Настоящее изобретение относится к способам выделения экспрессированных бакуловирусами вирусоподобных частиц (VLP) безоболочечных вирусов из клеток насекомых.
В настоящее время медицинская наука в значительной степени полагается на вакцины, когда дело доходит до борьбы с инфекционными заболеваниями. Такие вакцины могут представлять собой живые ослабленные вакцины или инактивированные вакцины, и те и другие с их собственными преимуществами и недостатками. Живые ослабленные вакцины точно имитируют естественную инфекцию, и, следовательно, они вызывают иммунный ответ, который сопоставим с иммунным ответом, вызываемым вирулентной формой вируса или микроорганизма. Однако они могут вызвать некоторые нежелательные побочные эффекты, и уровень ослабления является крайне важным. Инактивированные вакцины, с другой стороны, являются безопасными, но они в ряде случаев не так эффективны, как их живые аналоги.
В случае вирусных вакцин третий тип вакцины вызывает все больший интерес: так называемая вакцина на основе вирусоподобных частиц (VLP). Вирусоподобные частицы представляют собой частицы, которые морфологически очень похожи на вирус дикого типа. Они содержат все иммуногенные компоненты, которые обнаруживаются на вирусе дикого типа, и они содержат все указанные компоненты в их естественной организации и конформации. Однако они отличаются от вируса дикого типа тем, что они не способны производить инфекционное вирусное потомство, поскольку они не имеют вирусного генома.
Поэтому они являются привлекательной альтернативой как живых ослабленных, так и инактивированных вирусных вакцин.
VLP преимущественно продуцируются in vitro в клеточных системах. И во многих случаях они продуцируются в экспрессирующей системе бакуловирус/клетка насекомого. Указанные системы обладают тем преимуществом, что они способны продуцировать VLP в относительно больших количествах.
Белки, экспрессированные в экспрессирующей бакуловирусной векторной системе (BEVS), могут быть либо секретируемыми, либо несекретируемыми белками. Из этого следует, что несекретируемые VLP вначале необходимо выделить из клеток насекомых, прежде чем их можно дополнительно очистить. Стандартные способы выделения VLP включают замораживание-оттаивание, обработку ультразвуком, механические способы с высоким сдвиговым усилием, методы бусинок, "клеточную бомбу", осмотический шок, с использованием гипотонической среды, использование детергентов и ферментативных способов. Однако все указанные способы неизбежно приводят также к разрушению или лизису клеток и высвобождению всего содержимого клетки: РНК, ДНК, белков, клеточных органелл, клеточного дебриса, и в случае систем на основе бакуловируса, также живого бакуловируса в окружающую среду. Очевидно, что большинство, если не все указанные компоненты, должны быть удалены перед тем, как VLP будут готовы для использования в вакцине. Требуемый уровень очистки предполагает сложный многостадийный процесс, который включает в себя множество контролей качества.
Таким образом, существует явная необходимость в способах выделения несекретируемых VLP в культуральную жидкость, которые в то же время позволяют избежать выделения всей клеточной нуклеиновой кислоты и белкового содержимого. Целью настоящего изобретения является разработка способов выделения VLP, которые позволили бы уменьшить или даже избежать проблем, упомянутых выше. Способы согласно настоящему изобретению особенно подходят для выделения несекретируемых VLP безоболочечных вирусов, экспрессированных в экспрессирующей бакуловирусной векторной системе с использованием клеток насекомых. Неожиданно было обнаружено, что после того как клетки насекомых, содержащие безоболочечные VLP, смешивают с солью, так что раствор соли в воде, полученный в результате смешивания, содержит по меньшей мере 300 мосмоль/л соли, катион которой выбирают из группы, состоящей из щелочных металлов и щелочно-земельных металлов, и анион которой выбирают из группы, состоящей из CI-, Br- или I-, клетки высвобождают VLP без выделения всего содержимого клеток. Способ особенно подходит для клеток насекомых, когда их выращивают без носителя в суспензии.
Это действительно в высшей степени удивительный феномен: способ согласно настоящему изобретению, очевидно, оставляет большинство клеток интактными, как можно видеть, например, на фиг. 4. Механизм, лежащий в основе указанного выделения VLP, неясен.
Таким образом, первый вариант осуществления настоящего изобретения относится к способам выделения экспрессированных бакуловирусом вирусоподобных частиц (VLP) безоболочечных вирусов из клеток насекомых, отличающихся тем, что способ не включает стадий лизиса или разрушения клеток и включает стадию смешивания клеток насекомых с солью, где раствор соли в воде, полученный в результате смешивания, содержит по меньшей мере 300 мосмоль/л соли, катион которой выбирают из группы, состоящей из щелочных металлов и щелочноземельных металлов, и анион которой выбирают из группы, состоящей из CI-, Br- и I-.
В контексте этого способа понимают, что смешивание клеток насекомых с солью, таким образом, что раствор соли в воде, полученный в результате смешивания, содержит по меньшей мере 300 мосмоль/л соли, катион которой выбирают из группы, состоящей из щелочных металлов и щелочноземельных металлов, и анион которой выбирают из группы, состоящей из CI-, Br- и I-, включает следующие стадии: культуральную жидкость (клетки насекомых в окружающей их жидкости) смешивают с солью, катион которой выбирают из группы, состоящей из щелочных металлов и щелочноземельных металлов, и анион которой выбирают из группы, состоящей из CI-, Br- и I-, в таком объеме, что полученный солевой раствор после смешивания содержит по меньшей мере 300 мосмоль/л соли.
Само собой разумеется, что смешивание может быть осуществлено различными путями; например, путем добавления соли в твердом виде к культуральной жидкости или путем смешивания соли в водном растворе с культуральной жидкостью. Раствор соли в воде, который является результатом смешивания, как описано в настоящем изобретении, может содержать, например, по меньшей мере 150 мМ NaCl/л (при этом получают раствор соли в воде с осмолярностью по меньшей мере 300 мосмоль NaCl/л). Если солевой раствор, как описано в настоящем изобретении, создают на основе MgCl2, раствор соли в воде, полученный в результате смешивания, будет содержать по меньшей мере 100 мМ MgCl2/л (при этом получают раствор соли в воде с осмолярностью по меньшей мере 300 мосмоль MgCl2/л).
Следующие примеры служат просто в качестве иллюстрации; если 100 мл культуры клеток насекомых, содержащей клетки насекомых и культуральную среду, смешивают с 100 мл раствора 300 мМ NaCl/л, конечные 200 мл жидкости, которую получают в результате указанного действия, содержат 150 мМ NaCl/л. Конечный объем раствора соли, разумеется, не имеет отношения к полезным эффектам способа согласно изобретению. Если 75 мл культуры клеток насекомых, содержащей клетки насекомых и культуральную среду, смешивают с 25 мл раствора 1200 мМ NaCl/л, конечные 100 мл жидкости, которую получают в результате указанного действия, содержат 300 мМ NaCl/л.
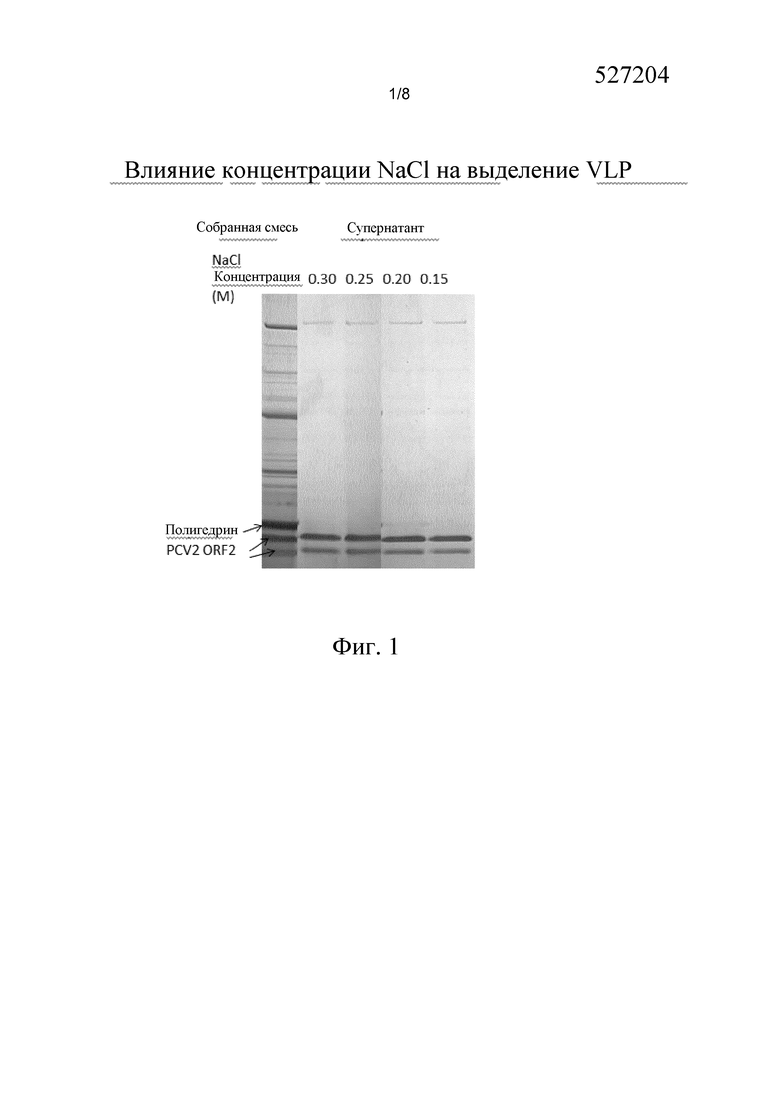
Визуальное отображение на геле неожиданного феномена по изобретению представлено на фиг. 1. Как можно видеть на фиг. 1, дорожки, соответствующие супернатанту клеток после обработки способом согласно изобретению, содержащему вырабатываемые VLP, являются исключительно чистыми (для подробностей смотреть раздел “Примеры”).
Хотя, как указано выше, конечный объем раствора соли не относится к способу как таковому, на практике целесообразно ввести стадию концентрации перед использованием культуральной жидкости на стадии смешивания согласно изобретению. Основной причиной использования стадии концентрации является то, что VLP-продукция в коммерческих целях требует относительно больших объемов культуры клеток. Не эффективно получать VLP непосредственно из указанных больших объемов; получение VLP из небольшого объема требует гораздо меньшей обработки.
Уровень концентрации на данной стадии не является критическим. Концентрация культуральной жидкости в объеме, составляющем половину исходного объема культуральной жидкости (т.е. до 50% исходного объема), уже будет давать значительное преимущество в простоте получения. Дальнейшая концентрация до 40%, 30%, 25%, 20%, 15%, 10% или даже 5% является предпочтительной в указанном порядке предпочтений.
Концентрацию можно осуществить различными путями, которые известны в данной области. Одним из наиболее часто используемых способов является ультрафильтрация, например, в виде тангенциального проточного диализа.
Другим часто используемым способом является низкоскоростное центрифугирование. Низкоскоростное центрифугирование с последующим удалением части или всей культуральной жидкости приводит к концентрации и в конечном счете к осаждению клеток, но клетки по-прежнему будут интактными и по-прежнему будут окружены небольшим количеством культуральной жидкости. Затем такие клеточные осадки можно легко смешать с солью в водном растворе с получением соли в водном растворе, как описано в изобретении.
По практическим соображениям самым подходящим способом является простое хранение культуральной жидкости, содержащей клетки, при котором клетки опускаются на дно сосуда под действием гравитации. Спустя примерно 1-18 часов, в зависимости от собранного объема и размера сосуда, большинство клеток, несомненно, будет находиться в нижних 10% культуральной жидкости, так что не содержащие клеток верхние 90%> культуральной жидкости можно удалить. Указанный способ позволяет избежать проблемы центрифугирования и/или ультрафильтрации больших объемов культуральной жидкости. VLP-содержащие клетки в нижних 10%> культуральной жидкости, полученные таким образом, можно сразу же использовать в стадии смешивания клеток насекомых с солью, как описано в изобретении.
Таким образом, предпочтительная форма этого варианта осуществления относится к способу согласно изобретению, отличающемуся тем, что стадии смешивания клеток насекомых с солью предшествует стадия концентрации, на которой клетки насекомых концентрируют. В принципе, катионы, выбранные из группы, состоящей из щелочных металлов и щелочноземельных металлов, могут быть использованы в способе согласно изобретению. Однако предпочтительными являются катионы, выбранные из группы, состоящей из Na+, Mg2+, Ca2+ и K+, поскольку они недороги и подходят для использования в медицинских целях.
Таким образом, другая предпочтительная форма этого варианта осуществления настоящего изобретения относится к способу согласно изобретению, отличающемуся тем, что катион выбирают из группы, состоящей из Na+, Mg2+, Ca2+ и K+.
Предпочтительным анионом является Cl-, поскольку соли, содержащие анион Cl-, недороги и подходят для использования в медицинских целях.
Таким образом, более предпочтительная форма этого варианта осуществления относится к способу согласно изобретению, отличающемуся тем, что соль выбирают из группы, состоящей из NaCl, KC1, CaCl2 и MgCl2.
Культуральная жидкость, содержащая клетки насекомых, которые несут VLP, дополнительно содержит множество компонентов, которые необходимы для культивирования клеток. Она также содержит продукты жизнедеятельности клеток.
Указанные продукты являются нежелательными побочными продуктами, которые могут быть вредны для конечной вакцины, содержащей VLP.
Такие продукты, соответственно, предпочтительно отделяют от клеток насекомых, содержащих VLP, перед применением способа согласно изобретению на клетках. Это можно легко выполнить путем осаждения клеток, удаления супернатанта, ресуспендирования клеток в растворе соли, как описано в изобретении. Если это делать аккуратно, то, несомненно, можно удалить большую часть супернатанта. В таком случае, раствор соли в воде, полученный в результате смешивания клеток и соли, по существу состоит из соли, как описано в изобретении, и воды. По существу в описании подразумевается, что раствор соли в воде, полученный в результате смешивания, содержит 10% культуральной жидкости. Предпочтительно, водный раствор, полученный в результате смешивания, содержит меньше 8%, меньше 6%, меньше 4% или даже меньше 2% культуральной жидкости.
Таким образом, предпочтительная форма этого варианта осуществления относится к способам согласно изобретению, отличающимся тем, что раствор соли в воде, полученный в результате смешивания, по существу состоит из соли, как описано в изобретении, и воды.
Способ по настоящему изобретению подходит для несекретируемых безоболочечных VLP, которые, в общем случае, экспрессируются в BEVS.
Таким образом, способ среди прочего подходит для BEVS-полученных VLP членов семейства Parvoviridae и Circoviridae; вирусных семейств, которые имеют несколько членов, для которых в настоящее время VLP создают на коммерческой основе.
Таким образом, другая предпочтительная форма этого варианта осуществления настоящего изобретения относится к способу согласно изобретению, отличающемуся тем, что безоболочечный вирус является членом семейства Parvoviridae или Circoviridae.
Примерами важных вакцин против вирусов Parvoviridae и Circoviridae, предназначенных для использования в ветеринарии, на основе безоболочечного вируса, который можно успешно вводить в форме VPL, произведенных в бакуловирусной системе экспрессии на основе клеток насекомых, являются вакцины против парвовируса свиней (PPV) и цирковируса свиней 2-го типа (PCV2).
Таким образом, более предпочтительная форма указанного варианта осуществления настоящего изобретения относится к способу согласно изобретению, отличающемуся тем, что безоболочечный вирус представляет собой парвовирус свиней (PPV) или цирковирус свиней 2-го типа (PCV2).
Продукция VLP в BEVS в принципе также приводит к продукции бакуловирусов. Такие бакуловирусы являются нежелательным побочным продуктом, и поэтому они должны быть удалены или по меньшей мере инактивированы.
Неожиданно обнаружили, если раствор соли в воде хранить при температуре 30-55°C, предпочтительно 35-50°C, более предпочтительно 40-45°C, количество живых бакуловирусов значительно уменьшается. Кроме того, количество вырабатываемого антигена также существенно увеличивается при повышении температуры.
Указанное снижение является очень полезным, поскольку количество вируса, который необходимо инактивировать после применения способа согласно изобретению, является относительно низким, и поэтому придется использовать более низкие количества инактивирующих химических препаратов.
Только в качестве примера; если раствор соли в воде хранить при температуре 30-40°C, можно легко получить снижение вирусного титра в количестве от 1000 до 10000 раз.
Время, в течение которого следует применять определенную температуру, зависит от выбранной температуры: при температуре приблизительно 40°C, снижение вирусного титра в 10 раз достигается через 1 час, тогда как при температуре приблизительно 35°C такое снижение достигается через 5-6 часов. В целом, хранение раствора соли в воде, полученного в результате смешивания, при определенной температуре в течение 18-24 часов подходит в любом случае. Два типа клеток насекомых, которые наиболее часто используют для VLP-экспрессии, представляют собой клетки SF9 или SF21. Способ, описанный в настоящем изобретении, может быть весьма успешно применен к обоим указанным типам клеток. Таким образом, другая предпочтительная форма указанного варианта осуществления настоящего изобретения относится к способу согласно изобретению, в котором клетки насекомых являются клетками SF9 или SF21. Описанный способ обычно является частью обычно применяемых способов очистки бакуловирус-экспрессированных вирусоподобных частиц (VLP) безоболочечных вирусов, которые включают в себя несколько стадий, в числе которых есть стадии a) инфицирования клеток насекомых рекомбинантным бакуловирусом, кодирующим VLP-белки, b) культивирования клеток насекомых, c) выделения VLP из клеток насекомых и d) отделения VLP.
Присутствует стадия c), стадия выделения VLP, которая была бы очень полезна в составе способа согласно изобретению.
Таким образом, другой вариант осуществления настоящего изобретения относится к способам очистки экспрессированных бакуловирусами вирусоподобных частиц (VLP) безоболочечных вирусов, где способ включает стадии
a) инфицирования клеток насекомых рекомбинантным бакуловирусом, кодирующим VLP-белки,
b) культивирования клеток насекомых,
c) выделения VLP из клеток насекомых,
d) отделения VLP,
при этом способ не включает стадий лизиса или разрушения клеток, и стадия c) включает способ по настоящему изобретению выделения VLP.
ПРИМЕРЫ
Пример 1. Продукция VLP PCV2
В этом примере использовали BEVS-произведенные VLP PCV2. Создание указанного рекомбинантного бакуловируса подробно описано ранее в патенте США US 2001/0064765. В дальнейшем вирус обозначается как вирус BacPCV-2-ORF-2.
Для получения продукта экспрессии в максимальных количествах осуществляли предварительные эксперименты с целью оптимизации условий получения рекомбинантных VLP PCV-2 ORF-2. Все эксперименты осуществляли с использованием клеток Spodoptera frugiperda 21 (Sf21) в суспензионной культуре при 28°C. Для инфицирования использовали вирус BacPCV-2-ORF-2 на уровне 4-го пассажа от исходного вакцинного вируса. Для оптимизированной продукции плотность клеток во время инфицирования составляла 1,4×106 клеток/мл, множественность заражения (MOI) составляла 0,01, и культуру поддерживали в течение 6 дней после инфицирования. Полученную смесь называли собранной смесью, содержащей продукта экспрессии.
Образцы культуральной жидкости до и после низкоскоростного центрифугирования подвергали денатурирующему электрофорезу в полиакриламидном геле в присутствии SDS по методу Лэммли (Laemmli, U.K. (1970). Nature 227, 680-685). Используемые 4-12% градиентные гели окрашивали кумасси бриллиантовым голубым. Все гели в Примерах также представляют собой денатурирующие SDS-полиакриламидные гели, полученные по методу Лэммли.
Пример 2. Выделение VLP из клеток насекомых
Клетки собирали в соответствии с Примером 1. На фиг. 1 дорожка, обозначенная “собранная смесь”, показывает содержание белка в собранной смеси клеточной культуры. Указанная дорожка показывает все белки, представленные в культуральной жидкости, и клетки, в том числе клеточные белки, бакуловирусные белки и белки, относящиеся к VLP.
После центрифугирования собранной смеси при 200 g, 90% бесклеточного супернатанта заменяли равным объемом раствора соли в воде, таким образом, что конечная концентрация после смешивания составляла 0,15, 0,20, 0,25 и 0,30 мМ NaCl. Указанные смеси выдерживали в течение 18 часов при 40°C и оставшиеся клетки затем осаждали центрифугированием при 3000 g. Супернатанты подвергали гель-электрофорезу.
Как можно видеть на фиг. 1, на дорожках, обозначенных "супернатант", супернатант содержит практически чистый белок VLP PCV2 (и следовые количества не-VLP белков).
Пример 3. Сравнение действия солей щелочных металлов и фосфатных буферов
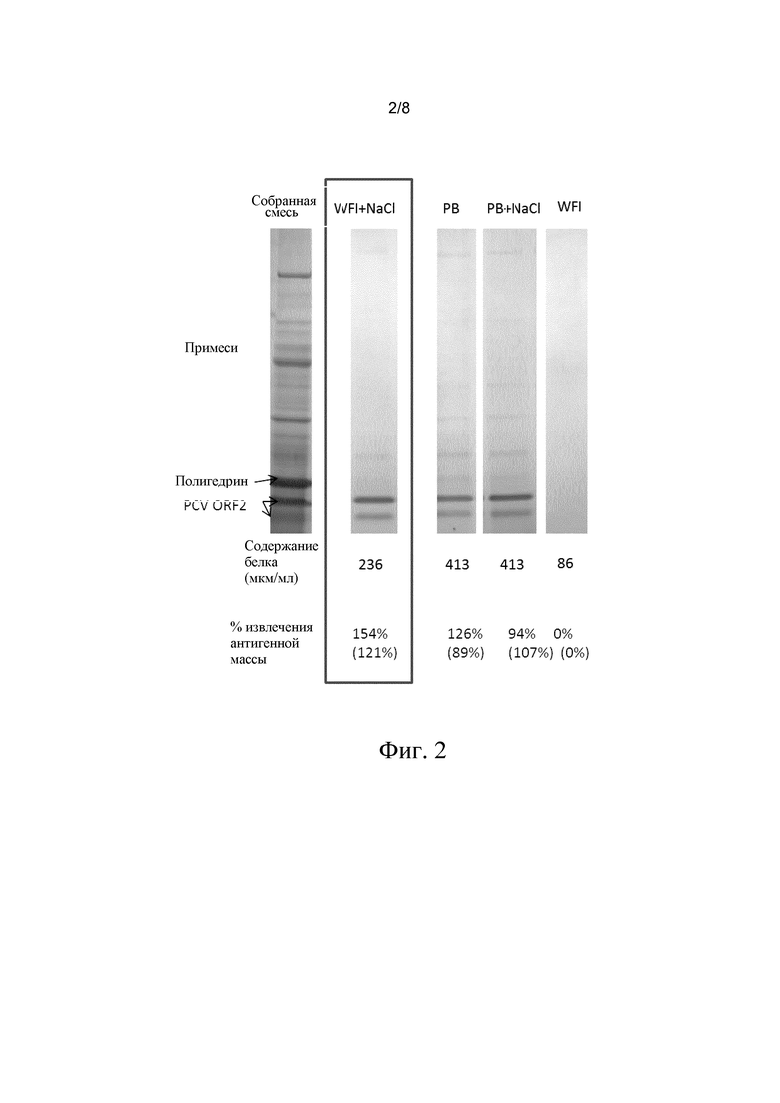
В этом эксперименте собирали культуральную жидкость, клетки осаждали при 200 g, 90% супернатанта удаляли и клетки ресуспендировали в равном объеме либо PB (250 мМ фосфатный буфер, pH 7,4), либо PB + 0,5 M NaCl, либо H2О (= WFI = вода-для-инъекций), либо в водном растворе 0,5 M NaCl, как описано в изобретении.
Инкубацию осуществляли при 40°C, в течение ночи. Затем клетки удаляли центрифугированием при 3000 g. На фиг. 2 можно видеть, что применение WFI в чистом виде не позволяет выделить VLP, тогда как применение PB+NaCl, NaCl в водном растворе и PB позволяет выделить VLP. Как следует из изображения, дорожке WFI + NaCl (= вода-для-инъекций + NaCl, = NaCl в водном растворе) соответствует самое большое количество чистых VLP и самое низкое фоновое содержание белков, не относящихся к VLP.
Антигенную массу (выраженную в Антигенных Единицах на мл - АЕ/мл) всех образцов определяли с помощью ELISA относительно стандартного образца с известной антигенной массой. Затем рассчитывали степени извлечения (в %) для всех тестируемых образцов относительно степени извлечения, установленной для образца собранной смеси. В качестве альтернативного способа интенсивность специфических белковых полос PCV2 на геле рассчитывали относительно интенсивности специфической белковой полосы PCV2 в образце собранной смеси.
Пример 4. Сравнение действия солей щелочных металлов и фосфатных буферов при повышенных температурах
В этом примере собирали культуральную жидкость, клетки осаждали при 200 g, 90% супернатанта удаляли и клетки ресуспендировали в равном объеме либо PB (250 мМ фосфата, pH 7,4), либо PB + 0,5 M NaCl, либо 0,01 M PBS (8 г NaCl/л, 0,2 г KC1/л, 1,44 г Na2HPО4/л, 0,2 г K2HPО4/л, pH 7,1), либо 0,5 M NaCl в водном растворе, как описано в изобретении.
Готовили несколько разведений, в целях сравнения эффекта в зависимости от уровня разведения культуральной жидкости. В одном эксперименте клетки в 30 мл собранной культуральной жидкости концентрировали в 10 раз и затем смешивали с раствором соли в воде до конечного объема 50 мл. Во втором эксперименте клетки в 30 мл собранной культуральной жидкости концентрировали в 10 раз и затем смешивали с раствором соли в воде до конечного объема 25 мл. В третьем эксперименте клетки в 30 мл собранной культуральной жидкости концентрировали в 10 раз и затем смешивали с раствором соли в воде до конечного объема 10 мл.
Инкубацию осуществляли при 40°C в течение ночи. Затем клетки удаляли центрифугированием при 3000 g. Из изображения геля на фиг. 3 можно видеть, что PB, PB+NaCl, PBS и NaCl в водном растворе (WFI + NaCl) все позволяют выделить VLP. Как следует из изображения, дорожке WFI + NaCl (= вода-для-инъекций + NaCl, = NaCl в водном растворе) соответствует самое большое количество чистых VLP и самое низкое фоновое содержание белков, не относящихся к VLP. Сходный эффект наблюдается для всех трех концентраций.
Кроме того, эксперимент с WFI + NaCl также осуществляли при 45°C. Как можно видеть на изображении дорожки "WFI + NaCl, 45°C", инкубация при данной температуре дает картину, сравнимую с обработкой WFI + NaCl при 40°C: VLP-белок со сравнительно низким количеством белковых примесей.
Пример 5. Поведение клеток в водно-солевой среде
В этом эксперименте были сделаны микрофотографии клеток насекомых в культуральной жидкости и клеток, которые обрабатывали по способу согласно изобретению. Клетки либо оставляли в виде необработанной культуральной жидкости, либо подвергали обработке согласно способу по изобретению, в данном случае смешиванию с раствором соли в воде, так что раствор соли в воде, полученный в результате смешивания, содержал 0,3 M NaCl/L. На фиг. 4 представлены результаты: на чертеже слева показаны клетки насекомых без обработки, тогда как на чертеже справа показаны клетки насекомых в растворе соли в воде, содержащем 0,3 M NaCl/л после смешивания. Как можно видеть, клетки на фотографии справа по-прежнему интактны, и по размеру они даже несколько меньше клеток на фотографии слева, из-за их нахождения в гиперосмотической среде раствора соли в воде, содержащего 0,3 мМ NaCl/L.
Пример 6. Влияние температуры инкубации на титр бакуловируса
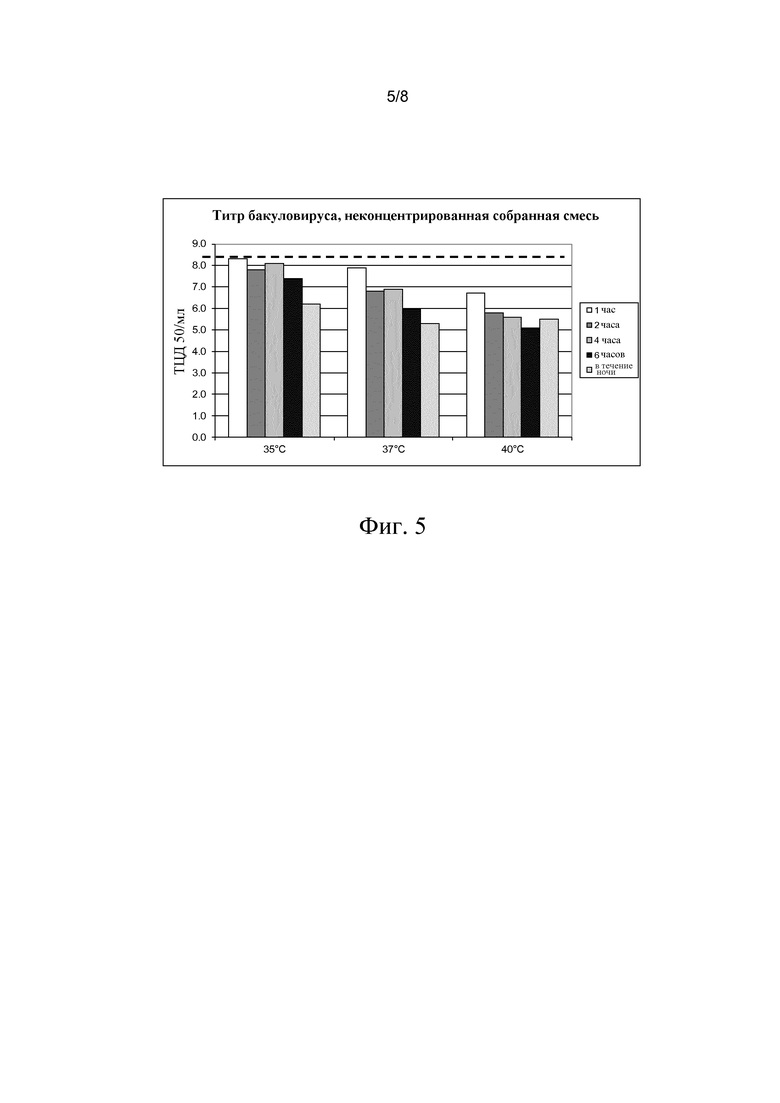
В этом примере клетки собирали в объеме 30 мл с помощью низкоскоростного центрифугирования, 90% бесклеточной культуральной жидкости удаляли и затем оставшийся клеточный осадок ресуспендировали в 25 мл раствора соли в воде до конечной концентрации 0,5 M NaCl/л.
Полученную таким образом суспензию подвергали в течение 1, 2, 4, 6 или 18 часов (в течение ночи) действию температур 35°C, 37°C и 40°C. Затем клетки удаляли центрифугированием при 3000 g. Как видно из фиг. 5, снижение титра бакуловируса примерно в 100 раз может быть получено в течение ночи при 35°C. При 40°C снижение титра в 100 раз достигается уже через 1-2 часа, тогда как снижение в 1000 раз достигается через 6-18 часов.
Пример 7. Сравнение действия различных концентраций нескольких щелочных металлов
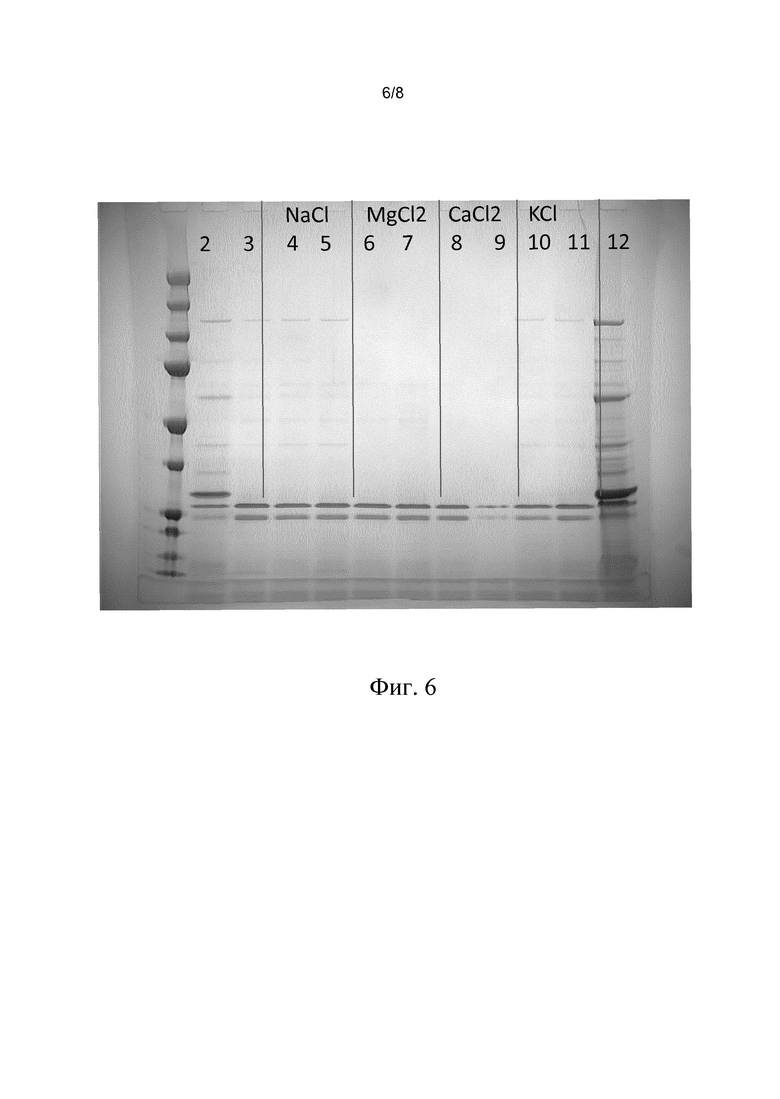
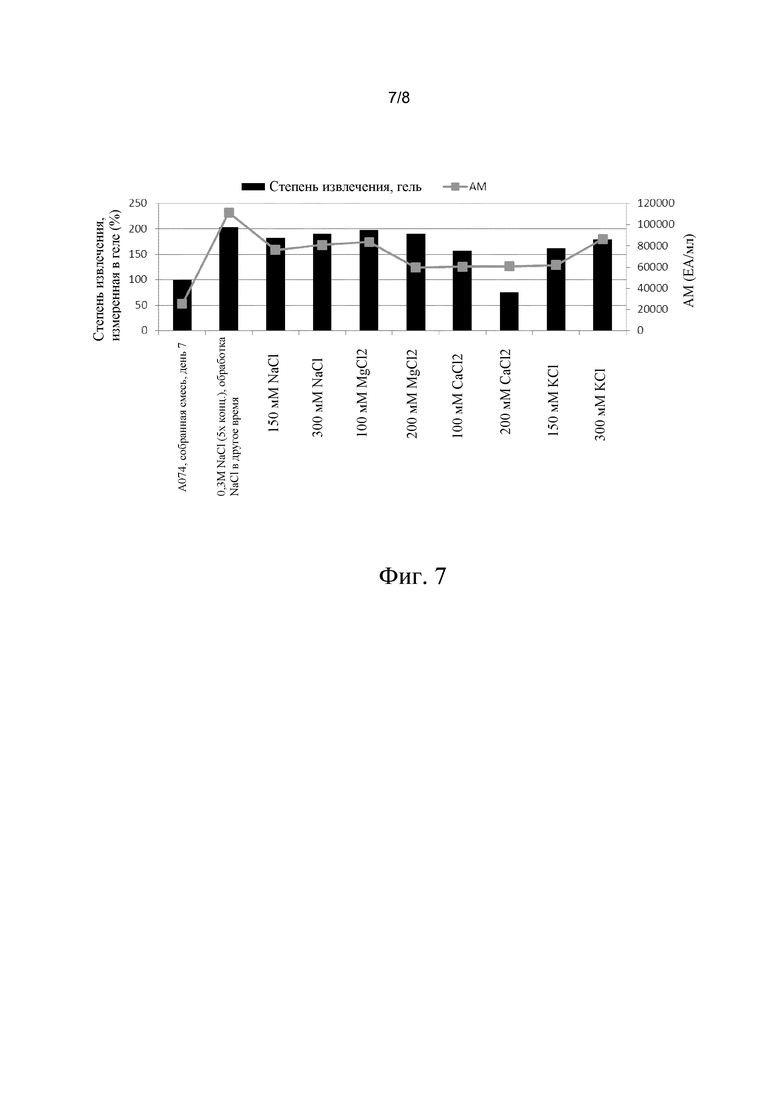
В этом эксперименте сравнивают выделение VLP с помощью разных солей. Клетки собирали, как описано, 90% бесклеточной культуральной жидкости удаляли. Затем добавляли растворы различных солей в воде, при этом объем растворов соли в воде всегда был равен объему концентрированной собранной смеси, для достижения конечной концентрации клеток в 5 раз. Тест осуществляли с использованием различных концентраций NaCl, KC1, MgCl2 и CaCl2. Затем клетки удаляли центрифугированием при 3000 g. Концентрация после смешивания показана в таблице 1 ниже.
Результаты показаны на фиг. 6. На дорожке 2 показаны белки, которые присутствуют в собранной смеси (после пятикратного разведения образцов (исключение: дорожка 12: неразведенный образец собранной смеси). Степень извлечения для образца собранной смеси, которую определяли в геле (таблица 1), указана в таблице как 100, относительная степень извлечения, которую определяли в геле, представлена во второй колонке справа таблицы 1. Выход антигенной массы представлен в правой колонке таблицы 1. Как видно из изображения геля на фиг. 6 и из таблицы, все виды обработки имеют более высокую степень извлечения, чем препарат на дорожке 2 (собранная смесь), и все виды обработки обеспечивают получение чистых препаратов VLP. (Относительно низкое значение степени извлечения, соответствующее дорожке 9, рассматривается как артефакт, поскольку определение антигенной массы препарата, который тестировали на дорожке 9, показало высокую степень извлечения, как и предполагалось). На фиг. 7 показано, что существует хорошая корреляция между степенью извлечения VLP, которую определяли в геле, и степенью извлечения VLP, которую определяли путем определения антигенной массы на основе стандартного теста ELISA.
Пример 8. Применение способа для клеток SF9 и SF21, продуцирующих парвовирус свиней
Эксперимент направлен на то, чтобы показать, что способ равным образом подходит для других безоболочечных несекретируемых VLP. В эксперименте выделяли BEVS-полученные VLP Parvoviridae: парвовируса свиней (PPV). Также в эксперименте использовали два различных вида широко используемых клеток насекомых: клетки SF9 и клетки SF21.
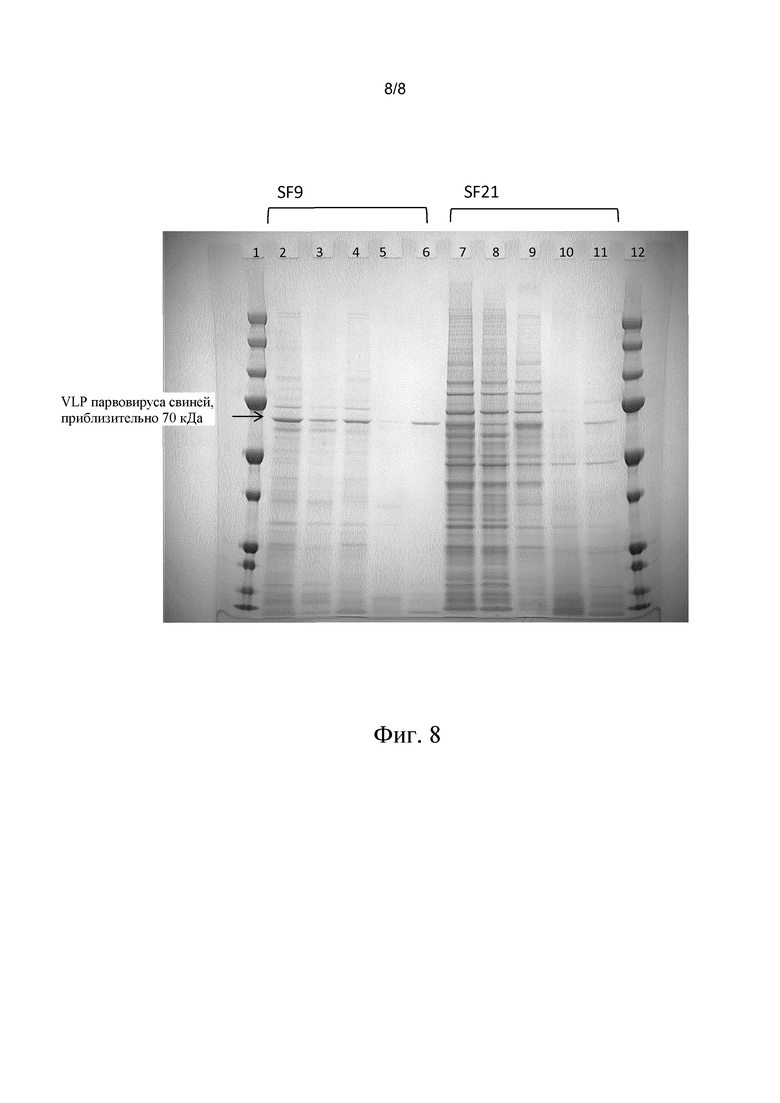
Клетки SF9 и SF21 инфицировали рекомбинантным бакуловирусом, экспрессирующим VLP парвовируса свиней (“собранная смесь”, фиг. 8, дорожки 2 и 7). Затем клетки осаждали при 200 g, 90% супернатанта удаляли (фиг. 8, дорожки 4 и 9) и клетки ресуспендировали либо в равном объеме PBS (фиг. 8, дорожки 3 и 8), либо в 0,3 M NaCl в водном растворе. После инкубации образцов, обработанных NaCl, в течение ночи при 40°C, образцы центрифугировали при 3000 g, супернатант собирали (фиг. 8, дорожки 6 и 11) и осадок ресуспендировали в равном объеме PBS (фиг. 8, дорожки 5 и 10).
На фиг. 8 представлены результаты, полученные при использовании способа в отношении PPV-VLP, произведенных в клетках SF9 и SF21. Дорожка 2: собранная смесь; дорожка 3: собранная смесь, осадок, ресуспендированный в PBS, дорожка 4: супернатант собранной смеси; дорожка 5: собранная смесь, 0,3 M NaCl в течение ночи, 40°C, осадок; дорожка 6: собранная смесь, 0,3 M NaCl в течение ночи 40°C, супернатант; дорожка 7: собранная смесь; дорожка 8: собранная смесь, осадок, ресуспендированный в PBS, дорожка 9: супернатант собранной смеси; дорожка 10: собранная смесь, 0,3 M NaCl в течение ночи 40°C, осадок; дорожка 11: собранная смесь, 0,3 M NaCl в течение ночи 40°C, супернатант.
Как видно из сравнения дорожки 5 и дорожки 3, обработка согласно способу по настоящему изобретению позволяет выделить практически все VLP из клеток насекомых: клетки, осажденные после обработки, как видно на дорожке 5, содержат крайне незначительное количество VLP или не содержат VLP. Как видно из сравнения дорожки 4 и дорожки 3 в отличие от картины, наблюдаемой в случае VLP PCV2, клетки насекомых, продуцирующие VLP CPV, уже выделяют часть VLP в супернатант. Причина данного явления неизвестна. Тем не менее, способ согласно изобретению также может быть применен подходящим образом для VLP CPV, поскольку без применения способа по настоящему изобретению значительное количество VLP CPV остается заключенным в клетках после сбора продукта.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг. 1. На фигуре показан эффект применения способа согласно изобретению с использованием различных концентраций NaCl в водном растворе, при 40°C в отношении клеток насекомых, которые содержат VLP.
Фиг. 2. На фигуре показан эффект применения способа согласно изобретению с использованием различных солей в водном растворе, в отношении клеток насекомых, которые содержат VLP.
Фиг. 3. На фигуре показан эффект применения способа согласно изобретению с использованием различных солей в водном растворе при повышенной температуре, в отношении клеток насекомых, которые содержат VLP.
Фиг. 4. На фигуре представлено сравнение клеток насекомых в их культуральной среде (левая картинка) и клеток насекомых после обработки по способу согласно изобретению (правая картинка).
Фиг. 5. На фигуре показано влияние пролонгированной инкубации при повышенной температуре на титр бакуловируса.
Фиг. 6. На фигуре показан эффект применения способа согласно изобретению с использованием NaCl, MgCl2, CaCl2 и KCl в качестве соли щелочного металла в растворе соли в воде, в отношении клеток насекомых, которые содержат VLP.
Фиг. 7. На фигуре показано отношение между количеством VLP, полученным с использованием способа согласно изобретению, которое определяли в геле, и которое определяли с помощью метода ELISA для антигенной массы.
Фиг. 8. На фигуре показан эффект применения способа согласно изобретению для двух различных клеточных линий насекомых, которые экспрессируют произведенные бакуловирусом VLP парвовируса свиней.
Дорожка 2, 7: собранная смесь. Дорожка 3, 8: собранная смесь, осадок, ресуспендированный в PBS. Дорожка 4, 9: супернатант собранной смеси. Дорожка 5, 10: собранная смесь, 0,3 M NaCl, в течение ночи, 40°C, осадок. Дорожка 6, 11: собранная смесь, 0,3 M NaCl в течение ночи, 40°C, супернатант.


Изобретения относятся к выделению экспрессированных бакуловирусами вирусоподобных частиц (VLP)безоболочечных вирусов из клеток насекомых и касаются способа выделения экспрессированных бакуловирусом VLP цирковируса свиней 2-го типа (PCV2) в клетках насекомых и способа получения экспрессированных бакуловирусами VLP PCV2. Представленный способ выделения экспрессированных бакуловирусом VLP PCV2 включает стадию смешивания клеток насекомых, выбранных из SF9 и SF21, с солью, где раствор соли в воде, полученный в результате смешивания, содержит от 300 мОсмоль/л до 600 мОсмоль/л соли, выбранной из группы, состоящей из NaCl, KCl, CaCl2 и MgCl2, и не включает стадий лизиса или разрушения клеток. Изобретения позволяют получать несекретируемые VLP в культуральную жидкость без необходимости выделения всей клеточной нуклеиновой кислоты и белкового содержимого. 2 н. и 4 з.п. ф-лы, 8 ил., 1 табл., 8 пр.
1. Способ выделения экспрессированных бакуловирусом вирусоподобных частиц (VLP) цирковируса свиней 2-го типа (PCV2) из клеток насекомых, выбранных из SF9 и SF21, отличающийся тем, что способ не включает стадий лизиса или разрушения клеток и включает стадию смешивания клеток насекомых с солью, где раствор соли в воде, полученный в результате смешивания, содержит от 300 мОсмоль/л до 600 мОсмоль/л соли, выбранной группы, состоящей из NaCl, KCl, CaCl2 и MgCl2.
2. Способ по п.1, отличающийся тем, что стадии смешивания клеток насекомых с солью предшествует стадия концентрации, во время которой клетки насекомых концентрируют.
3. Способ по п.1, отличающийся тем, что раствор соли в воде, полученный в результате смешивания, состоит по существу из соли согласно любому из пп. 1-2 и воды.
4. Способ по п.1, отличающийся тем, что раствор соли в воде хранят при температуре 35-50°C.
5. Способ по п.4, отличающийся тем, что раствор соли в воде хранят при температуре 40-45°C.
6. Способ получения экспрессированных бакуловирусами вирусоподобных частиц (VLP) цирковируса свиней 2-го типа (PCV2), включающий стадии
a) инфицирования клеток насекомых, выбранных из SF9 и SF21, рекомбинантным бакуловирусом, кодирующим VLP-белки,
b) культивирования клеток насекомых,
c) выделения VLP из клеток насекомых,
d) отделения VLP,
отличающийся тем, что способ не включает стадий лизиса или разрушения клеток и стадия c) включает способ по любому из пп. 1-5 для выделения VLP.
| LIU L.-J | |||
| et al, Efficient production of type 2 porcine circovirus-like particles by a recombinant baculovirus, Archives of Virology, 09.11.2008, 153, 2291-2295 | |||
| PALOMARES L.A | |||
| et al, Simultaneous expression of recombinant proteins in the insect cell-baculovirus system: production of virus-like particles, Methods, 28.01.2012, 56 (3), 389-395 | |||
| WO 03068993 A1, 21.08.2003 | |||
| WO 2010062757 A1, 03.06.2010 | |||
| ANDREWS BA et al, Purification of virus like particles from yeast cells using aqueous two-phase systems, Bioseparation, 04.1995, 5 (2), 105-112 | |||
| С | |||
| Н | |||
| БЕЛЖЕЛАРСКАЯ и др., Характеристика структурных белков и вирусоподобных частиц вируса гепатита С, синтезированных в клетках насекомых с помощью бакуловирусной системы экспрессии, Молекулярная биология, 2010, т.44., No.1, стр.107-119. |

