Изобретение относится к молекулам искусственной нуклеиновой кислоты, включающим открытую рамку считывания, элемент 3'-UTR и необязательно поли(А)-последовательность и/или сигнал полиаденилирования. Изобретение дополнительно относится к вектору, включающему элемент 3'-UTR, к фармацевтической композиции, включающей молекулу искусственной нуклеиновой кислоты, и к набору, включающему молекулу искусственной нуклеиновой кислоты, причем вектор и/или фармацевтическая композиция, включающие молекулу искусственной нуклеиновой кислоты, предпочтительно применяются в области генной терапии и/или генетической вакцинации.
Генная терапия и генетическая вакцинация принадлежат к наиболее многообещающим и быстро развивающимся методам современной медицины. С их помощью можно разработать высокоспецифичные и индивидуальные варианты лечения широкого спектра заболеваний. В частности, наследственные генетические заболевания, а также аутоиммунные заболевания, раковые или связанные с опухолями заболевания, а также воспалительные заболевания могут представлять собой объект для таких подходов к лечению. Предусматривается также предотвращение (раннего) проявления таких заболеваний с помощью этих подходов.
Главная рациональная концепция относительно генной терапии заключается в соответствующей модуляции нарушенной экспрессии генов, связанных с патологическими состояниями при конкретных заболеваниях. Патологически измененная экспрессия генов может привести к отсутствию или гиперпродукции важных продуктов генов, например, факторов сигнализации, таких как гормоны, факторы домашнего хозяйства, ферменты метаболизма, структурные белки или тому подобное. Измененная экспрессия генов может быть обусловлена не только неверной регуляцией транскрипции и/или трансляции, но также возникать в результате мутаций в пределах ORF, кодирующей конкретный белок. Патологические мутации могут быть вызваны, например, хромосомной аберрацией или более конкретными мутациями, такими как точечные мутации или мутации со сдвигом рамки считывания, причем все они приводят к ограниченному функционированию и потенциально к полной потере функции продукта гена. Однако нарушение регуляции транскрипции или трансляции может также возникать, если мутации возникают в генах, кодирующих белки, которые вовлечены в механизм транскрипции или трансляции в клетке. Такие мутации могут приводить к патологической позитивной или негативной регуляции генов, которые по существу являются функциональными. Гены, кодирующие генные продукты, которые имеют такие регуляторные функции, могут представлять собой, например, транскрипционные факторы, рецепторы сигнальных соединений, белки-посредники или тому подобное. Однако потеря функции таких генов, кодирующих регуляторные белки, может при определенных условиях быть возвращена искусственным введением других факторов, действующих далее по нижележащему пути относительно поврежденного генного продукта. Такие генные дефекты могут также компенсироваться генной терапией путем замены самого поврежденного гена.
Генетическая вакцинация позволяет вызвать желаемый иммунный ответ на выбранные антигены, такие как характерные компоненты поверхности бактерий, вирусных частиц, опухолевые антигены или тому подобное. В целом вакцинация является одним из главных достижений современной медицины. Однако эффективные вакцины в настоящее время доступны только для небольшого количества заболеваний. Соответственно, инфекции, которые не могут быть предотвращены вакцинацией, все еще поражают миллионы людей каждый год.
В целом вакцины могут быть подразделены на вакцины «первого», «второго» и «третьего» поколения. Вакцины «первого поколения» обычно представляют собой вакцины на основе целого организма. Они основаны либо на живых и ослабленных, либо на убитых патогенах, например, вирусах, бактериях или тому подобном. Главным недостатком живых и ослабленных вакцин является риск возвращения к вариантам, опасным для жизни. Таким образом, хотя и ослабленные, такие патогены могут все еще в действительности нести непредсказуемые риски. Убитые патогены могут быть не настолько эффективными, насколько это желательно для выработки специфического иммунного ответа. Для того чтобы минимизировать эти риски, были разработаны вакцины «второго поколения». Они обычно представляют собой субъединичные вакцины, состоящие из определенных антигенов или рекомбинантных белковых компонентов, которые происходят от патогенов.
Генетические вакцины, т.е. вакцины для генетической вакцинации, обычно обозначаются как вакцины «третьего поколения». Они обычно состоят из молекул нуклеиновой кислоты, созданных генно-инженерным способом, которые дают возможность экспрессироваться in vivo фрагментам пептида или белка (антигена), характерным для патогена или опухолевого антигена. Генетические вакцины экспрессируются при введении больному и захватываются компетентными клетками. Экспрессия введенных нуклеиновых кислот приводит к продукции кодируемых белков. В случае если эти белки узнаются иммунной системой больного как чужеродные, запускается иммунный ответ.
Как видно из представленного выше, оба метода, генная терапия и генетическая вакцинация, по существу основаны на введении молекул нуклеиновой кислоты больному и последующей транскрипции и/или трансляции кодируемой генетической информации. Альтернативно генетическая вакцинация или генная терапия может также включать методы, включающие выделение конкретных клеток из организма больного, подлежащего лечению, последующую трансфекцию in vitro таких клеток и повторное введение обработанных клеток больному.
В контексте генной терапии или генетической вакцинации в качестве молекул нуклеиновой кислоты для введения может быть использована ДНК, а также РНК. ДНК, как известно, является относительно стабильной и легкой для обработки. Однако использование ДНК несет риск нежелательной вставки введенных фрагментов ДНК в геном больного, потенциально ведущей к потере функции поврежденных генов. Дополнительным риском является нежелательная выработка антител против ДНК. Другим недостатком является ограниченный уровень экспрессии кодируемого пептида или белка, который может быть достигнут при введении ДНК и ее транскрипции/трансляции. Среди других причин, уровень экспрессии вводимой ДНК будет зависеть от присутствия специфических факторов транскрипции, которые регулируют транскрипцию ДНК. В отсутствие таких факторов транскрипция ДНК не будет давать удовлетворительные уровни РНК. В результате ограничивается уровень получаемого транслируемого пептида или белка.
При использовании для генной терапии или генетической вакцинации РНК вместо ДНК риск нежелательной интеграции в геном и выработки антител против ДНК сведен к минимуму или отсутствует. Однако РНК, как считается, являются относительно нестабильными типами молекул, которые могут легко деградировать под действием повсеместных РНКаз.
Деградация РНК in vivo вносит вклад в регуляцию периода полужизни РНК. Как предположено и доказано, этот эффект служит для тонкой настройки экспрессии генов эукариот (Friedel et al., Conserved principles of mammalian transcriptional regulation revealed by RNA half-life, Nucleic Acid Research, 2009, 1-12). Соответственно, каждая существующая в природе РНК имеет свой индивидуальный период полужизни в зависимости от гена, от которого происходит мРНК. Это вносит вклад в регуляцию уровня экспрессии этого гена. Нестабильные РНК важны для осуществления кратковременной экспрессии гена в отдельные точки времени. Однако долгоживущие РНК могут быть связаны с аккумуляцией конкретных белков или продолжительной экспрессией генов. Период полужизни мРНК in vivo может также зависеть от факторов окружающей среды, таких как лечение гормонами, как показано, например, для мРНК инсулиноподобного фактора роста I, актина и альбумина (Johnson et al., Newly synthesized RNA: Simultaneous measurement in intact cells of transcription rates and RNA stability of insulin-like growth factor I, actin, and albumin in growth hormone-stimulated hepatocytes, Proc. Natl. Acad. Sci., Vol. 88, pp. 5287-5291, 1991).
Для генной терапии и генетической вакцинации обычно желательна стабильная РНК. Это с одной стороны обусловлено тем фактом, что продукт, кодируемый последовательностью РНК, должен аккумулироваться in vivo. С другой стороны, РНК должна поддерживать свою структурную и функциональную целостность при получении в виде подходящей единицы лекарственной формы, в процессе ее хранения и при введении. Таким образом, существенное внимание уделялось предоставлению стабильных молекул РНК для генной терапии и генетической вакцинации для предохранения их как объекта ранней деградации или распада.
Сообщалось, что содержание G/C в молекулах нуклеиновой кислоты может влиять на их стабильность. Таким образом, нуклеиновые кислоты, включающие повышенное количество остатков гуанина (G) и/или цитозина (С), могут быть функционально более стабильными, чем нуклеиновые кислоты, содержащие большое количество нуклеотидов аденина (A), тимидина (T) или урацила (U). В этом контексте патент WO 02/098443 предлагает фармацевтическую композицию, содержащую мРНК, которая стабилизирована модификациями последовательности в транслируемой области. Такая модификация последовательности имеет преимущество в результате вырожденности генетического кода. Соответственно, кодоны, которые содержат менее благоприятное сочетание нуклеотидов (менее благоприятное в плане стабильности РНК), могут быть заменены альтернативными кодонами без изменения кодируемой аминокислотной последовательности. Этот метод стабилизации РНК ограничен резервами специфической последовательности нуклеотидов каждой отдельной молекулы РНК, которые не позволяют оставить пространство для желаемой аминокислотной последовательности. Этот подход ограничен также кодирующими областями РНК.
В качестве альтернативного варианта стабилизации мРНК, обнаружено, что природные молекулы мРНК эукариот содержат характерные стабилизирующие элементы. Например, они могут включать так называемые нетранслируемые области (UTR) на своем 5'-конце (5'-UTR) и/или на своем 3'-конце (3'-UTR), а также другие структурные характеристики, такие как структура в виде 5'-кэпа или 3'-поли(А)-хвоста. Как 5'-UTR, так и 3'-UTR обычно транскрибируются с геномной ДНК и являются, таким образом, элементами незрелой мРНК. Характерные структурные характеристики зрелой мРНК, такие как 5'-кэп и 3'-поли(А)-хвост (также называемый поли(А)-хвостом или поли(А)-последовательностью), обычно добавляются к транскрибированной (незрелой) мРНК при процессинге мРНК.
3'-поли(А)-хвост обычно представляет собой монотонный участок последовательности адениновых нуклеотидов, добавляемый к 3'-концу транскрибируемой мРНК. Он может включать приблизительно до 400 адениновых нуклеотидов. Обнаружено, что длина такого 3'-поли(А)-хвоста потенциально является решающим элементом для стабильности индивидуальной мРНК.
Показано также, что 3'-UTR мРНК α-глобина может быть важным фактором для хорошо известной стабильности мРНК α-глобина (Rodgers et al., Regulated α-globin mRNA decay is a cytoplasmic event proceeding through 3'-to-5' exosome-dependent decapping, RNA, 8, pp. 1526-1537, 2002). 3'-UTR мРНК α-глобина, очевидно, вовлечена в формирование конкретного рибонуклеопротеидного комплекса, α-комплекса, чье присутствие коррелирует со стабильностью мРНК in vitro (Wang et al., An mRNA stability complex functions with poly(A)-binding protein to stabilize mRNA in vitro, Molecular and Cellular biology, Vol 19, No. 7, July 1999, p. 4552-4560).
Независимо от факторов, влияющих на стабильность мРНК, эффективная трансляция вводимых молекул нуклеиновой кислоты клетками или тканями-мишенями имеет решающее значение для любого подхода с использованием молекул нуклеиновой кислоты для генной терапии или генетической вакцинации. Наряду с регуляцией стабильности, трансляция большинства мРНК также регулируется структурными особенностями, такими как UTR, 5'-кэп и 3'-поли(А)-хвост. В этом контексте сообщалось, что длина поли(А)-хвоста может также играть важную роль в эффективности трансляции. Стабилизирующие 3'-элементы, однако, могут также иметь ослабляющий эффект на трансляцию.
Целью настоящего изобретения является предоставление молекул нуклеиновой кислоты, которые могут быть пригодны для применения в генной терапии и/или генетической вакцинации. В частности, целью настоящего изобретения является обеспечение видами мРНК, которые стабилизированы относительно преждевременной деградации или распада без проявления значительной потери функциональности в плане эффективности трансляции. Другой целью настоящего изобретения является предоставление молекул нуклеиновой кислоты, кодирующих такие представленные выше виды мРНК, которые могут быть пригодны для использования при генной терапии и/или генетической вакцинации. Еще одной целью настоящего изобретения является создание фармацевтической композиции для применения при генной терапии и/или генетической вакцинации. Таким образом, целью настоящего изобретения является обеспечение улучшенными видами нуклеиновой кислоты, которые преодолевают описанные выше недостатки предшествующего уровня техники с помощью экономически эффективного и прямого подхода.
Цель, лежащая в основе настоящего изобретения, решается с помощью заявленного предмета изобретения.
Для ясности и понятности предоставляются следующие определения. Любой технический признак, упоминаемый в этих определениях, может быть истолкован в каждом без исключения варианте осуществления изобретения. Дополнительные определения и объяснения могут быть конкретно предоставлены в контексте этих вариантов осуществления.
Адаптивный иммунный ответ: Адаптивный иммунный ответ обычно понимается как ответ иммунной системы, специфичный для антигена. Антигенная специфичность позволяет генерировать ответы, которые адаптированы к конкретным патогенам или клеткам, инфицированным патогеном. Способность закреплять эти адаптивные ответы обычно поддерживается в организме «клетками памяти». Если патоген инфицировал организм более одного раза, эти конкретные клетки памяти используются для быстрого его устранения. В этом контексте первой стадией адаптивного иммунного ответа является активация наивных антигенспецифических Т-клеток или различных иммунных клеток, способных индуцировать антигенспецифический иммунный ответ антигенпрезентирующими клетками. Это происходит в лимфоидных тканях и органах, через которые постоянно проходят наивные Т-клетки. Три типа клеток, которые могут служить в качестве антигенпрезентирующих клеток, представляют собой дендритные клетки, макрофаги и В-клетки. Каждая из этих клеток имеет определенную функцию при индукции иммунных ответов. Дендритные клетки могут захватывать антигены путем фагоцитоза и макропиноцитоза и могут становиться стимулированными в результате контакта, например, с чужеродным антигеном, мигрировать в местную лимфоидную ткань, где они дифференцируются в зрелые дендритные клетки. Макрофаги поглощают частичковые антигены, такие как бактерии, и индуцируются инфекционными агентами или другими соответствующими стимулами для экспрессии молекул МНС. Уникальная способность В-клеток связывать и подвергать интернализации растворимые белковые антигены с помощью своих рецепторов также может быть важной для индукции Т-клеток. Молекулы MHC обычно отвечают за презентацию антигена Т-клеткам. Здесь презентация антигена на молекулах МНС приводит к активации Т-клеток, что вызывает их пролиферацию и дифференцировку в подготовленные к работе эффекторные Т-клетки. Наиболее важной функцией эффекторных Т-клеток является уничтожение инфицированных клеток с помощью CD8+ цитотоксических Т-клеток и активация макрофагов с помощью Th1-клеток, что вместе составляет клеточный иммунитет, а также активация В-клеток с помощью как Th2, так и Th1 клеток для продукции различных классов антител, и, таким образом, управления гуморальным иммунным ответом. Т-клетки распознают антиген своими Т-клеточными рецепторами, которые не распознают и не связывают антиген непосредственно, а вместо этого распознают короткие пептидные фрагменты, например, происходящих от патогена белковых антигенов, например, так называемые эпитопы, которые связаны с молекулами МНС на поверхности других клеток.
Адаптивная иммунная система: Адаптивная иммунная система по существу призвана прекратить или предотвратить рост патогенов. Она обычно регулирует адаптивный иммунный ответ, обеспечивая иммунную систему позвоночных способностью распознавать и запоминать конкретные патогены (для выработки иммунитета) и закреплять все более сильные атаки каждый раз при встрече с патогеном. Система является высоко адаптируемой из-за соматической гипермутации (процесса ускоренных соматических мутаций), и рекомбинации V(D)J (необратимой генетической рекомбинации сегментов гена рецептора антигена). Этот механизм позволяет небольшому количеству генов генерировать огромное количество рецепторов различных антигенов, которые затем уникально экспрессируются на каждом индивидуальном лимфоците. Поскольку перегруппировка генов приводит к необратимому изменению в ДНК каждой клетки, все потомство (потомки) такой клетки затем наследует гены, кодирующие рецептор с одной и той же специфичностью, включая В-клетки памяти и Т-клетки памяти, которые являются ключевыми для продолжительного специфического иммунитета.
Адъювант/компонент адъюванта: Адъювант или компонент адъюванта в самом широком смысле обычно представляет собой фармакологический и/или иммунологический агент, который может модифицировать, например, повышать действие других агентов, таких как лекарство или вакцина. Это следует интерпретировать в широком смысле, и это относится к широкому спектру веществ. Обычно эти вещества способны повысить иммуногенность антигенов. Например, адъюванты могут узнаваться врожденной иммунной системой и, например, могут вызывать врожденный иммунный ответ. «Адъюванты» обычно не вызывают адаптивный иммунный ответ. В этой связи «адъюванты» не могут рассматриваться в качестве антигенов. Их механизм действия отличается от эффектов, запускаемых антигенами, приводя к адаптивному иммунному ответу.
Антиген: В контексте настоящего изобретения «антиген» обычно относится к веществу, которое может узнаваться иммунной системой, предпочтительно адаптивной иммунной системой, и способно запускать антигенспецифический иммунный ответ, например, путем образования антител и/или антигенспецифических Т-клеток как части адаптивного иммунного ответа. Обычно антиген может представлять собой или может включать пептид или белок, который может презентироваться с помощью МНС Т-клеткам.
Молекула искусственной нуклеиновой кислоты: Молекула искусственной нуклеиновой кислоты может обычно пониматься как молекула нуклеиновой кислоты, например, ДНК или РНК, которая не существует в природе. Другими словами, молекула искусственной нуклеиновой кислоты может пониматься как неприродная молекула нуклеиновой кислоты. Такая молекула нуклеиновой кислоты может быть неприродной из-за своей индивидуальной последовательности (которая не встречается в природе) и/или за счет других модификаций, например, структурных модификаций нуклеотидов, которые не встречаются в природе. Молекула искусственной нуклеиновой кислоты может представлять собой молекулу ДНК, молекулу РНК или гибридную молекулу, включающую части ДНК и РНК. Обычно молекулы искусственной нуклеиновой кислоты могут быть сконструированы и/или созданы методами генной инженерии, чтобы соответствовать желаемой искусственной последовательности нуклеотидов (гетерологичная последовательность). В этом контексте искусственная последовательность обычно представляет собой последовательность, которая не может существовать в природе, т.е. она отличается от последовательности дикого типа по меньшей мере одним нуклеотидом. Термин «дикий тип» можно понимать как последовательность, существующую в природе. Кроме того, термин «молекула искусственной нуклеиновой кислоты» не ограничивается обозначением «одной единственной молекулы», но, обычно, понимается как включающий множество идентичных молекул. Соответственно, он может относиться к множеству идентичных молекул, содержащихся в аликвоте.
Бицистронная РНК, мультицистронная РНК: Бицистронная или мультицистронная РНК обычно представляет собой РНК, предпочтительно мРНК, которая обычно может иметь две (бицистронная) или более (мультицистронная) открытых рамок считывания (ORF). Открытая рамка считывания в данном контексте представляет собой последовательность кодонов, которая способна к трансляции в пептид или белок.
Носитель/полимерный носитель: Носитель в контексте настоящего изобретения обычно может представлять собой соединение, которое облегчает транспорт и/или комплексирование другого соединения (груза). Полимерный носитель обычно представляет собой носитель, который образован из полимера. Носитель может быть связан со своим грузом путем ковалентного или нековалентного взаимодействия. Носитель может транспортировать нуклеиновые кислоты, например, РНК или ДНК, в клетки-мишени. Носитель в некоторых вариантах осуществления может быть катионным компонентом.
Катионный компонент: Термин «катионный компонент» обычно относится к заряженной молекуле, которая является положительно заряженной (катионом) при величине рН обычно от 1 до 9, предпочтительно при величине рН 9 или ниже (например, от 5 до 9), 8 или ниже (например, от 5 до 8), 7 или ниже (например, от 5 до 7), наиболее предпочтительно при физиологическом рН, например, от 7,3 до 7,4. Таким образом, катионный компонент может представлять собой любое положительно заряженное соединение или полимер, предпочтительно катионный пептид или белок, который положительно заряжен в физиологических условиях, особенно в физиологических условиях in vivo. «Катионный пептид или белок» может содержать по меньшей мере одну положительно заряженную аминокислоту или более одной положительно заряженной аминокислоты, например, выбранных из Arg, His, Lys или Orn. Таким образом, «поликатионные» компоненты, имеющие более одного положительного заряда в данных условиях, также включены в объем изобретения.
5'-кэп: 5'-кэп является участком, обычно участком модифицированных нуклеотидов, который обычно «кэпирует» 5'-конец зрелой мРНК. 5'-кэп может обычно образовываться с помощью модифицированного нуклеотида, в частности, производного нуклеотида гуанина. Предпочтительно 5'-кэп связан с 5'-концом через 5'-5'-трифосфатную связь. 5'-кэп может быть метилированным, например, m7GpppN, где N представляет собой концевой 5'-нуклеотид нуклеиновой кислоты, несущей 5'-кэп, обычно 5'-конец РНК. Другие примеры структур 5'-кэпа включают глицерил, инвертированный дезоксиабазический остаток (часть), 4',5'-метиленнуклеотид, 1-(бета-D-эритрофуранозил)нуклеотид, 4'-тионуклеотид, карбоциклический нуклеотид, 1,5-ангидрогекситолнуклеотид, L-нуклеотиды, альфа-нуклеотид, модифицированное нуклеотидное основание, трео-пентофуранозилнуклеотид, ациклический 3',4'-секонуклеотид, ациклический 3,4-дигидроксибутилнуклеотид, ациклический 3,5-дигидроксипентилнуклеотид, 3'-3'-инвертированную нуклеотидную часть, 3'-3'-инвертированную абазическую часть, 3'-2'-инвертированную нуклеотидную часть, 3'-2'-инвертированную абазическую часть, 1,4-бутандиолфосфат, 3'-фосфороамидат, гексилфосфат, аминогексилфосфат, 3'-фосфат, 3'-фосфоротиоат, фосфородитиоат или связанную или не связанную мостиком метилфосфонатную часть.
Клеточный иммунитет/клеточный иммунный ответ: Клеточный иммунитет обычно относится к активации макрофагов, естественных клеток-киллеров (NK), антигенспецифических цитотоксических Т-лимфоцитов, а также к секреции различных цитокинов в ответ на антиген. В более общем плане клеточный иммунитет основан не на антителах, а на активации клеток иммунной системы. Обычно клеточный иммунный ответ может быть охарактеризован, например, активацией антигенспецифических цитотоксических Т-лимфоцитов, которые способны индуцировать апоптоз в клетках, например, в специфических иммунных клетках, таких как дендритные клетки или другие клетки, экспонирующие эпитопы чужеродных антигенов на своей поверхности. Такие клетки могут быть инфицированы вирусом или инфицированы внутриклеточными бактериями или являются раковыми клетками, экспонирующими опухолевые антигены. Другие характеристики могут представлять собой активацию макрофагов и естественных клеток-киллеров, что позволяет им уничтожить патогены и стимулировать клетки к секреции разнообразных цитокинов, которые влияют на функцию других клеток, вовлеченных в адаптивные иммунные ответы и врожденные иммунные ответы.
ДНК: ДНК представляет собой обычно сокращенное название дезоксирибонуклеиновой кислоты. Она представляет собой молекулу нуклеиновой кислоты, т.е. полимера, состоящего из нуклеотидов. Эти нуклеотиды обычно представляют собой мономеры дезоксиаденозинмонофосфата, дезокситимидинмонофосфата, дезоксигуанозинмонофосфата и дезоксицитидинмонофосфата, которые сами состоят из части сахара (дезоксирибозы), части основания и части фосфата, и полимеризуются в характерную структуру остова. Структура остова обычно образуется с помощью фосфодиэфирных связей между частью сахара, т.е. дезоксирибозой, первого нуклеотида и фосфатной частью второго соседнего мономера. Конкретный порядок мономеров, т.е. порядок оснований, связанных с сахаро/фосфатным остовом, называют последовательностью ДНК. ДНК может быть одноцепочечной или двухцепочечной. В двухцепочечной форме, нуклеотиды первой цепи обычно гибридизуются с нуклеотидами второй цепи, например, путем спаривания оснований A/T и спаривания оснований G/C.
Эпитоп: (также называемый «антигенной детерминантой») может быть подразделен на Т-клеточные эпитопы и B-клеточные эпитопы. Т-клеточные эпитопы или части белков в контексте настоящего изобретения могут включать фрагменты, предпочтительно имеющие в длину от приблизительно 6 до приблизительно 20 или даже более аминокислот, например, фрагменты, прошедшие процессинг и презентируемые молекулами класса I МНС, предпочтительно имеющие в длину от приблизительно 8 до приблизительно 10 аминокислот, например, 8, 9 или 10, (или даже 11 или 12 аминокислот), или фрагменты, прошедшие процессинг и презентируемые молекулами класса II МНС, предпочтительно имеющие в длину приблизительно 13 или более аминокислот, например, 13, 14, 15, 16, 17, 18, 19, 20 или даже более аминокислот, причем эти фрагменты могут быть выбраны из любой части аминокислотной последовательности. Эти фрагменты обычно узнаются Т-клетками в виде комплекса, состоящего из пептидного фрагмента и молекулы МНС, т.е. фрагменты обычно не узнаются в их нативной форме. В-клеточные эпитопы обычно представляют собой фрагменты, расположенные на наружной поверхности (нативных) белковых или пептидных антигенов, как определено в настоящем описании, предпочтительно содержащие от 5 до 15 аминокислот, более предпочтительно содержащие от 5 до 12 аминокислот, еще более предпочтительно содержащие от 6 до 9 аминокислот, которые могут узнаваться антителами, т.е. в их нативной форме.
Такие эпитопы белков или пептидов кроме того могут быть выбраны из любых упомянутых в данном документе вариантов таких белков или пептидов. В этом контексте антигенные детерминанты могут представлять собой конформационные или прерывистые эпитопы, которые включают сегменты белков или пептидов, как определено в настоящем описании, которые являются прерывистыми по аминокислотной последовательности белков или пептидов, как определено в настоящем описании, но собраны вместе в трехмерную структуру, или непрерывные или линейные эпитопы, которые состоят из одной полипептидной цепи.
Фрагмент последовательности: Фрагмент последовательности обычно может представлять собой более короткую часть полноразмерной последовательности, например, молекулы нуклеиновой кислоты или аминокислотной последовательности. Таким образом, фрагмент обычно состоит из последовательности, идентичной соответствующему участку в полноразмерной последовательности. Предпочтительный фрагмент последовательности в контексте настоящего изобретения состоит из непрерывного участка элементов, таких как нуклеотиды или аминокислоты, соответствующих непрерывному участку элементов в молекуле, из которой произошел фрагмент, который состоит из по меньшей мере 20%, предпочтительно по меньшей мере 30%, более предпочтительно по меньшей мере 40%, более предпочтительно по меньшей мере 50%, еще более предпочтительно по меньшей мере 60%, еще более предпочтительно по меньшей мере 70% и наиболее предпочтительно по меньшей мере 80% полной (т.е. полноразмерной) молекулы, из которой происходит этот фрагмент.
G/C модифицированная: G/С-модифицированная нуклеиновая кислота обычно может представлять собой нуклеиновую кислоту, предпочтительно молекулу искусственной нуклеиновой кислоты, как определено в настоящем описании, на основе модифицированной последовательности дикого типа, включающей предпочтительно повышенное количество гуанозиновых и/или цитозиновых нуклеотидов по сравнению с последовательностью дикого типа. Такое повышенное количество может быть создано путем замены кодонов, содержащих аденозиновые или тимидиновые нуклеотиды, кодонами, содержащими гуанозиновые или цитозиновые нуклеотиды. Если обогащенное содержание G/С присутствует в кодирующей области ДНК или РНК, пользуются вырожденностью генетического кода. Соответственно, замены кодонов предпочтительно не изменяют кодируемые аминокислотные остатки, а исключительно повышают содержание G/C в молекуле нуклеиновой кислоты.
Генная терапия: Генная терапия может обычно подразумевать способ лечения организма больного или выделенных элементов организма больного, например, выделенных тканей/клеток, с помощью нуклеиновых кислот, кодирующих пептид или белок. Он обычно может включать по меньшей мере одну из стадий: а) введения нуклеиновой кислоты, предпочтительно молекулы искусственной нуклеиновой кислоты, как определено в настоящем описании, непосредственно больному любым способом введения, или in vitro в выделенные клетки/ткани больного, что приводит к трансфекции клеток больного либо in vivo/ex vivo либо in vitro; б) транскрипции и/или трансляции введенной молекулы нуклеиновой кислоты; и, необязательно, с) повторного введения выделенных, трансфицированных клеток больному, если нуклеиновая кислота не была введена непосредственно больному.
Генетическая вакцинация: Генетическая вакцинация может обычно пониматься как вакцинация путем введения молекулы нуклеиновой кислоты, кодирующей антиген или иммуноген, или их фрагменты. Молекула нуклеиновой кислоты может быть введена в организм индивидуума или в выделенные клетки индивидуума. После трансфекции определенных клеток организма или трансфекции выделенных клеток антиген или иммуноген может экспрессироваться этими клетками, а затем презентироваться иммунной системе, вызывая адаптивный, т.е. антигенспецифический, иммунный ответ. Таким образом, генетическая вакцинация обычно включает по меньшей мере одну из стадий: а) введения нуклеиновой кислоты, предпочтительно молекулы искусственной нуклеиновой кислоты, как определено в настоящем описании, индивидууму, предпочтительно больному, или в выделенные клетки индивидуума, предпочтительно больного, что обычно приводит к трансфекции клеток индивидуума либо in vivo, либо in vitro, б) транскрипции и/или трансляции введенной молекулы нуклеиновой кислоты; и, необязательно, с) повторного введения выделенных, трансфицированных клеток индивидууму, предпочтительно больному, если нуклеиновая кислота не была введена непосредственно больному.
Гетерологичная последовательность: Две последовательности обычно считаются «гетерологичными», если они не происходят от одного и того же гена. Т.е., хотя гетерологичные последовательности могут происходить от одного и того же организма, они в естественных условиях (в природе) не присутствуют в одной и той же молекуле нуклеиновой кислоты, например, в одной и той же мРНК.
Гуморальный иммунитет/гуморальный иммунный ответ: Гуморальный иммунитет обычно относится к продукции антител и необязательно к дополнительным процессам, сопровождающим продукцию антител. Гуморальный иммунный ответ обычно может быть охарактеризован, например, активацией Th2 и продукцией цитокинов, формированием зародышевого центра и переключением изотипов, созреванием аффинности и генерацией клеток памяти. Гуморальный иммунитет также обычно может относиться к эффекторным функциям антител, которые включают нейтрализацию патогена и токсина, активацию классического комплемента и стимулируемый опсонином фагоцитоз и уничтожение патогенов.
Иммуноген: В контексте настоящего изобретения иммуноген может обычно пониматься как соединение, которое способно стимулировать иммунный ответ. Предпочтительно иммуноген представляет собой пептид, полипептид или белок. В особенно предпочтительном варианте осуществления иммуноген по настоящему изобретению представляет собой продукт трансляции предлагаемой молекулы нуклеиновой кислоты, предпочтительно молекулы искусственной нуклеиновой кислоты, как определено в настоящем описании. Обычно иммуноген вызывает, по меньшей мере, адаптивный иммунный ответ.
Иммуностимулирующая композиция: В контексте данного изобретения иммуностимулирующая композиция может обычно пониматься как композиция, содержащая по меньшей мере один компонент, который способен индуцировать иммунный ответ или из которой происходит компонент, который способен индуцировать иммунный ответ. Такой иммунный ответ может предпочтительно представлять собой врожденный иммунный ответ или сочетание адаптивного и врожденного иммунного ответа. Предпочтительно иммуностимулирующая композиция в контексте настоящего изобретения содержит по меньшей мере одну молекулу искусственной нуклеиновой кислоты, более предпочтительно РНК, например, молекулу мРНК. Иммуностимулирующий компонент, такой как мРНК, может комплексироваться с подходящим носителем. Таким образом, иммуностимулирующая композиция может включать комплекс мРНК/носитель. Кроме того, иммуностимулирующая композиция может включать адъювант и/или подходящий носитель для иммуностимулирующего компонента, такого как мРНК.
Иммунный ответ: Иммунный ответ может обычно представлять собой специфическую реакцию адаптивной иммунной системы на определенный антиген (так называемый специфический или адаптивный иммунный ответ) или неспецифическую реакцию врожденной иммунной системы (так называемый неспецифический или врожденный иммунный ответ) или их сочетание.
Иммунная система: Иммунная система может защищать организмы от инфекции. Если патогену удалось пройти через физический барьер организма и поступить в этот организм, врожденная иммунная система обеспечивает немедленный, но неспецифический ответ. Если патогены ускользают от этого врожденного ответа, позвоночные обладают вторым уровнем защиты, адаптивной иммунной системой. В этом случае иммунная система адаптирует свой ответ в процессе инфекции, чтобы улучшить свое узнавание патогена. Этот улучшенный ответ затем сохраняется после уничтожения патогена в виде иммунологической памяти и позволяет адаптивной иммунной системе закреплять более быстрые и более сильные атаки каждый раз при встрече с этим патогеном. В соответствии с этим, иммунная система состоит из врожденной и адаптивной иммунной системы. Каждая из этих двух частей обычно содержит так называемые гуморальные и клеточные компоненты.
Иммуностимулирующая РНК: Иммуностимулирующая РНК (isRNA) в контексте настоящего изобретения обычно может представлять собой РНК, которая способна индуцировать врожденный иммунный ответ. Она обычно не имеет открытой рамки считывания и, таким образом, не обеспечивает пептидом-антигеном или иммуногеном, но вызывает иммунный ответ, например, путем связывания с конкретным видом Toll-подобного рецептора (TLR) или другими подходящими рецепторами. Однако разумеется, мРНК, имеющие открытую рамку считывания и кодирующие пептид/белок, также могут вызвать врожденный иммунный ответ и, таким образом, могут представлять собой иммуностимулирующие РНК.
Врожденная иммунная система: Врожденная иммунная система, также известная как неспецифическая (или неспецифичная) иммунная система, обычно включает клетки и механизмы, которые защищают хозяина от инфицирования другими организмами неспецифическим способом. Это означает, что клетки врожденной системы могут распознавать и реагировать на патогены общим способом, но в отличие от адаптивной иммунной системы, она не создает у хозяина продолжительного или защитного иммунитета. Врожденная иммунная система может быть активирована, например, с помощью лигандов Toll-подобных рецепторов (TLR) или других вспомогательных соединений, таких как липополисахариды, TNF-альфа, лиганд CD40, или цитокины, лимфокины, монокины, интерлейкины или хемокины, IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11, IL-12, IL-13, IL-14, IL-15, IL-16, IL-17, IL-18, IL-19, IL-20, IL-21, IL-22, IL-23, IL-24, IL-25, IL-26, IL-27, IL-28, IL-29, IL-30, IL-31, IL-32, IL-33, IFN-альфа, IFN-бета, IFN-гамма, GM-CSF, G-CSF, M-CSF, LT-бета, TNF-альфа, факторы роста и hGH, лиганд Toll-подобного рецептора человека TLR1, TLR2, TLR3, TLR4, TLR5, TLR6, TLR7, TLR8, TLR9, TLR10, лиганд Toll-подобного рецептора мыши TLR1, TLR2, TLR3, TLR4, TLR5, TLR6, TLR7, TLR8, TLR9, TLR10, TLR11, TLR12 или TLR13, лиганд NOD-подобного рецептора, лиганд RIG-1-подобного рецептора, иммуностимулирующая нуклеиновая кислота, иммуностимулирующая РНК (isRNA), CpG-DNA, антибактериальный агент или противовирусный агент. Фармацевтическая композиция по настоящему изобретению может включать одно или более таких веществ. Обычно ответ врожденной иммунной системы включает рекрутирование иммунных клеток к местам инфекции путем продукции химических факторов, включая специализированные химические медиаторы, называемые цитокинами; активацию каскада комплемента; выявление и устранение чужеродных веществ, присутствующих в органах, тканях, крови и лимфе, специализированными лейкоцитами; активацию адаптивной иммунной системы; и/или действие в качестве физического и химического барьера для инфекционных агентов.
Сайт клонирования: Сайт клонирования обычно понимается как сегмент молекулы нуклеиновой кислоты, который подходит для вставки последовательности нуклеиновой кислоты, например, последовательности нуклеиновой кислоты, включающей открытую рамку считывания. Вставка может быть выполнена любым молекулярно-биологическим методом, известным специалисту в данной области техники, например, путем рестрикции и лигирования. Сайт клонирования обычно включает один или более сайтов рестрикции, узнаваемых ферментом (сайты рестрикции). Эти один или более сайтов рестрикции могут узнаваться ферментами рестрикции, которые расщепляют ДНК в этих сайтах. Сайт клонирования, который включает более одного сайта рестрикции, может также обозначаться как сайт множественного клонирования (MCS) или полилинкер.
Молекула нуклеиновой кислоты: Молекула нуклеиновой кислоты представляет собой молекулу, включающую, предпочтительно состоящую из компонентов нуклеиновой кислоты. Термин молекула нуклеиновой кислоты предпочтительно относится к молекулам ДНК или РНК. Он предпочтительно используется как синоним термину «полинуклеотид». Предпочтительно молекула нуклеиновой кислоты представляет собой полимер, включающий или состоящий из нуклеотидных мономеров, которые ковалентно связаны друг с другом с помощью фосфодиэфирных связей сахаро/фосфатного остова. Термин «молекула нуклеиновой кислоты» охватывает также молекулы модифицированных нуклеиновых кислот, например, молекулы ДНК или РНК с модифицированными основаниями, модифицированными сахарами или модифицированными остовами т.д.
Открытая рамка считывания: Открытая рамка считывания (ORF) в контексте изобретения может обычно представлять собой последовательность из нескольких триплетов нуклеотидов, которые могут транслироваться в пептид или белок. Открытая рамка считывания предпочтительно содержит инициирующий кодон, т.е. сочетание трех последовательных нуклеотидов, кодирующих обычно аминокислоту метионин (ATG), на ее 5'-конце и последующую область, которая обычно составляет в длину величину, кратную 3 нуклеотидам. ORF предпочтительно завершается стоп-кодоном (например, ТАА, TAG, TGA). Обычно это единственный стоп-кодон открытой рамки считывания. Таким образом, открытая рамка считывания в контексте настоящего изобретения предпочтительно представляет собой нуклеотидную последовательность, состоящую из количества нуклеотидов, которое можно разделить на три, которая начинается с инициирующего кодона (например, ATG) и которая предпочтительно заканчивается стоп-кодоном (например, ТАА, TGA или TAG). Открытая рамка считывания может быть отдельной, или она может быть включена в более длинную последовательность нуклеиновой кислоты, например, в вектор или мРНК. Открытая рамка считывания может обозначаться как «область, кодирующая белок».
Пептид: Пептид или полипептид обычно представляет собой полимер из аминокислотных мономеров, соединенных пептидными связями. Он обычно содержит менее 50 мономерных единиц. Тем не менее, термин пептид включает молекулы, имеющие более 50 мономерных единиц. Длинные пептиды называют также полипептидами, имеющими обычно от 50 до 600 мономерных единиц.
Фармацевтически эффективное количество: Фармацевтически эффективное количество в контексте настоящего изобретения обычно понимается как такое количество, которого достаточно, чтобы вызвать фармацевтический эффект, такой как иммунный ответ, изменение патологического уровня экспрессируемого пептида или белка или замена отсутствующего продукта гена, например, в случае патологического состояния.
Белок: Белок обычно включает один или более пептидов или полипептидов. Белок обычно имеет 3-х-мерную укладку, которая может требоваться, чтобы белок оказывал свое биологическое действие.
Поли(А)-последовательность: Поли(А)-последовательность, которая также называется поли(А)-хвост или 3'-поли(А)-хвост, обычно понимается как последовательность адениновых нуклеотидов, например, вплоть до приблизительно 400 адениновых нуклеотидов, например, от приблизительно 20 до приблизительно 400, предпочтительно от приблизительно 50 до приблизительно 400, более предпочтительно от приблизительно 50 до приблизительно 300, еще более предпочтительно от приблизительно 50 до приблизительно 250, наиболее предпочтительно от приблизительно 60 до приблизительно 250 адениновых нуклеотидов. Поли(А)-последовательность обычно расположена на 3'-конце мРНК. В контексте настоящего изобретения поли(А)-последовательность может быть расположена в мРНК или в любой другой молекуле нуклеиновой кислоты, такой как, например, вектор, например, вектор, служащий матрицей для продукции РНК, предпочтительно мРНК, например, путем транскрипции вектора.
Полиаденилирование: Полиаденилирование обычно понимается как добавление поли(А)-последовательности к молекуле нуклеиновой кислоты, такой как молекула РНК, например, незрелая мРНК. Полиаденилирование может быть индуцировано с помощью так называемого сигнала полиаденилирования. Этот сигнал предпочтительно локализован в пределах участка нуклеотидов на 3'-конце молекулы нуклеиновой кислоты, такой как молекула РНК, подлежащая полиаденилированию. Сигнал полиаденилирования обычно включает гексамер, состоящий из нуклеотидов аденина и урацила/тимина, предпочтительная последовательность гексамера AAUAAA. Возможны также другие последовательности для предпочтительных последовательностей гексамера. Полиаденилирование обычно происходит во время процессинга пре-мРНК (также называемой незрелой мРНК). Обычно созревание РНК (от пре-мРНК до зрелой мРНК) включает стадию полиаденилирования.
Сайт рестрикции: Сайт рестрикции, также называемый сайтом узнавания ферментами рестрикции, представляет собой нуклеотидную последовательность, узнаваемую ферментом рестрикции. Сайт рестрикции обычно представляет собой короткую, предпочтительно палиндромную нуклеотидную последовательность, например, последовательность, включающую от 4 до 8 нуклеотидов. Сайт рестрикции предпочтительно специфически узнается ферментом рестрикции. Фермент рестрикции обычно расщепляет нуклеотидную последовательность, включающую сайт рестрикции, в этом сайте. В двухцепочечной нуклеотидной последовательности, такой как двухцепочечная последовательность ДНК, фермент рестрикции обычно вызывает рестрикцию обеих цепей нуклеотидной последовательности.
РНК, мРНК: РНК представляет собой обычное сокращенное название рибонуклеиновой кислоты. Она представляет собой молекулу нуклеиновой кислоты, т.е. полимера, состоящую из нуклеотидов. Эти нуклеотиды обычно представляют собой мономеры аденозинмонофосфата, уридинмонофосфата, гуанозинмонофосфата и цитидинмонофосфата, которые соединены друг с другом в так называемом остове. Остов формируется с помощью фосфодиэфирных связей между сахаром, т.е. рибозой, первого и фосфатной частью второго прилегающего мономера. Специфический непрерывный ряд мономеров называется последовательностью РНК. Обычно РНК может быть получена путем транскрипции последовательности ДНК, например, внутри клетки. В эукариотных клетках транскрипция обычно осуществляется внутри ядра или митохондрии. In vivo транскрипция ДНК обычно приводит к так называемой незрелой РНК, процессинг которой должен привести к так называемой матричной РНК, обычно сокращенно называемой мРНК. Процессинг незрелой РНК, например, в эукариотных организмах, включает множество различных посттранскрипционных модификации, таких как сплайсинг, 5'-кэпирование, полиаденилирование, экспорт из ядра или митохондрии и тому подобное. Суммарно эти процессы называются также созреванием РНК. Зрелая матричная РНК обычно имеет нуклеотидную последовательность, которая может быть транслирована в аминокислотную последовательность конкретного пептида или белка. Обычно зрелая мРНК включает 5'-кэп, 5'-UTR, открытую рамку считывания, 3'-UTR и поли(А)-последовательность. Кроме матричной РНК существует несколько некодирующих типов РНК, которые могут быть вовлечены в регуляцию транскрипции и/или трансляции.
Последовательность молекулы нуклеиновой кислоты: Последовательность молекулы нуклеиновой кислоты обычно понимается как построенная в конкретном и индивидуальном порядке, т.е. в виде непрерывного ряда ее нуклеотидов. Последовательность белка или пептида обычно понимается как построенная в определенном порядке, т.е. в виде непрерывного ряда ее аминокислот.
Идентичность последовательностей: Две или более последовательности идентичны, если они демонстрируют одинаковую длину и порядок нуклеотидов или аминокислот. Процент идентичности обычно описывает степень, в которой две последовательности идентичны, т.е. он обычно описывает процент нуклеотидов, которые по их положению в последовательности соответствуют идентичным нуклеотидам эталонной последовательности. Для определения степени идентичности последовательности, подлежащие сравнению, рассматриваются как имеющие одинаковую длину, т.е. длину самой длинной последовательности из последовательностей, подлежащих сравнению. Это означает, что первая последовательность, состоящая из 8 нуклеотидов на 80% идентична второй последовательности, состоящей из 10 нуклеотидов, включающих первую последовательность. Другими словами, в контексте настоящего изобретения идентичность последовательностей предпочтительно относится к проценту нуклеотидов последовательности, которые имеют то же самое положение в двух или более последовательностях, имеющих одинаковую длину. Пробелы обычно рассматриваются как неидентичные положения, независимо от их фактического положения при выравнивании.
Стабилизированная молекула нуклеиновой кислоты: Стабилизированная молекула нуклеиновой кислоты представляет собой молекулу нуклеиновой кислоты, предпочтительно молекулу ДНК или РНК, которая модифицирована таким образом, что она более устойчива к распаду или деградации, например, под действием факторов окружающей среды или ферментативного гидролиза, например, деградации под действием экзо- или эндонуклеаз, чем молекула нуклеиновой кислоты без модификации. Предпочтительно стабилизированная молекула нуклеиновой кислоты в контексте настоящего изобретения стабилизируется в клетке, такой как прокариотная или эукариотная клетка, предпочтительно в клетке млекопитающего, такой как клетка человека. Эффект стабилизации также может быть вызван за пределами клетки, например, в буферном растворе и т.д., например, в процессе получения фармацевтической композиции, включающей стабилизированную молекулу нуклеиновой кислоты.
Трансфекция: Термин «трансфекции» относится к введению молекул нуклеиновой кислоты, таких как молекулы ДНК или РНК (например, мРНК), в клетки, предпочтительно в эукариотные клетки. В контексте настоящего изобретения термин «трансфекция» охватывает любой способ, известный специалисту в данной области техники, для введения молекул нуклеиновой кислоты в клетки, предпочтительно в эукариотные клетки, такие как клетки млекопитающих. Такие методы охватывают, например, электропорацию, липофекцию, например, на основе катионных липидов и/или липосом, преципитацию фосфатом кальция, трансфекцию на основе наночастиц, трансфекцию на основе вирусов или трансфекцию на основе катионных полимеров, таких как ДЭАЭ-декстран или полиэтиленимин и т.д. Предпочтительно введение осуществляется не на основе вирусов.
Вакцина: Вакцина обычно понимается как профилактический или терапевтический материал, обеспечивающий по меньшей мере одним антигеном, предпочтительно иммуногеном. Антиген или иммуноген может происходить от любого материала, который подходит для вакцинации. Например, антиген или иммуноген может происходить от патогена, такого как бактерии или вирусные частицы и т.д., или из опухоли или раковой ткани. Антиген или иммуноген стимулирует адаптивную иммунную систему организма, чтобы обеспечить адаптивный иммунный ответ.
Вектор: Термин «вектор» относится к молекуле нуклеиновой кислоты, предпочтительно к молекуле искусственной нуклеиновой кислоты. Вектор в контексте настоящего изобретения подходит для включения или переноса желаемой последовательности нуклеиновой кислоты, такой как последовательность нуклеиновой кислоты, содержащая открытую рамку считывания. Такие векторы могут представлять собой векторы хранения, экспрессионные векторы, клонирующие векторы, трансферные векторы и т.д. Вектор хранения представляет собой вектор, который обеспечивает удобное хранение молекулы нуклеиновой кислоты, например, молекулы мРНК. Таким образом, вектор может включать последовательность, соответствующую, например, желаемой последовательности мРНК или ее части, такую как последовательность, соответствующая открытой рамке считывания и 3'-UTR мРНК. Экспрессионный вектор может быть использован для получения продуктов экспрессии, таких как РНК, например мРНК, или пептиды, полипептиды или белки. Например, экспрессионный вектор может включать последовательности, необходимые для транскрипции участка последовательности вектора, такие как промоторная последовательность, например, промоторная последовательность РНК. Клонирующий вектор обычно представляет собой вектор, который содержит клонирующий сайт, который может быть использован для включения последовательностей нуклеиновой кислоты в вектор. Клонирующий вектор может представлять собой, например, плазмидный вектор или вектор на основе бактериофага. Трансферный вектор может представлять собой вектор, который пригоден для передачи молекул нуклеиновой кислоты в клетки или организмы, например, вирусный вектор. Вектор в контексте настоящего изобретения, может представлять собой, например, РНК-вектор или ДНК-вектор. Предпочтительно вектор представляет собой молекулу ДНК. Предпочтительно вектор по настоящей заявке включает в себя сайт клонирования, маркер селекции, такой как фактор устойчивости к антибиотику, и последовательность, подходящую для репликации вектора, такую как сайт инициации репликации. Предпочтительно вектор в контексте настоящей заявки представляет собой плазмидный вектор.
Носитель: Носитель обычно понимается как вещество, которое подходит для хранения, транспорта и/или введения соединения, такого как фармацевтически активное соединение. Например, это может быть физиологически приемлемая жидкость, которая подходит для хранения, транспорта и/или введения фармацевтически активного соединения.
3'-нетранслируемая область (3'-UTR): 3'-UTR обычно представляет собой часть мРНК, которая расположена между кодирующей белок областью (открытой рамкой считывания (ORF) или кодирующей последовательностью (CDS)) и поли(А)-последовательностью мРНК. 3'-UTR мРНК не транслируется в аминокислотную последовательность. Последовательность 3'-UTR обычно кодируется геном, который транскрибируется в соответствующую мРНК в процессе экспрессии гена. Геномная последовательность сначала транскрибируется в незрелую мРНК, которая включает факультативные интроны. Незрелая мРНК в процессе созревания проходит затем дополнительный процессинг до зрелой мРНК. Этот процесс созревания включает стадии 5'-кэпирования, сплайсинга незрелой мРНК для вырезания факультативных интронов и модификаций 3'-конца, таких как полиаденилирование 3'-конца незрелой мРНК и необязательное расщепление эндо- или экзонуклеазами и т.д. В контексте настоящего изобретения, 3'-UTR соответствует последовательности зрелой мРНК, которая примыкает с 3'-конца к стоп-кодону кодирующей белок области, предпочтительно непосредственно примыкает к 3'-концу стоп-кодона кодирующей белок области, и которая продолжается до 5'-конца поли(А)-последовательности, предпочтительно до нуклеотида, непосредственно примыкающего к 5'-концу поли(А)-последовательности. Термин «соответствует» означает, что последовательность 3'-UTR может представлять собой последовательность РНК, такую как последовательность мРНК, используемая для определения последовательности 3'-UTR, или последовательность ДНК, которая соответствует такой последовательности РНК. В контексте настоящего изобретения, термин «3'-UTR гена», такой как «3'-UTR гена альбумина», представляет собой последовательность, которая соответствует 3'-UTR зрелой мРНК, происходящей от этого гена, т.е. мРНК, полученной путем транскрипции гена и созревания незрелой мРНК. Термин «3'-UTR гена» охватывает 3'-UTR последовательности ДНК и последовательности РНК.
В первом аспекте настоящее изобретение относится к молекуле искусственной нуклеиновой кислоты, включающей
a. по меньшей мере одну открытую рамку считывания (ORF); и
b. по меньшей мере один элемент 3'-нетранслируемой области (элемент 3'-UTR), включающий или состоящий из последовательности нуклеиновой кислоты, которая происходит от 3'-UTR гена альбумина, или от варианта 3'-UTR гена альбумина.
Термин «элемент 3'-UTR» относится к последовательности нуклеиновой кислоты, которая включает или состоит из последовательности нуклеиновой кислоты, которая происходит от 3'-UTR или от варианта 3'-UTR. «Элемент 3'-UTR» предпочтительно обозначает последовательность нуклеиновой кислоты, которая представляет собой 3'-UTR последовательности искусственной нуклеиновой кислоты, такой как искусственная мРНК, или которая кодирует 3'-UTR молекулы искусственной нуклеиновой кислоты. Таким образом, по настоящему изобретению предпочтительно элемент 3'-UTR может представлять собой 3'-UTR мРНК, предпочтительно искусственной мРНК, или он может представлять собой транскрипционную матрицу для 3'-UTR мРНК. Таким образом, элемент 3'-UTR предпочтительно представляет собой последовательность нуклеиновой кислоты, которая соответствует 3'-UTR мРНК, предпочтительно 3'-UTR искусственной мРНК, такой как мРНК, полученная путем транскрипции векторного конструкта, созданного способом генной инженерии. Предпочтительно элемент 3'-UTR по настоящему изобретению функционирует как 3'-UTR или кодирует нуклеотидную последовательность, которая выполняет функцию 3'-UTR.
Предпочтительно по меньшей мере одна открытая рамка считывания и по меньшей мере один элемент 3'-UTR являются гетерологичными. Термин «гетерологичный» в данном контексте означает, что открытая рамка считывания и элемент 3'-UTR в этом сочетании не встречаются в природе (являются неприродными). Предпочтительно элемент 3'-UTR происходит от гена, отличного от гена открытой рамки считывания. Например, ORF может происходить от другого гена, чем элемент 3'-UTR, например, кодирующего другой белок или один и тот же белок, но разных видов и т.д. Предпочтительно, чтобы открытая рамка считывания не кодировала альбумин человека, предпочтительно, чтобы открытая рамка считывания не кодировала альбумин. Предпочтительно, чтобы открытая рамка считывания не кодировала репортерный белок, например, выбранный из группы, состоящей из белков глобинов (в частности, бета-глобина), белка люциферазы, белков GFP или их вариантов, например, вариантов, имеющих по меньшей мере 70% идентичность последовательности с белком глобином, белком люциферазой или белком GFP. В частности, в конкретном варианте осуществления предпочтительно, чтобы открытая рамка считывания не кодировала бета-глобин, или более конкретно бета-глобин кролика или их варианты, в частности, в том случае, когда элемент 3'-UTR происходит от 3'-UTR альбумина крысы или его вариантов. Кроме того, в конкретном варианте осуществления молекула искусственной нуклеиновой кислоты по изобретению не кодирует сигнальную последовательность (и, следовательно, не содержит сегмента, кодирующего такую сигнальную последовательность), которая является синонимом и также обозначается как сигнал локализации или направляющий сигнал. Такая сигнальная последовательность обычно располагается на 5'-конце кодируемой аминокислотной последовательности. В частности, искусственная нуклеиновая кислота по изобретению не кодирует белок, который (в искусственных условиях или в природе) содержит «сигнальную аминокислотную последовательность», в частности, не содержит сигнальную последовательность, направляющую кодируемый белок к полисомам, связанным с мембраной эндоплазматического ретикулума, и/или обеспечивающую транслокацию представляющего интерес белка, кодируемого нуклеиновой кислотой по изобретению, через мембрану эндоплазматического ретикулума. В частности, белок, кодируемый молекулой нуклеиновой кислоты по изобретению, не содержит сигнальной последовательности альбумина, более конкретно не содержит сигнальной последовательности альбумина крысы. Обычно белок, кодируемый молекулой искусственной нуклеиновой кислоты по изобретению, не содержит также сигнальную последовательность белка молока или гормона роста, в частности, если кодирующая область кодирует глобин, более конкретно бета-глобин, еще более конкретно бета-глобина кролика. В другом варианте осуществления последовательность искусственной нуклеиновой кислоты по изобретению не содержит последовательность 5'-UTR глобина, в частности, не содержит последовательность 5'-UTR бета-глобина, или более конкретно, не содержит последовательность 5'-UTR глобина кролика, в частности, если кодирующая область кодирует последовательность глобина, более конкретно, бета-глобина или его вариантов. Если молекула искусственной нуклеиновой кислоты содержит последовательность 5'-UTR, то эта последовательность 5'-UTR может быть выбрана таким образом, что она не представляет собой последовательность 5'-UTR гена альбумина, в частности, не происходит от гена альбумина крысы. В случае, если молекула искусственной нуклеиновой кислоты по изобретению содержит более одной 3'-UTR, другая(ие) 3'-UTR(s) предпочтительно не выбрана/выбраны из группы, состоящей из 3'-UTR глобина и 3'-UTR c-myc. В другом варианте осуществления 3'-UTR молекулы нуклеиновой кислоты по изобретению не соответствует 3'-UTR альбумина крысы, в частности, если кодирующая область кодирует глобин, более конкретно бета-глобин.
Кроме того, особенно предпочтительно, чтобы открытая рамка считывания не кодировала фактор IX человека или его варианты, например, варианты, имеющие по меньшей мере 70% идентичность последовательности с фактором IX человека. Молекула нуклеиновой кислоты не содержит промотор альбумина, в частности, промотор альбумина с точечной мутацией, более конкретно с точечной мутацией G52A, в частности, если кодирующая область молекулы нуклеиновой кислоты по изобретению кодирует фактор IX человека или его варианты, как описано выше.
В другом варианте осуществления молекула искусственной нуклеиновой кислоты по изобретению не соответствует элементу транспозона, например, транспозона плазмиды, или не содержит транспозон (в частности, не содержит транспозон Tn5 или не содержит мозаичные элементы Tn5), в частности, если кодирующая область кодирует ген устойчивости, в частности, ген устойчивости к неомицину. Молекула нуклеиновой кислоты по изобретению в таких условиях не может функционально взаимодействовать с транспозазой, в частности, с транспозазой Tn5. В функциональном отношении молекула нуклеиновой кислоты по изобретению обычно не должна образовывать комплекс между нуклеиновой кислотой (так как искусственная нуклеиновая кислота по изобретению не содержит транспозон) и транспозазой, специфичной для какого-либо транспозона. Кодирующая область (ORF) молекулы нуклеиновой кислоты по изобретению не кодирует siRNA, в частности, если молекула нуклеиновой кислоты по изобретению функционально взаимодействует с транспозазой.
В этом контексте в конкретном варианте осуществления особенно предпочтительно, чтобы открытая рамка считывания не содержала интрон, в частности, в случае, если открытая рамка считывания кодирует фактор IX человека или его варианты.
Кроме того, в этом контексте в конкретном варианте осуществления предпочтительно, чтобы открытая рамка считывания не кодировала фактор IX человека или его варианты, в частности, в том случае, когда элемент 3'-UTR происходит от 3'-UTR альбумина человека или его вариантов.
Молекула искусственной нуклеиновой кислоты по изобретению в конкретном варианте осуществления настоящего изобретения не является экспрессионной кассетой. Таким образом, молекула нуклеиновой кислоты по изобретению, например, не содержит 3'-промотор или 3'-конец промотора. В плане данного варианта осуществления молекула нуклеиновой кислоты по изобретению также не представляет собой «кассету для секреции», так как она не содержит сигнальную последовательность и предпочтительно не содержит 3'-промотор. В частности, молекула нуклеиновой кислоты по изобретению состоит из одной единой молекулы нуклеиновой кислоты, включающей ORF и область 3'-UTR, а также необязательно область 5'-UTR. Таким образом, молекула нуклеиновой кислоты по изобретению не соответствует кассете для секреции, которая состоит из более одного отдельного генетического элемента, в частности, не соответствует первому генетическому элементу, представляющему собой область выше кодирующей области (ORF), и отдельному второму генетическому элементу, представляющему собой область ниже ORF, которые предоставляются независимо, например, в виде частей набора.
Предпочтительно по меньшей мере один элемент 3'-UTR функционально связан с ORF. Это предпочтительно означает, что элемент 3'-UTR связан с ORF так, что он может оказывать влияние, такое как стабилизирующее влияние, на экспрессию ORF или стабилизирующее влияние на молекулу искусственной нуклеиновой кислоты. Предпочтительно ORF и элемент 3'-UTR связаны в 5'→3' направлении. Таким образом, предпочтительно молекула искусственной нуклеиновой кислоты включает структуру 5'-ORF-(необязательный)линкер-3'-UTR-элемент-3', где линкер может присутствовать или отсутствовать. Например, линкер может представлять собой один или более нуклеотидов, таких как участок из 1-50 или 1-20 нуклеотидов, например, включающий или состоящий из одного или более сайтов узнавания ферментом рестрикции (сайты рестрикции).
Предпочтительно по меньшей мере один элемент 3'-UTR включает или состоит из последовательности нуклеиновой кислоты, которая происходит от 3'-UTR гена альбумина позвоночных или его варианта, предпочтительно от 3'-UTR гена альбумина млекопитающих, такого как, например, 3'-UTR гена альбумина мыши, гена альбумина павиана анубис или гена альбумина человека или их варианта. Более предпочтительно, чтобы по меньшей мере один элемент 3'-UTR включал или состоял из последовательности нуклеиновой кислоты, происходящей от 3'-UTR гена альбумина приматов, в частности, от гена альбумина человека или от гена альбумина павиана анубис или от их варианта, еще более предпочтительно от 3'-UTR гена альбумина человека в соответствии с регистрационным номером GenBank NM_000477.5 или его варианта. В предпочтительном варианте осуществления элемент 3'-UTR не происходит от 3'-UTR гена альбумина Xenopus. Предпочтительно элемент 3'-UTR не включает поли(А)-лимитирующий элемент B (PLEB) 3'-UTR от гена альбумина Xenopus. Предпочтительно элемент 3'-UTR не состоит из PLEB 3'-UTR гена альбумина Xenopus.
Термин «последовательность нуклеиновой кислоты, которая происходит от 3'-UTR из […] гена альбумина» предпочтительно относится к последовательности нуклеиновой кислоты, которая основывается на последовательности 3'-UTR гена альбумина или на ее фрагменте или части. Этот термин включает последовательности, соответствующие полной последовательности 3'-UTR, т.е. полноразмерной последовательности 3'-UTR гена альбумина и последовательностям, соответствующим фрагменту последовательности 3'-UTR гена альбумина. Предпочтительно фрагмент 3'-UTR гена альбумина состоит из непрерывного участка нуклеотидов, соответствующих непрерывному участку нуклеотидов в полноразмерной 3'-UTR гена альбумина, который составляет по меньшей мере 20%, предпочтительно по меньшей мере 30%, более предпочтительно по меньшей мере 40%, более предпочтительно по меньшей мере 50%, еще более предпочтительно по меньшей мере 60%, еще более предпочтительно по меньшей мере 70%, еще более предпочтительно по меньшей мере 80% и наиболее предпочтительно по меньшей мере 90% полноразмерной 3'-UTR гена альбумина. Такой фрагмент по настоящему изобретению предпочтительно представляет собой функциональный фрагмент, как описано в настоящем описании. Термин «3'-UTR гена альбумина» предпочтительно обозначает 3'-UTR природного гена альбумина.
Термины «вариант 3'-UTR гена альбумина» и «его вариант» в контексте 3'-UTR гена альбумина относится к варианту 3'-UTR природного гена альбумина, предпочтительно к варианту 3'-UTR гена альбумина позвоночных, более предпочтительно к варианту 3'-UTR гена альбумина млекопитающих, такого как 3'-UTR гена альбумина мыши, еще более предпочтительно к варианту 3'-UTR гена альбумина приматов, особенно гена альбумина человека или гена альбумина павиана анубис, как описано выше. Такой вариант может представлять собой модифицированную 3'-UTR гена альбумина. Например, вариант 3'-UTR может иметь одну или более делеций нуклеотидов, вставок, добавлений и/или замен по сравнению с природной 3'-UTR, из которой произошел вариант. Предпочтительно вариант 3'-UTR гена альбумина идентичен по меньшей мере на 40%, предпочтительно по меньшей мере на 50%, более предпочтительно по меньшей мере на 60%, более предпочтительно по меньшей мере на 70%, еще более предпочтительно по меньшей мере на 80%, еще более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% природному варианту 3'-UTR, из которого он происходит. Предпочтительно вариант представляет собой функциональный вариант, как описано в настоящем описании.
Термин «последовательность нуклеиновой кислоты, которая происходит от варианта 3'-UTR гена альбумина» предпочтительно относится к последовательности нуклеиновой кислоты, которая основывается на варианте последовательности 3'-UTR гена альбумина, или на его фрагменте или части, как описано выше. Этот термин включает последовательности, соответствующие полной последовательности варианта 3'-UTR гена альбумина, т.е. полноразмерному варианту последовательности 3'-UTR гена альбумина, а также последовательности, соответствующие фрагменту варианта последовательности 3'-UTR гена альбумина. Предпочтительно фрагмент варианта 3'-UTR гена альбумина состоит из непрерывного участка нуклеотидов, соответствующих непрерывному участку нуклеотидов в полноразмерном варианте 3'-UTR гена альбумина, который составляет по меньшей мере 20%, предпочтительно по меньшей мере 30%, более предпочтительно по меньшей мере 40%, более предпочтительно по меньшей мере 50%, еще более предпочтительно по меньшей мере 60%, еще более предпочтительно по меньшей мере 70%, еще более предпочтительно по меньшей мере 80% и наиболее предпочтительно по меньшей мере 90% полноразмерного варианта 3'-UTR гена альбумина. Такой фрагмент варианта по настоящему изобретению предпочтительно представляет собой функциональный фрагмент варианта, как описано в настоящем описании.
Термины «функциональный вариант», «функциональный фрагмент» и «функциональный фрагмент варианта» (также называемый «функциональный вариант фрагмента») в контексте настоящего изобретения, означает, что фрагмент 3'-UTR, вариант 3'-UTR или фрагмент варианта 3'-UTR гена альбумина выполняет по меньшей мере одну, предпочтительно более одной функции природной 3'-UTR гена альбумина, из которой вариант, фрагмент или фрагмент варианта произошел. Такая функция может представлять собой, например, стабилизацию мРНК и/или стабилизацию и/или продление продукции белка с мРНК и/или увеличение экспрессии белка или общей продукции белка с мРНК предпочтительно в клетке млекопитающего, например, в клетке человека. Особенно предпочтительно, чтобы вариант, фрагмент и фрагмент варианта по настоящему изобретению выполнял функцию стабилизации мРНК, предпочтительно в клетке млекопитающего, такой как клетка человека, по сравнению с мРНК, включающей эталонную 3'-UTR или без 3'-UTR, и/или функцию стабилизации и/или продления получения белка с мРНК, предпочтительно в клетке млекопитающего, например, в клетке человека, по сравнению с мРНК, включающей эталонную 3'-UTR или без 3'-UTR, и/или функцию увеличения продукции белка с мРНК, предпочтительно в клетке млекопитающего, например, в клетке человека, по сравнению с мРНК, включающей эталонную 3'-UTR или без 3'-UTR. Эталонная 3'-UTR может представлять собой, например, природную 3'-UTR в сочетании с ORF. Кроме того, функциональный вариант, функциональный фрагмент или функциональный фрагмент варианта 3'-UTR гена альбумина предпочтительно не имеет существенно сниженного влияния на эффективность трансляции мРНК, которая включает такой вариант, фрагмент или фрагмент варианта 3'-UTR, по сравнению с 3'-UTR дикого типа, из которой вариант, фрагмент или фрагмент варианта произошел. Особенно предпочтительной функцией «функционального фрагмента», «функционального варианта» или «функционального фрагмента варианта» 3'-UTR гена альбумина в контексте настоящего изобретения является стабилизация и/или продление продукции белка в результате экспрессии мРНК, несущей функциональный фрагмент, функциональный вариант или функциональный фрагмент варианта, как описано выше.
Предпочтительно, чтобы эффективность одной или более функций, оказываемых функциональным вариантом, функциональным фрагментом или функциональным фрагментом варианта, таких как стабилизация эффективности продукции мРНК и/или белка и/или повышение эффективности продукции белка составляла по меньшей мере 40%, более предпочтительно по меньшей мере 50%, более предпочтительно по меньшей мере 60%, еще более предпочтительно по меньшей мере 70%, еще более предпочтительно по меньшей мере 80%, наиболее предпочтительно по меньшей мере 90% от стабилизации эффективности продукции мРНК и/или белка и/или повышения эффективности продукции белка, проявляемых природной 3'-UTR гена альбумина, от которого произошел вариант, фрагмент или фрагмент варианта.
В контексте настоящего изобретения фрагмент 3'-UTR гена альбумина или вариант 3'-UTR гена альбумина предпочтительно имеет длину по меньшей мере приблизительно 50 нуклеотидов, предпочтительно по меньшей мере приблизительно 75 нуклеотидов, более предпочтительно по меньшей мере приблизительно 100 нуклеотидов, еще более предпочтительно по меньшей мере приблизительно 125 нуклеотидов, наиболее предпочтительно по меньшей мере приблизительно 150 нуклеотидов. Предпочтительно такой фрагмент 3'-UTR гена альбумина или вариант 3'-UTR гена альбумина представляет собой функциональный фрагмент, как описано выше.
Предпочтительно, чтобы по меньшей мере один элемент 3'-UTR молекулы искусственной нуклеиновой кислоты по настоящему изобретению включал или состоял из «функционального фрагмента», «функционального варианта» или «функционального фрагмента варианта» 3'-UTR гена альбумина.
Предпочтительно, чтобы по меньшей мере один элемент 3'-UTR молекулы искусственной нуклеиновой кислоты по настоящему изобретению повышал устойчивость молекулы искусственной нуклеиновой кислоты, например, повышал стабильность мРНК по настоящему изобретению по сравнению с соответствующей мРНК (эталонной мРНК) без 3'-UTR или включающей эталонную 3'-UTR, такую как природная 3'-UTR в сочетании с ORF. Предпочтительно, чтобы по меньшей мере один элемент 3'-UTR молекулы искусственной нуклеиновой кислоты по настоящему изобретению повышал стабильность продукции белка с молекулы искусственной нуклеиновой кислоты по настоящему изобретению, например, с мРНК по настоящему изобретению, по сравнению с соответствующей мРНК без 3'-UTR или включающей эталонную 3-'UTR, такую как природная 3'-UTR в сочетании с ORF. Предпочтительно, чтобы по меньшей мере один элемент 3'-UTR молекулы искусственной нуклеиновой кислоты по настоящему изобретению продлевал продукцию белка с молекулы искусственной нуклеиновой кислоты по настоящему изобретению, например, с мРНК по настоящему изобретению по сравнению с соответствующей мРНК без 3'-UTR или включающей эталонную 3-'UTR, такую как природная 3'-UTR в сочетании с ORF. Предпочтительно, чтобы по меньшей мере один элемент 3'-UTR молекулы искусственной нуклеиновой кислоты по настоящему изобретению повышал экспрессию белка и/или суммарную продукцию белка с молекулы искусственной нуклеиновой кислоты по настоящему изобретению, например, с мРНК по настоящему изобретению по сравнению с соответствующей мРНК без 3'-UTR или включающей эталонную 3-'UTR, такую как природная 3'-UTR в сочетании с ORF. Предпочтительно, чтобы по меньшей мере один элемент 3'-UTR молекулы искусственной нуклеиновой кислоты по настоящему изобретению не влиял отрицательно на эффективность трансляции мРНК по сравнению с эффективностью трансляции соответствующей мРНК без 3'-UTR или включающей эталонную 3-'UTR, такую как природная 3'-UTR в сочетании с ORF. Термин «соответствующая мРНК» в данном контексте означает, что за исключением различных 3'-UTR эталонная мРНК сопоставима, предпочтительно идентична мРНК, включающей элемент 3'-UTR по изобретению.
Термин «стабилизация и/или продление продукции белка» с молекулы искусственной нуклеиновой кислоты, такой как искусственная мРНК, предпочтительно означает, что продукция белка с молекулы искусственной нуклеиновой кислоты, такой как искусственная мРНК, стабилизируется и/или продолжается по сравнению с продукцией белка с молекулы эталонной нуклеиновой кислоты, например, эталонной мРНК, например, включающей эталонную 3'-UTR или без 3'-UTR, предпочтительно в системе экспрессии млекопитающих, например, в клетках HeLa или HDF. Таким образом, белок, полученный с молекулы искусственной нуклеиновой кислоты, такой как искусственная мРНК, выявляется в течение более длительного периода времени, чем это может наблюдаться для белка, продуцируемого с молекулы эталонной нуклеиновой кислоты. Другими словами, количество белка, продуцируемое с молекулы искусственной нуклеиновой кислоты, такой как искусственная мРНК, измеряемое в зависимости от времени, снижается до пороговой величины в более поздний момент времени по сравнению с количеством белка, продуцируемым с молекулы эталонной нуклеиновой кислоты, такой как эталонная мРНК, измеряемым в зависимости от времени. Такая пороговая величина может представлять собой, например, количество белка, измеряемое в начальной фазе экспрессии, такой как 1, 2, 3, 4, 5 или 6 часов после инициации экспрессии, например, после трансфекции молекулы нуклеиновой кислоты (фиг. 17).
Например, продукция белка с молекулы искусственной нуклеиновой кислоты, такой как искусственная мРНК, в количестве, которое составляет, по меньшей мере, количество, наблюдаемое в начальной фазе экспрессии, такой как через 1, 2, 3, 4, 5 или 6 часов после инициации экспрессии, например, после трансфекции молекулы нуклеиновой кислоты, продлевается по меньшей мере приблизительно на 5 часов, предпочтительно по меньшей мере приблизительно на 10 часов, более предпочтительно по меньшей мере приблизительно на 24 часа по сравнению с продукцией белка с эталонной молекулы нуклеиновой кислоты, такой как эталонная мРНК, в системе экспрессии млекопитающих, например, в клетках млекопитающих, например, в клетках HeLa или HDF. Таким образом, молекула искусственной нуклеиновой кислоты по настоящему изобретению предпочтительно позволяет продлевать продукцию белка в количестве, которое представляет собой по меньшей мере количество, выявляемое в начальной фазе экспрессии, такой как через 1, 2, 3, 4, 5 или 6 часов после инициации экспрессии, например, после трансфекции по меньшей мере приблизительно на 5 часов, предпочтительно по меньшей мере приблизительно на 10 часов, более предпочтительно по меньшей мере приблизительно на 24 часа по сравнению с молекулой эталонной нуклеиновой кислоты без 3'-UTR или включающей эталонную 3'-UTR.
В предпочтительных вариантах осуществления продукция белка с молекулы искусственной нуклеиновой кислоты по настоящему изобретению продлевается по меньшей мере в 1,5 раза, предпочтительно по меньшей мере в 2 раза, более предпочтительно по меньшей мере в 2,5 раза по сравнению с продукцией белка с молекулы эталонной нуклеиновой кислоты без 3'-UTR или включающей эталонную 3'-UTR.
Этот эффект продления продукции белка может быть определен путем (i) измерения количеств белка, например, полученного путем экспрессии ORF, кодирующей репортерный белок, такой как люцифераза, предпочтительно в системе экспрессии млекопитающих, такой как клетки HeLa или HDF, в зависимости от времени, (ii) определения временной точки, при которой количество белка снижается по сравнению с количеством белка, наблюдаемым, например, через 1, 2, 3, 4, 5 или 6 часов после инициации экспрессии, например, через 1, 2, 3, 4, 5 или 6 часов после трансфекции молекулы искусственной нуклеиновой кислоты, и (iii) сравнения момента времени, при котором количество белка снижается по сравнению с количеством белка, наблюдаемым через 1, 2, 3, 4, 5 или 6 часов после инициации экспрессии, с указанным моментом времени, определенным для молекулы нуклеиновой кислоты без 3'-UTR или включающей эталонную 3'-UTR (фиг. 17).
Предпочтительно, чтобы это стабилизирующее и/или продлевающее влияние на продукцию белка достигалось, в то время как суммарное количество белка, продуцируемого с молекулы искусственной нуклеиновой кислоты по настоящему изобретению, например, в течение промежутка времени 48 или 72 часа, составляло, по меньшей мере, количество белка, продуцируемое с молекулы эталонной нуклеиновой кислоты без 3'-UTR или включающей эталонную 3'-UTR, такую как природная 3'-UTR с ORF молекулы искусственной нуклеиновой кислоты. Таким образом, в настоящем изобретении предлагается молекула искусственной нуклеиновой кислоты, которая дает возможность продолжительной и/или стабилизированной продукции белка в системе экспрессии млекопитающих, например, в клетках млекопитающих, например, в клетках HeLa или HDF, как указано выше, где общее количество белка, продуцируемого с указанной молекулы искусственной нуклеиновой кислоты, например, в течение промежутка времени 48 или 72 часа, составляет, по меньшей мере, суммарное количество белка, продуцируемого, например, в пределах указанного диапазона времени, с молекулы эталонной нуклеиновой кислоты без 3'-UTR, или включающей эталонную 3'-UTR, такую как природная 3'-UTR с ORF молекулы искусственной нуклеиновой кислоты.
Таким образом, «стабилизированная экспрессия белка» предпочтительно означает, что существует более равномерная продукция белка с молекулы искусственной нуклеиновой кислоты по настоящему изобретению в течение заранее определенного периода времени, например, более 24 часов, более предпочтительно более 48 часов, еще более предпочтительно более 72 часов по сравнению с молекулой эталонной нуклеиновой кислоты, например, мРНК, включающей эталонную 3'-UTR или без 3'-UTR. Соответственно, уровень продукции белка, например, в системе млекопитающих, с молекулы искусственной нуклеиновой кислоты, включающей элемент 3'-UTR по настоящему изобретению, например, с мРНК по настоящему изобретению, предпочтительно не падает до степени, наблюдаемой для эталонной молекулы нуклеиновой кислоты, такой как эталонная мРНК, как описано выше. Например, количество белка (кодируемого ORF), выявляемое через 6 часов после инициации экспрессии, например, через 6 часов после трансфекции молекулы искусственной нуклеиновой кислоты по настоящему изобретению в клетку, такую как клетка млекопитающего, может быть сравнимо с количеством белка, выявляемым через 48 часов после инициации экспрессии, например, через 48 часов после трансфекции. Таким образом, отношение количества белка, кодируемого ORF, такого как репортерный белок, например, люцифераза, выявляемого через 48 часов после инициации экспрессии, например, через 48 часов после трансфекции, к количеству белка, выявляемому через 6 часов после начала экспрессии, например, через 6 часов после трансфекции, составляет предпочтительно по меньшей мере приблизительно 0,4, более предпочтительно по меньшей мере приблизительно 0,5, более предпочтительно по меньшей мере приблизительно 0,6, еще более предпочтительно по меньшей мере приблизительно 0,7. Предпочтительно, чтобы отношение составляло между приблизительно 0,4 и приблизительно 4, предпочтительно между приблизительно 0,65 и приблизительно 3, более предпочтительно между приблизительно 0,7 и 2 для молекулы нуклеиновой кислоты по настоящему изобретению. Для соответствующей эталонной молекулы нуклеиновой кислоты, например, мРНК, включающей эталонную 3'-UTR или без 3'-UTR, указанное отношение может составлять, например, от приблизительно 0,05 до приблизительно 0,3.
Таким образом, в настоящем изобретении предлагается молекула искусственной нуклеиновой кислоты, включающая ORF и элемент 3'-UTR, как описано выше, где отношение количества (репортерного) белка, например, количества люциферазы, выявляемое через 48 часов после инициации экспрессии, к количеству (репортерного) белка, выявляемому через 6 часов после инициации экспрессии, предпочтительно в системе экспрессии млекопитающих, например, в клетках млекопитающих, например, в клетках HeLa, предпочтительно выше приблизительно 0,4, более предпочтительно выше приблизительно 0,5, более предпочтительно выше приблизительно 0,6, еще более предпочтительно выше приблизительно 0,7, например, от приблизительно 0,4 до приблизительно 4, предпочтительно между приблизительно 0,65 и приблизительно 3, более предпочтительно между приблизительно 0,7 и 2, где предпочтительно суммарное количество белка, продуцируемого с указанной молекулы искусственной нуклеиновой кислоты, например, в пределах промежутка времени 48 часов составляет, по меньшей мере, суммарное количество белка, продуцируемого, например, в пределах указанного промежутка времени с молекулы эталонной нуклеиновой кислоты без 3'-UTR или включающей эталонную 3'-UTR, такую как природная 3'-UTR с ORF молекулы искусственной нуклеиновой кислоты. В предпочтительном варианте осуществления в настоящем изобретении предлагается молекула искусственной нуклеиновой кислоты, включающая ORF и элемент 3'-UTR, как описано выше, где отношение количества (репортерного) белка, например, количества люциферазы, выявляемое через 72 часа после инициации экспрессии, к количеству (репортерного) белка, выявляемому через 6 часов после инициации экспрессии, предпочтительно в системе экспрессии млекопитающих, например, в клетках млекопитающих, например в клетках HeLa, предпочтительно выше приблизительно 0,4, более предпочтительно выше приблизительно 0,5, более предпочтительно выше приблизительно 0,6, еще более предпочтительно выше приблизительно 0,7, например, между приблизительно 0,4 и 1,15, предпочтительно между приблизительно 0,65 и приблизительно 1,15, более предпочтительно между приблизительно 0,7 и 1,0, где предпочтительно, чтобы суммарное количество белка, продуцируемое с указанной молекулы искусственной нуклеиновой кислоты, например, в течение промежутка времени 72 часа, составляет, по меньшей мере, суммарное количество белка, продуцируемого, например, в пределах указанного промежутка времени, с молекулы эталонной нуклеиновой кислоты без 3'-UTR или включающей эталонную 3'-UTR, такую как природная 3'-UTR с ORF молекулы искусственной нуклеиновой кислоты.
«Повышенная экспрессия белка» в контексте настоящего изобретения предпочтительно означает повышенную экспрессию белка в определенный момент времени после инициации экспрессии по сравнению с эталонной молекулой. Таким образом, уровень белка, выявляемый в определенный момент времени после инициации экспрессии, например, после трансфекции молекулы искусственной нуклеиновой кислоты по настоящему изобретению, например, после трансфекции мРНК по настоящему изобретению, например, через 48 или 72 часа после трансфекции, предпочтительно выше, чем уровень белка, выявляемый в тот же момент времени после инициации экспрессии, например, после трансфекции молекулы эталонной нуклеиновой кислоты, например, эталонной мРНК, включающей эталонную 3'-UTR или без 3'-UTR.
«Повышенная суммарная продукция белка» с молекулы искусственной нуклеиновой кислоты относится к повышенной продукции белка в течение периода времени, в который белок продуцируется с молекулы искусственной нуклеиновой кислоты, предпочтительно в системе экспрессии млекопитающих, например, в клетках млекопитающих, например в клетках HeLa или HDF. Таким образом, «суммарная продукция белка» предпочтительно относится к площади под кривой (AUC), представляющей продукцию белка в зависимости от времени.
Указанное увеличение стабильности молекулы искусственной нуклеиновой кислоты, указанное увеличение стабильности продукции белка, указанное продление продукции белка и/или указанное увеличение экспрессии белка и/или суммарной продукции белка предпочтительно определяют путем сравнения с соответствующей эталонной молекулой нуклеиновой кислоты без 3'-UTR, например, мРНК без 3'-UTR или с молекулой эталонной нуклеиновой кислоты, включающей эталонную 3'-UTR, например, природную 3'-UTR с ORF, как описано выше.
Стабилизирующее влияние на продукцию мРНК и/или белка и ее эффективность и/или на эффект увеличения продукции белка и ее эффективность вариантов, фрагментов и/или фрагментов вариантов 3'-UTR гена альбумина, а также стабилизирующее влияние на продукцию мРНК и/или белка и ее эффективность и/или эффект увеличения продукции белка и ее эффективность по меньшей мере одного элемента 3'-UTR молекулы искусственной нуклеиновой кислоты по настоящему изобретению могут быть определены с помощью любого способа, подходящего для этой цели, известного специалисту в данной области техники. Например, могут быть созданы молекулы искусственной мРНК, включающие кодирующую последовательность для репортерного белка, такого как люцифераза, без 3'-UTR, с 3'-UTR, происходящей от природного гена альбумина, 3'-UTR, происходящей от эталонного гена (т.е. эталонной 3'-UTR, такой как природная 3'-UTR с ORF), в виде 3'-UTR, варианта 3'-UTR гена альбумина, в виде фрагмента 3'-UTR природного гена альбумина или в виде 3'-UTR фрагмента варианта 3'-UTR гена альбумина. Такие мРНК могут быть созданы, например, путем транскрипции in vitro соответствующих векторов, таких как плазмидные векторы, например, включающие промотор Т7 и последовательность, кодирующую соответствующие последовательности мРНК. Созданные молекулы мРНК могут быть трансфицированы в клетки с помощью любого способа трансфекции, подходящего для трансфекции мРНК, например, они могут быть включены в клетки млекопитающих, такие как клетки HeLa, путем электропорации, и образцы могут быть проанализированы через определенные моменты времени после трансфекции, например, через 6 часов, 24 часа, 48 часов и 72 часа после трансфекции. Указанные образцы могут быть проанализированы на количество мРНК и/или количество белка с помощью методов, хорошо известных специалисту в данной области техники. Например, количество репортерной мРНК, присутствующей в клетках в момент времени взятия образцов, может быть определено количественными методами ПЦР. Количество репортерного белка, кодируемого соответствующими мРНК, может быть определено, например, с помощью методов ИФА или определения репортеров в зависимости от используемого репортерного белка, например, определения люциферазы. Эффект стабилизации экспрессии белка и/или продления экспрессии белка может быть проанализирован, например, с помощью определения отношения уровня белка, выявляемого через 48 часов после трансфекции, к уровню белка, выявляемому через 6 часов после трансфекции. Чем ближе указанная величина к 1, тем стабильнее экспрессия белка в пределах этого периода времени. Такие измерения могут, конечно, также осуществляться через 72 или более часов, и может быть определено отношение уровня белка, выявляемого через 72 часа после трансфекции, к уровню белка, выявляемому через 6 часов после трансфекции для определения стабильности экспрессии белка.
Предпочтительно по меньшей мере один элемент 3'-UTR молекулы искусственной нуклеиновой кислоты по настоящему изобретению включает или состоит из последовательности нуклеиновой кислоты, которая идентична по меньшей мере приблизительно на 40%, предпочтительно по меньшей мере приблизительно на 50%, предпочтительно по меньшей мере приблизительно на 60%, предпочтительно по меньшей мере приблизительно на 70%, более предпочтительно по меньшей мере приблизительно на 80%, более предпочтительно по меньшей мере приблизительно на 90%, еще более предпочтительно по меньшей мере приблизительно на 95%, еще более предпочтительно по меньшей мере приблизительно на 99%, наиболее предпочтительно на 100% последовательности нуклеиновой кислоты 3'-UTR гена альбумина, например, последовательности нуклеиновой кислоты в соответствии с SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34 или SEQ ID NO: 35, как показано ниже, где варианты последовательностей (например, идентичные по меньшей мере на 40%), предпочтительно представляют собой функциональные варианты, как описано выше:
CATCACATTT AAAAGCATCT CAGCCTACCA TGAGAATAAG AGAAAGAAAA TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT TGCCTCTTTT CTCTGTGCTT CAATTAATAA AAAATGGAAA GAATCT (SEQ ID NO: 1)
CAUCACAUUU AAAAGCAUCU CAGCCUACCA UGAGAAUAAG AGAAAGAAAA UGAAGAUCAA AAGCUUAUUC AUCUGUUUUU CUUUUUCGUU GGUGUAAAGC CAACACCCUG UCUAAAAAAC AUAAAUUUCU UUAAUCAUUU UGCCUCUUUU CUCUGUGCUU CAAUUAAUAA AAAAUGGAAA GAAUCU (SEQ ID NO: 2).
AAACATCACA ATTAAGAACA TCTCAGCCTA CCATGAGAAC AAGAGAAATA AAATGAAGAT CAAAAGCTTA TTCATCTGTT TTTCTTTTTC ATTGGTATAA AGCCAACACC CTGTCTAAAA AACTATAAAT TTCTTTAATC ATTTTGCCTC TTTTCTCTGT GCTTCAATTA ATAAAAAATG GAAAGAATCT AGATCTAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAA (SEQ ID NO: 32)
AAACAUCACA AUUAAGAACA UCUCAGCCUA CCAUGAGAAC AAGAGAAAUA AAAUGAAGAU CAAAAGCUUA UUCAUCUGUU UUUCUUUUUC AUUGGUAUAA AGCCAACACC CUGUCUAAAA AACUAUAAAU UUCUUUAAUC AUUUUGCCUC UUUUCUCUGU GCUUCAAUUA AUAAAAAAUG GAAAGAAUCU AGAUCUAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAA (SEQ ID NO: 33)
ACACATCACA ACCACAACCT TCTCAGGCTA CCCTGAGAAA AAAAGACATG AAGACTCAGG ACTCATCTTT TCTGTTGGTG TAAAATCAAC ACCCTAAGGA ACACAAATTT CTTTAAACAT TTGACTTCTT GTCTCTGTGC TGCAATTAAT AAAAAATGGA AAGAATCTAC AGATCTAAAA AAAA (SEQ ID NO: 34)
ACACAUCACA ACCACAACCU UCUCAGGCUA CCCUGAGAAA AAAAGACAUG AAGACUCAGG ACUCAUCUUU UCUGUUGGUG UAAAAUCAAC ACCCUAAGGA ACACAAAUUU CUUUAAACAU UUGACUUCUU GUCUCUGUGC UGCAAUUAAU AAAAAAUGGA AAGAAUCUAC AGAUCUAAAA AAAA (SEQ ID NO: 35)
По меньшей мере один элемент 3'-UTR молекулы искусственной нуклеиновой кислоты по настоящему изобретению может также включать или состоять из фрагмента с последовательностью нуклеиновой кислоты, которая идентична по меньшей мере приблизительно на 40%, предпочтительно по меньшей мере приблизительно на 50%, предпочтительно по меньшей мере приблизительно на 60%, предпочтительно по меньшей мере приблизительно на 70%, более предпочтительно по меньшей мере приблизительно на 80%, более предпочтительно по меньшей мере приблизительно на 90%, еще более предпочтительно по меньшей мере приблизительно на 95%, еще более предпочтительно по меньшей мере приблизительно на 99%, наиболее предпочтительно на 100% последовательности нуклеиновой кислоты 3'-UTR гена альбумина, например, с последовательностью нуклеиновой кислоты в соответствии с SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34 или SEQ ID NO: 35, где фрагмент предпочтительно представляет собой функциональный фрагмент или функциональный фрагмент варианта, как описано выше. Такой фрагмент предпочтительно имеет длину по меньшей мере приблизительно 50 нуклеотидов, предпочтительно по меньшей мере приблизительно 75 нуклеотидов, более предпочтительно по меньшей мере приблизительно 100 нуклеотидов, еще более предпочтительно по меньшей мере приблизительно 125 нуклеотидов, наиболее предпочтительно по меньшей мере приблизительно 150 нуклеотидов.
Например, такой фрагмент может иметь последовательность нуклеиновой кислоты согласно SEQ ID NO: 18-30, такую как
AAAAGCATCT CAGCCTACCA TGAGAATAAG AGAAAGAAAA TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT TGCCTCTTTT CTCTGTGCTT CAATT (SEQ ID NO: 18)
CATCACATTT AAAAGCATCT CAGCCTACCA TGAGAATAAG AGAAAGAAAA TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG (SEQ ID NO: 19)
AAAAGCATCT CAGCCTACCA TGAGAATAAG AGAAAGAAAA TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC (SEQ ID NO: 20)
CAGCCTACCA TGAGAATAAG AGAAAGAAAA TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT (SEQ ID NO: 21)
TGAGAATAAG AGAAAGAAAA TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT (SEQ ID NO: 22)
AGAAAGAAAA TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT TGCCTCTTTT (SEQ ID NO: 23)
TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT TGCCTCTTTT CTCTGTGCTT (SEQ ID NO: 24)
AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT TGCCTCTTTT CTCTGTGCTT CAATTAATAA (SEQ ID NO: 25)
ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT TGCCTCTTTT CTCTGTGCTT CAATTAATAA AAAATGGAAA (SEQ ID NO: 26)
CAGCCTACCA TGAGAATAAG AGAAAGAAAA TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT TGCCTCTTTT CTCTGTGCTT CAATTAATAA A (SEQ ID NO: 27)
TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT TGCCTCTTTT CTCTGTGCTT CAATTAATAA A (SEQ ID NO: 28)
CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT TGCCTCTTTT CTCTGTGCTT CAATTAATAA A (SEQ ID NO: 29)
AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC (SEQ ID NO: 30)
или соответствующую последовательность РНК, или последовательность нуклеиновой кислоты, которая является по меньшей мере на 40%, предпочтительно по меньшей мере приблизительно на 50%, предпочтительно по меньшей мере приблизительно на 60%, предпочтительно по меньшей мере приблизительно на 70%, более предпочтительно по меньшей мере приблизительно на 80%, более предпочтительно по меньшей мере приблизительно на 90%, еще более предпочтительно по меньшей мере приблизительно на 95%, еще более предпочтительно по меньшей мере приблизительно на 99% идентичной указанным последовательностям нуклеиновой кислоты или соответствующей последовательности РНК. Таким образом, по меньшей мере один элемент 3'-UTR молекулы искусственной нуклеиновой кислоты по настоящему изобретению может включать или состоять из фрагмента нуклеиновой кислоты, как описано выше. Очевидно, что тимидиновые нуклеотиды, включенные во фрагменты в соответствии с SEQ ID NO: 18-30, могут быть заменены уридиновыми нуклеотидами.
Предпочтительно указанные варианты, фрагменты или фрагменты вариантов представляют собой функциональные варианты, функциональные фрагменты или функциональные фрагменты их вариантов, как описано выше, проявляющие по меньшей мере одну функцию последовательности нуклеиновой кислоты в соответствии с SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34 или SEQ ID NO: 35, такую как стабилизация молекулы искусственной нуклеиновой кислоты по изобретению, стабилизированная и/или продолжительная экспрессия белка с молекулы искусственной нуклеиновой кислоты по настоящему изобретению, и/или увеличение продукции белка, предпочтительно с эффективностью по меньшей мере 40%, более предпочтительно по меньшей мере 50%, более предпочтительно по меньшей мере 60%, еще более предпочтительно по меньшей мере 70%, еще более предпочтительно по меньшей мере 80%, наиболее предпочтительно по меньшей мере 90% от эффективности стабилизации и/или повышения эффективности продукции белка, демонстрируемой последовательностью нуклеиновой кислоты согласно SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34 или SEQ ID NO: 35.
Предпочтительно по меньшей мере один элемент 3'-UTR молекулы искусственной нуклеиновой кислоты по настоящему изобретению имеет длину по меньшей мере приблизительно 50 нуклеотидов, предпочтительно по меньшей мере приблизительно 75 нуклеотидов, более предпочтительно по меньшей мере приблизительно 100 нуклеотидов, еще более предпочтительно по меньшей мере приблизительно 125 нуклеотидов, наиболее предпочтительно по меньшей мере приблизительно 150 нуклеотидов. Например, элемент 3'-UTR может иметь длину от приблизительно 50 до приблизительно 300 нуклеотидов, предпочтительно от приблизительно 100 до приблизительно 250 нуклеотидов, более предпочтительно от приблизительно 150 до приблизительно 200 нуклеотидов.
Кроме того, молекула искусственной нуклеиновой кислоты по настоящему изобретению может включать более одного элемента 3'-UTR, как описано выше. Например, молекула искусственной нуклеиновой кислоты по настоящему изобретению может включать один, два, три, четыре или более элементов 3'-UTR, где индивидуальные элементы 3'-UTR могут быть одинаковыми или они могут различаться. Например, молекула искусственной нуклеиновой кислоты по настоящему изобретению может включать два по существу идентичных элемента 3'-UTR, как описано выше, например, два элемента 3'-UTR, включающих или состоящих из последовательности нуклеиновой кислоты, которая происходит от 3'-UTR гена альбумина, или от варианта 3'-UTR гена альбумина, такого как последовательность нуклеиновой кислоты согласно SEQ ID NO: 1, 2, 32, 33, 34 или 35, от их функциональных вариантов, их функциональных фрагментов или их функциональных фрагментов вариантов, как описано выше.
Неожиданно авторы настоящего изобретения обнаружили, что молекула искусственной нуклеиновой кислоты, включающая 3'-UTR, как описано выше, может представлять собой или может обеспечить молекулой мРНК, которая позволяет продлить и/или стабилизировать продукцию белка. Таким образом, 3'-UTR, как описано в настоящем описании, может улучшить стабильность экспрессии белка с молекулы мРНК и/или улучшить эффективность трансляции.
Молекула искусственной нуклеиновой кислоты по настоящему изобретению может представлять собой РНК, такую как мРНК, ДНК, такую как ДНК вектора, или может представлять собой модифицированную молекулу РНК или ДНК. Она может быть предоставлена в виде двухцепочечной молекулы, имеющей смысловую цепь и антисмысловую цепь, например, в виде молекулы ДНК, имеющей смысловую цепь и антисмысловую цепь.
Искусственная нуклеиновая кислота по настоящему изобретению может дополнительно включать необязательные 5'-UTR и/или 5'-кэп. Необязательные 5'-кэп и/или 5'-UTR предпочтительно расположены к 5'-концу от ORF в пределах молекулы искусственной нуклеиновой кислоты по настоящему изобретению.
Предпочтительно молекула искусственной нуклеиновой кислоты по настоящему изобретению дополнительно включает поли(А)-последовательность и/или сигнал полиаденилирования. Предпочтительно необязательная поли(А)-последовательность расположена к 3'-концу от по меньшей мере одного элемента 3'-UTR, предпочтительно связана с 3'-концом элемента 3'-UTR. Связь может быть прямой или непрямой, например, через участок из 2, 4, 6, 8, 10, 20 и т.д. нуклеотидов, например, через линкер из 1-50, предпочтительно из 1-20 нуклеотидов, например, включающий или состоящий из одного или более сайтов рестрикции.
В одном варианте осуществления необязательный сигнал полиаденилирования расположен в элементе 3'-UTR. Предпочтительно сигнал полиаденилирования включает консенсусную последовательность NN(U/T)ANA, с N=A или U, предпочтительно AA(U/T)AAA или A(U/T)(U/T)AAA. Такая консенсусная последовательность может узнаваться большинством животных и бактериальных клеточных систем, например, по факторам полиаденилирования, таким как фактор специфичности расщепления/полиаденилирования (CPSF), работающий совместно с CstF, PAP, PAB2, CFI и/или CFII. Предпочтительно сигнал полиаденилирования, предпочтительно консенсусная последовательность NNUANA локализована менее чем приблизительно на 50 нуклеотидов, более предпочтительно менее чем приблизительно на 30 оснований, более предпочтительно менее чем приблизительно на 25 оснований, например, на 21 основание выше 3'-конца элемента 3'-UTR.
Транскрипция молекулы искусственной нуклеиновой кислоты по настоящему изобретению, например, искусственной молекулы ДНК, включающей сигнал полиаденилирования в элементе 3'-UTR, должна приводить к незрелой РНК, содержащей сигнал полиаденилирования в своем элементе 3'-UTR. Например, транскрипция молекулы ДНК, включающей элемент 3'-UTR в соответствии с SEQ ID NO: 1
CATCACATTT AAAAGCATCT CAGCCTACCA TGAGAATAAG AGAAAGAAAA TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT TGCCTCTTTT CTCTGTGCTT CAATTAATAA AAAATGGAAA GAATCT (SEQ ID NO: 1)
приведет к РНК, имеющей элемент 3-'UTR в соответствии с последовательностью
CAUCACAUUU AAAAGCAUCU CAGCCUACCA UGAGAAUAAG AGAAAGAAAA UGAAGAUCAA AAGCUUAUUC AUCUGUUUUU CUUUUUCGUU GGUGUAAAGC CAACACCCUG UCUAAAAAAC AUAAAUUUCU UUAAUCAUUU UGCCUCUUUU CUCUGUGCUU CAAUUAAUAA AAAAUGGAAA GAAUCU (SEQ ID NO: 2).
Использование подходящей системы транскрипции будет затем приводить к присоединению поли(А)-последовательности к незрелой РНК. Например, молекула искусственной нуклеиновой кислоты по изобретению может представлять собой молекулу ДНК, включающую элемент 3'-UTR, как описано выше, и сигнал полиаденилирования, который может приводить к полиаденилированию РНК при транскрипции этой молекулы ДНК. Таким образом, результирующая РНК может включать сочетание элемента 3'-UTR по изобретению с последующей поли(А)-последовательностью.
Потенциальные системы транскрипции представляют собой системы транскрипции in vitro или клеточные системы транскрипции и т.д. Соответственно, транскрипция молекулы искусственной нуклеиновой кислоты по изобретению, например, транскрипция молекулы искусственной нуклеиновой кислоты, включающая открытую рамку считывания, элемент 3'-UTR и сигнал полиаденилирования, может вести к молекуле мРНК, включающей открытую рамку считывания, элемент 3'-UTR и поли(А)-последовательность.
Таким образом, в изобретении предлагается также молекула искусственной нуклеиновой кислоты, которая представляет собой молекулу мРНК, включающую открытую рамку считывания, элемент 3'-UTR, как описано выше, и поли(А)-последовательность.
В одном варианте осуществления в настоящем изобретении предлагается молекула искусственной нуклеиновой кислоты, которая представляет собой молекулу искусственной ДНК, включающей открытую рамку считывания и последовательность в соответствии с SEQ ID NO: 1 или последовательность, идентичную по меньшей мере приблизительно на 40% или более SEQ ID NO: 1, или ее фрагмент, как описано выше. Кроме того, в изобретении предлагается молекула искусственной нуклеиновой кислоты, которая представляет собой молекулу искусственной РНК, включающей открытую рамку считывания и последовательность в соответствии с SEQ ID NO: 2 или последовательность, идентичную по меньшей мере приблизительно на 40% или более SEQ ID NO: 2, или ее фрагмент, как описано выше.
В одном дополнительном варианте осуществления в изобретении предлагается молекула искусственной нуклеиновой кислоты, которая представляет собой молекулу искусственной ДНК, включающую открытую рамку считывания и последовательность в соответствии с SEQ ID NO: 32 или последовательность, идентичную по меньшей мере приблизительно на 40% или более SEQ ID NO: 32, или ее фрагмент, как описано выше. Кроме того, в изобретении предлагается молекула искусственной нуклеиновой кислоты, которая представляет собой молекулу искусственной РНК, включающей открытую рамку считывания и последовательность в соответствии с SEQ ID NO: 33 или последовательность, идентичную по меньшей мере приблизительно на 40% или более SEQ ID NO: 33, или ее фрагмент, как описано выше.
Кроме того, в изобретении предлагается молекула искусственной нуклеиновой кислоты, которая представляет собой молекулу искусственной ДНК, включающей открытую рамку считывания и последовательность в соответствии с SEQ ID NO: 34 или последовательность, идентичную по меньшей мере приблизительно на 40% или более SEQ ID NO: 34, или ее фрагмент, как описано выше. Кроме того, в изобретении предлагается молекула искусственной нуклеиновой кислоты, которая представляет собой молекулу искусственной РНК, включающей открытую рамку считывания и последовательность в соответствии с SEQ ID NO: 35 или последовательность, идентичную по меньшей мере приблизительно на 40% или более SEQ ID NO: 35, или ее фрагмент, как описано выше.
Таким образом, в изобретении предлагается молекула искусственной нуклеиновой кислоты, которая может представлять собой матрицу для молекулы РНК, предпочтительно для молекулы мРНК, которая стабилизирована и оптимизирована в отношении эффективности трансляции. Другими словами, молекула искусственной нуклеиновой кислоты может представлять собой ДНК или РНК, которые могут быть использованы для получения мРНК. Полученная мРНК может, в свою очередь, транслироваться для продукции желаемого пептида или белка, кодируемого открытой рамкой считывания. Если молекула искусственной нуклеиновой кислоты представляет собой ДНК, она может быть использована, например, в виде двухцепочечной формы хранения для непрерывной и повторной продукции мРНК in vitro или in vivo.
В одном варианте осуществления молекула искусственной нуклеиновой кислоты по настоящему изобретению дополнительно включает поли(А)-последовательность. Длина поли(А)-последовательности может варьироваться. Например, поли(А)-последовательность может иметь длину от приблизительно 20 адениновых нуклеотидов до приблизительно 300 адениновых нуклеотидов, предпочтительно от приблизительно 40 до приблизительно 200 адениновых нуклеотидов, более предпочтительно от приблизительно 50 до приблизительно 100 адениновых нуклеотидов, например, приблизительно 60, 70, 80, 90 или 100 адениновых нуклеотидов.
Например, молекула искусственной нуклеиновой кислоты по настоящему изобретению может включать последовательность нуклеиновой кислоты, соответствующую последовательностям ДНК
CATCACATTT AAAAGCATCT CAGCCTACCA TGAGAATAAG AGAAAGAAAA TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT TGCCTCTTTT CTCTGTGCTT CAATTAATAA AAAATGGAAA GAATCTAGAT CTAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAA (SEQ ID NO: 3),
AAACATCACA ATTAAGAACA TCTCAGCCTA CCATGAGAAC AAGAGAAATA AAATGAAGAT CAAAAGCTTA TTCATCTGTT TTTCTTTTTC ATTGGTATAA AGCCAACACC CTGTCTAAAA AACTATAAAT TTCTTTAATC ATTTTGCCTC TTTTCTCTGT GCTTCAATTA ATAAAAAATG GAAAGAATCT AGATCTAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA (SEQ ID NO: 36)
или
ACACATCACA ACCACAACCT TCTCAGGCTA CCCTGAGAAA AAAAGACATG AAGACTCAGG ACTCATCTTT TCTGTTGGTG TAAAATCAAC ACCCTAAGGA ACACAAATTT CTTTAAACAT TTGACTTCTT GTCTCTGTGC TGCAATTAAT AAAAAATGGA AAGAATCTAC AGATCTAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA (SEQ ID NO: 38)
Транскрипция таких последовательностей может вести к молекулам искусственной нуклеиновой кислоты, включающим последовательности
CAUCACAUUU AAAAGCAUCU CAGCCUACCA UGAGAAUAAG AGAAAGAAAA UGAAGAUCAA AAGCUUAUUC AUCUGUUUUU CUUUUUCGUU GGUGUAAAGC CAACACCCUG UCUAAAAAAC AUAAAUUUCU UUAAUCAUUU UGCCUCUUUU CUCUGUGCUU CAAUUAAUAA AAAAUGGAAA GAAUCUAGAU CUAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAA (SEQ ID NO: 4),
AAACAUCACA AUUAAGAACA UCUCAGCCUA CCAUGAGAAC AAGAGAAAUA AAAUGAAGAU CAAAAGCUUA UUCAUCUGUU UUUCUUUUUC AUUGGUAUAA AGCCAACACC CUGUCUAAAA AACUAUAAAU UUCUTTAATC ATTTTGCCTC TTTTCTCTGT GCTTCAATTA ATAAAAAATG GAAAGAATCT AGATCTAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA (SEQ ID NO: 37)
или
ACACAUCACA ACCACAACCU UCUCAGGCUA CCCUGAGAAA AAAAGACAUG AAGACUCAGG ACUCAUCUUU UCUGUUGGUG UAAAAUCAAC ACCCUAAGGA ACACAAAUUU CUUUAAACAU UUGACUUCUU GUCUCUGUGC UGCAAUUAAU AAAAAAUGGA AAGAAUCUAC AGAUCUAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA (SEQ ID NO: 39)
Такие молекулы искусственной РНК, т.е. молекулы искусственной нуклеиновой кислоты, включающие последовательность в соответствии с SEQ ID NO: 4, 37 или 39, также могут быть получены in vitro обычными методами химического синтеза без необходимости транскрипции с прародительской ДНК.
В особенно предпочтительном варианте осуществления молекула искусственной нуклеиновой кислоты по настоящему изобретению представляет собой молекулу РНК, предпочтительно молекулу мРНК, включающую в направлении с 5'- к 3'-концу открытую рамку считывания, элемент 3'-UTR, как описано выше, и поли(А)-последовательность.
В предпочтительном варианте осуществления открытая рамка считывания не кодирует альбумин, особенно не кодирует альбумин человека, альбумин мыши или альбумин павиана анубис, учитывая, что элемент 3'-UTR идентичен 3'-UTR альбумина человека, альбумина мыши или альбумина бабуина анубис, соответственно. В некоторых других вариантах осуществления может быть предпочтительным, чтобы открытая рамка считывания не кодировала альбумин человека с соответствующим регистрационным номером GenBank NM_000477.5, при условии, что элемент 3'-UTR идентичен 3'-UTR альбумина человека. В некоторых других вариантах осуществления может быть предпочтительным, чтобы открытая рамка считывания не кодировала альбумин или его варианты, в котором последовательность элемента 3'-UTR идентична SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34 или SEQ ID NO: 35. Кроме того, в некоторых вариантах осуществления предпочтительно, чтобы открытая рамка считывания не кодировала фактор IX человека или репортерный белок, например, выбранный из группы, состоящей из белков глобина (в частности, бета-глобина), белков люциферазы, белков GFP или их вариантов, например, вариантов, имеющих последовательность, идентичную по меньшей мере на 70% белку глобину, белку люциферазы или белку GFP. Кроме того, в определенных вариантах осуществления предпочтительно, чтобы открытая рамка считывания не включала интрон, особенно в случае, если открытая рамка считывания кодирует фактор IX человека.
В одном варианте осуществления настоящее изобретение относится к молекуле искусственной ДНК, включающей открытую рамку считывания, предпочтительно открытую рамку считывания, кодирующую пептид или белок, отличный от альбумина; элемент 3'-UTR, включающий или состоящий из последовательности, идентичной по меньшей мере приблизительно на 60%, предпочтительно по меньшей мере приблизительно на 70%, более предпочтительно по меньшей мере приблизительно на 80%, более предпочтительно по меньшей мере приблизительно на 90%, еще более предпочтительно по меньшей мере приблизительно на 95%; еще более предпочтительно по меньшей мере на 99%; еще более предпочтительно на 100% последовательности с SEQ ID NO: 1, 32 или 34; и сигнал полиаденилирования и/или поли(А)-последовательность. Кроме того, в изобретении предлагается молекула искусственной ДНК, включающая открытую рамку считывания, предпочтительно открытую рамку считывания, которая кодирует любой пептид или белок, отличный от альбумина; элемент 3'-UTR, включающий или состоящий из последовательности, идентичной по меньшей мере приблизительно на 60%, предпочтительно по меньшей мере приблизительно на 70%, более предпочтительно по меньшей мере приблизительно на 80%, более предпочтительно по меньшей мере приблизительно на 90%, еще более предпочтительно по меньшей мере приблизительно на 95%; еще более предпочтительно по меньшей мере на 99%; даже более предпочтительно на 100% последовательности с SEQ ID NO: 3, 36 или 38.
Кроме того, настоящее изобретение относится к молекуле искусственной РНК, предпочтительно к молекуле искусственной мРНК или молекуле искусственной вирусной РНК, включающей открытую рамку считывания, предпочтительно открытую рамку считывания, кодирующую пептид или белок, отличный от альбумина; элемент 3'-UTR, включающий или состоящий из последовательности, идентичной по меньшей мере приблизительно на 60%, предпочтительно по меньшей мере приблизительно на 70%, более предпочтительно по меньшей мере приблизительно на 80%, более предпочтительно по меньшей мере приблизительно на 90%, еще более предпочтительно по меньшей мере приблизительно на 95%; еще более предпочтительно по меньшей мере на 99%; еще более предпочтительно на 100% последовательности с SEQ ID NO: 2, 33 или 35; и сигнал полиаденилирования и/или поли(А)-последовательность. Кроме того, в настоящем изобретении предлагается молекула искусственной РНК, предпочтительно молекула искусственной мРНК или молекула искусственной вирусной РНК, включающая открытую рамку считывания, предпочтительно открытую рамку считывания, кодирующую пептид или белок, отличный от альбумина; элемент 3'-UTR, включающий или состоящий из последовательности, идентичной по меньшей мере приблизительно на 60%, предпочтительно по меньшей мере приблизительно на 70%, более предпочтительно по меньшей мере приблизительно на 80%, более предпочтительно по меньшей мере приблизительно на 90%, еще более предпочтительно по меньшей мере приблизительно на 95%; еще более предпочтительно по меньшей мере на 99%; даже более предпочтительно на 100% последовательности с SEQ ID NO: 4, 37 или 39.
В изобретении предлагается молекула искусственной нуклеиновой кислоты, предпочтительно искусственной мРНК, которая может характеризоваться повышенной стабильностью и продолжительностью экспрессии кодируемого пептида или белка. Без связи с любой теорией повышенная стабильность экспрессии белка и в результате продолжительная экспрессия белка может быть результатом снижения деградации молекулы искусственной нуклеиновой кислоты, такой как молекула искусственной мРНК по настоящему изобретению. Таким образом, элемент 3'-UTR по изобретению может препятствовать деградации и распаду искусственной нуклеиновой кислоты.
В некоторых вариантах осуществления предпочтительно, чтобы элемент 3'-UTR не состоял из структуры стебель-петля гистона, предпочтительно не включал структуры стебель-петля гистона. В одном варианте осуществления молекула искусственной нуклеиновой кислоты по настоящему изобретению не включает структуры стебель-петля гистона. Тем не менее, в некоторых вариантах осуществления молекула искусственной нуклеиновой кислоты может включать структуру стебель-петля гистона в дополнение к последовательности нуклеиновой кислоты, происходящей от 3'-UTR гена альбумина. Такая молекула искусственной нуклеиновой кислоты по настоящему изобретению, например, может включать в направлении от 5'- к 3'-концу ORF, элемент 3'-UTR по изобретению, предпочтительно включающий сигнал полиаденилирования, необязательную структуру стебель-петля гистона и необязательную поли(А)-последовательность. Она может также включать в направлении от 5'- к 3'-концу ORF, элемент 3'-UTR по изобретению, например, включающий сигнал полиаденилирования, поли(А)-последовательность и необязательную структуру стебель-петля гистона.
В контексте настоящего изобретения, такая структура стебель-петля гистона обычно происходит от гена гистона и включает внутримолекулярное спаривание оснований двух соседних последовательностей, полностью или частично обратно комплементарных, что тем самым образует стебель петли. Структура стебель-петля может находиться в одноцепочечной ДНК или, более обычно, в РНК. Структура также известна как шпилька или петля шпильки и обычно состоит из стебля и (концевой) петли в последовательном порядке, где стебель образован двумя соседними последовательностями, полностью или частично обратно комплементарными, разделенными короткой последовательностью в виде варианта спейсера, который формирует петлю структуры стебель-петля. Две соседние последовательности, полностью или частично обратно комплементарные, могут быть определены, например, как элементы структуры стебель-петля stem1 и stem2. Структура стебель-петля образуется, когда эти две соседние последовательности, полностью или частично обратно комплементарные, например, элементы структуры стебель-петля stem1 и stem2 образуют друг с другом пары оснований, что приводит к двухцепочечной последовательности нуклеиновой кислоты, включающей неспаренную петлю на ее концевом участке, образованную короткой последовательностью, расположенной между элементами стебель-петля stem1 и stem2 на непрерывной последовательности. Неспаренная петля тем самым обычно представляет собой область нуклеиновой кислоты, которая не способна к спариванию оснований с любым из этих элементов стебля-петли. Результирующая структура в форме леденца на палочке является ключевым строительным блоком многих вторичных структур РНК. Формирование структуры стебель-петля, таким образом, зависит от стабильности получаемых областей стебля и петли, где первым необходимым условием обычно является присутствие последовательности, которая может создать свою обратную укладку с формированием спаренной двойной цепи. Стабильность спаренных элементов стебля-петли определяется длиной, количеством ошибочных спариваний или выпетливаний (небольшое количество ошибочных спариваний обычно приемлемо особенно в длинной двойной цепи), и составом оснований спаренной области. В контексте настоящего изобретения оптимальная длина петли составляет 3-10 оснований, более предпочтительно от 3 до 8, от 3 до 7, от 3 до 6 или даже более, предпочтительно от 4 до 5 оснований, и наиболее предпочтительно 4 основания.
Примером последовательности стебель-петля гистона является последовательность в соответствии с SEQ ID NO: 31 (CAAAGGCTCTTTTCAGAGCCACCA) или соответствующая последовательность РНК.
Таким образом, в некоторых вариантах осуществления молекула искусственной нуклеиновой кислоты по настоящему изобретению включает (a.) по меньшей мере одну открытую рамку считывания; (b.) по меньшей мере один элемент 3'-UTR, как описано в настоящем описании, и (d.) по меньшей мере одну структуру стебель-петля гистона, которая может, например, включать или состоять из последовательности, имеющей идентичность последовательности по меньшей мере приблизительно 75%, предпочтительно по меньшей мере приблизительно 80%, предпочтительно по меньшей мере приблизительно 85%, более предпочтительно по меньшей мере приблизительно 90%, еще более предпочтительно по меньшей мере приблизительно 95% с последовательностью SEQ ID NO: 31 или с соответствующей последовательностью РНК, где предпочтительно положения 6, 13 и 20 последовательности имеют идентичность последовательности по меньшей мере приблизительно на 75%, предпочтительно по меньшей мере приблизительно на 80%, предпочтительно по меньшей мере приблизительно на 85%, более предпочтительно по меньшей мере приблизительно на 90%, еще более предпочтительно по меньшей мере приблизительно на 95% с консервативной последовательностью SEQ ID NO: 31 или соответствующей последовательностью РНК, т.е. идентичны нуклеотидам в положениях 6, 13 и 20 SEQ ID NO: 31.
В некоторых вариантах осуществления молекула искусственной нуклеиновой кислоты включает дополнительные элементы, такие как 5'-кэп, поли(С)-последовательность и/или мотив IRES. 5'-кэп может быть добавлен к 5'-РНК после транскрипции. Кроме того, молекула искусственной нуклеиновой кислоты по изобретению, особенно, если нуклеиновая кислота находится в форме мРНК или кодирует мРНК, может быть модифицирована с помощью последовательности по меньшей мере из 10 цитидинов, предпочтительно по меньшей мере из 20 цитидинов, более предпочтительно по меньшей мере из 30 цитидинов (так называемая «поли(С)-последовательность»). В частности, молекула нуклеиновой кислоты по изобретению может содержать, особенно, если нуклеиновая кислота находится в форме мРНК или кодирует мРНК, поли(С)-последовательность обычно из приблизительно от 10 до 200 цитидиновых нуклеотидов, предпочтительно от приблизительно 10 до 100 цитидиновых нуклеотидов, более предпочтительно приблизительно от 10 до 70 цитидиновых нуклеотидов или еще более предпочтительно приблизительно от 20 до 50 или даже от 20 до 30 цитидиновых нуклеотидов. Таким образом, предпочтительно молекула искусственной нуклеиновой кислоты по настоящему изобретению включает, предпочтительно в направлении от 5'- к 3'-концу ORF, по меньшей мере один элемент 3'-UTR, как описано выше, поли(А)-последовательность или сигнал полиаденилирования и поли(С)-последовательность.
Последовательность участка внутренней посадки рибосомы (IRES) или мотив IRES может разделять несколько открытых рамок считывания, например, если молекула искусственной нуклеиновой кислоты кодирует два или более пептидов или белков. Последовательность IRES может быть особенно полезна, если мРНК представляет собой би- или мультицистронную РНК.
Кроме того, молекула искусственной нуклеиновой кислоты может включать дополнительные 5'-элементы, предпочтительно 5'-UTR, промотор или последовательность, содержащую 5'-UTR и промотор. Промотор может управлять транскрипцией или регулировать транскрипцию молекулы искусственной нуклеиновой кислоты по изобретению, например, молекулы искусственной ДНК по настоящему изобретению. Кроме того, 5'-UTR может взаимодействовать с элементом 3'-UTR по изобретению и, следовательно, может поддерживать стабилизирующий эффект элемента 3'-UTR по изобретению. Такие элементы могут затем поддерживать стабильность и эффективность трансляции. Соответственно, в некоторых вариантах осуществления, в изобретении предлагаются молекулы искусственной нуклеиновой кислоты, предпочтительно молекулы мРНК, включающие в направлении от 5'- к 3'-концу, по меньшей мере одну из следующих структур
5'-кэп - 5'-UTR - ORF - элемент 3'-UTR - стебель-петля гистона - поли(А)-последовательность
5'-кэп - 5'-UTR - ORF - элемент 3'-UTR - поли(А)-последовательность - стебель-петля гистона
5'-кэп - 5'-UTR - ORF - IRES - ORF - элемент 3'-UTR - стебель-петля гистона - поли(А)-последовательность
5'-кэп - 5'-UTR - ORF - IRES - ORF - элемент 3'-UTR - стебель-петля гистона - поли(А)-последовательность - поли(С)-последовательность
5'-кэп - 5'-UTR - ORF - IRES - ORF - элемент 3'-UTR - поли(А)-последовательность - стебель-петля гистона
5'-кэп - 5'-UTR - ORF - IRES - ORF - элемент 3'-UTR - поли(А)-последовательность - поли(С)-последовательность - стебель-петля гистона
5'-кэп - 5'-UTR - ORF - элемент 3'-UTR - поли(А)-последовательность, - поли(С)-последовательность
5'-кэп - 5'-UTR - ORF - элемент 3'-UTR - поли(А)-последовательность - поли(С)-последовательность - стебель-петля гистона
Предпочтительно молекула искусственной нуклеиновой кислоты по настоящему изобретению, предпочтительно открытая рамка считывания, по меньшей мере, частично модифицирована G/C. Таким образом, молекула искусственной нуклеиновой кислоты по изобретению может быть термодинамически стабилизирована путем модификации содержания G (гуанозина)/С (цитидина) молекулы. Содержание G/C в открытой рамке считывания молекулы искусственной нуклеиновой кислоты по настоящему изобретению может быть увеличено по сравнению с содержанием G/C в открытой рамке считывания соответствующей последовательности дикого типа, предпочтительно путем использования дегенерации генетического кода. Таким образом, аминокислотная последовательность, кодируемая молекулой нуклеиновой кислоты, предпочтительно не модифицируется в результате модификации G/C по сравнению с аминокислотной последовательностью, кодируемой конкретной последовательностью дикого типа. Кодоны кодирующей последовательности или вся молекула нуклеиновой кислоты, например, мРНК, поэтому могут варьироваться по сравнению с кодирующей последовательностью дикого типа так, что они включают повышенное количество G/C нуклеотидов, в то время как транслируемая аминокислотная последовательность сохраняется. В связи с тем, что несколько кодонов кодируют одну и ту же аминокислоту (так называемая вырожденность генетического кода), могут быть определены наиболее благоприятные кодоны для стабильности (так называемое альтернативное использование кодонов).
В зависимости от аминокислоты, подлежащей кодированию кодирующей областью молекулы нуклеиновой кислоты по изобретению, как определено в настоящем описании, существуют различные возможности для модификации последовательности нуклеиновой кислоты, например, открытой рамки считывания, по сравнению с ее кодирующей областью дикого типа. В случае аминокислот, которые кодируются кодонами, которые содержат исключительно G или С нуклеотиды, не требуется модификации кодона. Таким образом, кодоны Pro (CCC или CCG), Arg (CGC или CGG), Ala (GCC или GCG) и Gly (GGC или GGG) не требуют модификации, так как не присутствует А или U/T.
В отличие от этого, кодоны, которые содержат A и/или U/T нуклеотиды, могут быть модифицированы путем замены другими кодонам, которые кодируют те же аминокислоты, но не содержат A и/или U/T. Например,
кодоны Pro могут быть модифицированы с СС(U/T) или CCA до ССС или CCG;
кодоны Arg могут быть модифицированы с CG(U/T) или CGA, или AGA, или AGG до CGC или CGG;
кодоны Ala могут быть модифицированы с GC(U/T) или GCA до GCC или GCG;
кодоны Gly могут быть модифицированы с GG(U/T) или GGA до GGC или GGG.
В других случаях, хотя нуклеотиды A или (U/T) не могут быть исключены из кодонов, однако, можно уменьшить содержание А и (U/T) с помощью использования кодонов, которые включают нуклеотиды А и/или (U/T) в более низком количестве. Примерами этого являются:
кодоны Phe могут быть модифицированы с (U/T)(U/T)(U/T) до (U/T)(U/T)C;
кодоны Leu могут быть модифицированы с (U/T)(U/T)A, (U/T)(U/T)G, C(U/T)(U/T) или C(U/T)A до С(U/T)C или C(U/T)G;
кодоны Ser могут быть модифицированы с (U/T)C(U/T) или (U/T)СА или AG(U/T) до (U/T)CC, (U/T)CG или AGC;
кодон Tyr может быть модифицирован с (U/T)A(U/T) до (U/T)AC;
кодон Cys может быть модифицирован с (U/T)G(U/T) до (U/T)GC;
кодон His может быть модифицирован с CA(U/T) до CAC;
кодон Gln может быть модифицирован с CAA до CAG;
кодоны Ile могут быть модифицированы с А(U/T)(U/T) или A(U/T)А до А(U/T)C;
кодоны Thr могут быть модифицированы с AC(U/T) или ACA до ACC или ACG;
кодон Asn может быть модифицирован с АА(U/T) до AAC;
кодон Lys может быть модифицирован с ААА до AAG;
кодоны Val могут быть модифицированы с G(U/T)(U/T) или G(U/T)A до G(U/T)С или G(U/T)G;
кодон Asp может быть модифицирован с GA(U/T) до GAC;
кодон Glu может быть модифицирован с GAA до GAG;
стоп-кодон (U/Т)АA может быть модифицирован до (U/T)AG или (U/T)GA.
В случае кодонов Met (A(U/Т)G) и Trp ((U/Т)GG), с другой стороны, нет никакой возможности модификации последовательности без изменения кодируемой аминокислотной последовательности.
Замены, перечисленные выше, могут быть использованы либо индивидуально, либо во всех возможных сочетаниях, чтобы увеличить содержание G/C открытой рамки считывания последовательности нуклеиновой кислоты по изобретению, как определено в настоящем описании, по сравнению с этой конкретной открытой рамкой считывания дикого типа (т.е. исходной последовательностью). Таким образом, например, все кодоны Thr, существующие в последовательности дикого типа, могут быть модифицированы до ACC (или ACG).
Предпочтительно содержание G/C открытой рамки считывания молекулы искусственной нуклеиновой кислоты по изобретению, как определено в настоящем описании, увеличивается по меньшей мере на 7%, более предпочтительно по меньшей мере на 15%, особенно предпочтительно по меньшей мере на 20%, по сравнению с содержанием G/C кодирующей области дикого типа без изменения кодируемой аминокислотной последовательности, то есть с помощью использования вырожденности генетического кода. В соответствии с конкретным вариантом осуществления в открытой рамке считывания молекулы искусственной нуклеиновой кислоты по изобретению, или ее фрагменте, варианте или производном замещаются по меньшей мере 5%, 10%, 20%, 30%, 40%, 50%, 60%, более предпочтительно по меньшей мере 70%, еще более предпочтительно по меньшей мере 80% и наиболее предпочтительно по меньшей мере 90%, 95% или даже 100% замещаемых кодонов, увеличивая тем самым содержание G/C указанной открытой рамки считывания.
В этом контексте особенно предпочтительно увеличивать содержание G/C в открытой рамке считывания последовательности нуклеиновой кислоты по изобретению, как определено в настоящем описании, до максимума (т.е. до 100% замещаемых кодонов), по сравнению с открытой рамкой считывания дикого типа без изменения кодируемой аминокислотной последовательности.
Кроме того, открытая рамка считывания предпочтительно, по меньшей мере частично, имеет оптимизированные кодоны. Оптимизация кодонов основана на обнаружении того, что эффективность трансляции может быть определена по разной частоте присутствия транспортной РНК (тРНК) в клетках. Таким образом, если в кодирующей области молекулы искусственной нуклеиновой кислоты по изобретению присутствуют так называемые «редкие кодоны», как определено в настоящем описании, на повышенном уровне, трансляция соответствующей модифицированной последовательности нуклеиновой кислоты является менее эффективной, чем в случае, когда присутствуют кодоны, кодирующие относительно «частые» тРНК.
Таким образом, открытая рамка считывания последовательности нуклеиновой кислоты по изобретению предпочтительно модифицирована по сравнению с соответствующей кодирующей областью дикого типа, так что по меньшей мере один кодон последовательности дикого типа, который кодирует тРНК, которая является относительно редкой в клетке, заменяется на кодон, который кодирует тРНК, которая присутствует в клетке сравнительно часто и несет ту же аминокислоту, что и относительно редкая тРНК. С помощью этой модификации, открытая рамка считывания молекулы искусственной нуклеиновой кислоты по изобретению, как определено в настоящем описании, модифицируется таким образом, что кодоны, для которых часто встречающиеся тРНК доступны, могут заменять кодоны, которые соответствуют редким тРНК. Другими словами, в соответствии с изобретением с помощью такой модификации все кодоны открытой рамки считывания дикого типа, которые кодируют редкие тРНК, могут быть заменены на кодон, который кодирует тРНК, являющуюся более частой в клетке и несущую ту же аминокислоту что и редкая тРНК. Какие тРНК присутствуют относительно часто в клетке, и какие в отличие от этого присутствуют относительно редко, известно специалисту в этой области техники; сравни, например, Akashi, Curr. Opin. Genet. Dev. 2001, 11(6): 660-666. Соответственно, предпочтительно, чтобы открытая рамка считывания являлась кодон-оптимизированной, предпочтительно в отношении системы, в которой молекула нуклеиновой кислоты по настоящему изобретению подлежит экспрессии, предпочтительно в отношении системы, в которой молекула нуклеиновой кислоты по настоящему изобретению подлежит трансляции. Предпочтительно использование кодонов открытой рамки считывания является кодон-оптимизированным в соответствии с использованием кодонов млекопитающего, более предпочтительно в соответствии с использованием кодонов человека. Предпочтительно, чтобы открытая рамка считывания являлась кодон-оптимизированной и модифицированной по содержанию G/C.
Для дальнейшего улучшения устойчивости к деградации, например, устойчивости к деградации под действием экзо- или эндонуклеазы in vivo, и/или для дальнейшего улучшения стабильности экспрессии белка c молекулы искусственной нуклеиновой кислоты по настоящему изобретению, молекула искусственной нуклеиновой кислоты может дополнительно включать модификации, такие как модификации остова, модификации сахаров и/или модификации оснований, например, липидные модификации или тому подобное. Предпочтительно транскрипция и/или трансляция молекулы искусственной нуклеиновой кислоты по настоящему изобретению в результате указанных модификаций существенно не нарушается.
Нуклеотидные аналоги/модификации, которые могут быть использованы в контексте настоящего изобретения, могут быть выбраны, например, из 2-амино-6-хлорпуринрибозид-5'-трифосфата, 2-аминоаденозин-5'-трифосфата, 2-тиоцитидин-5'-трифосфата, 2-тиоуридин-5'-трифосфата, 4-тиоуридин-5'-трифосфата, 5-аминоаллилцитидин-5'-трифосфата, 5-аминоаллилуридин-5'-трифосфата, 5-бромоцитидин-5'-трифосфата, 5-бромоуридин-5'-трифосфата, 5-йодоцитидин-5'-трифосфата, 5-йодоуридин-5'-трифосфата, 5-метилцитидин-5'-трифосфата, 5-метилуридин-5'-трифосфата, 6-азацитидин-5'-трифосфата, 6-азауридин-5'-трифосфата, 6-хлорпуринрибозид-5'-трифосфата, 7-дезазааденозин-5'-трифосфата, 7-дезазагуанозин-5'-трифосфата, 8-азааденозин-5'-трифосфата, 8-азидоаденозин-5'-трифосфата, бензимидазолрибозид-5'-трифосфата, N1-метиладенозин-5'-трифосфата, N1-метилгуанозин-5'-трифосфата, N6-метиладенозин-5'-трифосфата, O6-метилгуанозин-5'-трифосфата, псевдоуридин-5'-трифосфата или пуромицин-5'-трифосфата, ксантозин-5'-трифосфата. Особое предпочтение отдается нуклеотидам с модификацией оснований, выбранным из группы нуклеотидов с модифицированными основаниями, состоящей из 5-метилцитидин-5'-трифосфата, 7-дезазагуанозин-5'-трифосфата, 5-бромцитидин-5'-трифосфата и псевдоуридин-5'-трифосфата.
Кроме того, молекулы искусственной нуклеиновой кислоты, модифицированные липидами, обычно могут включать по меньшей мере один линкер, который ковалентно связан с молекулой искусственной нуклеиновой кислоты и по меньшей мере один липид, который ковалентно связан с этим линкером. Альтернативно, молекула искусственной нуклеиновой кислоты, модифицированная липидами, может включать по меньшей мере одну молекулу искусственной нуклеиновой кислоты, как определено в настоящем описании, и по меньшей мере один предпочтительно бифункциональный липид, который ковалентно связан, предпочтительно без линкера, с этой молекулой искусственной нуклеиновой кислоты. Согласно третьей альтернативе, молекула искусственной нуклеиновой кислоты, модифицированная липидами, может включать молекулу искусственной нуклеиновой кислоты, как определено в настоящем описании, по меньшей мере один линкер, который ковалентно связан с этой молекулой искусственной нуклеиновой кислоты, по меньшей мере один липид, который ковалентно связан с этим линкером, и дополнительно, по меньшей мере один, предпочтительно бифункциональный липид, который ковалентно связан, предпочтительно без линкера, с молекулой искусственной нуклеиновой кислоты.
В еще одном аспекте, в настоящем изобретении предлагается вектор, включающий
a. открытую рамку считывания (ORF) и/или сайт клонирования, например, для вставки открытой рамки считывания или последовательности, включающей открытую рамку считывания; и
b. по меньшей мере один элемент 3'-нетранслируемой области (элемент 3'-UTR), включающий последовательность нуклеиновой кислоты, которая происходит от 3'-UTR гена альбумина, или от варианта 3'-UTR гена альбумина.
По меньшей мере, один элемент 3'-UTR и ORF представляют собой описанное выше для молекулы искусственной нуклеиновой кислоты по настоящему изобретению. Сайт клонирования может представлять собой любую последовательность, которая подходит для вставки открытой рамки считывания или последовательности, включающей открытую рамку считывания, например, один или более сайтов рестрикции. Таким образом, вектор, включающий сайт клонирования, предпочтительно подходит для вставки открытой рамки считывания в вектор, предпочтительно для вставки открытой рамки считывания к 5'-концу относительно элемента 3'-UTR. Предпочтительно сайт клонирования или ORF расположены к 5'-концу относительно элемента 3'-UTR, предпочтительно в непосредственной близости от 5'-конца элемента 3'-UTR. Например, сайт клонирования или ORF могут быть прямо соединены с 5'-концом элемента 3'-UTR, или они могут быть соединены через участок нуклеотидов, такой как участок из 2, 4, 6, 8, 10, 20 и т.д. нуклеотидов, как описано выше для молекулы искусственной нуклеиновой кислоты по настоящему изобретению.
Предпочтительно вектор по настоящему изобретению подходит для продукции молекулы искусственной нуклеиновой кислоты по настоящему изобретению, предпочтительно для продукции искусственной мРНК по настоящему изобретению, например, путем необязательной вставки в вектор открытой рамки считывания или последовательности, включающей открытую рамку считывания, и транскрипции вектора. Таким образом, предпочтительно, чтобы вектор включал элементы, необходимые для транскрипции, такие как промотор, например, промотор РНК-полимеразы. Предпочтительно, чтобы вектор подходил для транскрипции с использованием эукариотных, прокариотных, вирусных или фаговых систем транскрипции, таких как эукариотные клетки, прокариотные клетки или эукариотные, прокариотные, вирусные или фаговые системы транскрипции in vitro. Таким образом, например, вектор может включать промоторную последовательность, которая узнается полимеразой, такой как РНК-полимераза, например, эукариотной, прокариотной, вирусной или фаговой РНК-полимеразой. В предпочтительном варианте осуществления вектор включает промотор РНК-полимеразы фага, такого как Т7 или SP6, предпочтительно промотор Т7. Предпочтительно, чтобы вектор подходил для транскрипции in vitro с использованием фага на основе системы транскрипции in vitro, такой как система транскрипции in vitro на основе промотора Т7 РНК-полимеразы.
Вектор может дополнительно включать поли(А)-последовательность и/или сигнал полиаденилирования, как описано выше для молекулы искусственной нуклеиновой кислоты по настоящему изобретению.
Вектор может представлять собой РНК-вектор или ДНК-вектор. Предпочтительно вектор представляет собой ДНК-вектор. Вектор может представлять собой любой вектор, известный специалисту в данной области техники, такой как вирусный вектор или плазмидный вектор. Предпочтительно, чтобы вектор представлял собой плазмидный вектор, предпочтительно плазмидный ДНК-вектор.
В предпочтительном варианте осуществления вектор по настоящему изобретению включает молекулу искусственной нуклеиновой кислоты по настоящему изобретению.
Предпочтительно ДНК-вектор по настоящему изобретению включает последовательность в соответствии с SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 32, SEQ ID NO: 34, SEQ ID NO: 36 или SEQ ID NO: 38, или последовательность, идентичную по меньшей мере приблизительно на 40%, предпочтительно по меньшей мере приблизительно на 50%, предпочтительно по меньшей мере приблизительно на 60%, предпочтительно по меньшей мере приблизительно на 70%, более предпочтительно по меньшей мере приблизительно на 80%, более предпочтительно по меньшей мере приблизительно на 90%, еще более предпочтительно по меньшей мере приблизительно на 95%; еще более предпочтительно по меньшей мере приблизительно на 99% последовательности нуклеиновой кислоты согласно SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 32, SEQ ID NO: 34, SEQ ID NO: 36 или SEQ ID NO: 38, или ее фрагменту, как описано выше, предпочтительно ее функциональному фрагменту.
Предпочтительно РНК-вектор по настоящему изобретению включает последовательность в соответствии с SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 33, SEQ ID NO: 35, SEQ ID NO: 37 или SEQ ID NO: 39 или последовательность, идентичную по меньшей мере приблизительно на 40%, предпочтительно по меньшей мере приблизительно на 50%, предпочтительно по меньшей мере приблизительно на 60%, предпочтительно по меньшей мере приблизительно на 70%, более предпочтительно по меньшей мере приблизительно на 80%, более предпочтительно по меньшей мере приблизительно на 90%, еще более предпочтительно по меньшей мере приблизительно на 95%; еще более предпочтительно по меньшей мере приблизительно на 99% последовательности нуклеиновой кислоты согласно SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 33, SEQ ID NO: 35, SEQ ID NO: 37 или SEQ ID NO: 39 или ее фрагменту, предпочтительно ее функциональному фрагменту.
Предпочтительно, чтобы вектор представлял собой кольцевую молекулу. Предпочтительно, чтобы вектор представлял собой двухцепочечную молекулу, такую как двухцепочечная молекула ДНК. Такая кольцевая предпочтительно двухцепочечная молекула ДНК может быть удобно использована в качестве формы хранения молекулы искусственной нуклеиновой кислоты по изобретению. Кроме того, она может быть использована для трансфекции клеток, например, культивируемых клеток. Также она может быть использована для транскрипции in vitro для получения молекулы искусственной РНК по изобретению.
Предпочтительно вектор, предпочтительно кольцевой вектор, является линеаризуемым, например, путем расщепления ферментами рестрикции. В предпочтительном варианте осуществления вектор включает сайт расщепления, такой как сайт рестрикции, предпочтительно уникальный сайт расщепления, локализованный непосредственно к 3'-концу от элемента 3'-UTR, или, если она присутствует, локализованный к 3'-концу от поли(А)-последовательности или сигнала полиаденилирования, или, если она присутствует, локализованный к 3'-концу от поли(C)-последовательности, или, если она присутствует, локализованный к 3'-концу от системы стебель-петля гистона. Таким образом, предпочтительно, чтобы продукт, полученный с помощью линеаризованного вектора, заканчивался на 3'-конце 3'-концом элемента 3'-UTR, или, если она присутствует, 3'-концом поли(А)-последовательности или сигнала полиаденилирования, или, если она присутствует, 3'-концом поли(С)-последовательности. В том варианте осуществления, в котором вектор по настоящему изобретению включает молекулу искусственной нуклеиновой кислоты по настоящему изобретению, сайт рестрикции, предпочтительно уникальный сайт рестрикции, предпочтительно расположен непосредственно к 3'-концу от 3'-конца молекулы искусственной нуклеиновой кислоты.
В еще одном аспекте, настоящее изобретение относится к клетке, включающей молекулу искусственной нуклеиновой кислоты по настоящему изобретению или вектор по настоящему изобретению. Клетка может представлять собой любую клетку, такую как бактериальная клетка, клетка насекомых, клетка растений, клетка позвоночных, например, клетка млекопитающих. Такая клетка может быть использована, например, для репликации вектора по настоящему изобретению, например, в бактериальной клетке. Кроме того, клетка может быть использована для транскрипции молекулы искусственной нуклеиновой кислоты или вектора по настоящему изобретению и/или трансляции открытой рамки считывания молекулы искусственной нуклеиновой кислоты или вектора по настоящему изобретению. Например, клетка может быть использована для получения рекомбинантного белка.
Клетки по настоящему изобретению можно получать, например, с помощью стандартных методов переноса нуклеиновых кислот, таких как методы стандартной трансфекции, трансдукции или трансформации. Например, молекула искусственной нуклеиновой кислоты или вектор по настоящему изобретению могут быть перенесены в клетку с помощью электропорации, липофекции, например, на основе катионных липидов и/или липосом, преципитации фосфатом кальция, трансфекции на основе наночастиц, трансфекции на основе вирусов или на основе катионных полимеров, таких как DEAE-декстран или полиэтиленимин и т.д.
Предпочтительно клетка представляет собой клетку млекопитающего, такую как клетка человека, домашнего животного, лабораторного животного, такую как клетка мыши или крысы. Предпочтительно клетка является клеткой человека. Клетка может представлять собой клетку установленной клеточной линии, такой как СНО, ВНК, 293T, COS-7, HeLa, HEK и т.д., или клетка может представлять собой первичную клетку, такую как клетка HDF и т.д., предпочтительно клетка выделена из организма. В предпочтительном варианте осуществления клетка является выделенной клеткой млекопитающего, предпочтительно человека. Например, клетка может представлять собой иммунную клетку, такую как дендритная клетка, раковую или опухолевую клетку или любую соматическую клетку и т.д., предпочтительно от млекопитающего, предпочтительно от человека.
В еще одном аспекте настоящее изобретение относится к фармацевтической композиции, включающей молекулу искусственной нуклеиновой кислоты по настоящему изобретению, вектор по настоящему изобретению или клетку по настоящему изобретению. Фармацевтическая композиция по изобретению может быть использована, например, в качестве вакцины, например, для генетической вакцинации. Таким образом, ORF может, например, кодировать антиген для вакцинации, подлежащий введению больному. Таким образом, в предпочтительном варианте осуществления фармацевтическая композиция по настоящему изобретению представляет собой вакцину. Кроме того, фармацевтическая композиция по настоящему изобретению может быть использована, например, для генной терапии.
Предпочтительно фармацевтическая композиция дополнительно включает один или более фармацевтически приемлемых наполнителей, носителей, уплотнителей и/или разбавителей. В контексте настоящего изобретения фармацевтически приемлемый носитель обычно включает жидкость или нежидкую основу для фармацевтической композиции по изобретению. В одном варианте осуществления фармацевтическая композиция представлена в жидкой форме. В этом контексте предпочтительно, чтобы носитель основывался на воде, например, апирогенной воде, изотоническом физиологическом растворе или забуференных (водных) растворах, например, фосфатных, цитратных и т.д. буферных растворах. Буфер может быть гипертоническим, изотоническим или гипотоническим относительно конкретной эталонной среды, т.е. буфер может иметь более высокое, идентичное или более низкое содержание соли относительно конкретной эталонной среды, где могут быть использованы предпочтительно такие концентрации упомянутых выше солей, которые не приводят к повреждению клеток млекопитающих за счет осмоса или других концентрационных эффектов. Эталонными средами являются, например, жидкости, присутствующие в методах «in vivo», такие как кровь, лимфа, цитозольные жидкости или другие жидкости организма, или, например, жидкости, которые могут быть использованы в качестве эталонных сред в методах «in vitro», такие как обычные буферы или жидкости. Такие обычные буферы или жидкости известны специалисту в данной области техники. В качестве жидкой основы особенно предпочтительным является раствор Рингера-лактата.
В фармацевтической композиции по изобретению может быть использован также один или более совместимых твердых или жидких наполнителей или разбавителей или инкапсулирующих соединений, пригодных для введения больному. Термин «совместимые», используемый в настоящем описании, предпочтительно означает, что эти компоненты фармацевтической композиции по изобретению способны к смешиванию с нуклеиновой кислотой, вектором или клетками по изобретению, как определено в настоящем описании, таким образом, что не происходит никакого взаимодействия, которое могло бы существенно снизить фармацевтическую эффективность фармацевтической композиции по изобретению в типичных условиях применения.
Фармацевтическая композиция по настоящему изобретению может необязательно дополнительно включать один или более дополнительных фармацевтически активных компонентов. Фармацевтически активный компонент в этом контексте представляет собой соединение, которое проявляет терапевтический эффект в отношении излечения, облегчения или предотвращения конкретного симптома или заболевания. Такие соединения включают без какого-либо ограничения пептиды или белки, нуклеиновые кислоты, (терапевтически активные) низкомолекулярные органические или неорганические соединения (с молекулярной массой менее 5000, предпочтительно менее 1000), сахара, антигены или антитела, терапевтические агенты, уже известные в предшествующем уровне техники, антигенные клетки, антигенные клеточные фрагменты, клеточные фракции, компоненты клеточной стенки (например, полисахариды), модифицированные, ослабленные или инактивированные (например, химически или путем облучения) патогены (вирусы, бактерии и т.д.).
Кроме того, фармацевтическая композиция по изобретению может включать носитель для молекулы искусственной нуклеиновой кислоты или вектора. Такой носитель может быть пригоден для опосредования растворения в физиологически приемлемых жидкостях, транспорта и клеточного захвата фармацевтически активной молекулы искусственной нуклеиновой кислоты или вектора. Соответственно, такой носитель может представлять собой компонент, который может быть пригоден для депонирования и доставки молекулы искусственной нуклеиновой кислоты или вектора по изобретению. Такие компоненты могут представлять собой, например, катионные или поликатионные носители или соединения, которые могут служить в качестве агента для трансфекции или комплексирования.
Особенно предпочтительными агентами для трансфекции или комплексирования в этом контексте являются катионные или поликатионные соединения, включая протамин, нуклеолин, спермин или спермидин или другие катионные пептиды или белки, такие как поли-L-лизин (PLL), полиаргинин, основные полипептиды, проникающие в клетки пептиды (CPP), включая связывающие ВИЧ пептиды, HIV-1 Tat (HIV), происходящие от Tat пептиды, пенетратин, пептиды, происходящие от VP22 или аналогов, VP22 HSV (простого герпеса), MAP, KALA или домены трансдукции белков (PTD), PpT620, богатые пролином пептиды, богатые аргинином пептиды, богатые лизином пептиды, MPG-пептид(ы), Pep-1, L-олигомеры, пептид(ы) кальцитонина, пептиды, происходящие от Antennapedia (особенно от Drosophila Antennapedia), pAntp, plsI, FGF, лактоферрин, транспортан, буфорин-2, Bac715-24, SynB, SynB(1), pVEC, пептиды, происходящие от hCT, SAP или гистоны.
Кроме того, такие катионные или поликатионные соединения или носители могут быть катионными или поликатионными пептидами или белками, которые предпочтительно включают или дополнительно модифицируются, чтобы включать по меньшей мере одну группу -SH. Предпочтительно катионный или поликатионный носитель выбран из катионных пептидов, имеющих следующую общую формулу (I):
{(Arg)l;(Lys)m;(His)n;(Orn)o;(Хаа)х}; формула (I),
в которой l+m+n+o+х=3-100, и l, m, n или o, независимо друг от друга, представляют собой любое число, выбранное из 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21-30, 31-40, 41-50, 51-60, 61-70, 71-80, 81-90 и 91-100, предлагаемое так, что общее содержание Arg (аргинина), Lys (лизина), His (гистидина) и Orn (орнитина) составляет по меньшей мере 10% от всех аминокислот олигопептида; и Хаа обозначает любую аминокислоту, выбранную из природной (= существующей в природе) или неприродной аминокислоты за исключением Arg, Lys, His или Orn; и х представляет любое число, выбранное из 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21-30, 31-40, 41-50, 51-60, 61-70, 71-80, 81-90, предлагаемое так, что общее содержание Хаа не превышает 90% от всех аминокислот олигопептида. Любая из аминокислот Arg, Lys, His, Orn и Хаа может быть расположена в любом месте пептида. В этом контексте катионные пептиды или белки в диапазоне 7-30 аминокислот являются особенно предпочтительными.
Кроме того, катионный или поликатионный пептид или белок, который определен в соответствии с формулой {(Arg)l;(Lys)m;(His)n;(Orn)o;(Хаа)х} (формула (I)), как показано выше, и который включает или дополнительно модифицирован, чтобы включать по меньшей мере одну группу -SH, может быть без ограничения выбран из подформулы (Ia):
{(Arg)l;(Lys)m;(His)n;(Orn)o;(Хаа')х(Cys)y} подформула (Ia)
где (Arg)l;(Lys)m;(His)n;(Orn)o и х представляют собой определенное в данном описании, Хаа' обозначает любую аминокислоту, выбранную из природных (= существующих в природе) или неприродных аминокислот за исключением Arg, Lys, His, Orn или Cys, и y представляет собой любое число, выбранное из 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21-30, 31-40, 41-50, 51-60, 61-70, 71-80 и 81-90, предлагаемое так, что общее содержание Arg (аргинина), Lys (лизина), His (гистидина) и Orn (орнитина) составляет по меньшей мере 10% от всех аминокислот олигопептида. Кроме того, катионный или поликатионный пептид может быть выбран из подформулы (Ib):
Cys1{(Arg)l;(Lys)m;(His)n;(Orn)o;(Хаа)х}Cys2 подформула (Ib)
где эмпирическая формула {(Arg)l;(Lys)m;(His)n;(Orn)o;(Хаа)х} (формула (III)) представляет собой определенное в настоящем описании и образует остов аминокислотной последовательности в соответствии с (полуэмпирической) формулой (III) и где Cys1 и Cys2 представляют собой цистеины, расположенные проксимально или терминально от (Arg)l;(Lys)m;(His)n;(Orn)o;(Хаа)х.
Кроме того, предпочтительные катионные или поликатионные соединения, которые могут быть использованы в качестве агента трансфекции или комплексирования, могут включать катионные полисахариды, например, хитозан, полибрен, катионные полимеры, например, полиэтиленимин (PEI), катионные липиды, например, DOTMA: хлорид [1-(2,3-сиолеилокси)пропил)]-N,N,N-триметиламмония, DMRIE, ди-C14-амидин, DOTIM, SAINT, DC-Chol, BGTC, CTAP, DOPC, DODAP, DOPE: диолеилфосфатидилэтаноламин, DOSPA, DODAB, DOIC, DMEPC, DOGS: диоктадециламидоглицилспермин, DIMRI: димиристооксипропилдиметилгидроксиэтиламмония бромид, DOTAP: диолеоилокси-3-(триметиламмонио)пропан, DC-6-14: O,O-дитетрадеканоил-N-(α-триметиламмониоацетил)диэтаноламина хлорид, CLIP1: rac-[(2,3-диоктадецилоксипропил)(2-гидроксиэтил)]диметиламмония хлорид, CLIP6: rac-[2(2,3-дигексадецилоксипропилоксиметилокси)этил]триметиламмоний, CLIP9: rac-[2(2,3-дигексадецилоксипропилоксисукцинилокси)этил]триметиламмоний, олигофектамин, или катионные или поликатионные полимеры, например, модифицированные полиаминокислоты, такие как β-аминокислотные полимеры или обратные полиамиды и т.д., модифицированные полиэтилены, такие как PVP (поли(N-этил-4-винилпиридиния бромид)) и т.д., модифицированные акрилаты, такие как pDMAEMA (поли(диметиламиноэтил)метилакрилат) и т.д., модифицированные амидоамины, такие как pAMAM (поли(амидоамин)) и т.д., модифицированные полибетааминоэфиры (PBAE), такие как диамин и модифицированные полимеры 1,4-бутандиолдиакрилат-co-5-амино-1-пентанола и т.д., дендримеры, такие как дендримеры полипропиламина или дендримеры на основе pAMAM и т.д., полиимин(ы), такие как PEI: поли(этиленимин), поли(пропиленимин) и т.д., полиаллиламин, полимеры на основе сахарного остова, такие как полимеры на основе циклодекстрина, полимеры на основе декстрана, хитозан, и т.д., полимеры на основе силанового остова, такие как PMOXA-PDMS сополимеры и т.д., блокполимеры, состоящие из сочетания одного или более катионных блоков (например, выбранных из катионного полимера, как указано выше) и одного или более гидрофильных или гидрофобных блоков (например, полиэтиленгликоль) и т.д.
В этом контексте особенно предпочтительно, чтобы молекула искусственной нуклеиновой кислоты по изобретению или вектор по изобретению комплексировались, по меньшей мере, частично с катионным или поликатионным соединением, предпочтительно с катионными белками или пептидами. Отчасти это означает, что только часть молекулы искусственной нуклеиновой кислоты по изобретению или вектора по изобретению образует комплекс с катионным или поликатионным соединением, а остальные молекулы искусственной нуклеиновой кислоты по изобретению или вектор по изобретению находится в некомплексированной («свободной») форме. Предпочтительно отношение комплексированной нуклеиновой кислоты к свободной нуклеиновой кислоте выбрано из диапазона от приблизительно 5:1 (масс./масс.) до приблизительно 1:10 (масс./масс.), более предпочтительно из диапазона приблизительно 4:1 (масс./масс.) до приблизительно 1:8 (масс./масс.), еще более предпочтительно из диапазона от приблизительно 3:1 (масс./масс.) до приблизительно 1:5 (масс./масс.) или 1:3 (масс./масс.) и наиболее предпочтительно отношение комплексированной нуклеиновой кислоты к свободной нуклеиновой кислоте выбрано из отношения приблизительно 1:1 (масс./масс.).
Фармацевтическая композиция по настоящему изобретению может необязательно дополнительно включать один или более адъювантов, например, адъювантов для стимуляции врожденной иммунной системы или для повышения клеточного захвата молекулы искусственной нуклеиновой кислоты или вектора. В этом контексте адъювант может пониматься как любое соединение, которое подходит для инициации или повышения иммунного ответа врожденной иммунной системы, т.е. неспецифического иммунного ответа. Другими словами, при введении фармацевтическая композиция по изобретению предпочтительно вызывает врожденный иммунный ответ благодаря необязательно содержащемуся в ней адъюванту. Предпочтительно такой адъювант может представлять собой адъювант, поддерживающий индукцию врожденного иммунного ответа у млекопитающего. Такой адъювант может представлять собой, например, иммуностимулирующую нуклеиновую кислоту, т.е. нуклеиновую кислоту, которая может связываться с Toll-подобным рецептором или тому подобное, предпочтительно иммуностимулирующую РНК.
Такие адъюванты, предпочтительно такие иммуностимулирующие нуклеиновые кислоты, могут индуцировать врожденный, т.е. неспецифический, иммунный ответ, который может поддерживать специфический, т.е. адаптивный иммунный ответ на пептид или белок, т.е. на антиген, кодируемый молекулой искусственной нуклеиновой кислоты фармацевтической композиции, предпочтительно вакцины.
Фармацевтическая композиция по изобретению может также дополнительно включать любое дополнительное соединение, которое известно как иммуностимулирующее из-за своего связывающего сродства (в качестве лиганда) к Toll-подобным рецепторам человека TLR1, TLR2, TLR3, TLR4, TLR5, TLR6, TLR7, TLR8, TLR9, TLR10, или из-за своего связывающего сродства (в качестве лиганда) к Toll-подобным рецепторам мыши TLR1, TLR2, TLR3, TLR4, TLR5, TLR6, TLR7, TLR8, TLR9, TLR10, TLR11, TLR12 или TLR13.
Дополнительные добавки, которые могут быть включены в фармацевтическую композицию по изобретению, представляют собой, например, эмульгаторы, такие как, например, Твин®; смачивающие агенты, такие как, например, лаурилсульфат натрия; красители; придающие вкус агенты, фармацевтические носители; образующие таблетку агенты; стабилизаторы; антиоксиданты; консерванты и т.д.
Фармацевтическая композиция по настоящему изобретению предпочтительно включает «безопасное и эффективное количество» компонентов фармацевтической композиции, особенно последовательности нуклеиновой кислоты по изобретению, вектора и/или клеток, как определено в данном описании. При использовании в настоящем описании «безопасное и эффективное количество» означает количество, достаточное для существенной индукции положительного изменения заболевания или нарушения, как определено в данном описании. В то же время, тем не менее, «безопасное и эффективное количество» предпочтительно позволяет избежать серьезных побочных эффектов и обеспечивает целесообразное соотношение между преимуществом и риском. Определение этих пределов обычно входит в объем целесообразного медицинского заключения.
В еще одном аспекте настоящее изобретение относится к молекуле искусственной нуклеиновой кислоты по настоящему изобретению, вектору по настоящему изобретению, клетке по настоящему изобретению или фармацевтической композиции по настоящему изобретению для применения в качестве лекарства, например, в качестве вакцины (при генетической вакцинации), или в генной терапии.
Молекула искусственной нуклеиновой кислоты по настоящему изобретению, вектор по настоящему изобретению, клетка по настоящему изобретению или фармацевтическая композиция по настоящему изобретению особенно подходят для любого медицинского применения, когда необходимо использование терапевтического действия или эффекта пептидов, полипептидов или белков, или добавление определенного пептида или белка. Таким образом, в настоящем изобретении предлагается молекула искусственной нуклеиновой кислоты по настоящему изобретению, вектор по настоящему изобретению, клетка по настоящему изобретению или фармацевтическая композиция по настоящему изобретению для применения при лечении или профилактике заболевания или нарушения, поддающегося лечению с помощью терапевтического действия или эффекта пептидов, полипептидов или белков, или поддающегося лечению с помощью добавления конкретного пептида, полипептида или белка. Например, молекула искусственной нуклеиновой кислоты по настоящему изобретению, вектор по настоящему изобретению, клетка по настоящему изобретению или фармацевтическая композиция по настоящему изобретению могут быть использованы для лечения или профилактики генетических заболеваний, аутоиммунных заболеваний, раковых или опухолевых заболеваний, инфекционных заболеваний, хронических заболеваний и тому подобного, например, путем генетической вакцинации или генной терапии.
В частности, такое терапевтическое лечение, которое имеет преимущество из-за стабильного и длительного присутствия терапевтических пептидов, полипептидов или белков у индивидуума, подлежащего лечению, в контексте настоящего изобретения особенно подходит для медицинского применения, так как элемент 3'-UTR по изобретению обеспечивает стабильную и длительную экспрессию ORF молекулы нуклеиновой кислоты по изобретению. Таким образом, особенно пригодным медицинским применением молекулы искусственной нуклеиновой кислоты по настоящему изобретению, вектора по настоящему изобретению, клетки по настоящему изобретению или фармацевтической композиции по настоящему изобретению является вакцинация. Таким образом, в настоящем изобретении предлагается молекула искусственной нуклеиновой кислоты по настоящему изобретению, вектор по настоящему изобретению, клетка по настоящему изобретению или фармацевтическая композиция по настоящему изобретению для вакцинации индивидуума, предпочтительно индивидуума млекопитающих, более предпочтительно человека. Предпочтительное лечение вакцинированием представляет собой вакцинацию против инфекционных заболеваний, таких как бактериальные, протозойные или вирусные инфекции, и противоопухолевую вакцинацию. Такая вакцинация может быть профилактической или терапевтической.
В зависимости от заболевания, подлежащего лечению или профилактике, может быть выбрана ORF. Например, открытая рамка считывания может кодировать белок, который должен получать больной, страдающий полным отсутствием или, по меньшей мере, частичной потерей функции белка, такой как больной, страдающий генетическим заболеванием. Кроме того, открытая рамка считывания может быть выбрана из ORF, кодирующей пептид или белок, который благоприятно влияет на заболевание или состояние индивидуума. Кроме того, открытая рамка считывания может кодировать пептид или белок, который вызывает регуляторное снижение патологической гиперпродукции природного пептида или белка или гибель клеток, патологически экспрессирующих белок или пептид. Такие отсутствие, потеря функции или гиперпродукция могут, например, возникать в случае опухоли и неоплазии, аутоиммунных заболеваний, аллергий, инфекций, хронических заболеваний и тому подобного. Кроме того, открытая рамка считывания может кодировать антиген или иммуноген, например, эпитоп патогена или опухолевый антиген. Таким образом, в предпочтительных вариантах осуществления молекула искусственной нуклеиновой кислоты или вектор по настоящему изобретению включает ORF, кодирующую аминокислотную последовательность, включающую или состоящую из антигена или иммуногена, например, эпитопа патогена или связанного с опухолью антигена, элемент 3'-UTR, как описано выше, и необязательные дополнительные компоненты, такие как поли(А)-последовательность и т.д.
В контексте медицинского применения, в частности в контексте вакцинации, предпочтительно, чтобы молекула искусственной нуклеиновой кислоты по настоящему изобретению представляла собой РНК, предпочтительно мРНК, так как ДНК несет риск появления иммунного ответа против ДНК и имеет тенденцию к встраиванию в геномную ДНК. Однако в некоторых вариантах осуществления, например, если вирусный носитель для доставки, такой как аденовирусный носитель для доставки, используется для доставки молекулы искусственной нуклеиновой кислоты или вектора по настоящему изобретению, например, в контексте генных терапевтических процедур, может быть желательно, чтобы молекула искусственной нуклеиновой кислоты или вектор представляли собой молекулу ДНК.
Молекулу искусственной нуклеиновой кислоты по настоящему изобретению, вектор по настоящему изобретением, клетку по настоящему изобретению или фармацевтическую композицию по настоящему изобретению можно вводить перорально, парентерально, путем ингаляции, местно, ректально, интраназально, трансбуккально, вагинально или через имплантированный резервуар. Термин парентеральный, используемый в настоящем описании, включает методы подкожной, внутривенной, внутримышечной, внутрисуставной, внутрисиновиальной, внутримышечной, внутритекальной, внутрипеченочной инъекции или инфузии, инъекции или инфузии в очаг поражения, внутричерепной, трансдермальной, внутрикожной, внутрилегочной, внутрибрюшинной, внутрисердечной, внутриартериальной и подъязычной инъекции или инфузии.
Предпочтительно молекулу искусственной нуклеиновой кислоты по настоящему изобретению, вектор по настоящему изобретению, клетку по настоящему изобретению или фармацевтическую композицию по настоящему изобретению вводят парентерально, например, путем методов парентеральной инъекции, более предпочтительно путем подкожной, внутривенной, внутримышечной, внутрисуставной, внутрисиновиальной, интрастернальной, интратекальной, внутрипеченочной инъекции, инъекции в очаг поражения, внутричерепной, трансдермальной, внутрикожной, внутрилегочной, внутрибрюшинной, внутрисердечной, внутриартериальной, сублингвальной инъекции или с помощью инфузии. Особенно предпочтительным является внутрикожное и внутримышечное введение. Стерильные инъекционные формы фармацевтической композиции по настоящему изобретению могут представлять собой водную или масляную суспензию. Эти суспензии могут быть составлены в соответствии со способами, известными в данной области техники, с использованием подходящих диспергирующих или смачивающих агентов и суспендирующих агентов.
Молекулу искусственной нуклеиновой кислоты по настоящему изобретению, вектор по настоящему изобретению, клетку по настоящему изобретению или фармацевтическую композицию по настоящему изобретению можно также вводить перорально в любой перорально приемлемой лекарственной форме, включая, но не ограничиваясь этим, капсулы, таблетки, водные суспензии или растворы.
Молекулу искусственной нуклеиновой кислоты по настоящему изобретению, вектор по настоящему изобретению, клетку по настоящему изобретению или фармацевтическую композицию по настоящему изобретению также можно вводить местно, особенно когда мишень лечения включает области или органы, легкодоступные для местного применения, например, включая заболевания кожи или любой другой доступной эпителиальной ткани. Подходящие составы для местного применения легко получают для каждой из этих областей или органов. Для местного применения молекула искусственной нуклеиновой кислоты по настоящему изобретению, вектор по настоящему изобретению, клетка по настоящему изобретению или фармацевтическая композиция по настоящему изобретению могут быть составлены в виде подходящей мази, суспендированной или растворенной в одном или более носителях.
В одном варианте осуществления применение в качестве лекарственного средства включает стадию трансфекции клеток млекопитающих, предпочтительно трансфекции in vitro клеток млекопитающих, более предпочтительно трансфекции in vitro клеток, выделенных от индивидуума, подлежащего лечению с помощью лекарственного средства. Если применение включает трансфекцию in vitro выделенных клеток, применение в качестве лекарственного средства может дополнительно включать повторное введение трансфицированных клеток больному. Применение молекул искусственной нуклеиновой кислоты или вектора по изобретению в качестве лекарственного средства может дополнительно включать стадию отбора успешно трансфицированных выделенных клеток. Таким образом, это может быть удобно, если вектор дополнительно включает маркер селекции. Кроме того, применение в качестве лекарственного средства может включать трансфекцию in vitro выделенных клеток и очистку продукта экспрессии, т.е. кодируемого пептида или белка из этих клеток. Этот очищенный пептид или белок может в последующем вводиться нуждающемуся в этом индивидууму.
Настоящее изобретение также относится к способу лечения или профилактики заболевания или нарушения, как описано выше, включающему введение молекулы искусственной нуклеиновой кислоты по настоящему изобретению, вектора по настоящему изобретению, клетки по настоящему изобретению или фармацевтической композиции по настоящему изобретению нуждающемуся в этом индивидууму.
Кроме того, настоящее изобретение относится к способу лечения или профилактики заболевания или нарушения, включающему трансфекцию клетки молекулой искусственной нуклеиновой кислоты по настоящему изобретению или вектором по настоящему изобретению. Указанная трансфекция может быть выполнена in vitro или in vivo. В предпочтительном варианте осуществления трансфекция клетки выполняется in vitro и трансфицированные клетки вводят нуждающемуся в этом индивидууму, предпочтительно больному человеку. Предпочтительно клетка, которая должна быть трансфицирована in vitro, представляет собой клетку, выделенную от индивидуума, предпочтительно от больного человека. Таким образом, в настоящем изобретении предлагается способ лечения, включающий стадии выделения клетки от индивидуума, предпочтительно от больного человека, трансфекции выделенной клетки искусственной нуклеиновой кислотой по настоящему изобретению или вектором по настоящему изобретению, и введения трансфицированной клетки индивидууму, предпочтительно больному человеку.
Способ лечения или профилактики нарушения по настоящему изобретению предпочтительно представляет собой способ вакцинации и/или способ генной терапии, как описано выше.
Как описано выше, элемент 3'-UTR по изобретению способен стабилизировать молекулу мРНК и/или стабилизировать и/или продлевать продукцию белка с молекулы мРНК. Таким образом, в другом аспекте настоящее изобретение относится к способу стабилизации молекулы РНК, предпочтительно молекулы мРНК, включающему стадию связывания молекулы РНК, предпочтительно молекулы мРНК, или вектора, кодирующего молекулу РНК, с элементом 3'-UTR, включающим или состоящим из последовательности нуклеиновой кислоты, которая происходит от 3'-UTR гена альбумина или от варианта 3'-UTR гена альбумина, предпочтительно с элементом 3'-UTR, как описано выше.
Кроме того, настоящее изобретение относится к способу увеличения продукции белка с молекулы искусственной нуклеиновой кислоты или с вектора, предпочтительно с молекулы мРНК, и/или стабилизации и/или продления продукции белка с молекулы искусственной нуклеиновой кислоты или с вектора, предпочтительно с молекулы мРНК, причем способ включает стадию связывания молекулы искусственной нуклеиновой кислоты или вектора, предпочтительно молекулы мРНК, с элементом 3'-UTR, который включает или состоит из последовательности нуклеиновой кислоты, которая происходит от 3'-UTR гена альбумина или от варианта 3'-UTR гена альбумина, предпочтительно с элементом 3'-UTR, как описано выше.
Термин «связывание молекулы искусственной нуклеиновой кислоты или вектора с элементом 3'-UTR» в контексте настоящего изобретения предпочтительно означает функциональное связывание или функциональное объединение молекулы искусственной нуклеиновой кислоты или вектора с элементом 3'-UTR. Это означает, что молекула искусственной нуклеиновой кислоты или вектор и элемент 3'-UTR, предпочтительно элемент 3'-UTR, как описано выше, связаны или объединены так, что осуществляется функция элемента 3'-UTR, например, функция стабилизации продукции РНК и/или белка. Обычно это означает, что элемент 3'-UTR интегрирован в молекулу искусственной нуклеиновой кислоты или в вектор, предпочтительно в молекулу мРНК, к 3'-концу относительно открытой рамки считывания, предпочтительно непосредственно к 3'-концу открытой рамки считывания, предпочтительно между открытой рамкой считывания и поли(А)-последовательностью или сигналом полиаденилирования. Предпочтительно элемент 3'-UTR интегрирован в молекулу искусственной нуклеиновой кислоты или в вектор, предпочтительно в мРНК, в качестве 3'-UTR, то есть так, что элемент 3'-UTR представляет собой 3'-UTR молекулы искусственной нуклеиновой кислоты или вектора, предпочтительно мРНК, то есть так, что он расположен от 3'-конца открытой рамки считывания до 5'-конца поли(А)-последовательности или сигнала полиаденилирования, необязательно соединенных через короткий линкер, такой как последовательность, включающая или состоящая из одного или более сайтов рестрикции. Таким образом, предпочтительно термин «связывание молекулы искусственной нуклеиновой кислоты или вектора с элементом 3'-UTR» означает функциональную связь элемента 3'-UTR с открытой рамкой считывания, локализованной в молекуле искусственной нуклеиновой кислоты или в векторе, предпочтительно в молекуле мРНК. 3'-UTR и ORF, как описано выше, представляют собой молекулу искусственной нуклеиновой кислоты по настоящему изобретению, например, предпочтительно ORF и 3'-UTR являются гетерологичными, например, происходят от различных генов, как описано выше.
В еще одном аспекте настоящее изобретение относится к применению элемента 3'-UTR, предпочтительно элемента 3'-UTR, как описано выше, для повышения стабильности молекулы РНК, предпочтительно молекулы мРНК, где элемент 3'-UTR включает или состоит из последовательности нуклеиновой кислоты, которая происходит от 3'-UTR гена альбумина или от варианта 3'-UTR гена альбумина.
Кроме того, настоящее изобретение относится к применению элемента 3'-UTR, предпочтительно элемента 3'-UTR, как описано выше, для повышения продукции белка с молекулы искусственной нуклеиновой кислоты или вектора, предпочтительно с молекулы мРНК, и/или для стабилизации и/или продления продукции белка с молекулы искусственной нуклеиновой кислоты, или векторной молекулы, предпочтительно с молекулы мРНК, где элемент 3'-UTR включает или состоит из последовательности нуклеиновой кислоты, которая происходит от 3'-UTR гена альбумина или от варианта 3'-UTR гена альбумина, как описано выше.
Применение по настоящему изобретению предпочтительно включает связывание молекулы искусственной нуклеиновой кислоты, вектора или РНК с элементом 3-'UTR, как описано выше.
Соединения и ингредиенты фармацевтической композиции по изобретению могут быть также получены и проданы отдельно друг от друга. Таким образом, изобретение дополнительно относится к набору или к набору из частей, включающему молекулу искусственной нуклеиновой кислоты по изобретению, вектор по изобретению, клетку по изобретению и/или фармацевтическую композицию по изобретению. Предпочтительно такой набор или наборы из частей могут кроме того включать инструкции по применению, клетки для трансфекции, адъювант, средство для введения фармацевтической композиции, фармацевтически приемлемый носитель и/или фармацевтически приемлемый раствор для растворения или разбавления молекулы искусственной нуклеиновой кислоты, вектора, клеток или фармацевтической композиции.
Следующие фигуры, последовательности и примеры предназначены для дополнительной иллюстрации настоящего изобретения. Они не рассматриваются как ограничивающие предмет изобретения.
На фиг. 1 показано влияние 3'-UTR α-глобина человека, 3'-UTR альбумина человека и 3'-UTR β-глюкуронидазы человека на экспрессию люциферазы с искусственной мРНК. Здесь мРНК, включающая 3'-UTR альбумина человека, представляет собой мРНК по настоящему изобретению. Она включает открытую рамку считывания, кодирующую люциферазу Photinus pyralis, за которой следует в направлении от 5'- к 3'-концу элемент 3'-UTR в соответствии с SEQ ID NO: 2 и поли(А)-последовательность, имеющая в длину 64 аденина. Наблюдается заметное продление экспрессии белка с искусственной мРНК, содержащей 3'-UTR альбумина человека, соответствующую SEQ ID NO: 2.
Влияние 3'-UTR α-глобина человека, 3'-UTR альбумина человека или 3'-UTR β-глюкуронидазы человека на экспрессию люциферазы с мРНК было исследовано в сравнении с экспрессией люциферазы с мРНК без 3'-UTR. Таким образом, различные мРНК вводили электропорацией в клетки HeLa. Уровни люциферазы измеряли через 6, 24 и 48 часов после трансфекции. Уровень люциферазы с мРНК без 3'-UTR падает с 6 часов к 48 часам, через 48 часов остается 10% от сигнала через 6 часов. 3'-UTR α-глобина умеренно стабилизирует экспрессию люциферазы с мРНК. Удивительно, однако, что 3'-UTR альбумина человека по изобретению дополнительно заметно продлевает экспрессию люциферазы с мРНК. В отличие от этого 3'-UTR стабильной мРНК β-глюкуронидазы не продлевает экспрессию люциферазы в той степени, которая наблюдается для 3'-UTR альбумина, подтверждая, что 3'-UTR альбумина особенно эффективна для продления экспрессии белка с мРНК. Данные представлены как средние RLU ± SD (относительные световые единицы ± стандартное отклонение) для трех трансфекций. RLU суммированы в примере 5.1.
На фиг. 2 показано влияние 3'-UTR альбумина человека на экспрессию люциферазы с мРНК в сравнении с экспрессией люциферазы с мРНК, содержащей 3'-UTR α-глобина человека или 3'-UTR каждой из нескольких различных устойчивых мРНК. Таким образом, различные мРНК вводили электропорацией в клетки HeLa. Уровни люциферазы измеряли через 6, 24 и 48 часов после трансфекции. Уровень люциферазы с мРНК без 3'-UTR падает с 6 часов к 48 часам, через 48 часов остается 14% от сигнала через 6 часов. 3'-UTR α-глобина умеренно стабилизирует экспрессию люциферазы с мРНК. Удивительно, однако, что 3'-UTR альбумина человека по изобретению дополнительно заметно продлевает экспрессию люциферазы с мРНК. По сравнению с 3'-UTR альбумина 3'-UTR нескольких различных стабильных мРНК действительно влияют на экспрессию люциферазы с мРНК гораздо менее благоприятным образом: 3'-UTR atp5o и atp5l стабилизируют экспрессию люциферазы гораздо меньше, чем 3'-UTR альбумина. Кроме того, 3'-UTR atp5o и atp5l существенно снижают уровни люциферазы по сравнению с 3'-UTR альбумина. 3'-UTR стабильной мРНК ndufa1 действительно заметно стабилизирует экспрессию люциферазы. Однако 3'-UTR ndufa1 также существенно снижает уровни люциферазы. 3'-UTR альбумина является уникальной в отношении продления экспрессии белка при поддержании суммарной экспрессии белка. Данные представлены как средние RLU ± SD (относительные световые единицы ± стандартное отклонение) для трех трансфекций. RLU суммированы в примере 5.2.
На фиг. 3 показано влияние точечных мутаций для удаления либо HindIII и/или сайта рестрикции XbaI и/или сигнала терминации Т7 с 3'-UTR альбумина человека на экспрессию люциферазы с мРНК, содержащей 3'-UTR альбумин человека. Таким образом, различные мРНК вводили электропорацией в клетки HeLa. Уровни люциферазы измеряли через 6, 24, 48 и 72 часа после трансфекции. Сигнал PpLuc корректировали на эффективность трансфекции по сигналу котрансфицированной RrLuc. 3'-UTR α-глобина стабилизирует экспрессию люциферазы с мРНК только очень умеренно. В отличие от этого все варианты 3'-UTR альбумина заметно продлевают экспрессию люциферазы с мРНК. Данные представлены как средние RLU ± SD (относительные световые единицы ± стандартное отклонение) для трех трансфекций. RLU суммированы в примере 5.4.
На фиг. 4 показана последовательность мРНК PpLuc(GC) - A64 без 3'-UTR. На фиг. 5 показана последовательность мРНК PpLuc(GC) - альбумин - A64. 3'-UTR альбумина человека вставляли между ORF и поли(А). Последовательность была взята из Dugaiczyk et al. 1982; Proc Natl Acad Sci U S A. Jan; 79(1):71-5.
На фиг. 6 показана последовательность мРНК PpLuc(GC) - альбумин2 - A64. 3'-UTR альбумина человека с сигналом терминации фага Т7, удаленным с помощью одной точечной мутации, вставляли между ORF и поли(А).
На фиг. 7 показана последовательность мРНК PpLuc(GC) - альбумин3 - A64. 3'-UTR альбумина человека с сигналом терминации фага Т7, удаленным с помощью одной точечной мутации, вставляли между ORF и поли(А).
На фиг. 8 показана последовательность мРНК PpLuc(GC) - альбумин4 - A64. 3'-UTR альбумина человека с сигналом терминации фага Т7, удаленным с помощью одной точечной мутации, вставляли между ORF и поли(А).
На фиг. 9 показана последовательность мРНК PpLuc(GC) - альбумин5 - A64. 3'-UTR альбумина человека с сигналом терминации фага Т7, удаленным с помощью двух последовательных точечных мутаций, вставляли между ORF и поли(А).
На фиг. 10 показана последовательность мРНК PpLuc(GC) - альбумин6 - A64. 3'-UTR альбумина человека с сайтами рестрикции HindIII и XbaI, удаленными с помощью двух одиночных точечных мутаций, вставляли между ORF и поли(А).
На фиг. 11 показана последовательность мРНК PpLuc(GC) - альбумин7 - A64. 3'-UTR альбумина человека с сигналом терминации фага Т7, а также с сайтами рестрикции HindIII и XbaI, удаленными с помощью трех одиночных точечных мутаций, вставляли между ORF и поли(А).
На фиг. 12 показана последовательность мРНК PpLuc(GC) - ag - A64. Центр, участок 3'-UTR β-глобина человека, связывающий α-комплекс, вставляли между ORF и поли(А).
На фиг. 13 показана последовательность мРНК PpLuc(GC) - gusb - A64. 3'-UTR β-глюкуронидазы человека вставляли между ORF и поли(А).
На фиг. 14 показана последовательность мРНК PpLuc(GC) - atp5o - A64. 3'-UTR субъединицы O АТФ-синтазы человека вставляли между ORF и поли(А).
На фиг. 15 показана последовательность мРНК PpLuc(GC) - ndufa1 - A64. 3'-UTR субъединицы 1 подкомплекса 1α NADH-дегидрогеназы человека [убихинона] вставляли между ORF и поли(А).
На фиг. 16 показана последовательность мРНК PpLuc(GC) - atp5l - A64. 3'-UTR субъединицы g АТФ-синтазы человека вставляли между ORF и поли(А).
Фиг. 17 иллюстрирует в качестве примера действие элемента 3'-UTR по настоящему изобретению, стабилизирующее продукцию белка и/или пролонгирующее продукцию белка. Кривые представляют собой количество белка, продуцируемого с молекул нуклеиновой кислоты, например, в клетках млекопитающих, измеренное в зависимости от времени. Сплошная линия представляет собой продукцию белка с молекулы искусственной нуклеиновой кислоты по настоящему изобретению, например, искусственной мРНК, пунктирная линия представляет собой продукцию белка с молекулы эталонной нуклеиновой кислоты, например, с отсутствием 3'-UTR или включающей эталонную 3'-UTR, такую как природная 3'-UTR, с ORF, кодирующей репортерный белок. Непрерывная горизонтальная жирная линия представляет собой пороговое значение. Это может быть, например, количество белка, измеряемое через 1, 2, 3, 4, 5 или 6 часов после инициации экспрессии, например, после трансфекции молекулы нуклеиновой кислоты. Можно видеть, что количество белка, продуцируемое с молекулы эталонной нуклеиновой кислоты, пересекает пороговое значение приблизительно через 32 часа после инициации экспрессии, например, после трансфекции, тогда как количество белка, продуцируемого с молекулы искусственной нуклеиновой кислоты по настоящему изобретению, пересекает пороговое значение приблизительно через 68 часов после инициации экспрессии, например, после трансфекции. Суммарное количество продуцируемого белка равно площади под кривой (AUC). Предпочтительно, чтобы суммарное количество белка, продуцируемого с молекулы искусственной нуклеиновой кислоты по настоящему изобретению, составляло, по меньшей мере, суммарное количество белка, продуцируемого с молекулы эталонной нуклеиновой кислоты без 3'-UTR.
На фиг. 18 показано действие различных 3'-UTR альбумина приматов на экспрессию люциферазы с искусственной мРНК в сравнении с экспрессией люциферазы с мРНК без 3'-UTR. Здесь мРНК, включающая 3'-UTR альбумина человека (альбумин), и мРНК, включающая 3'-UTR альбумина павиана анубис (альбумин8), представляют собой мРНК по настоящему изобретению. Они включают открытую рамку считывания, кодирующую люциферазу Photinus pyralis, а затем в направлении с 5'- к 3'-концу элемент 3'-UTR в соответствии с SEQ ID NO: 2 или SEQ ID NO: 33 и поли(А)-последовательность, имеющую в длину 64 аденина. Наблюдается заметное продление экспрессии белка с искусственных мРНК, содержащих 3'-UTR альбумина, соответствующих SEQ ID NO: 2 или SEQ ID NO: 33.
Эффект 3'-UTR альбумина человека и 3'-UTR альбумина павиана анубис на экспрессию люциферазы с мРНК исследован в сравнении с экспрессией люциферазы с мРНК без 3'-UTR. Чтобы исследовать экспрессию люциферазы, различные мРНК вводили электропорацией в фибробласты дермы человека (HDF). Уровни люциферазы измеряли через 6, 24, 48 и 72 часа после трансфекции. Уровень люциферазы с мРНК без 3'-UTR падает с 6 часов к 72 часам, через 72 часа остается 5% от сигнала через 6 часов. Опять 3'-UTR альбумина человека по изобретению заметно продлевает экспрессию люциферазы с мРНК. 3'-UTR альбумина бабуина анубис (альбумин8) продлевает экспрессию люциферазы с мРНК в той же степени, что и последовательность 3'-UTR альбумина человека. 3'-UTR альбумина приматов, таким образом, особенно хорошо подходят для продления экспрессии белка с мРНК. Данные представлены в виде среднего RLU ± SD (относительные световые единицы ± стандартное отклонение) для трех трансфекций. RLU суммированы в примере 5.4, таблица 7.
На фиг. 19 показано действие различных 3'-UTR альбумина приматов на экспрессию люциферазы с мРНК в сравнении с экспрессией люциферазы с мРНК без 3'-UTR с использованием другого метода трансфекции. Для этого различные мРНК вводили липофекцией в фибробласты дермы человека (HDF). Уровни люциферазы измеряли через 6, 24, 48 и 72 часа после трансфекции. Уровень люциферазы с мРНК без 3'-UTR падает с 6 часов к 48 часам до 82% от сигнала через 6 часов. Опять 3'-UTR альбумина человека заметно продлевает экспрессию люциферазы с мРНК, причем сигнал через 48 часов выше, чем сигнал через 6 часов. 3'-UTR альбумина бабуина анубис (альбумин8) продлевает экспрессию люциферазы с мРНК в сходной степени, что и последовательность 3'-UTR альбумина человека. 3'-UTRs альбумина приматов, таким образом, особенно хорошо подходят для продления экспрессии белка с мРНК. Данные представлены в виде среднего RLU ± SD (относительные световые единицы ± стандартное отклонение) для трех трансфекций. RLU суммированы в примере 5.4, таблица 9.
На фиг. 20 показано действие различных 3'-UTR альбумина приматов на экспрессию люциферазы с мРНК в клетках мыши в сравнении с экспрессией люциферазы с мРНК без 3'-UTR. Для этого различные мРНК вводили липофекцией в клетки L-929, клеточную линию фибробластов мыши. Уровни люциферазы измеряли через 6, 24, 48 и 72 часа после трансфекции. Уровень люциферазы с мРНК без 3'-UTR падает с 6 часов к 48 часам, через 48 часов остается 23% от сигнала через 6 часов. Даже в клетках мыши 3'-UTR альбумина человека действительно заметно продлевает экспрессию люциферазы с мРНК. 3'-UTR альбумина бабуина анубис (альбумин8) в сходной степени продлевает экспрессию люциферазы с мРНК. 3'-UTR альбумина приматов, таким образом, особенно хорошо подходят для продления экспрессии белка с мРНК в клеточных типах млекопитающих. Данные представлены в виде среднего RLU ± SD (относительные световые единицы ± стандартное отклонение) для трех трансфекций. RLU суммированы в примере 5.5, таблица 11.
На фиг. 21 показано действие различных 3'-UTR альбумина млекопитающих на экспрессию люциферазы с мРНК в сравнении с экспрессией люциферазы с мРНК без 3'-UTR. Для этого различные мРНК вводили липофекцией в клетки HeLa. Уровни люциферазы измеряли через 6, 24, 48 и 72 часа после трансфекции. Уровень люциферазы с мРНК без 3'-UTR падает с 6 часов к 48 часам, через 48 часов остается 49% от сигнала через 6 часов. Опять 3'-UTR альбумина человека и 3'-UTR альбумина бабуина анубис (альбумин8) заметно продлевают экспрессию люциферазы с мРНК, причем сигнал через 48 часов выше, чем сигнал через 6 часов. Важно, что 3'-UTR альбумина мыши (альбумин9) также сходно продлевает экспрессию люциферазы с мРНК. 3'-UTR альбумина млекопитающих, таким образом, особенно хорошо подходят для продления экспрессии белка с мРНК. Данные представлены в виде среднего RLU ± SD (относительные световые единицы ± стандартное отклонение) для трех трансфекций. RLU суммированы в примере 5.6, таблица 13.
На фиг. 22 показана последовательность мРНК PpLuc(GC) - альбумин8 - A64. 3'-UTR альбумина бабуина анубис вставляли между ORF и поли(А). Последовательность взята из эталонной последовательности NCBI XM_003898783.1.
На фиг. 23 показана последовательность мРНК PpLuc(GC) - альбумин9 - A64. 3'-UTR альбумина мыши вставляли между ORF и поли(А). Последовательность взята из эталонной последовательности NCBI NM_009654.3.
Примеры
1. Получение ДНК-матриц
Для транскрипции in vitro конструировали вектор, содержащий промотор Т7, за которым следует GC-обогащенная последовательность, кодирующая люциферазу Photinus pyralis (PpLuc(GC)) и поли(А)-последовательность А64. За поли(А)-последовательностью сразу следовал сайт рестрикции, используемый для линеаризации вектора перед транскрипцией in vitro для того, чтобы получить мРНК, заканчивающуюся поли(А)-последовательностью А64. мРНК, полученную таким образом с этого вектора с помощью транскрипции in vitro, обозначали как «PpLuc(GC)-A64».
Этот вектор модифицировали для включения нетранслируемых 3'-последовательностей открытой рамки считывания (3'-UTR). Получены векторы, включающие следующие кодирующие последовательности мРНК (фигуры с 4 по 16 и фигуры 22 и 23):
мРНК, используемые в примерах, получали транскрипцией указанных векторов in vitro.
2. Транскрипция in vitro
ДНК-матрицу в соответствии с примером 1 линеаризовали и транскрибировали in vitro с использованием T7-полимеразы. ДНК-матрицу затем гидролизовали обработкой ДНКазой. Транскрипты мРНК содержали структуру 5'-кэпа, полученную путем добавления избытка N7-метилгуанозин-5'-трифосфат-5'-гуанозина в реакционную смесь для транскрипции. мРНК, полученную таким образом, очищали и ресуспендировали в воде.
3. Трансфекция клеток
3.1 Электропорация мРНК
Клетки обрабатывали трипсином и промывали в opti-MEM. 5×104 или 1×105 клеток в 200 мкл opti-MEM в каждом случае электропорировали 0,3 или 1 мкг PpLuc-кодирующих мРНК. В качестве контроля отдельно электропорировали мРНК, не кодирующую PpLuc. В некоторых экспериментах мРНК, кодирующую люциферазу Renilla reniformis (RrLuc), подвергали электропорации вместе с мРНК PpLuc для контроля эффективности трансфекции (0,1 мкг мРНК RrLuc). Электропорированные клетки высевали в 24-луночные планшеты в 1 мл среды. Через 6, 24 или 48 часов после трансфекции (или через 72 часов в некоторых экспериментах) среду аспирировали, и клетки лизировали в 200 мкл лизирующего буфера (25 мМ Трис, рН 7,5 (HCl), 2 мМ ЭДТА, 10% глицерин, 1% Тритон Х-100, 2 мМ ДТТ, 1 мМ PMSF или, альтернативно, пассивным буфером для лизиса, Promega). Лизаты хранили при -20°С до измерения активности люциферазы.
3.2 Липофекция мРНК
Клетки высевали в 96-луночные планшеты за три дня до трансфекции (2500 или 5000 клеток на лунку). Непосредственно перед липофекцией клетки промывали в opti-MEM. В клетки липофекцией вводили мРНК, кодирующую PpLuc, 25 нг на лунку, в комплексе с липофектамином 2000. мРНК, кодирующую люциферазу Renilla reniformis (RrLuc), котрансфицировали вместе с мРНК PpLuc для контроля эффективности трансфекции (2,5 нг мРНК RrLuc на лунку). Через 6, 24, 48 или 72 часа после трансфекции среду аспирировали, и клетки лизировали в 100 мкл лизирующего буфера (пассивного буфера для лизиса, Promega). Лизаты хранили при -80°С до измерения активности люциферазы.
4. Измерение активности люциферазы
Активность люциферазы измеряли в относительных световых единицах (RLU) в ридере для планшетов BioTek SynergyHT. Активность PpLuc измеряли через каждые 5 секунд времени, используя 50 мкл лизата и 200 мкл люциферинового буфера (75 мкМ люциферина, 25 мМ глицилглицина, рН 7,8 (NaOH), 15 мМ MgSO4, 2 мМ АТФ). Активность RrLuc измеряли через каждые 5 секунд времени, используя 50 мкл лизата и 200 мкл целентеразинового буфера (40 мкМ целентеразин, 2,2 мМ ЭДТА, 220 мМ KH2PO4/K2HPO4, рН 5,0, 1,1 М NaCl, 1,3 мМ NaN3, 0,44 г/л BSA).
Альтернативно, активность люциферазы измеряли в относительных световых единицах (RLU) в ридере для планшетов Hidex Chameleon. Активность PpLuc измеряли через каждые 2 секунды времени, используя 20 мкл лизата и 200 мкл люциферинового буфера (Beetle-Juice, PJK GmbH). Активность RrLuc измеряли через каждые 2 секунды времени, используя 20 мкл лизата и 100 мкл целентеразинового буфера (Renilla-Juice, PJK GmbH).
5. Результаты
5.1 3'-UTR альбумина продлевает экспрессию белка с мРНК заметно больше, чем хорошо известная 3'-UTR α-глобина
Для исследования действия 3'-нетранслируемых областей на экспрессию белка с мРНК синтезировали мРНК с различными 3'-UTR в соответствии с примерами 1-2: мРНК содержала либо центр, участок, связывающий α-комплекс 3'-UTR α-глобина человека (PpLuc(GC)-ag-A64 в соответствии с SEQ ID NO: 13), так как 3'-UTR α-глобина, как сообщалось, стабилизирует мРНК независимо от кодирующей области последовательности (Rodgers, N.D., Wang, Z. & Kiledjian, M., 2002. Regulated alpha-globin mRNA decay is a cytoplasmic event proceeding through 3'-to-5' exosome-dependent decapping. RNA, 8(12), S.1526-1537.). Альтернативно, мРНК содержала 3'-UTR альбумина человека (PpLuc (GC)-альбумин-A64 в соответствии с SEQ ID NO: 6). мРНК альбумина человека, как сообщалось, является стабильной (Johnson, T.R. et al., 1991. Newly synthesized RNA: simultaneous measurement in intact cells of transcription rates and RNA stability of insulin-like growth factor I, actin, and albumin in growth hormone-stimulated hepatocytes. Proceedings of the National Academy of Sciences, 88(12), S.5287-5291). Наконец, была использована мРНК, содержащая 3'-UTR β-глюкуронидазы человека (PpLuc(GC)-gusb-A64 в соответствии с SEQ ID NO: 14). Эта мРНК β-глюкуронидазы человека, также как сообщалось, является стабильной (Watson, G. & Paigen, K., 1987. Genetic variations in kinetic constants that describe beta-glucuronidase mRNA induction in androgen-treated mice. Molecular and Cellular Biology, 7(3), S.1085-1090.). Для сравнения использовали также мРНК без 3'-UTR (PpLuc (GC)-A64 согласно SEQ ID NO: 5). мРНК, кодирующие люциферазу, вводили электропорацией в клетки HeLa. Уровни люциферазы измеряли через 6, 24 и 48 часов после трансфекции (смотри ниже таблицу 1 и фигуру 1).
Уровень люциферазы с мРНК без 3'-UTR падал с 6 часов до 48 часов, через 48 часов оставалось 10% от сигнала через 6 часов. 3'-UTR α-глобина умеренно стабилизировала экспрессию люциферазы с мРНК. Удивительно, однако, что 3'-UTR альбумина человека дополнительно заметно продлевала экспрессию люциферазы с мРНК. В отличие от этого 3'-UTR β-глюкуронидазы не продлевала экспрессию люциферазы в той степени, которая наблюдалась для 3'-UTR альбумина.
Было рассчитано отношение уровня люциферазы через 48 часов к уровню через 6 часов, оказавшееся более высоким, что указывает на стабилизацию экспрессии белка. Эти данные, указывающие на то, насколько какая-либо 3'-UTR стабилизирует экспрессию белка во времени, суммированы в таблице 2.
3'-UTR альбумина стабилизировала экспрессию белка намного больше, чем хорошо известная 3'-UTR α-глобина, и больше, чем 3'-UTR стабильной мРНК β-глюкуронидазы. Этот результат показывает, что 3'-UTR альбумина особенно эффективна в отношении продления экспрессии белка с мРНК.
5.2 3'-UTR альбумина является уникальной в отношении продления экспрессии белка при поддержании суммарной экспрессии белка
Продление экспрессии белка с помощью 3'-UTR альбумина может отражать общее действие 3'-UTR стабильных мРНК (хотя даже очень ограниченное действие 3'-UTR α-глобина, наблюдаемое в примере 5.1, не подтверждает этот аргумент). Таким образом, синтезировали мРНК, содержащие 3'-UTR от стабильных мРНК (Friedel, C.C. et al., 2009. Conserved principles of mammalian transcriptional regulation revealed by RNA half-life. Nucleic Acids Research, 37(17), S.e115.), atp5o или ndufa1, или atp5l (субъединицы O АТФ-синтазы человека, субъединицы 1 подкомплекса 1α NADH-дегидрогеназы человека [убихинона] или субъединицы g АТФ-синтазы человека, соответственно). мРНК, кодирующие люциферазу, вводили электропорацией в клетки HeLa. Уровни люциферазы измеряли через 6, 24 и 48 часов после трансфекции (смотри ниже таблицу 3 и фигуру 2).
Уровень люциферазы с мРНК без 3'-UTR падал с 6 часов к 48 часам, через 48 часов оставалось 14% от сигнала через 6 часов. 3'-UTR α-глобина умеренно стабилизировала экспрессию люциферазы с мРНК. Удивительно, однако, что 3'-UTR альбумина человека дополнительно заметно продлевала экспрессию люциферазы с мРНК. По сравнению с действием 3'-UTR альбумина 3'-UTR нескольких различных стабильных мРНК влияют на экспрессию люциферазы с мРНК гораздо менее благоприятным образом: 3'-UTR atp5o и atp5l стабилизировали экспрессию люциферазы гораздо меньше, чем 3'-UTR альбумина. Кроме того, 3'-UTR atp5o и atp5l существенно снижали уровни люциферазы по сравнению с 3'-UTR альбумина. 3'-UTR стабильной мРНК ndufa1 действительно заметно стабилизирует экспрессию люциферазы. Однако 3'-UTR ndufa1 также существенно снижала уровни люциферазы.
Было рассчитано отношение уровня люциферазы через 48 часов к уровню через 6 часов, оказавшееся более высоким, что указывает на стабилизацию экспрессии белка. Эти данные, указывающие на то, насколько какая-либо 3'-UTR стабилизирует экспрессию белка во времени, суммированы в таблице 4.
3'-UTR альбумина являлась уникальной в отношении продления экспрессии белка при поддержании суммарной экспрессии белка. В наиболее поздний период времени 3'-UTR альбумина вызывала подъем до существенно более высокой экспрессии белка по сравнению с хорошо известной 3'-UTR α-глобина и 3'-UTR нескольких различных стабильных мРНК. Этот результат показывает, что 3'-UTR альбумина особенно подходит для продления экспрессии белка с мРНК.
5.3 Различные варианты 3'-UTR альбумина продлевают экспрессию белка с мРНК
3'-UTR альбумина человека содержит сайт рестрикции HindIII, сайт рестрикции XbaI и сигнал терминации T7. Таким образом, синтезировали мРНК, содержащую варианты 3'-UTR альбумина человека с сайтом рестрикции HindIII и/или XbaI и/или сигналом терминации Т7, удаленным(ми) с помощью точечной(ых) мутации(й). (PpLuc(GC)-альбумин2-7 в соответствии с SEQ ID NO: 17-12). мРНК, кодирующие люциферазу, вводили электропорацией в клетки HeLa. Уровни люциферазы измеряли через 6, 24, 48 и 72 часа после трансфекции. Сигнал PpLuc корректировали на эффективность трансфекции по сигналу котрансфицированной RrLuc. (смотри следующую таблицу 5 и фигуру 3).
Было рассчитано отношение уровня люциферазы через 72 часа к уровню через 6 часов, оказавшееся более высоким, что указывает на стабилизацию экспрессии белка. Эти данные, указывающие на то, насколько какая-либо 3'-UTR стабилизирует экспрессию белка во времени, суммированы в таблице 6.
Уровень люциферазы с мРНК без 3'-UTR падал с 6 часов к 72 часам, через 72 часа оставалось 5% от сигнала через 6 часов. 3'-UTR α-глобина только очень умеренно стабилизировала экспрессию люциферазы с мРНК. В отличие от этого, все варианты 3'-UTR альбумина заметно продлевали экспрессию люциферазы с мРНК.
5.4 3'-UTR альбумина приматов продлевают экспрессию белка с мРНК
Продление экспрессии белка под действием 3'-UTR альбумина может быть видоспецифичным. Сравнивали 3'-UTR альбумина различных приматов, 3'-UTR альбумина бабуина анубис была наименее гомологичной 3'-UTR альбумина человека (шимпанзе: 99% идентичности, карликовый шимпанзе: 99% идентичности, орангутанг Суматры: 99% идентичности, бабуин анубис: 96% идентичности). Таким образом, синтезировали мРНК, содержащую 3'-UTR гена альбумина бабуина анубис (PpLuc(GC)-альбумин8-A64 в соответствии с SEQ ID NO: 40). мРНК, кодирующие люциферазу, вводили электропорацией в фибробласты дермы человека (HDF). Уровни люциферазы измеряли через 6, 24, 48 и 72 часа после трансфекции. Сигнал PpLuc корректировали на эффективность трансфекции по сигналу котрансфицированной RrLuc. (смотри следующую таблицу 7 и фигуру 18).
Было рассчитано отношение уровня люциферазы через 72 часа к уровню через 6 часов, оказавшееся более высоким, что указывает на стабилизацию экспрессии белка. Эти данные, указывающие на то, насколько какая-либо 3'-UTR стабилизирует экспрессию белка во времени, суммированы в таблице 8.
Уровень люциферазы с мРНК без 3'-UTR падал с 6 часов к 72 часам, через 72 часа оставалось 5% от сигнала через 6 часов. 3'-UTR альбумина человека заметно продлевала экспрессию люциферазы с мРНК. 3'-UTR альбумина бабуина анубис продлевала экспрессию люциферазы с мРНК в той же степени, что и последовательность человека. Этот результат показывает, что 3'-UTR альбумина приматов особенно подходят для продления экспрессии белка с мРНК.
Продление экспрессии белка обеими 3'-UTR как человека, так и бабуина анубис наблюдалось также, если мРНК вводили липофекцией, а не электропорацией. мРНК, кодирующие люциферазу, вводили липофекцией в HDF. Уровни люциферазы измеряли через 6, 24, 48 и 72 часа после трансфекции. Сигнал PpLuc корректировали на эффективность трансфекции по сигналу котрансфицированной RrLuc. (смотри следующую таблицу 9 и фигуру 19).
Были рассчитаны отношения уровня люциферазы через 48 часов к уровню через 6 часов, оказавшиеся более высокими, что указывает на стабилизацию экспрессии белка. Эти данные, указывающие на то, насколько какая-либо 3'-UTR стабилизирует экспрессию белка во времени, суммированы в таблице 10.
При липофекции вместо электропорации, снова 3'-UTR как альбумина человека, так и 3'-UTR альбумина бабуина анубис заметно продлевали экспрессию люциферазы с мРНК. Этот результат подтверждает, что 3'-UTR альбумина приматов являются особенно пригодными для продления экспрессии белка с мРНК.
5.5 3'-UTR альбумина приматов продлевают экспрессию белка с мРНК в клетках мыши
Продление экспрессии белка под действием 3'-UTR альбумина может быть видоспецифичным. Таким образом, тестировали, будут ли 3'-UTR альбумина человека и 3'-UTR альбумина бабуина анубис продлевать экспрессию люциферазы с мРНК в клетках мыши. мРНК, кодирующие люциферазу, вводили липофекцией в клетки L-929, клеточную линию фибробластов мыши. Уровни люциферазы измеряли через 6, 24, 48 и 72 часа после трансфекции. Сигнал PpLuc корректировали на эффективность трансфекции по сигналу котрансфицированной RrLuc (смотри следующую таблицу 11 и фигуру 20).
Было рассчитано отношение уровня люциферазы через 48 часов к уровню через 6 часов, оказавшееся более высоким, что указывает на стабилизацию экспрессии белка. Эти данные, указывающие на то, насколько какая-либо 3'-UTR стабилизирует экспрессию белка во времени, суммированы в таблице 12.
Уровень люциферазы с мРНК без 3'-UTR падал с 6 часов к 48 часам, через 48 часов оставалось 23% от сигнала через 6 часов. 3'-UTR альбумина человека заметно продлевала экспрессию люциферазы с мРНК в клеточной линии мыши. 3'-UTR альбумина бабуина анубис также продлевала экспрессию люциферазы с мРНК в клеточной линии мыши. Этот результат показывает, что 3'-UTR альбумина приматов особенно подходят для продления экспрессии белка с мРНК в клеточных линиях млекопитающих.
5.6 3'-UTR альбумина млекопитающих продлевают экспрессию белка с мРНК
Продление экспрессии белка под действием 3'-UTR альбумина может быть видоспецифичным. Сравнивали 3'-UTR альбумина различных млекопитающих, 3'-UTR альбумина мыши была наименее гомологична 3'-UTR альбумина человека (лошадь: 86% идентичности, домашняя собака: 84% идентичности, крупный рогатый скот: 74% идентичности, крыса: 73% идентичности, мышь: 72% идентичности). Таким образом, синтезировали мРНК, содержащую 3'-UTR гена альбумина мыши (PpLuc(GC)-альбумин9-А64 в соответствии с SEQ ID NO: 41). мРНК, кодирующие люциферазу, вводили липофекцией в клетки HeLa. Уровни люциферазы измеряли через 6, 24, 48 и 72 часа после трансфекции. Сигнал PpLuc корректировали на эффективность трансфекции по сигналу котрансфицированной RrLuc (смотри ниже таблицу 13 и фигуру 21).
Было рассчитано отношение уровня люциферазы через 48 часов к уровню через 6 часов, оказавшееся более высоким, что указывает на стабилизацию экспрессии белка. Эти данные, указывающие на то, насколько какая-либо 3'-UTR стабилизирует экспрессию белка во времени, суммированы в таблице 14.
Уровень люциферазы с мРНК без 3'-UTR падал с 6 часов к 48 часам, через 48 часов оставалось 49% от сигнала через 6 часов. 3'-UTR альбумина человека и 3'-UTR альбумина бабуина анубис заметно продлевали экспрессию люциферазы с мРНК. Важно, что 3'-UTR альбумина мыши сходно продлевала экспрессию люциферазы с мРНК в клеточной линии человека HeLa. Этот результат показывает, что 3'-UTR альбумина млекопитающих особенно подходят для продления экспрессии белка с мРНК.
Последовательности:
SEQ ID NO: 1:
CATCACATTT AAAAGCATCT CAGCCTACCA TGAGAATAAG AGAAAGAAAA TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT TGCCTCTTTT CTCTGTGCTT CAATTAATAA AAAATGGAAA GAATCT
SEQ ID NO: 2:
CAUCACAUUU AAAAGCAUCU CAGCCUACCA UGAGAAUAAG AGAAAGAAAA UGAAGAUCAA AAGCUUAUUC AUCUGUUUUU CUUUUUCGUU GGUGUAAAGC CAACACCCUG UCUAAAAAAC AUAAAUUUCU UUAAUCAUUU UGCCUCUUUU CUCUGUGCUU CAAUUAAUAA AAAAUGGAAA GAAUCU
SEQ ID NO: 3:
CATCACATTT AAAAGCATCT CAGCCTACCA TGAGAATAAG AGAAAGAAAA TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT TGCCTCTTTT CTCTGTGCTT CAATTAATAA AAAATGGAAA GAATCTAGAT CTAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAA
SEQ ID NO: 4:
CAUCACAUUU AAAAGCAUCU CAGCCUACCA UGAGAAUAAG AGAAAGAAAA UGAAGAUCAA AAGCUUAUUC AUCUGUUUUU CUUUUUCGUU GGUGUAAAGC CAACACCCUG UCUAAAAAAC AUAAAUUUCU UUAAUCAUUU UGCCUCUUUU CUCUGUGCUU CAAUUAAUAA AAAAUGGAAA GAAUCUAGAU CUAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAA
SEQ ID NO: 5:
GGGAGAAAGCTTGAGGATGGAGGACGCCAAGAACATCAAGAAGGGCCCGGCGCCCTTCTA
CCCGCTGGAGGACGGGACCGCCGGCGAGCAGCTCCACAAGGCCATGAAGCGGTACGCCCT
GGTGCCGGGCACGATCGCCTTCACCGACGCCCACATCGAGGTCGACATCACCTACGCGGA
GTACTTCGAGATGAGCGTGCGCCTGGCCGAGGCCATGAAGCGGTACGGCCTGAACACCAA
CCACCGGATCGTGGTGTGCTCGGAGAACAGCCTGCAGTTCTTCATGCCGGTGCTGGGCGC
CCTCTTCATCGGCGTGGCCGTCGCCCCGGCGAACGACATCTACAACGAGCGGGAGCTGCT
GAACAGCATGGGGATCAGCCAGCCGACCGTGGTGTTCGTGAGCAAGAAGGGCCTGCAGAA
GATCCTGAACGTGCAGAAGAAGCTGCCCATCATCCAGAAGATCATCATCATGGACAGCAA
GACCGACTACCAGGGCTTCCAGTCGATGTACACGTTCGTGACCAGCCACCTCCCGCCGGG
CTTCAACGAGTACGACTTCGTCCCGGAGAGCTTCGACCGGGACAAGACCATCGCCCTGAT
CATGAACAGCAGCGGCAGCACCGGCCTGCCGAAGGGGGTGGCCCTGCCGCACCGGACCGC
CTGCGTGCGCTTCTCGCACGCCCGGGACCCCATCTTCGGCAACCAGATCATCCCGGACAC
CGCCATCCTGAGCGTGGTGCCGTTCCACCACGGCTTCGGCATGTTCACGACCCTGGGCTA
CCTCATCTGCGGCTTCCGGGTGGTCCTGATGTACCGGTTCGAGGAGGAGCTGTTCCTGCG
GAGCCTGCAGGACTACAAGATCCAGAGCGCGCTGCTCGTGCCGACCCTGTTCAGCTTCTT
CGCCAAGAGCACCCTGATCGACAAGTACGACCTGTCGAACCTGCACGAGATCGCCAGCGG
GGGCGCCCCGCTGAGCAAGGAGGTGGGCGAGGCCGTGGCCAAGCGGTTCCACCTCCCGGG
CATCCGCCAGGGCTACGGCCTGACCGAGACCACGAGCGCGATCCTGATCACCCCCGAGGG
GGACGACAAGCCGGGCGCCGTGGGCAAGGTGGTCCCGTTCTTCGAGGCCAAGGTGGTGGA
CCTGGACACCGGCAAGACCCTGGGCGTGAACCAGCGGGGCGAGCTGTGCGTGCGGGGGCC
GATGATCATGAGCGGCTACGTGAACAACCCGGAGGCCACCAACGCCCTCATCGACAAGGA
CGGCTGGCTGCACAGCGGCGACATCGCCTACTGGGACGAGGACGAGCACTTCTTCATCGT
CGACCGGCTGAAGTCGCTGATCAAGTACAAGGGCTACCAGGTGGCGCCGGCCGAGCTGGA
GAGCATCCTGCTCCAGCACCCCAACATCTTCGACGCCGGCGTGGCCGGGCTGCCGGACGA
CGACGCCGGCGAGCTGCCGGCCGCGGTGGTGGTGCTGGAGCACGGCAAGACCATGACGGA
GAAGGAGATCGTCGACTACGTGGCCAGCCAGGTGACCACCGCCAAGAAGCTGCGGGGCGG
CGTGGTGTTCGTGGACGAGGTCCCGAAGGGCCTGACCGGGAAGCTCGACGCCCGGAAGAT
CCGCGAGATCCTGATCAAGGCCAAGAAGGGCGGCAAGATCGCCGTGTAAGACTAGTAGAT
CTAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA
AAAAAA
SEQ ID NO: 6:
GGGAGAAAGCTTGAGGATGGAGGACGCCAAGAACATCAAGAAGGGCCCGGCGCCCTTCTA
CCCGCTGGAGGACGGGACCGCCGGCGAGCAGCTCCACAAGGCCATGAAGCGGTACGCCCT
GGTGCCGGGCACGATCGCCTTCACCGACGCCCACATCGAGGTCGACATCACCTACGCGGA
GTACTTCGAGATGAGCGTGCGCCTGGCCGAGGCCATGAAGCGGTACGGCCTGAACACCAA
CCACCGGATCGTGGTGTGCTCGGAGAACAGCCTGCAGTTCTTCATGCCGGTGCTGGGCGC
CCTCTTCATCGGCGTGGCCGTCGCCCCGGCGAACGACATCTACAACGAGCGGGAGCTGCT
GAACAGCATGGGGATCAGCCAGCCGACCGTGGTGTTCGTGAGCAAGAAGGGCCTGCAGAA
GATCCTGAACGTGCAGAAGAAGCTGCCCATCATCCAGAAGATCATCATCATGGACAGCAA
GACCGACTACCAGGGCTTCCAGTCGATGTACACGTTCGTGACCAGCCACCTCCCGCCGGG
CTTCAACGAGTACGACTTCGTCCCGGAGAGCTTCGACCGGGACAAGACCATCGCCCTGAT
CATGAACAGCAGCGGCAGCACCGGCCTGCCGAAGGGGGTGGCCCTGCCGCACCGGACCGC
CTGCGTGCGCTTCTCGCACGCCCGGGACCCCATCTTCGGCAACCAGATCATCCCGGACAC
CGCCATCCTGAGCGTGGTGCCGTTCCACCACGGCTTCGGCATGTTCACGACCCTGGGCTA
CCTCATCTGCGGCTTCCGGGTGGTCCTGATGTACCGGTTCGAGGAGGAGCTGTTCCTGCG
GAGCCTGCAGGACTACAAGATCCAGAGCGCGCTGCTCGTGCCGACCCTGTTCAGCTTCTT
CGCCAAGAGCACCCTGATCGACAAGTACGACCTGTCGAACCTGCACGAGATCGCCAGCGG
GGGCGCCCCGCTGAGCAAGGAGGTGGGCGAGGCCGTGGCCAAGCGGTTCCACCTCCCGGG
CATCCGCCAGGGCTACGGCCTGACCGAGACCACGAGCGCGATCCTGATCACCCCCGAGGG
GGACGACAAGCCGGGCGCCGTGGGCAAGGTGGTCCCGTTCTTCGAGGCCAAGGTGGTGGA
CCTGGACACCGGCAAGACCCTGGGCGTGAACCAGCGGGGCGAGCTGTGCGTGCGGGGGCC
GATGATCATGAGCGGCTACGTGAACAACCCGGAGGCCACCAACGCCCTCATCGACAAGGA
CGGCTGGCTGCACAGCGGCGACATCGCCTACTGGGACGAGGACGAGCACTTCTTCATCGT
CGACCGGCTGAAGTCGCTGATCAAGTACAAGGGCTACCAGGTGGCGCCGGCCGAGCTGGA
GAGCATCCTGCTCCAGCACCCCAACATCTTCGACGCCGGCGTGGCCGGGCTGCCGGACGA
CGACGCCGGCGAGCTGCCGGCCGCGGTGGTGGTGCTGGAGCACGGCAAGACCATGACGGA
GAAGGAGATCGTCGACTACGTGGCCAGCCAGGTGACCACCGCCAAGAAGCTGCGGGGCGG
CGTGGTGTTCGTGGACGAGGTCCCGAAGGGCCTGACCGGGAAGCTCGACGCCCGGAAGAT
CCGCGAGATCCTGATCAAGGCCAAGAAGGGCGGCAAGATCGCCGTGTAAGACTAGTGCAT
CACATTTAAAAGCATCTCAGCCTACCATGAGAATAAGAGAAAGAAAATGAAGATCAAAAG
CTTATTCATCTGTTTTTCTTTTTCGTTGGTGTAAAGCCAACACCCTGTCTAAAAAACATA
AATTTCTTTAATCATTTTGCCTCTTTTCTCTGTGCTTCAATTAATAAAAAATGGAAAGAA
TCTAGATCTAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA
AAAAAAAAAAAAA
SEQ ID NO: 7:
GGGAGAAAGCTTGAGGATGGAGGACGCCAAGAACATCAAGAAGGGCCCGGCGCCCTTCTA
CCCGCTGGAGGACGGGACCGCCGGCGAGCAGCTCCACAAGGCCATGAAGCGGTACGCCCT
GGTGCCGGGCACGATCGCCTTCACCGACGCCCACATCGAGGTCGACATCACCTACGCGGA
GTACTTCGAGATGAGCGTGCGCCTGGCCGAGGCCATGAAGCGGTACGGCCTGAACACCAA
CCACCGGATCGTGGTGTGCTCGGAGAACAGCCTGCAGTTCTTCATGCCGGTGCTGGGCGC
CCTCTTCATCGGCGTGGCCGTCGCCCCGGCGAACGACATCTACAACGAGCGGGAGCTGCT
GAACAGCATGGGGATCAGCCAGCCGACCGTGGTGTTCGTGAGCAAGAAGGGCCTGCAGAA
GATCCTGAACGTGCAGAAGAAGCTGCCCATCATCCAGAAGATCATCATCATGGACAGCAA
GACCGACTACCAGGGCTTCCAGTCGATGTACACGTTCGTGACCAGCCACCTCCCGCCGGG
CTTCAACGAGTACGACTTCGTCCCGGAGAGCTTCGACCGGGACAAGACCATCGCCCTGAT
CATGAACAGCAGCGGCAGCACCGGCCTGCCGAAGGGGGTGGCCCTGCCGCACCGGACCGC
CTGCGTGCGCTTCTCGCACGCCCGGGACCCCATCTTCGGCAACCAGATCATCCCGGACAC
CGCCATCCTGAGCGTGGTGCCGTTCCACCACGGCTTCGGCATGTTCACGACCCTGGGCTA
CCTCATCTGCGGCTTCCGGGTGGTCCTGATGTACCGGTTCGAGGAGGAGCTGTTCCTGCG
GAGCCTGCAGGACTACAAGATCCAGAGCGCGCTGCTCGTGCCGACCCTGTTCAGCTTCTT
CGCCAAGAGCACCCTGATCGACAAGTACGACCTGTCGAACCTGCACGAGATCGCCAGCGG
GGGCGCCCCGCTGAGCAAGGAGGTGGGCGAGGCCGTGGCCAAGCGGTTCCACCTCCCGGG
CATCCGCCAGGGCTACGGCCTGACCGAGACCACGAGCGCGATCCTGATCACCCCCGAGGG
GGACGACAAGCCGGGCGCCGTGGGCAAGGTGGTCCCGTTCTTCGAGGCCAAGGTGGTGGA
CCTGGACACCGGCAAGACCCTGGGCGTGAACCAGCGGGGCGAGCTGTGCGTGCGGGGGCC
GATGATCATGAGCGGCTACGTGAACAACCCGGAGGCCACCAACGCCCTCATCGACAAGGA
CGGCTGGCTGCACAGCGGCGACATCGCCTACTGGGACGAGGACGAGCACTTCTTCATCGT
CGACCGGCTGAAGTCGCTGATCAAGTACAAGGGCTACCAGGTGGCGCCGGCCGAGCTGGA
GAGCATCCTGCTCCAGCACCCCAACATCTTCGACGCCGGCGTGGCCGGGCTGCCGGACGA
CGACGCCGGCGAGCTGCCGGCCGCGGTGGTGGTGCTGGAGCACGGCAAGACCATGACGGA
GAAGGAGATCGTCGACTACGTGGCCAGCCAGGTGACCACCGCCAAGAAGCTGCGGGGCGG
CGTGGTGTTCGTGGACGAGGTCCCGAAGGGCCTGACCGGGAAGCTCGACGCCCGGAAGAT
CCGCGAGATCCTGATCAAGGCCAAGAAGGGCGGCAAGATCGCCGTGTAAGACTAGTGCAT
CACATTTAAAAGCATCTCAGCCTACCATGAGAATAAGAGAAAGAAAATGAAGATCAAAAG
CTTATTCGTCTGTTTTTCTTTTTCGTTGGTGTAAAGCCAACACCCTGTCTAAAAAACATA
AATTTCTTTAATCATTTTGCCTCTTTTCTCTGTGCTTCAATTAATAAAAAATGGAAAGAA
TCTAGATCTAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA
AAAAAAAAAAAAA
SEQ ID NO: 8:
GGGAGAAAGCTTGAGGATGGAGGACGCCAAGAACATCAAGAAGGGCCCGGCGCCCTTCTA
CCCGCTGGAGGACGGGACCGCCGGCGAGCAGCTCCACAAGGCCATGAAGCGGTACGCCCT
GGTGCCGGGCACGATCGCCTTCACCGACGCCCACATCGAGGTCGACATCACCTACGCGGA
GTACTTCGAGATGAGCGTGCGCCTGGCCGAGGCCATGAAGCGGTACGGCCTGAACACCAA
CCACCGGATCGTGGTGTGCTCGGAGAACAGCCTGCAGTTCTTCATGCCGGTGCTGGGCGC
CCTCTTCATCGGCGTGGCCGTCGCCCCGGCGAACGACATCTACAACGAGCGGGAGCTGCT
GAACAGCATGGGGATCAGCCAGCCGACCGTGGTGTTCGTGAGCAAGAAGGGCCTGCAGAA
GATCCTGAACGTGCAGAAGAAGCTGCCCATCATCCAGAAGATCATCATCATGGACAGCAA
GACCGACTACCAGGGCTTCCAGTCGATGTACACGTTCGTGACCAGCCACCTCCCGCCGGG
CTTCAACGAGTACGACTTCGTCCCGGAGAGCTTCGACCGGGACAAGACCATCGCCCTGAT
CATGAACAGCAGCGGCAGCACCGGCCTGCCGAAGGGGGTGGCCCTGCCGCACCGGACCGC
CTGCGTGCGCTTCTCGCACGCCCGGGACCCCATCTTCGGCAACCAGATCATCCCGGACAC
CGCCATCCTGAGCGTGGTGCCGTTCCACCACGGCTTCGGCATGTTCACGACCCTGGGCTA
CCTCATCTGCGGCTTCCGGGTGGTCCTGATGTACCGGTTCGAGGAGGAGCTGTTCCTGCG
GAGCCTGCAGGACTACAAGATCCAGAGCGCGCTGCTCGTGCCGACCCTGTTCAGCTTCTT
CGCCAAGAGCACCCTGATCGACAAGTACGACCTGTCGAACCTGCACGAGATCGCCAGCGG
GGGCGCCCCGCTGAGCAAGGAGGTGGGCGAGGCCGTGGCCAAGCGGTTCCACCTCCCGGG
CATCCGCCAGGGCTACGGCCTGACCGAGACCACGAGCGCGATCCTGATCACCCCCGAGGG
GGACGACAAGCCGGGCGCCGTGGGCAAGGTGGTCCCGTTCTTCGAGGCCAAGGTGGTGGA
CCTGGACACCGGCAAGACCCTGGGCGTGAACCAGCGGGGCGAGCTGTGCGTGCGGGGGCC
GATGATCATGAGCGGCTACGTGAACAACCCGGAGGCCACCAACGCCCTCATCGACAAGGA
CGGCTGGCTGCACAGCGGCGACATCGCCTACTGGGACGAGGACGAGCACTTCTTCATCGT
CGACCGGCTGAAGTCGCTGATCAAGTACAAGGGCTACCAGGTGGCGCCGGCCGAGCTGGA
GAGCATCCTGCTCCAGCACCCCAACATCTTCGACGCCGGCGTGGCCGGGCTGCCGGACGA
CGACGCCGGCGAGCTGCCGGCCGCGGTGGTGGTGCTGGAGCACGGCAAGACCATGACGGA
GAAGGAGATCGTCGACTACGTGGCCAGCCAGGTGACCACCGCCAAGAAGCTGCGGGGCGG
CGTGGTGTTCGTGGACGAGGTCCCGAAGGGCCTGACCGGGAAGCTCGACGCCCGGAAGAT
CCGCGAGATCCTGATCAAGGCCAAGAAGGGCGGCAAGATCGCCGTGTAAGACTAGTGCAT
CACATTTAAAAGCATCTCAGCCTACCATGAGAATAAGAGAAAGAAAATGAAGATCAAAAG
CTTATTCATCAGTTTTTCTTTTTCGTTGGTGTAAAGCCAACACCCTGTCTAAAAAACATA
AATTTCTTTAATCATTTTGCCTCTTTTCTCTGTGCTTCAATTAATAAAAAATGGAAAGAA
TCTAGATCTAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA
AAAAAAAAAAAAA
SEQ ID NO: 9:
GGGAGAAAGCTTGAGGATGGAGGACGCCAAGAACATCAAGAAGGGCCCGGCGCCCTTCTA
CCCGCTGGAGGACGGGACCGCCGGCGAGCAGCTCCACAAGGCCATGAAGCGGTACGCCCT
GGTGCCGGGCACGATCGCCTTCACCGACGCCCACATCGAGGTCGACATCACCTACGCGGA
GTACTTCGAGATGAGCGTGCGCCTGGCCGAGGCCATGAAGCGGTACGGCCTGAACACCAA
CCACCGGATCGTGGTGTGCTCGGAGAACAGCCTGCAGTTCTTCATGCCGGTGCTGGGCGC
CCTCTTCATCGGCGTGGCCGTCGCCCCGGCGAACGACATCTACAACGAGCGGGAGCTGCT
GAACAGCATGGGGATCAGCCAGCCGACCGTGGTGTTCGTGAGCAAGAAGGGCCTGCAGAA
GATCCTGAACGTGCAGAAGAAGCTGCCCATCATCCAGAAGATCATCATCATGGACAGCAA
GACCGACTACCAGGGCTTCCAGTCGATGTACACGTTCGTGACCAGCCACCTCCCGCCGGG
CTTCAACGAGTACGACTTCGTCCCGGAGAGCTTCGACCGGGACAAGACCATCGCCCTGAT
CATGAACAGCAGCGGCAGCACCGGCCTGCCGAAGGGGGTGGCCCTGCCGCACCGGACCGC
CTGCGTGCGCTTCTCGCACGCCCGGGACCCCATCTTCGGCAACCAGATCATCCCGGACAC
CGCCATCCTGAGCGTGGTGCCGTTCCACCACGGCTTCGGCATGTTCACGACCCTGGGCTA
CCTCATCTGCGGCTTCCGGGTGGTCCTGATGTACCGGTTCGAGGAGGAGCTGTTCCTGCG
GAGCCTGCAGGACTACAAGATCCAGAGCGCGCTGCTCGTGCCGACCCTGTTCAGCTTCTT
CGCCAAGAGCACCCTGATCGACAAGTACGACCTGTCGAACCTGCACGAGATCGCCAGCGG
GGGCGCCCCGCTGAGCAAGGAGGTGGGCGAGGCCGTGGCCAAGCGGTTCCACCTCCCGGG
CATCCGCCAGGGCTACGGCCTGACCGAGACCACGAGCGCGATCCTGATCACCCCCGAGGG
GGACGACAAGCCGGGCGCCGTGGGCAAGGTGGTCCCGTTCTTCGAGGCCAAGGTGGTGGA
CCTGGACACCGGCAAGACCCTGGGCGTGAACCAGCGGGGCGAGCTGTGCGTGCGGGGGCC
GATGATCATGAGCGGCTACGTGAACAACCCGGAGGCCACCAACGCCCTCATCGACAAGGA
CGGCTGGCTGCACAGCGGCGACATCGCCTACTGGGACGAGGACGAGCACTTCTTCATCGT
CGACCGGCTGAAGTCGCTGATCAAGTACAAGGGCTACCAGGTGGCGCCGGCCGAGCTGGA
GAGCATCCTGCTCCAGCACCCCAACATCTTCGACGCCGGCGTGGCCGGGCTGCCGGACGA
CGACGCCGGCGAGCTGCCGGCCGCGGTGGTGGTGCTGGAGCACGGCAAGACCATGACGGA
GAAGGAGATCGTCGACTACGTGGCCAGCCAGGTGACCACCGCCAAGAAGCTGCGGGGCGG
CGTGGTGTTCGTGGACGAGGTCCCGAAGGGCCTGACCGGGAAGCTCGACGCCCGGAAGAT
CCGCGAGATCCTGATCAAGGCCAAGAAGGGCGGCAAGATCGCCGTGTAAGACTAGTGCAT
CACATTTAAAAGCATCTCAGCCTACCATGAGAATAAGAGAAAGAAAATGAAGATCAAAAG
CTTATTCATCTCTTTTTCTTTTTCGTTGGTGTAAAGCCAACACCCTGTCTAAAAAACATA
AATTTCTTTAATCATTTTGCCTCTTTTCTCTGTGCTTCAATTAATAAAAAATGGAAAGAA
TCTAGATCTAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA
AAAAAAAAAAAAA
SEQ ID NO: 10:
GGGAGAAAGCTTGAGGATGGAGGACGCCAAGAACATCAAGAAGGGCCCGGCGCCCTTCTA
CCCGCTGGAGGACGGGACCGCCGGCGAGCAGCTCCACAAGGCCATGAAGCGGTACGCCCT
GGTGCCGGGCACGATCGCCTTCACCGACGCCCACATCGAGGTCGACATCACCTACGCGGA
GTACTTCGAGATGAGCGTGCGCCTGGCCGAGGCCATGAAGCGGTACGGCCTGAACACCAA
CCACCGGATCGTGGTGTGCTCGGAGAACAGCCTGCAGTTCTTCATGCCGGTGCTGGGCGC
CCTCTTCATCGGCGTGGCCGTCGCCCCGGCGAACGACATCTACAACGAGCGGGAGCTGCT
GAACAGCATGGGGATCAGCCAGCCGACCGTGGTGTTCGTGAGCAAGAAGGGCCTGCAGAA
GATCCTGAACGTGCAGAAGAAGCTGCCCATCATCCAGAAGATCATCATCATGGACAGCAA
GACCGACTACCAGGGCTTCCAGTCGATGTACACGTTCGTGACCAGCCACCTCCCGCCGGG
CTTCAACGAGTACGACTTCGTCCCGGAGAGCTTCGACCGGGACAAGACCATCGCCCTGAT
CATGAACAGCAGCGGCAGCACCGGCCTGCCGAAGGGGGTGGCCCTGCCGCACCGGACCGC
CTGCGTGCGCTTCTCGCACGCCCGGGACCCCATCTTCGGCAACCAGATCATCCCGGACAC
CGCCATCCTGAGCGTGGTGCCGTTCCACCACGGCTTCGGCATGTTCACGACCCTGGGCTA
CCTCATCTGCGGCTTCCGGGTGGTCCTGATGTACCGGTTCGAGGAGGAGCTGTTCCTGCG
GAGCCTGCAGGACTACAAGATCCAGAGCGCGCTGCTCGTGCCGACCCTGTTCAGCTTCTT
CGCCAAGAGCACCCTGATCGACAAGTACGACCTGTCGAACCTGCACGAGATCGCCAGCGG
GGGCGCCCCGCTGAGCAAGGAGGTGGGCGAGGCCGTGGCCAAGCGGTTCCACCTCCCGGG
CATCCGCCAGGGCTACGGCCTGACCGAGACCACGAGCGCGATCCTGATCACCCCCGAGGG
GGACGACAAGCCGGGCGCCGTGGGCAAGGTGGTCCCGTTCTTCGAGGCCAAGGTGGTGGA
CCTGGACACCGGCAAGACCCTGGGCGTGAACCAGCGGGGCGAGCTGTGCGTGCGGGGGCC
GATGATCATGAGCGGCTACGTGAACAACCCGGAGGCCACCAACGCCCTCATCGACAAGGA
CGGCTGGCTGCACAGCGGCGACATCGCCTACTGGGACGAGGACGAGCACTTCTTCATCGT
CGACCGGCTGAAGTCGCTGATCAAGTACAAGGGCTACCAGGTGGCGCCGGCCGAGCTGGA
GAGCATCCTGCTCCAGCACCCCAACATCTTCGACGCCGGCGTGGCCGGGCTGCCGGACGA
CGACGCCGGCGAGCTGCCGGCCGCGGTGGTGGTGCTGGAGCACGGCAAGACCATGACGGA
GAAGGAGATCGTCGACTACGTGGCCAGCCAGGTGACCACCGCCAAGAAGCTGCGGGGCGG
CGTGGTGTTCGTGGACGAGGTCCCGAAGGGCCTGACCGGGAAGCTCGACGCCCGGAAGAT
CCGCGAGATCCTGATCAAGGCCAAGAAGGGCGGCAAGATCGCCGTGTAAGACTAGTGCAT
CACATTTAAAAGCATCTCAGCCTACCATGAGAATAAGAGAAAGAAAATGAAGATCAAAAG
CTTATTCATCTGTTGGTCTTTTTCGTTGGTGTAAAGCCAACACCCTGTCTAAAAAACATA
AATTTCTTTAATCATTTTGCCTCTTTTCTCTGTGCTTCAATTAATAAAAAATGGAAAGAA
TCTAGATCTAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA
AAAAAAAAAAAAA
SEQ ID NO: 11:
GGGAGAAAGCTTGAGGATGGAGGACGCCAAGAACATCAAGAAGGGCCCGGCGCCCTTCTA
CCCGCTGGAGGACGGGACCGCCGGCGAGCAGCTCCACAAGGCCATGAAGCGGTACGCCCT
GGTGCCGGGCACGATCGCCTTCACCGACGCCCACATCGAGGTCGACATCACCTACGCGGA
GTACTTCGAGATGAGCGTGCGCCTGGCCGAGGCCATGAAGCGGTACGGCCTGAACACCAA
CCACCGGATCGTGGTGTGCTCGGAGAACAGCCTGCAGTTCTTCATGCCGGTGCTGGGCGC
CCTCTTCATCGGCGTGGCCGTCGCCCCGGCGAACGACATCTACAACGAGCGGGAGCTGCT
GAACAGCATGGGGATCAGCCAGCCGACCGTGGTGTTCGTGAGCAAGAAGGGCCTGCAGAA
GATCCTGAACGTGCAGAAGAAGCTGCCCATCATCCAGAAGATCATCATCATGGACAGCAA
GACCGACTACCAGGGCTTCCAGTCGATGTACACGTTCGTGACCAGCCACCTCCCGCCGGG
CTTCAACGAGTACGACTTCGTCCCGGAGAGCTTCGACCGGGACAAGACCATCGCCCTGAT
CATGAACAGCAGCGGCAGCACCGGCCTGCCGAAGGGGGTGGCCCTGCCGCACCGGACCGC
CTGCGTGCGCTTCTCGCACGCCCGGGACCCCATCTTCGGCAACCAGATCATCCCGGACAC
CGCCATCCTGAGCGTGGTGCCGTTCCACCACGGCTTCGGCATGTTCACGACCCTGGGCTA
CCTCATCTGCGGCTTCCGGGTGGTCCTGATGTACCGGTTCGAGGAGGAGCTGTTCCTGCG
GAGCCTGCAGGACTACAAGATCCAGAGCGCGCTGCTCGTGCCGACCCTGTTCAGCTTCTT
CGCCAAGAGCACCCTGATCGACAAGTACGACCTGTCGAACCTGCACGAGATCGCCAGCGG
GGGCGCCCCGCTGAGCAAGGAGGTGGGCGAGGCCGTGGCCAAGCGGTTCCACCTCCCGGG
CATCCGCCAGGGCTACGGCCTGACCGAGACCACGAGCGCGATCCTGATCACCCCCGAGGG
GGACGACAAGCCGGGCGCCGTGGGCAAGGTGGTCCCGTTCTTCGAGGCCAAGGTGGTGGA
CCTGGACACCGGCAAGACCCTGGGCGTGAACCAGCGGGGCGAGCTGTGCGTGCGGGGGCC
GATGATCATGAGCGGCTACGTGAACAACCCGGAGGCCACCAACGCCCTCATCGACAAGGA
CGGCTGGCTGCACAGCGGCGACATCGCCTACTGGGACGAGGACGAGCACTTCTTCATCGT
CGACCGGCTGAAGTCGCTGATCAAGTACAAGGGCTACCAGGTGGCGCCGGCCGAGCTGGA
GAGCATCCTGCTCCAGCACCCCAACATCTTCGACGCCGGCGTGGCCGGGCTGCCGGACGA
CGACGCCGGCGAGCTGCCGGCCGCGGTGGTGGTGCTGGAGCACGGCAAGACCATGACGGA
GAAGGAGATCGTCGACTACGTGGCCAGCCAGGTGACCACCGCCAAGAAGCTGCGGGGCGG
CGTGGTGTTCGTGGACGAGGTCCCGAAGGGCCTGACCGGGAAGCTCGACGCCCGGAAGAT
CCGCGAGATCCTGATCAAGGCCAAGAAGGGCGGCAAGATCGCCGTGTAAGACTAGTGCAT
CACATTTAAAAGCATCTCAGCCTACCATGAGAATAAGAGAAAGAAAATGAAGATCAATAG
CTTATTCATCTGTTTTTCTTTTTCGTTGGTGTAAAGCCAACACCCTGTCTAAAAAACATA
AATTTCTTTAATCATTTTGCCTCTTTTCTCTGTGCTTCAATTAATAAAAAATGGAAAGAA
CCTAGATCTAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA
AAAAAAAAAAAAA
SEQ ID NO: 12:
GGGAGAAAGCTTGAGGATGGAGGACGCCAAGAACATCAAGAAGGGCCCGGCGCCCTTCTA
CCCGCTGGAGGACGGGACCGCCGGCGAGCAGCTCCACAAGGCCATGAAGCGGTACGCCCT
GGTGCCGGGCACGATCGCCTTCACCGACGCCCACATCGAGGTCGACATCACCTACGCGGA
GTACTTCGAGATGAGCGTGCGCCTGGCCGAGGCCATGAAGCGGTACGGCCTGAACACCAA
CCACCGGATCGTGGTGTGCTCGGAGAACAGCCTGCAGTTCTTCATGCCGGTGCTGGGCGC
CCTCTTCATCGGCGTGGCCGTCGCCCCGGCGAACGACATCTACAACGAGCGGGAGCTGCT
GAACAGCATGGGGATCAGCCAGCCGACCGTGGTGTTCGTGAGCAAGAAGGGCCTGCAGAA
GATCCTGAACGTGCAGAAGAAGCTGCCCATCATCCAGAAGATCATCATCATGGACAGCAA
GACCGACTACCAGGGCTTCCAGTCGATGTACACGTTCGTGACCAGCCACCTCCCGCCGGG
CTTCAACGAGTACGACTTCGTCCCGGAGAGCTTCGACCGGGACAAGACCATCGCCCTGAT
CATGAACAGCAGCGGCAGCACCGGCCTGCCGAAGGGGGTGGCCCTGCCGCACCGGACCGC
CTGCGTGCGCTTCTCGCACGCCCGGGACCCCATCTTCGGCAACCAGATCATCCCGGACAC
CGCCATCCTGAGCGTGGTGCCGTTCCACCACGGCTTCGGCATGTTCACGACCCTGGGCTA
CCTCATCTGCGGCTTCCGGGTGGTCCTGATGTACCGGTTCGAGGAGGAGCTGTTCCTGCG
GAGCCTGCAGGACTACAAGATCCAGAGCGCGCTGCTCGTGCCGACCCTGTTCAGCTTCTT
CGCCAAGAGCACCCTGATCGACAAGTACGACCTGTCGAACCTGCACGAGATCGCCAGCGG
GGGCGCCCCGCTGAGCAAGGAGGTGGGCGAGGCCGTGGCCAAGCGGTTCCACCTCCCGGG
CATCCGCCAGGGCTACGGCCTGACCGAGACCACGAGCGCGATCCTGATCACCCCCGAGGG
GGACGACAAGCCGGGCGCCGTGGGCAAGGTGGTCCCGTTCTTCGAGGCCAAGGTGGTGGA
CCTGGACACCGGCAAGACCCTGGGCGTGAACCAGCGGGGCGAGCTGTGCGTGCGGGGGCC
GATGATCATGAGCGGCTACGTGAACAACCCGGAGGCCACCAACGCCCTCATCGACAAGGA
CGGCTGGCTGCACAGCGGCGACATCGCCTACTGGGACGAGGACGAGCACTTCTTCATCGT
CGACCGGCTGAAGTCGCTGATCAAGTACAAGGGCTACCAGGTGGCGCCGGCCGAGCTGGA
GAGCATCCTGCTCCAGCACCCCAACATCTTCGACGCCGGCGTGGCCGGGCTGCCGGACGA
CGACGCCGGCGAGCTGCCGGCCGCGGTGGTGGTGCTGGAGCACGGCAAGACCATGACGGA
GAAGGAGATCGTCGACTACGTGGCCAGCCAGGTGACCACCGCCAAGAAGCTGCGGGGCGG
CGTGGTGTTCGTGGACGAGGTCCCGAAGGGCCTGACCGGGAAGCTCGACGCCCGGAAGAT
CCGCGAGATCCTGATCAAGGCCAAGAAGGGCGGCAAGATCGCCGTGTAAGACTAGTGCAT
CACATTTAAAAGCATCTCAGCCTACCATGAGAATAAGAGAAAGAAAATGAAGATCAATAG
CTTATTCATCTCTTTTTCTTTTTCGTTGGTGTAAAGCCAACACCCTGTCTAAAAAACATA
AATTTCTTTAATCATTTTGCCTCTTTTCTCTGTGCTTCAATTAATAAAAAATGGAAAGAA
CCTAGATCTAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA
AAAAAAAAAAAAA
SEQ ID NO: 13:
GGGAGAAAGCTTGAGGATGGAGGACGCCAAGAACATCAAGAAGGGCCCGGCGCCCTTCTA
CCCGCTGGAGGACGGGACCGCCGGCGAGCAGCTCCACAAGGCCATGAAGCGGTACGCCCT
GGTGCCGGGCACGATCGCCTTCACCGACGCCCACATCGAGGTCGACATCACCTACGCGGA
GTACTTCGAGATGAGCGTGCGCCTGGCCGAGGCCATGAAGCGGTACGGCCTGAACACCAA
CCACCGGATCGTGGTGTGCTCGGAGAACAGCCTGCAGTTCTTCATGCCGGTGCTGGGCGC
CCTCTTCATCGGCGTGGCCGTCGCCCCGGCGAACGACATCTACAACGAGCGGGAGCTGCT
GAACAGCATGGGGATCAGCCAGCCGACCGTGGTGTTCGTGAGCAAGAAGGGCCTGCAGAA
GATCCTGAACGTGCAGAAGAAGCTGCCCATCATCCAGAAGATCATCATCATGGACAGCAA
GACCGACTACCAGGGCTTCCAGTCGATGTACACGTTCGTGACCAGCCACCTCCCGCCGGG
CTTCAACGAGTACGACTTCGTCCCGGAGAGCTTCGACCGGGACAAGACCATCGCCCTGAT
CATGAACAGCAGCGGCAGCACCGGCCTGCCGAAGGGGGTGGCCCTGCCGCACCGGACCGC
CTGCGTGCGCTTCTCGCACGCCCGGGACCCCATCTTCGGCAACCAGATCATCCCGGACAC
CGCCATCCTGAGCGTGGTGCCGTTCCACCACGGCTTCGGCATGTTCACGACCCTGGGCTA
CCTCATCTGCGGCTTCCGGGTGGTCCTGATGTACCGGTTCGAGGAGGAGCTGTTCCTGCG
GAGCCTGCAGGACTACAAGATCCAGAGCGCGCTGCTCGTGCCGACCCTGTTCAGCTTCTT
CGCCAAGAGCACCCTGATCGACAAGTACGACCTGTCGAACCTGCACGAGATCGCCAGCGG
GGGCGCCCCGCTGAGCAAGGAGGTGGGCGAGGCCGTGGCCAAGCGGTTCCACCTCCCGGG
CATCCGCCAGGGCTACGGCCTGACCGAGACCACGAGCGCGATCCTGATCACCCCCGAGGG
GGACGACAAGCCGGGCGCCGTGGGCAAGGTGGTCCCGTTCTTCGAGGCCAAGGTGGTGGA
CCTGGACACCGGCAAGACCCTGGGCGTGAACCAGCGGGGCGAGCTGTGCGTGCGGGGGCC
GATGATCATGAGCGGCTACGTGAACAACCCGGAGGCCACCAACGCCCTCATCGACAAGGA
CGGCTGGCTGCACAGCGGCGACATCGCCTACTGGGACGAGGACGAGCACTTCTTCATCGT
CGACCGGCTGAAGTCGCTGATCAAGTACAAGGGCTACCAGGTGGCGCCGGCCGAGCTGGA
GAGCATCCTGCTCCAGCACCCCAACATCTTCGACGCCGGCGTGGCCGGGCTGCCGGACGA
CGACGCCGGCGAGCTGCCGGCCGCGGTGGTGGTGCTGGAGCACGGCAAGACCATGACGGA
GAAGGAGATCGTCGACTACGTGGCCAGCCAGGTGACCACCGCCAAGAAGCTGCGGGGCGG
CGTGGTGTTCGTGGACGAGGTCCCGAAGGGCCTGACCGGGAAGCTCGACGCCCGGAAGAT
CCGCGAGATCCTGATCAAGGCCAAGAAGGGCGGCAAGATCGCCGTGTAAGACTAGTTATA
AGACTGACTAGCCCGATGGGCCTCCCAACGGGCCCTCCTCCCCTCCTTGCACCGAGATTA
ATAGATCTAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA
AAAAAAAAAAAA
SEQ ID NO: 14:
GGGAGAAAGCTTGAGGATGGAGGACGCCAAGAACATCAAGAAGGGCCCGGCGCCCTTCTA
CCCGCTGGAGGACGGGACCGCCGGCGAGCAGCTCCACAAGGCCATGAAGCGGTACGCCCT
GGTGCCGGGCACGATCGCCTTCACCGACGCCCACATCGAGGTCGACATCACCTACGCGGA
GTACTTCGAGATGAGCGTGCGCCTGGCCGAGGCCATGAAGCGGTACGGCCTGAACACCAA
CCACCGGATCGTGGTGTGCTCGGAGAACAGCCTGCAGTTCTTCATGCCGGTGCTGGGCGC
CCTCTTCATCGGCGTGGCCGTCGCCCCGGCGAACGACATCTACAACGAGCGGGAGCTGCT
GAACAGCATGGGGATCAGCCAGCCGACCGTGGTGTTCGTGAGCAAGAAGGGCCTGCAGAA
GATCCTGAACGTGCAGAAGAAGCTGCCCATCATCCAGAAGATCATCATCATGGACAGCAA
GACCGACTACCAGGGCTTCCAGTCGATGTACACGTTCGTGACCAGCCACCTCCCGCCGGG
CTTCAACGAGTACGACTTCGTCCCGGAGAGCTTCGACCGGGACAAGACCATCGCCCTGAT
CATGAACAGCAGCGGCAGCACCGGCCTGCCGAAGGGGGTGGCCCTGCCGCACCGGACCGC
CTGCGTGCGCTTCTCGCACGCCCGGGACCCCATCTTCGGCAACCAGATCATCCCGGACAC
CGCCATCCTGAGCGTGGTGCCGTTCCACCACGGCTTCGGCATGTTCACGACCCTGGGCTA
CCTCATCTGCGGCTTCCGGGTGGTCCTGATGTACCGGTTCGAGGAGGAGCTGTTCCTGCG
GAGCCTGCAGGACTACAAGATCCAGAGCGCGCTGCTCGTGCCGACCCTGTTCAGCTTCTT
CGCCAAGAGCACCCTGATCGACAAGTACGACCTGTCGAACCTGCACGAGATCGCCAGCGG
GGGCGCCCCGCTGAGCAAGGAGGTGGGCGAGGCCGTGGCCAAGCGGTTCCACCTCCCGGG
CATCCGCCAGGGCTACGGCCTGACCGAGACCACGAGCGCGATCCTGATCACCCCCGAGGG
GGACGACAAGCCGGGCGCCGTGGGCAAGGTGGTCCCGTTCTTCGAGGCCAAGGTGGTGGA
CCTGGACACCGGCAAGACCCTGGGCGTGAACCAGCGGGGCGAGCTGTGCGTGCGGGGGCC
GATGATCATGAGCGGCTACGTGAACAACCCGGAGGCCACCAACGCCCTCATCGACAAGGA
CGGCTGGCTGCACAGCGGCGACATCGCCTACTGGGACGAGGACGAGCACTTCTTCATCGT
CGACCGGCTGAAGTCGCTGATCAAGTACAAGGGCTACCAGGTGGCGCCGGCCGAGCTGGA
GAGCATCCTGCTCCAGCACCCCAACATCTTCGACGCCGGCGTGGCCGGGCTGCCGGACGA
CGACGCCGGCGAGCTGCCGGCCGCGGTGGTGGTGCTGGAGCACGGCAAGACCATGACGGA
GAAGGAGATCGTCGACTACGTGGCCAGCCAGGTGACCACCGCCAAGAAGCTGCGGGGCGG
CGTGGTGTTCGTGGACGAGGTCCCGAAGGGCCTGACCGGGAAGCTCGACGCCCGGAAGAT
CCGCGAGATCCTGATCAAGGCCAAGAAGGGCGGCAAGATCGCCGTGTAAGACTAGTGCAA
GACTGATACCACCTGCGTGTCCCTTCCTCCCCGAGTCAGGGCGACTTCCACAGCAGCAGA
ACAAGTGCCTCCTGGACTGTTCACGGCAGACCAGAACGTTTCTGGCCTGGGTTTTGTGGT
CATCTATTCTAGCAGGGAACACTAAAGGTGGAAATAAAAGATTTTCTATTATGGAAATAA
AGAGTTGGCATGAAAGTGGCTACTGAGATCTAAAAAAAAAAAAAAAAAAAAAAAAAAAAA
AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA
SEQ ID NO: 15:
GGGAGAAAGCTTGAGGATGGAGGACGCCAAGAACATCAAGAAGGGCCCGGCGCCCTTCTA
CCCGCTGGAGGACGGGACCGCCGGCGAGCAGCTCCACAAGGCCATGAAGCGGTACGCCCT
GGTGCCGGGCACGATCGCCTTCACCGACGCCCACATCGAGGTCGACATCACCTACGCGGA
GTACTTCGAGATGAGCGTGCGCCTGGCCGAGGCCATGAAGCGGTACGGCCTGAACACCAA
CCACCGGATCGTGGTGTGCTCGGAGAACAGCCTGCAGTTCTTCATGCCGGTGCTGGGCGC
CCTCTTCATCGGCGTGGCCGTCGCCCCGGCGAACGACATCTACAACGAGCGGGAGCTGCT
GAACAGCATGGGGATCAGCCAGCCGACCGTGGTGTTCGTGAGCAAGAAGGGCCTGCAGAA
GATCCTGAACGTGCAGAAGAAGCTGCCCATCATCCAGAAGATCATCATCATGGACAGCAA
GACCGACTACCAGGGCTTCCAGTCGATGTACACGTTCGTGACCAGCCACCTCCCGCCGGG
CTTCAACGAGTACGACTTCGTCCCGGAGAGCTTCGACCGGGACAAGACCATCGCCCTGAT
CATGAACAGCAGCGGCAGCACCGGCCTGCCGAAGGGGGTGGCCCTGCCGCACCGGACCGC
CTGCGTGCGCTTCTCGCACGCCCGGGACCCCATCTTCGGCAACCAGATCATCCCGGACAC
CGCCATCCTGAGCGTGGTGCCGTTCCACCACGGCTTCGGCATGTTCACGACCCTGGGCTA
CCTCATCTGCGGCTTCCGGGTGGTCCTGATGTACCGGTTCGAGGAGGAGCTGTTCCTGCG
GAGCCTGCAGGACTACAAGATCCAGAGCGCGCTGCTCGTGCCGACCCTGTTCAGCTTCTT
CGCCAAGAGCACCCTGATCGACAAGTACGACCTGTCGAACCTGCACGAGATCGCCAGCGG
GGGCGCCCCGCTGAGCAAGGAGGTGGGCGAGGCCGTGGCCAAGCGGTTCCACCTCCCGGG
CATCCGCCAGGGCTACGGCCTGACCGAGACCACGAGCGCGATCCTGATCACCCCCGAGGG
GGACGACAAGCCGGGCGCCGTGGGCAAGGTGGTCCCGTTCTTCGAGGCCAAGGTGGTGGA
CCTGGACACCGGCAAGACCCTGGGCGTGAACCAGCGGGGCGAGCTGTGCGTGCGGGGGCC
GATGATCATGAGCGGCTACGTGAACAACCCGGAGGCCACCAACGCCCTCATCGACAAGGA
CGGCTGGCTGCACAGCGGCGACATCGCCTACTGGGACGAGGACGAGCACTTCTTCATCGT
CGACCGGCTGAAGTCGCTGATCAAGTACAAGGGCTACCAGGTGGCGCCGGCCGAGCTGGA
GAGCATCCTGCTCCAGCACCCCAACATCTTCGACGCCGGCGTGGCCGGGCTGCCGGACGA
CGACGCCGGCGAGCTGCCGGCCGCGGTGGTGGTGCTGGAGCACGGCAAGACCATGACGGA
GAAGGAGATCGTCGACTACGTGGCCAGCCAGGTGACCACCGCCAAGAAGCTGCGGGGCGG
CGTGGTGTTCGTGGACGAGGTCCCGAAGGGCCTGACCGGGAAGCTCGACGCCCGGAAGAT
CCGCGAGATCCTGATCAAGGCCAAGAAGGGCGGCAAGATCGCCGTGTAAGACTAGTAAGT
GTTGGTTTTCTGCCATCAGTGAAAATTCTTAAACTTGGAGCAACAATAAAAAGCTTCCAG
AACAGATCAGATCTAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA
AAAAAAAAAAAAAAAAAA
SEQ ID NO: 16:
GGGAGAAAGCTTGAGGATGGAGGACGCCAAGAACATCAAGAAGGGCCCGGCGCCCTTCTA
CCCGCTGGAGGACGGGACCGCCGGCGAGCAGCTCCACAAGGCCATGAAGCGGTACGCCCT
GGTGCCGGGCACGATCGCCTTCACCGACGCCCACATCGAGGTCGACATCACCTACGCGGA
GTACTTCGAGATGAGCGTGCGCCTGGCCGAGGCCATGAAGCGGTACGGCCTGAACACCAA
CCACCGGATCGTGGTGTGCTCGGAGAACAGCCTGCAGTTCTTCATGCCGGTGCTGGGCGC
CCTCTTCATCGGCGTGGCCGTCGCCCCGGCGAACGACATCTACAACGAGCGGGAGCTGCT
GAACAGCATGGGGATCAGCCAGCCGACCGTGGTGTTCGTGAGCAAGAAGGGCCTGCAGAA
GATCCTGAACGTGCAGAAGAAGCTGCCCATCATCCAGAAGATCATCATCATGGACAGCAA
GACCGACTACCAGGGCTTCCAGTCGATGTACACGTTCGTGACCAGCCACCTCCCGCCGGG
CTTCAACGAGTACGACTTCGTCCCGGAGAGCTTCGACCGGGACAAGACCATCGCCCTGAT
CATGAACAGCAGCGGCAGCACCGGCCTGCCGAAGGGGGTGGCCCTGCCGCACCGGACCGC
CTGCGTGCGCTTCTCGCACGCCCGGGACCCCATCTTCGGCAACCAGATCATCCCGGACAC
CGCCATCCTGAGCGTGGTGCCGTTCCACCACGGCTTCGGCATGTTCACGACCCTGGGCTA
CCTCATCTGCGGCTTCCGGGTGGTCCTGATGTACCGGTTCGAGGAGGAGCTGTTCCTGCG
GAGCCTGCAGGACTACAAGATCCAGAGCGCGCTGCTCGTGCCGACCCTGTTCAGCTTCTT
CGCCAAGAGCACCCTGATCGACAAGTACGACCTGTCGAACCTGCACGAGATCGCCAGCGG
GGGCGCCCCGCTGAGCAAGGAGGTGGGCGAGGCCGTGGCCAAGCGGTTCCACCTCCCGGG
CATCCGCCAGGGCTACGGCCTGACCGAGACCACGAGCGCGATCCTGATCACCCCCGAGGG
GGACGACAAGCCGGGCGCCGTGGGCAAGGTGGTCCCGTTCTTCGAGGCCAAGGTGGTGGA
CCTGGACACCGGCAAGACCCTGGGCGTGAACCAGCGGGGCGAGCTGTGCGTGCGGGGGCC
GATGATCATGAGCGGCTACGTGAACAACCCGGAGGCCACCAACGCCCTCATCGACAAGGA
CGGCTGGCTGCACAGCGGCGACATCGCCTACTGGGACGAGGACGAGCACTTCTTCATCGT
CGACCGGCTGAAGTCGCTGATCAAGTACAAGGGCTACCAGGTGGCGCCGGCCGAGCTGGA
GAGCATCCTGCTCCAGCACCCCAACATCTTCGACGCCGGCGTGGCCGGGCTGCCGGACGA
CGACGCCGGCGAGCTGCCGGCCGCGGTGGTGGTGCTGGAGCACGGCAAGACCATGACGGA
GAAGGAGATCGTCGACTACGTGGCCAGCCAGGTGACCACCGCCAAGAAGCTGCGGGGCGG
CGTGGTGTTCGTGGACGAGGTCCCGAAGGGCCTGACCGGGAAGCTCGACGCCCGGAAGAT
CCGCGAGATCCTGATCAAGGCCAAGAAGGGCGGCAAGATCGCCGTGTAAGACTAGTGGAA
GCATTTTCCTGATTGATGAAAAAAATAACTCAGTTATGGCCATCTACCCCTGCTAGAAGG
TTACAGTGTATTATGTAGCATGCAATGTGTTATGTAGTGCTTAATAAAAATAAAATGAAA
AAAATGCAGATCTAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA
AAAAAAAAAAAAAAAAA
SEQ ID NO: 17:
GGGAGAAAGCTTGAGGATGGAGGACGCCAAGAACATCAAGAAGGGCCCGGCGCCCTTCTA
CCCGCTGGAGGACGGGACCGCCGGCGAGCAGCTCCACAAGGCCATGAAGCGGTACGCCCT
GGTGCCGGGCACGATCGCCTTCACCGACGCCCACATCGAGGTCGACATCACCTACGCGGA
GTACTTCGAGATGAGCGTGCGCCTGGCCGAGGCCATGAAGCGGTACGGCCTGAACACCAA
CCACCGGATCGTGGTGTGCTCGGAGAACAGCCTGCAGTTCTTCATGCCGGTGCTGGGCGC
CCTCTTCATCGGCGTGGCCGTCGCCCCGGCGAACGACATCTACAACGAGCGGGAGCTGCT
GAACAGCATGGGGATCAGCCAGCCGACCGTGGTGTTCGTGAGCAAGAAGGGCCTGCAGAA
GATCCTGAACGTGCAGAAGAAGCTGCCCATCATCCAGAAGATCATCATCATGGACAGCAA
GACCGACTACCAGGGCTTCCAGTCGATGTACACGTTCGTGACCAGCCACCTCCCGCCGGG
CTTCAACGAGTACGACTTCGTCCCGGAGAGCTTCGACCGGGACAAGACCATCGCCCTGAT
CATGAACAGCAGCGGCAGCACCGGCCTGCCGAAGGGGGTGGCCCTGCCGCACCGGACCGC
CTGCGTGCGCTTCTCGCACGCCCGGGACCCCATCTTCGGCAACCAGATCATCCCGGACAC
CGCCATCCTGAGCGTGGTGCCGTTCCACCACGGCTTCGGCATGTTCACGACCCTGGGCTA
CCTCATCTGCGGCTTCCGGGTGGTCCTGATGTACCGGTTCGAGGAGGAGCTGTTCCTGCG
GAGCCTGCAGGACTACAAGATCCAGAGCGCGCTGCTCGTGCCGACCCTGTTCAGCTTCTT
CGCCAAGAGCACCCTGATCGACAAGTACGACCTGTCGAACCTGCACGAGATCGCCAGCGG
GGGCGCCCCGCTGAGCAAGGAGGTGGGCGAGGCCGTGGCCAAGCGGTTCCACCTCCCGGG
CATCCGCCAGGGCTACGGCCTGACCGAGACCACGAGCGCGATCCTGATCACCCCCGAGGG
GGACGACAAGCCGGGCGCCGTGGGCAAGGTGGTCCCGTTCTTCGAGGCCAAGGTGGTGGA
CCTGGACACCGGCAAGACCCTGGGCGTGAACCAGCGGGGCGAGCTGTGCGTGCGGGGGCC
GATGATCATGAGCGGCTACGTGAACAACCCGGAGGCCACCAACGCCCTCATCGACAAGGA
CGGCTGGCTGCACAGCGGCGACATCGCCTACTGGGACGAGGACGAGCACTTCTTCATCGT
CGACCGGCTGAAGTCGCTGATCAAGTACAAGGGCTACCAGGTGGCGCCGGCCGAGCTGGA
GAGCATCCTGCTCCAGCACCCCAACATCTTCGACGCCGGCGTGGCCGGGCTGCCGGACGA
CGACGCCGGCGAGCTGCCGGCCGCGGTGGTGGTGCTGGAGCACGGCAAGACCATGACGGA
GAAGGAGATCGTCGACTACGTGGCCAGCCAGGTGACCACCGCCAAGAAGCTGCGGGGCGG
CGTGGTGTTCGTGGACGAGGTCCCGAAGGGCCTGACCGGGAAGCTCGACGCCCGGAAGAT
CCGCGAGATCCTGATCAAGGCCAAGAAGGGCGGCAAGATCGCCGTGTAAGACTAGTAGAC
CAATCTTTAACATCTGATTATATTTGATTTATTATTTGAGTGTTGTTGGACCATGTGTGA
TCAGACTGCTATCTGAATAAAATAAGATTTGTCAAAACTCAGTGTTTTCTCCATCAGACA
CTCCATGAAAGGTCACAATTTCTCTTGATATTAAGCTGGGTTGTCTTTAAACAACCCTAA
ATACACGTCTGTTTAGCCCGCAATTGGAAAGGATATATGTGGCAATATTAACCTGGTACA
TGAATATATGGGGATAACATTTTAATTTGAAGGTTTGGAATATATATATTTAAGCTTTAT
TTCCAGAACAGTGAGGGTTAGGTCTTGGGAAAACTATAACTTGCCAAAGTAGAAGAAATA
GTAGTACCATATGCCAAAGTGATAGAGATGAATCATGTCAGTAGTTAGAATAACATTTCA
ACTGTTTTCTTTGCTAAAATCACAGAAAGACCCTATTGACAACATCTATGTCTGTAAAAA
TGTTAGAGTACTTGTCATCTTGAATATAGCCTCCCCAAGAGAGAACAGGGTGGTATTCTA
AGTATGTTTCTTTGTAACATCTTTAGCAGTAGGACAGAGCCATACATGTGAAATCTGATT
TTTATGTGTGTTATTCGTTTGTCTGGTTTTACTACCTTTGCAAAAACAAAATACCCCAAA
GATATTTAAACAAGGTTATAATTTAGCATCTTCCCTGGATCTAAATAGTATATTATATCC
TGAAATAAATGAAATGATTGCTATAGATCTAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA
AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA
SEQ ID NO: 18:
AAAAGCATCT CAGCCTACCA TGAGAATAAG AGAAAGAAAA TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT TGCCTCTTTT CTCTGTGCTT CAATT
SEQ ID NO: 19:
CATCACATTT AAAAGCATCT CAGCCTACCA TGAGAATAAG AGAAAGAAAA TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG
SEQ ID NO: 20:
AAAAGCATCT CAGCCTACCA TGAGAATAAG AGAAAGAAAA TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC
SEQ ID NO: 21:
CAGCCTACCA TGAGAATAAG AGAAAGAAAA TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT
SEQ ID NO: 22:
TGAGAATAAG AGAAAGAAAA TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT
SEQ ID NO: 23:
AGAAAGAAAA TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT TGCCTCTTTT
SEQ ID NO: 24:
TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT TGCCTCTTTT CTCTGTGCTT
SEQ ID NO: 25:
AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT TGCCTCTTTT CTCTGTGCTT CAATTAATAA
SEQ ID NO: 26:
ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT TGCCTCTTTT CTCTGTGCTT CAATTAATAA AAAATGGAAA
SEQ ID NO: 27:
CAGCCTACCA TGAGAATAAG AGAAAGAAAA TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT TGCCTCTTTT CTCTGTGCTT CAATTAATAA A
SEQ ID NO: 28:
TGAAGATCAA AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT TGCCTCTTTT CTCTGTGCTT CAATTAATAA A
SEQ ID NO: 29:
CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC ATAAATTTCT TTAATCATTT TGCCTCTTTT CTCTGTGCTT CAATTAATAA A
SEQ ID NO: 30:
AAGCTTATTC ATCTGTTTTT CTTTTTCGTT GGTGTAAAGC CAACACCCTG TCTAAAAAAC
SEQ ID NO: 31:
CAAAGGCTCTTTTCAGAGCCACCA
SEQ ID NO: 32:
AAACATCACA ATTAAGAACA TCTCAGCCTA CCATGAGAAC AAGAGAAATA AAATGAAGAT CAAAAGCTTA TTCATCTGTT TTTCTTTTTC ATTGGTATAA AGCCAACACC CTGTCTAAAA AACTATAAAT TTCTTTAATC ATTTTGCCTC TTTTCTCTGT GCTTCAATTA ATAAAAAATG GAAAGAATCT AGATCTAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAA
SEQ ID NO: 33:
AAACAUCACA AUUAAGAACA UCUCAGCCUA CCAUGAGAAC AAGAGAAAUA AAAUGAAGAU CAAAAGCUUA UUCAUCUGUU UUUCUUUUUC AUUGGUAUAA AGCCAACACC CUGUCUAAAA AACUAUAAAU UUCUUUAAUC AUUUUGCCUC UUUUCUCUGU GCUUCAAUUA AUAAAAAAUG GAAAGAAUCU AGAUCUAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAA
SEQ ID NO: 34:
ACACATCACA ACCACAACCT TCTCAGGCTA CCCTGAGAAA AAAAGACATG AAGACTCAGG ACTCATCTTT TCTGTTGGTG TAAAATCAAC ACCCTAAGGA ACACAAATTT CTTTAAACAT TTGACTTCTT GTCTCTGTGC TGCAATTAAT AAAAAATGGA AAGAATCTAC AGATCTAAAA AAAA
SEQ ID NO: 35:
ACACAUCACA ACCACAACCU UCUCAGGCUA CCCUGAGAAA AAAAGACAUG AAGACUCAGG ACUCAUCUUU UCUGUUGGUG UAAAAUCAAC ACCCUAAGGA ACACAAAUUU CUUUAAACAU UUGACUUCUU GUCUCUGUGC UGCAAUUAAU AAAAAAUGGA AAGAAUCUAC AGAUCUAAAA AAAA
SEQ ID NO: 36:
AAACATCACA ATTAAGAACA TCTCAGCCTA CCATGAGAAC AAGAGAAATA AAATGAAGAT CAAAAGCTTA TTCATCTGTT TTTCTTTTTC ATTGGTATAA AGCCAACACC CTGTCTAAAA AACTATAAAT TTCTTTAATC ATTTTGCCTC TTTTCTCTGT GCTTCAATTA ATAAAAAATG GAAAGAATCT AGATCTAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA
SEQ ID NO: 37:
AAACAUCACA AUUAAGAACA UCUCAGCCUA CCAUGAGAAC AAGAGAAAUA AAAUGAAGAU CAAAAGCUUA UUCAUCUGUU UUUCUUUUUC AUUGGUAUAA AGCCAACACC CUGUCUAAAA AACUAUAAAU UUCUTTAATC ATTTTGCCTC TTTTCTCTGT GCTTCAATTA ATAAAAAATG GAAAGAATCT AGATCTAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA
SEQ ID NO: 38:
ACACATCACA ACCACAACCT TCTCAGGCTA CCCTGAGAAA AAAAGACATG AAGACTCAGG ACTCATCTTT TCTGTTGGTG TAAAATCAAC ACCCTAAGGA ACACAAATTT CTTTAAACAT TTGACTTCTT GTCTCTGTGC TGCAATTAAT AAAAAATGGA AAGAATCTAC AGATCTAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA
SEQ ID NO: 39:
ACACAUCACA ACCACAACCU UCUCAGGCUA CCCUGAGAAA AAAAGACAUG AAGACUCAGG ACUCAUCUUU UCUGUUGGUG UAAAAUCAAC ACCCUAAGGA ACACAAAUUU CUUUAAACAU UUGACUUCUU GUCUCUGUGC UGCAAUUAAU AAAAAAUGGA AAGAAUCUAC AGAUCUAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA
SEQ ID NO: 40:
GGGAGAAAGC TTGAGGATGG AGGACGCCAA GAACATCAAG AAGGGCCCGG CGCCCTTCTA
CCCGCTGGAG GACGGGACCG CCGGCGAGCA GCTCCACAAG GCCATGAAGC GGTACGCCCT
GGTGCCGGGC ACGATCGCCT TCACCGACGC CCACATCGAG GTCGACATCA CCTACGCGGA
GTACTTCGAG ATGAGCGTGC GCCTGGCCGA GGCCATGAAG CGGTACGGCC TGAACACCAA
CCACCGGATC GTGGTGTGCT CGGAGAACAG CCTGCAGTTC TTCATGCCGG TGCTGGGCGC
CCTCTTCATC GGCGTGGCCG TCGCCCCGGC GAACGACATC TACAACGAGC GGGAGCTGCT
GAACAGCATG GGGATCAGCC AGCCGACCGT GGTGTTCGTG AGCAAGAAGG GCCTGCAGAA
GATCCTGAAC GTGCAGAAGA AGCTGCCCAT CATCCAGAAG ATCATCATCA TGGACAGCAA
GACCGACTAC CAGGGCTTCC AGTCGATGTA CACGTTCGTG ACCAGCCACC TCCCGCCGGG
CTTCAACGAG TACGACTTCG TCCCGGAGAG CTTCGACCGG GACAAGACCA TCGCCCTGAT
CATGAACAGC AGCGGCAGCA CCGGCCTGCC GAAGGGGGTG GCCCTGCCGC ACCGGACCGC
CTGCGTGCGC TTCTCGCACG CCCGGGACCC CATCTTCGGC AACCAGATCA TCCCGGACAC
CGCCATCCTG AGCGTGGTGC CGTTCCACCA CGGCTTCGGC ATGTTCACGA CCCTGGGCTA
CCTCATCTGC GGCTTCCGGG TGGTCCTGAT GTACCGGTTC GAGGAGGAGC TGTTCCTGCG
GAGCCTGCAG GACTACAAGA TCCAGAGCGC GCTGCTCGTG CCGACCCTGT TCAGCTTCTT
CGCCAAGAGC ACCCTGATCG ACAAGTACGA CCTGTCGAAC CTGCACGAGA TCGCCAGCGG
GGGCGCCCCG CTGAGCAAGG AGGTGGGCGA GGCCGTGGCC AAGCGGTTCC ACCTCCCGGG
CATCCGCCAG GGCTACGGCC TGACCGAGAC CACGAGCGCG ATCCTGATCA CCCCCGAGGG
GGACGACAAG CCGGGCGCCG TGGGCAAGGT GGTCCCGTTC TTCGAGGCCA AGGTGGTGGA
CCTGGACACC GGCAAGACCC TGGGCGTGAA CCAGCGGGGC GAGCTGTGCG TGCGGGGGCC
GATGATCATG AGCGGCTACG TGAACAACCC GGAGGCCACC AACGCCCTCA TCGACAAGGA
CGGCTGGCTG CACAGCGGCG ACATCGCCTA CTGGGACGAG GACGAGCACT TCTTCATCGT
CGACCGGCTG AAGTCGCTGA TCAAGTACAA GGGCTACCAG GTGGCGCCGG CCGAGCTGGA
GAGCATCCTG CTCCAGCACC CCAACATCTT CGACGCCGGC GTGGCCGGGC TGCCGGACGA
CGACGCCGGC GAGCTGCCGG CCGCGGTGGT GGTGCTGGAG CACGGCAAGA CCATGACGGA
GAAGGAGATC GTCGACTACG TGGCCAGCCA GGTGACCACC GCCAAGAAGC TGCGGGGCGG
CGTGGTGTTC GTGGACGAGG TCCCGAAGGG CCTGACCGGG AAGCTCGACG CCCGGAAGAT
CCGCGAGATC CTGATCAAGG CCAAGAAGGG CGGCAAGATC GCCGTGTAAG ACTAGTAAAC
ATCACAATTA AGAACATCTC AGCCTACCAT GAGAACAAGA GAAATAAAAT GAAGATCAAA
AGCTTATTCA TCTGTTTTTC TTTTTCATTG GTATAAAGCC AACACCCTGT CTAAAAAACT
ATAAATTTCT TTAATCATTT TGCCTCTTTT CTCTGTGCTT CAATTAATAA AAAATGGAAA
GAATCTAGAT CTAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA
AAAAAAAAAA AAAAAA
SEQ ID NO: 41:
GGGAGAAAGC TTGAGGATGG AGGACGCCAA GAACATCAAG AAGGGCCCGG CGCCCTTCTA
CCCGCTGGAG GACGGGACCG CCGGCGAGCA GCTCCACAAG GCCATGAAGC GGTACGCCCT
GGTGCCGGGC ACGATCGCCT TCACCGACGC CCACATCGAG GTCGACATCA CCTACGCGGA
GTACTTCGAG ATGAGCGTGC GCCTGGCCGA GGCCATGAAG CGGTACGGCC TGAACACCAA
CCACCGGATC GTGGTGTGCT CGGAGAACAG CCTGCAGTTC TTCATGCCGG TGCTGGGCGC
CCTCTTCATC GGCGTGGCCG TCGCCCCGGC GAACGACATC TACAACGAGC GGGAGCTGCT
GAACAGCATG GGGATCAGCC AGCCGACCGT GGTGTTCGTG AGCAAGAAGG GCCTGCAGAA
GATCCTGAAC GTGCAGAAGA AGCTGCCCAT CATCCAGAAG ATCATCATCA TGGACAGCAA
GACCGACTAC CAGGGCTTCC AGTCGATGTA CACGTTCGTG ACCAGCCACC TCCCGCCGGG
CTTCAACGAG TACGACTTCG TCCCGGAGAG CTTCGACCGG GACAAGACCA TCGCCCTGAT
CATGAACAGC AGCGGCAGCA CCGGCCTGCC GAAGGGGGTG GCCCTGCCGC ACCGGACCGC
CTGCGTGCGC TTCTCGCACG CCCGGGACCC CATCTTCGGC AACCAGATCA TCCCGGACAC
CGCCATCCTG AGCGTGGTGC CGTTCCACCA CGGCTTCGGC ATGTTCACGA CCCTGGGCTA
CCTCATCTGC GGCTTCCGGG TGGTCCTGAT GTACCGGTTC GAGGAGGAGC TGTTCCTGCG
GAGCCTGCAG GACTACAAGA TCCAGAGCGC GCTGCTCGTG CCGACCCTGT TCAGCTTCTT
CGCCAAGAGC ACCCTGATCG ACAAGTACGA CCTGTCGAAC CTGCACGAGA TCGCCAGCGG
GGGCGCCCCG CTGAGCAAGG AGGTGGGCGA GGCCGTGGCC AAGCGGTTCC ACCTCCCGGG
CATCCGCCAG GGCTACGGCC TGACCGAGAC CACGAGCGCG ATCCTGATCA CCCCCGAGGG
GGACGACAAG CCGGGCGCCG TGGGCAAGGT GGTCCCGTTC TTCGAGGCCA AGGTGGTGGA
CCTGGACACC GGCAAGACCC TGGGCGTGAA CCAGCGGGGC GAGCTGTGCG TGCGGGGGCC
GATGATCATG AGCGGCTACG TGAACAACCC GGAGGCCACC AACGCCCTCA TCGACAAGGA
CGGCTGGCTG CACAGCGGCG ACATCGCCTA CTGGGACGAG GACGAGCACT TCTTCATCGT
CGACCGGCTG AAGTCGCTGA TCAAGTACAA GGGCTACCAG GTGGCGCCGG CCGAGCTGGA
GAGCATCCTG CTCCAGCACC CCAACATCTT CGACGCCGGC GTGGCCGGGC TGCCGGACGA
CGACGCCGGC GAGCTGCCGG CCGCGGTGGT GGTGCTGGAG CACGGCAAGA CCATGACGGA
GAAGGAGATC GTCGACTACG TGGCCAGCCA GGTGACCACC GCCAAGAAGC TGCGGGGCGG
CGTGGTGTTC GTGGACGAGG TCCCGAAGGG CCTGACCGGG AAGCTCGACG CCCGGAAGAT
CCGCGAGATC CTGATCAAGG CCAAGAAGGG CGGCAAGATC GCCGTGTAAG ACTAGTACAC
ATCACAACCA CAACCTTCTC AGGCTACCCT GAGAAAAAAA GACATGAAGA CTCAGGACTC
ATCTTTTCTG TTGGTGTAAA ATCAACACCC TAAGGAACAC AAATTTCTTT AAACATTTGA
CTTCTTGTCT CTGTGCTGCA ATTAATAAAA AATGGAAAGA ATCTACAGAT CTAAAAAAAA
AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAAAAAA AAAAAA

























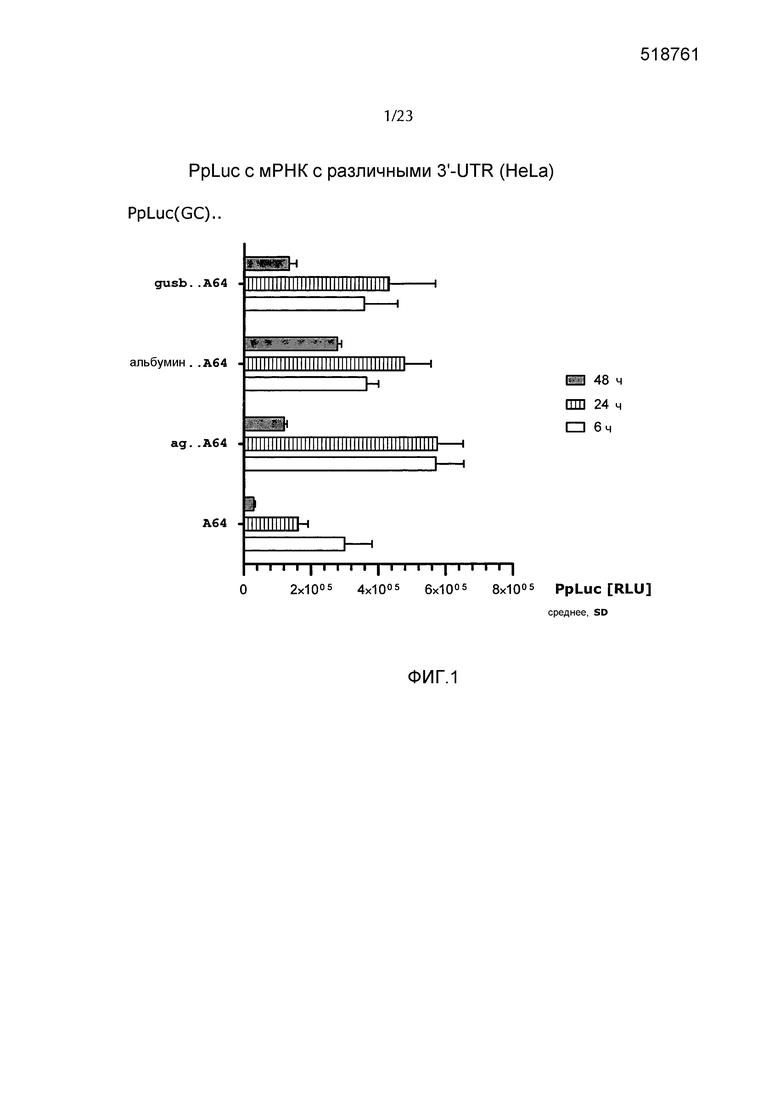
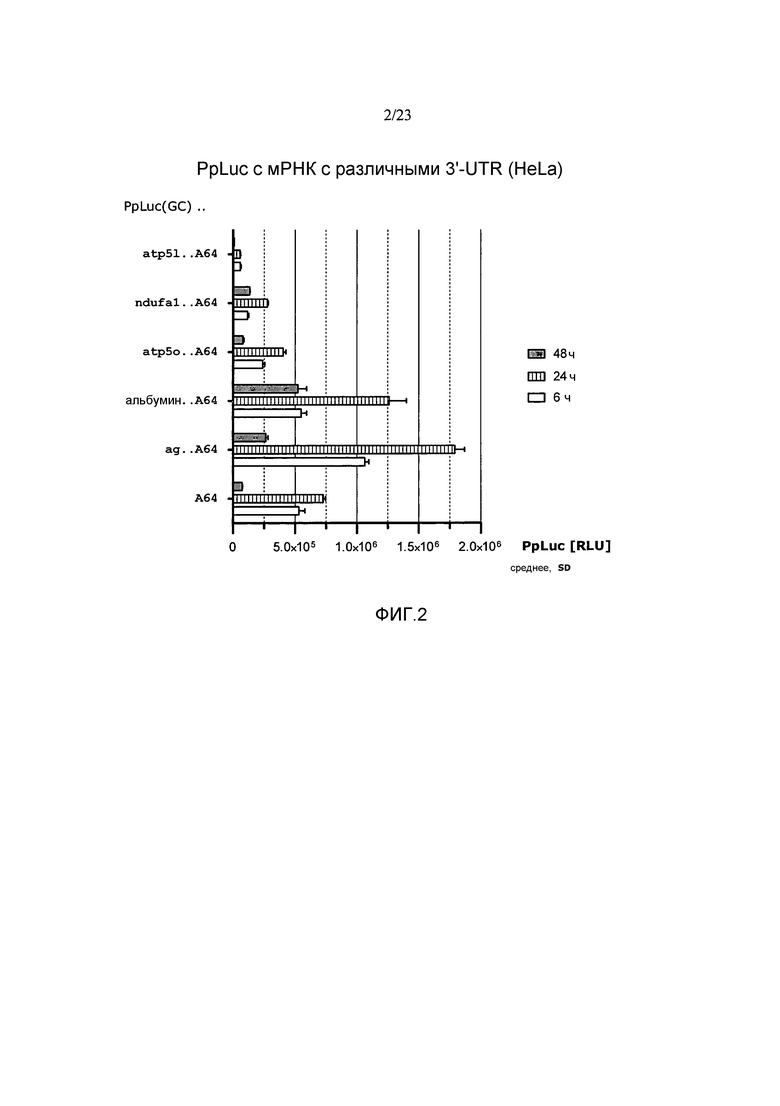
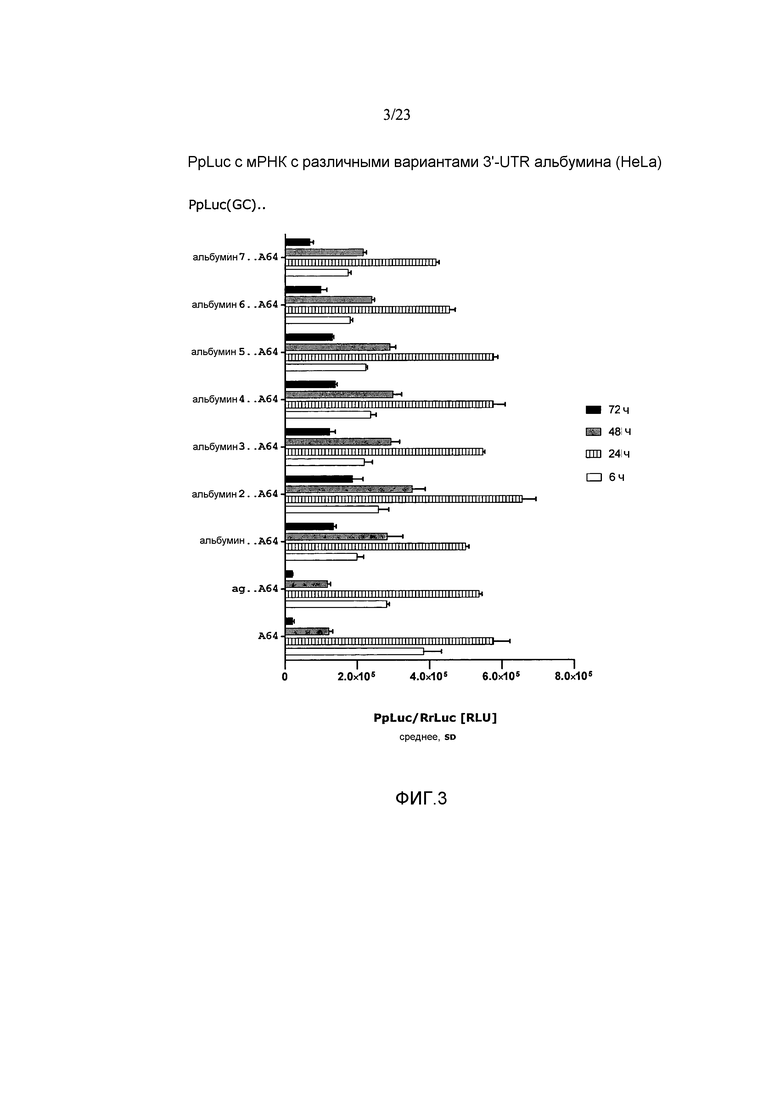













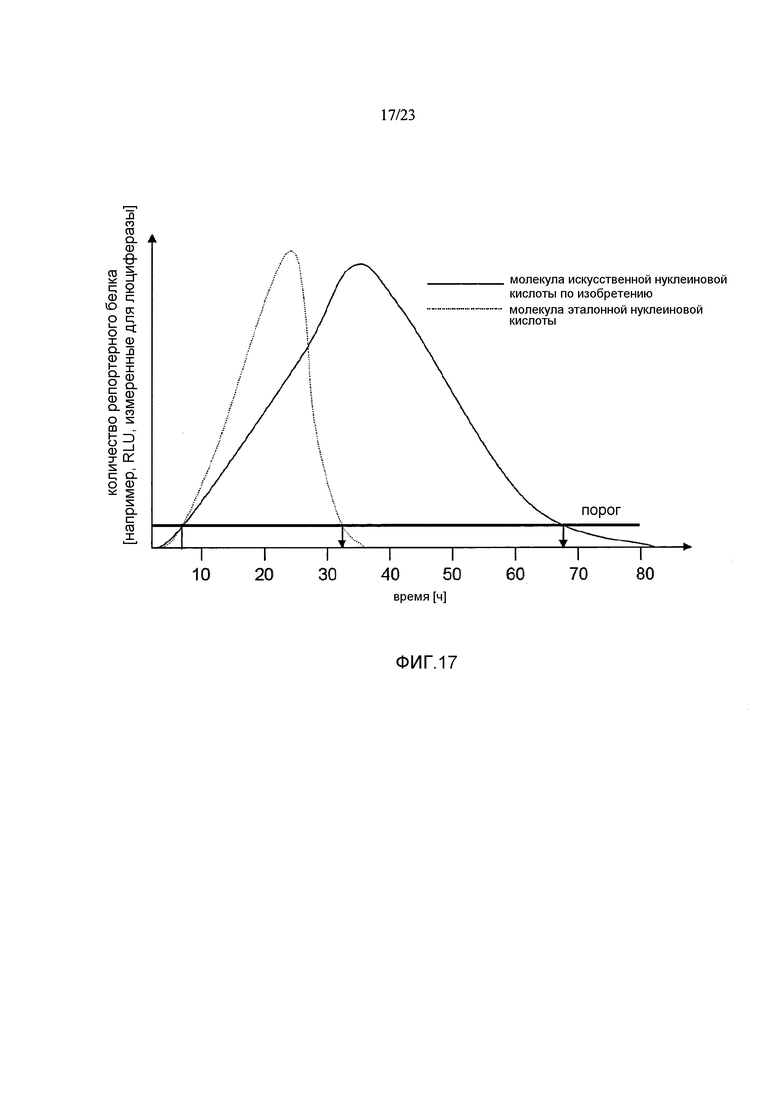
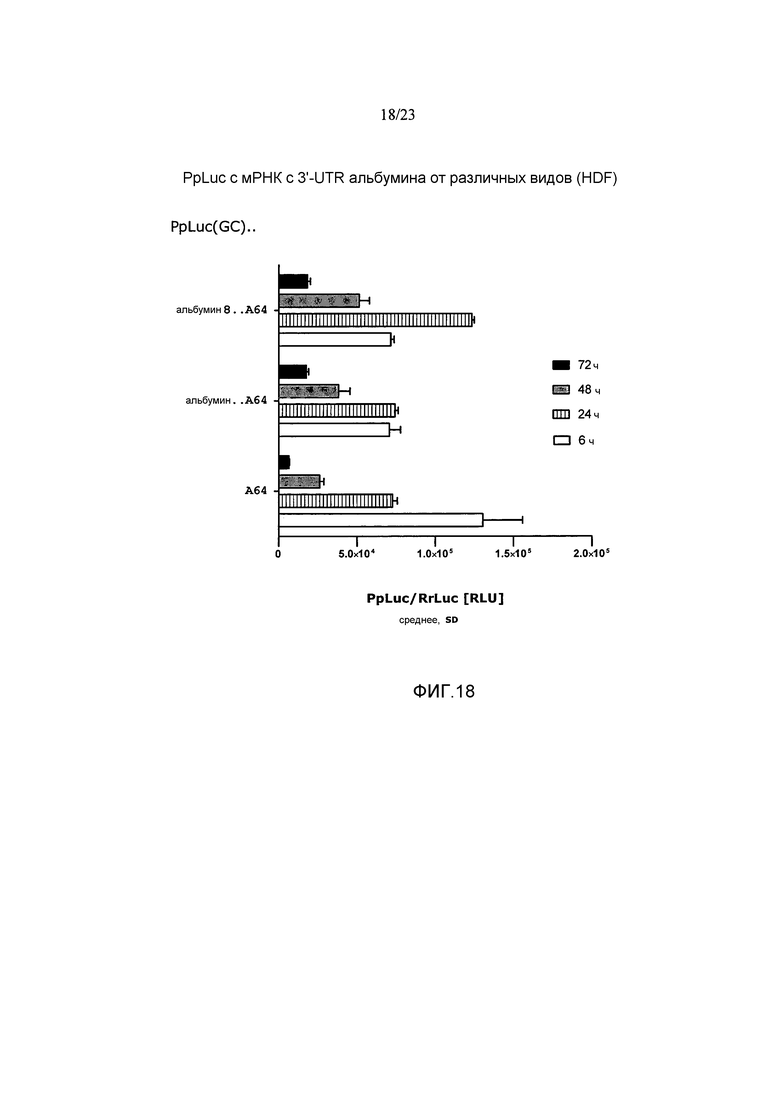
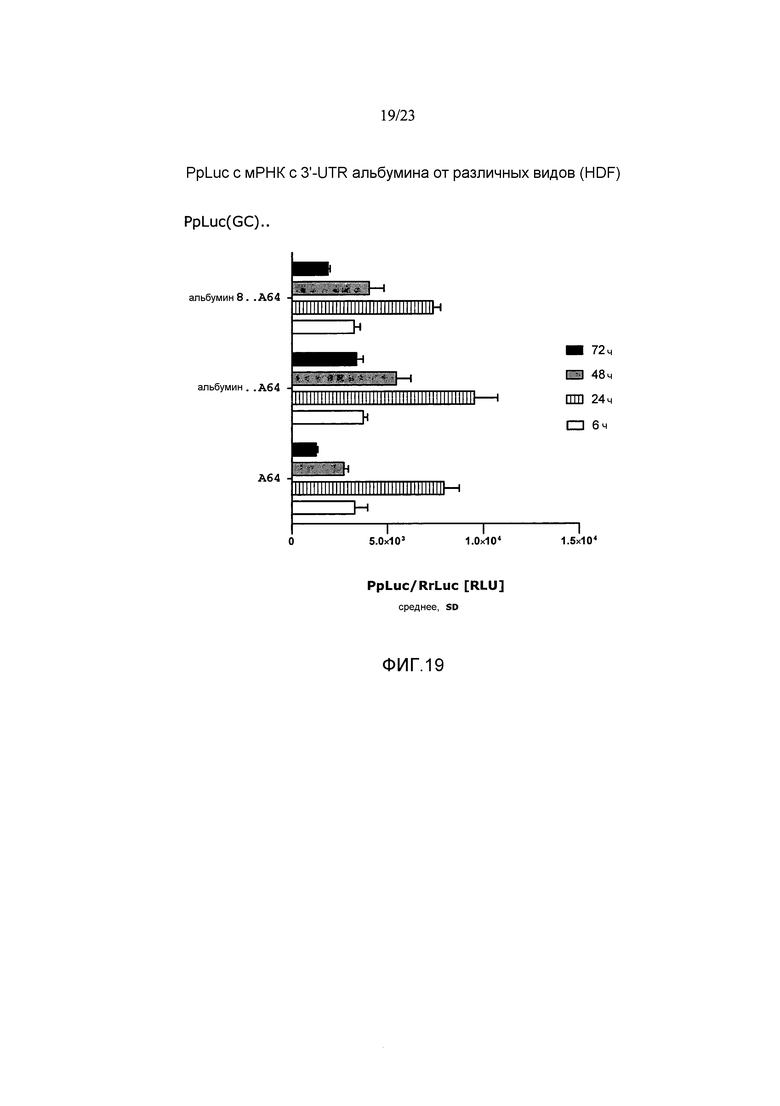
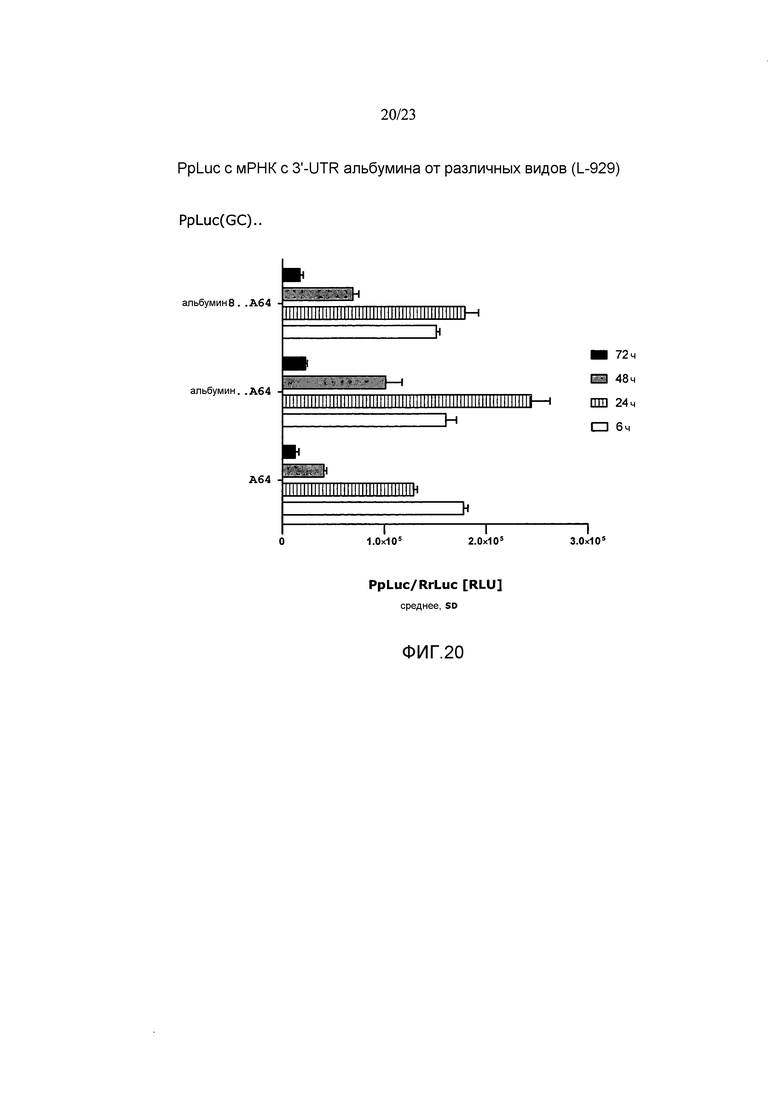
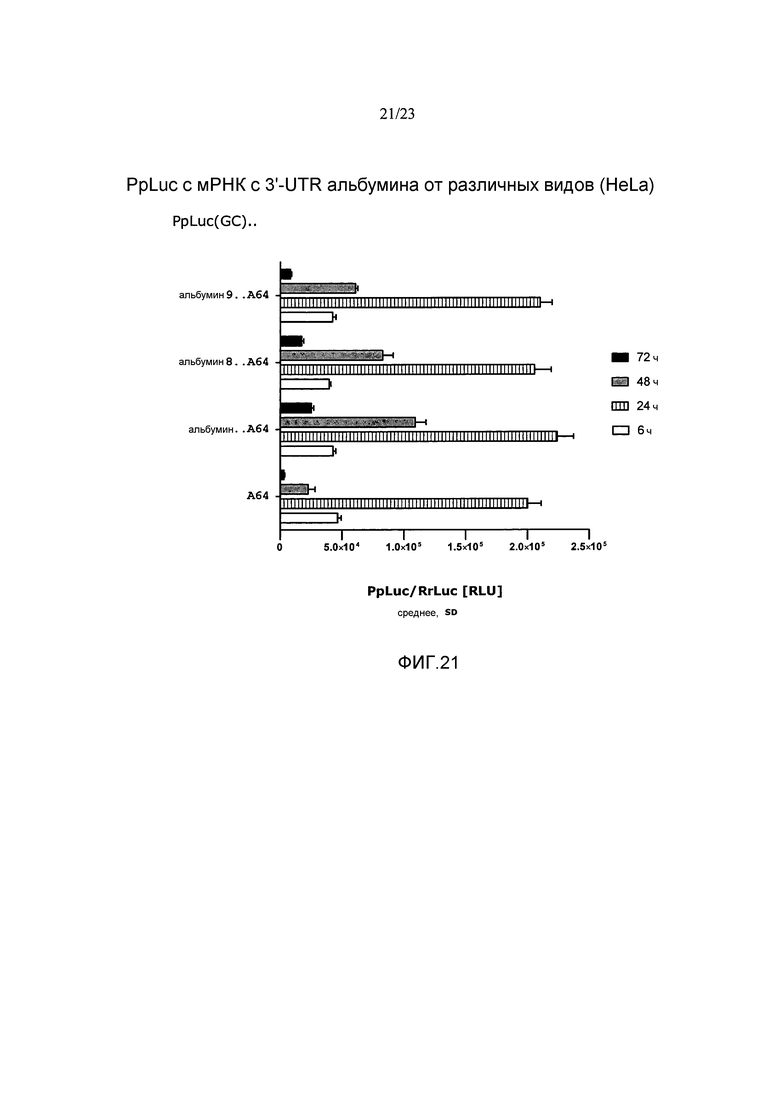


Изобретение относится к области биотехнологии, конкретно к молекуле искусственной нуклеиновой кислоты. Синергетический эффект, обусловленный наличием 3'-UTR гена альбумина и 3’-некодирующей области матричной РНК, уложенной во вторичную структуру типа «стебель-петля», в структуре искусственной нуклеиновой кислоты, позволяет повысить стабильность и продлить экспрессию кодируемого белка. Изобретение дополнительно относится к применению такой молекулы искусственной нуклеиновой кислоты, кодирующей терапевтические пептиды или белки в медицине при генной терапии и/или генетической вакцинации. 15 н. и 48 з.п. ф-лы., 23 ил., 14 табл., 5 пр.
1. Молекула искусственной мРНК для экспрессии представляющего интерес пептида или белка, где молекула искусственной нуклеиновой кислоты включает
a. по меньшей мере одну открытую рамку считывания (ORF), кодирующую представляющий интерес пептид или белок;
b. по меньшей мере один элемент 3'-нетранслируемой области (элемент 3'-UTR), включающий последовательность нуклеиновой кислоты, которая происходит от 3'-UTR гена альбумина или от варианта 3'-UTR гена альбумина, и
c. 3' -некодирующую область матричной РНК гистона, уложенную во вторичную структуру типа "стебель-петля".
2. Молекула искусственной мРНК по п. 1, где по меньшей мере один элемент 3'-UTR включает последовательность нуклеиновой кислоты, которая происходит от 3'-UTR гена альбумина позвоночных или от его варианта, предпочтительно от 3'-UTR гена альбумина млекопитающих, например от гена альбумина мыши или от его варианта, более предпочтительно от 3'-UTR гена альбумина приматов, особенно от гена альбумина человека или гена альбумина бабуина анубис или от их вариантов, еще более предпочтительно от 3'-UTR гена альбумина человека или его варианта.
3. Молекула искусственной мРНК по п. 1, где по меньшей мере один элемент 3'-UTR включает или состоит из последовательности нуклеиновой кислоты, которая идентична по меньшей мере приблизительно на 40%, предпочтительно по меньшей мере приблизительно на 50%, предпочтительно по меньшей мере приблизительно на 60%, предпочтительно по меньшей мере приблизительно на 70%, более предпочтительно по меньшей мере приблизительно на 80%, более предпочтительно по меньшей мере приблизительно на 90%, даже более предпочтительно по меньшей мере приблизительно на 95%, даже более предпочтительно по меньшей мере приблизительно на 99% последовательности нуклеиновой кислоты в соответствии с SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34 или SEQ ID NO: 35, или где по меньшей мере один элемент 3'-UTR включает или состоит из фрагмента последовательности нуклеиновой кислоты, который идентичен по меньшей мере приблизительно на 40%, предпочтительно по меньшей мере приблизительно на 50%, предпочтительно по меньшей мере приблизительно на 60%, предпочтительно по меньшей мере приблизительно на 70%, более предпочтительно по меньшей мере приблизительно на 80%, более предпочтительно по меньшей мере приблизительно на 90%, даже более предпочтительно по меньшей мере приблизительно на 95%, даже более предпочтительно по меньшей мере приблизительно на 99% последовательности нуклеиновой кислоты в соответствии с SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34 или SEQ ID NO: 35.
4. Молекула искусственной мРНК по п. 3, где фрагмент имеет длину по меньшей мере приблизительно 50 нуклеотидов, предпочтительно по меньшей мере приблизительно 75 нуклеотидов, более предпочтительно по меньшей мере приблизительно 100 нуклеотидов, даже более предпочтительно по меньшей мере приблизительно 125 нуклеотидов, наиболее предпочтительно по меньшей мере приблизительно 150 нуклеотидов.
5. Молекула искусственной мРНК по п. 1, где по меньшей мере один элемент 3'-UTR имеет длину по меньшей мере приблизительно 50 нуклеотидов, предпочтительно по меньшей мере приблизительно 75 нуклеотидов, более предпочтительно по меньшей мере приблизительно 100 нуклеотидов, еще более предпочтительно по меньшей мере приблизительно 125 нуклеотидов, наиболее предпочтительно по меньшей мере приблизительно 150 нуклеотидов.
6. Молекула искусственной мРНК по п. 1, дополнительно включающая поли(А)-последовательность и/или сигнал полиаденилирования.
7. Молекула искусственной мРНК по п. 6, где сигнал полиаденилирования локализован в пределах элемента 3'-UTR.
8. Молекула искусственной мРНК по п. 6 или 7, где сигнал полиаденилирования включает консенсусную последовательность NN(U/T)ANA с N=A или U, предпочтительно АА(U/T)ААА или A(U/T)(U/T)AAA.
9. Молекула искусственной мРНК по п. 6 или 7, где сигнал полиаденилирования, предпочтительно консенсусная последовательность NNUANA, локализован менее чем приблизительно на 50 нуклеотидов выше 3'-конца 3'-UTR.
10. Молекула искусственной мРНК по п. 6 или 7, где поли(А)-последовательность имеет длину от приблизительно 20 до приблизительно 300 адениновых нуклеотидов, предпочтительно от приблизительно 40 до приблизительно 200 адениновых нуклеотидов, более предпочтительно от приблизительно 50 до приблизительно 100 адениновых нуклеотидов, даже более предпочтительно от приблизительно 60 до приблизительно 70 адениновых нуклеотидов.
11. Молекула искусственной мРНК по п. 1, где открытая рамка считывания не кодирует альбумин, предпочтительно не кодирует альбумин человека.
12. Молекула искусственной мРНК по п. 1, дополнительно включающая структуру 5'-кэпа, поли(С)-последовательность и/или мотив IRES.
13. Молекула искусственной мРНК по п. 1, которая включает дополнительный 5'-элемент, предпочтительно 5'-UTR, промотор или 5'-UTR и последовательность, содержащую промотор.
14. Молекула искусственной мРНК по п. 1, в которой предпочтительно открытая рамка считывания является, по меньшей мере, частично G/C модифицированной, предпочтительно, где содержание G/C открытой рамки считывания повышено по сравнению с открытой рамкой считывания дикого типа.
15. Молекула искусственной мРНК по п. 1, где открытая рамка считывания включает кодон-оптимизированную область, предпочтительно, где открытая рамка считывания является кодон-оптимизированной.
16. Вектор для экспрессии представляющего интерес пептида или белка, где вектор включает молекулу искусственной мРНК по пп. 1-15.
17. Вектор по п. 16, где по меньшей мере один элемент 3'-UTR включает или состоит из последовательности нуклеиновой кислоты, которая происходит от 3'-UTR гена альбумина позвоночных или от его варианта, предпочтительно от 3'-UTR гена альбумина млекопитающих, например от гена альбумина мыши или от его варианта, более предпочтительно от 3'-UTR гена альбумина приматов, особенно от гена альбумина человека или от гена альбумина бабуина анубис или от их вариантов, еще более предпочтительно от 3'-UTR гена альбумина человека или от его варианта.
18. Вектор по п. 16, где по меньшей мере один элемент 3'-UTR включает или состоит из последовательности нуклеиновой кислоты, которая идентична по меньшей мере приблизительно на 40%, предпочтительно по меньшей мере приблизительно на 50%, предпочтительно по меньшей мере приблизительно на 60%, предпочтительно по меньшей мере приблизительно на 70%, более предпочтительно по меньшей мере приблизительно на 80%, более предпочтительно по меньшей мере приблизительно на 90%, даже более предпочтительно по меньшей мере приблизительно на 95%, даже более предпочтительно по меньшей мере приблизительно на 99% последовательности нуклеиновой кислоты в соответствии с SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34 или SEQ ID NO: 35, или где по меньшей мере один элемент 3'-UTR включает или состоит из фрагмента последовательности нуклеиновой кислоты, который идентичен по меньшей мере приблизительно на 40%, предпочтительно по меньшей мере приблизительно на 50%, предпочтительно по меньшей мере приблизительно на 60%, предпочтительно по меньшей мере приблизительно на 70%, более предпочтительно по меньшей мере приблизительно на 80%, более предпочтительно по меньшей мере приблизительно на 90%, даже более предпочтительно по меньшей мере приблизительно на 95%, даже более предпочтительно по меньшей мере приблизительно на 99% последовательности нуклеиновой кислоты в соответствии с SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34 или SEQ ID NO: 35.
19. Вектор по п. 18, где фрагмент имеет длину по меньшей мере приблизительно 50 нуклеотидов, предпочтительно по меньшей мере приблизительно 75 нуклеотидов, более предпочтительно по меньшей мере приблизительно 100 нуклеотидов, еще более предпочтительно по меньшей мере приблизительно 125 нуклеотидов, наиболее предпочтительно по меньшей мере приблизительно 150 нуклеотидов.
20. Вектор по п. 16, где по меньшей мере один элемент 3'-UTR имеет длину по меньшей мере приблизительно 50 нуклеотидов, предпочтительно по меньшей мере приблизительно 75 нуклеотидов, более предпочтительно по меньшей мере приблизительно 100 нуклеотидов, еще более предпочтительно по меньшей мере приблизительно 125 нуклеотидов, наиболее предпочтительно по меньшей мере приблизительно 150 нуклеотидов.
21. Вектор по п. 16, дополнительно включающий поли(А)-последовательность и/или сигнал полиаденилирования.
22. Вектор по п. 21, где сигнал полиаденилирования локализован в пределах элемента 3'-UTR.
23. Вектор по п. 21 или 22, где сигнал полиаденилирования включает консенсусную последовательность NN(U/T)ANA с N=A или U, предпочтительно AA(U/T)AAA или A(U/T)(U/T)AAA.
24. Вектор по п. 21 или 22, где сигнал полиаденилирования, предпочтительно консенсусная последовательность NNUANA, локализован менее чем приблизительно на 50 нуклеотидов выше 3'-конца элемента 3'-UTR.
25. Вектор по п. 21 или 22, где поли(А)-последовательность имеет длину от приблизительно 20 до приблизительно 300 адениновых нуклеотидов, предпочтительно от приблизительно 40 до приблизительно 200 адениновых нуклеотидов, более предпочтительно от приблизительно 50 до приблизительно 100 адениновых нуклеотидов, более предпочтительно от приблизительно 60 до приблизительно 70 адениновых нуклеотидов.
26. Вектор по п. 16, где открытая рамка считывания не кодирует альбумин человека, предпочтительно не кодирует альбумин.
27. Вектор по п. 16, где открытая рамка считывания не кодирует фактор IX человека или репортерный белок, предпочтительно не кодирует белок GFP, белок люциферазы, или белок глобина, или их варианты.
28. Вектор по п. 16, дополнительно включающий поли(С)-последовательность и/или мотив IRES.
29. Вектор по п. 16, дополнительно включающий 5'-элемент, предпочтительно 5'-UTR, промотор или 5'-UTR и последовательность, содержащую промотор.
30. Вектор по п. 16, который является, по меньшей мере, частично G/C модифицированным, предпочтительно, где открытая рамка считывания является, по меньшей мере, частично G/C модифицированной, предпочтительно, где содержание G/C открытой рамки считывания повышено по сравнению с открытой рамкой считывания дикого типа.
31. Вектор по п. 16, где открытая рамка считывания включает кодон-оптимизированную область, предпочтительно, где открытая рамка считывания является кодон-оптимизированной.
32. Вектор по п. 16, который представляет собой ДНК-вектор.
33. Вектор по п. 16, который представляет собой плазмидный вектор или вирусный вектор, предпочтительно плазмидный вектор.
34. Вектор по п. 16, который включает конструкцию молекулы искусственной нуклеиновой кислоты по любому из пп. 1-15.
35. Вектор по п. 16, который представляет собой кольцевую молекулу.
36. Вектор по п. 35, где за поли(А)-последовательностью или элементом 3'-UTR кодирующей цепи следует в направлении 5'→3' сайт рестрикции для линеаризации кольцевой молекулы вектора.
37. Клетка для экспрессии представляющего интерес пептида или белка, где клетка включает вектор по любому из пп. 16-36.
38. Клетка по п. 37, которая представляет собой клетку млекопитающего.
39. Клетка по п. 37 или 38, которая представляет собой клетку индивидуума-млекопитающего, предпочтительно выделенную клетку индивидуума-млекопитающего, предпочтительно человека.
40. Фармацевтическая композиция для экспрессии представляющего интерес пептида или белка, где фармацевтическая композиция включает конструкцию молекулы искусственной нуклеиновой кислоты по любому из пп. 1-15, вектор по любому из пп. 16-36 или клетку по любому из пп. 37-39 в эффективном количестве.
41. Фармацевтическая композиция по п. 40, дополнительно включающая один или более фармацевтически приемлемых разбавителей и/или наполнителей, и/или один или более адъювантов.
42. Применение конструкции молекулы искусственной нуклеиновой кислоты по любому из пп. 1-15 или вектора по любому из пп. 16-36 в генной терапии.
43. Применение конструкции молекулы искусственной нуклеиновой кислоты по любому из пп. 1-15 или вектора по любому из пп. 16-36 в генетической вакцинации.
44. Применение клетки по любому из пп. 37-39 в генной терапии.
45. Применение клетки по любому из пп. 37-39 в генетической вакцинации.
46. Применение фармацевтической композиции по п. 40 или 41 в генной терапии.
47. Применение фармацевтической композиции по п. 40 или 41 в генетической вакцинации.
48. Способ лечения или профилактики нарушения, включающий введение конструкции молекулы искусственной нуклеиновой кислоты по любому из пп. 1-15, вектора по любому из пп. 16-36, клетки по любому из пп. 37-39 или фармацевтической композиции по п. 40 или 41 нуждающемуся в этом индивидууму.
49. Способ лечения или профилактики нарушения, включающий трансфекцию клетки конструкцией молекулы искусственной нуклеиновой кислоты по любому из пп. 1-15 или вектором по любому из пп. 16-36.
50. Способ по п. 49, где трансфекция клетки осуществляется in vitro/ex vivo, и трансфицированная клетка вводится нуждающемуся в этом индивидууму, предпочтительно больному человеку.
51. Способ по п. 50, где клетка, подлежащая трансфекции in vitro, представляет собой выделенную клетку индивидуума, предпочтительно больного человека.
52. Способ по любому из пп. 48-51, который представляет собой способ вакцинации или способ генной терапии.
53. Способ стабилизации и/или продления продукции белка с молекулы мРНК или вектора, причем способ включает стадию связывания молекулы мРНК или вектора с элементом 3'-UTR, где элемент 3'-UTR включает или состоит из последовательности нуклеиновой кислоты, которая происходит от 3'-UTR гена альбумина или от варианта 3'-UTR гена альбумина и 3' -некодирующей области матричной РНК гистона, уложенная во вторичную структуру типа "стебель-петля".
54. Способ по п. 53, где элемент 3'-UTR включает или состоит из последовательности нуклеиновой кислоты, которая происходит от 3'-UTR гена альбумина позвоночных или от его варианта, предпочтительно от 3'-UTR гена альбумина млекопитающих, например от гена альбумина мыши или от его варианта, более предпочтительно от 3'-UTR гена альбумина приматов, особенно от гена альбумина человека или от гена альбумина бабуина анубис или от их вариантов, еще более предпочтительно от 3'-UTR гена альбумина человека или от его варианта.
55. Способ по п. 53, где элемент 3'-UTR включает или состоит из последовательности нуклеиновой кислоты, которая идентична по меньшей мере приблизительно на 40%, предпочтительно по меньшей мере приблизительно на 50%, предпочтительно по меньшей мере приблизительно на 60%, предпочтительно по меньшей мере приблизительно на 70%, более предпочтительно по меньшей мере приблизительно на 80%, более предпочтительно по меньшей мере приблизительно на 90%, даже более предпочтительно по меньшей мере приблизительно на 95%, даже более предпочтительно по меньшей мере приблизительно на 99% последовательности нуклеиновой кислоты в соответствии с SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34 или SEQ ID NO: 35, или где элемент 3'-UTR включает или состоит из фрагмента последовательности нуклеиновой кислоты, который идентичен по меньшей мере приблизительно на 40%, предпочтительно по меньшей мере приблизительно на 50%, предпочтительно по меньшей мере приблизительно на 60%, предпочтительно по меньшей мере приблизительно на 70%, более предпочтительно по меньшей мере приблизительно на 80%, более предпочтительно по меньшей мере приблизительно на 90%, даже более предпочтительно по меньшей мере приблизительно на 95%, даже более предпочтительно по меньшей мере приблизительно на 99% последовательности нуклеиновой кислоты в соответствии с SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 32. SEQ ID NO: 33, SEQ ID NO: 34 или SEQ ID NO: 35.
56. Способ no п. 55, где фрагмент имеет длину по меньшей мере приблизительно 50 нуклеотидов, предпочтительно по меньшей мере приблизительно 75 нуклеотидов, более предпочтительно по меньшей мере приблизительно 100 нуклеотидов, еще более предпочтительно по меньшей мере приблизительно 125 нуклеотидов, наиболее предпочтительно по меньшей мере приблизительно 150 нуклеотидов.
57. Способ по п. 53, где элемент 3'-UTR имеет длину по меньшей мере приблизительно 50 нуклеотидов, предпочтительно по меньшей мере приблизительно 75 нуклеотидов, более предпочтительно по меньшей мере приблизительно 100 нуклеотидов, еще более предпочтительно по меньшей мере приблизительно 125 нуклеотидов, наиболее предпочтительно по меньшей мере приблизительно 150 нуклеотидов.
58. Применение элемента 3'-UTR и 3' -некодирующей области матричной РНК гистона, уложенная во вторичную структуру типа "стебель-петля", для стабилизации и/или продления продукции белка с молекулы мРНК или вектора, где элемент 3'-UTR включает или состоит из последовательности нуклеиновой кислоты, которая происходит от 3'-UTR гена альбумина или от варианта 3'-UTR гена альбумина.
59. Применение по п. 58, где элемент 3'-UTR включает или состоит из последовательности нуклеиновой кислоты, которая происходит от 3'-UTR гена альбумина позвоночных или от его варианта, предпочтительно от 3'-UTR гена альбумина млекопитающих, например, от гена альбумина мыши или от его варианта, более предпочтительно от 3'-UTR гена альбумина приматов, особенно от гена альбумина человека или от гена альбумина бабуина анубис или от их вариантов, еще более предпочтительно от 3'-UTR гена альбумина человека или от его варианта.
60. Применение по п. 58, где элемент 3'-UTR включает или состоит из последовательности нуклеиновой кислоты, которая идентична по меньшей мере приблизительно на 40%, предпочтительно по меньшей мере приблизительно на 50%, предпочтительно по меньшей мере приблизительно на 60%, предпочтительно по меньшей мере приблизительно на 70%, более предпочтительно по меньшей мере приблизительно на 80%, более предпочтительно по меньшей мере приблизительно на 90%, даже более предпочтительно по меньшей мере приблизительно на 95%, даже более предпочтительно по меньшей мере приблизительно на 99% последовательности нуклеиновой кислоты в соответствии с SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34 или SEQ ID NO: 35, или где элемент 3'-UTR включает или состоит из фрагмента последовательности нуклеиновой кислоты, который идентичен по меньшей мере приблизительно на 40%, предпочтительно по меньшей мере приблизительно на 50%, предпочтительно по меньшей мере приблизительно на 60%, предпочтительно по меньшей мере приблизительно на 70%, более предпочтительно по меньшей мере приблизительно на 80%, более предпочтительно по меньшей мере приблизительно на 90%, даже более предпочтительно по меньшей мере приблизительно на 95%, даже более предпочтительно по меньшей мере приблизительно на 99% последовательности нуклеиновой кислоты в соответствии с SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34 или SEQ ID NO: 35.
61. Применение по п. 60, где фрагмент имеет длину по меньшей мере приблизительно 50 нуклеотидов, предпочтительно по меньшей мере приблизительно 75 нуклеотидов, более предпочтительно по меньшей мере приблизительно 100 нуклеотидов, еще более предпочтительно по меньшей мере приблизительно 125 нуклеотидов, наиболее предпочтительно по меньшей мере приблизительно 150 нуклеотидов.
62. Применение по п. 58, где элемент 3'-UTR имеет длину по меньшей мере приблизительно 50 нуклеотидов, предпочтительно по меньшей мере приблизительно 75 нуклеотидов, более предпочтительно по меньшей мере приблизительно 100 нуклеотидов, еще более предпочтительно по меньшей мере приблизительно 125 нуклеотидов, наиболее предпочтительно по меньшей мере приблизительно 150 нуклеотидов.
63. Набор для экспрессии представляющего интерес пептида или белка, включающий молекулу искусственной мРНК по любому из пп. 1-15, вектор по любому из пп. 16-36, клетку по любому из пп. 37-39 и/или фармацевтическую композицию по п. 40 или 41, где набор дополнительно включает инструкции по применению, клетки для трансфекции, адъювант, средство для введения фармацевтической композиции, фармацевтически приемлемый носитель и/или фармацевтически приемлемый раствор для растворения или разбавления конструкции молекулы искусственной нуклеиновой кислоты, ДНК-вектора или фармацевтической композиции.
| US 2010120152 A1, 13.05.2010 | |||
| WOODDELL et al., Sustained liver-specific transgene expression from the albumin promoter in mice following hydrodynamic plasmid DNA delivery, J.Gene Med., 2008, v | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| LEVY S | |||
| et al., Oligopyrimidine tract at the 5' end of mammalian ribosomal protein mRNAs is required for their translational control, PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCE, 1991, v | |||
| Шланговое соединение | 0 |
|
SU88A1 |
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| Центробежный насос | 1925 |
|
SU3319A1 |
| ULF A.O | |||
| et al., Micro-RNA-10a binds the 5'UTR of ribosomal protein mRNAs and enhances their translation, MOL.CELL, 2008, v | |||
| Способ обработки медных солей нафтеновых кислот | 1923 |
|
SU30A1 |
| PARTRIDGE K | |||
| et al., Competition between the signal sequence and a 3'UTR localisation singnal during redirection of beta-globin mRNA tj the endoplasmic reticulum: implications fr biotechnology, 1999, CYTOTECH., v | |||
| Способ обработки медных солей нафтеновых кислот | 1923 |
|
SU30A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Пишущая машина | 1922 |
|
SU37A1 |
| LEDDA M | |||
| et al., Effect of 3'utr length on the translational regulation of 5'-terminal oligopyrimidine mRNAs, GENE, 2005, v | |||
| Способ получения жидкой протравы для основных красителей | 1923 |
|
SU344A1 |
| Кулиса для фотографических трансформаторов и увеличительных аппаратов | 1921 |
|
SU213A1 |
| ПАРОВАЯ ФОРСУНКА | 1927 |
|
SU10056A1 |

