ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Настоящая заявка заявляет приоритет по предварительной заявке США № 61/739720, поданной 19 декабря 2012 года, которая включена в данный документ в полном объеме посредством ссылки.
ВКЛЮЧЕНИЕ СПИСКА ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Список последовательностей, который содержится в файле под названием "MONS323WOseq.txt", размером 345 Кб (измерено в Microsoft Windows®), и был создан 17 декабря 2013 года, подается при этом в электронной форме и включен в данный документ посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ
[0003] Настоящее изобретение относится к области молекулярной биологии растений, генной инженерии растений и молекулам ДНК, пригодным для модуляции экспрессии генов в растениях.
УРОВЕНЬ ТЕХНИКИ
[0004] Регуляторные элементы представляют собой генетические элементы, которые регулируют активность генов путем модуляции транскрипции функционально связанной транскрибируемой молекулы ДНК. Такие элементы включают промоторы, лидеры, энхансеры, интроны и 3’-нетранслируемые области и могут быть использованы в области молекулярной биологии растений и генетической инженерии растений.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0005] В изобретении предложены новые регуляторные элементы для использования в растениях и генетические конструкции, содержащие регуляторные элементы. В изобретении также предложены трансгенные клетки растений, растения, части растений и семена, содержащие регуляторные элементы. В одном варианте реализации изобретения в изобретении предложены регуляторные элементы, описанные в данном документе, функционально связанные с транскрибируемой молекулой ДНК. В некоторых вариантах реализации изобретения транскрибируемая молекула ДНК является гетерологической по отношению к последовательности регуляторного элемента, предложенного в данном документе. В данном документе также предложены способы получения и применения регуляторных элементов, описанных в данном документе, включая генетические конструкции, содержащие регуляторные элементы, и трансгенные растения, клетки растений, части растений и семена, содержащие регуляторные элементы, функционально связанные с транскрибируемой молекулой ДНК, гетерологической по отношению к регуляторному элементу.
[0006] Таким образом, в одном аспекте в изобретении предложена рекомбинантная молекула ДНК, содержащая последовательность ДНК, выбранную из группы, состоящей из: а) последовательности ДНК с по меньшей мере около 85 процентами идентичности последовательности к любой из SEQ ID №№: 1–98 и 168–171; б) последовательности ДНК, содержащей любую из SEQ ID №№: 1–98 и 168–171; и в) фрагмента любой из SEQ ID №№: 1–98 и 168–171, причем данный фрагмент обладает ген-регуляторной активностью; при этом последовательность ДНК функционально связана с гетерологической транскрибируемой молекулой ДНК. Под «гетерологической транскрибируемой молекулой ДНК» подразумевается, что транскрибируемая молекула ДНК является гетерологической по отношению к последовательности ДНК. В конкретных вариантах реализации изобретения рекомбинантная молекула ДНК содержит последовательность ДНК, имеющую по меньшей мере 90 процентов, по меньшей мере 91 процент, по меньшей мере 92 процента, по меньшей мере 93 процента, по меньшей мере 94 процента, по меньшей мере около 95 процентов, по меньшей мере 96 процентов, по меньшей мере 97 процентов, по меньшей мере 98 процентов или, по меньшей мере 99 процентов идентичности последовательности к последовательности ДНК любой из SEQ ID №№: 1–98 и 168–171. В конкретных вариантах реализации изобретения гетерологическая транскрибируемая молекула ДНК содержит ген, представляющий агрономический интерес, например, ген, способный придавать растениям устойчивость к гербицидам или вредителям. В еще других вариантах реализации изобретения в изобретении предложена генетическая конструкция, содержащая рекомбинантную молекулу ДНК, как предложено в данном документе.
[0007] В другом аспекте в данном документе предлагаются трансгенные клетки растений, содержащие рекомбинантную молекулу ДНК, включающую последовательность ДНК, выбранную из группы, состоящей из: а) последовательности ДНК с по меньшей мере около 85 процентами идентичности последовательности к любой из SEQ ID №№: 1–98 и 168–171; б) последовательности ДНК, содержащей любую из SEQ ID №№: 1–98 и 168–171; и в) фрагмента любой из SEQ ID №№: 1–98 и 168–171, причем данный фрагмент обладает ген-регуляторной активностью; при этом последовательность ДНК функционально связана с гетерологической транскрибируемой молекулой ДНК. В некоторых вариантах реализации изобретения трансгенная клетка растения представляет собой клетку однодольного растения. В других вариантах реализации изобретения трансгенная клетка растения представляет собой клетку двудольного растения.
[0008] В еще другом аспекте в данном документе ниже приведено трансгенное растение, или его часть, содержащее рекомбинантную молекулу ДНК, содержащую последовательность ДНК, выбранную из группы, состоящей из: а) последовательности ДНК с по меньшей мере около 85 процентами идентичности последовательности к любой из SEQ ID №№ 1–98 и 168–171; б) последовательности ДНК, содержащей любую из SEQ ID №№: 1–98 и 168–171; и в) фрагмента любой из SEQ ID №№: 1–98 и 168–171, причем данный фрагмент обладает ген-регуляторной активностью; при этом последовательность ДНК функционально связана с гетерологической транскрибируемой молекулой ДНК. В конкретных вариантах реализации изобретения трансгенное растение представляет собой дочернее растение любого поколения по отношению к исходному трансгенному растению и содержит молекулу рекомбинантной ДНК. Трансгенное семя, содержащее рекомбинантную молекулу ДНК, которое порождает такое трансгенное растение при выращивании, также предлагается в изобретении.
[0009] Еще в одном аспекте в изобретении предложен способ экспрессии транскрибируемой молекулы ДНК, например, гена, представляющего агрономический интерес, в трансгенном растении путем получения трансгенного растения, содержащего рекомбинантную молекулу ДНК по изобретению, и культивирования растения.
[00010] В данном документе также предложен способ, обеспечивающий трансгенное растение путем трансформации клетки растения рекомбинантной молекулой ДНК по изобретению, чтобы получить трансформированную клетку растения, и регенерации трансформированной клетки растения, чтобы получить трансгенное растение.
КРАТКОЕ ОПИСАНИЕ ФИГУР
[00011] Фиг. 1. Иллюстрирует выравнивание нескольких вариантов промоторов различных размеров, соответствующих промоторным элементам из Agrostis nebulosa. В частности, Фиг. 1 иллюстрирует выравнивание промотора длиной 2005 пар нуклеотидных оснований (п. н.) P-AGRne.Ubq1-1:1:5 (SEQ ID №: 2), включенного в группу регуляторных элементов экспрессии (EXP) EXP-AGRne.Ubq1:1:7 (SEQ ID №: 1), с вариантами промотора P-AGRne.Ubq1-1:1:5. Например, делеция на 5’-конце Р-AGRne.Ubq1-1:1:5 произвела промотор P-AGRne.Ubq1-1:1:4 (SEQ ID №: 6), последовательность длиной 999 п. н., которая включена в EXP-AGRne.Ubq1:1:8 (SEQ ID №: 5). Другой вариант промотора, проиллюстрированный на Фиг. 1, представляет собой P-AGRne.Ubq1-1:1:6 (SEQ ID №: 8), последовательность длиной 762 п. н., включенная в EXP-AGRne.Ubq1:1:9 (SEQ ID №: 7).
[00012] Фиг. 2. Иллюстрирует выравнивание нескольких вариантов промоторов различных размеров, соответствующих промоторным элементам из Arundo donax. В частности, Фиг. 2 иллюстрирует выравнивание промотора длиной 4114 п. н. P-ARUdo.Ubq1-1:1:4 (SEQ ID №: 10), включенного в группу регуляторных элементов экспрессии EXP-ARUdo.Ubq1:1:4 (SEQ ID №: 9), с вариантами промотора P-ARUdo.Ubq1-1:1:4. В выравнивание включены: промотор длиной 2012 п. н. P-ARUdo.Ubq1-1:1:5 (SEQ ID №: 14); промотор длиной 1000 п. н. P-ARUdo.Ubq1-1:1:6 (SEQ ID №: 17); и промотор длиной 755 п. н. P-ARUdo.Ubq1-1:1:8 (SEQ ID №: 22).
[00013] Фиг. 3. Иллюстрирует выравнивание нескольких вариантов промоторов различных размеров, соответствующих промоторным элементам из Arundo donax. В частности, Фиг. 3 иллюстрирует выравнивание промотора длиной 2033 п. н. P-ARUdo.Ubq2-1:1:4 (SEQ ID №: 24) с вариантами промотора P-ARUdo.Ubq2-1:1:4. В выравнивание включены: промотор длиной 2004 п. н. P-ARUdo.Ubq2-1:1:6 (SEQ ID №: 28); промотор длиной 1001 п. н. P-ARUdo.Ubq2-1:1:5 (SEQ ID №: 31); и промотор длиной 696 п. н. P-ARUdo.Ubq2-1:1:7 (SEQ ID №: 33).
[00014] Фиг. 4. Иллюстрирует выравнивание нескольких вариантов промоторов различных размеров, соответствующих промоторным элементам из Bouteloua gracilis. В частности, Фиг. 4 иллюстрирует выравнивание промотора длиной 2371 п. н. P-BOUgr.Ubq1-1:1:2 (SEQ ID №: 35) с вариантами промотора на 5’-конце P-BOUgr.Ubq1-1:1:2. В выравнивание включены: промотор длиной 1999 п. н. P-BOUgr.Ubq1-1:1:3 (SEQ ID №: 39); промотор длиной 1022 п. н. P-BOUgr.Ubq1-1:1:5 (SEQ ID №: 42); и промотор длиной 760 п. н. P-BOUgr.Ubq1-1:1:6 (SEQ ID №: 44).
[00015] Фиг. 5. Иллюстрирует выравнивание нескольких вариантов промоторов различных размеров, соответствующих промоторным элементам из Bouteloua gracilis. В частности, Фиг. 5 иллюстрирует выравнивание промоторного элемента длиной 2100 п. н. P-BOUgr.Ubq2-1:1:4 (SEQ ID №: 46) с вариантами промотора P-BOUgr.Ubq2-1:1:4. IВ выравнивание включены: промотор длиной 2043 п. н. P-BOUgr.Ubq2-1:1:7 (SEQ ID №: 50); промотор длиной 2002 п. н. P-BOUgr.Ubq2-1:1:5 (SEQ ID №: 53); промотор длиной 1024 п. н. P-BOUgr.Ubq2-1:1:6 (SEQ ID №: 56); и промотор длиной 749 п. н. P-BOUgr.Ubq2-1:1:8 (SEQ ID №: 61).
[00016] Фиг. 6. Иллюстрирует выравнивание нескольких вариантов промоторов различных размеров, соответствующих промоторным элементам из Miscanthus sinensis. В частности, Фиг. 6 иллюстрирует выравнивание промоторного элемента длиной 5359 п. н. P-MISsi.Ubq1-1:1:2 (SEQ ID №: 63) с вариантами промотора P-MISsi.Ubq1-1:1:2. В выравнивание включены: промотор длиной 2423 п. н. P-MISsi.Ubq1-1:1:11 (SEQ ID №: 67); промотор длиной 1447 п. н. P-MISsi.Ubq1-1:1:10 (SEQ ID №: 71); промотор длиной 899 п. н. P-MISsi.Ubq1-1:1:13 (SEQ ID №: 73); промотор длиной 691 п. н. P-MISsi.Ubq1-1:1:14 (SEQ ID №: 75); и промотор длиной 506 п. н. P-MISsi.Ubq1-1:1:9 (SEQ ID №: 77).
[00017] Фиг. 7. Иллюстрирует выравнивание нескольких вариантов промоторов различных размеров, соответствующих промоторным элементам из Schizachyium scoparium. В частности, Фиг. 7 иллюстрирует выравнивание промоторного элемента длиной 2831 п. н. P-SCHsc.Ubq1-1:1:12 (SEQ ID №: 79) с вариантами промотора P-SCHsc.Ubq1-1:1:12. В выравнивание включены: промотор длиной 2033 п. н. P-SCHsc.Ubq1-1:1:11 (SEQ ID №: 83); промотор длиной 1046 п. н. P-SCHsc.Ubq1-1:1:10 (SEQ ID №: 85); и промотор длиной 547 п. н. P-SCHsc.Ubq1-1:1:14 (SEQ ID №: 87).
[00018] Фиг. 8. Иллюстрирует выравнивание нескольких вариантов промоторов различных размеров, соответствующих промоторным элементам из Sorghastrum nutans. В частности, Фиг. 8 иллюстрирует выравнивание промоторного элемента длиной 2218 п. н. P-SORnu.Ubq1-1:1:4 (SEQ ID №: 89) с вариантами промотора P-SORnu.Ubq1-1:1:4. В выравнивание включены: промотор длиной 1964 п. н. P-SORnu.Ubq1-1:1:5 (SEQ ID №: 93); промотор длиной 1023 п. н. P-SORnu.Ubq1-1:1:6 (SEQ ID №: 96); и промотор длиной 724 п. н. P-SORnu.Ubq1-1:1:7 (SEQ ID №: 98).
[00019] Фиг. 9. Иллюстрирует конфигурации экспрессионных кассет по изобретению.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[00020] SEQ ID №№: 1, 5, 7, 9, 13, 16, 18, 19, 21, 23, 27, 30, 32, 34, 38, 41, 43, 45, 49, 52, 55, 58, 60, 62, 66, 70, 72, 74, 76, 78, 82, 84, 86, 88, 92, 95, 97, 99, 103, 106, 108, 110, 114, 116, 118, 120, 122, 126, 128, 132, 134, 138, 140, 144, 148, 150 и 168 представляют собой последовательности ДНК из групп регуляторных экспрессионных элементов (EXP), содержащих промоторную последовательность, функционально связанную с 5’-лидерной последовательностью, которая функционально связана с 5’-последовательностью интрона.
[00021] SEQ ID №№: 2, 6, 8, 10, 14, 17, 22, 24, 28, 31, 33, 35, 39, 42, 44, 46, 50, 53, 56, 61, 63, 67, 71, 73, 75, 77, 79, 83, 85, 87, 89, 93, 96, 98, 100, 104, 107, 109, 111, 117, 119, 121, 123, 129, 135, 141, 145, 151 и 169 представляют собой последовательности промотора.
[00022] SEQ ID №№: 3, 11, 25, 36, 47, 64, 68, 80, 90, 101, 112, 124, 130, 136, 142, 146, 152 и 170 представляют собой лидерные последовательности.
[00023] SEQ ID №№: 4, 12, 15, 20, 26, 29, 37, 40, 48, 51, 54, 57, 59, 65, 69, 81, 91, 94, 102, 105, 113, 115, 125, 127, 131, 133, 137, 139, 143, 147, 149, 153 и 171 представляют собой интронные последовательности.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
[00024] Изобретение относится к молекулам ДНК, имеющим ген-регуляторную активность в растениях. Нуклеотидные последовательности этих молекул ДНК приводятся в SEQ ID №№: 1–98 и 168–171. Эти молекулы ДНК, например, способны влиять на экспрессию функционально связанной транскрибируемой молекулы ДНК в тканях растений и, следовательно, регулировать экспрессию функционально связанного трансгена в трансгенных растениях. Изобретение также относится к способам их модификации, создания и применения. Изобретение также относится к композициям, которые включают трансгенные клетки растений, растения, части растений и семена, содержащие рекомбинантные молекулы ДНК по изобретению, и к способам их получения и применения.
[00025] Следующие определения и способы предлагаются для того, чтобы лучше обозначить изобретение и направлять к специалистам в данной области техники при применении изобретения. Если не указано иное, термины следует понимать в соответствии с общепринятым использованием специалистами в данной области техники.
Молекулы ДНК
[00026] Используемый в данном документе термин «ДНК» или «ДНК молекула» относится к двухцепочечной ДНК молекуле клеточного или синтетического происхождения, т. е. к полимеру дезоксирибонуклеотидных оснований. Используемый в данном документе термин «ДНК последовательность» относится к нуклеотидной последовательности ДНК молекулы. Номенклатура, использованная в данном документе, соответствует таковой по документу 37 Свода федеральных нормативных актов Соединенных Штатов § 1.822, и изложена в таблицах WIPO стандарта ST.25 (1998), приложение 2, таблицы 1 и 3.
[00027] Как используется в данном документе «рекомбинантная молекула ДНК» представляет собой молекулу ДНК, содержащую комбинацию молекул ДНК, которые бы не объединились естественным путем без вмешательства человека. Например, рекомбинантная молекула ДНК может быть молекулой ДНК, которая состоит по меньшей мере из двух гетерологических по отношению друг к другу молекул ДНК; молекулой ДНК, содержащей последовательность ДНК, которая отклоняется от существующих в природе последовательностей ДНК; или молекулой ДНК, которая была встроена в ДНК клетки-хозяина путем генетической трансформации.
[00028] Как используется в данном документе, термин «идентичность последовательностей» относится к степени, в которой две оптимально выровненные последовательности ДНК являются идентичными. Оптимальное выравнивание последовательностей создается путем выравнивания двух последовательностей ДНК вручную, например, референсной последовательности и другой последовательности ДНК, для создания максимального количества соответствий между нуклеотидами при выравнивании последовательностей с соответствующими внутренними нуклеотидными вставками, делециями или разрывами. Используемый в данном документе термин «референсная последовательность» относится к последовательности ДНК, приводимой в SEQ ID №№: 1–98 и 168–171.
[00029] Используемый в данном документе термин «процент идентичности последовательностей», или «процент идентичности», или «% идентичности» представляет собой долю идентичности, умноженную на 100. «Доля идентичности» для последовательности ДНК, оптимально выровненной с начальной последовательностью, представляет собой число соответствий между нуклеотидами при оптимальном выравнивании, деленное на общее число нуклеотидов в референсной последовательности, например, на общее число нуклеотидов во всей полноразмерной референсной последовательности. Таким образом, в одном варианте реализации изобретение относится к молекуле ДНК, содержащей последовательность ДНК, которая при оптимальном выравнивании с референсной последовательностью, представленной в данном документе как SEQ ID №№: 1–98 и 168–171 имеет по меньшей мере около 85 процентов идентичности, по меньшей мере около 86 процентов идентичности, по меньшей мере около 87 процентов идентичности, по меньшей мере около 88 процентов идентичности, по меньшей мере около 89 процентов идентичности, по меньшей мере около 90 процентов идентичности, по меньшей мере около 91 процента идентичности, по меньшей мере около 92 процентов идентичности, по меньшей мере около 93 процентов идентичности, по меньшей мере около 94 процентов идентичности, по меньшей мере около 95 процентов идентичности, по меньшей мере около 96 процентов идентичности, по меньшей мере около 97 процентов идентичности, по меньшей мере около 98 процентов идентичности, по меньшей мере около 99 процентов идентичности или, по меньшей мере около 100 процентов идентичности с референсной последовательностью.
Регуляторные элементы
[00030] Регуляторные элементы, такие как промоторы, лидеры, энхансеры, интроны и области терминации транскрипции (или 3´ UTR), играют существенную роль во всеобщей экспрессии генов в живых клетках. Термин «регуляторный элемент», используемый в данном документе, относится к молекулам ДНК, имеющим ген-регуляторную активность. Термин «ген-регуляторная активность», используемый в данном документе, относится к способности влияния на экспрессию функционально связанной транскрибируемой молекулы ДНК, например, путем воздействия на транскрипцию и/или трансляцию функционально связанной транскрибируемой молекулы ДНК. Регуляторные элементы, такие как промоторы, лидеры, энхансеры и интроны, функционирующие в растениях, являются, таким образом, полезными для модификации фенотипов растений с помощью генетической инженерии.
[00031] Используемая в данном документе «группа регуляторных элементов экспрессии» или «EXP» последовательность может относиться к группе функционально связанных регуляторных элементов, таких как энхансеры, промоторы, лидеры и интроны. Таким образом, группа регуляторных элементов экспрессии может состоять, например, из промотора, функционально связанного с 5’-лидерной последовательностью, которая, в свою очередь, функционально связана с 5’-последовательностью интрона.
[00032] Регуляторные элементы могут быть охарактеризованы по своему характеру экспрессии генов, например, по положительным и/или отрицательным воздействиям, таким как конститутивная экспрессия или временная, пространственная, зависящая от стадии развития, тканевая, зависящая от окружения, физиологическая, патологическая, клеточного цикла и/или химически чувствительная экспрессия, и любая из их комбинаций, а также по количественными или качественными показателям. Используемый в данном документе термин «характер экспрессии гена» представляет собой любую особенность транскрипции функционально связанной молекулы ДНК в транскрибируемую молекулу РНК. Транскрибируемая молекула РНК может быть транслирована для получения молекулы белка или может образовать антисмысловую или другую регуляторную молекулу РНК, такую как двухцепочечная РНК (дцРНК), транспортная РНК (тРНК), рибосомальная РНК (рРНК), микроРНК (микроРНК) и тому подобные.
[00033] Используемый в данном документе термин «экспрессия белка» представляет собой любую особенность трансляции транскрибируемой молекулы РНК в молекулу белка. Экспрессия белка может быть охарактеризована по своим временным, пространственным, относящимся к стадии развития или морфологическим качествам, а также по количественными или качественными показателям.
[00034] Промотор является полезным в качестве регуляторного элемента для модуляции экспрессии функционально связанной транскрибируемой молекулы ДНК. Как используется в данном документе, термин «промотор» относится в общем смысле к молекуле ДНК, которая участвует в распознавании и связывании РНК-полимеразы II и других белков, таких как транс-действующие факторы транскрипции, для инициации транскрипции. Промотор может происходить из 5’-нетранслируемой области (5´-UTR) гена. В альтернативном варианте промоторы могут быть получены синтетическим путем или в результате манипулирования молекулой ДНК. Промоторы также могут быть химерными. Химерные промоторы получают путем слияния двух или более гетерологических молекул ДНК. Промоторы, полезные в практической реализации изобретения, включают SEQ ID №№: 2, 6, 8, 10, 14, 17, 22, 24, 28, 31, 33, 35, 39, 42, 44, 46, 50, 53, 56, 61, 63, 67, 71, 73, 75, 77, 79, 83, 85, 87, 89, 93, 96, 98 и 169, включая их фрагменты или варианты. В конкретных вариантах реализации изобретения такие молекулы ДНК и любые их варианты или производные, описанные в данном документе, далее определяются как обладающие промоторной активностью, т. е. способны действовать в качестве промотора в клетке хозяина, такого как трансгенное растение. Еще в дополнительных конкретных вариантах реализации изобретения фрагмент может быть определен как проявляющий промоторную активность, которой обладает исходная молекула промотора, из которого он был получен, или фрагмент может содержать «минимальный промотор», обеспечивающий основной уровень транскрипции и включающий TATA бокс или эквивалентную последовательность ДНК для распознавания и связывания комплекса РНК-полимеразы II с целью инициации транскрипции.
[00035] В одном варианте реализации изобретения фрагменты снабжены промоторной последовательностью, описанной в данном документе. Фрагменты промоторов могут включать промоторную активность, как описано выше, и могут быть полезными по отдельности или в комбинации с другими промоторами и фрагментами промоторов, например, при создании химерных промоторов. В конкретных вариантах реализации изобретения предлагаются фрагменты промотора, содержащие, по меньшей мере около 50, по меньшей мере около 75, по меньшей мере около 95, по меньшей мере около 100, по меньшей мере около 125, по меньшей мере около 150, по меньшей мере около 175, по меньшей мере около 200, по меньшей мере около 225, по меньшей мере около 250, по меньшей мере около 275, по меньшей мере около 300, по меньшей мере около 500, по меньшей мере около 600, по меньшей мере около 700, по меньшей мере около 750, по меньшей мере около 800, по меньшей мере около 900 или по меньшей мере около 1000 последовательных нуклеотидов, или больше, молекулы ДНК, имеющей промоторную активность, описанную в данном документе. Способы получения таких фрагментов из исходной молекулы промотора хорошо известны в данной области техники.
[00036] Композиции, полученные из любых промоторов, представленных в SEQ ID №№: 2, 6, 8, 10, 14, 17, 22, 24, 28, 31, 33, 35, 39, 42, 44, 46, 50, 53, 56, 61, 63, 67, 71, 73, 75, 77, 79, 83, 85, 87, 89, 93, 96, 98 и 169, такие как внутренние или 5’-делеции, например, могут быть получены с использованием хорошо известных в данной области техники способов для улучшения или изменения экспрессии, включая удаление элементов, имеющих положительные или отрицательные воздействия на экспрессию; удвоение элементов, имеющих положительные или отрицательные воздействия на экспрессию; и/или удвоение или удаление элементов, имеющих ткане- или клеточно-специфические воздействия на экспрессию. Композиции, полученные из любых промоторов, представленных в SEQ ID №№: 2, 6, 8, 10, 14, 17, 22, 24, 28, 31, 33, 35, 39, 42, 44, 46, 50, 53, 56, 61, 63, 67, 71, 73, 75, 77, 79, 83, 85, 87, 89, 93, 96, 98 и 169, содержащие 3’-делеции, в которых элемент ТАТА-бокса или его эквивалентная последовательность ДНК и нижележащая последовательность удалены, могут быть использованы, например, для создания энхансерных элементов. Дополнительные делеции могут быть внесены для удаления любых элементов, имеющих положительные или отрицательные, тканево-специфичные, клеточно-специфичные или время-специфичные (такие как, но не ограничиваясь этим, суточные ритмы) воздействия на экспрессию. Любой из промоторов, представленный в SEQ ID №№: 2, 6, 8, 10, 14, 17, 22, 24, 28, 31, 33, 35, 39, 42, 44, 46, 50, 53, 56, 61, 63, 67, 71, 73, 75, 77, 79, 83, 85, 87, 89, 93, 96, 98 и 169, и фрагменты или энхансеры, полученные из них, могут быть использованы для получения композиций химерных регуляторных элементов, содержащих любой из промоторов, представленных в SEQ ID №№: 2, 6, 8, 10, 14, 17, 22, 24, 28, 31, 33, 35, 39, 42, 44, 46, 50, 53, 56, 61, 63, 67, 71, 73, 75, 77, 79, 83, 85, 87, 89, 93, 96, 98 и 169, и фрагменты или энхансеры, полученные из них, функционально связанные с другими энхансерами и промоторами.
[00037] В соответствии с настоящим изобретением, промотор или фрагмент промотора могут быть проанализированы на наличие известных промоторных элементов, т. е. характеристик последовательности ДНК, таких как ТАТА-бокс и других известных мотивов сайтов связывания транскрипционных факторов. Идентификация таких известных промоторных элементов может быть использована специалистом в данной области техники для разработки вариантов промотора, имеющих подобный характер экспрессии, как и исходный промотор.
[00038] Используемый в данном документе термин «лидер» относится к молекуле ДНК из 5’-нетранслируемой области (5’-UTR) гена и определяется в общем смысле как сегмент ДНК между сайтом инициации транскрипции (СИТ) и начальным сайтом последовательности, кодирующей белок. В альтернативном варианте, лидеры могут быть получены синтетическим путем или в результате манипулирования элементам ДНК. Лидер может быть использован в качестве 5’-регуляторного элемента для модуляции экспрессии функционально связанной транскрибируемой молекулы ДНК. Молекулы лидеров могут быть использованы с гетерологическим промотором или с их нативным промотором. Молекулы промоторов по изобретению, таким образом, могут быть функционально связанными с их нативным лидером или могут быть функционально связанными с гетерологическим лидером. Лидеры, полезные в практической реализации изобретения, включают SEQ ID №№: 3, 11, 25, 36, 47, 64, 68, 80, 90 и 170 или их фрагменты или варианты. В конкретных вариантах реализации изобретения такие последовательности ДНК могут быть определены как способные действовать в качестве лидера в клетке-хозяине, включая, например, трансгенную клетку растения. В одном варианте реализации изобретения такие последовательности ДНК могут быть расшифрованы как включающие лидерную активность.
[00039] Лидерные последовательности (5´-UTR), представленные в SEQ ID №№: 3, 11, 25, 36, 47, 64, 68, 80, 90 и 170, могут состоять из регуляторных элементов или могут принимать вторичные структуры, оказывающие влияние на транскрипцию или трансляцию функционально связанной молекулы ДНК. Лидерные последовательности, представленные в SEQ ID №№: 3, 11, 25, 36, 47, 64, 68, 80, 90 и 170, могут быть использованы, в соответствии с изобретением, для создания химерных регуляторных элементов, которые влияют на транскрипцию или трансляцию функционально связанной молекулы ДНК. В дополнение, лидерные последовательности представлены в SEQ ID №№: 3, 11, 25, 36, 47, 64, 68, 80, 90 и 170 могут быть использованы для создания химерных лидерных последовательностей, которые влияют на транскрипцию или трансляцию функционально связанной молекулы ДНК.
[00040] Используемый в данном документе термин «интрон» относится к молекуле ДНК, которая может быть выделена или идентифицирована из геномной копии гена и может быть определена в общем смысле как область, которая вырезается во время процессинга информационной РНК (мРНК) до трансляции. В альтернативном варианте, интрон может быть получен синтетическим путем или в результате манипулирования элементом ДНК. Интрон может содержать энхансерные элементы, влияющие на транскрипцию функционально связанных генов. Интрон может быть использован в качестве регуляторного элемента для модуляции экспрессии функционально связанной транскрибируемой молекулы ДНК. Генетическая конструкция может содержать интрон, и интрон может быть или не быть гетерологическим по отношению к транскрибируемой молекуле ДНК. Примеры интронов в данной области техники включают интрон актина риса и интрон HSP70 кукурузы.
[00041] По сравнению с генетическими конструкциями без интрона, включение некоторых интронов в генетические конструкции приводит к увеличению накопления мРНК и белка в растениях. Этот эффект был назван как «интрон опосредованное усиление» (IME) экспрессии генов. Интроны, стимулирующие экспрессию в растениях, были идентифицированы в генах кукурузы (например, tubA1, Adh1, Sh1, и Ubi1), в генах риса (например, tpi) и в генах двудольных растений, таких как петуния (например, rbcS), картофель (например, st-ls1) и Arabidopsis thaliana (например, ubq3 и pat1). Было показано, что делеции или мутации в пределах сайтов сплайсинга интрона снижают экспрессию генов, указывая, что сплайсинг может быть необходим для IME. Однако сплайсинг сам по себе не требуется, так как IME было показано для двудольных растений, с помощью точечных мутаций в пределах сайтов сплайсинга pat1 гена из A. thaliana. Было показано, что многократное использование одного и того же интрона в одном растении проявляет недостатки. В таких случаях необходимо иметь коллекцию основных элементов контроля для создания соответствующих рекомбинантных элементов ДНК.
[00042] Интроны, полезные в практической реализации изобретения, включают SEQ ID №№: 4, 12, 15, 20, 26, 29, 37, 40, 48, 51, 54, 57, 59, 65, 69, 81, 91, 94 и 171. Композиции, полученные из любых интронов, представленных в SEQ ID №№: 4, 12, 15, 20, 26, 29, 37, 40, 48, 51, 54, 57, 59, 65, 69, 81, 91, 94 и 171, могут состоять из внутренних делеций или дупликаций цис-регуляторных элементов; и/или изменения 5'- и 3'-последовательностей ДНК, содержащих интрон/экзон сплайсинговые сочленения, могут быть использованы для улучшения экспрессии или специфичности экспрессии, когда функционально связаны с промотором + лидером или химерным промотором + лидером и кодирующей последовательностью. При изменении граничных последовательностей интрон/экзон может быть полезным избегание использования нуклеотидной последовательности AT или нуклеотида А непосредственно перед 5’-концом сайта сплайсинга (GT) и нуклеотида G или нуклеотидной последовательности TG, соответственно, непосредственно после 3’-конца сайта сплайсинга (AG), для устранения потенциальных нежелательных стартовых кодонов, формирующихся во время процессинга информационной РНК в конечный транскрипт. Последовательность ДНК вблизи 5’- или 3’-концевых сайтов сплайсинговых сочленений интрона может быть модифицирована таким способом. Интроны и варианты интронов измененные, как описано в данном документе и с помощью способов, известных в данной области техники, могут быть проверены эмпирически, как описано в рабочих примерах, для определения влияния интрона на экспрессию функционально связанной молекулы ДНК.
[00043] Используемый в данном документе термин «3’-молекула терминации транскрипции», «3’-нетранслируемая область» или «3’-UTR» относится к молекуле ДНК, которая используется во время транскрипции с нетранслируемой области 3’- части молекулы мРНК. 3’-нетранслируемая область молекулы мРНК может быть получена путем специфического расщепления и 3’-полиаденилированием, также известным как поли(A)-хвост. 3’-UTR может быть функционально связанной с транскрибируемой молекулой ДНК и располагаться ниже от нее, и может включать сигнал полиаденилирования и другие регуляторные сигналы, способные воздействовать на транскрипцию, процессинг мРНК или экспрессию генов. Полагают, что поли(A)-хвосты выполняют функции стабилизации мРНК и инициации трансляции. Примеры 3’-молекул терминации транскрипции в данной области техники представляют собой 3’-область нопалинсинтазы, 3’-область hsp17 пшеницы, 3’-область малой субъединицы RuBisCO гороха, 3’-область E6 хлопка и 3’-UTR бусенника.
[00044] 3’-UTR, как правило, находят полезное применение для рекомбинантной экспрессии специфических молекул ДНК. Слабый 3’-UTR может генерировать сквозное прочитыванию, что может повлиять на экспрессию молекулы ДНК, расположенной в соседних экспрессионных кассетах. Подходящий контроль для терминации транскрипции может предотвратить сквозное прочитывание в последовательностях ДНК (например, других экспрессионных кассет), локализованных ниже, и может дополнительно обеспечить эффективное повторное использование РНК-полимеразы, для улучшения экспрессии генов. Эффективная терминация транскрипции (освобождение РНК-полимеразы II от ДНК) является предпосылкой для следующей инициации транскрипции и, таким образом, непосредственно влияет на общий уровень транскрипта. После терминации транскрипции зрелая мРНК освобождается от места синтеза, и матрица транспортируется в цитоплазму. Эукариотические мРНК накапливаются в формах поли(А) in vivo, что затрудняет обнаружение сайтов терминации транскрипции общепринятыми методами. Однако предсказание функциональных и эффективных 3’-UTR с помощью биоинформатических методов затруднено, так как отсутствуют консервативные последовательности ДНК, позволяющие легко предсказывать эффективную 3’-UTR.
[00045] С практической точки зрения, как правило, полезно, чтобы 3’-UTR, использованный в экспрессионной кассете, обладал следующими характеристиками. 3’-UTR должна быть способной рационально и эффективно завершать транскрипцию трансгена и предотвращать сквозное прочитывание транскрипта в любой из соседних последовательностей ДНК, которая может содержаться в другой экспрессионной кассете, как и в случае нескольких экспрессионных кассет, принадлежащих одной трансферной ДНК (Т-ДНК), или соседней хромосомной ДНК, в которую Т-ДНК была встроена. 3'-UTR не должна вести к снижению транскрипционной активности, наделяемой промотором, лидером, энхансерами и интронами, которые используются для контролирования экспрессии молекулы ДНК. В биотехнологии растений 3’-UTR часто используется для затравки реакций амплификации обратно транскрибируемой РНК, выделенной из трансформированных растений, и используется для: (1) оценки транскрипционной активности или экспрессии экспрессионной кассеты, интегрированной в хромосому растения; (2) оценки количества копий вставок в ДНК растения; и (3) оценки зиготности полученного семени после размножения. 3’-UTR также используется в реакциях амплификации ДНК, экстрагированной из трансформированных растений, для определения интактности встроенной кассеты.
[00046] Используемый в данном документе термин "энхансер" или "энхансерный элемент" относится к цис-действующему регуляторному элементу, называемому цис-элемент, который придает аспект общего характера экспрессии, но, как правило, недостаточный сам по себе для контролирования транскрипции, функционально связанной последовательности ДНК. В отличие промоторов, энхансерные элементы обычно не включают сайта инициации транскрипции (СИТ) или ТАТА-бокс или эквивалентую последовательность ДНК. Промотор или фрагмент промотора может по природе содержать один или более энхансерных элементов, влияющих на транскрипцию функционально связанной последовательности ДНК. Энхансерный элемент также может быть объединен с промотором для получения химерного промоторного цис-элемента, придающего аспект общей модуляции экспрессии генов.
[00047] Полагают, что многие энхансерные элементы промотора связывают ДНК-связывающие белки и/или влияют на топологию ДНК, производя локальные конформации, которые избирательно позволяют или ограничивают доступ РНК-полимеразы к ДНК-матрице или которые облегчают избирательное открытие двойной спирали на участке инициации транскрипции. Энхансерный элемент также может выполнять функцию связывания транскрипционных факторов, регулирующих транскрипцию. Некоторые энхансерные элементы связывают более чем один фактор транскрипции, и транскрипционные факторы могут взаимодействовать с различной аффинностью с более чем одним доменом энхансера. Энхансерные элементы могут быть идентифицированы с помощью ряда методов, включая анализ делеций, т. е. удаления одного или более нуклеотидов с 5'-конца или внутри промотора; анализ ДНК-связывающего белка, используя футпринтинг ДНКазы I, интерференцию метилирования, анализы сдвига электрофоретической мобильности, in vivo геномный футпринтинг путем лигирование-опосредованной полимеразной цепной реакции (ПЦР) и других общепринятых анализов; или путем анализа подобия последовательностей ДНК, используя известные мотивы цис-элементов или энхансерных элементов, как целевую последовательность или целевой мотив, с общепринятыми способами сравнения последовательностей ДНК, таких как BLAST. Тонкая структура домена энхансера может быть дополнительно исследована путем мутагенеза (или замещения) одного или более нуклеотидов или другими общепринятыми способами, известными в данной области техники. Энхансерные элементы могут быть получены путем химического синтеза или путем выделения из регуляторных элементов, которые включают такие элементы, и они могут быть синтезированы с дополнительными фланкирующими нуклеотидами, включающими применимые сайты рестрикции для облегчения последующих манипуляций. Таким образом, разработка, создание и использование энхансерных элементов, в соответствии со способами, описанными в данном документе, для модуляции экспрессии функционально связанных транскрибируемых молекул ДНК, охватываются по изобретению.
[00048] Используемый в данном документе термин «химерный» относится к отдельной молекуле ДНК, полученной путем слияния первой молекулы ДНК со второй молекулой ДНК, где ни первая, ни вторая молекула ДНК не будут по обыкновению содержаться в такой конфигурации, т. е. не будут объединены друг с другом. Химерная молекула ДНК представляет, таким образом, новую молекулу ДНК, иначе говоря, не содержащуюся обыкновенно в природе. Используемый в данном документе термин «химерный промотор» относится к промотору, полученному посредством такого манипулирования молекулами ДНК. Химерный промотор может объединять два или более фрагмента ДНК, например, слияние промотора с энхансерным элементом. Таким образом, разработка, создание и использование химерных промоторов, в соответствии со способами, описанными в данном документе, для модуляции экспрессии функционально связанных транскрибируемых молекул ДНК, охватываются по изобретению.
[00049] Используемый в данном документе термин «вариант» относится ко второй молекуле ДНК, такой как регуляторный элемент, которая аналогична по композиции, но не идентична первой молекуле ДНК, и причем вторая молекула ДНК все еще сохраняет общую функциональность, т. е. такой же или аналогичный характер экспрессии, например, через большую или меньшую, или эквивалентную транскрипционную или трансляционную активность по сравнению с первой молекулой ДНК. Вариант может быть укороченной или усеченной версией первой молекулы ДНК и/или измененной версией последовательности ДНК первой молекулы ДНК, такой как вариант с различными сайтами рестрикции и/или внутренними делециями, заменами и/или вставками. «Варианты» регуляторного элемента также охватывают варианты, возникающие в результате мутаций, которые происходят во время или в результате трансформации клеток бактерий и растений. В изобретении последовательность ДНК, приводимая в SEQ ID №№: 1–98 и 168–171, может быть использована для создания вариантов, которые аналогичны по композиции, но не идентичны последовательности ДНК исходного регуляторного элемента, хотя все еще сохраняют общую функциональность, т. е. такой же или аналогичный характер экспрессии, как и исходный регуляторный элемент. Получение таких вариантов по изобретению находится в пределах обычной квалификации в данной области техники в свете описания и осуществлено в пределах объема изобретения.
[00050] Химерные регуляторные элементы могут быть разработаны для включения различных составных элементов, которые могут быть функционально связаны различными способами, известными в данной области техники, такими как расщепление ферментами рестрикции и лигирование, клонирование независимое от лигирования, модульная сборка продуктов ПЦР в процессе амплификации или прямой химический синтез регуляторного элемента, а также другими способами, известными в данной области техники. Полученные различные химерные регуляторные элементы могут состоять из тех же или вариантов тех же составных элементов, но отличатся по последовательности ДНК, или последовательностей ДНК, включающих связывающую последовательность ДНК, или последовательностей, которые позволяют составным частям быть функционально связанными. В изобретении, последовательность ДНК, приводимая в SEQ ID №№: 1–98 и 168–171, может обеспечить референсную последовательность регуляторного элемента, в которой составные элементы, включающие референсную последовательность, могут быть соединены способами, известными в данной области техники, и могут включать замены, делеции и/или вставки одного или более нуклеотидов или мутации, которые по обыкновению встречаются при трансформации клеток бактерий и растений.
[00051] Действенность модификаций, дубликаций или делеций, описанных в данном документе, на желаемые аспекты экспрессии конкретного трансгена может быть проверена эмпирически в стабильных и транзиентных тестах растений, таких как те, которые описаны в рабочих примерах в данном документе, для того чтобы подтвердить результаты, которые могут варьировать в зависимости от произведенных изменений и цели изменения исходной молекулы ДНК.
Генетические конструкции
[00052] Используемый в данном документе термин «генетическая конструкция» означает любую рекомбинантную молекулу ДНК, такую как плазмида, космида, вирус, фаг или линейную или кольцевую молекулу ДНК или РНК, полученную из любого источника, способную к геномной интеграции или автономной репликации, включающую молекулу ДНК, где по меньшей мере одна молекула ДНК связана с другой молекулой ДНК функционально действующим способом, т. е. функционально связаны между собой. Используемый в данном документе термин «вектор» означает любую генетическую конструкцию, которая может быть использована для целей трансформации, т. е. введения гетерологической ДНК или РНК в клетку хозяина. Генетическая конструкция, как правило, включает одну или более экспрессионных кассет. Как используется в данном документе «экспрессионная кассета» относится к молекуле ДНК, содержащей по меньшей мере транскрибируемую молекулу ДНК, функционально связанную с одним или более регуляторных элементов, как правило, по меньшей мере промотором и 3’-UTR.
[00053] Используемый в данном документе термин «функционально связанный» относится к первой молекуле ДНК, соединенной со второй молекулой ДНК, причем первая и вторая молекулы ДНК расположены так, что первая молекула ДНК влияет на функцию второй молекулы ДНК. Две молекулы ДНК могут быть или не быть частью единой непрерывной молекулы ДНК и могут быть или не быть смежными. Например, промотор является функционально связанным с транскрибируемой молекулой ДНК, если промотор модулирует транскрипцию целевой транскрибируемой молекулы ДНК в клетке. Лидер, например, является функционально связанным с последовательностью ДНК, когда он способен воздействовать на транскрипцию или трансляцию последовательности ДНК.
[00054] Генетические конструкции по изобретению могут быть предложены в одном варианте реализации изобретения как генетические конструкции, состоящие из двух граничных областей опухолеиндуцирующей плазмиды, (Ti), имеющие области правого пограничного района (RB или AGRtu.RB) и левого пограничного района (LB или AGRtu.LB) Ti-плазмиды, изолированной из Agrobacterium tumefaciens, содержащей Т-ДНК, которая, наряду с переносом молекул, который обеспечивается клетками A. tumefaciens, позволяют интеграцию Т-ДНК в геном клетки растения (см., например, патент США 6603061). Генетические конструкции также могут содержать сегменты плазмидного каркаса ДНК, обеспечивающие функцию репликации и селекцию на антибиотике в клетках бактерий, например, ориджин репликации Escherichia coli, такой как ori322, ориджин репликации для широкого спектра хозяев, такой как oriV или oriRi, и кодирующую область селективного маркера, такого как Spec/Strp, кодирующего Tn7 аминогликозид аденилтрансферазу (aadA), придающую устойчивость спектиномицину или стрептомицину, или селективный маркерный ген гентамицина (Gm, Gent). Для трансформации растений часто используют бактериальный штамм хозяина ABI, C58 или LBA4404 A. tumefaciens; однако другие штаммы, известные специалистам в области трансформации растений, могут функционировать в изобретении.
[00055] В данной области техники известны способы для сборки и введения генетических конструкций в клетку таким образом, что транскрибируемая молекула ДНК транскрибируется в функциональную молекулу мРНК, которая транслируется и экспрессируется в виде белка. Для осуществления на практике изобретения, общепринятые композиции и способы получения и использования генетических конструкций и клеток хозяина хорошо известны специалистам в данной области техники. Типичные векторы, используемые для экспрессии нуклеиновых кислот в высших растениях, хорошо известны в данной области техники и включают векторы, полученные из Ti-плазмиды Agrobacterium tumefaciens и pCaMVCN контрольного вектора для перенесения.
[00056] Различные регуляторные элементы могут быть включены в генетическую конструкцию, включая любой из тех, что предложен в данном документе. Любые такие регуляторные элементы могут быть предложены в сочетании с другими регуляторными элементами. Такие комбинации могут быть разработаны или модифицированы для получения желаемых регуляторных функций. В одном варианте реализации изобретения генетические конструкции по изобретению содержат по меньшей мере один регуляторный элемент, функционально связанный с транскрибируемой молекулой ДНК, функционально связанной с 3’-UTR.
[00057] Генетические конструкции по изобретению могут включать любой промотор или лидер, предложенные в данном документе или известные в данной области техники. Например, промотор по изобретению может быть функционально связанным с гетерологическим нетранслируемым 5’-лидером, таким как один из полученных из гена белка теплового шока. В альтернативном варианте лидер по изобретению может быть функционально связанным с гетерологическим промотором, таким как 35S промоторный транскрипт вируса мозаики цветной капусты.
[00058] Экспрессионные кассеты также могут включать кодирующую последовательность транзитного пептида, которая кодирует пептид, полезный для внутриклеточного направления функционально связанного белка, в частности, к хлоропласту, лейкопласту или другой пластидной органелле, митохондрии, пероксисоме, вакуоле или внеклеточному пространству. Многие белки, локализованные в хлоропластах, экспрессируются генами ядра как предшественники и направляются в хлоропласт с помощью транзитного пептида хлоропласта (ТПХ). Примеры таких белков, изолированных из хлоропластов, включают, но не ограничиваются этими, белки, ассоциированные с малой субъединицей (МСЕ) рибулозо-1,5,-бифосфат карбоксилазы, ферредоксин, ферредоксин оксидоредуктазу, белок I и белок II фотосенсибилизирующего комплекса, тиоредоксин F и энолпирувил шикимат фосфатсинтазу (ЭПШФС). Транзитные пептиды хлоропластов описаны, например, в патенте США № 7193133. Было показано, что нехлоропластные белки могут быть направлены в хлоропласт при экспрессии гетерологического ТПХ, функционально связанного с трансгеном, кодирующим нехлоропластные белки.
Транскрибируемая молекула ДНК
[00059] Используемый в данном документе термин «транскрибируемая молекула ДНК» относится к любой молекуле ДНК, способной транскрибироваться в молекулу РНК, включая, но не ограничиваясь этим, те, которые имеют последовательности, кодирующие белок, и те, которые производят молекулы РНК, имеющие последовательности, полезные для супрессии гена. Тип молекулы ДНК может включать, но не ограничивается этими, молекулу ДНК из того же растения, молекулу ДНК из другого растения, молекулу ДНК из другого организма или синтетическую молекулу ДНК, такую как молекула ДНК, содержащая антисмысловую последовательность гена, или молекула ДНК, кодирующая искусственную, синтетическую или иным образом модифицированную версию трансгена. Типовая транскрибируемая молекула ДНК для включения в генетические конструкции по изобретению включает, например, молекулы ДНК или гены из другого вида, чем тот, в который молекула ДНК встроена, или гены, происходящие из или присутствующие в том же виде, но которые включены в клетки-реципиенты способами генной инженерии, а не классическими методами размножения.
[00060] «Трансген» относится к транскрибируемой молекуле ДНК гетерологической для клетки-хозяина по меньшей мере в отношении ее расположения в геноме клетки-хозяина и/или транскрибируемой молекуле ДНК искусственно включенной в геном клетки-хозяина в текущем или любом предыдущем поколении клеток.
[00061] Регуляторный элемент, такой как промотор по изобретению, может быть функционально связанным с транскрибируемой молекулой ДНК, которая гетерологическая по отношению к регуляторному элементу. Используемый в данном документе термин «гетерологический» относится к комбинации из двух или более молекул ДНК, когда такая комбинация по обыкновению не встречается в природе. Например, две молекулы ДНК могут быть получены из разных видов и/или две молекулы ДНК могут быть получены из разных генов, например, разных генов одного и того же вида или же одних и тех же генов из разных видов. Регуляторный элемент является, таким образом, гетерологическим по отношению к функционально связанной транскрибируемой молекуле ДНК, если такая комбинация по обыкновению не встречается в природе, т. е. транскрибируемая молекула ДНК по обыкновению не встречается функционально связанной с регуляторным элементом.
[00062] Транскрибируемая молекула ДНК в общем смысле может быть любой молекулой ДНК, для которой желательна экспрессия транскрипта. Такая экспрессия транскрипта может приводить к трансляции полученной молекулы мРНК и, соответственно, к экспрессии белка. В альтернативном варианте, например, транскрибируемая молекула ДНК может быть разработана, чтобы в итоге вызвать уменьшение экспрессии конкретного гена или белка. В одном варианте реализации изобретения это может быть достигнуто с помощью транскрибируемой молекулы ДНК, ориентированной в антисмысловом направлении. Специалист в данной области техники хорошо знаком с использованием такой антисмысловой технологии. Любой ген может негативно регулироваться таким способом, и в одном варианте реализации изобретения транскрибируемая молекула ДНК может быть разработана для супрессии конкретного гена через экспрессию дцРНК, миРНК или микроРНК молекулы.
[00063] Таким образом, в одном варианте реализации изобретения представляется рекомбинантная молекула ДНК, содержащая регуляторный элемент по изобретению, такой как те, что предложены в SEQ ID №№: 1–98 и 168–171, функционально связанный с гетерологической транскрибируемой молекулой ДНК для того, чтобы модулировать транскрипцию транскрибируемой молекулы ДНК на желаемом уровне или по желаемому характеру, когда генетическая конструкция интегрирована в геном клетки трансгенного растения. В одном варианте реализации изобретения транскрибируемая молекула ДНК содержит область гена, кодирующую белок, а в другом варианте реализации изобретения транскрибируемая молекула ДНК содержит антисмысловую область гена.
Гены, представляющие агрономический интерес
[00064] Транскрибируемая молекула ДНК может быть геном, представляющим агрономический интерес. Используемый в данном документе термин «ген, представляющий агрономический интерес» относится к транскрибируемой молекуле ДНК, которая при экспрессии в конкретной ткани растений, клетке или типе клетки придает желаемую характеристику. Продукт гена, представляющего агрономический интерес, может действовать внутри растения для того, чтобы влиять на морфологию растения, физиологию, рост, развитие, урожайность, композицию зерна, питательный профиль, устойчивость к болезням или вредителям и/или толерантность к факторам окружающей среды или химическим факторам, или может действовать в качестве пестицидного агента в рационе вредителя, который питается растением. В одном варианте реализации изобретения регуляторный элемент по изобретению включен в генетическую конструкцию таким образом, что регуляторный элемент функционально связан с транскрибируемой молекулой ДНК, которая является геном, представляющим агрономический интерес. В трансгенном растении, содержащем такую генетическую конструкцию, экспрессия гена, представляющего агрономический интерес, может придать полезную агрономическую черту. Полезная агрономическая черта может включать, но не ограничиваться этими, например, толерантность к гербицидам, борьбу с насекомыми, модифицирование урожайности, устойчивость к болезням, устойчивость к патогену, модифицирование роста и развития растения, модифицированное содержание крахмала, модифицированное содержание масла, модифицированное содержание жирных кислот, модифицированное содержание белка, модифицированное созревание плодов, улучшенное питание животных и человека, производства биополимеров, устойчивость к стрессовым факторам окружающей среды, фармацевтические пептиды, улучшенные свойства для переработки, улучшение вкуса, полезное производство гибридных семян, улучшенное производство волокна и желательное производство биотоплива.
[00065] Примеры генов, представляющих агрономический интерес, известных в данной области техники, включают устойчивость к гербицидам (патенты США №№ 6803501, 6448476, 6248876, 6225114, 6107549, 5866775, 5804425, 5633435 и 5463175), повышение урожайности (патенты США №№ USRE38446, 6716474, 6663906, 6476295, 6441277, 6423828, 6399330, 6372211, 6235971, 6222098 и 5716837), борьба с насекомыми (патенты США №№ 6809078, 6713063, 6686452, 6657046, 6645497, 6642030, 6639054, 6620988, 6593293, 6555655, 6538109, 6537756, 6521442, 6501009, 6468523, 6326351, 6313378, 6284949, 6281016, 6248536, 6242241, 6221649, 6177615, 6156573, 6153814, 6110464, 6093695, 6063756, 6063597, 6023013, 5959091, 5942664, 5942658, 5880275, 5763245 и 5763241), устойчивость к грибковым заболеваниям (патенты США №№ 6653280, 6573361, 6506962, 6316407, 6215048, 5516671, 5773696, 6121436, 6316407 и 6506962), устойчивость к вирусам (патенты США №№ 6617496, 6608241, 6015940, 6013864, 5850023 и 5304730), устойчивость к нематоде (патент США № 6228992), устойчивость к бактериальным заболеваниям (патент США № 5516671), рост и развитие растений (патенты США №№ 6723897 и 6518488), накопление крахмала (патенты США №№ 6538181, 6538179, 6538178, 5750876, 6476295), производство модифицированных масел (патенты США №№ 6444876, 6426447 и 6380462), повышенная продукция масел (патенты США №№ 6495739, 5608149, 6483008 и 6476295), модифицированное содержание жирных кислот (патенты США №№ 6828475, 6822141, 6770465, 6706950, 6660849, 6596538, 6589767, 6537750, 6489461 и 6459018), повышенная продукция белков (патент США № 6380466), созревание плодов (патент США № 5512466), улучшенное питание животных и человека (патенты США №№ 6723837, 6653530, 6541259; 5985605 и 6171640), биополимеры (патенты США №№ USRE37543, 6228623 и 5958745, и 6946588), устойчивость к стрессовым факторам окружающей среды (патент США № 6072103), фармацевтические пептиды и секретируемые пептиды (патенты США №№ 6812379, 6774283, 6140075 и 6080560), улучшенные свойства для переработки (патент США № 6476295), улучшенная усвояемость (патент США № 6531648), с низким содержанием рафинозы (патент США № 6166292), промышленное производство фермента (патент США № 5543576), улучшенный вкус (патент США № 6011199), фиксация азота (патент США № 5229114), производство гибридных семян (патент США № 5689041), производство волокна (патенты США №№ 6576818, 6271443, 5981834 и 5869720) и производство биотоплива (патент США № 5998700).
[00066] В альтернативном варианте ген, представляющий агрономический интерес, может повлиять на вышеупомянутые характеристики растения или фенотипы, посредством кодирования молекулы РНК, которая причиняет направленную модуляцию экспрессии эндогенного гена, например, через антисмысловые (см., например, патент США 5107065), ингибиторные РНК («РНК-интерференции», включающие модуляцию экспрессии генов с помощью микроРНК-, миРНК-, транс-действующих миРНК- и фазовых малых РНК-опосредованных механизмов, например, как описано в опубликованных заявках США 2006/0200878 и 2008/0066206 и в заявке на патент США 11/974469); или косупрессорно-опосредованные механизмы. РНК также может быть каталитической молекулой РНК (например, рибозимом или рибосвитчем, см., например, опубликованную заявку США 2006/0200878), разработаной для отщепления желаемого эндогенный продукта мРНК. В данной области техники известны способы для разработки и введения генетических конструкций в клетку, таким образом, что транскрибируемая молекула ДНК транскрибируется в молекулу, способную вызвать супрессию гена.
[00067] Экспрессия транскрибируемой молекулы ДНК в клетке растения также может быть использована для супрессии вредителей растений, питающихся клетками растения, например, композиции, выделенные из жесткокрылых насекомых- вредителей, и композиций, выделенных из нематодных вредителей. Вредители растений включают, но не ограничиваются этими, членистоногих вредителей, нематодных вредителей и грибковых или микробных вредителей.
Селективные маркеры
[00068] Селективные маркерные трансгены также могут быть использованы с регуляторными элементами по изобретению. Используемый в данном документе термин «селективный маркерный трансген» относится к любой транскрибируемой молекуле ДНК, чья экспрессия в трансгенном растении, ткани или клетке, или отсутствие таковых, может быть подвергнута скринингу или подсчитана каким-нибудь способом. Селективные маркерные гены и, связанные с ними, методы отбора и скрининга, предназначены для использования на практике изобретения, известны в данной области техники и включают, но не ограничиваются этими, транскрибируемые молекулы ДНК, кодирующие бета-глюкуронидазу (GUS), зеленый флуоресцентный белок (GFP), белки, придающие устойчивость к антибиотикам и белки, придающие устойчивость к гербицидам.
Трансформация клеток
[00069] Изобретение также направляется к способу получения трансформированных клеток и растений, содержащих один или более регуляторных элементов, функционально связанных с транскрибируемой молекулой ДНК.
[00070] Термин «трансформация» относится к введению молекулы ДНК в реципиентного хозяина. Используемый в данном документе термин «хозяин» относится к бактериями, грибами или растениям, включая любые клетки, ткани, органы, или к потомству бактерий, грибов или растений. Ткани и клетки растений, представляющие собой особый интерес, включают протопласты, каллус, корни, клубни, семена, стебли, листья, проростки, завязи и пыльцу.
[00071] Используемый в данном документе термин «трансформированный» относится к клетке, ткани, органу или организму, в который молекула чужеродной ДНК, такая как генетическая конструкция, была введена. Введенная молекула ДНК может быть интегрирована в геномную ДНК клетки, ткани, органа или организма реципиента так, что введенная молекула ДНК наследуется в дальнейшем потомстве. «Трансгенная» или «трансформированная» клетка или организм также может включать потомство клетки или организма и потомство, полученное в результате программы размножения с использованием такого трансгенного организма в качестве родителя при скрещивании и проявляющее измененный фенотип как результат присутствия чужеродных молекул ДНК. Введенная молекула ДНК также может быть временно введенной в клетку-реципиент так, что введенная молекула ДНК не наследуется в дальнейшем потомстве. Термин «трансгенный» относится к бактерии, грибу или растению, содержащим одну или более гетерологических молекул ДНК.
[00072] Существует много способов, хорошо известных специалистам в данной области техники, для введения молекулы ДНК в клетки растений. Процесс, как правило, включает этапы выбора подходящей клетки-хозяина, трансформации клетки-хозяина вектором и получения трансформированной клетки-хозяина. Методы и материалы для трансформации клеток растений, посредством введения растительной генетической конструкции в геном растений, в реализации на практике изобретения могут включать любой из известных и показанных способов. Подходящие способы включают, но не ограничиваются этими, бактериальную инфекцию (например, Agrobacterium ), бинарные БИХ векторы, прямой перенос ДНК (например, ПЭГ-опосредованную трансформацию, поглощение ДНК, опосредованное высушиванием/ингибированием, электропорацию, перемешивание с волокнами карбида кремния и ускорение частиц, покрытых ДНК) из числа других.
[00073] Клетки-хозяева могут представлять собой любую клетку или организм, например, клетку растения, клетку водоросли, водоросли, клетку гриба, грибы, бактериальную клетку или клетку насекомых. В конкретных вариантах реализации изобретения клетки-хозяева и трансформированные клетки могут включать клетки культурных растений.
[00074] Трансгенное растение впоследствии может быть регенерировано из клетки трансгенного растения по изобретению. С использованием общепринятых методов размножения или самоопыления можно получить семена из этого трансгенного растения. Такое семя и полученное в результате растение-потомок, выращенное из такого семени, будут содержать рекомбинантную молекулу ДНК по изобретению, и, следовательно, будут трансгенными.
[00075] Трансгенные растения по изобретению могут быть самоопылены, для предоставления семян гомозиготных трансгенных растений по изобретению (гомозиготных по рекомбинантной молекуле ДНК), или скрещены с нетрансгенными растениями или другими трансгенными растениями, для предоставления семян гомозиготных трансгенных растений по изобретению (гомозиготных по рекомбинантной молекуле ДНК). Как гомозиготные, так и гетерозиготные трансгенные растения упоминаются в данном документе как «растения-потомки». Растения потомки представляют собой трансгенные растения, произошедшие от исходного трансгенного растения и содержащие рекомбинантную молекулу ДНК по изобретению. Семена, полученные с использованием трансгенного растения по изобретению, могут быть собраны и использованы для выращивания поколений трансгенных растений, т. е. растений-потомков по изобретению, включающих генетическую конструкцию настоящего изобретения и экспрессию гена, представляющего агрономический интерес. Описания способов размножения, которые обычно используются для различных культур, могут быть найдены в одной из нескольких книжных ссылках, см., например, Allard, Principles of Plant Breeding, John Wiley & Sons, NY, U. of CA, Davis, CA, 50-98 (1960); Simmonds, Principles of Crop Improvement, Longman, Inc., NY, 369-399 (1979); Sneep and Hendriksen, Plant breeding Perspectives, Wageningen (ed), Center for Agricultural Publishing and Documentation (1979); Fehr, Soybeans: Improvement, Production and Uses, 2nd Edition, Monograph, 16:249 (1987); Fehr, Principles of Variety Development, Theory and Technique, (Vol. 1) и Crop Species Soybean (Vol. 2), Iowa State Univ., Macmillan Pub. Co., NY, 360-376 (1987).
[00076] Трансформированные растения могут быть проанализированы на наличие целевого гена или генов и по уровню экспрессии и/или профилю, предоставленному регуляторными элементами по изобретению. Специалистам в данной области техники известны многочисленные способы, доступные для анализа трансформированных растений. Например, методы анализа растений включают, но не ограничиваются этими, Саузерн блоттинги или нозерн блоттинги, подходы на основе ПЦР, биохимические анализы, способы фенотипического скрининга, полевые апробации и иммунодиагностические анализы. Экспрессия транскрибируемой молекулы ДНК может быть измерена с использованием реагентов TaqMan® (Applied Biosystems, Foster City, CA) и по способам, описанным производителем, и по количеству циклов ПЦР, определенному с использованием TaqMan® Testing Matrix. В альтернативном варианте реагенты Invader® (Third Wave Technologies, Madison, WI) и способы, описанные производителем, могут быть использованы для оценки экспрессии трансгена.
[00077] Изобретение также относится к частям растения по изобретению. Части растений включают, но не ограничиваются этими, листья, стебли, корни, клубни, семена, эндосперм, семязачаток и пыльцу. Части растений по изобретению могут быть жизнеспособными, нежизнеспособными, способными и/или не способными к регенерации. В изобретении также включены и предложены трансформированные клетки растения, содержащие молекулу ДНК по изобретению. Трансформированные клетки или клетки трансгенного растения по изобретению включают клетки способные и/или не способные к регенерации.
[00078] Изобретение может быть более легко понятным посредством ссылки на следующие примеры, которые предложены в качестве иллюстрации, и не подразумевают ограничения изобретения, если это не установлено. Специалистам в данной области техники должно быть понятно, что способы, описанные в следующих примерах, представляют собой способы, обнаруженные авторами изобретения, для нормального функционирования при применении изобретения. Однако специалистам в данной области техники должно быть понятно, в свете настоящего описания, что многие изменения могут быть совершены в конкретных описанных вариантах реализации изобретения, и по-прежнему получится похожий или аналогичный результат без отступления от сущности и объема изобретения, поэтому все имеющее значение размещается далее или показывается в прилагаемых графических материалах и должно быть интерпретировано в иллюстративном, а не в ограничивающем смысле.
ПРИМЕРЫ
Пример 1
Идентификация и клонирование регуляторных элементов
[00079] Новые регуляторные элементы убиквитина, или последовательности группы регуляторных элементов экспрессии (EXP), были выявлены и изолированы из геномной ДНК однодольных: полевицы тёмной (Agrostis nebulosa), арундо тростникового (Arundo donax), бутелоуи изящной (Bouteloua gracilis), мискантуса китайского (Miscanthus sinensis), шизахириума метельчатого (Schizachyrium scoparium), соргаструма поникающего (Sorghastrum nutans) и бусенника (Coix lacryma-jobi).
[00080] Последовательности транскрипта убиквитина 1 были идентифицированы в каждом из вышеуказанных видов. 5’-нетранслируемую область (5’-UTR) каждого из транскриптов убиквитина 1 использовали для разработки праймеров для амплификации соответствующих регуляторных элементов идентифицированного гена убиквитина, которая включает функционально связанные между собой промотор, лидер (5’-UTR) и первый интрон. Праймеры использовали с GenomeWalkerTM (Clontech Laboratories, Inc, Mountain View, CA) библиотеками, созданными в соответствии с протоколом производителя для клонирования 5’-области соответствующей последовательности геномной ДНК. Регуляторные элементы убиквитина также были выделены из однодольных Setaria italica, Setaria viridis и Zea mays subsp. Mexicana (Teosinte) с использованием GenomeWalkerTM библиотек, как описано выше. В дополнение, регуляторные элементы убиквитина также были выделены из однодольного Sorghum bicolor с использованием общедоступных последовательностей, которые представляют собой гомологи к генам убиквитинов 4, 6 и 7.
[00081] Используя идентифицированные последовательности, проводился биоинформатический анализ для выявления регуляторных элементов в пределах амплифицированной ДНК. Используя результаты этого анализа, регуляторные элементы были определены в пределах последовательностей ДНК и праймеров, разработанных для амплификации регуляторных элементов. Соответствующую молекулу ДНК для каждого регуляторного элемента амплифицировали с использованием стандартных условий полимеразной цепной реакции (ПЦР) с праймерами, содержащими уникальные сайты рестрикции, и геномной ДНК, выделенной из A. nebulosa, A donax, B. gracilis, M. sinensis, S. scoparium, S. nutans и C. lacryma-jobi. Полученные фрагменты ДНК были легированы в базовые растительные экспрессионные векторы и секвенированы. Затем проводили анализ сайта инициации транскрипции (СИТ) регуляторного элемента и интрон/экзон сплайсингового сочленения с помощью трансформированных протопластов растений. Вкратце, протопласты были трансформированы растительными экспрессионными векторами, содержащими клонированные фрагменты ДНК, функционально связанные с гетерологической транскрибируемой молекулой ДНК, и 5’-RACE-система для быстрой амплификации концов кДНК, версия 2.0 (Invitrogen, Carlsbad, California 92008) была использована для подтверждения СИТ регуляторного элемента и интрон/экзон сплайсингового сочленения путем анализа последовательности транскриптов информационной РНК (мРНК), полученных таким способом.
[00082] Последовательности ДНК, идентифицированных EXP, предложены в данном документе как SEQ ID №№: 1, 5, 7, 9, 13, 16, 18, 19, 21, 23, 27, 30, 32, 34, 38, 41, 43, 45, 49, 52, 55, 58, 60, 62, 66, 70, 72, 74, 76, 78, 82, 84, 86, 88, 92, 95, 97, 99, 103, 106, 108, 110, 114, 116, 118, 120, 122, 126, 128, 132, 134, 138, 140, 144, 148, 150 и 168, которые приведены ниже в таблице 1. Промоторные последовательности предложены в данном документе как SEQ ID №№: 2, 6, 8, 10, 14, 17, 22, 24, 28, 31, 33, 35, 39, 42, 44, 46, 50, 53, 56, 61, 63, 67, 71, 73, 75, 77, 79, 83, 85, 87, 89, 93, 96, 98, 100, 104, 107, 109, 111, 117, 119, 121, 123, 129, 135, 141, 145, 151 и 169. Лидерные последовательности предложены в данном документе как SEQ ID №№: 3, 11, 25, 36, 47, 64, 68, 80, 90, 101, 112, 124, 130, 136, 142, 146, 152 и 170. Интронные последовательности предложены в данном документе как SEQ ID №№: 4, 12, 15, 20, 26, 29, 37, 40, 48, 51, 54, 57, 59, 65, 69, 81, 91, 94, 102, 105, 113, 115, 125, 127, 131, 133, 137, 139, 143, 147, 149, 153 и 171.
Таблица 1. Группы регуляторных экспрессионных элементов («EXP»), промоторы, энхансеры, лидеры и интроны, изолированные из различных видов злаковых.
[00083] Как показано в таблице 1, например, регуляторная последовательность EXP обозначенная EXP-AGRne.Ubq1:1:7 (SEQ ID №: 1), с компонентами, изолированными из A. nebulosa, включает элемент промотора P-AGRne.Ubq1-1:1:5 (SEQ ID №: 2), функционально связанный с 5’-элементом лидера L-AGRne.Ubq1-1:1:1 (SEQ ID №: 3), функционально связанный с 5’-элементом интрона I-AGRne.Ubq1-1:1:3 (SEQ ID №: 4). Другие последовательности EXP связаны аналогично, как показано в таблице 1.
[00084] Как показано в таблице 1, списке последовательностей и на Фиг. 1-8, были разработаны варианты последовательностей промоторов из A. nebulosa, A donax, B. gracilis, M. sinensis, S. scoparium и S. nutans, содержащие укороченные фрагменты промоторов, например, P-AGRne.Ubq1-1:1:5 (SEQ ID №:2), P-ARUdo.Ubq1-1:1:4 (SEQ ID №:10), или других соответствующих промоторов из других видов, и в результате дали, например, P-AGRne.Ubq1-1:1:4 (SEQ ID №: 6) и P-ARUdo.Ubq1-1:1:5 (SEQ ID№: 14), а также другие фрагменты промоторов.
[00085] Также в таблицу 1 внесены три аллельных варианта, изолированные с использованием одних и тех же праймеров, разработанных для амплификации геномной ДНК из Z. mays subsp. Mexicana. Аллельные варианты последовательностей EXP Z. mays subsp. Mexicana содержат последовательности ДНК, имеющие определенную идентичность в пределах различных областей других последовательностей ДНК, но вставки, делеции и нуклеотидные несовпадения, также могут быть выявлены в пределах каждого промотора, лидера и/или интрона каждой из последовательностей EXP. Последовательности EXP, обозначенные EXP-Zm.UbqM1:1:6 (SEQ ID №: 122) и EXP-Zm.UbqM1:1:10 (SEQ ID №: 126) представляют собой первый аллель (аллель-1) Z. mays subsp. Mexicana в группе регуляторных элементов экспрессии с геном Ubq1, с единственным отличием между двумя последовательностями EXP, происходящим в последних 3’-нуклеотидах каждого соответствующего интрона, следующего за последовательностью 5’-AG-3’ из 3’-интрон сплайсингового сочленения. Последовательности EXP, обозначенные EXP-Zm.UbqM1:1:7 (SEQ ID №: 128) и EXP-Zm.UbqM1:1:12 (SEQ ID №: 132) представляют собой второй аллель (аллель-2) Z. mays subsp. Mexicana в группе регуляторных элементов экспрессии с геном Ubq1, с единственным отличием между двумя последовательностями EXP, происходящим в последних 3’-нуклеотидах каждого соответствующего интрона, следующего за последовательностью 5’-AG-3’ из 3’-интрон сплайсингового сочленения. EXP последовательности EXP-Zm.UbqM1:1:8 (SEQ ID №: 134) и EXP-Zm.UbqM1:1:11 (SEQ ID №: 138) представляют собой третий аллель (аллель-3) Z. mays subsp. Mexicana в группе регуляторных элементов экспрессии с геном Ubq1, с единственным отличием между двумя последовательностями EXP, происходящим в последних 3’-нуклеотидах каждого соответствующего интрона, следующего за последовательностью 5’-AG-3’ из 3’-интрон сплайсингового сочленения.
Пример 2
Анализ регуляторных элементов, контролирующих GUS в протопластах кукурузы, с использованием GUS экспрессионных кассет ампликонов
[00086] Протопласты листа кукурузы были трансформированы ампликонами ДНК, полученными из растительных экспрессионных векторов, содержащих последовательность EXP, которая контролирует экспрессию трансгена бета-глюкуронидазы (GUS), и сравнивались с протопластами листа, в которых экспрессия GUS была под контролем известных конститутивных промоторов, в серии экспериментов, представленных ниже.
[00087] В первой серии экспериментов протопласты клеток кукурузы, полученные из ткани листа, были трансформированы, как описано ранее, ампликонами, полученными в результате амплификации экспрессионных кассет с GUS геном, содержащих растительные экспрессионные векторы, для сравнения экспрессии трансгена (GUS) под контролем одной из последовательностей EXP-AGRne.Ubq1:1:7 (SEQ ID №: 1), EXP-AGRne.Ubq1:1:8 (SEQ ID №: 5), EXP-AGRne.Ubq1:1:9 (SEQ ID №: 7), EXP-ARUdo.Ubq1:1:8 (SEQ ID №: 13), EXP-ARUdo.Ubq1:1:9 (SEQ ID №: 18), EXP-ARUdo.Ubq1:1:11 (SEQ ID №: 21), EXP-ARUdo.Ubq2:1:8 (SEQ ID №: 27), EXP-ARUdo.Ubq2:1:9 (SEQ ID №: 30), EXP-ARUdo.Ubq2:1:10 (SEQ ID №: 32), EXP-BOUgr.Ubq1:1:6 (SEQ ID №: 38), EXP-BOUgr.Ubq1:1:7 (SEQ ID №: 41), EXP-BOUgr.Ubq1:1:8 (SEQ ID №: 43), EXP-BOUgr.Ubq2:1:14 (SEQ ID №: 52), EXP-BOUgr.Ubq2:1:16 (SEQ ID №: 58), EXP-BOUgr.Ubq2:1:17 (SEQ ID №: 60), EXP-MISsi.Ubq1:1:8 (SEQ ID №: 70), EXP-MISsi.Ubq1:1:10 (SEQ ID №: 72), EXP-MISsi.Ubq1:1:11 (SEQ ID №: 74), EXP-MISsi.Ubq1:1:7 (SEQ ID №: 76), EXP-SCHsc.Ubq1:1:9 (SEQ ID №: 78), EXP-SCHsc.Ubq1:1:7 (SEQ ID №: 84), EXP-SCHsc.Ubq1:1:10 (SEQ ID №: 86), EXP-SORnu.Ubq1:1:6 (SEQ ID №: 92), EXP-SORnu.Ubq1:1:7 (SEQ ID №: 95), EXP-SORnu.Ubq1:1:8 (SEQ ID №: 97), EXP-SETit.Ubq1:1:5 (SEQ ID №: 103), EXP-SETit.Ubq1:1:7 (SEQ ID №: 106), EXP-SETit.Ubq1:1:6 (SEQ ID №: 108), EXP-Sv.Ubq1:1:7 (SEQ ID №: 110), EXP-Sv.Ubq1:1:8 (SEQ ID №: 116), EXP-Sv.Ubq1:1:10 (SEQ ID №: 118), EXP-Zm.UbqM1:1:6 (SEQ ID №: 122), EXP-Zm.UbqM1:1:7 (SEQ ID №: 128), EXP-Zm.UbqM1:1:8 (SEQ ID №: 134), Exp-Sb.Ubq4:1:2 (SEQ ID №: 140) и Exp-Sb.Ubq6:1:2 (SEQ ID №: 144) с экспрессией под контролем известных конститутивных промоторов. Каждую последовательность EXP, содержащую матрицу амплификации, из которой экспрессионная кассета ампликона получена, клонировали с использованием способов, известных в данной области техники, в растительный экспрессионный вектор, показанный в таблице 2 ниже под заголовком «матрица ампликона». Полученные растительные экспрессионные векторы содержат последовательность EXP, функционально связанную с 5’-кодирующей последовательностью GUS, которая содержит либо процессируемый интрон («GUS-2», SEQ ID № 154), либо прилегающую кодирующую последовательность GUS («GUS-1», SEQ ID №: 153), функционально связанную с 5’- 3’-UTR T-AGRtu.nos-1:1:13 (SEQ ID №: 157) или T-Ta.Hsp17-1:1:1 (SEQ ID №: 158). Ампликоны были получены с использованием способов, известных специалистам в данной области техники с использованием матриц плазмидных конструкций, представленных в таблице 2 ниже. Вкратце, 5’-олигонуклеотидный праймер был разработан для отжига промоторной последовательности, а 3'- олигонуклеотидный праймер, который отжигается с 3’-конца 3’-UTR, был использован для амплификации каждой экспрессионной кассеты. Последовательные 5’-делеции были введены в промоторные последовательности, составляющие экспрессионные кассеты, давая начало различным последовательностям EXP, путем использования различных олигонуклеотидных праймеров, которые были разработаны для отжига при различных позициях в пределах промоторной последовательности, составляющей каждую матрицу ампликона.
Таблица 2. Ампликоны для экспрессии в растениях с последовательностью гена GUS и соответствующие матрицы плазмидных конструкций ампликонов, последовательность EXP, кодирующая последовательность GUS и 3’-UTR, использованные для трансформации протопластов листа кукурузы.
последовательность
[00088] Плазмидные конструкции, приведенные как матрицы ампликона в таблице 2, служили матрицами для амплификации экспрессионных кассет с трансгеном, содержащих перечисленные в таблице 2 последовательности EXP. Контрольные плазмиды, используемые для получения ампликонов с GUS трансгеном для сравнения, были созданы, как описано ранее, с конститутивными последовательностями EXP EXP-Os.Act1:1:9 (SEQ ID №: 162) и EXP-CaMV.35S-enh+Ta.Lhcb1+Os.Act1:1:1 (SEQ ID №: 161). Пустой вектор, не предназначенный для экспрессии трансгена, был использован в качестве отрицательного контроля для оценки фоновой экспрессии GUS и люциферазы.
[00089] Также были построены две плазмиды для использования при ко-трансформации и нормировании данных, с использованием способов, известных в данной области техники. Каждая плазмида содержала специфическую последовательность, кодирующую люциферазу, под контролем конститутивной последовательности EXP. Растительный вектор pMON19437 содержит экспрессионную кассету с конститутивным промотором, функционально связанным с 5’-интроном, (EXP-CaMV.35S-enh+Zm.DnaK:1:1, SEQ ID №: 163), функционально связанным с 5’-кодирующей последовательностью люциферазы светляка (Photinus pyralis) (LUCIFERASE:1:3, SEQ ID №: 156), функционально связанной с 5’-3’-UTR из гена нопалинсинтазы Agrobacterium tumefaciens (T-AGRtu.nos-1:1:13, SEQ ID №: 158). Растительный вектор pMON63934 содержит экспрессионную кассету с конститутивной последовательностью EXP (EXP-CaMV.35S-enh-Lhcb1, SEQ ID №: 164), функционально связанной с 5’-кодирующей последовательностью люциферазы морской губки (Renilla reniformis) (CR-Ren.hRenilla Lucife-0:0:1, SEQ ID №: 157), функционально связанной с 5’-3’-UTR из гена нопалинсинтазы Agrobacterium tumefaciens (T-AGRtu.nos-1:1:13, SEQ ID №: 158).
[00090] Протопласты листа кукурузы были трансформированы с использованием способа трансформации на основе ПЭГ, который хорошо известен в данной области техники. Клетки протопластов трансформировали плазмидными ДНК pMON19437 и pMON63934, и ампликонами, представленными в таблице 2, и инкубировали в течение ночи в полной темноте. Измерения как GUS, так и люциферазы проводили путем помещения аликвот лизированных препаратов клеток, трансформированных как описано ранее, в два различных планшета с мелкими лунками. Один планшет использовали для измерения GUS, а второй планшет использовали для выполнения двойного анализа люциферазы с использованием системы двойного анализа репортера люциферазы (Promega Corp., Madison, WI; см., например, Promega Notes Magazine, No: 57, 1996, p.02). Одна или две трансформации были проведены для каждой последовательности EXP и определены средние значения экспрессии для каждой последовательности EXP из нескольких образцов каждого эксперимента по трансформации. Измерения образцов были проведены с использованием четырех повторностей для каждой трансформации генетической конструкцией с последовательностью EXP, или в альтернативном варианте трех повторностей для каждого ампликона с последовательностью EXP в одном из двух экспериментах по трансформации. Значения уровней экспрессии GUS и люциферазы приводятся в таблице 3. В этой таблице, значения люциферазы светляка (например, в результате экспрессии pMON19437) приводятся в столбце, обозначенном «FLuc», а значения люциферазы Renilla приводятся в столбце, обозначенном «RLuc».
Таблица 3. Среднее значение активности GUS и люциферазы в протопластах клеток из листьев трансформированной кукурузы.
[00091] Для сравнения относительной активности каждой последовательности EXP, значения GUS были выражены как отношение активности GUS к активности люциферазы и нормированы по отношению к уровням экспрессии, наблюдаемым для EXP-Os.Act1:1:1 и EXP-CaMV.35S-enh+Ta.Lhcb1+Os.Act1:1:1. Таблица 4 ниже показывает GUS/RLuc отношения экспрессии, нормированные по отношению к экспрессии под контролем EXP-Os.Act1:1:1 и EXP-CaMV.35S-enh+Ta.Lhcb1+Os.Act1:1:1, в протопластах кукурузы. Таблица 5 ниже показывает GUS/FLuc отношения экспрессии, нормированные по отношению к экспрессии под контролем EXP-Os.Act1:1:1 и EXP-CaMV.35S-enh+Ta.Lhcb1+Os.Act1:1:1, в протопластах кукурузы.
Таблица 4. GUS/RLuc и GUS/FLuc отношения экспрессии, нормированные по отношению к EXP-CaMV.35S-enh+Ta.Lhcb1+Os.Act1:1:1 (SEQ ID №: 161) в протопластах кукурузы.
Os.Act1:1:1
Os.Act1:1:1
Таблица 5. GUS/RLuc и GUS/FLuc отношения экспрессии, нормированные по отношению к EXP-Os.Act1:1:9 (SEQ ID №: 162) в протопластах листьев кукурузы.
[00092] Как видно из таблиц 9 и 10, все последовательности EXP были способны проводить экспрессию GUS трансгена в клетках кукурузы. Среднее значение экспрессии GUS было выше для всех последовательностей EXP по сравнению с EXP-Os.Act1:1:9. EXP последовательности EXP-AGRne.Ubq1:1:7 (SEQ ID №: 1), EXP-AGRne.Ubq1:1:8 (SEQ ID №: 5), EXP-AGRne.Ubq1:1:9 (SEQ ID №: 7), EXP-ARUdo.Ubq1:1:8 (SEQ ID №: 13), EXP-ARUdo.Ubq1:1:9 (SEQ ID №: 18), EXP-ARUdo.Ubq1:1:11 (SEQ ID №: 21), EXP-ARUdo.Ubq2:1:8 (SEQ ID №: 27), EXP-ARUdo.Ubq2:1:9 (SEQ ID №: 30), EXP-ARUdo.Ubq2:1:10 (SEQ ID №: 32), EXP-BOUgr.Ubq1:1:6 (SEQ ID №: 38), EXP-BOUgr.Ubq1:1:7 (SEQ ID №: 41), EXP-BOUgr.Ubq2:1:14 (SEQ ID №: 52), EXP-BOUgr.Ubq2:1:16 (SEQ ID №: 58), EXP-BOUgr.Ubq2:1:17 (SEQ ID №: 60), EXP-MISsi.Ubq1:1:10 (SEQ ID №: 72), EXP-SCHsc.Ubq1:1:7 (SEQ ID №: 84), EXP-SCHsc.Ubq1:1:10 (SEQ ID №: 86), EXP-SORnu.Ubq1:1:7 (SEQ ID №: 95), EXP-SORnu.Ubq1:1:8 (SEQ ID №: 97), EXP-SETit.Ubq1:1:5 (SEQ ID №: 103), EXP-SETit.Ubq1:1:7 (SEQ ID №: 106), EXP-SETit.Ubq1:1:6 (SEQ ID №: 108), EXP-Sv.Ubq1:1:7 (SEQ ID №: 110), EXP-Sv.Ubq1:1:8 (SEQ ID №: 116), EXP-Sv.Ubq1:1:10 (SEQ ID №: 118), EXP-Zm.UbqM1:1:6 (SEQ ID №: 122), EXP-Zm.UbqM1:1:8 (SEQ ID №: 134), EXP-Sb.Ubq4:1:2 (SEQ ID №: 140) и EXP-Sb.Ubq6:1:2 (SEQ ID №: 144) показали уровни экспрессии GUS выше, чем EXP-CaMV.35S-enh+Ta.Lhcb1+Os.Act1:1:1.
[00093] Во второй серии экспериментов экспрессионную кассету ампликона с GUS геном, включающую последовательность EXP EXP-Zm.UbqM1:1:7 (SEQ ID №: 128) сравнивали с контрольными ампликонами, PCR0145942 (EXP-Os.Act1:1:9, SEQ ID №: 162) и PCR0145944 (EXP-CaMV.35S-enh+Zm.DnaK:1:1, SEQ ID №: 161) по отношению к экспрессии GUS. GUS экспрессия под контролем последовательности EXP EXP-Zm.UbqM1:1:7 была выше, чем в случае двух контролей. Таблица 6 ниже показывает средние значения GUS и люциферазы, определенные для каждого ампликона. Таблица 7 ниже показывает GUS/RLuc и GUS/FLuc отношения экспрессии, нормированные по отношению к экспрессии под контролем EXP-Os.Act1:1:9 и EXP-CaMV.35S-enh+Zm.DnaK:1:1, в протопластах кукурузы.
Таблица 6. Среднее значение активности GUS и люциферазы в протопластах клеток из листьев трансформированной кукурузы.
Таблица 7. GUS/RLuc и GUS/FLuc отношения экспрессии, нормированные по отношению к EXP-Os.Act1:1:9 (SEQ ID №: 161) и EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID №: 160) в протопластах листьев кукурузы.
[00094] Эффективность регуляторных элементов, контролирующих экспрессию GUS с ампликонов, может быть изучена подобным образом в протопластах листьев сахарного тростника. Например, протопласты сахарного тростника могут быть трансформированы ампликонами ДНК, полученными из растительных экспрессионных векторов, содержащих последовательность EXP, которая контролируют экспрессию трансгена GUS, и сравнены с протопластом листа, в котором экспрессия GUS контролируется известными конститутивными промоторами.
Пример 3
Анализ регуляторных элементов, контролирующих GUS в протопластах пшеницы, с использованием GUS экспрессионных кассет ампликонов
[00095] Протопласты листьев пшеницы были трансформированы ампликонами ДНК, полученными из растительных экспрессионных векторов, содержащих последовательность EXP, которая контролирует экспрессию трансгена GUS, и сравнивались с протопластом листа, в котором экспрессия GUS была под контролем известных конститутивных промоторов.
[00096] Клетки протопластов пшеницы, полученные из тканей листа, были трансформированы с использованием способов, известных в данной области техники, ампликонами, полученными в результате амплификации экспрессионных кассет с GUS геном, содержащих растительные экспрессионные векторы, для сравнения экспрессии трансгена (GUS) под контролем EXP последовательностей, перечисленных в таблице 3, с таковой под контролем известных конститутивных промоторов по методике, описанной в предыдущем примере (пример 2), используя такие же экспрессионные кассеты ампликонов с GUS геном, как были использованы ранее для анализа в кукурузе в примере 2. Ампликоны контрольной экспрессионной кассеты с GUS геном и плазмиды с геном люциферазы, используемые для трансформации протопластов пшеницы, были такими же, как представленые в предыдущем примере и приведенные ранее в таблице 3 примера 2. Подобным образом, отрицательные контроли использовались для определения фоновой активности GUS и люциферазы, как описано ранее. Протопласты листьев пшеницы были трансформированы с использованием способа трансформации на основе ПЭГ, как описано ранее в примере 2. В таблице 8 приведено среднее значение активности GUS и LUC, наблюдаемое в протопластах клеток из листьев трансформированной пшеницы, а таблицы 9 и 10 показывают нормированные GUS/FLuc и GUS/RLuc отношения экспрессии в протопласты пшеницы по отношению к экспрессии под конститутивными EXP контролями.
Таблица 8. Среднее значение активности GUS и люциферазы в протопластах клеток из листьев трансформированной пшеницы.
Таблица 9. GUS/RLuc и GUS/FLuc отношения экспрессии, нормированные по отношению к EXP-CaMV.35S-enh+Ta.Lhcb1+Os.Act1:1:1 (SEQ ID №: 161) в протопластах пшеницы.
Таблица 10. GUS/RLuc и GUS/FLuc отношения экспрессии, нормированные по отношению к EXP-Os.Act1:1:9 (SEQ ID №: 162) в протопластах листьев кукурузы.
[00097] Как видно из таблиц 9 и 10 выше, все последовательности EXP были способны проводить экспрессию трансгена GUS в клетках пшеницы. Все последовательностей EXP проводили экспрессию GUS на более высоком уровне, чем EXP-Os.Act1:1:9 в клетках пшеницы. EXP последовательности EXP-ARUdo.Ubq1:1:8 (SEQ ID №: 13), EXP-ARUdo.Ubq1:1:9 (SEQ ID №: 18), EXP-ARUdo.Ubq1:1:11 (SEQ ID №: 21), EXP-ARUdo.Ubq2:1:10 (SEQ ID №: 32), EXP-BOUgr.Ubq2:1:14 (SEQ ID №: 52), EXP-BOUgr.Ubq2:1:16 (SEQ ID №: 58), EXP-BOUgr.Ubq2:1:17 (SEQ ID №: 60), EXP-MISsi.Ubq1:1:10 (SEQ ID №: 72), EXP-SORnu.Ubq1:1:7 (SEQ ID №: 95), EXP-SETit.Ubq1:1:5 (SEQ ID №: 103), EXP-SETit.Ubq1:1:7 (SEQ ID №: 106), EXP-SETit.Ubq1:1:6 (SEQ ID №: 108), EXP-Sv.Ubq1:1:7 (SEQ ID №: 110), EXP-Sv.Ubq1:1:8 (SEQ ID №: 116), EXP-Sv.Ubq1:1:10 (SEQ ID №: 118), EXP-Zm.UbqM1:1:6 (SEQ ID №: 122) и EXP-Zm.UbqM1:1:8 (SEQ ID №: 134) показали уровни экспрессии GUS, равные или большие, чем уровни экспрессии GUS под контролем EXP-CaMV.35S-enh+Ta.Lhcb1+Os.Act1:1:1 в клетках пшеницы.
[00098] Во второй серии экспериментов, экспрессионную кассету ампликона с GUS геном, включающую EXP-ARUdo.Ubq1:1:11 (SEQ ID №: 21) сравнивали с контролями EXP-Os.Act1:1:9 (SEQ ID №: 162) и EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID №: 161). Таблица 11 ниже показывает средние значения GUS и люциферазы, определенные для каждого ампликона. Таблица 12 ниже показывает GUS/RLuc отношения экспрессии, нормированные по отношению к экспрессии под контролем EXP-Os.Act1:1:9 и EXP-CaMV.35S-enh+Zm.DnaK:1:1, в протопластах пшеницы.
Таблица 11. Среднее значение активности GUS и люциферазы в протопластах клеток из листьев трансформированной пшеницы.
Таблица 12. GUS/RLuc и GUS/FLuc отношения экспрессии, нормированные по отношению к EXP-Os.Act1:1:9 (SEQ ID №: 161) и EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID №: 160) в протопластах листьев пшеницы.
[00099] Как можно видеть ранее из таблицы 12, GUS экспрессия под контролем EXP-ARUdo.Ubq1:1:11 (SEQ ID №: 21) была выше, чем в случае двух конститутивных контролей EXP-Os.Act1:1:9 и EXP-CaMV.35S-enh+Zm.DnaK:1:1.
Пример 4
Анализ регуляторных элементов, контролирующих GUS в протопластах кукурузы и пшеницы
[000100] Протопласты листьев кукурузы и пшеницы трансформировали растительными экспрессионными векторами, содержащими последовательность EXP, которая контролирует экспрессию трансгена бета-глюкуронидазы (GUS), и сравнивали с экспрессией GUS в протопластах листьев, в которых экспрессия GUS контролировалась известными конститутивными промоторами.
[000101] Экспрессию трансгена под контролем EXP-Cl.Ubq10 (SEQ ID №: 168) сравнивали с экспрессией под контролем известных конститутивных промоторов. Вышеупомянутые последовательности EXP были клонированы в растительные экспрессионные векторы как показано в таблице 13 ниже, для получения векторов, в которых последовательность EXP функционально связана с 5’-GUS репортера, который содержал процессируемый интрон (именуемый GUS-2, SEQ ID №: 160), полученный из свето-индуцибильного тканеспецифического гена ST-LS1 картофеля (GenBank Accession: X04753), или прилегающую кодирующую последовательность GUS (GUS-1, SEQ ID №: 159), которая была функционально связана с 5’-3’-UTR из гена нопалинсинтазы A. tumefaciens (T-AGRtu.nos-1:1:13, SEQ ID №: 161) или гена пшеницы Hsp17 (T-Ta.Hsp17-1:1:1, SEQ ID №: 162).
Таблица 13. Растительная экспрессионная плазмидная конструкция с GUS геном и соответствующая последовательность EXP, кодирующая последовательность и 3’-UTR GUS, использованные для трансформации протопластов листа кукурузы. «SEQ ID №:» относится к данной последовательности EXP.
[000102] Также были построены две плазмиды для использования при ко-трансформации и нормировании данных, с использованием способов, известных в данной области техники. Каждая плазмида содержала специфическую последовательность, кодирующую люциферазу, под контролем конститутивной последовательности EXP. Растительный вектор pMON19437 содержит экспрессионную кассету с конститутивным промотором, функционально связанным с 5’-интроном, (EXP-CaMV.35S-enh+Zm.DnaK:1:1, SEQ ID №: 163), функционально связанным с 5’-кодирующей последовательностью люциферазы светляка (Photinus pyralis) (LUCIFERASE:1:3, SEQ ID №: 156), функционально связанной с 5’-3’-UTR из гена нопалинсинтазы Agrobacterium tumefaciens (T-AGRtu.nos-1:1:13, SEQ ID №: 158). Растительный вектор pMON63934 содержит экспрессионную кассету с конститутивной последовательностью EXP (EXP-CaMV.35S-enh-Lhcb1, SEQ ID №: 164), функционально связанной с 5’-кодирующей последовательностью люциферазы морской губки (Renilla reniformis) (CR-Ren.hRenilla Lucife-0:0:1, SEQ ID №: 157), функционально связанной с 5’-3’-UTR из гена нопалинсинтазы Agrobacterium tumefaciens (T-AGRtu.nos-1:1:13, SEQ ID №: 158).
[000103] Протопласты листа кукурузы были трансформированы с использованием способа трансформации на основе ПЭГ, который хорошо известен в данной области техники. Клетки протопластов трансформировали плазмидными ДНК pMON19437 и pMON63934, и плазмидами, представленными в таблице 13, и инкубировали в течение ночи в полной темноте. Измерения как GUS, так и люциферазы проводили подобным образом, как описано в примере 2 ранее. Одна или две трансформации были проведены для каждой последовательности EXP и определены средние значения экспрессии для каждой последовательности EXP из нескольких образцов каждого эксперимента по трансформации. Измерения образцов были проведены с использованием четырех повторностей для каждой трансформации генетической конструкцией с последовательностью EXP, или в альтернативном варианте трех повторностей для каждой генетической конструкции с последовательностью EXP в одном из двух экспериментах по трансформации. Значения уровней экспрессии GUS и люциферазы приводятся в таблице 14. В этой таблице, значения люциферазы светляка (например, в результате экспрессии pMON19437) приводятся в столбце, обозначенном «FLuc», а значения люциферазы Renilla приводятся в столбце, обозначенном «RLuc».
Таблица 14. Среднее значение активности GUS и люциферазы в протопластах клеток из листьев трансформированной кукурузы.
enh+Ta.Lhcb1+Os.Act1:1:1
[000104] Таблица 15 ниже показывает GUS/FLuc и GUS/RLuc отношения экспрессии, нормированные по отношению к экспрессии под контролем EXP-Os.Act1:1:9 и EXP-CaMV.35S-enh+Zm.DnaK:1:1, в протопластах кукурузы.
Таблица 15. GUS/FLuc и GUS/RLuc отношения экспрессии, нормированные по отношению к EXP-Os.Act1:1:9 (SEQ ID №: 161) и EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID №: 160) в протопластах листьев пшеницы.
[000105] Как можно видеть из таблицы 15 выше, EXP-Cl.Ubq10 (SEQ ID №: 168) был способен проводить экспрессию GUS, но на более низком уровне, чем в случае обоих конститутивных контролей.
[000106] Плазмиды, приведенные ранее в таблице 13, были также использованы для трансформации клеток протопластов листа пшеницы подобным образом, как и для протопластов листа кукурузы, описанных ранее. Средние значения GUS и люциферазы представлены в таблице 16 ниже. Таблица 17 ниже показывает GUS/FLuc и GUS/RLuc отношения экспрессии, нормированные по отношению к экспрессии под контролем EXP-CaMV.35S-enh+Zm.DnaK:1:1, в протопластах кукурузы.
Таблица 16. Среднее значение активности GUS и люциферазы в протопластах клеток из листьев трансформированной кукурузы.
Таблица 17. GUS/FLuc и GUS/RLuc отношения экспрессии, нормированные по отношению к EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID №: 160) в протопластах листьев пшеницы.
[000107] Как можно видеть из таблицы 17 выше, EXP-Cl.Ubq10 (SEQ ID №: 168) экспрессировал GUS на сходном уровне, что и EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID №: 160) в клетках протопластов пшеницы.
Пример 5
Анализ регуляторных элементов, контролирующих GUS в трансгенной кукурузе.
[000108] Растения кукурузы были трансформированы растительными экспрессионными векторами, содержащими последовательности EXP, которые контролируют экспрессию трансгена GUS, и полученные растения были проанализированы на экспрессию белка GUS. Последовательности EXP с убиквитином были клонированы в растительные бинарные трансформационные плазмидные конструкции с использованием способов, известных в данной области техники.
[000109] Полученные растительные экспрессионные векторы содержат правую граничную область из A. tumefaciens, первую экспрессионную кассету для анализа последовательности EXP, функционально связанной с кодирующей последовательностью GUS, которая имеет процессируемый интрон GUS-2, описанный ранее, функционально связанной с 5’-3’-UTR из гена белка переноса липидов риса (T-Os.LTP-1:1:1, SEQ ID №: 159); вторая кассета для селекции трансгена, используемая для отбора трансформированных клеток растения, придает устойчивость к гербициду глифосату (под контролем промотора актина 1 риса) и левой граничной области из A. tumefaciens. Полученные плазмиды были использованы для трансформации растений кукурузы. В таблице 18 приведены обозначения плазмид, последовательности EXP и SEQ ID №№, которые также описаны в таблице 1.
Таблица 18. Бинарные плазмиды для трансформации растений и ассоциированные последовательности EXP.
[000110] Растения были трансформированы путем Agrobacterium-опосредованной трансформации, например, как описано в публикации заявки на патент США 2009/0138985.
[000111] Гистохимический анализ GUS был использован для качественного анализа экспрессии трансформированных растений. Участки цельной ткани инкубировали с GUS окрашивающим раствором X-Gluc (5-бром-4-хлор-3-индолил-бета-глюкуронид) (1 мг/мл) в течение соответствующего периода времени, промывали и проверяли визуально по синему окрашиванию. GUS активность качественно определялась путем прямой визуальной проверки или проверки под микроскопом, используя выбранные органы и ткани растения. R0 растения были проверены на экспрессию в корнях и листьях, а также в пыльнике, шелке и развивающихся семенах и завязи, через 21 день после опыления (21 ДПО).
[000112] Для количественного анализа, общий белок экстрагировали из выбранных тканей трансформированных растений кукурузы. Один микрограмм общего белка был использован с флюорогенным субстратом 4-метилумбеллиферил-β-D-глюкуронидом (MUG) в общем объеме реакции 50 мкл. Продукт реакции, 4–метилумбеллиферон (4-MU), максимально флуоресцирует при высоких значениях рН, где гидроксильная группа ионизируется. Добавление щелочного раствора карбоната натрия одновременно останавливает реакцию и доводит рН, для количественного измерения флуоресцентного продукта. Флуоресценцию измеряли при длине волны возбуждения 365 нм, излучения – 445 нм с использованием Fluoromax-3 (Horiba; Киото, Япония) с Micromax Reader, с установленной шириной щели: 2 нм – на возбуждение и 3 нм – на излучение.
[000113] Средняя экспрессия GUS в R0, наблюдаемая для каждой трансформации, представлена в таблицах 19 и 20 ниже.
Таблица 19. Средняя экспрессия GUS в R0 в ткани корня и листа.
Таблица 20. Средняя экспрессия GUS в R0 в репродуктивных органах кукурузы (пыльник, шелк) и развивающихся семенах (завязь и эндосперм).
[000114] В R0 растениях кукурузы уровни экспрессии GUS в листе и корне отличалась среди последовательностей EXP с геном убиквитина. Несмотря на то, что все последовательности EXP показали способность контролировать экспрессию трансгена GUS в стабильно трансформированных растениях, каждая последовательность EXP продемонстрировала уникальный характер экспрессии в сравнении с другими. Например, EXP последовательности EXP-AGRne.Ubq1:1:7 (SEQ ID №: 1), EXP-AGRne.Ubq1:1:8 (SEQ ID №: 5), EXP-ARUdo.Ubq1:1:8 (SEQ ID №: 13), EXP-ARUdo.Ubq1:1:9 (SEQ ID №: 18), EXP-ARUdo.Ubq2:1:8 (SEQ ID №: 27), EXP-ARUdo.Ubq2:1:9 (SEQ ID №: 30), EXP-BOUgr.Ubq1:1:6 (SEQ ID №: 38), EXP-BOUgr.Ubq1:1:7 (SEQ ID №: 41), EXP-BOUgr.Ubq2:1:14 (SEQ ID №: 52), EXP-BOUgr.Ubq2:1:15 (SEQ ID №: 55), EXP-MISsi.Ubq1:1:7 (SEQ ID №: 76), EXP-SORnu.Ubq1:1:6 (SEQ ID №: 92), EXP-SORnu.Ubq1:1:7 (SEQ ID №: 95), EXP-SETit.Ubq1:1:10 (SEQ ID №: 99), EXP-Sv.Ubq1:1:11 (SEQ ID №: 114), EXP-Zm.UbqM1:1:12 (SEQ ID №: 132) и EXP-Sb.Ubq7:1:2 (SEQ ID №: 150) показали более низкие уровни экспрессии GUS в корне на V3 и V7 стадиях развития по отношению к EXP-Sv.Ubq1:1:12 (SEQ ID №: 120), EXP-Zm.UbqM1:1:10 (SEQ ID №: 126), EXP-Zm.UbqM1:1:11 (SEQ ID №: 138), EXP-Sb.Ubq4:1:2 (SEQ ID №: 140) и EXP-Sb.Ubq6:1:3 (SEQ ID №: 148). Более высокие уровни экспрессии GUS наблюдали на поздних стадиях развития корней (VT) для EXP-ARUdo.Ubq1:1:8 (SEQ ID №: 13), EXP-ARUdo.Ubq1:1:9 (SEQ ID №: 18), EXP-BOUgr.Ubq1:1:7 (SEQ ID №: 41), EXP-Zm.UbqM1:1:11 (SEQ ID №: 138) и EXP-Sb.Ubq6:1:3 (SEQ ID №: 148). Экспрессия в корнях под контролем EXP-Zm.UbqM1:1:10 (SEQ ID №: 140) показала отсутствие экспрессии на V3 стадии, но была высокой на V7 стадии, а затем снизилась на VT стадии. Экспрессия в корнях под контролем EXP-Zm.UbqM1:1:11 (SEQ ID №: 150) поддерживалась на сходном уровне на протяжении развития от стадий V3 и V7 вплоть до VT. Экспрессия GUS под контролем EXP-Cl.Ubq10 (SEQ ID №: 168) была относительно стабильной от V4 до V7 стадии, но снизилась примерно на половину на VT стадии по сравнению со стадиями V4 и V7.
[000115] Уровни экспрессии GUS также показали существенные различия в ткани листа. EXP последовательности EXP-Zm.UbqM1:1:10 (SEQ ID №: 126), EXP-Zm.UbqM1:1:12 (SEQ ID №: 132) и EXP-Zm.UbqM1:1:11 (SEQ ID №: 138) показали самый высокий уровень экспрессии GUS, наблюдаемый на всех трех стадиях развития (V3, V7 и VT). EXP последовательность EXP-Sb.Ubq4:1:2 (SEQ ID №: 140), показали снижение экспрессии на стадиях развития от V3 до VT. EXP последовательности EXP-ARUdo.Ubq1:1:8 (SEQ ID №: 13) и EXP-Sb.Ubq7:1:2 (SEQ ID №: 150) показали более высокие уровни экспрессии GUS на V3 и VT стадиях развития, с более низким уровнем экспрессии в середине роста на V7 стадии развития. EXP последовательность EXP-ARUdo.Ubq2:1:9 (SEQ ID №: 30), EXP-BOUgr.Ubq1:1:7 (SEQ ID №: 41) и EXP-MISsi.Ubq1:1:7 (SEQ ID №: 76) поддерживали экспрессию GUS на протяжении всех трех стадий, в то время как EXP-ARUdo.Ubq2:1:8 (SEQ ID №: 27), EXP-BOUgr.Ubq1:1:6 (SEQ ID №: 38) и EXP-BOUgr.Ubq2:1:15 (SEQ ID №: 55) показали небольшое снижение экспрессии на VT стадии. Экспрессия под контролем EXP-Cl.Ubq10 (SEQ ID №: 168) была сходная на V4 и VT стадиях, но уровень снижался на около половину на V7 стадии по сравнению со стадиями V4 и VT.
[000116] Подобным образом, по отношению к репродуктивной ткани (пыльнику и шелку) наблюдали различный характер экспрессии, уникальный для каждой последовательности EXP. Например, высокие уровни экспрессии наблюдали в пыльнике и шелке для последовательностей EXP: EXP-ARUdo.Ubq1:1:8 (SEQ ID №: 13), EXP-ARUdo.Ubq1:1:9 (SEQ ID №: 18), EXP-BOUgr.Ubq2:1:14 (SEQ ID №: 52), EXP-BOUgr.Ubq2:1:15 (SEQ ID №: 55), EXP-SORnu.Ubq1:1:7 (SEQ ID №: 95), EXP-Zm.UbqM1:1:10 (SEQ ID №: 126), EXP-Zm.UbqM1:1:12 (SEQ ID №: 132), EXP-Zm.UbqM1:1:11 (SEQ ID №: 138) и EXP-Sb.Ubq7:1:2 (SEQ ID №: 150). Экспрессия под контролем EXP последовательностей EXP-AGRne.Ubq1:1:7 (SEQ ID №: 1), EXP-SORnu.Ubq1:1:6 (SEQ ID №: 92), EXP-Sv.Ubq1:1:11 (SEQ ID №: 114), EXP-Sv.Ubq1:1:12 (SEQ ID №: 120), EXP-Sb.Ubq4:1:2 (SEQ ID №: 140), EXP-Sb.Ubq6:1:3 (SEQ ID №: 148) и EXP-Cl.Ubq10 (SEQ ID №: 168) была высокой в пыльнике, но более низкой в шелке, касательно каждой последовательности EXP, тогда как экспрессия под контролем EXP-BOUgr.Ubq1:1:6 (SEQ ID №: 38) была выше в шелке по сравнению с экспрессией в пыльнике.
[000117] Экспрессия в развивающихся семенах (21 ДПО завязь и эндосперм) отличалась среди последовательностей EXP. EXP последовательности EXP-Zm.UbqM1:1:10 (SEQ ID №: 126), EXP-Zm.UbqM1:1:12 (SEQ ID №: 132) и EXP-Zm.UbqM1:1:11 (SEQ ID №: 138) проводили высокую экспрессию GUS в тканях завязи и эндосперма развивающихся семян. Уровни экспрессии в эндосперме были около в два раза или более выше, чем в завязи, когда GUS находился под контролем последовательностей EXP: EXP-ARUdo.Ubq1:1:8 (SEQ ID №: 13), EXP-ARUdo.Ubq1:1:9 (SEQ ID №: 18), EXP-ARUdo.Ubq2:1:8 (SEQ ID №: 27), EXP-BOUgr.Ubq2:1:14 (SEQ ID №: 52), EXP-BOUgr.Ubq2:1:15 (SEQ ID №: 55), EXP-SORnu.Ubq1:1:6 (SEQ ID №: 92), EXP-SORnu.Ubq1:1:7 (SEQ ID №: 95), EXP-Sv.Ubq1:1:12 (SEQ ID №: 120), EXP-Sb.Ubq4:1:2 (SEQ ID №: 140) и EXP-Cl.Ubq10 (SEQ ID №: 168). Экспрессия GUS была в три раза выше в завязи, чем в эндосперме, под контролем EXP-Sb.Ubq7:1:2 (SEQ ID №: 150). Уровни экспрессии GUS были относительно эквивалентными в завязи и эндосперме, когда контролировались последовательностями EXP: EXP-AGRne.Ubq1:1:7 (SEQ ID №: 1), EXP-AGRne.Ubq1:1:8 (SEQ ID №: 5), EXP-BOUgr.Ubq1:1:6 (SEQ ID №: 38), EXP-BOUgr.Ubq1:1:7 (SEQ ID №: 41), EXP-MISsi.Ubq1:1:7 (SEQ ID №: 76), EXP-SETit.Ubq1:1:10 (SEQ ID №: 99) и EXP-Sb.Ubq6:1:3 (SEQ ID №: 148).
[000118] Каждая последовательность EXP показала способность контролировать экспрессию трансгена в стабильно трансформированных растениях кукурузы. Тем не менее, каждая последовательность EXP имела уникальный характер экспрессии для каждой ткани, дающий возможность выбрать последовательность EXP, которая наилучшим образом обеспечивает экспрессию конкретного трансгена в зависимости от стратегии экспрессии в ткани, необходимой для достижения желаемых результатов. Данный пример показывает, что последовательности EXP, выделенные из гомологичных генов, не обязательно ведут себя эквивалентно в трансформированном растении, и что экспрессия может быть определена только с помощью эмпирических исследований свойств каждой последовательности EXP и не может быть предсказана основываясь на гомологии гена, из которого промотор был получен.
Пример 6
Энхансеры, происходящие от регуляторных элементов.
[000119] Энхансеры происходят из промоторных элементов, предложенных в данном документе, таких, которые представлены в SEQ ID №№: 2, 6, 8, 10, 14, 17, 22, 24, 28, 31, 33, 35, 39, 42, 44, 46, 50, 53, 56, 61, 63, 67, 71, 73, 75, 77, 79, 83, 85, 87, 89, 93, 96, 98 и 169. Энхансерный элемент может содержать один или более цис регуляторных элементов, которые, в случае, когда они функционально связанны через 5' или 3' конец с промоторным элементом или функционально связаны через 5' или 3' конец с дополнительными энхансерными элементами, которые функционально связаны с промотором, могут повышать или модулировать экспрессию трансгена или обеспечить экспрессию трансгена в конкретном типе клеток или органе растения, или на конкретной временной точке в развитии или циркадного ритма. Энхансеры создают путем удаления ТАТА-бокса или функционально подобных элементов и любой нижележащей последовательности ДНК из промоторов, что позволяет инициировать транскрипцию с промоторов, описанных в данном документе, как описано выше, в том числе их фрагментов, в которых удалены ТАТА-бокс или функционально подобные элементы и последовательность ДНК, находящаяся ниже ТАТА-бокса.
[000120] Энхансерные элементы могут быть получены из промоторных элементов, описанных в данном документе, и клонированы с использованием способов, известных в данной области техники, таким образом, чтобы они были функционально связаны через 5' или 3' конец с промоторным элементом или функционально связаны через 5' или 3' конец с дополнительными энхансерными элементами которые функционально связаны с промотором. В альтернативном варианте энхансерные элементы клонируют, используя способы, известные в данной области техники, таким образом, чтобы они были функционально связаны с одной или более копий энхансерного элемента, который функционально связан через 5' или 3' конец с промоторным элементом или функционально связан через 5' или 3' конец с дополнительными энхансерными элементами, которые функционально связаны с промотором. Энхансерные элементы также могут быть клонированы таким образом, чтобы они были функционально связаны через 5' или 3' конец с промоторным элементом, полученным из организма другого рода, или функционально связаны через 5' или 3' конец с дополнительными энхансерными элементами, полученными из организмов другого рода или организма того же рода, которые функционально связаны с промотором, полученным из организма того же или другого рода, в результате чего образуется химерный регуляторный элемент. Экспрессионный вектор с геном GUS для трансформации растений создан с использованием способов, известных в данной области техники, подобно генетическим конструкциям, описанным в предыдущих примерах, в которых в итоговые растительные экспрессионные векторы содержат правую граничную область из A. tumefaciens, первую экспрессионную кассету для тестирования регуляторного или химерного регуляторного элемента, содержащую регуляторный или химерный регуляторный элемент, функционально связанный с интроном, полученным из белка теплового шока HSP70 из Z. mays (I-Zm.DnaK-1:1:1 SEQ ID №: 165) или любым из интронов, представленных в данном документе, или любым другим интроном, функционально связанным с кодирующей последовательностью GUS, которая либо имеет процессируемый интрон (GUS-2, SEQ ID №: 155) или не имеет интрона (GUS-1, SEQ ID №: 154), функционально связанной с 3’-UTR нопалинсинтазы из A. tumefaciens (T-AGRtu.nos-1:1:13, SEQ ID №: 158) или с 3'-UTR из гена белка переноса липидов риса (T-Os.LTP-1:1:1, SEQ ID №: 160); вторая кассета для селекции трансгена, используемая для отбора трансформированных клеток растения, придает устойчивость к гербициду глифосату (под контролем промотора актина 1 риса) или, в альтернативном варианте, к антибиотику канамицину (под контролем промотора актина 1 риса) и левой граничной области из A. tumefaciens. Полученные плазмиды используют для трансформации растений кукурузы или других растений этого рода с помощью способов, описанных ранее, или других способов Agrobacterium-опосредованной трансформации или бомбардировки микрочастицами, известными в данной области техники. В альтернативном варианте клетки протопластов, полученные из кукурузы или других растений рода, трансформируют с помощью способов, известных в данной области техники, для проведения анализа транзиентной экспрессии.
[000121] Экспрессия гена GUS под контролем регуляторного элемента, содержащего один или более энхансеров, оценивается при испытаниях растений со стабильной трансформацией или с транзиентной экспрессией для определения влияния энхансерного элемента на экспрессию трансгена. Модификации одного или более энхансерных элементов или дупликация одного или более энхансерных элементов осуществляется на основе эмпирических исследований и итоговой регуляции экспрессии генов, наблюдаемой при использовании композиции каждого регуляторного элемента. Изменение относительных позиций одного или более энхансеров в итоговом регуляторном или химерном регуляторном элементе может влиять на транскрипционную активность или специфичность регуляторного или химерного регуляторного элемента и определяется эмпирически для определения наилучших энхансеров для желаемого профиля экспрессии трансгена в пределах растения кукурузы или растения другого рода.
Пример 7
Анализ интронного усиления GUS активности с использованием протопластов растительного происхождения.
[000122] Интрон выбран на основании экспериментирования и сравнения с экспрессией контрольного безинтронного вектора для эмпирического выбора интрона и конфигурации в пределах расположения элементов переноса ДНК (Т-ДНК) вектора для оптимальной экспрессии трансгена. Например, при экспрессии гена устойчивости к гербициду, такому как CP4, который придает толерантность к глифосату, желательно получать экспрессию трансгена в репродуктивных тканях, а также в вегетативных тканях, для предотвращения потерю урожая после применения гербицида. Интрон, функционально связанный с конститутивным промотором, в данном случае будет выбран по его способности усиливать экспрессию устойчивости к гербициду, придаваемую трансгеном, в частности, в репродуктивных клетках и тканях трансгенного растения и, таким образом, обеспечивать как вегетативную, так и репродуктивную толерантность трансгенного растения при опрыскивании гербицидом. В большинстве генов убиквитина 5’-UTR состоит из лидера, который имеет последовательность интрона, встроенную в него. Поэтому, регуляторные элементы, полученные из таких генов, анализировали с использованием всей 5’-UTR, содержащей промотор, лидер и интрон. Для достижения различных профилей экспрессии или модулирования уровня экспрессии трансгена, интрон из такого регуляторного элемента может быть удален или замещен гетерологическим интроном.
[000123] Интроны, представленные в данном документе в SEQ ID №№: 4, 12, 15, 20, 26, 29, 37, 40, 48, 51, 54, 57, 59, 65, 69, 81, 91, 94 и 171, идентифицировали с использованием геномных ДНК контигов по сравнению с экспрессируемыми кластерами теговых последовательностей или кДНК контигов, для идентификации последовательностей экзонов и интронов в пределах геномной ДНК. В дополнение, 5’-UTR или лидерные последовательности также используются для определения интрон/экзон сплайсинговых сочленений одного или более интронов в условиях, когда последовательность гена кодирует лидерную последовательность, которая прерывается одним или более интроном. Интроны клонировали с использованием способов, известных в данной области техники, в вектор для трансформации растений так, чтобы были функционально связанными с 3'-регуляторным элементом и фрагментом лидера и функционально связанными с 5'- либо второго фрагмента лидера, либо кодирующими последовательностями, например, как показано в экспрессионных кассетах, представленных на фиг. 9.
[000124] Так, например, первая возможная экспрессионная кассета (конфигурация экспрессионной кассеты 1 на фиг. 9) состоит из промотора или химерного промоторного элемента [A], функционально связанного с 5’-лидерным элементом [B], функционально связанного с 5’-тестируемым интронным элементом [C], функционально связанного с кодирующей областью [D], которая функционально связана с 3'-UTR элементом [E]. В альтернативном варианте, вторая возможная экспрессионная кассета (конфигурация экспрессионной кассеты 2 на фиг. 9) состоит из промотора или химерного промоторного элемента [F], функционально связанного с 5’-первым лидерным элементом или фрагментом первого лидерного элемента [G], функционально связанного с 5’-тестируемым интронным элементом [H], функционально связанного с 5’-вторым лидерным элементом или вторым фрагментом первого лидерного элемента [I], функционально связанного с кодирующей областью [J], которая функционально связана с 3'-UTR элементом [K]. Дополнительно, третья возможная экспрессионная кассета (конфигурация экспрессионной кассеты 3 на фиг. 9) состоит из промотора или химерного промоторного элемента [L], функционально связанного с 5’-лидерным элементом [M], функционально связанного с 5’-первым фрагментом элемента кодирующей последовательности [N], функционально связанного с 5’-интронным элементом [O], функционально связанного с 5’-вторым фрагментом элемента кодирующей последовательности [P], которая функционально связана с 3'-UTR элементом [Q]. Конфигурация экспрессионной кассеты 3 разработана для обеспечения сплайсинга интрона таким способом, чтобы получить полную открытую рамку считывания без сдвига рамки между первым и вторым фрагментом кодирующей последовательности.
[000125] Как обсуждалось ранее, может быть предпочтительным избегание использования нуклеотидной последовательности AT или нуклеотида А непосредственно перед 5’-концом сайта сплайсинга (GT) и нуклеотида G или нуклеотидной последовательности TG, соответственно, непосредственно после 3’-конца сайта сплайсинга (AG), для устранения потенциальных нежелательных стартовых кодонов, формирующихся во время процессинга информационной РНК в конечный транскрипт. Последовательность ДНК вблизи 5’- или 3’-концевых сайтов сплайсинговых сочленений интрона, таким образом, может быть модифицирована.
[000126] Интроны тестировали на эффект усиления посредством возможности усиливать экспрессию при анализе транзиентной экспрессии или стабильной экспрессии в растении. Для анализа интронного усиления транзиентной экспрессии, сконструировали базовый растительный вектор с использованием способов, известных в данной области техники. Интрон клонировали в базовый растительный вектор, который содержит экспрессионную кассету, состоящую из конститутивного промотора, такого как промотор вируса мозаики цветной капусты P-CaMV.35S-enh-1:1:9 (SEQ ID №: 166), функционально связанного с 5’-лидерным элементом L-CaMV.35S-1:1:15 (SEQ ID №: 167), функционально связанным с 5’-тестируемым интронным элементом (например, один из SEQ ID №№: 4, 12, 15, 20, 26, 29, 37, 40, 48, 51, 54, 57, 59, 65, 69, 81, 91, 94 и 171), функционально связанным с кодирующей последовательностью GUS, которая либо имеет процессируемый интрон (GUS-2, SEQ ID №: 155) или не имеет интрона (GUS-1, SEQ ID №: 154), функционально связанной с 3’-UTR нопалинсинтазы из A. tumefaciens (T-AGRtu.nos-1:1:13, SEQ ID №: 158). Клеток протопластов, полученные из ткани растения кукурузы или другого рода, трансформировали базовым растительным вектором и контрольными векторами с геном люциферазы, как описано ранее в примере 2 выше, и анализировали в них активность. Для сравнения относительной способности интрона усиливать экспрессиию, значения GUS выражали как отношение активности GUS к активности люциферазы и сравнивали их с уровнями таковых, получаемых с помощью генетической конструкции, содержащей конститутивный промотор, функционально связанный с известным стандартом интрона, таким как интрон, полученный из HSP70 белка теплового шока Zea mays, I-Zm.DnaK-1:1:1 (SEQ ID №: 165), а также генетической конструкции, содержащей конститутивный промотор, но без интрона, функционально связанного с промотором.
[000127] Для анализа в стабильно трансформированных растениях интронов, представленных в SEQ ID №№: 4, 12, 15, 20, 26, 29, 37, 40, 48, 51, 54, 57, 59, 65, 69, 81, 91, 94 и 171, разработан экспрессионный вектор с геном GUS для трансформации растений аналогичный генетическим конструкциям, описанным в предыдущих примерах, в которых итоговые растительные экспрессионные векторы содержат правую граничную область из A. tumefaciens, первую экспрессионную кассету для тестирования интрона, состоящую из конститутивного промотора, такого как промотор вируса мозаики цветной капусты P-CaMV.35S-enh-1:1:9 (SEQ ID №: 166), функционально связанного с 5’-лидерным элементом L-CaMV.35S-1:1:15 (SEQ ID №: 167), функционально связанным с 5’-тестируемым интронным элементом, предложенным в данном документе, функционально связанным с кодирующей последовательностью GUS, которая либо имеет процессируемый интрон (GUS-2, SEQ ID №: 155) или не имеет интрона (GUS-1, SEQ ID №: 154), функционально связанной с 3’-UTR нопалинсинтазы из A. tumefaciens (T-AGRtu.nos-1:1:13, SEQ ID №: 158); вторая кассета для селекции трансгена, используемая для отбора трансформированных клеток растения, придает устойчивость к глифосату (под контролем промотора актина 1 риса) или, в альтернативном варианте, к антибиотику канамицину (под контролем промотора актина 1 риса) и левой граничной области из A. tumefaciens. Полученные плазмиды используют для трансформации растений кукурузы или других растений этого рода с помощью способов, описанных ранее, или Agrobacterium-опосредованными способами, известными в данной области техники. Однокопийные или низкокопийные трансформанты выбраны для сравнения с однокопийными или низкокопийными трансформированными растениями, трансформированными с помощью вектора для трансформации растений, идентичного тестируемому вектору, но без тестируемого интрона, чтобы определить, обеспечивает ли тестируемый интрон интрон-опосредованный эффект усиления.
[000128] Любой из интронов, представленных в SEQ ID №№: 4, 12, 15, 20, 26, 29, 37, 40, 48, 51, 54, 57, 59, 65, 69, 81, 91, 94 и 171, может быть модифицирован несколькими способами, такими как удаление фрагментов в пределах последовательности интрона, которое может уменьшать экспрессию, или дупликация фрагментов с интроном, которая может усиливать экспрессию. В дополнение, последовательности ДНК в пределах интрона, которые могут влиять на специфичность экспрессии либо в конкретных типах клеток, либо в тканях и органах, могут быть дуплицированы или изменены, или удалены, с целью повлиять на экспрессию и характеры экспрессии трансгена. В дополнение, интроны, предложенные в данном документе, могут быть модифицированы для удаления любых потенциальных стартовых кодонов (ATG), которые могут стать причиной непредусмотренных транскриптов, экспрессирующихся из неправильно сплайсированных интронов как отличающиеся, удлиненные или укороченные белки. После того, как интрон был эмпирически проверен или изменен на основании проведенного исследования, интрон используется для усиления экспрессии трансгена в стабильно трансформированных растениях, которые могут представлять собой однодольные или двудольные растения любого рода, при условии, что интрон обеспечивает усиление трансгена. Интрон также может быть использован для усиления экспрессии в других организмах, таких как водоросли, грибы или клетки животных, при условии, что интрон обеспечивает усиление или ослабление, или специфичность экспрессии трансгена, с которым он функционально связан.
[000129] Из иллюстраций и описания принципов изобретения специалистам в данной области техники будет очевидно, что форма и детали данного изобретения могут быть модифицированы без отступления от этих принципов. Мы заявляем все модификации, находящиеся в пределах сущности и объема формулы изобретения. Все публикации и опубликованные патентные документы, цитируемые в данном документе, включены посредством ссылки в той же степени, как если бы каждая отдельная публикация или заявка на патент конкретно и индивидуально указывались как включенные посредством ссылки.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2644205C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2800430C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2800425C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2800423C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2800424C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2799433C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2796556C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2675524C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2812893C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2639275C2 |









































































































































































































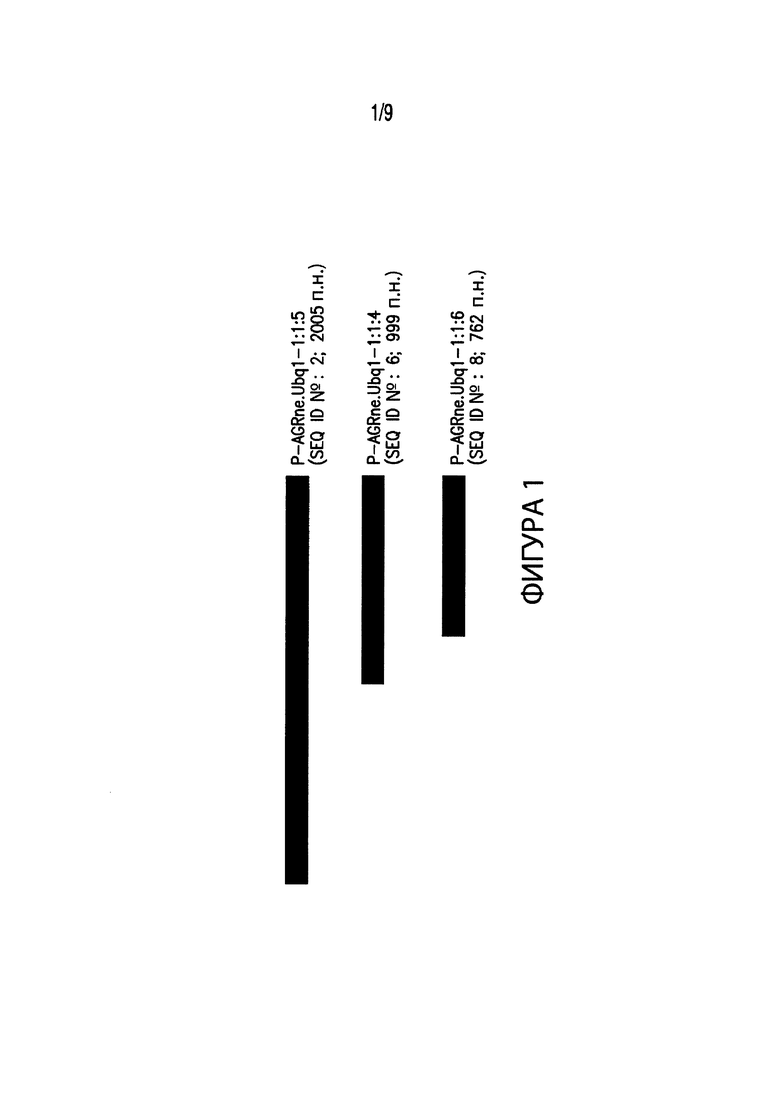
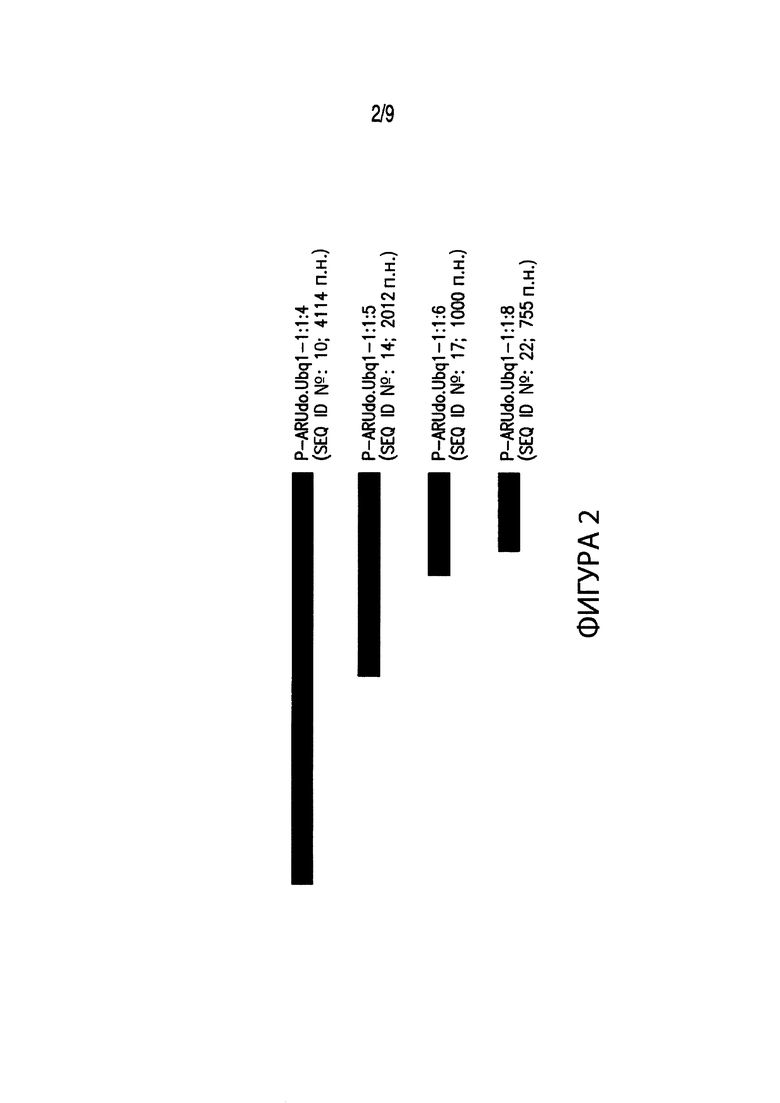
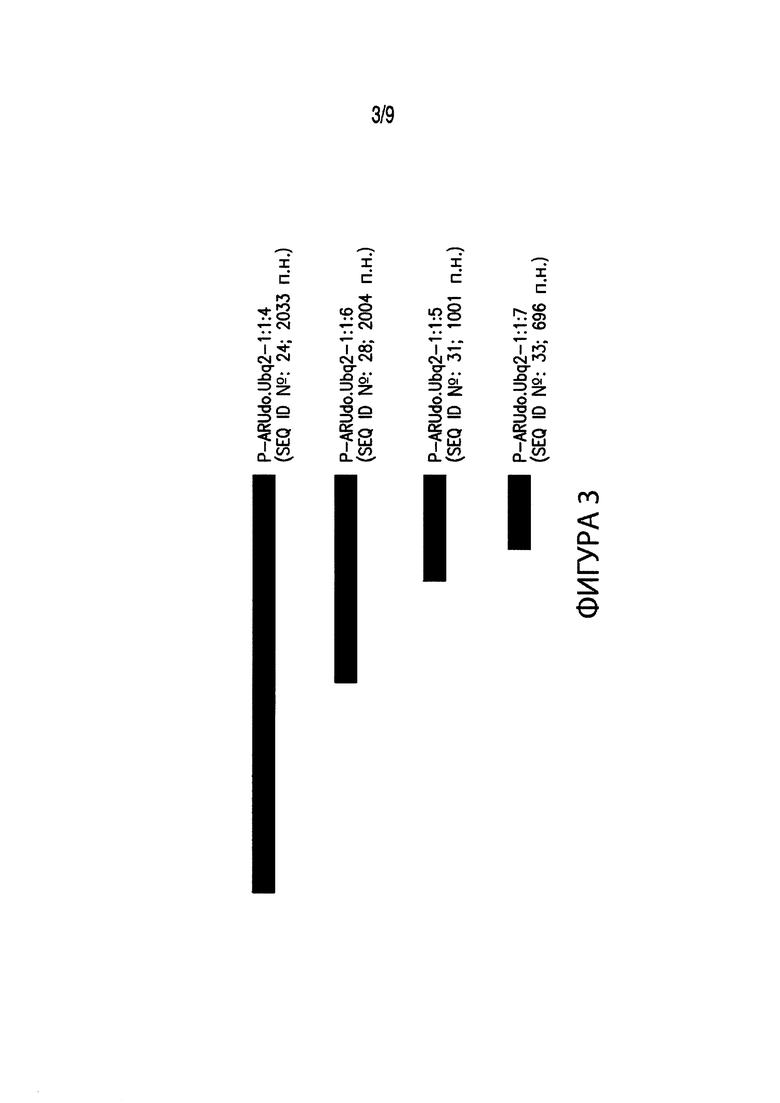
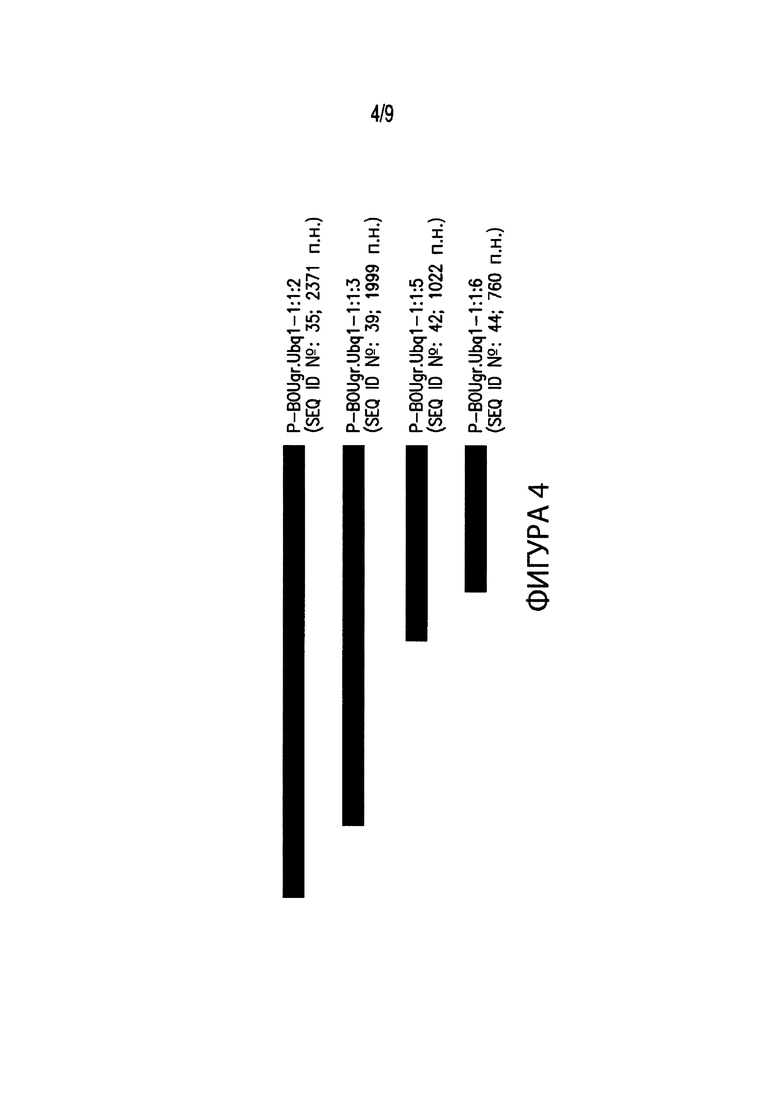
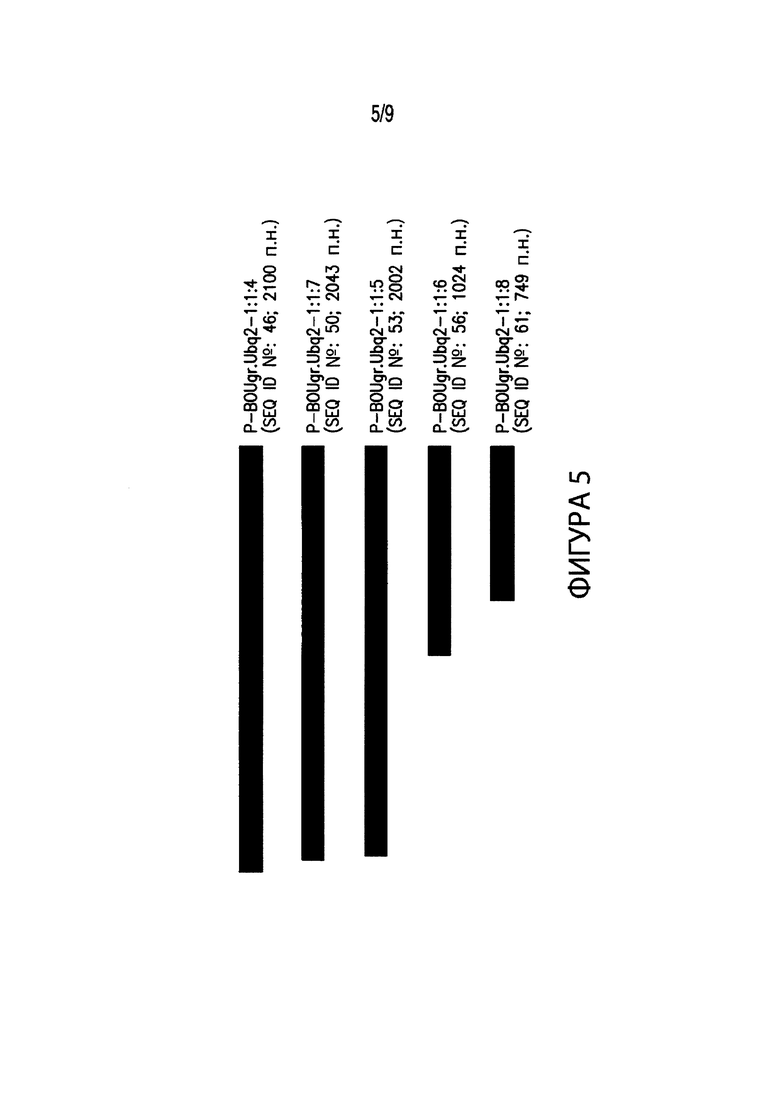
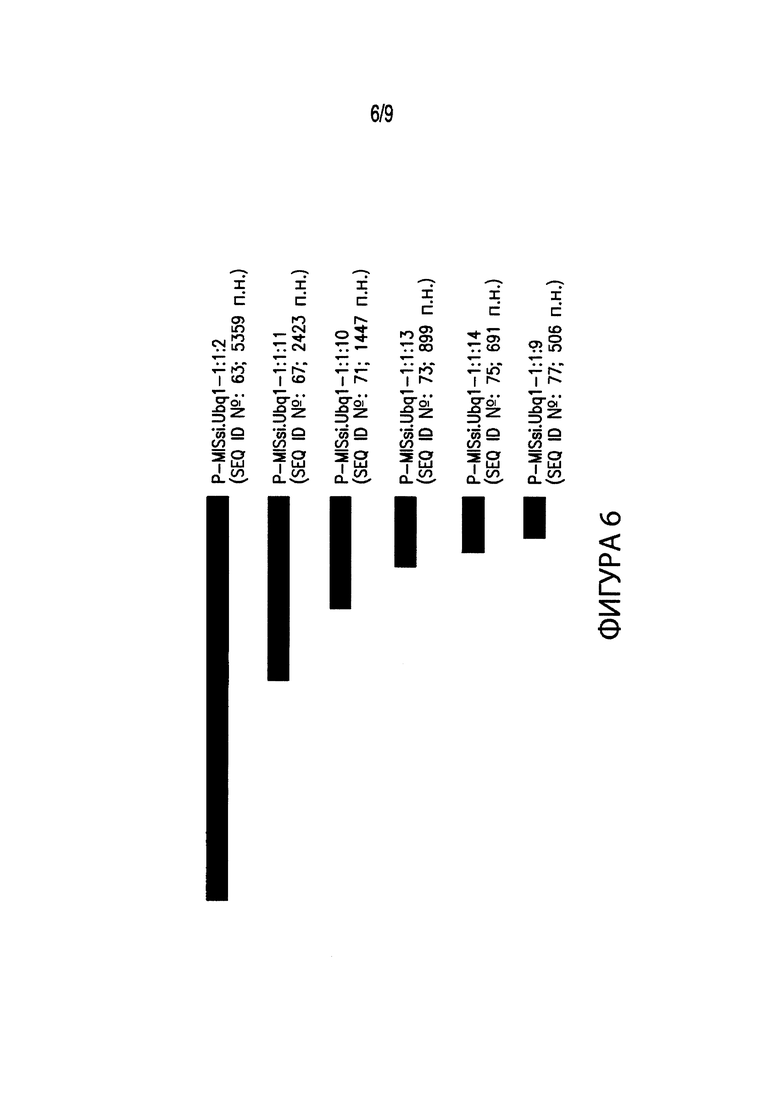
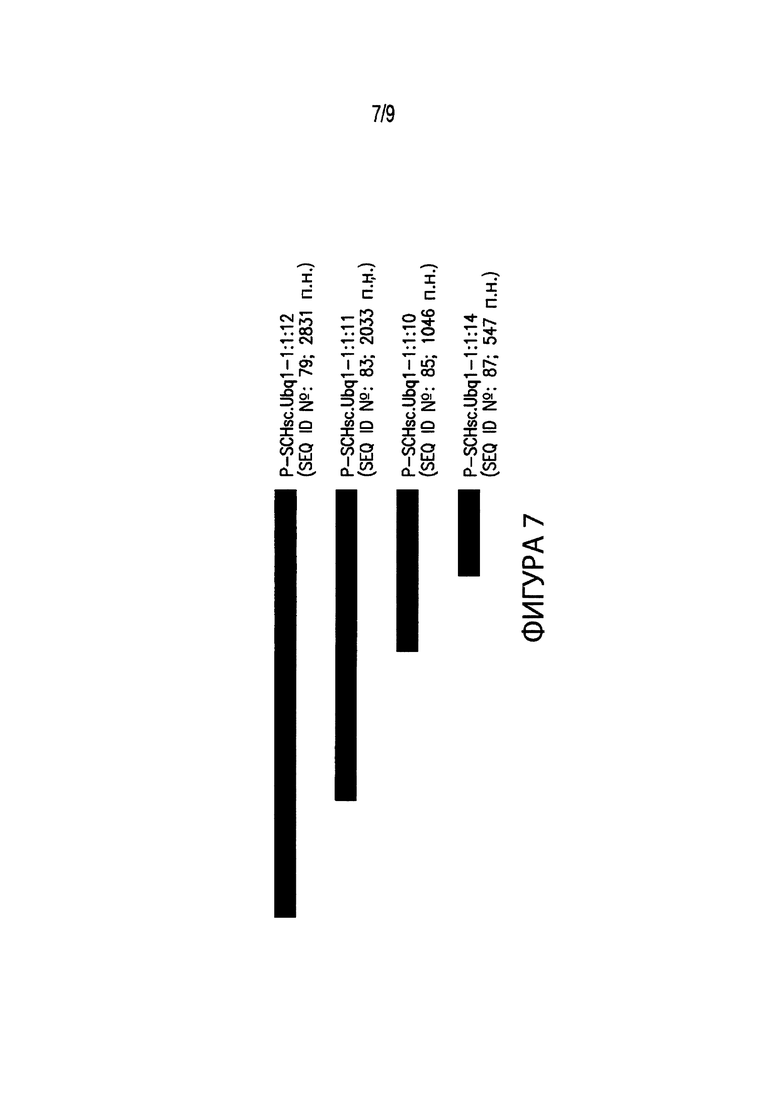
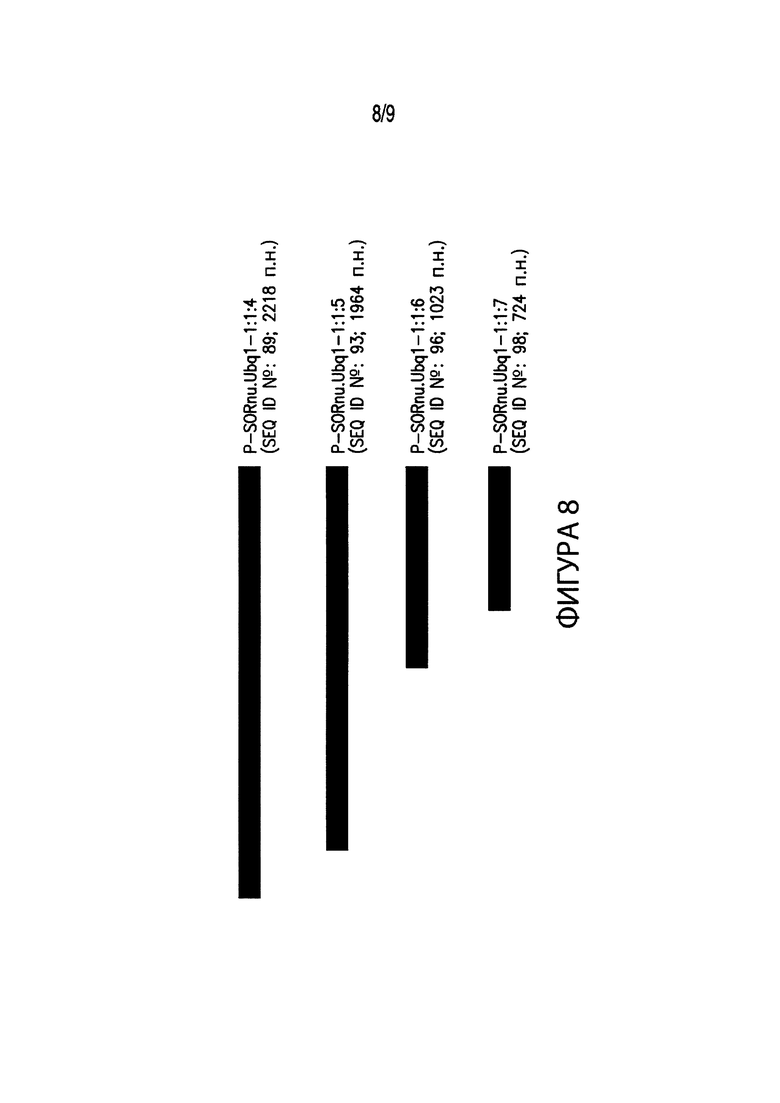
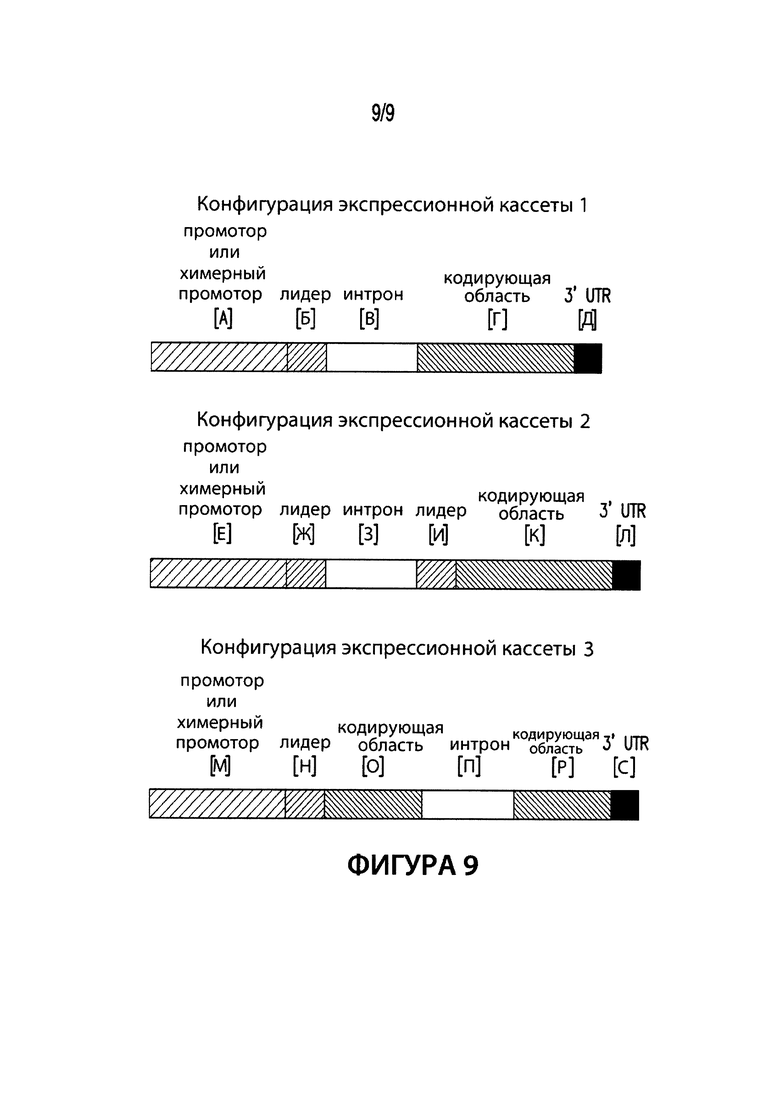
Изобретение относится к области биохимии, в частности к рекомбинантной молекуле ДНК для инициации транскрипции гетерологической транскрибируемой молекулы ДНК, а также к генетической конструкции для инициации транскрипции гетерологической транскрибируемой молекулы ДНК, ее содержащей. Также раскрыты клетка трансгенного растения, трансгенное растение, часть трансгенного растения, трансгенное семя, содержащие вышеуказанную молекулу ДНК. Изобретение также относится к способу экспрессии гетерологической транскрибируемой молекулы ДНК, а также к способу получения трансгенного растения с использованием вышеуказанной молекулы ДНК. Изобретение позволяет эффективно инициировать транскрипцию гетерологической транскрибируемой молекулы ДНК в растении. 8 н. и 5 з.п. ф-лы, 9 ил., 20 табл., 7 пр.
1. Рекомбинантная молекула ДНК для инициации транскрипции гетерологической транскрибируемой молекулы ДНК, содержащая последовательность ДНК, выбранную из группы, состоящей из:
а) последовательности ДНК с по меньшей мере 95 процентами идентичности полноразмерной последовательности к любой из SEQ ID №№: 16–18, где последовательность имеет промоторную активность; и
б) последовательности ДНК, содержащей любую из SEQ ID №№: 16–18;
причем указанная последовательность ДНК функционально связана с гетерологической транскрибируемой молекулой ДНК.
2. Молекула ДНК по п. 1, отличающаяся тем, что гетерологическая транскрибируемая молекула ДНК представляет собой ген, представляющий агрономический интерес.
3. Рекомбинантная молекула ДНК по п. 2, отличающаяся тем, что ген, представляющий агрономический интерес, обуславливает у растений толерантность к гербицидам.
4. Рекомбинантная молекула ДНК по п. 2, отличающаяся тем, что ген, представляющий агрономический интерес, обуславливает у растений устойчивость к вредителям.
5. Генетическая конструкция для инициации транскрипции гетерологической транскрибируемой молекулы ДНК, содержащая рекомбинантную молекулу ДНК по п. 1, где указанная молекула ДНК функционально связана с 3’UTR.
6. Клетка трансгенного растения, содержащая рекомбинантную молекулу ДНК, содержащую последовательность ДНК, выбранную из группы, состоящей из:
а) последовательности ДНК с по меньшей мере 95 процентами идентичности полноразмерной последовательности к любой из SEQ ID №№: 16–18, где последовательность имеет промоторную активность; и
б) последовательности ДНК, содержащей любую из SEQ ID №№: 16–18;
причем указанная последовательность ДНК функционально связана с гетерологической транскрибируемой молекулой ДНК, и
где указанная клетка растения имеет повышенную экспрессию указанной гетерологической транскрибируемой молекулы ДНК по сравнению с клеткой растения, не содержащей указанную рекомбинантную молекулу ДНК.
7. Клетка трансгенного растения по п. 6, отличающаяся тем, что указанная клетка трансгенного растения представляет собой клетку однодольного растения.
8. Клетка трансгенного растения по п. 6, отличающаяся тем, что указанная клетка трансгенного растения представляет собой клетку двудольного растения.
9. Трансгенное растение, содержащее рекомбинантную молекулу ДНК, содержащую последовательность ДНК, выбранную из группы, состоящей из:
а) последовательности ДНК с по меньшей мере 95 процентами идентичности полноразмерной последовательности к любой из SEQ ID №№: 16–18, где последовательность имеет промоторную активность; и
б) последовательности ДНК, содержащей любую из SEQ ID №№: 16–18;
причем указанная последовательность ДНК функционально связана с гетерологической транскрибируемой молекулой ДНК, и
где указанное трансгенное растение имеет повышенную экспрессию указанной гетерологической транскрибируемой молекулы ДНК по сравнению с растением, не содержащим указанную рекомбинантную молекулу ДНК.
10. Часть трансгенного растения, содержащая рекомбинантную молекулу ДНК, содержащую последовательность ДНК, выбранную из группы, состоящей из:
а) последовательности ДНК с по меньшей мере 95 процентами идентичности полноразмерной последовательности к любой из SEQ ID №№: 16–18, где последовательность имеет промоторную активность; и
б) последовательности ДНК, содержащей любую из SEQ ID №№: 16–18;
причем указанная последовательность ДНК функционально связана с гетерологической транскрибируемой молекулой ДНК, и
где указанная часть трансгенного растения имеет повышенную экспрессию указанной гетерологической транскрибируемой молекулы ДНК по сравнению с частью растения, не содержащей указанную рекомбинантную молекулу ДНК.
11. Трансгенное семя из трансгенного растения по п. 9 для получения растения, где данное семя содержит рекомбинантную молекулу ДНК по п.1, и где указанное растение имеет повышенную экспрессию указанной гетерологической транскрибируемой молекулы ДНК по сравнению с растением, не содержащим указанную рекомбинантную молекулу ДНК.
12. Способ экспрессии гетерологической транскрибируемой молекулы ДНК, включающий следующие этапы: получение трансгенного растения по п. 9 и культивирование указанного растения, причем транскрибируемая молекула ДНК экспрессируется.
13. Способ получения трансгенного растения, включающий следующие этапы:
а) трансформацию клетки растения рекомбинантной молекулой ДНК по п. 1 для получения трансформированной клетки растения; и
б) регенерацию трансгенного растения из трансформированной клетки растения;
где указанное трансгенное растение имеет повышенную экспрессию указанной гетерологической транскрибируемой молекулы ДНК по сравнению с растением, не содержащим указанную рекомбинантную молекулу ДНК.
| WO 2012134921 A2, 04.10.2012 | |||
| US 2010058495 A1, 04.03.2010 | |||
| WO 2011130894 A1, 27.10.2011 | |||
| US 2011023183 A1, 27.01.2011 | |||
| ИСКУССТВЕННЫЙ ПРОМОТОР ДЛЯ ЭКСПРЕССИИ ПОСЛЕДОВАТЕЛЬНОСТЕЙ ДНК В РАСТИТЕЛЬНЫХ КЛЕТКАХ | 2003 |
|
RU2326167C2 |

