Настоящее изобретение относится к последовательности нуклеиновых кислот, содержащей или кодирующей кодирующую область, кодирующую по меньшей мере один пептид или белок, содержащий опухолевый антиген или его фрагмент, вариант или производные по меньшей мере одну гистоновую структуру типа «стебель-петля» и поли(A)-последовательность или сигнал полиаденилирования. Кроме того, настоящее изобретение предоставляет применение нуклеиновой кислоты для увеличения экспрессии указанного кодируемого пептида или белка. Оно также раскрывает ее использование для подготовки фармацевтической композиции, особенно вакцины, например, для применения при лечении онкологических или опухолевых заболеваний. Настоящее изобретение далее описывает способ увеличения экспрессии пептида или белка, содержащего опухолевый антиген или его фрагмент, вариант или производное, с использованием нуклеиновой кислоты, содержащей или кодирующей гистоновую структуру типа «стебель-петля» и поли(A)-последовательность или сигнал полиаденилирования.
Помимо сердечно-сосудистых заболеваний и инфекционных заболеваний, возникновение опухолевых и онкологических заболеваний является одной из наиболее частых причин смерти в современном обществе и в большинстве случаев связано с большими затратами с точки зрения терапии и последующих реабилитационных мероприятий. Лечение опухолей и онкологических заболеваний в значительной степени зависит, например, от типа опухоли, которая имеет место, возраста, распространения раковых клеток у пациента, проходящего лечение, и др. Лечение рака в настоящее время обычно осуществляется путем применения лучевой терапии или химиотерапии, в дополнение к инвазивным операциям. Однако такие традиционные способы лечения, как правило, оказывают чрезвычайный стресс на иммунную систему и могут применяться в некоторых случаях лишь в ограниченной степени. Кроме того, большинство этих традиционных способов лечения требует длительных интервалов между отдельными процедурами лечения для обеспечения восстановления иммунной системы.
По этой причине, в дополнение к «традиционным способам лечения» в последние годы были исследованы дополнительные стратегии для того, чтобы избежать или хотя бы уменьшить воздействие таких способов лечения на иммунную систему. Один из таких дополнительных способов лечения, в частности, включает генно-терапевтические подходы или генетическую вакцинацию, которые уже оказались весьма перспективными для лечения или поддержки таких традиционных способов лечения.
Генная терапия и генетическая вакцинация представляют собой способы молекулярной медицины, которые уже доказали свою эффективность в терапии и профилактике заболеваний и, как правило, демонстрируют существенное влияние на повседневную медицинскую практику, в частности на лечение заболеваний, как упоминалось выше. Оба способа, генная терапия и генетическая вакцинация, основываются на введении нуклеиновых кислот в клетки или ткань пациента и последующей обработке информации, кодируемой нуклеиновой кислотой, которая была введена в клетки или ткань, то есть (белковой) экспрессии желательных полипептидов.
В подходах генной терапии, как правило, используется ДНК, хотя также известно об использовании РНК в последних разработках. Важно, что во всех этих подходах генной терапии мРНК функционирует в качестве мессенджера информации о последовательности кодируемого белка, независимо от того, используется ли ДНК, вирусная РНК или мРНК.
Вообще говоря, РНК считается нестабильной молекулой: РНКазы встречаются повсеместно, и их очень сложно инактивировать. Кроме того, РНК также является химически более лабильной, чем ДНК. Поэтому, наверное, является удивительным, что «состояние по умолчанию» мРНК в эукариотических клеток характеризуется относительной стабильностью и для ускорения распада отдельных мРНК требуются специальные сигналы. Основной причиной этого обнаружения, по-видимому, является то, что распад мРНК внутри клеток катализируется почти исключительно экзонуклеазами. При этом, концы эукариотических мРНК защищены от этих ферментов определенными концевыми структурами и ассоциированными с ними белками: m7GpppN CAP на 5'-конце и, как правило, поли(A)-последовательностью на 3'-конце. Удаление этих двух концевых модификаций, таким образом, рассматривается ограничивающим скорость распада мРНК. Несмотря на то, что стабилизирующий элемент был охарактеризован в 3' UTR мРНК альфа-глобина, последовательности РНК, влияющие на оборот эукариотических мРНК, обычно действуют в качестве катализатора распада, как правило, за счет ускорения деаденилирования (в качестве обзора, Meyer, S., C. Temme, et al. (2004), Crit Rev Biochem Mol Biol 39(4): 197-216.).
Как упоминалось выше, 5'-концы эукариотических мРНК, как правило, модифицируются посттранскрипционно, чтобы нести метилированную CAP-структуру, например, m7GpppN. Помимо ролей в сплайсинге, стабилизации и транспорте РНК, CAP-структура значительно увеличивает привлечение 40S-рибосомных субъединиц к 5'-концу мРНК во время инициации трансляции. Для последней функции требуется распознавание CAP-структуры комплексом эукариотического фактора инициации eIF4F. Поли(A)-последовательность дополнительно стимулирует трансляцию через усиление привлечения 40S-субъединиц к мРНК, эффект, который требует вмешательства поли(A)-связывающего белка (PABP). PABP, в свою очередь, как недавно был продемонстрировано, физически взаимодействует с eIF4G, который является частью CAP-связывающего eIF4F-комплекса. Соответственно, была постулирована модель замкнутой петли инициации трансляции на кэпированной, полиаденилированной мРНК (Michel, Y. M., D. Poncet, et al. (2000), J Biol Chem 275(41): 32268-76.).
Почти все эукариотические мРНК заканчиваются такой поли(A)-последовательностью, которая добавляется к 3'-концу вездесущим аппаратом расщепления/полиаденилирования. Присутствие поли(A)-последовательности на 3'-конце является одной из самых узнаваемых особенностей эукариотических мРНК. После расщепления большинство пре-мРНК, за исключением зависящих от репликации транскриптов гистонов, приобретают полиаденилированный хвост. В этом контексте, 3'-концевой процессинг представляет собой ядерный котранскрипционный процесс, который способствует транспорту мРНК из ядра в цитоплазму и влияет на стабильность и трансляцию мРНК. Формирование этого 3'-конца происходит за две стадии реакции, направляемой аппаратом расщепления/полиаденилирования, и зависит от наличия двух элементов последовательности в предшественниках мРНК (пре-мРНК): высококонсервативного гексануклеотида AAUAAA (сигнал полиаденилирования) и расположенной ниже по течению C/U-богатой последовательности. На первой стадии, пре-мРНК расщепляется между этими двумя элементами. На второй стадии, тесно связанной с первой стадией, вновь образованной 3'-конец удлиняется путем добавления поли(A)-последовательности, состоящей из 200-250 аденилатов, что впоследствии оказывает влияние на все аспекты метаболизма мРНК, включая экспорт, стабильность и трансляцию мРНК (Dominski, Z. and W. F. Marzluff (2007), Gene 396(2): 373-90.).
Единственным известным исключением из этого правила являются репликацие-зависимые гистоновые мРНК, которые заканчиваются гистоновыми структурами типа «стержень-петля», а не поли(A)-последовательностью. Типичные последовательности гистоновых структур типа «стебель-петля» описаны в Lopez et al. (Davila Lopez, M., & Samuelsson, T. (2008), RNA (New York, N.Y.), 14(1), 1-10. doi:10.1261/rna.782308.).
Структуры типа «стебель-петля» в пре-мРНК гистонов, как правило, следуют за пурин-богатой последовательностью, известной как гистоновый расположенный вниз по течению элемент (HDE). Эти пре-мРНК процессируются в ядре при помощи одного эндонуклеотического расщепления примерно 5 нуклеотидами ниже по течению структуры типа «стебель-петля», катализируемого U7 snRNP через спаривание оснований U7 snRNA с HDE. Последовательность 3'-UTR, включающую гистоновую структуру типа «стебель-петля», и расположенный вниз по течению элемент гистонов (HDE) (сайт связывания U7 snRNP) обычно называют гистоновым сигналом 3'-процессинга (см., например, Chodchoy, N., N. B. Pandey, et al. (1991). Mol Cell Biol 11(1): 497-509.).
Из-за необходимости упаковки вновь синтезированной ДНК в хроматин синтез гистонов регулируется согласовано с клеточным циклом. Увеличенный синтез гистоновых белков в процессе S-фазы достигается за счет активации транскрипции гистоновых генов, а также посттранскрипционной регуляции уровней гистоновых мРНК. Можно показать, что гистоновая структура типа «стебель-петля» имеет важное значение для всех посттранскрипционных стадий регуляции экспрессии гистонов. Это необходимо для эффективного процессинга, экспорта мРНК в цитоплазму, посадки на полирибосомы и регулирования стабильности мРНК.
В контексте вышесказанного, был идентифицирован 32 кДа белок, который ассоциирован с гистоновой структурой типа «стебель-петля» на 3'-конце гистоновых информационных РНК как в ядре, так и цитоплазме. Уровень экспрессии этого связывающегося со структурой типа «стебель-петля» белка (SLBP) регулируется клеточным циклом и является наибольшим в S-фазе, когда уровни гистоновых мРНК повышаются. SLBP необходим для эффективного 3'-концевого процессинга гистоновых пре-мРНК при помощи U7 snRNP. После завершения процессинга SLBP остается связанным со структурой типа «стебель-петля» на конце зрелых гистоновых мРНК и стимулирует их трансляцию в гистоновые белки в цитоплазме (Dominski, Z. and W. F. Marzluff (2007), Gene 396(2): 373-90). Интересно, что РНК-связывающий домен SLBP является консервативным у метазоа и простейших (Davila Lopez, M., & Samuelsson, T. (2008), RNA (New York, N.Y.), 14(1), 1-10. doi:10.1261/rna.782308), и может быть продемонстрировано, что его связывание с последовательностью гистоновой структуры типа «стебель-петля» зависит от устройства структуры типа «стебель-петля» и минимальный сайт связывания содержит по меньшей мере 3 нуклеотида с 5'- и 2 нуклеотида 3'-стороны структуры типа «стебель-петля» (Pandey, N. B., et al. (1994), Molecular and Cellular Biology, 14(3), 1709-1720 и Williams, A. S., & Marzluff, W. F., (1995), Nucleic Acids Research, 23(4), 654-662.).
Хотя гены гистонов, как правило, классифицируются как «репликацие-зависимые», дающие начало мРНК, которая заканчивается гистоновой структурой типа «стебель-петля», или «замещенного типа», дающие начало мРНК, несущей вместо этого поли-(A)-хвост, в некоторых очень редких случаях были идентифицированы природные мРНК, содержащие как гистоновую структуру типа «стебель-петля», так и поли(A) или олиго(A) на своем 3'-конце. Sanchez et al. исследовали влияние природных олиго(A)-хвостов, добавленных к 3'-концу гистоновый структуры типа «стебель-петля» гистоновой мРНК во время оогенеза Xenopus, с использованием люциферазы в качестве репортерного белка и обнаружили, что олиго(A)-хвост представляет собой активную часть механизма репрессии трансляции, который сайленсирует гистоновую мРНК во время оогенеза, и его удаление является частью механизма, который активирует трансляцию гистоновых мРНК (Sanchez, R. and W. F. Marzluff (2004), Mol Cell Biol 24(6): 2513-25).
Кроме того, требования регулирования репликацие-зависимых гистонов на уровне процессинга пре-мРНК и стабильности мРНК были исследованы при помощи искусственных конструкций, кодирующих маркерный белок альфа-глобин, пользуясь тем, что ген глобина содержит интроны, в отличие от интрон-несодержащих генов гистонов. Для этой цели были созданы конструкции, в которых за последовательностью, кодирующей альфа-глобин, следовал сигнал гистоновой структуры типа «стебель-петля» (гистоновая структура типа «стебель-петля» с последующим гистоновым расположенным вниз по течению элементом) и сигналом полиаденилирования (Whitelaw, E., et al. (1986). Nucleic Acids Research, 14(17), 7059-7070.; Pandey, N. B., & Marzluff, W. F. (1987). Molecular and Cellular Biology, 7(12), 4557-4559.; Pandey, N. B., et al. (1990). Nucleic Acids Research, 18(11), 3161 - 3170).
В другом подходе Lüscher et al. исследовали зависимую от клеточного типа регуляцию рекомбинантного гена гистона H4. Были созданы конструкции, в которых за кодирующей последовательностью H4 следовал сигнал гистоновой структуры типа «стебель-петля» и сигнал полиаденилирования, два сигналами процессинга, случайно разделенных кодирующей последовательностью галактокиназы (Lüscher, B. et al., (1985). Proc. Natl. Acad. Sci. USA, 82(13), 4389-4393).
Кроме того, Stauber et al. определили минимальную последовательность, необходимую для осуществления регуляции клеточного цикла на уровне мРНК гистона H4. Для этих исследований были использованы конструкции, включающие кодирующую последовательность для селективного маркера ксантин:гуанинфосфорибозилтрансферазы (GPT), предшествующую сигналу гистоновой структуры типа «стебель-петля», с последующим сигналом полиаденилирования (Stauber, C. et al., (1986). EMBO J, 5(12), 3297-3303).
При исследовании процессинга гистоновой пре-мРНК Wagner et al. выявили факторы, необходимые для расщепления гистоновых пре-мРНК, с помощью репортерной конструкции, в которой EGFP помещен между сигналом гистоновой структуры типа «стебель-петля» и сигналом полиаденилирования, так, что EGFP экспрессировался лишь в случае нарушения процессинга гистоновой пре-мРНК (Wagner, E. J. et al., (2007). Mol Cell 28(4), 692-9).
Следует отметить, что для трансляции полиаденилированной мРНК, как правило, требуется сближение 3' поли(A)- последовательности с 5' CAP. Это опосредуется через белок-белковое взаимодействие между поли(A)-связывающим белком и эукариотическим фактором инициации eIF4G. В отношении репликацие-зависимых гистоновых мРНК, аналогичный механизм не был раскрыт. В этом контексте, Gallie et al. показывают, что гистоновая структура типа «стебель-петля» функционально аналогична поли-(A)-последовательности в том, что она повышает эффективность трансляции и является со-зависимымой от 5'-CAP для установления эффективного уровня трансляции. Они показали, что гистоновая структура типа «стебель-петля» является достаточной и необходимой для увеличения трансляции репортерной мРНК в трансфицированных клетках яичника китайского хомячка, но должна располагаться на 3'-конце, чтобы оптимально функционировать. Поэтому, аналогично поли(А)-хвосту в других мРНК, 3'-конец этих гистоновых мРНК, по-видимому, необходимым для трансляции in vivo и функционально аналогичен поли(А)-хвосту (Gallie, D. R., Lewis, N. J., & Marzluff, W. F. (1 996), Nucleic Acids Research, 24(10), 1954-1962).
Кроме того, можно показать, что SLBP связан с цитоплазматической гистоновой мРНК и необходим для ее трансляции. Хотя SLBP не взаимодействует непосредственно с eIF4G, домен, необходимый для трансляции гистоновой мРНК, взаимодействует с недавно идентифицированным белком SLIP1. На следующей стадии, SLIP1 взаимодействует с eIF4G и обеспечивает циклизацию гистоновой мРНК и поддерживает эффективную трансляцию гистоновой мРНК с помощью механизма, аналогичного трансляции полиаденилированных мРНК.
Как упоминалось выше, в подходах генной терапии, как правило, используется ДНК для передачи закодированной информации в клетку, которая затем транскрибируется в мРНК, несущую естественные элементы мРНК, в частности, структуру 5'-CAP и 3' поли(A)-последовательность для обеспечения экспрессии кодируемого терапевтического или антигенного белка.
Однако во многих случаях система экспрессии, основанная на введении таких нуклеиновых кислот в клетки или ткань пациента и последующей экспрессии нужных полипептидов, кодируемых этими нуклеиновыми кислотами, не проявляет желаемого или даже необходимого уровня экспрессии, который может обеспечить эффективную терапию, независимо от того, используется ли ДНК или РНК.
В предшествующем уровне техники до сих пор предпринимались различные попытки увеличения уровня экспрессии кодируемого белка, в частности, путем применения улучшенных систем экспрессии, как in vitro, так и/или in vivo. Способы увеличения экспрессии, как правило, описываемые в предшествующем уровне техники, традиционно основываются на применении экспрессионных векторов или кассет, содержащих определенные промоторы и соответствующие регуляторные клетки. Поскольку эти экспрессионные векторы или кассеты обычно ограничиваются конкретными клеточными системами, эти системы экспрессии должны быть адаптированы для применения в различных клеточных системах. Такие адаптированные экспрессионные векторы или кассеты затем, как правило, трансфицируют в клетки и обычно обрабатывают в зависимости от конкретной клеточной линии. Таким образом, предпочтение отдается в первую очередь тем молекулам нуклеиновых кислот, которые способны экспрессировать кодируемые белки в клетке-мишени при помощи систем, внутренне присущих клетке, независимых от промоторов и регуляторных элементов, которые специфичны для отдельных типов клеток. В этом контексте можно проводить различие между стабилизирующими элементами мРНК и элементами, которые увеличивают эффективность трансляции мРНК.
мРНК, которые оптимизированы по своей кодирующей последовательности и которые, в основном, подходят для таких целей, описаны в заявке WO 02/098443 (CureVac GmbH). Например, WO 02/098443 описывает мРНК, которые стабилизируются в общем виде и оптимизированы для трансляции своих кодирующих областей. WO 02/098443 далее раскрывает способ определения модификаций последовательностей. WO 02/098443 дополнительно описывают возможности для замены нуклеотидов аденина и урацила в последовательностях мРНК в целях увеличения содержания гуанина/цитозина (G/C) последовательностей. В соответствии с WO 02/098443, такие замены и адаптации для увеличения G/C-содержания могут использоваться для генно-терапевтических применений, а также для генных вакцин при лечении онкологических или инфекционных заболеваний. В этом контексте, WO 02/098443, как правило, упоминает последовательности в качестве базовой последовательности для таких модификаций, в которой модифицированная мРНК кодирует по меньшей мере один биологически активный пептид или полипептид, который транслируется в проходящем лечение пациенте, например, либо не всех, либо неадекватно, либо с дефектами. Кроме того, WO 02/098443 предлагает мРНК, кодирующие антигены, например, опухолевые антигены или вирусных антигены в качестве базовых последовательностей для таких модификаций.
В другом подходе, для увеличения экспрессии кодируемого белка приложение WO 2007/036366 описывает положительный эффект длинных поли(A)-последовательностей (в частности, длиной более 120 п.н.) и комбинации по меньшей мере двух 3'-нетранслируемых областей бета-глобина на стабильность и трансляционную активность мРНК.
Однако, несмотря на то, что все эти недавние документы рассматриваемой области техники уже пытаются обеспечить достаточно эффективные инструменты для подходов генной терапии и дополнительно улучшают стабильность и трансляционную активность мРНК, по-прежнему остается проблема пониженной стабильности приложений на основе РНК против ДНК-вакцин и генно-терапевтических подходов на основе ДНК. Соответственно, в рассматриваемой области по-прежнему существует потребность в предоставлении улучшенных инструментов для подходов генной терапии и генной вакцинации, или таковых в качестве дополнительной терапии для традиционных способов лечения, как описано выше, которые позволяют лучше обеспечивать кодируемыми белками in vivo, например, путем дополнительного увеличения стабильности и/или трансляционной активности мРНК, предпочтительно для генной терапии и генной вакцинации.
Более того, несмотря на все успехи в рассматриваемой области, эффективная экспрессия кодируемого пептида или белка в бесклеточных системах, клетках или организмах (рекомбинантная экспрессия) до сих пор остается сложной проблемой.
Задача, лежащая в основе настоящего изобретения, представляет собой, следовательно, предоставление дополнительных и/или альтернативных способов увеличения экспрессии кодируемого белка, предпочтительно путем дополнительной стабилизации мРНК и/или увеличения эффективности трансляции такой мРНК по отношению к таким нуклеиновым кислотам, известным из предшествующего уровня техники, для применения в генной вакцинации при лечебном или профилактическом лечении онкологических или опухолевых заболеваний.
Эта задача решается содержанием прилагаемых пунктов формулы изобретения. В частности, задача, лежащая в основе настоящего изобретения, решается в соответствии с первым аспектом при помощи последовательности нуклеиновых кислот по изобретению, содержащую или кодирующую
a) кодирующую область, кодирующую по меньшей мере один пептид или белок, который содержит опухолевый антиген или его фрагмент, вариант или производное;
b) по меньшей мере, одну гистоновую структуру типа «стебель-петля» и
c) поли-(A)-последовательность или сигнал полиаденилирования,
предпочтительно для увеличения экспрессии указанного кодируемого пептида или белка.
Кроме того, любая соответствующая последовательность типа «стебель-петля», отличная от гистоновой последовательности типа «стебель-петля» (полученная из генов гистонов, в частности, генов гистонов семейств H1, H2A, H2B, H3 и H4)), может быть использована в настоящем изобретении во всех его аспектах и вариантах осуществления.
В этом контексте особенно предпочтительно, что нуклеиновую кислоту по изобретению в соответствии с первым аспектом изобретения продуцируют по меньшей мере частично путем синтеза ДНК или РНК, предпочтительно, как описано в настоящем документе, или она представляет собой выделенную нуклеиновую кислоту.
Настоящее изобретение основано на удивительном открытии авторов настоящего изобретения, что комбинация поли(A)-последовательности или сигнала полиаденилирования и по меньшей мере одной гистоновой структуры типа «стебель-петля», несмотря на то, что каждое из этого представляет альтернативные природные механизмы, действует синергически, поскольку эта комбинация увеличивает белковую экспрессию, многократно превышающую уровень, наблюдаемый с каждым из элементов по-отдельности. Синергетический эффект комбинации поли(A) и по меньшей мере одной гистоновой структуры типа «стебель-петля» наблюдается независимым от порядка поли(A) и гистоновой структуры типа «стебель-петля» и независимым от длины поли(A)-последовательности.
Поэтому особенно предпочтительно, что последовательность нуклеиновых кислот по изобретению включает или кодирует а) кодирующую область, кодирующую по меньшей мере один пептид или белок, который содержит опухолевый антиген или его фрагмент, вариант или производное; b) по меньшей мере, одну гистоновую структуру типа «стебель-петля» и c) поли(A)-последовательность или последовательность полиаденилирования; предпочтительно для увеличения уровня экспрессии указанного кодируемого пептида или белка. В некоторых вариантах осуществления может быть предпочтительно, если кодируемый белок не является гистоновым белком, в частности, отличным от гистонового белка из семейства гистонов H4, H3, Н2А и/или H2B или его фрагмента, производного или варианта, сохраняющего функцию гистона (гистоно-подобную), а именно формирование нуклеосомы. Кроме того, кодируемый белок, как правило, не соответствует гистоновому линкерному белку семейства H1-гистона. Молекула нуклеиновой кислоты по изобретению, как правило, не содержат каких-либо регуляторных сигналов (5'- и/или, в частности, 3'- из гистонового гена мыши, в частности, не содержит гена гистона Н2А мыши и, далее, наиболее предпочтительно, не содержит гена гистона H2A614 мыши. В частности, она не содержит гистоновой структуры типа «стебель-петля» и/или сигнала процессинга гистоновой структуры типа «стебель-петля» гена гистона мыши, в частности, гена гистона Н2А мыши и, наиболее предпочтительно, гена гистона H2A614 мыши.
Также, нуклеиновая кислота по изобретению, как правило, не предоставляет репортерный белок (например, люциферазу, GFP, EGFP, бета-гактозидазу, особенно EGFP) галактокиназу (galK) и/или маркерный или селективный белок (например, альфа-глобин, галактокиназу и ксантин-гуанинфосфорибозилтрансферазу (GPT)) или бактериальной репортерный белок, например, хлорамфениколацетил-трансферазу (CAT) или другие бактериальные белки устойчивости к антибиотикам, например, полученные с бактериальным геном nео в своем элементе(a).
Не подразумевается, что репортерный, маркерный или селективный белок, как правило, является опухолевым антигеном, в соответствии с изобретением. Репортерный, маркерный или селективный белок или лежащих в его основе ген обычно используется как инструмент для исследования микроорганизмов, клеточных культур, животных или растений. Они придают экспрессирующим их организмам (предпочтительно гетерологичным) легко идентифицируемую особенность, которая может определяться или которая позволяет осуществить селекцию. В частности, маркерный или селективный белок проявляет селектируемые функции. Как правило, такие селективные, маркерные или репортерные белки природно не встречаются в организме человека или других млекопитающих, но происходят из других организмов, в частности, бактерий и растений. Соответственно, белки с селективной, маркерной или репортерной функцией, происходящих из видов, отличных от видов млекопитающих, в частности, отличных от человека, предпочтительно исключают из рассмотрения в качестве белка, обладающего свойством действовать в качестве «опухолевого антигена», в соответствии с настоящим изобретением. В частности, селективный, маркерный или репортерный белок позволяет идентифицировать трансформированные клетки при помощи тестов in vitro, основанных, например, на флуоресцентных или других спектроскопических способах и устойчивости к антибиотикам. Селективные, маркерные или репортерные гены, придающие такие свойства трансформированным клеткам, поэтому обычно не рассматриваются в качестве белка, действующего в качестве опухолевого антигена, в соответствии с изобретением, in vivo.
В любом случае, репортерные, маркерные или селективные белки, как правило, не проявляют каких-либо опухолевых антигенных свойств и, следовательно, не оказывают иммунологического эффекта, который терапевтически позволяет лечить опухолевые заболевания. Если какой-либо отдельный репортерный, маркерный или селективный белок, тем не менее, будет так действовать (в дополнение к его репортерной, маркерной или селективной функции), такой репортерный, маркерный или селективный белок предпочтительно не рассматривается в качестве «опухолевого антигена» по смыслу настоящего изобретения.
В отличие от этого, белок или пептид, действующий в качестве опухолевого антигена (в частности, за исключением генов гистоновых семейств H1 (H2A, H2B, H3 и H4)) в соответствии с настоящим изобретением, как правило, не проявляет селективную, маркерную или репортерную функцию. Если какой-либо отдельный «опухолевый антиген», тем не менее, будет так действовать (в дополнение к его функции опухолевого антигена), то такой опухолевый антиген предпочтительно не рассматривается в качестве «селективного, маркерного или репортерного белка» по смыслу настоящего изобретения.
Наиболее предпочтительно рассматривается, что белок, действующий как опухолевый антиген в соответствии с изобретением, получен из млекопитающих, в частности людей, в частности, из опухолей млекопитающих, и не квалифицируется в качестве селективного, маркерного или репортерного белка. В частности, такие опухолевые антигены получают из опухолей млекопитающих, в частности, из опухолей человека. Эти опухолевые антигенные белки понимаются как антигенные, поскольку они предназначены для лечения индивидуума путем инициирования иммунный ответ у индивидуума так, что иммунная система индивидуума получает возможность бороться с опухолевыми клетками индивидуума при помощи иммунных реакций TH1 и/или TH2. Соответственно, такие антигенные опухолевые белки обычно представляют собой белки млекопитающих, в частности, белки человека, характеризующиеся типом рака индивидуума.
Соответственно, предпочтительно, что кодирующая область, а) кодирующая по меньшей мере один пептид или белок, гетерологична по меньшей мере (b) по меньшей мере, одной гистоновой структуре типа «стебель-петля», или в более широком смысле, любой соответствующей структуре типа «стебель-петля». Иными словами, «гетерологичный» в контексте настоящего изобретения означает, что по меньшей мере одна последовательность типа «стебель-петля» не встречается в природе как (регуляторная) последовательность (например, в 3'UTR) определенного гена, который кодирует (опухолевый антигенный) белок или пептид элемента (а) нуклеиновой кислоты по изобретению. Соответственно, (гистоновая) структура типа «стебель-петля» нуклеиновой кислоты по изобретению получена предпочтительно из 3' UTR гена, отличного от гена, включающего кодирующую область элемента (a) нуклеиновой кислоты по изобретению. Например, кодирующая область элемента (a) не должна кодировать гистоновый белок или его фрагмент, вариант или производное (при сохранении функции гистонового белка), если нуклеиновая кислота по изобретению является гетерологичной, но должна кодировать любой другой пептид или последовательность (того же или другого вида), который проявляет биологическую функцию, предпочтительно функцию опухолевого антигена, отличную от гистоново(-подобной) функции, например, должна кодировать белок (проявляющий функцию опухолевого антигена, например, в плане вакцинации против опухолей млекопитающих, в частности, опухолей человека), инициируя тем самым иммунологическую реакцию в отношении опухолевых клеток индивидуума, которые предпочтительно экспрессируют опухолевый антиген, кодируемый нуклеиновой кислотой по изобретению.
В этом контексте особенно предпочтительно, чтобы нуклеиновая кислота по изобретению включала или кодировала в направлении от 5' к 3':
a) кодирующую область, кодирующую по меньшей мере один пептид или белок, который содержит опухолевый антиген или его фрагмент, вариант или производное;
b) по меньшей мере, одну гистоновую структуру типа «стебель-петля», необязательно без гистонового расположенного ниже по течению элемента (HDE), 3' к «гистоновой структуре типа «стебель-петля»
c) поли(A)-последовательность или сигнал полиаденилирования.
Термин «гистоновый расположенный ниже по течению элемент (HDE)» относится к пурин-богатому полинуклеотидному участку примерно от 15 до 20 нуклеотидов, 3' к природным гистоновым структурам типа «стебель-петля», который представляет сайт связывания с U7 snRNA, вовлеченную в процессирование гистоновой пре-мРНК в зрелую гистоновую мРНК. Например, у морских ежей HDE представляет собой CAAGAAAGA (Dominski, Z. and W. F. Marzluff (2007), Gene 396(2): 373-90).
Кроме того, предпочтительно, чтобы нуклеиновая кислота по изобретению в соответствии с первым аспектом настоящего изобретения не содержала интрон.
В другом, особенно предпочтительном варианте осуществления, последовательность нуклеиновых кислот по изобретению в соответствии с первым аспектом настоящего изобретения содержит или кодирует в направлении от 5' к 3':
a) кодирующую область, предпочтительно кодирующую по меньшей мере один пептид или белок, который содержит опухолевый антиген или его фрагмент, вариант или производное;
c) поли(A)-последовательность; и
b) по меньшей мере, одну гистоновую структуру типа «стебель-петля».
Последовательность нуклеиновых кислот по изобретению, в соответствии с первым вариантом осуществления по настоящему изобретению, включает любую подходящую нуклеиновую кислоту, выбранную, например, из любой (одноцепочечной или двуцепочечной) ДНК, предпочтительно, без ограничения этим, например, геномной ДНК, плазмидной ДНК, молекул одноцепочечной ДНК, молекул двухцепочечной ДНК, или она может быть выбрана, например, из любой PNA (пептид-нуклеиновой кислоты), или она может быть выбрана, например, из любой (одноцепочечной или двуцепочечной) РНК, предпочтительно матричной РНК (мРНК); и др. Последовательность нуклеиновых кислот по изобретению может также включать вирусную РНК (вРНК). При этом последовательность нуклеиновых кислот по изобретению может не быть вирусной РНК или может не содержать вирусной РНК. Конкретнее, последовательность нуклеиновых кислот по изобретению может не содержать элементов вирусной последовательности, например, вирусных энхансеров или вирусных промоторов (например, отсутствие инактивированных вирусных промоторных элементов или элементов последовательности, конкретнее, не инактивированных при помощи стратегий замещения), или других элементов вирусной последовательности, или вирусных или ретровирусных последовательностей нуклеиновых кислот. Точнее говоря, последовательность нуклеиновых кислот по изобретению может не являться антиретровирусным или вирусным вектором, или модифицированным антиретровирусным или вирусным вектором.
В любом случае, последовательность нуклеиновых кислот по изобретению может содержать или может не содержать энхансерную и/или промоторную последовательность, которая может быть или может не быть модифицирована или которая может быть или может не быть активирована. Энхансер или промотор может экспрессироваться или может не экспрессироваться в растении, и/или может экспрессироваться или может не экспрессироваться в эукариотах, или может экспрессироваться или может не экспрессироваться в прокариотах. Последовательность нуклеиновых кислот по изобретению может содержать или может не содержать последовательность, кодирующую (самосплайсирующийся) рибозим.
В конкретных вариантах осуществления последовательность нуклеиновых кислот по изобретению может содержать или может не содержать самореплицирующуюся РНК (репликон).
Предпочтительно, последовательность нуклеиновых кислот по изобретению представляет собой плазмидную ДНК или РНК, в частности мРНК.
В конкретном варианте осуществления первого аспекта по настоящему изобретению, нуклеиновая кислота по изобретению представляет собой последовательность нуклеиновых кислот, содержащую нуклеиновую кислоту, подходящую для транскрипции in vitro, особенно в соответствующем векторе для транскрипции in vitro (например, плазмиде или линейной последовательности нуклеиновых кислот, содержащей специфические промоторы для транскрипции in vitro, например, промоторы T3, Т7 или Sp6).
В дополнительных конкретных предпочтительных вариантах осуществления первого аспекта настоящего изобретения, нуклеиновая кислота по изобретению включена в нуклеиновую кислоту, подходящую для транскрипции и/или трансляции в экспрессионной системе (например, в экспрессионном векторе или плазмиде), в частности, в прокариотической (например, бактерии наподобие E.coli) или эукариотической (например, клетки млекопитающих, наподобие клеток СНО, дрожжевых клеток или клеток насекомых или целых организмов, наподобие растений или животных) экспрессионной системе.
Термин «экспрессионная система» означает систему (культуру клеток или целые организмы), которая подходит для продуцирования пептидов, белков и РНК, особенно мРНК (рекомбинантная экспрессия).
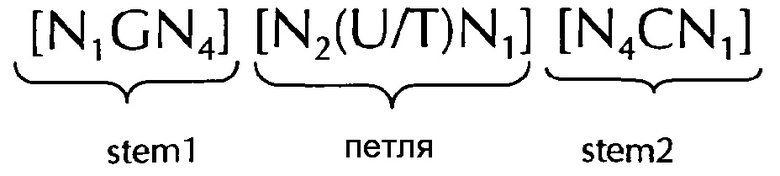
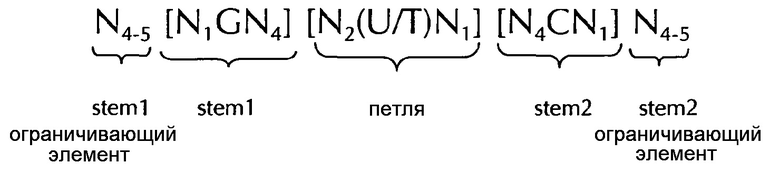
Последовательность нуклеиновых кислот по изобретению в соответствии с первым аспектом настоящего изобретения содержит или кодирует по меньшей мере одну гистоновую структуру типа «стебель-петля». В контексте настоящего изобретения, такая гистоновая структура типа «стебель-петля», в целом (независимо от того, является ли она гистоновой структурой типа «стебель-петля» или нет), как правило, получена из генов гистонов и включает внутримолекулярное спаривание оснований двух соседних полностью или частично обратно комплементарных последовательностей, образующих таким образом структуру типа «стебель-петля». Структура типа «стебель-петля» может иметь место в одноцепочечной ДНК, или, чаще, в РНК. Структура также известна как шпилька или шпилька-петля и, как правило, состоит из стебля и (терминальной) петли в пределах идущих подряд последовательностей, разделенных короткой последовательностью в виде спейсера, который образует петлю структуры типа «стебель-петля». Две соседние полностью или частично обратно комплементарные последовательности могут быть определены как, например, элементы структуры типа «стебель-петля» stem 1 и stem 2. Структура типа «стебель-петля» образуется, когда эти две соседние полностью или частично обратно комплементарные последовательности, например, элементы структуры типа «стебель-петля» stem 1 и stem 2, образуют пары оснований друг с другом, приводя к образованию участка двуцепочечной последовательности нуклеиновых кислот, содержащего неспаренную петлю на своем терминальном окончании, образованном короткой последовательностью, расположенной между элементами структуры типа «стебель-петля» stem 1 и stem 2 на консекутивной последовательности. Неспаренная петля, тем самым, как правило, представляет область нуклеиновой кислоты, которая не способна к образованию пар ни с одним из этих элементов структуры типа «стебель-петля». Образованная в результате структура, похожая на леденец на палочке, является ключевым строительным элементом многих вторичных структур РНК. Формирование структуры типа «стебель-петля», таким образом, зависит от стабильности полученных в результате областей стебля и петли, где первым условием, как правило, является наличие последовательности, которая может сворачиваться обратно на себя, чтобы образовывать спаренную двойную нить. Стабильность элементов спаренной двойной нити определяется длиной, количеством несовпадений или выпучиваний, которые она содержит (небольшое количество несовпадений обычно приемлемо, особенно в длинном двуцепочечном участке), и композицией оснований спаренной области. В контексте настоящего изобретения, возможной является петля длиной от 3 до 15 оснований, хотя более предпочтительная длина петли составляет от 3 до 10 оснований, более предпочтительно, от 3 до 8, от 3 до 7, от 3 до 6 или даже более предпочтительно, от 4 до 5 оснований, и наиболее предпочтительно, 4 основания. Последовательность стебля, образуемая двуцепочечной структурой, обычно составляет от 5 до 10 оснований в длину, более предпочтительно, от 5 до 8 оснований.
В контексте настоящего изобретения, гистоновую структуру типа «стебель-петля» получают, как правило, из гистоновых генов (например, гены гистоновых семейств H2A, H2B, H3, H4), и она содержит внутримолекулярный участок спаренных оснований двух соседних полностью или частично обратно комплементарных последовательностей, образующих таким образом структуру типа «стебель-петля». Как правило, гистоновая 3' UTR структура типа «стебель-петля» представляет собой РНК-элемент, участвующий в ядерно-цитоплазматическом транспорте гистоновых мРНК и в регуляции стабильности и эффективности трансляции в цитоплазме. В мРНК гистоновых генов метазоа отсутствует полиаденилирование и поли-А хвост, вместо этого 3'-концевой процессинг происходит в участке между этой высококонсервативной структурой типа «стебель-петля» и пурин-богатой областью примерно 20 нуклеотидами ниже по течению (гистоновый расположенный вниз по течению элемент, или HDE). Гистоновая структура типа «стебель-петля» связана с 31 кДа белком, связывающимся со структурой типа «стебель-петля» (SLBP - также называется связывающимся с гистоновой шпилькой белком, или НВР). Такие гистоновые структуры типа «стебель-петля» преимущественно используются в настоящем изобретении в комбинации с другими элементами последовательности и структурами, которые не встречаются природно (что означает: в нетрансформированных живых организмах/клетках) в гистоновых генах, но комбинируются, в соответствии с изобретением, для предоставления искусственной, гетерологичной нуклеиновой кислоты. Соответственно, настоящее изобретение, в частности, основывается на обнаружении о том, что искусственная (не природная) комбинация гистоновой структуры типа «стебель-петля» с другими гетерологичными элементами последовательности, которые не встречаются в гистоновых генах или гистоновых генах метазоа и выделены из областей операционных и/или регуляторных последовательностей (влияющих на транскрипцию и/или трансляцию) генов, кодирующих белки, отличных от гистонов, дает положительные эффекты. Соответственно, один из аспектов изобретения предоставляет комбинацию гистоновой структуры типа «стебель-петля» с поли(A)-последовательностью или последовательностью, представляющей сигнал полиаденилирования (3'-окончание кодирующей области), которая не присутствует в гистоновых генах простейших. В соответствии с другим предпочтительным аспектом изобретения, в настоящем документе предоставляется комбинация гистоновой структуры типа «стебель-петля» с кодирующей областью, кодирующей опухолевый антигенный белок, которая, предпочтительно, не присутствует в гистоновых генах простейших (кодирующая область и гистоновая последовательность структуры типа «стебель-петля» являются гетерологичными). Предпочтительно, если такие опухолевые антигенные белки присутствуют в клетках млекопитающих, предпочтительно людей, страдающих от опухолевых заболеваний. В еще более предпочтительном варианте осуществления, все элементы (a), (b) и (c) нуклеиновой кислоты по изобретению являются гетерологичными друг по отношению к другу и комбинируются искусственно из трех различных источников, например, (a) кодирующая область опухолевого антигенного белка гена человека, (b) гистоновая структура типа «стебель-петля» нетранслируемой области гистонового гена метазоа, например, млекопитающих, отличных от человека или человека, и (c) поли(A) последовательности или сигнала полиаденилирования, например, нетранслируемой области гена, отличного от гистонового гена и отличного от нетранслируемой области кодирующей области опухолевого антигена в соответствии с элементом (а) такой нуклеиновой кислоты по изобретению.
Поэтому гистоновая структура типа «стебель-петля» представляет собой структуру типа «стебель-петля», как описано в настоящем документе, которая, если, предпочтительно, функционально определено, проявляет/сохраняет свойство связывания со своим природным партнером по связыванию, белком, связывающимся со структурой типа «стебель-петля» (SLBP - также называемый связывающимся с гистоновой шпилькой белком, или НВР).
В соответствии с настоящим изобретением, последовательность гистоновый структура типа «стебель-петля» в соответствии с компонентом (b) пункта 1, может не быть получена из гена гистонового белка мыши. Конкретнее, последовательность гистоновой структуры типа «стебель-петля» не может быть получена из гистонового гена Н2А614 мыши. Кроме того, нуклеиновая кислота по изобретению не может ни содержать последовательность гистоновой структуры типа «стебель-петля» мыши, ни содержать гистонового гена H2A614 мыши. Далее, последовательность нуклеиновых кислот по изобретению может не содержать сигнал процессинга структуры типа «стебель-петля», конкретнее, сигнал процессинга гистона мыши и, в частности, не может содержать сигнал процессинга структуры типа «стебель-петля» H2kA614 мыши, даже если последовательность нуклеиновых кислот по изобретению может содержать по меньшей мере один гистоновый ген млекопитающего. При этом по меньшей мере один гистоновый ген млекопитающего может не представлять собой SEQ ID NO: 7 из WO 01/12824.
В соответствии с одним предпочтительным вариантом осуществления первого аспекта по изобретению, последовательность нуклеиновых кислот по изобретению включает или кодирует по меньшей мере одну последовательность гистоновой структуры типа «стебель-петля», предпочтительно в соответствии с по меньшей мере одной из следующих формул (I) или (II):
формула (I) (последовательность типа «стебель-петля» без ограничивающих элементов стебля):
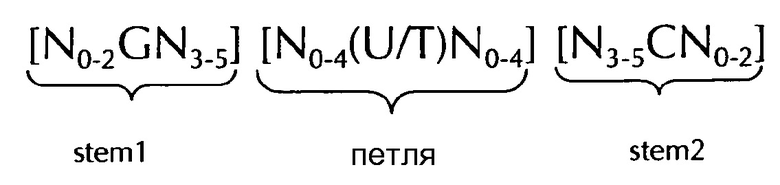
формула (II) (последовательность типа «стебель-петля» с ограничивающими элементами стебля):
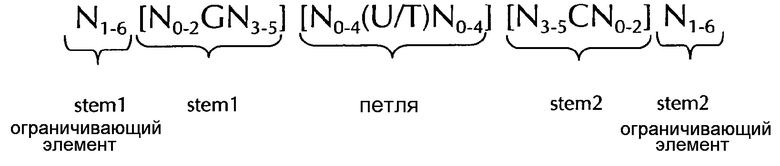
где
stem1- или stem2- ограничивающие элементы N1-6 представляет собой консекутивную последовательность от 1 до 6, предпочтительно от 2 до 6, более предпочтительно от 2 до 5, еще более предпочтительно, от 3 до 5, наиболее предпочтительно, от 4 до 5 или 5 N, где каждый N является независимо от другого выбранным из нуклеотида, выбранного из A, U, T, G и С или нуклеотидного аналога этого;
stem1 [N0-2GN3-5] представляет собой обратно комплементарную или частично обратно комплементарной элементу stem2, и представляет собой консекутивную последовательность длиной от 5 до 7 нуклеотидов;
где N0-2 представляет собой консекутивную последовательность длиной от 0 до 2 нуклеотидов, предпочтительно от 0 до 1, более предпочтительно, 1N, где каждый N является независимо от другого выбранным из нуклеотида, выбранного из A, U, T, G и С или нуклеотидного аналога этого;
где N3-5 представляет собой консекутивную последовательность длиной от 3 до 5, предпочтительно от 4 до 5, более предпочтительно, 4N, где каждый N является независимо от другого выбранным из нуклеотида, выбранного из A, U, T, G и С или нуклеотидного аналога этого;
и
где G представляет собой гуанозин или его аналог, и дополнительно может быть заменен цитидином или его аналогом, при условии, что его комплементарный нуклеотид цитидин в stem2 заменяется гуанозином;
последовательность петли [N0-4(U/T)N0-4] расположена между элементами stem1 и stem2 и представляет собой консекутивную последовательность от 3 до 5 нуклеотидов, более предпочтительно, 4 нуклеотида;
где каждый N0-4 является независимо от другого консекутивная последовательность длиной от 0 до 4, предпочтительно от 1 до 3, более предпочтительно от 1 до 2 нуклеотидов, где каждый N является независимо от другого выбранным из нуклеотида, выбранного из A, U, T, G и С или нуклеотидного аналога этого; и
где U/T представляет уридин, или, необязательно, тимидин;
stem2[N3-5СN0-2] представляет собой обратно комплементарную или частично обратно комплементарной элементу stem1, и представляет собой консекутивную последовательность длиной от 5 до 7 нуклеотидов;
где N3-5 представляет собой консекутивную последовательность длиной от 3 до 5, предпочтительно от 4 до 5, более предпочтительно, 4N, где каждый N является независимо от другого выбранным из нуклеотида, выбранного из A, U, T, G и С или нуклеотидного аналога этого;
где N0-2 представляет собой консекутивную последовательность длиной от 0 до 2 нуклеотидов, предпочтительно от 0 до 1, более предпочтительно, 1N, где каждый N является независимо от другого выбранным из нуклеотида, выбранного из A, U, T, G и С или нуклеотидного аналога этого;
и
где С представляет собой цитидин или его аналог, и дополнительно может быть заменен гуанозином или его аналогом, при условии, что его комплементарный нуклеотид гуанозин в stem1 заменяется цитидином;
где
stem1 и stem2 способны образовывать пары оснований друг с другом, образуя обратно комплементарную последовательность, где образование пар оснований может происходить между stem1 и stem2, например, за счет Уотсон-Криковского образования пар оснований нуклеотидов A и U/T или G и C, или за счет не-Уотсон-Криковского образования пар оснований, например, образования неустойчивых пар оснований, обратного Уотсон-Криковского образования пар оснований, образования пар оснований по Хугстену, обратного образования пар оснований по Хугстену, или способны образовывать пары оснований друг с другом, формируя частично обратно комплементарную последовательность, в пользу неполные базы, где неполное образование пар оснований может происходить между stem1 и stem2, на основании того, что одно или несколько оснований в одном стебле не имеют комплементарного основания в обратно комплементарной последовательности в другом стебле.
В контексте вышесказанного, образование неустойчивых пар оснований обычно не является Уотсон-Криковским образованием пар оснований между двумя нуклеотидами. Четыре основных неустойчивых пар оснований в данном контексте, который может быть использован, представляют собой гуанозин-уридин, инозин-уридин, инозин-аденозин, инозин-цитидин (G-U/T, I-U/T, I-A и I-C) и аденозин-цитидин (A-C).
Соответственно, в контексте настоящего изобретения, неустойчивое основание представляют собой основание, которое образует неустойчивую пару оснований с другим основанием, как описано выше. Поэтому не Уотсон-криковское образование пар оснований, например, образование неустойчивых пар оснований может происходить в стебле гистоновой структуры типа «стебель-петля» в соответствии с настоящим изобретением.
В контексте вышесказанного, частично обратно комплементарная последовательность содержит максимально 2, предпочтительно, только одно несовпадение в структуре стебля последовательности структуры типа «стебель-петля», сформированной образованием пар оснований stem1 и stem2. Иными словами, stem1 и stem2 предпочтительно способны к (полному) образованию пар оснований друг с другом на протяжении всей последовательности stem1 и stem2 (100% правильных Уотсон-Криковских или не Уотсон-Криковских образований пар оснований), тем самым формируя обратно комплементарную последовательность, где каждое основание имеет свое правильное Уотсон-Криковское или не Уотсон-Криковское дополнение основания в качестве комплементарного партнера связывания. Альтернативно, stem1 и stem2 предпочтительно способны к частичному образованию пар оснований друг с другом на протяжении всей последовательности stem1 и stem2, где по меньшей мере примерно 70%, 75%, 80%, 85%, 90% или 95% из 100% возможных правильных Уотсон-Криковских или не Уотсон-Криковских образований пар оснований занимают корректные Уотсон-Криковские или не Уотсон-Криковские образования пар оснований и, самое большее, примерно 30%, 25%, 20%, 15%, 10% или 5% оставшихся оснований являются неспаренными.
В соответствии с предпочтительным вариантом осуществления первого аспекта по изобретению по меньшей мере одна последовательность гистоновой структуры типа «стебель-петля» (с ограничивающими элементами стебля) последовательности нуклеиновых кислот по изобретению, как определено в настоящем документе, содержит примерно от 15 до примерно 45 нуклеотидов в длину, предпочтительно, примерно от 15 до примерно 40 нуклеотидов в длину, предпочтительно, примерно от 15 до примерно 35 нуклеотидов в длину, предпочтительно, примерно от 15 до примерно 30 нуклеотидов в длину и даже более предпочтительно, примерно от 20 до примерно 30 в длину и наиболее предпочтительно, примерно от 24 до примерно 28 нуклеотидов в длину.
В соответствии с дополнительным предпочтительным вариантом осуществления первого аспекта изобретения по меньшей мере одна последовательность гистоновой структуры типа «стебель-петля» (без ограничивающих элементов стебля) последовательности нуклеиновых кислот по изобретению, как определено в настоящем документе, содержит от примерно 10 до примерно 30 нуклеотидов в длину, предпочтительно от примерно 10 примерно 20 нуклеотидов в длину, предпочтительно от примерно 12 до примерно 20 нуклеотидов в длину, предпочтительно от примерно 14 до 20 нуклеотидов в длину и даже более предпочтительно от примерно 16 до примерно 17 в длину, и наиболее предпочтительно примерно 16 нуклеотидов в длину.
В соответствии с дополнительным предпочтительным вариантом осуществления первого аспект изобретения, последовательность нуклеиновых кислот по изобретению в соответствии с первым аспектом изобретения может содержать или кодировать по меньшей мере одну последовательность гистоновой структуры типа «стебель-петля» в соответствии с по меньшей мере одной из следующих конкретных формул (Iа) или (IIa):
Формула (Ia) (последовательность структуры типа «стебель-петля» без ограничивающих элементов стебля):
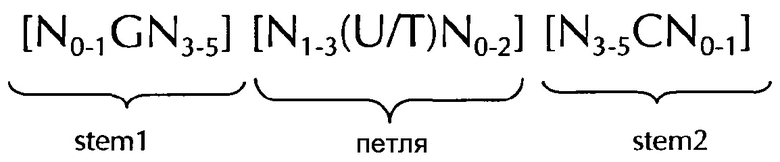
формула (IIa) (последовательность типа «стебель-петля» с ограничивающими элементами стебля):
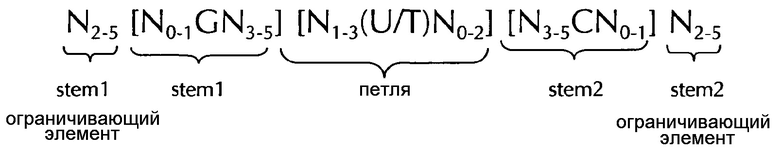
где:
N, C, G, T и U представляют собой то, как это определено выше.
В соответствии с дополнительным, еще более предпочтительным вариантом осуществления первого аспекта, последовательность нуклеиновых кислот по изобретению может включать или кодировать по меньшей мере одну последовательность гистоновой структуры типа «стебель-петля» в соответствии с по меньшей мере одной из следующих конкретных формул (Ib) или (IIb):
формула (Ib) (последовательность типа «стебель-петля» без ограничивающих элементов стебля):
формула (IIb) (последовательность типа «стебель-петля» с ограничивающими элементами стебля):
где:
N, C, G, T и U представляют собой то, как это определено выше.
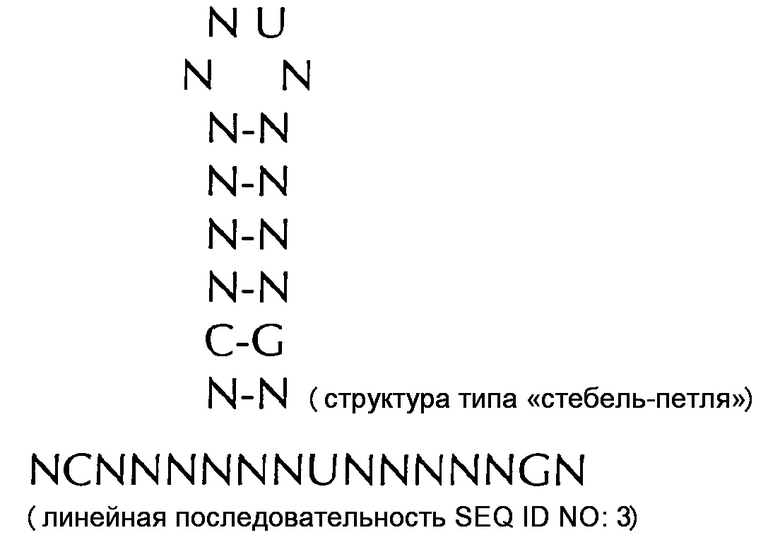
В соответствии с дополнительным еще более предпочтительным вариантом осуществления первого аспекта, последовательность нуклеиновых кислот по изобретению может включать или кодировать по меньшей мере одну последовательность гистоновой структуры типа «стебель-петля» в соответствии с по меньшей мере одной из следующих конкретных формул c (Ic) по (Ih) или с (IIc) по (IIh), показанных альтернативно в их структуре типа «стебель-петля» и в качестве линейной последовательности, представляющей гистоновые последовательности типа «стебель-петля», генерированные в соответствии с Примером 1:
формула (Ic): (консенсусная последовательность гистоновой структуры типа «стебель-петля» метазоа и простейших без ограничивающих элементов стебля):

формула (IIc): (консенсусная последовательность гистоновой структуры типа «стебель-петля» метазоа и простейших c ограничивающими элементами стебля):

формула (Id): (без ограничивающих элементов стебля):
формула (IId): (c ограничивающими элементами стебля):

формула (Ie): (консенсусная последовательность гистоновой структуры типа «стебель-петля» простейших без ограничивающих элементов стебля)

формула (IIe): (консенсусная последовательность гистоновой структуры типа «стебель-петля» простейших c ограничивающими элементами стебля)

формула (If): (консенсусная последовательность гистоновой структуры типа «стебель-петля» метазоа без ограничивающих элементов стебля)
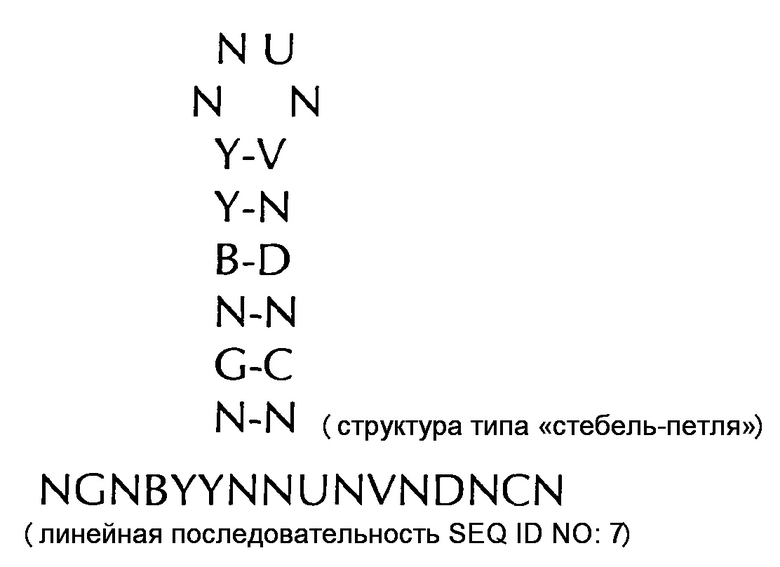
формула (IIf): (консенсусная последовательность гистоновой структуры типа «стебель-петля» метазоа c ограничивающими элементами стебля)

формула (Ig): (консенсусная последовательность гистоновой структуры типа «стебель-петля» позвоночных без ограничивающих элементов стебля)
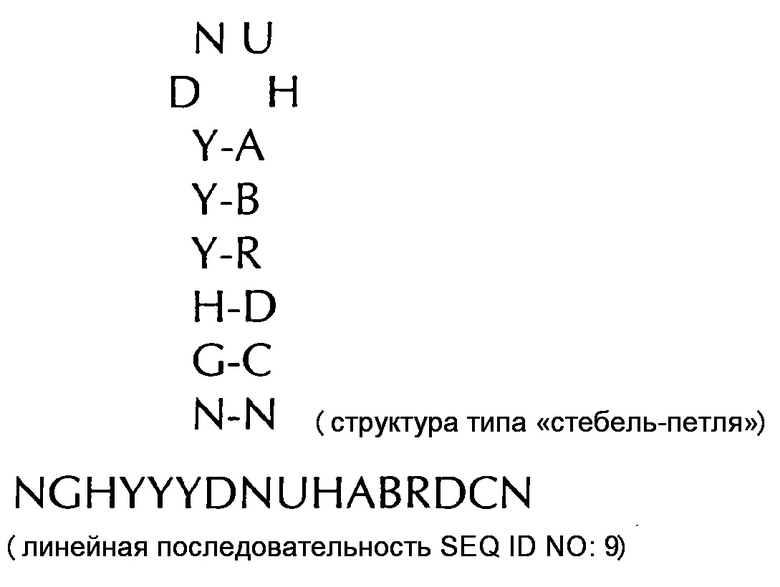
формула (IIg): (консенсусная последовательность гистоновой структуры типа «стебель-петля» позвоночных c ограничивающими элементами стебля)

формула (Ih): (консенсусная последовательность гистоновой структуры типа «стебель-петля» человека без ограничивающих элементов стебля)
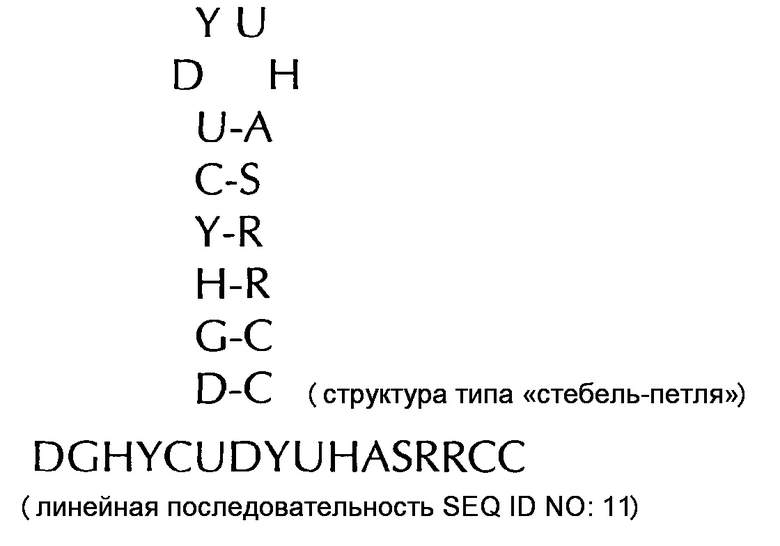
формула (IIh): (консенсусная последовательность гистоновой структуры типа «стебель-петля» человека c ограничивающими элементами стебля)

где в каждой из вышеприведенных формул с (Ic) по (Ih) или с (IIc) по (IIh):
N, C, G, A, T и U представляют собой то, как это определено выше;
каждый U может быть заменен на T;
каждый (высоко) консервативный G или C в элементах стебля 1 и 2 может быть заменены его комплементарным нуклеотидным основанием C или G, при условии, что его комплементарный нуклеотид в соответствующем стебле заменяется его комплементарным нуклеотидом в параллели; и/или
G, A, T, U, C, R, Y, M, K, S, W, H, B, V, D и N представляют собой нуклеотидные основания, как определено в следующей Таблице:
В этом контексте особенно предпочтительно, что последовательность гистоновой структуры типа «стебель-петля» в соответствии с по меньшей мере одной из формул от (I) или (Ia) до (Ih) или от (II) или (IIa) до (IIh) по настоящему изобретению выбирают из природной последовательности гистоновой структуры типа «стебель-петля», более предпочтительно, из последовательностей гистоновой структуры типа «стебель-петля» простейших или метазоа, и даже более особенно предпочтительно, из последовательностей гистоновой структуры типа «стебель-петля» позвоночных, и наиболее предпочтительно, из последовательностей гистоновой структуры типа «стебель-петля» млекопитающих, особенно из последовательностей гистоновой структуры типа «стебель-петля» человека.
В соответствии с особенно предпочтительным вариантом осуществления первого аспекта, последовательность гистоновой структуры типа «стебель-петля» в соответствии с по меньшей мере одной из конкретных формул с (I) или (Ia) до (Ih) или с (II) или (IIа) до (IIh) по настоящему изобретению является последовательностью гистоновой структуры типа «стебель-петля», содержащей в каждом нуклеотидном положении наиболее часто встречающийся нуклеотид, или либо наиболее часто, либо второй после наиболее часто встречающегося нуклеотида природных последовательностей гистоновой структуры типа «стебель-петля» метазоа и простейших (Фиг. 1), простейших (Фиг. 2), метазоа (Фиг. 3), позвоночных (Фиг. 4) и человека (Фиг. 5), как показано на Фиг. 1-5. В этом контексте особенно предпочтительно, что по меньшей мере 80%, предпочтительно по меньшей мере 85%, или наиболее предпочтительно по меньшей мере 90% всех нуклеотидов соответствуют наиболее часто встречающемуся нуклеотиду природной последовательности гистоновой структуры типа «стебель-петля».
В дополнительном конкретном варианте осуществления первого аспекта последовательность гистоновой структуры типа «стебель-петля» в соответствии с по меньшей мере одной из конкретных формул с (I) или (Ia) по (Ih) настоящего изобретения выбирается из следующих последовательностей гистоновой структуры типа «стебель-петля» (без ограничивающих элементов стебля), представляющих последовательности гистоновой структуры типа «стебель-петля», генерированных в соответствии с Примером 1:
VGYYYYHHTHRWRCB (SEQ ID NO: 13 в соответствии с формулой (Ic))
SGYYYTTYTMARRRCS (SEQ ID NO: 14 в соответствии с формулой (Ic))
SGYYCTTTTMAGRRCS (SEQ ID NO: 15, в соответствии с формуле (Ic))
DGNNNBNNTHVNNNCH (SEQ ID NO: 16 в соответствии с формулой (Ie))
RGNNNYHBTHRDNNCY (SEQ ID NO: 17 в соответствии с формулой (Ie))
RGNDBYHYTHRDHNCY (SEQ ID NO: 18 в соответствии с формулой (Ie))
VGYYYTYHTHRVRRCB (SEQ ID NO: 19 в соответствии с формулой (If))
SGYYCTTYTMAGRRCS (SEQ ID NO: 20 в соответствии с формулой (If))
SGYYCTTTTMAGRRCS (SEQ ID NO: 21 в соответствии с формулой (If))
GGYYCTTYTHAGRRCC (SEQ ID NO: 22 в соответствии с формулой (Ig))
GGCYCTTYTMAGRGCC (SEQ ID NO: 23 в соответствии с формулой (Ig))
GGCTCTTTTMAGRGCC (SEQ ID NO: 24 В соответствии с формулой (Ig))
DGHYCTDYTHASRRCC (SEQ ID NO: 25 в соответствии с формулой (Ih))
GGCYCTTTTHAGRGCC (SEQ ID NO: 26 в соответствии с формулой (Ih))
GGCYCTTTTMAGRGCC (SEQ ID NO: 27 в соответствии с формулой (Ih))
Кроме того, в этом контексте следующие последовательности гистоновой структуры типа «стебель-петля» (с ограничивающими элементами стебля), генерированные в соответствии с Примером 1, в соответствии с одной из конкретных формул с (II) или (IIa) до (IIh) являются особенно предпочтительными:
H*H*HHVVGYYYYHHTHRWRCBVHH*N*N* (SEQ ID NO: 28 в соответствии с формулой (IIc))
м*ч*MHMSGYYYTTYTMARRRCSMCH*H*H* (SEQ ID NO:29 в соответствии с формулой (IIc))
M*M*MMMSGYYCTTTTMAGRRCSACH*М*Н* (SEQ ID NO:30 в соответствии с формулой (IIc))
N*N*NNNDCNNNBNNTHVNNNCHNHN*N*N* (SEQ ID NO:31 в соответствии с формулой (IIе))
N*N*HHNRGNNNYHBTHRDNNCYDHH*N*N* (SEQ ID NO:32 в соответствии с формулой (IIе))
N*H*HHVRGNDBYHYTHRDHNCYRHH*H*H* (SEQ ID NO:33 в соответствии с формулой (IIе))
H*H*MHMVGYYYTYHTHRVRRCBVMH*Н*N* (SEQ ID NO:34 в соответствии с формулой (IIf))
M*M*MMMSGYYCTTYTMAGRRCSMCH*H*H* (SEQ ID NO:35 в соответствии с формулой (IIf))
M*M*MMMSGYYC TTTMAGRRCSACH*м*ч* (SEQ ID NO:36 в соответствии с формулой (IIf)
H*H*MAMGGYYCTTYTHAGRRCCVHN*N*M* (SEQ ID NO:37 в соответствии с формулой (IIg))
*H*AAMGGCYCTTYTMAGRGCCVCH*H*м* (SEQ ID NO:38 в соответствии с формулой (IIg))
M*M*AAMGGCTCTTTTMAGRGCCMCY*M*M* (SEQ ID NO:39 в соответствии с формулой (IIg))
N*H*AAHDGHYCTDYTHASRRCCVHB*N*H* (SEQ ID NO:40 в соответствии с формулой (IIh))
*H*AAMGGCYCTTTTHAGRGCCVMY*N*M* (SEQ ID NO:41 в соответствии с формулой (IIh))
H*м*AAAGGCYCTTTTMAGRGCCRMY*H*М* (SEQ ID NO:42 в соответствии с формулой (IIh))
В соответствии с дополнительным предпочтительным вариантом осуществления первого аспекта изобретения, последовательность нуклеиновых кислот по изобретению содержит или кодирует по меньшей мере одну последовательность гистоновой структуры типа «стебель-петля», демонстрирующую по меньшей мере примерно 80%, предпочтительно по меньшей мере примерно 85%, более предпочтительно по меньшей мере примерно 90% или даже более предпочтительно по меньшей мере примерно 95% идентичности последовательности с не до 100% консервативных нуклеотидов в последовательности гистоновой структуры типа «стебель-петля» в соответствии с по меньшей мере одной из конкретных формул с (I) или (Ia) до (Ih) или c (II) или (IIa) до (IIh) или с естественной последовательностью гистоновой структуры типа «стебель-петля».
В предпочтительном варианте осуществления последовательность гистоновой структуры типа «стебель-петля» не содержит последовательность петли 5'-UUUC-3'. Конкретнее, гистоновая структура типа «стебель-петля» не содержит последовательность stem1 5'-GGCUCU-3 и/или последовательность stem2 последовательности 5'-AGAGCC-3', соответственно. В другом предпочтительном варианте осуществления последовательность типа «стебель-петля» не содержит последовательность петли 5'-CCUGCCC-3' или последовательность петли 5'-UGAAU-3'. Конкретнее, структура типа «стебель-петля» не содержит последовательность stem1 5'-CCUGAGC-3' или не содержит последовательность stem1 5'-ACCUUUCUCCA-3 и/или последовательность stem2 5'-GCUCAGG-3 или 5'-UGGAGAAAGGU-3», соответственно. Также, поскольку изобретение специфически не ограничивает последовательности гистоновой структуры типа «стебель-петля», последовательности структуры типа «стебель-петля» предпочтительно не получены из 3'-нетранслируемой области рецептора инсулина млекопитающих. Также, предпочтительно, нуклеиновая кислота по изобретению не может содержать сигналы процессинга гистоновой структуры типа «стебель-петля», в частности, полученные из гена гистона H2A614 (H2kA614) мыши.
Последовательность нуклеиновых кислот по изобретению в соответствии с первым аспектом настоящего изобретения может необязательно включать или кодировать поли(A)-последовательность. При наличии такая поли(A)-последовательность включает последовательность примерно от 25 до примерно 400 нуклеотидов аденозина, предпочтительно, последовательность примерно от 30 или, более предпочтительно, примерно от 50 до примерно 400 нуклеотидов аденозина, более предпочтительно, последовательность примерно от 50 примерно до 300 нуклеотидов аденозина, даже более предпочтительно, последовательность примерно от 50 примерно до 250 нуклеотидов аденозина, наиболее предпочтительно, последовательность примерно от 60 примерно до 250 нуклеотидов аденозина. В этом контексте термин «примерно» означает отклонение в ±10% от значения (ий), к которому они относится. Соответственно, поли(A)-последовательность содержит по меньшей мере 25 или более чем 25, более предпочтительно по меньшей мере 30, более предпочтительно по меньшей мере 50 нуклеотидов аденозина. Поэтому, такая поли(A)-последовательность, как правило, не содержат менее 20 нуклеотидов аденозина. Конкретнее, она не содержит 10 и/или менее чем 10 нуклеотидов аденозина.
Предпочтительно, нуклеиновая кислота, в соответствии с настоящим изобретением, не содержат один или два, или по меньшей мере один или все, кроме одного или каждый из компонентов группы, состоящей из: последовательности, кодирующей рибозим (предпочтительно самоспайсирующийся рибозим), вирусной последовательности нуклеиновых кислот, сигнала процессинга гистоновой структуры типа «стебель-петля», в частности последовательности процессинга гистоновой структуры типа «стебель-петля», полученной из гистонового гена H2A614 мыши, гена Neo, последовательности инактивированного промотора и последовательности инактивированного энхансера. Даже более предпочтительно, нуклеиновая кислота, в соответствии с изобретением, не содержит рибозим, предпочтительно самоспайсирующийся рибозим, и один из группы, состоящей из: гена Neo, последовательности инактивированного промотора и последовательности инактивированного энхансера, сигнала процессинг гистоновой структуры типа «стебель-петля», в частности, последовательности процессинга гистоновой структуры типа «стебель-петля», полученной из гистонового гена H2A614 мыши. Соответственно, нуклеиновая кислота может, в предпочтительном способе, не содержать ни рибозим, предпочтительно самоспайсирующийся рибозим, ни ген Neo или, в качестве альтернативы, ни рибозим, предпочтительно самоспайсирующийся рибозим, ни ген устойчивости (например, обычно используемый для селекции). В другом предпочтительном способе, нуклеиновая кислота по изобретению может не содержать ни рибозим, предпочтительно самоспайсирующийся рибозим, ни сигнал процессинга гистоновой структуры типа «стебель-петля», в частности, последовательности процессинга гистоновой структуры типа «стебель-петля», полученной из гистонового гена H2A614 мыши.
Кроме того, в соответствии с первым аспектом настоящего изобретения, ядерная последовательность по изобретению необязательно включает сигнал полиаденилирования, который определяется в настоящем документе как сигнал, который передает полиаденилирование на (транскрибируемую) мРНК при помощи определенных белковых факторов (например, специфический фактор расщепления и полиаденилирования (CPSF), фактор стимуляции расщепления (CstF), факторы расщепления I и II (CF I и CF II), поли(A)-полимераза (PAP)). В этом контексте консенсусный сигнал полиаденилирования является предпочтительным, содержащим консенсусную последовательность NN(U/T)ANA. В определенном предпочтительном аспекте сигнал полиаденилирования содержит одну из следующих последовательностей: AA(U/T)ААА или A(U/T)(U/T)AAA (где уридин обычно присутствует в РНК и тимидин обычно присутствует в ДНК). В некоторых вариантах осуществления, сигнал полиаденилирования, используемый в нуклеиновой кислоте по изобретению, не соответствует U3 snRNA, U5, сигналу процессинга полиаденилирования гена G-CSF человека или сигнальной последовательности полиаденилирования SV40. В частности, вышеописанные сигналы полиаденилирования не комбинируется с любым геном антибиотикорезистентности (или любым другим репортерным, маркерным или селекционным геном), в частности, с геном устойчивости neo (неомицинфосфотрансфераза) (как ген кодирующей области в соответствии с элементом (a) нуклеиновой кислоты по изобретению в такой нуклеиновой кислоте по изобретению. И любой из вышеперечисленных сигналов полиаденилирования предпочтительно не комбинируется с гистоновой структурой типа «стебель-петля» или сигналом процессинга гистоновой структуру типа «стебель-петля» из гистонового гена H2A61 мыши в нуклеиновой кислоте по изобретению.
Последовательность нуклеиновых кислот по изобретению в соответствии с первым аспектом настоящего изобретения может, кроме того, кодировать белок или пептид, который содержит опухолевый антиген или его фрагмент, вариант или производное.
Опухолевые антигены предпочтительно расположены на поверхности (опухолевой) клетки, характеризуя опухоль млекопитающих, в частности, опухоль человека (например, при системных или твердоопухолевых заболеваниях). Опухолевые антигены также могут быть выбраны из белков, которые сверхэкспрессируются в опухолевых клетках по сравнению с нормальной клеткой. Кроме того, опухолевые антигены включают также антигены, экспрессируемые в клетках, которые не сами (или, первоначально, не сами) выродились (вырождаются), но связаны с предполагаемой опухолью. Антигены, которые связаны со снабжающими опухоль сосудами или их (пере)стройками, в частности, те антигены, которые связаны с неоваскуляризацией, например факторы роста, такие как VEGF, bFGF и т.д., также включены в настоящий документ. Антигены, связанные с опухолью, кроме того, включают антигены из клеток или тканей, как правило, заключающих в себе опухоль. Кроме того, некоторые вещества (обычно белки или пептиды) экспрессируются у пациентов, страдающих (осознанно или неосознанно) от онкологических заболеваний, и они проявляются в повышенной концентрации в жидкостях организма указанных пациентов. Эти вещества также называют «опухолевыми антигенами», однако они не являются антигенами в строгом значении вызывающего иммунный ответ вещества. Класс опухолевых антигенов может быть разбит далее на меньшие опухолевые специфические антигены (ТSA) и опухоле-ассоциированные антигены (ТАА). ТSA могут быть представлены только опухолевыми клетками и никогда нормальными «здоровыми» клетками. Они, как правило, являются следствием опухолевой специфической мутации. ТАА, которые являются более распространенными, как правило, представлены как опухолевыми, так и здоровыми клетками. Эти антигены распознаются, и антиген-представляющая клетка может быть уничтожена с помощью цитотоксических Т-клеток. Дополнительно, опухолевые антигены могут также иметь место на поверхности опухоли в виде, например, мутировавшего рецептора. В этом случае они могут распознаваться антителами.
Далее, опухоле-ассоциированные антигены могут быть классифицированы как ткане-специфические антигены, которые также называются меланоцит-специфические антигены, антигены рака яичек и опухолеве-специфические антигены. Под антигенами рака яичек, как правило, подразумевают пептиды и белки генов, ассоциированных с зародышевой линией, которые могут быть активированы в множестве опухолей. Антигены рака яичек человека можно далее подразделить на антигены, которые кодируются хромосомой X, так называемые антигены СТ-Х, и те антигены, которые не кодируются хромосомой X, так называемые (антигены non-СТ-Х). Антигены рака яичек, которые кодируются хромосомой X, включает в себя, например, семейство генов антигенов меланомы, так называемое MAGE-семейство. Гены MAGE-семейства могут характеризоваться общим гомологичным доменом MAGE (MHD). Каждый из этих антигенов, т.е. меланоцит-специфические антигены, антигены рака яичек и опухолево-специфические антигены, могут вызывать аутологичный клеточный и гуморальный иммунный ответ. Соответственно, опухолевый антиген, кодируемый последовательностью нуклеиновых кислот по изобретению, представляет собой, предпочтительно, меланоцит-специфический антиген, антиген рака яичек или опухолевые специфические антигены, предпочтительно, это может быть антиген СТ-Х, антигены non-СТ-Х, партнер по связыванию для антигена СТ-Х или партнер по связыванию для антигена non-СТ-Х или опухолевого специфического антигена, более предпочтительно антиген СТ-Х, партнер по связыванию для антигена non-СТ-Х или опухолевого специфического антигена.
Особенно предпочтительными являются опухолевые антигены, выбранные из перечня, состоящего из 5T4, 707-AP, 9D7, AFP, AlbZIP HPG1, альфа-5-бета-1-интегрина, альфа-5-бета-6-интегрина, альфа-актинина-4/m, альфа-метилацилкоэнзим-A-рацемазы, ART-4, ARTC1/m, B7H4, BAGE-1, BCL-2, bcr/abl, бета-катенина/m, BING-4, BRCA1/m, BRCA2/m, СA 15-3/СA 27-29, СA 19-9, CA72-4, CA125, калретикулина, CAMEL, CASP-8/m, катепсина B, катепсина L, CD19, CD20, CD22, CD25, CDE30, CD33, CD4, CD52, CD55, CD56, CD80, CDC27/m, CDK4/m, CDKN2A/m, CEA, CLCA2, CML28, CML66, COA-1/m, коактозин-подобного белка, collage XXIII, COX-2, СT-9/BRD6, Cten, циклина B1, циклина D1, cyp-B, CYPB1, DAM-10, DAM-6, DEK-CAN, EFTUD2/m, EGFR, ELF2/m, EMMPRIN, EpCam, EphA2, EphA3, ErbB3, ETV6-AML1, EZH2, FGF-5, FN, Frau-1, G250, GAGE-1, GAGE-2, GAGE-3, GAGE-4, GAGE-5, GAGE-6, GAGE7b, GAGE-8, GDEP, GnT-V, gp100, GPC3, GPNMB/m, HAGE, HAST-2, гепсина, Her2/neu, HERV-K-MEL, HLA-A*0201-R17I, HLA-A11/m, HLA-A2/m, HNE, гомеобоксa NKX3.1, HOM-TES-14/SCP-1, HOM-TES-85, HPV-E6, HPV-E7, HSP70-2M, HST-2, hTERT, iCE, IGF-1R, IL-13Ra2, IL-2R, IL-5, незрелого ламининового рецептора, калликреина-2, калликреина-4, Ki67, KIAA0205, KIAA0205/m, KK-LC-1, K-Ras/m, LAGE-A1, LDLR-FUT, MAGE-A1, MAGE-A2, MAGE-A3, MAGE-A4, MAGE-A6, MAGE-A9, MAGE-A10, MAGE-A12, MAGE-B1, MAGE-B2, MAGE-B3, MAGE-B4, MAGE-B5, MAGE-B6, MAGE-B10, MAGE-B16, MAGE-B17, MAGE-C1, MAGE-C2, MAGE-C3, MAGE-D1, MAGE-D2, MAGE-D4, MAGE-E1, MAGE-E2, MAGE-F1, MAGE-H1, MAGEL2, маммаглобина A, MART-1/melan-A, MART-2, MART-2/м, белка матриксa 22, MC1R, M-CSF, ME1/m, мезотелина, MG50/PXDN, MMP11, MN/СА IX-антигена, MRP-3, MUC-1, MUC-2, MUM-1/m, MUM-2/m, MUM-3/m, миозина класса I/m, NA88-A, N-ацетилглюкозаминилтрансферазы-V, Neo-PAP, Nео-PAP/m, NFYC/m, NGEP, NMP22, NPM/ALK, N-Ras/m, NSE, NY-ESO-B, NY-ESO-1, OA1, OFA-iLRP, OGT, OGT/m, OS-9, OS-9/m, остеокальцина, остеопонтина, р15, p190 минорного bcr-abl, p53, р53/m, PAGE-4, PAI-1, PAI-2, PAP, PART-1, PATE, PDEF, Pim-1-киназы, Pin-1, Pml/PARalpha, POTE, PRAME, PRDX5/m, простеина, протеиназы-3, PSA, PSCA, PSGR, PSM, PSMA, PTPRK/m, RAGE-1, RBAF600/m, RHAMM/CD168, RU1, RU2, S-100, SAGE, SART-1, SART-2, SART-3, SCC, SIRT2/m, Sp17, SSX-1, SSX-2/HOM-MEL-40, SSX-4, STAMP-1, STEAP-1, сурвивина, сурвивина-2B, SYT-SSX-1, SYT-SSX-2, ТА-90, TAG-72, TARP, ТEL-AML1, TGFбета, TGFбетаRII, TGM-4, TPI/m, TRAG-3, TRG, TRP-1, TRP-2/6b, TRP/INT2, TRP-p8, тирозиназы, UPA, VEGFR1, VEGFR-2/FLK-1 и WT1. Такие опухолевые антигены предпочтительно можно выбрать из группы, состоящей из р53, CA125, EGFR, Her2/neu, hTERT, PAP, MAGE-A1, MAGE-A3, мезотелина, MUC-1, GP100, MART-1, тирозиназы, PSA, PSCA, PSMA, STEAP-1, VEGF, VEGFR1, VEGFR2, Ras, CEA или WT1, и более предпочтительно из PAP, MAGE-A3, WT1 и MUC-1. Такие опухолевых антигены предпочтительно могут быть выбраны из группы, состоящей из MAGE-A1 (например, MAGE-A1 в соответствии с инв. номером M77481), MAGE-A2, MAGE-A3, MAGE-A6 (например, MAGE-A6 в соответствии с инв. номером NM_005363), MAGE-C1, MAGE-C2, мелана-А (например, мелан-А в соответствии с инв. номером NM_005511), GP100 (например, GP100 в соответствии с инв. номером M77348), тирозиназы (например, тирозиназы в соответствии с инв. номером NM_000372), сурвивина (например, сурвивин в соответствии с инв. номером AF077350), CEA (например, CEA в соответствии с инв. номером NM_004363), Her-2/neu (например, Her-2/neu в соответствии с инв. номером M11730), WT1 (например, WT1 в соответствии с инв. номером NM_000378), PRAME (например, PRAME в соответствии с инв. номером NM_0061 15), EGFRI (рецептор 1 эпидермального фактора роста) (например, EGFRI (рецептор 1 эпидермального фактора роста) в соответствии с инв. номером AF288738), MUC1, муцин-1 (например, муцин-1 в соответствии с инв. номером NMJD02456), SEC61G (например, SEC61G в соответствии с инв. номером NM_014302), hTERT (например, hTERT с инв. номером NM_198253), 5T4 (например, 5T4 в соответствии с инв. номером NM_006670), TRP-2 (например, TRP-2 в соответствии с инв. номером NM_001922), STEAP1, PCA, PSA, PSMA и др.
Кроме того, опухолевые антигены также могут включать в себя идиотипические антигены, связанные с онкологическим или опухолевым заболеванием, особенно лимфомой или связанным с лимфомой заболеванием, где указанный идиотипический антиген представляет собой идиотип иммуноглобулина лимфоидной клетки крови, или идиотип Т-клеточного рецептора лимфоидной клетки крови.
Опухолевые антигенные белки для лечения онкологических или опухолевых заболеваний, как правило, представляют собой белки происхождения из млекопитающих, предпочтительно человеческого происхождения. Их отбор для лечения индивидуума зависит от типа опухоли, предназначенной для лечения, и профиля экспрессии отдельной опухоли. Человека, страдающего от рака предстательной железы, например, предпочтительно лечить опухолевым антигеном, который обычно экспрессируется (или сверхэкспрессируется) в карциноме предстательной железы или специфически сверхэкспрессируется у проходящего лечение индивидуума, например, любой из PSМА, PSCA и/или PSA.
Кодирующая область нуклеиновой кислоты по изобретению в соответствии с первым аспектом настоящего изобретения может встречаться в виде моно-, ди- или даже мультицистронной нуклеиновой кислоты, т.е. нуклеиновой кислоты, которая несет кодирующие последовательности одного, двух или более белков или пептидов. Такие кодирующие последовательности в ди- или даже мультицистронных нуклеиновых кислотах могут быть разделены по меньшей мере одной последовательностью внутреннего рибосомного входного участка (IRES), например, как описано в настоящем документе, или сигнальными пептидами, которые вызывают расщепление полученного в результате полипептида, который включает в себя несколько белков или пептидов.
В соответствии с первым аспектом настоящего изобретения, последовательность нуклеиновых кислот по изобретению включает кодирующую область, кодирующую пептид или белок, который содержит опухолевый антиген или его фрагмент, вариант или производное. Предпочтительно, кодируемый опухолевый антиген не является гистоновым белком. В контексте настоящего изобретения такой гистоновый белок, как правило, является сильно основным белком, содержащимся в ядре эукариотической клетки, который упаковывает и направляет ДНК в структурные единицы, называемые нуклеосомы. Гистоновые белки являются главными белковыми компонентами хроматина, выступают в качестве катушки, вокруг которой наматывается ДНК, и играют важную роль в регуляции экспрессии генов. Без гистонов размотанная ДНК в хромосомах была бы очень длинной (отношение длины к ширине более 10 миллионов к одному у ДНК человека). Например, каждая человеческая клетка имеет примерно 1,8 метра ДНК, но при наматывании на гистоны она имеет примерно 90 миллиметров хроматина, который, при дубликации и конденсировании во время митоза приводит к образованию примерно 120 мкм хромосом. Более предпочтительно, в контексте настоящего изобретения, такой гистоновый белок обычно определяют как высоко консервативный белок, выбранный из одного из следующих пяти основных классов гистонов: H1/H5, H2A, H2B, H3 и H4, выбранных предпочтительно из гистонов млекопитающих, более предпочтительно, из гистонов человека или гистоновых белков. Такие гистоны или гистоновые белки, как правило, организованы в два суперкласса, определенных как основные гистоны, состоящие из гистонов H2A, H2B, H3 и H4, и линкерные гистоны, состоящие из гистонов Н1 и H5.
В этом контексте, линкерные гистоны, предпочтительно исключенные из сферы защиты находящегося в рассмотрении изобретения, предпочтительно линкерные гистоны млекопитающих, более предпочтительно, линкерные гистоны человека, обычно выбирают из H1, в том числе H1F, включая, в частности, H1FО, H1FNT, H1FОО, H1FX и H1H1, включая, в частности, HIST1H1А, HIST1H1B, HIST1H1C, HIST1H1D, HIST1H1E, HIST1H1T; и
Кроме того, основные гистоны, предпочтительно исключенные из сферы защиты находящегося в рассмотрении изобретения, предпочтительно основные гистоны млекопитающих, более предпочтительно, основные гистоны человека, обычно выбирают из Н2А, в том числе H2AF, включая, в частности, H2AFB1, H2AFB2, H2AFB3, H2AFJ, H2AFV, H2AFX, H2AFY, H2AFY2, H2AFZ и H2A1, включая, в частности, HIST1H2AA, HIST1H2AB, HIST1H2AC, HIST1H2AD, HIST1H2AE, HIST1H2AG, HIST1H2AI, HIST1H2AJ, HIST1Н2А, HIST1H2AL, HIST1H2AM и H2A2, включая, в частности, HIST2H2AA3, HIST2H2AC; H2B, в том числе H2BF, включая, в частности, H2BFM, H2BFO, H2BFS, H2BFWT, H2B1, включая, в частности, HIST1H2BA, HIST1H2BB, HIST1H2BC, HIST1H2BD, HIST1H2BE, HIST1H2BF, HIST1H2BG, HIST1H2BH, HIST1H2BI, HIST1H2BJ, HIST1H2BК, HIST1H2BL, HIST1H2BM, HIST1H2BN, HIST1H2BО, и H2B2, включая, в частности, HIST2H2BE; H3, в том числе H3A1, включая, в частности, HIST1H3A, HIST1H3B, HIST1H3C, HIST1H3D, HIST1H3E, HIST1H3F, HIST1H3G, HIST1H3H, HIST1H3I, HIST1H3J, и H3A2, включая, в частности, HIST2H3C, и H3A3, включая, в частности, HIST3H3; H4, в том числе H41, включая, в частности, HIST1H4A, HIST1H4B, HIST1H4C, HIST1H4D, HIST1H4E, HIST1H4F, HIST1H4C, HIST1H4H, HIST1H4I, HIST1H4J, HIST1H4K, HIST1H4L, и H44, включая, в частности, HIST4H4 и H5.
В соответствии с первым аспектом настоящего изобретения, последовательность нуклеиновых кислот по изобретению содержит кодирующую область, кодирующую пептид или белок, который содержит опухолевый антиген или его фрагмент, вариант или производное. Предпочтительно, кодируемый опухолевый антиген не является репортерным белком (например, люциферазой, зеленым флуоресцентным белком (GFP), усовершенствованным зеленым флуоресцентным белком (EGFP), бета-галактозидазой) и не является маркерным или селективным белком (например, альфа-глобином, галактокиназой и ксантин:гуанинфосфорибозилтрансферазой (GPT)). Предпочтительно, последовательность нуклеиновой кислоты по изобретению не содержит (бактериального) гена устойчивости к антибиотикам, в частности, последовательность гена neo (ген устойчивости к канамицину) или последовательность гена CAT (хлорамфениколацетилтрансфераза, ген устойчивости к хлорамфениколу).
Нуклеиновая кислота по изобретению, как определено выше, включает или кодирует а) кодирующую область, кодирующую пептид или белок, который содержит опухолевый антиген или его фрагмент, вариант или производные; b) по меньшей мере, одну гистоновую структуру типа «стебель-петля» и c) поли(A)-последовательность или сигнала полиаденилирования; предпочтительно для увеличения экспрессии указанного кодируемого пептида и белка, где кодируемый пептид или белок, предпочтительно не является гистоновым белком, не является репортерным белком и/или маркерным или селективным белком, как определено выше. Элементы с b) по c) нуклеиновой кислоты по изобретению могут располагаться в нуклеиновой кислоте по изобретению в любом порядке, т.е. клетки a), b) и c) могут располагаться в следующем порядке: а), b) и c) или a), c) и b) в направлении от 5' к 3' в последовательности нуклеиновых кислот по изобретению, где могут также содержаться дополнительные клетки, как описано в настоящем документе, такие как структура 5'-CAP, последовательность поли(C), стабилизирующие последовательности, последовательности IRES и т.д. Каждый из элементов с a) по c) нуклеиновой кислоты по изобретению, в частности, а) в ди- или мультицистронных конструкциях и/или каждый из элементов b) и c), более предпочтительно, элемент b) могут также повторяться по меньшей мере один раз, предпочтительно, два раза или более в нуклеиновой кислоте по изобретению. В качестве примера, нуклеиновая кислота по изобретению может представлять свои элементы последовательности a), b) и, при необходимости, c), например, в следующем порядке:
5' - кодирующая область - гистоновая структура типа «стебель-петля» - поли(А)-последовательность - 3'; или
5' - кодирующая область - гистоновая структура типа «стебель-петля» - сигнал полиаденилирования - 3'; или
5' - кодирующая область - поли(А)-последовательность -гистоновая структура типа «стебель-петля»-3'; или
5' - кодирующая область - сигнал полиаденилирования -гистоновая структура типа «стебель-петля» - 3'; или
5' - кодирующая область - кодирующая область - гистоновая структура типа «стебель-петля» - сигнал полиаденилирования - 3'; или
5' - кодирующая область - гистоновая структура типа «стебель-петля» - гистоновая структура типа «стебель-петля» - поли(А)-последовательность-3'; или
5'- кодирующая область - гистоновая структура типа «стебель-петля» - гистоновая структура типа «стебель-петля» -сигнал полиаденилирования - 3'; и др.
В этом контексте особенно предпочтительно, чтобы последовательность нуклеиновых кислот по изобретению включала или кодировала а) кодирующую область, кодирующую пептид или белок, который содержит опухолевый антиген или его фрагмент, вариант или производное; b) по меньшей мере, одну гистоновую структуру типа «стебель-петля» и c) поли(A)-последовательность или последовательность полиаденилирования; предпочтительно, для увеличения уровня экспрессии указанного кодируемого пептида и белка, где кодируемый белок предпочтительно не является гистоновым белком, не является репортерным белком (например, люциферазой, GFP, EGFP, бета-галактозидазой, особенно EGFP) и/или не является маркерым или селективным белком (например, альфа-глобином, галактокиназой и ксантин: гуанинфосфорибозилтрансферазой (GPT)).
В дополнительном предпочтительном варианте осуществления первого аспекта последовательность нуклеиновых кислот по изобретению, как определено в настоящем документе, может также находиться в виде модифицированной нуклеиновой кислоты.
В этом контексте, последовательность нуклеиновой кислоты по изобретению, как определено в настоящем документе, может быть модифицирована для предоставления «стабилизированной нуклеиновой кислоты», предпочтительно, стабилизированной РНК, более предпочтительно, РНК, которая, по существу, устойчива к деградации in vivo (например, экзо- или эндонуклеазой). Стабилизированная нуклеиновая кислота может, например, быть получена путем модификации G/C-состава кодирующей области последовательности нуклеиновых кислот по изобретению путем введения нуклеотидных аналогов (например, нуклеотиды с модификациями каркаса, модификациями сахара или модификациями основания) или путем введения стабилизирующих последовательностей в 3'- и 5'- нетранслируемую область последовательности нуклеиновых кислот по изобретению. Как уже упоминалось выше, последовательность нуклеиновых кислот по изобретению, как определено в настоящем документе, может содержать нуклеотидные аналоги/модификации, например, модификации каркаса, модификации сахара или модификации основания. Модификация каркаса в связи с настоящим изобретением является модификацией, при которой фосфаты каркаса нуклеотидов, содержащихся в последовательности нуклеиновых кислот по изобретению, как определено в настоящем документе, являются химически модифицированными. Модификация сахара в связи с настоящим изобретением является химической модификацией сахара нуклеотидов последовательности нуклеиновых кислот по изобретению, как определено в настоящем документе. Кроме того, модификация основания в связи с настоящим изобретением является химической модификацией части основания нуклеотидов молекулы нуклеиновой кислоты последовательности нуклеиновых кислот по изобретению. В этом контексте нуклеотидные аналоги или модификации предпочтительно выбирают из нуклеотидных аналогов, которые применимы для их транскрипции и/или трансляции.
В определенном предпочтительном варианте осуществления первого аспекта настоящего изобретения, определенные в настоящем документе нуклеотидные аналоги/модификации выбирают из модификаций оснований, которые дополнительно увеличивают экспрессию кодируемого белка и которые предпочтительно выбирают из 2-амино-6-хлорпуринрибозид-5'-трифосфата, 2-аминоаденозин-5'-трифосфата, 2-тиоцитидин-5'-трифосфата, 2-тиоуридин-5'-трифосфата, 4-тиоуридин-5'-трифосфата, 5-аминоаллилцитидин-5'-трифосфата, 5-аминоаллилуридин-5'-трифосфата, 5-бромоцитидин-5'-трифосфата, 5-бромоуридин-5'-трифосфата, 5-йодоцитидин-5'-аденозинтрифосфат, 5-йодоуридин-5'-трифосфата, 5-метилцитидин-5'-трифосфата, 5-метилуридин-5'-трифосфата, 6-азацитидин-5'-трифосфата, 6-азауридин-5'-трифосфата, 6-хлорпуринрибозид-5'-аденозинтрифосфата, 7-деазааденозин-5'-трифосфата, 7-деазагуанозин-5'-трифосфата, 8-азааденозин-5'-трифосфата, 8-азидоаденозин-5'-трифосфата, бензимидазолрибозид-5'-трифосфата, N1-метиладенозин-5'-трифосфата, N1-метилгуанозин-5'-трифосфата, N6-метиладенозин-5'-трифосфата, О6-метилгуанозин-5'-трифосфата, псевдоуридин-5'-трифосфата или пуромицин-5'-трифосфата, ксантозин-5'-трифосфата. Особое предпочтение отдается нуклеотидам для модификаций оснований, выбранным из группы нуклеотидов с модифицированными основаниями, состоящей из 5-метилцитидин-5'-трифосфата, 7-деазагуанозин-5'-трифосфата, 5-бромоцитидин-5'-трифосфата и псевдоуридин-5'-трифосфата.
В соответствии с дополнительным вариантом осуществления, последовательность нуклеиновых кислот по изобретению, как определено в настоящем документе, может содержать липидную модификацию. Такая липид-модифицированная нуклеиновая кислота, как правило, состоит из нуклеиновой кислоты, как определено в настоящем документе. Такая молекула липид-модифицированной нуклеиновой кислоты последовательность нуклеиновых кислот по изобретению, как определено в настоящем документе, как правило, дополнительно содержит по меньшей мере один линкер, ковалентно связанный с молекулой нуклеиновой кислоты, и по меньшей мере один липидов, ковалентно связанный с соответствующим линкером. Альтернативно, молекула липид-модифицированной нуклеиновой кислоты содержит по меньшей мере одну молекулу нуклеиновой кислоты, как определено в настоящем документе, и по меньшей мере один (бифункциональный) липид, ковалентно связанный (без линкера) с этой молекулой нуклеиновых кислот. В соответствии с третьей альтернативой, молекула липид-модифицированной нуклеиновой кислоты состоит из молекулы нуклеиновой кислоты, как определено в настоящем документе по меньшей мере одного линкера, ковалентно связанного с молекулой нуклеиновых кислот, и по меньшей мере одного липида, ковалентно связанного с соответствующим линкером, а также по меньшей мере одного (бифункционального) липида, ковалентно связанного (без линкера) с молекулой нуклеиновых кислот. В этом контексте особенно предпочтительно, что модификация липида присутствует на терминальных концах линейной последовательности нуклеиновых кислот по изобретению.
В соответствии с другим предпочтительным вариантом осуществления первого аспекта изобретения, последовательность нуклеиновых кислот по изобретению, как определено в настоящем документе, в особенности, если предоставляется в качестве (м)РНК, можно, тем самым, стабилизировать от расщепления РНКазами добавлением так называемой «5'-САР»-структуры.
В соответствии с дополнительным предпочтительным вариантом осуществления первого аспекта изобретения, последовательность нуклеиновых кислот по изобретению, как определено в настоящем документе, может быть модифицирована последовательностью по меньшей мере 10 цитидинов, предпочтительно по меньшей мере 20 цитидинов, более предпочтительно по меньшей мере 30 цитидинов (так называемая «поли(C)-последовательность»). В частности, последовательность нуклеиновых кислот по изобретению может содержать или кодировать для поли(C)-последовательность, как правило, примерно от 10 до 200 нуклеотидов цитидина, предпочтительно, примерно от 10 до 100 нуклеотидов цитидина, более предпочтительно, примерно от 10 до 70 нуклеотидов цитидина или даже более предпочтительно, примерно от 20 до 50 или даже от 20 до 30 нуклеотидов цитидина. Эта поли(C)-последовательность предпочтительно расположена с 3'-стороны от кодирующей области, содержащейся в нуклеиновой кислоте по изобретению в соответствии с первым аспектом настоящего изобретения.
В конкретном предпочтительном варианте осуществления настоящего изобретения, G/C-состав кодирующей области, кодирующей по меньшей мере один пептид или белок, который содержит опухолевый антиген или его фрагмент, вариант или производное последовательность нуклеиновых кислот по изобретению, как определено в настоящем документе, изменяют, в частности, увеличивают, по сравнению с G/C-составом конкретной кодирующей области дикого типа, т.е. немодифицированной кодирующей областью. Кодируемая аминокислотная последовательность кодирующей области предпочтительно не изменена по сравнению с кодируемой аминокислотной последовательностью конкретной кодирующей области дикого типа.
Модификация G/C-состава кодирующей области последовательности нуклеиновых кислот по изобретению, как определено в настоящем документе, основывается на том, что последовательность любой транслируемой области мРНК является важной для эффективной трансляции этой мРНК. Таким образом, композиция и последовательность разных нуклеотидов являются важными. В частности, последовательности мРНК, имеющие повышенное содержание G(гуанозин)/С(цитозин), более стабильны, чем последовательности мРНК, имеющих повышенное содержание А(аденозин)/U(урацил). В соответствии с изобретением, кодоны кодирующей области, следовательно, меняются по сравнению с ее кодирующей областью дикого типа, при сохранении транслированной аминокислотной последовательности, так что она включают повышенное количество G/C нуклеотидов. В отношении того, что некоторые кодоны кодируют одну и ту же аминокислоту (так называемая вырожденность генетического кода), могут быть определены наиболее благоприятные для стабильности кодоны (так называемые альтернативное использование кодонов).
В зависимости от аминокислоты, кодируемой кодирующей областью последовательность нуклеиновых кислот по изобретению, как определено в настоящем документе, существуют различные возможности для модификации последовательности нуклеиновых кислот, например, кодирующей области, по сравнению с кодирующей областью дикого типа. В случае аминокислот, которые кодируются кодонами, которые содержат исключительно нуклеотиды G или C, никакой модификации кодона не требуется. Таким образом, кодоны для Pro (ССС или CCG), Arg (CGC или CGG), Аlа (GCC или GCG) и Gly (GGC или GGG) не требуют модификации из-за отсутствия A или U.
В отличие от этого, кодоны, которые содержат нуклеотиды А и/или U, могут быть модифицированы за счет замещения другими кодонами, которые кодируют те же аминокислоты, но не содержат А и/или U. Их примерами являются:
кодоны для Pro могут быть изменены с CCU или CCA на ССС или CCG;
кодоны для Arg могут быть изменены с CGU или CGA или AGA или AGG на CGC или CGG;
кодоны для Аlа могут быть изменены с ССС или GCA на GCC или GCG;
кодоны для Gly могут быть изменены с GGU или GGA на GGC или GGG.
В других случаях, даже если нуклеотиды A или U не могут быть исключены из кодонов, при этом возможно уменьшение содержания A и U с помощью кодонов, которые включают меньшее содержание нуклеотидов А и/или U. Примерами этого являются:
кодоны для Phe могут быть изменены с UUU на UUC;
кодоны для Leu могут быть изменены с UUA, UUG, CUU или CUA на CUC или CUG;
кодоны для Ser могут быть изменены с UCU или UCA или AGU на UCC, UCG или AGG;
кодон для Tyr может быть изменен c UAU на UAC;
кодон для Cys может быть изменен c UGU на UGC;
кодон для His может быть изменен c CAU на САС;
кодон для Gln может быть изменен c CAA CAG;
кодон для Ile может быть изменен c AUU или АUА на AUC;
кодон для Thr может быть изменен c АCU или ACA на ACC или ACG;
кодон для Аsn может быть изменен с AAU на AAC;
кодон для Lys может быть изменен с ААА на АAG;
кодон для Val может быть изменен c GUU или GUA на GUC или GUG;
кодон для Asp может быть изменен c GAU GAC;
кодон для Glu может быть изменен c GAA на GAG;
стоп-кодон UAA может быть изменен с UAG или UGA.
В случае кодонов для Met (AUG) и Trp (UGG), с другой стороны, возможность модификации последовательности отсутствует.
Замены, перечисленные выше, могут использоваться либо по отдельности, либо во всех возможных комбинациях, чтобы увеличить G/C-состав кодирующей области последовательности нуклеиновых кислот по изобретению, как определено в настоящем документе, в частности, по сравнению с ее конкретной кодирующей областью дикого типа (т.е. исходной последовательностью). Так, например, все кодоны для Thr, содержащиеся в последовательности дикого типа, могут быть изменены на ACC (или АCG).
В контексте вышесказанного показаны кодоны мРНК. Соответственно, присутствующий в мРНК уридин может также присутствовать в виде тимидина в соответствующей ДНК, кодирующей конкретную мРНК.
Предпочтительно, G/C-состав кодирующей области последовательности нуклеиновых кислот по изобретению, как определено в настоящем документе, увеличивают по меньшей мере на 7%, более предпочтительно по меньшей мере на 15%, особенно предпочтительно по меньшей мере на 20% по сравнению с G/C-составом кодирующей области дикого типа. В соответствии с конкретным вариантом осуществления по меньшей мере 5%, 10%, 20%, 30%, 40%, 50%, 60%, более предпочтительно по меньшей мере 70%, даже более предпочтительно по меньшей мере 80% и наиболее предпочтительно по меньшей мере 90%, 95% или даже 100% замещаемых кодонов кодирующей области, кодирующей по меньшей мере один пептид или белок, который содержит опухолевый антиген или его фрагмент, вариант или производное, заменяют, увеличивая, таким образом, G/C-состав указанной кодирующей области.
В этом контексте, особенно предпочтительно увеличивать G/C-состав кодирующей области последовательности нуклеиновых кислот по изобретению, как определено в настоящем документе, до максимального (т.е. 100% заменяемых кодонов), по сравнению с кодирующей областью дикого типа.
В соответствии с изобретением, дополнительная предпочтительная модификация кодирующей области, кодирующей по меньшей мере один пептид или белок, который содержит опухолевый антиген или его фрагмент, вариант или производное последовательности нуклеиновых кислот по изобретению, как определено в настоящем документе, основывается на обнаружении, что эффективность трансляции также определяется различной частотой встречаемости тРНК в клетках. Таким образом, если так называемые «редкие кодоны» присутствуют в кодирующей области последовательности нуклеиновых кислот по изобретению, как определено в настоящем документе, в увеличенной степени, соответствующая модифицированная последовательность нуклеиновых кислот транслируется в значительно более слабой степени, чем в случае, когда присутствуют кодоны, кодирующие относительно «частые» тРНК.
В этом контексте кодирующую область последовательности нуклеиновых кислот по изобретению предпочтительно модифицируют по сравнению с соответствующей кодирующей областью дикого типа таким образом, чтобы по меньшей мере один кодон последовательности дикого типа, который кодирует тРНК, которая в клетке встречается относительно редко, заменяется на кодон, который кодирует тРНК, которая встречается относительно часто в клетке и несет ту же аминокислоту, что и сравнительно редкая тРНК. При помощи этой модификации, кодирующая область последовательности нуклеиновых кислот по изобретению, как определено в настоящем документе, модифицируется таким образом, что встраиваются кодоны, для которых доступны часто встречающиеся тРНК. Иными словами, в соответствии с изобретением, при помощи этой модификации все кодоны кодирующей области дикого типа, которые кодируют тРНК, которая встречается в клетке относительно редко, может быть в каждом случае заменен на кодон, который кодирует тРНК, которая относительно часто встречается в клетке и которая, в каждом случае, несет ту же аминокислоту, что и сравнительно редкая тРНК.
тРНК, которые встречаются относительно часто в клетке и который, напротив, встречаются сравнительно редко, известны специалистам в рассматриваемой области техники; Ср. например, Akashi, Curr. Opin. Genet. Dev. 2001, 11(6): 660-666. Кодоны, которые используются для определенной аминокислоты, тРНК, которая встречается наиболее часто, например, кодон для Gly, который использует тРНК, которая встречается наиболее часто в клетке человека, являются особенно предпочтительным.
В соответствии с изобретением особенно предпочтительно связать G/C-состав последовательности, который увеличивают, в частности максимизируют, в кодирующей области последовательности нуклеиновых кислот по изобретению, как определено в настоящем документе, с «частыми» кодонами без изменения аминокислотной последовательности пептида или белка, кодируемого кодирующей областью последовательности нуклеиновых кислот. Этот предпочтительный вариант осуществления позволяет предоставлять особенно эффективно транслируемую и стабилизированную (модифицированную) последовательность нуклеиновых кислот по изобретению, как определено в настоящем документе.
В соответствии с другим предпочтительным вариантом осуществления первого аспекта изобретения, последовательность нуклеиновых кислот по изобретению, как определено в настоящем документе, предпочтительно содержит дополнительно по меньшей мере одну 5'- и 3'-стабилизирующую последовательность. Эти стабилизирующие последовательности в 5'- и 3'-нетранслируемых областях обладают эффектом увеличения времени полужизни нуклеиновой кислоты, в частности мРНК в цитозоле. Эти стабилизирующие последовательности могут иметь 100% идентичности последовательности с природными последовательностями, которые присутствуют вирусах, бактериях и эукариотах, но также могут быть частично или полностью синтетическими. Нетранслируемые последовательности (UTR) гена (альфа)-глобина, например, из Homo sapiens или Xenopus laevis, могут быть упомянуты в качестве примера стабилизирующих последовательностей, которые могут быть использованы в настоящем изобретении для стабилизированных нуклеиновых кислот. Другой пример стабилизирующей последовательности имеет общую формулу (C/U)CCANxCCC(U/A)PyxUC(C/U)CC (SEQ ID NO:55), которая содержится в 3'-UTR очень стабильных РНК, которые кодируют (альфа)-глобин, тип(I)-коллаген, 15-липоксигеназу или тирозингидроксилазу (ср. Holcik et al., Proc. Natl. Acad. Sci. USA 1997, 94: 2410-2414). Такие стабилизирующие последовательности, конечно, могут быть использованы по отдельности или в комбинации друг с другом, а также в комбинации с другими стабилизирующими последовательностями, известными специалистам в рассматриваемой области техники. В этом контексте особенно предпочтительно, чтобы 3'-UTR последовательность гена альфа-глобина была расположена с 3'-стороны от кодирующей области, кодирующей по меньшей мере один пептид или белок, который содержит опухолевый антиген или его фрагмент, вариант или производное, входящей в последовательность нуклеиновых кислот по изобретению в соответствии с первым аспектом настоящего изобретения.
Замены, дополнения или удаления оснований предпочтительно проводят в последовательности нуклеиновых кислот по изобретению, как определено в настоящем документе, с помощью ДНК-матрицы для подготовки последовательности нуклеиновых кислот при помощи методик хорошо известного сайт-направленного мутагенеза или стратегии олигонуклеотидного лигирования (см., например, Maniatis et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, 3rd ed., Cold Spring Harbor, NY, 2001).
Любую из вышеперечисленных модификаций можно применить к последовательности нуклеиновых кислот по изобретению, как определено в настоящем документе, и дополнительно к любой нуклеиновой кислоте, как используется в контексте настоящего изобретения, и их можно, если применимо или при необходимости, объединить друг с другом в любой комбинации, при условии, что эти комбинации модификаций не мешают друг другу в соответствующей нуклеиновой кислоте. Специалист в данной области техники, соответственно, сможет сделать свой выбор.
Последовательности нуклеиновых кислот, используемые в соответствии с настоящим изобретением, как определено в настоящем документе, могут быть подготовлены с помощью любого способа, известного в рассматриваемой области техники, в том числе синтетических способов, таких как, например, твердофазного синтеза, а также способов in vitro, таких как реакции транскрипции in vitro или реакции in vivo, такие как размножение in vivo ДНК-плазмид в бактериях.
В таком процессе, для подготовки последовательности нуклеиновых кислот по изобретению, как определено в настоящем документе, особенно если нуклеиновая кислота находится в виде мРНК, соответствующая молекула ДНК может быть транскрибирована in vitro. Эта ДНК-матрицы предпочтительно включает подходящий промотор, например, промотор Т7 или SP6, для транскрипции in vitro, за которым следует нужная нуклеотидная последовательность молекулы нуклеиновых кислот, например, мРНК, которую нужно подготовить и сигнал терминации для транскрипции in vitro. Молекула ДНК, которая образует матрицу по меньшей мере одной представляющей интерес РНК, может быть подготовлена с помощью ферментативной пролиферации и последующего выделения как части плазмиды, который может реплицироваться в бактериях. Плазмиды, которые могут быть отмечены как подходящие для настоящего изобретения, представляют собой, например, плазмиды pT7Ts (инв. номер CenBank U26404; Lai et al., Development, 1995, 121: 2349-2360), серию pGEM®, например, pGEM®-1 (инв. номер GenBank X65300; Promega) и pSP64 (инв. номер GenBank X65327); См. также Mezei and Storts, Purification of PCR Products, in: Griffin and Griffin (ed.), PCR Technology: Current Innovation, CRC Press, Boca Raton, FL, 2001.
Последовательность нуклеиновых кислот по изобретению, как определено в настоящем документе, а также белки или пептиды, кодируемые с помощью этой последовательности нуклеиновых кислот, могут включать фрагменты или варианты таких последовательностей. Такие фрагменты или варианты могут, как правило, содержать последовательность, обладающую идентичностью последовательности с одной из вышеперечисленных последовательностей нуклеиновых кислот, или с одним из белков или пептидов и последовательностей, если кодируемая нуклеиновой кислотой по изобретению последовательностью по меньшей мере на 5%, 10%, 20%, 30%, 40%, 50%, 60%, предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 80%, в равной степени более предпочтительно по меньшей мере 85%, даже более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% или даже 97%, 98% или 99%, идентична со всей последовательностью дикого типа, либо на уровне нуклеиновой кислоты, либо на уровне аминокислоты.
«Фрагменты» белков или пептидов в контексте настоящего изобретения (например, как кодируется последовательностью нуклеиновых кислот по изобретению, как определено далее) могут включать последовательность белка или пептида, как определено в настоящем документе, который, с учетом его аминокислотной последовательности (или кодирующей его молекулой нуклеиновой кислоты), усеченной/укороченной с N-конца, C-конца и/или внутри последовательности по сравнению с аминокислотной последовательностью оригинального (нативного) белка (или кодирующей его молекулой нуклеиновой кислоты). Такое усечение, таким образом, может иметь место либо на аминокислотном уровне, либо, соответственно, на уровне нуклеиновых кислот. Идентичность последовательности в отношении такого фрагмента, как определено в настоящем документе, может поэтому предпочтительно относиться ко всему белку или пептиду, как определено в настоящем документе, или всей (кодирующей) молекуле нуклеиновых кислот подобного белка или пептида. Аналогичным образом, «фрагменты» нуклеиновых кислот в контексте настоящего изобретения могут содержать последовательность нуклеиновых кислот, как определено в настоящем документе, которая является, с учетом его молекулы нуклеиновой кислоты, 5'- и 3'- и/или внутри последовательности усеченной/сокращенной по сравнению с молекулой нуклеиновой кислоты исходной (нативной) молекулы нуклеиновой кислоты. Идентичность последовательности в отношении такого фрагмента, как определено в настоящем документе, может, поэтому предпочтительно относиться ко всей нуклеиновой кислоте, как определено в настоящем документе, и предпочтительный уровень идентичности последовательности, как правило, составляет такой, как указано в настоящем документе. Фрагменты имеют одну и ту же биологическую функцию или специфическую активность или по меньшей мере сохраняют активность природного полноразмерного белка по меньшей мере на 50%, более предпочтительно по меньшей мере на 70%, даже более предпочтительно по меньшей мере 90% (измеряется в соответствующем функциональном тесте, например, анализе, оценивающем антигенные свойства с помощью соответствующей системы тестирования, которая, например, определяет иммунологическую реакцию, например, экспрессию и/или секрецию соответствующего цитокина (в качестве индикатора иммунной реакции)) по сравнению с природным полноразмерным пептидом и белком, например, его специфическое антигенное или лечебное свойство. Соответственно, в предпочтительном варианте осуществления «фрагмент» представляет собой часть полноразмерного (природного) опухолевого антигенного белка, который оказывает опухолевые антигенные свойства на иммунную систему, как указано в настоящем документе.
Фрагменты белков или пептидов в контексте настоящего изобретения (например, как кодируется последовательностью нуклеиновых кислот по изобретению, как это определено в настоящем документе), кроме того, могут содержать последовательность белка или пептида, как определено в настоящем документе, который имеет длину от примерно 6 до примерно 20 или даже больше аминокислот, например, фрагменты, процессированные и представленные молекулами MHC (главного комплекса гистосовместимости) класса I, предпочтительно имеющие длину примерно от 8 до примерно 10 аминокислот, например, 8, 9 или 10, (или даже 6, 7, 11 или 12 аминокислот), или фрагменты, процессированные и представленные молекулами MHC класса II, предпочтительно имеющие длину примерно 13 или более аминокислот, например, 13, 14, 15, 16, 17, 18, 19, 20 или даже более аминокислот, причем эти фрагменты могут быть выбраны из любой части аминокислотной последовательности. Эти фрагменты, как правило, распознаются Т-клетками в виде комплекса, состоящего из пептидного фрагмента и молекулы MHC, т.е. фрагменты, как правило, не распознаются в их нативной форме. Фрагменты белков или пептидов, как определено в настоящем документе, могут содержать по меньшей мере один эпитоп этих белков или пептидов. Кроме того, домены белка, наподобие внеклеточного домена, внутриклеточного домена или трансмембранного домена и укороченных или усеченных версий белка также могут рассматриваться как содержащие фрагмент белка.
Фрагменты белков или пептидов, как определено в настоящем документе (например, кодируемые последовательность нуклеиновых кислот по изобретению, как определено в настоящем документе) могут также включать эпитопов этих белков или пептидов. Т-клеточные эпитопы или части белков в контексте настоящего изобретения могут включать фрагменты, предпочтительно, имеющие длину от 6 до 20 или даже больше аминокислот, например, фрагменты, процессированные и представленные молекулами MHC класса I, предпочтительно имеющие длину примерно от 8 до примерно 10 аминокислот, например, 8, 9 или 10, (или даже 11 или 12 аминокислот), или фрагменты, процессированные и представленные молекулами MHC класса II, предпочтительно имеющие длину примерно 13 или более аминокислот, например, 13, 14, 15, 16, 17, 18, 19, 20 или даже больше аминокислот, причем эти фрагменты могут быть выбраны из любого части аминокислотной последовательности. Эти фрагменты, как правило, распознаются Т-клетками в виде комплекса, состоящего из пептидного фрагмента и молекулы MHC, т.е. фрагменты, как правило, не распознаются в их нативной форме.
B-клеточные эпитопы обычно представляют собой фрагменты, расположенные на внешней поверхности (нативного) белковых или пептидных антигенов, как определено в настоящем документе, предпочтительно имеющие от 5 до 15 аминокислот, более предпочтительно, имеющие от 5 до 12 аминокислот, даже более предпочтительно, от 6 до 9 аминокислот, которые могут быть распознаны антителами, т.е. в их нативной форме.
Такие эпитопы белков или пептидов можно, кроме того, выбрать из любого из упомянутых в настоящем документе вариантов таких белков или пептидов. В этом контексте антигенные детерминанты могут быть конформационными или прерывающимися эпитопами, которые состоят из сегментов белков или пептидов, как определено в настоящем документе, которые являются прерывающимися по аминокислотной последовательности белков или пептидов, как определено в настоящем документе, но объединяются в трехмерную структуру, или непрерывными или линейными эпитопами, которые состоят из одной полипептидной цепи.
«Варианты» белков или пептидов, как определено в контексте настоящего изобретения, могут кодироваться последовательностью нуклеиновых кислот по изобретению, как определено в настоящем документе. Таким образом, можно создать белок или пептид, имеющий последовательность аминокислот, которая отличается от исходной последовательности одной или более (2, 3, 4, 5, 6, 7 или больше) мутацией(ями), например, одной или более замещенной, встроенной и/или удаленной аминокислотой (ами). Предпочтительный уровень идентичности последовательности «вариантов» в свете последовательности полноразмерного природного белка составляет, как правило, как указано в настоящем документе. Предпочтительно, варианты имеют одну и ту же биологическую функцию или специфическую активности или по меньшей мере сохраняют активность природного полноразмерного белка по меньшей мере на 50%, более предпочтительно по меньшей мере 70%, даже более предпочтительно по меньшей мере 90% (измеряется в соответствующем функциональном тесте, например, с помощью анализа оценки иммунологической реакции на опухолевый антиген на секрецию и/или экспрессию одного или нескольких цитокинов), по сравнению с полноразмерным нативным пептидом и белком, например, его специфические антигенные свойства. Соответственно, в предпочтительном варианте осуществления «вариант» представляет собой вариант опухолевого антигенного белка, который обладает опухолевыми антигенными свойствами в такой степени, как указано в настоящем документе.
«Варианты» белков или пептидов, как определено в контексте настоящего изобретения (например, как кодируемые нуклеиновой кислотой, как определено в настоящем документе) могут содержать консервативную аминокислотную замену(ы) по сравнению с их нативной, т.е. не мутированной физиологической последовательностью. Эти аминокислотные последовательности, а также кодирующие их нуклеотидные последовательности, в частности, подпадают под варианты терминов, как определено в настоящем документе. Замены, в которых аминокислоты, которые исходят из одного и того же класса, обмениваются друг к другу, называются консервативными заменами. В частности, такими являются аминокислоты, имеющие алифатические боковые цепи, положительно или отрицательно заряженные боковые цепи, ароматические группы в боковых цепях или аминокислоты, боковые цепи которых могут входить в водородные связи, например, боковые цепи, которые имеют гидроксильную функцию. Это означает, что, например, аминокислота, имеющая полярную боковую цепь, заменяется другой аминокислотой, также имеющей полярную боковую цепь, или, например, аминокислота, характеризующаяся гидрофобной боковой цепью, заменяется другой аминокислоты, также имеющей гидрофобную боковую цепь (например, серин (треонин) треонином (серином) или лейцин (изолейцин) изолейцином (лейцином)). Вставки и замены возможны, в частности, в тех положениях последовательности, которые не вызывают никакой модификации трехмерной структуры или не затрагивают область связывания. Модификации трехмерной структуры путем вставки(ок) или делеции(ий) можно легко определить, например, с помощью CD-спектров (спектры кругового дихроизма) (Urry, 1985, Absorption, Circular Dichroism and ORD of Polypeptides, in: Modern Physical Methods in Biochemistry, Neuberger eta/, (ed.), Elsevier, Амстердам).
Кроме того, варианты белков или пептидов, как определено в настоящем документе, которые могут кодироваться последовательностью нуклеиновых кислот по изобретению, как определено в настоящем документе, могут также содержать те последовательности, в которых нуклеотиды нуклеиновой кислоты обмениваются в соответствии с вырожденностью генетического кода, не приводя к изменению соответствующей аминокислотной последовательности белка или пептид, т.е. аминокислотная последовательность или по меньшей мере ее часть может не отличаться от исходной последовательности одной или несколькими мутациями(ей) в пределах вышеуказанного значения.
Для того чтобы определить процентное соотношение, с которым две последовательности идентичны, например, последовательности нуклеиновых кислот или аминокислотные последовательности, как определено в настоящем документе, предпочтительно, аминокислотные последовательности, кодируемые последовательностью нуклеиновых кислот по изобретению, как определено в настоящем документе, или аминокислотные последовательности сами по себе, последовательности могут быть выровнены для последующего выравнивания друг с другом. Соответственно, например, положение в первой последовательности может сравниваться с соответствующим положением во второй последовательности. Если положение в первой последовательности занимает такой же компонент, как в случае с положением во второй последовательности, то две последовательности идентичны по этому положению. Если это не так, последовательности отличаются в этом положении. Если имеют место вставки во второй последовательности по сравнению с первой последовательностью, пробелы могут быть встроены в первую последовательность, чтобы обеспечить дополнительное выравнивание. Если во второй последовательности имеют место делеции, по сравнению с первой последовательностью, во вторую последовательность могут быть вставлены пробелы, чтобы обеспечить дополнительное выравнивание. Процентное соотношение, при котором две последовательности идентичны, является тогда функцией количества одинаковых положений, деленного на общее число положений, включая те положения, которые заняты только в одной последовательности. Процентное соотношение, при котором две последовательности идентичны, может быть определено с помощью математического алгоритма. Предпочтительный, но не ограничивающий пример математического алгоритма, который может быть использован, представляет собой алгоритм Karlin et al. (1993), PNAS USA, 90:5873-5877 or Altschul et al. (1997), Nucleic Acids Res., 25:3389-3402. Такой алгоритм интегрирован в программу BLAST. Последовательности, которые идентичны последовательностям по настоящему изобретению, в определенной степени могут быть идентифицированы с помощью этой программы.
Последовательность нуклеиновых кислот по изобретению, как определено в настоящем документе, может кодировать производные пептида или белка. Такое производное пептида или белка представляет собой молекулу, которая получена из другой молекулы, такой как указанный пептид или белок. «Производное», как правило, содержит полноразмерную последовательность природного пептида или белка и дополнительные особенности последовательности, например, на N- или на C-конце, которые могут придавать дополнительную функцию природному полноразмерному пептиду/белка. Кроме того, такие производные имеют ту же биологическую функцию или специфическую активность или по меньшей мере сохраняют активность природного полноразмерного белка по меньшей мере на 50%, более предпочтительно по меньшей мере 70%, даже более предпочтительно по меньшей мере 90% (измеряется в соответствующем функциональном тесте, см. выше, например, как выражается при помощи экспрессии и/или секреции цитокинов в иммунологической реакции) по сравнению с полноразмерным пептидом и белком, например, его специфическое терапевтическое свойство. При этом, «производное» пептида или белка также включает в себя (химерное) слияние пептидов/белков, включающее пептид или белок, используемый в настоящем изобретении, или природный полноразмерный белок (или его вариант/фрагмент), слитый с отдельным пептидом/белком, присваивая, например, две или более биологических функций слитому пептиду/белку. Например, слияние включает в себя метку, такую как, например, эпитоп, например, FLAG-эпитоп или V5-эпитоп или HA-эпитоп. Например, эпитоп представляет собой FLAG-эпитоп. Такой тег полезен, например, для очистки слитого белка.
В этом контексте «вариант» белка или пептида может иметь по меньшей мере 70%, 75%, 80%, 85%, 90%, 95%, 98% или 99% аминокислотной идентичности на участке в 10, 20, 30, 50, 75 или 100 аминокислот такого белка или пептида. Аналогично, «вариант» последовательности нуклеиновых кислот или, в частности, фрагмент может иметь по меньшей мере 70%, 75%, 80%, 85%, 90%, 95%, 98% или 99% идентичности нуклеотидов на участке в 10, 20, 30, 50, 75 или 100 нуклеотидов такой последовательности нуклеиновых кислот; однако, как правило, имея в виду природные полноразмерные последовательности. В случае «фрагментов», как правило, идентичность последовательностей устанавливается для фрагмента с длиной (фрагмента) с частью полноразмерного белка (отражающей ту же длину, что и фрагмент), которая демонстрирует наивысший уровень идентичности последовательности.
В дополнительном предпочтительном варианте осуществления первого аспекта настоящего изобретения последовательность нуклеиновых кислот по изобретению связывают с носителем, трансфекцирующим или комплексообразующим агентом для увеличения эффективности трансфекции и/или иммуностимулирующих свойств последовательности нуклеиновых кислот по изобретению. Особенно предпочтительными агентами в этом контексте, подходящими для увеличения эффективности трансфекции, являются катионные или поликатионные соединения, в том числе протамин, нуклеолайн, спермин или спермидин, или другие катионные пептиды или белки, такие как поли-L-лизин (PLL), полиаргинин, основные полипептиды, проникающие в клетки пептиды (СРР), в том числе ВИЧ-связывающиеся пептиды, HIV-1 Tat (ВИЧ), Tat-производные пептиды, пенетратин, VP22-производные или аналоговые пептиды, HSV VP22 (Herpes simplex), MAP, KALA или домены белковой трансдукции (PTD), PpT620, пролин-богатые пептиды, аргинин-богатые пептиды, лизин-богатые пептиды, MPG-пептид(ы), Рер-1, L-олигомеры, пептид(ы) кальцитонина, пептиды-производные Antennapedia (в частности, из Drosophila antennapedia), pAntp, pIsl, FGF, лактоферрин, транспортан, буфорин-2, Bac715-24, SynB, SynB(1), pVEC, hCT-производные пептиды, SAP или гистоны. Кроме того, предпочтительные катионные или поликатионные белки или пептиды могут быть выбраны из следующих белков или пептидов, имеющих следующую общую формулу: (Arg)l;(Lys)m;(His)n;(Orn)o;(Хаа)x, где l+m+n+o+x=8-15, и l, m, n или o независимо друг от друга могут представлять собой любое число, выбранное из 0, 1 , 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15, при условии, что общее содержание Arg, Lys, His и Orn представляет по меньшей мере 50% всех аминокислот олигопептида; и Xaa может представлять собой любую аминокислоту, выбранную из нативной (= природной) или отличной от нативной аминокислоты, кроме Arg, Lys, His или Orn; и x может представлять собой любое число, выбранное из 0, 1, 2, 3 или 4, при условии, что общее содержание Xaa не превышает 50 % всех аминокислот олигопептида. Особенно предпочтительными катионными пептидами в этом контексте представляют собой, например, Arg7, Arg8, Arg9, H3R9, R9H3, H3R9H3, YSSR9SSY, (RKH)4, Y(RKH)2R и др. Другие предпочтительные катионные или поликатионные соединения, которые могут быть использованы в качестве средств для трансфекции, могут включать катионные полисахариды, например, хитозан, полибрен, катионные полимеры, например, полиэтиленимин (PEI), катионные липиды, например, DOTMA: [1-(2,3-сиолеилокси)пропил)]-N,N,N-триметиламмония хлорид, DMRIE, ди-C14-амидин, DOTIM, SAINT, DC-ChoI, BGTC, CTAP, DOPC, DODAP, DOPE: диолеилфосфатидилэтаноламин, DOSPA, DODAB, DOIC, DMEPC, DOGS: диоктадециламидоглицилспермин, DIMRI: димиристооксипропилдиметилгидроксиэтиламмония бромид, DOTAP: диолеоилокси-3-(триметиламмонио)пропан, DC-6-14: О,О- дитетрадеканоил-N-(α-триметиламмониоацетил)диэтаноламинхлорид, CLIP1:рац-[(2,3-диоктадецилоксипропил)(2-гидроксиэтил)]-диметиламмонийхлорид, CLIP6: рац-[2(2,3-дигексадецилоксипропил-оксиметилокси)этил]триметиламмония, CLIP9: рац-[2(2,3-дигексадецилоксипропил-оксисукцинилокси)этил]-триметиламмония, олигофектамин, или катионные или поликатионные полимеры, например, модифицированные полиаминокислоты, например, β-аминокислотные полимеры или обратные полиамиды и др., модифицированные полиэтилены, таких как PVP (поли(N-этил-4-винилпиридинийбромид)) и др., модифицированные акрилаты, такие как pDMAEMA (поли(диметиламиноэтилметилакрилат)) и др., модифицированные амидоамины, такие как рАМАМ (поли(амидоамин)) и др., модифицированный полибетааминоэфир (PBAE), такие как диаминовые концевые модифицированные 1,4 бутандиолдиакрилат-co-5-амино-1-пентанолполимеры и др., дендримеры, такие как полипропиленаминные дендримеры или дендримеры на основе рАМАМ и др., полиимин(ы), такие как PEI: поли(этиленимин), поли(пропиленимин)и др., полиаллиламин, полимеры на основе сахарного остова, такие как полимеры на основе циклодекстрина, полимеры на основе декстрана, хитозана и др., полимеры на основе силанового остова, такие как сополимеры PMOXA-PDMS и др., блокполимеры, состоящие из комбинации одного или более катионных блоков (например, выбранные из катионного полимера, как упоминалось выше) и одного или более гидрофильных или гидрофобных блоков (например, полиэтиленгликоль); и др.
В этом контексте особенно предпочтительно, что нуклеиновая кислота по изобретению образует комплекс по меньшей мере частично, с катионными или поликатионными соединениями, предпочтительно катионными белками или пептидами. «Частично» означает, что только часть нуклеиновой кислоты по изобретению образует комплекс с катионным или поликатионным соединением, а остаток нуклеиновой кислоты по изобретению находится в некомплексной форме («свободной»). Предпочтительно, соотношение нуклеиновой кислоты в комплексе: свободной нуклеиновой кислоте выбирают из диапазона от примерно 5:1 (масс/масс) до примерно 1:10 (масс/масс), что более предпочтительно, из диапазона от примерно 4:1 (масс/масс) до примерно 1:8 (масс/масс), даже более предпочтительно, от диапазона примерно 3:1 (масс/масс) до примерно 1:5 (масс/масс) или 1:3 (масс/масс), и наиболее предпочтительное соотношение нуклеиновой кислоты в комплексе к свободной нуклеиновой кислоте выбирается из соотношении примерно 1:1 (масс/масс)).
Предпочтительно, чтобы последовательность нуклеиновых кислот по изобретению предоставляется либо в свободном виде, либо в виде комплекса, например, с поликатионными соединениями любой химической структуры, предпочтительно поликатионными (поли)пептидами или синтетическими поликатионными соединениями. Предпочтительно, последовательность нуклеиновых кислот не предоставляется вместе с комплектующими клетками.
В дополнительном аспекте изобретение предоставляет композицию или набор или набор частей, содержащий множество или больше, чем одну, предпочтительно от 2 до 10, более предпочтительно от 2 до 5, наиболее предпочтительно от 2 до 4 последовательностей нуклеиновых кислот по изобретению, как определено в настоящем документе. Эти композиции по изобретению содержат более одной последовательности нуклеиновых кислот по изобретению, предпочтительно кодирующих различные пептиды и белки, которые содержат, предпочтительно, различные опухолевые антигены или их фрагменты, варианты или производные.
В предпочтительном варианте осуществления композиция или набор или набор частей по изобретению, содержащий множество (что означает, как правило, больше, чем 1, 2, 3, 4, 5, 6 или более 10 нуклеиновых кислот, например, от 2 до 10, предпочтительно от 2 до 5 нуклеиновых кислот) последовательностей нуклеиновых кислот по изобретению, в частности, для применения при лечении рака предстательной железы (РСа) содержит, по меньшей мере:
а) нуклеиновую кислоту по изобретению, кодирующую по меньшей мере один пептид и белок, где указанный кодируемый пептид или белок содержит опухолевый антиген PSA или его фрагмент, вариант или производное; и
b) нуклеиновую кислоту по изобретению, кодирующую по меньшей мере один пептид и белок, где указанный кодируемый пептид или белок содержит опухолевый антиген PSМA или его фрагмент, вариант или производное; и
c) нуклеиновую кислоту по изобретению, кодирующую по меньшей мере один пептид и белок, где указанный кодируемый пептид или белок содержит опухолевый антиген PSСA или его фрагмент, вариант или производное; и
d) нуклеиновую кислоту по изобретению, кодирующую по меньшей мере один пептид и белок, где указанный кодируемый пептид или белок содержит опухолевый антиген STEAP-1 или его фрагмент, вариант или производное.
В дополнительном предпочтительном варианте осуществления композиция или набор или набор частей по изобретению, содержащий множество (что означает, как правило, больше, чем 1, 2, 3, 4, 5, 6 или более 10 нуклеиновых кислот, например, от 2 до 10, предпочтительно от 2 до 5 нуклеиновых кислот) последовательностей нуклеиновых кислот по изобретению, в частности, для применения при лечении немелкоклеточного рака легкого (NSCLC), содержит, по меньшей мере:
a) последовательность нуклеиновых кислот, содержащую или кодирующую
i. кодирующую область, кодирующую по меньшей мере один пептид или белок, который содержит опухолевый антиген NY-ESO-1 или его фрагмент, вариант или производное,
ii. по меньшей мере, одну гистоновую структуру типа «стебель-петля» и
iii. поли(A)-последовательность или сигнал полиаденилирования;
b) нуклеиновую кислоту по изобретению, кодирующую по меньшей мере один пептид и белок, где указанный кодируемый пептид или белок содержит опухолевый антиген 5T4 или его фрагмент, вариант или производное; и
c) нуклеиновую кислоту по изобретению, кодирующую по меньшей мере один пептид и белок, где указанный кодируемый пептид или белок содержит опухолевый антиген сурвивин или его фрагмент, вариант или производное.
Кроме того, в качестве альтернативы, композиция или набор или набор частей по изобретению, содержащий множество (что означает, как правило, больше, чем 1, 2, 3, 4, 5, 6 или более 10 нуклеиновых кислот, например, от 2 до 10, предпочтительно от 2 до 5 нуклеиновых кислот) последовательностей нуклеиновых кислот по изобретению, в частности, для применения при лечении немелкоклеточного рака легкого (NSCLC), содержит, по меньшей мере:
a) последовательность нуклеиновых кислот, содержащую или кодирующую
i. кодирующую область, кодирующую по меньшей мере один пептид или белок, который содержит опухолевый антиген NY-ESO-1 или его фрагмент, вариант или производное,
ii. по меньшей мере, одну гистоновую структуру типа «стебель-петля» и
iii. поли(A)-последовательность или сигнал полиаденилирования;
b) нуклеиновую кислоту по изобретению, кодирующую по меньшей мере один пептид и белок, где указанный кодируемый пептид или белок содержит опухолевый антиген 5T4 или его фрагмент, вариант или производное; и
c) нуклеиновую кислоту по изобретению, кодирующую по меньшей мере один пептид и белок, где указанный кодируемый пептид или белок содержит опухолевый антиген сурвивин или его фрагмент, вариант или производное; и
d) нуклеиновую кислоту по изобретению, кодирующую по меньшей мере один пептид и белок, где указанный кодируемый пептид или белок содержит опухолевый антиген MAGE-C1 или его фрагмент, вариант или производное; и
e) нуклеиновую кислоту по изобретению, кодирующую по меньшей мере один пептид и белок, где указанный кодируемый пептид или белок содержит опухолевый антиген MAGE-C2 или его фрагмент, вариант или производное.
В соответствии с еще одним аспектом настоящее изобретение также предоставляет способ увеличения экспрессии кодируемого пептида или белка, включающий стадии, например, a) предоставление последовательности нуклеиновых кислот по изобретению, как определено в настоящем документе, или композиции по изобретению, содержащую множество (что означает, как правило, больше, чем 1, 2, 3, 4, 5, 6 или более 10 нуклеиновых кислот, например, от 2 до 10, предпочтительно от 2 до 5 нуклеиновые кислот) последовательностей нуклеиновых кислот по изобретению, как определено в настоящем документе, b) применение или введение последовательности нуклеиновых кислот по изобретению, как это определено в настоящем документе, или композиции по изобретению, содержащей множество последовательностей нуклеиновых кислот по изобретению, как определено в настоящем документе, в систему экспрессии, например, в бесклеточную систему экспрессии, клетку (например, экспрессионная клетка-хозяин или соматическая клетка), ткань или организм. Способ может применяться для лаборатории, для исследования, для диагностики, для коммерческого получения пептидов и белков и/или для терапевтических целей. В этом контексте, как правило, после подготовки последовательности нуклеиновых кислот по изобретению, как определено в настоящем документе, или композиции по изобретению, содержащей множество последовательностей нуклеиновых кислот по изобретению, как определено в настоящем документе, ее, как правило, применяют или вводят в бесклеточную систему экспрессии, клетку (например, экспрессионная клетка-хозяин или соматическая клетка), ткань или организм, например, в свободном виде или в виде комплекса, так и в виде фармацевтической композиции или вакцины, как описано в настоящем документе, предпочтительно через трансфекцию или с помощью любого из способов введения, как описано в настоящем документе. Способ можно проводить in vitro, in vivo и ex vivo. Кроме того, способ можно осуществлять в контексте лечения конкретного заболевания, в частности, при лечении онкологических или опухолевых заболеваний, предпочтительно, как определено в настоящем документе.
В этом контексте «in vitro» определяется в настоящем документе как трансфекция или трансдукция нуклеиновой кислоты по изобретению, как определено в настоящем документе, или композиции по изобретению, содержащей множество последовательностей нуклеиновых кислот по изобретению, как определено в настоящем документе, в клетки в культуре вне организма; «in vivo» определяется в настоящем документе, как трансфекция или трансдукция нуклеиновой кислоты по изобретению, как определено в настоящем документе, или композиции по изобретению, содержащей множество последовательностей нуклеиновых кислот по изобретению, в клетки путем применения нуклеиновой кислоты по изобретению или композиции по изобретению, в весь организм в целом или индивидуум, и «ex vivo» определяется здесь как трансфекция или трансдукция нуклеиновой кислоты по изобретению или композиции по изобретению, содержащей множество последовательностей нуклеиновых кислот по изобретению (что означает, как правило, больше, чем 1, 2, 3, 4, 5, 6 или более 10 нуклеиновых кислот, например, от 2 до 10, предпочтительно от 2 до 5 нуклеиновых кислот) в клетки вне организма или индивидуум, и последующее применение трансфицированных клеток к организму или индивидууму.
Точно также, в соответствии с другим аспектом, настоящее изобретение также предоставляет применение последовательности нуклеиновых кислот по изобретению, как определено в настоящем документе, или композиции по изобретению, содержащую множество последовательностей нуклеиновых кислот по изобретению, как определено в настоящем документе, предпочтительно для терапевтических или диагностических целей, для увеличения экспрессии кодируемого пептида и белка, например, путем применения либо введения последовательности нуклеиновых кислот по изобретению, как определено в настоящем документе, или композиции по изобретению, содержащую множество последовательностей нуклеиновых кислот по изобретению, как определено в настоящем документе, например, в бесклеточную систему экспрессии, клетку (например, экспрессионная клетка-хозяин или соматическая клетка), ткань или организм. Применение может применяться для лаборатории, для исследования, для диагностики, для коммерческого получения пептидов и белков, и/или для терапевтических целей. В этом контексте, как правило, после подготовки последовательность нуклеиновых кислот по изобретению, как определено в настоящем документе, или композиции по изобретению, содержащей множество последовательностей нуклеиновых кислот по изобретению, как определено в настоящем документе, ее, как правило, применяют или вводят в бесклеточную систему экспрессии, клетку (например, экспрессионная клетка-хозяин или соматическая клетка), ткань или организм, предпочтительно в свободном виде или в виде комплекса, или как фармацевтическую композицию или вакцину, как описано в настоящем документе, предпочтительно через трансфекцию или с помощью любого из способов введения, как это описано в настоящем документе. Применение может осуществляться in vitro, in vivo и ex vivo. Применение может быть, кроме того, осуществляется в контексте лечения конкретной болезни, особенно в лечении опухолевых заболеваний раком или, предпочтительно, как определено в настоящем документе.
В еще одном аспекте настоящее изобретение также относится к экспрессионной системе по изобретению, включающей последовательность нуклеиновых кислот по изобретению или вектор экспрессии или плазмиду в соответствии с первым аспектом настоящего изобретения. В этом контексте система экспрессии может представлять собой бесклеточную систему экспрессии (например, in vitro система транскрипции/трансляции), клеточную систему экспрессии (например, клетки млекопитающих наподобие клеток СНО, клеток насекомых, дрожжевых клеток, бактериальных клеток, таких как Е. coli) или организмы, используемые для экспрессии пептидов и белков (например, растения или животные, как коровы).
Кроме того, в соответствии с другим аспектом, настоящее изобретение также относится к применению нуклеиновой кислоты по изобретению, как определено в настоящем документе, или композиции по изобретению, содержащей множество последовательностей нуклеиновых кислот по изобретению, как определено в настоящем документе, для подготовки фармацевтической композиции для увеличения экспрессии кодируемого пептида и белка, например, для лечения онкологического или опухолевого заболевания, предпочтительно, как определено в настоящем документе, например, путем применения либо введения нуклеиновой кислоты по изобретению, как определено в настоящем документе, или композиции по изобретению, содержащей множество последовательностей нуклеиновых кислот по изобретению, как определено в настоящем документе, в клетку (например, экспрессионную клетку-хозяин или соматическую клетку), ткань или организм, предпочтительно, в свободном виде или в виде комплекса, как фармацевтическая композиция или вакцина, как описано в настоящем документе, более предпочтительно, с использованием любого из способов введения, как описано в настоящем документе.
Соответственно, в конкретном предпочтительном аспекте, настоящее изобретение также предоставляет фармацевтическую композицию, содержащую нуклеиновую кислоту по изобретению, как определено в настоящем документе, или композицию по изобретению, содержащую множество последовательностей нуклеиновых кислот по изобретению, как определено в настоящем документе, и, необязательно, фармацевтически приемлемый носитель и/или средство доставки.
В качестве первого ингредиента, фармацевтическая композиция по изобретению содержит по меньшей мере одну нуклеиновую кислоту по изобретению, как определено в настоящем документе.
В качестве второго ингредиента, фармацевтическая композиция по изобретению может необязательно включать по меньшей мере один дополнительный фармакологически активный компонент. Фармакологически активный компонент в этой связи представляет собой соединение, которое имеет терапевтический эффект исцеления, смягчения или предотвращения определенного показания или заболевания, как отмечено в настоящем документе, предпочтительно онкологических или опухолевых заболеваний. К таким соединениям относятся, без каких-либо ограничений, пептиды и белки, предпочтительно, как определено в настоящем документе, нуклеиновые кислоты, предпочтительно, как определено в настоящем документе, (терапевтически активные) низкомолекулярные органические или неорганические соединения (с молекулярной массой менее 5000, предпочтительно, менее 1000), сахара, антигены или антитела, предпочтительно, как определено в настоящем документе, терапевтические агенты, уже известные из уровня техники, антигенные клетки, антигенные клеточные фрагменты, клеточные фракции; компоненты клеточной стенки (например, полисахариды), модифицированные, ослабленные или дезактивированные (например, химически или облучением) патогены (вирусы, бактерии и др.), адъюванты, предпочтительно, как определено в настоящем документе, и др.
Кроме того, фармацевтическая композиция по изобретению может содержать фармацевтически приемлемый носитель и/или средство доставки. В контексте настоящего изобретения, фармацевтически приемлемый носитель, как правило, включает жидкую или нежидкую основу фармацевтической композиции по изобретению. Если фармацевтическая композиция по изобретению предоставляется в жидком виде, носителем, как правило, является апирогенная вода; изотонические солевые или буферизированные (водные) растворы, например фосфатный, цитратный и др. буферизированные растворы. Инъекционный буфер может быть гипертоническим, изотоническим или гипотоническим по отношению к определенной контрольной среде, то есть, буфер может иметь более высокое, идентичное или пониженное содержание соли по сравнению с определенной контрольной средой, причем предпочтительно могут использоваться такие концентрации упомянутых выше солей, которые не приводят к повреждению клеток за счет осмоса или других концентрационных эффектов. Контрольные среды представляют собой, например, жидкости, представленные в способах «in vivо», такие как кровь, лимфа, цитозольные жидкости или другие жидкости организма, или, например, жидкости, которые можно использовать, в способах «in vitrо», такие как общепринятые буферы или жидкости. Такие общепринятые буферы или жидкости известны специалистам в рассматриваемой области техники. Раствор лактата Рингера является особенно предпочтительным в качестве жидкой основы.
При этом одна или несколько совместимых жидких или твердых наполнителей или разбавителей или инкапсулирующих соединений также могут быть использованы для фармацевтической композиции по изобретению, которые подходят для введения пациенту, проходящему лечение. Термин «совместимость», как используется в настоящем документе, означает, что эти составляющие фармацевтической композиции по изобретению могут быть смешаны с нуклеиновой кислотой по изобретению, как определено в настоящем документе, таким способом, что не происходит взаимодействия, которое могло бы существенно снизить фармацевтическую эффективность фармацевтической композиции по изобретению при обычных условиях использования.
В соответствии с конкретным вариантом осуществления, фармацевтическая композиция по изобретению может содержать адъювант. В этом контексте, под адъювантом может подразумеваться любое соединение, которое подходит для инициации или усиления иммунного ответа врожденной иммунной системы, т.е. неспецифического иммунного ответа. Другими словами, фармацевтическая композиция по изобретению при введении предпочтительно вызывает врожденный иммунный ответ за счет адъюванта, который необязательно содержится в ней. Предпочтительно, такой адъювант может быть выбран из адъюванта, известного специалистам в рассматриваемой области техники и подходящего для настоящего случая, т.е. поддержка индукции врожденного иммунного ответа у млекопитающего, например, адъювантный белок, как это определено выше, или адъювант, как это определено далее.
Особенно предпочтительными в качестве адъювантов, подходящих для хранения и доставки, являются катионные или поликатионные соединения, как определено выше для последовательности нуклеиновых кислот по изобретению в качестве средства доставки, трансфекционного или комплексообразующего агента.
Фармацевтическая композиция по изобретению дополнительно может содержать один или несколько вспомогательных веществ в целях увеличения ее иммуногенности или иммуностимулирующей способности, при необходимости. Тем самым, предпочтительно, достигается синергическое действие последовательности нуклеиновых кислот по изобретению, как определено в настоящем документе, и вспомогательного вещества, которое может необязательно содержаться в фармацевтической композиции по изобретению. В зависимости от различных типов вспомогательных веществ, предметом рассмотрения в этом отношении могут стать различные механизмы. Например, соединения, которые позволяют созревать дендритным клеткам (ДК), например липополисахарида, TNF-альфа или CD40-лиганд образуют первый класс подходящих вспомогательных веществ. Как правило, можно использовать в качестве вспомогательного вещества любое средство, которое воздействует на иммунную систему в виде «сигнала опасности» (LPS, GP96 и др.) или цитокинов, таких как GM-CFS, которые позволяют иммунному ответу усиливаться и/или воздействовать целенаправленно. Особенно предпочтительными вспомогательными веществами являются цитокины, такие как монокины, лимфокинов, интерлейкины или хемокины, которые дополнительно стимулируют врожденный иммунный ответ, такие как IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-12, IL-13, IL-14, IL-15, IL-16, IL-17, IL-18, IL-19, IL-20, IL-21, IL-22, IL-23, IL-24, IL-25, IL-26, IL-27, IL-28, IL-29, IL-30, IL-31, IL-32, IL-33, IFN-альфа, IFN-бета, IFN-гамма, GM-CSF, C-CSF, M-CSF, LT-бета или TNF-Альфа, факторы роста, такие, как hGH.
Далее добавки, которые могут быть включены в фармацевтическую композицию по изобретению, представляют собой эмульгаторы, такие как, например, Tween®; смачивающие средства, такие, как, например, лаурилсульфат натрия; окрашивающие средства; придающие вкус средства, фармацевтические носители; средства таблетизирования; стабилизаторы; антиоксиданты; консерванты.
Фармацевтическая композиция по изобретению также может дополнительно содержать любое дополнительное соединение, которое, как известно, является иммуностимулирующим из-за его аффинности связывания (в качестве лигандов) с Toll-подобными рецепторами человека TLR1, TLR2, TLR3, TLR4, TLR5, TLR6, TLR7, TLR8, TLR9, TLR10, или из-за его аффинности связывания (в качестве лигандов) с Toll-подобными рецепторами мыши TLR1, TLR2, TLR3, TLR4, TLR5, TLR6, TLR7, TLR8, TLR9, TLR10, TLR11, TLR12 или TLR13.
Фармацевтическую композицию по изобретению можно вводить перорально, парентерально, при помощи спрея для ингаляции, местно, ректально, назально, трансбуккально, вагинально или с помощью имплантированной емкости. Термин «парентеральный», используемый в настоящем документе, включает в себя подкожный, внутривенный, внутримышечный, внутрисуставной, внутрисиновиальный, надчревный, интратекальный, внутрипеченочный, вводимый внутрь пораженных тканей, внутричерепной, накожный, внутрикожный, внутрилегочный, внутрибрюшинный, внутрисердечный, внутриартериальный и при помощи методик подъязычных инъекций или инфузий.
Предпочтительно, фармацевтическая композиция по изобретению может вводиться при помощи парентерального введения, более предпочтительно, в виде подкожного, внутривенного, внутримышечного, внутрисуставного, внутрисиновиального, надчревного, интратекального, печеновного, вводимого внутрь пораженной ткани, внутричерепного, накожного, внутрикожного, внутрилегочного, внутрибрюшинного, внутрисердечного, внутриартериального и сублингвального введения или при помощи инфузионных методик. Особенно предпочтительной является подкожная и внутримышечная инъекция. Стерильные инъекционные формы фармацевтических композиций по изобретению могут представлять собой водные или масляные суспензии. Эти суспензии могут быть составлены в соответствии со способами, известными в рассматриваемой области техники, с использованием подходящих диспергирующих или смачивающих средств и суспендирующих средств.
Фармацевтическую композицию по изобретению, как определено в настоящем документе, можно также вводить перорально, в любой пероральной приемлемой лекарственной формы, включая, в качестве неограничивающих примеров, капсулы, таблетки, водные суспензии или растворы.
Фармацевтическую композицию по изобретению также можно вводить местно, особенно когда цель лечения включает области или органы, легко доступные для местного применения, например, включая заболевания кожи или любых других доступных эпителиальных тканей. Подходящие местные составы легко подготавливают для каждого из этих областей или органов. Для местных применений, фармацевтическая композиция по изобретению может быть составлена в подходящей мази, содержащей нуклеиновую кислоту по изобретению, как определено в настоящем документе, суспендированную или растворенную в одном или нескольких носителях.
Фармацевтическая композиция по изобретению, как правило, включает «безопасное и эффективное количество» компонентов фармацевтической композиции по изобретению, в частности, последовательности(тей) нуклеиновых кислот по изобретению, как определено в настоящем документе. Как используется в настоящем документе, «безопасное и эффективное количество» означает количество последовательности(тей) нуклеиновых кислот по изобретению, как определено в настоящем документе, такое, которое достаточно для существенного стимулирования положительного изменения заболевания или расстройства, как определено в настоящем документе. В то же время, однако, «безопасное и эффективное количество» является достаточно малым для избежания серьезных побочных эффектов и для разрешения разумного отношения между преимуществом и риском. Определение этих ограничений, как правило, находится в пределах разумного медицинское освидетельствования.
Фармацевтическую композицию по изобретению можно применять для медицинских назначений у людей, а также для ветеринарных медицинских назначений, предпочтительно медицинских назначений у людей, в качестве фармацевтической композиции в целом или в качестве вакцины.
В соответствии с другим конкретным предпочтительным аспектом, фармацевтическая композиция по изобретению (или последовательность нуклеиновой кислоты по изобретению, как определено в настоящем документе, или композиция по изобретению, содержащая множество последовательностей нуклеиновых кислот по изобретению, как определено в настоящем документе) может предоставляться или применяться в качестве вакцины. Как правило, такая вакцина представляет собой то, как определено выше для фармацевтических композиций. Кроме того, такая вакцина, как правило, содержит нуклеиновую кислоту по изобретению, как определено в настоящем документе, или композицию по изобретению, содержащую множество последовательностей нуклеиновых кислот по изобретению, как определено в настоящем документе.
Вакцина по изобретению также может содержать фармацевтически приемлемый носитель, адъювант и/или средство доставки, как определено в настоящем документе для фармацевтической композиции по изобретению. В конкретном контексте вакцины по изобретению, выбор фармацевтически приемлемого носителя определяется, в принципе, способом, при помощи которого вакцину по изобретению вводят. Вакцину по изобретению можно вводить, например, системно или локально. Способы системного введения, как правило, включают в себя, например, трансдермальный, оральный, парентеральный способы, в том числе подкожный, внутривенный, внутримышечный, внутриартериальный, внутрикожный и интраперитонеально-инъекционный и/или интраназальный способы введения. Способы локального введения, как правило, включают в себя, например, способы местного введения, но также интрадермальный, трансдермальный, подкожный, внутримышечно-инъекционный или вводимый внутрь пораженной ткани, внутричерепной, внутрилегочный, внутрисердечный и сублингвальных инъекций. Более предпочтительно, вакцины можно вводить при помощи интрадермального, подкожного и внутримышечного способа. Вакцины по изобретению, поэтому предпочтительно составляют в жидкой (или иногда в твердой) форме.
Вакцина по изобретению может дополнительно содержать одно или более вспомогательных веществ для усиления иммуногенности или иммуностимуляционной способности, если требуется. Особенно предпочтительными являются адъюванты в качестве вспомогательных веществ или добавки, как определено для фармацевтической композиции.
Настоящее изобретение, кроме того, предоставляет несколько применений и использований последовательности нуклеиновой кислоты по изобретению, как определено в настоящем документе, композиции по изобретению, содержащей множество последовательностей нуклеиновых кислот по изобретению, как определено в настоящем документе, фармацевтической композиции по изобретению, вакцины по изобретению, каждая из которых содержит последовательность нуклеиновых кислот по изобретению, как определено в настоящем документе, или набор, содержащий это.
В соответствии с одним конкретным аспектом, настоящее изобретение направлено на первое медицинское применение последовательности нуклеиновой кислоты по изобретению, как определено в настоящем документе, или композиции по изобретению, содержащей множество последовательностей нуклеиновых кислот по изобретению, как определено в настоящем документе, в качестве лекарства, предпочтительно, в качестве вакцины, в частности, при лечении онкологических или опухолевых заболеваний.
В соответствии с другим аспектом, настоящее изобретение направлено на второе медицинское применение последовательности нуклеиновой кислоты по изобретению, как определено в настоящем документе, или композиции по изобретению, содержащей множество последовательностей нуклеиновых кислот по изобретению, как определено в настоящем документе, для лечения онкологических или опухолевых заболеваний, как определено в настоящем документе, предпочтительно для применения последовательности нуклеиновых кислот по изобретению, как определено в настоящем документе, композиции по изобретению, содержащей множество последовательности нуклеиновых кислот по изобретению, как определено в настоящем документе, фармацевтической композиции или вакцины, содержащей это, или наборов, содержащих это, для подготовки препарата лекарственного средства для профилактики, лечения и/или облегчения онкологических или опухолевых заболеваний, как определено в настоящем документе. Предпочтительно, с этой целью фармацевтическую композицию или вакцину применяют или вводят пациенту, нуждающемуся в этом.
Предпочтительно, заболевания, как упомянуто в настоящем документе, выбирают из онкологических или опухолевых заболеваний, которые предпочтительно включают, например, острый лимфобластный лейкоз, острый миелобластный лейкоз, адренокортикальную карциному, связанные со СПИДом раковые заболевания, связанную со СПИДом лимфому, рак прямой кишки, рак аппендикса, астроцитому, базально-клеточную карциному, рак желчных протоков, рак мочевого пузыря, рак костей, остеосаркому/злокачественную фиброзную гистиоцитому, глиому ствола мозга, опухоль головного мозга, астроцитому мозжечка, церебральную астроцитому/злокачественную глиому, эпендимому, медуллобластому, супратенториальные примитивные нейроэктодермальные опухоли, глиому зрительных проводящих путей и гипоталамуса, рак молочной железы, бронхиальные аденомы/карциноиды, лимфому Беркитта, карциноидную опухоль у детей, карциноидную опухоль желудочно-кишечного тракта, карциному неизвестной природы, первичную лимфому центральной нервной системы, детскую мозжечковую астроцитому, детскую мозжечковую астроцитому/злокачественную глиому, рак шейки матки, рак у детей, хронический лимфоцитарный лейкоз, хронический миелогенный лейкоз, хронические миелопролиферативные расстройства, рак кишечника, кожная T-клеточная лимфома, опухоль десмопластических малых круглых клеток, рак эндометрия, эпендимому, рак пищевода, саркома Юинга в семействе опухолей Юинга, экстракраниальная герминогенная опухоль у детей, внегонадная герминогенная опухоль, рак внепеченочных желчных путей, внутриглазную меланому, ретинобластому, рак желчного пузыря, рак желудка (живота), карциноидная опухоль желудочно-кишечного тракта, стромальная опухоль желудочно-кишечного тракта (GIST), экстракраниальная, внегонадная или овариальная герминогенная опухоль, гестационная трофобластическая опухоль, глиома ствола мозга, детская церебральная астроцитома, глиома зрительных проводящих путей и гипоталамуса у детей, карциноид желудка, волосатоклеточный лейкоз, рак головы и шеи, рак сердца, паренхиматозный (печеночный) рак, лимфома Ходжкина, гипофарингеальный рак, глиома гипоталамуса и зрительных проводящих путей у детей, внутриглазная меланома, карцинома островковых клеток (эндокринной поджелудочной железы), саркома Капоши, рак почки (почечно-клеточный рак), рак гортани, лейкозы, острый лимфобластный лейкоз, острый миелобластный лейкоз, хронический лимфоцитарный лейкоз, хронический миелогенный лейкоз, волосатоклеточный лейкоз, рак губ и ротовой полости, липосаркому, рак печени, немелкоклеточный рак легких, мелкоклеточный рак легких, лимфомы, связанная со СПИДом лимфому, лимфому Беркитта, кожную T-клеточную лимфому, лимфому Ходжкина, не-Ходжкинскую лимфому, лимфому первичной центральной нервной системы, макроглобулинемию Вальденстрема, злокачественную фиброзную гистиоцитому кости/остеосаркому, детскую медуллобластому, меланому, внутриглазную (глазную) меланому, Меркель-клеточную карциному, злокачественную мезотелиому у взрослых, детскую мезотелиому, метастатический плоскоклеточный рак шеи скрытой природы, рак рта, множественной эндокринной неоплазийный синдром у детей, множественную миелому/ новообразование плазматических клеток, грибовидный микоз, миелодиспластические синдромы, миелодиспластические/миелопролиферативные заболевания, хронический миелогенный лейкоз, острый миелоидный лейкоз у взрослых, детский острый миелоидный лейкоз, множественную миелому (рак костного мозга), хронические миелопролиферативные расстройства, рак полости носа и околоносовых пазух, карциному носоглотки, нейробластому, рак полости рта, рак ротоглотки, остеосаркому/злокачественную фиброзную гистиоцитому кости, рак яичников, эпителиальный рак яичников (поверхностная эпителиально-стромальная опухоль), овариальную герминогенную опухоль, овариальную низко злокачественную потенциальную опухоль, рак поджелудочной железы, рак островковых клеток поджелудочной железы, рак околоносовых пазух и носовой полости, рак паращитовидной железы, рак полового члена, рак глоточной железы, феохромоцитому, астроцитому шишковидной железы, герминому шишковидной железы, детскую пинеобластому и супратенториальные примитивные нейроэктодермальные опухоли, аденому гипофиза, неоплазию плазматических клеток/множественную миелому, плевролегочную бластому, первичную лимфому центральной нервной системы, рак простаты, рак прямой кишки, почечно-клеточную карциному (рак почки), рак почечной лоханки и мочеточника, ретинобластому, детскую рабдомиосаркома, рак слюнных желез, саркома Юингова семейства опухолей, саркома Капоши, саркома мягких тканей, саркома матки, синдром Sezary, рак кожи (отличный от меланомы), рак кожи (меланома), Меркель-клеточный рак кожи, рак тонкой кишки, плоскоклеточную карциному, метастатический плоскоклеточный рак шеи неустановленной природы, супратенториальную примитивную нейроэктодермальную опухоль у детей, тестикулярный рак, рак горла, детскую тимому, тимому и карциному тимуса, рак щитовидной железы, рак щитовидной железы у детей, переходно-клеточный рак почечной лоханки и мочеточника, гестационную трофобластическую опухоль, рак уретры, эндометриальный рак матки, саркома матки, рак влагалища, глиому зрительных проводящих путей и гипоталамуса у детей, рака вульвы, макроглобулинемию Вальденстрема и опухоль Вильмса у детей (рак почки).
В дополнительном предпочтительном аспекте, последовательность нуклеиновых кислот по изобретению, как определено в настоящем документе, или композиция по изобретению, содержащая множество последовательностей нуклеиновых кислот по изобретению, как определено в настоящем документе, может использоваться для подготовки фармацевтической композиции или вакцины, в частности, для целей, определенных в настоящем документе.
Фармацевтическая композиция по изобретению или вакцина может, кроме того, применяться для лечения заболевания или расстройства, предпочтительно онкологических или опухолевых заболеваний, как определено в настоящем документе.
В соответствии с последним аспектом, настоящее изобретение также предоставляет наборы, в частности наборы частей. Такие наборы, особенно наборы частей, как правило, содержат в качестве компонентов по отдельности или в комбинации с другими компонентами, как определено в настоящем документе по меньшей мере одну последовательность нуклеиновых кислот по изобретению, как определено в настоящем документе, фармацевтическую композицию по изобретению или вакцину, содержащую последовательность нуклеиновых кислот по изобретению. По меньшей мере, одна последовательность нуклеиновых кислот по изобретению, как определено в настоящем документе, необязательно находится в комбинации с другими компонентами, как определено в настоящем документе, тем самым по меньшей мере одна нуклеиновая кислота по изобретению предоставляется отдельно (первая часть набора) от по меньшей мере другой части набора, содержащего один или несколько компонентов. Фармацевтическая композиция по изобретению и/или вакцина по изобретению может, например, находиться в одной или разных частях набора. В качестве примера, например, по меньшей мере одна часть набора может содержать по меньшей мере одну последовательность нуклеиновых кислот по изобретению, как определено в настоящем документе, и по меньшей мере одну дополнительную часть набора по меньшей мере другого компонента, как определено в настоящем документе, например, по меньшей мере одна другая часть набора может содержать по меньшей мере одну фармацевтическую композицию или вакцину или их части, например, по меньшей мере одна часть набора может содержать последовательность нуклеиновых кислот по изобретению, как определено в настоящем документе по меньшей мере одну дополнительную часть набора по меньшей мере одного компонента, как определено в настоящем документе по меньшей мере еще одну часть набора по меньшей мере одного компонента фармацевтической композиции или вакцины по изобретению или фармацевтической композиции или вакцины по изобретению в целом, и по меньшей мере еще одну часть набора, например, по меньшей мере один фармацевтический носитель или средство доставки и др. В случае если набор или набор частей содержит множество последовательностей нуклеиновых кислот по изобретению, один компонент набора может содержать только одну, нескольких или все последовательности нуклеиновых кислот по изобретению, входящие в набор. В альтернативном вариант осуществления каждая/каждая последовательность нуклеиновых кислот по изобретению может состоять в другом /отдельном компоненте набора, так что каждый компонент образует часть набора. Кроме того, более чем одна нуклеиновая кислота может содержаться в первом компоненте как часть набора, в то время как один или несколько других (второй, третий и т.д.) компонентов (предоставляющих одну или несколько других частей набора) может содержать либо одну, либо более чем одну нуклеиновую кислоту по изобретению, которые могут быть идентичны или частично идентичны или отличаться от первого компонента. Набор или набор частей, кроме того, может содержать технические инструкции с информацией о приеме и дозировке последовательности нуклеиновых кислот по изобретению, фармацевтической композиции по изобретению или вакцины по изобретению или каких-либо его компонентов или частей, например, если набор подготовлен в виде набора частей.
Взятые вместе, изобретение предоставляет последовательность нуклеиновых кислот, включающую или кодирующую
а) кодирующую область, кодирующую по меньшей мере один пептид или белок;
b) по меньшей мере, одну гистоновую структуру типа «стебель-петля» и
c) поли(A)-последовательность или сигнал полиаденилирования;
где указанный пептид или белок содержит опухолевый антиген, фрагмент, вариант или производное указанного опухолевого антигена, предпочтительно, где опухолевый антиген представляет собой меланоцит-специфический антиген, антиген рака яичка или опухолевый специфический антиген, предпочтительно антиген CT-X, антиген non-CT-X, партнер по связыванию для антигена CT-X или партнер по связыванию для антигена non-CT-X или опухолевый специфический антиген, более предпочтительно антиген CT-X, партнер по связыванию для антигена non-CT-X или опухолевый специфический антиген или фрагмент, вариант или производное указанного опухолевого антигена.
В дополнительном предпочтительном варианте осуществления изобретение относится к композиции, содержащей по меньшей мере один тип последовательности нуклеиновых кислот, содержащей или кодирующей
а) кодирующую область, кодирующую по меньшей мере один пептид или белок;
b) по меньшей мере, одну гистоновую структуру типа «стебель-петля» и
c) поли(A)-последовательность или сигнал полиаденилирования;
где указанный пептид или белок содержит опухолевый антиген, фрагмент, вариант или производное указанного опухолевого антигена, предпочтительно, где опухолевый антиген представляет собой меланоцит-специфический антиген, антиген рака яичка или опухолевый специфический антиген, предпочтительно антиген CT-X, антиген non-CT-X, партнер по связыванию для антигена CT-X или партнер по связыванию для антигена non-CT-X или опухолевый специфический антиген, более предпочтительно антиген CT-X, партнер по связыванию для антигена non-CT-X или опухолевый специфический антиген или фрагмент, вариант или производное указанного опухолевого антигена; и где каждая из последовательностей нуклеиновых кислот кодирует отличный пептид или белок.
Композиция может дополнительно включать фармацевтически приемлемый носитель и/или фармацевтически приемлемые адъюванты, как определено в настоящем документе. Композиция может применяться в качестве вакцины или для лечения заболеваний, связанных с раком или опухолью.
В дополнительном предпочтительном варианте осуществления, изобретение предоставляет композицию, содержащую по меньшей мере две, предпочтительно, две или больше, более предпочтительно, множество последовательностей нуклеиновых кислот (что означает, как правило, больше, чем 1, 2, 3, 4, 5, 6 или более 10 нуклеиновых кислот, например, от 2 до 10, предпочтительно от 2 до 5 нуклеиновых кислот), последовательность, содержащую или кодирующую
а) кодирующую область, кодирующую по меньшей мере один пептид или белок;
b) по меньшей мере, одну гистоновую структуру типа «стебель-петля» и
c) поли(A)-последовательность или сигнал полиаденилирования;
где указанный пептид или белок содержит опухолевый антиген, фрагмент, вариант или производное указанного опухолевого антигена, предпочтительно, где опухолевый антиген представляет собой меланоцит-специфический антиген, антиген рака яичка или опухолевый специфический антиген, предпочтительно антиген CT-X, антиген non-CT-X, партнер по связыванию для антигена CT-X или партнер по связыванию для антигена non-CT-X или опухолевый специфический антиген, более предпочтительно антиген CT-X, партнер по связыванию для антигена non-CT-X или опухолевый специфический антиген или фрагмент, вариант или производное указанного опухолевого антигена; и где каждая из последовательностей нуклеиновых кислот кодирует отличный пептид или белок.
Композиция может дополнительно включать фармацевтически приемлемый носитель и/или фармацевтически приемлемые адъюванты, как определено в настоящем документе. Композиция может применяться в качестве вакцины или для лечения заболеваний, связанных с раком или опухолью.
В дополнительном предпочтительном варианте осуществления, изобретение предоставляет композицию, содержащую по меньшей мере две, предпочтительно, две или больше, более предпочтительно, множество (что означает, как правило, больше, чем 1, 2, 3, 4, 5, 6 или более 10 нуклеиновых кислот, например, от 2 до 10, предпочтительно от 2 до 5 нуклеиновых кислот) последовательностей нуклеиновых кислот, последовательность, содержащую или кодирующую
а) кодирующую область, кодирующую по меньшей мере один пептид или белок;
b) по меньшей мере, одну гистоновую структуру типа «стебель-петля» и
c) поли(A)-последовательность или сигнал полиаденилирования;
где указанный пептид или белок содержит опухолевый антиген, фрагмент, вариант или производное указанного опухолевого антигена, предпочтительно, где опухолевый антиген представляет собой меланоцит-специфический антиген, антиген рака яичка или опухолевый специфический антиген, предпочтительно антиген CT-X, антиген non-CT-X, партнер по связыванию для антигена CT-X или партнер по связыванию для антигена non-CT-X или опухолевый специфический антиген, более предпочтительно антиген CT-X, партнер по связыванию для антигена non-CT-X или опухолевый специфический антиген или фрагмент, вариант или производное указанного опухолевого антигена; и где каждая из последовательностей нуклеиновых кислот кодирует отличный пептид или белок; и, предпочтительно, где каждый тип последовательности нуклеиновых кислот кодирует отличный пептид или белок, предпочтительно на разных опухолевых антигенов.
Композиция может дополнительно включать фармацевтически приемлемый носитель и/или фармацевтически приемлемые адъюванты, как определено в настоящем документе. Композиция может применяться в качестве вакцины или для лечения заболеваний, связанных с раком или опухолью.
В дополнительном предпочтительном варианте осуществления, изобретение предоставляет композицию, содержащую по меньшей мере две, предпочтительно, две или больше, более предпочтительно, множество последовательностей нуклеиновых кислот, последовательность (что означает, как правило, больше, чем 1, 2, 3, 4, 5, 6 или более 10 нуклеиновых кислот, например, от 2 до 10, предпочтительно от 2 до 5 нуклеиновых кислот), содержащую или кодирующую
а) кодирующую область, кодирующую по меньшей мере один пептид или белок;
b) по меньшей мере, одну гистоновую структуру типа «стебель-петля» и
c) поли(A)-последовательность или сигнал полиаденилирования;
где указанный пептид или белок содержит опухолевый антиген, фрагмент, вариант или производное указанного опухолевого антигена, предпочтительно, где опухолевый антиген представляет собой меланоцит-специфический антиген, антиген рака яичка или опухолевый специфический антиген, предпочтительно антиген CT-X, антиген non-CT-X, партнер по связыванию для антигена CT-X или партнер по связыванию для антигена non-CT-X или опухолевый специфический антиген, более предпочтительно антиген CT-X, партнер по связыванию для антигена non-CT-X или опухолевый специфический антиген или фрагмент, вариант или производное указанного опухолевого антигена; и где каждая из последовательностей нуклеиновых кислот кодирует отличный пептид или белок; и, предпочтительно, где каждый тип последовательности нуклеиновых кислот кодирует отличный пептид и белок, предпочтительно отличный опухолевый антиген, более предпочтительно, где один тип содержащихся последовательностей нуклеиновых кислот кодирует PSА, PSМА, PSCA, STEAP-1, NY-ESO-1, 5T4, сурвивин, MAGE-C1 или MAGE-C2.
Композиция может дополнительно включать фармацевтически приемлемый носитель и/или фармацевтически приемлемые адъюванты, как определено в настоящем документе. Композиция может применяться в качестве вакцины или для лечения заболеваний, связанных с раком или опухолью.
В некоторых вариантах осуществления может быть предпочтительным, при условии, что композиция содержит только один тип последовательности нуклеиновых кислот, если последовательность нуклеиновых кислот не кодирует NY-ESO1, при условии, что композиция содержит только один тип последовательности нуклеиновых кислот.
В дополнительном предпочтительном варианте осуществления изобретение предоставляет композицию, содержащую по меньшей мере две последовательности нуклеиновых кислот, последовательность, содержащую или кодирующую
а) кодирующую область, кодирующую по меньшей мере один пептид или белок;
b) по меньшей мере, одну гистоновую структуру типа «стебель-петля» и
c) поли(A)-последовательность или сигнал полиаденилирования;
где указанный пептид или белок содержит опухолевый антиген, фрагмент, вариант или производное указанного опухолевого антигена, предпочтительно, где опухолевый антиген представляет собой меланоцит-специфический антиген, антиген рака яичка или опухолевый специфический антиген, предпочтительно антиген CT-X, антиген non-CT-X, партнер по связыванию для антигена CT-X или партнер по связыванию для антигена non-CT-X или опухолевый специфический антиген, более предпочтительно антиген CT-X, партнер по связыванию для антигена non-CT-X или опухолевый специфический антиген или фрагмент, вариант или производное указанного опухолевого антигена; и где каждая из последовательностей нуклеиновых кислот кодирует отличный пептид или белок; и где по меньшей мере одна из последовательностей нуклеиновых кислот кодирует 5T4, 707-AP, 9D7, AFP, AlbZIP HPG1, альфа-5-бета-1-интегрин, альфа-5-бета-6-интегрин, альфа-актинин-4/m, альфа-метилацилкоэнзим-A-рацемазe, ART-4, ARTC1/m, B7H4, BAGE-1, BCL-2, bcr/abl, бета-катенин/m, BING-4, BRCA1/m, BRCA2/m, СA 15-3/СA 27-29, СA 19-9, CA72-4, CA125, калретикулин, CAMEL, CASP-8/m, катепсин B, катепсин L, CD19, CD20, CD22, CD25, CDE30, CD33, CD4, CD52, CD55, CD56, CD80, CDC27/m, CDK4/m, CDKN2A/m, CEA, CLCA2, CML28, CML66, COA-1/m, коактозин-подобный белок, collage XXIII, COX-2, СT-9/BRD6, Cten, циклин B1, циклин D1, cyp-B, CYPB1, DAM-10, DAM-6, DEK-CAN, EFTUD2/m, EGFR, ELF2/m, EMMPRIN, EpCam, EphA2, EphA3, ErbB3, ETV6-AML1, EZH2, FGF-5, FN, Frau-1, G250, GAGE-1, GAGE-2, GAGE-3, GAGE-4, GAGE-5, GAGE-6, GAGE7b, GAGE-8, GDEP, GnT-V, gp100, GPC3, GPNMB/m, HAGE, HAST-2, гепсин, Her2/neu, HERV-K-MEL, HLA-A*0201-R17I, HLA-A11/m, HLA-A2/m, HNE, гомеобокс NKX3.1, HOM-TES-14/SCP-1, HOM-TES-85, HPV-E6, HPV-E7, HSP70-2M, HST-2, hTERT, iCE, IGF-1R, IL-13Ra2, IL-2R, IL-5, незрелый ламининовый рецептор, калликреин-2, калликреина-4, Ki67, KIAA0205, KIAA0205/m, KK-LC-1, K-Ras/m, LAGE-A1, LDLR-FUT, MAGE-A1, MAGE-A2, MAGE-A3, MAGE-A4, MAGE-A6, MAGE-A9, MAGE-A10, MAGE-A12, MAGE-B1, MAGE-B2, MAGE-B3, MAGE-B4, MAGE-B5, MAGE-B6, MAGE-B10, MAGE-B16, MAGE-B17, MAGE-C1, MAGE-C2, MAGE-C3, MAGE-D1, MAGE-D2, MAGE-D4, MAGE-E1, MAGE-E2, MAGE-F1, MAGE-H1, MAGEL2, маммаглобин A, MART-1/melan-A, MART-2, MART-2/м, белок матриксa 22, MC1R, M-CSF, ME1/m, мезотелин, MG50/PXDN, MMP11, MN/СА IX-антиген, MRP-3, MUC-1, MUC-2, MUM-1/m, MUM-2/m, MUM-3/m, миозин класса I/m, NA88-A, N-ацетилглюкозаминилтрансферазу-V, Neo-PAP, Nео-PAP/m, NFYC/m, NGEP, NMP22, NPM/ALK, N-Ras/m, NSE, NY-ESO-B, NY-ESO-1, OA1, OFA-iLRP, OGT, OGT/m, OS-9, OS-9/m, остеокальцин, остеопонтин, р15, p190 минорный bcr-abl, p53, р53/m, PAGE-4, PAI-1, PAI-2, PAP, PART-1, PATE, PDEF, Pim-1-киназу, Pin-1, Pml/PARalpha, POTE, PRAME, PRDX5/m, простеин, протеиназу-3, PSA, PSCA, PSGR, PSM, PSMA, PTPRK/m, RAGE-1, RBAF600/m, RHAMM/CD168, RU1, RU2, S-100, SAGE, SART-1, SART-2, SART-3, SCC, SIRT2/m, Sp17, SSX-1, SSX-2/HOM-MEL-40, SSX-4, STAMP-1, STEAP-1, сурвивин, сурвивин-2B, SYT-SSX-1, SYT-SSX-2, ТА-90, TAG-72, TARP, ТEL-AML1, TGFбета, TGFбетаRII, TGM-4, TPI/m, TRAG-3, TRG, TRP-1, TRP-2/6b, TRP/INT2, TRP-p8, тирозиназу, UPA, VEGFR1, VEGFR-2/FLK-1 и WT1 и иммуноглобулиновый идиотип лимфоидных клеток крови, или идиотип Т-клеточного рецептора лимфоидных клеток крови, или фрагмент, вариант или производное указанного опухолевого антигена; предпочтительно сурвивин или его гомолог, антиген из MAGE-семейства или его партнер по связыванию или фрагмент, вариант или производное указанного опухолевого антигена.
Композиция может дополнительно включать фармацевтически приемлемый носитель и/или фармацевтически приемлемые адъюванты, как определено в настоящем документе. Композиция может применяться в качестве вакцины или для лечения заболеваний, связанных с раком или опухолью.
В настоящем изобретении, если не указано иное, различные особенности альтернативных вариантов и вариантов осуществления могут комбинироваться друг с другом. Кроме того, термин «содержащий» не должен толковаться как означающий «состоящий из», если не оговорено отдельно. При этом, в контексте настоящего изобретения, термин «содержащий» может быть заменен на термин «состоящий из», где это применимо.
Фигуры:
Следующие Фигуры предназначены для дополнительной иллюстрации изобретения и не должны истолковываться как ограничивающие настоящее изобретение.
Фигура 1: показывает консенсусную последовательность гистоновой структуры типа «стебель-петля», генерируемую из последовательностей структур типа «стебель-петля» метазоа и простейших (как сообщили в Davila Lopez, M., & Samuelsson, T. (2008), RNA (New York, N.Y.), 14(1), 1-10. doi:10.1261/rna.782308). 4001 последовательность гистоновых структур типа «стебель-петля» из метазоа и простейших выравнивали и количество имеющих место нуклеотидов указывается для каждой позиции в последовательности структуры типа «стебель-петля». Генерированная консенсусная последовательность, представляющая все нуклеотиды, присутствующие в анализируемых последовательностях, представляется с использованием однобуквенного нуклеотидного кода. В дополнение к консенсусной последовательности, показаны последовательности, представляющие по меньшей мере 99%, 95% и 90% нуклеотидов, присутствующих в анализируемых последовательностях.
Фигура 2: показывает консенсусную последовательность гистоновой структуры типа «стебель-петля», генерируемую из последовательностей структур типа «стебель-петля» простейших (как сообщили в Davila Lopez, M., & Samuelsson, T. (2008), RNA (New York, N.Y.), 14(1), 1-10. doi:10.1261/rna.782308). 131 последовательность гистоновых структур типа «стебель-петля» из простейших выравнивали, и количество имеющих место нуклеотидов указывается для каждой позиции в последовательности структуры типа «стебель-петля». Генерированная консенсусная последовательность, представляющая все нуклеотиды, присутствующие в анализируемых последовательностях, представляется с использованием однобуквенного нуклеотидного кода. В дополнение к консенсусной последовательности, показаны последовательности, представляющие по меньшей мере 99%, 95% и 90% нуклеотидов, присутствующих в анализируемых последовательностях.
Фигура 3: показывает консенсусную последовательность гистоновой структуры типа «стебель-петля», генерируемую из последовательностей структур типа «стебель-петля» метазоа (как сообщили в Davila Lopez, M., & Samuelsson, T. (2008), RNA (New York, N.Y.), 14(1), 1-10. doi:10.1261/rna.782308). 3870 последовательность гистоновых структур типа «стебель-петля» из метазоа выравнивали, и количество имеющих место нуклеотидов указывается для каждой позиции в последовательности структуры типа «стебель-петля». Генерированная консенсусная последовательность, представляющая все нуклеотиды, присутствующие в анализируемых последовательностях, представляется с использованием однобуквенного нуклеотидного кода. В дополнение к консенсусной последовательности, показаны последовательности, представляющие по меньшей мере 99%, 95% и 90% нуклеотидов, присутствующих в анализируемых последовательностях.
Фигура 4: показывает консенсусную последовательность гистоновой структуры типа «стебель-петля», генерируемую из последовательностей структур типа «стебель-петля» позвоночных (как сообщили в Davila Lopez, M., & Samuelsson, T. (2008), RNA (New York, N.Y.), 14(1), 1-10. doi:10.1261/rna.782308). 1333 последовательности гистоновых структур типа «стебель-петля» из позвоночных выравнивали, и количество имеющих место нуклеотидов указывается для каждой позиции в последовательности структуры типа «стебель-петля». Генерированная консенсусная последовательность, представляющая все нуклеотиды, присутствующие в анализируемых последовательностях, представляется с использованием однобуквенного нуклеотидного кода. В дополнение к консенсусной последовательности, показаны последовательности, представляющие по меньшей мере 99%, 95% и 90% нуклеотидов, присутствующих в анализируемых последовательностях.
Фигура 5: показывает консенсусную последовательность гистоновой структуры типа «стебель-петля», генерируемую из последовательностей структур типа «стебель-петля» человека (Homo sapiens) (как сообщили в Davila Lopez, M., & Samuelsson, T. (2008), RNA (New York, N.Y.), 14(1), 1-10. doi:10.1261/rna.782308). 84 последовательность гистоновых структур типа «стебель-петля» из людей выравнивали, и количество имеющих место нуклеотидов указывается для каждой позиции в последовательности структуры типа «стебель-петля». Генерированная консенсусная последовательность, представляющая все нуклеотиды, присутствующие в анализируемых последовательностях, представляется с использованием однобуквенного нуклеотидного кода. В дополнение к консенсусной последовательности, показаны последовательности, представляющие по меньшей мере 99%, 95% и 90% нуклеотидов, присутствующих в анализируемых последовательностях.
Фигуры с 6 по 21: показывают мРНК от транскрипции in vitro.
Приводятся наименование и последовательность мРНК, полученных при транскрипции in vitro. Используются следующие сокращения:
ppLuc (GC): GC-богатая последовательность мРНК, кодирующая люциферазу Photinus pyralis
ag: 3'-нетранслируемая область (UTR) гена альфа-глобина
A64: поли(A)-последовательность с 64 аденилатами
A120: поли(A)-последовательность с 120 аденилатами
histoneSL: гистоновая структура типа «стебель-петля»
aCPSL: структура типа «стебель-петля», которая была отобрана из библиотеки по ее специфическому связыванию белка αCP-2KL
PolioCL: 5' структура клеверного листа из геномной РНК вируса полиомиелита
G30: поли(G)-последовательность с 30 гуанилатами
U30: поли(U) последовательности с 30 уридилатами
SL: неспецифическая/искусственная структура типа «стебель-петля»
N32: неспецифическая последовательность из 32 нуклеотидов
NY-ESO-1 (G/C): GC-богатая последовательность мРНК, кодирующая опухолевый антиген NY-ESO-1 человека
Survivin(G/C): GC-богатая последовательность мРНК, кодирующая опухолевый антиген сурвивин человека
MAGE-C1(G/C): GC-богатая последовательность мРНК, кодирующая опухолевый антиген MAGE-C1 человека
Внутри последовательностей выделены следующие элементы: кодирующая область (ORF) (ПРОПИСНЫМИ буквами), ag (жирным шрифтом), histoneSL (подчеркнуто), другие отдельные проверенные последовательности (курсив).
Фигура 6: показывает последовательность мРНК ppLuc(GC)-ag (SEQ ID NO: 43).
Путем линеаризации исходного вектора по сайту рестрикции сразу после 3'-UTR альфа-глобина (ag) получается мРНК с отсутствием поли(A)-последовательности.
Фигура 7: показывает последовательность мРНК ppLuc(GC)-ag-A64 (SEQ ID NO: 44).
Путем линеаризации исходного вектора по сайту рестрикции сразу после поли(A)-последовательность A64 получается мРНК, заканчивающаяся поли(A) последовательностью A64.
Фигура 8: показывает последовательность мРНК ppLuc(GC)-ag-histoneSL (SEQ ID NO: 45).
Поли(A)-последовательность A64 была заменена histoneSL. Гистоновая структура типа «стебель-петля», используемая в примерах, была получена от Cakmakci et al. (2008). Molecular and Cellular Biology, 28(3), 1182-1194.
Фигура 9: показывает последовательность мРНК ppLuc(GC)-ag-A64-histoneSL (SFO ID No: 46).
histoneSL была добавлена с 3' от poly(А) A64.
Фигура 10: показывает последовательность мРНК ppLuc(GC)-ag-А120 (SEQ ID NO: 47). Поли(A)-последовательность A64 была заменена на поли(A)-последовательность A120.
Фигура 11: показывает последовательность мРНК ppLuc(GC)-ag-A64-ag (SEQ ID NP: 48). Вторая 3'-UTR альфа-глобина была добавлена с 3' от A64 поли(А).
Фигура 12: показывает последовательность мРНК ppLuc(GC)-ag-A64-aCPSL (SEQ ID NО: 49).
Структура типа «стебель-петля» был добавлена с 3' от поли(А) A64. Структура типа «стебель-петля» была выбрана из библиотеки по ее специфическому связыванию белка αСР-2KL (Thisted et al., (2001), The Journal of Biological Chemistry, 276(20), 17484-1 7496). αСР-2KL представляет собой изоформу αCP-2, наиболее сильно экспрессируемую белок αCP (белок, связывающийся с поли(C) мРНК альфа-глобина) (Makeyev et al., (2000), Genomics, 67(3), 301 -316), группа РНК-связывающих белков, которые связываются с 3'-UTR альфа-глобина (Chkheidze et al., (1999), Molecular and Cellular Biology, 19(7), 4572-4581).
Фигура 13: показывает последовательность мРНК ppLuc(GC)-ag-A64-PolioCL (SEQ ID NP: 50).
5' структура «клеверного листа» из геномной РНК вируса полиомиелита была добавлена с 3' от поли(А) A64.
Фигура 14: показывает последовательность мРНК ppLuc(GC)-ag-A64-G30 (SEQ ID NО: 51). Участок из 30 гуанилатов был добавлен с 3' от поли(А) A64.
Фигура 15: показывает последовательность мРНК ppLuc(GC)-ag-A64-U30 (SEQ ID NО: 52). Участок из 30 уридилатов был добавлен с 3' от поли(А) A64.
Фигура 16: показывает последовательность мРНК ppLuc(GC)-ag-A64-SL (SEP ID NО: 53)
Структура типа «стебель-петля» была добавлена с 3' от поли(А) A64. Верхняя часть стебля и петли были взяты из (Babendure et al., (2006), RNA (New York, N.Y.), 12(5), 851-861). Структура типа «стебель-петля» состоит из 17 пар оснований в длину, CG-богатого стебля и петли длиной в 6 оснований.
Фигура 17: показывает ppLuc(GC)-ag-A64-N32 (SEQ ID NO: 54)
Путем линеаризации исходного вектора по альтернативному сайту рестрикции получена мРНК с 32 дополнительными нуклеотидами после поли(А).
Фигура 18: показывает последовательность мРНК NY-ESO-1(GС)-ag-A64-C30 (SEQ ID NО: 55)
Фигура 19: показывает последовательность мРНК NY-ESO-1(GС)-ag-A64-C30-histoneSL (SEQ ID NО: 56)
Фигура 20: показывает последовательность мРНК Survivin(GС)-ag-A64-C30-histoneSL (SEQ ID NP: 57)
Фигура 21: показывает последовательность мРНК MAGE-C1 (GC)-ag-A64-C30-histoneSL (SEQ ID NP: 58)
Фигура 22: показывает, что комбинация поли(A) и histoneSL увеличивает белковую экспрессию с мРНК синергетическим образом
Исследовался эффект поли(A)-последовательности, histoneSL и комбинации поли(A) и histoneSL на экспрессию люциферазы с мРНК. Соответственно, различные мРНК электропорировали в клетки HeLa. Уровни люциферазы измеряли через 6, 24 и 48 часов после трансфекции. Мало люциферазы экспрессируется с мРНК, не имеющей ни поли(A)-последовательности, ни histoneSL. Как поли(A)-последовательность, так и histoneSL увеличивают уровень люциферазы. Поразительно, однако, комбинация поли(A) и histoneSL дополнительного сильно увеличивает уровень люциферазы, многократно выше уровня, наблюдаемого с любым из отдельных элементов, тем самым действуя синергетически. Данные приводятся на графике, как среднее RLU±SD (относительные единицы освещенности ± стандартное отклонение) для трех повторностей трансфекций. Конкретные RLU приведены в Примере 14.2.
Фигура 23: показывает, что комбинация поли(A) и histoneSL увеличивает белковую экспрессию с мРНК независимо от их порядка.
Исследовался эффект поли(A)-последовательности, histoneSL, комбинации поли(A) и histoneSL и их порядка на экспрессию люциферазы с мРНК. Соответственно, различные мРНК липофецировали в клетки HeLa. Уровни люциферазы измеряли через 6, 24 и 48 часов после начала трансфекции. Как поли(A)-последовательность A64, так и histoneSL приводили к сравнимым уровням люциферазы. Увеличение длины поли(A)-последовательности с A64 до A120 или A300 увеличивает уровень люциферазы умеренно. В отличие от этого, комбинация поли(A) и histoneSL увеличивает уровень люциферазы гораздо сильнее, чем удлинение поли(A)-последовательности. Комбинация поли(A) и histoneSL действует синергически, так как увеличивается уровень люциферазы многократно выше уровня, наблюдавшегося с любым из отдельных элементов. Синергетический эффект комбинации поли(A) и histoneSL оказывается независимым от порядка поли(A) и histoneSL и независимым от длины поли(A) у мРНК с A64-histoneSL или histoneSL-A250. Данные приводятся на графике, как среднее RLU ± SD для трех повторностей трансфекций. Конкретные RLU приведены в Примере 14.3.
Фигура 24: показывает, что рост уровня белковой экспрессии при комбинации поли(A) и histoneSL является специфичным.
Исследовался эффект комбинации поли(A) и histoneSL или альтернативных последовательностей на экспрессию люциферазы с мРНК. Соответственно, различные мРНК были электропорированы в клетки HeLa. Уровни люциферазы измеряли через 6, 24 и 48 часов после трансфекции. Как поли(A)-последовательность, так и histoneSL приводили к сравнимым уровням люциферазы. Комбинация поли(A) и histoneSL сильно увеличивает уровень люциферазы, многократно выше уровня, наблюдавшегося с любым из отдельных элементов, тем самым действуя синергетически. В отличие от этого, комбинация поли(A) с любыми другими последовательностями не влияет на уровень люциферазы по сравнению с мРНК, содержащей только поли(A)-последовательность. Таким образом, комбинация поли(A) и histoneSL действует специфически и синергетически. Данные приводятся на графике, как среднее RLU ± SD для трех повторностей трансфекций. Конкретные RLU приведены в Примере 14.4.
Фигура 25: показывает, что комбинация поли(A) и histoneSL увеличивает белковую экспрессию с мРНК синергетическим образом in vivo.
Исследовался эффект поли(A)-последовательности, histoneSL и комбинации поли(A) и histoneSL на экспрессию люциферазы с мРНК in vivo. Соответственно, различные мРНК вводили внутрикожно в мышей. Мышей забивали через 16 часов после инъекции, и определяли уровни люциферазы в участках инъекции. Люцифераза экспрессируется с мРНК, имеющей либо histoneSL, либо поли(A)-последовательность. Поразительно, однако, комбинация поли(A) и histoneSL сильно увеличивает уровень люциферазы, многократно выше уровня, наблюдавшегося с любым из отдельных элементов, тем самым действуя синергетически. Данные приводятся на графике в качестве среднего RLU±SEM (относительные единицы освещенности ± стандартная ошибка среднего). Конкретные RLU суммированы в примере 14.5.
Фигура 26: показывает, что комбинация поли(A) и histoneSL увеличивает экспрессию белка NY-ESO-1 с мРНК.
Исследовался эффект поли(A)-последовательности и комбинации поли(A) и histoneSL на экспрессию NY-ESO-1 с мРНК. Соответственно, различные мРНК были электропорированы в клетки HeLa. Уровни NY-ESO-1 измеряли через 24 часа после трансфекции с помощью проточной цитометрии. NY-ESO-1 экспрессируется с мРНК, имеющей только поли(A)-последовательность. Поразительно, однако, комбинация поли(A) и histoneSL сильно увеличивает уровень NY-ESO-1, многократно выше уровня, наблюдаемого с поли(A)-последовательностью только. Данные приведены на графике, как зависимость счета от интенсивности флуоресценции. Средние интенсивности флуоресценции (MFI) приведены в примере 14.6.
Фигура 27: показывает, что комбинация поли(A) и histoneSL повышается уровень антител, достигаемый в результате вакцинации с мРНК.
Исследовался эффект поли(A)-последовательности и комбинации поли(A) и histoneSL на индукцию антител против NY-ESO-1, достигаемую в результате вакцинации с мРНК. Соответственно, мыши C57BL/6 были вакцинированы внутрикожно разных мРНК в комплексе с протамином. Уровень NY-ESO-1-специфических антител в вакцинированных и контрольных мышах анализировали способом ELISA с последовательным разведением сывороток. IgG2a[b] против NY-ESO-1 индуцируют мРНК, имеющей только поли(A)-последовательность. Поразительно, однако, комбинация поли(A) и histoneSL сильно увеличивает уровень IgG2a[b] против NY-ESO-1, многократно выше уровня, наблюдаемого с поли(A)-последовательностью только. Данные приведены на графике в качестве средних точечных титров. Средние точечные титры приведены в примере 14.7.
ПРИМЕРЫ:
Следующие примеры предназначены для дополнительной иллюстрации изобретения дальше и не должны истолковываться как ограничивающие изобретение.
1. Генерирование консенсусных последовательностей гистоновой структуры типа «стержень-петля»
Перед экспериментами, консенсусные последовательности гистоновой структуры типа «стержень-петля» были определены на основе последовательностей гистоновой структуры типа «стебель-петля» метазоа и простейших. Последовательности были взяты из приложения, предоставленного Lopez et al. (Davila Lopez, M., & Samuelsson, T. (2008), RNA (New York, N.Y.), 14(1), 1-10. doi:10.1261/rna.782308), которые выявили большое количество природных последовательностей гистоновых структур типа «стебель-петля» путем изучения геномных последовательностей и экспрессированных тегов последовательностей. Во-первых, все последовательности из метазоа и простейших (4001 последовательность), или все последовательности из простейших (131 последовательность) или, альтернативно, из метазоа (3870 последовательностей), или из позвоночных (1333 последовательности) или из человека (84 последовательности) были сгруппированы и выровнены. Затем, для каждого положения определялось количество представленных нуклеотидов. На основе таблиц, полученные таким образом, были генерированы консенсусные последовательности для 5 различных групп последовательностей, представляющие все нуклеотиды, присутствующие в анализируемых последовательностях. Кроме того, были получены более строгие консенсусные последовательности, все более подчеркивающие консервативные нуклеотиды.
2. Подготовка ДНК-матриц
Был сконструирован вектор для транскрипции in vitro, содержащий T7-промотор, за которым следует GC-богатая последовательность, кодирующая люциферазу Photinus pyralis (ppLuc(GC)), центральная часть 3' нетранслируемой область (UTR) альфа-глобина (ag) и поли(A)-последовательность. Поли(A)-последовательность следовала сразу за сайтом рестрикции, используемым для линеаризации вектора перед транскрипцией in vitro, для получения завершения мРНК поли(A)-последовательностью A64. мРНК, полученная с этого вектора в соответствии с транскрипцией in vitro, обозначена как «ppLuc(GC)-ag-A64».
Линеаризация этого вектора по альтернативному сайту рестрикции перед транскрипции in vitro позволило получить мРНК, либо удлиненную дополнительными нуклеотидами, расположенными с 3'-стороны от A64, либо утратившую A64. Кроме того, исходный вектор был модифицирован с целью включения альтернативных последовательностей. В целом, следующие мРНК были получены с этих векторов путем транскрипции in vitro (последовательности мРНК приведены на фигурах с 6 по 17):
ppLuc(CC)-ag (SEQ ID NO: 43)
ppLuc(GC) A64 (SEQ ID NO: 44)
ppLuc(CC)-ag-histoneSL (SEQ ID NO: 45)
ppLuc(CC)-ag-A64-histoneSL (SEQ ID NO: 46)
ppLuc(CC)-ag-A120 (SEQ ID NO: 47)
ppLuc(GC) A64-ag (SEQ ID NO: 48)
ppLuc(GC)-ag-A64-aCPSL (SEQ ID NO: 49)
ppLuc(CC)-ag-A64-PolioCL (SEQ ID NO: 50)
ppLuc(CC)-ag-A64-G30 (SEQ ID NO: 51 )
ppLuc(GC)-ag-A64-U30 (SEQ ID NO: 52)
ppLuc(GC)-ag-A64-SL (SEQ ID NO: 53)
ppLuc(GC)-ag-A64-N32 (SEQ ID No: 54)
Кроме того, последовательности плазмидной ДНК, кодирующие опухолевые антигены NY-ESO-1, сурвивин и MAGE-C1, были подготовлены соответствующим образом, как описано выше.
Таким образом, были получены следующие мРНК с этих векторов путем транскрипции in vitro (последовательности мРНК приведены на фигурах 18-21):
NY-ESO-1 (GC)-ag-A62-C30 (SEQ ID NO: 55)
NY-ESO-1 (GC)-ag-A62-C30-histoneSL (SEQ ID NO: 56)
Survivin(GC)-ag-A62-C30-histoneSL (SEQ ID NO: 57)
MAGE-C1 (GC)-ag-A64-C30-histoneSL (SEQ ID NO: 58)
Транскрипция in vitro
ДНК-матрицу в соответствии с примером 2 линеаризировали и транскрибировали in vitro при помощи T7-полимеразы. ДНК-матрицу затем расщепляют при помощи обработки ДНКазой. Все мРНК-транскрипты содержали 5'-CAP структуру, полученную путем добавления избытка N7-метилгуанозин-5'-трифосфат-5'-гуанозина в реакцию транскрипции. Полученные таким образом мРНК очищали и ресуспендировали в воде.
Ферментативное аденилирование мРНК
Две мРНК были ферментативно аденилированы:
ppLuc(GC)-ag-A64 и ppLuc(GC)-ag histoneSL.
C этой целью, РНК инкубировали с поли(A)-полимеразой E coli и ATP (Poly(A) Polymerase Tailing Kit, Epicentre, Мэдисон, США), следуя инструкциям производителя. мРНК с удлиненной поли(A)-последовательностью очищали и ресуспендировали в воде. Длину поли(A)-последовательности определяется путем агарозного гель-электрофореза. Начальные мРНК были удлинены примерно на 250 аденилатов, полученные мРНК обозначены как
ppLuc(GC)-ag-A300 и ppLuc(GC)-ag-histoneSL-A250, соответственно.
Экспрессия люциферазы при помощи электропорации мРНК
Клетки HeLa трипсинизировали и промывали в opti-МЕМ. Каждые из 1 x105 клеток в 200 мкл opti-МЕМ были электропорированы с 0,5 мкг ppLuc-кодирующей мРНК. В качестве контроля, мРНК, не кодирующую ppLuc, электропорировали отдельно. Электропорированные клетки засевали в 24-луночные планшеты в 1 мл среды RPMI 1640. Через 6, 24 или 48 часов после трансфекции, среду аспирировали и клетки лизировали в 200 мкл буфера для лизиса (25 мМ Трис, рН 7,5 (HCl), 2 мМ ЭДТА, 10% глицерина, 1% Triton X-100, 2 мМ DTT, 1 мМ PMSF). Лизаты хранили при -20°C до измерения активности ppLuc.
Экспрессия люциферазы при помощи липофекции мРНК
Клетки HeLa засевали в 96-луночные планшеты при плотности 2x104 клеток на лунку. На следующий день, клетки отмывали в opti-МЕМ, а затем трансфицировали 0,25 мкг ppLuc-кодирующей мРНК в комплексе с липофектином в 150 мкл opti-МЕМ. В качестве контроля, отдельно липофектировали мРНК, которая не кодировала ppLuc. В некоторых лунках, opti-МЕМ аспирировали и клетки лизировали в 200 мкл буфера для лизиса через 6 часов после начала трансфекции. В оставшихся лунках opti-МЕМ заменяли на среду RPMI 1640 в это время. В этих лунках среду аспирировали и клетки растворяли в 200 мкл буфера для лизиса через 24 или 48 часов после начала трансфекции. Лизаты хранили при -20 ºC до измерения активности ppLuc.
Определение люциферазы
Активность ppLuc определяли как относительные световые единицы (RLU) в планшет-ридере BioTek SynergyHT при измерении времени 5 секунд с помощью 50 мкл лизата и 200 мкл люциферинового буфера (25 мМ глицилглицина, рН 7,8 (NaOH), 15 мМ MgS04, 2 мМ АТФ, 75 мкМ люциферина). Конкретные RLU рассчитывали путем вычитания RLU контрольной РНК из общей RLU.
Экспрессия люциферазы путем внутрикожной инъекции мРНК (экспрессия люциферазы in vivo)
Мышей анестезировали смесью Rompun и Ketavet. Каждую ppLuc-кодирующую мРНК вводили внутрикожно (0,5 мкг мРНК в 50 мкл на инъекцию). В качестве контроля отдельно вводили мРНК, которая не кодировала ppLuc. Через 16 часов после инъекции мышей забивали и тканей собирали. Образцы тканей быстро замораживали в жидком азоте и лизировали в устройстве для лизиса ткани (Qiagen) в 800 мкл буфера для лизиса (25 мМ Трис, рН 7,5 (HCl), 2 мМ ЭДТА, 10% глицерина, 1% Triton X-100, 2 мМ DTT, 1 мМ PMSF).Затем образцы центрифугировали при 13500 об/мин при 4ºC в течение 10 минут. Лизаты хранили при -80ºC до определения ppLuc-активности (см. 7. определения люциферазы).
Экспрессия NY-ESO-1 при помощи электропорации мРНК
Клетки HeLa триптинизировали и промывали в opti-МЕМ. 2x105 клеток в 200 мкл opti-МЕМ электропорировали с 10 пг NY-ESO-1-кодирующей мРНК. Клетки из трех электропораций объединяли и засевали в 6-луночный планшет в 2 мл среды RPMI 1640. Через 24 часа после трансфекции, клетки собирали и переносили в 96-луночные планшеты с V-образным дном (2 лунки на мРНК). Клетки отмывали буферизованным фосфатом солевым раствором (PBS) и пермеализировали с 200 мкл на лунку Cytofix/Cytoperm (Becton Dickinson (BD)). Через 15 минут, клетки промывали PermWash (BD). Затем клетки инкубировали 1 ч при комнатной температуре либо с мышиными IgG1 против NY-ESO-1, либо с изотипными контрольными (20 мкг/ мл). Клетки снова промывали два раза PermWash. Далее клетки инкубировали 1 ч при 4ºC с Alexa-647-конъюгированными козлиными антимышиными IgG в разведении 1:500. Наконец, клетки промывали в два раза с PermWash. Клетки ресуспендировали в 200 мкл буфера (PBS, 2% FCS, 2 мМ ЭДТА, 0,01% азид натрия). Экспрессию NY-ESO-1 определяли количественно с помощью проточной цитометрии как среднюю интенсивность флуоресценции (МFI).
Индукция антител против NY-ESO-1 при помощи вакцинации с мРНК
Мышей линии C57BL/6 вакцинировали внутрикожно с NY-ESO-1-кодирующей мРНК в комплексе с протамином (5 раз в 14 дней). Контрольных мышей подвергали лечению буфером. Уровень NY-ESO-1-специфических антител у вакцинированных и контрольных мышей анализировали через 8 дней после последней вакцинации способом ELISA: 96-луночные планшеты для ELISA (Nunc) покрывали 100 мкл на лунку 10 мкг/мл рекомбинантного белка NY-ESO-1 в течение 16 часов при 4°C. Планшеты промывали по два раза промывочным буфером (PBS, 0,05% Tween-20). Для блокирования неспецифического связывания планшеты затем выдерживали в течение 2 ч при 37°C с блокирующим буфером (PBS, 0,05%, Tween-20, 1% BSA). После блокирования 100 мкл на лунку последовательно разведенных мышиных сывороток добавляли и инкубировали в течение 4 часов при комнатной температуре. Планшеты промывали три раза промывочным буфером. Далее, 100 мкл на лунку биотинилированных крысиных анти-мышиных детектирующих антител IgG2a[b] (BD Biosciences), разведенных 1:600 в блокирующем буфере, давали возможность связываться в течение 1 часа при комнатной температуре. Планшеты промывали снова три раза промывочным буфером, с последующей инкубацией в течение 30 минут при комнатной температуре с 100 мкл на лунку стрептавидина, конъюгированного с пероксидазой хрена. После четырех промывок промывочным буфером 100 мкл на лунку добавляли 3,3',5,5'-тетраметилбензидин (Thermo Scientific). После полученного в результате изменения цвета добавляли 100 мкл 20% серной кислоты. Абсорбцию определяли при 405 нм.
Индукция антител против сурвивина при помощи вакцинации с мРНК
Мышей линии C57BL/6 вакцинировали внутрикожно с сурвивина-кодирующей мРНК в комплексе с протамином (5 раз в 14 дней). Контрольных мышей подвергали лечению буфером. Уровень сурвивин-специфических антител у вакцинированных и контрольных мышей анализировали через 8 дней после последней вакцинации способом ELISA. 96-луночные планшеты для ELISA (Nunc) покрывали 100 мкл на лунку 10 мкг/мл рекомбинантного белка сурвивина в течение 16 часов при 4°C. Планшеты промывали по два раза промывочным буфером (PBS, 0,05% Tween-20). Для блокирования неспецифического связывания планшеты затем выдерживали в течение 2 ч при 37°C с блокирующим буфером (PBS, 0,05%, Tween-20, 1% BSA). После блокирования 100 мкл на лунку последовательно разведенных мышиных сывороток добавляли и инкубировали в течение 4 часов при комнатной температуре. Планшеты промывали три раза промывочным буфером. Далее, 100 мкл на лунку биотинилированных крысиных анти-мышиных детектирующих антител IgG2a[b] (BD Biosciences), разведенных 1:600 в блокирующем буфере, давали возможность связываться в течение 1 часа при комнатной температуре. Планшеты промывали снова три раза промывочным буфером, с последующей инкубацией в течение 30 минут при комнатной температуре с 100 мкл на лунку стрептавидина, конъюгированного с пероксидазой хрена. После четырех промывок промывочным буфером 100 мкл на лунку добавляли 3,3',5,5'-тетраметилбензидин (Thermo Scientific). После полученного в результате изменения цвета добавляли 100 мкл 20% серной кислоты. Абсорбцию определяли при 405 нм.
Индукция антител против MAGE-C1 с помощью вакцинации с мРНК
Мышей линии C57BL/6 вакцинировали внутрикожно с MAGE-C1-кодирующей мРНК в комплексе с протамином (5 раз в 14 дней). Контрольных мышей подвергали лечению буфером. Уровень MAGE-C1-специфических антител у вакцинированных и контрольных мышей анализировали через 8 дней после последней вакцинации способом ELISA. 96-луночные планшеты для ELISA (Nunc) покрывали 100 мкл на лунку 10 мкг/мл рекомбинантного белка MAGE-C1в течение 16 часов при 4°C. Планшеты промывали по два раза промывочным буфером (PBS, 0,05% Tween-20). Для блокирования неспецифического связывания планшеты затем выдерживали в течение 2 ч при 37°C с блокирующим буфером (PBS, 0,05%, Tween-20, 1% BSA). После блокирования 100 мкл на лунку последовательно разведенных мышиных сывороток добавляли и инкубировали в течение 4 часов при комнатной температуре. Планшеты промывали три раза промывочным буфером. Далее, 100 мкл на лунку биотинилированных крысиных анти-мышиных детектирующих антител IgG2a[b] (BD Biosciences), разведенных 1:600 в блокирующем буфере, давали возможность связываться в течение 1 часа при комнатной температуре. Планшеты промывали снова три раза промывочным буфером, с последующей инкубацией в течение 30 минут при комнатной температуре с 100 мкл на лунку стрептавидина, конъюгированного с пероксидазой хрена. После четырех промывок промывочным буфером 100 мкл на лунку добавляли 3,3',5,5'-тетраметилбензидин (Thermo Scientific). После полученного в результате изменения цвета добавляли 100 мкл 20% серной кислоты. Абсорбцию определяли при 405 нм.
Детектирование антиген-специфического клеточного иммунного ответа (Т-клеточного иммунный ответ) при помощи ELISPQT:
Мышей линии C57BL/6 вакцинировали внутрикожно с MAGE-C1-кодирующей мРНК в комплексе с протамином (5 раз в 14 дней). Контрольных мышей подвергали лечению буфером. Через 1 неделю после последней вакцинации мышей забивали, удаляли селезенки и выделяли спленоциты. Спленоциты повторно стимулировали в течение 7 дней в присутствии пептидов вышеописанного антигена (пептидная библиотека) или соинкубировали с дендритными клетками, генерированными из клеток костного мозга нативных сингенных мышей, которые электропорируют с мРНК, кодирующей антиген. Для детектирования антиген-специфического клеточного иммунного ответа определяли секрецию IFN-гамма после повторной стимуляции. Для детектирования IFN-гамма покрытые планшеты для мультискрининга (Millipore) инкубируют в течение ночи с покрывающим буфером (0,1 М карбонат-бикарбонатный буфер, рН 9,6, 10,59 г/л Na2CО3, 8,4 г/л NaHCО3), содержащим антитела против INFy (BD Pharmingen, Гейдельберг, Германия). Стимуляторы и эффекторные клетки инкубируют вместе в планшете в соотношении 1:20 в течение 24 часов. Планшет промывают 1 xPBS и инкубируют с биотин-связанным вторичным антителом. После промывки с 1xPBS/ 0,05% Tween-20 добавляют субстрат (5-бром-4-хлор-3-индолилфосфат/Nitro Blue Tetrazolium Liquid Substrate System от Sigma Aldrich, Taufkirchen, Германия) к планшету, и преобразование субстрата можно детектировать визуально.
РЕЗУЛЬТАТЫ
Последовательности гистоновых структур типа «стебель-петля»:
Для того, чтобы охарактеризовать последовательности гистоновых структур типа «стебель-петля», последовательности из метазоа и простейших (4001 последовательность), или из простейших (131 последовательность) или, альтернативно, из метазоа (3870 последовательностей) или из позвоночных (1333 последовательности) или из человека (84 последовательности) были сгруппированы и выровнены. Затем, количество представленных нуклеотидов определяли для каждого положения. На основе таблиц, полученных таким образом, были генерированы консенсусные последовательности для 5 различных групп последовательностей, представляющие все нуклеотиды, присутствующие в анализируемых последовательностях. В рамках объединенной консенсусной последовательности метазоа и простейших, 3 нуклеотида являются консервативными, T/U в петле и G и C в стебле, образуя пару оснований. Конструктивно, как правило, формируется стебель из 6 пар оснований и петля из 4 нуклеотидов. Однако отклоняющиеся структуры являются общими: из 84 гистоновых структур типа «стебель-петля» человека два содержат стебель только в 5 нуклеотидов, состоящий из 4 пар оснований и одного несоответствия. Другая гистоновая структура типа «стебель-петля» человека содержит стебель только в 5 пар оснований. Еще четыре гистоновых структуры типа «стебель-петля» человека содержат стебель длиной в 6 нуклеотидов, но и включает одно несоответствие в трех различных положениях, соответственно. Кроме того, четыре гистоновые структуры типа «стебель-петля» человека содержат одну неустойчивую пару оснований в двух разных положениях, соответственно. Что касается петли, длина в 4 нуклеотида, похоже, не является строго обязательной, так как петля из 5 нуклеотидов была идентифицирована в D. discoideum.
В дополнение к консенсусным последовательностям, представляющим все нуклеотиды, присутствующие в анализируемой последовательности, были получены более строгие консенсусные последовательности, все более подчеркивающие консервативные нуклеотиды.
В итоге были получены следующие последовательности:
(Cons): обозначает все присутствующие нуклеотиды
(99%): представляет по меньшей мере 99% всех присутствующих нуклеотидов
(95%): представляет по меньшей мере 95% всех присутствующих нуклеотидов
(90%): представляет по меньшей мере 90% всех присутствующих нуклеотидов
Результаты анализа последовательностей гистоновых структур типа «стебель-петля» приведены в следующих таблицах 1-5 (см. также Фигуры от 1 до 5):
Консенсусная последовательность гистоновой структуры типа «стебель-петля» метазоа и простейших: (на основе выравнивания 4001 последовательности гистоновых структур типа «стебель-петля» метазоа и простейших) (см. также Фигуру 1)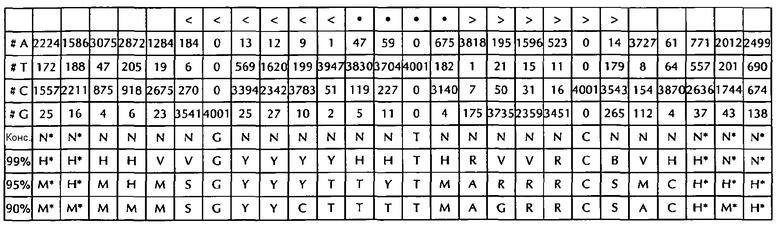
Консенсусная последовательность гистоновой структуры типа «стебель-петля» простейших: (на основе выравнивания 131 последовательности гистоновых структур типа «стебель-петля» простейших) (см. также Фигуру 2)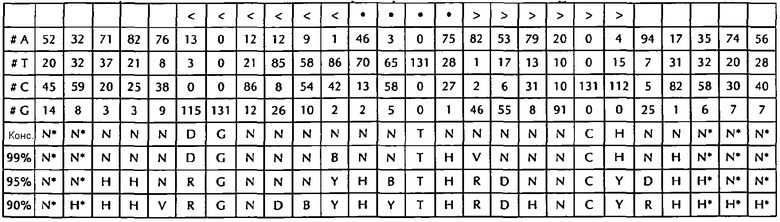
Консенсусная последовательность гистоновой структуры типа «стебель-петля» метазоа: (на основе выравнивания 3870 (в том числе 1333 последовательностей позвоночных) последовательностей гистоновых структур типа «стебель-петля» метазоа) (см. также Фигуру 3)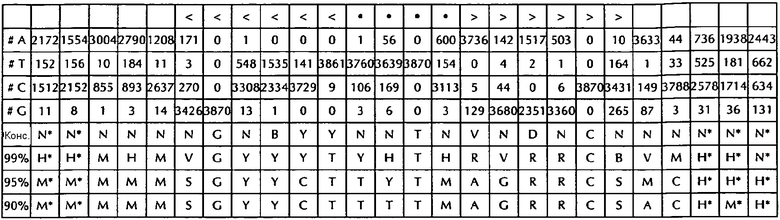
Консенсусная последовательность гистоновой структуры типа «стебель-петля» позвоночных: (на основе выравнивания 1333 последовательностей гистоновых структур типа «стебель-петля» позвоночных) (см. также Фигуру 4)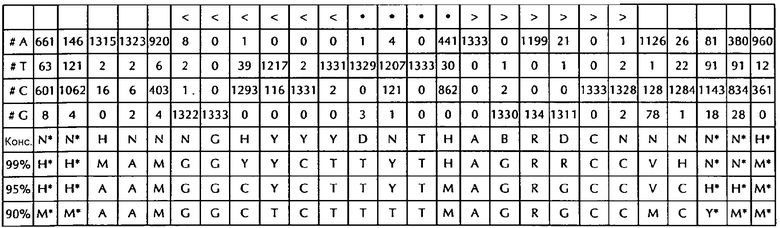
Консенсусная последовательность гистоновой структуры типа «стебель-петля» Homo sapiens: (на основе выравнивания 84 последовательностей гистоновых структур типа «стебель-петля» человека) (см. также Фигуру 5)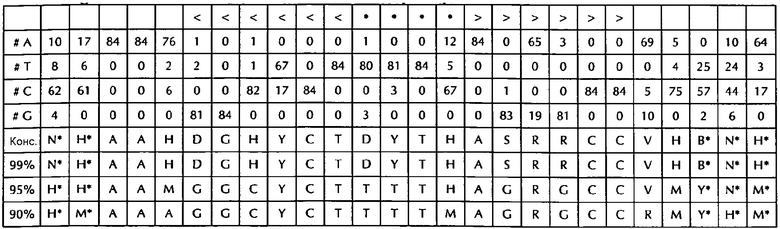
где использованные сокращения были определены следующим образом:
Комбинация поли(A) и histoneSL увеличивает экспрессию белков с мРНК синергетическим образом.
Для изучения эффекта комбинации поли(A) и histoneSL на экспрессию белков с мРНК, были синтезированы мРНК с различными последовательностями 3' альфа-глобина 3'-UTR: мРНК либо заканчивается сразу же на 3'-конце 3'-UTR, поэтому не содержит ни поли(A)-последовательности, ни histoneSL, или содержит либо поли(A)-последовательность A64, либо histoneSL вместо этого, или как поли(A) A64, так и histoneSL с 3' 3'-UTR. Кодирующие люциферазу мРНК или контрольная мРНК были электропорированы в клетки HeLa. Уровни люциферазы измеряли через 6, 24 и 48 часов после трансфекции (см. ниже Таблицу 6 и Фигуру 22).
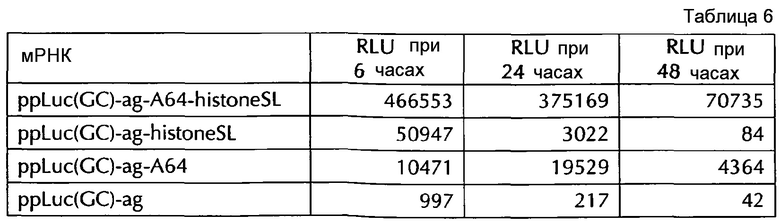
Мало люциферазы экспрессировалось с мРНК, не имеющей ни поли(A)-последовательности, ни histoneSL. Как поли(A)-последовательность, так и histoneSL увеличили уровень люциферазы в одной и той же степени. Либо мРНК приводила к уровню люциферазы, гораздо более высокому, чем для мРНК при отсутствии как поли(A), так и histoneSL. Поразительно, однако, комбинация поли(A) и histoneSL также сильно увеличивала уровень люциферазы, многократно выше уровня, наблюдавшегося с любым из отдельных элементов. Величина увеличения уровня люциферазы за счет комбинирования поли(A) и histoneSL в одной и той же мРНК показывает, что они действуют синергически.
Синергия между поли(A) и histoneSL были количественно определены путем деления сигнала от поли(A)-histoneSL мРНК (+/+) на сумму сигналов от histoneSL мРНК (-/+) плюс поли(A) мРНК (+/-) (см. следующую Таблицу 7).
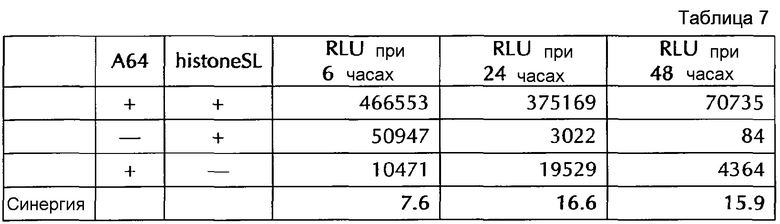
Коэффициент, рассчитанный таким образом, определяет, насколько выше уровень люциферазы с мРНК, комбинирующей поли(A) и histoneSL, чем можно было бы ожидать, если эффекты от поли(A) и histoneSL были чисто аддитивным. Уровень люциферазы с мРНК, комбинирующей поли(A) и histoneSL, был в 16,6 раза выше, чем если бы их эффекты были чисто аддитивные. Этот результат подтверждает, что комбинация поли(A) и histoneSL вызывает заметное синергидное увеличение белковой экспрессии.
Комбинация поли(A) и histoneSL увеличение белковую экспрессию с мРНК независимо от их порядка.
Эффект комбинации поли(A) и histoneSL может зависеть от длины поли(A)-последовательности и порядка поли(A) и histoneSL. При этом были синтезированы мРНК с увеличенной длиной поли(A)-последовательности и мРНК с поли(A) и histoneSL, расположенными в обратном порядке: Две мРНК, содержащие с 3'-стороны 3'-UTR либо поли(A) последовательность A120, либо A300. Еще одна мРНК содержала с 3'-стороны 3'-UTR сначала histoneSL, а затем поли(A)-последовательность A250. Кодирующие люциферазу мРНК или контрольная мРНК были липофектированы в клетки HeLa. Уровни люциферазы определяли через 6, 24 и 48 часов после начала трансфекции (см. нижеследующие Таблицу 8 и Фигуру 23).
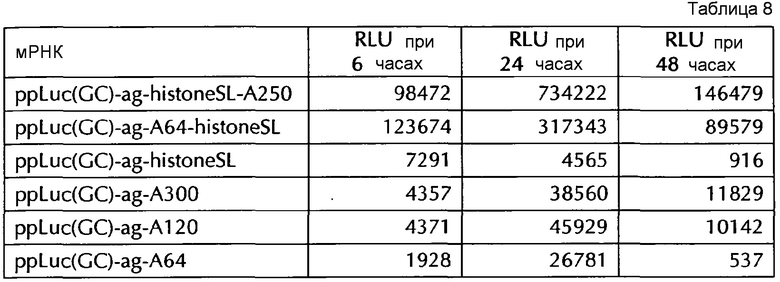
Как поли(A)-последовательность A64, так и histoneSL вызывали сопоставимые уровни люциферазы. В согласии с предыдущим экспериментом комбинация A64 и histoneSL сильно увеличивала уровень люциферазы, многократно выше уровня, наблюдавшегося с любым из отдельных элементов. Величина увеличения уровня люциферазы за счет комбинирования поли(A) и histoneSL в одной и той же мРНК показывает, что они действуют синергически. Синергия между A64 и histoneSL были количественно определены, как и прежде, на основе уровней люциферазы с A64-histoneSL, A64 и histoneSL мРНК (см. следующую таблицу 9). Уровень люциферазы с мРНК, комбинирующей A64 и histoneSL, был вплоть до 61,7 раза выше, чем в случае, если бы эффекты поли(A) и histoneSL были чисто аддитивными.
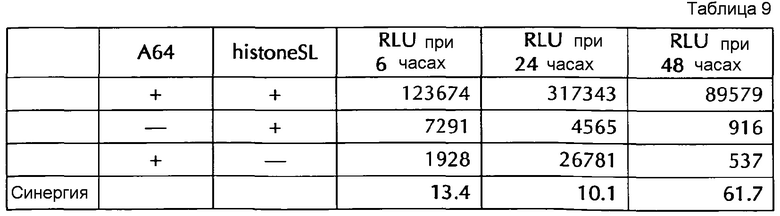
В отличие от этого, увеличение длины поли(A)-последовательности с A64 до A120 или до A300 увеличивал уровень люциферазы лишь незначительно (см. Таблицу 8 и Фигуру 19). мРНК с самой длинной поли(A)-последовательностью, A300, также сравнивали с мРНК, в которой поли(A)-последовательность аналогичной длины комбинировали с histoneSL, histoneSL-A250. В дополнение к обладанию длинной поли(A)-последовательностью, порядок histoneSL и поли(А) в этой мРНК является обратным по сравнению с A64-histoneSL мРНК. Комбинация A250 и histoneSL сильно увеличивала уровень люциферазы, многократно выше уровня, наблюдавшегося либо с histoneSL, либо с A300. Снова, была количественно определена синергия между A250 и histoneSL, как и прежде при сравнении RLU c histoneSL-A250 mRNA c RLU c A300 мРНК плюс histoneSL мРНК (см. следующую Таблицу 10). Уровень люциферазы с мРНК, комбинирующей A250 и histoneSL, был вплоть до 17,0 раз выше, чем если бы эффекты поли(A) и histoneSL были чисто аддитивными.
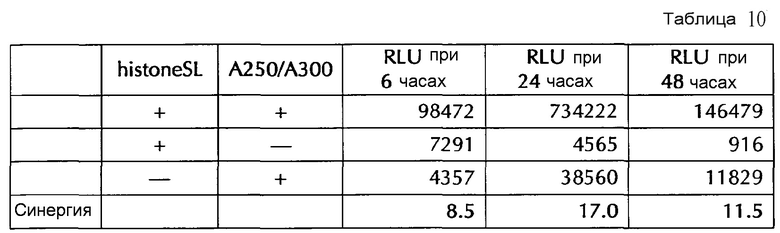
В заключение, был продемонстрирован высоко синергетический эффект комбинации histoneSL и поли(A) на белковую экспрессию с мРНК для существенно различных длин поли(A) независимо от порядка поли(A) и histoneSL.
Увеличение белковой экспрессии при помощи комбинации поли(A) и histoneSL является специфичным
Для исследования того, является ли специфичным эффект комбинации поли(A) и histoneSL на белковую экспрессию с мРНК, были синтезированы мРНК с альтернативными последовательностями в комбинации с поли(A). Эти мРНК содержали с 3'-стороны A64 одну из семи различных последовательностей, соответственно. Кодирующие люциферазу мРНК или контрольная мРНК были электропорированы в клетки HeLa. Уровни люциферазы определяли через 6, 24 и 48 часов после трансфекции (см. нижеследующие Таблицу 11 и Фигуру 24).
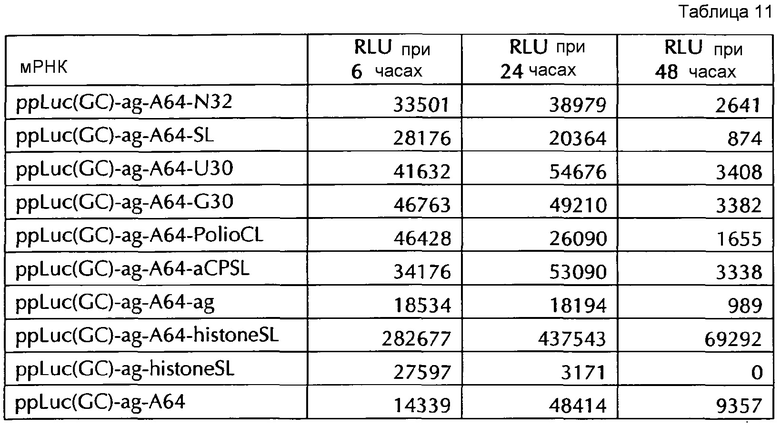
Как поли(A)-последовательность, так и histoneSL вызывали сопоставимые уровни люциферазы. Снова, комбинация поли(A) и histoneSL сильно увеличивает уровень люциферазы, многократно превышая уровень, наблюдающийся с любым из отдельных элементов, действуя, таким образом, синергетически. В отличие от этого, комбинирование поли(A) с любой из альтернативных последовательностей не оказывало влияния на уровень люциферазы по сравнению с мРНК, содержащие только поли(A)-последовательность. Таким образом, комбинация поли(A) и histoneSL увеличивает белковую экспрессию с мРНК синергичным образом, и этот эффект является специфическим.
Комбинация поли(A) и histoneSL увеличивает белковую экспрессию с мРНК синергичным образом in vivo.
Для изучения эффекта комбинации поли(A) и histoneSL на белковую экспрессию с мРНК in vivo, кодирующие люциферазу мРНК с различными последовательностями с 3'-стороны 3'-UTR альфа-глобина или контрольную мРНК инъецировали внутрикожно в мышей: мРНК, содержащие либо поли(A)-последовательность A64, либо histoneSL вместо этого, или как A64 поли(A), так и histoneSL 3' 3'-UTR. Уровни люциферазы определяли через 16 часов после инъекции (см. следующие Таблицу 12 и Фигуру 25).
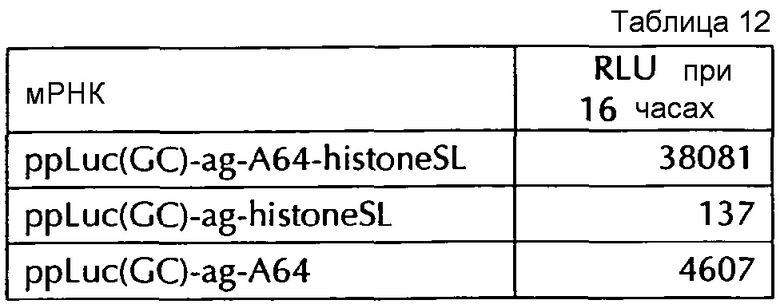
Люциферазу экспрессировали с мРНК, имеющую либо histoneSL, либо поли(A)-последовательность. Поразительно, однако, комбинация поли(A) и histoneSL дополнительного резко увеличивало уровень люциферазы, многократно выше уровня, наблюдавшегося с любым из отдельных элементов. Величина увеличения уровня люциферазы за счет комбинации поли(A) и histoneSL в одной и той же мРНК показывает, что они действуют синергически.
Синергию между поли(A) и histoneSL количественно определяли путем деления сигнала от поли(A)-histoneSL мРНК (+/+) на сумму сигналов от histoneSL мРНК (-/+) плюс поли(A) мРНК (+/-) (см. нижеследующую Таблицу 13).
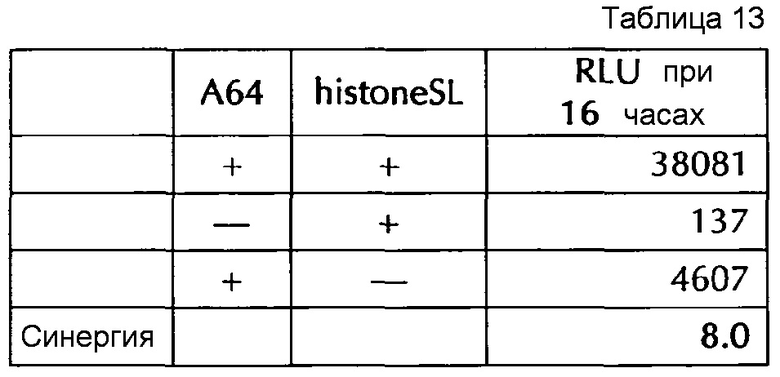
Коэффициент, рассчитанный таким образом определяет, насколько выше уровень люциферазы с мРНК, комбинирующей поли(A) и histoneSL, чем ожидалось бы, если бы эффекты поли(A) и histoneSL были чисто аддитивными. Уровень люциферазы с мРНК, комбинирующей поли(A) и histoneSL, был в 8 раз выше, чем если бы их эффекты были чисто аддитивными. Этот результат подтверждает, что комбинация поли(A) и histoneSL приводит к заметному синергичному увеличению белковой экспрессии in vivo.
Комбинация поли(A) и histoneSL увеличивает экспрессию белка NY-ESO-1 с мРНК.
Для изучения эффекта комбинации поли(A) и histoneSL на белковую экспрессию с мРНК, были синтезированы кодирующие NY-ESO-1 мРНК с различными последовательностями с 3'-стороны 3'-UTR альфа-глобина: мРНК, содержащие либо поли(A)-последовательность A64, либо как поли(A) A64, так и histoneSL с 3'-стороны 3'-UTR. NY-ESO-1-кодирующие мРНК были электропорированы в клетки HeLa. Уровни NY-ESO-1 определяли через 24 часа после трансфекции с помощью проточной цитометрии (см. следующие ниже Таблицу 14 и Фигуру 26).

NY-ESO-1 экспрессировали с мРНК, имеющей только поли(A)-последовательность. Поразительно, однако, комбинация поли(A) и histoneSL сильно увеличивала уровень NY-ESO-1, многократно выше уровня, наблюдаемого только с поли(A)-последовательностью.
Комбинация поли(A) и histoneSL увеличивает уровень антител, достигаемый при помощи вакцинации с мРНК.
Для изучения эффекта комбинации поли(A) и histoneSL на индукцию антител, достигаемую при помощи вакцинации с мРНК, мышей линии C57BL/6 вакцинировали внутрикожно NY-ESO-1-кодирующими мРНК в комплексе с протамином, с различными последовательностями 3' Альфа-глобина 3'-UTR. мРНК содержали либо поли(A)-последовательность A64, или как поли(A) A64, так и histoneSL 3' 3'-UTR. Уровень NY-ESO-1-специфических антител у вакцинированных и контрольных мышей анализировали способом ELISA с последовательными разведениями сывороток (см. следующие ниже Таблицу 15 и Фигуру 27).
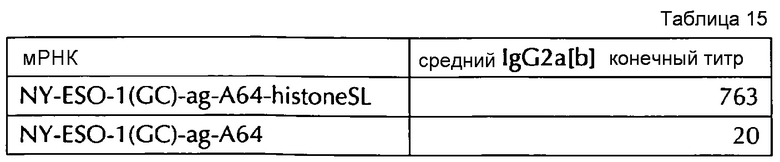
IgG2a[b] против NY-ESO-1 индуцировали при помощи мРНК, имеющей только поли(A)-последовательность. Поразительно, однако, комбинация поли(A) и histoneSL сильно увеличивала уровень IgG2a[b] против NY-ESO-1, многократно выше уровня, наблюдаемого только с поли(A)-последовательностью.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CUREVAC GMBH
<120> НУКЛЕИНОВАЯ КИСЛОТА, СОДЕРЖАЩАЯ ИЛИ КОДИРУЮЩАЯ ГИСТОНОВУЮ СТРУКТУРУ
ТИПА «СТЕБЕЛЬ-ПЕТЛЯ» И ПОЛИ(A)-ПОСЛЕДОВАТЕЛЬНОСТЬ ИЛИ СИГНАЛ
ПОЛИАДЕНИЛИРОВАНИЯ, ДЛЯ УВЕЛИЧЕНИЯ ЭКСПРЕССИИ КОДИРУЕМОГО
ОПУХОЛЕВОГО АНТИГЕНА
<130> CU01P128WO1
<140> PCT/EP2012/000674
<141> 2012-02-15
<160> 58
<170> PatentIn version 3.5
<210> 1
<211> 16
<212> РНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля» в
соответствии с формулой (Ic): консенсусная последовательность
гистоновой структуры типа «стебель-петля» метазоа и простейших без
ограничивающих элементов стебля
<220>
<221> misc_feature
<222> (1)..(1)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (3)..(8)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (10)..(14)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (16)..(16)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<400> 1
ngnnnnnnun nnnncn 16
<210> 2
<211> 26
<212> РНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля» в
соответствии с формулой (IIc): консенсусная последовательность
гистоновой структуры типа «стебель-петля» метазоа и простейших без
ограничивающих элементов стебля
<220>
<221> misc_feature
<222> (1)..(6)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (8)..(13)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (15)..(19)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (21)..(26)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<400> 2
nnnnnngnnn nnnunnnnnc nnnnnn 26
<210> 3
<211> 16
<212> РНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля» в
соответствии с формулой (Id): без ограничивающих элементов стебля
<220>
<221> misc_feature
<222> (1)..(1)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (3)..(8)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (10)..(14)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (16)..(16)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<400> 3
ncnnnnnnun nnnngn 16
<210> 4
<211> 26
<212> РНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля» в
соответствии с формулой (IId): с ограничивающими элементами стебля
<220>
<221> misc_feature
<222> (1)..(6)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (8)..(13)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (15)..(19)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (21)..(26)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<400> 4
nnnnnncnnn nnnunnnnng nnnnnn 26
<210> 5
<211> 16
<212> РНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля» в
соответствии с формулой (Iе): консенсусная последовательность
гистоновой структуры типа «стебель-петля» простейших без
ограничивающих элементов стебля
<220>
<221> misc_feature
<222> (3)..(8)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (10)..(14)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<400> 5
dgnnnnnnun nnnnch 16
<210> 6
<211> 26
<212> РНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля» в
соответствии с формулой (IIе): консенсусная последовательность
гистоновой структуры типа «стебель-петля» простейших с
ограничивающими элементами стебля
<220>
<221> misc_feature
<222> (1)..(5)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (8)..(13)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (15)..(19)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (22)..(26)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<400> 6
nnnnndgnnn nnnunnnnnc hnnnnn 26
<210> 7
<211> 16
<212> РНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля» в
соответствии с формулой (If): консенсусная последовательность
гистоновой структуры типа «стебель-петля» метазоа без ограничивающих
элементов стебля
<220>
<221> misc_feature
<222> (1)..(1)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (3)..(3)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (7)..(8)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (10)..(10)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (12)..(12)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (14)..(14)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (16)..(16)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<400> 7
ngnbyynnun vndncn 16
<210> 8
<211> 26
<212> РНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля» в
соответствии с формулой (IIf): консенсусная последовательность
гистоновой структуры типа «стебель-петля» метазоа с ограничивающими
элементами стебля
<220>
<221> misc_feature
<222> (1)..(6)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (8)..(8)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (12)..(13)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (15)..(15)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (17)..(17)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (19)..(19)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (21)..(26)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<400> 8
nnnnnngnby ynnunvndnc nnnnnn 26
<210> 9
<211> 16
<212> РНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля» в
соответствии с формулой (Ig): консенсусная последовательность
гистоновой структуры типа «стебель-петля» позвоночных без
ограничивающих элементов стебля
<220>
<221> misc_feature
<222> (1)..(1)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (8)..(8)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (16)..(16)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<400> 9
nghyyydnuh abrdcn 16
<210> 10
<211> 26
<212> РНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля» в
соответствии с формулой (IIg): консенсусная последовательность
гистоновой структуры типа «стебель-петля» позвоночных с
ограничивающими элементами стебля
<220>
<221> misc_feature
<222> (1)..(2)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (4)..(6)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (13)..(13)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (21)..(25)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<400> 10
nnhnnnghyy ydnuhabrdc nnnnnh 26
<210> 11
<211> 16
<212> РНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля» в
соответствии с формулой (Ih): консенсусная последовательность
гистоновой структуры типа «стебель-петля» человека (Homo sapiens)
без ограничивающих элементов стебля
<400> 11
dghycudyuh asrrcc 16
<210> 12
<211> 26
<212> РНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля» в
соответствии с формулой (IIh): консенсусная последовательность
гистоновой структуры типа «стебель-петля» человека (Homo sapiens)
с ограничивающими элементами стебля
<220>
<221> misc_feature
<222> (1)..(1)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (25)..(25)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<400> 12
nhaahdghyc udyuhasrrc cvhbnh 26
<210> 13
<211> 16
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(без ограничивающих элементов стебля) в соответствии с формулой (Iс)
<400> 13
vgyyyyhhth rvvrcb 16
<210> 14
<211> 16
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(без ограничивающих элементов стебля) в соответствии с формулой (Iс)
<400> 14
sgyyyttytm arrrcs 16
<210> 15
<211> 16
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(без ограничивающих элементов стебля) в соответствии с формулой (Iс)
<400> 15
sgyycttttm agrrcs 16
<210> 16
<211> 16
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(без ограничивающих элементов стебля) в соответствии с формулой (Ie)
<220>
<221> misc_feature
<222> (3)..(5)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (7)..(8)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (12)..(14)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<400> 16
dgnnnbnnth vnnnch 16
<210> 17
<211> 16
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(без ограничивающих элементов стебля) в соответствии с формулой (Ie)
<220>
<221> misc_feature
<222> (3)..(5)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (13)..(14)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<400> 17
rgnnnyhbth rdnncy 16
<210> 18
<211> 16
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(без ограничивающих элементов стебля) в соответствии с формулой (Ie)
<220>
<221> misc_feature
<222> (3)..(3)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (14)..(14)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<400> 18
rgndbyhyth rdhncy 16
<210> 19
<211> 16
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(без ограничивающих элементов стебля) в соответствии с формулой (If)
<400> 19
vgyyytyhth rvrrcb 16
<210> 20
<211> 16
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(без ограничивающих элементов стебля) в соответствии с формулой (If)
<400> 20
sgyycttytm agrrcs 16
<210> 21
<211> 16
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(без ограничивающих элементов стебля) в соответствии с формулой (If)
<400> 21
sgyycttttm agrrcs 16
<210> 22
<211> 16
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(без ограничивающих элементов стебля) в соответствии с формулой (Ig)
<400> 22
ggyycttyth agrrcc 16
<210> 23
<211> 16
<212> ДНК
<213> Искусственная
<220>
<223> histone stem-loop sequences (without stem-bordering elements)
according to formula (Ig)
<400> 23
ggcycttytm agrgcc 16
<210> 24
<211> 16
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(без ограничивающих элементов стебля) в соответствии с формулой (Ig)
<400> 24
ggctcttttm agrgcc 16
<210> 25
<211> 16
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(без ограничивающих элементов стебля) в соответствии с формулой (Ih)
<400> 25
dghyctdyth asrrcc 16
<210> 26
<211> 16
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(без ограничивающих элементов стебля) в соответствии с формулой (Ih)
<400> 26
ggcyctttth agrgcc 16
<210> 27
<211> 16
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(без ограничивающих элементов стебля) в соответствии с формулой (Ih)
<400> 27
ggcycttttm agrgcc 16
<210> 28
<211> 26
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(c ограничивающими элементами стебля) в соответствии с формулой (IIc)
<220>
<221> misc_feature
<222> (25)..(26)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<400> 28
hhhhvvgyyy yhhthrvvrc bvhhnn 26
<210> 29
<211> 26
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(c ограничивающими элементами стебля) в соответствии с формулой (IIc)
<400> 29
mhmhmsgyyy ttytmarrrc smchhh 26
<210> 30
<211> 26
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(c ограничивающими элементами стебля) в соответствии с формулой (IIc)
<400> 30
mmmmmsgyyc ttttmagrrc sachmh 26
<210> 31
<211> 26
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(c ограничивающими элементами стебля) в соответствии с формулой (IIe)
<220>
<221> misc_feature
<222> (1)..(5)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (8)..(10)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (12)..(13)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (17)..(19)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (22)..(22)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (24)..(26)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<400> 31
nnnnndgnnn bnnthvnnnc hnhnnn 26
<210> 32
<211> 26
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(c ограничивающими элементами стебля) в соответствии с формулой (IIe)
<220>
<221> misc_feature
<222> (1)..(2)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (5)..(5)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (8)..(10)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (18)..(19)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (25)..(26)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<400> 32
nnhhnrgnnn yhbthrdnnc ydhhnn 26
<210> 33
<211> 26
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(c ограничивающими элементами стебля) в соответствии с формулой (IIe)
<220>
<221> misc_feature
<222> (1)..(1)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (8)..(8)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (19)..(19)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<400> 33
nhhhvrgndb yhythrdhnc yrhhhh 26
<210> 34
<211> 26
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(c ограничивающими элементами стебля) в соответствии с формулой (IIf)
<220>
<221> misc_feature
<222> (26)..(26)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<400> 34
hhmhmvgyyy tyhthrvrrc bvmhhn 26
<210> 35
<211> 26
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(c ограничивающими элементами стебля) в соответствии с формулой (IIf)
<400> 35
mmmmmsgyyc ttytmagrrc smchhh 26
<210> 36
<211> 26
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(c ограничивающими элементами стебля) в соответствии с формулой (IIf)
<400> 36
mmmmmsgyyc ttttmagrrc sachmh 26
<210> 37
<211> 26
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(c ограничивающими элементами стебля) в соответствии с формулой (IIg)
<220>
<221> misc_feature
<222> (24)..(25)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<400> 37
hhmamggyyc ttythagrrc cvhnnm 26
<210> 38
<211> 26
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(c ограничивающими элементами стебля) в соответствии с формулой (IIg)
<400> 38
hhaamggcyc ttytmagrgc cvchhm 26
<210> 39
<211> 26
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(c ограничивающими элементами стебля) в соответствии с формулой (IIg)
<400> 39
mmaamggctc ttttmagrgc cmcymm 26
<210> 40
<211> 26
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(c ограничивающими элементами стебля) в соответствии с формулой (IIh)
<220>
<221> misc_feature
<222> (1)..(1)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<220>
<221> misc_feature
<222> (25)..(25)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<400> 40
nhaahdghyc tdythasrrc cvhbnh 26
<210> 41
<211> 26
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(c ограничивающими элементами стебля) в соответствии с формулой (IIh)
<220>
<221> misc_feature
<222> (25)..(25)
<223> n выбирают из нуклеотидов, выбранных из A,U,T,G и С или нуклеотидного
аналога этого
<400> 41
hhaamggcyc tttthagrgc cvmynm 26
<210> 42
<211> 26
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность гистоновой структуры типа «стебель-петля»
(c ограничивающими элементами стебля) в соответствии с формулой (IIh)
<400> 42
hmaaaggcyc ttttmagrgc crmyhm 26
<210> 43
<211> 1747
<212> РНК
<213> Искусственная
<220>
<223> Последовательность мРНК ppLuc(GC)-ag
<400> 43
gggagaaagc uugaggaugg aggacgccaa gaacaucaag aagggcccgg cgcccuucua 60
cccgcuggag gacgggaccg ccggcgagca gcuccacaag gccaugaagc gguacgcccu 120
ggugccgggc acgaucgccu ucaccgacgc ccacaucgag gucgacauca ccuacgcgga 180
guacuucgag augagcgugc gccuggccga ggccaugaag cgguacggcc ugaacaccaa 240
ccaccggauc guggugugcu cggagaacag ccugcaguuc uucaugccgg ugcugggcgc 300
ccucuucauc ggcguggccg ucgccccggc gaacgacauc uacaacgagc gggagcugcu 360
gaacagcaug gggaucagcc agccgaccgu gguguucgug agcaagaagg gccugcagaa 420
gauccugaac gugcagaaga agcugcccau cauccagaag aucaucauca uggacagcaa 480
gaccgacuac cagggcuucc agucgaugua cacguucgug accagccacc ucccgccggg 540
cuucaacgag uacgacuucg ucccggagag cuucgaccgg gacaagacca ucgcccugau 600
caugaacagc agcggcagca ccggccugcc gaagggggug gcccugccgc accggaccgc 660
cugcgugcgc uucucgcacg cccgggaccc caucuucggc aaccagauca ucccggacac 720
cgccauccug agcguggugc cguuccacca cggcuucggc auguucacga cccugggcua 780
ccucaucugc ggcuuccggg ugguccugau guaccgguuc gaggaggagc uguuccugcg 840
gagccugcag gacuacaaga uccagagcgc gcugcucgug ccgacccugu ucagcuucuu 900
cgccaagagc acccugaucg acaaguacga ccugucgaac cugcacgaga ucgccagcgg 960
gggcgccccg cugagcaagg aggugggcga ggccguggcc aagcgguucc accucccggg 1020
cauccgccag ggcuacggcc ugaccgagac cacgagcgcg auccugauca cccccgaggg 1080
ggacgacaag ccgggcgccg ugggcaaggu ggucccguuc uucgaggcca agguggugga 1140
ccuggacacc ggcaagaccc ugggcgugaa ccagcggggc gagcugugcg ugcgggggcc 1200
gaugaucaug agcggcuacg ugaacaaccc ggaggccacc aacgcccuca ucgacaagga 1260
cggcuggcug cacagcggcg acaucgccua cugggacgag gacgagcacu ucuucaucgu 1320
cgaccggcug aagucgcuga ucaaguacaa gggcuaccag guggcgccgg ccgagcugga 1380
gagcauccug cuccagcacc ccaacaucuu cgacgccggc guggccgggc ugccggacga 1440
cgacgccggc gagcugccgg ccgcgguggu ggugcuggag cacggcaaga ccaugacgga 1500
gaaggagauc gucgacuacg uggccagcca ggugaccacc gccaagaagc ugcggggcgg 1560
cgugguguuc guggacgagg ucccgaaggg ccugaccggg aagcucgacg cccggaagau 1620
ccgcgagauc cugaucaagg ccaagaaggg cggcaagauc gccguguaag acuaguuaua 1680
agacugacua gcccgauggg ccucccaacg ggcccuccuc cccuccuugc accgagauua 1740
auagauc 1747
<210> 44
<211> 1806
<212> РНК
<213> Искусственная
<220>
<223> Последовательность мРНК ppLuc(GC)-ag-A64
<400> 44
gggagaaagc uugaggaugg aggacgccaa gaacaucaag aagggcccgg cgcccuucua 60
cccgcuggag gacgggaccg ccggcgagca gcuccacaag gccaugaagc gguacgcccu 120
ggugccgggc acgaucgccu ucaccgacgc ccacaucgag gucgacauca ccuacgcgga 180
guacuucgag augagcgugc gccuggccga ggccaugaag cgguacggcc ugaacaccaa 240
ccaccggauc guggugugcu cggagaacag ccugcaguuc uucaugccgg ugcugggcgc 300
ccucuucauc ggcguggccg ucgccccggc gaacgacauc uacaacgagc gggagcugcu 360
gaacagcaug gggaucagcc agccgaccgu gguguucgug agcaagaagg gccugcagaa 420
gauccugaac gugcagaaga agcugcccau cauccagaag aucaucauca uggacagcaa 480
gaccgacuac cagggcuucc agucgaugua cacguucgug accagccacc ucccgccggg 540
cuucaacgag uacgacuucg ucccggagag cuucgaccgg gacaagacca ucgcccugau 600
caugaacagc agcggcagca ccggccugcc gaagggggug gcccugccgc accggaccgc 660
cugcgugcgc uucucgcacg cccgggaccc caucuucggc aaccagauca ucccggacac 720
cgccauccug agcguggugc cguuccacca cggcuucggc auguucacga cccugggcua 780
ccucaucugc ggcuuccggg ugguccugau guaccgguuc gaggaggagc uguuccugcg 840
gagccugcag gacuacaaga uccagagcgc gcugcucgug ccgacccugu ucagcuucuu 900
cgccaagagc acccugaucg acaaguacga ccugucgaac cugcacgaga ucgccagcgg 960
gggcgccccg cugagcaagg aggugggcga ggccguggcc aagcgguucc accucccggg 1020
cauccgccag ggcuacggcc ugaccgagac cacgagcgcg auccugauca cccccgaggg 1080
ggacgacaag ccgggcgccg ugggcaaggu ggucccguuc uucgaggcca agguggugga 1140
ccuggacacc ggcaagaccc ugggcgugaa ccagcggggc gagcugugcg ugcgggggcc 1200
gaugaucaug agcggcuacg ugaacaaccc ggaggccacc aacgcccuca ucgacaagga 1260
cggcuggcug cacagcggcg acaucgccua cugggacgag gacgagcacu ucuucaucgu 1320
cgaccggcug aagucgcuga ucaaguacaa gggcuaccag guggcgccgg ccgagcugga 1380
gagcauccug cuccagcacc ccaacaucuu cgacgccggc guggccgggc ugccggacga 1440
cgacgccggc gagcugccgg ccgcgguggu ggugcuggag cacggcaaga ccaugacgga 1500
gaaggagauc gucgacuacg uggccagcca ggugaccacc gccaagaagc ugcggggcgg 1560
cgugguguuc guggacgagg ucccgaaggg ccugaccggg aagcucgacg cccggaagau 1620
ccgcgagauc cugaucaagg ccaagaaggg cggcaagauc gccguguaag acuaguuaua 1680
agacugacua gcccgauggg ccucccaacg ggcccuccuc cccuccuugc accgagauua 1740
auaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1800
aaaaaa 1806
<210> 45
<211> 1772
<212> РНК
<213> Искусственная
<220>
<223> Последовательность мРНК ppLuc(GC)-ag-histoneSL
<400> 45
gggagaaagc uugaggaugg aggacgccaa gaacaucaag aagggcccgg cgcccuucua 60
cccgcuggag gacgggaccg ccggcgagca gcuccacaag gccaugaagc gguacgcccu 120
ggugccgggc acgaucgccu ucaccgacgc ccacaucgag gucgacauca ccuacgcgga 180
guacuucgag augagcgugc gccuggccga ggccaugaag cgguacggcc ugaacaccaa 240
ccaccggauc guggugugcu cggagaacag ccugcaguuc uucaugccgg ugcugggcgc 300
ccucuucauc ggcguggccg ucgccccggc gaacgacauc uacaacgagc gggagcugcu 360
gaacagcaug gggaucagcc agccgaccgu gguguucgug agcaagaagg gccugcagaa 420
gauccugaac gugcagaaga agcugcccau cauccagaag aucaucauca uggacagcaa 480
gaccgacuac cagggcuucc agucgaugua cacguucgug accagccacc ucccgccggg 540
cuucaacgag uacgacuucg ucccggagag cuucgaccgg gacaagacca ucgcccugau 600
caugaacagc agcggcagca ccggccugcc gaagggggug gcccugccgc accggaccgc 660
cugcgugcgc uucucgcacg cccgggaccc caucuucggc aaccagauca ucccggacac 720
cgccauccug agcguggugc cguuccacca cggcuucggc auguucacga cccugggcua 780
ccucaucugc ggcuuccggg ugguccugau guaccgguuc gaggaggagc uguuccugcg 840
gagccugcag gacuacaaga uccagagcgc gcugcucgug ccgacccugu ucagcuucuu 900
cgccaagagc acccugaucg acaaguacga ccugucgaac cugcacgaga ucgccagcgg 960
gggcgccccg cugagcaagg aggugggcga ggccguggcc aagcgguucc accucccggg 1020
cauccgccag ggcuacggcc ugaccgagac cacgagcgcg auccugauca cccccgaggg 1080
ggacgacaag ccgggcgccg ugggcaaggu ggucccguuc uucgaggcca agguggugga 1140
ccuggacacc ggcaagaccc ugggcgugaa ccagcggggc gagcugugcg ugcgggggcc 1200
gaugaucaug agcggcuacg ugaacaaccc ggaggccacc aacgcccuca ucgacaagga 1260
cggcuggcug cacagcggcg acaucgccua cugggacgag gacgagcacu ucuucaucgu 1320
cgaccggcug aagucgcuga ucaaguacaa gggcuaccag guggcgccgg ccgagcugga 1380
gagcauccug cuccagcacc ccaacaucuu cgacgccggc guggccgggc ugccggacga 1440
cgacgccggc gagcugccgg ccgcgguggu ggugcuggag cacggcaaga ccaugacgga 1500
gaaggagauc gucgacuacg uggccagcca ggugaccacc gccaagaagc ugcggggcgg 1560
cgugguguuc guggacgagg ucccgaaggg ccugaccggg aagcucgacg cccggaagau 1620
ccgcgagauc cugaucaagg ccaagaaggg cggcaagauc gccguguaag acuaguuaua 1680
agacugacua gcccgauggg ccucccaacg ggcccuccuc cccuccuugc accgagauua 1740
auagaucuca aaggcucuuu ucagagccac ca 1772
<210> 46
<211> 1835
<212> РНК
<213> Искусственная
<220>
<223> Последовательность мРНК ppLuc(GC)-ag-A64-histoneSL
<400> 46
gggagaaagc uugaggaugg aggacgccaa gaacaucaag aagggcccgg cgcccuucua 60
cccgcuggag gacgggaccg ccggcgagca gcuccacaag gccaugaagc gguacgcccu 120
ggugccgggc acgaucgccu ucaccgacgc ccacaucgag gucgacauca ccuacgcgga 180
guacuucgag augagcgugc gccuggccga ggccaugaag cgguacggcc ugaacaccaa 240
ccaccggauc guggugugcu cggagaacag ccugcaguuc uucaugccgg ugcugggcgc 300
ccucuucauc ggcguggccg ucgccccggc gaacgacauc uacaacgagc gggagcugcu 360
gaacagcaug gggaucagcc agccgaccgu gguguucgug agcaagaagg gccugcagaa 420
gauccugaac gugcagaaga agcugcccau cauccagaag aucaucauca uggacagcaa 480
gaccgacuac cagggcuucc agucgaugua cacguucgug accagccacc ucccgccggg 540
cuucaacgag uacgacuucg ucccggagag cuucgaccgg gacaagacca ucgcccugau 600
caugaacagc agcggcagca ccggccugcc gaagggggug gcccugccgc accggaccgc 660
cugcgugcgc uucucgcacg cccgggaccc caucuucggc aaccagauca ucccggacac 720
cgccauccug agcguggugc cguuccacca cggcuucggc auguucacga cccugggcua 780
ccucaucugc ggcuuccggg ugguccugau guaccgguuc gaggaggagc uguuccugcg 840
gagccugcag gacuacaaga uccagagcgc gcugcucgug ccgacccugu ucagcuucuu 900
cgccaagagc acccugaucg acaaguacga ccugucgaac cugcacgaga ucgccagcgg 960
gggcgccccg cugagcaagg aggugggcga ggccguggcc aagcgguucc accucccggg 1020
cauccgccag ggcuacggcc ugaccgagac cacgagcgcg auccugauca cccccgaggg 1080
ggacgacaag ccgggcgccg ugggcaaggu ggucccguuc uucgaggcca agguggugga 1140
ccuggacacc ggcaagaccc ugggcgugaa ccagcggggc gagcugugcg ugcgggggcc 1200
gaugaucaug agcggcuacg ugaacaaccc ggaggccacc aacgcccuca ucgacaagga 1260
cggcuggcug cacagcggcg acaucgccua cugggacgag gacgagcacu ucuucaucgu 1320
cgaccggcug aagucgcuga ucaaguacaa gggcuaccag guggcgccgg ccgagcugga 1380
gagcauccug cuccagcacc ccaacaucuu cgacgccggc guggccgggc ugccggacga 1440
cgacgccggc gagcugccgg ccgcgguggu ggugcuggag cacggcaaga ccaugacgga 1500
gaaggagauc gucgacuacg uggccagcca ggugaccacc gccaagaagc ugcggggcgg 1560
cgugguguuc guggacgagg ucccgaaggg ccugaccggg aagcucgacg cccggaagau 1620
ccgcgagauc cugaucaagg ccaagaaggg cggcaagauc gccguguaag acuaguuaua 1680
agacugacua gcccgauggg ccucccaacg ggcccuccuc cccuccuugc accgagauua 1740
auaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1800
aaaaaaugca ucaaaggcuc uuuucagagc cacca 1835
<210> 47
<211> 1869
<212> РНК
<213> Искусственная
<220>
<223> Последовательность мРНК ppLuc(GC)-ag-A120
<400> 47
gggagaaagc uugaggaugg aggacgccaa gaacaucaag aagggcccgg cgcccuucua 60
cccgcuggag gacgggaccg ccggcgagca gcuccacaag gccaugaagc gguacgcccu 120
ggugccgggc acgaucgccu ucaccgacgc ccacaucgag gucgacauca ccuacgcgga 180
guacuucgag augagcgugc gccuggccga ggccaugaag cgguacggcc ugaacaccaa 240
ccaccggauc guggugugcu cggagaacag ccugcaguuc uucaugccgg ugcugggcgc 300
ccucuucauc ggcguggccg ucgccccggc gaacgacauc uacaacgagc gggagcugcu 360
gaacagcaug gggaucagcc agccgaccgu gguguucgug agcaagaagg gccugcagaa 420
gauccugaac gugcagaaga agcugcccau cauccagaag aucaucauca uggacagcaa 480
gaccgacuac cagggcuucc agucgaugua cacguucgug accagccacc ucccgccggg 540
cuucaacgag uacgacuucg ucccggagag cuucgaccgg gacaagacca ucgcccugau 600
caugaacagc agcggcagca ccggccugcc gaagggggug gcccugccgc accggaccgc 660
cugcgugcgc uucucgcacg cccgggaccc caucuucggc aaccagauca ucccggacac 720
cgccauccug agcguggugc cguuccacca cggcuucggc auguucacga cccugggcua 780
ccucaucugc ggcuuccggg ugguccugau guaccgguuc gaggaggagc uguuccugcg 840
gagccugcag gacuacaaga uccagagcgc gcugcucgug ccgacccugu ucagcuucuu 900
cgccaagagc acccugaucg acaaguacga ccugucgaac cugcacgaga ucgccagcgg 960
gggcgccccg cugagcaagg aggugggcga ggccguggcc aagcgguucc accucccggg 1020
cauccgccag ggcuacggcc ugaccgagac cacgagcgcg auccugauca cccccgaggg 1080
ggacgacaag ccgggcgccg ugggcaaggu ggucccguuc uucgaggcca agguggugga 1140
ccuggacacc ggcaagaccc ugggcgugaa ccagcggggc gagcugugcg ugcgggggcc 1200
gaugaucaug agcggcuacg ugaacaaccc ggaggccacc aacgcccuca ucgacaagga 1260
cggcuggcug cacagcggcg acaucgccua cugggacgag gacgagcacu ucuucaucgu 1320
cgaccggcug aagucgcuga ucaaguacaa gggcuaccag guggcgccgg ccgagcugga 1380
gagcauccug cuccagcacc ccaacaucuu cgacgccggc guggccgggc ugccggacga 1440
cgacgccggc gagcugccgg ccgcgguggu ggugcuggag cacggcaaga ccaugacgga 1500
gaaggagauc gucgacuacg uggccagcca ggugaccacc gccaagaagc ugcggggcgg 1560
cgugguguuc guggacgagg ucccgaaggg ccugaccggg aagcucgacg cccggaagau 1620
ccgcgagauc cugaucaagg ccaagaaggg cggcaagauc gccguguaag acuaguuaua 1680
agacugacua gcccgauggg ccucccaacg ggcccuccuc cccuccuugc accgagauua 1740
auagaucuaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1800
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1860
aaaaaaaaa 1869
<210> 48
<211> 1858
<212> РНК
<213> Искусственная
<220>
<223> Последовательность мРНК ppLuc(GC)-ag-A64-ag
<400> 48
gggagaaagc uugaggaugg aggacgccaa gaacaucaag aagggcccgg cgcccuucua 60
cccgcuggag gacgggaccg ccggcgagca gcuccacaag gccaugaagc gguacgcccu 120
ggugccgggc acgaucgccu ucaccgacgc ccacaucgag gucgacauca ccuacgcgga 180
guacuucgag augagcgugc gccuggccga ggccaugaag cgguacggcc ugaacaccaa 240
ccaccggauc guggugugcu cggagaacag ccugcaguuc uucaugccgg ugcugggcgc 300
ccucuucauc ggcguggccg ucgccccggc gaacgacauc uacaacgagc gggagcugcu 360
gaacagcaug gggaucagcc agccgaccgu gguguucgug agcaagaagg gccugcagaa 420
gauccugaac gugcagaaga agcugcccau cauccagaag aucaucauca uggacagcaa 480
gaccgacuac cagggcuucc agucgaugua cacguucgug accagccacc ucccgccggg 540
cuucaacgag uacgacuucg ucccggagag cuucgaccgg gacaagacca ucgcccugau 600
caugaacagc agcggcagca ccggccugcc gaagggggug gcccugccgc accggaccgc 660
cugcgugcgc uucucgcacg cccgggaccc caucuucggc aaccagauca ucccggacac 720
cgccauccug agcguggugc cguuccacca cggcuucggc auguucacga cccugggcua 780
ccucaucugc ggcuuccggg ugguccugau guaccgguuc gaggaggagc uguuccugcg 840
gagccugcag gacuacaaga uccagagcgc gcugcucgug ccgacccugu ucagcuucuu 900
cgccaagagc acccugaucg acaaguacga ccugucgaac cugcacgaga ucgccagcgg 960
gggcgccccg cugagcaagg aggugggcga ggccguggcc aagcgguucc accucccggg 1020
cauccgccag ggcuacggcc ugaccgagac cacgagcgcg auccugauca cccccgaggg 1080
ggacgacaag ccgggcgccg ugggcaaggu ggucccguuc uucgaggcca agguggugga 1140
ccuggacacc ggcaagaccc ugggcgugaa ccagcggggc gagcugugcg ugcgggggcc 1200
gaugaucaug agcggcuacg ugaacaaccc ggaggccacc aacgcccuca ucgacaagga 1260
cggcuggcug cacagcggcg acaucgccua cugggacgag gacgagcacu ucuucaucgu 1320
cgaccggcug aagucgcuga ucaaguacaa gggcuaccag guggcgccgg ccgagcugga 1380
gagcauccug cuccagcacc ccaacaucuu cgacgccggc guggccgggc ugccggacga 1440
cgacgccggc gagcugccgg ccgcgguggu ggugcuggag cacggcaaga ccaugacgga 1500
gaaggagauc gucgacuacg uggccagcca ggugaccacc gccaagaagc ugcggggcgg 1560
cgugguguuc guggacgagg ucccgaaggg ccugaccggg aagcucgacg cccggaagau 1620
ccgcgagauc cugaucaagg ccaagaaggg cggcaagauc gccguguaag acuaguuaua 1680
agacugacua gcccgauggg ccucccaacg ggcccuccuc cccuccuugc accgagauua 1740
auaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1800
aaaaaaugca uccugcccga ugggccuccc aacgggcccu ccuccccucc uugcaccg 1858
<210> 49
<211> 1894
<212> РНК
<213> Искусственная
<220>
<223> Последовательность мРНК ppLuc(GC)-ag-A64-aCPSL
<400> 49
gggagaaagc uugaggaugg aggacgccaa gaacaucaag aagggcccgg cgcccuucua 60
cccgcuggag gacgggaccg ccggcgagca gcuccacaag gccaugaagc gguacgcccu 120
ggugccgggc acgaucgccu ucaccgacgc ccacaucgag gucgacauca ccuacgcgga 180
guacuucgag augagcgugc gccuggccga ggccaugaag cgguacggcc ugaacaccaa 240
ccaccggauc guggugugcu cggagaacag ccugcaguuc uucaugccgg ugcugggcgc 300
ccucuucauc ggcguggccg ucgccccggc gaacgacauc uacaacgagc gggagcugcu 360
gaacagcaug gggaucagcc agccgaccgu gguguucgug agcaagaagg gccugcagaa 420
gauccugaac gugcagaaga agcugcccau cauccagaag aucaucauca uggacagcaa 480
gaccgacuac cagggcuucc agucgaugua cacguucgug accagccacc ucccgccggg 540
cuucaacgag uacgacuucg ucccggagag cuucgaccgg gacaagacca ucgcccugau 600
caugaacagc agcggcagca ccggccugcc gaagggggug gcccugccgc accggaccgc 660
cugcgugcgc uucucgcacg cccgggaccc caucuucggc aaccagauca ucccggacac 720
cgccauccug agcguggugc cguuccacca cggcuucggc auguucacga cccugggcua 780
ccucaucugc ggcuuccggg ugguccugau guaccgguuc gaggaggagc uguuccugcg 840
gagccugcag gacuacaaga uccagagcgc gcugcucgug ccgacccugu ucagcuucuu 900
cgccaagagc acccugaucg acaaguacga ccugucgaac cugcacgaga ucgccagcgg 960
gggcgccccg cugagcaagg aggugggcga ggccguggcc aagcgguucc accucccggg 1020
cauccgccag ggcuacggcc ugaccgagac cacgagcgcg auccugauca cccccgaggg 1080
ggacgacaag ccgggcgccg ugggcaaggu ggucccguuc uucgaggcca agguggugga 1140
ccuggacacc ggcaagaccc ugggcgugaa ccagcggggc gagcugugcg ugcgggggcc 1200
gaugaucaug agcggcuacg ugaacaaccc ggaggccacc aacgcccuca ucgacaagga 1260
cggcuggcug cacagcggcg acaucgccua cugggacgag gacgagcacu ucuucaucgu 1320
cgaccggcug aagucgcuga ucaaguacaa gggcuaccag guggcgccgg ccgagcugga 1380
gagcauccug cuccagcacc ccaacaucuu cgacgccggc guggccgggc ugccggacga 1440
cgacgccggc gagcugccgg ccgcgguggu ggugcuggag cacggcaaga ccaugacgga 1500
gaaggagauc gucgacuacg uggccagcca ggugaccacc gccaagaagc ugcggggcgg 1560
cgugguguuc guggacgagg ucccgaaggg ccugaccggg aagcucgacg cccggaagau 1620
ccgcgagauc cugaucaagg ccaagaaggg cggcaagauc gccguguaag acuaguuaua 1680
agacugacua gcccgauggg ccucccaacg ggcccuccuc cccuccuugc accgagauua 1740
auaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1800
aaaaaaugca ucaauuccua cacgugaggc gcugugauuc ccuauccccc uucauucccu 1860
auacauuagc acagcgccau ugcauguagg aauu 1894
<210> 50
<211> 1909
<212> РНК
<213> Искусственная
<220>
<223> Последовательность мРНК ppLuc(GC)-ag-A64-PolioCL
<400> 50
gggagaaagc uugaggaugg aggacgccaa gaacaucaag aagggcccgg cgcccuucua 60
cccgcuggag gacgggaccg ccggcgagca gcuccacaag gccaugaagc gguacgcccu 120
ggugccgggc acgaucgccu ucaccgacgc ccacaucgag gucgacauca ccuacgcgga 180
guacuucgag augagcgugc gccuggccga ggccaugaag cgguacggcc ugaacaccaa 240
ccaccggauc guggugugcu cggagaacag ccugcaguuc uucaugccgg ugcugggcgc 300
ccucuucauc ggcguggccg ucgccccggc gaacgacauc uacaacgagc gggagcugcu 360
gaacagcaug gggaucagcc agccgaccgu gguguucgug agcaagaagg gccugcagaa 420
gauccugaac gugcagaaga agcugcccau cauccagaag aucaucauca uggacagcaa 480
gaccgacuac cagggcuucc agucgaugua cacguucgug accagccacc ucccgccggg 540
cuucaacgag uacgacuucg ucccggagag cuucgaccgg gacaagacca ucgcccugau 600
caugaacagc agcggcagca ccggccugcc gaagggggug gcccugccgc accggaccgc 660
cugcgugcgc uucucgcacg cccgggaccc caucuucggc aaccagauca ucccggacac 720
cgccauccug agcguggugc cguuccacca cggcuucggc auguucacga cccugggcua 780
ccucaucugc ggcuuccggg ugguccugau guaccgguuc gaggaggagc uguuccugcg 840
gagccugcag gacuacaaga uccagagcgc gcugcucgug ccgacccugu ucagcuucuu 900
cgccaagagc acccugaucg acaaguacga ccugucgaac cugcacgaga ucgccagcgg 960
gggcgccccg cugagcaagg aggugggcga ggccguggcc aagcgguucc accucccggg 1020
cauccgccag ggcuacggcc ugaccgagac cacgagcgcg auccugauca cccccgaggg 1080
ggacgacaag ccgggcgccg ugggcaaggu ggucccguuc uucgaggcca agguggugga 1140
ccuggacacc ggcaagaccc ugggcgugaa ccagcggggc gagcugugcg ugcgggggcc 1200
gaugaucaug agcggcuacg ugaacaaccc ggaggccacc aacgcccuca ucgacaagga 1260
cggcuggcug cacagcggcg acaucgccua cugggacgag gacgagcacu ucuucaucgu 1320
cgaccggcug aagucgcuga ucaaguacaa gggcuaccag guggcgccgg ccgagcugga 1380
gagcauccug cuccagcacc ccaacaucuu cgacgccggc guggccgggc ugccggacga 1440
cgacgccggc gagcugccgg ccgcgguggu ggugcuggag cacggcaaga ccaugacgga 1500
gaaggagauc gucgacuacg uggccagcca ggugaccacc gccaagaagc ugcggggcgg 1560
cgugguguuc guggacgagg ucccgaaggg ccugaccggg aagcucgacg cccggaagau 1620
ccgcgagauc cugaucaagg ccaagaaggg cggcaagauc gccguguaag acuaguuaua 1680
agacugacua gcccgauggg ccucccaacg ggcccuccuc cccuccuugc accgagauua 1740
auaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1800
aaaaaaugca ucaauucuaa aacagcucug ggguuguacc caccccagag gcccacgugg 1860
cggcuaguac uccgguauug cgguacccuu guacgccugu uuuagaauu 1909
<210> 51
<211> 1841
<212> РНК
<213> Искусственная
<220>
<223> Последовательность мРНК ppLuc(GC)-ag-A64-G30
<400> 51
gggagaaagc uugaggaugg aggacgccaa gaacaucaag aagggcccgg cgcccuucua 60
cccgcuggag gacgggaccg ccggcgagca gcuccacaag gccaugaagc gguacgcccu 120
ggugccgggc acgaucgccu ucaccgacgc ccacaucgag gucgacauca ccuacgcgga 180
guacuucgag augagcgugc gccuggccga ggccaugaag cgguacggcc ugaacaccaa 240
ccaccggauc guggugugcu cggagaacag ccugcaguuc uucaugccgg ugcugggcgc 300
ccucuucauc ggcguggccg ucgccccggc gaacgacauc uacaacgagc gggagcugcu 360
gaacagcaug gggaucagcc agccgaccgu gguguucgug agcaagaagg gccugcagaa 420
gauccugaac gugcagaaga agcugcccau cauccagaag aucaucauca uggacagcaa 480
gaccgacuac cagggcuucc agucgaugua cacguucgug accagccacc ucccgccggg 540
cuucaacgag uacgacuucg ucccggagag cuucgaccgg gacaagacca ucgcccugau 600
caugaacagc agcggcagca ccggccugcc gaagggggug gcccugccgc accggaccgc 660
cugcgugcgc uucucgcacg cccgggaccc caucuucggc aaccagauca ucccggacac 720
cgccauccug agcguggugc cguuccacca cggcuucggc auguucacga cccugggcua 780
ccucaucugc ggcuuccggg ugguccugau guaccgguuc gaggaggagc uguuccugcg 840
gagccugcag gacuacaaga uccagagcgc gcugcucgug ccgacccugu ucagcuucuu 900
cgccaagagc acccugaucg acaaguacga ccugucgaac cugcacgaga ucgccagcgg 960
gggcgccccg cugagcaagg aggugggcga ggccguggcc aagcgguucc accucccggg 1020
cauccgccag ggcuacggcc ugaccgagac cacgagcgcg auccugauca cccccgaggg 1080
ggacgacaag ccgggcgccg ugggcaaggu ggucccguuc uucgaggcca agguggugga 1140
ccuggacacc ggcaagaccc ugggcgugaa ccagcggggc gagcugugcg ugcgggggcc 1200
gaugaucaug agcggcuacg ugaacaaccc ggaggccacc aacgcccuca ucgacaagga 1260
cggcuggcug cacagcggcg acaucgccua cugggacgag gacgagcacu ucuucaucgu 1320
cgaccggcug aagucgcuga ucaaguacaa gggcuaccag guggcgccgg ccgagcugga 1380
gagcauccug cuccagcacc ccaacaucuu cgacgccggc guggccgggc ugccggacga 1440
cgacgccggc gagcugccgg ccgcgguggu ggugcuggag cacggcaaga ccaugacgga 1500
gaaggagauc gucgacuacg uggccagcca ggugaccacc gccaagaagc ugcggggcgg 1560
cgugguguuc guggacgagg ucccgaaggg ccugaccggg aagcucgacg cccggaagau 1620
ccgcgagauc cugaucaagg ccaagaaggg cggcaagauc gccguguaag acuaguuaua 1680
agacugacua gcccgauggg ccucccaacg ggcccuccuc cccuccuugc accgagauua 1740
auaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1800
aaaaaaugca uggggggggg gggggggggg gggggggggg g 1841
<210> 52
<211> 1841
<212> РНК
<213> Искусственная
<220>
<223> Последовательность мРНК ppLuc(GC)-ag-A64-U30
<400> 52
gggagaaagc uugaggaugg aggacgccaa gaacaucaag aagggcccgg cgcccuucua 60
cccgcuggag gacgggaccg ccggcgagca gcuccacaag gccaugaagc gguacgcccu 120
ggugccgggc acgaucgccu ucaccgacgc ccacaucgag gucgacauca ccuacgcgga 180
guacuucgag augagcgugc gccuggccga ggccaugaag cgguacggcc ugaacaccaa 240
ccaccggauc guggugugcu cggagaacag ccugcaguuc uucaugccgg ugcugggcgc 300
ccucuucauc ggcguggccg ucgccccggc gaacgacauc uacaacgagc gggagcugcu 360
gaacagcaug gggaucagcc agccgaccgu gguguucgug agcaagaagg gccugcagaa 420
gauccugaac gugcagaaga agcugcccau cauccagaag aucaucauca uggacagcaa 480
gaccgacuac cagggcuucc agucgaugua cacguucgug accagccacc ucccgccggg 540
cuucaacgag uacgacuucg ucccggagag cuucgaccgg gacaagacca ucgcccugau 600
caugaacagc agcggcagca ccggccugcc gaagggggug gcccugccgc accggaccgc 660
cugcgugcgc uucucgcacg cccgggaccc caucuucggc aaccagauca ucccggacac 720
cgccauccug agcguggugc cguuccacca cggcuucggc auguucacga cccugggcua 780
ccucaucugc ggcuuccggg ugguccugau guaccgguuc gaggaggagc uguuccugcg 840
gagccugcag gacuacaaga uccagagcgc gcugcucgug ccgacccugu ucagcuucuu 900
cgccaagagc acccugaucg acaaguacga ccugucgaac cugcacgaga ucgccagcgg 960
gggcgccccg cugagcaagg aggugggcga ggccguggcc aagcgguucc accucccggg 1020
cauccgccag ggcuacggcc ugaccgagac cacgagcgcg auccugauca cccccgaggg 1080
ggacgacaag ccgggcgccg ugggcaaggu ggucccguuc uucgaggcca agguggugga 1140
ccuggacacc ggcaagaccc ugggcgugaa ccagcggggc gagcugugcg ugcgggggcc 1200
gaugaucaug agcggcuacg ugaacaaccc ggaggccacc aacgcccuca ucgacaagga 1260
cggcuggcug cacagcggcg acaucgccua cugggacgag gacgagcacu ucuucaucgu 1320
cgaccggcug aagucgcuga ucaaguacaa gggcuaccag guggcgccgg ccgagcugga 1380
gagcauccug cuccagcacc ccaacaucuu cgacgccggc guggccgggc ugccggacga 1440
cgacgccggc gagcugccgg ccgcgguggu ggugcuggag cacggcaaga ccaugacgga 1500
gaaggagauc gucgacuacg uggccagcca ggugaccacc gccaagaagc ugcggggcgg 1560
cgugguguuc guggacgagg ucccgaaggg ccugaccggg aagcucgacg cccggaagau 1620
ccgcgagauc cugaucaagg ccaagaaggg cggcaagauc gccguguaag acuaguuaua 1680
agacugacua gcccgauggg ccucccaacg ggcccuccuc cccuccuugc accgagauua 1740
auaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1800
aaaaaaugca uuuuuuuuuu uuuuuuuuuu uuuuuuuuuu u 1841
<210> 53
<211> 1857
<212> РНК
<213> Искусственная
<220>
<223> Последовательность мРНК ppLuc(GC)-ag-A64-SL
<400> 53
gggagaaagc uugaggaugg aggacgccaa gaacaucaag aagggcccgg cgcccuucua 60
cccgcuggag gacgggaccg ccggcgagca gcuccacaag gccaugaagc gguacgcccu 120
ggugccgggc acgaucgccu ucaccgacgc ccacaucgag gucgacauca ccuacgcgga 180
guacuucgag augagcgugc gccuggccga ggccaugaag cgguacggcc ugaacaccaa 240
ccaccggauc guggugugcu cggagaacag ccugcaguuc uucaugccgg ugcugggcgc 300
ccucuucauc ggcguggccg ucgccccggc gaacgacauc uacaacgagc gggagcugcu 360
gaacagcaug gggaucagcc agccgaccgu gguguucgug agcaagaagg gccugcagaa 420
gauccugaac gugcagaaga agcugcccau cauccagaag aucaucauca uggacagcaa 480
gaccgacuac cagggcuucc agucgaugua cacguucgug accagccacc ucccgccggg 540
cuucaacgag uacgacuucg ucccggagag cuucgaccgg gacaagacca ucgcccugau 600
caugaacagc agcggcagca ccggccugcc gaagggggug gcccugccgc accggaccgc 660
cugcgugcgc uucucgcacg cccgggaccc caucuucggc aaccagauca ucccggacac 720
cgccauccug agcguggugc cguuccacca cggcuucggc auguucacga cccugggcua 780
ccucaucugc ggcuuccggg ugguccugau guaccgguuc gaggaggagc uguuccugcg 840
gagccugcag gacuacaaga uccagagcgc gcugcucgug ccgacccugu ucagcuucuu 900
cgccaagagc acccugaucg acaaguacga ccugucgaac cugcacgaga ucgccagcgg 960
gggcgccccg cugagcaagg aggugggcga ggccguggcc aagcgguucc accucccggg 1020
cauccgccag ggcuacggcc ugaccgagac cacgagcgcg auccugauca cccccgaggg 1080
ggacgacaag ccgggcgccg ugggcaaggu ggucccguuc uucgaggcca agguggugga 1140
ccuggacacc ggcaagaccc ugggcgugaa ccagcggggc gagcugugcg ugcgggggcc 1200
gaugaucaug agcggcuacg ugaacaaccc ggaggccacc aacgcccuca ucgacaagga 1260
cggcuggcug cacagcggcg acaucgccua cugggacgag gacgagcacu ucuucaucgu 1320
cgaccggcug aagucgcuga ucaaguacaa gggcuaccag guggcgccgg ccgagcugga 1380
gagcauccug cuccagcacc ccaacaucuu cgacgccggc guggccgggc ugccggacga 1440
cgacgccggc gagcugccgg ccgcgguggu ggugcuggag cacggcaaga ccaugacgga 1500
gaaggagauc gucgacuacg uggccagcca ggugaccacc gccaagaagc ugcggggcgg 1560
cgugguguuc guggacgagg ucccgaaggg ccugaccggg aagcucgacg cccggaagau 1620
ccgcgagauc cugaucaagg ccaagaaggg cggcaagauc gccguguaag acuaguuaua 1680
agacugacua gcccgauggg ccucccaacg ggcccuccuc cccuccuugc accgagauua 1740
auaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1800
aaaaaaugca uuauggcggc cguguccacc acggauauca ccguggugga cgcggcc 1857
<210> 54
<211> 1838
<212> РНК
<213> Искусственная
<220>
<223> ppLuc(GC)-ag-A64-N32
<400> 54
gggagaaagc uugaggaugg aggacgccaa gaacaucaag aagggcccgg cgcccuucua 60
cccgcuggag gacgggaccg ccggcgagca gcuccacaag gccaugaagc gguacgcccu 120
ggugccgggc acgaucgccu ucaccgacgc ccacaucgag gucgacauca ccuacgcgga 180
guacuucgag augagcgugc gccuggccga ggccaugaag cgguacggcc ugaacaccaa 240
ccaccggauc guggugugcu cggagaacag ccugcaguuc uucaugccgg ugcugggcgc 300
ccucuucauc ggcguggccg ucgccccggc gaacgacauc uacaacgagc gggagcugcu 360
gaacagcaug gggaucagcc agccgaccgu gguguucgug agcaagaagg gccugcagaa 420
gauccugaac gugcagaaga agcugcccau cauccagaag aucaucauca uggacagcaa 480
gaccgacuac cagggcuucc agucgaugua cacguucgug accagccacc ucccgccggg 540
cuucaacgag uacgacuucg ucccggagag cuucgaccgg gacaagacca ucgcccugau 600
caugaacagc agcggcagca ccggccugcc gaagggggug gcccugccgc accggaccgc 660
cugcgugcgc uucucgcacg cccgggaccc caucuucggc aaccagauca ucccggacac 720
cgccauccug agcguggugc cguuccacca cggcuucggc auguucacga cccugggcua 780
ccucaucugc ggcuuccggg ugguccugau guaccgguuc gaggaggagc uguuccugcg 840
gagccugcag gacuacaaga uccagagcgc gcugcucgug ccgacccugu ucagcuucuu 900
cgccaagagc acccugaucg acaaguacga ccugucgaac cugcacgaga ucgccagcgg 960
gggcgccccg cugagcaagg aggugggcga ggccguggcc aagcgguucc accucccggg 1020
cauccgccag ggcuacggcc ugaccgagac cacgagcgcg auccugauca cccccgaggg 1080
ggacgacaag ccgggcgccg ugggcaaggu ggucccguuc uucgaggcca agguggugga 1140
ccuggacacc ggcaagaccc ugggcgugaa ccagcggggc gagcugugcg ugcgggggcc 1200
gaugaucaug agcggcuacg ugaacaaccc ggaggccacc aacgcccuca ucgacaagga 1260
cggcuggcug cacagcggcg acaucgccua cugggacgag gacgagcacu ucuucaucgu 1320
cgaccggcug aagucgcuga ucaaguacaa gggcuaccag guggcgccgg ccgagcugga 1380
gagcauccug cuccagcacc ccaacaucuu cgacgccggc guggccgggc ugccggacga 1440
cgacgccggc gagcugccgg ccgcgguggu ggugcuggag cacggcaaga ccaugacgga 1500
gaaggagauc gucgacuacg uggccagcca ggugaccacc gccaagaagc ugcggggcgg 1560
cgugguguuc guggacgagg ucccgaaggg ccugaccggg aagcucgacg cccggaagau 1620
ccgcgagauc cugaucaagg ccaagaaggg cggcaagauc gccguguaag acuaguuaua 1680
agacugacua gcccgauggg ccucccaacg ggcccuccuc cccuccuugc accgagauua 1740
auaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1800
aaaaaaugca ucccccucua gacaauugga auuccaua 1838
<210> 55
<211> 747
<212> РНК
<213> Искусственная
<220>
<223> Последовательность мРНК NY-ESO-1(GC)-ag-A64-C30
<400> 55
gggagaaagc uuaccaugca ggccgagggc cgcggcaccg gcggcucgac cggcgacgcc 60
gacgggcccg gcggcccggg caucccggac ggcccgggcg ggaacgcggg cggcccgggc 120
gaggccggcg ccaccggcgg gcggggcccg cggggcgccg gcgccgcccg ggcgagcggc 180
cccggcgggg gcgccccgcg gggcccgcac ggcggcgccg ccagcggccu gaacgggugc 240
ugccggugcg gcgcccgcgg cccggagagc cggcuccugg aguucuaccu ggccaugccg 300
uucgcgaccc cgauggaggc cgagcuggcc cggcggagcc uggcccagga cgccccgccg 360
cugcccgugc cgggcgugcu ccugaaggag uucacgguga gcggcaacau ccugaccauc 420
cggcugaccg ccgcggacca ccggcagcug cagcugucga ucagcagcug ccuccagcag 480
cugagccugc ugauguggau cacccagugc uuccugccgg uguuccuggc ccagccgccc 540
agcggccagc gccggugacc acuaguuaua agacugacua gcccgauggg ccucccaacg 600
ggcccuccuc cccuccuugc accgagauua auaaaaaaaa aaaaaaaaaa aaaaaaaaaa 660
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaauauu cccccccccc cccccccccc 720
cccccccccc ucuagacaau uggaauu 747
<210> 56
<211> 761
<212> РНК
<213> Искусственная
<220>
<223> Последовательность мРНК NY-ESO-1(GC)-ag-A64-C30-histoneSL
<400> 56
gggagaaagc uuaccaugca ggccgagggc cgcggcaccg gcggcucgac cggcgacgcc 60
gacgggcccg gcggcccggg caucccggac ggcccgggcg ggaacgcggg cggcccgggc 120
gaggccggcg ccaccggcgg gcggggcccg cggggcgccg gcgccgcccg ggcgagcggc 180
cccggcgggg gcgccccgcg gggcccgcac ggcggcgccg ccagcggccu gaacgggugc 240
ugccggugcg gcgcccgcgg cccggagagc cggcuccugg aguucuaccu ggccaugccg 300
uucgcgaccc cgauggaggc cgagcuggcc cggcggagcc uggcccagga cgccccgccg 360
cugcccgugc cgggcgugcu ccugaaggag uucacgguga gcggcaacau ccugaccauc 420
cggcugaccg ccgcggacca ccggcagcug cagcugucga ucagcagcug ccuccagcag 480
cugagccugc ugauguggau cacccagugc uuccugccgg uguuccuggc ccagccgccc 540
agcggccagc gccggugacc acuaguuaua agacugacua gcccgauggg ccucccaacg 600
ggcccuccuc cccuccuugc accgagauua auaaaaaaaa aaaaaaaaaa aaaaaaaaaa 660
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaugca uccccccccc cccccccccc 720
cccccccccc ccaaaggcuc uuuucagagc caccaggaau u 761
<210> 57
<211> 646
<212> РНК
<213> Искусственная
<220>
<223> Последовательность мРНК Survivin(GC)-ag-A64-C30-histoneSL
<400> 57
gggagaaagc uuaccauggg cgcccccacc cugccgccgg ccuggcagcc guuccucaag 60
gaccaccgca ucucgaccuu caagaacugg ccguuccugg agggcugcgc gugcaccccg 120
gagcggaugg ccgaggccgg cuucauccac ugccccaccg agaacgagcc ggaccuggcc 180
cagugcuucu ucugcuucaa ggagcuggag ggcugggagc cggacgacga cccgaucgag 240
gagcacaaga agcacagcag cggcugcgcc uuccugagcg ugaagaagca guucgaggag 300
cugacgcucg gggaguuccu gaagcuggac cgggagcggg ccaagaacaa gaucgcgaag 360
gagaccaaca acaagaagaa ggaguucgag gagaccgcca agaaggugcg gcgggccauc 420
gagcagcugg ccgccaugga cugaccacua guuauaagac ugacuagccc gaugggccuc 480
ccaacgggcc cuccuccccu ccuugcaccg agauuaauaa aaaaaaaaaa aaaaaaaaaa 540
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaugcauccc cccccccccc 600
cccccccccc ccccccccaa aggcucuuuu cagagccacc agaauu 646
<210> 58
<211> 1813
<212> РНК
<213> Искусственная
<220>
<223> Последовательность мРНК MAGE-C1(GC)-ag-A64-C30-histoneSL
<400> 58
gggagaaagc uuaccaugca guccccgcug cagggcgagg aguuccagag cucccugcag 60
agccccgugu ccaucugcag cuccagcacc cccuccagcc ucccgcagag cuuccccgag 120
uccagccagu ccccccccga gggcccgguc cagagccccc ugcacucccc gcagagcccc 180
ccggagggga ugcacuccca gagcccccug cagucccccg agagcgcccc cgagggcgag 240
gacucccuca gcccgcugca gaucccccag uccccgcugg agggggagga cagccucucc 300
agccugcacu ucccccaguc cccgcccgag ugggaggaca gccugagccc ccuccacuuc 360
ccccaguucc cgccccaggg cgaggacuuc caguccagcc ugcagucccc cgugagcauc 420
ugcuccagcu ccacgagccu gucccucccc cagagcuucc cggagucccc ccagagcccg 480
cccgaggggc cggcgcaguc cccccugcag cgccccguga gcuccuucuu cagcuacacc 540
cuggccuccc uccugcagag cucccacgag agcccgcaga gcccgcccga gggccccgcc 600
caguccccgc ugcagagccc cgucuccagc uuccccucca gcaccuccag cucccucagc 660
caguccagcc ccguguccag cuucccgucc agcaccucca gcucccugag caagagcucc 720
cccgagagcc cccugcaguc ccccgugauc agcuucucca gcuccacgag ccucuccccg 780
uucagcgagg aguccagcuc ccccgucgac gaguacacca gcuccagcga cacccugcug 840
gaguccgaca gccucaccga cuccgagagc cugaucgaga gcgagccccu guucaccuac 900
acgcucgacg agaaggugga cgagcuggcc cgguuccugc uccugaagua ccaggugaag 960
cagcccauca ccaaggccga gaugcugacc aacgucaucu cccgcuacac cggcuacuuc 1020
ccggugaucu uccggaaggc gcgcgaguuc aucgagaucc ucuucgggau cagccugcgg 1080
gagguggacc ccgacgacuc cuacgucuuc gugaacacgc uggaccucac cagcgagggc 1140
ugccuguccg acgagcaggg gaugagccag aaccgccugc ucauccugau ccuguccauc 1200
aucuucauca agggcaccua cgccagcgag gaggucaucu gggacgugcu cuccgggauc 1260
ggcgugcggg ccggccgcga gcacuucgcc uucggggagc cccgggagcu gcugaccaag 1320
gucugggugc aggagcacua ccucgaguac cgcgaggugc ccaacagcuc cccgccccgg 1380
uacgaguucc uguggggccc ccgcgcccac agcgagguca ucaagcggaa ggugguggag 1440
uuccuggcga ugcucaagaa cacggucccc aucaccuucc cguccagcua caaggacgcc 1500
cugaaggacg uggaggagcg ggcccaggcc aucaucgaca ccaccgacga cuccacggcc 1560
accgagagcg cguccagcuc cgugaugagc cccagcuucu ccagcgagug accacuaguu 1620
auaagacuga cuagcccgau gggccuccca acgggcccuc cuccccuccu ugcaccgaga 1680
uuaauaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1740
aaaaaaaaau gcaucccccc cccccccccc cccccccccc cccccaaagg cucuuuucag 1800
agccaccaga auu 1813
<---
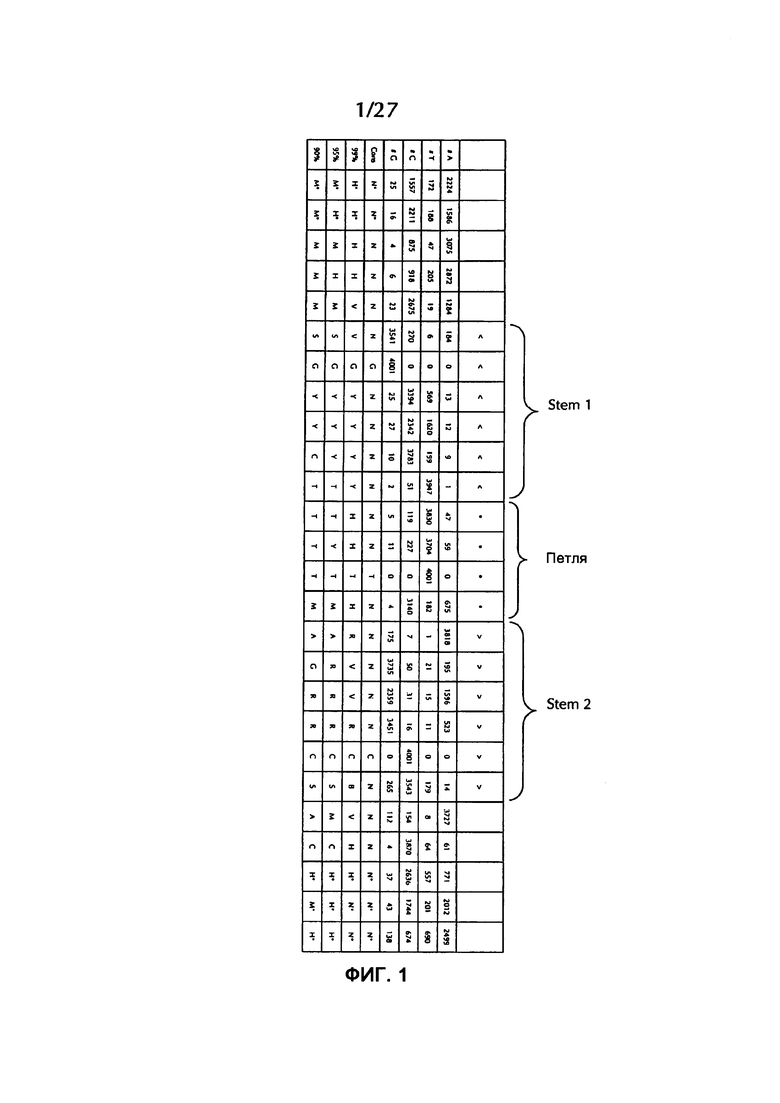
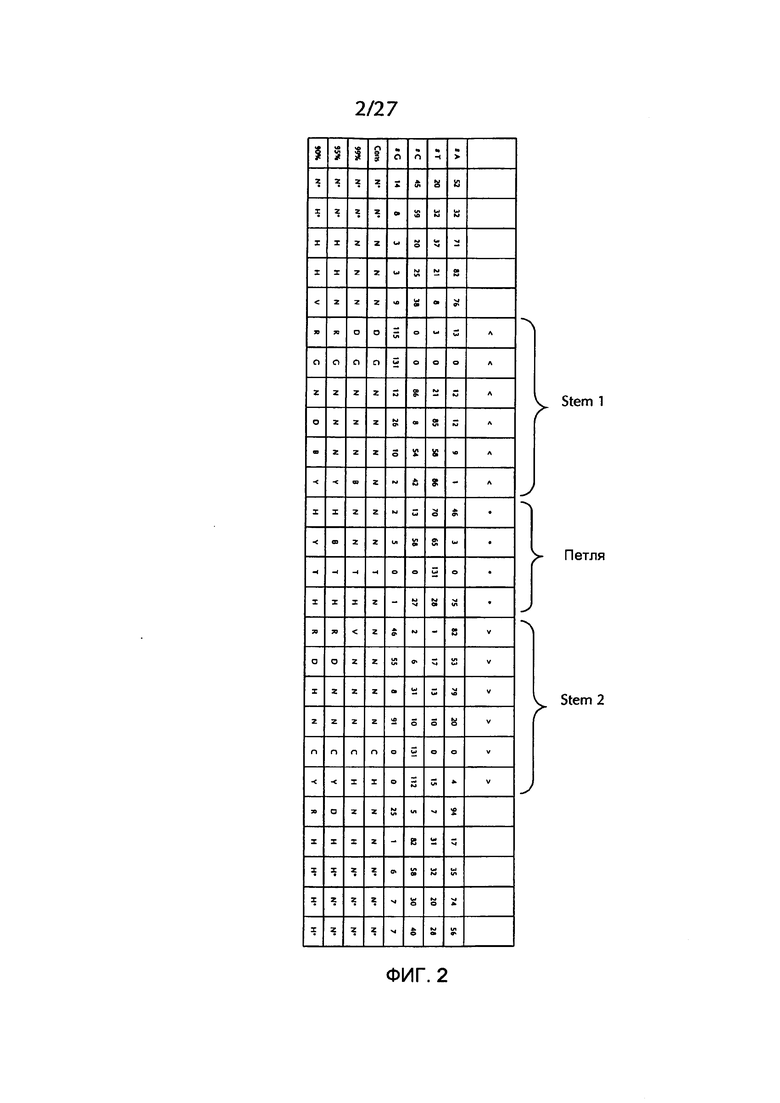
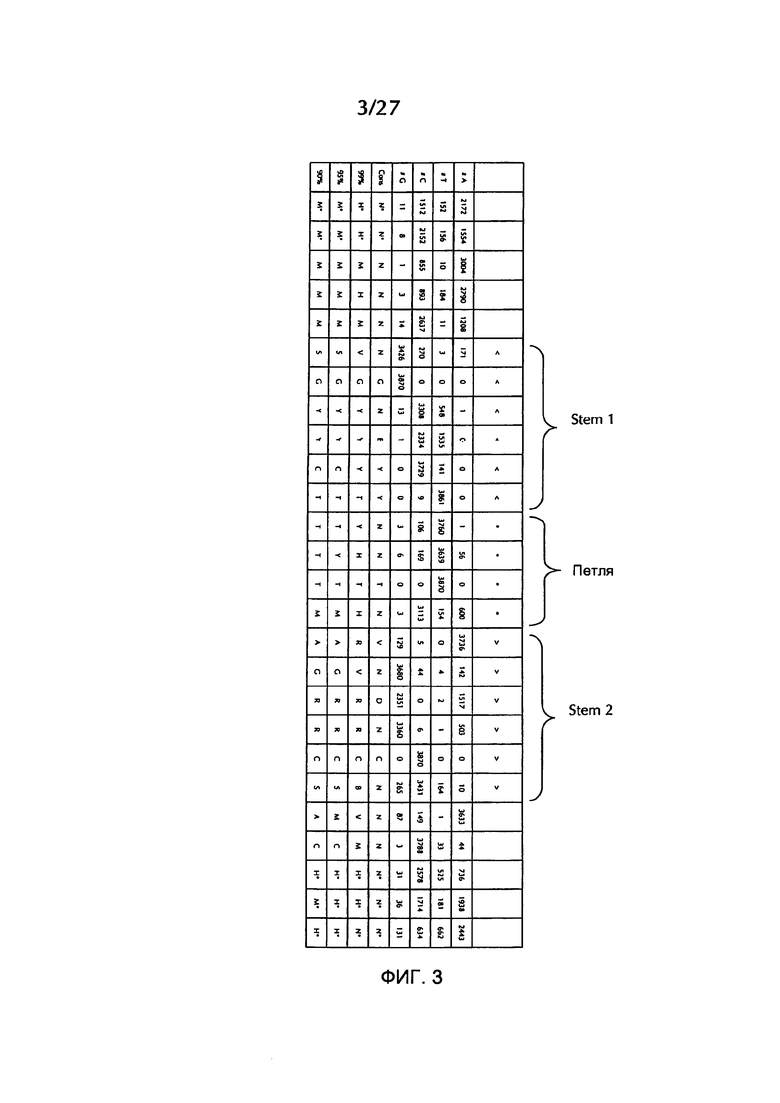
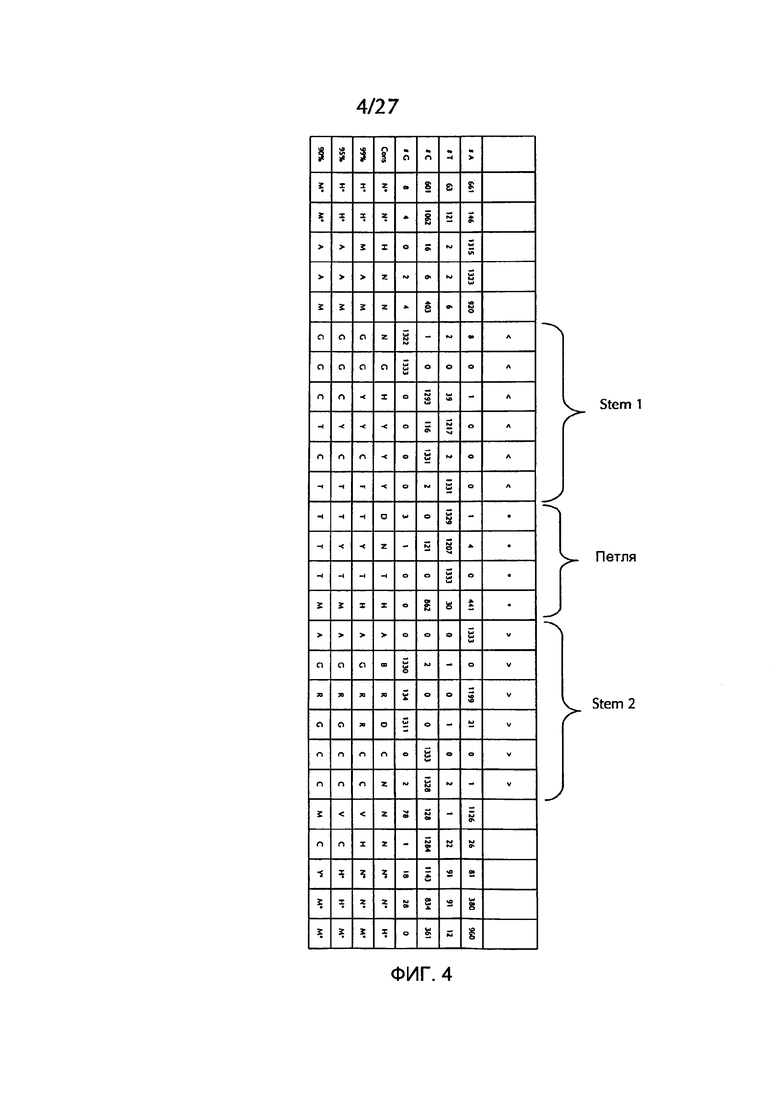
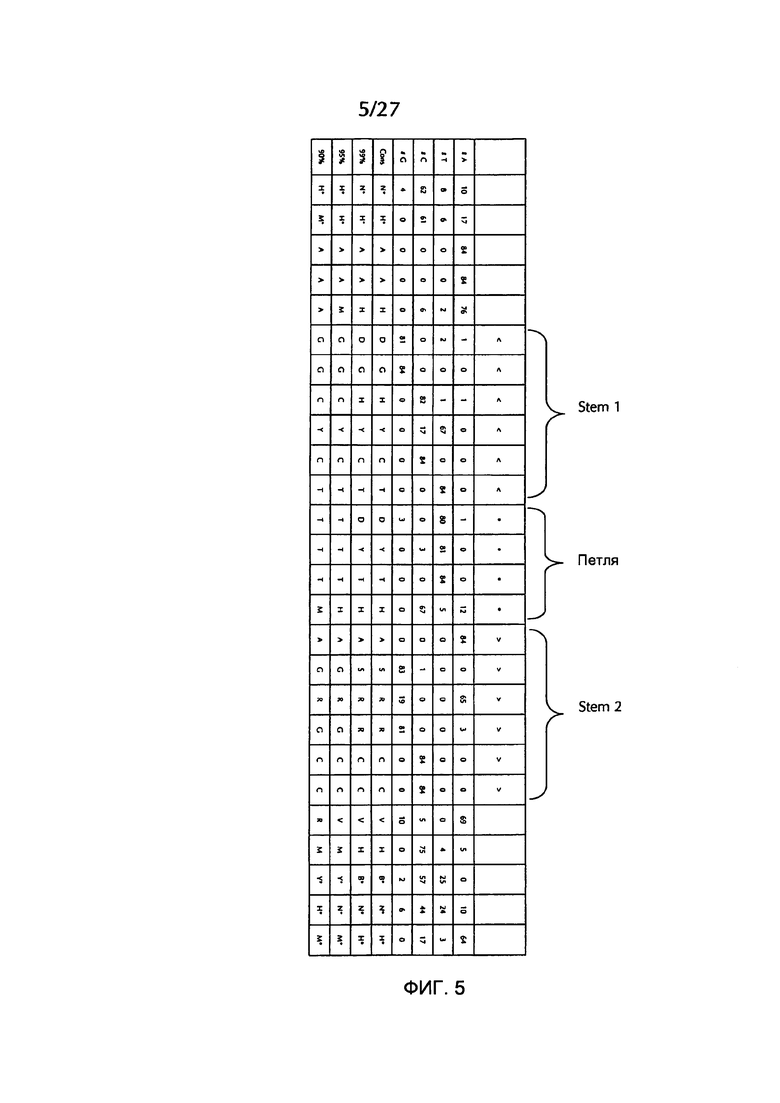
















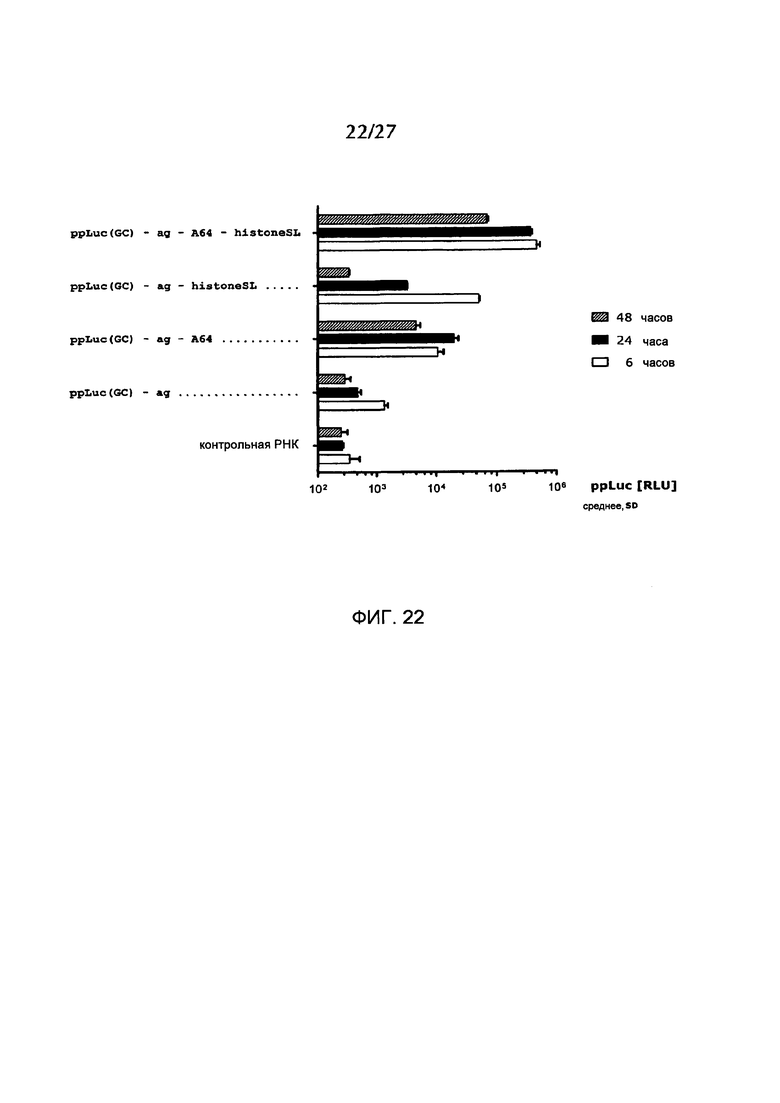
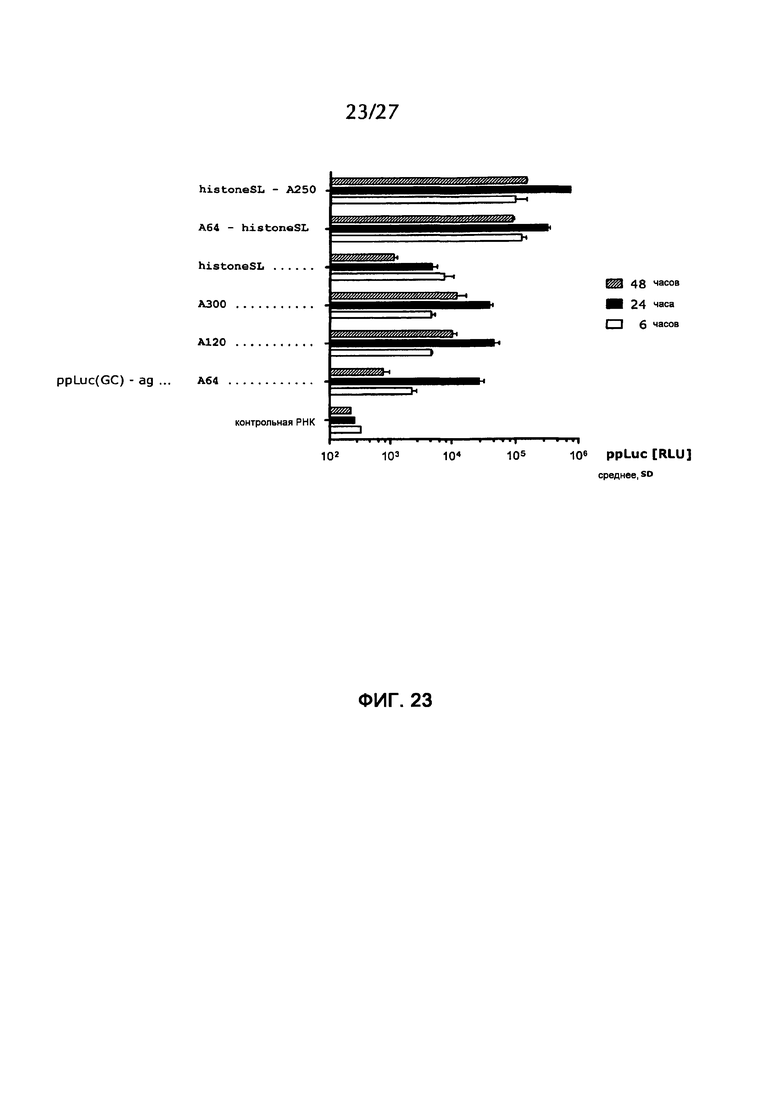
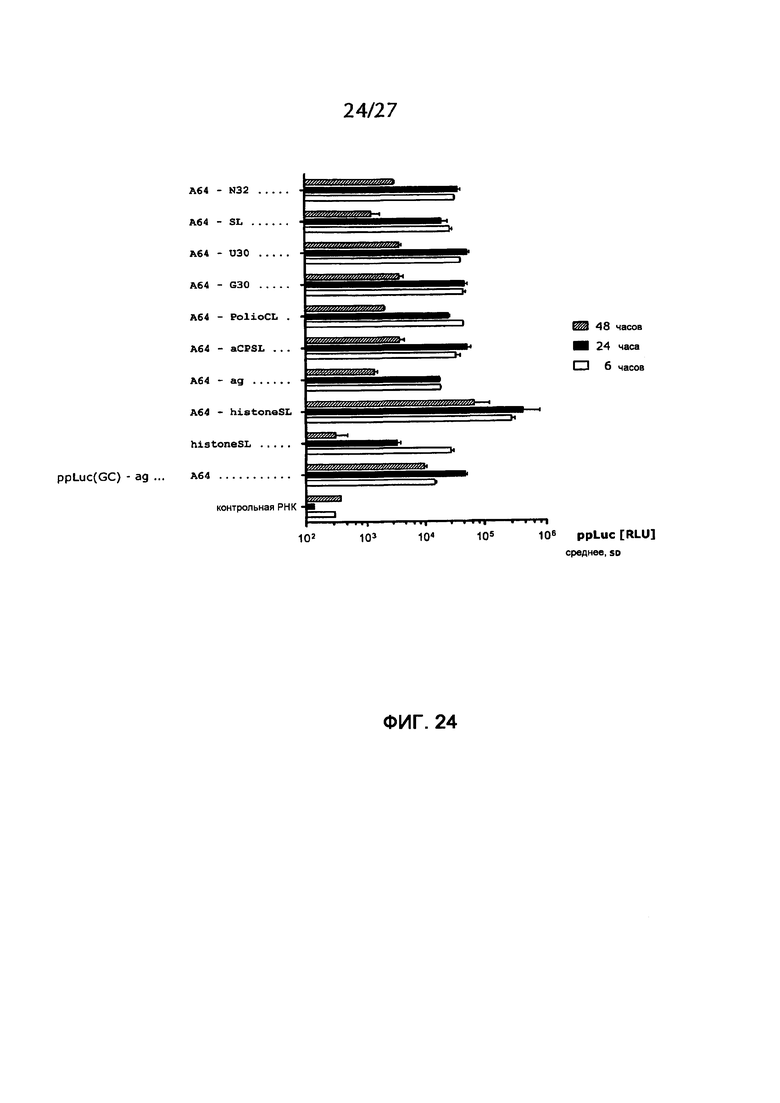
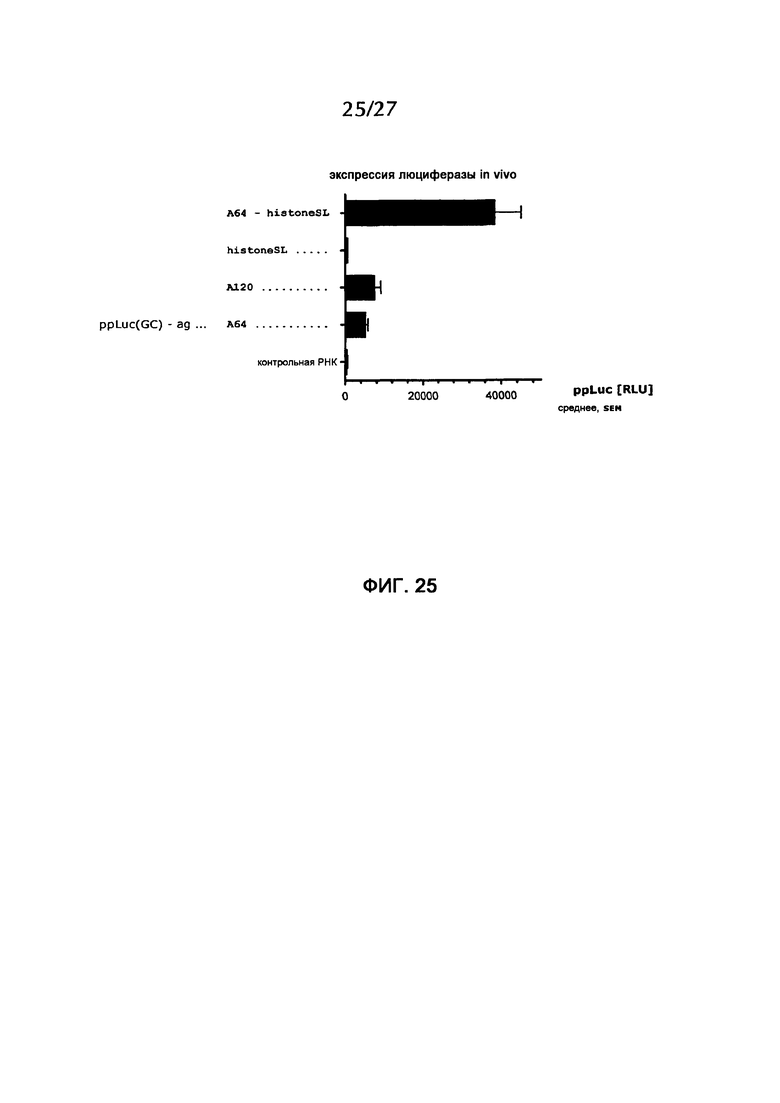
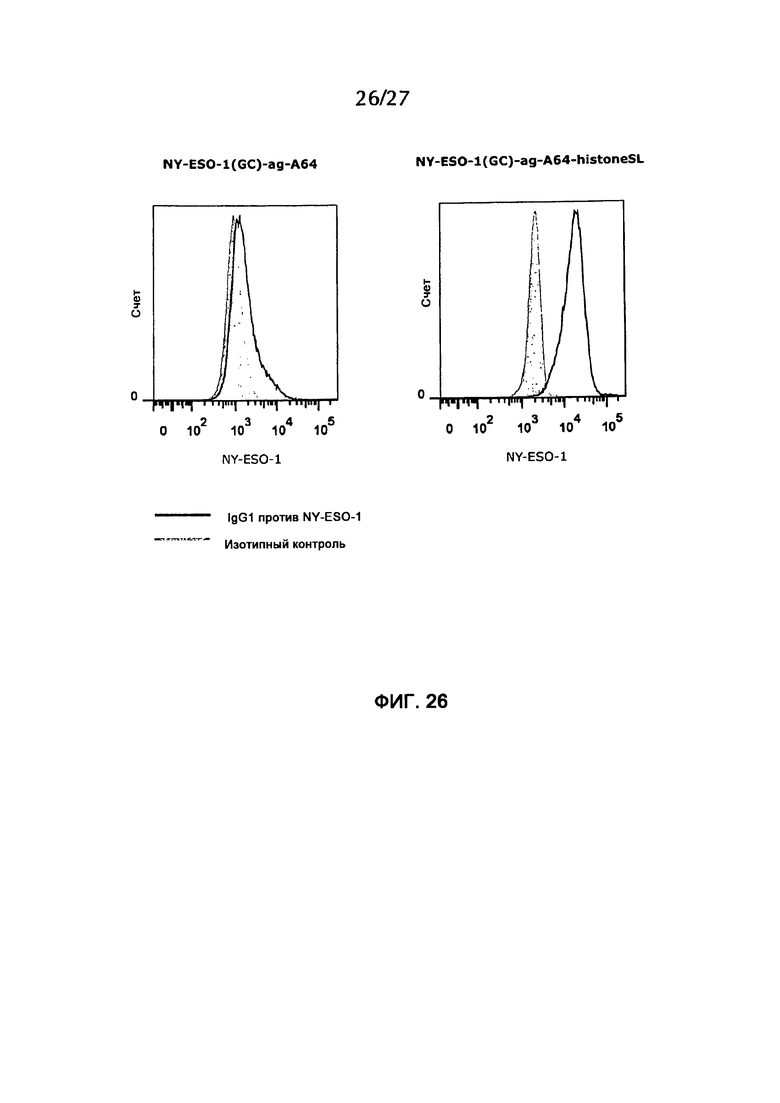

Изобретение относится к биотехнологии. Предложен набор, композиция, применение и способ для лечения опухолевых заболеваний. Изобретение может быть использовано в медицине. 7 н. и 20 з.п. ф-лы, 15 табл., 27 ил.
1. Набор для лечения опухолевых заболеваний, включающий:
А) нуклеиновую кислоту, включающую или кодирующую кодирующую область, которая кодирует по меньшей мере один пептид или протеин, который содержит опухолевый антиген NY-ESO-1 или его фрагмент; сохраняющий активность полноразмерного белка;
Б) нуклеиновую кислоту, включающую или кодирующую кодирующую область, которая кодирует по меньшей мере один пептид или протеин, содержащий опухолевый антиген 5Т4 или его фрагмент, сохраняющий активность полноразмерного белка;
В) нуклеиновую кислоту, включающую или кодирующую кодирующую область, которая кодирует по меньшей мере один пептид или протеин, содержащий опухолевый антиген сурвивин или его фрагмент, сохраняющий активность полноразмерного белка;
Г) нуклеиновую кислоту, включающую или кодирующую кодирующую область, которая кодирует по меньшей мере один пептид или протеин, содержащий опухолевый антиген MAGE-C1 или его фрагмент, сохраняющий активность полноразмерного белка; и
Д) нуклеиновую кислоту, включающую или кодирующую кодирующую область, которая кодирует по меньшей мере один пептид или протеин, содержащий опухолевый антиген MAGE-C2 или его фрагмент, сохраняющий активность полноразмерного белка;
где фрагмент NY-ESO-1, 5Т4, сурвивина, MAGE-C1 или MAGE-C2 имеет длину по меньшей мере шесть аминокислотных остатков;
где каждая нуклеиновая кислота А, Б, В, Г или Д в 5' - 3' направлении включает или кодирует
i)
а) кодирующую область,
б) по меньшей мере одну гистоновую структуру типа "стебель петля" и
в) поли(А) последовательность или
ii)
а) кодирующую область,
б) поли(А) последовательность и
в) по меньшей мере одну гистоновую структуру типа "стебель петля",
где каждая нуклеиновая кислота А, Б, В, Г или Д дополнительно включает или кодирует поли(С) последовательность, включающую по меньшей мере 10 цитидиновых нуклеотидов.
2. Набор по п. 1, где опухолевое заболевание представляет собой онкологическое заболевание.
3. Набор по п. 1, дополнительно включающий нуклеиновую кислоту, содержащую или кодирующую кодирующую область опухолевого антигена, где опухолевый антиген представляет собой меланоцит-специфический антиген, антиген рака яичка или опухолевый специфический антиген, выбранный из антигена CT-X, антигена non-CT-X, партнера по связыванию для антигена CT-X и партнера по связыванию для антигена non-CT-X, или фрагмент опухолевого антигена, где фрагмент имеет длину по меньшей мере шесть аминокислотных остатков.
4. Набор по п. 3, дополнительно включающий нуклеиновую кислоту, содержащую или кодирующую кодирующую область опухолевого антигена, где опухолевый антиген выбирается из перечня:
5T4, 707-AP, 9D7, AFP, AlbZIP HPG1, альфа-5-бета-1-интегрин, альфа-5-бета-6-интегрин, альфа-актинин-4/m, альфа-метилацилкоэнзим-A-рацемаза, ART-4, ARTC1/m, B7H4, BAGE-1, BCL-2, bcr/abl, бета-катенин/m, BING-4, BRCA1/m, BRCA2/m, СA 15-3/СA 27-29, СA 19-9, CA72-4, CA125, калретикулин, CAMEL, CASP-8/m, катепсин B, катепсин L, CD19, CD20, CD22, CD25, CDE30, CD33, CD4, CD52, CD55, CD56, CD80, CDC27/m, CDK4/m, CDKN2A/m, CEA, CLCA2, CML28, CML66, COA-1/m, коактозин-подобный белок, коллаж XXIII, COX-2, СT-9/BRD6, Cten, циклин B1, циклин D1, cyp-B, CYPB1, DAM-10, DAM-6, DEK-CAN, EFTUD2/m, EGFR, ELF2/m, EMMPRIN, EpCam, EphA2, EphA3, ErbB3, ETV6-AML1, EZH2, FGF-5, FN, Frau-1, G250, GAGE-1, GAGE-2, GAGE-3, GAGE-4, GAGE-5, GAGE-6, GAGE7b, GAGE-8, GDEP, GnT-V, gp100, GPC3, GPNMB/m, HAGE, HAST-2, гепсин, Her2/neu, HERV-K-MEL, HLA-A*0201-R17I, HLA-A11/m, HLA-A2/m, HNE, гомеобокс NKX3.1, HOM-TES-14/SCP-1, HOM-TES-85, HPV-E6, HPV-E7, HSP70-2M, HST-2, hTERT, iCE, IGF-1R, IL-13Ra2, IL-2R, IL-5, незрелый ламининовый рецептор, калликреин-2, калликреина-4, Ki67, KIAA0205, KIAA0205/m, KK-LC-1, K-Ras/m, LAGE-A1, LDLR-FUT, MAGE-A1, MAGE-A2, MAGE-A3, MAGE-A4, MAGE-A6, MAGE-A9, MAGE-A10, MAGE-A12, MAGE-B1, MAGE-B2, MAGE-B3, MAGE-B4, MAGE-B5, MAGE-B6, MAGE-B10, MAGE-B16, MAGE-B17, MAGE-C1, MAGE-C2, MAGE-C3, MAGE-D1, MAGE-D2, MAGE-D4, MAGE-E1, MAGE-E2, MAGE-F1, MAGE-H1, MAGEL2, маммаглобин A, MART-1/мелан-A, MART-2, MART-2/м, белок матриксa 22, MC1R, M-CSF, ME1/m, мезотелин, MG50/PXDN, MMP11, MN/СА IX-антиген, MRP-3, MUC-1, MUC-2, MUM-1/m, MUM-2/m, MUM-3/m, миозин класса I/m, NA88-A, N-ацетилглюкозаминилтрансфераза-V, Neo-PAP, Nео-PAP/m, NFYC/m, NGEP, NMP22, NPM/ALK, N-Ras/m, NSE, NY-ESO-B, NY-ESO-1, OA1, OFA-iLRP, OGT, OGT/m, OS-9, OS-9/m, остеокальцин, остеопонтин, р15, p190 минорный bcr-abl, p53, р53/m, PAGE-4, PAI-1, PAI-2, PAP, PART-1, PATE, PDEF, Pim-1-киназа, Pin-1, Pml/PARaльфа, POTE, PRAME, PRDX5/m, простеин, протеиназа-3, PSA, PSCA, PSGR, PSM, PSMA, PTPRK/m, RAGE-1, RBAF600/m, RHAMM/CD168, RU1, RU2, S-100, SAGE, SART-1, SART-2, SART-3, SCC, SIRT2/m, Sp17, SSX-1, SSX-2/HOM-MEL-40, SSX-4, STAMP-1, STEAP-1, сурвивин, сурвивин-2B, SYT-SSX-1, SYT-SSX-2, ТА-90, TAG-72, TARP, ТEL-AML1, TGFбета, TGFбетаRII, TGM-4, TPI/m, TRAG-3, TRG, TRP-1, TRP-2/6b, TRP/INT2, TRP-p8, тирозиназа, UPA, VEGFR1, VEGFR-2/FLK-1 и WT1 и иммуноглобулиновый идиотип лимфоидных клеток крови, или идиотип Т-клеточного рецептора лимфоидных клеток крови, или фрагмент указанного опухолевого антигена, где фрагмент имеет длину по меньшей мере шесть аминокислотных остатков.
5. Набор по любому из пп. 1-4, где по меньшей мере одна гистоновая структура типа «стебель-петля» является гетерологичной для кодирующей области, где кодирующая область не кодирует гистоновый белок или его фрагмент, производное или вариант, обладающий гистоновой или гистоноподобной функцией.
6. Набор по любому из пп. 1-5, где ее кодирующая область не кодирует репортерный, маркерный или селективный белок, причем репортерный, маркерный или селективный белок не является NY-ESO-1, 5Т4, сурвивином, MAGE-C1, MAGE-C2 или опухолевым антигеном, как определено в п. 3 или 4.
7. Набор по любому из пп. 1-6, где нуклеиновая кислота представляет собой мРНК.
8. Набор по любому пп. 1-7, где по меньшей мере одну гистоновую структуру типа «стебель-петля» выбирают из следующей формулы (I) или (II):
формула (I) (последовательность типа «стебель-петля» без ограничивающих элементов стебля)
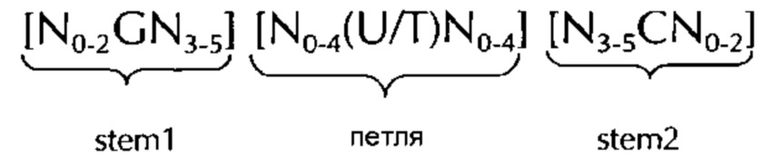
формула (II) (последовательность типа «стебель-петля» с ограничивающими элементами стебля)
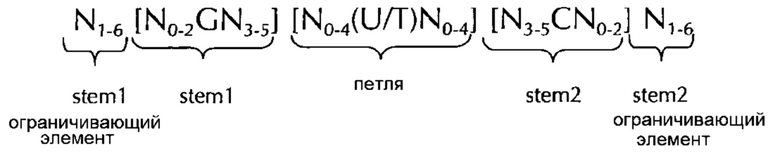
где stem1 или stem2 - ограничивающие элементы N1-6, представляют собой последовательность от 1 до 6 следующих друг за другом N, где каждый N является независимо от другого выбранным из нуклеотида, выбранного из A, U, T, G и С или нуклеотидного аналога этого;
stem1 [N0-2GN3-5] является обратно комплементарной или частично обратно комплементарной элементу stem2 и представляет собой последовательность длиной от 5 до 7 следующих друг за другом нуклеотидов;
где N0-2 представляет собой последовательность длиной от 0 до 2 следующих друг за другом N, где каждый N является независимо от другого выбранным из нуклеотида, выбранного из A, U, T, G и С или нуклеотидного аналога этого;
где N3-5 представляет собой последовательность длиной от 3 до 5 следующих друг за другом N, где каждый N является независимо от другого выбранным из нуклеотида, выбранного из A, U, T, G и С или нуклеотидного аналога этого; и
где G представляет собой гуанозин или его аналог и может быть дополнительно заменен цитидином или его аналогом при условии, что его комплементарный нуклеотид цитидин в stem2 заменяется гуанозином;
последовательность петли [N0-4(U/T)N0-4] расположена между элементами stem1 и stem2 и представляет собой последовательность длиной от 3 до 5 следующих друг за другом нуклеотидов,
где каждый N0-4 является независимо от другого последовательностью длиной от 0 до 4 следующих друг за другом N, где каждый N является независимо от другого выбранным из нуклеотида, выбранного из A, U, T, G и С или нуклеотидного аналога этого; и
где U/T представляет уридин или тимидин;
stem2[N3-5СN0-2] представляет собой обратно комплементарную или частично обратно комплементарную с элементом stem1 и представляет собой последовательность длиной от 5 до 7 следующих друг за другом нуклеотидов;
где N3-5 представляет собой последовательность длиной от 3 до 5 следующих друг за другом N, где каждый N является независимо от другого выбранным из нуклеотида, выбранного из A, U, T, G и С или нуклеотидного аналога этого;
где N0-2 представляет собой последовательность длиной от 0 до 2 следующих друг за другом N, где каждый N является независимо от другого выбранным из нуклеотида, выбранного из A, U, T, G и С или нуклеотидного аналога этого; и
где С представляет собой цитидин или его аналог и дополнительно может быть заменен гуанозином или его аналогом при условии, что его комплементарный нуклеотид гуанозин в stem1 заменяется цитидином;
где stem1 и stem2 способны образовывать пары оснований друг с другом,
образуя обратно комплементарную последовательность, где образование пар оснований может происходить между stem1 и stem2, или
образуя частично комплементарную последовательность, где образование неполных пар оснований может происходить между сегментом stem1 и stem2.
9. Набор по п. 8, где по меньшей мере одну гистоновую структуру типа «стебель-петля» выбирают из по меньшей мере одной из следующих формул (Iа) или (IIa):
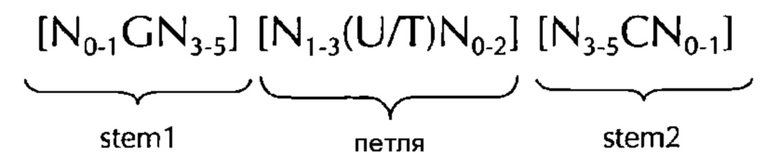
формула (Ia) (последовательность структуры типа «стебель-петля» без ограничивающих элементов стебля)
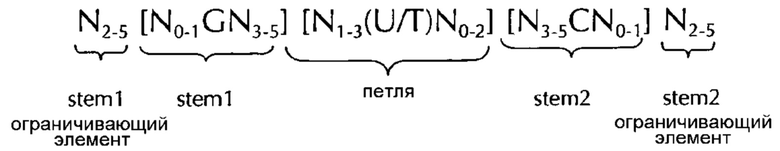
формула (IIa) (последовательность типа «стебель-петля» с ограничивающими элементами стебля).
10. Набор по любому из пп. 1-9, где поли(A)-последовательность включает последовательность от примерно 25 до примерно 400 нуклеотидов аденозина.
11. Набор по любому из пп. 1-10, где последовательность нуклеиновых кислот является модифицированной для стабилизации нуклеиновой кислотой.
12. Набор по любому из пп. 1-11, где
нуклеиновая кислота С состоит из нуклеотидной последовательности SEQ ID NO:57 и/или
нуклеиновая кислота D состоит из нуклеотидной последовательности SEQ ID NO:58.
13. Композиция для приготовления лекарственного средства для лечения опухолевых заболеваний, включающая эффективное количество нуклеиновых кислот, как они определены в любом из пп. 1-12 в отношении набора, наполнители, или разбавители, или инкапсулирующие соединения.
14. Композиция по п. 13, где опухолевое заболевание представляет собой онкологическое заболевание.
15. Применение набора по любому из пп. 1-12 для приготовления лекарственного средства для лечения опухолевого заболевания.
16. Применение набора по п. 15, где опухолевое заболевание представляет собой онкологическое заболевание.
17. Применение композиции по п. 13 для приготовления лекарственного средства для лечения опухолевого заболевания.
18. Применение композиции по п. 17, где опухолевое заболевание представляет собой онкологическое заболевание.
19. Применение набора по любому из пп. 1-12 для лечения опухолевого заболевания.
20. Применение набора по п. 19, где опухолевое заболевание представляет собой онкологическое заболевание.
21. Применение набора по п. 20, где онкологическое заболевание является немелкоклеточным раком легких.
22. Фармацевтическая композиция для лечения опухолевого заболевания, включающая нуклеиновые кислоты, как они определены в любом из пп. 1-12 в отношении набора, или композицию, как она определена в п. 13, и фармацевтически приемлемый носитель.
23. Фармацевтическая композиция по п. 22, где опухолевое заболевание представляет собой онкологическое заболевание.
24. Фармацевтическая композиция по п. 23, где онкологическое заболевание является немелкоклеточным раком легких.
25. Применение набора, который содержит нуклеиновую кислоту, кодирующую пептид или протеин, как она определена в любом из пп. 1-12, для увеличения экспрессии in vitro кодируемого пептида или протеина.
26. Применение композиции по п. 17 для увеличения экспрессии in vitro пептида или протеина, кодируемого входящей в ее состав нуклеиновой кислотой.
27. Способ увеличения экспрессии in vitro кодируемого пептида или протеина, кодируемого нуклеиновой кислотой, как она определена в любом из пп. 1-12, включающий этапы:
а) обеспечение набора по любому из пп. 1-12 или композиции по п. 17;
б) применение или введение нуклеиновых кислот, входящих в набор или композицию, в бесклеточную систему экспрессии, клетку или ткани.
| COLLART D | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| J Cell Biochem | |||
| Пуговица для прикрепления ее к материи без пришивки | 1921 |
|
SU1992A1 |
| WO 2010132867 A1, 18.11.2010 | |||
| Б | |||
| ГЛИК, ДЖ | |||
| ПАСТЕРНАК | |||
| Молекулярная биотехнология | |||
| Принципы и применение | |||
| Москва | |||
| Мир | |||
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |

