Область техники настоящего изобретения
Изобретение относится к областям молекулярной биологии и трансгенеза и представляет собой две линии животных с экспрессией антитромбина III человека в печени, а также способ их получения, включающий способ подготовки линейных двуцепочечных фрагментов ДНК для микроинъекций в зиготы с использованием фермента протеломеразы TelN из фага N15, а также рекомбинантные плазмиды для его осуществления.
Работа выполнена при финансовой поддержке Министерства Науки и Высшего образования Российской Федерации в рамках Соглашения № 075-15-2019-1661 от 31.10.2019 на базе Центра высокоточного редактирования и генетических технологий для биомедицины (ЦВРГТБ) ИБГ РАН.
Область техники
Ген SERPINC1 человека кодируют белок антитромбин III (АТIII), который играет важнейшую роль в регуляции системы гемостаза. Белок АТIII относится суперсемейству ингибиторов сериновых протеаз (серпинов) и действует за счет связывания и инактивации тромбина и факторов свертывания Xa и IXa [Mulder R, Croles FN, Mulder AB, Huntington JA, Meijer K, Lukens MV. SERPINC1 gene mutations in antithrombin deficiency. Br J Haematol. 2017Jul;178(2):279-285. doi: 10.1111/bjh.14658. Epub 2017 Mar 20. PMID: 28317092].
Наследственный дефицит АТIII у человека является одной из наиболее часто встречающихся тромбофилий с частотой в популяции 1:2000 - 1:5000 [Kumar R, Chan AK, Dawson JE, Forman-Kay JD, Kahr WH, Williams S. Clinical presentation and molecular basis of congenital antithrombin deficiency in children: a cohort study. Br J Haematol. 2014 Jul;166(1):130-9. doi:10.1111/bjh.12842. Epub 2014 Mar 29. PMID: 24684277]. Дефицит АТIII сопряжен прежде всего с развитием венозных тромбозов, угрожающих жизни и несущих риск инвалидизации [Luxembourg B, Delev D, Geisen C, Spannagl M, Krause M, Miesbach W, Heller C,Bergmann F, Schmeink U, Grossmann R, Lindhoff-Last E, Seifried E, Oldenburg J, Pavlova A. Molecular basis of antithrombin deficiency. Thromb Haemost. 2011 Apr;105(4):635-46. doi: 10.1160/TH10-08-0538. Epub 2011 Jan 25. PMID: 21264449]. Риск таких осложнений возрастает при травмах, оперативных вмешательствах [Ehrhardt JD Jr, Boneva D, McKenney M, Elkbuli A. Antithrombin Deficiency in Trauma and Surgical Critical Care. J Surg Res. 2020 Dec;256:536-542. doi:10.1016/j.jss.2020.07.010. Epub 2020 Aug 13. PMID: 32799002]. Для предотвращения тромбоэмболических осложнений у пациентов с дефицитом АТIII при оперативных вмешательствах или родах показано введение препаратов АТIII, который получают либо из плазма здоровых доноров, либо из молока трансгенных коз. В частности, препарат ATryn® был первым одобренным FDA лекарственным препаратом, получаемым из молока трансгенных животных [Lavine G. FDA approves first biological product derived from transgenic animal. Am J Health Syst Pharm. 2009 Mar 15;66(6):518. doi: 10.2146/news090023.PMID: 19265177].
Получение АТIII из плазмы доноров сопряжено с высокой стоимостью, необходимостью тестирования образцов на наличие вирусов человека и др. Поэтому получение рекомбинантных белков крови человека из молока трансгенных животных представляет собой один из актуальных и перспективных способов получения таких белков. В настоящий момент на рынке присутствуют три подобных препарата: ATryn ®, рекомбинантный антитромбин III, получаемый из молока трансгенных коз; Sevenfact®, рекомбинантный фактор свертывания крови VIIa, получаемый из молока трансгенных кроликов; и Ruconest ®, рекомбинантный ингибитор С1-эстеразы человека, получаемый из молока трансгенных кроликов.
Известно, что белки крови животных способны проникать в молоко, поэтому препараты рекомбинантных белков крови человека, получаемые из молока трансгенных животных, содержат примеси гомологичных белков крови животного. В частности, гомологичные белки животного проникают в молоко путем активного транспорта из крови [Monks J, Neville MC. Albumin transcytosis across the epithelium of the lactating mouse mammary gland. J Physiol. 2004 Oct 1;560(Pt 1):267-80. doi:10.1113/jphysiol.2004.068403. Epub 2004 Aug 5. PMID: 15297572] или за счет параклеточного транспорта при повышении проницаемости гемато-лактационного барьера в начале лактации или при мастите [ Wall SK, Gross JJ, Kessler EC, Villez K, Bruckmaier RM. Blood-derived proteins in milk at start of lactation: Indicators of active or passive transfer. J Dairy Sci. 2015 Nov;98(11):7748-56. doi: 10.3168/jds.2015-9440. Epub2015 Aug 20. PMID: 26298756]. Физико-химическое разделение белка человека и животного практически невозможно, в силу их высокой гомологии. Присутствие примесей белка крови животного в препарате рекомбинантного белка, получаемого из молока трансгенного животного, способно вызвать иммунную реакцию у пациента при введении препарата.
Для решения этой проблемы предложено получать гуманизированных животных-продуцентов, у которых инактивирован ген белка крови, а его функции замещаются за счет экспрессии гомологичного гена человека. Появление технологий CRISPR/Cas9-опосредованного геномного редактирования существенно упростило получение трансгенных животных с направленными модификациями генома [Shepelev MV, Kalinichenko SV, Deykin AV, Korobko IV. Production of Recombinant Proteins in the Milk of Transgenic Animals: Current State and Prospects. Acta Naturae. 2018 Jul-Sep;10(3):40-47. PMID: 30397525; PMCID: PMC6209402], в частности животных с нокаутом гена. Однако, нокаут гена Serpinc1 у мыши приводит к гибели эмбрионов на ранних стадиях развития [Ishiguro K, Kojima T, Kadomatsu K, Nakayama Y, Takagi A, Suzuki M, Takeda N, Ito M, Yamamoto K, Matsushita T, Kusugami K, Muramatsu T, Saito H. Complete antithrombin deficiency in mice results in embryonic lethality. J Clin Invest. 2000 Oct;106(7):873-8. doi: 10.1172/JCI10489. PMID: 11018075; PMCID: PMC517819], поэтому подобная стратегия не применима к гену, кодирующему ATIII.
Таким образом, получение гуманизированных по гену Serpinc1 животных, возможно за счет реализации двух стратегий. В первом случае, за счет CRISPR/Cas9-опосредованной таргетной встройки в локус гена Serpinc1 мыши генетической конструкции, кодирующей ATIII человека. В этом случае, ген мыши инактивируется за счет интеграции конструкции ATIII человека, которая и обеспечивает замещение функции ATIII мыши. Однако эффективность таргетной интеграции больших генетических конструкций в геном с помощью системы CRISPR/Cas9 остается достаточно низкой. Во втором случае, получают линию генно-модифицированных мышей с помощью трансгенеза со случайной встройкой в геном конструкции, кодирующей ATIII человека. Далее, в такой линии мышей на фоне экспрессии ATIII человека осуществляют нокаут гена Serpinc1 с помощью системы CRISPR/Cas9, что существенно проще и эффективней по сравнению со встройкой больших генетических конструкций.
Таким образом, одна из задач, решаемых настоящим изобретением, заключается в получении линии генно-модифицированной мышей, экспрессирующих ген ATIII человека, методом трансгенеза со случайной встройкой генетической конструкции в геном.
При получении трансгенных животных с помощью классического метода трансгенеза проводят микроинъекции линейных двуцепочечных фрагментов ДНК, которые получают с помощью гидролиза кольцевой плазмидной ДНК эндонуклеазами рестрикции с последующей очисткой фрагмента ДНК из агарозного геля. Недавно было предложен метод геномного редактирования, названный Tild-CRISPR, в котором в качестве матрицы для репарации двуцепочечных разрывов ДНК использовали линейные двуцепочечные фрагменты ДНК, получаемые с помощью эндонуклеаз рестрикции. Авторы заявляют о высокой эффективности нокина с помощью метода Tild-CRISPR при использовании линейных фрагментов ДНК в качестве матриц для репарации по сравнению с кольцевыми плазмидами и рядом других видов матриц для репарации [Yao X, Zhang M, Wang X, Ying W, Hu X, Dai P, Meng F, Shi L, Sun Y, Yao N,Zhong W, Li Y, Wu K, Li W, Chen ZJ, Yang H. Tild-CRISPR Allows for Efficient and Precise Gene Knockin in Mouse and Human Cells. Dev Cell. 2018 May 21;45(4):526-536.e5. doi: 10.1016/j.devcel.2018.04.021. PMID: 29787711]. Кроме того, авторы указывают на низкий уровень случайной интеграции таких линейных матриц в геном. Таким образом, разработка нового способа получения линейных двуцепочечных молекул ДНК имеет актуальность как для технологий геномного редактирования, так и для методов трансгенеза путем случайной встройки.
Считается, что фрагмент ДНК (трансген) не должен содержать последовательностей остова плазмиды или каких-то еще бактериальных нуклеотидных последовательностей. Однако далеко не всегда можно подобрать подходящие эндонуклеазы рестрикции для вырезания фрагмента ДНК строго по границам трансгена. Например, подходящие сайты рестрикции могут просто отсутствовать, либо генетическая конструкция для трансгенеза или матрица для репарации может содержать внутренние сайты эндонуклеаз рестрикции, не позволяющие вырезать нужный фрагмент. Для решения этой проблемы в данном изобретении был предложен подход для получения линейных двуцепочечных фрагментов ДНК с помощью фермента протеломеразы TelN. Преимущество этого метода основано на получении рекомбинантной плазмиды, в которой полилинкер фланкирован сайтами гидролиза протеломеразы TelN. Это позволяет решить задачу выбора ферментов для получения линейных фрагментов ДНК. Сайт узнавания TelN имеет длину 56 п.о., что исключает вероятность случайного присутствия таких сайтов в каких-либо генетических конструкциях. Кроме того, при расщеплении ДНК фермент TelN формирует ковалентные связи между комплементарными нуклеотидами на концах ДНК, что приводит к формированию шпилек на концах фрагмента ДНК. Такие структуры повышают устойчивость молекул ДНК к действию экзонуклеаз [Khan IM, Coulson JM. A novel method to stabilise antisense oligonucleotides against exonuclease degradation. Nucleic Acids Res. 1993 Jun 25;21(12):2957-8.doi: 10.1093/nar/21.12.2957. Erratum in: Nucleic Acids Res. 1993 Sep11;21(18):4433. PMID: 8392712; PMCID: PMC309707; Hirao I, Nishimura Y, Tagawa Y, Watanabe K, Miura K. Extraordinarily stable mini-hairpins: electrophoretical and thermal properties of the various sequence variants of d(GCGAAAGC) and their effect on DNA sequencing. Nucleic Acids Res.1992 Aug 11;20(15):3891-6. doi: 10.1093/nar/20.15.3891. PMID: 1508675; PMCID: PMC334063]. Таким образом, в рекомбинантную плазмиду клонируется фрагмент ДНК по сайтам эндонуклеаз рестрикции, расположенных в полилинкере. После этого, линейный фрагмент ДНК для микроинъекций получают с помощью фермента TelN.
Фермент протеломераза кодируется геном 29 фага N15, инфицирующего клетки Escherichia coli, и участвует в репликации фага N15, формируя из кольцевого генома линейную двуцепочечную молекулу профага с ковалентно замкнутыми концами. Белок TelN состоит из 630 аминокислот и имеет молекулярный вес 72.2 кДа. Последовательность-мишень для протеломеразы в геноме фага N15 называется tos (telomerase occupancy site). Сайт tos включает в себя шесть инвертированных повторов R1/L1, R2/L2 и R3/L3 и палиндромную последовательности telO в центре. Однако непосредственно для расщепления и линеаризации молекулы ДНК ферментом TelN достаточно наличия сайта-мишени telRL длиной 56 п.о., состоящего из инвертированных повторов R3/L3 длиной 14 п.о., фланкирующих палиндром telO длиной 22 п.о., который отделен от повторов последовательностями в 3 п.о. (Фиг.2). TelN расщепляет ДНК в середине палиндрома telO, что сопровождается формированием ковалентных связей между двумя комплементарными нуклеотидами (G и С) на конце молекулы ДНК и образованием шпильки из шести нуклеотидов (Фиг.2) [Deneke J, Ziegelin G, Lurz R, Lanka E. The protelomerase of temperate Escherichia coli phage N15 has cleaving-joining activity. Proc Natl Acad Sci USA. 2000 Jul 5;97(14):7721-6. doi:10.1073/pnas.97.14.7721. PMID: 10884403; PMCID: PMC16611].
Было показано, что формирование линейных молекулы ДНК с использованием TelN обеспечивает более высокий уровень экспрессии маркерного гена Egfp по сравнению с линейными фрагментами, полученными с помощью эндонуклеаз рестрикции. На мышиной модели формирования метастаз в легком при инъекции клеток меланомы B16F10 было показано, что полученная с помощью TelN линейная молекула ДНК, экспрессирующая IL-12, обеспечивает более высокий ингибирующий эффект на формирование метастазов по сравнению с исходной кольцевой плазмидой [Heinrich J, Schultz J, Bosse M, Ziegelin G, Lanka E, Moelling K. Linear closed mini DNA generated by the prokaryotic cleaving-joining enzyme TelN is functional in mammalian cells. J Mol Med (Berl). 2002 Oct;80(10):648-54. doi:10.1007/s00109-002-0362-2. PMID: 12395149]. Эти результаты указывают на то, что молекулы ДНК с ковалентно замкнутыми концами могут быть более стабильны в клетках млекопитающих.
Интересно отметить, что фермент TelN проявляет активность не только in vitro, но и в клетках млекопитающих [Liew PS, Chen Q, Ng AWR, Chew YC, Ravin NV, Sim EUH, Lee CW, Narayanan K. Phage N15 protelomerase resolves its tos recognition site into hairpin telomeres within mammalian cells. Anal Biochem. 2019 Oct 15;583:113361. doi:10.1016/j.ab.2019.113361. PMID: 31306622], а линейные молекулы ДНК содержащие tos, способны реплицироваться в клетках млекопитающих, и обеспечивать стабильную экспрессию трансгена [Liew PS, Tan TH, Wong YC, Sim EUH, Lee CW, Narayanan K. A Self-Replicating Linear DNA. ACS Synth Biol. 2020 Apr 17;9(4):804-813. doi:10.1021/acssynbio.9b00478. PMID: 32196315]. Кроме того, фермент TelN находит применение при молекулярном клонировании протяженных последовательностей ДНК [Cui YZ, Zhou JT, Li BZ, Yuan YJ. The TelN/tos-assisted precise targeting of chromosome segments (TAPE). J Adv Res. 2022 Nov;41:169-177. doi:10.1016/j.jare.2022.01.017. PMID: 36328746; PMCID: PMC9637730].
Таким образом полученные с помощью TelN фрагменты ДНК функциональны в клетках млекопитающих, а наличие ковалентно замкнутых шпилек на концах молекулы ДНК, по всей видимости, повышает их стабильность в клетках, в частности, за счет резистентности к действию экзонуклеаз. На основе этих данных нами был предложен новый метод получения линейных фрагментов ДНК для микроинъекций в зиготы мыши с помощью фермента протеломеразы TelN.
Раскрытие изобретения
Задачами заявляемого изобретения являются разработка универсального способа получения линейных двуцепочечных фрагментов ДНК с помощью фермента протеломеразы TelN из фага N15, пригодных для микроинъекций в зиготы млекопитающих, и создание линии генетически модифицированных мышей, экспрессирующих миниген антитромбина III человека в ткани печени.
Задача решается тем, что на основе вектора pBluescriptSK II (-) (Stratagene, США) конструируют рекомбинантную плазмиду pTelN (SEQ ID NO: 1), в которой полилинкер с двух сторон фланкирован сайтам расщепления протеломеразы TelN из фага N15 длиной 56 п.о. Кроме того, рекомбинантная плазмида TelN содержит промотор гена β-лактамазы и открытую рамку считывания β-лактамазы (ген устойчивости к ампициллину), высококопийную точку начала репликации ColE1/pMB1/pBR322/pUC, сайт связывания CAP (Catabolite activator protein), Lac промотор и Lac оператор, промоторы для РНК полимераз бактериофагов T3 и T7 и точку начала репликации для фага f1 (Фиг.1). Полученная рекомбинантная плазмида pTelN решает задачу по разработке универсального метода получения линейных двуцепочечных фрагментов ДНК за счет того, что в полилинкер pTelN, содержащий ряд уникальных сайтов рестрикции, клонируют фрагмент ДНК, который необходимо получить в виде линейного двуцепочечного фрагмента. Далее, плазмиду содержащую целевой фрагмент ДНК, обрабатывают ферментом протеломеразой TelN, получая линейный двуцепочечный фрагмент ДНК с ковалентно замкнутыми концами, несущий целевую последовательность ДНК.
Преимуществом предложенного в изобретении способа получения линейного двуцепочечного фрагмента ДНК для микроинъекций является использование фермента TelN, сайт распознавания которого составляет 56 п.о., что исключает случайное присутствие таких сайтов в каких-бы то ни было нуклеотидных последовательностях из геномов млекопитающих, таким образом устраняется необходимость в подборе уникальных эндонуклеаз рестрикции для вырезания фрагмента ДНК из плазмидного вектора, так как сайты рестриктаз как-правило имеют длину 6-8 п.о. и существенно чаще встречаются в геномах млекопитающих, и, соответственно, могут присутствовать в нуклеотидных последовательностях вырезаемых фрагментов ДНК, тем самым затрудняя подбор эндонуклеаз рестрикции для получения линейных фрагментов ДНК. Кроме того, фермент TelN после расщепления ДНК формирует ковалентные связи между нуклеотидами в прямой и обратной цепях ДНК на конце получаемых фрагментов ДНК с формированием шпилечных структур (Фиг.1). Это способствует повышению стабильности таких фрагментов ДНК в клетках млекопитающих за счет устойчивости к гидролизу экзонуклеазами.
Ранее нами был сконструирован миниген АТIII человека с оптимизированным паттерном сплайсинга путем минимизации протяженных интронов гена SERPINC1 и получена генетическая конструкция pmhATg7Spl, несущая миниген фланкированный плечами гомологии для локуса Serpinc1 мыши [Shepelev MV, Saakian EK, Kalinichenko SV, Korobko IV. Human Antithrombin III Minigene with an Optimized Splicing Pattern. Mol Biol (Mosk). 2019 May-Jun;53(3):411-420. doi: 10.1134/S0026898419030170. PMID: 31184606]. Подобная конструкция универсальна, и может быть использована как получения гуманизированных мышей с помощью CRISPR/Cas9-опосредованного нокина в локус Serpinc1, так и для трансгенеза с помощью случайной встройки. Далее, поставленная задача решается тем, что на основе плазмид pTelN и pmhATg7Spl конструируют рекомбинантную плазмиду pTelN-gAT3Spl, кодирующую миниген антитромбина III человека, и предназначенную для получения гуманизированных по гену Serpinc1 мышей. Плазмида pTelN-gAT3Spl включает миниген ATIII человека под транскрипционным контролем регуляторных элементов гена Serpinc1 мыши, сигнала полиаденилирования вируса SV40 и терминатора транскрипции из гена SKIV2L человека. Миниген АТIII человека фланкирован геномными последовательностями локуса Serpinc1 мыши для обеспечения возможности осуществления нокина минигена АТIII человека в локус Serpinc1 по механизму гомологичной рекомбинации с помощью системы CRISPR/Cas9. В частности, левое плечо гомологии включает 1965 п.о., первого интрона гена Serpinc1 и 102 п.о. второго экзона, вплоть до ATG кодона. Кроме того, мишень для гидовой РНК sg1 (см. ниже) расположена во втором экзоне в левом плече, поэтому для предотвращения расщепления плазмиды pTel-gAT3Spl левое плечо несет мутированный сайт PAM гидовой РНК sg1. Рекомбинантную плазмиду pTelN-gAT3Spl (SEQ ID NO:2) конструируют на основе вектора pGEM5Zа(+) (Promega, США) используя описанную ранее рекомбинантную плазмиду pmhATg7Spl, несущую миниген ATIII человека с оптимизированным паттерном сплайсинга [Shepelev MV, Saakian EK, Kalinichenko SV, Korobko IV. Human Antithrombin III Minigene with an Optimized Splicing Pattern. Mol Biol (Mosk). 2019 May-Jun;53(3):411-420. doi: 10.1134/S0026898419030170. PMID: 31184606].
Сконструированная плазмида pTel-gAT3Spl содержит элементы плазмиды pGEM5Zа(+), включающие промотор гена β-лактамазы и открытую рамку считывания β-лактамазы (ген устойчивости к ампициллину), высококопийную точку начала репликации ColE1/pMB1/pBR322/pUC, сайт связывания CAP (Catabolite activator protein), Lac промотор и Lac оператор, промоторы для РНК полимераз бактериофагов T3 и T7 и точку начала репликации для фага f1.
Кроме того, рекомбинантная плазмида pTelN-gAT3Spl содержит:
- нуклеотидную последовательность минигена ATIII человека, который включает в себя белоккодирующие экзоны гена SERPINC1 человека (со второго по седьмой), разделенные минимизированными вторым, пятым и шестым интронами и полноразмерными третьим и четвертым интронами гена SERPINC1 человека.
- левое и правое плечи гомологии для локуса Serpinc1 мыши, длиной 2076 и 1834 п.о., соответственно, необходимые для обеспечения возможности репарации двуцепочечного разрыва ДНК, вносимого Cas9, по механизму гомологичной рекомбинации для реализации стратегии гуманизации мышей с помощью CRISPR/Cas9-опосредованного нокина конструкции для экспрессии ATIII человека.
- левое плечо гомологии включает в себя 1965 п.о. первого интрона гена Serpinс1 и 102 п.о. второго экзона вплоть до ATG кодона. Наличие этих генетических элементов обеспечивает экспрессию транскрипта минигена при случайной встройке фрагмента ДНК в геном мыши.
- для предотвращения расщепления плазмиды pTelN-gAT3Spl нуклеазой Cas9 при осуществлении CRISPR/Cas9-опосредованного геномного редактирования для получения гуманизированных мышей по гену Serpinс1 левое плечо гомологии мыши содержит мутированный PAM сайт для выбранной гидовой РНК (sg1).
Для эффективной терминации транскрипции и полиаденилирования мРНК в 3'-области минигена помещен сигнал полиаденилирования из вируса SV40. Помимо этого, в 3'-области относительно минигена рекомбинантная плазмида содержит двунаправленный терминатор транскрипции из гена SKIV2L человека для блокирования возможной проходящей транскрипции минигена и предотвращения формирования аберрантных транскриптов.
Таким образом, еще одна поставленная задача решается тем, что путем трансгенеза методом случайного встраивания получают новую линию генно-модифицированных мышей с экспрессией антитромбина III человека в печени. Для этого предложенную рекомбинантную плазмиду pTelN-gAT3Spl обрабатывают ферментом TelN и получают линейный двуцепочечный фрагмент ДНК, несущий миниген АТIII, фланкированный плечами гомологии. Далее проводят микроинъекции такого фрагмента ДНК в оплодотворённые яйцеклетки мыши. В результате случайной встройки фрагмента в геном получают линию мышей с экспрессией минигена АТIII в ткани печени. Полученную линию генно-модифицированных мышей используют в качестве модели для разработки подхода к получению гуманизированных по гену Serpinc1 мышей с помощью комбинации трансгенеза со случайной встройкой и CRISPR/Cas9-опосредованнного нокаута гена Serpinc1. Для этого в полученной линии генно-модифицированных мышей с помощью системы CRISPR/Cas9 инактивируют ген Serpinc1. Таким образом, получают гуманизированных животных, у которых функция гена Serpinc1 компенсирована за счет экспрессии АТIII человека, кодируемого случайно встроенным в геном фрагментом ДНК, несущим минигена АТIII человека.
Кроме того, предложенная рекомбинантная плазмида pTelN-gAT3Spl может быть использована для получения гуманизированных по гену Serpinc1 мышей с помощью CRIPSR/Cas9-опосредованного нокина в локус Serpinc1. Для этого описанную в настоящем изобретении рекомбинантную плазмиду pTelN-gAT3Spl используют в качестве матрицы для репарации двуцепочечного разрыва ДНК, вносимого в геном мыши с помощью системы CRISPR/Cas9. Для этого подбирают гидовую РНК вносящую двуцепочечный разрыв во втором экзоне гена Serpinc1 мыши (sg1; последовательность в геноме мыши, комплементарная sg1: 5'-gagacgatcactccgaaaactgg -3'), получают препарат гидовой РНК с помощью транскрипции in vitro и проводят микроинъекции в зиготы мыши смеси гидовой РНК, мРНК нуклеазы Cas9 и рекомбинантной плазмиды pTelN-gAT3Spl в качестве матрицы для репарации разрыва ДНК. В случае корректной репарации разрыва ДНК по механизму гомологичной рекомбинации в геном мыши встраивается миниген АТIII человека таким образом, что в геноме мыши сохраняются эндогенные регуляторные последовательности гена Serpinc1, включающие промотор, первый экзон, первый интрон и часть второго экзона вплоть до кодона инициации трансляции белка АТIII, после которого следует миниген ATIII человека. Таким образом, за счет встройки минигена ATIII человека достигается инактивация гена Serpinc1 мыши, а функция белка ATIII мыши компенсируется за счет экспрессии белка ATIII человека, кодируемого минигеном.
Генотипирование трансгенных мышей проводят с помощью количественной ПЦР. Для этого выделяют геномную ДНК из тканей животных. Подбирают олигонуклеотидные праймеры для ПЦР, специфичные для нуклеотидных последовательностей трансгена, в частности для минигена ATIII человека и терминатора транскрипции SKIV2L из генома человека. Специфичность праймеров проверяют на образцах геномной ДНК из ткани животных дикого типа и геномной ДНК человека. Копийность трансгена определяют сходным образом с помощью количественной ПЦР. В качестве референсного гена для расчета копийности используют ген Bmp4 мыши, присутствующий в геноме в количестве двух копий.
Экспрессию минигена АТIII человека в печени трансгенных мышей проверяют с помощью ПЦР и количественной ПЦР. Для этого выделяют суммарную РНК из ткани печени трансгенных животных и проводят реакцию обратной транскрипции. Уровень экспрессии минигена АТIII человека определяют с помощью количественной ПЦР с олигонуклеотидными праймерами, специфичными для кДНК АТIII человека, используя для нормализации уровень транскрипта гена Gapdh мыши. Паттерн сплайсинга и экспрессию полноразмерного транскрипта минигена АТIII, определяют с помощью ПЦР. Для этого подбирают пары праймеров, позволяющие специфично амплифицировать полноразмерный транскрипта минигена АТIII человека и эндогенный транскрипт АТIII мыши. Специфичность соответствующих пар праймеров подтверждают на образцах первых цепей кДНК, полученных из ткани нетрансгенных мышей, у которых отсутствует экспрессия минигена АТIII человека.
Авторами предложен универсальный способ получения линейных фрагментов ДНК пригодных для микроинъекций в зиготы мыши с помощью фермента протеломеразы TelN из фага N15 и описана рекомбинантная плазмида pTelN для осуществления указанного способа. Авторами сконструирована рекомбинантная плазмида pTelN-gAT3Spl несущая миниген АТIII человека под транскрипционным контролем регуляторных элементов гена Serpinс1 мыши. Используя фрагмент ДНК, полученный описанным в данном изобретении способом, авторами впервые получена линия генно-модифицированных мышей, экспрессирующих миниген АТIII человека в ткани печени. Установлено, что миниген АТIII с оптимизированным паттерном сплайсинга эффективно сплайсируется в клетках печени мыши и не отмечается формирования аберрантных транскриптов. Изобретение решает задачу создания модели для получения гуманизированных по гену Serpinc1 животных. Кроме того, предложенная рекомбинантная плазмида pTelN-gAT3Spl может быть использована для получения трансгенных мышей, гуманизированных по гену Serping1, с помощью технологии CRIPSR/Cas9.
Краткое описание чертежей
На Фиг.1 показана нуклеотидная последовательность сайта telRL, являющегося мишенью для протеломеразы TelN. Сайт telRL имеет длину 56 п.о. и состоит из двух инвертированных повторов telR и telL длиной 14 п.о., отделенных 3 п.о. от палиндрома telO длиной 22 п.о.. Разрезание ДНК происходит в середине палиндрома telO (место разрезания показано треугольниками). После разрезания формируются ковалентные связи между комплементарными нуклеотидами С и G на концах фрагментов ДНК, что приводит также к формированию шпилек на свободных концах ДНК (показаны стрелкой).
На Фиг.2 показана генетическая карта плазмидной конструкции pTelN, имеющей размер 3037 п.о.
Показаны основные элементы плазмиды: “TelN” - сайт связывания протеломеразы TelN, длиной 56 п.о.; “telR” - часть сайта TelN, длиной 28 п.о., содержащая инвертированный повтор R3 длиной 14 п.о., и соответствующая правому плечу профага N15; “telL” - часть сайта TelN, длиной 28 п.о., содержащая инвертированный повтор L3 длиной 14 п.о., и соответствующая левому плечу профага N15; “AmpR promoter” - промотор гена устойчивости к ампициллину; “AmpR“ - ген устойчивости к ампициллину (β-лактамаза); “ColE1 ori” - точка начала репликации; “f1 ori” - точка начала репликации из бактериофага f1; “lac operator” - lac оператор; “lac promoter” - lac промотор; “CAP binding site” - cайт связывания белка CAP (Catabolite activator protein); “polylinker” - полилинкер плазмиды pTelN, содержащий ряд уникальных сайтов рестрикции (XhoI, NcoI, KpnI, NdeI, и др.).
На Фиг.3 показана генетическая карта плазмидной конструкции pTelN-gAT3Spl, кодирующей миниген АТIII человека, и имеющей размер 12185 п.о.
Показаны основные элементы плазмиды: “AmpR promoter” - промотор гена устойчивости к ампициллину; “AmpR“ - ген устойчивости к ампициллину (β-лактамаза); “ColE1 ori” - точка начала репликации; “f1 ori” - точка начала репликации из бактериофага f1; “lac operator” - lac оператор; “lac promoter” - lac промотор; “CAP binding site” - cайт связывания белка CAP (Catabolite activator protein); “Mm AT3 LHA” (left homology arm) - левое плечо гомологии локуса гена Serpinc1 мыши; “Mm AT3 RHA” (right homology arm) - правое плечо гомологии локуса гена Serpinc1 мыши;“AT3 minigene” - миниген АТ3 человека, состоящий из фрагментов гена SERPINС1, несущих второй (hAT3ex2), с третьего по пятый (hATex3-5), шестой (hAT3ex6) и седьмой (hAT3x7) экзоны, фланкированные интронными последовательностями; “SV40 polyA” - последовательность полиаденилирования из вируса SV40; “SKIV2L terminator” - терминатор транскрипции из гена SKIV2L человека; мутация PAM сайта для гидовой РНК sg1 - “PAM mut C->T”; ряд элементов плазмиды (кодоны инициации и терминации трансляции белка АТIII, гибридный первый экзон минигена) не показаны на карте из-за нехватки места; кроме того, показан ряд уникальных сайтов рестрикции (XhoI, NcoI, KpnI, NdeI, и др.).
На Фиг.4А показан пример анализа в 1%-ом ТАЕ агарозном геле 1.28 мкг плазмиды pTelN-gAT3Spl, гидролизованной ферментом протеломеразой TelN. Треугольником показаны положения полос маркера молекулярного веса длиной 10000 и 3000 п.о., определяющих положение в геле фрагмента ДНК длиной 9323 п.о., несущего миниген АТIII, и остова вектора pGEM5Zа(+) длиной 2862 п.о., на основе которого была создана плазмида pTelN-gAT3. Фрагмент длиной 9323 п.о. очищали из геля с помощью набора реагентов QiaexII (Qiagen).
На Фиг.4Б показан электрофоретический анализ очищенного фрагмента ДНК для микроинъекций, несущего миниген АТIII человека. Дорожка 1 -фрагмент ДНК фильтровали после очистки из геля, дорожка 2 - фрагмент центрифугировали после очистки из геля. Маркер молекулярного веса ДНК - O'GeneRuler 1 kb DNA Ladder (#SM1163, Thermo Scientific).
На Фиг.5 показан пример результатов генотипирования двух животных 4960 (трансген) и 4957 (нетрансген). Показаны кривые амплификации фрагментов минигена АТIII человека (hAT3), терминатора транскрипции SKIV2L (SKIV) и референсного гена Bmp4 мыши (Bmp4). Из рисунка видно, что фрагменты гена Bmp4 амплифицируются одинаково эффективно в обоих образцах (Bmp4, 4960, 4957), а значения порогового цикла Ct составляют около 20. В то же время, на матрице геномной ДНК из трансгенного животного 4960 отмечается эффективная амплификация фрагментов hAT3 и SKIV со значениями Ct около 16-17, что свидетельствует об интеграции в геном множественных копий генетической конструкции. Видно, что на матрице геномной ДНК из нетрансгенного животного 4957с праймерами на hAT3 и SKIV идет амплификация продуктов со значениями Ct около 32-33, что указывает на неспецифическую амплификацию фрагментов ДНК.
На Фиг.6 показаны результаты анализа экспрессии транскрипта минигена АТIII человека и транскрипта гена Serpinc1 мыши. Транскрипты амплифицировали с помощью ПЦР используя праймеры для амплификации полноразмерных транскриптов минигена (Mg) и гена Serpinc1 мыши (Ms) на матрице первых цепей кДНК из печени трансгенных животных поколения F1 трех независимых линий (животное 527 - линия 2032; животное 5607 - линия 2034 и животное 9538 - линия 2038) и нетрансгенных (wt) животных. Продукты амплификации разделяли в 1%-ом TAE-агарозном геле. RT+ - образцы реакции обратной транскрипции с добавлением обратной транскриптазы. RT- контрольные образцы реакции обратной транскрипции без добавления обратной транскриптазы. Стрелкой показано положение полосы маркера молекулярного веса ДНК (O'GeneRuler 1 kb DNA Ladder, #SM1163, Thermo Scientific) длиной 1500 п.о.
Примеры осуществления настоящего изобретения
Пример 1. Конструирование рекомбинантной плазмиды pTelN, несущей сайты протеломеразы TelN из фага N15
Вначале на основе вектора pBluescriptSK (-) (Stratagene, США) получают плазмиду pTelN (SEQ ID NO1), несущую сайты узнавания для фермента протеломеразы TelN из фага N15. Для этого, с использованием pBluescriptSK (-) в качестве матрицы амплифицируют остов плазмиды используя праймеры 5- actattgtgtgctgataggttaattgcgcgcttggcgtaatcatggtcatagctgtt-3 и 5- gcaattgtgtgctgatattacgcgcgctcactggccgtcgttttacaacg -3, затем полученный ПЦР продукт реамплифицируют с олигонуклеотидными праймерами TelN_dir 5-aaagagctctctagatatcagcacacaattgcccattatacgcgcgtataatggactattgtgtgctgataggttaattgcgcgcttggcg-3 и TelN_rev 5-acagggcccgctagctgcagtatcagcacacaatagtccattatacgcgcgtataatgggcaattgtgtgctgatattacgcgcgctcactggc-3, вносящими сайты протеломеразы и сайты рестрикции SacI, XbaI и EcoRV с 5'-конца ампликона и сайты PstI, NheI и ApaI с 3'-конца. Получающийся в результате ПЦР-продукт обрабатывают эндонуклеазами рестрикции SacI и ApaI и лигируют с фрагментом SacI-ApaI, вырезанным из вектора pBluescriptSK (-), и несущим полилинкер с рядом уникальных сайтов рестрикции.
Полученная рекомбинантная плазмида pTelN содержит следующие элементы: сайты распознавания протеломеразы TelN длиной 56 п.о. в позициях 50-105 и 225-280 п.о.; сайты telR, длиной 28 п.о., в позициях 50-77 и 225-252 п.о., содержащие инвертированный повтор L3 длиной 14 п.о., и соответствующие левому плечу профага N15; сайты telL, длиной 28 п.о., в позициях 78-105 и 253-280 п.о., содержащие инвертированный повтор L3 длиной 14 п.о., и соответствующие левому плечу профага N15; полилинкер длиной 199 п.о. в позиции 106-224 п.о.; промотор гена β-лактамазы длиной 105 п.о. в позиции 2331-2435 п.о.; открытая рамка считывания β-лактамазы длиной 861 п.о. в позиции 1470-2330 п.о.; точка начала репликации ColE1/pMB1/pBR322/pUC длиной 589 п.о. в позиции 711-1299 п.о.; точка начала репликации бактериофага f1 длиной 456 п.о. в позиции 2461-2916 п.о.; Lac оператор длиной 17 п.о. в позиции 333-349; Lac промотор длиной 31 п.о. в позиции 357-387 п.о.; сайт связывания белка CAP (Catabolite activator protein) длиной 22 п.о. в позиции 402-423 п.о..
Пример 2. Конструирование рекомбинантной плазмиды pTelN-gAT3Spl, кодирующей миниген антитромбина III человека
Рекомбинантную плазмиду pTelN-gAT3Spl (SEQ ID NO2), кодирующую миниген АТIII человека с оптимизированным паттерном сплайсинга, получают на основе ранее описанной плазмиды pmhATg7Spl [Shepelev MV, Saakian EK, Kalinichenko SV, Korobko IV. Human Antithrombin III Minigene with an Optimized Splicing Pattern. Mol Biol (Mosk). 2019 May-Jun;53(3):411-420. doi: 10.1134/S0026898419030170. PMID: 31184606.]. Вначале конструируют промежуточную рекомбинантную плазмиду pZTermRA12, несущую правое плечо гомологии локуса Serpinc1 мыши и ранее описанный эффективный терминатор транскрипции SKIV2L [Deykin A, Tikhonov M, Kalmykov V, Korobko I, Georgiev P, Maksimenko O. Transcription termination sequences support the expression of transgene product secreted with milk. Transgenic Res. 2019 Aug;28(3-4):401-410. doi:10.1007/s11248-019-00122-9. Epub 2019 Mar 28. PMID: 30919251].
Пример2а. Конструирование промежуточной рекомбинантной плазмиды pZTermRA12
Для получения плазмиды pmATRArow фрагмент мышиного генома длиной 1918 п.о. (правое плечо локуса Serpinc1 мыши) амплифицируют с праймерами 5'-atctctgtaggagcatcggc-3' и 5'-tccgtgacagatagcacagc-3' и клонируют в вектор pCR-Blunt (Invitrogen, США). Полученную плазмиду секвенируют, определяя полную нуклеотидную последовательность ПЦР-продукта. Далее, плазмиду pmATRArow используют в качестве матрицы в ПЦР-реакции и реамплифицируют последовательность правого плеча гомологии с праймерами 5'-aatctagaccctggggcaggaagtgg-3' и 5'-tccgtgacagatagcacagc-3, продукт ПЦР клонируют в вектор pCR-Blunt получая плазмиду pATRA12. Далее фрагмент XbaI-HindIII (сайт гидролиза рестриктазы HindIII обрабатывают фрагментом Кленова (Fermentas, EP0054) клонируют из pATRA12 в плазмиду pBl-KIV-DOM, несущую терминатор транскрипции из гена SKIV2L и полученную ранее [Deykin A, Tikhonov M, Kalmykov V, Korobko I, Georgiev P, Maksimenko O.Transcription termination sequences support the expression of transgene product secreted with milk. Transgenic Res. 2019 Aug;28(3-4):401-410. doi:10.1007/s11248-019-00122-9. Epub 2019 Mar 28. PMID: 30919251], по сайтам XbaI-Cfr42I (сайт гидролиза рестриктазы Cfr42I обрабатывают ферментом Т4 ДНК полимеразой, Thermo Scientific, EP0061) получая плазмиду pATermRA12. Далее, фрагмент SalI-SacI клонируют из плазмиды pATermRA12 по сайтам SalI-SaсI в плазмиду pGEM5Zа(+) (Promega, США) получая вектор pZTermRA12.
Пример2б. Конструирование рекомбинантной плазмиды pTelN-gAT3Spl
Далее получают плазмиду pgATHDR1Spl путем клонирования фрагмента NotI-SalI из плазмиды pmhATg7Spl в вектор pZTermRA12 по этим же сайтам. Затем из плазмиды pgATHDR1Spl клонируют фрагмент NotI-MluI (сайт гидролиза рестриктазой MluI обрабатывают фрагментом Кленова) в вектор pTelN по сайтам HincII-NotI получая рекомбинантную плазмиду pTelN-gAT3Spl.
Полученная рекомбинантная плазмида pTelN-gAT3Spl содержит следующие элементы: промотор гена β-лактамазы длиной 105 п.о. в позиции 603-707 п.о.; открытая рамка считывания β-лактамазы длиной 861 п.о. в позиции 708-1586 п.о.; точка начала репликации ColE1/pMB1/pBR322/pUC длиной 589 п.о. в позиции 1739-2327 п.о.; сайт связывания белка CAP (Catabolite activator protein) длиной 22 п.о. в позиции 2615-2636 п.о.; Lac промотор длиной 31 п.о. в позиции 2651-2681 п.о. ; Lac оператор длиной 17 п.о. в позиции 2689-2705; сайты распознавания протеломеразы TelN длиной 56 п.о. в позициях 2758-2813 и 12081-12136 п.о.; сигнал полиаденилирования из вируса SV40 длиной 262 п.о. в позиции 9093-9354 п.о. ; точка начала репликации бактериофага f1 в позиции 10428-10856 п.о. длиной 429 п.о.; терминатор транскрипции из гена SKIV2L человека в позиции 9386-10154 п.о. длиной 769 п.о. ; левое плечо гомологии локуса гена Serpinс1 мыши длиной 2076 п.о. в позиции 2838-4913 п.о. ; правое плечо гомологии локуса гена Serpinс1 мыши длиной 1834 п.о. в позиции 10200-12033 п.о. ; мутация PAM сайта в позиции 4851 п.о. длиной 1 п.о. ; миниген АТIII человека длиной 4151 п.о. в позиции 4905-9055 п.о. ; гибридный первый экзон минигена, состоящий из 5'-части второго экзона гена Serpinc1 мыши до ATG кодона и 3'-части первого экзона и части первого интрона гена SERPINC1 человека, длиной 266 п.о. в позиции 4803-5068 п.о.; фрагмент генома человека, несущий второй экзон гена SERPINC1, фланкированный интронными последовательностями, длиной 680 п.о. в позиции 5074-5753 п.о.; фрагмент генома человека, несущий с третьего по пятый экзоны гена SERPINC1, фланкированный интронными последовательностями, длиной 2700 п.о. в позиции 5760-8459 п.о.; фрагмент генома человека, несущий шестой экзон гена SERPINC1, фланкированный интронными последовательностями, длиной 276 п.о. в позиции 8466-8741 п.о.; фрагмент генома человека, несущий седьмой экзон гена SERPINC1, фланкированный интронными последовательностями, длиной 339 п.о. в позиции 8748-9086 п.о.; кодон инициации трансляции белка АТIII в позиции 4905-4907 п.о. длиной 3 п.о. и кодон терминации трансляции белка АТIII в позиции 9053-9055 п.о. длиной 3 п.о.
Пример 3. Получению линейного фрагмента ДНК для микроинъекций с помощью фермента протеломеразы TelN.
Плазмиду pTelN-gAT3Spl в количестве 2568 нг обрабатывают протеломеразой TelN (New England Biolabs, # M0651S) в реакции объемом 50 мкл.
Для этого смешивают:
Реакцию инкубируют при +30°С в течении 1 ч 30 мин, и затем 15 мин при +70°С для диссоциации TelN от молекул ДНК. Далее продукты реакции разделяют в 1%-ТАЕ агарозном геле (Фиг.4А), фрагмент длиной около 9350 п.о. вырезают из геля и очищают с помощью набора реактивов QiaexII (Qiagen, #20021). Концентрацию полученного фрагмента ДНК определяют с помощью спектрофотометра Nanophotometer ND120 (Implen). Концентрация составляет 30 нг/мкл. Для подготовки фрагмента ДНК для микроинъекций в зиготы, часть очищенного фрагмента центрифугируют при 16000g в течение 10 мин для осаждения возможных примесей частиц носителя для очистки фрагмента из геля; часть очищенного фрагмента фильтруют через фильтр Ultrafree-MC, 0.2 μm (Millipore, #UFC30LG25). На Фиг.4Б показан электрофоретический анализ полученных фрагментов ДНК для микроинъекций. Видно, что полученные фрагменты не содержат примесных полос, а также нет признаков деградации ДНК.
Пример 4. Получение трансгенных мышей, экспрессирующих миниген АТIII человека
Пример 4а. Получение яйцеклеток мышей.
Яйцеклетки для микроинъекций получали методом индукции суперовуляции. Для этого неполовозрелым самкам гибридам F1(CBA×C57BL/6) весом 12-13 г внутрибрюшинно вводили 8 ед. гонадотропина сыворотки жеребых кобыл (ГСЖК) и через 48 час - 8 ед. хорионического гонадотропина человека (ХгЧ). После такой обработки самок ссаживали с самцами-производителями линии F1(CBA×C57BL/6). Факт спаривания констатировали на следующее утро по наличию копулятивной пробки. Схема индукции суперовуляции включала: в 12:00 - введение ГСЖК, через 48 часов - введение ХгЧ. В 17:00 этого же дня - подсадка к самцам-производителям. Отбор доноров производили на следующий день в 9:00. Световой режим в виварии был установлен с 7:00 до 19:00. Отобранных самок-доноров умерщвляли, извлекали яйцеводы, затем вымывали яйцеклетки в среде HEPES-KSOM с добавлением гиалуронидазы. Процедуру проводили под бинокулярным микроскопом (Zeiss Stemi DV4) с увеличением в 32 раза. Для вымывания яйцеклеток использовали стеклянные капилляры с внутренним диаметром примерно 100 мкм, изготовленные на пуллере Narishige PC-10 (Япония) и микрокузнице Narishige MF-900 (Япония).
Пример 4б. Проведение микроинъекции в зиготы.
Полученные зиготы культивировали в течение двух часов при t=37°C и 5% СО2 в капле среды KSOM под минеральным маслом (Sigma, США), затем зиготы помещали в микроинъекционную камеру. Микроинъекции проводили в среде HEPES-KSOM под контролем микроскопа Zeiss Axiovert 200М при увеличении в 400-600 раз, используя микроманипуляторы Narishige. Для изготовления игл для микроинъекций использовали пуллер Sutter instrument Со Р-97 (США), для изготовления удерживающей пипетки использовали пуллер Narishige PC-10 и микрокузницу Narishige MF-900. После окончания микроинъекций выжившие зиготы переносили в каплю среды KSOM под минеральное масло (Sigma) и культивировали в течение 1 часа для выявления жизнеспособных эмбрионов.
Пример 4в. Получение самок-реципиентов.
Самок-реципиентов яйцеклеток получали следующим образом: половозрелых самок гибридной линии F1(CBA×C57BL/6) весом не менее 24 г ссаживали с вазэктомированными самцами той же линии. Через 18 часов псевдобеременных реципиентов отбирали по наличию копулятивных пробок. Выжившие после микроинъекции зиготы трансплантировали в яйцеводы псевдобеременной самки. Одной псевдобеременной самке пересаживали 6-10 эмбрионов. Операцию проводили под наркозом (смесь золетила и рометара), вводимого внутрибрюшинно.
Пример 4г. Операция вазэктомирования.
Операцию вазэктомирования проводили заранее под наркозом (смесь золетила и рометара, вводимого внутрибрюшинно). Через надрез в коже и брюшной стенке вытягивали из брюшной полости семенник, придатки семенника и семявыносящий проток, после чего раскаленным пинцетом разрушали семявыносящий проток. Органы возвращали в брюшную полость, и повторяли всю процедуру на другом семявыносящем протоке. На завершающем этапе на брюшную стенку и на кожу накладывали швы с последующей антисептической обработкой операционного поля.
Пример 4д. Получение новорожденных мышей.
На 19 день после пересадки микроинъецированных эмбрионов самку-реципиента умерщвляли путем цервикальной дислокации и проводили кесарево сечение, после чего выживших детенышей помещали к заранее подготовленной кормилице. Через 8-14 дней после рождения у мышат брали образец ткани, выделяли геномную ДНК и анализировали наличие целевой генетической конструкции в геноме с помощью количественной ПЦР.
Пример 4г. Проведение микроинъекций для получения линии генно-модифицированных мышей, экспрессирующих антитромбин III человека
В эксперименте, заявленном в настоящем изобретении, для получения трансгенных мышей, экспрессирующих миниген АТIII человека, в зиготы инъецировали смесь, содержащую 20 нг/мкл рибонуклеопротеинового комплекса (РНП) белка Cas9 D10A и гидовой РНК sg1, 20 нг/мкл РНП белка Cas9 D10A и гидовой РНК sg2 (последовательность мишень в геноме мыши: 5-gcatcggccatgtattcccctgg-3) и 5 нг/мкл фрагмента ДНК (gAT3-TelN), несущего миниген АТIII человека, и вырезанного из плазмиды pTelN-gAT3 с помощью фермента протеломеразы TelN (Фиг.4). Задача, решаемая в данном изобретении, заключается в том, что путем трансгенеза методом случайного встраивания получают новую линию генетически модифицированных мышей. Однако проводимый эксперимент по микроинъекциям в зиготы мыши, помимо трансгенеза, включал также и анализ активности РНП белка Cas9 D10A в комплексе с гидовыми РНК, вносящими разрывы в области второго экзона гена Serpinc1 мыши. Таким образом, в рамках одного эксперимента решались три технические задачи: (1) оценка активности РНП комплексов в рамках разработки стратегии получения гуманизированных по гену Serpinc1 мышей с помощью системы CRISPR/Cas9; (2) оценка пригодности фрагмента ДНК, полученного с помощью TelN, как матрицы для репарации разрывов ДНК, вносимых Cas9, в соответствии с методом Tild-CRISPR; а также (3), оценка эффективности случайной интеграции полученного линейного фрагмента ДНК с минигеном АТIII в геном мыши, что одновременно является и трансгенезом методом случайного встраивания.
В результате, в общей сложности, указанной смесью были инъецированы 721 зигота мыши, после инъекций выжило 259 клеток (36%), которые были пересажены в яйцеводы 21 псевдобеременного реципиента. В результате от 21 реципиента было получено потомство численностью 14 мышат. Был проведен анализ эффективности редактирования в локусе Serpinc1 с помощью анализа с T7 эндонуклеазой как описано в статье [Sakurai T, Watanabe S, Kamiyoshi A, Sato M, Shindo T. A single blastocyst assay optimized for detecting CRISPR/Cas9 system-induced indel mutations in mice. BMC Biotechnol. 2014; 14: 69. doi: 10.1186/1472-6750-14-69], который не выявил наличия инделов в целевом локусе, что указывает на неэффективность РНП Cas9 D10A в использованной концентрации (данные не показаны). Далее проводили генотипирование полученного потомства на предмет интеграции в геном фрагмента ДНК, несущего миниген АТIII человека, с помощью количественной ПЦР (см. ниже).
Пример 5. Генотипирование животных.
Пример 5а. Выделение геномной ДНК из тканей мышей.
Выделение геномной ДНК из тканей животных проводят в соответствии с протоколом [Green MR, Sambrook J. Preparation of Genomic DNA from Mouse Tails and Other Small Samples. Cold Spring Harb Protoc. 2017 Sep 1;2017(9):pdb.prot093518. doi:10.1101/pdb.prot093518. PMID: 28864567] с небольшими модификациями. В частности, образцы тканей животных лизируют при +56°С в течение 14-16 часов в 250 мкл буфера SNET (20 мМ Tris-HCl pH=8.0, 400 мМ NaCl, 5 мМ ЭДТА pH=8.0, 1% додецилсульфат натрия), содержащем 0.8 мкг/мкл протеиназы К (Sigma, #P2308). Далее, к образцам добавляют 125 мкл смеси хлороформа:изоамилового спирта в соотношении 24:1 и 125 мкл фенола, насыщенного 1М Tris-HCl pH=8.0. Образцы встряхивают на миксере и инкубируют 10-15 мин при постоянном перемешивании. Образцы центрифугируют в течении 5 мин при 16000g при комнатной температуре. Собирают верхнюю фазу и добавляют 250 мкл изопропанола, перемешивают образцы. Для осаждения геномной ДНК образцы центрифугируют в течении 15 мин при 16000g при +4°С. Затем осадок промывают 70% этанолом, высушивают и растворяют в 400 мкл буфера 10 мМ Трис-HCl, 0.5 мМ ЭДТА pH=9.0.
Пример 5б. Генотипирование животных с помощью количественной ПЦР.
Генотипирование животных, рожденных самками-реципиентами микроинъецированных зигот, проводят с помощью количественной ПЦР с праймерами, специфичными для нуклеотидных последовательностей минигена антитромбина III человека и терминатора транскрипции SKIV2L из генома человека. В качестве референсного гена для контроля качества геномной ДНК используют праймеры на ген Bmp4 мыши.
В частности, количественную ПЦР проводят, используя мастермикс iTaq™ Universal SYBR® Green Supermix (#1725121, Bio-Rad), реакции объемом 20 мкл смешивают в стрипованных пробирках 0.2 ml 8-Tube PCR Strips without Caps, low profile, clear (#TLS0801, Bio-Rad) и закрывают стрипованными оптически прозрачными крышками 0.2 ml Flat PCR Tube 8-Cap Strips, optical, ultraclear (#TCS0803, Bio-Rad). Одна реакция содержит 1-х кратный мастермикс, прямой и обратный праймеры в концентрации 200 нМ, и 1 мкл геномной ДНК в качестве матрицы для ПЦР. Амплификацию проводят в амплификаторе CFX96 (Bio-Rad) при следующих условиях: 95°C - 1 мин; 40 циклов: 95°С - 15 с, 60°C - 30 с, детекция флуоресценции; анализ кривой плавления от 60°С до 95°С с интервалом 0.5°С.
Фрагмент минигена ATIII человека длиной 108 п.о. амплифицируют с праймерами 5qhAT3 (5-ctgatggagagtcgtgttcagca-3, SEQ ID NO:3) и 3qhAT3 (5- catcacctttgaagggcaactca-3, SEQ ID NO:4). Фрагмент терминатора транскрипции SKIV2L человека длиной 107 п.о. амплифицируют с праймерами 5qSKIV (5- gctaagaagacagcgagagaa-3 SEQ ID NO:5) и 3qSKIV (5- caagactccctctcccaaatag- 3 SEQ ID NO:6). Фрагмент гена Bmp4 мыши длиной 89 п.о. амплифицируют с праймерами 5qBmp4 (5- cagggcttccaccgtataaac-3 SEQ ID NO:7) и 3qBmp4 (5-gtgtccagtagtcgtgtgatg-3 SEQ ID NO:8).
Из графиков амплификации определяют значения порогового цикла (Ct) для всех амплифицируемых фрагментов. Пример результатов генотипирования животных поколения F1 приведен на Фиг.5. Видно, что фрагменты референсного гена Bmp4 амплифицируются в обеих образцах (животные 4957 и 4960), тогда как фрагменты минигена АТIII и терминатора SKIV2L амплифицируются только на матрице геномной ДНК из ткани трансгенного животного 4960.
В результате генотипирования четырнадцати мышей поколения F0, родившихся у самок-реципиентов, было выявлено три животных (2032, 2034 и 2038), которые несли в геноме трансген (миниген АТIII). С помощью ПЦР было установлено, что конструкция не интегрировала в локус Serpinc1 (данные не показаны), что еще раз подтверждает неэффективность использованных в эксперименте РНП Cas9 D10A. Трансгенные мыши поколения F0 были использованы для последующего размножения и получения линий генно-модифицированных мышей, экспрессирующих минигена ATIII человека.
Кроме того, результат эксперимента, во-первых, указывает на то, что полученные с помощью фермента TelN линейные двуцепочечные фрагменты ДНК с ковалентно замкнутыми шпильками на концах эффективно интегрируют в геном, и могут использоваться для трансгенеза методом случайного встраивания. Во-вторых, учитывая неэффективность редактирования в локусе Serpinc1 при использовании РНП, невозможно оценить является ли линейный двуцепочечный фрагмент ДНК с ковалентно замкнутыми концами эффективной матрицей для репарации, как это указывается в методе Tild-CRISPR. В-третьих, важно отметить, что в методе Tild-CRISPR авторы использовали для микроинъекций в зиготы мыши линейные двуцепочечные фрагменты ДНК, полученные с помощью эндонуклеаз рестрикции, в концентрациях от 33 до 100 нг/мкл [Yao X, Zhang M, Wang X, Ying W, Hu X, Dai P, Meng F, Shi L, Sun Y, Yao N, Zhong W, Li Y, Wu K, Li W, Chen ZJ, Yang H. Tild-CRISPR Allows for Efficient and Precise Gene Knockin in Mouse and Human Cells. Dev Cell. 2018 May 21;45(4):526-536.e5. doi: 10.1016/j.devcel.2018.04.021. PMID: 29787711.]. При этом авторы указывают на отсутствие или низкий уровень случайной интеграции в геном мыши фрагментов ДНК, инъецированных в таких концентрациях. Результаты нашего эксперимента ставят под сомнение вывод данной работы, так как, уже при микроинъекции фрагмента ДНК в концентрации 5 нг/мл, нами были получены три животных со случайной встройкой фрагмента в геном. Таким образом, в одном эксперименте была проведена оценка эффективности ряда методов геномного редактирования, и одновременно была решена задача настоящего изобретения по получению линии генно-модифицированных мышей, экспрессирующих миниген АТIII человека.
Пример 6. Анализ экспрессии минигена, кодирующего антитромбин III человека, в ткани печени мыши
Пример 6а. Выделение суммарной РНК из ткани печени мыши
Cуммарную РНК из ткани печени мыши выделяют с помощью набора реагентов RNeasy mini kit (Qiagen, #74104), дополнительно обрабатывая РНК ферментом ДНКазой для удаления примесей геномной ДНК. Ткани печени хранят при -80°С, для выделения РНК около 30 мг ткани гомогенизируют в жидком азоте с помощью ступки и пестика в 600 мкл буфера RLT, содержащего 6 мкл β-меркаптоэтанола, и переносят лизат в стерильную микроцентрифужную пробирку. Для гомогенизации лизат пропускают 5 раз через иглу калибра 23G с помощью стерильного одноразового шприца объемом 2 мл. Полученные лизаты замораживают и хранят при -70°С. Для выделения РНК лизаты оттаивают при +37°С, добавляют равный объем 70%-го спирта, перемешивают пипетированием и наносят лизат на колонку из набора RNeasy mini kit, центрифугируют 30 с при скорости 16000g для связывания РНК с колонкой. Далее колонку промывают, используя 350 мкл буфера RW1, после чего обрабатывают РНК на колонке с помощью ДНКазы из набора RNase-free DNase set (Qiagen, #79254). Для этого смешивают 70 мкл буфера RDD и 10 мкл ДНКазы, наносят смесь на колонку и инкубируют 15 мин при комнатной температуре. Далее, колонку промывают, используя 350 мкл буфера RW1, удаляют проскок. Затем, колонку дважды промывают, используя 500 мкл буфера RPE, центрифугируют первый раз 15 с, удаляют проскок, второй раз - 2 мин для подсушивания РНК. Связанную с колонкой РНК элюируют с помощью 50 мкл стерильной воды свободной от РНКаз, концентрацию РНК определяют с помощью спектрофотометра NanoPhotometer N120 (Implen).
Пример 6б. Проведение реакции обратной транскрипции
Реакцию обратной транскрипции проводят с использованием обратной транскриптазы SuperScript IV (Thermo Fisher Scientific, #18090010) используя 500 нг суммарной РНК на одну реакцию. В стрипованных пробирках 0.2mL SnapStrip II PCR Tubes (SSIbio, #3245-00) смешивают 1000 нг РНК каждого из образцов (из расчета на две реакции: с добавлением обратной транскриптазы и контрольную без добавления обратной транскриптазы) со стерильной водой в объеме 23 мкл. В отдельной пробирке смешивают из расчета на один образец: 1 мкл 10 мМ дезоксинуклеотидтрифосфатов (дНТФ) и 0.5 мкл праймеров (смесь случайных гексамеров, Thermo Fisher Scientific #48190-011) в концентрации 300 нг/мкл. 3 мкл смеси праймеров и дНТФ добавляют к смеси РНК и воды, перемешивают, и 13 мкл переносят в отдельную стрипованную пробирку. Далее, образцы инкубируют при +65°С в течение 5 мин и сразу переносят в лед. Далее на льду к образцам добавляют 4 мкл 5-кратного реакционного буфера для SuperScript IV, 1 мкл 0.1 ДТТ, 0.5 мкл ингибитора РНКаз Ribolock 40 ед./мкл (Thermo Fisher Scientific #EO0381) и 0.5 мкл SuperScript IV 200 ед./мкл. В контрольные образцы вместо SuperScript IV добавляют 0.5 мкл стерильной деионизованной воды. Реакции аккуратно перемешивают, осаждают жидкость центрифугированием. Инкубируют в ПЦР-амплификаторе Dyad (Bio-Rad) при следующих условиях: +23°С - 10 мин, +50°С - 15 мин, +80°С - 10 мин. Затем добавляют 20 мкл стерильной воды для разведения образца и хранят при -80°С.
Пример 6в. Анализ паттерна экспрессии и сплайсинга полноразмерного транскрипта минигена АТIII с помощью ПЦР.
Экспрессию полноразмерного транскрипта минигена АТIII человека в печени трансгенных мышей анализируют с помощью ПЦР с детекцией продуктов реакции в агарозном геле используя мастермикс LongAmp® Hot Start Taq 2X Master Mix (NEB, #M0533S). Смешивают реакцию объемом 10 мкл, содержащую 1-х кратный LongAmp мастермикс, прямой и обратный праймеры в концентрации 500 нМ и 1 мкл реакции обратной транскрипции в качестве матрицы. Амплификацию проводят в амплификаторе Dyad (Bio-Rad) при следующих условиях: 94°С - 3 мин; 35 циклов -94°С - 30с, 60°С - 30 с, 65°С -1 мин 30 с; 65°С - 5 мин, 4°С - постоянно. Далее 4 мкл реакции ПЦР разделяют в 1%-ом ТАЕ-агарозном геле. Документацию результатов электрофореза проводят с помощью системы гель-документации ChemiDoc XRS (Bio-Rad).
Фрагмент транскрипта минигена АТIII человека длиной 1457 п.о. амплифицируют в ПЦР используя праймеры ATRAS (SEQ ID NO:9 5-atctctgtaggagcatcggc-3) и 3mhAT3 (SEQ ID NO:10 5-cgactctagaaataggaagagg -3), ожигающиеся на последовательности второго экзона гена Serpinc1 мыши в левом плече гомологии и седьмого экзона минигена ATIII человека.
В качестве контроля амплифицируют фрагмент транскрипта гена Serpinс1 мыши длиной 1438 п.о. используя праймеры ATRAS (SEQ ID NO:9) и Mm_AT3_R (SEQ ID NO:11 5-tgcaaagattaagaatattttagttc-3 ) ожигающиеся на последовательности первого и восьмого экзонов гена.
Как видно из Фиг.6, с праймерами на транскрипт гена мыши (панель Ms) амплифицируются фрагменты ожидаемого размера около 1500 п.о. на матрице первых цепей кДНК из ткани печени трансгенных (527, 5607 и 9538) и нетрансгенных животных (wt). В тоже время, с праймерами, специфичными для транскрипта минигена (панель Mg) амплифицируются фрагменты ожидаемого размера около 1500 п.о. только на матрице первых цепей кДНК из ткани печени трансгенных животных. При этом у животного 527 линии 2032 было обнаружена амплификация фрагмента длиной около 1000 п.о., что указывает на формирование аберрантного укороченного транскрипта минигена у животных, полученных от первичного трансгена линии 2032. Таким образом, у трансгенных мышей двух линий (2034 и 2038) амплифицируется фрагмент транскрипта минигена ожидаемого размера. Кроме того, продукт ПЦР животного 9538 очищали из агарозного геля, клонировали в вектор pJET1.2 и полностью секвенировали, что подтвердило корректность сплайсинга транскрипта минигена АТIII человека (данные не показаны). Важно отметить, что на матрице первых цепей кДНК из ткани печени трансгенных животных линий 2034 и 2038 не отмечалась амплификация продуктов, отличных по размеру от ожидаемого, что свидетельствует о прохождении корректного сплайсинга минигена АТIII человека в ткани печени трансгенных мышей.
Пример 7. Определение уровня транскрипта минигена АТIII человека с помощью количественной ПЦР.
Количественную ПЦР проводят, используя мастермикс iTaq™ Universal SYBR® Green Supermix (#1725121, Bio-Rad), реакции объемом 20 мкл смешивают в стрипованных пробирках 0.2 ml 8-Tube PCR Strips without Caps, low profile, clear (#TLS0801, Bio-Rad) и закрывают стрипованными оптически прозрачными крышками 0.2 ml Flat PCR Tube 8-Cap Strips, optical, ultraclear (#TCS0803, Bio-Rad). Одна реакция содержит 1-х кратный мастермикс, прямой и обратный праймеры в концентрации 200 нМ, и 2 мкл реакции обратной транскрипции в качестве матрицы для ПЦР. Амплификацию проводят в амплификаторе CFX96 (Bio-Rad) при следующих условиях: 95°C - 1 мин; 45 циклов: 95°С - 15 с, 60°C - 30 с, детекция флуоресценции; анализ кривой плавления от 60°С до 95°С с интервалом 0.5°С. Из графиков амплификации определяют значения порогового цикла (Ct) для всех транскриптов, и рассчитывают относительный уровень экспрессии по формуле 2-ΔΔCt, как описано в работе [Schmittgen TD, Livak KJ. Analyzing real-time PCR data by the comparative C(T) method. Nat Protoc. 2008;3(6):1101-8. doi: 10.1038/nprot.2008.73. PMID:18546601.]. В качестве референсного гена используют транскрипт гена Gapdh мыши.
Фрагмент транскрипта гена Serpinc1 мыши длиной 101 п.о. амплифицируют с праймерами 5qMmAT3(SEQ ID NO:14; 5- tcgatgggcagtcatgcccagtg -3) и 3qMmAT3 (SEQ ID NO:15; 5- cttgaagggcagctctagcacct -3).
Фрагмент транскрипта гена Gapdh мыши длиной 153 п.о. амплифицируют с праймерами 5qMmGAPD (SEQ ID NO:12; 5-catcactgccacccagaagactg-3) и 3qMmGAPD (SEQ ID NO:13; 5-atgccagtgagcttcccgttcag-3).
Фрагмент минигена ATIII человека длиной 108 п.о. амплифицируют с праймерами 5qhAT3 (SEQ ID NO:3) и 3qhAT3 (SEQ ID NO:4).
Как видно из Таблицы 1, наибольший уровень экспрессии транскрипта минигена ATIII человека детектируется у животных линий 2034 и 2038. Например, у животного № 5607 линии 2034 различия в уровнях экспрессии эндогенного транскрипта Serpinc1 и транскрипта минигена составляли 2.27±0.303 циклов. То есть уровень экспрессии минигена был ниже уровня экспрессии транскрипта Serpinc1 в 4.82 раза (Табл.1). Стоит отметить, что уровень экспрессии анализировали у гемизиготных животных, несущих трансген только в одной из двух хромосом. Получение гомозиготных животных позволит повысить уровень экспрессии минигена как минимум в два раза. Таким образом, можно ожидать что уровень экспрессии минигена АТIII у гомозиготных животных этой линии будет примерно в 2 раза ниже по сравнению с уровнем эндогенного транскрипта Serpinc1, что позволит компенсировать функции АТIII с гомозиготным нокаутом гена Serpinc1 и таким образом получить гуманизированных по этому гену животных.
Важно отметить, что плазмидная конструкция pTelN-gAT3 не содержит последовательностей какого -либо сильного промотора для активации транскрипции минигена. Это обусловлено универсальным дизайном генетической конструкции pTelN-gAT3, которая предназначена как для случайной встройки в геном, так и для CRISPR/Cas9-опосредованного нокина в локус Serpinc1. Данная плазмида содержит левое плечо гомологии локуса Serpinc1, которое включает в себя первый интрон гена Serpinc1 мыши, но не его промотор. Это обуславливает относительно низкий уровень экспрессии минигена. Стоит отметить, что наличие в конструкции сильного промотора могло бы привести к экспрессии минигена на уровне выше, чем физиологический для мыши, что чревато нарушением гемостаза у трансгенных животных. Таким образом, совокупность низкого уровня экспрессии минигена за счет отсутствия сильного промотора в генетической конструкции и встройка множественных копий трансгена в геном обеспечивают уровень экспрессии минигена примерно в пять раз ниже, чем уровень эндогенного антитромбина III. Получение гомозиготных животных позволит повысить уровень экспрессии минигена как минимум в два раза.
Пример 8. Определение копийности трансгена с помощью количественной ПЦР
Количество копий генетической конструкции, встроенной в геном трех линий генно-модифицированных мышей (линии 2032, 2034 и 2038), определяют с помощью количественной ПЦР с праймерами, специфичными для нуклеотидных последовательностей минигена антитромбина III человека и терминатора транскрипции SKIV2L из генома человека. В качестве референсного гена для расчета копийности используют ген Bmp4 мыши, присутствующий в диплоидном геноме мыши в количестве двух копий.
В частности, количественную ПЦР проводят, используя мастермикс 5X qPCRmix-HS SYBR (#PKL147L, Evrogen), реакции объемом 20 мкл смешивают в стрипованных пробирках 0.2 ml 8-Tube PCR Strips without Caps, low profile, clear (#TLS0801, Bio-Rad) и закрывают стрипованными оптически прозрачными крышками 0.2 ml Flat PCR Tube 8-Cap Strips, optical, ultraclear (#TCS0803, Bio-Rad). Одна реакция содержит 1-х кратный мастермикс, прямой и обратный праймеры в концентрации 200 нМ, и 1 мкл геномной ДНК в качестве матрицы для ПЦР. Амплификацию проводят в амплификаторе CFX96 (Bio-Rad) при следующих условиях: 95°C - 5 мин; 40 циклов: 95°С - 15 с, 60°C - 15 с, 72C - 30 c; детекция флуоресценции; анализ кривой плавления от 60°С до 95°С с интервалом 0.5°С. Для определения копийности используют те же праймеры, что и для генотипирования животных (см. пример 5б; SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:7 и SEQ ID NO:8).
Для расчета копийности анализируют по 2-3 образца геномной ДНК трансгенных животных каждой из независимых линий 2032, 2034 и 2038. Реакция проводят в дубликатах в трех повторах. Из графиков амплификации определяют значения порогового цикла (Ct) для всех амплифицируемых фрагментов. Копийность трансгенов рассчитывают по формуле 2-ΔCt, где ΔCt=Сt трансгена -Ct Bmp4 в соответствии с публикацией [Sommeregger W, Prewein B, Reinhart D, Mader A, Kunert R. Transgene copy number comparison in recombinant mammalian cell lines: critical reflection of quantitative real-time PCR evaluation. Cytotechnology. 2013 Oct;65(5):811-8.doi: 10.1007/s10616-013-9606-y. Epub 2013 Jun 27. PMID: 23807595; PMCID: PMC3967612]. Пример результатов расчета количества копий трансгена с использованием значений Ct для амплифицируемого фрагмента терминатора транскрипции SKIV2L приведен в Таблице 2.
Таким образом, установлено, что максимальное количество копий генетической конструкции (>150) содержит геном животных линии 2034. Это коррелирует с наивысшим уровнем экспрессии транскрипта минигена ATIII (пример 7). В свою очередь, наименьшее количество копий для животных линии 2032 (6-7 копий) коррелирует с наиболее низким уровнем экспрессии транскрипта минигена АТIII у этой линии животных (см. пример 7, Таблица 1). Полученные результаты указывают на то, что фрагмент для микроинъекций в зиготы, полученный с помощью фермента TelN, обеспечивает уровень экспрессии трансгена, коррелирующий с количеством копий трансгена в геноме.
--->
Информация о перечнях последовательностей:
Версия DTD: V1_3
Название файла: Мыши антитромбин III.xml
Название программного обеспечения: WIPO Sequence
Версия программного обеспечения: 2.1.2
Дата производства: 2022-12-27
Общая информация:
Текущая заявка / Ведомство ИС: RU
Текущая заявка / Номер дела заявителя: non
Имя заявителя: Федеральное государственное бюджетное учреждение
науки Институт биологии гена Российской академии наук
Имя заявителя / Язык: ru
Имя заявителя / Имя латиницей: Institute of Gene Biology Russian
Academy of Sciences
Название изобретения: Способ получения генно-модифицированных мышей,
экспрессирующих миниген антитромбина III человека, с помощью
микроинъекций TelN-линеаризованного фрагмента ДНК ( ru )
Общее количество последовательностей: 15
Последовательности:
Номер последовательности (ID): 1
Длина: 3037
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..3037
> mol_type, other DNA
> organism, synthetic construct
Остатки:
gttttcccag tcacgacgtt gtaaaacgac ggccagtgag cgcgcgtaat
atcagcacac 60
aattgcccat tatacgcgcg tataatggac tattgtgtgc tgatactgca
gctagcgggc 120
cccccctcga ggtcgacggt atcgataagc ttgatatcga attcctgcag
cccgggggat 180
ccactagttc tagagcggcc gccaccgcgg tggagctctc tagatatcag
cacacaattg 240
cccattatac gcgcgtataa tggactattg tgtgctgata ggttaattgc
gcgcttggcg 300
taatcatggt catagctgtt tcctgtgtga aattgttatc cgctcacaat
tccacacaac 360
atacgagccg gaagcataaa gtgtaaagcc tggggtgcct aatgagtgag
ctaactcaca 420
ttaattgcgt tgcgctcact gcccgctttc cagtcgggaa acctgtcgtg
ccagctgcat 480
taatgaatcg gccaacgcgc ggggagaggc ggtttgcgta ttgggcgctc
ttccgcttcc 540
tcgctcactg actcgctgcg ctcggtcgtt cggctgcggc gagcggtatc
agctcactca 600
aaggcggtaa tacggttatc cacagaatca ggggataacg caggaaagaa
catgtgagca 660
aaaggccagc aaaaggccag gaaccgtaaa aaggccgcgt tgctggcgtt
tttccatagg 720
ctccgccccc ctgacgagca tcacaaaaat cgacgctcaa gtcagaggtg
gcgaaacccg 780
acaggactat aaagatacca ggcgtttccc cctggaagct ccctcgtgcg
ctctcctgtt 840
ccgaccctgc cgcttaccgg atacctgtcc gcctttctcc cttcgggaag
cgtggcgctt 900
tctcatagct cacgctgtag gtatctcagt tcggtgtagg tcgttcgctc
caagctgggc 960
tgtgtgcacg aaccccccgt tcagcccgac cgctgcgcct tatccggtaa
ctatcgtctt 1020
gagtccaacc cggtaagaca cgacttatcg ccactggcag cagccactgg
taacaggatt 1080
agcagagcga ggtatgtagg cggtgctaca gagttcttga agtggtggcc
taactacggc 1140
tacactagaa ggacagtatt tggtatctgc gctctgctga agccagttac
cttcggaaaa 1200
agagttggta gctcttgatc cggcaaacaa accaccgctg gtagcggtgg
tttttttgtt 1260
tgcaagcagc agattacgcg cagaaaaaaa ggatctcaag aagatccttt
gatcttttct 1320
acggggtctg acgctcagtg gaacgaaaac tcacgttaag ggattttggt
catgagatta 1380
tcaaaaagga tcttcaccta gatcctttta aattaaaaat gaagttttaa
atcaatctaa 1440
agtatatatg agtaaacttg gtctgacagt taccaatgct taatcagtga
ggcacctatc 1500
tcagcgatct gtctatttcg ttcatccata gttgcctgac tccccgtcgt
gtagataact 1560
acgatacggg agggcttacc atctggcccc agtgctgcaa tgataccgcg
agacccacgc 1620
tcaccggctc cagatttatc agcaataaac cagccagccg gaagggccga
gcgcagaagt 1680
ggtcctgcaa ctttatccgc ctccatccag tctattaatt gttgccggga
agctagagta 1740
agtagttcgc cagttaatag tttgcgcaac gttgttgcca ttgctacagg
catcgtggtg 1800
tcacgctcgt cgtttggtat ggcttcattc agctccggtt cccaacgatc
aaggcgagtt 1860
acatgatccc ccatgttgtg caaaaaagcg gttagctcct tcggtcctcc
gatcgttgtc 1920
agaagtaagt tggccgcagt gttatcactc atggttatgg cagcactgca
taattctctt 1980
actgtcatgc catccgtaag atgcttttct gtgactggtg agtactcaac
caagtcattc 2040
tgagaatagt gtatgcggcg accgagttgc tcttgcccgg cgtcaatacg
ggataatacc 2100
gcgccacata gcagaacttt aaaagtgctc atcattggaa aacgttcttc
ggggcgaaaa 2160
ctctcaagga tcttaccgct gttgagatcc agttcgatgt aacccactcg
tgcacccaac 2220
tgatcttcag catcttttac tttcaccagc gtttctgggt gagcaaaaac
aggaaggcaa 2280
aatgccgcaa aaaagggaat aagggcgaca cggaaatgtt gaatactcat
actcttcctt 2340
tttcaatatt attgaagcat ttatcagggt tattgtctca tgagcggata
catatttgaa 2400
tgtatttaga aaaataaaca aataggggtt ccgcgcacat ttccccgaaa
agtgccacct 2460
aaattgtaag cgttaatatt ttgttaaaat tcgcgttaaa tttttgttaa
atcagctcat 2520
tttttaacca ataggccgaa atcggcaaaa tcccttataa atcaaaagaa
tagaccgaga 2580
tagggttgag tgttgttcca gtttggaaca agagtccact attaaagaac
gtggactcca 2640
acgtcaaagg gcgaaaaacc gtctatcagg gcgatggccc actacgtgaa
ccatcaccct 2700
aatcaagttt tttggggtcg aggtgccgta aagcactaaa tcggaaccct
aaagggagcc 2760
cccgatttag agcttgacgg ggaaagccgg cgaacgtggc gagaaaggaa
gggaagaaag 2820
cgaaaggagc gggcgctagg gcgctggcaa gtgtagcggt cacgctgcgc
gtaaccacca 2880
cacccgccgc gcttaatgcg ccgctacagg gcgcgtccca ttcgccattc
aggctgcgca 2940
actgttggga agggcgatcg gtgcgggcct cttcgctatt acgccagctg
gcgaaagggg 3000
gatgtgctgc aaggcgatta agttgggtaa cgccagg
3037
Номер последовательности (ID): 2
Длина: 12185
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..12185
> mol_type, other DNA
> organism, synthetic construct
Остатки:
cctggcgtta cccaacttaa tcgccttgca gcacatcccc ctttcgccag
ctggcgtaat 60
agcgaagagg cccgcaccga tcgcccttcc caacagttgc gcagcctgaa
tggcgaatgg 120
gacgcgccct gtagcggcgc attaagcgcg gcgggtgtgg tggttacgcg
cagcgtgacc 180
gctacacttg ccagcgccct agcgcccgct cctttcgctt tcttcccttc
ctttctcgcc 240
acgttcgccg gctttccccg tcaagctcta aatcgggggc tccctttagg
gttccgattt 300
agtgctttac ggcacctcga ccccaaaaaa cttgattagg gtgatggttc
acgtagtggg 360
ccatcgccct gatagacggt ttttcgccct ttgacgttgg agtccacgtt
ctttaatagt 420
ggactcttgt tccaaactgg aacaacactc aaccctatct cggtctattc
ttttgattta 480
taagggattt tgccgatttc ggcctattgg ttaaaaaatg agctgattta
acaaaaattt 540
aacgcgaatt ttaacaaaat attaacgctt acaatttagg tggcactttt
cggggaaatg 600
tgcgcggaac ccctatttgt ttatttttct aaatacattc aaatatgtat
ccgctcatga 660
gacaataacc ctgataaatg cttcaataat attgaaaaag gaagagtatg
agtattcaac 720
atttccgtgt cgcccttatt cccttttttg cggcattttg ccttcctgtt
tttgctcacc 780
cagaaacgct ggtgaaagta aaagatgctg aagatcagtt gggtgcacga
gtgggttaca 840
tcgaactgga tctcaacagc ggtaagatcc ttgagagttt tcgccccgaa
gaacgttttc 900
caatgatgag cacttttaaa gttctgctat gtggcgcggt attatcccgt
attgacgccg 960
ggcaagagca actcggtcgc cgcatacact attctcagaa tgacttggtt
gagtactcac 1020
cagtcacaga aaagcatctt acggatggca tgacagtaag agaattatgc
agtgctgcca 1080
taaccatgag tgataacact gcggccaact tacttctgac aacgatcgga
ggaccgaagg 1140
agctaaccgc ttttttgcac aacatggggg atcatgtaac tcgccttgat
cgttgggaac 1200
cggagctgaa tgaagccata ccaaacgacg agcgtgacac cacgatgcct
gtagcaatgg 1260
caacaacgtt gcgcaaacta ttaactggcg aactacttac tctagcttcc
cggcaacaat 1320
taatagactg gatggaggcg gataaagttg caggaccact tctgcgctcg
gcccttccgg 1380
ctggctggtt tattgctgat aaatctggag ccggtgagcg tgggtctcgc
ggtatcattg 1440
cagcactggg gccagatggt aagccctccc gtatcgtagt tatctacacg
acggggagtc 1500
aggcaactat ggatgaacga aatagacaga tcgctgagat aggtgcctca
ctgattaagc 1560
attggtaact gtcagaccaa gtttactcat atatacttta gattgattta
aaacttcatt 1620
tttaatttaa aaggatctag gtgaagatcc tttttgataa tctcatgacc
aaaatccctt 1680
aacgtgagtt ttcgttccac tgagcgtcag accccgtaga aaagatcaaa
ggatcttctt 1740
gagatccttt ttttctgcgc gtaatctgct gcttgcaaac aaaaaaacca
ccgctaccag 1800
cggtggtttg tttgccggat caagagctac caactctttt tccgaaggta
actggcttca 1860
gcagagcgca gataccaaat actgtccttc tagtgtagcc gtagttaggc
caccacttca 1920
agaactctgt agcaccgcct acatacctcg ctctgctaat cctgttacca
gtggctgctg 1980
ccagtggcga taagtcgtgt cttaccgggt tggactcaag acgatagtta
ccggataagg 2040
cgcagcggtc gggctgaacg gggggttcgt gcacacagcc cagcttggag
cgaacgacct 2100
acaccgaact gagataccta cagcgtgagc tatgagaaag cgccacgctt
cccgaaggga 2160
gaaaggcgga caggtatccg gtaagcggca gggtcggaac aggagagcgc
acgagggagc 2220
ttccaggggg aaacgcctgg tatctttata gtcctgtcgg gtttcgccac
ctctgacttg 2280
agcgtcgatt tttgtgatgc tcgtcagggg ggcggagcct atggaaaaac
gccagcaacg 2340
cggccttttt acggttcctg gccttttgct ggccttttgc tcacatgttc
tttcctgcgt 2400
tatcccctga ttctgtggat aaccgtatta ccgcctttga gtgagctgat
accgctcgcc 2460
gcagccgaac gaccgagcgc agcgagtcag tgagcgagga agcggaagag
cgcccaatac 2520
gcaaaccgcc tctccccgcg cgttggccga ttcattaatg cagctggcac
gacaggtttc 2580
ccgactggaa agcgggcagt gagcgcaacg caattaatgt gagttagctc
actcattagg 2640
caccccaggc tttacacttt atgcttccgg ctcgtatgtt gtgtggaatt
gtgagcggat 2700
aacaatttca cacaggaaac agctatgacc atgattacgc caagcgcgca
attaacctat 2760
cagcacacaa tagtccatta tacgcgcgta taatgggcaa ttgtgtgctg
atatctagag 2820
agctccaccg cggtggcggc cgcataaagg tgaggagaga accacgtcag
cagagttgtc 2880
ctctgaccct cacacatgct ggtgccaggg ttacccgtaa tcacacgcat
gtatcataca 2940
cacacacaca catccttaag tgtttgcttg tttgttgtgt atgatctcac
aaacatgcta 3000
cagtgcagat ggaggtcaga ggacatctgt gggagtcaga tttcttcttc
taccacgtgt 3060
gtcccaggga ccaaactcag gtcatcgggt tggtagcaga tgcctttacc
cactgaggcg 3120
ttgtgctgtc cctatcccca ctttttgtat gttggttaca tacatggctg
ctctctaacc 3180
catgcagatg cctgcagctc agaacatatc cctctattgg ctaaacaggt
gatgtttgtc 3240
tttagtcaag agtaatgatg ggcctttgag caacaaattc tggcatccat
ccttgaattg 3300
tcccgctcag tcaggcaaga cctttgctgt gacttcagac ctaatgagtt
cctgtctaga 3360
tagtgaagca ttccccacaa ttcctcacac agaatgctga acacagagca
atcctagtga 3420
ggaaaaccga gataaaggag cctcctatct ccacttagct tgaatttact
taacccccgc 3480
ccccagaact actaaaaata ttaataataa tatgtacaga cataatttgg
cttgttcgct 3540
ctttaagaaa tatttcgagc caggagttgg tggtgaatgc ctataatctc
agcagtctgg 3600
agcagaggca ggcagatctc ttgagttcag tgccagcctg atctacagag
tgagttccag 3660
gacagccagg gctgcacaga gaaaccctgt ctcaaaaaga ccttaaaaaa
aaaaaaaaaa 3720
agaggtgttt tgggttgtgc agagtcgcca ccatgagaaa ggccctcctg
aactgaactg 3780
aactgaactg tatggacaag aagttcttat tgaagtgaaa tggtggcaga
catgtccaag 3840
ggatgaatgg cgaatgatga gtgtggagag cagaagcaac actgagcaac
agagcaacct 3900
tgcaatggtg gtcgccaagg aaacagcatc atcaggcaga agccttggag
agagtctaaa 3960
caacggccct gcaccttcag caggagtgca cccaccccac ccccaacaca
cacacagggc 4020
ctgtctgact gtgggtgtag aactgggtca tggacatttt cataagaaac
tcttttgttt 4080
gtttggtttg gtttggttta gttttttgtt ttttgagaca gggtttctct
gtagacccct 4140
ggctgtcctg gaactcactc tgtagaccag ggtggcctcg aactcagaaa
tccgcctgcc 4200
tctgccttcc gaatgctggg attaaaggcg tgtgccacca tgcccagcaa
gaaacttatt 4260
tctaaataaa cttctgtgat actccaatat taaaaaaaga aaaaaatagg
agaggaaagg 4320
aaacgatttc ggtagttgct gctcttgcgc acagtaggcg aagcagatgc
agaggctcac 4380
acttgtaatg ccagcccttg ggaggtagag acaggagagc agatgtccaa
ggtcagcctt 4440
agatacataa caaacctgag gctagcccgt cagaggcagg cagggagcag
aacaagcagg 4500
tgttccccgt catcggcctc cgcgtctgca ctctcctgac ttgaggcacc
agggcttttc 4560
cccacagagc cctcttgagt cgggaacccc ctcagtggat ctgagtcctc
catgtgcccc 4620
ccttttcctt tcaaagaaga gagaagaatg gagggctgcg ggcaagaaga
gggggacaca 4680
ccttccctag gcctgagtga actttacctt gtctactaat taaacaacac
ggagttctgt 4740
gctttgctta accctgggaa ccggtcatca gcctttgacc tcagccctcc
tctcctgacc 4800
agctccctcc cccacctggc ctctggacct ctcagattta ggggaaagaa
tcagttttcg 4860
gagtgatcgt ctcagtcagc accatctctg taggagcatc ggccatgtat
tccaatgtga 4920
taggaactgt aacctctgga aaaaggtaag aggggtgagc tttccccttg
cctgccccta 4980
ctgggttttg tgacctccaa aggactcaca ggaatgacct ccaacacctt
tgagaagacc 5040
aggccctctc cctggtagtt acagtcaagg atctttctaa ttccaacatg
accacgtcca 5100
ggcaactgaa ctgtcccctg gcaagaagtc taggactgaa cctgtcccgg
gcccctgtac 5160
ttggttcaaa ggatttagcc tttctcttgg ccacaccagg tgggctggaa
tcctctgctt 5220
tactggggca accctgtggt gggcagtggg gctaggggtt gcagcctagc
ttaacttggc 5280
attttgtctc cttgcaggaa ggtttatctt ttgtccttgc tgctcattgg
cttctgggac 5340
tgcgtgacct gtcacgggag ccctgtggac atctgcacag ccaagccgcg
ggacattccc 5400
atgaatccca tgtgcattta ccgctccccg gagaagaagg caactgagga
tgagggctca 5460
gaacagaaga tcccggaggc caccaaccgg cgtgtctggg aactgtccaa
ggccaattcc 5520
cgctttgcta ccactttcta tcagcacctg gcagattcca agaatgacaa
tgataacatt 5580
ttcctgtcac ccctgagtat ctccacggct tttgctatga ccaagctggg
tgcctgtaat 5640
gacaccctcc agcaactgat ggaggtacga ccaaaggtct tctgcccagc
caccttgtta 5700
ggagcacctt tggggcttcc ataggcccaa gtccaatgat tcctcaacca
acactgcagt 5760
cccaggtact gtgcttgaag gagttaacaa ctgaggtggc tattagtcag
agactgacca 5820
gcatgtgctc accacccatg ttaactaggc agcccaccaa acccaccacc
attttttttt 5880
gacttctata ggtatttaag tttgacacca tatctgagaa aacatctgat
cagatccact 5940
tcttctttgc caaactgaac tgccgactct atcgaaaagc caacaaatcc
tccaagttag 6000
tatcagccaa tcgccttttt ggagacaaat cccttacctt caatgagacc
taccaggaca 6060
tcagtgagtt ggtatatgga gccaagctcc agcccctgga cttcaaggtg
agttgcagat 6120
gttacccctg acctccgagt tcttcctctc cactcagaga ttgaggaggt
ggagaaacag 6180
catccaaatt cacactgctt tgctgctgaa gactgctgga gggctgacta
aaagttagaa 6240
cccctgcaat agttattctt acttgaaacc tgagaaatca aaggtatcca
tgcttggatt 6300
gtagtgactg cccagaaaac atgaattaat aatcaattct tcattccatc
caccaacttc 6360
aaatatatac caaagggtgt tttgaagatg ccagttctac aagatatctt
acttaatttg 6420
aactgttatc atggtcaaat aaagttggta catgatgcat gttacattct
cctcttggag 6480
attcatgaag cacatgggcc tatgaaggtt ctgagaaact ctgcaacaaa
gaaatctgtt 6540
ggctttattc aatcggcatt cctcaaatgt atttgactgc atgggcattt
ctctcctcca 6600
tataacctgc caaccccata taacctgcca accccatata acctgcaatc
attcattgct 6660
tcccctggca catgccttgg aaattctact tttgtgagtt aaggttttcc
aaagtcagag 6720
aaaataatat tttatcttct ttttcccaga ctattttcgc cttccttctt
ttcatttatt 6780
tcttcctatt tctttttgtt ctttttcttc tgataatatt tattaactac
aggaaagatt 6840
catggaacta tattagatat gtgaggcttc cccaatttgg gttagagcaa
tggcttctta 6900
atcaaatggt gggaaaggac agagggatgg tgagaaaaat aaaatgctgc
ctgggaaaat 6960
ggagaagcca attgaatagc acaggtgagt aggtttattt tctgttctcc
tcaggaaaat 7020
gcagagcaat ccagagcggc catcaacaaa tgggtgtcca ataagaccga
aggccgaatc 7080
accgatgtca ttccctcgga agccatcaat gagctcactg ttctggtgct
ggttaacacc 7140
atttacttca aggtactcag aatggccctg gagagacccc agggacttcc
tcttgctctt 7200
cagcttaccc cctttttttt taaatggcga gaccgaagcc ctgagagggc
aaatggactg 7260
ccgaaagcta cacaggtaca ggtcagcagg gcaggtcaat ctattattta
tttatttatt 7320
tatttttgac agagtctcgc tctgtcgccc aggctggagt gcagtggcgt
gatctcggct 7380
cactgcaagc ttcgcctcct gggttcacgg cattctcctg cctcagcctc
ccaagtagct 7440
gggaatacag gcacccacca ccatgcctgg ctaatttttt ggtttttttt
tagtaaagac 7500
ggggtttcac cgtgttagcc aggatagtct tgatctcctg acctcgtgat
ctgcccacct 7560
cggcctccca aagtgctggg attacaggca tgagccaccg cgcccggcag
attggcttct 7620
ttcacctagt aaaatgcatt tactgttcct ttgtgttttc gtggctttgt
cactcatttc 7680
ttcttagcat ggaatagcat tacatttggt ctggatgtac cacagtctgt
ctattcatct 7740
actgaaggac attttggctg cttccaaggt ttgacagtta tgaataaacc
tactcataat 7800
tccatcattc tgacacagcc attgttaacc tttttgtgca tatcccgcca
gtcttttttc 7860
cgaataatta tatattaatg taacactata atatggatat gtcttgtgtc
aataactatc 7920
ctcctatgaa tgtttgtgtt cttactttgt gattctcttc cagggcctgt
ggaagtcaaa 7980
gttcagccct gagaacacaa ggaaggaact gttctacaag gctgatggag
agtcgtgttc 8040
agcatctatg atgtaccagg aaggcaagtt ccgttatcgg cgcgtggctg
aaggcaccca 8100
ggtgcttgag ttgcccttca aaggtgatga catcaccatg gtcctcatct
tgcccaagcc 8160
tgagaagagc ctggccaagg tagagaagga actcacccca gaggtgctgc
aagagtggct 8220
ggatgaattg gaggagatga tgctggtggt ccacatgccc cgcttccgca
ttgaggacgg 8280
cttcagtttg aaggagcagc tgcaagacat gggccttgtc gatctgttca
gccctgaaaa 8340
gtccaaactc ccaggtttgt ctaggaagga gtttcctccc ttctccaccc
gcaaggtagt 8400
ctgaccaaaa gtggaagagt tggagaaaga atagaaagga gcaacaagtc
aggactcctg 8460
gatctaggat ttcttcttat gtttcaaaaa gccccaaagg atctcttaat
ccaaactgaa 8520
ttcccatctg tgggttgaag ccaactttct cccatctcac aaagacttct
ccggtcttcc 8580
ttccaggtat tgttgcagaa ggccgagatg acctctatgt ctcagatgca
ttccataagg 8640
catttcttga ggtgagtaca ccttccccac tctcttaggg tacagaaagg
agatgcatga 8700
acagcaggaa cacgtggaaa aggcctgttt ccagtgttaa ggcatgcaac
ctgacccaaa 8760
tgtacttttt actggaaaac aaaagcattt gaggaattgc tgtgtctgtg
gatgatttac 8820
ctgccaaaaa tgaacggcag agtggctaat ttagttttat ttcccatgtg
acctgcaggt 8880
aaatgaagaa ggcagtgaag cagctgcaag taccgctgtt gtgattgctg
gccgttcgct 8940
aaaccccaac agggtgactt tcaaggccaa caggcctttc ctggttttta
taagagaagt 9000
tcctctgaac actattatct tcatgggcag agtagccaac ccttgtgtta
agtaaaatgt 9060
tcttattctt tgcacctctt cctatttcta gagtcggggc ggccggccgc
ttcgagcaga 9120
catgataaga tacattgatg agtttggaca aaccacaact agaatgcagt
gaaaaaaatg 9180
ctttatttgt gaaatttgtg atgctattgc tttatttgta accattataa
gctgcaataa 9240
acaagttaac aacaacaatt gcattcattt tatgtttcag gttcaggggg
aggtgtggga 9300
ggttttttaa agcaagtaaa acctctacaa atgtggtaaa atcgataagg
atccgtcgac 9360
ggtatcgata agcttgataa cggccgccca gagcacggtt gtccaggatg
accccaggtg 9420
aggcattcag ctctgtccct cccctctgga tcccaggatc cagcctctgg
ccctcaactg 9480
atgcctccat ctgcccccca ggctcgttca tctcttctct tgggagcctg
gcggcccagt 9540
aaccgtgtct gtacaccaag atgcacctta cgccttcctg cccatatggt
atgtggaagc 9600
tatgactcag gacctcccat caccccccaa gactccctct cccaaatagt
aatgctttag 9660
agggaggcag tcatatctct gtgtgcagat aataaaagca tatttctaag
aggttctctc 9720
gctgtcttct tagctgagtc atccctgtcc agcaactcaa gcacacaaca
gtgctttgct 9780
gttttatcat catgttttta catggggcat tcactgggtg tagaggctgg
ccgcaaatac 9840
gatgtcccgc cgtagcaagg tagccgctgt ctccatcttg gcacccagca
caggctctcc 9900
taccaggcgg gctgcccccc gcagtgagcg acacatctca gccaggcgct
gaatgcagcg 9960
gaccaccagg ccctcagggg tccctgagag ccctgccaac tcggagaagg
gctgtgggga 10020
aagtagggtg aggactggat gcactcagcc tgggcaggtt ctccccagcc
agccgtctgc 10080
aaaatcccaa acctcaggta ctcaccatgc cccgggccca ctcatataca
acctcaacca 10140
gcccaaaatt cagcggccgt atcgaattcc tgcagcccgg gggatccact
agttctagac 10200
cctggggcag gaagtggggc tgctggtgag aggtaagcgg tgggcttccc
tctgtcagct 10260
ctggctgggt tttctgacct ctgaaggact ctcaaggatg acctctgaca
tttttgagac 10320
gaccaggccc tcaccctagt agttagtcaa agaccttttg gaagccatca
cttcacctaa 10380
gccacacctg aggttcttag ctcctttaga agccaagggt gcttcccttg
ggaagggatg 10440
ggtaagccag agcgcacttc cagggcagcc tcctaatctg ctcctcttcc
tactcaactg 10500
ccccccacct gccactgtga cctccgacta cctcccaaag cctccttccc
ctttctccct 10560
tcttggtgta ggttgttgct gtaggttgct aaaaatgaag aggtagcctg
accattatgg 10620
ttcctgttta tacccagcag agcttaggac ccctcctgct tctattcaca
tttccagatg 10680
tgcccaagaa caaaatggga aagacaagac aacagcgatg gctctctctc
cccttcctca 10740
ctcctctctc cccactccct cttccctctt gtcactctac ctgttgatat
cagaccacac 10800
aaaaatccac gcatcctttg gttgccctta ccccccagtt agaccatctc
ccactgagca 10860
gcctacctgc ctgacttttt cttccagaag ggactggaaa ggcgcaatgc
ttagatacag 10920
actccctctc cttacccagg taccccagag cacctggagg gagagcattt
acttcttggt 10980
tattacttta ttgagtatcc acattctctt gccaatctca ctacttcatg
tttttaactg 11040
aggtataatg tagcccaggc tgccctcgtg cctgttacct tagcctccca
aatgctgaga 11100
ttaaggttgt aaccctcacc acacctggct actagctcag ttttttagaa
cgagatacca 11160
agctgggcat gatagtgaat gcctataatc ccagcacttg ggagctggag
gaaggaaagt 11220
cagaagttta aggtcatctt cagctacata gtaagaatgg ggcagcctgg
gctatccctg 11280
acaccctcaa agtgagatta cctttacttt tagtgaccct tgttagaagc
tattaaaaca 11340
tttcaggaga cagaaaataa tgagggcaga actaaggagg gggcagtaga
ggtcagaggg 11400
taaggaagga gaattcattg aatctgggat ctgactcaac caggaggggg
aggggaagga 11460
gctgggtgac ttctaggttt ctggactggt gtcaggatcc actgggttga
aggatagaag 11520
agcaggtgtg tgggggaggt gacagctcag catgtcacat ctcactatgg
aagtcgctgg 11580
ccttccagac cgaactgtcc ggccaatagt aagtctggct gtctgcgtgc
aggctggagc 11640
tgcagatctg gggccttctg tactcagctg ctccttcgtt gagttcacat
tctcttaccc 11700
attttcgcca aagccagtcc ttcaagattc aggctaaagt ctgtccttgt
ctggggtttt 11760
ctgacccctt caggtggctc ccttcacact ctcgactctc tctactgggc
tcatgtgggc 11820
gttccgttca tcaaaggcag cgttgcattc ttttttagcc tagcatggac
acacacacaa 11880
ctgtgtccag gcaaggagcc taggactggt tttgtcctag atggacaagg
atgcttagct 11940
caaacgtcaa ggccggaatc ctacactttc aggaataaga ctgtggtggt
cagtggggat 12000
ggggagtcat ggttctgtcc cttgtaggaa gctggtggag ctcccaacgc
ggacctcgag 12060
ggggggcccg ctagctgcag tatcagcaca caatagtcca ttatacgcgc
gtataatggg 12120
caattgtgtg ctgatattac gcgcgctcac tggccgtcgt tttacaacgt
cgtgactggg 12180
aaaac
12185
Номер последовательности (ID): 3
Длина: 23
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..23
> mol_type, other DNA
> organism, synthetic construct
Остатки:
ctgatggaga gtcgtgttca gca
23
Номер последовательности (ID): 4
Длина: 23
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..23
> mol_type, other DNA
> organism, synthetic construct
Остатки:
catcaccttt gaagggcaac tca
23
Номер последовательности (ID): 5
Длина: 21
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..21
> mol_type, other DNA
> organism, synthetic construct
Остатки:
gctaagaaga cagcgagaga a
21
Номер последовательности (ID): 6
Длина: 22
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..22
> mol_type, other DNA
> organism, synthetic construct
Остатки:
caagactccc tctcccaaat ag
22
Номер последовательности (ID): 7
Длина: 21
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..21
> mol_type, other DNA
> organism, synthetic construct
Остатки:
cagggcttcc accgtataaa c
21
Номер последовательности (ID): 8
Длина: 21
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..21
> mol_type, other DNA
> organism, synthetic construct
Остатки:
gtgtccagta gtcgtgtgat g
21
Номер последовательности (ID): 9
Длина: 20
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..20
> mol_type, other DNA
> organism, synthetic construct
Остатки:
atctctgtag gagcatcggc
20
Номер последовательности (ID): 10
Длина: 22
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..22
> mol_type, other DNA
> organism, synthetic construct
Остатки:
cgactctaga aataggaaga gg
22
Номер последовательности (ID): 11
Длина: 26
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..26
> mol_type, other DNA
> organism, synthetic construct
Остатки:
tgcaaagatt aagaatattt tagttc
26
Номер последовательности (ID): 12
Длина: 23
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..23
> mol_type, other DNA
> organism, synthetic construct
Остатки:
catcactgcc acccagaaga ctg
23
Номер последовательности (ID): 13
Длина: 23
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..23
> mol_type, other DNA
> organism, synthetic construct
Остатки:
atgccagtga gcttcccgtt cag
23
Номер последовательности (ID): 14
Длина: 23
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..23
> mol_type, other DNA
> organism, synthetic construct
Остатки:
tcgatgggca gtcatgccca gtg
23
Номер последовательности (ID): 15
Длина: 23
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..23
> mol_type, other DNA
> organism, synthetic construct
Остатки:
cttgaagggc agctctagca cct
23
END
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ конструирования минигенов млекопитающих и рекомбинантная плазмида pgC1HDR, кодирующая миниген ингибитора С1 эстеразы человека, предназначенная для получения гуманизированных по гену Serping1 мышей | 2022 |
|
RU2805177C1 |
| Способ получения генно-модифицированных кроликов с нокаутом гена LEPR с помощью системы CRISPR/Cas9 | 2023 |
|
RU2836438C1 |
| Способ получения генно-модифицированных лабораторных животных с индуцируемой системной и тканеспецифической экспрессией циклофилин А человека | 2022 |
|
RU2800264C1 |
| Способ получения мышиной модели для изучения миодистрофии Дюшенна и вариантов ее терапии | 2023 |
|
RU2815936C1 |
| СИСТЕМЫ CRISPR-CAS И СПОСОБЫ ИЗМЕНЕНИЯ ЭКСПРЕССИИ ПРОДУКТОВ ГЕНОВ | 2013 |
|
RU2687451C1 |
| Технология получения мышей с гуманизированным участком Gnao1 в области однонуклеотидного полиморфизма rs587777057 для тестирования РНК-терапии для GNAO1 c.607 G>A пациентов | 2021 |
|
RU2757121C1 |
| Способ получения мышиной модели для изучения синдрома Леша-Нихена путем внесения делеции p.Val8del в ген hprt1 | 2021 |
|
RU2768048C1 |
| Способ получения генно-модифицированных лабораторных животных с нуль-аллелем гена P2rx3 | 2022 |
|
RU2805173C1 |
| ЛИНИЯ ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫХ МЫШЕЙ ДЛЯ МОДЕЛИРОВАНИЯ БОЛЕЗНИ АЛЬЦГЕЙМЕРА, СПОСОБ ЕЕ ПОЛУЧЕНИЯ И ЕЕ ПРИМЕНЕНИЕ, sgPHK И КОМБИНАЦИЯ crRNA/tracrRNA, ИСПОЛЬЗУЕМЫЕ В СПОСОБЕ, И НАБОР ПРАЙМЕРОВ | 2024 |
|
RU2835497C1 |
| ГЕНЕТИЧЕСКИЕ КОНСТРУКЦИИ LTF3, LTF5, LTF7, LTF10, LTF11 ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ЛАКТОФЕРРИНА ЧЕЛОВЕКА (ВАРИАНТЫ) | 2011 |
|
RU2491343C1 |
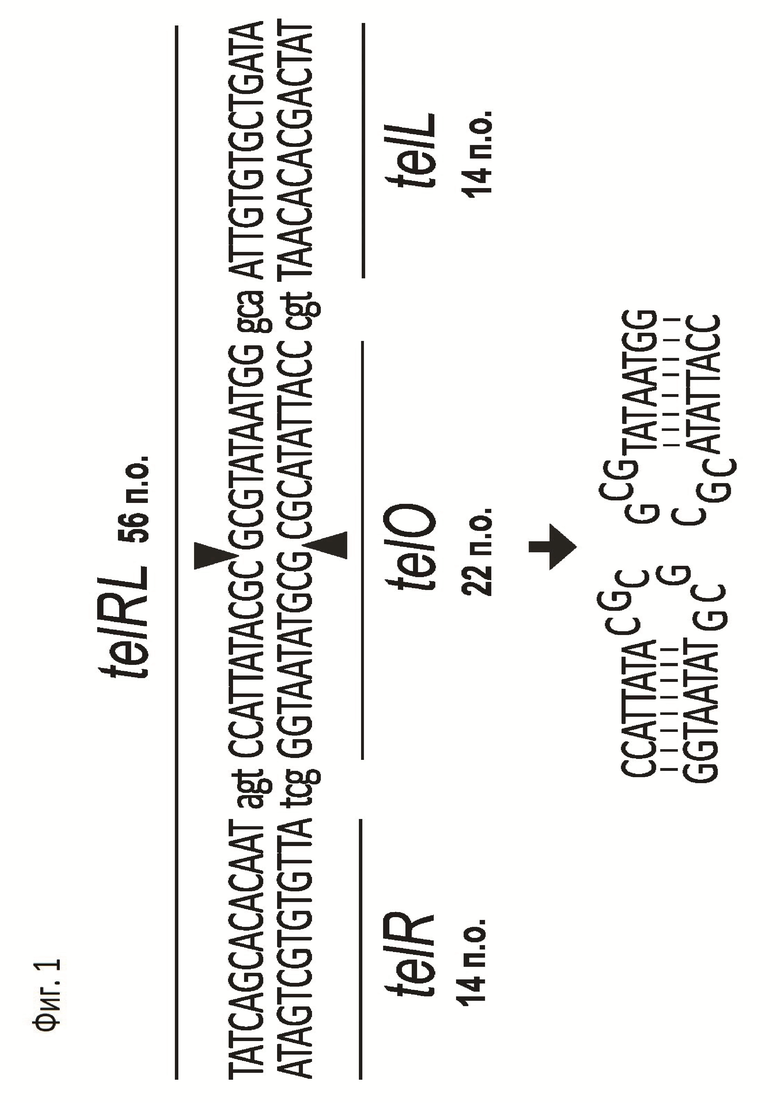

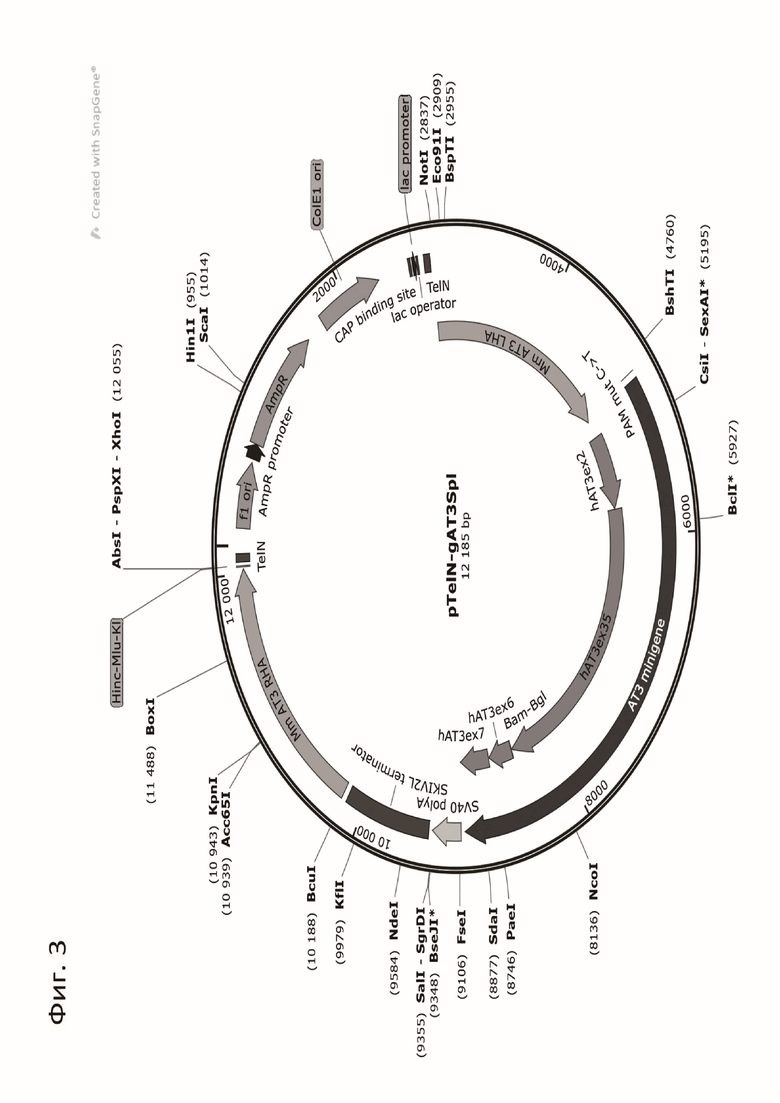
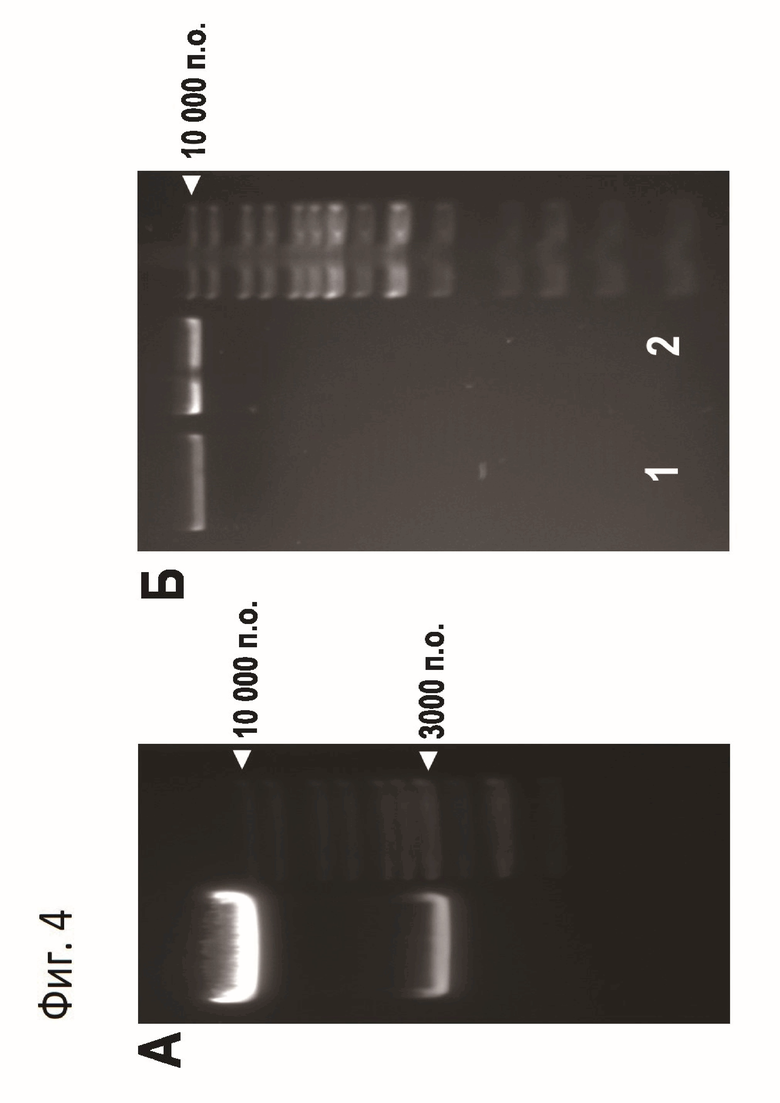


Изобретение относится к области биотехнологии, а именно к рекомбинантной плазмидной ДНК, кодирующей миниген антитромбина III человека под транскрипционным контролем регуляторных элементов гена Serpinc1 мыши, сигнала полиаденилирования вируса SV40 и терминатора транскрипции из гена SKIV2L человека, фланкированный плечами гомологии локуса Serpinс1 мыши с мутированным сайтом PAM. Также раскрыт способ получения трансгенных мышей, экспрессирующих миниген антитромбина III человека, предусматривающий использование указанной плазмидной ДНК. Изобретение эффективно для получения генно-модифицированных мышей, гуманизированных по гену Serpinс1. 2 н.п. ф-лы, 6 ил., 2 табл., 8 пр.
1. Рекомбинантная плазмидная ДНК последовательностью SEQ ID NO: 2 для получения генно-модифицированных мышей, гуманизированных по гену Serpinс1, кодирующая миниген антитромбина III человека под транскрипционным контролем регуляторных элементов гена Serpinc1 мыши, сигнала полиаденилирования вируса SV40 и терминатора транскрипции из гена SKIV2L человека, фланкированный плечами гомологии локуса Serpinс1 мыши с мутированным сайтом PAM.
2. Способ получения трансгенных мышей, экспрессирующих миниген антитромбина III человека, в котором:
1) обрабатывают плазмидную ДНК по п.1 ферментом протеломеразы TelN из фага N15 и получают линейный двуцепочечный фрагмент ДНК, несущий миниген АТIII;
2) проводят микроинъекции полученного фрагмента ДНК в оплодотворённые яйцеклетки мыши;
3) микроинъецированные яйцеклетки пересаживают в яйцеводы псевдобеременных самок-реципиентов, от которых получают потомство поколения F0;
4) из тканей животных поколения F0 выделяют геномную ДНК и проводят генотипирование с помощью количественной ПЦР с олигонуклеотидными праймерами, имеющими последовательности SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8, с детекцией результатов с помощью интеркалирующего красителя SYBR Green I или его аналогов.
| KAZUHIRO ISHIGURO et al | |||
| Complete antithrombin deficiency in mice results in embryonic lethality, J Clin Invest., 2000, 106(7), pp | |||
| Щелевая форсунка | 1925 |
|
SU873A1 |
| XUAN YAO et al | |||
| Tild-CRISPR Allows for Efficient and Precise Gene Knockin in Mouse and Human Cells, Dev Cell, 2018, 45(4), pp.526-536.e5 | |||
| RENE MULDER et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
