3-23/2-8/4, 3-23/3-3/4, 3-23/3-10/4, 3-23/6-6/4, 3-23/7-27/4, 3-30/1-1/4, 3-30/1-7/4, 3-30/3-3/4, 3-30/3-22/5, 3-30/5-5/2, 3-30/5-12/4, 3-30/6-6/1, 3-30/6-6/3, 3-30/6-6/4, 3-30/6-6/5, 3-30/6-13/4, 3-30/7-27/4, 3-30/7-27/5, 3-30/7-27/6, 4-59/3-16/3, 4-59/3-16/4, 4-59/3-22/3, 5-51/5-5/3, 1-69/6-6/5 и 1-69/6-13/4.
Согласно одному варианту осуществления реаранжированная область VL человека представляет собой последовательность Vκ3-20Jκ1 человека, и мышь экспрессирует обратную химерную легкую цепь, содержащую (i) домен VL, происходящий из реаранжированной последовательности VL/JL человека, и (ii) CL мыши; причем легкая цепь ассоциирована с обратной химерной тяжелой цепью, содержащей (i) CH мыши, и (ii) соматически мутированный VH человека, происходящий из генного сегмента VH человека, выбранного из генного сегмента VH человека 1-2, 1-8, 1-24, 1-69, 2-5, 3-7, 3-9, 3-11, 3-13, 3-15, 3-20, 3-23, 3-30, 3-33, 3-48, 3-53, 4-31, 4-39, 4-59, 5-51, 6-1 и их комбинации. Согласно одному варианту осуществления мышь экспрессирует легкую цепь, которая является соматически мутированной. Согласно одному варианту осуществления CL представляет собой Cκ мыши. Согласно конкретному варианту осуществления генный сегмент VH человека выбран из генного сегмента 3-30, 3-33, 3-53, 4-39 и 5-51. Согласно конкретному варианту осуществления соматически мутированный домен VH человека содержит последовательность, происходящую из сегмента DH, выбранного из 1-1, 1-7, 1-26, 2-15, 3-3, 3-16 и 6-13. Согласно конкретному варианту осуществления соматически мутированный домен VH человека содержит последовательность, происходящую из сегмента JH, выбранного из 3, 4, 5 и 6. Согласно конкретному варианту осуществления соматически мутированный домен VH человека кодируется реаранжированной последовательностью VH/DH/JH человека, выбранной из 3-30/1-1/4, 3-30/3-3/3, 3-33/1-7/4, 3-33/2-15/4, 4-39/1-26/3, 5-51/3-16/6, 5-51/6-13/5 и 3-53/1-1/4.
Согласно одному варианту осуществления мышь содержит как реаранжированную последовательность Vκ1-39Jκ5 человека, так и реаранжированную последовательность Vκ3-20Jκ1 человека, и мышь экспрессирует обратную химерную легкую цепь, содержащую (i) домен VL, происходящий из последовательности Vκ1-39Jκ5 человека или последовательности Vκ3-20Jκ1 человека, и (ii) CL мыши; причем легкая цепь ассоциирована с обратной химерной тяжелой цепью, содержащей (i) CH мыши, и (ii) соматически мутированный VH человека, происходящий из генного сегмента VH человека, выбранного из генного сегмента VH человека 1-2, 1-8, 1-24, 1-69, 2-5, 3-7, 3-9, 3-11, 3-13, 3-15, 3-20, 3-23, 3-30, 3-33, 3-48, 3-53, 4-31, 4-39, 4-59, 5-51, 6-1 и их комбинации. Согласно одному варианту осуществления мышь экспрессирует легкую цепь, которая является соматически мутированной. Согласно одному варианту осуществления CL представляет собой Cκ мыши.
Согласно одному варианту осуществления 90-100% эндогенных нереаранжированных генных сегментов VH мыши замещают по меньшей мере одним нереаранжированным генным сегментом VH человека. Согласно конкретному варианту осуществления все или по существу все эндогенные нереаранжированные генные сегменты VH мыши замещают по меньшей мере одним нереаранжированным генным сегментом VH человека. Согласно одному варианту осуществления замещение осуществляют с помощью по меньшей мере 18, по меньшей мере 39, по меньшей мере 80 или 81 нереаранжированного генного сегмента VH человека. Согласно одному варианту осуществления замещение осуществляют с помощью по меньшей мере 12 функциональных нереаранжированных генных сегментов VH человека, по меньшей мере 25 функциональных нереаранжированных генных сегментов VH человека или по меньшей мере 43 нереаранжированных генных сегментов VH человека.
Согласно одному варианту осуществления генетически модифицированная мышь представляет собой линию C57BL, согласно конкретному варианту осуществления выбранную из C57BL/A, C57BL/An, C57BL/GrFa, C57BL/KaLwN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr и C57BL/01a. Согласно конкретному варианту осуществления генетически модифицированная мышь представляет собой сочетание вышеупомянутой линии 129 и вышеупомянутой линии C57BL/6. Согласно другому конкретному варианту осуществления мышь представляет собой сочетание вышеупомянутых линий 129 или сочетание вышеупомянутых линий BL/6. Согласно конкретному варианту осуществления линия 129 сочетания представляет собой линию 129S6 (129/SvEvTac).
Согласно одному варианту осуществления мышь экспрессирует обратное химерное антитело, содержащее легкую цепь, которая содержит Cκ мыши и соматически мутированный домен VL человека, происходящий из реаранжированной последовательности Vκ1-39Jκ5 человека или реаранжированной последовательности Vκ3-20Jκ1 человека, и тяжелую цепь, которая содержит CH мыши и соматически мутированный домен VH человека, происходящий из генного сегмента VH человека, выбранного из генного сегмента VH человека 1-2, 1-8, 1-24, 1-69, 2-5, 3-7, 3-9, 3-11, 3-13, 3-15, 3-20, 3-23, 3-30, 3-33, 3-48, 3-53, 4-31, 4-39, 4-59, 5-51 и 6-1, причем мышь не экспрессирует полностью мышиное антитело и не экспрессирует полностью человеческое антитело. Согласно одному варианту осуществления мышь содержит локус κ легкой цепи, который содержит замещение эндогенных генных сегментов κ легкой цепи мыши реаранжированной последовательностью Vκ1-39Jκ5 человека или реаранжированной последовательностью Vκ3-20Jκ1 человека и содержит замещение всех или по существу всех эндогенных генных сегментов VH мыши полным или по существу полным репертуаром генных сегментов VH человека.
Согласно одному аспекту предусмотрена популяция антиген-специфических антител, полученных от описанной в настоящем документе мыши, причем антитела содержат ген легкой цепи, происходящий из реаранжировки Vκ1-39/Jκ5 человека или реаранжировки Vκ3-20/Jκ1 человека, и причем антитела содержат реаранжированный ген тяжелой цепи иммуноглобулина, происходящий из реаранжировки генного сегмента VH человека, выбранного из генного сегмента VH человека 1-2, 1-3, 1-8, 1-18, 1-24, 1-46, 1-58, 1-69, 2-5, 2-26, 2-70, 3-7, 3-9, 3-11, 3-13, 3-15, 3-16, 3-20, 3-21, 3-23, 3-30, 3-33, 3-43, 3-48, 3-53, 3-64, 3-72, 3-73, 4-31, 4-34, 4-39, 4-59, 5-51 и 6-1. Согласно одному варианту осуществления один или несколько генных сегментов VH человека реаранжированы с генным сегментом тяжелой цепи JH человека, выбранным из 1, 2, 3, 4, 5 и 6. Согласно конкретному варианту осуществления легкая цепь содержит 1, 2, 3, 4 или 5 или больше соматических гипермутаций.
Согласно одному варианту осуществления легкая цепь содержит 1, 2, 3, или 4 соматические гипермутации. Согласно одному варианту осуществления ген легкой цепи содержит 1 или 2 мутации. Согласно различным вариантам осуществления ген легкой цепи способен подвергаться множественным мутациям на всем протяжении своей последовательности.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ1-39/Jκ5 человека, и легкая цепь содержит по меньшей мере одну или не больше четырех соматических гипермутаций. Согласно одному варианту осуществления легкая цепь содержит по меньшей мере две соматических гипермутации. Согласно одному варианту осуществления легкая цепь содержит по меньшей мере три соматических гипермутации. Согласно одному варианту осуществления легкая цепь содержит по меньшей мере четыре соматические гипермутации. Согласно конкретному варианту осуществления мутации присутствуют в одной или нескольких каркасных областях (FW) легкой цепи. Согласно конкретному варианту осуществления мутации присутствуют в одной или нескольких определяющих комплементарность областях (CDR) легкой цепи. Согласно конкретному варианту осуществления мутации присутствуют в одной или нескольких FW и/или одной или нескольких CDR легкой цепи. Согласно различным вариантам осуществления каркасные области выбраны из каркасной области 1 (FW1), каркасной области 2 (FW2), каркасной области 3 (FW3) и/или их комбинации. Согласно различным вариантам осуществления CDR выбраны из CDR1, CDR2, CDR3 и/или их комбинации.
Согласно одному варианту осуществления тяжелая цепь содержит по меньшей мере одну мутацию в одной или нескольких FW или одной или нескольких CDR. Согласно одному варианту осуществления тяжелая цепь содержит по меньшей мере одну мутацию в одной или нескольких FW и одной или нескольких CDR. Согласно одному варианту осуществления тяжелая цепь содержит по меньшей мере две мутации в одной или нескольких FW и одной или нескольких CDR. Согласно одному варианту осуществления тяжелая цепь содержит по меньшей мере три мутации в одной или нескольких FW и одной или нескольких CDR. Согласно одному варианту осуществления тяжелая цепь содержит по меньшей мере четыре мутации в одной или нескольких FW и одной или нескольких CDR. Согласно одному варианту осуществления тяжелая цепь содержит по меньшей мере пять или больше пяти мутаций в одной или нескольких FW и одной или нескольких CDR; согласно конкретному варианту осуществления тяжелая цепь содержит по меньшей мере пять или больше пяти мутаций в двух FW; согласно конкретному варианту осуществления тяжелая цепь содержит по меньшей мере пять или больше пяти мутаций в одной FW и одной CDR.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ1-39/Jκ5 человека, и приблизительно 9% происходящих из Vκ1-39/Jκ5 легких цепей содержат по меньшей мере одну мутацию, присутствующую в FW1; согласно одному варианту осуществления по меньшей мере 9% легких цепей содержат одну мутацию, присутствующую в FW1. Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ1-39/Jκ5 человека, и приблизительно 25% происходящих из Vκ1-39/Jκ5 легких цепей содержат по меньшей мере одну или не больше двух мутаций, присутствующих в CDR1; согласно одному варианту осуществления по меньшей мере 19% легких цепей содержат одну мутацию, присутствующую в CDR1; согласно одному варианту осуществления по меньшей мере 5% легких цепей содержат две мутации, присутствующие в CDR1.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ1-39/Jκ5 человека, и приблизительно 20% происходящих из Vκ1-39/Jκ5 легких цепей содержат по меньшей мере одну или не больше трех мутаций, присутствующих в FW2; согласно одному варианту осуществления по меньшей мере 17% легких цепей содержат одну мутацию, присутствующую в FW2; согласно одному варианту осуществления по меньшей мере 1% легких цепей содержат две мутации, присутствующие в FW2; согласно одному варианту осуществления по меньшей мере 1% легких цепей содержат три мутации, присутствующие в FW2.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ1-39/Jκ5 человека, и приблизительно 10% происходящих из Vκ1-39/Jκ5 легких цепей содержат по меньшей мере одну или не больше двух мутаций, присутствующих в CDR2; согласно одному варианту осуществления по меньшей мере 10% легких цепей содержат одну мутацию, присутствующую в CDR2; согласно одному варианту осуществления по меньшей мере 1% легких цепей содержат две мутации, присутствующие в CDR2.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ1-39/Jκ5 человека, и приблизительно 29% происходящих из Vκ1-39/Jκ5 легких цепей содержат по меньшей мере одну или не больше четырех мутаций, присутствующих в FW3; согласно одному варианту осуществления по меньшей мере 21% легких цепей содержат одну мутацию, присутствующую в FW3; согласно одному варианту осуществления по меньшей мере 5% легких цепей содержат две мутации, присутствующие в FW3; согласно одному варианту осуществления по меньшей мере 2% легких цепей содержат три мутации, присутствующие в FW3; согласно одному варианту осуществления по меньшей мере 2% легких цепей содержат четыре мутации, присутствующие в FW3.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ1-39/Jκ5 человека, и приблизительно 37% происходящих из Vκ1-39/Jκ5 легких цепей содержат по меньшей мере одну или не больше четырех мутаций, присутствующих в CDR3; согласно одному варианту осуществления по меньшей мере 27% легких цепей содержат одну мутацию, присутствующую в CDR3; согласно одному варианту осуществления по меньшей мере 8% легких цепей содержат две мутации, присутствующие в CDR3; согласно одному варианту осуществления по меньшей мере 1% легких цепей содержат три мутации, присутствующие в CDR3; согласно одному варианту осуществления по меньшей мере 1% легких цепей содержат четыре мутации, присутствующие в CDR3.
Согласно одному варианту осуществления предусмотрена популяция антиген-специфических антител, полученных от описанной в настоящем документе мыши, причем антитела содержат легкую цепь, происходящую из реаранжировки Vκ1-39/Jκ5 человека, и приблизительно 9% происходящих из Vκ1-39/Jκ5 легких цепей содержат одну или несколько мутаций, присутствующих в FW1, приблизительно 25% происходящих из Vκ1-39/Jκ5 легких цепей содержат одну или несколько мутаций, присутствующих в CDR1, приблизительно 20% происходящих из Vκ1-39/Jκ5 легких цепей содержат одну или несколько мутаций, присутствующих в FW2, приблизительно 10% происходящих из Vκ1-39/Jκ5 легких цепей содержат одну или несколько мутаций, присутствующих в CDR2, приблизительно 29% происходящих из Vκ1-39/Jκ5 легких цепей содержат одну или несколько мутаций, присутствующих в FW3, и приблизительно 37% происходящих из Vκ1-39/Jκ5 легких цепей содержат одну или несколько мутаций, присутствующих в CDR3.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ1-39/Jκ5 человека, и приблизительно 35% тяжелых цепей содержат по меньшей мере одну мутацию, присутствующую в FW1; согласно одному варианту осуществления по меньшей мере 25% тяжелых цепей содержат одну мутацию, присутствующую в FW1; согласно одному варианту осуществления по меньшей мере 9% тяжелых цепей содержат две мутации, присутствующие в FW1; согласно одному варианту осуществления по меньшей мере 1% тяжелых цепей содержит три мутации, присутствующие в FW1; согласно одному варианту осуществления по меньшей мере 1% тяжелых цепей содержит больше пяти мутаций, присутствующих в FW1.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ1-39/Jκ5 человека, и приблизительно 92% тяжелых цепей содержат по меньшей мере одну или не больше четырех мутаций, присутствующих в CDR1; согласно одному варианту осуществления по меньшей мере 92% тяжелых цепей содержат по меньшей мере одну, по меньшей мере две, по меньшей мере три или по меньшей мере четыре мутации, присутствующие в CDR1; согласно одному варианту осуществления по меньшей мере 26% тяжелых цепей содержат одну мутацию, присутствующую в CDR1; согласно одному варианту осуществления по меньшей мере 44% тяжелых цепей содержат две мутации, присутствующие в CDR1; согласно одному варианту осуществления по меньшей мере 19% тяжелых цепей содержат три мутации, присутствующие в CDR1; согласно одному варианту осуществления по меньшей мере 3% тяжелых цепей содержат четыре мутации, присутствующие в CDR1.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ1-39/Jκ5 человека, и приблизительно 66% тяжелых цепей содержат по меньшей мере одну или не больше трех мутаций, присутствующих в FW2; согласно одному варианту осуществления по меньшей мере 66% тяжелых цепей содержат по меньшей мере одну, по меньшей мере две или по меньшей мере три мутации, присутствующие в FW2; согласно одному варианту осуществления по меньшей мере 35% тяжелых цепей содержат одну мутацию, присутствующую в FW2; согласно одному варианту осуществления по меньшей мере 23% тяжелых цепей содержат две мутации, присутствующие в FW2; согласно одному варианту осуществления по меньшей мере 8% тяжелых цепей содержат три мутации, присутствующие в FW2.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ1-39/Jκ5 человека, и приблизительно 70% тяжелых цепей содержат по меньшей мере одну или не больше четырех мутаций, присутствующих в CDR2; согласно одному варианту осуществления по меньшей мере 70% тяжелых цепей содержат по меньшей мере две, по меньшей мере три или по меньшей мере четыре мутации, присутствующие в CDR2; согласно одному варианту осуществления по меньшей мере 34% тяжелых цепей содержат одну мутацию, присутствующую в CDR2; согласно одному варианту осуществления по меньшей мере 20% тяжелых цепей содержат две мутации, присутствующие в CDR2; согласно одному варианту осуществления по меньшей мере 12% тяжелых цепей содержат три мутации, присутствующие в CDR2; согласно одному варианту осуществления по меньшей мере 5% тяжелых цепей содержат четыре мутации, присутствующие в CDR2.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ1-39/Jκ5 человека, и приблизительно 91% тяжелых цепей содержат по меньшей мере один или до пяти или больше мутаций, присутствующих в FW3; согласно одному варианту осуществления по меньшей мере 91% тяжелых цепей содержат по меньшей мере две, по меньшей мере три, по меньшей мере четыре или по меньшей мере пять или больше мутаций, присутствующих в FW3; согласно одному варианту осуществления по меньшей мере 19% тяжелых цепей содержат одну мутацию, присутствующую в FW3; согласно одному варианту осуществления по меньшей мере 33% тяжелых цепей содержат две мутации, присутствующие в FW3; согласно одному варианту осуществления по меньшей мере 22% тяжелых цепей содержат три мутации, присутствующие в FW3; согласно одному варианту осуществления по меньшей мере 11% тяжелых цепей содержат четыре мутации, присутствующие в FW3; согласно одному варианту осуществления по меньшей мере 7% тяжелых цепей содержат пять или больше мутаций, присутствующих в FW3.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ1-39/Jκ5 человека, и приблизительно 63% тяжелых цепей содержат по меньшей мере одну или не больше двух мутаций, присутствующих в CDR3; согласно одному варианту осуществления по меньшей мере 63% тяжелых цепей содержат одну мутацию, присутствующую в CDR3; согласно одному варианту осуществления по меньшей мере 54% тяжелых цепей содержат одну мутацию, присутствующую в CDR3; согласно одному варианту осуществления по меньшей мере 9% тяжелых цепей содержат две мутации, присутствующие в CDR3.
Согласно одному варианту осуществления предусмотрена популяция антиген-специфических антител, полученных от описанной в настоящем документе мыши, причем антитела содержат легкую цепь, происходящую из реаранжировки Vκ1-39/Jκ5 человека, и по меньшей мере 35% тяжелых цепей содержат одну или несколько мутаций, присутствующих в FW1, приблизительно 92% тяжелых цепей содержат одну или несколько мутаций, присутствующих в CDR1, приблизительно 66% тяжелых цепей содержат одну или несколько мутаций, присутствующих в FW2, приблизительно 70% тяжелых цепей содержат одну или несколько мутаций, присутствующих в CDR2, приблизительно 91% тяжелых цепей содержат одну или несколько мутаций, присутствующих в FW3, и приблизительно 63% тяжелых цепей содержат одну или несколько мутаций, присутствующих в CDR3.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ3-20/Jκ1 человека, и ген легкой цепи содержит по меньшей мере одну или не больше двух соматических гипермутаций; согласно одному варианту осуществления ген легкой цепи содержит по меньшей мере две, по меньшей мере три, по меньшей мере четыре или больше соматических гипермутаций. Согласно конкретному варианту осуществления мутации присутствуют в одной или нескольких каркасных областях легкой цепи. Согласно конкретному варианту осуществления мутации присутствуют в одной или нескольких областях CDR легкой цепи. Согласно конкретному варианту осуществления мутации присутствуют в одной или нескольких каркасных областях и/или одной или нескольких областях CDR легкой цепи. Согласно различным вариантам осуществления каркасные области выбраны из каркасной области 1 (FW1), каркасной области 2 (FW2), каркасной области 3 (FW3) и/или их комбинации.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ3-20/Jκ1 человека, и приблизительно 10% происходящих из Vκ3-20/Jκ1 легких цепей содержат по меньшей мере одну мутацию, присутствующую в FW1; согласно одному варианту осуществления по меньшей мере 10% легких цепей содержат одну мутацию в FW1.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ3-20/Jκ1 человека, и приблизительно 53% происходящих из Vκ3-20/Jκ1 легких цепей содержат по меньшей мере одну или не больше двух мутаций, присутствующих в CDR1; согласно одному варианту осуществления по меньшей мере 27% легких цепей содержат одну или несколько мутаций в CDR1; согласно одному варианту осуществления приблизительно 54% легких цепей содержат один или две мутации, присутствующие в CDR1.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ3-20/Jκ1 человека, и приблизительно 6% происходящих из Vκ3-20/Jκ1 легких цепей содержат по меньшей мере одну или не больше двух мутаций, присутствующих в FW2; согласно одному варианту осуществления по меньшей мере 6% легких цепей содержат по меньшей мере одну мутацию, присутствующую в FW2; согласно одному варианту осуществления по меньшей мере 3% легких цепей содержат одну мутацию, присутствующую в FW2; согласно одному варианту осуществления по меньшей мере 3% легких цепей содержат две мутации, присутствующие в FW2.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ3-20/Jκ1 человека, и по меньшей мере приблизительно 3% происходящих из Vκ3-20/Jκ1 легких цепей содержат по меньшей мере одну мутацию, присутствующую в CDR2; согласно одному варианту осуществления по меньшей мере 3% легких цепей содержат одну мутацию в CDR2.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ3-20/Jκ1 человека, и приблизительно 17% или больше происходящих из Vκ3-20/Jκ1 легких цепей содержат по меньшей мере одну или не больше двух мутаций, присутствующих в FW3; согласно одному варианту осуществления по меньшей мере 20% легкой цепи содержат одну мутацию, присутствующую в FW3; согласно одному варианту осуществления по меньшей мере 17% легких цепей содержат две мутации, присутствующие в FW3.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ3-20/Jκ1 человека, и по меньшей мере 43% происходящих из Vκ3-20/Jκ1 легких цепей содержат по меньшей мере одну мутацию, присутствующую в CDR3; согласно одному варианту осуществления по меньшей мере 43% легких цепей содержат одну мутацию в CDR3.
Согласно одному варианту осуществления предусмотрена популяция антиген-специфических антител, полученных от описанной в настоящем документе мыши, причем антитела содержат легкую цепь, происходящую из реаранжировки Vκ3-20/Jκ1 человека, и приблизительно 10% происходящих из Vκ3-20/Jκ1 легких цепей содержат одну или несколько мутаций, присутствующих в FW1, приблизительно 53% происходящих из Vκ3-20/Jκ1 легких цепей содержат одну или несколько мутаций, присутствующих в CDR1, приблизительно 6% происходящих из Vκ3-20/Jκ1 легких цепей содержат одну или несколько мутаций, присутствующих в FW2, приблизительно 3% происходящих из Vκ3-20/Jκ1 легких цепей содержат одну или несколько мутаций, присутствующих в CDR2, приблизительно 37% происходящих из Vκ3-20/Jκ1 легких цепей содержат одну или несколько мутаций, присутствующих в FW3, и приблизительно 43% происходящих из Vκ3-20/Jκ1 легких цепей содержат одну или несколько мутаций, присутствующих в CDR3.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ3-20/Jκ1 человека, и приблизительно 43% тяжелых цепей содержат по меньшей мере одну или не больше двух мутаций, присутствующих в FW1; согласно одному варианту осуществления по меньшей мере 41% тяжелых цепей содержат по меньшей мере одну мутацию, присутствующую в FW1; согласно одному варианту осуществления приблизительно 41% тяжелых цепей содержат одну мутацию, присутствующую в FW1; согласно одному варианту осуществления приблизительно 2% тяжелых цепей содержат две мутации, присутствующие в FW1.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ3-20/Jκ1 человека, и приблизительно 92% тяжелых цепей содержат по меньшей мере одну или не больше четырех мутаций, присутствующих в CDR1; согласно одному варианту осуществления по меньшей мере 43% тяжелых цепей содержат по меньшей мере одну мутацию, присутствующую в CDR1; согласно одному варианту осуществления по меньшей мере 25% тяжелых цепей содержат по меньшей мере две мутации, присутствующие в CDR1; согласно одному варианту осуществления по меньшей мере 15% тяжелых цепей содержат по меньшей мере 3 мутации, присутствующие в CDR1; согласно одному варианту осуществления по меньшей мере 10% тяжелых цепей содержат 4 или больше мутаций, присутствующих в CDR1.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ3-20/Jκ1 человека и приблизительно 46% тяжелых цепей содержат по меньшей мере одну или не больше трех мутаций, присутствующих в FW2; согласно одному варианту осуществления по меньшей мере 34% тяжелых цепей содержат по меньшей мере одну мутацию, присутствующую в FW2; согласно одному варианту осуществления по меньшей мере 10% тяжелых цепей содержат две или больше мутаций, присутствующих в FW2; согласно одному варианту осуществления по меньшей мере 2% тяжелых цепей содержат три или больше мутаций, присутствующих в FW2.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ3-20/Jκ1 человека, и приблизительно 84% тяжелых цепей содержат по меньшей мере одну или вплоть до пяти или больше пяти мутаций, присутствующих в CDR2; согласно одному варианту осуществления по меньшей мере 39% тяжелых цепей содержат одну или несколько мутаций, присутствующих в CDR2; согласно одному варианту осуществления по меньшей мере 18% тяжелых цепей содержат две или больше мутаций, присутствующих в CDR2; согласно одному варианту осуществления по меньшей мере 21% тяжелых цепей содержат три или больше мутаций, присутствующих в CDR2; согласно одному варианту осуществления по меньшей мере 3% тяжелых цепей содержат четыре или больше мутаций, присутствующих в CDR2; согласно одному варианту осуществления по меньшей мере 2% тяжелых цепей содержат пять или больше мутаций, присутствующих в CDR2.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ3-20/Jκ1 человека, и приблизительно 92% тяжелых цепей содержат по меньшей мере одну или вплоть до пяти или больше пяти мутаций, присутствующих в FW3; согласно одному варианту осуществления по меньшей мере 21% легких цепей содержат по меньшей мере одну мутацию, присутствующую в FW3; согласно одному варианту осуществления по меньшей мере 20% тяжелых цепей содержат по меньшей мере две мутации, присутствующие в FW3; согласно одному варианту осуществления по меньшей мере 13% тяжелых цепей содержат по меньшей мере три мутации, присутствующие в FW3; согласно одному варианту осуществления по меньшей мере 20% тяжелых цепей содержат по меньшей мере четыре мутации в FW3; согласно одному варианту осуществления по меньшей мере 18% тяжелых цепей содержат по меньшей мере 5 мутаций в FW3.
Согласно одному варианту осуществления легкая цепь происходит из реаранжировки Vκ3-20/Jκ1 человека, и приблизительно 7% тяжелых цепей содержат по меньшей мере одну мутацию, присутствующую в CDR3; согласно одному варианту осуществления приблизительно 7% тяжелых цепей содержат одну мутацию в CDR3.
Согласно одному варианту осуществления предусмотрена популяция антиген-специфических антител, полученных от описанной в настоящем документе мыши, причем антитела содержат легкую цепь, происходящую из реаранжировки Vκ3-20/Jκ1 человека, и приблизительно 43% тяжелых цепей содержат одну или несколько мутаций, присутствующих в FW1, приблизительно 92% тяжелых цепей содержат одну или несколько мутаций, присутствующих в CDR1, приблизительно 46% тяжелых цепей содержат одну или несколько мутаций, присутствующих в FW2, приблизительно 84% тяжелых цепей содержат одну или несколько мутаций, присутствующих в CDR2, приблизительно 92% тяжелых цепей содержат одну или несколько мутаций, присутствующих в FW3, и приблизительно 7% тяжелых цепей содержат одну или несколько мутаций, присутствующих в CDR3.
Согласно одному аспекту предусмотрена мышь, которая экспрессирует легкую цепь иммуноглобулина из реаранжированной последовательности легкой цепи иммуноглобулина, причем реаранжированная последовательность легкой цепи иммуноглобулина присутствует в зародышевой линии мыши, причем легкая цепь иммуноглобулина содержит вариабельную последовательность человека. Согласно одному варианту осуществления зародышевая линия мыши содержит реаранжированную последовательность легкой цепи иммуноглобулина, которая происходит из того же сегмента V и того же сегмента J, что и все несуррогатные последовательности легкой цепи, присутствующие в каждой В-клетке мыши, которая содержит реаранжированную последовательность легкой цепи.
Согласно одному варианту осуществления зародышевая линия мыши не содержит функциональный нереаранжированный генный сегмент V легкой цепи иммуноглобулина. Согласно одному варианту осуществления зародышевая линия мыши не содержит функциональный нереаранжированный генный сегмент J легкой цепи иммуноглобулина.
Согласно одному варианту осуществления зародышевая линия мыши содержит не больше одной, не больше двух или не больше трех реаранжированных последовательностей (V/J) легкой цепи.
Согласно одному варианту осуществления реаранжированная последовательность V/J содержит последовательность κ легкой цепи. Согласно конкретному варианту осуществления последовательность κ легкой цепи представляет собой последовательность κ легкой цепи человека. Согласно конкретному варианту осуществления последовательность κ легкой цепи выбрана из последовательности Vκ1-39/J человека, последовательности Vκ3-20/J человека и их комбинации. Согласно конкретному варианту осуществления последовательность κ легкой цепи представляет собой последовательность Vκ1-39/Jκ5 человека. Согласно конкретному варианту осуществления последовательность κ легкой цепи представляет собой последовательность Vκ3-20/Jκ1 человека.
Согласно одному варианту осуществления мышь дополнительно содержит в своей зародышевой линии последовательность, выбранную из κ интронного энхансера мыши 5' по отношению к реаранжированной последовательности легкой цепи иммуноглобулина, κ 3' энхансера мыши и их комбинации.
Согласно одному варианту осуществления мышь содержит нереаранжированный генный сегмент VH человека, нереаранжированный генный сегмент DH человека и нереаранжированный генный сегмент JH человека, причем указанные генные сегменты VH, DH и JH способны к реаранжировке для образования вариабельной генной последовательности тяжелой цепи иммуноглобулина, функционально связанной с константной генной последовательностью тяжелой цепи. Согласно одному варианту осуществления мышь содержит множество генных сегментов VH, DH и JH человека. Согласно конкретному варианту осуществления генные сегменты VH, DH и JH человека замещают эндогенные генные сегменты VH, DH и JH мыши на эндогенном локусе тяжелой цепи иммуноглобулина мыши. Согласно конкретному варианту осуществления мышь содержит замещение всех или по существу всех функциональных генных сегментов VH, DH и JH мыши всеми или по существу всеми функциональными генными сегментами VH, DH и JH человека.
Согласно одному варианту осуществления мышь экспрессирует легкую цепь иммуноглобулина, которая содержит константную последовательность мыши. Согласно одному варианту осуществления мышь экспрессирует легкую цепь иммуноглобулина, которая содержит константную последовательность человека.
Согласно одному варианту осуществления мышь экспрессирует тяжелую цепь иммуноглобулина, которая содержит последовательность мыши, выбранную из последовательности CH1, шарнирной последовательности, последовательности CH2, последовательности CH3 и их комбинации.
Согласно одному варианту осуществления мышь экспрессирует тяжелую цепь иммуноглобулина, которая содержит последовательность человека, выбранную из последовательности CH1, шарнирной последовательности, последовательности CH2, последовательности CH3 и их комбинации.
Согласно одному варианту осуществления реаранжированная последовательность легкой цепи иммуноглобулина в зародышевой линии мыши находится на эндогенном локусе легкой цепи иммуноглобулина мыши. Согласно конкретному варианту осуществления реаранжированная последовательность легкой цепи иммуноглобулина в зародышевой линии мыши замещает все или по существу все последовательности V и J легкой цепи мыши на эндогенном локусе легкой цепи иммуноглобулина мыши.
Согласно одному аспекту предусмотрена мышь, которая содержит популяцию В-клеток, характеризующуюся тем, что каждая В-клетка, которая содержит несуррогатную последовательность легкой цепи, содержит реаранжированный ген легкой цепи, который происходит из единственного сегмента V человека и единственного сегмента J человека, причем единственная вариабельная последовательность легкой цепи в зародышевой линии мыши представляет собой реаранжированную последовательность, происходящую из единственного сегмента V человека и единственного сегмента J человека, и причем каждая В-клетка, которая содержит реаранжированный ген легкой цепи, дополнительно содержит ген, кодирующий когнатный вариабельный домен тяжелой цепи человека, и причем реаранжированный ген легкой цепи содержит по меньшей мере одну, по меньшей мере две, по меньшей мере три или по меньшей мере четыре соматических гипермутации.
Согласно одному аспекту предусмотрена плюрипотентная, индуцированная плюрипотентная или тотипотентная клетка, полученная от описанной в настоящем документе мыши. Согласно конкретному варианту осуществления клетка представляет собой эмбриональную стволовую (ES) клетку мыши.
Согласно одному аспекту предусмотрена ткань, полученная от описанной в настоящем документе мыши. Согласно одному варианту осуществления ткань происходит из селезенки, лимфатического узла или костного мозга описанной в настоящем документе мыши.
Согласно одному аспекту предусмотрено ядро, полученное от описанной в настоящем документе мыши. Согласно одному варианту осуществления ядро происходит из диплоидной клетки, которая не является В-клеткой.
Согласно одному аспекту предусмотрена клетка мыши, которая выделена из описанной в настоящем документе мыши. Согласно одному варианту осуществления клетка представляет собой ES клетку. Согласно одному варианту осуществления клетка представляет собой лимфоцит. Согласно одному варианту осуществления лимфоцит представляет собой В-клетку. Согласно одному варианту осуществления В-клетка экспрессирует химерную тяжелую цепь, содержащую вариабельный домен, происходящий из генного сегмента человека; и легкую цепь, происходящую из реаранжированной последовательности Vκ1-39/J человека, реаранжированной последовательности Vκ3-20/J человека или их комбинации; причем вариабельный домен тяжелой цепи слит с константной областью мыши, и вариабельный домен легкой цепи слит с константной областью мыши или человека.
Согласно одному аспекту предусмотрена гибридома, причем гибридома образована из В-клетки описанной в настоящем документе мыши. Согласно конкретному варианту осуществления В-клетка получена от описанной в настоящем документе мышь, которая была иммунизирована иммуногеном, содержащим представляющий интерес эпитоп, и В-клетка экспрессирует связывающий белок, который связывает представляющий интерес эпитоп, связывающий белок содержит соматически мутированный домен VH человека и CH мыши, и содержит домен VL человека, происходящий из реаранжированной области Vκ1-39Jκ5 человека или реаранжированной области Vκ3-20Jκ1 человека, и CL мыши.
Согласно одному аспекту предусмотрен зародыш мыши, причем зародыш содержит донорную ES клетку, которая получена от описанной в настоящем документе мыши.
Согласно одному аспекту предусмотрен нацеливающий вектор, содержащий от 5' к 3' в направлении транскрипции относительно последовательностей 5' и 3' относящихся к мыши гомологичных плечей вектора 5' гомологичное плечо мыши, промотор иммуноглобулина человека или мыши, лидерную последовательность человека или мыши и область VL человека, выбранную из реаранжированной области Vκ1-39Jκ5 человека или реаранжированной области Vκ3-20Jκ1 человека, и 3' гомологичное плечо мыши. Согласно одному варианту осуществления 5' и 3' гомологичные плечи нацеливают вектор на последовательность 5' по отношению к энхансерной последовательности, которая присутствует 5' и проксимально к гену Cκ мыши. Согласно одному варианту осуществления промотор представляет собой промотор генного сегмента вариабельной области иммуноглобулина человека. Согласно конкретному варианту осуществления промотор представляет собой промотор Vκ3-15 человека. Согласно одному варианту осуществления лидерная последовательность представляет собой лидерную последовательность мыши. Согласно конкретному варианту осуществления лидерная последовательность мыши представляет собой лидерную последовательность Vκ3-7 мыши.
Согласно одному аспекту предусмотрен нацеливающий вектор, описанный выше, но вместо 5' гомологичного плеча мыши промотор человека или мыши фланкирован 5' сайтом распознавания сайт-специфической рекомбиназы (SRRS), и вместо 3' гомологичного плеча мыши область VL человека фланкирована 3' SRRS.
Согласно одному аспекту предусмотрено обратное химерное антитело, произведенное описанной в настоящем документе мышью, причем обратное химерное антитело содержит легкую цепь, содержащую VL человека и CL мыши, и тяжелую цепь, содержащую VH человека и CH мыши.
Согласно одному аспекту предусмотрен способ получения антитела, предусматривающий экспрессию в одной клетке (a) первой генной последовательности VH иммунизированной описанной в настоящем документе мыши, слитой с генной последовательностью CH человека; (b) генной последовательности VL иммунизированной описанной в настоящем документе мыши, слитой с генной последовательностью CL человека; и (c) содержание клетки при условиях, достаточных для экспрессии полностью человеческого антитела, и выделение антитела. Согласно одному варианту осуществления клетка содержит вторую генную последовательность VH второй иммунизированной описанной в настоящем документе мыши, слитую с генной последовательностью CH человека, первый генная последовательность VH кодирует домен VH, который распознает первый эпитоп, а вторая генная последовательность VH кодирует домен VH, который распознает второй эпитоп, причем первый эпитоп и второй эпитоп не являются идентичными.
Согласно одному аспекту предусмотрен способ получения эпитопсвязывающего белка, предусматривающий воздействие на описанную в настоящем документе мышь иммуногена, который содержит представляющий интерес эпитоп, содержание мыши при условиях, достаточных для того, чтобы мышь производила молекулу иммуноглобулина, которая специфически связывает представляющий интерес эпитоп, и выделение молекулы иммуноглобулина, которая специфически связывает представляющий интерес эпитоп; причем эпитопсвязывающий белок содержит тяжелую цепь, которая содержит соматически мутированный VH человека и CH мыши, ассоциированную с легкой цепью, содержащей CL мыши и VL человека, происходящий из реаранжированного сегмента Vκ1-39Jκ5 человека или реаранжированного сегмента Vκ3-20Jκ1 человека.
Согласно одному аспекту предусмотрена клетка, которая экспрессирует эпитопсвязывающий белок, причем клетка содержит: (а) нуклеотидную последовательность человека, кодирующую домен VL человека, который происходит из реаранжированного сегмента Vκ1-39Jκ5 человека или реаранжированного сегмента Vκ3-20Jκ1 человека, причем нуклеотидная последовательность человека слита (напрямую или через линкер) с последовательностью кДНК константного домена легкой цепи иммуноглобулина человека (например, последовательностью ДНК к константного домена человека); и (b) первую нуклеотидную последовательность VH человека, кодирующую домен VH человека, происходящий из первой нуклеотидной последовательности VH человека, причем первая нуклеотидная последовательность VH человека слита (напрямую или через линкер) с последовательностью кДНК константного домена тяжелой цепи иммуноглобулина человека; причем эпитопсвязывающий белок распознает первый эпитоп. Согласно одному варианту осуществления эпитопсвязывающий белок связывает первый эпитоп с константой диссоциации, составляющей ниже чем 10-6 M, ниже чем 10-8 M, ниже чем 10-9 М, ниже чем 10-10 М, ниже чем 10-11 M или ниже чем 10-12 М.
Согласно одному варианту осуществления клетка содержит вторую нуклеотидную последовательность человека, кодирующую второй домен VH человека, причем вторая последовательность человека слита (напрямую или через линкер) с последовательностью кДНК константного домена тяжелой цепи иммуноглобулина человека, и причем второй домен VH человека не распознает специфически первый эпитоп (например, проявляет константу диссоциации, составляющую, например, 10-6 M, 10-5 M, 10-4 M или выше), и причем эпитопсвязывающий белок распознает первый эпитоп и второй эпитоп, и причем каждая из первой и второй тяжелой цепи иммуноглобулина ассоциируется с идентичной легкой цепью согласно (а).
Согласно одному варианту осуществления второй домен VH связывает второй эпитоп с константой диссоциации, которая составляет ниже чем 10-6 М, ниже чем 10-7 М, ниже чем 10-8 М, ниже чем 10-9 М, ниже чем 10-10 М, ниже чем 10-11 M или ниже чем 10-12 М.
Согласно одному варианту осуществления эпитопсвязывающий белок содержит первую тяжелую цепь иммуноглобулина и вторую тяжелую цепь иммуноглобулина, каждая из которых ассоциирована с идентичной легкой цепью, происходящей из реаранжированной области VL человека, выбранной из Vκ1-39Jκ5 человека или Vκ3-20Jκ1 человека, причем первая тяжелая цепь иммуноглобулина связывает первый эпитоп с константой диссоциации в наномолярном - пикомолярном диапазоне, вторая тяжелая цепь иммуноглобулина связывает второй эпитоп с константой диссоциации в наномолярном - пикомолярном диапазоне, первый эпитоп и второй эпитоп не являются идентичными, первая тяжелая цепь иммуноглобулина не связывает второй эпитоп или связывает второй эпитоп с константой диссоциации, которая является слабее микромолярного диапазона (например, в миллимолярном диапазоне), вторая тяжелая цепь иммуноглобулина не связывает первый эпитоп или связывает первый эпитоп с константой диссоциации, которая является слабее микромолярного диапазона (например, в миллимолярном диапазоне), и одна или несколько из VL, VH первой тяжелой цепи иммуноглобулина и VH второй тяжелой цепи иммуноглобулина являются соматически мутированными.
Согласно одному варианту осуществления первая тяжелая цепь иммуноглобулина содержит связывающий белок А остаток, и вторая тяжелая цепь иммуноглобулина не содержит связывающий белок А остаток.
Согласно одному варианту осуществления клетка выбрана из СНО, COS, 293, HeLa и ретинальной клетки, экспрессирующей последовательность вирусной нуклеиновой кислоты (например, клетки PERC.6™).
Согласно одному аспекту предусмотрено обратное химерное антитело, содержащее VH человека и константный домен тяжелой цепи мыши, VL человека и константный домен легкой цепи мыши, причем антитело получено способом, предусматривающим иммунизацию описанной в настоящем документе мыши иммуногеном, содержащим эпитоп, и антитело специфически связывает эпитоп иммуногена, которым иммунизировали мышь. Согласно одному варианту осуществления домен VL является соматически мутированным. Согласно одному варианту осуществления домен VH является соматически мутированным. Согласно одному варианту осуществления как домен VL, так и домен VH являются соматически мутированными. Согласно одному варианту осуществления VL связан с доменом Cκ мыши.
Согласно одному аспекту предусмотрена мышь, содержащая генные сегменты VH человека, замещающие все или по существу все генные сегменты VH мыши на эндогенном локусе тяжелой цепи мыши; не больше одной или двух реаранжированных последовательностей VL/JL легкой цепи человека, выбранных из реаранжированной Vκ1-39/J и реаранжированной Vκ3-20/J или их комбинации, замещающих все генные сегменты легкой цепи мыши; причем вариабельные генные сегменты тяжелой цепи человека связаны с константным геном мыши, и реаранжированные последовательности легкой цепи человека связаны с константным геном человека или мыши.
Согласно одному аспекту предусмотрена ES клетка мыши, содержащая замещение всех или по существу всех вариабельных генных сегментов тяжелой цепи мыши вариабельными генными сегментами тяжелой цепи человека, и не больше одной или двух реаранжированных последовательностей легкой цепи VL/JL человека, причем вариабельные генные сегменты тяжелой цепи человека связаны с константным геном тяжелой цепи иммуноглобулина мыши, и реаранжированные последовательности VL/JL легкой цепи человека связаны с константным геном легкой цепи иммуноглобулина мыши или человека. Согласно конкретному варианту осуществления константный ген легкой цепи представляет собой константный ген мыши.
Согласно одному аспекту предусмотрен антигенсвязывающий белок, произведенный описанной в настоящем документе мышью. Согласно конкретному варианту осуществления антигенсвязывающий белок содержит вариабельную область тяжелой цепи иммуноглобулина человека, слитую с константной областью мыши, и вариабельную область легкой цепи иммуноглобулина человека, происходящую из генного сегмента Vκ1-39 или генного сегмента Vκ3-20, причем константная область легкой цепи представляет собой константную область мыши.
Согласно одному аспекту предусмотрен полностью человеческий антигенсвязывающий белок, произведенный из генной последовательности вариабельной области иммуноглобулина от описанной в настоящем документе мыши, причем антигенсвязывающий белок содержит полностью человеческую тяжелую цепь, содержащую вариабельную область человека, происходящую из последовательности описанной в настоящем документе мыши, и полностью человеческую легкую цепь, содержащую Vκ1-39 или Vκ3-20. Согласно одному варианту осуществления вариабельная область легкой цепи содержит от одной до пяти соматических мутаций. Согласно одному варианту осуществления вариабельная область легкой цепи представляет собой когнатную вариабельную область легкой цепи, которая спаривается в В-клетке мыши с вариабельной областью тяжелой цепи.
Согласно одному варианту осуществления полностью человеческий антигенсвязывающий белок содержит первую тяжелую цепь и вторую тяжелую цепь, причем первая тяжелая цепь и вторая тяжелая цепь содержат неидентичные вариабельные области, независимо происходящие от описанной в настоящем документе мыши, и причем каждая из первой и второй тяжелых цепей экспрессируется из клетки-хозяина ассоциированной с легкой цепью человека, происходящей из генного сегмента Vκ1-39 или генного сегмента Vκ3-20. Согласно одному варианту осуществления первая тяжелая цепь содержит первую вариабельную область тяжелой цепи, которая специфически связывает первый эпитоп первого антигена, и вторая тяжелая цепь содержит вторую вариабельную область тяжелой цепи, которая специфически связывает второй эпитоп второго антигена. Согласно конкретному варианту осуществления первый антиген и второй антиген являются различными. Согласно конкретному варианту осуществления первый антиген и второй антиген являются одинаковыми, и первый эпитоп и второй эпитоп не являются идентичными; согласно конкретному варианту осуществления связывание первого эпитопа первой молекулой связывающего белка не блокирует связывание второго эпитопа второй молекулой связывающего белка.
Согласно одному аспекту полностью человеческий связывающий белок, происходящий из последовательности иммуноглобулина человека описанной в настоящем документе мыши, содержит первую тяжелую цепь иммуноглобулина и вторую тяжелую цепь иммуноглобулина, причем первая тяжелая цепь иммуноглобулина содержит первую вариабельную область, которая не является идентичной вариабельной области второй тяжелой цепи иммуноглобулина, и причем первая тяжелая цепь иммуноглобулина содержит связывающую белок А детерминанту дикого типа, и вторая тяжелая цепь не содержит связывающую белок А детерминанту дикого типа. Согласно одному варианту осуществления первая тяжелая цепь иммуноглобулина связывает белок А при условиях выделения, и вторая тяжелая цепь иммуноглобулина не связывает белок А или связывает белок А по меньшей мере в 10 раз, в 100 раз или в 1000 раз слабее, чем первая тяжелая цепь иммуноглобулина связывает белок А при условиях выделения. Согласно конкретному варианту осуществления первая и вторая тяжелые цепи представляют собой изотипы IgG1, причем вторая тяжелая цепь содержит модификацию, выбранную из 95R (EU 435R), 96F (EU 436F) и их комбинации, и причем первая тяжелая цепь не содержит такой модификации.
Согласно одному аспекту предусмотрен способ получения биспецифического антигенсвязывающего белка, предусматривающий воздействие на первую описанную в настоящем документе мышь первого представляющего интерес антигена, который содержит первый эпитоп, воздействие на вторую описанную в настоящем документе мышь второго представляющего интерес антигена, который содержит второй эпитоп, предоставление каждой из первой и второй мыши возможности развить иммунные ответы на представляющие интерес антигены, идентификацию у первой мыши первой вариабельной области тяжелой цепи человека, которая связывает первый эпитоп первого представляющего интерес антигена, идентификацию у второй мыши второй вариабельной области тяжелой цепи человека, которая связывает второй эпитоп второго представляющего интерес антигена, получение первого полностью человеческого гена тяжелой цепи, который кодирует первую тяжелую цепь, которая связывает первый эпитоп первого представляющего интерес антигена, получение второго полностью человеческого гена тяжелой цепи, который кодирует вторую тяжелую цепь, которая связывает второй эпитоп второго представляющего интерес антигена, экспрессию первой тяжелой цепи и второй тяжелой цепи в клетке, которая экспрессирует единственную полностью человеческую легкую цепь, происходящую из генного сегмента Vκ1-39 человека или генного сегмента Vκ3-20 человека для образования биспецифического антигенсвязывающего белка, и выделение биспецифического антигенсвязывающего белка.
Согласно одному варианту осуществления первый антиген и второй антиген не являются идентичными.
Согласно одному варианту осуществления первый антиген и второй антиген являются идентичными, и первый эпитоп и второй эпитоп не являются идентичными. Согласно одному варианту осуществления связывание первой вариабельной области тяжелой цепи с первым эпитопом не блокирует связывание второй вариабельной области тяжелой цепи со вторым эпитопом.
Согласно одному варианту осуществления первый антиген выбран из растворимого антигена и антигена клеточной поверхности (например, опухолевого антигена), и второй антиген содержит рецептор клеточной поверхности. Согласно конкретному варианту осуществления рецептор клеточной поверхности представляет собой рецептор иммуноглобулина. Согласно конкретному варианту осуществления рецептор иммуноглобулина представляет собой рецептор Fc. Согласно одному варианту осуществления первый антиген и второй антиген представляют собой один и тот же рецептор клеточной поверхности, и связывание первой тяжелой цепи с первым эпитопом не блокирует связывание второй тяжелой цепи со вторым эпитопом.
Согласно одному варианту осуществления вариабельный домен легкой цепи легкой цепи содержит 2-5 соматических мутаций. Согласно одному варианту осуществления вариабельный домен легкой цепи представляет собой соматически мутированную когнатную легкую цепь, экспрессируемую в В-клетке первой или второй иммунизированной мыши или с первым, или со вторым вариабельным доменом тяжелой цепи.
Согласно одному варианту осуществления первая полностью человеческая тяжелая цепь содержит аминокислотную модификацию, которая снижает ее аффинность к белку А, и вторая полностью человеческая тяжелая цепь не содержит модификацию, которая снижает ее аффинность к белку А.
Согласно одному аспекту предусмотрено антитело или биспецифическое антитело, содержащее вариабельный домен тяжелой цепи человека, полученный согласно настоящему изобретению. Согласно другому аспекту предусмотрено применение описанной в настоящем документе мыши для получения полностью человеческого антитела или полностью человеческого биспецифического антитела.
Согласно одному аспекту генетически модифицированная мышь, зародыш или клетка, описанная в настоящем документе, содержит локус κ легкой цепи, который сохраняет эндогенные регуляторные или контролирующие элементы, например, κ интронный энхансер мыши, κ 3' энхансер мыши или как интронный энхансер, так и 3' энхансер, причем регуляторные или контролирующие элементы облегчают соматическую мутацию и созревание аффинности экспрессируемой последовательности локуса κ легкой цепи.
Согласно одному аспекту предусмотрена мышь, которая содержит В-клеточную популяцию, характеризующуюся наличием легких цепей иммуноглобулина, происходящих не более чем из одного или не более чем их двух реаранжированных или нереаранжированных генных сегментов V и J легкой цепи иммуноглобулина, причем мышь проявляет соотношение легких цепей κ:λ, которое является приблизительно таким же, как у мыши, которая содержит сегмент дикого типа, комплементарный генным сегментам V и J легкой цепи иммуноглобулина.
Согласно одному варианту осуществления легкие цепи иммуноглобулина происходят не более чем из одного или не более чем из двух реаранжированных генных сегментов V и J легкой цепи иммуноглобулина. Согласно конкретному варианту осуществления легкие цепи происходят не более чем из одного реаранжированного генного сегмента V и J легкой цепи иммуноглобулина.
Согласно одному аспекту предусмотрена описанная в настоящем документе мышь, которая экспрессирует легкую цепь иммуноглобулина, происходящую не более чем из одной или не более чем из двух последовательностей Vκ/Jκ человека, причем мышь содержит замещение всех или по существу всех эндогенных генных сегментов вариабельной области тяжелой цепи мыши одним или несколькими генные сегменты вариабельной области тяжелая цепь человека, и мышь проявляет соотношение (а) CD19+ В-клеток, которые экспрессируют иммуноглобулин, содержащий λ легкую цепь, к (b) CD19+ В-клеткам, которые экспрессируют иммуноглобулин, содержащий κ легкую цепь, составляющее приблизительно 1 к приблизительно 20.
Согласно одному варианту осуществления мышь экспрессирует единственную κ легкую цепь, происходящую из последовательности Vκ1-39Jκ5 человека, и соотношение CD19+ В-клеток, которые экспрессируют иммуноглобулин, содержащий λ легкую цепь, к CD19+ В-клеткам, которые экспрессируют иммуноглобулин, содержащий κ легкую цепь, составляет приблизительно 1 к приблизительно 20; согласно одному варианту осуществления соотношение составляет приблизительно 1 к по меньшей мере приблизительно 66; согласно конкретному варианту осуществления соотношение составляет приблизительно 1 к 66.
Согласно одному варианту осуществления мышь экспрессирует единственную κ легкую цепь, происходящую из последовательности Vκ3-20Jκ5 человека, и соотношение CD19+ В-клеток, которые экспрессируют иммуноглобулин, содержащий λ легкую цепь, к CD19+ В-клеткам, которые экспрессируют иммуноглобулин, содержащий κ легкую цепь, составляет приблизительно 1 к приблизительно 20; согласно одному варианту осуществления соотношение составляет приблизительно 1 к приблизительно 21. Согласно конкретным вариантам осуществления соотношение составляет 1 к 20 или 1 к 21.
Согласно одному аспекту предусмотрена генетически модифицированная мышь, которая экспрессирует единственную реаранжированную κ легкую цепь, причем мышь содержит функциональный локус λ легкой цепи, и причем мышь экспрессирует популяцию В-клеток, которая содержит Igκ+ клетки, которые экспрессируют κ легкую цепь, происходящую из этой же единственной реаранжированной κ легкой цепи. Согласно одному варианту осуществления процентное отношение Igκ+Igλ+ В-клеток у мыши является приблизительно таким же, как у мыши дикого типа. Согласно конкретному варианту осуществления процентное отношение Igκ+Igλ+ В-клеток у мыши составляет приблизительно 2 - приблизительно 6%. Согласно конкретному варианту осуществления процентное отношение Igκ+Igλ+ В-клеток у мыши, у которой единственная реаранжированная к легкая цепь происходит из последовательности Vκ1-39Jκ5, составляет приблизительно 2 - приблизительно 3; согласно конкретному варианту осуществления процентное отношение составляет приблизительно 2,6. Согласно конкретному варианту осуществления процентное отношение Igκ+Igλ+ В-клеток к мыши, у которой единственная реаранжированная к легкая цепь происходит из последовательности Vκ3-20Jκ1, составляет приблизительно 4 - приблизительно 8; согласно конкретному варианту осуществления приблизительно 6.
Согласно одному аспекту предусмотрена генетически модифицированная мышь, причем мышь экспрессирует единственную реаранжированную κ легкую цепь, происходящую из генного сегмента Vκ и Jκ человека, причем мышь экспрессирует популяцию В-клеток, которая содержит единственную κ легкую цепь, происходящую из единственной реаранжированной последовательности κ легкой цепи, причем генетически модифицированная мышь не становилась устойчивой к соматическим гипермутациям. Согласно одному варианту осуществления по меньшей мере 90% κ легких цепей, экспрессируемых на В-клетке мыши, проявляют от по меньшей мере одной до приблизительно пяти соматических гипермутаций.
Согласно одному аспекту предусмотрена генетически модифицированная мышь, которая модифицирована для экспрессии единственной κ легкой цепи, происходящей не более чем из одной или не более чем из двух реаранжированных последовательностей κ легкой цепи, причем мышь проявляет частоту использования κ легкой цепи, которая приблизительно в два раза или больше, по меньшей мере приблизительно в три раза или больше, или по меньшей мере приблизительно в четыре раза или больше выше, чем частота использования κ легкой цепи, проявляемая мышью дикого типа, или выше, чем частота использования κ легкой цепи, проявляемая мышью той же линии, которая содержит репертуар генных сегментов κ легкой цепи дикого типа. Согласно конкретному варианту осуществления мышь экспрессирует единственную κ легкую цепь не более чем из одной реаранжированной последовательности κ легкой цепи. Согласно более конкретному варианту осуществления реаранжированная последовательность κ легкой цепи выбрана из последовательности Vκ1-39Jκ5 и Vκ3-20Jκ1. Согласно одному варианту осуществления реаранжированная последовательность κ легкой цепи представляет собой последовательность Vκ1-39Jκ5. Согласно одному варианту осуществления реаранжированная последовательность κ легкой цепи представляет собой последовательность Vκ3-20Jκ1.
Согласно одному аспекту предусмотрена генетически модифицированная мышь, которая экспрессирует единственную κ легкую цепь, происходящую не более чем из одной или не более чем из двух реаранжированных последовательностей κ легкой цепи, причем мышь проявляет частоту использования κ легкой цепи, которая приблизительно в 100 раз или больше, по меньшей мере приблизительно в 200 раз или больше, по меньшей мере приблизительно в 300 раз или больше, по меньшей мере приблизительно 400 раз или больше, по меньшей мере приблизительно 500 раз или больше, по меньшей мере приблизительно 600 раз или больше, по меньшей мере приблизительно 700 раз или больше, по меньшей мере приблизительно 800 раз или больше, по меньшей мере приблизительно 900 раз или больше, по меньшей мере приблизительно 1000 раз или больше выше, чем частота использования такой же κ легкой цепи, проявляемая мышью, содержащей полный или по существу полный локус κ легкой цепи человека. Согласно конкретному варианту осуществления мышь, содержащая полный или по существу полный локус κ легкой цепи человека, не содержит функциональную нереаранжированную последовательность κ легкой цепи мыши. Согласно конкретному варианту осуществления мышь экспрессирует единственную κ легкую цепь не более чем из одной реаранжированной последовательности κ легкой цепи. Согласно одному варианту осуществления мышь содержит одну копию реаранжированной последовательности κ легкой цепи (например, гетерозигота). Согласно одному варианту осуществления мышь содержит две копии реаранжированной последовательности κ легкой цепи (например, гомозигота). Согласно более конкретному варианту осуществления реаранжированная последовательность κ легкой цепи выбрана из последовательности Vκ1-39Jκ5 и Vκ3-20Jk1. Согласно одному варианту осуществления реаранжированная последовательность κ легкой цепи представляет собой последовательность Vκ1-39Jκ5. Согласно одному варианту осуществления реаранжированная последовательность κ легкой цепи представляет собой последовательность Vκ3-20Jκ1.
Согласно одному аспекту предусмотрена генетически модифицированная мышь, которая экспрессирует единственную легкую цепь, происходящую не более чем из одной или не более чем из двух реаранжированных последовательностей легкой цепи, причем легкая цепь у генетически модифицированной мыши проявляет уровень экспрессии, который по меньшей мере в 10 раз - приблизительно в 1000 раз, в 100 раз - приблизительно в 1000 раз, в 200 раз - приблизительно в 1000 раз, в 300 раз - приблизительно в 1000 раз, в 400 раз - приблизительно в 1000 раз, в 500 раз - приблизительно в 1000 раз, в 600 раз - приблизительно в 1000 раз, в 700 раз - приблизительно в 1000 раз, в 800 раз - приблизительно в 1000 раз или в 900 раз - приблизительно в 1000 раз выше, чем экспрессия такой же реаранжированной легкой цепи, проявляемая мышью, содержащей полный или по существу полный локус легкой цепи. Согласно одному варианту осуществления легкая цепь содержит последовательность человека. Согласно конкретному варианту осуществления последовательность человека представляет собой к последовательность. Согласно одному варианту осуществления последовательность человека представляет собой λ последовательность. Согласно одному варианту осуществления легкая цепь представляет собой полностью человеческую легкую цепь.
Согласно одному варианту осуществления уровень экспрессии характеризуется количественным определением мРНК транскрибированной последовательности легкой цепи и сравнением ее с транскрибированной последовательностью легкой цепи мыши, содержащей полный или по существу полный локус легкой цепи.
Согласно одному аспекту предусмотрена генетически модифицированная мышь, которая экспрессирует единственную κ легкую цепь, происходящую не более чем из одной или не более чем из двух реаранжированных последовательностей κ легкой цепи, причем мышь, при иммунизации антигеном, проявляет сывороточный титр, который сопоставим с таковым у мыши дикого типа, иммунизированной таким же антигеном. Согласно конкретному варианту осуществления мышь экспрессирует единственную κ легкую цепь не более чем из одной реаранжированной последовательности κ легкой цепи. Согласно одному варианту осуществления сывороточный титр определяется как общий иммуноглобулин. Согласно конкретному варианту осуществления сывороточный титр определяется как IgM-специфический титр. Согласно конкретному варианту осуществления сывороточный титр определяется как IgG-специфический титр. Согласно более конкретному варианту осуществления реаранжированная последовательность κ легкой цепи выбрана из последовательности Vκ1-39Jκ5 и Vκ3-20Jκ1. Согласно одному варианту осуществления реаранжированная последовательность κ легкой цепи представляет собой последовательность Vκ1-39Jκ5. Согласно одному варианту осуществления реаранжированная последовательность κ легкой цепи представляет собой последовательность Vκ3-20Jκ1.
Согласно одному аспекту предусмотрена генетически модифицированная мышь, которая экспрессирует популяцию антиген-специфических антител, причем все легкие цепи иммуноглобулина популяции антиген-специфических антител содержат вариабельную область легкой цепи (VL) человека, происходящую из одного и того же единственного генного сегмента VL человека, и тяжелые цепи иммуноглобулина содержат вариабельную область тяжелой цепи (VH) человека, происходящую из одного из множества генных сегментов VH человека.
Согласно различным вариантам осуществления генные сегменты VH человека выбраны из VH1-2, VH1-3, VH1-8, VH1-18, VH1-24, VH1-46, VH1-58, VH1-69, VH2-5, VH2-26, VH2-70, VH3-7, VH3-9, VH3-11, VH3-13, VH3-15, VH3-20, VH3-21, VH3-23, VH3-30, VH3-33, VH3-43, VH3-48, VH3-53, VH3-64, VH3-72, VH3-73, VH4-31, VH4-34, VH4-39, VH4-59, VH5-51 и VH6-1.
Согласно различным вариантам осуществления один и тот же единственный генный сегмент VL человека выбран из генного сегмента Vκ1-39 человека и генного сегмента Vκ3-20 человека. Согласно различным вариантам осуществления все легкие цепи иммуноглобулина содержат генный сегмент J легкой цепи (JL) человека, выбранный из генного сегмента Jκ и Jλ. Согласно конкретному варианту осуществления генный сегмент JL человека выбран из генного сегмента Jκ1 и Jκ5 человека. Согласно различным вариантам осуществления мышь не содержит последовательность, выбранную из генного сегмента VL иммуноглобулина мыши, генного сегмента JL иммуноглобулина мыши и их комбинации. Согласно различным вариантам осуществления область VL человека функционально связана с константной областью легкой цепи (CL) иммуноглобулина человека, мыши или крысы. Согласно конкретному варианту осуществления область VL человека функционально связана с областью Cκ мыши. Согласно конкретному варианту осуществления область VL человека функционально связана с областью Cκ крысы.
Согласно различным вариантам осуществления область VL человека экспрессируется из эндогенного локуса легкой цепи иммуноглобулина. Согласно различным вариантам осуществления область VH человека функционально связана с константной областью тяжелой цепи (CH) иммуноглобулина человека, мыши или крысы. Согласно различным вариантам осуществления область (CH) включает в себя последовательность человека, выбранную из CH1, шарнира, CH2, CH3, CH4 и/или их комбинации. Согласно различным вариантам осуществления область VH человека экспрессируется из эндогенного локуса тяжелой цепи иммуноглобулина.
Согласно одному аспекту предусмотрена генетически модифицированная мышь, которая экспрессирует множество тяжелых цепей иммуноглобулина, ассоциированных с единственной легкой цепью. Согласно одному варианту осуществления тяжелая цепь содержит последовательность человека. Согласно различным вариантам осуществления последовательность человека выбрана из вариабельной последовательности, CH1, шарнира, CH2, CH3 и их комбинации. Согласно одному варианту осуществления единственная легкая цепь содержит последовательность человека. Согласно различным вариантам осуществления последовательность человека выбрана из вариабельной последовательности, константной последовательности и их комбинации. Согласно одному варианту осуществления мышь содержит недееспособный эндогенный локус иммуноглобулина и экспрессирует тяжелую цепь и/или легкую цепь из трансгена или внехромосомной эписомы. Согласно одному варианту осуществления мышь содержит замещение на эндогенном локусе мыши некоторых или всех эндогенных генных сегментов тяжелой цепи мыши (т.е. V, D, J), и/или некоторых или всех эндогенных константных последовательностей тяжелой цепи мыши (например, CH1, шарнира, CH2, CH3 или их комбинации), и/или некоторых или всех эндогенных последовательностей легкой цепи мыши (например, константных последовательностей V, J или их комбинации) одной или несколькими последовательностями иммуноглобулина человека.
Согласно одному аспекту предусмотрена мышь, подходящая для получения антител, которые содержат одинаковую легкую цепь, причем все или по существу все антитела, произведенные мышью, экспрессируются с одинаковой легкой цепью. Согласно одному варианту осуществления легкая цепь экспрессируется из эндогенного локуса легкой цепи.
Согласно одному аспекту предусмотрен способ получения легкой цепи из антитела человека, предусматривающий получение от описанной в настоящем документе мыши последовательности легкой цепи и последовательности тяжелой цепи, и использование последовательности легкой цепи и последовательности тяжелой цепи в получении антитела человека. Согласно одному варианту осуществления антитело человека представляет собой биспецифическое антитело.
Согласно одному аспекту предусмотрен способ идентификации вариабельного домена тяжелой цепи человека, который способен связывать представляющий интерес антиген с описанной в настоящем документе сконструированной легкой цепью, причем способ предусматривает получение вариабельного домена тяжелой цепи, происходящего из первого антитела, которое способно связывать антиген, повторное спаривание вариабельного домена тяжелой цепи с зародышевой последовательностью легкой цепи и трансфекцию клетки так, чтобы каждая экспрессировалась для образования второго антитела, воздействие антигена на второе антитело и измерение связывания второго антитела с антигеном.
Согласно одному варианту осуществления легкая цепь первого антитела содержит последовательность Vκ1-39 человека. Согласно одному варианту осуществления легкая цепь первого антитела содержит последовательность Vκ3-20 человека. Согласно одному варианту осуществления зародышевая последовательность легкой цепи содержит последовательность Vκ1-39 или Vκ3-20 человека. Согласно различным вариантам осуществления связывание второго антитела с антигеном определяют путем сравнения связывания первого антитела с антигеном.
Любой из описанных в настоящем документе вариантов осуществления и аспектов может использоваться в сочетании друг с другом, если иное не указано или не очевидно из контекста. Другие варианты осуществления станут очевидными специалистам в настоящей области техники при рассмотрении последующего описания.
Краткое описание графических материалов
На фиг. 1 показана стратегия нацеливания для замещения эндогенных генных сегментов вариабельной области легкой цепи иммуноглобулина мыши генной областью Vκ1-39Jκ5 человека.
На фиг. 2 показана стратегия нацеливания для замещения эндогенных генных сегментов вариабельной области легкой цепи иммуноглобулина мыши генной областью Vκ3-20Jκ1 человека.
На фиг. 3 показана стратегия нацеливания для замещения эндогенных генных сегментов вариабельной области легкой цепи иммуноглобулина мыши генной областью VpreB/Jλ5 человека.
На фиг. 4 показано процентное отношение CD19+ В-клеток (ось y) из периферической крови мышей дикого типа (WT), мышей, гомозиготных в отношении сконструированной реаранжированной области легкой цепи Vκ1-39Jκ5 человека (Vκ1-39Jκ5 НО) и мышей, гомозиготных в отношении сконструированной реаранжированной области легкой цепи Vκ3-20Jκ1 человека (Vκ3-20Jκ1 НО).
На фиг. 5А показана относительная экспрессия мРНК (ось y) происходящей из Vκ1-39 легкой цепи в анализе количественной ПЦР с использованием зондов, специфических для участка соединения сконструированной реаранжированной области легкой цепи Vκ1-39Jκ5 человека (зонд участка соединения Vκ1-39Jκ5) и генного сегмента Vκ1-39 человека (зонд Vκ1-39) у мыши, гомозиготной в отношении замещения эндогенных генных сегментов Vκ и Jκ генными сегментами Vκ и Jκ человека (Нκ), мыши дикого типа (WT) и мыши, гетерозиготной в отношении сконструированной реаранжированной области легкой цепи Vκ1-39Jκ5 человека (Vκ1-39Jκ5 НЕТ). Сигналы нормируют к экспрессии Cκ мыши. N.D.: не обнаружено.
На фиг. 5В показана относительная экспрессия мРНК (ось y) происходящей из Vκ1-39 легкой цепи в анализе количественной ПЦР с использованием зондов, специфических для участка соединения сконструированной реаранжированной области легкой цепи Vκ1-39Jκ5 человека (зонд участка соединения Vκ1-39Jκ5) и генного сегмента Vκ1-39 человека (зонд Vκ1-39) у мыши, гомозиготной в отношении замещения эндогенных генных сегментов Vκ и Jκ генными сегментами Vκ и Jκ человека (Нκ), мыши дикого типа (WT) и мыши, гомозиготной в отношении сконструированной реаранжированной области легкой цепи Vκ1-39Jκ5 человека (Vκ1-39Jκ5 НО). Сигналы нормируют к экспрессии Cκ мыши.
На фиг. 5С показана относительная экспрессия мРНК (ось y) происходящей из Vκ3-20 легкой цепи в анализе количественной ПЦР с использованием зондов, специфических для участка соединения сконструированной реаранжированной области легкой цепи Vκ3-20Jκ1 человека (зонд участка соединения Vκ3-20Jκ1) и генного сегмента Vκ3-20 человека (зонд Vκ3-20) у мыши, гомозиготной в отношении замещения эндогенных генных сегментов Vκ и Jκ генными сегментами Vκ и Jκ человека (Нκ), мыши дикого типа (WT) и мыши, гетерозиготной (НЕТ) и гомозиготной (НО) в отношении сконструированной реаранжированной области легкой цепи Vκ3-20Jκ1 человека. Сигналы нормируют к экспрессии Cκ мыши.
На фиг. 6А показан титр IgM (слева) и IgG (справа) у мышей дикого типа (WT; N=2) и мышей, гомозиготных в отношении сконструированной реаранжированной области легкой цепи Vκ1-39Jκ5 человека (Vκ1-39Jκ5 НО; N=2), иммунизированных β-галактозидазой.
На фиг. 6В показан общий титр иммуноглобулинов (IgM, IgG, IgA) у мышей дикого типа (WT; N=5) и мышей, гомозиготных в отношении сконструированной реаранжированной области легкой цепи Vκ3-20Jκ1 человека (Vκ3-20Jκ1 НО; N=5), иммунизированных β-галактозидазой.
Подробное описание настоящего изобретения
Настоящее изобретение не ограничивается конкретными способами и описанными экспериментальными условиями, поскольку такие способы и условия могут варьировать. Кроме того, следует понимать, что используемая в настоящем документе терминология предназначена только для цели описания конкретных вариантов осуществления и не предусматривается для ограничения, поскольку объем настоящего изобретения определяется его формулой
Если не указано иное, все используемые в настоящем документе термины и фразы, включают в себя те значения, которые подразумеваются под терминами и фразами в настоящей области техники, если иное прямо не указано или явно не следует из контекста, в котором используется термин или фраза. Хотя любые способы и материалы, подобные или эквивалентные описанным в настоящем документе, могут быть использованы на практике или при испытании настоящего изобретения, ниже будут описаны конкретные способы и материалы.
Используемый в настоящем документе термин "антитело" включает в себя молекулы иммуноглобулина, содержащие четыре полипептидных цепи, две тяжелых (H) цепи и две легких (L) цепи, взаимосвязанных дисульфидными связями. Каждая тяжелая цепь содержит вариабельную область (VH) тяжелой цепи и константную область (CH) тяжелой цепи. Константная область тяжелой цепи содержит три домена, CH1, CH2 и CH3. Каждая легкая цепь содержит вариабельную область (VL) легкой цепи и константную область (CL) легкой цепи. Области VH и VL могут быть дополнительно подразделены на области гипервариабельности, имеющие название определяющие комплементарность области (CDR), чередующиеся с областями, которые являются более консервативными, имеющими название каркасные области (FR). Каждая VH и VL содержит три CDR и четыре FR, расположенные от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4 (CDR тяжелой цепи могут иметь сокращение HCDR1, HCDR2 и HCDR3; CDR легкой цепи могут иметь сокращение LCDR1, LCDR2 и LCDR3). Термин антитело с "высокой аффинностью" относится к антителу, которое характеризуется KD по отношению к его целевому эпитопу, составляющей приблизительно 10-9 M или ниже (например, приблизительно 1×10-9 M, 1×10-10 M, 1×10-11 M или приблизительно 1×10-12 M). Согласно одному варианту осуществления KD измеряют с помощью поверхностного плазмонного резонанса, например, BIACORE™; согласно другому варианту осуществления KD измеряют с помощью ELISA.
Фраза "биспецифическое антитело" включает в себя антитело, способное к селективному связыванию двух или более эпитопов. Биспецифические антитела, как правило, содержат две неидентичных тяжелых цепи, причем каждая тяжелая цепь специфически связывает различный эпитоп либо на двух различных молекулах (например, различные эпитопы на двух различных иммуногенах), либо на одной молекуле (например, различные эпитопы на одном иммуногене). Если биспецифическое антитело способно к селективному связыванию двух различных эпитопов (первого эпитопа и второго эпитопа), то аффинность первой тяжелой цепи к первому эпитопу будет, как правило, по меньшей мере на один-два или три или четыре или более порядков величин ниже, чем аффинность первой тяжелой цепи ко второму эпитопу, и наоборот. Эпитопы, специфически связанные биспецифическим антителом, могут присутствовать на одинаковой или отличающейся мишени (например, на одинаковом или отличающемся белке). Биспецифические антитела могут быть получены, например, путем комбинации тяжелых цепей, которые распознают различные эпитопы одного иммуногена. Например, последовательности нуклеиновых кислот, кодирующие вариабельные последовательности тяжелой цепи, которые распознают различные эпитопы одного иммуногена, могут быть слиты с последовательностями нуклеиновых кислот, кодирующими одинаковые или различные константные области тяжелой цепи, и такие последовательности могут быть экспрессированы в клетке, которая экспрессирует легкую цепь иммуноглобулина. Типичное биспецифическое антитело содержит две тяжелых цепи, каждая из которых содержит три CDR тяжелой цепи, за которыми следуют (от N-конца к С-концу) домен CH1, шарнир, домен CH2 и домен CH3, и легкую цепь иммуноглобулина, которая либо не обеспечивает специфичности связывания с эпитопом, но которая может соединяться с каждой тяжелой цепью, либо может соединяться с каждой тяжелой цепью и которая может связывать один или несколько эпитопов, связанных связывающими эпитоп областями тяжелой цепи, или которая может соединяться с каждой тяжелой цепью и обеспечивает связывание или одной или обеих тяжелых цепей с одним или обоими эпитопами.
Термин "клетка" включает в себя любую клетку, которая является подходящей для экспрессии рекомбинантной последовательности нуклеиновой кислоты. Клетки предусматривают клетки прокариот и эукариот (одноклеточных или многоклеточных), бактериальные клетки (например, штаммы Е. coli, Bacillus spp., Streptomyces spp. и т.д.), клетки микобактерий, клетки грибов, дрожжевые клетки (например, S. cerevisiae, S. pombe, P. pastoris, P. methanolica и т.д.), растительные клетки, клетки насекомых (например, SF-9, SF-21, инфицированные бакуловирусом клетки насекомых, Trichoplusia ni и т.д.), клетки не относящихся к человеку животных, клетки человека или такие слияния клеток, как, например, гибридомы или квадромы. Согласно некоторым вариантам осуществления клетка представляет собой клетку человека, мартышки, человекообразной обезьяны, хомяка, крысы или мыши. Согласно некоторым вариантам осуществления клетка является эукариотической и выбрана из следующих клеток: СНО (например, СНО K1, DXB-11 СНО, Veggie -СНО), COS (например, COS-7), ретинальная клетка, Vero, CV1, почечная клетка (например, HEK293, 293 EBNA, MSR 293, MDCK, HaK, BHK), HeLa, HepG2, WI38, MRC 5, Colo205, НВ 8065, HL-60, (например, BHK21), Jurkat, Daudi, А431 (эпидермальная), CV-1, U937, 3T3, клетка L, клетка С127, SP2/0, NS-0, ММТ 060562, клетка Сертоли, клетка BRL 3А, клетка НТ1080, клетка миеломы, опухолевая клетка и клеточная линия, происходящая из вышеупомянутой клетки. Согласно некоторым вариантам осуществления клетка содержит один или несколько вирусных генов, например, ретинальная клетка, которая экспрессирует вирусный ген (например, клетка PER.C6™).
Фраза "определяющая комплементарность область", или термин "CDR", предусматривает аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты генов иммуноглобулина организма, которая в норме (т.е. у животного дикого типа) находится между двумя каркасными областями в вариабельной области легкой или тяжелой цепи молекулы иммуноглобулина (например, антитела или Т-клеточного рецептора). CDR может кодироваться, например, зародышевой последовательностью или реаранжированной или нереаранжированной последовательностью, и, например, наивной или зрелой В-клетки или Т-клетки. CDR может быть соматически мутировала (например, отличаться от последовательности, кодируемой в зародышевой линии животного), гуманизирована и/или модифицирована с помощью аминокислотных замен, вставок или делеций. В некоторых обстоятельствах (например, для CDR3) CDR могут кодироваться двумя или более последовательностями (например, зародышевыми последовательностями), которые не являются смежными (например, в нереаранжированной последовательности нуклеиновой кислоты), но являются смежными в последовательности нуклеиновой кислоты В-клетки, например, в результате сплайсинга или соединения последовательностей (например, V-D-J рекомбинации для формирования CDR3 тяжелой цепи).
Термин "консервативный" при использовании для описания консервативной аминокислотной замены предусматривает замену аминокислотного остатка другим аминокислотным остатком, содержащим R-группу боковой цепи со сходными химическими свойствами (например, заряд или гидрофобность). Как правило, консервативная аминокислотная замена не будет существенно изменять представляющие интерес функциональные свойства белка, например, способность вариабельной области специфически связывать целевой эпитоп с необходимой аффинностью. Примеры групп аминокислот, которые содержат боковые цепи со сходными химическими свойствами, включают в себя такие алифатические боковые цепи, как глицин, аланин, валин, лейцин и изолейцин; такие алифатическо-гидроксильные боковые цепи, как серин и треонин; такие содержащие амид боковые цепи, как аспарагин и глутамин; такие ароматические боковые цепи, как фенилаланин, тирозин и триптофан; такие основные боковые цепи, как лизин, аргинин и гистидин; такие кислотные боковые цепи, как аспарагиновая кислота и глутаминовая кислота; и, такие серосодержащие боковые цепи, как цистеин и метионин. Группы консервативных замен аминокислот включают в себя, например, валин/лейцин/изолейцин, фенилаланин/тирозин, лизин/аргинин, аланин/валин, глутамат/аспартат и аспарагин/глутамин. Согласно некоторым вариантам осуществления консервативная аминокислотная замена может представлять собой замену любого нативного остатка в белке аланином, как используется, например, сканирующем аланином мутагенезе. Согласно некоторым вариантам осуществления производится консервативная замена, которая характеризуется положительным значением в матрице логарифмического правдоподобия РАМ250, раскрытой в Gonnet et al. (1992) Exhaustive Matching of the Entire Protein Sequence Database, Science 256:1443-45. Согласно некоторым вариантам осуществления замена представляет собой умеренно консервативную замену, причем замена характеризуется неотрицательным значением в матрице логарифмического правдоподобия РАМ250.
Согласно некоторым вариантам осуществления положения остатков в легкой цепи или тяжелой цепи иммуноглобулина отличаются на одну или несколько консервативных аминокислотных замен. Согласно некоторым вариантам осуществления положения остатков в легкой цепи иммуноглобулина или ее функциональном фрагменте (например, фрагменте, который обеспечивает экспрессию и секрецию, например, из В-клетки) не идентичны легкой цепи, чья аминокислотная последовательность представлена в настоящем документе, но отличается на одну или несколько консервативных аминокислотных замен.
Фраза "эпитопсвязывающий белок" предусматривает белок, характеризующийся по меньшей мере одной CDR и который способен к селективному распознаванию эпитопа, например, способен к связыванию эпитопа с KD, составляющей приблизительно один микромоль или ниже (например, KD, составляющей приблизительно 1×10-6 M, 1×10-7 Μ, 1×10-9 Μ, 1×10-9 Μ, 1×10-10 Μ, 1×10-11 Μ или приблизительно 1×10-12 Μ). Для терапевтических эпитопсвязывающих белков (например, терапевтических антител) зачастую необходима KD, которая находится в наномолярном или пикомолярном диапазоне.
Фраза "функциональный фрагмент" предусматривает фрагменты эпитопсвязывающих белков, которые могут экспрессироваться, секретироваться и специфически связывать эпитоп с KD в микромолярном, наномолярном или пикомолярном диапазоне. Специфическое распознавание предусматривает наличие KD, которая находится по меньшей мере в микромолярном диапазоне, наномолярном диапазоне или пикомолярном диапазоне.
Термин "зародышевый" включает в себя ссылку на последовательность нуклеиновой кислоты иммуноглобулина в несоматически мутированной клетке, например, несоматически мутированной В-клетке или пре-В-клетке или гематопоэтической клетке.
Фраза "тяжелая цепь" или "тяжелая цепь иммуноглобулина" предусматривает последовательность константной области тяжелой цепи иммуноглобулина из любого организма. Вариабельные домены тяжелой цепи включают в себя три CDR тяжелой цепи и четыре FR области, если не указано иное. Фрагменты тяжелых цепей включают в себя CDR, CDR и FR и их комбинации. Типичная тяжелая цепь содержит после вариабельного домена (в направлении от N-конца к С-концу) домен CH1, шарнир, домен CH2 и домен CH3. Функциональный фрагмент тяжелой цепи содержит фрагмент, который способен к специфическому распознаванию эпитопа (например, распознаванию эпитопа с KD в микромолярном, наномолярном или пикомолярном диапазоне), который способен к экспрессии и секреции из клетки и который содержит по меньшей мере одну CDR.
Термин "идентичность" при использовании в отношении последовательности предусматривает идентичность, которая определяется с помощью ряда различных известных в настоящей области техники алгоритмов, которые могут использоваться для измерения идентичности нуклеотидных и/или аминокислотных последовательностей. Согласно некоторым вариантам осуществления описанные в настоящем документе идентичности определяют с использованием ClustalW версии 1.83 (медленного) выравнивания, использующего штраф за открытие гэпа, составляющий 10.0, штраф за продление гэпа, составляющий 0,1 и с использованием матрицы подобия Gonnet (MACVECTOR™ 10.0.2, MacVector Inc., 2008). Длина последовательностей, сравниваемых относительно идентичности последовательностей, будет зависеть от конкретных последовательностей, но в случае константного домена легкой цепи длина должна содержать последовательность достаточной длины для укладки в константный домен легкой цепи, который способен к самоассоциации для формирования канонического константного домена легкой цепи, например, способного к формированию двух складчатых бета-слоев, содержащих бета-тяжи и способного взаимодействовать по меньшей мере с одним доменом CH1 человека или мыши. В случае домена CH1 длина последовательности должна содержать последовательность достаточной длины для укладки в домен CH1, который способен к формированию двух складчатых бета-слоев, содержащих бета-тяжи и способен взаимодействовать по меньшей мере с одним константным доменом легкой цепи мыши или человека.
Фраза "молекула иммуноглобулина" включает в себя две тяжелых цепи иммуноглобулина и две легких цепи иммуноглобулина. Тяжелые цепи могут быть идентичными или различными, и легкие цепи могут быть идентичными или различными.
Фраза "легкая цепь" предусматривает последовательность легкой цепи иммуноглобулина из любого организма и если не указано иное, предусматривает κ и λ легкие цепи человека и VpreB, а также суррогатные легкие цепи. Вариабельные домены (VL) легкой цепи, как правило, содержат три CDR легкой цепи и четыре каркасные (FR) области, если не указано иное. Как правило, полноразмерная легкая цепь включает в себя от амино-конца к карбокси-концу домен VL, который содержит FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4, и константный домен легкой цепи. Легкие цепи включают в себя такие, например, которые не связывают селективно или первый, или второй эпитоп, селективно связанный связывающим эпитоп белком, в котором они имеют место. Легкие цепи также включают в себя те, которые связывают и распознают или содействуют тяжелой цепи в связывании и распознавании одного или нескольких эпитопов, селективно связанных связывающим эпитоп белком, в котором они имеют место. Общие легкие цепи представляют собой легкие цепи, происходящие из реаранжированной последовательности Vκ1-39Jκ5 человека или реаранжированной последовательности Vκ3-20Jκ1 человека, и включают в себя соматически мутированные версии (например, с созревшей аффинностью).
Фраза "микромолярный диапазон" подразумевает 1-999 микромоль; фраза "наномолярный диапазон" подразумевает 1-999 наномоль; фраза "пикомолярный диапазон" подразумевает 1-999 пикомоль.
Фраза "соматически мутированный" предусматривает ссылку на последовательность нуклеиновой кислоты из В-клетки, которая подверглась переключению класса, причем последовательность нуклеиновой кислоты вариабельной области иммуноглобулина (например, вариабельного домена тяжелой цепи или включающей в себя последовательность CDR или FR тяжелой цепи) в подвергшейся переключению класса В-клетке не идентична последовательности нуклеиновой кислоты в В-клетке перед переключением класса, так что, например, различие в последовательности нуклеиновой кислоты CDR или каркасной области между В-клеткой, которая не подверглась переключению класса и В-клеткой, которая подверглась переключению класса. "Соматически мутированный" предусматривает ссылку на последовательности нуклеиновых кислот из В-клеток с созревшей аффинностью, которые не являются идентичными соответствующим последовательностям вариабельной области иммуноглобулина в В-клетках с несозревшей аффинностью (т.е. последовательности в геноме гаметоцитов). Фраза "соматически мутированный" также предусматривает ссылку на последовательность нуклеиновой кислоты вариабельной области иммуноглобулина из В-клетки после воздействия на В-клетку представляющего интерес эпитопа, причем последовательность нуклеиновой кислоты отличается от соответствующей последовательности нуклеиновой кислоты перед воздействием на В-клетку представляющего интерес эпитопа. Фраза "соматически мутированный" относится к последовательностям из антител, которые были образованы у животного, например, мыши, содержащей последовательности нуклеиновых кислот вариабельной области иммуноглобулина человека, в ответ на иммунизцию иммуногеном, и которые являются результатом процессов отбора, наследственно функционирующих у такого животного.
Термин "нереаранжированный" в отношении последовательности нуклеиновой кислоты включает в себя последовательности нуклеиновых кислот, которые существуют в зародышевой линии клетки животного.
Фраза "вариабельный домен" предусматривает аминокислотную последовательность легкой или тяжелой цепи иммуноглобулина (модифицированной при необходимости), которая содержит следующие аминокислотные области, последовательно от N-конца к С-концу (если не указано иное): FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4.
Общая легкая цепь
Предыдущие попытки получить применимые мультиспецифические эпитопсвязывающие белки, например, биспецифические антитела, были затруднены различными проблемами, которые зачастую характеризовались общей парадигмой: in vitro селекция или манипуляция с последовательностями для рационального конструирования или для конструирования методом проб и ошибок подходящего формата для образования пары гетеродимерного биспецифического иммуноглобулина человека. К сожалению, большинство, если не все из подходов по конструированию in vitro обеспечивают преимущественно узкоспециализированные решения проблем, которые являются подходящими, если вообще ими являются, для отдельных молекул. С другой стороны, способы in vivo для использования сложных организмов для отбора соответствующих пар, которые способны привести к получению терапевтических средств для людей, не были реализованы.
Как правило, нативные последовательности мыши зачастую не представляют собой хороший источник терапевтических последовательностей человека. По меньшей мере по этой причине, создание вариабельных областей тяжелой цепи иммуноглобулина мыши, которые образуют пары с общей легкой цепью человека, находит ограниченную практическую применимость. Большее число попыток конструирования in vitro будет затрачено в методе проб и ошибок в попытке гуманизировать вариабельные последовательности тяжелой цепи мыши, при этом надеясь сохранить специфичность и аффинность эпитопа при поддержании способности соединяться с общей легкой цепью человека с неопределенным результатом. В конце такого процесса конечный продукт может сохранять некоторую специфичность и аффинность, и связываться с общей легкой цепью, но в конечном счете иммуногенность у человека, вероятно, будет сохранять значительную степень опасности.
Следовательно, подходящая мышь для получения терапевтических средств для людей будет содержать соответствующе большой репертуар генных сегментов вариабельной области тяжелой цепи человека вместо эндогенных генных сегментов вариабельной области тяжелой цепи мыши. Генные сегменты вариабельной области тяжелой цепи человека должны быть способны реаранжироваться и рекомбинироваться с эндогенным константным доменом тяжелой цепи мыши для образования обратной химерной тяжелой цепи (т.е. тяжелой цепи, содержащей вариабельный домен человека и константную область мыши). Тяжелая цепь должна быть способна к переключению класса и соматической гипермутации так, чтобы соответствующе большой репертуар вариабельных доменов тяжелой цепи был доступен мыши для выбора одного, который может связываться с ограниченным репертуаром вариабельные области легкой цепи человека.
Мышь, которая выбирает общую легкую цепь для множества тяжелых цепей, характеризуется практической применимостью. Согласно различным вариантам осуществления антитела, которые экспрессируются у мыши, которая может экспрессировать только общую легкую цепь, будут содержать тяжелые цепи, которые могут связываться и экспрессироваться с идентичной или по существу идентичной легкой цепью. Это особенно применимо в получении биспецифических антител. Например, такая мышь может быть иммунизирована первым иммуногеном для образования В-клетки, которая экспрессирует антитело, которое специфически связывает первый эпитоп. Мышь (или мышь, генетически аналогичная) может быть иммунизирована вторым иммуногеном для образования В-клетки, которая экспрессирует антитело, которое специфически связывает второй эпитоп. Вариабельные тяжелые области могут быть клонированы из В-клеток и экспрессированы с одинаковой константной областью тяжелой цепи и одинаковой легкой цепью и экспрессированы в клетке для образования биспецифического антитела, причем компонент легкой цепи биспецифического антитела был выбран мышью для связывания и экспрессии с компонентом легкой цепи.
Авторы настоящего изобретения сконструировали мышь для образования легких цепей иммуноглобулина, которые будут подходящим образом создавать пары с достаточно разнообразным семейством тяжелых цепей, включающим в себя тяжелые цепи, чьи вариабельные области отклоняются от зародышевых последовательностей, например, вариабельные области с созревшей аффинностью или соматически мутированные вариабельные области. Согласно различным вариантам осуществления мышь разработана для спаривания вариабельных доменов легкой цепи человека с вариабельными доменами тяжелой цепи человека, которые содержат соматические мутации, таким образом, обеспечивая путь к получению связывающих белков с высокой аффинностью, подходящих для применения в качестве терапевтических средств для людей.
Генетически сконструированная мышь посредством длительного и сложного процесса селекции антитела в пределах организма образует биологически приемлемые выборы в спаривании разнообразного набора вариабельных доменов тяжелой цепи человека с ограниченным числом вариантов легкой цепи человека. Для достижения этого мышь конструируют так, чтобы она представляла ограниченное число вариантов вариабельных доменов легкой цепи человека в сочетании с широким разнообразием вариантов вариабельных доменов тяжелой цепи человека. При сенсибилизации иммуногеном мышь максимально увеличивает число решений в ее репертуаре для развития антитела к иммуногену, ограниченное преимущественно или исключительно числом или вариантами легких цепей в ее репертуаре. Согласно различным вариантам осуществления это включает в себя предоставление мыши возможности достичь подходящих и совместимых соматических мутаций вариабельного домена легкой цепи, которые будут при этом совместимыми с относительно широким разнообразием вариабельных доменов тяжелой цепи человека, предусматривая, в частности, соматически мутированные вариабельные домены тяжелой цепи человека.
Для достижения ограниченного репертуара вариантов легкой цепи мышь конструируют для приведение в нефункциональное или по существу нефункциональное состояние ее способности образовывать, или реаранжировать, нативный вариабельный домен легкой цепи мыши. Этого можно достичь, например, путем удаления генных сегментов вариабельной области легкой цепи мыши. Эндогенный локус мыши может затем быть модифицирован с помощью экзогенного подходящего предпочтительного генного сегмента вариабельной области легкой цепи человека, функционально связанного с эндогенным константным доменом легкой цепи мыши, таким способом, чтобы экзогенные генные сегменты вариабельной области человека могли комбинироваться с эндогенным геном константной области легкой цепи мыши и формировать реаранжированный ген обратной химерной легкой цепи (вариабельная область человека, константная область мыши). Согласно различным вариантам осуществления вариабельная область легкой цепи способна быть соматически мутированной. Согласно различным вариантам осуществления для максимизации способности вариабельной области легкой цепи к достижению соматических мутаций у мыши сохраняют подходящий(е) энхансер(ы). Например, в модификации локуса κ легкой цепи мыши для замещения эндогенных генных сегментов κ легкой цепи мыши генными сегментами κ легкой цепи человека к интронный энхансер мыши и к 3' энхансер мыши являются функционально сохраненными или не разрушенными.
Предусматривается генетически сконструированная мышь, которая экспрессирует ограниченный репертуар обратных химерных (вариабельная область человека, константная область мыши) легких цепей, связанных с разнообразием обратных химерных (вариабельная область человека, константная область мыши) тяжелых цепей. Согласно различным вариантам осуществления эндогенные генные сегменты κ легкой цепи мыши подвергают делеции и замещают одной (или двумя) реаранжированной(ыми) областью(ями) легкой цепи человека, функционально связанной(ыми) с эндогенным геном Cκ мыши. Согласно вариантам осуществления для максимизации соматической гипермутации реаранжированной области легкой цепи человека сохраняют к интронный энхансер мыши и κ 3' энхансер мыши. Согласно различным вариантам осуществления мышь также содержит нефункциональный локус λ легкой цепи или его делецию или делецию, которая делает локус неспособным образовывать λ легкую цепь.
Предусматривается генетически сконструированная мышь, которая согласно различным вариантам осуществления содержит локус вариабельной области легкой цепи, не содержащий эндогенные генные сегменты VL и JL легкой цепи мыши и содержащий реаранжированную вариабельную область легкой цепи человека, согласно одному варианту осуществления реаранжированную последовательность VL/JL человека, функционально связанную с константной областью мыши, причем локус способен подвергаться соматической гипермутации и при этом локус экспрессирует легкую цепь, содержащую последовательность VL/JL человека, связанную с константной областью мыши. Таким образом, согласно различным вариантам осуществления локус содержит κ 3' энхансер мыши, который связан с нормальным уровнем соматической гипермутации, или уровнем дикого типа.
Генетически сконструированная мышь согласно различным вариантам осуществления, будучи иммунизированной представляющим интерес антигеном, образует В-клетки, которые проявляют разнообразие реаранжировок вариабельных областей тяжелой цепи иммуноглобулина человека, которые экспрессируют и функционируют с одной или двумя реаранжированными легкими цепями, предусматривая варианты осуществления, где одна или две легкие цепи содержат вариабельные области легкой цепи человека, которые содержат, например, 1-5 соматических мутаций. Согласно различным вариантам осуществления легкие цепи человека, экспрессированные таким образом, способны к ассоциации и экспрессии с любой вариабельной областью тяжелой цепи иммуноглобулина человека, экспрессированной у мыши.
Связывающие эпитоп белки, связывающие более чем один эпитоп
Описанные в настоящем документе композиции и способы могут использоваться для образования связывающих белков, которые связывают более чем один эпитоп с высокой аффинностью, например, биспецифических антител. Преимущества настоящего изобретения предусматривают способность выбирать подходящим образом высокую степень связывания тяжелых цепей иммуноглобулина (например, с созревшей аффинностью), каждая из которых будет ассоциироваться с одной легкой цепью.
Синтез и экспрессия биспецифических связывающих белков были проблематичными, частично вследствие ограничений, связанных с установлением подходящей легкой цепи, которая может связываться и экспрессироваться с двумя различными тяжелыми цепями, и частично вследствие проблем с выделением. Описанные в настоящем документе способы и композиции предусматривают генетически модифицированную мышь для выбора, посредством в остальном нормальных процессов, подходящей легкой цепи, которая может связываться и экспрессироваться более чем с одной тяжелой цепью, включая в себя тяжелые цепи, являющиеся соматически мутированными (например, с созревшей аффинностью). Последовательности VL и VH человека из подходящих В-клеток описанных в настоящем документе иммунизированных мышей, которые экспрессируют антитела с созревшей аффинностью, содержащие обратные химерные тяжелые цепи (т.е. вариабельная область человека и константная область мыши), могут быть установлены и клонированы в вектор экспрессии с подходящей генной последовательностью константной области человека (например, IgG1 человека). Могут быть получены два таких конструкта, причем каждый конструкт кодирует вариабельный домен тяжелой цепи человека, который связывает различный эпитоп. Одна из VL человека (например, Vκ1-39Jκ5 человека или Vκ3-20Jκ1 человека), в зародышевой последовательности или из В-клетки, причем последовательность была соматически мутирована, может быть слита внутри рамки считывания с подходящим геном константной области человека (например, геном κ константной области человека). Эти три полностью человеческих тяжелых и легких конструкта могут быть помещены в подходящую клетку для экспрессии. Клетка будет экспрессировать два основных вида: гомодимерную тяжелую цепь с идентичной легкой цепью и гетеродимерную тяжелую цепь с идентичной легкой цепью. Для обеспечения легкого разделения этих основных видов одну из тяжелых цепей модифицируют для исключения связывающей белок А детерминанты, получая в результате дифференциальную аффинность гомодимерного связывающего белка от гетеродимерного связывающего белка. Адресованные этому вопросу композиции и способы описаны в USSN 12/832838, поданной 25 июня 2010, имеющей название "Readily Isolated Bispecific Antibodies with Native Immunoglobulin Format", опубликованной в виде US 2010/0331527 A1.
Согласно одному аспекту предусматривается описанный в настоящем документе эпитопсвязывающий белок, причем последовательности VL и VH человека происходят от описанных в настоящем документе мышей, которые были иммунизированы содержащим представляющий интерес эпитоп антигеном.
Согласно одному варианту осуществления предусматривается эпитопсвязывающий белок, который содержит первый и второй полипептид, причем первый полипептид содержит, с N-конца к С-концу, связывающую первый эпитоп область, которая селективно связывает первый эпитоп, за которой следует константная область, которая содержит первую область CH3 IgG человека, выбранного из IgG1, IgG2, IgG4 и их комбинации; и второй полипептид содержит, с N-конца к С-концу, связывающую второй эпитоп область, которая селективно связывает второй эпитоп, за которой следует константная область, которая содержит вторую область CH3 человека IgG, выбранного из IgG1, IgG2, IgG4 и их комбинации, причем вторая область CH3 содержит модификацию, которая снижает или устраняет связывание второго домена CH3 с белком А.
Согласно одному варианту осуществления вторая область CH3 содержит модификацию H95R (по нумерации экзонов IMGT; H435R по нумерации EU). Согласно другому варианту осуществления вторая область CH3 дополнительно содержит модификацию Y96F (IMGT; Y436F по EU).
Согласно одному варианту осуществления вторая область CH3 происходит из модифицированного IgG1 человека и дополнительно содержит модификацию, выбранную из группы, состоящей из D16E, L18M, N44S, K52N, V57M и V82I (IMGT; D356E, L358M, N384S, K392N, V397M и V422I по EU).
Согласно одному варианту осуществления вторая область CH3 происходит из модифицированного IgG2 человека и дополнительно содержит модификацию, выбранную из группы, состоящей из N44S, K52N и V82I (IMGT; N384S, K392N и V422I по EU).
Согласно одному варианту осуществления вторая область CH3 происходит из модифицированного IgG4 человека и дополнительно содержит модификацию, выбранную из группы, состоящей из Q15R, N44S, K52N, V57M, R69K, E79Q и V82I (IMGT; Q355R, N384S, K392N, V397M, R409K, E419Q и V422I по EU).
Одним способом получения эпитопсвязывающего белка, который связывает более чем один эпитоп, является иммунизация первой мыши согласно настоящему изобретению антигеном, который содержит первый представляющий интерес эпитоп, причем мышь содержит эндогенный локус вариабельной области легкой цепи иммуноглобулина, который не содержит эндогенную VL мыши, которая способна к реаранжировке и формированию легкой цепи, причем на эндогенном локусе вариабельной области легкой цепи иммуноглобулина мыши находится единственная реаранжированная область VL человека, функционально связанная с эндогенным геном константной области легкой цепи мыши, и реаранжированная область VL человека выбрана из Vκ1-39Jκ5 человека и Vκ3-20Jκ1 человека, и эндогенные генные сегменты VH мыши были замещены полностью или частично генными сегментами VH человека, так чтобы тяжелые цепи иммуноглобулина, образованные мышью, представляли собой исключительно или по существу тяжелые цепи, которые содержат вариабельные домены человека и константные домены мыши. Будучи иммунизированной, такая мышь будет образовывать обратное химерное антитело, содержащее только один из двух вариабельных доменов легкой цепи человека (например, один из Vκ1-39Jκ5 человека или Vκ3-20Jκ1 человека). Как только определяют В-клетку, которая кодирует VH, который связывает представляющий интерес эпитоп, нуклеотидная последовательность VH (и необязательно VL) может быть извлечена (например, с помощью ПЦР) и клонирована в конструкт экспрессии внутри рамки считывания с подходящим константным доменом иммуноглобулина человека. Этот процесс может быть повторен для определения второго домена VH, который связывает второй эпитоп, и вторая генная последовательность VH может быть извлечена и клонирована в вектор экспрессии внутри рамки считывания со вторым подходящим константным доменом иммуноглобулина. Первый и второй константные домены иммуноглобулина могут принадлежать одному или различному изотипу, и один из константных доменов иммуноглобулина (но не оставшийся) может быть модифицирован, как описано в настоящем документе или в заявке на патент США №2010/0331527 А1, и эпитопсвязывающий белок может быть экспрессирован в подходящей клетке и выделен на основании его дифференциальной аффинности к белку А по сравнению с гомодимерным связывающим эпитоп белком, например, как описано в заявке на патент США №2010/0331527 А1.
Согласно одному варианту осуществления предусматривается способ получения биспецифического эпитопсвязывающего белка, предусматривающий определение первой нуклеотидной последовательности VH (VH1) с созревшей аффинностью (например, содержащей одну или несколько соматических гипермутаций) человека из описанной в настоящем документе мыши, определение второй нуклеотидной последовательности VH (VH2) с созревшей аффинностью (например, содержащей одну или несколько соматических гипермутаций) человека от описанной в настоящем документе мыши, клонирование VH1 внутри рамки считывания с тяжелой цепью человека, не содержащей модификацию детерминанты белка А, как описано в заявке на патент США №US 2010/0331527 А1, для образования тяжелой цепи 1 (НС1), клонирование VH2 внутри рамки считывания с тяжелой цепью человека, содержащей детерминанту белка А, как описано в заявке на патент США №2010/0331527 А1, для образования тяжелой цепи 2 (НС2), введение вектора экспрессии, содержащего НС1, и аналогичного или другого вектора экспрессии, содержащего НС2, в клетку, причем клетка также экспрессирует легкую цепь иммуноглобулина человека, которая содержит Vκ1-39 человека/Jκ5 человека или Vκ3-20 человека/Jκ1 человека, слитую с константным доменом легкой цепи человека, позволяя клетке экспрессировать биспецифический эпитопсвязывающий белок, содержащий домен VH, кодируемый VH1, и домен VH, кодируемый VH2, и выделение биспецифического эпитопсвязывающего белка на основании его дифференциальной способности связывать белок А по сравнению с моноспецифическим гомодимерным эпитопсвязывающим белком. Согласно конкретному варианту осуществления НС1 представляет собой IgG1 и НС2 представляет собой IgG1, который содержит модификацию H95R (IMGT; H435R по EU) и дополнительно содержит модификацию Y96F (IMGT; Y436F по EU). Согласно одному варианту осуществления домен VH, кодируемый VH1, домен VH, кодируемый VH2, или оба, являются соматически мутированными.
Гены VH человека, которые экспрессируются с общей VL человека
Разнообразные вариабельные области человека из антител с созревшей аффинностью, возникших к четырем различным антигенам, экспрессировали либо с их когнатной легкой цепью, либо по меньшей мере с одной из легких цепей человека, выбранной из Vκ1-39Jκ5 человека, Vκ3-20Jκ1 человека или VpreBJλ5 человека (смотрите пример 10). Для антител к каждому из антигенов соматически мутированные тяжелые цепи высокой аффинности из различных семейств генов успешно образовывали пары с реаранжированными зародышевыми областями Vκ1-39Jκ5 и Vκ3-20Jκ1 человека и секретировались из клеток, экспрессирующих тяжелые и легкие цепи. Для Vκ1-39Jκ5 и Vκ3-20Jκ1 благоприятно экспрессировались домены VH, происходящие из следующих семейств генов VH человека: 1-2, 1-8, 1-24, 2-5, 3-7, 3-9, 3-11, 3-13, 3-15, 3-20, 3-23, 3-30, 3-33, 3-48, 4-31, 4-39, 4-59, 5-51 и 6-1. Таким образом, мышь, которую конструировали для экспрессии ограниченного репертуара доменов VL человека из одной или обеих Vκ1-39Jκ5 и Vκ3-20Jκ1, будет образовывать разнообразную популяцию соматически мутированных доменов VH человека из локуса VH, модифицированного для замещения генных сегментов VH мыши генными сегментами VH человека.
Мыши, генетически сконструированные для экспрессии обратных химерных (вариабельная область человека, константная область мыши) тяжелых цепей иммуноглобулина, ассоциированных с единственной реаранжированной легкой цепью (например, Vκ1-39/J или Vκ3-20/J), будучи иммунизированной представляющим интерес антигеном, образовывали В-клетки, которые содержали разнообразие реаранжировок VH человека и экспрессировали разнообразие высокоаффинных антиген-специфических антител с разнообразными свойствами относительно их способности блокировать связывание антигена с его лигандом и относительно их способности связывать варианты антигена (смотрите примеры 5-10).
Таким образом, описанные в настоящем документе мыши и способы применимы в получении и селекции вариабельных доменов тяжелой цепи иммуноглобулина человека, включающих в себя соматически мутированные вариабельные домены тяжелой цепи человека, которые являются результатом разнообразия реаранжировок, которые проявляют широкий спектр аффинностей (предусматривая проявление KD, являющейся приблизительно наномолярной или меньше), широкий спектр специфичностей (предусматривая связывание с различными эпитопами одного антигена) и которые связываются и экспрессируются с одинаковой или по существу одинаковой вариабельной областью легкой цепи иммуноглобулина человека.
Следующие примеры представлены для того, чтобы описать специалистам в настоящей области техники получение и применение способов и композиций настоящего изобретения и не предусмотрены для ограничения объема того, что авторы настоящего изобретения рассматривают как свое изобретение. Были предприняты попытки соблюдения точности относительно используемых чисел (например, количеств, температуры и т.д.), но необходимо учитывать некоторые экспериментальные погрешности и отклонения. Если не указано иное, части представляют собой части по массе, молекулярная масса представляет собой среднюю молекулярную массу, температура указана в градусах Цельсия, и давление равно или близко атмосферному.
ПРИМЕРЫ
Следующие примеры представлены для того, чтобы описать специалистам в настоящей области техники получение и применение способов и композиций настоящего изобретения и не предусмотрены для ограничения объема того, что авторы настоящего изобретения рассматривают как собственное изобретение. Были предприняты попытки соблюдения точности относительно используемых чисел (например, количеств, температуры и т.д.), но необходимо учитывать некоторые экспериментальные погрешности и отклонения. Если не указано иное, температура указана в градусах Цельсия, давление равно или близко атмосферному, части представляют собой части по массе, и молекулярная масса представляет собой среднюю молекулярную массу.
Пример 1
Идентификация вариабельных областей тяжелой цепи человеке, которые ассоциируются с выбранными вариабельными областями легкой цепи человека
Конструировали систему экспрессии in vitro для определения того, может ли единственная реаранжированная зародышевая легкая цепь человека совместно экспрессироваться с тяжелыми цепями человека из антиген-специфических антител человека.
Известны способы получения антител человека у генетически модифицированных мышей (смотрите, например, патент США №6596541, Regeneran Pharmaceuticals, VELOCIMMUNE®). Технология VELOCIMMUNE® предусматривает создание генетически модифицированной мыши, характеризующейся геномом, содержащим вариабельные области тяжелой и легкой цепи человека, функционально связанные с эндогенными локусами константной области мыши так, чтобы мышь производила антитело, содержащее вариабельную область человека и константную область мыши в ответ на стимуляцию антигеном. ДНК, кодирующие вариабельные области тяжелых и легких цепей антител, производимых мышью VELOCIMMUNE®, являются полностью человеческими. Вначале выделяют химерные антитела с высокой аффинностью, характеризующиеся вариабельной областью человека и константной областью мыши. Как описано ниже, определяют характеристики антител и отбирают по необходимым характеристикам, включающим в себя аффинность, селективность, эпитоп и т.д. Константные области мыши замещают необходимой константной областью человека для получения полностью человеческого антитела, содержащего не относящийся к IgM изотип, например, IgG1, IgG2, IgG3 или IgG4 дикого типа или модифицированный IgG1, IgG2, IgG3 или IgG4. Наряду с тем что выбранная константная область может варьировать в соответствии с конкретным применением, характеристики высокоаффинного связывания с антигеном и целевой специфичности находятся в вариабельной области.
Мышь VELOCIMMUNE® иммунизировали с помощью фактора роста, который стимулирует ангиогенез (антиген С), и антиген-специфические человеческие антитела выделяли и секвенировали в отношении частоты использования генов V с использованием стандартных техник, принятых в настоящей области техники. Выбранные антитела клонировали в отношении константных областей тяжелой и легкой цепи человека и 69 тяжелых цепей выбирали для спаривания с одной из следующих трех легких цепей человека: (1) когнатная к легкая цепь, связанная с к константной областью человека, (2) реаранжированная зародышевая Vκ1-39Jκ5 человека, связанная с к константной областью человека, или (3) реаранжированная зародышевая Vκ3-20Jκ1 человека, связанная с κ константной областью человека. Каждую пару тяжелой цепи с легкой цепью котрансфектировали в клетки CHO-K1 с использованием стандартных техник. Присутствие антитела в супернатанте обнаруживали с помощью антитела к IgG человека в анализе ELISA. Титр антител (нг/мл) определяли для каждой пары тяжелой цепи с легкой цепью и титры с различными реаранжированными зародышевыми легкими цепями сравнивали с титрами, полученными с молекулой родительского антитела (т.е. тяжелая цепь, спаренная с когнатной легкой цепью) и рассчитывали процент нативного титра (таблица 1). VH: вариабельный ген тяжелой цепи. ND: при текущих экспериментальных условиях экспрессия не обнаружена.
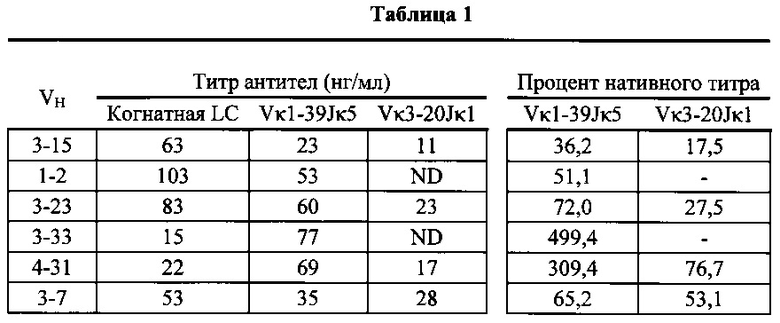
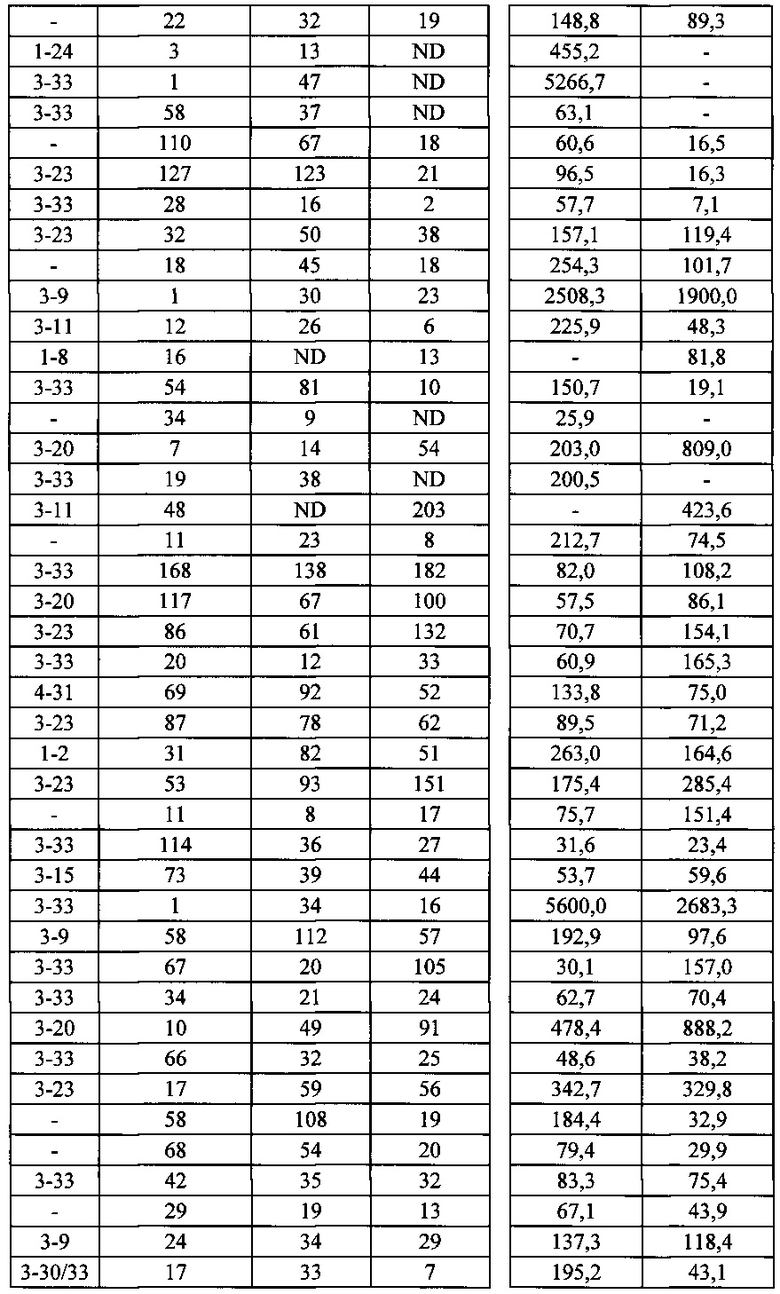
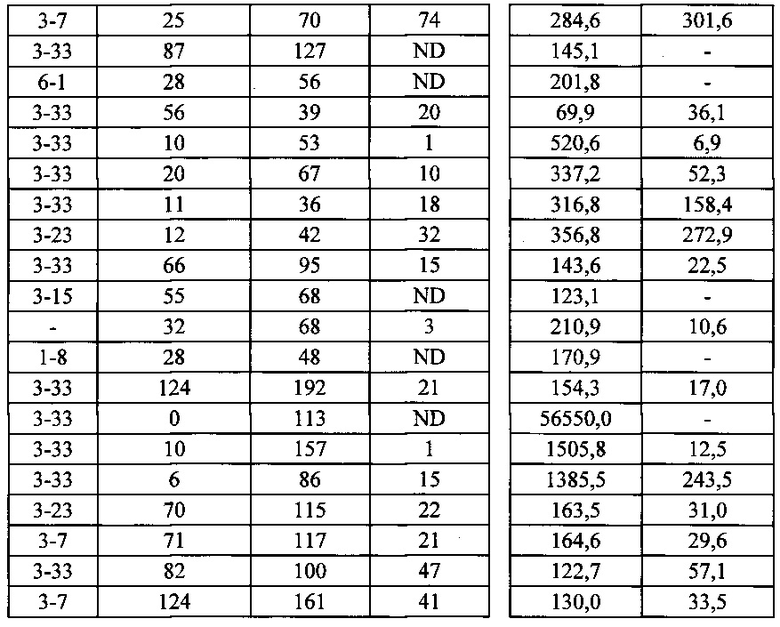
В аналогичном эксперименте мышей VELOCIMMUNE® иммунизировали с помощью нескольких различных антигенов и выбранные тяжелые цепи антиген-специфических антител человека исследовали в отношении их способности спариваться с различными реаранжированными зародышевыми легкими цепями человека (как описано выше). Используемые в настоящем эксперименте антигены включали в себя фермент, вовлеченный в гомеостаз холестерола (антиген А), гормон сыворотки, вовлеченный в регуляцию гомеостаза глюкозы (антиген В), фактор роста, который стимулирует ангиогенез (антиген С), и рецептор клеточной поверхности (антиген D). Антиген-специфические антитела выделяли из мышей каждой группы иммунизации и вариабельные области тяжелой цепи и легкой цепи клонировали и секвенировали. Из последовательности тяжелых и легких цепей определяли частоту использования генов V и выбранные тяжелые цепи спаривали или с их когнатной легкой цепью, или с реаранжированной зародышевой областью Vκ1-39Jκ5 человека. Каждую пару тяжелой цепи с легкой цепью котрансфектировали в клетки CHO-K1 и присутствие антитела в супернатанте обнаруживали с помощью антитела к IgG человека в анализе ELISA. Титр антител (мкг/мл) определяли для каждого спаривания тяжелой цепи с легкой цепью и титры с различными реаранжированными зародышевыми легкими цепями человека сравнивали с титрами, полученными с молекулой родительского антитела (т.е. тяжелая цепь, спаренная с когнатной легкой цепью) и рассчитывали процент нативного титра (таблица 2). VH: вариабельный ген тяжелой цепи. Vκ: вариабельный ген κ легкой цепи. ND: при текущих экспериментальных условиях экспрессия не обнаружена.
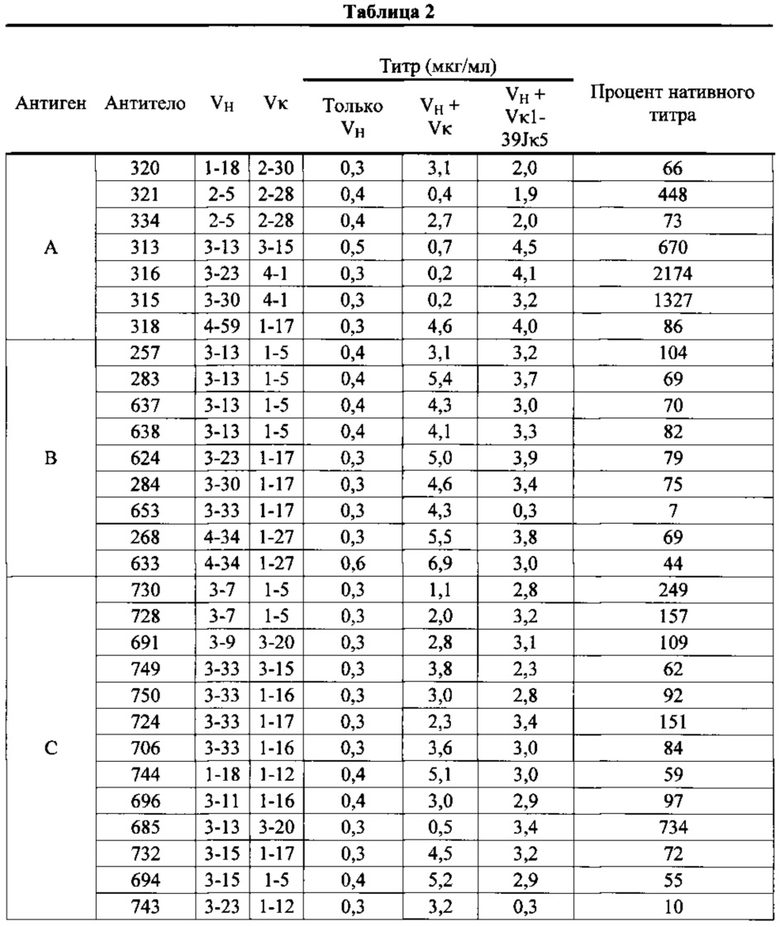
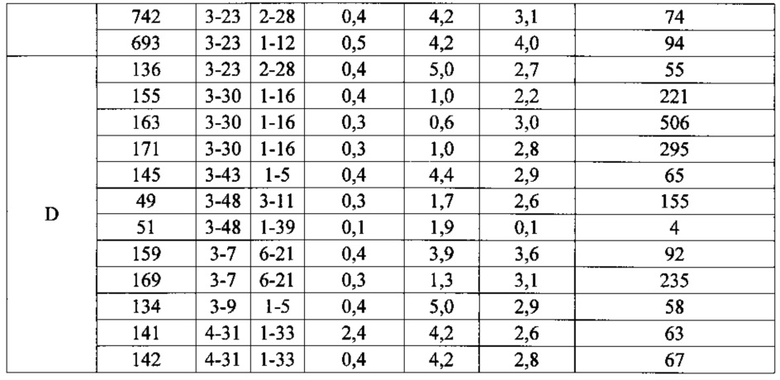
Полученные из настоящих экспериментов результаты показывают, что соматически мутированные, высокоаффинные тяжелые цепи из различных семейств генов способны спариваться с реаранжированными зародышевыми областями Vκ1-39Jκ5 и Vκ3-20Jκ1 человека и секретироваться из клетки в виде нормальной молекулы антитела. Как показано в таблице 1, титр антител увеличивался приблизительно для 61% (42 из 69) тяжелых цепей при спаривании с реаранжированной легкой цепью Vκ1-39Jκ5 человека и приблизительно для 29% (20 из 69) тяжелых цепей при спаривании с реаранжированной легкой цепью Vκ3-20Jκ1 человека по сравнению с когнатной легкой цепью родительского антитела. Приблизительно для 20% (14 из 69) тяжелых цепей обе реаранжированные зародышевые легкие цепи человека обеспечивали увеличение экспрессии по сравнению с когнатной легкой цепью родительского антитела. Как показано в таблице 2, реаранжированная зародышевая область Vκ1-39Jκ5 человека обеспечивала увеличение экспрессии нескольких тяжелых цепей, специфических для диапазона различных классов антигенов по сравнению с когнатной легкой цепью для родительских антител. Титр антител увеличивался более чем в два раза приблизительно для 35% (15/43) тяжелых цепей по сравнению с когнатной легкой цепью родительских антител. Для двух тяжелых цепей (315 и 316) увеличение являлось больше чем десятикратным по сравнению с родительским антителом. Во всех тяжелых цепях, которые показали увеличение экспрессии относительно когнатной легкой цепи родительского антитела, тяжелые цепи третьего семейства (VH3) были представлены в избытке по сравнению с другими генными семействами вариабельной области тяжелой цепи. Это демонстрирует благоприятное взаимодействие тяжелых цепей VH3 человека в спаривании с реаранжированными зародышевыми легкими цепями Vκ1-39Jκ5 и Vκ3-20Jκ1 человека.
Пример 2
Получение реаранжированного зародышевого локуса легкой цепи человека
Различные нацеливающие векторы реаранжированной зародышевой легкой цепи человека получали с использованием технологии VELOCIGENE® (смотрите, например, патент США №6586251 и Valenzuela et al. (2003) High-throughput engineering of the mouse genome coupled with high-resolution expression analysis, Nature Biotech. 21(6):652-659) для модификации мышиных геномных клонов на основе бактериальной искусственной хромосоме (ВАС) 302g12 и 254m04 (Invitrogen). С использованием этих двух ВАС клонов геномные конструкты конструировали так, чтобы они содержали единственную реаранжированную зародышевую область легкой цепи человека и вставляли в эндогенный локус κ легкой цепи, который предварительно модифицировали для удаления эндогенных κ вариабельных и соединяющих генных сегментов.
Конструкция нацеливающих векторов реаранжированной зародышевой легкой цепи человека. Три различных реаранжированных зародышевых области легкой цепи человека получали с использованием стандартных молекулярно-биологических техник, принятых в настоящей области техники. Вариабельные генные сегменты человека, используемые для конструкции этих трех областей, включали в себя реаранжированную последовательность Vκ1-39Jκ5 человека, реаранжированную последовательность Vκ3-20Jκ1 человека и реаранжированную последовательность VpreBJλ5 человека.
Сегмент ДНК, содержащий экзон 1 (кодирующий лидерный пептид) и интрон 1 гена Vκ3-7 мыши, получали с помощью синтеза ДНК de novo (Integrated DNA Technologies). Обеспечивали часть 5' нетранслируемой области около встречающегося в природе сайта рестрикционного фермента BlpI. Экзоны генов Vκ1-39 и Vκ3-20 человека амплифицировали с помощью ПЦР из геномных ВАС библиотек человека. Прямые праймеры содержали 5' удлинение, содержащее акцепторный сайт сплайсинга интрона 1 гена Vκ3-7 мыши. Обратный праймер, используемый для ПЦР последовательности Vκ1-39 человека, содержал удлинение, кодирующее Jκ5 человека, тогда как обратный праймер, используемый для ПЦР последовательности Vκ3-20 человека, содержал удлинение, кодирующее Jκ1 человека. Последовательность VpreBJλ5 человека получали с помощью синтеза ДНК de novo (Integrated DNA Technologies). Часть интрона Jκ-Cκ человека, содержащая донорный сайт сплайсинга, амплифицировали с помощью ПЦР из плазмиды pBS-296-HA18-PISceI. Прямой праймер ПЦР содержал удлинение, кодирующее часть любой из последовательностей Jκ5, Jκ1 или Jλ5 человека. Обратный праймер содержал сайт PI-SceI, который ранее сконструировали в интрон.
Vκ3-7 экзон 1/интрон 1 мыши, экзоны вариабельной легкой цепи человека и фрагменты Jκ-Cκ интрона человека сшивали вместе с помощью ПЦР перекрывающегося удлинения, обрабатывали с помощью BlpI и PI-SceI и лигировали в плазмиду pBS-296-HA18-PISceI, которая содержала промотор из вариабельного генного сегмента Vκ3-15 человека. Фланкированную lox кассету гигромицина в плазмиде pBS-296-HA18-PISceI замещали фланкированной FRT кассетой гигромицина, фланкированной сайтами NotI и AscI. Фрагмент NotI/PI-SceI этой плазмиды лигировали в модифицированную ВАС 254m04 мыши, которая содержала часть интрона Jκ-Cκ мыши, экзон Cκ мыши и приблизительно 75 т.п.н. геномной последовательности ниже по ходу транскрипции от κ локуса мыши, которые обеспечивали 3' гомологичное плечо для гомологичной рекомбинации в ES клетках мыши. Фрагмент NotI/AscI этой ВАС затем лигировали в модифицированную ВАС 302g12 мыши, которая содержала фланкированную FRT кассету неомицина и приблизительно 23 т.п.н. геномной последовательности выше против хода транскрипции от эндогенного κ локуса для гомологичной рекомбинации в ES клетках мыши.
Нацеливающий вектор реаранжированной зародышевой Vκ1-39Jκ5 человека (фиг. 1). Сайты рестрикционных ферментов вводили на 5' и 3' концы сконструированной вставки легкой цепи для клонирования в нацеливающий вектор: сайт AscI на 5' конец и сайт PI-SceI на 3' конец. В пределах 5' сайта AscI и 3' сайта PI-SceI нацеливающий конструкт в направлении 5'-3' содержал 5' гомологичное плечо, содержащее последовательность 5' по отношению к эндогенному локусу κ легкой цепи мыши, полученному из ВАС клона 302g12 мыши, фланкированный FRT ген устойчивости к неомицину, геномную последовательность, содержащую промотор Vκ3-15 человека, лидерную последовательность вариабельного генного сегмента Vκ3-7 мыши, последовательность интрона вариабельного генного сегмента Vκ3-7 мыши, открытую рамку считывания реаранжированной зародышевой области Vκ1-39Jk5 человека, геномную последовательность, содержащую часть интрона Jκ-Cκ человека и 3' гомологичное плечо, содержащее последовательность 3' относительно эндогенного генного сегмента Jκ5 мыши, полученного из ВАС клона 254m04 мыши (фигура 1, средняя часть). Гены и/или последовательности выше против хода транскрипции от эндогенного локуса κ легкой цепи мыши и ниже по ходу транскрипции от наиболее 3' генного сегмента Jκ (например, эндогенный 3' энхансер) не были модифицированы нацеливающим конструктом (смотрите фигуру 1). Последовательность сконструированного локуса Vκ1-39Jκ5 человека показана в SEQ ID ΝΟ: 1.
Нацеленную вставку реаранжированной зародышевой области Vκ1-39Jκ5 человека в ДНК ВАС подтверждали с помощью полимеразной цепной реакции (ПЦР) с использованием праймеров, расположенных на последовательностях в пределах реаранжированной зародышевой области легкой цепи человека. Кратко, последовательность интрона 3' по отношению к лидерной последовательности Vκ3-7 мыши подтверждали с помощью праймеров ULC-m1F (AGGTGAGGGT ACAGATAAGT GTTATGAG; SEQ ID NO: 2) и ULC-m1R (TGACAAATGC CCTAATTATA GTGATCA; SEQ ID NO: 3). Открытую рамку считывания реаранжированной зародышевой области Vκ1-39Jκ5 человека подтверждали с помощью праймеров 1633-h2F (GGGCAAGTCA GAGCATTAGC A; SEQ ID NO: 4) и 1633-h2R (TGCAAACTGG ATGCAGCATA G; SEQ ID NO: 5). Кассету неомицина подтверждали с помощью праймеров neoF (GGTGGAGAGG CTATTCGGC; SEQ ID NO: 6) и neoR (GAACACGGCG GCATCAG; SEQ ID NO: 7). Нацеленную ДНК ВАС затем использовали для электропорации ES клеток мыши для создания модифицированных ES клеток для получения химерных мышей, которые экспрессируют реаранжированную зародышевую область Vκ1-39Jκ5 человека.
Положительные ES клеточные клоны подтверждали с помощью скрининга и кариотипирования TAQMAN™ с использованием зондов, специфических для сконструированной области легкой цепи Vκ1-39Jκ5, вставленной в эндогенный локус. Кратко, зонд neoP (TGGGCACAAC AGACAATCGG CTG; SEQ ID NO: 8), который связывается в пределах маркерного гена неомицина, зонд ULC-m1P (CCATTATGAT GCTCCATGCC TCTCTGTTC; SEQ ID NO: 9), который связывается в пределах интронной последовательности 3' с лидерной последовательностью Vκ3-7 мыши, и зонд 1633h2P (ATCAGCAGAA ACCAGGGAAA GCCCCT; SEQ ID NO: 10), который связывается в пределах открытой рамки считывания реаранжированной зародышевой Vκ1-39Jκ5. Положительные ES клеточные клоны затем использовали для имплантации самкам мышей для получения помета детенышей, экспрессирующих зародышевую область легкой цепи Vκ1-39Jκ5.
Альтернативно, ES клетки, содержащие реаранжированную зародышевую область легкой цепи Vκ1-39Jκ5 человека, трансфектировали с помощью конструкта, который экспрессирует FLP для удаления фланкированной FRT кассеты неомицина, введенной нацеливающим конструктом. Необязательно, кассету неомицина удаляют путем скрещивания с мышами, которые экспрессируют рекомбиназу FLP (например, патент США №6774279). Необаятельно, кассету неомицина сохраняют у мышей.
Нацеливающий вектор реаранжированной зародышевой Vκ3-20Jκ1 человека (фиг. 2). Аналогичным образом, сконструированный локус легкой цепи, экспрессирующий реаранжированную зародышевую область Vκ3-20Jκ1 человека, получали с использованием нацеливающего конструкта, включающего в себя, в направлении 5'-3', 5' гомологичное плечо, содержащее последовательность 5' по отношению к эндогенному к локусу легкой цепи мыши, полученному из ВАС клона 302g12 мыши, фланкированный FRT ген устойчивости к неомицину, геномную последовательность, включающую в себя промотор Vκ3-15 человека, лидерную последовательность вариабельного генного сегмента Vκ3-7 мыши, последовательность интрона вариабельного генного сегмента Vκ3-7 мыши, открытую рамку считывания реаранжированной зародышевой области Vκ3-20Jκ1 человека, геномную последовательность, содержащую часть интрона Jκ-Cκ человека, и 3' гомологичное плечо, содержащее последовательность 3' относительно эндогенного генного сегмента Jκ5 мыши, полученного из ВАС клона 254m04 мыши (фигура 2, средняя часть). Последовательность сконструированного локуса Vκ3-20Jκ1 человека показана в SEQ ID NO: 11.
Нацеленную вставку реаранжированной зародышевой области Vκ3-20Jκ1 человека в ВАС ДНК подтверждали с помощью полимеразной цепной реакции (ПЦР) с использованием праймеров, расположенных на последовательностях в пределах реаранжированной зародышевой области легкой цепи Vκ3-20Jκ1 человека. Кратко, последовательность интрона 3' относительно мыши лидерной последовательности Vκ3-7 подтверждали с помощью праймеров ULC-m1F (SEQ ID NO: 2) и ULC-m1R (SEQ ID NO: 3). Открытую рамку считывания реаранжированной зародышевой области Vκ3-20Jκ1 человека подтверждали с помощью праймеров 1635-h2F (TCCAGGCACC CTGTCTTTG; SEQ ID NO: 12) и 1635-h2R (AAGTAGCTGC TGCTAACACT CTGACT; SEQ ID NO: 13). Кассету неомицина подтверждали с помощью праймеров neoF (SEQ ID NO: 6) и neoR (SEQ ID NO: 7). Нацеленную ДНК ВАС затем использовали для электропорации ES клеток мыши для создания модифицированных ES клеток для получения химерных мышей, которые экспрессируют, которые экспрессируют реаранжированную зародышевую легкую цепь Vκ3-20Jκ1 человека.
Положительные ES клеточные клоны подтверждали с помощью скрининга и кариотипирования TAQMAN™ с использованием зондов, специфических для сконструированной области легкой цепи Vκ3-20Jκ1, вставленной в эндогенный локус κ легкой цепи. Кратко, зонд neoP (SEQ ID NO: 8), который связывается в пределах маркерного гена неомицина, зонд ULC-m1P (SEQ ID NO: 9), который связывается в пределах лидерной последовательности Vκ3-7 мыши, и зонд 1635h2P (AAAGAGCCAC CCTCTCCTGC AGGG; SEQ ID NO: 14), который связывается в пределах открытой рамку считывания Vκ3-20Jκ1 человека. Положительные ES клеточные клоны затем использовали для имплантации самкам мышей для получения помета детенышей, экспрессирующих зародышевую область легкой цепи Vκ3-20Jκ1 человека.
Альтернативно, ES клетки, содержащие зародышевую область легкой цепи Vκ3-20Jκ1 человека, могут трансфектировать с помощью конструкта, который экспрессирует FLP, для удаления фланкированной FRT кассеты неомицина, введенной нацеливающим конструктом. Необязательно, кассету неомицина могут удалять путем скрещивания с мышами, которые экспрессируют рекомбиназу FLP (например, патент США №6774279). Необязательно, кассету неомицина оставляют у мышей.
Нацеливающий вектор реаранжированной зародышевой VpreBJλ5 человека (фиг. 3). Аналогичным образом, сконструированный локус легкой цепи, экспрессирующий реаранжированную зародышевую область VpreBJλ5 человека, получали с использованием нацеливающего конструкта, включающего в себя, в направлении 5'-3', 5' гомологичное плечо, содержащее последовательность 5' относительно эндогенного локуса κ легкой цепи мыши, полученного из ВАС клона 302g12 мыши, фланкированный FRT ген устойчивости к неомицину, геномную последовательность, включающую в себя промотор Vκ3-15 человека, лидерную последовательность вариабельного генного сегмента Vκ3-7 мыши, последовательность интрона вариабельного генного сегмента Vκ3-7 мыши, открытую рамку считывания реаранжированной зародышевой области VpreBJλ5 человека, геномную последовательность, содержащую часть интрона Jκ-Cκ человека, и 3' гомологичное плечо, содержащее последовательность 3' относительно эндогенного мыши Jκ5 генного сегмента, полученного из ВАС клона 254m04 мыши (фигура 3, средняя часть). Последовательность сконструированного локуса VpreBJλ5 человека показана в SEQ ID NO:15.
Нацеленную вставку реаранжированной зародышевой область VpreBJλ5 человека в ВАС ДНК подтверждали с помощью полимеразной цепной реакции (ПЦР) с использованием праймеров, расположенный на последовательностях в пределах реаранжированной зародышевой области легкой цепи VpreBJλ5 человека. Кратко, последовательность интрона 3' относительно лидерной последовательности Vκ3-7 мыши подтверждали с помощью праймеров ULC-m1F (SEQ ID NO: 2) и ULC-m1R (SEQ ID NO: 3). Открытую рамку считывания реаранжированной зародышевой области VpreBJλ5 человека подтверждали с помощью праймеров 1616-h1F (TGTCCTCGGC CCTTGGA; SEQ ID NO: 16) и 1616-h1R (CCGATGTCAT GGTCGTTCCT; SEQ ID NO: 17). Кассету неомицина подтверждали с помощью праймеров neoF (SEQ ID NO: 6) и neoR (SEQ ID NO: 7). Нацеленную ДНК ВАС затем использовали для электропорации ES клеток мыши для создания модифицированных ES клеток для получения химерных мышей, которые экспрессируют реаранжированную зародышевую легкую цепь VpreBJλ5 человека.
Положительные ES клеточные клоны подтверждают с помощью скрининга и кариотипирования TAQMAN™ с использованием зондов, специфических для сконструированной области легкой цепи VpreBJλ5, вставленной в эндогенный локус κ легкой цепи. Кратко, зонд neoP (SEQ ID NO: 8), который связывается в пределах маркерного гена неомицина, зонд ULC-m1P (SEQ ID NO: 9), который связывается в пределах лидерной последовательности IgVκ3-7 мыши, и зонд 1616h1P (ACAATCCGCC TCACCTGCAC ССТ; SEQ ID NO: 18), который связывается в пределах открытой рамки считывания VpreBJλ5 человека. Положительные ES клеточные клоны затем используют для имплантации самкам мышей для получения помета детенышей, экспрессирующих зародышевую область легкой цепи.
Альтернативно, ES клетки, содержащие реаранжированную зародышевую область легкой цепи линия VpreBJλ5 человека, трансфектировали с помощью конструкта, который экспрессирует FLP, для удаления фланкированной FRT кассеты неомицина, введенной нацеливающим конструктом. Необязательно, кассету неомицина удаляют путем скрещивания с мышами, которые экспрессируют рекомбиназу FLP (например, патент США №6774279). Необязательно, кассету неомицина сохраняют у мышей.
Пример 3
Получение мышей, экспрессирующих единственную реаранжированную легкую цепь человека
Описанный выше нацеленные ES клетки использовали в качестве донорных ES клеток и вводили в эмбрион мыши на стадии 8 клеток с помощью способа VELOCIMOUSE® (смотрите, например, патент США №7294754 и Poueymirou et al. (2007) F0 generation mice that are essentially fully derived from the donor gene-targeted ES cells allowing immediate phenotypic analyses, Nature Biotech. 25(1):91-99. VELOCIMICE®, несущих независимо сконструированную зародышевую область легкой цепи Vκ1-39Jκ5 человека, область легкой цепи Vκ3-20Jκ1 или область легкой цепи VpreBJλ5 определяли путем генотипирования с использованием модификации анализа аллелей (Valenzuela et al., ранее), который обнаруживает присутствие уникальной реаранжированной зародышевой области легкой цепи человека.
Детенышей генотипируют и детеныша, гетерозиготного или гомозиготного в отношении уникальной реаранжированной зародышевой области легкой цепи человека выбирают путем получения характеристик экспрессии реаранжированной зародышевой области легкой цепи человека.
Проточная цитометрия. Экспрессию реаранжированной области легкой цепи человека в репертуаре нормальных антител мышей с общей легкой цепью подтверждали с помощью анализа экспрессии κ и λ иммуноглобулина в спленоцитах и периферической крови мышей с общей легкой цепью. Суспензии клеток от собранных селезенок и периферической крови мышей дикого типа (n=5), мышей, гетерозиготных в отношении Vκ1-39Jκ5 общей легкой цепи (n=3), гомозиготных в отношении Vκ1-39Jκ5 общей легкой цепи (n=3), гетерозиготных в отношении Vκ3-20Jκ1 общей легкой цепи (n=2) и гомозиготных в отношении Vκ3-20Jκ1 общей легкой цепи (n=2), получали с использованием стандартных способов и окрашивали с помощью CD19+, Igλ+ и Igκ+ с использованием флуоресцентно меченых антител (BD Pharmigen).
Кратко, 1×106 клеток инкубировали с антителом к CD16/CD32 мыши (клон 2.4G2, BD Pharmigen) на льду в течение 10 минут с последующим мечением следующим коктейлем антител в течение 30 минут на льду: конъюгированное с АРС антитело к CD19 мыши (клон 1D3, BD Pharmigen), конъюгированное с PerCP-Cy5.5 антитело к CD3 мыши (клон 17А2, BioLegend), конъюгированное с FITC антитело к Igκ мыши (клон 187.1, BD Pharmigen), конъюгированное с РЕ антитело к Igλ мыши (клон RML-42, BioLegend). После окрашивания клетки отмывали и фиксировали в 2% формальдегиде. Сбор данных проводили на проточном питометре LSRII и анализировали с помощью FlowJo. Гейтирование: общие В-клетки (CD19+CD3-), Igκ+ В-клетки (Igκ+Igλ-CD19+CD3-), Igλ+ В-клетки (Igκ-Igλ+CD19+CD3-). Данные, собранные от образцов крови и спленоцитов, показали сходные результаты. В таблице 3 представлено процентное отношение положительных CD19+ В-клеток из периферической крови одной репрезентативной мыши из каждой группы, которые являлись Igλ+, Igκ+ или Igλ+Igκ+. Процентное отношение CD19+ В-клеток в периферической крови от мышей дикого типа (WT) и мышей, гомозиготных в отношении или Vκ1-39Jκ5, или Vκ3-20Jκ1 общей легкой цепи, показаны на фиг. 4.
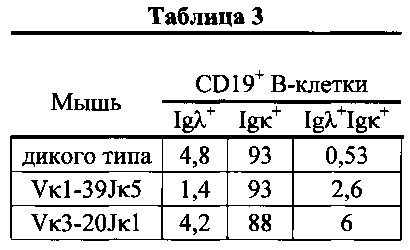
Экспрессия общей легкой цепи
Экспрессию каждой общей легкой цепи (Vκ1-39Jκ5 и Vκ3-20Jκ1) анализировали у гетерозиготных и гомозиготных мышей с использованием анализа количественной ПЦР (например, TAQMAN™).
Кратко, очищали CD19+ В-клетки из селезенок мышей дикого типа, мышей, гомозиготных в отношении замещения локусов вариабельной области тяжелой цепи и κ легкой цепи мыши соответствующими локусами вариабельной области тяжелой цепи и κ легкой цепи (Нκ) человека, а также мышей, гомозиготных и гетерозиготных в отношении каждой реаранжированной области легкой цепи человека (Vκ1-39Jκ5 или Vκ3-20Jκ1) с использованием микрогранул CD19 мыши (Miltenyi Biotec) в соответствии с инструкциями производителя. Общую РНК очищали от CD19+ В-клеток с использованием мининабора RNeasy (Qiagen) в соответствии с инструкциями производителя и геномную РНК удаляли с использованием обработки не содержащей РНазу ДНКазой на колонке (Qiagen). 200 нг мРНК обратно транскрибировали в кДНК с использованием набора First Stand cDNA Synthesis (Invitrogen) и полученную кДНК амплифицировали с использованием универсального мастер-микса для ПЦР Taqman (Applied Biosystems). Все реакции проводили с использованием системы обнаружения последовательности ABI 7900 (Applied Biosystems) с использованием праймеров и Taqman MGB зондов, охватывающих (1) участок соединения Vκ-Jκ для обеих общих легких цепей, (2) только ген Vκ (т.е. Vκ1-39 и Vκ3-20), и (3) область Cκ мыши. На таблице 13 представлены последовательности праймеров и зондов, используемых для настоящего анализа. Относительную экспрессию нормировали к экспрессии области Cκ мыши. Результаты показаны на фиг. 5А, 5В и 5С.

Антиген-специфические антитела с общей легкой цепью. Мышей с общей легкой цепью, содержащих или Vκ1-39Jκ5, или Vκ3-20Jκ1 общую легкую цепь на эндогенном локусе κ легкой цепи мыши, иммунизировали с помощью β-галактозидазы и измеряли титр антител.
Кратко, β-галактозидазу (Sigma) эмульгировали в адъюванте Titermax™ (Sigma), согласно инструкциям производителя. Мышей дикого типа (n=7), мышей, гомозиготных в отношении общей легкой цепи Vκ1-39Jκ5 (n=2) и гомозиготных в отношении общей легкой цепи Vκ3-20Jκ1 (n=5) иммунизировали с помощью подкожной инъекций в количестве 100 мкг β-галактозидазы/Titermax™. Мышей повторно иммунизировали с помощью подкожной инъекции два раза, с интервалом в 3 недели, в количестве 50 мкг β-галактозидазы/Titermax™. После второй повторной иммунизации кровь собирали от находящихся под анестезией мышей с использованием ретроорбитального кровотечения в пробирки для отделения сыворотки (BD Biosciences) согласно инструкциям производителя. Для измерения антител IgM или IgG к β-галактозидазе планшеты для ELISA (Nunc) покрывали 1 мкг/мл β-галактозидазы в течение ночи при 4°C. Избыток антигенов смывали перед блокированием с помощью PBS с 1% BSA в течение одного часа при комнатной температуре. Серийные разведения сыворотки добавляли к планшетам и инкубировали в течение одного часа при комнатной температуре перед отмывкой. Планшеты затем инкубировали с конъюгированным с HRP антителом к IgM (Southern Biotech) или антителом к IgG (Southern Biotech) в течение одного часа при комнатной температуре. После следующей отмывки планшеты проявляли с помощью субстрата ТМВ (BD Biosciences). Реакции останавливали с помощью 1 н. серной кислоты и считывали OD450 с использованием сканирующего спектрофотометра для прочтения планшетов Victor Х5 (Perkin Elmer). Данные анализировали с помощью GraphPad™ Prism и сигнал рассчитывали как разведение сыворотки, которое в два раза выше фонового значения. Результаты показаны на фиг. 6А и 6В.
Как показано в настоящем примере, отношение κ/λ В-клеток как в компартменте селезенки, так и периферических компартментах мышей с общей легкой цепью Vκ1-39Jκ5 и Vκ3-20Jκ1 показало паттерн приблизительно дикого типа (Таблица 3 и фиг. 4). Мыши с общей легкой цепью VpreBJλ5 тем не менее показали меньшее количество периферических В-клеток, из которых приблизительно 1-2% экспрессируют сконструированную область легкой цепи человека (данные не показаны). Уровни экспрессии реаранжированных областей легкой цепи человека Vκ1-39Jκ5 и Vκ3-20Jκ1 из эндогенного локуса κ легкой цепи повышались по сравнению с эндогенным локусом κ легкой цепи, содержащим полное замещение генных сегментов Vκ и Jκ мыши генными сегментами Vκ и Jκ человека (фиг. 5А, 5В и 5С). Уровни экспрессии VpreBJλ5 реаранжированной области легкой цепи человека демонстрировали сходную высокую экспрессию из эндогенного локуса κ легкой цепи как у гетерозиготных, так и гомозиготных мышей (данные не показаны). Это демонстрирует, что в прямой конкуренции с эндогенным локусом λ, κ легкой цепи мыши или обоими эндогенными локусами легкой цепи мыши, единственная реаранжированная последовательность VL/JL человека может давать в результате более высокие, чем уровень экспрессии дикого типа из эндогенного локуса κ легкой цепи и приводить к нормальному количеству В-клеток селезенки и крови. Кроме того, присутствие сконструированного κ локуса легкой цепи, содержащего или последовательность Vκ1-39Jκ5 человека, или последовательность Vκ3-20Jκ1 человека, хорошо переносилось мышами и, вероятно, функционировало способом дикого типа путем репрезентации значительной части репертуара легкой цепи в гуморальном компоненте иммунного ответа (фиг. 6А и 6В).
Пример 4
Скрещивание мышей, экспрессирующих единственную реаранжированную зародышевую легкую цепь человека
В настоящем примере описаны некоторые другие штаммы генетически модифицированных мышей, которые могут скрещиваться с какой-либо из описанных в настоящем документе мышей с общей легкой цепью для создания множественных штаммов генетически модифицированных мышей, характеризующихся множественными генетически модифицированными локусами иммуноглобулина.
Нокаут эндогенного Igλ (KO). Для оптимизации частоты использования сконструированного локуса легкой цепи, мышей, несущих одну из реаранжированных зародышевых областей легкой цепи человека, скрещивают с другой мышью, содержащей делецию в эндогенном локусе λ легкой цепи. Таким образом, полученное потомство будет экспрессировать, в качестве своей единственной легкой цепи, реаранжированную зародышевую область легкой цепи человека, как описано в примере 2. Скрещивание проводят с помощью стандартных техник, принятых в настоящей области техники, и, альтернативно, с помощью коммерческого производителя (например, Jackson Laboratory). Штаммы мышей, несущие сконструированный локус легкой цепи и делецию эндогенного локуса λ легкой цепи, подвергают скринингу в отношении присутствия уникальной области легкой цепи и отсутствия эндогенных λ легких цепей мыши.
Гуманизированный эндогенный локус тяжелой цепи. Мышей, несущих сконструированный зародышевый локус легкой цепи человека, скрещивают с мышами, которые содержат замещение эндогенного вариабельного генного локуса тяжелой цепи мыши вариабельным генным локусом тяжелой цепи человека (смотрите патент США №6596541; мышь VELOCIMMUNE®, Regeneran Pharmaceuticals, Inc.). Мышь VELOCIMMUNE® содержит геном, содержащий вариабельные области тяжелой цепи человека, функционально связанные с эндогенными локусами константной области мыши так, чтобы мышь производила антитела, содержащие вариабельную область тяжелой цепи человека и константную область тяжелой цепи мыши в ответ на антигенную стимуляцию. ДНК, кодирующая вариабельные области тяжелых цепей антител, выделяют и функционально связывают с ДНК, кодирующей константные области тяжелой цепи человека. ДНК затем экспрессируют в клетке, способной к экспрессии полностью человеческой тяжелой цепи антитела.
Получают мышей, содержащих замещение эндогенного локуса VH мыши с локусом VH человека и единственную реаранжированную зародышевую область VL человека на эндогенном локусе κ легкой цепи. Обратные химерные антитела, содержащие соматически мутированные тяжелые цепи (VH человека и CH мыши) с единственной легкой цепью человека (VL человека и CL мыши) получают при иммунизации с помощью представляющего интерес антигена. Определяют нуклеотидные последовательности VH и VL В-клеток, экспрессирующих антитела и полностью человеческие антитела получают путем слияния нуклеотидных последовательностей VH и VL с нуклеотидными последовательностями CH и CL человека в подходящей системе экспрессии.
Пример 5
Получение антител из мышей, экспрессирующих тяжелые цепи человека и реаранжированную зародышевую область легкой цепи человека
После скрещивания мышей, которые содержат сконструированную область легкой цепи человека с различными необходимыми штаммами, содержащими модификации и делеции других эндогенных локусов Ig (как описано в примере 4), выбранных мышей могут иммунизировать с помощью представляющего интерес антигена.
Как правило, мышь VELOCIMMUNE®, содержащую одну из единственных реаранжированных зародышевых областей легкой цепи человека, подвергают сенсибилизации с помощью антигена, и лимфатические клетки (такие как В-клетки) выделяют из сыворотки животных. Лимфатические клетки подвергают слиянию с миеломной клеточной линией для получения иммортализированных гибридомных клеточных линий, и такие гибридомные клеточные линии подвергают скринингу и селекции для определения гибридомных клеточных линий, которые производят антитела, содержащие вариабельные области тяжелой цепи человека и реаранжированные зародышевые легкие цепи человека, которые являются специфическими по отношению к используемому для иммунизация антигену. ДНК, кодирующую вариабельные области тяжелых цепей и легкой цепи, выделяют и связывают с необходимыми изотипическими константными областями тяжелой цепи и легкой цепи. Вследствие присутствия эндогенных последовательностей мыши и каких-либо дополнительных цис-действующих элементов, присутствующих в эндогенном локусе, единственная легкая цепь каждого антитела может быть соматически мутировала. Это вносит дополнительное разнообразие в антиген-специфический репертуар, содержащий единственную легкую цепь и разнообразные последовательности тяжелой цепи. Полученные клонированные последовательности антитела впоследствии экспрессируют в такой клетке, как клетка СНО. Альтернативно, ДНК, кодирующие антиген-специфические химерные антитела или вариабельные домены легких и тяжелых цепей, определяют непосредственно из антиген-специфических лимфоцитов.
Вначале, выделяют высокоаффинные химерные антитела, содержащие вариабельную область человека и константную область мыши. Как описано выше, определяют характеристики антител и проводят отбор антител в отношении необходимых характеристик, предусматривающих аффинность, селективность, эпитоп и т.д. Константные области мыши замещают необходимой константной областью человека для получения полностью человеческого антитела, содержащего соматически мутированную тяжелую цепь человека и единственную легкую цепь, происходящую из реаранжированной зародышевой области легкой цепи человека настоящего изобретения. Подходящие константные области человека включают в себя, например, IgG1 или IgG4 дикого типа или модифицированные IgG1 или IgG4.
Отдельные когорты мышей VELOCIMMUNE®, содержащие замещение эндогенный локус тяжелой цепи мыши генными сегментами VH, DH и JH человека и замещение эндогенного локуса κ легкой цепи мыши или сконструированной зародышевой областью легкой цепи Vκ1-39Jκ5 человека, или сконструированной зародышевой областью легкой цепи Vκ3-20Jκ1 человека (описанными выше), иммунизировали с помощью белка рецептора клеточной поверхности человека (антиген Е). Антиген Ε вводят непосредственно на подушечку стопы задней лапы мышей с помощью шести последовательных инъекций каждые 3-4 дня. 2-3 мкг антигена Ε смешивают с 10 мкг CpG олигонуклеотида (№ по кат. tlrl-modn - ODN1826 олигонуклеотид; InVivogen, Сан-Диего, Калифорния) и 25 мкг Adju-Phos (гелевый адъювант фосфата алюминия, № по кат. Н-71639-250; Brenntag Biosector, Фредерикссунд, Дания) перед инъекцией. В общем шесть инъекций вводят перед последней повторной иммунизацией антигеном, которую вводят за 3-5 дней до умерщвления. Образцы крови после 4-й и 6-й инъекции собирали и иммунный ответ антител подвергают мониторингу с помощью стандартного антиген-специфического иммуноанализа.
Когда достигается необходимый иммунный ответ, спленоциты собирают и подвергают слиянию с миеломными клетками мыши для сохранения их жизнеспособности и образования гибридомных клеточных линий. Гибридомные клеточные линии подвергают скринингу и селекции для определения клеточных линий, которые производят специфические для антигена Ε антитела с общей легкой цепью. С использованием этой техники получают некоторые специфические для антигена Ε антитела с общей легкой цепью (т.е. антитела, обладающие вариабельными доменами тяжелой цепи человека, одинаковым вариабельным доменом легкой цепи человека и константными доменами мыши).
Альтернативно, антитела с общей легкой цепью к антигену Ε выделяют непосредственно из антиген-положительных В-клеток без слияния с миеломными клетками, как описано в заявке на патент США №2007/0280945 A1. С использованием настоящего способа получала некоторые полностью человеческие антитела с общей легкой цепью к антигену Ε (т.е. антитела, обладающие вариабельными доменами тяжелой цепи человека, или сконструированной областью легкой цепи Vκ1-39Jκ5 человека, или сконструированной областью легкой цепи Vκ3-20Jκ1 человека и константными доменами человека).
Биологические свойства иллюстративных антител к антигену Ε с общей легкой цепью, полученных согласно способам настоящего примера, подробно описаны в разделах, представленных ниже.
Пример 6
Частота использования генного сегмента тяжелой цепи в антиген-специфических антителах с общей легкой цепью
Для анализа структуры произведенных человеческих антител к антигену Ε с общей легкой цепью нуклеиновые кислоты, кодирующие вариабельные области тяжелой цепи антитела, клонировали и секвенировали. Из последовательностей нуклеиновых кислот и предсказанных аминокислотных последовательностей антител частоту использования генов определяли для вариабельной области тяжелой цепи (HCVR) выбранных антител с общей легкой цепью, полученных из иммунизированных мышей VELOCIMMUNE®, содержащих или сконструированную область легкой цепи Vκ1-39Jκ5 человека, или сконструированную область легкой цепи Vκ3-20Jκ1 человека. Результаты показаны в таблицах 5 и 6, на которых показано, что мыши согласно настоящему изобретению образуют антиген-специфические антитела с общей легкой цепью из разнообразных генных сегментов тяжелой цепи человека, вследствие разнообразия реаранжировок, при использовании или мыши, которая экспрессирует легкую цепь только из легкой цепи, происходящей из Vκ1-39 человека или Vκ3-20 человека. Генные сегменты VH человека из 2, 3, 4 и 5 семейств реаранжировали с разнообразными сегментами человека DH и сегментами человека JH для получения антиген-специфических антител.
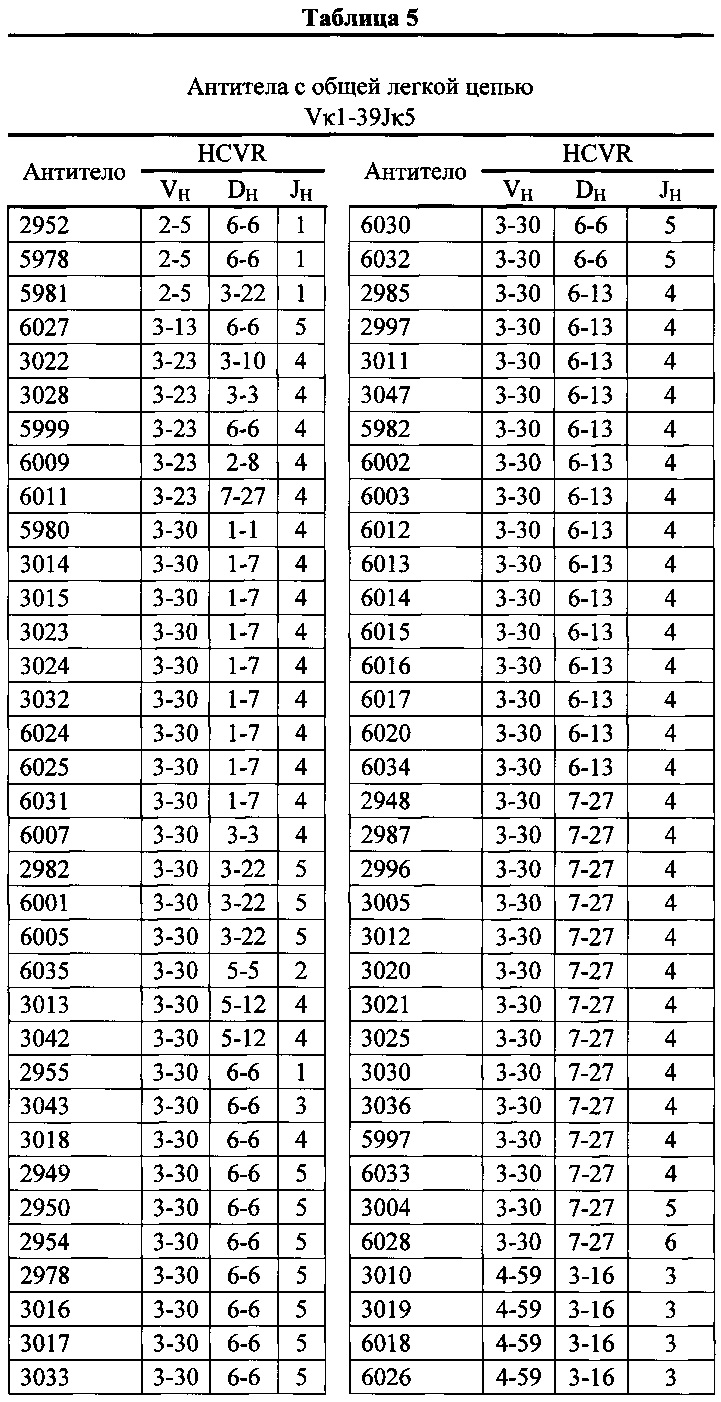
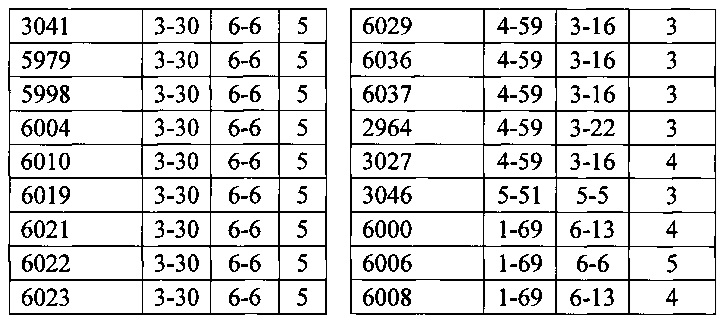
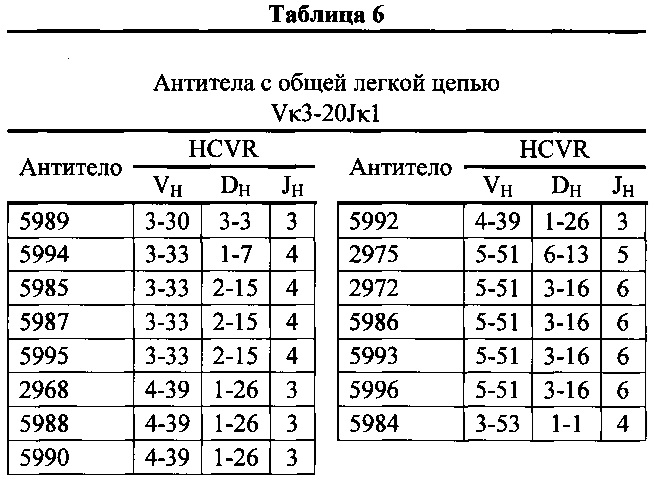
Пример 7
Определение блокирующей способности антиген-специфических антител с общей легкой цепью с помощью анализа LUMINEX™
Девяносто восемь антител с общей легкой цепью человека, возникших против антигена Е, анализировали в отношении их способности блокировать связывание естественного лиганда антигена Ε (лиганда Υ) с антигеном Ε в исследовании на основе изучения гранул.
Внеклеточный домен (ECD) антигена Ε конъюгировали с двумя метками туе эпитопа и 6Х гистидиновой меткой (антиген E-mmH) и связывали через аминогруппу с карбоксилированными микрогранулами в концентрации 20 мкг/мл в буфере MES. Смесь инкубировали в течение двух часов при комнатной температуре с последующей дезактивацией гранул с помощью 1 Μ Tris pH 8,0 с последующей отмывкой в PBS с 0,05% (объем/объем) Tween-20. Гранулы затем блокировали с помощью PBS (Irvine Scientific, Санта-Ана, Калифорния), содержащего 2% (вес/объем) BSA (Sigma-Aldrich Corp., Сент-Луис, Миссури). В 96-луночном планшете для фильтрации супернатанты, содержащие специфические для антигена Ε антитела с общей легкой цепью, разбавляли 1:15 в буфере. Получали отрицательный контроль, содержащий имитацию супернатанта с компонентами сред, аналогичными супернатанту антител. Меченные антигеном Ε гранулы добавляли к супернатантам и инкубировали в течение ночи при 4°C. Белок биотинилированного лиганда Y добавляли до конечной концентрации, составляющей 0,06 нМ, и инкубировали в течение двух часов при комнатной температуре. Обнаружение биотинилированного лиганда Y, связанного с меченными комплексом антиген E-myc-myc-6His гранулами, определяли с помощью R-фикоэритрина, конъюгированного с стрептавидином (Moss Inc, Пасадена, Мериленд) с последующим измерением на основанном на проточной цитометрии анализаторе LUMINEX™. Фоновую среднюю интенсивность флуоресценции (MFI) образца без лиганда Y отнимали от всех образцов. Процент блокирования рассчитывали путем деления вычтенного из фонового значения MFI каждого образца на скорректированное значение отрицательного контроля, умножая на 100 и вычитая полученное значение из 100.
Согласно аналогичному эксперименту такие же 98 антител с общей легкой цепью человека, возникших против антигена Е, анализировали в отношении их способности блокировать связывание антигена Ε с меченными лигандом Y гранулами.
Кратко, лиганд Y связывали через аминогруппу с карбоксилированными микрогранулами в концентрации 20 мкг/мл с разведением в буфере MES. Смесь инкубировали в течение двух часов при комнатной температуре с последующей дезактивацией гранул с помощью 1М Tris pH 8, затем отмывая в PBS с 0,05% (объем/объем) Tween-20. Гранулы затем блокировали с помощью PBS (Irvine Scientific, Санта-Ана, Калифорния), содержащего 2% (вес/объем) BSA (Sigma-Aldrich Corp., Сент-Луис, Миссури). В 96-луночном планшете для фильтрации супернатанты, содержащие специфические для антигена Ε антитела с общей легкой цепью, разбавляли 1:15 в буфере. Получали отрицательный контроль, содержащий имитацию супернатанта с компонентами сред, аналогичными супернатанту антител. Биотинилированный антиген E-mmH добавляли до конечной концентрации, составляющей 0,42 нМ, и инкубировали в течение ночи при 4°C. Меченные лигандом Y гранулы затем добавляли к смеси антитела с антигеном Ε и инкубировали в течение двух часов при комнатной температуре. Обнаружение биотинилированного антигена E-mmH, связанного с меченными лигандом Υ гранулами, определяли с помощью R-фикоэритрина, конъюгированного с стрептавидином (Moss Inc, Пасадена, Мериленд) с последующим измерением на основанном на проточной цитометрии анализаторе LUMINEX™. Фоновую среднюю интенсивность флуоресценции (MFI) образца без антигена Ε отнимали от всех образцов. Процент блокирования рассчитывали путем деления вычтенного из фонового значения MFI каждого образца на скорректированное значение отрицательного контроля, умножая на 100 и вычитая полученное значение из 100.
На таблицах 7 и 8 показано процент блокирования для всех 98 антител к антигену Ε с общей легкой цепью, исследованных в обоих анализах LUMINEX™. ND: не определено при текущих экспериментальных условиях.
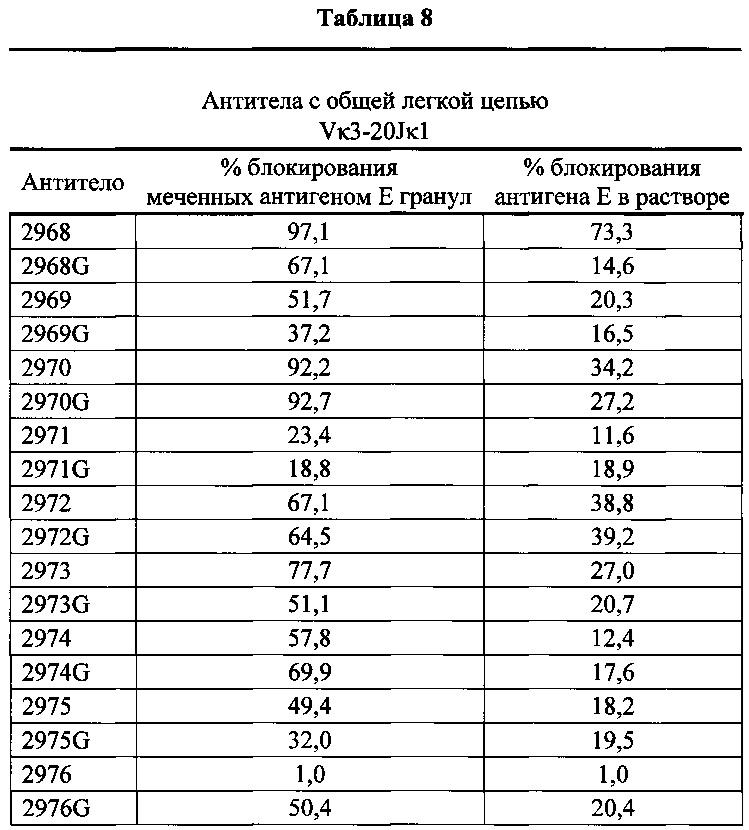
В первом описанном выше эксперименте LUMINEX™ 80 антител с общей легкой цепью, содержащих сконструированную легкую цепь Vκ1-39Jκ5, исследовали в отношении их способности блокировать связывание лиганда Y с меченными антигеном Ε гранулами. Из этих 80 антител с общей легкой цепью 68 продемонстрировали блокирование >50%, наряду с тем что 12 продемонстрировали блокирование <50% (6 при 25-50% блокировании и 6 при блокировании <25%). Для 18 антител с общей легкой цепью, содержащих сконструированную легкую цепь Vκ3-20Jκ1, 12 продемонстрировали блокирование >50%, наряду с тем что 6 продемонстрировали блокирование <50% (3 при 25-50% блокировании и 3 при блокировании <25%) связывания лиганда Y с меченными антигеном Ε гранулами.
Во втором описанном выше эксперименте LUMINEX™ те же 80 антител с общей легкой цепью, содержащих сконструированную легкую цепь Vκ1-39Jκ5, исследовали в отношении их способности блокировать связывание антигена Ε с меченными лигандом Y гранулами. Их этих 80 антител с общей легкой цепью 36 продемонстрировали блокирование >50%, наряду с тем что 44 продемонстрировали блокирование <50% (27 при 25-50% блокировании и 17 при блокировании <25%). Для 18 антител с общей легкой цепью, содержащих сконструированную легкую цепь Vκ3-20Jκ1, 1 продемонстрировало блокирование >50%, наряду с тем что 17 продемонстрировали блокирование <50% (5 при 25-50% блокировании и 12 при блокировании <25%) связывания антигена Ε с меченными лигандом Y гранулами.
Данные таблиц 7 и 8 показывают, что описанные в таблицах 5 и 6 реаранжировки создают специфические для антигена Ε антитела с общей легкой цепью, которые блокировали связывание лиганда Y с его когнатным рецептором антигеном Ε с переменными степенями эффективности, что находится в соответствии с антителами к антигену Ε с общей легкой цепью из таблиц 5 и 6, содержащих антитела со специфичностью к перекрывающимся и неперекрывающимся эпитопам по отношению к антигену Е.
Пример 8
Определение способности к блокированию антиген-специфических антител с общей легкой цепью с помощью ELISA
Человеческие антитела с общей легкой цепью, возникшие против антигена Е, исследовали в отношении их способности блокировать связывание антигена Ε с покрытой лигандом Y поверхностью в анализе ELISA.
Лигандом Y покрывали 96-луночные планшеты в концентрации 2 мкг/мл с разведением в PBS и инкубировали в течение ночи с последующей четырехкратной отмывкой в PBS с 0,05% Tween-20. Планшет затем блокировали с помощью PBS (Irvine Scientific, Санта-Ана, Калифорния), содержащего 0,5% (вес/объем) BSA (Sigma-Aldrich Corp., Сент-Луис, Миссури) в течение одного часа при комнатной температуре. В отдельном планшете супернатанты, содержащие антитела с общей легкой цепью к антигену Ε разбавляли 1:10 в буфере. Имитацию супернатанта с такими же компонентами антител использовали в качестве отрицательного контроля. Антиген Е-mmH (описанный выше) добавляли до конечной концентрации 0,150 нМ и инкубировали в течение одного часа при комнатной температуре. Смесь антитело/антиген E-mmH затем добавляли к планшету, содержащему лиганд Y, и инкубировали в течение одного часа при комнатной температуре. Обнаружение антигена E-mmH, связанного с лигандом Y, определяли с помощью пероксидазы хрена (HRP), конъюгированной с анти-пента-His антителом (Qiagen, Валенсия, Калифорния) и проявляли с помощью стандартной колориметрической реакции с использованием субстрата тетраметилбензидина (ТМВ) (BD Biosciences, Сан-Хосе, Калифорния), нейтрализованного серной кислотой. Поглощение считывали при OD450 в течение 0,1 сек. Фоновое поглощение образца без антигена Ε вычитали из всех образцов. Процент блокирования рассчитывали путем деления вычтенного из фонового значения MFI каждого образца на скорректированное значение отрицательного контроля, умножая на 100 и вычитая полученное значение из 100.
На таблицах 9 и 10 показан процент блокирования для всех 98 антител к антигену Ε с общей легкой цепью, исследованных в анализе ELISA. ND: не определено при текущих экспериментальных условиях.
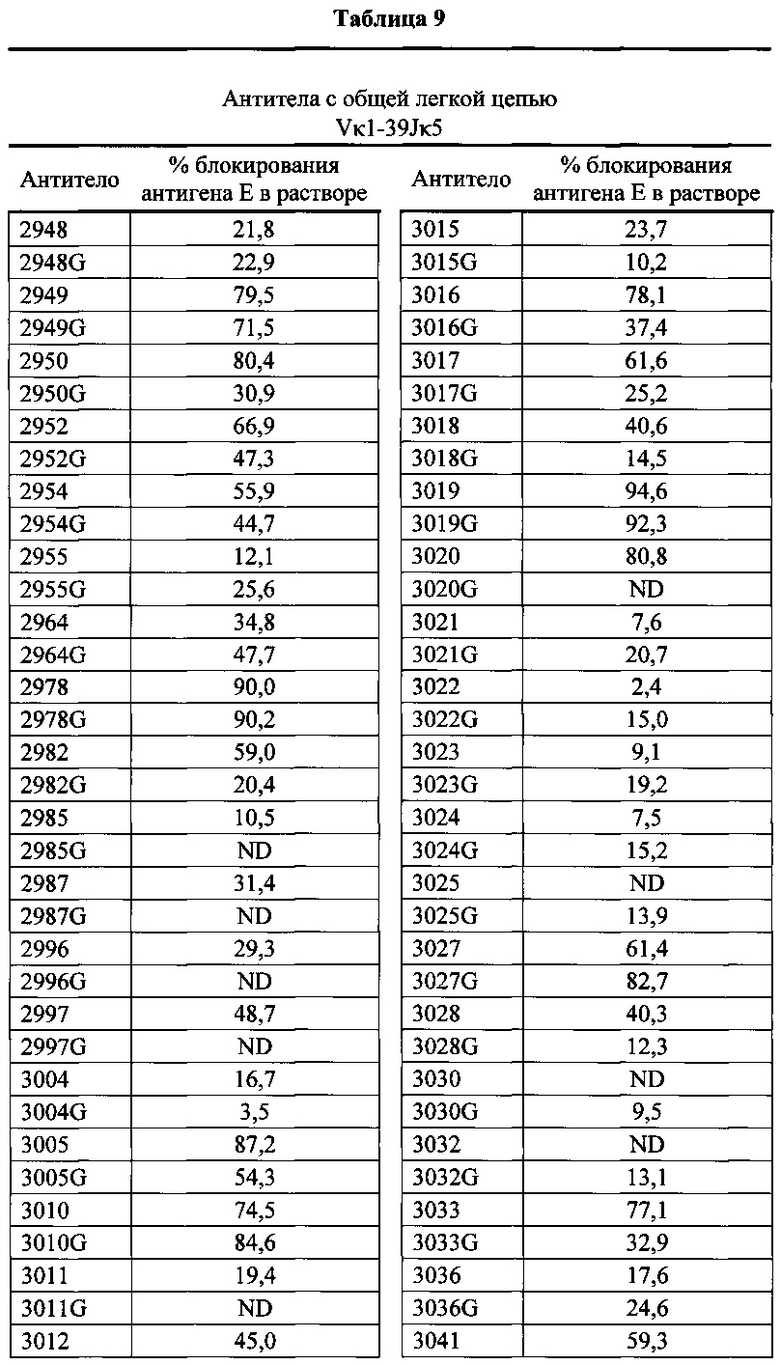
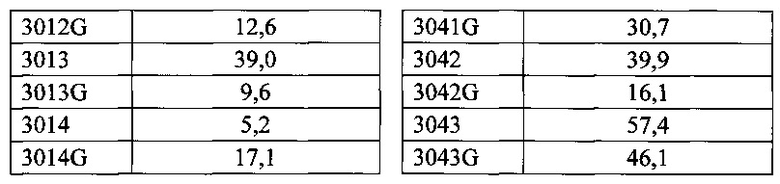
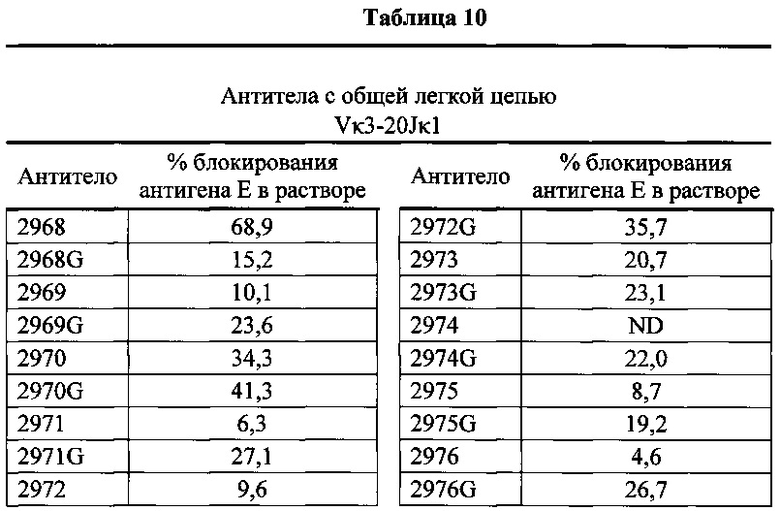
Как описано в настоящем примере, из 80 антител с общей легкой цепью, содержащих сконструированную легкую цепь Vκ1-39Jκ5, исследованных в отношении их способности блокировать связывание антигена Ε с покрытой лигандом Υ поверхностью, 22 продемонстрировали блокирование >50%, наряду с тем что 58 продемонстрировали блокирование <50% (20 при 25-50% блокировании и 38 при блокировании <25%). Для 18 антител с общей легкой цепью, содержащих сконструированную легкую цепь Vκ3-20Jκ1, одно продемонстрировало блокирование >50%, наряду с тем что 17 продемонстрировали блокирование <50% (5 при 25-50% блокировании и 12 при блокировании <25%) связывания антигена Ε с покрытой лигандом Υ поверхностью.
Эти результаты также согласуются с пулом антител с общей легкой цепью, специфических для антигена Е, содержащим антитела со специфичностью к перекрывающимся и неперекрывающимся эпитопам по отношению к антигену Е.
Пример 9
Определение аффинность с помощью BIACORE™ для антиген-специфических антител с общей легкой цепью
Равновесные константы диссоциации (KD) для выбранных супернатантов антител определяли с помощью SPR (поверхностного плазмонного резонанса) с использованием прибора BIACORE™ Τ100 (GE Healthcare). Все данные получали с использованием HBS-EP (10 мМ Hepes, 150 мМ NaCl, 0,3 мМ EDTA, 0,05% поверхностно-активного вещества Р20, pH 7,4) в качестве как подвижного буфера, так и буфера для образца, при 25°C. Антитела захватывали из сырых образцов супернатанта на поверхность сенсорного чипа СМ5, предварительно дериватизированную с помощью высокой плотности антител к Fc человека с использованием стандартного химического взаимодействия соединения через аминогруппу. В ходе стадии захвата супернатанты впрыскивали по всей поверхности антител к Fc человека с объемной скоростью потока, составляющей 3 мкл/мин в течение в общем 3 минут. За стадией захвата следовало введение или подвижного буфера, или аналита в концентрации 100 нМ в течение 2 минут при объемной скорости потока, составляющей 35 мкл/мин. Диссоциацию антигена от захваченного антитела отслеживали с течение 6 минут. Захваченное антитело удаляли кратковременным введением 10 мМ глицина, pH 1,5. Все сенсограммы дважды соотносили с эталонными значениями путем вычитания сенсограмм от введений буфера из сенсограмм аналита, тем самым удаляя артефакты, вызванные диссоциацией антитела от поверхности захвата. Данные связывания для каждого антитела согласовывали с 1:1 моделью связывания с переносом масс с использованием программного обеспечения для оценки BIAcore™ Τ100 v2.1. Результаты показаны в таблицах 11 и 12.
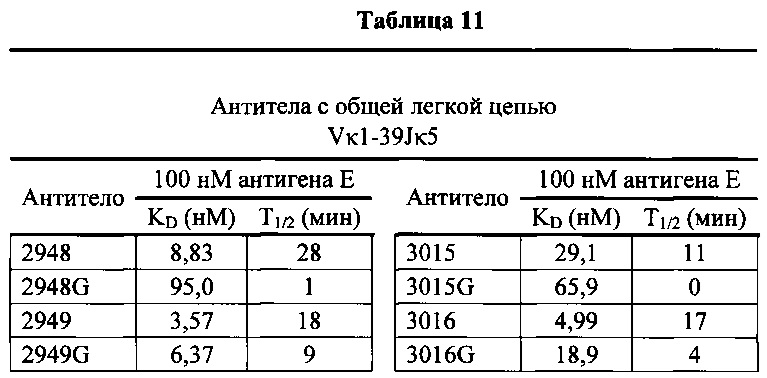
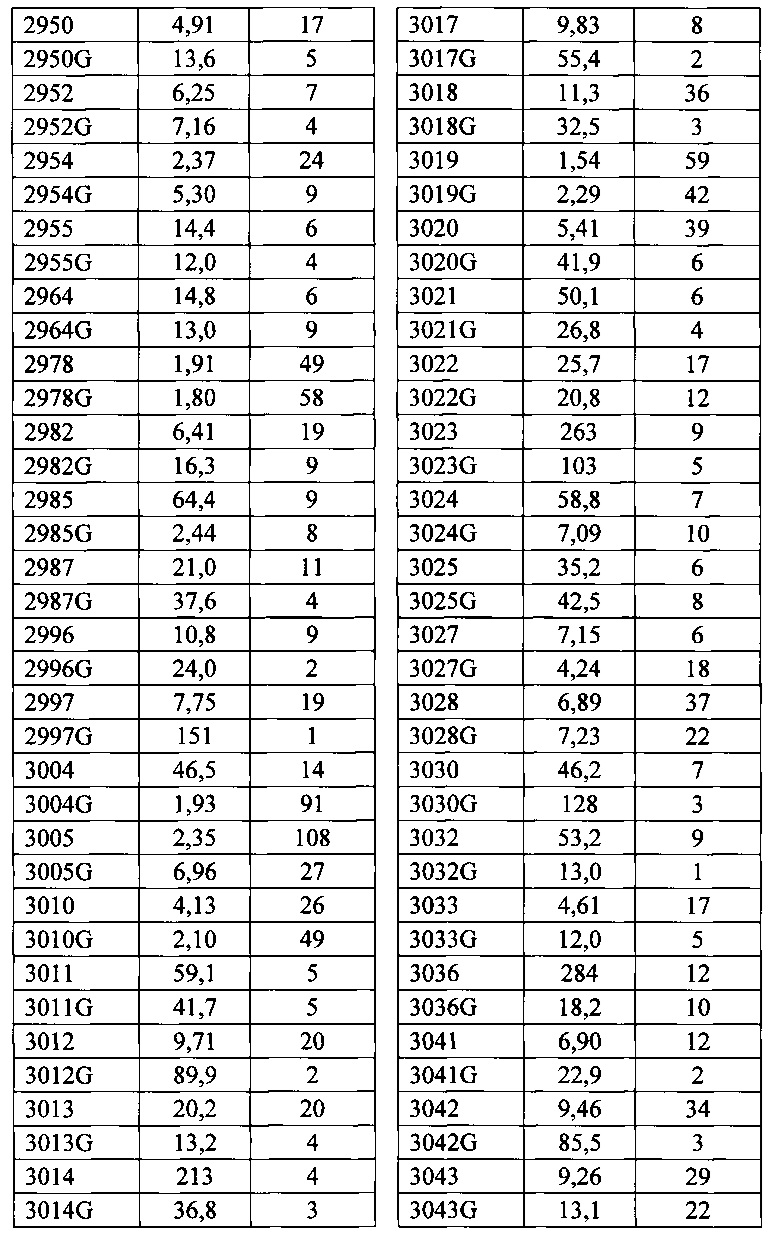
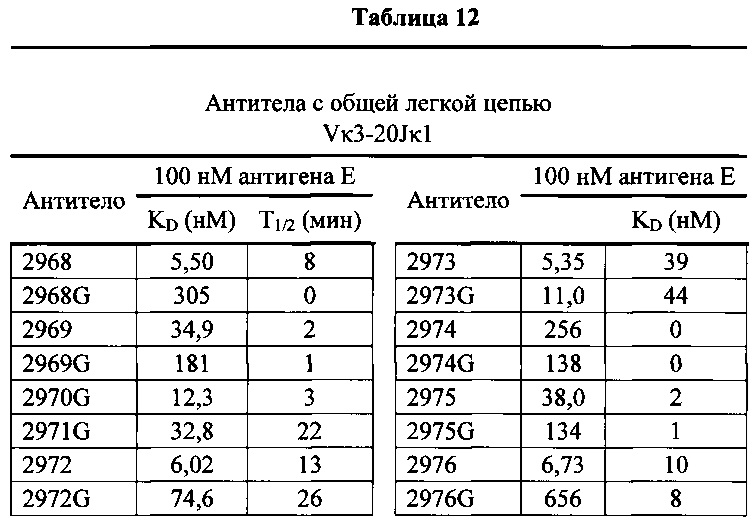
Аффинности связывания антител с общей легкой цепью, содержащих показанные в таблицах 5 и 6 реаранжировки, варьируют, причем почти все проявляют KD в наномолярном диапазоне. Данные аффинности согласуются с антителами с общей легкой цепью, полученными в результате комбинаторной ассоциации описанных в таблицах 5 и 6 реаранжированных вариабельных доменов, являющихся высокоаффинными, клонально-селективными и соматически мутированными. Объединяя c предварительно показанными данными, описанные в таблицах 5 и 6 антитела с общей легкой цепью содержат набор разнообразных, высокоаффинных антител, которые проявляют специфичность в отношении одного или нескольких эпитопов на антигене Ε
Пример 10
Определение специфичностей связывания антиген-специфических антител с общей легкой цепью с помощью анализа LUMINEX™
Выбранные антитела с общей легкой цепью к антигену Ε исследовали в отношении их способности связываться с ECD (внеклеточный домен) антигена Ε и вариантов ECD антигена Е, включающих в себя ортолог яванского макака (Mf антиген Е), который отличается от белка человека приблизительно на 10% его аминокислотных остатков; делеционный мутант антигена Е, у которого отсутствуют последние 10 аминокислот с С-конца ECD (антиген Е-ΔСТ); и два мутанта, содержащих аланиновую замену на предполагаемых положениях взаимодействия с лигандом Υ (антиген Е-Ala1 и антиген Е-Ala2). Белки антигена Ε получали в СНО клетках, и каждый содержал myc-myc-His С-концевую метку.
Для исследований связывания белок ECD антигена Ε или вариантный белок (описанный выше) из 1 мл культуральной среды захватывали путем инкубации в течение 2 часов при комнатной температуре с 1×106 микросферическими (LUMINEX™) гранулами, ковалентно покрытыми моноклональным антителом к myc (MAb 9Е10, гибридомная клеточная линия CRL-1729™; АТСС, Манассас, Виргиния). Гранулы затем отмывали с помощью PBS перед применением. Супернатанты, содержащие антитела с общей легкой цепью к антигену Е, разбавляли 1:4 в буфере и добавляли к 96-луночным планшетам для фильтрования. Имитацию супернатанта без антител использовали в качестве отрицательного контроля. Гранулы, содержащие захваченные белки антигена Е, затем добавляли к образцам антител (3000 гранул на лунку) и инкубировали в течение ночи при 4°C. На следующий день гранулы образца отмывали и связанное антитело с общей легкой цепью обнаруживали с помощью конъюгированного с R-фикоэритрином антитела к IgG человека. Интенсивность флуоресценции гранул (приблизительно 100 гранул, подсчитанных для связывания каждого образца антитела с каждым белком антигена Е) измеряли с помощью анализатора на основе проточной цитометрии LUMINEX™ и регистрировали среднюю интенсивность флуоресценции (MFI) по меньшей мере для 100 подсчитанных гранул на одно взаимодействие гранулы с антителом. Результаты показаны в таблицах 13 и 14.
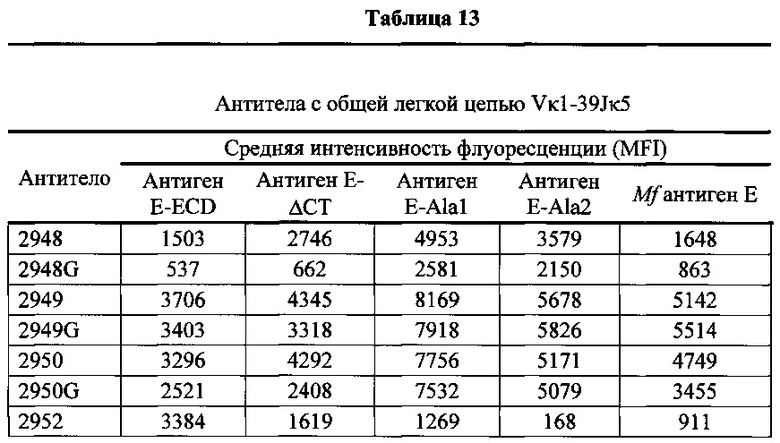
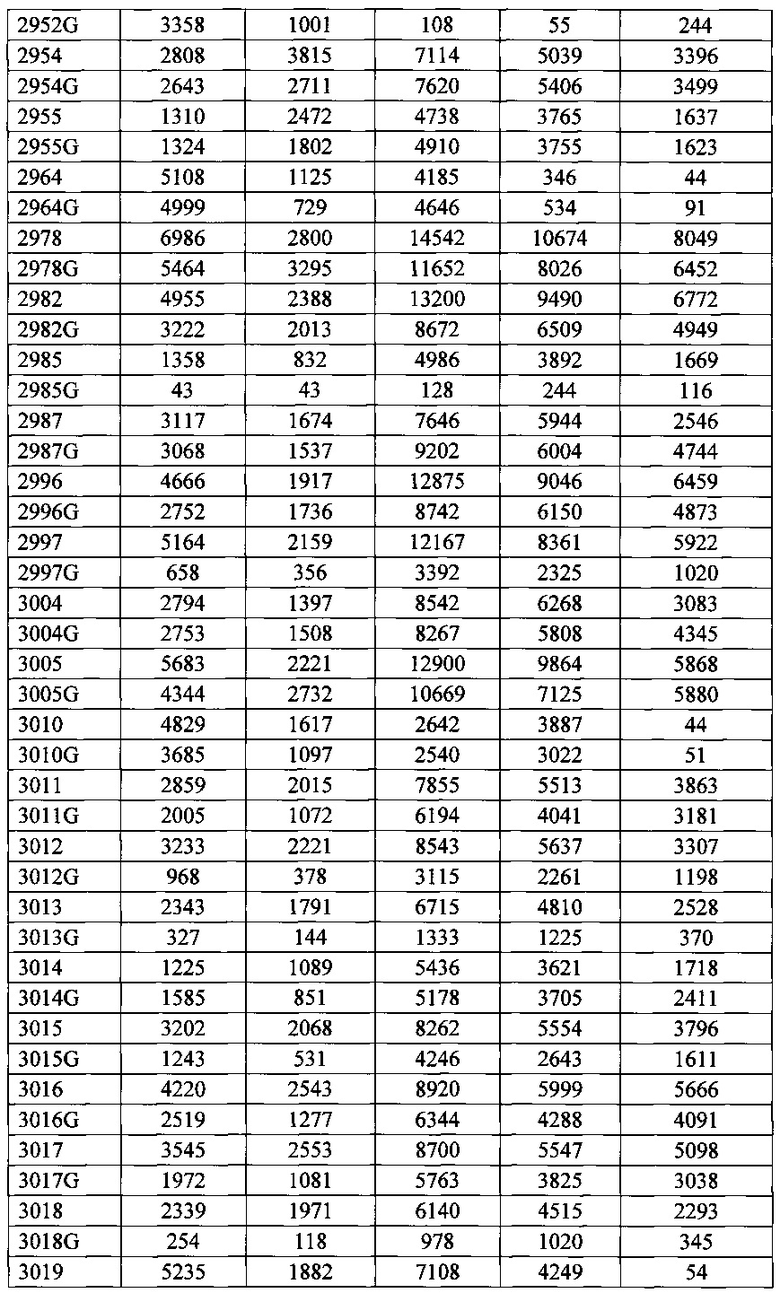
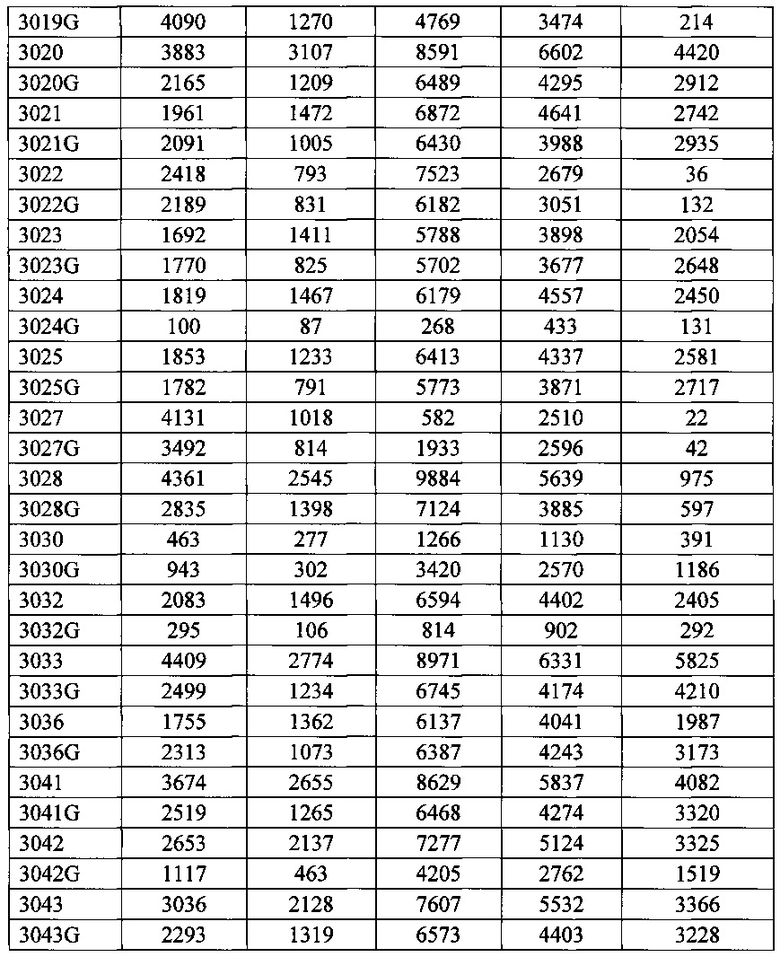

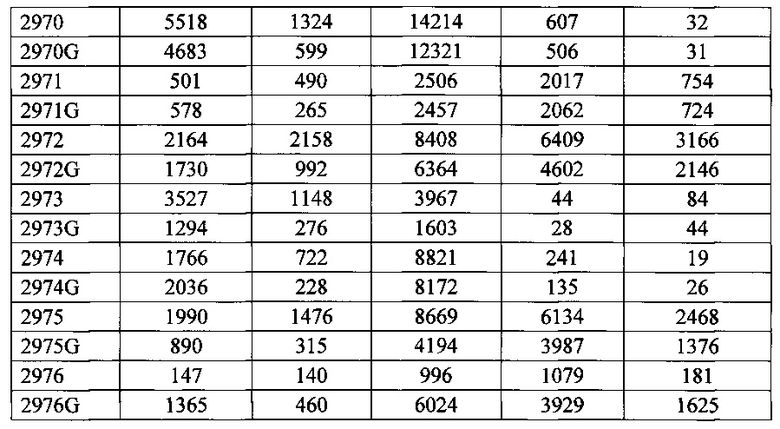
Супернатанты антител с общей легкой цепью к антигену Е проявляли высокоспецифическое связывание с гранулами, связанными с ECD антигена Е. Для этих гранул имитация супернатанта отрицательного контроля давала в результате пренебрежимо малый сигнал (<10 MFI) по сравнению с образцом гранул с ECD антигена, тогда как супернатанты, содержащие антитела с общей легкой цепью к антигену Е проявляли сильный сигнал связывания (средняя MFI, составляющая 2627, для 98 супернатанов антител; MFI>500 для 91 из 98 образцов антител).
В качестве измерения способности выбранных антител с общей легкой цепью к антигену Е определять различные эпитопы на ECD антигена Е, определяли относительное связывание антител с вариантами. Все четыре варианта антигена Е захватывали на гранулы к myc LUMINEX™, как описано выше для исследований связывания нативного ECD антигена Е, и определяли отношения относительного связывания (MFIвариант/MFIECD антигена E). Для 98 исследованных супернатантов антител с общей легкой цепью, показанных в таблицах 12 и 13, средние отношения (MFIвариант/MFTECD антигена E) отличалось для каждого варианта, вероятно, отражая различные захваченные количества белков на гранулах (средние отношения, составляющие 0,61, 2,9, 2,0 и 1,0 для антиген Е-ΔСТ, антиген Е-Ala1, антиген Е-Ala2 и Mf антиген Е, соответственно). Для каждого варианта белка связывание для подгруппы из 98 исследованных антител с общей легкой цепью показало значительно сниженное связывание, указывая на чувствительность к мутации, которая характеризовала изучаемый вариант. Например, 19 из образцов антител с общей легкой цепью связывались с Mf антигеном Е с отношением MFIвариант/MFIECD антигена E, составляющим <8%. Поскольку многие из этой группы содержали антитела с высокой или относительно высокой аффинностью (5 с KD<5 нМ, 15 с KD<50 нМ), возможно, что более низкий сигнал для этой группы является результатом чувствительности к различиям в последовательности (эпитопе) между нативным ECD антигена Ε и изучаемым вариантом, а не результатом более низких аффинностей.
Эти данные показывают, что описанные в таблицах 5 и 6 антитела с общей легкой цепью представляют разнообразную группу специфических для антигена Ε антител с общей легкой цепью, которые специфически распознают более чем один эпитоп на антигене Е.
Пример 11
Перестановка легких цепей в антителах с общей легкой цепью
Тяжелые цепи выбранных антиген-специфических антител с общей легкой цепью исследовали в отношении связывания с антигеном Ε после повторного спаривания тяжелых цепей или с зародышевой сконструированной легкой цепью Vκ1-39Jκ5, или с зародышевой сконструированной легкой цепью Vκ3-20Jκ1 (как описано в примере 1).
Кратко, 247 тяжелых цепей специфических к антигену Ε антител с общей легкой цепью (Vκ1-39Jκ5 и Vκ3-20Jκ1) трансфектировали с помощью или зародышевой Vκ1-39, или зародышевой Vκ3-20 сконструированной легкой цепи и подвергали повторному скринингу в отношении связывания с антигеном Ε с помощью анализа LUMINEX™ (описанного в примере 7 и примере 10). Связывание с антигеном Ε подтверждали с помощью BIACORE™ (описанного в примере 9). Результаты показаны в таблице 15.
Как показано в настоящем примере, 28 антител с общей легкой цепью, специфических к антигену Е, были способны связываться с антигеном Ε при повторном спаривании с зародышевой формой легкой цепи.

Пример 12
Частота использования гена тяжелой цепи и частота соматических гипермутаций в антителах с общей легкой цепью
Последовательности тяжелой и легкой цепей (>6000) антител, полученных у мышей VELCOIMMUNE® (например, патент США №№6596541 и 7105348), составляли с последовательностями тяжелой и легкой цепей (>600) антител с общей легкой цепью, полученных с помощью схемы мультиантигенной иммунизации с использованием мышей со сконструированной легкой цепью (описанных выше) для сравнения частоты использования генного сегмента тяжелой цепи и частот возникновения соматических гипермутаций цепей антител.
Частота использования гена тяжелой цепи. Последовательности тяжелой и легкой цепи, полученные от мышей VELOCIMMUNE®, содержащих замещение эндогенного локус тяжелой цепи мыши генными сегментами VH, DH и JH человека и замещение эндогенного локуса κ легкой цепи мыши или сконструированной зародышевой областью легкой цепи Vκ1-39Jκ5 человека, или сконструированной зародышевой областью легкой цепи Vκ3-20Jκ1 человека (как описано в примере 2), иммунизированных рецептором клеточной поверхности человека (антигеном Е), гетеродимером двух гликопротеинов клеточной поверхности человека (антигеном F), рецептором цитокина человека (антигеном G) и антигеном дифференциации опухоли человека (антигеном Н), анализировали в отношении частоты использования генного сегмента тяжелой цепи и регистрировали генные сегменты VH и JH. Результаты показаны в таблицах 16-18. Процентные отношения в таблицах 16-18 представляют округленные значения и в некоторых случаях могут не равняться 100% при суммировании.
На таблице 16 представлено процентное отношение частоты использования семейства тяжелых цепей для антител от мышей VELCOIMMUNE® (VI), антител от мышей VELCOIMMUNE®, содержащих когнатную легкую цепь Vκ1-39 (VI - Vκ1-39), антител от мышей со сконструированной легкой цепью Vκ1-39 (Vκ1-39), антител от мышей VELCOIMMUNE®, содержащих когнатную легкую цепь Vκ3-20 (VI - Vκ3-20), и антител от мышей со сконструированной легкой цепью Vκ3-20 (Vκ3-20). На таблице 17 представлено процентное отношение частоты использования генов VH и JH для антител от мышей VELCOIMMUNE® (VI), антител от мышей VELCOIMMUNE®, содержащих когнатную легкую цепь Vκ1-39 (VI - Vκ1-39), антител от мышей со сконструированной легкой цепью Vκ1-39 (Vκ1-39), антител от мышей VELCOIMMUNE®, содержащих когнатную легкую цепь Vκ3-20 (VI - Vκ3-20), и антител от мышей со сконструированной легкой цепью Vκ3-20 (Vκ3-20). На таблице 18 представлено процентное отношение частоты использования гена VH для антител от мышей со сконструированной легкой цепью Vκ1-39 (мышей Vκ1-39) из каждой группы иммунизации (антигены Е, F, G и Н) и процентное отношение частоты использования гена VH для антител от мышей со сконструированной легкой цепью Vκ3-20 (мышей Vκ3-20) из выбранных групп иммунизации (антигены Ε и G).
Как показано в настоящем примере, частота использования гена тяжелой цепи для антигенов, проанализированных у мышей со сконструированной легкой цепью Vκ1-39Jκ5, характеризовалась преимуществом III подгрупп семейства VH (VH3-7, VH3-9, VH3-11, VH3-13, VH3-20, VH3-23, VH3-30, VH3-33 и VH3-48). Значительная частота использования других подгрупп семейства VH характеризовалась частотой использования VH1-18, VH1-69, VH2-5, VH4-59 и VH6-1. Для антигенов, исследованных у мышей со сконструированной легкой цепью Vκ3-20Jκ1, частота использования гена тяжелой цепи характеризовалась преимуществом подгруппы III семейства VH, подгруппы IV семейства VH и подгруппы V семейства VH (VH3-11, VH3-30, VH3-33, VH4-39, VH4-59 и VH5-51). Значительная частота использования других подгрупп семейства VH характеризовалась частотой использования VH1-18, VH1-69, VH2-70 и VH6-1.
Частота соматических гипермутаций. Тяжелые и легкие цепи из антител, произведенных мышами VELCOIMMUNE® и мышами со сконструированной легкой цепью (описанными выше), выравнивали с зародышевыми последовательностями согласно частоте использования гена тяжелой и легкой цепи, продемонстрированной для каждой тяжелой и/или легкой цепи. Рассчитывали изменения в аминокислотах для каждой каркасной области (FW) и определяющей комплементарность области (CDR) как для тяжелой, так и легкой цепи каждой последовательности. Результаты показаны в таблицах 19-22. Процентные отношения в таблицах 21-24 представляют округленные значения и в некоторых случаях могут не равняться 100% при суммировании.
На таблице 19 представлено количество изменения аминокислот (АА), наблюдаемых в каждой FW и CDR области тяжелых цепей антител от мышей VELCOIMMUNE®, тяжелых цепей из антител от мышей со сконструированной легкой цепью Vκ1-39 (мышей Vκ1-39) и тяжелых цепей антител от мышей со сконструированной легкой цепью Vκ3-20 (мышей Vκ3-20). На таблице 20 представлено число изменений аминокислот (АА), наблюдаемых в каждой FW и CDR области легких цепей антител от мышей VELCOIMMUNE®, легкой цепи антител от сконструированных мышей Vκ1-39 (мышей Vκ1-39) и легкой цепи антител от сконструированных мышей Vκ3-20 (мышей Vκ3-20). На таблице 21 представлено число изменений аминокислот (АА), наблюдаемых в каждой FW и CDR области тяжелых цепей антител от мышей со сконструированной легкой цепью Vκ1-39 (мышей Vκ1-39) для выбранных групп иммунизации (антигены Е, F и Н). На таблице 22 представлено число изменений аминокислот (АА), наблюдаемых в каждой FW и CDR области тяжелых цепей антител от мышей со сконструированной легкой цепью Vκ3-20 (мышей Vκ3-20) для выбранных групп иммунизации (антигены Ε и G).
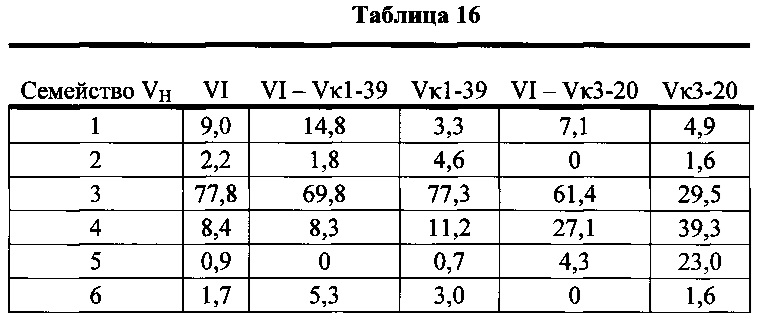
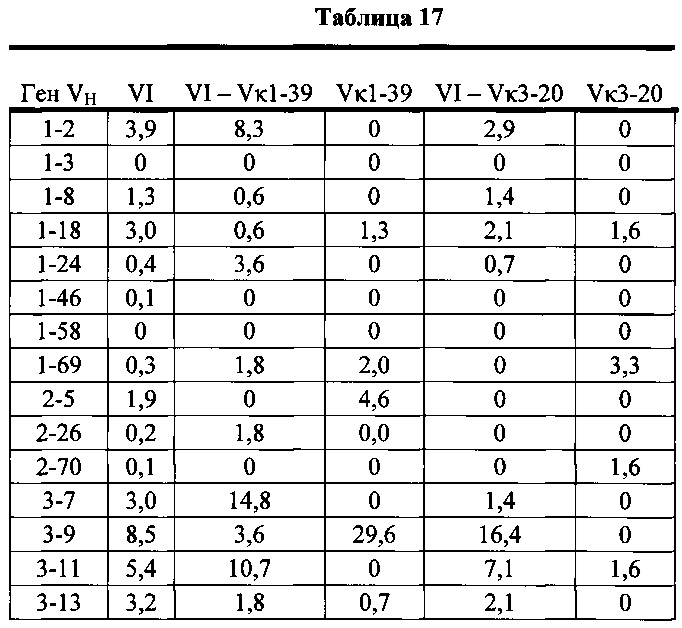
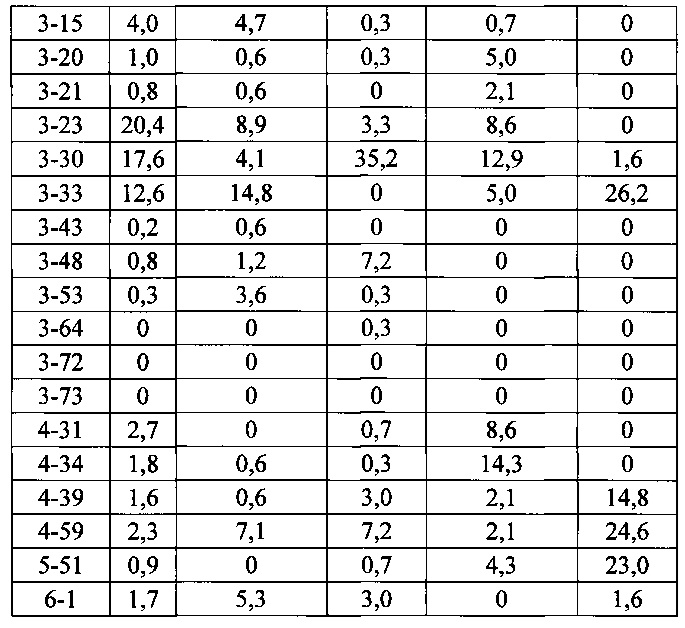
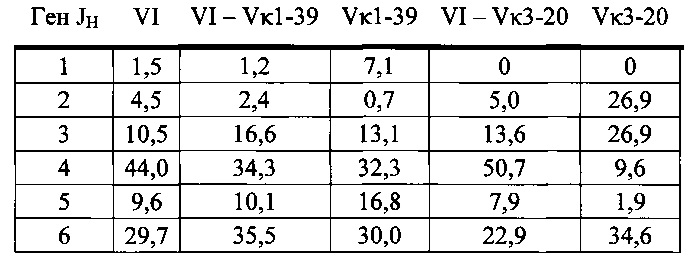
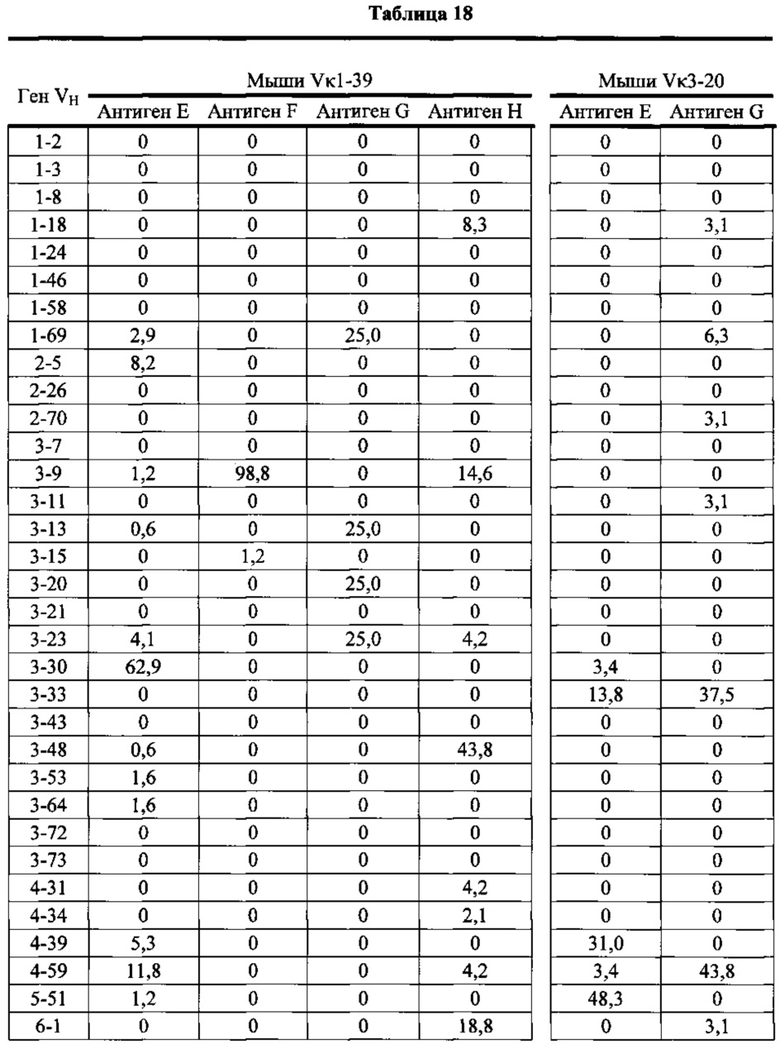
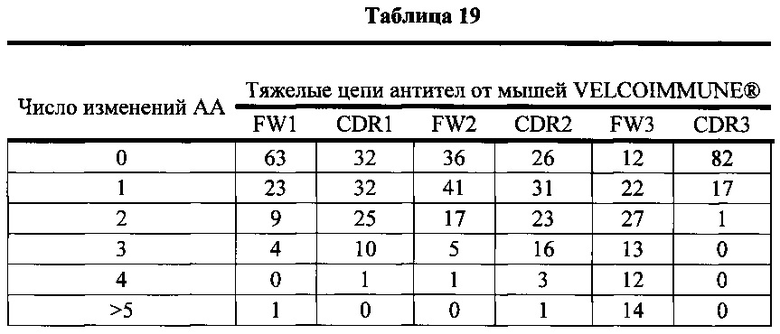
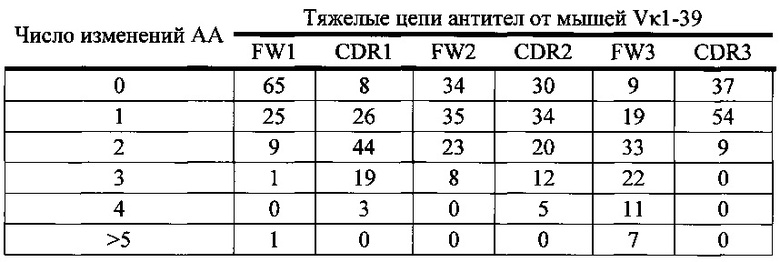
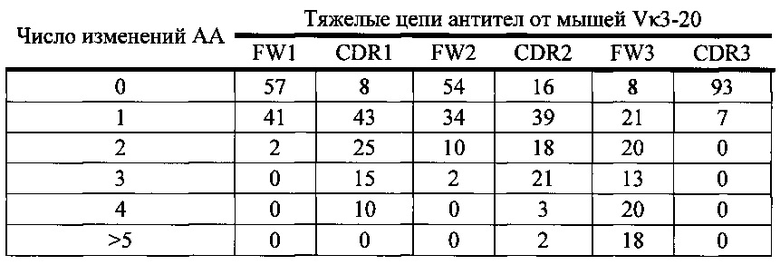
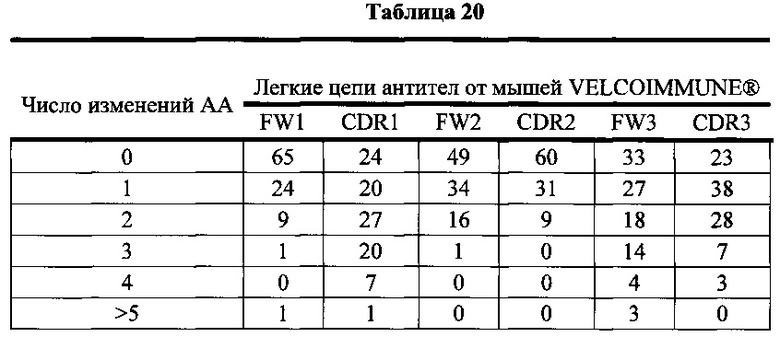
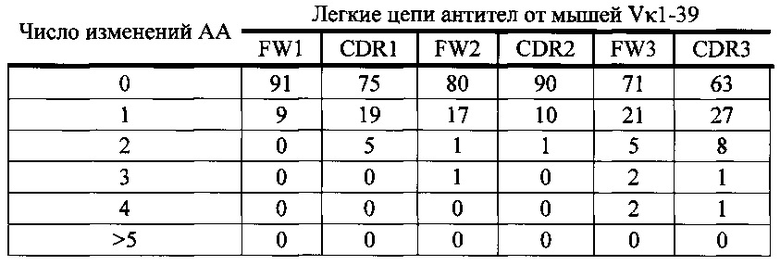
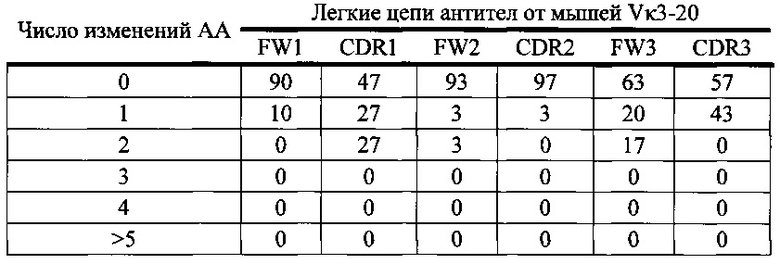
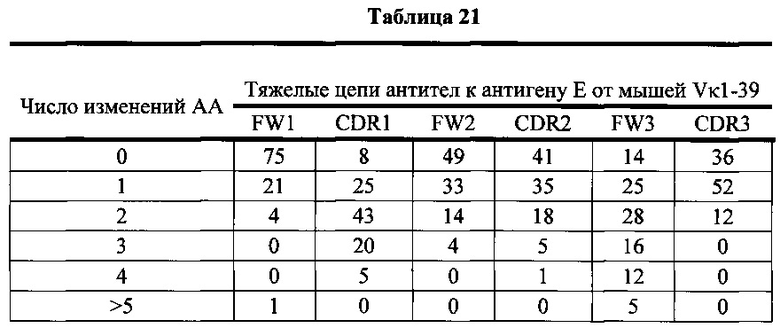

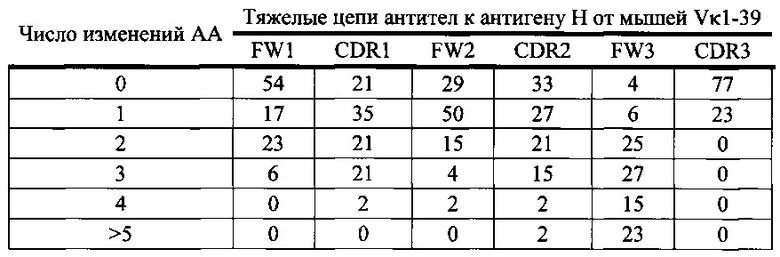
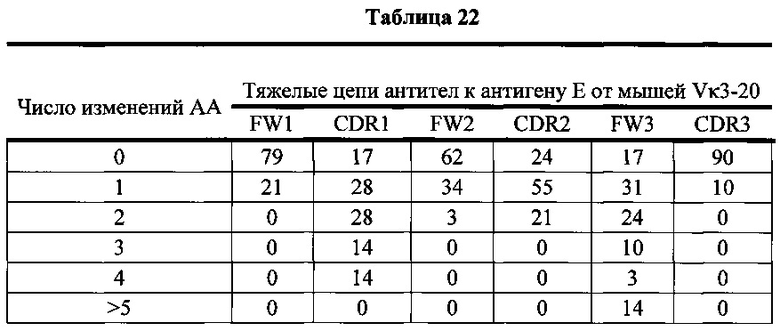
| название | год | авторы | номер документа |
|---|---|---|---|
| МЫШИ, ЭКСПРЕССИРУЮЩИЕ ОГРАНИЧЕННЫЙ РЕПЕРТУАР ЛЕГКИХ ЦЕПЕЙ ИММУНОГЛОБУЛИНА | 2014 |
|
RU2689664C2 |
| МЫШИ, ЭКСПРЕССИРУЮЩИЕ ОГРАНИЧЕННЫЙ РЕПЕРТУАР ЛЕГКИХ ЦЕПЕЙ ИММУНОГЛОБУЛИНА | 2014 |
|
RU2812814C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ УНИВЕРСАЛЬНОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2664232C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ УНИВЕРСАЛЬНОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2787781C2 |
| АНТИТЕЛА СО ВСТРОЕННЫМ В ЛЕГКИЕ ЦЕПИ ГИСТИДИНОМ И ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА ЖИВОТНЫЕ ДЛЯ ИХ ПОЛУЧЕНИЯ | 2013 |
|
RU2644684C2 |
| ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА ЖИВОТНЫЕ, ЭКСПРЕССИРУЮЩИЕ АНТИТЕЛА С ОБЩЕЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2614859C2 |
| МЫШЬ С ОБЩЕЙ ЛЕГКОЙ ЦЕПЬЮ | 2011 |
|
RU2571205C2 |
| АНТИТЕЛА СО ВСТРОЕННЫМ В ЛЕГКИЕ ЦЕПИ ГИСТИДИНОМ И ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА ЖИВОТНЫЕ ДЛЯ ИХ ПОЛУЧЕНИЯ | 2014 |
|
RU2694728C2 |
| МЫШЬ С ОБЩЕЙ ЛЕГКОЙ ЦЕПЬЮ | 2020 |
|
RU2826121C2 |
| МЫШЬ С ОБЩЕЙ ЛЕГКОЙ ЦЕПЬЮ | 2011 |
|
RU2724663C2 |














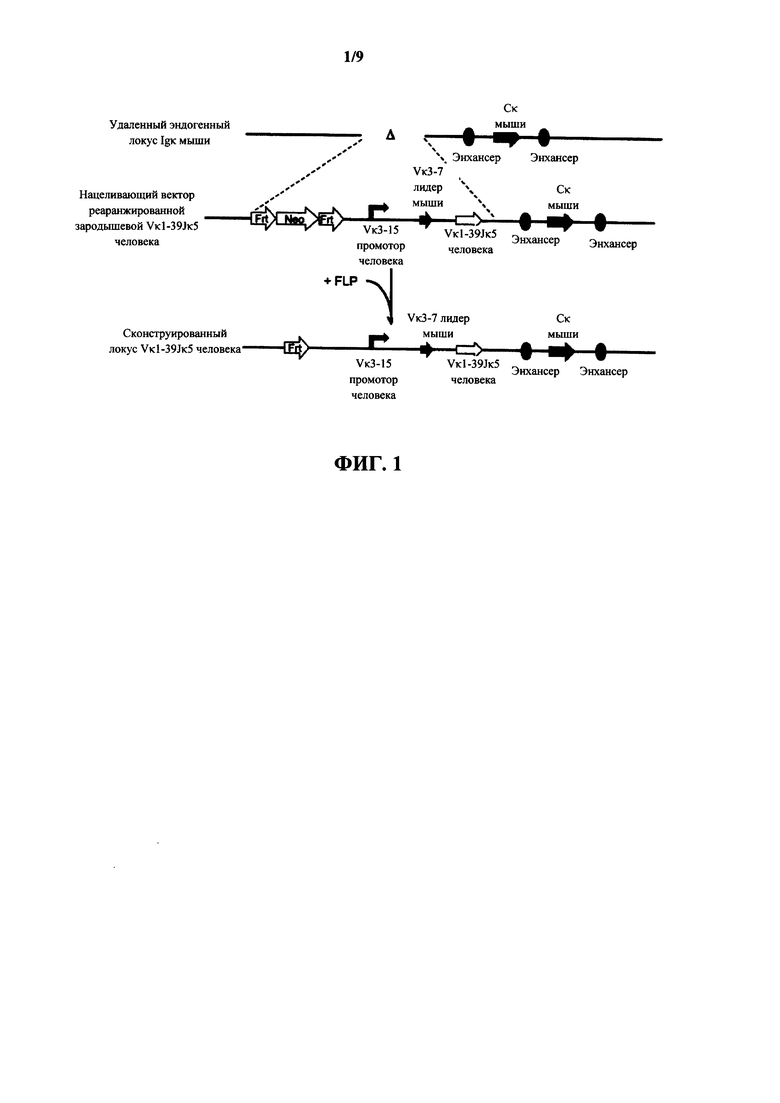
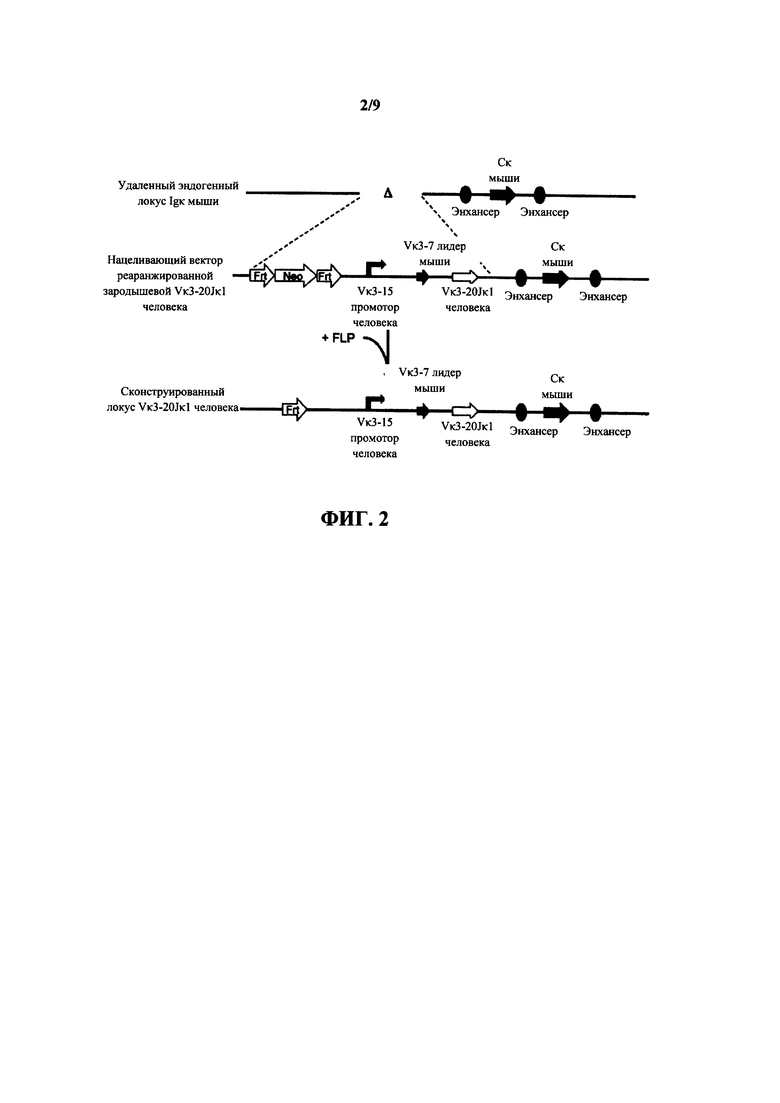
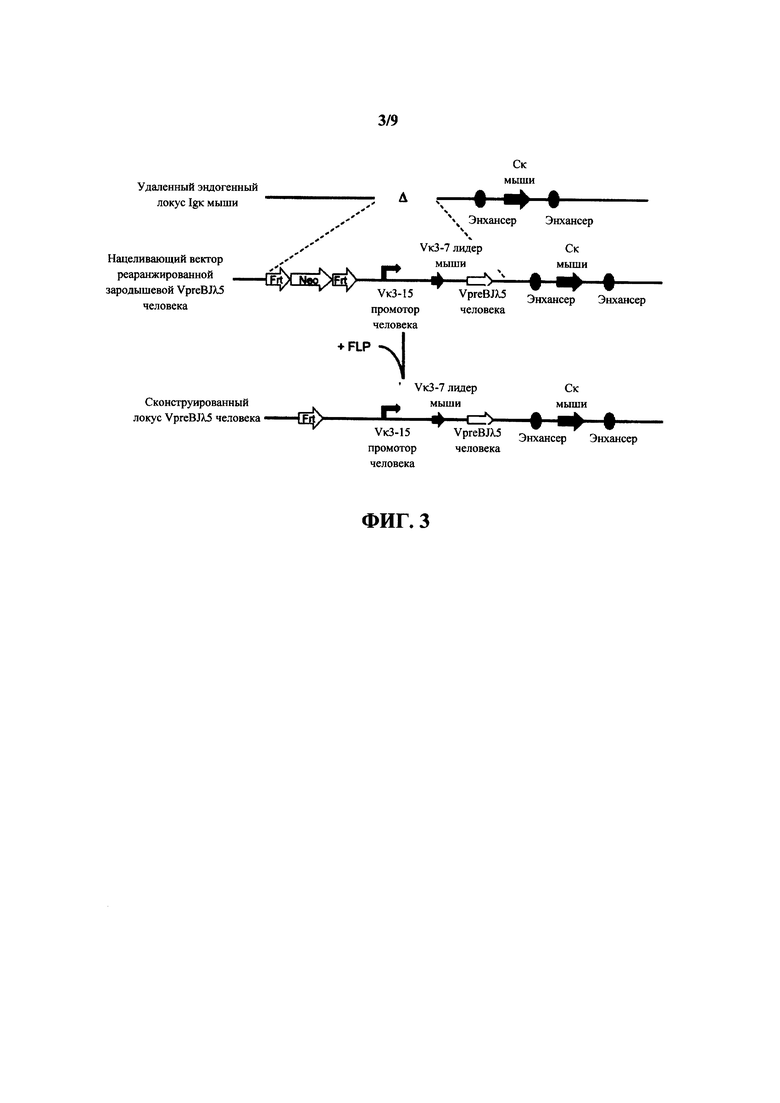
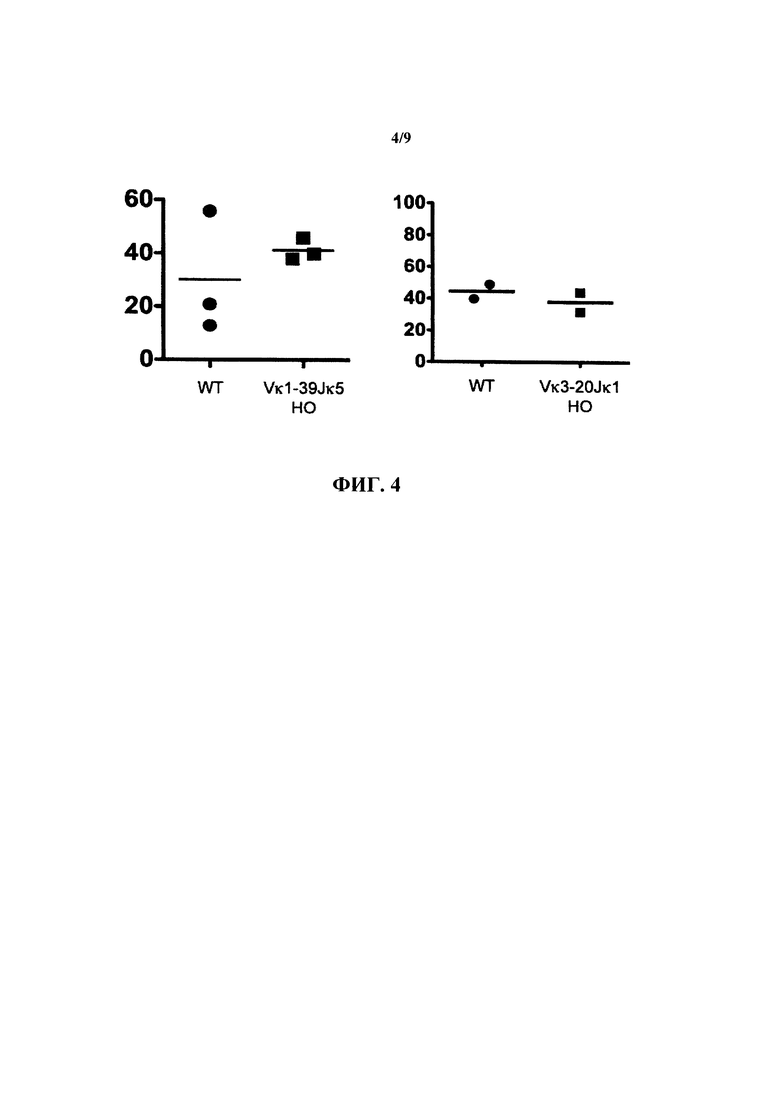
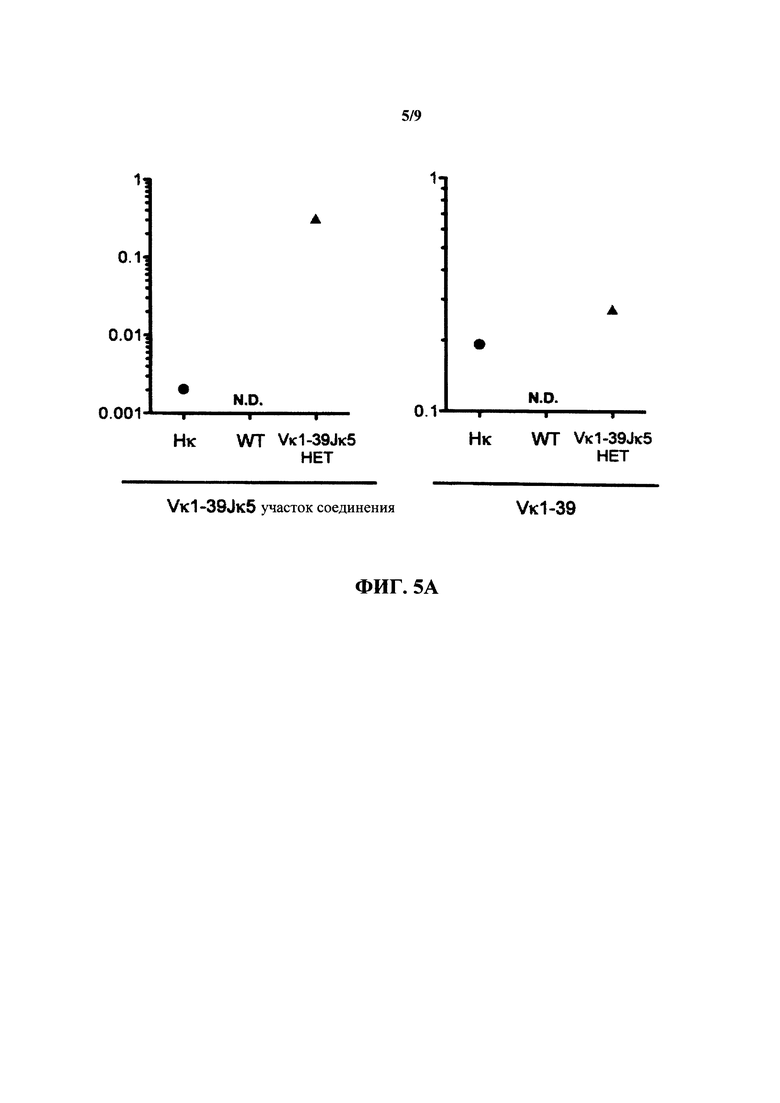
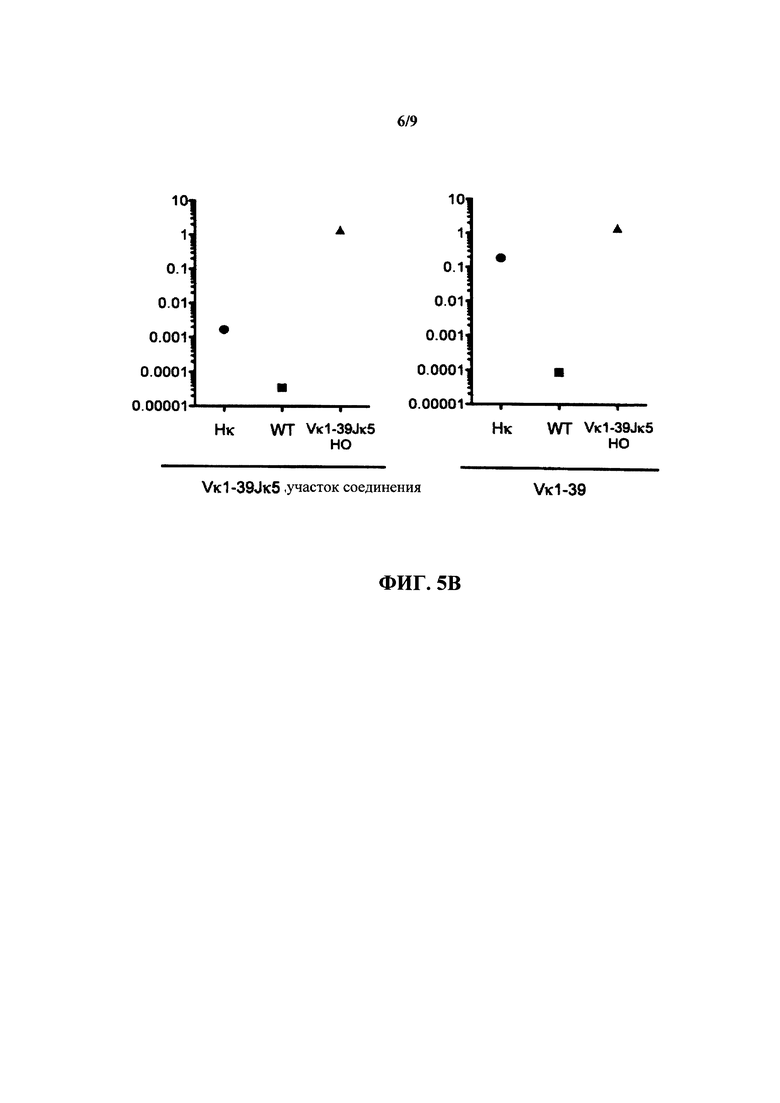
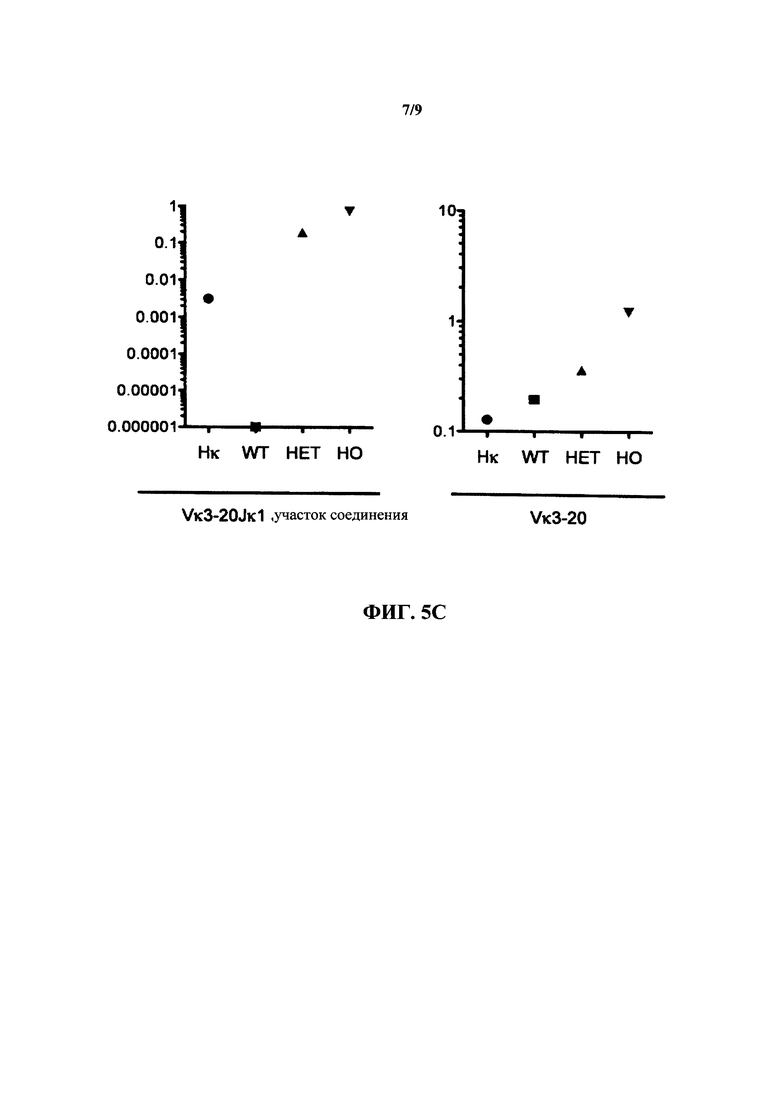
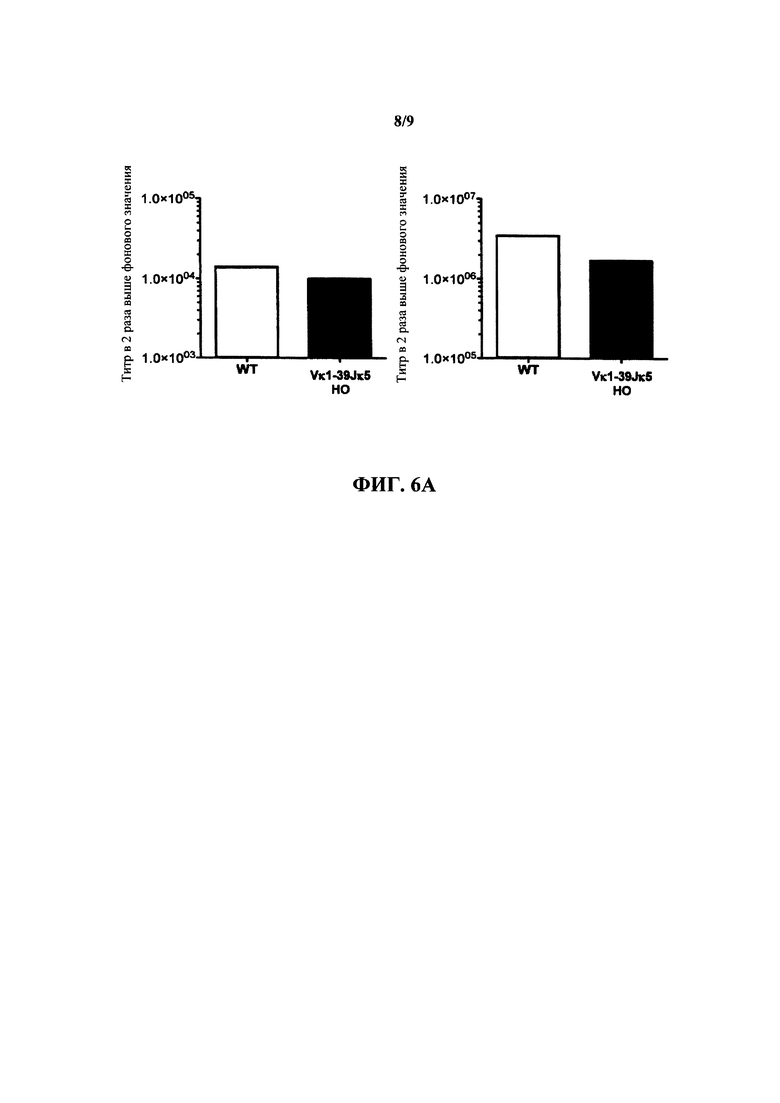
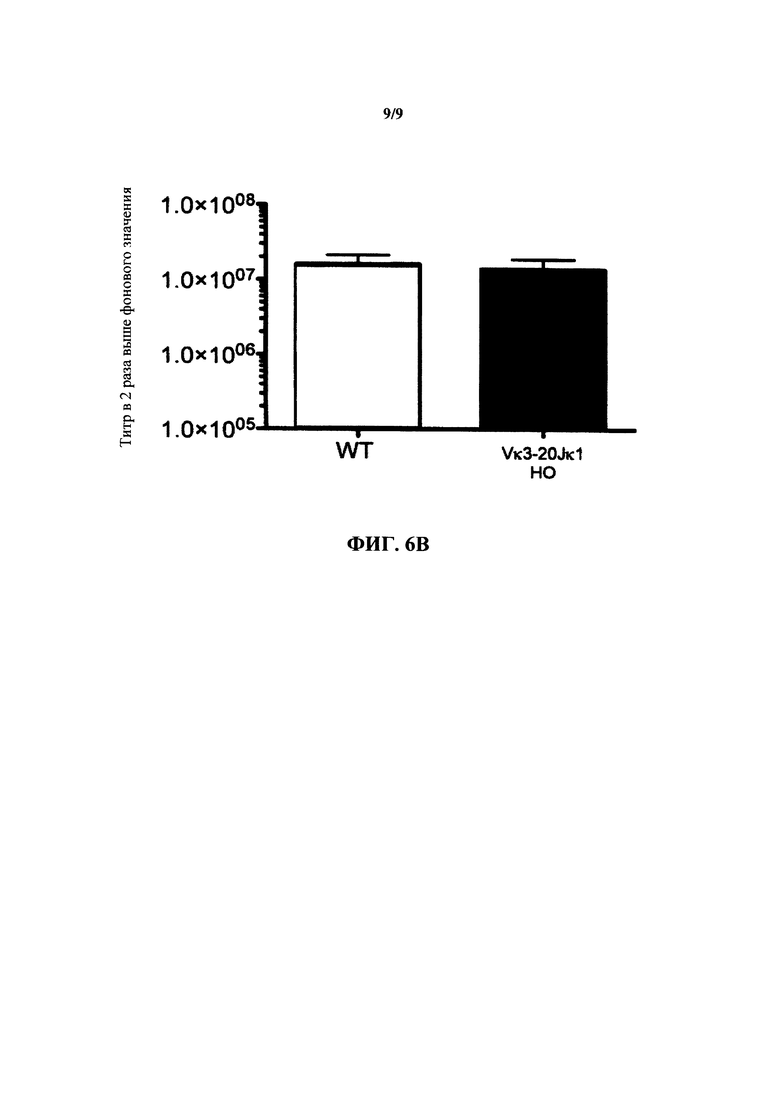
Изобретение относится к области биохимии, в частности к генетически модифицированной мыши для получения антитела, причем антитело содержит первую тяжелую цепь иммуноглобулина, содержащую первый вариабельный домен тяжелой цепи иммуноглобулина человека, кодируемый вариабельной областью тяжелой цепи иммуноглобулина человека, содержащей VH2-26, VH3-21, VH3-64 или их соматически гипермутированный вариант. Также раскрыто применение вышеуказанной генетически модифицированной мыши в получении антитела и в получении биспецифического антитела. Изобретение также относится к способу получения антитела к представляющему интерес антигену, причем антитело содержит тяжелую цепь иммуноглобулина, содержащую вариабельный домен тяжелой цепи иммуноглобулина человека, кодируемый последовательностью вариабельной области тяжелой цепи иммуноглобулина человека, содержащей VH2-26, VH3-21, VH3-64 человека или их соматически гипермутированный вариант, а также к способу получения полностью человеческого биспецифического антитела. Изобретение позволяет эффективно получать антитело в генетически модифицированной мыши. 7 н. и 17 з.п. ф-лы, 6 ил., 22 табл., 12 пр. 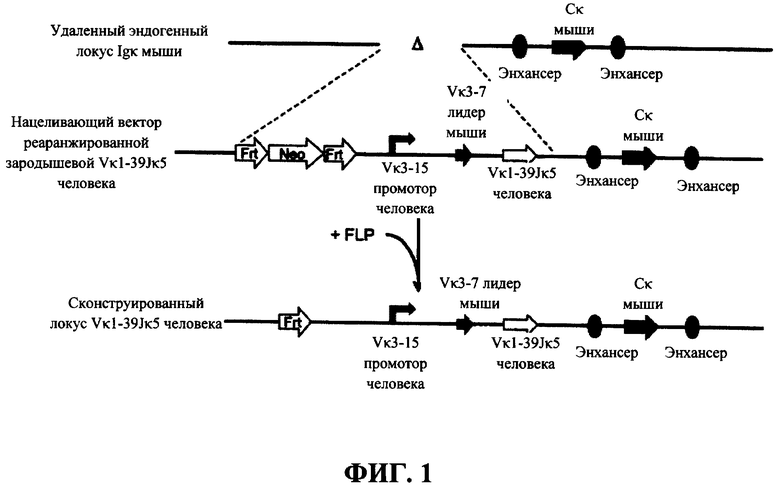
1. Генетически модифицированная мышь для получения антитела, причем антитело содержит первую тяжелую цепь иммуноглобулина, содержащую первый вариабельный домен тяжелой цепи иммуноглобулина человека, кодируемый вариабельной областью тяжелой цепи иммуноглобулина человека, содержащей VH2-26, VH3-21, VH3-64 или их соматически гипермутированный вариант,
причем генетически модифицированная мышь содержит популяцию В-клеток, каждая из которых экспрессирует легкую цепь иммуноглобулина, которая содержит вариабельный домен легкой цепи иммуноглобулина человека, кодируемый вариабельной областью легкой цепи иммуноглобулина человека, содержащей единственную реаранжированную последовательность Vκ1-39/Jκ человека, которая присутствует в зародышевой линии мыши, причем реаранжированная последовательность Vκ1-39/Jκ человека функционально связана с константной областью легкой цепи иммуноглобулина,
причем В-клетки популяции включают в себя по меньшей мере одну В-клетку, которая экспрессирует тяжелую цепь иммуноглобулина, которая содержит первый вариабельный домен тяжелой цепи иммуноглобулина человека, кодируемый вариабельной областью тяжелой цепи иммуноглобулина человека, содержащей генный сегмент VH человека группы, состоящей из VH2-26, VH3-21, VH3-64 и их соматически мутированных вариантов.
2. Мышь по п. 1, у которой В-клетки популяции вместе экспрессируют вариабельные домены тяжелой цепи иммуноглобулина человека, кодируемые вариабельными областями тяжелой цепи иммуноглобулина человека, содержащими:
(a) VH2-26 или его соматически гипермутированный вариант,
(b) VH3-21 или его соматически гипермутированный вариант, и
(c) VH3-64 или его соматически гипермутированный вариант.
3. Мышь по п. 1, у которой каждая из В-клеток в популяции экспрессирует вариабельный домен легкой цепи иммуноглобулина человека, кодируемый вариабельной областью легкой цепи иммуноглобулина человека, которая представляет собой соматически гипермутированный вариант реаранжированной последовательности Vκ1-39/Jκ человека.
4. Мышь по п. 1, у которой:
(a) реаранжированная последовательность Vκ1-39/Jκ человека представляет собой реаранжированную последовательность Vκ1-39/Jκ5 человека;
(b) реаранжированная последовательность Vκ1-39/Jκ человека функционально связана с областью Сκ мыши;
(c) вариабельная область тяжелой цепи иммуноглобулина человека, кодирующая первый вариабельный домен тяжелой цепи иммуноглобулина человека у мыши, функционально связана с последовательностью нуклеиновой кислоты константной (СН) области тяжелой цепи иммуноглобулина мыши, кодирующей СН1, шарнир, СН2, СН3 или их комбинацию; и/или
(d) вариабельная область тяжелой цепи иммуноглобулина человека, кодирующая первый вариабельный домен тяжелой цепи иммуноглобулина человека у мыши, экспрессирована из эндогенного локуса тяжелой цепи иммуноглобулина.
5. Мышь по п. 1, у которой реаранжированная последовательность Vκ1-39/Jκ человека:
(a) расположена так, что она замещает эндогенный нереаранжированный генный сегмент Vκ иммуноглобулина и эндогенный нереаранжированный генный сегмент Jκ иммуноглобулина;
(b) расположена так, что она замещает все функциональные эндогенные нереаранжированные генные сегменты Vκ иммуноглобулина и все функциональные эндогенные нереаранжированные генные сегменты Jκ иммуноглобулина; и/или
(c) функционально связана с константной областью легкой цепи иммуноглобулина мыши или константной областью легкой цепи иммуноглобулина крысы.
6. Мышь по п. 1, у которой антитело содержит легкую цепь иммуноглобулина, которая содержит вариабельный домен легкой цепи иммуноглобулина, кодируемый вариабельной областью легкой цепи иммуноглобулина человека, содержащей реаранжированную последовательность Vκ1-39/Jκ5 человека, которая расположена в геноме зародышевой линии так, что она замещает по меньшей мере один эндогенный нереаранжированный генный сегмент Vκ иммуноглобулина и по меньшей мере один эндогенный нереаранжированный генный сегмент Jκ иммуноглобулина.
7. Мышь по п. 1, у которой первая тяжелая цепь иммуноглобулина образует пару с легкой цепью иммуноглобулина, содержащей вариабельный домен легкой цепи иммуноглобулина человека, кодируемый вариабельной областью легкой цепи иммуноглобулина человека, которая содержит реаранжированную последовательность Vκ1-39/Jκ человека или ее соматически мутированный вариант.
8. Мышь по п. 1, у которой антитело содержит вторую тяжелую цепь иммуноглобулина, причем вторая тяжелая цепь иммуноглобулина содержит второй вариабельный домен тяжелой цепи иммуноглобулина человека, кодируемый вариабельной областью тяжелой цепи иммуноглобулина человека, содержащей VH2-26, VH3-21, VH3-64 человека или их соматически мутированный вариант.
9. Мышь по п. 1, у которой антитело содержит вторую тяжелую цепь иммуноглобулина, и одна или обе из первой тяжелой цепи иммуноглобулина и второй тяжелой цепи иммуноглобулина содержат вариабельный домен тяжелой цепи человека, кодируемый вариабельной областью тяжелой цепи иммуноглобулина человека, экспрессируемый В-клеткой мыши или выделенный из нее.
10. Применение генетически модифицированной мыши по п. 9 в получении антитела, предусматривающее: экспрессию первой тяжелой цепи иммуноглобулина, содержащей первый вариабельный домен тяжелой цепи иммуноглобулина человека, кодируемый вариабельной областью тяжелой цепи иммуноглобулина человека, и второй тяжелой цепи иммуноглобулина вместе в клетке, которая также экспрессирует легкую цепь иммуноглобулина, при этом последовательность первой вариабельной области тяжелой цепи иммуноглобулина человека была экспрессирована из указанной генетически модифицированной мыши или выделена из нее, и при этом легкая цепь иммуноглобулина содержит вариабельный домен легкой цепи иммуноглобулина, кодируемый вариабельной областью легкой цепи иммуноглобулина человека, содержащей реаранжированную Vκ1-39/Jκ человека или ее соматически мутированный вариант.
11. Применение генетически модифицированной мыши по п. 1 в получении антитела, предусматривающее:
(a) иммунизацию указанной генетически модифицированной мыши с помощью представляющего интерес антигена,
(b) выделение тяжелой цепи иммуноглобулина из указанной генетически модифицированной мыши, которая включает первый вариабельный домен тяжелой цепи иммуноглобулина человека,
(c) определение последовательности вариабельной области тяжелой цепи иммуноглобулина человека, кодирующей указанный первый вариабельный домен тяжелой цепи иммуноглобулина человека, и
(d) экспрессию в клетке последовательности, кодирующей тяжелую цепь, которая содержит вариабельную область тяжелой цепи иммуноглобулина человека и последовательности, кодирующей легкую цепь таким образом, что антитело формируется посредством спаривания экспрессированной первой тяжелой цепи иммуноглобулина человека с легкой цепью иммуноглобулина.
12. Применение генетически модифицированной мыши по п. 9 в получении антитела, при этом антитело представляет собой биспецифическое антитело, предусматривающее:
(a) идентификацию последовательности, кодирующей первую тяжелую цепь иммуноглобулина, причем первая тяжелая цепь иммуноглобулина содержит вариабельный домен тяжелой цепи иммуноглобулина человека, встречающийся в популяции В-клеток указанной генетически модифицированной мыши и выделение последовательности, кодирующей первый вариабельный домен тяжелой цепи иммуноглобулина из В-клетки, указанной генетически модифицированной мыши,
(b) идентификацию последовательности, кодирующей отличающуюся вторую тяжелую цепь иммуноглобулина, содержащую второй вариабельный домен тяжелой цепи иммуноглобулина человека, встречающийся в популяции В-клеток указанной генетически модифицированной мыши и выделение последовательности, кодирующей второй вариабельный домен тяжелой цепи иммуноглобулина из В-клетки, указанной генетически модифицированной мыши, и
(с) использование последовательности, кодирующей первый вариабельный домен тяжелой цепи иммуноглобулина и последовательности, кодирующей второй вариабельный домен тяжелой цепи иммуноглобулина для получения биспецифического антитела.
13. Применение генетически модифицированной мыши по п. 1 в получении биспецифического антитела, предусматривающее:
идентификацию последовательности, кодирующей первую тяжелую цепь иммуноглобулина, причем первая тяжелая цепь иммуноглобулина содержит вариабельный домен тяжелой цепи иммуноглобулина человека, встречающийся в популяции В-клеток указанной генетически модифицированной мыши и выделение последовательности, кодирующей первый вариабельный домен тяжелой цепи иммуноглобулина из В-клетки, указанной генетически модифицированной мыши, и
идентификацию последовательности, кодирующей отличающуюся вторую тяжелую цепь иммуноглобулина, причем вторая тяжелая цепь иммуноглобулина содержит второй вариабельный домен тяжелой цепи иммуноглобулина человека, встречающийся в популяции В-клетках, произведенных отличающейся второй генетически модифицированной мышью и выделение последовательности, кодирующей отличающийся второй вариабельный домен тяжелой цепи иммуноглобулина из В-клетки, указанной отличающейся второй генетически модифицированной мыши,
причем вторая популяция В-клеток указанной второй генетически модифицированной мыши представляет собой популяцию В-клеток, каждая из которых экспрессирует вариабельный домен легкой цепи иммуноглобулина человека, кодируемый вариабельной областью легкой цепи иммуноглобулина человека, содержащей реаранжированную последовательность Vκ1-39/Jκ человека или ее соматически гипермутированный вариант, причем реаранжированная последовательность Vκ1-39/Jκ человека присутствует в зародышевой линии второй мыши, и при этом реаранжированная последовательность Vκ1-39/Jκ человека функционально связана с константной областью легкой цепи иммуноглобулина, причем В-клетки второй популяции также включают в себя по меньшей мере одну В-клетку, которая экспрессирует второй вариабельный домен тяжелой цепи иммуноглобулина человека, кодируемый вариабельной областью тяжелой цепи иммуноглобулина человека, содержащей VH2-26, VH3-21, VH3-64 человека или их соматически мутированный вариант, и
использование указанной последовательности, кодирующей первый вариабельный домен тяжелой цепи иммуноглобулина и указанной последовательности, кодирующей второй вариабельный домен тяжелой цепи иммуноглобулина для получения антитела, при этом антитело представляет собой биспецифическое антитело.
14. Применение по любому из пп. 10-13, при котором зародышевая линия генетически модифицированной мыши не содержит эндогенный нереаранжированный генный сегмент Vκ иммуноглобулина, функциональный для экспрессии в легкой цепи иммуноглобулина, и зародышевая линия генетически модифицированной мыши не содержит эндогенный нереаранжированный генный сегмент Jκ иммуноглобулина, функциональный для экспрессии в легкой цепи иммуноглобулина.
15. Применение по любому из пп. 10-13, при котором антитело представляет собой полностью человеческое антитело.
16. Применение по любому из пп. 10-11, при котором антитело представляет собой биспецифическое антитело.
17. Способ получения антитела к представляющему интерес антигену, причем антитело содержит тяжелую цепь иммуноглобулина, содержащую вариабельный домен тяжелой цепи иммуноглобулина человека, кодируемый последовательностью вариабельной области тяжелой цепи иммуноглобулина человека, содержащей VH2-26, VH3-21, VH3-64 человека или их соматически гипермутированный вариант, причем способ предусматривает:
(а) идентификацию вариабельной области тяжелой цепи иммуноглобулина человека, из генетически модифицированной мыши и выделение вариабельной области тяжелой цепи иммуноглобулина из В-клетки, указанной генетически модифицированной мыши, причем указанная генетически модифицированная мышь отличается тем, что ее В-клетки производят популяцию антител, каждое из которых содержит:
(i) тяжелую цепь иммуноглобулина, содержащую вариабельный домен тяжелой цепи иммуноглобулина человека, кодируемый вариабельной областью тяжелой цепи иммуноглобулина человека, содержащей VH2-26, VH3-21, VH3-64 человека или их соматически мутированный вариант; и
(ii) легкую цепь иммуноглобулина, содержащую вариабельный домен легкой цепи иммуноглобулина человека, кодируемый вариабельной областью легкой цепи иммуноглобулина человека, содержащей реаранжированную последовательность Vκ1-39/Jκ человека; и
(b) использование указанной полученной в (а) вариабельной области тяжелой цепи иммуноглобулина человека для получения антитела, которое специфически связывает представляющий интерес антиген.
18. Способ получения полностью человеческого биспецифического антитела, причем способ предусматривает:
(а) экспрессию первой тяжелой цепи иммуноглобулина человека, содержащей последовательность, кодируемую первой вариабельной областью тяжелой цепи иммуноглобулина человека, происходящей из В-клетки генетически сконструированной мыши, причем мышь:
(i) характеризуется геномом зародышевой линии, который является гомозиготным или гетерозиготным в отношении реаранжированной последовательности Vκ1-39/Jκ человека, причем реаранжированная последовательность Vκ1-39/Jκ человека функционально связана с областью Сκ;
(ii) характеризуется геномом зародышевой линии, который включает в себя нереаранжированные генные сегменты VH человека, генные сегменты DH человека и генные сегменты JH человека, причем нереаранжированные генные сегменты VH человека функционально связаны с областью CH и включают, VH2-26 человека, VH3-21 человека, VH3-64 человека или их комбинацию; и
(iii) содержит популяцию В-клеток, каждая из которых экспрессирует легкую цепь иммуноглобулина, содержащую вариабельный домен легкой цепи иммуноглобулина человека, кодируемый вариабельной областью легкой цепи иммуноглобулина человека, из реаранжированной последовательности Vκ1-39/Jκ человека или ее соматически гипермутированного варианта, и популяция включает в себя по меньшей мере одну В-клетку, которая экспрессирует тяжелую цепь иммуноглобулина, содержащую вариабельный домен тяжелой цепи иммуноглобулина человека, кодируемый вариабельной областью тяжелой цепи иммуноглобулина человека, включая VH2-26, VH3-21, VH3-64 человека или их соматически гипермутированный вариант; и
(b) спаривание экспрессированной первой тяжелой цепи иммуноглобулина человека с легкой цепью иммуноглобулина человека и со второй тяжелой цепью иммуноглобулина человека, которая также образует пару с легкой цепью иммуноглобулина человека для получения полностью человеческого биспецифического антитела, причем вторая тяжелая цепь иммуноглобулина человека содержит последовательность, кодируемую второй вариабельной областью тяжелой цепи иммуноглобулина человека, причем одна или обе из первой вариабельной области тяжелой цепи иммуноглобулина человека и второй вариабельной области тяжелой цепи иммуноглобулина человека кодируются вариабельной областью тяжелой цепи иммуноглобулина человека, которая включает VH2-26, VH3-21, VH3-64 человека или их соматически гипермутированный вариант.
19. Способ по п. 18, при котором В-клетки популяции вместе экспрессируют вариабельные домены тяжелой цепи иммуноглобулина человека, кодируемые вариабельными областями тяжелой цепи иммуноглобулина человека, содержащими:
(a) VH2-26 или его соматически гипермутированный вариант,
(b) VH3-21 или его соматически гипермутированный вариант, и
(c) VH3-64 или его соматически гипермутированный вариант.
20. Способ по п. 18, при котором реаранжированная последовательность Vκ1-39/Jκ человека представляет собой Vκ1-39 зародышевой линии человека, реаранжированную с Jκ5 зародышевой линии человека.
21. Способ по п. 18, при котором антитело содержит вариабельный домен легкой цепи иммуноглобулина человека, кодируемый вариабельной областью легкой цепи иммуноглобулина человека, которая представляет собой реаранжированную последовательность Vκ1-39/Jκ человека или ее соматически мутированный вариант.
22. Способ по любому из пп. 18-21, причем способ предусматривает перед указанной стадией (а) стадию выделения первой вариабельной области тяжелой цепи иммуноглобулина человека из В-клетки генетически сконструированной мыши.
23. Способ по любому из пп. 18-21, при котором вторую вариабельную область тяжелой цепи иммуноглобулина человека выделяют из В-клетки генетически сконструированной мыши.
24. Способ по любому из пп. 18-21, при котором зародышевая линия генетически сконструированной мыши не содержит эндогенный нереаранжированный генный сегмент Vκ иммуноглобулина, функциональный для экспрессии в легкой цепи иммуноглобулина, и зародышевая линия генетически сконструированной мыши не содержит эндогенный нереаранжированный генный сегмент Jκ иммуноглобулина, функциональный для экспрессии в легкой цепи иммуноглобулина.
| WO 2011097603 A1, 11.08.2011 | |||
| DE KRUIF J | |||
| et al., Human immunoglobulin repertoires against tetanus toxoid contain a large and diverse fraction of high-affinity promiscuous VH genes, JOURNAL OF MOLECULAR BIOLOGY, ACADEMIC PRESS, UNITED KINGDOM, vol | |||
| Аппарат для получения газа под высоким давлением для работы в поршневом или турбинном двигателе | 1922 |
|
SU387A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| DE WILDT R.M | |||
| et al., Analysis of heavy and light chain pairings indicates that receptor editing shapes the human antibody repertoire, J Mol Biol., 1999, Vol.285, N.3, pp.895-901 | |||
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ МЫШЕЙ | 2009 |
|
RU2425880C2 |

