Изобретение относится к рыбному хозяйству на внутренних водоемах, в частности к генетическим способам селекции рыб, а именно к способу индуцированного; диплоидного гиногене- за у пеляди.
Целью изобретения является получение массовых количеств диплоидных гиногенетических личинок пеляди.
Сущность способа состоит в том, что спермии пеляди подвергают генетической инактивации УФ-облучением, осеменяют ими икру и последнюю на стадии ранней анафазы - II подвергают тепловому воздействию в течение . 30-40 мин при 15-17°С, начиная с 18- 22 мин после осеменения.
Параметры теплового воздействия наиболее существенны для достижения эффекта, так как тепловое воздейст- ,вие разрушает нити веретена деле-/ ния, в результате чего хромосомы второго редукционного тельца втягиваются в цитоплазму яйца и происходит восстановление диплоидности за счет обьединения женского пронуклеуса со вторым редукционным тельцем. В связи с этим особенно важным в способе является определение момента начала теплового воздействия.
С целью получения массовых гиногенетических партий пеляди производят узконапра.вленное непродолжительное тегшовое воздействие, которое захватьшает только стадию анафазы - II, которая поступает при темпера- i туре воды 2-6 С через 18-22 мин после осеменения (табл.1).
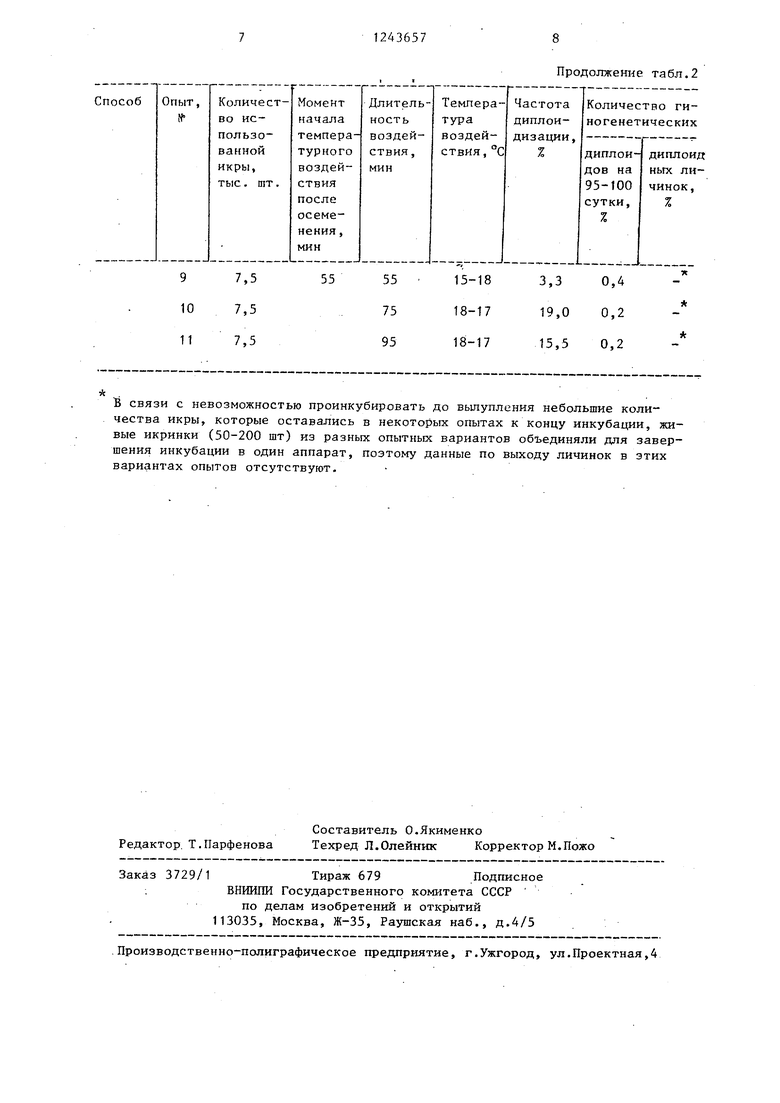
Подтверждением правильности предл .гаемых параметров являются результаты опытов, проведенных при запредельных режимах (табл.2). Из табл.2 видно, что более раннее начало шока (табл.2, опыты 1-5) или более позднее (табл.2 опыты 7-11) значительно .снижает вход диплоидных гиногенетических личинок. Изменение температуры шока выше 15-1У С и удлинение ил сокращение длительности шока хотя и приводят к значительному увеличению частоты диплоидизации (до 59,2% (табл.2, опыты 9-11), но резко снижают выживаемость икры в целом, что ведет к значительному сокращению числа гиногенетических диплоидов (д нескольких штук) уже на стадии пигментации глаз. Снижение температуры шока ниже оптимааьного значения так
2(36572
же приводит к уменьшению выхода .гиногенетических диплоидных личинок. Пример 1. Получили и перемешали спер от 10 самцов пеляди в
IJ возрасте 3+. Из полученной смеси отобрали 5 капель, которые суспендировали в ча.шке Петри в 12 мл раствора Гольтфретера двойной концентрации. Для генетической инактивации
)0 ядер сперматозоидов использовали ультрафиолетовые лучи от ртутной лампы типа БУВ-30 с длиной волны 257 мм. Лампа была откалибрована в институте Цитологии АН СССР. Для
15 облучения чашку Петри со спермой помещали под лампу на магнитную мешалку. Расстояние от лампы до поверхности раствора спермы было 45 см. При токе 0,4А в этом случае мощность до-,
зы
12 9 5
I jl. Xл
см
5
0
5
Экспозиция облучения
м
2 МИН. Доза облучения за две минуты составляла 15,0 мДж/см. Энергия дозы сохранялась на расстоянии +15 см от осей лампы. Подвижность спермы определяли до и после облучения при активации, их водой. Для чего в каплю воды.на предметном стекле, помещенном на предметном столике микроскопа, вносили спермиев препаровальной иглой, при этом наблюдали их активные вихревые движения. После облучения, как правило, подвижность спермиев несколько снижалась.
Икру получили от одной самки пеляди в возрасте 3+. Взяли в контроль и в опыт по 7,5 тыс. шт. икринок, - осеменили облученной спермой обычным сухим методом. Контрольную икру не подвергали температурным воздействиям, ее развитие происходило при температуре воды 1 - . Опытную икру через 20 мин после осеменения поместили 3 воду с температурой 16°С на 40 мин Для того, чтобы температура шока бьша постоянной, миску с опытной икрой поместили в термостатированную водяную баню. Температуру во время опыта контролировали при помо:щи термометра с точностью до О,1°С. После окончания срока теплового воздействия теплую воду слили, залили икру водой оптимальной для пеляди температуры 5°С и -инкубировали в стандартных условиях в аппара- тах Вейса. Наблюдения за развитием эмбрионов вели в течение всего инкубационного периода вплоть до вы- лупления личинок.
0
5
0
Число гиногенетических диплоидов высчитывали в процентах от количества приступивших к развитию эмбрионов и определяли на двух эмбриональных стадиях и на стадии личинок. Частоту диплоидизации определяли на стадии бластулы средних клеток (4 сут эмбрионального развития) , для чего икру фиксировали в уксуснокислом спирте (3:1). В контроле и опыте было исследовано по 50 эмбрионов. Окраску хромосом и приготовление давленых препаратов проводили по общепринятой методике. На препаратах каждый эмбрион располагали и исследовали отдельно. Определяли плоид- ностъ и процент клеток с аберрантными митозами. На стадии пигментации ,глаз проводились отбор и подсчет тиногенетических диплоидов, диплоиды отбирали по признакам размера и степени окраски глаз. На стадии личинок
подсчитывали число вьшупившихся нормальных диплоидных гиногенетических личинок. В результате теплового шока частота диплоидизации повысилась от 3,4% в контроле до 36,0% в опыте, количество гиногенетических диплоидов на 100-е сутки увеличилось от 0,1% в котроле до 10,6% в опыте, а число нормальных диплоидных гиногенетических личинок возросло от 0,03% в контроле до 6,4% в опыте. Таким образом, использование приведенных в этом примере параметров температурных воздействий позволило получить наилучшие результаты.
П.р и м е р 2. Получили и перемешали икру от 11 самок. Использовали в контроле 6,0 тыс. шт. икринок, в опыте 270,0 тыс.шт. Сперму получили от 10 самцов. Экспозиция облучения 12 мин. Дрза УФ-облучения спермы 90,0 мДж/см. Опытную партию
икры через 18 мин после осеменения
прогревали 30 мин при , в результате чего частота диплоидизации повысилась от 3,2% в контроле до 35,7% в опыте. Количество гиногенетических диплоидов на 100-е сутки
эмбрионального развития в опыте состав ляло 6,7%, а число гиногенетических личинок 3,5%.
Пример 3. Получили икру от
2самок -И сперму от 7 самцов. В контроле и в опыте использовали по
7,5 тыс. икринок. Экспозиция облуче- ния 12 мин. Доза облучения 90 ЬЩж/см. Опытную икру через 22 мин после oce-i
4
в воду
с тейпера10
15
20
25
30
35
менения поместили турой 17 С. Длительность прогрева 40 мин. В результате частота дипло- тизации 11,0%, количество гиногенетических диплоидов на 100-е сутки эмбрионального развития в контроле 1,2%, а в опыте 5,2%. Число гиногенетических личинок возросло от 0,08% в контроле до 3,4% опыте.
Согласно предлагаемому способу возможно получение массовых гиноге- нeт чecкиx партий нормально развивающихся диплоидных гиногенетических по- томко.д, пеляди, которые являются ценным селекционным и экспериментальным материалом.
Способ может найти применение при получении второго и последующих гиногенетических поколений пеляди, кроме того, при воспроизводстве гибрида пелчира в себе и получении триплоидных особей пеляди.
Получение диплоидных гиногенетических личинок пеляди в массовых количествах позволяет .внедрить метод индуцированного диплоидного гиноге- неза в практику селекционных работ с пелядью и ускорить в 2-3 раза процесс селекции. Использование этого эффективного генетического метода при разведении пеляди дает возможность проводить промышленные скрещивания гиногенетических линий и получать гетерозисный эффект при товарном выращивании пеляди в пру- довых и озерных хозяйствах северо- запада .
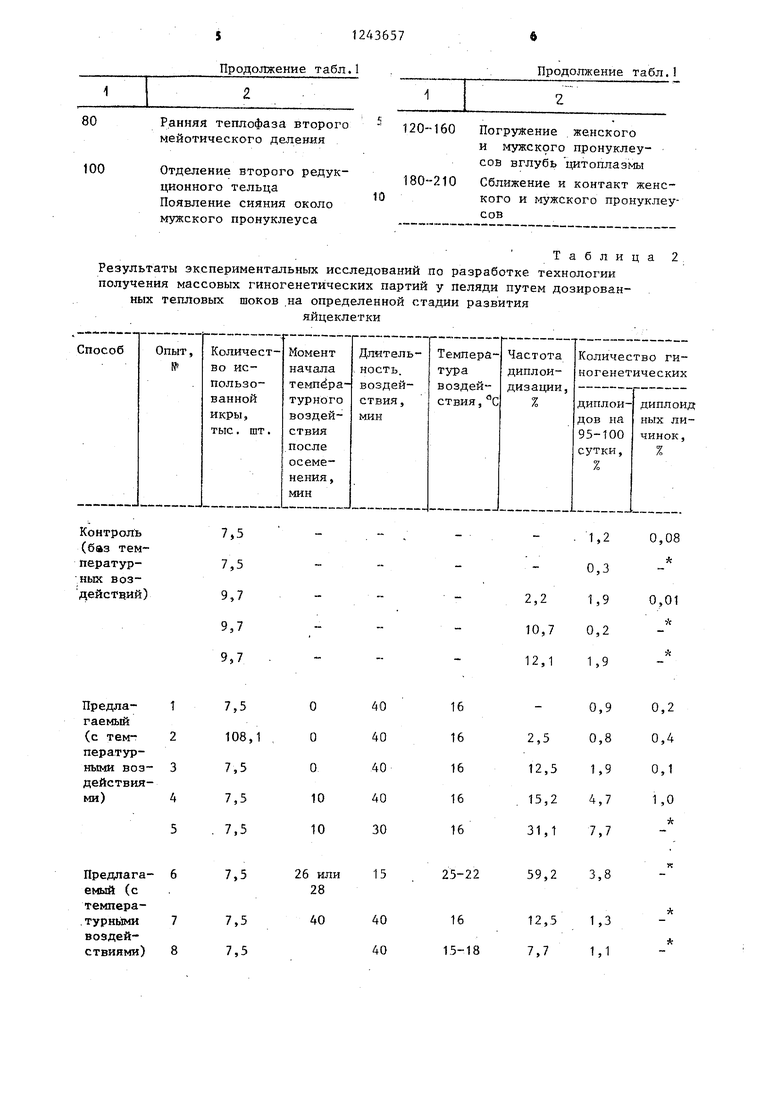
Таблица
Скорость прохождения второго мейотического деления в осемененной яйцеклетке пеляди при температуре воды (+2)-(+6)С,.
45
50
О Метафаза второго мейотического деления
20-40 Ранняя анафаза второго мейотического деления
60 Поздняя анафаза второго мейотического деления
Продолжение табл.1
Ранняя теплофаза второго мейотического деления
Отделение второго редукционного тельца Появление сияния около мужского пронуклеуса
Таблица 2
Результаты экспериментальных исследований по разработке технологии получения массовых гиногенетических партий у леляди путем дозированных тепловых шоков .на определенной стадии развития
яйцеклетки
Продолжение табл,1
120--160 Погружение женского
и мужского пронуклеу- сов вглубь цитоплазмы
180-210 Сближение и контакт женс- кого и мужского пронуклеуСОВ
В связи с невозможностью проинкубировать до вылупления небольшие количества икры, которые оставались в некоторых опытах к концу инкубации, живые икринки (50-200 шт) из разных опытных вариантов объединяли для завершения инкубации в один аппарат, поэтому данные по выходу личинок в этих вариантах опытов отсутствуют.
Редактор. Т.Парфенова
Составитель О.Якименко
Техред Л.Олейник Корректор М.Пожо
Заказ 3729/1Тираж 679Подписное
ВНИИПИ Государственного комитета СССР
по делам изобретений и открытий 113035, Москва, Ж-35, Раушская наб., д.4/5
Производственно-полиграфическое предприятие, г.Ужгород, ул.Проектная,4
Продолжение табл.2

| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ОДНОПОЛО-ЖЕНСКОГО ПОТОМСТВА У ОСЕТРОВЫХ РЫБ | 2005 |
|
RU2312495C2 |
| СПОСОБ ВОССТАНОВЛЕНИЯ ГЕНОТИПОВ РЫБ | 1992 |
|
RU2045896C1 |
| Способ получения диплоидного гиногенетического потомства у карповых рыб | 1974 |
|
SU738565A1 |
| СПОСОБ ВОСПРОИЗВОДСТВА СИГОВЫХ РЫБ | 2001 |
|
RU2202179C2 |
| Способ получения аллотетраплоидных гибридов серебряного карася с карпом | 1986 |
|
SU1364259A1 |
| СПОСОБ ПОЛУЧЕНИЯ ДНК ИЗ ЯЙЦЕКЛЕТОК РЫБ | 2015 |
|
RU2632649C2 |
| Способ искусственного оплодотворения икры рыб | 1983 |
|
SU1093311A1 |
| Способ стимуляции развития икры сиговых рыб | 1990 |
|
SU1803013A1 |
| Способ разведения рыб | 1990 |
|
SU1759356A1 |
| Способ повышения жизнестойкости оплодотворенной икры рыб | 1978 |
|
SU789063A1 |

| Черфас Н.Б., Иляс ова В.А | |||
| Рекомендации по получению диплоидного гиногенетического потомства у карпа | |||
| М.: ВНИИПРХ, 1979, с.32 | |||
| Refstic Т | |||
| Induction of diploid gynogenesis in Atlantic salmon and rainbow trouf using irradiated sperm and heat chock can | |||
| Гребенчатая передача | 1916 |
|
SU1983A1 |
| Мантельман И.И., Кайданова Т.Н | |||
| Применение химического способа инактивации спермиев при получении гино- генетических личинок пеляди | |||
| Известия ГосНИОРХа, т | |||
| Паровозный золотник (байпас) | 1921 |
|
SU153A1 |
