1.0 Предпосылки изобретения
1.1 Область изобретения
Данное изобретение относится в общем к трансгенным растениям, обладающим инсектицидными способностями, и к конструкциям ДНК, используемым для переноса генов, придающих устойчивость к насекомым, в геномы растений. Более конкретно, данное изобретение относится к способу экспрессии инсектицидных белков в растениях, трансформированных геном, кодирующим δ-эндотоксин В.thuringiensis, что приводит к эффективной защите от чувствительных вредителей-мишеней.
1.2 Описание уровня техники
1.2.1 Способы борьбы с инвазией насекомых в растениях
Грамположительная почвенная бактерия В.thuringiensis хорошо известна вследствие продуцирования ею белковых параспоральных кристаллов, или δ-эндотоксинов, которые являются токсичными для личинок многочисленных чешуекрылых, жесткокрылых и двукрылых насекомых. В.thuringiensis продуцирует во время споруляции кристаллические белки, которые являются специфически токсичными в отношении определенных видов насекомых. Было показано, что различные штаммы В.thuringiensis продуцируют инсектицидные кристаллические белки. Композиции, содержащие штаммы В.thuringiensis, которые продуцируют белки, имеющие инсектицидную активность, использовали коммерчески в качестве приемлемых для окружающей среды топических инсектицидов вследствие их токсичности для конкретного насекомого-мишени и отсутствия токсичности для растений и других не являющихся мишенью организмов.
Кристаллы δ-эндотоксина являются токсичными в отношении личинок насекомого при проглатывании. Растворение кристалла в средней кишке насекомого высвобождает протоксиновую форму δ-эндотоксина, которая в большинстве случаев процессируется затем в активный токсин протеазой средней кишки. Активированные токсины узнают щеточную каемку эпителия средней кишки насекомого через рецепторные белки. Несколько предположительных рецепторов кристаллических белков были выделены из личинок определенных насекомых (Knight et al., 1995; Gill et al., 1995; Masson et al., 1995). Связывание активных токсинов сопровождается интеркаляцией и агрегацией молекул токсина с образованием пор в эпителии средней кишки. Этот процесс приводит к осмотическому дисбалансу, набуханию, лизису клеток, выстилающих эпителий средней кишки, и в конце концов к гибели личинок.
1.2.2 Трансгенные δ-эндотоксины В.thuringiensis в качестве биопестицидов
Устойчивость растений и биологическая защита являются центральными тактическими линиями защиты в большинстве программ усовершенствования инсектицидов, применяемых к большинству разнообразных культур. С пришествием молекулярно-генетических способов были выделены гены различных δ-эндотоксинов и были определены их последовательности ДНК. Эти гены использовали для конструирования некоторых генетически сконструированных продуктов В.thuringiensis, которые были одобрены для коммерческого применения. Недавние развития разработали новые системы доставки δ-эндотоксинов, в том числе растения, которые содержат и экспрессируют генетически сконструированные гены δ-эндотоксинов. Экспрессия δ-эндотоксинов В.thuringiensis в растениях содержит в себе потенциал для эффективной борьбы с вредителями растений, пока могут быть преодолены определенные проблемы. Эти проблемы включают в себя развитие устойчивости насекомых к конкретному белку Cry, экспрессируемому в данном растении, и развитие морфологически отклоняющихся от нормы растений вследствие присутствия трансгена.
Было показано, что экспрессия δ-эндотоксинов В.thuringiensis в трансгенных хлопчатнике, кукурузе и картофеле является эффективным средством борьбы с сельскохозяйственно важными насекомыми-вредителями (Perlak et al., 1990; Koziel et al., 1993; Perlak et al., 1993). Трансгенные культуры, экспрессирующие δ-эндотоксины В.thuringiensis, позволяют растениеводам значительно уменьшить применение дорогих, токсичных и иногда неэффективных топических химических инсектицидов. Применение трансгенов, кодирующих δ-эндотоксины В.thuringiensis, является особенно предпочтительным, когда встраивание трансгена не оказывает отрицательного действия на урожай желаемого продукта из трансформированных растений. Наблюдения показали, что урожаи из сельскохозяйственных растений, экспрессирующих определенные δ-эндотоксины В.thuringiensis, такие как Cry1A или Сry3А, являются эквивалентными или более высокими, чем для остальных одинаковых нетрансгенных коммерческих сортов растений. Это указывает на то, что некоторые δ-эндотоксины В.thuringiensis не имеют значимого отрицательного влияния на рост и развитие растений. Однако это относится не ко всем δ-эндотоксинам В.thuringiensis, которые могут быть использованы для трансформации растений.
Применение топических произведенных из В.thuringiensis инсектицидов может также привести к развитию штаммов насекомых, устойчивых к этим инсектицидам. Устойчивость к δ-эндотоксинам Cry1A В.thuringiensis, наносимым в виде лиственных спреев, развивалась в по меньшей мере одном хорошо документированном случае (Shelton et al., 1993). Ожидается, что насекомые могут подобным образом развивать устойчивость к δ-эндотоксинам В.thuringiensis, экспрессируемым в трансгенных растениях. Такая устойчивость, если она станет распространенной, могла бы явно лимитировать коммерческую ценность зародышевой плазмы кукурузы, хлопчатника, картофеля и другой зародышевой плазмы, содержащей гены, кодирующие δ-эндотоксины В.thuringiensis. Один из возможных способов как увеличения эффективности инсектицида против вредителей-мишеней, так и уменьшения развития устойчивых к инсектицидам вредителей мог бы заключаться в гарантии того, что трансгенные культуры экспрессируют высокие уровни δ-эндотоксинов В.thuringiensis (McGaughey and Whalon, 1993; Roush, 1994).
Кроме получения трансгенного растения, которое экспрессирует δ-эндотоксины В.thuringiensis при высоких уровнях, коммерчески жизнеспособные гены В.thuringiensis должны удовлетворять нескольким дополнительным критериям. Например, экспрессия этих генов в трансгенных сельскохозяйственных культурах не должна уменьшать силы, жизнеспособности или фертильности этих растений, и не должна влиять на нормальную морфологию этих растений. Подобные вредные эффекты имеют два нежелательных результата: они могут препятствовать получению и размножению трансгенных растений; они могут также препятствовать развитию зрелых растений или придавать неприемлемые агрономические свойства.
Остается потребность в композициях и способах, применимых в получении трансгенных растений, которые экспрессируют δ-эндотоксины В.thuringiensis при уровнях, достаточно высоких для эффективной защиты от насекомых-вредителей растения-мишени, а также предупреждения развития инсектицид-резистентных штаммов вредителей. Способ, приводящий к высоким уровням экспрессии δ-эндотоксинов В.thuringiensis, обеспечит также преимущества более частого получения коммерчески жизнеспособных трансформированных линий растений и более эффективной защиты от заражения в течение всего вегетационного периода.
Остается также потребность в способе увеличения уровня экспрессии δ-эндотоксинов В.thuringiensis, который не приводит одновременно к морфологическим изменениям растения, мешающим оптимальному росту и развитию желательных тканей растения. Например, способ усиления экспрессии δ-эндотоксинов В.thuringiensis в кукурузе не должен приводить к растению кукурузы, которое не может оптимально развиваться для культивирования. Достижение этих задач, таких как высокие уровни экспрессии, а также получение морфологически нормальных растений, не удавалось, и поиск их решения был упорно продолжающимся и важным аспектом долгосрочной ценности инсектицидных продуктов растений.
2.0 Сущность изобретения
Описаны новые способы для экспрессии Сry2А δ-эндотоксинов В.thuringiensis, которые лишены значимой игибирующей двукрылых насекомых активности в трансформированных растениях. Этот способ выгодно приводит к повышенным уровням экспрессии δ-эндотоксинов В.thuringiensis, а также более высокой скорости восстановления морфологически нормальных растений.
Посредством достижения высоких уровней экспрессии данное изобретение влияет на другой недостаток предыдущего уровня: развитие устойчивости насекомых. Конкретно, данное изобретение обеспечивает превосходящую прежние способы стратегию задержки или элиминации развития устойчивости к Cry1A δ-эндотоксинам, белкам В.thuringiensis, наиболее обычно экспрессируемым трансгенными линиями. Описанные способы включают в себя экспрессию класса Сry2А δ-эндотоксинов В.thuringiensis и, в частности, тех, которые лишены ингибирующей двукрылых насекомых активности. δ-эндотоксины В.thuringiensis группы Сry2А не имеют значимой гомологии с δ-эндотоксинами Cry1A типа и проявляют отличающиеся характеристики связывания и порообразования (English et al., 1994), и ожидается, что, как таковые, они действуют на насекомых, которые становятся устойчивыми к δ-эндотоксинам Cry1A или на которые не действуют δ-эндотоксины Cry1A (Hofte and Whiteley, 1989).
В предпочтительных вариантах данное изобретение обеспечивает выделенную и очищенную конструкцию ДНК, содержащую кодирующий δ-эндотоксин Сry2А район, локализованный в пластиде или хлоропласте или локализованный в ядерном геноме клетки растения и функционально (оперативно) связанный с районом, кодирующим пластидный транзитный пептид (РТР). Предпочтительные конструкции ДНК данного изобретения включают в себя конструкции, которые кодируют δ-эндотоксины Cry2A, лишенные ингибирующей двукрылых насекомых активности, хотя полная инактивация в отношении двукрылых не является необходимой. В иллюстративном варианте конструкции ДНК данного изобретения кодируют δ-эндотоксин Cry2Ab, функционально связанный с сегментом (или последовательностью) ДНК, кодирующим пластидный транзитный пептид, который является одним из средств, делающим возможной локализацию δ-эндотоксина Cry2Ab в пластиде или хлоропласте. В некоторых вариантах δ-эндотоксин Cry2Ab содержит последовательность SEQ ID NO:2. Однако авторы изобретения предполагают, что любой δ-эндотоксин Cry2A, лишенный ингибирующей двукрылых насекомых активности, может быть использован в соответствии с данным изобретением, причем особенно предпочтительными являются δ-эндотоксины Cry2A, имеющие значимые гомологии относительно Cry2Ab.
В другом варианте конструкции ДНК данного изобретения используют сегменты нуклеиновой кислоты, кодирующие РТР, для потенциирования экспрессии δ-эндотоксина. Применение одного типа РТР, нацеливающего на хлоропласты пептида (СТР), в соединении с трансгеном Cry1A В.thuringiensis для активации экспрессии этого трансгена в трансформированном растении, описано в патенте США 5500365 (специально включенного здесь в качестве полной ссылки). Однако, когда наблюдается повышенная экспрессия, ее приписывают отчасти использованию новой 5'-нетранслируемой лидерной последовательности в экспрессирующем векторе.
В противоположность предыдущему уровню данной области данное изобретение описывает структурную последовательность ДНК, которая вызывает образование последовательности РНК, кодирующей целевой слитый белок, содержащий аминоконцевой пластидный транзитный пептид с δ-эндотоксином Cry2Ab; и 3’-нетранслируемую последовательность ДНК, которая функционирует в клетках растений, вызывая терминацию транскрипции и присоединение полиаденилированных нуклеотидов к 3’-концу указанной последовательности РНК. Неожиданно, эта конструкция ДНК приводит к увеличенным уровням экспрессии δ-эндотоксина Cry2A. Этот нацеленный слитый белок является неактивным для всех видов, но продуцируется как средство для локализации зрелого, инсектицидно активного белка δ-эндотоксина в хлоропласте, что дает удивительные и неожиданные выгодные агрономические эффекты.
Один вариант, представленный в данном изобретении, представляет собой введение гена, кодирующего δ-эндотоксин Cry2A, лишенный активности против двукрылых насекомых, в хлоропластный или пластидный геном. Альтернативно, ген, кодирующий δ-эндотоксин Cry2A, лишенный активности против двукрылых насекомых, мог бы экспрессироваться из автономно реплицирующегося эписомного элемента, локализованного в хлоропласте или пластиде.
В другом предпочтительном варианте данное изобретение обеспечивает трансгенные растения, которые были трансформированы выделенной и очищенной конструкцией ДНК, которая транслируется и экспрессируется при высоких уровнях данным растением. Как однодольные, так и двудольные растения могут быть трансформированы в соответствии с описанными здесь способами и с использованием описанных здесь конструкций ДНК. Растения, трансформированные согласно данному изобретению, могут быть получены в дополнительном предпочтительном варианте способом, включающим в себя получение выделенной и очищенной конструкции ДНК и затем трансформацию растения этой конструкцией таким образом, что данное растение экспрессирует белки, которые кодирует данная конструкция. Авторы изобретения наблюдали, что трансформация растений описанными способами приводит к увеличенной частоте трансформантов, которые экспрессируют данный трансген, а также к генерированию более морфологически нормальных растений из исходных трансформантов.
Предполагается, что увеличенные уровни экспрессии, наблюдаемые в описанном изобретении, позволят получить уменьшенное развитие устойчивости насекомых к δ-эндотоксинам В.thuringiensis. Это может быть достигнуто трансформацией растения предпочтительной конструкцией ДНК для получения только высоких уровней экспрессии Cry2A или посредством одновременного экспонирования насекомых-мишеней Cry1A и неактивным против двукрылых насекомых δ-эндотоксинам Cry2A, экспрессируемым в чувствительных растениях. Такие насекомые включают в себя Ostrina spp., Diatraea spp., Helicoverpa spp. и Spodoptera spp., в Zea mays; Heliothis virescens, Helicoverpa spp., Pectinophora spp., в Gossypium hirsutum; Anticarsia spp., Pseudoplusia spp., Epinotia spp., в Glycine max; и Scirpophaga incertulas в Oryza sativa.
Таким образом, предполагается, что способ, описанный данным изобретением, обеспечит многочисленные преимущества в сравнении с прежним уровнем, в том числе конкретно описанные выше. Эти преимущества включают в себя: достижение улучшенной борьбы в отношении чувствительных насекомых; минимизацию развития инсектицид-резистентности штаммов насекомых; получение большего числа коммерчески жизнеспособных устойчивых к насекомым линий растений; достижение сохраняющейся в течение всего вегетационного периода защиты от насекомых-патогенов и увеличение частоты встречаемости морфологически нормальных трансформированных растений. Дополнительным преимуществом данного изобретения является то, что будет требоваться получение уменьшенных количеств трансгенных линий для идентификации трансгенного события с нормальными характеристиками роста.
Таким образом, предполагается, что способ, описанный данным изобретением, обеспечит много преимуществ в сравнении с прежним уровнем, в том числе преимущества, конкретно очерченные выше. Эти преимущества включают в себя: достижение улучшенной борьбы в отношении чувствительных насекомых; минимизацию развития инсектицид-резистентности штаммов насекомых; получение большего числа коммерчески жизнеспособных устойчивых к насекомым линий растений; достижение сохраняющейся в течение всего вегетационного периода защиты от насекомых-патогенов и увеличение частоты встречаемости морфологически нормальных трансформированных растений. Дополнительным преимуществом данного изобретения является то, что будет требоваться получение уменьшенных количеств трансгенных линий для идентификации трансгенного события с нормальными характеристиками роста.
2.1 Композиции нуклеиновых кислот
В одном важном варианте данное изобретение обеспечивает выделенную и очищенную конструкцию нуклеиновой кислоты, содержащую кодирующий Cry2A район и кодирующий РТР район. Эти конструкции ДНК при перенесении в растение подвергаются клеточным процессам, приводящим к увеличенной экспрессии δ-эндотоксинов в трансгенном растении. δ-эндотоксины Cry2A данного изобретения являются предпочтительно неэффективными против двукрылых насекомых, хотя некоторые вредные действия в отношении двукрылых могут быть приемлемыми. В некоторых вариантах конструкция ДНК кодирует неактивный в отношении двукрылых насекомых δ-эндотоксин Cry2Ab, а в более предпочтительных вариантах δ-эндотоксин Cry2Ab имеет полипептидную последовательность SEQ ID NO:2 или последовательность, по существу гомологичную полипептидной последовательности SEQ ID NO:2. Такие нуклеотидные гомологи могут быть более чем на приблизительно 88% гомологичными, более чем на приблизительно 90% гомологичными, более чем на приблизительно 95% гомологичными и даже более, чем на 99% гомологичными с δ-эндотоксином Cry2Ab, представленным в SEQ ID NO:2. Примеры пептидов включают в себя пептиды, которые на приблизительно 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 и даже 99 или более процентов гомологичны δ-эндотоксину Cry2Ab, представленному в SEQ ID NO:2.
В даже более предпочтительных вариантах конструкция ДНК данного изобретения содержит кодирующий δ-эндотоксин Cry2Ab район с последовательностью нуклеиновой кислоты SEQ ID NO:1 или последовательностью со значительной гомологией относительно последовательности SEQ ID NO:1. Также рассматриваются как находящиеся в объеме данного изобретения конструкции ДНК, имеющие сегменты со значительными гомологиями относительно последовательности нуклеиновой кислоты, представленной в SEQ ID NO:1, такие как сегменты, которые могут быть приблизительно на 90% гомологичными или приблизительно на 95% гомологичными или даже на приблизительно 99% гомологичными. Более конкретно, гомологичные последовательности нуклеиновых кислот, включенные в данное изобретение, включают в себя последовательности, которые являются на приблизительно 90, 91, 92, 93, 94, 95, 96, 97, 98 и 99 процентов гомологичны последовательности нуклеиновой кислоты SEQ ID NO:1.
Конструкции ДНК, обеспеченные здесь, включают в себя также РТР-кодирующий район, расположенный выше по ходу транскрипции (слева) от района, кодирующего δ-эндотоксин Cry2A, и ниже по ходу транскрипции (справа) от промотора. Эти кодирующие пластидный транзитный пептид районы могут кодировать любой функциональный в растении РТР и могут функционировать для нацеливания кодируемых белков в определенные пластиды в клетке растений или для увеличения экспрессии δ-эндотоксина, кодируемого данной конструкцией ДНК. В предпочтительных вариантах данное изобретение может включать в себя РТР, выбранный из группы, состоящей из zmSSU, РТР1, РТР1Δ и РТР2, или любые другие функциональные в растении РТР. Более конкретно район, кодирующий пластидный транзитный пептид, кодирует пластидный транзитный пептид, имеющий аминокислотную последовательность SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8 или SEQ ID NO:10 или любую полипептидную последовательность, по существу гомологичную этим последовательностям. Даже более предпочтительно данное изобретение включает в себя район, кодирующий пластидный транзитный пептид, имеющий нуклеотидную последовательность SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7 или SEQ ID NO:9 или последовательность нуклеиновой кислоты, которая по существу гомологична этим последовательностям.
Авторы изобретения предполагают, что данное изобретение могло бы решить задачи увеличения патогенности для вредителей и получить уменьшенное развитие устойчивых к пестицидам насекомых, если конструкции ДНК, обеспеченные здесь, ко-экспрессировать вместе с другими пестицидными конструкциями, такими как другие белки. Таким образом, данное изобретение обеспечивает применение описанных конструкций ДНК, которые дополнительно содержат экспрессируемые в растении кодирующие районы других Cry-белков. В них могли бы быть включены кодирующие районы для Cry1-белков, таких как Cry1A, Cry1Ab, Cry1Bb или Cry1-химеры (см. находящиеся в процессе одновременного рассмотрения заявки на патент США с регистрационными номерами 08/754490 и 08/922505 и находящуюся в процессе одновременного рассмотрения РСТ заявку PCT/US97/17507 на основе заявки США с регистрационным номером 08/721259, каждая из которых включена здесь в полном виде в качестве ссылки).
В некоторых предпочтительных вариантах конструкция ДНК является экспрессионной кассетой, которая может быть вырезана и выделена из указанной плазмиды.
2.2 Дополнительные элементы композиции нуклеиновой кислоты
Полинуклеотидные композиции данного изобретения применимы в трансформации как однодольных, так и двудольных растений. Таким образом, конструкция ДНК данного изобретения может дополнительно содержать другие различные регуляторные элементы для содействия в экспрессии белков и для дополнительного облегчения введения данной конструкции ДНК в растение. Одним примером этого является включение в данную конструкцию ДНК интрона, расположенного в нетранслируемом лидере, выше по ходу транскрипции (слева) от района, кодирующего пластидный транзитный пептид. Одной из применимых лидерных последовательностей является хитшоковый белок (белок теплового шока) петунии. В различных альтернативных вариантах данный интрон может быть любым из следующих интронов: интроном 1 Adh, интроном сахарозосинтазы, омега-элементом TMV, хитшоковым белком 70 кукурузы (hsp 70) или интроном Act1 риса. В предпочтительных вариантах данный интрон является либо хитшоковым белком 70 кукурузы, либо хитшоковым белком 70 петунии.
В другом предпочтительном варианте данного изобретения обеспечена нуклеотидная последовательность, содержащая дополнительно кодирующий район по существу неактивного относительно двукрылых насекомых δ-эндотоксина Cry2A и кодирующий РТР район, расположенные под контролем функционального в растении промотора. Применение промотора необходимо для запуска клеточных процессов таким образом, чтобы максимизировалась экспрессия данного гена. Предпочтительные промоторы включают в себя следующие промоторы: CaMV 35S, гистона, CaMV 19S, nos, OCS, Adh, сахарозосинтазы, α-тубулина, актина, cab, ФЕП-карбоксилазы, ssRUBISCO (малой субъединицы РБФК), Act1, Famv, усиленного FMV, или промоторы, связанные с комплексом R-генов. В более предпочтительных вариантах промотор представляет собой усиленный или удвоенный промотор CaMV 35S (Кау et al., 1987). В дополнительных предпочтительных вариантах промотор является промотором FMV35S. Функциональные в хлоропластах или пластидах растений промоторы также входят в сферу действия данного изобретения.
Далее, данное изобретение рассматривает включение терминаторного района в конструкцию ДНК для содействия клеточным процессам, участвующим в экспрессии белка. В различных вариантах этот терминатор может быть любым из следующих терминаторов: терминатором гена нопалинсинтазы Agrobacterium tumefaciens, терминатором гена октопинсинтазы Agrobacterium tumefaciens и 3’-концом гена ингибитора протеазы I или II из картофеля или томата. В особенно предпочтительном варианте терминатором является терминатор гена нопалинсинтазы Agrobacterium tumefaciens.
2.3 Трансформирующие векторы
Поскольку конструкция ДНК данного изобретения предназначена прежде всего, хотя и не исключительно, для применения в трансформации растений, она в некоторых предпочтительных вариантах содержится в экспрессирующем векторе. Такие экспрессирующие векторы могут содержать множество регуляторных и других элементов, предназначенных для возможности оптимальной экспрессии желательных белков, которые кодирует данный экспрессирующий вектор. Эти дополнительные элементы могут включать в себя промоторы, терминаторы и интроны, описанные выше в разделе 2.2. Вектор, содержащий конструкцию ДНК и любые регуляторные или другие элементы, может быть выбран из группы, состоящей из искусственной хромосомы дрожжей, искусственной хромосомы бактерий, плазмиды или космиды.
Далее, сами экспрессирующие векторы могут быть в различных формах. Эти формы могут отличаться в силу разных причин и будут, вероятно, состоять из различных компонентов в зависимости от того, предназначены ли они для трансформации однодольного или двудольного растения. Например, фиг.1 иллюстрирует один возможный вариант, где экспрессирующий вектор однодольных растений содержит ген Cry2Ab в плазмиде, названной (SEQ ID NO:16). Кроме того, предполагается, что в качестве применимых трансформирующих конструкций будут применимы также другие экспрессирующие векторы, содержащие экспрессионные кассеты, представленные в этих плазмидных векторах, а также экспрессионные кассеты, содержащие по существу гомологичные последовательности. Таким образом, любой трансформирующий вектор, содержащий последовательность нуклеиновой кислоты от нуклеотида 1781 до нуклеотида 5869 SEQ ID NO:16, может быть использован.
Фиг.2 иллюстрирует один возможный экспрессирующий вектор двудольных растений. Он содержит ген Cry2Ab, представленный в плазмидах, названных pMON33827 (SEQ ID NO:13), pMON33828 (SEQ ID NO:14) и pMON33829 (SEQ ID NO:15). Как и в случае трансформирующих векторов однодольных растений авторы изобретения считают, что другие экспрессирующие векторы, содержащие экспрессионные кассеты, представленные в этих плазмидных векторах, или по существу гомологичные этим экспрессионным кассетам векторы, будут применимы в качестве трансформирующих конструкций двудольных растений. Предпочтительные экспрессионные кассеты двудольных растений включают в себя кассеты, представленные нуклеиновыми кислотами 17-3182 SEQ ID NO:13; нуклеиновыми кислотами 17-3092 SEQ ID NO:14 и нуклеиновыми кислотами 17-3155 SEQ ID NO:15. Иллюстративные варианты векторов, содержащих такие экспрессионные кассеты, описаны в последовательностях, обозначенных здесь как SEQ ID NO:13, SEQ ID NO:14 и SEQ ID NO:15.
Векторы, дополнительно рассматриваемые как находящиеся в сфере действия данного изобретения, включают в себя векторы, способные содержать как композиции нуклеиновых кислот неактивного к двукрылым насекомым Cry2A, описанные в разделе 2.1 выше, так и любые другие конструкции ДНК, которые дополнительно содержат экспрессируемые в растениях кодирующие районы для других белков Cry, таких как белок Cry1. Векторы, способные содержать обе из этих конструкций, могут также содержать множество различных цистронов, делающих их полицистронными или мультицистронными.
2.4 Трансформированные клетки-хозяева
Другой предпочтительный вариант данного изобретения включает в себя клетки, трансформированные конструкциями ДНК, описанными здесь в разделах 2.1 и 2.2, и с использованием трансформирующих векторов, описанных в разделе 2.3. Трансформированные клетки, рассматриваемые в данном изобретении, включают в себя как прокариотические, так и эукариотические клетки, экспрессирующие белки, кодируемые новыми конструкциями ДНК данного изобретения. Способ получения трансгенных клеток хорошо известен в данной области. В общих чертах данный способ предусматривает трансформацию подходящей клетки-хозяина сегментом ДНК, который содержит промотор, функционально (оперативно) связанный с кодирующим районом, который кодирует δ-эндотоксин В.thuringiensis. Такой кодирующий район обычно функционально связан с районом терминации транскрипции, вследствие чего данный промотор способен запускать транскрипцию данного кодирующего района в данной клетке и, следовательно, обеспечивать клетку способностью продуцировать δ-эндотоксин in vivo. Альтернативно, в случаях, когда желательно контролировать, регулировать или снижать количество конкретного δ-эндотоксина или эндотоксинов, экспрессируемых в конкретной трансгенной клетке, данное изобретение обеспечивает также экспрессию антисмысловой мРНК δ-эндотоксина; антисмысловой мРНК интрона; антисмысловой мРНК РТР или антисмысловой мРНК UTR. Применение антисмысловой мРНК как средства регуляции или уменьшения количества конкретного представляющего интерес белка в клетке хорошо известно в данной области.
В предпочтительном варианте данное изобретение включает в себя клетку растения, которая была трансформирована сегментом нуклеиновой кислоты или конструкцией ДНК данного изобретения и которая экспрессирует ген или сегмент гена, кодирующий один или несколько неактивных относительно двукрылых насекомых δ-эндотоксинов Cry2A В.thuringiensis, как описано здесь. В применении здесь термин “трансгенная клетка растения” относится к клетке растения, которая имеет включенные последовательности ДНК, в том числе, но не только, гены, которые, возможно, в норме не присутствуют, последовательности ДНК, которые в норме не транскрибируются в РНК или не транслируются (“не экспрессируются”) в белок, или любые другие гены или последовательности ДНК, которые желательно ввести в нетрансформированное растение, такие как гены, которые в норме присутствуют в нетрансформированном растении, но которые желательно сконструировать генетически или которые должны иметь измененную экспрессию.
Предполагается, что в некоторых случаях геном трансгенного растения данного изобретения будет увеличен посредством стабильного введения конструкций ДНК, кодирующих неактивный относительно двукрылых насекомых δ-эндотоксин Cry2A В.thuringiensis, как описано в разделах 2.1 и 2.2 выше. В некоторых случаях более одного трансгена будут включены в ядерный геном или в хлоропластный или пластидный геном трансформированной клетки-хозяина растения. Это имеет место, когда более чем один сегмент ДНК, кодирующий кристаллический белок, включен в геном такого растения. В некоторых ситуациях может быть желательно иметь один, два, три, четыре или даже больше кодирующих кристаллический белок В.thuringiensis полинуклеотидов (либо нативных, либо рекомбинантно-сконструированных), включенных и стабильно экспрессируемых в трансформированном трансгенном растении.
В предпочтительных вариантах введение трансгена в геном клетки растения приводит к стабильной интеграции, при которой потомство таких растений также содержит копию трансгена в их геноме. Наследуемость этого генетического элемента потомством растения, в которое исходно был введен данный ген, является предпочтительным аспектом данного изобретения. Предпочтительный ген, который может вводиться, включает в себя, например, ген δ-эндотоксина В.thuringiensis и, в частности, один или несколько из генов, описанных здесь.
Средства для трансформации клетки растения и получения трансгенной клеточной линии хорошо известны в данной области (примеры приведены в патентах США 5550318; 5508468; 5482852; 5384253; 5276269 и 5225341, все специально включены здесь в полном виде в качестве ссылок) и вкратце обсуждаются здесь. Векторы, плазмиды, космиды, YAC (искусственные хромосомы дрожжей) и сегменты ДНК для применения в трансформации таких клеток будут, конечно, обычно содержать либо опероны, гены, либо произведенные из генов последовательности данного изобретения, либо нативные, либо синтетически полученные, и, в частности, кодирующие описанные кристаллические белки. Эти конструкции ДНК могут дополнительно включать в себя структуры, такие как промоторы, энхансеры, полилинкеры или даже генные последовательности, которые имеют положительно или отрицательно регулирующую активность в отношении конкретных представляющих интерес генов, если это желательно. Сегмент ДНК или ген может кодировать либо нативный, либо модифицированный кристаллический белок, который будет экспрессироваться в полученных рекомбинантных клетках и/или который будет придавать улучшенный фенотип регенерированному растению.
Трансгенные клетки, конкретно рассматриваемые в данном изобретении, включают в себя трансгенные клетки растений. Особенно предпочтительные клетки растений включают в себя клетки, полученные из кукурузы, пшеницы, сои, дернообразующих трав, декоративных растений, плодового дерева, кустарников, овощей, зерновых, бобовых и т.п. или любого растения, в которое желательно введение трансгена δ-эндотоксина В.thuringiensis, неактивного в отношении двукрылых насекомых.
2.5 Трансформированные растения
В другом аспекте растения, трансформированные любой конструкцией ДНК данного изобретения, которая экспрессирует белки, кодируемые данной конструкцией, рассматриваются как часть данного изобретения. Таким образом, данное изобретение обеспечивает далее трансгенные растения, которые были трансформированы конструкцией ДНК, описанной здесь в разделах 2.1 и 2.2, и были трансформированы с использованием трансформирующих векторов, описанных в разделе 2.3. Агрономические, плодоовощные, декоративные и другие экономически или коммерчески ценные растения могут быть созданы в соответствии со способами, описанными здесь, для экспрессии δ-эндотоксинов В.thuringiensis при уровнях, достаточно высоких для придания устойчивости к насекомым-патогенам, причем эти растения остаются морфологически нормальными.
Такие растения могут ко-экспрессировать полипептид δ-эндотоксина вместе с другими противогрибковыми, антибактериальными или антивирусными связанными с патогенезом пептидами, полипептидами или белками; инсектицидными белками; белками, придающими устойчивость к гербицидам; и белками, участвующими в улучшении качества или количества продуктов растений или агрономической продуктивности растений. Одновременная ко-экспрессия множественных белков в растениях является выгодной потому, что она использует более одного способа действия для борьбы с патогенным повреждением растений. Это может минимизировать возможность развития устойчивых штаммов патогенов, расширить сферу устойчивости и потенциально привести к синергическому инсектицидному действию, усиливая тем самым способности растения противостоять инвазии насекомых (Intl. Patent Appl. Publ. No. WO 92/17591, 15 October 1992, включенная здесь в качестве полной ссылки).
Трансформированное растение данного изобретения может быть либо однодольным растением, либо двудольным растением. Если это растение является однодольным растением, оно может быть любым из многочисленных видов. Предпочтительные однодольные виды, включенные в данное изобретение, могут включать в себя кукурузу, рис, пшеницу, ячмень, овес, рожь, просо, сорго, сахарный тростник, спаржу, дернообразующие травы или любое из ряда других зерновых или злаковых растений. В предпочтительных вариантах однодольным растением является кукуруза.
Данное изобретение рассматривает также множество двудольных растений, таких как хлопчатник, соя, томаты, цитрусовое растение, табак, сахарная свекла, люцерна, кормовые (конские) бобы, горох, фасоль, яблоня, вишня, груша, земляника, малина или любое другое бобовое, клубневое или плодовое растение. В предпочтительных вариантах двудольным растением является растение сои, растение табака или растение хлопчатника.
Многие из растений, которые предназначены для трансформации в соответствии с данным изобретением, являются коммерческими сельскохозяйственными культурами. Коммерческой формой этих растений могут быть исходные растения или их потомство, которое наследовало желательные трансгены. Таким образом, растения, дополнительно рассматриваемые в сфере данного изобретения, включают в себя любое потомство растений, трансформированных любой из пермутаций конструкции ДНК, указанных в данной заявке. Конкретно, этот потомок может быть определен как трансгенное растение R0. Другие потомства трансформированного растения также включены в сферу действия данного изобретения, в том числе любое растение-потомок любой генерации трансформированного растения, когда это растение-потомок унаследовало конструкцию ДНК из любого растения R0.
При трансформации специфической конструкцией ДНК сегменты нуклеиновой кислоты или полинуклеотидные сегменты данной конструкции могут быть включены в различных частях в хромосому трансформанта. Таким образом, в другом варианте данное изобретение включает в себя любое растение или любую клетку растения, полученные с использованием описанной здесь конструкции ДНК. Такие растение или клетка, включенные в данное изобретение, включают в себя растение или клетку, полученные способом, который предусматривает следующие стадии: (1) получение конструкции ДНК, включающей в себя район, кодирующий δ-эндотоксин Cry2A В.thuringiensis, неактивный в отношении двукрылых насекомых, расположенный в рамке считывания и под контролем промотора, функционального (операбельного) в растении, и район, кодирующий пластидный транзитный пептид, расположенный выше по ходу транскрипции (слева) от кодирующего района δ-эндотоксина Cry2A В.thuringiensis и ниже по ходу транскрипции (справа) от промотора; и (2) трансформацию растения полученной конструкцией ДНК, так что данное растение экспрессирует δ-эндотоксин Cry2A В.thuringiensis. Растение может быть также трансформировано таким образом, что оно дополнительно включает в свой геном и экспрессирует другие δ-эндотоксины Cry.
В родственном аспекте данное изобретение включает в себя также семена, продуцируемые трансформированным растением, потомство таких семян и семена, продуцируемые потомством исходного трансгенного растения, полученного в соответствии с описанным выше способом. Такое потомство и такие семена будут иметь в качестве трансгена δ-эндотоксин В.thuringiensis, неактивный в отношении двукрылых насекомых, и такое потомство будет наследовать признаки, привнесенные введением стабильного трансгена, в соответствии с менделевским расщеплением. Все такие трансгенные растения, имеющие включенные в их геном трансгенные сегменты ДНК, кодирующие любую включенную в них конструкцию ДНК, в частности конструкции ДНК, описанные в разделах 2.1 и 2.2, являются аспектами данного изобретения.
Могли бы быть также получены рекомбинантные растения, клетки, семена и другие ткани, в которых были изменены только митохондриальная или хлоропластная ДНК включением молекул, рассматриваемых в данной заявке. Промоторы, которые функционируют в хлоропластах, известны в данной области (Hanley-Bowden et al., Trends in Biochemical Sciences 12:67-70, 1987). Способы и композиции для получения клеток, содержащих хлоропласты, в которые была введена гетерологичная ДНК, были описаны Daniell et al., U.S. Pat. No. 5693507 (1997).
2.6 Способы трансформации растений
2.6.1 Способ экспрессии δ-эндотоксина Cry2A в растении
В другом предпочтительном варианте данное изобретение обеспечивает способ экспрессии δ-эндотоксинов Cry2A В.thuringiensis, неактивных в отношении двукрылых насекомых, при высоких уровнях в трансгенных растениях. Описанные способы могут использовать любую из конструкций ДНК, описанных в разделах 2.1 и 2.2 выше, а также любой из трансформирующих векторов, описанных, например, в разделе 2.3 выше. Рассматриваемые способы позволяют δ-эндотоксинам Cry2A, альтернативе δ-эндотоксинов Cry1A В.thuringiensis для борьбы с некоторыми насекомыми-вредителями, экспрессироваться в растениях без негативного воздействия на получение агрономических качеств трансгенных растений. Изобретение, описанное здесь, делает также возможной экспрессию δ-эндотоксинов Cry2A при уровнях, в 25 раз более высоких чем уровни, достигаемые существующими способами.
Таким образом, описанный здесь способ позволяет растениям экспрессировать Cry2A для использования либо в качестве альтернативы, либо в качестве дополнения к растениям, экспрессирующим δ-эндотоксины типа Cry1A В.thuringiensis как для уничтожения, так и для преодоления устойчивости ключевых насекомых-вредителей, в том числе Ostrina sp., Diatraea sp., Helicoverpa sp., Spodoptera sp. в Zea mays; Heliothis virescens, Helicoverpa sp., Pectinophora sp. в Gossypium hirsutum; и Anticarsia sp., Pseudoplusia sp., Epinotia sp. в Glycine max. Предполагается также, что описанные способы могут быть использованы для драматического увеличения экспрессии δ-эндотоксинов В.thuringiensis, включающих в себя Cry2A и родственных Cry2A, с увеличением таким образом эффективности против насекомых-мишеней и уменьшением вероятности развития резистентности к этим белкам. В одном варианте данного изобретения экспрессируется δ-эндотоксин Сry2Ab. Насекомые-мишени этого белка и их обычные хозяева показаны ниже в таблице 1.

Способ экспрессии δ-эндотоксина Cry2A В.thuringiensis в растении, описанный здесь, включает в себя стадии: (1) получения сегмента нуклеиновой кислоты, содержащего промотор, функционально (оперативно) связанный с первой полинуклеотидной последовательностью, кодирующей пластидный транзитный пептид, и второй полинуклеотидной последовательностью, кодирующей δ-эндотоксин Cry2A В.thuringiensis, лишенный активности против двукрылых насекомых, с получением слитого белка, состоящего из аминоконцевого пластидного транзитного пептида и δ-эндотоксина Cry2A В.thuringiensis, лишенного активности против двукрылых насекомых, и (2) трансформации растения конструкцией ДНК стадии 1 таким образом, что данное растение экспрессирует данный слитый белок. В предпочтительном варианте сегмент нуклеиновой кислоты, используемый в стадии (1) данного способа, построен таким образом, что 5’-конец второй полинуклеотидной последовательности функционально связан в одной и той же трансляционной рамке считывания с 3’-концом первой полинуклеотидной последовательности.
Растение или клетка растения, трансформированные по описанному здесь способу, могут быть либо однодольным растением или двудольным растением. Если данное растение является однодольным растением, оно может быть любым из множества видов. Предпочтительные однодольные растения, включенные в данное изобретение, могут включать в себя кукурузу, рис, пшеницу, ячмень, овес, рожь, просо, сорго, сахарный тростник, спаржу, дернообразующие травы или любое из ряда других зерновых или злаковых растений. В предпочтительных вариантах однодольным растением является кукуруза.
Данное изобретение рассматривает также способ, при помощи которого трансформируют двудольные растения или клетки растений. Такие двудольные растения могут включать в себя такие растения, как хлопчатник, соя, томаты, картофель, цитрусовое растение, табак, сахарная свекла, люцерна, кормовые (конские) бобы, горох, фасоль, яблоня, вишня, груша, земляника, малина или любое другое бобовое, клубневое или плодовое растение. В предпочтительных вариантах двудольным растением является растение сои, растение или клетка табака или растение или клетка хлопчатника.
2.6.2 Способ экспрессии δ-эндотоксина Cry2Ab в потомстве растения
Как указывалось в отношении других вариантов, описанных в данном изобретении, многие растения, предназначенные для трансформации в соответствии с данным изобретением, являются коммерческими сельскохозяйственными растениями. Коммерческой формой этих растений могут быть исходные растения или их потомство, которое наследовало желательные трансгены. Таким образом, авторы изобретения дополнительно предполагают, что описанный здесь способ включает в себя способ получения трансгенного растения-потомка или клетки-растения-потомка. Способ получения такого потомства включает в себя следующее. Способ экспрессии δ-эндотоксина Cry2Ab В.thuringiensis в описанном здесь растении включает в себя стадии: (1) получения сегмента нуклеиновой кислоты, содержащей промотор, функционально (оперативно) связанный с первой полинуклеотидной последовательностью, кодирующей пластидный транзитный пептид, и второй полинуклеотидной последовательностью, кодирующей δ-эндотоксин Cry2A В.thuringiensis, лишенный активности против двукрылых насекомых; (2) получения второго растения и (3) скрещивания первого и второго растений с получением скрещенного трансгенного потомка-растения или потомка-клетки-растения, которые унаследовали нуклеиновые сегменты из первого растения. Данное изобретение конкретно включает в себя потомство, растение или семена потомства из любого из однодольных или двудольных растений, в том числе указанных в разделах 2.5 и 2.6.1 выше.
2.6.3 Способ ко-экспрессии Cry2Ab и других δ-эндотоксинов Cry В.thuringiensis в растении и растении-потомке
В другом предпочтительном варианте описанный здесь способ экспрессии неактивного в отношении Diptera δ-эндотоксина Cry2A В.thuringiensis включает в себя ко-экспрессию описанной конструкции ДНК в любом из ее описанных вариантов вместе с δ-эндотоксином Cry1 В.thuringiensis. Ожидается, что данный способ экспрессии вместе этих δ-эндотоксинов Cry В.thuringiensis позволит получить повышенные инсектицидные свойства в трансформированном растении через увеличенную экспрессию и пониженное развитие резистентности насекомых, которые являются желательными результатами, не присутствующими в существующих способах. Эта ко-экспрессия может иметь место в исходном трансформанте или в любом числе поколений потомства исходного трансформанта, которое унаследовало эти гены, для ко-экспрессии белков, кодируемых любой из конструкций ДНК, описанных здесь.
3.0 Краткое описание чертежей
Следующие чертежи образуют часть данного описания и включены для дополнительной демонстрации некоторых аспектов данного изобретения. Данное изобретение может быть лучше понято со ссылкой на один или несколько из этих чертежей в сочетании с подробным описанием характерных экспериментов, представленных здесь.
Фиг.1. Схематическая иллюстрация элементов экспрессирующих векторов Cry2Ab однодольных растений pMON30464, pMON30463 и pMON26800.
Фиг.2. Схематическая иллюстрация элементов экспрессирующих векторов Cry2Ab двудольных растений pMON33830, pMON33827, pMON33828 и pMON33829.
Фиг.3. Схематическая иллюстрация элементов экспрессирующих векторов Cry2Aа двудольных растений pMON33803, pMON33812, pMON33811 и pMON33806.
Фиг.4. Плазмида, названная pMON30464.
Фиг.5. Плазмида, названная pMON33827.
Фиг.6. Плазмида, названная pMON33828.
Фиг.7. Плазмида, названная pMON33829.
4.0. Описание иллюстративных вариантов
Следующее подробное описание данного изобретения обеспечено для помощи специалистам, квалифицированным в данной области, в применении на практике данного изобретения. Даже в данном случае следующее далее подробное описание не должно рассматриваться как чрезмерно ограничивающее данное изобретение, так как модификации и вариации в обсуждаемых здесь вариантах могут быть произведены специалистами с квалификацией в данной области без отклонения от идеи и сферы действия данного изобретательского открытия.
4.1. Идентификация последовательностей SEQ ID NO:1. Последовательность нуклеиновой кислоты гена Cry2Ab.
SEQ ID NO:2. Аминокислотная последовательность δ-эндотоксина Cry2Ab В. thuringiensis.
SEQ ID NO:3. Последовательность нуклеиновой кислоты пластидного транзитного пептида zmSSU.
SEQ ID NO:4. Аминокислотная последовательность пластидного транзитного пептида zmSSU.
SEQ ID NO:5. Последовательность нуклеиновой кислоты пластидного транзитного пептида 1 (РТР1).
SEQ ID NO:6. Аминокислотная последовательность РТР1.
SEQ ID NO:7. Последовательность нуклеиновой кислоты пластидного транзитного пептида 1Δ (РТР1Δ).
SEQ ID NO:8. Аминокислотная последовательность РТР1Δ.
SEQ ID NO:9. Последовательность нуклеиновой кислоты пластидного транзитного пептида 2 (РТР2).
SEQ ID NO:10. Аминокислотная последовательность РТР2.
SEQ ID NO:11. Последовательность нуклеиновой кислоты гена Cry2Aa.
SEQ ID NO:12. Аминокислотная последовательность полипептида Cry2Aa.
SEQ ID NO:13. pMON33827.
SEQ ID NO:14. pMON33828.
SEQ ID NO:15. pMON33829.
SEQ ID NO:16. pMON30464.
SEQ ID NO:17. Последовательность гена Cry2Ab Bacillus thuringiensis, UWGCG номер доступа М23724 (Widner and Whiteley).
SEQ ID NO:18. Аминокислотная последовательность cry2Ab Bacillus thuringiensis, транслируемая из SEQ ID NO:17.
4.2 Определения
Следующие слова и фразы имеют здесь приведенные ниже значения.
Биологические функциональные эквиваленты. В применении здесь такие эквиваленты, касающиеся инсектицидных белков данного изобретения, являются пептидами, полипептидами и белками, которые содержат сходство последовательности или части, представляющей последовательность, с новыми пептидами данного изобретения, такими как Cry2Ab, и которые проявляют одинаковые или сходные функциональные свойства со свойствами описанных здесь полипептидов, в том числе инсектицидную активность. Биологические эквиваленты включают в себя также пептиды, полипептиды и белки, которые взаимодействуют, т.е. специфически связываются с антителами, индуцированными против Cry2Ab, и которые проявляют одинаковую или сходную инсектицидную активность, в том числе как моноклональными, так и поликлональными антителами.
Термин локализованные в хлоропласте или пластиде в применении здесь относится к биологической молекуле, полинуклеотиду или полипептиду, которая расположена в хлоропласте или пластиде, так что эта молекула изолировна от клеточной цитоплазматической среды, и функционирует в цитоплазме хлоропласта или пластиды с обеспечением эффектов, заявленных в данном изобретении. Локализация биологической молекулы в хлоропласте или пластиде может происходить в случае полинуклеотидов с использованием искусственных механических средств, таких как электропорация, механическая микроинъекция, или бомбардировкой покрытыми полинуклеотидами микроснарядами, или в случае полипептидов с использованием секреторных или импортных средств, где используют природную, синтетическую или гетерологичную нацеливающую на пластиду или хлоропласт пептидную последовательность, которая функционируют для нацеливания, инсертирования, содействия переносу или локализации связанного полипептида в хлоропласт или пластиду.
Борьба или контроль в отношении повреждения насекомыми в сельскохозяйственном контексте относится к снижению повреждения сельскохозяйственной культуры, вызываемого заражением насекомым-вредителем. Более обычно эта фраза относится к уменьшению вредных эффектов, вызываемых присутствием нежелательного насекомого в любом конкретном местоположении.
Событие относится к трансгенному растению, полученному из введения чужеродной ДНК в один или несколько уникальных сайтов в ядерной геномной ДНК.
Экспрессия: Комбинация внутриклеточных процессов, в том числе транскрипции, трансляции и других функций процессинга и стабилизации внутриклеточного белка и РНК, которым подвергается кодирующая молекула ДНК, такая как структурный ген, для продуцирования полипептида.
Инсектицидный полипептид относится к полипептиду, имеющему инсектицидные свойства, например полипептиду, который ингибирует рост, развитие, жизнеспособность или плодовитость насекомых-вредителей, являющихся мишенями.
Функционально (оперативно) связанные: Кодирующие сегменты нуклеиновой кислоты, связанные в рамке считывания таким образом, что свойства одного влияют на экспрессию другого.
Экспрессируемые в растении кодирующие районы: Кодирующие районы, которые являются экспрессируемыми in planta, так как они содержат типичные для растений регуляторные элементы для облегчения экспрессии представляющего интерес гена.
Пластидный транзитный пептид: Любая аминокислотная последовательность, применимая в нацеливании или локализации связанной аминокислоты, такой как слитый белок, в субклеточный компартмент или органеллу, такую как пластида.
Потомство: “Потомство” включает в себя любого отпрыска или потомка трансгенного растения или любое последующее растение, которое имеет трансформант в его линии дифференцировки (родословной). Потомство не ограничивается одной генерацией, а охватывает потомков трансформанта до тех пор, пока они содержат или экспрессируют данный трансген. Семена, содержащие трансгенные эмбрионы, а также семена из трансгенных растений и их потомки или отпрыски также являются важными частями данного изобретения.
Промотор: Сайт узнавания на последовательности ДНК или группе последовательностей ДНК, который обеспечивает регуляторный элемент экспрессии для структурного гена и с которым специфически связывается РНК-полимераза и инициирует синтез РНК (транскрипцию) данного гена.
R0 обозначает первичное регенерированное растение, полученное из трансформации ткани или клеток растения в культуре. Последующее потомство или последующие генерации, полученные из данного R0, называют R1 (первая генерация), R2 (вторая генерация) и т.д.
Регенерация: Процесс выращивания растения из клетки растения (например, протопласта или эксплантата растения).
Стабильно сохраняемые в пластиде или хлоропласте растения относится к введению посредством электропорации, трансформации, трансдукции или мицелло- или липосомоподобного слияния полинуклеотида или нуклеиновой кислоты в хлоропласт или пластиду таким образом, что эта нуклеиновая кислота остается в реципиентных хлоропласте или пластиде и во всем последующем потомстве реципиентных хлоропласта или пластиды либо посредством включения рекомбинацией в геном хлоропласта или пластиды, либо в виде ковалентно связанного кольцевого репликона, находящегося в хлоропласте или пластиде, посредством выращивания любого растения, клетки растения или ткани растения, содержащей такие трансформированные хлоропласт или пластиду, и в присутствии химикалия или соединения, которое требует одного или нескольких генов, присутствующих на репликоне и экспрессируемых репликоном, для гарантии выживания трансформированных пластиды или хлоропласта и их потомства пластид или хлоропластов в растении, клетки растения или ткани растения.
Структурная кодирующая последовательность обозначает последовательность ДНК, которая кодирует пептид, полипептид или белок, который продуцируется клеткой после транскрипции структурной кодирующей последовательности в мессенджер-РНК (мРНК), с последующей трансляцией этой мРНК в желаемый пептидный, полипептидный или белковый продукт.
Структурный ген: Ген, который экспрессируется с образованием полипептида.
Существенная гомология: В применении здесь этот термин обозначает последовательности нуклеиновой кислоты или полипептида, которые являются на приблизительно 86% гомологичными, до приблизительно 90% гомологичными, до приблизительно 95% гомологичными, до приблизительно 99% гомологичными. Более конкретно, авторы считают, что существенные гомологи являются до приблизительно 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 и 99 процентов гомологичными ссылочной последовательности нуклеиновой кислоты полипептида.
Существенная временная или пространственная регуляция относится к экспрессии гена в растении или ткани растения от функционального в растении промотора. В случае временной регуляции промотор может регулироваться для экспрессии только во время специфических периодов времени во время роста и развития клетки растения или ткани растения или даже целого растения. Промотор, который активно экспрессирует один или несколько генов во время прорастания семян, может быть одним из примеров временной регуляции. Другой пример мог бы включать промоторы, которые активно экспрессируют один или несколько генов только в периоды времени, когда растение, клетка растения или ткань растения экспонированы определенным интенсивностям света или во время полной темноты. Существенная временная регуляция имеет ввиду промотор, который активно экспрессируется в определенное время, но который может быть или может не быть подавленным в другие периоды времени, так что экспрессия может все еще детектироваться мониторингом на присутствие какого-либо индикатора, такого как фермент, продуцируемый из кодирующей последовательности, связанной с таким промотором, или путем измерения увеличения или уменьшения некоторого продукта гена, такого как мРНК, продуцируемого в различные периоды времени во время всего роста, дифференцировки и развития растения и/или в ответ на различные стимулы окружающей среды. Существенная пространственная регуляция обозначает экспрессию гена, связанного с промотором, от которого экспрессия протекает только во время роста и развития определенных клеток или тканей в данном растении. Например, можно было бы ожидать, что тапетальный промотор является экспрессируемым только в клетках корня или тканях корня. Существенно пространственно регулируемый обозначает также уровень экспрессии от специфического промотора конкретной ткани в этой конкретной ткани в сравнении с уровнями экспрессии от этого или подобного промотора в других тканях, причем экспрессия может также детектироваться в тканях других, чем эта конкретная ткань, в которой экспрессия данного промотора является предпочтительной, но при значительно более низких уровнях экспрессии, как измерено по продуцированию фермента, продуцируемого из кодирующей последовательности, связанной с этим промотором, или по появлению какого-либо детектируемого генного продукта. Промоторы могут быть также существенно врменно или существенно пространственно регулируемыми вместе и одновременно координированно регулируемым образом.
Синтетический ген: Синтетические гены, кодирующие δ-эндотоксины В.thuringiensis данного изобретения, являются генами, получаемыми способом, включающим в себя любой тип генетического выделения или манипуляции. Это включает в себя выделение гена из природно встречающегося состояния, манипулирование этим геном, такое как модификация кодонов (как описано здесь) или сайт-специфический мутагенез (как описано здесь), укорочение гена или любой другой способ манипуляции или выделения.
Терминатор: 3’-концевая последовательность терминации транскрипции и полиаденилирования.
Трансформация: Способ введения экзогенной последовательности ДНК (например, вектора или рекомбинантной молекулы ДНК) в клетку или протопласт, в которых эта экзогенная ДНК включается в хромосому или способна к автономной репликации.
Трансформированная клетка: Клетка, которая была изменена введением одной или нескольких молекул ДНК в эту клетку.
Трансген: Генная конструкция или сегмент ДНК, содержащий ген, который желательно экспрессировать в реципиентных клетке, ткани или организме. Он может включать в себя целую плазмиду или другой вектор или может просто включать в себя функциональный кодирующий участок, район, домен или сегмент переносимой конструкции ДНК.
Трансгенная клетка: Любая клетка, полученная или регенерированная из трансформированной клетки или происходящая из трансгенной клетки. Примеры трансгенных клеток включают в себя каллусы растений, происходящие из трансформированной клетки растения, и конкретные клетки, такие как клетки листа, корня, стебля, например соматические клетки или репродуктивные (зародышевые) клетки, полученные из трансгенного растения.
Трансгенное событие: Растение или его потомство, полученное из введения чужеродной ДНК в ядерный геном клетки или протопласта растения.
Трансгенное растение: Растение или его потомство, которое было генетически модифицировано таким образом, что оно содержит и экспрессирует гетерологичные последовательности ДНК в виде белков. В качестве конкретного приведенного здесь примера трансгенное растение сои генетически модифицируют таким образом, что оно содержит и экспрессирует по меньшей мере одну гетерологичную последовательность ДНК, функционально связанную с регуляторными последовательностями транскрипции и находящуюся под регуляторным контролем регуляторных последовательностей транскрипции, которые функционируют в клетках или тканях растения или в целых растениях. Трансгенное растение может также называться трансформированным растением. Трансгенным растением также называют потомство исходного трансгенного растения, когда это потомство содержит и способно экспрессировать гетерологичную кодирующую последовательность под регуляторным контролем экспрессируемых в растении описанных здесь регуляторных последовательностей транскрипции.
Вектор: Молекула ДНК, способная реплицироваться в клетке-хозяине, и/или молекула ДНК, с которой может быть функционально связан другой сегмент ДНК таким образом, чтобы произошла репликация этого связанного сегмента. Примером вектора является плазмида.
4.3 Синтез и выделение сегмента нуклеиновой кислоты, кодирующего δ-эндотоксин B.thuringiensis и нацеливающие на пластиды последовательности
Данное изобретение описывает новые конструкции ДНК, содержащие полинуклеотидные последовательности, кодирующие δ-эндотоксины В.thuringiensis, а также нацеливающие на пластиды последовательности. Способы конструирования и экспрессии синтетических генов В.thuringiensis в растениях хорошо известны лицам с обычной квалификацией в данной области и описаны подробно в патенте США 5500365. Данное изобретение рассматривает применение генов Cry2A В.thuringiensis в трансформации как однодольных, так и двудольных растений. Для потенциирования экспрессии этих генов данное изобретение обеспечивает конструкции ДНК, содержащие полинуклеотидные сегменты, кодирующие нацеливающие на пластиды пептиды, расположенные выше по ходу транскрипции (слева) от полинуклеотидных последовательностей, кодирующих целевые δ-эндотоксины В.thuringiensis. В частности, рассматриваются последовательности, кодирующие δ-эндотоксины В.thuringiensis, лишенные по существу ингибиторной активности против видов Diptera.
4.4 Зонды и праймеры
В одном аспекте информация о нуклеотидных последовательностях, обеспеченная данным изобретением, позволяет получать относительно короткие последовательности ДНК, способные специфически гибридизоваться с последовательностями генов описанных здесь выбранных полинуклеотидов. В этих аспектах зонды нуклеиновых кислот подходящей длины готовят с учетом выбранных полинуклеотидных последовательностей, кодирующих полипептиды δ-эндотоксина СryА, например такой последовательности, которая показана в SEQ ID NO:1. Эти зонды нуклеиновых кислот могут быть также приготовлены с учетом выбранных полинуклеотидных последовательностей, кодирующих нацеливающий на пластиды пептид, таких как последовательности, показанные в SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7 и SEQ ID NO:9. Способность таких зондов нуклеиновых кислот специфически гибридизоваться с последовательностью гена, кодирующего полипептид δ-эндотоксина или последовательность нацеливающего на пластиды пептида, делает их конкретно применимыми в различных вариантах. Наиболее важно то, что эти зонды могут быть использованы в многочисленных тестах для детектирования присутствия комплементарной последовательности в конкретной пробе.
В некоторых вариантах предпочтительно использовать олигонуклеотидные праймеры. Последовательности таких праймеров сконструированы с использованием полинуклеотида данного изобретения для применения в детектировании, амплификации или мутации определенного сегмента гена кристаллического белка из В.thuringiensis с использованием ПЦРтм-технологии. Этот способ может быть использован для детектирования, амплификации или мутации определенного сегмента полинуклеотида, кодирующего нацеливающий на пластиды пептид. Сегменты генов, родственные полинуклеотидам, кодирующим полипептиды δ-эндотоксина и нацеливающие на пластиды пептиды, данного изобретения могут быть также амплифицированы при помощи ПЦР™ с использованием таких праймеров.
Для обеспечения некоторых преимуществ в соответствии с данным изобретением предпочтительная последовательность нуклеиновой кислоты, используемая для гибридизационных исследований или анализов, включает в себя последовательности, которые комплементарны по меньшей мере отрезку из 14-30 нуклеотидов или примерно такой длины полинуклеотидной последовательности, кодирующей кристаллический белок, такой как показанная в SEQ ID NO:1, или последовательности, которые комплементарны по меньшей мере отрезку из 14-30 нуклеотидов или примерно такой длины последовательности, кодирующей нацеливающий на пластиды пептид, такой как показанные в SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7 и SEQ ID NO:9.
Размер по меньшей мере 14 нуклеотидов в длину помогает гарантировать, что этот фрагмент будет фрагментом достаточной длины для образования дуплексной молекулы, которая является как стабильной, так и селективной. Молекулы, имеющие комплементарные последовательности на протяжении сегментов, больших чем 14 оснований в длину, обычно являются предпочтительными. Для увеличения стабильности и селективности гибрида и тем самым улучшения качества и степени полученных гибридных молекул, обычно предпочитают конструировать молекулы, имеющие комплементарные гену последовательности из 14-20 нуклеотидов или даже более длинные, если желательно. Такие фрагменты могут быть легко получены, например, прямым синтезом фрагмента химическими способами, с использованием технологии репродукции нуклеиновых кислот, такой как ПЦРтм-технология патентов США 4683195 и 4683202 (каждый из которых включен здесь в качестве ссылки), или вырезанием выбранных фрагментов ДНК из рекомбинантных плазмид, содержащих подходящие инсерты и подходящие сайты рестрикции.
4.5 Экспрессирующие векторы
Данное изобретение рассматривает также экспрессирующий вектор, содержащий полинуклеотид данного изобретения. Таким образом, в одном варианте экспрессирующий вектор является выделенной и очищенной молекулой ДНК, содержащей промотор, функционально связанный с кодирующей последовательностью, которая кодирует полипептид данного изобретения, причем кодирующий район функционально связан с районом терминации транскрипции, посредством чего этот промотор запускает транскрипцию данного кодирующего района. Кодирующий район может включать в себя сегмент, кодирующий δ-эндотоксин В.thuringiensis, и сегмент, кодирующий пластидный нацеливающий пептид. Молекула ДНК, содержащая экспрессирующий вектор, может также содержать функциональный интрон.
В применении здесь термины "функционально (оперативно) связанные" или "операбельно связанные" означают, что промотор связан с кодирующим районом таким образом, что транскрипция данного кодирующего района управляется и регулируется данным промотором. Способы функционального связывания промотора с кодирующим районом для регуляции как выше по ходу транскрипции, так и ниже по ходу транскрипции от кодирующего района хорошо известны в данной области.
Предпочтительные трансформирующие векторы растений включают в себя векторы, произведенные из Ti-плазмиды Agrobacterium tumefaciens, а также описанные, например, Herrera-Estrella (1983), Bevan (1983), Klee (1985) и Eur. Pat. Appl. No. EP 0120516 (все включены специально в качестве ссылки).
Промоторы, которые функционируют в бактериях, хорошо известны в данной области. Примеры и предпочтительные промоторы для кристаллических белков В.thuringiensis включают в себя промоторы генов sigA, sigE и sigK. Альтернативно могут быть использованы сами промоторы генов, кодирующих нативные, мутагенизированные, гетерологичные или рекомбинантные кристаллические белки.
При использовании экспрессирующего вектора данного изобретения для трансформации растения выбирают промотор, который способен запускать экспрессию в данном конкретном виде растения. Промоторы, которые функционируют в различных видах растений, также хорошо известны в данной области. Промоторы, применимые в экспрессии полипептидов в растениях, являются промоторами, которые являются индуцибельными, вирусными, синтетическими или конститутивными, как описано (Odell et al., 1985), и/или регулируемыми во времени, пространственно регулируемыми и пространственно и во времени регулируемыми. Предпочтительные промоторы включают в себя усиленные промоторы CaMV35S и промотор FMV35S.
4.5.1 Векторы с кодирующими нацеливающий на пластиды пептид сегментами
В соответствии с данным изобретением экспрессирующие векторы, сконструированные для специфического потенциирования экспрессии полипептида в трансформированном растении, могут включать в себя определенные районы, кодирующие нацеливающие на пластиды пептиды (РТР). Эти районы позволяют полностью использовать клеточные процессы, участвующие в транскрипции, трансляции и экспрессии кодируемого белка, при связи с определенными δ-эндотоксинами В.thuringiensis. Такие нацеливающие на пластиды пептиды функционируют различным образом, таким как, например, перенесением экспрессируемого белка в клеточную структуру, в которой он может эффективно функционировать, или перенесением экспрессируемого белка к зонам клетки, в которых сконцентрированы клеточные процессы, необходимые для его экспрессии.
Использование РТР может также увеличивать частоту восстановления морфологически нормальных растений и частоту, с которой могут быть получены трансгенные растения. При условии, что коммерчески жизнеспособная экспрессия δ-эндотоксинов как типа Cry1A, так и типа Сry3А В.thuringiensis достигалась экспрессией форм этих белков, которые остаются локализованными в цитозоле (т.е. в ненацеленной форме), экспрессию ненацеленных форм как Cry2Aа, так и Cry2Ab также сначала пытались использовать в трансгенных хлопчатнике, табаке и кукурузе.
В кукурузе ненацеленные экспрессирующие Cry2Ab трансформирующие векторы дали относительно мало трансгенных событий (т.е. независимых событий инсерции в геном кукурузы) с уровнями экспрессии Cry2Ab, достаточными для коммерчески приемлемого контроля насекомых. Кроме того, многие из трансформантов кукурузы, экспрессирующие ненацеленный Cry2Ab, проявляли явные дефекты роста, такие как серьезное уменьшение в росте (задержка роста) или сильное пожелтение листьев (хлороз), которые делали растения коммерчески неприемлемыми. Уровни экспрессии ненацеленного Cry2Ab в кукурузе были не выше приблизительно 15 м.д. от уровня, минимально необходимого для опосредованной Cry2Ab защиты от мотылька кукурузного (ЕСВ).
Хотя были описаны исследования, включающие в себя экспрессию нацеленных на пластиды δ-эндотоксинов типа Cry1A В.thuringiensis в трансгенных растениях (Wong et al., 1992), нацеливание негомологичных белков Cry2A или Cry2A не было ранее описано. Одно сообщение о нацеленной на пластиды экспрессии Cry1Ac показало, что нацеливание приводит к небольшому увеличению или к отсутствию увеличения экспрессии Cry1Ac (патент США № 5500365). Другое сообщение показало, что увеличение экспрессии нацеленной на пластиды формы Cry1Ac требовало включения новой 5’-нетранслируемой лидерной последовательности (Wong et al., 1992) и что действие лидерной и нацеливающей последовательностей на экспрессию сильно зависело от кодирующей последовательности структурного гена. Wong et al. сделали вывод, что как лидерная последовательность, так и пластидный трансзитный пептид увеличивали экспрессию Cry1Ac в 18 раз, но те же самые последовательности увеличивали экспрессию β-глюкуронидазы только в 6 раз. Наконец, ни одно из предыдущих сообщений не предполагало, что нацеливание на пластиды могло бы приводить к увеличенному выходу морфологически нормальных экспрессирующих В.thuringiensis растений.
Данное изобретение описывает, что трансгенные растения кукурузы, экспрессирующие δ-эндотоксины Cry2A, неактивные в отношении Diptera, такие как Cry2Ab, при уровнях до в 10 раз более высоких, чем требующиеся для борьбы с ЕСВ, получали морфологически при значительно более высоких частотах при использовании нацеленной на пластиды формы Cry2A. В случае Cry2Ab повышенная экспрессия является решающей в получении трансгенной кукурузы с защитой против ЕСВ, так как LC50 Cry2Ab против ЕСВ является значимо более высокой, чем LC50 ЕСВ Cry2Ab В. thuringiensis, используемого в настоящее время для борьбы с ЕСВ в трансгенной кукурузе (патент США 5338544, 1994; MacIntosh et al., 1990; Armstrong et al., 1995).
Увеличенная экспрессия является особенно ценной в связи с тем, что она обеспечивает дополнительную защиту против развития резистентности через стратегию высоких доз (McGaughey and Whalon, 1993; Roush, 1994). Экспрессия высокого уровня желательна, кроме того, потому, что она обеспечивает стойкую (длительную) защиту от насекомых в случаях, когда экспрессия инсектицидного гена снижается вследствие условий окружающей среды. Дополнительно и неожиданно растения кукурузы, трансформированные нацеленными на пластиды экспрессирующими Cry2Ab векторами, проявляли нормальные рост и развитие.
Значимым различием между нацеленной и ненацеленной (цитозольной) экспрессией Cry2Ab было драматическое увеличение уровней белка Cry2Ab в растениях, трансформированных нацеленным на пластиды Cry2Ab экспрессирующим вектором относительно растений, трансформированных цитозольным вектором Cry2Ab. Этот результат был очень неожиданным. Также в противоположность описаниям предыдущей работы данное изобретение выявило, что увеличенный выход фенотипически нормальных трансгенных растений может быть достигнут с использованием описанных способов нацеленной на пластиды экспрессии.
Примером нацеливающего на плазмиды пептида (РТР) является пептид хлоропластного нацеливания. Было обнаружено, что пептиды хлоропластного нацеливания применимы, в частности, в системе глифозат-резистентного селектируемого маркера. В этой системе растения, трансформированные для экспрессии белка, придающего устойчивость к глифозату, трансформируют РТР, который нацеливает этот пептид на хлоропласты клетки. Глифозат ингибирует пути шикимовой кислоты, которые ведут к биосинтезу ароматических соединений, в том числе аминокислот и витаминов. Конкретно, глифозат ингибирует превращение фосфоенолпировиноградной кислоты и 3-фосфошикимовой кислоты в 5-енолпирувил-3-фосфошикимовую кислоту путем ингибирования фермента синтазы 5-енолпирувил-3-фосфошикимовой кислоты (ЕРSР-синтазы или EPSPS). Дополнительная EPSPS, образовавшаяся через введение трансгена, кодирующего этот фермент, позволяет клетке противостоять действию глифозата. Таким образом, когда гербицид глифозат действует, убивая данную клетку путем прерывания биосинтеза ароматических аминокислот, в частности, в хлоропласте этой клетки, РТР позволяет увеличить устойчивость к этому гербициду путем сосредоточения экспрессии фермента устойчивости к глифозату в хлоропласте клетки, т.е. в органелле-мишени данной клетки. Примеры ферментов устойчивости к гербициду включают в себя ESPS, как отмечалось выше, глифозатоксидоредуктазу (GOX) и ген аroА (см. патент США № 4535060, специально включенный здесь в полном виде в качестве ссылки).
РТР могут нацеливать белки на хлоропласты и другие пластиды. Например, органеллой-мишенью может быть амилопласт. Предпочтительные РТР данного изобретения включают в себя РТР нацеливания как на хлоропласты, так и на другие пластиды. Характерные примеры предпочтительных РТР включают в себя РТР белка RUBISCO SSU кукурузы и функционально родственные пептиды, такие как РТР1, РТРΔ и РТР2. Примерами этих РТР являются полипептиды, показанные в SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8 и SEQ ID NO:10. Полинуклеотидные последовательности, кодирующие эти полипептиды, показаны в SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7 и SEQ ID NO:9.
Могли бы быть также получены рекомбинантные растения, клетки, семена и другие ткани растений, в которых только митохондриальная или хлоропластная ДНК изменена включением молекул, рассматриваемых в этой заявке. Промоторы, которые функционируют в хлоропластах, были известны в данной области (Hanley-Bowden et al., Trends in Biochemical Sciences 12:67-70, 1987). Способы и композиции для получения клеток, содержащих хлоропласты, в которые была встроена гетерологичная ДНК, были описаны Daniell et al., U.S. Pat. № 5693507 (1997). McBride et al., (WO 95/24492) описывают локализацию и экспрессию генов, кодирующих белок δ-эндотоксин Cry1A, в хлоропластных геномах растений табака. Как описано здесь, локализация Cry2Aа в хлоропласте или пластиде приводит к пониженным уровням экспрессии, как измерено по накоплению δ-эндотоксина Cry2Aа, что противоречит улучшенной экспрессии локализованного в хлоропласте или пластиде δ-эндотоксина Cry2Ab.
4.5.2 Применение промоторов в экспрессирующих векторах
Экспрессия гена, который существует в форме двухцепочечной ДНК, включает в себя транскрипцию мессенджер-РНК (мРНК) из кодирующей цепи этой ДНК РНК-полимеразным ферментом и последующий процессинг первичного транскрипта мРНК в ядре. Транскрипция ДНК в мРНК регулируется районом ДНК, обычно называемым "промотором". Район промотора содержит последовательность оснований, которая передает сигнал РНК-полимеразе для связывания с ДНК и инициации транскрипции мРНК с использованием одной из цепей этой ДНК в качестве матрицы для образования соответствующей цепи РНК. Конкретный выбранный промотор должен быть способен индуцировать достаточную экспрессию кодирующей фермент последовательности для получения продуцирования эффективного инсектицидного количества белка В.thuringiensis.
3’-нетранслируемый район химерных генов растений данного изобретения также содержит сигнал полиаденилирования, который функционирует в растениях, вызывая присоединение аденилатнуклеотидов к 3’-концу этой РНК. Примерами предпочтительных 3’-районов являются (1) 3’-транскрибируемые, нетранслируемые районы, содержащие сигнал полиаденилирования индуцирующих опухоль (Ti) плазмидных генов Agrobacterium, таких как ген нопалинсинтазы (NOS), и (2) 3’-концы генов растений, таких как ген Е9 ssRUBISCO гороха (Fischhoff et al., 1987).
Промотор выбирают по его способности направлять транскрипционную активность трансформированной клетки растения или трансгенного растения на кодирующий район для гарантии достаточной экспрессии кодирующей фермент последовательности для получения продуцирования инсектицидных количеств белка В.thuringiensis. Структурные гены могут запускаться различными промоторами в тканях растений. Промоторы могут быть почти конститутивными (т.е. они управляют транскрипцией трансгена во всех тканях), такие как промотор CaMV35S, или тканеспецифическими, или специфическими для определенной стадии развития промоторами, действующими в двудольных или однодольных растениях. Если промотор является почти конститутивным промотором, таким как CaMV35S или FMV35S, увеличения в экспрессии полипептида обнаруживаются в разнообразных трансформированных тканях растения и в большинстве органов растения (например, каллусе, листе, семени и корне). Усиленные или дупликатные версии промоторов CaMV35S и FMV35S являются особенно применимыми в использовании на практике данного изобретения (Кау et al., 1987; Rogers, U.S. Patent 5378619).
Специалистам с квалификацией в данной области будет понятно, что имеется ряд промоторов, которые активны в клетках растений, и они были описаны в литературе. Такие промоторы могут быть получены из растений или вирусов растений и включают в себя, но не ограничиваются ими, промоторы нопалинсинтазы (NOS) и октопинсинтазы (OCS) (которые находятся на индуцирующих опухоль плазмидах А. tumefaciens), промоторы 19S и 35S вируса мозаичной болезни цветной капусты (CaMV), индуцируемый светом промотор из малой субъединицы рибулозо-1,5-бисфосфаткарбоксилазы (ssRUBISCO, очень обильного полипептида растений), промотор ActI риса и промотор 35S вируса мозаичной болезни (FMV) норичника шишковатого. Все эти промоторы использовали для создания различных типов конструкций ДНК, которые экспрессировали в растениях (см., например, McElroy et al., 1990, U.S. Patent 5463175).
Кроме того, можно также предпочтительно вызвать экспрессию δ-эндотоксина В.thuringiensis в специфических тканях растения с использованием интегрирующихся в растение векторов, содержащих тканеспецифический промотор. Специфические ткани-мишени могут включать в себя лист, стебель, корень, клубень, семя, плод и т.д., а выбранный промотор должен иметь желательную тканеспецифичность или специфичность в отношении стадии развития. Таким образом, функция промотора должна быть оптимизирована выбором промотора с желательной способностью экспрессии в определенной ткани и соответствующей силой промотора и отбором трансформанта, который продуцирует желательную инсектицидную активность в этих тканях-мишенях. Этот подход селекции из пула трансформантов рутинно используется в экспрессии гетерологичных структурных генов в растениях, так как существует вариация между трансформантами, содержащими один и тот же гетерологичный ген вследствие сайта инсертирования гена в геноме растения (обычно называемая "эффектом положения"). Кроме промоторов, о которых известно, что они индуцируют транскрипцию (конститутивную или тканеспецифическую) ДНК в клетках растения, другие промоторы могут быть идентифицированы для использования в настоящем изобретении посредством скрининга библиотеки кДНК растений для генов, которые селективно или предпочтительно экспрессируются в тканях-мишенях, с последующим определением районов промоторов.
Примером тканеспецифического промотора является промотор лектина, который является специфическим для ткани семени. Лектиновый белок в семенах сои кодируется единственным геном (LeI), который экспрессируется только во время созревания семян и ответственен за приблизительно 2 - приблизительно 5% общей мРНК семян. Ген лектина и специфическиий для семян промотор были полностью охарактеризованы и использовались для запуска специфической для семян экспрессии в трансгенных растениях табака (Vodkin et al., 1983; Lindstrom et al., 1990). Экспрессирующий вектор, содержащий кодирующий район, который кодирует представляющий интерес пептид, может быть сконструирован таким образом, что он находится под контролем промотора лектина, и этот вектор может быть введен в растения с использованием, например, способа трансформации протопластов (Dhir et al., 1991). Затем экспрессия этого полипептида будет направляться специфически в семена данного трансгенного растения.
Трансгенное растение данного изобретения, полученное из клетки растения, трансформированной тканеспецифическим промотором, может быть скрещено со вторым трансгенным растением, полученным из клетки растения, трансформированной специфическим для другой ткани промотором, для получения гибридного трансгенного растения, которое обнаруживает эффекты трансформации в более чем одной специфической ткани.
Другими примерами тканеспецифических промоторов являются промоторы сахарозосинтазы 1 кукурузы (Yang et al., 1990), алкогольдегидрогеназы 1 кукурузы (Vogel et al., 1989), светособирающего комплекса кукурузы (Simpson, 1986), хитшокового белка кукурузы (Odell et al., 1985), малой субъединицы РБФ-карбоксилазы гороха (Poulsen et al., 1986; Cashmore et al., 1983), маннопинсинтазы Ti-плазмиды (McBride and Summerfelt, 1989), нопалинсинтазы Ti-плазмиды (Langridge et al., 1989), халконизомеразы петунии (Van Tunen et al., 1988), богатого глицином белка 1 фасоли (Keller et al., 1989), СаМV35S-транскрипта (Odell et al., 1985) и пататина картофеля (Wenzler et al., 1989). Предпочтительными промоторами являются промотор вируса мозаики цветной капусты (CaMV35S) и промотор малой субъединицы РБФ-карбоксилазы S-E9.
Промоторы, используемые в конструкциях ДНК данного изобретения, могут быть модифицированы, если желательно, для воздействия на их регуляторные характеристики. Например, промотор CaMV35S может быть лигирован с частью гена ssRUBISCO, который подавляет экспрессию ssRUBISCO в отсутствие света, для создания промотора, который является активным в листьях, но не в корнях. Полученный химерный промотор может быть использован, как описано здесь. Для целей данного описания фраза промотор "CaMV35S" включает в себя, следовательно, вариации промотора CaMV35S, например промоторы, полученные лигированием с операторными районами, случайным или регулируемым мутагенезом и т.д.. Кроме того, эти промоторы могут быть изменены для содержания ″энхансерных последовательностей″ для содействия повышению экспрессии гена. Примеры таких энхансерных последовательностей сообщались Кау et al., (1987). Хлоропластные и пластидные промоторы известны в данной области (Daniell et al., US Pat. № 5693507; включен здесь в качестве ссылки), например промоторы, полученные из хлоропластных генов, таких как ген psbA из шпината или гороха, rbcL и промоторный район atpB из кукурузы и промоторы рРНК. Любой функциональный в хлоропластах или пластидах промотор находится в рамках данного изобретения.
РНК, продуцируемая конструкцией ДНК данного изобретения, также содержит 5’-нетранслируемую лидерную последовательность. Эта последовательность может быть произведена из промотора, выбранного для экспрессии гена, и может быть специфически модифицирована для увеличения трансляции этой мРНК. 5’-нетранслируемые районы могут быть также получены из вирусных РНК, из подходящих эукариотических генов или из синтетической генной последовательности. Данное изобретение не ограничивается конструкциями, в которых нетранслируемый район произведен из 5’-нетранслируемой последовательности, которая сопутствует промоторной последовательности. Как показано ниже, лидерная последовательность гена растения, которая применима в данном изобретении, является лидером белка 70 теплового шока петунии (hsp70) (Winter et al., 1988).
Пример варианта данного изобретения включает в себя аминокислотную последовательность В.thuringiensis пластидного нацеливания или пластидной локализации. Последовательности пластидного нацеливания были выделены из многочисленных ядерно-кодируемых генов растений, и было показано, что они управляют импортом цитоплазматически синтезированных белков в пластиды (обзор в Keegstra and Olsen, 1989). Множество последовательностей пластидного нацеливания, хорошо известные в данной области, в том числе, но не только, ADPGPP, EPSP-синтаза или ssRUBISCO, могут быть использованы в применении на практике данного изобретения. В альтернативных предпочтительных вариантах последовательности пластидного нацеливания (пептид и нуклеиновая кислота) для однодольных культур могут состоять из геномного кодирующего фрагмента, содержащего последовательность интрона, а также удвоенный сайт протеолитического расщепления в кодируемых последовательностях пластидного нацеливания.
Наиболее предпочтительная последовательность нуклеиновой кислоты, называемая здесь zmSSU PTP (SEQ ID NO:3), которая состоит из геномного кодирующего фрагмента, содержащего последовательность интрона, а также удвоенный сайт протеолитического расщепления в кодируемых последовательностях пластидного нацеливания, была произведена из последовательности пластидного нацеливания zmSl (Russel et al., 1993). Прямые трансляционные слияния пептидной последовательности zmSSU PTP (SEQ ID NO:4) с аминоконцом этих последовательностей применимы в получении повышенных уровней этого полипептида в трансгенной кукурузе. Слияния в рамке считывания последовательности нуклеиновой кислоты zmSSU РТР (SEQ ID NO:3) с геном Cry2Ab (SEQ ID NO:1) могут выполняться лигированием сайта NcoI при 3’ (С-концевое кодирование) конце последовательности zmSSU РТР с 5’-NcoI-сайтом (N-концевое кодирование) последовательности Cry2Ab.
Предпочтительная последовательность для двудольных культур, называемая здесь РТР2 (SEQ ID NO:9), состоит из геномного кодирующего фрагмента, содержащего последовательность пептида хлоропластного нацеливания из гена EPSP-синтазы Arabidopsis thaliana, в которой сайт отщепления транзитного пептида ssRUBISCO РТР заменяет нативный сайт отщепления РТР EPSP-синтазы (Klee et al., 1987).
Как отмечалось выше, 3’-нетранслируемый район химерных генов растений данного изобретения содержит сигнал полиаденилирования, который функционирует в растениях, вызывая присоединение аденилатнуклеотидов к 3’-концу РНК. Примерами предпочтительных 3’-районов являются (1) 3’-транскрибируемые, нетранслируемые районы, содержащие сигнал полиаденилирования индуцирующих опухоль (Ti) плазмидных генов Agrobacterium, таких как ген нопалинсинтазы (NOS), и (2) гены растений, такие как ген Е9 ssRUBISCO гороха (Fischhoff et al., 1987).
4.5.3 Применение интронов в экспрессирующих векторах
Для оптимизированной экспрессии в однодольных растениях в экспрессионную конструкцию ДНК может быть включен также интрон. Такой интрон обычно расположен вблизи 5’-конца мРНК в нетранслируемой последовательности. Интрон мог бы быть получен из, но не только, ряда интронов, состоящих из интрона белка 70 теплового шока (HSP) (патент США 5424412; 1995), интрона Act1 риса (McElroy et al., 1990), интрона 1 Adh (Callis et al., 1987) или интрона сахарозосинтазы (Vasil et al., 1989). Как показано здесь, интрон HSP70 кукурузы применим в данном исследовании.
4.5.4 Применение терминаторов в экспрессирующих векторах
РНК-полимераза транскрибирует кодирующую последовательность ДНК ядерного генома через сайт, в котором имеет место полиаденилирование. Обычно последовательности ДНК, локализованные на несколько пар оснований ниже по ходу транскрипции (справа) от сайта полиаденилирования, служат для терминации транскрипции. Эти последовательности ДНК называют здесь районами терминации транскрипции. Эти районы требуются для эффективного полиаденилирования транскрибируемой мессенджер РНК (мРНК). Для кодирующих последовательностей, введенных в хлоропласт или пластиду, или в хлоропласт или пластидный геном, терминация транскрипции мРНК одинакова со способами, хорошо известными в области экспрессии бактериальных генов. Например, в полицистронной или моноцистронной последовательности транскрипция может терминироваться структурами типа "стебель-петля" или структурами, сходными с rho-зависимыми последовательностями.
Конструкции обычно будут включать в себя представляющий интерес ген вместе с 3’-концевой последовательностью ДНК, которая действует как сигнал терминации транскрипции и в конструкциях, предназначенных для экспрессии в ядерном геноме, делает возможным полиаденилирование полученной мРНК. Предполагается, что наиболее предпочтительными 3’-элементами являются элементы из гена нопалинсинтазы A.tumefaciens (3’-конец nos) (Bevan et al., 1983), терминатор для Т7-транскрипта из гена октопинсинтазы A. tumefaciens и 3’-конец генов ингибитора протеазы i и ii из картофеля и томата. Регуляторные элементы, такие как элемент Ω TMV (Gallie et al., 1989) могут быть дополнительно включены, если желательно.
4.5.5 Другие усиливающие экспрессию элементы
Другим типом элемента, который может регулировать экспрессию генов, является последовательность ДНК между сайтом инициации транскрипции и началом кодирующей последовательности, называемая нетранслируемой лидерной последовательностью. Лидерная последовательность может влиять на экспрессию генов. Могут быть сделаны компиляции лидерных последовательностей для прогнозирования оптимальных или субоптимальных последовательностей и генерирования "консенсусных" и предпочтительных лидерных последовательностей (Joshi, 1987). Предполагается, что предпочтительные лидерные последовательности включают в себя те последовательности, которые содержат последовательности, в отношении которых предсказано, что они управляют оптимальной экспрессией связанного структурного гена, т.е. включают в себя предпочтительную консенсусную лидерную последовательность, которая может увеличивать или поддерживать стабильность мРНК и предотвращать неподходящую инициацию трансляции. Выбор таких последовательностей будет известен для специалистов в данной области в свете данного описания. Последовательности, произведенные из генов, которые экспрессируются на высоком уровне в растениях, в частности в кукурузе, будут наиболее предпочтительными. Одним из особенно ценных лидеров может быть лидер HSP70 петунии.
Энхансеры транскрипции или дупликации энхансеров могли бы быть использованы для увеличения экспрессии. Эти энхансеры часто обнаруживаются 5’ (слева) от начала транскрипци в промоторе, функционирующем в эукариотических клетках, но часто могут быть встроены в прямой или обратной ориентации 5’ (слева) или 3’ (справа) от кодирующей последовательности. Примеры энхансеров включают в себя элементы из промотора CaMV 35S, генов октопинсинтазы (Ellis et al., 1987), гена актина риса и промотора из эукариот не-растений (например, дрожжей; Ма et al., 1988).
4.5.6 Мультигенные векторные конструкции и IRES
В некоторых вариантах данного изобретения использование элементов внутренних сайтов связывания рибосом (IRES) применяют для создания мультигенных или полицистронных матриц. IRES-элементы способны обходить модель рибосомного сканирования зависимой от 5’-метилированного кэпа трансляции и начинать трансляцию при внутренних сайтах (Pelletier and Sonenberg, 1988). Описаны IRES-элементы из двух членов семейства пикорнавирусов (полиомиелита и энцефаломиокардита) (Pelletier and Sonenberg, 1988), а также IRES из матрицы млекопитающего (Macejak and Sarnow, 1991). IRES-элементы могут быть связаны с гетерологичными открытыми рамками считывания. Множественные открытые рамки считывания могут транскрибироваться вместе, каждая из которых отделена посредством IRES, создавая полицистронные матрицы. Благодаря IRES-элементу каждая открытая рамка считывания доступна для рибосом для эффективной трансляции. Множественные гены могут эффективно экспрессироваться с использованием единственного промотора /энхансера для транскрибирования единственной матрицы.
Любая гетерологичная открытая рамка считывания может быть связана с IRES-элементами. Это охватывает гены для секретируемых белков, мультисубъединичных белков, кодируемых независимыми генами, внутриклеточных или мембраносвязанных белков и селектируемых маркеров. Таким путем экспрессия нескольких белков может быть одновременно сконструирована в клетке с использованием единственной конструкции и единственного селектируемого маркера.
Конструкции, предназначенные для экспрессии из использования в хлоропласте или пластиде специфического аппарата транскрипции и трансляции, могут содержать либо моноцистронные, либо полицистронные последовательности.
4.5.7 Конструирование экспрессирующего вектора
Выбор, какой экспрессирующий вектор использовать и, в конечном счете, с каким промотором следует связать функционально кодирующий полипептид район, зависит непосредственно от желательных функциональных свойств, т.е. локализации и тайминга экспрессии белка, и клетки-хозяина, которая должна быть трансформирована. Эти моменты являются хорошо известными ограничениями, присущими области конструирования рекомбинантных молекул ДНК. Однако вектор, применимый в использовании на практике данного изобретения, способен направлять экспрессию кодирующего полипептид района, с которым он функционально связан.
Типичные векторы, применимые для экспрессии генов в высших растениях, хорошо известны в данной области и включают в себя векторы, произведенные из описанной индуцирующей опухоль (Ti) плазмиды A.tumefaciens (Rogers et al., 1987). Однако известно, что несколько других интегрирующихся в растения векторных систем функционируют в растениях, в том числе описанный регулирующий транспорт вектор pCaMVCN (Fromm. et al., 1986). PCaMVCN (доступный из Pharmacia, Piscataway, NJ) включает в себя промотор CaMV35S.
В предпочтительных вариантах вектор, используемый для экспрессии полипептида, включает в себя селектируемый маркер, который эффективен в клетке растения, предпочтительно селективный маркер устойчивости к лекарственному средству.
Одним из предпочтительных маркеров устойчивости к лекарственному средству является ген, экспрессия которого приводит к устойчивости к канамицину, т.е. описанный химерный ген, содержащий промотор нопалинсинтазы, неомицинфосфотрансферазу II Tn5 (nptII) и 3’-нетранслируемый район нопалинсинтазы (Rogers et al., 1988).
Способы получения экспрессирующих векторов хорошо известны в данной области. Экспрессирующие (трансформирующие) векторы, используемые для трансформации растений, и способы приготовления этих векторов описаны в патентах США 4971908, 4940835, 4769061 и 4757011 (каждый из которых специально включен здесь в качестве ссылки). Эти векторы могут быть модифицированы для включения кодирующей последовательности в соответствии с данным изобретением.
Были разработаны различные способы для функционального связывания ДНК с векторами через комплементарные липкие концы или тупые концы. Например, комплементарные гомополимерные участки могут быть присоединены к сегменту ДНК, который должен быть инсертирован, и к векторной ДНК. Затем эти вектор и сегмент ДНК соединяются водородными связями между комплементарными гомополимерными хвостами с образованием рекомбинантных молекул ДНК.
Кодирующий район, который кодирует полипептид, способный придавать инсектицидную активность клетке, предпочтительно является полинуклеотидом, кодирующим δ-эндотоксин В.thuringiensis, или функциональным эквивалентом такого полинуклеотида. В соответствии с такими вариантами кодирующий район, содержащий последовательность ДНК SEQ ID NO:1, является также предпочтительным.
Специфические кодирующие полипептид δ-эндотоксина В.thuringiensis гены, которые, как было показано, успешно трансформируют растения в сочетании с генами, кодирующими пептид пластидного нацеливания, для экспрессии δ-эндотоксинов В.thuringiensis при высоких уровнях, являются генами, содержащимися в плазмидных векторах. Предпочтительные плазмиды, содержащие последовательность пластидного нацеливания, включают в себя pMON30464, pMON33827, pMON33828, pMON33829. Эти плазмиды кодируются последовательностями, показанными в SEQ ID NO:16, SEQ ID NO:13, SEQ ID NO:14, SEQ ID NO:15. Более конкретно, растения могут быть успешно трансформированы любым вектором, содержащим экспрессионные кассеты, включающие в себя нуклеотидные последовательности от нуклеотида 1781 до нуклеотида 5869 SEQ ID NO:16, от нуклеотида 17 до нуклеотида 3182 SEQ ID NO:13, от нуклеотида 17 до нуклеотида 3092 SEQ ID NO:14 или от нуклеотида 17 до нуклеотида 3155 SEQ ID NO:15.
Описанное здесь исследование идентифицировало способы потенциирования in planta экспрессии δ-эндотоксинов В.thuringiensis, которые сообщают устойчивость к насекомым-патогенам при включении в ядерный, пластидный или хлоропластный геном чувствительных растений. В патенте США 5500365 (специально включенном здесь в качестве ссылки) описан способ синтеза генов растений для оптимизации уровня экспрессии белка, кодируемого синтезированным геном. Этот способ относится к модификации последовательностей структурных генов экзогенного трансгена, чтобы сделать их более "растение-подобными" и, следовательно, имеющими большую вероятность трансляции и экспрессии растением. Подобный способ для усиления экспрессии трансгенов предпочтительно в однодольных растениях описан в патенте США 5689052 (специально включенном здесь в качестве ссылки). Агрономические, плодоовощные, декоративные и другие экономически или коммерчески ценные растения могут быть созданы в соответствии с описанными здесь способами для экспрессии δ-эндотоксинов В.thuringiensis при уровнях, достаточно высоких для придания устойчивости к насекомым-патогенам.
Такие растения могут ко-экспрессировать полипептид δ-эндотоксина В.thuringiensis вместе с другими противогрибковыми, антибактериальными или антивирусными связанными с патогенезом пептидами, полипептидами или белками; инсектицидными белками; белками, придающими устойчивость к гербицидам; и белками, участвующими в улучшении качества продуктов растений или агрономической продуктивности растений. Одновременная ко-экспрессия множественных белков в растениях является выгодной потому, что она использует более одного способа действия для борьбы с патогенным повреждением растений. Это может минимизировать возможность развития устойчивых штаммов патогенов, расширить сферу устойчивости и потенциально привести к синергическому инсектицидному действию, усиливая тем самым способность растения противостоять инвазии насекомых (WO 92/17591).
Специально для применения в соответствии с данным изобретением рассматриваются векторы, которые включают в себя энхансерный элемент ocs. Этот элемент был сначала идентифицирован в виде палиндромного энхансера 16 п.н. из гена октопинсинтазы (ocs) Agrobacterium (Ellis et al., 1987) и присутствует в по меньшей мере 10 других промоторах (Bouchez et al., 1989). Предполагается, что использование энхансерного элемента, такого как элемент ocs, и, в частности, множественных копий этого элемента может быть использовано для увеличения уровня транскрипции из смежных промоторов при применении в контексте трансформации однодольных растений.
Предполагается, что введение больших последовательностей ДНК, содержащих более одного гена, может быть желательным. Введение таких последовательностей может быть облегчено использованием бактериальных или дрожжевых искусственных хромосом (ВАС или YAC соответственно) или даже искусственных хромосом растений. Например, применение ВАС для опосредованной Agrobacterium трансформации описано Hamilton et al. (1996).
В конечном счете наиболее желательными сегментами ДНК для введения в геном однодольных растений могут быть гомологичные гены или семейства генов, кодирующие желательный признак (например, увеличенный урожай) и вводимые под контролем новых промоторов или энхансеров, и т.д., или, возможно, даже гомологичных или тканеспецифических (например, специфических для корневой шейки/корневого влагалища, мутовки, стебля, початка кукурузы, зерна или листа) промоторов или регуляторных элементов. В самом деле, предполагается, что конкретное использование данного изобретения может состоять в получении трансформантов, содержащих трансген, который нацелен тканеспецифическим образом. Например, гены устойчивости к насекомым могут экспрессироваться специфически в тканях мутовки и тканях корневой шейки/корневого влагалища, которые являются мишенями первого и второго потомства соответственно ЕСВ. Подобно этому гены, кодирующие белки с конкретной активностью против блошки длинноусой, могут быть нацелены в корневые ткани.
Векторы для применения в тканеспецифическом нацеливании генной экспрессии в трансгенных растениях обычно будут содержать тканеспецифические промоторы и также будут содержать другие тканеспецифические регуляторные элементы, такие как энхансерные последовательности. Промоторы, которые направляют специфическую или усиленную экспрессию в определенных тканях растений, будут известны лицам с квалификацией в данной области в свете данного описания.
Ожидается также, что тканеспецифическая экспрессия может функционально выполняться введением конститутивно экспрессируемого гена (во всех тканях) в комбинации с антисмысловым геном, который экспрессируется только в тех тканях, где продукт этого гена не является желательным. Например, ген, кодирующий кристаллический токсиновый белок из В.thuringiensis, может быть введен таким образом, что он экспрессируется во всех тканях с использованием промотора 35S из вируса мозаичной болезни цветной капусты. Альтернативно, промотор актина риса или промотор гистона из двудольных или однодольных видов мог бы быть также использован для конститутивной экспрессии гена. Кроме того, ожидается, что могут использоваться промоторы, комбинирующие элементы из более чем одного промотора. Например, в патенте США 5491288 описано комбинирование промотора вируса мозаичной болезни цветной капусты с промотором гистона. Таким образом, экспрессия антисмыслового транскрипта гена B.t. в зерне кукурузы с использованием, например, промотора зеина могла бы предотвращать накопление δ-эндотоксина в семенах. Таким образом, белок, кодируемый введенным геном, присутствовал бы во всех тканях, кроме зерна. Авторы изобретения специально обсуждают вопрос о том, что подобная стратегия могла бы использоваться с применением данного изобретения для управления экспрессией скринируемого или селектируемого маркера в ткани семени.
Альтернативно, может быть желательным получение новых тканеспецифических промоторных последовательностей для применения в соответствии с данным изобретением. Для достижения этого можно сначала выделить клоны кДНК из рассматриваемой ткани и идентифицировать клоны, которые экспрессируются специфически в этой ткани, например, при помощи Нозерн-блоттинга. Идеально, было бы желательно идентифицировать ген, который не присутствует с высоким числом копий, но продукт которого является относительно обильным в специфических тканях. Промоторные и регуляторные элементы соответствующих геномных клонов могут быть также локализованы с использованием способов молекулярной биологии, известных специалистам в данной области.
Предполагается, что экспрессия некоторых генов в трансгенных растениях будет желательной при указанных условиях. Например, предполагается, что экспрессия определенных генов, придающих устойчивость к стрессовым факторам окружающей среды, таким как засуха, будет желательной при фактических стрессовых условиях. Далее, предполагается, что экспрессия таких генов на протяжении всего развития растений может иметь вредные действия. Известно, что имеется большое число генов, которые отвечают на условия окружающей среды. Например, экспрессия некоторых генов, таких как rbcS, кодирующий малую субъединицу рибулозобисфосфаткарбоксилазы, регулируется светом через фитохром. Другие гены индуцируются вторичными стимулами. Например, синтез абсцизовой кислоты (АВА) индуцируется определенными факторами окружающей среды, в том числе, но не только, водным стрессом. Было показано, что ряд генов индуцируются АВА (Skriver and Mundy, 1990). Ожидается также, что экспрессия генов, сообщающих устойчивость к поеданию насекомыми, была бы желательной только в условиях действительного заражения насекомыми. Таким образом, для некоторых желательных признаков индуцибельная экспрессия генов в трансгенных растениях будет желательной.
Предполагается, что в некоторых вариантах данного изобретения экспрессия гена в трансгенном растении будет желательной только в определенный период времени во время развития данного растения. Связанный с развитием тайминг часто коррелирует с тканеспецифической экспрессией генов. Например, экспрессия запасных зеиновых белков инициируется в эндосперме спустя приблизительно 15 дней после опыления.
Предполагается также, что может быть полезным нацеливание самой ДНК клеткой. Например, может быть полезным нацелить вводимую ДНК в ядро, так как это может увеличить частоту трансформации. В самом ядре она была бы полезна для нацеливания гена для достижения сайт-специфической интеграции. Например, было бы полезно иметь ген, введенный посредством трансформации, для замены существующего в этой клетке гена.
4.6 Идентификация и выделение инсектицидных δ-эндотоксинов и генов В.thuringiensis
Предполагается, что описанный в данном изобретении способ мог бы быть использован для получения существенно улучшенной экспрессии ряда новых δ-эндотоксинов В.thuringiensis, выделенных, как описано ниже. Идентификация новых штаммов Bacillus thuringiensis, кодирующих кристаллические эндотоксины с инсектицидной активностью, была описана ранее (Donovan et al., 1992). Выделение эндотоксина В.thuringiensis с последующими секвенированием аминоконцевых аминокислот, обратной трансляцией аминокислотной последовательности для конструирования олигонуклеотидного зонда или использованием родственного гена В.thuringiensis в качестве зонда, с последующим клонированием гена, кодирующего этот эндотоксин, гибридизацией известны лицам с квалификацией в данной области и были описаны (см., например, Donovan et al., 1992; патент США 5264364, включенные здесь в качестве ссылки).
Улучшенная экспрессия неактивных против Diptera δ-эндотоксинов Cry2A В.thuringiensis в трансгенных растениях может быть достигнута посредством способов, описанных в данном изобретении. Одним белком, для которого получена улучшенная экспрессия, является Cry2Ab.
Предыдущая работа показала, что некоторые δ-эндотоксины Cry2A имеют более широкую специфичность в отношении круга хозяев, чем другие близкородственные δ-эндотоксины Cry2A, причем не только виды чешуекрылых насекомых, но и виды двукрылых насекомых, в частности, также чувствительны к очень низким дозам токсинов. В противоположность этому было показано, что близкородственные эндотоксины Cry2A, не обнаруживающие существенной ингибиторной активности против двукрылых насекомых, имели более узкую специфичность в отношении круга хозяев (Widner et al., 1989, J. Bacteriol, 171:965-974; Widner et al. (a), 1990, J. Bacteriol. 172:2826-2832). Эти работы показали, что Cry2Ab, используемый здесь, не полностью лишен ингибиторной активности против двукрылых насекомых, но просто является менее сильнодействующим, чем близкородственные δ-эндотоксины Cry2A В.thuringiensis. Эти работы показали, что Cry2A, в частности, был гораздо менее эффективен чем Cry2Aа и, следовательно, лишен активности против двукрылых насекомых при испытании против Aedes egyptii. He существовало ни одного приемлемого способа для различения между близкородственными δ-эндотоксинами, однако, как показано здесь, отбор на подходящие Cry2A мог бы выполняться с использованием одного способа или комбинации способов, включающих в себя, но не только, сравнения гомологии общей аминокислотной последовательности, сравнений близко сфокусированного сходства между отдельными Cry2A в районе аминокислотной последовательности 307-382, или на основе данных IC50. Widner et al. продемонстрировали, что в 50-100 раз больше Cry2Ab чем Cry2Aа требовалось для получения одинаковых IС50-эффектов на насекомых Diptera. Таким образом, диапазон чувствительности видов Diptera к белку Cry2A мог бы быть использован в качестве одного из средств измерения и установления различий чувствительности насекомых-мишеней между различными классами белков Cry2A. Например, величина IC50 в м.д., приблизительно в 3 раза более высокая, чем проявляемая Cry2Aа против Aedes egyptii, могла бы использоваться как признак для исключения определенных белков Cry2A как лишенных существенной ингибиторной активности против видов Diptera. Однако применение подхода на основе диапазонов ТС50-ингибиторной активности следует использовать острожно, так как эти величины зависят от ряда высоко вариабельных условий, в том числе, но не только, от способов и материалов, используемых для тестирования белков, и от физического кондиционирования тестируемых насекомых. Альтернативный способ отличения δ-эндотоксинов Cry2A, лишенных существенной активности против видов Diptera, от δ-эндотоксинов, которые не находятся в сфере действия данного изобретения, мог бы охватывать исключение белков Cry2A, которые имеют сходство аминокислотной последовательности более приблизительно 87% относительно аминокислотной последовательности Cry2Aа, или более предпочтительно исключение белков Cry2A, которые имеют сходство аминокислотной последовательности более приблизительно 90% относительно аминокислотной последовательности Cry2Aа. В частности, считается, что район Cry2Aа, соответствующий аминокислотным остаткам от приблизительно 307 до приблизительно 382, является критическим для ингибиторной активности против Diptera данного белка, и при замене этого комплементарного района несходства в Cry2Ab придает ингибиторную активность против Diptera белку Cry2Ab. Таким образом, дополнительный способ для отличения δ-эндотоксинов Cry2A, которые находятся в сфере действия данного изобретения, мог бы включать в себя сравнение сходства этого района белка без необходимости учета уровня гомологии при сравнении любых конкретных δ-эндотоксинов Cry2A с этим районом в Cry2Aа. При вариабельных аминокислотах в этом домене из 76 аминокислот δ-эндотоксинами Cry2A, которые находятся в рамках данного изобретения, были бы предпочтительно те, которые имеют более приблизительно 80 - приблизительно 99 процентов сходства с Cry2Aа в данной последовательности, или более предпочтительно те, которые имеют более приблизительно 60-79 процентов сходства с Cry2αА в данной последовательности, или те, которые имеют более приблизительно 40 - приблизительно 59 процентов сходства с Cry2Aа в данной последовательности, или даже более предпочтительно те, которые имеют более приблизительно 24 - приблизительно 39 процентов сходства с Cry2Aа в данной последовательности, или наиболее предпочтительно те δ-эндотоксины Cry2A, которые имеют более приблизительно 0 - приблизительно 23 процента сходства с Cry2Aа в данной последовательности.
4.7 Трансформированные клетки растения и трансгенные растения
Рассматривается также клетка растения, трансформированная экспрессирующим вектором данного изобретения. Трансгенное растение, полученное из такой трансформированной или трансгенной клетки, также обсуждается. Лицам с квалификацией в данной области будет понятно, что химерный ген растения, содержащий структурную кодирующую последовательность данного изобретения, может быть встроен в геном растения способами, хорошо известными в данной области. Такие способы для трансформации ДНК растительных клеток включают в себя опосредованную Agrobacterium трансформацию растений, применение липосом, трансформацию с использованием вирусов или пыльцы, электропорацию, трансформацию протопластов, перенос генов в пыльцу, инъекцию в репродуктивные органы, инъекцию в незрелые эмбрионы и бомбардировку частицами. Каждый из этих способов имеет отличающиеся преимущества и недостатки. Таким образом, конкретный способ введения генов в конкретную разновидность растения может не быть обязательно наиболее эффективным для другой разновидности растения, но хорошо известно, какие способы применимы для конкретной разновидности растения.
Имеется много способов для введения трансформирующих сегментов ДНК в клетки, но не все они пригодны для доставки ДНК в растительные клетки. Считается, что подходящие способы включают в себя фактически любой способ, при помощи которого ДНК может быть введена в клетку, такие как инфицирование А.tumefaciens и родственными штаммами Agrobacterium, прямая доставка ДНК, такая как, например, ПЭГ-опосредованная трансформация протопластов (Omirulleh et al., 1993), опосредованное высушиванием/ингибированием поглощение ДНК, трансформация электропорацией, перемешиванием с волокнами карбида кремния, ускорением покрытых ДНК частиц и т.д. В некоторых вариантах предпочтительными являются способы ускорения и они включают в себя, например, бомбардировку микроснарядами и т.п.
Технология введения ДНК в клетки хорошо известна в данной области. Описаны четыре основных способа доставки гена в клетки: (1) химические способы (Graham and van der Eb, 1973); (2) физические способы, такие как микроинъекция (Capecchi, 1980), электропорация (Wong and Neumann, 1982; Fromm et al., 1985) и генное ружье (Johnston and Tang, 1994; Fynan et al., 1993); (3) вирусные векторы (Clapp, 1993; Lu et al., 1993; Eglitis and Anderson, 1988a; 1988b); и (4) опосредованные рецептором механизмы (Curiel et al., 1991; Wagner et al., 1992).
4.7.1 Электропорация
Применение коротких электрических импульсов высокого напряжения к различным животным и растительным клеткам приводит к образованию пор размером порядка нанометров в плазматической мембране. ДНК поглощается непосредственно в цитоплазму клеток либо через эти поры, либо как следствие перераспределения мембранных компонентов, которое сопровождает закрытие этих пор. Электропорация может быть чрезвычайно эффективной и может быть использована как для временной (преходящей) экспрессии клонированных генов, так и для создания клеточных линий, которые несут интегрированные копии представляющих интерес генов. Электропорация в противоположность кальций-фосфат-опосредованной трансфекции и слиянию протопластов часто приводит к появлению клеточных линий, несущих один или самое большее несколько интегрированных копий чужеродной ДНК.
Введение ДНК посредством электропорации хорошо известно специалистам в данной области. Для выполнения электропорации можно использовать либо рыхлые ткани, такие как суспензионная культура клеток, либо эмбриогенный каллус, или альтернативно, можно трансформировать незрелые зародыши или другие организованные ткани непосредственно. Можно было бы частично деградировать клеточные стенки выбранных клеток выдерживанием их с пектин-деградирующими ферментами (пектолиазами) или механическим раневым повреждением контролируемым образом, что делает эти клетки более восприимчивыми к трансформации. Затем такие клетки были бы реципиентами для переноса ДНК электропорацией, которая может проводиться на этой стадии, и затем трансформированные клетки идентифицируют при помощи подходящего протокола отбора или скрининга в зависимости от характера новой включенной ДНК.
4.7.2 Бомбардировка микроснарядами
Другим предпочтительным способом доставки трансформирующих сегментов ДНК в растительные клетки является бомбардировка микроснарядами. В этом способе частицы могут быть покрыты нуклеиновыми кислотами и доставлены в клетки движущим усилием. Примеры частиц включают в себя частицы, состоящие из вольфрама, золота, платины и т.п. С использованием этих частиц ДНК проносится через клеточную стенку и в цитоплазму на поверхности небольших металлических частиц, как описано (Klein et al., 1987; Klein et al., 1988; Kawata et al., 1988). Металлические частицы проникают через несколько слоев клеток и таким образом делают возможной трансформацию в тканевых эксплантатах. Способ бомбардировки микроснарядами является предпочтительным для идентификации управляемых хлоропластами или пластидами событий трансформации.
Преимущество бомбардировки микроснарядами, кроме того, что она является эффективным средством воспроизводимо стабильной трансформации растительных клеток, заключается в том, что она не требует ни выделения протопластов (Cristou et al., 1988), ни восприимчивости к инфекции Agrobacterium. Иллюстративным вариантом способа доставки ДНК в растительные клетки путем ускорения является Biolistics Particle Delivery System, которая может быть использована для проталкивания частиц, покрытых ДНК, или клеток через сито, например сито из нержавеющей стали или Nytex, на поверхностть фильтра, покрытую культивируемыми в суспензии клетками растения. Сито диспергирует эти частицы таким образом, что они не доставляются к реципиентным клеткам в виде больших агрегатов. Считается, что сито, включенное между аппаратом для бомбардировки микроснарядами и клетками, подлежащими бомбардировке, уменьшает размер агрегатов микроснарядов и может способствовать более высокой частоте трансформации путем уменьшения повреждений, наносимых реципиентным клеткам микроснарядами, которые являются слишком большими.
Для бомбардировки клетки в суспензии являются предпочтительно сконцентрированными на фильтрах или твердой культуральной среде. Альтернативно незрелые эмбрионы или другие клетки-мишени могут быть размещены на твердой культуральной среде. Клетки, подлежащие бомбардировке, размещают на подходящем расстоянии ниже пластинки остановки микроснарядов. Если желательно, одно или несколько сит могут быть расположены между ускоряющим устройством и подлежащими бомбардировке клетками. Посредством использования описанного здесь способа можно получать до 1000 или более очагов клеток, временно экспрессирующих маркерный ген. Число клеток в очаге, которые экспрессируют экзогенный продукт через 48 часов после бомбардировки, часто находится в диапазоне от 1 до 10, в среднем 1-3.
В трансформации посредством бомбардировки можно оптимизировать условия культивирования перед бомбардировкой и параметры бомбардировки для получения максимальных чисел стабильных трансформантов. Как физические, так и биологические параметры для бомбардировки являются важными в этой технологии. Физическими факторами являются факторы, которые включают в себя манипулирование осадком ДНК/микроснаряды, или факторы, которые влияют на полет и скорость макро- или микрочастиц. Биологические факторы включают в себя все стадии, участвующие в манипуляции клетками до и непосредственно после бомбардировки, а также природа трансформирующей ДНК, такой как линеаризованная ДНК или интактные суперспиральные плазмиды. Считается, что манипуляции перед бомбардировкой особенно важны для успешной трансформации незрелых зародышей растений.
Таким образом, предполагается, что может быть желательным приспособление различных параметров бомбардировки к исследованиям малого масштаба для полной оптимизации условий. Могут быть желательными приспособления физических параметров, таких как расстояние промежутка, расстояние полета, расстояние до ткани и давление гелия. Можно также минимизировать факторы снижения травмы (TRF) модификацией условий, которые влияют на физиологическое состояние реципиентных клеток и которые могут, следовательно, влиять на эффективности трансформации и интеграции. Например, осмотическое состояние, гидратация ткани и стадия субкультуры или клеточного цикла реципиентных клеток могут быть приспособлены для оптимальной трансформации. Выполнение других рутинных приспособлений будет известно специалистам в данной области в свете данного изобретения.
Способы опосредованной частицами трансформации хорошо известны специалистам в данной области. В патенте США 5015580 (специально включенном здесь в качестве ссылки) описана трансформация сои с использованием такого способа.
4.7.3 Опосредованный Agrobacterium перенос генов
Опосредованный Agrobacterium перенос является широко применяемой системой для введения генов в растительные клетки, так как ДНК может быть введена в ткани целого растения, обходя тем самым необходимость регенерации интактного растения из протопласта. Применение опосредованных Agrobacterium интегрирующихся в растение векторов для введения ДНК в клетки растений хорошо известно в данной области. См., например, описанные способы (Fraley et al., 1985; Rogers et al., 1987). Генетическая инженерия растений хлопчатника с использованием опосредованного Agrobacterium переноса описана в патенте США 5004863 (специально включенном здесь в качестве ссылки); как трансформация растений латука описана в патенте США 5349124 (специально включенном здесь в качестве ссылки) и опосредованная Agrobacterium трансформация сои описана в патенте США 5416011 (специально включенном здесь в качестве ссылки). Далее, интеграция Ti-ДНК является относительно точным процессом, приводящим к малому числу реаранжировок. Район переносимой ДНК определяется граничными последовательностями, и промежуточную ДНК обычно встраивают в геном растений, как описано (Spielmann et al., 1986; Jorgensen et al., 1987).
Современные векторы трансформаци на основе Agrobacterium способны реплицироваться в Е. coli, а также в Agrobacterium, делая возможными подходящие описанные манипуляции (Klee et al., 1985). Кроме того, недавние технологические успехи в создании векторов для опосредованного Agrobacterium переноса генов улучшили аранжировку генов и сайтов рестрикции в этих векторах для облегчения конструирования векторов, способных экспрессировать различные кодирующие полипептиды гены. Описанные векторы (Rogers et al., 1987) имеют удобные мультилинкерные районы, фланкированные промотором и сайтом полиаденилирования, для прямой экспрессии встроенных кодирующих полипептиды генов и пригодны для целей данного изобретения. Кроме того, Agrobacterium, содержащая как “вооруженные”, так и “разоруженные” Ti-гены, может быть использована для трансформации. В разновидностях растений, в которых опосредованная Agrobacterium трансформация является эффективной, этот способ является лучшим способом вследствие легкой и определенной природы переноса генов.
Опосредованная Agrobacterium трансформация листовых дисков и других тканей, таких как семядоли и гипокотили, по-видимому, ограничивается растениями, которые Agrobacterium природно инфицирует. Опосредованная Agrobacterium трансформация является наиболее эффективной в двудольных растениях. Небольшое число однодольных растений, по-видимому, являются природными хозяевами для Agrobacterium, хотя трансгенные растения были получены в случае спаржи с использованием векторов Agrobacterium, как описано (Bytebier et al., 1987). Недавно другие однодольные растения были трансформированы Agrobacterium. В эту группу включены кукуруза (Ishida et al.) и рис (Cheng et al.).
Трансгенные растения, образованные с использованием способов трансформации с применением Agrobacterium, обычно содержат единственный ген на одной хромосоме. Такие трансгенные растения могут быть названы гетерозиготными в отношении добавленного гена. Однако, поскольку применение слова “гетерозиготные” обычно подразумевает присутствие комплементарного гена в том же самом локусе второй хромосомы пары хромосом и нет такого гена в растении, содержащем один добавленный ген, как в этом случае, считается, что более точное название для такого растения будет независимый сегрегант, так как этот добавленный экзогенный ген расщепляется независимо во время митоза и мейоза.
Независимый сегрегант может быть предпочтительным, когда растение ставится на коммерческую основу в виде гибрида, такого как кукуруза. В этом случае независимый сегрегант, содержащий этот ген, скрещивают с другим растением с получением гибридного растения, которое является гетерозиготным в отношении представляющего интерес гена.
Другим предпочтительным вариантом является трансгенное растение, которое является гомозиготным в отношении добавленного структурного гена; т.е. трансгенное растение, которое содержит два добавленных гена, один ген в одном и том же локусе на каждой хромосоме пары хромосом. Гомозиготное трансгенное растение может быть получено половым спариванием (самоопылением) независимого трансгенного растения-сегреганта, которое содержит единственный добавленный ген, проращиванием части полученных семян и анализом полученных растений на активность представляющего интерес гена и менделирующую наследственность, указывающую на гомозиготность относительно контроля (нативных, нетрансгенных растений) или трансгенное растение, являющееся независимым сегрегантом.
Два различных трансгенных растения могут быть скрещены с образованием потомства, которое содержит два независимо расщепляющихся добавленных экзогенных гена. Самоопыление подходящего потомка могут производить растения, которые являются гомозиготными для обоих добавленных экзогенных генов, кодирующих представляющий интерес полипептид. Обратное скрещивание с родительским растением и ауткроссинг с нетрансгенным растением также рассматриваются.
Трансформация протопластов растений может быть достигнута с использованием способов на основе кальций-фосфатного осаждения, обработки полиэтиленгликолем, электропорации и комбинации этих обработок (см., например, Potrykus et al., 1985; Lorz et al., 1985; Fromm et al., 1985; Uchimiya et al., 1986; Callis et al., 1987; Marcotte et al., 1988).
Применение этих систем для различных зародышевых плазм растений зависит от способности регенерировать данную конкретную разновидность растения из протопластов. Иллюстративные способы для регенерации зерновых из протопластов описаны (см., например, Fujimura et al., 1985; Toriyama et al., 1986; Yamada et al., 1986; Abdullah et al., 1986).
Для трансформации растительной зародышевой плазмы, которая не может успешно регенерировать из протопластов, могут быть использованы другие пути введения ДНК в интактные клетки или ткани. Например, регенерация зерновых из незрелых зародышей или эксплантатов может выполняться, как описано (Vasil, 1988).
4.8 Экспрессия генов в растениях
Немодифицированные бактериальные гены часто слабо экспрессируются в трансгенных растительных клетках. Использование кодонов в растениях более близко сходно с использованием кодонов человека и других высших организмов, чем одноклеточных организмов, таких как бактерии. В нескольких сообщениях были описаны способы улучшения экспрессии рекомбинантных генов в растениях (Murray et al., 1989; Diehn et al., 1996; Iannacone et al., 1997; Rouwendal et al., 1997; Futterer et al., 1997 и Futterer and Hohn, 1996). Эти сообщения описывают различные способы конструирования кодирующих последовательностей для представления последовательностей, которые более эффективно транслируются, на основе таблиц частоты встречаемости кодонов в растении, улучшений в неоднозначности основания третьего положения кодона, с использованием рекомбинантных последовательностей, которые избегают подозреваемого полиаденилирования или А/Т-богатых доменов или консенсусных последовательностией сплайсинга интронов. Хотя эти способы для конструирования синтетических генов являются известными, синтетические гены данного изобретения получали согласно способу Brown et al., (патент США № 5689052; 1997), который включен здесь в качестве полной ссылки. Таким образом, данное изобретение обеспечивает способ получения синтетических генов растений, которые экспрессируют in planta желаемый продукт при уровнях, значимо более высоких, чем гены дикого типа. Вкратце, согласно Brown et al. частоту редких и полуредких кодонов однодольных растений в полинуклеотидной последовательности, кодирующей желаемый белок, уменьшают и заменяют более предпочтительными кодонами однодольных растений. Повышенное накопление желаемого полипептида, кодируемого модифицированной полинуклеотидной последовательностью, в однодольном растении является результатом увеличения частоты встречаемости предпочтительных кодонов анализом кодирующей последовательности в последовательных шести нуклеотидных фрагментах и изменением этой последовательности на основе частоты встречаемости этих шестимеров относительно частоты встречаемости самых редких 284, 484 и 664 шестимеров в однодольных растениях. Кроме того. Brown et al. описывают увеличенную экспрессию рекомбинантного гена посредством способа уменьшения частоты редких кодонов способами для уменьшения встречаемости сигналов полиаденилирования и сайтов сплайсинга интронов в нуклеотидной последовательности, удаления самокомплементарных последовательностей в этой нуклеотидной последовательности и замены не-самокомплементарными нуклеотидами с сохранением структурного гена, кодирующего данный полипептид, и уменьшения частоты встречаемости динуклеотидных пар 5’-CG-3’ в этой нуклеотидной последовательности. Эти стадии выполняются последовательно и имеют кумулятивный эффект, приводящий к нуклеотидной последовательности, содержащей предпочтительное (преферентивное) использование более предпочтительных кодонов однодольных растений, для однодольных растений в отношении большинства аминокислот, присутствующих в желаемом полипептиде.
Работа, описанная здесь, идентифицировала способы потенциирования in planta экспрессии δ-эндотоксинов В.thuringiensis, которые придают устойчивость к насекомым-патогенам, при включении в ядерный, пластидный или хлоропластный геном чувствительных растений. В патенте США 5500365 (специально включенном здесь в качестве ссылки) описан способ синтеза генов растений для оптимизации уровня экспрессии белка, кодируемого синтезированным геном. Этот способ относится к модификации последовательностей структурного гена экзогенного трансгена, чтобы сделать их более "растение-подобными" и, следовательно, имеющими большую вероятность быть транслированными и экспрессируемыми растением, однодольным или двудольным. Однако способ, описанный в патенте США 5689052, обеспечивает усиленную экспрессию трансгенов, предпочтительно в однодольных растениях.
4.9 Получение устойчивых к насекомым трансгенных растений
Таким образом, количество гена, кодирующего представляющий интерес полипептид (т.е. бактериальный кристаллический белок или полипептид δ-эндотоксина и пептид нацеливания на пластиды) может быть увеличено в растениях трансформацией этих растений с использованием способов трансформации, таких как описанные в разделе 4.7. В частности, трансформация хлоропластов или пластид может приводить к желательным кодирующим последовательностям, которые присутствуют в количестве до приблизительно 10000 копий на клетку в ткани, содержащей эти субклеточные структуры-органеллы (McBride et al., Bio/Technology 13:362-365, 1995).
ДНК может быть также введена в растения прямым переносом ДНК в пыльцу, как описано (Zhou et al., 1983; Hess, 1987). Экспрессия кодирующих полипептид генов может быть получена инъекцией ДНК в репродуктивные органы, как описано (Реnа et al., 1987). ДНК может быть также введена инъекцией непосредственно в клетки незрелых зародышей и регидратацией высушенных зародышей, как описано (Neuhaus et al., 1987; Benbrook et al., 1986).
4.9.1 Отбор трансформированных клеток
После выполнения доставки экзогенной ДНК в реципиентные клетки следующая стадия получения трансгенного растения обычно состоит в идентификации трансформированных клеток для дальнейшего культивирования и регенерации растений. Как упоминалось здесь, для улучшения способности идентифицировать трансформанты может быть желательным применение селектируемого или скринируемого гена в качестве экспрессируемого гена или в дополнение к экспрессируемому представляющему интерес гену. В этом случае обычно можно было бы затем анализировать популяцию потенциально трансформированных клеток экспонированием клеток селективному агенту или агентам или скринировать эти клетки на желаемый признак маркерного гена.
Пример варианта способов для идентификации трансформированных клеток включает в себя экспонирование трансформированных клеток селективному агенту, такому как метаболический ингибитор, антибиотик, гербицид или т.п. Клетки, которые были трансформированы и имеют стабильно интегрированный маркерный ген, сообщающий устойчивость к данному селективному агенту, будут расти и делиться в культуре. Чувствительные клетки не будут поддаваться дальнейшему культивированию. Один пример предпочтительного маркерного гена придает устойчивость к глифозату. При использовании этого гена в качестве селектируемого маркера предположительно трансформированные клетки обрабатывают глифозатом. При обработке трансгенные клетки будут доступны для дальнейшего культивирования, тогда как чувствительные, или не-трансформированные клетки не будут пригодны для культивирования. Этот способ описан подробно в патенте США 5569834, специально включенном здесь в качестве ссылки. Другим примером предпочтительной селектируемой маркерной системы является система устойчивости к неомицинфосфотрансферазе (nptII), при помощи которой сообщается устойчивость к антибиотику канамицину, как описано в патенте США 5569834 (специально включенном здесь в качестве ссылки). Опять после трансформации с этой системой трансформированные клетки будут доступны для дальнейшего культивирования при обработке канамицином, тогда как не-трансформированные клетки не будут пригодны для культивирования. Еще одна предпочтительная система селектируемого маркера включает в себя использование генной конструкции, придающей устойчивость к паромомицину. Применение этого типа системы селектируемого маркера описано в патенте США 5424412 (специально включенном здесь в качестве ссылки).
Все рассматриваемые анализы являются недеструктивными, и трансформированные клетки могут культивироваться далее после идентификации. Другой скринируемый маркер, который может быть использован, представляет собой ген, кодирующий зеленый флуоресцентный белок.
Транспластономический отбор (отбор трансформационных событий пластид или хлоропластов) является упрощенным благодаря использованию преимущественной чувствительности хлоропластов или пластид к спектиномицину, ингибитору белкового синтеза пластид и хлоропластов, но не белкового синтеза кодируемыми ядерным геномом цитоплазматическими рибосомами. Спектиномицин предотвращает накопление хлоропластных белков, требующихся для фотосинтеза, и в результате устойчивые к спектиномицину трансформированные клетки растения могут быть отличены на основе их различия в цвете: устойчивые, трансформированные клетки являются зелеными, тогда как чувствительные клетки являются белыми вследствие ингибирования пластидного белкового синтеза. Трансформация хлоропластов или пластид подходящим бактериальным геном aad, или геном, кодирующим устойчивую к спектиномицину функциональную в пластиде или хлоропласте рибосомальную РНК, обеспечивает средство для отбора и поддержания транспластономических событий (Maliga, Trends in Biotechnology 11:101-106, 1993).
Далее, предполагается, что комбинации скринируемых и селектируемых маркеров будут применимы для идентификации трансформированных клеток. В некоторых типах клеток или тканей селективный агент, такой как глифозат или канамицин, может либо не обеспечивать достаточную убивающую активность для ясного узнавания трансформированных клеток, либо может индуцировать существенное неселективное ингибирование как трансформантов, так и равным образом не-трансформантов, делая этот способ отбора неэффективным. Предполагается, что отбор с использованием ингибирующего рост соединения, такого как глифозат, при концентрациях ниже концентраций, вызывающих 100%-ное ингибирование, с последующим скринингом растущей ткани на экспрессию скринируемого маркерного гена, такого как ген устойчивости к канамицину, позволило бы извлекать трансформанты из типов клеток и тканей, которые не поддаются только отбору. Предполагается, что комбинации отбора и скрининга могут сделать возможной идентификацию трансформантов в широком разнообразии типов клеток и тканей.
4.9.2 Регенерация трансформантов
Развитие или регенерация растений либо из отдельных протопластов растений, либо из различных эксплантатов хорошо известны в данной области (Weissbach and Weissbach, 1988). Этот процесс регенерации и роста обычно включает в себя стадии отбора трансформированных клеток, культивирования индивидуализованных клеток посредством обычных стадий развития эмбрионов через стадию укорененного проростка. Трансгенные зародыши и семена регенерируют подобным образом. Полученные трансгенные укорененные побеги высаживают после этого в подходящую среду для выращивания растений, такую как почва.
Развитие или регенерация растений, содержащих чужеродный экзогенный ген, кодирующий представляющий интерес пептид, введенный с использованием Agrobacterium, из листовых эксплантатов может быть достигнуто способами, хорошо известными в данной области, такими как описанные (Horsch et al., 1985). В этой процедуре трансформанты культивируют в присутствии агента селекции и в среде, которая индуцирует регенерацию побегов в разновидности трансформируемого растения, как описано (Fraley et al., 1983). В частности, патент США 5349124 (специально включенный здесь в качестве ссылки) описывает подробно создание генетически трансформированных клеток латука и происходящих из них растений, которые экспрессируют гибридные кристаллические белки, придающие инсектицидную активность против личинок чешуекрылых насекомых таким растениям.
Эта процедура обычно производит побеги в пределах двух-четырех месяцев, и затем эти побеги переносят на подходящую индуцирующую корнеобразование среду, содержащую селективный агент и антибиотик для предотвращения бактериального роста. Побеги, которые укореняются в присутствии селективного агента с образованием проростков, затем переносят в почву или другие среды, чтобы сделать возможным образование корней. Эти процедуры варьируют в зависимости от конкретного используемого вида растения, причем такие вариации хорошо известны в данной области.
Предпочтительно регенерированные растения самоопыляются, давая гомозиготные трансгенные растения, или пыльцу, полученную из регенерированных растений, скрещивают с выращенными из семян растениями агрономически важных, предпочтительно инбредных линий. Наоборот, пыльцу из растений этих важных линий используют для опыления регенерированных растений. Трансгенное растение данного изобретения, содержащее желательный полипептид, культивируют с использованием способов, хорошо известных специалистам в данной области.
Таким образом, трансгенное растение данного изобретения имеет увеличенное количество кодирующего района, кодирующего полипептид δ-эндотоксина В.thuringiensis и пептид пластидного нацеливания. Предпочтительное трансгенное растение является независимым сегрегантом и может передавать этот ген и его активность своему потомству. Более предпочтительное трансгенное растение является гомозиготным в отношении указанного гена и передает этот ген всем из его потомков при половом размножении. Семена из трансгенного растения могут выращиваться в поле или вегетационном домике, и полученные половозрелые трансгенные растения самоопыляются с образованием истинных селекционных растений. Потомство из этих растений становится истинными селекционными линиями, которые оценивают на увеличенную экспрессию трансгена В.thuringiensis.
4.10 Идентификация событий трансгенных растений с устойчивостью к насекомым
Для идентификации трансгенного растения, экспрессирующего высокие уровни представляющего интерес δ-эндотоксина, необходимо подвергнуть скринингу устойчивые к гербициду или антибиотику трансгенные регенерированные растения (генерацию R0) на инсектицидную активность и/или экспрессию представляющего интерес гена. Это может быть выполнено различными способами, хорошо известными специалистам в данной области, в том числе, но не только: 1) получением небольших проб ткани из трансгенного растения R0 и прямым анализом этой ткани на активность против чувствительных насекомых параллельно с тканью, происходящей из неэкспрессирующих растений отрицательного контроля. Например, трансгенные растения R0 кукурузы, экспрессирующие эндотоксины В.thuringiensis, такие как Cry2Ab, могут быть идентифицированы анализом ткани листа, полученной из таких растений, на активность против ЕСВ; 2) анализом белковых экстрактов при помощи твердофазного иммуноферментного анализа (ELISA), специфического для представляющего интерес гена (Cry2Ab); или 3) ПЦРтм с обратной транскриптазой (ОТ-ПЦР) для идентификации событий экспрессии представляющего интерес гена.
4.11 Выделение гомологичного гена и фрагментов гена
Гены и δ-эндотоксины в соответствии с данным изобретением включают в себя не только полноразмерные последовательности, описанные здесь, но также фрагменты этих последовательностей или слитые белки, которые сохраняют характерную инсектицидную активность последовательностей, приведенных здесь конкретно в качестве примеров.
Для лиц с квалификацией в данной области должно быть очевидно, что инсектицидные δ-эндотоксины могут быть идентифицированы и получены несколькими способами. Специфические гены или их части могут быть получены из депозитариев культур или сконструированы синтетически, например, с использованием автоматического синтезатора генов. Вариации этих генов могут быть легко сконструированы с использованием стандартных способов получения точковых мутаций. Фрагменты этих генов могут быть также получены с использованием коммерчески доступных экзонуклеаз или эндонуклеаз в соответствии со стандартными процедурами. Например, ферменты, такие как Ваl31, или сайт-направленный мутагенез могут быть использованы для систематического отрезания нуклеотидов от концов этих генов. Гены, кодирующие активные фрагменты, могут быть также получены с использованием многочисленных других рестрикционных ферментов. Протеазы могут быть использованы для прямого получения активных фрагментов этих δ-эндотоксинов.
Эквивалентные δ-эндотоксины и/или гены, кодирующие эти δ-эндотоксины, могут быть также выделены из штаммов Bacillus и/или библиотек ДНК с использованием обеспеченнного здесь руководства. Например, антитела к описанным и заявленным здесь δ-эндотоксинам могут быть использованы для идентификации и выделения других δ-эндотоксинов из смеси белков. Кокретно, антитела могут быть индуцированы к частям δ-эндотоксинов, которые являются наиболее константными и наиболее отличающимися от других δ-эндотоксинов В.thuringiensis. Затем эти антитела могут быть использованы для специфической идентификации эквивалентных δ-эндотоксинов с характерной инсектицидной активностью посредством иммунопреципитации, ферментного иммуносорбентного анализа (ELISA) или Вестерн-блоттинга.
Дополнительным способом для идентификации δ-эндотоксинов и генов данного изобретения является способ с использованием олигонуклеотидных зондов. Эти зонды являются нуклеотидными последовательностями, имеющими детектируемую метку. Как хорошо известно в данной области, если молекула зонда и проба нуклеиновой кислоты гибридизуются с образованием сильной связи между этими двумя молекулами, то можно резонно предположить, что данный зонд и проба являются по существу идентичными. Детектируемая метка зонда обеспечивает средство для определения известным образом, произошла ли гибридизация. Такой анализ с использованием зондов обеспечивает быстрый способ идентификации генов инсектицидных δ-эндотоксинов данного изобретения.
Нуклеотидные сегменты, которые используют в качестве зондов согласно данному изобретению, могут быть синтезированы при помощи синтезаторов ДНК с использованием стандартных процедур. В применении нуклеотидных сегментов в качестве зондов конкретный зонд метят любой подходящей меткой, известной специалистам в данной области, в том числе радиоактивной или нерадиоактивной метками. Типичные радиоактивные метки включают в себя 32P, 125I, 35S или т.п. Зонд, меченый радиоактивным изотопом, может быть сконструирован из нуклеотидной последовательности, комплементарной пробе ДНК, обычной реакцией ник-трансляции с использованием ДНКазы и полимеразы ДНК. Затем зонд и проба могут быть объединены в растворе буфера для гибридизации и выдержаны при подходящей температуре во время прохождения отжига. После этого мембрану промывают от посторонних материалов, оставляя пробу, и связанные молекулы зонда обычно детектируют и определяют количественно при помощи радиоавтографии и/или жидкостного сцинтилляционного счета.
Нерадиоактивные метки включают в себя, например, лиганды, такие как биотин или тироксин, а также ферменты, такие как гидролазы или пероксидазы, или различные хемилюминесцирующие молекулы, такие как люциферин, или флуоресцентные соединения, такие как флуоресцеин и его производные. Зонд может быть помечен на обоих концах различными типами меток для легкости отделения, например, с использованием изотопной метки на конце, как упомянуто выше, и биотиновой метки на другом конце.
Образование дуплекса и стабильность зависят от существенной комплементарности между двумя цепями гибрида, и, как отмечалось выше, может быть допустима некоторая степень ошибочного спаривания. Таким образом, зонды данного изобретения включают в себя мутации (как одиночные, так и множественные), делеции, инсерции описанных последовательностей и их комбинации, причем указанные мутации, инсерции и делеции позволяют образование стабильных гибридов с представляющим интерес полинуклеотидом-мишенью. Мутации, инсерции и делеции могут быть получены в конкретной полинуклеотидной последовательностии многочисленными путями, способами, известными в настоящее время специалисту с обычной квалификацией в данной области, и, возможно, другими способами, которые могут стать известными в будущем.
Потенциальные вариации в перечисленных зондах ответственны частично за избыточность генетического кода. Вследствие избыточности генетического кода более чем один кодирующий триплет нуклеотидов (кодон) может быть использован для большинства аминокислот, используемых в образовании белков. Таким образом, различные нуклеотидные последовательности могут кодировать конкретную аминокислоту. Таким образом, аминокислотные последовательности δ-эндотоксинов и пептидов В.thuringiensis и пептидов пластидного нацеливания и полинуклеотиды, кодирующие их, могут быть получены с использованием эквивалентных нуклеотидных последовательностей, кодирующих одну и ту же аминокислотную последовательность белка или пептида. Таким образом, данное изобретение включает в себя такие эквивалентные нуклеотидные последовательности. Также инвертированные или комплементарные последовательности являются аспектом данного изобретения и могут быть легко использованы лицом, квалифицированным в данной области. Кроме того, было показано, что белки идентифицированной структуры и функции могут быть сконструированы посредством изменения аминокислотной последовательности, если такие изменения не изменяют вторичную структуру белка (Kaiser and Kezdy, 1984). Таким образом, данное изобретение включает в себя мутанты аминокислотной последовательности, изображенной здесь, которые не изменяют вторичную структуру белка, или, если эта структура изменена, биологическая активность по существу сохраняется. Кроме того, данное изобретение включает в себя также мутанты организмов, являющихся хозяевами всего гена или части гена, кодирующего δ-эндотоксин, и гена, кодирующего пептид пластидного нацеливания, обсуждаемых в данном изобретении. Такие мутанты могут быть получены способами, хорошо известными лицам с квалификацией в данной области. Например, УФ-излучение может быть использовано для получения мутантов организмов-хозяев. Подобным образом, такие мутанты могут включать в себя аспорогенные клетки-хозяева, которые также могут быть получены процедурами, хорошо известными в данной области.
4.12 Сайт-специфический мутагенез
Сайт-специфический мутагенез является способом, применимым в получении индивидуальных пептидов или биологически функциональных эквивалентных белков или пептидов, посредством специфического мутагенеза основной ДНК. Описанный далее способ обеспечивает легкую возможность приготовления вариантов тест-последовательности, например, с учетом одного или нескольких предыдущих рассмотрении, введением одного или нескольких изменений нуклеотидной последовательности в ДНК. Сайт-специфический мутагенез позволяет получить мутанты посредством использования специфических олигонуклеотидных последовательностей, кодирующих последовательность ДНК желаемой мутации, а также достаточное число соседних нуклеотидов, для обеспечения праймерной последовательности достаточного размера и достаточной сложности последовательности для образования стабильного дуплекса на обеих сторонах пересекаемого делеционного сочленения. Обычно предпочтительным является праймер длиной приблизительно 17-25 нуклеотидов, с приблизительно 5-10 остатками на обеих сторонах сочленения изменяемой последовательности.
В общем, способ сайт-специфического мутагенеза хорошо известен в данной области, как видно из примеров многочисленных публикаций. Как должно быть понятно, этот способ обычно использует фаговый вектор, который существует как в одноцепочечной, так и в двухцепочечной форме. Типичные векторы, применимые в сайт-направленном мутагенезе, включают в себя такие векторы, как фаг М13. Эти фаги коммерчески легко доступны и их применение обычно хорошо известно специалистам в данной области. Двухцепочечные плазмиды также рутинно используются в сайт-направленном мутагенезе, что исключает стадию переноса представляющего интерес гена из плазмиды в фаг.
В общем сайт-направленный мутагенез в соответствии с данным изобретением выполняют получением сначала одноцепочечного вектора или расплавлением двух цепей двухцепочечного вектора, который содержит в его последовательности последовательность ДНК, кодирующую желаемый пептид. Олигонуклеотидный праймер, несущий желательную мутированную последовательность, готовят обычно синтетически. Затем этот праймер отжигают с одноцепочечным вектором и подвергают действию полимеризующих ДНК ферментов, таких как фрагмент Кленова полимеразы I E.coli, для завершения синтеза несущей мутацию цепи. Таким образом, образуется гетеродуплекс, в котором одна цепь кодирует исходную немутированную последовательность, а вторая цепь несет желаемую мутацию. Затем этот гетеродуплексный вектор используют для трансформации подходящих клеток, таких как клетки Е.coli, и отбирают клоны, которые включают в себя рекомбинантные векторы, несущие расположенную в них мутированную последовательность.
Получение вариантов последовательностей кодирующих выбранный пептид сегментов ДНК с использованием сайт-направленного мутагенеза обеспечено как средство получения потенциально ценных разновидностей и не предназначено для ограничения, так как существуют другие пути, в которых могут быть получены варианты последовательностей пептидов и кодирующих их последовательностей ДНК. Например, рекомбинантные векторы, кодирующие желательную пептидную последовательность, могут быть обработаны мутагенными агентами, такими как гидроксиламин, с получением вариантов последовательностей. Такие процедуры могут выгодно изменять биохимические и биофизические характеристики белка или способ его действия. Они включают в себя, но не ограничиваются ими: 1) улучшенное образование δ-эндотоксина, 2) улучшенную стабильность белка или уменьшенную деградацию протеазами, 3) улучшенное узнавание мембранного рецептора насекомого и связывание в ним, 4) улучшенную олигомеризацию или образование каналов в эндотелии средней кишки насекомого и 5) улучшенную инсектицидную активность или инсектицидную специфичность, обусловленную любой или всеми указанными выше причинами.
4.13 Биологические функциональные эквиваленты
Модификация и изменения могут быть сделаны в структуре пептидов данного изобретения и сегментов ДНК, кодирующих их, при получении все еще функциональной молекулы, которая кодирует белок или пептид с желаемыми характеристиками. Биологически функциональные эквивалентные пептиды, полипептиды и белки, обсуждаемые здесь, должны обладать приблизительно 80% или большим сходством последовательности, предпочтительно приблизительно 85% или большим сходством последовательности и наиболее предпочтительно приблизительно 90% или большим сходством последовательности с последовательностью, или соответствующей частью в ней, основной аминокислотной последовательности Cry2Ab.
Далее следует обсуждение, основанное на изменении аминокислот белка, для создания эквивалента или даже улучшенной молекулы второго поколения. В конкретных вариантах данного изобретения предполагается, что мутированные кристаллические белки применимы для увеличения инсектицидной активности данного белка и, следовательно, для увеличения инсектицидной активности и/или экспрессии рекомбинантного трансгена в растительной клетке. Замены аминокислот могут быть получены изменением кодонов последовательности ДНК в соответствии с кодонами, приведенными в таблице 3.
Например, определенные аминокислоты могут быть заменены другими аминокислотами в структуре белка без заметной потери интерактивной способности связывания в такими структурами, как, например, антигенсвязывающие районы антител или сайты связывания на молекулах субстрата. Поскольку именно эта интерактивная способность и природа белка определяет биологическую функциональную активность белка, определенные аминокислотные замены можно произвести в белковой последовательности и, конечно, его основной кодирующей последовательности ДНК, и тем не менее получить белок с такими же свойствами. Таким образом, авторы изобретения считают, что различные замены могут быть сделаны в пептидных последовательностях описанных композиций или соответствующих последовательностях ДНК, которые кодируют указанные пептиды, без ощутимой потери их биологической применимости или активности.
При выполнении таких замен может учитываться гидропатический индекс аминокислот. Важность гидропатического индекса аминокислот в придании интерактивной биологической функции белку обычно признается в данной области (Kyte and Doolittle, 1982, включено здесь в качестве ссылки). Признается, что относительный гидропатический характер аминокислоты вносит вклад во вторичную структуру полученного белка, что, в свою очередь, определяет взаимодействие этого белка с другими молекулами, например ферментами, субстратами, рецепторами, ДНК, антителами, антигенами и т.п.
Каждой аминокислоте был приписан гидропатический индекс на основе ее гидрофобности и характеристик заряда (Kyte and Doolittle, 1982), эти гидропатические индексы таковы: изолейцин (+4,5); валин (+4,2); лейцин (+3,8); фенилаланин (+2,8); цистеин/цистин (+2,5); метионин (+1,9); аланин (+1,8); глицин (-0,4); треонин (-0,7); серин (-0,8); триптофан (-0,9); тирозин (-1,3); пролин (-1,6); гистидин (-3,2); глутамат (-3,5); глутамин (-3,5); аспартат (-3,5); аспарагин (-3,5); лизин (-3,9) и аргинин (-4,5).
Известно в данной области, что определенные аминокислоты можно заменить другими аминокислотами, имеющими такой же гидропатический индекс или балл, и все же получить белок с такой же биологической активностью, т.е. все же получить биологический функционально эквивалентный белок. При проведении таких замен предпочтительна замена аминокислот, гидропатические индексы которых находятся в пределах ±2, особенно предпочтительна замена аминокислот, индексы которых находятся в пределах ±1, и даже более предпочтительна замена аминокислот, индексы которых находятся в пределах ±0,5.
В данной области признается также, что замена похожих аминокислот может выполняться эффективно на основе гидрофильности. В патенте США 4554101, включенном здесь в качестве ссылки, утверждается, что наивысшая локальная средняя гидрофильность белка, определяемая гидрофильностью его смежных аминокислот, коррелирует с биологическим свойством этого белка.
Как описано в патенте 4554101, следующие величины гидрофильности были приписаны аминокислотным остаткам: аргинин (+3,0); лизин (+3,0); аспартат (+3,0±1); глутамат (+3,0±1); серин (+0,3); аспарагин (+0,2); глутамин (+0,2); глицин (0); треонин (-0,4); пролин (-0,5±1); аланин (-0,5); гистидин (-0,5); цистеин (-1,0); метионин (-1,3); валин (-1,5); лейцин (-1,8); изолейцин (-1,8); тирозин (-2,3); фенилаланин (-2,5); триптофан (-3,4).
Понятно, что аминокислоту можно заменить другой, имеющей такую же величину гидрофильности, и все же получить биологически эквивалентный и, в частности, иммунологически эквивалентный белок. В случае таких замен предпочтительна замена аминокислот, величины гидрофильности которых находятся в пределах ±2, особенно предпочтительна замена аминокислот, величины гидрофильности которых находятся в пределах ±1, и даже более предпочтительна замена аминокислот, величины гидрофильности которых находятся в пределах ±0,5.
Таким образом, как указано выше, аминокислотные замены обычно основаны на относительном сходстве заместителей боковых цепей аминокислот, например их гидрофобности, гидрофильности, заряде, размере и т.п. Примеры замен, которые учитывают различные из предыдущих характеристик, хорошо известны специалистам в данной области и включают в себя: аргинин и лизин; глутамат и аспартат; серин и треонин; глутамин и аспарагин; валин, лейцин и изолейцин.
Специалистам в данной области известно, что полинуклеотиды, кодирующие δ-эндотоксины, произведенные из В.thuringiensis, слабо экспрессируются при включении в ядерную ДНК трансгенных растений (обзор Diehn et al., 1996). Предпочтительно нуклеотидную последовательность, кодирующую представляющий интерес δ-эндотоксин, конструируют по существу, как описано в патенте США 5500365 и патенте США 5689052 (каждый из которых включен здесь специально в качестве ссылки). Примеры нуклеотидных последовательностей, применимых для экспрессии, включают в себя, но не ограничиваются ею, Cry2Ab (SEQ ID NO:1).
Пептиды, полипептиды и белки, биологически функционально эквивалентные Cry2Ab, включают в себя аминокислотные последовательности, содержащие консервативные замены аминокислот в основной последовательности, показанной в SEQ ID NO:2. В таких аминокислотных последовательностях одна или несколько аминокислот в основной последовательности заменена (заменены) другой аминокислотой (другими аминокислотами), заряд и полярность которых являются одинаковыми с зарядом и полярностью нативной аминокислоты, т.е. произведена консервативная замена аминокислот, приводящая к молчащему изменению.
Замены для аминокислоты в основной полипептидной последовательности могут быть выбраны из других членов класса, к которому принадлежит природно встречающаяся аминокислота. Аминокислоты могут быть разделены на следующие четыре группы: (1) кислые аминокислоты; (2) основные аминокислоты; (3) нейтральные полярные аминокислоты и (4) нейтральные неполярные аминокислоты. Характерные аминокислоты в этих различных группах включают в себя, но не ограничиваются ими: (1) кислые (отрицательно заряженные) аминокислоты, такие как аспарагиновая кислота и глутаминовая кислота; (2) основные (положительно заряженные) аминокислоты, такие как аргинин, гистидин и лизин; (3) нейтральные полярные аминокислоты, такие как глицин, серин, треонин, цистеин, цистин, тирозин, аспарагин и глутамин; (4) нейтральные неполярные (гидрофобные) аминокислоты, такие как аланин, лейцин, изолейцин, валин, пролин, фенилаланин, триптофан и метионин.
Консервативные аминокислотные замены в основной полипептидной последовательности могут быть выполнены заменой одной аминокислоты в одной из этих групп другой аминокислотой в той же самой группе. Биологически функциональные эквиваленты Cry2Ab могут иметь 10 или менее консервативных аминокислотных замен, более предпочтительно семь или менее консервативных аминокислотных замен и наиболее предпочтительно пять или менее консервативных аминокислотных замен. Таким образом, кодирующая нуклеотидная последовательность (ген, плазмидная ДНК, кДНК или синтетическая ДНК) будет иметь соответствующие замены оснований, позволяющие ей кодировать биологически функциональные эквивалентные формы Cry2Ab.
5.0 Примеры
Следующие далее примеры включены для демонстрации предпочтительных вариантов данного изобретения. Специалистам в данной области должно быть понятно, что способы, описанные в нижеследующих примерах, представляют способы, которые описаны автором изобретения как функционирующие хорошо в применении на практике данного изобретения, и, следовательно, могут рассматриваться как предпочтительные способы его применения. Однако специалисты в данной области должны понимать в свете данного описания, что многочисленные изменения могут быть сделаны в конкретных описанных здесь вариантах, и при этом можно получать такой же или сходный результат без отклонения от идеи и сферы действия данного изобретения.
5.1 Пример 1 - Увеличенная экспрессия Cry2Ab нацеленными векторами
Экспрессию белка Cry2Ab в растениях кукурузы, трансформированных нацеленными и ненацеленными экспрессирующими Cry2Ab векторами, сравнивали, и она была значимо более высокой в растениях с нацеленным вектором. Ненацеленные растительные экспрессирующие векторы Cry2Ab pMON26800 и pMON30463 содержат экспрессионную кассету, составленную из усиленного промотора CaMV35S, интрона hsp70 кукурузы, синтетического гена Cry2Ab с кодонами инициации и терминации трансляции (SEQ ID NO:1), и сайт полиаденилирования нопалинсинтазы.
Нацеленный растительный экспрессирующий вектор pMON30464 (SEQ ID NO:16) содержит экспрессионную кассету, включающую в себя усиленный промотор CaMV35S, интрон hsp70 кукурузы, хлоропластный транзитный пептид ssRUBISCO кукурузы (SEQ ID NO:3), слитый в рамке считывания с синтетическим геном Cry2Ab, и сайт полиаденилирования нопалинсинтазы.
Все векторы (pMON26800, pMON30463 и pMON30464) содержат также кассету, сообщающую устойчивость к паромомицину трансформированной ткани растения. В случае pMON26800 эта кассета состоит из усиленного промотора CaMV35S, интрона hsp70 кукурузы, гена неомицинфосфотрансферазы с кодонами инициации и терминации трансляции, и сайта полиаденилирования нопалинсинтазы. В случае pMON30463 и pMON30464 эта кассета состоит из промотора CaMV35S, гена неомицинфосфотрансферазы с кодонами инициации и терминации трансляции, и сайта полиаденилирования нопалинсинтазы. Трансгенные растения кукурузы, устойчивые к паромомицину, получали по существу, как описано в патенте США 5424412 (специально включенном здесь в качестве ссылки).
Ткань листа из независимо трансформированных трансгенных событий в стадии R0 подвергали количественному анализу на уровни белка Cry2Ab при помощи количественного теста ELISA. В этом ELISA применяли прямой сэндвич-тест, в котором использовали моноклональное улавливающее антитело, индуцированное против Cry2Ab, другое моноклональное антитело против Cry2Aа, конъюгированное со щелочной фосфатазой, в качестве вторичного антитела и очищенный белок Cry2Aа в качестве стандарта.
Сравнение уровней экспрессии Cry2Ab в содержащих pMON30463 (ненацеленный) и pMON30464 (нацеленный) растениях кукурузы показывает, что экспрессия ненацеленного Cry2Ab не превышает 15 м.д., тогда как нацеленная экспрессия часто превышает 100 м.д. (таблица 4). Блот-анализы белка подтверждают, что увеличенный уровень перекрестно-реактивного материала, продуцируемого pMON30464 (нацеленным) обусловлен увеличенным накоплением белка приблизительно Mr 71000, который ко-мигрирует с Cry2Ab, продуцируемым pMON30463 (ненацеленным) и стандартом Cry2Aа из В.thuringiensis. Эти данные показывают, что нацеливающий пептид, слитый с N-концом белка Cry2Ab, эффективно процессировался или удалялся.
Увеличенную экспрессию Сry2Ab в векторах pMON30464 (нацеленных) относительно векторов pMON26800 (ненацеленных) наблюдали также в растениях потомства R1, полученных из исходных трансгенных событий R0, что указывает на то, что высокая экспрессия является наследуемой (таблица 5).

Для эффективной борьбы с насекомыми, которые питаются на различных тканях кукурузы, решающим является то, что инсектицидный белок должен экспрессироваться при высоких уровнях на всех потенциальных участках питания. Для определения, имеют ли место увеличения нацеленной экспрессии Cry2Ab в других тканях, независимые нацеленные и ненацеленные трансгенные события, представляющие линии с высокой экспрессией, полученные с соответствующими типами векторов, анализировали параллельно на уровни экспрессии Cry2Ab. Экспрессия Cry2Ab является увеличенной практически во всех тканях кукурузы, атакуемых вредителями, такими как Ostrina nubialis и Helicoverpa zea, посредством нацеленной экспрессии (таблица 6). Однородно высокий уровень экспрессии этого типа является особенно ценным потому, что маловероятно, что он допустит развитие устойчивости вредителей-мишеней через поведенческую (питание) адаптацию.

Дополнительные анализы показывают, что увеличенные уровни белка Cry2Ab, продуцируемые pMON30464, приводят к соответствующему увеличению уровня биоактивности, как измерено непосредственно в тестах с кормлением. Для оценки уровня продуцируемой инсектицидной активности ткань листьев кукурузы из контрольных (не-трансгенных), “нацеленных” (pMON30464) и “ненацеленных” (pMON30463) растений анализировали на активность против Heliothis virescens в исследованиях с наложением скармливаемой ткани (таблица 7). Две концентрации ткани (0,0016 и 0,0031%) подвергали биоанализу и одну и ту же пробу ткани, используемую в наложении корма, подвергали количественным ELISA-определениям уровней Cry2Ab. Увеличение в 7,5 раз уровней Cry2Ab в нацеленных (pMON30464) пробах относительно ненацеленных (pMON30463) проб явно коррелирует с соответствующим различием в 6 раз в среднем весе личинок, наблюдаемое при обоих уровнях концентраций. Таким образом, эти данные показывают, что увеличенные уровни Cry2Ab, продуцируемые pMON30464, приводят к соответствующим увеличениям уровней биоактивности.
5.2 Пластидное нацеливание Cry2Ab увеличивает частоту агрономически нормальных растений, полученных из трансформации
Для получения коммерчески жизнеспособного признака борьбы с насекомыми на основе трансгена решающим является получение события с нормальными характеристиками роста растений. В большинстве случаев вполне большое число независимых трансгенных событий успешно получали в полевых тестах для гарантии того, что событие, которое удовлетворяет всем ключевым критериям (таким как эффективная борьба с насекомыми, нормальное менделевское расщепление трансгена и нормальные характеристики роста или агрономические характеристики), будет идентифицировано. Способы, которые увеличивают частоту, с которой получают нормальные события, явно являются ценными, так как они увеличивают шансы идентификации события, которое может быть переведено на коммерческую основу. Полезно также увеличение размера пула перспективных событий для скрининга путем увеличения процента R0-событий (первично регенерируемых растений) со способностью к оплодотворению. Поскольку трансформация растений в лаборатории является напряженной (трудоемкой), любой способ, который уменьшает число R0-событий, которое должно быть произведено для получения трансгенного события с подходящей производительностью и подходящими характеристиками роста, также является ценным.
Большие популяции независимых инсерционных R0-событий трансгена ненацеленных векторов pMON26800 и pMON30463 и нацеленного вектора pMON30464 были получены и оценены в баллах на фертильность. Наблюдали, что более высокий процент R0-событий, генерируемых с нацеленным вектором, был фертильным (таблица 8). Потомство фертильных R0-событий вводили затем в полевые испытания, где они оценивались на устойчивость в отношении мотылька кукурузного (ЕСВ1) и нормальное расщепление.
Способы для определения оценок ЕСВ1 и величин расщепления были по существу такими, как описанные способы (Armstrong et al., 1995). События, которые удовлетворяли критериям ЕСВ1 и расщепления, оценивали затем на задержку роста и уменьшение высоты растений. В то время как 60% ненацеленных событий проявляли уменьшения высоты растений, только 3% нацеленных событий обнаружили задержку роста (таблица 8). Улучшенная способность к оплодотворению и уменьшенная задержка роста приводили к значительно улучшенному (37% против 8%) выходу ECB1-положительных событий с отсутствием задержки роста с нацеленным вектором Cry2Ab. В целом в 4 раза больше ненацеленных R0-событий должно продуцироваться и подвергаться скринингу для получения того же самого числа нормальных, ЕСВ+R0-событий, которое получено с нацеленным вектором Cry2Ab в исследовании по трансформации.

5.3 Пример 3 - Пластидное нацеливание Cry2Ab увеличивает частоту уничтожения высокого уровня мотылька кукурузного в трансгенной кукурузе
Ранее описанные популяции независимо трансформированных событий, происходящих как из нацеленного (pMON30464), так и ненацеленного (pMON30463 или pMON26800) экспрессирующих векторов Cry2Ab, подвергали также скринингу на устойчивость к заражению второй генерацией мотылька кукурузного (ЕСВ2). Для облегчения этих исследований коммерчески эффективное событие трансгенной кукурузы MON810 (Yieldgardтм), трансформированной геном Cry2Ab, включали в качестве положительного контроля. Эффективность против ЕСВ2 испытывали в полевых тестах по существу, как описано (Armstrong et al., 1995). В полевом испытании 1996 года 18 независимых ненацеленных событий pMON26800 сравнивали с MON810 (Cry1Ab). Из этих 18 событий только одно давало защиту от ЕСВ2, которая была статистически неотличимой от MON810 и была значимо меньше, чем не-трансгенный отрицательный контроль (событие UT1 в таблице 9). В полевом испытании 1997 года 18 независимых нацеленных событий (pMON30464) испытывали параллельно с 3 ненацеленными событиями (1 событие pMON30463 и два события pMON26800, полученные из испытаний 1996 года) и MON810 (таблица 10). Девять из восемнадцати нацеленных pMON30464 событий давали защиту от ЕСВ2, которая была статистически неотличимой от защиты от ЕСВ2, сообщаемой коммерчески эффективным событием Cry1Ab-экспрессирующего MON810 (Yieldgardтм), и все имели значительно меньшее повреждение ЕСВ2, чем не-трансгенный отрицательный контроль (таблица 10).
Эти данные показывают, что абсолютное число и частота коммерчески эффективных Cry2Ab-линий из нацеленного вектора pMON30464 являются гораздо большими, чем полученные из ненацеленного вектора pMON26800. В то время как 9 из 18 нацеленных Cry2Ab-событий (50%) давали защиту от ЕСВ2, которая была статистически неотличимой от коммерческого стандарта MON810 Cry1Ab и была значимо меньше, чем не-трансгенный отрицательный контроль, только 1 из 18 ненацеленных Cry2Ab-событий (6%) обнаруживали защиту от ЕСВ2, которая была статистически неотличимой от коммерческого стандарта MON810 Cry1Ab и была значимо меньше, чем не-трансгенный отрицательный контроль. Превосходство нацеленного экспрессирующего Cry2Ab вектора особенно очевидно, если учесть, что 9 коммерчески эффективных Cry2Ab-событий получали из общего числа 78 положительных в тесте ЕСВ с кормлением листовым диском R0-растений (частота выхода 11,5%), тогда как только 3 коммерчески эффективных Cry2Ab-события получали из общего числа 192 положительных в тесте ЕСВ с кормлением листовым диском R0-растений (частота выхода 1,6%) (Ro-ECB данные из таблицы 6).


5.4 Пример 4 - Пластидное нацеливание белка Cry2Ab приводит к увеличенной экспрессии в трансгенной каллусной ткани хлопчатника
Сравнивали уровни белка Cry2Ab в каллусной ткани хлопчатника, трансформированной нацеленным на пластиды и ненацеленным экспрессирующими Cry2Ab векторами. Уровни Cry2Ab были значимо более высокими в каллусе, который был трансформирован нацеленными на пластиды генами (таблица 11).
Экспрессирующий вектор растений pMON33830 содержал экспрессионную кассету Cry2Ab, состоящую из следующих генетических элементов, функционально связанных для продуцирования функционального белка Cry2Ab в клетках растений: усиленного промотора CaMV35S, 5’-нетранслируемого лидера hsp70 петунии, синтетического гена Cry2Ab с кодоном инициации трансляции (SEQ ID NO:1) и последовательностями терминации транскрипции и полиаденилирования из гена нопалинсинтазы (NOS) A.tumefaciens.
Экспрессирующие векторы растений pMON33827 (SEQ ID NO:13), pMON33828 (SEQ ID NO:14) и pMON33829 (SEQ ID NO:15) содержали экспрессионные кассеты Cry2Ab, похожие на содержащуюся в pMON33830, за исключением того, что в каждой из них отличающаяся последовательность хлоропластного нацеливания была трансляционно слита с N-концом синтетического гена cry2Ab. pMON33827 содержал кодирующую последовательность для РТР1 (SEQ ID NO:5), которая состоит из нацеливающей на хлоропласт последовательности ssRUBISCO (SSU) Arabidopsis thaliana и последовательностей, кодирующих первые 24 аминокислоты белка ssRUBISCO (SSU) (Wong et al., 1992). SEQ ID NO:6 представляет последовательность нацеливающего пептида РТР1. Этот пептид содержит полную нативную нацеливающую последовательность, включающую в себя сайт отщепления пептида пластидного нацеливания вместе с первыми 24 аминокислотами зрелого белка RUBISCO SSU, последовательно связанного с удвоенной последовательностью аминокислот (положения аминокислот 50-57 SEQ ID NO:6), содержащей сайт отщепления пептида пластидного нацеливания RUBISCO SSU (положения аминокислот 80-87 SEQ ID NO:6). Таким образом, РТР1 содержит удвоенный сайт отщепления пептида пластидного нацеливания. Полинуклеотидная кассета, содержащая эту РТР-кодирующую последовательность, связана на ее 3’-конце с сайтом рестрикции NcoI, который делает возможным встраивания кодирующих последовательностей, которые находятся трансляционно в рамке считывания с РТР-кодирующей последовательностью, например последовательностей, которые кодируют Cry2Ab, Cry2Aа, их варианты и другие кодирующие полезные полипептиды последовательности.
pMON33828 содержала кодирующую последовательность для РТР1Δ (SEQ ID NO:7), модификацию РТР1, в которой 24 аминокислоты SSU между двумя сайгами отщепления транзитного пептида были удалены расщеплением рестриктазой SphI, которая разрезает один раз в каждой копии сайта отщепления транзитного пептида, и повторным лигированием, что приводило к присутствию только одной части транзитного пептида РТР1, за которым следовали единственная копия сайта отщепления транзитного пептида и сайт NcoI. Пептидная последовательность для РТР1Δ названа SEQ ID NO:8.
pMON33829 содержала кодирующую последовательность для РТР2 (SEQ ID NO:9), последовательность транзитного пептида из гена EPSP-синтазы Arabidopsis thaliana. Пептидная последовательность для РТР2 была названа SEQ ID NO:10.
Все описанные выше экспрессирующие векторы трансформации растений содержали также кассету селектируемого маркерного гена, которая придает устойчивость к канамицину трансформированным клеткам растений.
Каллусную ткань хлопчатника из 12 случайно выбранных, независимых трансгенных событий из трансформации каждым из pMON33827, pMON33828, pMON33829 и pMON33830 подвергали количественному анализу уровней белка Cry2Ab с использованием количественного ELISA-анализа. В этом ELISA применяли прямой сэндвич-тест, в котором использовали моноклональное улавливающее антитело, индуцированное против Cry2Aа, другое моноклональное антитело против Cry2Aа, конъюгированное со щелочной фосфатазой, в качестве вторичного антитела и очищенный белок Cry2Aа в качестве стандарта. Сравнение уровней экспрессии Cry2Ab в нацеленной и не-нацеленной каллусной ткани показало значимое увеличение экспрессии при включении нацеливающей на хлоропласт последовательности (таблица 11). РТР1Δ давал значимо более высокий средний уровень экспрессии при сравнении с ненацеленным Cry2Ab, как определено с использованием t-критерия (t=2,31, р=0,03). РТР2 обеспечивал значимо более высокую вероятность получения каллусных линий, экспрессирующих более высокие уровни Cry2Ab, как определено с использованием G-критерия (G2/X2=5,6, р=0,02).
5.5 Пример 5 - Нацеливание белка Cry2Aа в пластиды приводит к уменьшенной экспрессии в трансгенной каллусной ткани хлопчатника
В противоположность примеру 4 выше и приводя пример, что увеличение экспрессии, полученной с использованием последовательностей пластидного нацеливания, является специфически для конкретных генов cry, авторы изобретения обнаружили, что те же самые последовательности пластидного нацеливания, описанные выше, РТР1, РТР1А и РТР2, приводили к значимо более низким уровням экспрессии близкородственного гена Cry2Aа в трансгенном каллусе хлопчатника (таблица 12). Экспрессирующий вектор растений pMON33803 содержал экспрессионную кассету Cry2Aа, состоящую из следующих генетических элементов, функционально связанных для продуцирования функционального белка Cry2Aа в клетках растений: промотора FMV35S, 5’-нетранслируемого лидера белка теплового шока HSP70 петунии, синтетического гена cry2Aa (SEQ ID NO:11) с кодоном инициации трансляции и сайтом рестрикции NcoI на 5’-конце и последовательностями терминации транскрипции и полиаденилирования из гена SSU E9 из гороха. Пептидная последовательность для белка Cry2Aa была названа SEQ ID NO:12. pMON33812, pMON33811 и pMON33806 содержали экспрессионные кассеты Cry2Aa, похожие на присутствующую в pMON33803, за исключением того, что в каждом случае отличающаяся последовательность хлоропластного нацеливания (РТР1, РТР1Δ и РТР2 соответственно) была трансляционно слита с N-концом синтетического гена cry2Aa. Все эти векторы содержали также кассету гена селектируемого маркера, придающую устойчивость к глифозату трансформированным клеткам растения.
Каллусную ткань хлопчатника из 10 случайно выбранных, независимых трансгенных событий из трансформаций каждым из pMON33803, pMON33812, pMON33811 и pMON33806 подвергали количественному анализу уровней белка Cry2Aа с использованием количественного ELISA-анализа для Сгу2. Сравнение уровней экспрессии Cry2Aа в нацеленной и ненацеленной каллусной ткани обнаружило значимое уменьшение экспрессии при включении последовательностей хлоропластного нацеливания (таблица 12). Ненацеленный ген Cry2Aа давал уровни экспрессии, которые отличались значимо от получаемых с использованием любого из трех нацеленных генов Cry2Aа, как определено с использованием HSD-критерия Tukey-Kramer (Тьюки-Крамера) (α=0,05).
5.6 Пример 6 - Нацеливание белка Cry2Aа в пластиды приводит к пониженной экспрессии и увеличенной фитотоксичности в трансгенных растениях табака
Трансформированные растения табака генерировали с использованием pMON33803, ненацеленного экспрессирующего Cry2Aа вектора растений и pMON33806, нацеленного на хлоропласт РТР2-Cry2Aа-экспрессирующего вектора растений.
Пробы ткани листьев эквивалентного веса из 48 содержащих pMON33803 и 41 содержащих pMON33806 растений экстрагировали в равных объемах буфера для экстракции и определяли относительные уровни Cry2Aа с использованием количественного ELISA (таблица 13). В этом ELISA применяли прямой сэндвич-тест, в котором использовали поликлональное улавливающее антитело, индуцированное против Cry2Aа, то же самое поликлональное антитело, конъюгированное со щелочной фосфатазой, в качестве вторичного антитела и очищенный белок Cry2Aа в качестве стандарта.
Доля общего числа растений, полученных из трансформации, которые экспрессировали ненацеленный Cry2Aа при высоких уровнях, была выше, чем доля полученных растений, которые экспрессировали нацеленный Cry2Aа при высоких уровнях. Наоборот, доля общего числа растений, полученных из трансформации, которые не экспрессировали детектируемый нацеленный Cry2Aа, была больше, чем доля полученных растений, которые не экспрессировали ненацеленный Cry2Aа. Все РТР2-Cry2Aа-содержащие растения, которые имели детектируемые уровни экспрессии Cry2Aа, проявляли сильно отклоняющийся от нормы фенотип; эти растения были сильно задержанными в росте, имели укороченные междоузлия, имели деформированные, сморщенные листья и были бесплодными. Все РТР2-Cry2Aа-растения, которые не имели экспрессии Cry2Aа, выглядели нормально. В противоположность этому только некоторые из растений, имеющих высокую экспрессию ненацеленного Cry2Aа, проявляли задержанный в росте фенотип.

5.7 Пример 7 - Трансформация хлоропласта табака геном Cry2Ab
Могут быть получены рекомбинантные растения, в которых только митохондриальная или хлоропластная ДНК была изменена включением молекул, рассматриваемых в данной заявке. Промоторы, которые функционируют в хлоропластах, известны в данной области (Hanley-Bowden et al., Trends in Biochemical Sciences 12:67-70, 1987). Способы и композиции для получения клеток, содержащих хлоропласты, в которые была встроена гетерологичная ДНК, были описаны, например, Daniell et al., (патент США № 5693507; 1997) и Maliga et al., (патент США № 5451513; 1995). Может быть сконструирован вектор, который содержит экспрессионную кассету, из которой мог бы быть получен белок Cry2A. Кассета могла бы содержать функциональную в хлоропласте промоторную последовательность, запускающую экспрессию гена кристаллического белка Cry2A, сконструированную очень сходным образом с конструированием других описанных здесь полинуклеотидов, с использованием термических методологий амплификации, расщепления рестриктазами и лигирования и т.д. Экспрессируемый в хлоропласте ген мог бы обеспечить промотор и 5’-нетранслируемый район из гетерологичного гена или хлоропластного гена, такого как psbA, которые обеспечили бы транскрипцию и трансляцию последовательности ДНК, кодирующей белок Cry2A в хлоропласте; последовательность ДНК, кодирующую белок Cry2A; и район терминации транскрипции и трансляции, такой как 3’-инвертированный повторяющийся район хлоропластного гена, который мог бы стабилизировать экспрессируемую мРНК Cry2A. Экспрессия из хлоропласта усилила бы накопление продукта гена Cry2A. Клетка-хозяин, содержащая хлоропласты или пластиды, может быть трансформирована этой экспрессионной кассетой, и затем полученная клетка, содержащая трансформированные хлоропласты, может выращиваться для экспрессии белка Cry2A. Кассета может содержать также ген устойчивости к антибиотику, ген устойчивости к гербициду или ген другого селектируемого маркера, кроме гена Cry2A. Экспрессионная кассета может быть фланкирована последовательностями ДНК, полученными из хлоропластной ДНК, которые могли бы облегчать стабильную интеграцию этой экспрессионной кассеты в хлоропластный геном, в частности гомологичной рекомбинацией. Альтернативно, экспрессионная кассета может не интегрироваться, но посредством включения начала репликации, полученного из хлоропластной ДНК, она могла бы быть способной обеспечивать репликацию гетерологичного гена Cry2A в хлоропласте. Растения могут быть генерированы из клеток, содержащих трансформированные хлоропласты, и могут затем выращиваться для получения семян, из которых могут быть получены дополнительные растения. Такие способы трансформации являются более выгодными, чем трансформация ядерного генома, в частности, когда трансформация хлоропластов выполняется интегрированием в хлоропластный геном, поскольку хлоропластные гены обычно наследуются по материнской линии. Это обеспечивает "более безопасные" трансгенные растения в отношении окружающей среды, фактически исключая возможность ускользания в окружающую среду. Кроме того, хлоропласты могут быть трансформированы многократно для получения функциональных хлоропластных геномов, которые экспрессируют множественные желаемые рекомбинантные белки, тогда как трансформация ядерного генома, как было показано, является скорее лимитированной в случае, когда желательно получение множественных генов. Таким образом, при использовании хлоропластной или пластидной трансформации можно избежать событий расщепления. В противоположность экспрессии растительного ядерного генома экспрессия в хлоропластах или пластидах может инициироваться из только одного промотора и продолжаться через полицистронный район, продуцируя множественные пептиды из единственной мРНК.
Эту экспрессионную кассету можно было бы получить во многом тем же самым путем, что и другие векторы трансформации растений. Функциональные в хлоропластах растений последовательности ДНК могут быть встроены в бактериальную плазмиду и связаны с последовательностями ДНК, экспрессирующими желательные генные продукты, такие как белки Cry2A, так что белок Cry2A продуцируется внутри хлоропласта, что исключает необходимость регуляции ядерных генов, кэпирования, сплайсинга или полиаденилирования регулируемых ядром генов или необходимость нацеливающих на хлоропласт или пластиду последовательностей. Экспрессионная кассета, содержащая ген Cry2A, который является либо синтетически сконструированным, либо нативным геном, полученным непосредственно из генома В.thuringiensis или из эписомного элемента В. thuringiensis, может быть встроена в сайт рестрикции в векторе, сконструированном для цели трансформации хлоропласта или пластиды. Кассета могла бы быть фланкирована выше по ходу транскрипции (слева) функциональным в хлоропласте или пластиде промотором, а ниже по ходу транскрипции (справа) функциональной в хлоропласте или пластиде последовательностью терминации транскрипции и трансляции. Полученная кассета могла бы быть включена в хлоропластный или пластидный геном с использованием хорошо известных способов гомологичной рекомбинации.
Альтернативно, трансформация хлоропластов или пластид могла бы быть достигнута с использованием автономно реплицирующейся плазмиды или другого вектора, способного размножаться в хлоропласте или пластиде. Одним из средств выполнения данного способа было бы использование части хлоропластного или пластидного генома, требующейся для инициации репликации хлоропласта или пластиды, как средства сохранения этих плазмиды или вектора в трансформированных хлоропласте или пластиде. Последовательность, способная к стабильной репликации эпигенетического элемента хлоропласта или пластиды, могла бы быть легко идентифицирована из случайного клонирования хлоропластного или пластидного генома в стандартный бактериальный вектор, который также содержит ген селектируемого маркера хлоропласта или пластиды, с последующей трансформацией хлоропластов или пластид и отбором на трансформированные клетки на подходящей селективной среде. Введение описанной здесь экспрессионной кассеты в эпигенетический элемент хлоропласта или пластиды обеспечило бы эффективное средство для локализации Cry2Aа δ-эндотоксина В.thuringiensis в хлоропласте или пластиде.
6.0 Ссылки
Следующие ссылки в той степени, в которой они обеспечивают примеры процедурных или других подробностей, дополняющих изложенные здесь детали, специально включены здесь в качестве ссылок.
U.S. Patent № 4535060; 1985.
U.S. Patent № 4554101; 1985.
U.S. Patent № 4683195; 1987.
U.S. Patent № 4683202; 1987.
U.S. Patent № 4757011; 1988.
U.S. Patent № 4769061; 1988.
U.S. Patent № 4940835; 1990.
U.S. Patent № 4971908; 1990.
U.S. Patent № 5004863; 1991.
U.S. Patent № 5015580; 1991.
U.S. Patent № 5023179; 1991.
U.S. Patent № 5225341; 1993.
U.S. Patent № 5264364; 1993.
U.S. Patent № 5276269; 1994.
U.S. Patent № 5322687; 1994.
U.S. Patent № 5338544; 1994.
U.S. Patent № 5349124; 1994.
U.S. Patent № 5378619; 1995.
U.S. Patent № 5384253; 1995.
U.S. Patent № 5416011; 1995.
U.S. Patent № 5424412; 1995.
U.S. Patent № 5451513; 1995.
U.S. Patent № 5463175, 1995.
U.S. Patent № 5482852; 1996.
U.S. Patent № 5491288; 1996.
U.S. Patent № 5500365; 1996.
U.S. Patent № 5508468; 1996.
U.S. Patent № 5569834; 1996.
U.S. Patent № 5689052, 1997.
U.S. Patent № 5693507; 1997.
EРО 0120516.
WO 92/17591.
WO 95/24492.
Armstrong et al., Plant Cell Rep., 9:335-339, 1990.
Armstrong et al., Crop Science, 35(2):550-557, 1995.
Barton et al., Plant Physiol., 85:1103-1109, 1987.
Benfey et al., EMBO J., 8:2195-2202, 1989.
Bevan et al., Nature, 304:184, 1983.
Callis et al., Genes Dev., 1:1183-1200; 1987.
Cheng et al., Proc. Natl. Acad. Sci. USA, 96(6):2767-2772, 1998.
Dhir et al., Plant Cell Rep., 10: 106-10; 1991.
Diehn et al., In: Genetic Engineering, Ed. J.K. Setlow, Plenum Press, New York, NY, 18:83-99, 1996.
Donovan et al., Mol. Gen. Genet., 214:365-372, 1988.
Donovan et al., Appl. Environ. Microbiol., 58:3921-3927, 1992.
English et al., Insect. Biochem. Molec. Bio., 24(10):1025-1026, (1994).
Feinberg and Vogelstein, Anal. Biochem., 132:6-13, 1983.
Fischhoff et al., Bio/Technology, 5:807-813, 1987.
Frischauf et al., Methods Enzymol., 153:103-115, 1987.
Fromm et al., Nature, 319:791-793, 1986.
Fromm et al., Bio/Technology, 8:833-839, 1990.
Gould et al., J. Cell Biol., 105:2923-2931, 1987.
Hanley-Bowden et al., Trends in Biochemical Sciences 12:67-70; 1987.
Herrera-Estrella et al., Nature, 303:209, 1983.
Hertig et al., Plant Mol. Biol., 16:171-174, 1991.
Hofte and Whitetey, Microbiol. Rev., 53(2):242-255, 1989.
Horsch et al., Science, 227:1229-1231, 1985.
Horton et al., Gene, 77:61-68, 1989.
Ishida et al., Nas. Biotechnol., 14(6):745-750, 1996.
Kay et al., Science, 236:1299-1302, 1987.
Keegstra and Olsen, Ann. Rev. Plant Physiol. Mol. Biol., 40:471-501, 1989.
Klee et al., Bio/Technology, 3:637-642, 1985.
Klee et al., Mol. Gen. Genet., 210:437-442, 1987.
Koehler and Ho, Plant Cell, 2:769-783, 1990.
Koziel et al., Bio/Technology, 11:194-200, 1993.
Lambert et al., Appl. Environ. Microbiol, 62:80-86, 1996.
Lee et al., Science, 239:1288-1291, 1988.
Lindstrom et al., Dev. Genet., 11:160-7; 1990.
Macejak and Sarnow, Nature, 353:90-94, 1991.
MacIntosh et al., J. Invert. Pathol., 56:258-266, 1990.
Maliga, Trends in Biotechnology, 11:101-106, 1993.
McElroy et al., Plant Cell, 2:163-171, 1990.
McGaughey and Whalon, Science, 258:1451-1455, 1993.
Nayak et al., Proc. Natl. Acad. Sci. USA, 94(6):2111-2116, 1997.
Odell et al., Nature, 313: 810-12; 1985.
Pelletier and Sonenberg, Nature, 334:320-325, 1988.
Perlak et al., Bio/technology, 8:939-943, 1990.
Perlak et al., Plant Molecular Biol., 22:313-321, 1993.
Roush, Biocontrol Sci. Technol., 4:501-516, 1994.
Russetl et al., Plant Cell Reports, 13:24-27, 1993.
Shelton et al., J. Econ. Entomol., 86:697-705, 1993.
Tang et al., Appl. Environ. Microbiol., 62:564-569, 1996.
Tillmann et al., EMBO J., 8(9):2463-2467, 1989.
Vaeck et al., Nature, 328:33-37, 1987.
Vasil et al., Plant Physiol., 91:1575-1579; 1989.
Vodkin et al., Cell, 34:1023-1031, 1983.
Widner et al., J. Bacteriol., 171:965-974; 1989.
Widner et al., (a), J. Bacteriol., 172:2826-2832; 1990.
Winter et al., Mol. Gen. Genet., 221(2):315-319, 1988.
Wong et al., Plant Molec. Biol., 20:81-93, 1992.
Xu et al., Plant Mol. Biol., 27:237-248, 1995.
Yamamoto et al., Plant Cell :371-382, 1991.
Все композиции и способы, описанные и заявленные здесь, могут быть приготовлены и осуществлены без чрезмерного экспериментирования в свете данного описания. Хотя композиции и способы данного изобретения были описаны в виде предпочтительных вариантов, специалистам в данной области должно быть ясно, что могут быть применены вариации к этим композициям и способам и в стадиях или последовательности стадий описанного здесь способа без отклонения от концепции, идеи и сферы действия данного изобретения. Более конкретно, должно быть очевидно, что некоторые агенты, которые являются как химически, так и физиологически родственными, могут быть применены вместо описанных здесь агентов с получением тех же самых или сходных результатов. Следует считать, что все такие сходные замены и модификации, очевидные для специалистов в данной области, находятся в рамках идеи, объема и концепции данного изобретения, определяемых прилагаемой формулой изобретения.
Список последовательностей приведен после формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СИНТЕТИЧЕСКИЕ ТРАНЗИТНЫЕ ПЕПТИДЫ ХЛОРОПЛАСТА ИЗ BRASSICA | 2013 |
|
RU2636035C2 |
| НОВЫЕ БЕЛКИ, ПРОЯВЛЯЮЩИЕ ИНГИБИРУЮЩУЮ АКТИВНОСТЬ ПО ОТНОШЕНИЮ К НАСЕКОМЫМ | 2016 |
|
RU2765310C2 |
| СИНТЕТИЧЕСКИЕ ПРОИСХОДЯЩИЕ ИЗ BRASSICA ТРАНЗИТНЫЕ ПЕПТИДЫ ХЛОРОПЛАСТОВ | 2013 |
|
RU2626535C2 |
| ТОКСИН AXMI277 ПРОТИВ НЕМАТОД И СПОСОБЫ ЕГО ПРИМЕНЕНИЯ | 2012 |
|
RU2706488C2 |
| СИНТЕТИЧЕСКИЕ ТРАНЗИТНЫЕ ПЕПТИДЫ ХЛОРОПЛАСТОВ | 2013 |
|
RU2632648C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ, ВЫДЕЛЕННЫЕ ИЗ ВИДОВ БАКТЕРИЙ Bacillus, И ИХ ПРИМЕНЕНИЕ | 2004 |
|
RU2382822C2 |
| КОМПОЗИЦИИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2016 |
|
RU2759224C2 |
| ГЕН АХМ1554, КОДИРУЮЩИЙ ДЕЛЬТА-ЭНДОТОКСИН, И СПОСОБЫ ЕГО ПРИМЕНЕНИЯ | 2016 |
|
RU2755282C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ | 2010 |
|
RU2613778C2 |
| УНИВЕРСАЛЬНЫЙ ВЕКТОР ДЛЯ СТАБИЛЬНОЙ ТРАНСФОРМАЦИИ ХЛОРОПЛАСТНОГО ГЕНОМА, СПОСОБ СТАБИЛЬНОЙ ТРАНСФОРМАЦИИ РАСТЕНИЯ-МИШЕНИ, СПОСОБ ПРИДАНИЯ УСТОЙЧИВОСТИ К ГЕРБИЦИДУ РАСТЕНИЮ-МИШЕНИ, СПОСОБ ОПРЕДЕЛЕНИЯ ТРАНСФОРМАЦИИ ХЛОРОПЛАСТА И СТАБИЛЬНО ТРАНСФОРМИРОВАННЫЙ ХЛОРОПЛАСТНЫЙ ГЕНОМ | 1998 |
|
RU2238324C2 |



















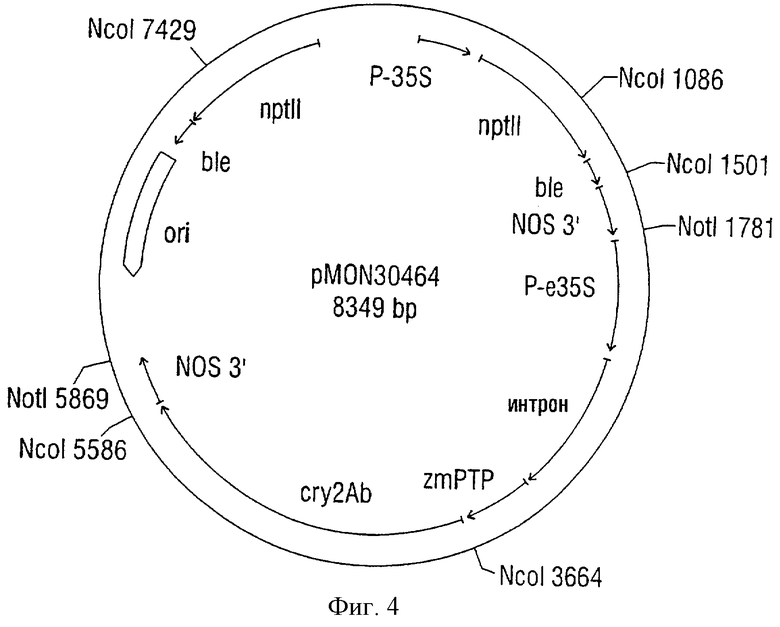
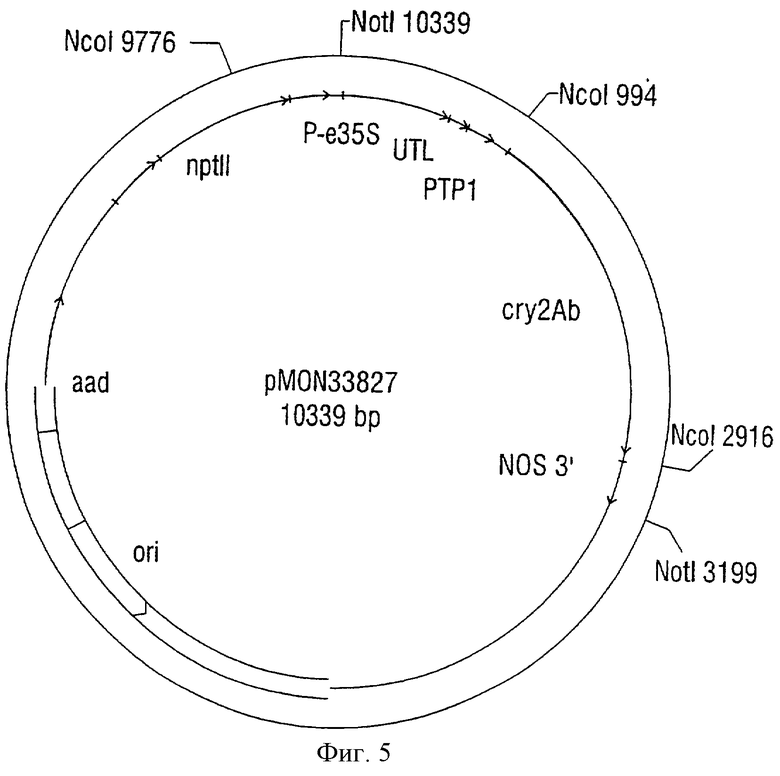
Изобретение относится к биотехнологии, в частности к созданию трансгенных растений, обладающих инсектицидными свойствами. Выделяют последовательность нуклеиновой кислоты (варианты), содержащую промотор, функционально связанный с первой полинуклеотидной последовательностью, кодирующей пластидный транзитный пептид, которая в свою очередь связана в рамке считывания со второй полинуклеотидной последовательностью, кодирующей белок Cry2Ab δ-эндотоксина Bacillus thuringiensis, причем экспрессия указанной последовательности нуклеиновой кислоты растительной клеткой позволяет получить слитый белок, содержащий аминоконцевой пластидный транзитный пептид, ковалентно связанный с указанным белком δ-эндотоксина, и причем слитый белок действует таким образом, что локализует экспрессию белка δ-эндотоксина в субклеточной органелле или субклеточном компартменте. Вводят в геном растения вышеуказанную последовательность нуклеиновой кислоты. Регенерируют растение из ткани растения, трансформированной вышеуказанной последовательностью нуклеиновой кислоты. Скрещивают два полученных трансгенных растения между собой с получением растения-потомка. Описанное изобретение позволяет снизить производственные затраты и повысить сбор сельскохозяйственной продукции. 5 с. и 43 з.п. ф-лы, 7 ил., 13 табл.
| US 5338544, 16.08.1994 | |||
| WO 9524492, 14.09.1995 | |||
| СПОСОБ ПОЛУЧЕНИЯ БИСАЗОМЕТИНОВ — ПРОИЗВОДНЫХ ДИФЕНИЛМЕТАНА И ТРИФЕНИЛМЕТАНА | 0 |
|
SU385962A1 |
| US 5500365, 19.03.1996 | |||
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК РВТN II, ОПРЕДЕЛЯЮЩАЯ СИНТЕЗ КРИСТАЛЛИЧЕСКОГО ИНСЕКТИЦИДНОГО ДЕЛЬТА-ЭНДОТОКСИНА CRY IIIA-ТИПА, СПОСОБ ЕЕ КОНСТРУИРОВАНИЯ И ШТАММ БАКТЕРИЙ PSEUDOMONAS PUTIDA, СОДЕРЖАЩИЙ РЕКОМБИНАНТНУЮ ПЛАЗМИДУ ДНК РВТN II-ПРОДУЦЕНТ КРИСТАЛЛИЧЕСКОГО ИНСЕКТИЦИДНОГО ДЕЛЬТА-ЭНДОТОКСИНА CRY IIIA-ТИПА, АКТИВНОГО ПРОТИВ НАСЕКОМЫХ ОТРЯДА COLEOPTERA | 1995 |
|
RU2103363C1 |
