ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННУЮ ЗАЯВКУ НА ПАТЕНТ
Настоящая заявка испрашивает приоритет и преимущество на основании заявки на патент США №62/837758, озаглавленной «Рекомбинантная вирусоподобная частица (VLP) капсида цирковируса: композиции, способы и применения», которая полностью включена в настоящую заявку посредством ссылки.
1. ОБЛАСТЬ ТЕХНИКИ
Настоящая заявка относится к композициям, содержащим цирковирус-2 свиней (PCV2) и, в частности, вирусоподобные частицы PCV2 (VLP).
2. УРОВЕНЬ ТЕХНИКИ
Цирковирусы представляют собой небольшие безоболочечные икосаэдрические вирусы, которые содержат небольшой кольцевой, ковалентно замкнутый геном одноцепочечной ДНК (оцДНК). Цирковирусы были идентифицированы у различных видов и, как известно, вызывают инфекции у птиц, водных и наземных животных (1, 2). Изменение морфологии капсида и организации генома недавно привели к классификации семейства Circoviridae на два отдельных рода, Circovirus и Cyclovirus (3). Род circovirus включает цирковирусы свиней типов 1 (PCV1), 2 (PCV2) и 3 (PCV3) (2). Последовательности генома, связанные с родом Cyclovirus, были связаны с несколькими видами позвоночных и беспозвоночных, хотя распознавание окончательных хозяев для этой группы все еще неясно (2). Инфекции PCV2 являются причиной значительной смертности среди свиней в качестве возбудителя заболевания, связанного с цирковирусом свиней (PCVAD), а также связаны с синдромом дерматита и нефропатии свиней (PDNS), а также с репродуктивными расстройствами свиней (1, 4).
Частица вириона имеет диаметр приблизительно 19 нм, а размер генома PCV составляет от 1,7 т.п.н. до 2,3 т.п.н. Циклическая природа генома привела к возникновению семейства вирусов circovirus. Были изучена история эволюции, и она позволила подробно описать филогенетические деревья и вариации в структуре поверхности капсида (5-8). Первоначальная реконструкция крио-ЭМ изображений нескольких нативных цирковирусов продемонстрировала, что капсид имеет икосаэдрическую симметрию Т=1 (9). Геном PCV кодирует один структурный капсидный белок (CP). Экспрессия и очистка CP белка PCV2b из E. coli продемонстрировали самосборку и имитацию общей морфологии для 70 инфекционных вирусов. Кристаллическая структура этой вирусоподобной частицы (VLP) визуализировала укладку CP как укладку в виде канонического вирусного «желейного рулета», состоящего из двух четырехнитевых β-листов (10, 11). Петли, соединяющие р-нити, образуют элементы на поверхности вируса и могут включать антигенные эпитопы. CP PCV2b также экспрессировали и очищали как VLP из клеток насекомых Trichoplasia ni (11). Реконструкция крио-ЭМ изображения этой VLP продемонстрировала, что N-конец расположен внутри капсида, и авторы пришли к выводу, что антигенные свойства, связанные с N-концом, вероятно, являются результатом того, что N-конец подвергается временной экстернализации из капсида посредством процесса, называемого вирусным «дыханием» (11-13). Экстернализация N-конца может играть важную роль в жизненном цикле вируса.
Аминокислотные последовательности записей PCV2 в GenBank были разделены на четыре различных генотипа (PCVa-d) (14). PCV2a был доминирующим глобальным генотипом до 2003 г., когда наблюдался сдвиг генотипа на PCV2b (15, 16). Генотип PCV2c, возможно, вымер, поскольку в GenBank присутствует всего три записи депонирования. В 2013 году Wei et al. сообщили и депонировали большое количество последовательностей PCV2d в GenBank (5). По результатам дополнительных отчетов и депонирования последовательностей PCV2d присутствует приблизительно 320 депонирований в GenBank (7). Увеличение количества депонирований последовательностей PCV2d может быть результатом того, что PCV2d становится новым и преобладающим генотипом в Азии, Европе, Северной и Южной Америке. Увеличение может быть результатом наличия мутантов, «ускользнувших» от вакцинации (17). Следовательно, PCV2d может представлять собой второй глобальный сдвиг генотипа, который может быть невосприимчивым к действующей в настоящее время программе вакцинации против PCV2 (7). Несмотря на значительное количество филогенетических исследований генотипов PCV2 и растущее значение PCV2d в мировой индустрии свиноводства, нет существует отчетов, описывающих структуру капсида PCV2d.
С появлением PCV2d необходимо более глубокое понимание генотипов PCV2 и их различий для разработки методов лечения заболеваний, вызываемых этими типами вирусов. Эти и другие проблемы решаются в настоящей заявке.
3. КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Согласно первому аспекту предложена система экспрессии у млекопитающих для получения рекомбинантных вирусоподобных частиц (VLP) цирковируса свиней типа 2 (PCV2). Система экспрессии включает клетку млекопитающего и плазмиду, содержащую ген PCV2, кодирующий капсидный белок. Ген PCV2 оптимизирован по кодонам, и клетка млекопитающего трансфицируется плазмидой. Система экспрессии продуцирует рекомбинантные VLP PCV2.
Согласно другому аспекту клетка млекопитающего представляет собой клетку эмбриональной почки человека -293 (HEK-293).
Согласно другому аспекту ген PCV2 включает сайт распознавания NheI, последовательность Козака и сайт распознавания NotI. Сайт распознавания NheI и последовательность Козака расположены выше стартового кодона гена PCV2, и сайт распознавания NotI встроен после кодона терминации гена PCV2.
Согласно другому аспекту большинство продуцируемых рекомбинантных VLP PCV2 присутствует в ядре клеток млекопитающего.
Согласно другому аспекту капсидный белок содержит аминокислотную последовательность SEQ ID NO: 2. Согласно другому аспекту капсидный белок кодируется нуклеотидной последовательностью SEQ ID NO: 1.
Согласно другому аспекту капсидный белок модифицирован сигнальной последовательностью секреции, введенной на NH2-конце капсидного белка. Согласно дополнительному аспекту капсидный белок содержит аминокислотную последовательность SEQ ID NO: 4. Согласно дополнительному аспекту капсидный белок кодируется нуклеотидной последовательностью SEQ ID NO: 3.
Согласно другому аспекту продуцируемые рекомбинантные VLP PCV2 выбраны из группы, состоящей из: VLP PCV2a, VLP PCV2b, VLP PCV2c, VLP PCV2d и VLP PCV2e. Согласно дополнительному аспекту продуцируемые рекомбинантные VLP PCV2 представляют собой VLP PVC2d.
Согласно другому аспекту плазмида представляет собой pcDNA3.4-PCV2. Согласно другому аспекту ген PCV2 оптимизирован по кодонам с использованием оптимизированной по кодонам аминокислотной последовательности, показанной на Фиг. 1А.
Согласно второму аспекту предложен способ получения вирусоподобных частиц (VLP) цирковируса свиней типа 2 (PCV2). В способе предложена суспензия культивируемых клеток млекопитающего. Клетки млекопитающего трансфицируют плазмидой, содержащей ген PCV2, кодирующий капсидный белок. Натриевую соль вальпроевой кислоты (VPA) добавляют к трансфицированным клеткам млекопитающего, и добавление натриевой соли VPA ингибирует пролиферацию клеток. Трансфицированные клетки млекопитающего центрифугируют и промывают. Затем центрифугированные клетки млекопитающего суспендируют в забуференном фосфатом физиологическом растворе (PBS). Для клеток млекопитающего осуществляют множественные циклы замораживания и оттаивания, а затем клетки млекопитающего обрабатывают ультразвуком в нескольких циклах. Затем проводят два последовательных цикла центрифугирования клеток млекопитающего для получения VLP PCV2, и большинство продуцируемых VLP PCV2 присутствует в ядре клеток млекопитающего.
Согласно другому аспекту способа стадия центрифугирования и промывки трансфицированных клеток млекопитающего включает: центрифугирование клеток млекопитающего при 2000×g в течение 15 мин,
промывку клеток млекопитающего раствором PBS и повторное центрифугирование клеток млекопитающего при 2000×g в течение 15 мин.
Согласно другому аспекту способа центрифугированные клетки млекопитающего замораживают при приблизительно -80°С и оттаивают при приблизительно 37°С во время циклов замораживания и оттаивания.
Согласно другому аспекту способа первый из двух последовательных циклов центрифугирования осуществляют при 2000×g в течение 15 минут, и второй из двух последовательных циклов центрифугирования осуществляют при 8000×g в течение 15 минут.
Согласно другому аспекту способа VLP PCV2 очищают ультрацентрифугированием.
Согласно другому аспекту способа клетки млекопитающего представляют собой клетки эмбриональной почки человека -293 (HEK-293).
Согласно другому аспекту способа ген PCV2 включает сайт распознавания NheI, последовательность Козака и сайт распознавания NotI. Сайт распознавания NheI и последовательность Козака расположены выше стартового кодона гена PCV2, и сайт распознавания NotI встроен после кодона терминации гена PCV2.
Согласно другому аспекту способа плазмида представляет собой pcDNA3.4-PCV2.
Согласно другому аспекту способа продуцируемые VLP PCV2 выбраны из группы, состоящей из: VLP PCV2a, VLP PCV2b, VLP PCV2c, VLP PCV2d и VLP PCV2e. Согласно дополнительному аспекту продуцируемые VLP PCV2 представляют собой VLP PVC2d.
Согласно третьему аспекту предложена VLP PCV2, полученная вышеуказанным способом.
4. КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг. 1А-1D. Экспрессия вирусоподобных частиц PCV2 в клетках млекопитающего. Фиг. 1А) Плазмида, созданная для экспрессии 404 капсидного белка PCV2d. Ген капсида PCV2d, оптимизированный по кодонам, синтезировали (Blue Heron Technologies, Bothell, WA) и клонировали в вектор экспрессии pcDNA3.4 (Fisher Scientific). Фиг. 1B) Экспрессию белка проводили во временно трансфицированных суспензионных культурах клеток Expi293 (Life Technologies). SDS-PAGE анализ очищенных VLP PCV2d (1 мкг белка) и окрашенных кумасси голубым. Фиг. 1С) SDS-PAGE анализ очищенных VLP PCV2d (0,5 мкг белка), перенесенных на нитроцеллюлозную мембрану и исследованных вестерн-блоттингом с помощью первичных кроличьих поликлональных капсидных антител против PCV2 (Cab 183908, Abeam, UK). Фиг. ID) Негативная электронно-микроскопическая микрофотография очищенных VLP, окрашенных уранилацетатом. Размеры частиц составляют примерно 19 нм в диаметре.
Фиг. 2А-2С. Структурное исследование VLP PCV2d. Фиг. 2А) Икосаэдрическая крио-ЭМ реконструкция изображения очищенных VLP PCV2d, окрашенная в соответствии с локальным разрешением. Градиентная цветная карта с левой стороны указывает разрешение для цветов. Фиг. 2В) Ленточный рисунок с координатами атомов с одной асимметричной единицей, окрашенной в голубой цвет. Фиг. 2С) Структурное наложение PCV2a (темно-синий), PCV2b (голубой) и PCV2d (желтый). Петли помечены в соответствии с соединяемыми ими р-нитями. Фигуры созданы с использованием UCSF ChimeraX (47).
Фиг. 3А-3D. Сравнение последовательностей 1377 записей капсидного белка PCV2, которые нанесены на координаты атомов PCV2d. Фиг. 3А) Выравнивание последовательностей сервером Clustal Omega было использовано для создания диаграммы WebLogo для демонстрации вариации последовательностей. Горизонтальная ось выравнивания указывает аминокислоту, а вертикальная ось указывает ее наблюдаемую частоту. Столбцы, соединяющие аминокислоты 77, 80, 99, 91, 190, 19 (черный) и 53, 215 (серый), представляют собой эволюционно связанные кластеры, показанные на панели D. Фиг. 3В) Модель заполнения пространства координат атомов PCV2d с модифицированной схемой цветового кодирования ConSurf. Цветная полоса внизу показывает степень консервативности, определенную сервером ConSurf. Желтый прямоугольник указывает на недостаточность данных, определенных сервером (1 указывает на плохую консервативность, а 9 указывает на высококонсервативные мутации). Верхний правый квадрант поверхности VLP был удален, чтобы показать консервативность последовательности внутри капсида. Изображение сделано с помощью UCSF ChimeraX и окрашено с использованием подсвечивания поверхности. Фиг. 3С) Участки высококонсервативных аминокислот на поверхности капсида (аминокислоты 82, 170, 188, 189 и 193 - зеленым, 55, 56, 51 и 73 - синим). Антитела, направленные против этих остатков, могут обладать широкой нейтрализующей способностью. Фиг. 3D) Ленточный рисунок субъединицы PCV2d. Остатки у больных эволюционно связаны друг с другом, как было определено с помощью plmc. MATLAB 2019 и программы EVzoom. Фигуры созданы с применением UCSF ChimeraX (47).
Фиг. 4А-4Н. Сайты нейтрализации антител. Вверху) Модель заполнения пространства атомных координат PCV2d с открытыми на поверхности аминокислотами, окрашенными в голубой цвет, аминокислотами с вариабельными последовательностями в фиолетовый цвет и аминокислотами, связывающими антитело, в желтый цвет. В центре) диаграмма WebLogo из 11 аминокислот, содержащих аминокислоты, подверженные воздействию растворителя. Внизу) Аминокислоты на поверхности капсида.
Фиг. 5. Изменение массы тела во время фазы исследования для каждой группы обработки: Т01 (PIGONE 1mL), Т02 (PIGONE 2mL), Т03_A (Huve-PCV2_А), Т03_В (Huve-PCV2_В), Т04 (HVP-DNA), Т05 (CIRCOFLEX [инактивированный бакулоэкспрессируемый ORf2 PCV2]) и Т06 (PBS). Ось Y представляет массу тела в кг, а ось X - дни исследования от SD0 до SD63.
Фиг. 6. Среднесуточный прирост массы тела во время периодов исследования для каждой группы обработки: Т01 (PIGONE 1mL), Т02 (PIGONE 2mL), Т03_A (Huve-PCV2_A), Т03 В (Huve-PCV2_В), T04 (HVP-DNA), T05 (CIRCOFLEX (Инактивированный бакулоэкспрессируемый ORf2 PCV2) и T06 (PBS). Ось Y представляет прибавку в массе тела в граммах, а ось X - периоды исследования SD0-SD35; SD35-SD63; SD0-SD63.
Фиг. 7. Динамика изменения серологии во время фазы исследования для каждой группы обработки: Т01 (PIGONE 1mL), Т02 (PIGONE 2mL), T03_A (Huve-PCV2_A), T03_B (Huve-PCV2_B), T04 (HVP-DNA), T05 (CIRCOFLEX [инактивированный бакулоэкспрессируемый PCV2 ORf2]) и T06 (PBS). Ось Y представляет значение S/P, а ось X - дни исследования от SD0 до SD63.
Фиг. 8. Процент поддающихся количественной оценке положительных и не поддающихся количественной оценке положительных животных во время фазы заражения для каждой группы обработки: Т01 (PIGONE 1mL), Т02 (PIGONE 2mL), T03_A (Huve-PCV2_A), Т03_В (Huve-PCV2_В), Т04 (HVP-DNA), Т05 (CIRCOFLEX (инактивированный бакулоэкспрессируемый ORf2 PCV2)) и Т06 (PBS). Ось Y представляет процент животных, а ось X - дни исследования от SD35 до SD63.
Фиг. 9. Средние вирусные нагрузки у поддающихся количественной оценке положительных животных (>4log10 копий/мл) во время фазы заражения для каждой группы обработки: Т01 (PIGONE 1mL), Т02 (PIGONE 2mL), Т03_A (Huve-PCV2_A), Т03_В (Huve-PCV2_В), T04 (HVP-DNA), T05 (CIRCOFLEX (инактивированный бакулоэкспрессируемый ORf2 PCV2)) и T06 (PBS). Ось Y представляет вирусную нагрузку, а ось X - дни исследования от SD41 до SD63.
Фиг. 10. Процент поддающихся количественной оценке положительных, не поддающихся количественной оценке положительных и ниже предела обнаружения животных во время фазы заражения для каждой группы обработки: Т01 (PIGONE 1mL), Т02 (PIGONE 2mL), Т03_A (Huve-PCV2_A), T03_B (Huve-PCV2_B), T04 (HVP-DNA), T05 (CIRCOFLEX (инактивированный бакулоэкспрессируемый ORf2 PCV2)) и T06 (PBS). Ось Y представляет процент животных, а ось X - дни исследования от SD35 до SD63.
Фиг. 11. Средние вирусные нагрузки всех животных (>2log10 копий/мл) во время фазы заражения для каждой
группы обработки: Т01 (PIGONE 1mL), Т02 (PIGONE 2mL), T03_A (Huve-PCV2_A), Т03_В (Huve-PCV2_В), Т04 (HVP-DNA), Т05 (CIRCOFLEX (инактивированный бакулоэкспрессируемый ORf2 PCV2)) и Т06 (PBS). Ось Y представляет вирусную нагрузку, а ось X - дни исследования от SD35 до SD63.
5. ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
На практике различных вариантов реализации настоящей заявки можно использовать, если не указано иное, обычные способы химии, биохимии, молекулярной биологии, иммунологии и фармакологии в пределах квалификации в данной области техники. Такие способы полностью объяснены в литературе. См., например, Remington's Pharmaceutical Sciences, 18th Edition (Easton, Pennsylvania: Mack Publishing Company, 1990); Methods In Enzymology (S. Colowick and N. Kaplan, eds., Academic Press, Inc.); и Handbook of Experimental Immunology, Vols. I-IV (D.M. Weir and С.C. Blackwell, eds., 1986, Blackwell Scientific Publications); Sambrook, et al., Molecular Cloning: A Laboratory Manual (2nd Edition, 1989); Short Protocols in Molecular Biology, 4th ed. (Ausubel et al. eds., 1999, John Wiley & Sons); Molecular Biology Techniques: An Intensive Laboratory Course, (Ream et al., eds., 1998, Academic Press); PCR (Introduction to Biotechniques Series), 2nd ed. (Newton & Graham eds., 1997, Springer Verlag); Fundamental Virology, Second Edition (Fields & Knipe eds., 1991, Raven Press, New York).
Все цитируемые в настоящем документе публикации, патенты и заявки на патенты включены в настоящее описание в качестве ссылки во всей полноте.
В настоящем описании и прилагаемой формуле изобретения, формы единственного числа включают ссылки на множественное число, если содержание явно не указывает иное. Таким образом, например, ссылка на «VLP» может включать смесь двух или более таких VLP.
Определения
Используемый в настоящем документе термин «вирусоподобная частица» или «VLP» относится к нереплицирующейся вирусной оболочке. VLP обычно состоят из одного или более вирусных белков, таких как, но не ограничиваясь ими, те белки, которые называются капсидными, оболочечными (coat), оболочечными (shell), поверхностными белками и/или белками оболочки, или образующих частицы полипептидов, полученных из этих белков. VLP могут образовываться спонтанно при экспрессии рекомбинантного белка в соответствующей системе экспрессии. Способы получения конкретных VLP обсуждаются более подробно ниже. Присутствие VLP после экспрессии рекомбинантных вирусных белков может быть обнаружено с использованием обычных методов, известных в данной области техники, таких как электронная микроскопия, биофизическая характеризация и тому подобное. См., например, Baker et al., Biophys. J. (1991) 60:1445-1456; Hagensee et al., J. Virol (1994) 68:4503-4505. Например, VLP могут быть выделены центрифугированием в градиенте плотности и/или идентифицированы по характерным полосам плотности (например, Примеры). В качестве альтернативы, криоэлектронная микроскопия может быть проведена на витрифицированных водных образцах рассматриваемого препарата VLP, а изображения записаны при соответствующих условиях воздействия. Дополнительные способы очистки VLP включают, но не ограничиваются ими, хроматографические методы, такие как методики на основе аффинности, ионного обмена, исключения по размеру и с применением обращенной фазы.
Используемый в настоящем документе термин «гибридный» или «химерный» относится к молекуле (например, белку или VLP), которая содержит части, по меньшей мере, из двух разных белков. Очевидно, что описанная в настоящем документе гибридная или химерная молекула может включать полноразмерные белки, слитые с дополнительными гетерологичными полипептидами (полная длина или их части), а также части белков, слитые с дополнительными гетерологичными полипептидами (полная длина или их части). Также будет очевидно, что гибридная или химерная молекула может включать последовательности дикого типа или мутантные последовательности в любом одном, некоторых или всех гетерологичных доменах.
«Антиген» относится к молекуле, содержащей один или более эпитопов (т.е. «антигенные эпитопы»), линейный, конформационный или оба, которые будут стимулировать иммунную систему хозяина для выработки гуморального и/или клеточного антиген-специфического ответа. В одном или более вариантах реализации эпитоп будет включать примерно от 7 до 15 аминокислот, например, 9, 10, 12 или 15 аминокислот. Термин включает полипептиды, которые включают модификации, такие как делеции, добавления и замены (обычно консервативные по природе) по сравнению с нативной последовательностью, при условии, что белок сохраняет способность вызывать иммунологический ответ, как определено в настоящем документе. Эти модификации могут быть намеренными, например, посредством сайт-направленного мутагенеза, или могут быть случайными, например, посредством мутаций хозяев, продуцирующих антигены.
«Иммуногенная композиция» представляет собой композицию, которая содержит антигенную молекулу, когда введение композиции субъекту приводит к развитию у субъекта гуморального и/или клеточного иммунного ответа на представляющую интерес антигенную молекулу.
«Очищенный» или «очистка» в целом относится к выделению вещества (соединения, полинуклеотида, белка, полипептида, полипептидной композиции) таким образом, что вещество составляет большую процентную часть образца, в котором оно находится. Обычно в образце «очищенный» компонент составляет 50%, предпочтительно 80-85% или более предпочтительно 90-95% образца. Способы очистки представляющих интерес полинуклеотидов и полипептидов хорошо известны в данной области техники и включают, например, ионообменную хроматографию, аффинную хроматографию и седиментацию в зависимости от плотности.
«Кодирующая последовательность» или последовательность, которая «кодирует» выбранный полипептид, представляет собой молекулу нуклеиновой кислоты, которая транскрибируется (в случае ДНК) и транслируется (в случае мРНК) в полипептид in vivo при помещении под контроль соответствующих регуляторных последовательностей (или «контрольных элементов»). Границы кодирующей последовательности определяются стартовым кодоном на 5' (амино) конце и стоп-кодоном трансляции на 3'(карбокси) конце. Кодирующая последовательность может включать, без ограничения, кДНК из вирусной, прокариотической или эукариотической мРНК, последовательности геномной ДНК из вирусной или прокариотической ДНК и даже синтетические последовательности ДНК. Последовательность терминации транскрипции может располагаться с 3' конца относительно кодирующей последовательности. Типичные «контрольные элементы» включают, но не ограничиваются ими, промоторы транскрипции, энхансерные элементы транскрипции, сигналы терминации транскрипции, последовательности полиаденилирования (расположенные с 3' конца от стоп-кодона трансляции), последовательности для оптимизации инициации трансляции (расположенные с 5' конца от кодирующей последовательности), и последовательности терминации трансляции, и/или элементы последовательности, контролирующие открытую структуру хроматина, см., например, McCaughan et al. (1995) PNAS USA 92:5431-5435; Kochetov et al. (1998) FEBS Letts. 440: 351-355.
Молекула «нуклеиновой кислоты» может включать, без ограничения, прокариотические последовательности, эукариотическую мРНК, кДНК из эукариотической мРНК, геномные последовательности ДНК из ДНК эукариот (например, млекопитающих) и даже синтетические последовательности ДНК. Термин также охватывает последовательности, которые включают любые известные аналоги оснований ДНК и РНК.
Термин «рекомбинантный», используемый в настоящем документе для описания молекулы нуклеиновой кислоты, означает полинуклеотид геномного, кДНК, полусинтетического или синтетического происхождения, который в силу своего происхождения или манипуляции: (1) не связан со всем или частью полинуклеотида, с которым он связан в природе; и/или (2) связан с полинуклеотидом, отличным от того, с которым он связан в природе. Термин «рекомбинантный», используемый в отношении белка или полипептида, означает полипептид, продуцируемый путем экспрессии рекомбинантного полинуклеотида. «Рекомбинантные клетки-хозяева», «клетки-хозяева», «клетки», «линии клеток», «культуры клеток» и другие подобные термины, обозначающие прокариотические микроорганизмы или линии эукариотических клеток, культивируемые как одноклеточные единицы, используются взаимозаменяемо и относятся к клеткам, которые могут использоваться или использовались в качестве реципиентов для рекомбинантных векторов или другой переносимой ДНК, и могут включать потомство исходной клетки, которая была трансфицирована. Понятно, что потомство единственной родительской клетки не обязательно может быть полностью идентичным по морфологии или геномной или полной ДНК-комплементарности исходному родителю вследствие случайной или намеренной мутации. Потомство родительской клетки, которое являются в достаточной степени сходным с родительской клеткой, чтобы характеризоваться соответствующим свойством, таким как наличие нуклеотидной последовательности, кодирующей желаемый пептид, включается в потомство, подразумеваемое этим определением, и охватывается вышеуказанными терминами.
«Оптимизация кодонов» или «оптимизированный кодон» обычно относится к подходам генной инженерии, которые используют синонимичные замены кодонов для увеличения продукции белка.
Методы определения «сходства» аминокислотных последовательностей хорошо известны в данной области техники. В общем, «сходство» означает точное сравнение аминокислоты с аминокислотой двух или более полипептидов в соответствующем месте, где аминокислоты идентичны или обладают аналогичными химическими и/или физическими свойствами, такими как заряд или гидрофобность. Затем можно определить так называемое «процентное сходство» между сравниваемыми полипептидными последовательностями. Методы определения идентичности последовательности нуклеиновой кислоты и аминокислотной последовательности также хорошо известны в данной области техники и включают определение нуклеотидной последовательности мРНК для этого гена (обычно через промежуточную кДНК) и определение аминокислотной последовательности, кодируемой им, и сравнение ее с второй аминокислотной последовательностью. В общем, «идентичность» относится к точному соответствию нуклеотида с нуклеотидом или аминокислоты с аминокислотой двух полинуклеотидных или полипептидных последовательностей соответственно.
Две или более полинуклеотидных последовательностей можно сравнить, определив их «процент идентичности». Две или более аминокислотных последовательностей аналогичным образом можно сравнить, определив их «процент идентичности». Процент идентичности двух последовательностей, будь то последовательности нуклеиновой кислоты или пептида, обычно описывается как количество точных совпадений между двумя выровненными последовательностями, деленное на длину более короткой последовательности и умноженное на 100. Приблизительное выравнивание последовательностей нуклеиновых кислот обеспечивается алгоритмом локальной гомологии согласно Smith and Waterman, Advances in Applied Mathematics 2:482-489 (1981). Этот алгоритм может быть расширен для использования с пептидными последовательностями с использованием оценочной матрицы, разработанной Dayhoff, Atlas of Protein Sequences and Structure, M.O. Dayhoff ed., 5 suppl. 3:353-358, National Biomedical Research Foundation, Washington, D.C., USA, и нормированной no Gribskov, Nucl. Acids Res. 14(6):6745-6763 (1986). Подходящие программы для расчета процента идентичности или сходства между последовательностями обычно известны в данной области техники.
«Вектор» способен переносить последовательности генов в клетки-мишени (например, бактериальные плазмидные векторы, вирусные векторы, невирусные векторы, носители в виде частиц и липосомы). Обычно «векторная конструкция», «вектор экспрессии» и «вектор для переноса гена» означают любую конструкцию нуклеиновой кислоты, способную управлять экспрессией одной или более представляющих интерес последовательностей в клетке-хозяине. Таким образом, термин включает носители для клонирования и экспрессии, а также вирусные векторы. Термин используется взаимозаменяемо с терминами «вектор экспрессии нуклеиновой кислоты» и «кассета экспрессии».
Используемый в настоящем документе термин «субъект» обычно относится к любому представителю подтипа хордовых, включая, без ограничения, людей и других приматов, включая не являющихся человеком приматов, таких как шимпанзе и другие виды человекообразных обезьян и обезьян; сельскохозяйственных животных, таких как крупный рогатый скот, овцы, свиньи, козы и лошади; домашних млекопитающих, таких как собаки и кошки; лабораторных животных, включая грызунов, таких как мыши, крысы и морские свинки; птиц, включая домашних, диких и промысловых птиц, таких как куры, индейки и другие куриные, утки, гуси и т.п. Термин не обозначает конкретный возраст. Таким образом, предполагается охватить как взрослых, так и новорожденных. Настоящие системы, описанные в данном документе, предназначены для использования у любого из вышеупомянутых видов позвоночных, поскольку иммунные системы всех этих позвоночных функцирнируют одинаково.
Под «фармацевтически приемлемым» или «фармакологически приемлемым» подразумевается материал, который не является нежелательным с биологической или иной точки зрения, т.е. материал можно вводить индивидууму в составе или композиции без каких-либо неприемлемых биологических эффектов или вредного взаимодействия с любым из компонентов состава, в котором он содержится.
В настоящем документе «лечение» относится к любому из (i) предотвращения инфекции или повторной инфекции, как в случае традиционной вакцины, (ii) уменьшения или устранения симптомов и (iii) существенного или полного устранения рассматриваемого патогена. Лечение можно проводить профилактически (до заражения) или терапевтически (после заражения).
Используемый в настоящем документе термин «адъювант» относится к соединению, которое при использовании в комбинации со специфическим иммуногеном (например, VLP) в составе будет усиливать или иным образом изменять или модифицировать полученный в результате иммунный ответ. Модификация иммунного ответа включает усиление или расширение специфичности одного или обоих антительного и клеточного иммунного ответов. Модификация иммунного ответа также может означать снижение или подавление определенных антиген-специфических иммунных ответов.
Используемый в настоящем документе термин «эффективная доза» обычно относится к тому количеству VLP одного или более вариантов реализации настоящей заявки, которое является достаточным для индукции иммунитета, предотвращения и/или облегчения инфекции или уменьшения по меньшей мере одного симптома инфекции и/или для повышения эффективности другой дозы VLP. Эффективная доза может относиться к количеству VLP, достаточному для отсрочки или минимизации начала инфекции. Эффективная доза может также относиться к количеству VLP, которое обеспечивает терапевтический эффект при лечении или контроле инфекции. Кроме того, эффективная доза представляет собой количество по отношению к VLP в одном или более вариантах реализации настоящей заявки отдельно или в комбинации с другими видами лечения, которое обеспечивает терапевтический эффект при лечении или контроле инфекции. Эффективная доза также может представлять собой количество, достаточное для усиления собственного иммунного ответа субъекта (например, человека) против последующего воздействия инфекционного агента. Уровни иммунитета можно контролировать, например, путем измерения количества нейтрализующих секреторных и/или сывороточных антител, например, путем нейтрализации бляшек, фиксации комплемента, иммуноферментного связывания или анализа микронейтрализации. В случае вакцины «эффективная доза» представляет собой дозу, которая предотвращает заболевание и/или снижает тяжесть симптомов.
Используемый в настоящем документе термин «эффективное количество» относится к количеству VLP, необходимому или достаточному для реализации желаемого биологического эффекта. Эффективным количеством композиции будет количество, при котором достигается выбранный результат, и такое количество может быть определено специалистом в рамках обычного эксперимента. Например, эффективное количество для предотвращения, лечения и/или облегчения инфекции может представлять собой такое количество, которое необходимо для активации иммунной системы, приводящей к развитию антигенспецифического иммунного ответа при воздействии на VLP согласно настоящей заявке. Этот термин также является синонимом термина «достаточное количество».
Используемый в настоящем документе термин «поливалентный» относится к VLP, которые содержат несколько антигенных белков против нескольких типов или штаммов инфекционных агентов.
Используемый в настоящем документе термин «защитный иммунный ответ» или «защитный ответ» относится к иммунному ответу, опосредованному антителами против инфекционного агента, который проявляется у позвоночного (например, человека), который предотвращает или облегчает инфекцию или снижает хотя бы один его симптом. VLP согласно настоящей заявке могут стимулировать продукцию антител, которые, например, нейтрализуют инфекционные агенты, блокируют проникновение инфекционных агентов в клетки, блокируют репликацию указанных инфекционных агентов и/или защищают клетки-хозяева от инфекции и разрушения. Термин также может относиться к иммунному ответу, опосредованному Т-лимфоцитами и/или другими лейкоцитами, против инфекционного агента, проявляемому позвоночным (например, человеком), который предотвращает или облегчает инфекцию гриппа или снижает по меньшей мере один его симптом.
Используемый в настоящем документе термин «вакцина» относится к составу, который содержит VLP согласно настоящей заявке, который находится в форме, которую можно вводить позвоночному, и который индуцирует защитный иммунный ответ, достаточный для индукции иммунитета для предотвращения и/или облегчения инфекции, и/или уменьшения по меньшей мере одного симптома инфекции и/или повышения эффективности другой дозы VLP. Обычно вакцина содержит среду обычного физиологического раствора или буферного водного раствора, в котором композиция (композиции) согласно настоящей заявке суспендирована или растворена. В этой форме композицию (композиции) согласно настоящей заявке можно удобно применять для предотвращения, ослабления или иного лечения инфекции. При введении хозяину вакцина способна вызывать иммунный ответ, включая, помимо прочего, продукцию антител и/или цитокинов и/или активацию цитотоксических Т-клеток, антигенпредставляющих клеток, Т-хелперных клеток, дендритных клеток и/или другие клеточные ответы.
В одном или более вариантах реализации настоящая заявка относится к композициям, содержащим вирусы семейства Circoviridae [например, цирковирус-2 свиней (PCV2), его различные генотипы и серотипы или другие члены этого семейства], вирусоподобные частицы (VLP), и к способам получения и применения этих VLP, включая создание и получение вакцин на основе вирусоподобных частиц (VLP) (например, моновалентных, поливалентных, состоящих из частиц одного вида универсальных или поливалентных, состоящих из частиц одного вида мозаичных или модифицированных химерных композиций), а также их применению для терапевтической доставки [(например, небольших молекул, нуклеиновых кислот, антител, ферментов (наноносителей, нанотел)] для диагностики, иммуномодулирующих функций и в случае терапевтических показаний. В частности, настоящее описание включает стратегии и способы, используемые для разработки новых моновалентных, поливалентных или универсальных вакцин против цирковируса свиней, которые способны защищать свиней от заражения одним или более серотипами, кладами или антигенными вариантами рода цирковирусов свиней. В настоящем документе также описаны способы получения VLP (например, системы секреции), которые продуцируют VLP, которые проявляют определенные антигенные конфигурации или модификации. Эти VLP содержат конформационные нативные или химерные эпитопы, необходимые для генерации усиленного нейтрализующего иммунного ответа на цирковирус свиней или другие вирусные агенты. Состоящие из частиц одного вида моновалентные, двухвалентные, поливалентные, универсальные или химерные (наприпер, различные серотипы и генотипы, такие как PCV2a, PCV2b, PCV2c, PCV2d и PCV2e) VLP подвергают сборке и применяют для приготовления вакцинных композиций, что позволяет осуществлять иммунизацию и последующую защиту против одного или более серотипов или антигенно отличных вирусов (например, серотипов из Азии, Европы или Северной Америки и т.д.) VLP с нативными, модифицированными или реконструированными мономерами капсида позволяют связывать/конъюгировать различные молекулярные единицы с внешней поверхностью частицы (малые или большие молекулярные единицы) или инкапсулировать такие молекулярные единицы в структуре частицы посредством разборки и повторной сборки VLP или альтернативных способов упаковки.
Кроме того, VLP также применяют для диагностики инфекции или по терапевтическим показаниям. Вакцины VLP могут быть получены в суспензионной культуре эукариотических клеток и сохранены в клетках или высвобождены в культуральную среду. После очистки, концентрирования и приготовления вакцину можно вводить любым подходящим путем, например, через слизистую оболочку или парентерально, и вызывать иммунный ответ, способный защищать от любого или всех серотипов цирковируса свиней, антигенных вариантов и т.д. Также предложены VLP, содержащие терапевтические средства, с иммуномодулирующими функциями и для диагностического применения.
Эти и другие аспекты предложенных композиций и способов ниже описаны более подробно со ссылкой на прилагаемые фигуры и примеры, на которых показаны один или более проиллюстрированных вариантов реализации и/или расположения VLP PCV2. Композиции и способы согласно настоящей заявке никоим образом не ограничиваются проиллюстрированными вариантами реализации и/или расположения. Следует понимать, что композиции и способы, показанные на сопровождающих фигурах, являются просто примерами композиций и способов согласно настоящей заявке, которые могут быть воплощены в различных формах, как это понятно специалисту в данной области техники.
Согласно одному или более вариантам реализации настоящей заявки предложена система экспрессии у млекопитающих для получения рекомбинантных вирусоподобных частиц (VLP) цирковируса свиней типа 2 (PCV2). Система экспрессии включает клетку млекопитающего и плазмиду, содержащую ген PCV2, кодирующий капсидный белок. Ген PCV2 оптимизирован по кодонам, и клетка млекопитающего трансфицируется плазмидой. Система экспрессии продуцирует рекомбинантные VLP PCV2. Согласно по меньшей мере одному варианту реализации клетка млекопитающего представляет собой клетку эмбриональной почки человека -293 (HEK-293).
Согласно одному или более вариантам реализации ген PCV2 системы экспрессии может включать сайт распознавания NheI, последовательность Козака и сайт распознавания NotI. Сайт распознавания NheI и последовательность Козака расположены выше стартового кодона гена PCV2, и сайт распознавания NotI встроен после кодона терминации гена PCV2.
Согласно по меньшей мере одному варианту реализации большинство продуцируемых рекомбинантных VLP PCV2 присутствует в ядре клеток млекопитающего.
Согласно одному или более вариантам реализации капсидный белок содержит аминокислотную последовательность SEQ ID NO: 2. Согласно по меньшей мере одному варианту реализации капсидный белок кодируется нуклеотидной последовательностью SEQ ID NO: 1.
Согласно по меньшей мере одному варианту реализации капсидный белок модифицирован сигнальной последовательностью секреции, введенной на NH2-KOHue капсидного белка.. Согласно одному или более другим вариантам реализации капсидный белок содержит аминокислотную последовательность SEQ ID NO: 4. Согласно одному или более другим вариантам реализации капсидный белок кодируется нуклеотидной последовательностью SEQ ID NO: 3.
Согласно одному или более вариантам реализации продуцируемые рекомбинантные VLP PCV2 могут представлять собой по меньшей мере одну из следующих: VLP PCV2a, VLP PCV2b, VLP PCV2c, VLP PCV2d и VLP PCV2e.
Согласно предпочтительному варианту реализации продуцируемые рекомбинантные VLP PCV2 представляют собой VLP PVC2d.
Согласно по меньшей мере одному варианту реализации плазмида представляет собой pcDNA3.4-PCV2. Согласно одному или более вариантам реализации ген PCV2 оптимизирован по кодонам с использованием оптимизированной по кодонам аминокислотной последовательности, показанной на Фиг. 1А.
Согласно одному или более вариантам реализации настоящей заявки предложен способ получения вирусоподобных частиц (VLP) цирковируса свиней типа 2 (PCV2). В способе предложена суспензия культивируемых клеток млекопитающего. Клетки млекопитающего трансфицируют плазмидой, содержащей ген PCV2, кодирующий капсидный белок. Натриевую соль вальпроевой кислоты (VPA) добавляют к трансфицированным клеткам млекопитающего, и добавление натриевой соли VPA ингибирует пролиферацию клеток. Трансфицированные клетки млекопитающего центрифугируют и промывают. Затем центрифугированные клетки млекопитающего суспендируют в забуференном фосфатом физиологическом растворе (PBS). Для клеток млекопитающего осуществляют по меньшей мере один цикл замораживания и оттаивания, и предпочтительно множество циклов замораживания и оттаивания, а затем клетки млекопитающего обрабатывают ультразвуком в нескольких циклах. Согласно одному или более вариантам реализации затем осуществляют два последовательных цикла центрифугирования клеток млекопитающего для получения VLP PCV2. Согласно по меньшей мере одному варианту реализации большинство продуцируемых VLP PCV2 присутствует в ядре клеток млекопитающего.
Согласно по меньшей мере одному варианту реализации стадия центрифугирования и промывки трансфицированных клеток млекопитающего может содержать: центрифугирование клеток млекопитающего при 2000×g в течение 15 мин, промывку клеток млекопитающего раствором PBS и повторное центрифугирование клеток млекопитающего при 2000×g в течение 15 мин.
Согласно одному или более вариантам реализации центрифугированные клетки млекопитающего можно замораживать при приблизительно -80°С и оттаивать при приблизительно 37°С во время циклов замораживания и оттаивания.
Согласно по меньшей мере одному варианту реализации первый из двух последовательных циклов центрифугирования осуществляют при 2000×g в течение 15 минут, и второй из двух последовательных циклов центрифугирования осуществляют при 8000×g в течение 15 минут.
Согласно по меньшей мере одному варианту реализации VLP PCV2 очищают ультрацентрифугированием.
Согласно по меньшей мере одному варианту реализации плазмида представляет собой pcDNA3.4-PCV2. Согласно одному или более вариантам способа продуцируемые VLP PCV2 выбраны из группы, состоящей из: VLP PCV2a, VLP PCV2b, VLP PCV2c, VLP PCV2d и VLP PCV2e. Согласно по меньшей мере одному предпочтительному варианту реализации продуцируемые VLP PCV2 представляют собой VLP PVC2d. Согласно по меньшей мере одному варианту реализации предложена VLP PCV2, полученная вышеуказанным способом.
Таким образом, в соответствии с одним или более вариантами реализации в настоящей заявке раскрыта система сборки VLP PCV2 у млекопитающего (например, VLP PCV2d). В настоящей заявке дополнительно раскрыт структурный анализ, основанный на реконструкции с помощью криоэлектронной микроскопии, который выявляет внешние и внутренние особенности VLP, которые имеют важное значение для разработки новых вакцин против PCV-2 и использования частиц в качестве нанотранспортеров для доставки различных молекулярных единиц для профилактического, терапевтического, иммунотерапевтического применения, иммунизации (гетерологичной, гомологичной, поливалентной), диагностического применения из различных вариантов применения. Эти анализы и другие примеры, относящиеся к VLP PCV2, более подробно описаны ниже.
6. ПРИМЕРЫ
6.1. Пример 1
МАТЕРИАЛЫ И СПОСОБЫ
Клетки, ген капсида, плазмида и антитело. Суспензионные культуры клеток человека Expi293F (Life Technologies, СА) выращивали в бессывороточной экспрессионной среде Expi293 (Life Technologies, СА) при 37°С в среде 5% CO2 и перемешивали при 150 об/мин в колбах Эрленмейера. Ген цирковируса свиней типа 2 (PCV2), кодирующий капсидный белок (штамм RQ3), был химически синтезирован с использованием оптимизированной по кодонам последовательности (аминокислотная последовательность, показанная на Фиг. 1А) компанией Blue Heron Technologies (Bothell, WA). Сайт распознавания NheI и последовательность Козака были добавлены непосредственно перед стартовым кодоном, и сайт распознавания NotI был встроен после кодона терминации. Синтезированный ген Сар выделяли из транспортной плазмиды путем двойного расщепления рестрикционными ферментами NheI и 289 NotI и субклонировали после очистки в геле в экспрессионную плазмиду млекопитающих pcDNA3.4, разрезанную теми же ферментами. Сшитую плазмиду трансформировали в клетки MAX Efficiency Stbl2 (Life Technologies), и правильный клон идентифицировали с помощью анализа рестрикционными ферментами и проверяли путем секвенирования.
Получение и очистка вирусоподобных частиц (VLP). VLP PCV2 продуцировали в суспензионной культуре клеток млекопитающего Expi293F после временной трансфекции плазмидой pcDNA3.4-PCV2 (Фиг. 1A-1D). Клетки Expi293F высевали в концентрации 2×106 клеток/мл и культивировали в течение 16 часов перед трансфекцией. Плазмидную ДНК (1 мкг/мл) разводили в объеме Opti-MEM, составляющем 5% от общего объема культуры. Отдельно готовили полиэтиленимин (PEI) в эквивалентном объеме Opti-MEM (4 мкг/мл). После 5 мин инкубации при комнатной температуре раствор PEI по каплям добавляли в пробирку, содержащую ДНК, и после 30 мин инкубации при комнатной температуре смесь добавляли по каплям к суспензии клеток. Через 24 часа после трансфекции к культуре клеток добавляли натриевую соль вальпроевой кислоты (VPA) до конечной концентрации 3,75 мМ для ингибирования пролиферации клеток. Через 72 часа после трансфекции клетки осаждали центрифугированием при 2000×g в течение 15 минут, и затем промывали один раз фосфатно-солевым буфером (PBS) и снова центрифугировали при 2000×g в течение 15 минут. Осадок клеток ресуспендировали в PBS и затем подвергали трем циклам замораживания (-80°С) и оттаивания (37°С). Впоследствии клетки дополнительно фрагментировали тремя циклами обработки ультразвуком и осветляли двумя последовательными центрифугированиями, сначала при 2000 × g в течение 15 минут, а затем при 8000 × g в течение 15 минут. Эта процедура высвободила часть VLP в цитоплазму, но большая часть частиц оказалась захваченной внутри ядра клетки (данные не показаны). VLP PCV2, содержащиеся в осветленном супернатанте, были дополнительно очищены ультрацентрифугированием в двухслойном градиенте плотности CsCl: нижний слой, 5 мл 1,4 г/мл CsCl и верхний слой, 10 мл 1,25 г/мл CsCl, оба приготовлены в 10 мМ Трис-HCl (рН 7,9). Образцы загружали в градиент и центрифугировали при 15°С в течение 4 часов при 140000 × g, используя ротор SW28 (Beckman Coulter, СА). VLP выглядели как непрозрачная полоса на границе слоев 1,25 и 1,4 г/мл CsCl, и были собраны путем прокалывания пробирки иглой 18G и шприцом. Собранный раствор смешивали с 37% CsCl в 10 мМ Трис-HCl (рН 7,9) до конечного объема 12 мл, а затем центрифугировали при 15°С в течение 16 часов при 155000×g, используя ротор SW 41Ti (Beckman Coulter, СА). VLP обнаруживали в нижней части пробирки и извлекали, как описано выше. Собранный материал VLP подвергали диализу против 10 мМ Трис-HCl рН 7,9 и 150 мМ NaCl при 10°С в течение ночи с использованием кассеты Slide-A-Lyzer. Очищенные VLP PCV2 концентрировали, и буфер заменяли на фосфатно-солевой буфер (PBS) с использованием центробежных фильтрующих устройств Amicon Utra-4 (Merck Millopore, MA). Образцы VLP PCV2 хранили в аликвотах по 50-100 мкл при -80°С.
Вестерн блоттинг и краситель Кумасси голубой. Очищенные VLP смешивали с загрузочным буфером, нагревали при 100°С в течение 5 минут и пропускали через 4-12% бис-трис-808-полиакриламидный гель (Life Technologies, СА). Количество загружаемых белков составляло 1 мкг для последующего окрашивания кумасси, и 0,5 мкг - для вестерн-блоттинга. После электрофоретического разделения гель окрашивали кумасси синим, или белки подвергали электропереносу на нитроцеллюлозную мембрану 0,45 мкм (Life Technologies LC2001). Затем мембрану блокировали 5% обезжиренным молоком в TBST (10 мМ Трис-HCl, 130 мМ NaCl и 0,05% Твин-20, рН 7,4) в течение 1 часа при (20°С) с последующей инкубацией в течение ночи при (20°С) в первичных антителах кролика против цирковируса свиней (Cab 183908, Abeam, UK), разбавленных блокирующим буфером. Мембраны промывали 3 раза посредством TBST, а затем инкубировали в течение 2 часов со вторичными антителами (антитела козы к IgG кролика, конъюгированные с HRP 332, 1: 1000), разведенными в блокирующем буфере. Наконец, мембраны 3 раза промывали посредством TBST и проявляли с помощью системы вестерн-блоттинга ECL (Life Technologies, СА) в соответствии с инструкциями производителя. Изображения окрашенного геля и иммуноблоттинга получали с помощью прибора FluorChem Imager (Protein Simple, СА). 337
Негативное окрашивание и ТЕМ исследование. 5 мкл образцов pCV2-2 VLP наносили на углеродную пленку CF200-CU с медными сетками 200 меш (Electron Microscopy Science) на 1 мин, затем сетки промывали посредством 200 мкл 50 мМ буфера какодилата натрия, а затем сразу же окрашивали с помощью 50 мкл 0,5% уранилацетата в течение 1 мин. Сетки исследовали с помощью просвечивающего электронного микроскопа JOEL 2100, работающего при 200 кВ, с CCD-матрицей Orius 2048 × 2048 пикселей (Gatan Inc., Pleasanton, СА).
Сбор данных крио-ЭМ. Замороженные гидратированные образцы VLP PCV2d были приготовлены на медных сетках Quantifoil R 2/2, 200 меш (Electron Microscopy Science). Предварительно просеянный образец VLP объемом 4 мкл наносили на сетку, которую промокали в течение 3 секунд, и мгновенно замораживали в жидком этане с использованием прибора FEI Vitrobot. Сетки хранили в жидком азоте до сбора данных. Данные собирали при сверхнизких температурах на FEI Titan Krios, работающем при 300 кВ, с камерой Gatan К2 с квантовым энергетическим фильтром GIF с шириной 15 эВ. Сбор данных производили с помощью программного пакета Leginon (36).
Реконструкция изображения. Пакет MotionCor2 использовали для коррекции движения частиц (37). Для коррекции использовали параметры по умолчанию и взвешивание дозы, за исключением опции patch 5, и первый кадр каждой записи отбрасывали во время совмещения. Частицы были выбраны автоматически с помощью Gautomatch v0.53, а оценка функции передачи контраста (CTF) была выполнена на совмещенных микрофотографиях с использованием Gctf v0.50 (38). Relion 3.0 применяли для извлечения 23358 частиц при 300×300 пикселей из микрофотографий взвешенной дозы с использованием координат, определенных Gautomatch. Нереференсная двухмерная классификация была выполнена с помощью Relion 3.0. Опции по умолчанию для этой стадии включали диаметр 250 Å, и было запрошено 128 классов (39). Первоначальная модель была сгенерирована с разрешением 60 Å с использованием кристаллической структуры PCV2 (запись PDB 3R0R) с функцией molmap UCSF Chimera (40). Трехмерная классификация была проведена на 7196 частицах с использованием Relion 3.0 с диаметром 250 Å, 3 классами и симметрией С1. Одиночный класс с 4442 частицами показал самое высокое разрешение. Эти частицы использовали для реконструкции изображений с высоким разрешением с помощью Relion 3.0. Опять же диаметр 250 Å и симметрию 11 использовали с остальными параметрами по умолчанию Relion 3.0. Бинарная маска была создана с помощью программы relion_mask_create в Relion 3.0. Бинарная маска для последующей обработки была создана следующим образом: 1) реконструкция изображения с высоким разрешением была обработана фильтром низких частот до разрешения 15Å с использованием relion_image_handler (Relion 3.0), 2) наименьший порог внешнего шума капсида PCV2 был идентифицирован для этого объема с использованием UCSF Chimera, 3) relion_mask_create (Relion 3.0) использовали для преобразования этого объема в бинарную маску с идентифицированным порогом, расширением маски 7 пикселей и сглаживанием края 2 пикселя. Полученная маска была затем проверена с помощью UCSF Chimera, чтобы убедиться в отсутствии внутренних полостей.
Локальное разрешение рассчитывали с помощью программы MonoRes. Для расчета использовали ту же бинарную маску, которую использовали во время последующей обработки с применением Relion. Использовали диапазон разрешения от 2,8 до 6,0 Å (41).
Уточнение структуры 376.
Координаты атомов для кристаллической структуры PCV2b (запись PDB 3R0R) были модифицированы с помощью Coot (42). Биологические матрицы, необходимые для создания вирусоподобной частицы, присутствуют в PDB, и были использованы Coot для создания VLP PCV2d. UCSF Chimera использовали для ручной стыковки VLP с симметризованными реконструкциями изображений. Полученные координаты итерационно уточняли с использованием phenix.real_refine из программного пакета Phenix с применением ограничений некристаллографической симметрии (NCS) и ручной подгонкой с помощью Coot (43).
Выравнивание последовательностей и эволюционная связь
При поиске белка Blast с последовательностью PCV2d и фильтром по общему названию организма «Porcine circovirus 2» (taxid: 85708) получили в общей сложности 1966 последовательностей PCV2 (44). Частичные последовательности, последовательности с названиями, содержащими «предполагаемый», «Р3», «неизвестный» и «Р27.9», были удалены вручную. Выравнивание последовательностей выполняли для остальных последовательностей с помощью MUSCLE с применением параметров по умолчанию (45). Последовательности, которые генерировали пробелы, содержали более десяти аминокислот с отличающейся последовательностью или не имели последовательностей, подобных им, удаляли вручную. Это выполняли для того, чтобы удалить случайные ошибки/артефакты, которые могли возникнуть в процессе секвенирования (т.е. искусственная рекомбинация во время ПЦР с Taq-полимеразой (46)). Для удаления идентичных последовательностей было выполнено несколько циклов выравнивания и делеции. В результате осталось 1377 последовательностей. Последний цикл выравнивания проводили с помощью Clustal Omega с применением параметров по умолчанию (19). Расчеты эволюционной связи не могли быть успешно выполнены с сервером EVcouplings 394 (evfold.org) вследствие ограниченного диапазона в значении Expect (Е), представляющем последовательность (т.е. последовательности слишком похожи). Следовательно, plmc использовали для генерации коэволюции и ковариации внутри последовательностей. Лямбда L2 для используемых полей и связей составляла 0,01 и 16,0 соответственно, и было выполнено максимальное количество итераций, составляющее 100. Результаты были преобразованы для визуализации с помощью EVzoom с использованием MATLAB 2019 и скриптов, предоставленных программой plmc (22). Полученная матрица была визуализирована с помощью EVzoom (22), а структурная ковариация была визуально подтверждена с помощью UCSF Chimera (40).
РЕЗУЛЬТАТЫ
Рекомбинантные VLP PCV2, собранные в клетках млекопитающего. Ген капсидного белка (CP) PCV2d (GenBank: АВХ71783.1) был субклонирован в экспрессионную плазмиду pcDNA 3.4 млекопитающих для получения рекомбинантных VLP PCV2d в клетках млекопитающего (Фиг. 1А). Трансфекция суспензионной культуры клеток млекопитающего HEK-293 плазмидой приводила к продукции CP и сборке VLP. На сегодняшний день VLP PCV2 продуцировали с использованием клеток насекомых, дрожжей или E. coli (11), таким образом, экспрессия VLP с использованием клеток млекопитающего обеспечивает субстрат, аналогичный субстрату, используемому при естественной инфекции PCV2. Во время очистки было определено, что большая часть VLP или CP находилась в ядре клетки, в то время как часть была обнаружена в цитоплазме инфицированной клетки. Это предполагает, что VLP могут выходить из ядра, возможно, с помощью скрытого ядерного сигнала экспорта, который не может быть достоверно идентифицирован с помощью инструментов биоинформатики (данные не показаны). Анализ очищенных VLP PCV2 и их белкового состава был подтвержден посредством SDS-PAGE и вестерн-блоттингом (Фиг. 1В и С). Дальнейшее исследование с помощью электронной микроскопии с негативным окрашиванием показало однородные сферические частицы с гладкими краями, слегка шероховатую поверхность диаметром ~ 19 нм (Фиг. 1D). CP, описанный в этом исследовании, обладает аминокислотной последовательностью, идентифицированной из ряда недавно выделенных и зарегистрированных записей генома вируса PCV2d в GenBank, таких как С/2013/3, выделенный на Тайване (AWD32058.1), W233-12, выделенный в Японии (ВВЕ28610.1), CN-FJC011, выделенный в Китае (AVZ66019.1), и England/15-P0222-09-14, выделенный в Англии (AATN97185.1).
Криоэлектронная микроскопия и реконструкция изображений VLP PCV2d. Реконструкция икосаэдрического крио-ЭМ изображения очищенных VLP PCV2d была определена с разрешением 3,3 Å (Фиг. 2А). Тщательный анализ реконструкции изображения показывает, что молекулярные оболочки боковых цепей имеют достаточное качество для моделирования координат атомов. Координаты из кристаллической структуры PCV2b были вручную введены в реконструкцию изображения, и были выполнены соответствующие модификации, чтобы отразить аминокислотную последовательность PCV2d. Подобранная модель была уточнена с помощью нескольких итераций автоматического уточнения с помощью Phenix и ручной корректировки с помощью программы Coot (Фиг. 2В). Окончательная статистика уточнения приведена в Таблице 1 ниже.

Субъединицы структур генотипа PCV2a, b и d накладывали друг на друга и генерировали графики среднеквадратичных отклонений эквивалентных атомов Сα (9, 18). Области, которые демонстрируют наибольшее разнообразие, соответствуют двум из открытых петель, которые состоят из аминокислот 85-91 (петля CD) и 188-194 (петля GH) (Фиг. 2С).
Консервативность последовательностей четырех генотипов может помочь в создании универсальной вакцины против PCV 2. Был опубликован ряд публикаций, в которых аминокислотную последовательность CP PCV2 сравнивали для понимания эволюции капсида PCV2 (6, 7). Однако авторам настоящего изобретения не было известно об исследовании, в котором информация о выравнивании последовательностей была нанесена на карту атомных координат капсида, чтобы понять, как наблюдаемые мутации коррелируют со структурой или «ускользанием» от иммунитета. Всего 1377 неизбыточных последовательностей CP PCV2 подвергали выравниванию последовательностей с помощью сервера Clustal Omega (19). Выравнивание последовательностей использовали для создания диаграммы WebLogo для наблюдения консервативности последовательности (Фиг. 3А). Горизонтальная ось выравнивания указывает аминокислоту, а вертикальная ось указывает наблюдаемую частоту аминокислоты. Последовательности с наибольшим разнообразием (записи GenBank: AVZ65995.1 и ALK0432.1) имеют 74,4% идентичности последовательностей. Хотя диаграмма WebLogo успешно демонстрирует частоту встречаемости наиболее популярных аминокислот в каждом положении, она снижает/устраняет частоту встречаемости менее популярных аминокислот. Действительно, мутации можно наблюдать внутри гидрофобного ядра белка, на границе раздела между субъединицами, и в остатках на внешней и внутренней поверхности капсида. Более того, более тщательный анализ выровненных последовательностей показывает, что только Met1 и Arg147 абсолютно консервативны. Наблюдаемое разнообразие последовательностей предполагает, что капсид способен переносить мутации почти в каждом положении, оставаясь при этом инфекционным вирусом. Для визуализации консервативности/изменчивости последовательности в каждой аминокислотной позиции информация о выравнивании последовательностей была нанесена на атомные координаты PCV2d с использованием сервера ConSurf (Фиг. 3В) (20). Фигура показывает значительную степень неконсервативных мутаций, произошедших в капсиде. Области, окрашенные в красный цвет, претерпевают наибольшие изменения, в то время как области, отмеченные синим цветом, претерпевают наименьшие изменения. По всей структуре размещены высококонсервативные остатки; однако два набора остатков образуют пятна на поверхности капсида. К ним относятся Tyr55, Thr56, Met71 и Arg73, а также Pro82, Thr170, Gln188, Thr189 и Val193 (Фиг. 3С). Присутствие этих пятен предполагает, что можно создать вакцины, способные нейтрализовать все доступные генотипы PCV2, если продуцирование антител может быть направлено на эти аминокислоты (Фиг. 3С).
Эволюционно связанные мутации дифференцируют генотипы PCV-2. Большое количество депонированных последовательностей CP в GenBank позволило авторам настоящего изобретения задать вопрос, является ли какое-либо из положений аминокислот эволюционно ограниченным (21). Для эволюционно ограниченных аминокислот мутация одной аминокислоты требует мутации другой аминокислоты. В простейшем случае это может быть связано с тем, что аминокислоты упаковываются друг против друга в структуре белка, поэтому мутация до более крупной аминокислоты в первом положении требует мутации до более мелкой аминокислоты во втором положении для правильной упаковки. Такую информацию можно использовать для прогнозирования укладки белка или идентификации функционально важных сайтов (21-23). Измерения эволюционной связи (ЕС), полученные на основе 1377 уникальных последовательностей, показывают, что два независимых участка в структуре демонстрируют связь. Первый участок состоит из аминокислот 53 и 215, а второй участок состоит из аминокислот 77, 80, 89, 90, 190 и 191 (Фиг. 3С). Удивительно, но различия последовательностей во втором участке ответственны за структурное разнообразие, наблюдаемое между PCV2a и PCV2b/d для петель CD и GH (Фиг. 2С) (9, 18). Результаты ЕС могут указывать на то, что связанные остатки являются функционально значимыми. Расчеты доступной для растворителя площади поверхности с помощью сервера GetArea (curie.utmb.edu) показывают, что остатки 77, 80, 89 и 90 углублены более чем на 75%, в то время как остатки 190 и 191 подвергаются воздействию растворителя (24). Остатки 190 и 191 помогают определить нейтрализующий эпитоп на поверхности капсида.
Изменение последовательности на поверхности капсида является ответом на нейтрализующие антитела. Координаты атомов PCV2d были использованы для идентификации аминокислот на поверхности VLP с боковыми цепями, подвергнутыми воздействию растворителя (53, 55-56, 58-64, 70-71, 73, 75, 77-78, 82-83, 85, 88-89, 102, 113, 123, 127, 131-137, 148, 155, 156, 158, 161, 166, 168-170, 188-191, 194, 204, 206-208, 210, 229-234). Эти аминокислоты могут быть антигенными детерминантами, поскольку боковые цепи обеспечивают поверхность для взаимодействия антител. Непрерывные последовательности, которые могут представлять линейные эпитопы, показаны на Фиг. 4А-4Н. Затем были идентифицированы наиболее вариабельные аминокислоты в этих областях (53, 57, 59, 60, 63, 75, 77, 88, 89, 131, 133, 134, 136, 169, 190, 191, 211, 215 и 232.). Вариация может быть результатом «ускользания» мутантов от нейтрализации антител. Наличие мутаций в качестве реакции «ускользания» от нейтрализации антител подтверждается экспериментами по выявлению подтипов антигенов, проведенными Lefebvre et al. и Saha et al., где панели нейтрализующих моноклональных антител тестировали на их способность связываться с различными штаммами PCV2. Исследования показали, что положения аминокислот 59, 63, 88, 89, 130, 133, 206 и 210 отвечают за дифференциацию связывания антител (25, 26). Исследование Franzo at al. при описании эволюции PCV2 до и после введения вакцинации выявило изменения в аминокислотах 59, 191, 206, 210, 228 и 232 (27). За исключением аминокислоты 130, которая углублена на границе раздела субъединица-субъединица, эти аминокислоты находятся на поверхности капсида и претерпевают мутации (Фиг. 4А-4Н).
Внутреннюю область PCV2 VLP можно использовать для упаковки материала для нанотехнологий. Срез реконструкции, близкий к центру, был извлечен из реконструкции крио-ЭМ изображения, и график плотности значений пикселей был рассчитан в горизонтальном и вертикальном направлениях. На основе центрального среза реконструкции внешний диаметр VLP PCV2 составляет приблизительно 18,5 нм, принимая для расчета примерно сферическую форму. Таким образом, внешний объем оценивается в 3,3×103 нм3. Внутренний диаметр с использованием той же оценки сферической формы имеет диаметр приблизительно 13 нм. Следовательно, объем внутренней области составляет примерно 1,2×103 нм3. Сканирование центрального среза трехмерного молекулярного объема показало, что внутренняя область занята. Плотность является более низкой, чем у капсидной оболочки, вероятно, представляющей кумулятивную неупорядоченную матрицу N-конца из 60 CP единиц 200, которые составляют всю VLP.
Обсуждение
Геном цирковируса свиней 2 кодирует четыре известных белка: репликазу (ORF1), отвечающую за репликацию генома, капсидный белок (ORF2), отвечающий за образование оболочки капсида, и ORF3 и ORF4, которые, как полагают, играют роль в регуляции клеточного апоптоза (28, 29). Филогенетический анализ последовательности CP PCV2 показывает, что существуют 4 генотипа, распределенных глобально (PCV2a-d) (14). В настоящее время считается, что PCV2b является доминантным генотипом; однако недавнее увеличение количества отложений CP PCV2d в GenBank позволяет предполагать, что может иметь место сдвиг генотипа с PCV2b на PCV2d (5). Увеличение числа записей PCV2d может быть результатом «ускользнувших» мутантов в ответ на вакцинацию (17). Чтобы учитывать потенциальный сдвиг в сторону генотипа PCV2d и возможность того, что этот новый генотип может быть устойчивым к вакцинам, представленным на рынке, в настоящей заявке раскрыта система экспрессии у млекопитающих для получения больших количеств вирусоподобных частиц PCV2d (VLP). В настоящей заявке предложена первая система, в которой клетки млекопитающих были использованы для генерации большого количества VLP. Система экспрессии у млекопитающих особенно благоприятна для E. coli или бакуловирусной экспрессии, поскольку она больше похожа на клетки, естественно инфицированные PCV2, и, таким образом, позволяет изучать детали капсида PCV2 в контексте цикла репликации вируса. Например, N-конец CP обладает множественными сигналами ядерной локализации, и ожидается, что сборка вируса происходит в ядре инфицированной клетки, где реплицируется геном оцДНК (30). Однако ранее было неизвестно, могут ли капсиды PCV2 выходить из интактных ядер и выходить из инфицированных клеток.
Следовательно, в одном или более вариантах реализации настоящая система экспрессии может помочь выяснить, способен ли собранный капсид выходить из ядра. В одном или более вариантах реализации преобладающая фракция VLP в настоящей системе экспрессии находится в ядре клетки. Это неожиданно, поскольку биоинформатический поиск не идентифицирует ядерные сигналы экспорта (NES) в последовательности CP (данные не показаны). Таким образом, капсид PCV2 может иметь скрытый NES, который может играть важную роль в жизненном цикле вируса. Ядерный цитоплазматический транспорт, однако, может быть уменьшен путем перенаправления трансляции капсидного белка в сторону секреторного пути путем введения последовательности сигнала секреции на NH2-конце капсидного белка. Эта генетическая модификация приводит к увеличению синтеза капсидного белка, сборке VLP и высвобождению частиц в культуральную среду.
Реконструкция крио-ЭМ изображения и его структурный анализ показывают, что ИР, экспрессируемые млекопитающими, бакуловирусом и E. coli, практически не различимы. Сравнение координат атомов PCV2a, b и d выявляет различия в конформации открытых петель, состоящих из аминокислот 86-91 (петля CD), 131-136 (петля EF) и 188-194 (петля GH) (Фиг. 2А-2С). Структуры PCV2b и PCV2d более похожи друг на друга, чем любой из них с PCV2a. Эти различия объясняются эволюционно связанными мутациями в этих петлях (Фиг. 3В) и обеспечивают структурное описание антигенного сдвига, наблюдаемого в 2003 г. от PCV2a к PCV2b (15, 16). Расширение симметрии с последующей целенаправленной классификацией субъединицы капсида не идентифицирует какие-либо классы, которые могут указывать на посттрансляционную модификацию капсида (данные не показаны). Хотя для дальнейшего рассмотрения возможности посттрансляционной модификации VLP PCV2, генерируемой из системы экспрессии у млекопитающих, может быть необходимо использовать масс-спектроскопию. Выравнивание 1377 уникальных последовательностей CP PCV2 указывает на то, что только две аминокислоты (Met1 и Arg147) являются абсолютно консервативными, а остальные положения в последовательности претерпели мутации. Это демонстрирует замечательную пластичность структуры капсида для мутации при сохранении инфекционного вируса и, таким образом, подчеркивает способность PCV2 избегать нейтрализации антител. Координаты атомов, полученные при реконструкции крио-ЭМ изображения, идентифицируют участки на поверхности капсида, которые могут быть потенциальными мишенями для нейтрализации антител. В настоящей заявке идентифицированы непрерывные области на поверхности капсида, которые могут служить эпитопами, которые потенциально могут вызывать нейтрализующие антитела против нескольких генотипов. Пептиды, которые демонстрируют вариацию последовательности, могут представлять собой мутанты, «ускользнувшие» от нейтрализации антител. Следовательно, описанный протокол экспрессии можно использовать для создания капсидов, собранных в виде поливалентной мозаики, которая одновременно отображает нейтрализующие эпитопы нескольких генотипов. Позиции, которые не продемонстрировали вариабельность, могут представлять области вирусного капсида, которые могут быть идеальными для дизайна универсальной вакцины.
Ответ на собранные в млекопитающих VLP может более точно имитировать иммунный ответ на реальную инфекцию. Система экспрессии VLP у млекопитающих может обеспечить рекомбинантную вакцину со значительной эффективностью. Например, было обнаружено, что антительный ответ свиней, инокулированных рекомбинантным белком Сар, полученным из бакуловируса, и свиней, испытавших естественную инфекцию, различается (31, 32). У свиней после рекомбинантной вакцины предпочтительно распознавался только самый крупный полипептидный фрагмент, CP (43-233), тогда как экспериментально инфицированные свиньи и свиньи с PDNS показали сильную реактивность против олигопептида CP, 169-180. Пептид меньшего размера является общим для подтипов PCV2a и PCV2b и может служить приманкой, которая отвлекает защитный ответ от более крупного пептида 43-233.
Используемые в настоящее время вакцины были получены с использованием капсида из генома PCV2a, и в нескольких исследованиях сообщалось о неудачах иммунизации в результате инфекции PCV2d (7, 33, 34). Кроме того, всегда существует вероятность низкой эффективности вакцины из-за сдвига генотипа, и этот фактор необходимо учитывать при разработке вакцины в будущем. G. Franzo и др. (27) изучали причины давления отбора, производимого вакциной, посредством вакцинации. Они сообщили о высокой частоте мутаций в положениях аминокислот 59, 191, 206 для PCV2a и 131, 228 для PCV2b, снижающих связывание антител, которые ранее связывались с капсидом; возможно, вызывая «ускользание» от иммунного ответа от защиты вакцины (Фиг. 4) (35). Следовательно, необходима платформа для экспрессии VLP PCV2d. Таким образом, в соответствии с по меньшей мере одним вариантом реализации в настоящей заявке раскрыта экспрессия для продукции VLP PVC2d в клетках млекопитающих.
В одном или более вариантах реализации система экспрессии VLP согласно настоящей заявке также имеет применение в области трансляционой медицины. N-концевые 40 аминокислот CP богаты аргинином, имеют высокий положительный заряд и предположительно взаимодействуют с геномом оцДНК во время морфогенеза вируса. Этот мотив расположен во внутреннем пространстве VLP и, по-видимому, доступен для связывания специфических меток (или просто для абсорбции отрицательно заряженных малых молекул или олигонуклеотидов), что позволяет использовать частицу в качестве наночастицы для переноса терапевтических или иммуномодулирующих фрагментов, направленных на определенные ткани. В качестве альтернативы, N-конец PCV2 может быть заменен на гидрофобные аминокислоты для создания внутренней гидрофобной среды для гидрофобных молекул. Такие наноструктуры можно использовать в качестве диагностического антигена или в составе вакцины. Кроме того, замена открытых аминокислот на тиолсодержащие остатки или неприродные аминокислоты может позволить конъюгацию малых, средних или больших молекул, полезных для гетерологичной вакцинации, доставки лекарств, терапевтического лечения, диагностики и т.д.
В соответствии с одним или более вариантами реализации система экспрессии согласно настоящей заявке может продуцировать большие количества VLP PVC2d в клетках млекопитающего (клетки эмбриональной почки человека: НЕК 293). Эта система позволяет быстро изучить жизненный цикл PCV2 в фоновом режиме, которая очень похожа на естественного хозяина PCV2. Как более подробно поясняется ниже, реконструкция крио-ЭМ изображения VLP была определена с разрешением 3,3 А и использовалась для идентификации потенциальных антигенных эпитопов, которые могут быть использованы при конструировании вакцины или в составах для доставки небольших терапевтических молекул. Сравнение 1377 уникальных записей CP PCV2 в GenBank показывает, что за исключением двух аминокислот в каждой позиции аминокислоты произошла мутация. Однако две группы аминокислот, которые образуют отдельные участки на поверхности капсида, демонстрируют ограниченное изменение последовательности. Вакцины, способные направлять антитела к этим участкам, могут служить универсальными вакцинами против PCV2.
6.2. Пример 2
Исследование введения вакцины для оценки эффективности вакцин против цирковируса 2 свиней.
ДИЗАЙН ЭКСПЕРИМЕНТА. Приблизительно в возрасте 1 недели на ферме были взяты образцы крови от 120 поросят мужского и женского пола. Эти образцы крови были проанализированы на наличие PCV-2 с помощью количественного анализа полимеразной цепной реакции в реальном времени (количественная ПЦР в реальном времени) для исключения любых положительных поросят. Эти образцы количественной ПЦР в реальном времени, отрицательные на PCV-2, были протестированы на наличие антител против PCV-2 с использованием коммерческого иммуноферментного анализа (ELISA). Затем были отобраны 72 двухнедельных поросенка, самцов и самок, которые продемонстрировали отрицательный результат на PCV-2 с помощью количественной ПЦР в реальном времени (взятые из помета со всеми протестированными поросятами с отрицательными результатами) и с самыми низкими уровнями антител к PCV-2, которые были отобраны и транспортированы в участок проведения испытаний на экспериментальной ферме. По прибытии в помещения животных взвешивали, и на следующий день случайным образом распределяли на шесть экспериментальных групп по двенадцать свиней в каждой на основании титров S/P PCV-2 (при скрининге), массы тела, пола и количества свиноматок. Впоследствии свиней помещали в четыре помещения на период акклиматизации в течение 10 дней.
В возрасте 21±3 дня (день исследования 0) свиней иммунизировали внутримышечно (в/м) в правую сторону шеи. Поросят из группы обработки Т01 иммунизировали посредством 1 мл вакцины PIGONE (цирковирус свиней типа 2 (PCV2), штамм VQ2610 1×109 - 5×109 копий ДНК, эквивалентных предварительной активации вируса). Поросят из группы обработки Т02 иммунизировали посредством 2 мл вакцины PIGONE (цирковирус свиней типа 2 (PCV2), штамм VQ2610 1×109 - 5×109 копий ДНК, эквивалентных предварительной активации вируса) (далее «PIGONE»). Животных из Т03 иммунизировали посредством 2 мл вакцины Huve-PCV2 (6 - Huve-PCV_2 А и 6 - Huve-PCV2_В). Huve-PCV2_А содержит белок ORf2 PCV2d, экспрессируемый эукариотической клеткой, a Huve-PCV2_B содержит клетку млекопитающего, экспрессирующую VLP PCV2d. Животных из группы обработки Т04 иммунизировали посредством 1 мл вакцины HVP-DNA (ДНК HVP включает ДНК-вакцину ORF2 PCV2b), а животных из группы обработки Т05 иммунизировали посредством 1 мл коммерческой вакцины Ingelvac CIRCOFLEX (инактивированный бакулоэкспрессируемый ORf2 PCV2) (Boehringer Ingelheim). Поросятам из группы обработки Т06 вводили физиологический раствор с фосфатным буфером (PBS) и использовали в качестве группы отрицательного контроля. В 21 день исследования (SD21), примерно в возрасте 6 недель, свиней иммунизировали внутримышечно (в/м) в левую сторону шеи. Свиней из группы обработки Т01 иммунизировали посредством 1 мл вакцины PIGONE, свиней из группы обработки Т02 иммунизировали посредством 2 мл вакцины PIGONE, животных из группы обработки Т03 иммунизировали посредством 2 мл вакцины Huve-PCV2 (6 посредством Huve-PCV2_A и 6 посредством Huve-PCV2_В), и свиней из группы обработки Т04 иммунизировали посредством 1 мл вакцины HVP-DNA. Группы Т05 и Т06 внутримышечно иммунизировали посредством 1 мл PBS. Перед каждой вакцинацией проводили клинический осмотр свиней.
Приблизительно в возрасте 8 недель (WOA) на 35 день исследования всем животным из шести экспериментальных групп интраназально (и/н) вводили 3 мл инокулята 104,95 TCID50/мл PCV-2b штамма Sp-10-7-54-13.
Образцы крови собирали еженедельно в течение периода исследования для серологического анализа (ELISA) и/или обнаружения PCV-2 в сыворотке (количественная ПЦР в реальном времени). Кроме того, мазки из носа и прямой кишки получали еженедельно после заражения для определения профиля выделения PCV-2 (при необходимости). Животных оценивали по клиническим признакам еженедельно после заражения, и массу тела регистрировали при первой вакцинации, контрольном заражении и вскрытии.
В возрасте 12 недель (SD63 и SD64) все свиньи из пяти обработанных групп были умерщвлены. При вскрытии были собраны миндалины, трахеобронхиальные лимфатические узлы, брыжеечные лимфатические узлы и поверхностные паховые лимфатические узлы для гистопатологического исследования и обнаружения PCV-2 в тканях (при необходимости). Схема эксперимента представлена в Таблице 2.
РАСПРЕДЕЛЕНИЕ ОБРАБОТКИ. Животные были разделены на 6 групп по 12 животных (Т01, Т02, Т03, Т04, Т05 или Т06) в соответствии с титрами антител против PCV-2 при скрининге, массой в день поступления, полом и количеством свиноматок. Переменные были ранжированы, затем отсортированы и использованы для отнесения свиней к шести группам обработки блоками по 6. Животных из групп обработки Т01, Т02, Т03, Т04 и Т05 случайным образом смешивали в 3 комнатах (комнаты 5, 6 и 7), помещая по четыре животных из каждой группы в комнату. Обработку Т06 проводили в отдельной комнате (комната 8).
Животных из группы Т03 (Huve-PCV2) разделяли на 2 подгруппы: Т03_A (Huve-PCV2_A) и Т03_В (Huve-PCV2_В). Это было сделано случайным образом на SD0, выбрав по два животных из Т03 из каждой комнаты для каждой подгруппы.
Порядок входа в комнаты был установлен следующим образом: фаза иммунизации: комната 8- комната 7- комната 6- комната 5; фаза заражения: комната 5- комната 6-комната 7- комната 8.
Комната, в которой размещалась группа PBS (Т06), была первой комнатой, в которую входили во время фазы иммунизации, и, таким образом, первой комнатой, в которой была проведена имитационная вакцинация и заражение. После заражения в комнаты с вакцинированными животными входили перед входом в комнату с контрольной группой PBS (Т06).

ПРЕДВАРИТЕЛЬНЫЕ РЕЗУЛЬТАТЫ
События здоровья. Животное №472 из группы Т01 (PIGONE 2 мл) было обнаружено мертвым на SD32. Проведено вскрытие трупа и описан фибринозный перикардит. Причина смерти, вероятно, была связана с бактериальной септицемией и не связана с испытуемым образцом.
Клинические наблюдения отдельных животных. Все свиньи были оценены на предмет депрессии, физического состояния и респираторного дистресс-синдрома в дни вакцинации (SD0 и SD21) перед вакцинацией. Кроме того, у животных также оценивали клинические признаки еженедельно после заражения, в дни исследования 35 до заражения, 41, 49, 56 и 63. Во время фазы исследования у исследуемых животных не было зарегистрировано никаких клинических признаков.
Масса тела. Массу тела регистрировали у всех свиней при поступлении на экспериментальную площадку в течение назначения лечения, при первой вакцинации (SD0), при заражении (SD35) и при вскрытии (SD63). Среднесуточный прирост массы тела (ADWG) рассчитывали для периодов: первая вакцинация - заражение, заражение - вскрытие и первая вакцинация - вскрытие. Изменение массы тела представлено на Фиг. 5 и ADWG на Фиг. 6.
Группа PBS (Т06) имела немного более высокую среднюю массу тела на SD35 и SD63 и более высокий ADWG в трех наблюдаемых периодах по сравнению с другими группами, но, вероятно, они статистически не отличаются.
Серология. Наличие антител против PCV-2 в крови в дни исследования SD0 до вакцинации, SD7 ± 1, SD13, SD20, SD29, SD35 до заражения, SD41, SD49, SD56 и SD63 тестировали с помощью коммерческого набора ELISA IngezimCirco IgG. Результаты были выражены как титры S/P.
Кинетика антител представлена на Фиг. 7. Представлено семь групп обработки, группа обработки Т03 (Huve-PCV2) разделена на две подгруппы: Huve-PCV2_А (Т03_А) и Huve-PCV2_В (Т03_В).
При SD0 (первая вакцинация) значения S/P всех исследуемых животных находились в диапазоне от 0,2 до 1, с общим средним значением 0,71.
Усиленный эффект второй вакцинации наблюдается в 3 группах на SD29: PIGONE 1 мл и 2 мл (Т01 и Т02) и Huve-PCV2_B (Т03).
В день заражения группы с CIRCOFLEX (инактивированный бакулоэкспрессиуемый ORf2 PCV2) (Т05), PBS (Т06), Huve-PCV2_А (Т03_А) и HVP-DNA (Т04) имели низкие значения S/P, между 0,38-0,4. После контрольного заражения, как правило, значения S/P снижались, и для некоторых вакцинированных групп наблюдался умеренный ответ на SD56 или SD63.
Единственной группой с высоким уровнем антител после второй вакцинации была Huve-PCV2_В (Т03_В).
Виремия. Коммерческий набор для количественной ПЦР в реальном времени LSI VetMAX Porcine Circovirus Type 2 - Quantification (Life Technologies, код ссылки QPCV) был использован для обнаружения и количественного определения ДНК PCV-2 в образцах сыворотки, полученных в дни исследования перед вакцинацией SD0, перед заражением SD35,
SD41, SD49, SD56 и SD63. Результаты выражали в виде числа копий PCV-2 log10/мл и классифицировали как отрицательные (<3 log10 копий/мл), не поддающиеся количественной оценке положительные (от 3log10копий/мл до 4 log10 копий/мл) или положительные (>4log10 копий/мл).
РЕЗУЛЬТАТЫ ВИРЕМИИ С УЧЕТОМ ВИРУСНЫХ НАГРУЗОК >3log10 копий/мл.
Результаты представлены на Фиг. 8 и 9, в виде процента поддающихся количественной оценке положительных и не поддающихся количественной оценке положительных животных и средних значений поддающихся количественной оценке положительных животных (>4log10 копий/мл).
Как правило, низкий процент положительных животных (>3log10 копий/мл) наблюдается во время фазы заражения. Обработка Т01 и Т02, вакцинированных посредством PIGONE (1 мл или 2 мл), показывает численно более высокий процент положительных животных по SD49 и SD56, но в любом случае достигает 50% положительных результатов (поддается количественной оценке + не поддается количественной оценке). CIRCOFLEX (инактивированный бакулоэкспрессируемый ORf2 PCV2) (Т04) не продемонстрировал положительных (поддающихся количественной оценке + не поддающихся количественной оценке) животных во время фазы заражения.
Huve-PCV2_В (Т03_В), CIRCOFLEX (инактивированный бакулоэкспрессируемый ORf2 PCV2) (Т05) и PBS (Т06) не продемонстрировали поддающихся количественной оценке положительных животных (>4log10 копий/мл) во время фазы заражения. В остальных группах было от 1 до 3 положительных животных по меньшей мере в первый день отбора проб во время фазы заражения, при этом SD56 представлял собой день с большим количеством групп с положительными животными. Средние значения положительных поддающихся количественной оценке животных находились в диапазоне от 4,12log10 до 5,34log10 копий PCV-2/мл.
РЕЗУЛЬТАТЫ ВИРЕМИИ С УЧЕТОМ ВИРУСНЫХ НАГРУЗОК >2log10 копий/мл.
Результаты представлены на Фиг. 10 и 11, в процентах от зараженных вирусом животных и как средние значения для всех животных (>2log10 копий/мл).
При рассмотрении всех животных, включая образцы ниже предела обнаружения, более высокий процент положительных результатов наблюдается во всех исследуемых группах. Группы PIGONE (Т01 или Т02) продолжают оставаться группами с численно более высоким процентом положительных результатов на SD49 (PIGONE 1 мл, Т01), SD56 и SD63 (PIGONE 2 мл, Т02), за которыми следует HVPDNA (Т04).
Средние вирусные нагрузки для всех животных со значениями количественной ПЦР в реальном времени находились в диапазоне 2,19-4,831log10копий/мл.
Группы Т01 (PIGONE 1 мл), Т02 (PIGONE 2 мл), Т03_В (Huve-PCV2_B) и Т04 (HVP-ДНК) представляют средние значения >2log10 копий/мл в течение четырех дней отбора проб после заражения. Группа Huve-PCV2_А (Т03_А) и контрольная группа PBS (Т06) имели положительные значения через три дня, от SD49 до SD63, в то время как CIRCOFLEX (инактивированный бакулоэкспрессируемый ORf2 PCV2) (Т05) продемонстрировал наличие зараженных вирусом животных через два дня, через одну неделю после заражения (SD41) и при вскрытии (SD63).
На SD35 одно животное из группы Т02 (PIGONE 2 мл) имело вирусную нагрузку 2,76log10 копий/мл. Это животное было отрицательным согласно количественной ПЦР в реальном времени при отборе животных, SD0, SD7, SD14 и SD21. Также после испытания на SD41 и SD49. Во время контрольного заражения его вирусная нагрузка на SD56 и SD63 составляла >3log10 копий/мл.
Серология. Значения S/P при SD0 были выше, чем в предыдущем исследовании, из-за материнских антител. Это не позволяет увидеть явную сероконверсию после вакцинации. PIGONE 1 мл и 2 мл (Т01, Т02) и Huve-PCV2_В (Т03_В), по-видимому, показывают сероконверсию после 2-й вакцинации по сравнению с введением PBS (Т06). CIRCOFLEX (инактивированный бакулоэкспрессируемый ORf2 PCV2) (Т05) и вакцина HVP-DNA (Т04) не проявляют ответа антител. Они имеют аналогичную кинетику антител, что и у группы, получившей PBS.
Отсутствие сероконверсии в группе CIRCOFLEX (инактивированный бакулоэкспрессируемый ORf2 PCV2) (Т05) ожидалось с учетом серологических значений во время вакцинации. Защита CIRCOFLEX (инактивированный бакулоэкспрессируемый ORf2 PCV2) основана на клеточном иммунитете. Обычно в полевых условиях не наблюдается серологического ответа у животных с материнскими антителами при вакцинации CIRCOFLEX (инактивированный бакулоэкспрессируемый ORf2 PCV2).
Подгруппы Huve-PCV2 нельзя сравнивать с другими группами, поскольку n составляет половину остальных (6 животных в подгруппе). Однако, сравнивая их, они имеют разные результаты, так как Т03_В имеет более высокий серологический ответ.
Виремия. На SD35 перед заражением одно животное из группы PIGONE 2 мл (Т02) было заражено вирусом, ниже предела обнаружения (вирусная нагрузка: 2,76 log10 копий/мл). Это животное было отрицательным согласно количественной ПЦР в реальном времени при отборе животных, SD0, SD7, SD14 и SD21. Также после испытания на SD41 и SD49. Была предпринята попытка секвенировать эту ДНК на SD35, но это не удалось из-за низкой вирусной нагрузки. Этому есть два возможных объяснения: 1) субклиническая инфекция полевым штаммом PCV-2 во время фазы иммунизации с очень низкой вирусной нагрузкой; и 2) обнаруживается заражение ДНК PCV-2, а не всего вируса Неизвестного происхождения.
Обычно инокуляция PCV-2 вызывала легкую субклиническую инфекцию, поскольку во время заражения наблюдалась низкая вирусная нагрузка. Таким образом, было определено, что следует учитывать все результаты количественной ПЦР в реальном времени, в том числе «ниже предела обнаружения» (2-3log10 копий/мл). Группы PIGONE 1 мл (Т01), PIGONE 2 мл (Т02) и HVP-DNA (Т04) показывают более высокий процент положительных животных. Группы Huve-PCV2_В (Т03_В), CIRCOFLEX (инактивированный бакулоэкспрессируемый ORf2 PCV2) (Т05) и PBS (Т06) демонстрировали отсутствие / очень низкий процент зараженных вирусом животных на протяжении фазы заражения. Средние значения составляют <3log10 копий/мл. При сравнении подгрупп Huve-PCV2 (Т03) среди них, подгруппа Т03_В имеет численно более низкий процент зараженных вирусом животных и среднюю вирусную нагрузку от SD49 до SD63.
Информация о дне заражения. Посевной материал готовили следующим образом: его размораживали, перемешивали и затем разделяли на 5 флаконов с прибл. 50 мл/флакон. Титрование в день заражения показало 104,95 TCID50. Флаконы были разделены между комнатами, т.е. начинали один флакон, и пока он не был закончен, новый не использовали, поэтому один и тот же флакон мог использоваться в двух помещениях. До использования флаконы с посевным материалом держали охлажденными.
Порядок заражения: сначала заражали группу PBS (Т06) (комната 8), а затем вакцинированные группы (комнаты 7, 6 и 5). Возможный эффект помещения: в комнате PBS (Т06) было меньше животных, чем в других трех комнатах с вакцинированными группами. 12 или 20 животных в комнате соответственно. 4 комнаты были равны по площади и условиям содержания (температура, влажность и т.д.).
Дополнительные варианты реализации
В соответствии с одним или более вариантами реализации в настоящей заявке раскрыта сборка рекомбинантных капсидов цирковируса свиней или вирусоподобных частиц (VLP), обладающих уникальными структурными и антигенными свойствами, где один или более составляющих мономеров капсида включают последовательности, придающие вакцине моновалентные или поливалентные свойства, и/или способность инкапсулировать или удерживать молекулярные единицы с различными биологическими активностями. По меньшей мере в одном варианте реализации рекомбинантный капсид или VLP включает последовательность сигнала секреции, генетически связанную с кодирующей последовательностью капсида, которая обеспечивает высвобождение и/или секрецию частиц из продуцирующих клеток в культуральную среду.
В одном или более вариантах реализации рекомбинантный капсид или последовательность VLP, кодирующая поверхностные мотивы и петли, мутированы, изменены или модифицированы, следовательно, антигенные свойства собранных VLP являются новыми, различными или химерными.
В одном или нескольких вариантах реализации рекомбинантного капсида VLP сборку и/или получение проводят с двумя или более различными мономерами капсидного белка, в результате чего получают VLP смешанного состава (мозаичного или химерного) с уникальными антигенными и биологическими свойствами. В одном или более вариантах реализации рекомбинантный капсид или VLP включает модифицированную поверхность, которая позволяет конъюгировать различные молекулярные объекты с поверхностью VLP.
В одном или более вариантах реализации рекомбинантный капсид или VLP включает NH2-концевые или внутренние аминокислоты мономеров капсида, которые мутированы, удалены или модифицированы, что позволяет включать небольшие молекулы, нуклеиновые кислоты или другие молекулярные фрагменты во время сборки в VLP-продуцирующих клетках или in vitro путем разборки и повторной сборки VLP.
В одном или более вариантах реализации полученные VLP нацелены на определенные ткани или клетки для доставки молекулярных единиц и проявляют биологическую активность.
По меньшей мере в одном варианте реализации в настоящей заявке также раскрыта конструкция ДНК, содержащая последовательности, кодирующие капсидный белок во всей его последовательности и форме, используемая для сборки VLP. Конструкция ДНК содержит последовательности, кодирующие различные формы мономеров капсида. В одном или более вариантах реализации в настоящей заявке дополнительно раскрыт способ получения VLP, причем способ включает введение в клетку-хозяина одной или более конструкций ДНК в условиях, при которых клетка продуцирует VLP. По меньшей мере в одном варианте реализации клетка-хозяин представляет собой эукариотическую клетку, выбранную из группы, состоящей из клеток млекопитающих, дрожжей, насекомых, растений, амфибий и птиц. В одном или более вариантах реализации конструкция ДНК вводится в клетки и интегрируется в клеточный геном или равномерно делится и сегрегируется на дочерние клетки, что приводит к непрерывному продуцированию VLP.
По меньшей мере, в одном варианте реализации в настоящей заявке раскрыта VLP, полученная указанным выше способом.
В одном или более вариантах реализации предложена иммуногенная композиция, которая содержит по меньшей мере одну VLP согласно любому из вышеуказанных вариантов реализации. По меньшей мере в одном варианте реализации иммуногенная композиция дополнительно содержит адъювант. В одном или более вариантах реализации предложен способ генерации иммунного ответа на один или более цирковирусов свиней у животного. Способ включает введение животному эффективного количества иммуногенной композиции. По меньшей мере в одном варианте реализации способа композицию вводят через слизистую, внутрикожно, подкожно, внутримышечно или перорально. По меньшей мере в одном варианте реализации иммунный ответ вакцинирует животное против нескольких серотипов или антигенных вариантов одного или более цирковирусов свиней. По меньшей мере в одном варианте реализации способа животное представляет собой свинью.
В одном или более вариантах реализации рекомбинантный капсид или VLP согласно настоящей заявке содержат молекулы с фармацевтической или биологической активностью. По меньшей мере в одном варианте реализации рекомбинантный капсид или VLP включает фармацевтическую композицию для доставки лекарственного средства VLP, гетерологичной иммунизации и/или иммуномодуляции.
В одном или более вариантах реализации способа генерации иммунного ответа на один или более цирковирусов свиней у животного иммуногенную композицию вводят субъекту в качестве терапевтического или профилактического лечения. Настоящие способы обеспечивают эффективное применение композиций согласно настоящей заявке у субъекта. По меньшей мере в одном варианте реализации субъектом является человек или животное.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO: 1. Нуклеотидная последовательность капсида PCV2 дикого типа.
SEQ ID NO: 2. Аминокислотная последовательность капсида PCV2 дикого типа.
SEQ ID NO: 3. Нуклеотидная последовательность модифицированного капсида PCV2 с сигнальной последовательностью секреции.
SEQ ID NO: 4. Аминокислотная последовательность модифицированного капсида PCV2 с сигнальной последовательностью секреции.
SEQ ID NO: 5. Нуклеотидная последовательность Pcdna3.4-PCV2. (Фиг. 1А).
SEQ ID NO: 1. Капсид штамма цирковируса-2 свиней PCV2
Длина: 705
Тип: ДНК

SEQ ID NO: 2. Капсид штамма цирковируса-2 свиней PCV2
Длина: 234
Тип: Аминокислота

SEQ ID NO: 3. Капсид модифицированного цирковируса-2 свиней (PCV2)
Длина: 750 Тип: ДНК

SEQ ID NO: 4. Капсид модифицированного штамма цирковируса-2 свиней PCV2
Длина: 249
Тип: Аминокислота

SEQ ID NO: 5. Pcdna3.4-PCV2. (Фиг. 1А).
Длина: 6778
Тип: ДНК


Объем настоящего изобретения не ограничивается описанными в настоящем документе конкретными вариантами реализации. Действительно, различные модификации настоящего изобретения в дополнение к описанным в настоящем документе станут очевидными для специалистов в данной области техники из предшествующего описания и сопровождающих фигур. Предполагается, что такие модификации входят в объем прилагаемой формулы изобретения.
Патенты, заявки на патенты и публикации цитируются по всей этой заявке, описания которых, в частности, включая все раскрытые химические структуры, включены в настоящий документ в качестве ссылки. Цитирование вышеупомянутых публикаций или документов не является признанием того, что любое из вышеперечисленного относится к предшествующему уровню техники, а также не является признанием содержания или даты этих публикаций или документов. Все цитируемые в настоящем документе ссылки включены в качестве ссылки в той же степени, как если бы каждая отдельная публикация, заявка на патент или патент были специально и индивидуально указаны для включения в качестве ссылки.
Вышеизложенное письменное описание считается достаточным для того, чтобы дать возможность специалисту в данной области техники применить настоящее изобретение на практике. Различные модификации изобретения в дополнение к тем, которые показаны и описаны в настоящем документе, станут очевидны специалистам в данной области техники из предшествующего описания и подпадают под объем прилагаемой формулы изобретения.
Примерные системы и способы изложены в следующих пунктах:
Пункт 1: Система экспрессии у млекопитающих для получения рекомбинантных вирусоподобных частиц (VLP) цирковируса свиней типа 2 (PCV2), при этом система экспрессии содержит:
клетку млекопитающего;
плазмиду, содержащую ген PCV2, кодирующий капсидный белок, при этом ген PCV2 оптимизирован по кодонам;
при этом клетка млекопитающего трансфицирована плазмидой; и
при этом система экспрессии продуцирует рекомбинантные VLP PCV2.
Пункт 2: Система экспрессии по п. 1, отличающаяся тем, что клетка млекопитающего представляет собой клетку эмбриональной почки человека-293 (HEK-293) человека.
Пункт 3: Система экспрессии по пп. 1-2, отличающаяся тем, что ген PCV2 включает сайт распознавания NheI, последовательность Козака и сайт распознавания NotI, и
причем сайт распознавания NheI и последовательность Козака расположены выше стартового кодона гена PCV2, и сайт распознавания NotI встроен после кодона терминации гена PCV2.
Пункт 4: Система экспрессии по пп. 1-3, отличающаяся тем, что большинство продуцируемых рекомбинантных VLP PCV2 присутствует в ядре клеток млекопитающего.
Пункт 5: Система экспрессии по пп. 1-4, отличающаяся тем, что капсидный белок содержит аминокислотную последовательность SEQ ID NO: 2.
Пункт 6: Система экспрессии по пп. 1-5, отличающаяся тем, что капсидный белок кодируется нуклеотидной последовательностью SEQ ID NO: 1.
Пункт 7: Система экспрессии по пп. 1-4, отличающаяся тем, что капсидный белок модифицирован сигнальной последовательностью секреции, введенной на NH2-конце капсидного белка.
Пункт 8: Система экспрессии по п. 7, отличающаяся тем, что капсидный белок содержит аминокислотную последовательность SEQ ID NO: 4.
Пункт 9: Система экспрессии по пп. 7-8, отличающаяся тем, что капсидный белок кодируется нуклеотидной последовательностью SEQ ID NO: 3.
Пункт 10: Система экспрессии по пп. 1-9, отличающаяся тем, что продуцируемые рекомбинантные VLP PCV2 выбраны из группы, состоящей из: VLP PCV2a, VLP PCV2b, VLP PCV2c, VLP PCV2d и VLP PCV2e.
Пункт 11: Система экспрессии по пп. 1-10, отличающаяся тем, что продуцируемые рекомбинантные VLP PCV2 представляют собой VLP PVC2d.
Пункт 12: Система экспрессии по пп. 1-11, отличающаяся тем, что плазмида представляет собой pcDNA3.4-PCV2.
Пункт 13: Система экспрессии по пп. 1-12, отличающаяся тем, что ген PCV2 оптимизирован по кодонам с применением оптимизированной по кодонам аминокислотной последовательности, показанной на Фиг. 1А.
Пункт 14: Способ получения вирусоподобных частиц (VLP) цирковируса свиней типа 2 (PCV2), включающий:
обеспечение суспензии культивируемых клеток млекопитающего;
трансфекцию клеток млекопитающего плазмидой, содержащей ген PCV2, кодирующий капсидный белок;
добавление натриевой соли вальпроевой кислоты (VPA) к трансфицированным клеткам млекопитающего, причем добавление натриевой соли VPA ингибирует пролиферацию клеток;
центрифугирование и промывание трансфицированных клеток млекопитающего;
суспендирование центрифугированных клеток млекопитающего в фосфатно-солевом буферном растворе (PBS);
осуществление нескольких циклов замораживания и оттаивания клеток млекопитающего;
обработку ультразвуком клеток млекопитающего в нескольких циклах; и
осуществление двух последовательных циклов центрифугирования клеток млекопитающего для получения VLP PCV2,
причем большинство продуцируемых VLP PCV2 присутствуют в ядре клеток млекопитающего.
Пункт 15: Способ по п. 14, согласно которому стадия центрифугирования и промывания трансфицированных клеток млекопитающего включает:
центрифугирование клеток млекопитающего при 2000×g в течение 15 мин;
промывание клеток млекопитающего раствором PBS; и
центрифугирование клеток млекопитающего снова при 2000×g в течение 15 мин.
Пункт 16: Способ по пп. 14-15, согласно которому центрифугированные клетки млекопитающего замораживают при приблизительно -80°С и оттаивают при приблизительно 37°С во время циклов замораживания и оттаивания.
Пункт 17: Способ по пп. 14-16, согласно которому первый из двух последовательных циклов центрифугирования осуществляют при 2000×g в течение 15 минут, и второй из двух последовательных циклов центрифугирования осуществляют при 8000×g в течение 15 минут.
Пункт 18: Способ по пп. 14-17, дополнительно включающий очистку VLP PCV2 с помощью ультрацентрифугирования.
Пункт 19: Способ по пп. 14-18, согласно которому клетки млекопитающего представляют собой клетки эмбриональной почки человека -19 (HEK-293).
Пункт 20: Способ по пп. 14-19, согласно которому ген PCV2 включает сайт распознавания NheI, последовательность Козака и сайт распознавания NotI, и
причем сайт распознавания NheI и последовательность Козака расположены выше стартового кодона гена PCV2, и сайт распознавания NotI встроен после кодона терминации гена PCV2.
Пункт 21: Способ по пп. 14-20, согласно которому плазмида представляет собой pcDNA3.4-PCV2.
Пункт 22: Способ по пп. 14-21, согласно которому продуцируемые VLP PCV2 выбраны из группы, состоящей из: PCV2a VLPs, PCV2b VLPs, PCV2c VLPs, PCV2d VLPs и PCV2e VLP.
Пункт 23: Способ по пп. 14-22, согласно которому продуцируемые VLP PCV2 представляют собой VLP PVC2d.
Пункт 24: VLP PCV2, созданная способом по пп. 14-23.
ССЫЛКИ
1. Ellis J. 2014. Porcine circovirus: a historical perspective. Vet Pathol 51:315-27. 455
2. Rosario K, Breitbart M, Harrach B, Segales J, Delwart E, Biagini P, Varsani A. 2017. Revisiting the taxonomy of the family Circoviridae: establishment of the genus Cyclovirus and removal of the genus Gyrovirus. Arch Virol 162:1447-1463. 458
3. Breitbart M, Delwart E, Rosario K, Segales J, Varsani A, Ictv Report C. 2017. ICTV Virus Taxonomy Profile: Circoviridae. J Gen Virol 98:1997-1998.
4. Palinski R, Pineyro P, Shang P, Yuan F, Guo R, Fang Y, Byers E, Hause BM. 2017. A Novel Porcine Circovirus Distantly Related to Known Circoviruses Is Associated with Porcine Dermatitis 462 and Nephropathy Syndrome and Reproductive Failure. J Virol 91.
5. Wei C, Zhang M, Chen Y, Xie J, Huang Z, Zhu W, Xu T, Cao Z, Zhou P, Su S, Zhang G. 2013. Genetic evolution and phylogenetic analysis of porcine circovirus type 2 infections in southern China from 2011 to 2012. Infect Genet Evol 17:87-92.
6. Franzo G, Cortey M, Olvera A, Novosel D, Castro AM, Biagini P, Segales J, Drigo M. 2015. Revisiting the taxonomical classification of Porcine Circovirus type 2 (PCV2): still a real challenge. 468 Virol J 12:131.
7. Xiao CT, Halbur PG, Opriessnig T. 2015. Global molecular genetic analysis of porcine circovirus type 2 (PCV2) sequences confirms the presence of four main PCV2 genotypes and reveals a rapid increase of PCV2d. J Gen Virol 96:1830-41.
8. Wang N, Zhan Y, Wang A, Zhang L, Khayat R, Yang Y. 2016. In silico analysis of surface structure variation of PCV2 capsid resulting from loop mutations of its capsid protein (Cap). J Gen Virol 97:3331-3344.
9. Liu Z, Guo F, Wang F, Li TC, Jiang W. 2016. 2.9 A Resolution Cryo-EM 3D Reconstruction of Close-Packed Virus Particles. Structure 24:319-28.
10. Khayat R, Johnson JE. 2011. Passthe jelly rolls. Structure 19:904-6. 478
11. Khayat R, Brunn N, Speir JA, Hardham JM, Ankenbauer RG, Schneemann A, Johnson JE. 2011. The 2.3-angstrom structure of porcine circovirus 2. J Virol 85:7856-62.
12. Guo L, Lu Y, Huang L, Wei Y, Liu C. 2011. Identification of a new antigen epitope in the nuclear localization signal region of porcine circovirus type 2 capsid protein. Intervirology 54:156-63.
13. Mahe D, Blanchard P, Truong C, Amauld C, Le Cann P, Cariolet R, Madec F, Albina E, Jestin A. 2000. Differential recognition of ORF2 protein from type 1 and type 2 porcine circoviruses and identification of immunorelevant epitopes. J Gen Virol 81:1815-24.
14. Guo LJ, Lu YH, Wei YW, Huang LP, Liu CM. 2010. Porcine circovirus type 2 (PCV2): genetic variation and newly emerging genotypes in China. Virol J 7:273.
15. Beach NM, Meng XJ. 2012. Efficacy and future prospects of commercially available and experimental vaccines against porcine circovirus type 2 (PCV2). Virus Res 164:33-42.
16. Patterson AR, Opriessnig T. 2010. Epidemiology and horizontal transmission of porcine circovirus type 2 (PCV2). Anim Health Res Rev 11:217-34.
17. Xiao CT, Halbur PG, Opriessnig T. 2012. Complete genome sequence of a novel porcine circovirus type 2b variant present in cases of vaccine failures in the United States. J Virol 86:12469.
18. Dhindwal S, Avila B, Feng S, Khayat R. 2019. Porcine circovirus 2 uses a multitude of weak binding sites to interact with heparan sulfate, and the interactions do not follow the symmetry of the capsid. Journal of Virology doi: 10.1128/jvi.02222-18.
19. Sievers F, Wilm A, Dineen D, Gibson TJ, Karplus K, Li W, Lopez R, McWilliam H, Remmert M, Soding J, Thompson JD, Higgins DG. 2011. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol Syst Biol 7:539.
20. Ashkenazy H, Abadi S, Martz E, Chay O, Mayrose I, Pupko T, Ben-Tal N. 2016. ConSurf 2016: an improved methodology to estimate and visualize evolutionary conservation in macromolecules. Nucleic Acids Res 44: W344-50.
22. Hopf ТА, Ingraham JB, Poelwijk FJ,  CPI, Springer M, Sander C, Marks DS. 2017. Mutation effects predicted from sequence co-variation. Nature Biotechnology 35:128.
CPI, Springer M, Sander C, Marks DS. 2017. Mutation effects predicted from sequence co-variation. Nature Biotechnology 35:128.
23. Shamsi Z, Moffett AS, Shukla D. 2017. Enhanced unbiased sampling of protein dynamics using evolutionary coupling information. Sci Rep 7:12700.
24. Fraczkiewicz R, Braun W. 1998. Exact and efficient analytical calculation of the accessible surface areas and their gradients for macromolecules. Journal of Computational Chemistry 19:319-333.
25. Saha D, Huang L, Bussalleu E, Lefebvre DJ, Fort M, Van Doorsselaere J, Nauwynck HJ. 2012. 513 Antigenic subtyping and epitopes' competition analysis of porcine circovirus type 2 using monoclonal antibodies. Vet Microbiol 157:13-22.
26. Lefebvre DJ, Costers S, Van Doorsselaere J, Misinzo G, Delputte PL, Nauwynck HJ. 2008. Antigenic differences among porcine circovirus type 2 strains, as demonstrated by the use of monoclonal antibodies. J Gen Virol 89:177-87.
27. Franzo G, Tucciarone CM, Cecchinato M, Drigo M. 2016. Porcine circovirus type 2 (PCV2) evolution before and after the vaccination introduction: A large scale epidemiological study. Sci Rep 6:39458.
28. Teras M, Viisileht E, Pahtma-Hall M, Rump A, Paalme V, Pata P, Pata I, Langevin C, Ruutel 522 Boudinot S. 2018. Porcine circovirus type 2 ORF3 protein induces apoptosis in melanoma cells. BMC Cancer 18:1237.
29. Pan Y, Li P, Jia R, Wang M, Yin Z, Cheng A. 2018. Regulation of Apoptosis During Porcine Circovirus Type 2 Infection. Front Microbiol 9:2086.
30. Shuai J, Wei W, Jiang L, Li XI, Chen N, Fang W. 2008. Mapping of the nuclear localization signals in open reading frame 2 protein from porcine circovirus type 1. Acta Biochimica et Biophysica Sinica 40:71-77.
31. Trible BR, Suddith AW, Kerrigan MA, Cino-Ozuna AG, Hesse RA, Rowland RR. 2012. Recognition of the different structural forms of the capsid protein determines the outcome following infection with porcine circovirus type 2. J Virol 86:13508-14.
32. Trible BR, Kerrigan M, Crossland N, Potter M, Faaberg K, Hesse R, Rowland RR. 2011. Antibody recognition of porcine circovirus type 2 capsid protein epitopes after vaccination, infection, and disease. Clin Vaccine Immunol 18:749-57.
33. Seo HW, Park C, Kang I, Choi K, Jeong J, Park SJ, Chae C. 2014. Genetic and antigenic characterization of a newly emerging porcine circovirus type 2b mutant first isolated in cases of vaccine failure in Korea. Arch Virol 159:3107-11.
34. Opriessnig T, Gerber PF, Xiao CT, Halbur PG, Matzinger SR, Meng XJ. 2014. Commercial PCV2a-based vaccines are effective in protecting naturally PCV2b-infected finisher pigs against experimental challenge with a 2012 mutant PCV2. Vaccine 32:4342-8.
35. Franzo G, Cecchinato M, Martini M, Ceglie L, Gigli A, Drigo M. 2014. Observation of high recombination occurrence of Porcine Reproductive and Respiratory Syndrome Virus in field condition. Virus Res 194:159-66.
36. Suloway C, Shi J, Cheng A, Pulokas J, Carragher B, Potter CS, Zheng SQ, Agard DA, Jensen GJ. 2009. Fully automated, sequential tilt-series acquisition with Leginon. J Struct Biol 167:11-8.
37. Zheng SQ, Palovcak E, Armache JP, Verba KA, Cheng Y, Agard DA. 2017. MotionCor2: anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nat Methods 14:331-332.
38. Zhang T, Gu Y, Fan H. 2016. Effect of impurities and post-experimental purification in SAD phasing with serial femtosecond crystallography data. Acta Crystallogr D Struct Biol 72:789-94.
39. Scheres SH. 2012. RELION: implementation of a Bayesian approach to cryo-EM structure determination. J Struct Biol 180:519-30.
40. Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, Meng EC, Ferrin ТЕ. 2004. UCSF Chimera--a visualization system for exploratory research and analysis. J Comput Chem 25:1605-12.
41. Vilas JL, Gomez-Bianco J, Conesa P, Melero R, Miguel de la Rosa-Trevin J, Oton J, Cuenca J, Marabini R, Carazo JM, Vargas J, Sorzano COS. 2018. MonoRes: Automatic and Accurate Estimation of Local Resolution for Electron Microscopy Maps. Structure 26:337-344 e4.
42. Emsley P, Cowtan K. 2004. Coot: model-building tools for molecular graphics. Acta Crystallogr D Biol Crystallogr 60:2126-32.
43. Afonine PA, Headd JJ, Terwilliger TC, Adams PC. 2013. Computational Crystallography Newsletter 4:43-44.
44. Johnson M, Zaretskaya I, Raytselis Y, Merezhuk Y, McGinnis S, Madden TL. 2008. NCBI BLAST: abetter web interface. Nucleic Acids Res 36: W5-9.
45. Edgar RC. 2004. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res 32:1792-7.
46. Martin DP, Lemey P, Lott M, Moulton V, Posada D, Lefeuvre P. 2010. RDP3: a flexible and fast computer program for analyzing recombination. Bioinformatics 26:2462-3.
47. Goddard TD, Huang CC, Meng EC, Pettersen EF, Couch GS, Morris JH, Ferrin ТЕ. 2018. UCSF ChimeraX: Meeting modem challenges in visualization and analysis. Protein Sci 27:14-25.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ЭКСПРЕССИЯ БЕЛКА ORF2 PCV2b В КЛЕТКАХ НАСЕКОМЫХ | 2018 |
|
RU2779423C2 |
| PCV2B ДИВЕРГЕНТНАЯ ВАКЦИННАЯ КОМПОЗИЦИЯ И СПОСОБЫ ЕЁ ПРИМЕНЕНИЯ | 2014 |
|
RU2662685C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ИММУНИЗАЦИИ СВИНЕЙ ПРОТИВ СВИНОГО ЦИРКОВИРУСА | 2008 |
|
RU2493254C9 |
| ВАКЦИНА, ВКЛЮЧАЮЩАЯ БЕЛОК ORF2 PCV2 ГЕНОТИПА 2B | 2018 |
|
RU2774926C2 |
| РЕКОМБИНАНТНЫЕ АНТИГЕНЫ ЦИРКОВИРУСА СВИНЕЙ ТИПА 2 (PCV-2) ДЛЯ КОМПОЗИЦИЙ ВАКЦИН, ДИАГНОСТИЧЕСКИЙ НАБОР И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2687001C2 |
| ИММУНОГЕННЫЕ КОМПОЗИЦИИ ДЛЯ ИММУНИЗАЦИИ СВИНЕЙ ПРОТИВ ЦИРКОВИРУСА ТИПА 3 И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2016 |
|
RU2744193C2 |
| ИММУНОГЕННЫЕ КОМПОЗИЦИИ ДЛЯ ИММУНИЗАЦИИ СВИНЕЙ ПРОТИВ ЦИРКОВИРУСА ТИПА 3 И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2022 |
|
RU2803427C1 |
| ИММУНОГЕННЫЕ КОМПОЗИЦИИ ДЛЯ ИММУНИЗАЦИИ СВИНЕЙ ПРОТИВ ЦИРКОВИРУСА ТИПА 3 И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2016 |
|
RU2771533C2 |
| Способ получения секреторного рекомбинантного белка ORF2 цирковируса свиней второго типа в дрожжах Komagataella phaffii BG11/ pPic-CV2-7 в форме вирусоподобных частиц | 2023 |
|
RU2832292C1 |
| ВАКЦИНЫ ПРОТИВ СВИНОГО ЦИРКОВИРУСА 3 ТИПА (PCV3), ИХ ПРОИЗВОДСТВО И ПРИМЕНЕНИЕ | 2020 |
|
RU2813989C2 |



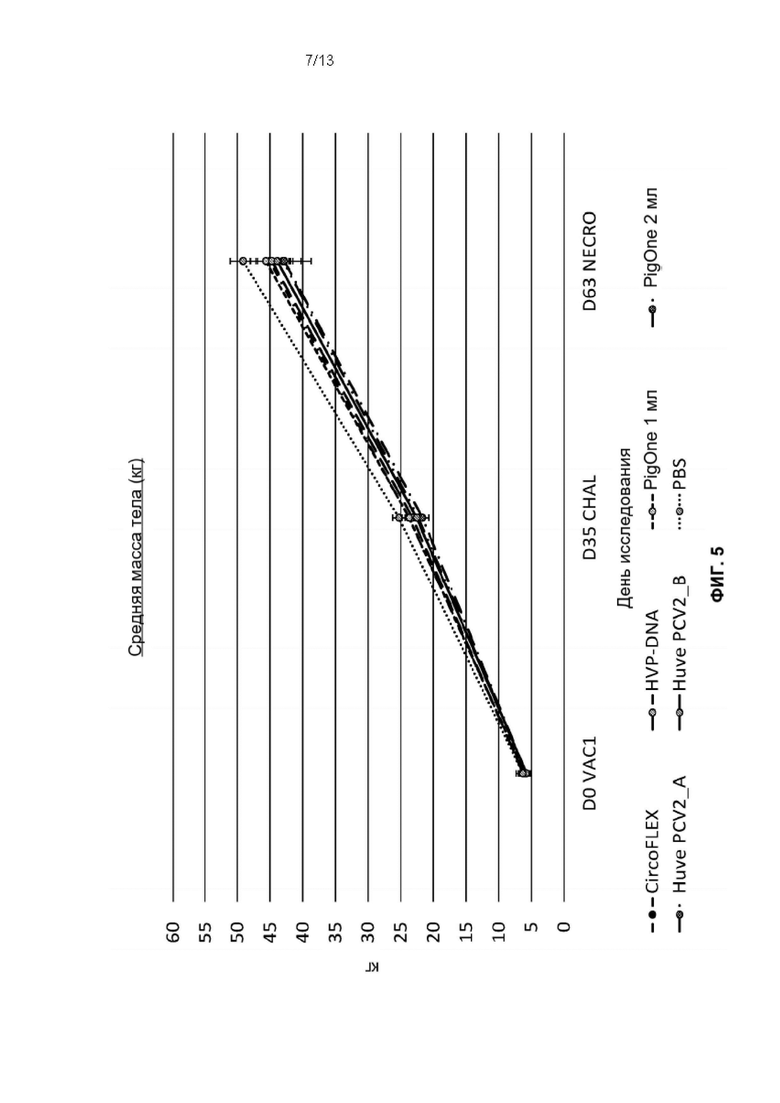
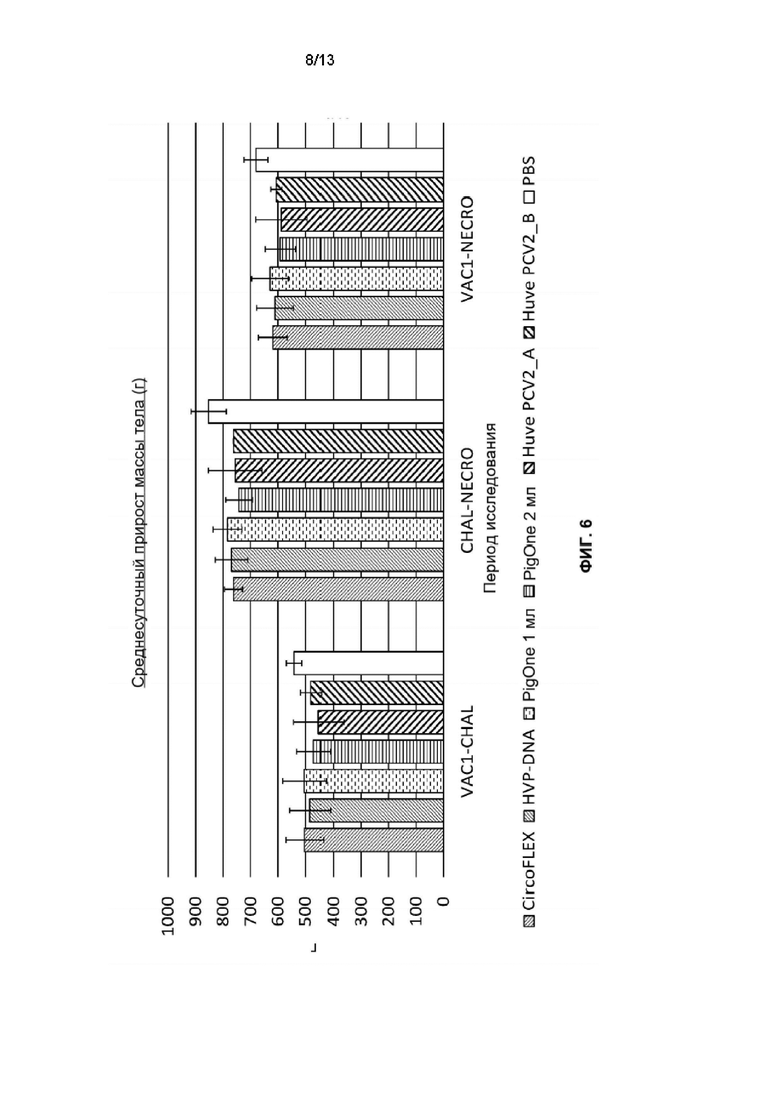
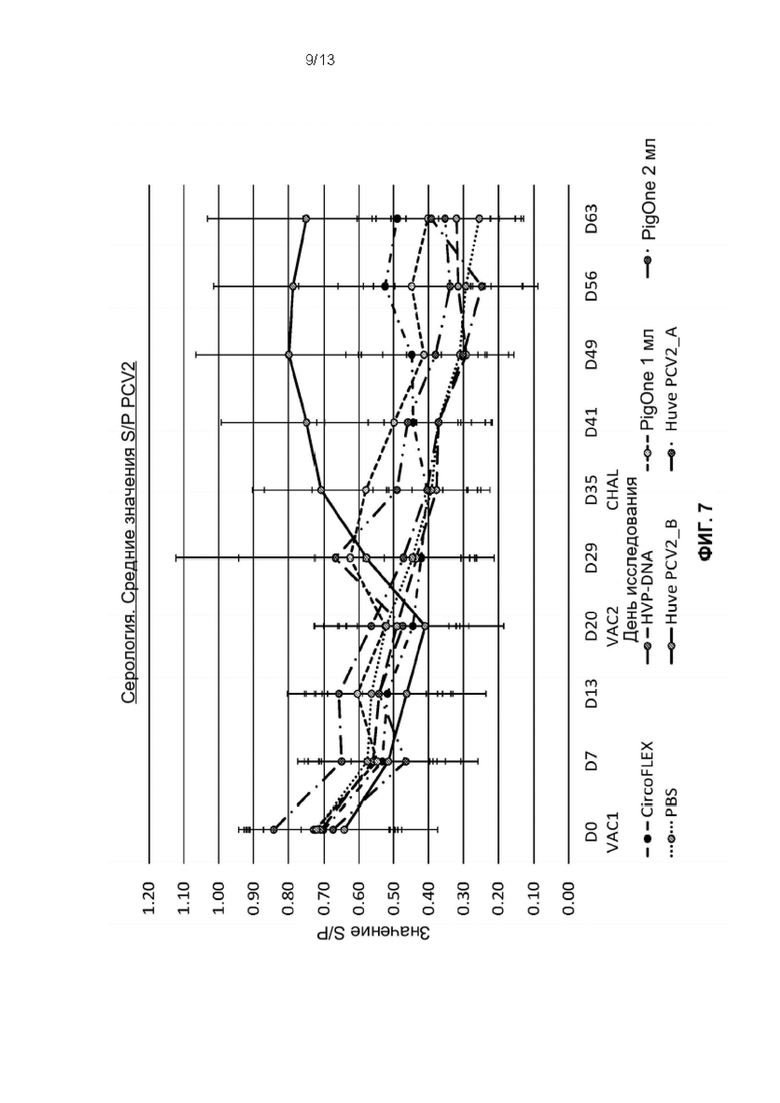

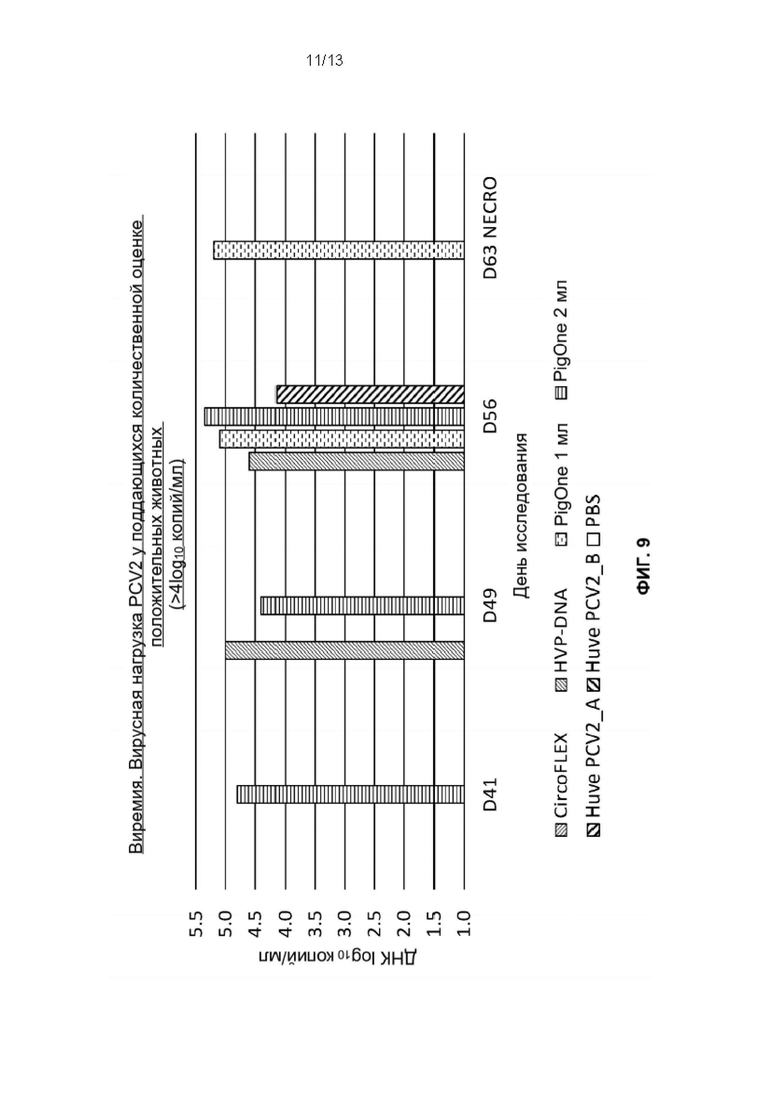
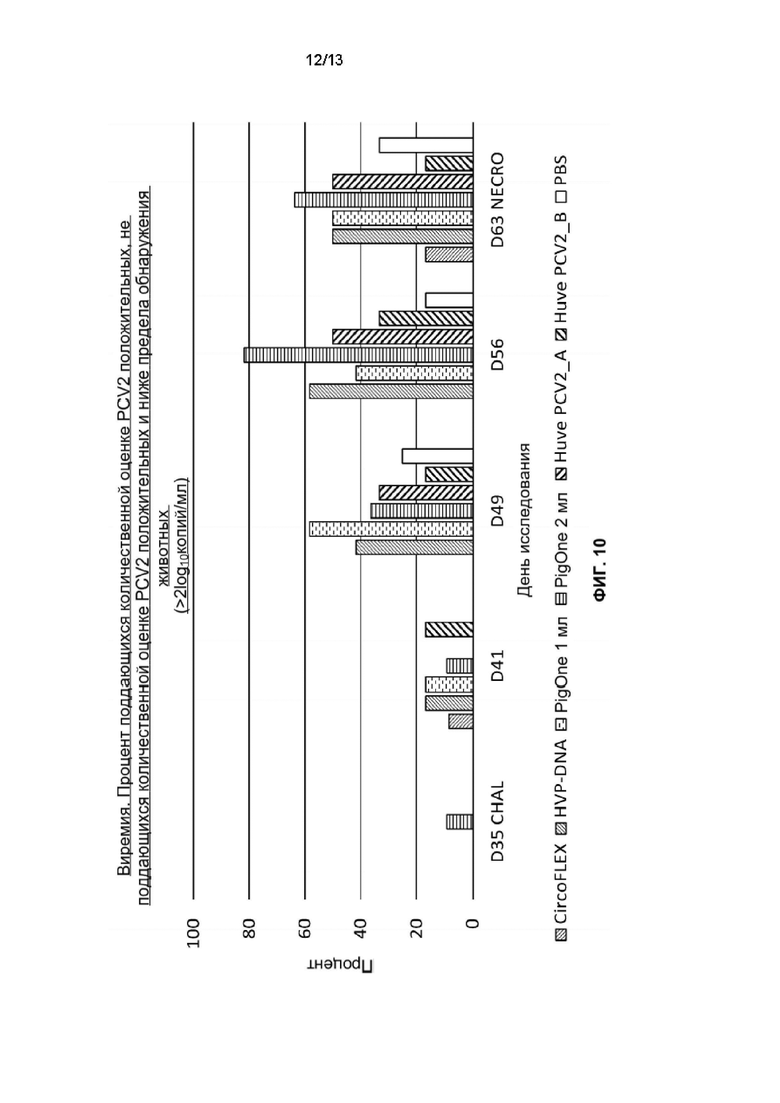
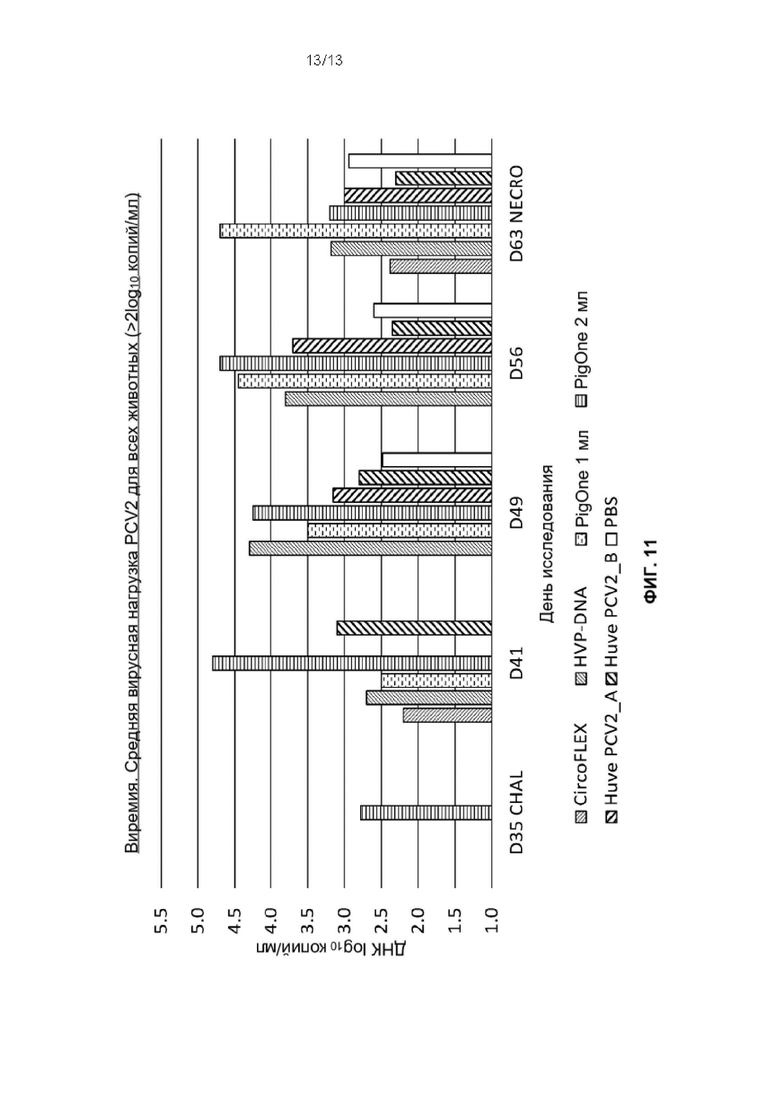
Изобретение относится к биотехнологии, вирусологии и ветеринарии. Предложена система экспрессии у млекопитающих для получения рекомбинантных вирусоподобных частиц (VLP) цирковируса свиней типа 2 (PCV2), при этом указанная система экспрессии содержит: клетку млекопитающего; плазмиду, содержащую ген PCV2, кодирующий капсидный белок, при этом указанный капсидный белок содержит аминокислотную последовательность SEQ ID NO: 4 и/или указанный капсидный белок кодируется нуклеотидной последовательностью SEQ ID NO: 3; при этом указанная клетка млекопитающего трансфицирована указанной плазмидой; и при этом указанная система экспрессии продуцирует рекомбинантные VLP PCV2, и причем указанный капсидный белок модифицирован сигнальной последовательностью секреции, введенной на NH2-конце указанного капсидного белка. Предложен способ получения вирусоподобных частиц (VLP) цирковируса свиней типа 2 (PCV2), включающий: обеспечение суспензии культивируемых клеток млекопитающего; трансфекцию указанных клеток млекопитающего плазмидой, содержащей ген PCV2, кодирующий капсидный белок, причем указанный капсидный белок содержит аминокислотную последовательность SEQ ID NO: 4 и/или указанный капсидный белок кодируется нуклеотидной последовательностью SEQ ID NO: 3, и причем указанный капсидный белок модифицирован сигнальной последовательностью секреции, введенной на NH2-конце указанного капсидного белка; добавление натриевой соли вальпроевой кислоты (VPA) к трансфицированным клеткам млекопитающего, причем добавление натриевой соли VPA ингибирует пролиферацию клеток; центрифугирование и промывание трансфицированных клеток млекопитающего; суспендирование центрифугированных клеток млекопитающего в фосфатно-солевом буферном растворе (PBS); осуществление нескольких циклов замораживания и оттаивания клеток млекопитающего; обработку ультразвуком клеток млекопитающего в нескольких циклах; и осуществление двух последовательных циклов центрифугирования клеток млекопитающего для получения VLP PCV2, причем большинство продуцируемых VLP PCV2 присутствуют в ядре клеток млекопитающего. Предложена рекомбинантная вирусоподобная частица цирковируса свиней типа 2 (VLP PCV2), причем капсидный белок, входящий в указанную VLP PCV2, модифицирован сигнальной последовательностью секреции, введенной на NH2-конце указанного капсидного белка, и при этом указанный капсидный белок содержит аминокислотную последовательность SEQ ID NO: 4 и/или указанный капсидный белок кодируется нуклеотидной последовательностью SEQ ID NO: 3. Изобретение позволяет расширить арсенал средств получения рекомбинантных вирусоподобных частиц для индукции иммунного ответа. 3 н. и 15 з.п. ф-лы, 2 табл., 25 ил., 2 пр.
1. Система экспрессии у млекопитающих для получения рекомбинантных вирусоподобных частиц (VLP) цирковируса свиней типа 2 (PCV2), при этом указанная система экспрессии содержит:
клетку млекопитающего;
плазмиду, содержащую ген PCV2, кодирующий капсидный белок, при этом указанный капсидный белок содержит аминокислотную последовательность SEQ ID NO: 4 и/или указанный капсидный белок кодируется нуклеотидной последовательностью SEQ ID NO: 3;
при этом указанная клетка млекопитающего трансфицирована указанной плазмидой; и
при этом указанная система экспрессии продуцирует рекомбинантные VLP PCV2, и причем указанный капсидный белок модифицирован сигнальной последовательностью секреции, введенной на NH2-конце указанного капсидного белка.
2. Система экспрессии по п. 1, отличающаяся тем, что указанная клетка млекопитающего представляет собой клетку эмбриональной почки человека-293 (HEK-293).
3. Система экспрессии по п. 1 или 2, отличающаяся тем, что ген PCV2 включает сайт распознавания NheI, последовательность Козака и сайт распознавания NotI, и
причем сайт распознавания NheI и последовательность Козака расположены выше стартового кодона гена PCV2, и сайт распознавания NotI встроен после кодона терминации гена PCV2.
4. Система экспрессии по любому из пп. 1-3, отличающаяся тем, что продуцируемые рекомбинантные VLP PCV2 выбраны из группы, состоящей из: VLP PCV2a, VLP PCV2b, VLP PCV2c, VLP PCV2d и VLP PCV2e.
5. Система экспрессии по п. 4, отличающаяся тем, что продуцируемые рекомбинантные VLP PCV2 представляют собой VLP PVC2d.
6. Система экспрессии по любому из пп. 1-5, отличающаяся тем, что указанная плазмида представляет собой pcDNA3.4-PCV2.
7. Способ получения вирусоподобных частиц (VLP) цирковируса свиней типа 2 (PCV2), включающий:
обеспечение суспензии культивируемых клеток млекопитающего;
трансфекцию указанных клеток млекопитающего плазмидой, содержащей ген PCV2, кодирующий капсидный белок, причем указанный капсидный белок содержит аминокислотную последовательность SEQ ID NO: 4 и/или указанный капсидный белок кодируется нуклеотидной последовательностью SEQ ID NO: 3, и причем указанный капсидный белок модифицирован сигнальной последовательностью секреции, введенной на NH2-конце указанного капсидного белка;
добавление натриевой соли вальпроевой кислоты (VPA) к трансфицированным клеткам млекопитающего, причем добавление натриевой соли VPA ингибирует пролиферацию клеток;
центрифугирование и промывание трансфицированных клеток млекопитающего;
суспендирование центрифугированных клеток млекопитающего в фосфатно-солевом буферном растворе (PBS);
осуществление нескольких циклов замораживания и оттаивания клеток млекопитающего;
обработку ультразвуком клеток млекопитающего в нескольких циклах; и
осуществление двух последовательных циклов центрифугирования клеток млекопитающего для получения VLP PCV2,
причем большинство продуцируемых VLP PCV2 присутствуют в ядре клеток млекопитающего.
8. Способ по п. 7, согласно которому стадия центрифугирования и промывания трансфицированных клеток млекопитающего включает:
центрифугирование клеток млекопитающего при 2000×g в течение 15 мин;
промывание клеток млекопитающего раствором PBS; и
центрифугирование клеток млекопитающего снова при 2000×g в течение 15 мин.
9. Способ по любому из пп. 7, 8, согласно которому центрифугированные клетки млекопитающего замораживают при приблизительно -80°C и оттаивают при приблизительно 37°C во время циклов замораживания и оттаивания.
10. Способ по любому из пп. 7-9, согласно которому первый из двух последовательных циклов центрифугирования осуществляют при 2000×g в течение 15 минут, и второй из двух последовательных циклов центрифугирования осуществляют при 8000×g в течение 15 минут.
11. Способ по любому из пп. 7-10, дополнительно включающий очистку VLP PCV2 с помощью ультрацентрифугирования.
12. Способ по любому из пп. 7-11, согласно которому клетки млекопитающего представляют собой клетки эмбриональной почки человека-293 (HEK-293).
13. Способ по любому из пп. 7-12, согласно которому ген PCV2 включает сайт распознавания NheI, последовательность Козака и сайт распознавания NotI, и
причем сайт распознавания NheI и последовательность Козака расположены выше стартового кодона гена PCV2, и сайт распознавания NotI встроен после кодона терминации гена PCV2.
14. Способ по любому из пп. 7-13, согласно которому указанная плазмида представляет собой pcDNA3.4-PCV2.
15. Способ по любому из пп. 7-14, согласно которому продуцируемые VLP PCV2 выбраны из группы, состоящей из: VLP PCV2a, VLP PCV2b, VLP PCV2c, VLP PCV2d и VLP PCV2e.
16. Способ по любому из пп. 7-15, согласно которому продуцируемые VLP PCV2 представляют собой VLP PVC2d.
17. Рекомбинантная вирусоподобная частица цирковируса свиней типа 2 (VLP PCV2), причем капсидный белок, входящий в указанную VLP PCV2, модифицирован сигнальной последовательностью секреции, введенной на NH2-конце указанного капсидного белка, и при этом указанный капсидный белок содержит аминокислотную последовательность SEQ ID NO: 4 и/или указанный капсидный белок кодируется нуклеотидной последовательностью SEQ ID NO: 3.
18. VLP PCV2 по п. 17, отличающаяся тем, что указанная вирусоподобная частица получена способом по любому из пп. 7-16.
| KHAYAT, R et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| bioRxiv | |||
| Способ изготовления электрических сопротивлений посредством осаждения слоя проводника на поверхности изолятора | 1921 |
|
SU19A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| Машина для добывания торфа и т.п. | 1922 |
|
SU22A1 |

