ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Данная заявка испрашивает приоритет относительно предварительной заявки на патент США №63/036902, поданной 9 июня 2020 года, и предварительной заявки на патент США №63/036910, поданной 9 июня 2020 года, содержимое каждой из которых включено в данный документ посредством ссылки в полном объеме.
ЗАЯВЛЕНИЕ О СПОНСИРОВАНИИ ИССЛЕДОВАНИЯ ФЕДЕРАЛВНЫМ ПРАВИТЕЛВСТВОМ США
Данное изобретение разработано при поддержке Правительства США в рамках гранта №1444514 от Национального научного фонда США. Правительство США имеет определенные права на данное изобретение.
ССЫЛКА НА ПЕРЕЧЕНВ ПОСЛЕДОВАТЕЛВНОСТЕЙ
Перечень последовательностей, поданный в виде текстового файла, озаглавленного «UGA 2020 139 03 РСТ ST25.txt», созданного 7 июня 2021 года и имеющего размер 20835 байт, настоящим включен в данный документ посредством ссылки согласно § 1.52(e)(5) Раздела 37 Свода федеральных правил США.
ОБЛАСТВ ТЕХНИКИ
Область техники данного изобретения в целом относится к линиям растений - индукторов гаплоидов и способам их применения для создания генетически модифицированных растений - удвоенных гаплоидов.
УРОВЕНВ ТЕХНИКИ
Селекционные компании по всему миру ежегодно создают десятки тысяч гаплоидных линий кукурузы в качестве предшественников для создания новых инбридов, которые в конечном итоге используются для создания гибридов для продажи. Индуцированные гаплоиды удваивают путем химической обработки и немедленно исследуют на предмет агрономической
производительности. Способствующей этому технологией стало открытие инбридного сорта кукурузы под названием Stock 6, который индуцирует гаплоиды при скрещивании в качестве растения мужского пола (Сое, et al., The American Naturalist, 93 (873): 381-82 (1959)). Около 3% потомства от этого скрещивания являются гаплоидными по материнскому геному. Теперь данная линия улучшена и отобрана для улучшенного формирования гаплоидов, которые могут составлять от 3-20% (Hu et al., Genetics, 202 (4): 1267-76 (2016); Prigge et al., Genetics, 190 (2): 781-93 (2012)). Bee способы индукции гаплоидов кукурузы происходят от данного основополагающего открытия и выведенным на его основе селекционным линиям. Соответствующая литература по данной теме была тщательно проанализирована (Chaikam et al., TAG. Theoretical and Applied Genetics. Theoretische Und Angewandte Genetik, 132 (12): 3227-43 (2019); Comai and Tan, Trends in Genetics. doi:10.1016/j.tig.2019.07.005 (2019); Kalinowska et al., TAG. Theoretical and Applied Genetics. Theoretische Und Angewandte Genetik, 132 (3): 593-605 (2019)). Ген, ответственный за индукцию гаплоидов на основе Stock 6 (Matrilineal, или mat1), представляет собой пататин-подобную фосфолипазу, экспрессируемую главным образом в пыльце (Kelliher et al., Nature, 542 (7639): 105-9 (2017); Liu et al., Molecular Plant, 10 (3): 520-22 (2017); Gilles et al., The EMBO Journal, 36 (6): 707-17 (2017)). Механизм его действия не понятен, но может включать в себя изменение свойств мембраны во время оплодотворения, что приводит к внезапной потере отцовского генотипа. Мутации в mat1 также индуцируют гаплоиды у риса (Yao et al., Nature Plants, 4 (8): 530-33 (2018)), указывая на то, что данный способ индукции гаплоидов можно широко применять для однодольных видов сельскохозяйственных культур.
Индукция гаплоидов сама по себе представляет широкий интерес. Также важна последующая демонстрация того, что mat1 можно применять для «невидимой» передачи кассеты CRISPR/Cas9 в любой инбридный фон (Kelliher et al., Nature Biotechnology, 37 (3):287-92 (2019)) и редактировать гены у больше чем 3% гаплоидного потомства. Полученные в результате гаплоиды затем можно удваивать для создания инбридов с гомозиготными мутациями быстрым способом без ГМО. Любой другой способ редактирования генов требует трансформации и регенерации; процесс, который сильно зависит от генотипа. Путем применения генотип-независимого индуктора гаплоидов данного лимитирующего процесса можно избежать. Однако данный способ является трудоемким, поскольку для него необходимо создать мутацию в mat1, прежде чем начинать редактирование гена в другом сайте. К тому же линии, содержащие мутацию mat1, нездоровы и их трудно размножать. Кроме того, поскольку механизм действия mat1 неизвестен, неясно, как Cas9 способен получить доступ к материнскому геному. Поскольку данный механизм неизвестен, усовершенствовать данную технологию сложно.
Другим способом индукции гаплоидов является опосредованная центромерой индукция гаплоидов (Ravi and Chan (Ravi, et al., Genetics, 186 (2): 461-71 (2010), Ravi, et al., PLoS Genet., 7 (6): e1002121 (2011) doi: 10.1371/journal.pgen.1002121, Ravi et al., Nature, 464 (7288): 615-18 (2011)). Было обнаружено, что нулевой мутант арабидопсиса cenh3-/-, дополненный модифицированной версией CENH3, называемой «GFP-tai1swap», может индуцировать отцовские гаплоиды с высокой частотой (~25%). GFP-tailswap представляет собой сложный трансген с заменой в гистоновом хвосте и большим фрагментом GFP, добавленным к N-концу. Другие формы tailswap, включающие в себя гены CENH3 от разных видов, с GFP или без него (Britt and Kuppu, Frontiers in Plant Science, 1 (April): 357, (2016)), и точечные мутации, которые приводят к изменениям одной аминокислоты в CENH3, также могут индуцировать гаплоиды с более низкой частотой (Karimi-Ashtiyani et al., PNAS, 112 (36): 11211-16 (2015); Kuppu et al., PLoS Genetics, 11 (9): e1005494 (2015), Kalinowska et al., TAG. Theoretical and Applied Genetics. Theoretische Und Angewandte Genetik, 132 (3): 593-605 (2019)).
Опосредованная центромерой индукция гаплоидов также была успешной у ряда других видов двудольных растений (Kalinowska et al., TAG. Theoretical and Applied Genetics. Theoretische Und Angewandte Genetik, 132 (3): 593-605 (2019)). Результаты у однодольных растений ограничены и ненадежны (Kelliher et al., Frontiers in Plant Science, 1 (March): 414 (2016)), что приводит к общему мнению о том, что для однодольных растений применяют систему Matrilineal, а для двудольных - опосредованную центромерой гаплоидную индукцию (Kalinowska et al., TAG. Theoretical and Applied Genetics. Theoretische Und Angewandte Genetik, 132 (3): 593-605 (2019)).
Следовательно, сохраняется потребность в улучшенных композициях и способах для индукции гаплоидов у однодольных растений и образованных из них гаплоидов, необязательно - в сочетании с одновременным редактированием генов для индукции одной или большего числа мутаций относительно фонового генома.
КРАТКОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Представлены однодольные растения, гетерозиготные по центромерному гистону 3 (CenH3), и их применение в способах эффективной опосредованной центромерой индукции гаплоидов у целевого растения. Однодольные растения - индукторы гаплоидов, как правило, состоят из диплоидных растительных клеток, имеющих только одну аллель CENH3, которая полностью функциональна. Диплоидные растительные клетки могут также включать в себя, например, одну аллель CenH3, кодирующую нефункциональный белок CENH3. В некоторых вариантах осуществления аллель, кодирующая нефункциональный белок CENH3, представляет собой белок-нулевую аллель, РНК-нулевую аллель или их комбинацию. В некоторых вариантах осуществления эндогенный локус CenH3 на первой диплоидной хромосоме мутирован или частично или полностью удален. В некоторых вариантах осуществления мутация представляет собой мутацию со сдвигом рамки, которая вводит стоп-кодон, заставляя ген экспрессировать усеченный нефункциональный белок. Как правило, эндогенные локусы CenH3 на второй (т.е. другой) диплоидной хромосоме остаются неповрежденными. Функциональный белок CENH3 может представлять собой белок CENH3 дикого типа.
Как правило, у растения отсутствует хромосомно интегрированный или внехромосомный трансген, кодирующий CenH3 дикого типа, варианты белка CENH3 и слитые белки. Следовательно, в растении, как правило, отсутствует конструкция, кодирующая слитый белок CENH3 - зеленый флуоресцентный белок (GFP), такая как GFP-tai1swap.В некоторых вариантах осуществления нулевую cenh3 используют отдельно или в комбинации с другими технологиями, использующими гаплоиды, такими как синтетический апомиксис или перенос сконструированных хромосом из одной линии в другую.
Однодольное растение - индуктор гаплоидов также необязательно может включать в себя механизм редактирования генов, такой как сайт-направленная нуклеаза и, необязательно, гидовая РНК, стабильно экспрессируемая клетками данного однодольного растения. Как правило, конструкции, экспрессирующие механизм редактирования генов (например, нуклеазу и, необязательно, гидовую РНК), стабильно экспрессируются однодольным растением. В некоторых вариантах осуществления сайт-направленная нуклеаза представляет собой систему на основе CRISPR, нуклеазу на основе эффектора, подобного активатору транскрипции (TALEN), или нуклеазу с «цинковыми пальцами» (ZFN), которые можно применять в виде цитидиндезаминазных или адениндезаминазных слитых белков. В некоторых вариантах осуществления в геном растения - индуктора гаплоидов интегрирована гетерологичная конструкция нуклеиновой кислоты, кодирующая нуклеазу.
В некоторых вариантах осуществления геном однодольного растения - индуктора гаплоидов включает в себя последовательность донорной нуклеиновой кислоты, которая вводится в геном целевого растения путем направляемой гомологией репарации (НГР) после расщепления нуклеазой.
Также представлены яйцеклетки и спермии, образованные растениями - индукторами гаплоидов, лишенные одной аллели, кодирующей функциональный белок CENH3, и экспрессирующие механизм редактирования генов. В некоторых вариантах осуществления яйцеклетки содержат не больше чем около 12,5% функционального белка CENH3 по сравнению с соответствующей яйцеклеткой, образованной растением, гомозиготным по CenH3. В некоторых вариантах осуществления клетки спермиев содержат не больше чем около 25% функционального белка CENH3 по сравнению с соответствующей клеткой спермия, образованной растением, гомозиготным по CenH3.
Также представлены способы индукции образования целевого однодольного гаплоидного растения. Такие способы, как правило, включают в себя опыление целевого родительского однодольного растения пыльцой от однодольного растения - индуктора гаплоидов или опыление однодольного растения - индуктора гаплоидов пыльцой от целевого родительского однодольного растения. Гаплоиды индуцируются, когда яйцеклетка или пыльца, несущие аллель cenh3 (т.е. нулевую аллель), с разбавленным количеством CENH3, оплодотворяются пыльцой от линии дикого типа. Затем отбирают гаплоидное потомство, полученное в результате опыления.
Также представлены способы модификации генома целевого однодольного растения. Такие способы, как правило, включают в себя индукцию образования целевого гаплоидного однодольного растения, экспрессирующего механизм редактирования генов, отбор гаплоидного потомства с геномом целевого однодольного растения, но не однодольного растения - индуктора гаплоидов, и при этом геном гаплоидного потомства модифицирован сайт-направленной нуклеазой и, необязательно, по меньшей мере одной гидовой РНК, доставляемой однодольным растением - индуктором гаплоидов.
Любой из данных способов может дополнительно включать в себя дополнительные этапы, например, удвоение хромосом выбранного гаплоидного потомства. Удвоение хромосом может быть спонтанным или индуцированным, например, индуцированным агентом для удвоения хромосом, необязательно выбранным из колхицина, пронамида, дитипира, трифлуралина или другого агента против микротрубочек.
Однодольное растение - индуктор гаплоидов может представлять собой, например, кукурузу, пшеницу, рис, сорго, ячмень, овес, тритикале, рожь, просо африканское, просо пальчатое, просо обыкновенное, просо итальянское, банан, бамбук, сахарный тростник, просо прутьевидное, мискантус, аспарагус, лук, чеснок, лук скороду или ямс. В предпочтительном варианте осуществления растение-индуктор представляет собой кукурузу.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На фиг.1А-1С проиллюстрировано создание в кукурузе мутации cenh3-нулъ с помощью CRISPR/Cas9. Фиг. 1А представляет собой схематическое изображение, показывающее конструкции векторов для редактирования генома у кукурузы. Ubi-Cas9 включает в себя кодон-оптимизированный Cas9, управляемый промотором полиубиквитина кукурузы. гРНК-ImmuneCENH3 включает в себя гидовую РНК (гРНК), нацеленную на четвертый экзон Cenh3, и нерасщепляемый ген ImmuneCenH3, управляемый нативным промотором Cenh3 длиной 2,1 тыс. п.о. TailswapCENH3 основан на ImmuneCENH3, но включает в себя модифицированный N-концевой хвост и GFP-метку. Фиг. 1В представляет собой схематическое изображение геномной структуры гена CENH3. Экзоны показаны рамками. Также показаны мотив, примыкающий к протоспейсеру (МПП), и целевая последовательность единой гидовой РНК (егРНК) длиной в 20 п.о. (SEQ ID NOS: 16), выровненные с комплементарной последовательностью/гибридизующимся сегментом егРНК (SEQ ID NO: 17). Фиг. 1С представляет собой хроматограмму последовательности из гетерозиготной линии, показывающую сдвиг рамки в мутации cenh3-нуль (SEQ ID NO: 18-19). Проиллюстрированы МПП, делеция и стоп-кодон.
На фиг.2A-2F проиллюстрированы результаты анализов, подтверждающие, что растения являются гаплоидами. Фиг. 2А и 2В представляют собой графики, отображающие анализ диплоидов и гаплоидов способом проточной цитометрии. Диплоидные растения (2А) демонстрируют пики при 2N и 4N, где 4N является результатом эндоредупликации в дифференцированных тканях. Гаплоидные растения имеют пики 1N и 2N (2В). Фиг. 2С и 2D представляют собой изображения хромосомных препаратов. Диплоиды кукурузы имеют 20 хромосом (2С), в то время как гаплоиды имеют 10 (2D). Фиг. 2Е и 2F представляют собой изображения растений: гаплоидные растения являются более низкими (2Е) и стерильными, без выступающих пыльников (2F).
Фиг. 3А и 3В представляют собой графики, иллюстрирующие молекулярные кариотипы анеуплоидов. На обеих панелях хромосомы показаны конец к концу в верхней части. На фиг.3А показаны анеуплоиды, полученные от скрещиваний g18. Анеуплоид_1 является трисомными по хромосоме 3, а анеуплоид_2 является моносомным по хромосомам 2 и 4, и трисомным по хромосоме 10. На фиг.3В показаны анеуплоиды, полученные от скрещиваний g11. Анеуплоид_3 и анеуплоид_4 являются моносомными по хромосоме 7, анеуплоид_5 является моносомным по хромосомам 3, 6 и 7, анеуплоид 6 является моносомным по всем хромосомам, кроме хромосомы 1, и анеуплоид_7 является моносомным по хромосоме 9. Охват в каждом образце был нормализован относительно охвата в соответствующем диплоиде от каждого скрещивания.
Фиг. 4А представляет собой диаграмму последовательности этапов, показывающую неудачную стратегию создания линии-индуктора Tailswap-CENH3 у кукурузы. ImmuneCenH3 заменяли версией конструкции GFP-tailswap для кукурузы и несколькими другими вариантами по Cenh3, чтобы посмотреть, будут ли данные линии индуцировать гаплоиды. Ни один из данных вариантов по CenH3 не дополнял нулевую аллель («неудача»). Фиг. 4В представляет собой диаграмму последовательности этапов, показывающую распределение CENH3 во время развития женской гаметы. В женском гаметофите происходит три клеточных деления, которые предшествуют образованию яйцеклетки. Яйцеклетка, несущая cenh3-нуль, может содержать не больше чем 12,5% белка CENH3.
На фиг.5A-5D проиллюстрировано применение cenh3-нуль в одновременной индукции гаплоидов и редактировании генов. Фиг. 5А представляет собой плазмидную карту конструкции CRISPR, применяемой для одновременной индукции гаплоидов и редактирования генов. Компоненты конструкций указаны. Фиг. 5В и 5С представляют собой фотографии фенотипа дикого типа (ДТ) (5В) и фенотипа «Young ear» (fea2) (5С) (адаптировано из работы Taguchi-Shiobara, et al., Genes Dev. 15: 2755-2766 (2001), в которой фенотип мутанта fea2 был описан впервые). Фиг. 5D представляет собой фотографию фенотипа «Young ear» у гаплоидного растения с отредактированным генотипом fea2, полученного с помощью cenh3-индуктора гаплоидов.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
I. Определения
Термин «около» предназначен для описания значений либо выше, либо ниже заявленного значения, в диапазоне приблизительно +/- 10%. Диапазоны предназначены для того, чтобы быть понятными в контексте, и никаких дополнительных ограничений не подразумевается. Употребление в контексте данного документа любых без исключения примеров или иллюстративных формулировок (например, «такой, как») предназначено только для лучшего освещения данного описания и не налагает ограничения на объем данного описания, если не заявлено иное.
Термин «растение» употребляется в наиболее широком смысле. Он включает в себя, но не ограничивается ими, любые виды древесных, орнаментальных или декоративных культур или злаков, а также фруктовых или овощных растений. Он также относится к множеству растительных клеток, которые в значительной степени дифференцированы в структуру, присутствующую на любой стадии развития растения. Такие структуры включают в себя, но не ограничиваются ими, плод, побег, стебель, лист, лепесток цветка ит.д.
Термин «растительная ткань» включает в себя дифференцированные и недифференцированные ткани растений, включительно с теми, которые присутствуют в корнях, побегах, листьях, пыльце, семенах и опухолях, а также клетки в культуре (например, одиночные клетки, протопласты, эмбрионы, каллюс и т.д.). Растительная ткань может находиться in planta, в культуре органов, культуре тканей или культуре клеток. При употреблении в контексте данного документа термин «часть растения» относится к структуре растения, органу растения или ткани растения.
Термин «растительный материал» относится к листьям, стеблям, корням, цветкам или цветочным частям, плодам, пыльце, яйцеклеткам, зиготам, семенам, черенкам, культурам клеток или тканей, или любой другой части или продукту растения.
Термин «орган растения» относится к отчетливо структурированной и дифференцированной части растения, такой как корень, стебель, лист, бутон цветка или эмбрион.
Термин «растительная клетка» относится к структурной и физиологической единице растения, содержащей протопласт и клеточную стенку. Растительная клетка может быть в виде выделенной одиночной клетки или культивируемой клетки, или как часть более высокоорганизованной единицы, такой как, например, растительная ткань, орган растения или целое растение.
Термин «культура растительных клеток» относится к культурам растительных единиц, таких как, например, протопласты, клетки клеточной культуры, клетки в тканях растений, пыльца, пыльцевые трубки, яйцеклетки, зародышевые мешки, зиготы и эмбрионы на различных стадиях развития.
Термин «трансгенное растение» относится к растению или дереву, которое содержит рекомбинантный генетический материал, обычно не встречающийся в растениях или деревьях данного типа, и который был введен в рассматриваемое растение (или в предшественников данного растения) путем манипуляций человека. Следовательно, растение, выращенное из растительной клетки, в которую путем трансформации введена рекомбинантная ДНК, является трансгенным растением, как и все потомки этого растения, которые содержат введенный трансген (независимо от того, получены они половым или бесполым путем). Следует понимать, что термин «трансгенное растение» охватывает все растение или дерево и части растения или дерева, например, зерна, семена, цветы, листья, корни, плоды, пыльцу, стебли ит.д.
Термин «конструкция» относится к рекомбинантной генетической молекуле, имеющей одну или большее число выделенных полинуклеотидных последовательностей. Генетические конструкции, используемые для экспрессии трансгена в организме хозяина, включают в себя, в направлении от 5' к 3': промоторную последовательность; последовательность, кодирующую ген, представляющий интерес; и последовательность терминации. Конструкция может также включать в себя селектируемый (-е) маркерный (-е) ген (-ы) и другие регуляторные элементы для экспрессии.
Термин «ген» относится к последовательности ДНК, которая кодирует через свою матрицу или информационную РНК последовательность аминокислот, характерных для конкретного пептида, полипептида или белка. Термин «ген» также относится к последовательности ДНК, которая кодирует РНК-продукт. Термин «ген» при употреблении по отношению к геномной ДНК включает в себя промежуточные, некодирующие области, а также регуляторные области, и может включать в себя 5'- и 3'-концы.
Термин «ортологичные гены», или «ортологи», относится к генам, которые имеют сходную последовательность нуклеиновых кислот, поскольку они были разделены видообразующим событием.
Термин «полипептид» в общем относится к пептидам и белкам, содержащим больше чем около десяти аминокислот. Полипептиды могут быть «экзогенными», что означает, что они являются «гетерологичными», т.е. чужеродными для используемой клетки-хозяина, такими как полипептид человека, продуцируемый бактериальной клеткой.
Термин «выделенное (-ый, -ая, -ые)» предназначен для описания соединения, представляющего интерес (например, нуклеиновых кислот), которое находится в среде, отличной от той, в которой данное соединение встречается в природе, например, отделено от его естественной окружающей среды, например, путем концентрирования пептида до концентрации, в которой он не встречается в природе. «Выделенное (-ый, -ая, -ые)» означает включающее (-ий, -ая, -ие) в себя соединения, находящиеся в образцах, существенно обогащенных в отношении соединения, представляющего интерес, и/или в образцах, в которых соединение, представляющее интерес, является частично очищенным или по существу очищенным. Выделенные нуклеиновые кислоты по меньшей мере на 60% свободны, предпочтительно - на 75% свободны, и наиболее предпочтительно - на 90% свободны от других ассоциированных с ними компонентов. «Выделенная» молекула нуклеиновой кислоты или полинуклеотида представляет собой молекулу нуклеиновой кислоты, которая идентифицирована и отделена по меньшей мере от одной загрязняющей молекулы нуклеиновой кислоты, с которой она обычно ассоциирована в природном источнике. Выделенная нуклеиновая кислота может быть, например, свободна от ассоциации со всеми компонентами, с которыми она естественным образом ассоциирована. Выделенная молекула нуклеиновой кислоты отлична от той формы или окружения, в которых она встречается в природе.
Термин «локус» относится к определенному положению вдоль хромосомы или последовательности ДНК. В зависимости от контекста локусом может быть ген, маркер, хромосомная полоса или определенная последовательность из одного или большего числа нуклеотидов.
Термин «аллель» относится к одной из двух или большего числа альтернативных форм гена.
Термин «вектор» относится к репликону, такому как плазмида, фаг или космида, в который может быть вставлен другой сегмент ДНК, чтобы вызвать репликацию данного вставленного сегмента. Векторы могут быть векторами экспрессии.
Термин «вектор экспрессии» относится к вектору, который включает в себя одну или большее число последовательностей контроля экспрессии.
Термин «последовательность контроля экспрессии» относится к последовательности ДНК, которая контролирует и регулирует транскрипцию и/или трансляцию другой последовательности ДНК. Последовательности контроля экспрессии, которые подходят для прокариот, например, включают в себя промотор, необязательно -последовательность оператора, сайт связывания с рибосомой и тому подобное. Известно, что эукариотические клетки используют промоторы, сигналы полиаденилирования и энхансеры.
Термин «промотор» относится к регуляторной последовательности нуклеиновой кислоты, как правило, расположенной «выше» (в направлении к 5') от гена или последовательности, кодирующей белок, которая в сочетании с различными элементами отвечает за регулирование экспрессии гена или последовательности, кодирующей белок. Промоторы, подходящие для использования в конструкциях согласно данному изобретению, являются функциональными в растениях и в организмах-хозяевах, используемых для экспрессии представленных в данном изобретении полинуклеотидов. Многие растительные промоторы широко известны. К ним относятся конститутивные промоторы, индуцируемые промоторы, ткане- и клеточноспецифические промоторы, и промоторы, регулируемые развитием. Иллюстративные промоторы и слитые промоторы описаны, например, в патенте США №6717 034, который включен в данный документ посредством ссылки во всей своей полноте.
Последовательность нуклеиновой кислоты или полинуклеотид являются «функционально связанными», когда они находятся в функциональной взаимосвязи с другой последовательностью нуклеиновой кислоты. Например, ДНК для предпоследовательности или секреторного лидера функционально связана с ДНК для полипептида, если она экспрессируется в виде препротеина, который участвует в секреции данного полипептида; промотор или энхансер функционально связаны с кодирующей последовательностью, если они влияют на транскрипцию данной последовательности; или сайт связывания рибосомы функционально связан с кодирующей последовательностью, если он расположен так, чтобы способствовать трансляции. В целом, «функционально связанные» означает, что связанные последовательности ДНК являются смежными и, в случае секреторного лидера, смежными и в рамке считывания. Связывания можно достичь путем лигирования в удобных сайтах рестрикции. Если такие сайты не существуют, в соответствии с общепринятой практикой применяют синтетические олигонуклеотидные адаптеры или линкеры.
Термины «трансформированный», «трансгенный», «трансфицированный» и «рекомбинантный» относятся к организму-хозяину, такому как бактерия или растение, в который была введена гетерологичная молекула нуклеиновой кислоты. Молекула нуклеиновой кислоты может быть стабильно интегрирована в геном хозяина или молекула нуклеиновой кислоты может также присутствовать в качестве внехромосомной молекулы. Такая внехромосомная молекула может быть аутореплицирующейся. Под трансформированными клетками, тканями или растениями понимают не только конечный продукт процесса трансформации, но и их трансгенное потомство. «Нетрансформированный», «нетрансгенный» или «нерекомбинантный» хозяин относится к организму дикого типа, например, бактерии или растению, который не содержит гетерологичную молекулу нуклеиновой кислоты.
Термин «эндогенная» в отношении нуклеиновой кислоты относится к нуклеиновым кислотам, обычно присутствующим в организме хозяина.
Термин «гетерологичный (-ая, -ое, -ые)» относится к элементам, встречающимся там, где они обычно не встречаются. Например, промотор может быть связан с гетерологичной последовательностью нуклеиновой кислоты, например, последовательностью, которая обычно не обнаруживается функционально связанной с данным промотором. При употреблении в контексте данного документа для описания промоторного элемента «гетерологичный» означает промоторный элемент, который отличается от того, который обычно обнаруживается в нативном промоторе, по последовательности, виду или количеству. Например, гетерологичный элемент контроля в последовательности промотора может быть контролирующим/регуляторным элементом другого промотора, добавленного для усиления промоторного контроля, или дополнительным контролирующим элементом того же промотора. Следовательно, термин «гетерологичный» может также охватывать «экзогенные» и «не нативные» элементы.
При употреблении в контексте данного документа термин «гомологичный» означает полученный из того же вида. Например, гомологичный признак - это любая характеристика организмов, происходящая от их общего предка. Гомологичные последовательности могут быть ортологичными или паралогичными. Гомологичные последовательности являются ортологичными, если они были разделены видообразующим событием: когда один вид расходится на два отдельных вида, расходящиеся копии одного гена в образованном в результате этого виде считаются ортологичными. Ортологи, или ортологичные гены, представляют собой гены у разных видов, которые похожи друг на друга, потому что они произошли от общего предка. Гомологичные последовательности являются паралогичными, если они были разделены событием дупликации гена: если ген в организме дублируется, занимая два разных положения в одном и том же геноме, то такие две копии являются паралогичными.
При употреблении в контексте данного документа термин «полипептид» в общем относится к пептидам и белкам, содержащим больше чем около десяти аминокислот. Полипептиды могут быть «экзогенными», что означает, что они являются «гетерологичными», т.е. чужеродными для используемой клетки-хозяина, такими как полипептид человека, продуцируемый бактериальной клеткой.
При употреблении в контексте данного документа «культивар» относится к культивируемому сорту.
При употреблении в контексте данного документа «гермоплазма» относится к одной или большему числу фенотипических характеристик, или к одному или большему числу генов, кодирующих указанные одну или большее число фенотипических характеристик, способным передаваться между поколениями.
При употреблении в контексте данного документа термин «прародитель» относится к любому из видов, сортов, культиваров или гермоплазме, из которых получено растение.
При употреблении в контексте данного документа термин «производный вид, производная гермоплазма или производный сорт» относится к любому виду растений, гермоплазме или сорту, которые получены с использованием указанного вида, сорта, культивара или гермоплазмы с помощью стандартных процедур половой гибридизации, технологии рекомбинантной ДНК, культуры ткани, мутагенеза или комбинации любого одного или большего числа из указанных процедур.
При употреблении в контексте данного документа термины «интрогрессия», «интрогрессированный» и «интрогрессировать» относятся как к естественному, так и к искусственному процессу, посредством которого гены одного вида, сорта или культивара перемещаются в геном другого вида, сорта или культивара путем скрещивания данных видов. Данный процесс необязательно может завершаться обратным скрещиванием к повторяющемуся родителю.
При употреблении в контексте данного документа термин «часть растения», или «растительная часть», может включать в себя, но не ограничивается ими, черенки, клетки, протопласты, культуры клеточной ткани, каллюс (каллюсы), скопления клеток, эмбрионы, тычинки, пыльцу, пыльники, пестики, семяпочки, цветки, семена, лепестки, листья, стебли и корни.
При употреблении в контексте данного документа «гибрид», как правило, происходит от одного или большего числа скрещиваний между разными сортами, гермоплазмами, популяциями, породами или культиварами в пределах одного вида, между разными подвидами в пределах вида или между разными видами в пределах рода. Как правило, гибриды между подвидами называются «внутривидовыми гибридами», а гибриды между разными видами в пределах рода называются «межвидовыми гибридами».
Приведенные в данном документе диапазоны значений предназначены только для того, чтобы служить сокращенным способом индивидуальной ссылки на каждое отдельное значение, попадающее в данный диапазон, если в данном документе не указано иное, и каждое отдельное значение включено в данный документ, как если бы оно было указано в данном документе индивидуально.
Применение термина «около» предназначено для описания значений либо выше, либо ниже заявленного значения, в диапазоне приблизительно +/- 10%; в других формах значения могут варьировать по значению либо выше, либо ниже заявленного значения, в диапазоне приблизительно +/- 5%; в других формах значения могут варьировать по значению либо выше, либо ниже заявленного значения, в диапазоне приблизительно +/- 2%; в других формах значения могут варьировать по значению либо выше, либо ниже заявленного значения, в диапазоне приблизительно +/-1%. Предшествующие диапазоны предназначены для того, чтобы быть понятными в контексте, и никаких дополнительных ограничений не подразумевается.
Представлены материалы, композиции и компоненты, которые можно применять для, можно применять в сочетании с, можно применять в процессе описанных в данном документе способе и композициях, или которые являются продуктами описанных в данном документе способа и композиций. Эти и другие материалы описаны в данном документе, и следует понимать, что когда описываются комбинации, подмножества, взаимодействия, группы и т.д. этих материалов, конкретно предусматривается и описывается в данном документе каждая (-ый) из них, хотя конкретные ссылки на каждую из различных индивидуальных и коллективных комбинаций и перестановок этих соединений могут не быть описаны явно. Например, если описываются и обсуждаются лиганд и ряд модификаций, которые могут быть внесены в ряд молекул, включая данный лиганд, конкретно предусматриваются каждая комбинация, перестановка лиганда и возможные модификации, если специально не указано иное. Следовательно, если описан класс молекул А, В и С, а также класс молекул D, Е и F, и описан пример комбинированной молекулы A-D, то даже если каждая из них не указана по отдельности, каждая предусмотрена индивидуально и коллективно. Следовательно, в данном примере каждая из комбинаций А-Е, A-F, B-D, В-Е, B-F, C-D, С-Е и C-F конкретно предусмотрена и должна считаться описанной в описании А, В и С; D, Е и F; и иллюстративной комбинации A-D. Подобным образом, любое их подмножество или комбинация из них также конкретно предусматриваются и описываются. Следовательно, например, подгруппы А-Е, B-F и С-Е конкретно предусмотрены и должны считаться описанными в описании А, В и С; D, Е и F; и иллюстративной комбинации A-D. Кроме того, каждый из материалов, композиций, компонентов и т.д., предусматриваемых и описанных, как указано выше, также может быть конкретно и независимо включен или исключен из любой группы, подгруппы, перечня, набора ит.д. таких материалов.
Данные концепции применимы ко всем аспектам данной заявки, включая, но не ограничиваясь ими, этапы в способах получения и применения композиций, представленных в данном документе. Следовательно, если существует множество дополнительных этапов, которые могут быть выполнены, следует понимать, что каждый из этих дополнительных этапов может быть выполнен с любым конкретным вариантом осуществления или комбинацией вариантов осуществления способов, представленных в данном документе, и что каждая такая комбинация конкретно предусмотрена и должна считаться описанной.
Все способы, описанные в данном документе, могут быть выполнены в любом подходящем порядке, если не указано иное или иное явно не противоречит контексту. Употребление в контексте данного документа любых без исключения примеров или иллюстративных формулировок (например, «такой, как») предназначено только для лучшего освещения вариантов осуществления данного изобретения и не налагает ограничения на объем вариантов осуществления данного изобретения, если не заявлено иное. Ни одна из языковых формулировок в данном документе не должна истолковываться как указывающая на какой-либо не заявленный элемент как на существенный для практики данного изобретения.
II. Растения
Гаплоидные растения широко используются для ускорения производства новых инбридных линий. Индукция гаплоидов включает в себя временное диплоидное состояние с последующей потерей одного из способствующих геномов. Данный механизм также позволяет встраивать механизмы редактирования генов независимым от генотипа способом без стабильной трансформации целевой линии.
В представленных способах используют простую нулевую мутацию гена центромерного гистона Н3. Результаты, представленные в примерах, приведенных ниже, показывают, что растение, гетерозиготное по селл3-нуль, образует гаплоиды с частотой 5% при скрещивании в качестве растения женского пола и 0,5% при скрещивании в качестве растения мужского пола. Механизм индукции гаплоидов включает в себя последовательную потерю хромосом в зиготе. Представленные растения и способы их применения также позволяют встраивать механизмы редактирования генов в любую линию простым, быстрым и не требующим ГМО способом.
Номенклатура генов кукурузы предусматривает, что мутантные аллели обозначают обычным шрифтом или выделенным курсивом не заглавным шрифтом, где первая буква является прописной (например, cenh3), гены дикого типа обозначают обычным шрифтом или выделенным курсивом шрифтом, где первая буква является заглавной (например, CenH3), а экспрессируемый белковый продукт обозначают всеми заглавными буквами (CENH3). Поскольку эксперименты, описанные ниже, проводили на кукурузе, в данном документе, как правило, употребляется номенклатура, характерная для кукурузы. Следует, однако, понимать, что композиции и способы, описанные в данном документе, также применимы к соответствующему гену в других однодольных растениях и, следовательно, также описаны для такого соответствующего гена, поэтому использование данной номенклатуры не должно толковаться как ограничение описанных композиций и способов только кукурузой.
Описанные в данном документе способы индукции гаплоидов, как правило, включают в себя скрещивание линии - индуктора гаплоидов и целевой линии, подлежащей индукции для создания гаплоидов.
Описанные в данном документе способы редактирования генов, как правило, включают в себя скрещивание линии - индуктора гаплоидов, включающей в себя механизм редактирования генов, и целевой линии, подлежащей индукции для создания гаплоидов, которая также является целевой для модификации генов. Процесс индукции гаплоидов in vivo разработан для введения механизма редактирования в целевую гермоплазму путем включения его в родительский индуктор гаплоидов. Как правило, механизм редактирования стабильно интегрирован в качестве трансгена. Одновременное редактирование плюс гаплоидную индукцию можно выполнять у различных однодольных растений с помощью широкого скрещивания или гаплоидной индукции de novo.
Как правило, одно или большее число растений, используемых в скрещиваниях, описанных в данном документе, и/или полученное от них потомство, являются растениями, не встречающимися в природе. «Растение, не встречающееся в природе» относится к растению, которое без вмешательства человека не встречается в природе. Растения, не встречающиеся в природе, включают в себя трансгенные растения, а также растения, полученные нетрансгенными способами, такими как селекция растений.
А. Целевые линии
Целевая линия, подлежащая индукции, как правило, является однодольным растением. Однодольные растения включают в себя одну из крупных таксономических единиц покрытосеменных растении (цветущие растения с семенами, защищенными внутри сосуда). Они являются травянистыми растениями, имеющими листья с параллельным жилкованием, и имеют зародыш с одной семядолей, в отличие от двудольных растений (Двудольные), которые имеют зародыш с двумя семядолями. В некоторых вариантах осуществления целевая линия представляет собой однодольное растение, выбранное из кукурузы, пшеницы, риса, сорго, ячменя, овса, тритикале, ржи, проса африканского, проса пальчатого, проса обыкновенного, проса итальянского, банана, бамбука, сахарного тростника, проса прутьевидного, мискантуса, аспарагуса, лука, чеснока, лука скороды или ямса.
Целевой линией, как правило, является та, в которой желательно наличие гаплоидного растения, как правило, для ускорения производства новых инбридных линий. В некоторых вариантах осуществления целевая линия представляет собой элитную инбридную линию, в которой уже была проведена обширная селекция, но необходимы генетические модификации для улучшения линии, чтобы, например, она была устойчива к болезням или вредителям, или лучше адаптировалась к различным условиям окружающей среды.
В. Линии-индукторы
Представлены линии растений - индукторов гаплоидов, необязательно экспрессирующие механизм редактирования генов. Линия-индуктор также является однодольной, например, представляет собой кукурузу, пшеницу, рис, сорго, ячмень, овес, тритикале, рожь, просо африканское, просо пальчатое, просо обыкновенное, просо итальянское, банан, бамбук, сахарный тростник, просо прутьевидное, мискантус, аспарагус, лук, чеснок, лук скороду или ямс.
Как целевая линия, так и линия-индуктор могут быть линиями растений, которые, как правило, размножаются половым путем. В других вариантах осуществления скрещивание представляет собой межвидовое и межродовое гибридное скрещивание родственных видов или родов, которые обычно не размножаются половым путем друг с другом. Данные скрещивания также можно назвать широкими скрещиваниями. У пшеницы, риса, ячменя, капусты и других культур путь к индукции гаплоидов может заключаться в использовании донора пыльцы, который индуцирует гаплоиды посредством широкого скрещивания. Например, можно было бы использовать пыльцу кукурузы на пшенице, пыльцу проса на пшенице, пыльцу ячменя на других видах ячменя или любой другой способ широкого скрещивания.
Как более подробно обсуждается ниже, растение-индуктор является гетерозиготным по cenh3-нуль и необязательно включает в себя механизм редактирования генов.
1. cenh3-нули
Растения-индукторы являются гетерозиготными cenh3-нулями.
Иллюстративные учетные номера GenBank, предоставляющие местоположения генов и последовательности для CenH3 в различных однодольных растениях, включают в себя: кукурузу, AF519807.2; рис, AY438639.1; пшеницу, JF969287.1; ячмень, JF419329.1; банан, КР878235.1, каждый из который включен в данный документ посредством ссылки во всей своей полноте, и предоставляют следующие аминокислотные и нуклеотидные (мРНК/кДНК) последовательности для CENH3:






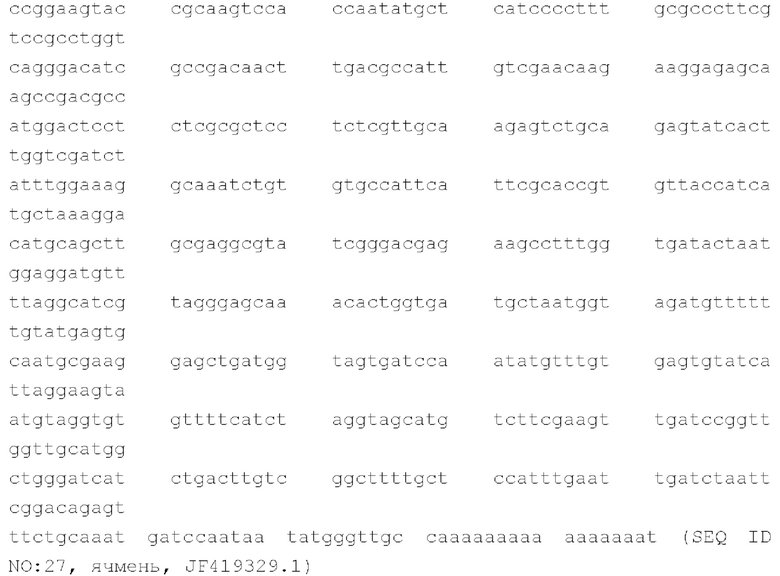

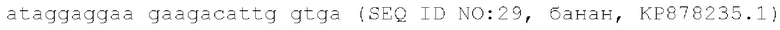
Ген Cenh3 является консервативным во всех растениях, грибах и животных, за редким исключением в некоторых линиях насекомых. Он выполняет принципиально важную роль определения границ функциональной центромеры, а также инициирования и организации кинетохора (Cheeseman and Desai, Nature Reviews Molecular Cell Biology, 9: 33-46 (2008). CENH3 дикого типа может представлять собой CENH3 однодольного растения-индуктора, например, кукурузы, пшеницы, риса, сорго, ячменя, овса, тритикале, ржи, проса африканского, проса пальчатого, проса обыкновенного, проса итальянского, банана, бамбука, сахарного тростника, проса прутьевидного, мискантуса, аспарагуса, лука, чеснока, лука скороды или ямса. В некоторых вариантах осуществления CENH3 дикого типа имеет аминокислотную последовательность любой из последовательностей SEQ ID NO: 20, 22, 24, 26 или 28, или аминокислотную последовательность, которая по меньшей мере на 75%, 80%, 85%, 90%, 95% или большее число процентов идентична им; кодируется нуклеиновой кислотой любой из последовательностей SEQ ID NO: 21, 23, 25, 27 или 29, нуклеиновой кислотой, которая по меньшей мере на 75%, 80%, 85%, 90%, 95% или большее число процентов идентична им; является гомологом, таким как ортолог или паралог, вышеуказанных последовательностей; или любая их комбинация.
Нулевая аллель представляет собой нефункциональную аллель, вызванную генетической мутацией. Такие мутации могут привести к полному отсутствию продуцирования соответствующего генного продукта или к продукту, который не функционирует должным образом; в любом случае, такая аллель считается нефункциональной. Например, белок CENH3 связывается с ДНК и рекрутирует все налагающиеся выше белки, которые образуют кинетохор, опосредующий сегрегацию хромосом в растительной клетке. Нефункциональный CENH3 не будет способствовать образованию центромеры, образованию кинетохора и/или сегрегации хромосом в растительной клетке. Его нулевая аллель, которая не кодирует функциональный белок, отличается от вариантов CENH3, таких как GFP-tailswap, или вариантов, которые продуцируют измененные или частично удаленные белки CENH3. См., например, работу Kuppu, et al., "A Variety of Changes, Including CRISPR/Cas9-mediated Deletions, in CENH3 Lead to Haploid Induction on Outcrossing", Plant Biotechnol J, 2020, doi: 10.1111/pbi.13365, которая конкретным способом включена в данный документ посредством ссылки во всей своей полноте. GFP-tailswap и вариантные формы могут замещать нативный CENH3 и сохранять достаточную функцию для организации кинетохора, пусть и несовершенно. Нулевые аллели представляют собой особую категорию мутаций, которые приводят к полной потере функции.
Мутантная аллель, которая не продуцирует транскрипт РНК, называется РНК-нулевой аллелью (выявляется нозерн-блоттингом, полным секвенированием РНК или секвенированием ДНК делеционной аллели), а та, которая не продуцирует белок, называется белок-нулевой аллелью (выявляется вестерн-блоттингом). Нулевые аллели часто вызываются мутациями со сдвигом рамки. Генетический код считывается в виде триплетов нуклеотидов, так что любая последовательность может быть прочитана в трех рамках, где только одна является верной. Мутации, которые вызывают небольшую делецию или добавление нуклеотидов, могут сместить рамку считывания в сторону бессмысленного белка и часто приводят к преждевременной остановке трансляции белка. Мутации со сдвигом рамки, которые вызывают преждевременную остановку, особенно когда большая часть прогнозируемого белка отсутствует, как правило, интерпретируются как нулевые аллели. Например, мутация со сдвигом рамки, которая приводит к стоп-кодону в N-концевом хвосте CENH3, кодирует слишком усеченный белок, у которого отсутствует способность связываться с ДНК или другими гистонами (фиг.1С). Генетическая нулевая или аморфная аллель имеет тот же фенотип при гомозиготности, что и при гетерозиготности с дефицитом, нарушающим рассматриваемый локус.Генетическая нулевая аллель может быть как белок-нулевой, так и РНК-нулевой, но также может экспрессировать нормальные уровни генного продукта, который нефункционален из-за мутации. Нулевая аллель cenh3 может быть делецией всего локуса или мутацией одного или большего числа нуклеотидов в нем, приводящей к отсутствию продуцирования продукта гена cenh3 или к продукту, который не функционирует должным образом. Нулевая аллель cenh3 может быть белок-нулевой, РНК-нулевой или обеими.
Нулевая аллель описываемых cenh3-нулей, как правило, отличается от конструкций GFP-tailswap и нескольких вариантов Cenh3, таких как те, которые описаны в патенте США №8618354, публикации заявки на патент США №2018/0116141, публикации заявки на патент США №2019/0343060 и в WO 2017/004375, которые создают варианты белков CENH3, являющиеся функциональными, хотя и со сниженной или измененной функцией или уровнем относительно аллели cenh3 дикого типа.
В некоторых вариантах осуществления спермий, несущий нулевой cenh3, содержит не больше чем 25% от нормального количества функционального (например, дикого типа) белка CENH3, а яйцеклетка, несущая нулевой cenh3, содержит больше чем 12,5% функционального (например, дикого типа) белка CENH3.
В некоторых вариантах осуществления гетерозиготная по cenh3-нуль линия-индуктор не включает в себя рекомбинантный ген, экспрессирующий мутантный или вариантный CENH3. Следовательно, в некоторых вариантах осуществления нулевая аллель не дополняется экспрессией неэндогенного CENH3. В некоторых вариантах осуществления количественные сокращения одного только CENH3 индуцируют опосредованную центромерой гаплоидную индукцию.
В некоторых вариантах осуществления единственный функциональный CENH3 в гетерозиготной по cenh3-нуль линии-индукторе экспрессируется из эндогенной аллели дикого типа растения, гетерозиготного по cenh3-нуль.
Неограничивающий способ создания cenh3-нуль описан ниже. Кратко, трансгенную линию, содержащую Cas9, скрещивали с другой трансгенной линией, содержащей гидовую РНК, нацеленную на первый экзон CenH3, а также интактный полноразмерный геномный клон CenH3, который содержит пять молчащих изменений нуклеотидов в сайте гидовой РНК (фиг.1А-1С; нерасщепляемый ген указан как ImmuneCenH3). Когда линии, содержащие данные конструкции, скрещивали вместе, Cas9 вызывал мутацию нативного cenh3, в то время как функция CenH3 обеспечивалась геном ImmuneCenH3. Следовательно, в некоторых вариантах осуществления, нулевая аллель имеет одну или большее число мутаций в первом экзоне, которые сводят у нулю экспрессию функционального белка из данной нулевой аллели. В некоторых вариантах осуществления нулевая аллель вызвана мутацией в последовательности, кодирующей N-концевой хвост, которая удаляет всю последовательность белка CENH3, взаимодействующую с ДНК или другими гистонами.
При каждом делении клетки CENH3 естественным образом разделяется поровну между реплицированными нитями ДНК в S-фазе и пополняется позже в G2 (Lermontova et al., The Plant Journal: For Cell and Molecular Biology, 68 (1): 40-50 (2006)). Это не может произойти в гаплоидной клетке, которая является cenh3-нулевой, и клеточный цикл вынужден протекать с вдвое меньшим количеством CENH3, чем присутствует обычно. В мужском гаметофите (пыльце) происходит два клеточных деления, которые предшествуют образованию спермия, а в женском гаметофите происходит три клеточных деления, которые предшествуют образованию яйцеклетки. Спермий, несущий cenh3-нуль, может содержать не больше чем 25% от нормального количества CENH3, а яйцеклетка, несущая cenh3-нуль, может содержать не больше чем 12,5% (фиг.4В). Результаты также показывают, что 0,5% потомства от растения мужского пола, несущего cenh3-нуль, являются гаплоидами, и 5,0% потомства от растения женского пола являются гаплоидами. Следовательно, индуктор можно применять либо в качестве растения мужского пола для скрещивания, либо в качестве растения женского пола для скрещивания. Растения-индукторы женского пола генерируют более высокий процент гаплоидов, следовательно, в некоторых вариантах осуществления индуктор гаплоидов предпочтительно представляет собой растение женского пола, но, в качестве альтернативы, способы можно выполнять с применением индуктора гаплоидов в качестве растения мужского пола.
2. Маркеры
Индуктор гаплоидов может иметь маркер, который может способствовать идентификации семян, являющихся гаплоидными. Например, индуктор гаплоидов может иметь доминантный ген пурпурного пигмента (например, R1-nj). Семена гаплоидных особей имеют фиолетовый алейрон, но в эндосперме (щитке) фиолетовый пигмент отсутствует, что указывает на то, что данная зародышевая линия не содержит хромосомы индуктора гаплоидов. Семена, которые имеют желтым эндосперм и фиолетовый алейрон, высаживают и выращивают как саженцы. У этих саженцев удваивают число хромосом с помощью колхицина или других способов, которые более подробно обсуждаются ниже. Гаплоиды с удвоенным набором хромосом выращивают в теплице и/или пересаживают в поле, и растения с удвоенным набором хромосом самоопыляют для получения семян растений - удвоенных гаплоидов, как более подробно обсуждается в другой части данного документа.
3. Механизм редактирования генов
Линия-индуктор также необязательно включает в себя механизм редактирования генов. Например, растение-индуктор может иметь закодированный в своей ДНК механизм, необходимый для выполнения редактирования генома целевого растения.
Нацеленный мутагенез (также известный как редактирование генов) является очень важной технологией для селекции сельскохозяйственных культур. В настоящее время существует множество способов редактирования конкретных целевых генов, включительно с CRISPR, TALEN, мегануклеазами и цинковыми пальцами. Эндонуклеазу можно сконструировать так, чтобы она нацеливалась практически на любую последовательность. Эндонуклеазу (-ы) можно сконструировать с использованием таких способов, но не ограничиваясь ими, как способы, которые описаны в работах Svitashev, et al., Plant Physiology, 169: 931-945 (2015), Lee, et al., Plant Biotechnology, 17 (2): 362-372 (2019)), Sander et al., Nature Met, 8 (1): 67-69, (2011), Cermak et al., Nucl Acids Res, 39 (17): 7879 (2011); with correction at Nucl Acids Res, 39:e82. doi: 10.1093/nar/gkr218, 2011); и Liang et al., et al., J Genet Genom, 41 (2): 63-68, (2014). Промотор, используемый для стимулирования экспрессии эндонуклеазы, может представлять собой промотор, который экспрессируется на протяжении всего развития или конкретно в яйцеклетках, или в период раннего развития эмбриона, и может быть эндогенным или экзогенным. Примерам промоторов являются 35S (CaMV d35S) или производные (например, двойной 35S) ZmUb1 (кукуруза), АРХ (рис), OsCc1 (рис) EIF5 (рис), R1G1B (рис), PGD1 (рис), Act1 (рис), SCP1 (рис).
Конструкция (-и) механизма редактирования генов может (могут) включать в себя селектируемый маркер (например, устойчивость к гербицидам) для содействия получению трансгена во время трансформации всего растения и последующего обратного скрещивания. В некоторых случаях одну или большее число (например, две или большее число, или три или большее число) эндонуклеаз и/или гидовых РНК CRISPR объединяют в единую конструкцию для нацеливания на одну или большее число последовательностей ДНК.
Одним из способов введения в растения механизма редактирования генов является применение к растительной ткани способа на основе агробактерий (такого как способ, описанный в работе Ishida et al., Nature Biotechnol, 146: 745-750 (1996)), или бомбардировки частицами (такой как способ, описанный в работе Gordon-Kainin et al., Plant Cell Online, 2 (7): 603-618 (1990)). Разработаны новые способы, охватывающие собой гены -регуляторы развития, которые позволяют трансформировать растения без обширной культуры тканей. См., например, работу Lowe, et al., The Plant Cell, 28: 1998-2015 (2016). При трансформации ДНК, кодирующую механизм редактирования (например, CAS9 и гидовую РНК), вводят в каллюс растения, семена или эмбриональную ткань. Стабильно трансформированные растения (события) затем выделяют, необязательно - с помощью селектируемого маркера.
В качестве альтернативы, линию, поддающуюся трансформации, сначала трансформируют механизмом редактирования генов, а затем полученную линию скрещивают с линией - индуктором гаплоидов. Полученное в результате этого поколение Flf являющееся гетерозиготным по cenh3, само становится индуктором гаплоидов. Дополнительного обратного скрещивания не требуется. В данном случае линия Fi - индуктор гаплоидов содержит трансген эндонуклеазы. Затем линию-индуктор, несущую (например, кодирующую, экспрессирующую и т.д.) механизм редактирования генов, можно опылять вторым растением, подлежащим
редактированию. В результате данного события опыления образуется потомство (например, эмбрионы или семена), по меньшей мере одно из которых будет гаплоидным семенем. Это гаплоидное семя будет содержать только хромосомы второго растения; хромосомы растения-индуктора исчезли (были устранены, потеряны или деградировали), но перед этим хромосомы растения-индуктора позволили экспрессировать механизм редактирования генов.
В качестве альтернативы, и не желая быть связанным теорией, растение-индуктор поставляет уже экспрессированный механизм редактирования при опылении через пыльцевую трубку. Или, в случае, когда линия - индуктор гаплоидов является растением женского пола в скрещивании, яйцеклетка данного растения, индуцирующего гаплоиды, содержит механизм редактирования, который присутствует и, возможно, уже экспрессируется при оплодотворении «диким типом» или пыльцевым зерном растения, не являющегося индуктором гаплоидов. С помощью любого из данных путей гаплоидное потомство, полученное в результате скрещивания, также подвергнется редактированию своего генома. В случае кукурузы, где можно произвести много скрещиваний с одним растением мужского пола, F1, содержащее cenh3-нуль и механизм редактирования генов, можно применять для редактирования нескольких линий путем выполнения нескольких отдельных скрещиваний.
Механизм редактирования генов, как правило, включает в себя элемент или элементы, которые индуцируют одноцепочечный или двухцепочечный разрыв в геноме целевой клетки. Например, механизмом редактирования может быть любой фермент, модифицирующий ДНК, но предпочтительно это сайт-направленная нуклеаза. Сайт-направленная нуклеаза предпочтительно основана на CRISPR, но также может представлять собой мегануклеазу, нуклеазу на основе эффектора, подобного активатору транскрипции (TALEN), или нуклеазу с «цинковыми пальцами». Нуклеаза, например, может представлять собой Cas9 или Cfp1/Cas12a. В одном аспекте нуклеаза предназначена для расщепления ДНК с целью создания небольших делеций или дупликаций в целевом сайте. Возникающие в результате этого небольшие делеций и дупликации могут нарушить функцию гена.
В другом аспекте фермент, модифицирующий ДНК, представляет собой сайт-направленный фермент редактирования оснований, такой как Cas9-цитидиндезаминаза или Cas9-адениндезаминаза, при этом у Cas9 может быть инактивирована одна или обе его нуклеазные активности, т.е. dCas9.
В еще одном варианте осуществления механизм редактирования генов может быть объединен с дополнительной матрицей репарации, так что за расщеплением следует направляемая гомологией репарация (НГР), приводящая к модификации или замещению ДНК в целевом сайте. Назначение индуктора гаплоидов в данном контексте заключается в том, чтобы быстро перенести механизм редактирования генов из трансформируемой линии в любую другую линию без прохождения фазы культивирования тканей или повторного обратного скрещивания.
Механизмы редактирования генов, которые можно применять, более подробно обсуждаются ниже.
а. Элементы, индуцирующие разрыв цепи
i. CRISPR/Cas
В предпочтительных вариантах осуществления элемент, который индуцирует одно- или двухцепочечный разрыв в геноме целевой клетки, представляет собой систему CRISPR/Cas. Как и в других системах на животных моделях, экспрессии Cas9 и егРНК в целевых клетках достаточно для модификации геномов растений (Deepa, et al., Front. Plant Sci., 9:985 (2018), doi:10.3389/fpis.2018.00985. В то время как Cas9 применяется широко, подобным образом можно применять любую систему на основе CRISPR/Cas, например, Cfp1/Cas12a (Tang, et al., Genome Biology, 19 (84) (2018), doi/10.1186/s13059-018-1458-5). Широко используемые промоторы РНК-полимеразы II (такие как 35S или ZmUb1) часто используются для экспрессии генов Cas, но промоторы, экспрессирующиеся в яйцеклетках, могут быть более применимы в данном изобретении. Специфичные для растений промоторы РНК-полимеразы III [AtU6 (арабидопсис); TaU6 (пшеница); OsU6 или OsU3 (рис)] были использованы для экспрессии егРНК в растительных системах. Другие варианты осуществления могут включать в себя мультиплексные системы гидовых РНК, управляемые другими промоторами (Lowder, et al., Plant Physiol., 169 (2): 971-985 (2015), He, et al., J Genet Genomics, 20; 44 (9): 469-472 (2017)). Гены Cas могут быть полностью функциональными и сконструированными для создания двухцепочечных разрывов, которые репарируются путем негомологичного соединения концов (НГСК), что приводит к мутациям, нарушающим функцию гена. В качестве альтернативы, гены Cas можно частично инактивировать, чтобы вызвать одноцепочечные надрезы (например, nCas9), или полностью инактивировать (например, dCas9), чтобы приводить к связыванию, но не расщеплению, и просто направлять другой фермент, такой как аденин- или цитидиндезаминаза, в желаемый сайт (Eid, et al., Biochem J, 475 (11): 1955-1964 (2018)). Существует несколько коммерчески доступных векторов для экспрессии Cas9 или вариантов Cas9 и гРНК в растительных системах, и они включают в себя пустые каркасы гРНК, имеющие промотор РНК-полимеразы III растений, и каркасы гРНК, в которые практик может вставить гРНК, представляющую интерес.Системы на основе CRISPR можно также адаптировать для изменения генов путем направляемой гомологией репарации, как описано ниже. Одним из ограничений является то, что в подходах на основе CRISPR используют последовательности, которые включают в себя короткие мотивы, примыкающие к протоспейсеру (сайты МПП).
Геном растения-индуктора может включать в себя одну или большее число нуклеиновых кислот, кодирующих фермент Cas, и гидовую РНК в качестве компонентов системы CRISPR. Геном растения-индуктора может необязательно включать в себя донорную полинуклеотидную последовательность, подлежащую рекомбинации в геном целевой клетки в целевом сайте или смежно с ним (например, сайт одно- или двухцепочечного разрыва, индуцированного Cas9).
Способы изготовления композиций для применения в редактировании генома с использованием систем CRISPR/Cas подробно описаны, например, в WO 2013/176772, WO 2014/018423, Cong, Science, 15: 339 (6121): 819-823 (2013), и Jinek, et al., Science, 337 (6096): 816-21 (2012).
В общем, «система CRISPR» относится в совокупности к транскриптам и другим элементам, вовлеченным в экспрессию или направляющим активность CRISPR-ассоциированных («Cas») генов, включительно с белком Cas или последовательностями, кодирующими ген Cas, последовательностью tracr (трансактивирующей CRISPR) (например, tracr-PHK или активная частичная tracr-PHK), последовательностью tracr-mate (включающая в себя «прямой повтор» и обработанный tracr-PHK частичный прямой повтор в контексте эндогенной системы CRISPR), гидовой последовательностью (также называемой «спейсером» в контексте эндогенной системы CRISPR), или другими последовательностями и транскриптами из локуса CRISPR. Одна или большее число последовательностей tracr-mate, функционально связанных с гидовой последовательностью (например, прямой повтор - спейсер -прямой повтор), также могут быть указаны как пре-cr-PHK (пре-CRISPR РНК) до процессинга или cr-РНК после процессинга нуклеазой.
В некоторых вариантах осуществления tracr-PHK и cr-РНК связаны и образуют химерный гибрид cr-РНК - tracr-PHK, в котором зрелая cr-РНК слита с частичной tracr-PHK через синтетическую структуру «стебель - петля» для имитации естественного дуплекса cr-РНК: tracr-PHK, как описано в работах Cong, Science, 15: 339 (6121): 819-823 (2013), и Jinek, et al., Science, 337 (6096): 816-21 (2012)). Одиночная слитая конструкция cr-РНК - tracr-PHK также упоминается в данном документе как гидовая РНК или гРНК (или единая гидовая РНК (егРНК)). В пределах егРНК часть, образованная из сг-РНК, может быть идентифицирована как «целевая последовательность», а часть, образованная из tracr-PHK, часто упоминается как «каркас».
В некоторых вариантах осуществления один или большее число элементов системы CRISPR происходят из системы CRISPR типа I, типа II или типа III. В некоторых вариантах осуществления один или большее число элементов системы CRISPR происходят из конкретного организма, включающего в себя эндогенную систему CRISPR, такого как Streptococcus pyogenes.
В общем, система CRISPR характеризуется элементами, которые способствуют образованию комплекса CRISPR в сайте целевой последовательности (также называемого протоспейсером в контексте эндогенной системы CRISPR). В контексте формирования комплекса CRISPR термин «целевая последовательность» относится к последовательности, для которой разработана комплементарная гидовая последовательность, при этом гибридизация между целевой последовательностью и гидовой последовательностью способствует образованию комплекса CRISPR. Целевой последовательностью может быть любой полинуклеотид, такой как полинуклеотиды ДНК или РНК. В некоторых вариантах осуществления целевая последовательность расположена в ядре или цитоплазме клетки.
В целевой нуклеиновой кислоте каждый протоспейсер ассоциирован с мотивом, примыкающим к протоспейсеру (МПП), распознавание которого является специфичным для конкретных систем CRISPR. В системе CRISPR/Cas из Streptococcus pyogenes МПП представляет собой нуклеотидную последовательность NGG. В системе CRISPR/Cas из Streptococcus thermophiles МПП представляет собой нуклеотидную последовательность NNAGAAW. Дуплекс tracr-PHK направляет Cas к целевой ДНК, состоящей из протоспейсера и необходимого МПП, посредством образования гетеродуплекса между спейсерной областью сг-РНК и протоспейсерной ДНК.
Доступно множество ресурсов, помогающих практикам определить подходящие целевые сайты после определения желаемой последовательности целевой ДНК. См., например, crispr.u-psud.fr/, инструмент, предназначенный для того, чтобы помогать ученым находить сайты нацеливания CRISPR у широкого спектра видов и генерировать соответствующую последовательность с-РНК.
В некоторых вариантах осуществления один или большее число полинуклеотидов, стимулирующих экспрессию одного или большего числа элементов системы CRISPR, вводят в геном растения-индуктора так, что экспрессия элементов системы CRISPR направляет образование комплекса CRISPR в одном или большем числе целевых сайтов. Например, фермент Cas и одна или большее число гидовых последовательностей, связанных с последовательностью tracr-mate, и каждая последовательность tracr могут быть функционально связаны с отдельными регуляторными элементами в отдельных конструкциях экспрессии (например, в егРНК). В качестве альтернативы, два или большее число элементов, экспрессируемых из одних и тех же или разных регуляторных элементов, могут быть объединены в единую конструкцию, при этом одна или большее число дополнительных конструкций обеспечивают любые компоненты системы CRISPR, не включенные в первую конструкцию. Элементы системы CRISPR, которые объединены в единую конструкцию, могут быть расположены в любой подходящей ориентации, например, один элемент может быть расположен в направлении к 5' от («выше») или в направлении к 3' от («ниже») второго элемента. Кодирующая последовательность одного элемента может быть расположена на той же или противоположной нити кодирующей последовательности второго элемента и ориентирована в том же или противоположном направлении. В некоторых вариантах осуществления одиночный промотор управляет экспрессией транскрипта, кодирующего фермент CRISPR, и одной или большим числом из гидовой последовательности, последовательности tracr-mate (необязательно функционально связанной с гидовой последовательностью) и последовательности tracr, встроенной в одну или большее число интронных последовательностей (например, каждая в другом интроне, две или большее число в по меньшей мере одном интроне или все в одном интроне). В некоторых вариантах осуществления фермент CRISPR, гидовая последовательность, последовательность tracr-mate и последовательность tracr функционально связаны и экспрессируются из одного и того же промотора.
В некоторых вариантах осуществления конструкция включает в себя регуляторный элемент, функционально связанный с кодирующей фермент последовательностью, кодирующей фермент CRISPR, такой как белок Cas.
Неограничивающие примеры белков Cas включают в себя Cas1, Cas1B, Cas2, Cas3, Cas4, Cas5, Cas6, Cas7, Cas8, Cas9 (также известен как Csn1 и Csx12), Cas10, Csy1, Csy2, Csy3, Cse1, Cse2, Csc1, Csc2, Csa5, Csn2, Csm2, Csm3, Csm4, Csm5, CsinG, Cmr1, Cmr3, Cmr4, Cmr5, Cmr6, Csb1, Csb2, Csb3, Csx17, Csx14, Csx10, Csx16, CsaX, Csx3, Csx1, Csx15, Csf1, Csf2, Csf3, Csf4, их гомологи или их модифицированные версии. В некоторых вариантах осуществления немодифицированный фермент CRISPR обладает активностью по расщеплению ДНК, такой как Cas9. В некоторых вариантах осуществления фермент CRISPR направляет расщепление одной или обеих цепей в местоположении целевой последовательности, например, внутри целевой последовательности и/или внутри последовательности, которая комплементарна целевой последовательности.
В некоторых вариантах осуществления конструкция кодирует фермент CRISPR, который мутирован по сравнению с соответствующим ферментом дикого типа так, что мутированный фермент CRISPR лишен способности расщеплять одну или обе цепи целевого полинуклеотида, содержащего целевую последовательность.
Например, замещение аспартата аланином (D10A) в каталитическом домене RuvC I Cas9 из S. pyogenes превращает Cas9 из нуклеазы, которая расщепляет обе цепи, в никазу (расщепляет одну цепь). Другие примеры мутаций, которые превращают Cas9 в никазу, включают в себя, но не ограничиваются ими, Н840А, N854A и N863A. В качестве дополнительного примера, можно мутировать два или большее число каталитических доменов Cas9 (RuvC I, RuvC II и RuvC III) для получения мутированного Cas9, по существу лишенного всей активности расщепления ДНК. В некоторых вариантах осуществления мутацию D10A комбинируют с одной или большим числом мутаций Н840А, N854A или N863A для получения фермента Cas9, по существу лишенного всей активности по расщеплению ДНК. В некоторых вариантах осуществления фермент CRISPR считается по существу лишенным всей активности по расщеплению ДНК, если активность по расщеплению ДНК мутированного фермента составляет меньше чем около 25%, 10%, 5%>, 1%>, 0,1%>, 0,01%, или меньше по сравнению с его немутированной формой.
В некоторых вариантах осуществления фермент-кодирующая последовательность, кодирующая фермент CRISPR, является кодон-оптимизированной для экспрессии в конкретных клетках. Разные виды демонстрируют конкретное предпочтение в отношении определенных кодонов конкретной аминокислоты. Предпочтение кодонов (различия в использовании кодонов между организмами) часто коррелирует с эффективностью трансляции информационной РНК (иРНК), которая, в свою очередь, как полагают, зависит, среди прочего, от свойств транслируемых кодонов и доступности конкретных молекул транспортной РНК (тРНК).
Преобладание выбранных тРНК в клетке, как правило, является отражением кодонов, наиболее часто используемых в синтезе пептидов. Соответственно, можно конструировать гены для оптимальной экспрессии генов в данном организме на основе оптимизации кодонов. Таблицы использования кодонов легко доступны, например, в «Базе данных использования кодонов» (англ. «Codon Usage Database»), и эти таблицы можно адаптировать несколькими способами. См. работу Nakamura, Y., et al., Nucl. Acids Res., 28: 2 92 (2000). Также доступны компьютерные алгоритмы для кодон-оптимизации конкретной последовательности для экспрессии в конкретной клетке-хозяине, например, Gene Forge (Aptagen; Джакобус, штат Пенсильвания, США). В некоторых вариантах осуществления один или большее число кодонов (например, 1, 2, 3, 4, 5, 10, 15, 20, 25, 50 или большее число, или все кодоны) в последовательности, кодирующей фермент CRISPR, соответствуют наиболее часто используемому кодону для конкретной аминокислоты.
Хотя специфика может варьировать в различных разработанных системах CRISPR, общая методология схожа. Практик, заинтересованный в использовании технологии CRISPR для нацеливания на последовательность ДНК (идентифицированную с помощью одного из многих доступных онлайн-инструментов), может вставить короткий фрагмент ДНК, содержащий целевую последовательность, в конструкцию экспрессии гидовой РНК. Конструкция экспрессии единой гидовой РНК (егРНК) содержит целевую последовательность (около 20 нуклеотидов), форму последовательности tracr-PHK (каркас), а также подходящий промотор и необходимые элементы для надлежащего процессинга в целевых клетках. В некоторых вариантах осуществления множественные гидовые РНК экспрессируются из одной конструкции либо путем соединения вместе многих кассет экспрессии с промоторами РНК-полимеразы III, либо путем создания длинных РНК, которые включают в себя тРНК или сайты самоотщепления рибозима, которые высвобождают множество функциональных егРНК, нацеленных на один или несколько генов (Cermak, et al., Plant Cell, 29 (6): 1196-1217 (2017), He, et al., J Genet Genomics, 44 (9): 469-472 (2017)). Векторы являются коммерчески доступными (см., например, Addgene). Многие из систем основаны на специально разработанных комплементарных олигосахаридах, которые отжигаются с образованием двухцепочечной ДНК, а затем клонируются в плазмиду экспрессии егРНК. Коэкспрессия егРНК и соответствующего фермента Cas из одних и тех же или отдельных конструкций в клетках приводит к одноцепочечному или двухцепочечному разрыву (в зависимости от активности фермента Cas) в желаемом целевом сайте. Основанные на CRISPR/Cas подходы к редактированию генов полностью совместимы с системой индукции гаплоидов cenh3, и эти две технологии можно с пользой комбинировать для облегчения независимого от генотипа редактирования генов.
ii. Нуклеазы с «цинковыми пальцами»
В некоторых вариантах осуществления элемент, который индуцирует одно- или двухцепочечный разрыв в геноме целевого растения, представляет собой конструкцию нуклеиновой кислоты или конструкции нуклеиновых кислот, кодирующие нуклеазы с «цинковыми пальцами» (ZFN). ZFN, как правило, представляют собой слитые белки, которые включают в себя ДНК-связывающий домен, полученный из белка с «цинковыми пальцами», связанного с доменом расщепления, таким как фермент Fok I типа IIS (Miller, et al., Nature Biotechnology, 25: 7 7 8-785(2007)). Fok I катализирует двухцепочечное расщепление ДНК в 9 нуклеотидах от ее сайта распознавания на одной нити и в 13 нуклеотидах от ее сайта распознавания на другой. См. также патенты США №5356802; №5436150 и №5487994; а также работы Li et al. Proc.r Natl. Acad. Sci. USA 89 (1992): 4275-4279; Li et al. Proc. Natl. Acad. Sci. USA, 90: 2764-2768 (1993); Kim et al. Proc. Natl. Acad. Sci. USA. 91: 883-887 (1994a); Kim et al. J. Biol. Chem. 269: 31,978-31,982 (1994b). Один или большее число из этих ферментов (или их ферментативно функциональных фрагментов) можно применять в качестве источника доменов расщепления.
Иллюстративные рестрикционные ферменты типа IIS описаны в международной публикации WO 07/014275. Дополнительные рестрикционные ферменты также содержат разделяемые домены связывания и расщепления. См., например, работу Roberts et al. Nucleic Acids Res., 31: 418-420 (2003). В определенных вариантах осуществления домен расщепления включает в себя один или большее число сконструированных полудоменов расщепления (также называемых мутантами домена димеризации), которые минимизируют или предотвращают гомодимеризацию, как описано, например, в патентных публикациях США №2005/0064474, №2006/0188987 и №2008/0131962. В определенных вариантах осуществления полудомен расщепления является мутантом полудомена расщепления из Fok I дикого типа. В некоторых вариантах осуществления полудомены расщепления модифицируют так, чтобы они включали в себя ядерные или другие сигналы локализации, пептидные метки или другие домены связывания.
ДНК-связывающий домен, который, в принципе, может быть сконструирован для нацеливания на любое представляющее интерес геномное местоположение, может представлять собой тандемный массив «цинковых пальцев» Cys2His2, каждый из которых, как правило, распознает от трех до четырех нуклеотидов в целевой последовательности ДНК. Соединяя вместе множество «пальцев» (количество варьирует: в опубликованных исследованиях на каждый мономер использовали от трех до шести «пальцев»), пары ZFN можно сконструировать так, чтобы они связывались с геномными последовательностями длиной 18-36 нуклеотидов.
Другой тип «цинковых пальцев», называемый «цинковыми пальцами» Cys2Cys2, который связывает цинк между 2 парами цистеинов, был обнаружен в ряде ДНК-связывающих белков.
ДНК-связывающий домен ZFN может состоять из двух-шести «цинковых пальцев». Как правило, считается, что каждый мотив «цинковых пальцев» распознает и связывается с последовательностью из трех пар оснований, и, как таковой, белок, включающий в себя большее число «цинковых пальцев», нацелен на более длинную последовательность и, следовательно, может обладать большей специфичностью и аффинностью к целевому сайту. Домены, связывающие «цинковые пальцы», могут быть «сконструированы» для связывания с заранее определенной нуклеотидной последовательностью. См., например, работы Beerli et al. Nature Biotechnol. 20: 135-141 (2002); Pabo et al. Ann. Rev. Biochem. 70: 313-340 (2001); Isalan et al., Nature Biotechnol. 19: 656-660 (2001); Segal et al. Curr. Opin. Biotechnol. 12: 632-637 (2001); Choo et al., Curr. Opin. Struct. Biol. 10: 41 1-416 (2000).
Стандартные ZFN соединяют домен расщепления с С-концом каждого домена «цинкового пальца». Чтобы позволить двум доменам расщепления димеризоваться и расщеплять ДНК, две отдельные ZFN должны связывать противоположные нити ДНК своими С-концами на определенном расстоянии друг от друга, как правило, от 5 до 7 п.о. Возможно как одноцепочечное расщепление, так и двухцепочечное расщепление, и двухцепочечное расщепление может произойти в результате двух разных событий одноцепочечного расщепления. Репарация индуцированных «цинковыми пальцами» двухцепочечных разрывов, как правило, происходит путем негомологичного соединения концов (НГСК) и приводит к мутациям, которые нарушают функцию гена. Системы нуклеаз с «цинковыми пальцами» можно также комбинировать со второй конструкцией, содержащей донорную молекулу, для вставки новых последовательностей ДНК путем направляемой гомологией репарации, как описано ниже. См. также работу Shukla, et al., Nature, 459, 437-441(2009).
См. также патенты США №6140081; №6453242; №6534261; №6610512; №6746838; №6866997; №7067617; опубликованные заявки на патент США №2 002/0165356; №2 004/0197892; №2007/0154989; №2007/0213269; и публикации международных патентных заявок № WO 98/53059 и WO 2003/016496, для дальнейшего обсуждения конструкций.
Сильная сторона нуклеаз с «цинковыми пальцами» заключается в том, что можно нацеливаться на любой сайт, и это не ограничивается сайтами МПП, которые необходимы для нацеливания Cas9. Основанные на нуклеазе с «цинковыми пальцами» подходы к редактированию генов полностью совместимы с системой индукции гаплоидов cenh3, и эти две технологии можно с пользой комбинировать для облегчения независимого от генотипа редактирования генов.
iii. Нуклеазы на основе эффекторов, подобных активаторам транскрипции
В некоторых вариантах осуществления элемент, который индуцирует одно- или двухцепочечный разрыв в геноме целевого растения, представляет собой конструкцию нуклеиновой кислоты или конструкции нуклеиновых кислот, кодирующие нуклеазы на основе эффекторов, подобных активаторам транскрипции (TALEN). TALEN имеют общую архитектуру, подобную архитектуре ZFN, с основным отличием в том, что ДНК-связывающий домен происходит от эффекторных белков TAL - факторов транскрипции из патогенных бактерий растений. ДНК-связывающий домен TALEN представляет собой тандемный массив аминокислотных повторов, каждый длиной около 34 остатков. Повторы очень похожи друг на друга; как правило, они различаются главным образом в двух положениях (аминокислоты 12 и 13, называемые вариабельной двухосновой повтора, или ВДП, англ. «RVD»). Каждая ВДП определяет преимущественное связывание с одним из четырех возможных нуклеотидов, что означает, что каждый повтор TALEN связывается с одной парой оснований, хотя известно, что ВДП NN связывает аденины в дополнение к гуанину. Подобно системам «цинковых пальцев» ВДП связаны друг с другом для придания им специфичности в отношении уникальных целевых сайтов и слиты с доменом расщепления, таким как Fokl (Cermak, et al., Nucleic Acids Research, 39 (12) (2011), Page e82, doi/10.1093/nar/gkr218).
Репарация TALEN-индуцированных двухцепочечных разрывов, как правило, происходит путем негомологичного соединения концов (НТСК) и приводит к мутациям, которые нарушают функцию гена. TALEN можно также комбинировать с донорной матрицей для НГР, как описано ниже. Сильная сторона подхода на основе TALEN заключается в том, что можно нацеливаться на любой сайт, и это не ограничивается сайтами МПП, которые необходимы для нацеливания Cas. Основанные на TALEN подходы к редактированию генов полностью совместимы с системой индукции гаплоидов cenh3, и эти две технологии можно с пользой комбинировать для облегчения независимого от генотипа редактирования генов.
См. также Cermak, et al, Nucl. Acids Res. 1-11 (2011), публикацию заявки на патент США №2 011/0145940, Miller et al., Nature Biotechnol 29: 143 (2011), для дальнейшего обсуждения конструкций TALEN.
b. Последовательности ДНК, изменяющие гены
Нуклеазная активность систем редактирования генома, описанных в данном документе, приводит к расщеплению целевой ДНК с образованием одноцепочечных или двухцепочечных разрывов в данной целевой ДНК. Двухцепочечные разрывы могут быть репарированы клеткой одним из по меньшей мере двух способов: негомологичным соединением концов (НГСК) и направляемой гомологией репарацией (НГР). При негомологичном соединении концов двухцепочечные разрывы репарируются путем прямого лигирования концов разрывов друг с другом. Как таковой, никакой новый материал нуклеиновой кислоты не вставляется в сайт, хотя некоторый материал нуклеиновой кислоты может быть потерян или получен в процессе репарации, что приводит к небольшим делециям или вставкам, которые могут нарушить функцию гена. При направляемой гомологией репарации донорный полинуклеотид, гомологичный расщепленной целевой последовательности ДНК, используется в качестве матрицы для репарации данной расщепленной целевой последовательности ДНК, что приводит к передаче генетической информации от донорного полинуклеотида к целевой ДНК. Следовательно, новый материал нуклеиновой кислоты может быть вставлен/скопирован в данный сайт.
Следовательно, в некоторых вариантах осуществления растение-индуктор необязательно включает в себя донорный полинуклеотид, например, в качестве сегмента генома растения-индуктора. Модификации ДНК целевого растения, обусловленные направляемой гомологией репарацией, можно применять для индуцирования коррекции генов, замены генов, мечения генов, вставки трансгена, делеций нуклеотидов, нарушения генов, мутации генов и т.д.
В данной заявке донорная полинуклеотидная последовательность, как правило, включает в себя области последовательности, гомологичные последовательности целевой ДНК, известные как «гомологичные плечи». Последовательность между гомологичными плечами может представлять собой естественную или сконструированную последовательность, которая вводит новые признаки, такие как метки, промоторные мотивы, экспрессируемые белковые мотивы или другие последовательности, представляющие интерес.Во время НГР гомологичные плечи используются для направления репарации двухцепочечного разрыва, что приводит к вставке новой последовательности в целевой сайт.
В применениях, в которых желательно вставить полинуклеотидную последовательность в последовательность целевой ДНК, донорная последовательность, подлежащая вставке, также предоставляется геномом растения-индуктора. Донорная последовательность с гомологичными плечами может быть в виде второй конструкции ДНК или компонента конструкции (Shi, et al., Plant Biotechnology Journal, 15: 207-216 (2017)), или конструкции, экспрессирующей РНК, которую можно использовать в качестве донорной матрицы для репарации (Li, et al., Nature Biotechnology, 37: 445-450 (2019)).
При введении донорной последовательности с целью изменения геномной последовательности целевого растения донорная последовательность, как правило, не идентична геномной последовательности, которую она замещает. Скорее, донорная последовательность может содержать по меньшей мере одно или большее число изменений одного основания, вставок, делеций, инверсий или перегруппировок по сравнению с геномной последовательностью, при условии, что присутствует достаточная гомология для поддержки направляемой гомологией репарации. В некоторых вариантах осуществления донорная последовательность включает в себя негомологичную последовательность, фланкированную двумя областями гомологии, так что направляемая гомологией репарация между целевой областью ДНК и двумя фланкирующими последовательностями приводит к вставке данной негомологичной последовательности в данную целевую область.
Донорная последовательность может включать в себя сайты рестрикции, нуклеотидные полиморфизмы, селектируемые маркеры (например, гены устойчивости к лекарственным препаратам, флуоресцентные белки, ферменты и т.д.) и т.д. по сравнению с геномной последовательностью, которые можно применять для оценки успешной вставки донорной последовательности в сайт расщепления или, в некоторых случаях, можно применять для других целей (например, для обозначения экспрессии в целевом геномном локусе). В некоторых случаях, если они расположены в кодирующей области, такие различия в последовательности нуклеотидов не изменят аминокислотную последовательность или внесут «молчащие» аминокислотные изменения (т.е. изменения, которые не влияют на структуру или функцию белка). В качестве альтернативы, эти различия в последовательностях могут включать в себя фланкирующие последовательности рекомбинации, такие как последовательности FLP, loxP или подобные, которые могут быть активированы в более позднее время для удаления маркерной последовательности.
Считается, что все применяемые в настоящее время способы редактирования генов, включительно с CRISPR/Cas9, «цинковыми пальцами», TALEN, их комбинации с редакторами оснований и их использование для прямой вставки измененных последовательностей ДНК с помощью НГР, совместимы с cenh3-системой индукции гаплоидов. Ценность cenh3-индуктора гаплоидов в данном контексте заключается в том, чтобы облегчить перенос механизмов редактирования генов из легко трансформируемой линии в элитную гермоплазму быстрым и эффективным способом.
III. Способы применения
А. Индукция гаплоидов
Представлены способы для индукции гаплоидов. Представленные способы индукции гаплоидов, как правило, включают в себя скрещивание линии, подлежащей индукции, с линией индуктора гаплоидов. Как обсуждалось выше, индуктор гаплоидов, как правило, представляет собой растение, гетерозиготное по cenh3-нуль.
Гаплоидные растения получают путем скрещивания индуктора гаплоидов почти с любой инбридной, гибридной или другой гермоплазмой, представляющей интерес.Гаплоиды образуются, когда хромосомы из растения - индуктора гаплоидов не сохраняются в течение первых клеточных делений эмбрионов. Полученный фенотип не является полностью проникающим, при этом некоторые семязачатки содержат гаплоидные эмбрионы, а другие содержат диплоидные эмбрионы, анеуплоидные эмбрионы, химерные эмбрионы или абортированные эмбрионы. После индукции гаплоидов гаплоидные эмбрионы или семена, как правило, отделяют от диплоидных и анеуплоидных эмбрионов или семян того же поколения с помощью скрининга фенотипических или генетических маркеров и выращивают в гаплоидные растения. Затем эти растения превращают с помощью химических манипуляций (например, с использованием агента против микротрубочек, такого как колхицин) в удвоенные гаплоидные («УГ») растения, которые затем продуцируют инбридные семена.
Селекция растений значительно облегчается за счет использования удвоенных гаплоидных (УГ) растений. Производство УГ-растений позволяет селекционерам получать инбридные линии без инбридинга нескольких поколений, тем самым сокращая время, необходимое для получения гомозиготных растений.
Индуктора гаплоидов скрещивают (либо в качестве растения мужского пола, либо в качестве растения женского пола) с целевой линией для получения гаплоидного потомства. В результате получаются гаплоидный эмбрион, растение или семя, которые содержат набор хромосом только от растения-родителя, не являющегося индуктором.
Если индуктор используется в качестве растения мужского пола, то полученное потомство будет иметь цитоплазму целевой линии. Если желательна цитоплазма индуктора (например, для получения мужской стерильной цитоплазмы), в качестве растения женского пола можно использовать индуктор гаплоидов.
Дополнительные этапы могут включать в себя методики идентификации гаплоидов и последующие методики удвоения хромосом, такие как, но не ограничиваясь ими, те, которые описаны в работе Prigge and Melchinger ("Production of Haploids and Doubled Haploids," in Maize Plant Cell Culture Protocols, Methods in Molecular Biology, Volume 877, pp.161-172, 2012) и другие, которые включают в себя, например, применение колхицина, пронамида, дитипира, трифлуралина или другого известного агента против микротрубочек, или другого ингибитора митоза. Затем эту линию можно непосредственно применять в последующих программах разведения.
Простота применения должна сделать его особенно универсальным в сочетании с другими технологиями, основанными на гаплоидах. Одним из таких применений является синтетический апомиксис, при котором растения конструируют так, чтобы они пропускали меиоз и производили диплоидные гаметы; когда такие линии скрещивают с индуктором гаплоидов, генотип получающегося в результате этого потомства идентичен родительскому генотипу (Marimuthu, et al., Science, 331 (6019): 876 (2011)). Другое применение - область хромосомной инженерии, в которой индуктор гаплоидов применяют для быстрого перемещения небольших, сконструированных или полностью синтетических хромосом из одной линии в другую (Birchler, et al., Current Opinion in Plant Biology, 19: 76-80 (2014).
В. Редактирование генов у однодольных
1. Способы редактирования генов
Также представлены способы одновременной индукции гаплоидов и редактирования генов. Представленные способы, как правило, включают в себя скрещивание линии, подлежащей индукции, с линией индуктора гаплоидов. Как представлено выше, индуктор гаплоидов, как правило, представляет собой растение, гетерозиготное по cenh3-нуль, а также кодирует механизмы редактирования генов типов CRISPR/Cas9, «цинковые пальцы», TALEN, либо отдельно, либо в комбинации с ферментами-редакторами оснований и/или молекулами-донорами, которые позволяют выполнять НГР и замену или модификацию генов.
При одновременном редактировании плюс индукции гаплоидов возможно быстрое и экономичное получение отредактированных культур и элитных линий без культивирования тканей. Линия, в которой проводят редактирование, может быть элитной гермоплазмой, и сам механизм редактирования будет устранен в процессе индукции гаплоидов. В то же время получаются отредактированные линии удвоенных гаплоидов.
Механизм редактирования генов доставляют посредством линии-индуктора. ДНК, РНК и белки, составляющие механизм редактирования генов, кодируются линией-индуктором и присутствуют в ней. Как правило, механизмы редактирования генов стабильно вставляются в индуктор, например, посредством бомбардировки или трансформации, опосредованной агробактериями. Трансгенные линии - индукторы гаплоидов, экспрессирующие механизмы редактирования, можно применять в качестве доноров или акцепторов пыльцы при межвидовых или межродовых скрещиваниях для индукции гаплоидов и одновременного редактирования генома.
Индуктора гаплоидов скрещивают (либо в качестве растения мужского пола, либо в качестве растения женского пола) с целевой линией для получения гаплоидного потомства. После оплодотворения механизмы редактирования вносят изменения в неиндуцированные целевые гены до или во время элиминации хромосом индуктора. Результатом являются гаплоидный эмбрион, растение или семя, которые содержат набор хромосом только от родительского растения, не являющегося индуктором, при этом данный набор хромосом содержит последовательности ДНК, которые были отредактированы.
Промотор (_ы), используемый (-е) в конструкции эндонуклеазы, как правило, приводит к экспрессии эндонуклеазы до оплодотворения, во время первой пары клеточных делений, или их комбинации. При использовании индуктора гаплоидов в качестве растения женского пола, если эндонуклеаза экспрессируется в яйцеклетке до опыления и на первых стадиях развития клетки, данная эндонуклеаза может немедленно начать мутирование целевой последовательности после опыления и продолжать мутирование целевой последовательности до того, как геном индуктора гаплоидов будет потерян из клетки. На первых стадиях митоза, до элиминации генома индуктора гаплоидов, целевая эндонуклеаза индуцирует нацеленный (-е) разрыв (-ы) цепей ДНК в ДНК целевой линии. Эти разрывы репарируются путем НГСК для создания небольших вставок или делеций, или исправляются путем НГР, в зависимости от применения редактирования генов.
Геномы гаплоидного потомства могут быть удвоены до или после скрининга потомства на наличие мутации (мутаций). Как только геномы этих гаплоидных особей удвоятся, их можно будет вырастить и самоопылить, чтобы получить семена удвоенных гаплоидов. Можно выполнять различные мутации и оценивать события мутаций, чтобы определить, дают ли полученные мутации желаемый результат. Представленные способы можно выполнять на всех (или многих) линиях однодольных, которые селекционер планирует использовать в качестве родителей для разведения. Если селекционер создает популяции, используя линии, которые имеют желаемую (-ые) мутацию (-и) во всех целевых локусах, популяции не расщепляются относительно желаемой (-ых) мутации (-й). Следовательно, усилия по разведению упрощаются за счет отсутствия необходимости отбора относительно желаемой (-ых) мутации (-й).
Представленные способы могут производить удвоенных гаплоидных особей с целевой (-ыми) мутацией (-ями) без затрат времени и средств на обратное скрещивание по желаемой целевой мутации в целевой линии. cenh3-индуктор гаплоидов является генетически доминантным. Когда растение с cenh3-нуль скрещивают с любой линией, полученное в результате F1 само становится индуктором гаплоидов. Растение F1, содержащее ген cenh3-нуль и механизм редактирования генов, можно скрещивать с сотнями или даже тысячами элитных линий, особенно тогда, когда в качестве растения мужского пола используется стоковая линия-индуктор. В некоторых вариантах осуществления удвоенные гаплоидные особи из нескольких элитных линий, все с целевой (-ыми) мутацией (-ями), производятся меньше чем за один год.
В некоторых вариантах осуществления линия-индуктор может быть нацелена на 2, 3, 4, 5 или большее число мутаций путем включения, например, множественных трансгенов эндонуклеаз и/или гРНК, и/или донорных последовательностей и т.д.
Полученные удвоенные гаплоидные особи могут иметь не все желаемые мутации. В некоторых вариантах осуществления, когда желательны множественные мутации, удвоенное гаплоидное потомство с единичными мутациями можно скрещивать друг с другом, и потомство F2 можно проверять на наличие особей, гомозиготных по всем желаемым мутациям.
Если индуктор используется в качестве растения мужского пола, то полученное потомство будет иметь цитоплазму целевой линии. Если желательна цитоплазма индуктора (например, для получения мужской стерильной цитоплазмы), в качестве растения женского пола можно использовать индуктор гаплоидов. Если желательна цитоплазма целевой линии, можно провести скрещивание между немутированной версией целевой линии (в качестве растения женского пола) и мутированной версией целевой линии (в качестве растения мужского пола).
2. Виды редактирования генов
Как было представлено выше, подходящие системы редактирования генов включают в себя, но не ограничиваются ими, те, которые вызывают мутации или изменения оснований, и целенаправленную вставку последовательности с помощью НГР. Считается, что любую форму процесса стабильного редактирования генов, будь то CRISPR/Cas9, «цинковые пальцы» или TALEN, можно комбинировать с cenh3-нуль для модификации генома целевой линии. Используя одну нуклеазу Cas9 и несколько гРНК, можно одновременно нацеливаться на и изменять больше чем один сайт.
Когда активный белок Cas нацелен на локус и не предоставляется донорная матрица, одним из возможных результатов являются небольшие делеций или вставки, которые создаются в процессе репарации путем НГСК. Если целевое местоположение выбрано правильно, мутация может создать сдвиг рамки в кодирующей последовательности и отменить функцию гена или изменить промотор, чтобы изменить паттерн экспрессии. Если цель состоит в том, чтобы подавить экспрессию, множественные гены часто нацеливают на несколько областей кодирующей последовательности. Если несколько гРНК нацелены на промотор, могут произойти значительные и полезные изменения в экспрессии генов (Rodriguez, et al., Cell, 171 (2): 470-80 (2017)).
Нацеленный мутагенез последовательности ДНК также может быть достигнут путем прямого преобразования одного основания ДНК в другое без необходимости двухцепочечных разрывов (ДЦР). Например, цитидиндезаминазу АРОВЕС1, адениндезаминазу и другие энхансерные компоненты, такие как урацил-ДНК-гликозилаза (UDG), можно объединить с никазой Cas9 (А840Н) или с Cas9, у которого инактивирована нуклеазная активность (dCa9), для прямого редактирования последовательности ДНК без введения двухцепочечных разрывов ДНК (Komor et al., Nature, 533: 420-424 (2016) doi:10.1038/naturel7946; Gaudelli et al., Nature, 551: 464-471 (2017) doi:10.1038/nature2 4 64 4; Komor et al., Science Advances, 3 (8) eaao4774 (2017), DOI: 10.1126/sciadv.aao4774). Данный вид механизма редактирования оснований также можно доставлять посредством линии - индуктора гаплоидов, чтобы напрямую индуцировать редактирование оснований в целевых последовательностях в других сортах.
НГР, которая является альтернативным средством восстановления ДЦР в хромосомах, представляет собой механизм для конструирования геномов растений, который способствует более тонким модификациям последовательности ДНК, включительно с коррекцией ДНК, нацеленным включением или замещением, или любым типом желаемой мутации. НГР происходит во время деления клеток (Ceccaldi, et al., Trends Cell Biol, 26 (1): 52-64 (2016). doi: 10.1016/j.tcb.2015.07.009) и может быть особенно активна во время быстрого деления клеток молодого эмбриона. В подтверждение этого представленные ниже данные указывают на то, что НГР может быть ответственна за мутацию в растении, полученную в результате одновременной индукции гаплоидов и сквозного редактирования генов в примере 2.
Следовательно, считается, что систему индукции гаплоидов in vivo можно применять не только для введения белка, РНК или ДНК для расщепления или преобразования целевой последовательности, ее также можно применять, вместе с подходящей матрицей репарации, для введения точных изменений последовательности в области, предназначенные для редактирования генов.
ДНК-матрицу можно вставлять в геном линии индуктора, несущей механизмы редактирования генома, такие как система CRISPR-Cas9, либо в том же трансгенном локусе, либо в другом локусе. Когда в индуцированных гаплоидных эмбрионах присутствуют как Cas9-erPHK, так и ДНК-матрица, расщепление целевой последовательности приведет к репарации хромосомного разрыва с использованием гомологичной последовательности трансгенной ДНК в качестве матрицы.
Трансгены можно вводить в ДЦР, если предоставленный шаблон содержит трансген, окруженный последовательностями, которые гомологичны последовательностям по обе стороны от данного ДЦР («гомологичные плечи», см. работу Shukla et al., Nature, 459: 437-441, (2009)). Трансгенное событие (например, для вставки гена, представляющего интерес) вносят в линию индуктора гаплоидов путем скрещивания. Ген эндонуклеазы можно использовать для определения относительного положения трансгена в нетрансгенной линии. Трансгенное событие, подлежащее вставке, должно быть фланкировано с обеих сторон последовательностями ДНК, гомологичными ДНК, фланкирующей целевой сайт. Когда индуктор гаплоидов скрещивают с целевой линией, эндонуклеаза вызывает двухцепочечный разрыв в целевом сайте. Если ДНК целевой линии репарируется с помощью НГР с использованием ДНК исходной линии индуктора гаплоидов (и трансгена) в качестве матрицы, ДНК целевой линии «репарирует» двухцепочечный разрыв, помещая последовательность трансгена в сайт двухцепочечного разрыва. Следовательно, представленные способы можно применять для размещения трансгенов в целевых линиях без необходимости обратного скрещивания.
Представленные композиции и способы можно более полно понять из пронумерованных параграфов, представленных ниже.
1. Однодольное растение - гаплоидный индуктор, гетерозиготное по центромерному гистону 3 (CenH3 - англ.: centromeric histone 3), содержащее диплоидные растительные клетки, содержащие только один аллель, кодирующий функциональный белок CENH3.
2. Однодольное растение по параграфу 1, отличающееся тем, что диплоидные растительные клетки содержат одну аллель CenH3, кодирующую нефункциональный белок CENH3.
3. Однодольное растение по параграфу 2, отличающееся тем, что аллель, кодирующая нефункциональный белок CENH3, представляет собой белок-нулевую аллель.
4. Однодольное растение по параграфу 2 или 3, отличающееся тем, что аллель, кодирующая нефункциональный белок CENH3, представляет собой РНК-нулевую аллель.
5. Однодольное растение по любому из параграфов 1-4, отличающееся тем, что аллель, кодирующая нефункциональный белок CENH3, вызвана мутацией со сдвигом рамки, которая создает стоп-кодон, отменяющий функцию.
6. Однодольное растение по любому из параграфов 1-5, отличающееся тем, что эндогенные локусы CenH3 на первой диплоидной хромосоме частично или полностью удалены.
7. Однодольное растение по любому из параграфов 1-6, отличающееся тем, что эндогенные локусы CenH3 на второй диплоидной хромосоме являются интактными.
8. Однодольное растение по любому из параграфов 1-7, отличающееся тем, что функциональный белок CENH3 представляет собой белок CENH3 дикого типа.
9. Однодольное растение по любому из параграфов 1-8, отличающееся тем, что у данного растения отсутствует хромосомно интегрированный или внехромосомный трансген, кодирующий CenH3 дикого типа.
10. Однодольное растение по любому из параграфов 1-9, отличающееся тем, что у данного растения отсутствует хромосомно интегрированный или внехромосомный трансген, кодирующий вариантный или слитый белок CENH3.
11. Однодольное растение по параграфу 10, отличающееся тем, что слитый белок, который отсутствует у растения, содержит зеленый флуоресцентный белок.
12. Однодольное растение по параграфу 10 или 11, отличающееся тем, что слитый белок, который отсутствует у растения, представляет собой GFP-tailswap.
13. Однодольное растение по любому из параграфов 1-12, отличающееся тем, что спермии или яйцеклетки содержат меньше CENH3, чем спермии или яйцеклетки растения дикого типа, гомозиготного по CENH3, необязательно отличающееся тем, что спермии или яйцеклетки содержат меньше чем 50% CENH3 по сравнению со спермиями или яйцеклетками растения дикого типа, предпочтительно отличающееся тем, что спермии или яйцеклетки содержат от около 50% до 10%, например, 50%, 25% или 12,5% CENH3 по сравнению со спермиями или яйцеклетками растения дикого типа.
14. Однодольное растение по любому из параграфов 1-13, отличающееся тем, что данное растение представляет собой кукурузу, пшеницу, рис, сорго, ячмень, овес, тритикале, рожь, просо африканское, просо пальчатое, просо обыкновенное, просо итальянское, банан, бамбук, сахарный тростник, просо прутьевидное, мискантус, аспарагус, лук, чеснок, лук скороду или ямс.
15. Однодольное растение по параграфу 14, отличающееся тем, что данное растение представляет собой кукурузу.
16. Однодольное растение по любому из параграфов 1-15, дополнительно содержащее экзогенную сайт-нацеленную нуклеазу, экспрессируемую клетками данного однодольного растения.
17. Однодольное растение по параграфу 16, отличающееся тем, что диплоидные растительные клетки содержат одну аллель CenH3, кодирующую нефункциональный белок CENH3.
18. Однодольное растение по параграфу 17, отличающееся тем, что аллель, кодирующая нефункциональный белок CENH3, представляет собой белок-нулевую аллель.
19. Однодольное растение по параграфу 17 или 18, отличающееся тем, что аллель, кодирующая нефункциональный белок CENH3, представляет собой РНК-нулевую аллель.
20. Однодольное растение по любому из параграфов 16-19, отличающееся тем, что аллель, кодирующая нефункциональный белок CENH3, вызвана мутацией со сдвигом рамки, которая создает стоп-кодон, отменяющий функцию.
21. Однодольное растение по любому из параграфов 16-20, отличающееся тем, что эндогенные локусы CenH3 на первой диплоидной хромосоме частично или полностью удалены.
22. Однодольное растение по любому из параграфов 16-21, отличающееся тем, что эндогенные локусы CenH3 на второй диплоидной хромосоме являются интактными.
23. Однодольное растение по любому из параграфов 16-22, отличающееся тем, что функциональный белок CENH3 представляет собой белок CENH3 дикого типа.
24. Однодольное растение по любому из параграфов 16-23, отличающееся тем, что у данного растения отсутствует хромосомно интегрированный или внехромосомный трансген, кодирующий CenH3 дикого типа.
25. Однодольное растение по любому из параграфов 16-24, отличающееся тем, что у данного растения отсутствует хромосомно интегрированный или внехромосомный трансген, кодирующий вариантный или слитый белок CENH3.
26. Однодольное растение по параграфу 25, отличающееся тем, что слитый белок, который отсутствует у растения, содержит зеленый флуоресцентный белок.
27. Однодольное растение по параграфу 25 или 26, отличающееся тем, что слитый белок, который отсутствует у растения, представляет собой GFP-tailswap.
28. Однодольное растение по любому из параграфов 16-27, отличающееся тем, что спермии или яйцеклетки содержат меньше CENH3, чем спермии или яйцеклетки растения дикого типа, гомозиготного по CENH3, необязательно отличающееся тем, что спермии или яйцеклетки содержат меньше чем 50% CENH3 по сравнению со спермиями или яйцеклетками растения дикого типа, предпочтительно отличающееся тем, что спермии или яйцеклетки содержат от около 50% до 10%, например, 50%, 25% или 12,5% CENH3 по сравнению со спермиями или яйцеклетками растения дикого типа.
29. Однодольное растение по любому из параграфов 26-28, отличающееся тем, что сайт-нацеленная нуклеаза стабильно экспрессируется клетками данного однодольного растения.
30. Однодольное растение по параграфу 2 9, отличающееся тем, что сайт-нацеленная нуклеаза представляет собой мегануклеазу (MN), нуклеазу с «цинковыми пальцами» (ZFN), нуклеазу на основе эффектора, подобного активатору транскрипции (TALEN), или нуклеазу на основе CRISPR, необязательно отличающееся тем, что нуклеазу выбирают из нуклеазы Cas9, нуклеазы Cfp1, dCas9-FokI, dCpf1-FokI, химерной Cas9-цитидиндезаминазы, химерной Cas9-адениндезаминазы, химерной FEN1-FokI, Mega-TAL, никазы Cas9 (nCas9), химерной нуклеазы dCas9 не FokI, нуклеазы dCpf1 не FokI, химерной Cpf1-цитидиндезаминазы и Cpf1-адениндезаминазы.
31. Однодольное растение по параграфу 29 или 30, отличающееся тем, что геном данного растения содержит гетерологичную конструкцию нуклеиновой кислоты, кодирующую нуклеазу.
32. Однодольное растение по любому из параграфов 16-31, дополнительно содержащее гидовую РНК, экспрессируемую клетками данного однодольного растения.
33. Однодольное растение по параграфу 29 или 30, отличающееся тем, что геном данного растения содержит гетерологичную конструкцию нуклеиновой кислоты, кодирующую гРНК.
34. Однодольное растение по любому из параграфов 16-33, отличающееся тем, что геном данного растения содержит донорную последовательность нуклеиновой кислоты, подлежащую вставке путем рекомбинации в сайт расщепления, индуцированный нуклеазой.
35. Однодольное растение по любому из параграфов 16-34, отличающееся тем, что данное растение представляет собой кукурузу, пшеницу, рис, сорго, ячмень, овес, тритикале, рожь, просо африканское, просо пальчатое, просо обыкновенное, просо итальянское, банан, бамбук, сахарный тростник, просо прутьевидное, мискантус, аспарагус, лук, чеснок, лук скороду или ямс.
36. Яйцеклетка, образованная растением по любому из параграфов 1-15, в которой отсутствует одна аллель, кодирующая функциональный белок CENH3, и которая содержит не больше чем около 12,5% функционального белка CENH3 по сравнению с соответствующей яйцеклеткой, образованной растением, гомозиготным по CenH3.
37. Спермий, образованный растением по любому из параграфов 1-15, спермий, в котором отсутствует одна аллель, кодирующая функциональный белок CENH3, и который содержит не больше чем около 25% функционального белка CENH3 по сравнению с соответствующим спермием, образованным растением, гомозиготным по CenH3.
38. Яйцеклетка, образованная растением по любому из параграфов 16-35, в которой отсутствует одна аллель, кодирующая функциональный белок CENH3, и которая содержит не больше чем около 12,5% функционального белка CENH3 по сравнению с соответствующей яйцеклеткой, образованной растением, гомозиготным по CenH3.
39. Спермий, образованный растением по любому из параграфов 16-35, спермий, в котором отсутствует одна аллель, кодирующая функциональный белок CENH3, и который содержит не больше чем около 25% функционального белка CENH3 по сравнению с соответствующим спермием, образованным растением, гомозиготным по CenH3.
40. Способ индукции образования целевого гаплоидного однодольного растения, включающий в себя опыление целевого родительского однодольного растения пыльцой от однодольного растения - индуктора гаплоидов по любому из параграфов 1-15; и отбор по меньшей мере одного гаплоидного потомка, полученного в результате данного опыления.
41. Способ индукции образования целевого гаплоидного однодольного растения, включающий в себя опыление однодольного растения - индуктора гаплоидов по любому из параграфов 1-15 пыльцой от целевого родительского однодольного растения; и отбор по меньшей мере одного гаплоидного потомка, полученного в результате данного опыления.
42. Способ по параграфу 40 или 41, дополнительно включающий в себя удвоение хромосом отобранного гаплоидного потомка.
43. Способ по параграфу 42, отличающийся тем, что удвоение хромосом является спонтанным или индуцированным, например, индуцированным агентом для удвоения хромосом, необязательно выбранным из колхицина, пронамида, дитипира, трифлуралина или другого агента против микротрубочек.
44. Способ по любому из параграфов 40-43, отличающийся тем, что целевое однодольное растение выбирают из кукурузы, пшеницы, риса, сорго, ячменя, овса, тритикале, ржи, проса африканского, проса пальчатого, проса обыкновенного, проса итальянского, банана, бамбука, сахарного тростника, проса прутьевидного, мискантуса, аспарагуса, лука, чеснока, лука скороды или ямса.
45. Способ модификации генома целевого однодольного растения, включающий:
индукцию образования целевого гаплоидного однодольного растения, включающую в себя опыление целевого родительского однодольного растения пыльцой от однодольного растения индуктора гаплоидов по любому из параграфов 16-35, и
отбор по меньшей мере одного гаплоидного потомка, полученного в результате опыления,
при этом гаплоидный потомок содержит геном целевого однодольного растения, но не однодольного растения - гаплоидного индуктора, и геном гаплоидного потомка был модифицирован сайт-направленной нуклеазой и, необязательно, по меньшей мере одной направляющей РНК, доставляемой однодольным растением гаплоидным индуктором.
46. Способ модификации генома целевого однодольного растения, включающий:
индукцию образования целевого гаплоидного однодольного растения, включающую в себя опыление однодольного растения индуктора гаплоидов по любому из параграфов 16-35 пыльцой от целевого родительского однодольного растения, и
отбор по меньшей мере одного гаплоидного потомка, полученного в результате опыления,
при этом гаплоидный потомок содержит геном целевого однодольного растения, но не однодольного растения - гаплоидного индуктора, и геном гаплоидного потомка был модифицирован сайт-направленной нуклеазой и, необязательно, по меньшей мере одной направляющей РНК, доставляемой однодольным растением гаплоидным индуктором.
47. Способ по параграфу 45 или 46, дополнительно включающий в себя удвоение хромосом отобранного гаплоидного потомка.
48. Способ по параграфу 47, отличающийся тем, что удвоение хромосом является спонтанным или индуцированным, например, индуцированным агентом для удвоения хромосом, необязательно выбранным из колхицина, пронамида, дитипира, трифлуралина или другого агента против микротрубочек.
49. Способ по любому из параграфов 45-48, отличающийся тем, что целевое однодольное растение выбирают из кукурузы, пшеницы, риса, сорго, ячменя, овса, тритикале, ржи, проса африканского, проса пальчатого, проса обыкновенного, проса итальянского, банана, бамбука, сахарного тростника, проса прутьевидного, мискантуса, аспарагуса, лука, чеснока, лука скороды или ямса.
Примеры
Пример 1: Получение растения кукурузы с мутацией cenh3-нуль и его применение для индукции гаплоидов.
Материалы и методы
Растительные материалы
Линии со вставками транспозонов g11, g18 и cenh3-mu1015598 были получены из Центра генетического сотрудничества по культурам кукурузы (Maize Genetics Cooperation Stock Center), г. Урбана, штат Иллинойс, США. Аллель cenh3-mu1015598 является одной из нескольких мутаций в стоковой линии UFMu-01386. Все растения выращивали в теплицах Факультета биологии растений Университета Джорджии.
Подготовка, конструкции и трансформация
Конструкция Ubi-Cas9 содержит 1991 п.о. промотора полиубиквитина кукурузы (GenBank: S94464.1), управляющего кодон-оптимизированной версией Cas9 кукурузы, оканчивающегося терминатором Nos.
Конструкция гРНК-ImmuneCENH3 содержит два компонента модуль гидовой РНК и ген ImmuneCENH3. Часть, образованная гидовой РНК, содержит промотор U6 кукурузы (Svitashev, et al., Plant Physiol. 169, 931-945 (2015)), управляющий гидовой РНК (TCCCGCAGCGCTACAGTCCC) (SEQ ID NO: 1), оканчивающийся терминатором PolIII TTTTTTTT. Часть, образованная геном ImmuneCENH3, содержит 6455 п.о. нативного гена CENH3 (координаты: хромосома 6: 166705239-166711693 в Zm-B73-REFERENCE-NAM-5.0), но имеет пять «молчащих» изменений кодонов в целевой области гРНК (CCAGGTACGGTCGCCCTGCGCGA) (SEQ ID NO: 2). Промотор включает в себя 2184 п.о. последовательности «выше» от ATG.
Чтобы создать конструкцию гРНК-TailswapCENH3, сохраняли естественный 5' UTR CENH3 и вставляли кодон-оптимизированную последовательность GFP в ATG ImmuneCENH3. За этим следовала последовательность линкера ATGGATGAACTATACAAGGGCGGAGGCGGTGGAGGCGTCGAC (SEQ ID NO: 3) и последовательность гена Н3. 3 кукурузы (Genbank NM_001294303.2), включительно с его интроном, слитым с нативным геном CENH3 на 3 п.о. «выше» от целевой области гидовой РНК. Трансген GFP-tailswap арабидопсиса также включает в себя часть, образованную Н3. 3. Данная конструкция основана на последовательности GFP-tailswap арабидопсиса, полученной из лаборатории Comai.
Данные три конструкции синтезировали с помощью GenScript (www.genscript.com) и клонировали в двоичный вектор pTF101.1 (Paz, et al., Euphytica 136, 167-179 (2004)). Для получения мутации cenh3 трансгенные линии, несущие Ubi-Cas9, скрещивали с линиями, несущими гРНК-ImmuneCENH3.
Экстрагирование ДНК, генотипирование и анализ последовательностей
Для стандартного генотипирования листьев геномную ДНК получали с использованием протокола СТАВ (Clarke, Cold Spring Harbor Protocols vol. 2009 db.prot5177-pdb.prot5177 (2009)). Ткань эндосперма собирали после того, как зерна проросли и можно было различить глянцевый фенотип. Зародыши и околоплодники удаляли пинцетом, а эндосперм измельчали в порошок с помощью ступки и пестика. ДНК эндосперма экстрагировали с помощью набора реактивов IBI Plant Genomic DNA Mini Kit (IBI Scientific IB47231).
Чтобы идентифицировать присутствие ImmuneCENH3 и Cas9 в трансгенных линиях, праймеры CENH3-F2 и CENH3-R3 использовали для амплификации ImmuneCENH3, а праймеры Cas9-F1 и Cas9-R1 использовали для амплификации Cas9 (таблица 1). Для идентификации исходной мутации cenh3 в растениях с Cas9 выполняли ПЦР с использованием набора Phusion High-Fidelity PGR Kit (New England Biolabs, г. Ипсвич, штат Массачусетс, США) с праймерами CENH3-F1 и CEMH3-R1, представленными в таблице 1 (SEQ ID NO: 4-14, в порядке убывания, как они указаны в данной таблице).
Продукты ПЦР либо напрямую секвенировали по Сэнгеру, либо клонировали с использованием набора для клонирования ТОРО ТА cloning kit (Thermo Fisher #К457501), а затем секвенировали по Сэнгеру.
В линиях, у которых отсутствует ImmuneCENH3, нулевую аллель сепЬЗ дифференцировали от нативной аллели CENH3 посредством ПЦР и расщепления рестрикционным ферментом. ПЦР амплифицирует ПЦР-продукт размером 4 96 п. о. при использовании праймеров CENH3-F2 и CENH3-R2. Когда данный продукт расщепляется рестрикционной эндонуклеазой AlwNI (New England Biolabs), аллель дикого типа расщепляется на две части размером 284 п.о. и 212 п.о., в то время как мутантная аллель cenh3 не расщепляется.
Аллель cenh3-mu1015598 оценивали с использованием праймеров CENH3-F4, CENH3-R4 и Mumix (смесь двух праймеров Mu1 и Mu2 в соотношении 1:1, представлено в таблице 1). Аллель дикого типа амплифицировали с помощью CENH3-F4 и CENH3-R4, в то время как аллель Ми амплифицировали с помощью CENH3-F4 и Mumix.
Оценка, плоидности
Потомство от скрещиваний +/cenh3 выращивали в помещении под лампами для выращивания в течение 10-13 суток и опрыскивали саженцы водой для определения глянцевого фенотипа. Все растения с глянцевым фенотипом впоследствии анализировали способом проточной цитометрии. Для каждого растения собирали около 1 г быстрозамороженных листьев или корней и измельчали в 1,5 мл предварительно охлажденного буфера для экстракции ядер (2 мМ ЭДТА, 15 мМ Трис-HCl рН 7,5, 20 мМ NaCl, 80 мМ KCl, 0,5 мМ спермина, 15 мМ 2-меркаптоэтанола, 0,1 мМ PMSF, 0,1% Тритон X-100). После измельчения смесь дважды фильтровали через сетчатое сито с ячейками 4 0 мкм. Ядра окрашивали 4,6-диамидино-2-фенилиндолом и загружали в проточные цитометры, размещенные в Лаборатории общих ресурсов цитометрии CTEGD Университета Джорджии.
Хромосомные препараты
Хромосомный анализ выполняли так, как описано в работе (Dawe, et al., Cell 173, 839-850.e18 (2018)). Кратко, концы корней собирали у гаплоидных и диплоидных растений, инкубировали в камере с закисью азота в течение трех часов и фиксировали 90%-й уксусной кислотой. Концы корней срезали лезвием бритвы и расщепляли в растворе фермента (1% пектолиаза Y-23, 2% целлюлаза Onozuka R-10) при 37°С течение 50 минут. Срез корня промывали в этаноле, затем погружали в 90%-ю уксусную кислоту. Концы корней раздавливали металлической киркой и по 10 мкл клеточной суспензии наносили на предметные стекла для микроскопии. Предметные стекла высушивали, наносили ProLong Gold с DAPI (Thermo Fisher, № по каталогу: Р36931) и накрывали покровными стеклами. Полученные препараты визуализировали с помощью флуоресцентного микроскопа Zeiss Axio Imager.M1 с 63-кратным масляным объективом Plan-APO Chromat, а для анализа данных использовали программное обеспечение Slidebook (Intelligent Imaging Innovations, г. Денвер, штат Колорадо, США).
Поверхностное секвенирование гаплоидов и анеуплоидов Для каждого образца по 12 нг/мкл ДНК обрабатывали ультразвуком в объеме 100 мкл с помощью соникатора Diagenode Bioruptor в течение семи минут при высоком режиме с 30-секундными интервалами включения-выключения, получая фрагменты длиной в среднем около 500 п.о. Библиотеки для секвенирования ДНК получали с использованием набора КАРА Hyperprep Kit (KK8502) с одноиндексными адаптерами КАРА (КК8700). По 600 нг обработанной ультразвуком ДНК использовали в качестве анализируемого материала для каждого образца, и для амплификации библиотек использовали 3 цикла ПЦР. 150-нуклеотидные считывания секвенирования от анализатора Illumina нормализовали для отсечения адаптеров и фильтровали по качеству с использованием Cutadapt версии 1.9.1 (Martin, et al., EMBnet.journal vol. 17 10 (2 011)) со следующими параметрами: «-q 20 -a AGATCGGAAGAGC -e.05 -О 1 -m 50» (SEQ ID NO: 15). Считывания выравнивали относительно Zm-B7 3-RE FERENCE-NAM-5.0 с использованием BWA-mem версии 0.7.15 в однонаправленном режиме с параметрами по умолчанию (Li, & Durbin, Bioinformatics vol. 25 1754-1760 (2009)). Покрытие считываний визуализировали с использованием IGVTools версии 2.3.98 (Thorvaldsdottir, et al., Brief. Bioinform. 14, 178-192 (2013)) с расчетом покрытия на интервалах по 25 млн. п.о.
Результаты
Выращивание удвоенных гаплоидов широко применяется для ускорения производства новых инбридных линий (Kalinowska, et al., Theor. Appl. Genet. 132, 593-605 (2019)). Один из распространенных подходов, используемых во всей отрасли селекции кукурузы, заключается в создании гаплоидов с мутантами, которые препятствуют оплодотворению (Kelliher, et al., Nature 542, 105-109 (2017), Liu, et al., Mol. Plant 10, 520-522 (2017), Gilles, et al., EMBO J. 36, 707-717 (2017), Yao, et al., Nat Plants 4, 530-533 (2018), Zhong, et al., Nat Plants 6, 466-472 (2020)). Совершенно другой способ индукции гаплоидов был впервые предложен Саймоном Чаном и его коллегами, которые показали, что при скрещивании линий арабидопсиса со структурно измененным белком центромерного гистона Н3 (CENH3) получаются гаплоиды и анеуплоиды с высокой частотой до 25-45% (Ravi and Chan, Nature 464, 615-618 (2010)). CENH3 представляет собой вариант гистона, который определяет местоположение центромеры и рекрутирует налагающиеся выше белки кинетохора (Cheeseman & Desai, Nat. Rev. Mol. Cell Biol. 9, 33-46 (2008), Black & Bassett, Curr. Opin. Cell Biol. 20, 91-100 (2008)). Первоначальное исследование включало в себя конструкцию под названием GFP-tailswap, в которой N-концевой хвост CENH3 был модифицирован с помощью GFP-метки, однако точечные мутации и небольшие делеций CENH3 также могут индуцировать гаплоиды с аналогичными частотами (Karimi-Ashtiyani, et al., Proc. Natl. Acad. Sci. U. S. A. 112, 11211-11216 (2015), Kuppu, et al., PLoS Genet. 11, el005494 (2015), Kuppu, et al., Plant Biotechnol. J. (2020) doi:10.1111/pbi.13365). Кроме арабидопсиса опосредованная центромерой индукция гаплоидов у других растений оказалась менее эффективной, как правило, образуя <1% гаплоидов (Kalinowska, et al., Theor. Appl. Genet. 132, 593-605 (2019)).
Данное исследование было разработано для изучения механизма опосредованной центромерой индукции гаплоидов у кукурузы, первоначально - с использованием способа с GFP-tailswap. Однако данный подход осложняется тем фактом, что для него требовался как мутант нативного cenh3, так и функциональный трансген GFP-tailswap, который дополняет данного мутанта. Другая группа уже продемонстрировала некоторый успех, используя существующий мутант кукурузы (cenh3-mu1015598), полученный вставкой мутатора Робертсона (Ми) в 5' UTR данного гена (Kelliher, et al., Front. Plant Sci. 7, 414 (2016), Feng, et al., Plant J. (2019) doi:10.1111/tpj.14606)). Путем скрещивания они внесли GFP-tailswap в генетический фон cenh3-mul015598 и наблюдали в среднем 0,86% гаплоидов при скрещивании в качестве растения мужского пола и отсутствие гаплоидов при скрещивании в качестве растения женского пола (Kelliher, et al., Front. Plant Sci. 7, 414 (2016)). Получили cenh3-mu1015598 и выполнили самокрещивания трех гетерозиготных растений. Генотипирование показало, что у двух початков с низкой частотой расщеплялись гомозиготные мутанты, которые выросли до различных состояний зрелости (таблица 2).

Получение гомозиготных мутантов указывает на то, что cenh3-mu1015598 не является нулевым, и что предыдущие результаты, возможно, были искажены низким уровнем экспрессии CENH3 дикого типа. Вариабельная пенетрантность аллели cenh3-mul015598 может объясняться тем фактом, что элементы Mu могут способствовать низким уровням экспрессии при вставке в области 5' UTR (Barkan & Martienssen, Proc. Natl. Acad. sci. U. S. A. 88, 3502-3506 (1991)).
Чтобы преодолеть отбор по истинным cenh3-нулевым аллелям, cenh3-иулъ был создан с использованием подхода CRISPR/Cas9 с двумя конструкциями. Одну линию трансформировали конструкцией, экспрессирующей Cas9, управляемый промотором убиквитина. Вторую трансформировали конструкцией, экспрессирующей гРНК, нацеленную на четвертый экзон нативного гена CENH3, и геном «Immune CENH3», который содержит полноразмерный нативный ген CENH3 с пятью «молчащими» изменениями нуклеотидов в области нацеливания гРНК (фиг.1А, 1В). После скрещивания двух данных линий друг с другом Cas9 вызвал мутации в нативном гене CENH3, но оставил трансген незатронутым. Была выбрана аллель cenh3 с делецией одного нуклеотида, которая вызывает немедленный стоп-кодон в N-концевом хвосте CENH3 (фиг.1С). В присутствии ImmuneCENH3 мутация cenh3 расщепляется как простой рецессивный менделевский признак (таблица 3).
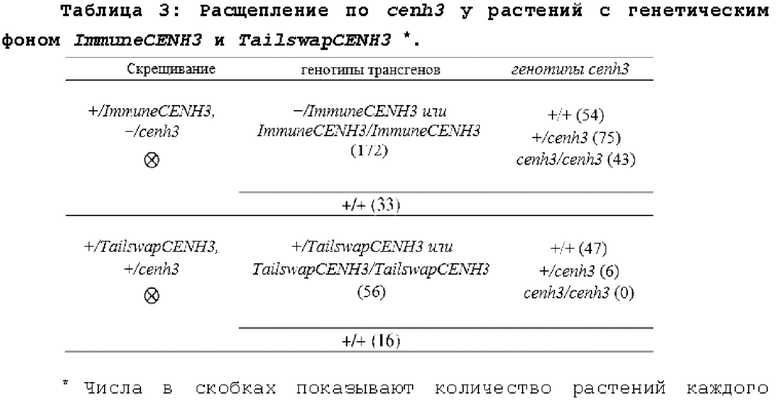
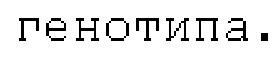
Затем создавали трансгены с TailswapCENH3 - близкой копией конструкции GFP-tailswap арабидопсиса, и скрещивали их с мутацией cenh3. Растения, которые содержали TallswapCENH3 и являлись гомозиготными по cenh3 (таблица 3) получены не были, что указывает на то, что данный трансген не дополняет истинную нулевую аллель (фиг.4А).
В ходе данных исследований было обнаружено, что сепЪЗ иногда передавался при отсутствии ImmuneCENH3. Путем скрещивания с линиями дикого типа получили линию с простым расщеплением по cenh3, в которой отсутствовали оба исходных трансгена. Среди самоскрещенного потомства от линии +/cenh3 было 163 +/+ растения дикого типа, 55 гетерозигот +/cenh3 и нуль гомозигот cenh3/cenh3, что указывает на то, что данный мутант является гомозиготно-летальным и плохо передается через гаметофиты. Также выполняли реципрокные скрещивания между гетерозиготами +/cenh3 и растениями дикого типа. Менделевский признак обычно передается 50% потомков при тестовом скрещивании, однако было замечено, что только 12,1% потомков получили cenh3 при скрещивании через растение мужского пола и 25% при скрещивании через растение женского пола (таблица 4).
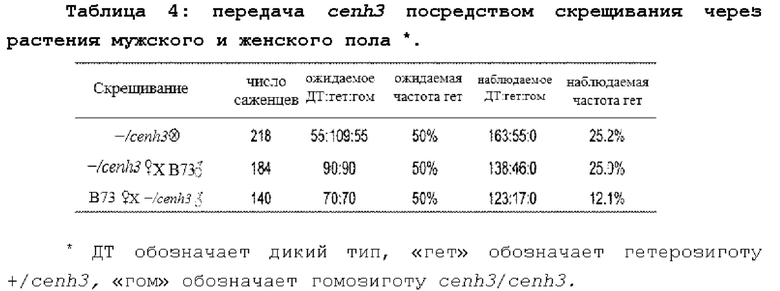
Снижение передачи может объясняться тем, что спермии и яйцеклетки переносятся внутри многоклеточных гаплоидных гаметофитов. Образованию спермия предшествуют два деления гаплоидных клеток, а образованию яйцеклетки предшествуют три деления гаплоидных клеток. Те гаметофиты, у которых присутствует аллель cenh3, должны использовать CENH3, перенесенный из фазы спорофита, в то время как он естественным образом разбавляется в каждом клеточном цикле (Lermontova, et al., Plant Cell 18, 2443-2451 (2006)). В данной модели спермий с cenh3 будет иметь около ¼ от нормального количества CENH3, а яйцеклетка, несущая cenh3, будет иметь около ⅛ по сравнению с растением, гетерозиготным по cenh3 (фиг.4В). Предполагая отсутствие компенсации дозы, эти значения будут дополнительно снижены на ½ по сравнению с нормальным гомозиготным родителем дикого типа. В результате этого спермии и яйцеклетки, несущие cenh3, могут иметь меньшие центромеры.
Чтобы проверить, способны ли гетерозиготные мутанты +/cenh3 индуцировать гаплоиды, гетерозиготы по cenh3 скрещивали с тестовыми линиями в обоих направлениях. В первом тесте растения дикого типа и растения +/cenh3 скрещивали с линией, гомозиготной по рецессивной мутации glossy8 (g18) на хромосоме 5, которая придает листьям саженцев блестящий вид (Xu, et al., Plant Physiology vol. 115, 501-510 (1997)). Было замечено, что 0,5% потомства имели глянцевый фенотип, когда гетерозиготы +/cenh3 скрещивали как растения мужского пола, и 5,0% потомства были гаплоидными, когда растения +/cenh3 скрещивали как растения женского (таблица 5).
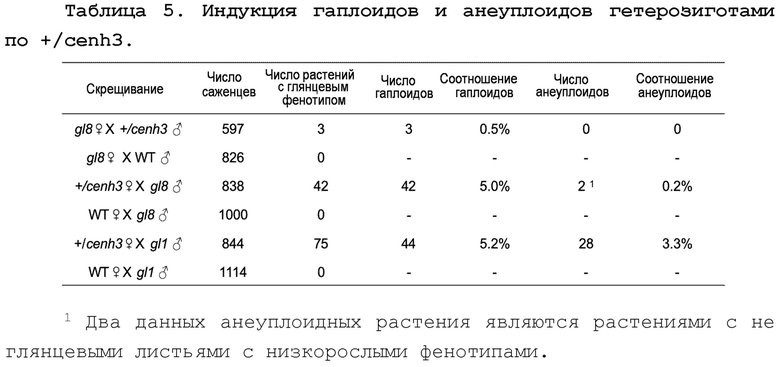
Анализ способом проточной цитометрии показал, что все растения с глянцевыми листьями были гаплоидами (фиг.2А-2В) интерпретация, которая была подтверждена подсчетом хромосом в клетках концов корней трех растений (фиг.2C-2D). При выращивании до зрелости гаплоидные растения были низкорослыми и стерильными (Chase, Bot. Rev. 35, 117-168 (1969)) (фиг.2E-2F). Также наблюдались два не глянцевых растения с низкорослыми фенотипами, которые, как полагали, были анеуплоидами. Эти два растения подвергали поверхностному секвенированию вместе с шестью гаплоидами. В то время как гаплоиды показали равномерный охват последовательностей, низкорослые растения этого не сделали; одно было трисомным по хромосоме 3, а другое было моносомным по хромосомам 2 и 4, и трисомным по хромосоме 10 (фиг.3А-3В).
Второй набор тестов проводили с использованием растений glossyl (g11), которые имеют аналогичный фенотип, но мутация находится в хромосоме 7 (Sturaro, et al., Plant Physiol. 138, 478-489 (2005)). В этих скрещиваниях также оценивали скорость прорастания, которая является косвенным показателем
кариотипической аномалии, широко используемым для оценки эффективности индукторов гаплоидов арабидопсиса (Kuppu, et al., PLoS Genet. 11, e1005494 (2015), Ravi, et al., Nature Communications vol. 5 (2014), Maheshwari, et al., PLoS Genet. 11, e1004970 (2015)). В скрещиваниях, в которых гетерозиготы+/cenh3 были растениями женского пола, 5,2% потомства демонстрировали глянцевый фенотип и были гаплоидными на основании результатов анализов способом проточной цитометрии. Еще 3,3% потомства демонстрировали глянцевый фенотип, но имели более высокое содержание ДНК, чем ожидалось для гаплоидов, и были оценены как анеуплоиды (таблица б).
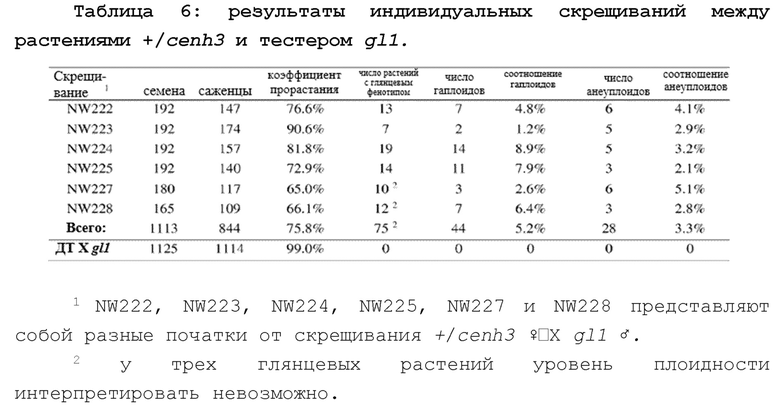
Разные скрещивания значительно различались по скорости прорастания (65-91%), частотам гаплоидов (1,2-8,9%) и анеуплоидов (2,1-5,1%) (таблица б). Данные о последовательностях пяти анеуплоидных растений подтвердили, что у всех, кроме одного, отсутствовала хромосома 7, иногда в сочетании с потерей других хромосом. У одного растения с глянцевым фенотипом, которое, по-видимому, имело две полные копии хромосомы 7, возможно, была небольшая интерстициальная делеция, которая не была обнаружена при поверхностном секвенировании (сегментарные анеуплоиды распространены у гибридов арабидопсиса с GFP-tailswap (Tan, et al., Elife 4, (2015))). Результаты тестовых скрещиваний g11 больше соответствуют тому, что наблюдалось у арабидопсиса, где любое данное скрещивание с GFP-tail swap, как правило, дает гаплоиды и анеуплоиды в одинаковых пропорциях (Ravi and Chan, Nature 464, 615-618 (2010), Ravi, et al., Nature Communications vol. 5 (2014), Maheshwari, et al., PLoS Genet. 11, el004 970 (2015)).
Если разбавление CENH3 является основным механизмом индукции гаплоидов, то только гаметы, несущие мутацию cenh3 от родителя +/cenh.3, должны индуцировать гаплоиды. К сожалению, невозможно оценить саженцы на наличие аллели cenh3, поскольку геном гаплоидного индуктора теряется. Однако данные от скрещиваний GFP-tailswap арабидопсиса показывают, что эндосперм редко демонстрирует полную элиминацию однородительского генома, когда саженец является гаплоидным (Ravi, et al., Nature Communications vol. 5 (2014)). Если это верно и для кукурузы, то генотип эндосперма можно было бы использовать для определения исходного генотипа саженца. Генотипировали остаточные эндоспермы из набора из одиннадцати гаплоидных растений, полученных в результате скрещивания a +/cenh3 X g18. Результаты показали, что все одиннадцать были гетерозиготными по аллели cenh3, что убедительно подтверждает интерпретацию о том, что индукция гаплоидов является результатом низких уровней CENH3 в гаметах с cenh3.
Одним из поразительных свойств опосредованной центромерой индукции гаплоидов является то, что она эффективна только у подмножества потомков. У некоторых особей теряются все хромосомы от родителя - индуктора гаплоидов, а у другого, гораздо большего подмножества, потеря хромосом не происходит.Относительно небольшой класс анеуплоидов представляет собой события «частичной индукции гаплоидов», когда некоторые хромосомы были потеряны, но другие выжили. Тот факт, что скрещивания g18 дали больше истинных гаплоидов, чем скрещивания g11, может быть связан с тем фактом, что первые были проведены летом, а вторые - зимой. Также возможно, что роль сыграла схема отбора.
Исследования с использованием линии кукурузы с делецией r-X1, которая вызывает моносомию с высокой частотой, продемонстрировали, что некоторые хромосомы восстанавливаются как моносомные с большей частотой, чем другие 28. Моносомики по хромосоме 5 (с g18) образуются редко, тогда как моносомики по хромосоме 7 (с g11) образуются гораздо чаще (в 17 раз чаще (Weber, Use of Maize Monosomies for Gene Localization and Dosage Studies, in The Maize Handbook (eds. Freeling, M. & Walbot, V.) 350-358 (Springer New York, 1994))). Действительно, два из пяти секвенированных анеуплоидов от скрещиваний g11 были моносомными только по хромосоме 7 (таблица 6). Эти данные могут указывать на то, что тестер g18 способствует образованию гаплоидов, в то время как тестер g11 приводит к более широкому спектру плоидности.
Считается, что вся предшествующая литература по опосредованной центромерой индукции гаплоидов описывает комплементацию нулевой аллели вариантом CENH3 или аллелями, которые продуцируют измененные или частично удаленные формы CENH3 (Ravi and Chan, Nature 464, 615-618 (2010), Karimi-Ashtiyani, et al., Proc. Natl. Acad. Sci. U. S. A. 112, 11211-11216 (2015), Kuppu, et al., Plant Biotechnol. J. (2020) doi:10.1111/pbi.13365, Maheshwari, et al., PLoS Genet. 11, e1004970 (2015), Ishii, et al., Annu. Rev. Plant Biol. 67, 421-438 (2016)). Эти данные послужили подтверждением первоначальной интерпретации, согласно которой индукция гаплоидов вызвана конкуренцией между двумя структурно различными формами CENH3 и окончательным отторжением измененных центромер механизмом устранения неправильной сборки (Ravi and Chan, Nature 464, 615-618 (2010), Britt and Kuppu, Front. Plant Sci. 7, 357 (2016), Kalinowska, et al., Theor. Appl. Genet. 132, 593-605 (2019), Kuppu, et al., Plant Biotechnol. J. (2020) doi:10.1111/pbi.13365, Maheshwari, et al., PLoS Genet. 11, e1004970 (2015), Copenhaver, & Preuss, Nat. Biotechnol. 28, 423-424 (2010)), по сравнению с другими возможными механизмами (Wang & Dawe, Molecular Plant vol. 11 398-406 (2018), Karimi-Ashtiyani, et al., Proc. Natl. Acad. Sci. U. S. A. 112, 11211-11216 (2015), Ravi, et al., PLoS Genet. 7,, el002121 (2011), Wang, et al., Plant Methods 15, 42 (2019), Sanei, et al., Proc. Natl. Acad. Sci. U. S. A. 108, E498-505 (2011), Tan, et al., Elife 4, (2015)).
Напротив, представленные в данном документе данные свидетельствуют о достижении высоких уровней индукции гаплоидов при использовании мутации cenh3 в N-концевом хвосте, которая удаляет всю последовательность, которая взаимодействует с ДНК или другими гистонами. Следовательно, количественные сокращения одного только CENH3 индуцируют опосредованную центромерой гаплоидную индукцию. Главное преимущество подхода, основанного на cenh3-нуль, заключается в том, что растения имеют высокую жизнеспособность, а сам процесс прост. Любая линия, скрещенная с cenh3, становится индуктором гаплоидов. Простота применения данного подхода должна сделать его особенно универсальным в сочетании с другими технологиями, основанными на гаплоидах, такими как синтетический апомиксис (Marimuthu, et al., Science, 331 (6019): 876 (2011), Wang, et al., aBIOTECH, 1: 15-20(2020)), перенос сконструированных хромосом из одной линии в другую (Birchler, et al., Current Opinion in Plant Biology, 19: 76-80 (2014)) и независимое от генотипа редактирование генов (Kelliher, et al., Nat. Biotechnol. 37, 287-292 (2019)).
Пример 2: Индукция гаплоидов с одновременным редактированием генов
Материалы и методы
Применение cenh3-индуктора гаплоидов включает в себя одновременную индукцию гаплоидов и редактирование генов. В примере такого применения растение cenh3-нуль сначала скрещивают с линией, содержащей конструкцию CRISPR (экспрессирующую как Cas9, так и одну или большее число гидовых РНК). Эту гибридную линию, содержащую компоненты cenh3 и CRISPR, затем скрещивают в качестве растений женского пола с мужской линией кукурузы дикого типа. Считается, что после оплодотворения около 5% потомков будут гаплоидными, и среди них около половины потомков получат конструкцию CRISPR, экспрессирующую Cas9 и видовую (-ые) РНК. Компоненты CRISPR экспрессируются в ранних делениях зиготы и при этом могут катализировать редактирование генов в отцовском геноме. Материнский геном будет быстро утерян во время индукции гаплоидов, удаляя компоненты CRISPR и оставляя только отцовский геном, часть которого будет подвергнута устойчивому редактированию генов.
Чтобы протестировать индукцию гаплоидов с одновременным редактированием генов, были разработаны эксперименты по скрещиванию cenh3-нулъ с конструкцией CRISPR, экспрессирующей Cas9 из промотора убиквитина и восемь видовых РНК, нацеленных на четыре гена, которые контролируют развитие растений: фасциированный початок-2 (fea2), фасциированный початок-3 (fea3), компактное растение-2 (ct2) и карликовое растение с толстой метелкой-1 (td1). Сначала конструкцию CRISPR ввели в инбридную линию под названием В104 путем Agrobacterium-опосредованной трансформации. Затем трансгенную линию В104 скрещивали с растением, гетерозиготным по cenh3-нуль. Потомков от данного скрещивания - растения как с конструкцией CRISPR, так и с cenh3-скрещивали с линией кукурузы, гомозиготной по рецессивной мутации, называемой luteus-1, для идентификации гаплоидов.
Результаты
Было выявлено, что из общего числа 192-х растений 7 были гаплоидами. Генотипировали в общей сложности 40 диплоидных растений и все 7 гаплоидных растений (путем ампликон-секвенирования по Сэнгеру) на предмет изменений, внесенных редактированием в каждый сайт гидовой РНК. Общая частота редактирования была низкой, что привело в общей сложности к пяти изменениям, внесенным редактированием, среди сорока диплоидных растений. Однако, что важно, одно из гаплоидных растений имело изменение в гене fea2. Изменение в гаплоидном растении дало фенотип фасциированного початка, соответствующий известному рецессивному фенотипу данного мутанта (Taguchi-Shiobara, et al., Genes Dev. 15: 2755-2766 (2001)). Частота редактирования, составляющая 1/7 у гаплоидов, примерно соответствует частоте редактирования, составляющей 5/40 у диплоидов, что указывает в данном небольшом, раннем пилотном эксперименте на то, что редактирование примерно одинаково эффективно у гаплоидов и диплоидов.
Дополнительно следует отметить тот факт, что единственное внесенное редактированием изменение относится к нестандартному типу. В то время как изменения в диплоидах соответствовали расщеплению и репарации путем негомологичного соединения концов (НГСК), гаплоидное растение содержало вставку, фланкированную короткой областью гомологии с обеих сторон. Данный тип редактирования может указывать на ошибочную направляемую гомологией репарацию (НГР) (Xue and Greene, Trends In Genetics, DOI: 10.1016/j.tig.2021.02.008. (2021)). Эти данные дают основания полагать, что НГР активна во время ранних делений зиготы, и что расщепление и репарация могут происходить в течение короткого времени, в течение которого присутствуют оба генома, до потери хромосом из линии - индуктора гаплоидов. НГР является важной формой репарации для многих применений CRISPR, включающих в себя замещение промоторов или генов новыми последовательностями (Zhang et al., Nat Plants, 5: 778-794 (2019)).
Результаты данных экспериментов проиллюстрированы на фиг.5A-5D и в таблицах 7 и 8. Фиг. 5А представляет собой плазмидную карту конструкции CRISPR, применяемой для одновременной индукции гаплоидов и редактирования генов. Компоненты конструкций указаны.
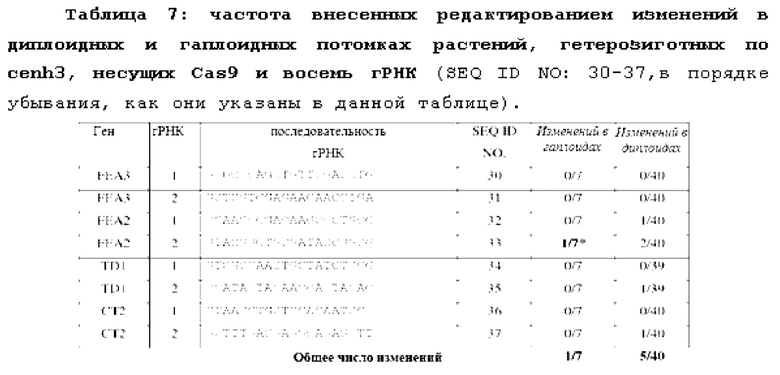


Внесенные редактированием изменения в диплоидных растениях находятся в пределах областей гРНК, что согласуется с расщеплением и репарацией путем НГСК. Внесенное редактированием изменение в гаплоидном растении представляет собой вставку из 81 п.о., что предполагает репарацию путем ошибочной НГР, предположительно опосредованной фланкирующей областью
микрогомологии. Обозначены следующие особенности: целевые области гидовых РНК, сайты МПП; удаленные основания (тире); область дупликации; фланкирующая гомологичная дупликация. ДТ = дикий тип.
Очень небольшой эксперимент с другой конструкцией (нацеленной на ZmGB1) не выявил никаких обнаруживаемых внесенных редактированием изменений в гаплоидах и только несколько - у диплоидов.
Если не определено иное, все технические и научные термины, употребляемые в данном документе, имеют такие же значения, которые обычно понимаются специалистом в области техники, к которой принадлежит данное изобретение. Публикации, цитируемые в данном документе, и материалы, на которые они ссылаются, конкретным образом включены в данный документ посредством ссылки.
Специалисты в данной области техники распознают или смогут установить, используя не более чем рутинные эксперименты, множество эквивалентов конкретным вариантам осуществления данного изобретения, описанным в данном документе. Предусмотрено, что такие эквиваленты охватываются следующими пунктами формулы изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> UNIVERSITY OF GEORGIA RESEARCH FOUNDATION, INC.
COLD SPRING HARBOR LABORATORY
DAWE, R. KELLY
JACKSON, DAVID
<120> ГЕТЕРОЗИГОТНЫЕ ПО CENH3 ОДНОДОЛЬНЫЕ РАСТЕНИЯ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ
ДЛЯ ИНДУКЦИИ ГАПЛОИДОВ И ОДНОВРЕМЕННОГО РЕДАКТИРОВАНИЯ ГЕНОМА
<130> UGA 2020-139-03
<150> U.S.S.N. 63/036902
<151> 09.06.2020
<150> U.S.S.N. 63/036910
<151> 09.06.2020
<160> 47
<170> PatentIn, версия 3.5
<210> 1
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 1
tcccgcagcg ctacagtccc 20
<210> 2
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 2
ccaggtacgg tcgccctgcg cga 23
<210> 3
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 3
atggatgaac tatacaaggg cggaggcggt ggaggcgtcg ac 42
<210> 4
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 4
tgcaagatga gggcgaatgt g 21
<210> 5
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 5
tacttcctga tctcccgcag c 21
<210> 6
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 6
ggctgctctt acttgcttgc 20
<210> 7
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 7
cgctctactt tcccgtttgt tac 23
<210> 8
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 8
tacttcctga tctcgcgcag ggcg 24
<210> 9
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 9
acgagaagta cccgacaatc tacc 24
<210> 10
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 10
tgatttgaag ttcggcgtca gg 22
<210> 11
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 11
cgctcgaact ggagcttctt 20
<210> 12
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 12
aggttggcag gtagccgtta 20
<210> 13
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 13
gcctctattt cgtcgaatcc g 21
<210> 14
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 14
gcctccattt cgtcgaatcc c 21
<210> 15
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 15
agatcggaag agc 13
<210> 16
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 16
ccagggactg tagcgctgcg gga 23
<210> 17
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 17
tcccgcagcg ctacagtccc tgg 23
<210> 18
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 18
gccaccgctg gcggccaggg actgtagcgc tgcgg 35
<210> 19
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 19
gccaccgctg gcggccaggg ctgtagcgct gcgg 34
<210> 20
<211> 157
<212> БЕЛОК
<213> Zea mays
<400> 20
Met Ala Arg Thr Lys His Gln Ala Val Arg Lys Thr Ala Glu Lys Pro
1 5 10 15
Lys Lys Lys Leu Gln Phe Glu Arg Ser Gly Gly Ala Ser Thr Ser Ala
20 25 30
Thr Pro Glu Arg Ala Ala Gly Thr Gly Gly Arg Ala Ala Ser Gly Gly
35 40 45
Asp Ser Val Lys Lys Thr Lys Pro Arg His Arg Trp Arg Pro Gly Thr
50 55 60
Val Ala Leu Arg Glu Ile Arg Lys Tyr Gln Lys Ser Thr Glu Pro Leu
65 70 75 80
Ile Pro Phe Ala Pro Phe Val Arg Val Val Arg Glu Leu Thr Asn Phe
85 90 95
Val Thr Asn Gly Lys Val Glu Arg Tyr Thr Ala Glu Ala Leu Leu Ala
100 105 110
Leu Gln Glu Ala Ala Glu Phe His Leu Ile Glu Leu Phe Glu Met Ala
115 120 125
Asn Leu Cys Ala Ile His Ala Lys Arg Val Thr Ile Met Gln Lys Asp
130 135 140
Ile Gln Leu Ala Arg Arg Ile Gly Gly Arg Arg Trp Ala
145 150 155
<210> 21
<211> 1145
<212> ДНК
<213> Zea mays
<400> 21
ctcccgtccc gagagttctg aatcgaaacc gtcggccacg agagcagtgc gaggcgccca 60
ccgcgatggc tcgaaccaag caccaggccg tgaggaagac ggcggagaag cccaagaaga 120
agctccagtt cgagcgctca ggtggtgcga gtacctcggc gacgccggaa agggctgctg 180
ggaccggggg aagagcggcg tctggaggtg actcagttaa gaagacgaaa ccacgccacc 240
gctggcggcc agggactgta gcgctgcggg agatcaggaa gtaccagaag tccactgaac 300
cgctcatccc ctttgcgcct ttcgtccgtg tggtgaggga gttaaccaat ttcgtaacaa 360
acgggaaagt agagcgctat accgcagaag ccctccttgc gctgcaagag gcagcagaat 420
tccacttgat agaactgttt gaaatggcga atctgtgtgc catccatgcc aagcgtgtca 480
caatcatgca aaaggacata caacttgcaa ggcgtatcgg aggaaggcgt tgggcatgat 540
atataatatc cattctgatt gcatcattct tgtgaatttg tttgtaggag ctagacatta 600
gtgttgttga atgctgcatg gttcctaatc cttttcgcag tctaacatct gtggagttag 660
tatgttacat ggcaacagct gaacatctgt ggactataac tatatggcaa cagccgaaga 720
ttgtgtctgt gggataactg gttgttttgg ttgctcttca gtagtttgtt tgcttcaggt 780
aaccatgctg cgaactatga tgttttcatt ctcggtttgc ttcagctaac cgagatcgat 840
tcagtctgca gtatatggac tatggagtaa actgcatgct gaaacccgaa ccactgctga 900
aacggcagtt gccaggatag caggagggcc ctttatgcac agtggaattg agtagagaac 960
tgagtaaacc atggttcttt ctccttttga actggaacac aaacacagtt ggatcttgtt 1020
tctcttctta ggccattgtc atcgtgtttc ttaggggtgt aaatggtatc tgtccgtatt 1080
cgaatttgat ctatctaaca aggctgaaat ccgaaaaaaa aaaaaaaaaa aaaaaaaaaa 1140
aaaaa 1145
<210> 22
<211> 164
<212> БЕЛОК
<213> Oryza sativa
<400> 22
Met Ala Arg Thr Lys His Pro Ala Val Arg Lys Ser Lys Ala Glu Pro
1 5 10 15
Lys Lys Lys Leu Gln Phe Glu Arg Ser Pro Arg Pro Ser Lys Ala Gln
20 25 30
Arg Ala Gly Gly Gly Thr Gly Thr Ser Ala Thr Thr Arg Ser Ala Ala
35 40 45
Gly Thr Ser Ala Ser Gly Thr Pro Arg Gln Gln Thr Lys Gln Arg Lys
50 55 60
Pro His Arg Phe Arg Pro Gly Thr Val Ala Leu Arg Glu Ile Arg Lys
65 70 75 80
Phe Gln Lys Thr Thr Glu Leu Leu Ile Pro Phe Ala Pro Phe Ser Arg
85 90 95
Leu Val Arg Glu Ile Thr Asp Phe Tyr Ser Lys Asp Val Ser Arg Trp
100 105 110
Thr Leu Glu Ala Leu Leu Ala Leu Gln Glu Ala Ala Glu Tyr His Leu
115 120 125
Val Asp Ile Phe Glu Val Ser Asn Leu Cys Ala Ile His Ala Lys Arg
130 135 140
Val Thr Ile Met Gln Lys Asp Met Gln Leu Ala Arg Arg Ile Gly Gly
145 150 155 160
Arg Arg Pro Trp
<210> 23
<211> 828
<212> ДНК
<213> Oryza sativa
<400> 23
acgccgcttc agtttgaaaa cccaccgcca cgtcgccgcc gccgccgccg ccgccgccga 60
cgccgagatg gctcgcacga agcacccggc ggtgaggaag tcgaaggcgg agcccaagaa 120
gaagctccag ttcgaacgct cccctcggcc gtcgaaggcg cagcgcgctg gtggcggcac 180
gggtacctcg gcgaccacga ggagcgcggc tggaacatcg gcttcaggga cgcctaggca 240
gcaaacgaag cagaggaagc cacaccgctt ccgtccaggc acagtggcac tgcgggagat 300
caggaaattt cagaaaacca ccgaactgct gatcccgttt gcaccatttt ctcggctggt 360
cagggagatc actgatttct attcaaagga tgtgtcacgg tggacccttg aagctctcct 420
tgcattgcaa gaggcagcag aataccactt agtggacata tttgaagtgt caaatctctg 480
cgccatccat gctaagcgtg ttaccatcat gcaaaaggac atgcaacttg ccaggcgtat 540
cggtgggcgg aggccatggt gaaaatttgt ttgcgagcca tgcagcatga tggacaagga 600
gcaacatgtg tcgttgatta acattttaga aagtagtgta gatgtatctt cacataggga 660
tcaacttacc cttcgttccc attctaattc agttgatgtt agtatttacc ttttgctcca 720
tttggattgg tcgaattcag gatttcatca aacagtcgat tgtgaaatgt gaaccaggaa 780
ttgttgtgtt gattgcaata atgggttcct ctcaaaaaaa aaaaaaaa 828
<210> 24
<211> 163
<212> БЕЛОК
<213> Triticum aestivum
<400> 24
Met Ala Arg Thr Lys His Pro Ala Val Arg Lys Thr Lys Ala Pro Pro
1 5 10 15
Lys Lys Gln Leu Gly Pro Arg Pro Ala Gln Arg Arg Gln Glu Thr Asp
20 25 30
Gly Ala Gly Thr Ser Ala Thr Pro Arg Arg Ala Gly Arg Ala Ala Ala
35 40 45
Pro Gly Gly Ala Gln Gly Ala Thr Gly Gln Pro Lys Gln Arg Lys Pro
50 55 60
His Arg Phe Arg Pro Gly Thr Val Ala Leu Arg Glu Ile Arg Arg Tyr
65 70 75 80
Gln Lys Ser Val Asp Phe Leu Ile Pro Phe Ala Pro Phe Val Arg Leu
85 90 95
Ile Lys Glu Val Thr Asp Phe Phe Cys Pro Glu Ile Ser Arg Trp Thr
100 105 110
Pro Gln Ala Leu Val Ala Ile Gln Glu Ala Ala Glu Tyr His Leu Val
115 120 125
Asp Val Phe Glu Arg Ala Asn His Cys Ala Ile His Ala Lys Arg Val
130 135 140
Thr Val Met Gln Lys Asp Ile Gln Leu Ala Arg Arg Ile Gly Gly Arg
145 150 155 160
Arg Leu Trp
<210> 25
<211> 492
<212> ДНК
<213> Triticum aestivum
<400> 25
atggcccgca ccaagcaccc ggccgtcagg aagaccaagg cgccgcccaa gaagcagctc 60
gggccccgcc ccgcgcagcg gcggcaggag acagatggcg cgggcacgtc ggcgacaccg 120
aggcgagccg ggcgggcggc ggccccaggg ggggctcaag gggcaactgg gcaacccaag 180
cagaggaaac cacaccggtt caggccaggc acggtggcac tgcgggagat caggaggtat 240
cagaagtcgg tcgactttct catcccgttt gcaccatttg tccgtctgat caaggaggtc 300
accgacttct tctgtcctga aatcagccgc tggactcccc aagcgctcgt cgcgattcaa 360
gaggctgcag agtatcacct cgtcgacgta tttgaaaggg caaatcactg tgccatccat 420
gcaaagcgtg ttaccgtcat gcaaaaggac atacagcttg caaggcgtat cggcgggagg 480
aggctttggt ga 492
<210> 26
<211> 139
<212> БЕЛОК
<213> Hordeum vulgare
<400> 26
Met Ala Arg Thr Lys Lys Thr Val Ala Ala Lys Glu Lys Arg Pro Pro
1 5 10 15
Cys Ser Lys Ser Glu Pro Gln Ser Gln Pro Lys Lys Lys Glu Lys Arg
20 25 30
Ala Tyr Arg Phe Arg Pro Gly Thr Val Ala Leu Arg Glu Ile Arg Lys
35 40 45
Tyr Arg Lys Ser Thr Asn Met Leu Ile Pro Phe Ala Pro Phe Val Arg
50 55 60
Leu Val Arg Asp Ile Ala Asp Asn Leu Thr Pro Leu Ser Asn Lys Lys
65 70 75 80
Glu Ser Lys Pro Thr Pro Trp Thr Pro Leu Ala Leu Leu Ser Leu Gln
85 90 95
Glu Ser Ala Glu Tyr His Leu Val Asp Leu Phe Gly Lys Ala Asn Leu
100 105 110
Cys Ala Ile His Ser His Arg Val Thr Ile Met Leu Lys Asp Met Gln
115 120 125
Leu Ala Arg Arg Ile Gly Thr Arg Ser Leu Trp
130 135
<210> 27
<211> 768
<212> ДНК
<213> Hordeum vulgare
<400> 27
gaacgaactc tatctctctc tctgcctgct ttccccacct gcgatggctc gcacgaagaa 60
aacggtggcg gcgaaggaga agcgcccccc ttgctccaag tcggagccgc agtcgcagcc 120
gaagaagaag gagaagcggg cgtaccggtt ccggccgggc acggtggcgc tgcgggagat 180
ccggaagtac cgcaagtcca ccaatatgct catccccttt gcgcccttcg tccgcctggt 240
cagggacatc gccgacaact tgacgccatt gtcgaacaag aaggagagca agccgacgcc 300
atggactcct ctcgcgctcc tctcgttgca agagtctgca gagtatcact tggtcgatct 360
atttggaaag gcaaatctgt gtgccattca ttcgcaccgt gttaccatca tgctaaagga 420
catgcagctt gcgaggcgta tcgggacgag aagcctttgg tgatactaat ggaggatgtt 480
ttaggcatcg tagggagcaa acactggtga tgctaatggt agatgttttt tgtatgagtg 540
caatgcgaag gagctgatgg tagtgatcca atatgtttgt gagtgtatca ttaggaagta 600
atgtaggtgt gttttcatct aggtagcatg tcttcgaagt tgatccggtt ggttgcatgg 660
ctgggatcat ctgacttgtc ggcttttgct ccatttgaat tgatctaatt cggacagagt 720
ttctgcaaat gatccaataa tatgggttgc caaaaaaaaa aaaaaaat 768
<210> 28
<211> 167
<212> БЕЛОК
<213> Musa acuminata
<400> 28
Met Ala Arg Thr Lys His Leu Ser Asn Arg Ser Ser Ser Arg Pro Arg
1 5 10 15
Lys Arg Phe His Phe Gly Arg Ser Pro Gly Gln Arg Thr Pro Ala Asp
20 25 30
Ala Asn Arg Pro Ala Thr Pro Ser Gly Ala Thr Pro Arg Thr Thr Ala
35 40 45
Thr Arg Ser Arg Asp Thr Pro Gln Gly Ala Pro Ser Gln Ser Lys Gln
50 55 60
Gln Pro Arg Arg Arg Arg Phe Arg Pro Gly Val Val Ala Leu Arg Glu
65 70 75 80
Ile Arg Asn Leu Gln Lys Thr Trp Asn Leu Leu Ile Pro Phe Ala Pro
85 90 95
Phe Val Arg Leu Val Arg Glu Ile Thr His Phe Tyr Ser Lys Glu Val
100 105 110
Asn Arg Trp Thr Pro Glu Ala Leu Val Ala Ile Gln Glu Ala Ala Glu
115 120 125
Thr His Met Ile Glu Met Phe Glu Asp Ala Tyr Leu Cys Ala Ile His
130 135 140
Ala Lys Arg Val Thr Leu Met Gln Lys Asp Ile His Leu Ala Arg Arg
145 150 155 160
Ile Gly Gly Arg Arg His Trp
165
<210> 29
<211> 504
<212> ДНК
<213> Musa acuminata
<400> 29
atggcgagaa cgaagcatct gtccaacagg tcctcctctc gccctcggaa gcgcttccat 60
ttcggtcggt ctccagggca gcgaaccccc gctgatgcga atcggcctgc gacgccatct 120
ggtgctactc ctagaaccac ggccaccaga tcgagggata cgcctcaagg ggcaccgagc 180
caatcaaaac agcagccgag gcggcgcagg tttaggccgg gggtggtggc gctacgcgag 240
atcaggaatt tgcagaagac gtggaatcta ttgatccctt tcgctccgtt tgtcagactt 300
gttcgggaga tcactcattt ctactcgaaa gaagtaaacc gatggacccc tgaagcttta 360
gttgcgattc aagaggcagc ggaaactcat atgatagaaa tgtttgaaga tgcatatctc 420
tgtgcaattc atgcaaaacg tgttaccctt atgcaaaaag atatccatct agcaaggcga 480
ataggaggaa gaagacattg gtga 504
<210> 30
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 30
gcgcccagct gttcgacctg 20
<210> 31
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 31
gctcgtggag aacaacctga 20
<210> 32
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 32
ggaaggcgag aagcgctgcg 20
<210> 33
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 33
gtacggctgc gataccggcg 20
<210> 34
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 34
gtcgccaact gctatctccg 20
<210> 35
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 35
ggatactaca accagtacag 20
<210> 36
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 36
gtaaggtgct ggagaatcg 19
<210> 37
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 37
gctttgacga ggcagagctt 20
<210> 38
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 38
cgcggaaggc gagaagcgct gcgcggtcgg tggaggcagg gactgcaggt tgggtcgccg 60
acgccaggac 70
<210> 39
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 39
cgcggaaggc gagaagtgcg cggtcggtgg aggcagggac tgcaggttgg gtcgccga 58
<210> 40
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 40
tccgtacggc tgcgataccg gcggggatct cgccggagaa gcggttgtgg gagaggtcga 60
<210> 41
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 41
tccgtacggc tgcgatacgc ggggatctcg ccggagaagc ggttgtggga gaggtcga 58
<210> 42
<211> 59
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 42
tccgtacggc tgcgataccg cggggatctc gccggagaag cggttgtggg agaggtcga 59
<210> 43
<211> 266
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 43
tccgtacggc tgcgataccg gcggggatct cgccggagaa gcggttgtgg gagaggtcga 60
ggaggaggag agcggagttg tcggggtcgg cgacgatccg cggcgggacg gcgccggaga 120
agcggttgtg ggagaggtcg aggaggagga gagcggagtt gtcggggtcg gcgacgatcc 180
gcggcgggac ggcgccggag atggcgttgc gggagagatc aagggcagcg aggcgcgcgg 240
ggaaggagag acgcggggag agcggg 266
<210> 44
<211> 61
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 44
gtcggatact acaaccagta cagcggcggg gtcccgcgcg agttcggcgc gctccagtcg 60
c 61
<210> 45
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 45
gtcggatact acaaccagtc agcggcgggg tcccgcgcga gttcggcgcg ctccagtcgc 60
<210> 46
<211> 61
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 46
ctggcttgac gaggcagagc ttaggagcta cacatcagtc atccatgcta atgtgtatca 60
g 61
<210> 47
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетический полинуклеотид
<400> 47
ctggcttgac gaggcagact taggagctac acatcagtca tccatgctaa tgtgtatcag 60
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЕ АЛЛЕЛИ CENH3 ПШЕНИЦЫ | 2019 |
|
RU2836060C2 |
| КОМПОЗИЦИИ ДЛЯ ИНДУКЦИИ ГАПЛОИДИИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2816649C1 |
| КОМПОЗИЦИИ ДЛЯ ИНДУКЦИИ ГАПЛОИДИИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2771141C2 |
| ОДНОВРЕМЕННОЕ РЕДАКТИРОВАНИЕ ГЕНОВ И ИНДУКЦИЯ ГАПЛОИДОВ | 2017 |
|
RU2788349C2 |
| СОЗДАНИЕ ГАПЛОИДНЫХ РАСТЕНИЙ И УСОВЕРШЕНСТВОВАНИЕ СЕЛЕКЦИИ РАСТЕНИЙ | 2010 |
|
RU2571927C2 |
| НОВЫЕ ФЕРМЕНТЫ И СИСТЕМЫ CRISPR | 2016 |
|
RU2777988C2 |
| ЭКСПРЕССИРУЮЩИЕ CAS МЫШИНЫЕ ЭМБРИОНАЛЬНЫЕ СТВОЛОВЫЕ КЛЕТКИ И МЫШИ И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2782358C2 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ ПШЕНИЦЫ С БИАЛЛЕЛЬНЫМИ МУТАЦИЯМИ В ПРОТОМОТОРНОЙ ОБЛАСТИ ГЕНА VRN-A1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ CRISPR/CAS9 | 2020 |
|
RU2772575C2 |
| ДОСТАВКА, КОНСТРУИРОВАНИЕ И ОПТИМИЗАЦИЯ СИСТЕМ, СПОСОБОВ И КОМПОЗИЦИЙ ДЛЯ МАНИПУЛЯЦИИ С ПОСЛЕДОВАТЕЛЬНОСТЯМИ И ПРИМЕНЕНИЯ В ТЕРАПИИ | 2013 |
|
RU2721275C2 |
| СПОСОБ СОЗДАНИЯ НОВЫХ МУТАЦИЙ В ОРГАНИЗМАХ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2833881C1 |
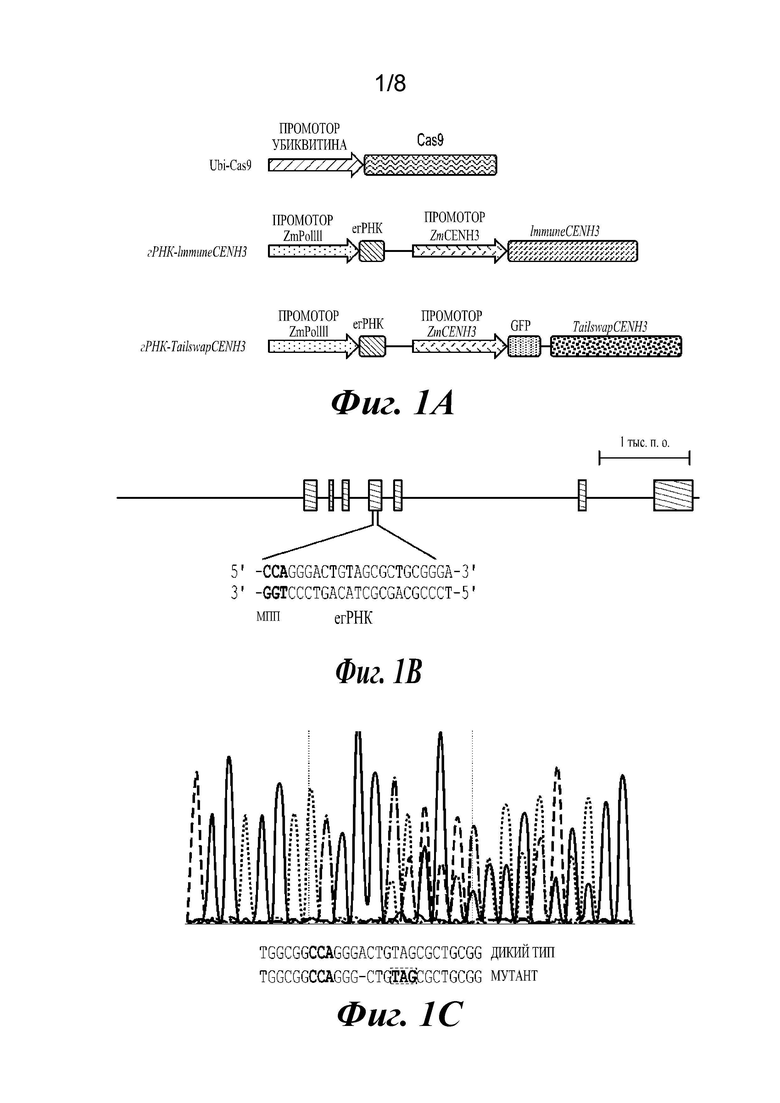
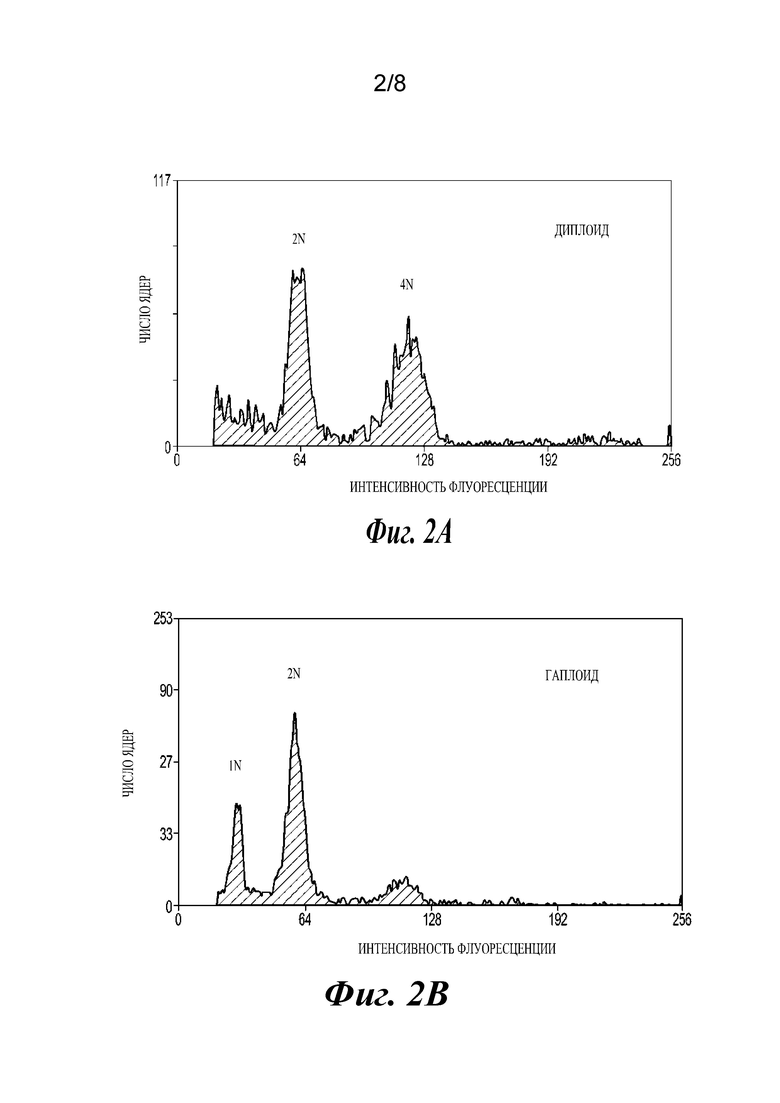
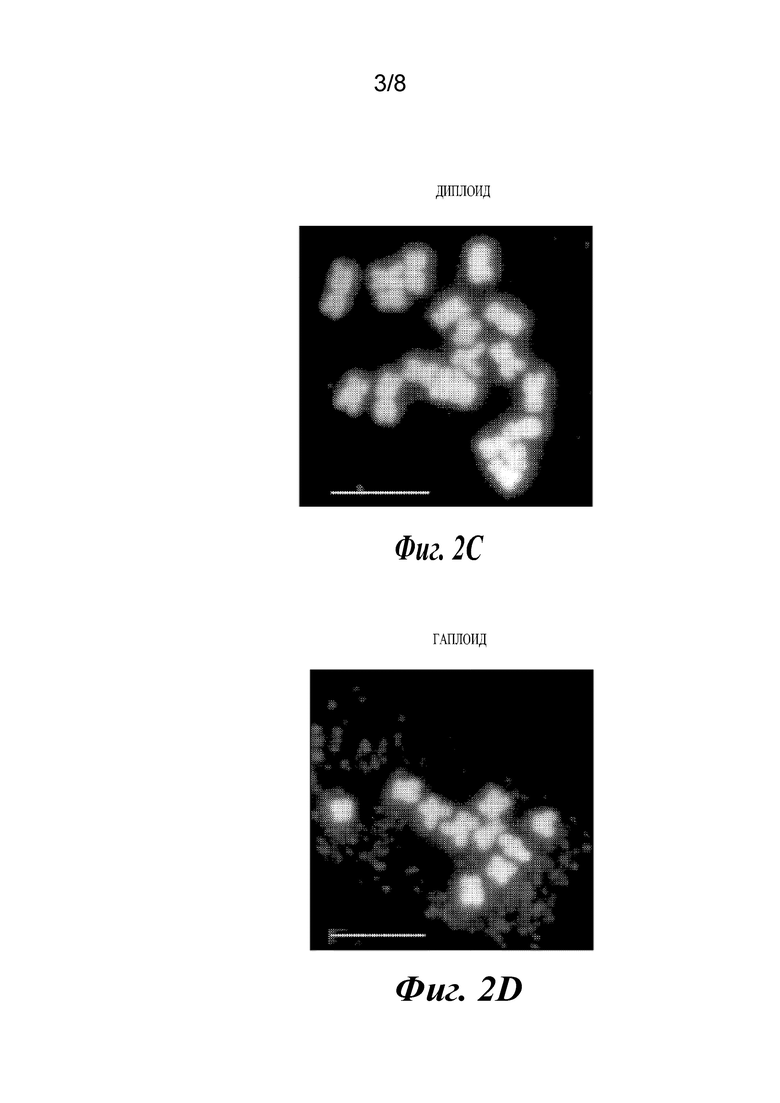
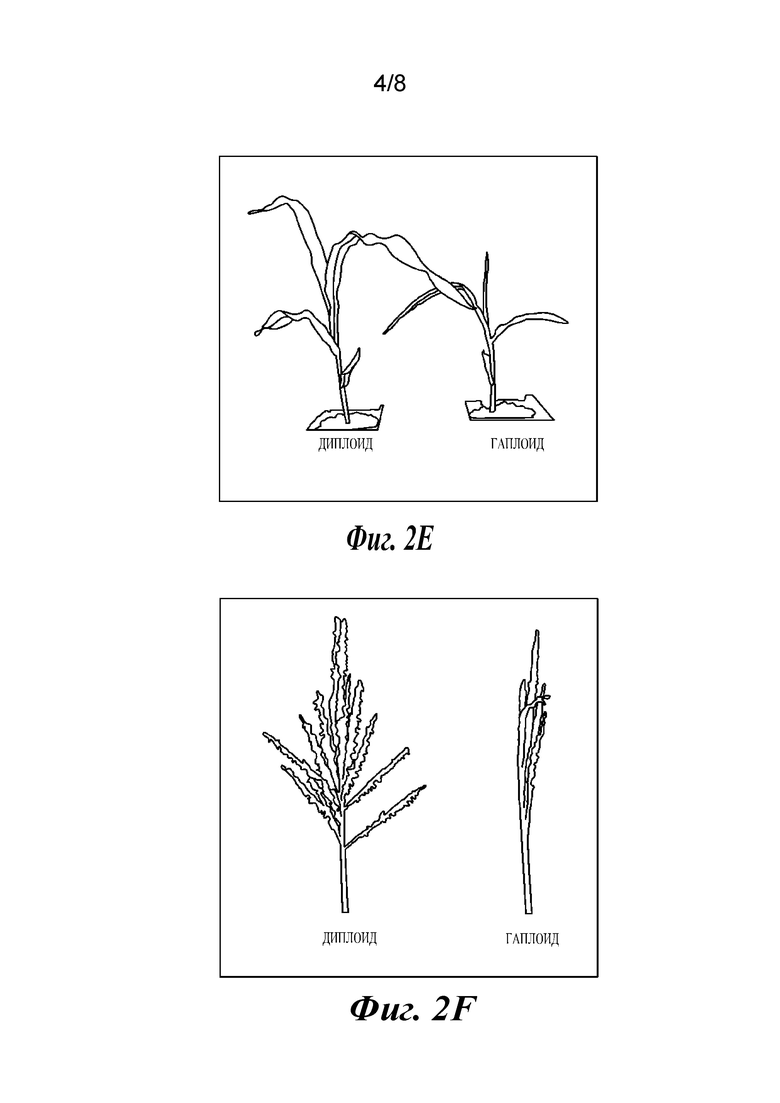
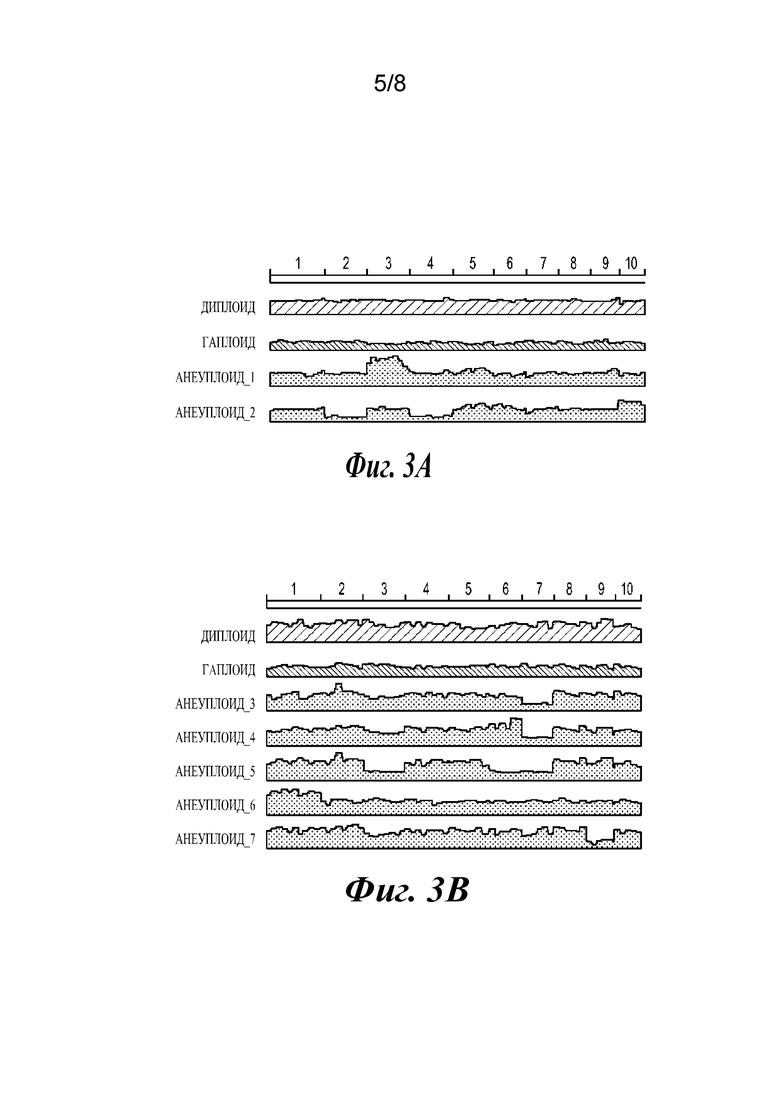
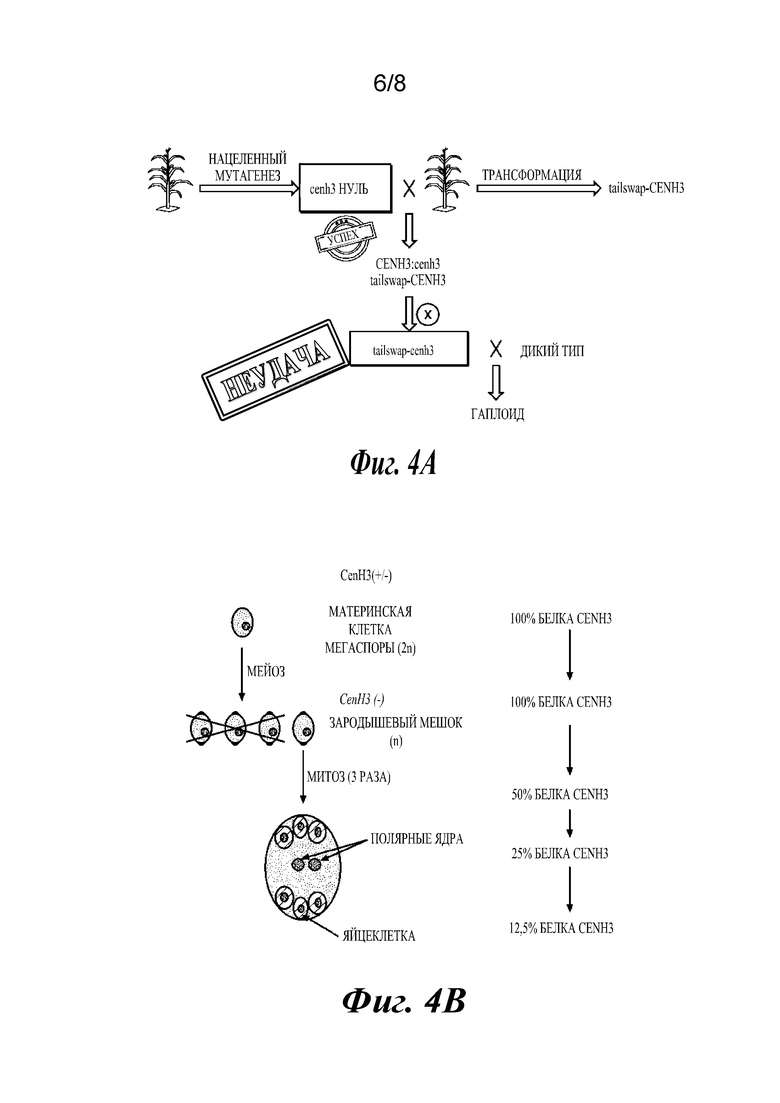
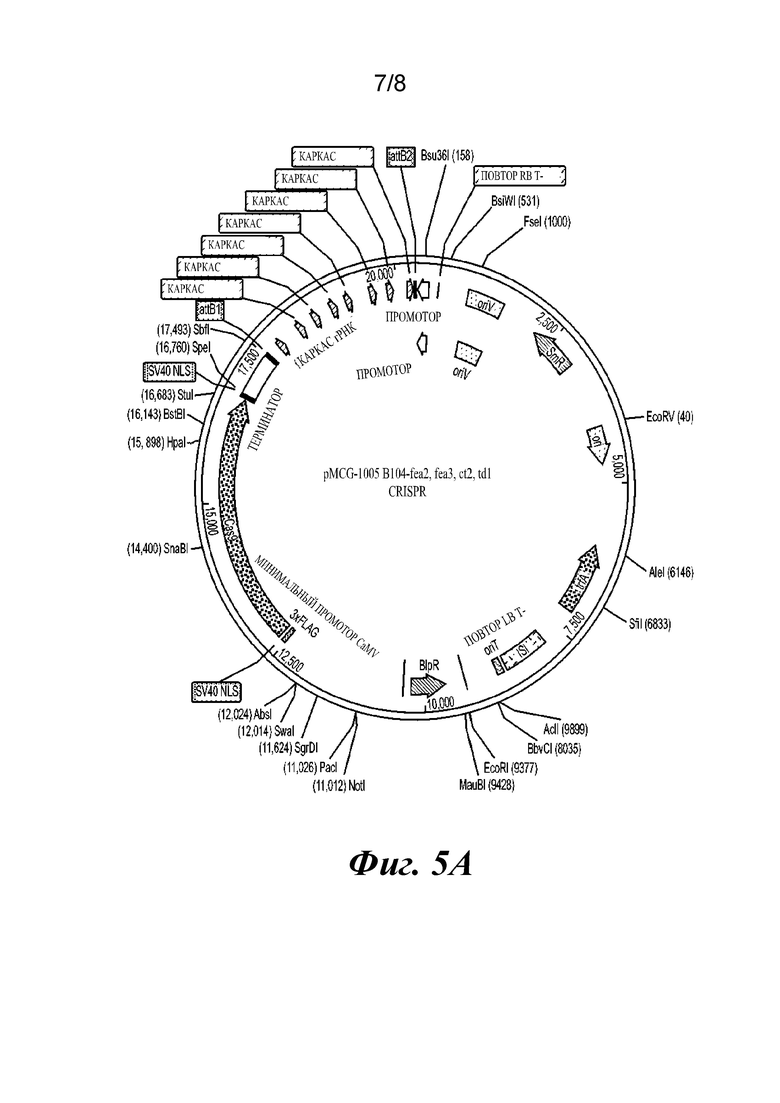
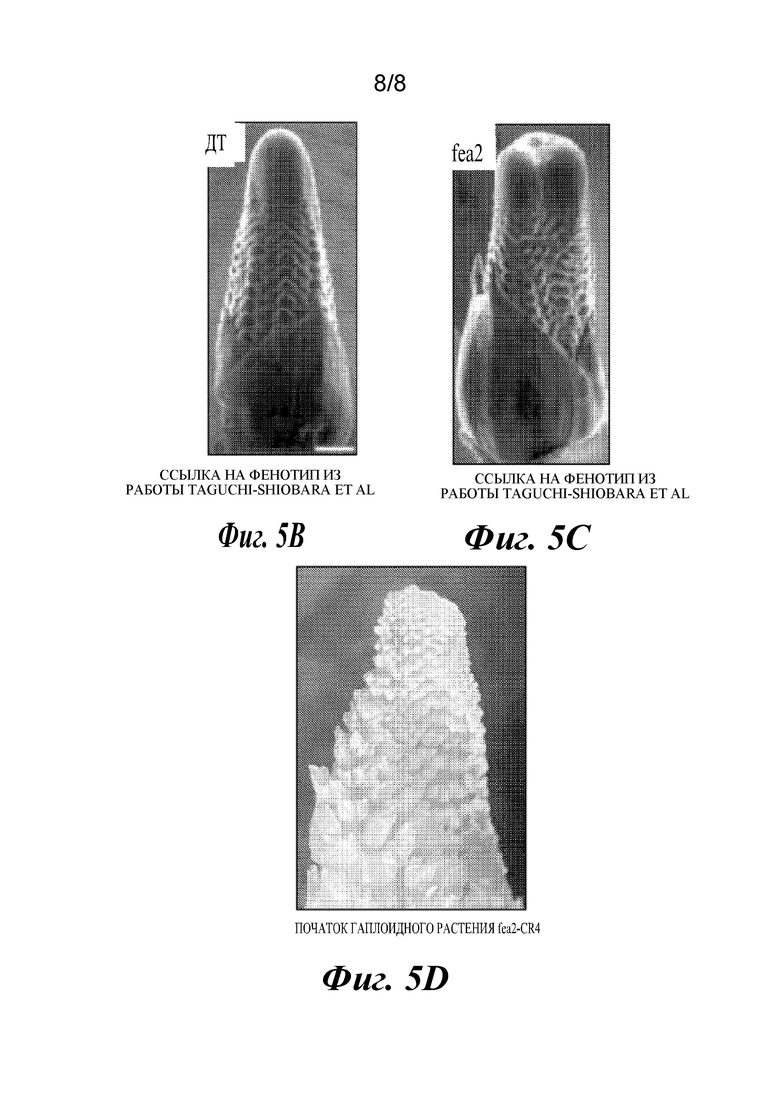
Изобретение относится к области биохимии, в частности к однодольному растению - гаплоидному индуктору, гетерозиготному по CenH3. Также раскрыты спермий и яйцеклетка указанного растения. Раскрыты способ индукции образования целевого гаплоидного однодольного растения; способ модификации генома целевого однодольного растения. Изобретение позволяет эффективно получать однодольное растение - гаплоидный индуктор, гетерозиготное по CenH3. 9 н. и 38 з.п. ф-лы, 5 ил., 8 табл., 2 пр.
1. Однодольное растение - гаплоидный индуктор, гетерозиготное по CenH3, содержащее диплоидные растительные клетки, содержащие только два локуса CenH3, где только один локус содержит аллель, кодирующий функциональный белок CENH3 дикого типа.
2. Однодольное растение по п. 1, отличающееся тем, что растение содержит один генетически модифицированный локус CenH3, содержащий аллель, кодирующий нефункциональный белок CENH3.
3. Однодольное растение по п. 2, отличающееся тем, что аллель, кодирующий нефункциональный белок CENH3, представляет собой белок-нулевой аллель.
4. Однодольное растение по п. 3, отличающееся тем, что аллель, кодирующий нефункциональный белок CENH3, представляет собой РНК-нулевой аллель.
5. Однодольное растение по п. 1, отличающееся тем, что аллель, кодирующий нефункциональный белок CENH3, вызван мутацией со сдвигом рамки, которая создает стоп-кодон, отменяющий функцию.
6. Однодольное растение по п. 1, отличающееся тем, что один из локусов CenH3 представляет собой эндогенный локус CenH3 на первой диплоидной хромосоме и его аллель представляет собой частично или полностью удаленную последовательность CenH3 дикого типа.
7. Однодольное растение по п. 3, отличающееся тем, что один из локусов CenH3 представляет собой эндогенный локус CenH3 на второй диплоидной хромосоме, его аллель представляет собой интактную последовательность CenH3, кодирующую функциональный белок CENH3.
8. Однодольное растение по п. 7, отличающееся тем, что функциональный белок CENH3 представляет собой белок CENH3 дикого типа, функция которого включает формирование и/или организацию кинетохора.
9. Однодольное растение по п. 8, отличающееся тем, что спермии или яйцеклетки содержат меньше CENH3, чем спермии или яйцеклетки растения дикого типа, гомозиготного по CENH3, необязательно отличающееся тем, что спермии или яйцеклетки содержат меньше чем 50% CENH3 по сравнению со спермиями или яйцеклетками растения дикого типа, предпочтительно отличающееся тем, что спермии или яйцеклетки содержат от около 50% до 10%, например 50%, 25% или 12,5% CENH3, по сравнению со спермиями или яйцеклетками растения дикого типа.
10. Однодольное растение по п. 1, отличающееся тем, что растение представляет собой кукурузу, пшеницу, рис, сорго, ячмень, овес, тритикале, рожь, просо африканское, просо пальчатое, просо обыкновенное, просо итальянское, банан, бамбук, сахарный тростник, просо прутьевидное, мискантус, аспарагус, лук, чеснок, лук скороду или ямс.
11. Однодольное растение по п. 10, отличающееся тем, что растение представляет собой кукурузу.
12. Однодольное растение по п. 1, дополнительно содержащее экзогенную сайт-нацеленную нуклеазу, экспрессируемую клетками однодольного растения.
13. Однодольное растение по п. 12, отличающееся тем, что растение содержит один генетически модифицированный локус, содержащий аллель, кодирующий нефункциональный белок CENH3.
14. Однодольное растение по п. 13, отличающееся тем, что аллель, кодирующий нефункциональный белок CENH3, представляет собой белок-нулевой аллель.
15. Однодольное растение по п. 14, отличающееся тем, что аллель, кодирующий нефункциональный белок CENH3, представляет собой РНК-нулевой аллель.
16. Однодольное растение по п. 12, отличающееся тем, что аллель, кодирующий нефункциональный белок CENH3, вызван мутацией со сдвигом рамки, которая создает стоп-кодон, отменяющий функцию.
17. Однодольное растение по п. 14, отличающееся тем, что один из локусов CenH3 представляет собой эндогенный локус CenH3 на первой диплоидной хромосоме и его аллель представляет собой частично или полностью удаленную последовательность CenH3 дикого типа.
18. Однодольное растение по п. 14, отличающееся тем, что один из локусов CenH3 представляет собой эндогенный локус CenH3 на второй диплоидной хромосоме и его аллель представляет собой интактную последовательность CenH3, кодирующую функциональный белок CENH3.
19. Однодольное растение по п. 18, отличающееся тем, что функциональный белок CENH3 представляет собой белок CENH3 дикого типа.
20. Однодольное растение по п. 19, отличающееся тем, что спермии или яйцеклетки содержат меньше CENH3, чем спермии или яйцеклетки растения дикого типа, гомозиготного по CENH3, необязательно отличающееся тем, что спермии или яйцеклетки содержат меньше чем 50% CENH3 по сравнению со спермиями или яйцеклетками растения дикого типа, предпочтительно отличающееся тем, что спермии или яйцеклетки содержат от около 50% до 10%, например 50%, 25% или 12,5% CENH3, по сравнению со спермиями или яйцеклетками растения дикого типа.
21. Однодольное растение по п. 12, отличающееся тем, что сайт-нацеленная нуклеаза стабильно экспрессируется клетками однодольного растения.
22. Однодольное растение по п. 12, отличающееся тем, что сайт-нацеленная нуклеаза представляет собой мегануклеазу (MN), нуклеазу с «цинковыми пальцами» (ZFN), нуклеазу на основе эффектора, подобного активатору транскрипции (TALEN), или нуклеазу на основе CRISPR, необязательно отличающееся тем, что нуклеаза выбрана из нуклеазы Cas9, нуклеазы Cfp1, dCas9-FokI, dCpf1-FokI, химерной Cas9-цитидиндезаминазы, химерной Cas9-адениндезаминазы, химерной FEN1-FokI, Mega-TAL, никазы Cas9 (nCas9), химерной нуклеазы dCas9, отличной отFokI, нуклеазы dCpf1, отличной от FokI, химерной Cpf1-цитидиндезаминазы и Cpf1-адениндезаминазы.
23. Однодольное растение по п. 12, отличающееся тем, что геном растения содержит гетерологичную конструкцию нуклеиновой кислоты, кодирующую нуклеазу.
24. Однодольное растение по п. 12, дополнительно содержащее направляющую РНК, экспрессируемую клетками однодольного растения.
25. Однодольное растение по п. 24, отличающееся тем, что геном растения содержит гетерологичную конструкцию нуклеиновой кислоты, кодирующую нРНК.
26. Однодольное растение по п. 25, отличающееся тем, что геном растения содержит донорную молекулу нуклеиновой кислоты.
27. Однодольное растение по п. 12, отличающееся тем, что растение представляет собой кукурузу, пшеницу, рис, сорго, ячмень, овес, тритикале, рожь, просо африканское, просо пальчатое, просо обыкновенное, просо итальянское, банан, бамбук, сахарный тростник, просо прутьевидное, мискантус, аспарагус, лук, чеснок, лук скороду или ямс.
28. Яйцеклетка, образованная растением по любому из пп. 1-11, где у яйцеклетки отсутствует один локус, содержащий аллель, кодирующий функциональный белок CENH3, и яйцеклетка содержит не больше чем около 12,5% функционального белка CENH3 по сравнению с соответствующей яйцеклеткой, образованной растением, гомозиготным по CenH3.
29. Спермий, образованный растением по любому из пп. 1-11, где у спермия, отсутствует один локус, содержащий аллель, кодирующий функциональный белок CENH3, и спермий содержит не больше чем около 25% функционального белка CENH3 по сравнению с соответствующей яйцеклеткой, образованной растением, гомозиготным по CenH3.
30. Яйцеклетка, образованная растением по любому из пп. 12-27, где у яйцеклетки отсутствует один локус, содержащий аллель, кодирующий функциональный белок CENH3, и яйцеклетка содержит не больше чем около 12,5% функционального белка CENH3 по сравнению с соответствующей яйцеклеткой, образованной растением, гомозиготным по CenH3.
31. Спермий, образованный растением по любому из пп. 12-27, где у спермия отсутствует один локус, содержащий аллель, кодирующий функциональный белок CENH3, и спермий содержит не больше чем около 25% функционального белка CENH3 по сравнению с соответствующей яйцеклеткой, образованной растением, гомозиготным по CenH3.
32. Способ индукции образования целевого гаплоидного однодольного растения, включающий опыление целевого родительского однодольного растения пыльцой от однодольного растения - гаплоидного индуктора - по любому из пп. 1-11, идентификацию гаплоидного потомка и отбор по меньшей мере одного гаплоидного потомка, полученного в результате опыления.
33. Способ по п. 32, дополнительно включающий удвоение хромосом отобранного гаплоидного потомка.
34. Способ по п. 33, отличающийся тем, что удвоение хромосом является спонтанным или индуцированным с помощью агента для удвоения хромосом, необязательно выбранного из колхицина, пронамида, дитипира, трифлуралина или другого антимикротрубочкового агента.
35. Способ по п. 32, отличающийся тем, что целевое однодольное растение выбрано из кукурузы, пшеницы, риса, сорго, ячменя, овса, тритикале, ржи, проса африканского, проса пальчатого, проса обыкновенного, проса итальянского, банана, бамбука, сахарного тростника, проса прутьевидного, мискантуса, аспарагуса, лука, чеснока, лука скороды или ямса.
36. Способ индукции образования целевого гаплоидного однодольного растения, включающий опыление однодольного растения - гаплоидного индуктора - по любому из пп. 1-11 пыльцой от целевого родительского однодольного растения, идентификацию гаплоидного потомка и отбор по меньшей мере одного гаплоидного потомка, полученного в результате опыления.
37. Способ по п. 36, дополнительно включающий удвоение хромосом отобранного гаплоидного потомка.
38. Способ по п. 37, отличающийся тем, что удвоение хромосом является спонтанным или индуцированным с помощью агента для удвоения хромосом, необязательно выбранного из колхицина, пронамида, дитипира, трифлуралина или другого антимикротрубочкового агента.
39. Способ по п. 36, отличающийся тем, что целевое однодольное растение выбрано из кукурузы, пшеницы, риса, сорго, ячменя, овса, тритикале, ржи, проса африканского, проса пальчатого, проса обыкновенного, проса итальянского, банана, бамбука, сахарного тростника, проса прутьевидного, мискантуса, аспарагуса, лука, чеснока, лука скороды или ямса.
40. Способ модификации генома целевого однодольного растения, включающий:
индукцию образования целевого гаплоидного однодольного растения, включающую опыление целевого родительского однодольного растения пыльцой от однодольного растения - гаплоидного индуктора - по любому из пп. 12-27, идентификацию гаплоидного потомка и отбор по меньшей мере одного гаплоидного потомка, полученного в результате опыления,
при этом гаплоидный потомок содержит геном целевого однодольного растения, но не однодольного растения - гаплоидного индуктора, и геном гаплоидного потомка был модифицирован сайт-направленной нуклеазой и, необязательно, по меньшей мере одной направляющей РНК, доставляемой однодольным растением - гаплоидным индуктором.
41. Способ по п. 40, дополнительно включающий удвоение хромосом отобранного гаплоидного потомка.
42. Способ по п. 41, отличающийся тем, что удвоение хромосом является спонтанным или индуцированным с помощью агента для удвоения хромосом, необязательно выбранного из колхицина, пронамида, дитипира, трифлуралина или другого антимикротрубочкового агента.
43. Способ по п. 40, отличающийся тем, что целевое однодольное растение выбрано из кукурузы, пшеницы, риса, сорго, ячменя, овса, тритикале, ржи, проса африканского, проса пальчатого, проса обыкновенного, проса итальянского, банана, бамбука, сахарного тростника, проса прутьевидного, мискантуса, аспарагуса, лука, чеснока, лука скороды или ямса.
44. Способ модификации генома целевого однодольного растения, включающий:
индукцию образования целевого гаплоидного однодольного растения, включающую опыление однодольного растения - гаплоидного индуктора - по любому из пп. 12-27 пыльцой от целевого родительского однодольного растения, идентификацию гаплоидного потомка и отбор по меньшей мере одного гаплоидного потомка, полученного в результате опыления,
при этом гаплоидный потомок содержит геном целевого однодольного растения, но не однодольного растения - гаплоидного индуктора, и геном гаплоидного потомка был модифицирован сайт-направленной нуклеазой и, необязательно, по меньшей мере одной направляющей РНК, доставляемой однодольным растением - гаплоидным индуктором.
45. Способ по п. 44, дополнительно включающий удвоение хромосом отобранного гаплоидного потомка.
46. Способ по п. 45, отличающийся тем, что удвоение хромосом является спонтанным или индуцированным с помощью агента для удвоения хромосом, необязательно выбранного из колхицина, пронамида, дитипира, трифлуралина или другого антимикротрубочкового агента.
47. Способ по п. 44, отличающийся тем, что целевое однодольное растение выбрано из кукурузы, пшеницы, риса, сорго, ячменя, овса, тритикале, ржи, проса африканского, проса пальчатого, проса обыкновенного, проса итальянского, банана, бамбука, сахарного тростника, проса прутьевидного, мискантуса, аспарагуса, лука, чеснока, лука скороды или ямса.
| US 20140090099 A1, 27.03.2014 | |||
| WO 2019136417 A2, 11.07.2019 | |||
| WO 2017200386 A1, 23.11.2017 | |||
| ГЕН И ВАРИАЦИИ, СВЯЗАННЫЕ С ФЕНОТИПОМ BM1, МОЛЕКУЛЯРНЫЕ МАРКЕРЫ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2617958C2 |

