Область изобретения
Настоящее изобретение относится к микроорганизму рода Corynebacterium, продуцирующему L-лизин, и способу получения L-лизина с использованием этого микроорганизма.
Предшествующий уровень техники
L-Лизин используют в кормах для животных и в производстве лекарственных средств или косметических средств для человека, и обычно его получают с помощью ферментации, используя штаммы рода Corynebacterium или штаммы рода Escherichia. Чтобы обеспечить получение L-лизина, проводят различные исследования с целью разработки штаммов с высокоэффективным продуцированием и технологий ферментационного процесса. Более конкретно, главным образом используют способы, направленные на дифференцированные подходы к желаемым веществам, такие как повышение экспрессии генов, кодирующих ферменты, связанные с биосинтезом L-лизина, или делетирование генов, не являющихся необходимыми для его биосинтеза (патент Кореи №10-0838038).
Для выявления эффективных параметров, которые могут повышать продуктивность по лизину, авторы настоящего изобретения в результате случайного введения эндогенного гена микроорганизма рода Corynebacterium обнаружили ген, ассоциированный с продуцированием высоких концентраций лизина, и в соответствии с указанным открытием, подтвердили, что когда экспрессия этого гена у микроорганизма рода Corynebacterium повышается, продуктивность по L-лизину может быть повышена, завершив тем самым настоящее изобретение.
Техническая задача
Задача настоящего изобретения заключается в предложении микроорганизма рода Corynebacterium, продуцирующего L-лизин, имеющего улучшенную активность белка, состоящего из аминокислотной последовательности SEQ ID NO: 1, по сравнению с эндогенной активностью.
Другая задача настоящего изобретение заключается в предложении способа получения L-лизина с использованием этого микроорганизма.
Техническое решение
Для выполнения указанных выше задач, в одном аспекте настоящего изобретения предложен микроорганизм рода Corynebacterium, продуцирующий L-лизин, имеющий улучшенную активность белка, состоящего из аминокислотной последовательности SEQ ID NO: 1, по сравнению с эндогенной активностью.
Ниже в данном документе настоящее изобретение будет описано подробно. Между тем, каждое из разъяснений и типичных воплощений, раскрытых в данном описании изобретения, может быть применено к другим разъяснениям и типичным воплощениям. Это означает, что все комбинации различных факторов, раскрытых здесь, входят в объем настоящего изобретения. Кроме того, объем настоящего изобретения не следует ограничивать конкретным описанием изобретения, предложенным ниже.
В настоящем изобретении термин "белок, состоящий из аминокислотной последовательности SEQ ID NO: 1," можно использовать взаимозаменяемо с термином "белок НМ1524". Дополнительно, его можно использовать взаимозаменяемо с термином "белок, кодированный геном НМ1524". Кроме того, выражение "белок, по существу состоящий из аминокислотной последовательности SEQ ID NO: 1" или "белок, состоящий из аминокислотной последовательности SEQ ID NO: 1" можно использовать взаимозаменяемо.
Дополнительно, белок может включать полипептид, имеющий гомологию с аминокислотной последовательностью SEQ ID NO: 1 по меньшей мере 80%, 90%, 95%, 97% или 99%. Например, очевидно, что белок, имеющий аминокислотную последовательность, в которой последовательность частично делетирована, модифицирована, замещена или имеет вставку, включен в объем настоящего изобретения, пока он имеет аминокислотную последовательность, имеющую указанную выше гомологию и проявляющую эффект, соответствующий таковому у белка, состоящего из аминокислотной последовательности SEQ ID NO: 1.
Кроме того, пока он имеет активность, соответствующую таковой у белка, состоящего из аминокислотной последовательности SEQ ID NO: 1, не исключается мутация, которая может иметь место в результате добавления не имеющей смысла последовательности выше или ниже нуклеиновокислотной последовательности аминокислотной последовательности SEQ ID NO: 1, или естественная мутация в ней или молчащая мутация в ней, и белок, имеющий аминокислотную последовательность SEQ ID NO: 1, также включен в объем настоящего изобретения.
При использовании в настоящем изобретении термин "гомология" относится к проценту идентичности между двумя фрагментами полинуклеотидов или полипептидов, означающему степень соответствия с данной аминокислотной последовательностью или нуклеотидной последовательностью, и может выражаться в процентах. В настоящем описании изобретения последовательность, гомологичная данной аминокислотной последовательности или нуклеотидной последовательности, имеющая активность, такую же или подобную активности данной аминокислотной последовательности или нуклеотидной последовательности, может быть указана в "% гомологии". Например, гомология может быть подтверждена с использованием стандартного программного обеспечения для вычисления параметров, таких как показатель сцепления генов, идентичность и подобие, более конкретно BLAST 2.0, или путем сравнения последовательностей посредством экспериментов саузерн-гибридизации в определенных жестких условиях, определенные подходящие условия гибридизации входят в релевантный технический диапазон и могут быть определены посредством способа, известного специалисту в данной области техники (например J. Sambrook et at., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; F.M. Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York).
Ген, кодирующий белок, состоящий из аминокислотной последовательности SEQ ID NO: 1, хотя не ограничивается этим, может представлять собой полинуклеотид, включающий нуклеотидную последовательность SEQ ID NO: 2, и может представлять собой полинуклеотид, имеющий гомологию с нуклеотидной последовательностью SEQ ID NO: 2 по меньшей мере 80%, 90%, 95%, 97% или 99%. Очевидно, что полинуклеотиды, которые, вследствие вырождения кодонов, могут быть транслированы в белок, состоящий из аминокислотной последовательности SEQ ID NO: 1, или белки, имеющие гомологию с ним, также могут быть включены. Альтернативно, зонд, который может быть получен из известной последовательности гена, например, любой последовательности, которая в жестких условиях гибридизуется с последовательностью, комплементарной всей или части нуклеотидной последовательности, для кодирования белка, имеющего активность белка, состоящего из аминокислотной последовательности SEQ ID NO: 1, может быть включен без ограничения. Термин "жесткие условия" относится к условиям, которые обеспечивают специфическую гибридизацию между полинуклеотидами. Такие условия более конкретно описаны в литературе (например J. Sambrook et al., infra). Например, жесткие условия могут включать условия, при которых гены, имеющие высокую гомологию, гомологию 80% или выше, более конкретно 90% или выше, еще более конкретно 95% или выше, намного более конкретно 97% или выше, и особенно более конкретно 99% или выше гибридизуются друг с другом, в то время как гены, имеющие гомологию более низкую, чем указанная выше гомология, не гибридизуются друг с другом; или могут включать обычные условия отмывки при Саузерн-гибридзациии, то есть отмывки один, более конкретно два или три раза при концентрации солей и температурах, соответствующих 60°С, 1×SSC и 0,1% SDS; более конкретно 60°С, 0,1×SSC и 0,1% SDS; и еще более конкретно 68°С, 0,1×SSC и 0,1% SDS.
Для гибридизации требуется, чтобы две нуклеиновые кислоты содержали комплементарные последовательности, хотя, в зависимости от жесткости гибридзации, могут быть возможны ошибки спаривания между нуклеотидами. Термин "комплементарный" используют для описания связи между нуклеотидами, которые могут гибридизоваться друг с другом. Например, что касается ДНК, аденозин комплементарен тимину и цитозин комплементарен гуанину. Следовательно, настоящее изобретение также может включать выделенный фрагмент нуклеиновых кислот, комплементарный целой последовательности, а также последовательность нуклеиновых кислот, по существу подобную ему.
Более конкретно, полинуклеотид, имеющий гомологию, может быть обнаружен с использованием условий гибридизации, включающих стадию гибридизации при значении Тпл 55°С в описанных выше условиях. Кроме того, значение Тm может представлять собой 60°С, 63°С или 65°С, но не ограничивается этим, и может подходящим образом регулироваться специалистом в данной области техники в зависимости от его цели.
Подходящая жесткость для гибридизации полинуклеотидов зависит от длины и степени комплементарно ста полинуклеотидов, и такие вариабельные величины хорошо известны в данной области техники.
Зонд, используемый в гибридизации, может представлять собой часть последовательности, комплементарной нуклеотидной последовательности. Такой зонд может быть изготовлен посредством ПЦР с использованием олигонуклеотида, полученного на основе известной последовательности в качестве праймера, и фрагмента гена, содержащего нуклеотидную последовательность в качестве матрицы. Фрагмент гена может представлять собой, например, по меньшей мере примерно 50 нуклеотидов, 60 нуклеотидов, 70 нуклеотидов, 80 нуклеотидов, 90 нуклеотидов или по меньшей мере 100 нуклеотидов. Дополнительно, при необходимости специалист в данной области техники может регулировать температуру и концентрацию солей в растворах для отмывки в зависимости от факторов, таких как длина зонда.
При использовании в настоящем изобретении термин "эндогенная активность" относится к конкретной активности белка, первоначально имеющейся у родительского штамма микроорганизма до его трансформации, в случае, когда свойство микроорганизма изменяется вследствие генетической модификации, вызванной естественными или искусственными факторами.
Как его используют в настоящем изобретении, термин "активность, которая улучшена по сравнению с эндогенной активностью" относится к повышению активности белка микроорганизма по сравнению с его эндогенной активностью или его активностью до модификации. Улучшение активности может включать как ввод чужеродного НМ1524, так и эндогенное повышение активности НМ1524.
Более конкретно, улучшение активности по настоящему изобретению может быть осуществлено посредством следующих способов:
1) способ увеличения числа копий полинуклеотида, кодирующего белок,
2) способ модификации последовательности регуляции экспрессии таким образом, чтобы экспрессия полинуклеотида повышалась,
3) способ модификации полинуклеотидной последовательности на хромосоме таким образом, чтобы активность белка повышалась,
4) способ введения модифицированного полинуклеотида, в котором кодоны чужеродного полинуклеотида или указанного выше полинуклеотида, проявляющего активность белка, были оптимизированы, или
5) способ модификации путем комбинации указанных выше способов для эффективного повышения и так далее; однако улучшение активности не ограничивается этим.
Увеличение числа копий полинуклеотида в способе (1) выше может быть осуществлено посредством функционального связывания с вектором, или может быть осуществлено посредством вставки в хромосому в клетке-хозяине, но не ограничивается конкретно этим. Более конкретно, оно может быть осуществлено посредством функционального связывания полинуклеотида, кодирующего белок по настоящему изобретению, с вектором, который может реплицироваться и функционировать независимо от клетки-хозяина, и ввода его в клетку-хозяина. Альтернативно, это может быть осуществлено посредством способа увеличения числа копий полинуклеотида в хромосоме в клетке-хозяине путем функционального связывания полинуклеотида с вектором, который может вставить полинуклеотид в хромосому в клетке-хозяине, и ввода его в клетку-хозяина.
Кроме того, в способе (2) выше модификация последовательности регуляции экспрессии таким образом, чтобы экспрессия полинуклеотида повышалась, может быть выполнена путем индукции модификации последовательности посредством делеции, вставки, или неконсервативной или консервативной замены в последовательности нуклеиновой кислоты, или посредством их комбинации с целью дополнительного повышения активности последовательности регуляции экспрессии, или путем замены последовательностью нуклеиновых кислот, имеющей более сильную активность, но не ограничивается конкретно этим. Последовательность регуляции экспрессии может включать промотор, операторную последовательность, последовательность, кодирующую сайт связывания рибосом, последовательность регуляции терминации транскрипции и трансляции и так далее, но не ограничивается конкретно этим.
Сильный гетер о логичный промотор может быть связан с участком выше по ходу транскрипции единицы экспрессии полинуклеотида в отличие от исходного промотора; примеры сильных промоторов представляют собой промотор CJ7 (патент Кореи №0620092 и международная публикация № WO 2006/065095), промотор lysCP1 (международная публикация № WO 2009/096689), промотор EF-Tu, промотор groEL, промотор асеА или асеВ и так далее, но сильный промотор не ограничивается этим. Кроме того, в способе (3) выше модификация полинуклеотидной последовательности на хромосоме может быть осуществлена путем индукции модификации в последовательности регуляции экспрессии посредством делеции, вставки, или неконсервативной или консервативной замены в последовательности нуклеиновой кислоты, или посредством их комбинации, чтобы дополнительно повысить активность полинуклеотидной последовательности, или она может быть выполнена путем замены полинуклеотидной последовательности на полинуклеотидную последовательность, модифицированную с целью получения дополнительной более высокой активности; однако модификация не ограничивается конкретно этим.
Дополнительно, в способе (4) выше введение чужеродной полинуклеотидной последовательности может быть выполнено путем ввдения в клетку-хозяина чужеродного полинуклеотида, кодирующего белок, который проявляет активность такую же или подобную активности белка, указанного выше, или модифицированного полинуклеотида, в котором кодоны чужеродного полинуклеотида были оптимизированы. Чужеродный полинуклеотид можно использовать без ограничения его происхождением или последовательностью, пока он проявляет активность такую же или подобную таковой у белка. Дополнительно, для оптимизированной транскрипции и трансляции чужеродного полинуклеотида в клетке-хозяине, он может быть введен в клетку-хозяин после оптимизации его кодонов. Введение может быть осуществлено специалистом в данной области техники путем подбора подходящего способа трансформации, известного в данной области техники, и посредством экспрессии введенного в клетку-хозяина полинуклеотида может продуцироваться белок, тем самым улучшая его активность.
Наконец, способ (5), который относится к модификации для эффективного повышения путем комбинации способов с (1) до (4), может быть осуществлен посредством комбинированного применения по меньшей мере одного из следующих: повышения числа копий полинуклеотида, кодирующего белок, модификации последовательности регуляции экспрессии таким образом, чтобы экспрессия полинуклеотида повышалась, модификации полинуклеотидной последовательности на хромосоме, и модификации чужеродного полинуклеотида, проявляющего активность белка или его кодон-оптимизированного модифицированного полинуклеотида.
При использовании здесь, термин "вектор" относится к ДНК-конструкции, включающей полинуклеотидную последовательность, кодирующую целевой белок, которую функционально связывают с подходящей регуляторной последовательностью так, чтобы целевой белок мог экспрессироваться в подходящем хозяине. Регуляторная последовательность включает промотор, способный инициировать транскрипцию, любую операторную последовательность для контроля транскрипции, последовательность, кодирующую подходящий мРНК-домен, связывающий рибосому, и последовательность, контролирующую терминацию транскрипции и трансляции. После введения в подходящую клетку-хозяина вектор может реплицироваться или функционировать независимо от генома хозяина, и он может интегрироваться в геном хозяина самостоятельно. В одном воплощении полинуклеотид, кодирующий целевой белок в хромосоме, может быть замещен модифицированным полинуклеотидом посредством вектора для вставки в хромосому. Вставка полинуклеотида в хромосому может быть выполнена посредством любого способа, известного в данной области техники, например гомологичной рекомбинации, но не ограничивается этим.
Вектор, используемый в настоящем изобретении, не ограничен конкретно, и можно использовать любой вектор, известный в данной области техники. Примеры обычно используемых векторов могут включать естественные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, в качестве фагового вектора или космидного вектора можно использовать pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A, Charon21A и так далее, и в качестве плазмидного вектора можно использовать векторы на основе pBR, pUC, pBluescriptII, pGEM, pTZ, pCL, pET и так далее. Более конкретно, можно использовать векторы pDZ, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCC1BAC и так далее.
При использовании здесь, термин "трансформация" относится к процессу введения вектора, включающего полинуклеотид, кодирующий целевой белок, в клетку-хозяина, обеспечивая тем самым экспрессию белка, кодируемого полинуклеотидом, в клетке-хозяине. Пока трансформированный полинуклеотид может экспрессироваться в клетке-хозяине, не имеет значения, вставлен ли он в хромосому клетки-хозяина и находится в ней или находится вне хромосомы, и могут быть включены оба случая. Дополнительно, полинуклеотид включает ДНК и РНК, которые кодируют целевой белок. Полинуклеотид может быть введен в любой форме, пока он может быть введен в клетку-хозяина и экспрессироваться в ней. Например, полинуклеотид может быть введен в клетку-хозяина в форме кассеты экспрессии, которая представляет собой конструкцию гена, включающую все элементы, необходимые для автоэкспрессии. Кассета экспрессии обычно может включать промотор, функционально связанный с полинуклеотидом, терминатор, сайт связывания рибосом и стоп-кодон. Кассета экспрессии может быть в форме вектора экспрессии, способного к саморепликации. Дополнительно, полинуклеотид может быть введен в клетку-хозяина как есть и функционально связан с последовательностью, необходимой для его экспрессии в клетке-хозяине, но ограничивается этим.
Дополнительно, как используется выше, термин "функционально связанный" относится к функциональной связи между указанной выше последовательностью гена и последовательностью промотора, который инициирует и опосредует транскрипцию полинуклеотида, кодирующего целевой белок по настоящему изобретению.
Способ трансформации вектора по настоящему изобретению включает любой способ введения нуклеиновых кислот в клетку и может быть выполнен путем подбора подходящей стандартной методики, известной в данной области техники, в зависимости от клетки-хозяина. Примеры способа включают электропорацию, осаждение фосфатом кальция (CaHPO4), осаждение хлоридом кальция (CaCl2), микроинъекцию, методику с полиэтиленгликолем (PEG), методику с диэтиламиноэтилдекстраном, методику с катионными липосомами, методику с ацетатом лития и ДМСО и так далее, но способ не ограничивается этими.
В качестве клетки-хозяина предпочтительно использовать хозяина, имеющего высокую эффективность введения ДНК и высокую эффективность экспрессии для введенной ДНК; например, клетка-хозяин может представлять собой микроорганизм рода Corynebacterium.
При использовании здесь, термин "L-лизин" относится к основной α-аминокислоте, которая, как незаменимая аминокислота, не может синтезироваться in vivo, и которая представляет собой L-аминокислоту с химической формулой NH2(CH2)4CH(NH2)COOH. Дополнительно, L-лизин может быть включен в объем настоящего изобретения даже когда находится в форме соли.
При использовании здесь, термин "продуцирующий L-лизин микроорганизм" относится к штамму микроорганизма, который может продуцировать L-лизин, исходя из задач настоящего изобретения, более конкретно относится к штамму, который может продуцировать L-лизин в высокой концентрации посредством операций в соответствии с настоящим изобретением. Соответственно, если микроорганизм способен продуцировать L-лизин, тип его родительского штамма не ограничен конкретно. То есть в настоящем изобретении родительский штамм может включать как штаммы с L-продуктивностью по лизину, так и штаммы без продуктивности по L-лизину. Продуктивность по L-лизину может включать как штаммы, которые встречаются в природе, так и штаммы, которые были созданы искусственно. Микроорганизм, имеющий искусственно созданную продуктивность по L-лизину, может быть модифицирован для получения продуктивности по L-лизину посредством вещества, индуцирующего мутацию, такого как нитрозогуанидин (NTG) и так далее, или уровень экспрессии или активность целевого белка могут регулироваться для получения продуктивности по L-лизину, но микроорганизм не ограничивается этим. Более конкретно, целевой белок может включать все белки, которые непосредственно/косвенным образом действуют в пути биосинтеза L-лизина, и путем повышения или снижения уровня экспрессии или его активности, микроорганизм может быть модифицирован для получения продуктивности по L-лизину; дополнительно, модификация также может быть индуцирована в аминокислотной последовательности или нуклеотидной последовательности такого белка для получения продуктивности по L-лизину. Искусственное создание микроорганизма, включая регулирование экспрессии целевого белка и индукцию случайной мутации с использованием NTG и так далее, описанные выше, может быть выполнено подходящим образом с помощью известного способа специалистом в данной области техники.
Более конкретно, микроорганизм может представлять собой микроорганизм рода Corynebacterium. Например, можно использовать Corynebacterium glutamicum, Corynebacterium ammoniagenes, Corynebacterium thermoaminogenes, Brevibacterium flavum или Brevibacterium fermentum и так далее, но микроорганизм не ограничивается этими. Например, в качестве микроорганизма рода Corynebacterium можно использовать Corynebacterium glutamicum. Однако микроорганизм не ограничивается этими примерами и можно использовать другие известные микроорганизмы рода Corynebacterium, имеющие продуктивность по L-лизину.
Примеры известных микроорганизмов рода Corynebacterium, имеющих продуктивность по L-лизину, представляют собой микроорганизмы, описанные в патенте Кореи №10-0397322 (или патентной публикации US №2003-0124688), патенте Кореи №10-0924065 (или патентной публикации US №2010-0143984), патенте Кореи №10-0073610 и/или Binder et al., Genome Biology 2012, 13:R40, и их содержание включено в данное описание изобретения посредством ссылки во всей их полноте.
В качестве другого аспекта, в настоящем изобретении предложен способ получения L-лизина, включающий: культивирование микроорганизма рода Corynebacterium, продуцирующего L-лизин, имеющего улучшенную активность белка, состоящего из аминокислотной последовательности SEQ ID NO: 1, по сравнению с эндогенной активностью, в среде; и выделение L-лизина из культивируемого микроорганизма или среды его культивирования.
Микроорганизм рода Corynebacterium, продуцирующий L-лизин, представляет собой такой, как описано выше.
При использовании здесь, термин "культивирование" означает выращивание микроорганизма в контролируемых подходящим образом условиях окружающей среды. Процесс культивирования по настоящему изобретению можно выполнять в соответствии с подходящими культуральными средами и условиями культивирования, известными в данной области техники. Такой процесс культивирования может быть легко скорректирован для применения специалистом в данной области техники в зависимости от штамма, подлежащего селекции. В способе, указанном выше, культивирование микроорганизма может быть выполнено посредством известного способа периодического культивирования, способа непрерывного культивирования, способа периодического культивирования с подпиткой и так далее, но не ограничивается этим. В частности, что касается условий культивирования, рН культивирования можно регулировать до подходящего рН (например рН от 5 до рН 9, более конкретно от рН 6 до рН 8 и наиболее конкретно рН 6,8) с использованием основного соединения (например, гидроксида натрия, гидроксида калия или аммиака) или кислого соединения (например, фосфорной кислоты или серной кислоты). Дополнительно, во время культивирования для предотвращения пенообразования можно добавлять пеногаситель, такой как полигликолевый эфир жирной кислоты; кроме того, можно вводить в культуральную среду кислород или кислородсодержащий газ для поддержания аэробного ее состояния или для поддержания анаэробного или микроаэробного состояния культуральной среды, можно вводить азот, водород или углекислый газ, или культивирование можно быть выполнено без ввода газа. Температуру культуральной среды можно поддерживать от 20°С до 45°С, более конкретно от 25°С до 40°С, и культивирование можно продолжать до получения желаемого количества полезного продуцируемого вещества, более конкретно в течение приблизительно от 10 до 160 часов. Однако культивирование не ограничивается указанным выше. L-лизин, полученный посредством культивирования, может выделяться в среду или может оставаться в клетках.
Дополнительно, в качестве источника углерода для культуральной среды, которая будет использоваться, можно использовать сахара и углеводы (например глюкозу, сахарозу, лактозу, фруктозу, мальтозу, мелассу, крахмал и целлюлозу), масла и жиры (например соевое масло, подсолнечное масло, арахисовое масло и кокосовое масло), жирные кислоты (например пальмитиновую кислоту, стеариновую кислоту и линолевую кислоту), спирты (например глицерин и этиловый спирт), органические кислоты (например уксусную кислоту) и так далее, по отдельности или в комбинации, но источник углерода не ограничивается этим. В качестве источника азота можно использовать, азотсодержащие органические соединения (например пептон, дрожжевой экстракт, мясной сок, солодовый экстракт, жидкий кукурузный экстракт, соевую муку и мочевину) или неорганические соединения (например сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония) и так далее, по отдельности или в комбинации, но источник азота не ограничивается этим. В качестве источника фосфора можно использовать однозамещенный фосфат калия, гидроортофосфат калия, соответствующие натрийсодержащие соли и так далее, по отдельности или в комбинации, но источника фосфора не ограничивается этим. Кроме того, в среде могут содержаться необходимые для стимуляции роста вещества, такие как соли других металлов (например сульфат магния или сульфат железа), аминокислоты, витамины и так далее.
В способе по настоящему изобретению для выделения L-лизина, продуцированного в ходе культивирования, целевая аминокислота может быть получена из культуральной жидкости с использованием подходящего способа, известного в данной области техники, в зависимости от способа культивирования. Например, можно использовать центрифугирование, фильтрацию, анионообменную хроматографию, кристаллизацию, ВЭЖХ и так далее, и целевой L-лизин может быть извлечен из среды или микроорганизма с использованием подходящего способа, известного в данной области техники. Дополнительно, стадия выделения может включать процесс очистки.
Полезные эффекты изобретения
L-Лизин может быть получен с высокой эффективностью с использованием микроорганизма по настоящему изобретению, который имеет продуктивность по L-лизину.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Далее настоящее изобретение будет описано подробно посредством типичных воплощений. Однако эти типичные воплощения предлагаются только для цели иллюстрации, и объем настоящего изобретения не ограничивается этими типичными воплощениями.
Пример 1: Получение библиотеки геномной ДНК дикого типа микроорганизма рода Corynebacterium
После выделения геномной ДНК из Corynebacterium glutamicum АТСС13032 геномную ДНК обрабатывали с помощью фермента рестрикции Sau3AI, и фрагменты ДНК разделяли по размерам путем электрофореза в агарозном геле и посредством этого получали селективно фрагменты ДНК от 3 т.о. до 4 т.о. После лигирования фрагментов с вектором pECCG117 (патент Кореи №10-0057684), который имеет сайт рестрикции BamHI, и введения в Е. coli DH5α, полученные в результате трансформанты высевали затем на твердую среду LB, содержащую канамицин (25 мг/л), и таким образом получали трансформированные колонии. ПЦР выполняли на 100 случайный колониях с использованием праймеров SEQ ID NOs: 3 и 4 и при этом было обнаружено, что соотношение колоний, содержащих вектор, в который были вставлены фрагменты целевой ДНК приблизительно от 3 т.о. до 4 т.о., равнялось 90% или выше. Все полученные колонии совместно культивировали после инокуляции в содержащую канамицин (25 мг/л) жидкую среду LB; затем выделяли плазмиды с использованием широко известного способа выделения плазмид, создав тем самым библиотеку геномной ДНК Corynebacterium glutamicum АТСС 13032.
Пример 2: Получение и оценка продуцирующего лизин микроорганизма с введенной библиотекой геномной ДНК дикого типа
С использованием способа электрического импульса библиотеку геномной ДНК, полученную в Примере 1, вводили в продуцирующий лизин штамм Corynebacterium glutamicum KCCM11016P (первоначально обозначенный как KFCC10881, микроорганизм был повторно депонирован в международном органе депонирования в соответствии с Будапештским договором и ему был присвоен номер доступа KCCM11016P; патент Кореи №10-0159812), и после посева на комплексную планшетную среду, содержащую канамицин (25 мг/л), и культивирования при 30°С в течение 24 часов получили приблизительно 2000 колоний.
Комплексная планшетная среда
20 г глюкозы, 50 г (NH4)2SO4, 10 г пептона, 5 г дрожжевого экстракта, 1,5 г мочевины, 5 г KH2PO4, 10 г K2HPO4, 0,5 г MgSO4⋅7H2O, 100 мкг биотина, 1000 мкг тиамина HCl, 2000 мкг пантотената кальция, 2000 мкг никотинамида, 20 г агара, 25 мг канамицин (на 1 л дистиллированной воды).
200 мкл комплексной жидкой среды вносили в каждую лунку 96-луночного планшета для культуры клеток и после инокуляции каждой из полученных колоний выполняли культивирование при встряхивании в течение 24 часов в условиях 30°С и 1200 об/мин. Клеточные тела и супернатант отделяли путем центрифугирования от бульонной культуры и 50 мкл супернатанта смешивали с раствором для реакции, содержащим лизиноксидазу.
Комплексная жидкая среда
20 г глюкозы, 10 г пептона, 5 г дрожжевого экстракта, 1,5 г мочевины, 4 г KH2PO4, 8 г K2HPO4, 0,5 г MgSO4⋅7H2O, 100 мкг биотина, 1000 мкг тиамина HCl, 2000 мкг пантотената кальция, 2000 мкг никотинамида, 25 мг канамицина (на 1 л дистиллированной воды)
Раствор для реакции
0,02 единиц лизиноксидазы (Sigma-Aldrich), 0,2 единиц пероксидазы (Sigma-Aldrich), 2 мг ABTS (на 1 мл раствора калий-фосфатного буфера).
После этого анализировали поглощение при OD405 в течение 30 минут и было отобрано 15 экспериментальных групп, показывающих более высокое поглощение, чем контрольная группа (KCCM11016P/pECCG117). Для подтверждения продуктивности по лизину каждого трансформанта каждый штамм инокулировали в 250 мл колбу с угловыми перегородками, содержащую 25 мл содержащей канамицин (25 мг/л) посевной среды, и культивировали при встряхивании в течение 20 часов в условиях 30°С и 200 об/мин. 1 мл посевной культуральной жидкости инокулировали в 250 мл колбу с угловыми перегородками, содержащую 24 мл содержащей канамицин (25 мг/л) продукционной среды, и культивировали при встряхивании в течение 96 часов при 37°С и 200 об/мин. После окончания культивирования анализировали концентрацию L-лизин с использованием ВЭЖХ (Таблица 1).
Посевная среда
20 г глюкозы, 5 г (NH4)2SO4, 10 г пептона, 5 г дрожжевого экстракта, 1,5 г мочевины, 4 г KH2PO4, 8 г K2HPO4, 0,5 г MgSO4⋅7H2O, 100 мкг биотина, 1000 мкг тиамина HCl, 2000 мкг пантотената кальция, 2000 мкг никотинамида (на 1 л дистиллированной воды)
Продукционная среда (рН 7,0)
100 г глюкозы, 40 г (NH4)2SO4, 2,5 г соевого белка, 5 г сухих веществ кукурузного экстракта, 3 г мочевины, 1 г KH2PO4, 0,5 г MgSO4⋅7H2O, 100 мкг биотина, 1000 мкг тиамина HCl, 2000 мкг пантотената кальция, 3000 мкг никотинамида, 30 г СаСО3 (на 1 л дистиллированной воды).
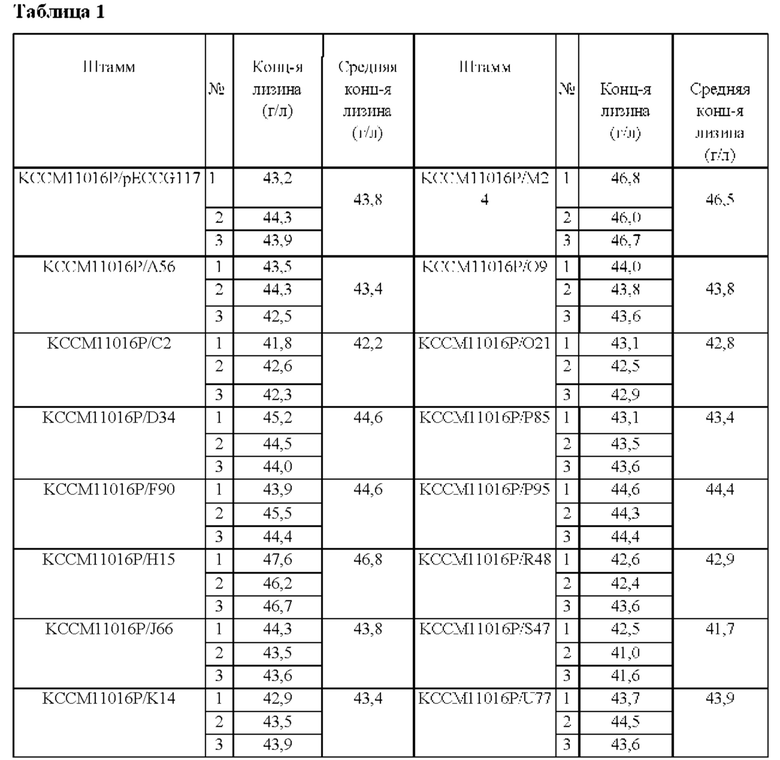
На основании приведенных выше результатов, были отобраны KCCM11016P/Н15 и KCCM11016P/М24, которые показывали эффект повышенной продуктивности по лизину по сравнению с контрольной группой, и выделяли плазмиды с использованием хорошо известного способа выделения плазмид. Плазмида, полученная из KCCM11016P/Н15, была названа рЕС-Н15, и плазмида, полученная из KCCM11016P/М24, была названа рЕС-М24. После этого выполняли анализ нуклеотидной последовательности с использованием праймеров SEQ ID NOs: 3 и 4. В результате было обнаружено, что плазмиды рЕС-Н15 и рЕС-М24 содержат нуклеотидные последовательности SEQ ID NO: 15 и 16, соответственно. Таким образом, было обнаружено, что обе указанные выше плазмиды содержат нуклеотидную последовательность SEQ ID NO: 2, которая кодирует аминокислотную последовательность SEQ ID NO: 1. Соответственно, ген, кодирующий аминокислотную последовательность SEQ ID NO: 1, был назван НМ1524, и далее в описании называется также.
Пример 3: Получение вектора сверхэкспрессии гена НМ1524
Для подтверждения эффекта НМ1524, обнаруженного в Примере 2, был получен вектор для сверхэкспрессии соответствующего гена.
На основе сведений о нуклеотидных последовательностях синтезировали праймеры, предназначенные для обеспечения вставки сайтов рестрикции XhoI в 5' и 3' концы (SEQ ID NO: 5 и 6, соответственно), с целью получения фрагмента ДНК, содержащего участок от примерно 200 п.н. выше инициирующего кодона НМ1524 до примерно 50 п.н. ниже его стоп-кодона, и выполняли ПЦР с использованием геномной ДНК Corynebacterium glutamicum в качестве матрицы. ПЦР выполняли путем начальной денатурации при 94°С в течение 5 минут; 30 циклов, состоящих из денатурации при 94°С в течение 30 секунд, отжига при 56°С в течение 30 секунд и полимеризации при 72°С в течение 90 секунд; и конечной полимеризации при 72°С в течение 7 минут.

После обработки указанного выше продукта ПЦР-амплификации с помощью фермента рестрикции XhoI, обрабатывали вектор pECCG117 с помощью XhoI, лигировали с полученными фрагментами ДНК и трансформировали в Е. coli DH5α, и полученный в результате трансформант высевали на твердую среду LB, содержащую канамицин (25 мг/л). После ПЦР-скрининга (с использованием SEQ ID NOs: 3 и 4) колоний, трансформированных с помощью вектора, в который был вставлен целевой ген, была получена плазмида с использованием хорошо известного способа извлечения плазмид, и эта плазмида была названа ECCG-HM1524.
Пример 4: Анализ продуктивности по лизину у штамма с введенным вектором сверхэкспрессии гена НМ1524
После ввода вектора pECCG-HM1524, полученного в Примере 3, в Corynebacterium glutamicum KCCM11016P (то есть продуцирующий лизин штамм) с использованием способа электрического импульса, полученные в результате трансформанты высевали на содержащую канамицин (25 мг/л) комплексную планшетную среду и после культивирования в течение 24 часов при 30°С были получены колонии. Полученный штамм назвали KCCM11016P/pECCG-HM1524, и концентрацию L-лизина в культуральной жидкости анализировали после культивирования трех партий в соответствии со способом культивирования в колбе Примера 2 (Таблица 2).
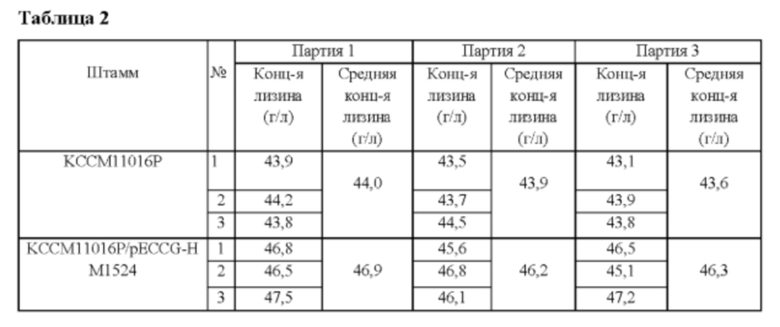
В результате было обнаружено, что продуктивность по лизину штамма KCCM11016P/pECCG-HM1524, у которого ген НМ1524 был сверхэкспрессирован, повышалась на 6% по сравнению с таковой у родительского штамма KCCM11016P.
Пример 5: Получение вектора для дополнительной вставки в хромосому гена НМ1524
Для подтверждения эффекта гена НМ1524, обнаруженного в Примере 4, был получен вектор для дальнейшей вставки гена в хромосому Corynebacterium. Для амплификации промотора Pcj7, полученного из Corynebacterium ammoniagenes (патент Кореи №10-0620092), синтезировали праймеры, которые были предназначены для обеспечения вставки сайта рестрикции EcoRI в 5'-конце и сайта рестрикции NdeI в 3'-конце промотора Pcj7 (SEQ ID NOs: 7 и 8, соответственно), и для обеспечения вставки сайта рестрикции SpeI в 5'-конце и сайта рестрикции SalI в 3'-конце промотора Pcj7 (SEQ ID NOs: 9 и 10, соответственно). В результате выполнения ПЦР с использованием синтезированных праймеров (SEQ ID NOs: 7 и 8, и SEQ ID NOs: 9 и 10) с геномном ДНК Corynebacterium ammoniagenes в качестве матрицы, были получены фрагменты ДНК гена НМ1524, содержащие сайты рестрикции EcoRI и NdeI в 5' и 3' концах, соответственно, а также сайты рестрикции SpeI и SalI в 5'- и 3'-концах, соответственно. ПЦР выполняли путем начальной денатурации при 94°С в течение 5 минут; 30 циклов, состоящих из денатурации при 94°С в течение 30 секунд, отжига при 56°С в течение 30 секунд и полимеризации при 72°С в течение 30 секунд; и конечной полимеризации при 72°С в течение 7 минут.
На основе нуклеотидной последовательности, о которой сообщалось, праймеры, предназначенные для обеспечения вставки сайта рестрикции NdeI в положение инициирующего кодона и сайта рестрикции SpeI ниже стоп-кодона (SEQ ID NOs: 11 и 12, соответственно), синтезировали для амплификации ORF (открытой рамки считывания) гена НМ1524. В результате выполнения ПЦР с использованием праймеров SEQ ID NOs: 11 и 12 с геномной ДНК Corynebacterium glutamicum АТСС13032 в качестве матрицы получали фрагменты ДНК гена НМ1524, содержащие сайты рестрикции NdeI и SpeI в позиции инициирующего кодона и ниже стоп-кодона, соответственно. ПЦР выполняли путем начальной денатурации при 94°С в течение 5 минут; 30 циклов, состоящих из денатурации при 94°С в течение 30 секунд, отжига при 56°С в течение 30 секунд, и полимеризации при 72°С в течение 90 секунд; и конечной полимеризации при 72°С в течение 7 минут.

После обработки каждого из указанных выше трех продуктов ПЦР-амплификации с помощью ферментов рестрикции, соответствующих сайтам рестрикции в каждом конце, лигировали вектор pDZ (патент Кореи №10-0924065) с фрагментами ДНК, которые были получены после обработки с помощью ферментов рестрикции EcoR и SalI, и таким образом получали вектор pDZ-Pcj7-HM1524.
Пример 6: Анализ продуктивности по лизину штамма с дополнительно вставленным в хромосому геном НМ1524
Вектор pDZ-Pcj7-HM1524, полученный в Примере 5, вводили в Corynebacterium glutamicum KCCM11016P с использованием способа электрического импульса, и среди колоний, трансформированных посредством гомологичной рекомбинации, отбирали те колонии, у которых ген НМ1524 был вставлен ниже стоп-кодона гена НМ1524 на хромосоме. Праймеры SEQ ID NOs: 13 и 14 использовали для скрининга колоний посредством ПЦР. Отобранный штамм назвали KCCM11016P::Pcj7-HM1524, и концентрацию L-лизина в культуральной жидкости анализировали после культивирования согласно способу культивирования в колбе Примера 2 (Таблица 3).
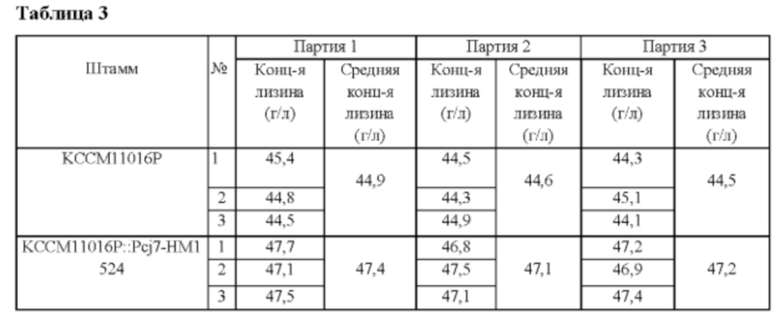
В результате было обнаружено, что продуктивность по лизину штамма KCCM11016P::Pcj7-HM1524, в который дополнительно был вставлен ген НМ1524, повышалась примерно на 6% по сравнению с таковой у родительского штамма KCCM11016P. Штамм KCCM11016P::Pcj7-HM1524 был назван СА01-2297 и депонирован в Корейском центре культур микроорганизмов (KCCM), международном органе депонирования, в соответствии с Будапештским договором, 2 августа 2016, с присвоенным номером доступа № KCCM11876P.
Пример 7: Получение лизина с использованием полученного из KCCM10770P микроорганизма, в который дополнительно вставлен ген НМ1524
Вектор pDZ-Pcj7-HM1524, полученный в Примере 5, трансформировали в Corynebacterium glutamicum KCCM10770P, продуцирующий лизин штамм (патент Кореи №10-0924065). Указанный выше Corynebacterium glutamicum KCCM10770P отличается тем, что 7 генов, ассоциированных с путем биосинтеза L-лизина, вставлены в его хромосому. Колонии селективно выделяли посредством ПЦР, и штамм с введенным геном НМ1524 был назван Corynebacterium glutamicum KCCM10770P::Pcj7-HM1524. После этого анализировали концентрацию L-лизина в культуральной жидкости после культивирования согласно способу культивирования в колбе Примера 2 (Таблица 4).
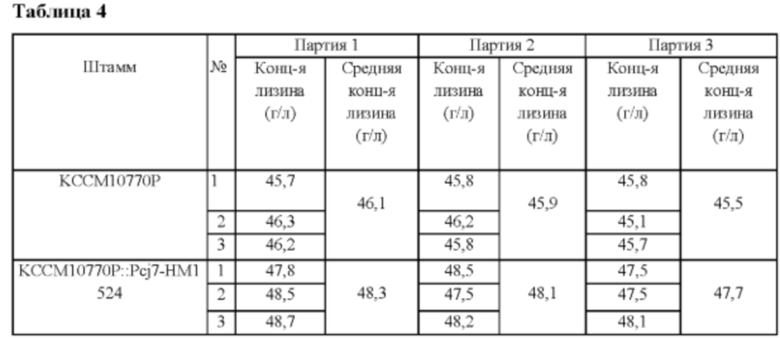
В результате было обнаружено, что продуктивность по лизину штамма Corynebacterium glutamicum KCCM10770P::Pcj7-HM1524 повышалась примерно на 5% по сравнению с таковой у родительского штамма.
Пример 8: Получение лизина с использованием полученного из CJ3P микроорганизма, в который дополнительно вставлен ген НМ1524
Вектор pDZ-Pcj7-HM1524, полученный в Примере 5, трансформировали в Corynebacterium glutamicum CJ3P, продуцирующий лизин штамм (Binder et al. Genome Biology 2012, 13:R40). Corynebacterium glutamicum CJ3P отличается тем, что 3 гена, ассоциированных с повышением продуктивности по L-лизину, вставлены в его хромосому. Колонии селективно выделяли посредством ПЦР и штамм с введенным геном НМ1524 назвали Corynebacterium glutamicum CJ3P::Pcj7-HM1524. Концентрацию L-лизина в культуральной жидкости анализировали после культивирования согласно способу культивирования в колбе Примера 2 (Таблица 5).
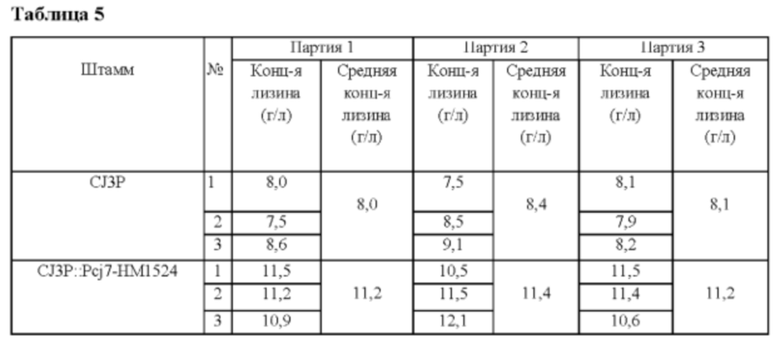
В результате было обнаружено, что продуктивность по лизину штамма Corynebacterium glutamicum CJ3P::Pcj7-HM1524 повышалась примерно на 38% по сравнению с таковой у родительского штамма.
Пример 9: Получение лизина с использованием полученного из KCCM11347P микроорганизма, в который дополнительно вставлен ген НМ1524
Вектор pDZ-Pcj7-HM1524, полученный в Примере 5, был трансформирован в Corynebacterium glutamicum KCCM11347P, продуцирующий лизин штамм (первоначально обозначенный как KFCC10750, микроорганизм был повторно депонирован в международном органе депонирования в соответствии с Будапештским договором и ему был присвоен номер доступа № KCCM11347P; патент Кореи №10-0073610). Corynebacterium glutamicum KCCM11347P отличается тем, что 3 гена, ассоциированных с повышением продуктивности по L-лизину, были вставлены в его хромосому. Колонии селективно выделяли посредством ПЦР, и штамм с введенным геном НМ1524 был назван Corynebacterium glutamicum KCCM11347P::Pcj7-HM1524. Концентрацию L-лизина в культуральной жидкости анализировали после культивирования согласно способу культивирования в колбе Примера 2 (Таблица 6).
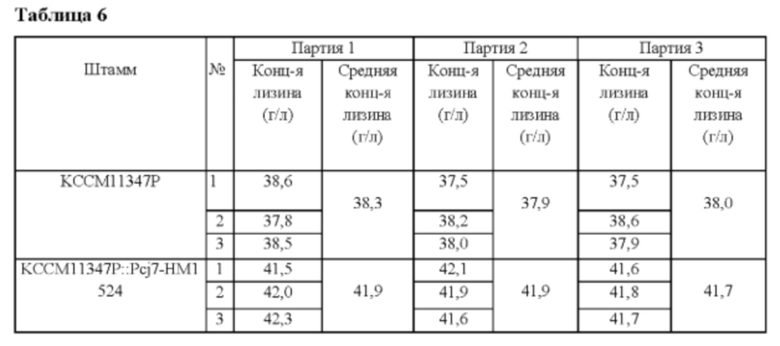
В результате было обнаружено, что продуктивность по лизину штамма Corynebacterium glutamicum KCCM11347P::Pcj7-HM1524 повышалась примерно на 10% по сравнению с таковой у родительского штамма.
В целом, приведенные выше результаты показывают, что для штаммов, имеющих повышенную активность гена НМ1524 по сравнению с его эндогенной активностью, продуктивность по лизину улучшалась, и дополнительно позволяют предположить, что лизин может продуцироваться в большом количестве посредством повышения активности белка, кодированного указанным выше геном, у микроорганизма.
Как видно из вышеизложенного, специалист в области техники, к которой принадлежит настоящее изобретение, сможет понять, что настоящее изобретение может быть осуществлено в других конкретных формах без модификации технических концепций или существенных признаков настоящего изобретения. В этом отношении типичные воплощения, раскрытые в данном описании изобретения, предназначены только для иллюстративных целей и не должны истолковываться как ограничивающие объем настоящего изобретения. Напротив, настоящее изобретение предназначено охватывать не только типичные воплощения, но также различные альтернативы, модификации, эквиваленты и другие воплощения, которые могут быть включены в сущность и объем настоящего изобретения, который определяется прилагаемой формулой изобретения.
Номер доступа
Название учреждения депонирования: Корейский центр культур микроорганизмов (KCCM) (Международный орган депонирования)
Номер доступа: KCCM11876P
Дата заявки: 2 августа 2016
--->
<110> CJ CheilJedang Corporation
<120> A microorganism of the genus Corynebacterium and a method for
producing L-lysine using the same
<130> OPA17196
<150> KR 10-2016-0152037
<151> 2016-11-15
<160> 16
<170> KopatentIn 2.0
<210> 1
<211> 418
<212> PRT
<213> Artificial Sequence
<220>
<223> HM1524 аминокислота
<400> 1
Met Arg Val Ala Met Ile Ser Met His Thr Ser Pro Leu Gln Gln Pro
1 5 10 15
Gly Thr Gly Asp Ser Gly Gly Met Asn Val Tyr Ile Leu Ser Thr Ala
20 25 30
Thr Glu Leu Ala Lys Gln Gly Ile Glu Val Asp Ile Tyr Thr Arg Ala
35 40 45
Thr Arg Pro Ser Gln Gly Glu Ile Val Arg Val Ala Glu Asn Leu Arg
50 55 60
Val Ile Asn Ile Ala Ala Gly Pro Tyr Glu Gly Leu Ser Lys Glu Glu
65 70 75 80
Leu Pro Thr Gln Leu Ala Ala Phe Thr Gly Gly Met Leu Ser Phe Thr
85 90 95
Arg Arg Glu Lys Val Thr Tyr Asp Leu Ile His Ser His Tyr Trp Leu
100 105 110
Ser Gly Gln Val Gly Trp Leu Leu Arg Asp Leu Trp Arg Ile Pro Leu
115 120 125
Ile His Thr Ala His Thr Leu Ala Ala Val Lys Asn Ser Tyr Arg Asp
130 135 140
Asp Ser Asp Thr Pro Glu Ser Glu Ala Arg Arg Ile Cys Glu Gln Gln
145 150 155 160
Leu Val Asp Asn Ala Asp Val Leu Ala Val Asn Thr Gln Glu Glu Met
165 170 175
Gln Asp Leu Met His His Tyr Asp Ala Asp Pro Asp Arg Ile Ser Val
180 185 190
Val Ser Pro Gly Ala Asp Val Glu Leu Tyr Ser Pro Gly Asn Asp Arg
195 200 205
Ala Thr Glu Arg Ser Arg Arg Glu Leu Gly Ile Pro Leu His Thr Lys
210 215 220
Val Val Ala Phe Val Gly Arg Leu Gln Pro Phe Lys Gly Pro Gln Val
225 230 235 240
Leu Ile Lys Ala Val Ala Ala Leu Phe Asp Arg Asp Pro Asp Arg Asn
245 250 255
Leu Arg Val Ile Ile Cys Gly Gly Pro Ser Gly Pro Asn Ala Thr Pro
260 265 270
Asp Thr Tyr Arg His Met Ala Glu Glu Leu Gly Val Glu Lys Arg Ile
275 280 285
Arg Phe Leu Asp Pro Arg Pro Pro Ser Glu Leu Val Ala Val Tyr Arg
290 295 300
Ala Ala Asp Ile Val Ala Val Pro Ser Phe Asn Glu Ser Phe Gly Leu
305 310 315 320
Val Ala Met Glu Ala Gln Ala Ser Gly Thr Pro Val Ile Ala Ala Arg
325 330 335
Val Gly Gly Leu Pro Ile Ala Val Ala Glu Gly Glu Thr Gly Leu Leu
340 345 350
Val Asp Gly His Ser Pro His Ala Trp Ala Asp Ala Leu Ala Thr Leu
355 360 365
Leu Asp Asp Asp Glu Thr Arg Ile Arg Met Gly Glu Asp Ala Val Glu
370 375 380
His Ala Arg Thr Phe Ser Trp Ala Ala Thr Ala Ala Gln Leu Ser Ser
385 390 395 400
Leu Tyr Asn Asp Ala Ile Ala Asn Glu Asn Val Asp Gly Glu Thr His
405 410 415
His Gly
<210> 2
<211> 1257
<212> DNA
<213> Artificial Sequence
<220>
<223> HM1524 нуклеотид
<400> 2
atgcgcgtag ctatgatttc catgcacacc tctccattgc agcagcccgg aactggtgat 60
tcaggcggca tgaacgtcta cattctttcg accgcgactg agctagcgaa acagggtatc 120
gaggtcgata tttacactcg tgccacgagg ccttctcagg gtgagatcgt gagagtagct 180
gagaatttgc gggtcattaa tatcgctgcg gggccgtatg aggggctttc caaagaggag 240
cttcctactc agttggcggc gtttaccggc ggaatgttgt cgtttacgcg ccgggagaag 300
gttacttatg atctgatcca ttctcactat tggctgtctg gtcaggtggg gtggttgctg 360
cgcgatttgt ggcggattcc ccttattcat acggcacaca ctttggcggc ggtgaagaat 420
tcttatcggg atgattcgga cactccggag tcggaggcgc gtcgcatttg tgagcagcag 480
ctggtggata acgctgacgt gttggcggtg aacactcagg aggagatgca ggatttgatg 540
catcactacg atgcggatcc ggatcggatt tctgtggtgt caccgggtgc ggacgtggaa 600
ctttatagcc ctggaaatga tcgcgcgacg gaacgttccc gtcgtgagct gggcattccg 660
ctgcacacaa aggtagtggc ttttgtgggt cggttgcagc cgtttaaggg cccgcaggtg 720
ctgatcaagg cggttgcggc gttgtttgat cgcgatccgg accgaaatct gcgcgtcatt 780
atttgtggcg gcccttctgg tccgaatgcg acaccggata cctataggca tatggcagag 840
gaactgggcg tcgaaaagcg aattcgcttt ttggacccgc gcccgccgag cgagctagtg 900
gccgtgtatc gggcggcgga catcgtggcc gtgccaagtt ttaatgagtc cttcggactc 960
gtcgccatgg aggcgcaagc cagcggcaca ccggtcattg cggcccgggt tggcggcctg 1020
cccatcgcag tcgcggaagg ggagacggga ttgcttgtcg acggccactc cccgcatgcc 1080
tgggccgacg ccttagccac actcttggac gatgacgaaa cgcgcatcag aatgggtgaa 1140
gacgccgtcg aacacgccag aacattctcc tgggcggcca ccgccgcaca gctatcgtcg 1200
ctgtacaacg acgctattgc caacgaaaat gtcgacggtg aaacgcatca cggctaa 1257
<210> 3
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> WT Библиотека геномной ДНК праймер-1
<400> 3
acgacgggat cagtaccga 19
<210> 4
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> WT библиотека геномной ДНК праймер-2
<400> 4
agctatctgt cgcagcgcc 19
<210> 5
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> 5' XhoI праймер
<400> 5
tcactcgagt gatggccagg ttgttgtc 28
<210> 6
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> 3' XhoI праймер
<400> 6
tcactcgagt tagtcatagg tactagttt 29
<210> 7
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> 5' EcoRI праймер
<400> 7
tcagaattct tccttcaggc taatctttt 29
<210> 8
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> 3' NdeI праймер
<400> 8
tcacatatgt gtttcctttc gttgggtac 29
<210> 9
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> 5' SpeI праймер
<400> 9
tcaactagtc ttccttcagg ctaatcttt 29
<210> 10
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> 3' SalI праймер
<400> 10
tcagtcgact gtttcctttc gttgggtac 29
<210> 11
<211> 26
<212> DNA
<213> Artificial Sequence
<220>
<223> s-NdeI праймер
<400> 11
tcacatatgc gcgtagctat gatttc 26
<210> 12
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> t-SpeI праймер
<400> 12
tcaactagtt tagccgtgat gcgtttcac 29
<210> 13
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> HM1524 селективный праймер-1
<400> 13
gtcgaacacg ccagaacatt 20
<210> 14
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> HM1524 селективный праймер-2
<400> 14
tactctcacg atctcaccct 20
<210> 15
<211> 4102
<212> DNA
<213> Artificial Sequence
<220>
<223> pEC-H15
<400> 15
gatcggatac ggctcagtcc actacgtttc ccacaccgga agggaaggcg attggtttca 60
gtgtggtttc agcccggcga agtccaaaat ctgcctgtat ggcctgaagg attcgcctcg 120
cggtgaggaa ttgctgcaga aacttggaaa atacaccgaa ggccgcggat gcgtgtacat 180
caataaaccg gaagacatcg atttggatgt tttagaggcc atgatcagcg agtcatgggc 240
cggccaaggc taggttgcaa atccccacca caagttgctc tgatcagcga ttttgtggtg 300
gatttttgcg tctccgccac ctgaaaccgc aaggattcac cacagattcg agttttcctt 360
tgaaacgtgg tggatccttg ccctgcaaac tttcaggaat cacaccagtc ccactggcca 420
caaatgggaa acccctcaga atcgcttctg aggggttatc tagcgccagt tggggtaagt 480
gcccatttgg gaaactcgac ctcttaaatc ggcgtctact tgccgagctt cttcaacgtc 540
cactcgtgcg gggcctgagt tgcaaatccc caccacaaat tgctctgatt agtggatttg 600
tggtggattt ttgcgtcttc gctatctgaa accgcgagga tccaccacag attcgagttt 660
tcctttgaaa cgtggttgat ccttgccctg gaatctttca ggaatcacat cagtcccact 720
ggttacaaag tggaaacccc tcagaatcgc ttctgggggg tgaactagcg ccagttgggg 780
taagtgccca tttgggaaac tcgacctctt aaatcggcgt ctacttgccg agcttcttca 840
acaactccgc ctgcacttca cgacgcctaa tcttgcccat ctgatcccgc ggcatctcct 900
caaagtggta gaaagtgcgc ggaaccttgt agcgggtgag gttcttgcgg gcgaattcct 960
tcaggccatc cggatccagc gctgcacctt ccaccaaagt gatggcagca acgacgtttt 1020
cggagccgtc ttcacgcggg ataccaacga ctgcggaatc ttcaatgtct gggtgctctg 1080
cgaggacttc ttcaacctca gctgggtaca cgttgaaacc gccagtgatg atgacttcct 1140
tgatgcgagc aactaggcgg atgaacccgt cttcttccat cactccgacg tcgccggtgc 1200
ggtaccactc gccgtggaag ctgttcttgg tggcttcttc ctggttgagg taacccttga 1260
acacctgtgg gcccttgact aggacttcgc cttcgctgcc gtcgggcatg gtttcgtcga 1320
ggttttctgg gtttgcgatg cgcacgatgg tgtcggggaa ggggattcct acgtagcctt 1380
ggcgtcggtg atcgctcatg gggttaccca cgatgatggg ggaggtttcg gtgaggccgt 1440
agccttcgac gaggcgtccg ccggtgtgct tttcccagcg ttcaacggtg cgctgggaga 1500
gtgtggatgc accggagaag gcgttgcgga ctcccttgat ggggattcct tctttttcgg 1560
aggcgtcgac gattttttcg taaagggtgg gcacgcctgg tagccaggtt ggggtgtgct 1620
ttttcattac gttcatgatc aggtcgatgc gtggggtggg aagtagcacc atttcgccac 1680
cgatgaacac ggacagtgtg ccgaccatgg tcagaccgta tgcgtggaac attggtaggg 1740
ctgcaagcat gcgttctggt ttgtctccga gacctggaac ccagtgcttt ccttggagga 1800
gattggagaa caggtttccg tgggtgagct gggcaccctt ggggcgtccg gtggtgccgg 1860
aggtgtagag gatcagcgcg acggattctt tggtcacggt gggttctgaa actacgtcgt 1920
cgccgtcgcc gcccattgct gcgctggtca gggtttcaaa aggaacggtg ttgggggctg 1980
cgccggagag ggattcgcgg ctcttgcgca gtgcagggat tgggagccga agtgctaggc 2040
gctggagtgg tggcatcgcg ttgatcatgt tgaccgacac gatggtttcc aactgggtct 2100
gtccacgtag ctgttcgacg gtgggggagg ctttgtccca gacgatggca acgcgggcac 2160
cgtggtcttt gaagggttcg agcagttcgt gggcggtgta gagcgggttg tgctcaatga 2220
ctactgcgcc gagtttcagc actgcgtaga aagctgcgat gtgctgtggg cagttgggga 2280
ggataatcgc tacgtgatcg ccggggcgga cacctagtgc gcgcaggcca gcggcagttt 2340
tgcggacttc tttgtccagt tcaccgtagg tttgtgaacg accgaaaaag taggtggctg 2400
gcttgtctgc gttgatggcc aggttgttgt cgtaaacgtc cagcagggtg gtgtcgccat 2460
attccagcga gtgtggcgtc cactctgggt agtgctggag ccattctttg gtttcgtatg 2520
ctgacatggt gtcccttcaa ctgcgttgct ttagtgccct ttagtatata gagacgtccc 2580
gctgctttct tcggcgatct agaatgtggg catgcgcgta gctatgattt ccatgcacac 2640
ctctccattg cagcagcccg gaactggtga ttcaggcggc atgaacgtct acattctttc 2700
gaccgcgact gagctagcga aacagggtat cgaggtcgat atttacactc gtgccacgag 2760
gccttctcag ggtgagatcg tgagagtagc tgagaatttg cgggtcatta atatcgctgc 2820
ggggccgtat gaggggcttt ccaaagagga gcttcctact cagttggcgg cgtttaccgg 2880
cggaatgttg tcgtttacgc gccgggagaa ggttacttat gatctgatcc attctcacta 2940
ttggctgtct ggtcaggtgg ggtggttgct gcgcgatttg tggcggattc cccttattca 3000
tacggcacac actttggcgg cggtgaagaa ttcttatcgg gatgattcgg acactccgga 3060
gtcggaggcg cgtcgcattt gtgagcagca gctggtggat aacgctgacg tgttggcggt 3120
gaacactcag gaggagatgc aggatttgat gcatcactac gatgcggatc cggatcggat 3180
ttctgtggtg tcaccgggtg cggacgtgga actttatagc cctggaaatg atcgcgcgac 3240
ggaacgttcc cgtcgtgagc tgggcattcc gctgcacaca aaggtagtgg cttttgtggg 3300
tcggttgcag ccgtttaagg gcccgcaggt gctgatcaag gcggttgcgg cgttgtttga 3360
tcgcgatccg gaccgaaatc tgcgcgtcat tatttgtggc ggcccttctg gtccgaatgc 3420
gacaccggat acctataggc atatggcaga ggaactgggc gtcgaaaagc gaattcgctt 3480
tttggacccg cgcccgccga gcgagctagt ggccgtgtat cgggcggcgg acatcgtggc 3540
cgtgccaagt tttaatgagt ccttcggact cgtcgccatg gaggcgcaag ccagcggcac 3600
accggtcatt gcggcccggg ttggcggcct gcccatcgca gtcgcggaag gggagacggg 3660
attgcttgtc gacggccact ccccgcatgc ctgggccgac gccttagcca cactcttgga 3720
cgatgacgaa acgcgcatca gaatgggtga agacgccgtc gaacacgcca gaacattctc 3780
ctgggcggcc accgccgcac agctatcgtc gctgtacaac gacgctattg ccaacgaaaa 3840
tgtcgacggt gaaacgcatc acggctaagt aaacgcgcgt cgtggaacat aaagtggcaa 3900
actagtacct atgactaacg gaaaattgat tcttcttcgt cacggtcaga gcgaatggaa 3960
cgcatccaac cagttcactg gatgggtcga cgtcaatctg accgaacagg gtgaggctga 4020
ggccaagcgc ggaggcgaac tcctcgtcga ggcaggcgtc ctcccaggcg ttgtatacac 4080
ctccttgctg cgtcgcgcga tc 4102
<210> 16
<211> 3181
<212> DNA
<213> Artificial Sequence
<220>
<223> pEC-M24
<400> 16
gatcgccggg gcggacacct agtgcgcgca ggccagcggc agttttgcgg acttctttgt 60
ccagttcacc gtaggtttgt gaacgaccga aaaagtaggt ggctggcttg tctgcgttga 120
tggccaggtt gttgtcgtaa acgtccagca gggtggtgtc gccatattcc agcgagtgtg 180
gcgtccactc tgggtagtgc tggagccatt ctttggtttc gtatgctgac atggtgtccc 240
ttcaactgcg ttgctttagt gccctttagt atatagagac gtcccgctgc tttcttcggc 300
gatctagaat gtgggcatgc gcgtagctat gatttccatg cacacctctc cattgcagca 360
gcccggaact ggtgattcag gcggcatgaa cgtctacatt ctttcgaccg cgactgagct 420
agcgaaacag ggtatcgagg tcgatattta cactcgtgcc acgaggcctt ctcagggtga 480
gatcgtgaga gtagctgaga atttgcgggt cattaatatc gctgcggggc cgtatgaggg 540
gctttccaaa gaggagcttc ctactcagtt ggcggcgttt accggcggaa tgttgtcgtt 600
tacgcgccgg gagaaggtta cttatgatct gatccattct cactattggc tgtctggtca 660
ggtggggtgg ttgctgcgcg atttgtggcg gattcccctt attcatacgg cacacacttt 720
ggcggcggtg aagaattctt atcgggatga ttcggacact ccggagtcgg aggcgcgtcg 780
catttgtgag cagcagctgg tggataacgc tgacgtgttg gcggtgaaca ctcaggagga 840
gatgcaggat ttgatgcatc actacgatgc ggatccggat cggatttctg tggtgtcacc 900
gggtgcggac gtggaacttt atagccctgg aaatgatcgc gcgacggaac gttcccgtcg 960
tgagctgggc attccgctgc acacaaaggt agtggctttt gtgggtcggt tgcagccgtt 1020
taagggcccg caggtgctga tcaaggcggt tgcggcgttg tttgatcgcg atccggaccg 1080
aaatctgcgc gtcattattt gtggcggccc ttctggtccg aatgcgacac cggataccta 1140
taggcatatg gcagaggaac tgggcgtcga aaagcgaatt cgctttttgg acccgcgccc 1200
gccgagcgag ctagtggccg tgtatcgggc ggcggacatc gtggccgtgc caagttttaa 1260
tgagtccttc ggactcgtcg ccatggaggc gcaagccagc ggcacaccgg tcattgcggc 1320
ccgggttggc ggcctgccca tcgcagtcgc ggaaggggag acgggattgc ttgtcgacgg 1380
ccactccccg catgcctggg ccgacgcctt agccacactc ttggacgatg acgaaacgcg 1440
catcagaatg ggtgaagacg ccgtcgaaca cgccagaaca ttctcctggg cggccaccgc 1500
cgcacagcta tcgtcgctgt acaacgacgc tattgccaac gaaaatgtcg acggtgaaac 1560
gcatcacggc taagtaaacg cgcgtcgtgg aacataaagt ggcaaactag tacctatgac 1620
taacggaaaa ttgattcttc ttcgtcacgg tcagagcgaa tggaacgcat ccaaccagtt 1680
cactggatgg gtcgacgtca atctgaccga acagggtgag gctgaggcca agcgcggagg 1740
cgaactcctc gtcgaggcag gcgtcctccc aggcgttgta tacacctcct tgctgcgtcg 1800
cgcgatccgc actgcaaaca tcgcactgaa cgctgcagac cgccactgga tcccagtgat 1860
ccgcgactgg cgcctcaacg agcgtcacta cggcgcactg cagggccttg acaaggctgc 1920
aaccaaggaa aaatacggcg acgaccagtt catggaatgg cgccgctcct acgacacccc 1980
accaccagag ctcgcggatg acgcagagta ctcccaggca aatgaccctc gttacgcgga 2040
cctcgacgta gttccacgca ccgaatgcct caaggacgtt gtggttcgtt ttgttcctta 2100
cttcgaggaa gaaatcctgc cacgcgcaaa gaagggcgaa accgtcctca tcgcagcaca 2160
cggcaactcc ctgcgtgcgc tggttaagca ccttgacggc atctccgatg ctgatatcgc 2220
agagctcaac atcccaaccg gcatcccact ggtctacgaa atcgccgaag acggttccgt 2280
agtaaaccca ggcggcacct acctcgatcc tgaggcagca gcagccggcg cagcagcagt 2340
agcaaaccag ggtaataagt agctatttgt aggtgagcac tcttcttgct ttcgtattgg 2400
gcgtggtcct catgggcctc gccctacctg cgtatacgaa aattaaagat cggatgcgtc 2460
gccacaagtc cgcggtcacc ctgtccgaaa accaggtcac cacggtgggg caggtcctcc 2520
acctggcgat tcaaggctcc ccaacgggaa tcacggttgt cgatcgcacc ggcgacgtca 2580
tcttatccaa cggccgcgcc cacgaattgg gcatcgtcca cgaaagatcc gtcgacggca 2640
acgtttggcg cgtcgcccag gaagccttcc aagaccaaga aacccactca ctcgacgtcc 2700
acccagaccg caatccgcgg cgcccgggta gtcgcatcac cgcagtgcag gcagtggtca 2760
agcctttaac gcttatcgac gatcgtttcg tgatcatcta tgcctccgac gaatccgaaa 2820
acgtgcgcat ggaatcggca cgccgagact tcgtcgcaaa cgtctcccac gaactgaaaa 2880
cccccgtcgg cggcatggca ctcctcgcgg aagccctcat ggaatcctcc gacgacccag 2940
aacaagtcga atacttcgga tccaggctcc accgcgaagc ccaccgcatg gccgacatga 3000
tcaacgaact gatctccctt tccaaacttc agggcgccga acgactccct gatatggaac 3060
ccgtccaggc tgacgacatc atcagcgaag ccatcgaacg cacccaactc gccgccgaca 3120
acgccaacat cgaaatcatt cgcggcgacc gcaccggcgt ttgggtagaa gccgatcgat 3180
c 3181
<---
Настоящее изобретение относится к микроорганизму, продуцирующему L-лизин, и способу получения L-лизина с использованием этого микроорганизма. Изобретение позволяет получать L-лизин с высокой степенью эффективности. 2 н. и 2 з.п. ф-лы, 6 табл., 9 пр.
1. Микроорганизм рода Corynebacterium, продуцирующий L-лизин, имеющий улучшенную активность белка, состоящего из аминокислотой последовательности SEQ ID NO:l, по сравнению с эндогенной активностью.
2. Микроорганизм рода Corynebacterium, продуцирующий L-лизин, по п. 1, где микроорганизм представляет собой Corynebacterium glutamicum.
3. Способ получения L-лизина, включающий культивирование в среде микроорганизма рода Corynebacterium, продуцирующего L-лизин, имеющего улучшенную активность белка, состоящего из аминокислотной последовательности SEQ ID NO:l, по сравнению с эндогенной активностью; и
выделение L-лизина из культивируемого микроорганизма или среды его культивирования.
4. Способ получения L-лизина по п. 3, где микроорганизм представляет собой Corynebacterium glutamicum.
| ФЕРМЕНТАЦИОННЫЙ СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА | 1994 |
|
RU2204605C2 |
| РЕКОМБИНАНТНАЯ ДНК И СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА | 2009 |
|
RU2451748C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА | 2000 |
|
RU2264461C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА С ИСПОЛЬЗОВАНИЕМ МИКРООРГАНИЗМОВ, ОБЛАДАЮЩИХ СПОСОБНОСТЬЮ ПРОДУЦИРОВАТЬ L-ЛИЗИН | 2012 |
|
RU2588665C2 |
| KR 101053429 B1, 03.08.2011. | |||

