Область техники
Изобретение относится к биотехнологии, молекулярной биофармакологии и может быть использовано для получения биологически активного белка теплового шока 70 (БТШ70) с использованием генно-инженерно модифицированных животных в качестве биореакторов. Белки теплового шока 70 играют важнейшую роль в развитии организма млекопитающих в нормальных условиях и при различных стрессовых воздействиях, и в перспективе могут быть использованы в качестве терапевтического средства для лечения некоторых форм нейродегенеративных заболеваний человека, связанных с неправильным синтезом и укладкой белков («протеинопатии»), а также при сепсисе (1, 2). Поэтому наработка эукариотического БТШ70 в количествах, достаточных для дальнейшего применения в терапии, является важной задачей.
Белки теплового шока играют важную роль для обеспечения гомеостаза организма на всех стадиях его развития, особенно в случае воздействия стресса, токсинов, радиации, сублетальной температуры, ацидоза, гипоксии или гипероксии, энергетического истощения клетки, а также при старении и микробных инфекциях.
Уровень техники
В настоящее время достаточно подробно изучены основные функции внутриклеточных стрессовых белков, которые обычно называют белками теплового шока (БТШ), связанные с участием этих молекул в процессах синтеза, фолдинга, транспорта, репарации внутриклеточных протеинов (шаперонные функции) и их важной ролью в системе защиты клеток от повреждающих воздействий внешней среды (протективные функции). Особо важную роль в гомеостазе клетки в норме и при различных вредных воздействиях играет древнее и весьма консервативное семейство белков теплового шока с молекулярной массой 70 килодальтон (БТШ70). В настоящее время известно более 10 представителей БТШ70 эукариот. Описаны также бактериальные ортологи БТШ70 (3). Структура и функции БТШ70 в настоящее время хорошо изучены. У этого семейства белков аминоконцевая часть молекулы содержит консервативный АТФазный домен (44 кДа), а в карбокситерминальной части молекулы локализован субстрат-связывающий домен (~25 кДа), необходимый для узнавания и связывания БТШ70 с полипептидами. Эти домены необходимы для эффективного связывания и транспортировки самых различных клеточных белков в норме и при различных патологиях.
Основными функциями белков семейства БТШ70 являются повышение клеточной термотолерантности к целому спектру повреждающих факторов, включая тепловой шок, гипоксию и пр. БТШ70 участвует в ренатурации и поддержании целостности структуры клеточных белков и обладает выраженным антиапоптотическим действием. Помимо этого он участвует в деградации белков с нарушенной структурой, препятствует образованию белковых агрегатов внутри клетки, а также играет важную роль в процессах ассоциации-диссоциации белков, а также их транслокации через мембраны внутриклеточных компартментов и органелл (4, 5).
Важнейшими функциями БТШ70 являются способность этих белков предотвращать образование агрегатов поврежденных и/или денатурированных белков, а также способность выступать в роли носителей иммуногенных пептидов. В частности, зарегистрирована поверхностная локализация БТШ70 у ряда линий опухолевых клеток, у вирус-инфицированных лимфоцитов, у активированных макрофагов и Т-лимфоцитов, а также обнаружено существование свободного БТШ70 в межклеточном пространстве (6). На основе иммуноадъювантных свойств БТШ70 интенсивно ведутся исследования по разработке вакцин для иммунотерапии злокачественных опухолей (7).
Рядом исследователей были разработаны продуценты для наработки рекомбинантных аналогов белков теплового шока БТШ70 патенты США №№6,524.825 (опубл. 25.02.2003 г.); 5,919,620 (опубл. 06.07.1999 г.).
Известна генетическая конструкция и метод для наработки БТШ70 в различных органах и в том числе в молочных железах (патент US 2005260746 А1). Данный патент отличается от заявленного тем, что в качестве трансгенного животного используется свинья, при этом структурные элементы конструкции включают в себя БТШ70 поставленный под контроль цитомегаловирусного энхансера и промотора. Полученный химерный ген содержит также сигнал полиаденилирования SV40. Трансгенные свиньи, содержащие такую конструкцию экспрессируют БТШ70 во всех органах, а не селективно в молоке, что обеспечивает полученная нами конструкция.
В своей работе мы использовали один из эффективных эукариотических векторов рВС1, предназначенный для создания экспрессионных конструкций для последующей продукции белков интереса в молоке трансгенных животных. Этот вектор и подробности работы с ним описаны в инструкции к киту («рВС1 Milk Expression Vector Kit for the Expression of Recombinant Proteins in the Milk of Transgenic Mice».
Использованная в настоящем патенте конструкция кроме рВС1 вектора содержит в своем составе сигнальный пептид лактоферрина человека, для секреции белка интереса (БТШ70) в молоко трансгенных животных.
Важным преимуществом полученной конструкции является то, что нам удалось клонировать сигнальную последовательность лактоферрина «встык», что позволяет получать высокий титр белка (БТШ70) без лишних аминокислот.
В патенте на изобретение РФ №2294210 (опубл. 13.05.2005 г.) нами предложено использовать рекомбинантный человеческий БТШ70 в качестве защиты от ряда опасных поражающих факторов.
В настоящее время большой интерес исследователей вызывают функции БТШ70 как молекул межклеточной сигнализации, способных вызывать продукцию цитокинов клетками иммунной системы путем рецептор-опосредуемых взаимодействий. Другие исследования показали, что иммунизация препаратами БТШ70, выделенными из опухолевых и инфицированных вирусами клеток, вызывает специфический антиопухолевый и антивирусный клеточный иммунный ответ. Специфичность такого ответа обусловлена способностью БТШ70 связывать различные белки и пептиды (в т.ч. онко- и вирусные пептиды) и доставлять их антиген-презентирующим клеткам. Возможность образования комплексов БТШ70 с белковыми антигенами in vitro позволила создавать на их основе противоопухолевые и противовирусные вакцины.
В последние десятилетия на клеточных и животных моделях (мыши, крысы) было показано, что рекомбинантный БТШ70 проявляет достоверные защитные свойства при сепсисе, вызываемом бактериальными токсинами, а также при различных видах нейродегенеративных заболеваний. Эти работы проводили на моделях грызунов (6, 7).
Природные белковые препараты БТШ70 млекопитающих были выделены из опухолевых и нормальных мышечных тканей. При этом накапливали и гомогенизировали ткани, экстрагировали целевой белок, а также фракционировался препарат с помощью нескольких хроматографических стадий, с очисткой на колонках с АТФ-сефарозой. Получение препарата белка БТШ70 из органов и тканей человека крайне не эффективно и опасно с точки зрения риска контаминации, в т.ч. вирусами, а также бактериальными токсинами.
Генно-инженерные методы позволяют в настоящее время нарабатывать аналитические количества БТШ70 в бактериях, необходимые для исследования различных функций белка и работы с модельными животными. Однако нарабатываемые в бактериях препараты БТШ70 не претерпевают пост-трансляционных модификаций, имеющих место в эукариотических клетках и могут содержать бактериальные токсины.
Отметим, что целевые белки и в том числе БТШ70 в принципе можно нарабатывать в различных эукариотических и прокариотических системах.
Известно, что в мире существует три основных стратегии получения рекомбинантных белков человека (посредством экспрессии а) в бактериальных системах, б) в культуре клеток животных, в) с использованием в качестве биореактора трансгенных животных. Каждый из этих подходов имеет свои преимущества и недостатки, но по экономической эффективности производство белков на основе трансгенных животных значительно превосходит другие методы.
При наработке белков в трансгенных животных, например, в молоке, ген данного белка клонируется в конструкцию под промотором генов казеина, лактоферрина или генов других «молочных» белков. Таким образом, данная конструкция экспрессируется только в молочной железе животного. Кроме того, целевой белок несет на своем С-конце слитый пептид, обеспечивающий его выход в молоко. Таким образом, трансгенные животные синтезируют целевой белок в молоко, из которого его далее можно очистить. Хотя белки, синтезируемые в молоко, обычно гликозилированы, ранее было показано, что это не влияет на свойства секретеруемых в норме белков, таких как лизоцим или лактоферрин.
Изобретательской задачей данной заявки на получение патента является разработка способа получения биологически активного белка человека БТШ70 в препаративных количествах для использования в качестве потенциального лекарственного средства.
Техническим результатом настоящего изобретения является получение биологически активного, рекомбинантного человеческого БТШ70, экспрессируемого в молоке трансгенных мышей.
Вывод о биологической активности полученного из молока препарата БТШ70 был сделан на основании ШАП-ИФА теста, позволяющего адекватно оценить шаперонные свойства полученного препарата БТШ70, а также на основании классического ЦТЛ (цитотоксических лимфоцитов) теста.
Осуществление изобретения
Получение конструкции для наработки функционально активного природного варианта белка БТШ70 при экспрессии в молочной железе трансгенных мышей.
Клонирование гена БТШ70 в плазмидный вектор для инъекций в животных.
Плазмида БТШ70 была создана на основе вектора pBC1 (Invitrogen) и геномной последовательности гена БТШ70 человека (Рис. 1). Стандартные условия выращивания in vitro, условия культивирования - линия Е. coli XL-2, среда 2xYT, ампициллин 50мкг/мл, 37°С, 18-20 часов.
Молекулярно-биологические свойства рекомбинантной плазмиды: размер конструкции: 16912 п.н., размер векторной части: 5877 п.н., маркерный признак: устойчивость к ампициллину.
Структурные элементы: тандемный повтор инсуляторов из бета-глобинового гена кур, бета-казеиновый промотор из генома коз, 5'-нетранслируемая область содержит два первых экзона бета-казеинового гена, кодирующая область содержит геномную последовательность сигнального пептида лактоферрина и кодирующую область гена БТШ70 человека, ген устойчивости к ампициллину, bla-промотор, ориджин репликации из плазмиды pBR322; сайт клонирования: NotI-SalI; размер кодируемого белка: 659 а.о. Устойчивость рекомбинантной плазмиды. Хранение в течение 2 лет (срок наблюдения) в виде плазмидной ДНК при температуре минус 20°С.
Аликвоту плазмидной ДНК извлекают из холодильника (температура хранения - минус 20,0±2,0°С) и проводят трансформацию культуры Е. coli методом электропорации. Проводят культивирование трансформированной культуры на среде 2xYT с добавлением 50 мкг/мл ампициллина. Продолжительность культивирования составляет от 18 до 20 часов при температуре +37,0±0,5°С.
Плазмидная ДНК выделяется методом щелочного лизиса.
Центрифугируют при ЦФ 4°С в течение 10-15', сливают среду; далее центрифугируют 4krpm в течение 1', удаляют остатки жидкости. Ресуспензируют в 10(5) ml раствора "I" (Глюкоза 50mM, Tris-HCl, рН 8.0 25mM, EDTA 10mM); + 1 ml свежеприготовленного раствора лизоцима (10mg/ml в р-ре "I"). Далее выдерживают при нормальной температуре в течение 15', добавляют 20 ml раствора "II" (NaOH 0.2N, SDS 1%), резко выливают, смешивают при 0°С в течение 5'; добавляют 10 ml холодного 10М AcONH4 (добавляют 5 ml пипеткой по каплям), смешивают. Выдерживают при 0°С в течение 5'.
Центрифугируют 5krpm, при 4°С в течение 10'; супернатант снимают, делят на 2 пробирки, добавляют к каждой по 12.5 мл изопропанола. Делают это на весах, чтобы пробирки имели равный вес; выдерживают при комнатной температуре в течение 10', далее центрифугируют 5krpm при 4°С в течение 10', сбросить супернатант. Центрифугируют при комнатной температуре в течение 3', отсасывают остатки жидкости.
Суспендируют осадок в 800μl 2 М AcONH4, объединяют в одной 2 ml пробирке; выдерживают при комнатной температуре в течение 5'; и центрифугируют при комнатной температуре в течение 10'. Супернатант переносят в пробирку с 800μl изопропанола; выдерживают при комнатной температуре в течение 5'; центрифугируют при комнатной температуре в течение 5'; далее споласкивают 70% EtOH; растворяют в 0.2-1 ml Н2О.
Выделенные и очищенные препараты плазмидной ДНК оценивают электрофоретически в 0,8% агарозном геле. Концентрацию измеряют на спектрофотометре и флуорометре. Проводится рестриктное картирование отдельно по EcoRI, HindIII, NotI-SalI. Рестриктные фрагменты разделяют электрофоретически на 0,8% агарозном геле и сравнивают с эталонными картами, представленными на фигурах 2, 3, 4.
Полученный препарат представляет собой очищенный препарат ДНК рекомбинантной плазмиды для экспрессии природного БТШ70, который может быть использован для подготовки фрагмента для микроинъекций.
Анализ трансгенных мышей
Создание трансгенных животных, получение потомства и анализ наличия трансгена проводили согласно описанным ниже методикам (Раздел «Создание трансгенных животных»).
С помощью ПЦР в режиме реального времени был оценено наличие трансгенов в молочной железе трансгенных мышей (Рис. 2). Таким образом, было показано, что трансгенные мыши, экспрессируют в молочной железе РНК, кодирующую природный вариант белка БТШ70.
Анализ свойств природного БТШ70, полученного в молочной железе трансгенных мышей.
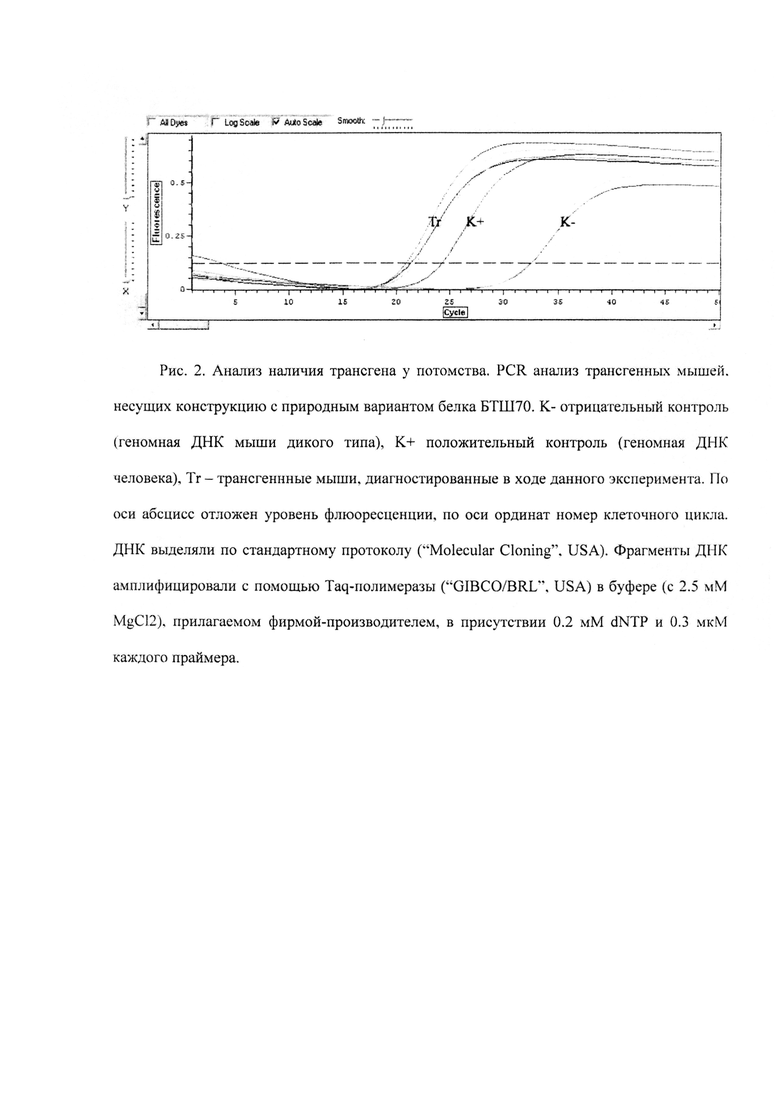
Далее было измерено примерное содержание БТШ70 (г/л) в молоке трансгенных мышей.). Молоко разбавляли в 5 раз буфером состава 20 мМ NaH2PO4/300 мМ NaCl рН 8.0, после чего разделяли методом диск-электрофореза в 10% полиакриламидном геле. После разделения белки переносили на мембрану Hybond ECL и обрабатывали моноклональными антителами к БТШ70 человека. Белок узнавался антителами, но его электрофоретическая подвижность была выше, чем у контрольного образца, что говорит о его посттрансляционной модификации (Рис. 3).
Создание трансгенных животных
Первичные трансгены были получены методом микроинъекций раствора ДНК в ТЕ-буфере (концентрация ДНК 5 нг/мкл) в мужской пронуклеус оплодотворенных яйцеклеток мышей гибридной линии (CBAxC57BL/6)F1 с последующей пересадкой выживших микроинъецированных зигот псевдобеременным реципиентам.
Яйцеклетки получали после индукции суперовуляции. Для этого неполовозрелым самкам (CBAxC57BL/6) F1 весом 12-13 г вводили внутрибрюшинно 8 ед. ГСЖК (гонадотропин сыворотки жеребых кобыл, фирма "МОСАГРОГЕН"), а затем, через 46 ч, внутрибрюшинно вводили 8 ед. Хгч (хорионический гонадотропин человека, Московский эндокринный завод). После такой обработки самок подсаживали к самцам-производителям гибридной линии (CBAxC57BL/6)F1. Факт спаривания констатировали на следующее утро по наличию копулятивной пробки. Схема получения суперовуляции: 13.00 ч - ГСЖК, через 46 ч в 11.00 ч - Хгч. В 17.00 ч этого же дня - подсадка к самцам-производителям. Отбор доноров производили на следующий день в 9.30 ч. Виварий освещали с 7.00 до 19.00 ч.
Самок с копулятивными пробками умерщвляли путем цервикальной дислокации и извлекали яйцеводы. Яйцеклетки вымывали из яйцеводов средой HEPES_KSOM с добавлением гиалуронидазы ("Sigma") под бинокуляром "Zeiss Stemi DV4" при увеличении 32х. Для вымывания использовали стеклянные капилляры с внутренним диаметром около 100 мкм, изготовленные на пуллере "Narishige PC-10" и микрокузнице "Narishige MF-900". Яйцеклетки культивировали в течение 2 ч (37°С, 5% CO2) в капле среды HEPES-KSOM или KSOM под минеральным маслом, затем помещали в микроинъекционную камеру. Микроинъекции проводили в среде HEPES-KSOM под микроскопом "Zeiss Axiovert 200М" при увеличении 400х-600х, используя микроманипуляторы "Narishige". Для изготовления игл для микроинъекций использовали пуллер "Sutter instrument Со Р-97". После окончания микроинъекций выжившие клетки переносили в каплю среды KSOM или HEPES-KSOM под минеральное масло и культивировали в течение 1 ч для выявления жизнеспособных клеток. Реципиентов получали следующим образом. Половозрелых самок (CBAxC57BL/6) F1 весом не менее 24 г подсаживали к вазэктомированным самцам той же линии. Псевдобеременных реципиентов отбирали на следующее утро по наличию копулятивных пробок.
Выжившие после микроинъекции зиготы трансплантировали в левый яйцевод псевдобеременной самки. В зависимости от числа клеток, выживших после микроинъекции, одной псевдобеременной самке пересаживали от 10 до 20 зигот. Для обездвиживания животных во время операции использовали авертин, который представляет собой 2.5% раствор в воде раствора 1 г 2,2,2 трибромэтанола в 1 мл 2_метил_2_бутанола. Авертин вводили внутрибрюшинно из расчета 15 мкл на 1 г веса животного.
Работы проводились на базе Центра коллективного пользования ИБГ РАН.
Получение потомства
При отсутствии естественных родов на 21-й день после пересадки микроинъецированных яйцеклеток реципиента умерщвляли путем цервикальной дислокации и проводили кесарево сечение, после чего выживших детенышей помещали к заранее подготовленной кормилице. Через 14-21 сут после рождения у мышат, рожденных после микроинъекций, отрезали кусочек хвоста, выделяли из них ДНК и анализировали наличие трансгена методом ПЦР.
Анализ наличия трансгена у потомства
ДНК выделяли по стандартному протоколу ("Molecular Cloning", USA). Фрагменты ДНК амплифицировали с помощью Taq-полимеразы ("GIBCO/BRL", USA) в буфере (с 2.5 мМ MgCl2), прилагаемом фирмой-производителем, в присутствии 0.2 мМ dNTP и 0.3 мкМ каждого праймера.
Для амплификации гена использовали следующие праймеры:
1 - 5'cgcaactccaccatccccaccaag
2 - 5'gccctctcgccctcgtacacctg
Получение молока, содержащего препарат БТШ70 из трансгенных мышей.
1. Пробы молока у трансгенных мышей получали на 2-20 день после родов не чаще 2 раз за лактацию. Взятие молока проводится у наркотизированной внутрибрюшинной инъекцией раствора Avertin (0,25%) мыши из расчета 10 мкл/г, для увеличения молокоотдачи мыши внутрибрюшинно вводится окситоцин (10 ед./мл) из расчета 10 мкл/г.
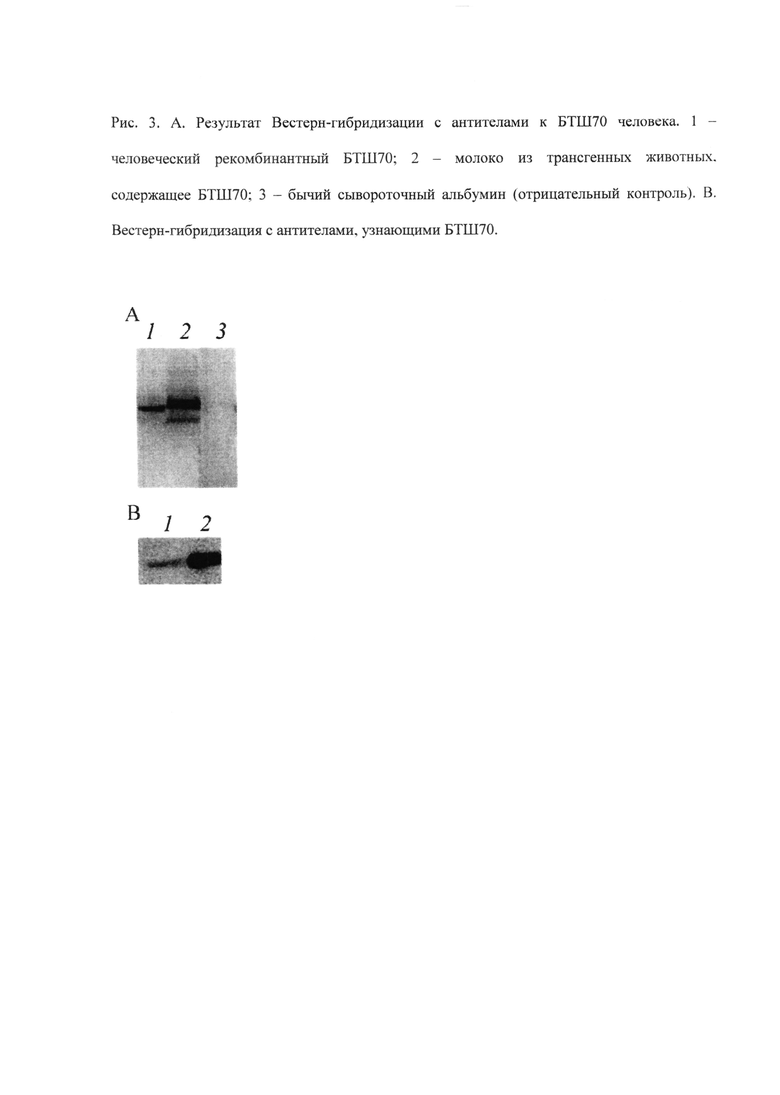
Молоко отбирается с помощью доильного аппарата, представляющего собой перистальтический насос, создающий разряженную атмосферу в пробирке для образцов; она соединяется со стеклянным капилляром, второй конец которого поочередно подносится к соскам мыши, и, за счет отрицательного давления в нем, молоко высасывается из протоков молочной железы и поступает в пробирку для образцов (рисунок 4).
Молоко разбавляли высокосолевым буфером Tris-HCl pH8.0/300mM NaCl в соотношении 1:10 (10 частей буфера на 1 часть молока), после чего центрифугировали 10 мин при ускорении не меньше 8000 g и отбрасывали жировую фракцию.
Анализ биологической активности молока, содержащего БТШ70
I. Тест ШАП-ИФА представляет собой систему, с помощью которой определяется способность активной части молекул шаперона, в данном случае БТШ70, узнавать и связывать субстрат, денатурированный белок. В данной модификации теста необратимо денатурированный белок, карбоксиметилированный лактальбумин, наносится на поверхность 96-луночного планшета для иммунологических реакций. После удаления неспецифического связывания в лунки вносится раствор БТШ70 в различных концентрациях и далее - поликлональные антитела RSIII, полученные в лаборатории ЗМК ИНЦ РАН. Тест основан на работах групп Wawrzynow-Liberek 1991 и Cheetham 1994 и подробно описан в работе Лазарев и др. 2011 (8).
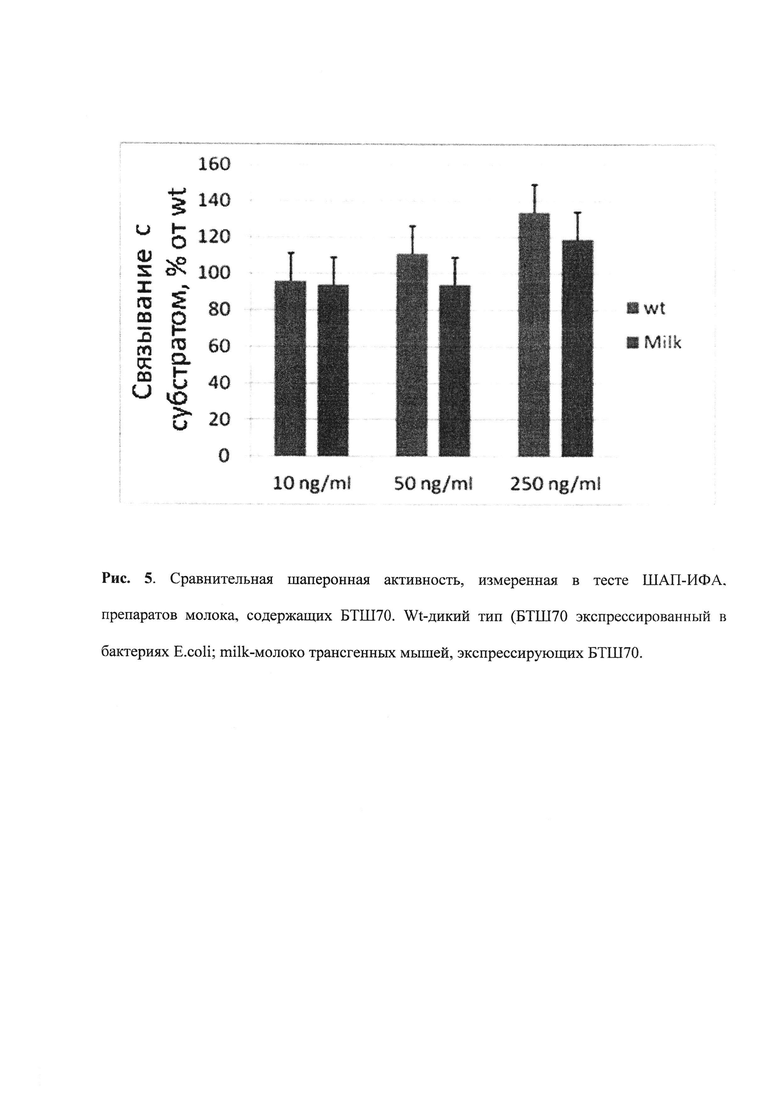
Очевидно, чем выше сигнал, тем большая часть молекул шаперона способна связывать субстрат. В сочетании с точным методом определения концентрации БТШ70 (r-Hu-Hsp70, HSPA1A, выделен из биомассы Е. coli), взятого в качестве эталона, можно вычислить относительную шаперонную активность (рисунок 5).
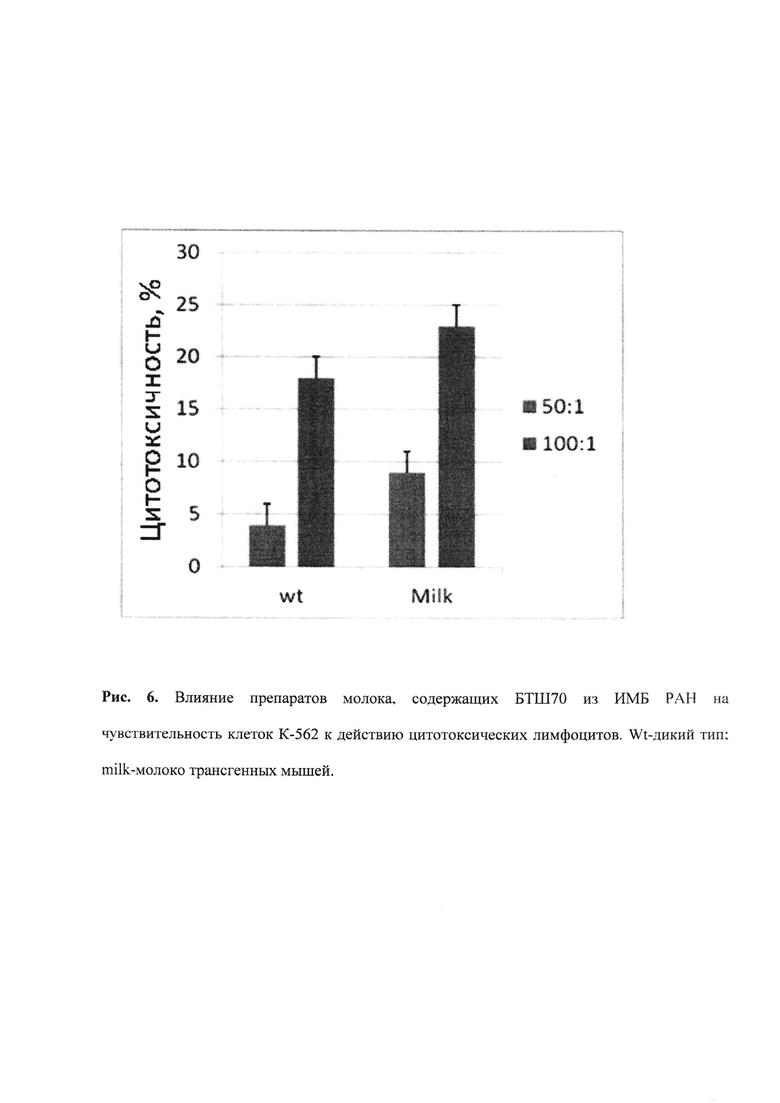
II. ЦТЛ (CTL) классический тест цитотоксических лимфоцитов, в котором последние должны узнавать и убивать опухолевые клетки. Тест является одним из ключевых в иммунологии при определении активности клеток врожденного и приобретенного иммунитета. В нашем случае в качестве эффекторов служат неактивированные спленоциты, выделенные из мышей С3Н в соответствии со стандартным протоколом; по данным проточной цитометрии 2-7% от результирующей популяции спленоцитов составляют натуральные киллеры. Мишенью для них служат клетки эритробластомы человека К-562, обработанные в течение 18 час препаратами БТШ70 (50 мкг/мл). Тест основан на том, что при инкубации раковых клеток-мишеней с функционально активным БТШ70 их чувствительность к натуральны киллерам увеличивается. Метод подробно описан в работах (9). После совместной инкубации в соотношении эффекторы (киллеры) : опухолевые клетки 50:1 и 100:1 клетки помещаются в лунки 96-луночной платы, где после введения необходимых составляющих определяется активность ЛДГ, вышедшего в среду из погибающих клеток с помощью тест-системы CytoTox-96 (Promega). Рисунок 6.
Краткое описание рисунков
Рис. 1. Карта конструкции (рБТШ70), содержащей ген нативного БТШ70 (Hsp70), для инъекции в животных. Структурные элементы конструкции: бета-казеиновый промотор из генома козы, сигнальная последовательность лактоферрина, тандемный повтор инсуляторов из бета-глобиного гена кур, ТАТА-бокс, ген устойчивости к ампициллину, ориджин репликации из плазмиды pBR322, кДНК копия человеческого гена БТШ70 (получена от Проф. Моримото GenBank Accession No M11717, аминокислотная последовательность соответствующего белка SwissProt Р08107 (SEQ ID NO:
Рис. 2. Анализ наличия трансгена у потомства. PCR анализ трансгенных мышей, несущих конструкцию с природным вариантом белка БТШ70. K- отрицательный контроль (геномная ДНК мыши дикого типа), K+ положительный контроль (геномная ДНК человека), Tr - трансгеннные мыши, диагностированные в ходе данного эксперимента. По оси абсцисс отложен уровень флюоресценции, по оси ординат номер клеточного цикла. ДНК выделяли по стандартному протоколу ("Molecular Cloning", USA). Фрагменты ДНК амплифицировали с помощью Taq-полимеразы ("GIBCO/BRL", USA) в буфере (с 2.5 мМ MgCl2), прилагаемом фирмой-производителем, в присутствии 0.2 мМ dNTP и 0.3 мкМ каждого праймера.
Рис. 3. А. Результат Вестерн-гибридизации с антителами к БТШ70 человека. 1 - человеческий рекомбинантный БТШ70; 2 - молоко из трансгенных животных, содержащее БТШ70; 3 - бычий сывороточный альбумин (отрицательный контроль). В. Вестерн-гибридизация с антителами, узнающими БТШ70.
Рисунок 4. Схема установки для доения мышей. А - Силиконовый шланг; В - Пробирка для образцов; С - Внешняя пробирка, внутри которой поддерживается разряженная атмосфера; D - Молоко попадает в пробирку для проб через систему «игла-силиконовый шланг-капилляр» (капилляр контактирует с соском молочной железы мыши); Е - Перистальтический насос; F - Внешний вид установки.
Рис. 5. Сравнительная шаперонная активность, измеренная в тесте ШАП-ИФА. препаратов молока, содержащих БТШ70. Wt-дикий тип (БТШ70 экспрессированный в бактериях E. coli; milk-молоко трансгенных мышей, экспрессирующих БТШ70.
Рис. 6. Влияние препаратов молока, содержащих БТШ70 из ИМБ РАН на чувствительность клеток К-562 к действию цитотоксических лимфоцитов. Wt-дикий тип; milk-молоко трансгенных мышей.
Источники информации
1. Kustanova GA, Murashev AN, Karpov VL, Margulis BA, Guzhova IV et al., (2006) Exogenous heat shock protein 70 mediates sepsis manifestations and decreases the mortality rate in rats. Cell Stress Chaperones 11:276-286.
2. Bobkova NV, Garbuz DG, Nesterova I, Medvinskaya N, Samokhin A et al., (2014) Therapeutic effect of exogenous hsp70 in mouse models of Alzheimer's disease. J Alzheimers Dis 38(2):425-435.
3. Action of the Hsp70 chaperone system observed with single proteins.unes JM, Mayer-Hartl M, Hartl FU, 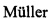 DJ.Nat Commun. 2015 Feb 17;6:6307. doi: 10.1038/ncomms7307.
DJ.Nat Commun. 2015 Feb 17;6:6307. doi: 10.1038/ncomms7307.
4. Kim YE, Hipp MS, Bracher A, Hayer-Hartl M, Hartl FU (2013) Molecular chaperone functions in protein folding and proteostasis. Annu Rev Biochem 82:323-355.
5. Guzhova I, Margulis В (2006) Hsp70 chaperone as a survival factor in cell pathology. IntRevCytol 254:101-149.
6. Ekimova IV, Nitsinskaya LE, Romanova IV, Pastukhov YF, Margulis BA et al., (2010) Exogenous protein Hsp70/Hsc70 can penetrate into brain structures and attenuate the severity of chemically-induced seizures. J Neurochem 115(4): 1035-1044.
7. Hansen JE1, Sohn W. Kim C, et al. Antibody-mediated Hsp70 protein therapy. Brain Res. 2006 May 9; 1088(1): 187-96.
8. Lazarev VF, Onokhin KV, Antimonova OI, Polonik SG, Guzhova IV, Margulis BA. Kinetics of chaperone activity of proteins Hsp70 and Hdj1 in human leukemia u-937 cells after preconditioning with thermal shock or compound u-133. Biochemistrv (Mosc). 2011 May; 76(5):590-5.
9. Shevtsov MA, Pozdnyakov AV, Mikhrina AL, et al., Effective immunotherapy of rat glioblastoma with prolonged intratumoral delivery of exogenous heat shock protein Hsp70. Int J Cancer. 2014 Nov 1; 135(9):2118-28.
| название | год | авторы | номер документа |
|---|---|---|---|
| Генетическая конструкция для экспрессии функционально-активного человеческого стресс-белка (БТШ70) с мутированными сайтами гликозилирования для наработки в эукариотических экспрессионных системах | 2016 |
|
RU2647570C1 |
| ГЕНЕТИЧЕСКИЕ КОНСТРУКЦИИ LTF3, LTF5, LTF7, LTF10, LTF11 ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ЛАКТОФЕРРИНА ЧЕЛОВЕКА (ВАРИАНТЫ) | 2011 |
|
RU2491343C1 |
| Способ получения кролика с нокаутом гена CSN2 | 2023 |
|
RU2834016C1 |
| СПОСОБ БИОПРОТЕКЦИИ, ОСНОВАННЫЙ НА ИСПОЛЬЗОВАНИИ ПРЕПАРАТА БТШ70, ПРЕДНАЗНАЧЕННЫЙ ДЛЯ СОЗДАНИЯ СРЕДСТВ И МЕТОДОВ НЕСПЕЦИФИЧЕСКОЙ ЗАЩИТЫ ОТ ОПАСНЫХ БИОЛОГИЧЕСКИХ ПОРАЖАЮЩИХ ФАКТОРОВ | 2005 |
|
RU2294210C1 |
| СПОСОБ ТЕРАПИИ ОПУХОЛЕВЫХ ЗАБОЛЕВАНИЙ | 2013 |
|
RU2597414C2 |
| ШТАММ БАКТЕРИЙ Escherichia coli - ПРОДУЦЕНТ БЕЛКА ТЕПЛОВОГО ШОКА 70 И СПОСОБ ПОЛУЧЕНИЯ ПРЕПАРАТА БЕЛКА ТЕПЛОВОГО ШОКА ЧЕЛОВЕКА | 2013 |
|
RU2564120C2 |
| Способ получения генно-модифицированных мышей, экспрессирующих миниген антитромбина III человека, с помощью микроинъекций TelN-линеаризованного фрагмента ДНК | 2022 |
|
RU2806568C1 |
| ТРАНСГЕН ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ПОЛИПЕПТИДА В МОЛОКЕ ТРАНСГЕННЫХ КОРОВ, СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННОЙ КОРОВЫ (ВАРИАНТЫ), МОЛОКО ОТ ТРАНСГЕННОЙ КОРОВЫ, ПИЩЕВОЙ СОСТАВ | 1990 |
|
RU2095414C1 |
| ГЕННО-ИНЖЕНЕРНАЯ КОНСТРУКЦИЯ pGoatcasGCSF, ОБЕСПЕЧИВАЮЩАЯ ПРОДУКЦИЮ ГРАНУЛОЦИТ-КОЛОНИЕСТИМУЛИРУЮЩЕГО ФАКТОРА ЧЕЛОВЕКА В МОЛОКО ТРАНСГЕННЫХ ЖИВОТНЫХ | 2010 |
|
RU2422529C1 |
| СПОСОБ РАЗДЕЛЕНИЯ ЛАКТОФЕРРИНОВ ЧЕЛОВЕКА И КОЗЫ С ПОМОЩЬЮ ДИФФЕРЕНЦИАЛЬНОЙ ИММУНОАФФИННОЙ ХРОМАТОГРАФИИ С ИСПОЛЬЗОВАНИЕМ ОДНОДОМЕННЫХ МИНИ-АНТИТЕЛ | 2014 |
|
RU2553515C1 |

Изобретение относится к биотехнологии, конкретно к рекомбинантному получению белка теплового шока человека 70 (БТШ70), и может быть использовано для получения БТШ70 в молоке трансгенных животных. На основе экспрессионного вектора рВС1 получают экспрессионную плазмиду рБТШ70 размером 16912 п. н., с картой, приведенной на рис. 1, которая содержит ген БТШ70 человека и кодирует сигнальную последовательность лактоферрина. Изобретение позволяет получить высокий титр человеческого БТШ70, обладающего шаперонными свойствами, характерными для нативного человеческого БТШ70, в молоке трансгенных животных. 6 ил.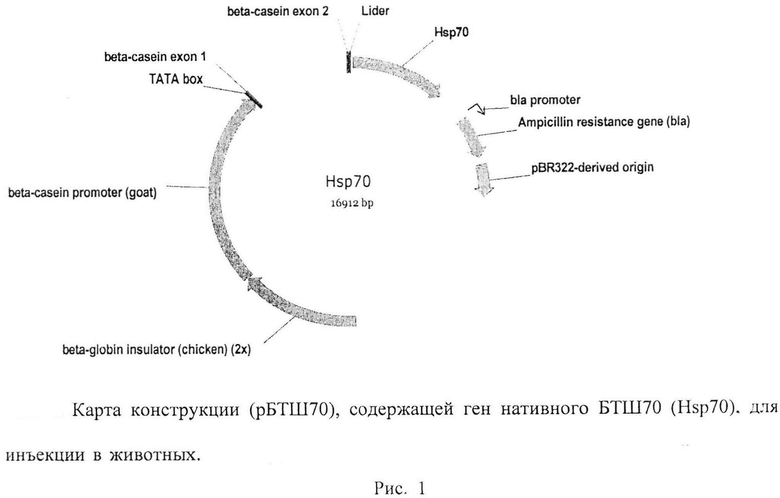
Генетическая конструкция рБТШ70, представляющая собой экспрессионную плазмиду размером 16912 п.н., с картой, приведенной на рис. 1, полученная на основе экспрессионного вектора рВС1, содержащая ген белка теплового шока человека 70 (БТШ70), и кодирующая сигнальную последовательность лактоферрина, предназначенная для продукции биологически активного рекомбинантного белка теплового шока человека 70 (БТШ70) в молоке трансгенных животных.
| US 2005260746 A1, 24.11.2005 | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Приспособление для уменьшения дымовой тяги паровоза | 1920 |
|
SU270A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| LIEDEL J.L | |||
| et al., Mother's milk-induced Hsp70 expression preserves intestinal epithelial barrier function in an immature rat pup model, Pediatr Res., 2011, v.69, n.5 Pt 1, p.395-400. | |||

