Изобретение относится к биотехнологии и рыбоводству, в частности к проведению молекулярно-генетического анализа рыб.
Молекулярно-генетические исследования в ихтиологии и рыбоводстве проводятся в течение многих лет для дифференцировки природных популяций рыб, идентификации и паспортизации пород и линий, а также для выявления различий между дикими и искусственно воспроизводимыми особями.
Успешное применение молекулярно-генетических методов анализа, самый эффективный из которых - полимеразная цепная реакция (ПЦР), в значительной степени зависит от получения достаточного количества высокомолекулярной ДНК с ненарушенной структурой, пригодной для постановки ПЦР.
У рыб наилучшим материалом для выделения ДНК, который можно взять прижизненно без причинения серьезных повреждений, являются фрагменты плавников и кровь. Однако нередки случаи, когда необходимо выделять ДНК из гамет, в том числе из отдельных яйцеклеток. Такая необходимость возникает, например, при изучении различий между соматической и гаметической ДНК у одной и той же особи, при разработке некоторых специальных методов хромосомных манипуляций, при поиске полоопределяющих факторов у видов с женской гетерогаметностью, в исследованиях элиминации трансгена в мейозе. Для этих целей необходимо проведение анализа ядерных генетических маркеров.
Методы выделения ДНК включают 2 основных этапа - лизис образцов и очистку ДНК лизата от примесей (протеинов, жиров, а также остатков тканей и клеток).
Для некоторых видов биологических образцов, а также для большинства фиксированных разными способами образцов необходима предварительная пробоподготовка.
Наиболее универсальный способ получения высокомолекулярой ДНК из биологических материалов, который с небольшими модификациями может быть использован для разных видов клеток и тканей, включает в себя пробоподготовку и выделение ДНК с помощью фенольно-хлороформной экстракции (Методические рекомендации по использованию полимеразной цепной реакции в животноводстве. - Дубровицы, 1998. - С. 5-7).
Предварительная пробоподготовка заключается в осаждении нужных фракций клеток из жидких биологических образцов, оттаивании, промывке и др. Выделение ДНК состоит в том, что сначала проводится лизис образца в лизирующем буфере с протеиназой К, далее очистка лизата фенолом или смесью фенол : хлороформ : изоамиловый спирт, затем смесью хлороформ : изоамиловый спирт. Из очищенного лизата суммарную ДНК осаждают этиловым или изопропиловым спиртом.
Однако использование данного метода для выделения ДНК из яйцеклеток рыб имеет ограничения из-за продолжительного и неполного лизиса, что обусловлено сложностью разрушения плотных яйцевых оболочек.
При неполном лизисе яйцеклеток, а также низком соотношении ядро (хромосомы)/цитоплазма (всего 1 гаплоидный набор хромосом на крупную клетку с высоким содержанием протеинов и липидов) возникает необходимость увеличивать число этапов очистки лизатов органическими растворителями для удаления белков, жиров и нелизированных остатков. В результате повышается вероятность потери ДНК и нарушения ее структуры.
Известен способ получения высокомолекулярой ДНК из разных клеток и тканей, включающий пробоподготовку биоматериала и выделение ДНК с помощью солевой экстракции (Методические рекомендации по использованию полимеразной цепной реакции в животноводстве. - Дубровицы, 1998. - С. 10-11). Его основное преимущество перед стандартным фенольно-хлороформным методом заключается в том, что не требуется экстракция фенолом, который является высокоопасным веществом (2-й класс опасности) и к тому же сильным ингибитором ПЦР.
В данном способе пробоподготовку проводят так же, как в предыдущем (промывка, осаждение клеток из жидких образцов, оттаивание и др.). Выделение ДНК проводят путем лизиса образца в гомогенизационном буфере с протеиназой К, встряхивания лизата с хлоридом натрия, очистки смесью хлороформ: изоамиловый спирт, осаждения из очищенного лизата суммарной ДНК этиловым или изопропиловым спиртом.
При использовании этого метода для выделения ДНК из яйцеклеток рыб ограничения те же, что и в предыдущем случае, т.е продолжительный и неполный лизис из-за сложности разрушения плотных яйцевых оболочек.
Наиболее близким по технической сущности и достигаемому результату является способ получения ДНК из икры рыб, включающий пробоподготовку и дальнейшее выделение ДНК с помощью модификации фенольно-хлороформной экстракции. (Fain S.R. DNA extraction from caviar with organic solvents. - US Department of the Interior, Fish and Wildlife Service, version 5.3.1998).
Пробоподготовка проводится следующим образом: небольшое количество икры промывают в солевом растворе для удаления жира и разъединения икринок, из них выбирают интактные икринки. Для выделения ДНК 1 икринку помещают в лизирующий раствор с протеиназой К, в который для повышения эффективности лизиса добавлен детергент Triton Х-100, и инкубируют в термостате 1 час при 65°С. Затем проводят очистку лизата смесью фенол: хлороформ, осаждение ДНК 95% этанолом, растворение ДНК в ТЕ-буфере. Этот способ позволяет получать из 1 икринки значительное количество митохондриальной ДНК, пригодной для ПЦР.
Недостатком данного метода является невозможность получения из икры (яйцеклеток) достаточного количества ядерной геномной (хромосомной) ДНК, пригодной для проведения молекулярно-генетического анализа (в частности, для использования в ПЦР), и в результате невозможность проведения анализа ядерных генетических маркеров.
Это связано с тем, что ядерная ДНК в гаплоидном яйце представлена только одной хромосомной копией, а сама яйцеклетка рыб содержит большое количество белков и жиров (желтка), составляющих запас питательных веществ для развития эмбриона. Трудно выделить такое малое количество ДНК, не повредив ее, и без потерь очистить от большого количества желтка, запасенного в яйцеклетке.
Использование фенола для очистки ДНК делает метод высокоопасным.
Технический результат предлагаемого изобретения заключается в облегчении выделения и увеличении количества выделенной из яйцеклеток ядерной (хромосомной) ДНК, пригодной для проведения молекулярно-генетического анализа, в частности использования в ПЦР при проведении анализа ядерных генетических маркеров, а также в снижении опасности процесса получения ДНК.
Указанный технический результат достигается тем, что в способе получения ДНК из яйцеклеток рыб, включающем пробоподготовку и выделение ДНК, особенность заключается в том, что на стадии пробоподготовки яйцеклетки осеменяют генетически инактивированной спермой для получения гиногенетических эмбрионов, из которых после вылупления выделяют ДНК. При этом выделение ДНК проводят методом солевой экстракции.
Получение на стадии пробоподготовки гиногенетических эмбрионов обусловлено тем, что при гиногенезе в развитии зародыша участвует только ядро яйцеклетки. В процессе развития гиногенетических эмбрионов хромосомный набор исходных яйцеклеток многократно копируется, что позволяет получить из них препараты, содержащие в достаточном количестве хромосомную ДНК, пригодную для использования в ПЦР при проведении анализа ядерных генетических маркеров.
При вылуплении эмбрионы освобождаются от плотных оболочек яйца, в результате чего при выделении ДНК из вылупившихся эмбрионов достигается их полный лизис, уменьшается его продолжительность и количество этапов очистки лизата органическими растворителями, и как следствие, снижается вероятность потери ДНК и нарушения ее структуры.
Выделение ДНК из гиногенетических эмбрионов методом солевой экстракции, в котором не используется высокотоксичный фенол, снижает опасность процесса получения ДНК.
Способ применим для получения ДНК из яйцеклеток практически любых видов рыб с внешним размножением.
Проведенный анализ уровня техники позволил установить, что не обнаружен источник, характеризующийся признаками, тождественными всем существенным признакам заявленного изобретения, следовательно, предлагаемое изобретение соответствует условию «новизна».
Дополнительный поиск известных решений показал, что заявленное изобретение не вытекает для специалиста явным образом из известного уровня техники. Применение известного метода индуцированного гиногинеза в заявленном изобретении приводит к получению нового технического результата - облегчению выделения и увеличению количества ядерной ДНК, пригодной для проведения молекулярно-генетического анализа, в частности, использования в ПЦР при проведении анализа ядерных генетических маркеров. Совокупность существенных признаков в заявленном способе делает возможным выделение ядерной ДНК из отдельной яйцеклетки и проведение молекулярно-генетического анализа для самых различных целей.
Таким образом, заявленное изобретение соответствует условию "изобретательский уровень".
Способ осуществляется следующим образом.
От нерестующих рыб получают зрелые половые продукты. Из икринок (яйцеклеток) с помощью индуцированного гиногенеза получают гиногенетические эмбрионы. Для этого икринки осеменяют генетически инактивированной посредством УФ-облучения спермой. После вылупления гиногенетических эмбрионов проводят их отбор (и фиксацию, если предусматривается хранение), затем выделяют из них ДНК методом солевой экстракции.
Лизис эмбрионов проводят в гомогенизационном буфере с протеиназой К, полученный лизат встряхивают с хлоридом натрия, очищают смесью хлороформ: изоамиловый спирт и осаждают из очищенного лизата суммарную ДНК этиловым или изопропиловым спиртом.
Примеры осуществления способа проиллюстрированы фотографиями (фиг. 1-4), где приведены электрофореграммы ДНК из икры и из гиногенетических эмбрионов, а также электрофореграммы продуктов ПЦР, полученных с этими препаратами ДНК.
Пример 1. Из полученной от самки карпа икры отбирали 2 небольшие порции.
Одну из них промывали в солевом растворе, выбирали интактные яйцеклетки для последующего выделения ДНК посредством фенольно-хлороформного метода в модификации для икры (в соответствии с прототипом). ДНК выделяли из 1, 10, 50, 90 и 270 яйцеклеток. Указанные количества яйцеклеток помещали в лизирующий раствор с протеиназой К и детергентом Triton Х-100 и инкубировали при 65°С. Время лизиса увеличивалось с увеличением числа лизируемых яйцеклеток (икринок). Лизаты очищали смесью фенол: хлороформ, число экстракций увеличивалось с увеличением числа используемых для выделения ДНК икринок. Затем проводили осаждение ДНК 95% этанолом. Осадок ДНК растворяли в бидистиллированной воде.
Другую порцию икры осеменяли генетически инактивированной спермой для получения гиногенетических эмбрионов.
Для облучения спермы в качестве источника коротковолнового УФ-излучения (300 Дж/м2) использовали ртутную бактерицидную лампу. Расстояние от лампы до объекта облучения составляло 45 см, продолжительность облучения - 11 мин. Перед облучением сперму разбавляли в соотношении 1:12 (6 капель спермы на 10 мл 0,9% раствора NaCl). 10 мл суспензии разбавленной спермы наливали в 9 см чашку Петри и облучали при постоянном перемешивании на магнитной мешалке. Гиногенетические эмбрионы (гаплоидные) инкубировались в чашках Петри при комнатной температуре. Для предотвращения фотореактивации УФ-повреждений спермиев начальный период инкубации проводили в затемненном помещении. Гиногенетических эмбрионов отбирали для выделения ДНК после вылупления.
Для последующего выделения ДНК из гиногенетических гаплоидных эмбрионов использовали метод солевой экстракции. 1 гаплоидный эмбрион помещали для лизиса в гомогенизационный буфер с протеиназой К и инкубировали при 62°С 1-2 часа. Лизат встряхивали с хлоридом натрия, очищали смесью хлороформ: изоамиловый спирт и осаждали из очищенного лизата ДНК этиловым спиртом. Осадок ДНК растворяли в бидистиллированной воде.
Полученную суммарную ДНК разделяли методом горизонтального электрофореза в 1,0% агарозном геле с бромистым этидием в буфере ТАЕ (трис-ЭДТА-ацетатная система) и визуализировали под УФ-светом в темной комнате AutoChemi на трансиллюминаторе 2UV. Регистрацию электрофореграмм проводили на компьютере с помощью системы AutoChemi для изучения изображения (UVP).
Длина фрагментов ДНК приводится в парах оснований (bp).
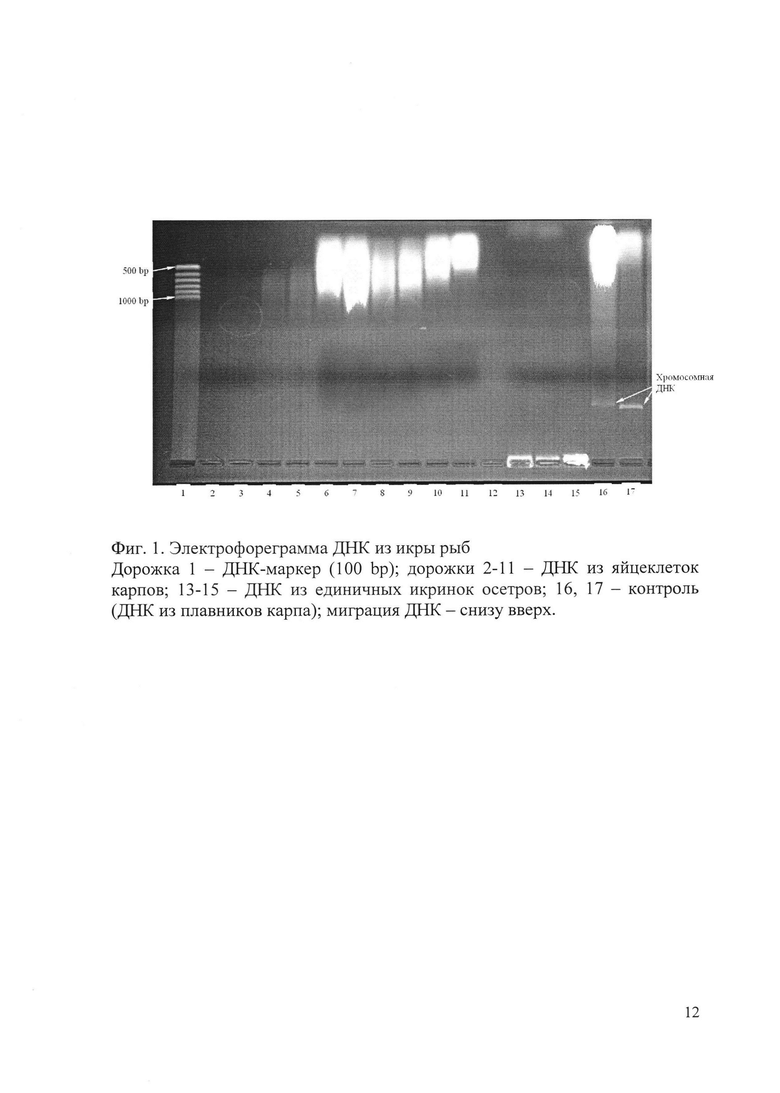
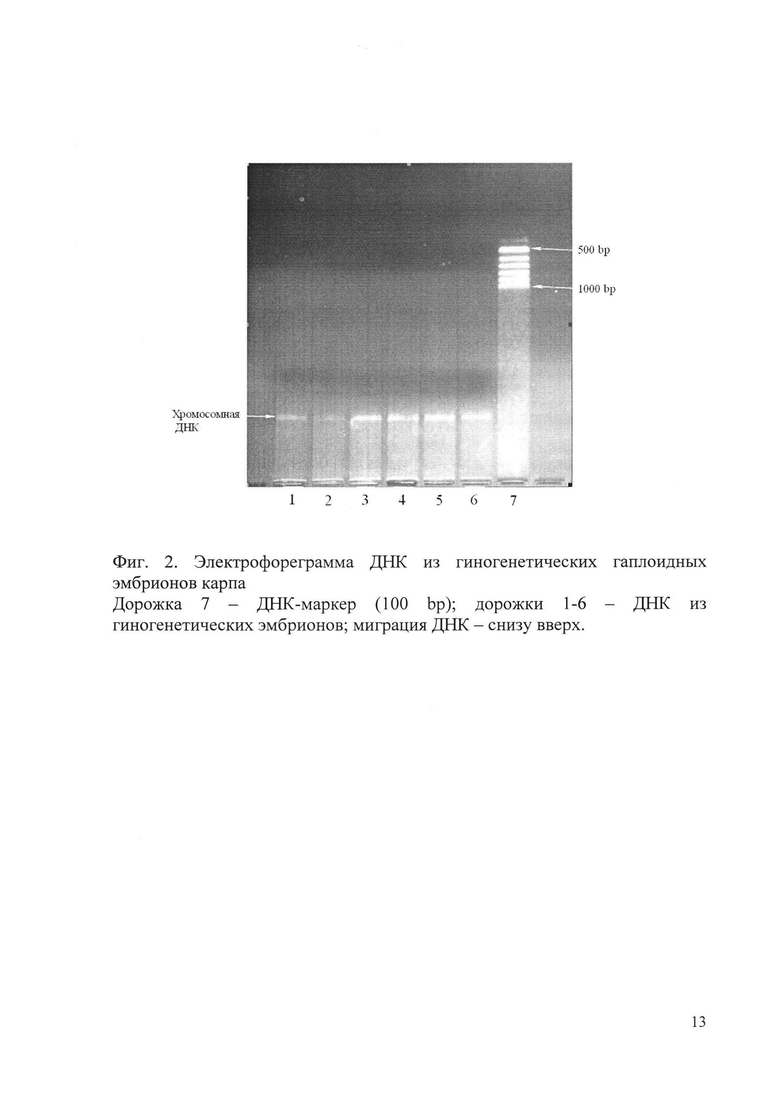
Результаты электрофореза экстрактов ДНК из яйцеклеток рыб представлены на фиг. 1, из гиногенетических гаплоидных эмбрионов - на фиг. 2. Миграция ДНК на всех электрофореграммах - снизу вверх.
На фиг. 1 на дорожке 1 - ДНК-маркер (100 bp); на дорожках 2, 3 - ДНК из 1 яйцеклетки карпов, 4, 5 - ДНК из 10 яйцеклеток карпов, 6, 7 - ДНК из 50 яйцеклеток карпов, 8, 9 - ДНК из 90 яйцеклеток карпов, 10, 11 - ДНК из 270 яйцеклеток карпов; 13-15 - ДНК из 1 икринки осетров; 16, 17 - контроль (экстракты из плавников карпа, содержащие хромосомную ДНК).
Как видно на фиг. 1, полоса, соответствующая высокомолекулярной хромосомной ДНК (размер порядка 10000 bp и более), наблюдается только на дорожках 16 и 17, на которых проводилось разделение контрольных препаратов ДНК из соматических тканей карпа. На дорожках 2-11, где проводилось разделение экстрактов ДНК из икры (яйцеклеток) карпа, полоса, соответствующая хромосомной ДНК, не визуализируется. Возможные причины ее отсутствия - потеря или разрушение ДНК в процессе выделения.
Полоса, соответствующая хромосомной ДНК, хорошо видна на дорожках 1-6 (фиг. 2), на которых проводилось разделение препаратов ДНК из гиногенетических гаплоидных эмбрионов карпа. Таким образом, можно сделать вывод о присутствии высокомолекулярной хромосомной ДНК во всех этих препаратах. Каждый препарат ДНК получен из 1 гиногенетического гаплоидного эмбриона.
Пример 2. Чтобы оценить пригодность препаратов ДНК, полученных из яйцеклеток и гиногенетических гаплоидных эмбрионов карпа, для проведения ПЦР, их использовали в амплификации целевого фрагмента ДНК длиной 540 bp (рекомбинантная конструкция ptMTa-scGH, встроенная в ядерный геном) со специфическими праймерами AL4 и G35.
ПЦР проводили в конечном объеме 25 мкл с 1×ПЦР-буфером, 2 мМ MgCl2, 0,2 мМ динуклеотидтрифосфатов, 10 пмол каждого праймера и 1,5 Ед Taq-полимеразы, используя 2,5 мкл экстракта ДНК.
Режим ПЦР: предварительная денатурация ДНК - 95°С - 13 мин; синтез ПЦР-продуктов: 5 циклов - 94°С - 40 сек, 62°С - 40 сек, 72°С - 40 сек; 5 циклов - 94°С - 40 сек, 60°С - 40 сек, 72°С - 40 сек; 35 циклов - 94°С - 30 сек, 58°С - 15 сек, 72°С - 15 сек; окончательная достройка цепей - 72°С - 10 мин.
Продукты ПЦР разделяли с помощью электрофореза в 1,0% агарозном геле с бромистым этидием в буфере ТАЕ (трис-ЭДТА-ацетатная система) и визуализировали под УФ-светом в темной комнате AutoChemi на трансиллюминаторе 2UV. Регистрацию электрофореграмм проводили на компьютере с помощью системы AutoChemi для изучения изображения (UVP).
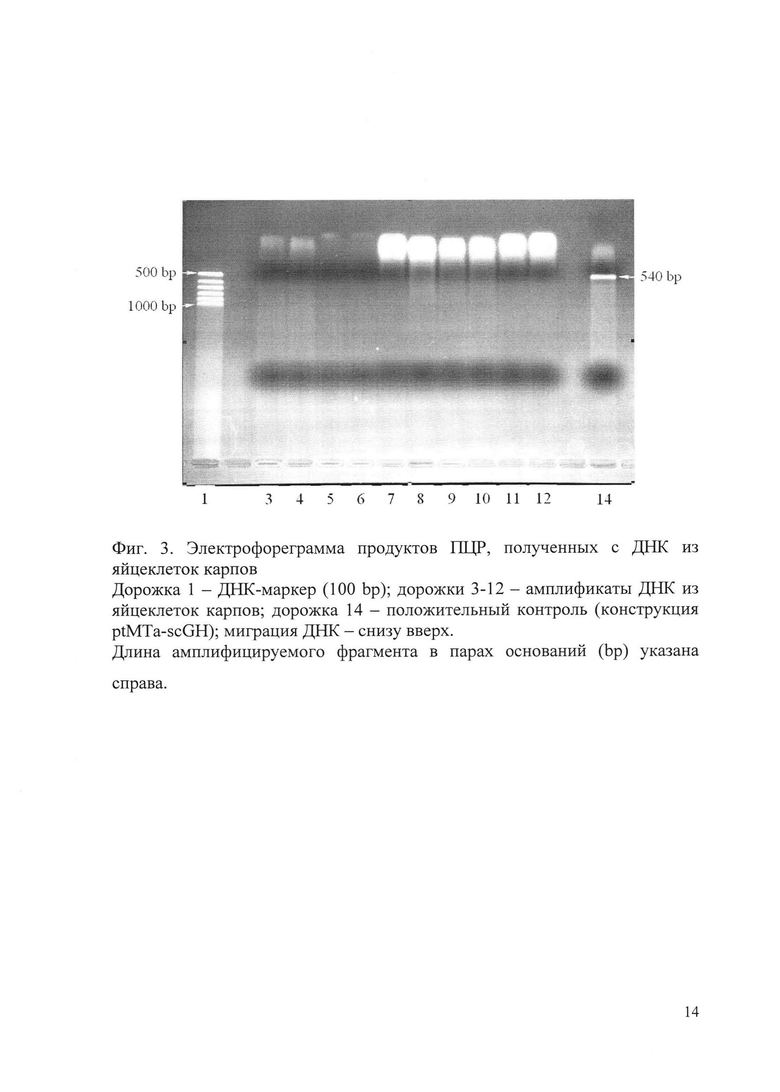
Результаты электрофореза продуктов ПЦР, полученных с ДНК из яйцеклеток карпа, представлены на фиг. 3, с ДНК из гиногенетических гаплоидных эмбрионов - на фиг. 4. Миграция ДНК на обеих электрофореграммах - снизу вверх.
В продуктах ПЦР, полученных с препаратами ДНК из яйцеклеток карпов, амплифицируемый целевой фрагмент длиной 540 bp отсутствует (дорожки 3-12 на фиг. 3). Это может быть связано с недостаточным количеством хромосомной ДНК для проведения ПЦР, а также с нарушением ее структуры в процессе экстракции.
Этот целевой фрагмент выявлен в продуктах ПЦР, полученных с 10 препаратами ДНК из отдельных гиногенетических гаплоидных эмбрионов карпа (дорожки 2-4, 6, 10, 12-15, 17 на фиг. 4). Его отсутствие в остальных 9 амплификатах (дорожки 1, 5, 7-9, 11, 16, 18, 19 на фиг. 4) связано с гемизиготностью самок, от которых получали икру, по этому фрагменту ДНК (конструкция ptMTa-scGH).
Приведенные примеры иллюстрируют, что предлагаемый способ позволяет получить высокомолекулярную хромосомную ДНК, пригодную для исследования с помощью ПЦР, в достаточном количестве, а также снизить опасность и трудоемкость процедуры выделения ДНК.
Таким образом, изложенные выше сведения свидетельствуют о выполнении при использовании заявленного изобретения следующей совокупности условий:
- способ получения ДНК из яйцеклеток рыб по заявленному изобретению предназначен для использования в рыбоводстве, в частности при получении ДНК для молекулярно-генетического анализа рыб;
- для заявленного способа в том виде, как он охарактеризован в независимом пункте изложенной формулы изобретения, подтверждена возможность его осуществления с помощью описанных в заявке средств и методов.
Следовательно, заявленное изобретение соответствует условию "промышленная применимость".
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ОДНОПОЛО-ЖЕНСКОГО ПОТОМСТВА У ОСЕТРОВЫХ РЫБ | 2005 |
|
RU2312495C2 |
| СПОСОБ ВОССТАНОВЛЕНИЯ ГЕНОТИПОВ РЫБ | 1992 |
|
RU2045896C1 |
| Способ получения аллотетраплоидных гибридов серебряного карася с карпом | 1986 |
|
SU1364259A1 |
| Способ индуцирования диплоидного гиногенеза у пеляди | 1984 |
|
SU1243657A1 |
| БИОЛОГИЧЕСКИЙ МАРКЕР ДЛЯ ОПРЕДЕЛЕНИЯ ПОРОДЫ РЫБ, НАБОР И СПОСОБ ОПРЕДЕЛЕНИЯ ПОРОДНОЙ ПРИНАДЛЕЖНОСТИ РЫБ | 2005 |
|
RU2294633C2 |
| Способ получения диплоидного гиногенетического потомства у карповых рыб | 1974 |
|
SU738565A1 |
| СПОСОБ КРИОКОНСЕРВАЦИИ ЯЙЦЕКЛЕТОК ОСЕТРОВЫХ РЫБ | 2010 |
|
RU2460284C2 |
| ГЕТЕРОЗИГОТНЫЕ ПО CENH3 ОДНОДОЛЬНЫЕ РАСТЕНИЯ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ ДЛЯ ИНДУКЦИИ ГАПЛОИДОВ И ОДНОВРЕМЕННОГО РЕДАКТИРОВАНИЯ ГЕНОМА | 2021 |
|
RU2832578C1 |
| Способ инкубации икры лососеобразных рыб в шестилуночных культуральных планшетах с использованием стимуляторов развития | 2021 |
|
RU2786438C1 |
| СПОСОБЫ ДОСТАВКИ АГЕНТА В ИКРИНКИ РЫБ | 2016 |
|
RU2741645C2 |

Изобретение относится к области биотехнологии и рыбоводства и касается способа получения ДНК из яйцеклеток рыб. Представленный способ включает пробоподготовку и выделение ДНК. В процессе пробоподготовки яйцеклетки осеменяют генетически инактивированной спермой для получения гиногенетических эмбрионов, из которых после вылупления выделяют ДНК. Перед инактивацией сперму разбавляют 0,9%-ным раствором хлорида натрия в соотношении 1:12, инактивацию проводят УФ-излучением в дозе 300 Дж/м2, а выделение ДНК осуществляют методом солевой экстракции путем лизиса образца в гомогенизационном буфере с протеиназой К с дальнейшим встряхиванием с хлоридом натрия, очисткой смесью хлороформ: изоамиловый спирт и осаждением ДНК из очищенного лизата этиловым спиртом. Изобретение позволяет облегчить выделение и увеличить количество ядерной ДНК, пригодной для проведения молекулярно-генетического анализа, в частности использования ПЦР при проведении анализа ядерных генетических материалов. 4 ил., 2 пр.
Способ получения ДНК из яйцеклеток рыб, предусматривающий пробоподготовку и выделение ДНК, отличающийся тем, что в процессе пробоподготовки яйцеклетки осеменяют генетически инактивированной спермой для получения гиногенетических эмбрионов, из которых после вылупления выделяют ДНК, при этом перед инактивацией сперму разбавляют 0,9%-ным раствором хлорида натрия в соотношении 1:12, инактивацию проводят УФ-излучением в дозе 300 Дж/м2, а выделение ДНК осуществляют методом солевой экстракции путем лизиса образца в гомогенизационном буфере с протеиназой К с дальнейшим встряхиванием с хлоридом натрия, очисткой смесью хлороформ: изоамиловый спирт и осаждением ДНК из очищенного лизата этиловым спиртом.
| IRENE E SAMONTE-PADILLA et al., Induction of diploid gynogenesis in an evolutionary model organism, the three-spined stickleback (Gasterosteus aculeatus), BMC Dev Biol., 2011, Vol.11, No.55, pp.1-11 | |||
| СПОСОБ ПОЛУЧЕНИЯ ОДНОПОЛО-ЖЕНСКОГО ПОТОМСТВА У ОСЕТРОВЫХ РЫБ | 2005 |
|
RU2312495C2 |

