Изобретение относится к биотехнологии сельскохозяйственных растений и может найти применение для нужд сельского хозяйства и растениеводства при диагностике трангенного растительного материала. Изобретение может быть использовано в пищевой промышленности при выявлении генноинженерно модифицированных организмов (ГМО).
Известен способ определения трансгенных растений [1, 2], сущность которого состоит в анализе нуклеотидных последовательностей методом полимеразной цепной реакции (ПЦР) с последующей гибридизацией ее продуктов на микрочипах. Основным недостатком этого способа является необходимость переноса амплификата после проведения ПЦР в ячейки биочипов, что создает предпосылку для загрязнения реактивов и лабораторного оборудования продуктами ПЦР и может привести к получению ложных результатов анализа.
Известен способ определения трансгенных растений [3], основанный на амплификации методом ПЦР в реальном времени последовательностей некоторых разновидностей промотора 35S и терминатора nos. Однако этот способ не позволяет выявить значительное количество трансгенных линий, содержащих отличающиеся последовательности данных маркеров.
ПЦР в режиме реального времени позволяет контролировать увеличение количества синтезируемого продукта в ходе реакции и не требует проведения электрофореза на последующих стадиях. Это стало возможным благодаря добавлению в реакционную среду красителей, обеспечивающих флуоресценцию, прямо пропорциональную количеству синтезированного ПЦР-продукта, - репортерную флуоресценцию.
ПЦР в режиме реального времени существует в двух основных модификациях, различающихся по способам генерации репортерной флуоресценции. Первая из них связана с применением интеркалирующих флуоресцентных агентов, свечение которых значительно возрастает при связывании с двуцепочечной ДНК, вторая связана с использованием меченных флуоресцентными агентами олигонуклеотидных проб, комплементарных участку ПЦР-продукта. Вторая модификация ПЦР является более предпочтительной для диагностических целей, поскольку дает дополнительную возможность контроля специфичности реакции.
Использование ПЦР в реальном времени позволяет свести к минимуму проблему контаминации продуктом реакции, ускорить процедуру анализа за счет отсутствия необходимости проведения электрофореза, позволяет получать количественные характеристики присутствия интересующей ДНК в пробе.
На настоящий момент на основе ПЦР технологии разработано огромное количество диагностических систем различного назначения, в том числе для нужд сельского хозяйства и растениеводства.
Известны аналогичные изобретения «Патент РФ 2386698, Кузнецов В.В., Абрамов Д.Д., Митрохин И.А., Цыдендамбаев В.Д. Способ идентификации трансгенных последовательностей днк в растительном материале и продуктах на его основе» [3].
Однако аналогичные наборы не позволяют выявить все разнообразие коммерческих трансгенных линий растений. Это связано с тем, что в генно-инженерных конструкциях использовано несколько разновидностей последовательностей терминатора nos и большое разнообразие последовательностей промоторов 35S.
По данным базы NCBI (ncbi.nlm.nih.gov) последовательности 35S варьируют по длине от 47 до 485 нуклеотидов, а также по нуклеотидному составу. Данные о последовательностях 35S приведены в приложениях 1 и 2.
Сущность изобретения поясняется фиг. 1 и 2.
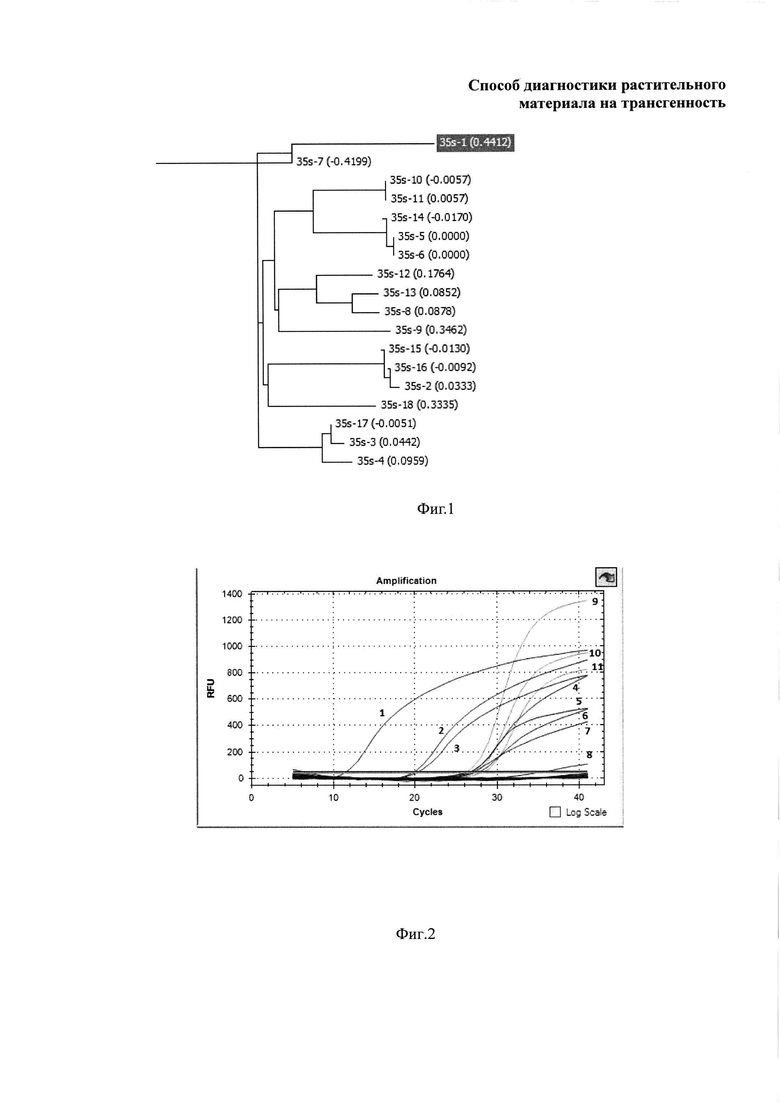
Фиг. 1 иллюстрирует нецелесообразность применения праймеров к промотору 35S для диагностики трансгенности в виду большой молекулярной дивергенции ДНК мишеней.
Древо, отражающее сходство последовательностей, построенное в программе Vector NTI, представлено на фиг. 1.
Как видно из фиг. 1, последовательности 35S существенно отличаются друг от друга и формируют не менее 6 групп сходства. При этом варьирование в пределах каждой группы может быть существенным. Эти обстоятельства делают невозможным применение данной последовательности в качестве универсальной мишени для подбора праймеров, детектирующих ГМО. Анализ последовательностей терминатора nos, напротив, показал, что существует 2 его основные вариации.
То есть данная мишень является пригодной для разработки тест-системы, но необходимо учитывать две разновидности последовательности терминатора nos. Анализ распространения селективного маркера nptII (таблица 1) показал, что он присутствует в более сотни линий ГМО и может быть использован как вспомогательный [4].
Заявителю известно, что работа над созданием праймеров, которые используются в подобных наборах, строится обычно следующим образом.
1) С помощью открытых и коммерческих баз данных нуклеотидных последовательностей генно-инженерных конструкций выбирается их консервативный участок, специфичный для как можно большего количества генно-инженерно модифицированных организмов.
2) На основании выбранного участка генома с помощью специального программного обеспечения подбирается последовательность олигонуклеотидов, используемых для проведения ПЦР-реакции (часто 2 праймера и зонд). На данном этапе работа заключается в создании выравнивания многих нуклеотидных последовательностей и выборе участка последовательности рекомбинантной ДНК с наименьшим полиморфизмом.
3) Изготовление праймеров.
4) С помощью практических экспериментов доказывается пригодность подобранных последовательностей для конкретных целей (например, для определения наличия/отсутствия генно-инженерно модифицированных растений в биоматериале).
Наиболее близким способом по достигаемому техническому результату является способ Диагностики ГМО [3], принятый в качестве прототипа. Сущность его состоит в проведении ПЦР с использованием специфических праймеров и флуоресцентномеченых зондов, комплиментарных трансгенным последовательностям ДНК (таблица 1), с последующей детекцией продуктов амплификации в закрытых амплификационных пробирках с помощью флуоресцентного ПЦР-детектора. Ключевым элементом заявляемого способа является использование флуоресцентномеченых зондов, представляющих собой олигонуклеотиды, меченные молекулами флуорофора и гасителя флуоресценции. Зонды добавляют в реакционную смесь наряду с праймерами и другими компонентами реакции. В ходе ПЦР флуоресцентномеченый зонд гибридизуется на специфических ампликонах и разрушается Taq-полимеразой, в результате чего флуорофор оказывается свободным от гасителя. Таким образом, уровень флуоресценции в амплификационных пробирках возрастает пропорционально количеству образовавшихся специфических продуктов ПЦР.
Недостатком известного способа, принятого в качестве прототипа, является невозможность выявить некоторые трансгенные линии с его помощью. Например, некоторые линии сои, устойчивой к раундапу (см. последовательности NCBI ##AJ783418.1, АВ209952.1, AF465641.1), кукурузы с геном Bt (см. последовательности NCBI ##EU363768.1 EU363766.1). Изобретение свободно от этих недостатков.
Технический результат изобретения состоит в расширении диапазона генотипов диагностируемых ГМО.
Указанный технический результат достигается тем, что в способе диагностики биоматериалов на наличие в них трансгенных растений, заключающемся в выделении из биоматериала ДНК и проведении на ней полимеразной цепной реакции в режиме реального времени зондами, меченными флуоресцентными красителями и гасителями флуоресценции, в соответствии с изобретением после выделения из образцов ДНК в концентрации 0,1-100 нг/мкл на ее основе составляют реакционную смесь, содержащую праймеры 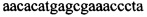 ,
, 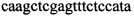 ,
, 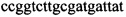 ,
, 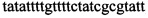 ,
, 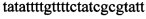 ,
, 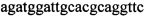 ,
, 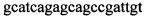 и зонды
и зонды 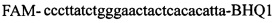 ,
, 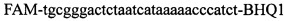 ,
, 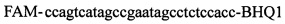 . Полимеразную цепную реакцию проводят в режиме реального времени с температурой отжига праймеров 60°C при 40 циклах, при этом осуществляют непрерывный контроль флуоресценции и по экспоненциальному ее нарастанию диагностируют трансгенность растений. Кроме этого, указанный технический результат достигается тем, что в качестве термостабильной полимеразы с активностью 0,1-0,3 ед. на 1 мкл смеси берут Taq. Вместе с тем указанный технический результат достигается тем, что в качестве термостабильной полимеразы с активностью 0,1-0,3 ед. на 1 мкл смеси берут Tth.
. Полимеразную цепную реакцию проводят в режиме реального времени с температурой отжига праймеров 60°C при 40 циклах, при этом осуществляют непрерывный контроль флуоресценции и по экспоненциальному ее нарастанию диагностируют трансгенность растений. Кроме этого, указанный технический результат достигается тем, что в качестве термостабильной полимеразы с активностью 0,1-0,3 ед. на 1 мкл смеси берут Taq. Вместе с тем указанный технический результат достигается тем, что в качестве термостабильной полимеразы с активностью 0,1-0,3 ед. на 1 мкл смеси берут Tth.
Сущность изобретения состоит в том, что для диагностики наличия в образце трансгенных растений или продуктов их переработки проводят ПЦР в реальном времени с праймерами и зондами к регуляторным последовательностям ДНК (терминатор nos) нескольких разновидностей и селективному маркеру nptII, используемым в большинстве линий трансгенных растений. Например, при сравнении уровня техники:
По патенту РФ 2386698 (RU):
Способ идентификации трансгенных последовательностей ДНК в растительном материале и продуктах на его основе (RU).
При схожей технологии детекции в известном патенте описывается детекция более узкого набора линий, содержащих трансгены.
Существенными признаками изобретения являются:
из анализируемого образца выделяют ДНК чистоты, достаточной для проведения ферментативных реакций, после этого составляют реакционную смесь, содержащую, по меньшей мере: праймеры 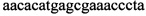 ,
, 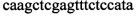 ,
, 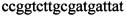 ,
, 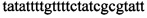 ,
, 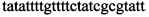 ,
, 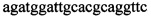 ,
, 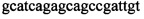 и зонды
и зонды 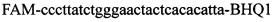 ,
, 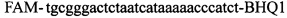 ,
,  , зонд может содержать и другие флуоресцентные красители или гасители флуоресценции;
, зонд может содержать и другие флуоресцентные красители или гасители флуоресценции;
Полимеразную цепную реакцию проводят в амплификаторе (термоциклире) при температуре отжига праймеров 60°C при 40 циклах, при этом осуществляют непрерывный контроль флуоресценции и по экспоненциальному ее нарастанию диагностируют трансгенность растений. Реакцию можно проводить в амплификаторе, обладающем возможностью детектировать изменение флуоресценции в режиме реального времени или в обычном амплификаторе без детектора (в этом случае о прохождении реакции судят по изменению уровня флуоресценции, измеренной перед началом реакции и после ее прохождения).
В случае, если прохождение реакции в виде увеличения уровня флуоресценции образца будет детектировано, делается вывод о наличии трансгенных растений в анализируемом образце.
Реализация заявленного способа.
Для обнаружения заявленным способом трансгенных растений или продуктов их переработки в образце требуется:
1. Выделить ДНК из образца СТАБ-методом. Для этого нужно гомогенезировать образец в СТАВ-буфере (NaCl 1,4М, Трис HCl (pH 8) 0,1М, ЭДТА (pH 8) 20 мМ, СТАВ 2% m/v), инкубировать полученную смесь 1 ч при 56°C, 40-60 мин экстрагировать хлороформом, разделить фракции центрифугированием, верхнюю (водную), содержащую ДНК фракцию переосадить этанолом, ДНК растворить в воде.
2. Приготовить реакционную смесь для ПЦР, которая должна содержать 1х буфер для полимеразы, катионы Mg 2+ в концентрации 0,1-0,3 мM, нуклеотиды в концентрации 2-10 мкМ, Taq полимеразу (или другую термостабильную полимеразу, обладающую 5' экзонуклеазной активностью), 0,1-0,3 ед. активности на 1 мкл смеси, ДНК матрицу исследуемого образца 0,1-100 нг/мкл, праймеры, следующего состава: 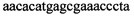 ,
, 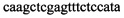 ,
, 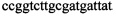 ,
, 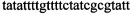 ,
, 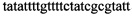 ,
, 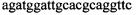 ,
, 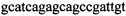 и зонды
и зонды 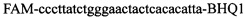 ,
, 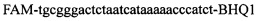 ,
, 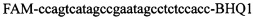 , зонд может содержать и другие флуоресцентные красители или гасители флуоресценции.
, зонд может содержать и другие флуоресцентные красители или гасители флуоресценции.
3. Провести полимеразную цепную реакцию в режиме реального времени с температурой отжига праймеров 60°C при 40 циклах. Пример программы: 95°C - 5 мин, 40 циклов (95°C - 18 с, 60°C - 30 с, 72°C - 30 с). Прохождение реакции контролируют посредством прибора для измерения уровня флуоресценции в ходе реакции, при прохождении реакции хотя бы в одной из реакционных смесей, наблюдается повышение уровня флуоресценции, делается вывод о наличии трансгенных растений в исследуемом образце.
Заявленный способ апробирован в реальном времени на лабораторной базе Санкт-Петербургского государственного университета и поясняется конкретными примерами.
Пример конкретной реализации.
Для обнаружения описываемым методом трансгенных растений и продуктов их переработки в образце требуется:
1. Выделить ДНК из образца СТАБ-методом (подготовительный этап).
2. Провести ПЦР в реальном времени (собственно анализ).
Выделение ДНК из образцов трансгенных корней каланхоэ (трансформированных штаммом А4 Agrobacterium rhizogenes), клубней картофеля семян сои и мясного фарша с добавлением сои.
Гомогенезировать образец в СТАВ-буфере (NaCl 1,4М, Трис HCl (pH 8) 0,1М, ЭДТА (pH 8) 20 мМ, СТАВ 2% m/v), инкубировать полученную смесь 1 ч при 56°C, 40-60 мин экстрагировать хлороформом, разделить фракции центрифугированием, верхнюю (водную), содержащую ДНК фракцию переосадить этанолом, ДНК растворить в воде.
ПОСТАНОВКА ПЦР
В ходе апробации заявленного способа были приготовлены 2 реакционные смеси для ПЦР. Первая смесь содержала 1х буфер для полимеразы, катионы Mg 2+ в концентрации 0,2 мМ, нуклеотиды в концентрации 2 мкМ, Taq полимеразу, 0,2 ед. активности на 1 мкл смеси, ДНК матрицу исследуемого образца 10-100 нг/мкл, праймеры, следующего состава: 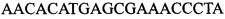 ,
, 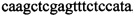 ,
, 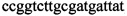 ,
, 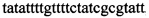 ,
, 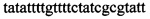 ,
, 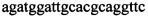 ,
, 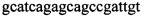 в концентрации 2 mM и зонды
в концентрации 2 mM и зонды 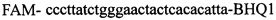 ,
, 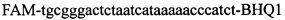 ,
, 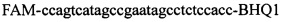
Полимеразную цепную реакцию в режиме реального времени проводили по программе:
95°C - 5 мин, 40 циклов (95°C - 18 с, 60°C - 30 с, 72°C - 30 с).
Прохождение реакции контролировали посредством прибора для измерения уровня флуоресценции в ходе реакции, например амплификатор АНК32 (испытания проводились на лабораторной базе Института Аналитического приборостроения РАН), или CFX96 (Biorad, США). Результаты реакции представлены в качестве примера на фиг. 2.
Фиг. 2 иллюстрирует прохождение реакции с разработанными авторами праймерами на трансгенном и контрольном растительном материале. ПЦР с разработанными авторами праймерами и зондами для детекции ГМО (графики 1-8) и праймерами к гену лектина сои (графики 9-11) на контрольных ДНК плазмиды pART27 (график 1), штаммов агробактерии Т37, С58 (графики 2-3), опытных ДНК трансгенной сои (графики 4, 9), трансгенного картофеля (графики 5-7), трансгенного каланхоэ (график 8), нетрансгенной сои (график 10), мясного фарша (график 11).
Как видно из фиг. 2, амплификация фрагмента гена лектина проходит только на ДНК сои, амплификация терминатора nos - только на трансгенных растениях. Значения пороговых циклов для лектина и nos отличаются на 1 у трансгенной сои, что может быть объяснено большей дозой генов с терминатором nos по сравнению с референсом. Аналогичные результаты получены на трансгенных линиях картофеля с референсным геном соланина и терминатором nos.
Таким образом, наблюдается прохождение реакции, свидетельствующей о работе апробированной диагностической системы. Можно сделать вывод об успешной работе диагностической системы при использовании ДНК заявленных концентраций.
Технико-экономическая эффективность изобретения состоит в том, что предлагаемый новый способ диагностики растительного материала на предмет трансгенности направлен на решение проблемы быстрой диагностики биологического материала растительного происхождения (растения и продукты их переработки), т.е. на эффективное и оперативное выявление наличия в исследуемых биоматериалах трансгенных растений. Заявленный способ по достигаемому техническому результату отличается от известных в данной области аналогов: позволяет расширить список детектируемых трансгенных линий растений.
Источники информации
1. Патент РФ №2453605, Грановский И.Э., Белецкий И.П., Шляпникова Е.А., Шляпников Ю.М., Гаврюшкин А.В., Бирюков С.В. Дифференцирующий и специфический олигонуклеотиды для идентификации последовательностей днк трансгенных растений в пищевых продуктах, способ идентификации трансгенных продуктов, биочип, комбинация олигонуклеотидов (варианты) и набор для осуществления этого способа.
2. Патент РФ 2270254, Мирзабеков А.Д., Грядунов Д.А., Михайлович В.М., Заседателев А.С., Романов Г.А., Кузнецов В.В., Цыденедамбаев В.Д., Митрохин И.А., Крылова Е.М. Способ идентификации трансгенных последовательностей днк в растительном материале и продуктах на его основе, набор олигонуклеотидов и биочип для осуществления этого способа.
3. Патент РФ 2386698, Кузнецов В.В., Абрамов Д.Д., Митрохин И.А., Цыдендамбаев В.Д. Способ идентификации трансгенных последовательностей ДНК в растительном материале и продуктах на его основе (RU 2386698) – прототип.
4. Navarro M.J., Natividad-Tome K.G. Gimutao K.A / From Monologue to Stakeholder Engagement: The Evolution of Biotech Communication ISAAA Briefs N48, 2012, 137. P.

| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ДИАГНОСТИКИ БИОМАТЕРИАЛОВ НА НАЛИЧИЕ В НИХ АГРОБАКТЕРИЙ | 2011 |
|
RU2458142C1 |
| ВЫСОКОСПЕЦИФИЧНЫЙ ДНК-МАРКЕР, ИСПОЛЬЗУЕМЫЙ В КАЧЕСТВЕ ЭНДОГЕННОГО РЕФЕРЕНСНОГО КОНТРОЛЯ ДЛЯ ОБНАРУЖЕНИЯ ГЕНОМНОЙ ДНК КАРТОФЕЛЯ В РАСТИТЕЛЬНОМ МАТЕРИАЛЕ И ПИЩЕВЫХ ПРОДУКТАХ, В ТОМ ЧИСЛЕ ПРИ ИДЕНТИФИКАЦИИ ГМО | 2013 |
|
RU2539756C1 |
| СПОСОБ ИДЕНТИФИКАЦИИ ТРАНСГЕННЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ ДНК В РАСТИТЕЛЬНОМ МАТЕРИАЛЕ И ПРОДУКТАХ НА ЕГО ОСНОВЕ, НАБОР ОЛИГОНУКЛЕОТИДОВ И БИОЧИП ДЛЯ ОСУЩЕСТВЛЕНИЯ ЭТОГО СПОСОБА | 2004 |
|
RU2270254C2 |
| Тест-система для обнаружения генома возбудителя ротовируса типа А у сельскохозяйственных животных с помощью мультиплексной полимеразной цепной реакции с флуоресцентной детекцией в режиме реального времени | 2018 |
|
RU2694501C1 |
| Способ выявления генома возбудителя ротовирусной инфекции у сельскохозяйственных животных | 2018 |
|
RU2689718C1 |
| СПОСОБ ИДЕНТИФИКАЦИИ ТРАНСГЕННЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ ДНК В РАСТИТЕЛЬНОМ МАТЕРИАЛЕ И ПРОДУКТАХ НА ЕГО ОСНОВЕ | 2008 |
|
RU2386698C1 |
| СПОСОБ ДЕТЕКЦИИ ОДНОНУКЛЕОТИДНОГО ПОЛИМОРФИЗМА ЛОКУСА PRL-RSAI С ПОМОЩЬЮ СПЕЦИФИЧЕСКИХ ПРАЙМЕРОВ В ПОЛИМЕРАЗНОЙ ЦЕПНОЙ РЕАКЦИИ В РЕАЛЬНОМ ВРЕМЕНИ | 2022 |
|
RU2807577C1 |
| НАБОР ОЛИГОНУКЛЕОТИДНЫХ ПРАЙМЕРОВ И ФЛУОРЕСЦЕНТНО-МЕЧЕНОГО ЗОНДА ДЛЯ ИДЕНТИФИКАЦИИ Burkholderia mallei И ДИФФЕРЕНЦИАЦИИ ЕГО ОТ Burkholderia pseudomallei | 2014 |
|
RU2551208C1 |
| Тест-система для обнаружения ДНК вируса африканской чумы свиней с помощью полимеразной цепной реакции в режиме реального времени | 2017 |
|
RU2645263C1 |
| Олигонуклеотидные праймеры и флуоресцентно-меченый зонд, способ и тест-система для выявления генома вируса заразного узелкового дерматита (нодулярного дерматита) КРС в реакции полимеразной цепной реакции в режиме реального времени | 2019 |
|
RU2714045C1 |
Изобретение относится к области биохимии, в частности к способу диагностики растительного материала на трансгенность. Способ включает выделение из биоматериала ДНК и проведение полимеразной цепной реакции в режиме реального времени с зондами, меченными флуоресцентными красителями и гасителями флуоресценции. При этом составляют реакционную смесь, содержащую праймеры aacacatgagcgaaacccta, caagctcgagtttctccata, ccggtcttgcgatgattat, tatattttgttttctatcgcgtatt, tatattttgttttctatcgcgtatt, agatggattgcacgcaggttc, gcatcagagcagccgattgt и зонды FAM-cccttatctgggaactactcacacatta-BHQ1, FAM-tgcgggactctaatcataaaaacccatct-BHQ1, FAM-ccagtcatagccgaatagcctctccacc-BHQ1. Полимеразную цепную реакцию проводят в режиме реального времени, при этом осуществляют непрерывный контроль флуоресценции и по экспоненциальному ее нарастанию диагностируют трансгенность растений. Изобретение позволяет эффективно диагностировать растительный материал на трансгенность. 2 ил., 1 табл.
Способ диагностики растительного материала на трансгенность, заключающийся в выделении из биоматериала ДНК и проведении на ней полимеразной цепной реакции в режиме реального времени с зондами, меченными флуоресцентными красителями и гасителями флуоресценции, отличающийся тем, что после выделения из образцов ДНК в концентрации 0,1-100 нг/мкл на основе ее составляют реакционную смесь, которая содержит праймеры aacacatgagcgaaacccta, caagctcgagtttctccata, ccggtcttgcgatgattat, tatattttgttttctatcgcgtatt, tatattttgttttctatcgcgtatt, agatggattgcacgcaggttc, gcatcagagcagccgattgt и зонды FAM-cccttatctgggaactactcacacatta-BHQ1, FAM-tgcgggactctaatcataaaaacccatct-BHQ1, FAM-ccagtcatagccgaatagcctctccacc-BHQ1, праймеры в составе реакционных смесей имеют концентрацию 0,2-2 мM, а зонды 0,1-1 мM, реакционные смеси содержат буфер для полимеразы, катионы Mg 2+ в концентрации 0,1-0,3 мM, нуклеотиды в концентрации 2-10 мкМ, Taq полимеразу 0,1-0,3 ед. активности на 1 мкл смеси, полимеразную цепную реакцию проводят в режиме реального времени с температурой отжига праймеров 60°С при 40 циклах, при этом осуществляют непрерывный контроль флуоресценции и по экспоненциальному ее нарастанию диагностируют трансгенность растений.
| СПОСОБ ИДЕНТИФИКАЦИИ ТРАНСГЕННЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ ДНК В РАСТИТЕЛЬНОМ МАТЕРИАЛЕ И ПРОДУКТАХ НА ЕГО ОСНОВЕ | 2008 |
|
RU2386698C1 |
| RU 2009123913 А, 27.12.2010 | |||
| US 2009239234 A1, 04.09.2009 | |||
| СПОСОБ ПОЛУЧЕНИЯ ДИЗАЩИЩЕННОГО 2,3-ГИДРОКСИМЕТИЛЦИТИЛЦИКЛОБУТАНОЛА | 1993 |
|
RU2118312C1 |

