Изобретение относится к молекулярной генетике, а именно к способам определения полиморфизма генов, а именно гена апоптотического протеаза-активирующего фактора 1 (APAF1), ассоциированного с гаплотипом фертильности голштинского скота НН1, и может быть использовано в селекции крупного рогатого скота.
Рецессивные генетические дефекты, ассоциированные с эмбриональной смертностью, рассматривают сегодня в качестве одной из важных причин снижения воспроизводительной способности коров [Моногенные наследственные дефекты и их роль в воспроизводстве / Н.А. Зиновьева, Н.И. Стрекозов, Г.В. Ескин, И.С. Турбина, И.Н. Янчуков, А.Н. Ермилов // Животноводство России. 2015. №6. С. 30-31]. Большой прогресс в их идентификации был достигнут благодаря разработке и использованию нового подхода, получившего название «картирования гомозиготности» [Highly effective SNP-based association mapping and management of recessive defects in livestock / C. Charlier, W. Coppieters, F. Rollin, D. Desmecht, JS. Agerholm et al. // Nat Genet. 2008. 40. Pp. 449-454]. Он основан на генотипировании десятков или сотен тысяч SNP с помощью ДНК-чипов средней и высокой плотности и последующей идентификации регионов хромосом, характеризующихся отсутствием одного из гомозиготных генотипов, что позволяет рассматривать соответствующие регионы в качестве кандидатов локализации генов, ответственных за проявление летальных генетических дефектов. Такой метод позволяет осуществлять идентификацию летальных генетических дефектов в течение очень короткого периода времени и при наличии ограниченного числа случаев их проявляется. Для обозначения выявленных этим способом дефектов был предложен термин «гаплотипы фертильности».
С использованием картирования гомозиготности удалось идентифицировать рецессивные генетические дефекты, ассоциированные с эмбриональной смертностью, практически во всех основных породах молочного скота. Так, по крайней мере, пять таких гаплотипов (НН1-НН5) выявлено в голштинской породе, два (JH1, JH2) - в джерсейской, два (ВН1, ВН2) - в бурой швицкой, два (МН1, МН2) - в монбельярдской, один (АН1) - в айрширской породе и четыре гаплотипа (FH1-FH4) - у немецкого симментальского скота [Harmful recessive effects on fertility detected by absence of homozygous haplotypes / P.M. VanRaden, K.M. Olson, D.J. Null, J.L. Hutchison // J. Dairy Sci. 2011. Is. 94. Pp. 6153-6161; Detection of Haplotypes Associated with Prenatal Death in Dairy Cattle and Identification of Deleterious Mutations in GART, SHBG and SLC37A2 / S. Fritz, A. Capitan, A. Djari, S.C. Rodriguez, A. Barbat, A. Baur, C. Grohs, B. Weiss, M. Boussaha,  C. Klopp, D. Rocha, D. Boichard // PLoS ONE. 2013. Vol. 8. Is. 6: e65550; Genomic evaluation of Ayrshire dairy cattle and new haplotypes affecting fertility and stillbirth in Holstein, Brown Swiss and Ayrshire breeds / T.A. Cooper, G.R. Wiggans, P.M. VanRaden, J.L. Hutchison, J.B. Cole, D.J. Null // Amer. Dairy Sci. Assoc. - Amer. Soc. Anim. Sci. joint annual meeting, Indianapolis, IN, July 9, 2013. Poster T206; Identification of a nonsense mutation in CWC15 associated with decreased reproductive efficiency in Jersey cattle / T.S. Sonstegard, J.B. Cole, P.M. VanRaden, C.P. Van Tassell, D.J. Null, S.G. Schroeder, D. Bickhart, M.C. McClure // PLoS. 2013. ONE 8: e54872; Homozygous haplotype deficiency reveals deleterious mutations compromising reproductive and rearing success in cattle / H. Pausch, H. Schwarzenbacher, J. Burgstaller, K. Flisikowski, C. Wurmser, S. Jansen, S. Jung, A. Schnieke, T. Wittek, R. Fries // BMC Genomics. 2015. 16:312. DOI: 10.1186/s12864-015-1483-7].
C. Klopp, D. Rocha, D. Boichard // PLoS ONE. 2013. Vol. 8. Is. 6: e65550; Genomic evaluation of Ayrshire dairy cattle and new haplotypes affecting fertility and stillbirth in Holstein, Brown Swiss and Ayrshire breeds / T.A. Cooper, G.R. Wiggans, P.M. VanRaden, J.L. Hutchison, J.B. Cole, D.J. Null // Amer. Dairy Sci. Assoc. - Amer. Soc. Anim. Sci. joint annual meeting, Indianapolis, IN, July 9, 2013. Poster T206; Identification of a nonsense mutation in CWC15 associated with decreased reproductive efficiency in Jersey cattle / T.S. Sonstegard, J.B. Cole, P.M. VanRaden, C.P. Van Tassell, D.J. Null, S.G. Schroeder, D. Bickhart, M.C. McClure // PLoS. 2013. ONE 8: e54872; Homozygous haplotype deficiency reveals deleterious mutations compromising reproductive and rearing success in cattle / H. Pausch, H. Schwarzenbacher, J. Burgstaller, K. Flisikowski, C. Wurmser, S. Jansen, S. Jung, A. Schnieke, T. Wittek, R. Fries // BMC Genomics. 2015. 16:312. DOI: 10.1186/s12864-015-1483-7].
Экономическая значимость летальных генетических дефектов обусловлена прежде всего влиянием на фертильность коров, чем собственно на гибель плода. Селекционное значение имеют те дефекты, носителями которых служат интенсивно используемые быки-производители, что приводит к постепенному росту частоты встречаемости скрытых носителей среди коров, а следовательно, и к повышению вероятности спаривания двух скрытых носителей и тем самым появлению с 25%-ной вероятностью плодов, несущих гомозиготный по летальному аллелю генотип [Молекулярные методы в диагностике заболеваний и наследственных дефектов сельскохозяйственных животных / Е.А. Гладырь, Н.А. Зиновьева, Л.К. Эрнст, О.В. Костюнина, А.С. Быкова, А.Д. Банникова, Е.П. Кудина, Г. Брем // Зоотехния. 2009. №8. С. 26-27; Роль ДНК-диагностики в контроле и элиминации рецессивных наследственных аномалий сельскохозяйственных животных / Н.А. Зиновьева, Е.А. Гладырь, В.Р. Харзинова, О.В. Костюнина, М.В. Покровская, Н.Г. Друшляк, Я.А. Кабицкая // Достижения науки и техники АПК. 2012. №11. С. 37-40].
С использованием данных о воспроизводительных качествах десятков тысяч животных североамериканской популяции голштинской породы, генотипированных с помощью Bovine SNP50 BeadChip (Illumina Inc., США), на хромосоме 5 в области 58-66 Mb (сборка генома UMD 3.0) был картирован гаплотип, ассоциированный с эмбриональной смертностью на различных сроках стельности, получивший название голштинского гаплотипа 1 - НН1 [Harmful recessive effects on fertility detected by absence of homozygous haplotypes / P.M. VanRaden, K.M. Olson, D.J. Null, J.L. Hutchison // J. Dairy Sci. 2011. Is. 94. Pp. 6153-6161]. Его наличие и локализацию позже подтвердили Fritz с соавторами [Detection of Haplotypes Associated with Prenatal Death in Dairy Cattle and Identification of Deleterious Mutations in GART, SHBG and SLC37A2 / S. Fritz, A. Capitan, A. Djari, S.C. Rodriguez, A. Barbat, A. Baur, C. Grohs, B. Weiss, M. Boussaha, C. Klopp, D. Rocha, D. Boichard // PLoS ONE. 2013. Vol. 8. Is. 6: е65550]. Было установлено, что причиной снижения фертильности, ассоциированной с НН1, служит нонсенс-мутация C→T в гене апоптотического протеаза-активирующего фактора 1, APAF1, приводящая к замене Q→Х в позиции 579 аминокислотной последовательности [Identification of a nonsense mutation in APAF1 that is causal for a decrease in reproductive efficiency in dairy cattle / H.A. Adams, T. Sonstegard, P.M. VanRaden, D.J. Null, C. Van Tassell, H. Lewin. // Proc. Plant Anim. Genome XX Conf., abstr. 2012. Р0555]. Функциональный пептид APAF1 необходим для нормального эмбрионального развития. Мутация в APAF1 в случае спаривания быков - скрытых носителей с коровами - скрытыми носителями мутации приводит к спонтанным абортам и, как следствие, к снижению степени стельности. Было показано, что в североамериканской и французской популяциях голштинов снижение стельности составило соответственно 3,1 и 4,3-6,7% [Harmful recessive effects on fertility detected by absence of homozygous haplotypes / P.M. VanRaden, K.M. Olson, D.J. Null, J.L. Hutchison // J. Dairy Sci. 2011. Is. 94. Pp. 6153-6161; Detection of Haplotypes Associated with Prenatal Death in Dairy Cattle and Identification of Deleterious Mutations in GART, SHBG and SLC37A2 / S. Fritz, A. Capitan, A. Djari, S.C. Rodriguez, A. Barbat, A. Baur, C. Grohs, B. Weiss, M. Boussaha,  C. Klopp, D. Rocha, D. Boichard // PLoS ONE. 2013. Vol. 8. Is. 6: e65550].
C. Klopp, D. Rocha, D. Boichard // PLoS ONE. 2013. Vol. 8. Is. 6: e65550].
Анализ происхождения выявленных скрытых носителей НН1 показал, что вероятный родоначальник мутаций - известный бык-производитель американской селекции USA 1427381 Pawnee Farm Arlinda Chief (1962 г.р.). Распространению мутации в популяциях коров способствовали его потомки, активно использующиеся в системе искусственного осеменения голштинского и голштинизированного скота: USA 1773417 Walkway Chief MARK (F1, 1978 г.р.), AN 382748 A Townson LINDY (F3, 1984 г.р.), USA 17378279 Ja-Bob JORDAN-RED (F4, 1997 г.р.), CAN 5279989 Shoremar MASON (F4, 1990 г.р.), USA 137332056 Glenn-Ann PALERMO (F6, 2006 г.р.). Частота встречаемости скрытых носителей НН1 среди быков-производителей в США составляет 1,9% [Identification of a nonsense mutation in APAF1 that is causal for a decrease in reproductive efficiency in dairy cattle / H.A. Adams, T. Sonstegard, P.M. VanRaden, D.J. Null, C. Van Tassell, H. Lewin. // Proc. Plant Anim. Genome XX Conf., abstr. 2012. Р0555]. Проведенный нами анализ родословных 560 быков-производителей, используемых в системе искусственного осеменения в России, показал, что в линиях предков 18-ти из них встречаются носители мутантного аллеля APAF1. Скрытые носители принадлежат к трем широко распространенным генеалогическим линиям - Рефлекшн Соверинга, Монтвик Чифтейна и Вис Бэк Айдиал.
Зачастую носители генетических дефектов - выдающиеся быки-производители, обладающие высоким генетическим потенциалом по продуктивным показателям, однако этот факт не следует рассматривать как причину для их незамедлительного исключения из воспроизводства. Основным инструментом в использовании быков - скрытых носителей должен стать подбор, исключающий их скрещивание с коровами и телками, потомками скрытых носителей аналогичного генетического дефекта, а также обязательная ДНК-диагностика коров быкопроизводящей группы и ремонтных бычков [Моногенные наследственные дефекты и их роль в воспроизводстве / Н.А. Зиновьева, Н.И. Стрекозов, Г.В. Ескин, И.С. Турбина, И.Н. Янчуков, А.Н. Ермилов // Животноводство России. 2015. №6. С. 30-31].
Анализ научно-технической, патентной и иной информации показал, что единственным применяемым сегодня способом диагностики НН1, используемым в качестве аналога, является использование кастомных (смоделированных пользователем) биочипов, используемых для полногеномного сканирования SNP. Однако проведение ДНК-диагностики данным способом требует наличия дорогостоящего оборудования и, кроме того, сопряжено с высокой стоимостью биочипов. Если для проведения комплексной ДНК-диагностики (геномная оценка + исследование на наличие нескольких генетических аномалий) такой способ является экономически оправданным, то для массового скрининга популяций по НН1 необходима разработка простого, относительно дешевого способа, не требующего использования дорогостоящего оборудования.
В качестве прототипа заявленного способа можно считать секвенирование фрагмента гена APAF1, в результате которого был выявлена нонсенс-мутация C→T, приводящая к замене Q→Х в позиции 579 аминокислотной последовательности [Identification of a nonsense mutation in APAF1 that is causal for a decrease in reproductive efficiency in dairy cattle / H.A. Adams, T. Sonstegard, P.M. VanRaden, D.J. Null, C. Van Tassell, H. Lewin. // Proc. Plant Anim. Genome XX Conf., abstr. 2012. Р0555].
Основной недостаток прототипа заключается в необходимости использования дорогостоящего оборудования и высокой стоимости исследований.
Задача нашего изобретения - создание дешевого способа идентификации полиморфизма в гене APAF1, ассоциированного с гаплотипом фертильности НН1, для использования в селекции крупного рогатого скота.
Технический результат изобретения достигается тем, что предложен способ диагностики полиморфизма гена APAF1, ассоциированного с гаплотипом фертильности НН1 крупного рогатого скота с использованием метода полимеразной цепной реакции с последующим анализом полиморфизма длин рестрицированных фрагментов (ПЦР-ПДРФ), позволяющим определять исключение сайта рестрикции эндонуклеазы BstC8I (последовательность узнавания GCN↓NGC) в области мутации и проводить идентификацию результатов с помощью метода электрофореза в агарозном геле без использования дорогостоящего оборудования, что обеспечивает относительно невысокую стоимость разрабатываемого способа.
Принцип действия разрабатываемого способа основан на использовании двух праймеров, фланкирующих фрагмент APAF1, содержащий диагностируемую мутацию. Принимая во внимание наличие второго неспецифического сайта рестрикции этой эндонуклеазы на расстоянии 10 п.о. «down stream» от места мутации, дизайн тест-системы предусматривает локализацию обратного праймера в области неспецифического сайта рестрикции BstC8I и введение в него нуклеотидной замены, приводящей к исключению указанного сайта в амплифицированном фрагменте. Исследуемая мутация C→T приводит к образованию сайта рестрикции BstC8I, поэтому нормальному аллелю Q соответствуют два фрагмента меньшей длины, образующихся в результате гидролиза, в то время как мутантному аллелю X, ассоциированному с НН1, - нерестрицированный фрагмент большей длины, что позволяет дифференцировать мутантные и немутантные аллели APAF1 методом электрофореза в агарозном геле.
Способ отличается тем, что с применением нескольких технически простых и не требующих дорогостоящих реактивов, оборудования, затрат сил и времени методов возможно выявление мутантного аллеля X гена APAF1, что позволит применить данный метод в селекции животных.
Сущность изобретения - определение полиморфизма гена APAF1, ассоциированного с гаплотипом фертильности НН1 крупного рогатого скота, методом ПЦР-ПДРФ.
Разрабатываемый способ базируется на определении нуклеотидной трансверсии C→T в позиции 63150400 гена APAF1 (UMD 3.1). С этой целью был выбран участок гена APAF1 с мутацией, ассоциированной с гаплотипом фертильности крупного рогатого скота НН1. В электронной базе была найдена нуклеотидная последовательность интересующего нас участка ДНК (номер GenBank AC_000162.1).
Для создания серии референтных образцов с известными генотипами по APAF1, ассоциированными с НН1 (n=80), из проб ткани (ушной выщип, n=40) и спермы (n=40) быков и коров голштинской и голштинизированной черно-пестрой породы выделяли ДНК: по 20 образцов каждого из видов биоматериала методом экстракции перхлоратом [Введение в молекулярную генную диагностику сельскохозяйственных животных / Н.А. Зиновьева, Е.А. Гладырь, Л.К. Эрнст, Г. Брем // Дубровицы: ВИЖ, 2002. 112 с.], по 10 образцов - с использованием магнитных частиц (ООО «Изоген», Россия) и по 10 образцов - с использованием колонок Nexttec (Nexttec Biotechnologie GmbH, Германия) в соответствии с рекомендациями производителей. Референтные генотипы генерировали посредством прямого определения последовательности в области мутации методом пиросеквенирования. С этой целью проводили амплификацию фрагмента длиной 172 п.о., содержащего мутацию, с использованием праймеров APAF1_1Pyro и APAF1_2Pyro_Bio (мечен биотином) с последующим отжигом зонда APAF1_Zond и пиросеквенированием. Для пиросеквенирования использовали базовую последовательность 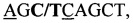 где
где 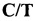 - мутируемый нуклеотид,
- мутируемый нуклеотид, 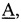
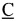 - контрольные нуклеотиды.
- контрольные нуклеотиды.
Краткое описание чертежей
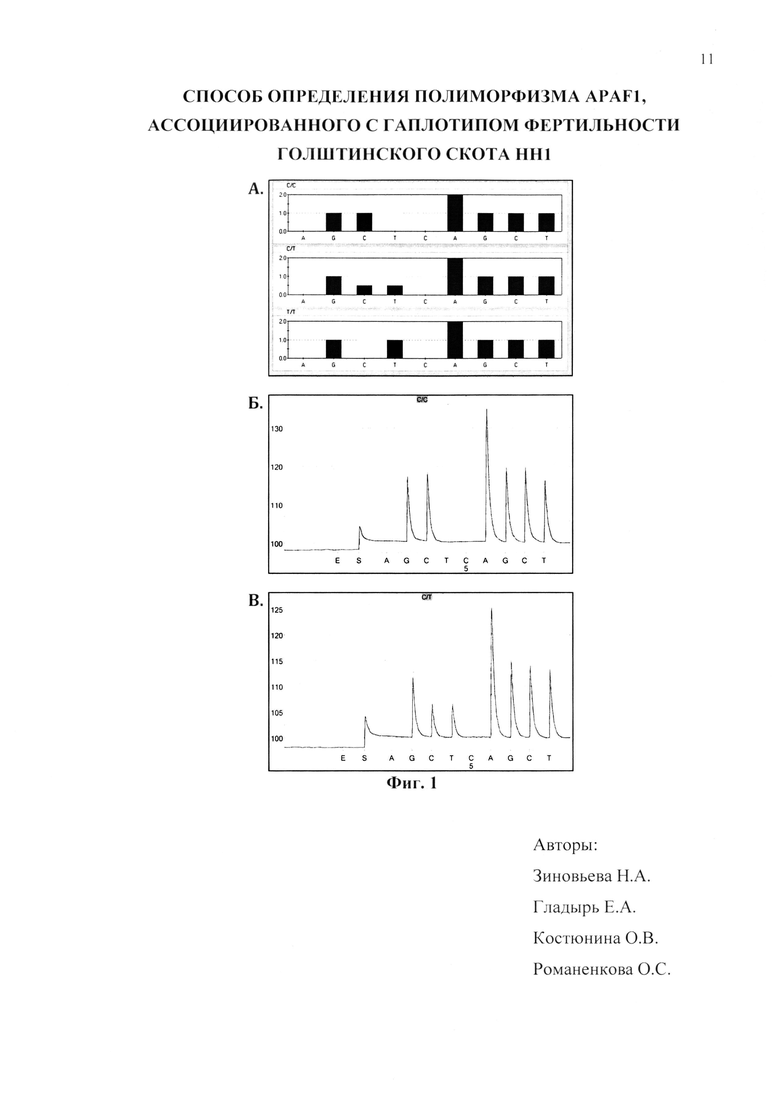
Фиг. 1 - теоретическая модель тест-системы определения полиморфизма APAF1 (гаплотипа НН1) на основе метода пиросеквенирования (А) и результаты генотипирования референтных образцов (Б и В).
Фиг. 2 - теоретическая модель тест-системы определения полиморфизма APAF1 (гаплотипа НН1) на основе ПЦР-ПДРФ (А) и результаты генотипирования референтных образцов (Б).
На фиг. 1А представлены теоретически смоделированные гистограммы возможных генотипов APAF1, где на верхнем рисунке гистограмма соответствует генотипу QQ (последовательность С/С), на среднем - генотипу QX (последовательность С/Т) и на нижнем - генотипу XX (последовательность Т/Т). На фиг. 1Б представлена результирующая пикограмма последовательности гена APAF1 в области исследуемой мутации, соответствующая генотипу QQ, на фиг. 1В - результирующая пикограмма последовательности гена APAF1 в области исследуемой мутации, соответствующая генотипу QX.
В результате проведенного генотипирования была создана серия референтных образцов (n=80), в том числе 14 образцов с генотипом QX (скрытый носитель НН1) и 66 образцов с генотипом QQ (неноситель НН1). Генотип XX является летальным и поэтому не может быть выявлен среди взрослых животных.
Определение полиморфизма APAF1 предложенным способом выполняли следующим образом.
1. Исходя из локализации мутации были подобраны два праймера, фланкирующие определяемую мутацию, при этом праймер APAF1-1 является прямым, а праймер APAF2 - обратным:
APAF1-1 - 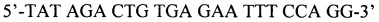
APAF1-2 - 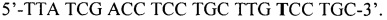
Принимая во внимание наличие второго неспецифического сайта рестрикции этой эндонуклеазы на расстоянии 10 п.о. «down stream» от места мутации, дизайн тест-системы предусматривал локализацию обратного праймера APAF1-2 в области неспецифического сайта рестрикции BstC8I и введение в него нуклеотидной замены, приводящей к исключению указанного сайта в амплифицированном фрагменте.
Продукт амплификации праймеров APAF1-1 и APAF1-2 имеет длину 156 п.о. Место исследуемой мутации помечено звездочкой. Аллель Q, содержащий сайт рестрикции BstC8I, характеризуется двумя фрагментами длиной 123 и 33 п.о., аллель X - одним, длиной 156 п.о. Фиг. 2А иллюстрирует описанный выше вариант настоящего изобретения.
2. Реакции проводили в 15 мкл реакционной смеси следующего состава: 1xПЦР буфер (16.6 мМ (NH4)2SO4, 67.7 мМ Трис-HCl, pH=8.8, 0.1% (v/v) Tween 20, 1.5 мМ MgCl2), 0,2 мМ дНТФ, 10 пмол каждого из праймеров, 2 мМ MgCl2, 1 Ед Taq-полимеразы и 1 мкл ДНК при следующем температурно-временном режиме: 1 цикл - 95°C 7 мин, 40 циклов последовательно - 94°C - 30 с, 59°C - 30 с, 72°C - 45 с, заключительная элонгация при 72°C - 7 мин;
3. По завершении ПЦР к реакционной смеси добавляли 0,2 мкл эндонуклеазы BstC8I (10 МЕ/мкл, ООО "СибЭнзим", Россия) и 1,5 мкл буфера W+BSA (ООО "СибЭнзим", Россия) и инкубировали при 37°C в течение 10-12 ч.
4. Определение аллелей APAF1 осуществляли методом гель-электрофореза, нанося по 5 мкл продуктов гидролиза в 2,5% агарозный гель, гель электрофоретически разделяли в 1х ТАЕ буфере 20 мин при 100 V и детектировали под ультрафиолетовым светом (УФ), при этом генотипу QQ (неносители НН1) соответствуют два фрагмента длиной 123 и 33 п.о., генотипу QX (скрытые носители НН1) - три фрагмента длиной 156, 123 и 33 п.о. (Фиг. 2А, 2Б). Генотипу XX (летальный, может быть выявлен только среди плодов на ранних этапах стельности) соответствует наличие одного фрагмента длиной 156 п.о. Длины фрагментов сравнивали в сопоставлении с М - маркером длины 100 п.о. (500×2), Биосан, Россия.
5. Результативность разработанной тест-системы оценивали посредством сравнения результатов генотипирования референтных образцов.
Пример. Контрольное использование предложенного способа определения полиморфизма APAF1 было апробировано на выборке племенного поголовья голштинского и голштинизированного черно-пестрого скота, в том числе 593 быков-производителей и 270 коров выявило наличие 39 скрытых носителей НН1 (генотип QX), в том числе 23 быков и 16 коров, что соответствует частотам встречаемости 3,9 и 5,9%.
Таким образом, разработанный способ может быть использована для выявления животных, являющихся скрытыми носителями мутации Т в гене APAF1, ассоциированной с гаплотипом фертильности НН1.
Предложенный способ может быть использован в генетике сельскохозяйственных животных для выявления полиморфизма в гене APAF1 крупного рогатого скота, ассоциированном с гаплотипом фертильности НН1, с целью последующего использования полученных результатов в популяционной генетике, разведении и селекции крупного рогатого скота.

Изобретение относится к молекулярной генетике. Описан способ определения полиморфизма генов, ассоциированных с фертильностью крупного рогатого скота, а именно гена апоптотического протеаза-активирующего фактора 1 (APAF1), связанного с гаплотипом фертильности голштинского скота НН1. Способ включает ПЦР-ПДРФ определение полиморфизма C→T в позиции 63150400 (сборка генома UMD 3.1) гена APAF1. При этом амплификацию фрагмента гена, содержащего мутацию, проводят с использованием двух праймеров. В обратный праймер вводится нуклеотидная замена, приводящая к исключению неспецифического сайта рестрикции эндонуклеазы BstC8I, расположенного на расстоянии 10 п.о. «down stream» от места исследуемой мутации, с последующим рестрикционным гидролизом продуктов ПЦР эндонуклеазой BstC8I. Фрагменты меньшей длины, образующиеся в результате рестрикции, соответствуют аллелю Q, в то время как нерестрицированный фрагмент большей длины соответствует аллелю X. Идентификация генотипов животных осуществляется по результатам электрофореза продуктов ПЦР-ПДРФ в агарозном геле. Способ может быть использован в генетике сельскохозяйственных животных для выявления полиморфизма в гене APAF1 крупного рогатого скота, ассоциированном с гаплотипом фертильности НН1, с целью последующего использования полученных результатов в популяционной генетике, разведении и селекции крупного рогатого скота. 2 ил.
Способ диагностики полиморфизма APAF1, ассоциированного с гаплотипом фертильности НН1 крупного рогатого скота, включающий ПЦР-ПДРФ анализ фрагмента гена APAF1, содержащего полиморфизм С→Т в позиции 63150400 (сборка генома UMD 3.1), и отличающийся тем, что в обратный праймер вводится нуклеотидная замена, приводящая к исключению неспецифического сайта рестрикции эндонуклеазы BstC8I, расположенного на расстоянии 10 п.о. «down stream» от места исследуемой мутации, с последующим гидролизом продуктов ПЦР эндонуклеазой BstC8I, при этом фрагменты меньшей длины, образующиеся в результате рестрикции, соответствуют аллелю Q (нуклеотид С), в то время как нерестрицированный фрагмент большей длины соответствует аллелю X (нуклеотид Т), а идентификация генотипов животных осуществляется по результатам электрофореза продуктов ПЦР-ПДРФ в агарозном геле.
| ADAMS et al, Identification of a nonsense mutation in APAF1 that is causal for a decrease in reproductive efficiency in dairy cattle, Proc | |||
| Plant Anim, 2012, реферат | |||
| RU 2008120600 А, 10.12.2009. |

