ОБЛАСТЬ ТЕХНИКИ
Настоящая заявка относится к микроорганизму рода Corynebacterium, продуцирующему L-лизин, и способу получения L-лизина с его помощью.
УРОВЕНЬ ТЕХНИКИ
L-лизин биосинтезируется из предшественника оксалоацетата посредством пути биосинтеза лизина и оксалоацетат распадается на пируват и двуокись углерода с помощью оксалоацетат-декарбоксилазы (odx).
Ранее Simon Klaffl и Bernhard J. Eikmanns просто продемонстрировали, что ген cg1458 кодирует оксалоацетат-декарбоксилазу, подтвердив, что сверхэкспрессия внутреннего гена cg1458 в Corynebacterium glutamicum повышает активность оксалоацетат-декарбоксилазы, а также уменьшает получение L-лизина, и активность оксалоацетат-декарбоксилазы уменьшается с помощью инактивации гена, однако они не продемонстрировали изменения при получении L-лизина с помощью инактивации cg1458 (Klaffl S, Eikmanns BJ, et al., J. Bacteriol., May 2010, 192(10), p. 2604-12). Тем не менее, авторы настоящего открытия предсказали, что при ингибировании активности эндогенной оксалоацетат-декарбоксилазы в L-лизин-продуцирующем штамме Corynebacterium увеличивается подвод оксалоацетата для пути биосинтеза L-лизина без потребления оксалоацетата в других путях, и они инактивируют ген оксалоацетат-декарбоксилазы в различных классах L-лизин-продуцирующих штаммов. В результате авторы настоящего открытия установили, что инактивация гена оксалоацетат-декарбоксилазы приводит к улучшению способности штамма продуцировать L-лизин, осуществляя тем самым настоящую заявку.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
ТЕХНИЧЕСКАЯ ЗАДАЧА
Соответственно целью настоящей заявки является обеспечение микроорганизма рода Corynebacterium, продуцирующего L-лизин.
Другой целью настоящей заявки является обеспечение способа получения L-лизина с помощью микроорганизма рода Corynebacterium.
ТЕХНИЧЕСКОЕ РЕШЕНИЕ
Для достижения вышеуказанных целей, один из аспектов настоящей заявки предусматривает микроорганизм рода Corynebacterium, продуцирующий L-лизин, в котором активность оксалоацетат-декарбоксилазы инактивирована.
Дополнительно, другой аспект настоящей заявки предусматривает способ получения L-лизина с помощью культивирования микроорганизма рода Corynebacterium.
ПРЕИМУЩЕСТВЕННЫЕ ЭФФЕКТЫ ИЗОБРЕТЕНИЯ
Настоящая заявка предусматривает рекомбинантный микроорганизм рода Corynebacterium с повышенным продуцированием L-лизина с помощью инактивации гена оксалоацетат-декарбоксилазы в L-лизин-продуцирующем штамме Corynebacterium. Рекомбинантный микроорганизм рода Corynebacterium способен продуцировать L-лизин с высоким выходом, вследствие чего имеет промышленно полезное применение.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
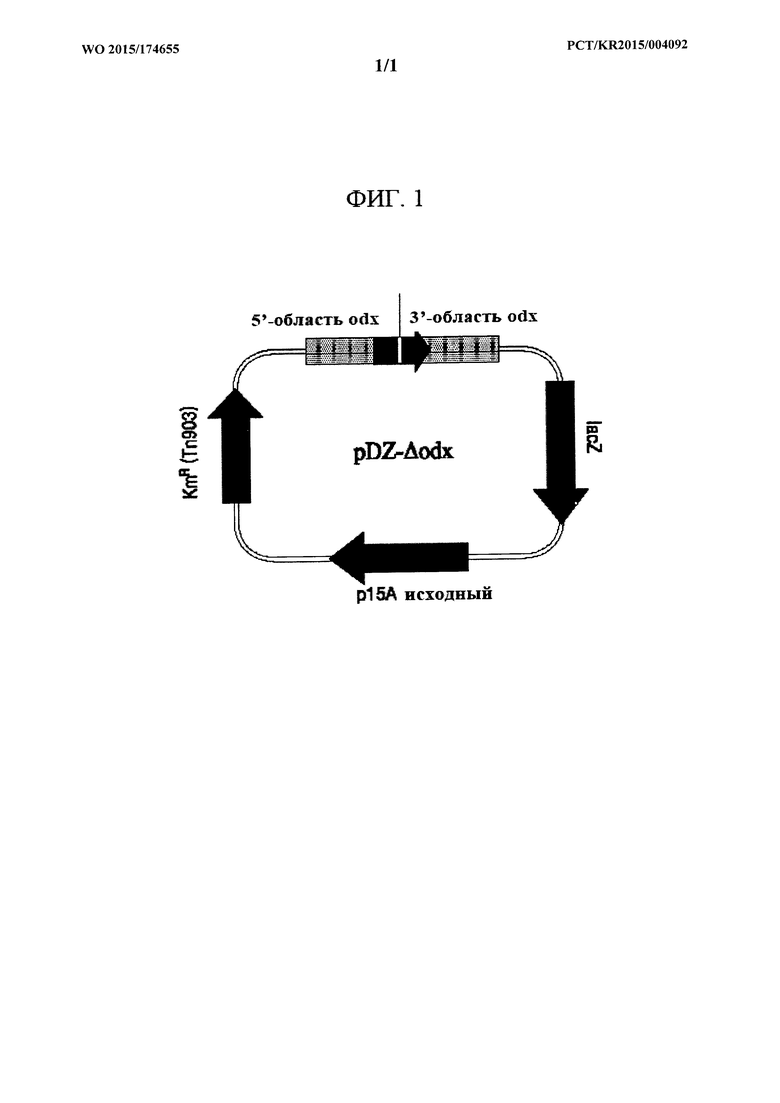
На ФИГ. 1 показан вектор pDZ-Δodx для удаления гена cdx в хромосоме Corynebacterium.
ПРИНЦИП ИЗОБРЕТЕНИЯ
Далее в данном документе настоящая заявка будет описана более подробно.
Один из аспектов настоящей заявки предусматривает микроорганизм рода Corynebacterium с повышенным продуцированием L-лизина с помощью инактивации оксалоацетат-декарбоксилазной активности.
Оксалоацетат-декарбоксилаза (odx) представляет собой фермент, который катализирует распад оксалоацетата на пируват и диоксид углерода, а также можно использовать фермент, полученный из любого микроорганизма, при условии, что фермент обладает вышеуказанной активностью. В частности, можно использовать фермент, полученный из микроорганизма Coryneform, а более конкретно фермент, полученный из Corynebacterium glutamicum, но не ограничиваясь ними.
Более конкретно, оксалоацетат-декарбоксилаза по настоящей заявке может иметь аминокислотную последовательность под SEQ ID NO: 1 и может быть закодирована геном odx, имеющим нуклеотидную последовательность под SEQ ID NO: 2, но не ограничиваться аминокислотной последовательностью или нуклеотидной последовательностью. В частности, белок с гомологией 80% или выше, а конкретно 90% или выше, 95% или выше, и более конкретно 97% или выше, с аминокислотной последовательностью под SEQ ID NO: 1 также включен в объем настоящей заявки при условии, что белок обладает оксалоацетат-декарбоксилазной активностью. Очевидно, что любая аминокислотная последовательность с делецией, модификацией, заменой или добавлением какой-нибудь последовательности может быть в пределах объема настоящей заявки при условии, что аминокислотная последовательность в пределах диапазона гомологии обладает биологической активностью, которая по существу идентична или соответствует белку под SEQ ID NO: 1. В настоящую заявку может быть включен также любой вариант вышеуказанных нуклеотидных последовательностей, кодирующих такие же аминокислотные последовательности, что является результатом вырождения генетического кода.
Термин "гомология", используемый в данном документе, относится к степени совпадения с данной аминокислотной последовательностью или нуклеотидной последовательностью, и гомология может быть выражена в виде процентов. В настоящей заявке гомологичная последовательность, обладающая активностью, которая является идентичной или аналогичной данной аминокислотной последовательности или нуклеотидной последовательности, выражается в виде "% гомологии". Гомологичную последовательность можно определять с помощью, например, алгоритма BLAST [см. Karlin and Altschul, Pro. Natl. Acad. Sci. USA, 90, 5873(1993)] или FASTA от Pearson [см. Methods Enzymol., 183, 63(1990)]. На основе данного алгоритма была разработана программа под названием BLASTN или BLASTX [см. http://www.ncbi.nlm.nih.gov]. Гомологичную последовательность можно идентифицировать также с помощью сравнения последовательностей в эксперименте Саузерн-гибридизации при жестких условиях, согласно определению, и определение подходящих условий гибридизации известно специалистам в данной области техники (см. например Sambrook et al., 1989, infra.), и можно определять способом, хорошо известным специалистам в данной области техники.
Термин "инактивация", используемый в данном документе, означает, что активность дополнительно ослаблена или выключена, по сравнению с активностью белка, который встречается в естественном состоянии или немодифицированном состоянии исходного микроорганизма, и инактивацию можно выполнять с помощью любого способа инактивации, известного в данной области техники. В настоящей заявке инактивация оксалоацетат-декарбоксилазы означает, что внутренний ген odx экспрессируется на низком уровне или не экспрессируется по сравнению с родительским штаммом или немодифицированным штаммом, или активность выключена или уменьшена, даже если ген экспрессируется.
В частности, внутреннюю активность оксалоацетат-декарбоксилазы можно инактивировать с помощью введения рекомбинантного вектора, содержащего часть гена, которая кодирует оксалоацетат-декарбоксилазу, в микроорганизм для того, чтобы удалить или мутировать внутренний ген оксалоацетат-декарбоксилазы, экспрессию белка можно уменьшать модификацией нуклеотидной последовательности промоторной области или 5'-UTR области гена, либо активность белка можно уменьшать введением мутации в ORF область соответствующего гена. Кроме того, введение гена в хромосому можно осуществлять любым способом, известным в данной области техники.
Термин "внутренняя активность", используемый в данном документе, относится к активности белка исходного микроорганизма в естественном состоянии или немодифицированном состоянии.
Более конкретно, инактивацию можно вызывать сайт-специфичным разрушением гена, но не ограничиваясь ним.
Термин "рекомбинантный вектор", используемый в данном документе, относится к ДНК-конструкции, содержащей последовательность полинуклеотида, кодирующего целевой белок, который функционально связан с соответствующей регуляторной последовательностью таким образом, что целевой белок можно экспрессировать в соответствующем хозяине. Регуляторная последовательность может включать в себя промотор, способный инициировать транскрипцию, любую операторную последовательность для регуляции транскрипции, последовательность, кодирующую подходящий мРНК-сайт связывания рибосомы и последовательность, регулирующую терминацию транскрипции и трансляции. Вектор, после трансформации в подходящую клетку-хозяина, может быть реплицирован, или он может функционировать независимо от генома хозяина, или он может быть интегрирован в сам геном. Вектор, используемый в настоящей заявке, не имеет особых ограничений при условии, что вектор является воспроизводимым в хозяине, и при этом можно использовать любой вектор, известный в данной области техники.
Термин "трансформация", используемый в данном документе, означает, что ген вводят в клетку-хозяина, обеспечивая тем самым возможность экспрессии гена в клетке-хозяине. Трансформированный ген может находится внутри или снаружи хромосомы клетки-хозяина при условии, что его можно экспрессировать в клетке-хозяине. Ген можно вводить в любой форме при условии, что его можно вводить в клетку-хозяина и экспрессировать там.
В качестве родительской клетки, в которую вводят рекомбинантный вектор, можно использовать любой микроорганизм без ограничения, при условии, что микроорганизм способен продуцировать L-лизин. В частности, можно использовать микроорганизм рода Corynebacterium или рода Brevibacterium, а более конкретно Corynebacterium glutamicum, но не ограничиваясь ними.
В конкретном варианте осуществления настоящей заявки рекомбинантный вектор конструировали с помощью вставки части гена odx в вектор pDZ (патент Кореи №0924065), который не реплицируется в Corynebacterium glutamicum, а трансформация рзкомбинантного вектора в микроорганизм и гомологичная рекомбинация рекомбинантного вектора в хромосому давали возможность получить микроорганизм рода Corynebacterium с повышенным продуцированием L-лизина, в котором оксалоацетат-декарбоксилаза инактивирована, однако настоящая заявка этим не ограничивается.
Все штаммы Corynebacterium glutamicum, полученные в варианте осуществления настоящей заявки, продемонстрировали увеличенное продуцирование L-лизина по сравнению с родительским штаммом, указывая, что они способны производить L-лизин ^'высоким выходом с помощью инактивации гена odx.
Другой аспект настоящей заявки предусматривает способ получения L-лизина с помощью культивирования микроорганизма рода Corynebacterium. Более подробно, предусматривается способ получения L-лизина, способ, включающий продуцирование L-лизина в культуре или клетках микроорганизма с помощью культивирования микроорганизма по настоящей заявке; и извлечение L-лизина из культуры или клеток микроорганизма.
В способе по настоящей заявке культивирование микроорганизма рода Corynebacterium можно проводить с помощью любых условий культивирования и способов, известных в данной области техники, не ограничиваясь следующими примерами.
Примером среды, которую можно использовать для культивирования микроорганизма рода Corynebacterium, может быть среда, описанная в Manual of Methods for General Bacteriology от the American Society for Bacteriology (Washington D.C., USA, 1981).
Источники углерода, используемые в среде, могут включать сахариды и углеводы, такие как глюкоза, сахароза, лактоза, фруктоза, мальтоза, крахмал и целлюлоза, масла и липиды, такие как соевое масло, подсолнечное масло, касторовое масло и кокосовое масло, жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота и линолевая кислота, спирты, такие как глицерин и этанол, а также органичные кислоты, такие как уксусная кислота. Эти материалы можно использовать по отдельности или в смеси.
Используемые источники азота могут включать пептон, экстракт дрожжей, мясной бульон, экстракт солода, жидкий кукурузный экстракт, соевую муку и мочевину, или неорганические соединения, например, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Данные источники азота можно использовать также по отдельности или в смеси.
Используемые источники фосфора могут включать вторичный кислый фосфат калия, дигидрофосфат калия, или соответствующие натрий-содержащие соли, а культуральная среда может включать необходимые для роста соли металлов, такие как сульфат магния или сульфат железа. В дополнение к таким материалам можно Использовать необходимые материалы для роста, такие как аминокислоты и витамины. Кроме того, в культуральную среду можно добавлять соответствующие предшественники. Эти материалы можно добавлять в культуру в ходе культивирования с помощью подходящего способа в периодическом или непрерывном режиме.
В ходе культивирования микроорганизма значение рН культуры можно регулировать основным соединением, таким как гидроксид натрия, гидроксид калия или аммиак, или кислотным соединением, таким как фосфорная кислота или серная кислота. Образование пены можно сдерживать с помощью антивспенивателя, такого как сложный полигликолевый эфир жирной кислоты. Для поддержания аэробных условий в культуру можно вводить кислород или кислородсодержащий газ (например воздух). Обычно температура культуры может составлять от 20°С до 45°С, в частности от 25°С до 40°С. Культивирование можно продолжать до тех пор, пока не будет получено желаемое количество L-лизина. В частности, время культивирования может составлять от 10 ч. до 160 ч.
В способе по настоящей заявке культивирование можно проводить в непрерывном или периодическом режиме, таком как периодический процесс, периодический процесс с подпиткой и повторяющийся периодический процесс с подпиткой. Способ культивирования хорошо известен в данной области техники, и при этом можно использовать любой способ.
Далее настоящая заявка будет описана более подробно со ссылками на примеры. Однако данные примеры приведены только в целях иллюстрации, и ограничение объема настоящей заявки данными примерами не подразумевается.
[Пример]
Пример 1. Конструирование рекомбинантного вектора для инактивации гена odx, полученного из Corynebacterium
Для того, чтобы инактивировать ген odx (SEQ ID NO: 2), кодирующий оксалоацетат-декарбоксилазу Corynebacterium glutamicum, имеющий аминокислотную последовательность, представленную SEQ ID NO: 1, с помощью сайт-специфичного разрушения гена, конструировали плазмидный вектор следующим способом. Во-первых, на основе NIH Genbank (США) получали нуклеотидную последовательность гена odx и на основе этой последовательности синтезировали праймеры для получения инактивированных фрагментов гена odx.
Проводили ПЦР [Sambrook et al, Molecular Cloning, a Laboratory Manual (1989), Cold Spring Harbor Laboratories] с использованием хромосомы дикого типа Corynebacterium glutamicum АТСС13032 в качестве матрицы и праймеров SEQ ID NO: 3 и SEQ ID NO: 4, SEQ ID NO: 5 и SEQ ID NO: 6. После денатурации при 95°C в течение 5 минут, повторяли 30 циклов денатурации при 95°С в течение 30 секунд, отжиг при 55°С в течение 30 секунд, и полимеризацию при 72°С в течение 30 секунд, и затем проводили полимеризацию при 72°С в течение 7 минут. В результате получали SEQ ID NO: 7 из 536 п. о. и SEQ ID NO: 8 из 534 п. о., включая 5'-конец и 3'-конец гена odx, соответственно.
SEQ ID NO:3: 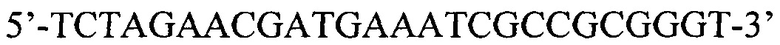
SEQ ID NO: 4: 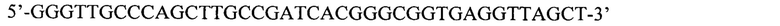
SEQ ID NO: 5: 
SEQ ID NO: 6: 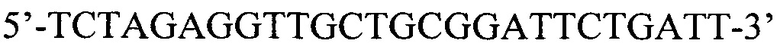
Проводили ПЦР с помощью амплифицированных SEQ ID NO: 7 и SEQ ID NO: 8 в качестве матрицы, а также SEQ ID NO: 3 и SEQ ID NO: 6. После денатурации при 95°C в течение 5 минут повторяли 30 циклов денатурации при 95°С в течение 30 секунд, отжиг при 55°С в течение 30 секунд, и полимеризацию при 72°С в течение 60 секунд, и затем проводили полимеризацию при 72°С в течение 7 минут. В результате амплифицировали SEQ ID NO: 9 (далее упоминаемый в данном документе как Δodx) из 1030 п. о., в котором 5'-конец и 3'-конец гена odx были связаны друг с другом.
Вектор pDZ (патент Кореи №0924065), который не реплицируется в Corynebacterium glutamicum, и амплифицированный фрагмент ДНК Δodx обрабатывали соответственно рестрикционным ферментом XbaI и затем лигировали друг с другом с помощью ДНК-лигазы. Для получения плазмиды проводили клонирование. Эта плазмида была обозначена как pDZ-Δodx (ФИГ. 1).
Пример 2. Инактивация odx в L-лизин-продуцирующем Corynebacterium и анализ продуцирования L-лизина
pDZ-Δodx, полученный в примере 1, трансформировали соответственно в L-лизин-продуцирующие штаммы Corynebacterium glutamicum КССМ11016Р и КССМ11347Р с помощью способа электрических импульсов (Appl. Microbiol. Biotechnol. (1999) 52:541-545), и трансформированные штаммы получали на селективной среде, содержащей 25 мг/л канамицина. Штаммы, содержащие инактивированный ген odx, получали с помощью фрагмента ДНК Δodx, вставленного в геном с помощью вторичной рекомбинации (кроссовер), и штаммы обозначали соответственно как KCCM11016P/Δodx и KCCM11347P/Δodx.
Для того, чтобы исследовать влияет ли на самом деле инактивация гена odx на увеличение продуцирования лизина, odx-инактивированные штаммы КССМ11016Р/Δodx и КССМ11347Р/Δodx культивировали с помощью следующих способов и исследовали их продуцирование.
Каждый из штаммов инокулировали в колбу с угловыми перегородками объемом 250 мл, содержащую 25 мл следующей среды для посева, с последующим культивированием при 37°С в течение 20 часов с перемешиванием при 200 об/мин. 1 мл культуры для посева инокулировали в колбу с угловыми перегородками объемом 250 мл, содержащую 24 мл следующей среды для продуцирования, с последующим культивированием при 30°С в течение 72 часов с перемешиванием при 200 об/мин.
* Состав среды для посева (рН 7,0):
20 г глюкозы, 10 г пептона, 10 г экстракта дрожжей, 5 г мочевины, 4 г КН2РО4, 8 г K2HPO4, 0,5 г MgSO4⋅7H2O, 100 мкг биотина, 1000 мкг тиамина HCl (в 1 литре технологической воды)
* Состав среды для продуцирования (рН 7,0):
80 г глюкозы, 20 г молассы или предварительно обработанной молассы (в качестве восстанавливающего сахара), 5 г жидкого кукурузного экстракта, 40 г (NH4)2SO4, 4 г мочевины, 1 г KH2PO4, 2,5 г NaCl, 1 г MgSO4*7H2O, 10 мг FeSO4⋅7H2O, 10 мг MnSO4⋅5H2O, 100 мкг биотина, 200 мкг тиамина HCl, 40 г СаСО3, если необходимо, 0,4 г L-лейцина, если необходимо, 0,1 г L-треонина, если необходимо, 0,1 г L-метионина (в 1 литре технологической воды)
После завершения культивирования измеряли степень продуцирования L-лизина с помощью HPLC. В результате, содержание L-лизина в культуре каждого штамма было таким, как показано в следующей таблице 1.
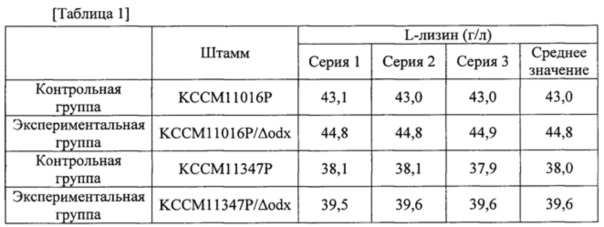
Как показано в таблице 1, odx-инактивированные штаммы KCCM11016P/Δodx и KCCM11347P/Δodx продемонстрировали приблизительно 4,2% повышение продуцирования L-лизина по сравнению с их родительскими штаммами КССМ11016Р и КССМ11347Р, соответственно.
Среди них штамм KCCM11016P/Δodx обозначили как СА01-2276 и 22 ноября 2013 года передали на депонирование под номером доступа КССМ11478Р в Международный депозитарий, Корейский центр культур микроорганизмов, который является дочерней Коллекцией культур Коллекции культур Корейской федерации, расположенной по адресу 361-221, Hongje-l-Dong, Seodaemungu-Gu, Сеул, Корея.
Пример 3. Инактивация odx в L-лизин-продуцирующем Corynebacterium с повышенной эффективностью пути биосинтеза L-лизина и анализ продуцирования лизина
pDZ-Δodx, полученный в примере 1, трансформировали в L-лизин-продуцирующий штамм с повышенной эффективностью пути биосинтеза L-лизина Corynebacterium glutamicum КССМ10770Р (патент Кореи №10-0924065) таким же способом, как и в примере 2, для того, чтобы получить трансформированный штамм, который был обозначен как KCCM10770P/Δodx.
Для того, чтобы исследовать влияет ли на самом деле инактивация гена odx на увеличение продуцирования L-лизина, odx-инактивированный штамм KCCM10770P/Δodx культивировали таким же способом, как и в примере 2, и исследовали его продуцирование. Содержание L-лизина в культуре каждого штамма было таким, как показано в следующей таблице 2.
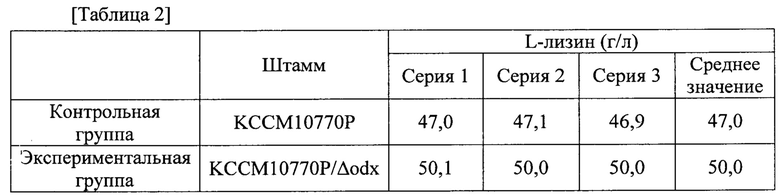
Как показано в таблице 2, odx-инактивированный штамм KCCM10770P/Δodx продемонстрировал приблизительно 6,4% повышение продуцирования L-лизина по сравнению с родительским штаммом КССМ10770Р.
Пример 4. Инактивация odx в полученном из дикого типа L-лизин-продуцирующем Corynebacterium и анализ продуцирования лизина
pDZ-Δodx, полученный в примере 1, трансформировали в полученный из дикого типа L-лизин-продуцирующий штамм Corynebacterium glutamicum CJ3P (Binder et al. Genome Biology 2012, 13:R40, pyc(P458S), hom(V59A), lysC(T311I)) таким же способом, как и в примере 2, для того, чтобы получить трансформированный штамм, который был обозначен как CJ3P/Δodx.
Для того, чтобы исследовать влияет ли на самом деле инактивация гена odx на увеличение продуцирования лизина, odx-инактивированный штамм CJ3P/Δodx культивировали таким же способом, как и в примере 2, и исследовали его продуцирование. Содержание L-лизина в культуре каждого штамма было таким, как показано в следующей таблице 3.
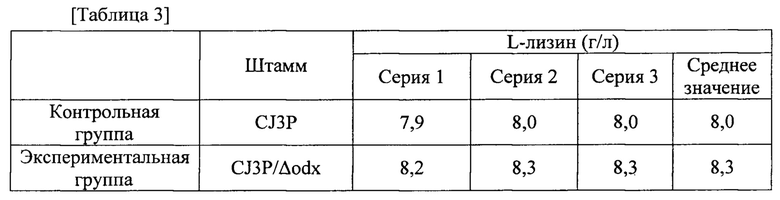
Как показано в таблице 3, при инактивации odx в родительском штамме CJ3P, который представляет собой полученный из дикого типа L-лизин-продуцирующий штамм, также наблюдали приблизительно 3,8% повышение продуцирования лизина.
Соответственно в настоящей заявке в примерах 2-4 продемонстрировано, что повышенное продуцирование лизина обычно наблюдали в разных классах микроорганизмов рода Corynebacterium с помощью инактивации гена odx, указывая на то, что инактивация гена odx является полезным свойством при получении L-лизина.




Группа изобретений относится к области биотехнологии. Предложен микроорганизм рода Corynebacterium, продуцирующий L-лизин, где оксалоацетат-декарбоксилаза с аминокислотной последовательностью SEQ ID NO: 1 инактивирована. Предложен способ получения L-лизина с использованием указанного микроорганизма. Группа изобретений позволяет повысить продукцию L-лизина в модифицированном штамме по сравнению с родительским штаммом. 2 н. и 1 з.п. ф-лы, 1 ил., 3 табл., 4 пр.
1. Микроорганизм рода Corynebacterium, продуцирующий L-лизин, где оксалоацетат-декарбоксилаза с аминокислотной последовательностью, изложенной в SEQ ID NO: 1, является инактивированной.
2. Микроорганизм рода Corynebacterium по п.1, где микроорганизм рода Corynebacterium представляет собой Corynebacterium glutamicum.
3. Способ получения L-лизина, при этом способ предусматривает:
культивирование микроорганизма по любому из пп.1 и 2 с получением культуры или клеток микроорганизма и
извлечение L-лизина из культуры или клеток микроорганизма.
| KR 101285945 B1, 12.07.2013 | |||
| KR 100789271 B1, 02.01.2008 | |||
| Chain A, Crystal Structures Of Cg1458, NCBI Accession: 4DBF_A; Найдено в Интернет по адресу: www.ncbi.nlm.nih.gov/protein/4dbf_a | |||
| БАКТЕРИЯ - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 2005 |
|
RU2316588C1 |

