Область изобретения
Настоящее изобретение относится к областям биотехнологии, генной инженерии и медицинской химии. В одном воплощении данное изобретение относится к области экспрессии гена. В другом воплощении данное изобретение относится к диацилгидразиновым лигандам и хиральным диацилгидразиновым лигандам для естественных и мутантных ядерных рецепторов и к их применению в индуцируемой системе генной экспрессии на основе ядерного рецептора, и к способам модуляции экспрессии гена в клетке-хозяине с использованием таких лигандов и индуцируемой системы экспрессии гена.
Предшествующий уровень техники
В данном описании изобретения упоминаются различные публикации, содержание которых включено посредством ссылки во всей их полноте. Однако цитирование любой ссылки в данном описании изобретения не следует истолковывать как признание того, что эта ссылка является "предшествующим уровнем техники" для настоящей заявки.
В области генной инженерии точное регулирование генной экспрессии является полезным средством для исследования, воздействия на развитие и регуляции развития и других физиологических процессов. Генная экспрессия является сложным биологическим процессом, включающим ряд специфических взаимодействий белок-белок. Для инициации генной экспрессии, так чтобы продуцировалась РНК, необходимая в качестве первой стадии белкового синтеза, активатор транскрипции нужно приблизить к промотору, который регулирует транскрипцию гена. Обычно сам активатор транскрипции ассоциирован с белком, имеющим по меньшей мере один ДНК-связывающий домен, который связывается с ДНК-связывающими сайтами, присутствующими в промоторных участках генов. Таким образом, для того, чтобы произошла экспрессия гена, белок, содержащий ДНК-связывающий домен и домен трансактивации, расположенный на должном расстоянии от ДНК-связывающего домена, необходимо привести в правильное положение в промоторном участке гена.
В традиционном трансгенном подходе используется специфический для данного типа клетки промотор для запуска экспрессии сконструированного трансгена. ДНК-конструкция, содержащая трансген, сначала встраивается в геном хозяина. При инициации посредством активатора транскрипции, в данном клеточном типе происходит экспрессия трансгена.
Другим средством регуляции экспрессии чужих генов в клетках являются индуцируемые промоторы. Примеры использования таких индуцируемых промоторов включают промотор PR1-a, прокариотические системы репрессор-оператор, системы иммуносупрессант-иммунофилин, и системы активации транскрипции высших эукариот, такие как системы рецепторов стероидных гормонов, и описаны ниже.
Промотор PR1-a из табака индуцируется в условиях системного ответа приобретенной устойчивости после патогенной атаки. Применение PR1-a может быть ограничено, так как он часто реагирует на эндогенные вещества и внешние факторы, такие как патогены, УФ-В излучение и загрязнения. Были описаны системы генной регуляции на основе промоторов, индуцируемые с помощью теплового шока, интерферона и тяжелых металлов (Wurn et al., Proc. Natl. Acad. Sci. USA 55: 5414-5418 (1986); Arnheiter et ah, Клетка 62: 51-61 (1990); Filmus et al, Nucleic Acids Research 20: 27550-27560 (1992)). Однако такие системы имеют ограничения из-за их воздействия на экспрессию генов-немишеней. Такие системы также являются ликовыми.
Прокариотические системы репрессор-оператор используют бактериальные репрессорные белки и уникальные операторные ДНК последовательности, с которым они связываются. Как тетрациклиновые ("Tet"), так и лактозные ("Lac") системы репрессор-оператор из бактерий Escherichia coli использовали в растениях и животных для контроля генной экспрессии. В Tet-системе тетрациклин связывается с репрессорным белком TetR, вызывая конформационное изменение, которое высвобождает репрессорный белок из оператора, что в результате позволяет произойти транскрипции. В Lac-системе lac-оперон активируется в ответ на присутствие лактозы или синтетических аналогов, таких как изопропил-b-O-тиогалактозид. К сожалению, использование таких систем ограничивается нестабильной химией лигандов, то есть тетрациклина и лактозы, их токсичностью, их естественным присутствием или относительно высоких уровней, требующихся для индукции или репрессии. По тем же причинам пригодность таких систем в животных ограничена.
Иммуносупрессивные молекулы, такие как FK506, рапамицин и циклоспорин А, могут связываться с иммунофилинами FKBP12, циклофилином и так далее. С использованием данной информации была разработана общая стратегия сближения любых двух белков просто путем помещения FK506 на каждый из двух белков или путем помещения FK506 на один и циклоспорина А на другой белок. Синтетический гомодимер FK506 (FK1012) или соединение, полученное путем слияния FK506-циклоспорина (FKCsA), затем можно использовать для индуцирования димеризации этих молекул (Spencer et al, Science 262: 1019-24 (1993); Belshaw et al, Proc Natl Acad Sci USA 93: 4604-7 (1996)). ДНК-связывающий домен Gal4, слитый с FKBP12, и активаторный домен VP16, слитый с циклофилином, и соединение FKCsA использовали, чтобы продемонстрировать гетеродимеризацию и активацию репортерного гена под контролем промотора, содержащего Gal4-связывающие сайты. К сожалению, такая система включает иммуносупрессанты, которые могут иметь нежелательные побочные эффекты и, вследствие этого, ограничивают ее использование для различных применений по переключению генов у млекопитающих.
Также были использованы системы активации транскрипции у высших эукариот, такие как системы рецепторов стероидных гормонов. Рецепторы стероидных гормонов являются членами суперсемейства ядерных рецепторов и обнаружены в клетках позвоночных и беспозвоночных. К сожалению, использование стероидных соединений, которые активируют рецепторы, для регуляции генной экспрессии, особенно у растений и млекопитающих, ограничено из-за их участия во многих других естественных биологических путях в таких организмах. С целью преодоления таких затруднений была разработана альтернативная система с использованием экдизоновых рецепторов насекомых (EcR).
Рост, линька и развитие насекомых регулируются стероидным гормоном экдизоном (гормоном линьки) и ювенильными гормонами (Dhadialla et al, Annu. Rev. Entomol. 43: 545-569 (1998)). Молекулярная мишень для экдизона у насекомых состоит по меньшей мере из экдизонового рецептор (EcR) и белок Ultraspiracle (USP). EcR является членом суперсемейства стероидных ядерных рецепторов, которые отличаются характерными ДНК- и лиганд-связывающими доменами, и доменом активации (Koelle et al, Клетка, 67: 59-71 (1991)). EcR-рецепторы восприимчивы к ряду стероидных соединений, таких как понастерон А и муристерон А. Недавно были описаны нестероидные соединения с экдистероидной агонистической активностью, включая имеющиеся в продаже инсектициды тебуфенозид и метоксифенозид, которые представляет на мировом рынке Rohm and Haas Company (смотри WO 96/27673 и US 5530028). Оба аналога имеют исключительные профили безопасности в других организмах.
Экдизоновые рецепторы насекомых (EcR) гетеродимеризуются с Ultraspiracle (USP), имеющимся у насекомых гомологом ретиноидного X-рецептора млекопитающих (RXR), и связывают экдистероиды и элементы ответа экдизоновых рецепторов и активируют транскрипцию реагирующих на экдизон генов. Комплексы EcR/USP/лиганд играют важные роли в развитии и размножении насекомого. EcR имеет пять модулярных доменов, домены A/B (трансактивация), С (ДНК-связывание, гетеродимеризация), D (шарнир, гетеродимеризация), E (связывание лиганда, гетеродимеризация и трансактивация) и F (трансактивация). Некоторые из этих доменов, такие как A/B, C и E, сохраняют свою функцию, когда они конденсированы с другими белками.
Строго регулируемые индуцируемые системы экспрессии гена или "переключатели гена" полезны для различных применений, таких как генная терапия, крупномасштабное продуцирование белков в клетках, высокопроизводительные клеточные скрининговые анализы, функциональная геномика и регулирование свойств трансгенных растений и животных.
В первом варианте генного переключателя на основе EcR использовали EcR Drosophila melanogaster (DmEcR) и RXR Mus musculus (MmRXR) и обнаружили, что эти рецепторы в присутствие стероида, понастерона A, трансактивируют репортерные гены в клеточных линиях млекопитающих и у трансгенных мышей (Christopherson et al, Proc. Natl. Acad. Sci. U.S.A. 59: 6314-6318 (1992); No et al., Proc. Natl. Acad. Sci. U.S.A. 95: 3346-3351 (1996)). Позже Suhr etal, Proc. Natl. Acad. Sci. 95: 7999-8004 (1998) показали, что нестероидный агонист экдизона, тебуфенозид, индуцировал высокий уровень трансактивации репортерных генов в клетках млекопитающих при помощи EcR Bombyx mori (BmEcR) в отсутствие экзогенного гетеродимерного партнера.
В WO 97/38117 и W099/58155 раскрыты способы модуляции экспрессии экзогенного гена, в которых ДНК-конструкция, содержащая экзогенный ген, и экдизоновый элемент ответа активируется с помощью второй ДНК-конструкции, содержащей экдизоновый рецептор, который в присутствие лиганда, вследствие этого, и возможно в присутствие рецептора, способного действовать в качестве молчащего партнера, связывается с экдизоновым элементом ответа для индуцирования экспрессии гена. Выбранные экдизоновые рецепторы выделяли из Drosophila melanogaster. Обычно такие системы требуют присутствия молчащего партнера, предпочтительно ретиноидного Х-рецептора (RXR), с целью обеспечения оптимальной активации. В клетках млекопитающих экдизоновый рецептор насекомых (EcR) гетеродимеризуются с ретиноидным X-рецептором (RXR) и регулирует экспрессию генов-мишеней лиганд-зависимым способом. В WO 99/02683 раскрыто, что экдизоновый рецептор, выделенный из шелкопряда Bombyx mori, функционирует в системах млекопитающих без необходимости в экзогенном димерном партнере.
В US 6265173 B1 раскрыто, что различные члены суперсемейства стероидных/тиреоидных рецепторов могут объединяться с рецептором Ultraspiracle из Drosophila melanogaster (USP) или его фрагментами, содержащими по меньшей мере домен димеризации USP, для использования в системе экспрессии гена. В US 5880333 раскрыта EcR Drosophila melanogaster и гетеродимерная система Ultraspiracle (USP), используемая в растениях, в которых домен трансактивации и ДНК-связывающий домен расположены на двух разных гибридных белках. К сожалению такие системы на основе USP являются конститутивными в животных клетках и, вследствие этого, являются неэффективными для регуляции экспрессии репортерного гена.
В каждом из данных случаев домен трансактивации и ДНК-связывающий домен (или в виде нативного EcR, как в WO 99/02683, или в виде модифицированного EcR, как в WO 97/38117) внедряли в одну молекулу, и другие гетеродимерные партнеры, или USP или RXR, использовали в их нативном состоянии.
Недостатки описанной выше системы регуляции гена, основанной на EcR, включают значительную фоновую активность в отсутствие лигандов и непригодность таких систем для использования как в растениях, так и у животных (смотри US 5880333). Следовательно, в данной области техники существует потребность в улучшенных системах на основе EcR для точного модуляции экспрессии экзогенных генов как в растениях, так и у животных. Такие улучшенные системы могли бы стать полезными для таких применений, как генная терапия, крупномасштабное продуцирование белков и антител, высокопроизводительные клеточные скрининговые анализы, функциональная геномика и регуляция свойств трансгенных животных. Для некоторых применений, таких как генная терапия, может быть желательно иметь индуцируемую систему экспрессии генов, которая хорошо реагирует на синтетические нестероидные лиганды и в то же время невосприимчива к естественным стероидам. Таким образом, улучшенные системы, которые являются простыми, компактными и зависят от лигандов, которые являются относительно недорогими, общедоступными и низкотоксичными для хозяина, будут полезны для регуляции биологических систем.
Недавно было показано, что индуцируемая система экспрессии гена на основе экдизонового рецептора, в которой домен трансактивации и ДНК-связывающий домен отделены друг от друга путем размещения их на двух разных белках, приводит к существенно сниженной фоновой активности в отсутствие лиганда и значительно повышенной активности по сравнению с фоном в присутствии лиганда (смотри WO 01/70816 A1, включенную в данное описание изобретения во всей ее полноте посредством ссылки). Такая двухгибридная система представляет собой значительно улучшенную индуцируемую систему модуляции экспрессии гена по сравнению с двумя системами, раскрытыми в заявках WO 97/38117 и WO 99/02683. В двухгибридной системе используется способность пары взаимодействующих белков приводить домен активации транскрипции в более благоприятное положение относительно ДНК-связывающего домена так, что когда ДНК-связывающий домен связывается с ДНК-связывающим сайтом гена, домен трансактивации более эффективно активирует промотор (смотри, например, US 5283173). Кратко, двухгибридная система экспрессии гена содержит две кассеты экспрессии гена; первую, кодирующую ДНК-связывающий домен, слитый с полипептидом ядерного рецептора, и вторую, кодирующую домен трансактивации, конденсированный с другим полипептидом ядерного рецептора. В присутствие лиганда взаимодействие первого полипептида со вторым полипептидом эффективно связывает ДНК-связывающий домен с доменом трансактивации. Фоновая активность в отсутствие лиганда существенно понижена, поскольку ДНК-связывающий домен и домен трансактивации находятся на двух разных молекулах.
Двухгибридная система также обеспечивает улучшенную чувствительность к нестероидным лигандам, например диацилгидразинам, по сравнению со стероидными лигандами, например понастероном А ("PonA") или муристероном A ("MurA"). Значит, по сравнению со стероидами, нестероидные лиганды обеспечивают более высокую активность при более низкой концентрации. Кроме того, так как трансактивация, основанная на переключателях генов EcR, часто является зависимой от клеточной линии, легче адаптировать системы генных переключателей для получения максимальной возможности трансактивации для каждого применения. Более того, в двухгибридной системе устранены некоторые побочные эффекты сверхэкспрессии RXR, которая часто имеет место при использовании в качестве партнера гетероди мерного рецептора немодифицированного RXR. В одной двухгибридной системе домен связывания нативной ДНК и домен трансактивации удалены из EcR или RXR, и в результате эти гибридные молекулы имеют меньшую возможность взаимодействовать с другими рецепторами стероидного гормона, присутствующего в клетке, что приводит к пониженным побочным эффектам. Дополнительные системы генных переключателей включают системы, описанные ниже, каждая из которых включена посредством ссылки: US 7091038; WO 2004078924; EP 1266015; US 20010044151; US 20020110861; US 20020119521; US 20040033600; US 20040197861; US 20040235097; US 20060020146; US 20040049437; US 20040096942; US 20050228016; US 20050266457; US 20060100416; WO 2001/70816; WO 2002/29075; WO 2002/066612; WO 2002/066613; WO 2002/066614; WO 2002/066615; WO 2005/108617; US 6258603; US 20050209283; US 20050228016; US 20060020146; EP 0965644; US 7304162; US 7304161; MX 234742; KR 10-0563143; AU 765306; AU 2002-248500; и AU 2002-306550.
С улучшением в системах регуляции генов на основе экдизоновых рецепторов происходит увеличение их использования для различных применений, что приводит к увеличению спроса на лиганды с более высокой активностью чем те, что имеются в наличие в настоящее время. В US 6258603 B1, US 2005/0209283 A1 и US 2006/0020146 A1 (и патентах, указанных здесь) раскрыты дибензоилгидразиновые лиганды. Однако существует потребность в дополнительных лигандах с улучшенными фармакологическими свойствами. Заявители обнаружили хиральные диацилгидразиновые лиганды, которые не были описаны ранее, и которые обладают неожиданной биологической активностью и способностью неожиданным образом модулировать экспрессию трансгенов.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В настоящем изобретении предложены диацилгидразиновые лиганды Формулы I и хиральные диацилгидразиновые лиганды Формул II или III для применения с индуцируемыми системами экспрессии гена на основе экдизоновых рецепторов, полезные для модуляции экспрессии представляющего интерес гена-мишени в клетке-хозяине. Хиральные диацилгидразиновые лиганды по изобретению являются энантиомерно обогащенными либо R- либо S-стереоизомером. Заявители обнаружили, что эти новые хиральные диацилгидразиновые лиганды удивительно эффективны. Таким образом, настоящее изобретение является полезным для таких применений как генная терапия, крупномасштабное продуцирование белков и антител, скрининговые клеточные анализы, функциональная геномика, протеомика, метоболомика и регуляция свойств трансгенных организмов, где является желательным контроль уровней экспрессии гена. Преимущество настоящего изобретения состоит в том, что в нем предлагаются средства регуляции генной экспрессии и адаптации уровней экспрессии для удовлетворения требованиям пользователя.
Настоящее изобретение относится к соединениям Формулы I
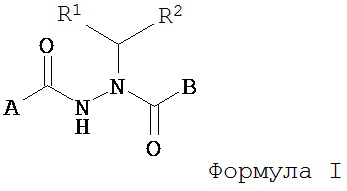
где
A представляет собой алкокси, арилалкилокси или арилокси;
B представляет собой возможно замещенный арил или возможно замещенный гетероарил; и
R1 и R2 независимо представляют собой возможно замещенный алкил, арилалкил, гидроксиалкил, галогеноалкил, возможно замещенный циклоалкил, возможно замещенный алкенил, возможно замещенный алкинил, возможно замещенный гетероцикл, возможно замещенный арил или возможно замещенный гетероарил;
или его фармацевтически приемлемые соли, гидраты, кристаллические формы или аморфные формы.
В другом воплощении настоящее изобретение относится к энантиомерно обогащенными соединениям Формулы II
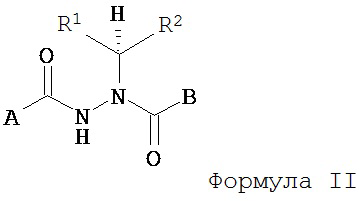
где
A представляет собой алкокси, арилалкилокси, арилокси, арилалкил, возможно замещенный арил или возможно замещенный гетероарил;
B представляет собой возможно замещенный арил или возможно замещенный гетероарил; и
R’ и R” независимо представляют собой возможно замещенный алкил, арилалкил, гидроксиалкил, галогеноалкил, возможно замещенный циклоалкил, возможно замещенный алкенил, возможно замещенный алкинил, возможно замещенный гетероцикл, возможно замещенный арил или возможно замещенный гетероарил;
при условии, что R1 отличается от R2;
где абсолютной конфигурацией асимметрического атома углерода, несущего R1 и R2, преимущественно является S;
или его фармацевтически приемлемые соли, гидраты, кристаллические формы или аморфные формы.
В другом воплощении настоящее изобретение относится к энантиомерно обогащенным соединениям Формулы III
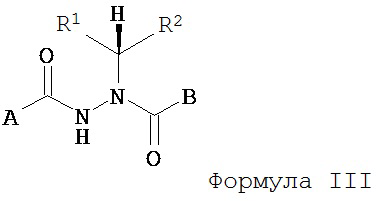
где
A представляет собой алкокси, арилалкилокси, арилокси, арилалкил, возможно замещенный арил или возможно замещенный гетероарил;
B представляет собой возможно замещенный арил или возможно замещенный гетероарил; и
R1 и R2 независимо представляют собой возможно замещенный алкил, арилалкил, гидроксиалкил, галогеноалкил, возможно замещенный циклоалкил, возможно замещенный алкенил, возможно замещенный алкинил, возможно замещенный гетероцикл, возможно замещенный арил или возможно замещенный гетероарил;
при условии, что R1 отличается от R2;
где абсолютной конфигурацией асимметрического атома углерода, несущего R1 и R2, преимущественно является R;
или его фармацевтически приемлемые соли, гидраты, кристаллические формы или аморфные формы.
В одном воплощении настоящее изобретение относится к N-(1-трет-бутил-бутил)-N’-(2-этил-3-метоксибензоил)гидразиду (R)-3,5-диметилбензойной кислоты, имеющему энантиомерный избыток по меньшей мере 95%, или к его фармацевтически приемлемой соли, гидрату, кристаллической форме или аморфной форме.
В другом воплощении настоящее изобретение относится к фармацевтической композиции, содержащей N-(1-трет-бутил-бутил)-N’-(2-этил-3-метоксибензоил)гидразид (R)-3,5-диметилбензойной кислоты, имеющий энантиомерный избыток по меньшей мере 95%, или его фармацевтически приемлемую соль, гидрат, его кристаллическую форму или аморфную форму.
Настоящее изобретение также относится к способу получения соединения Формулы IV
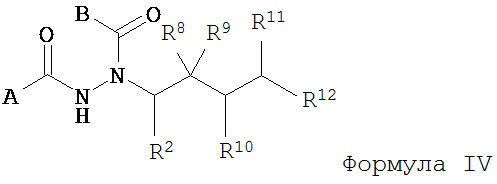
где
A представляет собой арилалкил, возможно замещенный арил или возможно замещенный гетероарил;
B представляет собой возможно замещенный арил или возможно замещенный гетероарил;
R2 представляет собой возможно замещенный алкил, арилалкил, гидроксиалкил, галогеноалкил, возможно замещенный циклоалкил, возможно замещенный алкенил, возможно замещенный алкинил, возможно замещенный гетероцикл, возможно замещенный арил или возможно замещенный гетероарил;
при условии, что R2 отличается от -CR8R9CHR10CR11R12; и
R8, R9, R10, R11 и R12 независимо выбраны из водорода, алкила, циклоалкила, гетероцикла, арила или гетероарила;
где асимметрический атом углерода, к которому присоединен R2, является энантиомерно обогащенным либо R- либо S-изомером;
включающий:
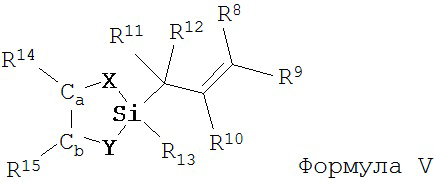
а) взаимодействие соединения Формулы V
с соединением Формулы VI
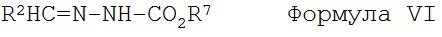
где
X и Y независимо представляют собой О или NR, при этом R представляет собой алкил или арил;
Ca и Cb независимо представляют собой асимметрический атом углерода в S-конфигурации или асимметрический атом углерода в R-конфигурации;
R14 и R15 независимо представляют собой алкил или арил;
R13 представляет собой галоген, водород, алкил, алкокси или OSO2CF3;
R7 представляет собой алкил, арилалкил или арил; и
R2, R8, R9, R10, R11 и R12 имеют значения, указанные выше;
с образованием соединения Формулы VII;
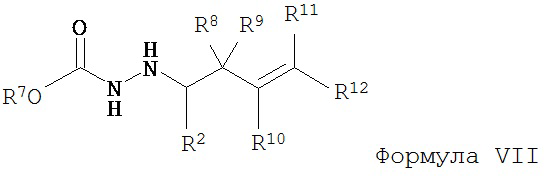
б) восстановление соединения Формулы VII с образованием соединения Формулы VIII;
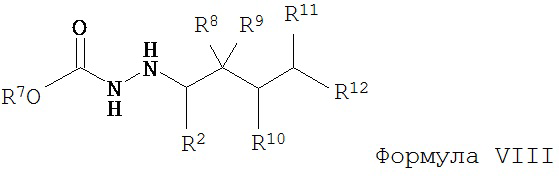
в) взаимодействие соединения Формулы VIII с соединением Формулы B-CO-LG, где LG представляет собой уходящую группу, с образованием соединения Формулы IX;
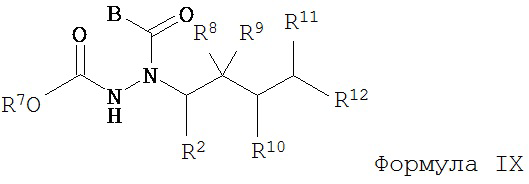
г) удаление группы R7CO2- из соединения Формулы IX с образованием соединения Формулы X; и
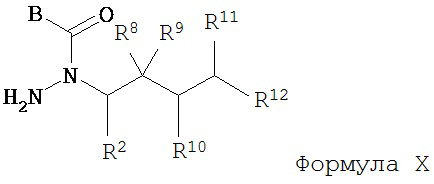
д) взаимодействие соединения Формулы Х с соединением Формулы A-CO-LG, где LG представляет собой уходящую группу, с образованием соединения Формулы IV.
Настоящее изобретение также относится к способам модуляции экспрессии гена в клетке-хозяине с использованием системы модуляции экспрессии гена с диацилгидразиновым лигандом Формулы I или хиральным диацилгидразиновым лигандом Формулы II или III.
В одном воплощении настоящее изобретение относится к применению диацилгидразинового лиганда Формулы I или хирального диацилгидразинового лиганда Формулы II или III в индуцируемой системе экспрессии гена, которая имеет пониженный уровень фоновой экспрессии гена и реагирует на субмикромолекулярные концентрации лиганда.
В другом воплощении изобретение относится к способу модуляции экспрессии гена-мишени в клетке-хозяине, где клетка-хозяин содержит первую кассету экспрессии гена, содержащую первый полинуклеотид, кодирующую первый полипептид, содержащую:
1) домен трансактивации;
2) ДНК-связывающий домен; и
3) лиганд-связывающий домен ядерного рецептора группы H;
и вторую кассету экспрессии гена, содержащую:
1) элемент ответа, способный связываться с указанным ДНК-связывающим доменом;
2) промотор, который активируется доменом трансактивации; и
3) указанный ген-мишень;
включающий приведение в контакт указанной клетки-хозяина с диацилгидразиновым лигандом Формулы I или хиральным диацилгидразиновым лигандом Формулы II или III; где экспрессия гена-мишени модулируется.
В другом воплощении изобретение относится к способу регуляции экспрессии эндогенного или гетерологичного гена в трансгенном субъекте, включающему приведение лиганда в контакт с экдизон-рецепторным комплексом в клетках субъекта, где клетки дополнительно содержат ДНК-связывающую последовательность для экдизон-рецепторного комплекса в комбинации с лигандом, и где образование комплекса «экдизон-рецепторный комплекс-лиганд-ДНК-связывающая последовательность» индуцирует экспрессию гена, и где лиганд представляет собой диацилгидразиновый лиганд Формулы I или хиральный диацилгидразиновый лиганд Формулы II или III; где экспрессию эндогенного или гетерологичного гена в трансгенном субъекте регулируется.
В другом воплощении изобретение относится к способу модуляции экспрессии гена-мишени в клетке-хозяине, включающему стадии:
а) введение в клетку-хозяина системы модуляции экспрессии гена, содержащей:
1) первую кассету экспрессии гена, которая способна экспрессироваться в клетке-хозяине, где указанная первая кассета экспрессии гена содержит полинуклеотидную последовательность, которая кодирует первый гибридный полипептид, содержащий:
(а) ДНК-связывающий домен, который распознает элемент ответа, ассоциированный с геном, экспрессия которого подлежит модулированию; и
(б) лиганд-связывающий домен экдизонового рецептора;
2) вторую кассету экспрессии гена, которая способна экспрессироваться в клетке-хозяине, где указанная вторая кассета экспрессии гена содержит полинуклеотидную последовательность, которая кодирует второй гибридный полипептид, содержащий:
(а) домен трансактивации; и
(б) лиганд-связывающий домен химерного ретиноидного X-рецептора; и
3) третью кассету экспрессии гена, которая способна экспрессироваться в клетке-хозяине, где указанная третья кассета экспрессии гена содержит полинуклеотидную последовательность, включающую:
(а) элемент ответа, распознаваемый с помощью ДНК-связывающего домена первого гибридного полипептида;
(б) промотор, который активируется доменом трансактивации второго гибридного полипептида; и
(в) ген, экспрессия которого подлежит модулированию; и
(б) введение в клетку-хозяина диацилгидразинового лиганда Формулы I или хирального диацилгидразинового лиганда Формулы II или III; где экспрессия гена в клетке-хозяине модулируется.
В другом воплощении настоящее изобретение относится к способу продуцирования полипептида, включающему стадии:
а) селекции клетки, которая является по существу нечувствительной к воздействию диацилгидразинового лиганда Формулы I или хирального диацилгидразинового лиганда Формулы II или III;
б) введения в клетку:
1) ДНК-конструкции, содержащей:
(а) экзогенный ген, кодирующий полипептид; и
(б) элемент ответа;
где ген находится под контролем элемента ответа; и
2) экдизон-рецепторного комплекса, содержащего:
(а) ДНК-связывающий домен, который связывается с элементом ответа;
(б) связывающий домен для указанного лиганда; и
(в) домен трансактивации; и
в) воздействие на клетку указанного лиганда; при котором продуцируется полипептид.
Данное воплощение изобретения обеспечивает преимущество контролируемого во времени продуцирования полипептида клеткой. Более того, в тех случаях, когда накопление такого полипептида может повредить клетку, экспрессия полипептида может быть ограничена короткими периодами воздействия на указанную клетку соединений по настоящему изобретению. Такой контроль является особенно важным, когда экзогенный ген представляет собой терапевтический ген. Терапевтические гены могут быть привлечены для продуцирования полипептидов, который контролирует необходимые функции, такие как продуцирование инсулина у пациентов с диабетом. Также они могут быть использованы для продуцирования повреждающих или даже летальных белков, таких как белки, летальные для раковых клеток. Также такой контроль может быть важным, когда уровни продуцируемого белка могут привести к метаболическому уменьшению роста или репродукции, например у трансгенных растений.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг.1: Схема конструкции-переключателя CVBE и репортерной конструкции 6XEcRE Lac Z. Фланкируют обе структуры длинные концевые повторы, G418 и пуромицин представляют собой селектируемые маркеры, CMV представляет собой цитомегаловирусный промотор, VBE представляет собой кодирующую последовательность для аминокислот 26-546 из EcR Bombyx mori, вставленную ниже домена трансактивации VP16, 6XEcRE представляет собой шесть копий экдизонового элемента ответа, lacZ кодирует репортерный фермента P-галактозидазу.
Фиг.2A-2B: Сравнение с помощью хиральной ВЭЖХ (A) рац-, (Б) (R)- и (B) (S)-3,5-диметилбензойной кислоты. N-(1-трет-бутил-бутил)-N’-(2,6-дихлорбензоил)гидразида.
Фиг.3: График, демонстрирующий сравнение in vivo рац-, (R)- и (S)-2-этил-3-метокси-бензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида при индукции экспрессии гена терапевтической системы RheoSwitch® у мышей. Черные кружки представляют собой S-энантиомер, черные треугольники представляют собой рацемат, и белые кружки представляют собой R-энантиомер.
Фиг.4: Таблица серий образцов или рацемата и R-энантиомера 2-этил-3-метокси-бензойной кислоты N’-(трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида. Образцы R-энантиомера, полученного посредством быстрой кристаллизации/осаждения из смеси либо метанол/вода или толуол/гептан, давали одинаковую картину дифракции рентгеновских лучей на порошке (данные не показаны), и имели по существу одинаковую точку плавления ([толуол/гептан] 166,2-167,1°C, [CH3OH/H2O] 166,5-167,4°C) по сравнению друг с другом и по сравнению со стандартом, полученным при кристаллизации из CH3OH (165,1-166,5°C). Образцы двух отдельных препаратов рацемата, полученных при выпаривании метанола, показывали одинаковую точку плавления (170-171°C, 169-170°C) в пределах погрешности эксперимента и незначительные варьирования по чистоте.
Фиг.5: Таблица, показывающая распределение по размерам частиц микронизированного R-энантиомера 2-этил-3-метокси-бензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида. D (об, 0,1): распределение по объему, 10% частиц имеют такой же размер или менее. D (об, 0,5): распределение по объему, 50% частиц имеют такой же размер или менее. D (об, 0.9): распределение по объему, 90% частиц имеют такой же размер или менее.
Фиг.6: Таблица, показывающая распределение по размерам частиц микронизированного рацемата 2-этил-3-метокси-бензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида. D (об, 0,1): распределение по объему, 10% частиц имеют такой же размер или менее. D (об, 0,5): распределение по объему, 50% частиц имеют такой же размер или менее. D (об, 0.9): распределение по объему, 90% частиц имеют такой же размер или менее.
Фиг.7: Таблица, показывающая результаты анализа насыпной плотности и плотности утряски микронизированного рацемата и R-энантиомера 2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида.
Фиг.8: Данные термогравиметрического анализа/дифференциального термического анализа (TGA/DTA) (термическая кривая) микронизированного рацемата и R-энантиомера 2-этил-3-метокси-бензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида демонстрируют разные кристаллические формы. Номера серий: REH-28-9-1 и REH-28-4-2; масса образца 7,71994 мг. Выход сигнала в единицах мкВ на мг образца (мкВ/мг); термогравиметрия - изменение массового процента образца (TG%). Оба вещества показали эндотермический эпизод в профиле DTA. Начальная температура для R-энантиомера (163,6°C) была значительно ниже, чем у рацемата (171,2°C). Теплота плавления R-энантиомера (59,8 мкВ.с/мг) также была значительно ниже чем у рацемата (80,8 мкВ.с/мг).
Фиг.9: Таблица, показывающая качественное исследование сравнительной растворимости рацемата и R-энантиомера 2-этил-3-метокси-бензойной кислоты N’-(трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида в фармацевтических эксципиентах.
Фиг.10: Таблица, показывающая результаты анализа термодинамического равновесия (90°C в течение 5 минут или указанного времени; обработка 2) с последующим охлаждением до комнатной температуры и затравки рацемата и R-энантиомера 2-этил-3-метокси-бензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида в фармацевтических эксципиентах. Обработка 1 представляет собой результат перемешивания при комнатной температуре в течение ≤2,5 ч.
Фиг.11: Таблица, показывающая растворимость (мкМ) рацемата и R-энантиомера 2-этил-3-метокси-бензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида в 20%-ном PEG 1000 в дистиллированной воде, рН 7,0.
Фиг.12: Таблица, показывающая растворимость рацемата и R-энантиомера N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида 2-этил-3-метокси-бензойной кислоты в водном полисорбате 80.
Фиг.13: Таблица, показывающая двустороннюю проникающую способность рацемата и R-энантиомера 2-этил-3-метокси-бензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида через клеточные монослои Caco-2. aКлассификация проходимости: (Papp A-B)<1,0×10-6 см/с = НИЗКАЯ; (Papp A-B)>1,0×10-6 см/с = ВЫСОКАЯ; bЗначительная утечка: утечка >3,0 и (Papp B-A)>1,0×10-6 см/с.
Фиг.14: Таблица, показывающая проникающую способность рацемата и R-энантиомера 2-этил-3-метокси-бензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида в клетки MDR1-MDCK. Классификация: A-B Papp>3,0 и соотношение утечки <3,0: высокое. А-В Papp>3,0 и 10 > отношение утечки >3,0: среднее. A-B Papp>3,0 и отношение утечки >10: низкое. A-B Papp<3,0: низкое.
Фиг.15: Таблица, показывающая стабильность рацемата и R-энантиомера 2-этил-3-метокси-бензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида в микросомах печени человека.
Фиг.16: Таблица, показывающая режим дозирования рацемата и R-энантиомера 2-этил-3-метокси-бензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида /препаратов Labrasol у мышей C57BL/6N:Crl. aBвeдeниe дозы в течение 9 суток (первые 18 самок/в группе,) или 12 суток (следующие 18 самок/в группе); bВключены в качестве возможных замен.
Фиг.17: Микроснимки немикронизированного и микронизированного образцов рацемата 2-этил-3-метокси-бензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида. Указана шкала отсчета 50 микрон.
Фиг.18: Таблица, показывающая уровни рацемата и R-энантиомера 2-этил-3-метокси-бензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида в сыворотке крови, введенного в Labrasol с 4 уровнями дозирования (3, 10, 30 и 50 мг/кг/сутки) через 9 суток после введения лекарственного средства и часов после суточной дозы.
Фиг.19: Таблица, показывающая уровни рацемата и R-энантиомера 2-зтил-3-метокси-бензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида в сыворотке крови, введенного в Labrasol с 4 уровнями дозирования (3, 10, 30 и 50 мг/кг/сутки) через 12 суток после введения лекарственного средства и часов после суточной дозы.
Фиг.20: Диаграммное изображение плазмиды pCMV/GEVY(DEF). Плазмида pCMV/GEV/Y(DEF) состоит из доменов D, Е и F из EcR Choristoneura fumiferana, содержащих мутации V390I/Y410E/E274V, слитых по направлению транскрипции с дрожжевой GAL4-DBD (aa 1-147) и помещенных под контроль промотора CMV и нижележащего SV40 сигнала полиаденилирования в векторе pBESfD (Promega Corporation, Madison, WI, USA). DEF-домены показанных EcR амплифицировали, используя праймеры, сконструированные на основе последовательностей из 20-25 нуклеотидов на 5’- и 3’-концах. Сайты ферментной рестрикции BamH I и Xba I были добавлены к праймерам 5’ и 3’ соответственно. Продукты ПЦР обрабатывали соответствующими ферментами рестрикции и клонировали в вектор pBIND.
Фиг.21: Диаграммное изображение вектора pCMV/VP16-Hs-LmRXR. Вектор pCMV/W16-Hs-LmRXR содержит химерный RXR из RXRP Homo sapiens (спираль 1-8 домена Е) и RXR Locusta migratoria (спираль 9-12 домена E), слитые по направлению транскрипции от домена активации VP16 и помещенные под контроль промотора CMV в вектор pBIND.
Фиг.22: Диаграммное изображение репортерного вектора p6xGAL4RE-TTR-SEAP. Индуцируемый репортерный вектор SEAP p6xGAL4RE-TTR-SEAP содержит репортерный ген щелочной фосфатазы, секретируемой человеком, помещенный под контроль индуцируемого промотора, состоящего из 6 копий элемент ответа Gal4, против направления транскрипции от транстиретинового промотора (TTR).
Фиг.23: Диаграммное изображение Ad-RTS-hlL-12, где участки E1 и E3 удалены, и компоненты RTS-IL12 компоненты замещают участок E1.
Фиг.24: Диаграммы, показывающие экспрессию mlL12p70 в опухоли и сыворотке после инъекции клеток В16, инфицированных плазмидой Ad. RheoSPI-mlL12, и введения R-энантиомера или рацемата диацилгидразина в дозировках 0, 3, 10, 30, 50 мг/кг/сутки. Контроли: мыши, несущие нетрансдуцированный B16, получающие лиганд только в количестве 50 мг/кг/сутки в течение 3 суток (Лиганд 50), и необработанные (без лиганда) мыши, несущие опухоль (TuControl). "L3d затем перерыв 3d": мышей обрабатывали лигандом в течение 3 суток, затем в течение еще 3 суток лиганд не предлагали.
Фиг.25: Диаграммы, показывающие экспрессию mIFN-y в опухоли и сыворотке после инъекции клеток B16, инфицированных плазмидой Ad. RheoSPI-mlL12, и введения R-энантиомера или рацемата диацилгидразина в дозах 0, 3, 10, 30, 50 мг/кг/сутки. Контроли: мыши, несущие нетрансдуцированный В16, получающие лиганд только в количестве 50 мг/кг/сутки в течение 3 суток (Лиганд 50), и необработанный (без лиганда) мыши, несущие опухоль (TuControl). "L3d затем перерыв 3d": мышей обрабатывали лигандом в течение 3 суток, затем в течение еще 3 суток лиганд не предлагали.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к соединениям Формулы I
где
A представляет собой алкокси, арилалкилокси или арилокси;
B представляет собой возможно замещенный арил или возможно замещенный гетероарил; и
R1 и R2 независимо представляют собой возможно замещенный алкил, арилалкил, гидроксиалкил, галогеноалкил, возможно замещенный циклоалкил, возможно замещенный алкенил, возможно замещенный алкинил, возможно замещенный гетероцикл, возможно замещенный арил или возможно замещенный гетероарил;
или их фармацевтически приемлемым солям, гидратам, кристаллическим формам или аморфным формам.
В другом воплощении настоящее изобретение относится к энантиомерно обогащенным соединениям Формулы II
где
A представляет собой алкокси, арилалкилокси, арилокси, арилалкил, возможно замещенный арил или возможно замещенный гетероарил;
B представляет собой возможно замещенный арил или возможно замещенный гетероарил; и
R1 и R2 независимо представляют собой возможно замещенный алкил, арилалкил, гидроксиалкил, галогеноалкил, возможно замещенный циклоалкил, возможно замещенный алкенил, возможно замещенный алкинил, возможно замещенный гетероцикл, возможно замещенный арил или возможно замещенный гетероарил;
при условии, что R1 отличается от R2;
где абсолютной конфигурацией асимметрического атома углерода, несущего R1 и R2, преимущественно является S;
или их фармацевтически приемлемым солям, гидратам, кристаллическим формам или аморфным формам.
В другом воплощении настоящее изобретение относится к энантиомерно обогащенным соединениям Формулы III
где
A представляет собой алкокси, арилалкилокси, арилокси, арилалкил, возможно замещенный арил или возможно замещенный гетероарил;
B представляет собой возможно замещенный арил или возможно замещенный гетероарил; и
R1 и R2 независимо представляют собой возможно замещенный алкил, арилалкил, гидроксиалкил, галогеноалкил, возможно замещенный циклоалкил, возможно замещенный алкенил, возможно замещенный алкинил, возможно замещенный гетероцикл, возможно замещенный арил или возможно замещенный гетероарил;
при условии, что R1 отличается от R2;
где абсолютная конфигурация асимметрического атома углерода, несущего R1 и R2, преимущественно представляет собой R;
или его фармацевтически приемлемым солям, гидратам, кристаллическим формам или аморфным формам.
В другом воплощении настоящее изобретение относится к энантиомерно обогащенным соединениям Формулы II или III, где
А представляет собой -OC(CH3)3, -OCH2Ph, возможно замещенный арил или возможно замещенный гетероарил;
B представляет собой возможно замещенный арил;
R1 представляет собой возможно замещенный (C1-C6)алкил, гидрокси(C1-C4)алкил или возможно замещенный (C2-C6)алкенил; и
R2 представляет собой возможно замещенный (C1-C6)алкил, арил(C1-C4)алкил, возможно замещенный (C3-C7)циклоалкил или возможно замещенный арил;
при условии, что R1 отличается от R2;
где
абсолютная конфигурация асимметрического атома углерода, несущего R1 и R2, преимущественно представляет собой S в Формуле II; и
абсолютная конфигурация асимметрического атома углерода, несущего R1 и R2, преимущественно представляет собой R в Формуле III;
или их фармацевтически приемлемым солям, гидратам, кристаллическим формам или аморфным формам.
В другом воплощении настоящее изобретение относится к энантиомерно обогащенным соединениям Формулы II или III, где
A представляет собой возможно замещенный арил или возможно замещенный гетероарил;
B представляет собой возможно замещенный арил;
R1 представляет собой возможно замещенный (C1-C6)алкил, гидрокси(C1-C4)алкил или возможно замещенный (C2-C6)алкенил; и
R2 представляет собой возможно замещенный (C1-C6)алкил, арил(C1-C4)алкил, возможно замещенный (C3-C7)циклоалкил или возможно замещенный арил;
при условии, что R1 отличается от R2;
где
абсолютная конфигурация асимметрического атома углерода, несущего R1 и R2, преимущественно представляют собой S в Формуле II; и
абсолютная конфигурация асимметрического атома углерода, несущего R1 и R2, преимущественно представляют собой R в Формуле III;
или их фармацевтически приемлемым солям, гидратам, кристаллическим формам или аморфным формам.
В другом воплощении настоящее изобретение относится к энантиомерно обогащенным соединениям Формулы II или III, где
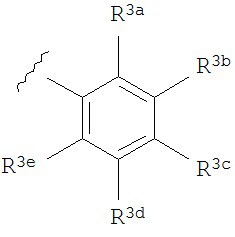
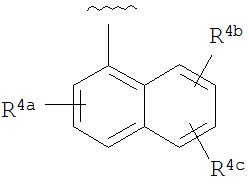
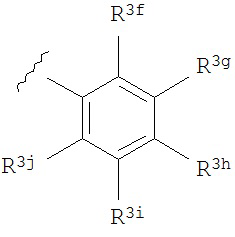
A представляет собой
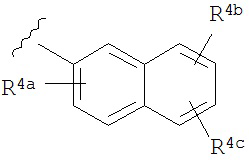
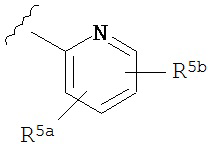
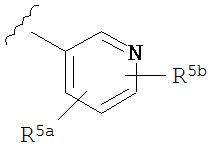
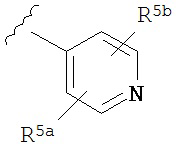
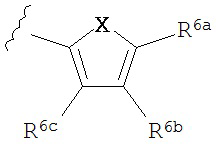 or
or 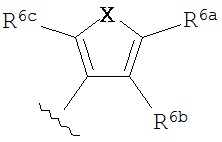
R3a, R3b, R3c, R3d и R3e независимо выбраны из водорода, галогено, нитро, циано, гидрокси, амино, возможно замещенного алкила, галогеноалкила, гидроксиалкила, арилалкила, возможно замещенного циклоалкила, возможно замещенного алкенила, возможно замещенного алкинила, возможно замещенного арила, возможно замещенного гетероарила, возможно замещенного гетероцикла, алкокси, арилокси, арилалкилокси, алкилтио, карбоксамидо, сульфонамидо, -CORc, -SO2Rd, -N(Re)CORf, -N(Re)SO2Rg или -N(Re)C=N(Rh)-амино; или
R3a и R3b взятые вместе с атомами углерода, к которым они присоединены, образуют пяти-, шести- или семичленное конденсированное циклоалкильное или гетероциклическое кольцо; или
R3d и R3c, взятые вместе с атомами углерода, к которым они присоединены, образуют пяти-, шести- или семичленное конденсированное циклоалкильное или гетероциклическое кольцо;
R4a, R4b, R4c, R5a, R5b, R6a, R6b и R6c независимо выбраны из водорода, галогено, нитро, циано, гидрокси, амино, возможно замещенного алкила, галогеноалкила, гидроксиалкила, арилалкила, возможно замещенного циклоалкила, возможно замещенный алкенила, возможно замещенного алкинила, возможно замещенного арила, возможно замещенного гетероарила, возможно замещенного гетероцикла, алкокси, арилокси, арилалкилокси, алкилтио, карбоксамидо, сульфонамидо, -CORc, -SO2Rd, -N(Re)CORf, -N(Re)SO2R9 или -N(Re)C=N(Rh)-амино;
X представляет собой O или S;
B представляет собой
и;
R3f, R3g, R3h, R3i и R3j независимо выбраны из водорода, галогено, нитро, циано, гидрокси, амино, возможно замещенного алкила, галогеноалкила, гидроксиалкила, арилалкила, возможно замещенного циклоалкила, возможно замещенного алкенила, возможно замещенного алкинила, возможно замещенный арила, возможно замещенного гетероарила, возможно замещенного гетероцикла, алкокси, арилокси, арилалкилокси, алкилтио, карбоксамидо, сульфонамидо, -CORc, -SO2Rd, -N(Re)CORf, -N(Re)SO2R9 или -N(Re)C=N(Rh)-амино; или
R3f и R3g, взятые вместе с атомами углерода, к которым они присоединены, образуют пяти-, шести- или семичленное конденсированное циклоалкильное или гетероциклическое кольцо; или
R3g и R3h, взятые вместе с атомами углерода, к которым они присоединены, образуют пяти-, шести- или семичленное конденсированное циклоалкильное или гетероциклическое кольцо;
где
Rc представляет собой водород, гидрокси, галогеноалкил, гидроксиалкил, арилалкил, возможно замещенный алкил, возможно замещенный циклоалкил, возможно замещенный алкенил, возможно замещенный алкинил, возможно замещенный гетероцикл, возможно замещенный арил, возможно замещенный гетероарил, алкокси, арилокси или арилалкилокси;
Rd представляет собой галогеноалкил, гидроксиалкил, арилалкил, возможно замещенный алкил, возможно замещенный циклоалкил, возможно замещенный алкенил, возможно замещенный алкинил, возможно замещенный гетероцикл, возможно замещенный арил или возможно замещенный гетероарил;
Re представляет собой водород, галогеноалкил, гидроксиалкил, арилалкил, возможно замещенный алкил, возможно замещенный циклоалкил, возможно замещенный алкенил, возможно замещенный алкинил, возможно замещенный гетероцикл, возможно замещенный арил или возможно замещенный гетероарил;
Rf представляет собой водород, галогеноалкил, гидроксиалкил, арилалкил, возможно замещенный алкил, возможно замещенный циклоалкил, возможно замещенный алкенил, возможно замещенный алкинил, возможно замещенный гетероцикл, возможно замещенный арил, возможно замещенный гетероарил, алкокси, арилокси, арилалкилокси или амино;
Rg представляет собой галогеноалкил, гидроксиалкил, арилалкил, возможно замещенный алкил, возможно замещенный циклоалкил, возможно замещенный алкенил, возможно замещенный алкинил, возможно замещенный гетероцикл, возможно замещенный арил, возможно замещенный гетероарил или амино;
Rh представляет собой водород, алкил, арил, циано или нитро;
R1 представляет собой возможно замещенный (C1-C6)алкил, гидрокси(C1-C4)алкил или возможно замещенный (C2-C6)алкенил; и
R2 представляет собой возможно замещенный (C1-C6)алкил, арил(C1-C4)алкил, возможно замещенный (C3-C7)циклоалкил или возможно замещенный арил;
при условии, что R1 отличается от R2;
где
абсолютная конфигурация асимметрического атома углерода, несущего R1 и R2, преимущественно представляет собой S в Формуле II; и
абсолютная конфигурация асимметрического атома углерода, несущего R1 и R2, преимущественно представляет собой R в Формуле III;
или их фармацевтически приемлемым солям, гидратам, кристаллическим формам или аморфным формам.
В другом воплощении настоящее изобретение относится к энантиомерно обогащенным соединениям Формулы II, где
A, B, R3a, R3b, R3c, R3d, R3e, R3f, R3g, R3h, R3i, R3j, R4a, R4b, R4c, R5a, R5b, R6a, R6b R6c и X имеют такие же значения, как указано выше;
R1 представляет собой (C1-C6)алкил, гидрокси(C1-C4)алкил или (C2-C4)алкенил; и
R2 представляет собой возможно замещенный (C3-C7)циклоалкил;
где абсолютная конфигурация асимметрического атома углерода, несущего R1 и R2, преимущественно представляет собой S;
или их фармацевтически приемлемым солям, гидратам, кристаллическим формам или аморфным формам.
В другом воплощении настоящее изобретение относится к энантиомерно обогащенным соединениям Формулы III, где
A, B, R3a, R3b, R3c, R3d, R3e, R3f, R3g, R3h, R3i, R3j, R4a, R4b, R4c, R5a, R5b, R6a, R6b R6c и X имеют такие же значения, как указано выше;
R1 представляет собой (C1-C6)алкил, гидрокси(C1-C4)алкил или (C2-C4)алкенил; и
R2 представляет собой возможно замещенный (C1-C6)алкил;
при условии, что R1 отличается от R2;
где абсолютная конфигурация асимметрического атома углерода, несущего R1 и R2, преимущественно представляет собой R;
или их фармацевтически приемлемым солям, гидратам, кристаллическим формам или аморфным формам.
В одном воплощении настоящее изобретение относится к энантиомерно обогащенным соединениям Формулы II, где
A представляет собой
B представляет собой
R3a, R3b, R3c, R3d, R3e, R3f, R3g, R3h, R3i и R3j независимо выбраны из водорода, галогена, (C1-C4)алкил или (C1-C4)алкокси;
R1 представляет собой (C1-C6)алкил, гидрокси(C1-C4)алкил или (C2-C4)алкенил;
R2 представляет собой возможно замещенный (C3-C7)циклоалкил;
где абсолютная конфигурация асимметрического атома углерода, несущего R1 и R2, преимущественно представляет собой S;
или их фармацевтически приемлемым солям, гидратам, кристаллическим формам или аморфным формам.
В другом воплощении настоящее изобретение относится к энантиомерно обогащенным соединениям Формулы III, где
A представляет собой
B представляет собой
R3a, R3b, R3c, R3d, R3e, R3f, R3g, R3h, R3i и R3j независимо выбраны из водорода, галогена, (C1-C4)алкила или (C1-C4)алкокси;
R1 представляет собой (C1-C6)алкил, гидрокси(C1-C4)алкил или (C2-C4)алкенил; и
R2 представляет собой возможно замещенный (C1-C6)алкил;
где абсолютная конфигурация асимметрического атома углерода, несущего R1 и R2, преимущественно представляет собой R;
или их фармацевтически приемлемым солям, гидратам, кристаллическим формам или аморфным формам.
В другом воплощении настоящее изобретение относится к энантиомерно обогащенным соединениям Формулы II, имеющим энантиомерный избыток S-изомера по меньшей мере 50%, 75%, 85%, 95% или >99%. В одном воплощении соединение Формулы II по существу состоит из S-изомера.
В другом воплощении настоящее изобретение относится к энантиомерно обогащенным соединениям Формулы III, имеющим энантиомерный избыток R-изомера по меньшей мере 50%, 75%, 85%, 95% или >99%. В одном воплощении соединение Формулы III состоит в основном из R-изомера.
В другом воплощении настоящее изобретение относится к фармацевтической композиции, содержащей соединение Формулы I, II или III.
В одном воплощении настоящее изобретение относится к N-(1-трет-бутил-бутил)-N’-(2-этил-3-метоксибензоил)гидразиду (R)-3,5-диметилбензойной кислоты, имеющему энантиомерный избыток по меньшей мере 95% или по меньшей мере 99%, или к его фармацевтически приемлемой соли, гидрату, кристаллической форме или аморфной форме. В одном воплощении соединение является энантиомерно чистым.
В другом воплощении настоящее изобретение относится к фармацевтической композиции, содержащей N-(1-трет-бутил-бутил)-N’-(2-этил-3-метоксибензоил)гидразид (R)-3,5-диметилбензойной кислоты, имеющий энантиомерный избыток по меньшей мере 95% или по меньшей мере 99%, или его фармацевтически приемлемой соли, гидрата, кристаллической форме или аморфной форме. В одном воплощении соединение является энантиомерно чистым.
Неожиданно было обнаружено, что энантиомерно чистый N-(1-трет-бутил-бутил)-N’-(2-этил-3-метоксибензоил)гидразид (R)-3,5-диметилбензойной кислоты обладает комбинацией полезных свойств, которые делают его исключительно пригодным для фармацевтического применения in vivo. Такие свойства включают более низкую точку плавления и более низкую теплоту плавления по сравнению с рацематом (Пример 68); лучшую растворимость во многих жидких фармацевтических эксципиентах по сравнению с рацематом (Пример 68); лучшую проникающую способность в некоторые клетки по сравнению с рацематом (Пример 69); вероятно лучше способен пройти гематоэнцефалический барьер чем рацемат (Пример 69); лучшую устойчивость к метаболизму гепатоцитов по сравнению с рацематом (Пример 70); при введении перорально в виде раствора Labrasol или суспензии достигает значительно более высоких уровней в плазме по сравнению с рацематом (Пример 71); и достигает более значительной экспрессии гена in vivo посредством активации переключателя генов на основе EcR по сравнению с рацематом (Пример 72).
Настоящее изобретение также относится к способу получения соединения Формулы IV,
где
A представляет собой арилалкил, возможно замещенный арил или возможно замещенный гетероарил;
B представляет собой возможно замещенный арил или возможно замещенный гетероарил;
R2 представляет собой возможно замещенный алкил, арилалкил, гидроксиалкил, галогеноалкил, возможно замещенный циклоалкил, возможно замещенный алкенил, возможно замещенный алкинил, возможно замещенный гетероцикл, возможно замещенный арил или возможно замещенный гетероарил;
при условии, что R2 не является -CR8R9CHR10CR11R12; и
R8, R9, R10, R11 и R12 независимо выбраны из водорода, алкила, циклоалкила, гетероцикла, арила или гетероарила;
где асимметрический атом углерода, к которому присоединен R2, является энантиомерно обогащенным либо R-, либо S-изомером;
включающему:
а) взаимодействие соединения Формулы V
с соединением Формулы VI
где
X и Y независимо представляют собой O или NR, где R представляет собой алкил или арил;
Ca и Cb независимо представляют собой асимметрический атом углерода в S-конфигурации или асимметрический атом углерода в R-конфигурации;
R14 и R15 независимо представляют собой алкил или арил;
R13 представляет собой галоген, водород, алкил, алкокси или OSO2CF3;
R7 представляет собой алкил, арилалкил или арил; и
R2, R8, R9, R10, R11 и R12 имеют значения, указанные выше;
с образованием соединения Формулы VII;
б) восстановление соединения Формулы VII с образованием соединения Формулы VIII;
в) взаимодействие соединения Формулы VIII с соединением Формулы B-CO-LG, где LG представляет собой уходящую группу, с образованием соединения Формулы IX;
г) удаление группы R7CO2- из соединения Формулы IX с образованием соединения Формулы X; и
д) взаимодействие соединения Формулы X с соединением Формулы A-CO-LG, где LG представляет собой уходящую группу, с образованием соединения Формулы IV.
В другом воплощении настоящее изобретение относится к способу получения соединения Формулы IV, где R8, R9, R10, R11 и R12 представляют собой водород.
В другом воплощении настоящее изобретение относится к способу получения соединения Формулы IV, где
R8, R9, R10, R11 и R12 представляют собой водород;
Х представляет собой NR, и R представляет собой метил;
Y представляет собой О;
R14 представляет собой метил;
R15 представляет собой фенил; и
каждый из Ca и Cb имеет R-конфигурацию.
В другом воплощении настоящее изобретение относится к способу получения соединения Формулы IV, где
R8, R9, R10, R11 и R12 представляют собой водород;
Х представляет собой NR, и R представляет собой метил;
Y представляет собой О;
R14 представляет собой метил;
R15 представляет собой фенил; и
каждый из Ca и Cb находятся в S-конфигурации.
В другом воплощении настоящее изобретение относится к способу получения соединения Формулы IV, где
R8, R9, R10, R11 и R12 представляют собой водород;
Х представляет собой NR, и R представляет собой метил;
Y представляет собой O;
R14 представляет собой метил;
R15 представляет собой фенил;
каждый из Ca и Cb находятся в S-конфигурации; и
В представляет собой 3,5-диметилфенил.
В другом воплощении настоящее изобретение относится к способу получения соединения Формулы IV, где
R8, R9, R10, R11 и R12 представляют собой водород;
X представляет собой NR и R представляет собой метил;
Y представляет собой О;
R14 представляет собой метил;
R15 представляет собой фенил;
каждый из Ca и Cb находятся в S-конфигурации; и
A представляет собой 2-этил-3-метоксифенил.
В другом воплощении настоящее изобретение относится к способу получения соединения Формулы IV, где
R8, R9, R10, R11 и R12 представляют собой водород;
X представляет собой NR, и R представляет собой метил;
Y представляет собой О;
R14 представляет собой метил;
R15 представляет собой фенил;
каждый из Ca и Cb находится в S-конфигурации; и
R2 представляет собой трет-бутил.
Настоящее изобретение также относится к способу получения соединения Формулы II или III, включающему:
а) взаимодействие ацилгидразина Формулы XI
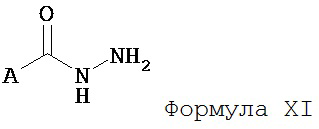
с кетоном Формулы XII
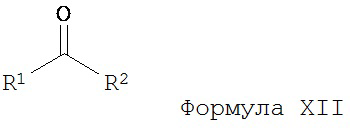
с образованием соединения Формулы XIII;

где R1 отличается от R2;
б) восстановление соединения Формулы XIII в присутствие хирального катализатора с образованием соединения Формулы S-XIV или R-XIV; и
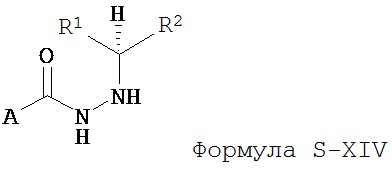
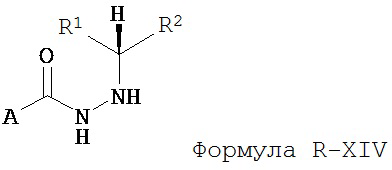
в) взаимодействие соединения Формулы S-XIV или R-XIV с соединением Формулы B-CO-LG, где LG представляет собой уходящую группу, с образованием соединения Формулы II или III.
Диацилгидразины Формулы I по изобретению включают, без ограничения ими: рац-N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразинкарбоновой кислоты бензиловый эфир; или рац-N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразинкарбоновой кислоты трет-бутиловый эфир.
Хиральные диацилгидразины Формулы II или III по настоящему изобретению включают, без ограничения ими:
(S)-3,5-диметилбензойной кислоты N-(1-циклогексил-бутил)-N’-(2-этил-3-метоксибензоил)гидразид;
(S)-3,5-диметилбензойной кислоты N-(1-циклогексил-бутил)-N’-(3-метоксибензоил)гидразид;
(S)-3,5-диметилбензойной кислоты N-(1-циклогексил-бутил)-N’-(4-метилбензоил)гидразид.
(R)-N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразинкарбоновой кислоты бензиловый эфир;
(R)-N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразинкарбоновой кислоты трет-бутиловый эфир;
(R)-N’-(1-трет-бутил-4-гидрокси-бутил)-N’-(3,5-диметилбензоил)гидразинкарбоновой кислоты бензиловый эфир;
(R)-N’-(1-трет-бутил-4-гидрокси-бутил)-N’-(3,5-диметилбензоил)гидразин-карбоновой кислоты бензиловый эфир;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2-этил-3-метоксибензоил)гидразид;
(R)-3,5-диметилбензойной кислоты N’-бензоил-N-(1-трет-бутил-бутил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2-метилбензоил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2-метоксибензоил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2-фтор-бензоил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2-хлор-бензоил)гидразид;
(R)-3,5-диметилбензойной кислоты N’-(2-бром-бензоил)-N-(1-трет-бутил-бутил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(3-метилбензоил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(3-метоксибензоил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(3-хлор-бензоил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(4-метилбензоил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(4-этилбензоил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(4-метоксибензоил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(4-хлор-бензоил)гидразид;
(R)-3,5-диметилбензойной кислоты (N-(1-трет-бутил-бутил)-N’-(2,6-дифтор-бензоил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2,6-дихлор-бензоил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(3,4-диметоксибензоил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(3,5-дифтор-бензоил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(3,5-диметокси-4-метилбензоил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(4-метилбензо[1,3]диоксол-5-карбонил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(5-метил-2,3-дигидробензо[1,4]диоксин-6-карбонил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(5-этил-2,3-дигидробензо[1,4]диоксин-6-карбонил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(нафталин-1-карбонил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(нафталин-2-карбонил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(тиофен-2-карбонил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2,5-диметил-фуран-3-карбонил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2-хлор-пиридин-3-карбонил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(6-хлор-пиридин-3-карбонил)гидразид;
(R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(3-метокси-2-метилбензоил)гидразид;
(R)-3,5-диметокси-4-метилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(3-метокси-2-метилбензоил)гидразид; и
(R)-3,5-диметилбензойной кислоты N’-(4-этилбензоил)-N-(1-фенетил-бут-3-енил)гидразид.
Хиральные диацилгидразины Формул II и III могут содержать хиральные центры в других положениях молекулы (то есть не на асимметрическом атоме углерода, содержащем R1 и R2). Подразумевается, что все возможные энантиомеры, диастереомеры и стереоизомеры заключены в объем настоящего изобретения.
Настоящее изобретение также относится к способам модуляции экспрессии гена в клетке-хозяине с использованием системы модуляции экспрессии гена с диацилгидразиновым лигандом Формулы I или хиральным диацилгидразиновым лигандом Формулы II или III.
В одном воплощении настоящее изобретение относится к применению диацилгидразинового лиганда Формулы I или хирального диацилгидразинового лиганда Формулы II или III в индуцируемой системе экспрессии гена, которая имеет пониженный уровень фоновой экспрессии гена в отсутствие лиганда и реагирует на субмикромолекулярные концентрации лиганда.
В другом воплощении настоящее изобретение относится к способу модуляции экспрессии гена-мишени в клетке-хозяине, где клетка-хозяин содержит первую кассету экспрессии гена, содержащую первый полинуклеотид, кодирующий первый полипептид, содержащую:
1) домен трансактивации;
2) ДНК-связывающий домен; и
3) лиганд-связывающий домен ядерного рецептора группы H;
и вторую кассету экспрессии гена, содержащую:
1) элемент ответа, способный связываться с указанным ДНК-связывающим доменом;
2) промотор, который активируется доменом трансактивации; и
3) указанный ген-мишень;
включающий приведение указанной клетки-хозяина в контакт с диацилгидразиновым лигандом Формулы I или хиральным диацилгидразиновым лигандом Формулы II или III; где экспрессия гена-мишени модулирована.
В другом воплощении настоящее изобретение относится к способу модуляции экспрессии интересующего гена в клетке-хозяине, где клетка-хозяин содержит первую рекомбинатную кассету экспрессии гена, содержащую первый полинуклеотид, кодирующий первый полипептид, содержащий:
1) ДНК-связывающий домен; и
2) лиганд-связывающий домен ядерного рецептора группы Н;
и вторую рекомбинатную кассету экспрессии гена, содержащую:
1) элемент ответа, способный связываться с указанным ДНК-связывающим доменом;
2) промотор; и
3) указанный интересующий ген;
включающий приведение указанной клетки-хозяина в контакт с диацилгидразиновым лигандом Формулы I или хиральным диацилгидразиновым лигандом Формулы II или III; где экспрессия интересующего гена модулирована.
В другом воплощении изобретение относится к способу регуляции экспрессии эндогенного или гетерологичного гена в трансгенных субъектах, включающему приведение лиганда в контакт с экдизон-рецепторным комплексом в клетках субъекта, где клетки дополнительно содержат ДНК-связывающую последовательность для экдизон-рецепторного комплекса, находящегося в комбинации с лигандом, и где образование комплекса «экдизон-рецепторный комплекс-лиганд-ДНК-связывающая последовательность» индуцирует экспрессию гена, и где лиганд представляет собой диацилгидразиновый лиганд Формулы I или хиральный диацилгидразиновый лиганд Формулы II или III, где экспрессия эндогенного или гетерологичного гена в трансгенном субъекте регулируется.
В одном воплощении клетки представляют собой аутологичные клетки, то есть клетки получают от субъекта и трансфицируют с помощью экдизон-рецепторного комплекса и ДНК-связывающей последовательности. Трансфицированные аутологичные клетки затем обратно имплантируют субъекту, которого затем обрабатывают лигандом. Аутологичные клетки можно имплантировать любым из ряда способов, включая инфузию или инъекцию, например прямую инъекцию. В одном воплощении аутологичные клетки имплантируют посредством внутриопухолевой инъекции.
В другом воплощении изобретение относится к способу регуляции экспрессии эндогенного или гетерологичного гена в трансгенном субъекте, включающему приведение лиганда в контакт с химерным экдизон-рецепторным комплексом в клетках субъекта, где клетки дополнительно содержат ДНК-связывающую последовательность, дополнительно содержащую промотор для экдизон-рецепторного комплекса, находящегося в комбинации с лигандом, и где образование комплекса «экдизон-рецепторный комплекс-лиганд-ДНК-связывающая последовательность» индуцирует экспрессию гена, и где лиганд представляет собой диацилгидразиновый лиганд Формулы I или хиральный диацилгидразиновый лиганд Формулы II или III, где экспрессия эндогенного или гетерологичного гена в трансгенном субъекте регулируется.
В другом воплощении трансгенный субъект представляет собой растение, насекомое, животное или млекопитающее, например человека или домашнее животное, такое как корова, свинья, овца, коза, лошадь, собака или кошка.
В другом воплощении изобретение относится к способу регуляции экспрессии трансгена у трансгенного субъекта, при котором указанный трансгенный субъект содержит рекомбинантный экдизон-рецепторный комплекс, способный регулировать экспрессию трансгена, включающему введение указанному субъекту эффективного количества диацилгидразинового лиганда Формулы I или хирального диацилгидразинового лиганда Формулы II или III.
В другом воплощении изобретение относится к способу модуляции экспрессии гена-мишени в клетке-хозяине, включающему стадии:
а) введение в клетку-хозяина системы модуляции экспрессии гена, содержащей:
1) первую кассету экспрессии гена, которая способна экспрессироваться в клетке-хозяине, где указанная первая кассета экспрессии гена, содержит полинуклеотидную последовательность, которая кодирует первый гибридный полипептид, содержащий:
(а) ДНК-связывающий домен, который распознает элемент ответа, ассоциированный с геном, экспрессию которого следует модулировать; и
(б) лиганд-связывающий домен экдизонового рецептора;
2) вторую кассету экспрессии гена, которая способна экспрессироваться в клетке-хозяине, где указанная вторая кассета экспрессии гена содержит полинуклеотидную последовательность, которая кодирует второй гибридный полипептид, содержащий:
(а) домен трансактивации домен; и
(б) лиганд-связывающий домен химерного ретиноидного рецептора X; и
3) третью кассету экспрессии гена, которая способна экспрессироваться в клетке-хозяине, где указанная третья кассета экспрессии гена содержит полинуклеотидную последовательность, содержащую:
(а) элемент ответа, распознаваемый ДНК-связывающим доменом первого гибридного полипептида;
(б) промотор, который активируется доменом трансактивации второго гибридного полипептида; и
(в) ген, экспрессию которого следует модулировать; и
б) введение в клетку-хозяина диацилгидразинового лиганда Формулы I или хирального диацилгидразинового лиганда Формулы II или III; где экспрессия гена в клетке-хозяине модулируется.
В другом воплощении настоящее изобретение относится к способу продуцирования полипептида, включающему стадии:
а) селекции клетки-хозяина, которая по существу нечувствительна к воздействию диацилгидразинового лиганда Формулы I или хирального диацилгидразинового лиганда Формулы II или III;
б) введение в клетку-хозяин:
1) ДНК-конструкции, содержащей:
(а) экзогенный гена, кодирующий полипептид; и
(б) элемента ответа;
где ген находится под контролем элемента ответа; и
2) экдизон-рецепторного комплекса, содержащего:
(а) ДНК-связывающий домен, который связывается с элементом ответа;
(б) связывающий домен для указанного соединения; и
(в) домен трансактивации; и
в) воздействия указанного соединения на клетку; при котором продуцируется полипептид.
В другом воплощении изобретение относится к способу, где экдизон-рецепторный комплекс или лиганд-связывающий домен экдизонового рецептора содержит мутацию.
Сокращения, используемые в данном описании изобретения, имеют свое общепринятое значение в химической и биологической областях техники, если не указано иное. Например: "ч" означает час(ы), "мин" означает минуту(ы), "сек" означает секунду(ы), "сут" означает сутки, "мкл" означает микролитр(ы), "мл" означает миллилитр(ы), "л" означает литр(ы), "мкМ" означает микромолярный, "мМ" означает миллимолярный, "M" означает молярный, "моль" означает моль, "ммоль" означает миллимоль, "мкг" означает микрограмм(ы), "мг" означает миллиграмм(ы), "A" означает аденин или аденозин, "T" означает тимин или тимидин, "G" означает гуанин или гуанозин, "C" означает цитидин или цитозин, "x g" означает кратность силы тяжести, "нт" означает нуклеотид(ы), "ак" означает аминокислоту(ы), "по" означает пару(ы) оснований, "кб" означает тысячу(и) нуклеотидов, "кг" означает кило, "мк" означает микро, "°C" означает градус Цельсия, "THF" означает тетрагидрофуран, "DME" означает диметоксиэтан, "DMF" означает диметилформамид, "ЯМР" означает ядерный магнитный резонанс, "psi" относится к фунтам на квадратный дюйм, и "TCX" означает тонкослойную хроматографию.
Термин "алкил", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к насыщенному алифатическому углеводороду с прямой или разветвленной цепью, имеющему от одного до десяти атомов углерода, или обозначенное число атомов углерода (C1-C10 означает от 1 до 10 атомов углерода). Типичные алкильные группы включают метил, этил, н-пропил, изопропил, н-бутил, sec-бутил, изобутил, трет-бутил, н-пентил, н-гексил, изогексил, н-гептил, 4,4-диметилпентил, н-октил, 2,2,4-триметилпентил, нонил, децил и тому подобное.
Термин "возможно замещенный алкил", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к алкилу, как он определен выше, который возможно замещен одним-тремя заместителями, независимо выбранными из нитро, циано, амино, возможно замещенного циклоалкила, возможно замещенного гетероарила, возможно замещенного гетероцикла, алкокси, арилокси, арилалкилокси, алкилтио, карбоксамидо, сульфонамидо, -CORc, -SO2Rd, -N(Re)CORf, -N(Re)SO2Rg или -N(Re)C=N(Rh)-амино. Типичные замещенные алкильные группы включают -CH2OCH3, -CH2CH2NH2, -CH2CH2CN, -CH2SO2CH3 и тому подобное.
Термин "галогеноалкил", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к алкилу, как он определен выше, имеющему от одного до шести галогеновых заместителей. Типичные галогеноалкильные группы включают трифторметил, -CH2CH2F и тому подобное.
Термин "гидроксиалкил", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к алкилу, как он определен выше, имеющему от одного до трех заместителей гидрокси, обычно один заместитель гидрокси. Типичные гидроксиалкильные группы включают гидроксиметил, гидроксиэтил, гидроксипропил и тому подобное.
Термин "арилалкил", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к алкилу, как он определен выше, имеющему от одного до трех арильных заместителей (указанные арильные заместители возможно могут быть замещены, как описано ниже), обычно один или два арильных заместителя. Типичные арилалкильные группы включают, например, бензил, фенилэтил, 4-фторфенилэтил, фенилпропил, дифенилметил и тому подобное.
Термин "циклоалкил", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к насыщенным и частично ненасыщенным (содержащим одну или две двойные связи) циклическим углеводородным группам, содержащим от одного до трех колец, имеющих от трех до двенадцати атомов углерода или указанное число атомов углерода. Типичные циклоалкильные группы включают циклопропил, циклобутил, циклопентил, циклогексил, циклогептил, циклооктил, норборнил, декалин, адамантил и тому подобное.
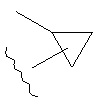
Термин "возможно замещенный циклоалкил", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к циклоалкилу, как он определен выше, который возможно замещен одним-тремя заместителями, независимо выбранными из галогено, нитро, циано, гидрокси, амино, возможно замещенного алкила, галогеноалкила, гидроксиалкила, арилалкила, возможно замещенного циклоалкила, возможно замещенного алкенила, возможно замещенного алкинила, возможно замещенного арила, возможно замещенного гетероарила, возможно замещенного гетероцикла, алкокси, арилокси, арилалкилокси, алкилтио, карбоксамидо, сульфонамидо, -CORc, -SO2Rd, -N(Re)CORf, -N(Re)SO2Rg или -N(Re)C=N(Rh)-амино. Типичные возможно замещенные циклоалкильные группы включают
и тому подобное. Возможно замещенный циклоалкил может быть конденсирован с арильной группой с получением возможно замещенного арила, как описано ниже.
Термин "алкенил", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к алкильной группе, как она определена выше, содержащей от одной до трех углерод-углеродных двойных связей, обычно одну или две двойных связи. Типичные алкенильные группы включают -CH=CH2, -CH2CH=CH2, -CH2CH2CH=CH2, -CH2CH2CH=CHCH3 и тому подобное.
Термин "возможно замещенный алкенил", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к алкенилу, как он определен выше, который возможно замещен одним-тремя заместителями, независимо выбранными из галогено, нитро, циано, гидрокси, амино, возможно замещенного алкила, галогеноалкила, гидроксиалкила, арилалкила, возможно замещенного циклоалкила, возможно замещенного алкенила, возможно замещенного алкинила, возможно замещенного арила, возможно замещенного гетероарила, возможно замещенного гетероцикла, алкокси, арилокси, арилалкилокси, алкилтио, карбоксамидо, сульфонамидо, -CORc, -SO2Rd, -N(Re)CORf, -N(Re)SO2Rg или -N(Re)C=N(Rh)-амино. Типичные возможно замещенные алкенильные группы включают -CH=CHPh, -CH2CH=CHPh и подобные.
Термин "алкинил", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к алкильной группе, как она определена выше, содержащей от одной до трех углерод-углеродных тройных связей, обычно одну или две тройные связи. Типичные алкинильные группы включают -C=CH, -C=CCH3, -CH2CH2C=CH и -CH2CH2C=CCH3.
Термин "возможно замещенный алкинил", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к алкинилу, как он определен выше, который возможно замещен одним-тремя заместителями, независимо выбранными из галогено, нитро, циано, гидрокси, амино, возможно замещенного алкила, галогеноалкила, гидроксиалкила, арилалкила, возможно замещенного циклоалкила, возможно замещенного алкенила, возможно замещенного алкинила, возможно замещенного арила, возможно замещенного гетероарила, возможно замещенного гетероцикла, алкокси, арилокси, арилалкилокси, алкилтио, карбоксамидо, сульфонамидо, -CORc, -SO2Rd, -N(Re)CORf, -N(Re)SO2Rg или -N(Re)C=N(Rh)-амино. Типичные возможно замещенные алкенильные группы включают -C=CPh, -CH2C=CPh и тому подобное.
Термин "арил", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к моноциклическим и бициклическим ароматическим кольцевым системам, имеющим от шести до двенадцати атомов углерода, таким как фенил (сокращено Ph), 1-нафтил и 2-нафтил.
Термин "возможно замещенный арил", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к арилу, как он определен выше, который возможно замещен одним-пятью заместителями, обычно одним-тремя заместителями, независимо выбранными из галогено, нитро, циано, гидрокси, амино, возможно замещенного алкила, галогеноалкила, гидроксиалкила, арилалкила, возможно замещенного циклоалкила, возможно замещенного алкенила, возможно замещенного алкинила, возможно замещенного арила, возможно замещенного гетероарила, возможно замещенного гетероцикла, алкокси, арилокси, арилалкилокси, алкилтио, карбоксамидо, сульфонамида -CORc, -SO2Rd, -N(Re)CORf, -N(Re)SO2Rg или -N(Re)C=N(Rh)-амино. Типичные возможно замещенные арильные группы включают 2-метилфенил, 2-метоксифенил, 2-фторфенил, 2-хлорфенил, 2-бромфенил, 3-метилфенил, 3-метоксифенил, 3-хлорфенил, 4-метилфенил, 4-этилфенил, 4-метоксифенил, 4-хлорфенил, 2,6-дифторфенил, 2,6-дихлорфенил, 2-метил, 3-метоксифенил, 2-этил, 3-метоксифенил, 3,4-диметоксифенил, 3,5-дифторфенил 3,5-диметилфенил и 3,5-диметокси, 4-метилфенил и тому подобное. Термин возможно замещенный арил предназначен для включения групп, имеющих конденсированные возможно замещенные циклоалкильные и конденсированные возможно замещенные гетероциклические кольца. Примеры включают
и тому подобное.
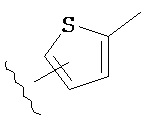
Термин "гетероарил", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к моноциклическим или бициклическим ароматическим кольцевым системам, имеющим от пяти до четырнадцати атомов углерода и один, два, три или четыре гетероатома, независимо выбранных из кислорода, азота и серы. Типичные гетероарильные группы включают 1-пирролил, 2-пирролил, 3-пирролил, 2-имидазолил, 4-имидазолил, пиразинил, 2-оксазолил, 4-оксазолил, 5-оксазолил, 3-изоксазолил, 4-изоксазолил, 5-изоксазолил, 2-тиазолил, 4-тиазолил, 5-тиазолил, 2-фурил, 3-фурил, 2-тиенил, 3-тиенил, 2-пиридил, 3-пиридил, 4-пиридил, 2-пиримидил, 4-пиримидил, 5-бензотиазолил, пуринил, 2-бензимидазолил, 4-бензимидазолил, 5-бензимидазолил, 2-бензотиазолил, 4-бензотиазолил, 5-бензотиазолил, 5-индолил, 5-индолил, 3-индазолил, 4-индазолил, 5-индазолил, 1-изохинолил, 5-изохинолил, 2-хиноксалинил, 5-хиноксалинил, 2-хинолил 3-хинолил, 6-хинолил и тому подобное. Подразумевается, что термин гетероарил включает возможные N-оксиды. Типичные N-оксиды включают пиридил N-оксид и тому подобное.
Термин "возможно замещенный гетероарил", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к гетероарилу, как он определен выше, который возможно замещен по любому доступному атому углерода одним-четырьмя заместителями, обычно одним или двумя заместителями, независимо выбранными из галогено, нитро, циано, гидрокси, амино, возможно замещенного алкила, галогеноалкила, гидроксиалкила, арилалкила, возможно замещенного циклоалкила, возможно замещенного алкенила, возможно замещенного алкинила, возможно замещенного арила, возможно замещенного гетероарила, возможно замещенного гетероцикла, алкокси, арилокси, арилалкилокси, алкилтио, карбоксамидо, сульфонамидо, -CORc, -SO2Rd, -N(Re)CORf, -N(Re)SO2Rg или -N(Re)C=N(Rh)-амино. Типичные замещенные гетероарильные группы включают
и тому подобное.
Конкретная химическая природа возможно замещенных арильных и возможно замещенных гетероарильных групп для концевых группировок A и B в ранее определенных хиральных диацилгидразиновых лигандах не является строго необходимой, и, как отмечено, предполагает широкий ряд заместителей. Предпочтительно заместители для возможно замещенного арила и возможно замещенного гетероарила выбраны таким образом, чтобы общее число атомов углерода и гетероатомов составляло не больше примерно двадцати, более предпочтительно не больше примерно пятнадцати.
Термин "гетероцикл", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к насыщенным и частично ненасыщенным (содержащим одну или две двойные связи) циклическим группам, содержащим одно-три кольца, имеющие от двух до двенадцати атомов углерода и один или два атома кислорода, серы или азота. Гетероцикл возможно может быть связан с остатком молекулы через атом углерода или атома азота. Типичные гетероциклические группы включают
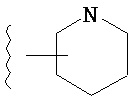
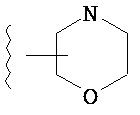
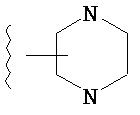

 и тому подобное.
и тому подобное.
Термин "возможно замещенный гетероцикл", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к гетероциклу, как он определен выше, который возможно замещен одним-четырьмя заместителями, обычно одним или двумя заместителями, независимо выбранными из галогено, нитро, циано, гидрокси, амино, возможно замещенного алкила, галогеноалкила, гидроксиалкила, арилалкила, возможно замещенного циклоалкила, возможно замещенного алкенила, возможно замещенного алкинила, возможно замещенного арила, возможно замещенного гетероарила, возможно замещенного гетероцикла, алкокси, арилокси, арилалкилокси, алкилтио, карбоксамидо, сульфонамидо, -CORc, -SO2Rd, -N(Re)CORf, -N(Re)SO2Rg или -N(Re)C=N(Rh)-амино. Типичные возможно замещенные гетероциклические группы включают
и тому подобное. Возможно замещенный гетероцикл может быть конденсирован с арильной группой с получением возможно замещенного арила, как описано выше.
Термин "алкокси", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к гидроксиалкилу, галогеноалкилу, арилалкилу, возможно замещенному алкилу, возможно замещенному циклоалкилу, возможно замещенному алкенилу или возможно замещенному алкинилу, присоединенному к концевому атому кислорода. Типичные алкоксигруппы включают метокси, трет-бутокси, -OCH2CH=CH2 и тому подобное.
Термин "арилокси", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к возможно замещенному арилу, присоединенному к концевому атому кислорода. Типичные группы арилокси включают фенокси и тому подобное.
Термин "арилалкилокси", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к арилалкилу, присоединенному к концевому атому кислорода. Типичные группы арилалкилокси включают бензилокси и тому подобное.
Термин "алкилтио", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к гидроксиалкилу, галогеноалкилу, арилалкилу, возможно замещенному алкилу, возможно замещенному алкенилу или возможно замещенному алкинилу, присоединенному к концевому атому серы. Типичные группы алкилтио включают -SCH3 и тому подобное.
Термин "галогено-" или "галоген", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к фтору, хлору, брому или йоду.
Термин "амино", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к радикалу формулы -NRaRb, где Ra и Rb независимо представляют собой водород, галогеноалкил, гидроксиалкил, арилалкил, возможно замещенный алкил, возможно замещенный циклоалкил, возможно замещенный гетероцикл, возможно замещенный арил или возможно замещенный гетероарил; или Ra и Rb, взятые вместе с атомом азота, к которому они присоединены, образуют четырех-семичленный гетероцикл. Типичные аминогруппы включают -NH2, -N(H)CH3, -N(CH3)2, -N(H)CH2Ph и тому подобное.
Термин "карбоксамидо", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к радикалу формулы -CO-амино. Типичные карбоксамидо-группы включают -CONH2, -CON(H)CH3, -CON(H)Ph и тому подобное.
Термин "сульфонамидо", при использовании в данном описании изобретения отдельно или как часть другой группы, относится к радикалу формулы -SO2-амино. Типичные сульфонамидо-группы включают -SO2NH2, -SO2N(H)CH3, -SO2N(H)Ph и тому подобное.
В отношении возможно замещенных групп, описанных выше, Rc представляет собой водород, гидрокси, галогеноалкил, гидроксиалкил, арилалкил, возможно замещенный алкил, возможно замещенный циклоалкил, возможно замещенный алкенил, возможно замещенный алкинил, возможно замещенный гетероцикл, возможно замещенный арил, возможно замещенный гетероарил, алкокси, арилокси или арилалкилокси; Rd представляет собой галогеноалкил, гидроксиалкил, арилалкил, возможно замещенный алкил, возможно замещенный циклоалкил, возможно замещенный алкенил, возможно замещенный алкинил, возможно замещенный гетероцикл, возможно замещенный арил или возможно замещенный гетероарил; Re представляет собой водород, галогеноалкил, гидроксиалкил, арилалкил, возможно замещенный алкил, возможно замещенный циклоалкил, возможно замещенный алкенил, возможно замещенный алкинил, возможно замещенный гетероцикл, возможно замещенный арил или возможно замещенный гетероарил; Rf представляет собой водород, галогеноалкил, гидроксиалкил, арилалкил, возможно замещенный алкил, возможно замещенный циклоалкил, возможно замещенный алкенил, возможно замещенный алкинил, возможно замещенный гетероцикл, возможно замещенный арил, возможно замещенный гетероарил, алкокси, арилокси, арилалкилокси или амино; Rg представляет собой галогеноалкил, гидроксиалкил, арилалкил, возможно замещенный алкил, возможно замещенный циклоалкил, возможно замещенный алкенил, возможно замещенный алкинил, возможно замещенный гетероцикл, возможно замещенный арил, возможно замещенный гетероарил или амино; и Rh представляет собой водород, алкил, арил, циано или нитро.
Во всем описании изобретения группы и возможные их заместители выбирают с получением стабильных группировок и соединений.
Соединения по настоящему изобретению могут содержать не встречающиеся в природе соотношения атомных изотопов одного или более атомов, образующих такие соединения. Например, соединения могут быть меченными радиоактивными изотопами, такими как, например, тритий (3H), иод-125 (125I) или углерод-14 (14C). Все изотопные варианты соединений по настоящему изобретению, как радиоактивные, так и нет, как предполагается, включены в объем настоящего изобретения.
Соединения по настоящему изобретению могут образовывать соли, которые также находятся в объеме данного изобретения. Следует понимать, что ссылка на соединение по настоящему изобретению в данном описании включает ссылку на его соли, если не указано иное. Термин "соль(и)", при использовании в данном описании изобретения, означает кислые и/или основные соли, образованные неорганическими и/или органическими кислотами и основаниями. Кроме того, если соединение Формулы I, II или III содержит как основную группировку, так и кислую группировку, могут быть образованы цвиттер-ионы ("внутренние соли") и они включены в термин "соль(и)" при использовании в данном описании изобретения. Предпочтительными являются фармацевтически приемлемые (то есть нетоксичные, физиологически приемлемые) соли, хотя другие соли также являются полезными, например на стадиях выделения или очистки, которые можно использовать во время получения. Соли соединений Формулы I, II или III могут быть образованы, например, посредством взаимодействия соединения с таким количеством кислоты или основания, как эквивалентное количество, в среде, такой как среда, в которой соль осаждается или в водной среде после лиофилизации.
Соединения по настоящему изобретению, которые содержат основную группировку, могут образовывать соли со множеством органических и неорганических кислот. Типичные соли присоединения кислоты включают ацетаты (например образованные с уксусной кислотой или тригалогеноуксусной кислотой, например трифторуксусной кислотой), адипаты, альгинаты, аскорбаты, аспартаты, бензоаты, бензолсульфонаты, бисульфаты, бораты, бутираты, цитраты, камфораты, камфорсульфонаты, циклопентанпропионаты, диглюконаты, додецилсульфаты, этансульфонаты, фумараты, глюкогептаноаты, глицерофосфаты, гемисульфаты, гептаноаты, гексаноаты, гидрохлориды (образованные соляной кислотой), гидробромиды (образованные с бромистым водородом), гидроиодиды, 2-гидроксиэтансульфонаты, лактаты, малеаты (образованные малеиновой кислотой), метансульфонаты (образованные метансульфоновой кислотой), 2-нафталинсульфонаты, никотинаты, нитраты, оксалаты, пектинаты, персульфаты, 3-фенилпропионаты, фосфаты, пикраты, пивалаты, пропионаты, салицилаты, сукцинаты, сульфаты (например образованные серной кислотой), сульфонаты (например упомянутые в данном описании изобретения), тартраты, тиоцианаты, толуолсульфонаты, такие кактозилаты, ундеканоаты и тому подобное.
Соединения по настоящему изобретению, которые содержат кислотную группировку, могут образовывать соли со множеством органических и неорганических оснований. Типичные соли оснований включают соли аммония, соли щелочных металлов, такие как соли натрия, лития и калия, соли щелочноземельных металлов, такие как соли кальция и мания, соли органических оснований (например органические амины), такие как бензатины, дициклогексиламины, гидробамины (образованные N,N-бис(дигидроабиэтил)этилендиамином), N-метил-D-глюкамины, N-метил-D-глюкамиды, трет-бутиламины, и соли с аминокислотами, такими как аргинин, лизин и тому подобное.
Стереохимические термины и условные обозначения, используемые в данном описании изобретения, соответствуют терминам и обозначениям, описанным в Pure & Appl. Chem 68: 2193 (1996), если не указано иное.
Термин "энантиомерный избыток" или "э.и." относится к показателю, насколько больше присутствует одного энантиомера, чем другого. Для смеси R- и S-энантиомеров процент энантиомерного избытка определяют как |R-S|*100, где R и S представляют собой соответствующие мольные или массовые фракции энантиомеров в смеси, так что R+S=1. Зная оптическое вращение хирального вещества, процентный энантиомерный избыток определяют как ([a]obs/[a]max)*100, где [a]obs представляет собой оптическое вращение смеси энантиомеров и [a]maX представляет собой оптическое вращение чистого энантиомера. Возможно определение энантиомерного избытка с использованием ряда аналитических методик, включая ЯМР-спектроскопию, хиральную хроматографию на колонке или оптическую поляриметрию.
Термины "энантиомерно чистый" или "энантиочистый" относятся к образцу хирального вещества, все молекулы которого (в рамках определения) имеют одинаковую хиральность.
Термины "энантиомерно обогащенный" или "энантиообогащенный" относятся к образцу хирального вещества, соотношение энантиомеров которого больше 50:50. Энантиомерно обогащенные соединения могут быть энантиомерно чистыми.
Термин "асимметрический атом углерода" относится к атому углерода в молекуле органического соединения, который соединен с четырьмя разными атомами или группами атомов.
Термин "преимущественно" означает в отношении больше чем 50:50.
Термин "уходящая группа" или "LG" относится к атому или к группе, которая отделяется от атома или группы, которая считается остаточной или главной частью субстрата в конкретном взаимодействии. В реакциях амидного сочетания типичные уходящие группы включают -F, -Cl, -Br, -OC6F5 и тому подобное.
Термин "выделенный" для целей настоящего изобретения означает вещество (например химическое соединение или биологическое вещество, такое как нуклеиновая кислота или белок), которое было извлечено из его первоначальной окружающей среды (окружающей среды, в которой оно присутствует естественным образом). Например, полинуклеотид, присутствующий в естественном состоянии в растении или животном, не выделен, однако тот же самый полинуклеотид, отделенный от соседних нуклеиновых кислот, в которых он присутствует в природе, считается "выделенным".
Термин "очищенный", при применении к биологическим веществам не требует присутствия вещества в форме с абсолютной чистотой, без присутствия других соединений. Это скорее относительное определение.
"Нуклеиновая кислота", "молекула нуклеиновой кислоты", "олигонуклеотид" и "полинуклеотид" используются взаимозаменяемо и относятся к полимерной форме фосфатного эфира рибонуклеозидов (аденозин, гуанозин, уридин или цитидин; "молекулы РНК") или дезоксирибонуклеотидов (дезоксиаденозин, дезоксигуанозин, дезокситимидин или дезоксицитидин; "молекулы ДНК"), или любым их фосфоэфирным аналогам, таким как фосфоротиоаты и тиоэфиры, либо в однонитевой форме, либо в двухнитевой спирали. Возможны двухнитевые спирали ДНК-ДНК, ДНК-РНК и РНК-РНК. Термин «молекула нуклеиновой кислоты» и в частности молекула ДНК или РНК, относится только к первичной и вторичной структуре молекулы, и не ограничивает ее какими-либо конкретными третичными формами. Таким образом, данный термин включает двухнитевую ДНК, обнаруженную, среди прочего, в линейных или кольцевых молекулах ДНК (например фрагменты рестрикции), плазмидах, сверхспиральной ДНК и хромосомах. При обсуждении структуры конкретных двухнитевых молекул ДНК, последовательности могут быть описаны в данном описании изобретения согласно стандартному правилу приведения последовательности только в направлении от 5’ к 3’ вдоль нетранскрибируемой нити ДНК (то есть нити, имеющей последовательность, гомологичную мРНК). "Рекомбинантная молекула ДНК" представляет собой молекулу ДНК, которая была подвергнута молекулярно-биологической манипуляции. ДНК включает кДНК, геномную ДНК, плазмидную ДНК, синтетическую ДНК и полусинтетическую ДНК, но не ограничивается ими.
Термин "фрагмент" относится к нуклеотидной последовательности с длиной, уменьшенной по сравнению с исходной нуклеиновой кислоты, и содержащей, в общей части, нуклеотидную последовательность, идентичную исходной нуклеиновой кислоты. Такой фрагмент нуклеиновой кислоты по изобретению может быть, по необходимости включен в более крупный полинуклеотид, составной частью которого он является. Такие фрагменты содержат, или альтернативно состоят из, олигонуклеотиды(ов), с длиной, меняющейся от по меньшей мере 6, 8, 9, 10, 12, 15, 18, 20, 21, 22, 23, 24, 25, 30, 39, 40, 42, 45, 48, 50, 51, 54, 57, 60, 63, 66, 70, 75, 78, 80, 90, 100, 105, 120, 135, 150, 200, 300, 500, 720, 900, 1000 или 1500 последовательных нуклеотидов нуклеиновой кислоты по изобретению.
При использовании в данном описании изобретения "выделенный фрагмент нуклеиновой кислоты" относится к полимеру РНК или ДНК, который является одно- или двухнитевым, возможно содержащему синтетические, неприродные или измененные нуклеотидные основания. Выделенный фрагмент нуклеиновой кислоты в форме полимера ДНК может состоять из одного или более сегментов кДНК, геномной ДНК или синтетической ДНК.
"Ген" относится к полинуклеотиду, содержащему нуклеотиды, которые кодируют функциональную молекулу (например полипептид или РНК), и включает кДНК и геномной ДНК. "Ген" также относится к фрагменту нуклеиновой кислоты, который экспрессирует конкретную РНК, белок или полипептид, включая регуляторные последовательности, предшествующие (5’ некодирующие последовательности) кодирующей последовательности и следующие за (3’ некодирующие последовательности) кодирующей последовательностью. "Нативный ген" относится к гену в виде, обнаруженном в природе, со своими собственными регуляторными последовательностями. "Химерный ген" относится к любому гену, который не является нативным геном, содержащему регуляторную и/или кодирующую последовательности, которые не обнаружены вместе в природе. Таким образом, химерный ген может содержать регуляторные последовательности и кодирующие последовательности, которые получены из разных источников, или регуляторные последовательности и кодирующие последовательности, полученные из того же источника, но расположенные другим образом, чем обнаруживается в природе. Химерный ген может содержать кодирующие последовательности, полученные из разных источников и/или регуляторные последовательности, полученные из разных источников. "Эндогенный ген" относится к нативному гену в его естественной локализации в геноме организма. "Чужой" ген или "гетерологичный" ген относится к гену не обычно не обнаруживаемому в организм-хозяине, но который вводят в организм-хозяин посредством переноса генов. Чужие гены могут содержать нативные гены, вставленные в ненативный организм, или химерные гены. "Трансген" представляет собой ген, который был введен в геном с помощью методики трансформации.
"Гетерологичная" ДНК относится к ДНК, в природе не локализованной в клетке, или в хромосомном сайте клетки. Предпочтительно гетерологичная ДНК включает ген, чужой для этой клетки.
Термин "геном" включает хромосомную, а также митохондриальную, хлоропластную и вирусную ДНК или РНК.
Молекула нуклеиновой кислоты является "гибридизуемой" в другую молекулу нуклеиновой кислоты, такую как кДНК, геномная ДНК или РНК, если однонитевую форму молекулы нуклеиновой кислоты можно отжечь в другую молекулу нуклеиновой кислоты в подходящих условиях температуры и ионной силы раствора (смотри Sambrook et al, 1989 ниже). Условия гибридизации и промывания хорошо известны и проиллюстрированы в Sambrook et al. Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor (1989), в частности Глава 11 и Таблица 11.1 (включен в данное описание изобретения во всей полноте посредством ссылки). Условия температуры и ионной силы определяют "жесткость" гибридизации.
Жесткие условия могут быть настроены для скрининга от относительно подобных фрагментов, таких как гомологичные последовательности из организмов с низкой степенью родства, до в высокой степени подобных фрагментов, таких как гены, которые дублируют функциональные ферменты близкородственных организмов. Для предварительного скрининга гомологичных нуклеиновых кислот можно использовать менее жесткие условия гибридизации, соответствующие Tm=55°, например 5x SSC, 0,1% SDS, 0,25% молоко, и без формамида; или 30%-ный формамид 5x SSC, 0,5% SDS. Умеренно жесткие условия гибридизации соответствуют более высокой Tm, например, 40%-ный формамид с 5x или 6x SCC. В высшей степени жесткие условия гибридизации соответствуют самой высокой Tm, например 50%-ный формамид, 5x или 6x SCC.
Для гибридизации требуется, чтобы две нуклеиновые кислоты содержали комплементарные последовательности, хотя и зависящие от жесткости гибридизации, при этом возможны несоответствия между основаниями. Термин "комплементарный" используют для описания отношения между нуклеотидными основаниями, которые способны гибридизироваться друг с другом. Например, в отношении ДНК, аденозин является комплементарным тимину, и цитозин является комплементарным гуанину. Таким образом, настоящее изобретение также включает выделенные фрагменты нуклеиновой кислоты, которые комплементарны полным последовательностям, как они раскрыты или использованы в данном описании изобретения, а также по существу аналогичным нуклеиновокислым последовательностям.
В одном воплощении изобретения полинуклеотиды определяют, используя условия гибридизации, включающих стадию гибридизации при Tm=55°C, и используя условия, как изложено выше. В другом воплощении Tm равна 60°C; в одном воплощении Tm равна 63°C; в другом воплощении Tm равна 65°C.
Постгибридизационные промывки также определяют жесткость условий. В одном наборе условий используют серии промывок, начиная с 6X SSC, 0,5% SDS при комнатной температуре в течение 15 минут (мин), затем повторяли с 2X SSC, 0,5% SDS при 45°C в течение 30 минут, и затем повторяли дважды с 0,2X SSC, 0,5% SDS при 50°C в течение 30 минут. В одном наборе жестких условий использовали более высокие температуры, где промывки идентичны промывкам выше за исключением того, что температура последних двух 30-минутных промывок в 0,2X SSC, 0,5% SDS, возрастала до 60°C. В другом наборе условий высокой степени жесткости используют две последние промывки в 0,1X SSC, 0,1% SDS при 65°C.
Подходящая жесткость для гибридизации нуклеиновых кислот зависит от длины нуклеиновых кислот и степени комплементации, переменных, хорошо известных в данной области техники. Чем выше степень подобия или гомологии между двумя нуклеотидными последовательностями, тем выше величина Tm для гибридов нуклеиновых кислот, имеющих эти последовательности. Относительная стабильность (соответствующая более высокой Tm) гибридизаций нуклеиновой кислоты снижается в следующем порядке: РНК:РНК, ДНК:РНК, ДНК:ДНК. Для гибридов из более чем 100 нуклеотидов в длину, были получены уравнения для расчета Tm (смотри Sambrook et al., выше, 9.50-0.51). Для гибридизации с более короткими нуклеиновыми кислотами, например олигонуклеотидами, положение несоответствий становится более важным, и длина олигонуклеотида определяет его специфичность (смотри Sambrook et al, выше, 11.7-11.8).
В одном воплощении изобретения полинуклеотиды определяют, используя условия гибридизации, включающие стадию гибридизации в менее чем 500 мМ соли и по меньшей мере при 37 градусах Цельсия, и стадию промывания в 2X SSPE при по меньшей мере 63 градусах Цельсия. В одном воплощении условия гибридизации включают менее 200 мМ соли и по меньшей мере 37 градусов Цельсия на стадии гибридизации. В другом воплощении условия гибридизации включают 2X SSPE и 63 градуса Цельсия для стадий как гибридизации, так и промывки.
В другом воплощении длина гибридизуемой нуклеиновой кислоты составляет по меньшей мере примерно 10 нуклеотидов. Предпочтительно минимальная длина гибридизуемой нуклеиновой кислоты составляет по меньшей мере примерно 15 нуклеотидов; более предпочтительно по меньшей мере примерно 20 нуклеотидов; и наиболее предпочтительно длина составляет по меньшей мере 30 нуклеотидов. Более того, специалисту в данной области техники понятно, что температура и концентрация соли в промывном растворе может быть при необходимости скорректирована согласно таким факторам, как длина зонда.
Термин "зонд" относится к однонитевой молекуле нуклеиновой кислоты, которая может спариваться основаниями с комплементарной однонитевой нуклеиновой кислотой-мишенью с образованием двухнитевой молекулы.
При использовании в данном описании изобретения, термин "олигонуклеотид" относится к короткой нуклеиновой кислоте, которая гибридизуется в молекулу геномной ДНК, молекулу кДНК, молекулу плазмидную ДНК или молекулу мРНК. Олигонуклеотиды могут быть меченными, например 32P-нуклеотидами или нуклеотидами, к которым была ковалентно присоединена метка, такая как биотин. Меченный олигонуклеотид можно использовать в качестве зонда для определения присутствия нуклеиновой кислоты. Олигонуклеотиды (один или оба из которых могут быть меченными) можно использовать в качестве праймеров для ПЦР, либо для клонирования полноразмерной нуклеиновой кислоты или фрагмента нуклеиновой кислоты, либо для определения присутствия нуклеиновой кислоты. Олигонуклеотид также можно использовать для образования тройной спирали с молекулой ДНК. В общем случае Олигонуклеотиды готовят синтетически, предпочтительно на синтезаторе нуклеиновых кислот. Таким образом, Олигонуклеотиды могут быть приготовлены с не встречающимися в природе аналогами фосфороэфирных связей, такими как тиоэфирные связи, и так далее.
"Праймер" относится к олигонуклеотиду, который гибридизуется с целевой последовательностью нуклеиновой кислоты с созданием двухнитевого участка нуклеиновой кислоты, который может служить в качестве участка инициации для синтеза ДНК в подходящих условиях. Такие праймеры можно использовать в полимеразной цепной реакции.
"Полимеразная цепная реакция", сокращенно PCR (ПЦР), относится к in vitro способу ферментного амплифицирования конкретных последовательностей нуклеиновых кислот. ПЦР включает повторяющиеся серии температурных циклов, где каждый цикл содержит три стадии:
денатурацию матричной нуклеиновой кислоты для разделения нитей целевой молекулы, отжиг однонитевого олигонуклеотидного праймера для ПЦР до матричной нуклеиновой кислоты, и удлинение отожженного(ых) праймера(ов) с помощью ДНК-полимеразы. ПЦР обеспечивает средства обнаружения присутствия целевой молекулы и, в количественных или полуколичественных условиях, определения относительного количества такой целевой молекулы в исходном пуле нуклеиновых кислот.
"Полимеразная цепная реакция с обратной транскрипцией" сокращенно обозначается RT-PCR (ОТ-ПЦР), и относится к in vitro способу ферментного продуцирования целевой молекулы или молекул кДНК из молекулы или молекул РНК молекулы, с последующей ферментной амплификацией конкретной последовательности или последовательностей нуклеиновой кислоты в рамках целевой молекулу или молекул кДНК, как описано в выше. ОТ-ПЦР также предлагает средство обнаружения присутствия целевой молекулы и, в количественных или полуколичественных условиях, определения относительного количества такой целевой молекулы в исходном пуле нуклеиновых кислот.
"Кодирующая последовательность" ДНК относится к двухнитевой последовательности ДНК, которая кодирует полипептид и может быть транскрибирована и транслирована в полипептид в клетке, in vitro или in vivo, при помещении под контроль соответствующих регуляторных последовательностей. Термин "подходящие регуляторные последовательности" относится к нуклеотидным последовательностям, расположенным выше против хода транскрипции (5’ некодирующие последовательности), в рамках или ниже по ходу транскрипции (3’ некодирующие последовательности) кодирующей последовательности, и которые оказывают влияние на транскрипцию, РНК-процессирование или стабильность, или трансляцию ассоциированной кодирующей последовательности. Регуляторные последовательности могут включать промоторы, трансляционные лидерные последовательности, интроны, последовательности распознавания полиаденилирования, сайт процессирования РНК, сайт, связывающий эффектор, и структуры типа стебель-петля. Границы кодирующей последовательность определены стартовым кодоном на 5’ (амино) конце и трансляцинным стоп-кодоном на 3’ (карбоксильном) конце. Кодирующая последовательность может включать, но не ограничена ими, прокариотические последовательности, кДНК от мРНК, последовательности геномной ДНК и даже синтетические последовательности ДНК. Если кодирующая последовательность предназначена для экспрессии в эукариотической клетке, сигнал подиаденилирования и последовательность окончания транскрипции обычно будут находиться 3’ относительно кодирующей последовательности.
"Открытая рамка считывания" сокращенно обозначается ORF и относится к длине последовательности нуклеиновой кислоты, либо ДНК, кДНК, либо РНК, которая содержит сигнал начала трансляции или инициирующий кодон, такой как ATG или AUG, и кодон окончания, и потенциально может быть транслирована в полипептидную последовательность.
Термин "голова-к-голове" используется в данном описании изобретения для описания ориентации двух полинуклеотидных последовательностей относительно друг друга. Два полинуклеотида расположены в ориентации "голова-к-голове", когда 5’-конец кодирующей нити одного полинуклеотида соседствует с 5’-концом кодирующей нити другого полинуклеотида, в результате чего транскрипция каждого полинуклеотида направлена от 5’-конца другого полинуклеотида. Термин "голова-к-голове" может быть сокращено обозначен как (5’)-к-(5’) и также может быть указан с помощью символов (←→) или (3’←5’5’→3’).
Термин "хвост-к-хвосту" используют в данном описании изобретения для описания ориентации двух полинуклеотидных последовательностей относительно друг друга. Два полинуклеотида расположены в ориентации «хвосту-к-хвосту», если 3’-конец кодирующей нити одного полинуклеотида соседствует с 3’-концом кодирующей нити другого полинуклеотида, в результате чего транскрипция каждого полинуклеотида направлена навстречу другому полинуклеотиду. Термин "хвост-к-хвосту" может быть сокращен (3’)-к-(3’) и также может быть указан с помощью символов (→←) или (5→3’3←5’).
Термин "голова-к-хвосту" используют в данном описании изобретения для описания ориентации двух полинуклеотидных последовательностей относительно друг друга. Два полинуклеотида расположены в ориентации голова-к-хвосту, если 5’-конец кодирующей нити одного полинуклеотида соседствует с 3’-концом кодирующей нити другого полинуклеотида, в результате чего транскрипция каждого полинуклеотида в том же самом направлении, что и у другого полинуклеотида. Термин "голова-к-хвосту" может быть обозначен сокращенно (5’)-к-(3’) и также может быть указан с помощью символов (→→) или (5’→3’5’→3’).
Термин "по ходу транскрипции" относится к нуклеотидной последовательности, которая расположена 3’ к эталонной нуклеотидной последовательности. В частности, нуклеотидные последовательности по ходу транскрипции в большинстве случаев относятся к последовательностям, которые следуют за начальной точкой транскрипции. Например, кодон, инициирующий трансляцию гена, размещен по ходу транскрипции от начального сайта транскрипции.
Термин "против хода транскрипции" относится к нуклеотидной последовательности, которая расположена 5’ по отношению к эталонной нуклеотидной последовательности. В частности, нуклеотидные последовательности против хода транскрипции в большинстве случаев относятся к последовательностям, которые расположены на 5’ стороне кодирующей последовательности или от начальной точки транскрипции. Например, большинство промоторов располагают против хода транскрипции от начального сайта транскрипции.
Термины "эндонуклеаза рестрикции" и "фермент рестрикции" используются взаимозаменяемо и относят к ферменту, который сшивает и разрезает в рамках конкретной нуклеотидной последовательности в двухнитевой ДНК.
"Гомологичная рекомбинация" относится к вставке последовательности чужой ДНК в другую молекулу ДНК, например вставке вектора в хромосому. Предпочтительно вектор нацелен на конкретный хромосомный сайт для гомологичной рекомбинации. Для конкретной гомологичной рекомбинации вектор будет содержать достаточно длинные участки гомологии с последовательностями хромосомы, чтобы обеспечить комплементарное связывание и введение вектора в хромосому. Более длинные участки гомологии и более значительные степени подобия последовательностей могут увеличивать эффективность гомологичной рекомбинации.
Некоторые способы, известные в данной области техники, можно использовать для увеличения количества полинуклеотида согласно изобретению. После того как установлена подходящая система-хозяин и условия роста, количество рекомбинантных экспрессирующих векторов можно увеличить и получить их в большом количестве. Как описано в данном описании изобретения, экспрессирующие векторы, которые можно использовать, включают, без ограничения ими, следующие векторы или их производные: вирусы человека или животных, такие как вирус коровьей оспы или аденовирус; вирусы насекомых, такие как бакуловирус; дрожжевые векторы; бактериофаговые векторы (например лямбда), и векторы плазмидной и космидной ДНК, и многие другие.
"Вектор" относится к любому носителю для клонирования и/или переноса нуклеиновой кислоты в клетку-хозяина. Вектор может представлять собой репликон, к которому может быть присоединен сегмент другой ДНК так, чтобы осуществить репликацию присоединенного сегмента. "Репликон" относится к любому генетическому элементу (например плазмиде, фагу, космиде, хромосоме, вирусу), который действует в качестве самостоятельной единицы ДНК-репликации in vivo, то есть способен реплицироваться под собственным контролем. Термин "вектор" включает как вирусные, так и невирусные носители для введения нуклеиновой кислоты в клетку in vitro, ex vivo или in vivo. Большое число векторов, известных в данной области техники, может быть использовано для манипулирования нуклеиновыми кислотами, введения элементов ответа и промоторов в гены и так далее. Возможные векторы включают, например, плазмиды или модифицированные вирусы, включая, например, бактериофаги, такие как лямбда-производные, или плазмиды, такие как производные плазмид pBR322 или pUC, или вектор Bluescript. Например, вставка фрагментов ДНК, соответствующих элементам ответа и промоторам, в подходящий вектор может быть осуществлена посредством лигирования соответствующих фрагментов ДНК в выбранный вектор, который имеет комплементарные "липкие" концы. Альтернативно, концы молекул ДНК могут быть модифицированы ферментом, или любой сайт может быть получен путем лигирования нуклеотидных последовательностей (линкеров) в концы ДНК. Такие векторы могут быть сконструированы так, чтобы содержать гены селектируемого маркера, которые обеспечивают селекцию клеток, у которых маркер введен в клеточный геном. Такие маркеры обеспечивают идентификацию и/или селекцию клеток-хозяев, которые включают и экспрессируют белки, кодируемые маркером.
Вирусные векторы, и особенно ретровирусные векторы, использовались в широком ряде применений генной доставки в клетках, а также живых животных субъектах. Вирусные векторы, которые могут быть использованы, включают, без ограничения ими, векторы ретровируса, адено-ассоциированного вируса, оспы, бакуловируса, коровьей оспы, простого герпеса, Эпштейна-Барр, аденовируса, геминивируса и колимовируса. Невирусные векторы включают плазмиды, липосомы, электрически заряженные липиды (цитофектины), комплексы ДНК-белок и биополимеры. Кроме нуклеиновой кислоты вектор может также содержать один или более чем один регуляторный участок, и/или селектируемые маркеры, полезные в селектировании, измерении, и мониторинге результатов переноса нуклеиновой кислоты (переноса в какие ткани, продолжительность экспрессии, и так далее).
Термин "плазмида" относится к внехромосомному элементу, часто несущему ген, который не является частью центрального метаболизма клетки, и обычно в форме кольцевых двухнитевых молекул ДНК. Такие элементы могут представлять собой автономно реплицирующиеся последовательности, последовательности, интегрирующиеся в геном, фаговые или нуклеотидные последовательности, линейные, кольцевые или сверхспиральные одно- или двухнитевые ДНК или РНК, полученные из любого источника, в которых несколько нуклеотидных последовательностей объединены или рекомбинированы в уникальную конструкцию, которая способна ввести фрагмент промотора и последовательность ДНК для селектируемого генного продукта вместе с соответствующей 3’ нетранслированной последовательностью в клетку.
"Клонирующий вектор" относится к "репликону", который представляет собой единичную длину нуклеиновой кислоты, предпочтительно ДНК, которая реплицируется последовательно, и который содержит начало репликации, такую как плазмида, фаг или космида, к которой может быть присоединен сегмент другой нуклеиновой кислоты, так чтобы осуществить репликацию присоединенного сегмента. Клонирующие векторы могут быть способны реплицироваться в одном типе клеток и экспрессироваться в другом ("челночный вектор").
Термин "экспрессирующий вектор" относится к вектору, плазмиде или носителю, предназначенному обеспечивать экспрессию введенной последовательности нуклеиновой кислоты после трансформации в хозяина. Клонированный ген, то есть введенную последовательность нуклеиновой кислоты, обычно помещают под контроль контролирующих элементов, таких как промотор, минимальный промотор, энхансер или подобное. Участки регуляции инициации или промоторы, которые полезны для управления экспрессией нуклеиновой кислоты в желательной клетке-хозяине, многочисленны и хорошо известны специалистам в данной области техники. Практически любой промотор, способный управлять такими генами, является пригодным для настоящего изобретения, включая, но без ограничения ими: вирусные промоторы, бактериальные промоторы, промоторы животных, промоторы млекопитающих, синтетические промоторы, конститутивные промоторы, тканеспецифические промоторы, специфические промоторы развития, индуцибельные промоторы, лиганд-регулируемые промоторы; CYC1, HIS3, GAL1, GAL4, GAL10, ADH1, PGK, PH05, GAPDH, ADC J, TRPI, URA3, LEU2, ENO, TPI, промоторы щелочной фосфатазы (полезные для экспрессии в Saccharomyces); промотор AOX1 (полезный для экспрессии в Pichia); промоторы β-лактамазы, lac, ara, tet, trp, IPL IPR, T7, tac, и trc (полезные для экспрессии в Escherichia coli); промоторы светорегулируемые, семя-специфичные, пыльца-специфичные, специфичные в отношении яичников, связанные с патогенезом или заболеванием, вируса мозаики цветной капусты 35S, минимальные CMV 35S, вируса мозаики маниоки (CsVMV), хлорофилл a/b связывающего белка, рибулоза-1,5-бифосфат-карбоксилазы, специфические в отношении ростков, специфические в отношении корней, хитиназы, стресс-индуцибельные, бациллярного вируса риса тунгро (tungro), суперпромоторы растений, лейцин-аминопептидазы картофеля, нитратредуктазы, маннопин-синтазы, нопалин-синтазы, убиквитина, зеинового белка, и антоцианиновые промоторы (полезные для экспрессии в растительных клетках); промоторы животных и млекопитающих, известные в данной области техники, включают, без ограничения ими, ранний (SV40e) промоторный участок SV40, промотор, содержащийся в 3’-длинном терминальном повторе (LTR) вируса саркомы Рауса (RSV), промоторы E1 A или главный поздний промотор (MLP) генов аденовирусов (Ad), ранний промотор цитомегаловируса (CMV), промотор тимидинкиназы (TK) вируса простого герпеса (HSV), промотор бакуловируса IE1, промотор фактора элонгации 1-альфа (EF1), промотор фосфоглицераткиназы (PGK), промотор убиквитина (Ubc), промотор альбумина, регуляторные последовательности промотора мышиного металлотионеина-L и участков транскрипционного контроля, универсальные промоторы (HPRT, виментин, α-актин, тубулин и тому подобное), промоторы промежуточных микрофиламентов (десмин, нейромикрофиламенты, кератин, GFAP и тому подобное), промоторы терапевтических генов (MDR, CFTR или фактора VIII типа и тому подобное), промоторы, связанные с патогенезом или заболеванием, и промоторы, которые проявляют тканевую специфичность и используются у трансгенных животных, такие как участок контроля гена эластазы I, который является активным в панкреатических ацинарных клетках; участок контроля гена инсулина, активный в панкреатических бета-клетках, участок контроля гена иммуноглобулина, активный в лимфоидных клетках, участок контроля вируса опухоли молочной железы мышей, активный в тестикулярных клетках, клетках молочной железы, лимфоидных и тучных клетках; участки контроля гена альбумина, Apo Al и Apo All, активные в печени, участок контроля гена альфа-фетопротеина, активный в печени, участок контроля гена альфа 1-антитрипсина, активный в печени, участок контроля гена бета-глобулина, активный в миелоидных клетках, участок контроля гена основного миелинового белка, активный в олигодендроцитных клетках мозга, участок контроля гена легкой цепи-2 миозина, активный в скелетных мышцах, и участок контроля гена гонадотроп-высвобождающего гормона, активный в гипоталамусе, промотор пируваткиназы, промотор виллина, промотор кишечного белка, связывающего жирные кислоты, промотор α-актина клеток гладких мышц и тому подобное. Кроме того, данные экспрессионные последовательности могут быть модифицированы путем добавления энхансера или регуляторных последовательностей и тому подобного.
Векторы могут быть введены в желательные клетки-хозяин способами, известными в данной области техники, например с помощью трансфекции, электропорации, микроинъекции, трансдукции, слияния клеток, DEAE (диэтиламиноэтилового) декстрана, осаждения с помощью фосфата кальция, липофекции (лизосомное слияние), использования генной пушки, или переносчика ДНК-вектора (смотри, например Wu et al, J. Biol. Chem. 267: 963-967 (1992); Wu et al, J. Biol. Chem. 263: 14621-14624 (1988); и Hartmut et al., канадская патентная заявка 2012311, поданная 15 марта 1990).
Полинуклеотид по изобретению может быть введен также in vivo с помощью липофекции. В последнее возросло использование липосом для инкапсуляции и трансфекции нуклеиновых кислот in vitro. Синтетические катионные липиды, разработанные, чтобы огранить трудности и опасности, встречающихся при опосредованной липосомами трансфекции, можно использовать для изготовления липосом для in vivo трансфекции гена, кодирующего маркер (Feigner et al, Proc. Natl Acad. Sci. USA. 84: 7413 (1987); Mackey et al, Proc. Natl. Acad. Sci. USA 55: 8027-8031 (1988); и Ulmer et al, Science 259: 1745-1748 (1993)). Использование катионные липидов может способствовать инкапсуляции отрицательно заряженных нуклеиновых кислот, а также способствовать слиянию с отрицательно заряженными клеточными мембранами (Feigner et al, Science 337: 387-388 (1989)). Особенно полезные липидные соединения и композиции для переноса нуклеиновых кислот описаны в WO 95/18863, WO 96/17823 и US 5459127. Использование липофекции для введения экзогенных генов в конкретные органы in vivo имеет определенные практические преимущества. Молекулярное нацеливание липосом на конкретные клетки является одним из преимуществ. Понятно, что направленная трансфекция в конкретные типы клеток будет особенно предпочтительной в ткани с клеточной гетерогенностью, такой как поджелудочная железа, печень, почки и мозг. Для нацеливания липиды могут быть химически связаны с другими молекулами (Mackey et al. 1988, выше). Нацеленные пептиды, например гормоны или нейромедиаторы, и белки, такие как антитела, или непептидные молекулы, могут быть связаны с липосомами химически.
Другие молекулы, такие как катионный олигопептид (например WO 95/21931), пептиды, полученные из ДНК-связывающих белков (например WO 96/25508), или катионный полимер (например из WO 95/21931), также полезны для облегчения трансфекции нуклеиновой кислоты in vivo.
Также возможно вводить вектор in vivo в виде плазмиды с "голой" ДНК (смотри патенты США 5693622, 5589466 и 5580859). Также можно использовать подход с опосредованной рецептором доставкой ДНК (Curiel et al, Hum. Gene Ther. 5: 147-154 (1992); и Wu et al, J. Biol Chem. 262: 4429-4432 (1987)).
Термин "трансфекция" относится к внедрению экзогенной или гетерологичной РНК или ДНК в клетку. Клетка "трансфицирована" экзогенной или гетерологичной РНК или ДНК, если такая РНК или ДНК введена внутрь клетки. Клетка "трансформирована" экзогенной или гетерологичной РНК или ДНК, если трансфицирующая РНК или ДНК осуществляет изменение фенотипа. Трансформирующие РНК или ДНК могут быть интегрированы (ковалентно связаны) в хромосомную ДНК, образуя геном клетки.
"Трансформация" относится к переносу фрагмента нуклеиновой кислоты в геном организма-хозяина, что приводит к генетически устойчивому наследованию. Организмы-хозяева, содержащие фрагменты трансформированной нуклеиновой кислоты называются "трансгенными" или "рекомбинантными" или "трансформированными" организмами.
Кроме того, рекомбинантный вектор, содержащий полинуклеотид по изобретению, может включать одну или более точек начала репликации в клетках-хозяевах, в которых требуется их амплификация или экспрессия, маркера или селектируемых маркера.
Термин "селектируемые маркеры" относится к фактору идентификации, обычно к гену антибиотической или химической устойчивости, который можно селектировать в отношении эффекта, основанного на маркерном гене, то есть устойчивости к антибиотику, устойчивости к гербициду, колориметрическим маркерам, ферментам, флуоресцентным маркерам и тому подобному, где эффект используется для отслеживания наследования интересующей нуклеиновой кислоты и/или для идентификации клетки или организма, которые унаследовали интересующую нуклеиновую кислоту. Примеры селектируемых маркерных генов, известных и используемых в данной области техники, включают: гены, обеспечивающие устойчивость к ампициллину, стрептомицину, гентамицину, канамицину, гигромицину, гербициду биалафосу, сульфонамиду и тому подобные; и гены, которые используют в качестве фенотипических маркеров, то есть антоцианиновые регуляторные гены, ген изопентанилтрансферазы и тому подобные.
Термин "репортерный ген" относится к нуклеиновой кислоте, кодирующей фактор идентификации, который можно идентифицировать на основании эффекта репортерного гена, где эффект используют для отслеживания наследования интересующей нуклеиновой кислоты для идентификации клетки или организма, который наследует интересующую нуклеиновую кислоту, и/или для измерения индукции экспрессии гена или транскрипции. Примеры репортерных генов, известные и используемые в данной области техники, включают гены: люциферазы (Luc), зеленого флуоресцентного белка (GFP), хлорамфеникола ацетилтрансферазы (CAT), β-галактозидазы (LacZ), β-глюкуронидазы (Gus), и тому подобные. Селектируемые маркерные гены также можно считать репортерными генами.
Промотор и "промоторная последовательность" используются взаимозаменяемо и относятся к последовательности ДНК, способной контролировать экспрессию кодирующей последовательности или функциональной РНК. В общем случае, кодирующая последовательность помещена 3’ к промоторной последовательности. Промоторы могут быть получены во всей их полноте из нативного гена, или могут быть составлены из разных элементов, полученных из разных промоторов, обнаруженных в природе, или даже содержать сегменты синтетической ДНК. Специалисту в данной области техники понятно, что разные промоторы могут управлять экспрессией гена в разных тканях или типах клеток, или на разных стадиях развития, или в ответ на разные условия окружающей среды или физиологические условия. Промоторы, которые чаще всего вызывают экспрессию гена в большинстве типов клеток, обычно называются "конститутивными промоторами." Промоторы, которые вызывают экспрессию гена в конкретном типе клеток, обычно называются "клеточноспецифическими промоторами" или "тканеспецифическими промоторами". Промоторы, которые вызывают экспрессию гена на конкретной стадии развития или клеточной дифференциации, обычно называют "промоторами, специфическими в отношении развития" или "промоторами, специфическими в отношении клеточной дифференциации". Промоторы, которые индуцируются и вызывают экспрессию гена после воздействия на клетку агента, биологической молекулы, химического вещества, лиганда, излучения или тому подобного, что индуцирует промотор, или обработки клетки ими, обычно называют "индуцируемыми промоторами" или "регулируемыми промоторами". Кроме того, признано, что поскольку в большинстве случаев точные границы регуляторных последовательностей не были полностью определены, фрагменты ДНК, имеющие разную длину, могут обладать идентичной промоторной активностью.
Промоторная последовательность обычно ограничена на ее 3’-конце сайтом инициации транскрипции и тянется в направлении против хода транскрипции (5’-направление) с включением минимального числа оснований или элементов, необходимых для начала транскрипции при уровнях, обнаруживаемых выше фонового. В промоторной последовательности может быть обнаружен сайт инициации транскрипции (удобно определяемый, например, с помощью картирования нуклеазой S1), а также белок-связывающие домены (консенсусные последовательности), ответственные за связывание РНК-полимеразы.
Кодирующая последовательность находится в клетке "под контролем" последовательностей транскрипционного и трансляционного контроля, когда РНК-полимераза транскрибирует кодирующую последовательность в мРНК, которая затем транс-РНК-сплайсируется (если кодирующая последовательность содержит интроны) и транслируется в белок, кодируемый кодирующей последовательностью.
"Последовательности транскрипционного и трансляционного контроля" относятся к регуляторным последовательностям ДНК, таким как промоторы, энхансеры, терминаторы и тому подобное, которые обеспечивают экспрессию кодирующей последовательности в клетке-хозяине. В эукариотических клетках контролирующими последовательностями являются сигналы полиаденилирования.
Термин "элемент ответа" относится к одному или более цис-действующим элементам ДНК, которые придают реактивность на опосредованное промотором прямое взаимодействие с ДНК-связывающими доменами фактора транскрипции, например ДНК-связывающим доменом первого гибридного белка. Данный элемент ДНК может быть или палиндромным (совершенным или несовершенным) в своей последовательности, или состоящим из мотивов последовательности или полусайтов, разделенных переменным числом нуклеотидов. Полусайты могут быть подобными или идентичными, и располагаться в виде или прямых или обратных повторов, или в виде единичного полусайта, или мультимеров соседних полусайтов в тандеме. Элемент ответа может содержать минимальный промотор, выделенный из разных организмов, в зависимости от природы клетки или организма, в который элемент ответа будет введен. ДНК-связывающий домен первого гибридного белка связывается, в присутствии или в отсутствие лиганда, с последовательностью ДНК элемента ответа для инициации или подавления транскрипции гена(ов), расположенного(ых) по ходу транскрипции, регулируемых данным элементом ответа. Примеры последовательностей ДНК для элементов ответа естественного экдизонового рецептора включают: RRGG/TTCANTGAC/ACYY (SEQ ID NO: 16) (смотри Cherbas et. al. Genes Dev. 5: 120-131 (1991)); AGGTCAN(n)AGGTCA, где N(n) может представлять собой один или более чем один спейсерный нуклеотид (SEQ ID NO: 17) (смотри D’Avino et al, Mol. Cell.. Endocrinol. 113: 1-9 (1995)); и GGGTTGAATGAATTT (SEQ ID NO: 18) (смотри Antoniewski et al, Mol. Cell. Biol. 74: 4465-4474 (1994)).
Термин "функционально связанный" относится к ассоциации последовательностей нуклеиновых кислот в одном фрагменте нуклеиновой кислоты так, что действие одного находится под влиянием другого. Например, промотор функционально связан с кодирующей последовательностью, если он способен влиять на экспрессию этой кодирующей последовательности (то есть эта кодирующая последовательность находится под транскрипционным контролем промотора). Кодирующие последовательности могут быть функционально связаны с регуляторными последовательностями в смысловой или антисмысловой ориентации.
Термин "экспрессия" при использовании в данном описании изобретения относится к транскрипции и устойчивому накоплению смысловой (мРНК) или антисмысловой РНК, полученных от нуклеиновой кислоты или полинуклеотида. Экспрессия может также относится к трансляции мРНК в белок или полипептид.
Термины "кассета", "кассета экспрессии" и "кассета экспрессии гена" относятся к сегменту ДНК, который может быть встроен в нуклеиновую кислоту или полинуклеотид в особых сайтах рестрикции или посредством гомологичной рекомбинации. Сегмент ДНК содержит полинуклеотид, который кодирует интересующий полипептид, и кассету и сайты рестрикции сконструированы так, чтобы для обеспечить вставку кассеты в надлежащую рамку считывания для транскрипции и трансляции. "Кассета трансформации" относится к конкретному вектору, содержащему полинуклеотид, который кодирует интересующий полипептид, и имеющему, в дополнение к полинуклеотиду, элементы, которые облегчают трансформацию конкретной клетки-хозяина. Кассеты, кассеты экспрессии, кассеты экспрессии гена и кассеты трансформации по изобретению также могут содержать элементы, которые обеспечивают улучшенную экспрессию полинуклеотида, кодирующего интересующий полипептид, в клетке-хозяине. Эти элементы могут включать, без ограничения ими: промотор, минимальный промотор, энхансер, элемент ответа, терминирующую последовательность, последовательность полиаденилирования и тому подобное.
Для целей данного изобретения термин "переключатель генов" относится к комбинации элемента ответа в ассоциации с промотором, и системе на основе EcR, которая в присутствии одного или более лиганда(ов), модулирует экспрессию гена, в который встроен элемент ответа и промотор.
Термины "модулировать" и "модулирует" означают индуцировать, снижать или ингибировать экспрессию нуклеиновой кислоты или гена, что приводит к соответствующим индукции, снижению или ингибированию продуцирования белка или полипептида.
Плазмиды или векторы по изобретению могут дополнительно содержать по меньшей мере один промотор, пригодный для стимулирования экспрессии гена в клетке-хозяине.
Энхансеры, которые можно использовать в воплощениях изобретения, включают: энхансер SV40, энхансер цитомегаловируса (CMV), энхансер фактора элонгации 1 (EF1), дрожжевые энхансеры, энхансеры вирусных генов и тому подобные, но не ограничиваются ими.
Участки контроля термининации, то есть терминаторные последовательности или последовательности полиаденилирования, также могут также быть получены из различных генов, присущих предпочтительным хозяевам. Возможно, терминирующий сайт может не являться необходимым, однако наиболее предпочтительно его включать. В одном воплощении изобретение участок контроля термининации может состоять или быть получен из синтетической последовательности, синтетического сигнала полиаденилирования, позднего сигнала полиаденилирования SV40, сигнала полиаденилирования SV40, сигнала полиаденилирования гормона роста крупного рогатого скота (BGH), вирусных терминаторных последовательностей или подобного.
Термины "3’-некодирующие последовательности" или "3’-нетранслируемый участок (UTR)" относятся к последовательностям ДНК, расположенным вниз по ходу транскрипции (3’) кодирующей последовательности и могут содержать последовательности распознавания полиаденилирования [poly(A)] и другие последовательности, кодирующие регуляторные сигналы, способные влиять на процессирование мРНК или экспрессию гена. Сигнал полиаденилирования обычно характеризуется влиянием добавления участков полиадениловой кислоты к 3’-концу предшественника мРНК.
"Регуляторный участок" относится к последовательности нуклеиновой кислоты, которая регулирует экспрессию второй последовательности нуклеиновой кислоты. Регуляторный участок может включать последовательности, которые по природе являются ответственными за экспрессирование конкретной нуклеиновой кислоты (гомологичный участок) или могут включать последовательности разного происхождения, которые являются ответственными за экспрессирование разных белков или даже синтетических белков (гетерологичный участок). В частности, последовательности могут представлять собой последовательности прокариотических, эукариотических или вирусных генов или производные последовательности, которые стимулируют или подавляют транскрипцию гена специфическим или неспецифическим образом и индуцируемым или неиндуцируемым образом. Регуляторный участок включает точки начала репликации, сайты сплайсинга РНК, промоторы, энхансеры, последовательности окончания транскрипции, и сигнальные последовательности, которые направляют полипептид в секреторные пути клетки-мишени.
Регуляторный участок из "гетерологичного источника" относится к регуляторному участку, который в природе не ассоциирован с экспрессируемой нуклеиновой кислотой. В гетерологичные регуляторные участки включают регуляторные участки от разных видов, регуляторные участки из разных генов, гибридные регуляторные последовательности и регуляторные последовательности, которые не встречаются в природе, но которые сконструированы специалистом в данной области техники.
"РНК-транскрипт" относится к продукту, полученному от транскрипции последовательности ДНК, катализируемой РНК-полимеразой. Если РНК-транскрипт является безупречной комплементарной копией последовательности ДНК, то его называют первичным транскриптом, или он может представлять собой последовательность РНК, полученную при посттранскрипционном процессинге первичного транскрипта, и называется зрелой РНК. "Матричная РНК (м-РНК)" относится к РНК без интронов и которая может быть транслирована клеткой в белок. "кДНК" относится к двухнитевой ДНК, которая является комплементарной м-РНК и произведена от нее. "Смысловая" РНК относится к РНК-транскрипту, который включает м-РНК и таким образом может быть транслирована клеткой в белок. "Антисмысловая РНК" относится к РНК-транскрипту, который является комплементарным всему или части первичного транскрипта-мишени или м-РНК, и который блокирует экспрессию гена-мишени.
Комплементарность антисмысловой РНК может быть с любой частью конкретного транскрипта гена, то есть по 5’ некодирующей последовательности, 3’ некодирующей последовательности или кодирующей последовательности. "Функциональная РНК" относится к антисмысловой РНК, рибозимной РНК или другой РНК, которая, не транслируясь, оказывает воздействие на клеточные процессы.
"Полипептид", "пептид" и "белок" используются взаимозаменяемо и относят к полимерному соединению, состоящему из ковалентно связанных аминокислотных остатков. Аминокислоты имеют следующую общую структуру:

Термин "выделенный полипептид," "выделенный пептид" или "выделенный белок" относится к полипептиду или белку, который является по существу не содержащим тех соединений, которые обычно ассоциированы с ним в его естественном состоянии (например других белков или полипептидов, нуклеиновых кислот, углеводов, липидов). "Выделенный" не означает исключение искусственных или синтетических смесей с другими соединениями, или присутствия примесей, которые не препятствуют биологической активности, и которые могут присутствовать, например, из-за неполной очистки, добавления стабилизаторов, или приготовления фармацевтически приемлемого препарата.
"Полипептид с мутацией в виде замены" или "мутант с заменой" следует понимать как означающий мутантный полипептид, содержащий замену по меньшей мере одной (1) аминокислоты дикого типа или природной на другую аминокислоту, относящуюся к полипептиду дикого типа или природному. Мутантный полипептид с заменой может содержать только одно (1) замену аминокислоты дикого типа или естественной и может называться "точечным мутантным" или "одноточечным мутантным" полипептидом. Альтернативно, мутантный полипептид с заменой может содержать замещение двух (2) или более аминокислот дикого типа или природных на 2 или более аминокислоты, относящиеся к полипептиду дикого типа или природному. Согласно изобретению, полипептид лиганд-связывающего домена ядерных рецепторов группы H, содержащий мутацию в виде замены, содержит замену по меньшей мере одной (1) аминокислоты дикого типа или природной на другую аминокислоту, относящуюся к полипептиду дикого типа или природному полипептиду лиганд-связывающего домена ядерных рецепторов группы H.
Если полипептид с мутацией в виде замены содержит замену двух (2) или более аминокислот дикого типа или природных, такая замена может содержать либо эквивалентное количество аминокислот дикого типа или природных, удаленных для замены, то есть 2 аминокислоты дикого типа или природные, замененные 2 аминокислотами не дикого типа или неприродными, либо неэквивалентное количество аминокислот дикого типа, удаленных для замены, то есть 2 аминокислоты дикого типа, замененные 1 аминокислотой не дикого типа (мутация замена + делеция), или 2 аминокислоты дикого типа, замененные 3 аминокислотами не дикого типа (мутация замена + вставка).
Мутанты с заменой могут быть описаны с использованием сокращенной терминологической системы для указания замененного аминокислотного остатка и номера, в рамках указанной полипептидной последовательности, и нового замещающего аминокислотного остатка. Например, мутант с заменой, в которой замещен двадцатый (20-й) аминокислотный остаток полипептида, может быть сокращенно обозначен как "x20z", где "x" представляет собой аминокислоту, подлежащую замене, "20" представляет собой положение аминокислотного остатка или номер в полипептиде, и "z" представляет собой новую заменяющую аминокислоту. Следовательно, для мутанта с заменой, сокращенно взаимозаменяемо обозначенного как "E20A" или "Glu20Ala", указано, что мутант содержит остаток аланина (обычно сокращенно обозначаемой в данной области техники как "A" или "Ala") вместо глутаминовой кислоты (обычно сокращенно обозначаемой в данной области техники как "E" или "Glu") в положении 20 полипептида.
Мутация в виде замены может быть создана посредством любого метода мутагенеза, известного в данной области техники, включая сайт-направленный мутагенез in vitro (Hutchinson et al, J. Biol. Chem. 253: 6551 (1978); Zoller et al, DNA 3: 479-488 (1984); Oliphant et al, Gene 44: 111 (1986); Hutchinson et al, Proc. Natl Acad. Sci. USA 83: 110 (1986)), использование линкеров TAB® (Pharmacia), расщепление эндонуклеазой рестрикции/делеция фрагмента и замену, опосредованный PCR/олигонуклеотид-направленный мутагенез, и тому подобное, но не ограничиваясь ими. Методы на основе PCR являются предпочтительными для сайт-направленного мутагенеза (смотри Higuchi, 1989, "Using PCR to Engineer DNA", in PCR Technology: Principles and Applications for DNA Amplification, H. Erlich, ed., Stockton Press, Chapter 6, стр.61-70).
"Фрагмент" полипептида по изобретению относится к полипептиду, аминокислотная последовательность которого короче последовательности эталонного полипептида, и который содержит, по сравнению со всей частью этих эталонных полипептидов идентичную аминокислотную последовательность. Такие фрагменты могут быть, в необходимых случаях, включены в больший полипептид, частью которого они являются. Такие фрагменты полипептида по изобретению могут иметь длину по меньшей мере 2, 3, 4, 5, 6, 8, 10, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 25, 26, 30, 35, 40, 45, 50, 100, 200, 240 или 300 аминокислот.
"Вариант" полипептида или белка относится к любому аналогу, фрагменту, производному или мутанту, который получают из полипептида или белка, и который сохраняет по меньшей мере одно биологическое свойство полипептида или белка. В природе могут существовать разные варианты полипептида или белка. Такие варианты могут представлять собой аллельные вариации, отличающиеся различиями в нуклеотидных последовательностях структурного гена, кодирующего белок, или могут включать отличную сплайсинговую или посттрансляционную модификацию. Специалист в данной области техники может получить варианты, имеющие одиночные или множественные аминокислотные замены, делеции, присоединения или замены. Такие варианты могут включать, среди прочего: (а) варианты, где один или более чем один аминокислотный остаток замещен консервативными или неконсервативными аминокислотами, (б) варианты, где одна или более чем одна аминокислота добавлена к полипептиду или белку, (в) варианты, где одна или более чем одна аминокислота включают заменяющую группу, и (г) варианты, где полипептид или белок слит с другим полипептидом, таким как сывороточный альбумин. Способы получения этих вариантов, включая генетические (супрессии, удаления, мутации, так далее), химические и ферментные способы, известны специалистам в данной области техники. Вариантный полипептид предпочтительно содержит по меньшей мере примерно 14 аминокислот.
Термин "гомология" относится к проценту идентичности между двумя полинуклеотидными или двумя полипептидными группами. Соответствие между последовательностями от одной группы к другой может быть определено способами, известными в данной области техники. Например, гомологию можно определить путем прямого сравнения информации о последовательностях двух полипептидных молекул посредством выравнивания информации о последовательностях и используя легкодоступные компьютерные программы. Альтернативно, гомология может быть определена при помощи гибридизации полинуклеотидов при условиях, в которых образуются устойчивые дуплексы между гомологичными участками, с последующей деструкцией нуклеазой(ами), специфичной(ых) к одной нити, и определения размера расщепленных фрагментов.
При использовании в данном описании изобретения, термин "гомологичный" во всех его грамматических формах и вариантах написания относится к родству между белками, которые обладают "общим эволюционным происхождением," включая белки из суперсемейств ("например суперсемейства иммуноглобулинов) и гомологичные белки из разных видов ("например легкая цепь миозина и так далее) (Reeck et al, Cell 50: 667 (1987)). Такие белки (и кодирующие их гены) обладают гомологией последовательностей, что отражается в их высокой степени подобия последовательностей. Однако, в общем употреблении и в настоящей заявке термин "гомологичный" с модификацией наречием, таким как "в высшей степени," может относиться к подобию последовательностей, а не общему эволюционному происхождению.
Таким образом, термин "подобие последовательностей" во всех его грамматических формах относится к степени идентичности или соответствия между последовательностями нуклеиновых кислот или аминокислотными последовательностями белков, которые могут иметь или могут не иметь общее эволюционное происхождение (смотри Reeck et al, Cell 50: 667 (1987)). В одном воплощении две последовательности ДНК являются "по существу гомологичными" или "по существу подобными", когда по меньшей мере примерно 50% (предпочтительно по меньшей мере примерно 75%, и наиболее предпочтительно по меньшей мере примерно 90 или 95%) нуклеотидов совпадают на определенной длине последовательности ДНК. Последовательности, которые являются по существу гомологичными, могут быть идентифицированы путем сравнения последовательностей, с использованием стандартного программного обеспечения, имеющегося в банках данных о последовательностях, или в эксперименте Саузерн-гибридизации, например, при жестких условиях, как они определены для такой конкретной системы. Определение подходящих условий гибридизации находится в рамках квалификации специалиста в данной области техники (смотри например Sambrook et al., 1989, выше.).
При использовании в данном описании изобретения "по существу подобный" относится к фрагментам нуклеиновой кислоты, где изменения одного или более нуклеотидных оснований приводят к замене одной или более аминокислот, но не сказываются на функциональных свойствах белка, кодируемого последовательностью ДНК. "По существу подобный" также относится к фрагментам нуклеиновой кислоты, где изменения одного или более нуклеотидных оснований не сказывается на способности фрагмента нуклеиновой кислоты опосредовать изменение экспрессии гена с помощью антисмысловой или со-супрессионной технологии. "По существу подобный" также относится к модификациям фрагментов нуклеиновой кислоты по настоящему изобретению, таким как делеция или вставка одного или более нуклеотидных оснований, которые по существу не сказываются на функциональных свойствах полученного транскрипта. Таким образом, понятно, что изобретение включает больше чем конкретные типичные последовательности. Каждая из предложенных модификаций находится в рамках обычной практики специалиста в данной области техники, как и определение сохранения биологический активности кодируемых продуктов.
Более того, специалисту в данной области техники понятно, что по существу подобные последовательности, включенные в данное изобретение, также определяют по их способности гибридизироваться, в жестких условиях (0,1X SSC, 0,1% SDS, 65°C и промывали 2X SSC, 0,1% SDS, затем 0,1X SSC, 0,1% SDS), с последовательностями, приведенными в качестве примера в данном описании изобретения. По существу подобные фрагменты нуклеиновой кислоты по настоящему изобретению представляют собой те фрагменты нуклеиновой кислоты, последовательности ДНК которых по меньшей мере на 70% идентичны последовательности ДНК фрагментов нуклеиновой кислоты, представленных в данном описании изобретения. Фрагменты нуклеиновой кислоты по настоящему изобретению включают фрагменты нуклеиновой кислоты, последовательности ДНК которых являются по меньшей мере на 80% идентичными последовательности ДНК фрагментов нуклеиновой кислоты, представленных в данном описании изобретения. Другие фрагменты нуклеиновой кислоты по меньшей мере на 90% идентичны ДНК последовательности фрагментов нуклеиновой кислоты, представленных в данном описании изобретения. Другие фрагменты нуклеиновой кислоты по меньшей мере на 95% идентичны последовательности ДНК фрагментов нуклеиновой кислоты, представленных в данном описании изобретения.
Две аминокислотные последовательности являются "по существу гомологичными" или "по существу подобными", если более чем примерно 40% аминокислот являются идентичными, или более 60% являются подобными (функционально идентичными). Предпочтительно, подобные или гомологичные последовательности идентифицируют путем выравнивания используя, например, программу наложения GCG (Genetics Computer Group, Program Manual for the GCG Package, Version 7, Madison, Wisconsin).
Термин "соответствующий" используется в данном описании изобретения в отношении подобных или гомологичных последовательностей, независимо от того является ли точное положение идентичным или отличным от молекулы, подобие или гомологию с которой определяют. Выравнивание последовательностей нуклеиновых кислот или аминокислот может включать промежутки. Таким образом, термин "соответствующий " относится к подобию последовательностей, а не к нумерации аминокислотных остатков или нуклеотидных оснований.
"Существенная часть" аминокислотной или нуклеотидной последовательности содержит достаточно большую аминокислотную последовательность полипептидной или нуклеотидной последовательности гена, чтобы предположительно идентифицировать такой полипептид или ген, либо посредством ручной оценки последовательности специалистом в данной области техники, или посредством компьютерно-автоматизированного сравнения последовательностей и идентификации с использованием алгоритмов, таких как BLAST (Basic Local Alignment Search Tool; Altschul et al, J. Mol. Biol. 275: 403-410 (1993)); смотри также www.ncbi.nlm.nih.gov/BLAST/). В общем случае, последовательность из десяти или более соседних аминокислот или тридцати или более нуклеотидов необходима для предполагаемой идентификации полипептидной или нуклеиновокислой последовательности как гомологичных известному белку или гену. Кроме того, в отношении нуклеотидных последовательностей, ген-специфичные олигонуклеотидные зонды, содержащие 20-30 соседних нуклеотидов, можно использовать в зависимых от последовательности методах идентификации гена (например Саузерн-гибридизация) и выделения (например гибридизация бактериальных колоний или бляшек бактериофага in situ). Кроме того, короткие олигонуклеотиды из 12-15 оснований можно использовать в качестве праймеров для амплификации в ПЦР с целью получения конкретного фрагмента нуклеиновой кислоты, содержащего праймеры. Таким образом, "существенная часть" нуклеотидной последовательности содержит достаточную часть последовательности, чтобы конкретно идентифицировать и/или выделить фрагмент нуклеиновой кислоты, содержащий последовательность.
Термин "процент идентичности", как известно в данной области техники, представляет собой отношение между двумя или более полипептидными последовательностями или двумя или более полинуклеотидными последовательностями, определенное путем сравнения последовательностей. В данной области техники "идентичность" также означает степень родства последовательностей у полипептидных или полинуклеотидных последовательностями, соответственно, при определении посредством совпадения нитей таких последовательностей. "Идентичность" и "подобие" могут быть легко рассчитаны известными способами, включая описанные в: Computational Molecular Biology (Lesk, A.M., ed.) Oxford University Press, New York (1988); Biocomputing: Informatics and Genome Projects (Smith, D. W., ed.) Academic Press, New York (1993); Computer Analysis of Sequence Data, Part I (Griffin, A.M., and Griffin, H.G., eds.) Humana Press, New Jersey (1994); Sequence Analysis in Molecular Biology (von Heinje, G., ed.) Academic Press (1987); и Sequence Analysis Primer (Gribskov, M. and Devereux, J., eds.) Stockton Press, New York (1991), но не ограничивается этим. Методы определения идентичности разрабатывают с целью достижения лучшего совпадения между тестируемыми последовательностями. Методы определения идентичности и подобия кодифицированы в общедоступных компьютерных программах. Выравнивание последовательностей и расчеты процента идентичности можно выполнять, используя программу Megalign из пакета обработки данных для биоинформатики LASERGENE (DNASTAR Inc., Madison, WI). Множественное выравнивание последовательностей может быть выполнено с использованием метода выравнивания Clustal (Higgins et al, CAB/OS. 5: 151-153 (1989)) с параметрами по умолчанию (GAP PENALTY-10, GAP LENGTH PENALTY=10). Параметры по умолчанию для парных выравниваний с использованием метода Clustal могут быть выбраны: KTUPLE 1, GAP PENALTY = 3, WINDOW = 5 и DIAGONALS SAVED = 5.
Термин "программное обеспечение анализа последовательности" относится к любому компьютерному алгоритму или программе, который полезен для анализа нуклеотидных или аминокислотных последовательностей. "Программное обеспечение для анализа последовательностей" может иметься в продаже или быть независимо разработанным. Типичное программное обеспечение для анализа последовательности включает пакет программ GCG (Wisconsin Package Version 9.0, Genetics Computer Group, (GCG), Madison, Wl), BLASTP, BLASTN, BLASTX (Altschul et al, J. Mol. Biol. 275: 403-410 (1990)), и DNASTAR (DNASTAR, Inc. 1228 S. Park St. Madison, Wl 53715 USA), но не ограничивается этим. В контексте данной заявки следует понимать, что когда для анализа используется программное обеспечение для анализа последовательностей, результаты анализа будут основаны на "значениях по умолчанию" указанных программ, если не указано иное. При использовании в данном описании изобретения "значения по умолчанию" будут означать любой набор значений или параметров, которые изначально загружаются с программной при первом запуске.
"Химически синтезированный" в отношении последовательности ДНК, означает, что компонентные нуклеотиды были собраны in vitro. Выполняемый вручную химический синтез ДНК можно осуществлять, используя традиционные способы, или автоматизированный химический синтез можно выполнить, используя один из ряда имеющиеся в продаже приборов. Таким образом, гены могут быть адаптированы для оптимальной экспрессии гена на основе оптимизации нуклеотидной последовательности на отражение кодонной склонности клетки-хозяина. Специалист в данной области техники оценит вероятность успешной экспрессии гена, если использование кодона склоняется к кодонам, поддерживаемым хозяином. Определение предпочтительных кодонов может быть основано на исследовании генов, произведенных от клетки-хозяина, когда информация о последовательности доступна.
При использовании в данном описании изобретения две или более индивидуально действующие системы генной регуляции называются "ортогональными", когда а) модуляция каждой из данных систем соответствующим лигандом, в выбранной концентрации, приводит к измеряемому изменению величины экспрессии гена такой системы, и б) изменение статистически значимо различается от изменения экспрессии всех других систем, одновременно действующих в клетке, ткани или организме, в не зависимости от одновременности или последовательности фактической модуляции. Предпочтительно, модуляция каждой индивидуально действующей системы генной регуляции приводит к изменению экспрессии гена по меньшей мере в 2 раза большему, чем ото всех других действующих систем в клетке, ткани или организме. Более предпочтительно, если изменение больше по меньшей мере в 5 раз. Еще более предпочтительно, если изменение больше по меньшей мере в 10 раз. Еще более предпочтительно, если изменение больше по меньшей мере в 100 раз. Еще более предпочтительно, изменение больше по меньшей мере в 500 раз. Идеально, модуляция каждой из данных систем соответствующим ей лигандом в выбранной концентрации приводит к измеримому изменению величины экспрессии гена такой системой и к неизмеримому изменению экспрессии всеми другими системами, действующими в клетке, ткани или организме. В таких случаях множественную индуцируемую систему генной регуляции называют "полностью ортогональной". Настоящее изобретение является полезным для поиска ортогональных лигандов и ортогональных рецепторных систем экспрессии гена, таких как описанные в US 2002/0110861 A1, который включен в данное описание изобретения посредством ссылки во всей его полноте.
Термин "модулировать" означает способность данного комплекса лиганд/рецептор индуцировать или подавлять трансактивацию экзогенного гена.
Термин "экзогенный ген" означает ген, чужой для субъекта, то есть ген, который вводят субъекту посредством процесса трансформации, немутированную версию эндогенного мутированного гена или мутированную версию эндогенного немутированного гена. Метод трансформации не является критичным для данного изобретения и может представлять собой любой подходящий для субъекта метод, известный специалистам в данной области техники. Например, трансгенные растения получают посредством восстановления из трансформированных клеток. Многочисленные методы трансформации известны из литературы, такие как агроинфекция с использованием Agrobacterium tumefaciens или ее плазмиды Ti, электропорация, микроинъекции растительных клеток и протопластов и трансформации микропульками. Комплементарные методы известны для трансформации клеток животных и восстановления таких трансформированных клеток в трансгенных животных. Экзогенные гены могут представлять собой либо естественные, либо синтетические гены и терапевтические гены, которые вводят в субъект в форме ДНК или РНК, которые могут действовать через промежуточную ДНК, например посредством обратной транскриптазы. Такие гены могут быть введены в клетки-мишени, непосредственно введены субъекту, или опосредовано введены субъекту путем переноса трансформированных клеток, например аутологичных клеток. Термин "терапевтический ген" означает ген, который придает выгодную функцию клетке-хозяину, в которой такой ген экспрессируется.
Термин "экдизон-рецепторный комплекс" обычно относится к гетеродимерному белковому комплексу, состоящему из двух членов семейства стероидных рецепторов, экдизонового рецептора ("EcR") и белков Ultraspiracle ("USP") (смотри Yao et al, Nature 366: 476-479 (1993)); Yao et al., Cell 71: 63-72 (1992)). Функциональный комплекс экдистероидного рецептора может также включать дополнительный(е) белок(и), такой(ие) как иммунофилин(ы). Дополнительные члены семейства белков стероидных рецепторов, известные в качестве факторов транскрипции (такие как DHR38, betaFTZ-1 или другие гомологи насекомых), также могут являться лиганд-зависимыми или независимыми партнерами EcR и/или USP. Экдизон-рецепторный комплекс может также представлять собой гетеродимер белка экдизонового рецептора и гомолога белка Ultraspiracle позвоночных, белка Х-рецептора ретиновой кислоты ("RXR"). Гомодимерные комплексы белка экдизонового рецептора или USP могут также быть функциональными при определенных обстоятельствах.
Комплекс экдистероидного рецептора может быть активирован при помощи активного экдистероидного или нестероидного лиганда, связанного с одним из белков комплекса, включая EcR, но не исключая другие белки комплекса.
Экдизон-рецепторный комплекс включает белки, которые являются членами суперсемейства стероидных рецепторов, все члены которого характеризуются присутствием аминоконцевого домена трансактивации, ДНК-связывающего домена ("DBD") и лиганд-связывающего домена ("LBD"), разделенных шарнирным участком. Некоторые члены семейства также могут иметь другой домен трансактивации на карбоксиконцевой стороне LBD. DBD характеризуется присутствием двух богатых цистеином доменов «цинковые пальцы», между которыми находятся два аминокислотных мотива, P-box и D-box, которые придают специфичность к экдизоновым элементам ответа. Эти домены могут быть или нативными, модифицированными, или химерными из разных доменов гетерологичных рецепторных белков.
Последовательности ДНК, образующие экзогенный ген, элемент ответа и экдизон-рецепторный комплекс, могут быть введены в архибактерии, прокариотические клетки, такие как Escherichia coli, Bacillus subtilis, или другие энтеробактерии, или эукариотические клетки, такие как растительные или животные клетки. Однако, так как много белков, экспрессируемых при помощи гена, процессируются неточно в бактериях, предпочтительными являются эукариотические клетки. Клетки могут находиться в форме простых клеток или многоклеточных организмов. Нуклеотидные последовательности экзогенного гена, элемента ответа и рецепторного комплекса также могут быть введены в виде молекул РНК, предпочтительно в форме функциональных вирусных РНК, таких как вирус табачной мозаики. Из эукариотических клеток предпочтительными являются клетки позвоночных, так как у них в природе отсутствуют молекулы, которые обеспечивают ответы экдизонового рецептора на лиганды по изобретению. В результате они являются "по существу нечувствительными" к лигандам по данному изобретению. Таким образом, лиганды по данному изобретению будут оказывать незначительные физиологические или другие эффекты на трансформированные клетки или на организм в целом. Следовательно, клетки могут расти и экспрессировать желательный продукт, по существу не зависимо от присутствия самого лиганда.
Термин "субъект" означает целое насекомое, растение или животное или клетку насекомого, растения или животного. Также ожидается, что лиганды будут действовать одинаково хорошо, когда субъект представляет собой грибы или дрожжи. Если субъект представляет собой целое животное, предпочтительно животное представляет собой позвоночное, наиболее предпочтительно млекопитающее.
Диацилгидразиновые лиганды Формулы I и хиральные диацилгидразиновые лиганды Формулы II или III по настоящему изобретению при использовании с экдизон-рецепторным комплексом, который, в свою очередь, связан с элементом ответа, связанным с экзогенным геном, обеспечивают средства для внешней временной регуляции экспрессии экзогенного гена. Порядок, в котором различные компоненты связываются друг с другом, то есть лиганд с рецепторным комплексом и рецепторный комплекс с элементом ответа, не является критичным. Обычно модуляция экспрессии экзогенного гена происходит в ответ на связывание экдизон-рецепторного комплекса со специфическим контролирующим или регуляторным элементом ДНК. Белок экдизонового рецептора, подобно другим членам семейства стероидных рецепторов, имеет по меньшей мере три домена, домен трансактивации, ДНК-связывающий домен и лиганд-связывающий домен. Данный рецептор, подобно субпопуляции семейства стероидных рецепторов, также имеет меньше четко определенных участков, ответственных за свойства гетеродимеризации. Связывание лиганда с лиганд-связывающим доменом белка экдизонового рецептора, после гетеродимеризации с белком USP или RXR, обеспечивает возможность связывания ДНК-связывающих доменов гетеродимерных белков с элементом ответа в активированной форме, таким образом вызывая экспрессию или подавление экзогенного гена. Данный механизм не исключает возможности связывания лиганда или с EcR или с USP, и образования в результате этого активных гомодимерных комплексов (например EcR+EcR или USP+USP). Предпочтительно один или более доменов рецептора могут варьироваться, продуцируя переключатель химерных генов. Обычно один или более из трех доменов могут быть выбраны из источника, отличного от источника других доменов, так что химерный рецептор оптимизирован в выбранной клетке-хозяине или организме в отношении трансактивационной активности, комплементарного связывания лиганда и распознавания специфического элемента ответа. Кроме того, сам элемент ответа может быть модифицирован или замещен элементами ответа для других доменов ДНК-связывающего белка, таких как белок GAL-4 из дрожжей (смотри Sadowski et al, Nature 335: 563-564 (1988)) или белок LexA из E. coli (смотри Brent et al, Cell 43: 729-736 (1985)), для адаптации комплексов химерного экдизонового рецептора. Другим преимуществом химерных систем является то, что они допускают выбор промотора, используемого для управления экзогенным геном, в соответствии с желательным конечным результатом. Такой двойной контроль может быть особенно важным в областях генной терапии, особенно когда продуцируются цитотоксические белки, так как можно контролировать и время экспрессии, и клетки, где экспрессия происходит. Когда экзогенные гены, функционально связанные с подходящим промотором, внедряют в клетки субъекта, экспрессия экзогенных генов контролируется присутствием лиганда по изобретению. Промоторы могут быть конститутивно или индуцируемо регулируемыми, или могут быть ткане-специфическими (то есть, экспрессирующимися только в определенном типе клетки) или специфичными к конкретным стадиям развития организма.
Многочисленные нуклеиновокислые последовательности геномной и кДНК, кодирующие множество полипептидов, хорошо известны в данной области техники. Экзогенный генетический материал, полезный с лигандами по данному изобретению, включает гены, которые кодируют интересующие биологически активные белки, такие как, например, секреторные белки, которые могут высвобождаться из клетки; ферменты, которые могут метаболизировать субстрат из токсичного вещества в нетоксичное вещество, или из неактивного вещества в активное вещество; регуляторные белки; рецепторы клеточной поверхности; и тому подобное. Полезные гены также включают гены, которые кодируют факторы свертываемости крови, гормоны, такие как инсулин, паратиреоидный гормон, высвобождающий фактор лютеинизирующего гормона, альфа- и бета- семинальные ингибины, и гормон роста человека; ген, которые кодируют белки, такие как ферменты, отсутствие которых приводит к возникновению аномального состояния; гены, кодирующие цитокины или лимфокины, такие как интерфероны, гранулоцитарно-макрофагальный колониестимулирующий фактор, колониестимулирующий фактор-1, фактор некроза опухоли, и эритропоэтин; гены, кодирующие ингибирующие вещества, такие как альфагантитрипсин, гены, кодирующие вещества, которые функционируют в качестве лекарственных средств, таких как токсины дифтерии и холеры; и тому подобное. Полезные гены также включают гены, полезные для терапии рака и для лечения генетических расстройств. Специалист в данной области техники имеет доступ к информации о последовательностях нуклеиновых кислот практически для всех известных генов и может либо получить молекулу нуклеиновой кислоты непосредственно из общедоступного депозитария, организации, которая опубликовала последовательность, или использовать обычные способы получения молекулы.
В одном воплощении экзогенный ген представляет собой ген hIL-12 под контролем терапевтической системы RheoSwitch™ (RTS), ex vivo трансфицированный аденовирусом в аутологичных дендритных клетках.
Для использования в генной терапии лиганды, описанные в данном описании изобретения, можно вводить отдельно или как часть фармацевтической композиции, содержащей фармацевтически приемлемый носитель. В одном воплощении фармацевтическая композиция представлена в форме растворов, суспензий, таблеток, капсул, мазей, эликсиров или инъецируемых композиций.
Фармацевтически приемлемые носители включают наполнители, такие как сахариды, например лактозу или сахарозу, маннит или сорбит, препараты целлюлозы и/или фосфаты кальция, например трикальция фосфат или гидрофосфат кальция, а также связывающие вещества, такие как крахмальная паста, используя, например, кукурузный крахмал, пшеничный крахмал, рисовый крахмал, картофельный крахмал, желатин, трагакант, метилцеллюлозу, гидроксипропилметилцеллюлозу, натрия карбоксиметилцеллюлозу и/или поливинилпирролидон. При желании могут быть добавлены разрыхлители, такие как вышеуказанные крахмалы, а также карбоксиметил-крахмал, сшитый поливинилпирролидон, агар, или альгиновая кислота или ее соль, такая как натрия альгинат. Вспомогательными веществами являются регулирующие текучесть агенты и смазывающие вещества, например диоксид кремния, тальк, стеариновая кислота или ее соли, такие как стеарат магния или стеарат кальция, и/или полиэтиленгликоль. В одном воплощении ядра драже получают вместе с подходящими покрывающими веществами, которые, при желании, являются устойчивыми к желудочным сокам. Для этой цели можно использовать концентрированные растворы сахаридов, которые возможно могут содержать аравийскую камедь, тальк, поливинилпирролидон, полиэтиленгликоль и/или диоксид титана, лакирующие растворы и подходящие органические растворители или смеси растворителей. Для получения оболочек, устойчивых к желудочному соку, используют растворы подходящих препаратов целлюлозы, таких как фталат ацетилцеллюлозы или фталат гидроксипропилметилцеллюлозы. Красящие вещества или пигменты можно добавлять к покрывающим веществам таблеток или драже, например для идентификации или с целью отличить комбинации доз активного соединения.
Другие фармацевтические препараты, которые можно использовать перорально, включают плотно заполненные капсулы, изготовленные из желатина, а также мягкие, герметичные капсулы, изготовленные из желатина и пластификатора, такого как глицерин или сорбит. Плотно заполненные капсулы могут содержать активные соединения в форме гранул или наночастиц, которые возможно могут быть смешаны с наполнителями, такими как лактоза, связывающими веществами, такими как крахмалы, и/или смазывающими веществами, такими как тальк или стеарат магния и, возможно, стабилизаторами. В одном воплощении их растворяют или суспендируют в подходящих жидкостях, таких как жирные масла, или жидкие парафины, возможно со стабилизаторами.
Жирные масла могут содержать моно-, ди- или триглицериды. Моно-, ди- и триглицериды включают те, которые являются производными C6, C8, C10, C12, C14, C16, Q18, C20 и C22 кислот. Типичные диглицериды включают, в частности, диолеин, дипальмитолеин, и смешанные диглицериды каприлиновой/каприновой кислот. Предпочтительные триглицериды включают растительные масла, рыбьи жиры, животные жиры, гидрированные растительные масла, частично гидрированные растительные масла, синтетические триглицериды, модифицированные триглицериды, фракционированные триглицериды, средне- и длинноцепочечные триглицериды, структурированные триглицериды, и их смеси. Типичные триглицериды включают: миндальное масло; масло бабассу; масло бораго; масло из семян черной смородины; каноловое масло; касторовое масло; кокосовое масло; кукурузное масло; масло из семян хлопчатника; масло из энотеры; масло из виноградных косточек; арахисовое масло; масло из семян горчицы; оливковое масло; пальмовое масло; масло пальмового ядра; арахисовое масло; рапсовое масло; сафлоровое масло; кунжутное масло; масло из акульей печени; соевое масло; подсолнечное масло; гидрированное касторовое масло; гидрированное кокосовое масло; гидрированное пальмовое масло; гидрированное соевое масло; гидрированное растительные масло; гидрированное хлопковое и касторовое масло; частично гидрированное соевое масло; частично соевое и хлопковое масло; глицерилтрикапроат; глицерилтрикаприлат; глицерилтрикапрат; глицерилтриундеканоат; глицерилтрилаурат; глицерилтриолеат; глицерилтрилинолеат; глицерилтрилинолеат; глицерилтрикаприлат/капрат; глицерилтрикаприлат/капрат/лаурат; глицерилтрикаприлат/капрат/линолеат; и глицерилтрикаприлат/капрат/стеарат.
В одном воплощении триглицерид представляет собой среднецепочечный триглицерид, доступный под коммерческим названием LABRAFAC CC. Другие триглицериды включают нейтральные масла, например нейтральные растительные масла, в частности фракционированные кокосовые масла, такие как известные и имеющиеся в продаже под коммерческим названием MIGLYOL, включая продукты: MIGLYOL 810; MIGLYOL 812; MIGLYOL 818; и САРТЕХ 355. Другие триглицериды представляют собой триглицериды каприловой-каприновой кислот, например известные и имеющиеся в продаже под коммерческим названием MYRITOL, включая продукт MYRITOL 813. Кроме того, триглицеридами этого класса являются CAPMUL МСТ, САРТЕХ 200, САРТЕХ 300, САРТЕХ 800, NEOBEE M5 и MAZOL 1400.
Фармацевтические композиции, содержащие триглицериды, могут дополнительно содержать липофильные и/или гидрофильные поверхностно-активные вещества, которые могут образовывать прозрачные растворы при растворении в водном растворителе. Одно такое поверхностно-активное вещество представляет собой токоферола полиэтиленгликоля 1000 сукцинат (витамин Е TPGS). Примеры таких композиций описаны в патенте США 6267985.
В другом воплощении фармацевтически приемлемый носитель содержит LABRASOL (Gattefosse SA), который представляет собой PEG-8 глицериды каприловой/каприновой кислот. В другом воплощении фармацевтически приемлемый носитель содержит PL90G, витамин Е TPGS, и Miglyol 812N. Компоненты таких препаратов показаны в Таблице 1.
Возможные фармацевтические препараты, которые могут быть использованы ректально, включают, например, суппозитории, которые состоят из комбинации одного или более лигандов с суппозиторной основой. Пригодные суппозиторные основы представляют собой, например, натуральные или синтетические триглицериды, или парафиновые углеводы. Кроме того, также можно использовать желатиновые ректальные капсулы, которые состоят из комбинации лиганда с основанием. Возможные вещества основы включают, например, жидкие триглицериды, полиэтиленгликоли или парафиновые углеводы.
Пригодные препараты для парентерального введения включают водные растворы лиганда в водорастворимой форме, например в виде водорастворимых солей и щелочных растворов. Кроме того, можно вводить суспензии лиганда в виде подходящих масляных инъекционных суспензий. Пригодные липофильные растворители или средства доставки включают жирные масла, например кунжутное масло, или синтетические эфиры жирных кислот, например этилолеат, или триглицериды, или полиэтиленгликоль-400. Водные инъекционные суспензии могут содержать вещества, которые увеличивают вязкость суспензии, включая, например, натрия карбоксиметилцеллюлозу, сорбитол и/или декстран. Возможно суспензия может также содержать стабилизаторы.
Местные композиции могут быть приготовлены в виде масел, кремов, лосьонов, мазей и тому подобного посредством выбора соответствующих носителей. Подходящие носители включают растительные или минеральные масла, белый вазелин (белый мягкий парафин), жиры или масла с разветвленной цепью, животные жиры и высокомолекулярный спирт (более C12). Также могут быть включены эмульгаторы, стабилизаторы, увлажнители и антиоксиданты, а также, при необходимости, агенты, придающие цвет или аромат. Кроме того, в таких местных препаратах можно использовать энхансеры проникновения через кожу. Примеры таких энхансеров можно обнаружить в US 3989816 и 4444762.
Кремы можно приготовить в виде препарата из смеси минерального масла, самоэмульгирующегося пчелиного воска и воды, с которой перемешивают лиганд, растворенный в небольшом количестве масла, такого как миндальное масло. Типичным примером такого крема является крем, который включает примерно 40 частей воды, примерно 20 частей пчелиного воска, примерно 40 частей минерального масла и примерно 1 часть миндального масла.
Мази могут быть приготовлены в виде препарата путем смешивания суспензии лиганда в растительном масле, таком как миндальное масло, с теплым мягким парафином, после чего оставляют смесь охлаждаться. Типичный пример такой мази представляет собой мазь, которая включает примерно 30% миндального масло и примерно 70% белого мягкого парафина по массе.
Лосьоны могут быть удобно изготовлены путем приготовления суспензии лиганда в подходящем высокомолекулярном спирте, таком как пропиленгликоль или полиэтиленгликоль.
Примеры антиоксидантов, которые можно добавлять к фармацевтическим композициям, включают ВНА и ВНТ.
В одном воплощении фармацевтическая композиция содержит 30 мг лиганда на мл LABRASOL в твердой желатиновой капсуле. В другом воплощении капсула содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95 или 100 мг лиганда.
Фармацевтические композиции могут содержать от 0,01% до 99% по массе лиганда. Композиции могут находиться либо в однодозовой, либо во многодозовой формах. Количество лиганда в любой конкретной фармацевтической композиции будет зависеть от эффективной дозы, то есть дозы, требующейся для достижения желательной экспрессии или супрессии гена. В одном воплощении субъекту вводят от 0,1 до 7,5 мг/кг. В другом воплощении субъекту вводят от 0,1 до 3 мг/кг. В другом воплощении вводят от 0,1 до 3 мг/кг.
Подходящие пути введения фармацевтических композиций включают пероральный, ректальный, местный (включая дермальный, буккальный и сублингвальный), вагинальный, парентеральный (включая подкожный, внутримышечный, внутривенный, внутридермальный, интратекальный, внутриопухолевый и эпидуральный) и посредством назо-гастрального зонда. Специалисту в данной области техники понятно, что предпочтительный путь введения будет зависеть от состояния, подлежащего лечению, и может варьироваться в зависимости от факторов, таких как состояние пациента. Фармацевтические композиции можно вводить один или более раз в сутки. В одном воплощении фармацевтическую композицию вводят ежесуточно, начиная за 24 часа перед введением клеток, содержащих экдизон-рецепторный комплекс и ДНК-связывающую последовательность.
Диацилгидразиновые лиганды Формулы 1 и хиральные диацилгидразиновые лиганды Формулы II или III по настоящему изобретению, описанные в данном описании изобретения, также можно вводить вместе с другими фармацевтически активными соединениями. Специалисту в данной области техники будет понятно, что фармацевтически активные соединения для применения в комбинации с лигандами, описанными в данном описании изобретения, выбирают с целью избежать вредных воздействий на реципиента или нежелательных взаимодействий между соединениями. Примеры других фармацевтически активных соединений, которые можно использовать в комбинации с лигандами включают, например, химиотерапевтические агенты против СПИД, производные аминокислот, анальгетики, анестетики, аноректальные продукты, антациды и ветрогонные средства, антибиотики, антикоагулянты, антидоты, антифибринолитические агенты, антигистамины, противовоспалительные агенты, противоопухолевые средства, противопаразитические средства, антипротозойные средства, антипиретики, антисептики, противосудорожные и антихолинергические, противовирусные средства, препараты для подавления аппетита, лечебные средства для артрита, модификаторы биологического ответа, регуляторы метаболизма костей, средства для опорожнения кишечника, сердечно-сосудистые агенты, стимуляторы центральной нервной системы, церебральные метаболические энхансеры, церуменолитики, инибиторы холинэстеразы, препараты для простуды и кашля, колониестимулирующие факторы, контрацептивы, цитопротекторные агенты, стоматологические препараты, дезодоранты, дерматологические средства, агенты для детоксикации, диабетические агенты, диагностические средства, средства для лечения диареи, агонисты дофаминового рецептора, электролиты, ферменты и средства, стимулирующие пищеварение, препараты спорыньи, агенты фертильности, волоконные добавки, противогрибковые агенты, инибиторы галактореи, инибиторы секреции желудочной кислоты, желудочно-кишечные прокинетические агенты, инибиторы гонадотропина, стимуляторы роста волос, гематиники, геморрагические агенты, гемостатики, антагонисты гистаминового рецептора-H2, гормоны, гипергликемические агенты, гиполипидемики, иммуносупрессанты, слабительные, лепростатики, вспомогательные средства для лейкофереза, легочные сурфактанты, препараты против мигрени, муколитики, антагонисты мышечных релаксантов, мышечные релаксанты, наркотические антагонисты, назальные спреи, назальные лекарственные средства, нуклеозидные аналоги, диетические добавки, препараты для остеопороза, родостимулирующие вещества, парасимпатолики, парасимпатомиметики, лекарственные средства против паркинсонизма, пенициллиновые адъюванты, фосфолипиды, инибиторы тромбоцитов, агенты против порфирии, аналоги простагландина, простагландины, ингибиторы протонного насоса, противозудные лекарственные средства, психотропики, хинолоны, стимуляторы дыхания, слюнные стимуляторы, заменители соли, склерозирующие агенты, препараты для кожных ран, вспомогательные средства для прекращения курения, сульфонамиды, симпатолитики, тромболитики, агенты против синдрома Туретта, препараты против тремора, препараты против туберкулеза, средства, способствующие выведению мочевой кислоты, агенты для мочевого тракта, маточные контрактанты, маточные релаксанты, вагинальные препараты, агенты против головокружения, аналоги витамина D, витамины и контрастные среды для медицинской визуализации. В некоторых случаях лиганды могут быть полезны в качестве вспомогательного средства к лекарственной терапии, например для «выключения» гена, который продуцирует фермент, метаболизирующий конкретное лекарственное средство.
Для сельскохозяйственных применений, в дополнение к применениям, описанным выше, лиганды по данному изобретению также можно использовать для контроля экспрессии пестицидных белков, таких как токсин Bacillus thuringiensis (Bt). Такая экспрессия может быть специфической по отношению к ткани или растению. Кроме того, особенно при необходимости контролирования также вредителей растений, один или более пестицидов можно объединять с лигандами, описанными в данном описании изобретения, обеспечивая таким образом дополнительные преимущества и эффективность, включая меньшее общее количество использований, чем при раздельном использовании этих пестицидов. Если используются смеси с пестицидами, относительные соотношения каждого компонента в композиции будут зависеть от относительной эффективности и желательной скорости использования каждого пестицида относительно зерновых, вредителей, и/или сорняков, подлежащих обработке. Специалисту в данной области техники понятно, что смеси пестицидов могут обеспечивать такие преимущества, как более широкий спектр активности, по сравнению с использованием одного пестицида. Примеры пестицидов, которые могут быть объединены в композиции с лигандами, описанными в данном описании изобретения, включают фунгициды, гербициды, инсектициды, акарициды и бактерициды.
Диацилгидразиновые лиганды Формулы I и хиральные диацилгидразиновые лиганды Формулы II или III по настоящему изобретению, описанные в данном описании изобретения, можно использовать для листвы растений в виде водных спреев посредством обычно используемых способов, таких как обычные многолитровые гидравлические спреи, малолитровые спреи, сильная струя воздуха и воздушные спреи. Разведение и норма внесения будут зависеть от типа используемого оборудования, способа и частоты желаемого использования, и скорости нанесения лиганда. Может быть целесообразно включать дополнительные вспомогательные средства в резервуар опрыскивателя. Такие вспомогательные средства включают поверхностно-активные вещества, диспергаторы, спредеры, клеящие вещества, противопенные агенты, эмульгаторы и другие подобные вещества, описанные в McCutcheon’s Emulsifiers and Detergents, McCutcheon’s Emulsifiers and Detergents/Functional Materials, and McCutcheon’s Functional Materials, все публикуются ежегодно в McCutcheon Division из MC Publishing Company (Нью-Джерси). Такие лиганды можно также смешивать с удобрениями или удобряющими веществами перед их использованием. Такие лиганды и твердые удобряющие вещества также можно смешивать в оборудовании для смешивания или перемешивания, или они могут быть внесены вместе с минеральными удобрениями в гранулированные композиции. Можно использовать любое относительное соотношение удобрения, которое пригодно для зерновых культур и сорняков, подлежащих обработке. Лиганды, описанные в данном описании изобретения, обычно содержат от 5% до 50% удобряющей композиции. Такие композиции представляют удобряющие вещества, которые способствуют быстрому росту целевых растений, и в то же время контролируют экспрессию гена.
КЛЕТКИ-ХОЗЯЕВА И ОРГАНИЗМЫ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА
Как описано выше, диацилгидразиновые лиганды Формулы и хиральные диацилгидразиновые лиганды Формулы II или III можно использовать для модуляции экспрессии гена в клетке-хозяине. Экспрессия в трансгенных хозяйских клетках может быть полезна для экспрессии различных интересующих генов. В настоящем изобретении предлагаются лиганды для модуляции экспрессии гена в прокариотических и эукариотических клетках-хозяевах.
Экспрессия в трансгенных хозяйских клетках полезна для экспрессии различных интересующих полипептидов, включая антигены, продуцируемые в растениях в качестве вакцин; ферменты, подобные альфа-амилазе, фитазе, гликанам и ксиланазе; гены резистентности к насекомым, нематодам, грибам, бактериям, вирусам, и абиотическим стрессам у растений; антигены; нутрицевтики; фармацевтические средства; витамины; гены для модификации аминокислотного содержания, гербицидной устойчивости, холодо-, засухо- и жароустойчивости; промышленные продукты; масла, белок, углеводы; антиоксиданты; мужские стерильные растения; цветы; топливо; другие полезные характеристики; терапевтические полипептиды или продукты, которые можно использовать для лечения состояния, заболевания, расстройства, дисфункции или генетического дефекта, такие как моноклональные антитела, ферменты, протеазы, цитокины, интерфероны, инсулин, эритропоэтин, факторы свертывания крови, другие факторы крови или компоненты; вирусные векторы для генной терапии; вирус для вакцин; мишени для поиска новых лекарств, функциональной геномики, и протеомических анализов и использования: промежуточные соединения в каскаде пути для модуляции каскадов путей, уже существующих у хозяина; промежуточные соединения каскада пути для синтеза новых продуктов с использованием хозяина, ранее не имевшего такой возможности; клеточные анализы; анализы функциональной геномики; протеомические анализы и тому подобное, но не ограничиваясь ими. Кроме того генные продукты могут быть полезными в для получения с большей скоростью выходов у хозяина или чтобы сделать возможной альтернативную форму роста для использования.
Таким образом, в настоящем изобретении предлагаются диацилгидразиновые лиганды Формулы 1 и хиральные диацилгидразиновые лиганды Формулы II или III для модуляции экспрессии гена в выделенной клетке-хозяине. В одном воплощении выделенная клетка-хозяин является прокариотической клеткой-хозяином или эукариотической клеткой-хозяином. В другом воплощении выделенная клетка-хозяин представляет собой клетку-хозяина беспозвоночных или клетку-хозяина позвоночных. Предпочтительно, клетку-хозяина выбирают из группы, состоящей из бактериальной клетки, грибной клетки, дрожжевой клетки, клетки нематоды, клетки насекомого, клетки рыбы, растительной клетки, клетки птиц, животной клетки и клетки млекопитающего. Более предпочтительно клетка-хозяин представляет собой дрожжевую клетку, клетку нематоды, клетку насекомого, растительную клетку, клетку данио, клетку цыпленка, клетку хомяка, клетку мыши, клетку крысы, клетку кролика, клетку кошки, клетку собаки, клетку крупного рогатого скота, клетку козы, клетку коровы, клетку свиньи, клетку лошади, клетку овцы, клетку обезьяноподобных, клетку обезьяны, клетку шимпанзе или клетку человека. Примеры хозяйских клеток включают, виды грибов или дрожжей, такие как Aspergillus, Trichoderma, Saccharomyces, Pichia, Candida, Hansenula, или бактериальные виды, такие как виды главным образом Synechocystis, Synechococcus, Salmonella, Bacillus, Acinetobacter, Rhodococcus, Streptomyces, Eschehchia, Pseudomonas, Methylomonas, Methylobacter, Alcaligenes, Anabaena, Thiobacillus, Methanobactenum и Klebsiella; растительные виды выбирают из группы, состоящей из яблока, Arabidopsis, проса, банана, ячменя, бобовых, свеклы, blackgram, нуга, чили, огурца, баклажана, бобов, кукурузы, дыни, сорго, золотистой фасоли, овса, окры, Panicum, папайи, арахиса, гороха, перца, голубиного гороха, ананаса, Phaseolus, картофеля, тыквы, риса, сорго, сои, крупноплодной тыквы, сахарного тростника, сахарной свеклы, подсолнечника, батата, чая, томата, табака, арбуза и пшеницы; животное; и клетки-хозяева млекопитающих, но не ограничиваются ими.
В другом воплощении клетка-хозяин представляет собой дрожжевую клетку, выбранную из группы, состоящей из Saccharomyces, Pichia, и клетки-хозяина Candida.
В другом воплощении клетка-хозяин представляет собой клетку нематоды Caenorhabditis elegans.
В другом воплощении клетка-хозяин представляет собой клетку насекомого.
В другом воплощении клетка-хозяин представляет собой растительную клетку, выбранную из группы, состоящей из клетки яблока, Arabidopsis, проса, банана, ячменя, бобовых, свеклы, ниссы, нуга, чилийского перца, огурца, баклажана, яванской фасоли, кукурузы, арбуза, пшена, золотистой фасоли, овса, окры, Panicum, папайи, арахиса, гороха, перца, голубиного гороха, ананаса, Phaseolus, картофеля, тыквы, риса, сорго, сои, крупноплодной тыквы, сахарного тростника, сахарной свеклы, подсолнечника, батата, чая, томата, табака, арбуза и пшеницы.
В другом воплощении клетка-хозяин представляет собой клетку данио.
В другом воплощении клетка-хозяин представляет собой клетку цыпленка.
В другом воплощении клетка-хозяин представляет собой клетку млекопитающего, выбранную из группы, состоящей из клетки хомячка, клетки мыши, клетки крысы, клетки кролика, клетки кошки, клетки собаки, клетки крупного рогатого скота, клетки козы, клетки коровы, клетки свиньи, клетки лошади, клетки овцы, клетки обезьяны, клетки шимпанзе и клетки человека.
Трансформация клетки-хозяина хорошо известна в данной области техники и может быть осуществлена посредством множества способов, включая электропорацию, вирусную инфекции, трансфекцию плазмида/вектор, трансфекцию, опосредованную невирусным вектором, трансформацию, опосредованную Agrobacterium, бомбардировку частицами и тому подобное, но не ограничиваясь ими. Экспрессия желательных генных продуктов включает культивирование трансформированной клетки-хозяина в подходящих условиях и индуцирование экспрессии трансформированного гена. Условия выращивания и протоколы экспрессии гена в прокариотических и эукариотических клетках хорошо известны в данной области техники. Клетки могут быть собраны и генные продукты выделены согласно протоколам, специфическим для генного продукта.
Кроме того, можно выбрать клетку-хозяин, которая модулирует экспрессию встроенного полинуклеотида, или изменяет и процессирует полипептидный продукт конкретным требуемым способом. Разные клетки-хозяева имеют характерные и специфические механизмы трансляционного и пост-трансляционного процессинга и модификации Снапример гликозилирование, расщепление (например сигнальной последовательности)) белков. Подходящие клеточные линии или хозяйские системы могут быть выбраны для обеспечения требуемой модификации и процессинга чужих экспрессированных белков. Например, экспрессию в бактериальной системе можно использовать для продуцирования негликозилированного продукта коревого белка. Однако полипептид, экспрессируемый в бактериях, может быть неправильно уложенным. Экспрессия в дрожжах может продуцировать гликозилированный продукт. Экспрессия в эукариотических клетках может увеличивать вероятность "нативного" гликозилирования и укладку гетерологичного белка. Кроме того, экспрессия в клетках млекопитающего может обеспечить средство восстановления или установления полипептидной активности. Кроме того, разные системы экспрессии вектор/хозяин могут в разной степени влиять на реакции процессинга, такие как протеолитические расщепления. Диацилгидразиновые лиганды Формулы I и хиральные диацилгидразиновые лиганды Формулы II или III по настоящему изобретению можно использовать в организме, отличном от человеческого, содержащем выделенную клетку-хозяина. В одном воплощении организм, отличный от человеческого, представляет собой прокариотический организм или эукариотический организм. В другом воплощении организм, отличный от человеческого, представляет собой организм беспозвоночных или организм позвоночных.
Предпочтительно организм, отличный от человеческого, выбран из группы, состоящей из бактерии, гриба, дрожжей, нематоды, насекомого, рыбы, растения, птицы, животного и млекопитающего. Более предпочтительно организм, отличный от человеческого организма, представляет собой дрожжи, нематоду, насекомое, растение, данио, курицу, хомяка, мышь, крысу, кролика, кошку, собаку, быка, козу, корову, свинью, лошадь, овцу, обезьяноподобное, обезьяну или шимпанзе.
В другом воплощении организм, отличный от человеческого организма, представляет собой дрожжи, выбранные из группы, состоящей из Saccharomyces, Pichia и Candida.
В другом воплощении организм, отличный от человеческого, представляет собой нематоду Caenorhabditis elegans.
В другом воплощении организм, отличный от человеческого, представляет собой растение, выбранное из группы, состоящей из яблока, Arabidopsis, проса, банана, ячменя, бобовых, свеклы, ниссы, нуга, чилийского перца, огурца, баклажана, яванской фасоли, кукурузы, арбуза, пшена, золотистой фасоли, овса, окры, Panicum, папайи, арахиса, гороха, перца, голубиного гороха, ананаса, Phaseolus, картофеля, тыквы, риса, сорго, сои, крупноплодной тыквы, сахарного тростника, сахарной свеклы, подсолнечника, батата, чая, томата, табака, арбуза обыкновенного и пшеницы.
В другом воплощении организм, отличный от человеческого, представляет собой мышь Mus musculus.
СИСТЕМЫ МОДУЛЯЦИИ ЭКСПРЕССИИ ГЕНА
Настоящее изобретение относится к диацилгидразиновым лигандам Формулы I и хиральным диацилгидразиновым лигандам Формулы II или III, которые являются полезными в индуцируемой системе экспрессии гена на основе экдизонового рецептора. Такие диацилгидразиновые лиганды обеспечивают улучшенную индуцируемую систему экспрессии гена как в прокариотических, так и в эукариотических хозяйских клетках. Таким образом, настоящее изобретение относится к диацилгидразиновым лигандам Формулы I и хиральным диацилгидразиновым лигандам Формулы II или III, которые являются полезными для модуляции экспрессии генов. В частности, настоящее изобретение относится к диацилгидразиновым лигандам Формулы I и хиральным диацилгидразиновым лигандам Формулы II или III, обладающим способностью трансактивировать систему модуляции экспрессии гена, содержащую по меньшей мере одну кассету экспрессии гена, то есть способным экспрессироваться в клетке-хозяине, содержащей полинуклеотид, который кодирует полипептид, содержащий лиганд-связывающий домен ядерного рецептора группы Н. В одном воплощении лиганд-связывающий домен ядерного рецептора группы Н происходит от экдизонового рецептора, универсального рецептора, рецептора-сироты 1, NER-1, ядерного рецептора 1 стероидного гормона, белка-15, взаимодействующего с ретиноидным X-рецептором, X-рецептора печени β, белка, подобного рецептору стероидного гормона, X-рецептора печени, X рецептора печени α, фарнезоидного X-рецептора, рецептор-взаимодействующего белка-14 или рецептора фарнезола. В другом воплощении лиганд-связывающий домен ядерного рецептора группы Н происходит от экдизонового рецептора.
В другом воплощении система модуляции экспрессии гена содержит кассету экспрессии гена, содержащую полинуклеотид, который кодирует полипептид, содержащий домен трансактивации, ДНК-связывающий домен, который распознает элемент ответа, ассоциированный с геном, экспрессию которого следует модулировать; и лиганд-связывающий домен ядерного рецептора группы Н, содержащий мутацию в виде замены. Система модуляции экспрессии гена может дополнительно содержать вторую кассету экспрессии гена, содержащую: 1) элемент ответа, распознаваемый при помощи ДНК-связывающего домена кодируемого полипептида первой кассеты экспрессии гена; 2) промотор, который активируется доменом трансактивации кодируемого полипептида первой кассеты экспрессии гена; и 3) ген, экспрессию которого следует модулировать.
В другом воплощении система модуляции экспрессии гена содержит кассету экспрессии гена, содержащую а) полинуклеотид, который кодирует полипептид, содержащий домен трансактивации, ДНК-связывающий домен, который распознает элемент ответа, ассоциированный с геном, экспрессию которого следует модулировать; и лиганд-связывающий домен ядерного рецептора группы H, содержащий мутацию в виде замены, и б) лиганд-связывающий домен второго ядерного рецептора, выбранный из группы, состоящей из лиганд-связывающего домена ретиноидного Х-рецептора позвоночных, лиганд-связывающего домена ретиноидного Х-рецептора беспозвоночных, лиганд-связывающего домена белка Ultraspiracle, и химерного лиганд-связывающего домена, содержащего два полипептидных фрагмента, где первый полипептидный фрагмент происходит от лиганд-связывающего домена ретиноидного Х-рецептора позвоночных, лиганд-связывающего домена ретиноидного Х-рецептора беспозвоночных, или лиганд-связывающего домена белка Ultraspiracle, и второй полипептидный фрагмент происходит от другого лиганд-связывающего домена ретиноидного Х-рецептора позвоночных, лиганд-связывающего домена ретиноидного Х-рецептора беспозвоночных или лиганд-связывающего домена белка Ultraspiracle. Система модуляции экспрессии гена может дополнительно включать вторую кассету экспрессии гена, содержащую: 1) элемент ответа, распознаваемый ДНК-связывающим доменом кодируемого полипептида первой кассеты экспрессии гена; 2) промотор, который активируется доменом трансактивации кодируемого полипептида первой кассеты экспрессии гена; и 3) ген, экспрессию которого следует модулировать.
В другом воплощении система модуляции экспрессии гена содержит первую кассету экспрессии гена, содержащую полинуклеотид, который кодирует первый полипептид, содержащий ДНК-связывающий домен, который распознает элемент ответа, ассоциированный с геном, экспрессию которого следует модулировать, и лиганд-связывающий домен ядерного рецептора, и вторую кассету экспрессии гена, содержащую полинуклеотид, который кодирует второй полипептид, содержащий домен трансактивации, и лиганд-связывающий домен ядерного рецептора, где один из лиганд-связывающих доменов ядерного рецептора представляет собой лиганд-связывающий домен ядерного рецептора группы Н, содержащий мутацию в виде замены. В одном воплощении первый полипептид по существу не содержит домен трансактивации и второй полипептид по существу не содержит ДНК-связывающий домен. Для целей изобретения "по существу не содержит" означает, что рассматриваемый белок не содержит достаточную последовательность рассматриваемого домена, чтобы обеспечить активацию или связывающую активность. Система модуляции экспрессии гена может дополнительно содержать третью кассету экспрессии гена, содержащую: 1) элемент ответа, распознаваемый ДНК-связывающим доменом первого полипептида первой кассеты экспрессии гена; 2) промотор, который активируется доменом трансактивации второго полипептида второй кассеты экспрессии гена; и 3) ген, экспрессию которого следует модулировать.
Там где только один лиганд-связывающий домен ядерного рецептора представляет собой лиганд-связывающий домен группы H, содержащий мутацию в виде замены, другой лиганд-связывающий домен ядерного рецептора может быть от любого другого ядерного рецептора, который образует димер с лиганд-связывающим доменом группы H, содержащим мутацию в виде замены. Например, если лиганд-связывающий домен ядерного рецептора группы H, содержащий мутацию в виде замены, представляет собой лиганд-связывающий домен экдизонового рецептора, содержащий мутацию в виде замены, другой лиганд-связывающий домен ядерного рецептора ("партнер") может происходить от экдизонового рецептора, ретиноидного X-рецептора (RXR) позвоночных, RXR беспозвоночных, белка Ultraspiracle (USP) или химерного ядерного рецептора, содержащего по меньшей мере два разных полипептидных фрагмента лиганд-связывающего домена ядерного рецептора, выбранных из группы, состоящей из RXR позвоночных, RXR беспозвоночных, и USP (смотри WO 01/70816 A2, международная заявка на патент PCT/US02/05235 и US 2004/0096942 A1, включенные в данное описание изобретения посредством ссылки во всей их полноте). "Партнерский" лиганд-связывающий домен ядерного рецептора может дополнительно содержать мутацию виде усечения, делеционную мутацию, мутацию в виде замены или другую модификацию.
В одном воплощении лиганд-связывающий домен RXR позвоночных представляет собой RXR человека Homo sapiens, мыши Mus musculus, крысы Rattus norvegicus, цыпленка Gallus gallus, свиньи Sus scrota domestica, лягушки Xenopus laevis, данио Danio rerio, туникаты Polyurocarpa misakiensis, или медузы Tripedalia cysophora.
В одном воплощении лиганд-связывающий домен RXR беспозвоночных представляет собой полипептид Ultraspiracle саранчи Locusta migratoria ("LmUSP"), гомолог 1 RXR иксодового клеща Amblyomma americanum ("AmaRXRI"), гомолог 2 RXR иксодового клеща Amblyomma americanum ("AmaRXR2"), гомолог RXR манящего краба Celuca pugilator ("CpRXR"), гомолог RXR жука Tenebrio molitor ("TmRXR"), гомолог RXR медоносной пчелы Apis mellifera ("AmRXR"), гомолог RXR тли Myzus persicae ("MpRXR"), или гомолог RXR недвукрылых/нечешуекрылых.
В одном воплощении химерный лиганд-связывающий домен RXR содержит по меньшей мере два полипептидных фрагмента, выбранных из группы, состоящей из полипептидного фрагмента RXR позвоночных видов, полипептидного фрагмента RXR беспозвоночных видов, и гомологичного полипептидного фрагмента RXR недвукрылых/нечешуекрылых беспозвоночных видов. Химерный лиганд-связывающий домен RXR для применения в настоящем изобретении может содержать полипептидные фрагменты RXR по меньшей мере двух разных видов, или, когда виды являются одинаковыми, два или более полипептидных фрагмента могут представлять собой две или более разные изоформы видов полипептидного фрагмента RXR.
В одном воплощении химерный лиганд-связывающий домен RXR содержит по меньшей мере один полипептидный фрагмент RXR позвоночных видов и один полипептидный фрагмент RXR беспозвоночных видов.
В другом воплощении химерный лиганд-связывающий домен RXR содержит по меньшей мере один полипептидный фрагмент RXR позвоночных видов и один гомологичный полипептидный фрагмент RXR недвукрылых/нечешуекрылых беспозвоночных видов.
В другом воплощении ген, экспрессию которого следует модулировать, является гомологичным геном в отношении клетки-хозяина. В другом воплощении ген, экспрессию которого следует модулировать, является гетерологичным геном в отношении клетки-хозяина.
Диацилгидразиновые лиганды Формулы I и хиральные диацилгидразиновые лиганды Формулы II или III для применения по настоящему изобретению, как описано ниже, при комбинировании с лиганд-связывающим доменом ядерного рецептора(ов), который в свою очередь соединяют с элементом ответа, связанным с геном, обеспечивают средства для внешней временной регуляции экспрессии гена. Механизм связывания или порядок, в котором различные компоненты по данному изобретению связываются друг с другом, то есть, например, лиганд с лиганд-связывающим доменом, ДНК-связывающий домен с элементом ответа, домен трансактивации с промотором, так далее, не является критичным.
Экдизоновый рецептор является членом суперсемейства ядерных рецепторов и классифицирован в подсемейство 1, группу H (упоминаемую в данном описании изобретения как "ядерные рецепторы группы H"). Члены каждой группы имеют 40-60%-ное аминокислотное подобие в домене E (лиганд-связывающем) (Laudet et al., Cell 97: 161-163 (1999)). В дополнение к экдизоновому рецептору, другие члены данной группы Н субсемейства 1 ядерных рецепторов включают: универсальный рецептор (UR), рецептор-сироту 1 (OR-1), ядерный рецептор 1 стероидного гормона (NER-1), взаимодействующий белок-15 ретиноидного Х-рецептора (RIP-15), X-рецептор β из печени (LXRβ), белок, подобный рецептору стероидного гормона (RLD-1), Х-рецептор печени (LXR), Х-рецептор α из печени (LXRα), фарнезоидный X-рецептор (FXR), рецептор-взаимодействующий белок-14 (RIP-14), и рецептор фарнезола (HRR-1).
В конкретном примере связывание лиганда с лиганд-связывающим доменом ядерного рецептора группы H и его партнерским лиганд-связывающим доменом ядерного рецептора, делает возможной экспрессию или супрессию гена. Такой механизм не исключает возможность связывания лиганда с ядерным рецептором группы H (GHNR) или его партнером, и образования в результате активных гомодимерных комплексов (например GHNR+GHNR или партнер + партнер). Предпочтительно один или более доменов рецептора варьируются, продуцируя гибридный генный переключатель. Обычно один или более из трех доменов, DBD, LBD и домена трансактивации, может быть выбран из источника, отличного от источника других доменов, так, что гибридные гены и полученные гибридные белки были оптимизированы в выбранной клетке-хозяине или организме в отношении активности к трансактивации, комплементарного связывания лиганда и распознавания конкретного элемента ответа. Кроме того, сам элемент ответа может быть модифицирован или замещен элементами ответа для других доменов ДНК-связывающего белка, такими как белок GAL-4 из дрожжей (смотри Sadowski et al, Nature 355: 563-564 (1988)) или белок LexA из Eschenchia coli (смотри Brent et al., Cell 43: 129-136 (1985)) или синтетические элементы ответа, специфичные для нацеленных взаимодействий с белками, сконструированными, модифицированные и выбранными для таких специфических взаимодействий (смотри, например, Kim etal., Proc. Natl. Acad. Sci. USA, 24: 3616-3620 (1997)) для накопления гибридных рецепторов. Другое преимущество двухгибридных систем состоит в том, что они обеспечивают выбор промотора, используемого для управления экспрессией гена в соответствии с необходимым конечным результатом. Такой двойной контроль может быть особенно важным в областях генной терапии, особенно при продуцировании цитотоксических белков, так как можно контролировать как время экспрессии, так и клетки, в которых происходит экспрессия. Когда гены, функционально связанные с подходящим промотором, вводят в клетки субъекта, экспрессия экзогенных генов контролируется присутствием системы по данному изобретению. Промоторы могут быть конститутивно или индуцируемо регулируемыми или могут быть тканеспецифическими (то есть экспрессирующимися только в конкретном типе клеток) или специфическими к конкретным стадиям развития организма.
В частности, в данном описании изобретения описаны диацилгидразиновые лиганды Формулы I и хиральные диацилгидразиновые лиганды Формулы II или III, полезные в системе модуляции экспрессии гена, содержащей лиганд-связывающий домен ядерного рецептора группы H, содержащий мутацию в виде замены. Данная система экспрессии гена может представлять собой систему экспрессии гена на основе "одиночного переключателя", где домен трансактивации, ДНК-связывающий домен и лиганд-связывающий домен находятся на одном кодируемом полипептиде. Альтернативно, система модуляции экспрессии гена может представлять собой систему модуляции экспрессии гена на основе "двойного переключателя" или "двухгибридной", где домен трансактивации и ДНК-связывающий домен располагаются на двух разных кодируемых полипептидах.
Система модуляции экспрессии гена на основе экдизонового рецептора для применения в настоящем изобретении может быть либо гетеродимерной, либо гомодимерной. Функциональный комплекс EcR в большинстве случаев относится к гетеродимерному белковому комплексу, состоящему из двух членов семейства стероидных рецепторов, экдизонового рецепторного белка, полученного из различных насекомых, и белка Ultraspiracle (USP), или гомолога USP позвоночных, белка ретиноидного Х-рецептора, (смотри Yao et al, Nature 366: 476-479 (1993) и Yao et al, Cell 77: 63-72 (1992)). Однако комплекс также может представлять собой гомодимер, как подробно описано ниже. Функциональный экдистероидный рецепторный комплекс также может включать дополнительный(ые) белок(и), такой(ие) как иммунофилины. Дополнительные члены семейства белков стероидных рецепторов, известные как факторы транскрипции (такие как DHR38 или бетаFTZ-1), могут также являться лиганд-зависимыми или независимые партнерами EcR, USP и/или RXR. Кроме того, могут требоваться другие кофакторы, например белки главным образом известные как со-активаторы (также называемые адаптерами или медиаторами). Такие белки не связываются с ДНК специфически к последовательности и не вовлечены в базальную транскрипцию. Они могут оказывать эффект на активацию транскрипции посредством различных механизмов, включая стимулирование ДНК-связывания активаторов, путем воздействия на хроматиновую структуру или опосредуя комплексные взаимодействия активатор-инициация. Примеры таких со-активаторов включают RIP 140, TIF1, RAP46/Bag-1, ARA70, SRC-1/NCoA-I, TIF2/GRIP/NCoA-2, ACTR/AIBI/RAC3/pCIP, а также смешанный со-активатор C, элемент ответа B-связывающий белок, CBP/p300 (обзор смотри в Glass et al, Curr. Opin. Cell Biol. 9: 222-232 (1997)). Также белковые кофакторы, общеизвестные как со-репрессоры (также известные как репрессоры, сайленсеров, или подавляющих медиаторов) могут требоваться для эффективного подавления активации транскрипции в отсутствие лиганда. Эти со-репрессоры могут взаимодействовать с нелигандированным экдизоновым рецептором для подавления активности элемента ответа. Имеющиеся данные позволяют предположить, что связывание лиганда изменяет конформацию рецептора, что приводит к высвобождению со-репрессора и рекрутменту вышеописанных со-активаторов, в результате чего ликвидируется их подавляющую активность. Примеры со-репрессоров включают N-CoR и SMRT (обзор смотри в Horwitz et al, Mol Endocrinol. 10: стр.1167-1177 (1996)). Эти кофакторы могут быть либо эндогенными в клетке или организме, или могут быть добавлены экзогенно в качестве трансгенов, для экспрессии либо регулируемым, либо нерегулируемым образом. Гомодимерные комплексы экдизонового рецепторного белка, USP или RXR могут также быть функциональными в некоторых случаях.
Экдизон-рецепторный комплекс обычно включает белки, которые являются членами суперсемейства ядерных рецепторов, где все члены, как правило, отличаются присутствием аминоконцевого домена трансактивации, ДНК-связывающего домена ("DBD") и лиганд-связывающего домена ("LBD"), отделенного от DBD шарнирным участком. При использовании в данном описании изобретения, термин "ДНК-связывающий домен" включает в себя минимальную полипептидную последовательность ДНК-связывающего белка, вплоть до полной длины ДНК-связывающего белка, при условии что ДНК-связывающий домен функционирует с целью ассоциации с конкретным элементом ответа. Члены суперсемейства ядерных рецепторов также характеризуются присутствием четырех или пяти доменов: A/B, C, D, E, и, у некоторых, F (смотри US 4981784 и Evans, Science 240:889-895 (1988)). "A/B" домен соответствует домену трансактивации, "C" соответствует ДНК-связывающему домену, "D" соответствует шарнирному участку и "E" соответствует лиганд-связывающему домену. Некоторые члены этого семейства также могут иметь другой домен трансактивации на карбоксилконцевой стороне LBD, соответствующий "F".
DBD характеризуется присутствием двух богатых цистеином доменов «цинковые пальцы», между которыми находятся два аминокислотных мотива, P-box и D-box, которые обеспечивают специфичность к экдизоновым элементам ответа. Такие домены могут быть либо нативными, модифицированными, либо химерами разных доменов гетерологичных рецепторных белков. Рецептор EcR, подобно подгруппе семейства стероидных рецепторов, также имеет менее четко определенные участки, ответственные за свойства гетеродимеризации. Так как домены ядерных рецепторов являются модульными по природе, LBD, DBD и домены трансактивации могут быть взаимозаменяемыми.
Известно, что системы переключателя генов включают компоненты из экдизон-рецепторного комплекса. Однако, в данных известных системах, во всех случаях, когда используется EcR, он ассоциируется с нативными или модифицированными ДНК-связывающими доменами и доменами трансактивации на той же молекуле. USP или RXR обычно используют в качестве молчащих партнеров. Ранее было показано, что если ДНК-связывающие домены и домены трансактивации находятся на одной молекуле, то фоновая активность в отсутствие лиганд будет высокой, и что такая активность резко снижается, когда ДНК-связывающие домены и домены трансактивации находятся на разных молекулах, то есть на каждом из двух партнеров гетеродимерного или гомодимерного комплекса (смотри PCT/US01/09050).
СПОСОБ МОДУЛЯЦИИ ЭКСПРЕССИИ ГЕНА
Настоящее изобретение также относится к способу модуляции экспрессии гена в клетке-хозяине с использованием системы модуляции экспрессии гена и лигандов по настоящему изобретению. Конкретно, в настоящем изобретении предлагается способ модуляции экспрессии гена в клетке-хозяине, включающий стадии: а) введения в клетку-хозяина системы модуляции экспрессии гена; и б) введения в клетку-хозяина диацилгидразинового лиганда Формулы I или хирального диацилгидразинового лиганда Формулы II или III; где ген, который следует модулировать, является компонентом кассеты экспрессии гена, содержащей: 1) элемент ответа, содержащий домен, распознаваемый ДНК-связывающим доменом системы экспрессии гена; 2) промотор, который активируется доменом трансактивации системы экспрессии гена; и 3) ген, экспрессию которого следует модулировать, при этом после введения диацилгидразинового лиганда Формулы I или хирального диацилгидразинового лиганда Формулы II или III в клетку-хозяина экспрессия гена модулируется.
В настоящем изобретении также предлагается способ модуляции экспрессии гена в клетке-хозяине, включающий стадии: а) введения в клетку-хозяина системы модуляции экспрессии гена; б) введения в клетку-хозяина кассеты экспрессии гена, где кассета экспрессии гена содержит 1) элемент ответа, содержащий домен, распознаваемый ДНК-связывающим доменом из системы экспрессии гена; 2) промотор, который активируется доменом трансактивации система экспрессии гена; и 3) ген, экспрессию которого следует модулировать; и в) введения в клетку-хозяина диацилгидразинового лиганда Формулы I или хирального диацилгидразинового лиганда Формулы II или III; при этом после введения диацилгидразинового лиганда Формулы I или хирального диацилгидразинового лиганда Формулы II или III в клетку-хозяина экспрессия гена модулируется.
В настоящем изобретении также предлагается способ модуляции экспрессии гена в клетке-хозяине, содержащей кассету экспрессии гена, содержащую элемент ответа, содержащий домен, к которому присоединяется ДНК-связывающий домен из первого гибридного полипептида системы модуляции экспрессии гена; промотор, который активируется доменом трансактивации второго гибридного полипептида системы модуляции экспрессии гена; и ген, экспрессию которого следует модулировать; где способ включает стадии: а) введения в клетку-хозяина системы модуляции экспрессии гена; и б) введения в клетку-хозяина диацилгидразинового лиганда Формулы I или хирального диацилгидразинового лиганда Формулы II или III; при этом после введения лиганда в хозяина экспрессия гена модулируется.
Гены, представляющие интерес в отношении экспрессии в клетке-хозяине, с использованием способов, раскрытых в данном описании изобретения, могут представлять собой эндогенные гены или гетерологичные гены. Информация о нуклеиновокислой или аминокислотной последовательности нужного гена или белка может быть размещена в одном из многих баз данных открытого доступа, например GENBANK, EMBL, Swiss-Prot и PIR, или во многих связанных с биологией журнальных публикациях. Таким образом, специалист в данной области техники имеет доступ к информации о последовательности нуклеиновой кислоты практически для всех известных генов. Такую информацию затем можно использовать для создания требуемых конструкций для вставки интересующего гена в кассеты экспрессии гена с использованием способов, описанных в данном описании изобретения.
ОЦЕНКА ГЕННОЙ ЭКСПРЕССИИ/ТРАНСКРИПЦИИ
Одной полезной оценкой способов по изобретению является оценка транкрипционного состояния клетки, включая идентичности и количества РНК, предпочтительно разновидностей м-РНК. Такие оценки удобно производить, определяя количество кДНК любым из нескольких существующих технологий экспрессии гена.
Технология матрицы нуклеиновых кислот является полезным методом определения дифференциальной экспрессии м-РНК. Такая технология включает, например, олигонуклеотидные чипы и микроматрицы ДНК. Такие методы основаны на фрагментах ДНК или олигонуклеотидов, которые соответствуют разным генам или кДНК, которые иммобилизованы на твердой подложке и гибридизирующихся с зондами, полученными из общих пулов м-РНК, экстрагированных из клеток, тканей или целых организмов, и превращены в кДНК. Олигонуклеотидные чипы представляют собой матрицы олигонуклеотидов, синтезированных на субстрате с использованием фотолитографические методик. Были получены чипы, которые могут быть использованы для анализа вплоть до 1700 генов. Микроматрицы ДНК представляют собой матрицы образцов ДНК, обычно продуктов ПЦР, которые с помощью робота отпечатывают на предметное стекло. Каждый ген анализируют с помощью целевой последовательности ДНК полной или неполной длины. В настоящее время микроматрицы с генами в количестве вплоть до 10000 обычно получают в промышленных масштабах. Основное различие между этими двумя методами заключается в том, что в олигонуклеотидных чипах обычно используются 25-мерные олигонуклеотиды, которые обеспечивают фракционирование коротких молекул ДНК, тогда как более крупные ДНК-мишени микроматриц, приблизительно 1000 пар оснований, могут обеспечивать большую чувствительность в фракционировании сложных смесей ДНК.
Другой полезной оценкой способов по изобретению является определение трансляционного состояния клетки путем измерения количества составляющих белковых видов, присутствующих в клетке, с использованием способов, хорошо известных в данной области техники.
Когда желательна идентификация генов, ассоциированных с различными физиологическим функциям, можно использовать анализ, в котором измеряют изменения таких функций, как рост клетки, апоптоз, старение, дифференциация, адгезия, связывание со специфическими молекулами, связывание с другой клеткой, клеточная организация, органогенез, внутриклеточный транспорт, облегчение транспорта, превращение энергии, метаболизм, миогенез, нейрогенез и/или кроветворение.
Кроме того, селектируемый маркер или репортерный ген экспрессии можно использовать для оценки модуляции экспрессии гена с использованием настоящего изобретения.
Другие способы обнаружения продуктов экспрессии гена хорошо известны в данной области техники и включают анализы Саузерн-блоттинг (определение ДНК), дот- или слот-блоттинг (ДНК, РНК), нозерн-блоттинг (РНК), RT-PCR (РНК), вестерн-блоттинг (определение полипептида) и ELISA (полипептид). Хотя и менее предпочтительно, меченные белки можно использовать для определения конкретной нуклеиновокислой последовательности, с которой происходит гибридизация.
В некоторых случаях необходимо увеличивать количество нуклеиновокислой последовательности. Это можно выполнить, используя один или более из ряда подходящих способов, включающих, например, полимеразную цепную реакцию ("PCR"), лигазную цепную реакцию ("LCR"), амплификацию с вытеснением цепи ("SDA"), амплификацию на основе транскрипции и тому подобное. ПЦР выполняют в соответствии с известными методиками, где, например, образец нуклеиновой кислоты обрабатывают в присутствие термостабильной ДНК-полимеразы, в условиях гибридизации, с одной парой олигонуклеотидных праймеров, с одним праймером, гибридизирующимся с одной нитью (матричной) конкретной последовательности, подлежащей определению. Праймеры являются достаточно комплементарными для каждой матричной нити конкретной последовательности для гибридизации с ними. Синтезируется продукт удлинения каждого праймера, и он является комплементарным матричной нити нуклеиновой кислоты, с которую он гибридизируется. Продукт удлинения, синтезированный из каждого праймера может также служить в качестве матрицы для дополнительного синтеза продуктов удлинения с использованием тех же самых праймеров. После значительного количества циклов синтеза продуктов удлинения образец можно проанализировать, как описано выше, чтобы оценить: присутствует(ют) ли последовательность или последовательности, подлежащие определению.
ОБЩИЕ СПОСОБЫ
Стандартные методы рекомбинантной ДНК и молекулярного клонирования, используемые в данном описании изобретения, хорошо известны в данной области техники и описаны в Sambrook, J., Fritsch, E.F. и Maniatis, Т. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, N.Y. (1989) (Maniatis) и Т.J. Silhavy, M.L. Bennan, и L.W. Enquist, Experiments with Gene Fusions, Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. (1984) и в Ausubel et al., Current Protocols in Molecular Biology, Greene Publishing Assoc. and Wiley-lnterscience (1987).
Вещества и способы, подходящие для поддерживания и роста бактериальных культур, хорошо известны в данной области техники. Способы, пригодные для использования в следующих примерах, могут быть найдены в изложении в Manual of Methods for General Bacteriology (Phillipp Gerhardt, R.G.E. Murray, Ralph N. Costilow, Eugene W. Nester, Willis A. Wood, Noel R. Krieg и G. Briggs Philips, eds), American Society for Microbiology, Washington, DC. (1994)) или в Thomas D. Brock in Biotechnology: A Textbook of Industrial Microbiology, Second Edition, Sinauer Associates, Inc., Sunderland, MA (1989). Все реагенты, ферменты рестрикции и вещества, используемые для роста и поддержания хозяйских клеток, получали от Aldrich Chemical (Milwaukee, WI), DIFCO Laboratories (Detroit, MI), GIBCO/BRL (Gaithersburg, MD) или Sigma Chemical Company (St. Louis, МО), если не указано иное.
Манипуляции с генетическими последовательностями могут быть осуществлены с использованием пакета программ, доступных от Genetics Computer Group, Inc. (Wisconsin Package Version 9.0, Genetics Computer Group, (GCG), Madison, WI). Если используют программу GCG "Pileup", может быть использована величина создания бреши 12, заданная по умолчанию, и величина удлинения бреши 4, заданная по умолчанию. Если используют программу CGC "Gap" или "Bestfit", то может быть использован штраф создания бреши 50, заданный по умолчанию, и штраф по умолчанию удлинения бреши 3. В любом случае, когда параметры программы GCG не запрашиваются, в этих или любых других программах GCG, можно использовать величины, заданные по умолчанию.
Точки плавления измеряли в стеклянных капиллярных трубках с помощью устройства Electrothermal® и не корректировали. Оптическое вращение ([α]25589) определяли при комнатной температуре в кварцевой кювете длиной 10 см, используя поляриметр Perkin Elmer Model 341. Концентрации приведены в г/100 мл. Спектры 1H ЯМР записывали при 300 МГц или 400,13 МГц, используя Bruker NMR. Если не указано иное, внутренним эталоном был растворитель. Спектры 13C ЯМР записывали при 100,6 МГц с помощью Bruker NMR. Если не указано иное, внутренний эталон представлял собой растворитель. Анализ LC-MS выполняли, используя пакет Agilent 1100 LC вместе с одноквадропульным масс-спектрометром Agilent. Растворители представляли собой (А) H2O/0,1% муравьиная кислота и (Б) ACN/0,1% муравьиная кислота в градиенте от T=0-15% B до T=10-98% В и время остановки 20 мин, на колонке 75 мм×2,1 мм C18 со скоростью потока = 0,2 мл/мин. Точные анализы массы выполняли путем прямого нагнетания в масс-спектрометр Agilent ESI/TOF. Рентгенокристаллографические определения выполняли, используя Bruker SMART6000. Элементный анализ выполняли на CHN-анализаторе Perkin Elmer 2400. Аналитическую тонкослойную хроматографию или ТСХ выполняли, используя Macherey-Nagel Polygram® на 0,2 мм пластинках Sil G/UV254. Большинство пластинок визуализировали с помощью УФ-излучения. Некоторые проявляли, используя йод или фосфомолибденовую кислоту. Хроматографию на силикагеле выполняли, используя силикагель Aldrich (230-400 меш, 60 A) в стеклянных колонках под давлением N2 или напором аргона приблизительно 30 фунтов на квадратный дюйм (0,21 МПа). Аналитическую ВЭЖХ выполняли, используя систему ВЭЖХ Gilson, оснащенную динамическим миксером 811C, детектором 306 UV/VIS 155 и устройством подачи жидкости 215. УФ-поглощение измеряли при 220 нм и 254 нм и интегрирования выполняли при 254 нм. Скорости потока обычно поддерживали при 1 мл/мин. Хроматографию на нормальной фазе выполняли на колонке 4,6 мм×25 см DuPont Zorbax ODS. Хроматографию на обращенной фазе выполняли, используя 5 микрон Alltech Adsorbosphere, на колонке C18 4,6 мм×25 см. Хиральную ВЭЖХ выполняли, используя 5 микрон CHIRALCEL®, колонку OD-H 4,6 мм×25 см, 5 микрон CHIRAKPAK®, колонку AD-H 4,6 мм×25 см, или 5 микрон Regis Rexchrom (S,S) колонку ULMO, 100 A, 4,6 мм×25 см. Растворители являлись химически чистыми, если не указано иное. Безводные растворители использовали в покупном виде.
ОБЩИЕ СПОСОБЫ СИНТЕЗА
Энантиомерно обогащенные соединения Формулы II или III, где A, B и R2 определены, как указано выше, и R1 представляет собой алкенил, можно получать посредством асимметрического синтеза, как описано в Общей Схеме 1.
Общая Схема 1
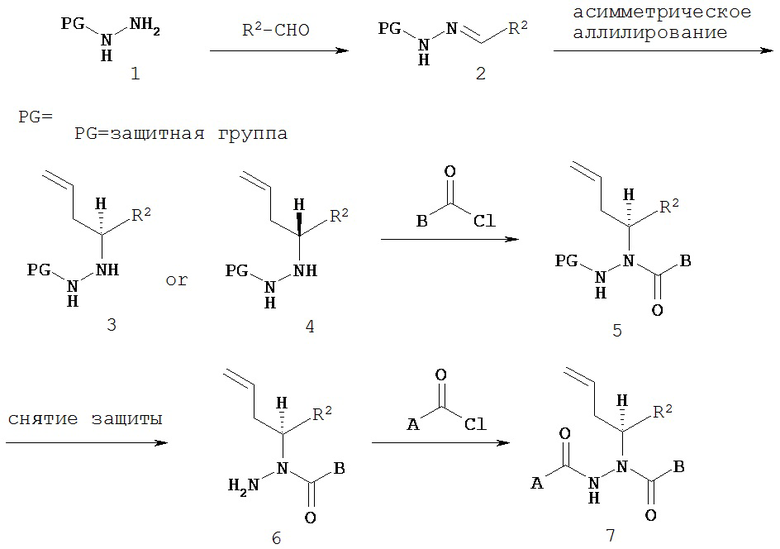
Взаимодействие защищенного гидразина 1 (например бензилкарбазата или трет-бутилкарбазата) с альдегидом R2CHO дает 2. Асимметрическое аллилирование 2 дает или 3 или 4 в энантиомерно обогащенной форме, в зависимости, среди прочего, от селекции хирального реагента и R2. Способы осуществления реакций асимметрического аллилирования известны в данной области техники и могут быть использованы для получения соединений по изобретению. Синтез хиральных реагентов, особенно полезных в реакциях асимметрического аллилирования, описан в Leighton et al., J. Am. Chem. Soc. 125: 9596 (2003) и WO 03/074534. Взаимодействие 3 с хлорангидридом карбоновой кислоты B-COCl дает 5. Карбоновую кислоту B-CO2H можно также соединять с 3 с получением 5. Удаление защитной группы из 5 дает 6. Специалисту в данной области техники известно множество способов снятия защиты с гидразина в зависимости от природы защитной группы. Пригодные стратегии защиты/снятия защиты обсуждаются в "Protective Groups in Organic Synthesis", Т.W. Greene и P.G.M. Wilts (1999). Взаимодействие 7 с хлорангидридом карбоновой кислоты A-COCl или с карбоновой кислотой А-CO2H дает 8, соединение Формулы II. Аналогично, взаимодействие 4 приводит к соединению Формулы III.
Энантиомерно обогащенные соединения Формулы II или III, где A, B, R1 и R2 определены, как указано выше, также можно получить путем асимметрического восстановления, как описано в Общей Схеме 2.
Общая Схема 2

Взаимодействие ацилгидразина 1 с кетоном R1COR2 дает гидразон 2. Асимметрическое восстановление 2 дает 3 или 4 в энантиомерно обогащенной форме, в зависимости, среди прочего, от селекции восстановителя, R1 и R2. В US 5250731 описано асимметрическое каталитическое гидрирование ацилгидразонов с получением оптически активных ацилгидразинов в присутствие каталитического комплекса хирального фосфолана. Специалисту в данной области техники понятно, что можно использовать также другие хиральные лиганды. Более того, специалисту в данной области техники ясно, что можно использовать дополнительные способы асимметрического восстановления. В данном примере взаимодействие 3 с BCOCl дает 5 и соединение Формулы II. Аналогично, взаимодействие 4 приводит к соединению Формулы III.
Кроме того, энантиомерно обогащенные соединения Формулы II или III можно получать посредством хирального разделения, или комбинации асимметрического синтеза и хирального разделения, или комбинации асимметрического гидрирования и хирального разделения. Как описано на Общей Схеме 3, в результате хиральной хроматографии ВЭЖХ рацемических диацилгидразинов получают энантиомерно обогащенные соединения Формулы II или III. Подходящие хиральные колонки для использования в хиральных разделениях включают, например, хиральные колонки Daicel CHIRALCEL® OD-Н, Daicel CHIRAKPAK® AD-H и Regis Technologies ULMO. Также возможны другие способы хирального разделения. Рацемические диацилгидразины для использования в способах хирального разделения могут быть получены с использованием методики, описанной в US 2005/0209283 Al и US 2006/0020146 A1.
Общая Схема 3
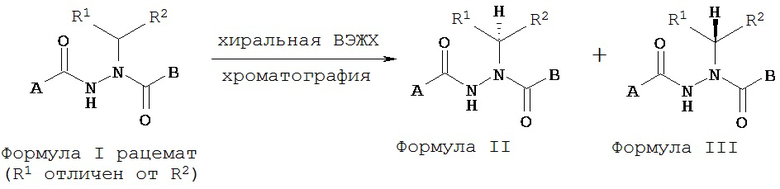
Энантиомерно обогащенные соединения Формулы II или III, где R1 представляет собой алкенил, затем могут быть обработаны с использованием стандартных химических трансформаций. Как описано в Общей Схеме 4 для энантиомерно обогащенного соединения Формулы III, алкенил можно превратить в алкил, гидроксиалкил, алкил, возможно замещенный циклоалкилом, алкил, возможно замещенный гетероциклом, алкил, замещенный алкокси, галогеноалкил и другие возможно замещенные алкильные группы. Специалисту в данной области техники понятен набор одно- или многостадийных химических превращений, которые можно использовать для превращения алкена в другие группы по изобретение.
Общая Схема 4
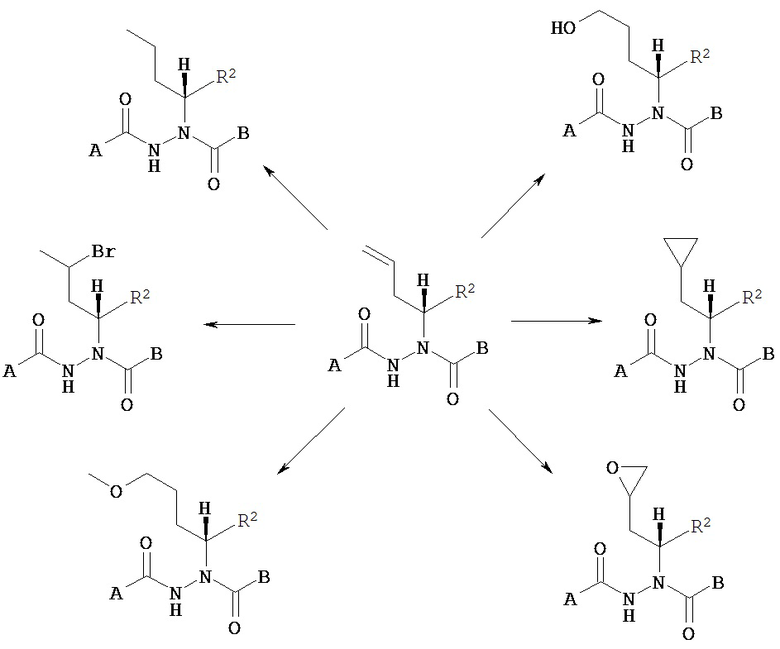
ПРИМЕРЫ
Настоящее изобретение может быть лучше понято со ссылкой на следующие общие синтетические способы, предложенные выше, и неограничивающие примеры, предложенные ниже, которые представлены как типичные примеры изобретения.
Пример 1
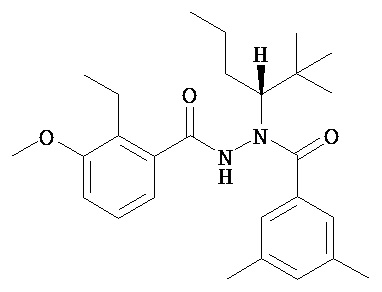
(R)-3,5-Диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2-этил-3-метоксибензоил)гидразид
СИНТЕЗ
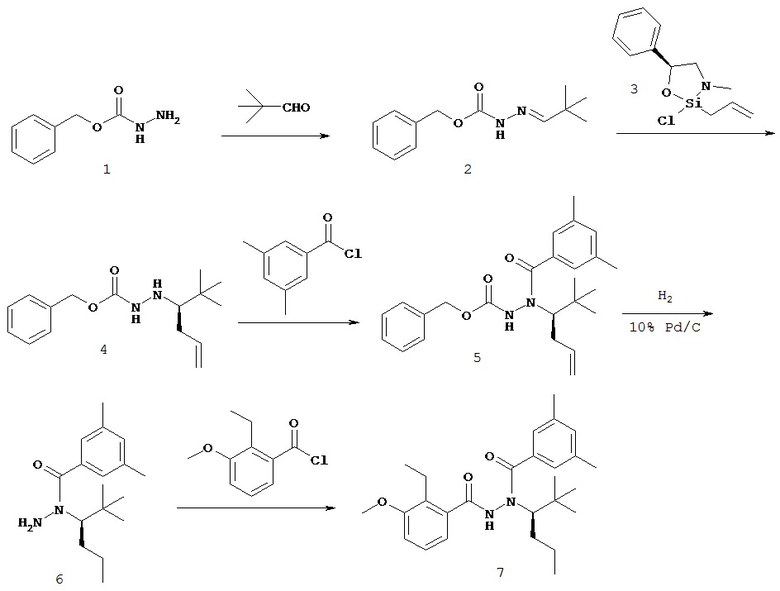
Соединение 1: Бензилкарбазат имеется в продаже.
Соединение 2: Бензилкарбазат (300 г, 1,81 молей) растворяли в 1,2 л метилового спирта в 3 л колбе с четырьмя горловинами, оснащенной обратным холодильником, термометром, магнитной мешалкой и 500 мл капельной воронкой. Ледяную уксусную кислоту (5 мл) добавляли к смеси, которую затем доводили до 45°C. Пивальдегид (248,8 г раствора, 2,17 молей, приблизительно 75% в mpem-бутаноле) добавляли порциями в течение 30 минут. Реакционную смесь нагревали с обратным холодильником в течение 30 минут, при контроле посредством ТСХ. Смесь оставляли охлаждаться почти до комнатной температуры и концентрировали в роторном испарителе до конечного объема приблизительно 700 мл. Добавляли приблизительно 7 г активированного угля приблизительно при 30°C. Смесь перемешивали в течение ночи и фильтровали через слой целита с размером пор "C" в стеклянной фриттированной воронке Бюхнера. Растворитель удаляли в вакууме с получением светло-желтого масла, которое наливали в чашку-кристаллизатор, и вызывали кристаллизацию манипулируя шпателем. Вещество гранулировали, и оставшийся растворитель оставляли испаряться на воздухе при комнатной температуре в течение нескольких суток, с получением 420,4 г неочищенного продукта. После перекристаллизации при комнатной температуре из первоначально кипящей смеси 300 мл этилацетата и 1 л гексанов получали 363,2 г бензилового эфира (E)-N’-(2,2-диметилпропилиден)гидразинкарбоновой кислоты в виде не совсем белого твердого вещества. Вторую порцию 43,1 г и идентичную точку плавления получали из смеси приблизительно 150 мл гексанов и 45 мл этилацетата. Общий выход: 96,1%. Rf=0,32 (2:1 гексан : этилацетат); (mp) точка плавления = 74-74,5°C; 1H ЯМР (400 МГц, CDCl3) δ 8,0 (1H, br s), 7.4-7.3 (5H, m), 7.1 (1H, s), 5.20 (2H, s), 1.1 (9H, s).
Соединение 3: (S,S)-2-Аллил-2-хлор-3,4-диметил-5-фенил-[1,3,2]оксазацилолидин получали, используя модифицированную методику из Leighton et al, J. Am. Chem. Soc. 125: 9596 (2003) и WO 03/074534. Все методику выполняли в безводной закрытой системе с минимизированными и кратковременными моментами воздействия со стороны атмосферы. В высушенную в сушильном шкафу 5 л колбу с 4 горловинами с подводом газа, верхним перемешиванием и капельной воронкой загружали сначала аллилтрихлорсилан (419,5 г, 2,39 моль), а затем 2 л безводного CH2Cl2. Раствор охлаждали в атмосфере аргона до 0°C в бане лед/соль. Триэтиламин (485 г, 4,79 молей) добавляли с помощью капельной воронки, поддерживая температуру 0°C. При этом значении реакционная смесь имела прозрачный янтарный цвет. S,S-псевдоэфедрин (359 г, 2,17 молей) добавляли порциями, используя капельную воронку для твердого вещества, в течение 2 ч, поддерживая внутреннюю температуру ниже 15°C. Капельную воронку промывали 200 мл CH2Cl2 в реакционную смесь. Постепенно образовались пастообразные полугранулированные твердые вещества (Et3NCl). Светло-коричневую вязкую суспензию перемешивали в течение приблизительно 2 ч на льду, затем ледяную баню убирали, позволяя реакционной смеси достичь комнатной температуре за несколько часов. Через приблизительно 12 ч большую часть CH2Cl2 удаляли путем перегонки, поддерживая при этом систему в атмосфере аргона. Добавляли 1 л безводного гексана, и растворитель снова отгоняли. Затем добавляли 1 л безводного пентана, и смесь перемешивали при комнатной температуре в течение приблизительно 1 ч в атмосфере аргона. Пастообразный твердый остаток, который ранее образовался в CH2Cl2, становился более гранулированным, сначала при добавлении гексана, и затем особенно после добавления пентана. Раствор имел светлый янтарно-коричневый цвет. По частям и используя слабое давление аргона, пентан и растворимый продукт перемещали из реакционной колбы через фильтр из стекловаты и трубку из PFA (перфторалкокси) во вторую сухую 1 л круглодонную колбу с 3 горловинами, оснащенную насадкой для перегонки и магнитной мешалкой. Попеременно с переносом раствора, пентан отгоняли из этой второй колбы, с получением концентрированного маслянистого продукта. Этот процесс повторяли, дважды промывая остаток Et3NCl, каждый раз 500 мл пентана, и переносили промывки во вторую колбу в атмосфере аргона. Продукт собирали при помощи перегонки фракций под вакуумом приблизительно 10 торр (1,333 кПа). Приблизительно 56,3 г (9,7%) собрали в головной фракции, которая вероятно содержала вплоть до приблизительно 3% аллилтрихлорсилана. Главную фракцию, очищенный (S,S)-2-аллил-2-хлор-3,4-диметил-5-фенил-[1,3,2]оксазасилолидин, получали в количестве 413,3 г (71%), bp=140-144°C, приблизительно 10 торр (1,33 КПа), в виде слабовязкой жидкости светло-желтой во время отбора, но которая становилась оранжевой через несколько суток при замораживании и плотной герметизации мембраной из парафильма. Вещество хранили при замораживании, не характеризовали, и использовали без дополнительной очистки в последующей реакции аллилирования.
Соединение 4: Колбу 5 л с 4 горловинами, оснащенную термометром и магнитной мешалкой, сушили в сушильном шкафу и поддерживали в атмосфере азота во время добавления 2 л безводного CH2Cl2, с последующим добавлением бензилового эфира N’-(2,2-диметил-пропилиден)-гидразинкарбоновой кислоты (220 г, 939 ммоля). Смесь охлаждали и перемешивали в большой, приблизительно 10 гал (37,85 л) ледяной-солевой бане при 2-3°C. (S,S)-2-аллил-2-хлор-3,4-диметил-5-фенил-[1,3,2]оксазацилолидин (363 г, 1,355 моля) добавляли в колбу в течение приблизительно 30 мин, используя канюлю и поддержку слабым давлением аргона. Изначально бледно-желтый раствор сделался прозрачный светло-оранжевым, в то время как температура сохранялась равной 2-3°C. Реакционной смеси оставляли нагреться до комнатной температуре самому по себе и наблюдали с помощью гашения небольшой аликвотой в CH3OH и анализирования посредством ТСХ (2:1 гексан : EtOAc). Через 36 часов после начала анализ ТСХ показал приблизительно 90%-ное завершение. Смесь охлаждали до 5°C, добавляли дополнительные 48 г (0,179 ммоль) оксазацилолидина, и смесь оставляли нагреваться до комнатной температуры. Через приблизительно 8 часов реакционную смесь опять охлаждали до 5°C и гасили предварительно охлажденным раствором 100 г K2CO3 в 100 мл деионизированной воды в течение приблизительно 1 ч. Во время гашения, экзотермический эффект повышал температуру до 17°C. Добавляли дополнительно приблизительно 100 мл деионизированной воды. Раствор становился прозрачным голубовато-зеленым. Ни разу во время взаимодействия или гашения не наблюдалось какого-либо видимого выделения газа. После 4 суток перемешивания при комнатной температуре, раствор оставался светло-желтого цвета, и через 5 суток органическая фаза загустела, но сохранила светло-желтый цвет. Смесь охлаждали до приблизительно 15°C (позднее стало очевидным, что это не требуется), добавляли приблизительно 1,25 л гексана, и гель частично разрушали и разбивали на мелкие части при помощи верхнего перемешивания. Надосадочную жидкость откачивали сифоном и фильтровали через стекловату, при этом поочередно добавляя приблизительно 1 л порции к белому гелю, остающемуся в колбе. Дополнительные гексаны также добавляли к надосадочной жидкости для осаждения псевдоэфедрина в виде белого твердого вещества и для разрушения какого-либо оставшегося геля. Гексановые экстракты объединяли и растворитель удаляли в вакууме. В целом, для экстракций геля, вызванного гексаном осаждения псевдоэфедрина, и фильтраций через стекловату и стеклянную фриттированную воронку Бюхнера, использовали приблизительно 5-6 л гексана для получения неочищенного аллилгидразида в виде желтого масла. Партии данного неочищенного вещества обрабатывали приблизительно тремя объемами гексана, оставлять стоять при комнатной температуре, декантировали с осажденного псевдоэфедрина, и переносили в делительную воронку. Раствор продукта отделяли от остаточного псевдоэфедрина, промывая три раза, приблизительно 25 мл 1,4 н. HCl каждый раз (большую часть окраски также удалили), с последующим промыванием 10%-ным K2CO3 и рассолом. Гексановый раствор неочищенного продукта сушили над твердым безводным Na2SO4. и растворитель удаляли в вакууме с получением светло-желтого вязкого масла. ТСХ (2:1 гексан : EtOAc) показала целевой продукт Rf=0,4; две примеси при 0,25 и 0,31 (5-10% каждой, одна из них, возможно, исходное вещество) и немного менее заметных примесей выше Rf=0,4. Псевдоэфедрин, ранее определенный на исходном уровне, теперь отсутствовал. В итоге, после удаления следов растворителя вакуумным насосом, выделили 196,8 г (75,8%) бензилового эфира (R)-N’-(1-трет-бутил-бут-3-инил)-гидразинкарбоновой кислоты в виде янтарного масла. Первоначальный гель, теперь отчасти уплотненный и гранулированный, экстрагировали приблизительно 2 л кипящих гексанов. Твердые вещества отфильтровали, гексаны отделяли, и масло декантировали с осажденного псевдоэфедрина, с получением приблизительно 18 г.Это вещество объединяли с остатками порции первичного продукта выше (приблизительно 3-4 г) и промывали 1,4 н. HCl, водным K2CO3 и рассолом, как ранее. Гексаны отделяли с выходом 20,6 г дополнительного продукта в виде янтарного масла, которое согласно ТСХ оказалось идентичным первой порции. Порцию 10,1 г бензилового эфира (R)-N’-(1-трет-бутил-бут-3-инил)-гидразинкарбоновой кислоты количественно хроматографировали на силикагеле с выходом 8,88 г (88%-ный выход для хроматографии) чистого продукта в виде прозрачного, не совсем белого масла. Rf=0,43 (2:1 гексан : этилацетат); 1H ЯМР (400 МГц, CDCl3) δ 7,4 (5H, br s), 6.2 (1H, br s), 6.0 (1H, br s), 5.15 (2H, s), 5.1 (2H, s), 4.1 (1H, br), 2.7 (1H, m), 2.4 (1H, br d), 2.0 (1H, m), 0.95 (9H, s); э.и. = 98.1%, AD-H column; [a]25589-42.5° (с 2.18, CH3OH).
Соединение 5: Бензиловый эфир (R)-N’-(1-трет-бутил-бут-3-инил)-гидразинкарбоновой кислоты (110 г, 398 ммоль) и 500 мл метиленхлорида добавляли в 2 л круглодонную колбу с термометром и магнитной мешалкой. Потом добавляли раствор K2CO3 (82,51 г, 597 ммоль в 200 мл деионизированной воды), и колбу охлаждали приблизительно до 10°C. Затем медленно добавляли чистый 3,5-диметилбензоилхлорид (73,82 г, 437,8 ммоль) в течение 20 мин. Для смывания остаточного хлорангидрида в колбу и для разбавления реакционной смеси использовали 1 л метиленхлорида. Во время добавления температуре не позволяли повышаться выше 15°C. Реакционную смесь перемешивали в течение ночи, сначала в ледяной бане и затем при комнатной температуре. ТСХ-Анализ приблизительно через 16 часов показал, что взаимодействие с оставшимся хлорангидридом закончено; смесь перемешивали в сумме в течение приблизительно 40 часов. Реакционную смесь наливали в 2 л делительную воронку. Органический слой собирали и объединяли с небольшой CH2Cl2 промывкой водного слоя. Органическую фазу промывали еще раз 10%-ным K2CO3, промывали рассолом, и затем сушили над твердым MgSO4/Na2SO4. Смесь отфильтровали от солей, и растворитель удаляли в вакууме с получением гранулированного светло-бежевого твердого вещества. Неочищенный продукт суспендировали и перемешивали в 300 мл смеси 2:1 гексан : эфир и фильтровали на воронке Бюхнера для удаления по существу остаточного 3,5-диметилбензоилхлорида. Собранные твердые вещества промывали еще четыре раза в сумме приблизительно 1200 мл смеси 2:1 гексан:эфир, получая белое гранулированное твердое вещество. Продукт собирали и оставляли сушиться на воздухе с получением 144,55 г бензилового эфира (R)-N’-(1-трет-бутил-бут-3-инил)-N’-(3,5-диметил-бензоил)-гидразинкарбоновой кислоты, выход = 88,9%. 30 г образца растворяли в 205 мл кипящего метанола и оставляли кристаллизоваться при комнатной температуре. Полученное флокулирующее вещество промывали на воронке Бюхнера 50 мл CH3OH с получением 22,2 г, Т.пл.=146°C, (э.и. >99,9%). Аналитический образец бензилового эфира (R)-N’-(1-трет-бутил-бут-3-инил)-N’-(3,5-диметилбензоил)гидразинкарбоновой кислоты получали путем двойной перекристаллизации данного вещества из кипящего CH3OH с последующей молекулярной перегонки в аппарате Кугельрора (Kugelrohr) и сбором в виде твердого вещества с получением чистого белого вещества: Т.пл.=145,5-147°C. Rf=0,12 (10:1 гексан : этилацетат); 1H ЯМР (400 МГц, CDCl3): δ 7.5-7.2 (m, 9H, ϕ+NH), 7.2-6.8 (m), 6.4 (S), 6.3 (S), 5.95 (br, 5H, бензиловый + C=C), 5.4-4.6 (m, 9 H, аллиловый N-CH, ϕ-CH3), 3.65 (br), 2.5 (m), 2.35 (S), 2.3 (S), 1.2-0.8 (m, 9H, -C(CH3)3); [а]25589-12,8° (с 2,01, CHCl3).
Соединение 6: Бензиловый эфир (R)-N’-(1-трет-бутил-бут-3-инил)-N’-(3,5-диметил-бензоил)-гидразинкарбоновой кислоты (13,5 г, 33 ммоль) суспендировали в 100 мл ледяной уксусной кислоты при комнатной температуре. Добавляли 10%-ный палладий на угле (0,24 г) в виде суспензии в 4,4 г уксусной кислоты. Серую суспензию встряхивали в течение 90 минут при давлении 10-30 фунтов на квадратный дюйм (69-207 КПа) в автоклаве для гидрирования Парра, наблюдая за реакцией при помощи ТСХ. Катализатору позволяли осесть, и реакционный раствор удаляли пипеткой и пропускали через бумажный складчатый фильтр (Schleicher & Schuell 597 1/2, 125 мм диаметром). Остаток катализатора промывали уксусной кислотой и промывки пропускали через фильтровальную бумагу с получением уксусной кислоты и продукта с объединенной общей массой 156 г. Смесь охлаждали на льду и добавляли приблизительно 600 мл деионизированной воды. За 30 минут продукт освобождали от масла и затем кристаллизовали. Его отфильтровывали через бумагу, сушили на воздухе, и собирали на бумаге с получением 7,9 г светло-желтого твердого вещества. Через несколько часов 0,52 г дополнительного продукта осаждалось из фильтрата с получением общего выхода 8,42 г (92,2%) N-(1-трет-бутил-бутил)-гидразида (R)-3,5-диметил-бензойной кислоты; Rf=0,5 (1:1 гексан : этилацетат); [a]25589+7,63 (с 1,98, CH3OH), 1H ЯМР (400 МГц, CDCl3) δ 7,05 (1H, s), 7.02 (2H, s), 4.6+3.5 (1H, 2d), 4.1 (2H, s, NH2), 2.38+2.37 (6H, 2s). 1.1-2.1 (4H, m), 1.0+0.9 (9H, 2s), 1.0+0.98 (3H, 2t), 2 разных конформационных изомера.
Соединение 7 (указанное в заголовке соединение): (R)-3,5-диметил-бензойной кислоты N-(1-трет-бутил-бутил)гидразид (35 г, 126,6 ммоль) растворяли в 100 мл метиленхлорида в 500 мл круглодонной колбе с магнитной мешалкой. Добавляли водный K2CO3 (26,25 г, 189,9 ммоль, раствор в 75 мл деионизированной воды), и смесь перемешивали на льду. Добавляли твердый 2-этил-3-метокси-бензоилхлорид (27,67 г, 139,3 ммоль) и смывали в колбу, используя 25 мл CH2Cl2. Смесь перемешивали на льду в течение приблизительно 40 ч, позволяя бане нагреться до комнатной температуры. Добавляли дополнительно метиленхлорид и воду по мере необходимости для содействия обработке. Органический слой отделяли и сушили над MgSO4. Добавляли приблизительно 5 г активированного древесного угля, и соль и уголь удаляли фильтрацией через бумагу. Растворитель выпаривали в вакууме, сначала на роторном испарителе и затем в глубоком вакууме, получая 57,8 г неочищенного продукта, содержащего следы CH2Cl2. Неочищенное вещество растирали сначала с 3×100 мл порциями эфира (2% этанол) и затем с 1×100 мл порцией смеси 1:1 гексан : эфир во фриттированной стеклянной воронке Бюхнера (размер пор ASTM 40-60 C, Pyrex). Затем продукт промывали 2×200 мл деионизированной водой с тщательным перемешиванием и оставляли сушиться на воздухе. (R)-3,5-Диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2-этил-3-метокси-бензоил)гидразид (также называемый (R)-2-этил-3-метокси-бензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметил-бензоил)-гидразид) собирали в виде блестящего белого порошкообразного твердого вещества (36,51 г, выход 65,7%, довольно чистый и очень высокий э.и. согласно ВЭЖХ и хиральной ВЭЖХ, одно пятно согласно ТСХ, 1:1 гексан : эфир). После упаривания объединенных органических промывок получали приблизительно 20 г вещества, которое растирали сначала с 200 мл смеси 2:1 гексан : эфир и затем с 2×100 мл смеси 2:1 гексан : эфир в фриттированной стеклянной колбе Бюхнера, размер пор ASTM 40-60°C. Вещество промывали 3×100 мл деионизированной воды с тщательным перемешиванием, и оставляли сушиться на воздухе. Вторую порцию собирали в виде белого порошка (12,98 г, выход 23,4%, довольно чистый и высокий э.и. согласно ВЭЖХ и хиральной ВЭЖХ). Третью порцию получали аналогичным образом в виде не совсем белого твердого вещества (1,52 г, 2,7%, чистота 98%, э.и. >99%). Конечный образец получали путем растворения в теплом метаноле и фильтрации через шприцевой фильтр 0,45 микрон в большую чашку-кристаллизатор. Вещество собирали, дробили шпателем, и нагревали при 50-58°C в вакуумном сушильном шкафу в сущности вплоть до постоянной массы. Анализ ВЭЖХ показал химическую чистоту 99,3% и э.и. >99,9% Rf=0,28 (1:1 гексан : этилацетат); Т.пл.=162,2-162,8°C; [a]25589+12.9 (с 2.04, CH3OH); 1H ЯМР (400 МГц, CD3OD) δ 7.0-7.1 (4H, m), 6.9 (1H, d), 6.1-6.4 (1H, 2d), 4.5-4.7 (1H, 2d), 3.78 (3H, s), 2.3 (6H, s), 2.3 (1H, m), 1.9 (2H, m), 1.55 (3H, m), 1.1-1.15 (9H, 2s), 0.9-1.0 (6H, m).
Примеры 2-29 получали, используя методику, описанную в Примере 1. Процент энантиомерного избытка (э.и.) определяли с помощью хиральной ВЭЖХ. В Таблице 2 представлены аналитические данные для примеров 2-29.
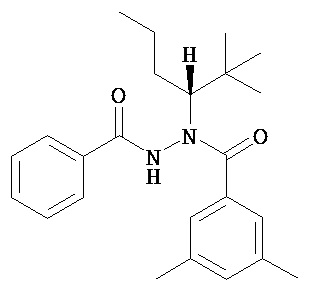
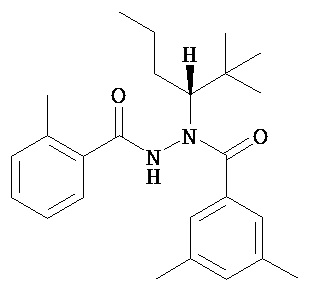
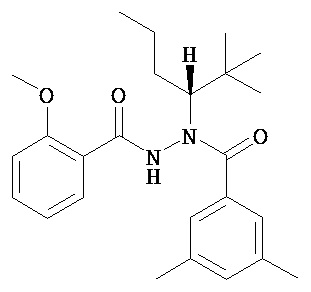
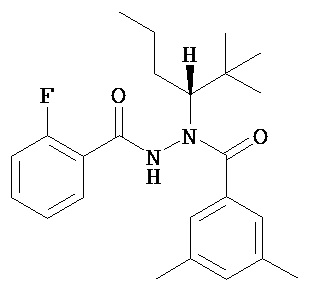
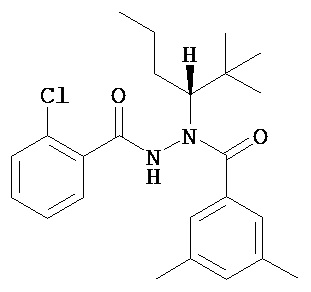
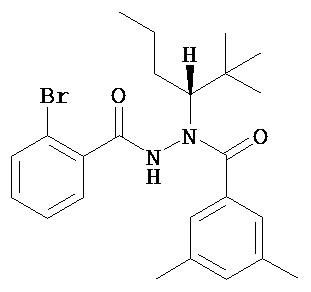
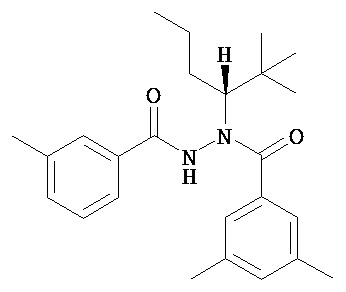
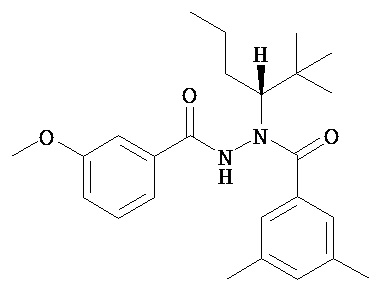
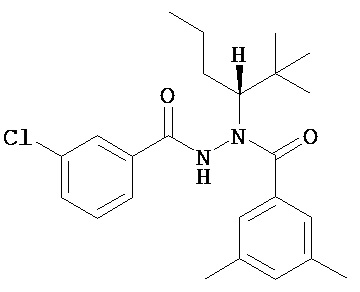
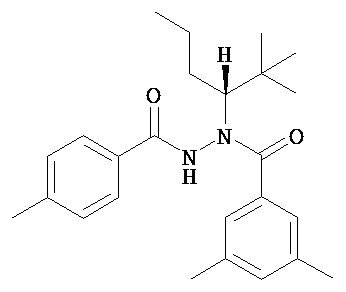
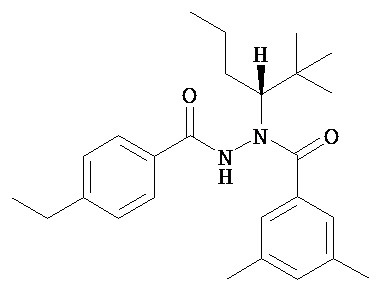
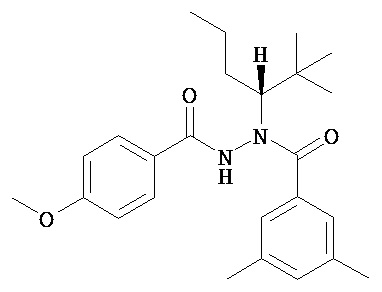
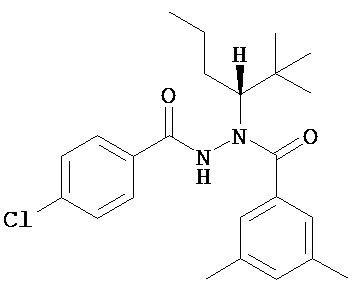
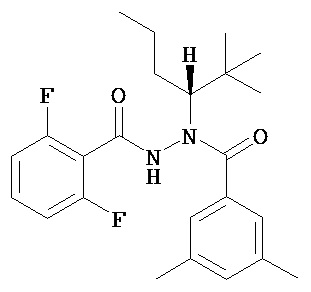
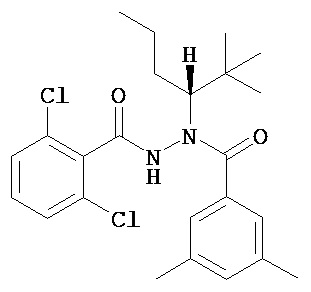
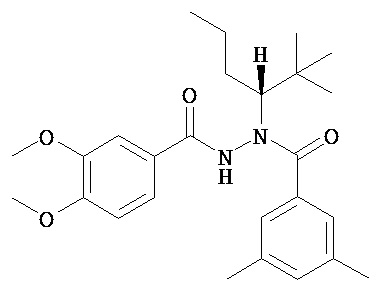
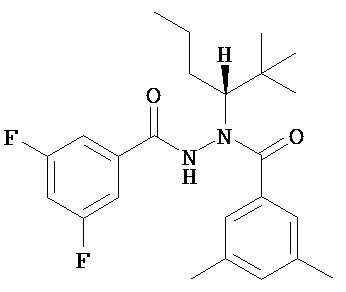
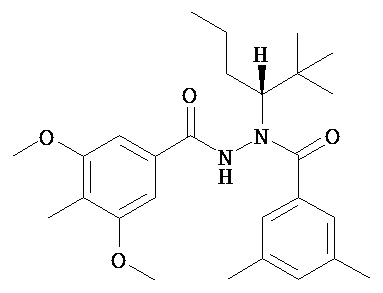
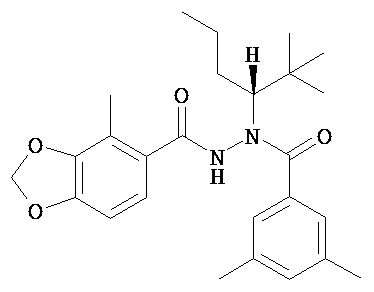
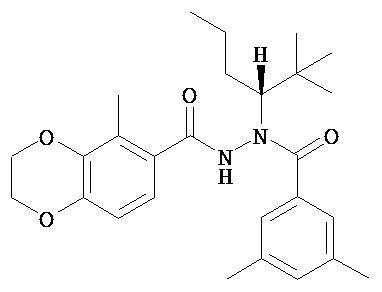
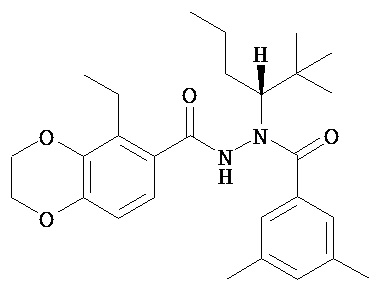
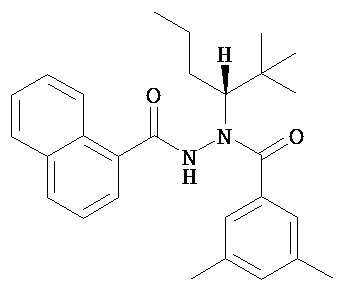
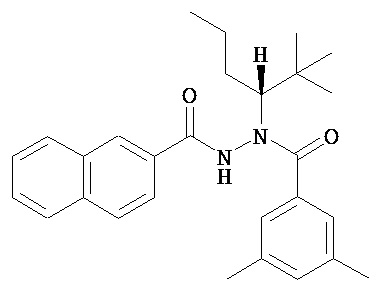
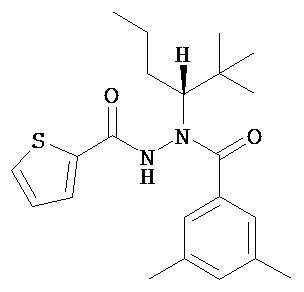
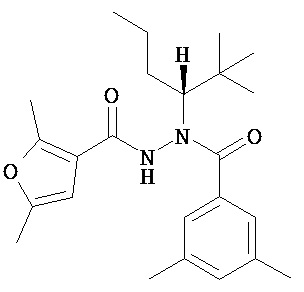
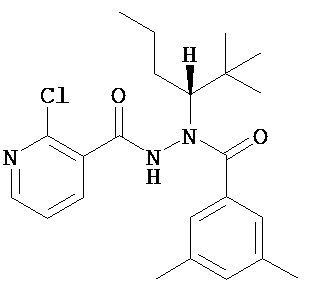

Пример 30
(R)-3,5-Диметил-бензойной кислоты N-(1-трет-бутил-бутил)-N’-(S)-[2-(6-метокси-нафталин-2-ил)-пропионил]-гидразид
СИНТЕЗ
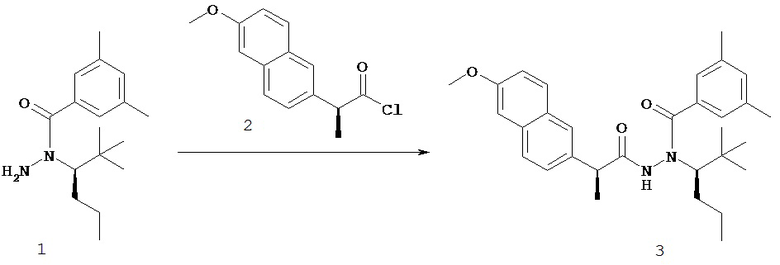
Соединение 1: (R)-3,5-диметил-бензойной кислоты N-(1-трет-бутил-бутил)-гидразид получали, как описано в Примере 1.
Соединение 2: Соединение 2 получали из соответствующей карбоновой кислоты. Тионилхлорид (1,24 г, 10,42 ммоль) добавляли к раствору (S)-(+)-2-(6-метоксинафталин-2-ил)-пропионовой кислоты (напроксен, 2 г, 8,69 ммоль) в 8,2 мл хлороформа в круглодонной колбе, оснащенной магнитной мешалкой. Добавляли каплю DMF и смесь нагревали в колбе с обратным холодильником в течение 3 часов. Хлороформ и избыток тионилхлорида отгоняли из реакционной смеси, куда добавляли метиленхлорид. В результате упаривания остаточного растворителя получали (S)-2-(6-метоксинафталин-2-ил)пропионилхлорид (1,918 г, выход 88,8%). Rf=0,2 (1:1 гексан : этиловый эфир). 1H ЯМР (400 МГц, CDCl3) δ 9,98 (br, 1H), 7.78 (t, 2H), 7.36 (d, 1H), 7.18 (d, 1H), 7.14 (m, 2H), 4.25 (q, 1H), 3.93 (s, 3H), 1.68 (d, 3H).
Соединение З (указанное в заголовке соединение): (R)-3,5-Диметилбензойной кислоты N-(1-трет-бутил-бутил)-гидразид (э.и. по оценкам >99% на основе предшественника, 1,1 г, 3,98 ммоль) растворяли в 10 мл метиленхлорида. Водный раствор K2CO3 (0,907 г, 6,57 ммоль в 2,5 мл) добавляли к реакционной смеси. Добавляли (S)-2-(6-метоксинафталин-2-ил)-пропионилхлорид (1,09 г, 4,38 ммоль), и реакционную смесь перемешивали в течение 24 часов и наблюдали посредством ТСХ. Дополнительно добавляли метиленхлорид и воду по необходимости для ускорения манипуляций. Органический слой отделяли, сушили над Na2SO4, фильтровали, и растворитель удаляли в вакууме с получением неочищенного продукта: Т.пл.=98°C (стекло) - 123°C. Неочищенное вещество растирали три раза с помощью смеси 2:1 гексан : эфир с получением 1,554 г, выход 80,1% (R)-3,5-диметилбензойной кислоты N-(трет-бутил-бутил)-N’-(S)-[2-(6-метоксинафталин-2-ил)пропионил]гидразид, Т.пл.=98°C (стекло) - 122°C. Растертое вещество кристаллизовали из изопропанола при 35°C с получением кристаллического продукта. Спектры 1H ЯМР из всех трех партий были идентичными. Rf=0,15 (1:1 гексан : этиловый эфир); Т.пл.=158-160°C (нагревание начинали при 150°C); Т.пл.=156-164°C (нагревание начинали при 25°C); 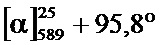
С целью определения стереохимического течения реакции асимметрического аллилирования в Примере 1, определяли рентгеновскую кристаллическую структуру (R)-3,5-диметилбензойной кислоты (N-(1-трет-бутил-бутил)-N’-(S)-2-(6-метоксинафталин-2-ил)пропионил]гидразида. В данном эксперименте установлено, что абсолютная конфигурация атома углерода, несущего трет-бутильную группу и н-пропильную группу, представляет собой R-конфигурацию.
Пример 31
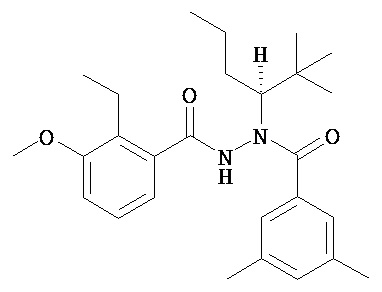
(S)-3,5-Диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2-этил-3-метоксибензоил)гидразид
СИНТЕЗ
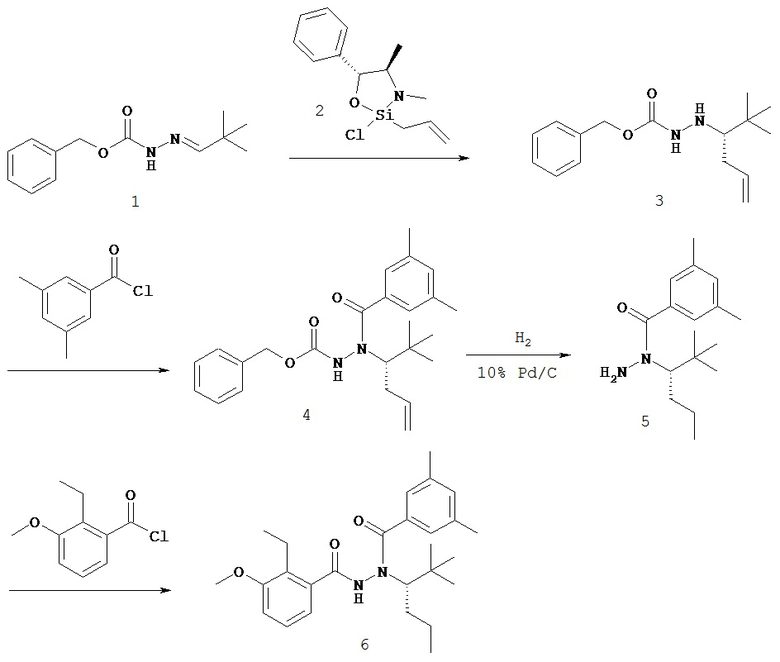
Соединение 1: бензиловый эфир (E)-N’-(2,2-диметилпропилиден)-гидразинкарбоновой кислоты получали, как описано в Примере 1.
Соединение 2: (R,R)-2-аллил-2-хлор-3,4-диметил-5-фенил-[1,3,2]оксазацилолидин получали, используя методику, описанную в Примере 1, используя R,R-псевдоэфедрин.
Соединение 3: бензиловый эфир (S)-N’-(1-трет-бутил-бут-3-инил)гидразинкарбоновой кислоты получали, используя методику, описанную в Примере 1. Аналитические данные: Rf=0,46 (2:1 гексан : этилацетат); Т.пл.=69-71°C; 1H ЯМР (400 МГц, CDCl3) δ 7,4 (br s, 5H), 6.2 (br s, 1H), 6.0 (br s, 1H), 5.15 (s, 2H), 5.1 (s, 2H),4.1 (br, 1H), 2.7 (m, 1H), 2.4 (br d, 1H), 2.0 (m, 1H), 0.95 (s, 9H); э.и.=98,2%, колонка ADH; 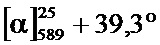
Соединение 4: бензиловый эфир (S)-N’-(1-трет-бутил-бит-3-инил)-N’-(3,5-диметилбензоил)гидразинкарбоновой кислоты получали, используя методику, описанную в Примере 1. Аналитические данные: Rf=0,43 (2:1 гексан : этилацетат); Т.пл.=150-151,5°C; 1H ЯМР (400 МГц, CDCl3) δ 7,45-6,88 (m, 7H), 6.44+6.28 (2s, 1H), 5.96 (br, 1H), 5,38-4,68 (m, 5H), 3.69 (br, 1H), 2.68-2.43 (m, 2H), 2.36+2.29 (2s, 6H), 1.13+1.02+0.94 (3s, 9H); 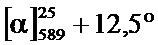
Соединение 5: (S)-3,5-Диметилбензойной кислоты N-(1-трет-бутил-бутил)гидразид получали, используя методику, описанную в Примере 1. Аналитические данные: Rf=0,5 (1:1 гексан : этилацетат); Т.пл.=112-112,8°C; 1H ЯМР (400 МГц, CDCl3) δ 7.05 (s, 1H), 7.02 (s, 2H), 4.6+3.5 (2d, 1H), 2.38+2.37 (2s, 6H), 1.2-2.1 (m, 4H), 1.1+1.0 (2s, 9H), 1.05+0.98 (2t, 3H) 2 различных изомера; 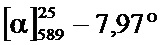
Соединение 6 (указанное в заголовке соединение): (S)-3,5-Диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2-этил-3-метокси-бензоил)-гидразид получали, используя методику, описанную в Примере 1. Аналитические данные: Rf=0,28 (1:1 гексан : этилацетат); Т.пл.=156,5-157,5°C; 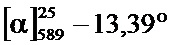
Примеры 32-47 получали, используя методику, описанную в Примере 31. Процент энантиомерного избытка (э.и.) определяли с помощью хиральной ВЭЖХ.
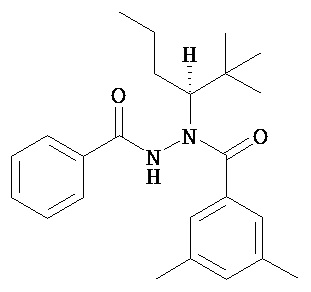
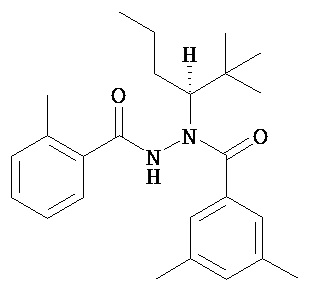
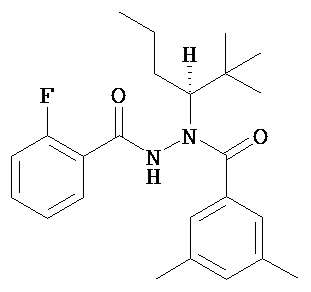
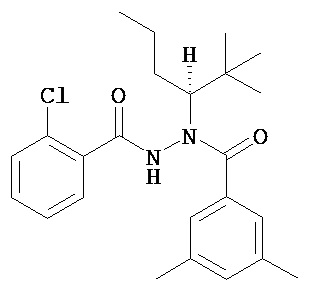
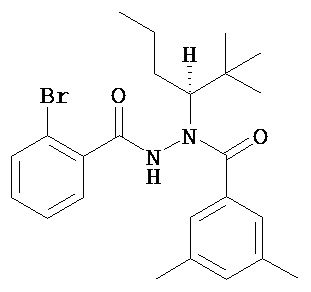
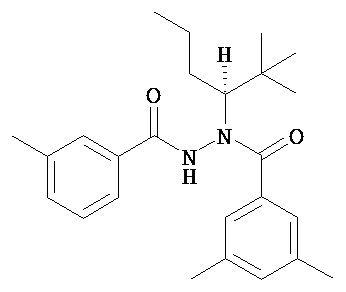
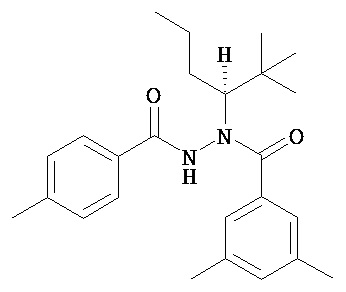
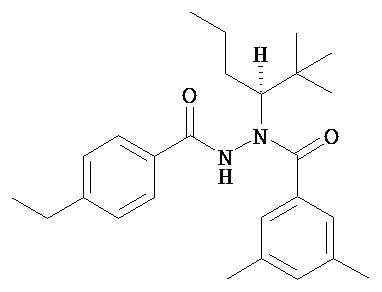
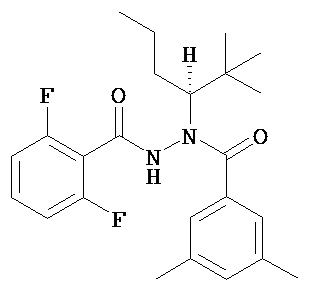
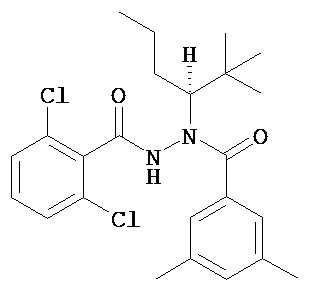
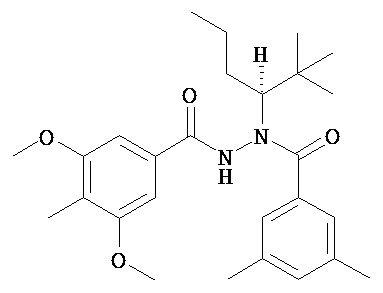
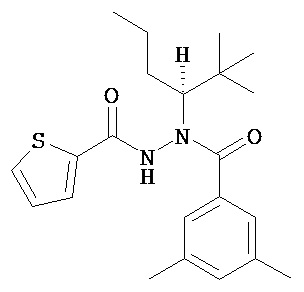
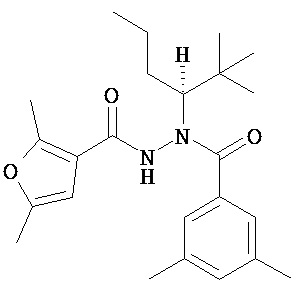
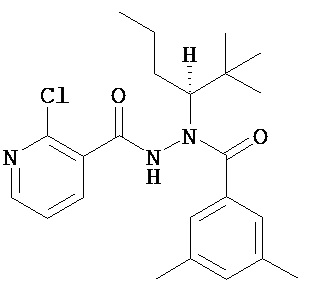

Пример 48
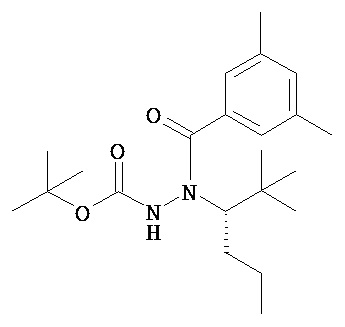
Трет-бутиловый эфир (S)-N’-(1-трет-бутил-бутил)-N’-(3,5-диметил-бензоил)гидразинкарбоновой кислоты
СИНТЕЗ
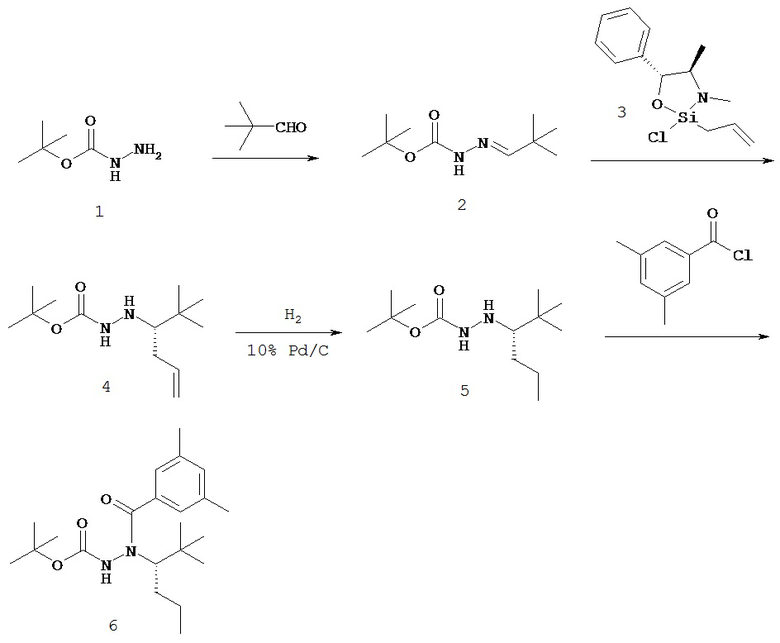
Соединение 1: трет-бутилкарбазат имеется в продаже.
Соединение 2: Пивальдегид (52,1 г, 454 ммоль, приблизительно 75%-ный раствор в трет-бутаноле) растворяли в 120 мл метанола в 1 л, круглодонной колбе с 3 горловинами и с обратным холодильником, термометром и капельной воронкой. Добавляли ледяную уксусную кислоту (1 мл) с последующим регулированным добавлением трет-бутилкарбазата (50 г, 378 ммоль), без отвода теплоты от реакционной смеси. Смесь перемешивали в течение 4 часов, наблюдая посредством ТСХ. По окончании взаимодействия, растворитель удаляли в вакууме, остаток переносили в пентан, и пентан выпаривали. Полученное масло кристаллизовалось при стоянии с получением mpem-бутилового эфира N’-(2,2-диметилпропилиден)гидразинкарбоновой кислоты (75,7 г, 93%). Rf=0,42 (2:1 гексан : этилацетат, 1% уксусная кислота); 1H ЯМР (400 МГц, CDCl3) δ 7.6 (br s, 1H), 7.1 (s, 1H), 1.5 (s, 9H), 1.1 (s, 9H) млн-1.
Соединение 3: (R,R)-2-Аллил-2-хлор-3,4-диметил-5-фенил-[1,3,2]оксазацилолидин получали, используя методику, описанную в Примере 1, с использованием R,R-псевдоэфедрина.
Соединение 4: трет-Бутиловый эфир N’-(2,2-диметил-пропилиден)-гидразинкарбоновой кислоты (12 г, 59,9 ммоль) загружали в высушенную 3-хгорлую 1 л круглодонную колбу с термометром, магнитной мешалкой и атмосферой азота. Используя катетер, в колбу загружали безводный хлористый метилен (200 мл). Смесь охлаждали до 0°C с помощью ледяной бани. Затем в колбу добавляли (R,R)-2-аллил-2-хлор-3,4-диметил-5-фенил-[1,3,2]оксазацилолидин (24,07 г, 89,87 ммоль) в нейтральных условиях. За считанные минуты, изначально светло-желтый раствор превратился в прозрачный темно-желтый. Реакционную смесь перемешивали в течение 6 ч при 0°C, затем оставляли нагреваться до комнатной температуры и перемешивали в течение ночи, наблюдая с помощью ТСХ. Реакционную смесь гасили путем добавления приблизительно 25 мл метанола, что приводило к осветлению цвета. Раствор концентрировали и остаток разбавляли 100 мл этилацетата и 100 мл воды. Фазы разделяли, и водный слой дважды экстрагировали этилацетатом. Органические фазы объединяли, промывали рассолом, сушили над Na2SO4 и концентрировали. Полученное масло очищали при помощи колоночной хроматографии, используя 10%-ный этилацетат в гексанах. Очищенный трет-бутиловый эфир (S)-N’-(1-трет-бутил-бут-3-инил)-гидразинкарбоновой кислоты получали в виде масла (2,87 г, выход 24,4%). Содержащую примеси фракцию также собирали (3,17 г). Rf=0,44 (5:1 гексан : этилацетат, I2-визуализация); 1H ЯМР (400 МГц, CDCl3) δ 6.07 (br, 1H, NH), 5.85 (m, 1H), 5.0 (dd, 2H), 2.6 (m, 1H), 2.33 (m, 1H), 2.3 (m, 1H), 1.4 (s, 9H), 0.9 (s, 9H).
Соединение 5: трет-Бутиловый эфир (S)-N’-(1-трет-бутил-бут-3-инил)-гидразинкарбоновой кислоты (2,34 г, 9,65 ммоль) растворяли в метаноле (50 мл) в сосуде Парра. Добавляли 10%-ный Pd/C (70 мг) в виде суспензии в воде (1,6 мл), и смесь встряхивали в гидрогенизаторе Парра в течение 4,5 ч с начальным давлением 55 фунтов на квадратный дюйм (366,8×103 Па). Снижение давления отслеживали в качестве оценки развития реакции, и оно показывало, что взаимодействие через 30 минут вероятно завершилось. Катализатору позволяли по существу осесть, и надосадочную жидкость удаляли с помощью пипетки, пропускали через шприцевой фильтр 0,45 микрон, и анализировали с помощью ТСХ. Растворитель удаляли в вакууме с выходом 2,19 г (92,7%) трет-бутилового эфира (S)-N’-(1-трет-бутил-бутил)-гидразинкарбоновой кислоты в виде бледно-палево-желтого масла. Rf=0,48 (5:1 гексан : этилацетат, I2-визуализация), 1H ЯМР (400 МГц, CDCl3) δ 6.0 (br, 1H), 4.0 (br, 1H), 2.5 (d, 1H), 1.5 (s, 9H), 1.1 (m, 2H), 1.6 (br, 2H), 0.94 (s, 9H), 0.94 (t, 3H).
Соединение 6 (указанное в заголовке соединение): mpem-бутиловый эфир (S)-N’-(1-трет-бутил-бутил)-гидразинкарбоновой кислоты (1 г, 4,09 ммоль) растворяли приблизительно в 3 мл метиленхлорида во флаконе при помощи магнитной мешалки. Добавляли раствор 0,8 г K2CO3 (5,79 ммоль) приблизительно в 2 мл воде, затем 0,81 г (4,80 ммоль) 3,5-диметилбензоилхлорида. Реакционную смесь перемешивали в течение ночи, сначала на льду, и затем оставляли нагреваться до комнатной температуры. Затем добавляли воду и метиленхлорид, каждого по несколько мл, для растворения всех твердых веществ. Водный слой удаляли, и органический слой сушили над твердым MgSO4. TCX показала, что реакция закончилась приблизительно на 95%. Смесь фильтровали через небольшое количество стекловаты во флакон. Растворитель выпаривали в струе N2, вводя смесь 3:1 гексан : эфир и пентан. Остаток обрабатывали с помощью шпателя, чтобы инициировать кристаллизацию, и после полного затвердевания, растирали трижды с 2-3 мл пентана в небольшой фриттированной стеклянной воронке. После сушки на воздухе получали 300 мг (19,5%) трет-бутилового эфира (S)-N’-(1-трет-бутил-бутил)-N’-(3,5-диметил-бензоил)-гидразинкарбоновой кислоты, достаточно чистой по показаниям TCX, хиральная ВЭЖХ показала э.и.=93,3%. Продукт сушили в вакуумном сушильном шкафу при 50°C для определения точки плавления. Rf=0,42 (4:1 гексан : этилацетат, UV (сильная), I2 (слабая) визуализация); Т.пл.=115,5-117,5°C; 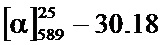
Пример 49
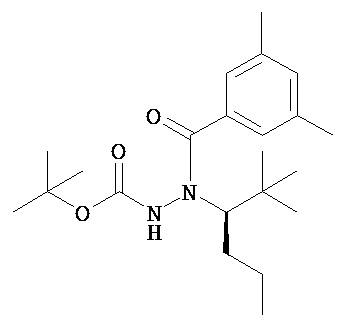
Трет-бутиловый эфир (R)-N’-(1-трет-бутил-бутил)-N’-(3,5-диметил-бензоил)гидразинкарбоновой кислоты
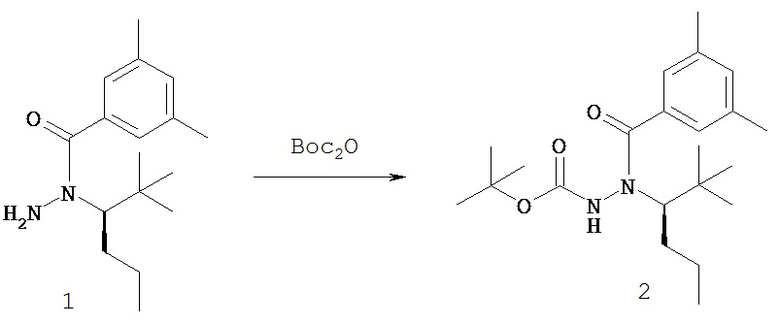
Соединение 1: (R)-N’-3,5-диметил-бензойной кислоты N-(1-трет-бутил-бут-3-инил)гидразид получали, как описано в Примере 1.
Соединение 2 (указанное в заголовке соединение): К раствору (R)-3,5-диметил-бензойной кислоты N-(1-трет-бутил-бутил)-гидразида (400 мг, 1,44 ммоль) в THF (10 мл) добавляли t-boc(трет-бутоксикарбонил)-ангидрид (629 мг, 2,88 ммоль). Реакционную смесь нагревали с обратным холодильником в течение 40 ч, охлаждали, разбавляли эфиром (20 мл), промывали H2O и сушили над безводным Na2SO4. Растворитель выпаривали в вакууме, и остаток очищали с помощью флэш-хроматографии и тонкослойной хроматографии с получением трет-бутилового эфира (R)-N’-(1-трет-бутил-бутил)-N’-(3,5-диметил-бензоил)-гидразинкарбоновой кислоты (45,7 мг, выход 4,1%) в виде белого твердого вещества. Rf=0,39 (1:3 EtOAc: н-гексан); 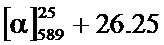
Пример 50
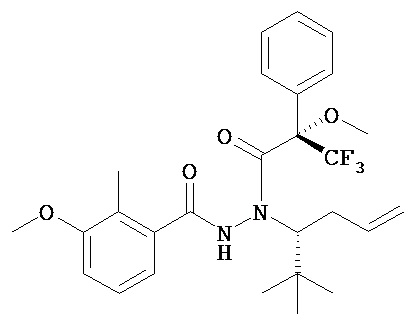
(R)-3-Метокси-2-метилбензойной кислоты N’-(1-трет-бутил-бут-3-инил)-N’-((S)-3,3,3-трифтор-2-метокси-2-фенилпропионил)гидразид
СИНТЕЗ
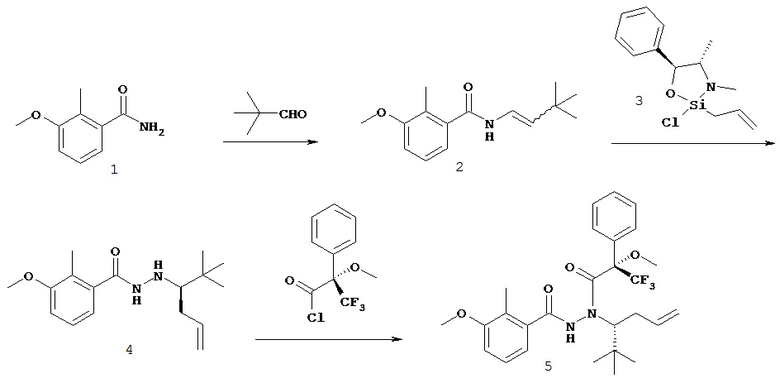
Соединение 1: гидразид 3-метокси-2-метилбензойной кислоты получали, используя методику, описанную в US 2005/0209283 A1.
Соединение 2: Пивальдегид (13,45 г, 156 ммоль) растворяли в 300 мл метанола в круглодонной колбе. Добавляли гидразид 3-метокси-2-метилбензойной кислоты (26,6 г, 147,6 ммоль) и 150 капель ледяной уксусной кислоты, и смесь нагревали с обратным холодильником в течение приблизительно 8 часов, наблюдая с помощью ТСХ. По окончании реакции растворитель удаляли в вакууме, и продукт суспендировали в холодных гексанах и фильтровали на воронке Бюхнера с получением 24,4 г (66%) (2,2-диметилпропилиден)гидразида 3-метокси-2-метилбензойной кислоты. Rf=0,19, (2:1 гексан : этилацетат); 1H ЯМР (400 МГц, DMSO-d6) δ 7.42+7.41 (3s, 1H), 7.1 (t, 1H), 6.9 (d, 1H), 6.82 (d, 1H), 3.78 (s, 3H), 2.2+2.1 (2s, 3H), 1.07+1.0 (3s, 9H), многочисленные конформации, возможно смесь E/Z.
Соединение 3: (S,S)-2-Аллил-2-хлор-3,4-диметил-5-фенил-[1,3,2]оксазацилолидин получали, как описано в Примере 1.
Соединение 4: 3-метокси-2-метилбензойной кислоты (2,2-диметил-пропилиден)гидразид (10,84 г, 43,31 ммоль) загружали в высушенную 1 л круглодонную колбу, находящуюся в атмосфере азота. Добавляли 350 мл безводного метиленхлорида через катетер. Колбу охлаждали примерно до 5°C в ледяной бане. Добавляли (S,S)-аллил-2-хлор-3,4-диметил-5-фенил-[1,3,2]оксазацилолидин (17,4 г, 65 ммоль) с помощью шприца. За считанные минуты изначально светлая смесь превратилась в темно-желтую прозрачную, и ее оставляли перемешиваться в атмосфере азота приблизительно при 5°C. Через 4 ч ТСХ показала окончание реакции. Реакционную смесь гасили, добавляя приблизительно 25 мл метанола при осторожном перемешивании. Темный цвет рассеивался. Раствор концентрировали, и остаток разбавляли этилацетатом (100 мл) и водой (100 мл). Фазы разделяли, и водный слой экстрагировали этилацетатом (2×100 мл). Объединенные органические фазы промывали один раз рассолом, сушили над MgSO4, декантировали и концентрировали с выходом густого желтого масла, которое кристаллизовалось при стоянии (13,66 г, выход 72,4%). Неочищенное вещество кристаллизовали из смеси 4:1 гексан:этилацетат, отделяя от нерастворимого порошковидного остатка, возможно псевдоэфедрина. После одной кристаллизации очищенное вещество показывало э.и. приблизительно 80%, предпочтительность R-изомера, как определено анализом Мошера. После 3 кристаллизации было получено 6,65 г (R)-3-метокси-2-метилбензойной кислоты N’-(1-трет-бутил-бут-3-енил)гидразида (35,2%) в виде кристаллического вещества, но без дальнейшего улучшения э.и. Rf=0,38 (2:1 гексан : этилацетат); 1H ЯМР (400 МГц, CDCl3) δ 7.22 (t, 1H),7.11 (br s, 1H), 6.95 (d, 1H), 6.15 (m, 1H),5.2 (d, 1H), 5.15 (d, 1H), 3.89 (s, 3H), 2.80 (d, 1H), 2.50 (dm, 1H), 2.33 (s, 3H), 2.2 (dt, 1H), 1.08 (s, 9H).
Соединение 5 (указанное в заголовке соединение): (R)-3-метокси-2-метил-бензойной кислоты N’-(1-трет-бутил-бут-3-инил)-гидразид (100 мг, 0,344 ммоль) и (R)-(+)-альфа-метил-альфа-(трифторметил)фенилацетилхлорид (104 мг, 0,413 ммоль) растворяли в 1,5 мл метиленхлорида во флаконе. Добавляли раствор K2CO3 (95 мг, 0,69 ммоль) приблизительно в 0,5 мл воды, и реакционную смесь перемешивали при комнатной температуре в течение ночи и наблюдали при помощи ТСХ. Фазы разделяли, добавляя дополнительный метиленхлорид и/или воду, как это требовалось, чтобы содействовать манипуляциям. Слой метиленхлорида сушили над MgSO4 или Na2SO4, и растворитель удаляли в вакууме с получением неочищенного продукта в виде маслянистого твердого вещества. Этот остаток растирали со смесью гексан/эфир, после чего начиналась кристаллизация. Кристаллизацию оставляли развиваться без дополнительных помех. Выделение из маточного раствора давало 83 мг (16,4%) кристаллического N’-(1-трет-бутил-бут-3-инил)-N’-((S)-3,3,3-трифтор-2-метокси-2-фенил-пропионил)-гидразида (R)-3-метокси-2-метил-бензойной кислоты, из которого получали рентгеновскую кристаллическую структуру. Т.пл.=128-129°C; 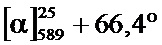
Для определения стереохимического течения реакции асимметрического аллилирования в Примере 50 определяли рентгеновскую кристаллическую структуру (N’-(1-трет-бутил-бут-3-инил)-N’-((3)-3,3,3-трифтор-2-метокси-2-фенил-припионил)-гидразида (R)-3-метокси-2-метил-бензойной кислоты. В данном эксперименте определили, что абсолютная конфигурация атома углерода, несущего трет-бутильную группу и н-пропильную группу представляет собой R-конфигурацию.
Пример 51
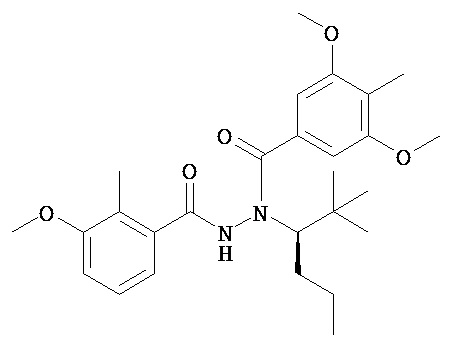
(R)-3,5-диметокси-4-метилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(3-метокси-2-метил-бензоил)-гидразид
СИНТЕЗ
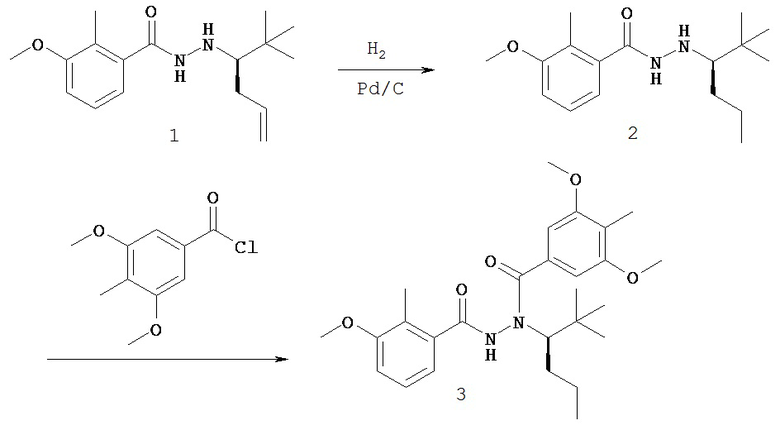
Соединение 1: (R)-3-Метокси-2-метил-бензойной кислоты N’-(1-трет-бутил-бут-3-инил)-гидразид получали, как описано в Примере 50.
Соединение 2: (R)-3-Метокси-2-метил-бензойной кислоты N’-(1-трет-бутил-бут-3-инил)-гидразид (3,05 г, 10,5 ммоль) растворяли в 100 мл метанола. Осторожно добавляли палладий на угле (1%, 240 мг), и смесь встряхивали в гидрогенизаторе Парра в течение 2,5 часов при начальном давлении 55 фунтов на квадратный дюйм (366,8 кПа). За снижением давления наблюдали в качестве оценки развития реакции; через 1,5 часа поглощение водорода прекратилось. Смесь фильтровали через слой целита, и растворитель удаляли в вакууме. Анализ посредством ТСХ с использованием нескольких растворителей значительно не преуспел в разделении двух гидразидных пятен, хотя 3:1 гексан : этилацетат (Rf=0,28) было немного более успешным. Неочищенный продукт кристаллизовали дважды из смеси 2:1 пентан : гексан с получением >0,6 г N’-(1-трет-бутил-бутил)-гидразида (R)-3-метокси-2-метил-бензойной кислоты. 1H ЯМР (400 МГц, CDCl3) δ 7.05 (t, 1H), 6.79 (d, 1H), 6.76 (d, 1H), 3.7 (s, 3H), 2.37 (m, 1H), 2.12 (s, 3H), 1.7-1.1 (m, 4H), 0.90 (s, 9H), 0.82 (t, 3H).
Соединение 3 (указанное в заголовке соединение): N’-(1-трет-Бутил-бутил)-гидразид (R)-3-метокси-2-метилбензойной кислоты (100 мг, 0,34 ммоль) и 3,5-диметокси-4-метил-бензоилхлорид (90 мг, 0,41 ммоль) растворяли в 1 мл метиленхлорида. Добавляли 1,5 экв. приблизительно 25%-ного K2CO3, и реакционную смесь перемешивали при комнатной температуре и наблюдали при помощи ТСХ. По завершении фазы разделяли, дополнительно добавляя CH2Cl2 и/или воду, как требовалось, чтобы способствовать манипуляциям. Фазу CH2Cl2 сушили над MgSO4 или Na2SO4, и растворитель удаляли в вакууме с получением маслянистого твердого вещества. Его растирали со смесью 1:1 гексан : эфир и оставляли сушиться на воздухе с получением N-(1-трет-бутил-бутил)-N’-(3-метокси-2-метил-бензоил)-гидразида (R)-3,5-диметокси-4-метил-бензойной кислоты. Rf=0,29 (2:1 гексан : этилацетат); 1H ЯМР (400 МГц, CDCl3) δ 7.1 (t, 1H), 7.0 (s, NH 1H), 6.9 (d, 1H), 6.7 (s, 2H), 6.3 (d, 1H), 4.78 (m, 1H), 3.95 (s, 3H), 3.85 (2s, 6H), 2.2 (s, 3H), 2.05 (m, 1H), 1.95 (s, 3H), 1.8 (m, 1H), 1.6 (m, 2H), 1.2+1.1+1.05 (3s, 9H), 1.05 (m, 3H); HRMS (ESI) m/z рассчитано для C27H39N2O5 [M+H]+ 471,2859, найдено 471,2850, рассчитано для C27H38NaN2O5 [M+Na]+ 493,2670, найдено 493,2678.
Пример 52
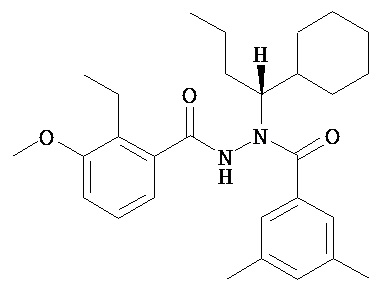
(R)-3,5-диметилбензойной кислоты N-(1-циклогексил-бутил)-N’-(2-этил-3-метоксибензоил)гидразид
СИНТЕЗ
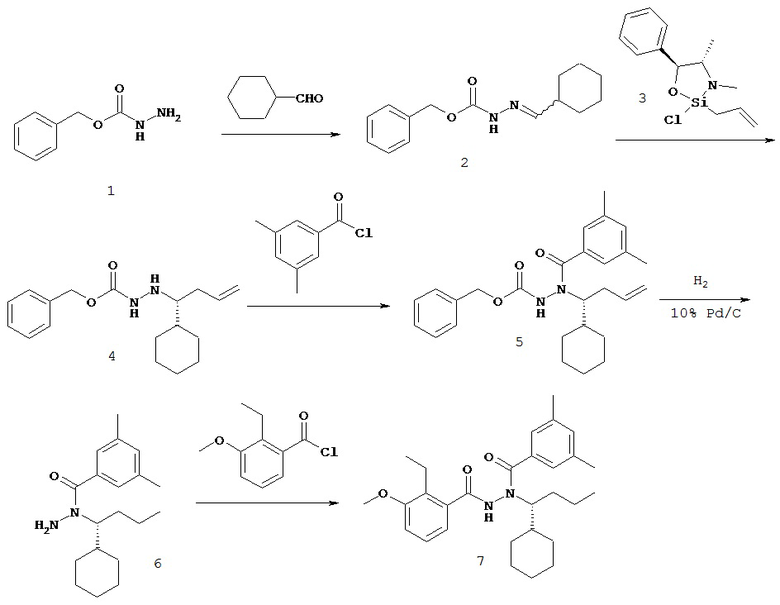
Соединение 1: Бензилкарбазат имеется в продаже.
Соединение 2: Бензилкарбазат (25 г, 150,44 ммоль) растворяли в 200 мл этанола в атмосфере азота в круглодонной колбе, оснащенной магнитным перемешиванием. Добавляли к раствору циклогексанкарбальдегид (17,7 г, 158 ммоль) и уксусную кислоту (1 мл), и смесь перемешивали при комнатной температуре в течение 8 ч. Полученный твердый осадок отфильтровывали, промывали гексаном, кристаллизовали из этилацетата и сушили в течение ночи на открытом лабораторном столе с получением 28,87 г (выход 73,7%) белого кристаллического бензилового эфира N’-циклогексилметилен-гидразинкарбоновой кислоты. Rf=0,67 (1:3 гексан:этилацетат), 1H ЯМР (400 МГц, CDCl3) δ 7.8 (1H, br s), 7.4 (5H, m), 7.1 (1H, br s), 5.3 (2H, s), 2.4 (1H, m), 1.7-1.9 (4H, m), 1.3 (6H, m); HRMS (ESI) m/z рассчитано для C15H21N2O2 [M+H]+ 261,1603, найдено 261,1592, рассчитано для C15H20NaN2O2 [M+Na]+ 283,1426, найдено 283,1417.
Соединение 3: (S,S)-2-Аллил-2-хлор-3,4-диметил-5-фенил-[1,3,2]оксазацилолидин получали, как описано в Примере 1.
Соединение 4: Круглодонную колбу с магнитным перемешиванием и атмосферой N2 наполняли 5 г (19,21 ммоль) бензилового эфира N’-циклогексилметилен-гидразинкарбоновой кислоты в 75 мл безводного CHCl3. Смесь охлаждали до 0°C, и добавляли к раствору (S,S)-оксазасилолидин (5,66 г, 21,1 ммоль), используя шприц. Ледяную баню убирали через приблизительно 30 минут, и реакционную смесь перемешивали при комнатной температуре в течение ночи. Реакционную смесь гасили водным Na2CO3. Органический слой удаляли, и водную фазу экстрагировали CHCl3. Объединенные органические фазы промывали несколько раз водой, сушили над безводным Na2SO4, и освобождали от растворителя, используя роторный испаритель. Полученный неочищенный продукт очищали с помощью колоночной хроматографии с получением бензилового эфира (R)-N’-(1-циклогексил-бут-3-инил)-гидразинкарбоновой кислоты в виде светло-желтого вязкого масла, 2,31 г, выход 39,8%. Rf=0,41 (3:1 гексан : этилацетат); 1H ЯМР (400 МГц, CDCl3) δ 7.4 (5H, m), 6.3 (1H, br), 5.9 (1H, br s), 5.25 (2H, s), 5.2 (2H, br s), 2.95 (1H, br s), 2.35 (1H, br d), 2.17 (1H, br m), 1.8 (2H, br m), 1.75 (2H, br d), 1.55 (1H, br t), 1.0-1.4 (6H, m) млн-1; HRMS (ESI) m/z рассчитано для C18H27N2O2 [M+H]+ 303,2072, найдено 303,2079, рассчитано для C18H26NaN2O2 [M+Na]+ 325,1892, найдено 325,1899.
Соединение 5: Бензиловый эфир R-N’-(1-циклогексил-бут-3-инил)-гидразинкарбоновой кислоты (2,20 г, 7,27 ммоль) растворяли в 2 мл метиленхлорида во флаконе с магнитным перемешиванием. Добавляли водный раствор K2CO3 (1,51 г, 10,91 ммоль в 4 мл), и смесь охлаждали на льду. Медленно добавляли чистый хлорангидрид, и приблизительно 1 мл метиленхлорида использовали для очистки и промывки. Смесь перемешивали в течение ночи сначала на льду, затем при комнатной температуре, наблюдая посредством ТСХ. Воду и/или CH2Cl2 добавляли к реакционной смеси, чтобы способствовать манипуляциям, и органический слой собирали. Водный слой экстрагировали один раз с помощью CH2Cl2, и органические слои объединяли и сушили над твердым Na2SO4. Растворитель удаляли в вакууме, и остаток растирали со смесью 2:1 гексан : эфир с выходом 2,93 г (92,7%) бензилового эфира (R)-N’-(1-циклогексил-бут-3-инил)-N’-(3,5-диметил-бензоил)-гидразинкарбоновой кислоты в виде белого твердого вещества. Rf=0,33 (3:1 гексан : этилацетат); 1H ЯМР (400 МГц, DMSO-d6) δ 9.8+9.65 (0.5H, NH, 2s), 7.1-7.5 (3.5H, m), 6.9-7.1 (5H, m), 6.05 (0.5H, m), 5.9 (0.5H, m), 5.2 (1H, m), 5.0 (2H, m), 4.8 (1H, m), 4.35 (1H, m), 2.35 (2H, br m), 2.25 (6H, s), 0.8-2.0 (11H, m); HMS (ESI) m/z рассчитано для C27H35N2O3 [M+H]+ 435,2647, найдено 435,2659, рассчитано для C27H34NaN2O3 [M+Na]+ 457,2467, найдено 457,2470.
Соединение 6: Бензиловый эфир (R)-N’-(1-циклогексил-бут-3-инил)-N’-(3,5-диметил-бензоил)-гидразинкарбоновой кислоты (2,91 г, 6,7 ммоль) суспендировали в приблизительно 50 мл ледяной уксусной кислоты. Добавляли 102 мг 10%-ного палладия на угле в виде суспензии в нескольких мл уксусной кислоты. Смесь разбавляли уксусной кислотой до 75 мл и встряхивали на гидрогенизаторе Парра в течение 90 минут при 20-30 фунтов на квадратный дюйм (133,4×103 Па - 200,1×103 Па). Суспензию оставляли стоять в течение двух суток, чтобы обеспечить оседание угля. Надосадочную жидкость фильтровали через шприцевой фильтр 0,45 микрон в чашку-кристаллизатор, и растворитель выпаривали с получением 2,06 г (102% массовый баланс) (R)-3,5-диметил-бензойной кислоты N-(1-циклогексил-бутил)гидразида в виде красновато-коричневого вязкого масла. Rf=0,24 (2:1 гексан : этилацетат); 1H ЯМР (400 МГц, CDCl3) δ 7.05 (1H, s), 7.0 (2H, s), 4.55+3.35 (1H, 2t), 2.4 (6H, s), 0.8-1.9 (15H, m), 0.9 (3H, t); HRMS (ESI) m/z рассчитано для C19H31N2O [M+H]+ 303,2436, найдено 303,2441, рассчитано для C19H30N2NaO [M+Na]+ 325,2256, найдено 325,2262.
Соединение 7 (указанное в заголовке соединение): N-(1-циклогексил-бутил)-гидразид (R)-3,5-диметилбензойной кислоты (0,14 г, 0,46 ммоль) растворяли в 1 мл метиленхлорида во флаконе с магнитным перемешиванием. Добавляли водный раствор K2CO3 (0,1 г, 0,69 ммоль, 2,77 мл). Колбу охлаждали на льду, и добавляли чистый хлорангидрид. Приблизительно 0,8 мл дополнительного метиленхлорида использовании для очистки и промывания. Смесь перемешивали в течение ночи, сначала на льду и затем при комнатной температуре, в то же время наблюдая за взаимодействием посредством ТСХ. Воду и CH2Cl2 добавляли к реакционной смеси, чтобы способствовать манипуляциям, и водный слой экстрагировали один раз с помощью CH2Cl2. Органические слои объединяли и сушили над твердым Na2SO4. Растворитель удаляли в вакууме, и неочищенный продукт очищали с помощью хроматографии с получением 143 мг (выход 67,9%) (R)-3,5-диметил-бензойной кислоты N-(1-циклогексил-бутил)-N’-(2-этил-3-метокси-бензоил)-гидразида в виде красновато-коричневого вязкого масла. 1H ЯМР (400 МГц, DMSO-d6) δ 10.4 (1H, 2s), 7.0-7.25 (5H, m), 6.2 (IH, m), 4.45 (IH, m), 3.8 (3H, s), 2.35 (6H, s), 2.0-2.2 (2H, m), 0.9-2.0 (15H, m), 0.9 (6H, m).
Примеры 53-55 получали, используя методику, описанную в Примере 52. Процент энантиомерного избытка (э.и.) определяли при помощи хиральной ВЭЖХ.
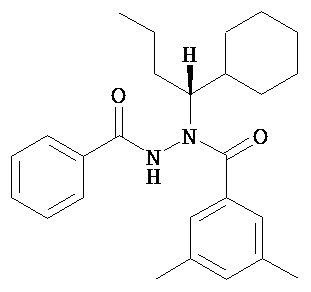
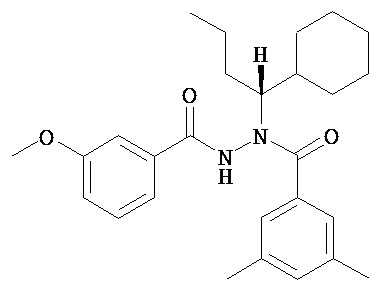
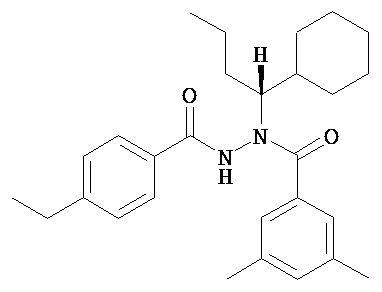
Пример 56
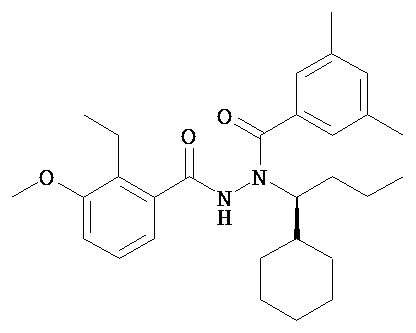
(R)-3,5-Диметилбензойной кислоты N-(1-циклогексилбутил)-N’-(2-этил-3-метокси-бензоил)гидразид
СИНТЕЗ
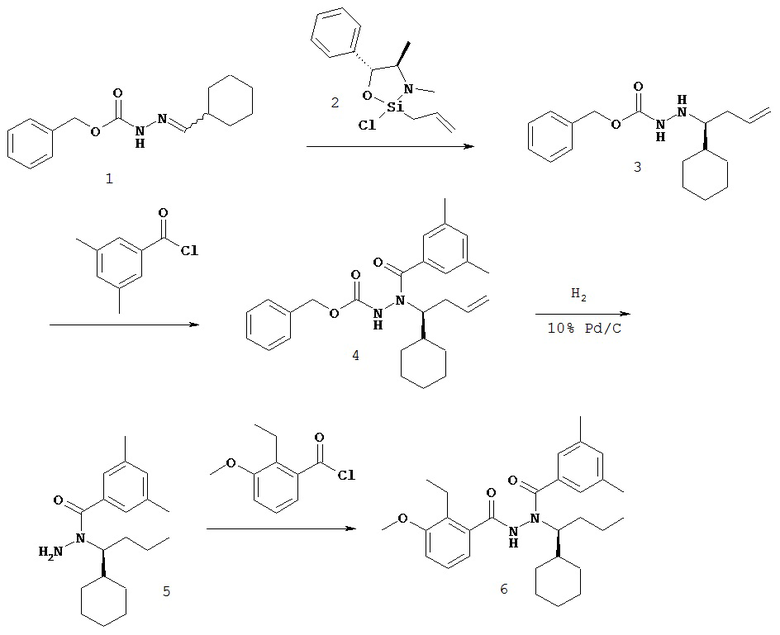
Соединение 1: N’-циклогексилметилен-гидразинкарбоновой кислоты бензиловый эфир получали, как описано в Примере 52.
Соединение 2: (R,R)-Аллил-2-хлор-3,4-диметил-6-фенил-[1,3,2]оксазацилолидин получали, используя методику, описанную в Примере 1, используя R,R-псевдоэфедрин.
Соединение 3: бензиловый эфир (S)-N’-(1-циклогексил-бут-3-инил)-гидразинкарбоновой кислоты получали с выходом 69% в виде светло-желтого вязкого масла, используя методику, описанную в Примере 52. Аналитические данные: Rf=0,41 (3:1 гексан : этилацетат); 1H ЯМР (400 МГц, DMSO-d6) δ 8.65 (1H, br s), 7.4 (5H, m), 5.85 (1H, br), 5.1 (4H, m), 4.2 (1H, s), 2.7 (1H, br s), 2.1 (1H, br m), 2.0 (1H, br m), 1.8 (2H, br m), 1.65 (2H, br m), 1.45 (1H, br m) 1.0-1.2 (6H, br m); ‘Н ЯМР (400 МГц, CDCl3) 5 7.4 (5H, m), 6.3 (1H, br), 5.9 (1H, br s), 5.25 (2H, s), 5.2 (2H, br s), 2.95 (1H, br s), 2.35 (1H, br d), 2.17 (1H, br m), 1.8 (2H, br m), 1.75 (2H, br d), 1.55 (IH, br t), 1.0-1.4 (6H, m); HRMS (ESI) m/z рассчитано для C18H27N2O2 [M+H]+ 303,2072, найдено 303,2057, рассчитано для C18H26NaN2O2 [M+Na]+ 325,1892, найдено 325,1873.
Соединение 4: бензиловый эфир (S)-N’-(1-циклогексил-бут-3-инил)-N’-(3,5-диметил-бензоил)-гидразинкарбоновой кислоты получали с выходом 84,8% в виде белого твердого вещества, используя методику, описанную в Примере 52. Аналитические данные: э.и. ≥80%. Rf=0,34 (3:1 гексан : этилацетат); 1H ЯМР (400 МГц, DMSO-d6) δ 9.8+9.65 (0.5H, NH, 2s), 7.1-7.5 (3.5H, m), 6.9-7.1 (5H, m), 6.05 (0.5H, m), 5.9 (0.5H, m), 5.2 (1H, m), 5.0 (2H, m), 4.8 (1H, m), 4.35 (1H, m), 2.35 (2H, br m), 2.25 (6H, s), 0.8-2.0 (11H, m); HRMS (ESI) m/z рассчитано для C27H35N2O3 [M+H]+ 435,2647, найдено 435,2655, рассчитано для C27H34NaN2O3 [M+Na]+ 457,2467, найдено 457,2469.
Соединение 5: (S)-3,5-Диметилбензойной кислоты N-(1-циклогексил-бутил)гидразид получали, используя методику, описанную в Примере 52. Аналитические данные: Rf=0,24 (2:1 гексан : этилацетат); 1H ЯМР (400 МГц, DMSO-d6) δ 7.05+6.95 (1H, s), 7.0+6.85 (2H, 2s), 4.55+4.05 (2H, 2s), 4.3+3.15 (1H, 2t), 2.3 (6H, 2s), 0.6-1.9 (15H, m), 0.9+0.8 (3H, 2t); (400 МГц, CDCl3) δ 7.05 (1H, s), 7.0 (2H, s), 4.55+3.35 (1H, 2t), 2.4 (6H, s), 0.8-1.9 (15H, m), 0.9 (3H, t); HRMS (ESI) m/z рассчитано для C19H31N2O [M+H]+ 303,2436, найдено 303,2440, рассчитано для C19H30N2NaO [M+Na]+ 325,2256, найдено 325,2260.
Соединение 6 (указанное в заголовке соединение): N-(1-циклогексил-бутил)-N’-(2-этил-3-метокси-бензоил)гидразид (S)-3,5-диметил-бензойной кислоты получали, используя методику, описанную в Примере 52 с э.и. 37%.
Примеры 57-59 получали, используя методику, описанную в Примере 56. Процент энантиомерного избытка (э.и.) определяли с помощью хиральной ВЭЖХ. В Таблице 3 приведены аналитические данные для примеров 57-59.
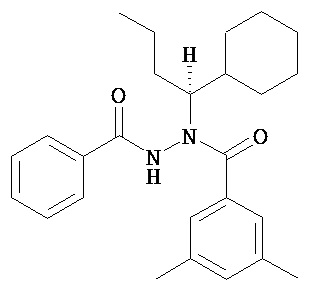
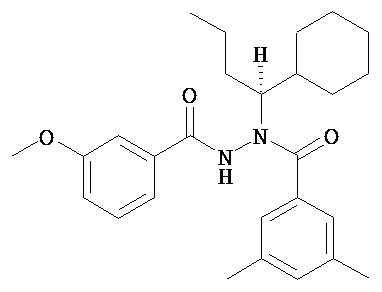
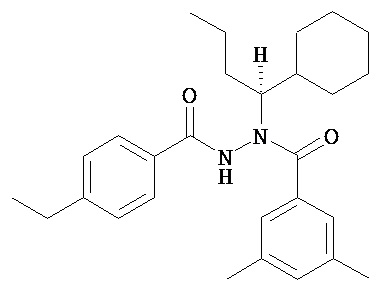
Пример 60
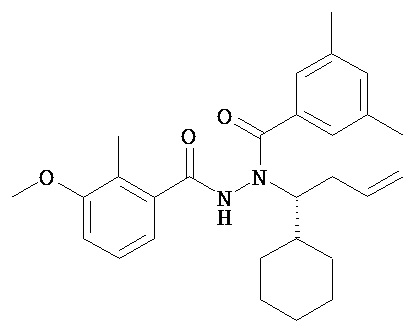
(R)-3,5-Диметилбензойной кислоты N-(1-циклогексил-бут-3-инил)-N’-(3-метокси-2-метилбензоил)гидразид
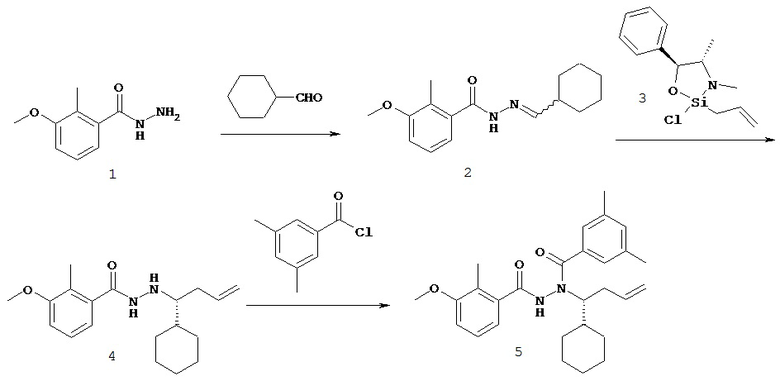
Соединение 1: Гидразид 3-метокси-2-метилбензойной кислоты получали, как описано в Примере 50.
Соединение 2: 10,0 г (55,5 ммоль) гидразида 3-метокси-2-метилбензойной кислоты растворяли в 200 мл абсолютного этилового спирта. Добавляли циклогексанкарбальдегид (6,85 г, 61,0 ммоль) и ледяную уксусную кислоту (3 мл), и реакционную смесь перемешивали в течение 18 ч, наблюдая посредством ТСХ. Осадок собирали фильтрацией и промывали гексаном. Продукт, 3-метокси-2-метил-бензойной кислоты циклогексилметилен-гидразид, получали в виде белого твердого вещества (9,36 г, выход 61%): Rf=0,20 (3:1 EtOAc : н-гексан); 1H ЯМР (400 МГц, CDCl3) δ 11.17 (s, 1H), 7.91 (s, 1H), 7.50 (d, J=4,8 Гц, 1H), 7.25 (t, J=6,4 Гц, 1H), 6.99 (t, J=5,6 Гц, 1H), 3.89 (s, 3H), 2.28 (s, 3H), 1.92-1.62 (m, 5H), 1.38-1.14 (m, 6H); HRMS (ESI) m/z рассчитано для C16H23N2O2 [M+H]+ 275,1760, найдено 275,1752, рассчитано для C16H22N2NaO2 [M+Na]+ 297,1579, найдено 297,1568.
Соединение 3: (S,S)-2-Аллил-2-хлор-3,4-диметил-5-фенил-[1,3,2]оксазацилолидин получали, как описано в Примере 1.
Соединение 4: В круглодонную колбу с мешалкой и подводом азота добавляли 3-метокси-2-метилбензойной кислоты циклогексилметилен-гидразид (2,8 г, 10,2 ммоль) и сухой CHCl3 (50 мл). Добавляли по каплям (S,S)-оксазацилолидин (3,28 г, 12,20 ммоль). Реакционную смесь перемешивали при 50°C в течение 6 ч, затем при комнатной температуре в течение 18 ч. Насыщенный раствор NaHCO3 (30 мл) добавляли для гашения реакции. Слой CHCl3 отделяли, и водный слой экстрагировали CHCl3 (2×30 мл). Раствор CHCl3 объединяли и сушили над безводным Na2SO4. Растворитель выпаривали в вакууме, и неочищенную смесь очищали при помощи флэш-хроматографии с получением N’-(1-циклогексил-бут-3-инил)гидразида (R)-3-метокси-2-метилбензойной кислоты в виде белого твердого вещества (1,74 г, выход 54%). 1H ЯМР (400 МГц, CDCl3) δ 9.69 (s, 1Н), 7.24 (t, J 8,0 Гц, 1H), 7.05 (d, J=8,0 Гц, 1H), 6.88 (d, J=7,6 Гц, 1H), 5.92 (m, 1H), 5.15 (dd, J=1,6, 17,2 Гц, 1H), 5.08 (dd, J=1,6, 10,4 Гц, 1H), 4.91 (dd, J=3,6, 6,8 Гц, 1H), 3.83 (s, 3H), 2.75 (m, 1H), 2.26-2.22 (m, 1H), 2.16 (s, 3H), 2.14-2.08 (m, 1H), 1.81-1.08 (m, 11H); HRMS (ESI) m/z рассчитано для C19H29N2O2 [M+H]+ 317,2229, найдено 317,2221, m/z рассчитано для C19H29N2O2 [M+Na]+ C19H28N2NaO2 339,2048, найдено 339,2037.
Соединение 5 (указанное в заголовке соединение): К раствору (R)-3-метокси-2-метилбензойной кислоты N’-(1-циклогексил-бут-3-инил)гидразида (1,3 г, 4,11 ммоль) в CH2Cl2 (5 мл), добавляли K2CO3 (2,27 г, 16,44 ммоль), H2O (5 мл) и наконец 3,5-диметилбензоилхлорид (727 мг, 4,31 ммоль). Реакционную смесь перемешивали при комнатной температуре в течение 24 ч. Добавляли дополнительно CH2Cl2 и H2O для облегчения манипуляций, слой CH2Cl2 отделяли, и водный слой экстрагировали при помощи CH2Cl2. Растворы CH2Cl2 объединяли и сушили над безводным Na2SO4. Растворитель выпаривали в вакууме, и остаток очищали при помощи флэш-хроматографии с получением (R)-3,5-диметил-бензойной кислоты N-(1-циклогексил-бут-3-енил)-N’-(3-метокси-2-метил-бензоил)-гидразида в виде белого твердого вещества (1,45 г, выход 78%). Rf=0,21 (3:1 EtOAc : н-гексан); 1H ЯМР (400 МГц, DMSO-d6) δ 10.47+10.39 (2s, 1H), 7.20-7.01 (m, 5H), 6.37 (d, J=7,2 Гц, 1H), 6.22-5.89 (m, 1H), 5.18 (dd, 1H), 5.02 (dd, 1H), 4.46 (m, 1H), 3.79 (s, 3H), 2.44-2.34 (m, 2H), 2.29 (s, 3H), 2.02-1.53 (m, 8H), 1.26-1.02 (m, 3H); HRMS (ESI) m/z рассчитано для C28H37N2O3 [M+H]+ 449,2804, найдено 449,2793, рассчитано для C28H36N2NaO3 [M+Na]+ 471,2624, найдено 471,2615.
Пример 61
(R)-4-Метокси-3,5-диметилбензойной кислоты N-(1-циклогексил-бут-3-инил)-N’-(3-метокси-2-метилбензоил)гидразид
СИНТЕЗ
(R)-4-Метокси-3,5-диметилбензойной кислоты N-(1-циклогексил-бут-3-инил)-N’-(3-метокси-2-метил-бензоил)-гидразид получали с выходом 71%, используя методику, описанную в Примере 60. Аналитические данные: Rf=0,14 (1:3 EtOAc : н-гексан); 1H ЯМР (400 МГц, DMSO-d6) δ 10.46+10.38 (2s, 1H), 7.21-6.42 (m, 5H), 6.16-5.93 (m, 1H), 5.23 (dd, 1H), 5.01 (dd, 1H), 4.45 (m, 1H), 3.79 (s, 3H), 3.77 (s, 6H), 2.42-2.28 (m, 2H), 2.04 (s, 3H), 1.88-1.05 (m, 14H); HRMS (ESI) m/z рассчитано для C29H38N2O5 [M+H]+ 495,2859, найдено 495,2848, рассчитано для C29H38N2NaO5 [M+Na]+ 517,2678, найдено 517,2667.
Пример 62
(R)-3,5-диметилбензойной кислоты N’-(4-этилбензоил)-N-(1-фенетил-бут-3-инил)гидразид
СИНТЕЗ
Соединение 1: гидразид 4-этил-бензойной кислоты имеется в продаже.
Соединение 2: гидразид 4-этил-бензойной кислоты (10 г, 60,9 ммоль) растворяли в 32 мл метанола. Добавляли уксусную кислоту (1 мл), смесь доводили до кипения с обратным холодильником, и затем медленно добавляли гидрокоричный альдегид (5,77 г, 67 ммоль). Реакционную смесь нагревали с обратным холодильником в течение 10 ч и наблюдали при помощи ТСХ. Осадки собирали посредством фильтрации и промывали гексанами. 4-Этил-бензойной кислоты (3-фенилпропилиден)гидразид получали в виде белого твердого вещества (9,34 г, выход 54,7%). Rf=0,36 (1:1 EtOAc : н-гексан); 1H ЯМР (400 МГц, CDCl3) δ 8.87 (s, 1H), 7.72-7.24 (m, 10Н), 2.87 (t, J=15,2 Гц, 2Н), 2.79-2.57 (m, 4Н), 1.24 (t, J=8 Гц, 3H).
Соединение 3: (R)-(+)-Метил-пара-толил сульфоксид (0,5 г, 3,21 ммоль) и тетрабутиламмония трифенил-дифторсиликат (ТВАТ) (0,12 г, 0,214 ммоль) растворяли в 0,038 мл 2-метил-2-бутена и 2,4 мл безводного метиленхлорида в атмосфере азота и при помощи магнитного перемешивания.
Аллилтрихлорсилан (0,23 мл, 1,61 ммоль) добавляли при -78°C, и реакционную смесь перемешивали в течение 15 минут.(1-Метил-3-фенил-пропилиден)гидразид 4-этилбензойной кислоты (0,3 г, 1,07 ммоль) растворяли в 11,6 мл безводного метиленхлорида и добавляли к реакционной смеси в течение 30 минут. Реакционную смесь перемешивали в течение 5 часов в атмосфере азота и поддерживали при температуре от -78°C до -70°C, наблюдая при помощи ТСХ. Реакцию гасили 1,5 мл триэтиламина и 3 мл безводного метанола при -78°C. Затем добавляли рассол, и смесь оставляли нагреваться до комнатной температуры. Органический слой удаляли, объединяли с тремя экстрактами метиленхлорида и сушили над сульфатом натрия. Растворитель выпаривали, и неочищенную смесь очищали с помощью флэш-хроматографии, используя ступенчатый градиент 2:1 и 1:1 пентан:этилацетат. Очищенный (R)-4-этил-бензойной кислоты N’-(1-фенетил-бут-3-инил)-гидразид получали в виде прозрачного масла (0,145 г, выход 41,4%, э.и. 57,6%, хиральная ВЭЖХ на Chiralcel ADH). Rf=0,50 (1:1 EtOAc : н-гексан); 1H ЯМР (400 МГц, CDCl3) δ 7.64-7.13 (m, 9H), 5.91 (m, 1H), 5.15 (t, J=28,0 Гц, 2H), 3.12 (m, 1H), 2.76-2.63 (m, 4H), 2.38-2.24 (m, 2H), 1.88-1.74 (m, 2H), 1.23 (t, J=8,0 Гц, 3H).
Соединение 4 (указанное в заголовке соединение): (R)-4-Этил-бензойной кислоты N’-(1-фенетил-бут-3-инил)гидразид (0,1 г, 0,31 ммоль) растворяли в 0,47 мл метиленхлорида с магнитным перемешиванием. К смеси добавляли бикарбонат калия (0,064 г, 0,37 ммоль) в 0,7 мл деионизированной воды и 3,5-диметилбензоилхлорид (0,063 г, 0,37 ммоль). Реакционную смесь сначала перемешивали на льду и затем оставляли нагреваться до комнатной температуры в течение выходных, наблюдая с помощью ТСХ. Органический слой удаляли, и водный слой экстрагировали метиленхлоридом три раза и сушили над сульфатом натрия. Растворитель выпаривали, и неочищенную смесь очищали с помощью флэш-хроматографии, используя смесь 2:1 гексаны:этилацетат в качестве элюента. Очищенный (R)-3,5-диметилбензойной кислоты N-(4-этилбензоил)-N-(1-фенетил-бут-3-инил)-гидразид получали в виде желтого масла (0,102 г, выход 72,9%, э.и. 51,2%, хиральная ВЭЖХ на Chiracel ADH). Rf=0,34 (2:1 гексан : EtOAc); 1H ЯМР (400 МГц, CDCl3) δ 7.8-7.02 (m, 12H), 5.99 (br, 1H), 5.22 (d, J=24,0 Гц, 2H), 2.73 (d, J=8 Гц, 2H), 2.31 (s, 6H), 2.12-2.03 (m, 2H), 1.87 (s, 1H), 1.61 (s, 2H), 1.29 (t, J=8,0 Гц, 3H); HRMS (ESI) m/z рассчитано для C30H35N2O2 [M+H]+ 455,2699, найдено 455,2685, рассчитано для C30H34N2NaO2 [M+Na]+ 477,2518, найдено 477,2505.
Пример 63
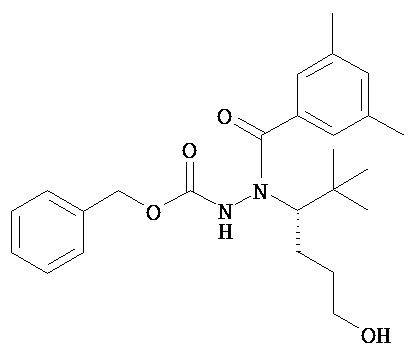
Бензиловый эфир (S)-N’-(1-трет-бутил-4-гидроксибутил)-N’-(3,5-диметилбензоил)гидразина карбоновой кислоты
СИНТЕЗ
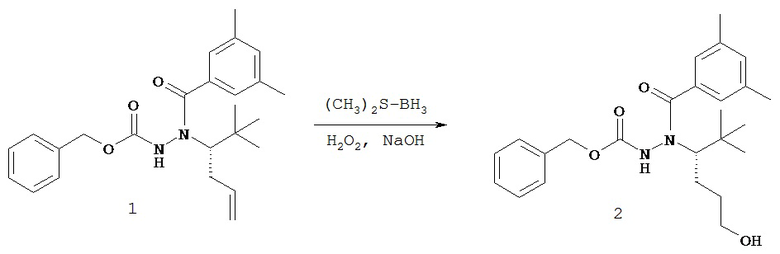
Соединение 1: Соединение 1 получали, как описано в Примере 31.
Соединение 2 (указанное в заголовке соединение): К раствору (S)-N’-(1-трет-бутил-бут-3-енил)-N’-(3,5-диметилбензоил)гидразинкарбоновой кислоты бензилового эфира (200 мг, 0,49 ммоль) в THF (3 мл), добавляли боран-диметилсульфид (2 М в THF, 125 мкл, 0,25 ммоль) при 0°C. Смесь перемешивали при комнатной температуре в течение 16 ч. Избыток борана и THF удаляли в вакууме, и добавляли дополнительно THF (3 мл), затем H2O2 (61 микролитр, 0,54 ммоль) и 3 н. NaOH (90 микролитров, 0,27 ммоль). После перемешивания при 50-55°C в течение 1 ч, опять добавляли воду, смесь экстрагировали эфиром, и эфирный слой сушили над безводным Na2SO4. Растворитель удаляли в вакууме, и остаток очищали при помощи флэш-хроматографии с получением бензилового эфира (S)-N’-(1-трет-бутил-4-гидрокси-бутил)-N’-(3,5-диметил-бензоил)-гидразинкарбоновой кислоты в виде белого твердого вещества (128 мг, выход 61%). Rf=0,30 (1:1 EtOAc : н-гексан). 1H ЯМР (400 МГц, DMSO-d6) δ 9.71+9.63 (2s, 1H), 7.44-6.88 (m, 8H), 5.06 (dd, J=3,6; 12,8 Гц, 1H), 4.79 (dd, J=12,44; 25,2 Гц, 1H), 4.43 (d, J=10,8 Гц, 1H), 4.28 (t, J=5,2 Гц, 1H), 3.47 (m, 2H), 2.27 (s, 6H), 1.86-1.21 (m, 4H), 1.01 (s, 9H); HRMS (ESI) m/z рассчитано для C25H35N2O4 [M+H]+ 427,2597, найдено 427,2587, рассчитано для C25H34N2NaO4 [M+Na]+ 449,2416, найдено 449,2407.
Пример 64
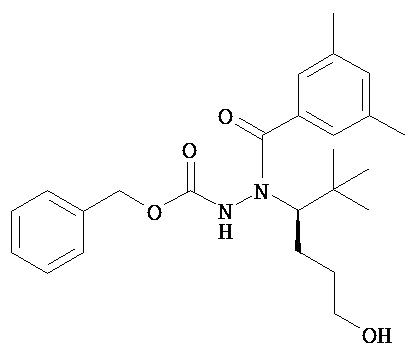
Бензиловый эфир (R)-N’-(1-трет-бутил-4-гидрокси-бутил)-N’-(3,5-диметилбензоил)гидразинкарбоновой кислоты
СИНТЕЗ
Бензиловый эфир (R)-N’-(1-трет-бутил-4-гидрокси-бутил)-N’-(3,5-диметил-бензоил)-гидразинкарбоновой кислоты получали с э.и. >99%, используя методику, описанную в Примере 63.
Пример 65
В данном примере проиллюстрирован аналитический способ, используемый для определения энантиомерного избытка (э.и.) соединений по изобретению.
Как показано на Фиг.2A-C, хроматограммы хиральной ВЭЖХ для рац-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2,6-дихлорбензоил)-гидразида (Пример 16), (R)-3,5-диметил-бензойной кислоты N-(1-трет-бутил-бутил)-N’-(2,6-дихлорбензоил)гидразида и (S)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2,6-дихлорбензоил)-гидразида (Пример 41) иллюстрируют способ, используемый для определения энантиомерного избытка. рац-3,5-Диметил-бензойной кислоты N-(1-трет-бутил-бутил)-N’-(2,6-дихлорбензоил)гидразид содержит 50% R-изомера (пик 1, приблизительно 16 минут) и 50% S-изомера (пик 2, приблизительно 19 минут). Энантиомерно обогащенный образец (R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2,6-дихлорбензоил)-гидразида по существу не содержит S-изомер с э.и. >99%. Энантиомерно обогащенный образец (S)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2,6-дихлорбензоил)гидразида по существу не содержит R-изомер с э.и. >99%. В этом эксперименте данные получали, используя колонку 4,6 мм×25 см Regis Rexchrom (S,S) ULMO при скорости потока 2,0 мл/мин, используя 2% метанол в гексане в качестве растворителя и инъецируемый объем 20 мкл. Энантиомерный избыток других соединений Формулы II или III определяли аналогичным образом, если не указано иное.
Пример 66
В данном примере проиллюстрировано in vitro испытание соединений по изобретению.
Кассета экспрессии гена
GAL4 DBD (1-147)-CfEcR(DEF)/VP16AD-βRXREF-LmUSPEF: Домены дикого типа D, E, и F из EcR листовертки еловой Choristoneura fumiferana ("CfEcR-DEF"; SEQ ID NO: 1) сливали с ДНК-связывающим доменом GAL4 ("GaI4DBD 1-147”; SEQ ID NO: 2) и помещали под контроль промотора фосфоглицераткиназы ("PGK"; SEQ ID NO: 3). Спирали 1-8 включительно доменов EF из RXRβ Homo sapiens ("HsRXRp-EF"; нуклеотиды 1-465 из SEQ ID NO: 4) и спирали 9-12 включительно доменов EF белка Ultraspiracle из Locusta migratoria ("LmUSP-EF"; нуклеотиды 403-630 из SEQ ID NO: 5) сливали с доменом трансактивации из VP16 ("VP16AD"; SEQ ID NO: 6) и помещали под контроль промотора фактора элонгации-1α ("EF-1α"; SEQ ID NO: 7). Пять консенсусных сайтов связывания элементов ответа GAL4 ("5XGAL4RE"; содержащая 5 копий GAL4RE, включающих SEQ ID NO: 8) сливали с синтетическим минимальным промотором ТАТА (SEQ ID NO: 9) и помещали в направлении против хода трансляции гена-репортера люциферазы (SEQ ID NO: 10).
Стабильная клеточная линия
Клетки CHO (яичников китайского хомячка) случайным образом трансфицировали транскрипционными кассетами для GAL4 DBD (1-147) CfEcR(DEF) и для VP16AD βRXREF-Z/wUSPEF, контролируемыми универсально активными клеточными промоторами (PGK и EF-1α, соответственно) на одной плазмиде. Стабильно трансфицированные клетки селектировали по стабильности к Зеоцину. Отдельно выделенные клоны CHO клеток случайным образом трансфицировали репортером RE-люциферазы GAL4 (pFR Luc). Клон 27-63 селектировали, используя гигромицин.
Обработка хиральным диацилгидразиновым лигандом
Клетки трипсинизировали и разбавляли до концентрации 2,5×104 клеток на мл. 100 мкл клеточной суспензии помещали в каждую лунку 96-луночного планшета и инкубировали при 37°C в 5%-ном CO2 в течение 24 ч. Получали исходные растворы лиганда в DMSO и разбавляли в 300 раз для всех обработок. Испытание доза-ответ состояло из 8 концентраций, в интервале от 33 мкМ до 0,01 мкМ.
Анализ гена-репортера
Экспрессию гена-репортера люциферазы измеряли через 48 ч после обработки клеток, используя систему анализа люциферазы Bright-Glo™ от Promega (E2650). Люминесценцию определяли при комнатной температуре, используя люминометр для микротитровальных планшетов Dynex MLX.
Стабильная клеточная линия
Получали популяцию стабильно трансформированных клеток, содержащих CVBE и 6XEcRE, как описано в Suhr et al, Proc. Natl. Acad. Sci. USA 95: 7999-804 (1998). Клетки человеческой почки 293, также называемые клетками HEK-293, последовательно инфицировали ретровирусными векторами, кодирующей сначала конструкцию переключателя CVBE, и затем репортерную конструкцию 6XEcRE Lac Z. Конструкция переключателя содержала кодирующую последовательность для аминокислот 26-546 из EcR (BE) из Bombyx mori (latrou), вставленную в рамку и по ходу транскрипции от домена (VBE) трансактивации VP16. Синтетический стартовый кодон ATG помещали под контроль ближайшего раннего промотора цитомегаловируса (CVBE) и фланкировали длинными концевыми повторами (LTR). Репортерная конструкция содержала шесть копий сайта связывания экдизонового элемента ответа (EcRE), помещенного против хода транскрипции LacZ, и фланкированного с обеих сторон последовательностями LTR (6XEcRE).
Клонирование серийным разбавлением использовали для изоляции отдельных клонов. Клоны селектировали, используя 450 мкг/мл G418 и 100 нг/мл пуромицина. Отдельные клоны оценивали на основе их ответа в присутствии и в отсутствие тестируемых лигандов. Клон Z3 отобрали для скрининга и SAR.
Клетки человеческой почки 293, стабильно трансформированные CVBE и 6XEcRE LacZ, поддерживали в минимальной эссенциальной среде (Mediates, 10-010-CV), содержащей 10% FBS (Life Technologies, 26140-087), 450 камеди G418 (Mediates, 30-234-CR), и 100 gnome promising (Sigma, P-7255), при 37°C в атмосфере, содержащей 5% CO2, и пересевали при достижении 75%-ного слияния.
Обработка хиральным диацилгидразиновым лигандом
Производили посев клеток Z3 в 96-луночные плашки для тканевых культур в концентрации 2,5×103 клеток на лунку и инкубировали при 37°C в 5% CO2 в течение двадцати четырех часов. Исходные растворы лиганда получали в DMSO. Исходные растворы лиганда разбавляли 100-кратно в среде, и 50 мкл данного разбавленного раствора лиганда (33 мкМ) добавляли к клеткам. Поддерживали 0,03%-ную конечную концентрацию DMSO как в контролях, так и в обработках.
Анализ гена-репортера
Экспрессию гена-репортера оценивали через 48 часов после обработки клеток. Активность (3-галактозидазы измеряли, используя биолюминесцентную систему анализа гена-репортера Gal Screen™ от Tropix (GSY1000). Показатели кратности индукции рассчитывали путем деления относительных световых единиц ("RLU") в клетках, обработанных лигандом, на RLU в клетках, обработанных DMSO. Люминесценцию определяли при комнатной температуре, используя люминометр для микротитровальных планшетов Dynex MLX.
Схема конструкции переключателя CVBE и конструкции репортера 6XEcRE Lac Z показана на Фиг.1. Фланкируют обе конструкции длинные концевые повторы, G418 и пуромицин представляют собой селектируемые маркеры, CMV представляет собой цитомегаловирусный промотор, VBE представляет собой кодирующую последовательность для аминокислот 26-546 из EcR Bombyx mori, вставленную по ходу транскрипции относительно домена трансактивации VP16, 6Х EcRE представляет собой шесть копий экдизононового элемента ответа, lacZ кодирует репортерный фермента β-галактозидазу.
Кассета экспрессии гена
GAL4 DBD-CfEcR(DEF)VP16AD-MMRXRE: Домены дикого типа D, E, и F EcR из листовертки еловой Choristoneura fumiferana ("CfEcR-DEF"; SEQ ID NO: 1) сливали с ДНК-связывающим доменом GAL4 ("Gal4DBDI-147"; SEQ ID NO: 2) и помещали под контроль промотора SV40e из вектора pM (PT3119-5, Clontech, Palo Alto, СА). Домены D и Е из RXR Mus Musculus ("МмRXR-DE"; SEQ ID NO: 11) сливали с доменом трансактивации из VP16 ("VP16AD"; SEQ ID NO: 6) и помещали под контроль промотора промотора SV40e из вектора pVP16 (PT3127-5, Clontech, Palo Alto, СА).
Стабильная клеточная линия
Клетки CHO случайным образом трансфицировали кассетами транскрипции для GAL4 DBD-CfEcR(DEF) и для VP16AD-MмRXRE, контролируемыми промоторами SV40e. Стабильно трансфицированные клетки селектировали, используя гигромицин. Отдельно выделенные клоны клеток CHO случайным образом трансфицировали репортером RE-люциферазы GAL4 (pFR-Luc, Stratagene, La Jolla, СА). Клон 13B3 селектировали, используя Зеоцин.
Обработка хиральным диацилгидразиновым лигандом
Клетки трипсинизировали и разбавляли до концентрации 2,5×104 клеток в мл. 100 мкл клеточной суспензии помещали в каждую лунку 96-луночного планшета и инкубировали при 37°C в 5%-ном CO2 в течение 24 ч. Стандартные растворы лиганда получали в DMSO и разбавляли в 300 раз для всех обработок. Испытание зависимости доза-ответ состояло из 8 концентраций в интервале от 33 мкМ до 0,01 мкМ.
Анализ переключателя генов - сконструированный EcR: системы USP/PXR
Анализы клеточного переключателя генов выполняли путем трансфицирования случайным образом следующих конструкций в фибробластные клетки мышиных эмбрионов (NTH3T3). Домены дикого типа D, E и F из EcR Choristoneura fumiferana или его мутанта V390I/Y410E/E274V сливали с GAL4-DBD и помещали под контроль промотора CMV в векторе pBIND (Promega Corporation, Madison, WI, USA). Домены DEF указанных EcRs амплифицировали, используя праймеры, созданные на основе 20-25-нуклеотидных последовательностей на 5’ и 3’ концах. Сайты фермента рестрикции EcoR I/BamH I и Xba I добавляли к праймерам 5’ и 3’ соответственно. Продукты ПЦР расщепляли соответствующими ферментами рестрикции и клонировали в вектор pBIND. EcR LBDs, клонированный в вектор pBIND верифицировали секвенированием. Химерный RXR из RXRp Homo sapiens и RXR Locusta migratoria, конденсированный с VP16-AD и помещенный под контроль промотора SV40e, конструировали, как описано ранее (Kumar P & Katakam A in Gene Transfer, Friedmann T и Rossi J, eds, стр.643-651 (2007), Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY). Репортерная плазмида индуцируемой люциферазы pFRLuc (Stratagene Cloning System, La Jolla, CA, USA) содержит пять копий элемента ответа GAL4 и синтетический минимальный промотор.
Клетки NIH3T3 поддерживали при 37°c и 5% CO2 в среде Игла, модифицированной Дулбекко (DMEM), с добавлением 10% плодной сыворотки телят, обе из которых были получены от Mediatech Inc., Manassas, VA. Клетки сеяли в 96-луночный планшет с плотностью 2500 клеток/лунка в 50 мкл среды для выращивания. На следующий день клетки сначала обрабатывали 35 мкл не содержащей сыворотки DMEM, содержащей диметилсульфоксид (DMSO; контроль), или раствором DMSO, содержащим лиганд. Клетки затем трансфицировали 15 мкл не содержащей сыворотки DMEM, содержащей 0,04 мкг конструкции EcR, 0,04 мкг конструкции RXR и 0,16 мкг конструкции репортера люциферазы на лунку, используя реагент трансфекции SuperFect™ (Qiagen Inc., Valencia, CA, USA) согласно инструкциям производителя. Лиганды испытывали в 8 дозах от 0,01 до 33 мкМ, и конечная концентрация DMSO составляла 0,33% как в контроле, так и в обработанных лунках. Через 48 часов после обработки и инкубирования трансфекции клетки анализировали на люциферазную активность, используя систему анализа люциферазы Bright-Glo™ (Promega Corporation, Madison, Wl, USA), следуя инструкциям производителя. Значения ec50 измеряли как минимум в двух повторах.
Анализ гена-репортера
Экспрессию гена-репортера люциферазы измеряли через 48 ч после обработки клеток, используя систему анализа люциферазы Bright-Glo™ от Promega (E2650). Люминесценцию определяли при комнатной температуре, используя люминометр для микротитровальных планшетов Dynex MLX.
Каждый анализ производили в двух отдельных лунках, и усредняли два значения. Кратность индукций рассчитывали путем деления относительных световых единиц ("RLU") в клетках, обработанных лигандом, на RLU в клетках, обработанных DMSO. RC50 рассчитывали из данных доза-ответ, используя трехпараметрическую логистическую модель. Относительный Max FI определяли как максимальную кратность индукции тестируемого лиганда (воплощение по изобретению), наблюдаемую при любой концентрации, относительно максимальной кратности индукции лиганда GS™-E (3,5-диметилбензойной кислоты N-трет-бутил-N’-(2-этил-3-метоксибензоил)гидразид), наблюдаемой при любой концентрации.
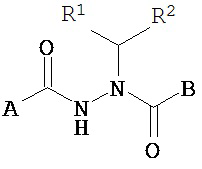
Пример 67
В данном примере проиллюстрировано in vivo тестирование соединений по изобретению. Индукцию in vivo репортерного фермента хиральными диацилгидразиновыми лигандами по настоящему изобретению оценивали на модельной системе мышей C57BL/6, содержащей переключатель генов.
Кассеты экспрессии гена
Домены дикого типа D, Е, и F из EcR листовертки еловой Choristoneura fumiferana ("CfficR-DEF"; SEQ ID NO: 1) подвергали мутированию [V107 (gtt)→1107 (att) и Y127 (tac)→E127 (gag)] и сливали с ДНК-связывающим доменом GAL4 ("Gal4DBDI-147"; SEQ ID NO: 2). Спирали 1-8 включительно доменов EF из RXRβ Homo sapiens ("HsRXRp-EF"; нуклеотиды 1-465 из SEQ ID NO: 4) и спирали 9-12 включительно доменов EF из белка Ultraspiracle Locusta migratoria ("LmUSP-EF"; нуклеотиды 403-630 из SEQ ID NO: 5) сливали с доменом трансактивации из VP16 ("VP16AD"; SEQ ID NO: 6), который регулирует репортерный ген человека, секретирующий щелочную фосфатазу ("SEAP", SEQ ID NO: 12), который помещали под контроль элемента ответа 6xGAL4 (SEQ ID NO: 13) и транстиретиновый промотор (SEQ ID NO: 14). Каждый элемент переключателя генов находился на отдельной плазмиде. Экспрессия рецептора находилась под контролем промотора CMV (SEQ ID NO: 15). Индукцию оценивали по количеству репортерного белка, экспрессируемого в присутствии лиганда.
Электропорация переключателя генов
Экспрессию SEAP в сыворотке мыши оценивали после электропорации переключателя генов в четырехглавую мышцу мыши. Мышей анестезировали с помощью 2 мкл/г смеси кетамина (100 мг/мл) и ксилазина (20 мг/мл). Животных затем обривали, векторы ДНК инъецировали в мышцу в объеме 2×50 мкл полиглютаминовой кислоты (12 мг/мл воды), прикладывали гель проводимости электрода, и калибр электрода (1 см×1 см; модель 384) помещали на заднюю конечность. Мышцу электропорировали 200 В/см, 8 раз, в течение 20 миллисекунд/импульс, с временными интервалами 1 сек. Направление поперечного электрического поля изменяли на обратное после того как животные получали половину импульсов. Электропорацию выполняли при помощи электропоратора ЕСМ 830 от ВТХ Molecular Delivery System.
Обработка хиральным диацилгидразиновым лигандом
В некоторых экспериментах мыши получали внутрибрюшинную инъекцию (IP) 2,6 мкмоль лиганда в 50 мкл DMSO через 3 суток после электропорации переключателя генов. В других экспериментах концентрацию лиганда снижали до 26 нмоль/50 мкл DMSO/мышь. Экспрессию SEAP оценивали через 2-11 суток после введения лиганда. В других экспериментах лиганд вводили в корм грызунов. Корм готовили путем растворения 2 г лиганда в 20 мл ацетона и добавления ее к 1 кг автоклавируемого корма LabDiet 5010 от Purina Mills. Все тщательно перемешивали в миксере Hobart и затем смешивали в течение еще 15 мин в миксере Cross Blend. Животные получали корм ad libitum (по своему желанию) в течение 1, 2 или 3 суток. Все значения представляют собой среднюю величину от четырех животных. Фоновое значение SEAP в сыворотках животных, обработанных одним вектором без добавления лиганда составляло 0-11 нг/мл сыворотки.
Репортерный анализ
Мышиную сыворотку получали путем центрифугирования крови, полученной путем ретроорбитального кровоизвлечения с небольшой стеклянной капиллярной пробиркой. Количественное выражение SEAP определяли, используя хемилюминесцентный набор Clontech Great Escape и сравнивая со стандартом SEAP Clontech.
Для оценки относительной эффективности in vivo (R)-2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)-гидразида (Пример 1) в индуцировании экспрессии гена RheoSwitch® Therapeutic System (RTS), двенадцать мышей C57BL6 электропорировали следующими плазмидами: pCMV/GEVY(DEF), pCMV/VP16-Hs-LmRXR и p6XGAL4RE-TTR-SEAP. Плазмида pCMV/GEVY (DEF) состоит из доменов D, E и F EcR из Choristoneura fumiferana, имеющих мутации V390I/Y410E/E274V, слитых по ходу транскрипции от дрожжевой GAL4-DBD (aa 1-147) и помещенных под контроль CMV-промотора и по ходу транскрипции от сигнала полиаденилирования SV40 в векторе pBIND (Promega Corporation, Madison, WI, USA). DEF-Домены указанных EcR амплифицировали, используя праймеры, сконструированные на основе 20-25-нуклеотидных последовательностях на 5’ и 3’ концах. Сайты фермента рестрикции BamH I и Xba I добавляли к 5’ и 3’ праймерам соответственно. Продукты ПЦР расщепляли соответствующими ферментами рестрикции и клонировали в вектор pBIND (Фиг.20). Вектор pCMV/VP16-Hs-LmRXR содержит химерный RXR из RXRβ Homo sapiens (спирали 1-8 домена Е) и RXR Locusta migratoria (спирали 9-12 домена E), слитые по ходу транскрипции от домена активации VP16 и помещенные под контроль промотора CMV в векторе pBDSfD (Фиг.21). Индуцируемый SEAP репортерный вектор p6xGAL4RE-TTR-SEAP содержит репортерный ген щелочной фосфатазы, секретируемой человеком, помещенный под контроль индуцируемого промотора, состоящего из 6 копий элемента ответа Gal4 расположенного против хода транскрипции от промотора транстиретина (TTR) (Фиг.22). Через трое суток после электропорации плазмидной ДНК в четырехглавую мышцу мышей, три лигандных препарата вводили мышам посредством внутрибрюшинной инъекции в количестве 26 нмоль/мышь (11,4 мкг/мышь, приблизительно 570 мкг/кг). Экспрессию щелочной фосфатазы, секретируемой человеком (SEAP), в мышиной сыворотке затем наблюдали в течение семнадцати суток после введения лиганда. Для сравнения оценивали относительную эффективность in vivo (S)-2-этил-3-метоксибензойной кислоты (N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида (Пример 31) и рац-2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида. Как показано на Фиг.3, (S)-2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)-гидразид (●-●) не индуцирует экспрессию репортерного гена RTS. N’-(1-трет-рац-2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразид индуцирует умеренную экспрессию репортерного гена RTS (▲-▲). (R)-2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)-гидразид (O-O) индуцирует существенную экспрессию репортерного гена RTS.
Пример 68
Физические свойства R-энантиомера и рацемата 2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметил-бензоил)-гидразида:
Клетки человеческой почки 293, стабильно трансформированные CVBE и 6XEcRE LacZ, поддерживали в минимальной эссенциальной среде (Mediates, 10-010-CV), содержащей 10% FBS (Life Technologies, 26140-087), 450 камеди G418 (Mediates, 30-234-CR), и 100 gnome promising (Sigma, P-7255), при 37°C в атмосфере, содержащей 5% CO2, и пересевали при достижении 75%-ного слияния.
Обработка хиральным диацилгидразиновым лигандом
Производили посев клеток Z3 в 96-луночные плашки для тканевых культур в концентрации 2,5×103 клеток на лунку и инкубировали при 37°C в 5% CO2 в течение двадцати четырех часов. Исходные растворы лиганда получали в DMSO. Исходные растворы лиганда разбавляли 100-кратно в среде, и 50 мкл данного разбавленного раствора лиганда (33 мкМ) добавляли к клеткам. Поддерживали 0,03%-ную конечную концентрацию DMSO как в контролях, так и в обработках.
Анализ гена-репортера
Экспрессию гена-репортера оценивали через 48 часов после обработки клеток. Активность β-галактозидазы измеряли, используя биолюминесцентную систему анализа гена-репортера Gal Screen™ от Tropix (GSY1000). Показатели кратности индукции рассчитывали путем деления относительных световых единиц ("RLU") в клетках, обработанных лигандом, на RLU в клетках, обработанных DMSO. Люминесценцию определяли при комнатной температуре, используя люминометр для микротитровальных планшетов Dynex MLX.
Схема конструкции переключателя CVBE и конструкции репортера 6XEcRE Lac Z показана на Фиг.1. Фланкируют обе конструкции длинные концевые повторы, G418 и пуромицин представляют собой селектируемые маркеры, CMV представляет собой цитомегаловирусный промотор, VBE представляет собой кодирующую последовательность для аминокислот 26-546 из EcR Bombyx топ, вставленную по ходу транскрипции относительно домена трансактивации VP16, 6XEcRE представляет собой шесть копий экдизононового элемента ответа, lacZ кодирует репортерный фермента β-галактозидазу.
Кассета экспрессии гена
GAL4 DBD-CfEcR(DEF)VP16AD-MMRXRE: Домены дикого типа D, E, и F EcR из листовертки еловой Choristoneura fumiferana ("CfEcR-DEF"; SEQ ID NO: 1) сливали с ДНК-связывающим доменом GAL4 ("Gal4DBDI-147"; SEQ ID NO: 2) и помещали под контроль промотора SV40e из вектора рМ (РТ3119-5, Clontech, Palo Alto, СА). Домены D и E из RXR Mus Musculus ("MмRXR-DE"; SEQ ID NO: 11) сливали с доменом трансактивации из VP16 ("VP16AD"; SEQ ID NO: 6) и помещали под контроль промотора промотора SV40e из вектора pVP16 (PT3127-5, Clontech, Palo Alto, CA).
Стабильная клеточная линия
Клетки CHO случайным образом трансфицировали кассетами транскрипции для GAL4 DBD-CfEcR(DEF) и для VP16AD-MMRXRE, контролируемыми промоторами SV40e. Стабильно трансфицированные клетки селектировали, используя гигромицин. Отдельно выделенные клоны клеток CHO случайным образом трансфицировали репортером RE-люциферазы GAL4 (pFR-Luc, Stratagene, La Jolla, CA). Клон 13B3 селектировали, используя Зеоцин.
Обработка хиральным диацилгидразиновым лигандом
Клетки трипсинизировали и разбавляли до концентрации 2,5×104 клеток в мл. 100 мкл клеточной суспензии помещали в каждую лунку 96-луночного планшета и инкубировали при 37°C в 5%-ном CO2 в течение 24 ч. Стандартные растворы лиганда получали в DMSO и разбавляли в 300 раз для всех обработок. Испытание зависимости доза-ответ состояло из 8 концентраций в интервале от 33 мкМ до 0,01 мкМ.
Анализ переключателя генов - сконструированный EcR: системы USP/PXR
Анализы клеточного переключателя генов выполняли путем трансфицирования случайным образом следующих конструкций в фибробластные клетки мышиных эмбрионов (NTH3T3). Домены дикого типа D, E и F из EcR Choristoneura fumiferana или его мутанта V390I/Y410E/E274V сливали с GAL4-DBD и помещали под контроль промотора CMV в векторе pBIND (Promega Corporation, Madison, WI, USA). Домены DEF указанных EcRs амплифицировали, используя праймеры, созданные на основе 20-25-нуклеотидных последовательностей на 5’ и 3’ концах. Сайты фермента рестрикции EcoR I/BamH I и Xba I добавляли к праймерам 5’ и 3’ соответственно. Продукты ПЦР расщепляли соответствующими ферментами рестрикции и клонировали в вектор pBIND. EcR LBDs, клонированный в вектор pBIND верифицировали секвенированием. Химерный RXR из RXRp Homo sapiens и RXR Locusta migratoria, конденсированный с VP16-AD и помещенный под контроль промотора SV40e, конструировали, как описано ранее (Kumar P & Katakam A in Gene Transfer, Friedmann Т и Rossi J, eds, стр.643-651 (2007), Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY). Репортерная плазмида индуцируемой люциферазы pFRLuc (Stratagene Cloning Sytem, La Jolla, CA, USA) содержит пять копий элемента ответа GAL4 и синтетический минимальный промотор.
Клетки NIH3T3 поддерживали при 37°C и 5% CO2 в среде Игла, модифицированной Дулбекко (DMEM), с добавлением 10% плодной сыворотки телят, обе из которых были получены от Mediatech Inc., Manassas, VA. Клетки сеяли в 96-луночный планшет с плотностью 2500 клеток/лунка в 50 мкл среды для выращивания. На следующий день клетки сначала обрабатывали 35 мкл не содержащей сыворотки DMEM, содержащей диметилсульфоксид (DMSO; контроль), или раствором DMSO, содержащим лиганд. Клетки затем трансфицировали 15 мкл не содержащей сыворотки DMEM, содержащей 0,04 мкг конструкции EcR, 0,04 мкг конструкции RXR и 0,16 мкг конструкции репортера люциферазы на лунку, используя реагент трансфекции SuperFect™ (Qiagen Inc., Valencia, CA, USA) согласно инструкциям производителя. Лиганды испытывали в 8 дозах от 0,01 до 33 мкМ, и конечная концентрация DMSO составляла 0,33% как в контроле, так и в обработанных лунках. Через 48 часов после обработки и инкубирования трансфекции клетки анализировали на люциферазную активность, используя систему анализа люциферазы Bright-Glo™ (Promega Corporation, Madison, WI, USA), следуя инструкциям производителя. Значения EC50 измеряли как минимум в двух повторах.
Анализ гена-репортера
Экспрессию гена-репортера люциферазы измеряли через 48 ч после обработки клеток, используя систему анализа люциферазы Bright-Glo™ от Promega (E2650). Люминесценцию определяли при комнатной температуре, используя люминометр для микротитровальных планшетов Dynex MLX.
Каждый анализ производили в двух отдельных лунках, и усредняли два значения. Кратность индукций рассчитывали путем деления относительных световых единиц ("RLU") в клетках, обработанных лигандом, на RLU в клетках, обработанных DMSO. EC50 рассчитывали из данных доза-ответ, используя трехпараметрическую логистическую модель. Относительный Max FI определяли как максимальную кратность индукции тестируемого лиганда (воплощение по изобретению), наблюдаемую при любой концентрации, относительно максимальной кратности индукции лиганда GS™-E (3,5-диметилбензойной кислоты N-трет-бутил-N’-(2-этил-3-метоксибензоил)гидразид), наблюдаемой при любой концентрации.
Пример 67
В данном примере проиллюстрировано in vivo тестирование соединений по изобретению. Индукцию in vivo репортерного фермента хиральными диацилгидразиновыми лигандами по настоящему изобретению оценивали на модельной системе мышей C57BL/6, содержащей переключатель генов.
Кассеты экспрессии гена
Домены дикого типа D, Е, и F из EcR листовертки еловой Choristoneura fumiferana ("CfficR-DEF"; SEQ ID NO: 1) подвергали мутированию [V107 (gtt)→1107 (att) и Y127 (tac)→E127 (gag)] и сливали с ДНК-связывающим доменом GAL4 ("Gal4DBDI-147"; SEQ ID NO: 2). Спирали 1-8 включительно доменов EF из RXRβ Homo sapiens ("HsRXRp-EF"; нуклеотиды 1-465 из SEQ ID NO: 4) и спирали 9-12 включительно доменов EF из белка Ultraspiracle Locusta migratoria ("LmUSP-EF"; нуклеотиды 403-630 из SEQ ID NO: 5) сливали с доменом трансактивации из VP16 ("VP16AD"; SEQ ID NO: 6), который регулирует репортерный ген человека, секретирующий щелочную фосфатазу ("SEAP", SEQ ID NO: 12), который помещали под контроль элемента ответа 6xGAL4 (SEQ ID NO: 13) и транстиретиновый промотор (SEQ ID NO: 14). Каждый элемент переключателя генов находился на отдельной плазмиде. Экспрессия рецептора находилась под контролем промотора CMV (SEQ ID NO: 15). Индукцию оценивали по количеству репортерного белка, экспрессируемого в присутствии лиганда.
Электропорация переключателя генов
Экспрессию SEAP в сыворотке мыши оценивали после электропорации переключателя генов в четырехглавую мышцу мыши. Мышей анестезировали с помощью 2 мкл/г смеси кетамина (100 мг/мл) и ксилазина (20 мг/мл). Животных затем обривали, векторы ДНК инъецировали в мышцу в объеме 2×50 мкл полиглютаминовой кислоты (12 мг/мл воды), прикладывали гель проводимости электрода, и калибр электрода (1 см×1 см; модель 384) помещали на заднюю конечность. Мышцу электропорировали 200 В/см, 8 раз, в течение 20 миллисекунд/импульс, с временными интервалами 1 сек. Направление поперечного электрического поля изменяли на обратное после того как животные получали половину импульсов. Электропорацию выполняли при помощи электропоратора ECM 830 от BTX Molecular Delivery System.
Обработка хиральным диацилгидразиновым лигандом
В некоторых экспериментах мыши получали внутрибрюшинную инъекцию (IP) 2,6 мкмоль лиганда в 50 мкл DMSO через 3 суток после электропорации переключателя генов. В других экспериментах концентрацию лиганда снижали до 26 нмоль/50 мкл DMSO/мышь. Экспрессию SEAP оценивали через 2-11 суток после введения лиганда. В других экспериментах лиганд вводили в корм грызунов. Корм готовили путем растворения 2 г лиганда в 20 мл ацетона и добавления ее к 1 кг автоклавируемого корма LabDiet 5010 от Purina Mills. Все тщательно перемешивали в миксере Hobart и затем смешивали в течение еще 15 мин в миксере Cross Blend. Животные получали корм ad libitum (по совему желании) в течение 1, 2 или 3 суток. Все значения представляют собой среднюю величину от четырех животных. Фоновое значение SEAP в сыворотках животных, обработанных одним вектором без добавления лиганда составляло 0-11 нг/мл сыворотки.
Репортерный анализ
Мышиную сыворотку получали путем центрифугирования крови, полученной путем ретроорбитального кровоизвлечения с небольшой стеклянной капиллярной пробиркой. Количественное выражение SEAP определяли, используя хемилюминесцентный набор Clontech Great Escape и сравнивая со стандартом SEAP Clontech.
Для оценки относительной эффективности in vivo (R)-2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)-гидразида (Пример 1) в индуцировании экспрессии гена RheoSwitch® Therapeutic System (RTS), двенадцать мышей C57BL6 электропорировали следующими плазмидами: pCMV/GEVY(DEF), pCMV/VP16-Hs-LmRXR и p6XGAL4RE-TTR-SEAP. Плазмида pCMV/GEVY (DEF) состоит из доменов D, E и F EcR из Choristoneura fumiferana, имеющих мутации V390I/Y410E/E274V, слитых по ходу транскрипции от дрожжевой GAL4-DBD (aa 1-147) и помещенных под контроль CMV-промотора и по ходу транскрипции от сигнала полиаденилирования SV40 в векторе pBIND (Promega Corporation, Madison, WI, USA). DEF-Домены указанных EcR амплифицировали, используя праймеры, сконструированные на основе 20-25-нуклеотидных последовательностях на 5’ и 3’ концах. Сайты фермента рестрикции BamH I и Xba I добавляли к 5’ и 3” праймерам соответственно. Продукты ПЦР расщепляли соответствующими ферментами рестрикции и клонировали в вектор pBIND (Фиг.20). Вектор pCMV/VP16-Hs-LmRXR содержит химерный RXR из RXRβ Homo sapiens (спирали 1-8 домена E) и RXR Locusta migratoria (спирали 9-12 домена Е), слитые по ходу транскрипции от домена активации VP16 и помещенные под контроль промотора CMV в векторе pBDSfD (Фиг.21). Индуцируемый SEAP репортерный вектор p6xGAL4RE-TTR-SEAP содержит репортерный ген щелочной фосфатазы, секретируемой человеком, помещенный под контроль индуцируемого промотора, состоящего из 6 копий элемента ответа Gal4 расположенного против хода транскрипции от промотора транстиретина (TTR) (Фиг.22). Через трое суток после электропорации плазмидной ДНК в четырехглавую мышцу мышей, три лигандных препарата вводили мышам посредством внутрибрюшинной инъекции в количестве 26 нмоль/мышь (11,4 мкг/мышь, приблизительно 570 мкг/кг). Экспрессию щелочной фосфатазы, секретируемой человеком (SEAP), в мышиной сыворотке затем наблюдали в течение семнадцати суток после введения лиганда. Для сравнения оценивали относительную эффективность in vivo (S)-2-этил-3-метоксибензойной кислоты (N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида (Пример 31) и рац-2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида. Как показано на Фиг.3, (S)-2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)-гидразид (●-●) не индуцирует экспрессию репортерного гена RTS. N’-(1-трет-рац-2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразид индуцирует умеренную экспрессию репортерного гена RTS (▲-▲). (R)-2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)-гидразид (O-O) индуцирует существенную экспрессию репортерного гена RTS.
Пример 68
Физические свойства R-энантиомера и рацемата 2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметил-бензоил)-гидразида:
Морфологическая форма: партии твердых образцов представлены на Фиг.4. В отношении R-энантиомера, образцы, полученные посредством быстрой кристаллизации/осаждения из смеси либо метанол/вода или толуол/гептан, имеют одинаковые картины дифракции рентгеновских лучей на порошке (данные не показаны), и имеют практически одинаковую точку плавления ([толуол/гептан] 166,2-167,1°C, [CH3CH/H2O] 166,5-167,4°C) по сравнению друг с другом и по сравнению со стандартом, полученным при кристаллизации из CH3OH (165,1-166,5°C). В отношении рацемата, два отдельных препарата, полученных путем выпаривания метанола, имели одинаковую точку плавления (170-171°C, 169-170°C) в рамках погрешности эксперимента и незначительные отклонения в чистоте. Морфологические формы рацемата и R-энантиомера 2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)-гидразида оказались типичными.
Микронизация и размер частиц: Чтобы нормализовать по размеру частиц, каждый из R-энантиомера и рацемата 2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида предварительно просеивали через 20 меш и микронизировали. Размер частиц определяли путем отвешивания примерно 20 мг испытуемого вещества в 15 мл пробирку Фалькона. В порошок добавляли 10 мл деионизированной воды. Суспензию перемешивали, обрабатывали ультразвуком в течение 20 минут и затем сразу же анализировали на размер частиц с помощью дифракции лазерного излучения, используя устройство Malvern Mastersizer. Распределение по размеру показано на Фиг.5 и 6. Хотя оказалось, что рацемат имеет немного больший общий размер частиц, чем R-энантиомер, два образца являлись совершенно похожими.
Анализ объемной плотности и плотности после уплотнения: Объемные плотности микронизированных R-энантиомера и рацемата 2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)-гидразида рассчитывали, используя измеренную массу и объем в 10 мл мерном цилиндре. Плотность после уплотнения микронизированного вещества рассчитывали, используя массу вещества и объем после уплотнения. Объем после уплотнения измеряли, используя анализатор плотности после уплотнения (модель: Stampfvolumeter, STAV2003, JEL), объем после уплотнения фиксировали, когда он становился постоянным (Фиг.7). R-энантиомер имеет немного более низкую объемную плотность и плотность после уплотнения. Оба вещества являются хлопьевидными (низкая насыпная плотность).
Термогравиметрический анализ/Дифференциальный термический анализ (TGA/DTA):
Микронизированные R-энантиомер и рацемат 2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида анализировали с помощью термогравиметрического анализа/дифференциального термического анализа. Термогравиметрическая кривая показана в Фиг.8. Оба вещества показали эндотермическое явление на профиле DTA. Начальная температура для R-энантиомера (163,6°C) была значительно ниже температуры рацемата (171,2°C). Теплота плавления для R-энантиомера (59,8 мкВ с/мг) также была значительно ниже, чем для рацемата (80,8 мкВ⋅с/мг). Это показывает, что два вещества представляют собой разные кристаллические формы. Более низкая точка плавления и более низкая теплота плавления R-энантиомера отражаются в различных свойствах растворимости.
При нагревании выше 150°C R-энантиомер терял 0,4% общей массы и рацемат терял 0,8% общей массы (Фиг.8). Влагосодержание (влага, удаляемая теплом) двух веществ слегка отличается.
Анализ динамической сорбции пара: При воздействии влагопоглощающих условий (20% RH (относительная влажность)), рацемат не показал никакой значительной потери массы, тогда как R-энантиомер потерял 0,9% от общей массы (данные не показаны), что может соответствовать свободно ассоциированной влаге, приобретенной во время переноса или хранения образца. После дегидратации во влагопоглощающих условиях, два вещества показали сравнимые увеличения массы при воздействии низкой (30-40% RH), средней (50-60% RH) и высокой (70-80% RH) влажности. При 70%-ной RH увеличение массы составляет 6,3% для R-энантиомера и 6,2% для рацемата (данные не показаны), указывая на умеренную гигроскопичность обоих веществ.
Растворимость в фармацевтических эксципиентах: Анализ 1: Качественно оценивали растворимость микронизированных R-энантиомера и рацемата 2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)-гидразида в имеющихся в продаже эксципиентах (Фиг.9). Исходные растворы/суспензии 10 мг/г (тестируемое соединение/эксципиент) в стеклянных флаконах получали при помощи обработки ультразвуком при комнатной температуре в течение 15 минут с закручиваем в течение 30 секунд с 5 минутными перерывах, с последующим нагреванием в алюминиевом термоблоке при 50°C в течение 12 минут с завихрением в течение 30 секунд с 3 минутными перерывами. На основании внешнего вида данного исходного образца эксципиента 10 мг/г анализировали 2-4 точечное множество концентраций при помощи селекции проб вверх или вниз в нескольких из следующих точек: 5, 10, 15, 20, 30, 50 мг/г (тестируемое соединение/эксципиент). R-энантиомер был более растворимым, чем рацемат во всех тестируемых фармацевтических эксципиентах, за исключением чистого полисорбата 80, в котором R-энантиомер и рацемат показывают сравнимую растворимость (смотри Фиг.9). Никакого изменения цвета не наблюдалось для любого из образцов. Были сделаны фотоснимки некоторых препаратов в прозрачных стеклянных флаконах (не показаны).
Анализ 2: Для каждого эксципиента в анализе, показанном на Фиг.9, устанавливали самую низкую концентрацию, при которой R-энантиомер или рацемат 2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида является нерастворимым. Суспензии обоих веществ в этой же концентрации готовили во флаконах. Каждый образец перемешивали при комнатной температуре в течение ≤2,5 ч (обработка 1). В качестве отдельного эксперимента, тестируемые комбинации соединение/эксципиент нагревали в алюминиевом блоке при 90°C в течение 5 минут, или, при необходимости, дольше; показано в Фиг.10, и оставляли охлаждаться до комнатной температуры, и вносили затравку минимального количества такого же микронизированного вещества (обработка 2). Физический внешний вид регистрировали в 72 ч или в указанное время (Фиг.10). Во всех случаях не наблюдалось изменения цвета. Данный анализ растворимости действует, начиная от состояния полного растворения, в котором исходный размер частиц и морфологическая форма являются несущественными. Наблюдение за осаждением/кристаллизацией при затравливании перенасыщенных растворов рацемата 2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида любой морфологической формой кристаллического рацемата, в то время как R-энантиомер остается полностью растворенным (особенно без нагревания), указывают на различие растворимости рацемата и R-энантиомера в тестируемой концентрации. Сделаны фотоснимки препаратов в прозрачных стеклянных флаконах (R-энантиомер в условиях анализа 1 без нагревания до 90°C и рацемат после обработки при 90°C, охлаждения и затравки кристалла, не показаны).
Анализ 3: R-энантиомер и рацемат 2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида смешивали с 20%-ным PEG 1000 в дистиллированной в воде, рН 7,0 в концентрации 1-2 мг порошка на мл. Смесь инкубировали при 37°C, перемешивали на вортексе при комнатной температуре в течение приблизительно 2 часов и оставляли отстаиваться в течение еще 12 часов при комнатной температуре. Раствор фильтровали или центрифугировали, и концентрации определяли количественно при помощи удвоенных инжекций LC/MS/MS, используя общий способ LC/MS/MS с минимум 4-точечной калибровочной кривой (Фиг.11). Смотри Lobenberg R, et al., Solubility as a limiting factor to drug absorption, In: Oral Drug Absorption, J.B. Dressman (Ed), Marcel Dekker, NY, 2000 и Mader WJ, Grady LT, Determination of solubility, Chapter 5 in Techniques of Chemistry, Physical Methods in Chemistry, Vol.1, Part V, Weissberger, V.A., Rossiter, B.W., Eds., Wiley, New York, 1971.
Анализ 4: Равновесная растворимость в эксципиентах композиции и биологических жидкостях влияет на биодоступность лекарственного средства, хотя кинетика растворения также может являться ограничивающим фактором в определении биодоступности. Растворимости R-энантиомера и рацемата 2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида количественно устанавливали в водных препаратах фармацевтического поверхностно-активного полисорбата 80 и затем получали действительную скорость растворения уплотненного тестируемого вещества. Смеси рацемата и R-энантиомера в 0,5%, 1%, и 1,5% растворах полисорбата 80 получали путем добавления 20+/-1 мг тестируемого вещества в 2 мл флаконе Эппендорфа, с последующим добавлением 500 мг раствора полисорбата. Смесь разбивали бусинками в течение 120 секунд, встряхивали при 25°C в течение 16-24 часов, и фильтровали, используя Spin-X (0,2 мкМ). Регистрировали рН фильтрата. Концентрацию тестируемого вещества в фильтрате определяли с помощью количественного анализа ВЭЖХ (Фиг.12). R-энантиомер является в 2-3 раза более растворимым в 1,0 и 1,5% растворах полисорбата 80. В деионизированной воде и 0,5%-ном полисорбате 80 растворимость рацемата и R-энантиомера сравнимы.
Затем профили действительного растворения R-энантиомера и рацемата 2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида определяли, используя спрессованные гранулы (диаметр 7 мм, 100 мг) в 1%-ном Полисорбате 80 в дистиллированной воде при 37°C, согласно следующим условиям испытания:
Емкость: 1000 мл
Среда: 1%-ный Полисорбат 80 в дистиллированной воде (вакуум-фильтрация через фильтр 0,8 мкм)
Объем среды: 1000 мл
Скорость: 100 об/мин
Температура: 37°C
Точки отбора проб: 15, 30, 45, 60, 90 и 120 мин
Объем проб: 1 мл (фильтровали через 30 микрон)
Анализ: ВЭЖХ (получают стандартные растворы от каждой партии)
Размер образца: n=2 гранулы/тестируемое вещество.
Высвобождение рацемата и R-энантиомера было ниже предела детектирования с помощью ВЭЖХ вплоть до 120 минут. Через 120 минут было замечено, что гранулы обоих веществ сохранились в среде в неизмененном виде.
Пример 69
Свойства проницаемости через клеточной мембраны R-энантиомера и рацемата 2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида:
Двухнаправленная проходимость через клеточные монослои Caco-2 (смотри Sambuy Y, et al., The Caco-2 cell line as a model of the intestinal barrier: influence of cell and culture-related factors on Caco-2 cell functional characteristics. Cell Biol Toxicol. 2005 Jan; 21(I):I-26 и Artursson P, et al., Caco-2 monolayers in experimental и theoretical predictions of drug transport. Adv Drug Deliv Rev. 2001 Mar 1; 46(I-3):27-43):
Данные показаны в Фиг.13. Тестируемые соединения готовили в виде 5 мкМ в HBSSg с максимальной концентрацией DMSO≤1%. Готовили 21-28 суточные слитые монослои клеток Caco-2. Приемную лунку готовили с добавлением 1%-ного BSA в модифицированном буфере Хэнкса HBSSg, и апикальные и базолатеральные стороны клеток Caco-2 поддерживали при рН 7,4±0,2. Два монослоя дозировали в каждом направлении (N=2): апикальную сторону дозировали для оценки (A→B) и базолатеральную сторону дозировали для оценки (B→A). Концентрацию тестируемого соединения измеряли как для апикальной так и для базолатеральной стороны в момент времени 120 минут, используя общий способ LC/MS/MS с минимум 4-точечной калибровочной кривой. Значения на Фиг.13 представляют собой: процент извлечения тестируемого соединения из лунок Transwell®, содержащих клеточные монослои Caco-2; видимая проницаемость (Papp) в обоих направлениях; отношение истечения (Papp B→A)/(Papp A→B); абсорбционный потенциал тестируемого соединения, классифицируемый или как Низкий или Высокий; классификация истечения является значительным при истечении ≥3,0 и Papp (B→A)≥1,0×10-6 см/с. R-энантиомер обладает лучшей проникающей способностью, чем рацемат, в обоих направлениях. Также, скорость истечения для R-энантиомера ниже.
Определение потенциала проницаемости через гематоэнцефалический барьер, с использованием клеток MDR1-MDCK (смотри Taub ME, et al., Fubctional assessment of multiple P-glycoprotein (P-gp) probe substrates: Influence of cell line и modulator concentration on P-gp activity. Drug Metab Dispos. 2005 Nov; 33(II): 1679-87 и Wang Q, et al., Evaluation of the MDR-MDCK cell line as a permeability screen for the кровь-brain barrier. Int J Pharm. 2005 Jan 20; 288(2): 349-59. (скорректирован из Absorption Systems Inc. bulletin):
Данные показаны на Фиг.14. Тестируемые соединения получали в виде 5 мкМ раствора в HBSSg с максимальной концентрацией DMSO≤1%. Готовили 7-11 суточные слитые монослои клеток MDR1-MDCK. Приемную лунку готовили с добавлением 1%-ного BSA в модифицированном буфере Хэнкса (HBSSg), и апикальные и базолатеральные стороны поддерживали при рН 7,4±0,2. Два монослоя дозировали в каждом направлении (N=2): апикальную сторону дозировали для оценки (A→B) и базолатеральную сторону дозировали для оценки (B→A). Концентрацию тестируемого соединения измеряли как для апикальной так и для базолатеральной стороны в момент времени 120 минут, используя общий способ LC/MS/MS с минимум 4-точечной калибровочной кривой. Зарегистрированные значения на Фиг.14: процент извлечения тестируемого соединения из лунок Transwell®, содержащих клеточные монослои MDR1-MDCK; видимая проходимость (Papp) в обоих направлениях; отношение истечения (Papp B→A)/(Papp A→B); и классификация потенциала проникновения через гематоэнцефалический барьер. R-энантиомер 2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида обладает большей проницающей способностью, чем рацемат, в обоих направлениях A→B и B→A в клетках MDR1-MDCK. Также, R-энантиомер демонстрирует меньшее истечение. R-энантиомер классифицируют как умеренно проникающий через гематоэнцефалический барьер, в то время как рацемат классифицируют как слабо проникающий. Более высокое проникновение через гематоэнцефалический барьер является значительным преимуществом для медицинских показаний, там, где требуется проникновение в центральную нервную систему (CNS).
Пример 70
Микросомальный метаболизм R-энантиомера и рацемата 2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)-гидразида (смотри Jeffrey P, et al., Utility of metabolic stability screening: comparison of in vitro и in vivo clearance. Xenobiotica. 2001 Aug-Sep; 31(8-9): 591-8 и Lin JH, et al., Role of pharmacokinetics и metabolism in drug discovery и development. Pharmacol Rev. 1997 Dec; 49(4): 403-49)
Тестируемые соединения получали в виде 5 мкМ растворов в DMSO (концентрация менее 0,25%), метаноле или ацетонитриле. Объединяли микросомы печени людей разного пола от ≥10 доноров и инкубировали с 1 мМ NADPH. Тестируемое соединение инкубировали при 37°C в буфере, содержащем 1,0 мг/мл микросомального белка с 1 мМ NADPH. В момент времени 0 и 60 минут от смеси отбирали пробы и наблюдали при помощи LC/MS/MS за площадью пика, соответствующего тестируемому соединению, с диапазоном 10-100%. Анализ выполняли дважды (N=2 отдельных инкубирования). Данные, представленные в Фиг.15, представляют собой процент сохранения тестостеронового стандарта и процент сохранения тестируемого соединения. R-энантиомер является немного более стабильным к метаболизму в микросомах печени человека, чем рацемат.
Пример 71
Биодоступность R-энантиомера и рацемата 2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида.
Соединения в Labrasol вводили мышам C57BL/6N:Crl (дикий тип) (Charles River Laboratories) при помощи перорального чреззондового питания, предполагаемого пути введения человеку. Дозировки установили в 3, 10, 30, или 50 мг/кг/сутки и 2,5 мл/кг/доза (Фиг.16). Дозы вводили в течение 9 суток (первые 18 самок/группа) или 12 суток (вторые 18 самок/группа) (Фиг.16). Кровь отбирали в точки времени 9 суток и 12 суток. Концентрации количественно определяли при помощи ВЭЖХ на обращенной фазе.
Для каждой концентрации образцы получали путем добавления Labrasol к твердому R-энантиомеру или рацемату в лабораторном стакане, обработки ультразвуком для разрушения видимых агрегатов и магнитного перемешивания вплоть до достижения однородного внешнего вида. Смеси переносили в мерный цилиндр и его дополняли до необходимого объема, используя промывку лабораторного стакана Labrasol. Содержимое цилиндра перемешивали путем переворачивания и перемешивали магнитной мешалкой вплоть до дозирования.
В данных дозированных препаратах Labrasol использовали микронизированный R-энантиомер, но немикронизированный рацемат. Для изучения вероятности того, что образец немикронизированного рацемата может оказывать влияние на скорость своего растворения в Labrasol, выполняли независимый эксперимент. В данном эксперименте исследовали растворимость и внешний вид 20 мг/мл препаратов микронизированного и немикронизированного рацемата. Каждый из микронизированного рацемата (партия REH-28-9-1/PYAP-2-8-2-2M), немикронизированного рацемата (партия #PYAP-2-8-2-2) и микронизированного R-энантиомера (партия #REH-28-4-2) суспендировали в Labrasol, размешивали вручную и затем обрабатывали ультразвуком в ультразвуковом аппарате настольного типа Branson 2100 при 25-28°C в течение 3×5 мин интервалов и интервала 1×10 мин, попеременно с размешиванием вручную. R-энантиомер легко растворялся, в то время как смеси и микронизированного и немикронизированного рацемата оставались мутными с суспендированными мелкими частицами (фотоснимки не показаны). При визуальном контроле количество нерастворенного вещества в образцах микронизированного и немикронизированного рацемата было по существу одинаковым. Микроскопирование (Фиг.17) показало, что большая часть частиц в суспензиях как микронизированного, так и немикронизированного рацемата была ниже 25 микрон (Фиг.17). Несколько крупных кристаллических фрагментов сохранялось в немикронизированном образце. Так как и микронизированные и немикронизированные образцы рацемата в результате имели аналогичные количества нерастворенного вещества, по-видимому главным различием между препаратами R-энантиомера и рацемата с Labrasol является действительная растворимость, а не исходный размер частиц растворенного вещества.
Уровни в сыворотке мышиной крови в виде функции от времени определяли при помощи количественной ВЭЖХ. R-энантиомер в Labrasol достигает значительно более высоких титров сыворотки крови при более высоких дозах 30 и 50 мг/кг/сутки (12 и 20 мг/мл введенного Labrasol). Уровни рацемата в сыворотке крови остаются существенно более низкими (<4-5 раз AUC) (Фиг.18 и 19). Различия в исходном размере частиц рацемата и R-энантиомера могут являться причиной части различий в сывороточном уровне. Также, более высокая действительная растворимость R-энантиомера по сравнению с рацематом в Labrasol может быть причиной части различий в сывороточном уровне. Кроме того, различия в сывороточном уровне могут быть следствием различий в мембранной проницаемости (эксперименты на клетках Caco-2 и клетках MDCK; Фиг.13 и 14), абсорбции, распределения или выделения.
Пример 72
Сравнение R-энантиомера и рацемата 2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида в отношении in vivo индукции продуцирования трансгена IL-12 из генномодифицированных опухолевых клеток.
Трансген, тестируемый в данном примере, представляли собой мышиный IL-12 под контролем RTS. Клетки меланомы мыши (B16) трансдуцировали аденовирусным вектором Ad-RTS-mIL-12, который несет мышиный трансген IL-12 под контролем RTS. Эти клетки инъецировали подкожно (п.к.) мышам C57BL/6 (гистосовместимых с клетками B16), и мышей обрабатывали разными дозами R-энантиомера и рацемического 2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразида в течение трех суток. Контрольные группы включали мышей, получавших нетрансдуцированные клетки B16 и более высокую дозу лиганда, и другую группу, получавшую трансдуцированные клетки B16 в отсутствие лиганда. Для оценки трансгенной экспрессии, непосредственно индуцированной лигандами, опухоли и сыворотку собирали от половины мышей из каждой группы через 72 часа после подкожной инъекции. Опухолевые экстракты и сыворотку готовили и анализировали в отношении IL-12 при помощи ELISA.
Остальных мышей использовали для тестирования выключения трансгена, когда введение лиганда прерывают в течение еще 72 часов. Результаты показывают различие экспрессии трансгена IL-12 в опухоли и сывороточных титров у мышей, обработанных рацематом и R-энантиомером лиганда. Прерывание введение лиганда приводило к выключению трансгена во всех группах аналогичным образом.
Материалы и методы: клетки В16 инфицировали Ad.-RTS-mIL-12 (MOI=100) in vitro. Через 48 ч, 5×105 (B16, правый бок) клеток инъецировали п.к. мышам C57BL/6 (10 мышей/группа). Лиганды, R-энантиомер и рацемат, растворяли в Labrasol посредством нагревания 10 мг/мл смесь при 50°C в течение 5 минут. Оба лиганда, согласно визуальному контролю, растворялись в Labrasol. Активирующие лиганды обеспечивали при помощи перорального кормления через зонд в следующих дозах (0 мг/кг; 3 мг/кг; 10 мг/кг; 30 мг/кг и 50 мг/кг) ежесуточно в течение 3 суток. Дополнительные контроли включали B16-немущих мышей, получавших только лиганд (50 мг/кг/сутки в течение 3 суток и необработанных опухоль-несущих мышей). Пять мышей/группа отбирали для выделения опухолевых поражений и периферической крови. Ткань гомогенизировали в общем объеме 1 мл фосфатно-солевого буферного раствора. Лизат/сыворотку криоконсервировали при -80°C. Других 5 мышей/группу оставляли сидеть в течение еще 72 ч в отсутствие дополнительного введения лиганда. Этих последних 5 мышей/группу забирали для выделения опухолевых поражений и периферической крови. Ткань гомогенизировали, как указано выше, и лизат/сыворотку криоконсервировали при -80°C. Образцы размораживали при 37°C в водяной бане и анализировали в отношении mlL-12p70 и также mIFN-y, дополнительного IL-12-зависимого цитокина, важного для CTL миграции в опухоли, используя специальные ELISA (BD-Pharmingen).
Результаты: Трансгенная экспрессия IL-12 в опухоли, а также уровни в сыворотке соответствуют дозозависимой диаграмме для обоих лигандов (Фиг.24 и 25). Однако, уровни экспрессии были в 2,5-3 раза выше у мышей, обработанных R-энантиомером 2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)-гидразида. Данную диаграмму наблюдали в опухолевых экстрактах и в сыворотке. Контрольные группы, в которых мыши получали или нетрансдуцированные клетки B16 плюс максимальную дозу лиганда, или трансдуцированные клетки B16 без введения лиганда, не демонстрировали никакой экспрессии IL-12. В группах, в которых обработку лигандом прерывали на 3 суток, уровни экспрессии снижались в равной степени во всех обработанных группах, указывая на то, что лиганд выводится, и активация RTS является обратимой, когда лиганд отменяют. При анализе уровней IFN-γ в опухоли и сыворотке, группа R-энантиомера отчетливо продемонстрировала усиленную экспрессию.
Пример 73
Фармацевтическая композиция, содержащая 2-этил-3-метоксибензойной кислоты N-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразид В Таблице 1 приведена фармацевтическая композиция, содержащая 2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразид, PL90G, витамин Е TPGS, ВНТ, и Miglyol 812N. PL90G представляет собой высокоочищенный фосфатидилхолин сои. Производителем PL90G является PHOSPHOLIPID GmbH, филиал Lipoid. Витамин Е TPGS представляет собой соединение GRAS (обычно признаваемый безопасным) и соответствует всем требованиям National Formulary (NF) (Национального формуляра). Miglyol 812N содержит триглицериды фракционированных растительных C8- и C10-жирных кислот. Жирные кислоты, используемые для получения Miglyol 812N, классифицируют как GRAS и они отвечают требованиям современных NF и ЕР для среднецепочечных триглицеридов. BHT [MeC6H2(CMe3)2OH] представляет собой органическое соединение, которое является липофильным (жирорастворимым) фенолом, который широко используется в качестве антиокисляющей пищевой добавки. Композицию распределяют в твердые желатиновые капсулы.
Пример 74
Эффективность в животных моделях заболевания Мышиные или человеческие дендритные клетки (DC), трансдуцированные аденовирусным вектором, кодирующим RTS и mIL-12 или hIL-12 соответственно, экспрессируют цитокин in vitro в присутствии (R)-2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)-гидразида дозозависимым способом. В отсутствие (R)-2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)-гидразида экспрессия IL-12 находится на нижнем уровне.
Сравнительный анализ влияния различных путей введения и доз (R)-2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)-гидразида на противоопухолевую эффективность доставляемых внутрь опухоли DC, трансдуцированных аденовирусным вектором, кодирующим мышиный интерлейкин-12p70 (DC-SPI(IL-12p70)), выполняли на мышиной модели меланомы B16. Результаты показали, что противоопухолевая эффективность была продемонстрирована при терапии (DC-SPI(IL-12p70)) в комбинации с (R)-2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразидом, введенным при помощи внутрибрюшинной (в.б.) инъекции, перорального кормления через зонд или диетической смеси. Оптимальные противоопухолевые эффекты имели место при дозе >30 мг/кг N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)-гидразида (R)-2-этил-3-метоксибензойной кислоты, введенной с терапией (DC-SPI(IL-12p70)).
В модели мышиной меланомы B16, в которой мышей инокулировали опухолью перед началом обработки (мышам C57/BL6 вводили подкожные инъекции клеток B16 для образования опухолей), трансдуцированные AdDC, содержащие мышиные гены IL-12, инъецированные в опухоль, опосредовали полную регрессию опухоли и пролонгированное выживание животного, которое зависело от системного введения N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)-гидразида (R)-2-этил-3-метоксибензойной кислоты в течение 48 часов после инъекции AdDC. Такой зависящий от лекарственного средства эффект был связан с: трансгенной экспрессией в опухоли и DLN; пролонгированным выживанием Ad-DC в опухолевой микросреде; миграцией и жизнестойкостью AdDC в DLN; и индукцией анти-B16 CD8+ T-клеток.
В указанной выше мышиный модели меланомы B16 внутриопухолевая терапия, состоящая из инъекции 107 AdDC в комбинации с 13-суточной обработкой (R)-2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразидом, демонстрировала защитный иммунитет, который был опухоль-специфическим. Когда животным без опухоли делали провокацию, на 50 сутки (после первоначальной инокуляции опухоли), эти животные были не восприимчивы к меланоме B16, но не к развитию карциномы толстого кишечника MC38.
Пример 75
Репликационно-некомпетентный рекомбинантный аденовирус, содержащий RTS и hIL-12
Фиг.23 представляет собой схематическое изображение Ad-RTS-hIL-12, где участки E1 и E3 делегированы, и компоненты RTS-IL12 замещают участок E1 (Фиг.23 не масштабирована). Термины, используемые на Фиг.23 имеют следующие значения:
RTS-IL-12: Человеческий IL-12 под контролем терапевтической системы RheoSwitch®
Preg: Промотор, стимулирующий экспрессию IL12, регулируемый активированным лекарственным средством
hIL-12: Кодирующие последовательности для субъединиц p40 и p35 человеческого EL-12, разделенные последовательностью IRES
polyA: Сигнал полиаденилирования
PUbC: Убиквитин C-промотор
VP16-RXR: Слияние домена активации транскрипции VP16 и химерного RXR
IRES: Внутренний сайт вхождения в рибосому
GaI4-EcR: Слитый белок из ДНК-связывающего домена GaI4 и лиганд-связывающего домена экдизонового рецептора
Ad: Основа аденовируса-5, содержащая генные делеции E1 и E3.
Рекомбинантный аденовирусный вектор Ad-RTS-hlL-12 получают, как описано в Anderson et al, Gene Therapy, 4: 1034-1038 (2000). Вектор Ad-RTS-hlL-12 продуцируют следующим способом. Кодирующие последовательности для рецепторных слитых белков, VP16-RXR и GaI4-EcR, разделенных EMCV-IRES (внутренний сайт вхождения в рибосому), вставляют в аденовирусный челночный вектор под контролем человеческого убиквитин C-промотора (конститутивного промотора). Затем кодирующие последовательности для субъединиц p40 и p35 hlL-12, разделенных IRES, помещенные под контроль синтетического индуцируемого промотора, вставляют против хода транскрипции от убиквитин C-промотора и рецепторных последовательностей. Челночный вектор содержит последовательности серотипа 5 аденовируса с левого конца до mu16, из которых последовательности E1 делегированы и замещены последовательностями RTS и IL-12 (RTS-hlL-12). Челночный вектор, несущий RTS-hlL-12, тестируют посредством неустойчивой трансфекции в клетки HT-1080 для лиганд-зависимой экспрессии IL-12. Челночный вектор затем рекомбинируют с E3-делетированной аденовирусной основой посредством котрансфекции в клетки HEK 293 с получением рекомбинантного аденовируса Ad-RTS-hIL-12. Аденовирусный вектор клинической чистоты продуцируют в cGMP аппаратуре.
Пример 76
Трансдукция аутологичных незрелых дендритных клеток аденовирусом, содержащим трансген hIL-12 и терапевтическую систему RheoSwitch®
Получение PBMC с помощью лейкафереза: Субъектов подвергали 90-120 минутному лейкаферезу в Hillman амбулаторном CTRC. Методика лейкафереза включает отвод крови из вены в одной руке; пропускание крови через центрифугу (клеточный сепаратор), где ее компоненты разделяют и один или более компонентов удаляют; и возвращение оставшихся компонентов в вену субъекта в той же самой или другой руке. Не более 15% от общего объема крови субъекта извлекают за один раз, в то время как кровь пропускают через сепаратор для клеток. В клеточном сепараторе кровь разделяют на плазму, тромбоциты, лейкоциты и эритроциты. Лейкоциты (WBC) извлекают, и все другие компоненты возвращают в кровоток субъекта. Каждую попытку выполняют с использованием двух периферических IV линий для данной процедуры. Если это невозможно, может быть необходима центральная линия. Субъект должен быть рекомендован терапевтом для лейкафереза, и его обычно проверяют в отношении основных показателей состояния организма (включая кровяное давление) перед процедурой.
Процессинг: После сбора, лейкопак доставляют вручную к CPL, и сразу же обрабатывают посредством ценрифужной сепарации в ELUTRA™. Она представляет собой замкнутую систему, утвержденную для клинического использования. Фракцию моноцитов извлекают, и после установления извлечения и жизнеспособности клеток, их переносят в картридж Aastrom для 6-суточного культивирования в присутствии IL-4 и GM-CSF. Все процедуры процессинга и промывания выполняют в стерильных условиях.
Первичный посев: Моноциты, извлеченные из одного лейкопака, подсчитывают в присутствии красителя трипанового синего для определения количества жизнеспособных клеток. Моноциты оценивают в отношении чистоты с помощью проточной цитометрии. Моноциты ресуспендируют в не содержащей сыворотки и антибиотиков среде CellGenix, содержащей 1000 МЕ/мл IL-4 и 1000 МЕ/мл GM-CSF на SOP-CPL-0166, и помещают в картридж Aastrom. Для инокулирования кассеты требуется минимальный загрузочный объем 50 мл и минимальное количество клеток.
Культивирование: Картридж Aastrom помещают в инкубатор в Replicell System, полностью замкнутое, cGMP-совместимое автоматизированное устройство для культивирования для генерации незрелых DC.
Сбор незрелых DC: На 6 сутки картридж Aastrom вынимают из инкубатора и собирают незрелые DC. Клетки извлекают центрифугированием при 1500 об/мин, промывают в среде CellGenix, подсчитывают в присутствии красителя трипанового синего и проверяют морфологические и фенотипические характеристики.
Жизнеспособность: Ее определяют, осуществляя гемоцитометрический подсчет клеток в присутствии трипанового синего. Обычно >95% собранных клеток является жизнеспособными, то есть, не включают краситель трипановый синий. Если жизнеспособность менее 70%, незрелые DC отбрасывают.
Фенотипирование: Клетки, полученные в культуре, подсчитывают при помощи микроскопического исследования на гемоцитометре, и получают предварительное определение лейкоцитарной формулы (DC против лимфоцитов), используя краситель трипановый синий. Подтверждение лейкоцитарной формулы получают при помощи проточной цитометрии, устанавливая DC против лимфоцитов и используя свойства интенсивного прямого и бокового рассеяния света незрелыми DC в качестве критерия их идентификации. Незрелые DC обычно содержат >80% клеток с дендритной клеточной морфологией и имеют фенотип DC.
Анализ эффективности IL-12p70: Было установлено, что зрелые DC (mDC) обладают способностью продуцировать IL-12p70 спонтанно или при активации посредством CD40L при добавлении или без добавления сигналов наследственного иммунитета (например LPS). Стандартизированный анализ продуцирования IL-12p70 в последнее время установлен и применим к небольшим образцам или крупным партиям DC-вакцин, созданных в различных условиях. Современный анализ эффективности состоит из двух разных стадий, первая из которых включает совместное инкубирование иммунокомпетентных DC с клетками лимфомы J588, стабильно трансфицированных геном лиганда CD40 человека, в качестве стимуляторов. Вторая стадия включает тестирование надосадочных жидкостей из данных совместных культур на уровни IL-12p70, секретируемых DC, стимулированными J558/CD40L+/-LPS в системе Luminex. Данный анализ эффективности имеет CV (коэффициент вариативности) анализов 18,5% (n=30) и широкий динамический диапазон, что облегчает определение различных DC-продуктов, характеризующихся очень различными уровнями продуцирования IL-12p70. Стандартный установленный диапазон для анализа с использованием DC-продуктов, генерируемых от моноцитов 13 обычных доноров, составлял 8-999 пг/мл, со средним значением 270 пг/мл.
Пример 77
Аденовирусная трансдукция незрелых DC
Незрелые DC собирают, подсчитывают и тестируют на жизнеспособность. Приблизительно 6-7×107 DC трансдуцируют аденовирусным вектором при оптимальной множественности инфекции (оптимальная MOI, между 500 и 1000, подлежит уточнению) в течение оптимального времени адсорбции вируса (между 2 ч и 4 ч, подлежит уточнению). После трансдукции клетки многократно промывали для удаления любых неадсорбированных вирусных частиц. Отделяют аликвоты для анализа стерильности, эндотоксина, микоплазмы, фенотипа DC и индукции трансгена IL-12. Целевую дозу 5×107 клеток поддерживают при 4°C в солевом растворе вплоть до завершения испытания при выпуске.
Трансдуцированные DC (AdDC), которые прошли все испытания при выпуске, доставляют в клинику для введения пациенту, как описано ниже.
Для получения результатов испытания in vitro индукции трансгена IL-12 с помощью трансдуцированных AdDC требуется как минимум двое суток, и следовательно продуцирование IL-12 в ответ на (R)-2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)гидразид не будет выпускающим критерием для внутриопухолевой инъекции AdDC. Однако, на основе предварительных исследований с использованием AdDC, полученных от трех здоровых добровольцев, индуцированной экспрессии IL-12 под влиянием (R)-2-этил-3-метоксибензойной кислоты N’-(1-трет-бутил-бутил)-N’-(3,5-диметилбензоил)-гидразида был достигнут в рамках целевого диапазона от 80 до 300 нг IL-12 на миллион клеток за 24 часа (уровень экспрессии 80 нг/миллион клеток/24 ч наблюдали в 2 из 3 препаратов AdDC). Следовательно, результаты индукции IL-12 in vitro используют для анализа результата обработки. Как таковые, трансдуцированные AdDC используют для инъекции после вирусной трансдукции и промывания. Любые оставшиеся нетрансдуцированные и трансдуцированные DC криоконсервируют для будущего анализа по мере необходимости.
Введение AdDC пациентам: Однократная внутриопухолевая инъекция AdDC в 0,9%-ном солевом растворе, который содержит приблизительно 5×107 клеток на инъекцию (или приблизительно 90% выхода DC от субъекта испытания) в объеме 150 микролитров. Субъектам следует дозировать максимально возможную дозу AdDC, но не более 5×107 клеток на инъекцию.
Пример 78
Введение людям трансдуцированных аденовирусом аутологичных дендритных клеток, сконструированных для экспрессии hIL-12 под контролем терапевтической системы RheoSwitch у субъектов с меланомой III и IV стадий.
Группа 1 состоит из 12 субъектов и группа 2 состоит из 28 субъектов, все из которых получают одну внутриопухолевую инъекцию трансдуцированных аутологичных AdDC приблизительно через 24 часа после первой дозы (R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2-этил-3-метоксибензоил)гидразида. Субъектов группы 1 помещают в 4 группы по 3 субъекта в каждую (A, B, C, D) для получения возрастающих доз (R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2-этил-3-метоксибензоил)гидразида. Субъекты группы 2 получают максимальную переносимую пероральную дозу (R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2-этил-3-метоксибензоил)гидразида (которую определяли на основании группы 1). Сразу же после инъекции AdDC субъектов обрабатывают тринадцатью дополнительными суточными дозами (R)-3,5-диметил-бензойной кислоты N-(1-трет-бутил-бутил)-N’-(2-этил-3-метоксибензоил)гидразида. Безопасность, переносимость и функцию трансгена оценивают для всех субъектов в каждой группе группы 1 в пределах до одного месяца после инъекции AdDC до регистрации субъектов для получения следующей более высокой дозы (R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2-этил-3-метоксибензоил)гидразида. Через тридцать суток группа 1 завершает фазу лечения в стационаре, и через по меньшей мере шесть-восемь недель после внутриопухолевой инъекции трансдуцированных аутологичных дендритных клеток, если субъект желает повторную обработку и субъект отвечает критериям для повторной обработки, можно вводить дополнительную(ые) инъекцию(и) DC с 14 последовательными суточными дозами (R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2-этил-3-метоксибензоил)гидразида. На основании группы 1 доза (R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2-этил-3-метоксибензоил)гидразида (0,01 мг/кг, 0,1 мг/кг, 1,0 мг/кг или 10,0 мг/кг) представляет собой максимальную переносимую дозу.
Пациентов оценивают с помощью физических осмотров (включая статус показателей ECOG), основных показателей состояния организма, сывороточной химии, анализа мочи, гематологии, побочных эффектов, антител и клеточного иммунного ответа на аденовирусный вектор и компоненты терапевтической системы RheoSwitch®. Также оценивают однократную дозу и фармокинетику в стационарном состоянии/ADME перорального (R)-3,5-диметилбензойной кислоты N-(1-трет-бутил-бутил)-N’-(2-этил-3-метоксибензоил)гидразида и его главных метаболитов, анализ функции трансгена путем измерения уровней hIL-12 в биопсиях целевой опухоли и/или дренирующих лимфатических узлов, оценки иммунологических эффектов с помощью измерения клеточного иммунного ответа (T-клетки) в биопсиях целевой опухоли, дренирующих лимфатических узлов и периферического кровообращения, и цитокинового профиля в сыворотке.
Следует понимать, что вышеизложенные воплощения и примеры не предназначены для ограничения в каком-либо отношении объема изобретения, и что представленная здесь формула изобретения, как предполагается, включает все воплощения и примеры, как подробно представленные, так и не представленные в данном описании изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ХИРАЛЬНЫЕ ДИАЦИЛГИДРАЗИНОВЫЕ ЛИГАНДЫ ДЛЯ МОДУЛЯЦИИ ЭКСПРЕССИИ ЭКЗОГЕННЫХ ГЕНОВ С ПОМОЩЬЮ ЭКДИЗОН-РЕЦЕПТОРНОГО КОМПЛЕКСА | 2008 |
|
RU2490253C2 |
| МУТАНТНЫЕ РЕЦЕПТОРЫ И ИХ ИСПОЛЬЗОВАНИЕ В СИСТЕМЕ ЭКСПРЕССИИ ИНДУЦИБЕЛЬНОГО ГЕНА НА ОСНОВЕ ЯДЕРНОГО РЕЦЕПТОРА | 2005 |
|
RU2539036C2 |
| БОРСОДЕРЖАЩИЕ ДИАЦИЛГИДРАЗИНЫ | 2014 |
|
RU2637946C2 |
| СКОНСТРУИРОВАННЫЕ КЛЕТКИ, ЭКСПРЕССИРУЮЩИЕ МНОЖЕСТВЕННЫЕ ИММУНОМОДУЛЯТОРЫ, И ИХ ПРИМЕНЕНИЯ | 2009 |
|
RU2573912C2 |
| КРИСТАЛЛИЧЕСКИЙ ДИАЦИЛГИДРАЗИН И ЕГО ПРИМЕНЕНИЕ | 2012 |
|
RU2629113C2 |
| ВЕКТОРЫ, УСЛОВНО ЭКСПРЕССИРУЮЩИЕ ТЕРАПЕВТИЧЕСКИЕ БЕЛКИ, КЛЕТКИ-ХОЗЯЕВА, СОДЕРЖАЩИЕ УКАЗАННЫЕ ВЕКТОРЫ, И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2612788C2 |
| НИЗКОМОЛЕКУЛЯРНЫЕ ИНГИБИТОРЫ N-КОНЦЕВОЙ АКТИВАЦИИ РЕЦЕПТОРА АНДРОГЕНОВ | 2009 |
|
RU2519948C2 |
| СОЕДИНЕНИЯ, ИНГИБИРУЮЩИЕ (БЛОКИРУЮЩИЕ) ГОРЬКИЙ ВКУС, СПОСОБЫ ИХ ПРИМЕНЕНИЯ И ПОЛУЧЕНИЯ | 2008 |
|
RU2522456C2 |
| МОДИФИЦИРОВАННЫЕ ДЕНДРИТНЫЕ КЛЕТКИ И ИХ ПРИМЕНЕНИЕ В ЛЕЧЕНИИ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ | 2013 |
|
RU2644210C2 |
| МОДИФИЦИРОВАННЫЕ ДЕНДРИТНЫЕ КЛЕТКИ И ИХ ПРИМЕНЕНИЕ В ЛЕЧЕНИИ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ | 2017 |
|
RU2711606C2 |









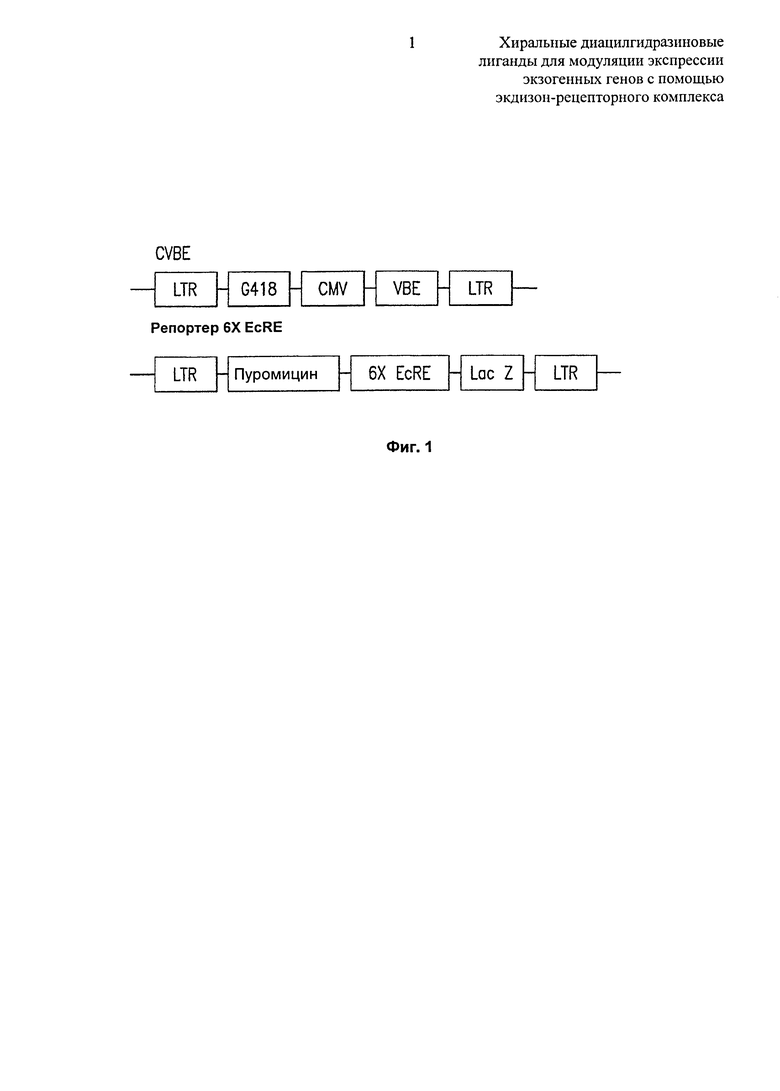
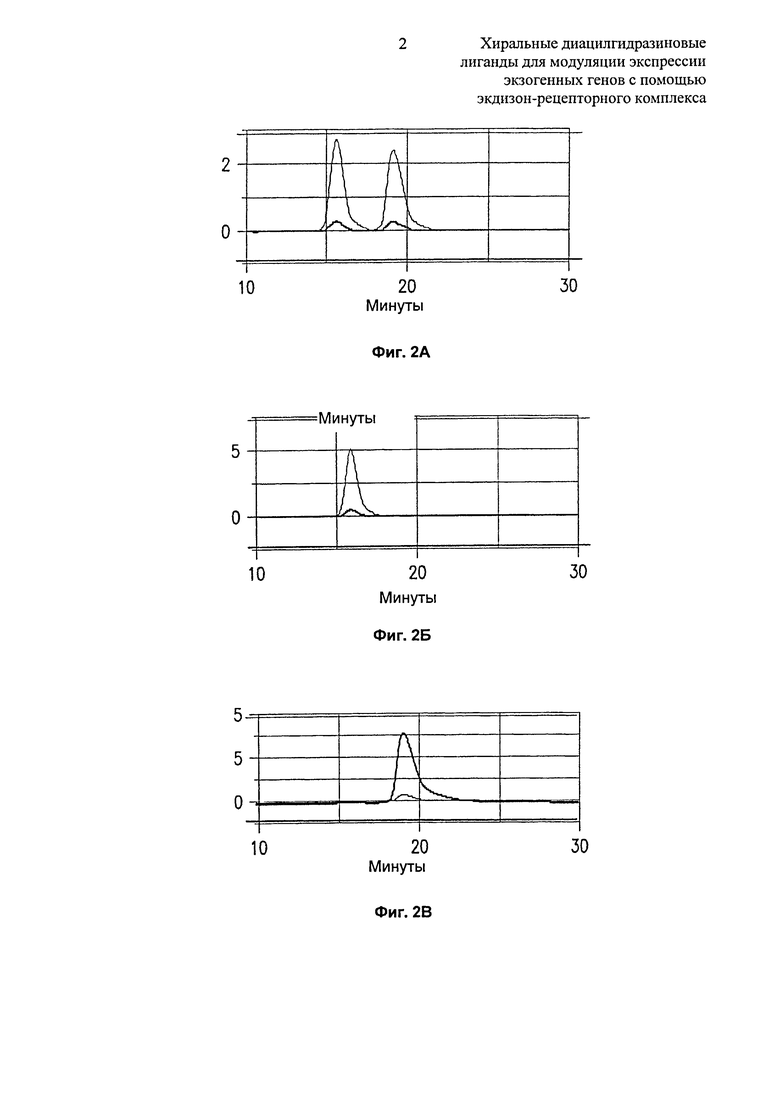
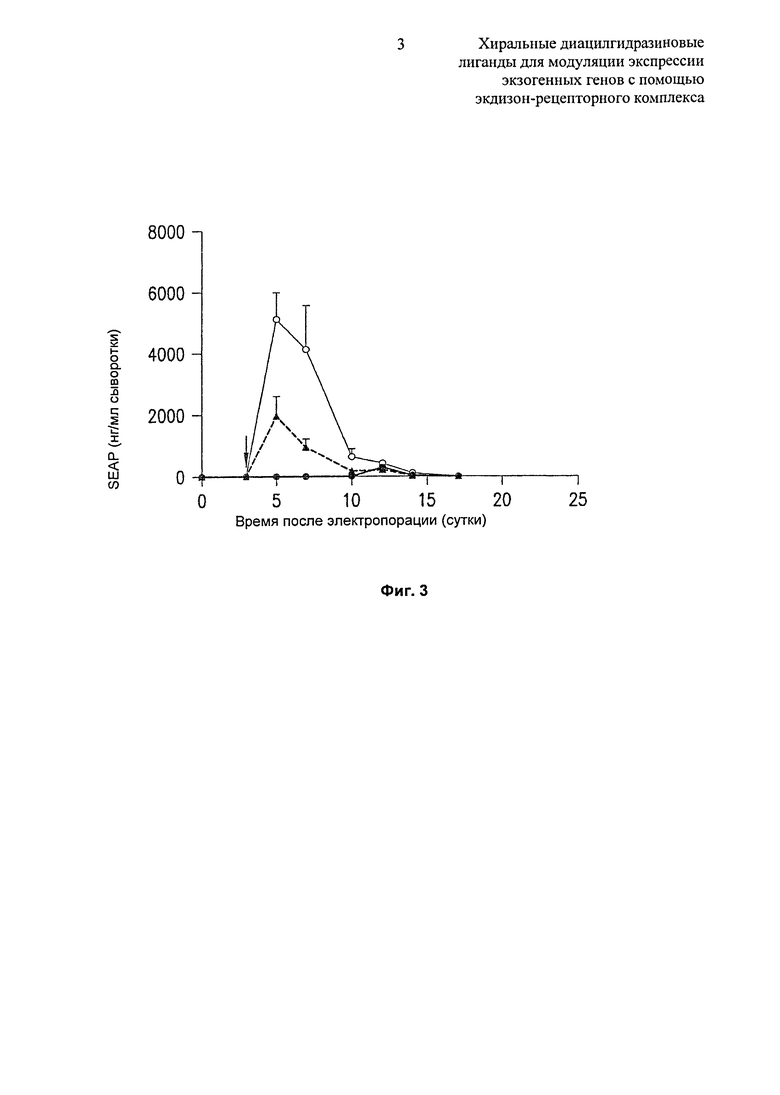
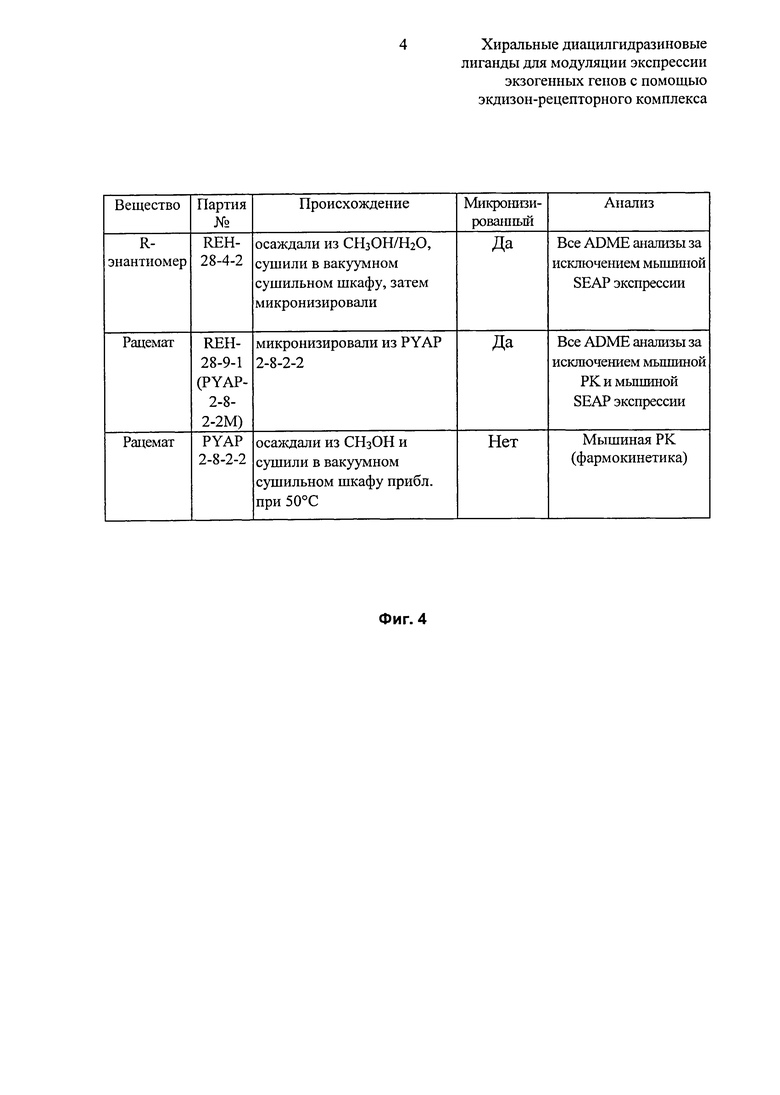
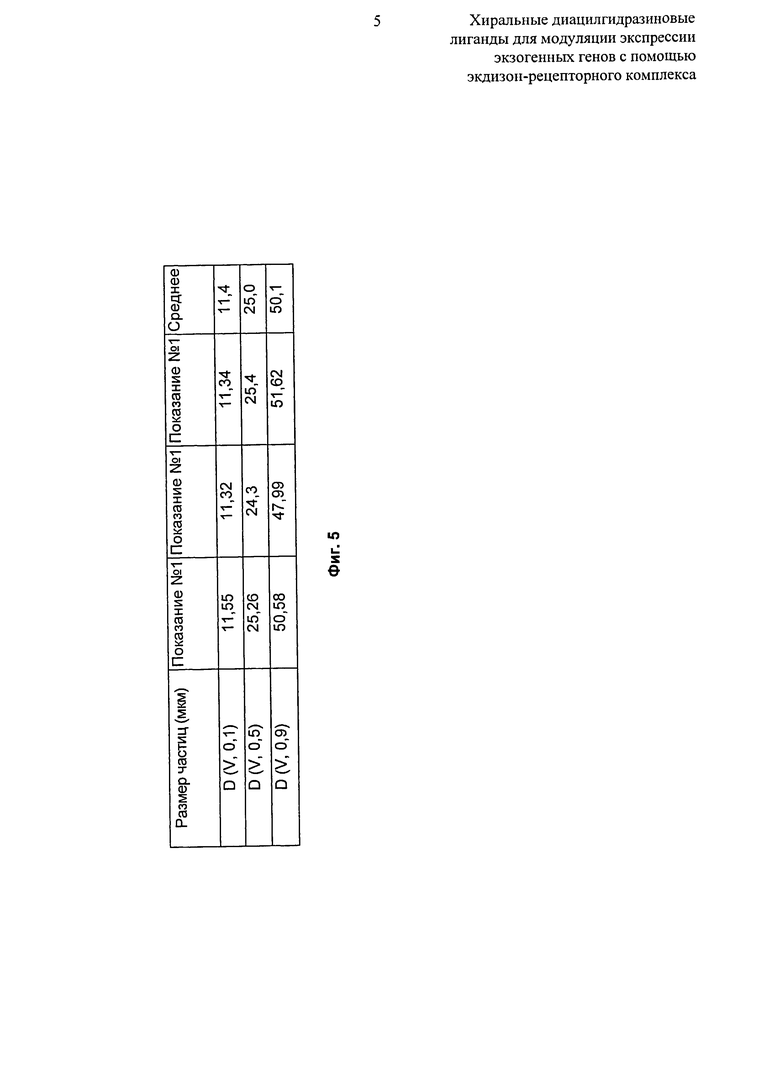
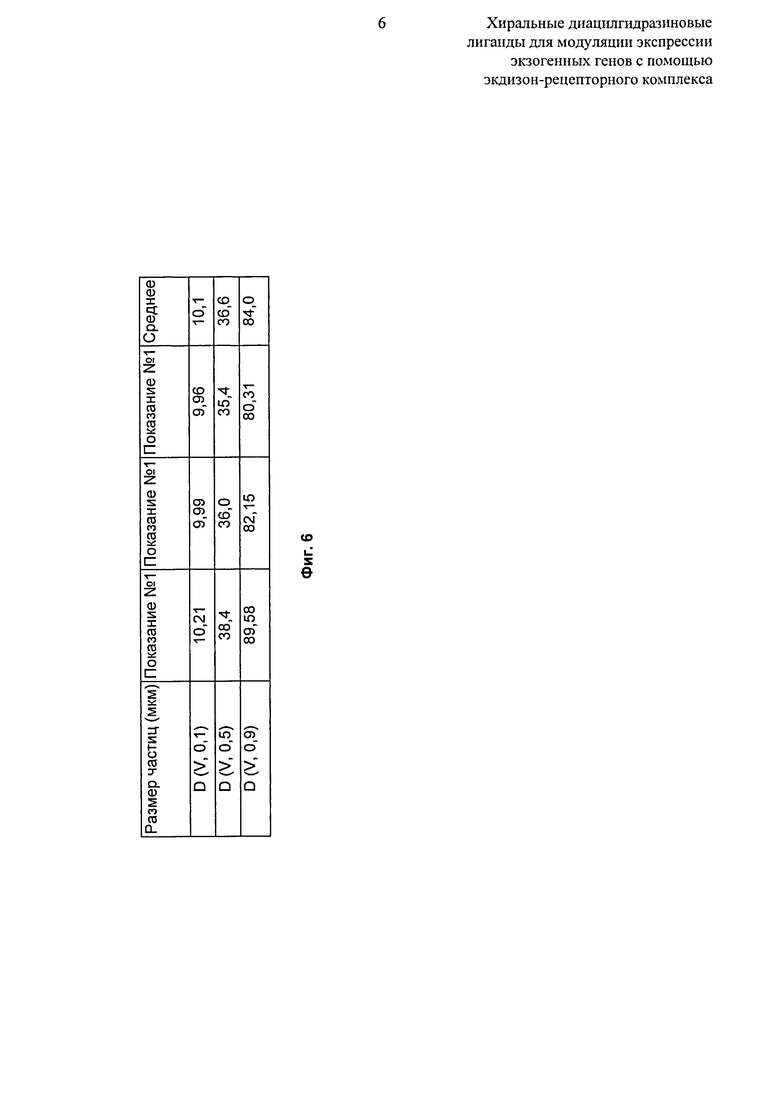
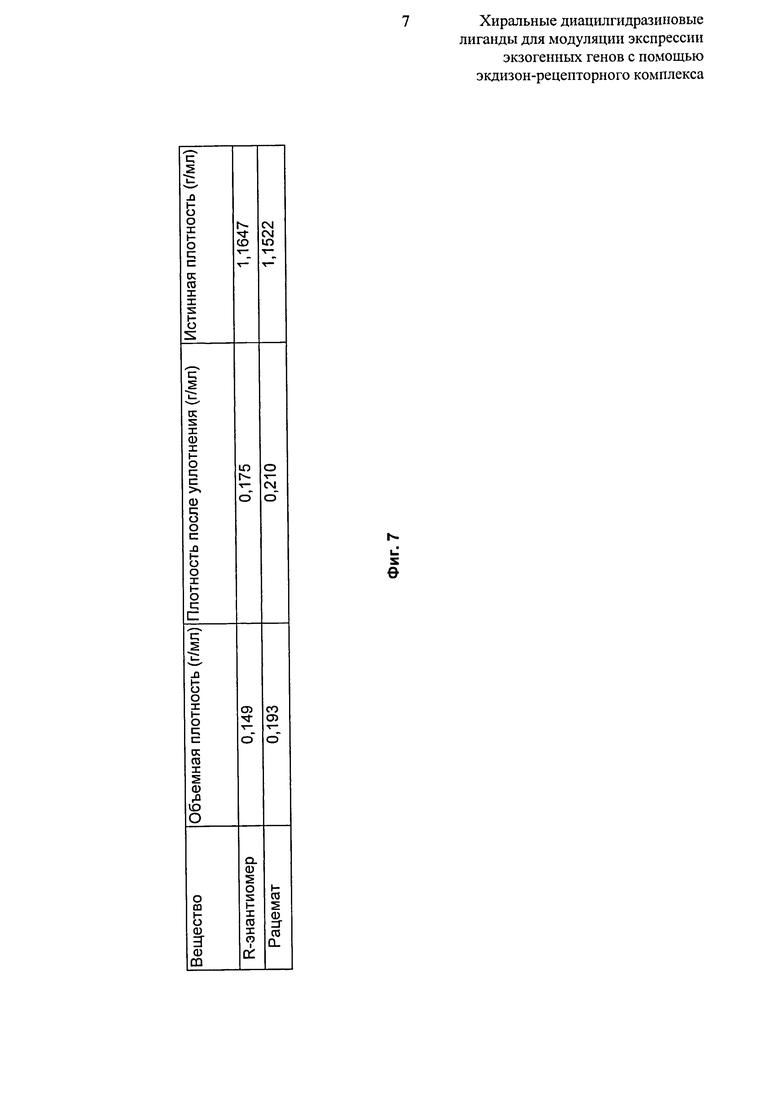
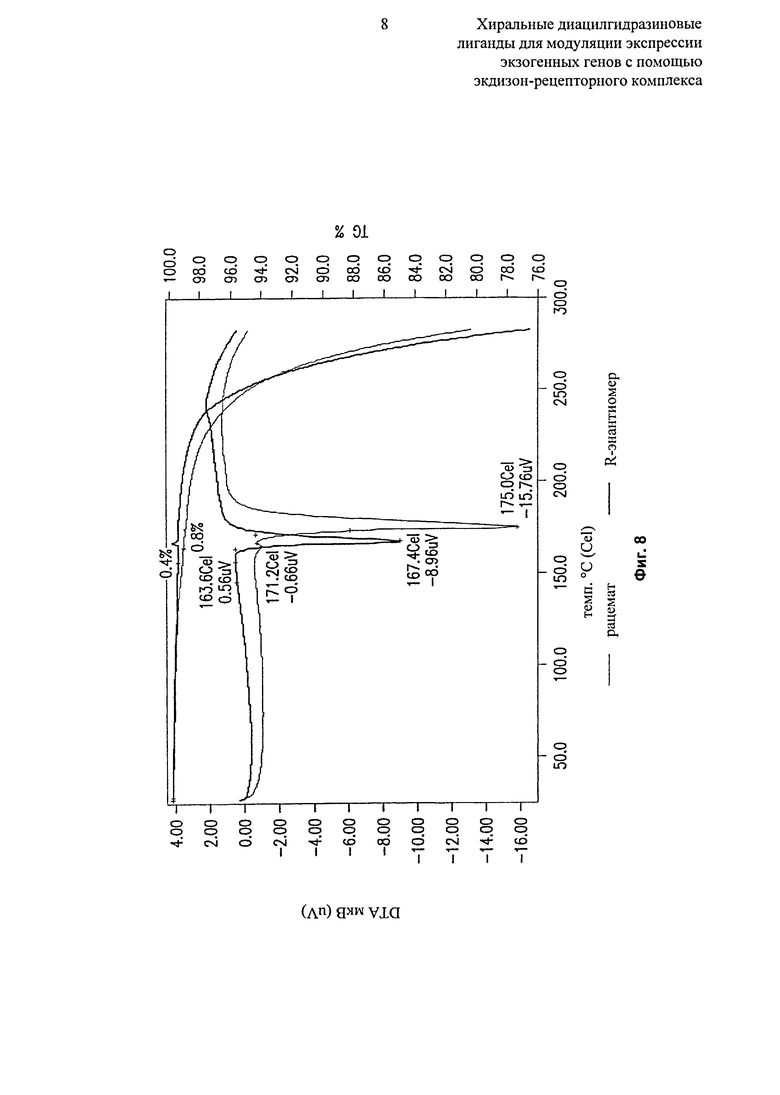
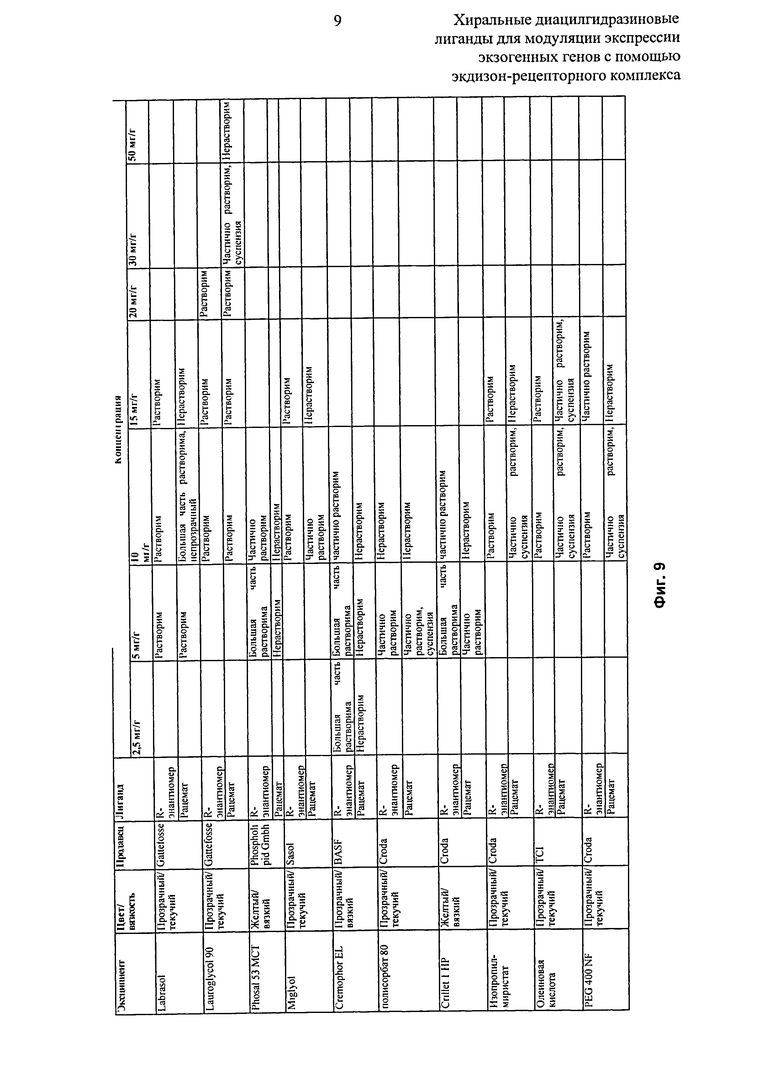
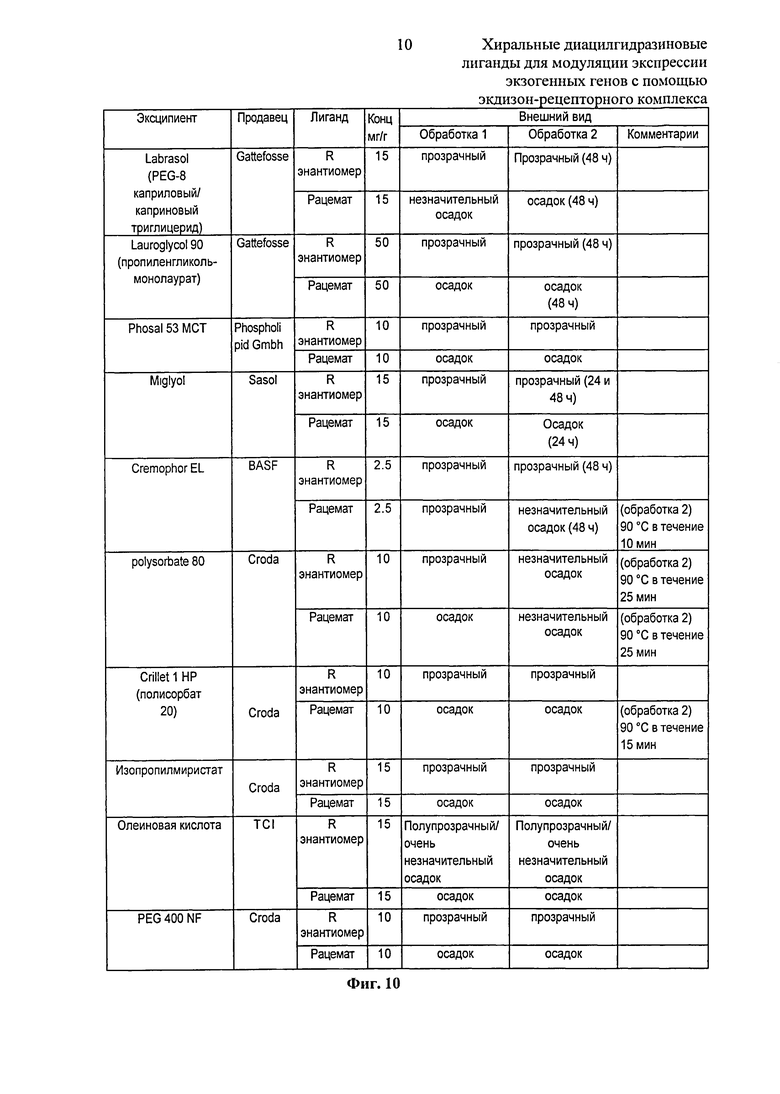
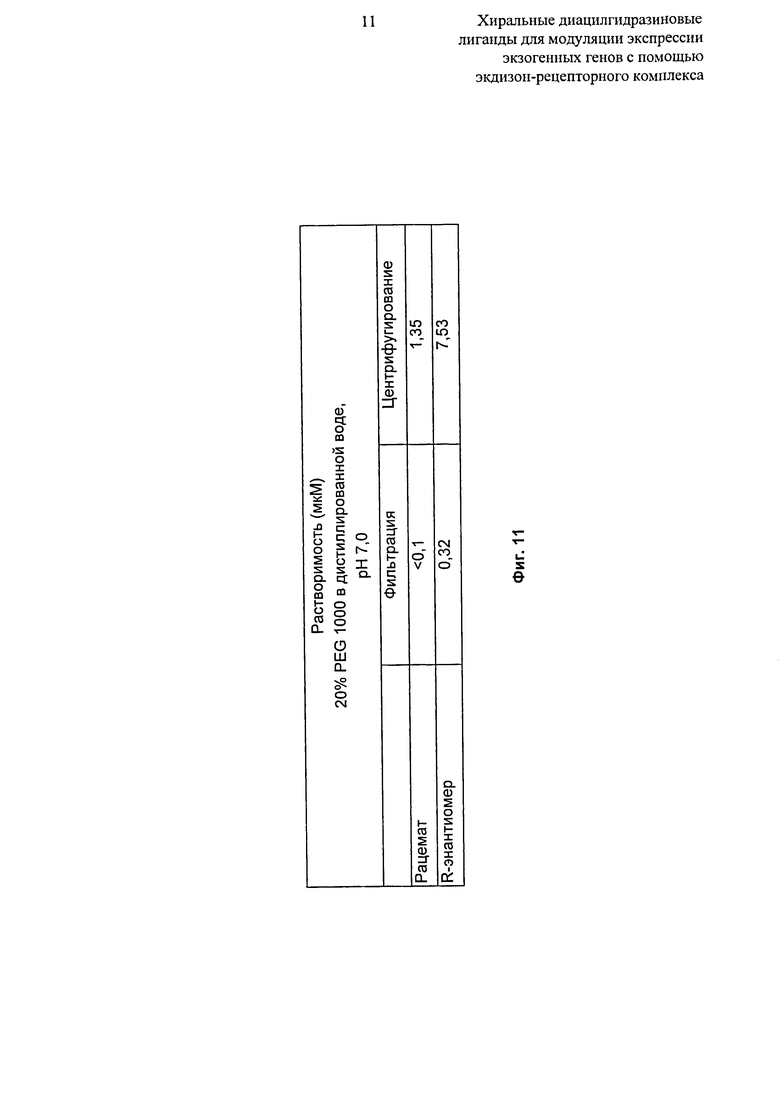
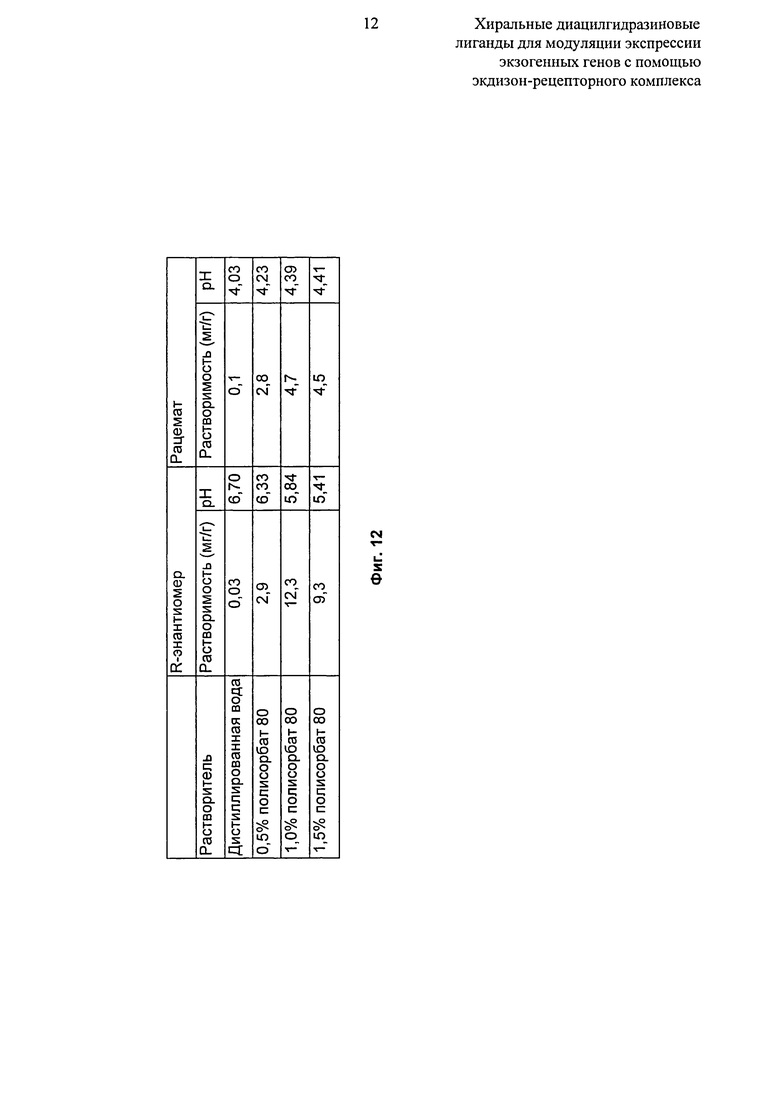
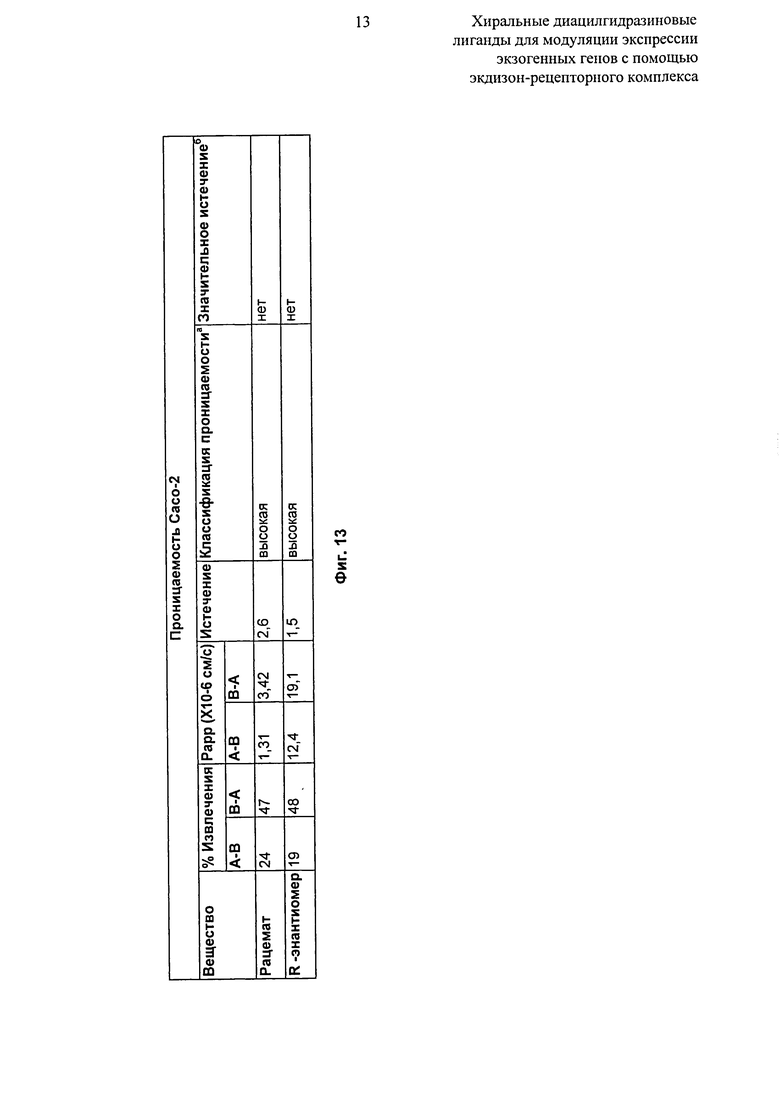
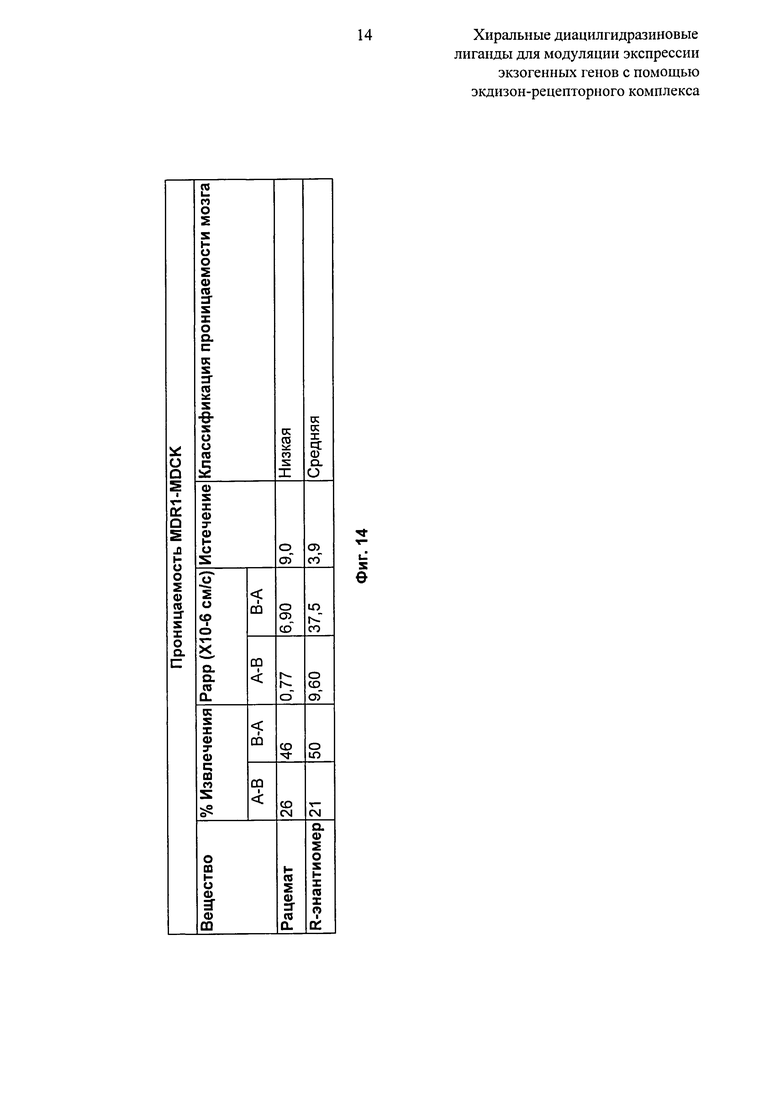
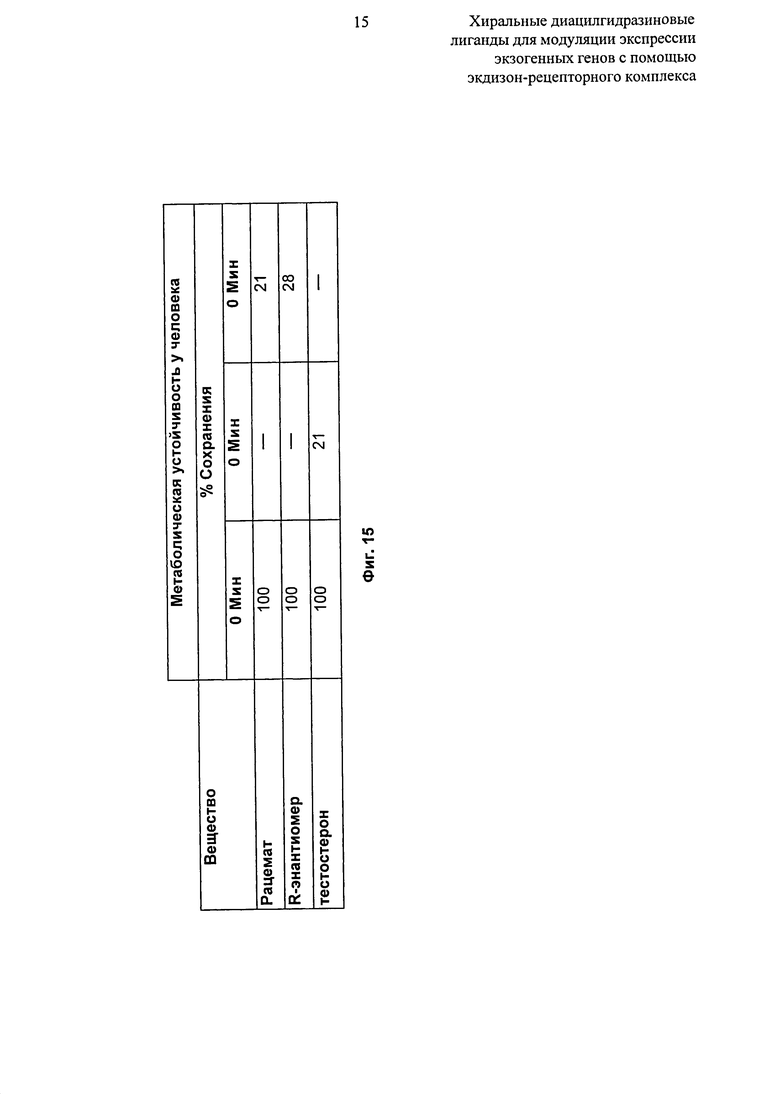
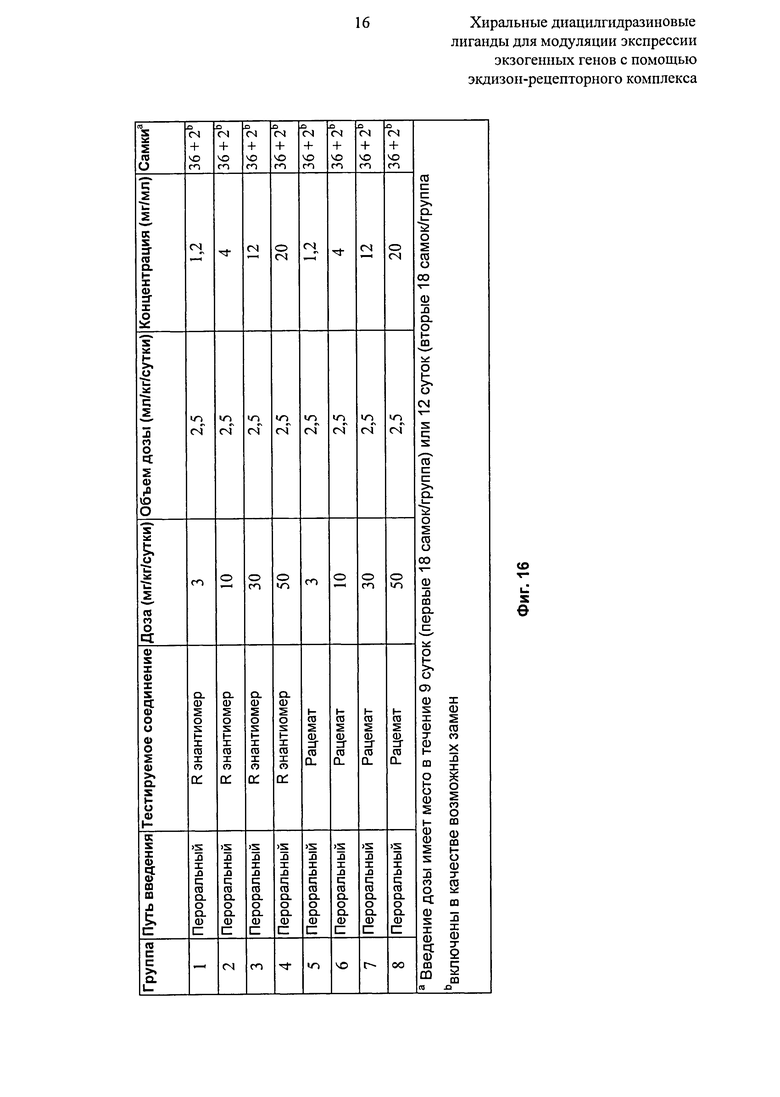

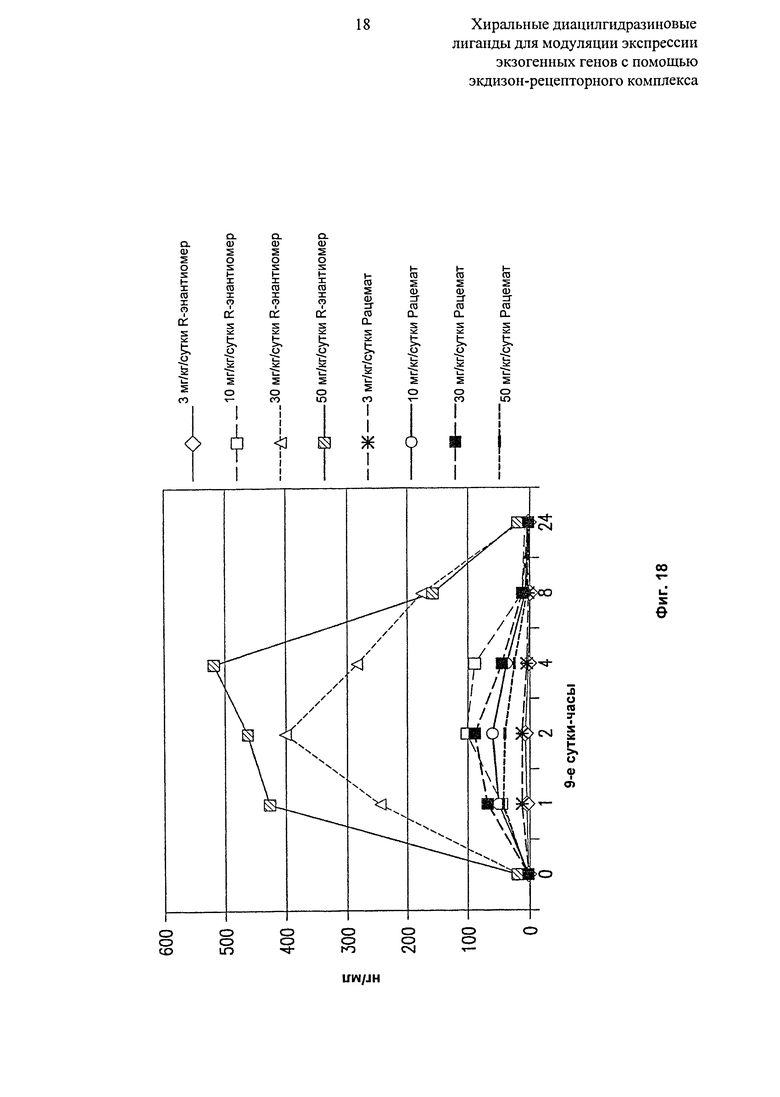
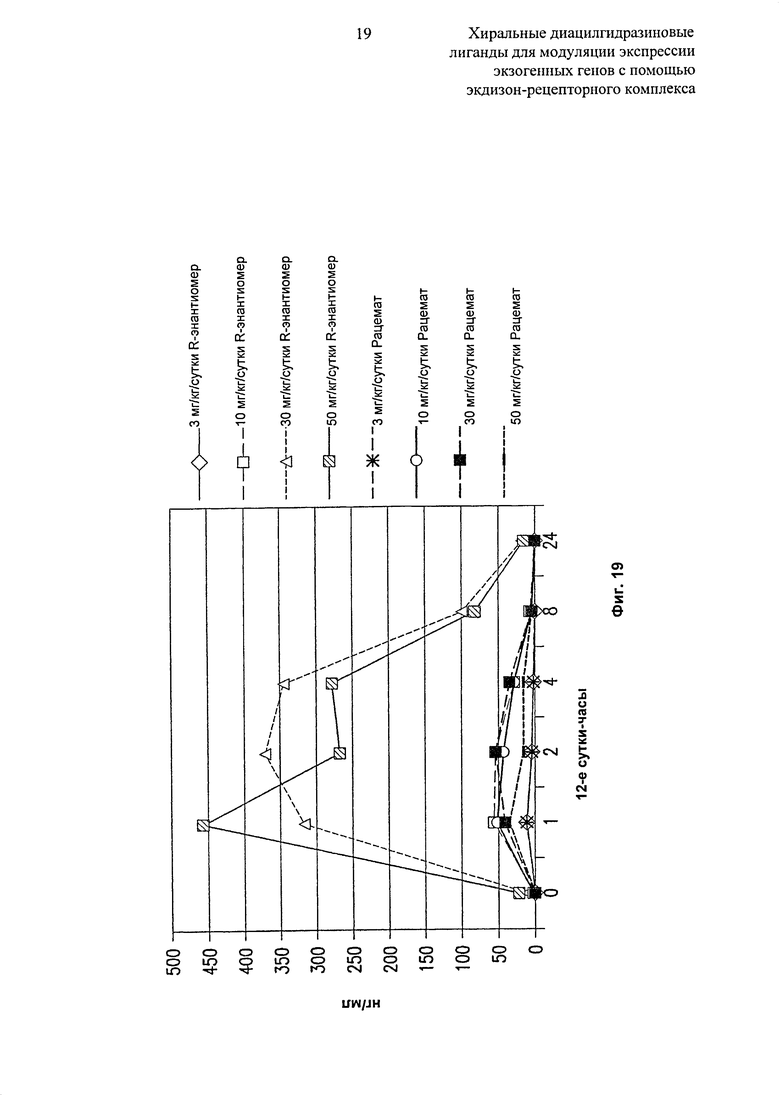
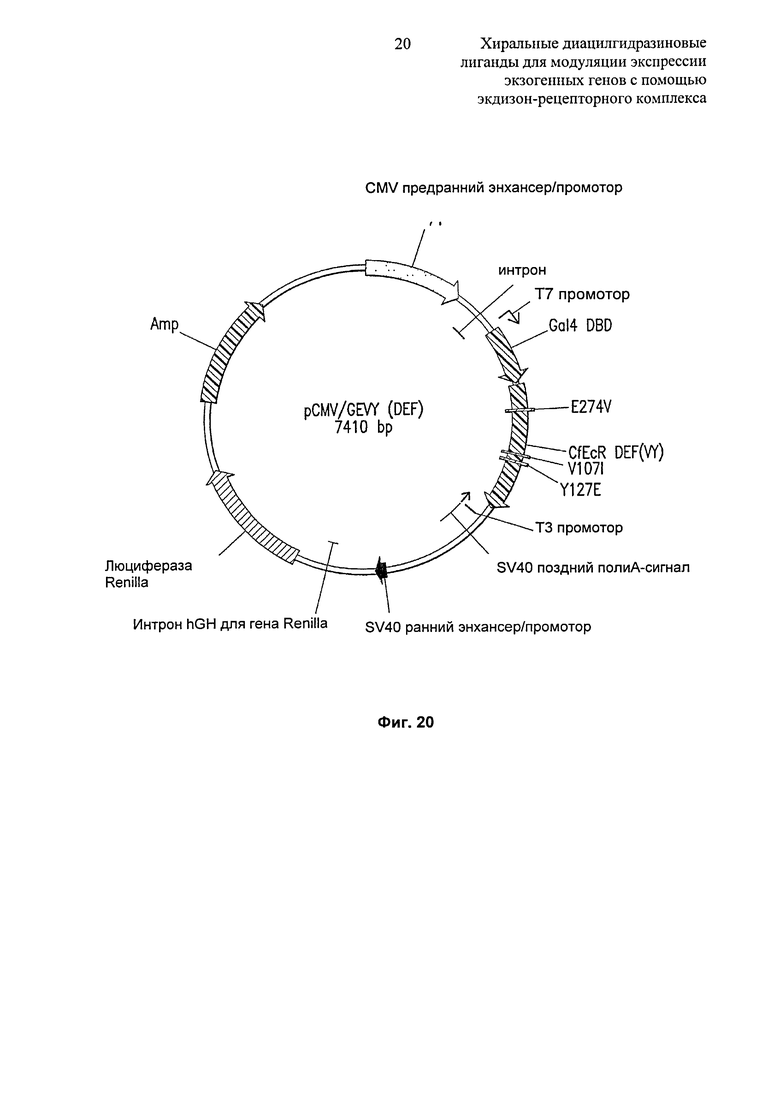
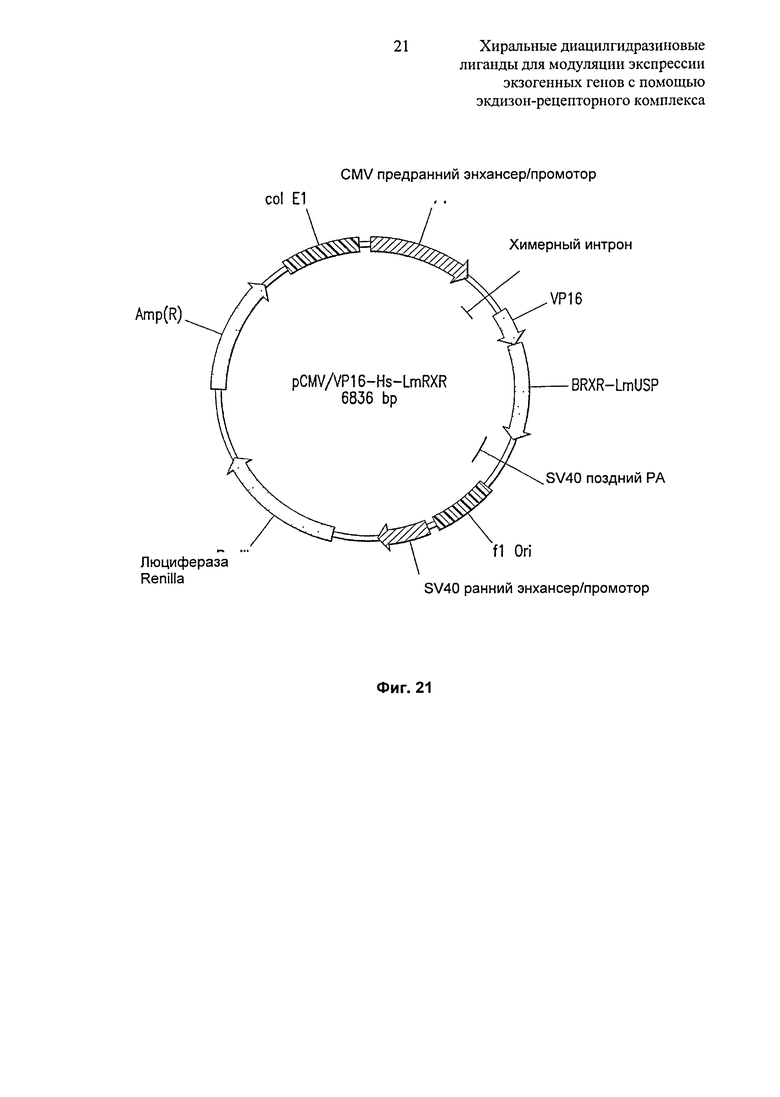
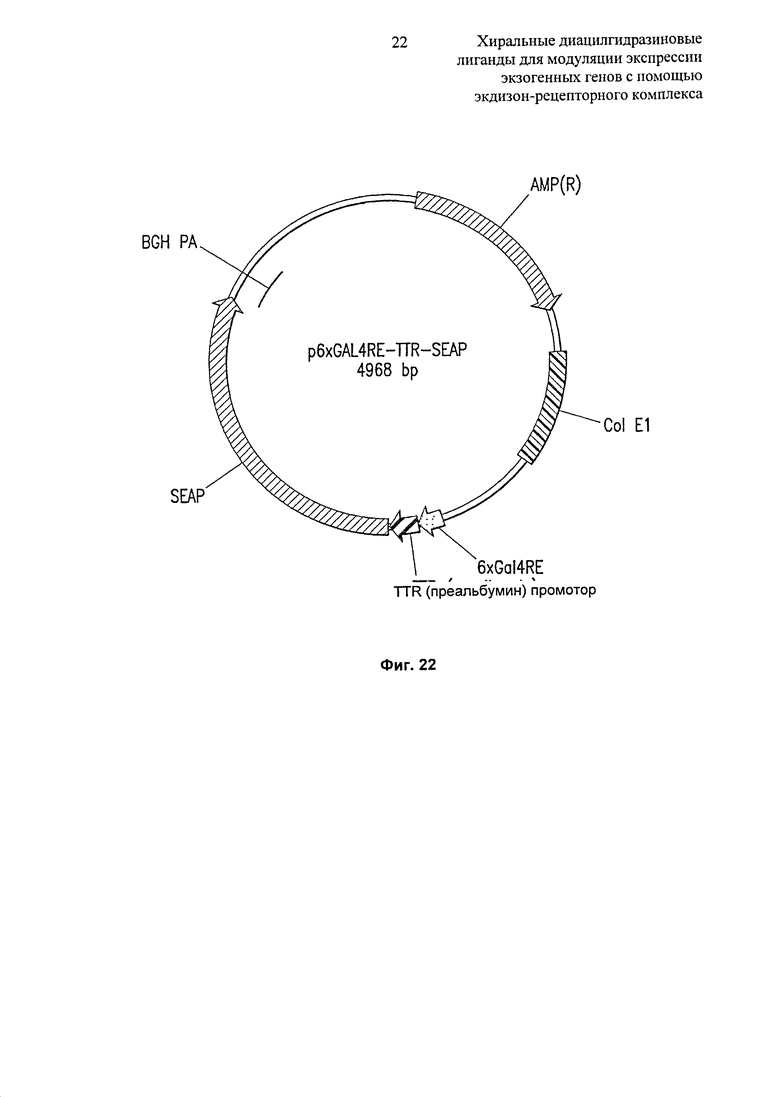
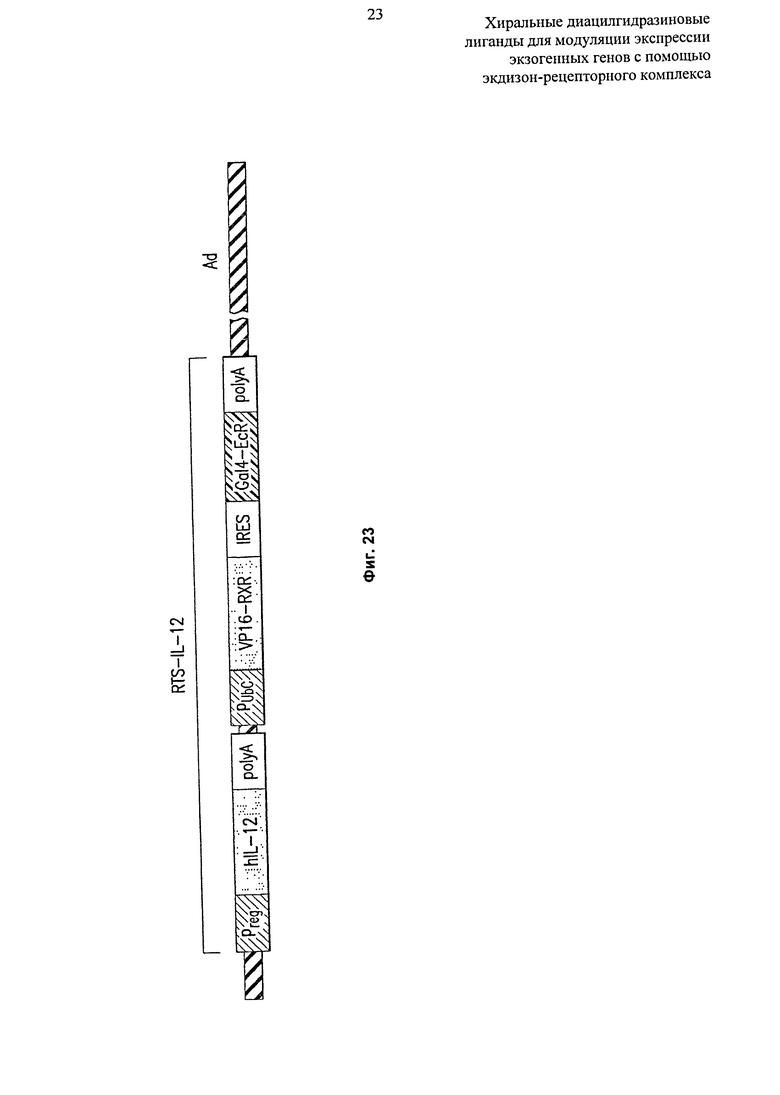
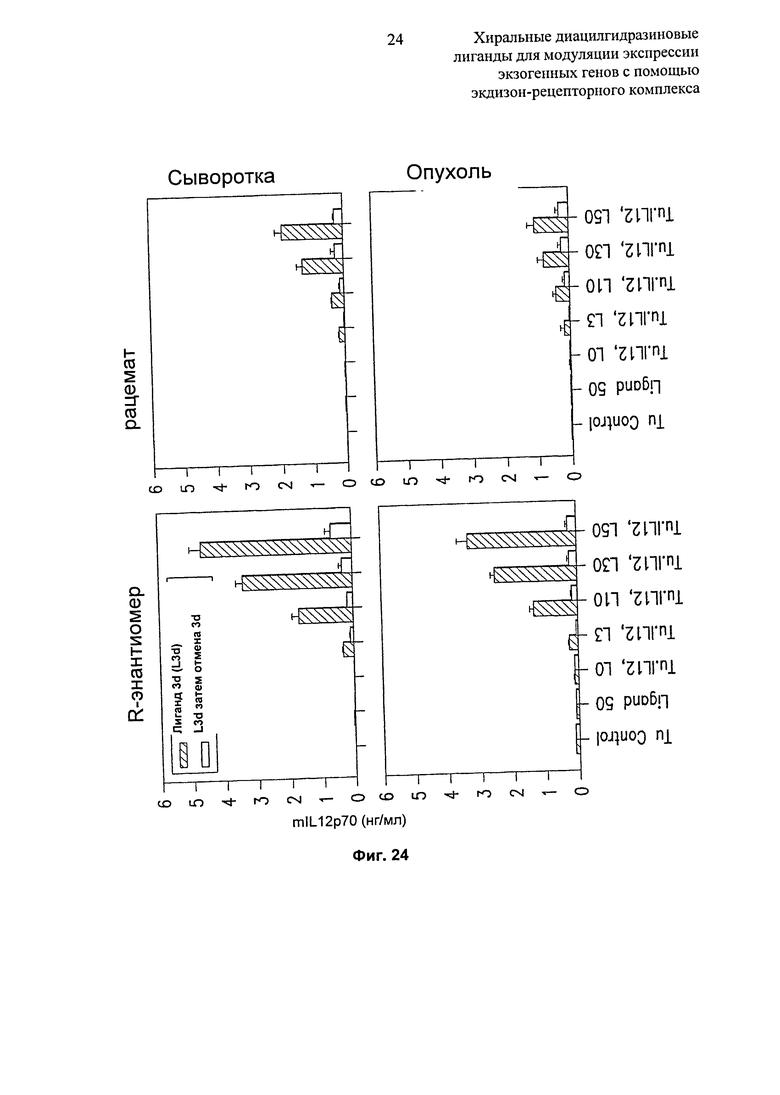
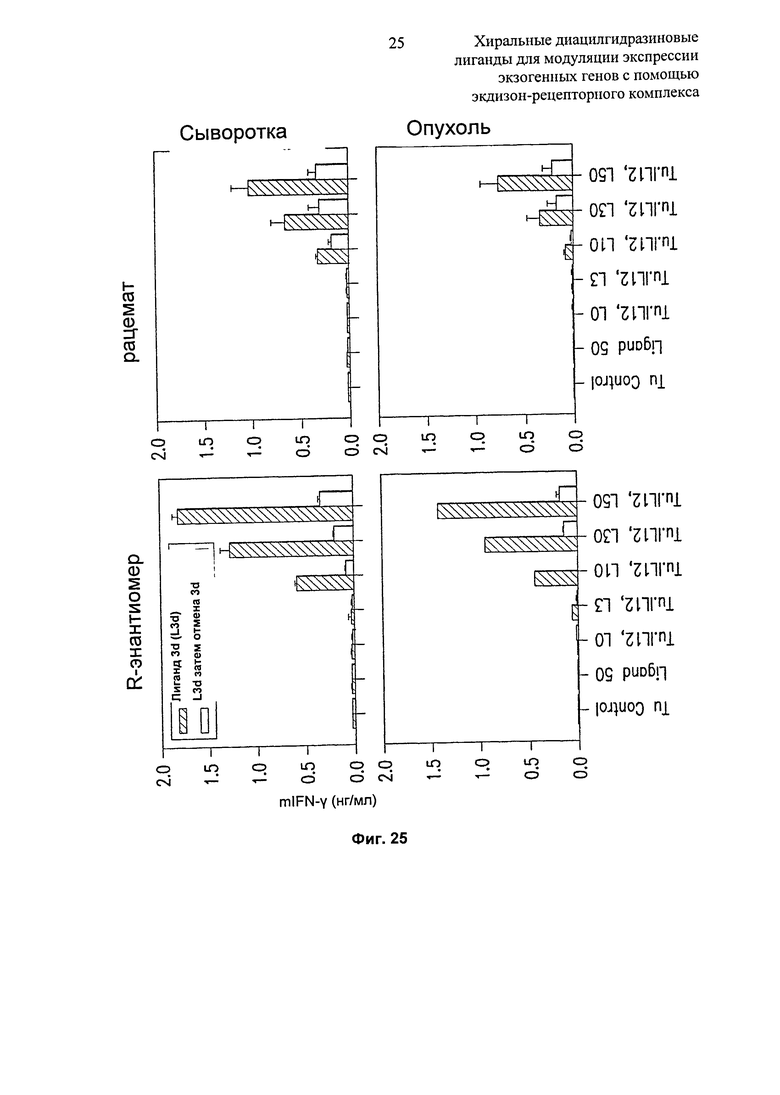
Изобретение относится к способу получения энантиомерно обогащенного соединения, имеющего Формулу III, где А представляет собой (С1-С6)алкил-O-, фенил-(С1-С6)алкил-O-; арил, выбранный из фенила, нафтила, бензо[1,3]диоксола, 2,3-бензо[1,4]диоксина, который возможно замещен 1-3 заместителями, где заместители выбраны из (С1-С6)алкила, (С3-С7)циклоалкила, (С1-С6)алкил-O-, гидрокси, амино и галогено; или гетероарил, имеющий четыре или пять атомов углерода и один гетероатом, выбранный из кислорода, азота и серы, который возможно замещен 1-3 заместителями, где заместители выбраны из (С1-С6)алкила, (С3-С7)циклоалкила, (С1-С6)алкил-O-, гидрокси, амино и галогено; В представляет собой фенил, возможно замещенный 1-3 заместителями, где заместители выбраны из (С1-С6)алкила, (С3-С7)циклоалкила, (С1-С6)алкил-O-, гидрокси, амино и галогено; и R1 и R2 независимо представляют собой (С1-С6)алкил, фенил-(С1-С6)алкил-, гидрокси-(С1-С6)алкил, (С3-С7)циклоалкил, (С2-С6)алкенил или (С2-С6)алкинил; при условии, что R1 отличается от R2; где абсолютной конфигурацией асимметрического атома углерода, несущего R1 и R2, является R-конфигурация; включающему (а) взаимодействие ацилгидразина Формулы XI с кетоном Формулы XII с образованием соединения Формулы XIII, где R1 отличается от R2, (б) восстановление соединения Формулы XIII в присутствии хирального катализатора с образованием соединения Формулы R-XIV и (в) взаимодействие соединения Формулы R-XIV с соединением Формулы B-CO-LG, где LG представляет собой уходящую группу, с образованием соединения, имеющего Формулу III. Технический результат - хиральные диацилгидразиновые лиганды для модуляции генов. 5 з.п. ф-лы, 25 ил., 11 табл.
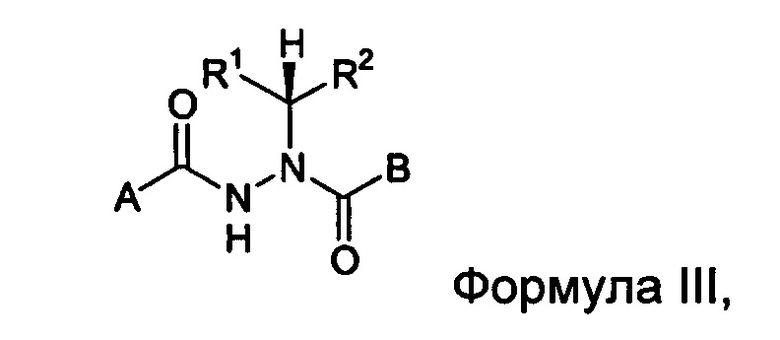
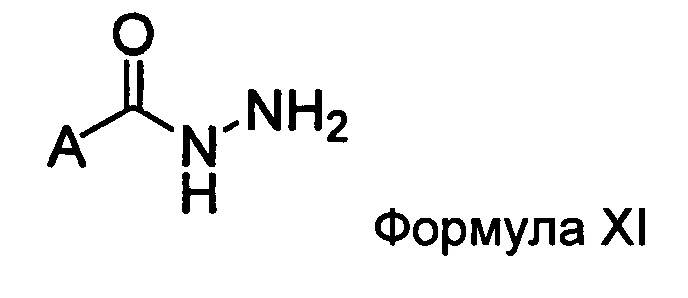 ,
, 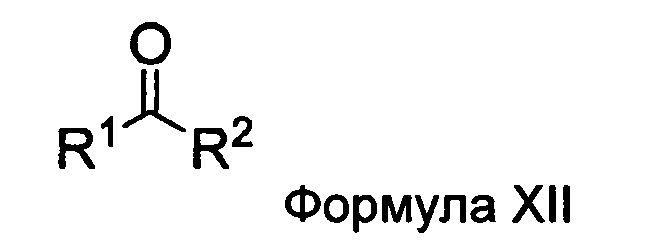 ,
, 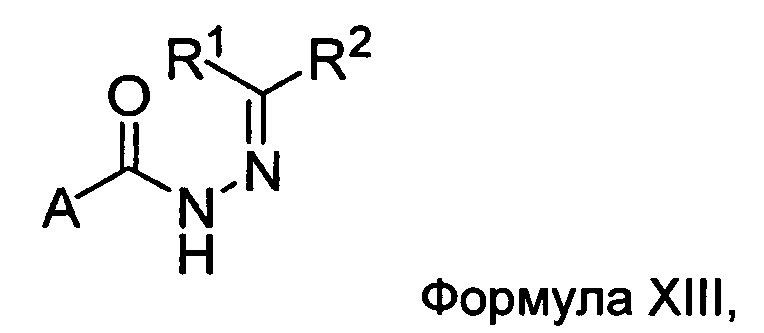
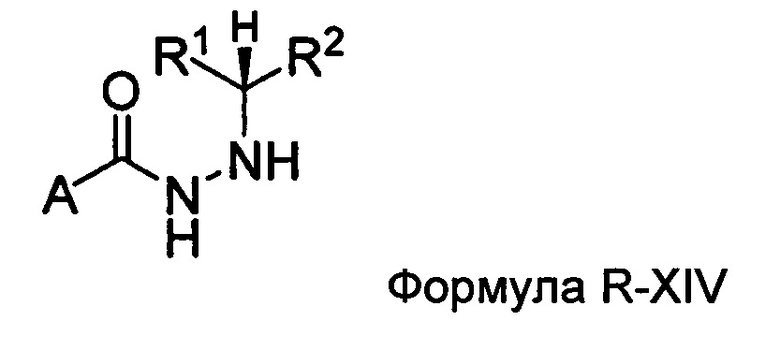
1. Способ получения энантиомерно обогащенного соединения, имеющего Формулу III
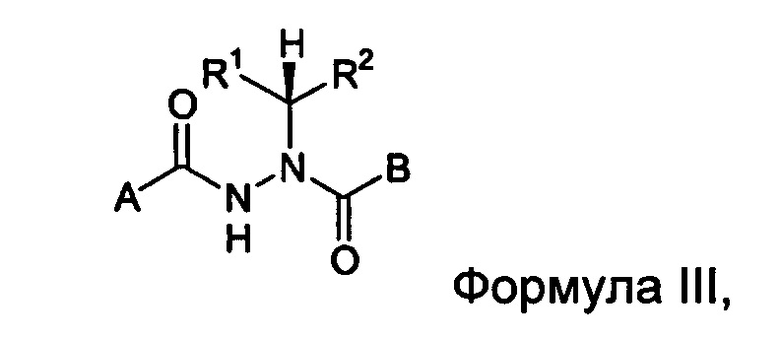
где
А представляет собой (С1-С6)алкил-O-, фенил-(С1-С6)алкил-O-; арил, выбранный из фенила, нафтила,  и
и  , который возможно замещен 1-3 заместителями, где заместители выбраны из (С1-С6)алкила, (С3-С7)циклоалкила, (С1-С6)алкил-O-, гидрокси, амино и галогено; или гетероарил, имеющий четыре или пять атомов углерода и один гетероатом, выбранный из кислорода, азота и серы, который возможно замещен 1-3 заместителями, где заместители выбраны из (С1-С6)алкила, (С3-С7)циклоалкила, (С1-С6)алкил-O-, гидрокси, амино и галогено;
, который возможно замещен 1-3 заместителями, где заместители выбраны из (С1-С6)алкила, (С3-С7)циклоалкила, (С1-С6)алкил-O-, гидрокси, амино и галогено; или гетероарил, имеющий четыре или пять атомов углерода и один гетероатом, выбранный из кислорода, азота и серы, который возможно замещен 1-3 заместителями, где заместители выбраны из (С1-С6)алкила, (С3-С7)циклоалкила, (С1-С6)алкил-O-, гидрокси, амино и галогено;
В представляет собой фенил, возможно замещенный 1-3 заместителями, где заместители выбраны из (С1-С6)алкила, (С3-С7)циклоалкила, (С1-С6)алкил-O-, гидрокси, амино и галогено; и
R1 и R2 независимо представляют собой (С1-С6)алкил, фенил-(С1-С6)алкил-, гидрокси-(С1-С6)алкил, (С3-С7)циклоалкил, (С2-С6)алкенил или (С2-С6)алкинил;
при условии, что R1 отличается от R2;
где абсолютной конфигурацией асимметрического атома углерода, несущего R1 и R2, является R-конфигурация;
включающий:
а) взаимодействие ацилгидразина Формулы XI
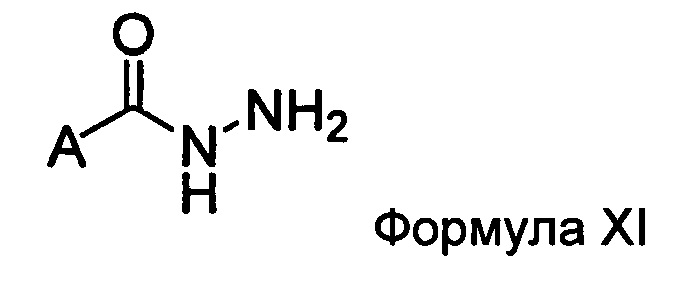
с кетоном Формулы XII
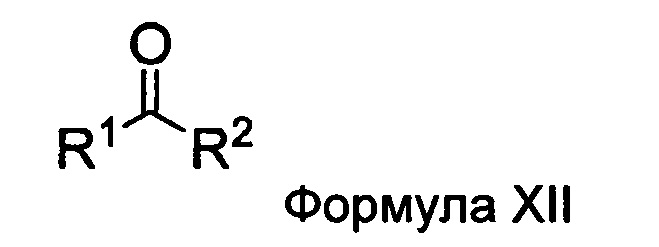
с образованием соединения Формулы XIII
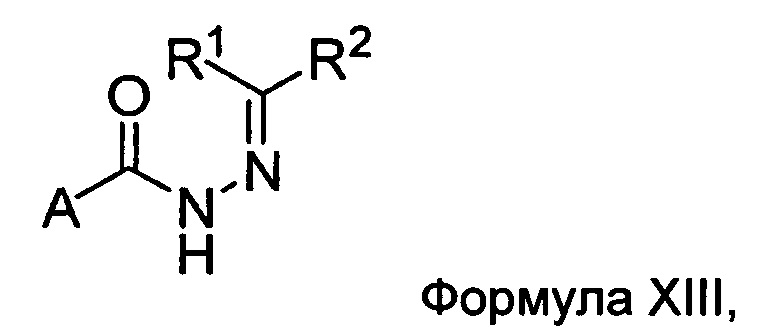
где R1 отличается от R2,
б) восстановление соединения Формулы XIII в присутствии хирального катализатора с образованием соединения Формулы R-XIV
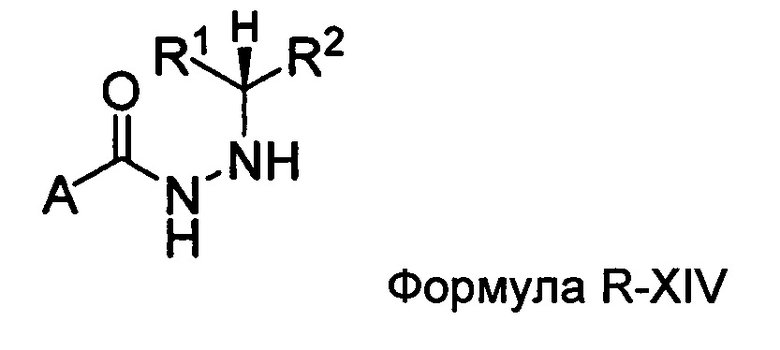
и
в) взаимодействие соединения Формулы R-XIV с соединением Формулы B-CO-LG, где LG представляет собой уходящую группу, с образованием соединения, имеющего Формулу III.
2. Способ по п. 1, где В представляет собой 3,5-диметилфенил.
3. Способ по п. 1, где R2 представляет собой трет-бутил.
4. Способ по п. 1, где А представляет собой 2-этил-3-метоксифенил.
5. Способ по п. 1, где соединение, имеющее Формулу III, выбрано из группы, состоящей из:
N-(1-трет-бутил-бутил)-N'-(2-этил-3-метокси-бензоил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N'-бензоил-N-(1-трет-бутил-бутил)-гидразида (R)-(3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(2-метил-бензоил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(2-метокси-бензоил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(2-фтор-бензоил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(2-хлор-бензоил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N'-(2-бром-бензоил)-N-(1-трет-бутил-бутил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(3-метил-бензоил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(3-метокси-бензоил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(3-хлор-бензоил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(4-метил-бензоил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(4-этил-бензоил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(4-метокси-бензоил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(трет-бутил-бутил)-N'-(4-хлор-бензоил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(2,6-дифтор-бензоил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(2,6-дихлор-бензоил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(3,4-диметокси-бензоил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(3,5-дифтор-бензоил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(3,5-диметокси-4-метил-бензоил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(4-метил-бензо[1,3]диоксол-5-карбонил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(5-метил-2,3-дигидро-бензо[1,4]диоксин-6-карбонил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(5-этил-2,3-дигидро-бензо[1,4]диоксин-6-карбонил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(нафталин-1-карбонил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(нафталин-2-карбонил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(тиофен-2-карбонил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(2,5-диметил-фуран-3-карбонил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(2-хлор-пиридин-3-карбонил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(6-хлор-пиридин-3-карбонил)-гидразида (R)-3,5-диметил-бензойной кислоты;
N-(1-трет-бутил-бутил)-N'-(3-метокси-2-метил-бензоил)-гидразида (R)-3,5-диметил-бензойной кислоты и
N-(1-трет-бутил-бутил)-N'-(3-метокси-2-метил-бензоил)-гидразида (R)-3,5-диметокси-4-метил-бензойной кислоты.
6. Способ по п. 5, где соединение, имеющее Формулу III, представляет собой N-(1-трет-бутил-бутил)-N'-(2-этил-3-метокси-бензоил)-гидразид (R)-3,5-диметил-бензойной кислоты.
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| US 4985461 A, 15.01.1991 | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| US 6326403 B1, 04.12.2001 | |||
| RU 2051902 C1, 10.01.1996. | |||

