2420-539339RU/019
РАСТЕНИЯ, ЭКСПРЕССИРУЮЩИЕ ФЕРМЕНТЫ, ДЕГРАДИРУЮЩИЕ КЛЕТОЧНУЮ СТЕНКУ, И ВЕКТОРЫ ЭКСПРЕССИИ
По настоящей заявке испрашивается приоритет временной заявки США No. 61/280635, поданной 6 ноября 2009 года и временной заявки США No. 61/398589, поданной 28 июня 2010 года, обе из которых включены в настоящее описание в качестве ссылок в полном объеме. Также данная заявка является частичным продолжением заявки США No. 12/590444, поданной 6 ноября 2009 года, которая включена в настоящее описание в качестве ссылки в полном объеме.
Список последовательностей, поданный в электронной форме вместе с данной заявкой под названием "Список последовательностей", который был создан 5 ноября 2010 года и имеет размер 2215456 байт, включен в настоящее описание в качестве ссылки в полном объеме.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Изобретение, раскрытое в настоящем описании, относится к растениям, экспрессирующим ферменты, деградирующие клеточную стенку, векторам, нуклеиновым кислотам, белкам, связанным с ними способам и их применениям.
УРОВЕНЬ ТЕХНИКИ
Гидролитические ферменты имеют важное промышленное и сельскохозяйственное применение, однако их экспрессия и продуцирование могут быть связаны с неблагоприятными фенотипическими эффектами, зависящими от экспрессирующего хозяина. В частности, экспрессия ферментов, деградирующих клеточную стенку, таких как целлюлазы, ксиланазы, лигниназы, эстеразы, пероксидазы и другие гидролитические ферменты, часто связана с неблагоприятными эффектами на рост, физиологические и агрономические характеристики при экспрессии в растениях. Некоторые из этих ферментов также могут плохо экспрессироваться в микробных хозяевах вследствие их гидролитической активности.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В одном из аспектов изобретение относится к трансгенному растению, включающему нуклеиновую кислоту, кодирующую аминокислотную последовательность, которая по меньшей мере на 90% идентична последовательности, выбранной из SEQ ID NO:44-115.
В одном из аспектов изобретение относится к трансгенному растению, включающему первую нуклеиновую кислоту, которая способна гибридизоваться в условиях умеренной жесткости со второй нуклеиновой кислотой, состоящей из нуклеотидной последовательности, выбранной из SEQ ID NO:116-187, или комплементарной ей последовательности.
В одном из аспектов изобретение относится к вектору, включающему первую нуклеиновую кислоту, способную гибридизоваться в условиях низкой, умеренной или высокой жесткости со второй нуклеиновой кислоты, состоящей из любой последовательности SEQ ID NO:116-187.
В одном из аспектов изобретение относится к вектору, включающему нуклеиновую кислоту, имеющую последовательность, которая по меньшей мере на 90% идентична эталонной последовательности, выбранной из SEQ ID NO:188-283.
В одном из аспектов изобретение относится к способу переработки растительной биомассы. Способ включает предварительную обработку растения или его части путем смешения растения или его части с жидкостью с образованием смеси, имеющей отношение жидкости к твердому веществу, меньшее или равное 15. Предварительная обработка также включает предоставление условий для поддержания смеси при температуре, меньшей или равной 100°C. Также способ включает предоставление одного или нескольких ферментов для модификации по меньшей мере одного компонента растения или его части.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Файл патента или заявки содержит по меньшей мере один чертеж, выполненный в цвете. Копии этой публикации патента или патентной заявки с цветным чертежом(ами) будут предоставлены ведомством при запросе и оплате необходимой пошлины.
Представленное ниже подробное описание предпочтительного варианта осуществления настоящего изобретения будет лучше понятным при прочтении совместно с прилагаемыми чертежами. Для цели иллюстрации изобретения на чертежах представлены варианты осуществления, которые в настоящее время являются предпочтительными. Однако понятно, что изобретение не ограничивается точными представленными схемами и инструментарием. На чертежах:
На фиг. 1 проиллюстрирована карта вектора pSB11.
На фиг. 2A проиллюстрирована карта вектора AG1000.
На фиг. 2B проиллюстрирована карта вектора pAG1001.
На фиг. 2C проиллюстрирована карта вектора pAG1002.
На фиг. 3A проиллюстрирована карта вектора pAG1003.
На фиг. 3B проиллюстрирована карта вектора pAG2000.
На фиг. 3C проиллюстрирована карта вектора pAG2004.
На фиг. 4 проиллюстрирована карта вектора pAG2014.
На фиг. 5 проиллюстрирована карта вектора pBSK:OsUbi3P:XmaI:AvrII:NosT.
На фиг. 6 проиллюстрирована карта вектора pBSK:OsUbi3P:XmaI:AvrII:NosT:L1.
На фиг. 7 проиллюстрирована удельная активность трех ксиланаз с регистрационными номерами P40942, P77853 и О30700.
На фиг. 8 проиллюстрирована активность различных образцов трансгенных растений, экспрессирующих ксиланазу P77853.
На фиг. 9 проиллюстрированы анализ температурной стабильности для О30700, P77853 и P40942.
На фиг. 10 проиллюстрирована технологическая схема процесса в макромасштабе.
На фиг. 11 проиллюстрирована технологическая схема процесса в микромасштабе.
На фиг. 12 представлены выходы глюкозы и ксилозы (процент от массы биомассы) после ферментативного гидролиза предварительно обработанной кукурузной соломы (2015.05 и 2004.8.4).
На фиг. 13 представлены выходы глюкозы и ксилозы (процент от массы биомассы) после ферментативного гидролиза предварительно обработанной кукурузной соломы (2004.8.4, 2063.13 и 2063.17).
На фиг. 14 представлены выходы глюкозы и ксилозы (процент от массы биомассы) после ферментативного гидролиза предварительно обработанной кукурузной соломы (2015.05 и 2004.8.4).
На фиг. 15 представлены выходы глюкозы и ксилозы (процент от массы биомассы) после ферментативного гидролиза предварительно обработанной кукурузной соломы (2064.17 и 2004.8.4).
На фиг. 16 проиллюстрирован выход глюкозы (процент от массы биомассы) после ферментативного гидролиза предварительно обработанной кукурузной соломы (2042.02, 2042.03, 2042.06 и 2004.8.4).
На фиг. 17A проиллюстрировано трансгенное растение, полученное с pAG3000.
На фиг. 17B проиллюстрировано трансгенное растение, полученное с pAG3001.
На фиг. 18A проиллюстрировано трансгенное растение, полученное с pAG2004.
На фиг. 18B проиллюстрирован початок из трансгенного растения, полученного с pAG2004.
На фиг. 18C проиллюстрирован початок из трансгенного растения, полученного с pAG2004.
На фиг. 19A проиллюстрировано трансгенное растение, полученное с pAG2005.
На фиг. 19B проиллюстрировано трансгенное растение, полученное с pAG2005.
На фиг. 20 проиллюстрировано измерение восстанавливающих сахаров в трансгенном растении события #15, трансформированном pAG2004.
На фиг. 21A проиллюстрировано трансгенное растение, полученное с pAG2016.
На фиг. 21B проиллюстрирован початок из трансгенного растения, полученного с pAG2016.
На фиг. 22 проиллюстрировано измерение восстанавливающих сахаров из трансгенных растений.
На фиг. 23 проиллюстрировано измерение ферментативной активности из высушенных увядающих образцов кукурузной соломы.
На фиг. 24 проиллюстрировано измерение активности фермента из образцов ткани листьев трансгенных растений, полученных с pAG2015, pAG2014 или pAG2004.
На фиг. 25A проиллюстрировано трансгенное растение, полученное с pAG2014.
На фиг. 25B проиллюстрировано трансгенное растение, полученное с pAG2014.
На фиг. 25C проиллюстрирован початок из трансгенного растения, полученного с pAG2014.
На фиг. 26A проиллюстрировано трансгенное растение, полученное с pAG2015.
На фиг. 26B проиллюстрировано трансгенное растение, полученное с pAG2015.
На фиг. 26C проиллюстрирован початок из трансгенного растения, полученного с pAG2015.
На фиг. 26D проиллюстрирован початок из трансгенного растения, полученного с pAG2015.
На фиг. 27A проиллюстрировано трансгенное растение, полученное с pAG2020.
На фиг. 27B проиллюстрировано трансгенное растение, полученное с pAG2020.
На фиг. 27C проиллюстрирован початок из трансгенного растения, полученного с pAG2020.
На фиг. 28A проиллюстрировано трансгенное растение, полученное с pAG2025.
На фиг. 28B проиллюстрировано трансгенное растение, полученное с pAG2025.
На фиг. 28C проиллюстрировано трансгенное растение, полученное с pAG2025.
На фиг. 29A проиллюстрировано трансгенное растение, полученное с pAG2017.
На фиг. 29B проиллюстрировано трансгенное растение, полученное с pAG2017.
На фиг. 29C проиллюстрирован початок из трансгенного растения, полученного с pAG2017.
На фиг. 29D проиллюстрирован початок из трансгенного растения, полученного с pAG2017.
На фиг. 30A проиллюстрировано трансгенное растение, полученное с pAG2019.
На фиг. 30B проиллюстрировано трансгенное растение, полученное с pAG2019, в сравнении с растением дикого типа.
На фиг. 31 проиллюстрировано трансгенное растение, полученное с pAG2019 или pAG2027, в сравнении с растением дикого типа. Левые три растения были получены с pAG2019. Правые три растения были получены с pAG2027.
На фиг. 32A проиллюстрировано два трансгенных растения, полученных с pAG2018, слева и два не экспрессирующих гидролазу растения справа.
На фиг. 32B проиллюстрировано трансгенное растение, полученное с pAG2018.
На фиг. 32C проиллюстрировано трансгенное растение, полученное с pAG2018.
На фиг. 33A проиллюстрировано трансгенное растение, полученное с pAG2026.
На фиг. 33B проиллюстрировано трансгенное растение, полученное с pAG2026.
На фиг. 33C проиллюстрировано трансгенное растение, полученное с pAG2026.
На фиг. 34A проиллюстрировано трансгенное растение, полученное с pAG2021.
На фиг. 34B проиллюстрировано трансгенное растение, полученное с pAG2021.
На фиг. 34C проиллюстрирован початок из трансгенного растения, полученного с pAG2021.
На фиг. 34D проиллюстрирован початок из трансгенного растения, полученного с pAG2021.
На фиг. 35A проиллюстрировано трансгенное растение, полученное с pAG2022.
На фиг. 35B проиллюстрировано трансгенное растение, полученное с pAG2022.
На фиг. 35C проиллюстрирован початок из трансгенного растения, полученного с pAG2022.
На фиг. 36A проиллюстрировано трансгенное растение, полученное с pAG2023.
На фиг. 36B проиллюстрировано трансгенное растение, полученное с pAG2023.
На фиг. 36C проиллюстрировано трансгенное растение, полученное с pAG2023.
На фиг. 37A проиллюстрировано трансгенное растение, полученное с pAG2024.
На фиг. 37B проиллюстрировано трансгенное растение, полученное с pAG2024.
На фиг. 37C проиллюстрировано трансгенное растение, полученное с pAG2024.
На фиг. 38 проиллюстрированы данные об активности для некоторых из событий pAG2021, вместе с измерениями для событий pAG2004 (отрицательные контроли для активности ксиланазы) и событий pAG20014 (положительный контроль для активности ксиланазы).
ПОДРОБНОЕ ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНЫХ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
Определенную терминологию используют в дальнейшем описании только для удобства и не для ограничения. Слова "правый", "левый", "верх" и "низ" обозначают направления на чертежах или в конкретных примерах, на которые приводится ссылка.
Форму единственного числа, как используют в формуле изобретения и в соответствующих частях описания, определяют как включающую один или несколько из упоминаемых объектов, если нет иных конкретных указаний. Выражение "по меньшей мере один" с последующим перечнем двух или более объектов, таким как "A, B или C" означает любой отдельный из A, B или C, а также любую их комбинацию.
Несмотря на потенциальные вредоносные эффекты ферментов на экспрессирующего хозяина, продуцирование ферментов в растениях, микроорганизмах и других организмах может создать большую экономическую пользу при продукции топлива, волокна, химических реагентов, сахара, текстиля, пульпы, бумаги и кормов для животных. В некоторых случаях существуют экономические преимущества для продукции ферментов в растениях, несмотря на агрономический или фенотипический эффект. Кроме того, некоторые фенотипические эффекты могут быть преодолены с использованием различных стратегий, которые защищают растение от активности фермента. Варианты осуществления, представленные в настоящем описании, включают но не ограничиваются ими, эти стратегии.
Стратегии для экспрессируемых в растениях ферментов могут быть зависимыми от культуры. Конкретный фермент может быть не важен, может иметь небольшую значимость или может быть значимым для экспресии в одной культуре, но значительную ценность или пользу для экспрессии в другой культуре. Значит, свойства модифицированного способами инженерии растения могут зависеть не только от конкретного фермента, но также от конкретного растения, которое экспрессирует фермент. Например, экспрессия ферментов ксиланаз в растениях может способствовать гидролизу гемицеллюлозы клеточной стенки растений и растительного волокна до ферментируемых сахаров (для продуцирования топлив и химических реагентов) или усвояемых сахаров (для кормов для животных и производства мяса). Однако конкретные ферменты ксиланазы также снижают выход зерна и могут вызывать бесплодие при экспрессии в кукурузе, препятствуя использованию этой культуры в качестве хозяина для экспрессии фермента. Несмотря на отрицательные эффекты на выход зерна и фертильность в кукурузе, которые могут снижать суммарную экономическую ценность полученного способами инженерии растения относительно не модифицированного способами инженерии растения, экспрессия идентичных ксиланаз в другой культуре, такой как просо, мискантус, сахарный тростник или сорго, в действительности может быть полезной, поскольку бесплодие в этих культурах будет препятствовать ауткроссингу гена ксиланазы и можно будет получать имеющие коммерческое значение количества ростков растений с использованием культуры ткани или вегетативного размножения. Хотя снижение фертильности, выхода зерна или сухого материала биомассы в кукурузе может препятствовать или снижать ценность экспрессии определенных ферментов ксиланаз, которые в ином случае были бы ценными в промышленности химической переработки и кормов для животных, экспрессия идентичных ферментов в просе, мискантусе, сорго или сахарном тростнике может не только обеспечить экономическую ценность, обеспечиваемую ферментом, но также может быть полезной с точки зрения нормативных актов и безопасности.
Аналогично, ценность фермента, экспрессируемого в одной ткани культуры, может отличаться при экспрессии в другой ткани или при экспрессии в той же ткани в другой культуре. Различная польза является следствием того, что конкретные ткани культуры (такие как зерно, семена, листья, стебли, корни, цветки, пыльца, и т.д.) могут иметь различную ценность, в зависимости от культуры и новых свойств, обеспечиваемых экспрессируемым ферментом. Конкретные ферменты ксиланазы и целлюлазы имеют существенные агрономические и фенотипические последствия при конститутивной экспрессии в кукурузе. Конститутивная экспрессия этих ферментов, по отдельности или в комбинации, часто приводит к недоразвитым растениям, бесплодным растениям или растением с более низким выходом и агрономическими характеристиками. Однако специфическая для семян экспрессия конкретных ферментов ксиланазы и целлюлазы может снизить или устранить какой-либо вредоносный агрономический эффект или снижение выхода или устранение какого-либо вредоносного агрономического эффекта или снижения выхода, все еще обеспечивая высокие уровни фермента. Это может быть полезным в зернах кукурузы. Продуцирование тех же ферментов в просе, мискантусе, сорго медвяном или сорго сахарном, или сахарном тростнике, где выход зерна на акр может быть значительно более низким по сравнению кукурузой, может приводить к отличающемуся профилю специфической для семян экспрессии ксиланазы или целлюлазы. Варианты осуществления включают экспрессию CWDE специфичным для семян образом в любом типе трансгенного растения. В зависимости от применения, такого как производство кормов для животных, мясной или молочной продукции, птицеводство, производство бумаги или производство ферментируемых сахаров, где содержащее фермент зерно может быть смешено с другим собранным в качестве урожая сырьем (предварительно обработанным или не обработанным предварительно), это может быть высоко эффективным путем предоставления благоприятных доз фермента в зернах кукурузы или в других зернах и семенах.
Суммарная экономическая ценность экспрессируемого в растении фермента может отличаться, в зависимости от того, где предполагается локализация и накопление фермента, и куда он нацелен. Например, конкретные ферменты ксиланазы и целлюлозы могут иметь фенотипические и агрономические эффекты при нацеливании на клеточную стенку растения, но небольшой эффект или не иметь эффекта при поддержании внутриклеточно или нацеливании в вакуоль. Это может обеспечивать экономическую пользу предоставления содержащегося внутриклеточно источника фермента для применений, где является желательным смешение фермента с субстратом. Напротив, хотя те же самые ферменты могут обеспечить ценность при применении в качестве добавки, таком как корма для животных или переработка предварительно обработанной биомассы, эти ферменты могут обеспечить небольшую ценность или не обеспечить ценности при применении для самопереработки, где нацеливание на клеточную стенку растения является предпочтительным для получения ферментируемых или усвояемых сахаров, но является проблематичным, поскольку оно приводит к фенотипическим или агрономическим эффектам.
Как описано выше, экзогенный фермент может экспрессироваться в конкретном растении, органе растения, ткани растения, клетки растения или субклеточной области или компартменте растения. Варианты осуществления также включают растение, в том числе экспрессирующее экзогенный фермент, где экзогенный фермент может быть во всем растении или он может быть локализован в области растения, в органе растения, в тканях растения или в субклеточной области или компартменте растения. Могут предусматриваться трансгенные растения, адаптированные к экзогенному CWDE или имеющие цитоплазматическое накопление экзогенного CWDE. Замысел, учитывающий, где в растении или в каком растении экспрессируется экзогенный фермент, может представлять собой, но не ограничиваться ими, замысел, в котором учитываются фенотипические, связанные с безопасностью, экономические и регуляторные вопросы, указанные выше.
В вариантах осуществления, представленных в настоящем описании, предусмотрены векторы для экспрессии белков в растениях. Белки могут представлять собой ферменты и ферменты могут представлять собой, но не ограничиваться ими, ферменты, деградирующие клеточную стенку. Предусмотрен ряд растений, предназначенных для экспрессии конкретных ферментов, деградирующих клеточную стенку. Растения могут иметь промышленное и/или сельскохозяйственное применения. Предусматриваются способы и материалы для получения векторов экспрессии и получения растений. Также предусматриваются способы, для которых можно использовать растения в промышленных и сельскохозяйственных применениях.
Предусматриваются векторы для экспрессии в растениях либо фермента, деградирующего клеточную стенку (или CWDE), либо варианта в виде модифицированного интеином CWDE. В одном из вариантов осуществления вектор пригоден для трансформации двудольного растения. В одном из вариантов осуществления вектор подходит для трансформации однодольного растения. CWDE, из которых может быть выбран CWDE в векторе или растении может быть выбран из, но не ограничиваясь ими, ксиланазы, целлюлазы, целлобиогидролазы, глюкозидазы, ксилозидазы, арабинофуранозидазы и эстеразы феруловой кислоты. В одном из вариантов осуществления кодирующая CWDE последовательность прерывается вследствие встраивания последовательности интеина. Встроенная последовательность интеина может инактивировать функцию соответствующего CWDE. В одном из вариантов осуществления созданный вектор позволяет встраивание по меньшей мере от трех до четырех кассет для экспрессии гена и/или подавления гена. Каждая кассета может включать CWDE или модифицированный интеином CWDE.
В одном из вариантов осуществления генетические элементы, используемые в векторе, представленном в настоящем описании, или в его конструкции, могут обеспечить по меньшей мере одно из следующих свойств: возможность выбрать трасгенные события после трансформации растения, возможность влиять на оптимальный уровень экспрессии генов в клетках или влиять на желаемое субклеточное нацеливание фермента. Векторы могут содержать селективный маркер, который может представлять собой, но не ограничиваться ими, ген фосфоманнозоизомеразы (PMI) E. coli. Другие селективные маркеры, которые могут быть включены в дополнение к маркеру PMI или вместо него, представляют собой маркеры, известные в данной области (такие как, но не ограничиваясь ими, EPSPS, BAR, npt-II, GUS и т.д.). Векторы также могут включать один или несколько промоторов. Промоторы могут представлять собой конститутивные или глобальные, тканеспецифические, специфические для семян, специфические для листьев, специфические для органов, специфические для субклеточной области или компартмента, или специфические для стадии развития промоторы. Предпочтительные промоторы включают промотор гена убиквитина 3 риса (OsUbi3P) с первым интроном (регистрационный номер No. AY954394, SEQ ID NO:1) или промотор гена актина 1 риса (регистрационный номер No. S44221, SEQ ID NO:2). Также можно использовать другие конститутивные промоторы, такие как, но не ограничиваясь ими, промотор убиквитина маиса (SEQ ID NO:3), и ими можно заменять промотор OsUbi3P или промотор актина 1 риса. Промоторы убиквитина 3 и гена актина 1 риса являются конститутивными и глобальными промоторами, которые можно использовать для обеспечения экспрессии генов в трансгенных растениях. Также в векторах может быть предусмотрен промотор глутелина из гена GluB-4 риса (регистрационный номер No. AY427571, SEQ ID NO:4) с его собственной сигнальной последовательностью. Промотор глутелина представляет собой специфический для семян промотор. В векторах могут присутствовать другие специфические для семян промоторы (такие как, но не ограничиваясь ими, промотор Zc2 maize zein, SEQ ID NO:5). Для доставки ферментов к их соответствующим субстратом или в места для высокого уровня накопления фермента, такие как вакуоли, в векторе могут быть предусмотрены различные нацеливающие сигнальные последовательности. Нацеливающие сигнальные последовательности, которые могут присутствовать в CWDE или векторе, кодирующем CWDE, включают, но не ограничиваются ими, PR1a (SEQ ID NO:6, кодируемая последовательностью нуклеиновой кислоты SEQ ID NO:7), BAASS (SEQ ID NO:8, кодируемая последовательностью нуклеиновой кислоты SEQ ID NO:9), и алеураин ячменя (SEQ ID NO:10, кодируемая нуклеиновой кислотой SEQ ID NO:11). Другие нацеливающие последовательности, которые могут быть включены, включают, но не ограничиваются ими, последовательность удержания в эндоплазматической сети (ER) SEKDEL (SEQ ID NO:12, кодируемая нуклеиновой кислотой SEQ ID NO:13), и укороченную последовательность KDEL (SEQ ID NO:10, кодируемую нуклеиновой кислотой SEQ ID NO:16). Ферменты могут быть предоставлены без нацеливающей последовательности. Ферменты могут быть предоставлены так, чтобы они накапливались в цитоплазме. Может быть предусмотрен терминатор транскрипции. В примерах экспрессирующих кассет генов, представленных в настоящем документе, используют последовательность эффективного терминатора транскрипции из гена нопалинсинтазы Agrobacterium tumefaciens.
В одном из вариантов осуществления предусмотрено трансгенное растение, включающее нуклеиновую кислоту, кодирующую CWDE или CWDE, модифицированный по меньшей мере одним из сигнальной последовательности или интеина. Последовательность нуклеиновой кислоты, кодирующая CWDE, может кодировать любую аминокислотную последовательность CWDE. Последовательность нуклеиновой кислоты, кодирующая CWDE, модифицированный по меньшей мере одним из сигнальной последовательности или интеина, может кодировать любую аминокислотную последовательность CWDE и по меньшей мере одно из сигнальной последовательности или любого интеина. Нуклеиновая кислота может кодировать белок, который по меньшей мере на 70, 72, 75, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен последовательности, выбранной из SEQ ID NO:44-115. Нуклеиновая кислота может кодировать белок, который по меньшей мере на 70, 72, 75, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен последовательности, выбранной из SEQ ID NO:44-45, 49-54, 57-59, 85-86, 94-96, 104-109 и 113-115. Нуклеиновая кислота может кодировать белок, который по меньшей мере на 70, 72, 75, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен последовательности, выбранной из SEQ ID NO:47 и 55. Нуклеиновая кислота может кодировать белок, который по меньшей мере на 70, 72, 75, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен последовательности, выбранной из SEQ ID NO:46, 48 и 56. Нуклеиновая кислота может кодировать белок, который по меньшей мере на 70, 72, 75, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен последовательности, выбранной из SEQ ID NO:60-67, 70 и 75. Нуклеиновая кислота может кодировать белок, который по меньшей мере на 70, 72, 75, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен последовательности, выбранной из SEQ ID NO:68-69, 71-74, 76-77 и 112. Нуклеиновая кислота может кодировать белок, который по меньшей мере на 70, 72, 75, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен последовательности, выбранной из SEQ ID NO:78-84. Нуклеиновая кислота может кодировать белок, который по меньшей мере на 70, 72, 75, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен последовательности, выбранной из SEQ ID NO:97-103. Нуклеиновая кислота может кодировать белок, который по меньшей мере на 70, 72, 75, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен последовательностт, выбранной из SEQ ID NO:87-93 и 110-111. Нуклеиновая кислота может кодировать белок, который по меньшей мере на 70, 72, 75, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен последовательности, выбранной из SEQ ID NO:44, 45, 49 и 54. Нуклеиновая кислота может кодировать белок, который по меньшей мере на 70, 72, 75, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен последовательности, выбранной из SEQ ID NO:45, 87, 104-106 и 113. Нуклеиновая кислота может кодировать белок, который по меньшей мере на 70, 72, 75, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен последовательности, выбранной из SEQ ID NO:50-53, 57-59, 94-96, 104-109 и 113-115. Нуклеиновая кислота может кодировать белок, который по меньшей мере на 70, 72, 75, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен последовательности, выбранной из SEQ ID NO:54-56 и 60-65. Любая из указанных выше нуклеиновых кислот, которые кодируют белок, обладающий менее чем 100% идентичностью с цитированной эталонной последовательностью, может кодировать белок, обладающий той же или по существу той же активностью, что и белок, обладающий 100% идентичностью с цитированной эталонной последовательностью. Активность можно оценивать с помощью анализов, известных в данной области для любого конкретного белка. Активность можно оценивать способом, указанным в примере или его части в настоящем описании. По существу та же активность будет известна в данной области. В одном из вариантов осуществления по существу та же активность находится в пределах 20% от активности белка, обладающего 100% идентичностью с цитированной эталонной последовательностью. В одном из вариантов осуществления по существу та же активность находится в пределах 15% от активности белка, обладающего 100% идентичностью с цитированной эталонной последовательностью. В одном из вариантов осуществления по существу та же активность находится в пределах 10% от активности белка, обладающего 100% идентичностью с цитированной эталонной последовательностью. В одном из вариантов осуществления по существу та же активность находится в пределах 5% от активности белка, обладающего 100% идентичностью с цитированной эталонной последовательностью. В одном из вариантов осуществления по существу та же активность находится в пределах 1% от активности белка, обладающего 100% идентичностью с цитированной эталонной последовательностью. Упомянутые выше нуклеиновые кислоты могут быть предоставлены в вариантах осуществления, представленных в настоящем документе, отдельно, в качестве части другой нуклеиновой кислоты, в качестве части вектора или, как указано выше, в качестве части трансгенного растения. Идентичность можно измерять с помощью алгоритма Смита-Ватермана (Smith TF, Waterman MS (1981), "Identification of Common Molecular Subsequences", Journal of Molecular Biology 147: 195-197, которая включена в настоящее описание в качестве ссылки в полном объеме). В одном из вариантов осуществления трансгенное растение может происходить из одного из кукурузы, проса, мискантуса, сахарного тростника или сорго. Трансгенное растение может быть получено посредством опосредуемой agrobacterium трансформации с использованием плазмиды, имеющей нуклеотидную последовательность, как указано выше. Плазмида обладает последовательностью, которая по меньшей мере на 70, 72, 75, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентична последовательности, выбранной из SEQ ID NO:188-283. Плазмида состоит по существу из последовательности, которая по меньшей мере на 70, 72, 75, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентична последовательности, выбранной из SEQ ID NO:188-283. Плазмида состоит из последовательности, которая по меньшей мере на 70, 72, 75, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентична последовательности, выбранной из SEQ ID NO:188-283.
В одном из вариантов осуществления предусмотрено трансгенное растение, включающее нуклеиновую кислоту, гибридизующуюся с эталонной нуклеиновой кислотой, кодирующей CWDE или CWDE, модифицированный по меньшей мере одной из сигнальной последовательности или интеина. Эталонная последовательность нуклеиновой кислоты, кодирующая CWDE, может кодировать любую аминокислотную последовательность CWDE. Эталонная последовательность нуклеиновой кислоты, кодирующая CWDE, модифицированный по меньшей мере одним из сигнальной последовательности или интеина, может кодировать любую аминокислотную последовательность CWDE и по меньшей мере одно из любой сигнальной последовательности или любого интеина. Нуклеиновая кислота, включенная в трансгенное растение, может быть обозначена как первая нуклеиновая кислота. Первая нуклеиновая кислота может быть способна гибридизоваться в условиях низкой жесткости со второй нуклеиновой кислотой, состоящей из нуклеотидной последовательности, выбранной из SEQ ID NO:116-187 или комплементарной им последовательности. Первая нуклеиновая кислота может быть способна гибридизоваться в условиях умеренной жесткости со второй нуклеиновой кислотой, состоящей из нуклеотидной последовательности, выбранной из SEQ ID NO:116-187, или комплементарной ей последовательности. Первая нуклеиновая кислота может быть способна гибридизоваться в условиях высокой жесткости со второй нуклеиновой кислотой, состоящей из нуклеотидной последовательности, выбранной из SEQ ID NO:116-187 или комплементарной ей последовательности. Первая нуклеиновая кислота может быть способна гибридизоваться в условиях низкой, умеренной или высокой жесткости со второй нуклеиновой кислотой, состоящей из нуклеотидной последовательности, выбранной из SEQ ID NO:116-117, 121-126, 129-131, 157-158, 166-168, 176-181 и 185-187, или комплементарной ей последовательности. Первая нуклеиновая кислота может быть способна гибридизоваться в условиях низкой, умеренной или высокой жесткости со второй нуклеиновой кислотой, состоящей из нуклеотидной последовательности, выбранной из SEQ ID NO:119 и 127, или комплементарной ей последовательности. Первая нуклеиновая кислота может быть способна гибридизоваться в условиях низкой, умеренной или высокой жесткости со второй нуклеиновой кислотой, состоящей из нуклеотидной последовательности, выбранной из SEQ ID NO:118, 120 и 128, или комплементарной ей последовательности. Первая нуклеиновая кислота может быть способна гибридизоваться в условиях низкой, умеренной или высокой жесткости со второй нуклеиновой кислотой, состоящей из нуклеотидной последовательности, выбранной из SEQ ID NO:132-139, 142 и 147, или комплементарной ей последовательности. Первая нуклеиновая кислота может быть способна гибридизоваться в условиях низкой, умеренной или высокой жесткости со второй нуклеиновой кислотой, состоящей из нуклеотидной последовательности, выбранной из SEQ ID NO:140-141, 143-146, 148-149 и 184, или комплементарной ей последовательности. Первая нуклеиновая кислота может быть способна гибридизоваться в условиях низкой, умеренной или высокой жесткости со второй нуклеиновой кислотой, состоящей из нуклеотидной последовательности, выбранной из SEQ ID NO:150-156, или комплементарной ей последовательности. Первая нуклеиновая кислота может быть способна гибридизоваться в условиях низкой, умеренной или высокой жесткости со второй нуклеиновой кислотой, состоящей из нуклеотидной последовательности, выбранной из SEQ ID NO:169-175, или комплементарной ей последовательности. Первая нуклеиновая кислота может быть способна гибридизоваться в условиях низкой, умеренной или высокой жесткости со второй нуклеиновой кислотой, состоящий из нуклеотидной последовательности, выбранной из SEQ ID NO:159-165 и 182-183, или комплементарной ей последовательности. Первая нуклеиновая кислота может быть способна гибридизоваться в условиях низкой, умеренной или высокой жесткости со второй нуклеиновой кислотой, состоящей из нуклеотидной последовательности, выбранной из SEQ ID NO:116, 117, 121 и 126, или комплементарной ей последовательности. Первая нуклеиновая кислота может быть способна гибридизоваться в условиях низкой, умеренной или высокой жесткости со второй нуклеиновой кислотой, состоящей из нуклеотидной последовательности, выбранной из SEQ ID NO:117, 159, 176-178 и 185, или комплементарной ей последовательности. Первая нуклеиновая кислота может быть способна гибридизоваться в условиях низкой, умеренной или высокой жесткости со второй нуклеиновой кислотой, состоящей из нуклеотидной последовательности, выбранной из SEQ ID NO:122-125, 129-131, 166-168, 176-181 и 185-187, или комплементарной ей последовательности. Первая нуклеиновая кислота может быть способна гибридизоваться в условиях низкой, умеренной или высокой жесткости со второй нуклеиновой кислотой, состоящей из нуклеотидной последовательности, выбранной из SEQ ID NO:126-128 и 132-137, или комплементарной ей последовательности. Примеры протоколов гибридизации и способов оптимизации протоколов гибридизации описаны в следующих книгах: Molecular Cloning, T. Maniatis, E.F. Fritsch, and J. Sambrook, Cold Spring Harbor Laboratory, 1982; и Current Protocols in Molecular Biology, F.M. Ausubel, R. Brent, R.E. Kingston, D.D. Moore, J.G. Seidman, J.A. Smith, K. Struhl, Volume 1, John Wiley & Sons, 2000, каждая из которых включена в настоящий документ в качестве ссылки в полном объеме. В качестве неограничивающего примера, методики для условий гибридизации умеренной жесткости являются следующими: фильтры, содержащие ДНК, предварительно обрабатывают в течение 2-4 ч при 68°C в растворе, содержащем 6X SSC (Amresco, Inc., Solon, OH), 0,5% SDS (Amersco, Inc., Solon, OH), 5X раствор Денхардта (Amersco, Inc., Solon, OH), и 100 мкг/мл денатурированной ДНК спермы лосося (Invitrogen Life Technologies, Inc., Carlsbad, CA). На квадратный сантиметр используемой мембраны используют приблизительно 0,2 мл раствора для предварительной обработки. Гибридизацию проводят в том же растворе со следующими модификациями: можно использовать 0,01 M EDTA (Amersco, Inc., Solon, OH), 100 мкг/мл ДНК спермы лосося, и 5-20×106 cpm 32P-меченных или флуоресцентно меченных зондов. Фильтры инкубируют в смеси для гибридизации в течение 16-20 ч при 68°C, а затем промывают в течение 15 минут при комнатной температуре (в пределах пяти градусов от 25°C) в растворе, содержащем 2X SSC и 0,1% SDS, при осторожном встряхивании. Раствор для промывания заменяют раствором, содержащим 0,1X SSC и 0,5% SDS и инкубируют в течение дополнительных 2 ч при 68°C при осторожном встряхивании. Фильтры подвергают блоттингу в сухом виде и экспонируют для проявления в устройстве для визуализации или путем радиоавтографии. Если необходимо, фильтры промывают в третий раз и подвергают повторному экспонированию для проявления. В качестве неограничивающего примера, низкая жесткость относится к условиям гибридизации, в которых используется низкая температура для гибридизации, например, температуры от 37°C до 60°C. В качестве неограничивающего примера, высокая жесткость относится к условиям гибридизации, указанным выше, но с модификацией посредством использования высоких температур, например, температур гибридизации свыше 68°C. Любые из нуклеиновых кислот, указанных выше, которые обладают менее чем 100% идентичностью с цитируемой эталонной последовательностью, могут кодировать белок, имеющий ту же или по существу ту же активность, что и белок, кодируемый последовательностью нуклеиновой кислоты, обладающей 100% идентичностью с цитированной эталонной последовательностью. Активность можно оценивать с помощью анализов, известных в данной области для любого конкретного белка. Активность можно оценивать способом, указанном в примере или его части в настоящем описании. По существу та же активность может быть известна в данной области. В одном из вариантов осуществления по существу та же активность находится в пределах 20% от активности белка, кодируемого последовательностью нуклеиновой кислоты, обладающей 100% идентичностью с цитированной эталонной последовательностью. В одном из вариантов осуществления по существу та же активность находится в пределах 15% от активности белка, кодируемого последовательностью нуклеиновой кислоты, обладающей 100% идентичностью с цитированной эталонной последовательностью. В одном из вариантов осуществления по существу та же активность находится в пределах 10% от активности белка, кодируемого последовательностью нуклеиновой кислоты, обладающей 100% идентичностью с цитированной эталонной последовательностью. В одном из вариантов осуществления, по существу та же активность находится в пределах 5% от активности белка, кодируемого последовательностью нуклеиновой кислоты, обладающей 100% идентичностью с цитированной эталонной последовательностью. В одном из вариантов осуществления по существу та же активность находится в пределах 1% от активности белка, кодируемого последовательностью нуклеиновой кислоты, обладающей 100% идентичностью с цитированной эталонной последовательностью. Трансгенное растение может происходить из одного из кукурузы, проса, мискантуса, сахарного тростника или сорго. Трансгенное растение можно получать с помощью опосредуемой agrobacterium трансформации с использованием плазмиды, включающей любую из указанных выше нуклеиновых кислот.
В одном из вариантов осуществления предусмотрен вектор, включающий нуклеиновую кислоту, кодирующую CWDE или CWDE, модифицированный по меньшей мере одним из сигнальной последовательности или интеина. Последовательность нуклеиновой кислоты, кодирующая CWDE, может кодировать любую аминокислотную последовательность CWDE. Последовательность нуклеиновой кислоты, кодирующая CWDE, модифицированный по меньшей мере одним из сигнальной последовательности или интеина, может кодировать любую аминокислотную последовательность CWDE и по меньшей мере одно из любой сигнальной последовательности или любого интеина. Нуклеиновая кислота может кодировать белок, обладающий по меньшей мере 70, 72, 75, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичностью с последовательностью, выбранной из SEQ ID NO:44-115. Последовательность нуклеиновой кислоты может гибридизоваться в условиях низкой жесткости с эталонной нуклеиновой кислотой, состоящей из последовательности одного из SEQ ID NO:116-187 или комплементарной ей последовательности. Последовательность нуклеиновой кислоты может гибридизоваться в условиях умеренной жесткости с эталонной нуклеиновой кислотой, состоящей из любой последовательности SEQ ID NO:116-187 или комплементарной ей последовательности. Последовательность нуклеиновой кислоты может гибридизоваться в условиях высокой жесткости с эталонной нуклеиновой кислотой, состоящей из любой последовательности SEQ ID NO:116-187, или комплементарной ей последовательности. Вектор может включать последовательность, обладающую 70, 72, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичностью с последовательностью, выбранной из SEQ ID NO:188-283. Вектор может по существу состоять из последовательности, обладающей 70, 72, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичностью с последовательностью, выбранной из SEQ ID NO:188-283. Вектор может состоять из последовательности, обладающей 70, 72, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичностью с последовательностью, выбранной из SEQ ID NO:188-283.
В одном из вариантов осуществления выделенную нуклеиновую кислоту, полинуклеотид или олигонуклеотид, кодирующие по меньшей мере часть любой из аминокислотных последовательностей согласно SEQ ID NO:44-115, можно использовать в качестве зонда для гибридизации или праймера. В одном из вариантов осуществления в качестве зонда или праймера можно использовать последовательность, комплементарную указанной выделенной нуклеиновой кислоте, полинуклеотиду или олигонуклеотиду. В одном из вариантов осуществления в качестве зонда для гибридизации или праймера можно использовать выделенную нуклеиновую кислоту, имеющую последовательность, которая гибридизуется в условиях низкой, умеренной или высокой жесткости по меньшей мере с частью нуклеиновой кислоты, имеющей любую последовательность SEQ ID NO:116-187 и 188-283 или комплементарную ей последовательность. Эти выделенные нуклеиновые кислоты, полинуклеотиды или олигонуклеотиды не ограничены, но они могут иметь длину в диапазоне 10-100, 10-90, 10-80, 10-70, 10-60, 10-50, 10-40, 10-35, 10-30, 10-25, 10-20 или 10-15 нуклеотидов, или 20-30 нуклеотидов, или они могут иметь длину 25 нуклеотидов. Диапазон длин нуклеотидных последовательностей, приведенных в настоящем описании, включает каждую длину нуклеотидной последовательности в этом диапазоне, включая граничные значения. Указанная длина нуклеотидов может начинаться в любом отдельном положении в эталонной последовательности, где после отдельного положения следует достаточно нуклеотидов для включения указанной длины. В одном из вариантов осуществления зонд для гибридизации или праймер на 85-100%, 90-100%, 91-100%, 92-100%, 93-100%, 94-100%, 95-100%, 96-100%, 97-100%, 98-100%, 99-100%, или 100% комплементарны нуклеиновой кислоте с той же длиной, что и зонд или праймер, и имеют последовательность, выбранную из длины нуклеотидов, соответствующей длине зонда или праймера в нуклеиновой кислоте, кодирующей один из белков SEQ ID NO:44-115, или в последовательности, комплементарной указанной нуклеиновой кислоте. В одном из вариантов осуществления зонд для гибридизации или праймер на 85-100%, 90-100%, 91-100%, 92-100%, 93-100%, 94-100%, 95-100%, 96-100%, 97-100%, 98-100%, 99-100% или на 100% комплементарны нуклеиновой кислоте с той же длиной, что и зонд или праймер, и имеют последовательность, выбранную из длины нуклеотидов, соответствующей длине зонда или праймера в нуклеиновой кислоте с любой последовательностью SEQ ID NO:116-283. В одном из вариантов осуществления зонд для гибридизации или праймер гибридизуются вдоль их длины с соответствующей длиной нуклеиновой кислоты, кодирующей любую последовательность SEQ ID NO:44-115 или последовательность, комплементарную указанной нуклеиновой кислоте. В одном из вариантов осуществления зонд для гибридизации или праймер гибридизуется вдоль его длины с соответствующей длиной нуклеиновой кислоты, имеющей любую последовательность SEQ ID NO:116-187 или комплементарную ей последовательность. В одном из вариантов осуществления гибридизация может происходить в условиях низкой жесткости. В одном из вариантов осуществления гибридизация может происходить в условиях умеренной жесткости. В одном из вариантов осуществления гибридизация может происходить в условиях высокой жесткости.
Выделенные нуклеиновые кислоты, полинуклеотиды или олигонуклеотиды согласно вариантам осуществления, описанным в настоящем описании, могут включать природные нуклеотиды, природные аналоги нуклеотидов или синтетические аналоги нуклеотидов. Нуклеиновые кислоты, полинуклеотиды или олигонуклеотиды согласно вариантам осуществления, описанным в настоящем описании, могут представлять собой нуклеиновую кислоту любого типа, включая дезоксирибонуклеиновую кислоту (ДНК), рибонуклеиновую кислоту (РНК) или пептидную нуклеиновую кислоту (PNA). Последовательности нуклеиновых кислот, приведенные в настоящем документе, приведены в качестве последовательностей ДНК, но в качестве вариантов осуществления, описанных в настоящем документе, предусмотрены другие нуклеиновые кислоты, включая последовательности РНК, где U заменяет T.
Хотя в вариантах осуществления, описанных в настоящем описании, можно использовать немеченые зонды для гибридизации или праймеры, зонды для гибридизации или праймеры могут быть мечены поддающейся детекции меткой и их можно использовать для детекции последовательности или синтеза нуклеиновых кислот. Характерные метки включают, но не ограничиваются ими, радионуклиды, поглощающие свет химические группы, красители и флуоресцентные группы. Метка может представлять собой флуоресцентную группу, такую как 6-карбоксифлуоресцеин (FAM), 6-карбокси-4,7,2',7'-тетрахлорфлуоресцеин (TET), родамин, JOE (2,7-диметокси-4,5-дихлор-6-карбоксифлуоресцеин), HEX (гексахлор-6-карбоксифлуоресцеин) или VIC.
В одном из вариантов осуществления предусмотрен способ переработки растительной биомассы. Способ может включать предварительную обработку растения или его части путем смешения растения или его части с жидкостью с образованием смеси, имеющей отношение жидкости к твердому веществу, меньшее или равное 15. Предварительная обработка может включать предоставление условий для поддержания смеси при температуре, меньшей или равной 100°C. Способ может включать предоставление одного или нескольких ферментов. Растительная биомасса может представлять собой любое растение или его часть или она может происходить из них. Растительная биомасса может представлять собой или может происходить из любого трансгенного растения или его части, описанных, проиллюстрированных или заявленных в настоящем документе. Способ может включать растение или часть растения, отличные от любого трансгенного растения или его части, описанных, проиллюстрированных или заявленных в настоящем документе, и комбинирование его с любым трансгенным растением или его частью, описанными, проиллюстрированными или заявленными в настоящем документе. Отношение жидкости к твердому веществу в смеси может представлять собой величину, меньшую или равную 25, 24, 23, 22, 21, 20, 19, 18, 17, 16, 15, 14, 13, 12, 11, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1. Отношение жидкости к твердому веществу может составлять 8 или менее. Отношение жидкости к твердому веществу может составлять 8. Стадия предварительной обработки может включать поддержание температуры, меньшей или равной 100°C, в течение по меньшей мере четырех часов. Стадия предварительной обработки может включать поддержание температуры от 40°C до 90°C. Жидкость, предоставляемая для получения смеси, может представлять собой любую жидкость. В одном из вариантов осуществления жидкость представляет собой воду. В одном из вариантов осуществления жидкость включает воду, бисульфит аммония и карбонат аммония. Бисульфит аммония может быть в любой подходящей концентрации. В одном из вариантов осуществления концентрация бисульфита аммония представляет собой величину 8%-38% (конечные значения включаются) в расчете на масс./масс. с растением или его частью. Карбонат аммония может быть при любом подходящем pH. В одном из вариантов осуществления pH карбоната аммония представляет собой pH в диапазоне от 7,6 до 8,5, включая конечные значения. Концентрация карбоната аммония может представлять собой любую подходящую концентрацию. В одном из вариантов осуществления концентрация карбоната аммония представляет собой величину от 4% до 19% (включая конечные значения) в расчете на масс./масс. с растением или его частью. Стадия предоставления одного или нескольких ферментов может включать предоставление любого фермента, пригодного для переработки растительной биомассы. В одном из вариантов осуществления один или несколько ферментов включают по меньшей мере один фермент, способный гидролизовать лигноцеллюлозный материал. В одном из вариантов осуществления один или несколько ферментов включают по меньшей мере один из эндоглюканазы, β-глюкозидазы, целлобиогидролазы или ксиланазы. В одном из вариантов осуществления один или несколько ферментов включают по меньшей мере один из ксиланазы, целлюлазы, целлобиогидролазы, глюкозидазы, ксилозидазы, арабинофуранозидазы или эстеразы феруловой кислоты. В одном из вариантов осуществления способ включает стадию предоставления одного или нескольких ферментов, где один или несколько ферментов не являются ксиланазой, а затем добавления ксиланазы в качестве дополнительной стадии.
Любой отдельный вариант осуществления, описанный в настоящем документе, может быть дополнен одним или несколькими элементами из любого одного или нескольких других вариантов осуществления, описанных в настоящем документе.
Примеры - Следующие неограничивающие примеры предоставлены для иллюстрации конкретных вариантов осуществления. Варианты осуществления, представленные в описании, могут быть дополнены одной или несколькими деталями из любого одного или нескольких примеров ниже.
Пример 1 - pSB11
Ссылаясь на фиг. 1, вектор согласно одному варианту осуществления, описанному в настоящем описании, может быть основан на промежуточной плазмиде pSB11 (производное pBR322). pSB11 доступа от Japan Tobacco. Плазмида pSB11 подходит для клонирования и она может легко поддерживаться в E. coli. Конъюгаты pSB11 с "супер-бинарным" акцепторным вектором pSB1 (обезвреженная плазмида Ti) могут поддерживаться в штамме LB4404 Agrobacterium tumefaciens посредством гомологичной рекомбинации с использованием участков cos и ori, присутствующих в обоих векторах. Встраивающийся продукт представляет собой гибридный вектор, который впоследствии можно использовать для трансформации растений. pSB1 содержит гены вирулентности, такие как virB, virC и virG, требуемые для процессинга T-ДНК и доставки ее в растительную клетку. pSB11 имеет множественный участок клонирования, содержащий уникальные участки для распознавания ферментом рестрикции для клонирования экспрессирующих кассет с последовательностями генов-мишеней.
Пример 2 - pAG1000
Ссылаясь на фиг. 2A, pAG1000 создавали путем модификации pSB11 для обеспечения акцептирования ей нескольких экспрессирующих гены кассет. Изначально, исходную экспрессирующую кассету, содержащую ген положительного селективного маркера manA, кодирующий фосфоманнозоизомеразу (PMI), управляемый промотором вируса скручивания желтых листьев цеструма (CMPS), клонировали из плазмиды pNOV2819 (Syngenta Biotechnology) в pSB11 в качестве фрагмента HindIII-KpnI с получением pAG1000.
Пример 3 - pAG1001, pAG1002 и pAG1003
pAG1000 далее модифицировали путем удаления участка EcoRI (нуклеотидное положение #7) с получением pAG1001 (фиг. 2B), а затем участка KpnI (нуклеотидное положение #1) с получением pAG1002 (фиг. 2C). Эти модификации делали участки EcoRI и KpnI доступными для последующего клонирования экспрессирующих кассет с представляющими интерес генами (GOI). Ссылаясь на фиг. 3A, последовательность участка множественного клонирования (MCS), представленную ниже, содержащую участки PacI, XhoI, SnaBI, NcoI, KpnI, XmaI, AvrII, EcoRI синтезировали способом ПЦР в качестве фрагмента PmeI-HindIII размером 249 п.н. и клонировали в участки PmeI-HindIII в pAG1002 с получением вектора pAG1003.
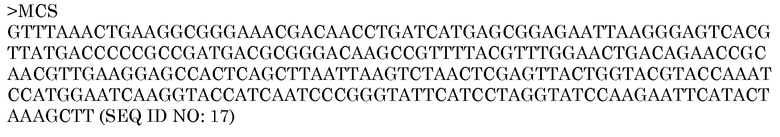
Пример 4 - pAG2000
Ссылаясь на фиг. 3B, более высокие уровни экспрессии могут быть обеспечены путем замены вирусного промотора CMPS в pAG1003 на промотор убиквитина 3 риса (SEQ ID NO:1), который представляет собой широко исследованный промотор с продемонстрированной эффективностью для экспрессии генов в однодольных растениях. OsUbi3P клонировали из плазмиды pRESQ101. pRESQ101 была описана E. Sivamani, J.D. Starmer, R. Qu, "Sequence analysis of rice rubi3 promoter gene expression cassettes for improved transgene expression", Plant Science, 177(6): 549-556, 2009, которая включена в настоящий документ в качестве ссылки в полном объеме. Проводили следующие модификации OsUbi3P для целей клонирования: 1) Участок EcoRI вносили на 5'-конце с помощью подхода ПЦР; 2) участок XmaI удаляли, в то время как добавляли участок BamHI на 3'-конце. Частичную последовательность OsUbi3P собирали в качестве фрагмента ApaI-BamHI в pBluescript, а затем клонировали в качестве полной промоторной области HindIII-BamHI, включающей первый интрон убиквитина, слитый с PMI в pAG1003, расщепленной HindIII-SpeI. Последнее клонирование приводило к вектору pAG2000.
Пример 5 - pAG2004 и pAG2005
Вектор pAG2000 далее модифицировали для разработки клонирующего вектора, способного включить в себя экспрессирующие кассеты GOI, при одновременном обеспечении усиленной экспрессии селективного маркера PMI для трансформации растений. Оптимизация экспрессии PMI включала замену исходной последовательности области соединения, соединяющей интрон OsUbi3 с начальным кодоном гена PMI в pAG2000 (представленной в SEQ ID NO:18, ниже), новой последовательностью из 9 нуклеотидов. Исходная последовательность области соединения подчеркнута и старт-кодон выделен полужирным шрифтом в версии SEQ ID NO:18, представленной ниже. Новая последовательность из 9 нуклеотидов представлена в качестве заключенной в рамку в версии SEQ ID NO:19, представленной ниже. Заключенная в рамку последовательность была подтверждена в качестве эффективной последовательности в отношении обеспечения высокого уровня временной экспрессии GUS в pRESQ48 в E. Sivamani and R. Qu (2006), которая включена в настоящий документ в качестве ссылки в полном объеме. Эта последовательность из 9 нуклеотидов соответствует трем начальным кодонам гена убиквитина 3 риса, где старт-кодон ATG модифицирован в ATC для устранения дополнительного участка инициации трансляции. Для обеспечения этой модификации фрагмент BglII-XcmI в pAG2000 (нуклеотидные положения 9726-105) заменяли синтезированным способом ПЦР фрагментом, который содержал требуемую последовательность области соединения из 9 нуклеотидов и был получен в последовательных реакциях с использованием праймеров P64/P68, P64/P66 и P64/P67.
>BglII-XcmI (9726-105) в pAG2000

>BglII-XcmI, синтезированный способом ПЦР фрагмент для конструирования pAG2004
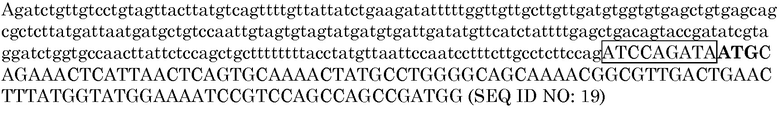

Ссылаясь на фиг. 3C, модификации, указанные выше, приводят к вектору pAG2004, который является вариантом осуществления, представленным в настоящем описании. Затем вектор pAG2004 использовали для конъюгации с pSB1 в штамме LBA4404 Agrobacterium tumefaciens и для трансформации незрелых эмбрионов маиса с использованием методики трансформации Japan Tobacco (Japan Tobacco Operating Manual for plasmid pSB1, Version 3.1, June 5, 2006; Komari, T., et. al., "Binary Vectors and Super-binary Vectors", Methods in Molecular Biology, Volume 343: Agrobacterium Protocols, pages 15-41, Humana Press, которая включена в настоящий документ в качестве ссылки в полном объеме). Эффективность трансформации маиса для pAG2004 и ее производного pAG2005, которое содержит промотор OsUbi3, клонированный в качестве KpnI-XmaI в pAG2004 MCS, может находиться в диапазоне 20-60%, в то время как pAG1003 с исходной экспрессирующей кассетой PMI из pNOV2819, где экспрессия manA запускается вирусным промотором CMPS, может обеспечивать вплоть до 15% эффективности трансформации.
Последовательность pAG2005 приведена в SEQ ID NO:24, которая представлена ниже:




Пример 5 - Генетические элементы, используемые в разработке векторов
Промоторы
Получали векторы, включающие последовательность промотора гена убиквитина 3 риса с первым интроном размером 2014 п.н. (OsUbi3P, регистрационный номер # AY954394, SEQ ID NO:1, представленная ниже) для конститутивной или "глобальной" экспрессии генов. Последовательность первого интрона OsUbi3P представлена строчными буквами в представлении SEQ ID NO:1 ниже. Векторы, представленные в настоящем описании, могут включать отличающиеся или дополнительные промоторы. Получали векторы, включающие промотор гена актина 1 риса с первым интроном гена (OsAct1P, регистрационный номер No. S44221, SEQ ID NO:2), который является конститутивным промотором. Промотор гена актина 1 риса можно использовать для экспрессии гена PMI в векторах, представленных в настоящем описании. Например, векторы pAG3000-pAG3003 включают промотор гена актина 1 риса с первым интроном гена. Некоторые векторы конструировали так, чтобы они включали промотор гена глутелина B-4 риса размером 1474 п.н. (OsGluB4P, регистрационный номер # AY427571, SEQ ID NO:4), который можно использовать для специфической для семян экспрессии генов и который используют для экспрессии ферментов и модифицированных интеином ферментов.

(SEQ ID NO:1), последовательность промотора представлена прописными буквами (SEQ ID NO:25), последовательность первого интрона представлена строчными буквами (SEQ ID NO:26)


Промотор гена убиквитина 3 риса клонировали из pRESQ101, как описано выше, одновременно синтезируя промоторы генов Act1 и GluB-4 риса. В случае слияния промотора гена Act1 риса с селективным маркером PMI, наблюдали вплоть до 23% эффективности трансформации при стабильной трансформации маиса с использованием селективной среды с маннозой в процессе культивирования тканей растений.
Сигнальные последовательности
Сигнальные последовательности могут быть включены с последовательностью CWDE (с дальнейшей модификацией или без нее; например с интеином) или в векторе для направления ферментов, экспрессируемых в растениях, в конкретные области внутри растительной клетки или вне ее. В некоторых примерах, описанных ниже, в CWDE или векторы, представленные в настоящем описании, были включены сигнальные последовательности PR1a табака (нацеливание на амилопласт) и альфа-амилазы BAASS ячменя (нацеливание на клеточную стенку). Эти сигнальные последовательности могут направлять ферменты в их соответствующие области для нацеливания. В некоторых примерах, описанных ниже, были включены сигнальные последовательности алеураина HvAleSP ячменя (нацеливание на вакуоль), GluB4 риса (экспрессия в семенах) и для удержания в ER (SEKDEL), и эти последовательности могут обеспечить локализацию белка в соответствующих клеточных компартментах или конкретных тканях. Целью такого нацеливания может быть достижение высоких уровней накопления белка, одновременно избегая потенциальных неблагоприятных эффектов на рост и развитие растения. Сигнальные последовательности, используемые в примерах, представленных в настоящем описании, и их соответствующие кодирующие нуклеотидные последовательности, представлены ниже:
Последовательность белка PR1a
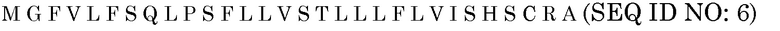
Нуклеотидная последовательность PR1a
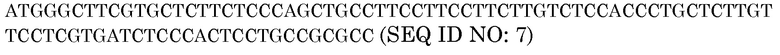
Последовательность белка BAASS
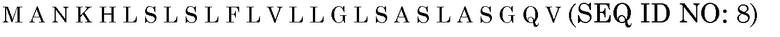
Нуклеотидная последовательность BAASS
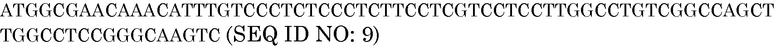
Последовательность белка HvAle
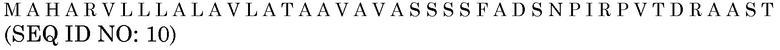
Нуклеотидная последовательность HvAle
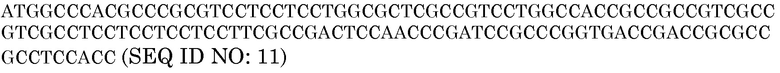

Последовательность белка GluB4SP
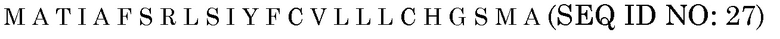
Нуклеотидная последовательность GluB4SP

Последовательности для нацеливания могут быть модифицированы из их исходных версий, чтобы они отражали частоту использования кодонов для оптимальной экспрессии генов в однодольных растениях. В одном из вариантов осуществления частоты использования кодонов хозяина представляют собой частоты для маиса. Каждую сигнальную последовательность можно синтезировать способом ПЦР с использованием специфических праймеров и связывать с 3'-концами последовательности; например, либо промотором OsUbi3, либо промотором OsGluB4, с использованием подхода ПЦР со слиянием.
Терминатор транскрипции
В векторы, представленные в настоящем описании, может быть включен терминатор транскрипции. В одном из вариантов осуществления в экспрессирующих гены кассетах, клонированных в векторах для трансформации растений, используют последовательность эффективного терминатора транскрипции (NosT) из гена нопалинсинтазы Agrobacterium tumefaciens. Последовательность представлена ниже:
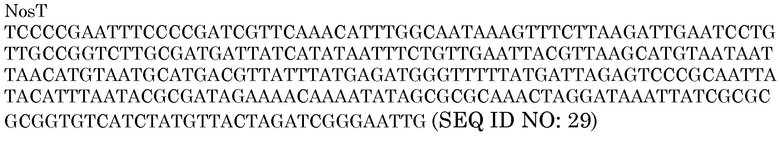
Эта последовательность встречается дважды в pAG2005 (SEQ ID NO:24). Второй раз она появляется в положениях 12034-12288 после второго промотора OsUbi3 вместе с последовательностью интрона и участком XmaI, и за ней следует участок рестрикции EcoRI (GAATTC, положения 12310-5 SEQ ID NO:24). Последовательность терминатора Nos можно амплифицировать способом ПЦР из pNOV2819 в качестве фрагмента из 276 п.н. Другие терминаторы транскрипции, известные в данной области, могут заменять терминатор Nos и использоваться вместо него. Другим терминатором, который можно использовать вместо терминатора Nos, является терминатор 35S.
Пример 6 - Разработка вектора для сверхэкспрессии ксиланазы P77853 дикого типа
Ссылаясь на фиг. 4, конструирование вектора pAG2014 обеспечивает пример типичного подхода для клонирования генов, кодирующих CWDE, таких как гены ксиланаз, целлюлаз и другие гены, представляющие особый интерес для разработки трансгенных однодольных растений, включая, но не ограничиваясь ими, маис, просо, сорго, мискантус и сахарный тростник.
Связь сигнальной последовательности с кодирующей областью зрелого фермента
Область соединения сигнальная последовательность -представляющий интерес белок можно определять экспериментально или с помощью моделей. Для этого примера для предсказания наилучшей области связывания сигнального пептида и фермента ксиланазы P77853 дикого типа использовали сервер SignalP 3.0, общедоступный через Center for Biological Sequence Analysis of the Technical University of Denmark (http://www.cbs.dtu.dk/index.shtml). Способ, используемый в SignalP 3.0, включает предсказание участков расщепления и предсказание сигнальный пептид/несигнальный пептид на основе комбинации нескольких искусственных нейронных сетей и скрытых моделей Маркова. Выходные данные программы обеспечивают показатель достоверности для отщепления сигнального пептида от зрелого белка. Оценивали три варианта области соединения; первый - с прямой связью между BAASS и P77853 (...GQV QTS...), второй - с удалением одной аминокислоты с С-конца BAASS (...GQ QTS...), и третий - с удалением одной аминокислоты с С-конца BAASS и удалением одной аминокислоты с N-конца P77853 (...GQ TS...). Вариант с наивысшим показателем отбирали для молекулярного клонирования. Последовательности BASS, P77853 и первой, второй и третьей областей соединения представлены ниже, причем область соединения подчеркнута:
BAASS из альфа-амилазы ячменя (регистрационный номер #X15226), 78 п.н.
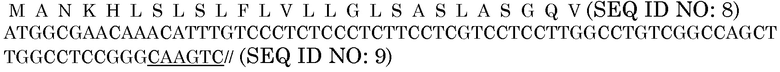

1 вариант связывания BAASS:P77853
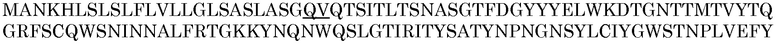
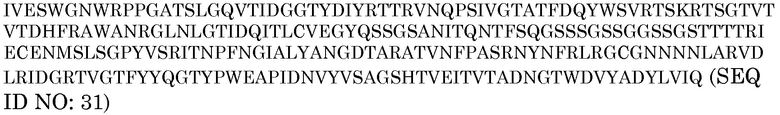
Предсказание сервером SignalP3.0: Сигнальный пептид
Наиболее вероятный участок расщепления между положениями 24 и 25: ASG-QV
Вероятность сигнального пептида: 1,000
Максимальная вероятность участка расщепления: 0,740 между положениями 24 и 25
2 вариант связывания BAASS:P77853

Предсказание сервером SignalP3.0: Сигнальный пептид
Наиболее вероятный участок расщепления между положениями 24 и 25: ASG-QQ
Вероятность сигнального пептида: 1,000
Максимальная вероятность участка расщепления: 0,768 между положениями 24 и 25
3 вариант связывания BAASS:P77853

Предсказание сервером SignalP3.0: Сигнальный пептид
Наиболее вероятный участок расщепления между положениями 24 и 25: ASG-QT
Вероятность сигнального пептида: 1,000
Максимальная вероятность участка расщепления: 0,582 между положениями 24 и 25
В этом примере 2 вариант связывания между BAASS и P77853 (...GQ QTS...) был выбран для разработки вектора pAG2014, исходя из выходных данных о максимальной вероятности участка расщепления из ServerP 3.0.
Отдельные генетические элементы для конструирования pAG2014 собирали в первичных реакциях ПЦР, как описано ниже. Первую реакцию ПЦР (ПЦР-1) использовали для амплификации 372 п.н. с 3'-конца первого интрона гена убиквитина 3 риса (представленного строчными буками), начиная с его собственного участка BglII (подчеркнуто). Этот фрагмент связывали с последовательностью из 9 нуклеотидов (показанную заглавными буквами, выделенными курсивом), соответствующей модифицированным трем начальным кодонам гена убиквитина 3 риса (подробное описание представлено выше), BAASS (показанной заглавными буквами) и последовательностью из 27 нуклеотидов (в рамке) с 5'-конца кодирующей области зрелого белка P77853. Вторую реакцию ПЦР (ПЦР-2) проводили для амплификации всей кодирующей области зрелого белка P77853, слитой со стоп-кодоном TAG, за которой следовал участок рестрикции AvrII (подчеркнут).
1. ПЦР-1 для амплификации 372 п.н. с 3'-конца первого интрона гена убиквитина 3 риса, области соединения из 9 п.н., BAASS и 5'-конца P77853:
Продукт ПЦР-1

Праймеры

2. ПЦР-2 для амплификации кодирующей области из 1017 п.н. зрелого белка P77853:
Продукт ПЦР-2

Праймеры
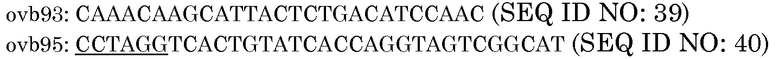
Последующий подход "ПЦР со слиянием" (Yon and Fried, 1989) использовали для "сшивания" генетических элементов, полученных в ПЦР-1 и ПЦР-2. Этот подход обеспечивал ожидаемую последовательность BglII-AvrII из 1362 п.н., состоящую из 261 п.н. с 3'-конца первого интрона гена убиквитина 3 риса с ее нативным 3'-концевым участком BglII, последовательностью для присоединения из 9 нуклеотидов между интроном и кодоном ATG сигнальной последовательности BAASS из 75 п.н. и кодирующей областью зрелой ксиланазы P77853 из 1011 п.н., заканчивающейся стоп-кодоном TGA, который фланкируется участком рестрикции AvrII:
3'OsUbi3Pint:BAASS:P77853 в качестве BglII-AvrII

Затем слитый продукт ПЦР вырезали из геля, очищали из геля с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (каталожный номер #28706) и лигировали с вектором pPCR-Blunt II TOPO. Слитый продукт ПЦР полностью секвенировали с использованием праймеров, специфичных к вектору и специфичных к гену. Слитый фрагмент ПЦР с подтвержденной последовательности высвобождали из вектора pPCR-Blunt II TOPO расщеплением BglII-AvrII и клонировали в pBluescript, который подготавливали с помощью следующих манипуляций:
1. Ссылаясь на фиг.5, фрагмент KpnI-EcoRI из pAG2005 размером 2362 п.н., который включает промотор OsUbi3, слитый с последовательностью CCCGGGTATTCATCCTAGG (SEQ ID NO:42) с участками XmaI (подчеркнут) и AvrII (заключен в рамку) и терминатором Nos, первоначально клонировали в pBluescript с получением вектора pBSK:OsUbi3P:XmaI:AvrII:NosT.
2. Ссылаясь на фиг.6, клонирование линкера L1 GAATTCTTACATTAGCACTAGAGCTC (SEQ ID NO:43) в участки EcoRI-SacI pBSK:OsUbi3P:XmaI:AvrII:NosT удаляло дополнительный участок XmaI и приводило к образованию "челночного" вектора pBSK:OsUbi3P:XmaI:AvrII:NosT:L1.
pBSK:OsUbi3P:XmaI:AvrII:NosT:L1 легко акцептирует расщепленные BglII-AvrII фрагменты ДНК. Таким образом, клонирование слитых продуктов ПЦР, сходное с клонированием, описанным в указанном выше примере, может привести к восстановлению полной экспрессирующей кассеты для представляющего интерес гена. Например, расщепленный BglII-AvrII слитый продукт ПЦР размером 1362 п.н., описанный выше для P77853, встраивали в расщепленный BglII-AvrII pBSK:OsUbi3P:XmaI:AvrII:NosT:L1 с получением экспрессирующей кассеты OsUbi3P:BAASS:P77853:NosT.
Полную экспрессирующую кассету OsUbi3P:BAASS:P77853:NosT далее вырезали в качестве фрагмента KpnI-EcoRI с использованием ферментов рестрикции и клонировали в pAG2005 с получением pAG2014. Вектор pAG2014 можно использовать для экспрессии ксиланазы P77853 дикого типа в трансгенных растениях из промотора гена убиквитина 3 риса, и нацеливания экспрессированного фермента на клеточную стенку растения с помощью сигнальной последовательности альфа-амилазы ячменя (BAASS). С использованием того же процесса получали векторы из следующего списка. Представленный ниже список также включает pAG1000, 1002, 1003, 1004, 1005, 2000, 2004. Представленные ниже векторы можно использовать для трансформации растений и экспрессии трансгенов.
1. pAG1000 – pAG1002 (SEQ ID NO:188-190, соответственно) представляют собой CMPSP:PMI в pSB11 с удалением различных участков рестрикции.
2. pAG1003 (SEQ ID NO:191) представляет собой pAG1002 с MCS.
3. pAG1004 представляет собой pAG1003 с GUS-int в MCS.
4. pAG1005 (SEQ ID NO:192) представляет собой pAG1003 с CPMSP:PMI, где PMI является кодон-оптимизированным и оптимизированным по экспрессии для маиса.
5. pAG2000 (SEQ ID NO:193) представляет собой pAG1003, где CMPSP:PMI заменен одним соединением промотора Ubi3 риса и PMI в HindIII-SpeI.
6. pAG2001 (SEQ ID NO:194) представляет собой pAG2000 с промотором Ubi3 риса в MCS.
7. pAG2002 (SEQ ID NO:195) представляет собой pAG2001 с промотором Ubi3 риса и терминатором Nos в MCS.
8. pAG2003 (SEQ ID NO:196) представляет собой pAG2000 со вторым соединением между промотором Ubi3 риса и PMI.
9. pAG2004 (SEQ ID NO:197) представляет собой pAG2000 с третьим соединением между промотором Ubi3 риса и PMI.
10. pAG2005 (SEQ ID NO:198) представляет собой pAG2004 с добавленным промотором Ubi3 риса и терминатором Nos из pAG2002 в MCS.
11. pAG2006 (SEQ ID NO:199) представляет собой pAG2005 с GUS между промотором Ubi3 риса и терминатором Nos, с использованием одного соединения между OsUbi3P и GUS.
12. pAG2007 (SEQ ID NO:200) представляет собой pAG2005 с GUS между промотором Ubi3 риса и терминатором Nos, с использованием второго соединения между OsUbi3P и GUS.
13. pAG2009 (SEQ ID NO:201) представляет собой pAG2005 с GUS, слитым с сигнальной последовательностью локализации во внутриклеточном пространстве PR1a (с использованием одного соединения) и между промотором Ubi3 риса и терминатором Nos.
14. pAG2010 (SEQ ID NO:202) представляет собой pAG2005 с GUS, слитым с сигнальной последовательностью локализации во внутриклеточном пространстве PR1a (с использованием второго соединения) и между промотором Ubi3 риса и терминатором Nos.
15. pAG2011 (SEQ ID NO:203) представляет собой pAG2005 с GUS, слитым с сигнальной последовательностью для нацеливания на клеточную стеку BAASS, и находящимся между промотором Ubi3 риса и терминатором Nos.
16. pAG2012 (SEQ ID NO:204) представляет собой pAG2007 с GUS между промотором глутелина GluB-4 риса и терминатором Nos.
17. pAG2013 (SEQ ID NO:205) представляет собой pAG2005 с GUS, слитым с сигнальной последовательностью для нацеливания на клеточную стенку HvExoI и находящимся между промотором Ubi3 риса и терминатором Nos.
18. pAG2014 (SEQ ID NO:206) представляет собой pAG2005 с WT P77853, слитым с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящимся между промотором Ubi3 риса и терминатором Nos.
19. pAG2015 (SEQ ID NO:207) представляет собой pAG2005 с WT P77853 между промотором Ubi3 риса и терминатором Nos.
20. pAG2016 (SEQ ID NO:208) представляет собой pAG2005 с GUS, слитым с сигнальной последовательностью локализации во внутриклеточном пространстве PR1a (оптимизированной для экспрессии в маисе) и находящимся между последовательностью промотора Ubi3 риса и терминатора Nos.
21. pAG2017 (SEQ ID NO:209) представляет собой pAG2005 с WT P40942 с сигнальной последовательностью локализации во внутриклеточном пространстве PR1a (оптимизированной для экспрессии в маисе) и между последовательностью промотора Ubi3 риса и терминатором Nos.
22. pAG2018 (SEQ ID NO:210) представляет собой pAG2005 с WT О30700, слитым с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящимся между промотором Ubi3 риса и терминатором Nos.
23. pAG2019 (SEQ ID NO:211) представляет собой pAG2005 с WT P40942, слитым с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящимся между промотором Ubi3 риса и терминатором Nos.
24. pAG2020 (SEQ ID NO:212) представляет собой pAG2005 с WT P77853, слитым с сигналом локализации во внутриклеточном пространстве PR1a (оптимизированной для экспрессии в маисе) и находящимся между последовательностью промотора Ubi3 риса и терминатором Nos.
25. pAG2021 (SEQ ID NO:213) представляет собой pAG2005 с P77853m3, слитым с сигналом локализации во внутриклеточном пространстве PR1a (оптимизированным для экспрессии в маисе) и находящимся между последовательностью промотора Ubi3 риса и терминатором Nos.
26. pAG2022 (SEQ ID NO:214) представляет собой pAG2005 с P77853m3:SEKDEL, слитым с сигнальной последовательностью локализации во внутриклеточном пространстве PR1a (оптимизированным для экспрессии в маисе) и находящимся между промотором Ubi3 риса и терминатором Nos.
27. pAG2023 (SEQ ID NO:215) представляет собой pAG2005 с P77853m3, слитым с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящимся между промотором Ubi3 риса и терминатором Nos.
28. pAG2024 (SEQ ID NO:216) представляет собой pAG2005 с P77853m3:SEKDEL, слитым с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящимся между промотором Ubi3 риса и терминатором Nos.
29. pAG2025 (SEQ ID NO:217) представляет собой pAG2012 с WT P77853, слитым с сигнальной последовательностью GluB-4 и находящимся между промотором глутелина GluB-4 риса и терминатором Nos.
30. pAG2026 (SEQ ID NO:218) представляет собой pAG2012 с WT O30700, слитым с сигнальной последовательностью GluB-4 и находящимся между промотором глутелина GluB-4 риса и терминатором Nos.
31. pAG2027 (SEQ ID NO:219) представляет собой pAG2012 с WT P40942, слитым с сигнальной последовательностью GluB-4 и находящимся между промотором глутелина GluB-4 риса и терминатором Nos.
32. pAG2028 (SEQ ID NO:220) представляет собой pAG2005 с P77853T134-195, слитым с сигнальной последовательностью для локализации во внутриклеточном пространстве PR1a (оптимизированной для экспрессии в маисе) и находящимся между промотором Ubi3 риса и терминатором Nos.
33. pAG2029 (SEQ ID NO:221) представляет собой pAG2005 с P77853T134-195, слитым с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящимся между промотором Ubi3 риса и терминатором Nos.
34. pAG2030 (SEQ ID NO:222) представляет собой pAG2005 с P77853m3 между промотором Ubi3 риса и терминатором Nos.
35. pAG2031 (SEQ ID NO:223) представляет собой pAG2012 с WT P54583, слитым с сигнальной последовательностью GluB-4 и находящимся между промотором глутелина GluB-4 риса и терминатором Nos.
36. pAG2032 (SEQ ID NO:224) представляет собой pAG2012 с WT P54583:SEKDEL, слитым с сигнальной последовательностью GluB-4 и находящимся между промотором глутелина GluB-4 риса и терминатором Nos.
37. pAG2033 (SEQ ID NO:225) представляет собой pAG2005 с WT P54583 между промотором Ubi3 риса и терминатором Nos.
38. pAG2034 (SEQ ID NO:226) представляет собой pAG2005 с WT P54583:SEKDEL между промотором Ubi3 риса и терминатором Nos.
39. pAG2035 (SEQ ID NO:227) представляет собой pAG2005 с WT P54583, слитым с сигналом локализации во внутриклеточном пространстве PR1a (оптимизированным для экспрессии в маисе) и находящимся между последовательностью промотора Ubi3 риса и терминатором Nos.
40. pAG2036 (SEQ ID NO:228) представляет собой pAG2005 с WT P54583:SEKDEL, слитым с сигнальной последовательностью локализации во внутриклеточном пространстве PR1a (оптимизированной для экспрессии в маисе), находящимся между промотором Ubi3 риса и терминатором Nos.
41. pAG2037 (SEQ ID NO:229) представляет собой pAG2005 с WT P54583, слитым с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящимся между промотором Ubi3 риса и терминатором Nos.
42. pAG2038 (SEQ ID NO:230) представляет собой pAG2005 с WT P54583:SEKDEL, слитым с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящимся между промотором Ubi3 риса и терминатором Nos.
43. pAG2039 (SEQ ID NO:231) представляет собой pAG2005 с GUS, слитым с HvAleSP и находящимся между промотором Ubi3 риса и терминатором Nos.
44. pAG2040 (SEQ ID NO:232) представляет собой pAG2005 с WT NtEGm, слитым с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящимся между промотором Ubi3 риса и терминатором Nos.
45. pAG2042 (SEQ ID NO:234) представляет собой pAG2005 с WT P54583, слитым с сигнальной последовательностью для нацеливания в вакуоль HvAleSP и находящимся между промотором Ubi3 риса и терминатором Nos.
46. pAG2043 (SEQ ID NO:235) представляет собой pAG2005 с WT NtEGm между промотором Ubi3 риса и терминатором Nos.
47. pAG2044 (SEQ ID NO:236) представляет собой pAG2005 с WT NtEGm, слитым с сигнальной последовательностью локализации во внутриклеточном пространстве PR1a (оптимизированной для экспрессии в маисе) и находящимся между промотором Ubi3 риса и терминатором Nos.
48. pAG2045 (SEQ ID NO:237) представляет собой pAG2005 с WT NtEGm:SEKDEL, слитым с сигнальной последовательностью локализации во внутриклеточном пространстве PR1a (оптимизированной для экспрессии в маисе) и находящимся между промотором Ubi3 риса и терминатором Nos.
49. pAG2046 (SEQ ID NO:238) представляет собой pAG2005 с WT NtEGm:SEKDEL, слитым с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящимся между промотором Ubi3 риса и терминатором Nos.
50. pAG2047 (SEQ ID NO:239) представляет собой pAG2005 с WT P54583:SEKDEL, слитым с сигнальной последовательностью для нацеливания на вакуоль HvAleSP и находящимся между промотором Ubi3 риса и терминатором Nos.
51. pAG2048 (SEQ ID NO:240) представляет собой pAG2005 с WT NtEGm между промотором Ubi3 риса, слитым с сигнальной последовательностью для нацеливания в вакуоль HvAleSP, и терминатором Nos.
52. pAG2049 (SEQ ID NO:241) представляет собой pAG2005 с WT NtEGm:SEKDEL, слитым с сигнальной последовательностью для нацеливания в вакуоль HvAleSP и находящимся между промотором Ubi3 риса и терминатором Nos.
53. pAG2050 (SEQ ID NO:242) представляет собой pAG2005 с WT P26222, находящимся между промотором Ubi3 риса и терминатором Nos.
54. pAG2051 (SEQ ID NO:243) представляет собой pAG2005 с WT P26222, слитым с сигнальной последовательностью локализации во внутриклеточном пространстве PR1a (оптимизированной для экспрессии в маисе) и находящимся между промотором Ubi3 риса и терминатором Nos.
55. pAG2052 (SEQ ID NO:244) представляет собой pAG2005 с WT P26222:SEKDEL, слитым с сигнальной последовательностью локализации во внутриклеточном пространстве PR1a (оптимизированной для экспрессии в маисе) и между промотором Ubi3 риса и терминатором Nos.
56. pAG2053 (SEQ ID NO:245) представляет собой pAG2005 с WT P26222, слитым с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящимся между промотором Ubi3 риса и терминатором Nos.
57. pAG2054 (SEQ ID NO:246) представляет собой pAG2005 с WT P26222:SEKDEL, слитым с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящимся между промотором Ubi3 риса и терминатором Nos.
58. pAG2055 (SEQ ID NO:247) представляет собой pAG2005 с WT P26222, слитым с сигнальной последовательностью для нацеливания в вакуоль HvAleSP и находящимся между промотором Ubi3 риса и терминатором Nos.
59. pAG2056 (SEQ ID NO 248) представляет собой pAG2005 с WT P26222:SEKDEL, слитым с сигнальной последовательностью для нацеливания в вакуоль HvAleSP и находящимся между промотором Ubi3 риса и терминатором Nos.
60. pAG2057 (SEQ ID NO:249) представляет собой pAG2005 с WT P77853:SEKDEL, слитым с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящимся между промотором Ubi3 риса и терминатором Nos.
61. pAG2058 (SEQ ID NO:250) представляет собой pAG2005 с WT P77853:SEKDEL, слитым с сигнальной последовательностью локализации во внутриклеточном пространстве PR1a (оптимизированной для экспрессии в маисе) и находящимся между промотором Ubi3 риса и терминатором Nos.
62. pAG2059 (SEQ ID NO:251) представляет собой pAG2005 с WT O43097 между промотором Ubi3 риса и терминатором Nos.
63. pAG2060 (SEQ ID NO:252) представляет собой pAG2005 с WT O43097 с сигнальной последовательностью локализации во внутриклеточном пространстве PR1a (оптимизированной для экспрессии в маисе) и находящимся между промотором Ubi3 риса и терминатором Nos.
64. pAG2061 (SEQ ID NO:253) представляет собой pAG2005 с WT O43097:SEKDEL, слитым с сигнальной последовательностью локализации во внутриклеточном пространстве PR1a (оптимизированной для экспрессии в маисе) и находящимся между промотором Ubi3 риса и терминатором Nos.
65. pAG2062 (SEQ ID NO:254) представляет собой pAG2005 с WT O43097, слитым с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящимся между промотором Ubi3 риса и терминатором Nos.
66. pAG2063 (SEQ ID NO:255) представляет собой pAG2005 с WT O43097:SEKDEL, слитым с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящимся между промотором Ubi3 риса и терминатором Nos.
67. pAG2064 (SEQ ID NO:256) представляет собой pAG2005 с WT O43097, слитым с сигнальной последовательностью для нацеливания в вакуоль HvAleSP и находящимся между промотором Ubi3 риса и терминатором Nos.
68. pAG2065 (SEQ ID NO:257) представляет собой pAG2005 с WT O43097:SEKDEL, слитым с сигнальной последовательностью для нацеливания в вакуоль HvAleSP и находящимся между промотором Ubi3 риса и терминатором Nos.
69. pAG2066 (SEQ ID NO:258) представляет собой pAG2005 с модифицированной интеином ксиланазой P77853-S158-2, слитой с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящимся между промотором Ubi3 риса и терминатором Nos.
70. pAG2067 (SEQ ID NO:259) представляет собой pAG2005 с модифицированной интеином ксиланазой P77853-S158-19, слитой с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящимся между промотором Ubi3 риса и терминатором Nos.
71. pAG2068 (SEQ ID NO:260) представляет собой pAG2005 с модифицированной интеином ксиланазой P77853-T134-1, слитой с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящимся между промотором Ubi3 риса и терминатором Nos.
72. pAG2069 (SEQ ID NO:261) представляет собой pAG2005 с WT О68438, находящимся между промотором Ubi3 риса и терминатором Nos.
73. pAG2070 (SEQ ID NO:262) представляет собой pAG2005 с WT О68438, слитым с сигналом локализации во внутриклеточном пространстве PR1a (оптимизированным для экспрессии в маисе) и находящимся между последовательностью промотора Ubi3 риса и терминатором Nos.
74. pAG2071 (SEQ ID NO:263) представляет собой pAG2005 с WT О68438:SEKDEL, слитым с сигнальной последовательностью локализации во внутриклеточном пространстве PR1a (оптимизированной для экспрессии в маисе) и находящимся между промотором Ubi3 риса и терминатором Nos.
75. pAG2072 (SEQ ID NO:264) представляет собой pAG2005 с WT О68438, слитым с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящимся между промотором Ubi3 риса и терминатором Nos.
76. pAG2073 (SEQ ID NO:265) представляет собой pAG2005 с WT О68438:SEKDEL, слитым с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящимся между промотором Ubi3 риса и терминатором Nos.
77. pAG2074 (SEQ ID NO:266) представляет собой pAG2005 c WT О68438, слитым с сигнальной последовательность для нацеливания в вакуоль HvAleSP и находящимся между промотором Ubi3 риса и терминатором Nos.
78. pAG2075 (SEQ ID NO:267) представляет собой pAG2005 с WT О68438:SEKDEL, слитым с сигнальной последовательность для нацеливания в вакуоль HvAleSP и находящимся между промотором Ubi3 риса и терминатором Nos.
79. pAG2076 (SEQ ID NO:268) представляет собой pAG2005 с модифицированной интеином ксиланазой P77853-S158-2 между промотором Ubi3 риса и терминатором Nos.
80. pAG2077 (SEQ ID NO:269) представляет собой pAG2005 с модифицированной интеином ксиланазой P77853-S158-19 между промотором Ubi3 риса и терминатором Nos.
81. pAG2078 (SEQ ID NO:270) представляет собой pAG2005 с модифицированной интеином ксиланазой P77853-T134-1 между промотором Ubi3 риса и терминатором Nos.
82. pAG2079 (SEQ ID NO:271) представляет собой pAG2005 с модифицированной интеином ксиланазой P77853-S158-2:SEKDEL, слитой с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящейся между промотором Ubi3 риса и терминатором Nos.
83. pAG2080 (SEQ ID NO:272) представляет собой pAG2005 с модифицированной интеином ксиланазой P77853-S158-19:SEKDEL, слитой с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящейся между промотором Ubi3 риса и терминатором Nos.
84. pAG2081 (SEQ ID NO:273) представляет собой pAG2005 с модифицированной интеином ксиланазой P77853-T134-1:SEKDEL, слитой с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящимся между промотором Ubi3 риса и терминатором Nos.
85. pAG3000 (SEQ ID NO:280) представляет собой pAG1003 с промотором Act1 риса, регулирующим PMI вместо CMPSP:PMI, с использованием одного соединения между OsAct1P и PMI (неполная консенсусная последовательность эукариотического участка инициации трансляции).
86. pAG3001 (SEQ ID NO:281) представляет собой pAG1003 с промотором Act1 риса, регулирующим PMI вместо CMPS-PMI, с использованием второго соединения между OsAct1P и PMI (полная консенсусная последовательность эукариотического участка инициации трансляции).
87. pAG3002 (SEQ ID NO:282) представляет собой pAG3000 с GUS между промотором Ubi3 риса, слитым с сигнальной последовательностью для нацеливания на клеточную стенку BAASS, и терминатором Nos.
88. pAG3003 (SEQ ID NO:283) представляет собой pAG3001 с GUS, слитым с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящимся между промотором Ubi3 риса и терминатором Nos.
89. pAG2041 (SEQ ID NO:233) представляет собой pAG2004 с NosT, клонированным в участки AvrII-EcoRI.
90. pAG2082 (SEQ ID NO:274) представляет собой pAG2005 с WT O43097, слитым с сигнальным пептидом глутелина B-4 и находящимся между промотором глутелина B-4 риса и терминатором Nos.
91. pAG2083 (SEQ ID NO:275) представляет собой pAG2005 с WT O43097:SEKDEL, слитым с сигнальным пептидом глутелина B-4 и находящимся между промотором глутелина B-4 риса и терминатором Nos.
92. pAG2084 (SEQ ID NO:276) представляет собой pAG2005 с WT NtEGm, слитым с сигнальным пептидом глутелина B-4 и находящимся между промотором глутелина B-4 риса и терминатором Nos.
93. pAG2085 (SEQ ID NO:275) представляет собой pAG2005 с модифицированной интеином ксиланазой P77853-T145-307 между промотором Ubi3 риса и терминатором Nos.
94. pAG2086 (SEQ ID NO:278) представляет собой pAG2005 с модифицированной интеином ксиланазой P77853-T145-307, слитой с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящейся между промотором Ubi3 риса и терминатором Nos.
95. pAG2087 (SEQ ID NO:279) представляет собой pAG2005 с модифицированной интеином ксиланазой P77853-T145-307:SEKDEL, слитой с сигнальной последовательностью для нацеливания на клеточную стенку BAASS и находящейся между промотором Ubi3 риса и терминатором Nos.
Аминокислотная последовательность белка, кодируемого каждый из приведенных выше векторов 18-19, 21-84 и 89-95, и нуклеиновой кислоты, кодирующей белок, представлена в таблице 1 ниже.
Варианты осуществления, представленные в настоящем документе, включают, но не ограничиваются ими, последовательности генов под заголовком "нуклеотидная последовательность" в таблице 1 ниже, аминокислотные последовательности под заголовком "последовательность белка" в таблице 1, растения, включающие последовательности генов из таблицы 1, векторы, включающие последовательности генов из таблицы 1, векторы под заголовком "вектор pAG" в таблице 1, растения, включающие векторы из таблицы 1, растения, включающие белки, кодируемые нуклеотидными последовательностями из таблицы 1, и растения, включающие последовательности белка из таблицы 1. Для векторов в таблице 1 каждая запись под заголовком "вектор pAG" включает номер. Номер добавлен к "pAG" для дополнения названия вектора. Например, указание на "2014" означает вектор pAG2014.






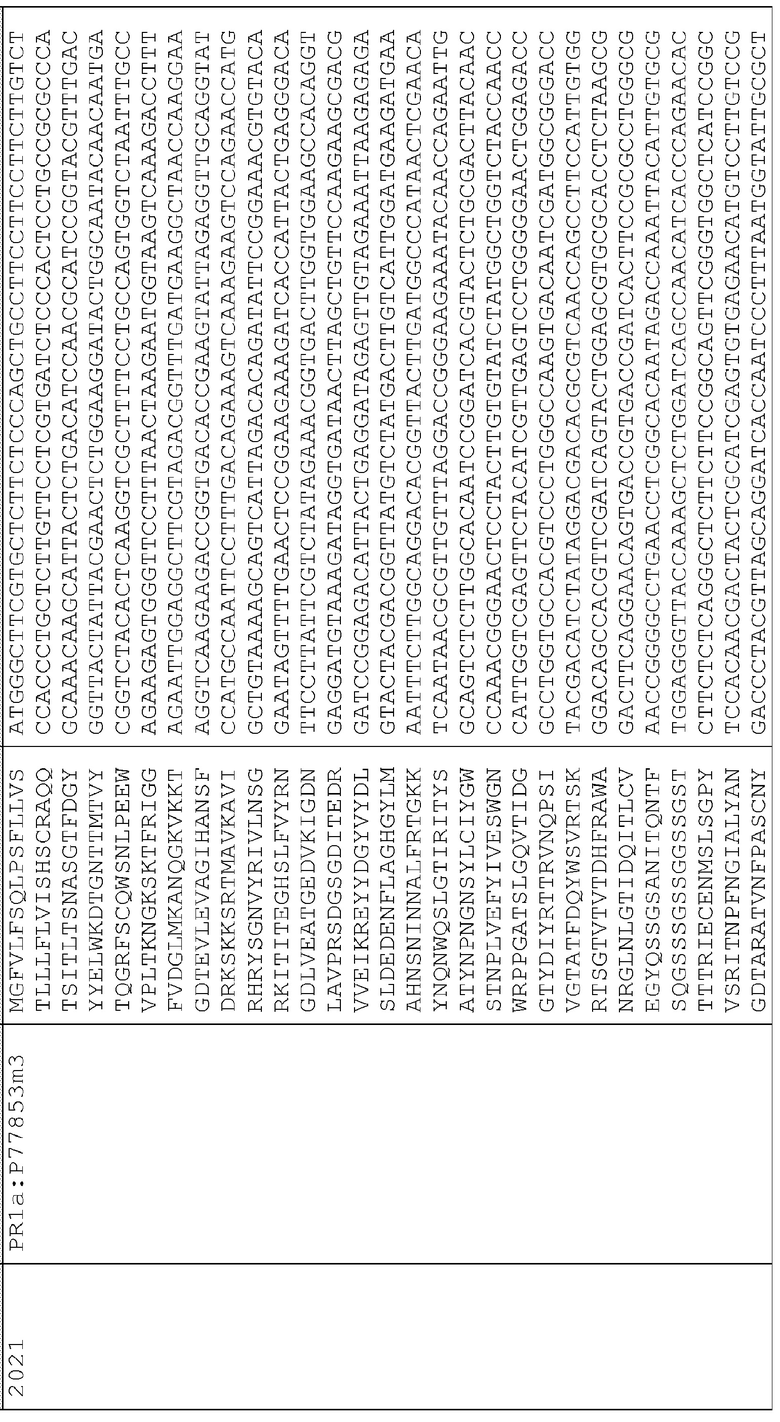

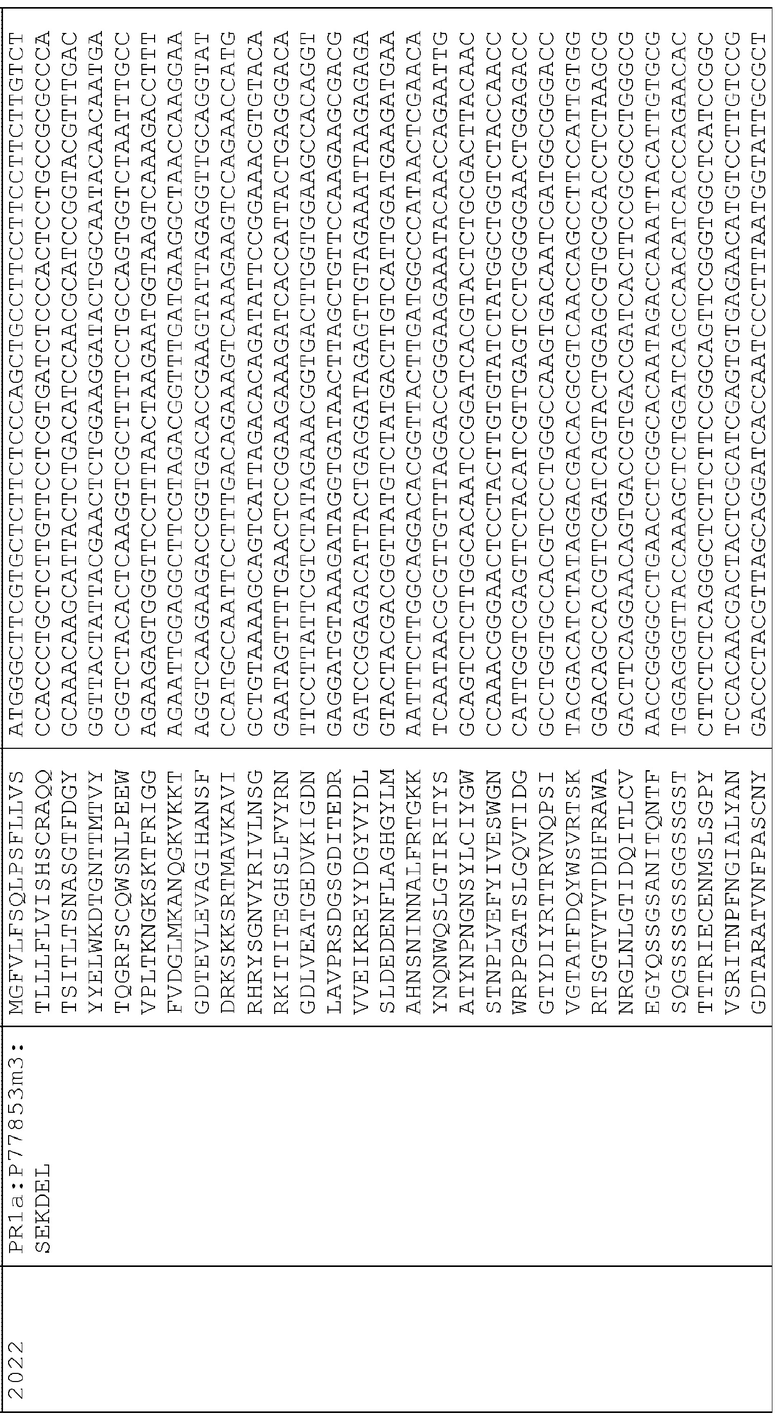




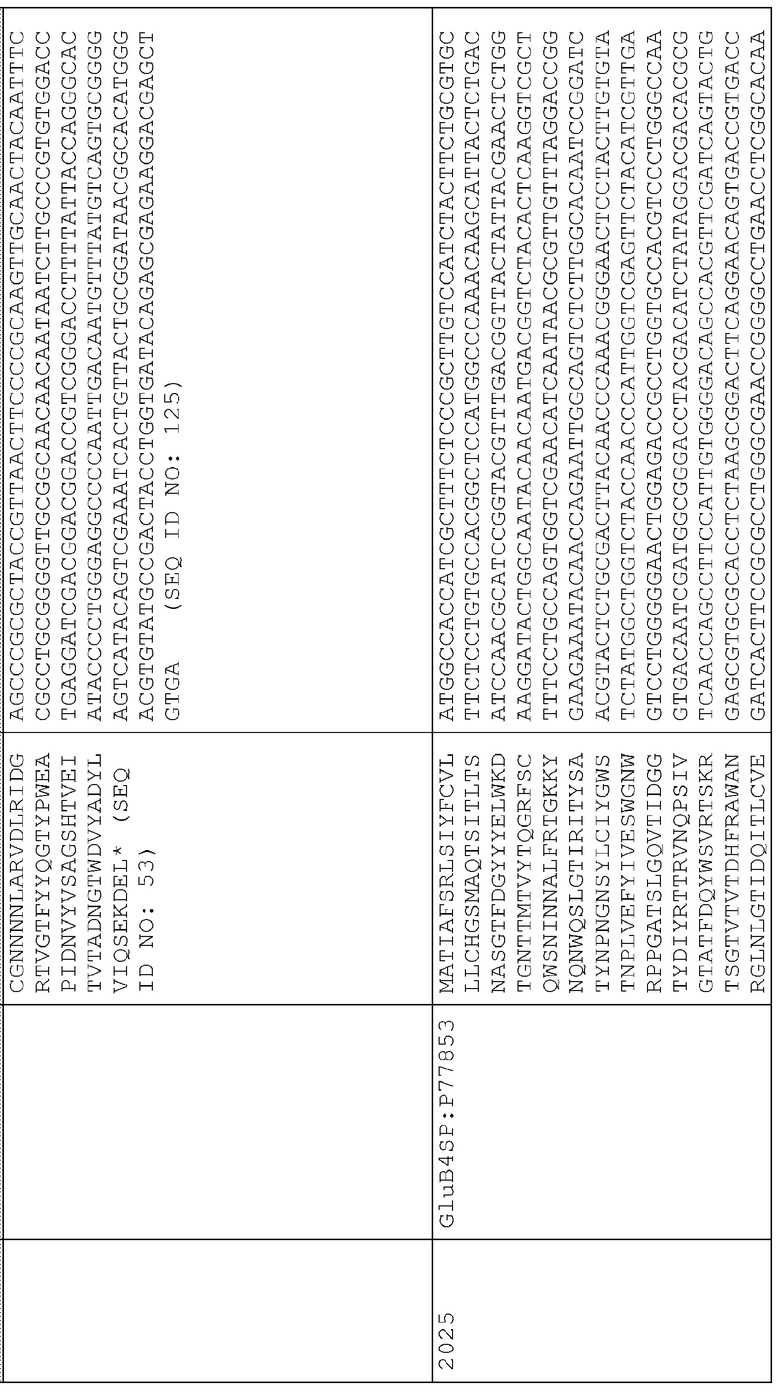
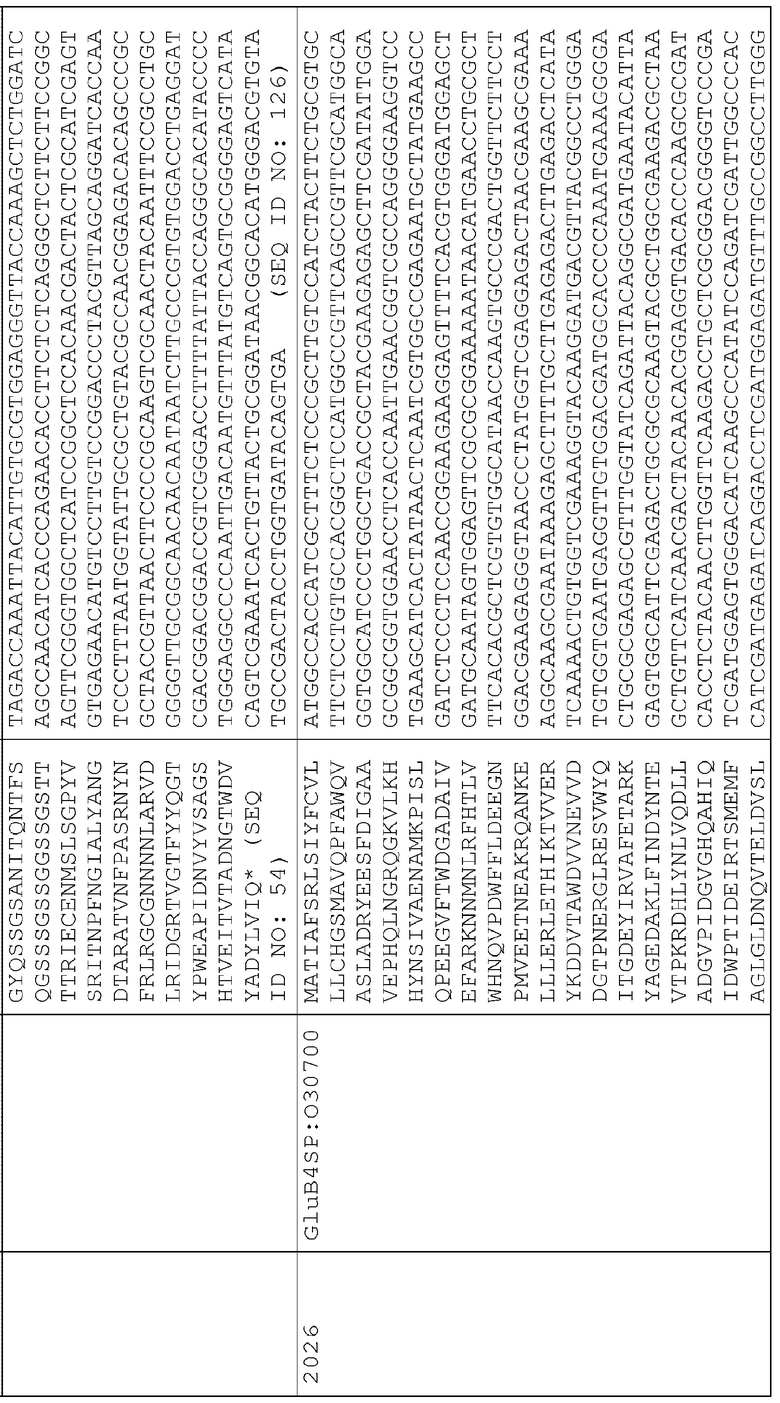
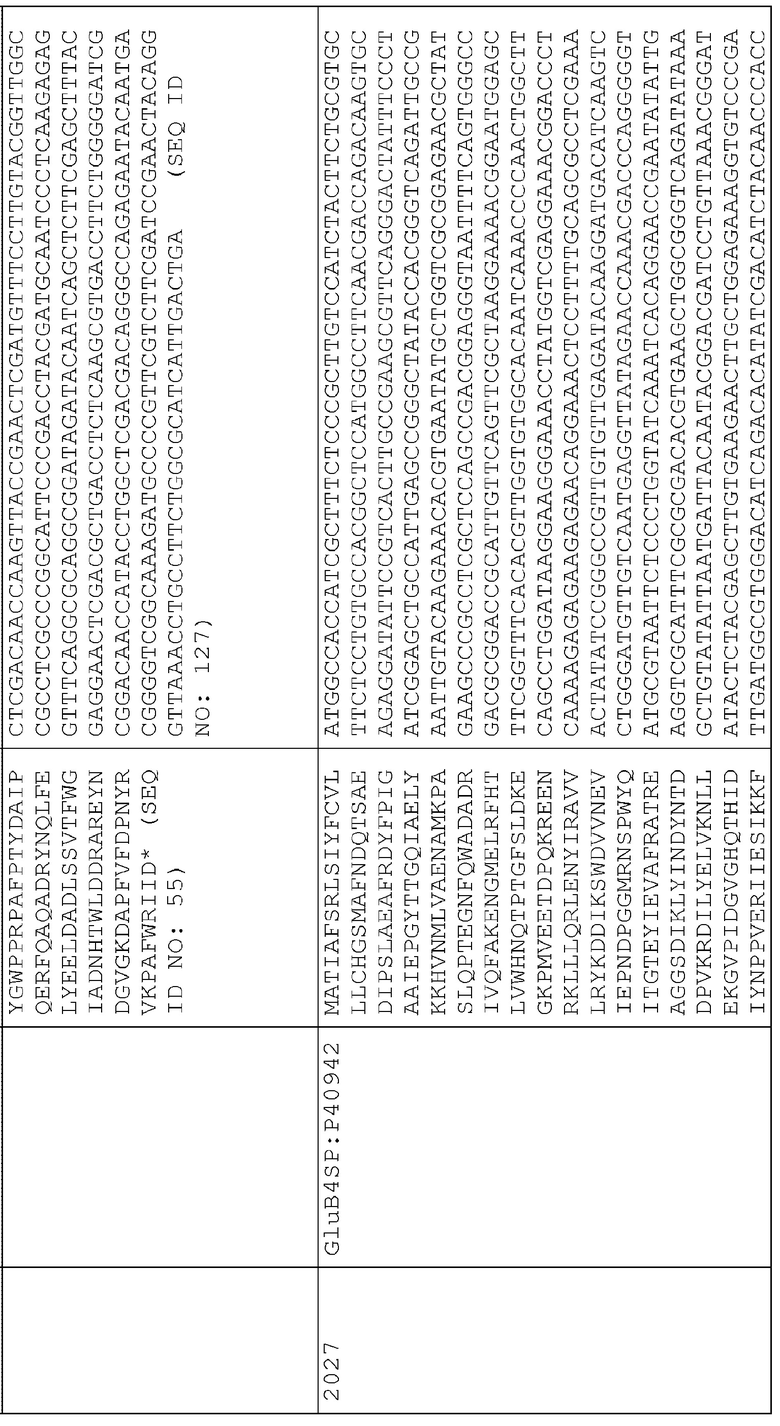









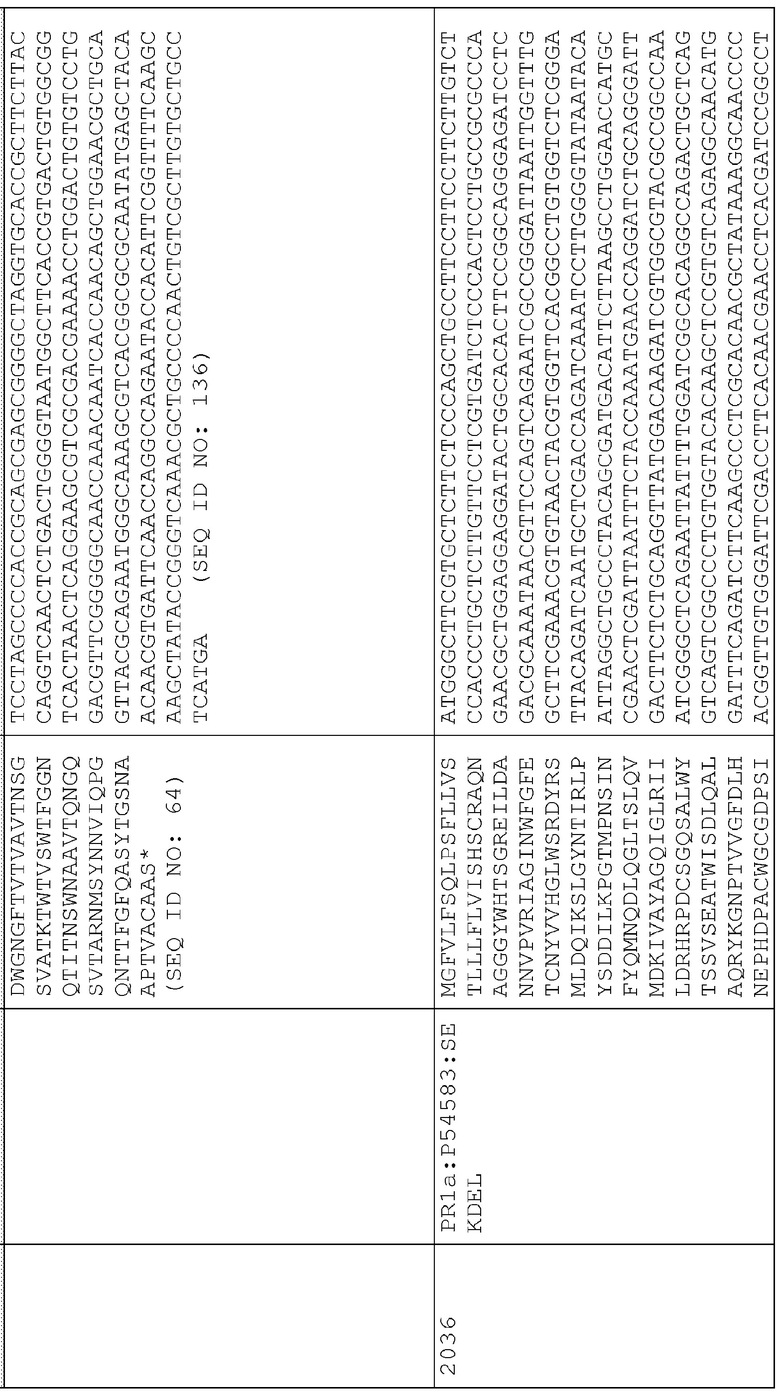






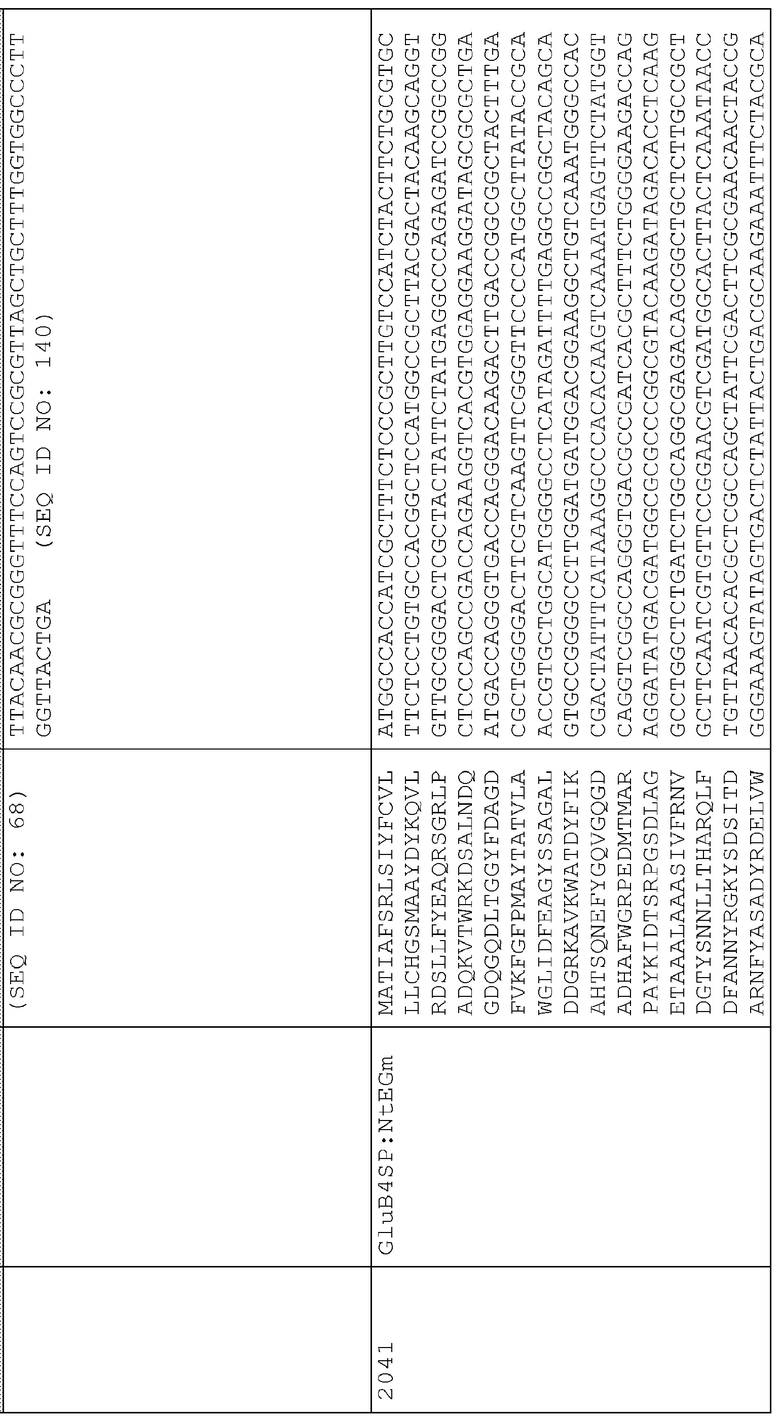












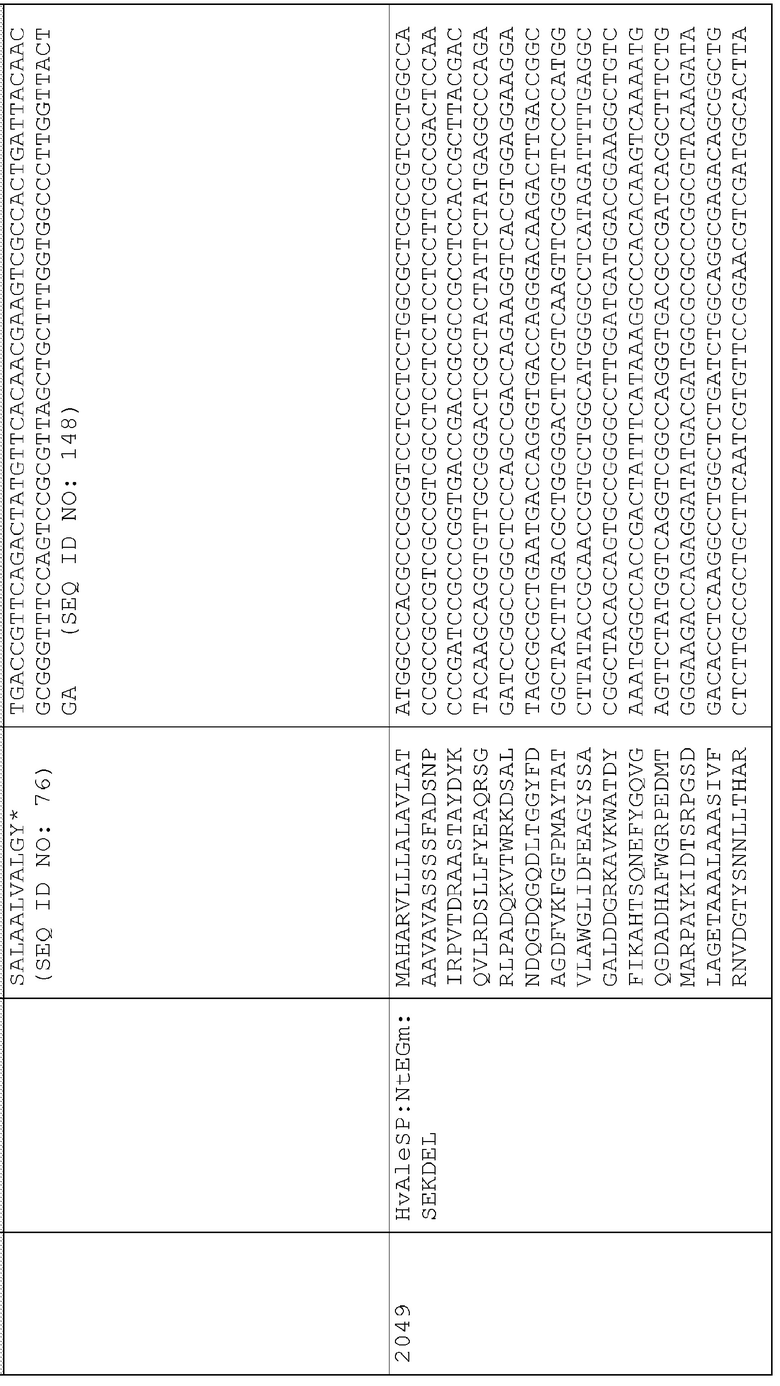




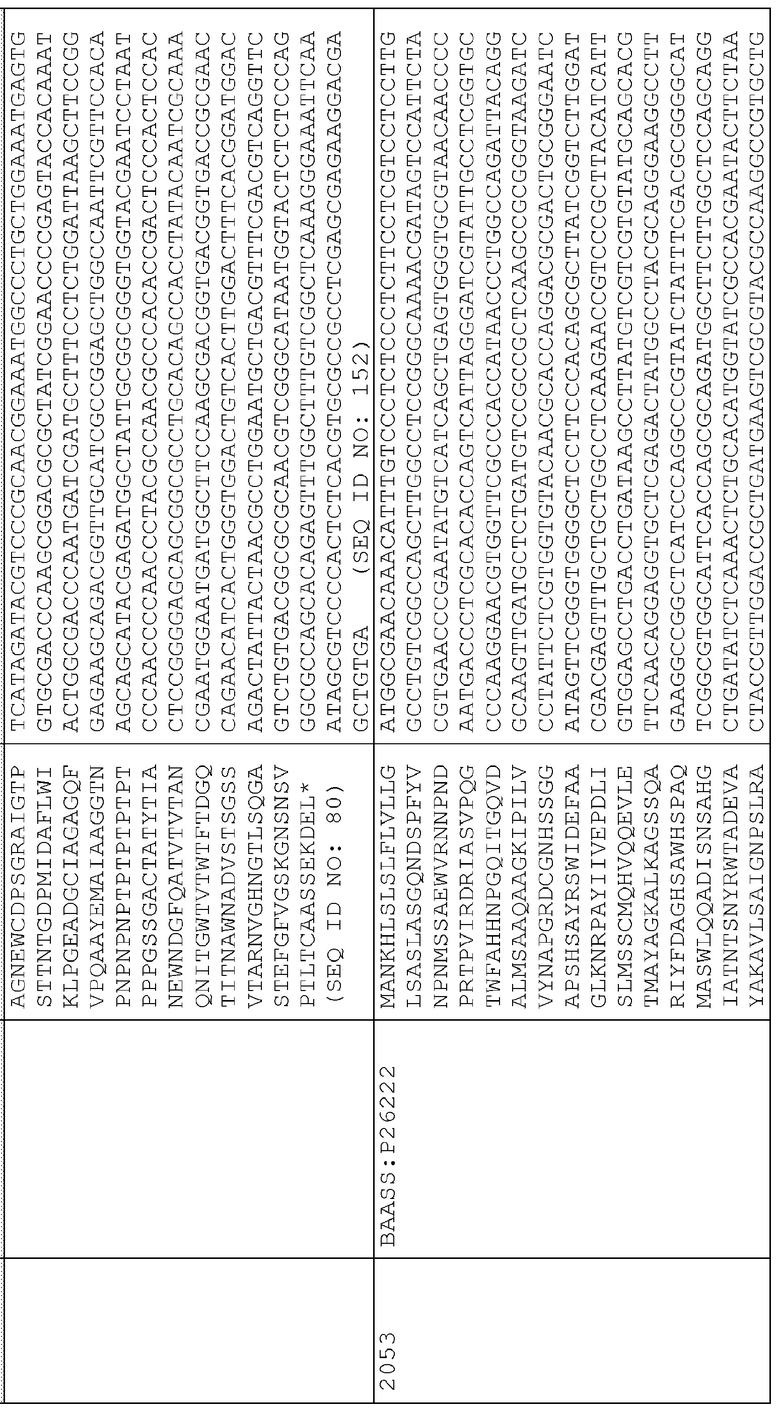





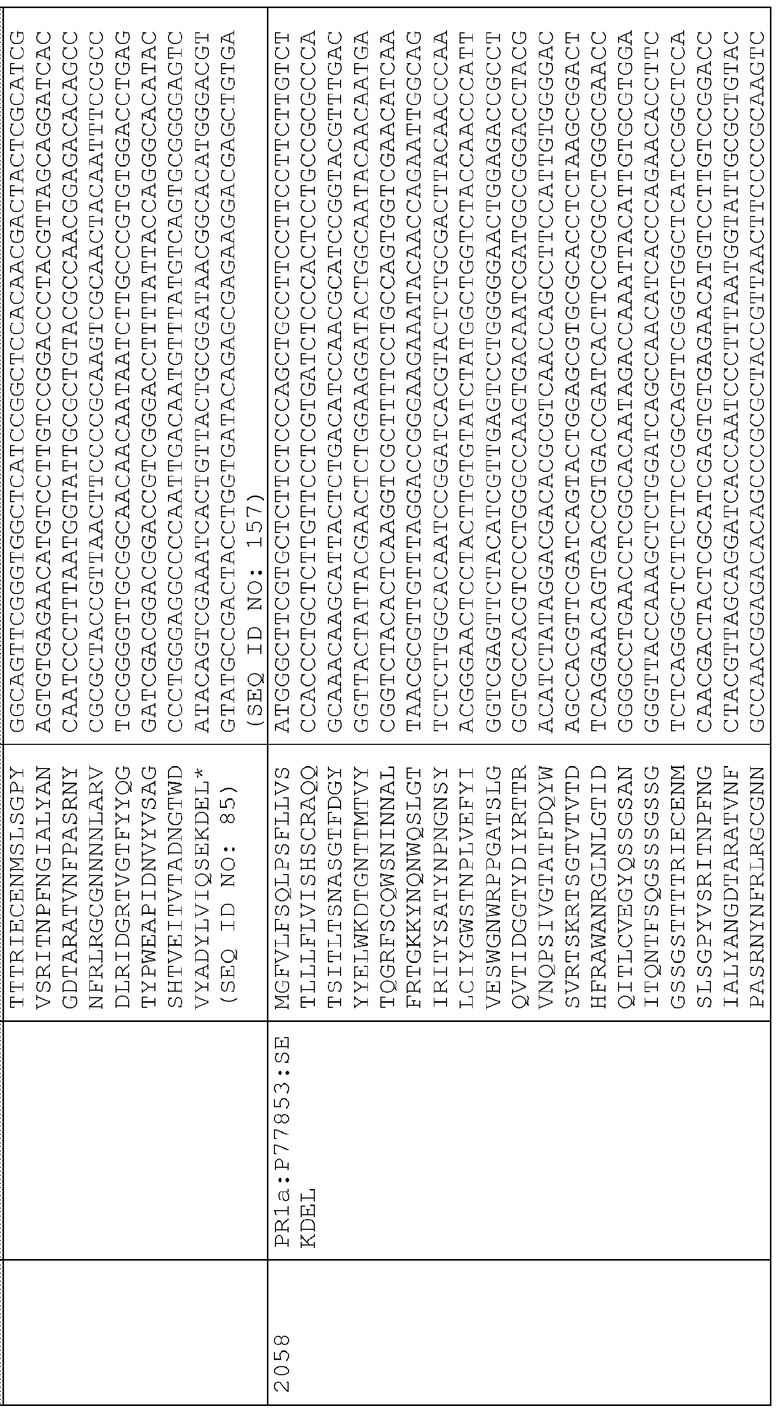
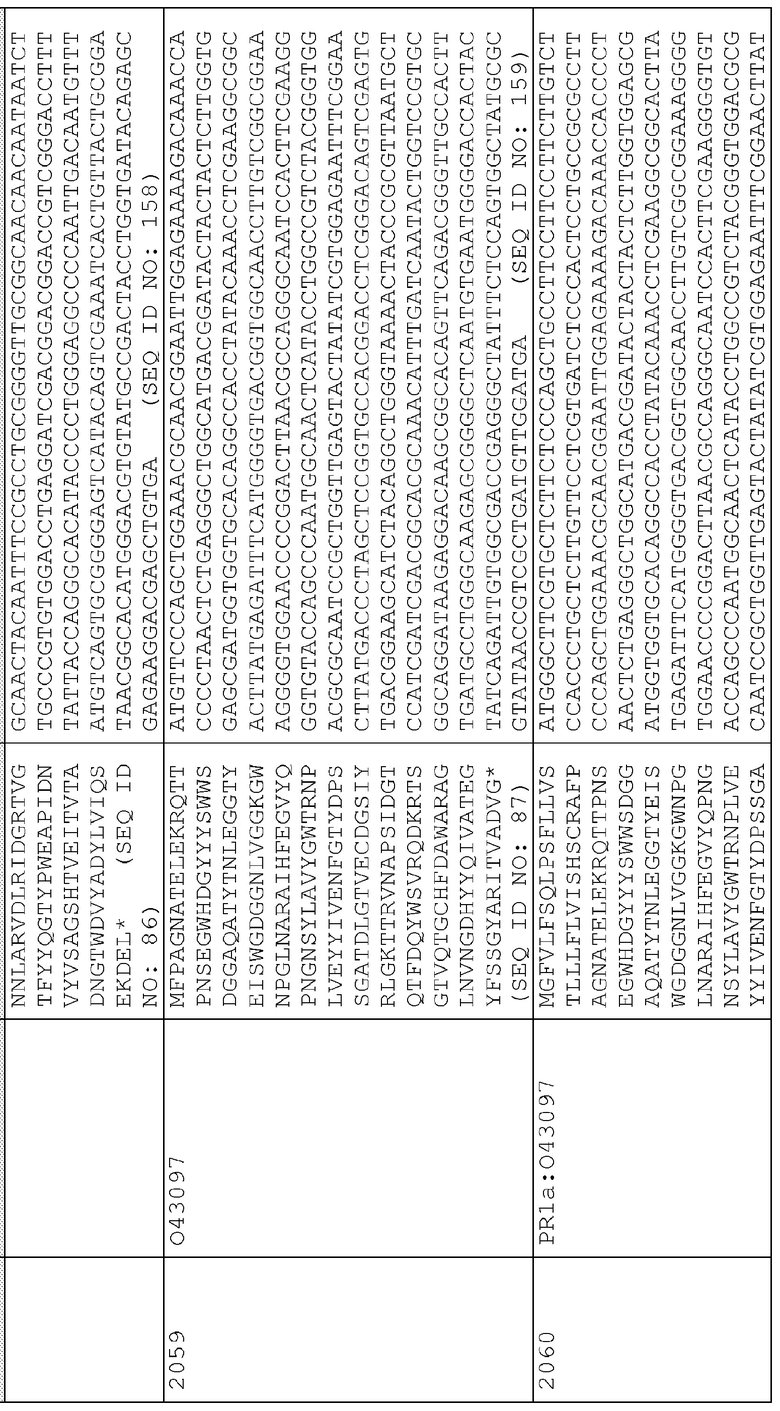

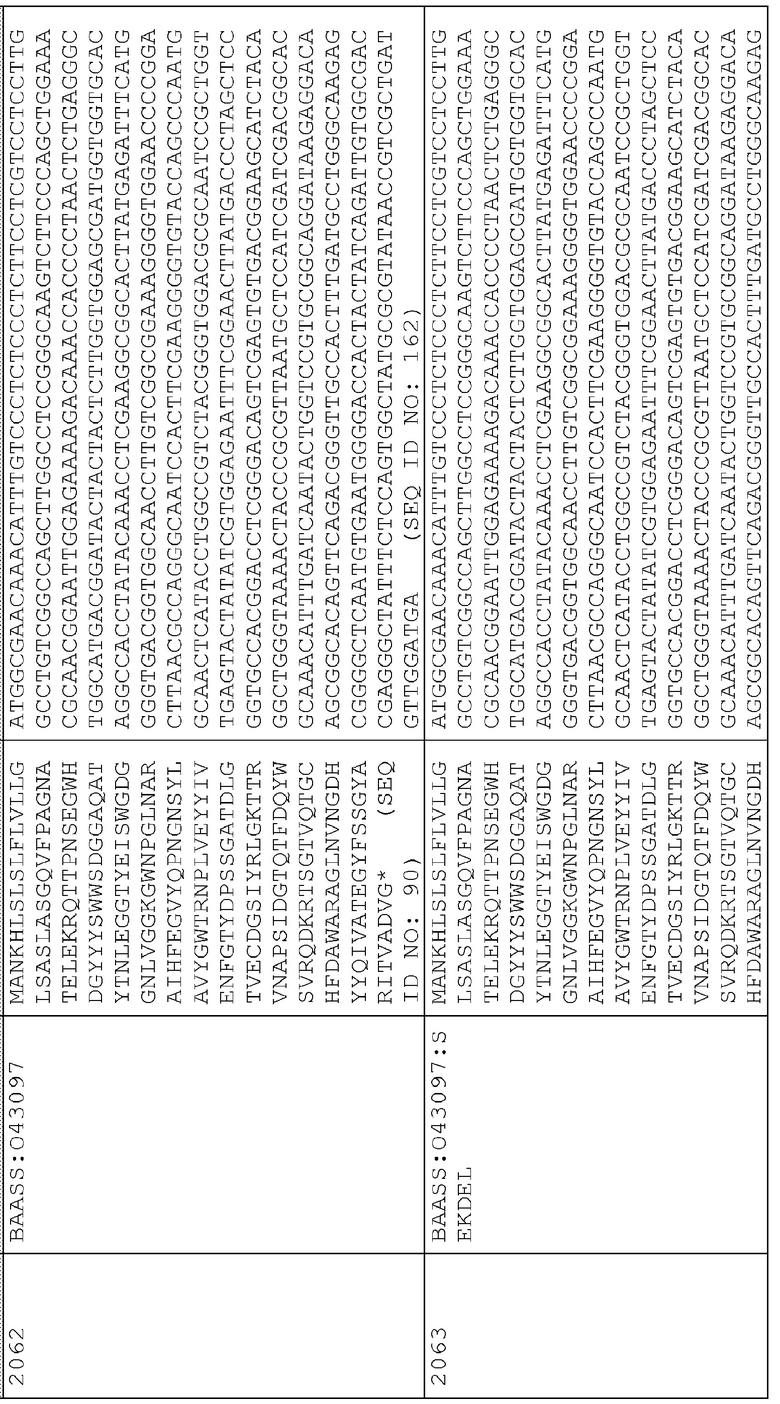











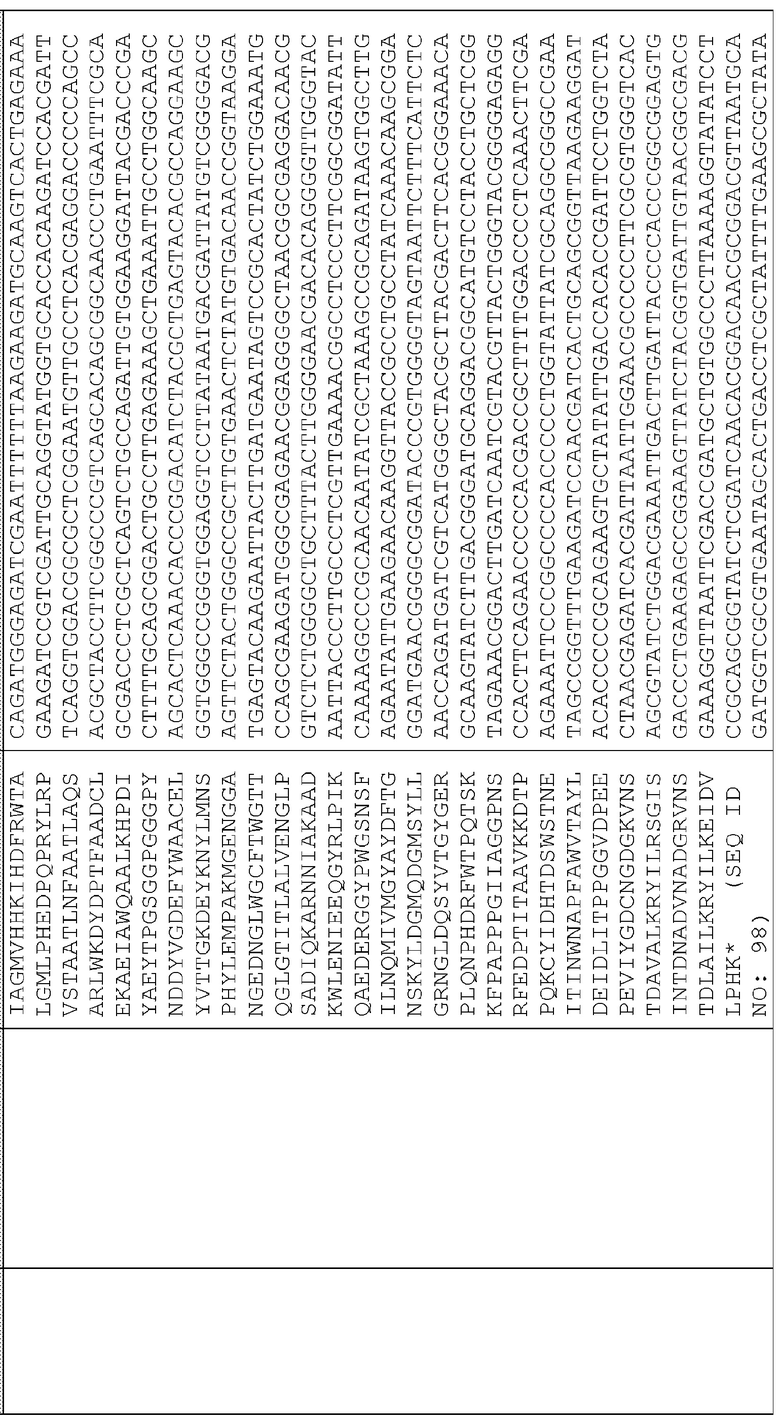







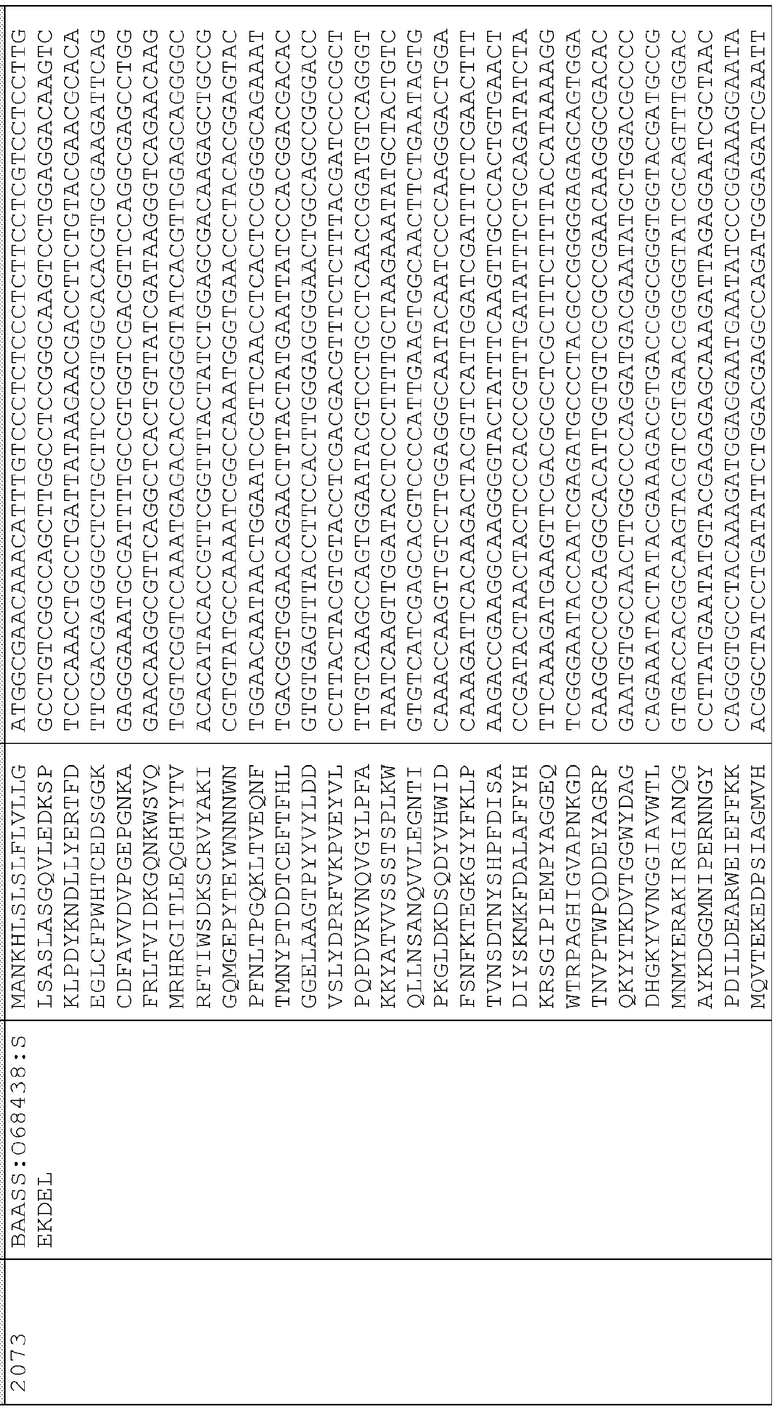




















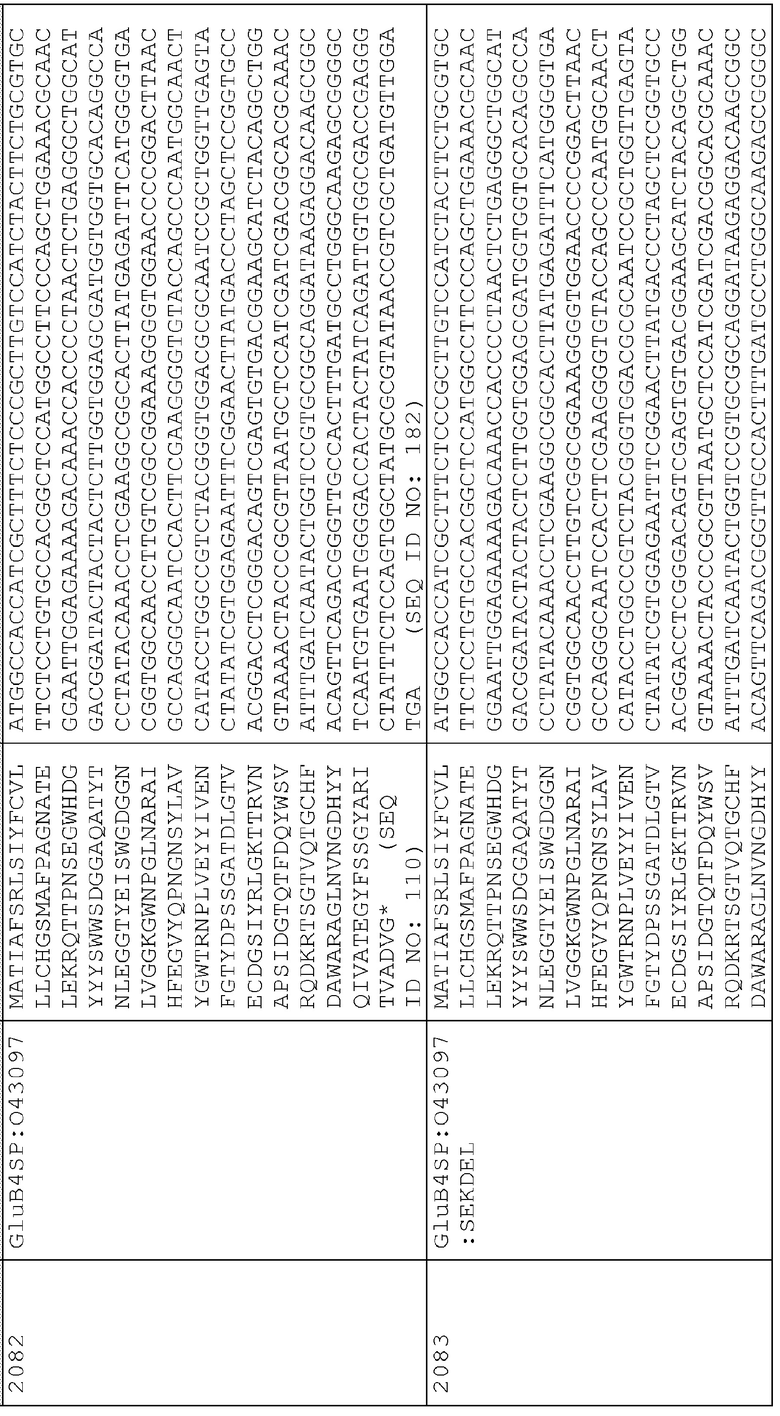







Пример 6 - Трансформация растений
Трансформация маиса
Опосредуемую Agrobacterium трансформацию незрелых эмбрионов маиса проводили, как описано в Negrotto et al., (2000) Plant Cell Reports 19: 798-803, которая включена в настоящий документ в качестве ссылки в полном объеме. Плазмиды для трансформации и гены селективных маркеров, используемые для трансформации, клонировали в вектор серии pAG, подходящий для трансформации однодольных растений, как описано выше. Векторы, используемые для этого примера, содержали ген фосфоманнозоизомеразы (PMI) (Negrotto et al. (2000) Plant Cell Reports 19: 798-803) в качестве селективного маркера, однако можно было использовать другие маркеры, обладающие такой же способностью.
Вектор для трансформации и штаммы Agrobacterium
Векторы для трансформации Agrobacterium tumefaciens конструировали с использованием стандартных молекулярных способов, известных в данной области, как описано выше. Плазмиды встраивали в штаммы Agrobacterium LBA4404+pSB1 (Ishida et al. (1996) Nature Biotechnology 14:745-750, которая включена в настоящий документ в качестве ссылки в полном объеме).
Ночные культуры штамма Agrobacterium, содержащего плазмиду, выращивали в течение двух суток на планшетах с твердой средой YP в течение от 2 до 4 суток при 28°C, содержавшей 100 мг/л спектиномицина и 10 мг/л тетрациклина.
Agrobacterium ресуспендировали в среде LS-inf, дополненной 100 мМ ацетосирингоном (As) (среда LSAs) (Negrotto et al., (2000) Plant Cell Rep 19: 798-803, которая включена в настоящий документ в качестве ссылки в полном объеме) пока клетки Agrobacterium не диспергировались однородно в суспензии. Затем суспензию Agrobacterium разбавляли до OD660 в диапазоне от 0,5 до 0,8 и встряхивали на устройстве для встряхивания в течение 15 секунд.
Инфицирование и одновременное культивирование незрелых эмбрионов маиса
Исходные растения маиса (Zea maize, культивары Hill, A188 или B73) выращивали в теплице при дневном свете в течение 16 часов при 28°C. Незрелые початки собирали от 7 до 15 суток после опыления и стерилизовали погружением в 20% хлорсодержащий отбеливатель (доступный под зарегистрированной торговой маркой CHLOROX®) в течение 15-20 минут. Затем стерилизованные початки тщательно промывали стерильной водой.
Незрелые зиготические эмбрионы выделяли из зерен и собирали в стерильную пробирку eppendorf, содержавшую жидкую среду LS-inf+100 plM As (LSAs). Эмбрионы встряхивали на устройстве для встряхивания в течение 5 секунд и промывали один раз свежей средой для инфицирования. Среду для инфицирования удаляли, добавляли раствор с Agrobacterium и эмбрионы встряхивали на устройстве для встряхивания в течение 30 секунд и позволяли им осесть с бактериями в течение 5 минут.
После инокуляции незрелые эмбрионы переносили стороной щитка вверх в среду LSAs и культивировали в темноте в течение от двух до трех суток при 22°С.
Выделение, селекция трансформированной эмбриогенной ткани маиса и регенерация растения
После одновременного культивирования незрелые эмбрионы переносили в среду LSDc, дополненную 200 мг/л тиментина и 1,6 мг/л нитрата серебра (Negrotto et al. 2000). Планшеты инкубировали в течение от 5 до 15 суток при 28°C в темноте.
Эмбрионы, образовавшие эмбриогенный каллюс, переносили в среду LSD1M0.5S (LSDc с 5 мг/л Dicamba, 10 г/л маннозы, 5 г/л сахарозы). Культуры подвергали селекции на этой среде в течение 6 недель с 3-недельными интервалами между пересевами. Выжившие культуры переносили либо в среду LSD1M0.5S для наращивания, либо в среду Reg1 (как описано в Negrotto et al., 2000). После культивирования на свету (режим 16 часов на свету/8 часов в темноте), зеленые ткани переносили в среду Reg2 без регуляторов роста (как описано в Negrotto et al., 2000) и инкубировали в течение 1-2 недель. Хорошо развитые проростки с листьями и корнями переносили в среду Reg3 (как описано в Negrotto et al., 2000) и выращивали на свету.
Собирали образцы листьев для анализа способом ПЦР в целях идентификации трансгенных растений, содержащих ген селективного маркера согласно Negrotto et al. (2000) и представляющий интерес ген. Положительные в ПЦР и пустившие корни растения промывали водой, чтобы смыть среду с агаром, и переносили в почву и выращивали в теплице для получения семян.
Трансформация проса
Среды, используемые для разработки протокола опосредуемой Agrobacterium трансформации, используемого для трансформации растения проса, получали с использованием стандартных способов, известных специалисту в данной области. В примерах, описанных в здесь, использовали следующие среды.
Среда для индукции соматических эмбрионов (SEI)
Среду SEI получали с использованием 4,3 г базальной солевой смеси MS, витаминов B5 (100 мг миоинозитола, 1 мг никотиновой кислоты, 1 мг пиридоксина HCl и 10 мг тиамина HCl), 30 г сахарозы, 5 мг 2,4-D и 10 мг BAP, 1,2 г/л Gelrite (Sigma, St. Louis, MO, США). Эти реагенты смешивали в стерильной воде, которую доводили до конечного объема 1 л. Перед автоклавированием pH доводили до 5,8.
Среда для регенерации
Среду для регенерации получали с использованием 4,3 г базальной солевой смеси MS, витаминов MS (100 мг миоинозитола, 1 мг никотиновой кислоты, 1 мг пиридоксина HCl и 10 мг тиамина HCl), 30 г сахарозы, и 1,2 г Gelrite (Sigma, St. Louis, MO, США). Эти реагенты смешивали в стерильной воде и доводили до конечного объема 1 л. Перед автоклавированием pH доводили до 5,8.
Среда для инокуляции (SW-1)
Среду SW-1 получали с использованием 4,3 г солей MS, витаминов B5 (100 мг миоинозитола, 1 мг никотиновой кислоты, 1 мг пиридоксина HCl и 10 мг тиамина HCl), 68,5 г сахарозы, 36 г глюкозы и 1 г казаминовых кислот. Эти реагенты смешивали в стерильной воде и доводили до конечного объема 1 л. Перед автоклавированием pH доводили до 5,8.
Среда для обновременного культивирования (SW-2)
Среду SW-2 приготавливали с использованием 4,3 г солей MS, витаминов B5 (100 мг миоинозитола, 1 мг никотиновой кислоты, 1 мг пиридоксина HCl и 10 мг тиамина HCl), 0,7 г L-пролина, 10 мг BAP, 5 мг 2,4-D, 0,5 г MES, 20 г сахарозы, 10 г глюкозы и 1,2 г Gelrite. Эти реагенты смешивали в стерильной воде и доводили до конечного объема 1 л. Перед автоклавированием pH доводили до 5,8.
Среда для покоящегося состояния (SW-3)
Среду SW-3 приготавливали с использованием 4,3 г солей MS, витаминов B5 (100 мг миоинозитола, 1 мг никотиновой кислоты, 1 мг пиридоксина HCl и 10 мг тиамина HCl), 10 мг BAP, 5 мг 2,4-D, 30 г сахарозы и 1,2 г Gelrite. Эти реагенты смешивали в стерильной воде и доводили до конечного объема 1 л. Перед автоклавированием pH доводили до 5,8.
Среде для селекции 1 (S1)
Среду S1 получали с использованием 4,3 г солей MS, витаминов B5 (100 мг миоинозитола, 1 мг никотиновой кислоты, 1 мг пиридоксина HCl и 10 мг тиамина HCl), 10 мг BAP, 5 мг 2,4-D, 5 г сахарозы, 10 г маннозы и 1,2 г Gelrite. Эти реагенты смешивали в стерильной воде и доводили до конечного объема 1 л. Перед автоклавированием pH доводили до 5,8.
Среда для регенерации (R1)
Среду R1 приготавливали с использованием 4,3 г солей MS, витаминов B5 (100 мг миоинозитола, 1 мг никотиновой кислоты, 1 мг пиридоксина HCl и 10 мг тиамина HCl), 30 г сахарозы и 1,2 г Gelrite. Эти реагенты смешивали в стерильной воде и доводили до конечного объема 1 л. Перед автоклавированием pH доводили до 5,8.
Инициация культур эмбриогенного каллюса
Зрелые семена проса (Panicum virgatum, cv. Alamo) приготавливали для трансформации путем удаления их оболочки с использованием наждачной бумаги. После удаления оболочки семян отдельные семена отбирали для стерилизации. Семена проса стерилизовали погружением в 20% хлорсодержащий отбеливатель (доступный под зарегистрированной торговой маркой CHLOROX®) в течение 5-10 минут. Затем стерилизованные семена тщательно промывали стерильной водой. Стерильные семена помещали на среду для индукции соматических эмбрионов (SEI) и инкубировали при 28°C в темноте в течение приблизительно 3-4 недель. Полученные культуры эмбриогенного каллюса переносили в свежую среду SEI и культивировали в течение дополнительных 6 недель с интервалами между пересевами 3 недели при 28°C в темноте.
Вектор для трансформации и штаммы Agrobacterium
Векторы для трансформации Agrobacterium tumefaciens конструировали, как описано выше, с использованием стандартных молекулярных способов, известных в данной области. Плазмиды вводили в штаммы Agrobacterium LBA4404+pSB1 (Ishida et al. (1996) Nature Biotechnology 14:745-750).
Ночные культуры штамма Agrobacterium, содержащего плазмиду, выращивали в течение двух суток на планшетах со средой YP, содержавшей 100 мг/л спектиномицина и 10 мг/л тетрациклина.
Приготовление Agrobacterium для трансформации
Культуру Agrobacterium инициировали еженедельно из исходной культуры в глицерине, хранящейся при -80°C, на полутвердой среде YP, содержавшей соответствующие антибиотики, и выращивали при 28°C в инкубаторе.
Agrobacterium наносили штрихами на свежую среду YP, содержавшую соответствующие антибиотики, за сутки до инокуляции и выращивали в инкубаторе при 28°C. Для использования в трансформации растений Agrobacterium собирали из чашки с использованием одноразовой пластмассовой петли для инокуляции и суспендировали в жидкой среде для инокуляции, такой как SW1, в стерильной 15-мл одноразовой полипропиленовой пробирке для центрифугирования. Agrobacterium ресуспендировали в пробирке путем встряхивания на устройстве для встряхивания в течение приблизительно 3-5 минут до тех пор, пока клетки Agrobacterium не диспергировались однородно в суспензии. Затем суспензию Agrobacterium разбавляли до OD660 в диапазоне 0,5-0,8 и встряхивали на устройстве для встряхивания в течение приблизительно 15 секунд.
Инфицирование и одновременного культивирование культур эмбриогенного каллюса проса
Многократные кластеры эмбриогенного каллюса проса типа II диаметром от 2 мм до 3 мм инфицировали Agrobacterium путем смешения эксплантатов с бактериальной суспензией, приготовленной, как описано выше, и встряхивали на устройстве для встряхивания в течение 30 с. Смесь инкубировали с приготовленными эксплантатами в течение приблизительно от 3 до 15 минут при комнатной температуре.
После инфицирования эксплантаты Agrobacterium помещали на среду для одновременного культивирования (SW-2) в чашках Петри размером 100×15 мм и инкубировали в течение от 2 до 3 суток при 22°C в темноте.
Регенерация и селекция трансгенных растений
После одновременного культивирования эксплантаты переносили на среду для выделения с антибиотиками для уничтожения Agrobacterium или для ингибирования роста Agrobacterium, без агента для селекции, такую как среда для выделения (SW3), дополненная 200 мг/л тиментина. Чашки инкубировали в течение от 5 до 15 суток при 28°C в темноте. Затем эксплантаты переносили на твердую среду S1 10 г/л маннозы и 5 г/л сахарозы), дополненную антибиотиками, на приблизительно от 14 до 21 суток. Затем эксплантаты переносили в свежую среду S1 (10 г/л маннозы и 5 г/л сахарозы) на приблизительно от 14 до 21 суток. Устойчивые клоны переносили в среду для дифференцировки эмбриона R1 (5 г/л маннозы и 10 г/л сахарозы) и инкубировали при 28°C в темноте в течение приблизительно от 2 до 3 недель.
Дифференцировавшиеся ткани растений переносили в свежую среду для дифференцировки эмбрионов R1 (5 г/л маннозы и 10 г/л сахарозы) и инкубировали при 26°C на свету в течение приблизительно от 2 до 3 недель.
Хорошо развившиеся проростки с листьями и корнями переносили в среду для прорастания корней. Образцы листьев брали для анализа ПЦР в целях идентификации трансгенных растений, содержащих ген селективного маркера согласно Negrotto et al. (2000) и представляющий интерес ген. Положительные в ПЦР и пустившие корни растения промывали водой, чтобы смыть среду с агаром, и переносили в почву и выращивали в теплице для получения семян.
Трансформация соматической эмбриогенной культуры сорго
Материалы и способы
Среды, используемые для протокола опосредуемой Agrobacterium трансформации, используемого для разработки трансформированных растений сорго, получали с использованием стандартных способов, известных специалисту в данной области. В примерах, описанных в настоящем описании, использовали следующие среды.
Среда для индукции соматических эмбрионов (SGWT-SEI)
4,3 г базальной смеси солей MS, витамины B5 (100 мг миоинозитола, 1 мг никотиновой кислоты, 1 мг пиридоксина HCl и 10 мг тиамина HCl), 1,2 г KH2PO4, 2,0 г L-пролина, 0,9 г L-аспарагина, 30 г сахарозы, 1,5 мг 2,4-D и 8 г агара (Sigma, St. Louis, MO, США) объединяли в стерильной воде. Конечный объем смеси доводили до 1 л с использованием стерильной воды. Перед автоклавированием pH доводили до 5,8.
Среда для регенерации (SGWT-R)
4,3 г базальной смеси солей MS, витамины B5 (100 мг миоинозитола, 1 мг никотиновой кислоты, 1 мг пиридоксина HCl и 10 мг тиамина HCl), 1,2 г KH2PO4, 2,0 г L-пролина, 0,9 г L-аспарагина, 30 г сахарозы, 1,0 мг IAA, 0,5 мг киннетина и 2,4 г Gelrite (Sigma, St. Louis, MO, США) объединяли в стерильной воде. Конечный объем смеси доводили до 1 л с использованием стерильной воды. Перед автоклавированием pH доводили до 5,8.
Среда для инокуляции (SGI-1)
4,3 г солей MS, витамины B5 (100 мг миоинозитола, 1 мг никотиновой кислоты, 1 мг пиридоксина HCl и 10 мг тиамина HCl), 68,5 г сахарозы, 36 г глюкозы, 1,0 г казаминовых кислот и 1,5 мг 2,4-D объединяли в стерильной воде. Конечный объем смеси доводили до 1 л с использованием стерильной воды. Перед автоклавированием pH доводили до 5,2.
Среда для одновременного культивирования (SGC-2)
4,3 г базальной смеси солей MS, витамины B5 (100 мг миоинозитола, 1 мг никотиновой кислоты, 1 мг пиридоксина HCl и 10 мг тиамина HCl), 1,2 г KH2PO4, 2,0 г L-пролина, 0,9 г L-аспарагина, 20 г сахарозы, 10 г глюкозы, 0,5 г MES, 1,5 мг 2,4-D, 40 мг ацетосирингона и 8 г агара объединяли в стерильной воде. Смесь доводили до конечного объема 1 л с использованием стерильной воды. pH доводили до 5,8.
Среда для индукции соматичесих эмбрионов (SGCI-3)
4,3 г базальной смеси солей MS, витамины B5 (100 мг миоинозитола, 1 мг никотиновой кислоты, 1 мг пиридоксина HCl и 10 мг тиамина HCl), 1,2 г KH2PO4, 2,0 г L-пролина, 0,9 г L-аспарагина, 30 г сахарозы, 1,5 мг 2,4-D и 8 г агара (Sigma, St. Louis, MO, США) объединяли в стерильной воде. Конечный объем смеси доводили до 1 л с использованием стерильной воды. pH доводили до 5,8. После автоклавирования добавляли тиментин до конечной концентрации 200 мг/л.
Среда для селекции 1 (SGS1-4)
4,3 г базальной смеси солей MS, витамины B5 (100 мг миоинозитола, 1 мг никотиновой кислоты, 1 мг пиридоксина HCl и 10 мг тиамина HCl), 1,2 г KH2PO4, 2,0 г L-пролина, 0,9 г L-аспарагина, 5 г сахарозы, 10 г маннозы, 1,5 мг 2,4-D и 8 г агара (Sigma, St. Louis, MO, США) объединяли в стерильной воде. Конечный объем смеси доводили до 1 л с использованием стерильной воды. pH доводили до 5,8. После автоклавирования добавляли тиментин до конечной концентрации 200 мг/л.
Среда для селекции 2 (SGS2-5)
4,3 г базальной смеси солей MS, витамины B5 (100 мг миоинозитола, 1 мг никотиновой кислоты, 1 мг пиридоксина HCl и 10 мг тиамина HCl), 1,2 г KH2PO4, 2,0 г L-пролина, 0,9 г L-аспарагина, 5 г сахарозы, 9,0 г маннозы, 1,5 мг 2,4-D и 8 г агара (Sigma, St. Louis, MO, США) объединяли в стерильной воде. Конечный объем смеси доводили до 1 л с использованием стерильной воды. pH доводили до 5,8. После автоклавирования добавляли тиментин до конечной концентрации 200 мг/л.
Среда для регенерации (SGR1-6)
4,3 г базальной смеси солей MS, витамины B5 (100 мг миоинозитола, 1 мг никотиновой кислоты, 1 мг пиридоксина HCl и 10 мг тиамина HCl), 1,2 г KH2PO4, 2,0 г L-пролина, 0,9 г L-аспарагина, 20 г сахарозы, 5,0 г маннозы, 1,0 мг IAA, 0,5 мг кинетин и 2,4 г Gelrite (Sigma, St. Louis, MO, USA) объединяли в стерильной воде. Конечный объем смеси доводили до 1 л с использованием стерильной воды. После автоклавирования добавляли тиментин до конечной концентрации 200 мг/л.
Инициация соматических эмбриогенных культур из незрелых зиготических эмбрионов
Незрелую зерновку сорго (Sorghum bicolor (L.) Moench) стерилизовали погружением в 20% хлорсодержащий отбеливатель (CHLOROX®) на 20 минут. Затем стерилизованную зерновку тщательно промывали стерильной водой.
Незрелые эмбрионы выделяли из зерновки и помещали в среду для индукции соматических эмбрионов (SGWT-SEI). Чашки инкубировали при 26-28°C в темноте в течение приблизительно 2-4 недель. Полученные соматические эмбриогенные кластеры использовали для экспериментов по трансформации или переносили в свежую среду SEI и культивировали в течение дополнительных от 3 до 6 недель с интервалами между пересевом 3 недели при 28°C в темноте перед применением в экспериментах по трансформации.
Вектор для трансформации и штаммы Agrobacterium
Векторы для трансформации Agrobacterium tumefaciens конструировали, как описано выше, с использованием стандартных молекулярных способов, известных в данной области. Плазмиды вводили в штаммы Agrobacterium LBA4404+pSB1 (Ishida et al. (1996) Nature Biotechnology 14:745-750).
Ночные культуры штамма Agrobacterium, содержащего плазмиду, выращивали в течение двух суток на планшетах со средой YP, содержащей 100 мг/л спектиномицина и 10 мг/л тетрациклина.
Приготовление Agrobacterium для трансформации
Культуру Agrobacterium инициировали еженедельно из исходной культуры в глицерине, хранящейся при -80°C, на полутвердой среде YP, содержавшей соответствующие антибиотики, и выращивали при 28°C в инкубаторе.
Agrobacterium наносили штрихами на свежую среду YP, содержавшую соответствующие антибиотики, за сутки до инокуляции и выращивали в инкубаторе при 28°C. Для использования в трансформации растений Agrobacterium собирали из чашки с использованием одноразовой пластмассовой петли для инокуляции и суспендировали в жидкой среде для инокуляции, такой как SW1, в стерильной 15-мл одноразовой полипропиленовой пробирке для центрифугирования. Agrobacterium ресуспендировали в пробирке путем встряхивания на устройстве для встряхивания в течение приблизительно 3-5 минут до тех пор, пока клетки Agrobacterium не диспергировались однородно в суспензии. Затем суспензию Agrobacterium разбавляли до OD660 в диапазоне от 0,5 до 0,8 и встряхивали на устройстве для встряхивания в течение приблизительно 15 секунд.
Инфицирование и одновременное культивирование соматических эмбриогенных культур сорго
Соматические эмбриогенные кластеры сорго инфицировали Agrobacterium путем смешения эксплантатов с бактериальной суспензией, приготовленной, как описано выше, и встряхивали на устройстве для встряхивания в течение 30 с. Смесь инкубировали с приготовленными эксплантатами в течение приблизительно от 3 до 15 минут при комнатной температуре.
После инфицирования эксплантаты Agrobacterium помещали на среду для одновременного культивирования (SGC-2) в чашках Петри размером 100×15 мм и инкубировали в течение от 2 до 3 суток при 22°C в темноте.
Регенерация и селекция трансгенных растений
После одновременного культивирования эксплантаты переносили на среду для выделения с антибиотиками для уничтожения Agrobacterium или для ингибирования роста Agrobacterium, без агента для селекции, такую как среда для выделения (SGCI-3), дополненная 200 мг/л тиментина. Чашки инкубировали в течение от 5 до 15 суток при 28°C в темноте.
Затем эксплантаты переносили на твердую среду SGS1-4 10 г/л маннозы и 5 г/л сахарозы), дополненную антибиотиками, на приблизительно от 14 до 21 суток.
Затем эксплантаты переносили в свежую среду SGS2-5 (10 г/л маннозы и 5 г/л сахарозы) на приблизительно 14-21 суток.
Устойчивые клоны переносили в среду для дифференцировки эмбриона SGR-6 (5 г/л маннозы и 10 г/л сахарозы) и инкубировали при 28°C в темноте в течение приблизительно от 2 до 3 недель.
Дифференцировавшиеся ткани растений переносили в свежую среду для дифференцировки эмбрионов R1 (5 г/л маннозы и 10 г/л сахарозы) и инкубировали при 26°C на свету в течение приблизительно от 2 до 3 недель.
Хорошо развившиеся проростки с листьями и корнями переносили в среду для прорастания корней.
Образцы листьев брали для анализа ПЦР в целях идентификации трансгенных растений, содержащих ген селективного маркера согласно Negrotto et al. (2000) и представляющий интерес ген. Положительные в ПЦР и пустившие корни растения промывали водой, чтобы смыть среду с агаром, и переносили в почву и выращивали в теплице для получения семян.
Пример 7 - Анализ трансгенных растений
Микробное продуцирование ферментов
В качестве части анализа трансгенных растений для получения стандартов ферментов можно использовать микробное продуцирование. Хотя продуцированные микробами ферменты могут обладать отличающимися паттернами гликозилирования или другими посттрансляционными модификациями, чем белок, экспрессируемый в растениях, микробный белок является приемлемым стандартом для получения антител, для измерения в анализах и для вестерн-блоттинга.
Пример 8 - Продуцирование ксиланаз с использованием P. pastoris
Гены, кодирующие представляющие интерес ферменты, клонировали в векторы экспрессии и трансформировали в пригодных экспрессирующих хозяев. Экспрессию в Pichai pastoris проводили в среде YPD при 30°C и 300 об./мин. Культуральные супернатанты собирали после экспрессии в течение от трех до пяти суток, что соответствовало моменту времени наиболее высокой активности фермента на мл очищенного супернатанта. Супернатант концентрировали проточной фильтрацией вдоль потока с 10-кДа мембраной MWCO и тщательно проводили замену буфера соответствующим буфером для реакции.
Количество фермента, присутствующего в концентрированных культуральных супернатантах, определяли путем обработки 10 мкл образца посредством PNGaseF (NEB) согласно протоколу изготовителя для удаления N-связанных гликанов из белка-мишени. Образец подвергали серийному разведению и 10 мкл каждого разведения фракционировали с помощью SDS-PAGE и окрашивали красителем Simply Blue Safe (Invitrogen) согласно инструкциям изготовителя. Концентрацию образца обозначали как наиболее высокий коэффициент разведения, при котором белок-мишень все еще поддавался детекции после окрашивания.
Получение антисыворотки кролика
Антитела, которые перекрестно реагируют с определенными белками, получали в New England Peptide. Представляющие интерес белки экспрессировали в Pichia pastoris. Полученный культуральный супернатант концентрировали проточной фильтрацией вдоль потока с использованием фильтра с 10-кДа MWCO (Millipore) и в некоторых случаях далее очищали колоночной хроматографией. Концентрат образца далее очищали с использованием устройства для фильтрации centricon с 10-кДа MWCO (Millipore), а затем фракционировали с помощью SDS-PAGE. Белковую полосу, соответствующую предсказанной молекулярной массе белка-мишени, вырезали из геля с использованием лезвия бритвы и отправляли в New England Peptide для получения антисыворотки. После получения специфичность каждой антисыворотки подтверждали вестерн-блоттингом, разделяли на аликвоты и хранили при 4°C или -20°C. Вестерн-блот анализ проводили в стандартных условиях, известных в данной области.
Пример 9 - определение активности ксиланазы с помощью измерения восстанавливающего сахара
Активность ксиланазы определяли с использованием ксилана березы в качестве субстрата и измеряя продуцирования концов с восстанавливающим сахаром с помощью микроанализа восстанавливающих сахаров Nelson-Somogyi (Green et al. 1989, Adaptation of the Nelson-Somogyi reducing-sugar assay to a microassay using microtiter plates, Anal Biochem. 1989 Nov 1;182(2):197-9, которая включена в настоящий документ в качестве ссылки в полном объеме). 2% (масс./об.) раствор субстрата приготавливали путем растворения ксилана березы (Sigma) в кипящей воде. В качестве консерванта добавляли 0,02% азид (конечная концентрация). Реагенты для анализа восстанавливающих сахаров Nelson-Somogyi приготавливали, как описано ранее (Green et al. 1989). Концентрации белка определяли с использованием набора для анализа белка BCA protein assay kit (Thermo Scientific) или предоставляли в качестве коэффициента разведения, как описано выше.
Анализируемая смесь состояла из 250 мкл 2% ксилана березы, 250 мкл буфера и различных объемов препарата ксиланазы (или стандартов ксиланазы, используемых для получения стандартной кривой) в общем объеме реакции один миллилитр. Анализы проводили при 60°C в течение 20 минут, затем помещали на лед для остановки реакции. Из каждой реакционной смеси 50 мкл каждой реакционной смеси анализировали в отношении присутствия восстанавливающих сахаров с использованием анализа восстанавливающих сахаров Nelson-Somogyi, как описано ранее. Единицы активности ксиланазы определяли из результатов, соответствующих линейному диапазону анализа. Удельную активность препаратов фермента вычисляли по следующему уравнению:
Удельная активность = (мМ образовавшихся восстанавливающих концов)/(концентрация в виде коэффициента разведения). Ссылаясь на фиг. 7, идентифицировали удельную активность трех ксиланаз с регистрационными номерами P40942, P77853 и O30700. Как показано, удельная активность O30700 в 5 раз превышает удельную активность P40942 и P77853, когда в качестве субстрата используют ксилан березы.
Пример 10 - анализ материала трансгенных растений
Трансгенные растения анализировали для определения уровней накопленного активного фермента. Для этих анализов образцы замороженной в жидком азоте ткани листьев растирали в ступке пестиком и растертый материал собирали. 10 мг растертого материала замороженных листьев распределяли в каждую лунку микропланшета для титрования. В каждую лунку добавляли 200 мкл 100-мМ буфера и реакционные смеси перемешивали пипетированием. Планшеты закрывали и помещали во вращающий инкубатор (200 об./мин.) при 55°C в течение 16 часов. После инкубации каждую реакционную смесь наносили на фильтрационный планшет Multiscreen HTS filterplate с 1,2-мкм фильтр из стекловолокна (Millipore, Billerica MA) и фильтровали центрифугированием при 500 x g в течение 3 минут. Активность фермента оценивали путем анализа 50 мкл полученного фильтрата с использованием анализа восстанавливающих сахаров Nelson-Somogyi, как описано ранее. Экстрагированный белок определяли с использованием набора для анализа белка BCA protein assay kit (Thermo). Уровни активности были представлены в качестве мМ концов восстанавливающих сахаров, образовавшихся на мг экстрагированного белка.
Ссылаясь на фиг. 8, представлена активность различных образцов трансгенных растений, экспрессирующих ксиланазу P77853. Образцы, обозначенные AG2014 и AG2015, трансформировали плазмидами pAG2014 и pAG2015, соответственно, и AG2004 представляет собой контроль. Продуцирование восстанавливающих сахаров в образцах трансгенных растений по сравнению с образцом дикого типа показало накопление активной ксиланазы в ткани трансгенного растения.
Пример 11 - определение активности против конъюгированных с pNP гликозидов
Для охарактеризации диапазона ферментативной активности конкретных ксиланаз проводили несколько анализов с использованием конъюгированных с п-нитрофенолом (pNP) гликозидов. Одномолярные исходные растворы субстратов получали в диметилсульфоксиде. Реакционные смеси состояли из 5-мМ (конечная концентрация) субстрата, 100-мМ буфера в 50 мкл и 1-10 мкл препарата фермента. Затем полученные реакционные смеси инкубировали при 60°C в течение одного часа. Реакции останавливали и проявляли добавлением 100 мкл 0,1-M карбонатного буфера, pH 10,5. Гидролиз субстрата, на который указывало образование NP, выявляли в качестве увеличения поглощения при 400 нм.
Субстраты для эндогидролиза полисахаридов также определяли с использованием источников конъюгированных с AZCL субстратов (Megazyme) и использовали согласно стандартному протоколу изготовителя. В кратком изложении, 250 мкл определенного буфера смешивали со 100 мкл препарата фермента и 150 мкл воды. Реакционную смесь помещали в инкубатор с водяной баней, установленный на желаемую температуру (обычно от 37°C до 70°C) на пять минут, после чего добавляли одну таблетку либо xylazyme AX, либо cellazyme C. Реакционную смесь инкубировали в течение 10 минут, затем удаляли из инкубатора и останавливали с помощью 10 мл 2% (масс./об.) Tris Base (Sigma®). На эндогидролиз полисахаридного субстрата указывало высвобождение растворимого синего красителя. Количество высвобожденного красителя количественно определяли путем измерения поглощения реакционного супернатанта при 590 нм. Контроли для этих реакций включали экстракты белка из штамма дикого типа и продуцирующего рекомбинантный фермент штамма P. pastoris или E. coli.
В таблице 1 ниже представлена выявленная активность нескольких ксиланаз. Как указано, активность эндоксиланазы выявляли для образцов P77853, О30700 и P40942. Активность целлобиогидролазы и β-глюкозидазы была определена в образцах, содержащих P40942, что указывает на то, что этот фермент способен к эндогидролизу ксилана и экзогидролизу целлюлозы и целлобиозы.
Пример 12 - определение термической стабильности
Термическую стабильность ферментов оценивали по восстановлению ферментативной активности после инкубации при повышенных температурах. В кратком изложении, препараты ксиланазы P77, O30 или O40 инкубировали при 4°C, 50°C, 60°C, 70°C или 80°C в течение одного часа, а затем анализировали с использованием субстрата xylazyme AX, как описано выше. Ссылаясь на фиг. 9, ксиланазы O30700 и P77853 сохраняют практически 100% активность после инкубации в течение одного часа при вплоть до 60°C, однако имеют сниженную активность при воздействии температурной обработки при 70°C и 80°C. Ксиланаза P40942 сохраняет практически 100% активность после одного часа при температурах вплоть до 70°C, в то время как при 80°C ее активность была снижена относительно воздействия при более низких протестированных температурах.
Термическая стабильность фермента является одной характеристикой, которая может повлиять на его применимость в различных применениях. Например, при переработке лигноцеллюлозной биомассы; например, которая происходит из кукурузы (соломы), проса, мискантуса, сорго или сахарного тростника, если трансгенный материал биомассы подлежит обработке при 70°C в течение одного часа, P40942 может быть лучшим ферментом для доставки обеспечения ксиланазы, чем O30700 или P77853, вследствие его увеличенной стабильности при этой температуре. Напротив, если трансгенное зерно; например, из трансгенной кукурузы или сорго, намереваются использовать для составления рациона на корме для животных, где корм растирают и перемешивают при температуре 50°C, тогда любой из этих ферментов может быть в достаточной степени термически стабильным. Однако это применение конкретных ферментов не исключает других применений тех же конкретных ферментов.
Пример 13 - материалы и способы для оценки трансгенных растений и способы их предварительной обработки и ферментативного гидролиза
Для переработки биомассы и определенных тканей растений можно использовать различные конфигурации способа. Одна из конфигураций способа называется макромасштабным способом, масштаб которого можно увеличивать, и он описан непосредственно ниже. Другая конфигурация способа называется микромасштабным способом, который можно использовать для оценки растений, и он подроблено описан ниже после описания макромасштабного способа.
Пример 13a - Макромасштабный способ - макромасштабная последовательная низкотемпературная хемимеханическая предварительная обработка (CMPT) и одностадийный ферментативный гидролиз:
Ссылаясь на фиг. 10, конверсию биомассы в ферментируемые сахара с помощью макромасштабного способа использовали с несколькими типами сырья. На фиг. 10 проиллюстрирована принципиальная схема для макромасштабного способа.
Получение субстрата биомассы:
Кукурузную солому трансформировали указанной плазмидой, содержащей любую из β-глюкозидазы, эндоглюканазы, целлобиогидролазы, FAE или ксиланазы, или комбинацию ферментов. Используемый вектор может представлять собой любой вектор, кодирующий CWDE или его производное, включая любой один или несколько из векторов, описанных в настоящем описании. В этом примере вектор представлял собой pAG2015, pAG2042 и pAG2063. Солому сушили в циркуляторе воздуха при 37°C в течение приблизительно 2 недель. Высушенную кукурузную солому 1010 нарезали до длины 1,0-1,5 дюйма (2,5-3,8 см).
Предварительная обработка:
Нарезанную высушенную кукурузную солому 1010 предварительно обрабатывали на стадии 1020 с использованием либо чистой воды, либо комбинации 8%-38% (масс./масс. в расчете на кукурузную солому) бисульфита аммония и 4%-19% (масс./масс. в расчете на кукурузную солому) карбоната аммония (pH 7,6-8,5). Биомассу добавляли в колбу с раствором для предварительной обработки при отношении жидкости к твердому веществу (L/S), равном 8. Смесь встряхивали при температурах 40°C-90°C в течение от четырех до 19 часов. Предварительно обработанный материал фильтровали с использованием фильтровальный бумаги VWR категории 415, и материал 1025 собирали для дальнейшего анализа.
Очистка:
Предварительно обработанную биомассу очищали на стадии 1030 в смеси с DI-водой при 40°C-90°C. После перемешивания биомассу фильтровали с использованием фильтровальной бумаги VWR категории 415. Очищенную биомассу (пульпу), которая не проходила, промывали DI-водой при 40°C-90°C. Пульпу 1035 хранили при 4°C для уравновешивания влаги и дальнейшего ферментативного гидролиза.
Ферменты:
Использовали фермент AccelleraseTM 1000 (Genencor International, Rochester, NY). Активность эндоглюканазы составляла 2500 CMC Е/г (минимум). Активность бета-глюкозидазы составляла 400 pNPG Е/г (минимум). Внешний вид представлял собой коричневую жидкость. Значение pH составляло 4,8-5,2.
Альтернативно использовали коктейль ферментов, который содержал: эндоглюканазу (C8546), β-глюкозидазу (49291) и ксиланазу (X2753), все приобретенные от Sigma (St. Louis, MO), и целлобиогидролазу (E-CBHI), которую приобретали от Megazyme (Wicklow, Ирландия).
Ферментативный гидролиз:
Следовали стандартному протоколу NREL (LAP-009). На стадии 1040 предварительно обработанную и очищенную солому гидролизовали в 0,1 M цитрате натрия (pH 5,0) при содержании твердого материала биомассы 6,0% с нагрузкой ферментом 0,2-0,4 мл на г кукурузной соломы для высвобождения сахара 1045. Реакцию проводили в 250 мл колбе erhlenmeyer при 250 об./мин. в течение периода 0-48 ч при 45°C-55°CВ зависимости от смеси ферментов и фермента, экспрессируемого в растении, pH варьировали от 5 до 9. Предпочтительное значение pH для этих смесей ферментов обычно составляло 5.
Необязательно в реакционную смесь для гидролиза можно добавлять тетрациклин или эквивалентный антибиотик для предотвращения роста какой-либо потенциальной микробной контаминации.
Анализ ферментируемых сахаров:
Образцы гидролизата нагревали при 95°C в течение 20 мин, а затем центрифугировали при 9000 x g, после чего супернатанты очищали пропусканием через 0,20-мкм фильтры PVDF (каталожный номер #: 09-910-13, Fisher Scientific, Pittsburg, PA). Концентрации моносахаридов и дисахаридов определяли высокоэффективной жидкостной хроматографией (ВЭЖХ) с использованием двойного насоса Shimadzu LC-20 AD с программным обеспечением LC solutions (Shimadzu, Kyoto, Япония). Концентрации сахара определяли с использованием колонки для сахаров Aminex HPX-87P (Bio-Rad Laboratories, Hercules, CA), действующей при 0,6 мл/мин и 85°C с дегазированной водой в качестве подвижной фазы. Области пиков для всех образцов, проанализированных с помощью детектора RI (RID 10AD), интегрировали и величины сравнивали со стандартными кривыми для количественного определения.
Результаты макромасштабной переработки
1 - кукурузная солома из растений AxB дикого типа. Для кукурузной соломы теоретический выход сахара составляет 33,5% (масс./масс.) глюкозы и 16,3% (масс./масс.) ксилозы.
Предварительная обработка: проводили, как описано выше либо с 8% бисульфатом аммоний и 4% карбонатом аммония, либо с 38% бисульфатом аммония, 19% карбонатом аммония при температуре 70°C в течение 4 ч.
Ферментативный гидролиз: проводили, как описано выше в течение 24 или 48 ч.
Результаты представлены в таблице 2, ниже. Выходов глюкозы 54,5% (24 часов) и 62,3% (48 часов), а также выходов ксилозы 20% (24 часов) и 27,5% (48 часов) можно достигнуть при ферментативном гидролизе в течение одних суток и двух суток после предварительной обработки разбавленным химическим реагентом. Результаты демонстрируют эффективность низкотемпературной CMPT в отношении ферментативного гидролиза.
Выход глюкозы и ксилозы после ферментативного гидролиза предварительно обработанной кукурузной соломы (АхВ)
4% Карбонат
19% Карбонат
2 - Солома. Высушенную в печи кукурузную солому AxB дикого типа тестировали и сравнивали против смеси соломы из девяти трансгенных растений кукурузы с pAG2015 (называемых в настоящем документе "2015M").
Предварительная обработка: проводили, как описано выше, с 16% бисульфатом аммония и 8% карбонатом аммония (pH 7,6) при 70°C в течение 4 ч.
Ферментативный гидролиз: проводили, как описано выше в течение 0 или 24 ч.
Результаты представлены в таблице 3, ниже. Лучшую эффективность в выражении выхода сахара наблюдали из трансгенных растений кукурузы с pAG2015, чем из растений AxB дикого типа.
Выход глюкозы и ксилозы после ферментативного гидролиза предварительно обработанной кукурузной соломы (АхВ и 2015М)
Пример 13b - Микромасштабный способ: Упрощенные низкотемпературная хемимеханическая предварительная обработка (CMPT) и ферментативный гидролиз
Ссылаясь на фиг. 11, микромасштабный способ осахаривания использовали для скрининга нескольких типов сырья биомассы в отношении конверсии в ферментируемые сахара с использованием либо одностадийного, либо двухстадийного ферментативного гидролиза.
Приготовление субстрата биомассы:
Получали кукурузную солому 1110 из кукурузы, трансформированной желаемым вектором, содержащим любую из бета-глюкозидазы, эндоглюканазы, целлобиогидролазы, FAE или ксиланазы, или комбинацией ферментов. Солому сушили в циркуляторе воздуха при 37°C в течение приблизительно 2 недель. После высушивания кукурузную солому нарезали до длины 1,0-1,5 дюйма (2,5-3,8 см). Солому измельчали на стадии 1120 с использованием мельницы UDY (Model 014, UDY Corporation, Fort Collins, Co) с 0,5-мм ситом.
Предварительная обработка:
Измельченную кукурузную солому предварительно обрабатывали на стадии 1130 с использованием либо чистой воды, либо химических реагентов. Биомассу добавляли в 2-мл пробирки с раствором для предварительной обработки в соотношении жидкости и твердого вещества 10. Можно было использовать 20 мг биомассы. Смесь встряхивали при температуре 40°C-90°C в течение 15-19 ч. Предварительно обработанный материал подвергали ферментативному гидролизу без промывания между стадиями.
Ферменты:
Все из эндоглюканазы (C8546), бета-глюкозидазы (49291) и ксиланазы (X2753) приобретали от Sigma® (St. Louis, MO). Целлобиогидролазу (E-CBHI) приобретали от Megazyme® (Wicklow, Ирландия).
Ферментативный гидролиз:
Способ основан на стандартном протоколе NREL (LAP-009).
Одностадийный гидролиз:
Измельченную предварительно обработанную солому суспендировали при нагрузке 2% (масс./об.) глюканом в polybuffer (50 мМ цитрат Na, 20 мМ фосфат K, двухосновный, 17 мМ аргинин, 40 мМ глицин, 25 мМ EPPS, 20 мМ HEPES, 0,02% азид натрия) с величинами pH в диапазоне от 3,5 до 5,0. Использованное значение pH было основано на конечном pH суспендированной предварительно обработанной соломы. Нагрузка ферментами коктейля была основана на экспериментах с использованием 10 мг соломы и она приведена в таблице 4 ниже. Анализ проводили на биомассе без добавления ферментов (без коктейля), и с коктейлем без ксиланазы, эндоглюканазы или других ферментов, которые экспрессировались в растении (коктейль минус ксиланаза или эндоглюканаза, в зависимости от фермента, экспрессируемого в растениях), при гидролизе. Его проводили для оценки эффекта в растениях экспрессируемых ферментов на гидролиз. Образцы гидролизовали при 40°C или 50°C в течение 48-96 ч при 200 об./мин. (реакционный объем 1 мл).
В смесь для гидролиза необязательно можно добавлять тетрациклин или эквивалентный антибиотик для предупреждения роста какой-либо потенциальной микробной контаминации.
Нагрузка ферментом для полного коктейля
Двухстадийный гидролиз:
Ферментативному гидролизу первой стадии давали название, в зависимости ферментов, экспрессируемых в растении (например, "гидролиз ксиланазой" или "гидролиз глюканазой"). Последующий ферментативный гидролиз второй стадии, был назван "гидролизом коктейлем ферментов".
Для первой стадии измельченную предварительно обработанную солому суспендировали при нагрузке 3% (масс./об.) глюканом в polybuffer с pH в диапазоне от 5,0 до 8,4. Использованное значение pH был основано на оптимальном pH для экспрессируемого в растении фермента. Этот гидролиз проводили при 55°C, 300 об./мин. в течение 24-48 ч.
Для гидролиза коктейлем ферментов pH при необходимости доводили до 5,0 с использованием концентрированной HCl. Затем к образцам добавляли ферменты коктейля, как указано для одностадийного ферментативного гидролиза, с получением образцов без коктейля, с полным коктейлем или с коктейлем минус ксиланаза или эндоглюканаза. Добавляли Polybuffer, pH 5,0, для конечного содержания твердых веществ 2%. Образцы гидролизовали при 50°C при 200 об./мин. в течение 48-96 ч.
Необязательно в реакционную смесь для гидролиза можно добавлять тетрациклин или эквивалентный антибиотик для предотвращения роста какой-либо потенциальной микробной контаминации.
Анализ ферментируемых сахаров:
Образцы гидролизата инкубировали при 95°C в течение 20 мин, а затем центрифугировали при 9000 x g, после чего супернатанты очищали пропусканием через 0,20-мкм фильтры PVDF. Концентрации моносахаридов и дисахаридов определяли высокоэффективной жидкостной хроматографией (ВЭЖХ) с использованием двойного насоса Shimadzu LC-20 AD с программным обеспечением LC solutions (Shimadzu, Kyoto, Япония). Концентрации сахара определяли с использованием колонки для сахаров Aminex HPX-87P (Bio-Rad Laboratories, Hercules, CA), действующей при 0,6 мл/мин и 85°C с дегазированной водой в качестве подвижной фазы. Области пиков для всех образцов, проанализированных с помощью детектора RI (RID 10AD), интегрировали и величины сравнивали со стандартными кривыми для количественного определения.
Результаты микромасштабной переработки
1 - Одностадийный ферментативный гидролиз, pAG2015. Анализированная растительная солома: для получения соломы использовали трансгенное растение кукурузы, обозначенное как 2015.05 (полученное трансформацией кукурузы посредством pAG2015, которая экспрессирует ксиланазу). Контрольное растение: для получения контрольной соломы использовали трансгенное растение кукурузы, обозначенное как 2004.8.4 (растение поколения T1, происходящее из исходного растения, которое было получено трансформацией кукурузы посредством pAG2004, которая не кодирует фермент ксиланазу). Теоретический выход сахара: 2015.05: 33,35% глюкозы, 18,69% ксилозы; 2004.8.4: 2015.05: 34,68% глюкозы, 20,6% ксилозы.
Предварительная обработка: проводили, как описано выше с 1:19 (об./об.) 15% NH4OH, 20% NH4Cl при 40°C или 60°C в течение 15 ч, 300 об./мин.
Одностадийный ферментативный гидролиз: как описано выше для 0,02% азида натрия при 50°C в течение 48 часов, 250 об./мин.
На фиг. 12 проиллюстрирован выход глюкозы и ксилозы (процент от массы биомассы) после ферментативного гидролиза предварительно обработанной кукурузной соломы (2015.05 и 2004.8.4). Как представлено на фиг. 12, 2015.05 демонстрирует лучшую эффективность гидролиза, исходя как из общего выхода гидролиза, так и из эффекта ксиланазы на гидролиз в растении (как показано с помощью обработки "коктейль-Xyl"). На фиг. 12, используют следующие обозначения: 40C PT: предварительную обработку проводили при 40°C; 60C PT: предварительную обработку проводили при 60°C. "Коктейль-Xyl" обозначает одностадийный ферментативный гидролиз, который проводили без ксиланазы в коктейле внешних ферментов. Каждый обозначенный образец на фиг. 12 демонстрирует результаты для отсутствия коктейля, полного коктейля и коктейля-Xyl слева направо.
2 - Одностадийный ферментативный гидролиз, pAG2063. Анализированная растительная солома: для получения соломы использовали трансгенные растения, обозначенные как 2063.13 и 2063.17 (полученные трансформацией кукурузы посредством pAG2063, которая экспрессирует ксиланазу). Для получения контрольной соломы использовали контрольное растение, обозначенное как 2004.8.4 (трансгенное растение, полученное трансформацией кукурузы посредством pAG2004; без экспрессии фермента ксиланазы).
Предварительная обработка: проводили, как описано выше, с 1:19 (масс./масс.) 15% NH4OH, 20% NH4Cl, при либо 40°C, либо 60°C, в течение 15 ч, 300 об./мин.
Одностадийный ферментативный гидролиз: проводили, как описано выше, с 1,0 мг/мл тетрациклина при 50°C в течение 48 часов, 250 об./мин.
На фиг. 13 проиллюстрирован выход глюкозы и ксилозы (процент от массы биомассы) после ферментативного гидролиза предварительно обработанной кукурузной соломы (2004.8.4, 2063.13 и 2063.17). Как представлено на фиг. 13, трансгенное растение 2063.17 демонстрирует лучшую эффективность гидролиза, чем контрольное растение и 2063.13, исходя как из общего выхода гидролиза, так и из эффекта ксиланазы на гидролиз в растении (как показано посредством обработки "коктейль-Xyl"). На фиг. 13 использовали следующие обозначения: 40C PT: предварительную обработку проводили при 40°C; 60C PT: предварительную обработку проводили при 60°C. "Коктейль-Xyl" обозначает одностадийный ферментативный гидролиз, который проводили без включения ксиланазы в коктейль внешних ферментов. Каждый обозначенный образец на фиг. 13 демонстрирует результаты для коктейля-Xyl и полного коктейля справа на лево. Образцы, где являются видимыми три столбца, демонстрируют результаты без коктейля слева от результатов для полного коктейля.
3 - Двухстадийный ферментативный гидролиз, pAG2014. Анализированная растительная солома: для получения соломы использовали трансгенное растение 2015.05; и для получения контрольной соломы использовали контрольное растение 2004.8.4. Как используют в настоящем изобретения, растение TO представляет собой 1 поколение; и растение T1 представляет собой 2 поколение, полученное из семян растений T0.
Предварительная обработка: проводили, как описано, с DI-водой при 55°C в течение 16 ч, 300 об./мин.
Ферментативный гидролиз первой стадии (гидролиз ксиланазы): проводили, как описано ранее, при 55°C, в течение 24 ч с 0,02% азидом натрия, 250 об./мин.
Гидролиз второй стадии (гидролиз коктейлем ферментов): проводили, как описано, при 50°C с использованием коктейля в течение 48 ч.
На фиг. 14 представлены выходы глюкозы и ксилозы (процент от массы биомассы) после ферментативного гидролиза предварительно обработанной кукурузной соломы (2015.05 и 2004.8.4). Растения 2015.05 как T0, так и T1, демонстрируют лучшую эффективность гидролиза, исходя из общего выхода гидролиза и эффекта ксиланазы на гидролиз в растении (см. фиг. 14, обработка "Ct-xyl"). На фиг. 14 использовали следующие обозначения: "N Ct": без коктейля, "F Ct": полный коктейль, "Ct-xyl": коктейль без ксиланазы. Каждый меченый образец на фиг. 14 демонстрирует результаты для отсутствия коктейля, полного коктейля и коктейля-Xyl слева направо.
4 - Двухстадийный ферментативный гидролиз, pAG2063. Анализированная растительная солома: Для получения соломы использовали трансгенное растение, обозначенное как 2063.17 (полученное трансформацией кукурузы посредством pAG2063). Для получения контрольной соломы использовали контрольное растение, обозначенное как 2004.8.4 (полученной трансформацией внутренней части посредством pAG2004).
Предварительная обработка: проводили, как описано, с DI-водой при 55°C в течение 16 ч, 300 об./мин.
Ферментативный гидролиз первой стадии (гидролиз ксиланазой): проводили, как описано ранее, при 55°C, в течение 24 ч с 0,02% азидом натрия, 250 об./мин.
Гидролиз второй стадии (гидролиз коктейлем ферментов): проводили, как описано, при 50°C с использованием коктейля в течение 96 ч.
На фиг. 15 представлены выходы глюкозы и ксилозы (процент от массы биомассы) после ферментативного гидролиза предварительно обработанной кукурузной соломы (2064.17 и 2004.8.4). Как представлено на фиг. 15, выходы как глюкозы, так и ксилозы для 2063.17 последовательно превышают выход для 2004.8.4 на протяжении курса предварительной обработки, гидролиза ксиланазой 1 стадии и гидролиза коктейлем ферментов 2 стадии. Выход ксилозы для 2063.17 возрастает на протяжении курсов, указывая на положительный эффект ксиланазы в растении на гидролиз ксилана.
На фиг. 15 использовали следующие обозначения: PT: уровни после предварительной обработки; PT-XH: уровни после гидролиза ксиланазой, 48 ч: уровни после второй стадии в течение 48 ч; 96 ч: уровни после второй стадии в течение 96 ч. "Коктейль-Xyl" обозначает одностадийный ферментативный гидролиз, который проводили без включения ксиланазы в коктейль внешних ферментов. Образцы 2004.8.4, PT2004.8.4 PT-XH, 2063.17 и PT2063.17 PT-XH демонстрируют только результаты без коктейля. Остальные образцы демонстрируют результаты для отсутствия коктейля, полного коктейля и коктейля минус ксиланаза слева направо.
5 - Одностадийный ферментативный гидролиз, pAG2042. Анализированная растительная солома: для получения соломы использовали трансгенные растения, обозначенные как 2042.2, 2042.3 и 2042.6 (полученные путем трансформации кукурузы посредством pAG2042). Для получения контрольной соломы использовали контрольное растение кукурузы 2004.8.4.
Предварительная обработка: проводили, как описано выше, с 0,3 M бисульфитом аммония/0,34 M карбонатом аммония при температурах либо 40°C, либо 60°C, в течение 19 ч, при 300 об./мин.
Одностадийный ферментативный гидролиз: проводили, как описано выше, с 1,0 мг/мл тетрациклина при 50°C в течение 48 часов, 250 об./мин.
На фиг. 16 представлен выход глюкозы (процент от массы биомассы) после ферментативного гидролиза предварительно обработанной кукурузной соломы (2042.02, 2042.03, 2042.06 и 2004.8.4). Как представлено на фиг. 16, выход глюкозы из 2042.3 значимо превышает выход глюкозы из других двух трансгенных растений (2042.2 и 2042.6), а также контрольного растения (2004.8.4). На фиг.16 использовали следующие обозначения: 40C PT: предварительную обработку проводили при 40°C; 60C PT: предварительную обработку проводили при 60°C. Каждый меченый образец на фиг. 16 демонстрирует результаты для отсутствия коктейля, полного коктейля и коктейля минус эндоглюканаза слева направо.
Пример 14 - Определение высвобождения восстанавливающего сахара в случае трансгенного растительного материала
Ссылаясь на фиг. 8, трансгенные растения анализировали для определения уровней накопленного активного фермента. Для этих анализов, замороженные в жидком азоте образцы ткани листьев растирали с помощью ступки и пестика и полученные растертые образцы собирали. 10 мг замороженных растертых образцов листьев отмеряли и помещали в лунку микропланшета для титрования. В каждую ленку добавляли 200 мкл 100 мМ натрий-фосфатного буфера (pH 6,5) и реакционные смеси перемешивали пипетированием. Планшеты закрывали фольгой и помещали во вращающий инкубатор (200 об./мин.) при 55°C в течение 16 часов. После инкубации каждую реакционную смесь наносили на фильтрационный планшет Multiscreen HTS с 1,2-мкм фильтром из стекловолокна (Millipore, Billerica MA) и фильтровали центрифугированием при 500 x g в течение 3 минут. Ферментативную активность оценивали путем анализа 50 мкл полученного фильтрата с использованием анализа восстанавливающих сахаров Nelson-Somogyi, как описано ранее. Экстрагированный белок определяли с использованием набора для анализа белка BCA protein assay kit (Thermo Scientific). Уровни активности были представлены в качестве мМ концов восстанавливающих сахаров, образовавшихся на мг экстрагированного белка. Продуцирование восстанавливающих сахаров в образцах трансгенных растений (AG2014 и AG2015) по сравнению с контрольным образцом трансгенного растения, экспрессирующего не ксиланазу (AG2004), продемонстрировало накопление активной ксиланазы в ткани трансгенного растения.
Пример 15 - детекция активности аутолизиса в трансгенной кукурузной соломе
Десять миллиграмм (+/-1 мг) растертого образца помещали в 1,5-мл микроцентрифужную пробирку. Растертый образец ресуспендировали в 1 мл 100-мМ натрий-фосфатного буфера, содержащего 40 мкг тетрациклина и 30 мкг циклогексимида. Реакционные смеси инкубировали в течение 64 часов при 60°C при перемешивании вращением с донышка на крышку (18 об./мин.). Супернатант реакционной смеси собирали и анализировали в отношении присутствия восстанавливающих сахаров с использованием анализа восстанавливающих сахаров Nelson-Somogyi. Результаты этого анализа были представлены в качестве мМ образовавшихся восстанавливающих концов эквивалентов ксилозы/мг соломы путем сравнения со стандартной кривой ксилозы.
Пример 16 - Трансгенные растения и трансгенные растения, экспрессирующие ферменты, деградирующие клеточную стенку
Как правило, для каждого вектора для трансформации, проводили по меньшей мере 20 событий. В некоторых случаях проводили больше (вплоть до 90) трансгенных событий и все события использовали для оценки эффекта процесса трансформации и экспрессии генов.
Трансгенные растения, сконструированные с использованием pAG3000 и pAG3001
Ссылаясь на фиг. 17A и 17B, растения T0 регенерировали после протокола трансформации, описанного выше, с использованием pAG3000 и pAG3001. Векторы для трансформации растений pAG3000 и pAG3001 описаны выше. Эти векторы имеют промотор актина 1 риса, регулирующий ген фосфоманнозоизомеразы (PMI) E. coli, который можно использовать для селекции трансгенных растений или для других целей. Отличия между pAG3000 и pAG3001 состоят в области соединения между промотором актина 1 риса и геном PMI. В pAG3000 использовали неполную консенсусную последовательность эукариотической области инициации трансляции, в то время как в pAG3001, использовали полную эукариотическую область инициации трансляции. Эмбрионы маиса трансформировали посредством pAG3000 и pAG3001, как описано выше.
Трансгенные растения, экспрессирующие pAG3000 и pAG3001, регенерировали, как описано выше. Исходя из экспериментальных результатов и следуя методикам, описанным выше, были отобраны трансгенные растения, имеющие pAG3000 и pAG3001 на среднем уровне 22,6% и 12,3%, соответственно, в маисе. В других видах эффективность трансформации (как определяют по количеству трансгенных растений, деленных на число мишеней для трасформации, где может быть достигнуто не более одного трансгенного события на мишень) не легко вычислить, поскольку целевые каллюсы не легко пересчитать в качестве отдельных мишеней. Максимальная наблюдаемая эффективность, наблюдаемая в любом отдельном эксперименте, составляла 28% для pAG3000 и 14% для pAG3001. Исходя из этих данных использование неполной консенсусной последовательности эукариотической области инициации трансляции обеспечило увеличенную эффективность трансформации по сравнению с полной эукариотической последовательностью инициации трансляции. Хотя промотор актина 1 риса считается относительно сильным конститутивным промотором, эффективность трансформации, полученная путем связывания его с PMI, была неизвестна, и было неясно, насколько они могли быть улучшенными относительно конструкции CMPS:PMI, полученной первоначально. Исходя из этих результатов средняя эффективность трансформации с селекцией при использовании CMPS:PMI составляла 1,5%, с максимум 14%, однако в отдельных экспериментах наблюдали эффективность 0%, 2%, 3%, 6%, 7%, 13% и 14%. На диапазоны эффективности трансформации может влиять качество материала мишени для трансформации, однако эти средние значения и диапазоны помогают определить, что можно было бы ожидать от трансформации с использованием этих конструкций. Исходя из этих результатов, связывание PMI с промотором актина 1 риса увеличивало эффективность трансформации PMI при использовании методик, описанных выше. Более того, использование области соединения между промотором актина 1 риса и PMI в pAG3000 увеличивало среднюю эффективность трансформации выше уровня увеличения при использовании области соединения, применяемой в pAG3001.
Как представлено на фиг.17A и 17B, трансгенные растения с pAG3000 (фиг. 17A) и pAG3001 (фиг. 17B) являются фенотипически нормальными для трансгенных растений на этой стадии развития. Трансгенную природу этих растений подтверждали с использованием ПЦР.
Пример 17 - Трансгенные растения, сконструированные с использованием pAG2004 и pAG2005
Ссылаясь на фиг. 18A, 18B, 18C, 19A и 19B, кукурузу трансформировали векторами для трансформации растений pAG2004 (фиг. 18A, 18B и 18C) и pAG2005 (фиг. 19A и 19B). Эти векторы имеют промотор убиквитина 3 риса, регулирующий ген фосфоманнозоизомеразы (PMI) E. coli, который можно использовать для селекции трансгенных растений или для других целей. Отличие между pAG2004 и pAG2005 состоит в том, что pAG2005 содержит дополнительную пустую экспрессирующую кассету, в которую можно клонировать другие представляющие интерес гены. С точки зрения выбора трансгенных событий pAG2004 и pAG2005 имеют идентичный промотор убиквитина 3 риса и кассету для селекции PMI. Эти два вектора вместе дают среднюю эффективность трансформации 20%. В отдельных экспериментах эти векторы обеспечивали эффективность трансформации 0%, 4%, 7%, 10%, 11%, 12%, 13%, 14%, 15%, 17%, 18%, 24%, 28%, 29%, 30%, 31%, 32%, 40%, 50%, 53% и 64%. На диапазоны эффективности трансформации может влиять качество материала мишени для трансформации, однако эти средние значения и диапазоны помогают определить, что можно было бы ожидать от трансформации с использованием этих конструкций.
Промотор убиквитина 3 риса, слитый с PMI, значительно увеличивал эффективность трансформации, которую наблюдали относительно CMPS:PMI, с использованием способа, описанного выше. Более того, средняя эффективность трансформации была выше, чем при использовании pAG3001, и сходной с эффективностью, наблюдаемой с использованием pAG3000. Поскольку максимальная эффективность, полученная с использованием pAG2004 и pAG2005 превышала эффективность, полученную с использованием pAG3000, кассеты для селекции pAG2004 и pAG2005 использовали для дальнейшей разработки трансгенных растений, как описано выше.
На фиг. 18A, 18B, 18C, 19A и 19B проиллюстрированы растения TO, регенерированные после протокола трансформации, описанного выше. На фиг. 18A показано, что почти стареющее трансгенное растение pAG2004 является фенотипически нормальным. На фиг. 18B и 18C показано, что початки трансгенного растения pAG2004 также являются фенотипически нормальными. На фиг. 19A и 19B показано, что трансгенные растения pAG2005 являются фенотипически нормальными. Трансгенную природу этих растений подтверждали с использованием ПЦР.
На фиг. 20 проиллюстрировано измерение восстанавливающих сахаров из трансгенного растения события #15, трансформированного pAG2004. На фиг. 20, образец буфера соответствует фоновому уровню анализа, где для измерения использовали 1 мг буфера. Поскольку pAG2004 не экспрессирует фермент, деградирующий клеточную стенку, измерение для него восстанавливающих сахаров соответствует отрицательному контролю для сравнения против него других растений и также соответствует нетрансгенным растениям дикого типа.
Пример 18 - Трансгенные растения, сконструированные с использованием pAG2016
Вектор для трансформации pAG2016 использовали при трансформации для регенерации трансгенных растений. Этот вектор для трансформации происходит из pAG2005 и он содержит экспрессирующую кассету для продуцирования бета-глюкуронидазы (GUS). В этой экспрессирующей кассете GUS слит с кодон-оптимизированным сигнальным пептидом PR1a, который направляет GUS в межклеточное пространство апопласта. Эффективность трансформации этого вектора имеет среднее значение 16%, и она находилась в ожидаемом диапазоне для используемой кассеты для селекции PMI.
Ссылаясь на фиг. 21A и 21B, трансгенные растения и початки с pAG2016 TO являются фенотипически нормальными. Растения регенерировали после протокола трансформации, описанного выше. Трансгенную природу этих растений подтверждали с использованием ПЦР. Эти растения демонстрируют, что трансген может эффективно экспрессироваться с экспрессирующей кассеты, содержащейся в pAG2005. Трансгенные растения также демонстрируют, что сигнальный пептид PR1a, который был слит с GUS в pAG2016, не снижает существенно эффективность трансформации или не ухудшает фенотип трансгенного растения.
Пример 19 - Трансгенные растения, сконструированные с использованием pAG2014, pAG2015, pAG2020, pAG2025
Векторы для трансформации pAG2014, pAG2015, pAG2020, pAG2025 использовали в трансформации для регенерации трансгенных растений. Векторы для трансформации pAG2014, pAG2015 и pAG2020 происходят из pAG2005 и каждый из них содержит экспрессирующую кассету для продуцирования ксиланазы (регистрационный номер P77853). В pAG2014 ген P77853 слит с сигнальной последовательностью альфа-амилазы ячменя (BAASS) для нацеливания на клеточную стенку. В pAG2015 ген P77853 не слит с каким-либо сигнальным пептидом и, таким образом, она должна накапливаться в цитоплазме клеток. В pAG2020, P77853 слита с сигнальным пептидом PR1a для нацеливания фермента в апопласт. Напротив, pAG2025 происходит из pAG2012, в котором используется промотор глутелина GluB-4 риса и сигнальная последовательность GluB-4 для обеспечения специфической для ткани семян экспрессии P77853. Средняя эффективность трансформации для pAG2014 составила 30%, для pAG2015 она составила 34%, для pAG2020 она составила 24%, и для pAG2025 она составила 10%. Все из этих эффективностей были в ожидаемом диапазоне эффективности трансформации при использовании промотора убиквитина 3 риса и кассеты для селекции PMI.
Измерение активности проводили для трансгенных событий, осуществленных с использованием способов, описанных выше. На представленных ниже фигурах продемонстрированы результаты измерения активности.
Ссылаясь на фиг. 22, для трансгенных растений проводили измерения восстанавливающих сахаров. На фиг. 22 представлено продуцирование восстанавливающих сахаров из трансгенных растений, имеющих pAG2014 (левый образец) или pAG2004 (средние образцы), и контроль в виде буфера (правый образец). Трансгенное растение события #5 (левый образец), полученное с pAG2014, которое экспрессирует ксиланазу P77853, продуцирует значительно большее количество восстанавливающих сахаров при инкубации при 60°C, чем растения, полученные с pAG2004.
Ссылаясь на фиг. 23, измерение ферментативной активности проводили из образцов сухой стареющей кукурузной соломы. Первые шесть образцов слева на фиг. 23 представляют собой отличающиеся трансгенные растения, имеющие pAG2014. Седьмой образец представляет собой отрицательный контроль из трансгенного растения, имеющего pAG2004. Трансгенным растениям, полученным с pAG2014, позволяли состариться, а затем высушивали в инкубаторе до абсолютно сухих уровней. Уровень сухости может составлять менее 1% влажности. Образцы соломы измельчали и анализировали, как описано выше. Как показано, ферментативная активность является стабильной, даже несмотря на процессы старения, высушивания и измельчения. В этих данных был получен диапазон активности с низких уровней (близких к контролю, экспрессирующему не ксиланазу (2004.15)) вплоть до свыше 8 мкг эквивалентов RBB/ мг соломы.
Ссылаясь на фиг. 24, измерения ферментативной активности проводили из образцов тканей листьев трансгенных растений, полученных с pAG2015, pAG2014 или pAG2004. Образец для pAG2014 представлен седьмым справа. Образец для pAG2004 представлен последним. Все другие образцы соответствуют отличающимся трансгенным событиям для растений pAG2015. Как видно, получают диапазон уровней активности, поскольку встраивание гена в геном растения является в высокой степени вариабельным и в значительной степени влияет на профили экспрессии. В целом, для данного вектора может быть достигнут максимальный уровень активности, и также возможна любая активность ниже этого уровня.
Как представлено на фиг. 24, pAG2015 (цитоплазматическая P77853) и pAG2014 (BAASS:P77853) обеспечивают значительные уровни активности. Активность для pAG2015 является значительной при экспрессии в растениях и взятии образцов из зеленых тканей и из стареющей кукурузной соломы, однако, таким образом, анализы показали, что pAG2014 обеспечивает более высокий уровень продуцирования восстанавливающих сахаров при анализе в соломе из стареющей кукурузы. Напротив, pAG2025 не обеспечивает активности в протестированных зеленых тканях (данные не представлены на фиг. 24), как можно было бы ожидать, учитывая специфическую для семян природу экспрессирующую кассету с трансгеном pAG2025.
На фиг. 25A и 25B проиллюстрированы трансгенные растения, полученные с pAG2014, на фиг. 25C проиллюстрирован початок из трансгенного растения, полученного с pAG2014. На фиг. 26A и 26B проиллюстрированы трансгенные растения, полученные с pAG2015, и на фиг. 26C и 26D проиллюстрированы початки из трансгенных растений, полученных с pAG2015. На фиг. 27A и 27B проиллюстрированы трансгенные растения, полученные с pAG2020, и на фиг. 27C проиллюстрирован початок из трансгенного растения, полученного с pAG2020. Ссылаясь на фиг. 28A, 28B и 28C, проиллюстрированы трансгенные растения, полученные с pAG2025. Эти растения демонстрируют, что ксиланаза P77853 может быть эффективно экспрессирована из экспрессирующей кассеты, содержащейся в pAG2005. Трансгенные растения также демонстрируют, что сигнальные пептиды BAASS и PR1a, которые слиты с P77853 в pAG2014 и pAG2020, соответственно, не препятствуют эффективности трансформации, но влияют на фенотип в отношении цитоплазматического накопления. Фенотипы этих растений были в высокой степени интересными и неожиданными. Отсутствуют известные работы, в которых показана экспрессия ферментов ксиланаз в кукурузе, просе, сорго или сахарном тростнике. На основе результатов, представленных в настоящем описании, ферменты ксиланазы обеспечивают определенные фенотипы, однако они в высокой степени зависят от конкретного используемого фермента, сигнального пептида и промотора, а также от присутствия сигнала удержания в ER, SEKDEL.
Ксиланаза P77853 является интересной, поскольку все трансгенные растения маиса, полученные с использованием pAG2014, pAG2015, pAG2020 и pAG2025, имели нормальные фенотипы роста, однако некоторые имели отличающиеся фенотипы семян. То, что растения развиваются нормально, в некоторой степени неожиданно, поскольку ксиланаза гидролизует ксилан в гемицеллюлозном компоненте клеточной стенки растений.
Ссылаясь на фиг. 25A, 25B и 25C, во многих из трансгенных событий для pAG2014 (BAASS:P77853) наблюдали несколько сморщенных зерен. Эти растения имели нормальный рост и развитие, однако во многих растениях наблюдали сегрегирующий фенотип семян. См. сморщенные семена 2510 на фиг. 25C. Сморщенные семена случайным образом отбирали совместно с нормальными семенами и тестировали в отношении увеличения активности ксиланазы (указывающей на присутствие фермента P77853). Среди протестированных семян, все сморщенные семена имели значительное увеличение активности ксиланазы, в то время как нормальные семена имели не поддающуюся детекции активность ксиланазы, также как и семена из растения дикого типа. Кроме того, из случайного початка было отобрано двенадцать сморщенных семян и посеяно рядом с 12 нормально выглядящими семенами. Среди посеянных семян только одно из 12 сморщенных семян проросло (его тестировали способом ПЦР, и было показано, что оно имеет ген P77853), в то время как из 12 нормальных семян проросли девять. Среди девяти нормальных семян, которые проросли, восемь не имели ген P77853, в то время как один имел P77853, при определении способом ПЦР. Это указывает на то, что P77853, при экспрессии в качестве слитой конструкции с сигнальной последовательностью BAASS, приводит к семенам, которые имеют сниженную фертильность относительно нетрансгенных семян, и что уровень неплодородности может зависеть от уровня экспрессии P77853. В то время как сморщенные семена и неплодородность могут обеспечить значительный коммерческий ущерб в кукурузе, они могут быть преимущественными в просе, сорго, мискантусе и сахарном тростнике, где стерильность растений может быть благоприятной с точки зрения перспективы разрешения контролирующего ведомства. Более того, многолетние культуры, такие как просол и сахарный тростник, можно клонально получать с помощью культуры ткани с использованием способов, известных в данной области, и вегетативно размножать. В этих культурах сниженная фертильность может быть в меньшей степени значима и она может быть преимущественной для изолирования гена. Таким образом, хотя неблагоприятный фенотип семян P77853 в кукурузе и других зерновых культурах является вредоносным, в кормовых, сахараносных и незерновых культурах, используемых в качестве сырья для кормов для животных или сырья для ферментации, P77853 может обеспечить существенную пользу для усвоения волокон, гидролиза и сниженной фертильности. Трансгенные события в просе, осуществленные с использованием pAG2014, были фенотипически нормальными.
Ссылаясь на фиг. 26A, 26B, 26C и 26D, для pAG2015, который не имел сигнального пептида и, таким образом, накапливал P77853 в цитоплазме растений, не наблюдали неблагоприятного фенотипа. Некоторые семена маиса были немного более нестандартного цвета, чем семена WT, однако до настоящего времени не наблюдали никакого другого аномального фенотипа (см. фиг. 26D). Эти растения накапливают значительные уровни активности ксиланазы, которая в среднем по меньшей мере равна, и в большинстве случаев несколько превышает, активности ксиланазы, выявленной в событиях pAG2014. То, что два растения не обладают общим фенотипом семян примечательно и указывает на то, что сигнальная последовательность BAASS для нацеливания на клеточную стенку, которую использовали в векторе pAG2014, вносит вклад в фенотип семян, наблюдаемый при событиях pAG2014. Поскольку эти растения накапливают высокие уровни активности ксиланазы, они могут быть пригодны в качестве источника фермента ксиланазы, в качестве сырья, которое может осуществлять аутогидролиз гемицеллюлозных компонентов для применения в промышленных процессах, таких как ферментация, в качестве корма для животного или добавки в корм для животного, и в качестве зернового корма для животного или добавки в корм. В отличие от трансгенных событий, осуществленных с использованием pAG2014, события, осуществленные с использованием pAG2015, не имели аномального фенотипа семян и могут оказаться полезными в зерновых культурах, таких как кукуруза, (зерновое) сорго, пшеница, ячмень и другие.
Ссылаясь на фиг. 27A, 27B и 27C, для событий pAG2020 (PR1a:P77853) как растения, так и початки, выглядели нормально и не имели значительного наблюдаемого фенотипа. Это является особенно неожиданным, поскольку PR1a нацеливает слитую ксиланазу P77853 в апопласт, где, как можно ожидать, она имеет эффект, сходный с эффектом для событий pAG2014. Неизвестно, вызывает ли сигнальный пептид PR1a более низкую экспрессию, более низкое накопление фермента или является ли он менее эффективным в отношении нацеливания белка P77853, однако отсутствие фенотипа семян в этих трансгенных растениях является неожиданным, учитывая результаты, полученные для pAG2014. Поскольку эти растения накапливают активность ксиланазы, они также могут быть полезны в качестве источника фермента ксиланазы, в качестве сырья, которое может осуществлять аутогидролиз компонентов гемицеллюлозы для применения в промышленных процессах, таких как ферментация, в качестве корма для животных или добавки в корм для животных. В отличие от трансгенных событий, осуществленных с использованием pAG2014, события, осуществленные с использованием pAG2020, не имели аномального фенотипа семян и могут оказаться полезными в зерновых культурах, таких как кукурузы, (зерновое) сорго, пшеница, ячмень и другие.
Ссылаясь на фиг. 28A, 28B и 28C, для событий pAG2025 (GluB4:P77853) все растения выглядели фенотипически нормальными.
Пример 20 - Трансгенные растения, сконструированные с использованием pAG2017, pAG2019 и pAG2027
Векторы для трансформации pAG2017, pAG2019 и pAG2027 использовали при трансформации для регенерации трансгенных растений. Векторы для трансформации pAG2017 и pAG2019 происходили из pAG2005, и каждый из них содержал экспрессирующую кассету для продуцирования ксиланазы (регистрационный номер P40942). Вектор pAG2027 происходит из pAG2012 и экспрессирует ксиланазу P40942 из промотора GluB-4, который экспрессируется преимущественно в семенах. В pAG2017 ксиланаза P40942 слита с сигнальным пептидом PR1a для нацеливания фермента в апопласт. В pAG2019 ген P40942 слит с сигнальной последовательностью альфа-амилазы ячменя (BAASS) для нацеливания на клеточную стенку. Средняя эффективность трансформации для pAG2017 составила 16%, для pAG2019 она составила 13% и для pAG2027 она составила 29%.
В противоположность трансгенным растениям, экспрессирующим P77853, все из которых были фенотипически нормальными, за исключением описанных выше аномалий семян, растения, экспрессирующие ксиланазу P40942, были в высокой степени низкорослыми, за исключением растений, полученных с pAG2027. Ссылаясь на фиг. 29A, 29B, 29C и 29D, растения, трансформированные pAG2017 (PR1a:P40942) являются в высокой степени низкорослыми и никогда не вырастают до той же высоты, что и растения дикого типа или растения, трансформированные pAG2020 (PR1a:P77853). На фиг. 29A представлено низкорослое трансгенное растение pAG2017. На фиг. 29B представлено низкорослое трансгенное растение с pAG2017 рядом с растением дикого типа справа. На фиг. 29C и 29D представлены початки из трансгенного растения с pAG2017 с частично сморщенными семенами, имеющими аномальную окраску. Результаты с pAG2017 были неожиданными, учитывая, что P77853 и P40942 имеют приблизительно одинаковую удельную активность при измерении in vitro на ксилане березы (см. выше). Также P40942 имеет некоторую активность целлобиогидролазы (CBH), так что возможно, что эта активность вносить вклад в наблюдаемый фенотип, однако другие группы имеют экспрессированные ферменты CBH в маисе с очевидно не наблюдаемым фенотипом роста. Значительные отличия в фенотипе роста между трансгенными растениями, полученными с pAG2017 и pAG2020, являются совершенно неожиданными и в высокой степени непредвиденными.
В дополнение к фенотипу роста в растениях pAG2017, семена из этих растений или полученные при ауткроссинге этих растений с нетрансгенными растениями AxB, также продемонстрировали сморщенный фенотип, сходный с фенотипом, наблюдаемым в семенах трансгенных растений, полученных с pAG2014, также демонстрируя некоторое обесцвечивание семян. Приблизительно 20 сморщенных семян было собрано из растений pAG2017 и все протестированные семена были положительными по активности ксиланазы, в то время как выполненные семена не имели поддающегося детекции увеличения активности ксиланазы при определении с использованием способов, описанных выше.
Ссылаясь на фиг. 30A и 30B, трансгенные растения, полученные с pAG2019 (BASS:P40942) также обладали низкорослым фенотипом, аналогично трансгенным растениям, полученным с pAG2017. Это было неожиданным, учитывая, что трансгенные растения, полученные с pAG2014 (BASS:P77853), не имели фенотипа роста, и в то же время ксиланазы P40942 и P77853 имеют приблизительно ту же удельную активность при измерении на ксилане березы. На фиг. 30A представлено низкорослое трансгенное растение, полученное с pAG2019, и на фиг. 30B представлено низкорослое трансгенное растение, полученное с pAG2019, вместе с растением дикого типа слева.
Ссылаясь на фиг. 31, трансгенные растения, полученные с pAG2027, которые экспрессируют P40942 с промотора GlutB риса, являются фенотипически нормальными в отношении роста. Левые три растения на фиг. 31 были получены с pAG2019. Правые три растения были получены с pAG2027. Результат с pAG2027 был противоположен трансгенным растениям, полученным с pAG2017 и pAG2019, и является неожиданным, поскольку P40942, экспрессированная с промотора убиквитина риса с использованием сигнальных последовательностей либо PR1a, либо BAASS, вызывала низкорослость. Однако этот результат согласуется с наблюдением, что растения, полученные с pAG2025 (промотор убиквитина 3 риса, регулирующий P77853), не являются низкорослыми и растут нормально. Учитывая отличия в фенотипах, наблюдаемые между векторами, экспрессирующими P77853 и P40942, нельзя было предсказать, каким будет результат для pAG2027. Поскольку промотор GlutB, главным образом, экспрессирует фермент в семенах, может быть, что ни один из ферментов, экспрессированных с промотора GluB, не продемонстрирует фенотип роста или связанный с зеленой тканью фенотип и продемонстрирует только фенотипы семян, сходные с фенотипами, наблюдаемыми в растениях, полученных с pAG2014 и pAG2017.
Пример 21 - Трансгенные растения, сконструированные с использованием pAG2018 и pAG2026
Векторы для трансформации pAG2018 и pAG2026 использовали для трансформации для регенерации трансгенных растений. Вектор pAG2018 происходит из pAG2005 и содержит экспрессирующую кассету для продуцирования ксиланазы (регистрационный номер О30700), слитой с сигнальной последовательностью BAASS. Вектор pAG2026 происходит из pAG2012 и экспрессирует ксиланазу O30700 с промотора GluB-4, который экспрессируется преимущественно в семенах. Средняя эффективность трансформации для pAG2018 составила 13% и для pAG2026 она составила 18%.
Как описано выше, трансгенные растения, экспрессирующие P77853, были фенотипически нормальными, за исключением описанных выше аномалий семян. Напротив, ссылаясь на фиг. 32A, 32B и 32C, трансгенные растения, полученные с pAG2018 и экспрессирующие ксиланазу O30700, были в высокой степени низкорослыми и никогда не вырастали до той же высоты, что и растения дикого типа или растения, трансформированные pAG2014. На фиг. 32A представлены два трансгенных растения, полученных с pAG2018, слева и два экспрессирующих не гидролазу растения справа. На каждой из фиг. 32B и 32C представлено трансгенное растение, полученное с pAG2018. Эти результаты были неожиданными, учитывая что как P77853, так и O30700 являются эндоксиланазными ферментами, и, в противоположность P40942, O30700 не имеет какой-либо активности CBH. Фенотип роста, наблюдаемый в случае O30700, был в высокой степени сходным с низкорослостью, наблюдаемой для растений с pAG2017 и pAG2019.
В противоположность трансгенным растениям, полученным с pAG2018, трансгенные растения, полученные с pAG2026, которые экспрессируют O30700 с промотора GlutB риса, являются фенотипически нормальными в отношении роста. См. фиг. 33A, 33B и 33C, на которых проиллюстрированы три различных трансгенных растения, полученных с pAG2026. Эти результаты являются неожиданными, поскольку O30700, экспрессированная с промотора убиквитина риса и слитая с сигнальной последовательностью BAASS, вызывала низкорослость. Напротив, этот результат согласуется с наблюдением, что растения, полученные с pAG2025 (промотор убиквитина 3 риса, регулирующий P77853), не являются низкорослыми и растут нормально, однако, учитывая отличия в фенотипах, наблюдаемые между векторами, экспрессирующими P77853 и O30700, нельзя было предсказать, каким будет этот результат. Поскольку промотор GlutB, главным образом, экспрессирует фермент в семенах, возможно, что ни один из ферментов, экспрессируемых с промотора GluB, не продемонстрирует фенотип роста или фенотип, связанный с зеленой тканью, и продемонстрирует только фенотипы семян, сходные с фенотипами, наблюдаемыми в растениях, полученных с pAG2014 и pAG2017.
Пример 22 - Трансгенные растения, сконструированные с использованием pAG2021, pAG2023 (P77853m3), pAG2022, pAG2024
Векторы для трансформации pAG2021, pAG2023, pAG2022 и pAG2024 использовали при трансформации для регенерации трансгенных растений. Все из этих векторов происходят из pAG2005 и содержат экспрессирующую кассету для продуцирования модифицированной интеином ксиланазы (обозначаемой как P77853m3). В векторах для трансформации pAG2021 и pAG2022, модифицированный интеином белок P77853m3 был слит с сигнальным пептидом PR1a, в то время как в pAG2023 и pAG2024, P77853m3 был слит с сигнальным пептидом BAASS. Векторы pAG2022 и pAG2024 также имеют последовательность удержания в эндоплазматической сети SEKDEL, присоединенную к P77853m3, в то время как pAG2021 и pAG2023 лишены последовательности SEKDEL. Средняя эффективность трансформации для pAG2021 составляла 19%, для pAG2022 она составляла 21%, для pAG2023 она составляла 24%, и для pAG2024 она составляла 38%.
Ни одно из трансгенных растений, полученных с pAG2021, pAG2022, pAG2023 и pAG2024, не имело аномальный фенотип. См. фиг. 34A, 34B, 34C и 34D для результатов pAG2021. Трансгенные растения, полученные с pAG2021, росли нормально, достигали нормального роста и имели нормальный набор семян. См. фиг. 35A, 35B и 35C для результатов для pAG2022. Трансгенные растения, полученные с pAG2022, также росли нормально, достигали нормальной высоты и имели нормальный набор семян. См. фиг. 36A, 36B и 36C для результатов для pAG2023. На этих фигурах показано, что трансгенные растения, полученные с pAG2023, росли нормально и достигали нормальной высоты. См. фиг. 37A, 37B и 37C для результатов для pAG2024. На этих фигурах показано, что трансгенные растения, полученные с pAG2024, также росли нормально и достигали нормальной высоты. Здесь продемонстрировано, что модификация интеином фермента, деградирующего клеточную стенку, может защитить растением от любого фенотипа, который может обеспечиваться не модифицированным интеином ферментом. В этом примере использовали цис-сплайсируемый интеин (mini-Psp-pol M1L4 m3), разработанный так, чтобы он имел чувствительную к температуре активность сплайсинга. Поскольку растения выращивали при температурах отсутствия сплайсинга, не наблюдали активности и связанных с ней фенотипов роста или семян. При некоторых температурах интеин может сплайсироваться в некоторой степени и проявлять активный фермент. Поскольку растения имеют нормальный фенотип, экспрессия модифицированных интеином белков является способом обеспечения включенной активности деградации клеточной стенки в растениях, которая впоследствии может быть вновь достигнута, но не имеет фенотипического эффекта на растение.
Ссылаясь на фиг. 38, оценивали ферментативную активность выбранных трансгенных событий. На этой фигуре представлены данные об активности для некоторых из событий pAG2021, совместно с измерениями для событий pAG2004 (отрицательные контроли для активности ксиланазы) и события pAG20014 (положительный контроль для активности ксиланазы). Для этого анализа образцы высушенной кукурузной соломы из стареющих растений анализировали с использованием способов, описанных выше. Образцы растений обозначали согласно номеру вектора, который использовали для их получения. Измерения для 2014.5 (трансгенное событие для маиса, осуществленное с pAG2014 и обозначенное как 2014.5) соответствует положительному контролю для активности ксиланазы, в то время как показатели активности для 2004.# (трансгенные события для маиса, осуществленные с pAG2004) соответствуют соломе отрицательного контроля по ксиланазе. Как показано, два из трансгенных растений, полученных с pAG2021, обеспечивают значительные уровни активности ксилиназы, однако растения были фенотипически нормальными, в отличие от событий pAG2014, которые продемонстрировали фенотип семян.
Варианты осуществления, представленные в настоящем описании, включают, но не ограничиваются ими, растения, описанные выше и/или проиллюстрированные на чертежах или их частях, векторы, кодирующие любую аминокислотную последовательность, представленную в настоящем описании, векторы, включающие любую последовательность нуклеиновой кислоты, представленную в настоящем описании, любую аминокислотную последовательность, представленную в настоящем описании, любую нуклеиновую кислоту, представленную в настоящем описании, любое растение, включающее вектор, представленный в настоящем описании, любое растение, включающее нуклеиновую кислоту, представленную в настоящем описании, любое растение, включающее аминокислотную последовательность, представленную в настоящем описании, и любой способ применения любого растения, части растения, вектора, аминокислотной последовательности или последовательности белка, представленные в настоящем описании.
Последовательность pAG2015 представляет собой:



Ссылки, цитированные на протяжении этой заявки, включены для всех целей, представленных в настоящем описании и в сами ссылки, как если бы каждая ссылка была полностью представлена. Для демонстрации, конкретные из этих ссылок цитированы в одном или нескольких конкретных местах настоящего описания. Цитирование ссылки в конкретном месте указывает на порядок(порядки), в котором включены идеи ссылки. Однако цитирование ссылки в конкретном месте не ограничивает порядок, в котором все из идей цитированной ссылки включены для любых целей.
Таким образом, понятно, что это изобретение не ограничивается конкретными описанными вариантами осуществления, а охватывает все модификации, которые находятся в пределах сущности и объема изобретения, как определено в прилагаемой формуле изобретения, в представленном выше описании и/или показано на прилагаемых чертежах.
| название | год | авторы | номер документа |
|---|---|---|---|
| РАСТЕНИЯ, ЭКСПРЕССИРУЮЩИЕ ФЕРМЕНТЫ, ДЕГРАДИРУЮЩИЕ КЛЕТОЧНУЮ СТЕНКУ, И ВЕКТОРЫ ЭКСПРЕССИИ | 2010 |
|
RU2606766C2 |
| СПОСОБЫ ТРАНСФОРМАЦИИ РАСТЕНИЙ ДЛЯ ЭКСПРЕССИИ ДЕЛЬТА-ЭНДОТОКСИНОВ BACILLUS THURINGIENSIS | 1999 |
|
RU2234531C2 |
| АУТОПРОЦЕССИРУЮЩИЕСЯ РАСТЕНИЯ И ЧАСТИ РАСТЕНИЙ | 2002 |
|
RU2312144C2 |
| РЕГУЛЯТОРНЫЕ МОЛЕКУЛЫ НУКЛЕИНОВЫХ КИСЛОТ ДЛЯ УСИЛЕНИЯ СЕМЯ-СПЕЦИФИЧНОЙ И/ИЛИ СЕМЯ-ПРЕДПОЧТИТЕЛЬНОЙ ГЕННОЙ ЭКСПРЕССИИ В РАСТЕНИЯХ | 2010 |
|
RU2559534C2 |
| КОНСТРУКТ И СПОСОБ ДЛЯ ЭКСПРЕССИИ ТРАНСГЕНОВ С ИСПОЛЬЗОВАНИЕМ ДВУНАПРАВЛЕННОГО КОНСТИТУТИВНОГО ПРОМОТОРА BRASSICA | 2013 |
|
RU2619178C2 |
| РАСТЕНИЯ С ИЗМЕНЕННЫМ УРОВНЕМ РАСТИТЕЛЬНОГО КРАХМАЛА | 2011 |
|
RU2565828C2 |
| Белок устойчивости к гербицидам, кодирующий его ген и их применение | 2016 |
|
RU2692553C2 |
| Белок устойчивости к гербицидам, кодирующие его гены и их применение | 2016 |
|
RU2681162C1 |
| СКОНСТРУИРОВАННАЯ СПОСОБАМИ ИНЖЕНЕРИИ ПЛАТФОРМА ДЛЯ ВСТРАИВАНИЯ ТРАНСГЕНА (ETIP) ДЛЯ НАЦЕЛИВАНИЯ ГЕНОВ И СТЭКИНГА ПРИЗНАКОВ | 2013 |
|
RU2666916C2 |
| РЕГУЛИРУЮЩИЙ ВЫСОТУ РАСТЕНИЙ ГЕН И ЕГО ПРИМЕНЕНИЯ | 2008 |
|
RU2458132C2 |
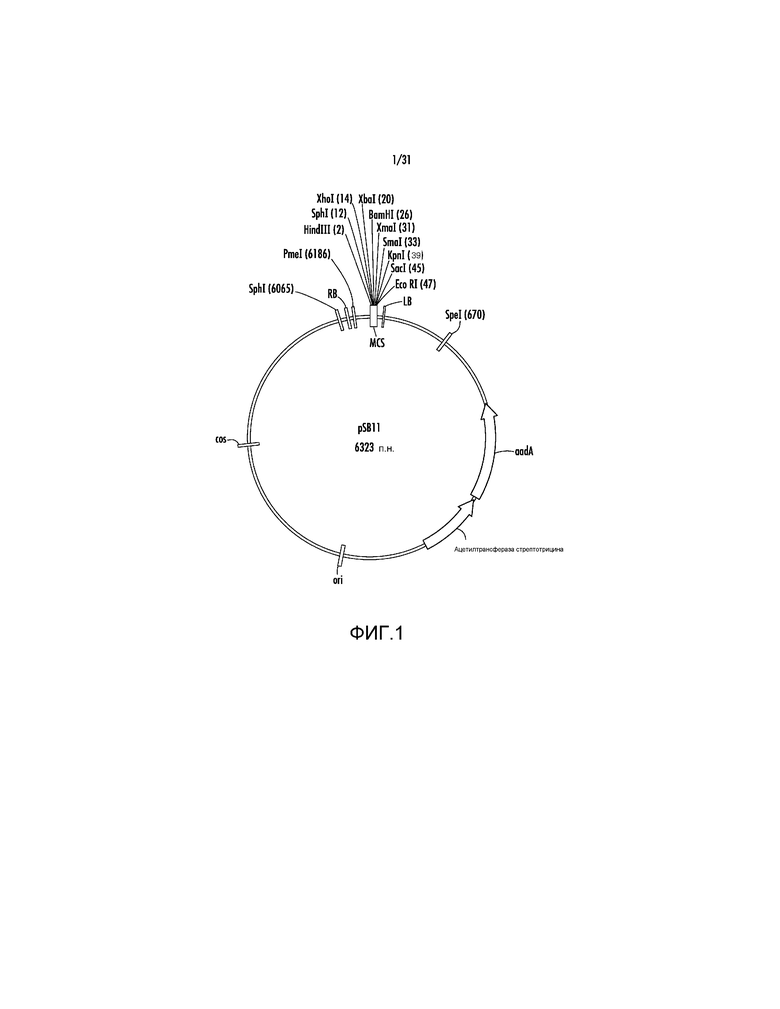
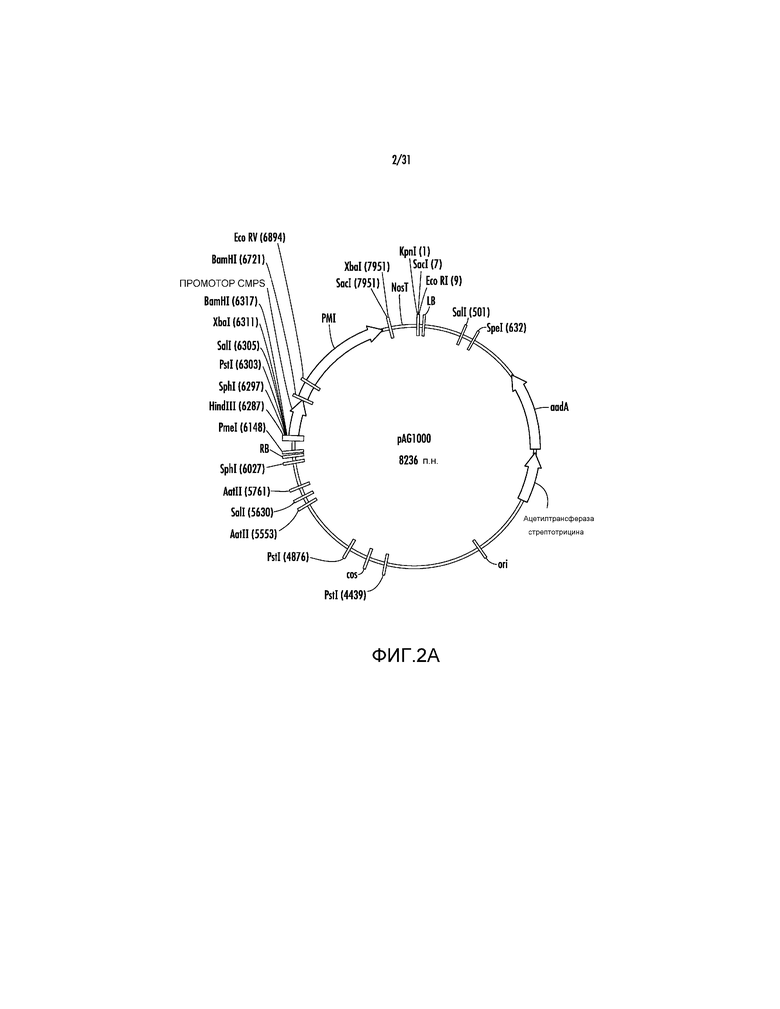
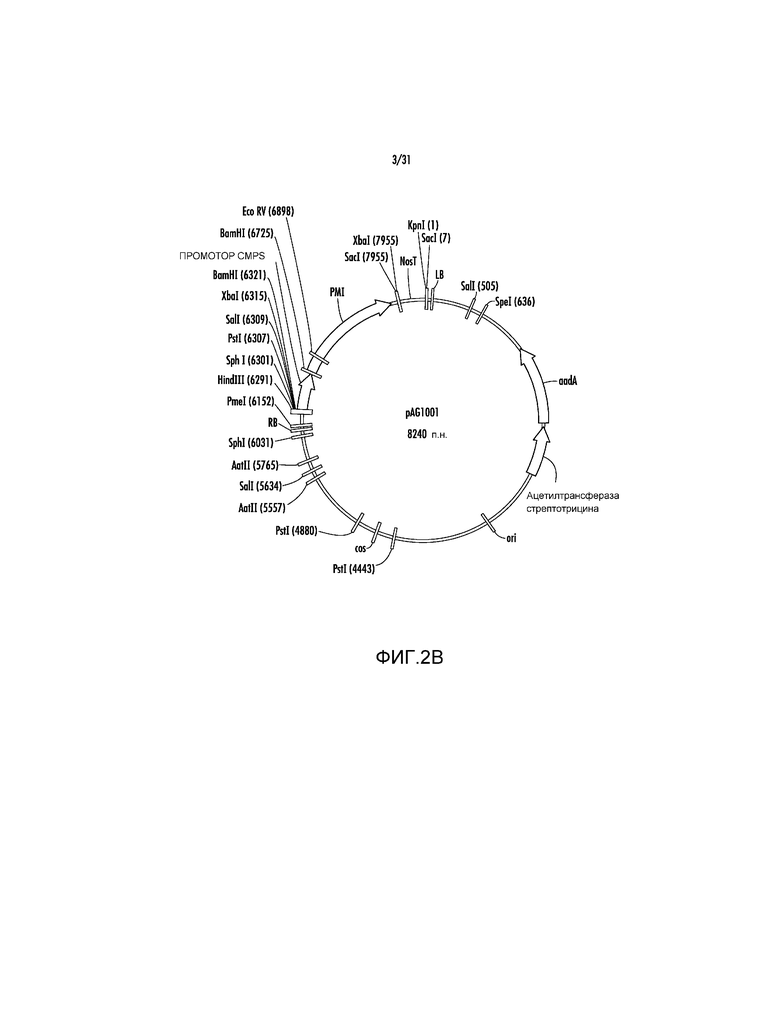
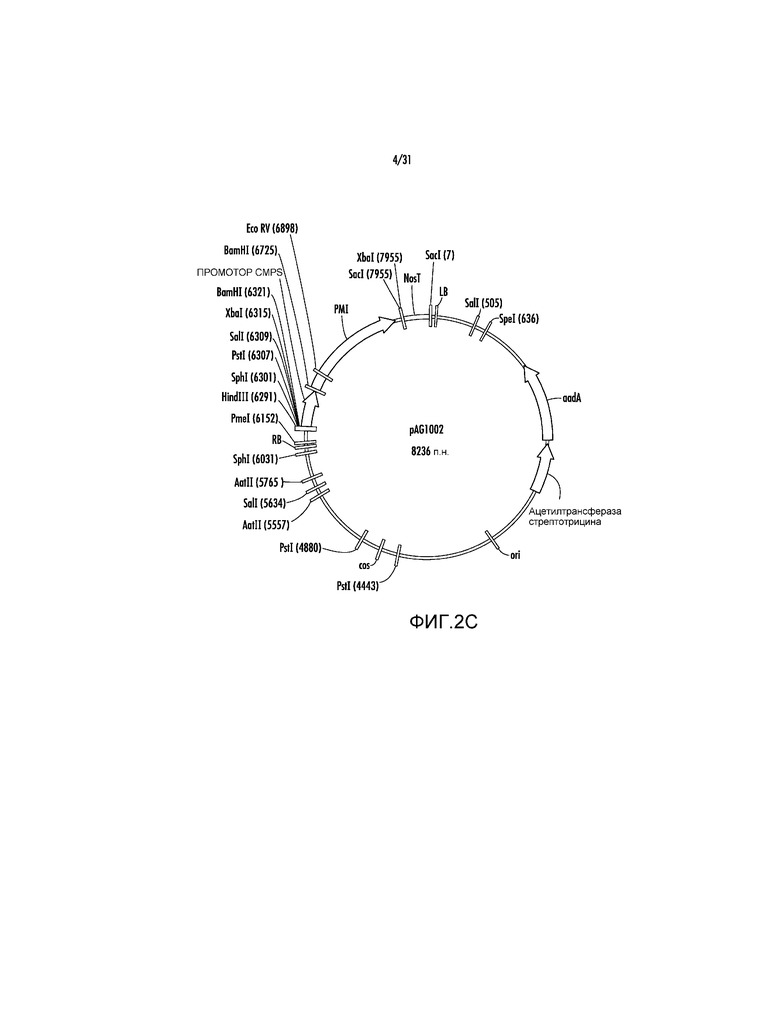
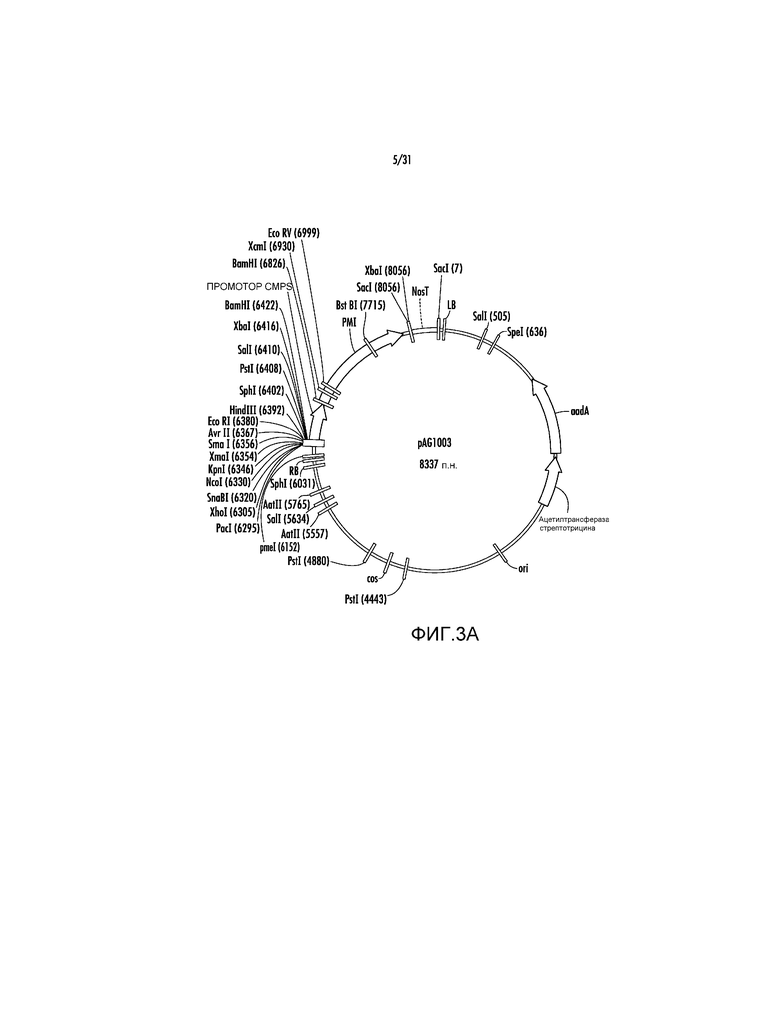

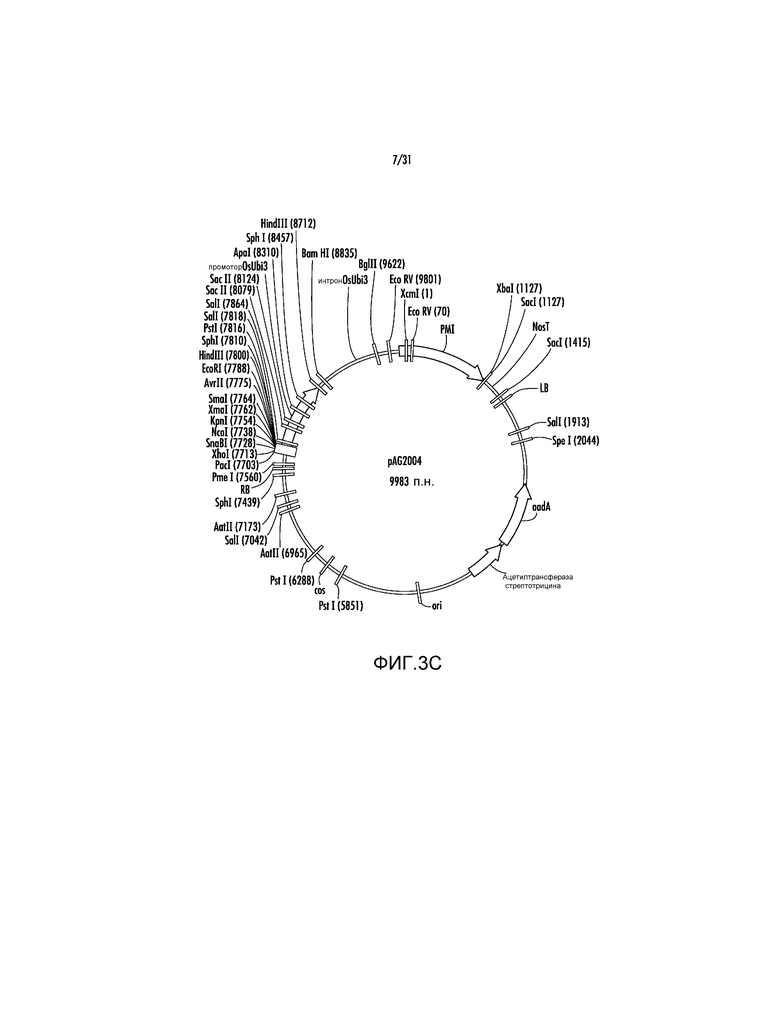
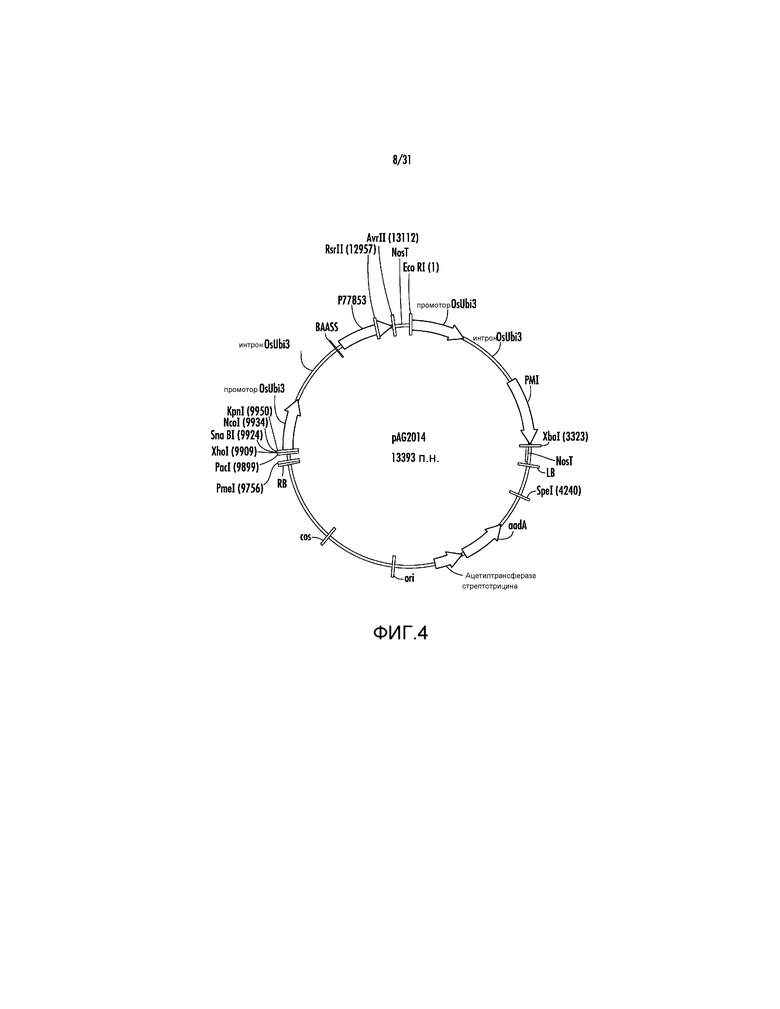
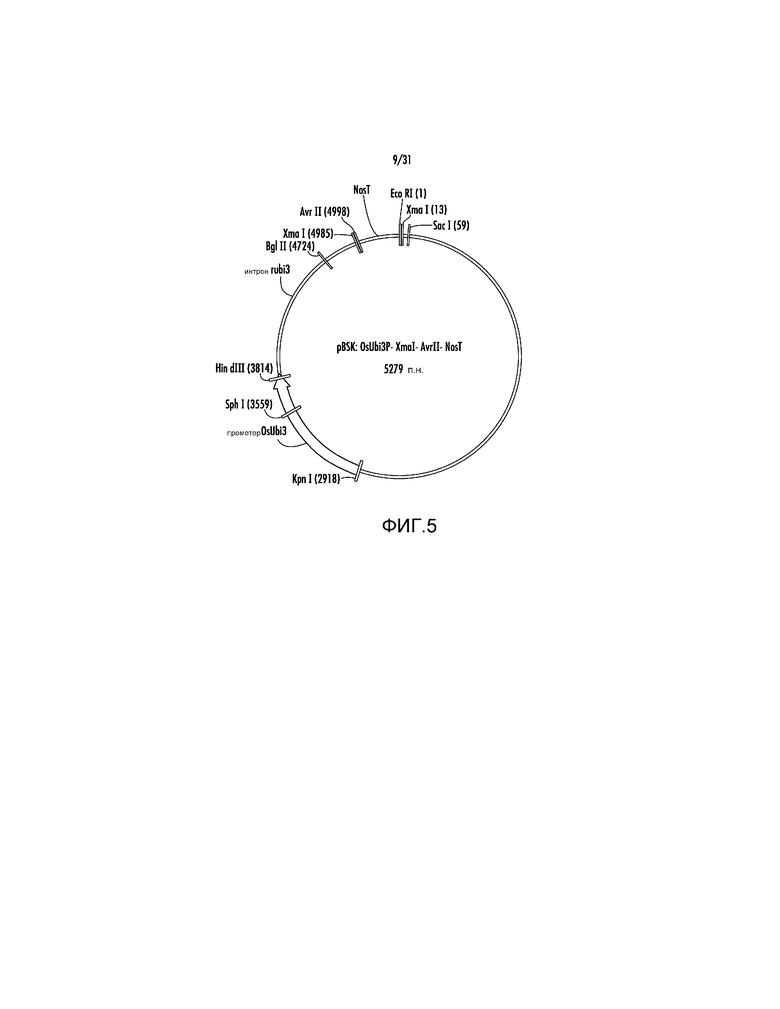
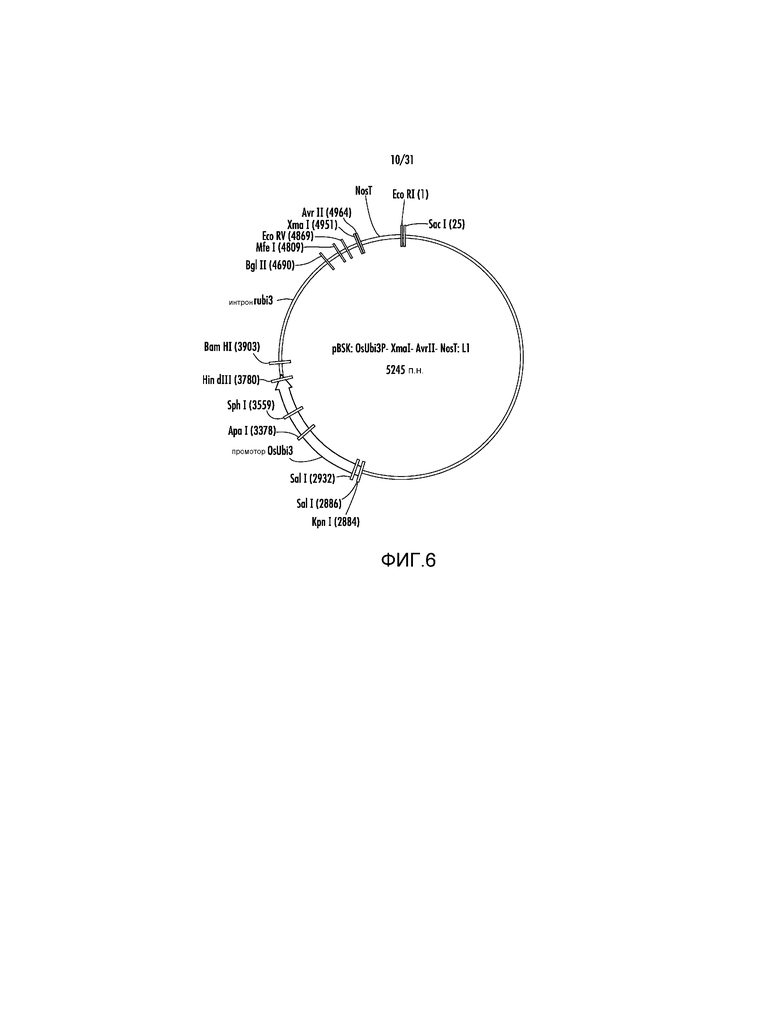
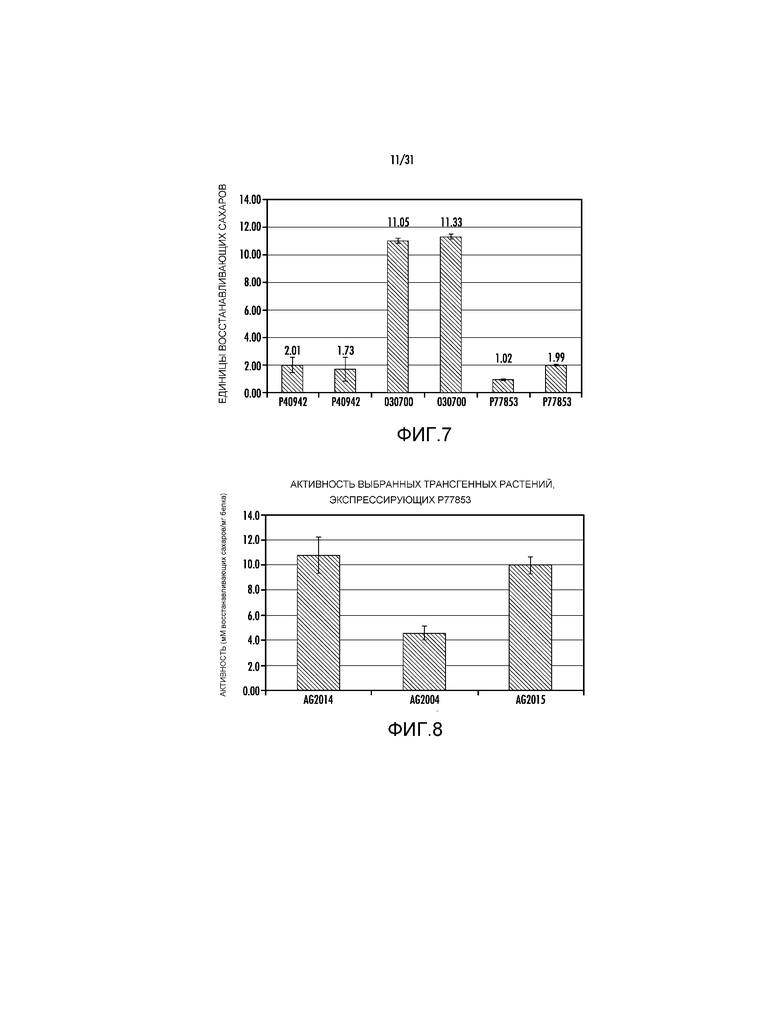
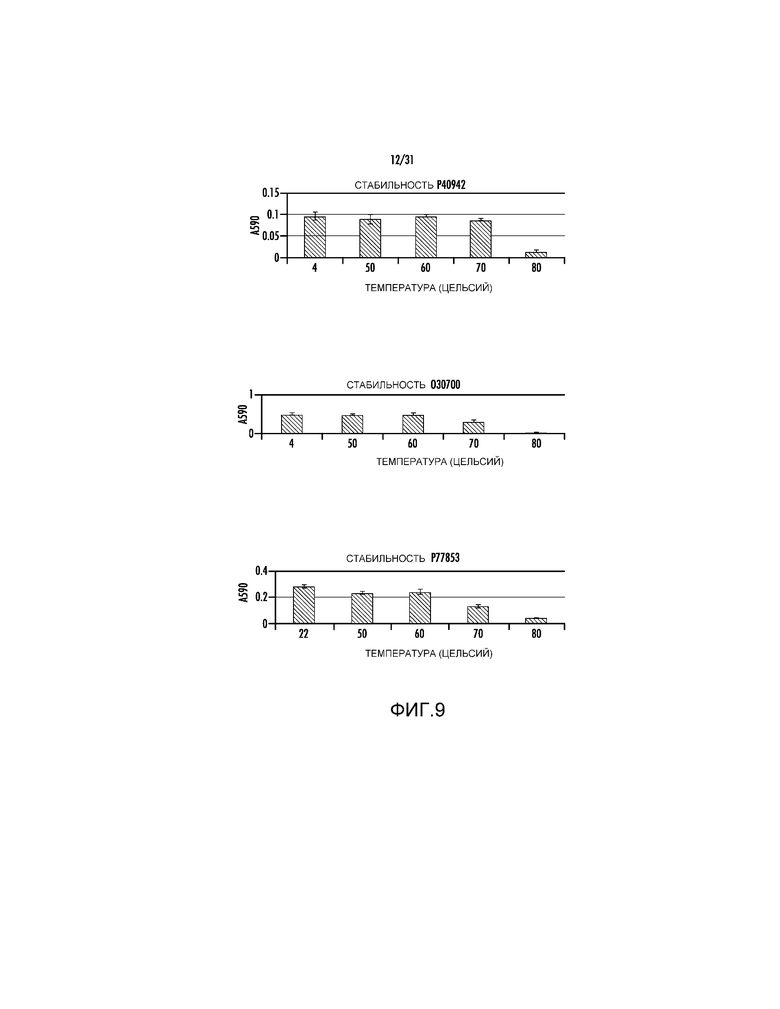

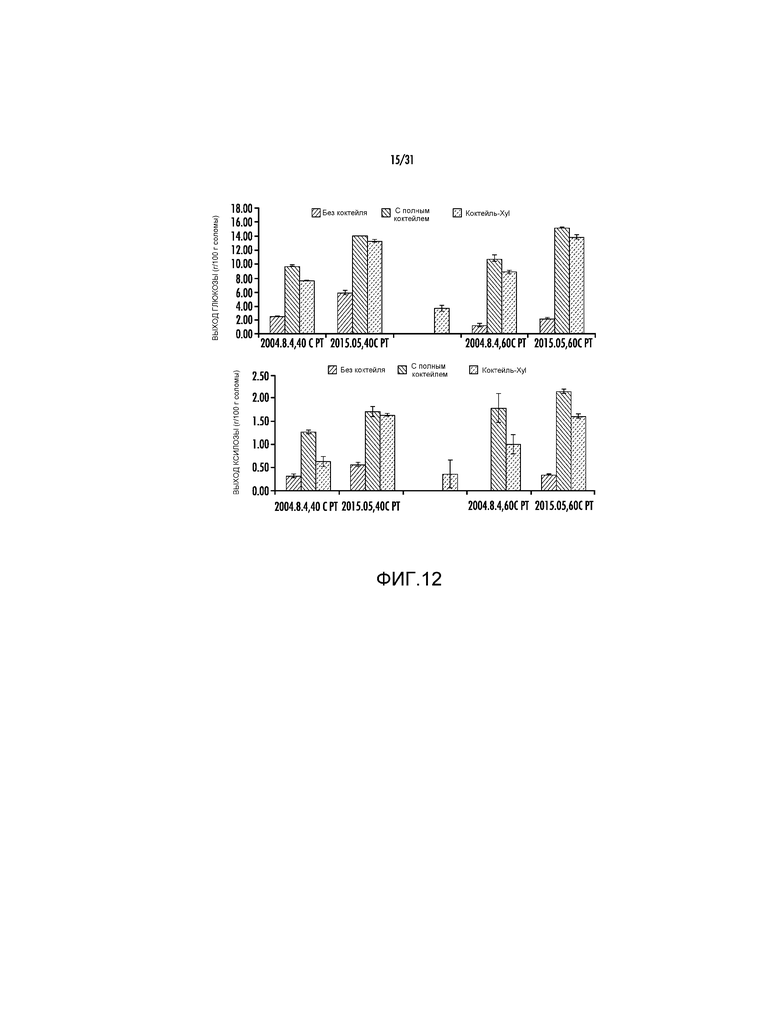
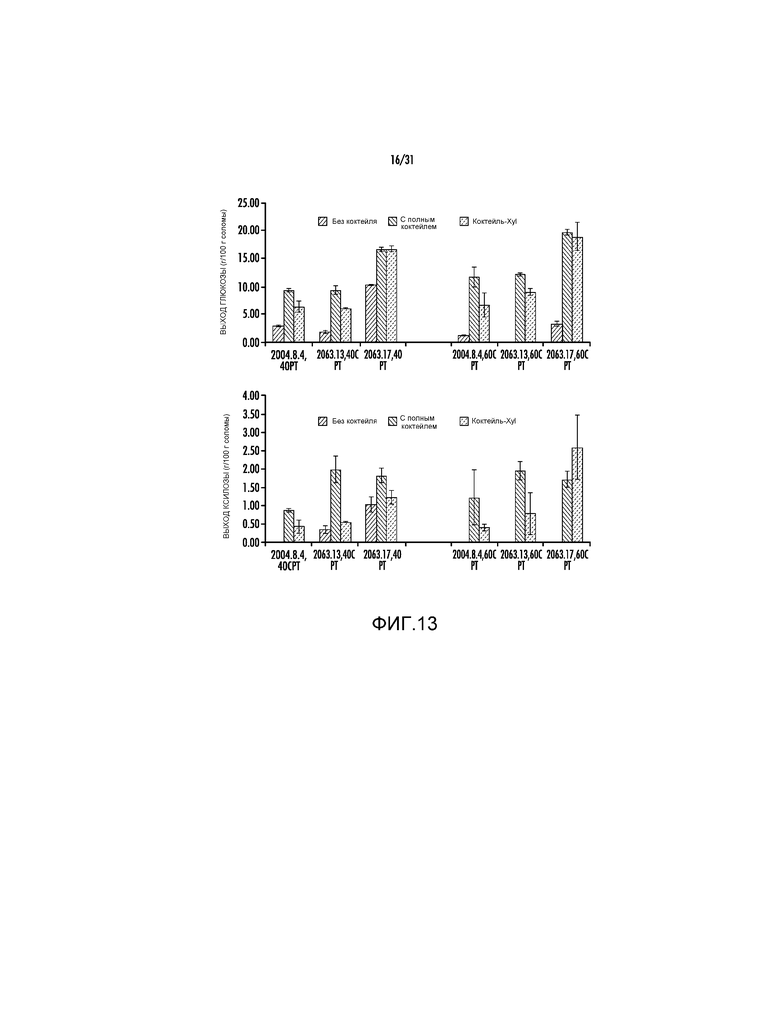
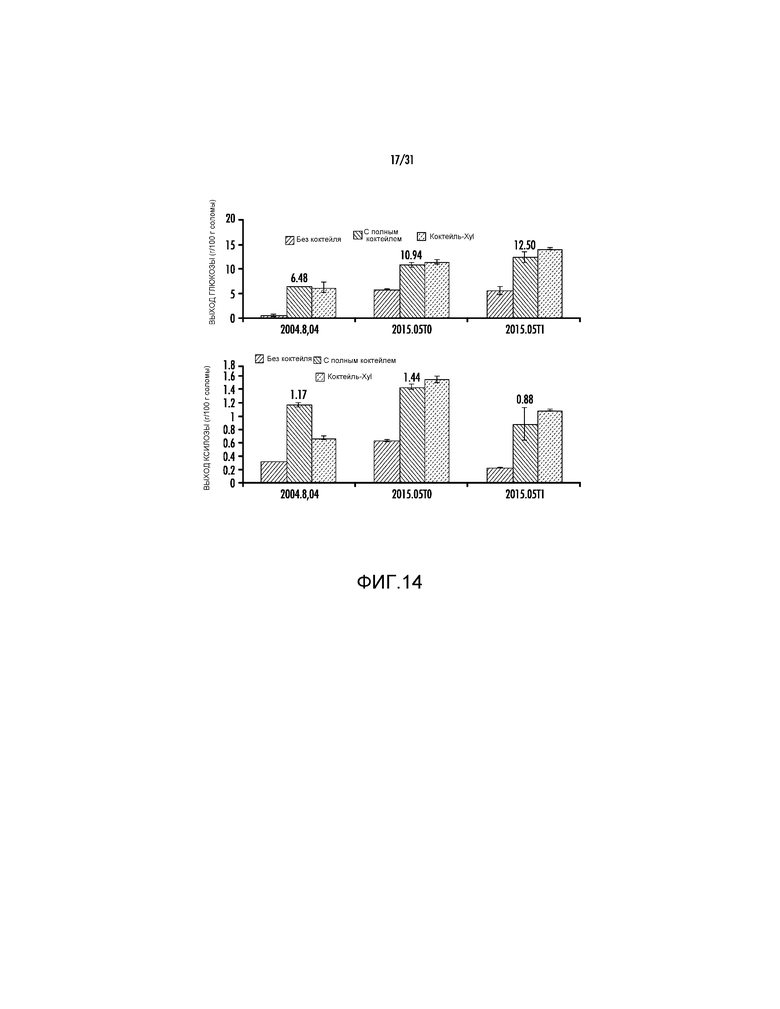
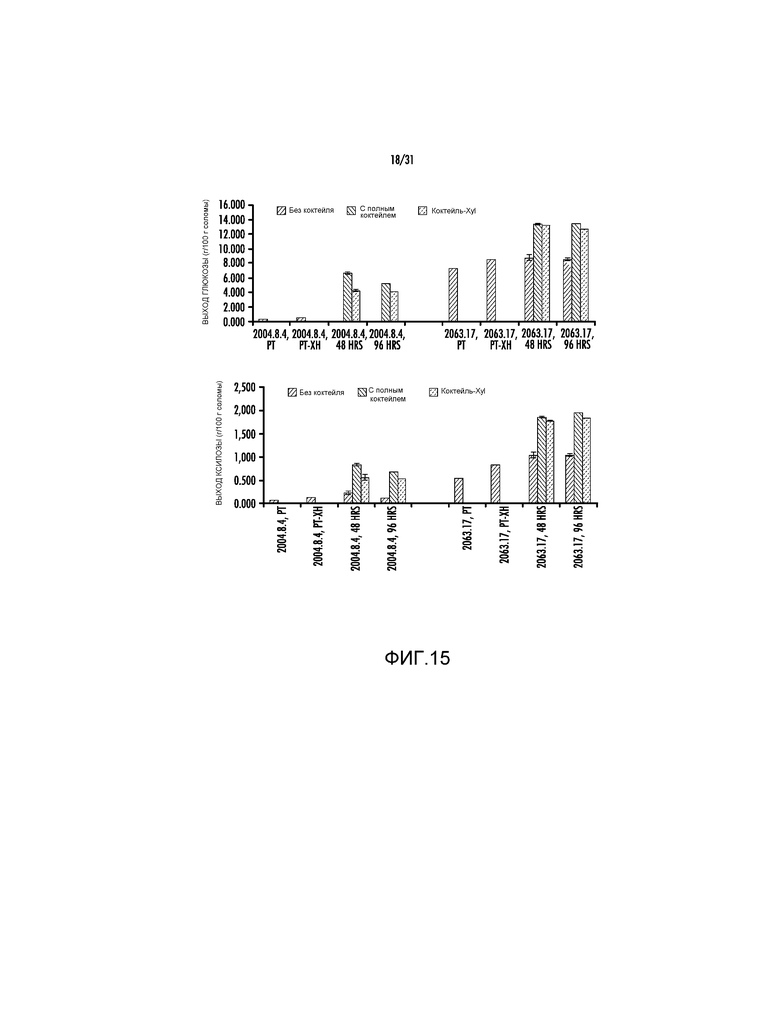
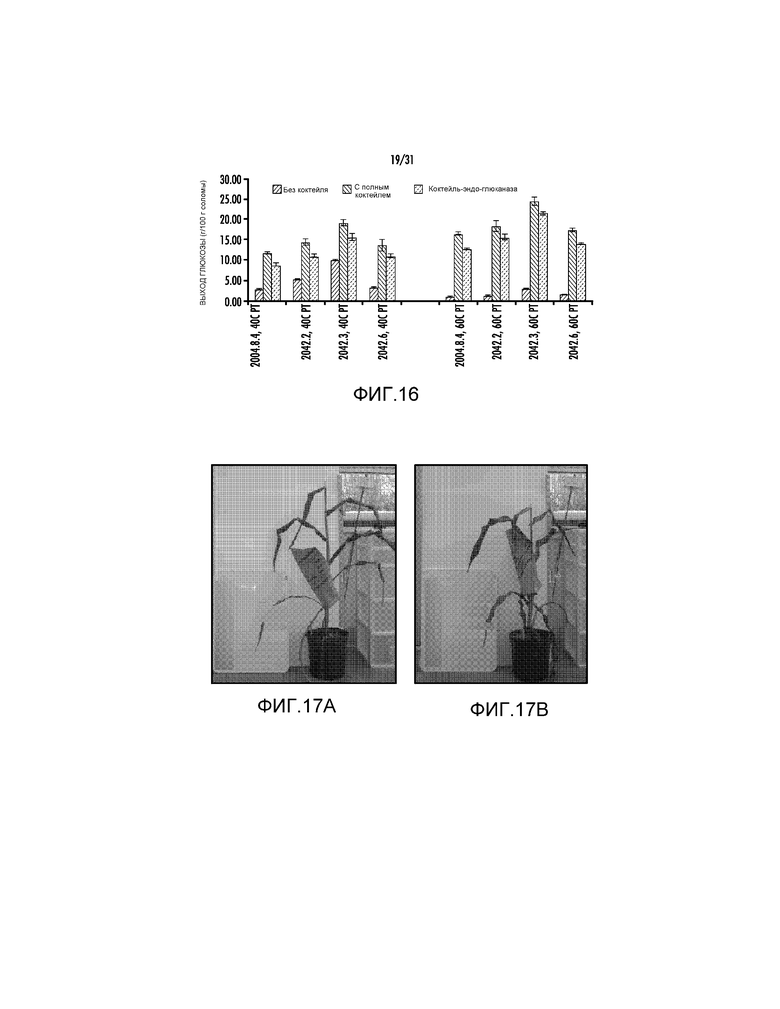

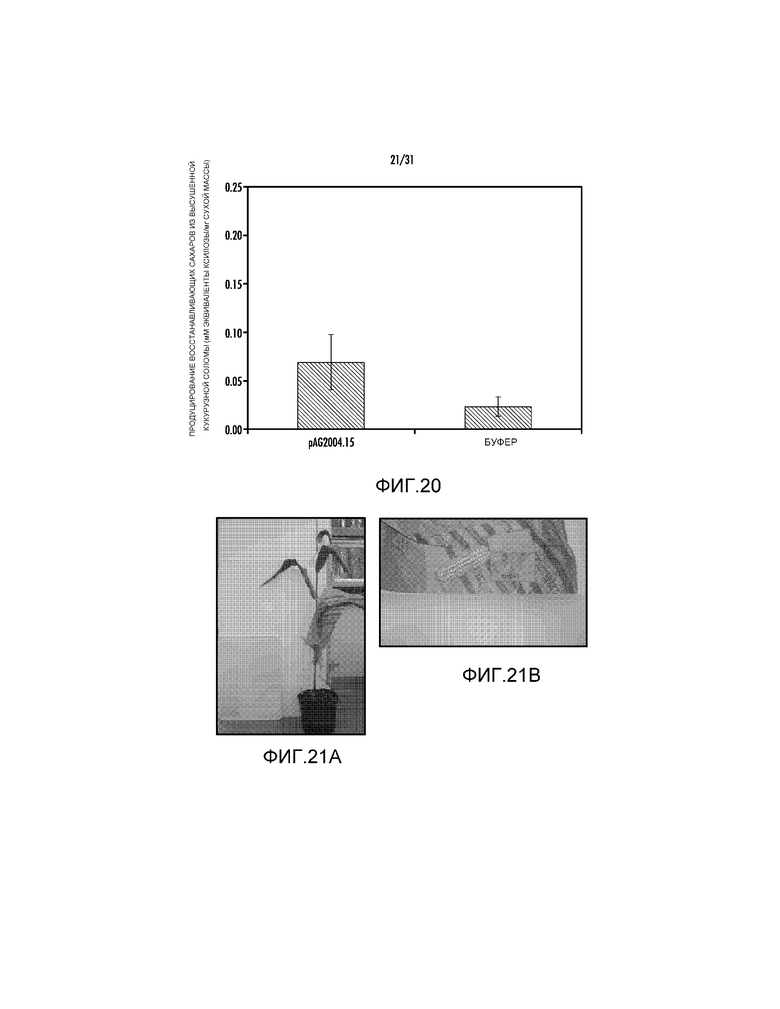
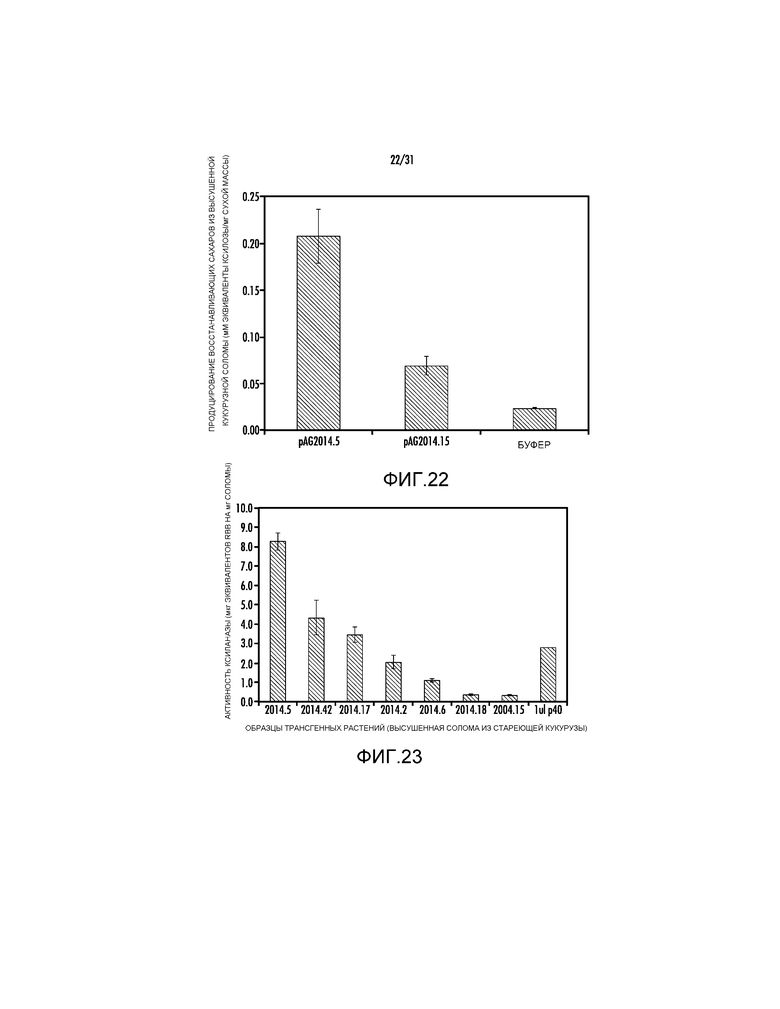
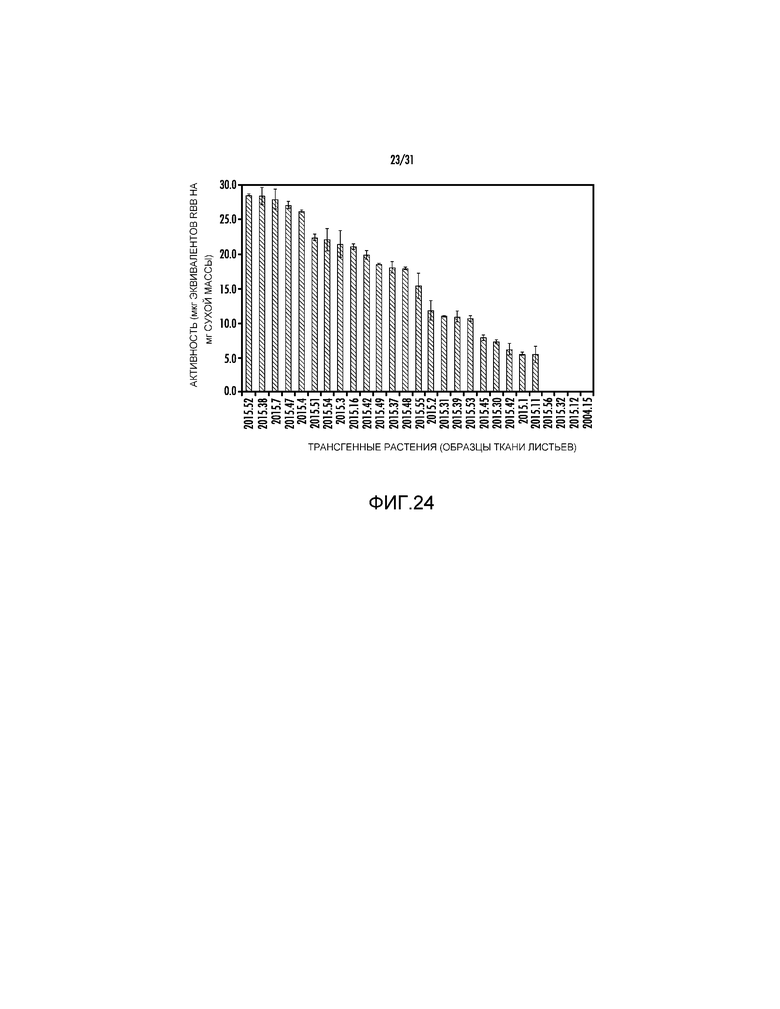
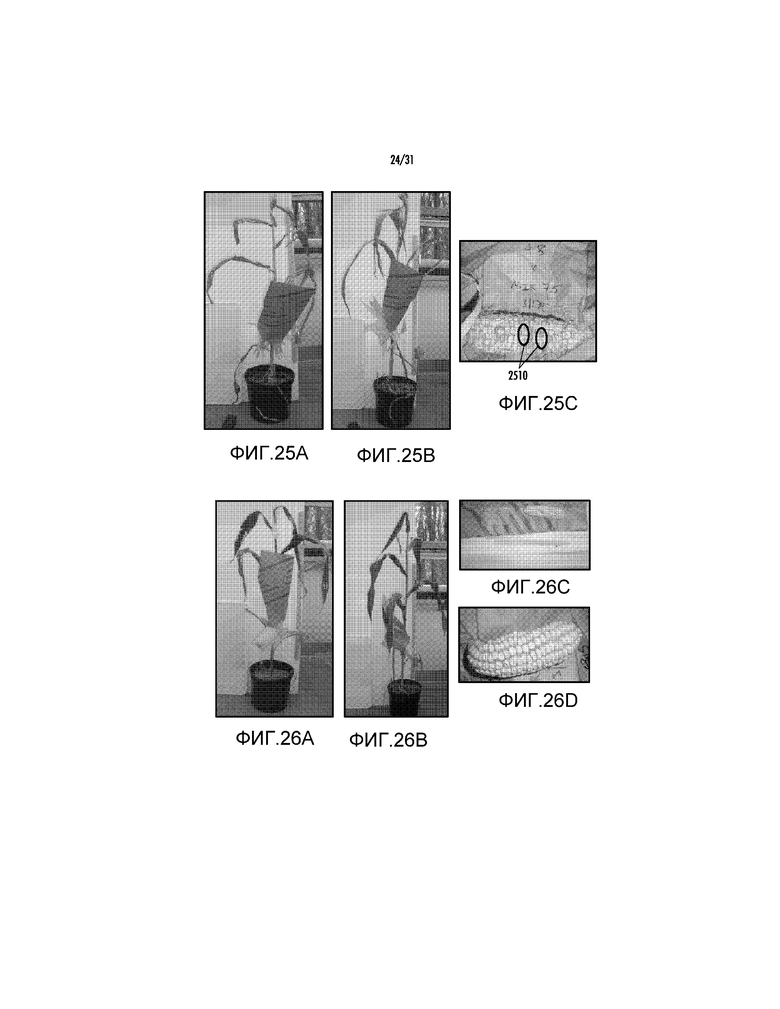



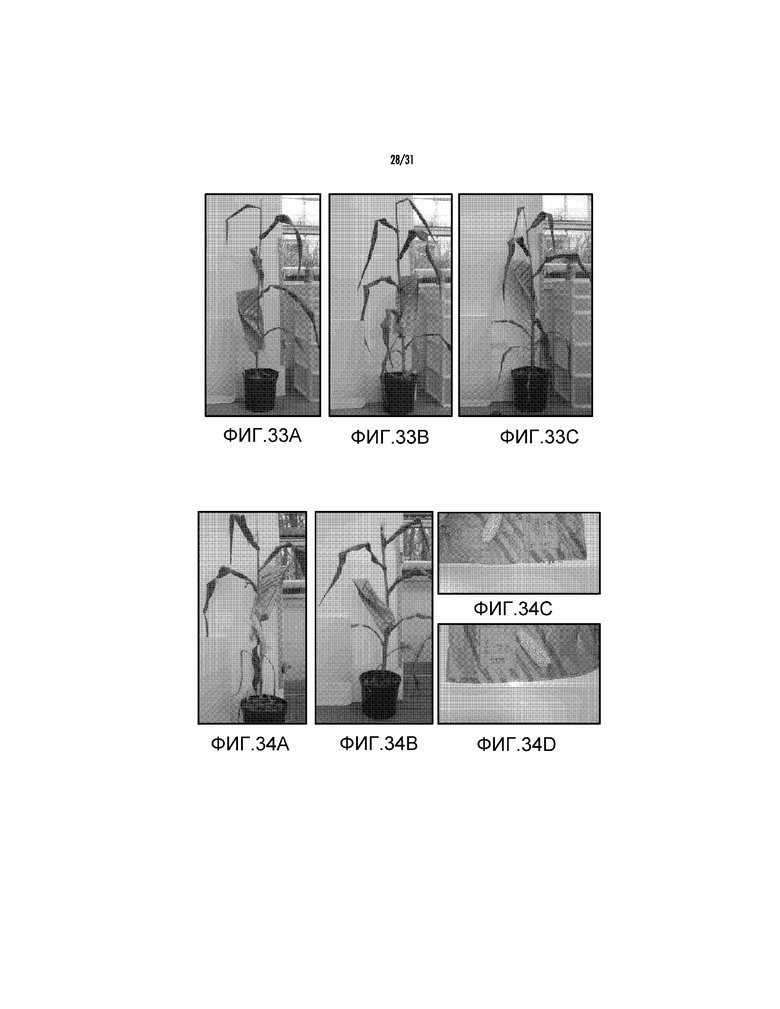


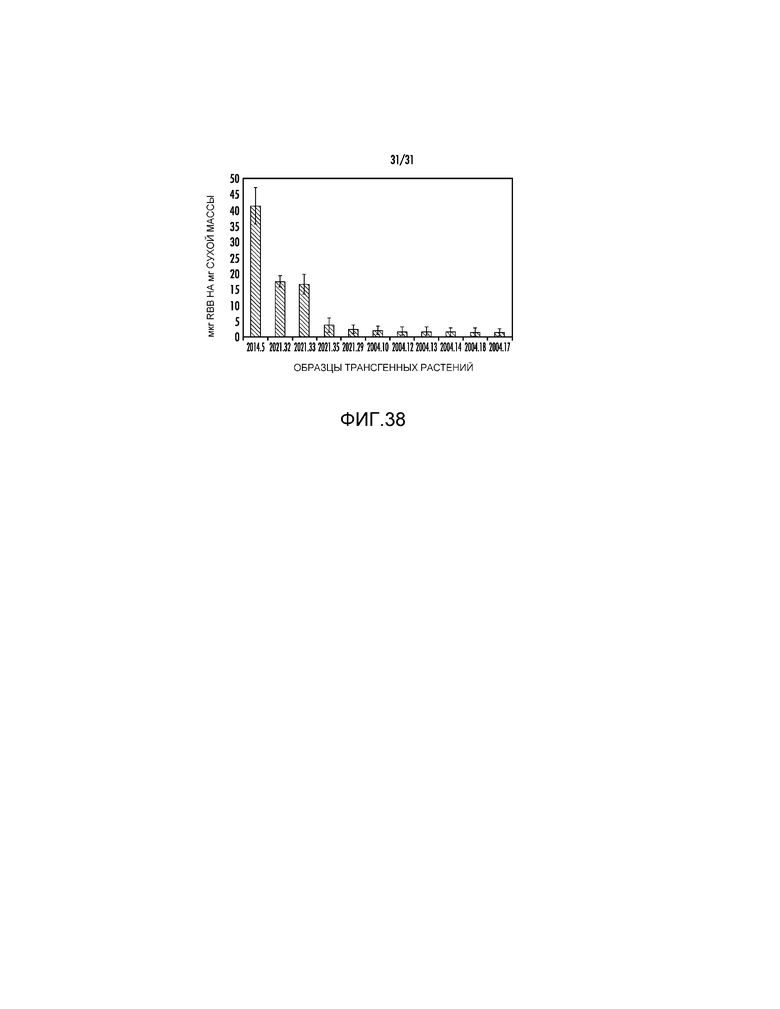
Изобретение относится к области биохимии, в частности к корму для животных, содержащему трансгенное растение или его часть для получения усвояемых сахаров с помощью фермента, деградирующего клеточную стенку, присутствующего в нем. Изобретение позволяет эффективно получать корм для животных, содержащий усвояемые сахара. 2 н. и 18 з.п. ф-лы, 38 ил., 5 табл., 22 пр.
1. Корм для животных, содержащий трансгенное растение или его часть для получения усвояемых сахаров с помощью фермента, деградирующего клеточную стенку, присутствующего в нем, причем трансгенное растение или его часть содержит нуклеиновую кислоту, кодирующую фермент, деградирующий клеточную стенку, где фермент, деградирующий клеточную стенку, содержит аминокислотную последовательность, имеющую по меньшей мере 90% идентичность с последовательностью, выбранной из SEQ ID NO:44-56, 59-67, 70, 75, 85-93 и 110-111, причем фермент, деградирующий клеточную стенку, имеет ту же самую или по существу ту же активность, что и фермент, деградирующий клеточную стенку, состоящий из последовательности, имеющей 100% идентичность с одной из SEQ ID NO:44-56, 59-67, 70, 75, 85-93 и 110-111.
2. Корм для животных по п.1, где аминокислотная последовательность представляет собой одну из имеющих по меньшей мере 90% идентичность с последовательностью, выбранной из SEQ ID NO:45, 49-54, 59 и 85.
3. Корм для животных по п.1, где аминокислотная последовательность представляет собой одну из имеющих по меньшей мере 90% идентичность с последовательностью, выбранной из SEQ ID NO:47 и 55.
4. Корм для животных по п.1, где аминокислотная последовательность представляет собой одну из имеющих по меньшей мере 90% идентичность с последовательностью, выбранной из SEQ ID NO:46, 48 и 56.
5. Корм для животных по п.1, где аминокислотная последовательность представляет собой одну из имеющих по меньшей мере 90% идентичность с последовательностью, выбранной из SEQ ID NO:60-67, 70 и 75.
6. Корм для животных по п.1, где аминокислотная последовательность представляет собой одну из имеющих по меньшей мере 90% идентичность с последовательностью, выбранной из SEQ ID NO:87-93 и 110-111.
7. Корм для животных по п.1, где аминокислотная последовательность представляет собой одну из имеющих по меньшей мере 90% идентичность с последовательностью, выбранной из SEQ ID NO:44, 45, 49 и 54.
8. Корм для животных по п.1, где аминокислотная последовательность представляет собой одну из имеющих по меньшей мере 90% идентичность с последовательностью, выбранной из SEQ ID NO:45 и 87.
9. Корм для животных по п.1, где аминокислотная последовательность представляет собой одну из имеющих по меньшей мере 90% идентичность с последовательностью, выбранной из SEQ ID NO:50-53 и 59.
10. Корм для животных по п.1, где трансгенное растение представляет собой одно из: кукурузы, проса, мискантуса, сахарного тростника или сорго.
11. Корм для животных по п.10, где трансгенное растение представляет собой кукурузу.
12. Корм для животных по п.10, где трансгенное растение представляет собой просо.
13. Корм для животных по любому из пп.1-12, дополнительно содержащий сырье из другого источника, чем трансгенное растение или его часть.
14. Корм для животных по любому из пп.1-12, где аминокислотная последовательность фермента, деградирующего клеточную стенку, кодируется нуклеиновой кислотой, имеющей 100% идентичности с одной из последовательностей, выбранной из SEQ ID NOS: 44-56, 59-67, 70, 75, 85-93 и 110-111.
15. Корм для животных по п.14, дополнительно содержащий сырье из другого источника, чем трансгенное растение или его часть.
16. Корм для животных, содержащий трансгенное растение или его часть для получения усвояемых сахаров с помощью фермента, деградирующего клеточную стенку, присутствующего в нем, причем трансгенное растение или его часть содержит нуклеиновую кислоту, которая способна гибридизоваться в условиях высокой жесткости с комплементом второй нуклеиновой кислоты, состоящей из нуклеотидной последовательности, выбранной из SEQ ID NO:116-128, 131-139, 142, 147, 157-165 и 182-183, где первая нуклеиновая кислота кодирует фермент, деградирующий клеточную стенку, способный гидролизовать компонент клеточной стенки трансгенного растения.
17. Корм для животных по п.16, где трансгенное растение представляет собой одно из: кукурузы, проса, мискантуса, сахарного тростника или сорго.
18. Корм для животных по п.16, где трансгенное растение представляет собой кукурузу.
19. Корм для животных по п.16, где трансгенное растение представляет собой просо.
20. Корм для животных по любому из пп.16-19, дополнительно содержащий сырье из другого источника, чем трансгенное растение или его часть.
| WO2007041419 A1, 12.04.2007 | |||
| US2007192900 A1, 16.08.2007 | |||
| WO2007146944 A2, 21.12.2007 | |||
| US2007250961 A1, 25.10.2007 | |||
| ПОЛИПЕПТИД И КОМПОЗИЦИЯ, ОБЛАДАЮЩИЕ АКТИВНОСТЬЮ КСИЛАНАЗЫ, ПРИМЕНЕНИЕ ПОЛИПЕПТИДА, ПОЛИНУКЛЕОТИД, КОДИРУЮЩИЙ ПОЛИПЕПТИД, ВЕКТОР ЭКСПРЕССИИ, СОДЕРЖАЩИЙ ПОЛИНУКЛЕОТИД, СПОСОБ ПОЛУЧЕНИЯ ПОЛИНУКЛЕОТИДА, СПОСОБ ОБРАБОТКИ РАСТИТЕЛЬНОГО ИЛИ КСИЛАНСОДЕРЖАЩЕГО МАТЕРИАЛА | 2000 |
|
RU2291901C2 |

