Ссылка на перечень последовательностей
Данная заявка включает перечень последовательностей в машиночитаемой форме, включенный сюда посредством ссылки.
Предшествующий уровень техники
Область изобретения
Настоящее изобретение относится к вариантам антимикробного пептида, полинуклеотидам, кодирующим указанные варианты, способам получения указанных вариантов и способам применения указанных вариантов.
Описание предшествующего уровня техники
В литературе описаны несколько классов антимикробных пептидов (AMP), примеры которых включают дефензины и альфа-спиральные пептиды.
Согласно настоящему изобретению предложены варианты антимикробного пептида, выделенного из Arenicola marina и описанного в WO 2007/023163.
Варианты антимикробного пептида по настоящему изобретению обладают лучшей антимикробной активностью по сравнению с исходным антимикробным пептидом. В частности, указанные варианты обладают улучшенной антимикробной активностью в присутствии белков сыворотки и крови. Другим преимуществом вариантов пептида по настоящему изобретению является пониженное связывание с белками, например белками сыворотки и крови, что приводит к улучшенной биодоступности по сравнению с исходным антимикробным пептидом.
Краткое описание сущности изобретения
Настоящее изобретение относится к выделенным вариантам антимикробного пептида, имеющего аминокислотную последовательность SEQ ID NO: 2, содержащим замену в одном или более (нескольких) из положений 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 15, 17, 19 и 21 зрелого пептида с SEQ ID NO: 2, обладающим антимикробной активностью.
Настоящее изобретение также относится к выделенным полинуклеотидам, кодирующим указанные варианты; конструкциям нуклеиновых кислот, векторам и клеткам-хозяевам, содержащим указанные полинуклеотиды; и способам получения указанных вариантов.
Настоящее изобретение также относится к способу лечения микробной инфекции с использованием вариантов по изобретению; и применению вариантов для изготовления лекарственного средства для лечения микробной инфекции.
Подробное описание изобретения
Настоящее изобретение относится к выделенным вариантам антимикробного пептида, имеющего аминокислотную последовательность SEQ ID NO: 2, содержащим замену в одном или более (нескольких) из положений 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 15, 17, 19 и 21 зрелого пептида с SEQ ID NO: 2, обладающим антимикробной активностью.
Определения
Антимикробная активность. Термин «антимикробная активность» определен здесь как способность уничтожать микробные клетки или ингибировать их рост. Подразумевают, что в контексте настоящего изобретения термин «антимикробный» означает наличие бактерицидного, и/или бактериостатического, и/или фунгицидного, и/или фунгистатического эффекта и/или вирулицидного эффекта, где термин «бактерицидный» следует понимать как способный уничтожать бактериальные клетки. Термин «бактериостатический» следует понимать как способный ингибировать бактериальный рост, то есть ингибировать рост бактериальных клеток. Термин «фунгицидный» следует понимать как способный уничтожать грибковые клетки. Термин «фунгистатический» следует понимать как способный ингибировать грибковый рост, то есть ингибировать рост грибковых клеток. Термин «вирулицидный» следует понимать как способный инактивировать вирус. Термин «микробные клетки» обозначает бактериальные или грибковые клетки (включая дрожжи).
Подразумевают, что в контексте настоящего изобретения термин «ингибирование роста микробных клеток» означает, что эти клетки не находятся в состоянии роста, то есть что они не способны к размножению.
В предпочтительном воплощении термин «антимикробная активность» определен как бактерицидная и/или бактериостатическая активность. Более предпочтительно, «антимикробная активность» определена как бактерицидная и/или бактериостатическая активность против Escherichia, предпочтительно Escherichia coli.
Для задач настоящего изобретения антимикробная активность может быть определена способом, описанным Lehrer et al., Journal of Immunological Methods, Vol.137 (2) pp.167-174 (1991). Альтернативно, антимикробная активность может быть определена в соответствии с руководствами Национального комитета по клиническим и лабораторным стандартам (National Committee for Clinical Laboratory Standards, NCCLS) от CLSI (Института клинических и лабораторных стандартов (Clinical and Laboratory Standards Institute), ранее известного как Национальный комитет по клиническим и лабораторным стандартам).
Пептиды, обладающие антимикробной активностью, могут быть способны уменьшать число жизнеспособных клеток Escherichia coli (DSM 1576) до 1/100 через 8 часов (предпочтительно через 4 часа, более предпочтительно через 2 часа, наиболее предпочтительно через 1 час и, в частности, через 30 минут) инкубации при 37°C в соответствующем субстрате для микробного роста при концентрации пептидов, обладающих антимикробной активностью, 500 мкг/мл, предпочтительно 250 мкг/мл, более предпочтительно 100 мкг/мл, еще более предпочтительно 50 мкг/мл, наиболее предпочтительно 25 мкг/мл и, в частности, 10 мкг/мл.
Пептиды, обладающие антимикробной активностью, могут также быть способны ингибировать рост Escherichia coli (DSM 1576) в течение 8 часов при 37°C в соответствующем субстрате для микробного роста при их добавлении в концентрации 500 мкг/мл, предпочтительно при их добавлении в концентрации 250 мкг/мл, более предпочтительно при их добавлении в концентрации 100 мкг/мл, еще более предпочтительно при их добавлении в концентрации 50 мкг/мл, наиболее предпочтительно при их добавлении в концентрации 10 мкг/мл и, в частности, при их добавлении в концентрации 5 мкг/мл.
Пептидные варианты по настоящему изобретению обладают улучшенной антимикробной активностью по сравнению с антимикробным пептидом с SEQ ID NO: 2. В одном воплощении пептидные варианты по настоящему изобретению обладают более чем 100% антимикробной активностью пептида с SEQ ID NO: 2 в присутствии сыворотки крови.
Вариант. Термин «вариант» означает пептид, обладающий антимикробной активностью, содержащий изменение, то есть замену, вставку и/или делецию, в одном или более (нескольких) положениях. Замена означает замещение аминокислоты, расположенной в том или ином положении, другой аминокислотой; делеция означает удаление аминокислоты, расположенной в том или ином положении; и вставка означает добавление 1-3 аминокислот рядом с аминокислотой, расположенной в том или ином положении.
Мутант. Термин «мутант» означает полинуклеотид, кодирующий вариант.
Антимикробный пептид дикого типа. Термин антимикробный пептид «дикого типа» означает антимикробный пептид, экспрессированный встречающимся в природе микроорганизмом, таким как бактерия, дрожжи или нитчатый гриб, обнаруживаемые в природе.
Исходный пептид или исходный антимикробный пептид. Термин «исходный пептид» или «исходный антимикробный пептид» означает антимикробный пептид, который подвергают изменению для получения ферментных вариантов по настоящему изобретению. Исходный пептид может представлять собой пептид, встречающийся в природе (пептид дикого типа), или его вариант.
Выделенный вариант. Термин «выделенный вариант» означает вариант, модифицированный человеком. В одном аспекте степень чистоты варианта составляет по меньшей мере 1%, например по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 40%, по меньшей мере 60%, по меньшей мере 80% и по меньшей мере 90%, как определено электрофорезом в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE).
По существу чистый вариант. Термин «по существу чистый вариант» означает препарат, содержащий не более 10%, не более 8%, не более 6%, не более 5%, не более 4%, не более 3%, не более 2%, не более 1% и не более 0,5% по массе другого пептидного вещества, ассоциированного естественным или рекомбинантным образом. Предпочтительно, степень чистоты варианта составляет по меньшей мере 92%, например по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99%, по меньшей мере 99,5% и 100% по массе всего пептидного вещества, присутствующего в препарате. Предпочтительно, варианты по настоящему изобретению представлены в по существу чистой форме. Это может быть обеспечено, например, получением варианта хорошо известными рекомбинантными способами или классическими способами очистки.
Зрелый пептид. Термин «зрелый пептид» означает пептид в его конечной форме после трансляции и любых посттрансляционных модификаций, таких как N-концевой процессинг, C-концевой процессинг, гликозилирование, фосфорилирование и так далее.
Последовательность, кодирующая зрелый пептид. Термин «последовательность, кодирующая зрелый пептид», означает полинуклеотид, кодирующий зрелый пептид, обладающий антимикробной активностью.
Идентичность последовательностей. Родство двух аминокислотных последовательностей или двух нуклеотидных последовательностей описывают параметром «идентичность последовательностей».
Для задач настоящего изобретения степень идентичности последовательностей между двумя аминокислотными последовательностями определяют с использованием алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48: 443-453), воплощенного в программе Needle программного пакета EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000, Trends Genet. 16: 276-277), предпочтительно версии 3.0.0 или более поздней версии. Используемые опционные параметры представляют собой штраф за начало разрыва 10, штраф за продолжение разрыва 0,5 и подстановочную матрицу EBLOSUM62 (EMBOSS-версию BLOSUM62). В качестве процента идентичности используют результат Needle, отмеченный как «наиболее продолжительная идентичность» (полученный с использованием опции -nobrief) и вычисленный следующим образом:
(идентичные остатки × 100)/(длина выравнивания - общее число разрывов при выравнивании).
Для задач настоящего изобретения степень идентичности последовательностей между двумя дезоксирибонуклеотидными последовательностями определяют с использованием алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, выше), воплощенного в программе Needle программного пакета EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000, supra), предпочтительно версии 3.0.0 или более поздней версии. Используемые опционные параметры представляют собой штраф за начало разрыва 10, штраф за продолжение разрыва 0,5 и подстановочную матрицу EDNAFULL (EMBOSS-версию NCBI NUC4.4). В качестве процента идентичности используют результат Needle, отмеченный как «наиболее продолжительная идентичность» (полученный с использованием опции - nobrief) и вычисленный следующим образом:
(идентичные дезоксирибонуклеотиды × 100)/(длина выравнивания - общее число разрывов при выравнивании).
Фрагмент. Термин «фрагмент» означает пептид, имеющий одну или более (несколько) аминокислот, удаленных с амино- и/или карбоксильного конца зрелого пептида, где указанный фрагмент обладает антимикробной активностью. В одном аспекте фрагмент содержит по меньшей мере 15 аминокислотных остатков, например, по меньшей мере 17 и по меньшей мере 19 аминокислотных остатков (например, аминокислотные остатки 1-20 в SEQ ID NO: 2).
Подпоследовательность. Термин «подпоследовательность» означает полинуклеотид, имеющий один или более (несколько) нуклеотидов, удаленных с 5'- и/или 3'-конца последовательности, кодирующей зрелый пептид, где подпоследовательность кодирует фрагмент, обладающий антимикробной активностью.
Аллельный вариант. Термин «аллельный вариант» означает любые две или более альтернативных форм гена, расположенных в одном и том же локусе хромосомы. Аллельное разнообразие появляется естественным образом в результате мутаций и может приводить к полиморфизму в популяциях. Мутации в генах могут быть молчащими (без изменения кодируемого пептида) или могут кодировать пептиды, имеющие измененные аминокислотные последовательности. Аллельный вариант пептида представляет собой пептид, кодируемый аллельным вариантом гена.
Выделенный полинуклеотид. Термин «выделенный полинуклеотид» означает полинуклеотид, модифицированный человеком. В одном аспекте степень чистоты выделенного полинуклеотида составляет по меньшей мере 1%, например по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 40%, по меньшей мере 60%, по меньшей мере 80%, по меньшей мере 90% и по меньшей мере 95%, как определено электрофорезом в агарозном геле. Полинуклеотиды могут быть геномного происхождения, иметь происхождение от комплементарной ДНК (кДНК), РНК, быть полусинтетического или синтетического происхождения, или представлять собой любые их комбинации.
По существу чистый полинуклеотид. Термин «по существу чистый полинуклеотид» означает полинуклеотидный препарат, свободный от других чужеродных или нежелательных нуклеотидов и представленный в форме, подходящей для применения в системах продуцирования генетически сконструированных пептидов. Таким образом, по существу чистый полинуклеотид содержит не более 10%, не более 8%, не более 6%, не более 5%, не более 4%, не более 3%, не более 2%, не более 1% и не более 0,5% по массе другого полинуклеотидного вещества, ассоциированного естественным или рекомбинантным образом. Тем не менее, по существу чистый полинуклеотид может включать встречающиеся в природе 5'- и 3'-нетранслируемые области, такие как промоторы и терминаторы. Предпочтительно, степень чистоты по существу чистого полинуклеотида составляет по меньшей мере 90%, например по меньшей мере 92%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% и по меньшей мере 99,5% по массе. Предпочтительно, полинуклеотиды по настоящему изобретению представлены в по существу чистой форме.
Кодирующая последовательность. Термин «кодирующая последовательность» означает полинуклеотид, непосредственно определяющий аминокислотную последовательность его пептидного продукта. Границы кодирующей последовательности обычно определены открытой рамкой считывания, которая обычно начинается с инициирующего кодона ATG или альтернативных инициирующих кодонов, таких как GTG и TTG, и заканчивается стоп-кодоном, таким как ТАА, TAG и TGA. Кодирующая последовательность может представлять собой ДНК, кДНК, синтетический или рекомбинантный полинуклеотид.
кДНК. Термин «кДНК» означает молекулу ДНК, которая может быть получена обратной транскрипцией зрелой сплайсированной молекулы матричной РНК (мРНК), полученной из эукариотической клетки. В кДНК отсутствуют последовательности интронов, которые могут присутствовать в соответствующей геномной ДНК. Исходный первичный РНК-транскрипт является предшественником мРНК, который проходит процессинг из нескольких стадий, включая сплайсинг, перед образованием зрелой сплайсированной мРНК.
Конструкция нуклеиновой кислоты. Термин «конструкция нуклеиновой кислоты» означает одно- или двухцепочечную молекулу нуклеиновой кислоты, которая выделена из встречающегося в природе гена или модифицирована для содержания сегментов нуклеиновых кислот таким образом, которым они в противном случае не существовали бы в природе, либо которая является синтетической. Термин «конструкция нуклеиновой кислоты» является синонимом термина «кассета экспрессии», когда конструкция нуклеиновой кислоты содержит контрольные последовательности, необходимые для экспрессии кодирующей последовательности по настоящему изобретению.
Контрольные последовательности. Термин «контрольные последовательности» означает все компоненты, необходимые для экспрессии полинуклеотида, кодирующего вариант по настоящему изобретению. Каждая контрольная последовательность может быть нативной или чужеродной по отношению к полинуклеотиду, кодирующему вариант, либо контрольные последовательности могут быть нативными или чужеродными по отношению друг к другу. Такие контрольные последовательности включают, без ограничения, лидерную последовательность, последовательность полиаденилирования, последовательность пропептида, промотор, последовательность сигнального пептида и терминатор транскрипции. По меньшей мере, контрольные последовательности включают промотор и сигналы окончания транскрипции и трансляции. Контрольные последовательности могут быть снабжены линкерами с целью включения специфичных сайтов рестрикции, облегчающих лигирование контрольных последовательностей с кодирующей областью полинуклеотида, кодирующего вариант.
Функционально связанный. Термин «функционально связанный» означает конфигурацию, в которой контрольная последовательность расположена в подходящем положении относительно кодирующей последовательности полинуклеотида, так что контрольная последовательность управляет экспрессией кодирующей последовательности.
Экспрессия. Термин «экспрессия» включает любую стадию, входящую в продуцирование варианта, включая, без ограничения, транскрипцию, посттранскрипционную модификацию, трансляцию, посттрансляционную модификацию и секрецию.
Вектор экспрессии. Термин «вектор экспрессии» означает линейную или кольцевую молекулу ДНК, содержащую полинуклеотид, кодирующий вариант, и функционально связанную с дополнительными нуклеотидами, обеспечивающими ее экспрессию.
Клетка-хозяин. Термин «клетка-хозяин» означает клетку любого типа, подходящую для трансформации, трансфекции, трансдукции и тому подобного с использованием конструкции нуклеиновой кислоты или вектора экспрессии, содержащего полинуклеотид по настоящему изобретению. Термин «клетка-хозяин» включает любое потомство исходной клетки, не идентичное исходной клетке в виду мутаций, которые возникают при репликации.
Условные обозначения вариантов
Для задач настоящего изобретения зрелый пептид, раскрытый в SEQ ID NO: 2, используют для определения соответствующего аминокислотного остатка в другом антимикробном пептиде. Аминокислотную последовательность другого антимикробного пептида выравнивают со зрелым пептидом, раскрытым в SEQ ID NO: 2, и на основании этого выравнивания определяют номер аминокислотного положения, соответствующего любому аминокислотному остатку зрелого пептида, раскрытого в SEQ ID NO: 2, с использованием алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48:443-453), воплощенного в программе Needle пакета EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000, Trends Genet. 16:276-277), предпочтительно версии 3.0.0 или более поздней версии.
Идентификация соответствующего аминокислотного остатка в другом антимикробном пептиде может быть подтверждена выравниванием последовательностей нескольких пептидов с использованием "ClustaIW" (Larkin et al., 2007, Bioinformatics 23:2947-2948).
Когда другой фермент отличается от зрелого пептида с SEQ ID NO: 2 настолько, что традиционное сравнение последовательностей не позволяет выявить их родство (Lindahl and Elofsson, 2000, J. Mol. Biol. 295:613-615), могут быть использованы другие алгоритмы попарного сравнения последовательностей. Большей чувствительности при поиске по последовательности можно достичь с использованием поисковых программ, в которых использованы вероятностные представления пептидных семейств (профилей) для поиска в базах данных. Например, программа PSI-BLAST генерирует профили способом повторного поиска в базах данных и позволяет выявлять отдаленные гомологи (Atschul et al., 1997, Nucleic Acids Res. 25:3389-3402). Еще большей чувствительности можно достичь, если семейство или суперсемейство данного пептида имеет одного или более представителей в базах данных структур белков. Такие программы, как GenTHREADER (Jones, 1999, J. Mol. Biol. 287:797-815; McGuffin and Jones, 2003, Bioinformatics 19:874-881), используют информацию из множества источников (PSI-BLAST, предсказание вторичной структуры, профили структурного выравнивания и потенциалы сольватации) в качестве входных данных для нейронной сети, предсказывающей структурные свойства запрашиваемой последовательности. Сходным образом, способ Gough et al., 2000, J. Mol. Biol. 313:903-919, может быть использован для выравнивания последовательности с неизвестной структурой с моделями суперсемейств в базе данных SCOP. Эти выравнивания могут, в свою очередь, быть использованы для получения моделей гомологии для данного пептида, и точность таких моделей можно оценивать с использованием множества инструментов, разработанных для этой цели.
Для белков с известной структурой доступно несколько инструментов и ресурсов для получения и генерирования структурных выравниваний. Например, было проведено структурное выравнивание SCOP-суперсемейств белков, и эти выравнивания доступны для просмотра и загрузки. Выравнивание двух или более белковых структур может быть проведено с использованием ряда алгоритмов, таких как дистанционная матрица выравнивания (Holm and Sander, 1998, Proteins 33:88-96) или комбинаторное расширение (Shindyalov and Bourne, 1998, Protein Engineering 11:739-747), и воплощения этих алгоритмов могут быть дополнительно использованы для поиска интересующей структуры в базах данных по структурам для обнаружения возможных структурных гомологов (например, Holm and Park, 2000, Bioinformatics 16:566-567).
При описании вариантов антимикробных пептидов по настоящему изобретению описанную ниже номенклатуру адаптируют для простоты использования. Используют принятые Международным союзом теоретической и прикладной химии (IUPAC) однобуквенные или трехбуквенные сокращения аминокислот.
Замены. Для аминокислотной замены используют следующую номенклатуру: исходная аминокислота, положение, замещенная аминокислота. Соответственно, замену треонина на аланин в положении 226 обозначают как «Thr226Ala» или «Т226А». Несколько мутаций разделяют знаками сложения («+»), например, «Gly205Arg + Ser411Phe» или «G205R + S411F», что означает замены в положениях 205 и 411 глицина (G) на аргинин (R) и серина (S) на фенилаланин (F), соответственно.
Делеции. Для аминокислотной делеции используют следующую номенклатуру: исходная аминокислота, положение, *. Соответственно, делецию глицина в положении 195 обозначают «Gly195*» или «G195*». Несколько делеции разделяют знаками сложения («+»), например, «Gly195* + Ser411*» или «G195* + S411*».
Вставки. Для аминокислотной вставки используют следующую номенклатуру: исходная аминокислота, положение, исходная аминокислота, встроенная аминокислота. Соответственно, вставку лизина после глицина в положении 195 обозначают «Gly195GlyLys» или «G195GK». Вставку нескольких аминокислот обозначают следующим образом: исходная аминокислота, положение, исходная аминокислота, встроенная аминокислота №1, встроенная аминокислота №2 и так далее. Например, вставку лизина и аланина после глицина в положении 195 обозначают «Gly195GlyLysAla» или «G195GKA».
В таких случаях встроенный(е) аминокислотный(е) остаток(ки) нумеруют путем добавления строчных букв к номеру положения аминокислотного остатка, предшествующего встроенному(ым) аминокислотному(ым) остатку(ам). Таким образом, в приведенном выше примере последовательность будет следующей.
Несколько замен. Варианты, содержащие несколько замен, разделяют знаками сложения («+»), например, «Arg170Tyr + Gly195Glu» или «R170Y + G195E», что обозначает замену аргинина и глицина тирозином и глутаминовой кислотой в положениях 170 и 195, соответственно.
Разные замены. Когда разные замены могут быть произведены в каком-либо положении, эти разные замены разделяют запятой, например, «Arg170Tyr,Glu» или «R170Y,E» обозначают замену аргинина тирозином или глутаминовой кислотой в положении 170. Так, «Tyr167Gly,Ala + Arg170Gly,Ala» или «Y167G,A + R170G,A» обозначает следующие варианты: «Tyr167Gly + Arg170Gly», «Tyr167Gly + Arg170Ala», «Tyr167Ala + Arg170Gly» и «Tyr167Ala + Arg170Ala».
Исходные антимикробные пептиды
Исходный антимикробный пептид представляет собой: (а) пептид, последовательность которого по меньшей мере на 60% идентична зрелому пептиду с SEQ ID NO: 2; (б) пептид, кодируемый полинуклеотидом, который гибридизуется в условиях средней строгости с (1) последовательностью SEQ ID NO: 1, кодирующей зрелый пептид, или (2) полноразмерной комплементарной цепью (1); или (в) пептид, кодируемый полинуклеотидом, последовательность которого по меньшей мере на 60% идентична последовательности SEQ ID NO: 1, кодирующей зрелый пептид.
В первом аспекте последовательность исходного пептида по меньшей мере на 60%, например по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична последовательности зрелого пептида SEQ ID NO: 2, обладающего антимикробной активностью. В одном аспекте аминокислотная последовательность исходного пептида отличается от зрелого пептида с SEQ ID NO: 2 не более чем на десять аминокислот, например на пять аминокислот, на четыре аминокислоты, на три аминокислоты, на две аминокислоты и на одну аминокислоту.
Предпочтительно, исходный пептид содержит аминокислотную последовательность SEQ ID NO: 2 или состоит из нее.
В одном воплощении исходный пептид представляет собой фрагмент пептида с SEQ ID NO: 2, содержащий по меньшей мере 15 аминокислотных остатков, например по меньшей мере 17 и по меньшей мере 19 аминокислотных остатков.
В другом воплощении исходный пептид представляет собой аллельный вариант пептида с SEQ ID NO: 2.
Во втором аспекте исходный пептид представляет собой пептид, кодируемый полинуклеотидом, который гибридизуется в условиях средней строгости, условиях средневысокой строгости, условиях высокой строгости или условиях очень высокой строгости с (1) последовательностью SEQ ID NO: 1, кодирующей зрелый пептид, или (2) полноразмерной комплементарной цепью (1) (J. Sambrook, E.F. Fritsch, and Т. Maniatis, 1989, Molecular Cloning, A Laboratory Manual, 2-я редакция, Cold Spring Harbor, New York).
Полинуклеотид с SEQ ID NO: 1 или его подпоследовательность, а также аминокислотная последовательность SEQ ID NO: 2 или ее фрагмент, могут быть использованы для конструирования нуклеиновокислотных зондов для идентификации и клонирования ДНК, кодирующей исходный пептид, из штаммов разных родов или видов способами, хорошо известными в данной области техники. В частности, такие зонды могут быть использованы для гибридизации с геномной или кДНК интересующего рода или вида после стандартных процедур Саузерн-блоттинга с целью идентификации и выделения его соответствующего гена. Такие зонды могут быть значительно короче полноразмерной последовательности, но их длина должна составлять по меньшей мере 14, например по меньшей мере 25 или по меньшей мере 35 нуклеотидов. Могут быть использованы как ДНК-, так и РНК-зонды. Зонды обычно включают метку для выявления соответствующего гена (например, 32P, 3H, 35S, биотин или авидин). Такие зонды охватываются настоящим изобретением.
Можно проводить скрининг библиотек геномной ДНК или кДНК, полученной из таких других организмов, на предмет ДНК, которая гибридизуется с описанными выше зондами и кодирует исходный пептид. Геномная или другая ДНК из таких других организмов может быть разделена электрофорезом в агарозном или полиакриламидном геле или другими методиками разделения. ДНК из библиотек или разделенная ДНК может быть перенесена и иммобилизована на нитроцеллюлозе или другом подходящем веществе-носителе. Для идентификации клона или ДНК, гомологичных SEQ ID NO: 1 или ее подпоследовательности, это вещество-носитель используют в Саузерн-блоттинге.
Для задач настоящего изобретения гибридизация показывает, что полинуклеотид гибридизуется с меченным нуклеотидным зондом, соответствующим полинуклеотиду, показанному в SEQ ID NO: 1, его комплементарной цепи или его подпоследовательности в условиях от низкой до очень высокой строгости. Молекулы, с которыми гибридизуется зонд, можно выявлять с использованием, например, рентгеновской пленки или любого другого способа выявления, известного в данной области техники.
В одном воплощении нуклеиновокислотный зонд представляет собой SEQ ID NO: 1.
Для длинных зондов длиной по меньшей мере 60 нуклеотидов условия от очень низкой до очень высокой строгости определяют как предварительную гибридизацию и гибридизацию при 42°C в 5Х SSPE, 0,3% додецилсульфате натрия (SDS), 200 микрограмм/мл ДНК гомогенизированных и денатурированных молок лососевых и 25% формамид для условий очень низкой и низкой строгости, 35% формамид для условий средней и средневысокой строгости или 50% формамид для условий высокой и очень высокой строгости, после стандартных процедур Саузерн-блоттинга, оптимально в течение 12-24 часов. Наконец, вещество-носитель отмывают три раза по 15 минут каждый с использованием 2Х SSC, 0,2% SDS при 45°C (условия очень низкой строгости), 50°C (условия низкой строгости), 55°C (условия средней строгости), 60°C (условия средневысокой строгости), 65°C (условия высокой строгости) или 70°C (условия очень высокой строгости).
Для коротких зондов длиной от приблизительно 15 нуклеотидов до приблизительно 60 нуклеотидов условия строгости определяют как предварительную гибридизацию и гибридизацию при температуре на приблизительно 5°C-приблизительно 10°C ниже расчетной температуры плавления (Tm), вычисленной способом Bolton и McCarthy (1962, Proc. Natl. Acad. Sci. USA 48:1390), в 0,9 M NaCI, 0,09 M трис-HCl, pH 7,6, 6 мМ этилендиаминтетрауксусной кислоте (EDTA), 0,5% NP-40, 1X растворе Денхардта, 1 мМ пирофосфате натрия, 1 мМ дигидрофосфате натрия, 0,1 мМ АТР и 0,2 мг дрожжевой РНК на мл после стандартных процедур Саузерн-блоттинга, оптимально в течение 12-24 часов. Наконец, вещество-носитель отмывают один раз в 6Х SCC с 0,1% SDS в течение 15 минут и два раза по 15 минут каждый с использованием 6Х SSC при температуре на 5°C-10°C ниже расчетной Tm.
В третьем аспекте исходный пептид представляет собой пептид, кодируемый полинуклеотидом, последовательность которого по меньшей мере на 60%, например по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична последовательности SEQ ID NO: 1, кодирующей пептид, обладающий антимикробной активностью.
Исходный пептид может быть получен из микроорганизмов любого рода. Для задач настоящего изобретения термин «получен из» при использовании здесь в связи с данным источником означает, что исходный пептид, кодируемый полинуклеотидом, продуцируется этим источником или клеткой, в которую был введен указанный полинуклеотид из источника. В одном аспекте исходный пептид секретируется во внеклеточную среду.
Исходный пептид может представлять собой бактериальный антимикробный пептид. Например, исходный пептид может представлять собой пептид грамположительной бактерии, такой как антимикробный пептид Bacillus, Clostridium, Enterococcus, Geobacillus, Lactobacillus, Lactococcus, Oceanobacillus, Staphylococcus, Streptococcus или Streptomyces, или пептид грамотрицательной бактерии, такой как антимикробный пептид Campylobacter, Е. coli, Flavobacterium, Fusobacterium, Helicobacter, llyobacter, Neisseria, Pseudomonas, Salmonella или Ureaplasma.
В одном аспекте исходный пептид представляет собой антимикробный пептид Bacillus alkalophilus, Bacillus amyloliquefaciens, Bacillus brevis, Bacillus circulans, Bacillus clausii, Bacillus coagulans, Bacillus firmus, Bacillus lautus, Bacillus lentus, Bacillus licheniformis, Bacillus megaterium, Bacillus pumilus, Bacillus stearothermophilus, Bacillus subtilis или Bacillus thuringiensis.
В другом аспекте исходный пептид представляет собой антимикробный пептид Streptococcus equisimilis, Streptococcus pyogenes, Streptococcus uberis или Streptococcus equi, subsp. zooepidemicus.
В другом аспекте исходный пептид представляет собой антимикробный пептид Streptomyces achromogenes, Streptomyces avermitilis, Streptomyces coelicolor, Streptomyces griseus или Streptomyces lividans.
Исходный пептид может представлять собой грибковый антимикробный пептид. Например, исходный пептид может представлять собой дрожжевой антимикробный пептид, такой как антимикробный пептид Candida, Kluyveromyces, Pichia, Saccharomyces, Schizosaccharomyces или Yarrowia. Например, исходный пептид может представлять собой антимикробный пептид нитчатого гриба, такой как антимикробный пептид Acremonium, Agaricus, Alternaria, Aspergillus, Aureobasidium, Botryospaeria, Ceriporiopsis, Chaetomidium, Chrysosporium, Claviceps, Cochliobolus, Coprinopsis, Coptotermes, Corynascus, Cryphonectria, Cryptococcus, Diplodia, Exidia, Filibasidium, Fusarium, Gibberella, Holomastigotoides, Humicola, Irpex, Lentinula, Leptospaeria, Magnaporthe, Melanocarpus, Meripilus, Mucor, Myceliophthora, Neocallimastix, Neurospora, Paecilomyces, Penicillium, Phanerochaete, Piromyces, Poitrasia, Pseudoplectania, Pseudotrichonympha, Rhizomucor, Schizophyllum, Scytalidium, Talaromyces, Thermoascus, Thielavia, Tolypocladium, Trichoderma, Trichophaea, Verticillium, Volvariella или Xylaria.
В другом аспекте исходный пептид представляет собой антимикробный пептид Saccharomyces carlsbergensis, Saccharomyces cerevisiae, Saccharomyces diastaticus, Saccharomyces douglasii, Saccharomyces kluyveri, Saccharomyces norbensis или Saccharomyces oviformis.
В другом аспекте исходный пептид представляет собой антимикробный пептид Acremonium cellulolyticus, Aspergillus aculeatus, Aspergillus awamori, Aspergillus foetidus, Aspergillus fumigatus, Aspergillus japonicus, Aspergillus nidulans, Aspergillus niger, Aspergillus oryzae, Chrysosporium inops, Chrysosporium keratinophilum, Chrysosporium lucknowense, Chrysosporium merdarium, Chrysosporium pannicola, Chrysosporium queenslandicum, Chrysosporium tropicum, Chrysosporium zonatum, Fusarium bactridioides, Fusarium cerealis, Fusarium crookwellense, Fusarium culmorum, Fusarium graminearum, Fusarium graminum, Fusarium heterosporum, Fusarium negundi, Fusarium oxysporum, Fusarium reticulatum, Fusarium roseum, Fusarium sambucinum, Fusarium sarcochroum, Fusarium sporotrichioides, Fusarium sulphureum, Fusarium torulosum, Fusarium trichothecioides, Fusarium venenatum, Humicola grisea, Humicola insolens, Humicola lanuginosa, Irpex lacteus, Mucor miehei, Myceliophthora thermophila, Neurospora crassa, Penicillium funiculosum, Penicillium purpurogenum, Phanerochaete chrysosporium, Thielavia achromatica, Thielavia albomyces, Thielavia albopilosa, Thielavia australeinsis, Thielavia fimeti, Thielavia microspora, Thielavia ovispora, Thielavia peruviana, Thielavia setosa, Thielavia spededonium, Thielavia subthermophila, Thielavia terrestris, Trichoderma harzianum, Trichoderma koningii, Trichoderma longibrachiatum, Trichoderma reesei или Trichoderma viride.
В другом аспекте исходный пептид представляет собой антимикробный пептид Arenicola marina, например антимикробный пептид с SEQ ID NO: 2.
Следует понимать, что для упомянутых выше видов изобретение включает как совершенные, так и несовершенные стадии, а также другие таксономические эквиваленты, например анаморфы, независимо от видового названия, под которым они известны. Специалисты в данной области техники легко распознают идентичность подходящих эквивалентов.
Штаммы этих видов легко- и общедоступны в нескольких коллекциях культур, таких как Американская коллекция типовых культур (АТСС), Германская коллекция микроорганизмов и клеточных культур (DSM), Центральное бюро плесневых культур (CBS) и Коллекция запатентованных культур Службы сельскохозяйственных исследований, Северный региональный исследовательский центр (NRRL).
Исходный пептид может быть идентифицирован и получен из других источников, включая микроорганизмы, выделенные из природы (например, почвы, компостов, воды и так далее), или образцы ДНК, полученные непосредственно из природы (например, почвы, компостов, воды и так далее) с использованием упомянутых выше зондов. Методики выделения микроорганизмов и ДНК непосредственно из природных сред хорошо известны в данной области техники. Полинуклеотид, кодирующий исходный пептид, может затем быть получен сходным образом путем скрининга библиотеки геномной ДНК или кДНК другого микроорганизма или смешанного образца ДНК. После выявления полинуклеотида, кодирующего исходный пептид, с использованием зонда(ов), полинуклеотид может быть выделен или клонирован с применением методик, известных специалистам в данной области техники (см., например, Sambrook et al., 1989, выше).
Исходный пептид может представлять собой гибридный пептид, где часть одного пептида слита с N-концом или C-концом части другого пептида.
Исходный пептид может также представлять собой слитый пептид или расщепляемый слитый пептид, в котором один пептид слит с N-концом или C-концом другого пептида. Слитый пептид получают путем слияния полинуклеотида, кодирующего один пептид, с полинуклеотидом, кодирующим другой пептид. Методики получения слитых пептидов известны в данной области техники и включают лигирование кодирующих последовательностей, кодирующих указанные пептиды, так чтобы они располагались в одной рамке и чтобы экспрессия слитого пептида контролировалась тем(и) же промотором(ами) и терминатором. Слитые белки могут также быть сконструированы с использованием интеиновой методики, где слитые конструкции получают после трансляции (Cooper et al., 1993, EMBO J. 12:2575-2583; Dawson et al., 1994, Science 266:776-779).
Слитый белок может также содержать сайт расщепления между двумя пептидами. При секреции такого слитого белка происходит расщепление сайта с высвобождением двух пептидов. Примеры сайтов расщепления включают, без ограничения, сайты, раскрытые в Martin et al., 2003, J. Ind. Microbiol. Biotechnol. 3:568-576; Svetina et al., 2000, J. Biotechnol. 76:245-251; Rasmussen-Wilson et al., 1997, Appl. Environ. Microbiol. 63:3488-3493; Ward et al., 1995, Biotechnology 13:498-503; и Contreras et al., 1991, Biotechnology 9:378-381; Eaton et al., 1986, Biochemistry 25:505-512; Collins-Racie et al., 1995, Biotechnology 13:982-987; Carter et al., 1989, Proteins: Structure, Function, and Genetics 6:240-248; и Stevens, 2003, Drug Discovery World 4:35-48.
Получение вариантов
Настоящее изобретение также относится к способам получения варианта, обладающего антимикробной активностью, включающим: (а) введение в исходный антимикробный пептид замены в одном или более (нескольких) положениях, соответствующих положениям 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 15, 17, 19 и 21 зрелого пептида с SEQ ID NO: 2, где указанный вариант обладает антимикробной активностью; и (б) выделение указанного варианта.
Варианты могут быть получены с использованием любого способа мутагенеза, известного в данной области техники, такого как сайт-направленный мутагенез, конструирование синтетических генов, конструирование полусинтетических генов, случайный мутагенез, перетасовка и так далее.
Сайт-направленный мутагенез представляет собой методику, при которой одну или более (несколько) мутаций создают в одном или более определенных сайтах полинуклеотида, кодирующего исходный пептид.
Сайт-направленный мутагенез может быть проведен in vitro полимеразной цепной реакцией (ПЦР), включающей использование олигонуклеотидных праймеров, содержащих желаемую мутацию. Сайт-направленный мутагенез может также быть проведен in vitro кассетным мутагенезом, включающим расщепление рестриктазой в сайте плазмиды, содержащей полинуклеотид, кодирующий исходный пептид, и последующее лигирование олигонуклеотида, содержащего мутацию в полинуклеотиде. Обычно рестриктазы, расщепляющие плазмиду и олигонуклеотид, одинаковы, что позволяет получить липкие концы плазмиды и вставки для их лигирования друг с другом. См., например, Scherer and Davis, 1979, Proc. Natl. Acad. Sci. USA 76:4949-4955; и Barton etal., 1990, Nucleic Acids Res. 18:7349-4966.
Сайт-направленный мутагенез может также быть проведен in vivo способами, известными в данной области техники. См., например, публикацию заявки на патент США №2004/0171154; Storici etal., 2001, Nature Biotechnol. 19:773-776; Kren et al., 1998, Nat. Med. 4:285-290; и Calissano and Macino, 1996, Fungal Genet. Newslett. 43:15-16.
В настоящем изобретении может быть использован любой способ сайт-направленного мутагенеза. Доступно множество коммерческих наборов, которые могут быть использованы для получения вариантов.
Конструирование синтетических генов включает in vitro синтез разработанной полинуклеотидной молекулы, кодирующей интересующий пептид. Синтез генов может быть проведен с использованием ряда методик, таких как мультиплексная технология на основе микрочипов, описанная Tian et al. (2004, Nature 432:1050-1054), и подобные технологии, где синтез и сборку олигонуклеотидов проводят на фотопрограммируемых микрожидкостных чипах.
Одну или несколько аминокислотных замен, делеций и/или вставок можно провести и оценить, применяя известные способы мутагенеза, рекомбинации и/или перетасовки с последующим подходящим способом скрининга, таким как описанные Reidhaar-Olson and Sauer, 1988, Science 241:53-57; Bowie and Sauer, 1989, Proc. Natl. Acad. Sci. USA 86:2152-2156; в WO 95/17413 или WO 95/22625. Другие способы, которые могут быть использованы, включают ПЦР с внесением ошибок, фаговый дисплей (например, Lowman et al., 1991, Biochemistry 30:10832-10837; патент США №5223409; WO 92/06204) и область-направленный мутагенез (Derbyshire et al., 1986, Gene 46:145, Ner et al., 1988, DNA 7:127).
Способы мутагенеза/перетасовки можно сочетать со способами высокоэффективного автоматизированного скрининга для определения активности клонированных мутированных пептидов, экспрессированных клетками-хозяевами (Ness et al., 1999, Nature Biotechnology 17:893-896). Мутированные молекулы ДНК, которые кодируют активные пептиды, могут быть выделены из клеток-хозяев и быстро секвенированы с использованием стандартных способов данной области техники. Эти способы позволяют быстро определить важность отдельных аминокислотных остатков пептида.
Конструирование полусинтетических генов проводят, комбинируя аспекты конструирования синтетических генов, и/или сайт-направленного мутагенеза, и/или случайного мутагенеза, и/или перетасовки. Типичным примером конструирования полусинтетических генов является способ с использованием синтетических полинуклеотидных фрагментов в комбинации с ПЦР-методиками. Таким образом, определенные области генов могут быть синтезированы de novo, в то время как другие области могут быть амплифицированы с использованием сайт-специфичных мутагенных праймеров, в то же время остальные области могут быть подвергнуты амплификации с использованием ПЦР с внесением ошибок или ПЦР без внесения ошибок. Затем может быть проведена перетасовка полинуклеотидных подпоследовательностей.
Варианты
Согласно настоящему изобретению также предложены варианты исходного антимикробного пептида, содержащие замену в одном или более (нескольких) положениях, соответствующих положениям 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 15, 17, 19 и 21 (предпочтительно положениям 1, 2, 4, 5, 6, 8, 9, 12, 13, 15, 17 и 21; более предпочтительно положениям 4, 5, 6, 8, 9, 12, 13, 15 и 17), обладающие антимикробной активностью. В одном воплощении вариант обладает улучшенной антимикробной активностью по сравнению с пептидом с SEQ ID NO: 2, предпочтительно в присутствии крови или сыворотки. В другом воплощении вариант демонстрирует меньшее связывание с белками по сравнению с пептидом с SEQ ID NO: 2. Предпочтительно, варианты антимикробного пептида демонстрируют связывание с белками сыворотки не более 99%. Варианты антимикробного пептида также демонстрируют улучшенную биодоступность. Предпочтительно, биодоступность при подкожном введении составляет по меньшей мере 30%, более предпочтительно по меньшей мере 40%, еще более предпочтительно по меньшей мере 50% и наиболее предпочтительно по меньшей мере 60%.
В одном воплощении последовательность варианта по меньшей мере на 60% идентична, например по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99%, но менее чем на 100% идентична аминокислотной последовательности исходного антимикробного пептида.
В другом воплощении последовательность варианта по меньшей мере на 60% идентична, например по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% и по меньшей мере на 99%, но менее чем на 100% идентична последовательности зрелого пептида SEQ ID NO: 2.
В одном аспекте число замен в вариантах по настоящему изобретению составляет 1-11, например, 1-10 замен, 1-9 замен, 1-8 замен, 1-7 замен, 1-6 замен, 1-5 замен, 1-4 замены, 1-3 замены и 1-2 замены.
В одном аспекте вариант содержит аминокислотную последовательность, показанную как SEQ ID NO: 3-SEQ ID NO: 548, или состоит из нее.
Подразумевают, что термин «SEQ ID NO: 3-SEQ ID NO: 548» обозначает любую из SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20, SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25, SEQ ID NO: 26, SEQ ID NO: 27, SEQ ID NO: 28, SEQ ID NO: 29, SEQ ID NO: 30, SEQ ID NO: 31, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34, SEQ ID NO: 35, SEQ ID NO: 36, SEQ ID NO: 37, SEQ ID NO: 38, SEQ ID NO: 39, SEQ ID NO: 40, SEQ ID NO: 41, SEQ ID NO: 42, SEQ ID NO: 43, SEQ ID NO: 44, SEQ ID NO: 45, SEQ ID NO: 46, SEQ ID NO: 47, SEQ ID NO: 48, SEQ ID NO: 49, SEQ ID NO: 50, SEQ ID NO: 51, SEQ ID NO: 52, SEQ ID NO: 53, SEQ ID NO: 54, SEQ ID NO: 55, SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, SEQ ID NO: 69, SEQ ID NO: 70, SEQ ID NO: 71, SEQ ID NO: 72, SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, SEQ ID NO: 79, SEQ ID NO: 80, SEQ ID NO: 81, SEQ ID NO: 82, SEQ ID NO: 83, SEQ ID NO: 84, SEQ ID NO: 85, SEQ ID NO: 86, SEQ ID NO: 87, SEQ ID NO: 88, SEQ ID NO: 89, SEQ ID NO: 90, SEQ ID NO: 91, SEQ ID NO: 92, SEQ ID NO: 93, SEQ ID NO: 94, SEQ ID NO: 95, SEQ ID NO: 96, SEQ ID NO: 97, SEQ ID NO: 98, SEQ ID NO: 99, SEQ ID NO: 100, SEQ ID NO: 101, SEQ ID NO: 102, SEQ ID NO: 103, SEQ ID NO: 104, SEQ ID NO: 105, SEQ ID NO: 106, SEQ ID NO: 107, SEQ ID NO: 108, SEQ ID NO: 109, SEQ ID NO: 110, SEQ ID NO: 111, SEQ ID NO: 112, SEQ ID NO: 113, SEQ ID NO: 114, SEQ ID NO: 115, SEQ ID NO: 116, SEQ ID NO: 117, SEQ ID NO: 118, SEQ ID NO: 119, SEQ ID NO: 120, SEQ ID NO: 121, SEQ ID NO: 122, SEQ ID NO: 123, SEQ ID NO: 124, SEQ ID NO: 125, SEQ ID NO: 126, SEQ ID NO: 127, SEQ ID NO: 128, SEQ ID NO: 129, SEQ ID NO: 130, SEQ ID NO: 131, SEQ ID NO: 132, SEQ ID NO: 133, SEQ ID NO: 134, SEQ ID NO: 135, SEQ ID NO: 136, SEQ ID NO: 137, SEQ ID NO: 138, SEQ ID NO: 139, SEQ ID NO: 140, SEQ ID NO: 141, SEQ ID NO: 142, SEQ ID NO: 143, SEQ ID NO: 144, SEQ ID NO: 145, SEQ ID NO: 146, SEQ ID NO: 147, SEQ ID NO: 148, SEQ ID NO: 149, SEQ ID NO: 150, SEQ ID NO: 151, SEQ ID NO: 152, SEQ ID NO: 153, SEQ ID NO: 154, SEQ ID NO: 155, SEQ ID NO: 156, SEQ ID NO: 157, SEQ ID NO: 158, SEQ ID NO: 159, SEQ ID NO: 160, SEQ ID NO: 161, SEQ ID NO: 162, SEQ ID NO: 163, SEQ ID NO: 164, SEQ ID NO: 165, SEQ ID NO: 166, SEQ ID NO: 167, SEQ ID NO: 168, SEQ ID NO: 169, SEQ ID NO: 170, SEQ ID NO: 171, SEQ ID NO: 172, SEQ ID NO: 173, SEQ ID NO: 174, SEQ ID NO: 175, SEQ ID NO: 176, SEQ ID NO: 177, SEQ ID NO: 178, SEQ ID NO: 179, SEQ ID NO: 180, SEQ ID NO: 181, SEQ ID NO: 182, SEQ ID NO: 183, SEQ ID NO: 184, SEQ ID NO: 185, SEQ ID NO: 186, SEQ ID NO: 187, SEQ ID NO: 188, SEQ ID NO: 189, SEQ ID NO: 190, SEQ ID NO: 191, SEQ ID NO: 192, SEQ ID NO: 193, SEQ ID NO: 194, SEQ ID NO: 195, SEQ ID NO: 196, SEQ ID NO: 197, SEQ ID NO: 198, SEQ ID NO: 199, SEQ ID NO: 200, SEQ ID NO: 201, SEQ ID NO: 202, SEQ ID NO: 203, SEQ ID NO: 204, SEQ ID NO: 205, SEQ ID NO: 206, SEQ ID NO: 207, SEQ ID NO: 208, SEQ ID NO: 209, SEQ ID NO: 210, SEQ ID NO: 211, SEQ ID NO: 212, SEQ ID NO: 213, SEQ ID NO: 214, SEQ ID NO: 215, SEQ ID NO: 216, SEQ ID NO: 217, SEQ ID NO: 218, SEQ ID NO: 219, SEQ ID NO: 220, SEQ ID NO: 221, SEQ ID NO: 222, SEQ ID NO: 223, SEQ ID NO: 224, SEQ ID NO: 225, SEQ ID NO: 226, SEQ ID NO: 227, SEQ ID NO: 228, SEQ ID NO: 229, SEQ ID NO: 230, SEQ ID NO: 231, SEQ ID NO: 232, SEQ ID NO: 233, SEQ ID NO: 234, SEQ ID NO: 235, SEQ ID NO: 236, SEQ ID NO: 237, SEQ ID NO: 238, SEQ ID NO: 239, SEQ ID NO: 240, SEQ ID NO: 241, SEQ ID NO: 242, SEQ ID NO: 243, SEQ ID NO: 244, SEQ ID NO: 245, SEQ ID NO: 246, SEQ ID NO: 247, SEQ ID NO: 248, SEQ ID NO: 249, SEQ ID NO: 250, SEQ ID NO: 251, SEQ ID NO: 252, SEQ ID NO: 253, SEQ ID NO: 254, SEQ ID NO: 255, SEQ ID NO: 256, SEQ ID NO: 257, SEQ ID NO: 258, SEQ ID NO: 259, SEQ ID NO: 260, SEQ ID NO: 261, SEQ ID NO: 262, SEQ ID NO: 263, SEQ ID NO: 264, SEQ ID NO: 265, SEQ ID NO: 266, SEQ ID NO: 267, SEQ ID NO: 268, SEQ ID NO: 269, SEQ ID NO: 270, SEQ ID NO: 271, SEQ ID NO: 272, SEQ ID NO: 273, SEQ ID NO: 274, SEQ ID NO: 275, SEQ ID NO: 276, SEQ ID NO: 277, SEQ ID NO: 278, SEQ ID NO: 279, SEQ ID NO: 280, SEQ ID NO: 281, SEQ ID NO: 282, SEQ ID NO: 283, SEQ ID NO: 284, SEQ ID NO: 285, SEQ ID NO: 286, SEQ ID NO: 287, SEQ ID NO: 288, SEQ ID NO: 289, SEQ ID NO: 290, SEQ ID NO: 291, SEQ ID NO: 292, SEQ ID NO: 293, SEQ ID NO: 294, SEQ ID NO: 295, SEQ ID NO: 296, SEQ ID NO: 297, SEQ ID NO: 298, SEQ ID NO: 299, SEQ ID NO: 300, SEQ ID NO: 301, SEQ ID NO: 302, SEQ ID NO: 303, SEQ ID NO: 304, SEQ ID NO: 305, SEQ ID NO: 306, SEQ ID NO: 307, SEQ ID NO: 308, SEQ ID NO: 309, SEQ ID NO: 310, SEQ ID NO: 311, SEQ ID NO: 312, SEQ ID NO: 313, SEQ ID NO: 314, SEQ ID NO: 315, SEQ ID NO: 316, SEQ ID NO: 317, SEQ ID NO: 318, SEQ ID NO: 319, SEQ ID NO: 320, SEQ ID NO: 321, SEQ ID NO: 322, SEQ ID NO: 323, SEQ ID NO: 324, SEQ ID NO: 325, SEQ ID NO: 326, SEQ ID NO: 327, SEQ ID NO: 328, SEQ ID NO: 329, SEQ ID NO: 330, SEQ ID NO: 331, SEQ ID NO: 332, SEQ ID NO: 333, SEQ ID NO: 334, SEQ ID NO: 335, SEQ ID NO: 336, SEQ ID NO: 337, SEQ ID NO: 338, SEQ ID NO: 339, SEQ ID NO: 340, SEQ ID NO: 341, SEQ ID NO: 342, SEQ ID NO: 343, SEQ ID NO: 344, SEQ ID NO: 345, SEQ ID NO: 346, SEQ ID NO: 347, SEQ ID NO: 348, SEQ ID NO: 349, SEQ ID NO: 350, SEQ ID NO: 351, SEQ ID NO: 352, SEQ ID NO: 353, SEQ ID NO: 354, SEQ ID NO: 355, SEQ ID NO: 356, SEQ ID NO: 357, SEQ ID NO: 358, SEQ ID NO: 359, SEQ ID NO: 360, SEQ ID NO: 361, SEQ ID NO: 362, SEQ ID NO: 363, SEQ ID NO: 364, SEQ ID NO: 365, SEQ ID NO: 366, SEQ ID NO: 367, SEQ ID NO: 368, SEQ ID NO: 369, SEQ ID NO: 370, SEQ ID NO: 371, SEQ ID NO: 372, SEQ ID NO: 373, SEQ ID NO: 374, SEQ ID NO: 375, SEQ ID NO: 376, SEQ ID NO: 377, SEQ ID NO: 378, SEQ ID NO: 379, SEQ ID NO: 380, SEQ ID NO: 381, SEQ ID NO: 382, SEQ ID NO: 383, SEQ ID NO: 384, SEQ ID NO: 385, SEQ ID NO: 386, SEQ ID NO: 387, SEQ ID NO: 388, SEQ ID NO: 389, SEQ ID NO: 390, SEQ ID NO: 391, SEQ ID NO: 392, SEQ ID NO: 393, SEQ ID NO: 394, SEQ ID NO: 395, SEQ ID NO: 396, SEQ ID NO: 397, SEQ ID NO: 398, SEQ ID NO: 399, SEQ ID NO: 400, SEQ ID NO: 401, SEQ ID NO: 402, SEQ ID NO: 403, SEQ ID NO: 404, SEQ ID NO: 405, SEQ ID NO: 406, SEQ ID NO: 407, SEQ ID NO: 408, SEQ ID NO: 409, SEQ ID NO: 410, SEQ ID NO: 411, SEQ ID NO: 412, SEQ ID NO: 413, SEQ ID NO: 414, SEQ ID NO: 415, SEQ ID NO: 416, SEQ ID NO: 417, SEQ ID NO: 418, SEQ ID NO: 419, SEQ ID NO: 420, SEQ ID NO: 421, SEQ ID NO: 422, SEQ ID NO: 423, SEQ ID NO: 424, SEQ ID NO: 425, SEQ ID NO: 426, SEQ ID NO: 427, SEQ ID NO: 428, SEQ ID NO: 429, SEQ ID NO: 430, SEQ ID NO: 431, SEQ ID NO: 432, SEQ ID NO: 433, SEQ ID NO: 434, SEQ ID NO: 435, SEQ ID NO: 436, SEQ ID NO: 437, SEQ ID NO: 438, SEQ ID NO: 439, SEQ ID NO: 440, SEQ ID NO: 441, SEQ ID NO: 442, SEQ ID NO: 443, SEQ ID NO: 444, SEQ ID NO: 445, SEQ ID NO: 446, SEQ ID NO: 447, SEQ ID NO: 448, SEQ ID NO: 449, SEQ ID NO: 450, SEQ ID NO: 451, SEQ ID NO: 452, SEQ ID NO: 453, SEQ ID NO: 454, SEQ ID NO: 455, SEQ ID NO: 456, SEQ ID NO: 457, SEQ ID NO: 458, SEQ ID NO: 459, SEQ ID NO: 460, SEQ ID NO: 461, SEQ ID NO: 462, SEQ ID NO: 463, SEQ ID NO: 464, SEQ ID NO: 465, SEQ ID NO: 466, SEQ ID NO: 467, SEQ ID NO: 468, SEQ ID NO: 469, SEQ ID NO: 470, SEQ ID NO: 471, SEQ ID NO: 472, SEQ ID NO: 473, SEQ ID NO: 474, SEQ ID NO: 475, SEQ ID NO: 476, SEQ ID NO: 477, SEQ ID NO: 478, SEQ ID NO: 479, SEQ ID NO: 480, SEQ ID NO: 481, SEQ ID NO: 482, SEQ ID NO: 483, SEQ ID NO: 484, SEQ ID NO: 485, SEQ ID NO: 486, SEQ ID NO: 487, SEQ ID NO: 488, SEQ ID NO: 489, SEQ ID NO: 490, SEQ ID NO: 491, SEQ ID NO: 492, SEQ ID NO: 493, SEQ ID NO: 494, SEQ ID NO: 495, SEQ ID NO: 496, SEQ ID NO: 497, SEQ ID NO: 498, SEQ ID NO: 499, SEQ ID NO: 500, SEQ ID NO: 501. SEQ ID NO: 502, SEQ ID NO: 503, SEQ ID NO: 504, SEQ ID NO: 505, SEQ ID NO: 506, SEQ ID NO: 507, SEQ ID NO: 508, SEQ ID NO: 509, SEQ ID NO: 510, SEQ ID NO: 511, SEQ ID NO: 512, SEQ ID NO: 513, SEQ ID NO: 514, SEQ ID NO: 515, SEQ ID NO: 516, SEQ ID NO: 517, SEQ ID NO: 518, SEQ ID NO: 519, SEQ ID NO: 520, SEQ ID NO: 521, SEQ ID NO: 522, SEQ ID NO: 523, SEQ ID NO: 524, SEQ ID NO: 525, SEQ ID NO: 526, SEQ ID NO: 527, SEQ ID NO: 528, SEQ ID NO: 529, SEQ ID NO: 530, SEQ ID NO: 531, SEQ ID NO: 532, SEQ ID NO: 533, SEQ ID NO: 534, SEQ ID NO: 535, SEQ ID NO: 536, SEQ ID NO: 537, SEQ ID NO: 538, SEQ ID NO: 539, SEQ ID NO: 540, SEQ ID NO: 541, SEQ ID NO: 542, SEQ ID NO: 543, SEQ ID NO: 544, SEQ ID NO: 545, SEQ ID NO: 546, SEQ ID NO: 547 и/или SEQ ID NO: 548.
В одном аспекте вариант содержит замену в одном или более (нескольких) положениях, соответствующих положениям 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 15, 17, 19 и 21; предпочтительно положениям 1, 2, 4, 5, 6, 8, 9, 12, 13, 15, 17 и 21; и более предпочтительно положениям 4, 5, 6, 8, 9, 12, 13, 15 и 17. В другом аспекте вариант содержит замену в двух положениях, соответствующих любым из положений 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 15, 17, 19 и 21; предпочтительно положениям 1, 2, 4, 5, 6, 8, 9, 12, 13, 15, 17 и 21; и более предпочтительно положениям 4, 5, 6, 8, 9, 12, 13, 15 и 17. В другом аспекте вариант содержит замену в трех положениях, соответствующих любым из положений 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 15, 17, 19 и 21; предпочтительно положениям 1, 2, 4, 5, 6, 8, 9, 12, 13, 15, 17 и 21; и более предпочтительно положениям 4, 5, 6, 8, 9, 12, 13, 15 и 17. В другом аспекте вариант содержит замену в четырех положениях, соответствующих любым из положений 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 15, 17, 19 и 21; предпочтительно положениям 1, 2, 4, 5, 6, 8, 9, 12, 13, 15, 17 и 21; и более предпочтительно положениям 4, 5, 6, 8, 9, 12, 13, 15 и 17. В другом аспекте вариант содержит замену в пяти положениях, соответствующих любым из положений 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 15, 17, 19 и 21; предпочтительно положениям 1, 2, 4, 5, 6, 8, 9, 12, 13, 15, 17 и 21; и более предпочтительно положениям 4, 5, 6, 8, 9, 12, 13, 15 и 17. В другом аспекте вариант содержит замену в шести положениях, соответствующих любым из положений 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 15, 17, 19 и 21; предпочтительно положениям 1, 2, 4, 5, 6, 8, 9, 12, 13, 15, 17 и 21; и более предпочтительно положениям 4, 5, 6, 8, 9, 12, 13, 15 и 17.
В другом аспекте вариант содержит замену G1A, D, F, H, I, K, M, Q, R, S, T, V, W, Y зрелого пептида с SEQ ID NO: 2. В другом аспекте вариант содержит замену F2A, G, H, I, L, M, P, S, V, W, Y зрелого пептида с SEQ ID NO: 2. В другом аспекте вариант содержит замену C3L зрелого пептида с SEQ ID NO: 2. В другом аспекте вариант содержит замену W4A, E, F, G, I, L, M, N, Q, S, T, V, Y зрелого пептида с SEQ ID NO: 2. В другом аспекте вариант содержит замену Y5E, F, G, H, K, N, R, S, W зрелого пептида с SEQ ID NO. 2. В другом аспекте вариант содержит замену V6A, C, E, F, G, H, I, L, M, N, Q, R, S, T, W, Y зрелого пептида с SEQ ID NO: 2. В другом аспекте вариант содержит замену C7V зрелого пептида с SEQ ID NO: 2. В другом аспекте вариант содержит замену V8A, F, G, H, I, L, N, S, T, W, Y зрелого пептида с SEQ ID NO: 2. В другом аспекте вариант содержит замену Y9A, D, F, G, H, I, K, M, Q, R, S, T, V, W зрелого пептида с SEQ ID NO: 2. В другом аспекте вариант содержит замену R10K, P, S, T зрелого пептида с SEQ ID NO: 2. В другом аспекте вариант содержит замену N11A, G, H, Q, R, S, Y зрелого пептида с SEQ ID NO: 2. В другом аспекте вариант содержит замену G12A, D, E, F, H, K, N, R, S, Y зрелого пептида с SEQ ID NO: 2. В другом аспекте вариант содержит замену V13A, C, F, G, H, K, L, P, Q, R, S, T, W, Y зрелого пептида с SEQ ID NO: 2. В другом аспекте вариант содержит замену V15A, C, F, G, H, I, K, L, M, N, P, Q, R, S, T, W, Y зрелого пептида с SEQ ID NO: 2. В другом аспекте вариант содержит замену Y17C, F, G, H, I, K, L, M, N, Q, R, S, T, V, W зрелого пептида с SEQ ID NO: 2. В другом аспекте вариант содержит замену R19D, H, K, M, T, Y зрелого пептида с SEQ ID NO: 2. В другом аспекте вариант содержит замену N21A, C, F, G, H, I, K, L, M, P, Q, R, S, T, W, Y зрелого пептида с SEQ ID NO: 2.
В одном аспекте вариант содержит замену в положении 1. В другом аспекте аминокислота в положении 1 заменена на Ala, Arg, Asn, Asp, Cys, GIn, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val, предпочтительно на Ala, Asp, Phe, His, IlE, Lys, Met, Gln, Arg, Ser, Thr, Val, Trp или Tyr. В другом аспекте вариант содержит замену G1A, D, F, H, I, K, M, Q, R, S, T, V, W, Y зрелого пептида с SEQ ID NO: 2.
В одном аспекте вариант содержит замену в положении 2. В другом аспекте аминокислота в положении 2 заменена на Ala, Arg, Asn, Asp, Cys, Gin, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val, предпочтительно на Ala, Gly, His, Ile, Leu, Met, Pro, Ser, Val, Trp или Tyr. В другом аспекте вариант содержит замену F2A, G, H, I, L, M, P, S, V, W, Y зрелого пептида с SEQ ID NO: 2.
В одном аспекте вариант содержит замену в положении 3. В другом аспекте аминокислота в положении 3 заменена на Ala, Arg, Asn, Asp, Cys, GIn, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val, предпочтительно на Leu. В другом аспекте вариант содержит замену C3L зрелого пептида с SEQ ID NO: 2.
В одном аспекте вариант содержит замену в положении 4. В другом аспекте аминокислота в положении 4 заменена на Ala, Arg, Asn, Asp, Cys, Gin, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val, предпочтительно на Ala, Glu, Phe, Gly, Ile, Leu, Met, Asn, Gin, Ser, Thr, Val или Tyr. В другом аспекте вариант содержит замену W4A, E, F, G, I, L, M, N, Q, S, T, V, Y зрелого пептида с SEQ ID NO: 2.
В другом аспекте вариант содержит замену в положении 5. В другом аспекте аминокислота в положении 5 заменена на Ala, Arg, Asn, Asp, Cys, Gin, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val, предпочтительно на Glu, Phe, Gly, His, Lys, Asn, Arg, Ser или Trp. В другом аспекте вариант содержит замену Y5E, F, G, H, K, N, R, S, W зрелого пептида с SEQ ID NO: 2.
В другом аспекте вариант содержит замену в положении 6. В другом аспекте аминокислота в положении 6 заменена на Ala, Arg, Asn, Asp, Cys, Gin, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val, предпочтительно на Ala, Cys, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Gin, Arg, Ser, Thr, Trp или Tyr. В другом аспекте вариант содержит замену V6A, C, E, F, G, H, I, L, M, N, Q, R, S, T, W, Y зрелого пептида с SEQ ID NO: 2.
В одном аспекте вариант содержит замену в положении 7. В другом аспекте аминокислота в положении 7 заменена на Ala, Arg, Asn, Asp, Cys, Gin, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val, предпочтительно на Val. В другом аспекте вариант содержит замену C7V зрелого пептида с SEQ ID NO: 2.
В другом аспекте вариант содержит замену в положении 8. В другом аспекте аминокислота в положении 8 заменена на Ala, Arg, Asn, Asp, Cys, Gin, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val, предпочтительно на Ala, Phe, Gly, His, Ile, Leu, Asn, Ser, Thr, Trp или Tyr. В другом аспекте вариант содержит замену V8A, F, G, H, I, L, N, S, T, W, Y зрелого пептида с SEQ ID NO: 2.
В другом аспекте вариант содержит замену в положении 9. В другом аспекте аминокислота в положении 9 заменена на Ala, Arg, Asn, Asp, Cys, Gin, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val, предпочтительно на Ala, Asp, Phe, Gly, His, Ile, Lys, Met, Gin, Arg, Ser, Thr, Val или Trp. В другом аспекте вариант содержит замену Y9A, D, F, G, H, I, K, M, Q, R, S, T, V, W зрелого пептида с SEQ ID NO: 2.
В одном аспекте вариант содержит замену в положении 10. В другом аспекте аминокислота в положении 10 заменена на Ala, Arg, Asn, Asp, Cys, Gin, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val, предпочтительно на Lys, Pro, Ser или Thr. В другом аспекте вариант содержит замену R10K, P, S, T зрелого пептида с SEQ ID NO: 2.
В одном аспекте вариант содержит замену в положении 11. В другом аспекте аминокислота в положении 11 заменена на Ala, Arg, Asn, Asp, Cys, Gin, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val, предпочтительно на Ala, Gly, His, Gin, Arg, Ser или Tyr. В другом аспекте вариант содержит замену N11A, G, H, Q, R, S, Y зрелого пептида с SEQ ID NO: 2.
В другом аспекте вариант содержит замену в положении 12. В другом аспекте аминокислота в положении 12 заменена на Ala, Arg, Asn, Asp, Cys, Gin, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val, предпочтительно на Ala, Asp, Glu, Phe, His, Lys, Asn, Arg, Ser или Tyr. В другом аспекте вариант содержит замену G12A, D, E, F, H, K, N, R, S, Y зрелого пептида с SEQ ID NO: 2.
В одном аспекте вариант содержит замену в положении 13. В другом аспекте аминокислота в положении 13 заменена на Ala, Arg, Asn, Asp, Cys, Gin, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val, предпочтительно на Ala, Cys, Phe, Gly, His, Lys, Leu, Pro, Gin, Arg, Ser, Thr, Trp или Tyr. В другом аспекте вариант содержит замену V13A, C, F, G, H, K, L, P, Q, R, S, T, W, Y зрелого пептида с SEQ ID NO: 2.
В другом аспекте вариант содержит замену в положении 15. В другом аспекте аминокислота в положении 15 заменена на Ala, Arg, Asn, Asp, Cys, Gin, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val, предпочтительно на Ala, Cys, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gin, Arg, Ser, Thr, Trp или Tyr. В другом аспекте вариант содержит замену V15A, C, F, G, H, I, K, L, M, N, P, Q, R, S, T, W, Y зрелого пептида с SEQ ID NO: 2.
В одном аспекте вариант содержит замену в положении 17. В другом аспекте аминокислота в положении 17 заменена на Ala, Arg, Asn, Asp, Cys, Gin, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val, предпочтительно на Cys, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gin, Arg, Ser, Thr, Val или Trp. В другом аспекте вариант содержит замену Y17C, F, G, H, I, K, L, M, N, Q, R, S, T, V, W зрелого пептида с SEQ ID NO: 2.
В одном аспекте вариант содержит замену в положении 19. В другом аспекте аминокислота в положении 19 заменена на Ala, Arg, Asn, Asp, Cys, Gin, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val, предпочтительно на Asp, His, Lys, Met, Thr или Tyr. В другом аспекте вариант содержит замену R19D, H, K, M, T, Y зрелого пептида с SEQ ID NO: 2.
В одном аспекте вариант содержит замену в положении 21. В другом аспекте аминокислота в положении 21 заменена на Ala, Arg, Asn, Asp, Cys, Gin, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val, предпочтительно на Ala, Cys, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gin, Arg, Ser, Thr, Trp или Tyr. В другом аспекте вариант содержит замену N21A, C, F, G, H, I, K, L, M, P, Q, R, S, T, W, Y зрелого пептида с SEQ ID NO: 2.
В другом аспекте вариант содержит замену в положениях, соответствующих положениям 5 и 17, такую как описанные выше.
В другом аспекте вариант содержит замены в положениях, соответствующих положениям 5 и 9, такие как описанные выше.
В другом аспекте вариант содержит замены в положениях, соответствующих положениям 5 и 15, такие как описанные выше.
В другом аспекте вариант содержит замены в положениях, соответствующих положениям 17 и 9, такие как описанные выше.
В другом аспекте вариант содержит замены в положениях, соответствующих положениям 17 и 15, такие как описанные выше.
В другом аспекте вариант содержит замены в положениях, соответствующих положениям 9 и 15, такие как описанные выше.
В другом аспекте вариант содержит замены в положениях, соответствующих положениям 5, 17 и 9, такие как описанные выше.
В другом аспекте вариант содержит замены в положениях, соответствующих положениям 5, 17 и 15, такие как описанные выше.
В другом аспекте вариант содержит замены в положениях, соответствующих положениям 5, 9 и 15, такие как описанные выше.
В другом аспекте вариант содержит замены в положениях, соответствующих положениям 17, 9 и 15, такие как описанные выше.
В другом аспекте вариант содержит замены в положениях, соответствующих положениям 5, 17, 9 и 15, такие как описанные выше.
В другом аспекте вариант содержит одну или более (несколько) замен, выбранных из группы, состоящей из:
G1A, G1D, G1F, G1H, G1I, G1K, G1M, G1Q, G1R, G1S, G1T, G1V, G1W, G1Y,
F2A, F2G, F2H, F2I, F2L, F2M, F2P, F2S, F2V, F2W, F2Y,
C3L,
W4A, W4E, W4F, W4G, W4I, W4L, W4M, W4N, W4Q, W4S, W4T, W4V, W4Y,
Y5E, Y5F, Y5G, Y5H, Y5K, Y5N, Y5R, Y5S, Y5W,
V6A, V6C, V6E, V6F, V6G, V6H, V6I, V6L, V6M, V6N, V6Q, V6R, V6S, V6T, V6W, V6Y,
C7V,
V8A, V8F, V8G, V8H, V8I, V8L, V8N, V8S, V8T, V8W, V8Y,
Y9A, Y9D, Y9F, Y9G, Y9H, Y9I, Y9K, Y9M, Y9Q, Y9R, Y9S, Y9T, Y9V, Y9W,
R10K, R10P, R10S, R10T,
N11A, N11G, N11H, N11Q, N11R, N11S, N11Y,
G12A, G12D, G12E, G12F, G12H, G12K, G12N, G12R, G12S, G12Y,
V13A, V13C, V13F, V13G, V13H, V13K, V13L, V13P, V13Q, V13R, V13S, V13T, V13W, V13Y,
V15A, V15C, V15F, V15G, V15H, V15I, V15K, V15L, V15M, V15N, V15P, V15Q, V15R, V15S, V15T, V15W, V15Y,
Y17C, Y17F, Y17G, Y17H, Y17I, Y17K, Y17L, Y17M, Y17N, Y17Q, Y17R, Y17S, Y17T, Y17V, Y17W,
R19D, R19H, R19K, R19M, R19T, R19Y,
N21A, N21C, N21F, N21G, N21H, N21I, N21K, N21L, N21M, N21P, N21Q, N21R, N21S, N21T, N21W и N21Y;
предпочтительно W4A, Y5H, Y5N, Y5R, V6A, V6F, V8A, Y9K, Y9R, G12R, G12K, V13A, V15I, V15S и Y17H.
В другом аспекте вариант содержит замены Y5N + Y17H зрелого пептида c SEQ ID NO: 2.
В другом аспекте вариант содержит замены Y5N + Y9R зрелого пептида с SEQ ID NO: 2.
В другом аспекте вариант содержит замены Y5N + Y9K зрелого пептида с SEQ ID NO: 2.
В другом аспекте вариант содержит замены Y17H + Y9R зрелого пептида c SEQ ID NO: 2.
В другом аспекте вариант содержит замены Y17H + Y9K зрелого пептида c SEQ ID NO: 2.
В другом аспекте вариант содержит замены Y5N+Y17H + Y9R зрелого пептида с SEQ ID NO: 2.
В другом аспекте вариант содержит замены Y5N+Y17H + Y9K зрелого пептида с SEQ ID NO: 2.
В другом аспекте вариант содержит замены Y5N + V6A + Y9K, или V8A + Y9R + V13A, или Y5N + Y9R + Y17H, или Y9K + V15S, или W4A + Y5R + Y9K, или Y5N + G12R + Y17H, или Y5N + V6F + Y17H, или Y5N + V15I + Y17H, или Y5H + V8A + Y9R, или Y5N + G12K + Y17H зрелого пептида с SEQ ID NO: 2.
Важные аминокислоты исходного пептида могут быть идентифицированы способами, известными в данной области техники, такими как сайт-направленный мутагенез или аланиновый сканирующий мутагенез (Cunningham and Wells, 1989, Science 244:1081-1085). В последней методике проводят отдельные аланиновые мутации по каждому остатку молекулы и исследуют антимикробную активность полученных мутантных молекул для идентификации аминокислотных остатков, необходимых для активности молекулы. См. также Hilton et al., 1996, J. Biol. Chem. 271:4699-4708. Важные аминокислоты могут также быть определены анализом идентичности с использованием пептидов, родственных по отношению к исходному пептиду.
Полинуклеотиды
Настоящее изобретение также относится к полинуклеотидам, кодирующим любые из вариантов по настоящему изобретению.
Конструкции нуклеиновых кислот
Настоящее изобретение также относится к конструкциям нуклеиновых кислот, содержащим полинуклеотид, кодирующий вариант по настоящему изобретению, функционально связанный с одной или более (несколькими) контрольными последовательностями, управляющими экспрессией кодирующей последовательности в подходящей клетке-хозяине в условиях, совместимых с контрольными последовательностями.
Для обеспечения экспрессии варианта полинуклеотидом можно подвергать его воздействию ряда способов. В зависимости от вектора экспрессии может быть желательна или необходима обработка полинуклеотида перед его введением в вектор. Методики модификации полинуклеотидов с использованием способов рекомбинантных ДНК хорошо известны в данной области техники.
Контрольная последовательность может представлять собой промоторную последовательность, распознаваемую клеткой-хозяином для экспрессии полинуклеотида. Промоторная последовательность содержит последовательности контроля транскрипции, опосредующие экспрессию варианта. Промотор может представлять собой любую последовательность нуклеиновой кислоты, демонстрирующую транскрипционную активность в клетке-хозяине, включая мутантные, процессированные и гибридные промоторы, и может быть получена из генов, кодирующих внеклеточные или внутриклеточные пептиды, гомологичные или гетерологичные по отношению к клетке-хозяину.
Примерами подходящих промоторов для управления транскрипцией конструкций нуклеиновых кислот по настоящему изобретению в бактериальной клетке-хозяине являются промоторы, полученные из гена альфа-амилазы Bacillus amyloliquefaciens (amyQ), гена альфа-амилазы Bacillus licheniformis (amyL), гена пенициллиназы Bacillus licheniformis (penP), гена мальтогенной амилазы Bacillus stearothermophilus (amyM), гена левансахаразы Bacillus subtilis (sacB), генов xylA и xylB Bacillus subtilis, lac-оперона E.coli, гена агаразы Streptomyces coelicolor (dagA) и прокариотического гена бета-лактамазы (Villa-Kamaroff et al., 1978, Proc. Natl. Acad. Sci. USA 75:3727-3731), а также tac-промотора (DeBoeref a/., 1983, Proc. Natl. Acad. Sci. USA 80:21-25). Другие промоторы описаны в разделе «Полезные белки рекомбинантных бактерий» ("Useful proteins from recombinant bacteria") в Gilbert et al., 1980, Scientific American 242:74-94, и в Sambrook et al., 1989, выше.
Примерами подходящих промоторов для управления транскрипцией конструкциями нуклеиновых кислот по настоящему изобретению в клетке-хозяине, представляющей собой клетку нитчатого гриба, являются промоторы, полученные из генов ацетамидазы Aspergillus nidulans, нейтральной альфа-амилазы Aspergillus niger, кислотоустойчивой альфа-амилазы Aspergillus niger, глюкоамилазы Aspergillus niger или Aspergillus awamori (glaA), ТАКА-амилазы Aspergillus oryzae, щелочной протеазы Aspergillus oryzae, триозофосфатизомеразы Aspergillus oryzae, трипсиноподобной протеазы Fusarium oxysporum (WO 96/00787), амилоглюкозидазы Fusarium venenatum (WO 00/56900), Daria Fusarium venenatum (WO 00/56900), Quinn Fusarium venenatum (WO 00/56900), липазы Rhizomucor miehei, аспарагиновой протеиназы Rhizomucor miehei, бета-глюкозидазы Trichoderma reesei, целлобиогидролазы I Trichoderma reesei, целлобиогидролазы II Trichoderma reesei, эндоглюканазы I Trichoderma reesei, эндоглюканазы II Trichoderma reesei, эндоглюканазы III Trichoderma reesei, эндоглюканазы IV Trichoderma reesei, эндоглюканазы V Trichoderma reesei, ксиланазы I Trichoderma reesei, ксиланазы II Trichoderma reesei, бета-ксилозидазы Trichoderma reesei, а также промотор NA2-tpi (модифицированный промотор, содержащий ген, кодирующий нейтральную альфа-амилазу аспергилл, где нетранслируемая лидерная последовательность была заменена нетранслируемой лидерной последовательностью гена, кодирующего триозофосфатизомеразу аспергилл; неограничивающие примеры включают модифицированные промоторы, включающие ген, кодирующий нейтральную альфа-амилазу Aspergillus niger, где нетранслируемая лидерная последовательность была заменена нетранслируемой лидерной последовательностью гена, кодирующего триозофосфатизомеразу Aspergillus nidulans или Aspergillus oryzae); и их мутантные, процессированные и гибридные промоторы.
У дрожжевого хозяина полезные промоторы получены из генов энолазы Saccharomyces cerevisiae (ENO-1), галактокиназы Saccharomyces cerevisiae (GAL1), алкогольдегидрогеназы/глицеральдегид-3-фосфатдегидрогеназы Saccharomyces cerevisiae (ADH1, ADH2/GAP), триозофосфатизомеразы Saccharomyces cerevisiae (TPI), металлотионеина Saccharomyces cerevisiae (CUP1) и 3-фосфоглицераткиназы Saccharomyces cerevisiae. Другие полезные промоторы для дрожжевых клеток-хозяев описаны Romanos et al., 1992, Yeast 8:423-488.
Контрольная последовательность может также представлять собой подходящую последовательность терминации транскрипции, распознаваемую клеткой-хозяином для завершения транскрипции. Последовательность терминации функционально связана с 3'-концом полинуклеотида, кодирующего данный вариант. Может быть использован любой терминатор, функционирующий в данной клетке-хозяине.
Предпочтительные терминаторы для клеток-хозяев, представляющих собой клетки нитчатых грибов, получены из генов антранилатсинтазы Aspergillus nidulans, альфа-глюкозидазы Aspergillus niger, глюкоамилазы Aspergillus niger, ТАКА-амилазы Aspergillus oryzae и трипсиноподобной протеазы Fusarium oxysporum.
Предпочтительные терминаторы для дрожжевых клеток-хозяев получены из генов энолазы Saccharomyces cerevisiae, цитохрома С (CYC1) Saccharomyces cerevisiae и глицеральдегид-3-фосфатдегидрогеназы Saccharomyces cerevisiae. Другие полезные терминаторы для дрожжевых клеток-хозяев описаны Romanos et al., 1992, выше.
Контрольная последовательность может также представлять собой подходящую лидерную последовательность, нетранслируемую область матричной РНК (мРНК), важную для трансляции клеткой-хозяином. Лидерная последовательность функционально связана с 5'-концом полинуклеотида, кодирующего вариант. Может быть использована любая лидерная последовательность, функционирующая в данной клетке-хозяине.
Предпочтительные лидерные последовательности для клеток-хозяев, представляющих собой клетки нитчатых грибов, получены из генов ТАКА-амилазы Aspergillus oryzae и триозофосфатизомеразы Aspergillus nidulans. Походящие лидерные последовательности для дрожжевых клеток-хозяев получены из генов энолазы Saccharomyces cerevisiae (ENO-1), 3-фосфоглицераткиназы Saccharomyces cerevisiae, альфа-фактора Saccharomyces cerevisiae и алкогольдегидрогеназы/глицеральдегид-3-фосфатдегидрогеназы Saccharomyces cerevisiae (ADH2/GAP).
Контрольная последовательность может также представлять собой последовательность полиаденилирования, последовательность, функционально связанную с 3'-концом последовательности, кодирующей вариант, при транскрипции распознаваемую клеткой-хозяином как сигнал для присоединения полиаденозиновых остатков к транскрибированной мРНК. Может быть использована любая последовательность полиаденилирования, функционирующая в данной клетке-хозяине.
Предпочтительные последовательности полиаденилирования для клеток-хозяев, представляющих собой клетки нитчатых грибов, получены из генов антранилатсинтазы Aspergillus nidulans, глюкоамилазы Aspergillus niger, альфа-глюкозидазы Aspergillus niger, ТАКА-амилазы Aspergillus oryzae и трипсиноподобной протеазы Fusarium oxysporum.
Полезные последовательности полиаденилирования для дрожжевых клеток-хозяев описаны Guo and Sherman, 1995, Mol. Cellular Biol. 15:5983-5990.
Контрольная последовательность может также представлять собой область, кодирующую сигнальный пептид, которая кодирует сигнальный пептид, связанный с N-концом варианта и направляет этот вариант в секреторный метаболический путь клетки. 5'-Конец кодирующей последовательности полинуклеотида может сам по себе содержать область, кодирующую сигнальный пептид, естественным образом связанную с сегментом кодирующей области, кодирующим вариант, в трансляционной рамке считывания. Альтернативно, 5’-конец кодирующей последовательности может содержать область, кодирующую сигнальный пептид, чужеродную по отношению к кодирующей последовательности. Чужеродная область, кодирующая сигнальный пептид, может быть необходима там, где кодирующая последовательность не содержит область, кодирующую сигнальный пептид, естественным образом. Альтернативно, чужеродная область, кодирующая сигнальный пептид, может просто заменять естественную область, кодирующую сигнальный пептид, для усиления секреции варианта. Тем не менее, может быть использована любая область, кодирующая сигнальный пептид, направляющая экспрессированный вариант в секреторный метаболический путь клетки-хозяина.
Эффективные последовательности, кодирующие сигнальные пептиды, для бактериальных клеток-хозяев представляют собой последовательности, кодирующие сигнальные пептиды, полученные из генов мальтогенной амилазы Bacillus NCIB 11837, субтилизина Bacillus licheniformis, бета-лактамазы Bacillus licheniformis, альфа-амилазы Bacillus stearothermophilus, нейтральных протеаз Bacillus stearothermophilus (nprT, nprS, nprM) и prsA Bacillus subtilis. Другие сигнальные пептиды описаны Simonen and Palva, 1993, Microbiological Reviews 57:109-137.
Эффективные последовательности, кодирующие сигнальные пептиды, для клеток-хозяев, являющихся клетками нитчатых грибов, представляют собой последовательности, кодирующие сигнальные пептиды, полученные из генов нейтральной амилазы Aspergillus niger, глюкоамилазы Aspergillus niger, ТАКА-амилазы Aspergillus oryzae, целлюлазы Humicola insolens, эндоглюканазы V Humicola insolens, липазы Humicola lanuginosa и аспарагиновой протеиназы Rhizomucor miehei.
Полезные сигнальные пептиды для дрожжевых клеток-хозяев получены из генов альфа-фактора Saccharomyces cerevisiae и инвертазы Saccharomyces cerevisiae. Другие полезные последовательности, кодирующие сигнальные пептиды, описаны Romanos et al., 1992, выше.
Контрольная последовательность может также представлять собой область, кодирующую пропептид, которая кодирует пропептид, расположенный на N-конце варианта. Полученный пептид известен как профермент или пропептид (или, в некоторых случаях, зимоген). Пропептид обычно неактивен и может быть превращен в активный пептид каталитическим или аутокаталитическим отщеплением пропептида от пропептида. Область, кодирующая пропептид, может быть получена от генов щелочной протеазы Bacillus subtilis (aprE), нейтральной протеазы Bacillus subtilis (nprT), лакказы Myceliophthora thermophila (WO 95/33836), аспарагиновой протеиназы Rhizomucor miehei и альфа-фактора Saccharomyces cerevisiae.
В случаях, когда на N-конце варианта присутствуют и область сигнального пептида, и область пропептида, указанная область пропептида расположена у N-конца варианта и указанная область сигнального пептида расположена у N-конца области пропептида.
Может также быть желательным добавление регуляторных последовательностей, позволяющих регулировать экспрессию варианта относительно роста клетки-хозяина. Примерами регуляторных систем являются системы, позволяющие включать или выключать экспрессию гена в ответ на химический или физический стимул, включая присутствие регуляторного соединения. Регуляторные системы в прокариотических системах включают системы операторов lac, tac и trp. У дрожжей может быть использована система ADH2 или система GAL1. У нитчатых грибов может быть использован промотор глюкоамилазы Aspergillus niger, промотор ТАКА-альфа-амилазы Aspergillus oryzae и промотор глюкоамилазы Aspergillus oryzae. Другими примерами регуляторных последовательностей являются последовательности, позволяющие амплифицировать гены. В эукариотических системах эти регуляторные последовательности включают ген дигидрофолатредуктазы, амплифицируемый в присутствии метотрексата, и гены металлотионеинов, амплифицируемые в присутствии тяжелых металлов. В этих случаях полинуклеотид, кодирующий вариант, функционально связан с регуляторной последовательностью.
Векторы экспрессии
Настоящее изобретение также относится к рекомбинантным векторам экспрессии, содержащим полинуклеотид по настоящему изобретению, промотор, сигналы окончания транскрипции и трансляции. Различные нуклеотидные и контрольные последовательности могут быть соединены друг с другом для получения рекомбинантного вектора экспрессии, который может содержать один или более (несколько) удобных сайтов рестрикции для обеспечения возможности введения или замены полинуклеотида, кодирующего вариант, в таких сайтах. Альтернативно, возможна экспрессия полинуклеотида посредством введения полинуклеотида или конструкции нуклеиновой кислоты, содержащей полинуклеотид, в подходящий вектор для экспрессии. При создании вектора экспрессии кодирующую последовательность располагают в векторе таким образом, чтобы кодирующая последовательность была функционально связана с подходящими контрольными последовательностями для экспрессии.
Рекомбинантный вектор экспрессии может представлять собой любой вектор (например, плазмиду или вирус), который можно удобно использовать в способах рекомбинантных ДНК и который способен приводить к экспрессии полинуклеотида. Выбор вектора будет обычно зависеть от совместимости вектора с клеткой-хозяином, в которую следует ввести вектор. Вектор может представлять собой линейную или замкнутую кольцевую плазмиду.
Вектор может представлять собой автономно реплицирующийся вектор, то есть вектор, существующий в форме внехромосомной структуры, репликация которой не зависит от репликации хромосом, например, плазмиду, внехромосомный элемент, минихромосому или искусственную хромосому. Вектор может включать любое средство обеспечения саморепликации. Альтернативно, вектор может представлять собой вектор, при введении которого в клетку-хозяина происходит его интеграция в геном и репликация вместе с хромосомой(ами), в которую он интегрирован. Кроме того, может быть использован один вектор или плазмида либо два или более векторов или плазмид, которые вместе содержат всю ДНК, которую следует ввести в геном клетки-хозяина, или транспозон.
Предпочтительно, вектор содержит один или более (несколько) селектируемых маркеров, которые позволяют проводить простую селекцию трансформированных, трансфицированных, трансдуцированных или тому подобных клеток. Селектируемый маркер представляет собой ген, продукт которого обеспечивает устойчивость к биоцидам или вирусам, устойчивость к тяжелым металлам, прототрофность ауксотрофам и тому подобное.
Примерами бактериальных селектируемых маркеров являются гены dal Bacillus licheniformis или Bacillus subtilis, или маркеры, придающие устойчивость к антибиотикам, такую как устойчивость к ампициллину, хлорамфениколу, канамицину или тетрациклину. Подходящими маркерами для дрожжевых клеток-хозяев являются ADE2, HIS3, LEU2, LYS2, МЕТ3, TRP1 и URA3. Селектируемые маркеры для использования в клетке-хозяине, являющейся клеткой нитчатого гриба, включают, без ограничения, amdS (ацетамидазу), argB (орнитинкарбамоилтрансферазу), bar (фосфинотрицинацетилтрансферазу), hph (гигромицинфосфотрансферазу), niaD (нитратредуктазу), pyrG (оротидин-5'-фосфатдекарбоксилазу), sC (сульфатаденилтрансферазу) и trpC (антранилатсинтазу), а также их эквиваленты. Для использования в клетках Aspergillus предпочтительны гены amdS и pyrG Aspergillus nidulans или Aspergillus oryzae и ген bar Streptomyces hygroscopicus.
Предпочтительно, вектор содержит элемент(ы), обеспечивающий(е) возможность интеграции вектора в геном клетки-хозяина или автономной репликации вектора в клетке независимо от генома.
Интеграция вектора в геном клетки-хозяина может быть основана на последовательности полинуклеотида, кодирующей вариант, или любом другом элементе вектора для интеграции в геном гомологичной или негомологичной рекомбинацией. Альтернативно, вектор может содержать дополнительные нуклеотидные последовательности для направления интеграции гомологичной рекомбинацией в геном клетки-хозяина в определенное(ые) положение(я) хромосом(ы). Для увеличения вероятности интеграции в определенном положении интеграционные элементы должны содержать достаточное количество нуклеиновых кислот, как например от 100 до 10000 пар оснований, от 400 до 10000 пар оснований и от 800 до 10000 пар оснований, с высокой степенью идентичности соответствующей целевой последовательности для увеличения вероятности гомологичной рекомбинации. Интеграционные элементы могут представлять собой любую последовательность, гомологичную целевой последовательности в геноме клетки-хозяина. Кроме того, интеграционные элементы могут представлять собой некодирующие или кодирующие нуклеотидные последовательности. С другой стороны, вектор может быть интегрирован в геном клетки-хозяина негомологичной рекомбинацией.
Для автономной репликации вектор может дополнительно содержать точку начала репликации, позволяющую вектору автономно реплицироваться в рассматриваемой клетке-хозяине. Точка начала репликации может представлять собой любой плазмидный репликатор, опосредующий автономную репликацию и функционирующий в клетке. Термин «точка начала репликации» или «плазмидный репликатор» означает нуклеотидную последовательность, позволяющую плазмиде или вектору реплицироваться in vivo.
Примерами бактериальных точек начала репликации являются точки начала репликации плазмид pBR322, pUC19, pACYC177 и pACYC184, обеспечивающие возможность репликации в Е. coli, и pUB110, рЕ194, pTA1060 и рАМβ1, обеспечивающие возможность репликации у Bacillus.
Примерами точек начала репликации для использования в дрожжевой клетке-хозяине являются 2-микронная точка начала репликации, ARS1, ARS4, комбинация ARS1 и CEN3 и комбинация ARS4 и CEN6.
Примерами точек начала репликации, полезных в клетке нитчатого гриба, являются АМА1 и ANS1 (Gems et al., 1991, Gene 98:61-67; Cullen et al., 1987, Nucleic Acids Res. 15:9163-9175; WO 00/24883). Выделение гена AMA1 и конструирование плазмид или векторов, содержащих данный ген, может быть проведено способами, раскрытыми в WO 00/24883.
Для усиления продуцирования варианта в клетку-хозяин можно ввести более одной копии полинуклеотида по настоящему изобретению. Увеличение числа копий полинуклеотида может быть обеспечено интеграцией по меньшей мере одной дополнительной копии последовательности в геном клетки-хозяина или включением амплифицируемого селектируемого маркерного гена в полинуклеотид, где клетки, содержащие амплифицированные копии селектируемого маркерного гена и, таким образом, дополнительные копии полинуклеотида могут быть селектированы посредством культивирования клеток в присутствии подходящего агента для селекции.
Способы, применяемые для лигирования элементов, описанных выше, с конструированием рекомбинантных векторов экспрессии по настоящему изобретению для получения по существу чистых вариантов хорошо известны специалисту в данной области техники (см., например, Sambrook et al., 1989, выше).
Клетки-хозяева
Настоящее изобретение также относится к рекомбинантным клеткам-хозяевам, содержащим полинуклеотид по настоящему изобретению, функционально связанный с одной или более (несколькими) контрольными последовательностями, управляющими продуцированием варианта по настоящему изобретению. Конструкцию или вектор, содержащие полинуклеотид, вводят в клетку-хозяина, таким образом, что конструкция или вектор сохраняются в интегрированной в хромосому форме или в форме самореплицирующегося внехромосомного вектора, как описано выше. Термин «клетка-хозяин» включает любое потомство исходной клетки, не идентичное исходной клетке в виду мутаций, происходящих при репликации. Выбор клетки-хозяина будет в значительной степени зависеть от гена, кодирующего вариант, и его источника.
Клетка-хозяин может представлять собой любую клетку, полезную в рекомбинантном продуцировании варианта, например, прокариотическую или эукариотическую клетку.
Прокариотическая клетка-хозяин может представлять собой любую грамположительную или грамотрицательную бактерию. Грамположительные бактерии включают, без ограничения, Bacillus, Clostridium, Enterococcus, Geobacillus, Lactobacillus, Lactococcus, Oceanobacillus, Staphylococcus, Streptococcus и Streptomyces. Грамотрицательные бактерии включают, без ограничения, Campylobacter, Е. coli, Flavobacterium, Fusobacterium, Helicobacter, Ilyobacter, Neisseria, Pseudomonas, Salmonella и Ureaplasma.
Бактериальная клетка-хозяин может представлять собой любую клетку Bacillus, включая, без ограничения, клетки Bacillus alkalophilus, Bacillus amyloliquefaciens, Bacillus brevis, Bacillus circulans, Bacillus clausii, Bacillus coagulans, Bacillus firmus, Bacillus lautus, Bacillus lentus, Bacillus licheniformis, Bacillus megaterium, Bacillus pumilus, Bacillus stearothermophilus, Bacillus subtilis и Bacillus thuringiensis.
Бактериальная клетка-хозяин может также представлять собой любую клетку Streptococcus, включая, без ограничения, клетки Streptococcus equisimilis, Streptococcus pyogenes, Streptococcus uberis и Streptococcus equi, subsp.zooepidemicus.
Бактериальная клетка-хозяин может также представлять собой любую клетку Streptomyces, включая, без ограничения, клетки Streptomyces achromogenes, Streptomyces avermitilis, Streptomyces coelicolor, Streptomyces griseus и Streptomyces lividans.
Введение ДНК в клетку Bacillus может, например, быть проведено трансформацией протопластов (см., например, Chang and Cohen, 1979, Mol. Gen. Genet. 168:111-115), использованием компетентных клеток (см., например, Young and Spizizen, 1961, J. Bacteriol. 81:823-829, или Dubnau and Davidoff-Abelson, 1971, J. Mol. Biol. 56:209-221), электропорацией (см., например, Shigekawa and Dower, 1988, Biotechniques 6:742-751) или конъюгацией (см., например, Koehler and Thorne, 1987, J. Bacteriol. 169:5271-5278). Введение ДНК в клетку Е. coli может, например, быть проведено трансформацией протопластов (см., например, Hanahan, 1983, J. Mol. Biol. 166:557-580) или электропорацией (см., например, Dower et al., 1988, Nucleic Acids Res. 16:6127-6145). Введение ДНК в клетку Streptomyces может, например, быть проведено трансформацией протопластов и электропорацией (см., например, Gong et al., 2004, Folia Microbiol. (Praha) 49:399-405), конъюгацией (см., например, Mazodier et al., 1989, J. Bacteriol. 171:3583-3585) или трансдукцией (см., например, Burke et al., 2001, Proc. Natl. Acad. Sci. USA 98:6289-6294). Введение ДНК в клетку Pseudomonas может, например, быть проведено электропорацией (см., например, Choi et al., 2006, J. Microbiol. Methods 64:391-397) или конъюгацией (см., например, Pinedo and Smets, 2005, Appl. Environ. Microbiol. 71:51-57). Введение ДНК в клетку Streptococcus может, например, быть проведено с использованием естественной компетентности (см., например, Perry and Kuramitsu, 1981, Infect. Immun. 32:1295-1297), трансформацией протопластов (см., например, Catt and Jollick, 1991, Microbios 68:189-2070), электропорацией (см., например, Buckley et al., 1999, Appl. Environ. Microbiol. 65:3800-3804) или конъюгацией (см., например, Clewell, 1981, Microbiol. Rev. 45:409-436). Тем не менее, может быть использован любой способ введения ДНК в клетку-хозяина, известный в данной области техники.
Клетка-хозяин может также представлять собой эукариотическую клетку, такую как клетка млекопитающего, насекомого, растения или гриба.
Клетка-хозяин может представлять собой клетку гриба. При использовании здесь «грибы» включают таксономические группы Ascomycota, Basidiomycota, Chytridiomycota и Zygomycota, а также Oomycota и все митоспоровые грибы (как определено Hawksworth et al., в Ainsworth and Bisby’s Dictionary of The Fungi, 8th edition, 1995, CAB International, University Press, Cambridge, UK).
Клетка-хозяин, являющаяся клеткой гриба, может представлять собой дрожжевую клетку. При использовании здесь «дрожжи» включают аскоспорогенные дрожжи (Endomycetales), базидиоспорогенные дрожжи и дрожжи, являющиеся несовершенными грибами (Fungi Imperfecti) (Blastomycetes). Поскольку в будущем возможны изменения классификации дрожжей, для задач данного изобретения дрожжи следует определять как описано в Biology and Activities of Yeast (Skinner, F.A., Passmore, S.M., and Davenport, R.R., eds, Soc. App. Bacteriol. Symposium Series No. 9, 1980).
Дрожжевая клетка-хозяин может представлять собой клетку Candida, Hansenula, Kluyveromyces, Pichia, Saccharomyces, Schizosaccharomyces или Yarrowia, такую как клетка Kluyveromyces lactis, Saccharomyces carlsbergensis, Saccharomyces cerevisiae, Saccharomyces diastaticus, Saccharomyces douglasii, Saccharomyces kluyveri, Saccharomyces norbensis, Saccharomyces oviformis или Yarrowia lipolytica.
Клетка-хозяин, являющаяся клеткой гриба, может представлять собой клетку нитчатого гриба. «Нитчатые грибы» включают все нитчатые формы таксономических групп Eumycota и Oomycota (как определено Hawksworth et al., 1995, supra). Для нитчатых грибов обычно характерна мицелиальная стенка, состоящая из хитина, целлюлозы, глюкана, хитозана, маннана и других сложных полисахаридов. Вегетативный рост происходит удлинением гиф, и углеродный катаболизм облигатно аэробен. В отличие от этого, вегетативный рост дрожжей, таких как Saccharomyces cerevisiae, происходит почкованием одноклеточного таллома, и катаболизм углерода может быть ферментативным.
Клетка-хозяин, являющаяся клеткой нитчатого гриба, может представлять собой клетку Acremonium, Aspergillus, Aureobasidium, Bjerkandera, Ceriporiopsis, Chrysosporium, Coprinus, Coriolus, Cryptococcus, Filibasidium, Fusarium, Humicola, Magnaporthe, Mucor, Myceliophthora, Neocallimastix, Neurospora, Paecilomyces, Penicillium, Phanerochaete, Phlebia, Piromyces, Pleurotus, Schizophyllum, Talaromyces, Thermoascus, Thielavia, Tolypocladium, Trametes или Trichoderma.
Например, клетка-хозяин, являющаяся клеткой нитчатого гриба, может представлять собой клетку Aspergillus awamori, Aspergillus foetidus, Aspergillus fumigatus, Aspergillus japonicus, Aspergillus nidulans, Aspergillus niger, Aspergillus oryzae, Bjerkandera adusta, Ceriporiopsis aneirina, Ceriporiopsis caregiea, Ceriporiopsis gilvescens, Ceriporiopsis pannocinta, Ceriporiopsis rivulosa, Ceriporiopsis subrufa, Ceriporiopsis subvermispora, Chrysosporium inops, Chrysosporium keratinophilum, Chrysosporium lucknowense, Chrysosporium merdarium, Chrysosporium pannicola, Chrysosporium queenslandicum, Chrysosporium tropicum, Chrysosporium zonatum, Coprinus cinereus, Coriolus hirsutus, Fusarium bactridioides, Fusarium cerealis, Fusarium crookwellense, Fusarium culmorum, Fusarium graminearum, Fusarium graminum, Fusarium heterosporum, Fusarium negundi, Fusarium oxysporum, Fusarium reticulatum, Fusarium roseum, Fusarium sambucinum, Fusarium sarcochroum, Fusarium sporotrichioides, Fusarium sulphureum, Fusarium torulosum, Fusarium trichothecioides, Fusarium venenatum, Humicola insolens, Humicola lanuginosa, Mucor miehei, Myceliophthora thermophila, Neurospora crassa, Penicillium purpurogenum, Phanerochaete chrysosporium, Phlebia radiata, Pleurotus eryngii, Thielavia terrestris, Trametes villosa, Trametes versicolor, Trichoderma harzianum, Trichoderma koningii, Trichoderma longibrachiatum, Trichoderma reesei или Trichoderma viride.
Клетки грибов можно трансформировать способом, включающим образование протопластов, трансформацию протопластов и регенерацию клеточной стенки способом, который известен сам по себе. Подходящие способы трансформации клеток-хозяев Aspergillus и Trichoderma описаны в EP 238023 и Yelton et al., 1984, Proc. Natl. Acad. Sci. USA 81:1470-1474. Подходящие способы трансформации для видов Fusarium описаны Malardier et al., 1989, Gene 78:147-156 и в WO 96/00787. Дрожжи можно трансформировать с использованием способов, описанных Becker and Guarente, в Abelson, J.N. and Simon, M.I., editors, Guide to Yeast Genetics and Molecular Biology, Methods in Enzymology, Volume 194, pp 182-187, Academic Press, Inc., New York; Ito et al., 1983, J. Bacteriol. 153:163; и Hinnen et al., 1978, Proc. Natl. Acad. Sci. USA 75:1920.
Способы получения
Настоящее изобретение также относится к способам получения варианта, включающим: (а) культивирование клетки-хозяина по настоящему изобретению в условиях, подходящих для экспрессии указанного варианта; и (б) выделение указанного варианта.
Клетки-хозяева культивируют в питательной среде, подходящей для продуцирования варианта, с использованием способов, известных в данной области техники. Например, клетку можно культивировать во встряхиваемой колбе или мелкомасштабной или крупномасштабной ферментацией (включая непрерывную, периодическую ферментацию, ферментацию с подпиткой или твердофазную ферментацию) в лабораторных или промышленных ферментаторах, проводимой в подходящей среде и в условиях, обеспечивающих экспрессию и/или выделение пептида. Культивирование проводят в подходящей питательной среде, содержащей источники углерода и азота и неорганические соли, с использованием способов, известных в данной области техники. Подходящие среды доступны от коммерческих поставщиков или могут быть приготовлены в соответствии с опубликованными составами (например, в каталогах Американской коллекции типовых культур). При секреции варианта в питательную среду вариант можно выделять непосредственно из среды. Если клетки не секретируют вариант, его можно выделять из лизатов клеток.
Вариант можно выявлять с использованием способов, известных в данной области техники, специфичных в отношении вариантов. Эти способы выявления могут включать применение специфичных антител, образование продукта фермента или исчезновение субстрата фермента. Например, ферментный анализ может быть использован для определения активности варианта.
Вариант может быть выделен способами, известными в данной области техники. Например, вариант может быть выделен из питательной среды обычными способами, включая, без ограничения, сбор, центрифугирование, фильтрацию, экстракцию, распылительную сушку, выпаривание или преципитацию.
Для получения по существу чистых вариантов вариант может быть очищен множеством способов, известных в данной области техники, включая, без ограничения, хроматографию (например ионообменную, аффинную, гидрофобную хроматографию, хроматофокусирование и гель-фильтрацию), электрофоретические способы (например препаративное изоэлектрическое фокусирование), дифференциальную растворимость (например фракционирование сульфатом аммония), SDS-PAGE или экстракцию (см., например, Protein Purification, J.-С. Janson and Lars Ryden, editors, VCH Publishers, New York, 1989).
В альтернативном аспекте вариант не выделяют, а клетку-хозяина по настоящему изобретению, экспрессирующую вариант, используют в качестве источника варианта.
Синтез in vitro
Полипептиды по изобретению могут также быть получены синтезом in vitro с использованием обычных способов, известных в данной области техники. Доступно множество коммерческих устройств для синтеза, например, автоматические устройства для синтеза от Applied Biosystems Inc., Beckman и так далее. С использованием устройств для синтеза встречающиеся в природе аминокислоты могут быть заменены искусственными аминокислотами, в частности D-изомерами (или D-формами), например D-аланином или D-изолейцином, диастереомерами, боковыми цепями другой длины или функциональности, и тому подобным. Конкретную последовательность и способ получения будут определять, исходя из требований удобства, экономических факторов, степени чистоты и тому подобного.
Можно обеспечить химическое связывание с различными пептидами или белками, содержащими подходящие функциональные группы для связывания, такие как аминогруппы для образования амидов или замещенных аминов, например, восстановительное аминирование, тиольные группы для образования тиоэфиров или дисульфидов, карбоксильные группы для образования амидов и тому подобное.
Если желательно, при синтезе или при экспрессии в пептид могут быть введены различные группы, позволяющие связывать его с другими молекулами или с поверхностью. Таким образом, могут быть использованы цистеины для получения тиоэфиров, гистидины для связывания с металлическим ионным комплексом, карбоксильные группы для образования амидов или эфиров, аминогруппы для образования амидов, и тому подобное.
Полипептиды могут также быть выделены и очищены обычными способами рекомбинантного синтеза. Может быть получен лизат экспрессирующего хозяина, и полученный лизат может быть очищен с использованием высокоэффективной жидкостной хроматографии (HPLC), гель-хроматографии, электрофореза в геле, аффинной хроматографии или другой методики очистки. По большей части, используемые композиции будут содержать по меньшей мере 20% желаемого продукта по массе, чаще по меньшей мере 75% по массе, предпочтительно по меньшей мере 95% по массе, и для терапевтических задач обычно по меньшей мере 95,5% по массе по отношению к примесям, связанным со способом получения продукта и его очистки. Обычно проценты будут основаны на общем количестве белка.
Растения
Настоящее изобретение также относится к растениям, например трансгенному растению, части растения или растительной клетке, содержащим полинуклеотид по настоящему изобретению, таким образом, что они экспрессируют и продуцируют вариант в выделяемых количествах. Вариант может быть выделен из растения или части растения. Альтернативно, растение или часть растения, содержащие вариант, могут быть использованы сами по себе для улучшения качества пищи или корма, например, улучшения пищевой ценности, вкусовых качеств и реологических свойств или для устранения непитательного фактора.
Трансгенное растение может быть двудольным или однодольным. Примерами однодольных растений являются травы, такие как мятлик луговой (мятлик, Роа), кормовая трава, такая как Festuca, Lolium, травы умеренной климатической зоны, такие как Agrostis, и злаки, например, пшеница, овес, рожь, ячмень, рис, сорго и маис (кукуруза).
Примерами двудольных растений являются табак, бобовые, такие как люпины, картофель, сахарная свекла, горох, фасоль и соя, и крестоцветные растения (семейство Brassicaceae), такие как цветная капуста, рапс и близкородственный модельный организм Arabidopsis thaliana.
Примерами частей растений являются стебель, каллюс, листья, корень, плоды, семена и клубни, а также отдельные ткани, входящие в состав этих частей, например, эпидермис, мезофилл, паренхима, сосудистые ткани, меристема. Определенные компартменты растительных клеток, такие как хлоропласты, апопласты, митохондрии, вакуоли, пероксисомы и цитоплазма также считают частями растений. Кроме того, любую растительную клетку, независимо от тканевого происхождения, считают частью растения. Сходным образом, части растений, такие как определенные ткани и клетки, например, зародыши, эндосперм, алейрон и семенные оболочки, также считают частями растений.
В объем настоящего изобретения также входит потомство таких растений, частей растений и растительных клеток.
Трансгенные растение или растительная клетка, экспрессирующие вариант, могут быть сконструированы способами, известными в данной области техники. Кратко, растение или растительную клетку конструируют введением одной или более (нескольких) экспрессионных конструкций, кодирующих вариант, в геном растения-хозяина или геном хлоропласта и размножением полученных в результате модифицированных растения или растительной клетки с получением трансгенного растения или растительной клетки.
Экспрессионная конструкция обычно представляет собой конструкцию нуклеиновой кислоты, содержащую полинуклеотид, кодирующий вариант, функционально связанный с подходящими регуляторными последовательностями, необходимыми для экспрессии полинуклеотида в выбранных растении или части растения. Кроме того, экспрессионная конструкция может содержать селектируемый маркер, полезный для идентификации растительных клеток, в которые интегрирована данная экспрессионная конструкция, и последовательностей ДНК, необходимых для введения конструкции в рассматриваемое растение (последнее зависит от применяемого способа введения ДНК).
Выбор регуляторных последовательностей, таких как промоторные и терминаторные последовательности и возможно сигнальные или транзитные последовательности, зависит, например, от того, когда, где и как желательно экспрессировать вариант. Например, экспрессия гена, кодирующего вариант, может быть конститутивной или индуцибельной или может быть специфической для стадии развития или ткани, и продукт гена может направляться в определенную ткань или часть растения, такие как семена или листья. Регуляторные последовательности описаны, например, в Tague et al., 1988, Plant Physiology 86:506.
Для конститутивной экспрессии можно использовать промотор 35S-CaMV, кукурузного убиквитина 1 и рисового актина 1 (Franck et al., 1980, Cell 21:285-294; Christensen et al., 1992, Plant Mol. Biol. 18:675-689; Zhang et al., 1991, Plant Cell 3:1155-1165). Органоспецифическими промоторами могут быть, например, промотор из запасающих тканей, таких как семена, клубни картофеля и плоды (Edwards and Coruzzi, 1990, Ann. Rev. Genet. 24:275-303), или метаболических тканей, таких как меристема (Ito et al., 1994, Plant Mol. Biol. 24:863-878), семяспецифичный промотор, такой как промоторы рисовых глютелина, проламина, глобулина или альбумина (Wu et al., 1998, Plant Cell Physiol. 39:885-889), промотор легумина В4 Vicia faba и гена неизвестного семенного белка Vicia faba (Conrad et al., 1998, J. Plant Physiol. 152:708-711), промотор белка масляного тельца семени (Chen et al., 1998, Plant Cell Physiol. 39:935-941), промотор запасного белка napA Brassica napus или любой другой семяспецифичный промотор, известный в данной области техники, например, как описано в WO 91/14772. Кроме того, промотор может представлять собой промотор, специфичный для листа, такой как промотор rbcs риса или томата (Kyozuka et al., 1993, Plant Physiol. 102:991-1000), промотор гена аденинметилтрансферазы вируса хлореллы (Mitra and Higgins, 1994, Plant Mol. Biol. 26:85-93), промотор гена aldP риса (Kagaya et al., 1995, Mol. Gen. Genet. 248:668-674) или промотор, индуцируемый повреждением, такой как промотор pin2 картофеля (Xu et al., 1993, Plant Mol. Biol. 22:573-588). Сходным образом, промотор может быть индуцирован абиотической обработкой, такой как температура, сухость или изменения содержания солей, или индуцирован экзогенными веществами, активирующими промотор, например, этанолом, эстрогенами, гормонами растений, такими как этилен, абсцизовая кислота и гибберелловая кислота, и тяжелыми металлами.
Можно также использовать энхансерный элемент промотора для достижения большей экспрессии варианта у растения. Например, энхансерным элементом промотора может быть интрон, расположенный между промотором и полинуклеотидом, кодирующим вариант. Например, в Xu et al., 1993, выше, раскрыто использование первого интрона гена актина 1 риса для усиления экспрессии.
Селектируемый маркерный ген и любые другие части экспрессионной конструкции могут быть выбраны из генов и любых других частей экспрессионной конструкции, доступных в данной области техники.
Конструкцию нуклеиновой кислоты включают в геном растения обычными методиками, известными в данной области, включая трансформацию, опосредованную Agrobacterium, вирус-опосредованную трансформацию, микроинъекцию, бомбардировку частицами, биолистическую трансформацию и электропорацию (Gasser et al., 1990, Science 244:1293; Potrykus, 1990, Bio/Technology 8:535; Shimamoto etal., 1989, Nature 338:274).
В настоящее время способом выбора для получения трансгенных двудольных растений является перенос генов, опосредованный Agrobacterium tumefaciens (см. обзор в Hooykas and Schilperoort, 1992, Plant Mol. Biol. 19: 15-38), и он может также быть использован для трансформации однодольных растений, хотя другие способы трансформации часто используют для этих растений. В настоящее время способом выбора для получения трансгенных однодольных растений является бомбардировка зародышевых каллюсов или развивающихся зародышей частицами (микроскопическими частицами золота или вольфрама, покрытыми трансформирующей ДНК) (Christou, 1992, Plant J. 2: 275-281; Shimamoto, 1994, Curr. Opin. Biotechnol. 5: 158-162; Vasil et al., 1992, Bio/Technology 10: 667-674). Альтернативный способ трансформации однодольных растений основан на трансформации протопластов, как описано в статье Omirulleh et al., 1993, PlantMol. Biol. 21: 415-428. Дополнительные способы трансформации для применения в соответствии с настоящим описанием включают способы, описанные в патентах США №№6395966 и 7151204 (каждый из которых полностью включен сюда посредством ссылки).
После трансформации проводят селекцию трансформантов с включенной экспрессионной конструкцией и их регенерацию с получением целых растений способами, хорошо известными в данной области техники. Часто способ трансформации разработан для селективного удаления селектируемых генов, либо в процессе регенерации, либо в последующих поколениях, с использованием, например, котрансформации двумя отдельными Т-ДНК-конструкциями или сайт-специфичного удаления селектируемого гена специфичной рекомбиназой.
Кроме прямой трансформации генотипа определенного растения конструкцией, полученной по настоящему изобретению, трансгенные растения могут быть получены скрещиванием растения, обладающего такой конструкцией, с растением без такой конструкции. Например, конструкция, кодирующая вариант, может быть введена в определенное множество растений скрещиванием, без необходимости когда-либо проводить прямую трансформацию растения данного множества. Таким образом, настоящее изобретение включает не только растение, полученное прямой регенерацией из клеток, трансформированных по настоящему изобретению, но также потомство таких растений. При использовании здесь потомство может относиться к потомству исходного растения, полученного по настоящему изобретению, любого поколения. Такое потомство может содержать ДНК-конструкцию, полученную по настоящему изобретению, или часть ДНК-конструкции, полученной по настоящему изобретению. Скрещивание приводит к введению трансгена в линию растений посредством перекрестного опыления исходной линии донорской линией растений. Неограничивающие примеры таких стадий описаны более подробно в патенте США №7151204.
Растения могут быть получены способом конверсии обратным скрещиванием. Например, растения включают растения, называемые генотипом, линией, инбредным растением или гибридом, полученными конверсией обратным скрещиванием.
Генетические маркеры могут быть использованы для облегчения интрогрессии одного или более трансгенов по изобретению из одного генетического фона в другой. Селекция с использованием маркеров обеспечивает преимущества по сравнению с обычной селекцией, так как она может быть использована во избежание ошибок, обусловленных фенотипической изменчивостью. Кроме того, генетические маркеры позволяют получить данные об относительном содержании элитной зародышевой плазмы в отдельном потомстве определенного скрещивания. Например, при скрещивании растения с желаемым признаком, но обладающего в остальном агрономически нежелательным генетическим фоном, с элитным родителем, генетические маркеры могут быть использованы для селекции потомства, обладающего не только интересующим признаком, но также относительно большой долей желаемой зародышевой плазмы. Таким способом минимизируют число поколений, необходимое для интрогрессии одного или более признаков в определенный генетический фон.
Настоящее изобретение также относится к способам получения варианта по настоящему изобретению, включающим: (а) культивирование трансгенного растения или растительной клетки, содержащих полинуклеотид, кодирующий вариант, в условиях, подходящих для продуцирования указанного варианта; и (б) выделение указанного варианта.
Способы и применения
Настоящее изобретение также относится к способам применения полипептидов, обладающих антимикробной активностью. Антимикробные полипептиды обычно полезны в любом месте, подверженном микробной контаминации. Обычно это места в водных системах, таких как системы охлаждения воды, в которых необходимо уничтожать микроорганизмы или контролировать их рост. Тем не менее, настоящее изобретение может также быть использовано во всех применениях, для которых полезны известные антимикробные композиции, таких как защита древесины, латекса, клеев, бумаги, картона, тканей, кожи и корма.
Другие применения включают сохранение продуктов питания, напитков, косметики, такой как лосьоны, кремы, гели, мази, мыла, шампуни, кондиционеры, антиперспиранты, дезодоранты, жидкостей для полоскания рта, продуктов для контактных линз или пищевых ингредиентов.
В целом, подразумевают, что антимикробные полипептиды по настоящему изобретению полезны для очистки, дезинфекции или ингибирования микробного роста на любой поверхности. Примерами поверхностей, с которыми можно полезным образом приводить в контакт антимикробные полипептиды по изобретению, являются поверхности используемого технологического оборудования, например, в молочном хозяйстве, установках для химической или фармацевтической промышленности. Антимикробные полипептиды по изобретению следует использовать в количестве, эффективном для очистки, дезинфекции или ингибирования микробного роста на рассматриваемой поверхности.
Кроме того, антимикробные полипептиды по изобретению могут быть использованы для очистки поверхностей и посуды для приготовления пищевых продуктов на предприятиях пищевой промышленности и в любой области, где готовят и подают пищу, такой как больницы, дома престарелых и рестораны.
Изобретение также относится к применению антимикробного пептида или композиции по изобретению в качестве лекарственного средства. Кроме того, антимикробный пептид или композиция по изобретению могут также быть использованы для изготовления лекарственного средства для контроля или борьбы с микроорганизмами, такими как грибы или бактерии, предпочтительно грамотрицательные бактерии.
Композиция и антимикробный пептид по изобретению могут быть использованы в качестве терапевтического или профилактического агента для применения в ветеринарии или у людей. Таким образом, композиция и антимикробный пептид по изобретению могут быть использованы в изготовлении терапевтических агентов или профилактических агентов для применения в ветеринарии или у людей для лечения микробных инфекций, таких как бактериальные или грибковые инфекции, предпочтительно инфекции, вызванные грамположительными бактериями. В частности, микробные инфекции могут быть связаны с заболеваниями легких, включая, без ограничения, туберкулез, пневмонию и муковисцидоз; кожные инфекции и инфекции глаза или полости рта; и заболевания, передающиеся половым путем, включая, без ограничения, гонорею и хламидиоз. Изобретение также относится к композициям или продуктам для заживления ран, таким как повязки, медицинским устройствам, таким как, например, катетеры.
Композиция по изобретению содержит эффективное количество антимикробного полипептида по изобретению.
Подразумевают, что при использовании здесь термин «эффективное количество» означает количество антимикробных полипептидов по изобретению, достаточное для ингибирования роста рассматриваемых микроорганизмов.
Композиции антимикробных полипептидов по изобретению вводят хозяину, страдающему микробной инфекцией или предрасположенному к ней. Введение может быть местным, локальным или системным, в зависимости от конкретного микроорганизма, предпочтительно, оно будет локальным. Обычно доза антимикробных полипептидов по изобретению будет достаточной для уменьшения микробной популяции по меньшей мере на 50%, обычно по меньшей мере на 1 log (логарифм) и, возможно, на 2 или более log уничтожения. Соединения по настоящему изобретению вводят в дозе, уменьшающей микробную популяцию при минимальных побочных эффектах. Предполагают, что для применения in vivo композицию будут получать и использовать под руководством врача. Антимикробные полипептиды по изобретению особенно полезны для уничтожения грамотрицательных бактерий, включая Pseudomonas aeruginosa и Chlamydia trachomatis; и грамположительных бактерий, включая стрептококков, таких как Streptococcus pneumonia, S. uberis, S. hyointestinalis, S. pyogenes и S. agalactiae; и стафилококков, таких как Staphylococcus aureus, S. epidermidis, S. simulans, S. xylosus и S. carnosus.
Композиции антимикробных полипептидов по изобретению могут быть введены хозяину, страдающему микробной инфекцией легких или предрасположенному к микробной инфекции легких, такой как пневмония; или к микробной раневой инфекции, такой как бактериальная раневая инфекция.
Композиции антимикробных полипептидов по изобретению могут также быть введены хозяину, страдающему кожной инфекцией или предрасположенному к кожной инфекции, такой как акне, атопический дерматит или себорейный дерматит; предпочтительно, кожная инфекция представляет собой бактериальную кожную инфекцию, например, вызванную Staphylococcus epidermidis, Staphylococcus aureus, Propionibacterium acnes, Pityrosporum ovale или Malassezia furfur.
Антимикробные полипептиды по изобретению также полезны для in vitro-композиций для уничтожения микробов, в частности там, где нежелательно применение обычных антибиотиков. Например, антимикробные полипептиды по изобретению могут быть добавлены в пищевые продукты для животных и/или людей; или они могут быть включены в качестве добавки в культуры клеток in vitro для предотвращения чрезмерного роста микробов в тканевой культуре.
Чувствительность определенного микроба к уничтожению антимикробными полипептидами по изобретению может быть определена исследованием in vitro, как подробно описано в экспериментальном разделе. Обычно в культуру микроба добавляют антимикробный полипептид в различных концентрациях на протяжении периода времени, достаточного для действия белка, обычно от одного часа до одних суток. Затем проводят подсчет жизнеспособных микробов и определяют уровень уничтожения.
Представляющие интерес микробы включают, без ограничения, грамотрицательные бактерии, например, Citrobacter sp.; Enterobacter sp.; Escherichia sp., например, E. coli; Klebsiella sp.; Morganella sp.; Proteus sp.; Providencia sp.; Salmonella sp., например, S. typhi, S. typhimurium; Serratia sp.; Shigella sp.; Pseudomonas sp., например, P. aeruginosa; Yersinia sp., например, Y. pestis, Y. pseudotuberculosis, Y. enterocolitica; Franciscella sp.; Pasturella sp.; Vibrio sp., например, V. cholerae, V. parahemolyticus; Campylobacter sp., например, С. jejuni; Haemophilus sp., например, H. influenzae, H. ducreyi; Bordetella sp., например, В. pertussis, В. bronchiseptica, В. parapertussis; Brucella sp., Neisseria sp., например, N. gonorrhoeae, N. meningitidis, и так далее. Другие интересующие бактерии включают Legionella sp., например, L. pneumophila; Listeria sp., например, L. monocytogenes; Mycoplasma sp., например, M. hominis, M. pneumoniae; Mycobacterium sp., например, M. tuberculosis, M. leprae; Treponema sp., например, Т. pallidum; Borrelia sp., например, В. burgdorferi; Leptospirae sp.; Rickettsia sp., например, R. rickettsii, R. typhi; Chlamydia sp., например, С.trachomatis, С.pneumoniae, С.psittaci; Helicobacter sp., например, H. pylori; и так далее.
Представляющие интерес небактериальные патогены включают грибковые и протозойные патогены, например, Plasmodia sp., например, Р. falciparum, Trypanosoma sp., например, Т. brucei; шистосомы; Entaemoeba sp., Cryptococcus sp., Candida sp., например, С.albicans; и так далее.
Могут быть использованы различные способы введения. Полипептидную композицию можно давать перорально или можно вводить внутрисосудистой, подкожной, внутрибрюшинной инъекцией, в форме аэрозоля, глазным способом введения, в мочевой пузырь, местно и так далее. Например, способы ингаляционного введения хорошо известны в данной области техники. Доза терапевтической композиции будет варьировать в широких пределах в зависимости от конкретного вводимого антимикробного полипептида, природы заболевания, частоты введения, способа введения, клиренса используемого агента из организма хозяина и тому подобного. Начальная доза может быть более высокой с последующими более низкими поддерживающими дозами. Дозу можно вводить с частотой один раз в неделю или один раз в две недели, или делить на меньшие дозы и вводить их один или несколько раз в сутки, два раза в неделю и так далее для поддержания эффективного уровня дозы. Во многих случаях для перорального введения будет необходима более высокая доза, чем для внутривенного введения. Амидные связи, а также N- и C-концы могут быть модифицированы для повышения стабильности при пероральном введении. Например, C-конец может быть амидирован.
Композиции
Соединения по данному изобретению могут быть включены во множество композиций для терапевтического введения. Более конкретно, соединения по настоящему изобретению могут быть включены в фармацевтические композиции в сочетании с подходящими фармацевтически приемлемыми носителями или разбавителями и могут быть включены в препараты в твердой, полутвердой, жидкой или газообразной формах, таких как капсулы, порошки, гранулы, мази, кремы, пены, растворы, суппозитории, инъекции, формы для ингаляционного применения, гели, микросферы, лосьоны и аэрозоли. Как таковое, введение соединений может быть осуществлено различными способами, включая пероральное, трансбуккальное, ректальное, парентеральное, внутрибрюшинное, внутрикожное, чрескожное, внутритрахеальное введение и так далее. Антимикробные полипептиды по изобретению могут распределяться системно после введения или могут быть локализованы с использованием имплантата или другой композиции, удерживающей активную дозу в месте имплантации.
Соединения по настоящему изобретению могут быть введены сами по себе, в комбинации друг с другом, или они могут быть использованы в комбинации с другими известными соединениями (например перфорином, противовоспалительными агентами, антибиотиками и так далее). В фармацевтических лекарственных формах соединения могут быть введены в форме их фармацевтически приемлемых солей. Следующие способы и эксципиенты приведены лишь в качестве примеров и никоим образом не являются ограничивающими.
Для препаратов для перорального введения соединения могут быть использованы сами по себе или в комбинации с подходящими добавками для изготовления таблеток, порошков, гранул или капсул, например, с обычными добавками, такими как лактоза, маннит, кукурузный крахмал или картофельный крахмал; со связывающими агентами, такими как кристаллическая целлюлоза, производные целлюлозы, аравийская камедь, кукурузный крахмал или желатины; с разрыхлителями, такими как кукурузный крахмал, картофельный крахмал или карбоксиметилцеллюлоза натрия; со смазывающими агентами, такими как тальк или стеарат магния; и, если желательно, с разбавителями, буферными агентами, увлажняющими агентами, консервантами и корригентами.
Соединения могут быть включены в композиции для инъекций путем их растворения, суспендирования или эмульгирования в водном или неводном растворителе, таком как растительные или другие подобные масла, синтетические глицериды алифатических кислот, эфиры высших алифатических кислот или пропиленгликоля; и, если желательно, с обычными добавками, такими как солюбилизаторы, изотонические агенты, суспендирующие агенты, эмульгаторы, стабилизаторы и консерванты.
Соединения могут быть использованы в аэрозольной композиции для ингаляционного введения. Соединения по настоящему изобретению могут быть включены в приемлемые пропелленты под давлением, такие как дихлордифторметан, пропан, азот и тому подобное.
Кроме того, соединения могут быть включены в суппозитории смешиванием с множеством основ, таких как эмульгирующие основы или водорастворимые основы. Соединения по настоящему изобретению могут быть введены ректально с использованием суппозитория. Суппозиторий может содержать наполнители, такие как масло какао, карбоваксы и полиэтиленгликоли, расплавляющиеся при температуре тела, но твердые при комнатной температуре.
Могут быть изготовлены стандартные лекарственные формы для перорального или ректального введения, такие как сиропы, эликсиры и суспензии, где каждая единица дозы, например, чайная ложка, столовая ложка, таблетка или суппозиторий, содержит предопределенное количество композиции, содержащей одно или более соединений по настоящему изобретению. Сходным образом, стандартные лекарственные формы для инъекции или внутривенного введения могут содержать соединение по настоящему изобретению в композиции в форме раствора в стерильной воде, нормальном физиологическом растворе или другом фармацевтически приемлемом носителе.
Имплантаты для длительного высвобождения композиций хорошо известны в данной области техники. Имплантаты изготавливают в форме микросфер, пластинок и так далее с биодеградируемыми или не являющимися биодеградируемыми полимерами. Например, полимеры молочной и/или гликолевой кислот образуют деградируемый полимер, хорошо переносимый хозяином. Имплантат, содержащий антимикробные полипептиды по изобретению, располагают близко к очагу инфекции, так чтобы локальная концентрация активного агента была повышенной по сравнению с остальными областями тела.
При использовании здесь термин «стандартная лекарственная форма» относится к физически дискретным единицам, подходящим для использования в качестве однократных доз для субъектов людей и животных, при этом каждая единица содержит предопределенное количество соединений по настоящему изобретению, которого, согласно вычислениям, достаточно для оказания желаемого эффекта, совместно с фармацевтически приемлемым разбавителем, носителем или наполнителем. Описания стандартных лекарственных форм по настоящему изобретению зависят от конкретного используемого соединения, и эффекта, который должен быть достигнут, и фармакодинамики используемого соединения у хозяина.
Фармацевтически приемлемые эксципиенты, такие как наполнители, адъюванты, носители или разбавители, общедоступны. Кроме того, общедоступны фармацевтически приемлемые вспомогательные вещества, такие как агенты для регулирования pH и буферные агенты, агенты для регулирования тоничности, стабилизаторы, смачивающие агенты и тому подобное.
Типичные дозы для системного введения варьируют от 0,1 пг до 100 миллиграмм на кг массы тела субъекта на одно введение. Типичная доза может представлять собой одну таблетку для приема от двух до шести раз в сутки или одну капсулу или таблетку с длительным высвобождением для приема один раз в сутки с пропорционально более высоким содержанием активного ингредиента. Эффект длительного высвобождения может быть обусловлен материалами, из которых изготовлена капсула, растворяющимися при различных значениях pH, капсулами, обеспечивающими медленное высвобождение под воздействием осмотического давления или любым другим известным способом контролируемого высвобождения.
Специалистам в данной области техники будет ясно, что уровни доз могут варьировать в зависимости от конкретного соединения, тяжести симптомов и предрасположенности субъекта к побочным эффектам. Некоторые из конкретных соединений обладают большей активностью, чем другие. Предпочтительные дозы данного соединения могут быть легко определены специалистами в данной области техники множеством способов. Предпочтительным способом является измерение физиологической активности данного соединения.
Один из интересующих способов представляет собой применение липосом в качестве наполнителя для доставки. Липосомы сливаются с клетками целевой области и обеспечивают доставку содержимого липосом внутрь клеток. Контакт липосом с клетками поддерживают в течение времени, достаточного для слияния, с использованием различных способов поддержания контакта, таких как выделение, связывающие агенты и тому подобное. В одном аспекте изобретения липосомы разработаны для получения аэрозоля для легочного введения. Липосомы могут быть изготовлены с очищенными белками или пептидами, опосредующими слияние мембран, такими как вирус Сендай или вирус гриппа и так далее. Липиды могут представлять собой любую полезную комбинацию известных липидов, образующих липосомы, включая катионные или цвиттер-ионные липиды, такие как фосфатидилхолин. Остальные липиды будут обычно нейтральными или кислыми липидами, такими как холестерин, фосфатидилсерин, фосфатидилглицерин и тому подобное.
Для получения липосом может быть использован способ, описанный Kato et al. (1991) J. Biol. Chem. 266:3361. Кратко, липиды и композицию для включения в липосомы, содержащую пептиды, смешивают в подходящей водной среде, подходящим образом в солевой среде, где общее содержание твердых веществ будет находиться в диапазоне приблизительно 1-10 масс.%. После интенсивного перемешивания в течение коротких периодов времени, приблизительно 5-60 сек, пробирку помещают в теплую водяную баню при приблизительно 25-40°C и этот цикл повторяют приблизительно 5-10 раз. Затем композицию обрабатывают ультразвуком на протяжении подходящего периода времени, обычно приблизительно 1-10 сек, и, возможно, дополнительно перемешивают на вихревой мешалке. Затем объем увеличивают добавлением водной среды, обычно увеличивая объем в приблизительно 1-2 раза, с последующим взбалтыванием и охлаждением. Способ позволяет включать в липосомы молекулы с высокой молекулярной массой.
Композиции с другими активными агентами
Для применения в рассматриваемых способах антимикробные полипептиды по изобретению могут быть включены в композиции с другими фармацевтически активными агентами, в частности другими антимикробными агентами. Другие интересующие агенты включают широкий спектр антибиотиков, известных в данной области техники. Классы антибиотиков включают пенициллины, например пенициллин G, пенициллин V, метициллин, оксациллин, карбенициллин, нафциллин, ампициллин и так далее; пенициллины в комбинации с ингибиторами бета-лактамазы; цефалоспорины, например цефаклор, цефазолин, цефуроксим, моксалактам и так далее; карбапенемы; монобактамы; аминогликозиды; тетрациклины; макролиды; линкомицины; полимиксины; сульфонамиды; хинолоны; хлорамфеникол; метронидазол; спектиномицин; триметоприм; ванкомицин; и так далее.
Также полезны противогрибковые агенты, включая полиены, например амфотерицин B, нистатин, 5-флукозин; и азолы, например миконазол, кетоконазол, итраконазол и флуконазол. Противотуберкулезные лекарственные средства включают изониазид, этамбутол, стрептомицин и рифампин. В композицию антимикробных полипептидов по изобретению могут также быть включены цитокины, например интерферон-гамма, фактор некроза опухоли-альфа, интерлейкин-12 и так далее.
Далее настоящее изобретение описано следующими примерами, которые не следует толковать как ограничивающие объем изобретения.
ПРИМЕРЫ
NZ17074 представляет собой антимикробный пептид с SEQ ID NO: 70.
ПРИМЕР 1
Выделение вариантов SEQ ID NO: 2, обладающих улучшенной антимикробной активностью
Проводили слияние кДНК, кодирующей SEQ ID NO: 2, с прообластью плектазина (см. Mygind et al; Nature, 437, 2005) и альфа-лидерной последовательностью фактора спаривания (Mating Factor) Saccharomyces cerevisiae, вводили ее в индуцибельный вектор экспрессии S. cerevisiae, pYES2, и трансформировали им S. cerevisiae. В этой системе использован промотор GAL1, репрессируемый глюкозой и активируемый галактозой.
Для получения вариантов полинуклеотида с SEQ ID NO: 1 использовали несколько способов. Полученные библиотеки клонировали и экспрессировали в S. cerevisiae. Проводили скрининг трансформированных клонов в чашечном анализе, включающем питательную среду, дополненную 1,5% галактозы, и 0,5% глюкозы, и либо лошадиной кровью (2,5-5%), либо сывороткой (5%), с верхним слоем организма-мишени, E. coli АТСС 10536 (см. Raventos et al Comb Chem High Throughput Screen. 2005; 8:219-33).
Условия чашечного анализа полностью ингибировали активность антимикробного пептида с SEQ ID NO: 2 (исходного антимикробного пептида). Варианты, демонстрирующие улучшенную антимикробную активность (приводящие к зонам просветления) в присутствии 2,5% крови, 5% крови или 5% сыворотки, были отобраны и секвенированы и показаны в Таблице 1.
Способ скрининга чашечным анализом
Приблизительно 300 колоний Saccharomyces cerevisiae, экспрессирующих варианты ареницина, распределяли по чашкам для скрининга, содержащим лошадиную кровь (2,5-5%) или 5% лошадиной сыворотки (см. содержимое чашек ниже). Чашки инкубировали 3 часа при 30 градусах по Цельсию, позволяя им сохнуть. Затем в чашки добавляли 25 мл верхних сред, темперированных при 42 градусах по Цельсию. После затвердевания сред чашки инкубировали 3 суток при 30 градусах по Цельсию.
На 4 сутки в чашки добавляли предварительно нагретые при 42 градусах по Цельсию верхние среды, содержащие либо 2,5% или 5% лошадиной крови, либо 5% лошадиной сыворотки и бактерии-мишени, Е. coli АТСС 10536 (см. подробный состав сред ниже). После затвердевания верхних сред чашки инкубировали 16 часов при 37 градусах по Цельсию. На следующие сутки чашки окрашивали, добавляя 10 мл 1,5 мМ МТТ в чашки, и инкубировали их при комнатной температуре в течение 30 минут. Клоны, приводившие к зонам просветления, отбирали и секвенировали.
Содержимое чашек и состав сред
При скрининге использовали три разных типа чашек для скрининга: (а), (б) и (в).
а) Чашки с 2,5% лошадиной крови
Нижний слой содержит 50 мл 1,5% агарозы + ¼ среды SC + 2,5% крови + 1,5% галактозы + 0,5% глюкозы. Первый верхний слой содержит 25 мл 1,5% агарозы + ¼ среды SC + 2,5% крови + 1,5% галактозы + 0,5% глюкозы. Самый верхний слой содержит 25 мл 0,2% МНВ (#212322; BD) + 1% агарозы (Sigma А-4718) + 2,5% лошадиной крови и 1,25×106 колониеобразующих единиц (КОЕ) Е. coli АТСС 10536.
б) Чашки с 5% лошадиной крови
Нижний слой содержит 50 мл 1,5% агарозы + ¼ среды SC + 5% крови + 1,5% галактозы + 0,5% глюкозы. Первый верхний слой содержит 25 мл 1,5% агарозы + ¼ среды SC + 5% крови + 1,5% галактозы + 0,5% глюкозы. Самый верхний слой содержит 25 мл 0,2% МНВ (#212322; BD) + 1% агарозы (Sigma А-4718) + 5% лошадиной крови и 1,25×106 колониеобразующих единиц (КОЕ) Е. coli АТСС 10536.
в) Чашки с 5% лошадиной сыворотки
Нижний слой содержит 50 мл 1,5% агарозы + ¼ среды SC + 5% сыворотки + 1,5% галактозы + 0,5% глюкозы. Первый верхний слой содержит 25 мл 1,5% агарозы + ¼ среды SC + 5% сыворотки + 1,5% галактозы + 0,5% глюкозы. Самый верхний слой содержит 25 мл 0,2% МНВ (#212322; BD) + 1% агарозы (Sigma А-4718) + 5% лошадиной сыворотки и 1,25×106 колониеобразующих единиц (КОЕ) E. coli АТСС 10536.
Состав среды SC
Значение pH доводили до 6, среду автоклавировали и разводили в соотношении ¼ при приготовлении чашек с кровью и в соотношении ½ при приготовлении чашек с сывороткой.
МТТ: бромид 3-(4,5-диметилтиазол-2-ил)-2,5-дифенил-2H-тетразолия, Sigma 13,503-8.
Определение связывания с белками
Анализы связывания с белками проводили следующим образом. Очищенные пептиды смешивали с 90% сывороткой и центрифугировали через 30 кДа фильтр. Ультрафильтрат и непрофильтрованные образцы сыворотки исследовали количественно посредством HPLC и затем вычисляли связывание с белками.
Антимикробный пептид с SEQ ID NO: 2 (исходный антимикробный пептид) демонстрировал связывание с белками 99,5% в данном анализе. Как показано в Таблице 1, все приведенные варианты демонстрируют меньшее связывание с белками, чем антимикробный пептид с SEQ ID NO: 2.
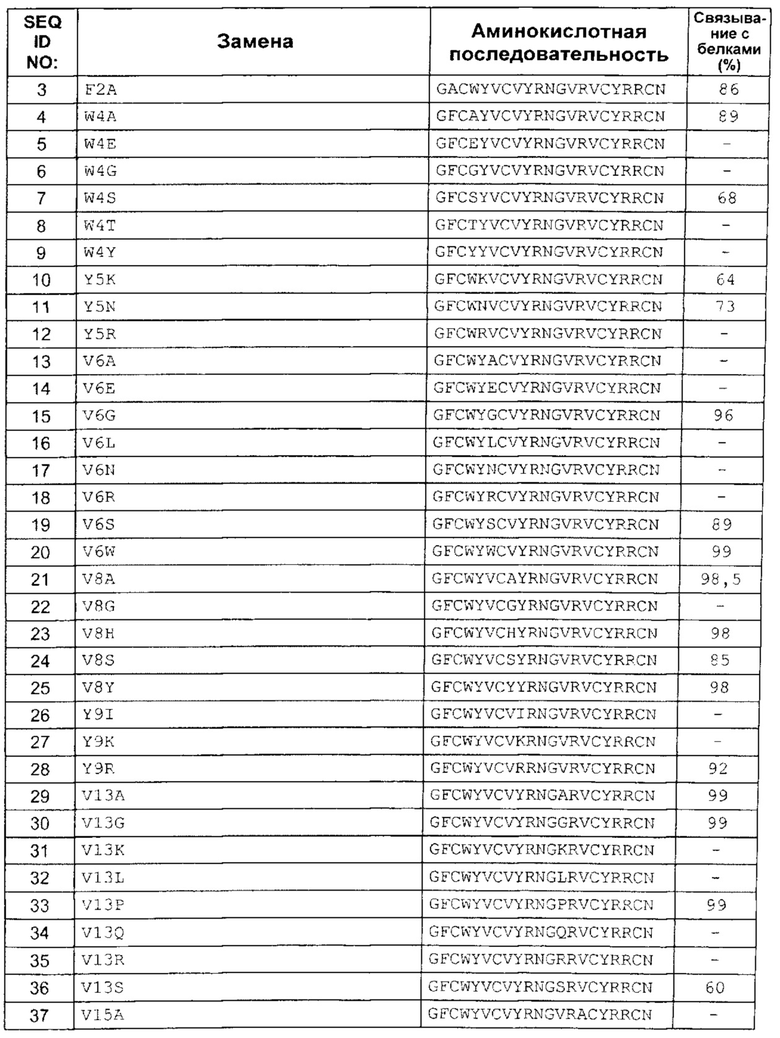
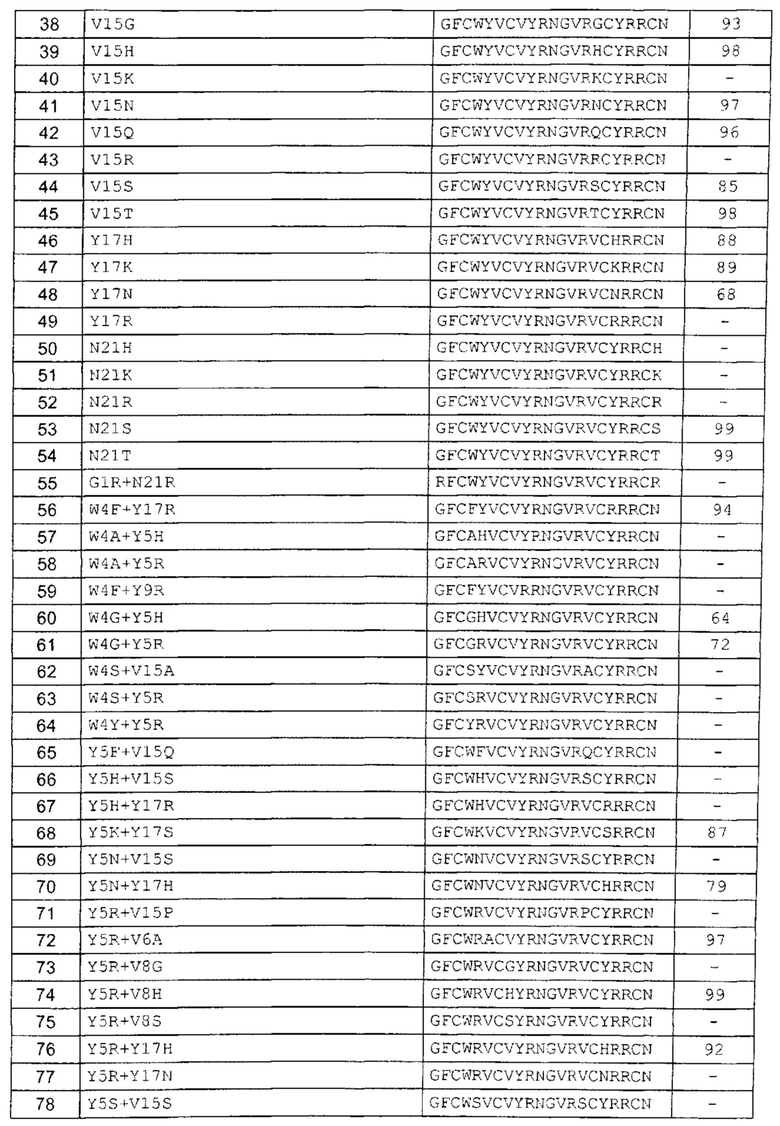
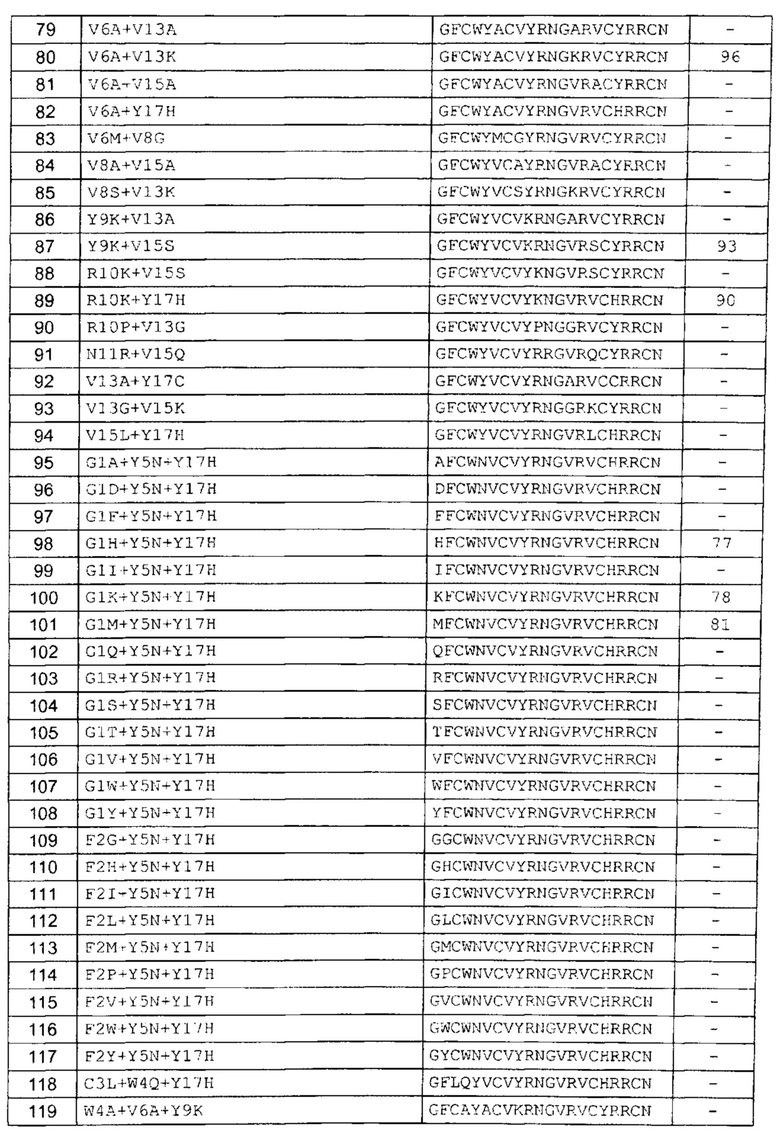
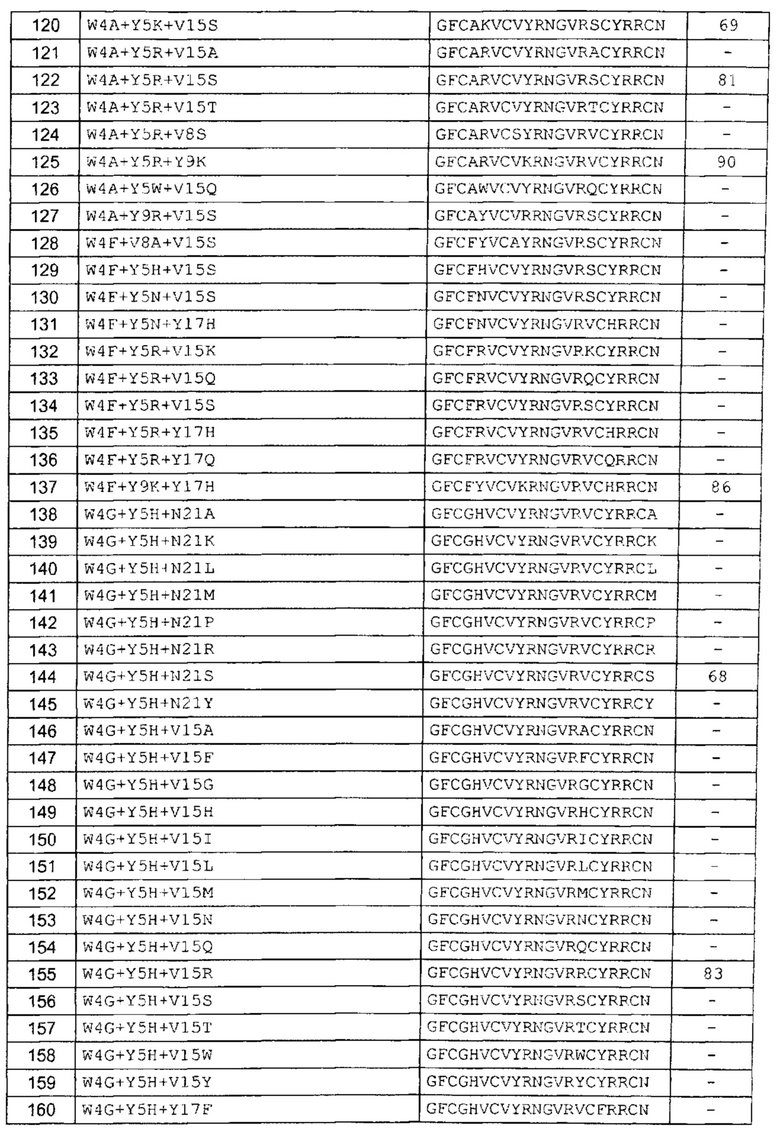
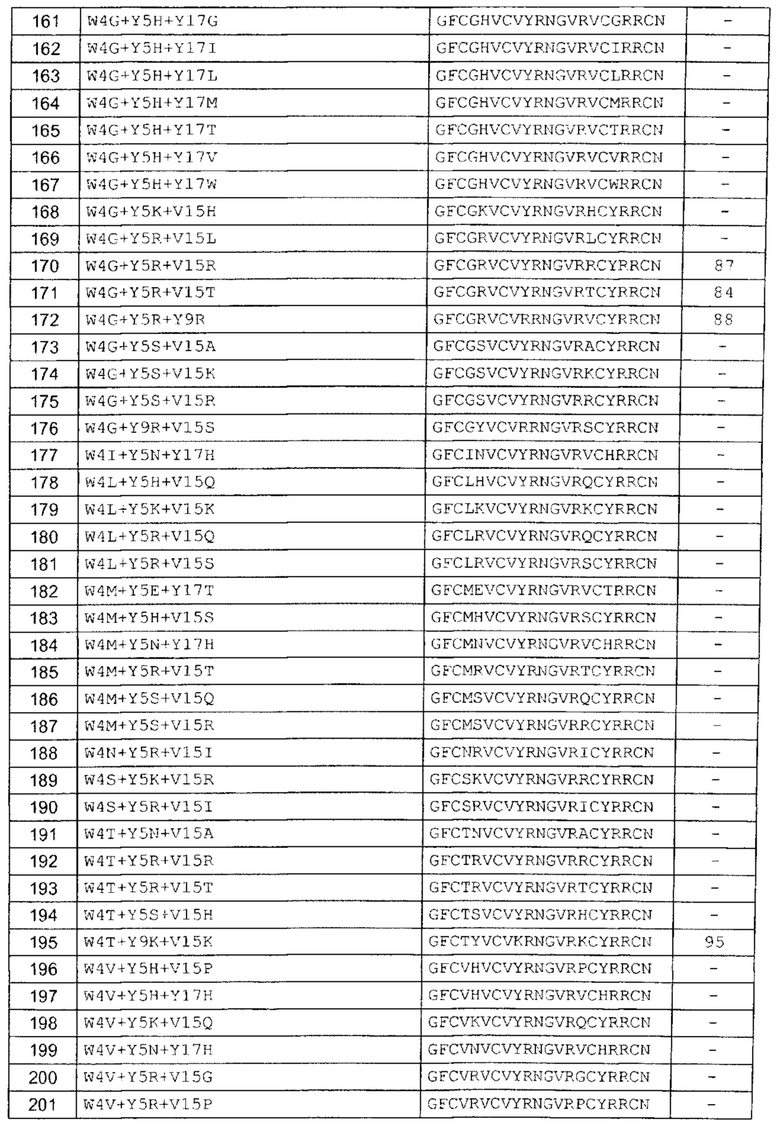
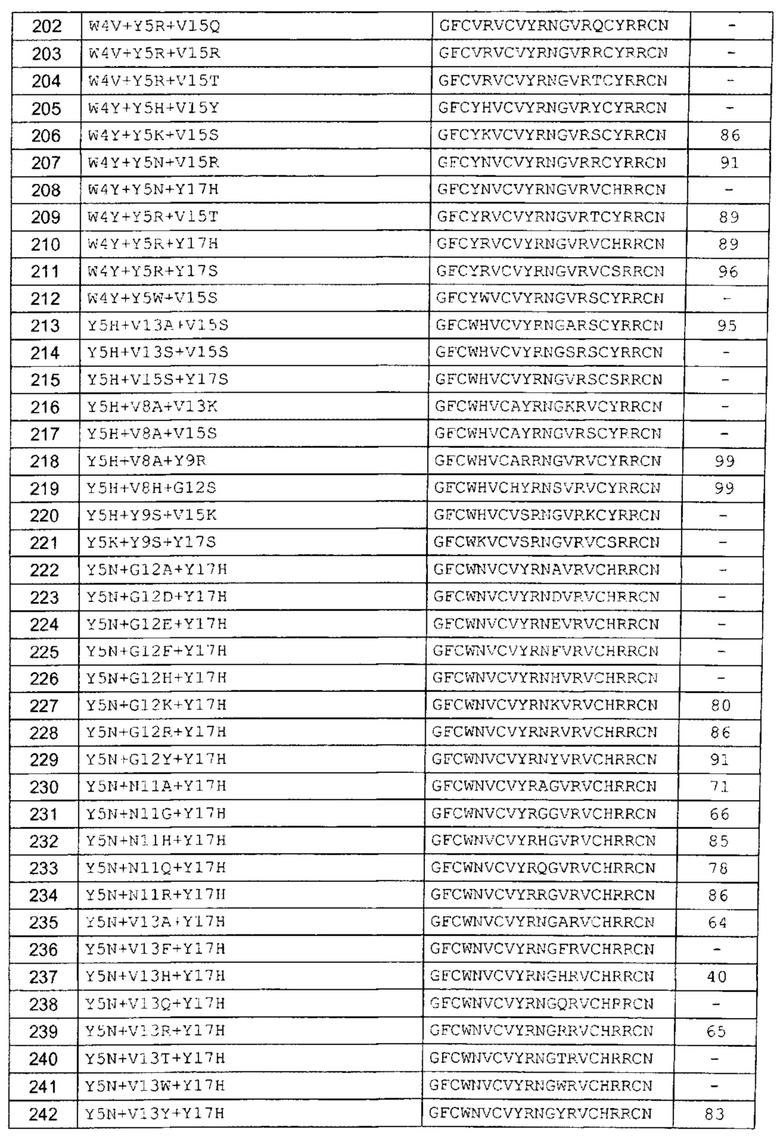
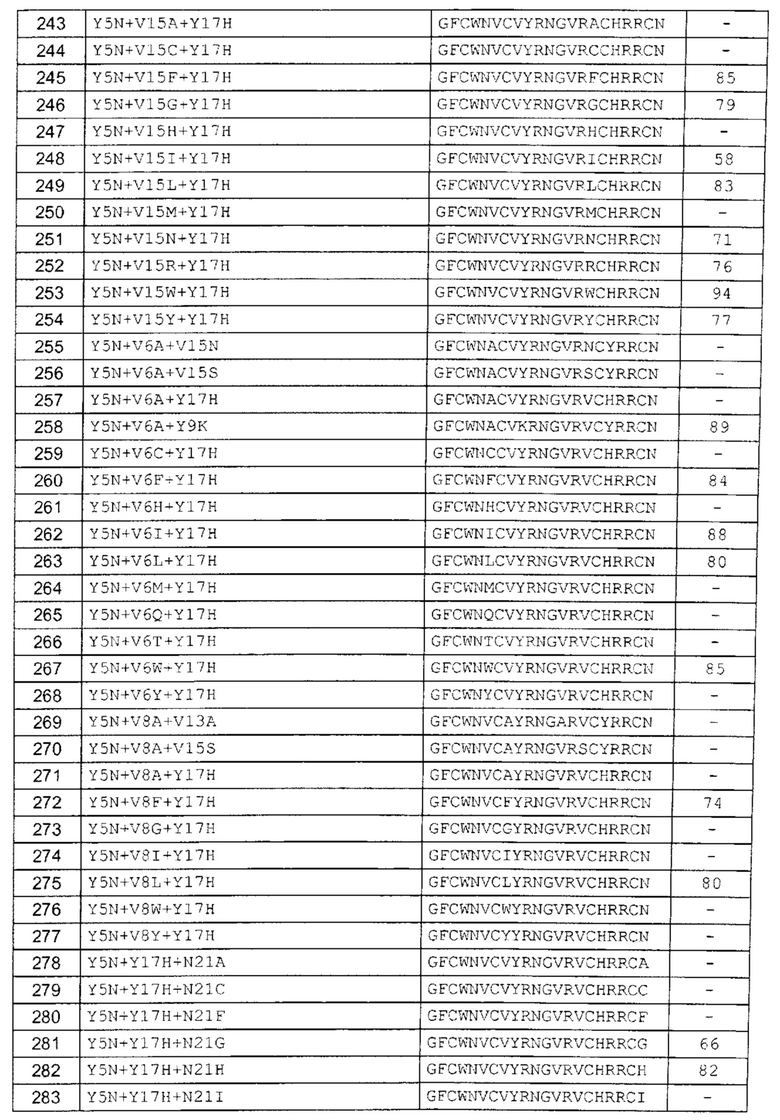
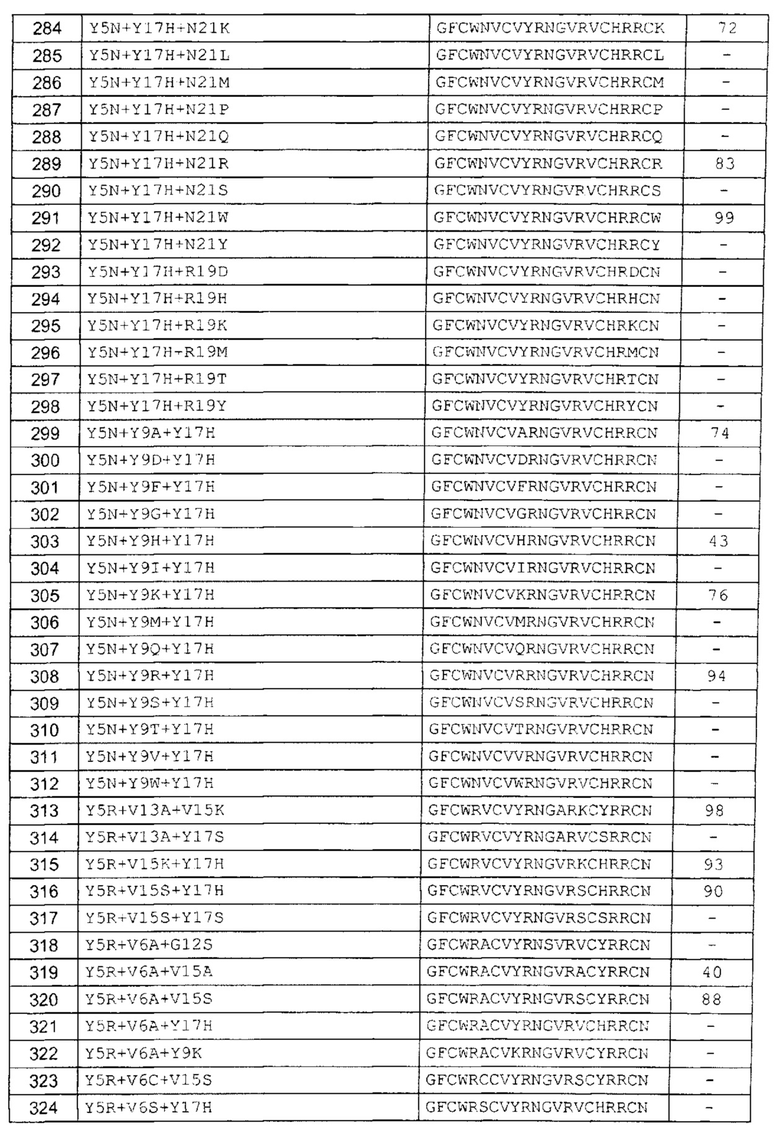
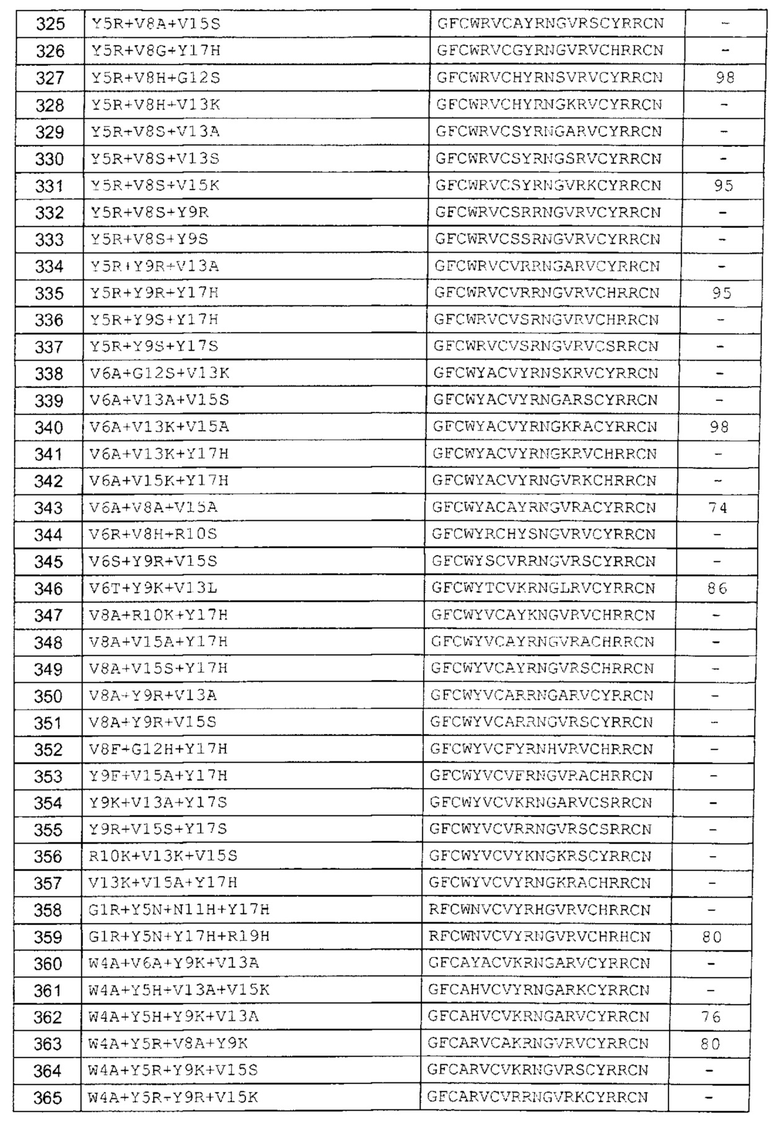
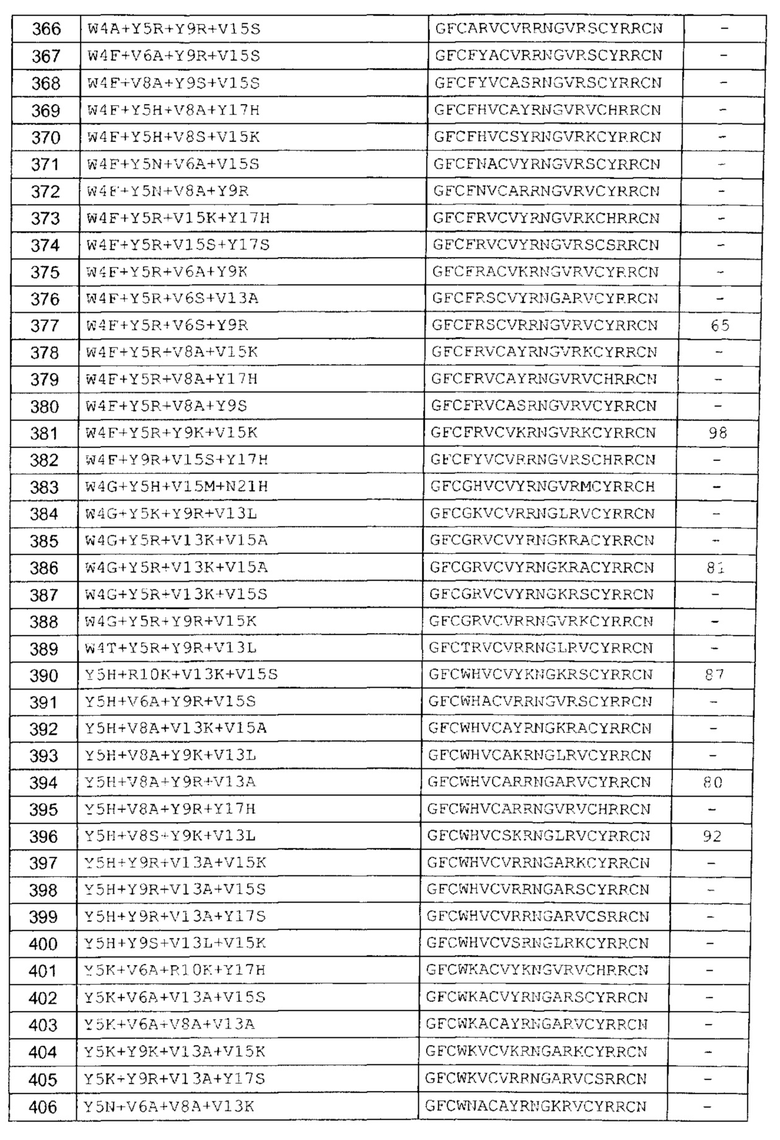
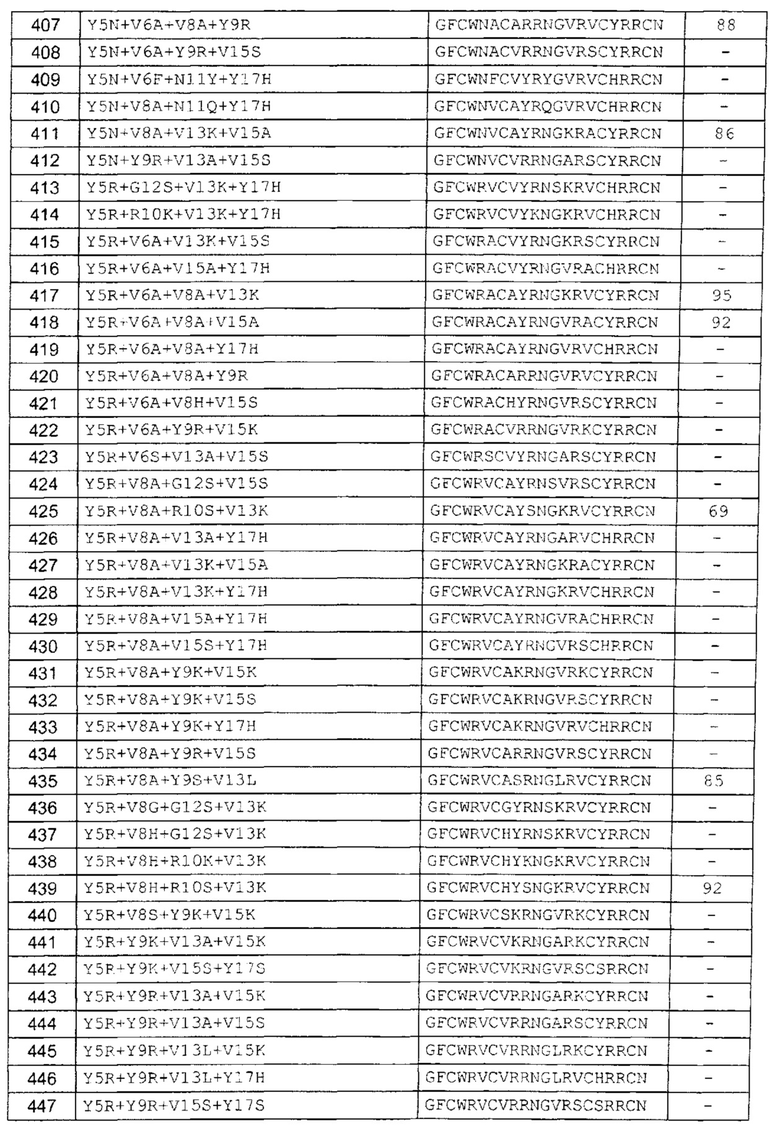
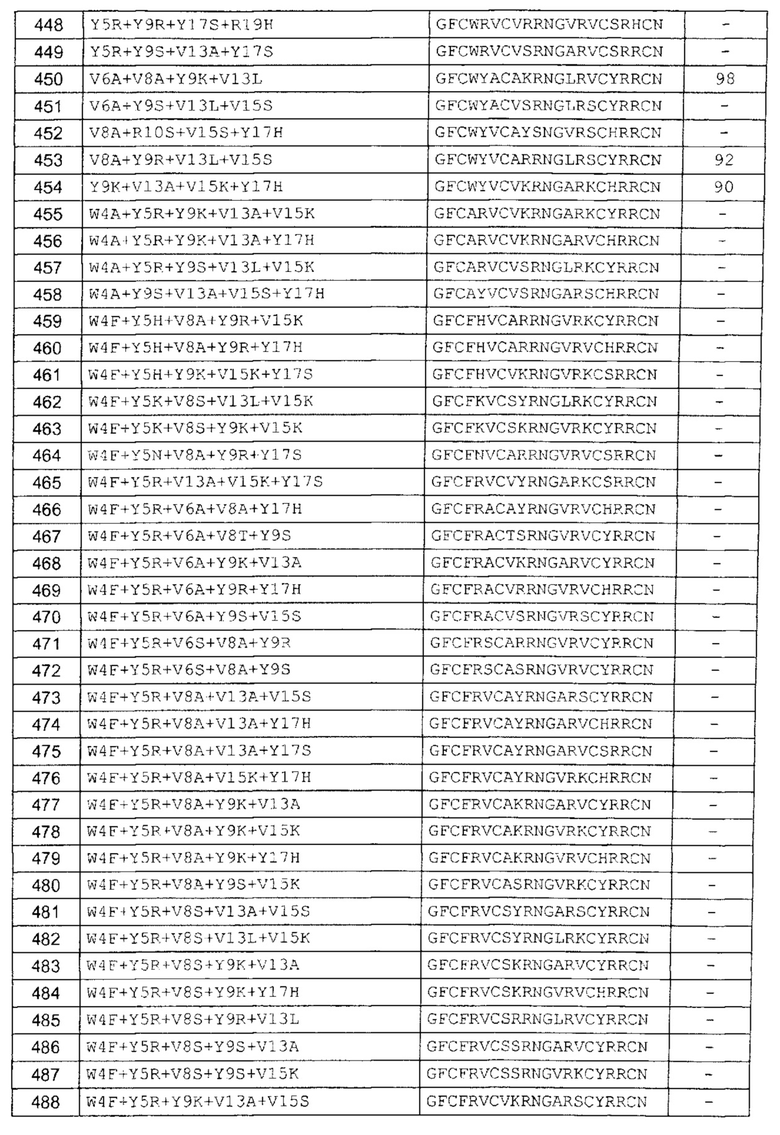
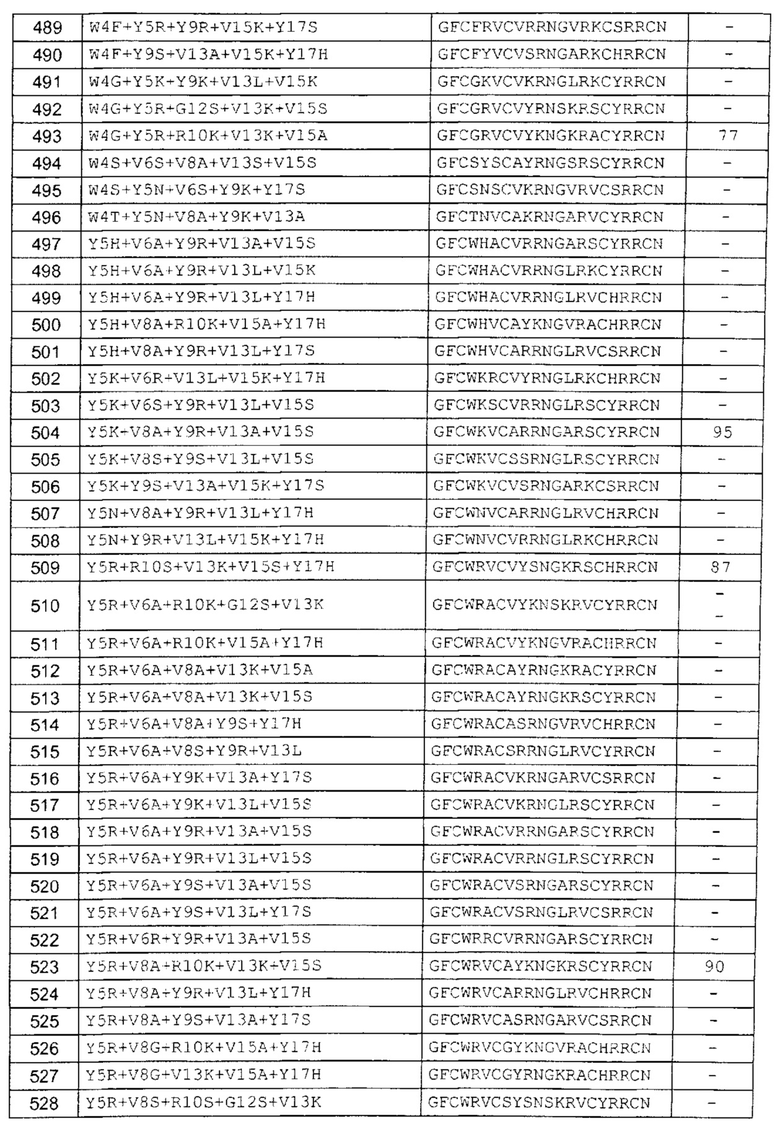

ПРИМЕР 2
Эффективность NZ17074 против Escherichia coli AID#172 в модели нейтропенического перитонита/сепсиса у мышей и оценка средней эффективной дозы (ED50)
Введение
Задачей данного исследования было исследование зависимости доза-ответ после внутривенного (в/в) введения однократной дозы NZ17074 в диапазоне 0,16-12 мг/кг. Эффект исследовали против Е. coli AID#172 в модели нейтропенического перитонита. Введение меропенема в дозе 40 мг/кг включали в качестве группы положительного контроля. Количество колоний в крови и перитонеальной жидкости определяли через 5 ч после введения.
Модель перитонита/сепсиса у мышей является хорошо известной моделью для исследований антимикробной активности, как описано N. Frimodt-M∅ller и J.D. Knudsen в Handbook of Animal Models of Infection (1999), ed. by O. Zak & M.A. Sande, Academic Press, San Diego, US.
Материалы и методы
- 30 самок беспородных мышей NMRI, 25-30 грамм (Harlan Scandinavia).
- Escherichia coli AID#172 от Statens Serum Instute, Copenhagen, Denmark. Клинический изолят из человеческой раны от 2003 г.с множественной лекарственной устойчивостью (к ампициллину, цефтазидиму, азтреонаму, гентамицину, ципрофлоксацину).
- NZ17074 в ацетате Рингера, pH 6, 1,2 мг/мл, 6,0 мл. Раствор хранили при 4°C до использования. Анализы использованных композиций для введения проводили по окончании фазы исследования, проводимой на живых животных, и в этих анализах были получены следующие результаты.
- Наполнитель (ацетат Рингера, pH 6). Раствор хранили при 4°C до использования.
- Meronem® (AstraZeneca, 500 мг вещества для инфузии, меропенем). Номер партии: 09466С. Срок годности: август 2013 г.
- Стерильная вода.
- Стерильный 0,9% физиологический раствор.
- Циклофосфамид, Apodan® (A-Pharma, 1 г). Номер партии: 928491. Срок годности: май 2012 г.
- Чашки с агаром и 5% лошадиной крови.
- Чашки с агаром, бромтимоловым синим и лактозой.
Лабораторный виварий и содержание мышей
Температуру и влажность в виварии регистрировали ежедневно. Температура составляла 21°C +/- 2°C, и ее можно было регулировать нагреванием и охлаждением. Влажность составляла 55 +/- 10%. Смена воздуха происходила приблизительно 10-20 раз в час, и период света/темноты был в 12-часовом интервале 06:00-18:00/18:00-06:00.
У мышей был свободный доступ к питьевой воде для домашних животных и пище (2016, Harlan). Мышей содержали в макролоновых клетках 3 типа по 3 мыши в клетке. В качестве подстилки использовали Aspen Wood от Tapvei. Кроме того, животным давали бумажные полоски Sizzle-nest в качестве материала для гнезд. Мышам метили хвосты для различения мышей в клетке. Мышей взвешивали за сутки до введения.
Приготовление растворов NZ17074
Раствор с концентрацией 1,2 мг/мл дополнительно разводили в PBS-наполнителе следующим образом.
Приготовление раствора меропенема
Введение меропенема в дозе 40 мг/кг было включено в качестве группы положительного контроля.
В общей сложности 500 мг меропенема (одну ампулу) растворяли в 10 мл воды в концентрации примерно 50 мг/мл. Этот исходный раствор разводили далее до 4 мг/мл (0,4 мл, 50 мг/мл + 4,6 мл физиологического раствора).
Приготовление циклофосфамида
В общей сложности 1 г циклофосфамида (одну ампулу Apodan 1 г) растворяли в 50 мл воды, примерно 20 мг/мл, на каждые сутки его использования. Этот исходный раствор разводили далее до 11 мг/мл (16,5 мл, 20 мг/мл + 13,5 мл физиологического раствора) для использования на -4 сутки или до 5 мг/кг (8,25 мл 20 мг/мл + 21,75 мл физиологического раствора) для использования на -1 сутки.
Введение циклофосфамида мышам
У мышей индуцировали нейтропению инъекцией 0,5 мл раствора циклофосфамида внутрибрюшинно за 4 суток (200 мг/кг) и 1 сутки (100 мг/кг) до заражения.
Заражение мышей
Свежие колонии Е. coli AID#172, полученные в течение ночи в чашке с агаром и 5% лошадиной крови, суспендировали и разводили в стерильном физиологическом растворе приблизительно до 2×106 КОЕ/мл. За один час до начала введения (момент времени -1 ч) проводили внутрибрюшинное заражение мышей 0,5 мл суспензии Е. coli в латеральный нижний квадрант живота. Приблизительно через 0,5-1 ч после введения мышам перорально вводили 45 мкл нейрофена (20 мг ибупрофена на мл, что соответствовало 30 мг/кг) в качестве болеутоляющего средства.
Введение препаратов мышам
Мышам проводили однократное в/в введение NZ17074, меропенема или наполнителя в латеральную хвостовую вену на протяжении приблизительно 30 секунд в объеме 10 мл/кг в момент времени 0 ч (см. Таблицу 1). Определение дозы было основано на средней массе тела 30 г. Мышам с массой тела 28-32 г вводили 0,30 мл раствора. Мышам с массой тела 27-28 г вводили 0,25 мл раствора и мышам с массой тела 32,1-36 г вводили 0,35 мл раствора.
T указывает время относительно введения. Числа в столбцах забора образцов представляют собой идентификационные номера мышей.
Клиническая балльная оценка мышей
Мышей наблюдали в течение исследования и присваивали им баллы от 0 до 5 в зависимости от их поведения и клинических признаков.
Балл 0: здоровые.
Балл 1: минимальные клинические признаки инфекции и воспаления, например, наблюдение минимальных признаков расстройства или изменения активности.
Балл 2: отчетливые признаки инфекции, такие как социальная самоизоляция, снижение любопытства, измененное положение тела, пилоэрекция или изменения характера движений.
Балл 3: выраженные признаки инфекции, такие как тугоподвижность, снижение любопытства, измененное положение тела, пилоэрекция, боль или изменения характера движений.
Балл 4: сильная боль, и мышь сразу умерщвляли для минимизации страданий животного.
Балл 5: смерть мыши.
Забор образцов
Количество колоний определяли в крови и перитонеальной жидкости на 0 и 5 ч. Мышей анестезировали CO2+O2 и забор крови производили из разреза в подмышечной области в пробирки типа Эппендорф с покрытием этилендиаминтетрауксусной кислотой (EDTA). Мышей умерщвляли сразу после забора крови, и в общей сложности 2 мл стерильного физиологического раствора вводили внутрибрюшинно и проводили мягкий массаж живота до его вскрытия и забора образцов жидкости пипеткой. Затем каждый образец разводили в 10 раз в физиологическом растворе и капли объемом 20 мкл наносили на чашки с синим агаром. Все чашки с агаром инкубировали 18-22 ч при 35°C на воздухе.
Результаты
Количество колоний определяли в начале введения и через 5 ч после введения. Количество КОЕ и клинические показатели мышей показаны в Таблице 3. Перед проведением вычислений проводили log10-трансформацию количества КОЕ.
КОЕ/мл в заражающем материале было определено как 6,29 log10. В начале введения среднее log10 КОЕ/мл составляло 5,76 в перитонеальной жидкости и 5,13 в крови, и уровни КОЕ сохранялись на сходном уровне в группе наполнителя (5,72 и 4,65 log10 КОЕ/мл в перитонеальной жидкости и крови, соответственно) через 5 ч после введения. Несколько сниженные уровни КОЕ наблюдали в крови и перитонеальной жидкости после введения NZ17074 в дозе 0,16-3,0 мг/кг. Введение NZ17074 в дозе 6 и 12 мг/кг приводило к значительному снижению уровней КОЕ (p<0,001) по сравнению с введением наполнителя, как в перитонеальной жидкости, так и в крови (Таблица 3). Введение меропенема в дозе 40 мг/кг также приводило к значимому снижению по сравнению с мышами, которым вводили наполнитель, как в крови (p<0,05), так и в перитонеальной жидкости (p<0,01).
Кривые доза-ответ (данные не показаны) вычисляли в GraphPad Prism с использованием сигмовидной кривой доза-ответ (вариабельный угол наклона). Определенные по этим кривым значения ED50 составили 3,09±2,07 мг/кг в перитонеальной жидкости и 3,17±0,53 мг/кг в крови.
Максимальный эффект NZ17074, Emax, определяли как разность log КОЕ при отсутствии ответа и при максимальном ответе. Отсутствие ответа характеризовали как количество колоний на уровне, определенном у мышей, которым вводили наполнитель. Emax, вычисленный как разность между «Верхним плато» ("Top plateau") и «Нижним плато» ("Bottom plateau") в GraphPad Prism с использованием сигмовидной кривой доза-ответ, составил 4,72 log10 КОЕ для перитонеальной жидкости и 3,15 log10 КОЕ для крови.
Кроме того, с использованием GraphPad Prism оценивали 1, 2 и 3 log уничтожения, определяемые как доза, необходимая для снижения бактериальной нагрузки на 1, 2 или 3 log по сравнению с началом лечения. 1, 2 и 3 log уничтожения для NZ17074 составляли 1,11 мг/кг, 2,95 мг/кг и 4,73 мг/кг, соответственно, в перитонеальной жидкости и 0,25 мг/кг, 2,75 мг/кг и 3,78 мг/кг, соответственно, в крови.
Во всех группах введения наблюдали нулевые или невысокие клинические баллы (Таблица 3).
Обсуждение и заключение
Задачей данного исследования было исследование зависимости доза-ответ после внутривенного (в/в) введения однократной дозы NZ17074 в диапазоне 0,16-12 мг/кг. Эффект исследовали против Е. coli AID#172 в модели нейтропенического перитонита/сепсиса.
Было определено, что значения ED50 для NZ17074 составили 3,09±2,07 мг/кг в перитонеальной жидкости и 3,17±0,53 мг/кг в крови. Согласно оценкам 1 log уничтожения составил 1,11 мг/кг в перитонеальной жидкости и 0,25 мг/кг в крови. Согласно оценкам 2 log уничтожения составил 2,95 мг/кг в перитонеальной жидкости и 2,76 мг/кг в крови. Согласно оценкам 3 log уничтожения составил 4,73 мг/кг в перитонеальной жидкости и 3,78 мг/кг в крови.
Предел выявления 1,4 log10 КОЕ/мл. Образцы без обнаруженных бактерий представлены как 1,0 log10 КОЕ/мл.
ПРИМЕР 3
Модель перитонита/сепсиса: эффект NZ17074 в дозе 7.5 мг/кг с течением времени против Escherichia coli AID#172 у мышей NMRI с нейтропенией
Введение
Задачей данного исследования было исследование эффективности NZ17074 in vivo после внутривенного (в/в) введения однократной дозы 7,5 мг/кг. Эффект исследовали против Escherichia coli AID#172 в модели перитонита у мышей NMRI с нейтропенией во избежание использования муцина, который обычно используют в модели перитонита у мышей. У мышей индуцировали нейтропению инъекциями циклофосфамида. Введение меропенема в дозе 40 мг/кг включали в качестве группы положительного контроля и введение наполнителя включали в качестве группы отрицательного контроля. Количество колоний в перитонеальной жидкости и крови определяли через 2 и 5 ч после введения.
Материалы и методы
- 30 самок беспородных мышей NMRI, 28-32 грамма (Harlan Scandinavia).
- Escherichia coli AID#172 от Statens Serum Instute, Copenhagen, Denmark. Клинический изолят из человеческой раны от 2003 г. с множественной лекарственной устойчивостью (к ампициллину, цефтазидиму, азтреонаму, гентамицину, ципрофлоксацину).
- NZ17074 в ацетате Рингера, pH 6, 1,2 мл, 0,75 мг/мл. Анализы вводимых композиций, проведенный после исследования, показал концентрацию приблизительно 0,78 мг/мл.
- Наполнитель (ацетат Рингера, pH 6), 3 мл.
- Meronem® (AstraZeneca, 500 мг вещества для инфузии, меропенем). Номер партии: 09466C. Срок годности: август 2013 г.
- Apodan® (A-Pharma, 1 г циклофосфамида). Номер партии: 928491. Срок годности: май 2012 г.
- Стерильная вода.
- Стерильный 0,9% физиологический раствор.
- Чашки с агаром и 5% лошадиной крови.
- Чашки с агаром, бромтимоловым синим и лактозой.
Лабораторный виварий и содержание мышей
Температуру и влажность в виварии регистрировали ежедневно. Температура составляла 21°C +/- 2°C, и ее можно было регулировать нагреванием и охлаждением. Влажность составляла 55 +/- 10%. Смена воздуха происходила приблизительно 10-20 раз в час, и период света/темноты был в 12-часовом интервале 06:00-18:00/18:00-06:00. У мышей был свободный доступ к питьевой воде для домашних животных и пище (2016, Harlan). Мышей содержали в макролоновых клетках 3 типа по 3 мыши в клетке. В качестве подстилки использовали Aspen Wood от Tapvei. Кроме того, животным давали бумажные полоски Sizzle-nest в качестве материала для гнезд. Мышам метили хвосты для различения мышей в клетке.
Раствор NZ17074
Раствор каждого исследуемого соединения с концентрацией 0,75 мг/мл хранили при +4°C до одного часа перед инъекцией, после этого при комнатной температуре.
Приготовление раствора меропенема
В общей сложности 500 мг меропенема (одну ампулу) растворяли в 10 мл воды, примерно 50 мг/мл, на сутки его использования. Этот исходный раствор разводили далее до 4 мг/мл (0,4 мл, 50 мг/мл + 4,6 мл физиологического раствора).
Приготовление циклофосфамида
В общей сложности 1 г циклофосфамида (одну ампулу Apodan) растворяли в 50 мл воды, примерно 20 мг/мл, на каждые сутки его использования. Этот исходный раствор разводили далее до 11 мг/мл (16,5 мл, 20 мг/мл + 13,5 мл физиологического раствора) для использования на -4 сутки или до 5 мг/кг (8,25 мл 20 мг/мл + 21,75 мл физиологического раствора) для использования на -1 сутки.
Введение циклофосфамида мышам
У мышей индуцировали нейтропению инъекцией 0,5 мл раствора циклофосфамида внутрибрюшинно за 4 суток (200 мг/кг) и 1 сутки (100 мг/кг) до заражения.
Заражение мышей
Свежие колонии Е. coli AID#172, полученные в течение ночи в чашке с агаром и 5% лошадиной крови, суспендировали и разводили в стерильном физиологическом растворе приблизительно до 2×106 КОЕ/мл.
За один час до начала введения (момент времени -1 ч) проводили внутрибрюшинное заражение мышей 0,5 мл суспензии Е. coli в латеральный нижний квадрант живота.
Через 2,5 ч после введения при существенных клинических признаках инфекции мышам перорально вводили 45 мкл нейрофена (20 мг ибупрофена на мл, что соответствовало 30 мг/кг) в качестве болеутоляющего средства.
Балльная оценка мышей
При каждом заборе образцов у мышей проводили балльную оценку клинических признаков инфекции.
Балл 0: здоровые.
Балл 1: минимальные клинические признаки инфекции и воспаления, например, наблюдение минимальных признаков расстройства или изменения активности.
Балл 2: отчетливые признаки инфекции, такие как социальная самоизоляция, снижение любопытства, измененное положение тела, пилоэрекция или изменения характера движений.
Балл 3: выраженные признаки инфекции, такие как тугоподвижность, снижение любопытства, измененное положение тела, пилоэрекция, боль или изменения характера движений.
Балл 4: сильная боль, и мышь сразу умерщвляли для минимизации страданий животного.
Балл 5: смерть мыши.
Введение препаратов мышам
Мышам проводили однократное в/в введение NZ17074, меропенема или наполнителя в латеральную хвостовую вену на протяжении приблизительно 30 секунд в момент времени 0 ч (см. Таблицу 1). Определение дозы было основано на средней массе тела 30 г. Мышам с массой тела 28-32 г вводили 0,30 мл раствора. Мышам с массой тела 27-28 г вводили 0,25 мл раствора и мышам с массой тела 32,1-36 г вводили 0,35 мл раствора. Мыши 17 случайно ввели 0,35 мл, несмотря на то, что ее масса тела составляла 29,5 г. По-видимому, это не повлияло на результаты, поскольку уровни КОЕ у этой мыши были очень сходны с двумя другими мышами данной группы.
Забор образцов
Количество колоний определяли в крови и перитонеальной жидкости на 0, 2 и 5 ч после введения согласно Таблице 1.
Мышей анестезировали CO2+O2 и забор крови проводили из разреза в подмышечной области. Мышей умерщвляли цервикальной дислокацией и в общей сложности 2 мл стерильного физиологического раствора вводили внутрибрюшинно и проводили мягкий массаж живота, затем его вскрывали и проводили забор образцов жидкости пипеткой. Каждый образец разводили в 10 раз в физиологическом растворе и капли объемом 20 мкл наносили на чашки с кровяным агаром. Все чашки с агаром инкубировали 18-22 ч при 35°C на воздухе.
Результаты
Количество колоний и клинические показатели мышей показаны в Таблице 2. Перед проведением вычислений проводили log10-трансформацию количества КОЕ для получения нормального распределения.
КОЕ/мл в заражающем материале было определено как 6,30 log10. В начале введения среднее log10 КОЕ/мл составляло 3,57 в перитонеальной жидкости и 3,54 в крови, и уровень КОЕ повышался до 5,43 и 4,58 в перитонеальной жидкости и крови, соответственно, через 2 ч у животных, которым вводили наполнитель, и до 5,72 и 4,74 в перитонеальной жидкости и крови, соответственно, через 5 ч у мышей, которым вводили наполнитель, чего и следовало ожидать.
Через 2 ч после введения NZ17074 наблюдали значимо сниженные уровни КОЕ, как в крови, так и в перитонеальной жидкости, по сравнению с введением наполнителя (p<0,001).
Дополнительное снижение уровней КОЕ, как в крови, так и в перитонеальной жидкости, наблюдали через 5 ч после введения NZ17074 (p<0,001 по сравнению с контролем наполнителем). Уровни КОЕ были более чем на 3 log10 КОЕ/мл ниже, чем после введения наполнителя.
Введение меропенема также приводило к значимому (p<0,01) снижению уровней КОЕ по сравнению с введением наполнителя в перитонеальной жидкости через 2 и 5 ч после введения, но в крови только через 5 ч после введения. Отсутствие значимого снижения в крови через 2 ч после введения может отражать скорее выраженную вариабельность в группе наполнителя, а не слабый эффект меропенема.
Различия уровней КОЕ после введения NZ17074 или меропенема по сравнению с введением наполнителя были следующими.
У всех мышеи были незначительные симптомы инфекции или не было симптомов инфекции (Таблица 2).
Обсуждение и заключение
Задачей данного исследования было исследование эффективности NZ17074 после внутривенного (в/в) введения однократной дозы 7,5 мг/кг в модели нейтропенического перитонита у мышей NMRI. Для NZ17074 наблюдали значимое (p<0,001) снижение более чем на 3 log10 КОЕ/мл по сравнению с введением наполнителя в крови и перитонеальной жидкости через 5 ч после введения. Кроме того, через 2 ч после введения NZ17074 наблюдали значимое снижение (p<0,001) как в крови, так и в перитонеальной жидкости. Меропенем продемонстрировал значимое снижение по сравнению с группой наполнителя (p<0,01) как в крови, так и в перитонеальной жидкости через 5 ч, но через 2 ч после введения только в перитонеальной жидкости.
ПРИМЕР 4
Модель инфекции бедра с нейтропенией: эффективность NZ17074 против Escherichia coli AID#172 и оценка ED50
Введение
Задачей данного исследования было исследование зависимости доза-ответ после внутривенного (в/в) введения однократной дозы NZ17074 в диапазоне 0,16-12 мг/кг. Эффект исследовали против Е. coli AID#172 в модели инфекции бедра с нейтропенией. Введение меропенема в дозе 40 мг/кг включали в качестве группы положительного контроля. Количество колоний в бедрах определяли через 5 ч после введения.
Модель инфекции бедра является хорошо известной моделью для исследований антимикробного эффекта и проникновения в ткани, как описано S. Gudmundsson & Н. Erlensdottir: Handbook of Animal Models of Infection (1999), ed. by O. Zak & M.A. Sande, Academic Press, San Diego, US, и в некоторых других публикациях. См. обзор D. Andes & С. Craig: Animal model pharmacokinetics and pharmacodynamics: a critical review. International Journal of Antimicrobial Agents, 19(4): 261-268.
Материалы и методы
- 40 самок беспородных мышей NMRI, 25-30 грамм (Harlan Scandinavia).
- Escherichia coli AID#172 от Statens Serum Instute, Copenhagen, Denmark. Клинический изолят из человеческой раны от 2003 г. с множественной лекарственной устойчивостью (к ампициллину, цефтазидиму, азтреонаму, гентамицину, ципрофлоксацину).
- NZ17074 в ацетате Рингера, pH 6, 1,2 мг/мл, 6,0 мл. Раствор хранили при 4°C до использования. Анализы использованных композиций для введения проводили по окончании фазы исследования, проводимой на живых животных, и в этих анализах были получены следующие результаты.
- Наполнитель (ацетат Рингера, pH 6). Раствор хранили при 4°C до использования.
- Meronem® (AstraZeneca, 500 мг вещества для инфузии, меропенем). Номер партии: 09466C. Срок годности: август 2013 г.
- Стерильная вода.
- Стерильный 0,9% физиологический раствор.
- Sendoxan® (циклофосфамид, Baxter, 1 г). Номер партии: 0A671C. Срок годности: январь 2013 г.
- Чашки с агаром и 5% лошадиной крови.
- Чашки с агаром, бромтимоловым синим и лактозой.
Лабораторный виварий и содержание мышей
Температуру и влажность в виварии регистрировали ежедневно. Температура составляла 21°C +/- 2°C, и ее можно было регулировать нагреванием и охлаждением. Влажность составляла 55 +/- 10%. Смена воздуха происходила приблизительно 10-20 раз в час, и период света/темноты был в 12-часовом интервале 06:00-18:00/18:00-06:00.
У мышей был свободный доступ к питьевой воде для домашних животных и пище (2016, Harlan). Мышей содержали в макролоновых клетках 3 типа по 4 мыши в клетке. В качестве подстилки использовали Aspen Wood от Tapvei. Кроме того, животным давали бумажные полоски Sizzle-nest в качестве материала для гнезд. Мышам метили хвосты для различения мышей в клетке. Мышей взвешивали за сутки до введения.
Приготовление растворов NZ17074
Раствор с концентрацией 1,2 мг/мл дополнительно разводили в PBS-наполнителе следующим образом.
Приготовление раствора меропенема
Введение меропенема в дозе 40 мг/кг было включено в качестве группы положительного контроля.
В общей сложности 500 мг меропенема (одну ампулу) растворяли в 10 мл воды, примерно 50 мг/мл. Этот исходный раствор разводили далее до 4 мг/мл (0,4 мл, 50 мг/мл + 4,6 мл физиологического раствора).
Приготовление циклофосфамида
В общей сложности 1 г циклофосфамида (одну ампулу Sendoxan® 1 г) растворяли в 50 мл воды, примерно 20 мг/мл, на каждые сутки его использования. Этот исходный раствор разводили далее до 11 мг/мл (16,5 мл, 20 мг/мл + 13,5 мл физиологического раствора) для использования на -4 сутки или до 5 мг/кг (8,25 мл 20 мг/мл + 21,75 мл физиологического раствора) для использования на -1 сутки.
Введение циклофосфамида мышам
У мышей индуцировали нейтропению инъекцией 0,5 мл раствора циклофосфамида внутрибрюшинно за 4 суток (200 мг/кг) и 1 сутки (100 мг/кг) до заражения.
Заражение мышей
Свежие колонии Е. coli AID#172, полученные в течение ночи в чашке с агаром и 5% лошадиной крови, суспендировали и разводили в стерильном физиологическом растворе приблизительно до 2×107 КОЕ/мл. За один час до начала введения (момент времени -1 ч) проводили внутримышечное заражение мышей 0,05 мл суспензии Е. coli в левую заднюю лапу. Приблизительно через 0,5 ч после введения мышам перорально вводили 45 мкл нейрофена (20 мг ибупрофена на мл, что соответствовало 30 мг/кг) в качестве болеутоляющего средства.
Введение препаратов мышам
Мышам проводили однократное в/в введение NZ17074, меропенема или наполнителя в латеральную хвостовую вену на протяжении приблизительно 30 секунд в объеме 10 мл/кг в момент времени 0 ч (см. Таблицу 1). Определение дозы было основано на средней массе тела 30 г. Мышам с массой тела 28-32 г вводили 0,30 мл раствора. Мышам с массой тела 27-28 г вводили 0,25 мл раствора и мышам с массой тела 32,1-36 г вводили 0,35 мл раствора.
Клиническая балльная оценка мышей
Мышей наблюдали в течение исследования и присваивали им баллы от 0 до 5 в зависимости от их поведения и клинических признаков.
Балл 0: здоровые.
Балл 1: минимальные клинические признаки инфекции и воспаления, например, наблюдение минимальных признаков расстройства или изменения активности.
Балл 2: отчетливые признаки инфекции, такие как социальная самоизоляция, снижение любопытства, измененное положение тела, пилоэрекция или изменения характера движений.
Балл 3: выраженные признаки инфекции, такие как тугоподвижность, снижение любопытства, измененное положение тела, пилоэрекция, боль или изменения характера движений.
Балл 4: сильная боль, и мышь сразу умерщвляли для минимизации страданий животного.
Балл 5: смерть мыши.
Забор образцов
Количество колоний определяли в бедрах на 0 и 5 ч. Мышей анестезировали CO2+O2 и умерщвляли. Сразу после этого убирали кожу, получали заднюю левую лапу и замораживали ее при -70°C. После оттаивания бедра гомогенизировали с использованием Dispomix® Drive. Затем каждый образец разводили в 10 раз в физиологическом растворе и капли объемом 20 мкл наносили на чашки с синим агаром. Все чашки с агаром инкубировали 18-22 ч при 35°C на воздухе.
Результаты
Количество колоний определяли в начале введения и через 5 ч после введения. Количество КОЕ показано в Таблице 3. Перед проведением вычислений проводили log10-трансформацию количества КОЕ.
КОЕ/мл в заражающем материале было определено как 7,35 log10, что соответствовало 6,05 log10 КОЕ/мышь. Наблюдаемая высокая вариабельность может быть обусловлена субоптимальным заражением некоторых мышей, приводящим к слишком низким показателям КОЕ. По этой причине наименьшее значение в каждой группе исключали из графиков и вычислений (см. Таблицу 3). В начале введения среднее log10 КОЕ/мл составляло 4,93 и повышалось до 6,49 log10 КОЕ/мл в группе наполнителя через 5 ч после введения. Несколько сниженные уровни КОЕ наблюдали после введения NZ17074 в дозе 0,16-3,0 мг/кг. После введения NZ17074 в дозе 6 мг/кг (p<0,05) и 12 мг/кг (p<0,01) наблюдали значительное снижение уровней КОЕ по сравнению с введением наполнителя (Таблица 3). Введение меропенема в дозе 40 мг/кг приводило к определенному, но незначимому снижению по сравнению с мышами, которым вводили наполнитель.
Кривые доза-ответ (не показаны) вычисляли в GraphPad Prism с использованием сигмовидной кривой доза-ответ (вариабельный угол наклона). Определенное по этим кривым значение ED50 составило 5,9 мг/кг. Тем не менее, нижнее плато получено не было, и, следовательно, это значение может быть заниженным.
Максимальный эффект NZ17074, Emax, определяли как разность log КОЕ при отсутствии ответа и при максимальном ответе. Отсутствие ответа характеризовали как количество колоний на уровне, определенном у мышей, которым вводили наполнитель. Emax, вычисленный как разность между «Верхним плато» и «Нижним плато» в GraphPad Prism с использованием сигмовидной кривой доза-ответ, составил 2,4 log10 КОЕ/мл. Кроме того, 1 log уничтожения, определяемый как доза, необходимая для снижения бактериальной нагрузки на 1 log по сравнению с началом лечения, определенный с использованием GraphPad Prism, составил 6,1 мг/кг. 2 и 3 log уничтожения получены не были.
Ни у одной мыши ни в одной временной точке не наблюдали клинических признаков инфекции.
Предел выявления 1,4 log10 КОЕ/мл.
Объем описанного и заявленного здесь изобретения не следует ограничивать конкретными аспектами, раскрытыми здесь, поскольку эти аспекты являются лишь иллюстрацией некоторых аспектов изобретения. Предполагают, что любые эквивалентные аспекты входят в объем данного изобретения. Действительно, различные модификации изобретения, в дополнение к показанным и описанным здесь, будут очевидны специалистам в данной области техники из предшествующего описания. Предполагают, что такие модификации также входят в объем прилагаемой формулы изобретения. В случае конфликта следует руководствоваться настоящим описанием, включая определения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ АНТИМИКРОБНОЙ АКТИВНОВНОСТЬЮ, И ПОЛИНУКЛЕОТИДЫ, КОДИРУЮЩИЕ ИХ | 2006 |
|
RU2512525C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ АНТИМИКРОБНОЙ АКТИВНОСТЬЮ, И ПОЛИНУКЛЕОТИДЫ, КОДИРУЮЩИЕ ИХ | 2006 |
|
RU2415150C2 |
| РЕКОМБИНАНТНАЯ ЭКСПРЕССИЯ ДЕФЕНСИНОВ В МИЦЕЛИАЛЬНЫХ ГРИБАХ | 2006 |
|
RU2404990C2 |
| ПОЛИПЕПТИДЫ С ПРОТИВОМИКРОБНОЙ АКТИВНОСТЬЮ И КОДИРУЮЩИЕ ИХ ПОЛИНУКЛЕОТИДЫ | 2005 |
|
RU2393224C2 |
| СПОСОБЫ УСИЛЕНИЯ ДЕГРАДАЦИИ ИЛИ ПРЕВРАЩЕНИЯ ЦЕЛЛЮЛОЗНОГО МАТЕРИАЛА | 2006 |
|
RU2441912C2 |
| ПРОТИВОМИКРОБНЫЕ ПОЛИПЕПТИДЫ ИЗ PSEUDOPLECTANIA NIGRELLA | 2002 |
|
RU2336278C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ ЭНДОПЕПТИДАЗНОЙ АКТИВНОСТЬЮ, И КОДИРУЮЩИЕ ИХ ПОЛИНУКЛЕОТИДЫ | 2011 |
|
RU2583293C2 |
| ПОЛИПЕПТИДЫ С ЛИЗОЦИМНОЙ АКТИВНОСТЬЮ И ПОЛИНУКЛЕОТИДЫ, КОДИРУЮЩИЕ ИХ | 2012 |
|
RU2619051C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ ПРОТИВОМИКРОБНЫМ ДЕЙСТВИЕМ, И КОДИРУЮЩИЕ ИХ ПОЛИНУКЛЕОТИДЫ | 2006 |
|
RU2403260C2 |
| СПОСОБЫ ПОВЫШЕНИЯ УСИЛИВАЮЩЕЙ ЦЕЛЛЮЛОЛИТИЧЕСКОЙ АКТИВНОСТИ ПОЛИПЕПТИДА | 2008 |
|
RU2510417C2 |
















































































































































Настоящее изобретение касается вариантов антимикробного пептида, выделенного полинуклеотида, вектора экспрессии, клетки-хозяина и способа получения таких вариантов. Представлен выделенный вариант антимикробного пептида, имеющего аминокислотную последовательность SEQ ID NO: 2, где указанный вариант обладает антимикробной активностью в отношении грамотрицательных бактерий и содержит аминокислотную последовательность любую из SEQ ID NO: 70, SEQ ID NO: 72, SEQ ID NO: 87, SEQ ID NO: 218, SEQ ID NO: 248, SEQ ID NO: 308, SEQ ID NO: 350, SEQ ID NO: 435 или SEQ ID NO: 450. Способ получения пептида включает культивирование клетки-хозяина, содержащей полинуклеотид, кодирующий описанный пептид, в условиях, подходящих для экспрессии, и выделение указанного варианта пептида. Представленные изобретения могут быть использованы для лечения инфекции, вызванной грамотрицательными бактериями. 6 н. и 2 з.п. ф-лы, 8 табл., 4 пр.
1. Выделенный вариант антимикробного пептида, имеющего аминокислотную последовательность SEQ ID NO: 2, где указанный вариант обладает антимикробной активностью в отношении грамотрицательных бактерий и содержит аминокислотную последовательность любую из SEQ ID NO: 70, SEQ ID NO: 72, SEQ ID NO: 87, SEQ ID NO: 218, SEQ ID NO: 248, SEQ ID NO: 308, SEQ ID NO: 350, SEQ ID NO: 435 или SEQ ID NO: 450.
2. Выделенный полинуклеотид, кодирующий вариант по п. 1, обладающий антимикробной активностью в отношении грамотрицательных бактерий.
3. Конструкция нуклеиновой кислоты, кодирующая вариант по п. 1, обладающий антимикробной активностью в отношении грамотрицательных бактерий, содержащая полинуклеотид по п. 2, функционально связанный с одной или более (несколькими) контрольными последовательностями, управляющими экспрессией кодирующей последовательности.
4. Вектор экспрессии, содержащий полинуклеотид по п. 2.
5. Клетка-хозяин, содержащая полинуклеотид по п. 2, для продуцирования варианта антимикробного пептида, имеющего аминокислотную последовательность SEQ ID NO: 2.
6. Способ получения варианта антимикробного пептида, имеющего аминокислотную последовательность SEQ ID NO: 2, как он определен в п. 1, включающий:
а) культивирование клетки-хозяина по п. 5 в условиях, подходящих для экспрессии указанного варианта; и
б) выделение указанного варианта.
7. Вариант по п. 1 для применения в качестве лекарственного средства против грамотрицательных бактерий.
8. Вариант по п. 1 для применения в изготовлении лекарственного средства для лечения инфекции, вызванной грамотрицательными бактериями.
| Втулочный затвор для бочек | 1924 |
|
SU12569A1 |
| WO 2007023163 A1, 01.03.2007 | |||
| WO 2006097110 A2, 21.09.2006 | |||
| OVCHINNIKOVA TV et al., Recombinant expression, synthesis, purification, and solution structure of arenicin, Biochem Biophys Res Commun., 2007 Aug 17, Vol.360, No.1, pp.156-62. | |||

