Настоящее изобретение относится к искусственным молекулам нуклеиновой кислоты, которые содержат открытую рамку считывания, элемент 3'-нетранслируемой области (3'-UTR-элемент) и необязательно поли(А)-последовательность и/или сигнал полиаденилирования. Изобретение относится также к вектору, содержащему 3'-UTR-элемент, к клетке, содержащей искусственную молекулу нуклеиновой кислоты или вектор, к фармацевтической композиции, содержащей искусственную молекулу нуклеиновой кислоты или вектор, и к набору, содержащему искусственную молекулу нуклеиновой кислоты, вектор и/или фармацевтическую композицию, предпочтительно предназначенным для применения в области генной терапии и/или генетической вакцинации.
Генная терапия и генетическая вакцинация относятся к наиболее перспективным и быстро развивающимся методам современной медицины. Они позволяют разрабатывать высокоспецифические и индивидуальные подходы для терапии широкого разнообразия заболеваний. Мишенью для таких подходов к лечению могут являться, прежде всего, врожденные генетические заболевания, злокачественные или связанные с опухолью заболевания, а также воспалительные заболевания. Подразумевается, что с помощью указанных подходов можно предупреждать раннее начало таких заболеваний.
Основная концептуальная идея генной терапии заключается в осуществлении соответствующей модуляции нарушенной генной экспрессии, ассоциированной с патологическими состояниями при конкретных заболеваниях. Патологически измененная генная экспрессия может приводить к недостатку или сверхпродуцированию имеющих важное значение генных продуктов, например, сигнальных факторов, таких как гормоны, факторы, обусловленные генами «домашнего хозяйства», метаболические ферменты, структурные белки или т.п. Измененная генная экспрессия может быть обусловлена не только нарушением регуляции транскрипции и/или трансляции, но также и мутациями в ОРС, кодирующей конкретный белок. Патологические мутации могут вызываться, например, хромосомной аберрацией или более специфическими мутациями, такими как точечные мутации или мутации, приводящие к сдвигу рамки считывания, которые все приводят к ограничению функциональности и возможно к полной потере функции генного продукта. Однако нарушение регуляции транскрипции или трансляции может иметь место также в том случае, если мутации воздействуют на гены, кодирующие белки, которые участвуют в механизме транскрипции или трансляции в клетке. Такие мутации могут приводить к патологической повышающей или понижающей регуляции генов, которые сами по себе являются функциональными. Гены, кодирующие генные продукты, которые обладают такими регуляторными функциями, могут представлять собой, например, факторы транскрипции, сигнальные рецепторы, белки-мессенджеры или т.п. Однако потеря функции таких генов, кодирующих регуляторные белки, при определенных обстоятельствах может быть восстановлена путем искусственной интродукции других факторов, функционирующих в прямом направлении относительно нарушенного генного продукта. Такие дефекты генов можно компенсировать также с помощью генной терапии посредством замены самого поврежденного гена.
Генетическая вакцинация позволяет вызывать требуемый иммунный ответ на выбранные антигены, такие как обладающие определенными характеристиками компоненты бактериальных поверхностей, вирусные частицы, опухолевые антигены или т.п. В целом, вакцинация представляет собой одно из наиболее важных достижений современной медицины. Однако, в настоящее время эффективные вакцины доступны только для ограниченного количества заболеваний. Поэтому инфекции, которые нельзя предупреждать путем вакцинации, все еще поражают каждый год миллионы людей.
Обычно вакцины можно подразделять на вакцины «первого», «второго» и «третьего» поколения. Вакцины «первого поколения», как правило, представляют собой вакцины, содержащие цельные организмы. Они основаны либо на живых и ослабленных, либо на убитых патогенах, например, вирусах, бактериях или т.п. Основным недостатком живых и ослабленных вакцин является риск реверсии с образованием угрожающих жизни вариантов. Таким образом, указанные патогены, хотя они и являются ослабленными, все еще могут нести присущие им непредсказуемые риски. Убитые патогены могут не являться достаточно эффективными для создания специфического иммунного ответа. Для того чтобы минимизировать указанные риски, были разработаны вакцины «второго поколения». Как правило, они представляют собой субъединичные вакцины, состоящие из компонентов определенных антигенов или рекомбинантных белков, происходящих из патогенов.
Генетические вакцины, т.е. вакцины для генетической вакцинации, как правило, называют вакцинами «третьего поколения». Они, как правило, состоят из созданных методами генной инженерии молекул нуклеиновой кислоты, которые обеспечивают экспрессию in vivo фрагментов пептида или белка (антигена), характерных для патогена или опухолевого антигена. При введении пациенту экспрессия генетических вакцин происходит после поглощения клетками-мишенями. Экспрессия введенных нуклеиновых кислот приводит к производству кодируемых белков. В том случае, когда эти белки распознаются иммунной системой пациента как чужие, запускается иммунный ответ.
Как следует из вышеизложенного, оба метода, и генная терапия, и генетическая вакцинация, по существу основаны на введении молекул нуклеиновой кислоты пациенту и последующей транскрипции и/или трансляции кодируемой генетической информации. Альтернативно этому, генетическая вакцинация или генная терапия может включать также методы, которые предусматривают выделение специфических клеток из организма пациента, подлежащего лечению, последующую трансфекцию ex vivo указанных клеток и повторное введение обработанных клеток пациенту.
В контексте генной терапии или генетической вакцинации в качестве молекул нуклеиновой кислоты можно вводить как ДНК, так и РНК. Известно, что ДНК является относительно стабильной и с ней проще манипулировать. Однако применение ДНК сопряжено с риском нежелательного встраивания введенных ДНК-фрагментов в геном пациента, что может приводить к мутагенным действиям, таким как потеря функции нарушенных генов. Другим риском является то, что происходит нежелательное возникновение антител к ДНК. Еще одним недостатком является ограниченный уровень экспрессии кодируемого пептида или белка, достижимый при введении ДНК, поскольку ДНК должна проникнуть в ядро для того, чтобы произошла ее транскрипция перед тем, как могла осуществляться трансляция полученной в результате этого мРНК. Уровень экспрессии введенной ДНК может зависеть, среди прочего, от присутствия специфических факторов транскрипции, которые регулируют транскрипцию ДНК. В случае отсутствия таких факторов транскрипция ДНК не приводит к получению ДНК в достаточных количествах. В результате достигаемый уровень транслированного пептида или белка является ограниченным.
При использовании РНК вместо ДНК для генной терапии или генетической вакцинации риск нежелательной интеграции в геном и образования антител к ДНК сводится к минимуму или устраняется. Однако считается, что РНК являться относительно нестабильной молекулой, которая может легко расщепляться под действием повсеместно присутствующих РНКаз.
Расщепление РНК in vivo вносит вклад в регулирование времени полужизни РНК. Этот эффект был изучен и было установлено, что он строго регулирует экспрессию эукариотических генов (Friedel и др., Conserved principles of mammalian transcriptional regulation revealed by RNA half-life, Nucleic Acid Research 37 (17), 2009, cc. 1-12). Таким образом, каждая встречающаяся в естественных условиях мРНК имеет свое индивидуальное время полужизни, зависящее от гена, из которого происходит мРНК, и от типа клетки, в которой происходит экспрессия. Оно вносит вклад в регуляцию уровня экспрессии рассматриваемого гена. Нестабильные РНК важны для реализации кратковременной генной экспрессии в определенные моменты времени. Однако РНК с продолжительным временем жизни могут ассоциироваться с накоплением различных белков или с непрерывной экспрессией генов. Время полужизни мРНК in vivo может зависеть также от факторов окружающей среды, таких как обработка гормонами, что продемонстрировано, например, для мРНК инсулин-подобного фактора роста I, актина, и альбумина (Johnson и др., Newly synthesized RNA: Simultaneous measurement in intact cells of transcription rates and RNA stability of insulin-like growth factor I, actin, and albumin in growth hormone-stimulated hepatocytes, Proc. Natl. Acad. Sci., Vol. 88, 1991, cc. 5287-5291).
Для генной терапии и генетической вакцинации, как правило, требуется стабильная РНК. Это обусловлено, с одной стороны, тем фактом, что, как правило, требуется, чтобы in vivo происходило накопление продукта, кодируемого последовательностью РНК. С другой стороны, РНК должна сохранять свою структурную и функциональную целостность при приготовлении пригодной лекарственной формы, в процессе ее хранения и при введении. Таким образом, предпринимались усилия для создания стабильных молекул РНК для генной терапии или генетической вакцинации с целью предупреждения их от раннего разложения или расщепления.
Опубликованы данные о том, что содержание G/C в молекулах нуклеиновой кислоты может оказывать влияние на ее стабильность. Так, нуклеиновые кислоты, содержащие в большем количестве остатки гуанина (G) и/или цитозина (С), могут быть функционально более стабильными, чем нуклеиновые кислоты, содержащие большое количество адениновых (А) и тиминовых (Т) или урациловых (U) нуклеотидов. В этом контексте в WO 02/098443 предложена фармацевтическая композиция, содержащая мРНК, которая стабилизирована посредством модификаций последовательности в кодирующей области. Такая модификация является возможной благодаря вырожденности генетического кода. В соответствии с этим кодоны, содержащие менее предпочтительную комбинацию нуклеотидов (менее предпочтительную с точки зрения стабильности РНК), быть заменены на альтернативные кодоны без изменения кодируемой аминокислотной последовательности. Такой метод стабилизации РНК ограничен тем, что для каждой отдельной молекулы РНК нужно получать конкретную нуклеотидную последовательность, которая не приводит к получению аминокислотной последовательности, отклоняющейся от требуемой. Указанный подход ограничен также тем, что его применяют к кодирующим областям РНК.
При поиске альтернативного подхода к стабилизации мРНК было установлено, что встречающиеся в естественных условиях молекулы эукариотической мРНК содержат характерные стабилизирующие элементы. Например, они могут содержать так называемые нетранслируемые области (UTR) на своем 5'-конце (5'-UTR) и/или на своем 3'-конце (3'-UTR), а также другие структурные особенности, такие как 5'-кэп-структура или 3'-поли(А)-хвост. Как 5'-UTR, так и 3'-UTR, как правило, транскрибируются с геномной ДНК и, следовательно, представляют собой элемент незрелой мРНК. Характерные структурные особенности зрелой мРНК, такие как 5'-кэп и 3'-поли(А)-хвост (которые обозначают также, как поли(А)-хвост или поли(А)-последовательность), как правило, добавляют к транскрибированной (незрелой) мРНК в ходе процессинга мРНК.
3'-поли(А)-хвост, как правило, представляет собой монотонный участок последовательности, который состоит из аденозиновых нуклеотидов, добавленный к 3'-концу транскрибированной мРНК. Он может содержать вплоть до примерно 400 аденозиновых нуклеотидов. Установлено, что длина такого 3'-поли(А)-хвоста может являться элементом, имеющим решающее значение для стабильности индивидуальной мРНК.
Было продемонстрировано также, что 3'-UTR мРНК α-глобина может являться важным фактором, обеспечивающим хорошо известную стабильность мРНК α-глобина (Rodgers и др., Regulated α-globin mRNA decay is a cytoplasmic event proceeding through 3'-to-5' exosome-dependent decapping, RNA, 8, 2002, cc. 1526-1537). 3'-UTR мРНК α-глобина, по-видимому, участвует в формировании специфического рибонуклеопротеинового комплекса, α-комплекса, присутствие которого коррелирует со стабильностью мРНК in vitro (Wang и др., An mRNA stability complex functions with poly(A)-binding protein to stabilize mRNA in vitro, Molecular and Cellular biology, том 19, №7, июль 1999 г., cc. 4552-4560).
Кроме того, для UTR в мРНК рибосомального белка была выявлена важная регуляторная функция: хотя 5'-UTR мРНК рибосомального белка контролирует ассоциированную с ростом трансляцию мРНК, строгость указанной регулировки обеспечивается соответствующей 3'-UTR в мРНК рибосомального белка (Ledda и др., Effect of the 3'-UTR length on the translational regulation of 5'-terminal oligopyrimidine mRNAs, Gene, том 344, 2005, cc. 213-220). Этот механизм оказывает влияние на специфическую схему экспрессии рибосомальных белков, которые, как правило, транскрибируются постоянным образом, вследствие чего мРНК некоторых рибосомальных белков, таких как рибосомальный белок S9 или рибосомальный белок L32, называют генами «домашнего хозяйства» (Janovick-Guretzky и др., Housekeeping Gene Expression in Bovine Liver is Affected by Physiological State, Feed Intake, and Dietary Treatment, J. Dairy Sci., том 90, 2007, cc. 2246-2252). Таким образом, ассоциированная с ростом схема экспрессии рибосомальных белков в основном обусловлена регуляцией на уровне трансляции.
Вне зависимости от факторов, влияющих на стабильность мРНК, эффективность трансляции введенных молекул нуклеиновой кислоты клетками-мишенями или тканью имеет решающее значение для любого подхода с использованием молекул нуклеиновой кислоты для генной терапии или генетической вакцинации. Как следует из процитированных выше примеров, структурные особенности, такие как UTR, 5'-кэп и 3'-поли(А)-хвост, наряду с регуляцией стабильности, регулируют также трансляцию большинства мРНК. В этом контексте, опубликованы данные о том, что длина поли(А)-хвоста может играть важную роль также и для эффективности трансляции. Однако стабилизирующие 3'-элементы могут оказывать также ослабляющее действие на трансляцию.
В основу настоящего изобретения была положена задача создать молекулы нуклеиновой кислоты, которые можно применять для генной терапии и/или генетической вакцинации. Задача изобретения заключалась, прежде всего, в том, чтобы создать виды мРНК, которые стабилизированы для защиты от преждевременного разложения или расщепления, но которые не характеризуются значительной функциональной потерей с точки зрения эффективности трансляции. Задачей изобретения являлось также создание искусственной молекулы нуклеиновой кислоты, предпочтительно мРНК, отличающейся повышенной экспрессией соответствующего белка, кодируемого указанной молекулой нуклеиновой кислоты. Одна из конкретных задач изобретения заключалась в создании мРНК, позволяющей повышать эффективность трансляции соответствующего кодируемого белка. Другая задача настоящего изобретения заключалась в том, чтобы создать молекулы нуклеиновой кислоты, кодирующие такие обладающие преимуществами виды мРНК, которые могут быть пригодными для применения в генной терапии и/или генетической вакцинации. Следующая задача настоящего изобретения заключалась в том, чтобы создать фармацевтическую композицию для применения в генной терапии и/или генетической вакцинации. В целом, задача настоящего изобретения заключалась в создании усовершенствованных видов нуклеиновой кислоты, которые позволяют преодолевать указанные выше недостатки существующего уровня техники с использованием экономичного и простого подхода.
Задачи, положенные в основу настоящего изобретения, решаются с помощью объектов изобретения, представленных в формуле изобретения.
Настоящее изобретение было создано при поддержке правительства в рамках соглашения № HR0011-11-3-0001, подписанного DARPA (Агентство передовых оборонных исследовательских проектов). Правительство обладает определенными правами на изобретение.
Для целей ясности и удобочитаемости ниже даны следующие определения. Все технические характеристики, указанные в этих определениях, применимы к каждому и всем вариантам осуществления изобретения. В контексте этих вариантов осуществления изобретения могут быть даны дополнительные определения и пояснения.
Адаптивный иммунный ответ: Как правило, под адаптивным иммунным ответом понимают антигенспецифический ответ иммунной системы. Специфичность в отношении антигена позволяет вырабатывать ответы, приспособленные к специфическим патогенам или инфицированным патогеном клеткам. Способность создавать такие приспособленные ответы, как правило, поддерживается в организме «клетками памяти». Если патоген инфицирует организм более одного раза, то указанные специфические клетки памяти используются для его быстрой элиминации. В этом контексте первая стадия адаптивного иммунного ответа представляет собой активацию наивных антигенспецифических Т-клеток или других иммунных клеток, способных индуцировать антигенспецифический иммунный ответ, антигенпрезентирующими клетками. Это происходит в лимфоидных тканях и органах, через которые постоянно проходят наивные Т-клетки. Три типа клеток, которые могут служить в качестве антигенпрезентирующих клеток, представляют собой дендритные клетки, макрофаги и В-клетки. Каждая из указанных клеток выполняет отдельную функцию при вызывании иммунных ответов. Дендритные клетки могут поглощать антигены посредством фагоцитоза и макропиноцитоза, и они могут стимулироваться при контакте, например, с чужим антигеном, к миграции в местную лимфоидную ткань, где может происходить их дифференцировка в зрелые дендритные клетки. Макрофаги поглощают находящиеся в форме частиц антигены, такие как бактерии, и могут индуцироваться инфекционными агентами или другими соответствующими стимулами и экспрессировать в результате этого молекулы ГКГС. Уникальная способность В-клеток к связыванию и интернализации растворимых белковых антигенов посредством своих рецепторов также может иметь важное значение для индукции Т-клеток. ГКГС-молекулы, как правило, ответственны за презентацию антигена Т-клеткам. При этом презентация антигена на молекулах ГКГС приводит к активации Т-клеток, что индуцирует их пролиферацию и дифференцировку в «вооруженные» эффекторные Т-клетки. Наиболее важной функцией эффекторных Т-клеток является уничтожение (цитолиз) инфицированных клеток цитотоксическими СВ8+-Т-клетками и активация макрофагов Тh1-клетками, что в совокупности создает опосредованный клетками (клеточный) иммунитет, а активация В-клеток как Th2-, так и Th1-клетками, приводящая к образованию различных классов антител, запускает гуморальный иммунный ответ. Т-клетки распознают антиген посредством своих Т-клеточных рецепторов, которые не распознают антиген и не связываются с ним непосредственно, но вместо этого распознают короткие пептидные фрагменты, например, выведенных из патогена белковых антигенов, например, так называемые эпитопы, которые связываются с молекулами ГКГС на поверхностях других клеток.
Адаптивная иммунная система: Адаптивная иммунная система в основном предназначена для устранения или предупреждения роста патогенов. Как правило, она регулирует адаптивный иммунный ответ, придавая иммунной системе позвоночных способность распознавать и запоминать специфические патогены (для создания иммунитета) и организовывать более сильную атаку в каждом случае при обнаружении патогена. Система обладает высокой способностью к адаптации вследствие соматической гипермутации (процесс ускоренных соматических мутаций) и V(D)J-рекомбинации (необратимая генетическая рекомбинация сегментов гена рецептора антигена). Этот механизм позволяет небольшому количеству генов создавать огромное количество различных рецепторов антигенов, которые затем уникальным образом экспрессируются на каждом индивидуальном лимфоците. Поскольку реаранжировка гена приводит к необратимому изменению ДНК каждой клетки, то все потомство такой клетки должно затем наследовать гены, кодирующие ту же самую рецепторную специфичность, включая В-клетки памяти и Т-клетки памяти, которые имеют решающее значение для долговременного специфического иммунитета.
Адъювант/адъювантный компонент: Адъювант или адъювантный компонент в наиболее широком смысле, как правило, представляет собой фармакологический и/или иммунологический агент, который может модифицировать, например, усиливать, действие других агентов, таких как лекарственное средство или вакцина. Это понятие следует рассматривать в широком смысле, и оно относится к широкому спектру субстанций. Как правило, указанные субстанции обладают способностью повышать иммуногенность антигенов. Например, адъюванты могут распознаваться врожденными иммунными системами и, например, могут вызывать врожденный иммунный ответ. «Адъюванты», как правило, не вызывают адаптивный иммунный ответ. Таким образом, «адъюванты» нельзя классифицировать как антигены. Механизм их действия отличен от механизма действий, запускаемых антигенами, которые приводят к адаптивному иммунному ответу.
Антиген: В контексте настоящего изобретения понятие «антиген» относится, как правило, к субстанции, которая может распознаваться иммунной системой, предпочтительно адаптивной иммунной системой, и которая обладает способностью запускать антигенспецифический иммунный ответ, например, посредством образования антител и/или антигенспецифических Т-клеток, в качестве компонента адаптивного иммунного ответа. Как правило, антиген может представлять собой или содержать пептид или белок, который может презентоваться ГКГС Т-клеткам. В контексте настоящего изобретения антиген может представлять собой продукт трансляции предложенной молекулы нуклеиновой кислоты, предпочтительно мРНК, представленной в настоящем описании. В этом контексте под антигенами подразумевают также фрагменты, варианты и производные пептидов и белков, содержащие по меньшей мере один эпитоп. В контексте настоящего изобретения наиболее предпочтительными являются опухолевые антигены и патогенные антигены.
Искусственная молекула нуклеиновой кислоты: Под искусственной молекулой нуклеиновой кислоты, как правило, понимают молекулу нуклеиновой кислоты, например, ДНК или РНК, которая не встречается в естественных условиях. Иными словами, искусственную молекулу нуклеиновой кислоты можно рассматривать как молекулу нуклеиновой кислоты, не встречающуюся в естественных условиях. Такая молекула нуклеиновой кислоты может не встречаться в естественных условиях из-за своей индивидуальной последовательности (которая не встречается в естественных условиях) и/или из-за других модификаций, например, структурных модификаций нуклеотидов, которые не встречаются в естественных условиях. Искусственная молекула нуклеиновой кислоты может представлять собой молекулу ДНК, молекулу РНК или гибридную молекулу, содержащую участки ДНК и РНК. Как правило, искусственные молекулы нуклеиновой кислоты можно конструировать и/или создавать методами генетической инженерии так, чтобы они соответствовали требуемой искусственной последовательности нуклеотидов (гетерологичной последовательности). В этом контексте искусственная последовательность, как правило, представляет собой последовательность, которая не может встречаться в естественных условиях, т.е. отличается от последовательности дикого типа по меньшей мере одним нуклеотидом. Можно считать, что понятие «дикий тип» относится к последовательности, встречающейся в природе. Кроме того, не следует понимать, что понятие «искусственная молекула нуклеиновой кислоты» обозначает «одну единственную молекулу», как правило, оно включает группу идентичных молекул. Следовательно, оно может относиться к множеству идентичных молекул, содержащихся в аликвоте.
Бицистронная РНК, полицистронная РНК: Бицистронная или полицистронная РНК, как правило, представляет собой РНК, предпочтительно мРНК, которая, как правило, может иметь две (бицистронная) или большее количество (полицистронная) открытых рамок считывания (ОРС). В этом контексте открытая рамка считывания представляет собой последовательность, состоящую из нескольких кодонов, которые могут транслироваться в пептид или белок.
Носитель/полимерный носитель: В контексте изобретения носитель, как правило, может представлять собой соединение, которое облегчает транспорт и/или включение в комплекс другого соединения (карго, полезный груз). Полимерный носитель, как правило, представляет собой носитель, образующий полимер. Носитель может быть связан со своим полезным грузом посредством ковалентного или нековалентного взаимодействия. Носитель может транспортировать нуклеиновые кислоты, например, РНК или ДНК, к клеткам-мишеням. Носитель может (в некоторых вариантах осуществления изобретения) представлять собой катионный компонент.
Катионный компонент: Понятие «катионный компонент», как правило, относится к заряженной молекуле, которая является положительно заряженной (катион) при значении рН, составляющем, как правило, от 1 до 9, предпочтительно при значении рН, равном или ниже 9 (например, от 5 до 9), равном или ниже 8 (например, от 5 до 8), равном или ниже 7 (например, от 5 до 7), наиболее предпочтительно при физиологическом значении рН, например, от 7,3 до 7.4. Таким образом, катионный компонент может представлять собой любое положительно заряженное соединение или полимер, предпочтительно катионный пептид или белок, который является положительно заряженным в физиологических условиях, прежде всего в физиологических условиях in vivo. «Катионный пептид или белок» может содержать по меньшей мере одну положительно заряженную аминокислоту, или более чем одну положительно заряженную аминокислоту, например, выбранную из Arg, His, Lys или Orn. Следовательно, «поликатионные» соединения также подпадают под указанное определение, если они несут более одного положительного заряда в указанных условиях.
5'-Кэп: 5'-Кэп представляет собой структуру, как правило, модифицированную нуклеотидную структуру, которая обычно «завершает» 5'-конец зрелой мРНК. 5'-кэп может, как правило, быть образован модифицированным нуклеотидом, прежде всего производным гуанинового нуклеотида. Предпочтительно 5'-кэп связан с 5'-концом через 5'-5'-трифосфатную связь. 5'-кэп может быть метилирован, например, может представлять собой m7GpppN, где N обозначает концевой 5'-нуклеотид нуклеиновой кислоты, несущей 5'-кэп, как правило, 5'-конец РНК. Другими примерами структур 5'-кэпа являются глицерил, инвертированная дезоксигруппа (фрагмент), лишенная азотистого основания, 4',5'-метиленовой нуклеотид, 1-(бета-D-эритрофуранозильный) нуклеотид, 4'-тионуклеотид, карбоциклический нуклеотид, 1,5-ангидрогекситольный нуклеотид, L-нуклеотиды, альфа-нуклеотид, нуклеотид с модифицированным основанием, треопентофуранозильный нуклеотид, ациклический 3',4'-секонуклеотид, ациклический 3,4-дигидроксибутильный нуклеотид, ациклический 3,5-дигидроксипентильный нуклеотид, 3'-3'-инвертированный нуклеотидный фрагмент, 3'-3'-инвертированный лишенный азотистого основания фрагмент, 3'-2'-инвертированный нуклеотидный фрагмент, 3'-2'-инвертированный лишенный азотистого основания фрагмент, 1,4-бутандиолфосфатный, 3'-фосфороамидатный, гексилфосфатный, аминогексилфосфатный, 3'-фосфатный, 3'-фосфоротиоатный, фосфородитиоатный или связанный мостиком или несвязанный мостиком метилфосфонатный фрагмент.
Клеточный иммунитет/клеточный иммунный ответ: Клеточный иммунитет относится, как правило, к активации макрофагов, естественных клеток-киллеров (NK), антигенспецифических цитотоксических Т-лимфоцитов и высвобождению различных цитокинов в ответ на антиген. В более общем смысле клеточный иммунитет основан не на антителах, а на активации клеток иммунной системы. Как правило, клеточный иммунный ответ может характеризоваться, например, активацией антигенспецифических цитотоксических Т-лимфоцитов, которые способны индуцировать апоптоз клеток, например, специфических иммунных клеток типа дендритных клеток или других клеток, экспонирующих эпитопы чужих генов на своей поверхности. Такие клетки могут представлять собой инфицированные вирусом или инфицированные внутриклеточными бактериями, или раковые клетки, экспонирующие опухолевые антигены. Другими характеристиками могут являться активация макрофагов и естественных клеток-киллеров, позволяющая им разрушать патогены, и стимуляция клеток к секреции различных цитокинов, которые влияют на функцию других клеток, участвующих в адаптивных иммунных ответах и врожденных иммунных ответах.
ДНК: ДНК является общепринятым сокращением дезоксирибонуклеиновой кислоты. Она представляет собой молекулу нуклеиновой кислоты, т.е. полимер, состоящий из нуклеотидов. Указанные нуклеотиды, как правило, представляют собой мономеры дезоксиаденозинмонофосфата, дезокситимидинмонофосфата, дезоксигуанозинмонофосфата и дезоксицитидинмонофосфата, которые сами состоят из сахарного фрагмента (дезоксирибоза), фрагмента, представляющего собой основание, и фосфатного фрагмента, и образуют полимеры с характерной каркасной структурой. Каркасная структура, как правило, образована с помощью фосфодиэфирных связей между сахарным фрагментом нуклеотида, т.е. дезоксирибозы, первого мономера и фосфатным фрагментом второго смежного мономера. Специфический порядок мономеров, т.е. порядок оснований, сцепленных с сахарным/фосфатным каркасом, называют ДНК-последовательностью. ДНК может быть одноцепочечной или двухцепочечной. В двухцепочечной форме нуклеотиды первой цепи, как правило, гибридизуются с нуклеотидами второй цепи, например, путем спаривания оснований А/Т и спаривания оснований G/C.
Эпитоп: Эпитопы (которые называют также «антигенными детерминантами») можно подразделить на Т-клеточные эпитопы и В-клеточные эпитопы. В контексте настоящего изобретения Т-клеточные эпитопы или части белков могут содержать фрагменты, предпочтительно имеющие длину от примерно 6 до примерно 20 аминокислот или даже более, например, фрагменты, процессируемые и презентируемые молекулами ГКГС класса I, предпочтительно имеют длину от примерно 8 до примерно 10 аминокислот, например, 8, 9 или 10, (или даже 11 или 12 аминокислот), или фрагменты, процессируемые и презентируемые молекулами ГКГС класса II, предпочтительно имеют длину примерно 13 аминокислот или более, например, 13, 14, 15, 16, 17, 18, 19, 20 или даже большее количество аминокислот, при этом указанные фрагменты можно выбирать из любой части аминокислотной последовательности. Указанные фрагменты, как правило, распознаются Т-клетками в форме комплекса, состоящего из пептидного фрагмента и молекулы ГКГС, т.е. фрагменты, как правило, не распознаются в их нативной форме. В-клеточные эпитопы, как правило, представляют собой фрагменты, локализованные на внешней поверхности (нативных) белковых или пептидных антигенов, указанных в настоящем описании, предпочтительно состоящих из 5-15 аминокислот, более предпочтительно состоящих из 5-12 аминокислот, еще более предпочтительно состоящих из 6-9 аминокислот, которые могут распознаваться антителами, т.е. в их нативной форме.
Такие эпитопы белков или пептидов можно, кроме того, выбирать из любых указанных вариантов таких белков или пептидов. В этом контексте антигенные детерминанты могут представлять собой конформационные или прерывистые эпитопы, состоящие из сегментов указанных в настоящем описании белков или пептидов, которые расположены с перерывами в аминокислотной последовательности белков или пептидов, указанных в настоящем описании, но которые находятся вблизи друг от друга в трехмерной структуре, или непрерывные или линейные эпитопы, состоящие из одной полипептидной цепи.
Фрагмент последовательности: фрагмент последовательности, как правило, представляет собой укороченный участок полноразмерной последовательности, например, молекулы нуклеиновой кислоты или аминокислотной последовательности. Следовательно, фрагмент, как правило, состоит из последовательности, которая идентична соответствующему сегменту полноразмерной последовательности. В контексте настоящего изобретения предпочтительный фрагмент последовательности состоит из непрерывного сегмента, содержащего такие элементы, как нуклеотиды или аминокислоты, который соответствует непрерывному состоящему из указанных элементов сегменту молекулы, из которой происходит фрагмент, длина которого составляет по меньшей мере 30%, более предпочтительно по меньшей мере 40%, более предпочтительно по меньшей мере 50%, еще более предпочтительно по меньшей мере 60%, еще более предпочтительно по меньшей мере 70%, и наиболее предпочтительно по меньшей мере 80% от всей (т.е. полноразмерной) молекулы, из которой происходит фрагмент.
G/C-модифицированная: G/C-модифицированная нуклеиновая кислота, как правило, может представлять собой нуклеиновую кислоту, предпочтительно молекулу РНК, указанную в настоящем описании, созданную на основе модифицированной последовательности дикого типа, которая содержит предпочтительно повышенное количество гуанозиновых и/или цитозиновых нуклеотидов по сравнению с последовательностью дикого типа. Для получения указанного повышенного количества можно осуществлять замену кодонов, содержащих аденозиновые или тимидиновые нуклеотиды, на кодоны, содержащие гуанозиновые или цитозиновые нуклеотиды. При повышенном содержании G/C в кодирующей области ДНК или РНК, при заменах можно использовать вырожденность генетического кода. Таким образом, замены кодонов предпочтительно не изменяют кодируемые аминокислотные остатки, но исключительно увеличивают содержание G/C в молекуле нуклеиновой кислоты.
Генная терапия: Генная терапия, как правило, может представлять собой лечение организма пациента или выделенных элементов организма пациента, например, выделенных тканей/клеток, с помощью нуклеиновых кислот, кодирующих пептид или белок. Как правило, она может включать по меньшей мере одну из следующих стадий а) введение нуклеиновой кислоты, предпочтительно молекулы РНК, указанной в настоящем описании, непосредственно пациенту - любым путем введения - либо in vitro в выделенные клетки/ткани пациента, что приводит к трансфекции клеток пациента, либо in vivo/ex vivo, либо in vitro; б) транскрипция и/или трансляция интродуцированной молекулы нуклеиновой кислоты; и необязательно в) повторное введение выделенных трансфектированных клеток пациенту, если нуклеиновую кислоту не вводили непосредственно пациенту.
Генетическая вакцинация: Генетическая вакцинация, как правило, может представлять собой вакцинацию, осуществляемую путем введения молекулы нуклеиновой кислоты, кодирующей антиген или иммуноген или его фрагмент. Молекулу нуклеиновой кислоты можно вводить в организм индивидуума или в выделенные клетки индивидуума. После трансфекции определенных клеток организма или после трансфекции выделенных клеток антиген или иммуноген может экспрессироваться указанными клетками и затем презентоваться иммунной системе, вызывая адаптивный, т.е. антигенспецифический иммунный ответ. Таким образом, генетическая вакцинация, как правило, включает по меньшей мере одну из следующих стадий: а) введение нуклеиновой кислоты, предпочтительно искусственной молекулы нуклеиновой кислоты, указанной в настоящем описании, индивидууму, предпочтительно пациенту, или в выделенные клетки индивидуума, предпочтительно пациента, что обычно приводит к трансфекции клеток индивидуума in vivo или in vitro; б) транскрипцию и/или трансляцию интродуцированной молекулы нуклеиновой кислоты; и необязательно в) повторное введение выделенных трансфектированных клеток индивидууму, предпочтительно пациенту, если нуклеиновую кислоту не вводили непосредственно пациенту.
Гетерологичная последовательность: Как правило, считается, что две последовательности являются «гетерологичными», если они не происходят из одного и того же гена. Это означает, что, хотя гетерологичные последовательности могут происходить из одного и того же организма, они в естественных условиях (в природе) не встречаются в одной и той же молекуле, например, в одной и той же мРНК.
Гуморальный иммунитет/гуморальный иммунный ответ: Гуморальный иммунитет, как правило, относится к производству антител и необязательно к второстепенным процессам, которые могут сопровождать его. Гуморальный иммунный ответ, как правило, может характеризоваться, например, активацией Th2 и производством цитокинов, образованием зародышевого центра и переключением изотипа, созреванием аффинности и образованием клеток памяти. Гуморальный иммунитет, как правило, может относиться также к эффекторным функциям антител, включая нейтрализацию патогенов и токсинов, классическую активацию комплемента и стимулирование опсонинами фагоцитоза и элиминации патогенов.
Иммуноген: В контексте настоящего изобретения под иммуногеном, как правило, понимают соединение, которое обладает способностью стимулировать иммунный ответ. Предпочтительно иммуноген представляет собой пептид, полипептид или белок. В наиболее предпочтительном варианте осуществления изобретения иммуноген в контексте настоящего изобретения представляет собой продукт трансляции применяемой молекулы нуклеиновой кислоты, предпочтительно искусственной молекулы нуклеиновой кислоты, указанной в настоящем описании. Как правило, иммуноген вызывает по меньшей мере адаптивный иммунный ответ.
Иммуностимулирующая композиция: В контексте изобретения иммуностимулирующая композиция, как правило, может представлять собой композицию, содержащую по меньшей мере один компонент, который обладает способностью индуцировать иммунный ответ или из которого может происходить компонент, обладающий способностью индуцировать иммунный ответ. Такой иммунный ответ предпочтительно может представлять собой врожденный иммунный ответ или комбинацию адаптивного и врожденного иммунного ответа. В контексте настоящего описания иммуностимулирующая композиция предпочтительно содержит по меньшей мере одну искусственную молекулу нуклеиновой кислоты, более предпочтительно РНК, например, молекулу мРНК. Иммуностимулирующий компонент, такой как мРНК, может находиться в составе комплекса с пригодным носителем. Таким образом, иммуностимулирующая композиция может содержать комплекс мРНК/носитель. Кроме того, иммуностимулирующая композиция может содержать адъювант и/или пригодный наполнитель для иммуностимулирующего компонента, такого как мРНК.
Иммунный ответ: Иммунный ответ может, как правило, представлять собой либо специфическую реакцию адаптивной иммунной системы на конкретный антиген (так называемый специфический или адаптивный иммунный ответ), либо неспецифическую реакцию врожденной иммунной системы (так называемый неспецифический или врожденный иммунный ответ).
Иммунная система: Иммунная система может защищать организмы от инфекции. Если патогену удалось преодолеть физический барьер организма и попасть в указанный организм, то врожденная иммунная система обеспечивает немедленный, но неспецифический ответ. Если патоген ускользает от этого врожденного ответа, у позвоночных имеется второй уровень защиты, представляющий собой адаптивную иммунную систему. В этом случае иммунная система адаптирует свой ответ в процессе инфекции для улучшения распознавания патогена. Затем этот улучшенный ответ сохраняется после элиминации патогена в форме иммунологической памяти и позволяет адаптивной иммунной системе организовывать более быстрые и сильные атаки каждый раз при проникновении указанного патогена. Таким образом, иммунная система включает врожденную и адаптивную иммунную систему. Каждая из этих двух частей, как правило, включает так называемые гуморальный и клеточный компоненты.
Иммуностимулирующая РНК: Иммуностимулирующая РНК (isPHK) в контексте изобретения, как правило, может представлять собой РНК, которая обладает способностью индуцировать врожденный иммунный ответ. Обычно она не имеет открытой рамки считывания и, таким образом, не кодирует пептид-антиген или иммуноген, но вызывает иммунный ответ, например, посредством связывания со специфическим типом Толл-подобного рецептора (TLR) или другими приемлемыми рецепторами. Однако, естественно, также и мРНК, имеющие открытую рамку считывания и кодирующие пептид/белок, могут индуцировать врожденный иммунный ответ и, следовательно, могут являться иммуностимулирующими РНК.
Врожденная иммунная система: Врожденная иммунная система, которую называют также неспецифической иммунной системой, как правило, включает клетки и механизмы, которые неспецифически защищают хозяина от заражения другими организмами. Это означает, что клетки врожденной системы могут распознавать патогены и реагировать на них обычным путем, но, в отличие от адаптивной иммунной системы, она не обеспечивает долговременный или защитный иммунитет хозяину. Врожденная иммунная система может, например, активироваться лигандами патоген-ассоциированных распознающих молекулярные паттерны (РАМР) рецепторов, например, Толл-подобных рецепторов (TLR), или другими вспомогательными субстанциями, такими как липополисахариды, TNF-альфа, лиганд CD40 или цитокины, монокины, лимфокины, интерлейкины или хемокины, IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11, IL-12, IL-13, IL-14, IL-15, IL-16, IL-17, IL-18, IL-19, IL-20, IL-21, IL-22, IL-23, IL-24, IL-25, IL-26, IL-27, IL-28, IL-29, IL-30, IL-31, IL-32, IL-33, IFN-альфа, IFN-бета, IFN-гамма, GM-CSF, G-CSF, M-CSF, LT-бета, TNF-альфа, факторы роста и hGH, лиганд человеческого Толл-подобного рецептора TLR1, TLR2, TLR3, TLR4, TLR5, TLR6, TLR7, TLR8, TLR9, TLR10, лиганд мышиного Толл-подобного рецептора TLR1, TLR2, TLR3, TLR4, TLR5, TLR6, TLR7, TLR8, TLR9, TLR10, TLR11, TLR12 или TLR13, лиганд NOD-подобного рецептора, лиганд RIG-I-подобного рецептора, иммуностимулирующая нуклеиновая кислота, иммуностимулирующая РНК (isPHK), CpG-ДНК, антибактериальный агент или противовирусный агент. Как правило, ответ врожденной иммунной системы включает рекрутинг иммунных клеток к областям заражения посредством производства химических факторов, включая специализированные химические медиаторы, которые называют цитокинами; активацию каскада комплемента; идентификацию и удаление чужих субстанций, присутствующих в органах, тканях, крови и лимфе, специализированными лейкоцитами; активацию адаптивной иммунной системы посредством процесса, известного как презентация антигена; и/или функционирование в качестве физического и химического барьера для инфекционных агентов.
Сайт клонирования: Под сайтом клонирования, как правило, понимают сегмент молекулы нуклеиновой кислоты, пригодный для встраивания нуклеотидной последовательности, например, нуклеотидной последовательности, содержащей открытую рамку считывания. Встраивание можно осуществлять с помощью любого метода молекулярной биологии, известного специалисту в данной области, например, посредством рестрикции и лигирования. Сайт клонирования, как правило, содержит один или несколько сайтов, распознаваемых рестриктазами (сайты рестрикции). Указанные один или несколько сайтов рестрикции могут распознаваться рестриктазами, расщепляющими ДНК в этих сайтах. Сайт клонирования, который содержит более одного сайта рестрикции, можно обозначать также как множественный сайт клонирования (MCS) или полилинкер.
Молекула нуклеиновой кислоты: Молекула нуклеиновой кислоты представляет собой молекулу, которая содержит, предпочтительно состоит из компонентов нуклеиновой кислоты. Понятие «молекула нуклеиновой кислоты» предпочтительно относится к молекулам ДНК или РНК. Его предпочтительно применяют в качестве синонима понятия «полинуклеотид». Предпочтительно молекула нуклеиновой кислоты представляет собой полимер, содержащий или состоящий из нуклеотидных мономеров, которые ковалентно связаны друг с другом фосфодиэфирными связями сахарного/фосфатного каркаса. Понятие «нуклеиновая кислота» включает также модифицированные молекулы нуклеиновой кислоты, такие как молекулы ДНК или РНК с модифицированным основанием, модифицированным сахарным фрагментом или модифицированным каркасом.
Открытая рамка считывания: Открытая рамка считывания (ОРС) в контексте изобретения, как правило, может представлять собой состоящую из нескольких нуклеотидных триплетов последовательность, которая может транслироваться в пептид или белок. Открытая рамка считывания предпочтительно содержит на своем 5'-конце стартовый кодон, т.е. комбинацию трех последовательно расположенных нуклеотидов, кодирующих, как правило, аминокислоту метионин (ATG или AUG), и следующую за ним область, длина которой, как правило, кратна 3 нуклеотидам. ОРС предпочтительно оканчивается стоп-кодоном (например, ТАА, TAG, TGA). Как правило, в открытой рамке считывания присутствует только один стоп-кодон. Таким образом, открытая рамка считывания в контексте настоящего изобретения предпочтительно представляет собой нуклеотидную последовательность, состоящую из кратного трем количества нуклеотидов, которая начинается стартовым кодоном (например, ATG или AUG), и которая предпочтительно заканчивается стоп-кодоном (например, ТАА, TGA или TAG, или UAA, UAG, UGA соответственно). Открытая рамка считывания может быть выделена или она может быть встроена в более длинную нуклеотидную последовательность, например, в вектор или мРНК. Открытую рамку считывания можно называть также «кодирующей белок областью».
Пептид: Пептид или полипептид, как правило, представляет собой полимер, состоящий из аминокислотных мономеров, связанных пептидными связями. Он, как правило, содержит менее 50 мономерных звеньев. Однако понятие «пептид» не исключает молекулы, имеющие более 50 мономерных звеньев. Длинные пептиды называют также полипептидами, как правило, они имеют от 50 до 600 мономерных звеньев.
Фармацевтически эффективное количество: В контексте изобретения под фармацевтически эффективным количеством, как правило, понимают количество, достаточное для того, чтобы индуцировать фармацевтическое действие, такое как иммунный ответ, измененный патологический уровень экспрессируемого пептида или белка, или замену недостающего генного продукта, например, в случае наличия патологической ситуации.
Белок: Белок, как правило, содержит один или несколько пептидов или полипептидов. Белок, как правило, уложен с образованием 3-мерной структуры, которая может требоваться для проявления биологической функции белка.
Поли(А)-последовательность: Поли(А)-последовательность, которую называют также поли(А)-хвостом или 3'-поли(А)-хвостом, как правило, представляет собой последовательность адениновых нуклеотидов, содержащую, например, вплоть до примерно 400 аденозиновых нуклеотидов, например, от примерно 25 до примерно 400, предпочтительно от примерно 50 до примерно 400, более предпочтительно от примерно 50 до примерно 300, еще более предпочтительно от примерно 50 до примерно 250, наиболее предпочтительно от примерно 60 до примерно 250 аденозиновых нуклеотидов. Поли(А)-последовательность, как правило, локализована на 3'-конце мРНК. В контексте настоящего изобретения поли(А)-последовательность может быть локализована внутри мРНК или любой другой молекулы нуклеиновой кислоты, например, внутри вектора, например, вектора, служащего в качестве матрицы для создания РНК, предпочтительно мРНК, например, при транскрипции вектора.
Полиаденилирование: Под полиаденилированием, как правило, понимают добавление поли(А)-последовательности к молекуле нуклеиновой кислоты, такой как молекула РНК, например, незрелая мРНК. Полиаденилирование может индуцироваться так называемым сигналом полиаденилирования. Указанный сигнал предпочтительно локализован в сегменте нуклеотидов на 3'-конце молекулы нуклеиновой кислоты, такой как молекула РНК, подлежащая полиаденилированию. Сигнал полиаденилирования, как правило, содержит гексамер, состоящий из адениновых и/или урациловых/тиминовых нуклеотидов, предпочтительно последовательность гексамера AAUAAA. Также возможны другие последовательности, предпочтительно гексамерные последовательности. Полиаденилирование, как правило, имеет место во время процессинга пре-мРНК (которую называют также незрелой мРНК). Как правило, созревание РНК (превращение пре-мРНК в зрелую мРНК) включает стадию полиаденилирования.
Сайт рестрикции: Сайт рестрикции, который называют также как сайт, распознаваемый рестриктазой, представляет собой нуклеотидную последовательность, распознаваемую рестрктазой. Сайт рестрикции, как правило, представляет собой короткую, предпочтительно палиндромную нуклеотидную последовательность, например, последовательность, содержащую от 4 до 8 нуклеотидов. Сайт рестрикции предпочтительно специфически распознается рестриктазой. Рестриктаза, как правило, расщепляет нуклеотидную последовательность, содержащую сайт рестрикции, в этом сайте. В двухцепочечной нуклеотидной последовательности, такой как двухцепочечная последовательность ДНК, рестриктаза, как правило, расщепляет обе цепи нуклеотидной последовательности.
РНК, мРНК: РНК является общепринятым сокращением для рибонуклеиновой кислоты. Она представляет собой молекулу нуклеиновой кислоты, т.е. полимер, состоящий из нуклеотидов. Указанные нуклеотиды представляют собой, как правило, мономеры аденозинмонофосфата, уридинмонофосфата, гуанозинмонофосфата и цитидинмонофосфата, соединенные друг с другом вдоль так называемого каркаса. Каркас формируется с помощью фосфодиэфирных связей между сахаром, т.е. рибозой, первого мономера и фосфатным фрагментом второго смежного мономера. Конкретную последовательность мономеров называют РНК-последовательностью. Как правило, РНК можно получать путем транскрипции ДНК-последовательности, например, внутри клетки. В эукариотических клетках транскрипция, как правило, происходит в ядре или в митохондрии. In vivo транскрипция ДНК, как правило, приводит к образованию так называемой незрелой РНК, которая в результате процессинга должна превратиться в так называемую матричную (информационную) РНК, которую сокращенно обозначают как мРНК. Процессинг незрелой РНК, например, в эукариотических организмах, включает ряд различных пост-транскрипционных модификаций, таких как сплайсинг, 5'-кэпирование, полиаденилирование, экспорт из ядра или митохондрии и т.п. Совокупность указанных процессов называют также созреванием РНК. Зрелая матричная РНК, как правило, представляет собой нуклеотидную последовательность, которая может транслироваться в аминокислотную последовательность конкретного пептида или белка. Как правило, зрелая мРНК содержит 5'-кэп, 5'UTR, открытую рамку считывания, необязательно 3'UTR и поли(А)-последовательность. Помимо матричной РНК существует несколько некодирующих типов РНК, которые могут участвовать в регуляции транскрипции и/или трансляции.
Последовательность молекулы нуклеиновой кислоты: Под последовательностью молекулы нуклеиновой кислоты, как правило, понимают конкретный и индивидуальный порядок расположения, т.е. последовательный ряд входящих в нее нуклеотидов. Под последовательностью белка или пептида, как правило, понимают порядок расположения, т.е. последовательный ряд входящих в него аминокислот.
Идентичность последовательности: Две или большее количество последовательностей являются идентичными, если они имеют одинаковую длину и порядок расположения нуклеотидов или аминокислот. Процент идентичности, как правило, характеризует степень идентичности двух последовательностей, т.е. он, как правило, характеризует процент нуклеотидов, которые соответствуют по своему положению идентичным нуклеотидам референс-последовательности. При определении степени идентичности считается, что подлежащие сравнению последовательности имеют одну и ту же длину, т.е. длину наиболее длинной последовательности из подлежащих сравнению последовательностей. Это означает, что первая последовательность, состоящая из 8 нуклеотидов, идентична на 80% второй последовательности, состоящей из 10 нуклеотидов, которая содержит первую последовательность. Другими словами, в контексте настоящего изобретения идентичность последовательностей предпочтительно относится к проценту нуклеотидов в последовательности, которые находятся в одних и тех же положениях в двух или большем количестве последовательностей, имеющих одинаковую длину. При сравнительном анализе первичной структуры последовательностей бреши обычно рассматривают как неидентичные положения безотносительно к их действительному положению.
Стабилизированная молекула нуклеиновой кислоты: Стабилизированная молекула нуклеиновой кислоты представляет собой молекулу нуклеиновой кислоты, предпочтительно молекулу ДНК или РНК, модифицированную таким образом, чтобы она обладала большей стабильностью к расщеплению или разрушению, например, факторами окружающей среды, или ферментативному расщеплению, такому как расщепление экзо- или эндонуклеазами, чем молекула нуклеиновой кислоты без модификации. Предпочтительно стабилизированная молекула нуклеиновой кислоты в контексте настоящего изобретения является стабилизированной в клетке, такой как прокариотическая или эукариотическая клетка, предпочтительно в клетке млекопитающего, такой как человеческая клетка. Стабилизирующее действие можно оказывать также вне клеток, например, в буферном растворе и т.д., например, в процессе производства фармацевтической композиции, которая содержит стабилизированную молекулу нуклеиновой кислоты.
Трансфекция: Понятие «трансфекция» относится к интродукции молекул нуклеиновой кислоты, таких как молекулы ДНК или РНК (например, мРНК), в клетки, предпочтительно в эукариотические клетки. В контексте настоящего изобретения под понятие «трансфекция» подпадает любой известный специалисту в данной области метод интродукции молекул нуклеиновой кислоты в клетки, предпочтительно в эукариотические клетки, такие как клетки млекопитающих. Такие методы включают, например, электропорацию, липофекцию, например, на основе катионных липидов и/или липосом, осаждение фосфатом кальция, трансфекцию на основе наночастиц, трансфекцию на основе вирусов или трансфекцию на основе катионных полимеров, таких как ДЭАЭ-декстран или полиэтиленимин, и т.д. Предпочтительно интродукцию осуществляют без использования вирусов.
Вакцина: Под вакциной, как правило, понимают предназначенный для профилактики или терапии продукт, содержащий по меньшей мере один антиген, предпочтительно иммуноген. Антиген или иммуноген может иметь происхождение из любого материала, пригодного для вакцинации. Например, антиген или иммуноген может происходить из патогена, такого как бактерии или вирусные частицы и т.д., или из опухолевой или раковой ткани. Антиген или иммуноген стимулирует адаптивную иммунную систему организма вырабатывать адаптивный иммунный ответ.
Вектор: Понятие «вектор» относится к молекуле нуклеиновой кислоты, предпочтительно к искусственной молекуле нуклеиновой кислоты. В контексте настоящего изобретения вектор можно применять для встраивания или хранения требуемой нуклеотидной последовательности, такой как нуклеотидная последовательность, содержащая открытую рамку считывания. Таким векторы могут представлять собой векторы для хранения, экспрессионные векторы, клонирующие векторы, векторы для переноса и т.д. Вектор для хранения представляет собой вектор, который позволяет удобно хранить молекулу нуклеиновой кислоты, например, молекулу мРНК. Так, вектор может содержать последовательность, соответствующую, например, требуемой последовательности мРНК или ее части, такой как последовательность, соответствующая открытой рамке считывания и 3'-UTR мРНК. Экспрессионный вектор можно применять для производства продуктов экспрессии, таких как РНК, например, мРНК, или пептиды, полипептиды или белки. Например, экспрессионный вектор может содержать последовательности, необходимые для транскрипции сегмента последовательности вектора, такой как промоторная последовательность, например, промоторная последовательность РНК-полимеразы. Клонирующий вектор, как правило, представляет собой вектор, который содержит сайт клонирования, который можно применять для встраивания нуклеотидных последовательностей в вектор. Клонирующий вектор может представлять собой, например, плазмидный вектор или вектор на основе бактериофага. Вектор для переноса может представлять собой вектор, который пригоден для переноса молекул нуклеиновой кислоты в клетки или организмы, например, вирусные векторы. В контексте настоящего изобретения вектор может представлять собой, например, РНК-вектор или ДНК-вектор. Предпочтительно вектор представляет собой молекулу ДНК. В контексте настоящего описания предпочтительно вектор содержит сайт клонирования, маркер для селекции, такой как фактор, обусловливающий устойчивость к антибиотикам, и последовательность, пригодную для размножения вектора, такую как сайт инициации репликации. В контексте настоящего описания предпочтительно вектор представляет собой плазмидный вектор.
Наполнитель: Под наполнителем, как правило, понимают материал, пригодный для хранения, транспортирования и/или введения соединения, такого как фармацевтически активное соединение. Он может представлять собой, например, физиологически приемлемую жидкость, пригодную для хранения, транспортирования и/или введения соединения, такого как фармацевтически активное соединение.
3'-нетранслируемая область (3'UTR): 3'UTR, как правило, является частью искусственной молекулы нуклеиновой кислоты, которая локализована с 3'-стороны (т.е. в прямом направлении) относительно открытой рамки считывания и которая не транслируется в белок. Как правило, 3'UTR представляет собой часть мРНК, которая локализована между кодирующей белок областью (открытой рамкой считывания (ОРС) или кодирующей последовательностью (CDS)) и поли(А)-последовательностью мРНК. В контексте изобретения понятие «3'UTR» может включать также элементы, которые не кодируются в матрице, с которой транскрибируется РНК, но которые добавляются после транскрипции в процессе созревания, например, поли(А)-последовательность. 3'UTR мРНК не транслируется в аминокислотную последовательность. Последовательность 3'UTR, как правило, кодируется геном, который транскрибируется в соответствующую мРНК в процессе экспрессии гена. Геномная последовательность сначала транскрибируется в незрелую мРНК, которая содержит необязательные интроны. Незрелая мРНК затем дополнительно процессируется с образованием зрелой мРНК в процессе созревания. Указанный процесс созревания включает стадии 5'-кэпирования, сплайсинга незрелой мРНК с вырезанием необязательных интронов и модификаций 3'-конца, таких как полиаденилирование 3'-конца незрелой мРНК, и необязательные расщепления эндо-/или экзонуклеазами и т.д. В контексте настоящего изобретения 3'UTR соответствует последовательности зрелой мРНК, которая локализована между стоп-кодоном кодирующей области белка, предпочтительно непосредственно примыкает с 3'-стороны к стоп-кодону кодирующей белок области, и поли(А)-последовательностью мРНК. Понятие «соответствует» означает, что последовательность 3'UTR может представлять собой последовательность РНК, такую как в последовательности мРНК, указанной в определении последовательности 3'UTR, или последовательность ДНК, которая соответствует указанной последовательности РНК. В контексте настоящего изобретения понятие «3'UTR гена», например, «3'UTR гена рибосомального белка», означает последовательность, которая соответствует 3'UTR зрелой мРНК, полученной из указанного гена, т.е. мРНК, полученной путем транскрипции гена и созревания незрелой мРНК. Понятие «3'UTR гена» включает последовательность ДНК и последовательность РНК (как смысловую, так и антисмысловую цепи и как зрелые, так и незрелые последовательности) 3'UTR.
5'-нетранслируемая область (5'UTR): Под 5'UTR, как правило, понимают конкретный сегмент матричной РНК (мРНК). Она локализована с 5'-стороны относительно открытой рамки считывания мРНК. Как правило, 5'UTR начинается с сайта инициации транскрипции и заканчивается за один нуклеотид до стартового кодона открытой рамки считывания. 5'UTR может содержать элементы, предназначенные для контроля генной экспрессии, так называемые регуляторные элементы. Указанные регуляторные элементы могут представлять собой, например, сайты связывания рибосом. 5'UTR может подвергаться посттранскрипционным модификациям, например, путем добавления 5'-кэпа. В контексте настоящего изобретения 5'UTR соответствует последовательности зрелой мРНК, локализованной между 5'-кэпом и стартовым кодоном. Предпочтительно 5'UTR соответствует последовательности, которая простирается от нуклеотида, расположенного с 3'-стороны 5'-кэпа, предпочтительно от расположенного с 3'-стороны нуклеотида, непосредственно примыкающего к 5'-кэпу, до нуклеотида, расположенного с 5'-стороны стартового кодона кодирующей белок области, предпочтительно до нуклеотида, непосредственно примыкающего с 5'-стороны к стартовому кодону кодирующей белок области. Нуклеотид, непосредственно примыкающий с 3'-стороны к 5'-кэпу зрелой мРНК, как правило, соответствует сайту инициации транскрипции. Понятие «соответствует» означает, что последовательность 5'UTR может представлять собой последовательность РНК, такую как в последовательности мРНК, указанной при определении последовательности 5'UTR, или последовательность ДНК, которая соответствует указанной последовательности РНК. В контексте настоящего изобретения понятие «5'UTR гена», например, «5'UTR ТОР-гена», обозначает последовательность, которая соответствует 5'UTR зрелой мРНК, полученной из этого гена, т.е. мРНК, полученной путем транскрипции гена и созревания незрелой мРНК. Понятие «5'UTR гена» включает последовательность ДНК и последовательность РНК 5'UTR. В вариантах осуществления изобретения указанная 5'-UTR может находиться на 5'-конце ОРС. Ее длина, как правило, составляет менее 500, 400, 300, 250 или менее 200 нуклеотидов. В других вариантах осуществления изобретения ее длина может находиться в пределах по меньшей мере от 10, 20, 30 или 40, предпочтительно вплоть до 100 или 150 нуклеотидов.
5'-концевой олигопиримидиновый тракт (ТОР): 5'-концевой олигопиримидиновый тракт (ТОР), как правило, обозначает сегмент пиримидиновых нуклеотидов, локализованный в 5'-концевой области молекулы нуклеиновой кислоты, например, в 5'-концевой области некоторых молекул мРНК или в 5'-концевой области функционального элемента, например, транскрибируемой области некоторых генов. Последовательность начинается с цитидина, который, как правило, соответствует сайту инициации транскрипции, и за которым следует сегмент, как правило, состоящий примерно из 3-30 пиримидиновых нуклеотидов. Например, ТОР может содержать 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30 или даже большее количество нуклеотидов. Пиримидиновый сегмент и, следовательно, 5'-ТОР, заканчивается одним нуклеотидом, расположенным в 5'-направлении относительно первого пуринового нуклеотида, локализованного в прямом направлении относительно ТОР. Матричную РНК, которая содержит 5'-концевой олигопиримидиновый тракт, часто обозначают как ТОР-мРНК. Таким образом, гены, из которых получают указанные матричные РНК, обозначают как ТОР-гены. Последовательности ТОР, например, обнаружены в генах и мРНК, кодирующих пептидные факторы элонгации и рибосомальные белки.
ТОР-мотив: В контексте настоящего изобретения ТОР-мотив обозначает нуклеотидную последовательность, которая соответствует 5'-ТОР, описанному выше. Таким образом, ТОР-мотив в контексте настоящего изобретения предпочтительно обозначает состоящий из пиримидиновых нуклеотидов сегмент длиной 3-30 нуклеотидов. Предпочтительно ТОР-мотив состоит по меньшей мере из 3 пиримидиновых нуклеотидов, предпочтительно по меньшей мере 4 пиримидиновых нуклеотидов, предпочтительно по меньшей мере 5 пиримидиновых нуклеотидов, более предпочтительно по меньшей мере 6 нуклеотидов, более предпочтительно по меньшей мере 7 нуклеотидов, наиболее предпочтительно по меньшей мере 8 пиримидиновых нуклеотидов, при этом сегмент пиримидиновых нуклеотидов предпочтительно начинается на его 5'-конце с цитозинового нуклеотида. В ТОР-генах и ТОР-мРНК ТОР-мотив предпочтительно начинается на его 5'-конце с сайта инициации транскрипции и заканчивается на расстоянии одного нуклеотида в 5'-направлении относительно первого пуринового остатка в указанном гене или мРНК. В контексте настоящего изобретения ТОР-мотив предпочтительно локализован на 5'-конце последовательности, которая представляет собой 5'UTR, или на 5'-конце последовательности, которая кодирует 5'UTR. Таким образом, в контексте настоящего изобретения сегмент из 3 или большего количества пиримидиновых нуклеотидов предпочтительно называют «ТОР-мотивом», если сегмент локализован на 5'-конце соответствующей последовательности, такой как мРНК, предлагаемая в изобретении, 5'UTR-элeмeнт мРНК, предлагаемой в изобретении, или нуклеотидная последовательность, которая происходит из 5'UTR ТОР-гена, указанного в настоящем описании. Другими словами, сегмент, состоящий из 3 или большего количества пиримидиновых нуклеотидов, который не локализован на 5'-конце 5'UTR или 5'UTR-элемента, но локализован где-либо внутри 5'UTR или 5'UTR-элемента, предпочтительно не обозначают как «ТОР-мотив».
ТОР-ген: ТОР-гены, как правило, характеризуются присутствием 5'-концевого олигопримидинового тракта. Кроме того, большинство ТОР-генов характеризуются связанной с ростом регуляцией трансляции. Однако известны также ТОР-гены с тканеспецифической регуляцией трансляции. Как указано выше, 5'UTR ТОР-гена соответствует последовательности 5'UTR зрелой мРНК, происходящей из ТОР-гена, которая предпочтительно простирается от нуклеотида, примыкающего с 3'-стороны к 5'-кэпу, до нуклеотида, примыкающего с 5'-стороны к стартовому кодону. 5'UTR ТОР-гена, как правило, не содержит никаких стартовых кодонов, предпочтительно не содержит расположенных в обратном направлении AUG (uAUG) или расположенных в обратном направлении открытых рамок считывания (иОРС). В данном контексте под расположенными в обратном направлении AUG и расположенными в обратном направлении открытыми рамками считывания, как правило, понимают AUG и открытые рамки считывания, которые находятся в 5'-направлении относительно стартового кодона (AUG) открытой рамки считывания, которая должна транслироваться. 5'UTR ТОР-генов, как правило, являются относительно короткими. Длины 5'UTR ТОР-генов могут варьироваться от 20 нуклеотидов вплоть до 500 нуклеотидов и, как правило, они содержат менее чем примерно 200 нуклеотидов, предпочтительно менее чем примерно 150 нуклеотидов, более предпочтительно менее чем примерно 100 нуклеотидов. Примерами 5'UTR ТОР-генов в контексте настоящего изобретения являются нуклеотидные последовательности, простирающиеся от нуклеотида в положении 5 до нуклеотида, непосредственно примыкающего к 5'-концу стартового кодона (например, ATG), указанные последовательности представлены в SEQ ID NO: 1-1363, SEQ ID NO: 1395, SEQ ID NO: 1421 и SEQ ID NO: 14221-1363 заявки на патент PCT/EP 2012/002448 WO 2013/143700 или представляют собой их гомологи или варианты, описание указанных документов включено в настоящее описание в качестве ссылки. В этом контексте наиболее предпочтительным фрагментом 5'UTR ТОР-гена является 5'UTR ТОР-гена без 5'ТОР-мотива. Понятие «5'UTR ТОР-гена» предпочтительно относится к 5'UTR встречающегося в естественных условиях ТОР-гена.
Первый объект настоящего изобретения относится к искусственной молекуле нуклеиновой кислоты, содержащей
а. по меньшей мере одну открытую рамку считывания (ОРС); и
б. по меньшей мере один элемент 3'-нетранслируемой области (3'-UTR-элемент), содержащий или состоящий из нуклеотидной последовательности, которая происходит из 3'-UTR гена рибосомального белка.
Понятие «3'-UTR-элeмeнт» относится к нуклеотидной последовательности, которая содержит или состоит из нуклеотидной последовательности, происходящей из 3'-UTR или из варианта 3'-UTR. Понятие «3'-UTR-элeмeнт» предпочтительно относится к нуклеотидной последовательности, которая представляет собой 3'-UTR искусственной нуклеотидной последовательности, такой как искусственная мРНК, или которая кодирует 3'-UTR искусственной молекулы нуклеиновой кислоты. Следовательно, в контексте настоящего изобретения 3'-UTR-элeмeнт предпочтительно может представлять собой 3'-UTR мРНК, предпочтительно искусственной мРНК, или может представлять собой матрицу для транскрипции 3'-UTR мРНК. Так, 3'-UTR-элeмeнт предпочтительно представляет собой нуклеотидную последовательность, которая соответствует 3'-UTR мРНК, предпочтительно 3'-UTR искусственной мРНК, такой как мРНК, полученная в результате транскрипции созданной генетическими методами векторной конструкции. Предпочтительно 3'-UTR-элемент в контексте настоящего изобретения функционирует в качестве 3'-UTR или кодирует нуклеотидную последовательность, которая выполняет функцию 3'-UTR.
Понятие «ген рибосомального белка», как правило, относится к гену, кодирующему рибосомальный белок. В контексте настоящего описания это понятие относится к любому гену рибосомального белка независимо от вида, из которого он происходит. Конкретно, понятие относится к гену эукариотического рибосомального белка. Кроме того, в контексте изобретения понятие «ген рибосомального белка» может относиться также к гену, сходному с геном рибосомального белка либо с точки зрения структуры, либо с точки зрения функции. В частности, понятие включает «подобные гену рибосомального белка», псевдогены и гены, обладающие характеристиками последовательности или структуры, прежде всего в их 3'-UTR-oблacти, присущими гену рибосомального белка. Предпочтительно понятие относится к гену рибосомального белка позвоночных, более предпочтительно к гену рибосомального белка млекопитающих, еще более предпочтительно к гену рибосомального белка приматов, прежде всего к гену рибосомального белка человека. Кроме того, понятие «ген рибосомального белка» включает также ген рибосомального белка грызунов, прежде всего ген мышиного рибосомального белка. Примерами генов рибосомального белка в контексте изобретения являются (но, не ограничиваясь только ими) гены рибосомального белка L9 (RPL9), рибосомального белка L3 (RPL3), рибосомального белка L4 (RPL4), рибосомального белка L5 (RPL5), рибосомального белка L6 (RPL6), рибосомального белка L7 (RPL7), рибосомального белка L7a (RPL7A), рибосомального белка L11 (RPL11), рибосомального белка L12 (RPL12), рибосомального белка L13 (RPL13), рибосомального белка L23 (RPL23), рибосомального белка L18 (RPL18), рибосомального белка L18a (RPL18A), рибосомального белка L19 (RPL19), рибосомального белка L21 (RPL21), рибосомального белка L22 (RPL22), рибосомального белка L23a (RPL23A), рибосомального белка L17 (RPL17), рибосомального белка L24 (RPL24), рибосомального белка L26 (RPL26), рибосомального белка L27 (RPL27), рибосомального белка L30 (RPL30), рибосомального белка L27a (RPL27A), рибосомального белка L28 (RPL28), рибосомального белка L29 (RPL29), рибосомального белка L31 (RPL31), рибосомального белка L32 (RPL32), рибосомального белка L35a (RPL35A), рибосомального белка L37 (RPL37), рибосомального белка L37a (RPL37A), рибосомального белка L38 (RPL38), рибосомального белка L39 (RPL39), рибосомального белка большой субъединицы, Р0 (RPLP0), рибосомального белка большой субъединицы, Р1 (RPLP1), рибосомального белка, большой субъединицы, Р2 (RPLP2), рибосомального белка S3 (RPS3), рибосомального белка S3A (RPS3A), рибосомального белка S4, Х-сцепленного (RPS4X), рибосомального белка S4, Y-сцепленного 1 (RPS4Y1), рибосомального белка S5 (RPS5), рибосомального белка S6 (RPS6), рибосомального белка S7 (RPS7), рибосомального белка S8 (RPS8), рибосомального белка S9 (RPS9), рибосомального белка S10 (RPS10), рибосомального белка S11 (RPS11), рибосомального белка S12 (RPS12), рибосомального белка S13 (RPS13), рибосомального белка S15 (RPS15), рибосомального белка S15a (RPS15A), рибосомального белка S16 (RPS16), рибосомального белка S19 (RPS19), рибосомального белка S20 (RPS20), рибосомального белка S21 (RPS21), рибосомального белка S23 (RPS23), рибосомального белка S25 (RPS25), рибосомального белка S26 (RPS26), рибосомального белка S27 (RPS27), рибосомального белка S27a (RPS27a), рибосомального белка S28 (RPS28), рибосомального белка S29 (RPS29), рибосомального белка L15 (RPL15), рибосомального белка S2 (RPS2), рибосомального белка L14 (RPL14), рибосомального белка S14 (RPS14), рибосомального белка L10 (RPL10), рибосомального белка L10a (RPL10A), рибосомального белка L35 (RPL35), рибосомального белка L13a (RPL13A), рибосомального белка L36 (RPL36), рибосомального белка L36a (RPL36A), рибосомального белка L41 (RPL41), рибосомального белка S18 (RPS18), рибосомального белка S24 (RPS24), рибосомального белка L8 (RPL8), рибосомального белка L34 (RPL34), рибосомального белка S17 (RPS17), рибосомального белка SA (RPSA), продукта 1 слияния: остаток убиквитина А-52-рибосомальный белок (UBA52), повсеместно экспрессируемому гену (FAU) вируса мышиной саркомы Финкеля-Бискиса-Рейли (FBR-MuSV), гену рибосомального белка L22-подобного 1 (RPL22L1), рибосомального белка S17 (RPS17), рибосомального белка L39-подобного (RPL39L), рибосомального белка L10-подобного (RPL10L), рибосомального белка L36а-подобного (RPL36AL), рибосомального белка L3-подобного (RPL3L), рибосомального белка S27-подобного (RPS27L), рибосомального белка L26-подобного 1 (RPL26L1), рибосомального белка L7-подобного 1 (RPL7L1), псевдогена рибосомального белка L13a (RPL13AP), псевдогена 8 рибосомального белка L37a (RPL37AP8), псевдогена 5 рибосомального белка S10 (RPS10P5), псевдогена рибосомального белка 11 S26 (RPS26P11), псевдогена 5 рибосомального белка L39 (RPL39P5), псевдогена 6 рибосомального белка большой субъединицы Р0 (RPLP0P6) и псевдогена 14 рибосомального белка L36 (RPL36P14). Предпочтительно понятие «ген рибосомального белка» относится к одному из вышеуказанных генов, которые происходят из млекопитающих, предпочтительно из Homo sapiens или Mus muscuius.
Предпочтительно по меньшей мере одна открытая рамка считывания и по меньше мере один 3'-UTR-элемент являются гетерологичными. В контексте настоящего описания понятие «гетерологичный» означает, что два элемента последовательности, содержащиеся в искусственной молекуле нуклеиновой кислоты, такие как открытая рамка считывания и 3'-UTR-элeмeнт, не встречаются в естественных условиях (в природе) в рассматриваемой комбинации. Предпочтительно 3'-UTR-элeмeнт происходит из гена, отличного от гена, из которого происходит открытая рамка считывания. Например, ОРС может происходить из гена, отличного от гена, из которого происходит 3'-UTR-элемент, например, кодирующего другой белок или такой же белок, но из другого вида, и т.д. Предпочтительно открытая рамка считывания не кодирует рибосомальный белок. В предпочтительном варианте осуществления изобретения ОРС не кодирует человеческий рибосомальный белок или растительный рибосомальный белок (прежде всего, из Arabidopsis), прежде всего человеческий рибосомальный белок S6 (RPS6), белок, подобный человеческому рибосомальному белку L36a (RPL36AL) или рибосомальный белок S16 Arabidopsis (RPS16). В другом предпочтительном варианте осуществления изобретения открытая рамка считывания (ОРС) не кодирует рибосомальный белок S6 (RPS6), белок, подобный рибосомальному белку L36a (RPL36AL) или рибосомальный белок S16 (RPS16).
В конкретных вариантах осуществления изобретения предпочтительно открытая рамка считывания не кодирует репортерный белок, например, выбранный из группы, состоящей из глобиновых белков (прежде всего, бета-глобина), белка люциферазы, белков GFP или их вариантов, например, вариантов, последовательности которых идентичны по меньшей мере на 70% последовательности белка глобина, белка люциферазы или белка GFP. В наиболее предпочтительном варианте осуществления изобретения открытая рамка считывания (ОРС) не кодирует репортерный ген или не происходит из репортерного гена, где репортерный ген предпочтительно не выбирают из группы, состоящей из (генов) глобиновых белков (прежде всего бета-глобина), белка люциферазы, бета-глюкуронидазы (GUS) и белков GFP или их вариантов, предпочтительно не выбирают из EGFP или из вариантов любого из вышеуказанных генов, последовательности которых, как правило, идентичны по меньшей мере на 70% последовательности любого из вышеуказанных репортерных генов, предпочтительно генов белка глобина, белка люциферазы или белка GFP.
Еще более предпочтительно 3'-UTR-элемент является гетерологичным любому другому элементу, содержащемуся в искусственной нуклеиновой кислоте, представленной в настоящем описании. Например, если искусственная нуклеиновая кислота, предлагаемая в изобретении, содержит 3'-UTR-элемент из рассматриваемого гена, то она предпочтительно не содержит никакой другой нуклеотидной последовательности, прежде всего никакой функциональной нуклеотидной последовательности (например, элемента кодирующей или регуляторной последовательности) из этого же гена, включая его регуляторные последовательности на 5'- и 3'-конце ОРС гена. В наиболее предпочтительном варианте осуществления изобретения искусственная молекула нуклеиновой кислоты содержит ОРС, 3'UTR и 5'-UTR, которые все являются гетерологичными друг другу, например, они являются рекомбинантными, поскольку каждый из указанных элементов происходит из различных генов (и их 5'- и 3'-UTR). В другом предпочтительном варианте осуществления изобретения 3'-UTR не происходит из 3'-UTR вирусного гена или происходит не из вируса.
Предпочтительно по меньшей мере один 3'-UTR-элeмeнт функционально связан с ОРС. Это означает, что предпочтительно 3'-UTR-элeмeнт ассоциирован с ОРС таким образом, что он может осуществлять функцию, такую как функция повышения или стабилизации экспрессии кодируемого пептида или белка, или функцию стабилизации искусственной молекулы нуклеиновой кислоты. Предпочтительно ОРС и 3'-UTR-элемент ассоциированы в направлении 5'→3'. Таким образом, предпочтительно искусственная молекула нуклеиновой кислоты содержит структуру 5'-ОРС-(необязательно)-линкер-3'-UTR-элемент-3', в которой линкер может присутствовать или отсутствовать. Например, линкер может состоять из одного или нескольких нуклеотидов, например, представлять собой сегмент, состоящий из 1-50 или 1-20 нуклеотидов, например, содержащий или состоящий из одного или нескольких сайтов, распознаваемых рестриктазами (сайтов рестрикции).
Предпочтительно по меньшей мере один 3'-UTR-элемент содержит нуклеотидную последовательность, происходящую из 3'-UTR гена эукариотического рибосомального белка, предпочтительно из 3'-UTR гена рибосомального белка позвоночных, более предпочтительно из 3'-UTR гена рибосомального белка млекопитающих, еще более предпочтительно из 3'-UTR гена рибосомального белка приматов, прежде всего гена рибосомального белка человека, или из 3'-UTR гена рибосомального белка грызунов, прежде всего гена мышиного рибосомального белка.
В предпочтительном варианте осуществления изобретения по меньшей мере один 3'-UTR-элeмeнт содержит или соответствует нуклеотидной последовательности, которая происходит из последовательности 3'-UTR транскрипта, выбранного из группы, состоящей из NM_000661.4, NM_001024921.2, NM_000967.3, NM_001033853.1, NM_000968.3, NM_000969.3, NM_001024662.1, NM_000970.3, NM_000971.3, NM_000972.2, NM_000975.3, NM_001199802.1, NM_000976.3, NM_000977.3, NM_033251.2, NM_001243130.1, NM_001243131, NM_000978.3, NM_000979.3, NM_001270490.1, NM_000980.3, NM_000981.3, NM_000982.3, NM_000983.3, NM_000984.5, NM_000985.4, NM_001035006.2, NM_001199340.1, NM_001199341.1, NM_001199342.1, NM_001199343.1, NM_001199344.1, NM_001199345.1, NM_000986.3, NM_000987.3, NM_000988.3, NM_000989.3, NM_000990.4, NM_001136134.1, NM_000991.4, NM_001136135.1, NM_001136136.1, NM_001136137.1, NM_000992.2, NM_000993.4, NM_001098577.2, NM_001099693.1, NM_000994.3, NM_001007073.1, NM_001007074.1, NM_000996.2, NM_000997.4, NM_000998.4, NM_000999.3, NM_001035258.1, NM_001000.3, NM_001002.3, NM_053275.3, NM_001003.2, NM_213725.1, NM_001004.3, NM_001005.4, NM_001256802.1, NM_001260506.1, NM_001260507.1, NM_001006.4, NM_001267699.1, NM_001007.4, NM_001008.3, NM_001009.3, NM_001010.2, NM_001011.3, NM_001012.1, NM_001013.3, NM_001203245.2, NM_001014.4, NM_001204091.1, NM_001015.4, NM_001016.3, NM_001017.2, NM_001018.3, NM_001030009.1, NM_001019.4, NM_001020.4, NM_001022.3, NM_001146227.1, NM_001023.3, NM_001024.3, NM_001025.4, NM_001028.2, NM_001029.3, NM_001030.4, NM_002954, NM_001135592.2, NM_001177413.1, NM_001031.4, NM_001032.4, NM_001030001.2, NM_002948.3, NM_001253379.1, NM_001253380.1, NM_001253382.1, NM_001253383.1, NM_001253384.1, NM_002952.3, NM_001034996.2, NM_001025071.1, NM_001025070.1, NM_005617.3, NM_006013.3, NM_001256577.1, NM_001256580.1, NM_007104.4, NM_007209.3, NM_012423.3, NM_001270491.1, NM_033643.2, NM_015414.3, NM 021029.5, NM_001199972.1, NM_021104.1, NM_022551.2, NM_033022.3, NM_001142284.1, NM_001026.4, NM_001142285.1, NM_001142283.1, NM_001142282.1, NM_000973.3, NM_033301.1, NM_000995.3, NM_033625.2, NM_001021.3, NM_002295.4, NM_001012321.1, NM_001033930.1, NM_003333.3, NM_001997.4, NM_001099645.1, NM_001021.3, NM_052969.1, NM_080746.2, NM_001001.4, NM_005061.2, NM_015920.3, NM_016093.2, NM_198486.2, NG_011172.1, NG_011253.1, NG_000952.4, NR_002309.1, NG_010827.2, NG_009952.2, NG_009517.1, NM_052835.3, NM_011287.2, NM_001162933.1, NM_009076.3, NM_009438.5, NM_025974.2, NM_025586.3, NM_001002239.3, NM_009077.2, NM_029751.4, NM_009078.2, NM_019647.6, NM_009079.3, NM_022891.3, NM_024218.4, NM_011975.3, NM_009081.2, NM_009082.2, NM_009083.4, NM_053257.3, ENSMUST00000081840 (NM_172086.2), NM_026724.2, NM_025592.3, NM_025589.4, NM_026069.3, NM_009084, NM_026055.1. NM_026594.2, NM_001163945.1, NM_024212.4, NM_016980.2, NM_011290.5, NM_011291.5, ENSMUST00000102898 (NM_013721.3), NM_025433.3, NM_012053,2. NM_011292.2, NM_007475.5, NM_018853.3, NM_026020.6, NM_025963.3, NM_013725.4, NM_011295.6, NM_020600.4, NM_009091.2, NM_170669.2, NM_013647.2, NM_009092.3, NM_008503.5, NM_026147.5 / ENSMUST00000138502, NM_207635.1, NM_024266.3, NM_013765.2, NM_024277.2, NM_026467.3, NM_012052.2, NM_016959.4, ENSMUST00000071745, NM_009095.2, NM_009096.3, NM_011300.3, NM_011029.4, NM_018860.4, NM_001277113.1, NM_001277114.1, NM_001271590.1, NM_007990, NM_025919, NM_016738, NM_026517, NM_207523, NM_009080, NM_011289, NM_013762, NM 021338, NM_018730, NM_019865, NM_023372.2, NM_026533.3, NM_009092, NM_011296, NM_023133, ENSMUST00000059080 (NM_025587.2), NM_024175, NM_027015, NM_016844, NM_009093.2, NM_009094, NM_009098, NM_029767 и NM_019883.
Фраза «нуклеотидная последовательность, которая происходит из 3'-UTR гена рибосомального белка» предпочтительно относится к нуклеотидной последовательности, основой которой является последовательность 3'-UTR гена рибосомального белка или ее фрагмент или участок. Указанная фраза включает последовательности, соответствующие полной последовательности 3'-UTR, т.е. полноразмерной последовательности 3'-UTR гена рибосомального белка, и последовательности, соответствующие фрагменту последовательности 3'-UTR гена рибосомального белка. Предпочтительно фрагмент 3'-UTR гена рибосомального белка состоит из непрерывного сегмента нуклеотидов, соответствующего непрерывному сегменту нуклеотидов полноразмерной последовательности 3'-UTR гена рибосомального белка, который включает по меньшей мере 5%, 10%, 20%, предпочтительно по меньшей мере 30%, более предпочтительно по меньшей мере 40%, более предпочтительно по меньшей мере 50%, еще более предпочтительно по меньшей мере 60%, еще более предпочтительно по меньшей мере 70%, еще более предпочтительно по меньшей мере 80% и наиболее предпочтительно по меньшей мере 90% полноразмерной последовательности 3'-UTR гена рибосомального белка. В контексте настоящего изобретения такой фрагмент предпочтительно представляет собой функциональный фрагмент, представленный в настоящем описании. Предпочтительно фрагмент сохраняет функцию регулирования трансляции ОРС, сцепленной с 3'-UTR или его фрагментом. Понятие «3'-UTR гена рибосомального белка» предпочтительно относится к 3'-UTR встречающегося в естественных условиях гена рибосомального белка.
Понятия «вариант 3'-UTR гена рибосомального белка» и «его вариант» в контексте 3'-UTR гена рибосомального белка относятся к варианту 3'-UTR встречающегося в естественных условиях гена рибосомального белка, предпочтительно к варианту 3'-UTR гена рибосомального белка позвоночных, более предпочтительно к варианту 3'-UTR гена рибосомального белка млекопитающих, еще более предпочтительно к варианту 3'-UTR варианту 3'-UTR гена рибосомального белка приматов, прежде всего гена рибосомального белка человека, представленного выше. Такой вариант может представлять собой модифицированный 3'-UTR гена рибосомального белка. Например, вариант 3'-UTR может иметь одну или несколько делеций, инсерций, добавлений и/или замен нуклеотидов по сравнению с встречающейся в естественных условиях 3'-UTR, из которой происходит вариант. Предпочтительно вариант 3'-UTR гена рибосомального белка по меньшей мере на 40%, предпочтительно по меньшей мере на 50%, более предпочтительно по меньшей мере на 60%, более предпочтительно по меньшей мере на 70%, еще более предпочтительно по меньшей мере на 80%, еще более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% идентичен встречающейся в естественных условиях 3'-UTR, из которой происходит вариант. Предпочтительно вариант представляет собой функциональный вариант, представленный в настоящем описании.
Фраза «нуклеотидная последовательность, которая происходит из варианта 3'-UTR гена рибосомального белка» предпочтительно относится к нуклеотидной последовательности, основой которой является вариант последовательности 3'-UTR гена рибосомального белка или его фрагмент или участок, представленный выше. Фраза включает последовательности, соответствующие полной последовательности варианта 3'-UTR гена рибосомального белка, т.е. полноразмерной последовательности варианта 3'-UTR гена рибосомального белка, и последовательности, соответствующие фрагменту варианта последовательности 3'-UTR гена рибосомального белка. Предпочтительно фрагмент варианта 3'-UTR гена рибосомального белка состоит из непрерывного сегмента нуклеотидов, соответствующего непрерывному сегменту нуклеотидов полноразмерного варианта последовательности 3'-UTR гена рибосомального белка, который включает по меньшей мере 20%, предпочтительно по меньшей мере 30%, более предпочтительно по меньшей мере 40%, более предпочтительно по меньшей мере 50%, еще более предпочтительно по меньшей мере 60%, еще более предпочтительно по меньшей мере 70%, еще более предпочтительно по меньшей мере 80%, и наиболее предпочтительно по меньшей мере 90% полноразмерного варианта 3'-UTR гена рибосомального белка. В контексте настоящего изобретения такой фрагмент варианта предпочтительно представляет собой функциональный фрагмент варианта, представленного в настоящем описании.
В контексте настоящего описания понятия «функциональный вариант» «функциональный фрагмент» и «функциональный фрагмент варианта» (который обозначают также как «функциональный вариантный фрагмент») означают, что фрагмент 3'-UTR, вариант 3'-UTR или фрагмент варианта 3'-UTR гена рибосомального белка выполняет по меньшей мере одну, предпочтительно более чем одну функцию встречающейся в естественных условиях 3'-UTR гена рибосомального белка, из которой происходит вариант, фрагмента или фрагмент варианта. Такая функция может заключаться, например, в стабилизации мРНК и/или увеличении, стабилизации и/или пролонгировании производства белка с мРНК, и/или увеличении экспрессии белка или общего производства белка с мРНК, предпочтительно в клетке млекопитающего, такой как человеческая клетка. Предпочтительно функция 3'-UTR касается трансляции белка, кодируемого ОРС. Более предпочтительно функция включает повышение эффективности трансляции ОРС, связанной с 3'-UTR или ее фрагментом или вариантом. В контексте настоящего изобретения наиболее предпочтительно, чтобы вариант, фрагмент и вариант фрагмента выполнял функцию стабилизации мРНК, предпочтительно в клетке млекопитающего, такой как человеческая клетка, по сравнению с мРНК, которая содержит референс-3'-UTR или в которой отсутствует 3'-UTR, и/или функцию увеличения, стабилизации и/или пролонгирования производства белка с мРНК, предпочтительно в клетке млекопитающего, такой как человеческая клетка, по сравнению с мРНК, которая содержит референс-3'-UTR или в которой отсутствует 3'-UTR, и/или функцию увеличения производства белка с мРНК, предпочтительно в клетке млекопитающего, такой как человеческая клетка, по сравнению с мРНК, которая содержит референс-3'-UTR или в которой отсутствует 3'-UTR. Референс-3'-UTR может представлять собой, например, 3'-UTR, встречающуюся в естественных условиях в комбинации с ОРС. Кроме того, функциональный вариант, функциональный фрагмент или функциональный вариант фрагмента 3'-UTR гена рибосомального белка предпочтительно не оказывает существенного понижающего действия на эффективность трансляции мРНК, которая содержит такой вариант, фрагмент или вариант фрагмента 3'-UTR, по сравнению с 3'-UTR дикого типа, из которой происходит вариант, фрагмент или вариант фрагмента. В контексте настоящего изобретения наиболее предпочтительная функция «функционального варианта», «функционального фрагмента» или «функционального варианта фрагмента» 3'-UTR гена рибосомального белка заключается в увеличении, стабилизации и/или пролонгировании производства белка посредством экспрессии мРНК, несущей функциональный вариант, функциональный фрагмент или функциональный вариант фрагмента, представленный выше.
Предпочтительно эффективность одной или нескольких функций, осуществляемых функциональным вариантом, функциональным фрагментом или функциональным вариантом фрагмента, такую как эффективность стабилизации производства мРНК и/или белка, и/или эффективность увеличения производства белка, повышают по меньшей мере на 5%, более предпочтительно по меньшей мере на 10%, более предпочтительно по меньшей мере на 20%, более предпочтительно по меньшей мере на 30%, более предпочтительно по меньшей мере на 40%, более предпочтительно по меньшей мере на 50%, более предпочтительно по меньшей мере на 60%, еще более предпочтительно по меньшей мере 70%, еще более предпочтительно по меньшей мере 80%, наиболее предпочтительно по меньшей мере на 90% по сравнению с эффективностью стабилизации производства мРНК и/или белка, и/или эффективностью увеличения производства белка, обеспечиваемой встречающейся в естественных условиях 3'-UTR гена рибосомального белка, из которого происходит вариант, фрагмент или вариант фрагмента.
В контексте настоящего изобретения фрагмент 3'-UTR гена рибосомального белка или вариант 3'-UTR гена рибосомального белка предпочтительно имеет длину по меньшей мере примерно 3 нуклеотида, предпочтительно по меньшей мере примерно 5 нуклеотидов, более предпочтительно по меньшей мере примерно 10, 15, 20, 25 или 30 нуклеотидов, еще более предпочтительно по меньшей мере примерно 50 нуклеотидов, наиболее предпочтительно по меньшей мере примерно 70 нуклеотидов. Предпочтительно такой фрагмент 3'-UTR гена рибосомального белка или вариант 3'-UTR гена рибосомального белка представляет собой функциональный фрагмент, описанный выше. В предпочтительном варианте осуществления изобретения 3'-UTR гена рибосомального белка или его фрагмент или вариант имеет длину от 3 до примерно 500 нуклеотидов, предпочтительно от 5 до примерно 150 нуклеотидов, более предпочтительно от 10 до 100 нуклеотидов, еще более предпочтительно от 15 до 90, наиболее предпочтительно от 20 до 70 нуклеотидов.
Предпочтительно по меньшей мере один 3'-UTR-элeмeнт искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, содержит или состоит из «функционального фрагмента», «функционального варианта» или «функционального фрагмента варианта» 3'-UTR гена рибосомального белка.
В предпочтительном варианте осуществления изобретения по меньшей мере один 3'-UTR-элeмeнт искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, повышает стабильность искусственной молекулы нуклеиновой кислоты, например, повышает стабильность мРНК, предлагаемой в настоящем изобретении, по сравнению с соответствующей нуклеиновой нуклеиновой референс-кислотой, в которой отсутствует 3'-UTR или которая содержит референс-3'-UTR, такую как 3'-UTR, встречающаяся в естественных условиях в комбинации с ОРС. Предпочтительно, по меньшей мере один 3'-UTR-элeмeнт искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, повышает стабильность производства белка с искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, например, с мРНК, предлагаемой в настоящем изобретении, по сравнению с соответствующей нуклеиновой кислотой, в которой отсутствует 3'-UTR или которая содержит референс-3'-UTR, такую как 3'-UTR, встречающаяся в естественных условиях в комбинации с ОРС. Предпочтительно по меньшей мере один 3'-UTR-элeмeнт искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, пролонгирует производство белка из искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, например, с мРНК, предлагаемой в настоящем изобретении, по сравнению с соответствующей нуклеиновой кислотой, в которой отсутствует 3'-UTR или которая содержит референс-3'-UTR, такую как 3'-UTR, встречающаяся в естественных условиях в комбинации с ОРС. Предпочтительно по меньшей мере один 3'-UTR-элeмeнт искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, повышает экспрессию белка и/или общее производство белка с искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, например, с мРНК, предлагаемой в настоящем изобретении, по сравнению с соответствующей нуклеиновой кислотой, в которой отсутствует 3'-UTR или которая содержит референс-3'-UTR, такую как 3'-UTR, встречающаяся в естественных условиях в комбинации с ОРС. Предпочтительно по меньшей мере один 3'-UTR-элeмeнт искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, не оказывает отрицательного влияния на эффективность трансляции нуклеиновой кислоты по сравнению с эффективностью трансляции соответствующей нуклеиновой кислоты, в которой отсутствует 3'-UTR или которая содержит референс-3'-UTR, такую как 3'-UTR, встречающаяся в естественных условиях в комбинации с ОРС. Еще более предпочтительно с помощью 3'-UTR повышают эффективность трансляции по сравнению с эффективностью трансляции белка, кодируемого соответствующей ОРС в естественных условиях.
Понятие «соответствующая молекула нуклеиновой кислоты» или «референс-молекула нуклеиновой кислоты» в указанном контексте означает, что референс-молекула нуклеиновой кислоты, за исключением различий в 3'-UTR, сопоставима, предпочтительно идентична предлагаемой в настоящем изобретении искусственной молекуле нуклеиновой кислоты, содержащей 3'-UTR-элемент.
Понятие «стабилизирует и/или пролонгирует производство белка» с искусственной молекулы нуклеиновой кислоты, такой как искусственная мРНК, предпочтительно означает, что производство белка с искусственной молекулы нуклеиновой кислоты, такой как искусственная мРНК, стабилизируется и/или пролонгируется по сравнению с производством белка с референс-молекулы нуклеиновой кислоты, такой как референс-мРНК, например, которая содержит референс-3'-UTR или в которой отсутствует 3'-UTR, предпочтительно в экспрессионной системе млекопитающих, такой как HeLa- или HDF-клетки. Так, белок, продуцируемый с искусственной молекулы нуклеиновой кислоты, такой как искусственная мРНК, поддается обнаружению в течение более длительного периода времени, чем время, в течение которого его можно обнаружить в случае белка, продуцируемого с референс-молекулы нуклеиновой кислоты. Иными словами, количество белка, продуцированного с искусственной молекулы нуклеиновой кислоты, такой как искусственная мРНК, измеряемое в зависимости от времени, снижается до уровня, находящегося ниже порогового значения, в более поздний момент времени, чем количество белка, продуцированного с референс-молекулы нуклеиновой кислоты, такой как референс-мРНК, измеряемое в зависимости от времени. Такое пороговое значение может представлять собой, например, количество белка, измеренное на начальной фазе экспрессии, например, через 1, 2, 3, 4, 5 или 6 ч после инициации экспрессии, например, после трансфекции молекулой нуклеиновой кислоты.
Например, производство белка с искусственной молекулы нуклеиновой кислоты, такой как искусственная мРНК, в количествах, равных по меньшей мере количеству, обнаруженному на начальной фазе экспрессии, например, через 1, 2, 3, 4, 5 или 6 ч после инициации экспрессии, например, после трансфекции молекулой нуклеиновой кислоты, пролонгируется по меньшей мере примерно на 5 ч, предпочтительно по меньшей мере примерно на 10 ч, более предпочтительно по меньшей мере примерно на 24 ч по сравнении с производством белка с референс-молекулы нуклеиновой кислоты, такой как референс-мРНК, в экспрессионной системе, такой как клетки млекопитающих, например, в HeLa- или HDF-клетках. Так, искусственная молекула нуклеиновой кислоты, предлагаемая в настоящем изобретении, предпочтительно позволяет пролонгировать производство белка в количестве, которое по меньшей мере равно количеству, обнаруженному на начальной фазе экспрессии, например, через 1, 2, 3, 4, 5 или 6 ч после инициации экспрессии, например, после трансфекции молекулой нуклеиновой кислоты, по меньшей мере примерно на 5 ч, предпочтительно по меньшей мере примерно на 10 ч, более предпочтительно по меньшей мере примерно на 24 ч по сравнении с производством белка с референс-молекулы нуклеиновой кислоты, в которой отсутствует 3'-UTR или которая содержит референс-3'-UTR.
В предпочтительных вариантах осуществления изобретения период производства белка с искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, удлиняют по меньшей мере в 1,5 раза, предпочтительно по меньшей мере в 2 раза, более предпочтительно по меньшей мере в 2,5 раза по сравнению с периодом производства белка с референс-молекулы нуклеиновой кислоты, в которой отсутствует 3'-UTR или которая содержит референс-3'-UTR.
Указанный эффект пролонгирования производства белка можно оценивать на основе (I) измерения количеств белка, например, полученных в результате экспрессии кодируемого репортерного белка, такого как люцифераза, предпочтительно в экспрессионной системе млекопитающих, такой как HeLa-или HDF-клетки, в зависимости от времени, (II) определения момента времени, в который количество белка становится ниже количества белка, обнаруженного, например, через 1, 2, 3, 4, 5 или 6 ч после инициации экспрессии, например, через 1, 2, 3, 4, 5 или 6 ч после трансфекции искусственной молекулой нуклеиновой кислоты, и (III) сравнения момента времени, который количество белка становится ниже количества белка, обнаруженного, например, через 1, 2, 3, 4, 5 или 6 ч после инициации экспрессии, с указанным моментом времени, определенным для молекулы нуклеиновой кислоты, в которой отсутствует 3'-UTR или которая содержит референс-3'-UTR.
Предпочтительно указанное стабилизирующее и/или пролонгирующее действие на производство белка достигают, когда общее количество белка, продуцированное с искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, например, в течение периода времени, составляющего 48 или 72 ч, по меньшей мере равно количеству белка, продуцированного с референс-молекулы нуклеиновой кислоты, в которой отсутствует 3'-UTR или которая содержит референс-3'-UTR, такую как 3'-UTR, встречающаяся в естественных условиях с ОРС искусственной молекулы нуклеиновой кислоты. Так, в настоящем изобретении предложена искусственная молекула нуклеиновой кислоты, которая обеспечивает пролонгированное и/или стабилизированное производство белка в экспрессионной системе млекопитающих, такой как клетки млекопитающих, например, в HeLa- или HDF-клетках, как указано выше, при этом общее количество белка, продуцированного с указанной искусственной молекулы нуклеиновой кислоты, например, в течение периода времени, составляющего 48 или 72 ч, по меньшей мере равно общему количеству белка, продуцированного, например, в течение указанного периода времени, с референс-молекулы нуклеиновой кислоты, в которой отсутствует 3'-UTR или которая содержит референс-3'-UTR, такую как 3'-UTR, встречающаяся в естественных условиях с ORF искусственной молекулы нуклеиновой кислоты.
Так, «стабилизированная экспрессия белка» предпочтительно означает, что имеет место более равномерное производство белка с искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, в течение заранее определенного периода времени, например, в течение 24 ч, более предпочтительно в течение 48 ч, еще более предпочтительно в течение 72 ч, по сравнению с референс-молекулой нуклеиновой кислоты, например, которая содержит референс-3'-UTR или в которой отсутствует 3'-UTR. Следовательно, уровень производства белка, например, в системе млекопитающих, с искусственной молекулы нуклеиновой кислоты, содержащей 3'-UTR-элeмeнт, предлагаемый в настоящем изобретении, например, с мРНК, предлагаемой в настоящем изобретении, предпочтительно не снижается до уровня, обнаруженного для референс-молекулы нуклеиновой кислоты, такой как референс-мРНК, указанная выше. Например, количество белка (кодируемого ОРС), обнаруженное через 6 ч после инициации экспрессии, например, через 6 ч после трансфекции искусственной молекулой нуклеиновой кислоты, предлагаемой в настоящем изобретении, клетки, такой как клетка млекопитающего, может быть сопоставимым с количеством белка, обнаруженного через 48 ч после инициации экспрессии, например, через 48 ч после трансфекции. Так, отношение количества белка, кодируемого ОРС, такого как репортерный белок, например, люцифераза, обнаруженное через 48 ч после инициации экспрессии, например, через 48 после трансфекции, к количеству белка, обнаруженному через 6 ч после инициации экспрессии, например, через 6 ч после трансфекции, предпочтительно составляет по меньшей мере примерно 0,4, более предпочтительно по меньшей мере примерно 0,5, более предпочтительно по меньшей мере примерно 0,6, еще более предпочтительно по меньшей мере примерно 0,7. Для молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении отношение предпочтительно составляет от примерно 0,4 до примерно 4, предпочтительно от примерно 0,65 до примерно 3, более предпочтительно от примерно 0,7 до 2. Для соответствующей референс-молекулы нуклеиновой кислоты, например, мРНК, которая содержит референс-3'-UTR или в которой отсутствует 3'-UTR, указанное отношение может составлять, например, от примерно 0,05 до примерно 0,3.
Так, в настоящем изобретении предложена искусственная молекула нуклеиновой кислоты, содержащая ОРС и 3'-UTR-элeмeнт, описанный выше, при этом отношение количества (репортерного) белка, например, количество люциферазы, обнаруженное через 48 ч после инициации экспрессии, к количеству (репортерного) белка, обнаруженному через 6 ч после инициации экспрессии, предпочтительно в экспрессионной системе млекопитающих, такой как клетки млекопитающих, например, в HeLa-клетках, предпочтительно превышает примерно 0,4, более предпочтительно превышает примерно 0,5, более предпочтительно превышает примерно 0,6, еще более предпочтительно превышает примерно 0,7, например, составляет от примерно 0,4 до примерно 4, предпочтительно от примерно 0,65 до примерно 3, более предпочтительно от примерно 0,7 до 2, при этом предпочтительно общее количество белка, продуцированного с указанной искусственной молекулы нуклеиновой кислоты, например, в течение периода времени, составляющего 48 ч, по меньшей мере равно общему количеству белка, продуцированного, например, в течение указанного периода времени, с референс-молекулы нуклеиновой кислоты, в которой отсутствует 3'-UTR или которая содержит референс-3'-UTR, такую как 3'-UTR, встречающаяся в естественных условиях с ОРС искусственной молекулы нуклеиновой кислоты. В предпочтительном варианте осуществления настоящего изобретения предложена искусственная молекула нуклеиновой кислоты, содержащая ОРС и 3'-UTR-элeмeнт, описанный выше, при этом отношение количества (репортерного) белка, например, количество люциферазы, обнаруженное через 72 ч после инициации экспрессии, к количеству (репортерного) белка, обнаруженному через 6 ч после инициации экспрессии, предпочтительно в экспрессионной системе млекопитающих, такой как клетки млекопитающих, например, в HeLa-клетках, предпочтительно превышает примерно 0,4, более предпочтительно превышает примерно 0,5, более предпочтительно превышает примерно 0,6, еще более предпочтительно превышает примерно 0,7, например, составляет от примерно 0,4 до 1,5, предпочтительно от примерно 0,65 до примерно 1,15, более предпочтительно от примерно 0,7 до 1.0, при этом предпочтительно общее количество белка, продуцированного с указанной искусственной молекулы нуклеиновой кислоты, например, в течение периода времени, составляющего 72 ч, по меньшей мере равно общему количеству белка, продуцированному, например, в течение указанного периода времени, с референс-молекулы нуклеиновой кислоты, в которой отсутствует 3'-UTR или которая содержит 3'-UTR, такую как 3'-UTR, встречающаяся в естественных условиях с ОРС искусственной молекулы нуклеиновой кислоты.
«Увеличенная экспрессия белка» или «повышенная экспрессия белка» в контексте настоящего изобретения предпочтительно обозначает увеличенную/повышенную экспрессию белка в один из моментов времени после инициации экспрессии или увеличенное/повышенное общее количество экспрессированного белка, по сравнению с экспрессией, индуцированной референс-молекулой нуклеиновой кислоты. Так, уровень белка, обнаруженный в определенный момент времени после инициации экспрессии, например, после трансфекции искусственной молекулой нуклеиновой кислоты, предлагаемой в настоящем изобретении, например, после трансфекции мРНК, предлагаемой в настоящем изобретении, например, через 6, 12, 24, 48 или 72 ч после трансфекции, предпочтительно превышает уровень белка, обнаруженный в этот же самый момент времени после инициации экспрессии, например, после трансфекции референс-молекулой нуклеиновой кислоты, такой как референс-мРНК, которая содержит референс-3'-UTR или в которой отсутствует 3'-UTR. В предпочтительном варианте осуществления изобретения максимальное количество белка (определенное, например, по активности или массе белка), экспрессированного с искусственной молекулы нуклеиновой кислоты, увеличивают по сравнению с количеством белка, экспрессированного с нуклеиновой референс-кислоты, которая содержит референс-3'-UTR или в которой отсутствует 3'-UTR. Пиковые уровни экспрессии предпочтительно достигаются в пределах 48 ч, более предпочтительно в пределах 24 ч и еще более предпочтительно в пределах 12 ч после, например, трансфекции.
В одном из вариантов осуществления изобретения «увеличенное общее производство белка» или «повышенное общее производство белка» с искусственной молекулы нуклеиновой кислоты, предлагаемой в изобретении, относится к увеличенному/повышенному производству белка за период времени, в течение которого белок продуцируется с искусственной молекулы нуклеиновой кислоты, предпочтительно в экспрессионной системе млекопитающих, такой как клетки млекопитающих, например, в HeLa- или HDF-клетках, по сравнению с референс-молекулой нуклеиновой кислоты, в которой отсутствует 3'-UTR или которая содержит референс-3'-UTR. Согласно предпочтительному варианту осуществления изобретения кумулятивное количество белка, экспрессированного в течение периода времени, увеличивается, если применяют искусственную молекулу нуклеиновой кислоты, предлагаемую в изобретении.
В изобретении предложена искусственная молекула нуклеиновой кислоты, которая отличается увеличенной экспрессией кодируемого белка по сравнению с соответствующей молекулой нуклеиновой кислоты, в которой отсутствует по меньшей мере один 3'-UTR-элeмeнт или которая содержит 3'-UTR («нуклеиновая референс-кислота»), содержащую нуклеотидную последовательность, происходящую из 3'-UTR гена рибосомального белка или из варианта 3'-UTR гена рибосомального белка. Для оценки in vivo производства белка с предлагаемой в изобретении искусственной молекулы нуклеиновой кислоты, определяют экспрессию кодируемого белка после инъекции/трансфекции предлагаемой в изобретении искусственной молекулой нуклеиновой кислоты клеток-/ткани-мишени и сравнивают с экспрессией белка, индуцированной нуклеиновой референс-кислотой. В данной области известны количественные методы определения экспрессии белка (например, вестерн-блоттинг, FACS, ELISA, масс-спектрометрия). В рассматриваемом контексте наиболее пригодным является определение экспрессии репортерных белков типа люциферазы, зеленого флуоресцентного белка (GFP) или секретированной щелочной фосфатазы (SEAP). Для этого искусственную нуклеиновую кислоту, предлагаемую в изобретении, или нуклеиновую референс-кислоту интродуцируют в ткань-мишень или клетку-мишень, например, посредством трансфекции или инъекции. Через несколько часов или несколько дней (например, через 6, 12, 24, 48 или 72 ч) после инициации экспрессии или после интродукции молекулы нуклеиновой кислоты берут образец клеток-мишеней и проводят измерения с помощью FACS и/или лизируют. После этого лизаты можно использовать для обнаружения экспрессированного белка (и определения тем самым эффективности экспрессии белка) с помощью нескольких методов, например, вестерн-блоттинга, FACS, ELISA, масс-спектрометрии или путем измерения флуоресценции или люминесценции.
Следовательно, если экспрессию белка с искусственной молекулы нуклеиновой кислоты, предлагаемой в изобретении, сравнивают с экспрессией белка с референс-молекулы нуклеиновой кислоты в определенные моменты времени (например, через 6, 12, 24, 48 или 72 ч после инициации экспрессии или после интродукции молекулы нуклеиновой кислоты), то обе молекулы нуклеиновой кислоты интродуцируют по отдельности в ткани-/клетки-мишени, берут образец ткани/клеток в определенный момент времени, приготавливают белковые лизаты согласно конкретному протоколу, скорректированному для конкретного метода обнаружения (например, для вестерн-блоттинга, ELISA и т.д., как это известно в данной области) и проводят обнаружение белка с помощью выбранного метода обнаружения. В качестве альтернативы измерениям количеств экспрессируемого белка в клеточных лизатах, или в дополнение к измерениям количеств экспрессируемого белка в клеточных лизатах, с помощью FACS-анализа можно определять количества белка также до лизиса собранных клеток или параллельно с использованием аликвот.
Если нужно измерять общее количество белка в течение определенного периода времени, то можно собирать ткань или клетки в несколько моментов времени после интродукции искусственной молекулы нуклеиновой кислоты (например, через 6, 12, 24, 48 и 72 ч после инициации экспрессии или после интродукции молекулы нуклеиновой кислоты; как правило, взятой из организма других подопытных животных), и определять количество белка в каждый момент времени как описано выше. Для расчета кумулятивного количества белка можно применять математический метод определения общего количества белка, например, можно определять площадь под кривой (AUC) согласно следующей формуле:
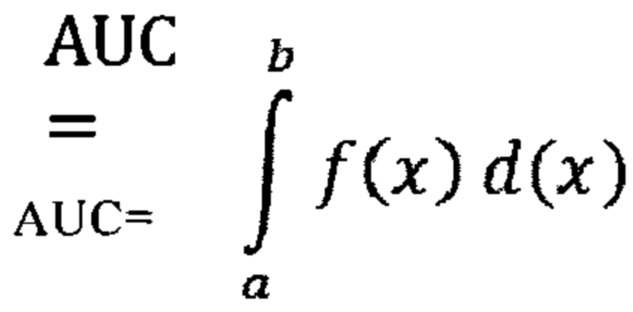
Для расчета площади под кривой для общего количества белка вычисляют интеграл уравнения кривой экспрессии между каждыми конечными точками (a и b).
Таким образом, понятие «общее производство белка» предпочтительно относится к площади под кривой (AUC), описывающей производство белка в зависимости от времени.
Указанное увеличение стабильности искусственной молекулы нуклеиновой кислоты, указанное увеличение стабильности производства белка, указанное пролонгирование производства белка и/или указанное увеличение/повышение экспрессии белка и/или общего производства белка предпочтительно определяют путем сравнения с соответствующей референс-молекулой нуклеиновой кислоты, в которой отсутствует 3'-UTR, например, мРНК, в которой отсутствует 3'-UTR, или референс-молекулой нуклеиновой кислоты, которая содержит референс-3'-UTR, такую как 3'-UTR, встречающуюся в естественных условиях с ОРС, как описано выше.
Стабилизирующее действие на мРНК и/или производство белка и эффективность, и/или повышающее производство белка действие и эффективность вариантов, фрагментов и/или фрагментов вариантов 3'-UTR гена рибосомального белка, а также стабилизирующее действие на мРНК и/или производство белка и эффективность, и/или повышающее производство белка действие и эффективность по меньшей мере одного 3'-UTR-элeмeнтa искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, можно определять с помощью любого пригодного для этой цели метода, известного специалисту в данной области. Например, можно создавать искусственные молекулы мРНК, содержащие кодирующую последовательность/открытую рамку считывания (ОРС) для репортерного белка, такого как люцифераза, и не содержащие 3'-UTR, содержащие 3'-UTR, происходящую из встречающегося в естественных условиях гена рибосомального белка, 3'-UTR, происходящую из референс-гена (т.е. референс-3'-UTR, такую как 3'-UTR, встречающаяся в естественных условиях с ОРС), в качестве 3'-UTR вариант 3'-UTR гена рибосомального белка, в качестве 3'-UTR фрагмент гена встречающегося в естественных условиях рибосомального белка, или в качестве 3'-UTR фрагмент варианта 3'-UTR гена рибосомального белка. Такие мРНК можно создавать, например, путем транскрипции in vitro соответствующих векторов, таких как плазмидные векторы, например, содержащие промотор Т7 и последовательность, кодирующую соответствующие последовательности мРНК. Созданными молекулами мРНК можно трансфектировать клетки с помощью метода трансфекции, пригодного для трансфекции мРНК, например, их можно интродуцировать путем электропорации в клетки млекопитающих, такие как HELA-клетки, и можно анализировать образцы в определенные моменты времени после трансфекции, например, через 6, 24, 48 и 72 ч после трансфекции. Указанные образцы можно анализировать для оценки количеств мРНК и/или количеств белка с помощью методов, хорошо известных специалистам в данной области. Например, количества репортерной мРНК, присутствующей в клетках в моменты времени взятия образцов, можно определять с помощью методов количественной ПЦР. Количества репортерного белка, кодируемого соответствующими мРНК, можно определять, например, с помощью вестерн-блоттинга, ELISA, FACS-анализа или репортерных анализов, таких как анализы люциферазы, в зависимости от используемого репортерного белка. Эффект стабилизации экспрессии белка и/или пролонгирования экспрессии белка можно анализировать, например, путем определения отношения уровня белка, обнаруженного через 48 ч после трансфекции и уровня белка, обнаруженного через 6 ч после трансфекции. Предпочтительно величина такого отношения превышает 1, т.е. экспрессия белка в более поздний момент времени больше, чем экспрессия белка в более ранний момент времени. Если величина такого отношения меньше 1, то белок тем более стабилен, чем ближе указанная величина к 1. Такие измерения можно, конечно, проводить также и через 72 ч или более, и для определения стабильности экспрессии белка можно определять отношение уровня белка, обнаруженного через 72 ч после трансфекции к уровню белка, обнаруженному через 6 ч после трансфекции.
В предпочтительном варианте осуществления изобретения 3'-UTR-элемент искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, происходит из 3'-UTR-области гена, кодирующего рибосомальный белок, предпочтительно из 3'-UTR-области гена рибосомального белка L9 (RPL9), рибосомального белка L3 (RPL3), рибосомального белка L4 (RPL4), рибосомального белка L5 (RPL5), рибосомального белка L6 (RPL6), рибосомального белка L7 (RPL7), рибосомального белка L7a (RPL7A), рибосомального белка L11 (RPL11), рибосомального белка L12 (RPL12), рибосомального белка L13 (RPL13), рибосомального белка L23 (RPL23), рибосомального белка L18 (RPL18), рибосомального белка L18a (RPL18A), рибосомального белка L19 (RPL19), рибосомального белка L21 (RPL21), рибосомального белка L22 (RPL22), рибосомального белка L23a (RPL23A), рибосомального белка L17 (RPL17), рибосомального белка L24 (RPL24), рибосомального белка L26 (RPL26), рибосомального белка L27 (RPL27), рибосомального белка L30 (RPL30), рибосомального белка L27a (RPL27A), рибосомального белка L28 (RPL28), рибосомального белка L29 (RPL29), рибосомального белка L31 (RPL31), рибосомального белка L32 (RPL32), рибосомального белка L35a (RPL35A), рибосомального белка L37 (RPL37), рибосомального белка L37a (RPL37A), рибосомального белка L38 (RPL38), рибосомального белка L39 (RPL39), рибосомального белка, большой субъединицы, Р0 (RPLP0), рибосомального белка, большой субъединицы, Р1 (RPLP1), рибосомального белка, большой субъединицы, Р2 (RPLP2), рибосомального белка S3 (RPS3), рибосомального белка S3A (RPS3A), рибосомального белка S4, Х-сцепленного (RPS4X), рибосомального белка S4, Y-сцепленного 1 (RPS4Y1), рибосомального белка S5 (RPS5), рибосомального белка S6 (RPS6), рибосомального белка S7 (RPS7), рибосомального белка S8 (RPS8), рибосомального белка S9 (RPS9), рибосомального белка S10 (RPS10), рибосомального белка S11 (RPS11), рибосомального белка S12 (RPS12), рибосомального белка S13 (RPS13), рибосомального белка S15 (RPS15), рибосомального белка S15а (RPS15A), рибосомального белка S16 (RPS16), рибосомального белка S19 (RPS19), рибосомального белка S20 (RPS20), рибосомального белка S21 (RPS21), рибосомального белка S23 (RPS23), рибосомального белка S25 (RPS25), рибосомального белка S26 (RPS26), рибосомального белка S27 (RPS27), рибосомального белка S27a (RPS27a), рибосомального белка S28 (RPS28), рибосомального белка S29 (RPS29), рибосомального белка L15 (RPL15), рибосомального белка S2 (RPS2), рибосомального белка L14 (RPL14), рибосомального белка S14 (RPS14), рибосомального белка L10 (RPL10), рибосомального белка L10a (RPL10A), рибосомального белка L35 (RPL35), рибосомального белка L13a (RPL13A), рибосомального белка L36 (RPL36), рибосомального белка L36a (RPL36A), рибосомального белка L41 (RPL41), рибосомального белка S18 (RPS18), рибосомального белка S24 (RPS24), рибосомального белка L8 (RPL8), рибосомального белка L34 (RPL34), рибосомального белка S17 (RPS17), рибосомального белка SA (RPSA) или рибосомального белка S17 (RPS17). В альтернативном варианте осуществления изобретения 3'-UTR-элемент может происходить из гена, кодирующего рибосомальный белок, или из гена, выбранного из генов продукта 1 слияния остаток убиквитина А-52-рибосомальный белок (UBA52), повсеместно экспрессируемого (FAU) вируса мышиной саркомы Финкеля-Бискиса-Рейли, белка 1, подобного рибосомальному белку L22 (RPL22L1), белка, подобного рибосомальному белку L39 (RPL39L), белка, подобного рибосомальному белку L10 (RPL10L), белка, подобного рибосомальному белку L36a (RPL36AL), белка, подобного рибосомальному белку L3 (RPL3L), белка, подобного рибосомальному белку S27 (RPS27L), белка 1, подобного рибосомальному белку L26 1 (RPL26L1), белка 1, подобного рибосомальному белку L7 (RPL7L1), псевдогена рибосомального белка L13а (RPL13AP), псевдогена 8 рибосомального белка L37a (RPL37AP8), псевдогена 5 рибосомального белка S10 (RPS10P5), псевдогена 11 рибосомального белка S26 (RPS26P11), псевдогена 5 рибосомального белка L39 (RPL39P5), псевдогена 6 рибосомального белка, большой субъединицы, Р0 (RPLP0P6) и псевдогена 14 рибосомального белка L36 (RPL36P14). Кроме того, 3'-UTR-элемент искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, предпочтительно происходит из 3'-UTR-oблacти гена, выбранного из группы, состоящей из генов белка, подобного рибосомальному белку S4 (RPS41), предполагаемой субъединицы 60S рибосомального белка L13а, предполагаемой субъединицы 60S белка, подобного рибосомальному белку L37a, предполагаемой субъединицы 40S белка, подобного рибосомальному белку S10, предполагаемой субъединицы 40S белка 1, подобного рибосомальному белку S26, предполагаемой субъединицы 60S белка 5, подобного рибосомальному белку L39, или субъединицы 60S белка, подобного кислому рибосомальному белку Р0.
В наиболее предпочтительном варианте осуществления изобретения 3'-UTR-элемент искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, происходит из 3'-UTR-oблacти гена, кодирующего рибосомальный белок, предпочтительно из 3'-UTR-oблacти гена рибосомального белка L3 (RPL3), рибосомального белка L11 (RPL11), рибосомального белка L13 (RPL13), рибосомального белка L23 (RPL23), рибосомального белка L23a (RPL23A), рибосомального белка L26 (RPL26), рибосомального белка L27 (RPL27), рибосомального белка L35a (RPL35A), рибосомального белка L38 (RPL38), рибосомального белка S4, Х-сцепленного (RPS4X), рибосомального белка S8 (RPS8), рибосомального белка S9 (RPS9), рибосомального белка S13 (RPS13), рибосомального белка S19 (RPS19), рибосомального белка S21 (RPS21), рибосомального белка S23 (RPS23), рибосомального белка S27 (RPS27), рибосомального белка S28 (RPS28), рибосомального белка S29 (RPS29), рибосомального белка L36 (RPL36), рибосомального белка L36a (RPL36A), рибосомального белка S18 (RPS18) или рибосомального белка S17 (RPS17). В другом предпочтительном варианте осуществления изобретения 3'-UTR-элeмeнт может происходить из гена, кодирующего рибосомальный белок, или из гена, выбранного из генов продукта 1 слияния остаток убиквитина А-52-рибосомальный белок (UBA52), повсеместно экспрессируемого (FAU) вируса мышиной саркомы Финкеля-Бискиса-Рейли или белка 1, подобного рибосомальному белку L22 (RPL22L1).
Предпочтительно по меньшей мере один 3'-UTR-элeмeнт искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, содержит или состоит из нуклеотидной последовательности, идентичной по меньшей мере примерно на 1, 2, 3, 4, 5, 10, 15, 20, 30 или 40%, предпочтительно по меньшей мере примерно на 50%, предпочтительно по меньшей мере примерно на 60%, предпочтительно по меньшей мере примерно на 70%, более предпочтительно по меньшей мере примерно на 80%, более предпочтительно по меньшей мере примерно на 90%, еще более предпочтительно по меньшей мере примерно на 95%, еще более предпочтительно по меньшей мере примерно на 99%, наиболее предпочтительно на 100% нуклеотидной последовательности 3'-UTR гена рибосомального белка, такой как нуклеотидная последовательность, представленная в SEQ ID NO: 10-115 или соответствующей последовательности РНК:
Рибосомальный белок L9 (RPL9) Homo sapiens

(SEQ ID NO: 10)
Рибосомальный белок L3 (RPL3) Homo sapiens
(SEQ ID NO: 11)
Рибосомальный белок L4 (RPL4) Homo sapiens

(SEQ ID NO: 12)
Рибосомальный белок L5 (RPL5) Homo sapiens
(SEQ ID NO: 13)
Рибосомальный белок L6 (RPL6) Homo sapiens
(SEQ ID NO: 14)
Рибосомальный белок L7 (RPL7) Homo sapiens
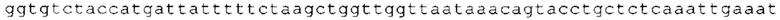
(SEQ ID NO: 15)
Рибосомальный белок L7a (RPL7A) Homo sapiens
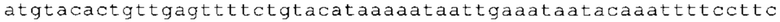
(SEQ ID NO: 16)
Рибосомальный белок L11 (RPL11) Homo sapiens
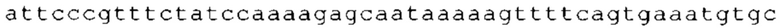
(SEQ ID NO: 17)
Рибосомальный белок L12 (RPL12) Homo sapiens
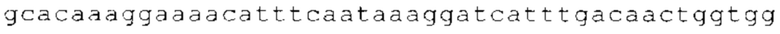
(SEQ ID NO: 18)
Рибосомальный белок L13 (RPL13) Homo sapiens


(SEQ ID NO: 19)
Рибосомальный белок L23 (RPL23) Homo sapiens
(SEQ ID NO: 20)
Рибосомальный белок L18 (RPL18) Homo sapiens
(SEQ ID NO: 21)
Рибосомальный белок L18a (RPL18A) Homo sapiens
(SEQ ID NO: 22)
Рибосомальный белок L19 (RPL19) Homo sapiens
(SEQ ID NO: 23)
Рибосомальный белок L21 (RPL21) Homo sapiens
(SEQ ID NO: 24)
Рибосомальный белок L22 (RPL22) Homo sapiens


(SEQ ID NO: 25)
Рибосомальный белок L23a (RPL23A) Homo sapiens

(SEQ ID NO: 26)
Рибосомальный белок L17 (RPL17) Homo sapiens
(SEQ ID NO: 27)
Рибосомальный белок L24 (RPL24) Homo sapiens
(SEQ ID NO: 28)
Рибосомальный белок L26 (RPL26) Homo sapiens
(SEQ ID NO: 29)
Рибосомальный белок L27 (RPL27) Homo sapiens
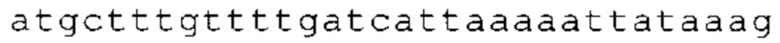
(SEQ ID NO: 30)
Рибосомальный белок L30 (RPL30) Homo sapiens

(SEQ ID NO: 31)
Рибосомальный белок L27a (RPL27A) Homo sapiens


(SEQ ID NO: 32)
Рибосомальный белок L28 (RPL28) Homo sapiens


(SEQ ID NO: 33)
Рибосомальный белок L28 (RPL28) Homo sapiens

(SEQ ID NO: 34)
Рибосомальный белок L28 (RPL28) Homo sapiens

(SEQ ID NO: 35)
Рибосомальный белок L29 (RPL29) Homo sapiens

(SEQ ID NO: 36)
Рибосомальный белок L31 (RPL31) Homo sapiens

(SEQ ID NO: 37)
Рибосомальный белок L31 (RPL31) Homo sapiens

(SEQ ID NO: 38)
Рибосомальный белок L32 (RPL32) Homo sapiens

(SEQ ID NO: 39)
Рибосомальный белок L35a (RPL35A) Homo sapiens
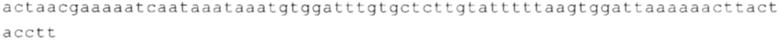
(SEQ ID NO: 40)
Рибосомальный белок L37 (RPL37) Homo sapiens

(SEQ ID NO: 41)
Рибосомальный белок L37a (RPL37A) Homo sapiens
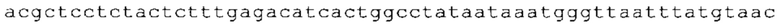
(SEQ ID NO: 42)
Рибосомальный белок L38 (RPL38) Homo sapiens
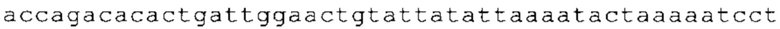
(SEQ ID NO: 43)
Рибосомальный белок L39 (RPL39) Homo sapiens

(SEQ ID NO: 44)
Рибосомальный белок большой субъединицы, Р0 (RPLP0) Homo sapiens

(SEQ ID NO: 45)
Рибосомальный белок большой субъединицы, P1 (RPLP1) Homo sapiens
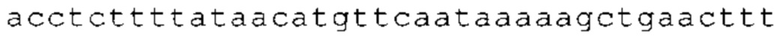
(SEQ ID NO: 46)
Рибосомальный белок большой субъединицы, Р2 (RPLP2)

(SEQ ID NO: 47)
Рибосомальный белок S3 (RPS3) Homo sapiens

(SEQ ID NO: 48)
Рибосомальный белок S3 (RPS3) Homo sapiens

(SEQ ID NO: 49)
Рибосомальный белок S3A (RPS3A) Homo sapiens
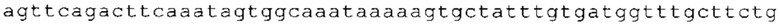
(SEQ ID NO: 50)
Рибосомальный белок S3A (RPS3A) Homo sapiens


(SEQ ID NO: 51)
Рибосомальный белок S4, Х-сцепленный (RPS4X) Homo sapiens
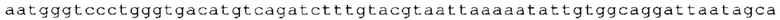
(SEQ ID NO: 52)
Рибосомальный белок S4, Y-сцепленный 1 (RPS4Y1) Homo sapiens
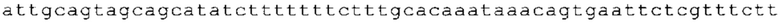
(SEQ ID NO: 53)
Рибосомальный белок S5 (RPS5) Homo sapiens
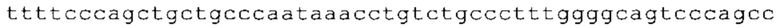
(SEQ ID NO: 54)
Рибосомальный белок S6 (RPS6) Homo sapiens
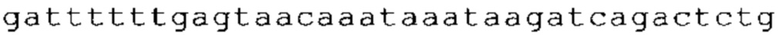
(SEQ ID NO: 55)
Рибосомальный белок S7 (RPS7) Homo sapiens
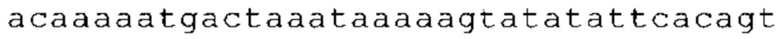
(SEQ ID NO: 56)
Рибосомальный белок S8 (RPS8) Homo sapiens
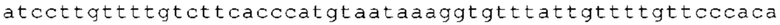
(SEQ ID NO: 57)
Рибосомальный белок S9 (RPS9) Homo sapiens
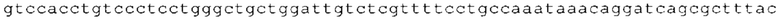
(SEQ ID NO: 58)
Рибосомальный белок S10 (RPS10) Homo sapiens
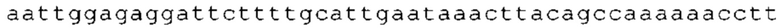
(SEQ ID NO: 59)
Рибосомальный белок S11 (RPS11) Homo sapiens
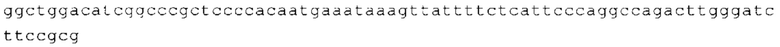
(SEQ ID NO: 60)
Рибосомальный белок S12 (RPS12) Homo sapiens
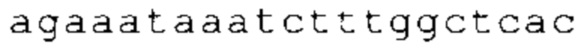
(SEQ ID NO: 61)
Рибосомальный белок S13 (RPS13) Homo sapiens
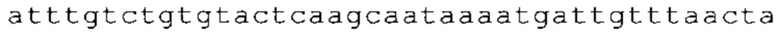
(SEQ ID NO: 62)
Рибосомальный белок S15 (RPS15) Homo sapiens
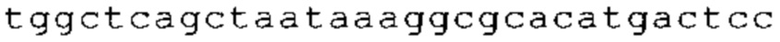
(SEQ ID NO: 63)
Рибосомальный белок S15a (RPS15A) Homo sapiens
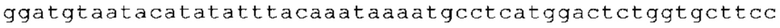
(SEQ ID NO: 64)
Рибосомальный белок S16 (RPS16) Homo sapiens
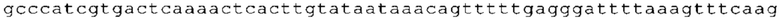
(SEQ ID NO: 65)
Рибосомальный белок S19 (RPS19) Homo sapiens
(SEQ ID NO: 66)
Рибосомальный белок S20 (RPS20) Homo sapiens

(SEQ ID NO: 67)
Рибосомальный белок S20 (RPS20) Homo sapiens

(SEQ ID NO: 68)
Рибосомальный белок S21 (RPS21) Homo sapiens
(SEQ ID NO: 69)
Рибосомальный белок S23 (RPS23) Homo sapiens

(SEQ ID NO: 70)
Рибосомальный белок S25 (RPS25) Homo sapiens
(SEQ ID NO: 71)
Рибосомальный белок S26 (RPS26) Homo sapiens
(SEQ ID NO: 72)
Рибосомальный белок S27 (RPS27) Homo sapiens
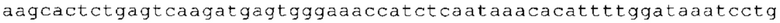
(SEQ ID NO: 73)
Рибосомальный белок S27a (RPS27a) Homo sapiens

(SEQ ID NO: 74)
Рибосомальный белок S28 (RPS28) Homo sapiens

(SEQ ID NO: 75)
Рибосомальный белок S29 (RPS29) Homo sapiens

(SEQ ID NO: 76)
Рибосомальный белок L15 (RPL15) Homo sapiens


(SEQ ID NO: 77)
Рибосомальный белок L15 (RPL15) Homo sapiens

(SEQ ID NO: 78)
Рибосомальный белок S2 (RPS2) Homo sapiens
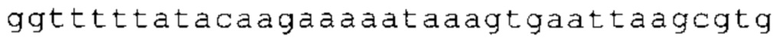
(SEQ ID NO: 79)
Рибосомальный белок L14 (RPL14) Homo sapiens

(SEQ ID NO: 80)
Рибосомальный белок S14 (RPS14) Homo sapiens
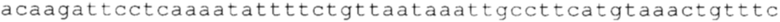
(SEQ ID NO: 81)
Рибосомальный белок L10 (RPL10) Homo sapiens

(SEQ ID NO: 82)
Рибосомальный белок L10a (RPL10A) Homo sapiens
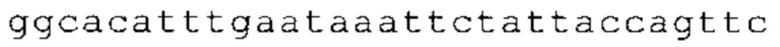
(SEQ ID NO: 83)
Рибосомальный белок L35 (RPL35) Homo sapiens
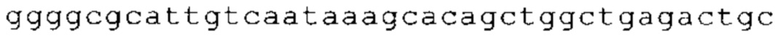
(SEQ ID NO: 84)
Рибосомальный белок L13a (RPL13A) Homo sapiens

(SEQ ID NO: 85)
Рибосомальный белок L36 (RPL36) Homo sapiens
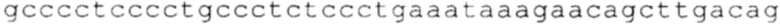
(SEQ ID NO: 86)
Homo sapiens ribosomal protein L36a (RPL36A) Homo sapiens

(SEQ ID NO: 87)
Рибосомальный белок L36a (RPL36A) Homo sapiens

(SEQ ID NO: 88)
Рибосомальный белок L41 (RPL41) Homo sapiens

(SEQ ID NO: 89)
Рибосомальный белок S18 (RPS18) Homo sapiens
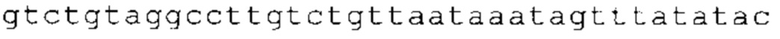
(SEQ ID NO: 90)
Рибосомальный белок S24 (RPS24) Homo sapiens
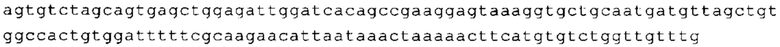
(SEQ ID NO: 91)
Рибосомальный белок S24 (RPS24) Homo sapiens

(SEQ ID NO: 92)
Рибосомальный белок L8 (RPL8) Homo sapiens
(SEQ ID NO: 93)
Рибосомальный белок L34 (RPL34) Homo sapiens

(SEQ ID NO: 94)
Рибосомальный белок L34 (RPL34) Homo sapiens

(SEQ ID NO: 95)
Рибосомальный белок S17 (RPS17) Homo sapiens
(SEQ ID NO: 96)
Рибосомальный белок SA (RPSA) Homo sapiens

(SEQ ID NO: 97)
Продукт 1 слияния: остаток А-52-рибосомальный белок 1 (UBA52)


(SEQ ID NO: 98)
Белок повсеместно экспрессируемого (FAU) вируса мышиной саркомы Финкеля-Бискиса-Рейли (FBR-MuSV)
(SEQ ID NO: 99)
Белок 1, подобный рибосомальному белку L22 (RPL22L1) Homo sapiens


(SEQ ID NO: 100)
Рибосомальный белок S17 (RPS17) Homo sapiens
(SEQ ID NO: 101)
Белок, подобный рибосомальному белку L39 (RPL39L) Homo sapiens
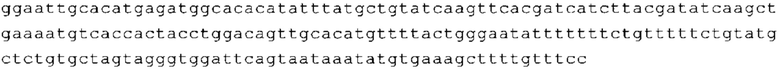
(SEQ ID NO: 102)
Белок, подобный рибосомальному белку L10 (RPL10L) Homo sapiens
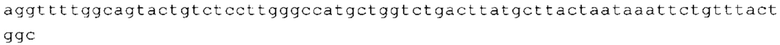
(SEQ ID NO: 103)
Белок, подобный рибосомальному белку L36a (RPL36AL) Homo sapiens

(SEQ ID NO: 104)
Белок, подобный рибосомальному белку L3 (RPL3L) Homo sapiens

(SEQ ID NO: 105)
Белок, подобный рибосомальному белку S27 (RPS27L) Homo sapiens

(SEQ ID NO: 106)
Белок 1, подобный рибосомальному белку L26 (RPL26L1) Homo sapiens

(SEQ ID NO: 107)
Белок 1, подобный рибосомальному белку L7 (RPL7L1) Homo sapiens

(SEQ ID NO: 108)
Псевдоген рибосомального белка L13a (RPL13AP) Homo sapiens

(SEQ ID NO: 109)
Псевдоген 8 рибосомального белка L37a 8 (RPL37AP8) Homo sapiens

(SEQ ID NO: 110)
Псевдоген 5 рибосомального белка S10 (RPS10P5) Homo sapiens
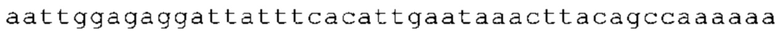
(SEQ ID NO: 111)
Псевдоген 11 рибосомального белка S26 (RPS26P11) Homo sapiens
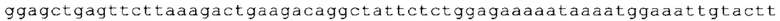
(SEQ ID NO: 112)
Псевдоген 5 рибосомального белка L39 (RPL39P5) Homo sapiens

(SEQ ID NO: 113)
Псевдоген 6 рибосомального белка Р0 большой субъединицы (RPLP0P6) Homo sapiens

(SEQ ID NO: 114)
Псевдоген 14 рибосомального белка P0 большой субъединицы L36 (RPL36P14) Homo sapiens
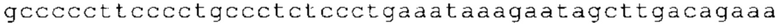
(SEQ ID NO: 115)
Кроме того, предпочтительно по меньшей мере один 3'-UTR-элемент искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, содержит или состоит из нуклеотидной последовательности, которая идентична по меньшей мере примерно на 1, 2, 3, 4, 5, 10, 15, 20, 30 или 40%, предпочтительно по меньшей мере примерно на 50%, предпочтительно по меньшей мере примерно на 60%, предпочтительно по меньшей мере примерно на 70%, более предпочтительно по меньшей мере примерно на 80%, более предпочтительно по меньшей мере примерно на 90%, еще более предпочтительно по меньшей мере примерно на 95%, еще более предпочтительно по меньшей мере примерно на 99%, наиболее предпочтительно на 100%) нуклеотидной последовательности 3'-UTR гена рибосомального белка, такой как нуклеотидная последовательность, представленная в SEQ ID NO: 116-205 или соответствующей последовательности РНК:
Рибосомальный белок L9 (RPL9) Mus musculus
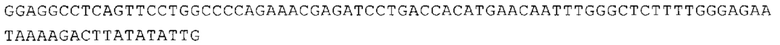
(SEQ ID NO: 116)
Рибосомальный белок L3 (RPL3) Mus musculus
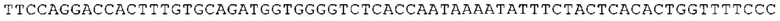
(SEQ ID NO: 117)
Рибосомальный белок L4 (RPL4) Mus musculus
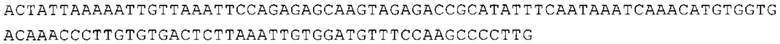
(SEQ ID NO: 118)
Рибосомальный белок L5 (RPL5) Mus musculus


(SEQ ID NO: 119)
Рибосомальный белок L6 (RPL6) Mus musculus

(SEQ ID NO: 120)
Рибосомальный белок L7 (RPL7) Mus musculus
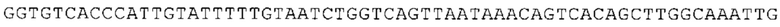
(SEQ ID NO: 121)
Рибосомальный белок L7a (RPL7A) Mus musculus
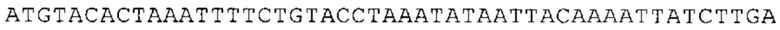
(SEQ ID NO: 122)
Рибосомальный белок L11 (RPL11) Mus musculus
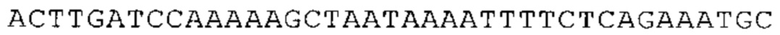
(SEQ ID NO: 123)
Рибосомальный белок L12 (RPL12) Mus musculus
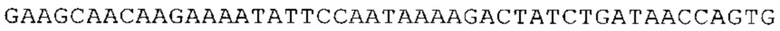
(SEQ ID NO: 124)
Рибосомальный белок L13 (RPL13) Mus musculus
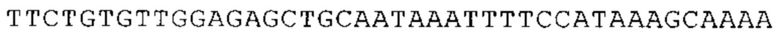
(SEQ ID NO: 125)
Рибосомальный белок L23 (RPL23) Mus musculus
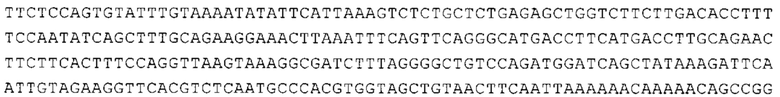
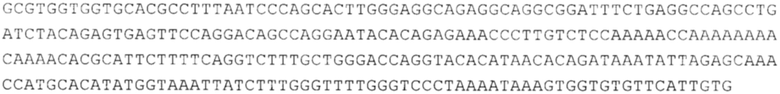
(SEQ ID NO: 126)
Рибосомальный белок L18 (RPL18) Mus musculus
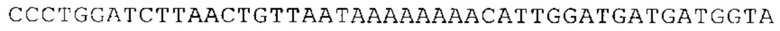
(SEQ ID NO:127)
Рибосомальный белок L18a (RPL18A) Mus musculus

(SEQ ID NO: 128)
Рибосомальный белок L19 (RPL19) Mus musculus
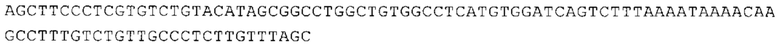
(SEQ ID NO: 129)
Рибосомальный белок L21 (RPL21) Mus musculus

(SEQ ID NO: 130)
Рибосомальный белок L22 (RPL22) Mus musculus

(SEQ ID NO: 131)
Рибосомальный белок L22 (RPL22) Mus musculus


(SEQ ID NO: 132)
Рибосомальный белок L23a (RPL23A) Mus musculus
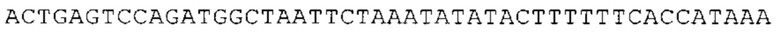
(SEQ ID NO: 133)
Рибосомальный белок L17 (RPL17) Mus musculus
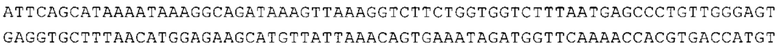
(SEQ ID NO: 134)
Рибосомальный белок L24 (RPL24) Mus musculus
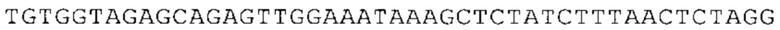
(SEQ ID NO: 135)
Рибосомальный белок L26 (RPL26) Mus musculus
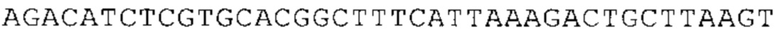
(SEQ ID NO: 136)
Рибосомальный белок L27 (RPL27) Mus musculus
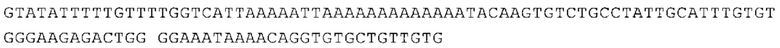
(SEQ ID NO: 137)
Рибосомальный белок L30 (RPL30) Mus musculus
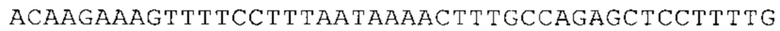
(SEQ ID NO: 138)
Рибосомальный белок L27a (RPL27A) Mus musculus

(SEQ ID NO: 139)
Рибосомальный белок L28 (RPL28) Mus musculus
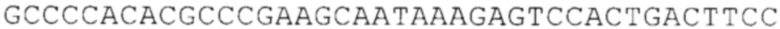
(SEQ ID NO: 140)
Рибосомальный белок L29 (RPL29) Mus musculus
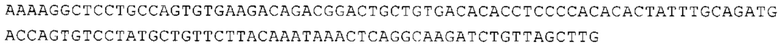
(SEQ ID NO: 141)
Рибосомальный белок L31 (RPL31) Mus musculus

(SEQ ID NO: 142)
Рибосомальный белок L32 (RPL32) Mus musculus
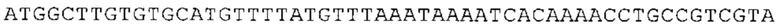
(SEQ ID NO: 143)
Рибосомальный белок L35a (RPL35A) Mus musculus
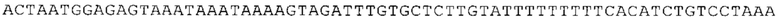
(SEQ ID NO: 144)
Рибосомальный белок L37 (RPL37) Mus musculus
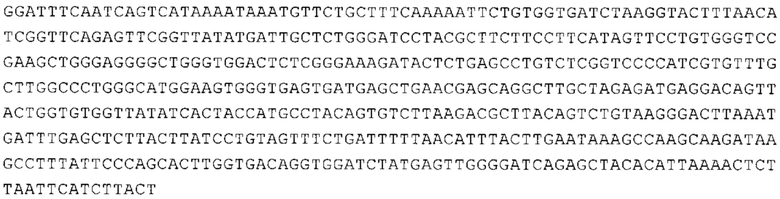
(SEQ ID NO: 145)
Рибосомальный белок (RPL37) Mus musculus

(SEQ ID NO: 146)
Рибосомальный белок L37a (RPL37A) Mus musculus

(SEQ ID NO: 147)
Рибосомальный белок L38 (RPL38) Mus musculus
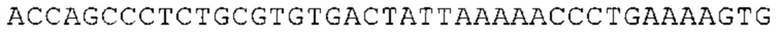
(SEQ ID NO: 148)
Рибосомальный белок L39 (RPL39) Mus musculus

(SEQ ID NO: 149)
Рибосомальный белок большой субъединицы Р0 (RPLP0) Mus musculus

(SEQ ID NO: 150)
Рибосомальный белок большой субъединицы P1 (RPLP1) Mus musculus
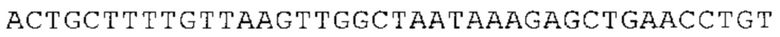
(SEQ ID NO: 151)
Рибосомальный белок большой субъединицы Р2 (RPLP2) Mus musculus
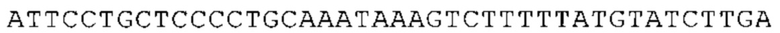
(SEQ ID NO: 152)
Рибосомальный белок S3 (RPS3) Mus musculus


(SEQ ID NO: 153)
Рибосомальный белок S3A (RPS3A) Mus musculus
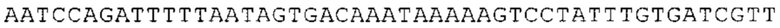
(SEQ ID NO: 154)
Рибосомальный белок S4, Х-сцепленный (RPS4X) Mus musculus
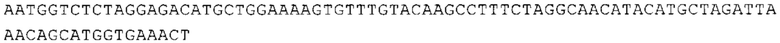
(SEQ ID NO: 155)
Рибосомальный белок 34-подобный (RPS41) Mus musculus
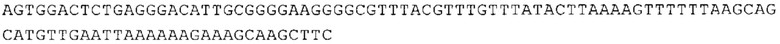
(SEQ ID NO: 156)
Рибосомальный белок S5 (RPS5) Mus musculus
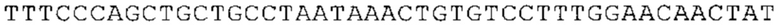
(SEQ ID NO: 157)
Рибосомальный белок S6 (RPS6) Mus musculus

(SEQ ID NO: 158)
Рибосомальный белок S7 (RPS7) Mus musculus
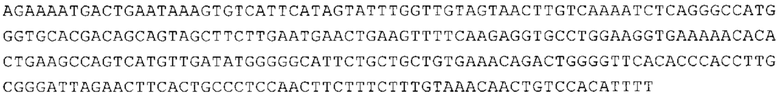
(SEQ ID NO: 159)
Рибосомальный белок S8 (RPS8) Mus musculus
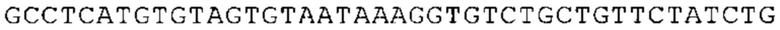
(SEQ ID NO: 160)
Рибосомальный белок S9 (RPS9) Mus musculus
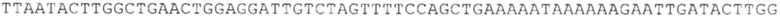
(SEQ ID NO: 161)
Рибосомальный белок S10 (RPS10) Mus musculus
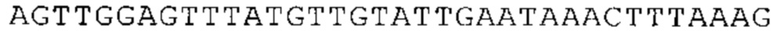
(SEQ ID NO: 162)
Рибосомальный белок S11 (RPS11) Mus musculus
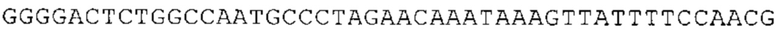
(SEQ ID NO: 163)
Рибосомальный белок S12 (RPS12) Mus musculus
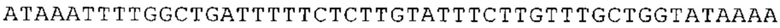
(SEQ ID NO: 164)
Рибосомальный белок S13 (RPS13) Mus musculus
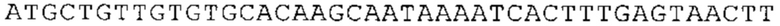
(SEQ ID NO: 165)
Рибосомальный белок S15 (RPS15) Mus musculus
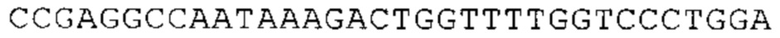
(SEQ ID NO: 166)
Рибосомальный белок S15a (RPS15A) Mus musculus


(SEQ ID NO: 167)
Mus musculus ribosomal protein S16 (RPS16) Mus musculus
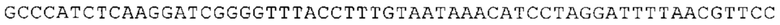
(SEQ ID NO: 168)
Рибосомальный белок S19 (RPS19) Mus musculus
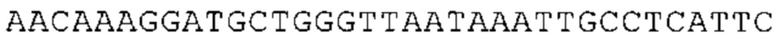
(SEQ ID NO: 169)
Рибосомальный белок S20 (RPS20) Mus musculus


(SEQ ID NO: 170)
Рибосомальный белок S21 (RPS21)
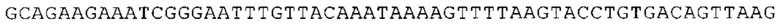
(SEQ ID NO: 171)
Рибосомальный белок S23 (RPS23) Mus musculus
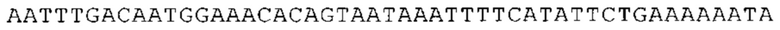
(SEQ ID NO: 172)
Рибосомальный белок S25 (RPS25) Mus musculus

(SEQ ID NO: 173)
Рибосомальный белок S26 (RPS26) Mus musculus
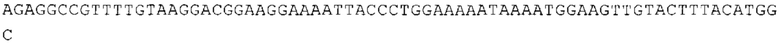
(SEQ ID NO: 174)
Рибосомальный белок S27 (RPS27) Mus musculus
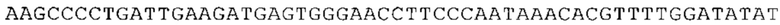
(SEQ ID NO: 175)
Рибосомальный белок S27a (RPS27a) Mus musculus
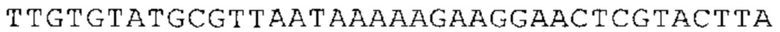
(SEQ ID NO: 176)
Рибосомальный белок S28 (RPS28) Mus musculus
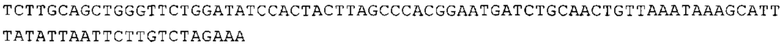
(SEQ ID NO: 177)
Рибосомальный белок S29 (RPS29) Mus musculus
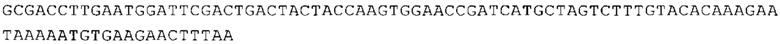
(SEQ ID NO: 178)
Рибосомальный белок L15 (RPL15) Mus musculus

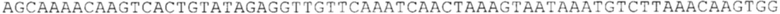
(SEQ ID NO: 179)
Рибосомальный белок S2 (RPS2) Mus musculus
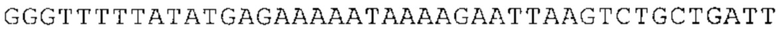
(SEQ ID NO: 180)
Рибосомальный белок L14 (RPL14) Mus musculus

(SEQ ID NO: 181)
Рибосомальный белок S14 (RPS14) Mus musculus
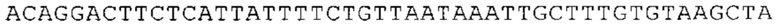
(SEQ ID NO: 182)
Рибосомальный белок L10 (RPL10) Mus musculus
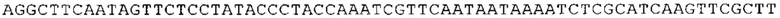
(SEQ ID NO: 183)
Рибосомальный белок L10a (RPL10A) Mus musculus
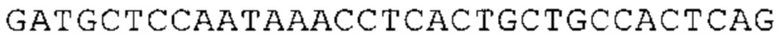
(SEQ ID NO: 184)
Рибосомальный белок L35 (RPL35) Mus musculus
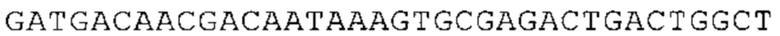
(SEQ ID NO: 185)
Рибосомальный белок L13a (RPL13A) Mus musculus
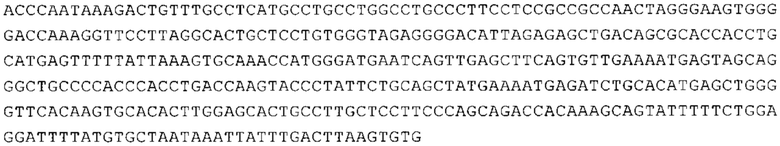
(SEQ ID NO: 186)
Рибосомальный белок L36 (RPL36) Mus musculus
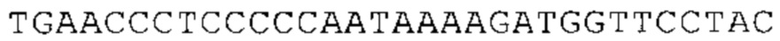
(SEQ ID NO: 187)
Рибосомальный белок L36a (RPL36A) Mus musculus
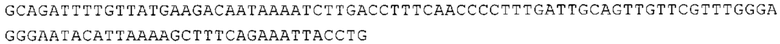
(SEQ ID NO: 188)
Рибосомальный белок L41 (RPL41) Mus musculus
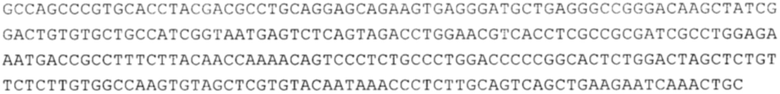
(SEQ ID NO: 189)
Рибосомальный белок S18 (RPS18) Mus musculus
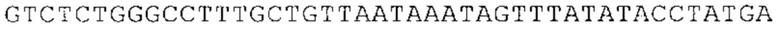
(SEQ ID NO: 190)
Рибосомальный белок S24 (RPS24) Mus musculus
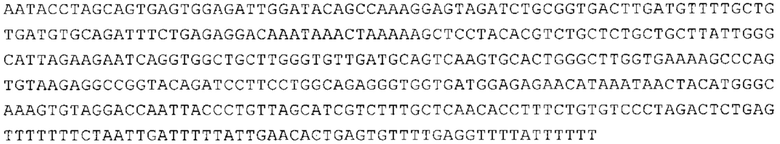
(SEQ ID NO: 191)
Рибосомальный белок L8 (RPL8) Mus musculus
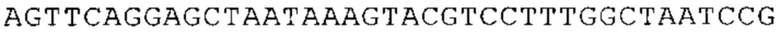
(SEQ ID NO: 192)
Рибосомальный белок L34 (RPL34) Mus musculus
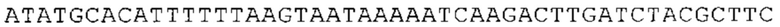
(SEQ ID NO: 193)
Рибосомальный белок S17 (RPS17) Mus musculus
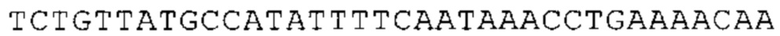
(SEQ ID NO: 194)
Рибосомальный белок SA (RPSA) Mus musculus

(SEQ ID NO: 195)
Продукт 1 слияния остаток убиквитина А-52-рибосомальный белок (UBA52) Mus musculus
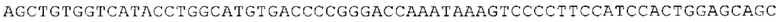
(SEQ ID NO: 196)
Повсеместно экспрессируемый белок (FAU) вируса мышиной саркомы Финкеля-Бискиса-Рейли (FBR-MuSV)
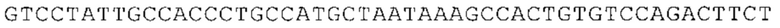
(SEQ ID NO: 197)
Рибосомальный белок L22-подобный 1 (RPL22L1) Mus musculus

(SEQ ID NO: 198)
Рибосомальный белок S17 (RPS17) Mus musculus
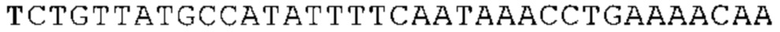
(SEQ ID NO: 199)
Рибосомальный белок L39-подобный (RPL39L) Mus musculus
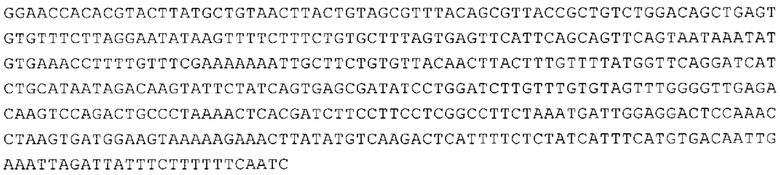
(SEQ ID NO: 200)
Рибосомальный белок L10-подобный (RPL10L) Mus musculus
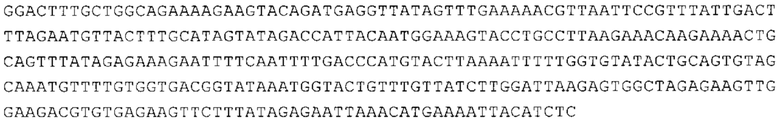
(SEQ ID NO: 201)
Рибосомальный белок L36а-подобный (RPL36AL) Mus musculus
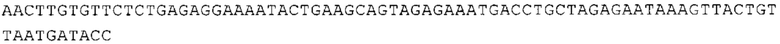
(SEQ ID NO: 202)
Рибосомальный белок L3-подобный (RPL3L) Mus musculus
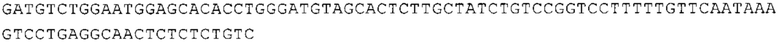
(SEQ ID NO: 203)
Рибосомальный белок S27-подобный (RPS27L) Mus musculus
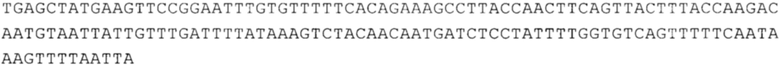
(SEQ ID NO: 204)
Рибосомальный белок L7-подобный 1 (RPL7L1) Mus musculus

(SEQ ID NO: 205)
В предпочтительном варианте осуществления изобретения по меньшей мере один 3'-UTR-элемент искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, содержит или состоит из нуклеотидной последовательности, которая идентична по меньшей мере примерно на 40%, предпочтительно по меньшей мере примерно на 50%, предпочтительно по меньшей мере примерно на 60%, предпочтительно по меньшей мере примерно на 70%, более предпочтительно по меньшей мере примерно на 80%, более предпочтительно по меньшей мере примерно на 90%, еще более предпочтительно по меньшей мере примерно на 95%, еще более предпочтительно по меньшей мере примерно на 99%, наиболее предпочтительно на 100% последовательности 3'-UTR гена рибосомального белка малой субъединицы 9 (RPS9). Наиболее предпочтительно по меньшей мере один 3'-UTR-элемент искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, содержит или состоит из нуклеотидной последовательности, которая идентична по меньшей мере примерно на 40%, предпочтительно по меньшей мере примерно на 50%, предпочтительно по меньшей мере примерно на 60%, предпочтительно по меньшей мере примерно на 70%, более предпочтительно по меньшей мере примерно на 80%, более предпочтительно по меньшей мере примерно на 90%, еще более предпочтительно по меньшей мере примерно на 95%, еще более предпочтительно по меньшей мере примерно на 99%, наиболее предпочтительно на 100% SEQ ID NO: 1 или SEQ ID NO: 2:
SEQ ID NO: 1
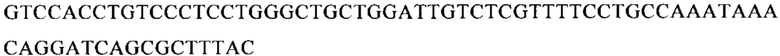
SEQ ID NO: 2
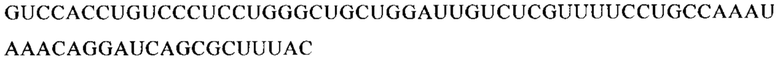
По меньшей мере один 3'-UTR-элeмeнт искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, может также содержать или состоять из фрагмента нуклеотидной последовательности, который идентичен по меньшей мере примерно на 40%, предпочтительно по меньшей мере примерно на 50%, предпочтительно по меньшей мере примерно на 60%, предпочтительно по меньшей мере примерно на 70%, более предпочтительно по меньшей мере примерно на 80%, более предпочтительно по меньшей мере примерно на 90%, еще более предпочтительно по меньшей мере примерно на 95%, еще более предпочтительно по меньшей мере примерно на 99%, наиболее предпочтительно на 100% нуклеотидной последовательности 3'-UTR гена рибосомального белка, такой как 3'-UTR, имеющая последовательность, представленную в SEQ ID NO: 10-205, где фрагмент предпочтительно представляет собой функциональный фрагмент или функциональный фрагмент варианта, описанный выше. Такой фрагмент предпочтительно имеет длину, составляющую по меньшей мере примерно 3 нуклеотида, предпочтительно по меньшей мере примерно 5 нуклеотида, более предпочтительно по меньшей мере примерно 10, 15, 20, 25 или 30 нуклеотидов, еще более предпочтительно по меньшей мере примерно 50 нуклеотидов, наиболее предпочтительно по меньшей мере примерно 70 нуклеотидов. В предпочтительном варианте осуществления изобретения фрагмент или его вариант имеет длину от 3 до примерно 500 нуклеотидов, предпочтительно от 5 до примерно 150 нуклеотидов, более предпочтительно от 10 до 100 нуклеотидов, еще более предпочтительно от 15 до 90, наиболее предпочтительно от 20 до 70 нуклеотидов.
Предпочтительно указанные варианты, фрагменты или фрагменты вариантов представляют собой функциональные варианты, функциональные фрагменты или функциональные фрагменты вариантов, описанные выше, которые обладают по меньшей мере одной функцией, присущей нуклеотидной последовательности, представленной в SEQ ID NO: 10-205, такой как стабилизация искусственной молекулы нуклеиновой кислоты, предлагаемой в изобретении, стабилизация и/или пролонгирование экспрессии белка с искусственной молекулы нуклеиновой кислоты, предлагаемой в изобретении, и/или увеличение производства белка, предпочтительно с эффективностью, составляющей по меньшей мере 40%, более предпочтительно по меньшей мере 50%, более предпочтительно по меньшей мере 60%, еще более предпочтительно по меньшей мере 70%, еще более предпочтительно по меньшей мере 80%, наиболее предпочтительно по меньшей мере 90% от эффективности стабилизации и/или эффективности увеличения производства белка с искусственной молекулы нуклеиновой кислоты, содержащей нуклеотидную последовательность, представленную в SEQ ID NO: 1 или SEQ ID NO: 2.
Предпочтительно по меньшей мере один 3'-UTR-элемент искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, имеет длину, составляющую по меньшей мере примерно 3 нуклеотида, предпочтительно по меньшей мере примерно 5 нуклеотида, более предпочтительно по меньшей мере примерно 10, 15, 20, 25 или 30 нуклеотидов, еще более предпочтительно по меньшей мере примерно 50 нуклеотидов, наиболее предпочтительно по меньшей мере примерно 70 нуклеотидов. Верхний предел длины 3'-UTR-элемента может составлять 500 нуклеотидов или менее, например, 400, 300, 200, 150 или 100 нуклеотидов. В других вариантах осуществления изобретения верхний предел может быть выбран из диапазона от 50 до 100 нуклеотидов. Например, фрагмент или вариант может иметь длину от 3 до примерно 500 нуклеотидов, предпочтительно от 5 до примерно 150 нуклеотидов, более предпочтительно от 10 до 100 нуклеотидов, еще более предпочтительно от 15 до 90, наиболее предпочтительно от 20 до 70 нуклеотидов.
Кроме того, искусственная молекула нуклеиновой кислоты, предлагаемая в настоящем изобретении, может содержать более одного 3'-UTR-элемента, описанного выше. Например, искусственная молекула нуклеиновой кислоты, предлагаемая в настоящем изобретении, может содержать один, два, три четыре или большее количество 3'-UTR-элементов, при этом индивидуальные 3'-UTR-элементы могут быть одинаковыми или могут быть различными. Например, искусственная молекула нуклеиновой кислоты, предлагаемая в настоящем изобретении, может содержать два практически идентичных 3'-UTR-элемента, описанных выше, например, два 3'-UTR-элемента, содержащих или состоящих из нуклеотидной последовательности, происходящей из 3'-UTR гена рибосомального белка, например, из последовательности, представленной в SEQ ID NO: 10-205, или из фрагмента или варианта 3'-UTR гена рибосомального белка, имеющего нуклеотидную последовательность, представленную в SEQ ID NO: 1 или 2, его функциональных вариантов, его функциональных фрагментов или функциональных фрагментов вариантов, описанных выше.
При создании изобретения неожиданно было установлено, что искусственная молекула нуклеиновой кислоты, содержащая 3'-UTR-элeмeнт, описанный выше, может представлять собой или позволяет создавать молекулу мРНК, которая обеспечивает повышенное, пролонгированное и/или стабилизированное производство белка. Так, 3'-UTR-элемент, представленный в настоящем описании, может повышать стабильность экспрессии белка с молекулы мРНК и/или повышать эффективность трансляции.
Искусственная молекула нуклеиновой кислоты, предлагаемая в настоящем изобретении, может представлять собой РНК, такую как мРНК, ДНК, такую как векторная ДНК, или может представлять собой модифицированную молекулу РНК или ДНК. Она может находиться в виде двухцепочечной молекулы, имеющей смысловую цепь и антисмысловую цепь, например, в виде молекулы ДНК, имеющей смысловую цепь и антисмысловую цепь.
Искусственная молекула нуклеиновой кислоты, предлагаемая в настоящем изобретении, кроме того, может необязательно содержать 5'-UTR и/или 5'-кэп. Необязательные 5'-кэп и/или 5'-UTR предпочтительно расположены с 5'-стороны относительно ОРС в искусственной молекуле нуклеиновой кислоты, предлагаемой в настоящем изобретении.
Предпочтительно искусственная молекула нуклеиновой кислоты, предлагаемая в настоящем изобретении, содержит также поли(А)-последовательность и/или сигнал полиаденилирования. Предпочтительно необязательная поли(А)-последовательность расположена с 3'-стороны относительно по меньшей мере одного 3'-UTR-элeмeнтa, предпочтительно необязательная поли(А)-последовательность присоединена к 3'-концу 3'-UTR-элемента. Соединение может осуществляться непосредственно или косвенным образом посредством сегмента, состоящего из 2, 4, 6, 8, 10, 20 и т.д. нуклеотидов, такой как линкер, состоящий из 1-50, предпочтительно из 1-20 нуклеотидов, например, содержащий или состоящий из одного или нескольких сайтов рестрикции.
В одном из вариантов осуществления изобретения необязательный сигнал полиаденилирования расположен в прямом направлении с 3'-стороны 3'-UTR-элемента. Предпочтительно сигнал полиаденилирования содержит консенсусную последовательность NN(U/T)ANA, в которой N обозначает А или U, предпочтительно AA(U/T)AAA или A(U/T)(U/T)AAA. Такая консенсусная последовательность может распознаваться большинством систем на основе клеток животных и бактериальных клеток, например, факторами полиаденилирования, такими как фактор специфичности расщепления/полиаденилирования (CPSF), в сочетании с CstF, PAP, РАВ2, CFI и/или CFII. Предпочтительно сигнал полиаденилирования, предпочтительно представляющий собой консенсусную последовательность NNUANA, расположен на расстоянии менее чем примерно 50 нуклеотидов, более предпочтительно менее чем примерно 30 оснований, наиболее предпочтительно менее чем примерно 25 оснований, например, на расстоянии 21 основания, в прямом направлении относительно 3'-конца 3'-UTR-элемента.
Транскрипция искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, например, искусственной молекулы ДНК, содержащей сигнал полиаденилирования, расположенный в прямом направлении относительно 3'-UTR-элемента, должна приводить к получению незрелой РНК, содержащей сигнал полиаденилирования, в прямом направлении относительно ее 3'-UTR-элемента. Например, транскрипция молекулы ДНК, содержащей 3'-UTR-элемент, имеющий SEQ ID NO: 1, должна приводить к получению РНК, несущей 3'-UTR-элемент, имеющий последовательность SEQ ID NO: 2.
Затем при использовании соответствующей системы транскрипции должно происходить присоединение поли(А)-последовательности к незрелой РНК. Например, предлагаемая в изобретении искусственная молекула нуклеиновой кислоты может представлять собой молекулу ДНК, содержащую 3'-UTR-элемент, описанный выше, и сигнал полиаденилирования, который может приводить к полиаденилированию РНК после транскрипции указанной молекулы ДНК. Следовательно, образовавшаяся РНК может содержать комбинацию предлагаемого в изобретении 3'-UTR-элемента, за которым располагается поли(А)-последовательность.
Возможными системами транскрипции являются системы транскрипции in vitro или клеточные системы транскрипции и т.д. Таким образом, транскрипция искусственной молекулы нуклеиновой кислоты, предлагаемой в изобретении, например, транскрипции искусственной молекулы нуклеиновой кислоты, содержащей открытую рамку считывания, 3'-UTR-элемент и сигнал полиаденилирования, может приводить к получению молекулы мРНК, содержащей открытую рамку считывания, 3'-UTR-элемент и поли(А)-последовательность.
Таким образом, в изобретении предложена также искусственная молекула нуклеиновой кислоты, которая представляет собой молекулу мРНК, содержащую открытую рамку считывания, 3'-UTR-элемент, описанный выше, и поли(А)-последовательность.
Один из вариантов осуществления изобретения относится к искусственной молекуле нуклеиновой кислоты, которая представляет собой искусственную молекулу ДНК, содержащую открытую рамку считывания и последовательность, представленную в SEQ ID NO: 1 или последовательность, идентичную по меньшей мере примерно на 40% или более SEQ ID NO: 1, или ее фрагмент, описанный выше. Кроме того, в изобретении предложена искусственная молекула нуклеиновой кислоты, которая представляет собой искусственную молекулу РНК, содержащую открытую рамку считывания и последовательность, представленную в SEQ ID NO: 2 или последовательность, идентичную по меньшей мере примерно на 40% или более SEQ ID NO: 2, или ее фрагмент, описанный выше.
Таким образом, в изобретении предложена также искусственная молекула нуклеиновой кислоты, которая может служить в качестве матрицы для молекулы РНК, предпочтительно для молекулы мРНК, которая стабилизирована и оптимизирована в отношении эффективности трансляции. Иными словами, искусственная молекула нуклеиновой кислоты может представлять собой ДНК, которую можно применять в качестве матрицы для получения мРНК. мРНК, которую можно получать таким путем, может, в свою очередь, транслироваться с получением требуемого пептида или белка, кодируемого открытой рамкой считывания. Если искусственная молекула нуклеиновой кислоты представляет собой ДНК, то ее можно, например, использовать в качестве двухцепочечной формы хранения для продолжения и повторения производства мРНК in vitro или in vivo.
В другом варианте осуществления изобретения 3'-UTR искусственной молекулы нуклеиновой кислоты, предлагаемая в изобретении, не содержит сигнал полиаденилирования или поли(А)-последовательность. Кроме того, предпочтительно искусственная молекула нуклеиновой кислоты, предлагаемая в изобретении, не содержит сигнал полиаденилирования или поли(А)-последовательность. Более предпочтительно 3'-UTR искусственной молекулы нуклеиновой кислоты или сама предлагаемая в изобретении искусственная молекула нуклеиновой кислоты не содержит сигнал полиаденилирования, прежде всего не содержит сигнал полиаденилирования AAU/TAAA.
В одном из вариантов осуществления изобретения искусственная молекула нуклеиновой кислоты, предлагаемая в настоящем изобретении, содержит также поли(А)-последовательность. Например, молекула ДНК, содержащая ОРС, за которой следует рибосомальная 3'-UTR, может содержать состоящий из тимидиновых нуклеотидов сегмент, который может транскрибироваться в поли(А)-последовательность в образующейся мРНК. Длина поли(А)-последовательности может варьироваться. Например, поли(А)-последовательность может иметь длину от примерно 20 адениновых нуклеотидов вплоть до 300 адениновых нуклеотидов, предпочтительно от примерно 40 до примерно 200 адениновых нуклеотидов, более предпочтительно от примерно 50 до примерно 100 адениновых нуклеотидов, например, примерно 60, 70, 80, 90 или 100 адениновых нуклеотидов. Наиболее предпочтительно предлагаемая в изобретении нуклеиновая кислота содержит поли(А)-последовательность, содержащую от примерно 60 до примерно 70 нуклеотидов, наиболее предпочтительно 64 адениновых нуклеотида.
Например, искусственная молекула нуклеиновой кислоты, предлагаемая в настоящем изобретении, может содержать нуклеотидную последовательность, соответствующую последовательности ДНК:
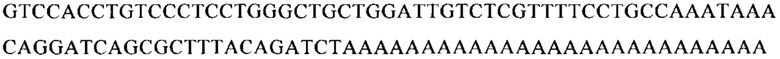
 (SEQ ID NO: 3).
(SEQ ID NO: 3).
Транскрипция указанных последовательностей может приводить к получению искусственных молекул нуклеиновой кислоты, содержащих последовательность:
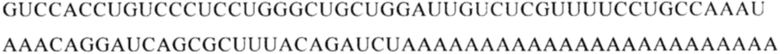
 (SEQ ID NO: 4).
(SEQ ID NO: 4).
Такие искусственные молекулы РНК, т.е. искусственные молекулы нуклеиновой кислоты, содержащие последовательность, представленную в SEQ ID NO: 4, можно также получать in vitro с помощью обычных методов химического синтеза, при этом не требуется осуществлять транскрипцию с ДНК-предшественника.
В наиболее предпочтительном варианте осуществления изобретения искусственная молекула нуклеиновой кислоты, предлагаемая в настоящем изобретении, представляет собой молекулу РНК, предпочтительно молекулу РНК, содержащую в 5→3'-направлении открытую рамку считывания, 3'-UTR-элемент, описанный выше, и поли(А)-последовательность.
Предпочтительно открытая рамка считывания не кодирует рибосомальный белок. В предпочтительном варианте осуществления изобретения открытая рамка считывания не кодирует рибосомальный белок, из которого происходит 3'-UTR-элемент предлагаемой в изобретении нуклеиновой кислоты, прежде всего не кодирует рибосомальный белок млекопитающих, при условии, что 3'-UTR-элемент идентичен 3'-UTR гена рибосомального белка млекопитающих. В некоторых других вариантах осуществления изобретения открытая рамка считывания не кодирует RPS9 или его варианты, при условии, что 3'-UTR-элемент имеет последовательность, идентичную SEQ ID NO: 1 или SEQ ID NO: 2.
В предпочтительном варианте осуществления изобретения ОРС не кодирует рибосомальные белки человека или растения, прежде всего Arabidopsis, прежде всего не кодирует человеческий рибосомальный белок S6 (RPS6), белок, подобный человеческому рибосомальному белку L36a (RPL36AL), или рибосомальный белок Arabidopsis S16 (RPS16). В другом предпочтительном варианте осуществления изобретения открытая рамка считывания (ОРС) не кодирует рибосомальный белок S6 (RPS6), белок, подобный рибосомальному белку L36a (RPL36AL) или рибосомальный белок S16 (RPS16) вне зависимости от его происхождения.
Один из вариантов осуществления изобретения относится к искусственной молекул ДНК, содержащей открытую рамку считывания, предпочтительно открытую рамку считывания, которая кодирует пептид или белок, отличный от рибосомального белка, из которого происходит 3'-UTR; 3'-UTR-элeмeнт, содержащий или состоящий из последовательности, идентичной по меньшей мере примерно на 60%, предпочтительно по меньшей мере примерно на 70%, более предпочтительно по меньшей мере примерно на 80%, более предпочтительно по меньшей мере примерно на 90%, еще более предпочтительно по меньшей мере примерно на 95%; еще более предпочтительно по меньшей мере на 99%; еще более предпочтительно на 100% последовательности, представленной в SEQ ID NO: 1; сигнал полиаденилирования и/или поли(А)-последовательность. Кроме того, в изобретении предложена искусственная молекула ДНК, содержащая открытую рамку считывания, предпочтительно открытую рамку считывания, которая кодирует любой пептид или белок, отличный от рибосомального белка, из которого происходит 3'-UTR; 3'-UTR-элeмeнт, содержащий или состоящий из последовательности, идентичной по меньшей мере примерно на 60%, предпочтительно по меньшей мере примерно на 70%, более предпочтительно по меньшей мере примерно на 80%, более предпочтительно по меньшей мере примерно на 90%, еще более предпочтительно по меньшей мере примерно на 95%; еще более предпочтительно по меньшей мере на 99%; еще более предпочтительно на 100% последовательности, представленной в SEQ ID NO: 3.
Кроме того, в изобретении предложена искусственная молекула РНК, предпочтительно искусственная молекула мРНК или искусственная вирусная молекула РНК, содержащая открытую рамку считывания, предпочтительно открытую рамку считывания, которая кодирует пептид или белок, отличный от рибосомального белка, из которого происходит 3'-UTR; 3'-UTR-элeмeнт, содержащий или состоящий из последовательности, идентичной по меньшей мере примерно на 60%, предпочтительно по меньшей мере примерно на 70%, более предпочтительно по меньшей мере примерно на 80%, более предпочтительно по меньшей мере примерно на 90%, еще более предпочтительно по меньшей мере примерно на 95%; еще более предпочтительно по меньшей мере на 99%; еще более предпочтительно на 100% последовательности, представленной в SEQ ID NO: 2; и сигнал полиаденилирования и/или поли(А)-последовательность. Кроме того, искусственная молекула РНК, предпочтительно искусственная молекула мРНК или искусственная вирусная молекула РНК, содержащая открытую рамку считывания, предпочтительно открытую рамку считывания, которая кодирует пептид или белок, отличный от рибосомального белка, из которого происходит 3'-UTR; 3'-UTR-элeмeнт, содержащий или состоящий из последовательности, идентичной по меньшей мере примерно на 60%, предпочтительно по меньшей мере примерно на 70%, более предпочтительно по меньшей мере примерно на 80%, более предпочтительно по меньшей мере примерно на 90%, еще более предпочтительно по меньшей мере примерно на 95%; еще более предпочтительно по меньшей мере на 99%; еще более предпочтительно на 100% последовательности, представленной в SEQ ID NO: 4.
В изобретении предложена искусственная молекула нуклеиновой кислоты, предпочтительно искусственная мРНК, которая может отличаться тем, что обеспечивает повышенную стабильность и пролонгирование экспрессии кодируемого пептида или белка. Не выдаваясь в какую-либо конкретную теорию, можно считать, что повышенная стабильность экспрессии белка и также пролонгированная экспрессия белка могут быть обусловлены уменьшением разложения искусственной молекулы нуклеиновой кислоты, такой как искусственная молекула мРНК, предлагаемая в настоящем изобретении. Таким образом, предлагаемый в изобретении 3'-UTR-элeмeнт может предупреждать разложение и расщепление искусственной нуклеиновой кислоты.
В некоторых вариантах осуществления изобретения искусственная молекула нуклеиновой кислоты может содержать гистоновую структуру типа «стебель-петля» в дополнение к нуклеотидной последовательности, происходящей из 3'-UTR гена рибосомального белка. Такая искусственная молекула нуклеиновой кислоты, предлагаемая в настоящем изобретении, может содержать, например, расположенные в 5'→3'-направлении ОРС, 3'-UTR-элемент, необязательную последовательность гистоновой структуры типа «стебель-петля», необязательную поли(А)-последовательность или сигнал полиаденилирования и необязательную поли(С)-последовательность. Она может содержать также расположенные в 5'→3'-направлении ОРС, 3'-UTR-элемент, необязательную поли(А)-последовательность, необязательную поли(С)-последовательность и необязательную последовательность гистоновой структуры типа «стебель-петля».
В предпочтительном варианте осуществления изобретения искусственная молекула нуклеиновой кислоты, предлагаемой в изобретении, содержит по меньшей мере одну последовательность гистоновой структуры типа «стебель-петля».
Последовательность гистоновой структуры типа «стебель-петля», которую можно применять в настоящем изобретении, предпочтительно выбирают по меньшей мере из одной из следующих формул (I) или (II):
формула (I) (последовательность структуры типа «стебель-петля» без пограничных элементов стебля):
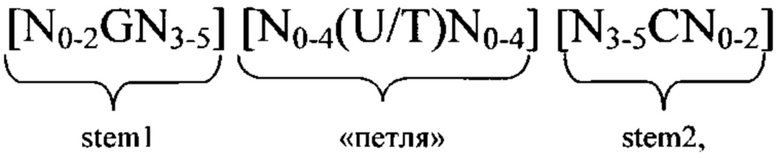
формула (II) (последовательность структуры типа «стебель-петля» с пограничными элементами стебля):
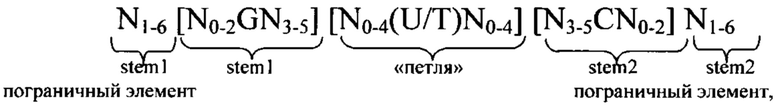
в которых:
где
у элементов stem1 и stem2 может происходить спаривание оснований друг с другом с образованием обратно комплементарной последовательности, где спаривание оснований может иметь место между steml и stem2, например, посредством спаривания оснований по Уотсону-Крику нуклеотидов А и U/T или G и С, или посредством спаривания оснований не по Уотсону-Крику, например, посредством спаривания «качающихся» оснований («качающегося» спаривания оснований), обратного спариванию оснований по Уотсону-Крику, посредством хугстиновского спаривания оснований, посредством обратного хугстиновскому спаривания оснований, или у них может происходить спаривание оснований друг с другом с образованием частично обратно комплементарной последовательности, при этом неполное спаривание оснований может иметь место между stem1 и stem2, вследствие того, что для одного или нескольких оснований в одном стебле не имеется комплементарного основания в обратно комплементарной последовательности другого стебля.
Согласно другому предпочтительному варианту осуществления изобретения мРНК, предлагаемая в настоящем изобретении, может содержать или кодировать по меньшей мере одну последовательность гистоновой структуры типа «стебель-петля», соответствующую по меньшей мере одной из следующих конкретных формул (Iа) или (IIа):
формула (Iа) (последовательность структуры типа «стебель-петля» без пограничных элементов стебля):
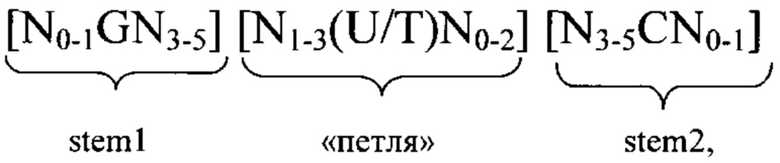
формула (IIа) (последовательность структуры типа «стебель-петля» с пограничными элементами стебля):
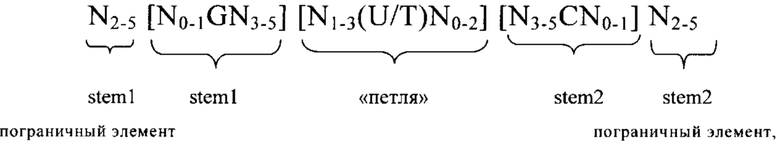
в которой:
N, С, G, Т и U имеют указанные выше значения.
Согласно другому предпочтительному варианту осуществления изобретения мРНК, предлагаемая в настоящем изобретении, может содержать или кодировать по меньшей мере одну последовательность гистоновой структуры типа «стебель-петля», соответствующую по меньшей мере одной из следующих конкретных формул (Ib) или (IIb):
формула (Ib) (последовательность структуры типа «стебель-петля» без пограничных элементов стебля):
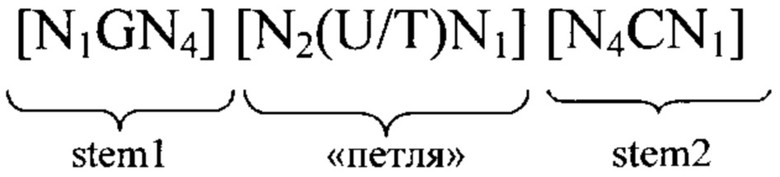
формула (IIb) (последовательность структуры типа «стебель-петля» с пограничными элементами стебля):
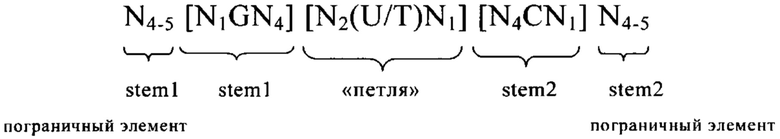
в которой:
N, С, G, Т и U имеют указанные выше значения.
Наиболее предпочтительной последовательностью гистоновой структуры типа «стебель-петля» является последовательность, представленная в SEQ ID NO: 5 CAAAGGCTCTTTTCAGAGCCACCA, или более предпочтительно последовательность РНК, соответствующая нуклеотидной последовательности, представленной в SEQ ID NO: 5.
Например, отдельные элементы могут присутствовать в искусственной молекуле нуклеиновой кислоты в следующем порядке:
5'-кэп - 5'-UTR - ОРС - 3'-UTR-элемент - гистоновая структура типа «стебель-петля» - поли(А)/(С)-последовательность;
5'-кэп - 5'-UTR - ОРС - 3'-UTR-элeмeнт - поли(А)/(С)-последовательность - гистоновая структура типа «стебель-петля»;
5'-кэп - 5'-UTR - ОРС - IRES - ОРС - 3'-UTR-элемент - гистоновая структура типа «стебель-петля» - поли(А)/(С)-последовательность;
5'-кэп - 5'-UTR - ОРС - IRES - ОРС - 3'-UTR-элемент - гистоновая структура типа «стебель-петля» - поли(А)/(С)-последовательность - поли(А)/(С)-последовательность;
5'-кэп - 5'-UTR - ОРС - IRES - ОРС - 3'-UTR-элемент - поли(А)/(С)-последовательность - гистоновая структура типа «стебель-петля»;
5'-кэп - 5'-UTR - ОРС - IRES - ОРС - 3'-UTR-элемент - поли(А)/(С)-последовательность - поли(А)/(С)-последовательность - гистоновая структура типа «стебель-петля»;
5'-кэп - 5'-UTR - ОРС - 3'-UTR-элeмeнт - поли(А)/(С)-последовательность - поли(А)/(С)-последовательность;
5'-кэп - 5'-UTR - ОРС - 3'-UTR-элeмeнт - поли(А)/(С)-последовательность - поли(А)/(С)-последовательность - гистоновая структура типа «стебель-петля»; и т.д.
В некоторых вариантах осуществления изобретения искусственная молекула нуклеиновой кислоты содержит дополнительные элементы, такие как 5'-кэп, поли(С)-последовательность и/или IRES-мотив. 5'-кэп может быть добавлен в процессе транскрипции или после транскрипции к 5'-концу РНК. Кроме того, предлагаемую в изобретении искусственную молекулу нуклеиновой кислоты, прежде всего в том случае, когда нуклеиновая кислота находится в форме мРНК или кодирует мРНК, можно модифицировать с помощью последовательности, состоящей по меньшей мере из 10 цитозинов, предпочтительно по меньшей мере из 20 цитозинов, более предпочтительно по меньшей мере из 30 цитозинов (так называемая «поли(С)-последовательность»). В частности, предлагаемая в изобретении искусственная молекула нуклеиновой кислоты может содержать, прежде всего в том случае, когда нуклеиновая кислота находится в форме мРНК или кодирует мРНК, поли(С)-последовательность которая, как правило, содержит примерно от 10 до 200 цитозиновых нуклеотидов, предпочтительно примерно от 10 до 100 цитидиновых нуклеотидов, более предпочтительно примерно от 10 до 70 цитидиновых нуклеотидов или еще более предпочтительно примерно от 20 до 50 или даже от 20 до 30 цитидиновых нуклеотидов. Наиболее предпочтительно, предлагаемая в изобретении нуклеиновая кислота содержит поли(С)-последовательность, состоящую из 30 цитидиновых оснований. Так, предпочтительно искусственная молекула нуклеиновой кислоты, предлагаемая в настоящем изобретении, содержит, предпочтительно в 5'→3' направлении ОРС, по меньшей мере один 3'-UTR-элeмeнт, описанный выше, поли(А)-последовательность или сигнал полиаденилирования и поли(С)-последовательность.
Последовательность внутреннего сайта связывания рибосом (IRES) или IRES-мотив может разделять несколько открытых рамок считывания, например в том случае, когда искусственная молекула нуклеиновой кислоты кодирует два или большее количество пептидов или белков. IRES-последовательность может быть наиболее полезной в том случае, когда искусственная молекула нуклеиновой кислоты представляет собой би- или полицистронную молекулу нуклеиновой кислоты.
Кроме того, искусственная молекула нуклеиновой кислоты может содержать дополнительные 5'-элементы, предпочтительно 5'-UTR, промотор или последовательность, содержащую 5'-UTR и промотор. Промотор может управлять транскрипцией или регулировать транскрипцию искусственной молекулы нуклеиновой кислоты предлагаемой в настоящем изобретении, например, искусственной молекулы ДНК, предлагаемой в настоящем изобретении. Кроме того, 5'-UTR может состоять или содержать 5'-UTR гена, представленную в настоящем описании. Кроме того, 5'-UTR может взаимодействовать с предлагаемым в изобретении 3'-UTR-элeмeнтoм и тем самым может поддерживать стабилизирующее действие предлагаемого в изобретении 3'-UTR-элeмeнтa. Такие элементы могут дополнительно поддерживать стабильность и эффективность трансляции. Следовательно, в некоторых вариантах осуществления изобретения предложены искусственные молекулы нуклеиновой кислоты, предпочтительно молекулы мРНК, содержащие в 5'→3'-направлении по меньшей мере одну из следующих структур
5'-кэп - 5'-UTR - ORF - 3'-UTR-элeмeнт - гистоновая структура типа «стебель-петля» - поли(А)/(С)-последовательность;
5'-кэп - 5'-UTR - ОРС - 3'-UTR-элeмeнт - поли(А)/(С)-последовательность - гистоновая структура типа «стебель-петля»;
5'-кэп - 5'-UTR - ОРС - IRES - ОРС - 3'-UTR-элемент - гистоновая структура типа «стебель-петля» - поли(А)/(С)-последовательность;
5'-кэп - 5'-UTR - ОРС - IRES - ОРС - 3'-UTR-элемент - гистоновая структура типа «стебель-петля» - поли(А)/(С)-последовательность - поли(А)/(С)-последовательность;
5'-кэп - 5'-UTR - ОРС - IRES - ОРС - 3'-UTR-элемент - поли(А)/(С)-последовательность - гистоновая структура типа «стебель-петля»;
5'-кэп - 5'-UTR - ОРС - IRES - ОРС - 3'-UTR-элемент - поли(А)/(С)-последовательность - поли(А)/(С)-последовательность - гистоновая структура типа «стебель-петля»;
5'-кэп - 5'-UTR - ОРС - 3'-UTR-элeмeнт - поли(А)/(С)-последовательность - поли(А)/(С)-последовательность;
5'-кэп - 5'-UTR - ОРС - 3'-UTR-элeмeнт - поли(А)/(С)-последовательность - поли(А)/(С)-последовательность - гистоновая структура типа «стебель-петля».
В наиболее предпочтительном варианте осуществления настоящего изобретения искусственная молекула нуклеиновой кислоты содержит по меньшей мере один элемент 5'-нетранслируемой области (5'UTR-элeмeнт) длиной менее чем 500, 400, 300, 250, 200, 150 или 100 нуклеотидов и/или более чем 20, 30, 40, 50 или 60 нуклеотидов, который предпочтительно содержит или состоит из нуклеотидной последовательности, которая происходит из 5'UTR ТОР-гена, или которая происходит из фрагмента, гомолога или варианта 5'UTR ТОР-гена.
Наиболее предпочтительно 5'UTR-элeмeнт не содержит ТОР-мотив или 5'-ТОР, указанный выше.
Нуклеотидная последовательность, которая происходит из 5'UTR ТОР-гена, происходит из эукараотического ТОР-гена, предпочтительно ТОР-гена растения или животного, более предпочтительно ТОР-гена хордовых, еще более предпочтительно ТОР-гена позвоночных, наиболее предпочтительно ТОР-гена млекопитающих, такого как ТОР-ген человека.
Например, 5'UTR-элемент предпочтительно выбирают из 5'-UTR-элементов, содержащих или состоящих из нуклеотидной последовательности, которая происходит из нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID NO: 1-1363, SEQ ID NO: 1395, SEQ ID NO: 1421 и SEQ ID NO: 1422, представленных в заявке на патент WO 2013/143700, описание которой включено в настоящее описание в качестве ссылки, из гомологов SEQ ID NO: 1-1363, SEQ ID NO: 1395, SEQ ID NO: 1421 и SEQ ID NO: 1422, представленных в заявке на патент WO 2013/143700, из ее варианта или предпочтительно из соответствующей последовательности РНК. Понятие «гомологи SEQ ID NO: 1-1363, SEQ ID NO: 1395, SEQ ID NO: 1421 и SEQ ID NO: 1422, представленные в заявке на патент WO 2013/143700» относится к последовательностям из видов кроме человека, которые гомологичны последовательностям SEQ ID NO: 1-1363, SEQ ID NO: 1395, SEQ ID NO: 1421 и SEQ ID NO: 1422, представленным в заявке на патент WO 2013/143700.
В предпочтительном варианте осуществления изобретения 5'UTR-элемент содержит или состоит из нуклеотидной последовательности, которая происходит из нуклеотидной последовательности, простирающейся от нуклеотидного положения 5 (т.е. нуклеотида, находящегося в последовательности в положении 5) до нуклеотида, непосредственно примыкающего с 5'-стороны к стартовому кодону (находящемуся на 3'-конце последовательностей), например, до нуклеотидного положения, непосредственно примыкающего с 5'-стороны к последовательности ATG нуклеотидной последовательности, выбранной из SEQ ID NO: 1-1363, SEQ ID NO: 1395, SEQ ID NO: 1421 и SEQ ID NO: 1422, представленных в заявке на патент WO 2013/143700, из гомологов SEQ ID NO: 1-1363, SEQ ID NO: 1395, SEQ ID NO: 1421 и SEQ ID NO: 1422, представленных в заявке на патент WO 2013/143700, из ее варианта, или соответствующей последовательности РНК. Наиболее предпочтительно 5'UTR-элемент происходит из нуклеотидной последовательности, простирающейся от нуклеотидного положения, непосредственно примыкающего с 3'-стороны к 5'-ТОР, до нуклеотидного положения, непосредственно примыкающего с 5'-стороны к стартовому кодону (находящемуся на 3'-конце последовательностей), например, до нуклеотидного положения, непосредственно примыкающего с 5'-стороны к последовательности ATG нуклеотидной последовательности выбранной из SEQ ID NO: 1-1363, SEQ ID NO: 1395, SEQ ID NO: 1421 и SEQ ID NO: 1422, представленных в заявке на патент WO 2013/143700, из гомологов SEQ ID NO: 1-1363, SEQ ID NO: 1395, SEQ ID NO: 1421 и SEQ ID NO: 1422, представленных в заявке на патент WO 2013/143700, из ее варианта, или соответствующей последовательности РНК.
В наиболее предпочтительном варианте осуществления изобретения 5'-UTR-элемент содержит или состоит из нуклеотидной последовательности, происходящей из 5'-UTR ТОР-гена, кодирующего рибосомальный белок, или из варианта 5'-UTR ТОР-гена, кодирующего рибосомальный белок. Например, 5'-UTR-элемент содержит или состоит из нуклеотидной последовательности, происходящей из 5'-UTR нуклеотидной последовательности, представленной в любой из SEQ ID NO: 170, 232, 244, 259, 1284, 1285, 1286, 1287, 1288, 1289, 1290, 1291, 1292, 1293, 1294, 1295, 1296, 1297, 1298, 1299, 1300, 1301, 1302, 1303, 1304, 1305, 1306, 1307, 1308, 1309, 1310, 1311, 1312, 1313, 1314, 1315, 1316, 1317, 1318, 1319, 1320, 1321, 1322, 1323, 1324, 1325, 1326, 1327, 1328, 1329, 1330, 1331, 1332, 1333, 1334, 1335, 1336, 1337, 1338, 1339, 1340, 1341, 1342, 1343, 1344, 1346, 1347, 1348, 1349, 1350, 1351, 1352, 1353, 1354, 1355, 1356, 1357, 1358, 1359 или 1360, представленных в заявке на патент WO 2013/143700, соответствующей последовательности РНК, ее гомолога или ее варианта, представленного в настоящем описании, предпочтительно без 5'-ТОР-мотива. Как описано выше, последовательность, простирающаяся от положения 5 до нуклеотида, непосредственно примыкающего с 5'-стороны к ATG (который находится на 3'-конце последовательностей), соответствует 5'-UTR указанных последовательностей.
Предпочтительно 5'UTR-элемент содержит или состоит из нуклеотидной последовательности, которая происходит из 5'-UTR ТОР-гена, кодирующего рибосомальный белок большой субъединицы (RPL), или из гомолога или варианта 5'-UTR ТОР-гена, кодирующего рибосомальный белок большой субъединицы (RPL). Например, 5'-UTR-элeмeнт содержит или состоит из нуклеотидной последовательности, происходящей из 5'-UTR нуклеотидной последовательности, представленной в любой из SEQ ID NO: 67, 259, 1284-1318, 1344, 1346, 1348-1354, 1357, 1358, 1421 и 1422, описанных в заявке на патент WO 2013/143700, соответствующей последовательности РНК, ее гомолога или ее варианта, представленного в настоящем описании, предпочтительно без 5'-ТОР-мотива.
В наиболее предпочтительном варианте осуществления изобретения 5'-UTR-элемент содержит или состоит из нуклеотидной последовательности, происходящей из 5'-UTR гена рибосомального белка большой субъединицы 32, предпочтительно из гена рибосомального белка большой субъединицы 32 (L32) позвоночных, более предпочтительно гена рибосомального белка большой субъединицы 32 (L32) млекопитающих, наиболее предпочтительно из гена рибосомального белка большой субъединицы 32 (L32) человека, или из варианта 5'-UTR гена рибосомального белка большой субъединицы 32, предпочтительно из гена рибосомального белка большой субъединицы 32 (L32) позвоночных, более предпочтительно из гена рибосомального белка большой субъединицы 32 (L32) млекопитающих, наиболее предпочтительно из гена рибосомального белка большой субъединицы 32 (L32) человека, где предпочтительно 5'-UTR-элемент не содержит 5'-ТОР указанного гена.
Следовательно, в наиболее предпочтительном варианте осуществления изобретения, 5'-UTR-элемент содержит или состоит из нуклеотидной последовательности, идентичной по меньшей мере примерно на 40%, предпочтительно по меньшей мере примерно на 50%, предпочтительно по меньшей мере примерно на 60%, предпочтительно по меньшей мере примерно на 70%, более предпочтительно по меньшей мере примерно на 80%, более предпочтительно по меньшей мере примерно на 90%, еще более предпочтительно по меньшей мере примерно на 95%, еще более предпочтительно по меньшей мере примерно на 99% нуклеотидной последовательности, представленной в SEQ ID NO: 6 (5'-UTR рибосомального белка большой субъединицы 32 человека без 5'-концевого олигопиримидинового тракта: GGCGCTGCCTACGGAGGTGGCAGCCATCTCCTTCTCGGCATC; соответствующая SEQ ID NO: 1368, представленной в заявке на патент WO 2013/143700) или предпочтительно соответствующей последовательности РНК, или где по меньшей мере один 5'UTR-элемент содержит или состоит из фрагмента нуклеотидной последовательности, идентичного по меньшей мере примерно на 40%, предпочтительно по меньшей мере примерно на 50%, предпочтительно по меньшей мере примерно на 60%, предпочтительно по меньшей мере примерно на 70%, более предпочтительно по меньшей мере примерно на 80%, более предпочтительно по меньшей мере примерно на 90%, еще более предпочтительно по меньшей мере примерно на 95%, еще более предпочтительно по меньшей мере примерно на 99% нуклеотидной последовательности, представленной в SEQ ID NO: 6, или более предпочтительно соответствующей последовательности РНК, где предпочтительно фрагмент представляет собой описанный выше фрагмент, непрерывный сегмент нуклеотидов, представляющий собой по меньшей мере 20% и т.д. полноразмерной 5'-UTR. Предпочтительно фрагмент имеет длину по меньшей мере примерно 20 нуклеотидов или более, предпочтительно по меньшей мере примерно 30 нуклеотидов или более, более предпочтительно по меньшей мере примерно 40 нуклеотидов или более. Предпочтительно фрагмент представляет собой функциональный фрагмент, представленный в настоящем описании.
В некоторых вариантах осуществления изобретения искусственная молекула нуклеиновой кислоты содержит 5'-UTR-элемент, который содержит или состоит из нуклеотидной последовательности, происходящей из 5'-UTR ТОР-гена позвоночных, например, млекопитающих, например, ТОР-гена человека, выбранного из RPSA, RPS2, RPS3, RPS3A, RPS4, RPS5, RPS6, RPS7, RPS8, RPS9, RPS10, RPS11, RPS12, RPS13, RPS14, RPS15, RPS15A, RPS16, RPS17, RPS18, RPS19, RPS20, RPS21, RPS23, RPS24, RPS25, RPS26, RPS27, RPS27A, RPS28, RPS29, RPS30, RPL3, RPL4, RPL5, RPL6, RPL7, RPL7A, RPL8, RPL9, RPL10, RPL10A, RPL11, RPL12, RPL13, RPL13A, RPL14, RPL15, RPL17, RPL18, RPL18A, RPL19, RPL21, RPL22, RPL23, RPL23A, RPL24, RPL26, RPL27, RPL27A, RPL28, RPL29, RPL30, RPL31, RPL32, RPL34, RPL35, RPL35A, RPL36, RPL36A, RPL37, RPL37A, RPL38, RPL39, RPL40, RPL41, RPLPO, RPLP1, RPLP2, RPLP3, RPLPO, RPLP1, RPLP2, EEF1A1, EEF1B2, EEF1D, EEF1G, EEF2, EIF3E, EIF3F, EIF3H, EIF2S3, EIF3C, EIF3K, EIF3EIP, EIF4A2, РАВРС1, HNRNPA1, ТРТ1, TUBB1, UBA52, NPM1, ATP5G2, GNB2L1, NME2, UQCRB или из его гомолога или варианта, где предпочтительно 5'-UTR-элeмeнт не содержит ТОР-мотив или 5'-ТОР указанных генов, и где необязательно 5'-UTR-элeмeнт начинается на его 5'-конце с нуклеотида, находящегося в положении 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 в прямом направлении относительно 5'-концевого олигопиримидинового тракта (ТОР) и где необязательно также 5'-UTR-элeмeнт, происходящий из 5'-UTR ТОР-гена, заканчивается на его 3'-конце нуклеотидом, находящимся в положении 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 в обратном направлении относительно стартового кодона (A(U/T)G) гена, из которого он происходит.
Предпочтительно в искусственной молекуле нуклеиновой кислоты, предлагаемой в настоящем изобретении, предпочтительно в открытой рамке считывания, модифицируют по меньшей мере содержание G/C. Так, предлагаемую в изобретении искусственную молекулу нуклеиновой кислоты можно стабилизировать с точки зрения ее термодинамических характеристик путем модификации содержания G (гуанозин)/С (цитидин) в молекуле. Содержание G/C в открытой рамке считывания искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, можно повышать по сравнению с содержанием G/C в открытой рамке считывания соответствующей последовательности дикого типа, предпочтительно на основе вырожденности генетического кода. Так, аминокислотную последовательность, кодируемую искусственной молекулой нуклеиновой кислоты, предпочтительно не модифицируют посредством модификации содержания G/C по сравнению с конкретной кодируемой аминокислотной последовательностью дикого типа. Таким образом, можно варьировать кодоны кодирующей последовательности или всей искусственной молекулы нуклеиновой кислоты, например, мРНК, по сравнению с кодирующей последовательностью дикого типа таким образом, чтобы они содержали большее количество G/C-нуклеотидов, сохраняя при этом транслируемую аминокислотную последовательность. Благодаря тому факту, что несколько кодонов кодируют одну и ту же аминокислоту (так называемая вырожденность генетического кода), можно изменять кодоны, не изменяя кодируемую последовательность пептида/белка (использование так называемых альтернативных кодонов). Следовательно, можно специально интродуцировать определенные кодоны (осуществляя обмен соответствующих кодонов дикого типа, кодирующих ту же самую аминокислоту), которые являются более предпочтительными с точки зрения стабильности РНК и/или наиболее часто встречающихся кодонов в организме индивидуума (так называемая оптимизация кодонов).
В зависимости от аминокислоты, которая должна кодироваться кодирующей областью предлагаемой в изобретении искусственной молекулы нуклеиновой кислоты, представленной в настоящем описании, существуют различные возможности для модификации нуклеотидной последовательности, например, открытой рамки считывания, по сравнению с ее кодирующей областью дикого типа. В случае аминокислот, которые кодируются кодонами, содержащими только нуклеотиды G или С, модификации кодонов не требуется. Таким образом, кодоны для Pro (ССС или CCG), Arg (CGC или CGG), Ala (GCC или GCG) и Gly (GGC или GGG) не требуют модификации, поскольку в них отсутствуют А или U/T.
В противоположность этому, кодоны, которые содержат нуклеотиды А и/или U, можно модифицировать путем замены на другие кодоны, которые кодируют эти же аминокислоты, но не содержат А и/или U.
Например,
кодирующие Pro кодоны СС(U/T) или ССА можно модифицировать на ССС или CCG;
кодирующие Arg кодоны CG(U/T) или CGA, или AGA, или AGG можно модифицировать на CGC или CGG;
кодирующие Ala кодоны GC(U/T) или GCA можно модифицировать на GCC или GCG;
кодирующие Gly кодоны GG(U/T) илиr GGA можно модифицировать на GGC или GGG.
В других случаях, когда нуклеотиды А или U нельзя элиминировать из кодонов, можно снижать содержание А и U с использованием кодонов, которые содержат нуклеотиды А и/или U в более низком количестве. Примерами указанных колонов являются:
кодирующие Phe кодоны (U/T)(U/T)(U/T) можно модифицировать на (U/T) (U/T)C;
кодирующие Leu кодоны (U/T) (U/T)A, (U/T) (U/T)G, C(U/T) (U/T) или C(U/T)A можно модифицировать на C(U/T)C или C(U/T)G;
кодирующие Ser кодоны (U/T)C(U/T) или (U/T)CA, или AG(U/T) можно модифицировать на (U/T)CC, (U/T)CG или AGC;
кодирующий Tyr кодон (U/T)A(U/T) можно модифицировать на (U/T)AC;
кодирующий Cys кодон (U/T)G(U/T) можно модифицировать на (U/T)GC;
кодирующий His кодон CA(U/T) можно модифицировать на САС;
кодирующий Gln кодон САА можно модифицировать на CAG;
кодирующие Ile кодоны A(U/T)(U/T) или A(U/T)A можно модифицировать на A(U/T)C
кодирующие Thr кодоны AC(U/T) или АСА можно модифицировать на АСС или ACG
кодирующий Asn кодон AA(U/T) можно модифицировать на ААС;
кодирующий Lys кодоны AAA можно модифицировать на AAG;
кодирующие Val кодоны G(U/T)(U/T) или G(U/T)A можно модифицировать на G(U/T)C или G(U/T)G;
кодирующий Asp кодон GA(U/T) можно модифицировать на GAC;
кодирующий Glu кодон GAA можно модифицировать на GAG;
стоп-кодон (U/T)AA можно модифицировать на (U/T)AG или (U/T)GA.
С другой стороны, в случае кодонов Met (AUG) и Trp (UGG), отсутствует возможность модификации последовательности без изменения кодируемой аминокислотной последовательности.
Перечисленные выше замены можно использовать либо индивидуально, либо в любых возможных комбинациях для повышения содержания G/C в открытой рамке считывания в предлагаемой в настоящем изобретении искусственной молекуле нуклеиновой кислоты, представленной в настоящем описании, по сравнению с конкретной открытой рамкой считывания дикого типа (т.е. исходной последовательности). Так, например, все кодоны Thr, встречающиеся в последовательности дикого типа, можно модифицировать на АСС (или ACG).
Предпочтительно содержание G/C в открытой рамке считывания в предлагаемой в настоящем изобретении искусственной молекуле нуклеиновой кислоты, представленной в настоящем описании, повышают по меньшей мере на 7%, более предпочтительно по меньшей мере на 15%, наиболее предпочтительно по меньшей мере на 20%, по сравнению с содержанием G/C в кодирующей области мРНК дикого типа без изменения кодируемой аминокислотной последовательности, т.е. используя вырожденность генетического кода. В конкретном варианте осуществления изобретения заменяют по меньшей мере 5%, 10%, 20%, 30%, 40%, 50%, 60%, более предпочтительно по меньшей мере 70%, еще более предпочтительно по меньшей мере 80% и наиболее предпочтительно по меньшей мере 90%, 95% или даже 100% пригодных для замены кодонов в открытой рамке считывания в предлагаемой в изобретении искусственной молекуле нуклеиновой кислоты или ее фрагменте, варианте или производном, повышая тем самым содержание G/C в указанной открытой рамке считывания.
В этом контексте наиболее предпочтительно повышать содержание G/C в открытая рамке считывания в предлагаемой в изобретении искусственной молекуле нуклеиновой кислоты, представленной в настоящем описании, до максимума (т.е. до 100% пригодных для замещения кодонов) по сравнению с открытой рамкой считывания дикого типа без изменения кодируемой аминокислотной последовательности.
Кроме того, в открытой рамке считывания предпочтительно осуществляют по меньшей мере частично оптимизацию кодонов. Оптимизация кодонов основана на открытии того факта, что эффективность трансляции может определяться также различной частотой встречаемости транспортных РНК (тРНК) в клетках. Таким образом, если в кодирующей области, предлагаемой в изобретении искусственной молекуле нуклеиновой кислоты, представленной в настоящем описании, присутствуют в повышенном количестве так называемые «редкие кодоны», то уровень трансляции соответствующей модифицированной нуклеотидной последовательности является более низким, чем в том случае, когда присутствуют кодоны, кодирующие относительно «часто встречающуюся» тРНК.
Так, открытую рамку считывания предлагаемой в изобретении искусственной молекуле нуклеиновой кислоты предпочтительно модифицируют по сравнению с соответствующей кодирующей областью дикого типа таким образом, чтобы по меньшей мере один кодон в последовательности дикого типа, который кодирует тРНК, которая является относительно редкой в клетке, был заменен на кодон, который кодирует тРНК, которая встречается в клетке относительно часто и «несет» такую же аминокислоту, что и относительно редко встречающаяся тРНК. Посредством такой модификации открытую рамку считывания предлагаемой в изобретении искусственной молекулы нуклеиновой кислоты, представленной в настоящем описании, модифицируют таким образом, чтобы кодонами, для которых доступны часто встречающиеся тРНК, заменять кодоны, которые соответствуют редким тРНК. Другими словами, согласно изобретению с помощью такой модификации все кодоны в открытой рамке считывания дикого типа, которые кодируют тРНК, являющуюся относительно редкой в клетке, можно заменять на кодоны, которые кодируют тРНК, относительно часто встречающуюся в клетке, и которая в каждом случае «несет» такую же аминокислоту, что и относительно редкая тРНК. тРНК, встречающиеся относительно часто в клетке, и наоборот, встречающиеся относительно редко, известны специалистам в данной области, см., например, Akashi, Curr. Opin. Genet. Dev., 11 (6), 2001, cc. 660-666. Таким образом, предпочтительно открытая рамка считывания имеет оптимизированные кодоны, предпочтительно применительно к системе, в которой должна экспрессироваться искусственная молекула нуклеиновой кислоты, предлагаемая в настоящем изобретении, предпочтительно применительно к системе, в которой должна транслироваться искусственная молекула нуклеиновой кислоты, предлагаемая в настоящем изобретении. Предпочтительно, оптимизацию кодонов для наиболее часто встречающихся кодонов в открытой рамке считывания осуществляют в соответствии с наиболее часто встречающимися кодонами млекопитающих, более предпочтительно в соответствии с наиболее часто встречающимися кодонами человека. Предпочтительно, в открытой рамке считывания осуществляют и оптимизацию кодонов, и модификацию содержания G/C.
Для дополнительного повышения устойчивости к расщеплению, например, устойчивости к расщеплению in vivo экзо- или эндонуклеазой, и/или для дополнительного повышения стабильности экспрессии белка с искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, можно осуществлять дополнительные модификации искусственной молекулы нуклеиновой кислоты, такие как модификации каркаса, модификации сахаров и/или модификации оснований, например, липидные модификации или т.п. Предпочтительно, такие модификации не приводят к значимому ухудшению транскрипции и/или трансляции искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении.
Как правило, искусственная молекула нуклеиновой кислоты, предлагаемая в настоящем изобретении, может содержать любой нативный (встречающийся в естественных условиях) нуклеотид, например, гуанозин, урацил, аденозин и/или цитозин, или их аналоги. В этом контексте нуклеотидные аналоги классифицируют как встречающиеся в естественных условиях и не встречающиеся в естественных условиях варианта встречающихся в естественных условиях нуклеотидов аденозина, цитозина, тимидина, гуанозина и уридина. Таким образом, аналоги представляют собой, например, химически модифицированные нуклеотиды, несущие не встречающиеся в естественных условиях функциональные группы, которые предпочтительно добавляют или изымают путем делеции из встречающегося в естественных условиях нуклеотида, или которыми заменяют встречающиеся в естественных условиях функциональные группы нуклеотида. Следовательно, можно модифицировать каждый компонент встречающегося в естественных условиях нуклеотида, а именно, Компонент, представляющий собой основание, компонент, представляющий собой сахар (рибоза) компонент и/или компонент, представляющий собой фосфат, формирующий каркас (см. выше) последовательности РНК. Аналоги гуанозина, урацила, аденозина и цитозина включают (но, не ограничиваясь только ими) любой встречающийся в естественных условиях или не встречающийся в естественных условиях гуанозин, урацил, аденозин, тимидин или цитозин, который был изменен химически, например, ацетилированием, метилированием, гидроксилированием и т.д., включая, например, 1-метиладенозин, 1-метилгуанозин, 1-метилинозин, 2,2-диметилгуанозин, 2,6-диаминопурин, 2'-амино-2'-дезоксиаденозин, 2'-амино-2'-дезоксицитидин, 2'-амино-2'-дезоксигуанозин, 2'-амино-2'-дезоксиуридин, 2-амино-6-хлорпуринорибозид, 2-аминопуринрибозид, 2'-арааденозин, 2'-арацитидин, 2'-арауридин, 2'-Азидо-2'-дезоксиаденозин, 2'-азидо-2'-дезоксицитидин, 2'-азидо-2'-дезоксигуанозин, 2'-азидо-2'-дезоксиуридин, 2-хлораденозин, 2'-фтор-2'-дезоксиаденозин, 2'-фтор-2'-дезоксицитидин, 2'-фтор-2'-дезоксигуанозин, 2'-фтор-2'-дезоксиуридин, 2'-фтортимидин, 2-метиладенозин, 2-метилгуанозин, 2-метилтио-N6-изопенениладенозин, 2'-O-метил-2-аминоаденозин, 2'-O-метил-2'-дезоксиаденозин, 2'-Ометил-2'-дезоксицитидин, 2'-O-метил-2'-дезоксигуанозин, 2'-O-метил-2'-дезоксиуридин, 2'-O-метил-5-метилуридин, 2'-O-метилинозин, 2'-O-метилпсевдоуридин, 2-тиоцитидин, 2-тиоцитозин, 3-метилцитозин, 4-ацетилцитозин, 4-тиоуридин, 5-(карбоксигидроксиметил)урацил, 5,6-дигидроуридин, 5-аминоаллилцитидин, 5-аминоаллилдезоксиуридин, 5-бромуридин, 5-карбоксиметиламинометил-2-тиоурацил, 5-карбоксиметиламинометилурацил, 5-хлор арацитозин, 5-фторуридин, 5-йодуридин, 5-метоксикарбонилметилуридин, 5-метоксиуридин, 5-метил-2-тиоуридин, 6-азацитидин, 6-азауридин, 6-хлор-7-деазагуанозин, 6-хлорпуринорибозид, 6-меркаптогуанозин, 6-метилмеркаптопуринрибозид, 7-деаза-2'-дезоксигуанозин, 7-деазааденозин, 7-метилгуанозин, 8-азааденозин, 8-бромаденозин, 8-бромгуанозин, 8-меркапторгуанозин, 8-оксогуанозин, бензимидазолрибозид, бета-D-маннозилквеозин, дигидроурацил, инозин, N1-метиладенозин, N6-([6-аминогексил]карбамоилметил)аденозин, N6-изопентениладенозин, N6-метиладенозин, N7-метилксантозин, метиловый эфир N-урацил-3-оксиуксусной кислоты, пуромицин, квеозин, урацил-5-оксиуксусная кислота, метиловый эфир урацил-5-оксиуксусной кислоты, вибутоксозин, ксантозин и ксилоаденозин. Получение таких аналогов известно специалисту в данной области, например, описано в US 4373071, US 4401796, US 4415732, US 4458066, US 4500707, US 4668777, US 4973679, US 5047524, US 5132418, US 5153319, US 5262530 и 5700642. Касательно аналогов, описанных выше, наиболее предпочтительными согласно некоторым вариантам осуществления изобретения могут быть те аналоги, которые увеличивают белковую экспрессию кодируемого пептида или белка, или повышают иммуногенность искусственной молекулы нуклеиновой кислоты, предлагаемой в изобретении, и/или не взаимодействуют с другой интродуцированной модификацией мРНК.
Согласно конкретному варианту осуществления изобретения мРНК в композиции, предлагаемой в настоящем изобретении, может содержать липидную модификацию.
В предпочтительном варианте осуществления изобретения искусственная молекула нуклеиновой кислоты содержит, предпочтительно в 5'→3'-направлении, следующие элементы:
5'-UTR;
по меньшей мере одну открытую рамку считывания (ОРС), где ОРС предпочтительно содержит по меньшей мере одну модификацию по сравнению с последовательностью дикого типа;
3'-UTR, происходящую из 3'-UTR рибосомального белка, предпочтительно из нуклеотидной последовательности, представленной в любой из SEQ ID NO: 10-115, более предпочтительно 3'-UTR RPS9, более предпочтительно 3'-UTR RPS9 человека;
поли(А)-последовательность, предпочтительно содержащую 64 аденилата;
поли(С)-последовательность, предпочтительно содержащую 30 цитидилатов;
гистоновую структуру типа «стебель-петля».
В другом предпочтительном варианте осуществления изобретения искусственная молекула нуклеиновой кислоты содержит или состоит из нуклеотидной последовательности, представленной в SEQ ID NO: 7 (см. фиг. 3) или комплементарной последовательности ДНК.
В наиболее предпочтительном варианте осуществления изобретения искусственная молекула нуклеиновой кислоты, предлагаемая в изобретении, может содержать также одну или несколько описанных ниже модификаций:
Химические модификации:
В контексте настоящего описания понятие «модификация» применительно к искусственной молекуле нуклеиновой кислоты может относиться к химическим модификациям, включающим модификации каркаса, а также модификации сахара или модификации основания.
В этом контексте искусственная молекула нуклеиновой кислоты, предпочтительно молекула РНК, представленная в настоящем описании, может содержать нуклеотидные аналоги/модификации, например, модификации каркаса, модификации сахара или модификации основания. Модификация каркаса согласно настоящему изобретению представляет собой модификацию, при которой химически модифицируют фосфаты каркаса нуклеотидов, содержащихся в молекуле нуклеиновой кислоты, представленной в настоящем описании. Модификация сахара согласно настоящему изобретению представляет собой химическую модификацию сахара нуклеотида в молекуле нуклеиновой кислоты, представленной в настоящем описании. Кроме того, модификация основания согласно настоящему изобретению представляет собой химическую модификацию оснóвного фрагмента нуклеотидов в молекуле нуклеиновой кислоты. В этом контексте нуклеотидные аналоги или модификации предпочтительно выбирают из нуклеотидных аналогов, пригодных для транскрипции и/или трансляции.
Модификации сахара:
Модифицированные нуклеозиды и нуклеотиды, которые могут быть включены в искусственную молекулу нуклеиновой кислоты, предпочтительно РНК, представленную в настоящем описании, могут быть модифицированы в сахарном фрагменте. Например, 2'-гидроксигруппу (ОН) в молекуле РНК можно модифицировать или заменять многочисленными «окси»- или «дезокси»-заместителями. Примерами модификаций, включающих «окси»-2'-гидроксигруппы, являются (но, не ограничиваясь только ими) алкоксигруппы или арилоксигруппы (-OR, например, R=Н, алкил, циклоалкил, арил, аралкил, гетероарил или сахар); полиэтиленгликоли (PEG), -O(CH2CH2O)nCH2CH2OR; «замкнутые» нуклеиновые кислоты (LNA), в которых 2'-гидроксил присоединен, например, с помощью метиленового мостика к 4'-углероду в том же самом сахарном фрагменте, представляющем собой рибозу; и аминогруппы (-О-аминогруппы, в которых аминогруппа, например, NRR, может представлять собой алкиламиногруппу, диалкиламиногруппу, гетероциклил, ариламиногруппу, диариламиногруппу, гетероариламиногруппу или дигетероариламиногруппу, этилендиамин, полиаминогруппу) или алкоксигруппу.
«Дезокси»-модификация включает водород, аминогруппу (например, NH2; алкиламиногруппу, диалкиламиногруппу, гетероциклил, ариламиногруппу, диариламиногруппу, гетероариламиногруппу, дигетероариламиногруппу или аминокислоту); или аминогруппу можно присоединять к сахару через линкер, где линкер содержит один или несколько атомов С, N и О.
Сахарная группа может содержать также один или несколько атомов углерода, которые могут находиться в стереохимической конфигурации, отличной от конфигурации соответствующего атома углерода в рибозе. Так, модифицированная молекула нуклеиновой кислоты может включать, например, в качестве сахара арабинозу.
Модификации каркаса:
Можно модифицировать также фосфатный каркас в модифицированных нуклеозидах и нуклеотидах, которые могут быть встроены в искусственную молекулу нуклеиновой кислоты, предпочтительно РНК, представленную в настоящем описании. Фосфатные группы в каркасе можно модифицировать посредством замены одного или нескольких атомов кислорода на различные заместители. Кроме того, в модифицированных нуклеозидах и нуклеотидах немодифицированный фосфатный фрагмент может быть полностью заменен на модифицированный фосфат, представленный в настоящем описании. Примеры модифицированных фосфатных групп включают (но, не ограничиваясь только ими) фосфоротиоаты, фосфороселенаты, боранофосфаты, сложные боранофосфатные эфиры, первичные фосфонаты, фосфороамидаты, алкил- или арилфосфонаты и сложные фосфотриэфиры. В фосфородитиоатах оба несвязывающих атома кислорода заменены серой. Фосфатный линкер также можно модифицировать путем замены связывающего атома кислорода на азот (мостиковые фосфороамидаты), серу (мостиковые фосфоротиоаты) и углерод (мостиковые метиленфосфонаты).
Модификации основания:
Модифицированные нуклеозиды и нуклеотиды, которые можно встраивать в искусственную молекулу нуклеиновой кислоты, предпочтительно молекулу РНК, представленную в настоящем описании, можно модифицировать также в фрагменте, представляющем собой нуклеооснование. Примерами нуклеооснований, входящих в РНК, являются (но, не ограничиваясь только ими) аденин, гуанин, цитозин и урацил. Например, нуклеозиды и нуклеотиды, представленные в настоящем описании, можно химически модифицировать на поверхности большой бороздки. В некоторых вариантах осуществления изобретения химические модификации главной бороздки включают аминогруппу, тиольную групп алкильную группу или галогруппу.
В наиболее предпочтительных вариантах осуществления настоящего изобретения нуклеотидные аналоги/модификации выбирают из модификаций основания, которые предпочтительно выбирают из 2-амино-6-хлорпуринрибозид-5'-трифосфат, 2-аминоаденозин-5'-трифосфат, 2-тиоцитидин-5'-трифосфат, 2-тиоуридин-5'-трифосфат, 4-тиоуридин-5'-трифосфат, 5-аминоаллилцитидин-5'-трифосфат, 5-аминоаллилуридин-5'-трифосфат, 5-бромцитидин-5'-трифосфат, 5-бромуридин-5'-трифосфат, 5-йодцитидин-5'-трифосфат, 5-йодуридин-5'-трифосфат, 5-метилцитидин-5'-трифосфат, 5-метилуридин-5'-трифосфат, 6-азацитидин-5'-трифосфат, 6-азауридин-5'-трифосфат, 6-хлорпуринрибозид-5'-трифосфат, 7-деазааденозин-5'-трифосфат, 7-деазагуанозин-5'-трифосфат, 8-азааденозин-5'-трифосфат, 8-азидоаденозин-5'-трифосфат, бензимидазолрибозид-5'-трифосфат, N1-метиладенозин-5'-трифосфат, N1-метилгуанозин-5'-трифосфат, N6-метиладенозин-5'-трифосфат, 06-метилгуанозин-5'-трифосфат, псевдоуридин-5'-трифосфат или пуромицин-5'-трифосфат, ксантозин-5'-трифосфат. Наиболее предпочтительные нуклеотиды для модификаций оснований выбирают из группы нуклеотидов с модифицированными основаниями, включающей 5-метилцитидин-5'-трифосфат, 7-деазагуанозин-5'-трифосфат, 5-бромцитидин-5'-трифосфат и псевдоуридин-5'-трифосфат.
В некоторых вариантах осуществления изобретения модифицированные нуклеозиды включают пиридин-4-оновый рибонуклеозид, 5-азауридин, 2-тио-5-азауридин, 2-тиоуридин, 4-тиопсевдоуридин, 2-тиопсевдоуридин, 5-гидроксиуридин, 3-метилуридин, 5-карбоксиметилуридин, 1-карбоксиметилпсевдоуридин, 5-пропинилуридин, 1-пропинилпсевдоуридин, 5-тауринометилуридин, 1-тауринометилпсевдоуридин, 5-тауринометил-2-тиоуридин, 1-тауринометил-4-тиоуридин, 5-метилуридин, 1-метилпсевдоуридин, 4-тио-1-метилпсевдоуридин, 2-тио-1-метилпсевдоуридин, 1-метил-1-деазапсевдоуридин, 2-тио-1-метил-1-деазапсевдоуридин, дигидроуридин, дигидропсевдоуридин, 2-тиодигидроуридин, 2-тиодигидропсевдоуридин, 2-метоксиуридин, 2-метокси-4-тиоуридин, 4-метоксипсевдоуридин и 4-метокси-2-тиопсевдоуридин.
В некоторых вариантах осуществления изобретения модифицированные нуклеозиды включают 5-азацитидин, псевдоизоцитидин, 3-метилцитидин, N4-ацетилцитидин, 5-формилцитидин, N4-метилцитидин, 5-гидроксиметилцитидин, 1-метилпсевдоизоцитидин, пирролоцитидин, пирролопсевдоизоцитидин, 2-тиоцитидин, 2-тио-5-метилцитидин, 4-тиопсевдоизоцитидин, 4-тио-1-метилпсевдоизоцитидин, 4-тио-1-метил-1-деазапсевдоизоцитидин, 1-метил-1-деазапсевдоизоцитидин, зебуларин, 5-азазебуларин, 5-метилзебуларин, 5-аза-2-тиозебуларин, 2-тиозебуларин, 2-метоксицитидин, 2-метокси-5-метилцитидин, 4-метоксипсевдоизоцитидин и 4-метокси-1-метилпсевдоизоцитидин.
В других вариантах осуществления изобретения модифицированные нуклеозиды включают 2-аминопурин, 2, 6-диаминопурин, 7-деазааденин, 7-деаза-8-азааденин, 7-деаза-2-аминопурин, 7-деаза-8-аза-2-аминопурин, 7-деаза-2,6-диаминопурин, 7-деаза-8-аза-2,6-диаминопурин, 1-метиладенозин, N6-метиладенозин, N6-изопентениладенозин, N6-(цис-гидроксиизопентенил)аденозин, 2-метилтио-N6-(цис-гидроксиизопентенил) аденозин, N6-глицинилкарбомоиладенозин, N6-треонилкарбамоиладенозин, 2-метилтио-N6-треонилкарбамоиладенозин, N6,N6-диметиладенозин, 7-метиладенин, 2-метилтиоаденин и 2-метоксиаденин.
В других вариантах осуществления изобретения модифицированные нуклеозиды включают инозин, 1-метилинозин, виозин, вибутозин, 7-деазагуанозин, 7-деаза-8-азагуанозин, 6-тиогуанозин, 6-тио-7-деазагуанозин, 6-тио-7-деаза-8-азагуанозин, 7-метилгуанозин, 6-тио-7-метилгуанозин, 7-метилинозин, 6-метоксигуанозин, 1-метилгуанозин, N2-метилгуанозин, N2,N2-диметилгуанозин, 8-оксогуанозин, 7-метил-8-оксогуанозин, 1-метил-6-тиогуанозин, N2-метил-6-тиогуанозин и N2,N2-диметил-6-тиогуанозин.
В некоторых вариантах осуществления изобретения нуклеотид можно модифицировать на поверхности главной бороздки и модификация может включать замену атома водорода на С-5 урацила метильной группой или галогруппой.
В конкретных вариантах осуществления изобретения модифицированный нуклеозид представляет собой 5'-O-(1-тиофосфат)-аденозин, 5'-O-(1-тиофосфат)-цитидин, 5'-O-(1-тиофосфат)-гуанозин, 5'-O-(1-тиофосфат)-уридин или 5'-O-(1-тиофосфат)-псевдоуридин.
В других конкретных вариантах осуществления изобретения искусственная молекула нуклеиновой кислоты, предпочтительно молекула РНК, может содержать нуклеозидные модификации, выбранные из 6-азацитидина, 2-тиоцитидина, альфа-тиоцитидина, псевдоизоцитидина, 5-аминоаллилуридина, 5-йодуридина, N1-метилпсевдоуридина, 5,6-дигидроуридина, альфа-тиоуридина, 4-тиоуридина, 6-азауридина, 5-гидроксиуридина, дезокситимидина, 5-метилуридина, пирролоцитидина, инозина, альфа-тиогуанозина, 6-метилгуанозина, 5-метилцитидина, 8-оксогуанозина, 7-деазагуанозина, N1-метиладенозина, 2-амино-6-хлорпурина, N6-метил-2-аминопурина, псевдоизоцитидина, 6-хлорпурина, N6-метиладенозин, альфа-тиоаденозина, 8-азидоаденозина, 7-деазааденозина.
Липидная модификация
Согласно следующему варианту осуществления изобретения искусственная молекула нуклеиновой кислоты, предпочтительно РНК, представленная в настоящем описании, может содержать липидную модификацию. Такая модифицированная липидами РНК, как правило, содержит РНК, представленную в настоящем описании. Такая модифицированная липидами молекула РНК, представленная в настоящем описании, как правило, дополнительно содержит по меньшей мере один линкер, ковалентно связанный с указанной молекулой РНК, и по меньшей мере один липид, ковалентно связанный с соответствующим линкером. Альтернативно этому, модифицированная липидами молекула РНК содержит (по меньшей мере одну) молекулу РНК, указанную в настоящем описании, и по меньшей мере один (бифункциональный) липид, ковалентно связанный (без линкера) с указанной молекулой РНК. Согласно третьей альтернативе модифицированная липидами молекула РНК содержит искусственную молекулу нуклеиновой кислоты, предпочтительно молекулу РНК, указанную в настоящем описании, по меньшей мере один линкер, ковалентно связанный с указанной молекулой РНК, и по меньшей мере один липид, ковалентно связанный с соответствующим линкером, и также по меньшей мере один (бифункциональный) липид, ковалентно связанный (без линкера) с указанной мРНК. В этом контексте наиболее предпочтительно липидную модификацию осуществляют на концах линейной последовательности РНК.
Модификация 5'-конца модифицированной РНК
Согласно другому предпочтительному варианту осуществления изобретения искусственную молекулу нуклеиновой кислоты, предпочтительно молекулу РНК, представленную в настоящем описании, можно модифицировать путем добавления так называемой «5'-кэп»-структуры.
5'-кэп представляет собой структуру, как правило, структуру, состоящую из модифицированных нуклеотидов, которая, как правило, завершает («кэпирует») 5'-конец зрелой мРНК. 5'-кэп, как правило, может быть сформирован модифицированным нуклеотидом, прежде всего производным гуанинового нуклеотида. Предпочтительно 5'-кэп связывают с 5'-концом с помощью 5'-5'-трифосфатной связи. 5'-кэп может быть метилирован, например, m7GpppN), где N представляет собой концевой 5'-нуклеотид нуклеиновой кислоты, несущей 5'-кэп, как правило, 5'-конец РНК. m7GpppN представляет собой 5'-кэп-структуру, которая встречается в естественных условиях в мРНК транскрибируемой полимеразой II, и, следовательно, не рассматривается как модификация, содержащаяся в модифицированной РНК, предлагаемой в изобретении. Это означает, что искусственная молекула нуклеиновой кислоты, предпочтительно молекула РНК, предлагаемая в настоящем изобретении, может содержать m7GpppN в качестве 5'-кэпа, но при этом дополнительно искусственная молекула нуклеиновой кислоты, предпочтительно молекула РНК, содержит по меньшей мере одну дополнительную модификацию, представленную в настоящем описании.
Другими примерами 5'-кэп-структур являются глицерил, инвертированная дезоксигруппа (фрагмент), лишенная азотистого основания, 4',5'-метиленовой нуклеотид, 1-(бета-D-эритрофуранозильный) нуклеотид, 4'-тионуклеотид, карбоциклический нуклеотид, 1,5-ангидрогекситольный нуклеотид, L-нуклеотиды, альфа-нуклеотид, нуклеотид с модифицированным основанием, треопентофуранозильный нуклеотид, ациклический 3',4'-секонуклеотид, ациклический 3,4-дигидроксибутильный нуклеотид, ациклический 3,5-дигидроксипентильный нуклеотид, 3'-3'-инвертированный нуклеотидный фрагмент, 3'-3'-инвертированный лишенный азотистого основания фрагмент, 3'-2'-инвертированный нуклеотидный фрагмент, 3'-2'-инвертированный лишенный азотистого основания фрагмент, 1,4-бутандиолфосфатный, 3'-фосфороамидатный, гексилфосфатный, аминогексилфосфатный, 3'-фосфатный, 3'-фосфоротиоатный, фосфородитиоатный или связанный мостиком или несвязанный мостиком метилфосфонатный фрагмент. Эти модифицированные 5'-кэп-структуры рассматриваются в качестве по меньшей мере одной модификации, содержащейся в искусственной молекуле нуклеиновой кислоты, предпочтительно в молекуле РНК, предлагаемой в настоящем изобретении.
Наиболее предпочтительными модифицированными 5'-кэп-структурами являются САР1 (метилирование рибозы соседнего с m7GpppN нуклеотида), САР2 (метилирование рибозы 2-ого нуклеотида, расположенного в прямом направлении относительно m7GpppN), САР3 (метилирование рибозы 3-его нуклеотида, расположенного в прямом направлении относительное m7GpppN), САР4 (метилирование рибозы 4-ого нуклеотида, расположенного в прямом направлении относительно m7GpppN), ARCA (аналог кэп-структуры с правильной ориентацией, модифицированный ARCA (например, модифицированный фосфотиоатом ARCA), инозин, N1-метилгуанозин, 2'-фторгуанозин, 7-деазагуанозин, 8-оксогуанозин, 2-аминогуанозин, ЗНК-гуанозин и 2-азидогуанозин.
В предпочтительном варианте осуществления изобретения по меньшей мере одна открытая рамка считывания кодирует терапевтический белок или пептид. В другом варианте осуществления изобретения по меньшей мере одной открытой рамкой считывания кодируется антиген, такой как патогенный антиген, опухолевый антиген, аллергенный антиген или аутоиммунный антиген. Таким образом, введение искусственной молекулы нуклеиновой кислоты, кодирующей антиген, применяют в подходе, заключающемся в генетической вакцинации против заболевания, в котором участвует указанный антиген.
В альтернативном варианте осуществления изобретения по меньшей мере одной открытой рамкой считывания искусственной молекулы нуклеиновой кислоты, предлагаемой в изобретении, кодируется антитело.
Антигены
Патогенные антигены
Искусственная молекула нуклеиновой кислоты, предлагаемой в настоящем изобретении, может кодировать белок или пептид, который содержит патогенный антиген или его фрагмент, вариант или производное. Такие патогенные антигены происходят из патогенных организмов, прежде всего, бактерий, вирусов или простейших (многоклеточных) патогенных организмов, которые вызывают иммунологическую реакцию у индивидуума, прежде всего у млекопитающего, более предпочтительно у человека. Более конкретно, патогенные антигены предпочтительно представляют собой поверхностные антигены, например, белки (или фрагменты белков, например, находящуюся снаружи часть поверхностного антигена), локализованные на поверхности вируса или бактерии или простейшего.
Патогенные антигены представляют собой пептидные или белковые антигены, предпочтительно происходящие из патогенов, ассоциированных с инфекционным заболеванием, которые предпочтительно выбирают из антигенов, происходящих из патогенов Acinetobacter baumannii, рода Anaplasma, Anaplasma phagocytophilum, Ancylostoma braziliense, Ancylostoma duodenale, Arcanobacterium haemolyticum, Ascaris lumbricoides, рода Aspergillus, Astroviridae, рода Babesia, Bacillus anthracis, Bacillus cereus, Bartonella henselae, вируса BK, Blastocystis hominis, Blastomyces dermatitidis, Bordetella pertussis, Borrelia burgdorferi, рода Borrelia, Borrelia spp, рода Brucella, Brugia malayi, семейства Bunyaviridae, Burkholderia cepacia и других видов Burkholderia, Burkholderia mallei, Burkholderia pseudomallei, семейства Caliciviridae, рода Campylobacter, Candida albicans, Candida spp, Chlamydia trachomatis, Chlamydophila pneumoniae, Chlamydophila psittaci, приона CJD, Clonorchis sinensis, Clostridium botulinum, Clostridium difficile, Clostridium perfringens, Clostridium perfringens, Clostridium spp, Clostridium tetani, Coccidioides spp, коронавирусов, Corynebacterium diphtheriae, Coxiella burnetii, вируса геморрагической лихорадки Крым-Конго, Cryptococcus neoformans, рода Cryptosporidium, Cytomegalovirus (CMV), вирусов Денге (DEN-1, DEN-2, DEN-3 и DEN-4), Dientamoeba fragilis, вируса Эбола (EBOV), рода Echinococcus, Ehrlichia chaffeensis, Ehrlichia ewingii, рода Ehrlichia, Entamoeba histolytica, рода Enterococcus, рода Enterovirus, энтеровирусов, прежде всего вируса Коксаки А и энтеровируса 71 (EV71), Epidermophyton spp, вируса Эпштейна-Барра (EBV), Escherichia coli O157:Н7, O111 и О104:Н4, Fasciola hepatica и Fasciola gigantica, приона FFI, суперсемейства Filarioidea, флавивирусов, Francisella tularensis, рода Fusobacterium, Geotrichum candidum, Giardia intestinalis, Gnathostoma spp, приона GSS, вируса Гуанарито, Haemophilus ducreyi, Haemophilus influenzae, Helicobacter pylori, генипавируса (вирус Гендра и вирус Нипа), вируса гепатита А, вируса гепатита В (HBV), вируса гепатита С (HCV), вируса гепатита D, вируса гепатита, вируса герпеса простого 1 и 2 (HSV-1 и HSV-2), Histoplasma capsulatum, HIV (вирус иммунодефицита человека), Hortaea werneckii, человеческого бокавируса (HBoV), человеческого вируса герпеса 6 (HHV-6) и человеческого вируса герпеса 7 (HHV-7), человеческого вируса метапенвмонии (hMPV), вируса папилломы человека (HPV), человеческих вирусов парагриппа (HPIV), вируса японского энцефалита, вируса JC, вируса Джунин, Kingella kingae, Klebsiella granulomatis, приона Куру, вируса Ласса, Legionella pneumophila, рода Leishmania, рода Leptospira, Listeria monocytogenes, вируса лимфоцитарного хориоменингита (LCMV), вируса Мачупо, Malassezia spp, вируса Марбурга, вируса кори, Metagonimus yokagawai, Microsporidia phylum, вируса контагиозного моллюска (MCV), вируса паротита, Mycobacterium leprae и Mycobacterium lepromatosis, Mycobacterium tuberculosis, Mycobacterium ulcerans, Mycoplasma pneumoniae, Naegleria fowleri, Necator americanus, Neisseria gonorrhoeae, Neisseria meningitidis, Nocardia asteroides, Nocardia spp, Onchocerca volvulus, Orientia tsutsugamushi, семейства Orthomyxoviridae (грипп), Paracoccidioides brasiliensis, Paragonimus spp, Paragonimus westermani, парвовируса В19, рода Pasteurella, рода Plasmodium, Pneumocystis jirovecii. полиовируса, вируса бешенства, респираторно-синтициального вируса (RSV), риновируса, риновирусов, Rickettsia akari, рода Rickettsia, Rickettsia prowazekii, Rickettsia rickettsii, Rickettsia typhi, вируса долины Рифта, ротавируса, вируса краснухи, вируса Сабиа, рода Salmonella, Sarcoptes scabiei, коронавируса SARS, рода Schistosoma, рода Shigella, вируса Син Номбре, хантавируса, Sporothrix schenckii, рода Staphylococcus, Streptococcus agalactiae, Streptococcus pneumoniae, Streptococcus pyogenes, Strongyloides stercoralis, рода Taenia, Taenia solium, вируса клещевого энцефалита (TBEV), Toxocara canis или Toxocara cati, Toxoplasma gondii, Treponema pallidum, Trichinella spiralis, Trichomonas vaginalis, Trichophyton spp, Trichuris trichiura, Trypanosoma brucei, Trypanosoma cruzi, Ureaplasma urealyticum, вируса ветряной оспы (VZV), Variola major или Variola minor, приона vCJD, вируса венесуэльского энцефалита лошадей, Vibrio cholerae, вируса Западного Нила, вируса западного энцефалита лошадей, Wuchereria bancrofti, вируса желтой лихорадки, Yersinia enterocolitica, Yersinia pestis и Yersinia pseudotuberculosis.
В этом контексте наиболее предпочтительными являются антигены из патогенов, выбранных из вируса гриппа, респираторно-синтициального вируса (RSV), вируса герпеса простого (HSV), вируса папилломы человека (HPV), вируса иммунодефицита человека (HIV), Plasmodium, Staphylococcus aureus, вируса Денге, Chlamydia trachomatis, цитомегаловируса (CMV), вируса гепатита В (HBV), Mycobacterium tuberculosis, вируса бешенства и вируса желтой лихорадки.
Опухолевые антигены
В другом варианте осуществления изобретения искусственная молекула нуклеиновой кислоты, предлагаемой в настоящем изобретении, может кодировать белок или пептид, который содержит пептид или белок, содержащий опухолевый антиген, фрагмент, вариант или производное указанного опухолевого антигена, где предпочтительно, опухолевый антиген представляет собой меланоцит-специфический антиген, раково-тестикулярный антиген или опухоль-специфический антиген, предпочтительно СТ-Х-антиген, не-Х-СТ-антиген, партнер, связывающий СТ-Х-антиген или партнер, связывающий не-Х-СТ-антиген, или опухоль-специфический антиген, более предпочтительно СТ-Х-антиген, партнер, связывающий не-Х-СТ-антиген или опухоль-специфический антиген, или фрагмент, вариант или производное указанного опухолевого антигена; и где каждая из нуклеотидных последовательностей кодирует отличный от других пептид или белок; и где по меньшей мере одна из нуклеотидных последовательностей кодирует 5Т4, 707-АР, 9D7, AFP, AlbZIP HPG1, альфа-5-бета-1-интегрин, альфа-5-бета-6-интегрин, альфа-актинин-4/m, альфа-метилацил-кофермент А-рацемазу, ART-4, ARTC1/m, В7Н4, BAGE-1, BCL-2, bcr/abl, бета-катенин/m, BING-4, BRCA1/m, BRCA2/m, CA 15-3/CA 27-29, CA 19-9, CA72-4, CA125, калретикулин, CAMEL, CASP-8/m, катепсин В, катепсин L, CD19, CD20, CD22, CD25, CDE30, CD33, CD4, CD52, CD55, CD56, CD80, CDC27/m, CDK4/m, CDKN2A/m, CEA, CLCA2, CML28, CML66, COA-1/m, коактозин-подобный белок, коллаген XXIII, COX-2, CT-9/BRD6, Cten, циклин B1, циклин D1, cyp-B, CYPB1, DAM-10, DAM-6, DEK-CAN, EFTUD2/m, EGFR, ELF2/m, EMMPRIN, EpCam, EphA2, EphA3, EphВ3, ETV6-AML1, EZH2, FGF-5, FN, Frau-1, G250, GAGE-1, GAGE-2, GAGE-3, GAGE-4, GAGE-5, GAGE-6, GAGE7b, GAGE-8, GDEP, GnT-V, gp100, GPC3, GPNMB/m, HAGE, HAST-2, гепсин, Her2/neu, HERV-K-MEL, HLA-A*0201-R17I, HLA-A11/m, HLA-A2/m, HNE, гомеобокс NKX3.1, HOM-TES-14/SCP-1, HOM-TES-85, HPV-E6, HPV-E7, HSP70-2M, HST-2, hTERT, iCE, IGF-1R, IL-13Ra2, IL-2R, IL-5, незрелый ламининовый рецептор, каллекреин-2, каллекреин-4, Ki67, KIAA0205, KIAA0205/m, KK-LC-1, K-Ras/m, LAGE-A1, LDLR-FUT, MAGE-A1, MAGE-A2, MAGE-A3, MAGE-A4, MAGE-A6, MAGE-A9, MAGE-A10, MAGE-A12, MAGE-B1, MAGE-B2, MAGE-B3, MAGE-B4, MAGE-B5, MAGE-B6, MAGE-B10, MAGE-B16, MAGE-B17, MAGE-C1, MAGE-C2, MAGE-C3, MAGE-D1, MAGE-D2, MAGE-D4, MAGE-E1, MAGE-E2, MAGE-F1, MAGE-H1, MAGEL2, маммоглобин A, MART-1/мелан-А, MART-2, MART-2/m, матриксный белок 22, MC1R, M-CSF, ME1/m, мезотелин, MG50/PXDN, MMP11, MN/CA IХ-антиген, MRP-3, MUC-1, MUC-2, MUM-1/m, MUM-2/m, MUM-3/m, миозин класса I/m, NA88-A, N-ацетилглюкозаминилтрансферазу-V, Neo-PAP, Neo-PAP/m, NFYC/m, NGEP, NMP22, NPM/ALK, N-Ras/m, NSE, NY-ESO-1, NY-ESO-B, OA1, OFA-iLRP, OGT, OGT/m, OS-9, OS-9/m, остеокальцин, остеопонтин, pi5, pi90 минорный bcr-abl, p53, p53/m, PAGE-4, PAI-1, PAI-2, PART-1, PATE, PDEF, Pim-1-киназу, Pin-1, Pml/PARaльфa, POTE, PRAME, PRDX5/m, простеин, протеиназу-3, PSA, PSCA, PSGR, PSM, PSMA, PTPRK/m, RAGE-1, RBAF600/m, RHAMM/CD168, RU1, RU2, S-100, SAGE, SART-1, SART-2, SART-3, SCC, SIRT2/m, Sp17, SSX-1, SSX-2/HOM-MEL-40, SSX-4, STAMP-1, STEAP, сурвивин, сурвивин-2 В, SYT-SSX-1, SYT-SSX-2, TA-90, TAG-72, TARP, TEL-AML1, TGFбета, TGFбетаRII, TGM-4, TPI/m, TRAG-3, TRG, TRP-1, TRP-2/6b, TRP/INT2, TRP-p8, тирозиназу, UPA, VEGF, VEGFR-2/FLK-1, WT1 и иммуноглобулиновый идиотип лимфоидной кровяной клетки или идиотип Т-клеточного рецептора, или фрагмент, вариант или производное указанного опухолевого антигена; предпочтительно сурвивин или его гомолог, антиген семейства MAGE или его связывающий партнер или фрагмент, вариант или производное указанного опухолевого антигена. Наиболее предпочтительными в этом контексте являются опухолевые антигены NY-ESO-1, 5Т4, MAGE-C1, MAGE-C2, сурвивин, Muc-1, PSA, PSMA, PSCA, STEAP и РАР.
В предпочтительном варианте осуществления изобретения искусственная молекула нуклеиновой кислоты кодирует белок или пептид, который содержит терапевтический белок или его фрагмент, вариант или производное.
В контексте настоящего описания терапевтические белки представляют собой пептиды или белки, которые оказывают полезное действие при лечении любого врожденного или приобретенного заболевания или которые улучшают состояние индивидуума. В частности, терапевтические белки имеют важное значение для создания терапевтических средств, которые могут помимо осуществления других функций модифицировать и осуществлять репарацию генетических ошибок, разрушать раковые клетки или инфицированные патогеном клетки, лечить нарушения иммунной системы, лечить метаболические или эндокринные нарушения. Например, эритропоэтин (ЕРО), белковый гормон, можно применять для лечения пациентов с дефицитом эритроцитов, который является обычной причиной почечных осложнений. Кроме того, под понятие «терапевтические белки» подпадают адъювантные белки, терапевтические антитела, а также гормонзаместительная терапия, которую применяют, например, при лечении женщин в период менопаузы. В более современных подходах используют соматические клетки пациента для перепрограммирования их в плюрипотентные стволовые клетки, что является заменой спорной терапии на основе стволовых клеток. Такие белки, применяемые для перепрограммирования соматических клеток или применяемые для дифференцировки стволовых клеток, также рассматриваются как терапевтические белки в контексте настоящего описания. Кроме того, терапевтические белки можно применять и для других целей, например, для заживления ран, регенерации тканей, ангиогенеза и т.д. Кроме того, в контексте настоящего описания антигенпрезентирующие В-клеточные рецепторы и их фрагменты и варианты рассматриваются как терапевтические белки.
Таким образом, терапевтические белки можно применять для различных целей, включая лечение различных заболеваний, таких, например, как инфекционные заболевания, неоплазмы (например, раковые или опухолевые заболевания), заболевания крови или кроветворных органов, эндокринных, связанных с питанием или метаболических заболеваний, заболеваний нервной системы, заболеваний системы кровообращения, заболеваний дыхательной системы, заболеваний пищеварительной системы, заболеваний кожи и подкожной ткани, заболеваний скелето-мышечной системы и соединительной ткани, и заболеваний мочеполовой системы, независимо от того, являются ли они врожденными или приобретенными.
В этом контексте наиболее предпочтительными терапевтическими белками, которые можно применять, среди прочего, для лечения метаболических или эндокринных нарушений, являются (в скобках указано конкретное заболевание, для лечения которого применяют терапевтический белок): кислая сфингомиелиназа (болезнь Нимана-Пика), адипотид (ожирение), агалзидаза-бета (человеческая галактозидаза А) (болезнь Фабри; препятствует накоплению липидов, которое может приводить к почечным и сердечно-сосудистым осложнениям), алглукозидаза (болезнь Помпе (гликогеноз (накопление гликогена) типа II)), альфа-галактозидаза А (альфа-GAL А, агалзидаза-альфа) (болезнь Фабри), альфа-глюкозидаза (гликогеноз (GSD), болезнь Помпе), альфа-L-идуронидаза (мукополисахаридоз (MPS), синдром Гурлер, синдром Шейе), альфа-N-ацетилглюкозаминидаза (синдром Санфилиппо), амфирегулин (рак, метаболическое нарушение), ангиопоэтин ((Ang1, Ang2, Ang3, Ang4, ANGPTL2, ANGPTL3, ANGPTL4, ANGPTL5, ANGPTL6, ANGPTL7) (ангиогенез, стабилизирует сосуды), бетацеллюлин (метаболическое нарушение), бета-глюкуронидаза (синдром Слая), костный морфогенетический белок BMP (ВМР1, ВМР2, ВМР3, ВМР4, ВМР5, ВМР6, ВМР7, ВМР8а, BMP8b, ВМР10, ВМР15) (регенеративное действие, состояния, связанные с костной тканью, хроническое заболевание почек (CKD)), белок CLN6 (болезнь, вызванная мутациями в локусе CLN6 - атипичный поздний инфантилизм, вариант с поздним началом, ранний ювенильный, нейрональный цероид-липофусциноз (NCL)), эпидермальный фактор роста (EGF) (заживление ран, регуляция роста, пролиферации и дифференцировки клеток), эпиген (метаболическое нарушение), эпирегулин (метаболическое нарушение), фактор роста фибробластов (FGF, FGF-1, FGF-2, FGF-3, FGF-4, FGF-5, FGF-6, FGF-7, FGF-8, FGF-9, FGF-10, FGF-11, FGF-12, FGF-13, FGF-14, FGF-16, FGF-17, FGF-17, FGF-18, FGF-19, FGF-20, FGF-21, FGF-22, FGF-23) (заживление ран, ангиогенез, эндокринные нарушения, регенерация ткани), галсульфаза (мукополисахаридоз типа VI), грелин (синдром раздраженного кишечника (IBS), ожирение, синдром Прадера-Вилли, сахарный диабет типа II), глюкоцереброзидаза (болезнь Гоше), GM-CSF (регенеративное действие, производство лейкоцитов, рак), гепаринсвязывающий EGF-подобный фактор роста (HB-EGF) (заживление ран, гипертрофия сердца и развитие и функция сердца), фактор роста гепатоцитов HGF (регенеративное действие, заживление ран), гепсидин (нарушения метаболизма железа, бета-талассемия), человеческий альбумин (пониженное производство альбумина (гипопротеинемия), повышенная потеря альбумина (нефротический синдром), гиповолемия, гипербилирубинемия), идурсульфаза (идуронат-2-сульфатаза) (мукополисахаридоз типа II (синдром Хантера)), интегрины αVβ3, αVβ5 и α5β1 (связывают матриксные макромолекулы и протеиназы, ангиогенез), идуронатсульфатаза (синдром Хантера), ларонидаза (мукополисахаридоз типа I, формы Гурлер и Гурлер-Шайе), N-ацетилгалактозамин-4-сульфатаза (rhASB; галсульфаза, арилсульфатаза A (ARSA), арилсульфатаза В (ARSB)) (дефицит арилсульфатазы В, синдром Марото-Лами, мукополисахаридоз типа VI), N-ацетилглюкозамин-6-сульфатаза (синдром Санфилиппо), фактор роста нервов (NGF, мозговой нейротрофический фактор (BDNF), нейротрофин-3 (NT-3) и нейротрофин 4/5 (NT-4/5) (регенеративное действие, сердечно-сосудистые заболевания, коронарный атеросклероз, ожирение, диабет типа 2, метаболический синдром, острые коронарные синдромы, деменция, депрессия, шизофрения, аутизм, синдром Ретта, нервная анорексия, нервная булимия, заживление ран, кожные язвы, изъязвления роговицы, болезнь Альцгеймера), нейрегулин (NRG1, NRG2, NRG3, NRG4) (метаболическое нарушение, шизофрения), нейропилин (NRP-1, NRP-2) (ангиогенез, управление аксонами, выживание, миграция клеток), обестатин (синдром раздраженного кишечника (IBS), ожирение, синдром Прадера-Вилли, сахарный диабет типа II), фактор роста тромбоцитов (PDGF (PDFF-A, PDGF-B, PDGF-C, PDGF-D) (регенеративное действие, заживление ран, нарушение ангиогенеза, артериосклероз, фиброз, рак), рецепторы TGF-бета (эндоглин, рецептор TGF-бета 1, рецептор TGF-бета 2, рецептор TGF-бета 3) (почечный фиброз, заболевание почек, диабет, терминальная стадия почечной болезни (ESRD), ангиогенез), тромбопоэтин (ТНРО) (фактор роста и развития мегакариоцитов (MGDF)) (тромбоцитарные нарушения, тромбоциты для донорской сдачи, восстановление уровня тромбоцитов после миелосупрессорной химиотерапии), трансформирующий фактор роста (TGF (TGF-альфа, TGF-бета (TGFбета1, TGFбета2 и TGFбета3))) (регенеративное действие, заживление ран, иммунитет, рак, заболевание сердца, диабет, синдром Марфана, синдром Лоеса-Дитца), VEGF (VEGF-A, VEGF-B, VEGF-C, VEGF-D, VEGF-E, VEGF-F и PIGF) (регенеративное действие, ангиогенез, заживление ран, рак, проницаемость), несритид (острая декомпенсированная застойная сердечная недостаточность), трипсин (декубитальная язва, варикозная язва, санация струпа, спонтанно раскрывающаяся рана, солнечный ожог, мекониевая непроходимость кишечника), адренокортикотропный гормон (АСТН) (болезнь Аддисона, мелкоклеточная саркома, адренолейкодистрофия, врожденная гиперплазия надпочечников, синдром Кушинга, синдром Нельсона, детские спазмы), предсердный натрийуретический пептид (ANP) (эндокринные нарушения), холецистокинин (различной длины), гастрин (гипогастринемия), лептин (диабет, гиперглицинемия, ожирение), окситоцин (стимулирует грудное кормление, ненаступление родов), соматостатин (симптоматическое лечение карционоидного синдрома, острое варикозное кровотечение и акромегалия, поликистозные заболевания печени и почек, акромегалия и симптомы, вызываемые нейроэндокринными опухолями), вазопрессин (антидиуретический гормон) (несахарный диабет), кальцитонин (постменопаузальный остеопороз, гиперкальциемия, болезнь Пэджета, метастазы в костную ткань, фантомная боль к конечностях, спинальный стеноз), экзенатид (диабет типа 2, устойчивый к обработке метформином и сульфонилмочевиной), гормон роста (GH), соматотропин (нарушение роста вследствие дефицита GH или хронической печеночной недостаточности, синдром Прадера-Вилли, синдром Турнера, обусловленное СПИДом истощение или кахексия при осуществлении противовирусной терапии), инсулин (сахарный диабет, диабетический кетоацидоз, гиперкалиемия), инсулиноподобный фактор роста 1 IGF-1 (нарушение роста у детей с делецией гена GH или серьезным первичным дефицитом IGF1, нейродегенеративное заболевание, сердечно-сосудистые заболевания, сердечная недостаточность), мекасермин ринфабат, аналог IGF-1 (нарушение роста у детей с делецией гена GH или серьезным первичным дефицитом IGF1, нейродегенеративное заболевание, сердечно-сосудистые заболевания, сердечная недостаточность), мегасермин, аналог IGF-1 (нарушение роста у детей с делецией гена GH или серьезным первичным дефицитом IGF1, нейродегенеративное заболевание, сердечно-сосудистые заболевания, сердечная недостаточность), пэгвисомант (акромегалия), прамлинтид (сахарный диабет, в комбинации с инсулином), терипаратид (человеческий паратиреоидный гормон, остатки 1-34) (серьезный остеопороз), бекаплермин (вспомогательное средство при санации диабетических язв), диботермин-альфа (костный морфогенный белок 2) (хирургическая операция по закреплению позвоночника (Spinal Fusion), восстановление кости при повреждении), гистрелина ацетат (гонадотропин-рилизинг гормон; GnRH) (преждевременное половое созревание), октреотид (акромегалия, симптоматическое ослабление VIP-секретирующей аденомы и метастатических карциноидных опухолей) ипалифермин (фактор роста кератиноцитов; KGF) (серьезный оральный мукозит у пациентов, подвергающихся химиотерапии, заживление ран).
Указанные и другие белки рассматриваются как терапевтические, поскольку они предназначены для лечения индивидуума путем возмещения нарушенного эндогенного производства в его организме функционального белка в достаточных количествах. Следовательно, такие терапевтические белки, как правило, представляют собой белки млекопитающих, прежде всего человеческие белки.
Для лечения нарушений крови, заболеваний системы кровообращения, заболеваний дыхательной системы, раковых или опухолевых заболеваний, инфекционных заболеваний или иммунодефицитов можно применять следующие терапевтические белки: альтерплаза (активатор тканевого плазминогена; tPA) (легочная эмболия, инфаркт миокарда, острый ишемический инсульт («удар»), окклюзия устройств центрального венозного доступа), анистреплаза (тромболизис), антитромбин III (АТ-III) (наследственный дефицит АТ-III, тромбоэмболия), бивалирудин (снижает риск свертывания крови при коронарной ангиопластике и индуцированной гепарином тромбоцитопении), дарбепоэтин-альфа (лечение анемии у пациентов с хронической почечной недостаточностью и хроническим почечным нарушением (+/- диализ)), дротрекогин-альфа (активированный белок С) (серьезный сепсис с высоким риском смерти), эритропоэтин, эпоэтин-альфа, эритропоэтин, эритропоетин (анемия при хроническом заболевании, миелодисплазия, анемия при почечной недостаточности или при осуществлении химиотерапии, предоперационная подготовка), фактор IX (гемофилия В), фактор VIIa (кровоизлияние у пациентов с гемофилией А или В и при применении ингибиторов фактора VIII или фактора IX), фактор VIII (гемофилия А), лепирудин (индуцируемая гепарином тромбоцитопения), концентрат белка С (венозный тромбоз, молниеносная пурпура), ретеплаза (делеция мутеина в tPA) (лечение острого инфаркта миокарда, улучшение функции желудочков), стрептокиназа (остро развивающийся трансмуральный инфаркт миокарда, легочная эмболия, тромбоз глубоких вен, артериальный тромбоз или эмболия, окклюзия артериовенозной канюли), тенестеплаза (острый инфаркт миокарда), урокиназа (легочная эмболия), ангиостатин (рак), анти-СD22 иммунотоксин (рецидивирующий острый миелоидный СD33+-лейкоз), денилейкин дифтитокс (кожная Т-клеточная лимфома (CTCL)), иммуноцианин (рак мочевого пузыря и предстательной железы), MPS (металлопанстимулин) (рак), афлиберцепт (немелкоклеточный рак легкого (NSCLC), метастатический колоректальный рак (mCRC), гормон-устойчивый метастатический рак предстательной железы, влажная форма дегенерации желтого пятна), эндостатин (рак, воспалительные заболевания типа ревматоидного артрита, а также болезнь Крона, диабетическая ретинопатияпсориаз и эндометриоз), коллагеназа (санация хронических кожных язв и областей серьезных ожогов, контрактура Дюпюитрена, болезнь Пейрони), человеческая дезоксирибонуклеаза I, дорназа (муковисцидоз; уменьшение инфекций респираторного тракта у выбранных пациентов с FVC, превышающим более чем на 40% предсказанную величину), гиалуронидаза (применяется в качестве адъюванта для повышения абсорбции и диспергирования инъецированных лекарственных средств, прежде всего анестезирующих средств при глазной хирургии и определенных визуализирующих агентов), папаин (санация некротической ткани или разжижение отторгающихся некротических масс при острых и хронических повреждениях, таких как пролежни, варикозные и диабетические язвы, ожоги, послеоперационные раны, пилонидальные кистозные раны, карбункулы и другие раны), L-аспарагиназа (острый лимфоцитарный лейкоз, требующий экзогенного аспарагина для пролиферации), пэг-аспарагиназа (острый лимфоцитарный лейкоз, требующий экзогенного аспарагина для пролиферации), расбуриказа (пациенты детского возраста с лейкозом, лимфомой и солидными опухолями, подвергающиеся противораковой терапии, которая может вызывать синдром лизиса опухоли), человеческий хорионный гонадотропин (HCG) (улучшает репродуктивную функцию), человеческий фолликулостимулирующий гормон (FSH) (улучшает репродуктивную функцию), лутропин-альфа (бесплодие, сопровождающееся дефицитом лютениизирующего гормона), пролактин (гипопролактинемия, дефицит сывороточного пролактина, дисфункция яичника у женщин, состояние страха, артериогенная эректильная дисфункция, преждевременное семяизвержение, олигозооспермия, астеноспермия, пониженная функция семенных пузырьков, гипоандрогенизм у мужчин), ингибитор альфа-1-протеиназы (врожденный дефицит антитрипсина), лактаза (газообразование, вздутие, спазмы и диарея вследствие неспособности расщеплять лактозу), панкреатические ферменты (липаза, амилаза, протеиназа) (муковисцидоз, хронический панкреатит, панкреатическая недостаточность, состояние после операции желудочного шунтирования по Билльрот типа II (Billroth II), обструкция протоков поджелудочной железы, стеаторрея, недостаточное пищеварение, газообразование, вздутие), аденозиндеаминаза (бычья пегадемаза, PEG-ADA) (заболевания с серьезным комбинированным иммунодефицитом вследствие дефицита аденозиндеаминазы), абатецепт (ревматоидный артрит (прежде всего, рефракторный в отношении ингибирования TNF-альфа)), алефацепт (бляшковидный псориаз), анакинра (ревматоидный артрит), этанецепт (ревматоидный артрит, многосуставной юношеский ревматоидный артрит, псориатический артрит, анкилоизирующий спондилит, бляшковидный псориаз, анкилоизирующий спондилит), анти\агонист рецептора интерлейкина-1 (IL-1), анакинра (воспаление и деградация хряща, ассоциированная с ревматоидным артритом), тимулин (нейродегенеративные заболевания, ревматизм, нервная анорексия), антагонист TNF-альфа (аутоиммунные заболевания, такие как ревматоидный артрит, анкилоизирующий спондилит, болезнь Крона, псориаз, гнойный гидраденит, рефрактерная астма), энфувиртид (HIV-1-инфекция) и тимозин α1 (гепатит В и С)
(в скобках указано конкретное заболевание, для лечения которого применяют терапевтический белок)
Следующим объектом настоящего изобретения является вектор, содержащий
а. открытую рамку считывания (ОРС) и/или сайт клонирования, например, предназначенный для встраивания открытой рамки считывания или последовательности, содержащей открытую рамку считывания; и
б. по меньшей мере один элемент 3'-нетранслируемой области (3'-UTR-элемент), содержащий нуклеотидную последовательность, которая происходит из 3'-UTR гена рибосомального белка.
По меньшей мере один 3'-UTR-элeмeнт и ОРС представляют собой конструкции, описанные выше для искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении. Сайт клонирования может представлять собой любую последовательность, пригодную для интродукции открытой рамки считывания или последовательности, содержащей открытую рамку считывания, такую как одни или несколько сайтов рестрикции. Так, вектор, содержащий сайт клонирования, предпочтительно является пригодным для встраивания открытой рамки считывания в вектор, предпочтительно для встраивания открытой рамки считывания с 5'-стороны относительно 3'-UTR-элемента. Предпочтительно, сайт клонирования или ОРС располагают с 5'-стороны относительно 3'-UTR-элeмeнта, предпочтительно в непосредственной близости от 5'-конца 3'-UTR-элeмeнтa. Например, сайт клонирования или ОРС можно непосредственно присоединять к 5'-концу 3'-UTR-элeмeнтa или их можно присоединять посредством сегмента нуклеотидов, например, сегмента, состоящего из 2, 4, 6, 8, 10, 20 и т.д. нуклеотидов, как описано выше для искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении.
Предпочтительно вектор, предлагаемой в настоящем изобретении, можно применять для производства искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, предпочтительно производства искусственной мРНК, предлагаемой в настоящем изобретении, например, необязательно путем встраивания открытой рамки считывания или последовательности, содержащей открытую рамку считывания, в вектор и транскрибирования вектора. Так, предпочтительно вектор содержит элементы, необходимые для транскрипции, такие как промотор, например, промотор РНК-полимеразы. Предпочтительно вектор пригоден для транскрипции с использованием эукариотической, прокариотической, вирусной или фаговой систем транскрипции, таких как эукариотические клетки, прокариотические клетки или эукариотические, прокариотические, вирусные или фаговые системы транскрипции in vitro. Так, например, вектор может содержать промоторную последовательность, которая распознается полимеразой, такой как РНК-полимераза, например, эукариотической, прокариотической, вирусной или фаговой РНК-полимеразой. В предпочтительном варианте осуществления изобретения вектор содержит промотор РНК-полимеразы фага, такой как промотор SP6, Т3 или Т7, предпочтительно промотор Т7. Предпочтительно вектор пригоден для транскрипции in vitro с использованием системы транскрипции in vitro на основе фага, такой как система транскрипции in vitro на основе РНК-полимеразы Т7.
В другом предпочтительном варианте осуществления изобретения вектор можно применять непосредственно для экспрессии кодируемого пептида или белка в клетках или ткани. Для этой цели вектор содержит конкретные элементы, необходимые для экспрессии в указанных клетках/ткани, например, конкретные последовательности промотора, такого как промотор CMV.
Вектор может содержать также поли(А)-последовательность и/или сигнал полиаденилирования, описанные выше для искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении.
Вектор может представлять собой РНК-вектор или ДНК-вектор. Предпочтительно вектор представляет собой ДНК-вектор. Вектор может представлять собой любой вектор, известный специалисту в данной области, такой как вирусный вектор или плазмидный вектор. Предпочтительно вектор представляет собой плазмидный вектор, предпочтительно плазмидный ДНК-вектор.
В предпочтительном варианте осуществления изобретения вектор, предлагаемый в настоящем изобретении, содержит искусственную молекулу нуклеиновой кислоты, предлагаемую в настоящем изобретении.
В одном из вариантов осуществления изобретения ДНК-вектор, предлагаемый в изобретении, содержит нуклеотидную последовательность, идентичную по меньшей мере примерно на 1, 2, 3, 4, 5, 10, 15, 20, 30 или 40%, предпочтительно по меньшей мере примерно на 50%, предпочтительно по меньшей мере примерно на 60%, предпочтительно по меньшей мере примерно на 70%, более предпочтительно по меньшей мере примерно на 80%, более предпочтительно по меньшей мере примерно на 90%, еще более предпочтительно по меньшей мере примерно на 95%, еще более предпочтительно по меньшей мере примерно на 99%, наиболее предпочтительно на 100% нуклеотидной последовательности 3'-UTR гена рибосомального белка, такой как нуклеотидная последовательность, представленная в SEQ ID NO: 10-115.
Предпочтительно ДНК-вектор, предлагаемый в настоящем изобретении, содержит последовательность, представленную в SEQ ID NO: 1, SEQ ID NO: 3, последовательность, комплементарную SEQ ID NO: 7 или последовательности, идентичные по меньшей мере примерно на 40%, предпочтительно по меньшей мере примерно на 50%, предпочтительно по меньшей мере примерно на 60%, предпочтительно по меньшей мере примерно на 70%, более предпочтительно по меньшей мере примерно на 80%, более предпочтительно по меньшей мере примерно на 90%, еще более предпочтительно по меньшей мере примерно на 95%; еще более предпочтительно по меньшей мере примерно на 99% нуклеотидной последовательности, представленной в SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 7, или ее фрагменту, описанному выше, предпочтительно ее функциональному фрагменту.
Предпочтительно РНК-вектор, предлагаемый в настоящем изобретении, содержит последовательность, представленную в SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 7 или последовательность, идентичную по меньшей мере примерно на 40%, предпочтительно по меньшей мере примерно на 50%, предпочтительно по меньшей мере примерно на 60%, предпочтительно по меньшей мере примерно на 70%, более предпочтительно по меньшей мере примерно на 80%, более предпочтительно по меньшей мере примерно на 90%, еще более предпочтительно по меньшей мере примерно на 95%; еще более предпочтительно по меньшей мере примерно на 99% нуклеотидной последовательности, представленной в SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 7 или ее фрагменту, предпочтительно ее функциональному фрагменту.
Предпочтительно вектор представляет собой кольцевую молекулу. Предпочтительно вектор представляет собой двухцепочечную молекулу, такую как двухцепочечная молекула ДНК. Такую кольцевую, предпочтительно двухцепочечную молекулу ДНК можно использовать в качестве удобной формы хранения предлагаемой в изобретении искусственной молекулы нуклеиновой кислоты. Кроме того, ее можно применять для трансфекции клеток, например, культивируемых клеток. Ее можно использовать также для транскрипции in vitro для получения искусственной молекулы РНК, предлагаемой в изобретении.
Предпочтительно вектор, предпочтительно кольцевой вектор, можно линеаризовать, например, путем расщепления рестриктазой. В предпочтительном варианте осуществления изобретения вектор содержит сайт расщепления, такой как сайт рестрикции, предпочтительно уникальный сайт расщепления, непосредственно примыкающий с 3'-стороны к 3'-UTR-элементу, или с 3'-стороны к поли(А)-последовательности или сигналу полиаденилирования, если она/он присутствует, или с 3'-стороны к подподпоследовательности, если она присутствует, или с 3'-стороны к гистоновой структуре типа «стебель-петля», если она присутствует. Так, предпочтительно продукт, полученный в результате линеаризации вектора, заканчивается на 3'-конце 3'-концом 3'-UTR-элемента или 3'-концом поли(А)-последовательности или сигналом полиаденилирования, если она/он присутствует, или 3'-концом поли(С)-последовательности, если она присутствует. В варианте осуществления изобретения, в котором вектор, предлагаемый в настоящем изобретении, содержит искусственную молекулу нуклеиновой кислоты, предлагаемую в настоящем изобретении, сайт рестрикции, предпочтительно уникальный сайт рестрикции, предпочтительно примыкающий непосредственно с 3'-стороны к 3'-концу искусственной молекулы нуклеиновой кислоты.
Следующий объект настоящего изобретения относится к клетке, содержащей искусственную молекулу нуклеиновой кислоты, предлагаемую в настоящем изобретении, или вектор, предлагаемый в настоящем изобретении. Клетка может представлять собой любую клетку, такую как бактериальная клетка, клетка насекомого, клетка растения, клетка позвоночного, например, клетка млекопитающего. Такую клетку можно применять, например, для репликации вектора, предлагаемого в настоящем изобретении, например, в клетке бактерии. Кроме того, клетку можно применять для осуществления транскрипции искусственной молекулы нуклеиновой кислоты или вектора, предлагаемой/предлагаемого в настоящем изобретении, и/или для трансляции открытой рамки считывания искусственной молекулы нуклеиновой кислоты или вектора, предлагаемой/предлагаемого в настоящем изобретении. Например, клетку можно применять для производства рекомбинантного белка.
Согласно настоящему изобретению клетки можно получать, например, с помощью стандартных методов переноса нуклеиновой кислоты, таких как стандартные методы трансфекции, трансдукции или трансформации. Например, искусственную молекулу нуклеиновой кислоты или вектор, предлагаемую/предлагаемый в настоящем изобретении, можно переносить в клетку путем электропорации, липофекции, например, с использованием катионных липидов и/или липосом, осаждения фосфатом кальция, трансфекции на основе наночастиц, трансфекции, основанной на применении вирусов или на основе катионных полимеров, таких как DEAE-декстран или полиэтиленимин и т.д.
Предпочтительно клетка представляет собой клетку млекопитающего, такую как клетка человека, домашнего животного, лабораторного животного, такую как клетка мыши или крысы. Предпочтительно клетка представляет собой человеческую клетку. Клетка может представлять собой клетку культивируемой клеточной линии, такой как СНО, ВНК, 293Т, COS-7, HELA, НЕК и т.д. или клетка может представлять собой первичную клетку, такую как человеческая кожная фибробластная (HDF) клетка и т.д., предпочтительно клетку, выделенную из организма. В предпочтительном варианте осуществления изобретения клетка представляет собой выделенную клетку млекопитающего, предпочтительно человека. Например, клетка может представлять собой иммунную клетку, такую как дендритная клетка, раковую или опухолевую клетку, или любую соматическую клетку и т.д., предпочтительно млекопитающего, предпочтительно человека.
Следующим объектом настоящего изобретения является фармацевтическая композиция, содержащая искусственную молекулу нуклеиновой кислоты, предлагаемую в настоящем изобретении, вектор, предлагаемый в настоящем изобретении, или клетку, предлагаемую в настоящем изобретении. Фармацевтическую композицию, предлагаемую в изобретении, можно применять, например, в качестве вакцины, например, для генетической вакцинации. Так, ОРС может, например, кодировать антиген, предназначенный для введения пациенту с цель вакцинации. Так, в предпочтительном варианте осуществления изобретения фармацевтическая композиция, предлагаемая в настоящем изобретении, представляет собой вакцину. Кроме того, фармацевтическую композицию, предлагаемую в настоящем изобретении, можно применять, например, для генной терапии.
Предпочтительно фармацевтическая композиция содержит также один или несколько фармацевтически приемлемых наполнителей, разбавителей и/или эксципиентов и/или один или несколько адъювантов. В контексте настоящего изобретения фармацевтически приемлемый наполнитель, как правило, включает жидкую или нежидкую основу предлагаемой в изобретении фармацевтической композиции. Если предлагаемая в изобретении фармацевтическая композиция находится в жидкой форме, но носитель, как правило, может представлять собой свободную от пирогенов воду; изотонический соляной раствор или забуференные (водные) растворы, например, растворы, забуференные фосфатом, цитратом и т.д. Предназначенный для инъекции буфер может быть гипертоническим, изотоническим или гипотоническим относительно конкретной референс-среды, т.е. содержание в буфере солей может быть выше, идентичным или ниже их содержания в конкретной референс-среде, при этом предпочтительно можно применять такие концентрации вышеуказанных солей, которые не приводят к повреждению клеток в результате осмоса или других связанных с концентрацией воздействий. Референс-среды могут представлять собой, например, жидкости, применяемые в методах «in vivo», такие как кровь, лимфа, цитозольные жидкости или другие жидкости организма, или, например, жидкости, которые можно использовать в качестве референс-сред в методах «in vitro», такие как обычные буферы или жидкости. Указанные обычные буферы или жидкости известны специалисту в данной области. Наиболее предпочтительной жидкой основой является лактированный раствор Рингера.
В предлагаемой в изобретении фармацевтической композиции можно использовать также один или несколько совместимых твердых или жидких наполнителей или разбавителей или капсулирующих соединений, которые можно применять для введения пациенту, подлежащему лечению. Понятие «совместимые» в контексте настоящего описания означает, что указанные компоненты предлагаемой в изобретении фармацевтической композиции можно смешивать с предлагаемой в изобретении нуклеиновой кислотой, указанной в настоящем описании, таким образом, чтобы не происходило взаимодействия, которое могло бы существенно снижать фармацевтическую эффективность предлагаемой в изобретении фармацевтической композиции в обычных условиях применения.
Фармацевтическая композиция, предлагаемая в настоящем изобретении, может необязательно содержать также один или несколько дополнительных фармацевтически активных компонентов. В данном контексте фармацевтически активный компонент представляет собой соединение, которое обладает терапевтическим действием в отношении лечения, облегчения или предупреждения конкретного показания или заболевания. Такие соединения включают (но, не ограничиваясь только ими) пептиды или белки, нуклеиновые кислоты, (терапевтически активные) низкомолекулярные органические или неорганические соединения (с молекулярной массой менее 5000, предпочтительно менее 1000), сахара, антигены или антитела, терапевтические агенты, уже известные из существующего уровня техники, антигенные клетки, антигенные клеточные фрагменты, клеточные фракции; компоненты клеточной оболочки (например, полисахариды), модифицированные, ослабленные или дезактивированные (например, химическим путем или путем облучения) патогены (вирусы, бактерии и т.д.), адъюванты, предпочтительно указанные в настоящем описании, и т.д.
Кроме того, предлагаемая в изобретении фармацевтическая композиция может содержать носитель для искусственной молекулы нуклеиновой кислоты или вектора. Такой носитель может быть пригоден для облегчения растворения в физиологически приемлемых жидкостях, транспортировки и поглощения клеткой фармацевтически активной искусственной молекулы нуклеиновой кислоты или вектора. Следовательно, такой носитель может представлять собой компонент, который может быть пригоден для хранения и доставки искусственной молекулы нуклеиновой кислоты или вектора, предлагаемой/предлагаемого в изобретении. Такие компоненты могут представлять собой, например, катионные или поликатионные носители или соединения, которые могут служить в качестве агентов для трансфекции или комплексообразования.
В этом контексте наиболее предпочтительными агентами для трансфекции или комплексообразования являются катионные или поликатионные соединения, включая протамин, нуклеолин, спермин или спермидин, или другие катионные пептиды или белки, такие как поли-L-лизин (PLL), полиаргинин,  полипептиды, обладающие способностью проникать в клетку пептиды (СРР), такие как ВИЧ-связывающие пептиды, Tat ВИЧ-1 (ВИЧ), выведенные из Tat пептиды, пенетратин, полученные из VP22 пептиды или аналоги, VP22 HSV (вирус герпеса простого), MAP, KALA или домены трансдукции белков (PTD), РрТ620, богатые пролином пептиды, богатые аргинином пептиды, богатые лизином пептиды, MPG-пептид(ы), Рер-1, L-олигомеры, пептид(ы) кальцитонина, выведенные из Antennapedia пептиды (прежде всего, выведенные из Drosophila antennapedia), pAntp, pIsl, FGF, лактоферрин, транспортан, буфорин-2, Вас715-24, SynB, SynB(1), pVEC, выведенные из hCT пептиды, SAP или гистоны.
полипептиды, обладающие способностью проникать в клетку пептиды (СРР), такие как ВИЧ-связывающие пептиды, Tat ВИЧ-1 (ВИЧ), выведенные из Tat пептиды, пенетратин, полученные из VP22 пептиды или аналоги, VP22 HSV (вирус герпеса простого), MAP, KALA или домены трансдукции белков (PTD), РрТ620, богатые пролином пептиды, богатые аргинином пептиды, богатые лизином пептиды, MPG-пептид(ы), Рер-1, L-олигомеры, пептид(ы) кальцитонина, выведенные из Antennapedia пептиды (прежде всего, выведенные из Drosophila antennapedia), pAntp, pIsl, FGF, лактоферрин, транспортан, буфорин-2, Вас715-24, SynB, SynB(1), pVEC, выведенные из hCT пептиды, SAP или гистоны.
Кроме того, такие катионные или поликатионные соединения могут представлять собой катионные или поликатионные пептиды или белки, которые предпочтительно содержат или дополнительно модифицированы так, чтобы они содержали по меньшей мере одну -SH-группу. Предпочтительно катионный или поликатионный носитель выбирают из катионных пептидов, имеющих следующую общую формулу (I):
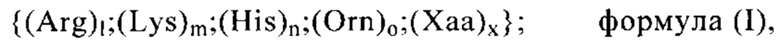
в которой l+m+n+o+x=3-100 и l, m, n или о независимо друг от друга обозначают любое число, выбранное из 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21-30, 31-40, 41-50, 51-60, 61-70, 71-80, 81-90 и 91-100, при условии, общее содержание Arg (аргинин), Lys (лизин), His (гистидин) и Orn (орнитин) составляет по меньшей мере 10% от общего содержания аминокислот в олигопептиде; а Хаа может обозначать любую аминокислоту, выбранную из нативных (т.е. встречающихся в естественных условиях) или не встречающихся в естественных условиях аминокислот за исключением Arg, Lys, His или Orn; и x может обозначать любое число, выбранное из 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21-30, 31-40, 41-50, 51-60, 61-70, 71-80, 81-90, при условии, что общее содержание Хаа не превышает 90% от общего содержания аминокислот в олигопептиде. Любую из аминокислот Arg, Lys, His, Orn и Хаа можно помещать в любое положение пептида. В этом контексте наиболее предпочтительными являются катионные пептиды или белки, содержащие 7-30 аминокислот.
Кроме того, катионный или поликатионный пептид или белок, определяемый формулой {(Arg)l;(Lys)m;(His)n;(Orn)o;(Xaa)x} (формула (I)), представленной выше, который содержит или дополнительно модифицирован так, что он содержал по меньшей мере одну -SH-группу, можно выбирать (но, не ограничиваясь только этим) из соединений подформулы (Iа):
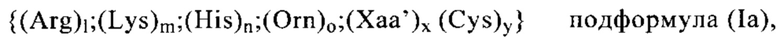
в которой (Arg)l;(Lys)m;(His)n;(Orn)o; и х имеют указанные выше значения, Хаа' обозначает любую аминокислоту, выбранную из нативных (т.е. встречающихся в естественных условиях) или не встречающихся в естественных условиях аминокислот за исключением Arg, Lys, His, Orn или Cys, и y обозначает любое число, выбранное из 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21-30, 31-40, 41-50, 51-60, 61-70, 71-80 и 81-90, условии, что общее содержание Arg (аргинин), Lys (лизин), His (гистидин) и Orn (оринитин) составляет по меньшей мере 10% от общего содержания аминокислот в олигопептиде. Кроме того, катионный или поликатионный пептид можно выбирать из соединений подформулы (Iб):
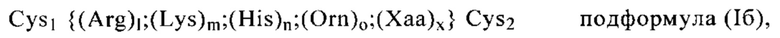
в которой эмпирическая формула {(Arg)l;(Lys)m;(His)n;(Orn)o;(Xaa)x} (формула (III)) представлена в настоящем описании и образует ядро аминокислотной последовательности, представленной (полуэмприрической) формулой (III), и в которой Cys1 и Cys2 обозначают остатки цистеина, расположенные проксимально или терминально относительно (Arg)l;(Lys)m;(His)n;(Orn)o;(Xaa)x.
Другие предпочтительные катионные или поликатионные соединения, которые можно применять в качестве агентов для трансфекции, могут представлять собой катионные полисахариды, например, хитозан, полибрен, катионные полимеры, например, полиэтиленимин (PEI), катионные липиды, например, DOTMA: хлорид [1-(2,3-сиолеилокси)пропил)]-N,N,N-триметиламмония, DMRIE, ди-С14-амидин, DOTIM, SAINT, DC-Chol, BGTC, СТАР, DOPC, DODAP, DOPE: диолеилфосфатидилэтаноламин, DOSPA, DODAB, DOIC, DMEPC, DOGS: диоктадециламидоглицилспермин, DIMRI: бромид димиристооксипропилдиметилгидроксиэтиламмония, DOTAP: диолеоилокси-3-(триметиламмонио)пропан, DC-6-14: хлорид O,O-дитетрадеканоил-N-(α-триметиламмониоацетил)диэтаноламина, CLIP1: хлорид рац-[(2,3-диоктадецилоксипропил)(2-гидроксиэтил)]диметиламмония, CLIP6: рац-[2(2,3-дигексадецилоксипропилоксиметилокси)этил]триметиламмоний, CLIP9: рац-[2(2,3-дигексадецилоксипропилоксисукцинилокси)этил]триметиламмоний, олигофектамин, или катионные или поликатионные полимеры, например, модифицированные полиаминокислоты, такие как полимеры β-аминокислот, или обратимые полиамиды и т.д., модифицированные полиэтилены, такие как PVP (бромид поли(N-этил-4-винилпиридиния)) и т.д. модифицированные акрилаты, такие как pDMAEMA (поли(диметиламиноэтилметилакрилат)) и т.д., модифицированные амидоамины, такие как рАМАМ (поли(амидоамин)) и т.д., модифицированный сложный полибетааминоэфир (РВАЕ), например, с модифицированным диаминовым концом сополимеры 1,4-бутандиолдиакрилата и 5-амино-1-пентанола, и т.д. дендримеры, например, дендримеры полипропиламина или дендримеры на основе рАМАМ, и т.д. полиимин(ы), например, PEI: поли(этиленимин), поли(пропиленимин) и т.д., полиаллиламин, полимеры на основе сахарного каркаса, такие как полимеры на основе циклодекстрина, полимеры на основе декстрана, хитозан и т.д. полимеры на основе силанового каркаса, например, сополимеры PMOXA-PDMS и т.д., блок-сополимеры, состоящие из комбинации одного или несколько катионных блоков (например, выбранных из катионного полимера, указанного выше) и одного или нескольких гидрофильных или гидрофобных блоков (например, полиэтиленгликоль); и т.д.
Согласно другому варианту осуществления изобретения фармацевтическая композиция, предлагаемая в изобретении, может содержать адъювант для повышения иммуностимулирующих свойств фармацевтической композиции. В данном контексте под адъювантом можно понимать любое соединение, которое пригодно для облегчения введения и доставки компонентов, таких как искусственная молекула нуклеиновой кислоты или вектор, которая/который содержится в фармацевтической композиции, предлагаемой в изобретении. Кроме того, такой адъювант может (но, не ограничиваясь только этим) инициировать или усиливать иммунный ответ врожденной иммунной системы, т.е. неспецифический иммунный ответ. Иными словами, после введения фармацевтическая композиция, предлагаемая в изобретении, как правило, инициирует адаптивный иммунный ответ, направленный на антиген, кодируемый искусственной молекулой нуклеиновой кислоты. Кроме того, фармацевтическая композиция, предлагаемая в изобретении, может вызывать (поддерживающий) врожденный иммунный ответ благодаря добавлению адъюванта, указанного в настоящем описании, к фармацевтической композиции, предлагаемой в изобретении.
Такой адъювант можно выбирать из любого адъюванта, известного специалисту в данной области и пригодного для рассматриваемого случая, т.е. для поддержания индукции иммунного ответа у млекопитающего. Предпочтительно адъювант можно выбирать из группы, включающей (но, не ограничиваясь только ими) TDM, MDP, мурамилдипептид, плюроники, раствор квасцов, гидроксид алюминия, ADJUMER™ (полифосфазен); гель фосфата алюминия; глюканы, выделенные из водорослей; алгаммулин; гель гидроксида алюминия (квасцы); высокоадсорбирующий белки гель гидроксида алюминия; обладающий низкой вязкостью гель гидроксида алюминия; AF или SPT (эмульсия, содержащая сквалан (5%), Твин 80 (0.2%), плюроник L121 (1,25%), забуференный фосфатом физиологический раствор, рН 7,4); AVRIDINE™ (пропандиамин); BAY R1005™ ((N-(2-дезокси-2-L-леуциламиноб-D-глюкопиранозил)-N-октадециламида гидроацетат); CALCITRIOL™ (1-альфа,25-дигидрокси-витамин D3); гель фосфата кальция; САР™ (наночастицы фосфата кальция); холерный голотоксин, слитый белок холерный токсин-А1-белок-A-D-фрагмент, субъединица В холерного токсина; CRL 1005 (блок-сополимер Р1205); содержащие цитокин липосомы; DDA (диметилдиоктадециламмония бромид); DHEA (дегидроэпиандростерон); DMPC (димиристоилфосфатидилхолин); DMPG (димиристоилфосфатидилглицерин); комплекс DOC/квасцы (натриевая соль дезоксихолиновой кислоты); полный адъювант Фрейнда; неполный адъювант Фрейнда; инулин-гамма; адъювант Гербу (смесь, содержащая: I) N-ацетилглюкозаминил-(Р1-4)-N-ацетилмурамоил-L-аланил-D35 глутамин (GMDP), II) диметилдиоктадециламмония хлорид (DDA), III) комплекс цинк-соль L-пролина (ZnPro-8); GM-CSF); GMDP (N-ацетилглюкозаминил-(b1-4)-N-ацетилмурамил-L47 аланил-D-изоглутамин); имиквимод (1-(2-метилпропил)-1Н-имидазо[4,5-с]хинолин-4-амин); ImmTher™ (N-ацетилглюкозаминил-N-ацетилмурамил-L-Ala-D-изоGlu-L-Ala-глицерина дипальмитат); DRV (иммунолипосомы, приготовленные из везикул, полученных методом гидратации-регидратации); интерферон-гамма; интерлейкин-1бета; интерлейкин-2; интерлейкин-7; интерлейкин-12; ISCOMSTM; ISCOPREP 7.0.3. ТМ; липосомы; LOXORIBINE™ (7-аллил-8-оксогуанозин); оральный адъювант LT 5 (лабильный энтеротоксин-протоксин из E. coli); микросферы и микрочастицы любого состава; MF59TM; (водная эмульсия сквалена); MONTANIDE ISA 51™ (очищенный неполный адъювант Фрейнда); MONTANIDE ISA 720™ (метаболизируемый масляный адъювант); MPL™ (3-Q-дезацил-4'-монофосфорил-липид А); липосомы МТР-РЕ и МТР-РЕ ((N-ацетил-L-аланил-D-изоглутаминил-L-аланин-2-(1,2-дипальмитоил-sn-глицеро-3-(гидроксифосфорилокси))этиламид, однонатриевая соль); MURAMETIDE™ (Nac-Mur-L-Ala-D-Gln-ОСН3); MURAPALMITINE™ и DMURAPALMITINE™ (Мас-Mur-L-Thr-D-изоGIn-sn-глицериндипальмитоил); NAGO (нейраминидаза-галактозооксидаза); наносферы или наночастицы любого состава; NISV (везикулы из неионогенного поверхностно-активного вещества); PLEURAN™ β-глюкан); PLGA, PGA и PLA (гомо- и сополимеры молочной кислоты и гликолевой кислоты; микросферы/наносферы); ПЛЮРОНИК L121™; РММА (полиметилметакрилат); PODDS™ (протеноидные микросферы); полиэтиленкарбаматные производные; поли-rA:поли-rU (комплекс полиадениловая кислота-полиуридиновая кислота); полисорбат 80 (Твин 80); белковые кохлеаты (фирма Avanti Polar Lipids, Inc., Алабастер, шт. Алабама); STIMULON™ (QS-21); Quil-A (сапонин Quil-A); S-28463 (4-аминоотекдиметил-2-этоксиметил-1Н-имидазо[4,5-с]хинолин-1-этанол); SAF-1™ («композиция адъювантов фирмы Syntex»); протеолипосомы вируса Сендаи и липидные матрицы, содержащие вирус Сендаи; Спан-85 (сорбитантриолеат); Specol (эмульсия Marcol 52, Спан 85 и Твин 85); сквален или Robane® (2,6,10,15,19,23-гексаметилтетракозан и 2,6,10,15,19,23-гексаметил-2,6,10,14,18,22-тетракозагексан); стеарилтирозин (октадецилтирозина гидрохлорид); Theramid® (N-ацетилглюкозаминил-N-ацетилмурамил-L-Ala-D-isoGlu-L-Ala-дипальмитоксипропиламид); Theronyl-MDP (Termurtide™ или [thr 1]-MDP; N-ацетилмурамил-L-треонил-D-изоглутамин); Ту-частицы (Ту-ВПЧ или вирусоподобные частицы); липосомы Уолтера-Рида (липосомы, содержащие липид А, адсорбированный на гидроксиде алюминия) и липопептиды, включая Pam3Cys, прежде всего соли алюминия, такие как адъю-фос (Adju-phos), алгидрогель, регидрагель и т.д., эмульсии, такие как CFA, SAF, IFA, MF59, провакс, TiterMax, монтанид, ваксфектин и т.д., сополимеры, такие как оптивакс (Optivax, CRL1005), L121, полоксамер 4010) и т.д., липосомы, такие как «липосомы-невидимки (Stealth)» и т.д., кохлеаты, такие как BIORAL, и т.д., адъюванты растительного происхождения, такие как QS21, Quil A, Iscomatrix, ISCOM; адъюванты, пригодные для костимуляции, включая томатин, биополимеры, такие как PLG, РММ, инулин и т.д., адъюванты микробного происхождения, такие как ромуртид, DETOX, MPL, CWS, манноза, нуклеотидные последовательности CpG, CpG7909, лиганды человеческого TLR 1-13, ISS-1018, 35 IC31, имидазохинолины, амплиген, Ribi529, IMOxine, IRIV, ВПЧ, холерный токсин, термолабильный токсин, Pam3Cys, флагеллин, GPI (гликозилфосфатидилинозитол)-якорь, LNFPIII/Lewis X, противомикробные пептиды, UC-1V150, слитый белок RSV, cdiGMP; и адъюванты, пригодные в качестве антагонистов, включая нейропептид CGRP.
Пригодные адъюванты можно выбирать также из катионных или поликатионных соединений, при этом адъювант предпочтительно получают путем образования комплекса, включающего искусственную молекулу нуклеиновой кислоты или вектор, входящую/входящий в фармацевтическую композицию, и катионное или поликатионное соединение. Ассоциация или образование комплекса, включающего искусственную молекулу нуклеиновой кислоты или вектор входящую/входящий в фармацевтическую композицию, и катионные или поликатионные соединения, указанные в настоящем описании, предпочтительно обеспечивает присущие адъюванту свойства и оказывает стабилизирующее действие на искусственную молекулу нуклеиновой кислоты или вектор, входящую/входящий в фармацевтическую композицию. Наиболее предпочтительно такие катионные или поликатионные соединения выбирают из катионных или поликатионных пептидов или белков, включая протамин, нуклеолин, спермин или спермидин, или другие катионные пептиды или белки, такие как поли-L-лизин (PLL), полиаргинин,  полипептиды, обладающие способностью проникать в клетку пептиды (СРР), такие как ВИЧ-связывающие пептиды, Tat ВИЧ-1 (ВИЧ), выведенные из Tat пептиды, пенетратин, полученные из VP22 пептиды или аналоги, VP22 HSV (вирус герпеса простого), MAP, KALA или домены трансдукции белков (PTD), РрТ620, богатые пролином пептиды, богатые аргинином пептиды, богатые лизином пептиды, МРG-пептид(ы), Рер-1, L-олигомеры, пептид(ы) кальцитонина, выведенные из Antennapedia пептиды (прежде всего, выведенные из Drosophila antennapedia), pAntp, pIsl, FGF, лактоферрин, транспортан, буфорин-2, Вас715-24, SynB, SynB(1), pVEC, выведенные из hCT пептиды, SAP или гистоны. Другими предпочтительными катионными или поликатионными соединениями могут являться катионные полисахариды, например хитозан, полибрен, катионные полимеры, например, полиэтиленимин (PEI), катионные липиды, например, DOTMA: хлорид [1-(2,3-сиолеилокси)пропил)]-N,N,N-триметиламмония, DMRIE, ди-С14-амидин, DOTIM, SAINT, DC-Chol, BGTC, СТАР, DOPC, DODAP, DOPE: диолеилфосфатидилэтаноламин, DOSPA, DODAB, DOIC, DMEPC, DOGS: диоктадециламидоглицилспермин, DIMRI: бромид димиристооксипропилдиметилгидроксиэтиламмония, DOTАР: диолеоилокси-3-(триметиламмонио)пропан, DC-6-14: хлорид O,O-дитетрадеканоил-N-(α-триметиламмониоацетил)диэтаноламина, CLIP1: хлорид рац-[(2,3-диоктадецилоксипропил)(2-гидроксиэтил)]диметиламмония, CLIP6: рац-[2(2,3-дигексадецилоксипропилоксиметилокси)этил]триметиламмоний, CLIP9: рац-[2(2,3-дигексадецилоксипропилоксисукцинилокси)этил]триметиламмоний, олигофектамин, или катионные или поликатионные полимеры, например, модифицированные полиаминокислоты, такие как полимеры β-аминокислот, или обратимые полиамиды и т.д., модифицированные полиэтилены, такие как PVP (бромид поли(N-этил-4-винилпиридиния)) и т.д. модифицированные акрилаты, такие как pDMAEMA (поли(диметиламиноэтилметилакрилат)) и т.д., модифицированные амидоамины, такие как рАМАМ (поли(амидоамин)) и т.д., модифицированный сложный полибетааминоэфир (РВАЕ), например, с модифицированным диаминовым концом сополимеры 1,4-бутандиолдиакрилата и 5-амино-1-пентанола, и т.д. дендримеры, например, дендримеры полипропиламина или дендримеры на основе рАМАМ, и т.д. полиимин(ы), например, PEI: поли(этиленимин), поли(пропиленимин) и т.д., полиаллиламин, полимеры на основе сахарного каркаса, такие как полимеры на основе циклодекстрина, полимеры на основе декстрана, хитозан и т.д. полимеры на основе силанового каркаса, например, сополимеры PMOXA-PDMS и т.д., блок-сополимеры, состоящие из комбинации одного или несколько катионных блоков (например, выбранных из катионного полимера, указанного выше) и одного или нескольких гидрофильных или гидрофобных блоков (например, полиэтиленгликоль); и т.д.
полипептиды, обладающие способностью проникать в клетку пептиды (СРР), такие как ВИЧ-связывающие пептиды, Tat ВИЧ-1 (ВИЧ), выведенные из Tat пептиды, пенетратин, полученные из VP22 пептиды или аналоги, VP22 HSV (вирус герпеса простого), MAP, KALA или домены трансдукции белков (PTD), РрТ620, богатые пролином пептиды, богатые аргинином пептиды, богатые лизином пептиды, МРG-пептид(ы), Рер-1, L-олигомеры, пептид(ы) кальцитонина, выведенные из Antennapedia пептиды (прежде всего, выведенные из Drosophila antennapedia), pAntp, pIsl, FGF, лактоферрин, транспортан, буфорин-2, Вас715-24, SynB, SynB(1), pVEC, выведенные из hCT пептиды, SAP или гистоны. Другими предпочтительными катионными или поликатионными соединениями могут являться катионные полисахариды, например хитозан, полибрен, катионные полимеры, например, полиэтиленимин (PEI), катионные липиды, например, DOTMA: хлорид [1-(2,3-сиолеилокси)пропил)]-N,N,N-триметиламмония, DMRIE, ди-С14-амидин, DOTIM, SAINT, DC-Chol, BGTC, СТАР, DOPC, DODAP, DOPE: диолеилфосфатидилэтаноламин, DOSPA, DODAB, DOIC, DMEPC, DOGS: диоктадециламидоглицилспермин, DIMRI: бромид димиристооксипропилдиметилгидроксиэтиламмония, DOTАР: диолеоилокси-3-(триметиламмонио)пропан, DC-6-14: хлорид O,O-дитетрадеканоил-N-(α-триметиламмониоацетил)диэтаноламина, CLIP1: хлорид рац-[(2,3-диоктадецилоксипропил)(2-гидроксиэтил)]диметиламмония, CLIP6: рац-[2(2,3-дигексадецилоксипропилоксиметилокси)этил]триметиламмоний, CLIP9: рац-[2(2,3-дигексадецилоксипропилоксисукцинилокси)этил]триметиламмоний, олигофектамин, или катионные или поликатионные полимеры, например, модифицированные полиаминокислоты, такие как полимеры β-аминокислот, или обратимые полиамиды и т.д., модифицированные полиэтилены, такие как PVP (бромид поли(N-этил-4-винилпиридиния)) и т.д. модифицированные акрилаты, такие как pDMAEMA (поли(диметиламиноэтилметилакрилат)) и т.д., модифицированные амидоамины, такие как рАМАМ (поли(амидоамин)) и т.д., модифицированный сложный полибетааминоэфир (РВАЕ), например, с модифицированным диаминовым концом сополимеры 1,4-бутандиолдиакрилата и 5-амино-1-пентанола, и т.д. дендримеры, например, дендримеры полипропиламина или дендримеры на основе рАМАМ, и т.д. полиимин(ы), например, PEI: поли(этиленимин), поли(пропиленимин) и т.д., полиаллиламин, полимеры на основе сахарного каркаса, такие как полимеры на основе циклодекстрина, полимеры на основе декстрана, хитозан и т.д. полимеры на основе силанового каркаса, например, сополимеры PMOXA-PDMS и т.д., блок-сополимеры, состоящие из комбинации одного или несколько катионных блоков (например, выбранных из катионного полимера, указанного выше) и одного или нескольких гидрофильных или гидрофобных блоков (например, полиэтиленгликоль); и т.д.
Кроме того, предпочтительные катионные или поликатионные белки или пептиды, которые можно применять в качестве адъюванта путем образования комплекса с искусственной молекулой нуклеиновой кислоты или вектором, предпочтительно РНК, включенной/включенным в композицию, можно выбирать из представленных ниже белков или пептидов, имеющих следующую общую формулу (I): (Arg)l;(Lys)m;(His)n;(Orn)o;(Xaa)x, в которой l+m+n+o+х=8-15 и l, m, n или о независимо друг от друга могут обозначать любое число, выбранное из 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15, при условии, что общее содержание Arg, Lys, His и Orn составляет по меньшей мере 50% от общего содержания аминокислот в олигопептиде; и Хаа может обозначать любую аминокислоту, выбранную из нативных (т.е. встречающихся в естественных условиях) или ненативных аминокислот за исключением Arg, Lys, His или Orn; и x может обозначать любое число, выбранное из 0, 1, 2, 3 или 4, при условии, что общее содержание Хаа не должно превышать 50% от всех аминокислот в олигопептиде. Наиболее предпочтительными олигоаргининами в данном контексте являются, например, Arg7, Arg8, Arg9, Arg7, H3R9, R9H3, H3R9H3, YSSR9SSY, (RKH)4, Y(RKH)2R и т.д.
Соотношение между искусственной нуклеиновой кислотой или вектором и катионным или поликатионным соединением можно рассчитывать на основе отношения содержания азота к содержанию фосфата (N/P-отношение) во всем включающем нуклеиновую кислоту комплексе. Например, 1 мкг РНК, как правило, содержит примерно 3 нмоля остатков фосфата, при условии, что для РНК характерно статистическое распределение оснований. Кроме того, 1 мл пептида, как правило, содержит примерно х нмолей остатков азота в зависимости от молекулярной массы и количества  аминокислот. Если в качестве примера сделать расчет для (Arg)9 (молекулярная масса 1424 г/моль, 9 атомов азота), то 1 мкг (Arg)9 содержит примерно 700 пмолей (Arg)9 и, следовательно, 700×9=6300 пмолей аминокислот, т.е. 6,3 нмолей атомов азота. Можно рассчитать, что для массового соотношения PHK/(Arg)9, составляющего примерно 1:1, отношение N/P должно составлять примерно 2. Если в качестве примера сделать расчет для протамина (молекулярная масса примерно 4250 г/моль, 21 атом азота, если применяют протамин из лосося) при массовом соотношении, составляющем примерно 2:1, то для 2 мкг РНК РНК дает 6 нмолей фосфата; 1 мкг протамина содержит примерно 235 пмолей молекул протамина и, следовательно, 235×21=4935 пмолей атомов азота, т.е. 4,9 нмолей атомов азота. Можно рассчитать, что для массового соотношения РНК/протамин, составляющего примерно 2:1, отношение N/P должно составлять примерно 0,81. Можно рассчитать, что для массового соотношения РНК/протамин, составляющего примерно 8:1, отношение N/P должно составлять примерно 0,2. В контексте настоящего изобретения касательно соотношения нуклеиновая кислота : пептид в комплексе N/P-отношение предпочтительно находится примерно в пределах 0,1-10, предпочтительно примерно в пределах 0,3-4, и наиболее предпочтительно примерно в пределах 0,5-2 или 0,7-2, и наиболее предпочтительно примерно в пределах 0,7-1,5.
аминокислот. Если в качестве примера сделать расчет для (Arg)9 (молекулярная масса 1424 г/моль, 9 атомов азота), то 1 мкг (Arg)9 содержит примерно 700 пмолей (Arg)9 и, следовательно, 700×9=6300 пмолей аминокислот, т.е. 6,3 нмолей атомов азота. Можно рассчитать, что для массового соотношения PHK/(Arg)9, составляющего примерно 1:1, отношение N/P должно составлять примерно 2. Если в качестве примера сделать расчет для протамина (молекулярная масса примерно 4250 г/моль, 21 атом азота, если применяют протамин из лосося) при массовом соотношении, составляющем примерно 2:1, то для 2 мкг РНК РНК дает 6 нмолей фосфата; 1 мкг протамина содержит примерно 235 пмолей молекул протамина и, следовательно, 235×21=4935 пмолей атомов азота, т.е. 4,9 нмолей атомов азота. Можно рассчитать, что для массового соотношения РНК/протамин, составляющего примерно 2:1, отношение N/P должно составлять примерно 0,81. Можно рассчитать, что для массового соотношения РНК/протамин, составляющего примерно 8:1, отношение N/P должно составлять примерно 0,2. В контексте настоящего изобретения касательно соотношения нуклеиновая кислота : пептид в комплексе N/P-отношение предпочтительно находится примерно в пределах 0,1-10, предпочтительно примерно в пределах 0,3-4, и наиболее предпочтительно примерно в пределах 0,5-2 или 0,7-2, и наиболее предпочтительно примерно в пределах 0,7-1,5.
В заявке на патент WO 2010/037539, содержание которой включено в настоящее описание в качестве ссылки, описана иммуностимулирующая композиция и методы получения иммуностимулирующей композиции. Так, в предпочтительном варианте осуществления изобретения композицию получают, осуществляя две отдельные стадии для достижения и эффективного иммуностимулирующего действия, и эффективной трансляции искусственной молекулы нуклеиновой кислоты, предлагаемой в изобретении. Для этого на первой стадии получают так называемый «адъювантный компонент» путем образования комплекса, включающего искусственную молекулу нуклеиновой кислоты или вектор, предпочтительно РНК, входящую в адъювантный компонент, и катионное или поликатионное соединение в конкретном соотношении для формирования стабильного комплекса. В этом контексте важно, чтобы в адъювантном компоненте отсутствовало или сохранялось лишь в ничтожно малых количествах свободное катионное или поликатионное соединение после образования комплекса с нуклеиновой кислотой. Таким образом, соотношение между нуклеиновой кислотой и катионным или поликатионным соединением в адъювантном компоненте, как правило, выбирают в таком диапазоне, чтобы нуклеиновая кислота была полностью включена в комплекс и отсутствовало или сохранялось лишь в ничтожно малых количествах свободное катионное или поликатионное соединение. Предпочтительно соотношение в адъювантном компоненте, т.е. соотношение между мРНК и катионным или поликатионным соединением, выбирают из диапазона, составляющего от примерно 6:1 (мас./мас.) до примерно 0,25:1 (мас./мас), более предпочтительно от примерно 5:1 (мас./мас.) до примерно 0,5:1 (мас./мас), еще более предпочтительно от примерно 4:1 (мас./мас.) до примерно 1:1 (мас./мас.) или от примерно 3:1 (мас./мас.) до примерно 1:1 (мас./мас.) и наиболее предпочтительно от примерно 3:1 (мас./мас.) до примерно 2:1 (мас./мас.).
Согласно предпочтительному варианту осуществления изобретения искусственную молекулу нуклеиновой кислоты или вектор, предпочтительно молекулу РНК, предлагаемую/предлагаемый в изобретении, добавляют на второй стадии к входящей в комплекс молекуле нуклеиновой кислоты, предпочтительно РНК, адъювантного компонента для получения (иммуностимулирующей) композиции, предлагаемой в изобретении. При этом искусственную молекулу нуклеиновой кислоты или вектор, предпочтительно молекулу РНК, предлагаемую/предлагаемый в изобретении, добавляют в виде свободной нуклеиновой кислоты, т.е. нуклеиновой кислоты, которая не входит в комплекс с другими соединениями. Перед добавлением свободную искусственную молекулу нуклеиновой кислоты или вектор не включают в комплекс и предпочтительно после добавления адъювантного компонента она/он не должна/должен подвергаться какой-либо поддающейся обнаружению или значимой реакции комплексообразования.
Приемлемые адъюванты можно выбирать также из нуклеиновых кислот, имеющих формулу (II): GlXmGn, в которой: G обозначает гуанозин, урацил или аналог гуанозина или урацила; X обозначает гуанозин, урацил, аденозин, тимидин, цитозин или аналог вышеуказанных нуклеотидов; 1 обозначает целое число от 1 до 40, при этом, когда l=1, то G представляет собой гуанозин или его аналог, когда l>1, то по меньшей мере 50% нуклеотидов представляют собой гуанозин или его аналог; m обозначает целое число и обозначает по меньшей мере 3; при этом, когда m=3, то X представляет собой урацил или его аналог, когда m>3, то по присутствуют меньшей мере 3 последовательно расположенных урацилов или аналогов урацила; n обозначает целое число от 1 до 40, при этом, когда n=1, то G представляет собой гуанозин или его аналог, когда n>1, то по меньшей мере 50% нуклеотидов представляет собой гуанозин или его аналог.
Другие приемлемые адъюванты можно выбирать также из нуклеиновых кислот, имеющих формулу (III): ClXmCn, в которой: С обозначает цитозин, урацил или аналог цитозина или урацила; X обозначает гуанозин, урацил, аденозин, тимидин, цитозин или аналог вышеуказанных нуклеотидов; 1 обозначает целое число от 1 до 40, при этом, когда l=1, то С представляет собой цитозин или его аналог, когда l>1, то по меньшей мере 50% нуклеотидов представляют собой цитозин или его аналог; m обозначает целое число и обозначает по меньшей мере 3; когда m=3, X представляет собой урацил или его аналог, когда m>3, то присутствуют по меньшей мере 3 последовательно расположенных урацила или аналогов урацила; n обозначает целое число от 1 до 40, при этом, когда n=1, то С представляет собой цитозин или его аналог, когда n>1, то по меньшей мере 50% нуклеотидов представляют собой цитозин или его аналог.
Предлагаемая в настоящем изобретении фармацевтическая композиция предпочтительно содержит в «безопасном и эффективном количестве» компоненты фармацевтической композиции, прежде всего предлагаемую в изобретении искусственную молекулу нуклеиновой кислоты, вектор и/или клетки, представленные в настоящем описании. В контексте настоящего описания понятие «безопасное и эффективное количество» обозначает количество, достаточное для значимой индукции положительной модификации заболевания или нарушения, указанного в настоящем описании. Однако в то же время «безопасное и эффективное количество» должно быть таким, чтобы избегать серьезных побочных действий и обеспечивать разумное соотношение между преимуществом и риском. Определение этих пределов, как правило, находится в компетенции осуществляющего лечение врача.
Следующим объектом настоящего изобретения является искусственная молекула нуклеиновой кислоты, предлагаемая в настоящем изобретении, вектор, предлагаемый в настоящем изобретении, клетка, предлагаемая в настоящем изобретении, или фармацевтическая композиция, предлагаемая в настоящем изобретении, предназначенная/предназначенный для применения в качестве лекарственного средства, например, в качестве вакцины (для генетической вакцинации), или в генной терапии.
Искусственная молекула нуклеиновой кислоты, предлагаемая в настоящем изобретении, вектор, предлагаемый в настоящем изобретении, клетка, предлагаемая в настоящем изобретении, или фармацевтическая композиция, предлагаемая в настоящем изобретении, наиболее пригодны для любого медицинского применения, в котором используется терапевтическое действие или эффект пептидов, полипептидов или белков, или в котором требуется добавление конкретного пептида или белка. Таким образом, в настоящем изобретении предложены искусственная молекула нуклеиновой кислоты, предлагаемая в настоящем изобретении, вектор, предлагаемый в настоящем изобретении, клетка, предлагаемая в настоящем изобретении, или фармацевтическая композиция, предлагаемая в настоящем изобретении, предназначенные для применения при лечении или предупреждении заболеваний или нарушений, поддающихся лечению посредством терапевтического действия или эффекта пептидов, полипептидов или белков, или поддающихся лечению путем добавления пептида, полипептида или белка. Например, искусственную молекулу нуклеиновой кислоты, предлагаемую в настоящем изобретении, вектор, предлагаемый в настоящем изобретении, клетку, предлагаемую в настоящем изобретении, или фармацевтическую композицию, предлагаемую в настоящем изобретении, можно применять для лечения или предупреждения генетических заболеваний, аутоиммунных заболеваний, раковых или связанных с опухолью заболеваний, инфекционных заболеваний, хронических заболеваний или т.п., например, путем генетической вакцинации или генной терапии.
В частности, такие терапевтические обработки, польза от которых определяется стабильным и пролонгированным присутствием терапевтических пептидов, полипептидов или белков в организме индивидуума, подлежащего лечению, являются наиболее пригодными в качестве медицинского подхода в контексте настоящего изобретения, поскольку предлагаемый в изобретении 3'-UTR-элемент обеспечивает стабильную и пролонгированную экспрессию пептида или белка, кодируемого предлагаемой в изобретении искусственной молекулой нуклеиновой кислоты или предлагаемым в изобретении вектором. Так, наиболее пригодным медицинским применением искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, вектора, предлагаемого в настоящем изобретении, клетки, предлагаемой в настоящем изобретении, или фармацевтической композиции, предлагаемой в настоящем изобретении, является вакцинация. Таким образом, в настоящем изобретении предложены искусственная молекула нуклеиновой кислоты, предлагаемая в настоящем изобретении, вектор, предлагаемый в настоящем изобретении, клетка, предлагаемая в настоящем изобретении, или фармацевтическая композиция, предлагаемая в настоящем изобретении, для вакцинации индивидуума, предпочтительно млекопитающего, более предпочтительно человека. Предпочтительными видами вакцинации являются вакцинация против инфекционных заболеваний, таких как бактериальные, вызываемые простейшими или вирусные инфекции, и противоопухолевая вакцинация. Такие виды вакцинации можно осуществлять в профилактических или терапевтических целях.
Можно выбирать ОРС в зависимости от заболевания, подлежащего лечению. Например, открытая рамка считывания может кодировать белок, который требуется поставлять в организм пациента, страдающего от полной потери или по меньшей мере от частичной потери функции белка, например, в организм пациента, страдающего генетическим заболеванием. Кроме того, открытую рамку считывания можно выбирать из ОРС, кодирующих пептид или белок, который оказывает благоприятное действие на заболевание или состояние пациента. Кроме того, открытая рамка считывания может кодировать пептид или белок, который осуществляет понижающую регуляцию патологического сверхпроизводства встречающегося в естественных условиях пептида или белка или элиминацию клеток, патологически экспрессирующих белок или пептид. Такое отсутствие, потеря функции или сверхпроизводство могут иметь место, например, в случае опухоли или неоплазии, при аутоиммунных заболеваниях, аллергиях, инфекциях или хронических заболеваниях, или т.п. Кроме того, открытая рамка считывания может кодировать антиген или иммуноген, например, эпитоп патогена или опухолевого антигена. Так, в предпочтительных вариантах осуществления изобретения искусственная молекула нуклеиновой кислоты или вектор, предлагаемая/предлагаемый в настоящем изобретении, содержит ОРС, кодирующую аминокислотную последовательность, содержащую или состоящую из последовательности антигена или иммуногена, например, эпитопа патогена или опухоль-ассоциированного антигена, 3'-UTR-элемент, указанный выше, и необязательно другие компоненты, такие как поли(А)-последовательность и т.п.
В контексте медицинского применения, прежде всего в контексте вакцинации, предпочтительно, чтобы искусственная молекула нуклеиновой кислоты, предлагаемая в настоящем изобретении, представляла собой РНК, предпочтительно мРНК, поскольку ДНК несет риск вызывания иммунного анти-ДНК-ответа и имеет тенденцию к встраиванию в геномную ДНК. Однако в некоторых вариантах осуществления изобретения, например, когда применяют вирусный носитель для доставки, такой как аденовирусный носитель для доставки, для доставки искусственной молекулы нуклеиновой кислоты или вектора, предлагаемой/предлагаемого в настоящем изобретении, например, в контексте различных видов генного терапии, может оказаться желательным, чтобы искусственная молекула нуклеиновой кислоты или вектор представляла/представлял собой молекулу ДНК.
Искусственную молекулу нуклеиновой кислоты, предлагаемую в настоящем изобретении, вектор, предлагаемый в настоящем изобретении, клетку, предлагаемую в настоящем изобретении, или фармацевтическую композицию, предлагаемую в настоящем изобретении, можно вводить орально, парентерально, путем ингаляции с использованием спрея, местно, ректально, интраназально, трансбуккально, вагинально, с использованием имплантированного резервуара или посредством струйной инъекции. В контексте настоящего описания понятие «парентерально» включает методы подкожной, внутривенной, внутримышечной, внутрисуставной, интрасиновиальной, надчревной, внутриоболочечной, внутрипеченочной, осуществляемой в область повреждения, внутричерепной, трансдермальной (чрескожной), внутрикожной, внутрилегочной, внутрибрюшинной, внутрисердечной, внутриартериальной и подъязычной инъекции или инфузии. В предпочтительном варианте осуществления изобретения искусственную молекулу нуклеиновой кислоты, предлагаемую в настоящем изобретении, вектор, предлагаемый в настоящем изобретении, клетку, предлагаемую в настоящем изобретении, или фармацевтическую композицию, предлагаемую в настоящем изобретении, вводят посредством безыгольной инъекции (например, струйной инъекции).
Предпочтительно искусственную молекулу нуклеиновой кислоты, предлагаемую в настоящем изобретении, вектор, предлагаемый в настоящем изобретении, клетку, предлагаемую в настоящем изобретении, или фармацевтическую композицию, предлагаемую в настоящем изобретении, вводят парентерально, например, путем парентеральной инъекции, более предпочтительно с помощью методов подкожной, внутривенной, внутримышечной, внутрисуставной, интрасиновиальной, надчревной, внутриоболочечной, внутрипеченочной, осуществляемой в область повреждения, внутричерепной, трансдермальной (чрескожной), внутрикожной, внутрилегочной, внутрибрюшинной, внутрисердечной, внутриартериальной и подъязычной инъекции или инфузии. Наиболее предпочтительными являются внутрикожная и внутримышечная инъекции. Стерильные инъекционные формы предлагаемой в изобретении фармацевтической композиции могут представлять собой водную или масляную суспензию. Такие суспензии можно приготавливать с помощью методов, известных в данной области, с применением пригодных диспергирующих или смачивающих средств. Предпочтительно, растворы или суспензии вводят посредством безыгольной инъекции (например, струйной инъекции).
Искусственную молекулу нуклеиновой кислоты, предлагаемую в настоящем изобретении, вектор, предлагаемый в настоящем изобретении, клетку, предлагаемую в настоящем изобретении, или фармацевтическую композицию, предлагаемую в настоящем изобретении, можно вводить также орально в любой пригодной для орального введения лекарственной форме, включая (но, не ограничиваясь только ими) капсулы, таблетки, водные суспензии или растворы.
Искусственную молекулу нуклеиновой кислоты, предлагаемую в настоящем изобретении, вектор, предлагаемый в настоящем изобретении, клетку, предлагаемую в настоящем изобретении, или фармацевтическую композицию, предлагаемую в настоящем изобретении, можно вводить также местно, прежде всего в тех случаях, когда мишень для лечения включает области или органы, легко доступные для местного нанесения, включая, например, заболевания кожи или любой другой доступной эпителиальной ткани. Для каждой из таких областей или органов можно легко приготавливать пригодные для местной обработки композиции. Для местного нанесения искусственную молекулу нуклеиновой кислоты, предлагаемую в настоящем изобретении, вектор, предлагаемый в настоящем изобретении, клетку, предлагаемую в настоящем изобретении, или фармацевтическую композицию, предлагаемую в настоящем изобретении, можно приготавливать в виде пригодной мази, суспендированной или растворенной в одном или нескольких носителях.
В одном из вариантов осуществления изобретения применение в качестве лекарственного средства включает стадию трансфекции клеток млекопитающих, предпочтительно трансфекцию in vitro или ex vivo клеток млекопитающих, более предпочтительно трансфекцию in vitro клеток, выделенных из организма пациента, подлежащего лечению лекарственным средством. Если применение включает трансфекцию in vitro выделенных клеток, то применение может предусматривать также повторное введение трансфектированных клеток пациенту. Применение предлагаемых в изобретении искусственных молекул нуклеиновой кислоты или вектора в качестве лекарственного средства может включать также стадию отбора успешно трансфектированных выделенных клеток. Таким образом, может оказаться целесообразным, чтобы вектор содержал также маркер для селекции. Применение в качестве лекарственного средства может включать также трансфекцию in vitro выделенных клеток и очистку продукта экспрессии, т.е. кодируемого пептида или белка из указанных клеток. Этот очищенный пептид или белок можно затем вводить индивидууму, нуждающемуся в этом.
В настоящем изобретении предложен также способ лечения или предупреждения заболевания или нарушения, указанного выше, заключающийся в том, что вводят искусственную молекулу нуклеиновой кислоты, предлагаемую в настоящем изобретении, вектор, предлагаемый в настоящем изобретении, клетку, предлагаемую в настоящем изобретении, или фармацевтическую композицию, предлагаемую в настоящем изобретении, индивидууму, нуждающемуся в этом.
Кроме того, предложен способ лечения или предупреждения заболевания или нарушения, заключающийся в том, что осуществляют трансфекцию клетки искусственной молекулой нуклеиновой кислоты, предлагаемой в настоящем изобретении, или вектором, предлагаемым в настоящем изобретении. Указанную трансфекцию можно осуществлять in vitro, ex vivo или in vivo. В предпочтительном варианте осуществления изобретения трансфекцию клетки осуществляют in vitro и трансфектированную клетку вводят индивидууму, нуждающемуся в этом, предпочтительно больному человеку. Предпочтительно клетка, которую требуется трансфектировать in vitro, представляет собой клетку, выделенную из организма индивидуума, предпочтительно больного человека. Таким образом, в настоящем изобретении предложен способ лечения, заключающийся в том, что осуществляют стадии, на которых выделяют клетку из организма индивидуума, предпочтительно из организма больного человека, трансфектируют выделенную клетку искусственной нуклеиновой кислотой, предлагаемой в настоящем изобретении, или вектором, предлагаемым в настоящем изобретении, и вводят трансфектированную клетку индивидууму, предпочтительно больному человеку.
Способ лечения или предупреждения нарушения, предлагаемый в настоящем изобретении, предпочтительно представляет собой способ вакцинации или способ генной терапии, описанный выше.
Как описано выше, предлагаемый в изобретении 3'-UTR-элемент обладает способностью стабилизировать молекулу мРНК и/или усиливать, стабилизировать и/или пролонгировать производство белка с молекулы мРНК. Таким образом, следующий объект настоящего изобретения относится к способу стабилизации молекулы РНК, предпочтительно молекулы мРНК, заключающийся в том, что выполняют стадию, на которой осуществляют ассоциацию молекулы РНК, предпочтительно молекулы мРНК, или вектора, кодирующего молекулу РНК, с 3'-UTR-элементом, содержащим или состоящим из нуклеотидной последовательности, происходящей из 3'-UTR гена рибосомального белка или из варианта 3'-UTR гена рибосомального белка, предпочтительно с 3'-UTR-элeмeнтoм, описанным выше.
Кроме того, настоящее изобретение относится к способу повышения, стабилизации и/или пролонгирования производства белка с искусственной молекулы нуклеиновой кислоты или с вектора, предпочтительно с молекулы мРНК, и/или стабилизации и/или пролонгирования производства белка с искусственной молекулы нуклеиновой кислоты или с вектора, предпочтительно с молекулы мРНК, заключающемуся в том, что выполняют стадию, на которой осуществляют ассоциацию искусственной молекулы нуклеиновой кислоты или вектора, предпочтительно молекулы мРНК, с 3'-UTR-элeмeнтoм, который содержит или состоит из нуклеотидной последовательности, происходящей из 3'-UTR гена рибосомального белка или из варианта 3'-UTR гена рибосомального белка, предпочтительно с 3'-UTR-элементом, описанным выше.
Выражение «ассоциация искусственной молекулы нуклеиновой кислоты или вектора с 3'-UTR-элeмeнтoм» в контексте настоящего изобретения предпочтительно означает функциональную ассоциацию или функциональное объединение искусственной молекулы нуклеиновой кислоты или вектора с 3'-UTR-элементом. Это означает, что осуществляют ассоциацию или связывание искусственной молекулы нуклеиновой кислоты или вектора с 3'-UTR-элементом, предпочтительно 3'-UTR-элементом, описанным выше, таким образом, чтобы осуществлялась функция 3'-UTR-элeмeнтa, например, функция стабилизации производства РНК и/или белка. Как правило, это означает, что 3'-UTR-элемент интегрируют в искусственную молекулу нуклеиновой кислоты или вектор, предпочтительно в молекулу мРНК, с 3'-стороны относительно открытой рамки считывания, предпочтительно так, чтобы он непосредственно примыкал с 3'-стороны к открытой рамке считывания, предпочтительно располагался между открытой рамкой считывания и поли(А)-последовательностью или сигналом полиаденилирования. Предпочтительно 3'-UTR-элeмeнт интегрируют в искусственную молекулу нуклеиновой кислоты или вектор, предпочтительно в мРНК, в качестве 3'-UTR, т.е. так, чтобы 3'-UTR-элeмeнт представлял собой 3'-UTR искусственной молекулы нуклеиновой кислоты или вектора, предпочтительно мРНК, т.е. так, чтобы он простирался от 3'-стороны открытой рамки считывания до 5'-стороны поли(А)-последовательности или сигнала полиаденилирования, необязательно был присоединен посредством короткого линкера, такого как последовательность, содержащая или состоящая из одного или нескольких сайтов рестрикции. Таким образом, предпочтительно выражение «ассоциация искусственной молекулы нуклеиновой кислоты или вектора с 3'-UTR-элементом» означает функциональную ассоциацию 3'-UTR-элeмeнтa с открытой рамкой считывания, локализованной в искусственной молекуле нуклеиновой кислоты или векторе, предпочтительно в молекуле мРНК. 3'-UTR и ОРС представляют собой конструкции, описанные выше для искусственной молекулы нуклеиновой кислоты, предлагаемой в настоящем изобретении, например, предпочтительно ОРС и 3'-UTR являются гетерологичными, например, происходят из различных генов, как описано выше.
Следующим объектом настоящего изобретения является применение 3'-UTR-элемента, предпочтительно 3'-UTR-элемента, описанного выше, для повышения стабильности молекулы РНК, предпочтительно молекулы мРНК, где 3'-UTR-элемент содержит или состоит из нуклеотидной последовательности, происходящей из 3'-UTR гена рибосомального белка или из варианта 3'-UTR гена рибосомального белка.
Кроме того, в настоящем изобретении предложено применение 3'-UTR-элемента, предпочтительно 3'-UTR-элемента, описанного выше, для повышения производства белка с искусственной молекулы нуклеиновой кислоты или вектора, предпочтительно с молекулы мРНК, и/или для стабилизации и/или пролонгирования производства белка с искусственной молекулы нуклеиновой кислоты или векторной молекулы, предпочтительно с молекулы мРНК, где 3'-UTR-элемент содержит или состоит из нуклеотидной последовательности, происходящей из 3'-UTR гена рибосомального белка или из варианта 3'-UTR гена рибосомального белка.
Варианты применения, предлагаемые в настоящем изобретении, предпочтительно включают осуществление ассоциации искусственной молекулы нуклеиновой кислоты, вектора или РНК с 3'-UTR-элeмeнтoм, описанным выше.
Соединения и ингредиенты предлагаемой в изобретении фармацевтической композиции можно также производить и продавать по отдельности друг от друга. Таким образом, изобретение относится также к набору или набору компонентов, содержащему искусственную молекулу нуклеиновой кислоты, предлагаемую в изобретении, вектор, предлагаемый в изобретении, клетку, предлагаемую в изобретении, и/или фармацевтическую композицию, предлагаемую в изобретении. Предпочтительно такой набор или набор компонентов может дополнительно содержать инструкции по применению, клетки для трансфекции, адъювант, средства для введения фармацевтической композиции, фармацевтически приемлемый носитель и/или фармацевтически приемлемый раствор для растворения или разведения искусственной молекулы нуклеиновой кислоты, вектора, клеток или фармацевтической композиции.
Представленные ниже чертежи, последовательности и примеры предназначены для дополнительной иллюстрации изобретения. Они не направлены на ограничение сущности рассматриваемого изобретения.
На чертежах показано:
На фиг. 1-3 - последовательности, кодирующие мРНК, которые можно получать путем транскрипции in vitro. Используются следующие сокращения:
rpl32: 5'-UTR человеческого рибосомального белка большой субъединицы 32, в которой отсутствует 5'-концевой олигопиримидиновый тракт;
PpLuc (GC): последовательность мРНК с высоким содержанием GC (GC-обогащенная), кодирующая люциферазу Photinus pyralis;
- А64: поли(А)-последовательность, содержащая 64 аденилата;
- ag: центральная, α-комплекс-связывающая часть 3'-UTR человеческого α-глобина;
- rps9: 3'-UTR-элeмeнт, происходящий из 3'-UTR человеческого рибосомального белка малой субъединицы 9;
- С30: поли(С)-последовательность, содержащая 30 цитидилатов;
- thctohSL: последовательность гистоновой структуры типа «стебель-петля», взятая из публикации Cakmakci, Lerner, Wagner, Zheng и William F Marzluff, Mol. Cell. Biol. 28 (3), 2008, cc. 1182-1194;
альбумин: 3'-UTR человеческого альбумина;
на фиг. 1 - представлена последовательность, кодирующая мРНК rpl32 - PpLuc(GC) - А64 - С30 - гистонSL (SEQ ID NO: 8);
на фиг. 2 - представлена последовательность, кодирующая мРНК rpl32 - PpLuc(GC) - ag - А64 - С30 - гистонSL (SEQ ID NO: 9). Центральная, α-комплекс-связывающая часть 3'-UTR человеческого α-глобина встроена между ОРС и поли(А). ОРС PpLuc(GC) выделена курсивом. 3'-UTR-элeмeнт, происходящий из α-глобина, подчеркнут;
на фиг. 3 - последовательность мРНК rpl32 - PpLuc(GC) - rps9 - А64 - С30 - hSL (SEQ ID NO: 7). 3'-UTR человеческого рибосомального белка малой субъединицы 9 встроена между ОРС и поли(А). ОРС PpLuc(GC) выделена курсивом, 3' UTR-элемент, происходящий из rps9, подчеркнут;
на фиг. 4 - данные, демонстрирующие, что 3'-UTR рибосомального белка малой субъединицы 9 существенно повышает экспрессию белка с мРНК;
Влияние предлагаемой в изобретении 3'-UTR человеческого рибосомального белка малой субъединицы 9 на экспрессию люциферазы с мРНК изучали в сравнении с экспрессией люциферазы с мРНК, в которой отсутствует 3'-UTR или которая содержит 3'-UTR человеческого α-глобина. Для этого различными мРНК трансфектировали путем липофекции человеческие кожные фибробласты (HDF). Уровни люциферазы измеряли через 6, 24, 48 и 72 ч после трансфекции. Очевидно, что имела место экспрессия люциферазы с мРНК, в которой отсутствовала 3'-UTR. При этом экспрессия люциферазы не возрастала при включении хорошо известной 3'-UTR человеческого α-глобина. В отличие от этого, 3'-UTR рибосомального белка малой субъединицы 9 приводила к существенному повышению экспрессии люциферазы. Данные представлены на графике в виде средней величины для трех трансфекции, выраженной в относительных световых единицах (RLU) ± SEM (стандартное отклонение). Результаты измерений в RLU обобщены в примере 5.1;
на фиг. 5 - последовательность, кодирующая мРНК rpl32 - PpLuc(GC) - альбумин - А64 - С30 - гистонSL (SEQ ID NO: 206). 3'-UTR человеческого альбумина встраивали между ОРС и поли(А). ОРС PpLuc(GC) выделена курсивом. 3'-UTR-элeмeнт, происходящий из альбумина, подчеркнут;
на фиг. 6 - данные, демонстрирующие, что 3'-UTR, происходящие из генов мышиного рибосомального белка rps21, rps29, rps9, rps27, rps28, rps19, rpl35a, rpl13, rpl36 и rpl23a повышают экспрессию белка с мРНК в HDF-клетках (человеческие кожные фибробласты) по меньшей мере в той же степени, что и мРНК, содержащая 3'-UTR гена альбумина, которая, как уже было продемонстрировано, повышает экспрессию белка с мРНК (WO 2013/143698);
на фиг. 7 - данные, демонстрирующие, что 3'-UTR, происходящие из генов мышиного рибосомального белка rps21, rps29, rps9, rps27, rps28, rps19, rpl35a, rpl13, rpl36 и rpl23a, повышают экспрессию белка с мРНК в HeLa-клетках по меньшей мере в той же степени, что и мРНК, содержащая 3'-UTR гена альбумина, которая, как уже было продемонстрировано, повышает экспрессию белка с мРНК (WO 2013/143698);
на фиг. 8 - данные, демонстрирующие, что 3'-UTR, происходящие из генов мышиного рибосомального белка rpl23, uba52, rpl2211, rpl36a, rps4x, rpl27, rpl3, rps23, rpsl3, rpl26, rps17, rps18, rps8, Fau, rps13, rpl11 и rpl38 повышают экспрессию белка с мРНК в HDF-клетках (человеческие кожные фибробласты) по меньшей мере в той же степени, что и мРНК, содержащая 3'-UTR гена альбумина, которая, как уже было продемонстрировано, повышает экспрессию белка с мРНК (WO 2013/143698);
на фиг. 9 - данные, демонстрирующие, что 3'-UTR, происходящие из генов мышиного рибосомального белка rpl23, uba52, rpl2211, rpl36a, rps4x, rpl27, rpl3, rps23, rps13, rpl26, rps17, rps18, rps8, Fau, rps13, rpl11 и rpl38, повышают экспрессию белка с мРНК в HeLa- по меньшей мере в той же степени, что и мРНК, содержащая 3'-UTR гена альбумина, которая, как уже было продемонстрировано, повышает экспрессию белка с мРНК (WO 2013/143698).
Примеры
1. Получение ДНК-матриц
Конструировали вектор для транскрипции in vitro, содержащий промотор Т7 и GC-обогащенную последовательность, кодирующую люциферазу Photinus pyralis (PpLuc(GC)). 5'-нетранслируемую область (5'-UTR) рибосомального белка большой субъединицы 32 встраивали с 5'-стороны относительно PpLuc(GC). Содержащую 64 аденина (А64) поли(А)-последовательность, за которой следовала последовательность, состоящая из 30 цитозинов (С30), и последовательность структуры типа «стебель-петля», встраивали с 3'-стороны относительно PpLuc(GC). За последовательностью структуры типа «стебель-петля» находился сайт рестрикции, который использовали для линеаризации вектора перед осуществлением транскрипции in vitro. мРНК, полученную с этого вектора после транскрипции in vitro обозначили как «rpl32 - PpLuc(GC) - А64 - С30 - гистонSL».
Указанный вектор модифицировали так, чтобы он включал нетранслируемые последовательности с 3'-стороны относительно открытой рамки считывания (3'-UTR). В целом, создавали векторы, которые содержали следующие кодирующие последовательности мРНК (некоторые из кодирующих последовательностей мРНК представлены в качестве примеров на фиг. 1-3):
rpl32 - PpLuc(GC) - А64 - С30 - гистонSL (SEQ ID NO: 8, фиг. 1);
rpl32 - PpLuc(GC) - ag - A64 - C30 - гистонSL (SEQ ID NO: 9, фиг. 2);
rpl32 - PpLuc(GC) - rps9 - A64 - C30 - гистонSL (SEQ ID NO: 7, фиг. 3);
rpl32 - PpLuc(GC) - rps21 - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rps29 - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rps9 - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rps27 - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rps28 - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rpsl9 - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rpl35a - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rpl13 - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rpl36 - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rpl23a - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rpl23 - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - uba52 - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rpl2211 - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rpl36a - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rps4x - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rpl27 - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rpl3 - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rps23 - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rpsl3 - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rpl26 - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rpsl7 - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rpsl8 - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - Fau - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rpsl3 - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rpl11 - A64 - C30 - гистонSL;
rpl32 - PpLuc(GC) - rpl38 - A64 - C30 - гистонSL.
2. Транскрипция in vitro
ДНК-матрицу, полученную согласно методу, описанному в примере 1, линеаризовали и транскрибировали in vitro с использованием Т7-РНК-полимеразы. Затем ДНК-матрицу расщепляли путем обработки ДНКазой. мРНК-транскрипты, содержащие 5'-кэп-структуру, получали путем добавления избытка N7-метил-гуанозин-5'-триптофосфат-5'-гуанозина в реакционную смесь для транскрипции. Полученную в результате этого мРНК очищали и ресуспендировали в воде.
3. Экспрессия люциферазы после липофекции мРНК
Человеческие кожные фибробласты (HDF) или HeLa-клетки высевали в 24-луночные планшеты за три дня до трансфекции с плотностью 3×104 клеток на лунку в среду (среда RPMI 1640, содержащая L-глутамин и 25 мМ Hepes (фирма Lonza, Базель, Швейцария), в которую добавляли 10% FCS, 1% Pen/Strep, 1% глутамина). Непосредственно перед осуществлением липофекции клетки промывали в среде Opti-MEM. Клетки подвергали липофекции с использованием 25 нг мРНК, кодирующей PpLuc, на лунку, включенной в комплекс с липофектамином 2000. Для контроля эффективности трансфекции наряду с мРНК PpLuc (2,5 нг мРНК RrLuc на лунку) осуществляли трансфекцию мРНК, кодирующей люциферазу Renilla reniformis (RrLuc). Через 90 мин после начала трансфекции среду заменяли на Opti-MEM. Через 6, 24, 48 и 72 ч после трансфекции среду удаляли путем аспирации и клетки лизировали в 100 мкл буфера для лизиса (буфер для пассивного лизиса, фирма Promega). Лизаты хранили при -80° до осуществления измерений люциферазной активности.
4. Измерение люциферазы
Люциферазную активность измеряли в относительных световых единицах (RLU) с помощью ридера для планшетов Hidex Chameleon. Активность PpLuc измеряли в течение периода измерения, составлявшего 2 с, с использованием 20 мкл лизата и 50 мкл содержащего люциферин буфера (буфер Beetle-Juice, фирма PJK GmbH). Активность RrLuc измеряли в течение периода измерения, составлявшего 2 с, с использованием 20 мкл лизата и 50 мкл содержащего коэлентеразин буфера (буфер Renilla-Juice, фирма PJK GmbH).
5. Результаты
5.1 3'-UTR генов рибосомального белка повышает экспрессию белка.
Для изучения воздействия 3'-UTR генов рибосомального белка на экспрессию белка с мРНК, синтезировали мРНК, содержащие различные UTR: мРНК либо не содержали 3'-UTR, либо содержали центральную связывающую α-комплекс часть 3'-UTR человеческого α-глобина (ag), либо содержали 3'-UTR человеческого рибосомального белка малой субъединицы 9 (rps9). мРНК, кодирующими люциферазу, трансфектировали человеческие кожные фибробласты (HDF). Уровни люциферазы измеряли через 6, 24, 48 и 72 ч после трансфекции. На основе этих данных рассчитывали общее количество экспрессированного белка в течение периода времени с 0 до 72 ч в виде площади под кривой (AUC) (см. представленную ниже таблицу 1 и фиг. 4).
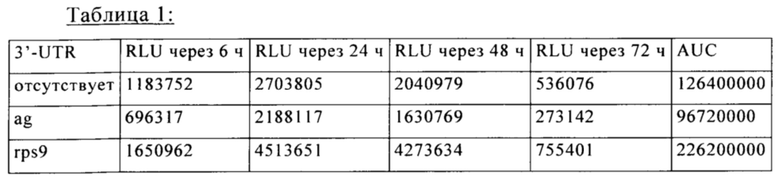
Очевидно, что имела место экспрессия люциферазы с мРНК, в которой отсутствует 3'-UTR. При этом экспрессия люциферазы не повышалась при наличии хорошо известной 3'-UTR α-глобина. В отличие от этого, 3'-UTR рибосомального белка малой субъединицы 9 существенно повышала экспрессию люциферазы.
5.2 3'-UTR генов мышиного рибосомального белка повышают экспрессию белка.
Для изучения воздействия 3'-UTR генов мышиного рибосомального белка на экспрессию белка с мРНК, синтезировали мРНК, содержащие различные UTR: мРНК, содержащие 3'-UTR различных мышиных рибосомальных белков (rps21, rps29, rps9, rps27, rps28, rps19, rpl35a, rpl13, rpl36 и rpl23a) и для сравнения 3'-UTR альбумина, которая, как известно, повышает экспрессию белка с мРНК (WO 2013/143698). мРНК, кодирующими люциферазу, трансфектировали человеческие кожные фибробласты (HDF). Уровни люциферазы измеряли через 24, 48 и 72 ч после трансфекции (см. представленную ниже таблицу 2 и фиг. 6).
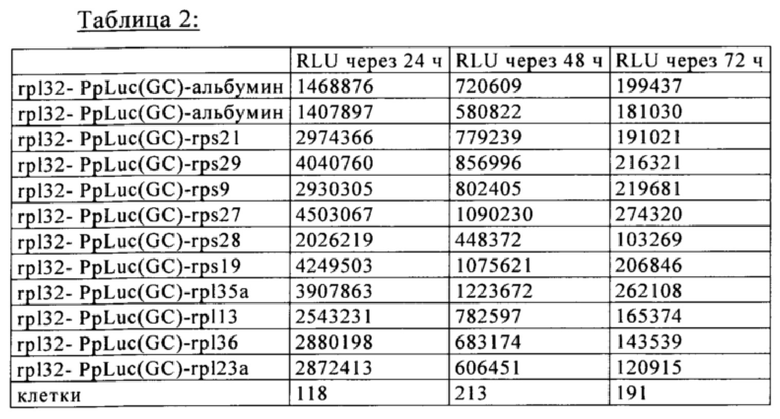
Результаты свидетельствуют о том, что с мРНК, содержащей 3'-UTR мышиных рибосомальных белков, люцифераза экспрессировалась в больших количествах, чем с мРНК, содержащей 3'-UTR альбумина, которая, как было описано, повышает производство белка.
5.3 3'-UTR генов мышиного рибосомального белка повышает экспрессию белка.
Для изучения воздействия 3'-UTR генов мышиного рибосомального белка на экспрессию белка с мРНК, синтезировали мРНК, содержащие различные UTR: мРНК, содержащие 3'-UTR различных мышиных рибосомальных белков (rps21, rps29, rps9, rps27, rps28, rps19, rpl35a, rpl13, rpl36 и rpl23a) и для сравнения 3'-UTR альбумина. мРНК, кодирующими люциферазу, трансфектировали HeLa-клетки. Уровни люциферазы измеряли через 24, 48 и 72 ч после трансфекции (см. представленную ниже таблицу 3 и фиг. 7).
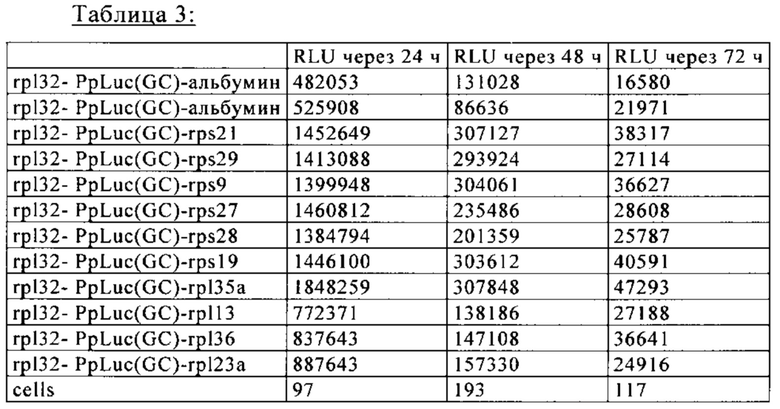
Результаты свидетельствуют о том, что с мРНК, содержащей 3'-UTR мышиных рибосомальных белков, люцифераза экспрессировалась в больших количествах, чем с мРНК, содержащей 3'-UTR альбумина, которая, как было описано, повышает производство белка.
5.4 3'-UTR генов мышиного рибосомального белка повышает экспрессию белка.
Для изучения воздействия 3'-UTR генов мышиного рибосомального белка на экспрессию белка с мРНК, синтезировали мРНК, содержащие различные UTR: мРНК, содержащие 3'-UTR различных мышиных рибосомальных белков (rpl23, uba52, rpl2211, rpl36a, rps4x, rpl27, rpl3, rps23, rpsl3, rpl26, rps17, rps18, rps8, Fau, rps13, rpl11 и rpl38) и для сравнения 3'-UTR альбумина. мРНК, кодирующими люциферазу, трансфектировали человеческие кожные фибробласты (HDF). Уровни люциферазы измеряли через 24, 48 и 72 ч после трансфекции (см. представленную ниже таблицу 4 и фиг. 8).
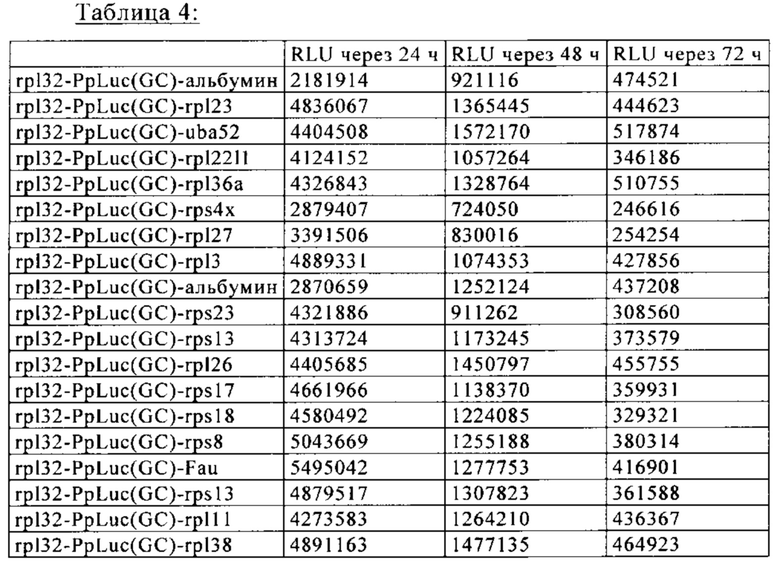
Результаты свидетельствуют о том, что с мРНК, содержащей 3'-UTR мышиных рибосомальных белков, люцифераза экспрессировалась в больших количествах, чем с мРНК, содержащей 3'-UTR альбумина, которая, как было описано, повышает производство белка.
5.5 3'-UTR генов мышиного рибосомального белка повышает экспрессию белка.
Для изучения воздействия 3'-UTR генов мышиного рибосомального белка на экспрессию белка с мРНК, синтезировали мРНК, содержащие различные UTR: мРНК, содержащие 3'-UTR различных мышиных рибосомальных белков (rpl23, uba52, rpl2211, rpl36a, rps4x, rpl27, rpl3, rps23, rps13, rpl26, rps17, rps18, rps8, Fau, rps13, rpl11 и rpl38) и для сравнения 3'-UTR альбумина. мРНК, кодирующими люциферазу, трансфектировали HeLa-клетки. Уровни люциферазы измеряли через 24, 48 и 72 ч после трансфекции (см. представленную ниже таблицу 5 и фиг. 9).
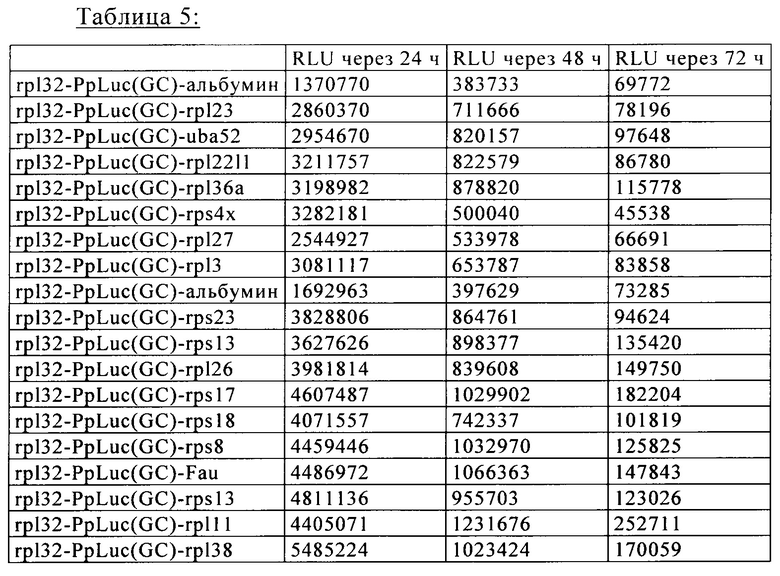
Результаты свидетельствуют о том, что с мРНК, содержащей 3'-UTR мышиных рибосомальных белков, люцифераза экспрессировалась в больших количествах, чем с мРНК, содержащей 3'-UTR альбумина, которая, как было описано, повышает производство белка.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> КуреВак ГмбХ
<120> Искусственные молекулы нуклеиновой кислоты
<130> CU01P155WO1
<160> 206
<170> PatentIn, версия 3.5
<210> 1
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3-UTR-элемент рибосомального белка S9 (RPS9) Homo sapiens
<400> 1
gtccacctgt ccctcctggg ctgctggatt gtctcgtttt cctgccaaat aaacaggatc 60
agcgctttac 70
<210> 2
<211> 70
<212> РНК
<213> Искусственная последовательность
<220>
<223> 3-UTR-элемент рибосомального белка S9 (RPS9) Homo sapiens
<400> 2
guccaccugu cccuccuggg cugcuggauu gucucguuuu ccugccaaau aaacaggauc 60
agcgcuuuac 70
<210> 3
<211> 140
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3-UTR-элемент рибосомального белка S9 (RPS9) Homo sapiens + поли-А
<400> 3
gtccacctgt ccctcctggg ctgctggatt gtctcgtttt cctgccaaat aaacaggatc 60
agcgctttac agatctaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 120
aaaaaaaaaa aaaaaaaaaa 140
<210> 4
<211> 140
<212> РНК
<213> Искусственная последовательность
<220>
<223> 3-UTR-элемент рибосомального белка S9 (RPS9) Homo sapiens + поли-А
<400> 4
guccaccugu cccuccuggg cugcuggauu gucucguuuu ccugccaaau aaacaggauc 60
agcgcuuuac agaucuaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 120
aaaaaaaaaa aaaaaaaaaa 140
<210> 5
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> наиболее предпочтительная последовательность гистоновой структуры типа «стебель-петля»
<400> 5
caaaggctct tttcagagcc acca 24
<210> 6
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5-UTR человеческого рибосомального белка большой субъединицы 32 без 5-концевого олигопиримидинового тракта
<400> 6
ggcgctgcct acggaggtgg cagccatctc cttctcggca tc 42
<210> 7
<211> 1918
<212> ДНК
<213> Искусственная последовательность
<220>
<223> rpl32 PpLuc(GC) rps9 A64 C30 гистонSL
<400> 7
ggggcgctgc ctacggaggt ggcagccatc tccttctcgg catcaagctt gaggatggag 60
gacgccaaga acatcaagaa gggcccggcg cccttctacc cgctggagga cgggaccgcc 120
ggcgagcagc tccacaaggc catgaagcgg tacgccctgg tgccgggcac gatcgccttc 180
accgacgccc acatcgaggt cgacatcacc tacgcggagt acttcgagat gagcgtgcgc 240
ctggccgagg ccatgaagcg gtacggcctg aacaccaacc accggatcgt ggtgtgctcg 300
gagaacagcc tgcagttctt catgccggtg ctgggcgccc tcttcatcgg cgtggccgtc 360
gccccggcga acgacatcta caacgagcgg gagctgctga acagcatggg gatcagccag 420
ccgaccgtgg tgttcgtgag caagaagggc ctgcagaaga tcctgaacgt gcagaagaag 480
ctgcccatca tccagaagat catcatcatg gacagcaaga ccgactacca gggcttccag 540
tcgatgtaca cgttcgtgac cagccacctc ccgccgggct tcaacgagta cgacttcgtc 600
ccggagagct tcgaccggga caagaccatc gccctgatca tgaacagcag cggcagcacc 660
ggcctgccga agggggtggc cctgccgcac cggaccgcct gcgtgcgctt ctcgcacgcc 720
cgggacccca tcttcggcaa ccagatcatc ccggacaccg ccatcctgag cgtggtgccg 780
ttccaccacg gcttcggcat gttcacgacc ctgggctacc tcatctgcgg cttccgggtg 840
gtcctgatgt accggttcga ggaggagctg ttcctgcgga gcctgcagga ctacaagatc 900
cagagcgcgc tgctcgtgcc gaccctgttc agcttcttcg ccaagagcac cctgatcgac 960
aagtacgacc tgtcgaacct gcacgagatc gccagcgggg gcgccccgct gagcaaggag 1020
gtgggcgagg ccgtggccaa gcggttccac ctcccgggca tccgccaggg ctacggcctg 1080
accgagacca cgagcgcgat cctgatcacc cccgaggggg acgacaagcc gggcgccgtg 1140
ggcaaggtgg tcccgttctt cgaggccaag gtggtggacc tggacaccgg caagaccctg 1200
ggcgtgaacc agcggggcga gctgtgcgtg cgggggccga tgatcatgag cggctacgtg 1260
aacaacccgg aggccaccaa cgccctcatc gacaaggacg gctggctgca cagcggcgac 1320
atcgcctact gggacgagga cgagcacttc ttcatcgtcg accggctgaa gtcgctgatc 1380
aagtacaagg gctaccaggt ggcgccggcc gagctggaga gcatcctgct ccagcacccc 1440
aacatcttcg acgccggcgt ggccgggctg ccggacgacg acgccggcga gctgccggcc 1500
gcggtggtgg tgctggagca cggcaagacc atgacggaga aggagatcgt cgactacgtg 1560
gccagccagg tgaccaccgc caagaagctg cggggcggcg tggtgttcgt ggacgaggtc 1620
ccgaagggcc tgaccgggaa gctcgacgcc cggaagatcc gcgagatcct gatcaaggcc 1680
aagaagggcg gcaagatcgc cgtgtaagac tagtgtccac ctgtccctcc tgggctgctg 1740
gattgtctcg ttttcctgcc aaataaacag gatcagcgct ttacagatct aaaaaaaaaa 1800
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaatgcatc 1860
cccccccccc cccccccccc cccccccccc aaaggctctt ttcagagcca ccagaatt 1918
<210> 8
<211> 1848
<212> ДНК
<213> Искусственная последовательность
<220>
<223> rpl32 PpLuc(GC) A64 C30 гистонSL
<400> 8
ggggcgctgc ctacggaggt ggcagccatc tccttctcgg catcaagctt gaggatggag 60
gacgccaaga acatcaagaa gggcccggcg cccttctacc cgctggagga cgggaccgcc 120
ggcgagcagc tccacaaggc catgaagcgg tacgccctgg tgccgggcac gatcgccttc 180
accgacgccc acatcgaggt cgacatcacc tacgcggagt acttcgagat gagcgtgcgc 240
ctggccgagg ccatgaagcg gtacggcctg aacaccaacc accggatcgt ggtgtgctcg 300
gagaacagcc tgcagttctt catgccggtg ctgggcgccc tcttcatcgg cgtggccgtc 360
gccccggcga acgacatcta caacgagcgg gagctgctga acagcatggg gatcagccag 420
ccgaccgtgg tgttcgtgag caagaagggc ctgcagaaga tcctgaacgt gcagaagaag 480
ctgcccatca tccagaagat catcatcatg gacagcaaga ccgactacca gggcttccag 540
tcgatgtaca cgttcgtgac cagccacctc ccgccgggct tcaacgagta cgacttcgtc 600
ccggagagct tcgaccggga caagaccatc gccctgatca tgaacagcag cggcagcacc 660
ggcctgccga agggggtggc cctgccgcac cggaccgcct gcgtgcgctt ctcgcacgcc 720
cgggacccca tcttcggcaa ccagatcatc ccggacaccg ccatcctgag cgtggtgccg 780
ttccaccacg gcttcggcat gttcacgacc ctgggctacc tcatctgcgg cttccgggtg 840
gtcctgatgt accggttcga ggaggagctg ttcctgcgga gcctgcagga ctacaagatc 900
cagagcgcgc tgctcgtgcc gaccctgttc agcttcttcg ccaagagcac cctgatcgac 960
aagtacgacc tgtcgaacct gcacgagatc gccagcgggg gcgccccgct gagcaaggag 1020
gtgggcgagg ccgtggccaa gcggttccac ctcccgggca tccgccaggg ctacggcctg 1080
accgagacca cgagcgcgat cctgatcacc cccgaggggg acgacaagcc gggcgccgtg 1140
ggcaaggtgg tcccgttctt cgaggccaag gtggtggacc tggacaccgg caagaccctg 1200
ggcgtgaacc agcggggcga gctgtgcgtg cgggggccga tgatcatgag cggctacgtg 1260
aacaacccgg aggccaccaa cgccctcatc gacaaggacg gctggctgca cagcggcgac 1320
atcgcctact gggacgagga cgagcacttc ttcatcgtcg accggctgaa gtcgctgatc 1380
aagtacaagg gctaccaggt ggcgccggcc gagctggaga gcatcctgct ccagcacccc 1440
aacatcttcg acgccggcgt ggccgggctg ccggacgacg acgccggcga gctgccggcc 1500
gcggtggtgg tgctggagca cggcaagacc atgacggaga aggagatcgt cgactacgtg 1560
gccagccagg tgaccaccgc caagaagctg cggggcggcg tggtgttcgt ggacgaggtc 1620
ccgaagggcc tgaccgggaa gctcgacgcc cggaagatcc gcgagatcct gatcaaggcc 1680
aagaagggcg gcaagatcgc cgtgtaagac tagtagatct aaaaaaaaaa aaaaaaaaaa 1740
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaatgcatc cccccccccc 1800
cccccccccc cccccccccc aaaggctctt ttcagagcca ccagaatt 1848
<210> 9
<211> 1914
<212> ДНК
<213> Искусственная последовательность
<220>
<223> rpl32 PpLuc(GC) ag A64 C30 гистонSL
<400> 9
ggggcgctgc ctacggaggt ggcagccatc tccttctcgg catcaagctt gaggatggag 60
gacgccaaga acatcaagaa gggcccggcg cccttctacc cgctggagga cgggaccgcc 120
ggcgagcagc tccacaaggc catgaagcgg tacgccctgg tgccgggcac gatcgccttc 180
accgacgccc acatcgaggt cgacatcacc tacgcggagt acttcgagat gagcgtgcgc 240
ctggccgagg ccatgaagcg gtacggcctg aacaccaacc accggatcgt ggtgtgctcg 300
gagaacagcc tgcagttctt catgccggtg ctgggcgccc tcttcatcgg cgtggccgtc 360
gccccggcga acgacatcta caacgagcgg gagctgctga acagcatggg gatcagccag 420
ccgaccgtgg tgttcgtgag caagaagggc ctgcagaaga tcctgaacgt gcagaagaag 480
ctgcccatca tccagaagat catcatcatg gacagcaaga ccgactacca gggcttccag 540
tcgatgtaca cgttcgtgac cagccacctc ccgccgggct tcaacgagta cgacttcgtc 600
ccggagagct tcgaccggga caagaccatc gccctgatca tgaacagcag cggcagcacc 660
ggcctgccga agggggtggc cctgccgcac cggaccgcct gcgtgcgctt ctcgcacgcc 720
cgggacccca tcttcggcaa ccagatcatc ccggacaccg ccatcctgag cgtggtgccg 780
ttccaccacg gcttcggcat gttcacgacc ctgggctacc tcatctgcgg cttccgggtg 840
gtcctgatgt accggttcga ggaggagctg ttcctgcgga gcctgcagga ctacaagatc 900
cagagcgcgc tgctcgtgcc gaccctgttc agcttcttcg ccaagagcac cctgatcgac 960
aagtacgacc tgtcgaacct gcacgagatc gccagcgggg gcgccccgct gagcaaggag 1020
gtgggcgagg ccgtggccaa gcggttccac ctcccgggca tccgccaggg ctacggcctg 1080
accgagacca cgagcgcgat cctgatcacc cccgaggggg acgacaagcc gggcgccgtg 1140
ggcaaggtgg tcccgttctt cgaggccaag gtggtggacc tggacaccgg caagaccctg 1200
ggcgtgaacc agcggggcga gctgtgcgtg cgggggccga tgatcatgag cggctacgtg 1260
aacaacccgg aggccaccaa cgccctcatc gacaaggacg gctggctgca cagcggcgac 1320
atcgcctact gggacgagga cgagcacttc ttcatcgtcg accggctgaa gtcgctgatc 1380
aagtacaagg gctaccaggt ggcgccggcc gagctggaga gcatcctgct ccagcacccc 1440
aacatcttcg acgccggcgt ggccgggctg ccggacgacg acgccggcga gctgccggcc 1500
gcggtggtgg tgctggagca cggcaagacc atgacggaga aggagatcgt cgactacgtg 1560
gccagccagg tgaccaccgc caagaagctg cggggcggcg tggtgttcgt ggacgaggtc 1620
ccgaagggcc tgaccgggaa gctcgacgcc cggaagatcc gcgagatcct gatcaaggcc 1680
aagaagggcg gcaagatcgc cgtgtaagac tagttataag actgactagc ccgatgggcc 1740
tcccaacggg ccctcctccc ctccttgcac cgagattaat agatctaaaa aaaaaaaaaa 1800
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa tgcatccccc 1860
cccccccccc cccccccccc ccccccaaag gctcttttca gagccaccag aatt 1914
<210> 10
<211> 109
<212> ДНК
<213> Homo sapiens
<400> 10
gatctaagag ttacctggct acagaaagaa gatgccagat gacacttaag acctacttgt 60
gatatttaaa tgatgcaata aaagacctat tgatttggac cttcttctt 109
<210> 11
<211> 57
<212> ДНК
<213> Homo sapiens
<400> 11
tgccaggaac agattttgca gttggtgggg tctcaataaa agttattttc cactgac 57
<210> 12
<211> 91
<212> ДНК
<213> Homo sapiens
<400> 12
actcttaaat ttgattattc cataaaggtc aaatcatttt ggacagcttc ttttgaataa 60
agacctgatt atacaggcag tgagaaacat g 91
<210> 13
<211> 58
<212> ДНК
<213> Homo sapiens
<400> 13
acccagcaat tttctatgat tttttcagat atagataata aacttatgaa cagcaact 58
<210> 14
<211> 33
<212> ДНК
<213> Homo sapiens
<400> 14
atgtcttaag aacctaatta aatagctgac tac 33
<210> 15
<211> 67
<212> ДНК
<213> Homo sapiens
<400> 15
ggtgtctacc atgattattt ttctaagctg gttggttaat aaacagtacc tgctctcaaa 60
ttgaaat 67
<210> 16
<211> 59
<212> ДНК
<213> Homo sapiens
<400> 16
atgtacactg ttgagttttc tgtacataaa aataattgaa ataatacaaa ttttccttc 59
<210> 17
<211> 50
<212> ДНК
<213> Homo sapiens
<400> 17
attcccgttt ctatccaaaa gagcaataaa aagttttcag tgaaatgtgc 50
<210> 18
<211> 46
<212> ДНК
<213> Homo sapiens
<400> 18
gcacaaagga aaacatttca ataaaggatc atttgacaac tggtgg 46
<210> 19
<211> 3787
<212> ДНК
<213> Homo sapiens
<400> 19
agccctcctg gggacttgga atcagtcggc agtcatgctg ggtctccacg tggtgtgttt 60
cgtgggaaca actgggcctg ggatggggct tcactgctgt gacttcctcc tgccagggga 120
tttggggctt tcttgaaaga cagtccaagc cctggataat gctttacttt ctgtgttgaa 180
gcactgttgg ttgtttggtt agtgactgat gtaaaacggt tttcttgtgg ggaggttaca 240
gaggctgact tcagagtgga cttgtgtttt ttctttttaa agaggcaagg ttgggctggt 300
gctcacagct gtaatcccag cactttgagg ttggctggga gttcaagacc agcctggcca 360
acatgtcaga actactaaaa ataaagaaat cagccatgct tggtgctgca cacttgtagt 420
tgcagctcct gggaggcaga ggtgagggat cacttaaccc aggaggcaga ggctgcactg 480
agccaggatc acgccactgc actctagcct gggcaacagt gagactgtct caaaaaaaaa 540
aaaagagaca gggtcttcgg cacccaggct ggagtacagt gccacaatca tggctcactg 600
cagtcttgaa ctcatggcct caagcagtcc tccctcagcc tcccaagtag aggggtttat 660
aggcacgaga ccctgcaccc aacctagagt tgcctttttt aagcaaagca gtttctagtt 720
aatgtagcat cttggacttt ggggcgtcat tcttaagctt gttgtgcccg gtaaccatgg 780
tcctcttgct ctgattaacc cttccttcaa tgggcttctt cacccagaca ccaaggtatg 840
agatggccct gccaagtgtc ggcctctcct gttaaacaaa aacattctaa agccattgtt 900
cttgcttcat ggacaagagg cagccagaga gagtgccagg gtgccctggt ctgagctggc 960
atccccatgt cttctgtgtc cgagggcagc atggtttctc gtgcagtgct cagacacagc 1020
ctgccctagt cctaccagct cacagcagca cctgctctcc ttggcagcta tggccatgac 1080
aaccccagag aagcagcttc agggaccgag tcagattctg ttttgtctac atgcctctgc 1140
cgggtgccgg tattgaggca cccagggagc tgttactggc gtggaaatag gtgatgctgc 1200
tacctctgct gctgcactca cagccacact tgatacacga tgacaccttg cttgtttgga 1260
aacatctaaa catctagtag atgacttgca ggctgttggc taccagtttc ctgtctgagg 1320
tgtatatgtt aacttcgtga tcagtttgta tgtttgggac tcttgtccta tgtaaagtta 1380
aggtgggccg ggtgcagtgg ctcacgcctg taatcctaac actgggaggc cgaggcgggt 1440
ggatcacctg atggtgaaac ctcatctcta ctgaaaatac aaaaattagc tgagtggtga 1500
cacacgcctg taatcccagc tacttggtag gcttgaaccc aggaggcaga gattgcagtg 1560
agccgagctg caccactgtg ctccagcctg ggtgacagcg agactcagtc tcaaaaaaag 1620
ttgtacaagg tggatggttg gaagcttgag cctaggctcg aatccctctc acgtgagagg 1680
gcctgaagat ttctggtgga ttccaacctg gctgaagact ggccgtgggg ggtgcagggg 1740
tctccagcgc tctgccctcc agcctgcttc ctccctgccc acaccgcact aggggaaggg 1800
cctttcctgc tgcctgcggg gccgcacctg gagtaggtaa tgccatgtgg tgacgtgaat 1860
ggagcagagg tctgtgcccc atcacaccgc cttgctgttt ttactgtggg acaaaagcac 1920
tctgatctgc gtgttccggg ggccctccta ccagccgact tgacgggaag tcagggttca 1980
ggtatcatct gtgcacctgg ggcggggtag tctgcactga acctgccaga gtcccctcct 2040
catttcactg aaagtcacag tctccagggc tgtgttgcta accttacgtt ctctccgttt 2100
gcttaatcta ttaagagccc taacaggaga ggatgggctt tctctgttgt ctggggccct 2160
gctgttggcc ggtgctctta gcaagaggtc atttttctag gttgcgctgg gacattgtga 2220
gtttggtgag ggtcatggat gtgggctggg ctgggctggg ctgggccggg ctgcctgctg 2280
cctgctgctc ccctacctga aatgcagcta gtgcggctct gcccttcctg gggctgagga 2340
aggcttctgc aggatagctg gggggctggg caggtgggtg aggcagcctc cctgctgaca 2400
ctcagtcctt gtagctggag caagatctcc tgatccaggt acgggcctgt ctgctccaag 2460
aaagactctg ccaccagatg caaaggggcc ctttgtttta acttagtccc tggggaccgc 2520
ctgattcagc acctgtcggc ccaggatacc ccgctggtgg ggacaagtgc ctgagtgtgg 2580
gccgtgcccg agtgtggcca tccctgagtg gggccgtcct gactaggaag tggcttttca 2640
gttgtgatgt gtgggcctga cctagggggc gctgtggaac ccgggctgga accagccctc 2700
tgtgccaggc cgcagacagg ttccgccggc cctgaggggc agctgccatg gcgtgggtca 2760
ctgggagctg agaggaaggg cccccaccgc acctcaggca aagcggctct gggaacacct 2820
tgatttcgtc catgtgagcc gtcccaggga gggcagccaa gctgtgaagc ctgagaaact 2880
gacctgtgtg ccacgagctt gtggtctgct gcccggtgga ggaagtgcag gtgcgcccag 2940
gctcctcatt ccgttttgca ggattccttc ggggtgtgag catttcctat tcagcctgtc 3000
gcccccgggg agcacgggct ggctctgtgg tgcccgtggc cttttgtaga agcgttggtt 3060
ttacggcagg ttcatctctg gggcagcctc ccacagtggg tggggctttg ccagcagtgc 3120
ccacgggggt catggggcca ggcgcgctcc ggcgcctgca gaactgatcg gggatagtct 3180
caggaggcgc tagtcacgtg ccccggtgat cggggatagt ctcagaaggc gctagtctcc 3240
tgccccggtg atcggggata gtctcaggag gcacgagtcg cctgcctcgg tgatgcaccg 3300
tttctcacac cggctgctct ggcccgagct aaaggggaag acgtgtgcgg ataggagctg 3360
cacacaattt tcctccatgt attgtttatt ttgctttttc ttttggctag acattaggaa 3420
tttcagtttt cccaagttgt atttttcctt ttctatttta aaattatcat gcagggctgg 3480
gtgaggtcgc tcacgcctat agtctcaaaa ctttgggagg ctgagggggg aggatggcat 3540
gagcccagga gtttaaggct gcagtgagcc gagatcgctc cactgtcctc cagcctgcat 3600
gacagagcga gaccctatct caggaaaaaa aaaaacaaaa ctattatgca gtagtttcga 3660
ccctggaaga cgagtgtgca tctttgagtt gtaacacgtg tacctcgccc atccaggcgt 3720
agtttcattt ggaatctggt tatcctgtag ttgctttgtt aaaaatatat gtaattgcaa 3780
atcattt 3787
<210> 20
<211> 63
<212> ДНК
<213> Homo sapiens
<400> 20
ttctccagta tatttgtaaa aaataaaaaa aaaaactaaa cccattaaaa agtatttgtt 60
tgc 63
<210> 21
<211> 45
<212> ДНК
<213> Homo sapiens
<400> 21
ccctggatcc tactctctta ttaaaaagat ttttgctgac agtgc 45
<210> 22
<211> 61
<212> ДНК
<213> Homo sapiens
<400> 22
gtgcagggcc ctcgtccggg tgtgccccaa ataaactcag gaacgccccg gtgctcgccg 60
c 61
<210> 23
<211> 79
<212> ДНК
<213> Homo sapiens
<400> 23
aacctcccac tttgtctgta catactggcc tctgtgatta catagatcag ccattaaaat 60
aaaacaagcc ttaatctgc 79
<210> 24
<211> 38
<212> ДНК
<213> Homo sapiens
<400> 24
taggtgttaa aaaaaaaaat aaaggacctc tgggctac 38
<210> 25
<211> 1651
<212> ДНК
<213> Homo sapiens
<400> 25
atttcattta tctggaaaat tttgtatgag ttcttgaata aaacttggga accaaaatgg 60
tggtttatcc ttgtatctct gcagtgtgga ttgaacagaa aattggaaat catagtcaaa 120
gggcttccct tggttcgcca ctcatttatt tgtaacttga cttctttttt tttctgctta 180
aaaatttcaa ttctcgtggt aataccagag tagaaggaga gggtgacttt accgaactga 240
cagccattgg ggaggcagat gcgggtgtgg aggtgtgggc tgaaggtagt gactgtttga 300
ttttaaaaag tgtgactgtc agttgtatct gttgcttttc tcaatgattc agggatacaa 360
atgggcttct ctcattcatt aaaagaaaac gcgacatctt tctaagattc tctgtgggaa 420
aatgactgtc aataaaatgc gggtttctgg gccattcgtc ttactttcat tttttgatta 480
caaatttctc ttgacgcaca caattatgtc tgctaatcct cttcttccta gagagagaaa 540
ctgtgctcct tcagtgttgc tgccataaag gggtttgggg aatcgattgt aaaagtccca 600
ggttctaaat taactaaatg tgtacagaaa tgaacgtgta agtaatgttt ctacaggtct 660
ttgcaacaaa ctgtcacttt cgtctccagc agagggagct gtaggaatag tgcttccaga 720
tgtggtctcc cgtgtggggc ccagcaatgg gggcccctga tgccaagagc tctggaggtt 780
cttgaaagag gggacacgaa ggaggagtga ctgggaagcc tcccatgcca aggaggtggg 840
aggtgccctg gaaatagctg cctcatgcca cttaggccat gactggattt aatgtcagtg 900
gtgtgccaca gtgcagaggc tagacaactg aaaggggcta ccaaggctgg gaaaaaaatg 960
caattgttgc tgtgagtgac tttgaaagac tctggtgcct tgtggtgccc ttctgaaatt 1020
caaacagtaa tgcaaaagtg tctgcattag aatttacggt gtctaaaatt catgttttta 1080
aaagagcttg cctacagatg gtttccacac ttgaaattgt gccctgcgag ttgcatagct 1140
ggaagttcaa tgctcagtcc taccttggct cccattaaac atttggtgct ctgtggattg 1200
agttgaacgt gttgaggctt tgcaatttca cttgtgttaa aggctctggc atttttccat 1260
ttctatgcaa atttctttga agcagaattg cttgcatatt tcttctctgc cgtcacagaa 1320
agcagagttt ctttcaaact tcactgaggc atcagttgct ctttggcaat gtcccttaac 1380
catgattatt aactaagttt gtggcttgag tttacaaatt ctacttgttg cattgatgtt 1440
cccatgtagt aagtcatttt tagtttggtt gtgaaaaaac cctgggctga agttggcatt 1500
tcagttaaaa gaaaaaaaga aactagtccc agatttgaaa acttgtaata aaattgaaac 1560
tcactggttt tctatgtctt tttgaactct tgtaatcgag ttttgatcat attttctatt 1620
aaagtggcta acacctggct actcttactg t 1651
<210> 26
<211> 474
<212> ДНК
<213> Homo sapiens
<400> 26
actgagtcca gctgcctaat tctgaatata tatatatata tatcttttca ccatatacat 60
gcctgtctgt caatttctgg ttgggctggg aggccacaca cacacactga catgacaggg 120
cttgggcaag actcctgttc tacttatcct tttgaaatac ctcaccctgc cactccacca 180
tgtatgatca ttccagagat ctttgtgact agagttagtg tcctaggaaa accagaactc 240
agaacttgcc tccatggttg agtaacaagc tgtacaagaa ccccttttat ccctggaaga 300
ggctgtgtat gaaaccaatg cccagggttt gaagggtgtt agcatccatt tcaggggagt 360
gtggattggc tggctctctg gtagcatttt gtcctcacac acccatctac tatgtccaac 420
cggtctgtct gcttccctca ccccttgccc aataaaggac aaggacttca gagg 474
<210> 27
<211> 36
<212> ДНК
<213> Homo sapiens
<400> 27
attcagcatt aaaataaatg taattaaaag gaaaag 36
<210> 28
<211> 45
<212> ДНК
<213> Homo sapiens
<400> 28
actggcagat tagattttta aataaagatt ggattataac tctag 45
<210> 29
<211> 48
<212> ДНК
<213> Homo sapiens
<400> 29
agtaatctta tatacaagct ttgattaaaa cttgaaacaa agagcctg 48
<210> 30
<211> 32
<212> ДНК
<213> Homo sapiens
<400> 30
atgctttgtt ttgatcatta aaaattataa ag 32
<210> 31
<211> 90
<212> ДНК
<213> Homo sapiens
<400> 31
accttttcac ctacaaaatt tcacctgcaa accttaaacc tgcaaaattt tcctttaata 60
aaatttgctt gttttaaaaa cattgtatct 90
<210> 32
<211> 4065
<212> ДНК
<213> Homo sapiens
<400> 32
agccacatgg agggagtttc attaaatgct aactactttt tccttgtggt gtgagtgtag 60
gttcttcagt ggcacctcta catcctgtgt gcattgggag cccaggttct agtacttagg 120
gtatgaagac atggggtcct ctcctgactt ccctcaaata tatggtaaac gtaagaccaa 180
cacagacgtt ggccagttaa acatttctgt ttataaagtc agaataatac ctgttgatca 240
ctgaaaggcc tgcatgtatt gtactctgaa ttttacagtg aatgagagaa tgtaccctaa 300
ttgttcaaca gggctcaaaa ggaaagattc cattttgatg ggtcacattc taaagagggg 360
cagtgtgata ggaatgagat ggtcctttag gacttaagtt ctcagcccaa ggtttttcca 420
cgtggccccc tcatcttttt ttttttttta aacggagtct ctcttgccag gctggagtgc 480
agtggcacga tctcggctca ctgcagcctc cgcctcccag gttaagcgat tctcctgcct 540
cagcttcctg actaactggg attacaggcg cccaccacca tgcccagcta atttttgtat 600
tttcagtaga gatggggttt caccatgttg gccatgctgg tctctaactc ctaacctcaa 660
gtgatctgcc cacatcggcc tccaaaagtt ctgggattat agtgtgagcc actgcgcccg 720
gccatggctc cttaatcttg atccaaatta ttgttacatc cagaatgtga tgaatcaaaa 780
tctcgagatg ggggtccagc aatctgaaat ttcagtatgc cagggctttt ctgtatgtca 840
aagtgggttt gaaatagtta atttttcttc tagtctgaaa tgtatcggga aaatttggaa 900
atcctgaagg ctggaaattg aaataagttt ttctaggatt tgtgtctctt gctattggaa 960
aactgatggt gaccaattca tgtttacaaa taagatcctc atagatctcg gtaaattata 1020
atttgctaca gttttatggt tcttcctgtg attttgagct ttttttgacc caaaataata 1080
cagtctaaaa ctatagacaa ataagatggc acttagactc ctgggtttta gttagtggag 1140
gtttccttag tgcactgtgg ggtcataata agccgagaac catggctgtc tatgggacac 1200
atctgtcagg acaaccttta gaggatgttg gggatcaaat agaaggcaca gagaagcact 1260
gaattggctt acataagaat aggctagaat tacaagtagt gaaacctcga ttcagctgga 1320
caattttaaa caaatgtatc atttggcttg tatcttctgt tgtgctggag aagttagaaa 1380
taagggctct ccagaccagc ctgaccaacc tggagaaacc ttgtctctac taaatacaca 1440
aaattagcca ggcgtggtgg cacatgcctg taatcccagc tactttggag gctgagccag 1500
gagaatctcc aggaggcgga ggttgctgtg agccgagatc gtgccattgc actccagctt 1560
gggcaacaag agtgaaactc tgtccacccc ccccaaaaaa agtaagggct ctccattagg 1620
gcccatagag gacttgtaat atggaacctg aatccaagga tcccacaata agtggtcagt 1680
agttcatgat gaattaaaag actcaatatt tggtcttcac ccaatacctg tgtgactttt 1740
agtcctaatt tcctcatctt taaaatttca gtgaaagtgc ctacctgagg attgtgtaga 1800
ttaaaatgga aaccgtgcac ttaatttttt gttttgtttt gagacggagt ctcgctctgt 1860
cgcccaggct ggagtgcagt ggtgcgatct cagatcactg caagctccgc ctcctaggtt 1920
cagaccattc tcctgcctca gcttcccaag tagctgggac tacaggcgcc cgccactgcg 1980
cccggctaat tttttgcatt tttagtagag acagggtttc accgtgttag ccaggatggt 2040
ctcgatctcc tgatctgccc gcctcagcct cccaaagtgc tgggattaca ggcatgagcc 2100
accgcgcccg gcccaggcac ttaatttttg tgtttgactt agtaacttaa gtgcaaacta 2160
ttacgggagc agatggagtc aattggcctt catgtgattg tcagtgggaa attggtccaa 2220
gcagagggaa tactggttca ggaaactggt ttgggaaggt taggcaaacg ggaagtgcta 2280
tggtggagag aaagattact ctggccgggc tgtaaaggac ggctacaatg ggaggctgaa 2340
ggcagaacca agaaaatggg agtgagtatg gaaaaggtac gattcagacg gcataatgga 2400
cgggacttgg agactgaatt gtagtgggcc gaccacaaaa tgataaggca tggaaggaag 2460
tagagtttgg ggggaaggat ccctagtccc ttaatggcta ccttcttccc caggagttgt 2520
taggccatcc gatcccctgg cctgggaaag aaacactgat ttcgttgctg gcttgttcac 2580
tcaccagaag ctacagctac taacagttct aaaaactgtt tcatgtgatg aggaacagac 2640
gaaaatagtt ttgagcccta agtccgccga ttccagtgct ttcttgaacc cgcatttact 2700
aaaatatttt catgactgcc aagctttgaa tagcctgctg tgttcatgga ggctcatact 2760
ggcgatctct agtggctggc taaagcttga attgcaaaag atctaatttc tggtctaatg 2820
tatatatgcc ttaaatatag ttgcgttcaa acgtgggagc tgcaggtgca acttgatttt 2880
atgacaaatg gctgccacat aatttgcaca agcagtgctc gtcaagggca gctaaatcag 2940
gcgagctttc aatcaaaata aatgtactac taaaccctac ttagcggcta actagcccaa 3000
gagcagacag cccacggacg gactgcaagt cggaagcgcg ggcggaagct gtgcagcgcc 3060
cacctggtgg ctccatcggc cgcgttcatc agtcagcacg acccgacctc agtggcgtcc 3120
tcacaacaca gaccggacct tgggtcttac cccggcacct gagaaccact tccggtgagt 3180
agcttctact tccggagacg atgactcccc cgcgtcccag accggaagaa gcccggcgga 3240
gaccggcctc gctcggccac ttccggcaag ggcggagccg gccagtggtg cgcgagcgca 3300
gataactccc ctggagaggc gggatgttca actccacccc tggtccttgg gcggccgtgg 3360
gtccccttcg aagcggagga atggccaacc tcgccgcact tcgagcccct ttagggtgcg 3420
tttaagaaca gtgggcgtgg cctttacgta aatcttcgag atgggaacct ccagaatttg 3480
tctcaattgt ctaaaaggta atgagcgtca gcgacattca agggcacttt gggctaaaaa 3540
agaaagtgct tgtacacgga tggaaatatt ctagaagaac ataaaaggaa tttcctctta 3600
ggaggttagg gaaatgagca cgaagtatgt tttggtgcag ttttttgttc aacccaatgc 3660
gtattttcat attgagaggc aatataaatg gagcgaaagt atcttgagaa aaaaaaaaaa 3720
actaccagaa cttgccgttg ctgaaaagta atattttctc tttcgagagt tttcatggcc 3780
ttttaaatta cacccccacc tccacaggca aataaatttg ttttggaatg cataccacat 3840
catctggctc tagaaacgta ttttgtgtag ctcccctagc aagaatatag gttaaagcgt 3900
aaatttaatt cctggctcta ttttacatcc caatttttat tttcctctca ttcccacttt 3960
acgttgtttc aaataaccta gtttgtgtat ccctgtaagt cattttggta taaagtaggt 4020
tataagtgta catgcgaaaa gatgttttta acaaaaatgt aactg 4065
<210> 33
<211> 3865
<212> ДНК
<213> Homo sapiens
<400> 33
gccttgctct gctcccccgc ccccaggcag ccatccgcag ggccagcgcc atcctgcgca 60
gccagaagcc tgtgatggtg aagaggaagc ggacccgccc caccaagagc tcctgagccc 120
cctgccccca gagcaataaa gtcagctggc tttctcacct gcctcgactg ggcctccctt 180
tttgaaacgc tctggggagc tctggccctg tgtgttgtca ttcaggccat gtcatcaaaa 240
ctctgcatgt caccttgtcc atctggaggt gatgtcaatg gctggccatg caggaggggt 300
ggggtagctg ccttgtccct ggtgagggca agggtcactg tcttcacaga aaaagtttgc 360
tgacttgtga ttgagaccta ctgtcccatt gtgaggtggc ctgaagaatc ccagctgggg 420
cagtggcttc cattcagaag aagaaaggcc ttttctagcc cagaagggtg caggctgagg 480
gctgggccct gggccctggt gctgtagcac ggtttgggga cttggggtgt tcccaagacc 540
tgggggacga cagacatcac gggaggaaga tgagatgact tttgcatcca gggagtgggt 600
gcagccacat ttggagggga tgggctttac ttgatgcaac ctcatctctg agatgggcaa 660
cttggtgggt ggtggcttat aactgtaagg gagatggcag ccccagggta cagccagcag 720
gcattgagca gccttagcat tgtcccccta ctcccgtcct ccaggtgtcc ccatccctcc 780
cctgtctctt tgagctggct cttgtcactt aggtctcatc tcagtggccg ctcctgggcc 840
accctgtcac ccaagctttc ctgattgccc agccctcttg tttcctttgg cctgtttgct 900
ccctagtgtt tattacagct tgtgaggcca ggagtttgag accatcctag gcaacataat 960
gagacaccgt ctctaaaata aaattagctg ggtgtggtgg tgcaccgcct gtggtcccag 1020
ctcctcagag gttgagtaga ggctgaggtg agcggagcac ttgagccaag agtatgaggc 1080
tgcagtgagc ccatgagccc caccactaca ctccagcctg gaagacacca tgacacacag 1140
tgaggcctgg atggggaaag agtcctgctg ttgatcctca catgtttcct gggcacctaa 1200
ctctgtcagc cactgccagg gaccaaggat ccagcatcca tggcacccct ggttcctgcc 1260
atcctggggt acccgattca aagaaggact ctgctccctg tctgagacca cccccggctc 1320
tgactgagag taaggggact gtcagggcct cgacttgcca ttggttgggg tcgtacgggg 1380
ctgggagccc tgcgttttga ggcagaccac tgcccttccg acctcagtcc tgtctgctcc 1440
agtcttgccc agctcgaagg agagcagatc tgaccacttg ccagcccctg tctgctgtga 1500
attaccattt cctttgtcct tcccttagtt gggtctatta gctcagattg agaggtgttg 1560
ccttaaaact gagttgggtg acttggtacc tgctcaggac cccccgcact gtcccaatcc 1620
cactcaggcc cacctccagc tggcctcact ccgctggtga cttcgtacct gctcaggagc 1680
ccccactgtc ccagtcccac tcaggcccat ctctggctgg cctcactgcg ctgggactcc 1740
gccttcataa ggagagctca ctgctcacgt tagtagatgg ccccttctcg tgaggcctct 1800
cccctggcac ctgcttcagt tgtcctccac agcactgatt tgcagcccac aagctggcag 1860
gtttatctgt ctcatgtttg tcttgtgctg gtgggcaagg ggtttgtcta gcacaccagc 1920
atataatgag atgcttgatg aatggtgcat attgaatgta taaagcccac cggtcctgag 1980
agtttgctca ctggagactt tctggagatg gagtctcgct ctgttgccca ggctggcgag 2040
tgcaatggcg cgatcttggc tcactgcagc ctccacctcc tgggttcaag cgattctcct 2100
gcctcagcct cccgagtagc tgggattaca ggtgggtgtc accacaccca gctcagtatt 2160
gtatttttag cagagatggg gtttcaccat tttgcccagg ctggtttgga actcctgact 2220
tcaaattacc cacctgcctc agcctcccaa agtgctggca ttacaggcgc tcgaggcttt 2280
ctgatgtggc tgctgctgct cagaaggcct tgtccttaac cacctccttg cctgccctgg 2340
aggcttgtgc ctctaggccc caccccctgt ggagtcctgc tggctttctc catccctatc 2400
tgaatcctcc ctgctgtgtg gcctcccctg gtctcatccg taacacagcc cagcttagtg 2460
ggcctctgtt cctgcgggtg gccagcctgt ctgtgtggct gggctgggga ggccacgtct 2520
ggtatctgaa tgctatcggt gggttggggt ggaggaacca ggagagggct ggagggaggg 2580
agatggtctc agccccacag agtttggagt cctcagtgtg ctgagcaaac gtggagacac 2640
catttccctc ctctagacct catcttggag agagagatgt tggatggggc catctattcc 2700
agctttattc acacaaatca tgtctgttgg cctggaaatt ggaaaaccag ttaaaccaaa 2760
aacatgatat taagaaaaca ggcaggctca ccatagtaaa aatgctgaaa gccaaagaca 2820
aaattgggag aacaaaagaa aagcgtcttg tcacatacag aaggtccctg ataaagttag 2880
tagctgccct catcagaaac caggcccagg cagtggggac acatccagag tgctgaaaga 2940
acctccccca ggtcatccta tccccaagag tgatgcccgg cagcattccc agctcagggc 3000
taatggttca cggaagccag gaatcaaact gcctgggttc cagtcccagc tctgccagtt 3060
atgcccagct gtggggactt gggcagctcg tttagtagca ccgtgcctca gtttcccata 3120
tgtaaaaggc cattttgagt gcctttcaca gccctgcata aggcaggtgt ctcagtgttc 3180
actgctgtct ctccagctct tagtccagta gctgcatggt gagtgagcgt agggcgcacc 3240
ctggaaggct gccaagccca aagttgtgca gagcgctggg gactccagac tccccacagc 3300
agcagagact cgggactgag gcatcctctg ttcacaggac atgctggcat ctactgggtc 3360
agggctctgc tgctcggtgg ctgtgcaacc ttgggcaagt tcctcaacct ctctgtgtct 3420
tcgtaccctc atctgtaaca tgcgtgtcga tagaccctac tactcagggt tgatgagaag 3480
attaaatgtg caaaacctgc ttgactgtgc ccacaaatcc tgattgtagg aataaattaa 3540
tgacttttta taaatatttt gatcagatgg actcatgatc acagatgtct tcacatgcct 3600
atgactaatt tgtacacaaa ctaatgctcg tgtttcccaa gcacctggaa gacatgccag 3660
atccatgtgc agtaatgcct ggtggctcca ggtctgcccc gccgtcctgt ggggctgtga 3720
gctttcccag cctcctgccc gtgtttgtga atatcattct gtcctcagct gcatttccag 3780
cccaggctgt ttggcgctgc ccaggaatgg tatcaattcc cctgtttctc ttgtagccag 3840
ttactagaat aaaatcatct acttt 3865
<210> 34
<211> 288
<212> ДНК
<213> Homo sapiens
<400> 34
ttttttactg tcaggcagga agagcggtaa ctgccatcgc ggcgggcatc cctggcgcca 60
gggtgttggt ctgggtaccg gcttccctct cggccgactt gtcagctctg tgagccgcgc 120
gcgtctgagc ccgtgtcctc acctgtaaag tggagaaatg aaaaaggacc tgaacttcct 180
cggtggttgt tgagagttaa ggcacggggt tgatgttttc agatgaaatt ctcaaagcaa 240
gtcagggtgg ggatggatgg tttcatccca caggtgggaa gattgagg 288
<210> 35
<211> 430
<212> ДНК
<213> Homo sapiens
<400> 35
gtttttctca ggtccttgat tggaactgcc tcagagccaa gggtcctttt actcagtggc 60
agcaacaaac gcagtctgtt ggctagtgat cctcctgtct cagggacacg tagtccaggg 120
agcagccaat tgcttggcac ttggggaccc cgttctgggg agtcctgaaa gctttcacct 180
cttggattgc cgaatacatg ggtggccctt cctagactaa gggactggcc tgagtgaggc 240
tgggcctctc agccaagctg atgttgaacc actgctgtgg ggatgggcct ggggttcctg 300
ggaagctgtt catacccatt gccaggagcg tgggctctgg ctggacctgg atcagatcct 360
aactgaagcg gcagctttct ggcatgagaa aggagtgttt tcatggtgga cagaattggg 420
ctatgagtgt 430
<210> 36
<211> 120
<212> ДНК
<213> Homo sapiens
<400> 36
atatctctgc caacatgagg acagaaggac tggtgcgacc ccccaccccc gcccctgggc 60
taccatctgc atggggctgg ggtcctcctg tgctatttgt acaaataaac ctgaggcagg 120
<210> 37
<211> 655
<212> ДНК
<213> Homo sapiens
<400> 37
agggagccct cctggaagtg gatgaggcct tgggtctcgg ctcttcattg cttcctgagc 60
tgcagcagat gcctttacaa ccaagctcac cgaggacgtc tgtctcccat attaccctgg 120
cagagggcca ggcctgttct acacggccgg ggtttcaaca aggtactgat gtcttctgcc 180
cttgcctctt cgacaggcaa gtaataagac ttaagtgaag agaattcttt aggcacacaa 240
attcacattt gatgtaatct cattatactt cctgatctgt gattgaaaac tttcatttcg 300
taactagtat gtctgtccca cctttaaaaa gtttttcatt atgaaagtaa gtatttgtta 360
gaattaagtc tatttaaatg aaaaaaactt agatatgagt ctgcatggcc tcaggaaaat 420
gatgttttaa aatagagatt ttaggttgtc tgcactctag cttttttgtc gttttcttaa 480
ggctttttta actgcatcaa aaattcagat acgaaacata cactaaaaaa taatacatca 540
tatcttaatt tccactgaac ttgatttaaa ttcagagtta cacagtatga atatcacaat 600
cagatatgtt caaaaaggtc tgaacaattg attttctgaa accatgaagg actac 655
<210> 38
<211> 331
<212> ДНК
<213> Homo sapiens
<400> 38
agccatttaa attcattaga aaaatgtcct tacctcttaa aatgtgaatt catctgttaa 60
gctaggggtg acacacgtca ttgtaccctt tttaaattgt tggtgtggga agatgctaaa 120
gaatgcaaaa ctgatccata tctgggatgt aaaaaggttg tggaaaatag aatgcccaga 180
cccgtctaca aaaggttttt agagttgaaa tatgaaatgt gatgtgggta tggaaattga 240
ctgttacttc ctttacagat ctacagacag tcaatgtgga tgagaactaa tcgctgatcg 300
tcagatcaaa taaagttata aaattgcctt c 331
<210> 39
<211> 1152
<212> ДНК
<213> Homo sapiens
<400> 39
gcagctcatg tgcacgtttt ctgtttaaat aaatgtaaaa actgccatct ggcatcttcc 60
ttccttgatt ttaagtcttc agcttcttgg ccaacttagt ttgccacaga gattgttctt 120
ttgcttaagc ccctttggaa tctcccattt ggaggggatt tgtaaaggac actcagtcct 180
tgaacagggg aatgtggcct caagtgcaca gactagcctt agtcatctcc agttgaggct 240
gggtatgagg ggtacagact tggccctcac accaggtagg ttctgagaca cttgaagaag 300
cttgtggctc ccaagccaca agtagtcatt cttagccttg cttttgtaaa gttaggtgac 360
aagttattcc atgtgatgct tgtgagaatt gagaaaatat gcatggaaat atccagatga 420
atttcttaca cagattctta cgggatgcct aaattgcatc ctgtaacttc tgtccaaaaa 480
gaacaggatg atgtacaaat tgctcttcca ggtaatccac cacggttaac tggaaaagca 540
ctttcagtct cctataaccc tcccaccagc tgctgcttca ggtataatgt tacagcagtt 600
tgccaaggcg gggacctaac tggtgacaat tgagcctctt gactggtact cagaatttag 660
tgacacgtgg tcctgatttt ttttggagac ggggtcttgc tctcacccag gctgggagtg 720
cagtggcaca ctgactacag ccttgacctc cccaggctca ggtgatcttc ccacctcagc 780
cttccaagta gctgggacta cagatgcaca cctccaaacc tgggtagttt ttgaagtttt 840
tttgtagagg tggtctagcc atgttgccta ggctcccgaa ctcctgagct caagcaatcc 900
tgcttcagcc tcccaaagta ctgggattac aggcatcttc tgtagtatat aggtcatgag 960
ggatatggga tgtggtactt atgagacaga aatgcttaca ggatgttttt ctgtaaccat 1020
cctggtcaac ttagcagaaa tgctgcgctg ggtataataa agcttttcta cttctagtct 1080
agacaggaat cttacagatt gtctcctgtt caaaacctag tcataaatat ttataatgca 1140
aactggtcct tc 1152
<210> 40
<211> 75
<212> ДНК
<213> Homo sapiens
<400> 40
actaacgaaa aatcaataaa taaatgtgga tttgtgctct tgtattttta agtggattaa 60
aaaacttact acctt 75
<210> 41
<211> 1176
<212> ДНК
<213> Homo sapiens
<400> 41
gaatgtcaac gattagtcat gcaataaatg ttctggtttt aaaaaataca tatctggttt 60
tggtaaggta tttttaatca attaggcttg tagtatcagt gaaatactgt aggtttaggg 120
actgggctag cttcatatca gatttacttg ttaagtgact gttttggaat gtttactttt 180
ggactgggtt tgtaacacgg ttaaaggcaa tgagaaacaa gcagaattcc aggagtcctt 240
gaagcagagg gcactggaag acaatatagc agattaaaat agcacagctc atgtggcata 300
ggtgggtatt ttagatgttt gagtaaattt gaaagagtat gatgtttaaa ttacctttag 360
caacatgttc atctgctatg ctgtcatgac tagggggatg attattagtc acatagagct 420
tgggagtacc actggaaacg tatgggtagg agtttaggtg gcttctgttt ttcaaaagat 480
gatcttatcc tagtatctgt aatgctcact tggcacacct gacttgtggg ctgtgtgtaa 540
ggtggctagc taagtgaaaa aagcctgcta ggtgtgagtc aacttaagaa tatgtaaata 600
ggtttgagaa aaagtagggc ttgggtgcaa gtaaagattg agcaggaaat aaaggaaaat 660
caagtataat ccctgagatt tgtagactaa aggcaatgat gtgggactac ttggtcgaat 720
ttttttagcc ctcaacttgg taattgggtg tttctgtgtt aaagcactga aacttgctgt 780
cgtgccttcc tagttttcgt ggtttattga cagggttggg ggtttttttt gtttttttaa 840
aatgaaggga caaagtcaac tggactgctg agtgagaggg caggggcagt tgaagggaac 900
atgaattgct ggaacagcta cataaaatag tgatgtagcc aagtcatgct atttaaatta 960
taattctcca ctgtgtttag aataacatct gaggttctta acctggcctt ggaagggtat 1020
cacttttact tgtaacctgg aatggcttta taatgtgcta gctaattgct actctcatct 1080
tgtattttaa ctcctaattt acccttcagg tctcagcttc agaacattca cttataaaga 1140
aaccctgctg attaaatctc tcttgggctt cctccc 1176
<210> 42
<211> 59
<212> ДНК
<213> Homo sapiens
<400> 42
acgctcctct actctttgag acatcactgg cctataataa atgggttaat ttatgtaac 59
<210> 43
<211> 49
<212> ДНК
<213> Homo sapiens
<400> 43
accagacaca ctgattggaa ctgtattata ttaaaatact aaaaatcct 49
<210> 44
<211> 178
<212> ДНК
<213> Homo sapiens
<400> 44
ggaattgcac atgagatggc acacatattt atgctgtctg aaggtcacga tcatgttacc 60
atatcaagct gaaaatgtca ccactatctg gagatttcga cgtgttttcc tctctgaatc 120
tgttatgaac acgttggttg gctggattca gtaataaata tgtaaggcct ttcttttt 178
<210> 45
<211> 73
<212> ДНК
<213> Homo sapiens
<400> 45
tcaccaaaaa gcaaccaact tagccagttt tatttgcaaa acaaggaaat aaaggcttac 60
ttctttaaaa agt 73
<210> 46
<211> 38
<212> ДНК
<213> Homo sapiens
<400> 46
acctctttta taacatgttc aataaaaagc tgaacttt 38
<210> 47
<211> 40
<212> ДНК
<213> Homo sapiens
<400> 47
attcctgctc ccctgcaaat aaagcctttt tacacatctc 40
<210> 48
<211> 1301
<212> ДНК
<213> Homo sapiens
<400> 48
cagggtctcc ttggcagctg tattctggag tctggatgtt gctctctaaa gacctttaat 60
aaaattttgt acaaagacac aaggtctgac tagactgttc agtattcaga ctgaggggca 120
tgttggcctc tggagcatta catatcttct tggttttaac catacttgtg gtatttgcaa 180
gggccagaac agtaagaccc aagcagagcc aaccagagaa ataatatttg tgtgatagag 240
aaggctgata gcaagcaagg cagcaccttg attcgttgtc ctgtagttca ggattgtagg 300
tttagaagag ggatatgttt gagtttttcc tatgcataag gcgatccacg ttgcacatag 360
aaagtgaata taaatggcca ttatattttg tgtcatgctg tgctctaagt gttctttaca 420
tatgtactcg ttaatcaacc tctctaaagt gtaaaggaaa tttgcttgca ccactgaagg 480
cacataaggc tcagaagtaa atttgcctaa gcagtataaa gctatcatta gaatccacat 540
tcctaagttg tgttctctta ggggatcatg gaaccagtca ttggtactac aggctattat 600
gttctggaga actgtgaaga acatttaaat tgtctctgat tttatctatc aatgttttga 660
agtattttct accagtgtct gtacttcaca agaaattcgg cactattttt tcaggcaaaa 720
ctagtgaggg acaggttggc ttgaaaatca tgagactgtt gttaaatcag atgctggttg 780
atcacagagg ggacttccag ggaaagctgt tatcaggtgg ctgcttcctg gtgatgcagc 840
ctggctgatg agataaccct ggctccacag atggcttagc aggtgctgtg atgatttggt 900
tttcttctca attagactga gctgcacatg gtgtttatat tgcttggcac atggtaaggg 960
cttaatattt gaggtaatta tgtagggcgt acactgacaa gtatctgacc cccccttcct 1020
ttttgactca taaattggtc atcttaacca tttaagtgta cacttctata gtgacagagt 1080
tagccctctg tccaagggat ttgcatctgt ggattcaacc aactttgggt caaaaataat 1140
caaaaaggat ggttgtgtgt gtattgaaca tgtagactta tttttcttat tttcaaaata 1200
ctatattttc ttgtcactta ttttcttgta cactgcagtt gtaacagcta tgtagcatgt 1260
acattaggta ttaaaagtaa tccagtgaag attgaaagtc t 1301
<210> 49
<211> 576
<212> ДНК
<213> Homo sapiens
<400> 49
cagcctcttc catgagtggg gagcccgctg cttgtctcca gctcctagca gtgagtcctg 60
ataatctcaa atttaaggac agtaactttg tctgggatga gtgtgggaaa ggatgtgttt 120
gggaacagac gcgagcctgc agaggtgttt gtaaccatct ctttctaagt ggtgggaagc 180
agacatttta ttctttaact gttaatatat atagtgtgtg ttttttatgc atgaaatatt 240
ttatagtttt taaaaatgcc cacactacta ttttgaaagt aaatgaggta atgtatgtgt 300
cagaacccaa tacccaaagc gatcgtagta agaggtgggg cctttgggaa ggcattaaat 360
tgcttaggga atgagggtgg aaccctcatg aatgagatta gagccttata ggagaggttg 420
gagggagttg cctggcctcc ctctcccatg tgaagactca gcaagaaaac attatttagg 480
aagcagagag ccctcatcaa acaccagatc tgctggccac ctgatctggc actttccagc 540
cttcagaact gtgagaaata aatttctgtt gtctat 576
<210> 50
<211> 57
<212> ДНК
<213> Homo sapiens
<400> 50
agttcagact tcaaatagtg gcaaataaaa agtgctattt gtgatggttt gcttctg 57
<210> 51
<211> 1324
<212> ДНК
<213> Homo sapiens
<400> 51
agctcacgtt gatgtcaaga ctaccgatgg ttacttgctt cgtctgttct gtgttggttt 60
tactaaaaaa cgcaacaatc agatacggaa gacctcttat gctcagcacc aacaggtccg 120
ccaaatccgg aagaagatga tggaaatcat gacccgagag gtgcagacaa atgacttgaa 180
agaagtggtc aataaattgt aagtgtttct ttgcttcctc acacaacaca accttgagta 240
ttggattatt cctgagatga gagaacgcat atgagacaag gtaaaggtct gttgaaatcc 300
tgtctgtgaa tccttctagc tatatctctt taagtgaaag agtgttaagt actcagtaaa 360
tatgattatt attactatta ttatttgagt cagagtcttg ctctgttgcc caggctcgag 420
tgcagtattg tgatcctcct tggctcactg taaccactgc ttcctgggtt caagcagttc 480
ttgagcctca gcctcctgag tatctgggaa tacaggggac tgccaccata cccagctaat 540
ttttttaaat ttttagtaga gatggggttt catcatgttg gccaggctgg tcttgaactc 600
ctgacttcag gtgatctgcc agtactctaa atgataacag ttttttcgtg tttatttatt 660
ttgaatgaag ctgtctcaca gtagatggag ttgaaggaca ggaaatgttt ttcccctact 720
tggaaaatac actgaataag ttgagtgggg tgggatgtgc ctggagtccc agctactcag 780
gaggctgagg tggtaggatt gtttgagccc aggagtttga ggccagcctg ggcaatatag 840
ggagaccctg tcccaaaaaa taaaaaatat acgtatatat atatacacac acaaagaaaa 900
aatacactga atagacaaaa cctttcatga ttaatgatgc acgggaataa gtgatgaaaa 960
aagtttcggt cccagatgat ggccagtgat aacaacattt ttctgatgtt cccatgcaat 1020
atacagttag ctaagagggt gtaatggaaa aagcataagg cttggactca gaagactcta 1080
ctaactttgc cactagctag ctatgtaatt cagatcatct atcctttaca tgtgaaaggt 1140
aaataatggc ttatcttaac aggaggattt atgcaggtta aatgaggtag gtgttatgtg 1200
taggtttatt ccaaggcttc tctactttta aaggaaatgg cttatatctg agaactagga 1260
cttttagaaa aaaatttact gttactggtt tgcaggattc cagacagcat tggaaaagac 1320
atag 1324
<210> 52
<211> 68
<212> ДНК
<213> Homo sapiens
<400> 52
aatgggtccc tgggtgacat gtcagatctt tgtacgtaat taaaaatatt gtggcaggat 60
taatagca 68
<210> 53
<211> 60
<212> ДНК
<213> Homo sapiens
<400> 53
attgcagtag cagcatatct ttttttcttt gcacaaataa acagtgaatt ctcgtttctt 60
<210> 54
<211> 53
<212> ДНК
<213> Homo sapiens
<400> 54
ttttcccagc tgctgcccaa taaacctgtc tgccctttgg ggcagtccca gcc 53
<210> 55
<211> 37
<212> ДНК
<213> Homo sapiens
<400> 55
gattttttga gtaacaaata aataagatca gactctg 37
<210> 56
<211> 36
<212> ДНК
<213> Homo sapiens
<400> 56
acaaaaatga ctaaataaaa agtatatatt cacagt 36
<210> 57
<211> 55
<212> ДНК
<213> Homo sapiens
<400> 57
atccttgttt tgtcttcacc catgtaataa aggtgtttat tgttttgttc ccaca 55
<210> 58
<211> 70
<212> ДНК
<213> Homo sapiens
<400> 58
gtccacctgt ccctcctggg ctgctggatt gtctcgtttt cctgccaaat aaacaggatc 60
agcgctttac 70
<210> 59
<211> 50
<212> ДНК
<213> Homo sapiens
<400> 59
aattggagag gattcttttg cattgaataa acttacagcc aaaaaacctt 50
<210> 60
<211> 77
<212> ДНК
<213> Homo sapiens
<400> 60
ggctggacat cggcccgctc cccacaatga aataaagtta ttttctcatt cccaggccag 60
acttgggatc ttccgcg 77
<210> 61
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 61
agaaataaat ctttggctca c 21
<210> 62
<211> 41
<212> ДНК
<213> Homo sapiens
<400> 62
atttgtctgt gtactcaagc aataaaatga ttgtttaact a 41
<210> 63
<211> 31
<212> ДНК
<213> Homo sapiens
<400> 63
tggctcagct aataaaggcg cacatgactc c 31
<210> 64
<211> 53
<212> ДНК
<213> Homo sapiens
<400> 64
ggatgtaata catatattta caaataaaat gcctcatgga ctctggtgct tcc 53
<210> 65
<211> 64
<212> ДНК
<213> Homo sapiens
<400> 65
gcccatcgtg actcaaaact cacttgtata ataaacagtt tttgagggat tttaaagttt 60
caag 64
<210> 66
<211> 37
<212> ДНК
<213> Homo sapiens
<400> 66
aacaaaccat gctgggttaa taaattgcct cattcgt 37
<210> 67
<211> 386
<212> ДНК
<213> Homo sapiens
<400> 67
ctgcattctc ctccgccaaa aaagtgacca agcagagtct ttctctgtca cccaggctgg 60
agtgcaatgg cgtgatctca gctcactgca acctctgcct cctgggttca agtgattctc 120
gtgtctcagc ctcctgagta gctgagacta caggtgtgca ccagtgttcc cagctgattt 180
ttgtatttta tgtagagatg gggttatgcc attttggcca ggctagtctc gaactcctga 240
gctcaggtga tacacacacc tcagcaaatc ttttaaatta tacattctgt gatatttcct 300
tgactttctt atccagcact tgtattgatt atttttcatt ttgataatgt tgggttttta 360
aaaactcctt tatgatggaa aatttc 386
<210> 68
<211> 281
<212> ДНК
<213> Homo sapiens
<400> 68
gtcaactatt ttaataaatt gatgaccagt tgttaacttc tgttggtttt tattcagaat 60
actggcagat tttaggaata taaaggtgta ctatgagact tccacttttc aggtggaata 120
tatgggtatc ttagagtggt ctatcctgtt ttcgttgtcg tttgagtcat ttgaaaactg 180
gattccgtta actacataat atgtgagacc tgactggttt tattggacac tggcagttta 240
taactttggc atactctaga taaattctga ttggtatggg g 281
<210> 69
<211> 53
<212> ДНК
<213> Homo sapiens
<400> 69
ctggagagaa tcacagatgt ggaatatttg tcataaataa ataatgaaaa cct 53
<210> 70
<211> 2785
<212> ДНК
<213> Homo sapiens
<400> 70
atattaatgg tgaaaacact gtagtaataa attttcatat gccaaaaaat gtttgtatct 60
tactgtcccc tgttctcacc acgaagatca tgttcattac caccaccacc cccccttatt 120
ttttttatcc taaaccagca aacgcaggac ctgtaccaat tttaggagac aataagacag 180
ggttgtttca ggattctcta gagttaataa catttgtaac ctggcacagt ttccctcatc 240
ctgtggaata agaaaatggg atagatctgg aataaatgtg cagtattgta gtattacttt 300
aagaacttta agggaacttc aaaaactcac tgaaattcta gtgagatact ttctttttta 360
ttcttggtat tttccatatc gggtgcaaca cttcagttac caaatttcat tgcacataga 420
ttatcttagg tacccttgga aatgcacatt cttgtatcca tcttacaggg gcccaagatg 480
ataaatagta aactcaaaat tgctccccac tctgtttatt atttaaaggt gtcaggatct 540
gtgttgtaat gtgtctacat taatgtgttt aggagaatac aggcattgga tcatttagtt 600
gatggaagta tatgccaggc aagggagata aggtatacga caagactgat gttttcagta 660
tcttctcatg aggttgtcag agaccttcat gtcttcaaag actagtcagc aaatgaagtg 720
gtttagtgta gagacaagat tggttgtgtt ttgataattt aagctaggta ttgagtacat 780
gtggattttg ctgtccacaa atacttgttt cagagttttc atggatacag tggcatggtt 840
gaaatgaagc tgtgagcctt ctgctttaaa tctgatgtaa gaaactcctg ttaacaaata 900
gtaagtatgg gttaattagc cctttgatca aagcctagct ttacattgtt taggatcttt 960
ggaaaacaat tggtttggtt gcccactttc cgtaggatca agagcagaac ctttcacatg 1020
gcacagaaga acccaggttg cgcttcatac ctgcatattc cagccttagc ctgccatttc 1080
tctccttggc actttgtgct ccagcaacac tggtctcagt tggtcatcct caaacttggg 1140
ttccatatcc agcctcagga cctctgttcc tgttactatg gttccttgca tgtcgcctgc 1200
tcttactaaa gagctcgtgt gttttccagc acacttcggt ttatctcttg atgatgatgc 1260
tagtctctcc ctccgcaagg gcggaaaggc tgcctgttgg tttgtaccag tgtttcctaa 1320
cgtgtagctg cagtcagtat ttggctaagc tgttcccagg ggctcaacag atgctttcgg 1380
atgagcctta actgacccaa tcctttgtga tgcgggagag attgctaggc ctcgctcacc 1440
tggccagaac cagggaaaga ggccgcggtt gcagcgcgat tccaggccct gggcgtcagg 1500
cgcggggtgg gcagctctcc ccgggcggtg gggcccttgt gaccgcgagg cggggcgcac 1560
caggaaggga gtgggacagc gcgggcgccc agggatgtgg cctggttacc tgccttctct 1620
gatacgtcaa gacaccttca acaatggctt gcagctgtac cctgttggct gcacccagga 1680
cgcccttttc actgctaagc agtcctacct gaggcccagg ggctgccaga ttgacccata 1740
aataatctcc ggcgcctcag atccagaagc tgctgagcct gatcttagtg ccttctcctt 1800
tctctgtgtg gccccccagc ccctttcccc actgccttgt gtccaaggcc ctttccttca 1860
tgtatccatg gaggagagac aaaaatacac atcaataaaa taagataggg aatccataaa 1920
tagacattca gaagtatggc caacggattt atcttaaaac caatggagga agaagagttt 1980
caataaatgt tgtggacttc catttgtcaa agaccaaaac aaaggaaccc caaccttaca 2040
tgtaatacaa acttaactca aaatggatca tatatctaaa tgtaaaatgg aaagctataa 2100
aactgaaaac agactatctt tacaacctag gcgtaggtat agtttttaga cattacacca 2160
aaagcacatg ccgtaaaaga aaaaatagat aaattggtgg atttcattaa aattaaaaaa 2220
ctttttctct ctgaaaaatc ctgttaagct gggcgctgtg gttcatgcct gtaatcccag 2280
cactttggga ggctgagttg ggaagaaatt aatagcttga ggccaggagt tcaagatcat 2340
cctgggcagc aaagtcatac actcttgagg gaagagagag accttctcat attgttttat 2400
attgttttat actcagtacc tgttttaaga aaaaaacaag gaagtgaaat caaagacagg 2460
cagcccggca ccaggcctga aaccagccct gggcctgcct ggcctaaacc tagtagttaa 2520
aaatcaactt acgacttaga acctgatgtt atccgtagat tccaagcatt gtataaaaaa 2580
attgtgaaac tccctgttgt gttctgtacc agtgcatgaa acccctgtca catatcccct 2640
agattgctca atcaatcacg accctttcat gtgaaatctt tagtgttgtg agcccttaaa 2700
agggacagaa attgtgcact tgaggagctc agattttaag gctgtagctt gccgatgctc 2760
ccagctgaat aaagcccttc cttct 2785
<210> 71
<211> 48
<212> ДНК
<213> Homo sapiens
<400> 71
ataggtccaa ccagctgtac atttggaaaa ataaaacttt attaaatc 48
<210> 72
<211> 67
<212> ДНК
<213> Homo sapiens
<400> 72
ggagctgagt tcttaaagac tgaagacagg ctattctctg gagaaaaata aaatggaaat 60
tgtactt 67
<210> 73
<211> 62
<212> ДНК
<213> Homo sapiens
<400> 73
aagcactctg agtcaagatg agtgggaaac catctcaata aacacatttt ggataaatcc 60
tg 62
<210> 74
<211> 276
<212> ДНК
<213> Homo sapiens
<400> 74
ctgtatgagt taataaaaga catgaactaa catttattgt tgggttttat tgcagtaaaa 60
agaatggttt ttaagcacca aattgatggt cacaccattt ccttttagta gtgctactgc 120
tatcgctgtg tgaatgttgc ctctggggat tatgtgaccc agtggttctg tatacctgcc 180
aggtgccaac cacttgtaaa ggtcttgata ttttcaattc ttagactacc tatactttgg 240
cagaagttat atttaatgta agttgtctaa atataa 276
<210> 75
<211> 139
<212> ДНК
<213> Homo sapiens
<400> 75
gcttggctgc tcgctgggtc ttggatgtcg ggttcgacca cttggccgat gggaatggtc 60
tgtcacagtc tgctcctttt ttttgtccgc cacacgtaac tgagatgctc ctttaaataa 120
agcgtttgtg tttcaagtt 139
<210> 76
<211> 94
<212> ДНК
<213> Homo sapiens
<400> 76
atgctcttcc ttcagaggat tatccggggc atctactcaa tgaaaaacca tgataattct 60
ttgtatataa aataaacatt tgaaaaaacc cttc 94
<210> 77
<211> 1355
<212> ДНК
<213> Homo sapiens
<400> 77
tataagtaaa gtttgtaaaa ttcatactta ataaacaatt taggacagtc atgtctgctt 60
acaggtgtta tttgtctgtt aaaactagtc tgcagatgtt tcttgaatgc tttgtcaaat 120
taagaaagtt aaagtgcaat aatgtttgaa gacaataagt ggtggtgtat cttgtttcta 180
ataagataaa cttttttgtc tttgctttat cttattaggg agttgtatgt cagtgtataa 240
aacatactgt gtggtataac aggcttaata aattctttaa aaggagagaa ctgaaactag 300
ccctgtagat ttgtctggtg catgtgatga aacctgcagc tttatcggag tgatggcaat 360
gctctgctgg tttattttca agtggctgcg ttttttttag tttggcaggt gtagactttt 420
taagttgggc tttagaaaat ctgggttagc ctgaagaaaa ttgcctcagc ctccacagta 480
ccattttaaa ttcacataaa aggtgaaagc tcctggttca gtgccatggc ttcatggcat 540
tcagtgatta gtggtaatgg taaacactgg tgtgttttga agttgaatgt gcgataaaat 600
tattagcctt aagattggta agctagcaat gaatgctagg gtgggaagct ggtgagccag 660
tggccattag ataaatacct ttcaagtgtg agcttagacg tcaaccctaa aatacttaac 720
cgtaatgcta attgtgatca ttatgaatcc cttcagtcac attaggggga aagtagttgg 780
ctataagtac gtcattctta gtccagtcag tcttaaaaac atcttgggtt acccactctg 840
tccactccca taggctacag aaaaagtcac aagcgcatgg tttccaacca tatgtgtttt 900
ctgcagttat ttctcttgtt ctggccaaac aaccctaaaa atccttacca ttccacaaag 960
ttggaccatc acttgtgcac ccactttgac tatgagtata ccaccacatt gcatttctgt 1020
ttgcaccatg tcttccagga gactagacta ctgttgtcca gggtcaattt gagtgtaaag 1080
aaaatgtaga caaggaattg cccaatttta aattctgact ttgctgactt aatttaaatg 1140
ctcgttctga accaattttc tcctatcttc tctaggggtt tcaaaagact cagttaattg 1200
atttccagga agtactcata gcaagttcat aaaagttctt gagacctaaa tttcttcaca 1260
aaaaaagaaa agatcttaag tcatacattt taattgtgta gaggttgttc aactgaagga 1320
ataaatgtct attaaactaa aacaaatgga ccttc 1355
<210> 78
<211> 2017
<212> ДНК
<213> Homo sapiens
<400> 78
gcaattcttc tgcctcggcc tcccaaatag ccaggactac aggcgcacac tgccatgccc 60
agctaagttt tgtattttta gtagagactg ggtttcacta tgttggccag gctggtctcg 120
aactcctgac ctcaagtgat ccacctgcct tggcctccca aagtgctggg attacaggcg 180
tgagccacca cccccagccc aatttttatt ttttgtacag acaggatctc actatgttgc 240
ccaggttggt ctcaaactac tggcctcaag caatcctgcc ttggcctccc aaagtgctgg 300
aattatagga atgagccacc acaccgggcc caaatttact ttagtaataa caacaattgg 360
ctgggtgcgg tggctcacgc ctgcaatccc aacactttcg gtaaccaagg tgggcttgag 420
ctcatgagtt agagagcagc ctgagcaacg tggtgagagc ccatctcaca aaaaataaca 480
aatcagctgg gcatggtgtt gcacgcctgt agtctccgaa atcacaccac tgcactccca 540
tcttgggtga tagagccaga acttgtctca aaaataacaa ttggtttctt acaatcccaa 600
aaggtgcagt tactagtatt aatccttttt tgccaatgag gaaacacaaa gatgaagcaa 660
cttgctcaaa gtcatacagt gacagtctga attcaaatcc tatacactta aagtttattt 720
gttttgtttt ggtttttttt gagatggagt ctcactgtgt cgcaaggctg gagtgcagtg 780
gcacgatctc agctcactgc aacccgggtt caagcgattc tcctgcctca gcctcccgag 840
tagctgggac tacaggcacg caccaccaca cccagctaat ttttgtattt ttagtagaga 900
cggtttcacc atgttggcca ggatggtctc gagctcctga cctcaggtga tcctcccgcc 960
ttggcctccc aaagtgccgg gattacaggt gtcagccact gcacgtggcc aacttaaagt 1020
ttttgataga taatacatta acgttaaaaa ttcaaaagat aagtataggc tctacagtac 1080
aaacccttct gcctcctagt tcctctccct ggaggcaagg tgatcagttt aacaatattt 1140
ttttattttg agacagggtc tcactgttgc ccaggctgga gtgtagtggc gcgttcacaa 1200
cttactgtag cctcaacctc ctggctcaag caatcctccc acctcagcct gtcgagtagc 1260
tggaaccaca ggtgcacacc accatgccag gctaattttt gtattttttg tagagacagg 1320
gtttcaccat gttgttcagg ctggtctcaa agtcctgggc tcaagcaatc ttcctgtctc 1380
tgcttcccaa agtgctggga ttacagatgt gggccacggt gcctggccta catatgtatt 1440
ttttcctttt cttccccaag tggtaggata tgatacacat tgttgatttt tttgtttagt 1500
tatgtatctc agagcttatt ctttatcagc tcatgaggaa cttcattttt tttttttttt 1560
ttgagatgta gttttgctct tatagcccag gttggagtac agtaacacaa tcttggctcg 1620
cagcaacttc tgcctcccag gttcaagcga ttctcctgcc tcagcctccg agtagctagg 1680
attacaggtg cctgccacta catccagcta tttttgtatt ttcagtagag acggggtttc 1740
accattttgg ccaagctggt ctcgaactcc tgacctcagg tgatccgccc atctcagcct 1800
cccaaagtag tgggattaca ggcatgagca accgtgcccg gctggaactt cattcttttg 1860
gtataactgc atggtatccc atcatgtgga tgtaccatga ttcattggat gtggaccctc 1920
ctgatggaca tttaaatttc ttccaatctg ttgctattac aaaaagaaaa atgtgtgcat 1980
acatctttat tcatctgtag aataaattct tagaagt 2017
<210> 79
<211> 37
<212> ДНК
<213> Homo sapiens
<400> 79
ggtttttata caagaaaaat aaagtgaatt aagcgtg 37
<210> 80
<211> 140
<212> ДНК
<213> Homo sapiens
<400> 80
gtggcaatca taaaaagtaa taaaggttct ttttgacctg ttgacaaatg tatttaagcc 60
tttggattta aagcctgttg aggctggagt taggaggcag attgatagta ggattataat 120
aaacattaaa taatcagttc 140
<210> 81
<211> 55
<212> ДНК
<213> Homo sapiens
<400> 81
acaagattcc tcaaaatatt ttctgttaat aaattgcctt catgtaaact gtttc 55
<210> 82
<211> 1485
<212> ДНК
<213> Homo sapiens
<400> 82
gggcttccaa tgtgctgccc ccctcttaat actcaccaat aaattctact tcctgtccac 60
ctatgtcttt gtatctacat tcttgacggg gaaggaactt cctctgggaa cctttgggtc 120
attgcccttt cacttcagaa acaggttgac aactcagccc tgctcatgag gcagcaaacc 180
ctgcaaaggg ctgggactgg tggccttatg tcagttgtct actctggagc ttgacttgga 240
cctccccagg tcctaggcag taggttgaaa aacactgaag tgcttttcat gaagcacagc 300
tgcagcaaag ccttgcaatc ccaggctggg gtcagcctac agttgtgttg cttattacaa 360
cacatgcgga ccaagagggg cttgtgggct agaggctgac cagcagcgtt tatttagcaa 420
gggtaggtgt gcatcacatt gggcttgttc tcacccatct ggtttggcca ttcctccttg 480
gtgggaatca tccaggtact gctgaggtca cctgcgattt gccccatttc ctatctctag 540
caacctcctg ggccccatgc ccccacccct tctagaacct gcattcccag ggccttcacc 600
acctgaccaa aggtctaggc taacctttgg tcatttgtaa caagacctcg gaacagacac 660
gtgtgtggca tggtttggcc tggggatctt agatgtctga cctgaactat tgtagaacag 720
cgctggcttt tgggggagca gcaaaaatga gaggagtgct aggtgggtgg cctgagcatc 780
tgtatccagg gacaggactc caaaggcttt tggtcccaga gctggggtat gttggcccca 840
gcccccagcc tgtggctccc aaaaggcctc tggttttttg taatctcagt ttacagccat 900
ttcttaggtt tttaattacc tttattttat tttgccaaac atacctggga atacctttta 960
tttttttttt accttggggt gatggttcca aaccataaat gtgattatag ttaacacatg 1020
acccttctag cgtcccagcc agtgtttttc ctgacctctg ttctttggag aggaggatgg 1080
aagggagggg tccggcacgc tgctggcatt ttgctgtgtc ctgcagcccc tttccgggac 1140
acctgggttc acacagcttt ttagcttaca taactggtgc agattttctg tgtggagatg 1200
ttgccttgac cagccttggc tggactttac caggcatgca gaagcctgta ccaacacaga 1260
ctacagcacc caggaggtgc gagtgtggct gctcagcggt tataacaggc ctgactgcat 1320
tgttcaccgg attataatga gccaaaatgt ttcccggtgt ttgctggttt cagggaagga 1380
gtttgatata gcagattaac caccctcctt gtagctattg gggcttaatg gtttcctggt 1440
gattcttacc aatccacaat aaacatggcc cattggcata tctgc 1485
<210> 83
<211> 31
<212> ДНК
<213> Homo sapiens
<400> 83
ggcacatttg aataaattct attaccagtt c 31
<210> 84
<211> 39
<212> ДНК
<213> Homo sapiens
<400> 84
ggggcgcatt gtcaataaag cacagctggc tgagactgc 39
<210> 85
<211> 486
<212> ДНК
<213> Homo sapiens
<400> 85
gcccaataaa gactgttaat tcctcatgcg ttgcctgccc ttcctccatt gttgccctgg 60
aatgtacggg acccaggggc agcagcagtc caggtgccac aggcagccct gggacatagg 120
aagctgggag caaggaaagg gtcttagtca ctgcctcccg aagttgcttg aaagcactcg 180
gagaattgtg caggtgtcat ttatctatga ccaataggaa gagcaaccag ttactatgag 240
tgaaagggag ccagaagact gattggaggg ccctatcttg tgagtggggc atctgttgga 300
ctttccacct ggtcatatac tctgcagctg ttagaatgtg caagcacttg gggacagcat 360
gagcttgctg ttgtacacag ggtatttcta gaagcagaaa tagactggga agatgcacaa 420
ccaaggggtt acaggcatcg cccatgctcc tcacctgtat tttgtaatca gaaataaatt 480
gctttt 486
<210> 86
<211> 44
<212> ДНК
<213> Homo sapiens
<400> 86
gcccctcccc tgccctctcc ctgaaataaa gaacagcttg acag 44
<210> 87
<211> 406
<212> ДНК
<213> Homo sapiens
<400> 87
gtgtcatctt ttattatgaa gacaataaaa tcttgagttt atgttcactt catttgtttg 60
ctgttcatct tttgggaggg aataagctag agccatcaat acaattccgc ttgtggggaa 120
atttatgcct cttactggta ctacttgttt tgcattgaag ctgactggtt gagttcacat 180
catatgttgc aattttctaa tttggcactt caatcactag gggccttatg aggcagtttg 240
tcattatgca atggttattg gttatcatgt gagtagacac atttcaggct aatagggaga 300
agtcagtaac acattcatag tgaatatgag atgtctttgc taagagttaa gtgtcagatc 360
tttgttataa cagttaattt aataaagaat tttggcattg ttcttc 406
<210> 88
<211> 1994
<212> ДНК
<213> Homo sapiens
<400> 88
ttgccgtaag gatatgcact tgtctctagt ccacacactt catgatatag gtatagcgtt 60
agtttagcga agttttcact gcactgatat atctagtagg tgatggagct gggaatgcaa 120
ctcatgtctg actagtccac aatactgcac tatttcagtg tttacgattt tttatccttt 180
cccttctgaa gaggcaaaaa attgaggaat gtgccctgct ttcctaagaa ctgaagtgtg 240
agtacactgg taaatccttt catttgcctt gttccttatc tgtcaatatg tctgaatcct 300
cgcttgttgg ttgcactaag aattgttctg ttgtttctca tcacagaaat ctgcagtcaa 360
ctacctgttc tcgtgaagtc ttaaaactct tatagaatag ccatttaggc ctttctgcta 420
gcctcctgaa ttctgtattc tcaggctgag cgagtttctg tttactctca aaccttaggt 480
gatttggcta actcttaaag taattagcac gatgattgga acggagcatt ctctccaaca 540
cagcatttct tttggcactt tgcttcttgt gcagtttagc tccagaaagt attaaggaat 600
gactttagtg ctcatttgga tgcagtaagt ggtttgatct cagggtggca aaaagaatgc 660
tttttttata ccttttcaca ttcggataac ttgtttagaa gacagaggtt ctaactaggt 720
tttggcctat taagaactgc aaactagcag cagcagaact ctggctaaag gggcaagctt 780
attaggaaat tgagtattta aaagttgagc taccatatga tccaacaatc ccactgctgg 840
gtatataccc agaagaaaat cggtatatca aagagatatc tgcactccta tgtttgttgt 900
agcactgttt ataatagcta agatttagaa gcaaccttag tgtccatcgg gatgaatgga 960
taaagaaaat gtacctatac gcggccaggc acggtggctt gtgcctagca ctttggaaag 1020
ccgaggcggg tggatcacct gaggtcagga gttcgagacc agcctggcca agatagtgaa 1080
accccgtctc tagtaaaaat acaaaaatta gccgggcttg tggtgtgggc ctgtaatctc 1140
agccacccgg gaggctgagg caggagaatc gctggaacct gggaggcaga ggctgcagtg 1200
agccgagatc acgccactgt actccagcct gggcgacaga gcaagactcc atctcaaaaa 1260
aaaaaaaaaa aaaaagggaa aaagaaaatg cacctataca cagtggtact attcagccat 1320
aaaaagaatg agatccagtc atttacaaca acatgggtgg aactggagat cgttatgtta 1380
agtgaaatag gcacacaaag acaagcatca catgttcttg tttgtgggat ctaaaaatca 1440
aaacaagtgg acttgtcata tagagagtag aaggatggtt accagaagct gagaacttct 1500
ggtggcggga ggtggggatg gttaatgggt acaaaaagaa aaaagaatga attagaccaa 1560
ctatttgata gcacgacagc gtgactaaag tcaataactt agttacatat tttaaaataa 1620
cttagagtgt aattggattg tttgtacctc aaagaaaaaa tgcaataaaa ctttacagtg 1680
gagaaaccta acaagcacta cctcagccag gtaatcaagg ttaacatcaa cagtcacgag 1740
tcatgttgat atataccctt gataaggtgt gatgaaaatg acacttaaac ctaaaaatcc 1800
ataaccctat ctaatgagaa aaataacaaa tcccaagagg ggcattttac aaaatacttg 1860
accagtagtg cggaaattgt caaggtcatc aaaaaagtct gagaaattgc cacagccaaa 1920
ggagtctaga gacatgatga ctaaatgtta ggtggtgtcc tgcgtggggt cctagaacag 1980
aaaaaggaca ttag 1994
<210> 89
<211> 307
<212> ДНК
<213> Homo sapiens
<400> 89
accgctagct tgttgcaccg tggaggccac aggagcagaa acatggaatg ccagacgctg 60
gggatgctgg tacaagttgt gggactgcat gctactgtct agagcttgtc tcaatggatc 120
tagaacttca tcgccctctg atcgccgatc acctctgaga cccaccttgc tcataaacaa 180
aatgcccatg ttggtcctct gccctggacc tgtgacattc tggactattt ctgtgtttat 240
ttgtggccga gtgtaacaac catataataa atcacctctt ccgctgtttt agctgaagaa 300
ttaaatc 307
<210> 90
<211> 38
<212> ДНК
<213> Homo sapiens
<400> 90
gtctgtaggc cttgtctgtt aataaatagt ttatatac 38
<210> 91
<211> 133
<212> ДНК
<213> Homo sapiens
<400> 91
agtgtctagc agtgagctgg agattggatc acagccgaag gagtaaaggt gctgcaatga 60
tgttagctgt ggccactgtg gatttttcgc aagaacatta ataaactaaa aacttcatgt 120
gtctggttgt ttg 133
<210> 92
<211> 1802
<212> ДНК
<213> Homo sapiens
<400> 92
tgtcactgcc atggccgcct tgctgcattt ctgaggatgc ttcatctctc caccttcttc 60
tccactcagc agccagcagg gcactgtgga aatcggagtc acatgagctg gcacctctgt 120
tcagaaccct ccagggctcc acatctctct cacccaaatg ccaaagacct ccccacgccc 180
ccacaatccc ccacgacctg gccactggcc tcccaccacc ttccagctcc agcggctcct 240
accacattta aggctttcct tcctagtttt aatttttcct cgtcagcagt tgattttatt 300
attttcttgt ttattggtat tttcccacta gaaatgaagc tgcgtgaagt tagagatttt 360
tttttttggt ctgtgttcct aattagctca ttgctatacc cctggcgccc agaacaatgc 420
cttggacaca gtacgcagta gactaaataa atacttgttg aatgactgac tgacggaatg 480
acggctgtgt ggggagtgga ttgggtcgtg aggcagaggc tgcggtggaa actcaggcag 540
gaggtgatgg tggttcttgg ggctgcggaa tgccaagttt agaagctctt cctctgctgt 600
ggcacatgaa ccggtcactc gagaaggctt ttagatttac tttgcctaat cccctcttag 660
tgcatgtggg gaaactgagg tacacaaaag gaattcccca ccaagttagg ggcagaacct 720
agcccccttg tctcccagat ggatatcttc tttttttttt gagacggagt cttgctctgt 780
tgcccaggct ggagtgcagt ggtaccatct tggctcactg caacctctgc ttcccaggtt 840
caagcgattc tcctgcctca gcctcctgag tgtctgcgat tacaggtgca cacaaccacg 900
cctggctaat ttttgtattt ttagtagaga cggggtttca ccgtgttggt cagggtgacc 960
tcaaactcct gacctcatga tccacccagc tcagcctccc aacgtgctgg gattacaggc 1020
atgagccacc gtgcctggct ggacatcttg ttattaaagc ttcttctctc tttgtagggg 1080
agggggagat gcctctggtg gagaagacca gtgtggcagt gactgtgtct gttagtgaac 1140
ctggtggctg gttgagggtc tgtcgtggtg actgaggaca catacaaagt gcttttctca 1200
gtggtcacct tggtgttggt gaataagggt cagaagatgg ctcctgtcct agggcactgc 1260
cagtcggttt ggaagctgaa atgcctgctt agcagtttga ggaaacacag accttggagg 1320
atcttctggt tgcctcttca agaattcatt ctattcccct tctgctcccc aaatttgctt 1380
ttcttggggt gggtcttggt tggcctaagc caagaaagta tggcatctac tccttccata 1440
gcaatagctc aggaataggc agtgacccag acctgaacca atcagtgcat ggaattaccc 1500
ctggccaaag tggttgattg aggctgggtg caagcagagt tgtgagaagg ctcccatttg 1560
gtggttggag agatcgcact tgctccagag gtcataatgt gcagatctga ggcttggaac 1620
tgctgcagac attttgctac cacaagtgaa gccaccctga cgacacagtt gacaatttgg 1680
agcagggcag agctgagaga acagcaggga aacagccaga gtcttgctca agcctccctg 1740
aagtatctat acccctggac tctagttatg ggggctaata aatgttatat actgtttaag 1800
gt 1802
<210> 93
<211> 34
<212> ДНК
<213> Homo sapiens
<400> 93
tgctgagggc ctcaataaag tttgtgttta tgcc 34
<210> 94
<211> 502
<212> ДНК
<213> Homo sapiens
<400> 94
aaaaatgaaa cttttttgag taataaaaat gaaaagacgc tgtccaatag aaaaagttgg 60
tgtgctggag ctacctcacc tcagcttgag agagccagtt gtgtgcatct ctttccagtt 120
ttgcatccag tgacgtctgc ttggcatctt gagattgtta tggtgagagt atttacacct 180
cagcaaatgc tgcaaaatcc tgttttcccc cagagagctg gaggttaaat actaccagca 240
catccctaga tactactcaa gttacagtat atgatcacta atatagtatg ctcttggtac 300
caggagctct gatatatatc tggtacatgt ttgataatga cttgattgtt attataagta 360
cttattaata cttcgattct gtaaagagtt tagggtttga ttttataaaa tccaaaatga 420
gccttttatt gaatccagtt ctctatgtga ccagttctct gtatgaatgg aagggaaaag 480
aattaaaaat cttgcaaagg gg 502
<210> 95
<211> 502
<212> ДНК
<213> Homo sapiens
<400> 95
aaaaatgaaa cttttttgag taataaaaat gaaaagacgc tgtccaatag aaaaagttgg 60
tgtgctggag ctacctcacc tcagcttgag agagccagtt gtgtgcatct ctttccagtt 120
ttgcatccag tgacgtctgc ttggcatctt gagattgtta tggtgagagt atttacacct 180
cagcaaatgc tgcaaaatcc tgttttcccc cagagagctg gaggttaaat actaccagca 240
catccctaga tactactcaa gttacagtat atgatcacta atatagtatg ctcttggtac 300
caggagctct gatatatatc tggtacatgt ttgataatga cttgattgtt attataagta 360
cttattaata cttcgattct gtaaagagtt tagggtttga ttttataaaa tccaaaatga 420
gccttttatt gaatccagtt ctctatgtga ccagttctct gtatgaatgg aagggaaaag 480
aattaaaaat cttgcaaagg gg 502
<210> 96
<211> 48
<212> ДНК
<213> Homo sapiens
<400> 96
attttttctg tagtgctgta ttattttcaa taaatctggg acaacagc 48
<210> 97
<211> 171
<212> ДНК
<213> Homo sapiens
<400> 97
gctgttcttg cataggctct taagcagcat ggaaaaatgg ttgatggaaa ataaacatca 60
gtttctaaaa gttgtcttca tttagtttgc tttttactcc agatcagaat acctgggatt 120
gcatatcaaa gcataataat aaatacatgt ctcgacatga gttgtacttc t 171
<210> 98
<211> 2309
<212> ДНК
<213> Homo sapiens
<400> 98
ggtggttctt tccttgaagg gcagcctcct gcccaggccc cgtggccctg gagcctcaat 60
aaagtgtccc tttcattgac tggagcagca attggtgtcc tcatggctga tctgtccagg 120
gaggtggctg aagagtgggc atctccctta gggactctac tcagcactcc attctgtgcc 180
acctgtgggg tcttctgtcc tagattctgt cacatcggca ttggtccctg ccctatgccc 240
ctgactctgg atttgtcatc tgtaaaactg gagtaaaaac ctcagtcgtg taattggtgg 300
gactgaggat cagttttgtc attgctggga tcctgtcagg cactttgagg tgtccctcag 360
gccttggccc tgaagtgtct aggtgtgtgg agatgggtag aaaattaggt acacccaatg 420
gtgtagaacg ttgattctca aattttttta ttttatacaa atggggtctc actatgttgt 480
ccaggctggt cttgaactcc tgggctcaag ccatccgccc atctcagccc ctcaaagtgt 540
tgggattaca agcaagaact gccatgcctg acccagttct cagttttttg tttgtttgtt 600
tgtttgtttg ttttgagacg gagtcttgct ctgtcgccca ggctggagtg cagtggcgca 660
gtctcggctt actacaacct ctgcctccgg ggttcacatc cttctcctgc ctcagcctcc 720
cgagtagctg ggactacagg tgcccgccac aactcctggc taattttttg tatttttagt 780
agagacgggg tttcactggg ttagccaggt tggtctcgat ctcctgacct tgtgatccat 840
tcgccttggc ctcccagaat gctggtatta caggcgtgag ccagcacgcc tggcccagtt 900
actcagtttt gaatctgagg ccgtgacatc actcatggtc tgcagtcagt gctctgcccc 960
tgagctgtac cctctcctat gataatcact cttaagaagg gcaacccttg gtgttttccc 1020
cttaaggtca cccaggctgg aatgcagtgg tgtggtcatg gctccctgta ccctggaact 1080
caggcttggg tgatcctctc tcctttgcct ccgaagtagc caggactaca ggtgtgcacc 1140
caccaccaca ctcagataat tgctttggtg tttttaaagc ttgtaatgat cagtaggctg 1200
aggtgggcaa atcataaggt caagagtttt ttagatgggg tgagcacaga ccaattcctg 1260
ttttatttac tgatttaaaa ttttgagaca gtctcactgt cacccaggtt ggggtgcagt 1320
ggtaggatca tagcttgctg cagccttgat ctcccaggat cttgcctcag cctcccgagt 1380
agctgggact gcatgcttgt gccaccacac tcggttaata ttttgtagag atggggtctt 1440
gctatgttgc ccaggctggc ttcaaactcc tgaacttaaa agcctcctgt ttagttttgg 1500
ttttttatca cttttttttt ttttttttga gatggagcct tgctcccatc gtgcaggctg 1560
gagtgcggtg gcgcagtctc ggctcactgc agcttctgcc tctcgggttc aagcgattct 1620
cctttctcag cctcttgagt agctggaatt accagtgtgc gccaccacca ccacgcctgg 1680
ctagtttttc tgtttttagt agagacaggg ttttgctatg ttggccaggc tggtcttgaa 1740
ctactgacct cttgtgatct acctgtcttg gccttccaaa gtgctaggat tacaagcgta 1800
agccacagcg cctggccttg ctacattttt tttttttttt ttttttttac agacatggtc 1860
tcgctatgtt gcccagaatg gttttgcact gggtccaagc agttctgccg cagcctccca 1920
aagtgctggg attacagggg tgaggcacct tgctggcccc tgttttgatt agggtgcagt 1980
gctggtgaag ccggtgcacg aggccagtga tgcatcctaa tgaggggtgg agttggcggg 2040
acttcctggg ccagtttggg gactttcaca aaagaccccc atgactcagg gttttgagtt 2100
cttaactgat cgaatgaagg attcaaaatt aaccactcca aggggggatt gaaggaagaa 2160
ccactcttaa tggacaaaaa gaaagaaagg ggagggagta acagggatat gagctctagc 2220
cgcccaagct agcaatggca acccttctgg gtccccttcc agcatgtgga agctttcctt 2280
tcgcttcatt caataaacag ctgctgctc 2309
<210> 99
<211> 58
<212> ДНК
<213> Homo sapiens
<400> 99
gtcttttgta attctggctt tctctaataa aaaagccact tagttcagtc atcgaaaa 58
<210> 100
<211> 1502
<212> ДНК
<213> Homo sapiens
<400> 100
gcaaaggctc cccttacagg gctttgctta ttaataaaat aaatgaagta tacatgagaa 60
ataccaagaa attggctttt agtttatcag tgaataaaaa atattatact cttgaacttt 120
tgtctcattt ttttgagtat gctgtttata tgattttgat ttccctctga taactatcaa 180
cagtatttaa atagcttata gctggtataa ttttttccca cgatttccaa aatcttttat 240
gtactcaggt aaaagtagcg ttatatagga aatctttttt ttagacactc tcgttctgtc 300
acccaggctg gagtgcagtg actcagcttc ctaaatagct ggaattacag gtgtgagcca 360
ccatgcccgg ctaatttttt gtacttttag tagagtaggg tttggccatg ttggccaggc 420
tggtttcaaa ctcctgacct caagtgatct acccacctcg gcttcccaaa gtgctgatta 480
tagctgtgaa ccaccatgcc cggccaggaa atcttactgt agaacaattt tttatatagc 540
tgtataaaat gtatatgatt gtcttgacag tctcaaatac tgtttttaat agcttgtaaa 600
tgtaatctca agtgcttaga acagttctta catataagtt gctctgtagt ttgctcttat 660
agttagccca aagactctgg gtgtgaggcc tgctgtaaac caatgttaaa ctgcttatta 720
gaaagcccta accacctgct ttgtaggcac cagaaactca aaaccaaatc tcaactcagc 780
tacagaatct actgtggtcc ttgtctgaaa aaattagttc actcggttgg aatcttgtct 840
cagagcatcc tcatctcttt ctcaaaagcc cctaccccaa caccggcgtg ttggttgtct 900
attgaaactt acaagtggat ggaccctttc tcccgaataa actggccttt gaaagctcta 960
atcgaaatgg tttggcaaaa tccatactgc aggagattag ggaggacaag aatgatgtgc 1020
ctttttgtac tgctgagcct gatggtggtg ccactacttc aggtacttag atgagtcttg 1080
atgctaatag aattgtgtcg ccaaacatat ctggacagtt acaacctaat ctatgcatta 1140
attggtttgg gaattgcttg aaattattgt ttaattcaat gttttaattc gttttcctaa 1200
aaatttaagt gcccccatca tcgtgcaata cctcagtgca gcaactcctt gattcttgga 1260
tgactgaact tcctaacttg gctctgcccc attgttccca tttttcatgt ttttcacaaa 1320
tagttaacca ggtacctact actgtgcacc gctgcagagc attgaggatg tatgtgatga 1380
gtaaaaacac ccagcctgct ctgctgtgtt agtattatga cggaaactga tcaaatcaca 1440
tgtgaacaaa tttactgcta caaaagggag ggcttaataa aaggaatttc atctgggaag 1500
gc 1502
<210> 101
<211> 48
<212> ДНК
<213> Homo sapiens
<400> 101
attttttctg tagtgctgta ttattttcaa taaatctggg acaacagc 48
<210> 102
<211> 193
<212> ДНК
<213> Homo sapiens
<400> 102
ggaattgcac atgagatggc acacatattt atgctgtatc aagttcacga tcatcttacg 60
atatcaagct gaaaatgtca ccactacctg gacagttgca catgttttac tgggaatatt 120
tttttctgtt tttctgtatg ctctgtgcta gtagggtgga ttcagtaata aatatgtgaa 180
agcttttgtt tcc 193
<210> 103
<211> 73
<212> ДНК
<213> Homo sapiens
<400> 103
aggttttggc agtactgtct ccttgggcca tgctggtctg acttatgctt actaataaat 60
tctgtttact ggc 73
<210> 104
<211> 93
<212> ДНК
<213> Homo sapiens
<400> 104
actttgggat atttttcttc aattttgaag agaaaatggt gaagccatag aaaagttacc 60
cgagggaaaa taaatacagt gatattctta cgc 93
<210> 105
<211> 257
<212> ДНК
<213> Homo sapiens
<400> 105
gctgtgtggg gtggatgaac cctgaagcgc accgcactgt ctgccccaat gtctaacaaa 60
ggccggaggc gactcttcct gcgaggtctc agagcgctgt gtaaccgccc aaggggttca 120
ccttgcctgc tgcctagaca aagccgattc attaagacag gggaattgca atagagaaag 180
agtaattcac acagagctgg ctgtgcggga gaccggagtt ttatgtttta ttattactca 240
aatcgatctc tttgagc 257
<210> 106
<211> 692
<212> ДНК
<213> Homo sapiens
<400> 106
tgattcaaac agcttcctga attttaattt tgtgttgtct cacagaaagc cttatcataa 60
attccataat tctaattaat ttaccaagat aatgtaatta catttggttt tgtaaggtat 120
acagcagtaa tctcctattt tggtgtcagt ttttcaataa agttttgatt atgggcaaat 180
cccctctttt tcttttttta aaatatattt gagtatgcca tacatttata tatatggtgt 240
atatgaattt ggtttaaaca ttttaaaatt tattctgatt agtttgtgtc tttttttttt 300
tttttgagag agagagtcct gctctgtcac tcaagctgga gtgcagtggt gcgatctcgg 360
ctcactgcaa cctccgcctc ccaggtccaa gcaattctct tgccttgtcc tcccaagtag 420
ctgggattat aggcacacac caccatgcct ggctaatttg tgtctcattt tcaagagtag 480
aaaccctaaa tattttattt tcattccttt tccaaattgc tatgaatggg attaaaggat 540
tacagatgta aagtctatta tttgtgaatt ctaaatgtag ttctgctgtt gtacctgtgg 600
aaacatctta aagaagtaca tattttgcac gtcctgcacg tgtaccccag aacttaaact 660
ataattaaaa agaatagttt caaaaaaata ca 692
<210> 107
<211> 228
<212> ДНК
<213> Homo sapiens
<400> 107
atagaacctg ttgtgcaacc acggtttaac cggagatttt gaggctaggg tgtgtttctt 60
tcgaactttt cggaatgtct ggaacatttc atttcctgtt ttgttacctg tgcctctgta 120
aatctacttt tgcaatttta agtaataatt ttatgaataa aaatgggaaa tgcttcctaa 180
ttccacatag tatttgcatt gttttataaa taaattccac ttactatc 228
<210> 108
<211> 529
<212> ДНК
<213> Homo sapiens
<400> 108
acccaggtga ggcagggctg aaaactgccc ttgggctgac ttttgatagg ccatgccttg 60
ccactttaca agttcttttt gcatttacta gtatttaaga gtaaccttga gattgggagg 120
aatagaggag gctggtacaa atagatggag acctgctggg atcagtgaat gcctgattag 180
gacatggggc tatgcatagc ctaagagtta taggcttaaa gatgtcgagt aactaaaaac 240
tgtattgctg gccgggcgcg gtggctcacg cctgtaatcc cagcactttg ggaggccaag 300
gcgggcagac catgaggtca ggagattgag accatcctgg ccaacatggt gaaaccctgt 360
ctctactaaa aatacaaaaa tgagctgggt gtggtggcac gtgcctgtag tcccagctac 420
tcgagaggct aaggcaggaa aatcgcttga acccaggagg cagagattgc agtgagccaa 480
gattgcacca gtgcactcca gctgggcgac agagcgagac tccatctcg 529
<210> 109
<211> 174
<212> ДНК
<213> Homo sapiens
<400> 109
gtggaaaaga acatgaaaaa gaaaactgac aaatacacac aggtctcctc aagatccatg 60
gacttctggt ctgagcctaa taaagactgt ttgtttattc ctcaaaaaca aacaaacaaa 120
aaaaaaccct ctgtattata aattattctg tgtaatggtg tgttaccata catt 174
<210> 110
<211> 156
<212> ДНК
<213> Homo sapiens
<400> 110
atgctcctct actctttgag acatctctgg cctataacaa atgggttaat ttatgttaaa 60
aaaaaaaaaa gagagagaga gtgaaacaac aatctacaca atcagagaaa atatttgcaa 120
atcttatatc tgattagaaa ttagtatctg gaacat 156
<210> 111
<211> 46
<212> ДНК
<213> Homo sapiens
<400> 111
aattggagag gattatttca cattgaataa acttacagcc aaaaaa 46
<210> 112
<211> 67
<212> ДНК
<213> Homo sapiens
<400> 112
ggagctgagt tcttaaagac tgaagacagg ctattctctg gagaaaaata aaatggaaat 60
tgtactt 67
<210> 113
<211> 294
<212> ДНК
<213> Homo sapiens
<400> 113
ggaattgaac atgagatggc acacatattt atgctgtcta aaggtcacaa tcatgttacc 60
atatcaagct gaaaatgtca ccactatctg gacagttgga catgtttttt tgggaatata 120
ctttttctct ctgaatctgt taggaacttt ctggttggct gggttccgta ataaatacat 180
gagacctttc atttcaaaaa aaagaaaaat aggcctcctt cccaggggct ccggatttca 240
tcagccttct gtgcatgccc agccatacaa accacgcagg gatggctcca agtg 294
<210> 114
<211> 171
<212> ДНК
<213> Homo sapiens
<400> 114
tcaccaaaaa gcaaccaact tagccagctt tatttgcaaa acaaggaaat aaaggcttac 60
ttctttaaaa aataaataaa taaataaata aataaataat aaataaataa ataaataaat 120
aaatagataa ataaataaaa agttttctac tcacactgaa gtgacgaagt c 171
<210> 115
<211> 49
<212> ДНК
<213> Homo sapiens
<400> 115
gcccccttcc cctgccctct ccctgaaata aagaatagct tgacagaaa 49
<210> 116
<211> 88
<212> ДНК
<213> Mus musculus
<400> 116
ggaggcctca gttcctggcc ccagaaacga gatcctgacc acatgaacaa tttgggctct 60
tttgggagaa taaaagactt atatattg 88
<210> 117
<211> 69
<212> ДНК
<213> Mus musculus
<400> 117
ttccaggacc actttgtgca gatggtgggg tctcaccaat aaaatatttc tactcacact 60
ggttttccc 69
<210> 118
<211> 118
<212> ДНК
<213> Mus musculus
<400> 118
actattaaaa attgttaaat tccagagagc aagtagagac cgcatatttc aataaatcaa 60
acatgtggtg acaaaccctt gtgtgactct taaattgtgg atgtttccaa gccccttg 118
<210> 119
<211> 1008
<212> ДНК
<213> Mus musculus
<400> 119
agcagttttc tatgaagatt ttttcataaa gacaataaac atattgatca agcagctttt 60
tctgtgttaa gctgttatta atgagactat aggaaatagt gtgaaattac aaaagcaaag 120
aagtagatag ttatttaatt aattaaatta attttacctt ttgtgttgca ccataaccta 180
ccactggtgg gattaagggc aagtattacc atgcctagct gagagtcttt ctccaggaaa 240
aaccagctta catgggttcc tgcaaatctc atgagtgttt cttgggtttc tagtcttcct 300
gggaggtgtc cttatctttc agattttcag atctggtaat tagcatgatc atcaggacat 360
ttattacaaa caaattgatt agtgggaaga aagtatctca aggtcaatct tggaagtgaa 420
caactggtgc taatccatgg ctttaaagat ttgagaacaa cggtgaaatt tggtttgagg 480
agaagggggt gtctaggacg tttcattttt atggtacatg ccagacatga atgtacatag 540
gaaaataact tgaaagggtc aaatattaaa ccttgaatat caggttcact tgggaaagca 600
ttaggtgctt atgcctctta gtaaatagcc cttcatccca gaaggagcaa gaattgtctt 660
cctgacttaa tccagtctta gctgaggtgc tgtgcatctt tatcatcttt gccttgcctc 720
acagtgtcag gctctgtggt actggggcta cacaggtcag gtaaacagtt aactgcttac 780
ctacatcccc agcaaagata atgtgacgat actaagatga acctatcaga gcttaaagat 840
aatgagtttc agtcacagtg ataactgcat gctaacttca gcatgtagaa tatatgccga 900
agctaaaagc cattccacag ttgactccat ctgaagttaa agtgtgtaag tacacagtaa 960
atcatgctat attaactgaa ctttttaata aatgagtcat ttgaattt 1008
<210> 120
<211> 306
<212> ДНК
<213> Mus musculus
<400> 120
attgttaacc taattaaaca gcttcatagg ttcttttggt gtcctttttg tgtgttgtgt 60
gtgcacatgt ttgttgggtg ggtgttttgc tggtgtcttt tcctctgtgt cttcctctgg 120
ccctttctgg aaagacctgc ttaatctgaa gcatgtgagc taggctagtc cactgggtcc 180
tgctctctgc ccatccccag ctggctttgg attagaggca catacactgc catggctgcc 240
ttttactgtg gctgtggttt tgcccttttt ttttaagcaa atagaaaatg ctgctgacta 300
tactgg 306
<210> 121
<211> 63
<212> ДНК
<213> Mus musculus
<400> 121
ggtgtcaccc attgtatttt tgtaatctgg tcagttaata aacagtcaca gcttggcaaa 60
ttg 63
<210> 122
<211> 49
<212> ДНК
<213> Mus musculus
<400> 122
atgtacacta aattttctgt acctaaatat aattacaaaa ttatcttga 49
<210> 123
<211> 39
<212> ДНК
<213> Mus musculus
<400> 123
acttgatcca aaaagctaat aaaattttct cagaaatgc 39
<210> 124
<211> 47
<212> ДНК
<213> Mus musculus
<400> 124
gaagcaacaa gaaaatattc caataaaaga ctatctgata accagtg 47
<210> 125
<211> 42
<212> ДНК
<213> Mus musculus
<400> 125
ttctgtgttg gagagctgca ataaattttc cataaagcaa aa 42
<210> 126
<211> 556
<212> ДНК
<213> Mus musculus
<400> 126
ttctccagtg tatttgtaaa atatattcat taaagtctct gctctgagag ctggtcttct 60
tgacaccttt tccaatatca gctttgcaga aggaaactta aatttcagtt cagggcatga 120
ccttcatgac cttgcagaac ttcttcactt tccaggttaa gtaaaggcga tctttagggg 180
ctgtccagat ggatcagcta taaagattca attgtagaag gttcacgtct caatgcccac 240
gtggtagctg taacttcaat taaaaaacaa aaacagccgg gcgtggtggt gcacgccttt 300
aatcccagca cttgggaggc agaggcaggc ggatttctga ggccagcctg atctacagag 360
tgagttccag gacagccagg aatacacaga gaaacccttg tctccaaaaa ccaaaaaaaa 420
caaaacacgc attcttttca ggtctttgct gggaccaggt acacataaca cagataaata 480
ttagagcaaa ccatgcacat atggtaaatt atctttgggt tttgggtccc taaaataaag 540
tggtgtgttc attgtg 556
<210> 127
<211> 47
<212> ДНК
<213> Mus musculus
<400> 127
ccctggatct taactgttaa taaaaaaaac attggatgat gatggta 47
<210> 128
<211> 693
<212> ДНК
<213> Mus musculus
<400> 128
acacagagac ccactgaata aaaacttgag actgtccttg cttgtttgct tctatgtccc 60
tggagaggtc ccagttggtc ccgtccctaa caacatgcta gccctgctca cctgcctgtc 120
agccttgctc agtggcatct ttccataggt gtgtatcccc ttagattagc ttcagcccca 180
ctacgatttg tctaggacat agcctgagcc ctgcctgtga cactgagggg tagcagtctg 240
tttctggact ccagggtgct gctgtctcag gcctaagaat tccagacatg actataatcc 300
aagcctgggg acctggttga gctttttatc ctgctggctc taagcttcag ctaggtggaa 360
atgaggccag ccaagcccca cagtgagctt gcaagcttta gatggggaca gggttacgct 420
ttggtgaatg atggaggaaa catgggggtt ccttttgttg ggtgcagcca gcacggcatc 480
atcatggtgc ccaatcttga aagggcacag gcctgaagct tcctgggact gttctgtcac 540
agggaggaac ctactgcagt tgcctacaat tgctacctct gagggacttg cctctggccc 600
cttgtagaca tttccatgtc tacacatggc ccagagtact ttcagggata gcaatgtgtg 660
aatggcactt agaagataac atgtgaaagc cat 693
<210> 129
<211> 98
<212> ДНК
<213> Mus musculus
<400> 129
agcttccctc gtgtctgtac atagcggcct ggctgtggcc tcatgtggat cagtctttaa 60
aataaaacaa gcctttgtct gttgccctct tgtttagc 98
<210> 130
<211> 1077
<212> ДНК
<213> Mus musculus
<400> 130
tgtacacaaa gaaataaaat accagcacca ggactgtgaa gtgttttcct tagactgtag 60
tgtggggttt gctcattggc tttcttgttc agattttact aattgttcta aatgatacag 120
cttagtgtgc agaaaatatc ctcttgattg gaaaatagcc aaatatttac aaaacaggta 180
tactagtttg aagaggctct atatgggggg agggggtgct ggaaaacatt agtgggtgac 240
cagtaatggt ggtacagctc taactcctag caccaggagt ccgaggcagg gggatcttca 300
ggctgcatga tgtgcatagt agcatgctgg tattagggag tgggtatctg tggttcccac 360
tttgaaaata accaaaattc tccaaagtgg gcagacctaa gccaggagag ggctggccac 420
agacatttgg tactgcttgc tgagaaagca ctgattgttt tcctaaccta aagattatat 480
atggcccacc ataccatctt tgaaacaatg tgtactggcc tttggttcac ctttcttgtc 540
tttgaagttg tacttggtgg gtgcatttaa ccttgccaca gagtggggag gatagagtct 600
aatggacctt aagtggtctc tggtggccat gtcggcagtg cttaggttgt agcccagggt 660
tggagtcggc agtgacaagc aaataactat attcttgctt gctgtggcag ctatacagaa 720
atttacggta taggtaagag ggttctctag aagtactacc tgtcttagtg aaagagattg 780
cttggttaac atcctgttat gtagtggggc tacttttaaa ctgtgtgaag tccccattag 840
ccacctccat aggcaatgga gctaacattc ttgctacagt ggccgcagct cattaacacc 900
taatgatgtg tttaacatgt gtccacatgg tgtgaatgtg ggtacgcatg tgcccagtat 960
tcagttcaca gaattgtcat catcttccat catgtcttca gtgagggact ctgcagatgc 1020
ccaccccagt ccttggttgt ggtgattctg ttagcattaa atgcactgga gagcttc 1077
<210> 131
<211> 1658
<212> ДНК
<213> Mus musculus
<400> 131
gacacattgg tctgcaatgt tttgtattaa ttcataaata aaatttagga acaaaaccgg 60
tggtttatcc ttgcatctct gcagtgtgga ttggacagga agttggaaat gacagggact 120
ttaactgggc tgctgctcct ttgtatatag acactttttt cctgctcaga aacttgagtt 180
ctccagtagc aatggccaaa cagaagaacc aggctagggg gctgcatctg acagagcaag 240
tagacgagag gctgggtggt gggctccggc cagcccgagt cttagagctg gtggttggtt 300
atatctggtg cctgtctcga ggagggcttg agacacagtg tggtgctcct cagaagcaga 360
caggtgattt ctttgtgtga tttttctttt cccctgggac aatgacagtc agtaagacag 420
gtttcaggga cttttgtgtc caggtctgag cactagtcgc tcacagttgt gtgtactaac 480
cttcttcctt cctattgaaa tggcaggggt ctttgagtct cactgctgca tgttctgcct 540
tcatagggat ctgtaagtat gctgggcatc tgggctttta gggggctctc tatagggtgt 600
ctgagataga ggtcaacaag ggcttataga caactcaaac aaagcccatg gcttgagcaa 660
gtctgcaaca agctgtttgt ctagcctcca gcagagggcg agggagacag cttccagatg 720
ttcccagtag gtggagcccc tccaagccca gggctcagga ggcttacagg gtgggaactc 780
caatactggt ggagggagga gggcgtttga tgggaagata gggaagttgc tgcttcctaa 840
actgtcacaa ctgggcttgg ataggagtca tagtctggga ccacagccct gtggtagaat 900
gctagcctgg tgtgctccag gtttaatctc catcactgca gaaatgagtc caagctgtgt 960
gtacctccag ggcactgggc atggggttcc cttgccattg tgtgtgcccg gagaactggc 1020
aggcgggaaa tgtctttatc aagggttacc ttggaagagg tcccaacact gtagggtgct 1080
cctgttgtca aaacctatgc agaggcatct gcttgctctc taataacagt atgcaatgct 1140
aaagggctcg cttacagccg gtggccacac tggaggcctg cacatcaggt ggccacaagt 1200
tctgctgctg cgcctccgag gaaacacttg gtcctccgat cgattttaac ctgttgaggc 1260
tttgcaatcc ccctgtggca aaggctccag tgttttctat ttctatgcaa atttcttgaa 1320
gcagaactgt tactgtcttt ctcctctgcc ctgggaggag gcgctagcgt ttccttccaa 1380
cttcaggtgc agcccccctc gtggttagcg gtcttaagtt cgtgacttgg gtttgcagat 1440
cttttttgtt acatcgccgg accatgtggt ggtctttagc tgtaaacaac attaaccctg 1500
ggttgattag catatgcttc taaaagatgg tcccagattc tgcgacttgt aataaaatgg 1560
aaacttgctg gtttttatgc ctttctaact cttgtatttg aatgaatgtt gatcactttt 1620
tgtattaaag tggctgacac atggctactg tcactgtg 1658
<210> 132
<211> 1795
<212> ДНК
<213> Mus musculus
<400> 132
aatatctcac caaaaaatat ttgaagaaga acaacctccg agactggctg cgtgttgtcg 60
ccaacagcaa agagagttac gagctgcgtt acttccagat taaccaggat gaagaggagg 120
aggaagacga ggattaggac acattggtct gcaatgtttt gtattaattc ataaataaaa 180
tttaggaaca aaaccggtgg tttatccttg catctctgca gtgtggattg gacaggaagt 240
tggaaatgac agggacttta actgggctgc tgctcctttg tatatagaca cttttttcct 300
gctcagaaac ttgagttctc cagtagcaat ggccaaacag aagaaccagg ctagggggct 360
gcatctgaca gagcaagtag acgagaggct gggtggtggg ctccggccag cccgagtctt 420
agagctggtg gttggttata tctggtgcct gtctcgagga gggcttgaga cacagtgtgg 480
tgctcctcag aagcagacag gtgatttctt tgtgtgattt ttcttttccc ctgggacaat 540
gacagtcagt aagacaggtt tcagggactt ttgtgtccag gtctgagcac tagtcgctca 600
cagttgtgtg tactaacctt cttccttcct attgaaatgg caggggtctt tgagtctcac 660
tgctgcatgt tctgccttca tagggatctg taagtatgct gggcatctgg gcttttaggg 720
ggctctctat agggtgtctg agatagaggt caacaagggc ttatagacaa ctcaaacaaa 780
gcccatggct tgagcaagtc tgcaacaagc tgtttgtcta gcctccagca gagggcgagg 840
gagacagctt ccagatgttc ccagtaggtg gagcccctcc aagcccaggg ctcaggaggc 900
ttacagggtg ggaactccaa tactggtgga gggaggaggg cgtttgatgg gaagataggg 960
aagttgctgc ttcctaaact gtcacaactg ggcttggata ggagtcatag tctgggacca 1020
cagccctgtg gtagaatgct agcctggtgt gctccaggtt taatctccat cactgcagaa 1080
atgagtccaa gctgtgtgta cctccagggc actgggcatg gggttccctt gccattgtgt 1140
gtgcccggag aactggcagg cgggaaatgt ctttatcaag ggttaccttg gaagaggtcc 1200
caacactgta gggtgctcct gttgtcaaaa cctatgcaga ggcatctgct tgctctctaa 1260
taacagtatg caatgctaaa gggctcgctt acagccggtg gccacactgg aggcctgcac 1320
atcaggtggc cacaagttct gctgctgcgc ctccgaggaa acacttggtc ctccgatcga 1380
ttttaacctg ttgaggcttt gcaatccccc tgtggcaaag gctccagtgt tttctatttc 1440
tatgcaaatt tcttgaagca gaactgttac tgtctttctc ctctgccctg ggaggaggcg 1500
ctagcgtttc cttccaactt caggtgcagc ccccctcgtg gttagcggtc ttaagttcgt 1560
gacttgggtt tgcagatctt ttttgttaca tcgccggacc atgtggtggt ctttagctgt 1620
aaacaacatt aaccctgggt tgattagcat atgcttctaa aagatggtcc cagattctgc 1680
gacttgtaat aaaatggaaa cttgctggtt tttatgcctt tctaactctt gtatttgaat 1740
gaatgttgat cactttttgt attaaagtgg ctgacacatg gctactgtca ctgtg 1795
<210> 133
<211> 48
<212> ДНК
<213> Mus musculus
<400> 133
actgagtcca gatggctaat tctaaatata tacttttttc accataaa 48
<210> 134
<211> 140
<212> ДНК
<213> Mus musculus
<400> 134
attcagcata aaataaaggc agataaagtt aaaggtcttc tggtggtctt taatgagccc 60
tgttgggagt gaggtgcttt aacatggaga agcatgttat taaacagtga aatagatggt 120
tcaaaaccac gtgaccatgt 140
<210> 135
<211> 46
<212> ДНК
<213> Mus musculus
<400> 135
tgtggtagag cagagttgga aataaagctc tatctttaac tctagg 46
<210> 136
<211> 40
<212> ДНК
<213> Mus musculus
<400> 136
agacatctcg tgcacggctt tcattaaaga ctgcttaagt 40
<210> 137
<211> 109
<212> ДНК
<213> Mus musculus
<400> 137
gtatattttt gttttggtca ttaaaaatta aaaaaaaaaa aatacaagtg tctgcctatt 60
gcatttgtgt gggaagagac tggggaaata aaacaggtgt gctgttgtg 109
<210> 138
<211> 45
<212> ДНК
<213> Mus musculus
<400> 138
acaagaaagt tttcctttaa taaaactttg ccagagctcc ttttg 45
<210> 139
<211> 640
<212> ДНК
<213> Mus musculus
<400> 139
aagccacacc ggaggttaat taaatgctaa cattttccat gtggtctttg catccttcct 60
tgtctgcatg ttggaaatct gcctaacatt ctaggaagag gtgaggtgtg ggcccttgag 120
agtcagtctg tgggaataag tgtagcccaa ctatgcacag ttgtaaattc ctacatcccc 180
gtgtgtattg gtcttgatat tcaaagaatt gatgaatgcc attactttca gtccaaagtg 240
aagaaacctg gtctcaaaaa atcccgagga ccagaaatga gatgggtttt cctgaaaatc 300
taaagttctt gaaaaacctt gccatccaga ttgctagcaa ctgcctagct ttgtaagctt 360
actgtgatgg acaggtagct caggacgact ggtcacttaa tactggacag attagcatgg 420
aaaacttaag gggaggagga ggtagtaggt tccatccagc ttcgctttgt tggtggcatc 480
taggtgttgt tccaagggag catgcctacc tgcaacagga catcactggt tgggaatact 540
gtagaaccag agctgtgacc tttgaactac tagaaagatg aaattttatg taaagagtac 600
cttggagtaa ataaataaag cccaagatcc tgattgtcta 640
<210> 140
<211> 40
<212> ДНК
<213> Mus musculus
<400> 140
gccccacacg cccgaagcaa taaagagtcc actgacttcc 40
<210> 141
<211> 124
<212> ДНК
<213> Mus musculus
<400> 141
aaaaggctcc tgccagtgtg aagacagacg gactgctgtg acacacctcc ccacacacta 60
tttgcagatg accagtgtcc tatgctgttc ttacaaataa actcaggcaa gatctgttag 120
cttg 124
<210> 142
<211> 575
<212> ДНК
<213> Mus musculus
<400> 142
cctgctcgtg tcaaataaag ttgcagaact gccttcaggg tttggttttc ctttctgttg 60
tctgcctcat gggtggaatt tttgggtcta cagggtgttg gaaattaatc tgagaatctc 120
tgttctgggt acatgggaaa ttagaaatac gtgaaacatt cttttcacag aagtcacttt 180
attaggattg tggatttggg ttggttttga aacagggttt cttgtggcac tgcttgttct 240
atagaatagg gtggccttga actcagaaat ccacctgcct tttcctccct agtattggca 300
attaaatgcc cagcttgttt gtaagctctc attttcagtt ccaggtttat gtgtgagcct 360
aagattaggt aaagattgag gttataactt aaacgtactg aattaactta tgttgtgtgg 420
gtcccaggaa ttggacctgg gacatcaact ctgcctttcc agccatcttt gccaaccagt 480
agctcatctc tgggatgtgt ctgccctcaa aatgacattt taaaaaagtc agtacaaaag 540
aacgattttt attaaaaacc ttgaggacaa acatt 575
<210> 143
<211> 54
<212> ДНК
<213> Mus musculus
<400> 143
atggcttgtg tgcatgtttt atgtttaaat aaaatcacaa aacctgccgt cgta 54
<210> 144
<211> 68
<212> ДНК
<213> Mus musculus
<400> 144
actaatggag agtaaataaa taaaagtaga tttgtgctct tgtatttttt tttcacatct 60
gtcctaaa 68
<210> 145
<211> 504
<212> ДНК
<213> Mus musculus
<400> 145
ggatttcaat cagtcataaa ataaatgttc tgctttcaaa aattctgtgg tgatctaagg 60
tactttaaca tcggttcaga gttcggttat atgattgctc tgggatccta cgcttcttcc 120
ttcatagttc ctgtgggtcc gaagctggga ggggctgggt ggactctcgg gaaagatact 180
ctgagcctgt ctcggtcccc atcgtgtttg cttggccctg ggcatggaag tgggtgagtg 240
atgagctgaa cgagcaggct tgctagagat gaggacagtt actggtgtgg ttatatcact 300
accatgccta cagtgtctta agacgcttac agtctgtaag ggacttaaat gatttgagct 360
cttacttatc ctgtagtttc tgatttttaa catttacttg aataaagcca agcaagataa 420
gcctttattc ccagcacttg gtgacaggtg gatctatgag ttggggatca gagctacaca 480
ttaaaactct taattcatct tact 504
<210> 146
<211> 306
<212> ДНК
<213> Mus musculus
<400> 146
ggatttcaat cagtcataaa ataaatgttc tgctttcaaa aaaaaaaaaa attaatcctc 60
tgtgatggcc agcagttaac attcaacagt ttctctctag gctcttgatt ctctgactat 120
tgtagggatt cgatcagcac tcgcatacca gaagtgtgag atggtccgtc ctttttcaag 180
acaagatttc tctgtgtagc cctggctgtc ctggaaccca ctctgtagac caggctggcc 240
ttgaatttac agagatcccc ttgcctccgc ttgctgagtg ctaggattaa aggcatgcgc 300
actatg 306
<210> 147
<211> 246
<212> ДНК
<213> Mus musculus
<400> 147
aagccctgct gtctgagact tgcctagcct gcaataaacg ggttatttac gtaacttttt 60
tttttttgcc ttgtttgtgg ttaattaaaa catttggtgt gtgttctatt ttttattttc 120
gaaagatgct tgttttgaga catactgtgt gaccctggct ggccttgagt gcctggttct 180
ccttacaagt gtagatacat ctggcttaag attttagtct ttcagaaata aaaatgttgc 240
taagac 246
<210> 148
<211> 41
<212> ДНК
<213> Mus musculus
<400> 148
accagccctc tgcgtgtgac tattaaaaac cctgaaaagt g 41
<210> 149
<211> 181
<212> ДНК
<213> Mus musculus
<400> 149
ggattcacac aatggcaaga ctgaggattt atactgaatt gtcatcaatc agtcctacca 60
gatggatttc aacatttaaa cctggagact cttcgtgtct tgaattagga tgtttgtcca 120
gtaataaaat atagaacctt tcaaaatgct tttctggttt ataaagtact gaattgccct 180
t 181
<210> 150
<211> 285
<212> ДНК
<213> Mus musculus
<400> 150
tcccgccaaa gcaaccaagt cagcctgctt aatttgagaa agatggaaat aaaggcttac 60
ttctcttaaa actccggtct ggatttattt agtttgttca cttaagcagg atgaaaaagc 120
aaaaccgcta ctgtttactt tgtgttggca tctttgtttc taaaattaaa gctcctagtg 180
tttttgtggg ctttgtttgt tttttgagac agtctcttga cttggtgcca tagctagtct 240
gggacaaaga ttttccaggt gtgaattaaa ggtgtatgtc atcaa 285
<210> 151
<211> 40
<212> ДНК
<213> Mus musculus
<400> 151
actgcttttg ttaagttggc taataaagag ctgaacctgt 40
<210> 152
<211> 42
<212> ДНК
<213> Mus musculus
<400> 152
attcctgctc ccctgcaaat aaagtctttt tatgtatctt ga 42
<210> 153
<211> 1059
<212> ДНК
<213> Mus musculus
<400> 153
cagggtcttg gcagctgcat ctggaggcat ttaataaaat aaagacattt aataaaatct 60
tgaacaaaga caaggcctga ctggattgtg tccagtattc aactgagtta tgttgtctat 120
ggagccatgc ttattctgtt ggtttaagct ggagggcatg agcagagctg accagagaag 180
tcatgaagtt ggtgaccctg tgttgaacaa ttgagggtta gaagagcagt ttggttttgg 240
tgctcttgat ggaacccagg tgcttggaca tagtaagcac acataagaca gagtaagact 300
gctgtgtctc tggcctggag tagtctttct tgcttttttt tttttttttt tttttctcta 360
gaatgaaagc agatggccca gcgagttagg tgcttcctat gaaagcatgt gtgctggttt 420
gtcatgcaca cagccctgca ggagagagta tggcaacaca gccgctcagc atcccaagat 480
aaaaagggag tttctactgc cattttgagc ttgggagttt gaaatgtaaa gcctgtccat 540
atgttttaag gatccatgta tttctgtttt gtttgttttt caaaacaggg tttctctgta 600
gccctggctg tcctggaacc cactctgtat gtagaccagg ctggccttga actcagaaat 660
ccacctgcct ctggcgatcc atatatttct aagtcctgta cttagacgct gttttggaaa 720
attcattttg gaagcattta ctgttggtgt gttttgtggg gaatgaatga tagcttggga 780
attcttttct gtttggtgag agtgaagctg tcagcccggt tgtagcctgg ctggtgctca 840
aaggctttct ctcattgtct tcacctacgt agctttacgt ggggtaagga cttaagttac 900
ttaagttggg tgcacactga ccatgtccac aacctgttaa ccaactctac atgatgagta 960
cagatgtacc tttttagaaa gtgttaatgt gtagccctgg ctggcctctg cctcagggta 1020
ttatgaataa agtgtgcaac cttcatctgg ttgattaaa 1059
<210> 154
<211> 49
<212> ДНК
<213> Mus musculus
<400> 154
aatccagatt tttaatagtg acaaataaaa agtcctattt gtgatcgtt 49
<210> 155
<211> 87
<212> ДНК
<213> Mus musculus
<400> 155
aatggtctct aggagacatg ctggaaaagt gtttgtacaa gcctttctag gcaacataca 60
tgctagatta aacagcatgg tgaaact 87
<210> 156
<211> 100
<212> ДНК
<213> Mus musculus
<400> 156
agtggactct gagggacatt gcggggaagg ggcgtttacg tttgtttata cttaaaagtt 60
ttttaagcag catgttgaat taaaaaagaa agcaagcttc 100
<210> 157
<211> 45
<212> ДНК
<213> Mus musculus
<400> 157
tttcccagct gctgcctaat aaactgtgtc ctttggaaca actat 45
<210> 158
<211> 603
<212> ДНК
<213> Mus musculus
<400> 158
gtctttaaga gcaacaaata aataatgacc ttgaatcttt cattggcttt cattaatagt 60
gtaactagat aaatgatggg aaagatgaga cagaagaagg aatacatcta taggactgct 120
agaatatggg gagagtgatt attttcaaat taatatgtat cgagcttcta ccccaaggta 180
aataaataac atttggagac cattaaaatg taggatggca tagaagaggc ctttactaag 240
attaataatt aaagaaacac agcctttaaa gtaaaaaaca cactgtgcct ttgaaacttg 300
ctaaaaagat taacttctgt cccaaaaggt atcagccatg cgctaccagc ctccctgccc 360
ctacagtggc agtggctgca ttcttggtga atggtagtgg aagggattaa acctaggcct 420
cagtcatgct tcccagtcac tggtactgat ttgtatgcac ccgcttaggt gtgaaggtag 480
ttttggtgtg tatcacaagt tagcctgtgt agcgaagaca ggttttctcc accgtgtttt 540
ttgttacaca tgactattca caaatgtgct gcagacagta aaatgagaaa tacccttcca 600
agg 603
<210> 159
<211> 269
<212> ДНК
<213> Mus musculus
<400> 159
agaaaatgac tgaataaagt gtcattcata gtatttggtt gtagtaactt gtcaaaatct 60
cagggccatg ggtgcacgac agcagtagct tcttgaatga actgaagttt tcaagaggtg 120
cctggaaggt gaaaaacaca ctgaagccag tcatgttgat atgggggcat tctgctgctg 180
tgaaacagac tggggttcac acccaccttg cgggattaga acttcactgc cctccaactt 240
ctttctttgt aaacaactgt ccacatttt 269
<210> 160
<211> 42
<212> ДНК
<213> Mus musculus
<400> 160
gcctcatgtg tagtgtaata aaggtgtctg ctgttctatc tg 42
<210> 161
<211> 69
<212> ДНК
<213> Mus musculus
<400> 161
ttaatacttg gctgaactgg aggattgtct agttttccag ctgaaaaata aaaaagaatt 60
gatacttgg 69
<210> 162
<211> 36
<212> ДНК
<213> Mus musculus
<400> 162
agttggagtt tatgttgtat tgaataaact ttaaag 36
<210> 163
<211> 47
<212> ДНК
<213> Mus musculus
<400> 163
ggggactctg gccaatgccc tagaacaaat aaagttattt tccaacg 47
<210> 164
<211> 50
<212> ДНК
<213> Mus musculus
<400> 164
ataaattttg gctgattttt ctcttgtatt tcttgtttgc tggtataaaa 50
<210> 165
<211> 43
<212> ДНК
<213> Mus musculus
<400> 165
atgctgttgt gtgcacaagc aataaaatca ctttgagtaa ctt 43
<210> 166
<211> 34
<212> ДНК
<213> Mus musculus
<400> 166
ccgaggccaa taaagactgg ttttggtccc tgga 34
<210> 167
<211> 3272
<212> ДНК
<213> Mus musculus
<400> 167
acgtaaagca taaataaaaa gcctttgtgg actgtgctca gggtcagtcc ttttgaatct 60
ctgcagcaga gtagctggct gtgctgactg gtgacacttc tggtgatgct cagctgtgag 120
gttttatgta gatattgaaa gcatgaccat tgtcttcact tcacctccag cttgggttgt 180
atgccagtaa catcagcata aggtggttaa tgacaggatg gtcccttgag tgtgcagtga 240
gtctggttta tttgccaatg agaagcacag gcctcctgta tgggtctttg cctacagccc 300
cctttcatca cccagacttg gtagacttac attctgtcac actgttggct cttaatctca 360
gccctgaaaa atgccatttc ttgggtatca aggctagtct agattcagaa accatataaa 420
ggttgacagc tggtttaaaa aaaaaaaggc ttggagcttg agttgggttc gcaggttatt 480
ccagggtatc tgtctgcact ttgtctccca gatttaaagg taagtgccac catgcctagc 540
atggtgactt attagctttg ttgctgtgga acatacatca aatgaaacat tggtatggct 600
ctggtttcac tgtccatggt tagtatctgg tgggtggaca cctggtggaa ccaagctgct 660
catcccagaa ctaaggagcc attccccaga gacttgtctt cctactaagt tccatcccta 720
cagcttctag tagtagcttc agaggttgtg aattgtggac ttagtctgcc ataacattta 780
aaataggtat taaattcaag tcatttggtc actcagcacc cacgtggctc ttcagaacaa 840
cagaagcccc tcggacttgt ctgttggaaa aaccagtttg aaataatgta cctgctttag 900
ttgagaaaac gctacaactg gtgctgtgtc ctgccatgct gatgagctct gctctgcgac 960
ctgccgaact tgggggatct ctacccccag actttgctca gatctgttga tgatttgtcc 1020
atgcaggaaa gtttacaagg tctctgtgtg tctactactt actagttgct gtgacaaaaa 1080
tactgaaagt gtttactgtg tgtgaggcac aaagtttgtg ggaagctgta ccctccctca 1140
ttttggtgct gctcctgcct tgactgacag gatgagctgc cccaaccatc gttgccatct 1200
tccataagaa gcaggtggct ctattgagtt ccctggaggt gatcccaagg gaaggaggag 1260
cctgggaaag tggatctcaa gtcccctagt ctggcagttg gctgtttagg aaagtccagt 1320
gtcagtgttt gatatgttgt aaggaaacaa attcagtttt atttagctta ttggctctgg 1380
ggaaatggca gttcccatta attggtgctg tctttctctt tgaggatcaa aattagcttc 1440
ctgttcagtt gttaagcata tttcatagtc aaataatcct cttatcttta caagtgaggt 1500
tttccttcga gtggatatca gagtccctcc cacggcttct atccctccta tccttgtgta 1560
ggaagttaag cttgctcatt tgtagattag tggtcggtta cactgcaatt tagagtatca 1620
tgtgtactct acacatgatt gattctaagc ccccttccct ttccatgtcc ttcaaaaatt 1680
tttttattct gagacagtgt taggattttt cctgggctag ccaaatgcaa gaagtgttgg 1740
tcccagactt gtaatctttc tgccttagcc tcccaaattc taggattata gatttatgtc 1800
atgtgatgag tggtctttta aagattatct tttatttttt ttgagatggg gtttttctgt 1860
gtagttttgg ctttcttggg acttgctctg tagacccagg atggtcttta ctcagatctg 1920
cttgcctctg cctcccaact gctaggatta aagcatgagc ctgagccttc atgcctggct 1980
gacaggtgaa ttctcaatac ctacctagcc atagggaaag tgattgtgtc cccttcctca 2040
tagaggggca tagtcaccca ggccatacct ttagcttggg cttttggtca gtgaagaagt 2100
atgaagggac aagaacacta tcaagagcct aatgtgctct ggcctggatg gtcagcacaa 2160
aatgaataga cttaccaaat tctgctgtct ccttggtaga tgtgaagttg ttggaagagt 2220
cctaaattta gcagactata ctgtcagcct atcagactat aggctgccgg agggcaagtc 2280
tgctacctat ttccatctca tgcctgcatt gttcatcccc ctatgtaacc cacctacctg 2340
tcactcattc ctccatccaa aaactattgt aggttcagtg gaaatttcaa gcttgcctgt 2400
ctcagcatct ttcttacctt acccctaagg atggcatctc tcttggctac atctttggtt 2460
tatctggaga tccttgatta atttgaacaa gagctacctt gggttatgca gtttatgcct 2520
ccagtgtccc agagaccggc atttgagaga tccctgatag caaacccata gggtggcctt 2580
tttttcatcc accccattct ccctcccacc tccctctttt gaccttgagt cctcaccaga 2640
gagaaaccag gcccacttaa ttagttctac atgtgtacac tacatgggtg cagtgcccaa 2700
gcaggagagg tgttagattc ccttttaact atagttaata gacagttttt aagccccagt 2760
ggatgctggg aagcaaacct acatcctcta gaagagcagc tatttctctg agccatttct 2820
ccagcaccat tttcccctct tttaaaagca ggtcttgcag tgtggcctag tctggccccc 2880
tgaggtgttt gcattgcatg gcaggcatgt ccacaggaac accatagttc tcaccactcg 2940
tacagcacag caagtggggt gccgcagggg attatcactt gagtataaaa taagggttgc 3000
tttagattga ataggataac cacgcgttct cagaacaatc aaggaaggct ggggtgagcc 3060
agcaccgacc ttaattgttt acttagtaaa ctactaaatg tatgcacgtg taagcttttg 3120
ccttgattga ggtcaagctg tcgagaaatg gttctcttta cagtggatcc agtcaggatt 3180
ggcagcaagc acctttgcct gctgagccta catgttgtat agaatggcaa cgttgtgtag 3240
aatggcagta cattaaatgg gtttttcatt ta 3272
<210> 168
<211> 59
<212> ДНК
<213> Mus musculus
<400> 168
gcccatctca aggatcgggg tttacctttg taataaacat cctaggattt taacgttcc 59
<210> 169
<211> 35
<212> ДНК
<213> Mus musculus
<400> 169
aacaaaggat gctgggttaa taaattgcct cattc 35
<210> 170
<211> 3167
<212> ДНК
<213> Mus musculus
<400> 170
gacaactgaa taaatcgtct taatggtcaa attttgctgg cttttgttca ggtttttttt 60
ttttaattca tgtttatgag tgtaaatgtg tgtgtcacag gggtgtcaga tgttctttga 120
accaccatgt aggtgctgga aacccaacct gcatcctcgg agagaacagg ttccttaacc 180
actgagcaag tactgaagca ttaaactgct tttaaaaatg aaggtgtgct aacagattgg 240
tcaggtgaaa aagagacgtt aggtttcctg cagggggcgc taagccaatt taaagactaa 300
gttgggttag aaaagagcag attgcatcct tgatctttta agcctgggga ttttgttttg 360
ttttgggata gggtcccaac acagaacagg ctgacctcat taaatatcaa tcttatttga 420
ttgcctctgc tcccagagta ctggaattaa aggcagggac cactgtatta gccattctga 480
gttatttgaa atggactctg caggccatac ttggtcaaaa ttctgccttc tcaattacag 540
gcatgagcca ctatgcctgg tttacttact aatagatgtc caaagactag tgtatgaaaa 600
ttttgctttt ccaggtgatt tgtgaaaggc agggtggcct ctcccatgtc acactacttg 660
ggttactcat gttgcaacat atctgcaact ttaggttgag gggatttgag cctgcatgtg 720
ccactttggc caactgaact aatctttaat tccatctaaa acttttaaat ctcagtcatg 780
tgttcaactg gaaaataacc tagagtgtgc tatgttgact tcaggtacac atcaaagcag 840
gttttagtga tgtagaagct gtgtttgagt tgaactagtg ttgaggctag gcttaggtac 900
catagaactt tggtttttca agacagggtt tctctgtgta gccctggctg tcctggaact 960
cactgtagac caggctggcc ttgaactcag aaatctgcct gcctgtgcct cccaacacac 1020
ccagctctag ctttaaattc cttgcaccaa gagatgcttt atccctccgc tgagaataca 1080
ggtgcatgtc agcatgctaa actctagata aaatttcatc ttgtttgaaa ggacaataat 1140
ataagaaaag tgtatttgca ctgtatacca tgcccttttg tgtttaaagt taaactggca 1200
acagtgtccc atagaggttc cagaagaaac tgcttctaag gggaggacca taaagggaaa 1260
tgcttaccat agaacttttt aaatgttcct acaggttgta ctctggatag gtatatgaac 1320
acctttctat tagaacagtt ttatagtagt acttagtgaa tatgtaaata atactatttg 1380
tgaaatagtt tgaggtttct ctatagtcct ggctggcttg agctctatac cagattggct 1440
tctaaatcag aaatctgcct ctatctcctg aatggttatg atagagatgc aagttactta 1500
atttcttaca tgaattgcac tttgtacatg cttttggata tgggcctagg cttttgcttt 1560
tggattagac tgtctttatt acatttactg gcttgatact ttacagtctt aagcccactt 1620
gcctgggtgt gatgcacacc tttaatccca gcactttggg atttctgagt tccaggacag 1680
ccagggctac gcagagaaac cctgtcttgg aggggaaaaa aagaaacttt ggaaatcaaa 1740
acttcttgga aagccacttt tagagacttg aatctaagga taactaacca ggtagtaacc 1800
acgagtcatt gattctgtga atcttgtatg agtgggttac aggcagaaat taacttcctc 1860
tgagagcact gctgttttag aaatgcgacc tagtaattac caaaggcatt gaagccactt 1920
gactacagtc tcaggtttct gcatctgaca ttgctgggac tgtgtggggt ttatgggtgt 1980
aaaataaaac aggcgaaagg atgttgagtg gaaagctttg gcttcagaat caaattccaa 2040
atcaattacc agatcaaatc aaatgccaga tcaaattaca agacttcctt atgactaaat 2100
tcttcagtga cttaggatac taatagtatc tgcctcaaaa gtgtatgtgg ttatttttgt 2160
cccagtgaag ccaagattca catgctatgg gcatggattt cctgaggaca tgctagccta 2220
tggtatgagt tacttgaaag gactctgaga aatcttagtc cccagctgtg aggtttttaa 2280
agatcatgct caatggaaag tggggcagta atttgagagc atggcacaaa tgaggtttta 2340
tcttttgaaa ctaagtgaat ctatgtcctt ggactagcat attttaaatc acacatcaaa 2400
tgaaatttct gttcaattcc tataaacagt ttatttcata ttttgtagtt accattttca 2460
ttaacagcat acacccttca attgtgttgc taaaactgag tcacattatt ctgtaagaac 2520
ttactccagt atcacaactt ggtgctcatc caatattttt attttcattc tattttccct 2580
gtctagccac gtatggccct attctctctt cttggattga ccctagcttc aacacaagaa 2640
agcttgcagt attttttttt tgcgcctggt tcattgtttt gtcctcaagt tctatccatg 2700
tgtccagaaa ttcgaggatt ttttaaaatt tacttttctg cctaaatact ggtatgtgga 2760
tagtctgtta tttttactgt tggttgcata tctgtagcat atttctgtat ttgcaatgac 2820
tacattgaat gtcccttagt taacctcatt cttcttaact attttgtagg cgtttgctat 2880
tcaaagcaca atctcaatta aaatgttttt aatagcatct ttccacttgg atgtgtaaag 2940
aaagtatttc tagaagtctg aatttttgtt gcattgtaga tgtgtacaca atagggctgg 3000
gaaagtagct cacagtgggt aaaagccatt tgttgccaat cctaacagcc tgagttcaat 3060
cccagaatct ataaggtaga agaaaaaaac cccagctccc acaagttatc tggccctctg 3120
cacacatata aatagtgcaa taaaaattaa ccatttaaaa atataaa 3167
<210> 171
<211> 58
<212> ДНК
<213> Mus musculus
<400> 171
gcagaagaaa tcgggaattt gttacaaata aaagttttaa gtacctgtga cagttaag 58
<210> 172
<211> 49
<212> ДНК
<213> Mus musculus
<400> 172
aatttgacaa tggaaacaca gtaataaatt ttcatattct gaaaaaata 49
<210> 173
<211> 264
<212> ДНК
<213> Mus musculus
<400> 173
acaggttcaa tcagctgtac atttggaaaa ataaaacttt attgaatcaa atgaatgggt 60
gcatctgttt cctaaggcag ccggggagga tttggtctta ggaataatag ctggaattgg 120
tttgttggcc atgaagtcag atgcaattgc gcctgggaac cttcagcttt tccctttacg 180
tggttgcttg cttcttgttg cagcttcggt tttgaattga tgcctgaaag aaaataaaaa 240
cttagcaaga ctaatggtaa atct 264
<210> 174
<211> 71
<212> ДНК
<213> Mus musculus
<400> 174
agaggccgtt ttgtaaggac ggaaggaaaa ttaccctgga aaaataaaat ggaagttgta 60
ctttacatgg c 71
<210> 175
<211> 56
<212> ДНК
<213> Mus musculus
<400> 175
aagcccctga ttgaagatga gtgggaacct tcccaataaa cacgttttgg atatat 56
<210> 176
<211> 38
<212> ДНК
<213> Mus musculus
<400> 176
ttgtgtatgc gttaataaaa agaaggaact cgtactta 38
<210> 177
<211> 92
<212> ДНК
<213> Mus musculus
<400> 177
tcttgcagct gggttctgga tatccactac ttagcccacg gaatgatctg caactgttaa 60
ataaagcatt tatattaatt cttgtctaga aa 92
<210> 178
<211> 91
<212> ДНК
<213> Mus musculus
<400> 178
gcgaccttga atggattcga ctgactacta ccaagtggaa ccgatcatgc tagtctttgt 60
acacaaagaa taaaaatgtg aagaacttta a 91
<210> 179
<211> 1187
<212> ДНК
<213> Mus musculus
<400> 179
tacacgtgat atttgtaaaa ttcatatcca ataaacaatt taggacagtc atttctgctt 60
aaaggtgtta tttgttaaaa ctagtctaca gattgtcatg agtgttctgt gaaaatgtag 120
aagttaagtg caataattga aaactgcagg tgatggcata tcttgtttct gatgtacttt 180
gcattatctg cttatgagat taagtgtata tagtgtttgt gccaagtggt gttctgtgtg 240
taagaccctg taagaggtaa aaagtcctga aactgaccct ggatgtgttg gtgcatgaga 300
tagaatctac agctttacga tggcatcttt tggttcactg aagtggctgc ttgggagttt 360
gatgagtaca aactttatag agttggattt tgcttagaaa tgtgtaggaa gagagggttt 420
caaaacctgt tttgtgcata gaaaagtgag atcaactata acccacattt tgagaattga 480
atccagtgtt attttcagaa gaccaggtaa atttgttgga agaggtttca ctttctgttg 540
gatctagagt atggatttgg agatgaaggt tgaacattgg atgtagtagg gcttatttag 600
ggcagattat ttccattaga tttgtaaact tggttgtggt tgcaagttaa tatcaaccaa 660
gcaaatataa gtttgattaa gcttgcagac attaagcttt tctagcagct ggtgtgtgca 720
gaggcagatg gctctctggt tccaggacta cctacaatat agagaaattt tctctattcc 780
agtcaattca tcatgggtaa gaatagtcca ttttggtagt gttatttcat cgtttacaat 840
ctacctatgg gtagtggctg gtaactgcct ggatgttgac atttcacaaa ggccatactt 900
aaccacactt ttatttctat ttgtgcgatg ccttggagta gtttcccaaa gtgattttga 960
gtgtggaaga aatggtattg tccccgaaca gctggcttgg tctcaaaatc taattgatgc 1020
ttttattaaa ttggttttcc tttggagatt ttaaaaggat gatttgattt ccagaaaata 1080
ctagactcaa aatcttgata gctaaaattc ttttctattc agcaaaacaa gtcactgtat 1140
agaggttgtt caaatcaact aaagtaataa atgtcttaaa caagtgg 1187
<210> 180
<211> 43
<212> ДНК
<213> Mus musculus
<400> 180
gggtttttat atgagaaaaa taaaagaatt aagtctgctg att 43
<210> 181
<211> 259
<212> ДНК
<213> Mus musculus
<400> 181
ggctacacag aataataaag gttctttttg acggtggtaa atctcatgtg tggactctaa 60
gcttgtcgcc aagtgggaaa tagactggtg ggattgtaga taggatgggc tacttaaact 120
cattctaccc aggccttagt acttagcata cagccagagt caaactgatc ctttatacag 180
ggggtaccat gacagtacaa cagtgtcgtt aaccctaaca aataaatttc ccaccaacgg 240
gtggaattcc ttcattttg 259
<210> 182
<211> 48
<212> ДНК
<213> Mus musculus
<400> 182
acaggacttc tcattatttt ctgttaataa attgctttgt gtaagcta 48
<210> 183
<211> 67
<212> ДНК
<213> Mus musculus
<400> 183
aggcttcaat agttctccta taccctacca aatcgttcaa taataaaatc tcgcatcaag 60
ttcgctt 67
<210> 184
<211> 33
<212> ДНК
<213> Mus musculus
<400> 184
gatgctccaa taaacctcac tgctgccact cag 33
<210> 185
<211> 36
<212> ДНК
<213> Mus musculus
<400> 185
gatgacaacg acaataaagt gcgagactga ctggct 36
<210> 186
<211> 388
<212> ДНК
<213> Mus musculus
<400> 186
acccaataaa gactgtttgc ctcatgcctg cctggcctgc ccttcctccg ccgccaacta 60
gggaagtggg gaccaaaggt tccttaggca ctgctcctgt gggtagaggg gacattagag 120
agctgacagc gcaccacctg catgagtttt tattaaagtg caaaccatgg gatgaatcag 180
ttgagcttca gtgttgaaaa tgagtagcag ggctgcccca cccacctgac caagtaccct 240
attctgcagc tatgaaaatg agatctgcac atgagctggg gttcacaagt gcacacttgg 300
agcactgcct tgctccttcc cagcagacca caaagcagta tttttctgga ggattttatg 360
tgctaataaa ttatttgact taagtgtg 388
<210> 187
<211> 32
<212> ДНК
<213> Mus musculus
<400> 187
tgaaccctcc cccaataaaa gatggttcct ac 32
<210> 188
<211> 103
<212> ДНК
<213> Mus musculus
<400> 188
gcagattttg ttatgaagac aataaaatct tgacctttca acccctttga ttgcagttgt 60
tcgtttggga gggaatacat taaaagcttt cagaaattac ctg 103
<210> 189
<211> 277
<212> ДНК
<213> Mus musculus
<400> 189
gccagcccgt gcacctacga cgcctgcagg agcagaagtg agggatgctg agggccggga 60
caagctatcg gactgtgtgc tgccatcggt aatgagtctc agtagacctg gaacgtcacc 120
tcgccgcgat cgcctggaga aatgaccgcc tttcttacaa ccaaaacagt ccctctgccc 180
tggacccccg gcactctgga ctagctctgt tctcttgtgg ccaagtgtag ctcgtgtaca 240
ataaaccctc ttgcagtcag ctgaagaatc aaactgc 277
<210> 190
<211> 44
<212> ДНК
<213> Mus musculus
<400> 190
gtctctgggc ctttgctgtt aataaatagt ttatatacct atga 44
<210> 191
<211> 404
<212> ДНК
<213> Mus musculus
<400> 191
aatacctagc agtgagtgga gattggatac agccaaagga gtagatctgc ggtgacttga 60
tgttttgctg tgatgtgcag atttctgaga ggacaaataa actaaaaagc tcctacacgt 120
ctgctctgct gcttattggg cattagaaga atcaggtggc tgcttgggtg ttgatgcagt 180
caagtgcact gggcttggtg aaaagcccag tgtaagaggc cggtacagat ccttcctggc 240
agagggtggt gatggagaga acataaataa ctacatgggc aaagtgtagg accaattacc 300
ctgttagcat cgtctttgct caacaccttt ctgtgtccct agactctgag tttttttcta 360
attgattttt attgaacact gagtgttttg aggttttatt tttt 404
<210> 192
<211> 39
<212> ДНК
<213> Mus musculus
<400> 192
agttcaggag ctaataaagt acgtcctttg gctaatccg 39
<210> 193
<211> 48
<212> ДНК
<213> Mus musculus
<400> 193
atatgcacat tttttaagta ataaaaatca agacttgatc tacgcttc 48
<210> 194
<211> 36
<212> ДНК
<213> Mus musculus
<400> 194
tctgttatgc catattttca ataaacctga aaacaa 36
<210> 195
<211> 875
<212> ДНК
<213> Mus musculus
<400> 195
gctgctgtgc aggtgcctga gcaaagggaa aaaagatgga aggaaaataa agttgctaaa 60
agctgtctta tggtcctcac tgcagactgt acctggattg gcatttggct atacaacaga 120
ggcatggtcc tactgacatg ttttgtgttg aatacttaag catgtgaaca catgggtttt 180
tttttttttt aatgtaaaat gtagtaacac aatgtttagg tggctttggt gttagctctg 240
gagacttcat gtgtcatcta ggtgaggtgt tctttaacac agggtctctg ttctgtcatg 300
cctcatagat ccttctacct ccagattgga gagggaaaag gcttatgtca ctgaacctgg 360
ccagattggg attttgtgtc ccaggaacaa agttaatgct aaaaagttaa tgccttggtg 420
agactgatag tctgatggtg tgaattcaca gtaagtggtt gggattgcca gatggaattc 480
cctgagctgc cgtgacaggt ggcattgcag aagtgaagga ttcaggaatt tgagtgttgg 540
gtgggggcct gtgaatagca cttgggctgg gaggggagac tgctgcccct gaatgtcctg 600
gaattcaagg acagtacctg gttaaatgtt tttctagctt ttctaaaaag tttgttaggc 660
ctggcattgg cggcgcacac ctttaatccc agcacttggg agtcaggcag gtggatttct 720
ggcctggtct acagagagtt ccaggatagc cagggataca cagaaatcct gcctcaggaa 780
aaaaccaaaa agaagtttgc taaaaataag catttttgct tgatggtatt gaagattgta 840
agacattaaa ttgtgtcatt acttctccag gtact 875
<210> 196
<211> 67
<212> ДНК
<213> Mus musculus
<400> 196
agctgtggtc atacctggca tgtgaccccg ggaccaaata aagtcccctt ccatccactg 60
gagcagc 67
<210> 197
<211> 50
<212> ДНК
<213> Mus musculus
<400> 197
gtcctattgc caccctgcca tgctaataaa gccactgtgt ccagacttct 50
<210> 198
<211> 84
<212> ДНК
<213> Mus musculus
<400> 198
ttggcctctg cttgtaatac aatgaaagta ttctaaagaa atataaaatt ggactttatg 60
agaaaataaa agtcatttca ctct 84
<210> 199
<211> 36
<212> ДНК
<213> Mus musculus
<400> 199
tctgttatgc catattttca ataaacctga aaacaa 36
<210> 200
<211> 446
<212> ДНК
<213> Mus musculus
<400> 200
ggaaccacac gtacttatgc tgtaacttac tgtagcgttt acagcgttac cgctgtctgg 60
acagctgagt gtgtttctta ggaatataag ttttctttct gtgctttagt gagttcattc 120
agcagttcag taataaatat gtgaaacctt ttgtttcgaa aaaaattgct tctgtgttac 180
aacttacttt gttttatggt tcaggatcat ctgcataata gacaagtatt ctatcagtga 240
gcgatatcct ggatcttgtt tgtgtagttt ggggttgaga caagtccaga ctgccctaaa 300
actcacgatc ttccttcctc ggccttctaa atgattggag gactccaaac ctaagtgatg 360
gaagtaaaaa gaaacttata tgtcaagact cattttctct atcatttcat gtgacaattg 420
aaattagatt atttcttttt tcaatc 446
<210> 201
<211> 332
<212> ДНК
<213> Mus musculus
<400> 201
ggactttgct ggcagaaaag aagtacagat gaggttatag tttgaaaaac gttaattccg 60
tttattgact ttagaatgtt actttgcata gtatagacca ttacaatgga aagtacctgc 120
cttaagaaac aagaaaactg cagtttatag agaaagaatt ttcaattttg acccatgtac 180
ttaaaatttt tggtgtatac tgcagtgtag caaatgtttt gtggtgacgg tataaatggt 240
actgtttgtt atcttggatt aagagtggct agagaagttg gaagacgtgt gagaagttct 300
ttatagagaa ttaaacatga aaattacatc tc 332
<210> 202
<211> 80
<212> ДНК
<213> Mus musculus
<400> 202
aacttgtgtt ctctgagagg aaaatactga agcagtagag aaatgacctg ctagagaata 60
aagttactgt taatgatacc 80
<210> 203
<211> 93
<212> ДНК
<213> Mus musculus
<400> 203
gatgtctgga atggagcaca cctgggatgt agcactcttg ctatctgtcc ggtccttttt 60
gttcaataaa gtcctgaggc aactctctct gtc 93
<210> 204
<211> 152
<212> ДНК
<213> Mus musculus
<400> 204
tgagctatga agttccggaa tttgtgtttt tcacagaaag ccttaccaac ttcagttact 60
ttaccaagac aatgtaatta ttgtttgatt ttataaagtc tacaacaatg atctcctatt 120
ttggtgtcag tttttcaata aagttttaat ta 152
<210> 205
<211> 1841
<212> ДНК
<213> Mus musculus
<400> 205
agcaggagca ggttttccaa aagcacccct cggaagtgtt tttgtcgtcg tttaaaatta 60
tcaagtatct tcagagaaga ttattttctg ccttcagaaa ctgaaggaag gcttgggcct 120
agagaacgac agtaaggtgc gagcaccgga gacacttaac acagctcagt ccatggaagg 180
acgagttccc tcattggctg cctgtctcga aatccacgca agctgtggag gaaagaatta 240
ccctgctcat cctgccttct atcttggtgt ttaatgttgg gtgggcaaca agcacaaacc 300
tccctcccac cccctccaag actgttagag cagtgggcca gaccaagcgg cgcacttgaa 360
catggatcaa gagggtcccg gttttacttt ttatttttgt cagggtaggc agtcttgtgt 420
ttgctttgtt caaagcaggg tctccctgtt ggccctggct ggccttatac tccacagcag 480
tcctgcctcc tcctcctagg tgctgggatt aaaggcgtgc gccaccacgc ccggctacag 540
cctgcatttt tatgcacatt ggtctgttaa gctagttgca ttctgtgcta ccggagggga 600
ctgaagttta atcacttgtc ttctattaaa aactagtgtt tgcctgggcc tggtgtgtat 660
acctttagtt gcagcgcttg ggaggcagag gcaggcagac ttcatgagtt cagggacagg 720
caagcctgct ctacagattt ccaggatacc cagggctaca catagagaaa atgttaaaga 780
taaacaaaaa gctggacagt gggggagcac acctttaatc ccagcactcg ggaggcagag 840
gcaggcggat ttctgagttc gaggccagcc tggtctacag agtgagttcc aggacagcca 900
ggactacaca gagaaaccct gtctcagaaa aaaaacaaaa caaaaaccaa tgcagtgata 960
gattgttgtt tcctaaacca catgtaccca ggaaatgccc actaaatttc acctggatca 1020
gtgttaactg atcattggga aatgaggtga ccaaaaatgc atcgcaacct tggacaaaca 1080
gcatggctat ttaacattct gggatcctgc agaatcctgc atcttcctaa gtagggaagc 1140
actgtagcat tggagagagg cctgggcgag cagagctaag gcttccattt ctggcttgct 1200
tggaatttaa aacaagcttt tttctatata gtaaaagatt gtttttaaga tttttgcgtg 1260
tgagtacatg ccaaagtagc caggaagtgt cacttgccct ggagctagaa ttactggcaa 1320
atgaaggctc agaggtggat gctgggacca attctaggcc ctctgagaaa gcaggtgcac 1380
ttggcttgtg cctccagccc caaaggcgat ggcttattgt gagcctgagg ccagccaggg 1440
ttacagagac tcaagaaaca agtggggttg tccatgttgc tggagatgac ccaggtctat 1500
taggaccttg actacatgga tagacattct ggcagctagt atactgccat tggggcctat 1560
ggagaagtag ccaccgagcc tatttagaaa gaaggactgc tggcaagctt ggtgtcacta 1620
tgaaggcaga caaagatcga tgtattaacg accccgactc caaaaacact cgagggggcc 1680
caaggtgggc tcagtggtta agagccgttc gcccaggggc tggagagttg gctcagtggt 1740
accacatggt ggctcacaac catctgtaat gagatctgac gccctcttct ggtgtatctg 1800
aagacagcta cagtgtactt acataaaata aataaatctt t 1841
<210> 206
<211> 2035
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность, кодирующая мРНК rpl32 - PpLuc(GC) - альбумин - A64 - C30 - гистонSL
<400> 206
ggggcgctgc ctacggaggt ggcagccatc tccttctcgg catcaagctt gaggatggag 60
gacgccaaga acatcaagaa gggcccggcg cccttctacc cgctggagga cgggaccgcc 120
ggcgagcagc tccacaaggc catgaagcgg tacgccctgg tgccgggcac gatcgccttc 180
accgacgccc acatcgaggt cgacatcacc tacgcggagt acttcgagat gagcgtgcgc 240
ctggccgagg ccatgaagcg gtacggcctg aacaccaacc accggatcgt ggtgtgctcg 300
gagaacagcc tgcagttctt catgccggtg ctgggcgccc tcttcatcgg cgtggccgtc 360
gccccggcga acgacatcta caacgagcgg gagctgctga acagcatggg gatcagccag 420
ccgaccgtgg tgttcgtgag caagaagggc ctgcagaaga tcctgaacgt gcagaagaag 480
ctgcccatca tccagaagat catcatcatg gacagcaaga ccgactacca gggcttccag 540
tcgatgtaca cgttcgtgac cagccacctc ccgccgggct tcaacgagta cgacttcgtc 600
ccggagagct tcgaccggga caagaccatc gccctgatca tgaacagcag cggcagcacc 660
ggcctgccga agggggtggc cctgccgcac cggaccgcct gcgtgcgctt ctcgcacgcc 720
cgggacccca tcttcggcaa ccagatcatc ccggacaccg ccatcctgag cgtggtgccg 780
ttccaccacg gcttcggcat gttcacgacc ctgggctacc tcatctgcgg cttccgggtg 840
gtcctgatgt accggttcga ggaggagctg ttcctgcgga gcctgcagga ctacaagatc 900
cagagcgcgc tgctcgtgcc gaccctgttc agcttcttcg ccaagagcac cctgatcgac 960
aagtacgacc tgtcgaacct gcacgagatc gccagcgggg gcgccccgct gagcaaggag 1020
gtgggcgagg ccgtggccaa gcggttccac ctcccgggca tccgccaggg ctacggcctg 1080
accgagacca cgagcgcgat cctgatcacc cccgaggggg acgacaagcc gggcgccgtg 1140
ggcaaggtgg tcccgttctt cgaggccaag gtggtggacc tggacaccgg caagaccctg 1200
ggcgtgaacc agcggggcga gctgtgcgtg cgggggccga tgatcatgag cggctacgtg 1260
aacaacccgg aggccaccaa cgccctcatc gacaaggacg gctggctgca cagcggcgac 1320
atcgcctact gggacgagga cgagcacttc ttcatcgtcg accggctgaa gtcgctgatc 1380
aagtacaagg gctaccaggt ggcgccggcc gagctggaga gcatcctgct ccagcacccc 1440
aacatcttcg acgccggcgt ggccgggctg ccggacgacg acgccggcga gctgccggcc 1500
gcggtggtgg tgctggagca cggcaagacc atgacggaga aggagatcgt cgactacgtg 1560
gccagccagg tgaccaccgc caagaagctg cggggcggcg tggtgttcgt ggacgaggtc 1620
ccgaagggcc tgaccgggaa gctcgacgcc cggaagatcc gcgagatcct gatcaaggcc 1680
aagaagggcg gcaagatcgc cgtgtaagac tagtgcatca catttaaaag catctcagcc 1740
taccatgaga ataagagaaa gaaaatgaag atcaatagct tattcatctc tttttctttt 1800
tcgttggtgt aaagccaaca ccctgtctaa aaaacataaa tttctttaat cattttgcct 1860
cttttctctg tgcttcaatt aataaaaaat ggaaagaacc tagatctaaa aaaaaaaaaa 1920
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa atgcatcccc 1980
cccccccccc cccccccccc cccccccaaa ggctcttttc agagccacca gaatt 2035
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ИСКУССТВЕННАЯ МОДИФИКАЦИЯ ГЕНОМА ДЛЯ РЕГУЛЯЦИИ ЭКСПРЕССИИ ГЕНА | 2018 |
|
RU2767201C2 |
| ИСКУССТВЕННЫЕ МОЛЕКУЛЫ НУКЛЕИНОВОЙ КИСЛОТЫ | 2016 |
|
RU2772349C2 |
| ПОСЛЕДОВАТЕЛЬНОСТИ 3'-UTR ДЛЯ СТАБИЛИЗАЦИИ РНК | 2016 |
|
RU2783165C1 |
| МОЛЕКУЛЫ ИСКУССТВЕННОЙ НУКЛЕИНОВОЙ КИСЛОТЫ, СОДЕРЖАЩИЕ 5'UTR ГЕНА TOP | 2013 |
|
RU2660565C2 |
| ПОСЛЕДОВАТЕЛЬНОСТИ 3'-UTR ДЛЯ СТАБИЛИЗАЦИИ РНК | 2016 |
|
RU2720934C2 |
| СЛИТЫЙ БЕЛОК НА ОСНОВЕ АНТИТЕЛА К ЕТА И TGF-β TRAP, СОДЕРЖАЩАЯ ЕГО ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2832190C2 |
| ИСКУССТВЕННЫЕ МОЛЕКУЛЫ НУКЛЕИНОВЫХ КИСЛОТ ДЛЯ УЛУЧШЕННОЙ ЭКСПРЕССИИ БЕЛКОВ И ПЕПТИДОВ | 2013 |
|
RU2750261C2 |
| ИСКУССТВЕННЫЕ МОЛЕКУЛЫ НУКЛЕИНОВЫХ КИСЛОТ ДЛЯ УЛУЧШЕННОЙ ЭКСПРЕССИИ БЕЛКОВ ИЛИ ПЕПТИДОВ | 2013 |
|
RU2658490C2 |
| ВАРИАНТЫ HSD17B13 И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2760851C2 |
| ДНК С ЗАМКНУТЫМИ КОНЦАМИ (зкДНК) И ПРИМЕНЕНИЕ В СПОСОБАХ УМЕНЬШЕНИЯ ИММУННОГО ОТВЕТА, СВЯЗАННОГО С ГЕННОЙ ТЕРАПИЕЙ ИЛИ ТЕРАПИЕЙ НУКЛЕИНОВОЙ КИСЛОТОЙ | 2020 |
|
RU2829367C2 |



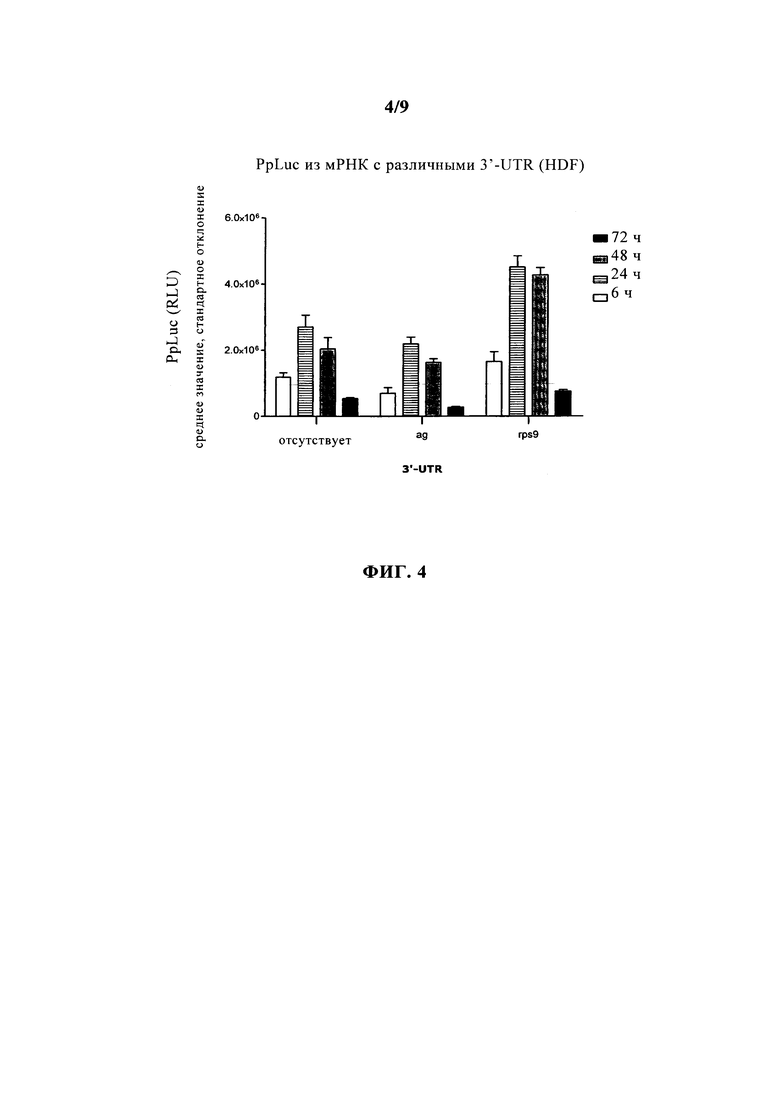

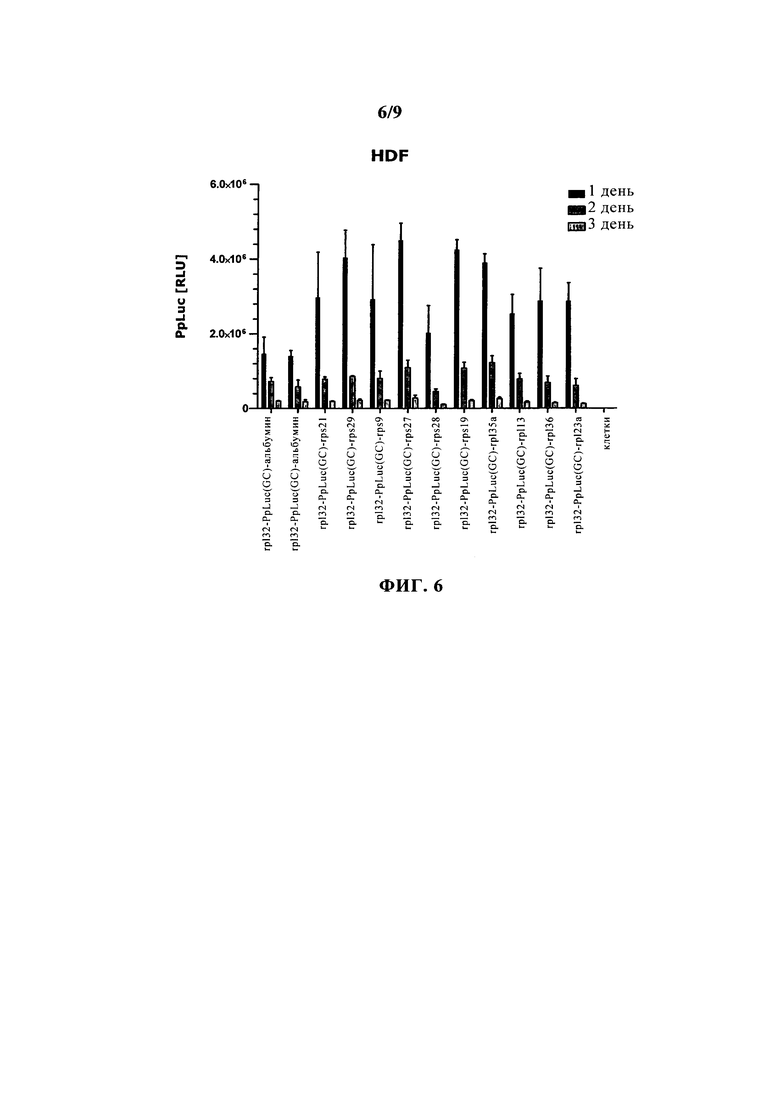
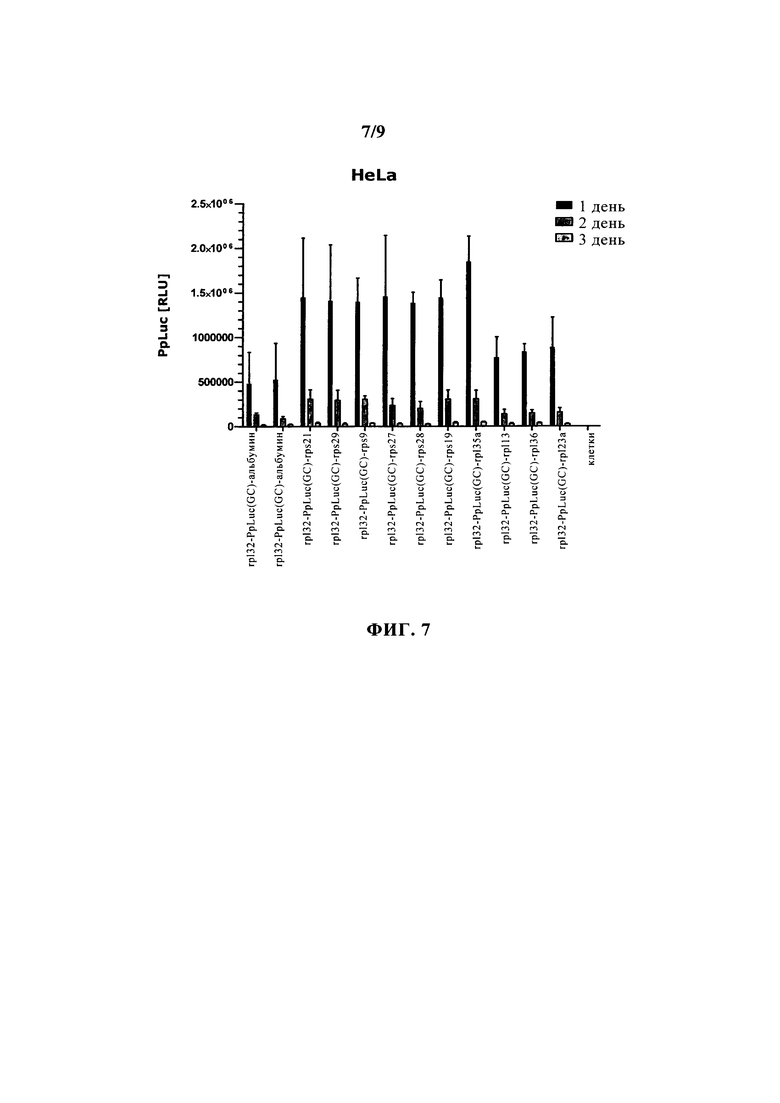
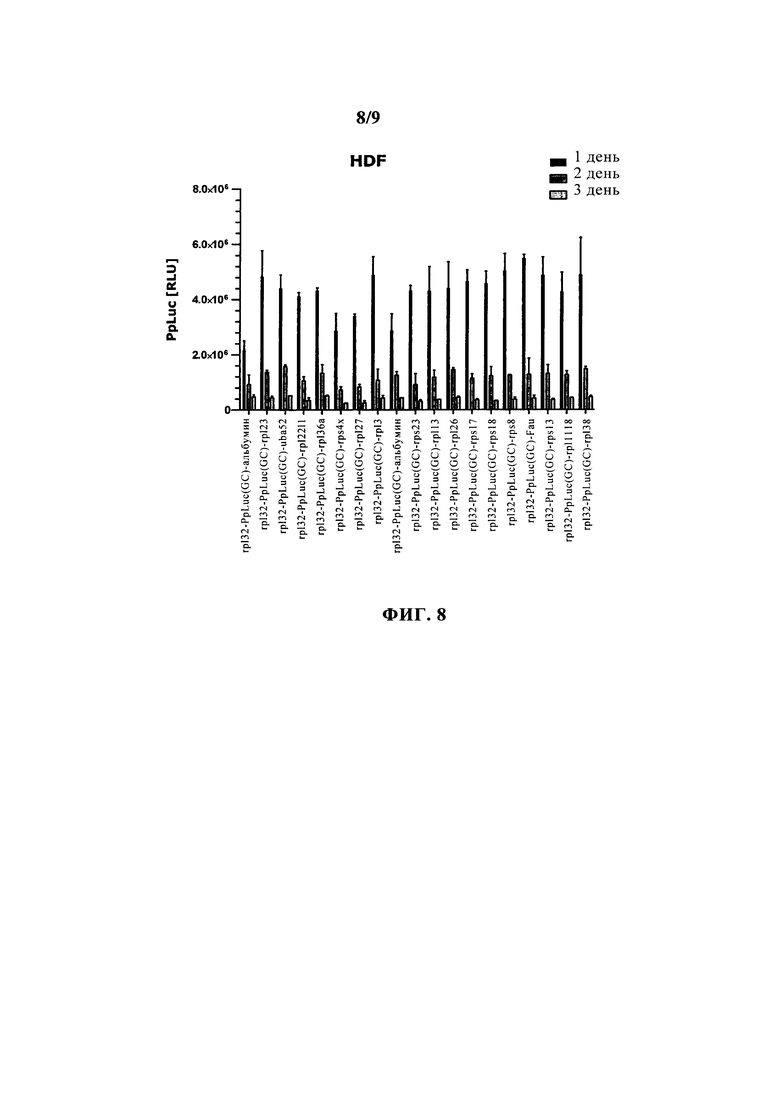
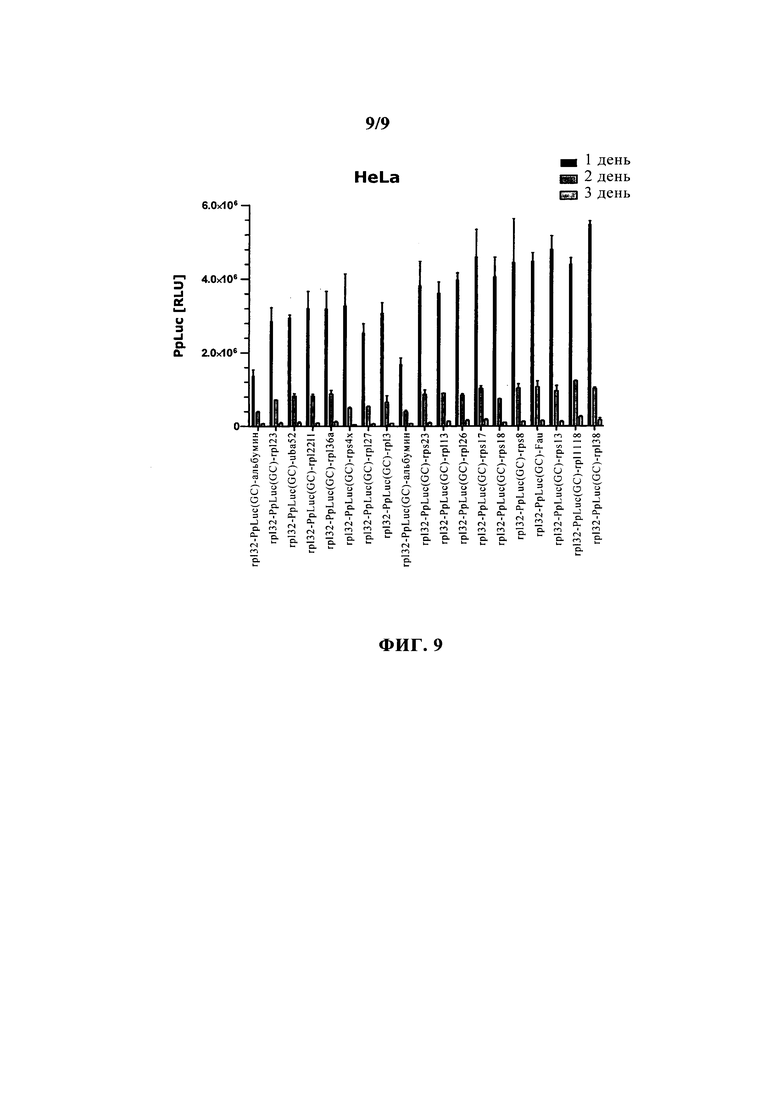
Изобретение относится к биотехнологии. Описана искусственная молекула нуклеиновой кислоты для генетической терапии или генетической вакцинации, содержащая по меньшей мере одну открытую рамку считывания (ОРС), кодирующую терапевтический пептид; и по меньшей мере один элемент 3'-нетранслируемой области (3'-UTR), который содержит нуклеотидную последовательность, происходящую из 3'-UTR гена рибосомального белка или из функционального варианта 3'-UTR гена рибосомального белка, в котором указанный функциональный вариант по меньшей мере на 80% идентичен указанному 3'-UTR гену рибосомального белка; и 5'-нетранслируемая область (5'-UTR), как дополнительного 5'-конце элемента, где 5'-UTR - это 5'TOP-UTR, который не содержит 5'-ТОР мотив. В заявке описана искусственная молекула нуклеиновой кислоты, содержащая по меньшей мере одну открытую рамку считывания и по меньшей мере один элемент 3'-нетранслируемой области (3'-UTR), который содержит нуклеотидную последовательность, происходящую из 3'-UTR гена рибосомального белка. Изобретение может быть использовано в генной терапии и/или генетической вакцинации. 9 н. и 15 з.п. ф-лы, 9 ил., 5 табл., 1 пр.
1. Искусственная молекула нуклеиновой кислоты для генетической терапии или генетической вакцинации, содержащая
а. по меньшей мере одну открытую рамку считывания (ОРС), кодирующую терапевтический пептид; и
б. по меньшей мере один элемент 3'-нетранслируемой области (3'-UTR), который содержит нуклеотидную последовательность, происходящую из 3'-UTR гена рибосомального белка или из функционального варианта 3'-UTR гена рибосомального белка, в котором указанный функциональный вариант по меньшей мере на 80% идентичен указанному 3'-UTR гену рибосомального белка; и
в. 5'-нетранслируемая область (5'-UTR), как дополнительного 5'-конце элемента, где 5'-UTR - это 5'TOP-UTR, который не содержит 5'-ТОР мотив.
2. Искусственная молекула нуклеиновой кислоты по п. 1, в которой по меньшей мере один 3'-UTR-элемент повышает, стабилизирует и/или пролонгирует получение белка с указанной искусственной молекулы нуклеиновой кислоты.
3. Искусственная молекула нуклеиновой кислоты по п. 1 или 2, в которой 3'-UTR гетерологична ОРС.
4. Искусственная молекула нуклеиновой кислоты по любому из пп. 1-3, в которой по меньшей мере один 3'-UTR-элемент содержит нуклеотидную последовательность, происходящую из 3'-UTR гена эукариотического рибосомального белка, предпочтительно из 3'-UTR гена рибосомального белка позвоночного, более предпочтительно из 3'-UTR гена рибосомального белка млекопитающего, еще более предпочтительно из 3'-UTR гена рибосомального белка примата, в частности гена рибосомального белка человека, или гена рибосомального белка грызуна, в частности гена рибосомального белка мыши.
5. Искусственная молекула нуклеиновой кислоты по любому из пп. 1-4, в которой ОРС не кодирует репортерный ген или не происходит из репортерного гена, где репортерный ген предпочтительно не выбирают из группы, состоящей из белков глобина, прежде всего бета-глобина, белка люциферазы, белков бета-глюкуронидазы (GUS) и GFP или их вариантов, предпочтительно не из EGFP, или вариантов, последовательности которых идентичны по меньшей мере на 70% последовательности белка глобина, белка люциферазы или белка GFP;
ОРС не кодирует рибосомальный белок, предпочтительно не эукариотический рибосомальный белок, где рибосомальный белок предпочтительно не выбран из рибосомального белка S6 (RPS6), рибосомального белка L36a-подобного (RPL36AL) или рибосомального белка S16 (RPS16); или
3'-UTR не происходит из вируса.
6. Искусственная молекула нуклеиновой кислоты по любому из пп. 1-5, в которой 3'-UTR, предпочтительно искусственная молекула нуклеиновой кислоты, не содержит поли(А)-последовательность или сигнал полиаденилирования, или
3'-UTR, предпочтительно искусственная молекула нуклеиновой кислоты, содержит поли(А)-последовательность или сигнал полиаденилирования, где
поли(А)-последовательность или сигнал полиаденилирования предпочтительно локализован на 3'-конце 3'-UTR-элемента.
7. Искусственная молекула нуклеиновой кислоты по любому из пп. 1-6, в которой по меньшей мере один 3'-UTR-элемент содержит нуклеотидную последовательность, происходящую из 3'-UTR последовательности, выбранной из группы, которая состоит из рибосомального белка L9 (RPL9), рибосомального белка L3 (RPL3), рибосомального белка L4 (RPL4), рибосомального белка L5 (RPL5), рибосомального белка L6 (RPL6), рибосомального белка L7 (RPL7), рибосомального белка L7a (RPL7A), рибосомального белка L11 (RPL11), рибосомального белка L12 (RPL12), рибосомального белка L13 (RPL13), рибосомального белка L23 (RPL23), рибосомального белка L18 (RPL18), рибосомального белка L18a (RPL18A), рибосомального белка L19 (RPL19), рибосомального белка L21 (RPL21), рибосомального белка L22 (RPL22), рибосомального белка L23a (RPL23A), рибосомального белка L17 (RPL17), рибосомального белка L24 (RPL24), рибосомального белка L26 (RPL26), рибосомального белка L27 (RPL27), рибосомального белка L30 (RPL30), рибосомального белка L27a (RPL27A), рибосомального белка L28 (RPL28), рибосомального белка L29 (RPL29), рибосомального белка L31 (RPL31), рибосомального белка L32 (RPL32), рибосомального белка L35a (RPL35A), рибосомального белка L37 (RPL37), рибосомального белка L37a (RPL37A), рибосомального белка L38 (RPL38), рибосомального белка L39 (RPL39), рибосомального белка, большой субъединицы, Р0 (RPLP0), рибосомального белка, большой субъединицы, P1 (RPLP1), рибосомального белка, большой субъединицы, Р2 (RPLP2), рибосомального белка S3 (RPS3), рибосомального белка S3A (RPS3A), рибосомального белка S4, X-сцепленного (RPS4X), Y-сцепленного 1 (RPS4Y1), рибосомального белка S5 (RPS5), рибосомального белка S6 (RPS6), рибосомального белка S7 (RPS7), рибосомального белка S8 (RPS8), рибосомального белка S9 (RPS9), рибосомального белка S10 (RPS10), рибосомального белка S11 (RPS11), рибосомального белка S12 (RPS12), рибосомального белка S13 (RPS13), рибосомального белка S15 (RPS15), рибосомального белка S15a (RPS15A), рибосомального белка S16 (RPS16), рибосомального белка S19 (RPS19), рибосомального белка S20 (RPS20), рибосомального белка S21 (RPS21), рибосомального белка S23 (RPS23), рибосомального белка S25 (RPS25), рибосомального белка S26 (RPS26), рибосомального белка S27 (RPS27), рибосомального белка S27a (RPS27a), рибосомального белка S28 (RPS28), рибосомального белка S29 (RPS29), рибосомального белка L15 (RPL15), рибосомального белка S2 (RPS2), рибосомального белка L14 (RPL14), рибосомального белка S14 (RPS14), рибосомального белка L10 (RPL10), рибосомального белка L10a (RPL10A), рибосомального белка L35 (RPL35), рибосомального белка L13a (RPL13A), рибосомального белка L36 (RPL36), рибосомального белка L36a (RPL36A), рибосомального белка L41 (RPL41), рибосомального белка S18 (RPS18), рибосомального белка S24 (RPS24), рибосомального белка L8 (RPL8), рибосомального белка L34 (RPL34), рибосомального белка S17 (RPS17), рибосомального белка SA (RPSA), продукта 1 слияния остаток убиквитина А-52-рибосомальный белок (UBA52), повсеместно экспрессируемого (FAU) вируса мышиной саркомы Финкеля-Бискиса-Рейли (FBR-MuSV), белка 1, подобного рибосомальному белку L22 (RPL22L1), подобного рибосомальному белку L39 (RPL39L), белка, подобного рибосомальному белку L10 (RPL10L), белка, подобного рибосомальному белку L36a (RPL36AL), белка, подобного рибосомальному белку L3 (RPL3L), белка, подобного рибосомальному белку S27 (RPS27L), белка 1, подобного рибосомальному белку L26 1 (RPL26L1), белка 1, подобного рибосомальному белку L7 (RPL7L1), псевдогена рибосомального белка L13a (RPL13AP), псевдогена 8 рибосомального белка L37a (RPL37AP8), псевдогена 5 рибосомального белка S10 (RPS10P5), псевдогена 11 рибосомального белка S26 (RPS26P11), псевдогена 5 рибосомального белка L39 (RPL39P5), псевдогена 6 рибосомального белка, большой субъединицы, Р0 (RPLP0P6) и псевдогена 14 рибосомального белка L36 (RPL36P14).
8. Искусственная молекула нуклеиновой кислоты по любому из пп. 1-7, в которой по меньшей мере один 3,-UTR-элемент содержит или состоит из нуклеотидной последовательности, идентичной по меньшей мере примерно на 80%, более предпочтительно по меньшей мере примерно на 90%, еще более предпочтительно по меньшей мере примерно на 95%, еще более предпочтительно по меньшей мере примерно на 99% нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID NO: 10-205.
9. Искусственная молекула нуклеиновой кислоты по любому из пп. 1-8, содержащая 5'-кэп-структуру, поли(С)-последовательность, гистоновую структуру типа «стебель-петля» и/или IRES-мотив.
10. Искусственная молекула нуклеиновой кислоты по п. 9, в которой гистоновая структура типа «стебель-петля» содержит последовательность SEQ ID NO: 5.
11. Искусственная молекула нуклеиновой кислоты по любому из пп. 1-10, где искусственная молекула нуклеиновой кислоты, предпочтительно ОРС, имеет по меньшей мере частично модифицированное содержание G/C, где предпочтительно содержание G/C в ОРС увеличено по сравнению с ОРС дикого типа, и/или ОРС содержит область с оптимизированными кодонами, в которой предпочтительно ОРС имеет оптимизированные кодоны.
12. Искусственная молекула нуклеиновой кислоты по любому из пп. 1-11, которая представляет собой РНК, предпочтительно молекулу мРНК.
13. Вектор экспрессии, содержащий искусственную молекулу нуклеиновой кислоты по любому из пп. 1-12.
14. Вектор по п. 13, представляющий собой плазмидный вектор или вирусный вектор, предпочтительно плазмидный вектор.
15. Клетка для получения искусственной молекулы нуклеиновой кислоты любой из пп. 1-12 или кодируемого таким образом пептида, содержащая вектор п. 13 или 14.
16. Фармацевтическая композиция для генетической терапии или генетической вакцинации, содержащая эффективное количество искусственной молекулы нуклеиновой кислоты по одному из пп. 1-12, вектора по п. 13 или 14, или клетки по п. 15 и один или несколько фармацевтически приемлемых наполнителей, разбавителей и/или эксципиентов, и/или один или несколько адъювантов.
17. Применение искусственной молекулы нуклеиновой кислоты по любому из пп. 1-12, вектора по п. 13 или 14, клетки по п. 15 или фармацевтической композиции по п. 16 для получения лекарственного средства для генетической терапии или генетической вакцинации.
18. Применение искусственной молекулы нуклеиновой кислоты по любому из пп. 1-12, вектора по п. 13 или 14, клетки по п. 15 или фармацевтической композиции по п. 16 для генетической терапии или генетической вакцинации.
19. Способ увеличения продукции белка из искусственной молекулы нуклеиновой кислоты in vitro, включающий стадию ассоциации молекулы нуклеиновой кислоты с 3'-UTR-элементом, как определено в любом из пп. 1-8.
20. Способ по п. 19, в котором искусственная молекула нуклеиновой кислоты представляет собой молекулу мРНК.
21. Применение 3'-UTR-элемента, как определено в любом из пп. 1-8, для увеличения продукции белка из молекулы нуклеиновой кислоты или вектора in vitro.
22. Применение по п. 21, в котором искусственная молекула нуклеиновой кислоты представляет собой молекулу мРНК.
23. Набор для генетической терапии или генетической вакцинации, содержащий искусственную молекулу нуклеиновой кислоты по любому из пп. 1-12, вектор по п. 13 или 14, клетку по п. 15 и фармацевтическую композицию по п. 16.
24. Набор по п. 23, содержащий инструкции по применению, клетки для трансфекции, адъювант, средства для введения фармацевтической композиции, фармацевтически приемлемый носитель и/или фармацевтически приемлемый раствор для растворения или разведения искусственной молекулы нуклеиновой кислоты, вектора, клеток или фармацевтической композиции.
| LEDDA M., Effect of 3'UTR length on the translational regulation of 5'-terminal oligopyrimidine mRNAs, Gene, 2005, v.344, p.213-220 | |||
| WO 2007068265 A1, 21.06.2007 | |||
| WO 2001007590 A2, 01.02.2001 | |||
| ПАРОВАЯ ФОРСУНКА | 1927 |
|
SU10056A1 |

