ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[001] Данная заявка заявляет приоритет Предварительной заявки Соединенных штатов № 61/467875, поданной 25 марта 2011 года, которая включена в настоящее изобретение посредством ссылки в ее полном виде.
ВКЛЮЧЕНИЕ СПИСКА ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[002] Список последовательностей, который содержится в файле, названном "MONS282WO_seq.txt", который равен 347 килобайтам (как измерено в Microsoft Windows®) и создан 21 марта 2012 года, зарегистрирован в настоящем изобретении электронным представлением и включен в данное описание посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ДАННОЕ ИЗОБРЕТЕНИЕ
[003] Настоящее изобретение относится к области молекулярной биологии растений и генетической инженерии растений и ДНК-молекулам, применимым для модуляции экспрессии генов в растениях.
УРОВЕНЬ ТЕХНИКИ
[004] Регуляторные элементы являются генетическими элементами, которые регулируют активность модуляции транскрипции функционально связанной транскрибируемой полинуклеотидной молекулы. Такие элементы включают в себя промоторы, лидеры, интроны и 3’-нетранслируемые районы и применимы в области молекулярной биологии растений и генетической инженерии растений.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[005] Настоящее изобретение обеспечивает новые регуляторные элементы генов для применения в растениях. Настоящее изобретение обеспечивает также ДНК-конструкты, содержащие эти регуляторные элементы. Настоящее изобретение обеспечивает также трансгенные клетки растений, растения и семена, содержащие эти регуляторные элементы. Эти последовательности могут быть функционально связаны с транскрибируемой полинуклеотидной молекулой. В одном варианте осуществления, эта транскрибируемая полинуклеотидная молекула может быть гетерологичной относительно обеспеченной настоящим изобретением регуляторной последовательности. Таким образом, последовательность регуляторных элементов, обеспеченная настоящим изобретением, может быть, в конкретных вариантах осуществления, определена как функционально связанная с гетерологичной транскрибируемой полинуклеотидной молекулой. Настоящее изобретение обеспечивает также способы изготовления и применения этих регуляторных элементов, ДНК-конструктов, содержащих эти регуляторные элементы, и трансгенных клеток растений, растений и семян, содержащих эти регуляторные элементы, функционально связанные с транскрибируемой полинуклеотидной молекулой.
[006] Таким образом, в одном аспекте, это изобретение обеспечивает молекулу ДНК, содержащую последовательность ДНК, выбранную из группы, состоящей из: a) последовательности по меньшей мере с 85% идентичностью последовательности с любой из SEQ ID NO: 1-158 и 180-183; b) последовательности, содержащей любую из SEQ ID NO: 1-158 и 180-183; и c) фрагмент любой из последовательностей SEQ ID NO: 1-158 и 180-183, где этот фрагмент имеет ген-регуляторную активность; где эта последовательность функционально связана с гетерологичной транскрибируемой полинуклеотидной молекулой. В конкретных вариантах осуществления, эта молекула ДНК содержит по меньшей мере приблизительно 90-процентную, по меньшей мере приблизительно 95-процентную, по меньшей мере приблизительно 98-процентную или по меньшей мере приблизительно 99-процентную идентичность последовательности с последовательностью ДНК любой из SEQ ID NO: 1-158 и 180-183. В некоторых вариантах этой молекулы ДНК, эта последовательность ДНК содержит регуляторный элемент. В некоторых вариантах осуществления этот регуляторный элемент содержит промотор. В конкретных вариантах осуществления, эта гетерологичная транскрибируемая полинуклеотидая молекула содержит представляющий агрономический интерес ген, такой как ген, способный обеспечивать устойчивость к гербицидам в растениях, или ген, способный обеспечивать устойчивость к вредителям в растениях.
[007] Настоящее изобретение обеспечивает также клетку трансгенного растения, содержащую гетерологичный ДНК-конструкт, обеспечиваемый изобретением, включающий в себя последовательность любой из SEQ ID NO: 1-158 и 180-183, или ее фрагмент или вариант, где указанная последовательность функционально связана с гетерологичной транскрибируемой полинуклеотидной молекулой. В некоторых вариантах осуществления, эта клетка трансгенного растения является клеткой однодольного растения. В других вариантах осуществления, эта клетка трансгенного растения является клеткой однодольного растения.
[008] Кроме того, настоящее изобретение обеспечивает трансгенное растение, или его часть, содержащие обеспеченную здесь ДНК-молекулу, включающую в себя последовательность ДНК, выбранную из группы, состоящей из: a) последовательности по меньшей мере с 85% идентичностью последовательности с любой из SEQ ID NO: 1-158 и 180-183; b) последовательности, содержащей любую из SEQ ID NO: 1-158 и 180-183; и c) фрагмент любой из последовательностей SEQ ID NO: 1-158 и 180-183, где этот фрагмент имеет ген-регуляторную активность; где эта последовательность функционально связана с гетерологичной транскрибируемой полинуклеотидной молекулой. В конкретных вариантах осуществления, это трансгенное растение может быть растением-потомком любой генерации, которое содержит молекулу ДНК, родственную с первоначальным трансгенным растением, содержащим эту молекулу ДНК. Кроме того, обеспечены трансгенные семена, содержащие молекулу ДНК настоящего изобретения.
[009] Еще в одном аспекте, настоящее изобретение обеспечивает способ получения товарного продукта, предусматривающий получение трансгенного растения или его части в соответствии с изобретением и получение из них товарного продукта. В одном варианте осуществления, товарным продуктом настоящего изобретения является концентрат белка, изолят белка, зерно, крахмал, семена, мучка, мука, биомасса или масло из семян растений. В другом аспекте, настоящее изобретение обеспечивает товарный продукт, обеспечиваемый вышеуказанным способом. Например, в одном варианте осуществления настоящего изобретения, изобретение обеспечивает товарный продукт, содержащий молекулу ДНК, обеспеченную здесь, включающую в себя последовательность ДНК, выбранную из группы, состоящей из: a) последовательности по меньшей мере с 85% идентичностью последовательности с любой из SEQ ID NO: 1-158 и 180-183; b) последовательности, содержащей любую из SEQ ID NO: 1-158 и 180-183; и c) фрагмент любой из последовательностей SEQ ID NO: 1-158 и 180-183, где этот фрагмент имеет ген-регуляторную активность; где эта последовательность функционально связана с гетерологичной транскрибируемой полинуклеотидной молекулой.
[0010] Еще в одном аспекте, настоящее изобретение обеспечивает способ экспрессии транскрибируемой полинуклеотидной молекулы, который предусматривает получение трансгенного растения в соответствии с настоящим изобретением, такого как растение, содержащее молекулу ДНК, представленную в настоящем описании, и культивирование растения, где в этой молекуле ДНК экспрессируется транскрибируемый полинуклеотид.
[0011] Должно быть понятно, что во всем этом описании и в формуле изобретения, если нет других указаний, слово "содержат" и его вариации, такие как "содержит" и "содержащий", должны пониматься как включение указанных композиции, стадии и/или величины или их групп, а не как исключение любых других композиции, стадии и/или величины или их групп.
КРАТКОЕ ОПИСАНИЕ ФИГУР
[0012] Фигуры 1a-1h изображают сопоставление вариантов размеров промоторов, соответствующих промоторным элементам, выделенным из вида травянистого растения Andropogon gerardii. В частности, фиг. 1а-1h показывают сопоставление промоторной последовательности 2603 п.н. P-ANDge.Ubq1-1:1:11 (SEQ ID NO: 2), обнаруженной в группе транскрипционного регуляторного элемента экспрессии EXP-ANDge.Ubq1:1:9 (SEQ ID NO: 1), с промоторными последовательностями, полученными с использованием делеционного анализа P-ANDge.Ubql-1:1:11. Делеция, например, 5’-конца P-ANDge.Ubq1-l:l:11, продуцировала промотор P-ANDge.Ubq1-1:l:9 (SEQ ID NO: 6), последовательность 2114 п.н. которого была обнаружена в EXP-ANDge.Ubq1-:1:7 (SEQ ID NO: 5). Другие промоторные последовательности на фиг. 1 включают в себя P-ANDge.Ubq1-1:1:10 (SEQ ID NO: 9), последовательность 1644 п.н. содержащуюся в EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8); P-ANDge.Ubq1-1:1:12 (SEQ ID NO: 11), последовательность 1472 п.н., содержащуюся в EXP-ANDge.Ubq1:1:10 (SEQ ID NO: 10); P-ANDge.Ubq1-1:1:8 (SEQ ID NO: 13), последовательность 1114 п.н., содержащуюся в EXP-ANDge.Ubq1:1:6 (SEQ ID NO: 12); P-ANDge.Ubq1-1:1:13 (SEQ ID NO: 15), последовательность 771 п.н., содержащуюся в EXP-ANDge.Ubq1:1:11 (SEQ ID NO: 14); и P-ANDge.Ubq1-1:1:14 (SEQ ID NO: 17), последовательность 482 п.н., содержащуюся в EXP-ANDge.Ubq1:1:12 (SEQ ID NO: 16).
[0013] Фигуры 2a-2g изображают сопоставление вариантов промоторов, выделенных из вида травянистого растения Saccharum ravennae (Erianthus ravennae). В частности, фигуры 2a-2g показывают сопоставление промоторной последовательности 2536 п.н. P-ERIra.Ubq1-1:1:10 (SEQ ID NO: 19) (обнаруженной, например, в группе транскрипционного регуляторного элемента экспрессии, например, EXP-ERIra.Ubq1 (SEQ ID NO: 18)) с промоторными последовательностями, полученными из делеционного анализа P-ERIra.Ubq1-1:1:10; промоторной последовательностью 2014 п.н. P-ERIra.Ubq1-1:1:9 (SEQ ID NO: 23); промоторной последовательностью 1525 п.н. P-ERIra.Ubq1-1:1:11 (SEQ ID NO: 26); промоторной последовательностью 1044 п.н. P-ERIra.Ubq1-1:1:8 (SEQ ID NO: 28); последовательностью 796 п.н. P-ERIra.Ubq1-1:1:12 (SEQ ID NO: 30); и последовательностью 511 п.н. P-ERIra.Ubq1-1:1:13 (SEQ ID NO: 32).
[0014] Фигуры 3a-3c изображают сопоставление вариантов размеров промоторов, соответствующих промоторным элементам, выделенным из вида травянистого растения Setaria viridis. В частности, фигуры 3a-3c показывают сопоставление промоторной последовательности 1493 п.н., P-Sv.Ubq1-1:1:1 (SEQ ID NO: 34) с промоторами, полученными из делеционного анализа 5’-конца P-Sv.Ubq1-1:1:1; промотором с размером 1035 п.н. P-Sv.Ubq1-1:1:2 (SEQ ID NO: 38); и промоторной последовательностью 681 п.н. P-Sv.Ubq1-1:1:3 (SEQ ID NO: 40).
[0015] Фигуры 4a-4e изображают сопоставление вариантов групп транскрипционных регуляторных элементов экспрессии, полученных из подвида травянистого растения Zea mays subsp. mexicana. В частности, фигуры 4a-4e сравнивают группу транскрипционных регуляторных элементов экспрессии 2005 п.н., названную EXP-Zm.UbqM1:1:2 (SEQ ID NO: 49), с аллельным вариантом EXP-Zm.UbqM1:1:5 (SEQ ID NO: 53), а также с вариантами размеров EXP-Zm.UbqM1:1:1 (SEQ ID NO: 41), который имеет длину 1922 п.н., и EXP-Zm.UbqM1:1:4 (SEQ ID NO: 45), который имеет длину 1971 п.н.
[0016] Фигуры 5a-5b изображают сопоставление вариантов размеров промоторов, выделенных из травянистого растения Sorghum bicolor. В частности, фигуры 5a-5b показывают сопоставление промоторного элемента с размером 791 п.н., P-Sb.Ubq6-1:1:2 (SEQ ID NO: 60), содержащегося в группе транскрипционных регуляторных элементов экспрессии EXP-Sb.Ubq6 (SEQ ID NO: 59), с промоторным элементом 855 п.н. P-Sb.Ubq6-1:1:1 (SEQ ID NO: 64), содержащимся в EXP-Sb.Ubq6:1:1 (SEQ ID NO: 63).
[0017] Фигуры 6a-6с изображают сопоставление вариантов размеров промоторов, выделенных из травянистого растения Setaria italica. В частности, Фигуры 6a-6c показывают сопоставление варианта промотора 1492 п.н. P-SETit.Ubq1-1:1:1 (SEQ ID NO: 70) с вариантом промотора 1492 п.н. P-SETit.Ubq1-1:1:4 (SEQ ID NO: 74), промоторным элементом 1034 п.н. P-SETit.Ubq1-1:1:2 (SEQ ID NO: 76) и промоторным элементом 680 п.н. P-SETit.Ubq1-1:1:3 (SEQ ID NO: 78).
[0018] Фигуры 7a-7b изображают варианты размеров промоторов и энхансерного элемента, соответствующего промоторным элементам, выделенным из вида травянистого растения Coix lachryma-jobi. В частности, фигуры 7a и 7b показывают сопоставление варианта промотора 837 п.н., P-Cl.Ubq1-1:1:l (SEQ ID NO: 80), обнаруженного в группе транскрипционных регуляторных элементов экспрессии EXP-Cl.Ubq1:1:1 (SEQ ID NO: 79), с энхансерным фрагментом, полученным из P-Cl.Ubq1-1:1:1, названным E-Cl.Ubq1: 1:1 (SEQ ID NO: 89), который имеет длину 798 п.н., а также с тремя 5’-концевыми делеционными вариантами P-Cl.Ubq1-1:1:1: элементом 742 п.н. P-Cl.Ubq1-1:1:4 (SEQ ID NO: 84); элементом 401 п.н. P-Cl.Ubq1-1:1:3 (SEQ ID NO: 86); и минимальным промоторным элементом 54 п.н. P-Cl.Ubq1-1:1:5 (SEQ ID NO: 88).
[0019] Фигура 8 изображает конфигурации трансгенных кассет настоящего изобретения.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0020] SEQ ID NO: 1, 5, 8, 10, 12, 14, 16, 18, 22, 25, 27, 29, 31, 33, 37, 39, 41, 45, 49, 53, 55, 59, 63, 65, 69, 73, 75, 77, 79, 83, 85, 87, 90, 93, 95, 97, 98, 99, 100, 102, 104, 106, 108, 110, 112, 114, 115, 116, 117, 119, 121, 123, 124, 125, 126, 128, 130, 132, 133, 134, 136, 137, 139, 141, 143, 145, 147, 149, 151, 153, 155, 157, 180, 181 и 183 являются последовательностями групп транскрипционных регуляторных элементов экспрессии или EXP-последовательностями, содержащими промоторную последовательность, функционально связанную 5’ (слева) от лидерной последовательности, которая функционально связана 5’ (слева) от последовательности интрона.
[0021] SEQ ID NO: 2, 6, 9, 11, 13, 15, 17, 19, 23, 26, 28, 30, 32, 34, 38, 40, 42, 46, 50, 56, 60, 64, 66, 70, 74, 76, 78, 80, 84, 86, 88, 91, 96 и 135 являются промоторными последовательностями.
[0022] SEQ ID NO: 3, 20, 35, 43, 47, 51, 57, 61, 67, 71 и 81 являются лидерными последовательностями.
[0023] SEQ ID NO: 4, 7, 21, 24, 36, 44, 48, 52, 54, 58, 62, 68, 72, 82, 92, 94, 101, 103, 105, 107, 109, 111, 113, 118, 120, 122, 127, 129, 131, 138, 140, 142, 144, 146, 148, 150, 152, 154, 156, 158 и 182 являются последовательностями интронов.
[0024] SEQ ID NO: 89 является последовательностью энхансера.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0025] Описанное изобретение обеспечивает полинуклеотидные молекулы, имеющие выгодную ген-регуляторную активность, полученные из растительных источников. Изобретение обеспечивает дизайн, конструирование и использование этих полинуклеотидных молекул. Нуклеотидные последовательности этих полинуклеотидных молекул обеспечены среди SEQ ID NO: 1-158 и 180-183. Эти полинуклеотидные молекулы способны, например, влиять на экспрессию функционально связанной транскрибируемой молекулы в тканях растений и, следовательно, регулировать экспрессию гена, или активность кодируемого геном продукта, в трансгенных растениях. Настоящее изобретение обеспечивает также способы их модификации, получения и применения. Настоящее изобретение обеспечивает также композиции, трансформированные клетки-хозяева, трансгенные растения и семена, содержащие эти промоторы, и/или другие описанные нуклеотидные последовательности и способы их получения и применения.
[0026] Следующие определения и способы обеспечены для лучшего определения настоящего изобретения и помощи специалистам с обычной квалификацией в данной области в применении на практике настоящего изобретения. Если нет других указаний, термины должны пониматься в соответствии с общепринятым применением их специалистами с обычной квалификацией в релевантной области.
Молекулы ДНК
[0027] В данном контексте, термин "ДНК" или "молекула ДНК" относится к двухцепочечной молекуле ДНК геномного или синтетического происхождения, т.е. к полимеру дезоксирибонуклеотидных оснований, или к молекуле полинуклеотида, считываемой от 5’ (левого) конца к 3’ (правому) концу. В данном контексте, термин "последовательность ДНК" относится к нуклеотидной последовательности молекулы ДНК. Используемая здесь номенклатура соответствует номенклатуре Title 37 of the United States Code of Federal Regulations § 1.822, и представлена в таблицах в WIPO Standard ST.25 (1998), Appendix 2, Tables 1 and 3.
[0028] В этом контексте, термин “выделенная молекула ДНК" относится к молекуле ДНК, по меньшей мере частично отделенной от других молекул, обычно ассоциированных с ней в ее нативном или природном состоянии. В одном варианте осуществления, термин "выделенная” молекула ДНК относится к молекуле ДНК, которая по меньшей мере частично отделена от некоторых нуклеиновых кислот, которые обычно фланкируют эту молекулу ДНК в ее нативном или природном состоянии. Таким образом, молекулы ДНК, слитые с регуляторными или кодирующими последовательностями, с которыми они обычно не ассоциированы, например, в результате рекомбинантных способов, считаются здесь выделенными. Такие молекулы считаются выделенными при интеграции в хромосому клетки-хозяина или при присутствии в растворе нуклеиновых кислот с другими молекулами ДНК, в том смысле, что они не находятся в их нативном состоянии.
[0029] Любое количество способов, хорошо известных квалифицированным в данной области специалистам, могут быть использованы для выделения и манипуляции молекулы ДНК, или ее фрагмента, описанных в настоящем изобретении. Например, технология ПЦР (полимеразной цепной реакции) может быть использована для амплификации конкретной исходной молекулы ДНК и/или получения вариантов этой первоначальной молекулы. Молекулы ДНК или их фрагмент могут быть также получены другими способами, такими как непосредственный синтез этого фрагмента химическим способом, как обычно практикуется с использованием автоматизированного синтезатора олигонуклеотидов.
[0030] В данном контексте, термин "идентичность последовательности" относится к степени, до которой являются идентичными две оптимально сопоставленные полинуклеотидные последовательности или две оптимально сопоставленные полипептидные последовательности. Оптимальное сопоставление последовательностей создается ручным сопоставлением двух последовательностей, например, ссылочной последовательности и другой последовательности, для максимизации количества нуклеотидных совпадений в этом сопоставлении последовательностей с подходящими внутренними нуклеотидными инсерциями, делециями или гэпами. В данном контексте, термин "ссылочная последовательность" относится к последовательности, обеспеченной в качестве полинуклеотидных последовательностей SEQ ID NO: 1-158 и 180-183.
[0031] В данном контексте, термин "процентная идентичность последовательности" или “процентная идентичность" или "%-ная идентичность" обозначает умножение полученной идентичности в виде дроби на 100. Эта "дробь идентичности" для последовательности, оптимально сопоставленной со ссылочной последовательностью, является числом совпадений нуклеотидов в оптимальном сопоставлении, деленным на общее число нуклеотидных совпадений в ссылочной последовательности, например, общее число нуклеотидов в полной длине всей ссылочной последовательности. Таким образом, одним вариантом настоящего изобретения является молекула ДНК, содержащая последовательность, которая при оптимальном сопоставлении со ссылочной последовательностью, обеспеченной в виде SEQ ID NO: 1-158 и 180-183, имеет по меньшей мере приблизительно 85-процентную идентичность, по меньшей мере приблизительно 90-процентную идентичность, по меньшей мере приблизительно 95-процентную идентичность, по меньшей мере приблизительно 96-процентную идентичность, по меньшей мере приблизительно 97-процентную идентичность, по меньшей мере приблизительно 98-процентную идентичность или по меньшей мере приблизительно 99-процентную идентичность со ссылочной последовательностью. В конкретных вариантах такие последовательности могут быть определены как имеющие ген-регуляторную активность.
Регуляторные элементы
[0032] Одним регуляторным элементом является молекула ДНК, имеющая ген-регуляторную активность, т.е. молекула ДНК, которая имеет способность влиять на транскрипцию и/или трансляцию функционально связанной транскрибируемой полинуклеотидной молекулы. Таким образом, термин "ген-регуляторная активность” относится к способности влиять на характер экспрессии функционально связанной транскрибируемой полинуклеотидной молекулы влиянием на транскрипцию и/или трансляцию этой функционально связанной транскрибируемой полинуклеотидной молекулы. В данном контексте, группа транскрипционных регуляторных элементов экспрессии или "EXP"-последовательность, может состоять из элементов экспрессии, таких как энхансеры, промоторы, лидеры и интроны, функционально связанные. Таким образом, группа элементов транскрипционной регуляторной экспрессии может состоять, например, из промотора, функционально связанного 5’ (слева) от лидерной последовательности, которая, в свою очередь, функционально связана 5’ (слева) от интронной последовательности. Эта интронная последовательность может содержать последовательность, начинающуюся в точке первой интрон/экзонной границы сплайсинга нативной последовательности, и может дополнительно состоять из малого лидерного фрагмента, содержащего вторую интрон/экзонную границу сплайсинга, чтобы обеспечить должный процессинг интрона/экзона для облегчения транскрипции и должный процессинг полученного транскрипта. Лидеры и интроны могут положительно влиять на транскрипцию функционально связанной транскрибируемой полинуклеотидной молекулы, а также на трансляцию полученной транскрибируемой РНК. Эта пре-процессированная молекула РНК содержит лидеры и интроны, которые могут влиять на посттранскрипционный процессинг транскрибируемой РНК и/или экспорт этой молекулы транскрибируемой РНК из ядра клетки в цитоплазму. После посттранскрипционного процессинга этой транскрибируемой молекулы РНК, лидерная последовательность может удерживаться в виде части конечной мессенджер-РНК и может положительно влиять на трансляцию этой молекулы мессенджер-РНК.
[0033] Регуляторные элементы, такие как промоторы, лидеры, интроны и районы терминации транскрипции (или 3’-UTR) являются молекулами ДНК, которые имеют ген-регуляторную активность и играют интегральную (существенную) роль во всей экспрессии генов в живых клетках. Термин "регуляторный элемент" относится к молекуле ДНК, имеющей ген-регуляторную активность, т.е. она способна влиять на транскрипцию и/или трансляцию функционально связанной транскрибируемой полинуклеотидной молекулы. Поэтому, выделенные регуляторные элементы, такие как промоторы и лидеры, которые функционируют в растениях, применимы для модификации фенотипов растений посредством способов генетической инженерии.
[0034] Регуляторные элементы могут характеризоваться по их характеру (паттерну) действий на экспрессию (количественно и/или качественно), например, положительных или отрицательных действий и/или других действий, например, по их временному, пространственному, относящемуся к развитию, тканевому, относящемуся к окружающей среде, физиологическому, патологическому, относящемуся к клеточному циклу и/или химически реагировать характеру экспрессии, и любой их комбинации, а также по количественным или качественным показателям. Промотор применим в качестве регуляторного элемента для модификации экспрессии функционально связанной транскрибируемой полинуклеотидной молекулы.
[0035] В данном контексте, "паттерн экспрессии гена" является любым паттерном транскрипции функционально связанной молекулы ДНК в транскрибируемую молекулу РНК. Эта транскрибируемая молекула РНК может быть транслирована для получения молекулы белка или может обеспечивать антисмысловую или другую регуляторную молекулу РНК, такую как dsRNA, tRNA, rRNA, miRNA и т.п.
[0036] В данном контексте, термин "экспрессия белка" обозначает любой паттерн трансляции транскрибируемой молекулы РНК в молекулу белка. Экспрессия белка может характеризоваться по его временным, пространственным, относящимся к развитию или морфологическим качествам, а также по количественным или качественным показателям.
[0037] В данном контексте, термин "промотор" относится обычно к молекуле ДНК, которая участвует в узнавании и связывании РНК-полимеразой II или другими белками (транс-активирующими факторами транскрипции) для инициации транскрипции. Промотор может быть сначала выделенным из 5’-нетранслируемого района (5’-UTR) геномной копии гена. Альтернативно, промоторы могут быть полученными синтетически или подвергнутыми модуляции молекулами ДНК. Промоторы могут быть также химерными, т.е. промотором, полученным через слияние двух или более гетерологичных молекул ДНК. Промоторы, применимые в практике настоящего изобретения, включают в себя SEQ ID NO: 2, 6, 9, 11, 13, 15, 17, 19, 23, 26, 28, 30, 32, 34, 38, 40, 42, 46, 50, 56, 60, 64, 66, 70, 74, 76, 78, 80, 84, 86, 88, 91, 96 и 135, или их фрагменты или варианты. В конкретных вариантах осуществления изобретения, такие молекулы и их любые варианты или производные, описанные здесь, дополнительно определяются как содержащие промоторную активность, т.е. способные действовать в качестве промотора в клетке-хозяине, например, в трансгенном растении. В других конкретных вариантах осуществления, фрагмент может быть определен как проявляющий промоторную активность, которой обладала исходная молекула промотора, из которого он получен, или фрагмент может содержать "минимальный промотор", который обеспечивает фоновый уровень транскрипции и состоит из TATA-бокса или эквивалентной последовательности для узнавания и связывания комплекса РНК-полимеразы II для инициации транскрипции.
[0038] В одном варианте осуществления, обеспечены фрагменты описанной здесь промоторной последовательности. Фрагменты промотора могут иметь промоторную активность, как описано выше, и могут быть применимы по отдельности или в комбинации с другими промоторами и фрагментами промоторов, например, в конструировании химерных промоторов. В конкретных вариантах осуществления, обеспечены фрагменты промотора, содержащие по меньшей мере приблизительно 50, 95, 150, 250, 500, 750 или по меньшей мере приблизительно 1000 смежных нуклеотидов, или более длинные, полинуклеотидной молекулы, имеющей промоторную активность, описанную в данном документе.
[0039] Композиции, произведенные из любого из промоторов, представленных как SEQ ID NO: 2, 6, 9, 11, 13, 15, 17, 19, 23, 26, 28, 30, 32, 34, 38, 40, 42, 46, 50, 56, 60, 64, 66, 70, 74, 76, 78, 80, 84, 86, 88, 91, 96 и 135, такие как внутренние или 5’-делеции, например, могут быть получены с использованием способов, известных в данной области, для улучшения или изменения экспрессии, в том числе удалением элементов, которые имеют либо положительные, либо отрицательные действия на экспрессию; дупликацией элементов, которые имеют положительные или отрицательные действия на экспрессию; и/или дупликацией или удалением элементов, которые имеют тканеспецифические или клеткоспецифические действия на экспрессию. Композиции, произведенные из любого из промоторов, представленных как SEQ ID NO: 2, 6, 9, 11, 13, 15, 17, 19, 23, 26, 28, 30, 32, 34, 38, 40, 42, 46, 50, 56, 60, 64, 66, 70, 74, 76, 78, 80, 84, 86, 88, 91, 96 и 135, состоящих из 3’-делеций, в которых элемент ТАТА-бокс или его эквивалентная последовательность и последовательность слева удалены, могут быть использованы, например, для получения энхансерных элементов. Кроме того, делеции могут быть получены для удаления любых элементов, которые являются положительными или отрицательными; тканеспецифическими; клеткоспецифическими; или специфическими в отношении тайминга (но не ограничиваемые циркадным ритмом) действиями на экспрессии. Любой из промоторов, представленных как SEQ ID NO: 2, 6, 9, 11, 13, 15, 17, 19, 23, 26, 28, 30, 32, 34, 38, 40, 42, 46, 50, 56, 60, 64, 66, 70, 74, 76, 78, 80, 84, 86, 88, 91, 96 и 135, и фрагментов или энхансеров, произведенных из них, может быть использован для получения химерных композиций транскрибируемых регуляторных элементов, состоящих из любого из промоторов, представленных как SEQ ID NO: 2, 6, 9, 11, 13, 15, 17, 19, 23, 26, 28, 30, 32, 34, 38, 40, 42, 46, 50, 56, 60, 64, 66, 70, 74, 76, 78, 80, 84, 86, 88, 91, 96 и 135, и произведенных из них фрагментов или энхансеров, функционально связанных с другими энхансерами и промоторами. Эффективность этих модификаций, дупликаций или делеций, описанных здесь, на желаемых аспектах экспрессии конкретного трансгена могут быть тестированы эмпирически в стабильных и транзиторных анализах растений, таких как анализы, описанные в рабочих примерах здесь, для валидизации результатов, которые могут варьироваться в зависимости от произведенных изменений и цели этого изменения в исходной молекуле.
[0040] В данном контексте, термин "лидер" относится к молекуле ДНК, выделенной из нетранслированного 5’-района (5’-UTR) геномной копии гена, и определяется обычно как нуклеотидный сегмент между стартовым сайтом транскрипции (TSS) и стартовым сайтом кодирующей белок последовательности. Альтернативно, лидеры могут быть получены синтетически или манипуляцией элементами ДНК. Молекулы лидера могут быть использованы для модуляции экспрессии функциональной транскрибируемой полинуклеотидной молекулы. Молекулы лидера могут быть использованы с гетерологичным промотором или с их природным промотором. Таким образом, промоторные молекулы настоящего изобретения могут быть функционально связаны с их природным лидером или могут быть функционально связаны с гетерологичным лидером. Лидеры, применимые в практике настоящего изобретения, включают в себя SEQ ID NO: 3, 20, 35, 43, 47, 51, 57, 61, 67, 71 и 81 или их фрагменты или варианты. В конкретных вариантах осуществления, могут быть обеспечены такие последовательности, определяемые как последовательности, способные действовать в качестве лидера в клетке-хозяине, в том числе, например, трансгенной клетке растения. В одном варианте осуществления, такие последовательности декодируются как содержащие лидерную активность.
[0041] Эти лидерные последовательности (5’-UTR), представленные как SEQ ID NO: 3, 20, 35, 43, 47, 51, 57, 61, 67, 71 и 81, могут состоять из регуляторных элементов или могут принимать вторичные структуры, которые могут влиять на транскрипцию или трансляцию трансгена. Лидерные последовательности, представленные как SEQ ID NO: 3, 20, 35, 43, 47, 51, 57, 61, 67, 71 и 81, могут быть использованы в соответствии с данным изобретением для получения химерных регуляторных элементов, которые влияют на транскрипцию или трансляцию трансгена. Кроме того, лидерные последовательности, представленные как SEQ ID NO: 3, 20, 35, 43, 47, 51, 57, 61, 67, 71 и 81, могут быть использованы для получения химерных лидерных последовательностей, которые влияют на транскрипцию или трансляцию трансгена.
[0042] Введение чужеродного гена в новое растение-хозяин не всегда приводит к высокой экспрессии получаемого гена. Кроме того, при рассмотрении комплексных признаков, иногда необходимо модулировать несколько генов, с пространственно или транзиторно отличающимися паттернами (характерами) экспрессии. Такую модуляцию могут обеспечивать в основном интроны. Однако, показано, что множественное применение одного и того же интрона обнаруживает недостатки. В этих случаях, необходимо иметь коллекцию интронов, известную в данной области, с усиливающими экспрессию свойствами. Поскольку доступная коллекция интронов, известная в данной области, с усиливающими экспрессию свойствами, является ограниченной, необходимы альтернативы.
[0043] Композиции, произведенные из любых из интронов, представленных как SEQ ID NO: 4, 7, 21, 24, 36, 44, 48, 52, 54, 58, 62, 68, 72, 82, 92, 94, 101, 103, 105, 107, 109, 111, 113, 118, 120, 122, 127, 129, 131, 138, 140, 142, 144, 146, 148, 150, 152, 154, 156, 158 и 182, могут состоять из внутренних делеций или дупликаций цис-регуляторных элементов; и/или изменений 5’- и 3’-последовательностей, содержащих границы сплайсинга интрон/экзон, могут быть использованы для улучшения экспрессии или специфичности экспрессии при функциональном связывании с промотором + лидером или химерным промотором + лидером и кодирующей последовательностью. Изменения 5’- и 3’-районов, содержащих границу сплайсинга интрон/экзон, могут быть также произведены для уменьшения потенциала для введения фальстарта и стоп-кодонов, продуцирующихся в полученном транскрипте, после процессинга и сплайсинга этой мессенджер-РНК. Эти интроны могут быть тестированы эмпирически, как описано в рабочих примерах, для определения действия интрона на экспрессию трансгена.
[0044] В соответствии с изобретением, промотор или фрагмент промотора могут быть анализированы на присутствие известных промоторных элементов, т.е. такие характеристики последовательности ДНК, как ТАТА-бокс и другие известные мотивы сайта связывания факторов транскрипции. Идентификация таких известных промоторных элементов может быть использована квалифицированным в данной области специалистом для конструирования вариантов промотора, имеющих сходный паттерн (характер) экспрессии с исходным промотором.
[0045] В данном контексте, термин "энхансер" или "энхансерный элемент" относится к цис-действующему транскрипционному регуляторному элементу, a.k.a. цис-элементу, который придает аспект общего паттерна экспрессии, но обычно является недостаточным в отдельности для управления транскрипцией, функционально связанной полинуклеотидной последовательности. В отличие от промоторов, энхансерные элементы обычно не включают в себя стартовый сайт транскрипции (TSS) или TATA-бокс или эквивалентную последовательность. Промотор может природно содержать один или несколько энхансерных элементов, которые влияют на транскрипцию функционально связанной полинуклеотидной последовательности. Выделенный энхансерный элемент может быть также слит с промотором для продуцирования химерного промоторного цис-элемента, который придает аспект общей модуляции транскрипци гена. Промотор или фрагмент промотора может содержать один или несколько энхансерных элементов, которые влияют на транскрипцию функционально связанных генов. Считается, что многие энхансерные элементы промотора связывают ДНК-связывающие белки и/или влияют на топологию ДНК, производя локальные конформации, которые селективно позволяют или ограничивают доступ РНК-полимеразы к ДНК-матрице или, которые облегчают селективное открывание двойной спирали в сайте инициации транскрипции. Некоторые энхансерные элементы связывают более одного фактора транскрипции, и факторы транскрипции могут взаимодействовать с различными аффинностями с более чем одним энхансерным доменом. Энхансерные элементы могут быть идентифицированы рядом способов, включающих в себя делеционный анализ, т.е. делеции одного или нескольких нуклеотидов из 5’-конца или внутренних относительно промотора; анализ ДНК-связывающего белка с использованием футпринтинга с ДНКазой I, интерференцию метилирования, анализы смещения мобильности в электрофорезе, in vivo геномный футпринтинг с использованием опосредованной лигированием ПЦР и другие общепринятые анализы; или анализ сходства последовательности ДНК с использованием известных мотивов цис-элементов или энхансерных элементов в качестве последовательности-мишени или мотива-мишени с общепринятыми способами сравнения последовательностей ДНК, такими как BLAST. Тонкая структура энхансерного домена может быть дополнительно исследована мутагенезом (или заменой) одного или нескольких нуклеотидов или другими общепринятыми способами. Энхансерные элементы могут быть получены химическим синтезом или выделением из регуляторных элементов, которые включают в себя такие элементы, и они могут быть синтезированы с дополнительными фланкирующими нуклеотидами, которые содержат применимые сайты рестрикционных ферментов для облегчения манипуляции субпоследовательности. Таким образом, дизайн, конструирование и применение энхансерных элементов в соответствии со способами, описанными здесь, для модуляции экспрессии функционально связанных полинуклеотидных молекул включены в настоящее изобретение.
[0046] В растениях, включение некоторых интронов в генные конструкты приводит к увеличенному накапливанию мРНК и белка относительно конструктов, лишенных этого интрона.
[0047] Этот эффект был назван "опосредуемым интроном усилением" (IME) экспрессии генов (Mascarenhas et al., (1990) Plant Mol. Biol. 15:913-920). Интроны, о которых известно, что они стимулируют экспрессию в растениях, были идентифицированы в генах кукурузы {например, tubA1, Adh1, Shi, Ubi1 (Jeon et al. (2000) Plant Physiol. 123: 1005-1014; Callis et al. (1987) Genes Dev. 1:1183-1200; Vasil et al. (1989) Plant Physiol. 91:1575-1579; Christiansen et al. (1992) Plant Mol. Biol. 18:675-689) и генах риса (e.g. salt, tpi: McElroy et al., Plant Cell 2:163-171 (1990); Xu et al., Plant Physiol. 106:459-467 (1994)). Подобным образом, было обнаружено, что интроны из генов двудольных растений, такие как интроны из петунии (например, rbcS), картофеля (например, st-ls1) и из Arabidopsis thaliana (например, ubq3 и pat1) повышают скорости экспрессии генов (Dean et al. (1989) Plant Cell 1:201-208; Leon et al. (1991) Plant Physiol. 95:968-972; Norris et al. (1993) Plant Mol Biol 21:895-906; Rose and Last (1997) Plant J.11:455-464). Было показано, что сплайсинг может быть необходимым для некоторых IME (Mascarenhas et al. (1990) Plant Mol Biol. 15:913-920; Clancy and Hannah (2002) Plant Physiol. 130:918-929). Однако было показано с использованием точковых мутаций в сайтах сплайсинга гена pat1 из A. thaliana, что сплайсинг сам по себе, не является необходимым для некоторых IME в двудольных растениях (Rose and Beliakoff (2000) Plant Physiol. 122:535-542).
[0048] Усиление экспрессии генов интронами не является общим феноменом, так как некоторые инсерции интронов в рекомбинантные экспрессионные кассеты не усиливали экспрессию (например, интроны из генов двудольных растений (гена rbcS из гороха, гена фазеолина из фасоли и гена stls-1 из Solarium tuberosum) и интроны из генов кукурузы (гена adh1 девятого интрона, гена hsp81 первого интрона)) (Chee et al. (1986) Gene 41:47-57; Kuhlemeier et al. (1988) Mol Gen Genet 212:405-411; Mascarenhas et al. (1990) Plant Mol. Biol. 15:913-920; Sinibaldi and Mettler (1992) В WE Cohn, K Moldave, eds, Progress in Nucleic Acid Research and Molecular Biology, Vol 42. Academic Press, New York, pp 229-257; Vancanneyt et al. 1990 Mol. Gen. Genet. 220:245-250). Таким образом, не каждый интрон может быть использован для манипуляции уровня экспрессии неэндогенных генов или эндогенных генов в трансгенных растениях. В известном уровне техники невозможно предсказать, какие характеристики или признаки конкретных последовательностей должны присутствовать в интронной последовательности для усиления экспрессии конкретного гена и, следовательно, на основании известного уровня техники невозможно предсказать, будет ли конкретный интрон растения, при использовании в качестве гетерологичного гена, вызывать усиление экспрессии на уровне ДНК или на уровне транскрипта (IME).
[0049] В данном контексте, термин "химерная" относится к единой молекуле ДНК, произведенной слиянием первой молекулы ДНК со второй молекулой ДНК, где ни первая молекула ДНК, ни вторая молекула ДНК в норме не обнаруживается в этой конфигурации, т.е. слитыми друг с другом. Таким образом, эта химерная молекула ДНК является новой молекулой ДНК, в противном случае не обнаруживаемой в природе. В данном контексте, термин "химерный промотор" относится к промотору, произведенному посредством такой манипуляции молекул ДНК. Химерный промотор может объединять два или более фрагментов ДНК; одним примером может быть слияние промотора с энхансерным элементом. Таким образом, дизайн, конструирование и применение функционально связанных транскрибируемых полинуклеотидных молекул включено в настоящее изобретение.
[0050] В данном контексте, термин "вариант" относится ко второй молекуле ДНК, которая находится в композиции, сходной, но не идентичной первой молекуле ДНК, и эта вторая молекула ДНК все еще сохраняет общую функциональность, т.е. тот же самый или сходный паттерн (характер) экспрессии первой молекулы ДНК. Вариант может быть более короткой или усеченной версией последовательности первой молекулы ДНК, и/или измененной версией последовательности первой молекулы ДНК, например, молекулой ДНК, такой как молекула ДНК, с отличающимися сайтами рестрикционных ферментов (рестриктаз) и/или внутренними делециями, заменами и/или инсерциями. "Вариант" может также включать в себя регуляторный элемент, имеющий нуклеотидную последовательность, содержащую замену, делецию и/или инсерцию одного или нескольких нуклеотидов ссылочной последовательности, где этот произведенный регуляторный элемент имеет большую или меньшую или эквивалентную транскрипционную или трансляционную активность, чем соответствующая исходная регуляторная молекула. "Варианты" регуляторного элемента будут также включать в себя варианты, возникающие из мутаций, которые встречаются в природе в трансформации бактериальных клеток и клетках растений. В настоящем изобретении, полинуклеотидная последовательность, обеспеченная как SEQ ID NO: 1-158 и 180-183, может быть использована для создания вариантов, которые являются по составу сходными, но не идентичными полинуклеотидной последовательности первоначального регуляторного элемента, хотя еще могут сохранять общую функциональность, т.е. тот же самый или сходный паттерн (характер) экспрессии, первоначального регуляторного элемента. Получение таких вариантов настоящего изобретения находится вполне в рамках обычной квалификации в данной области в свете этого описания и включено в объем настоящего изобретения. “Варианты” химерного регуляторного элемента включают в себя те же самые составляющие элементы, что и ссылочная последовательность, но эти составляющие элементы, содержащие химерный регуляторный элемент, могут быть функционально связаны различными способами, известными в данной области, такими как расщепление рестрикционными ферментами и лигирование, независимое от лигирования клонирование, модулярная сборка ПЦР-продуктов во время амплификации или прямой химический синтез регуляторного элемента, а также другие способы, известные в данной области. Полученный “вариант” химерного регуляторного элемента может состоять из тех же самых, или вариантов тех же самых, составляющих элементов ссылочной последовательности, но отличается по последовательности и/или последовательностям, которые содержат связывающую последовательность или последовательности, которые позволяют составляющим частям функционально связываться. В настоящем изобретении, полинуклеотидные последовательности, представленные как SEQ ID NO: 1-158 и 180-183, обеспечивают ссылочную последовательность, где составляющие элементы, которые содержат эту ссылочную последовательность, могут быть соединены способами, известными в данной области, и могут содержать замены, делеции и/или инсерции одного или нескольких нуклеотидов или мутаций, которые встречаются в природе в трансформации бактериальных клеток и клеток растений.
Конструкты
[0051] В данном контексте, термин "конструкт" обозначает любую рекомбинантую полинуклеотидную молекулу, такую как полимераза, космида, вирус, автономно реплицирующуюся полинуклеотидную молекулу, фаг или линейную или кольцевую одноцепочечную или двухцепочечную молекулу ДНК или РНК, полученную из любого источника, способную к геномной интеграции или автономной репликации, содержащей полинуклеотидную молекулу, где одна или несколько полинуклеотидных молекул были связаны функционально действующим образом, т.е. функционально связаны. В данном контексте, термин "вектор" обозначает любой рекомбинантный полинуклеотидный конструкт, который может быть использован для целей трансформации, т.е. введения гетерологичной ДНК в клетку-хозяина. Этот термин включает в себя экспрессионные кассеты, выделенные из любой из вышеуказанных молекул.
[0052] В данном контексте, термин "функционально связанные" относится к первой молекуле, соединенной со второй молекулой, где эти молекулы помещены таким образом, что эта первая молекула влияет на функцию этой второй молекулы. Эти две молекулы могут быть или могут не быть частью единой смежной молекулы и могут быть или могут не быть смежными. Например, промотор функционально связан с транскрибируемой полинуклеотидной молекулой, если этот промотор модулирует транскрипцию этой, представляющей интерес, транскрибируемой полинуклеотидной молекулы в клетке. Лидер, например, функционально связан с кодирующей последовательностью, когда он может служить в качестве лидера для полипептида, кодируемого этой кодирующей последовательностью.
[0053] Конструкты настоящего изобретения могут быть обеспечены, в одном варианте осуществления, в виде двойных конструктов Ti плазмида-граничная ДНК, которые имеют районы правой границы (RB или AGRtu.RB) и левой границы (LB или AGRtu.LB) Ti-плазмиды, выделенные из Agrobacterium tumefaciens, содержащие T-ДНК, которые вместе с транспортными молекулами, обеспечиваемыми клетками A. tumefaciens, позволяют интеграцию этой Т-ДНК в геном клетки растения {см., например, Патент США 6603061). Эти конструкты могут также содержать сегменты ДНК скелета плазмиды, которые обеспечивают функцию репликации и селекции с использованием антибиотика в бактериальных клетках, например, сайт инициации репликации Escherichia coli, точка начала репликации, такая как ori322, широкий набор точек репликации, такой как oriV или oriRi, и кодирующий район для селектируемого маркера, такого как Spec/Strp, который кодирует Tn7-аминогликозидаденилтрансферазу (aadA), придающую устойчивость к спектиномицину или стрептомицину, или селектируемый с использованием гентамицина (Gm, Gent) маркерный ген. Для трансформации растений, бактериальным штаммом-хозяином является часто A. tumefaciens ABI, C58 или LBA4404; однако, и другие штаммы, известные специалистам в области трансформации растений, могут функционировать в данном изобретении.
[0054] В данной области известны способы для сборки и введения конструктов в клетку таким образом, что транскрибируемая полинуклеотидная молекула транскрибируется в функциональную молекулу мРНК, которая транслируется и экспрессируется в виде белкового продукта. Для применения на практике настоящего изобретения, общепринятые композиции и способы получения и использования конструктов и клеток-хозяев, хорошо известны квалифицированному в данной области специалисту, см., например, Molecular Cloning: A Laboratory Manual, 3rd edition Volumes 1, 2, and 3 (2000) J. Sambrook, D.W. Russell, and N. Irwin, Cold Spring Harbor Laboratory Press. Способы для получения рекомбинантных векторов, особенно подходящих для трансформации растений, включают в себя, без ограничения, способы, описанные в Патентах США № 4971908; 4940835; 4769061 и 4757011 в их полном виде. Эти типы векторов были описаны в виде обзоров в научной литературе {см., например, Rodriguez, et al, Vectors: A Survey of Molecular Cloning Vectors and Their Uses, Butterworths, Boston, (1988) and Glick, et al., Methods in Plant Molecular Biology and Biotechnology, CRC Press, Boca Raton, FL. (1993)). Типичные векторы, применимые для экспрессии нуклеиновых кислот в высших растениях, хорошо известны в данной области и включают в себя векторы, произведенные из индуцирующей опухоли (Ti) плазмиды Agrobacterium tumefaciens (Rogers, et al, Methods in Enzymology 153: 253-277 (1987)). Другие рекомбинантные векторы, применимые для трансформации растений, включающие в себя транспортный контрольный вектор pCaMVCN, были также описаны в научной литературе (см., например, Fromm, et al., Proc. Natl. Acad. Sci. USA 82: 5824-5828 (1985)).
[0055] Различные регуляторные элементы могут быть включены в конструкт, включающие в себя любые предусмотренные в данном описании. Любые такие регуляторные элементы могут быть обеспечены в комбинации с другими регуляторными элементами. Такие комбинации могут быть сконструированы или модифицированы для получения желаемых регуляторных признаков. В одном варианте осуществления, конструкты настоящего изобретения содержат по меньшей мере один регуляторный элемент, функционально связанный с транскрибируемой полинуклеотидной молекулой, функционально связанной с 3’-UTR.
[0056] Конструкты настоящего изобретения могут включать в себя любой промотор или лидер, обеспеченный здесь или известный в данной области. Например, промотор настоящего изобретения может быть функционально связан с гетерологичным нетранслируемым 5’-лидером, таким как лидер, произведенный из гена белка теплового шока (см., например, Патенты США с номерами 5659122 и 5362865). Альтернативно, лидер настоящего изобретения может быть функционально связан с гетерологичным промотором, таким как промотор транскрипта 35S вируса мозаики цветной капусты (см. Патент США № 5352605).
[0057] В данном контексте, термин "интрон" относится к молекуле ДНК, которая может быть выделена или идентифицирована из геномной копии гена и может быть определена в общем как район, сплайсированный наружу во время процессинга мРНК перед трансляцией. Альтернативно, интрон может быть полученным синтетически или манипулированным элементом ДНК. Интрон может содержать энхансерные элементы, которые влияют на транскрипцию функционально связанных генов. Интрон может быть использован в качестве регуляторного элемента для модуляции экспрессии функционально связанной транскрибируемой полинуклеотидной молекулы. Конструкт ДНК может содержать интрон, и этот интрон может быть или может не быть гетерологичным относительно последовательности транскрибируемой полинуклеотидной молекулы. Примеры интронов в данной области включают в себя интрон актина риса (Патент США № 5641876) и интрон HSP70 кукурузы (Патент США № 5859347). Интроны, применимые в практике настоящего изобретения, включают в себя SEQ ID NO: 4, 7, 21, 24, 36, 44, 48, 52, 54, 58, 62, 68, 72, 82, 92, 94, 101, 103, 105, 107, 109, 111, 113, 118, 120, 122, 127, 129, 131, 138, 140, 142, 144, 146, 148, 150, 152, 154, 156, 158 и 182. Далее, при модификации интрон/экзонного сочленения, может быть предпочтительным избежание использования нуклеотидной последовательности АТ или нуклеотида А непосредственно перед 5’-концом сайта сплайсинга (GT) и нуклеотида G или нуклеотидной последовательности TG, соответственно, непосредственно после 3’-конца сайта сплайсинга (AG) для элиминирования потенциала нежелательных стартовых кодонов от образования во время процессинга этой мессенджер-РНК в конечном транскрипте. Последовательность около 5’- или 3’- концевых сайтов границ сплайсинга этого интрона может быть модифицирована таким образом.
[0058] В данном контексте, термин "3’-молекула терминации транскрипции" или "3’-UTR" относится к молекуле ДНК, которая используется во время транскрипции для продуцирования 3’-нетранслируемого района (3’-UTR) молекулы мРНК. 3’-нетранслируемый район молекулы мРНК может быть генерирован специфическим расщеплением и 3’-полиаденилированием, a.k.a. с использованием полиА-хвоста. 3’-UTR может быть функционально связан и помещен справа от транскрибируемой полинуклеотидной молекулы и может включать в себя полинуклеотиды, которые обеспечивают сигнал полиаденилирования и другие регуляторные сигналы, способные влиять на транскрипцию, процессинг мРНК или экспрессию гена. Предполагается, что полиА-хвосты функционируют в стабильности мРНК и в инициации трансляции. Примерами 3’-молекул терминации транскрипции в данной области являются 3’-район нопалинсинтазы {см. Fraley, et al., Proc. Natl. Acad. Sci. USA, 80: 4803-4807 (1983)); 3’-район hspl7 пшеницы; 3’-район малой субъединицы rubisco (рибулозобисфосфаткарбоксилазы) гороха; 3’-район E6 хлопчатника (Патент США № 6096950); 3’-районы, описанные в WO0011200A2; и 3’-UTR соиксина (Патент США № 6635806).
[0059] 3-UTR обычно находят полезное использование для рекомбинантной экспрессии конкретных генов. В системах животных, аппарат 3’-UTR был хорошо определен (например, Zhao et al., Microbiol Mol Biol Rev 63:405-445 (1999); Proudfoot, Nature 322:562-565 (1986); Kim et al., Biotechnology Progress 19: 1620-1622 (2003); Yonaha and Proudfoot, EMBO J. 19:3770-3777 (2000); Cramer et al, FEBS Letters 498: 179-182 (2001); Kuerstem and Goodwin, Nature Reviews Genetics 4:626-637 (2003)). Эффективная терминация транскриптов РНК является необходимой для предотвращения нежелательной транскрипции несвязанных с признаком (расположенных слева) последовательностей, которые могут интерферировать с генерированием признака. Аранжировка кассет экспрессии множественных генов в локальной близости друг с другом (например, в одной Т-ДНК) может вызывать супрессию экспрессии гена из одного или нескольких генов в указанном конструкте в сравнении с независимыми инсертами. (Padidam and Cao, BioTechniques 31:328-334 (2001). Это может мешать достижению адекватных уровней экспрессии, например, в случаях, когда была желательной сильная экспрессия генов из всех кассет.
[0060] В растениях неизвестны явно определенные последовательности сигнала полиаденилирования. Hasegawa et al, Plant J. 33: 1063-1072, (2003)) не смогли идентифицировать консервативные последовательности сигнала полиаденилирования в системах как in vitro, так и in vivo в Nicotiana sylvestris и определить фактическую длину первичного (не-полиаденилированного) транскрипта. Слабый 3’-UTR имеет потенциал генерирования сквозного прочитывания, который может влиять на экспрессию генов, локализованных в соседних экспрессионных кассетах (Padidam and Cao, BioTechniques 31:328-334 (2001)). Подходящий контроль терминации транскрипции может предотвращать сквозное прочитывание в последовательности {например, другие экспрессионные кассеты), локализованные справа, и может дополнительно позволять эффективный рециклинг РНК-полимеразы для улучшения экспрессии гена. Эффективная терминация транскрипции (высвобождение РНК-полимеразы II из этой ДНК) является предпосылкой для повторной ре-инициации транскрипции и посредством этого непосредственно влияет на общий уровень транскриптов. После терминации транскрипции, зрелая мРНК высвобождается из сайта синтеза и матрицы в цитоплазму. Эукариотические мРНК накапливаются в виде poly(A)-форм in vivo, так что трудно детектировать сайты терминации транскрипции общепринятыми способами. Однако, предсказание функциональных и эффективных 3’-UTR способов биоинформатики является трудным в том смысле, что не имеются консервативные последовательности, которые могли бы позволить легкое предсказание эффективных 3’-UTR.
[0061] С практической точки зрения, обычно является полезным, чтобы 3’-UTR, используемый в кассете трансгена, имел следующие характеристики. 3’-UTR должен быть способен продуктивно и эффективно терминировать транскрипцию трансгена и предотвращать сквозное прочитывание транскрипта в любую соседнюю последовательность ДНК, которая может состоять из другой кассеты трансгена, как в случае множественных кассет, находящихся в одной Т-ДНК, или в соседнюю хромосомную ДНК, в которую была инсертирована эта Т-ДНК. 3-UTR не должен вызывать уменьшения в транскрипционной активности, придаваемой промотором, лидером и интронами, которые используются для запуска экспрессии этого трансгена. В биотехнологии растений, 3’-UTR часто используется для праймирования реакций амплификации обратно транскрибируемой РНК, экстрагируемой из трансформированного растения и используемой для (1) оценивания транскрипционной активности или экспрессии трансгенной кассеты после интегрирования в хромосому растения; (2) оценивания числа копий инсерций в ДНК растения и (3) оценивания зиготности полученных семян после скрещивания. 3’-UTR используется также в реакциях амплификации ДНК, экстрагированной из трансформированного растения для характеристики интактности этой инсертированной кассеты.
[0062] 3-UTR, применимые в обеспечении экспрессии трансгена в растениях, могут быть идентифицированы на основе экспрессии маркеров экспрессирующейся последовательности (EST) в кДНК-библиотеках, созданных из мессенджер-РНК, выделенной из семян, цветков и других тканей, полученных из бородача большого (Andropogon gerardii), эриантуса (Saccharum ravennae (Erianthus ravennae)), щетинника зеленого (Setaria viridis), теосинте (Zea mays subsp. mexicana), проса итальянского (Setaria italica) или коикса (Coix lacryma-jobi). Библиотеки кДНК готовят из тканей, выделенных из выбранных видов растений с использованием способов, известных специалистам, квалифицированным в данной области, из ткани цветков, семян, листа и корня. Полученные кДНК секвенируют с использованием различных способов секвенирования, известных в данной области. Полученные EST собирают в кластеры с использованием программного обеспечения биоинформатики, такого как clc_ref_assemble_complete version 2.01.37139 (CLC bio USA, Cambridge, Massachusetts 02142). Число копий транскриптов каждого кластера определяют подсчетом числа считываний кДНК для каждого кластера. Идентифицированные 3’-UTR могут состоять из последовательности, произведенной из последовательности кДНК, а также последовательности, произведенной из геномной ДНК. Эту последовательность кДНК используют для создания праймеров, которые затем используют с библиотеками GenomeWalker™ (Clontech Laboratories, Inc, Mountain View, CA), сконструированными согласно протоколу производителя, для клонирования 3’-района соответствующей геномной последовательности ДНК для обеспечения более длинной последовательности терминации. Анализ относительного числа копий транскрипта либо непосредственным подсчетом, либо нормализованным подсчетом считываний наблюдаемой последовательности для библиотеки каждой ткани может быть использован для заключения о свойствах относительно паттернов экспрессии. Например, некоторые 3’-UTR могут быть обнаружены в высоком числе копий в транскриптах, наблюдаемых в высоком числе копий в ткани корня в сравнении с листом. Это предполагает, что этот транскрипт высоко экспрессируется в корне и что свойства экспрессии в корне могут быть отнесены к транскрипционной регуляции промотора, лидера, интронов или 3’-UTR. Эмпирическое тестирование 3-UTR, идентифицированного свойствами экспрессии в конкретных типах органов, тканей или клеток, может приводить к идентификации 3’-UTR, которые усиливают экспрессию в этих конкретных типах органов, тканей или клеток.
[0063] Конструкты и векторы могут также включать в себя кодирующую транзитный пептид последовательность, которая экспрессирует связанный пептид, который применим для нацеливания белкового продукта, в частности, в хлоропласт, лейкопласт или другую пластидную органеллу; митохондрии; пероксисому; вакуоль или внеклеточное местоположение. В отношении описания применения хлоропластных транзитных пептидов, см. Патент США № 5188642 и Патент США № 5728925. Многие локализованные в хлоропластах белки экспрессируются из ядерных генов в виде предшественников и нацеливаются на хлоропласт хлоропластным транзитным пептидом (СТР). Примеры таких выделенных хлоропластных белков включают в себя, но не ограничиваются ими, белки, ассоциированные с малой субъединицей (SSU) рибулозо-1,5-бисфосфаткарбоксилазы, ферредоксином, ферредоксин-оксидоредуктазой, белком I и белком II светособирающего комплекса, тиоредоксином F, енолпирувилшикимат-фосфатсинтазой (EPSPS) и транзитными пептидами, описанными в Патенте США № 7193133. Было продемонстрировано in vivo и in vitro, что нехлоропластные белки могут быть нацелены в хлоропласт с использованием слитых белков с гетерологичным CTP, и что этот CTP является достаточным для нацеливания белка в хлоропласт. Было показано, что включение подходящего хлоропластного транзитного пептида, такого как EPSPS CTP (CTP2) Arabidopsis thaliana {См., Klee et al, Mol. Gen. Genet. 210:437-442 (1987)) или EPSPS CTP (CTP4) Petunia hybrida) {См., della-Cioppa et al, Proc. Natl. Acad. Sci. USA 83:6873-6877 (1986)), нацеливает гетерологичные последовательности белка EPSPS в хлоропласты в трансгенных растениях {См., Патенты США с номерами 5627061; 5633435 и 5312910 и EP 0218571; EP 189707; EP 508909 и EP 924299).
Транскрибируемые полинуклеотидные молекулы
[0064] В данном контексте, термин "транскрибируемая полинуклеотидная молекула" относится к любой молекуле ДНК, способной транскрибироваться в молекулу РНК, включающей в себя, но не ограничивающейся ими, молекулы, имеющие кодирующие белок последовательности, и продуцирующие РНК молекулы, имеющие последовательности, применимые для супрессии генов. "Трансген" относится к транскрибируемой полинуклеотидной молекуле, гетерологичной в отношении клетки-хозяина по меньшей мере в отношении ее местоположения в геноме, и/или транскрибируемой полинуклеотидной молекуле, искусственно включенной в геном клетки-хозяина в существующей или любой предыдущей генерации этой клетки.
[0065] Промотор настоящего изобретения может быть функционально связан с транскрибируемой полинуклеотидной молекулой, которая является гетерологичной в отношении молекулы этого промотора. В данном контексте, термин "гетерологичный" относится к комбинации двух или более полинуклеотидных молекул, когда такая комбинация не обнаруживается обычно в природе. Например, эти две молекулы могут быть произведены из разных видов и/или эти две молекулы могут быть произведены из разных генов, например, разных генов из одного и того же вида, или одних и тех же генов из разных видов. Таким образом, промотор является гетерологичным в отношении функционально связанной транскрибируемой полинуклеотидной молекулы, если такая комбинация не обнаруживается обычно в природе, т.е. эта транскрибируемая полинуклеотидная молекула является, не встречающейся в природе, функционально связанной в комбинации с этой промоторной молекулой.
[0066] Эта транскрибируемая полинуклеотидная молекула может быть обычно любой молекулой ДНК, для которой является желаемой экспрессия транскрипта РНК. Такая экспрессия транскрипта РНК может приводить к трансляции полученной молекулы мРНК и, следовательно, к экспрессии белка. Альтернативно, например, эта транскрибируемая полинуклеотидная молекула может быть сконструирована для вызывания, в конечном счете, уменьшенной экспрессии конкретного гена или белка. В одном варианте осуществления, это может выполняться с использованием транскрибируемой полинуклеотидной молекулы, которая ориентирована в антисмысловом направлении. Квалифицированный в данной области специалист знаком с использованием такой антисмысловой технологией. Вкратце, когда транскрибируется антисмысловая транскрибируемая полинуклеотидая молекула, продукт РНК гибридизуется с комплементарной молекулой РНК или изолирует комплементарную молекулу РНК внутри этой клетки. Эта дуплексная молекула РНК не может транслироваться в белок аппаратом трансляции клетки и деградируется в этой клетке. Любой ген может быть отрицательно регулирован таким образом.
[0067] Таким образом, одним вариантом осуществления настоящего изобретения является регуляторный элемент настоящего изобретения, такой как регуляторные элементы, обеспеченные как SEQ ID NO: 1-158 и 180-183, функционально связанные с транскрибируемой полинуклеотидной молекулой при желаемом уровне или в желаемом паттерне, когда этот конструкт интегрируется в геном клетки растения. В одном варианте осуществления, эта транскрибируемая полинуклеотидная молекула содержит кодирующий белок район гена, и этот промотор влияет на транскрипцию молекулы РНК, которая транслируется и экспрессируется в виде белкового продукта. В одном варианте осуществления, эта транскрибируемая полинуклеотидная молекула содержит антисмысловой район гена, и промотор влияет на транскрипцию антисмысловой молекулы РНК, двухцепочечной РНК или другой подобной ингибиторной молекулы РНК для ингибирования экспрессии конкретной, представляющей интерес молекулы РНК в клетке-хозяине, являющейся мишенью.
Представляющие агрономический интерес гены
[0068] Транскрибируемые полинуклеотидные молекулы могут быть представляющими агрономический интерес генами. В данном контексте, термин "представляющий агрономический интерес ген" относится к транскрибируемой полинуклеотидной молекуле, которая при экспрессии в конкретной ткани, клетке или в конкретном типе клеток растения придает желаемые характеристики, такие как ассоциированная с растением морфология, физиология, рост, развитие, урожайность, продукты, питательный профиль, болезни или вредители, и/или устойчивость к химикатам. Представляющие интерес гены включают в себя, но не ограничиваются ими, гены, кодирующие получаемый белок, устойчивый к стрессу белок, относящийся к развитию контрольный белок, относящийся к дифференцировке ткани белок, меристемный белок, белок сбрасывания, белок источника, осаждающийся белок, контрольный белок цветков, белок семян, белок с устойчивостью к гербицидам, белок с устойчивостью к заболеваниям, фермент биосинтеза жирных кислот, фермент биосинтеза токоферола, фермент биосинтеза аминокислот, пестицидный белок, или любой другой агент, такой как антисмысловая или RNAi молекула, поражающая конкретный ген для супрессии. Продукт представляющего агрономический интерес гена может действовать в растении для вызывания влияния на физиологию или метаболизм растения или может действовать в качестве пестицидного агента в диете вредителя, который питается на этом растении.
[0069] В одном варианте осуществления настоящего изобретения, промотор настоящего изобретения включают в конструкт, так что этот промотор функционально связан с транскрибируемой полинуклеотидной молекулой, которая является представляющим агрономический интерес геном. Экспрессия этого представляющего агрономический интерес гена является желаемой для придания агрономически полезного признака. Например, агрономически полезным признаком может быть, но не ограничивается ими, устойчивость к гербицидам, контроль насекомых, модифицированный выход, устойчивость к грибковым болезням, устойчивость к вирусам, устойчивость к нематодам, устойчивость к бактериальным болезням, рост и развитие растений, продуцирование крахмала, продуцирование модифицированных масел, высокое продуцирование масла, модифицированное содержание жирных кислот, высокое продуцирование белка, созревание плодов, усиленное питание животных и человека, биополимеры, устойчивость к стрессу, вызываемому окружающей средой, фармацевтические пептиды и секретируемые пептиды, признаки улучшенного процессинга, улучшенная перевариваемость, продуцирование ферментов, вкус, фиксация азота, продуцирование гибридных семян, продуцирование волокна и получение биотоплива. Примеры представляющих агрономический интерес генов, известные в данной области, включают в себя гены для устойчивости к гербицидам (Патенты США с номерами 6803501; 6448476; 6248876; 6225114; 6107549; 5866775; 5804425; 5633435; и 5463175), увеличенного выхода (Патенты США с номерами USRE38446; 6716474; 6663906; 6476295; 6441277; 6423828; 6399330; 6372211; 6235971; 6222098; и 5716837), контроля насекомых (Патенты США с номерами 6809078; 6713063; 6686452; 6657046; 6645497; 6642030; 6639054; 6620988; 6593293; 6555655; 6538109; 6537756; 6521442; 6501009; 6468523; 6326351; 6313378; 6284949; 6281016; 6248536; 6242241; 6221649; 6177615; 6156573; 6153814; 6110464; 6093695; 6063756; 6063597; 6023013; 5959091; 5942664; 5942658, 5880275; 5763245; и 5763241), устойчивости к грибковым болезням (Патенты США с номерами 6653280; 6573361; 6506962; 6316407; 6215048; 5516671; 5773696; 6121436; 6316407; и 6506962), устойчивости к вирусам (Патенты США с номерами 6617496; 6608241; 6015940; 6013864; 5850023 и 5304730), устойчивости к нематодам (Патент США № 6228992), устойчивости к бактериальным болезням (Патент США № 5516671), роста и развития растений (Патенты США с номерами 6723897 и 6518488), получения крахмала (Патенты США с номерами 6538181; 6538179; 6538178; 5750876; 6476295), продуцирования модифицированных масел (Патенты США с номерами 6444876; 6426447 и 6380462), высокого продуцирования масла (Патенты США с номерами 6495739; 5608149; 6483008 и 6476295), модифицированного содержания жирных кислот (Патенты США с номерами 6828475; 6822141; 6770465; 6706950; 6660849; 6596538; 6589767; 6537750; 6489461 и 6459018), высокого получения белка (Патент США № 6380466), созревания плодов (Патент США № 5512466), усиленного питания животных и человека (Патенты США с номерами 6723837; 6653530; 6541259; 5985605 и 6171640), биополимеров (Патенты США с номерами USRE37543; 6228623 и 5958745 и 6946588), устойчивости к стрессу, вызываемому окружающей средой (U.S. Patent No. 6072103), фармацевтических пептидов и секретируемых пептидов (Патенты США с номерами 6812379; 6774283; 6140075 и 6080560), улучшенных признаков процессинга (Патент США № 6476295), улучшенной перевариваемости (Патент США № 6531648), низкой раффинозы (Патент США № 6166292), продуцирования промышленного фермента (Патент США № 5543576), улучшенного фактора (Патент США № 6011199), фиксации азота (Патент США № 5229114), продуцирования гибридных семян (Патент США № 5689041), получения волокон (Патенты США с номерами 6576818; 6271443; 5981834 и 5869720) и получения биотоплива (Патент США № 5998700).
[0070] Альтернативно, представляющий агрономический интерес ген может влиять на вышеупомянутые характеристики или фенотип растений кодированием молекулы РНК, которая вызывает нацеленную модуляцию экспрессии генов эндогенного гена, например, посредством антисмысловой последовательности (см., например, Патент США № 5107065); ингибиторной РНК ("RNAi", включающей в себя модуляцию экспрессии гена с использованием опосредуемых miRNA, siRNA, трансактивирующей siRNA и фазовой sRNA механизмов, например, как описано в опубликованных заявках US 2006/0200878 и US 2008/0066206, и в Заявке на патент США 11/974469); или опосредуемых косупрессией механизмов. Эта РНК могла бы также быть каталитической молекулой РНК (такой как, например, рибозим или рибосвитч; см., например, Патент США 2006/0200878), сконструированной для расщепления желаемого эндогенного продукта мРНК. Таким образом, любая транскрибируемая полинуклеотидная молекула, которая кодирует транскрибируемую молекулу РНК, которая влияет на агрономически важный фенотип или представляющее интерес морфологическое изменение, может быть использована для осуществления на практике настоящего изобретения. В данной области известны способы для конструирования и введения конструктов в клетку таким образом, что эта транскрибируемая полинуклеотидная молекула транскрибируется в молекулу, которая способна вызывать супрессию генов. Например, посттранскрипционная супрессия генов с использованием конструкта с антисмысловой ориентацией транскрибируемой полинуклеотидной молекулы для регуляции экспрессии генов в клетках растений, описана в Патентах США № 5107065 и 5759829, и посттранскрипционная супрессия генов с использованием конструкта со смысловой ориентацией транскрибируемой полинуклеотидной молекулы для регуляции экспрессии генов в растениях, описана в Патентах США с номерами 5283184 и 5231020. Экспрессия транскрибируемого полинуклеотида в клетке растения может быть также использована для супрессии питания вредителей растений на клетках растений, например, композиций, выделенных из вредителей Coleoptera (Публикация Патента США № US20070124836), и композиций, выделенных из нематодных вредителей (Публикация Патента США № US20070250947). Вредители растений включают в себя, но не ограничиваются ими, членистоногих вредителей, нематодных вредителей и грибковых или микробных вредителей. Примерные транскрибируемые полинуклеотидые молекулы для включения в конструкты настоящего изобретения включают в себя, например, молекулы или гены ДНК из вида, другого, чем виды- или гены-мишени, которые появляются с некоторыми видами или присутствуют на некоторых видах в том же самом виде, но являются включенными в клетки-реципиенты способами генной инженерии, а не классическими способами репродукции или селекции. Тип полинуклеотидной молекулы может включать в себя, но не ограничивается ими, полинуклеотидную молекулу, которая уже присутствует в клетке растения, полинуклеотидную молекулу из другого растения, полинуклеотидную молекулу из другого организма или полинуклеотидную молекулу, генерированную вне растения, такую как полинуклеотидная молекула, содержащая антисмысловую матрицу гена, или полинуклеотидную молекулу, кодирующую искусственную, синтетическую или иным образом модифицированную версию трансгена.
Селектируемые маркеры
[0071] В данном контексте, термин "маркер" относится к любой транскрибируемой полинуклеотидной молекуле, экспрессия которой, или отсутствие ее, может быть подвергнуто скринингу или оценено в баллах некоторым образом. Гены маркеров для применения в практике настоящего изобретения включают в себя, но не ограничиваются ими, транскрибируемые полинуклеотидные молекулы, кодирующие β-глюкуронидазу (GUS, описанную в Патенте США № 5599670), зеленый флуоресцентный белок и его варианты (GFP, описанный в Патентах США с номерами 5491084 и 6146826), белки, которые придают устойчивость к антибиотикам, или белки, которые придают устойчивость к гербицидам. Применимые маркеры устойчивости к антибиотикам, включающие в себя маркеры, кодирующие белки, придающие устойчивость к канамицину (nptII), гигромицину В (aph IV), стрептомицину или спектиномицину (aad, spec/strep) и гентамицину (aac3 и aacC4), известны в данной области. Гербициды, для которых была продемонстрирована устойчивость трансгенного растения, и может быть применен способ настоящего изобретения, включают в себя, но не ограничиваются ими: аминометилфосфоновую кислоту, глифозат, глюфозинат, сульфонилмочевины, имидазолиноны, бромоксинил, далапон, дикамба, циклогександион, ингибиторы оксидазы протопорфириногена и изоксасфлутоловые гербициды. Транскрибируемые полинуклеотидные молекулы, кодирующие белки, участвующие в устойчивости к гербицидам, известны в данной области и включают в себя, но не ограничиваются ими, транскрибируемую полинуклеотидную молекулу, кодирующую 5-енолпирувилшикимат-3-фосфатсинтазу (EPSPS для устойчивости к глифозату, описанную в Патентах США с номерами 5627061; 5633435; 6040497 и 5094945); транскрибируемую полинуклеотидную молекулу, кодирующую глифозат-оксидоредуктазу и глифозат-N-ацетилтрансферазу (GOX, описанную в Патенте США № 5463175; GAT, описанную в Патенте США № 20030083480, и монооксигеназу дикамба Патент США № 20030135879); транскрибируемую полинуклеотидную молекулу, кодирующую бромоксинилнитрилазу (Bxn для устойчивости к бромоксинилу, описанную в Патенте США № 4810648); транскрибируемую полинуклеотидную молекулу, кодирующую фитоендесатуразу (crtI), описанную в Misawa, et al., Plant Journal 4:833-840 (1993) и Misawa, et al., Plant Journal 6:481-489 (1994) для устойчивости к норфлуразону; транскрибируемую полинуклеотидную молекулу, кодирующую синтазу ацетогидроксикислоты (AHAS, aka ALS), описанную в Sathasiivan, et al., Nucl. Acids Res. 18:2188-2193 (1990) для устойчивости к гербицидам, содержащим сульфонилмочевину; и ген bar, описанный в DeBlock, et al., EMBO Journal 6:2513-2519 (1987) для устойчивости к глюфосфинату и устойчивости к биалафосу. Промоторные молекулы настоящего изобретения могут экспрессировать связанные транскрибируемые полинуклеотидные молекулы, которые кодируют фосфинотрицин-ацетилтрансферазу, устойчивую к глифозату EPSPS, аминогликозид-фосфотрансферазу, гидроксифенилпируватдегидрогеназу, гигромицинфосфотрансферазу, неомицинфосфотрансферазу, далапондегалогеназу, устойчивую к бромоксинилу нитрилазу, антралинатсинтазу, арилоксиалканоатдиоксигеназы, ацетил-CoA-карбоксилазу, глифозатоксидоредуктазу и глифозат-N-ацетилтрансферазу.
[0072] В термине "селектируемые маркеры" включены также гены, которые кодируют секретируемый маркер, секреция которого может быть детектирована как способ идентификации или селекции для трансформированных клеток. Примеры включают в себя маркеры, которые кодируют секретируемый антиген, который может быть идентифицирован взаимодействием с антителом, или даже секретируемые ферменты, которые могут быть детектированы каталитически. Селектируемые секретируемые маркерные белки попадают в ряд классов, включающих в себя малые диффундируемые белки, которые являются детектируемыми (например, при помощи ELISA), малые активные ферменты, которые являются детектируемыми во внеклеточном растворе (например, альфа-амилаза, бета-лактамаза, фосфинотрицинтрансфераза, или белки, которые инсертированы или уловлены в клеточной стенке (такие как белки, которые включают в себя лидерную последовательность, такую как лидерная последовательность, которая обнаружена в экспрессионной единице выступающего конца или связанных с патогенезом табака белках, также известных как PR-S табака). Другие возможные гены селектируемых маркеров будут очевидны квалифицированным специалистам в данной области, и включены в настоящее изобретение.
Трансформация клеток
[0073] Изобретение относится также к способу получения трансформированных клеток и растений, которые содержат промотор, функционально связанный с транскрибируемой полинуклеотидной молекулой.
[0074] Термин "трансформация" относится к введению нуклеиновой кислоты в реципиента-хозяина. В данном контексте, термин "хозяин" относится к бактериям, грибам или растениям, включающим в себя любые клетки, ткани и органы, или потомство этих бактерий, грибов или растений. Представляющие особый интерес ткани и клетки растений включают в себя протопласты, каллусы, корни, клубни, семена, стебли, листья, проростки, эмбрионы и пыльцу.
[0075] В данном контексте, термин "трансформированные" относится к клетке, ткани, органу или организму, в которые была введена чужеродная полинуклеотидная молекула, такая как конструкт. Эта введенная полинуклеотидная молекула может быть интегрирована в геномную ДНК реципиентных клетки, ткани, органа или организма, так что эта введенная полинуклеотидная молекула наследуется следующим потомством. "Трансгенные" или "трансформированные" клетка или организм также включают в себя потомство этих клетки и организма и потомство, полученное из программы селекции, использующей такой трансгенный организм в качестве родителя в скрещивании, и проявляющее измененный фенотип, происходящий из присутствия чужеродной полинуклеотидной молекулы. Термин "трансгенные" относится к бактериям, грибам или растениям, содержащим одну или несколько гетерологичных молекул полинуклеиновой кислоты.
[0076] Имеется много способов для введения молекулы полинуклеиновой кислоты в клетки растений. Этот способ обычно предусматривает стадии селекции подходящей клетки-хозяина, трансформацию этой клетки-хозяина рекомбинантным вектором, и получение трансформированной клетки-хозяина. Подходящие способы включают в себя бактериальную инъекцию (например, Agrobacterium), бинарные бактериальные векторы искусственной хромосомы, прямую доставку ДНК (например, посредством PEG-опосредованной трансформации, опосредованное высушиванием/ингибированием поглощение ДНК, электропорацию, встряхивание с силикон-карбидными волокнами и акселерацию покрытых ДНК частиц, и т.д. (обзор в Potrykus, et al., Ann. Rev. Plant Physiol. Plant Mol. Biol. 42: 205 (1991)).
[0077] Технология для введения молекул ДНК хорошо известна квалифицированным специалистам в данной области. Способы и материалы для трансформации клеток растений введением конструкта ДНК растения в геном растения в практике настоящего изобретения может включать в себя любой из хорошо известных и демонстрированных способов. Любые способы трансформации могут быть использованы для трансформации клетки-хозяина с одним или несколькими промоторами и/или имеющимися конструктами. Клетками-хозяевами могут быть любая клетка или любой организм, такие как клетка растения, клетка водоросли, водоросли, грибная клетка, грибы, бактериальная клетка или клетка насекомого. Предпочтительные хозяева и трансформированные клетки включают в себя клетки из: растений, Aspergillus, дрожжей, насекомых, бактерий и водорослей.
[0078] Регенерированные трансгенные растения могут быть самоопыляемыми для обеспечения гомозиготных трансгенных растений. Альтернативно, пыльца, полученная из регенерированных трансгенных растений, может быть скрещена с нетрансгенными растениями, предпочтительно инбредными линиями агрономически важных видов. Описания способов селекции, которые обычно используются для различных признаков и культур, могут быть найдены в нескольких ссылочных книгах, см., например, Allard, Principles of Plant Breeding, John Wiley & Sons, NY, U. of CA, Davis, CA, 50-98 (1960); Simmonds, Principles of crop improvement, Longman, Inc., NY, 369-399 (1979); Sneep and Hendriksen, Plant breeding perspectives, Wageningen (ed), Center for Agricultural Publishing and Documentation (1979); Fehr, Soybeans: Improvement, Production and Uses, 2nd Edition, Monograph, 16:249 (1987); Fehr, Principles of variety development, Theory and Technique, (Vol. 1) and Crop Species Soybean (Vol 2), Iowa State Univ., Macmillan Pub. Co., NY, 360-376 (1987). Напротив, пыльца из нетрансгенных растений может быть использована для опыления регенерированных трансгенных растений.
[0079] Эти трансформированные растения могут быть анализированы на присутствие представляющих интерес генов и уровень экспрессии и/или профиль, придаваемый регуляторными элементами настоящего изобретения. Квалифицированные специалисты в данной области знакомы с многочисленными способами, доступными для анализа трансформированных растений. Например, способы для анализа растений включают в себя, но не ограничиваются ими, Саузерн-блоттинг или Нозерн-блоттинг, подходы на основе ПЦР, биохимические анализы, фенотипические способы скрининга, полевые оценивания и иммунодиагностические анализы. Экспрессия транскрибируемой полинуклеотидной молекулы может быть измерена с использованием реагентов и способов TaqMan® (Applied Biosystems, Foster City, CA), как описано изготовителем, и количества циклов ПЦР могут быть определены с использованием TaqMan® Testing Matrix. Альтернативно, могут быть использованы реагенты и способы Invader® (Third Wave Technologies, Madison, WI), как описано изготовителем, для трансгенной экспрессии.
[0080] Семена растений изобретения могут быть собраны из фертильных трансгенных растений, и использованы для выращивания генераций потомков трансформированных растений этого изобретения, в том числе гибридных линий растений, содержащих конструкт этого изобретения и экспрессирующих ген, представляющий агрономический интерес.
[0081] Настоящее изобретение обеспечивает также части растений настоящего изобретения. Части растений, без ограничения, включают в себя листья, стебли, корни, клубни, семена, эндосперм, семяпочку и пыльцу. Настоящее изобретение включает в себя, и обеспечивает также трансформированные клетки растений, которые содержат молекулу нуклеиновой кислоты настоящего изобретения.
[0082] Трансгенное растение может переходить вместе с трансгенной полинуклеотидной молекулой в его потомство. Потомство включает в себя любую регенерированную часть растения или семена, содержащие трансген, произведенный из растения-предка. Это трансгенное растение предпочтительно является гомозиготным в отношении трансформированной полинуклеотидной молекулы, и переносит эту последовательность во все потомство, как результат полового размножения. Потомство может быть выращено из семян, продуцируемых этим трансгенным растением. Эти дополнительные растения могут затем самоопыляться для генерирования точной генеалогической линии растений. Потомство из этих растений оценивают, среди прочего, на экспрессию генов. Эта экспрессия генов может детектироваться несколькими обычными способами, такими как вестерн-блоттинг, нозерн-блоттинг, иммунопреципитация и ELISA.
Товарные продукты
[0083] Настоящее изобретение обеспечивает товарный продукт, содержащий молекулы ДНК изобретения. В данном контексте, термин "товарный продукт" относится к любой композиции или продукту, которые состоят из материала, произведенного из растения, семян, клетки растения или части растения, содержащих молекулу ДНК этого изобретения. Товарные продукты могут продаваться потребителям и могут быть жизнеспособными или нежизнеспособными. Нежизнеспособные товарные продукты включают в себя, но не ограничиваются ими, нежизнеспособные семена и зерно; обработанные семена, части семян и части растений; дегидратированную ткань растения, замороженную ткань растения и обработанную ткань растения; семена и части растений, обработанные для кормления животных для потребления наземными и/или водными животными, масло, мучку, муку, хлопья, отруби, волокна, молоко, сыр, бумагу, сливки, вино и любые другие пищевые продукты для потребления человеком; и биомассы и топливные продукты. Жизнеспособные товарные продукты включают в себя, но не ограничиваются ими, семена и клетки растений. Таким образом, растения, содержащие молекулу ДНК в соответствии с настоящим изобретением, могут быть использованы для изготовления любого товарного продукта, обычно получаемого из растений или их частей.
[0084] После общего описания настоящего изобретения, то же самое будет более легко пониматься с использованием ссылки на следующие примеры, которые обеспечены посредством иллюстрации и без ограничения настоящего изобретения, если нет иных указаний. Специалистам в данной области должно быть понятно, что способы, описанные в следующих примерах, представляют способы, раскрытые авторами изобретения, для хорошего функционирования в применении на практике изобретения. Однако, квалифицированные в данной области специалисты должны, в свете данного описания, понимать, что многие изменения могут быть произведены в конкретных вариантах осуществления, которые описаны, с получением все еще подобного или сходного результата, без отклонения от идеи и объема изобретения, и, следовательно, весь материал, представленный или показанный в сопутствующих фигурах, должен интерпретироваться как иллюстративный, и не в смысле ограничения.
ПРИМЕРЫ
Пример 1: Идентификация и клонирование регуляторных элементов
[0085] Новые убиквитиновые транскрипционные регуляторные элементы, или транскрипционные регуляторные последовательности группы элементов экспрессии (EXP) идентифицировали и выделяли из геномной ДНК однодольных видов бородача большого (Andropogon gerardii), эриантуса (Saccharum ravennae (Erianthus ravennae)), щетинника зеленого (Setaria viridis), теосинте (Zea mays subsp. mexicana), проса итальянского (Setaria italica) или коикса (Coix lacryma-jobi).
[0086] Последовательности транскрипта убиквитина 1 идентифицировали из каждого из этих видов. 5-нетранслируемый район (5’-UTR) каждого из транскриптов убиквитина 1 использовали для конструирования праймеров для амплификации соответствующих транскрипционных регуляторных элементов для идентифицированного гена Убиквитина, который содержит функционально связанные промотор, лидер (5’-UTR) и первый интрон. Эти праймеры использовали с применением библиотек GenomeWalker™ (Clontech Laboratories, Inc, Mountain View, CA), сконструированных согласно протоколу изготовителя для клонирования 5’-района соответствующей геномной последовательности ДНК. Транскрипционные регуляторные элементы Убиквитина выделяли также из однодольного Sorghum bicolor с использованием публично доступных последовательностей, которые являются гомологами генов Убиквитина 4, 6 и 7 Zea mays.
[0087] С использованием идентичных последовательностей проводили биоинформатический анализ для идентификации регуляторных элементов в этой амплифицированной ДНК. С использованием результатов этого анализа регуляторные элементы были определены в последовательностях ДНК и праймерах, сконструированных для амплификации этих регуляторных элементов. Соответствующую молекулу ДНК для каждого регуляторного элемента амплифицировали с использованием стандартных условий полимеразной цепной реакции с праймерами, содержащими уникальные сайты рестрикционных ферментов (рестриктаз) и геномной ДНК, выделенной из A. gerardii, S. ravennae, S. viridis, Z. mays subsp. mexicana, S. italica, C. lacryma-jobi и S. bicolor. Полученные фрагменты ДНК лигировали в базовые экспрессирующие векторы растений и секвенировали. Затем выполняли анализ регуляторного элемента TSS и интрон/экзон границ сплайсинга с использованием трансформированных протопластов растений. Вкратце, эти протопласты трансформировали экспрессирующими векторами растений, содержащими клонированные фрагменты ДНК, функционально связанные с гетерологичной транскрибируемой полинуклеотидной последовательностью, и использовали 5’-RACE System for Rapid Amplification of cDNA Ends, Version 2.0 (Invitrogen, Carlsbad, California 92008) для подтверждения регуляторного элемента TSS и интрон/экзон-границ сплайсинга анализом последовательности мРНК-транскриптов, получаемых посредством этого.
[0088] Последовательности идентифицированных групп транскрипционных регуляторных элементов экспрессии ("EXP") обеспечены здесь как SEQ ID NO: 1, 5, 8, 10, 12, 14, 16, 18, 22, 25, 27, 29, 31, 33, 37, 39, 41, 45, 49, 53, 55, 59, 63, 65, 69, 73, 75, 77, 79, 83, 85, 87, 90, 93, 95, 97, 98, 99, 100, 102, 104, 106, 108, 110, 112, 114, 115, 116, 117, 119, 121, 123, 124, 125, 126, 128, 130, 132, 133, 134, 136, 137, 139, 141, 143, 145, 147, 149, 151, 153, 155, 157, 180, 181 и 183, как показано в Таблице 1 ниже. Промоторные последовательности обеспечены здесь как SEQ ID NO: 2, 6, 9, 11, 13, 15, 17, 19, 23, 26, 28, 30, 32, 34, 38, 40, 42, 46, 50, 56, 60, 64, 66, 70, 74, 76, 78, 80, 84, 86, 88, 91, 96 и 135. Лидерные последовательности обеспечены здесь как SEQ ID NO: 3, 20, 35, 43, 47, 51, 57, 61, 67, 71 и 81. Интронные последовательности обеспечены здесь как SEQ ID NO: 4, 7, 21, 24, 36, 44, 48, 52, 54, 58, 62, 68, 72, 82, 92, 94, 101, 103, 105, 107, 109, 111, 113, 118, 120, 122, 127, 129, 131, 138, 140, 142, 144, 146, 148, 150, 152, 154, 156, 158 и 182. Энхансерная последовательность обеспечена как SEQ ID NO: 89.
Группы транскрипционных регуляторных элементов экспрессии ("EXP”), промоторов, энхансеров, лидеров и интронов, выделенных из различных видов травянистых растений
PCR42
ID NO:20); I-ERIra.Ubq1-1:1:2 (SEQ ID NO:24).
ID NO: 94)
ID NO: 43); I-Zm.UbqM1-1:1:13 (SEQ ID NO: 138)
ID NO: 51); I-Zm.UbqM1-1:1:20 (SEQ ID NO: 182)
[0089] Как показано в Таблице 1, например, эта транскрипционная регуляторная EXP-последовательность, обозначенная как EXP-ANDge.Ubq1:l:9 (SEQ ID NO: 1), с компонентами, выделенными из A. gerardii, содержит промоторный элемент P-ANDge.Ubq1-1:1:11 (SEQ ID NO: 2), функционально связанный 5’ (слева) от лидерного элемента, L-ANDge.Ubq1-1:1:2 (SEQ ID NO: 3), функционально связанный 5’ (слева) от интронного элемента, I-ANDge.Ubq1-1:1:3 (SEQ ID NO: 4). Другие EXP связаны сходным образом, как описано в Таблице 1.
[0090] Как показано в Таблице 1, списке последовательностей и фигурах 1-7, были сконструированы варианты промоторных последовательностей из видов A. gerardii, E. ravennae, Z. mays subsp. mexicana, S. bicolor, C. lacryma-jobi, S. italica, и S. viridis, которые содержат более короткие фрагменты промоторов, например, P-ANDge.Ubql-1:1:11 (SEQ ID NO:2), P-ERIra.Ubq1-1:1:10 (SEQ ID NO: 19) или другие соответствующие промоторы из других видов, и, например, приводящие к P-ANDge.Ubq1-1:1:9 (SEQ ID NO: 6), P-ERIra.Ubq1-1:1:9 (SEQ ID NO: 23), P-Cl.Ubq1-1:1:10 (SEQ ID NO: 96), P-SETit.Ubq1-1:1:2 (SEQ ID NO: 76) и P-Sv.Ubq1-1:1:2 (SEQ ID NO: 38), а также другие промоторные фрагменты. P-SETit.Ubq1-1:1:4 (SEQ ID NO: 74) содержит единственное изменение нуклеотида относительно P-SETit.Ubq1-1:1:1 (SEQ ID NO: 70). Подобным образом, P-Sv.Ubq1-1:1:3 (SEQ ID NO: 40) содержит единственное изменение нуклеотида относительно P-Sv.Ubq1-1:1:4 (SEQ ID NO: 135).
[0091] В некоторых случаях, варианты конкретных интронов создавали изменением последних 3 нуклеотидов каждого соответствующего интрона после последовательности 5’-AG-3’ 3’-интрон-границы сплайсинга. Эти варианты интронов показаны в Таблице 2 ниже.
3’-концевая последовательность вариантов интронов
[0092] В Таблице 1 перечислены также три аллельных варианта, выделенные с использованием тех же самых наборов праймеров, сконструированных для амплификации геномной ДНК из Z. mays subsp. mexicana. Аллельные варианты этих последовательностей EXP состоят из последовательности, которая имеет некоторую идентичность в различных районах других последовательностей, но инсерции, делеции и ошибочные спаривания нуклеотидов могут быть также обнаружены в каждом промоторе, лидере и/или интроне каждой из этих EXP-последовательностей. Эта EXP-последовательность, обозначенная как EXP-Zm.UbqM1:1:1 (SEQ ID NO: 41), представляет первый аллель (Аллель 1) группы транскрипционных регуляторных элементов экспрессии гена Udq1 Z. mays subsp. mexicana. EXP-последовательности, обозначенные как EXP-Zm.UbqM1:1:6 (SEQ ID NO: 137) и EXP-Zm.UbqM1:1:10 (SEQ ID NO: 139), представляют первый аллель (Аллель-1), только с одним различием между двумя EXP, встречающимися в последних 3’-нуклеотидах каждого соответствующего интрона после последовательности 5’-AG-3’ 3’-интрон-границы сплайсинга. EXP-последовательность, обозначенная как EXP-Zm.UbqM1:1:4 (SEQ ID NO: 45), представляет второй аллель (Аллель 2) группы транскрипционных регуляторных элементов экспрессии гена Ubq1 Z. mays subsp. mexicana. EXP-последовательности, обозначенные как EXP-Zm.UbqM1:1:7 (SEQ ID NO: 141) и EXP-Zm.UbqM1:1:12 (SEQ ID NO: 143) представляют второй аллель (Аллель-2) только с одним различием между двумя EXP, встречающимся в последних 3’-нуклеотидах каждого соответствующего интрона после последовательности 5’-AG-3’ 3’-интрон-границы сплайсинга. EXP-последовательности EXP-Zm.UbqM1:1:2 (SEQ ID NO: 49) и EXP-Zm.UbqM1:1:5 (SEQ ID NO: 53) представляет третий аллель (Аллель-3) группы транскрипционных регуляторных элементов экспрессии гена Ubq1 Z. mays subsp. mexicana и содержат единственное различие нуклеотидов в положении 1034 в их соответствующих интронах (G вместо I-Zm.UbqM1-1:1:1l, SEQ ID NO: 52 и T вместо I-Zm.UbqM1-1:1:12, SEQ ID NO: 54). EXP-последовательности, обозначенные как EXP-Zm.UbqM1:1:8 (SEQ ID NO: 145), EXP-Zm.UbqM1:1:9 (SEQ ID NO: 147), EXP-Zm.UbqM1:1:11 (SEQ ID NO: 149) и EXP-Zm.UbqM1:1:13 (SEQ ID NO: 181) также представляют третий аллель (Аллель 3). Интрон EXP-Zm.UbqMe1:1:9, 1-Zm.UbqM1-1:1:16 (SEQ ID NO: 148) содержит остаток тимина в положении 1034, тогда как интроны EXP-Zm.UbqM1:1:8, EXP-Zm.UbqM1:1:11 и EXP-Zm.UbqM1:1:13 (I-Zm.UbqM1-1:1:15, SEQ ID NO: 146; I-Zm.UbqM1-1:1:18, SEQ ID NO: 11 и I-Zm.UbqMl-1:1:20, SEQ ID NO: 182), каждый, содержит остаток гуанина в положении 1034. Кроме того, последние 3, 3’-концевые нуклеотиды EXP-Zm.UbqM1:1:8 (SEQ ID NO: 145) и EXP-Zm.UbqM1:1:9 (SEQ ID NO: 147) отличаются от нуклеотидов EXP-Zm.UbqM1:1:11 (SEQ ID NO: 149) и EXP-Zm.UbqM1:1:13 (SEQ ID NO: 181).
Пример 2: Анализ регуляторных элементов, управляющих GUS в протопластах кукурузы.
[0093] Протопласты листьев кукурузы трансформировали экспрессирующими векторами растений, содержащими последовательность EXP, управляющую экспрессией трансгена β-глюкуронидазы, и сравнивали с экспрессией GUS в протопластах листьев, в которых экспрессия GUS управляется известными конститутивными промоторами.
[0094] Экспрессию трансгена, управляемую EXP-ANDge.Ubq1:1:7 (SEQ ID NO: 5), EXP-ANDge.Ubq1:1:6 (SEQ ID NO: 12), EXP-ERIra.Ubq1:1:9 (SEQ ID NO: 22) или EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), сравнивали с экспрессией от известных конститутивных промоторов. Эти вышеупомянутые EXP-последовательности клонировали в экспрессирующие векторы растений, как показано в Таблице 3 ниже, для получения векторов, в которых последовательность EXP функционально связана 5’ (слева) с репортером β-глюкуронидазы (GUS), который содержал процессируемый интрон (называемый GUS-2, SEQ ID NO: 160), произведенный из светоиндуцируемого тканеспецифического гена ST-LS 1 картофеля (GenBank Accession: X04753), или кодирующую последовательность соседнего GUS (GUS-1, SEQ ID NOS: 159), которая была функционально связана 5’ (слева) с 3’-UTR, произведенную из гена нопалинсинтазы A. tumefaciens (T-AGRtu.nos-1:1:13, SEQ ID NO: 161) или гена Hspl7 пшеницы (T-Ta.Hspl7-1:1:1, SEQ ID NO: 162).
Конструкт экспрессирующей плазмиды GUS растения и соответствующие последовательность EXP, GUS-кодирующая последовательность и 3’-UTR, используемые для трансформации протопластов листьев кукурузы. “SEQ ID NO:” относится к конкретной EXP-последовательности
[0095] Контрольные плазмиды (pMON19469, pMON65328, pMON25455 и pMON122605), используемые для сравнения, конструировали, как описано выше, и они содержали известную EXP-последовательность: EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID NO: 170), EXP-CaMV.35S-enh+Ta.Lhcbl+Os.Act1:1:1(SEQ ID NO: 163), EXP-Os.Act1:1:9 (SEQ ID NO: 179) или EXP-Os.TubA-3:1:1 (SEQ ID NO: 165), соответственно, функционально связанные 5’ (слева) с кодирующей GUS последовательностью и 3’-UTR. Три дополнительных контроля были обеспечены для оценивания базовой экспрессии GUS и люциферазы: без контроля ДНК, пустого вектора, который не конструировали для трансгенной экспрессии, и экспрессирующего вектора, используемого для экспрессии зеленого флуоресцентного белка (GFR).
[0096] Две плазмиды, для применения в котрансформации и нормализации данных, также конструировали с использованием способов, известных в данной области. Каждая плазмида содержала специфическую кодирующую люциферазу последовательность, которая управлялась конститутивной EXP-последовательностью. Вектор растения pMON19437 содержит трансгенную кассету с конститутивным промотором, функционально связанным 5’ (слева) с интроном, (EXP-CaMV.35S-enh+Zm.DnaK:1:1, SEQ ID NO: 170), функционально связанным 5’ (слева) с кодирующей последовательностью люциферазы светляка (Photinus pyralis) (LUCIFERASE:1:3, SEQ ID NO: 166), функционально связанной 5’ (слева) с 3’-UTR из гена нопалинсинтазы Agrobacterium tumefaciens (T-AGRtu.nos-1:1:13, SEQ ID NO: 161). Вектор растений pMON63934 содержит трансгенную кассету с конститутивной последовательностью EXP (EXP-CaMV.35S-enh-Lhcb1, SEQ ID NO: 168), функционально связанной 5’ (слева) c кодирующей последовательностью люциферазы морской фиалки (Renilla reniformis) (CR-Ren.hRenilla Lucife-0:0:1, SEQ ID NO: 167), функционально связанной 5’ (слева) с 3’-UTR гена нопалинсинтазы Agrobacterium tumefaciens (T-AGRtu.nos-1:1:13, SEQ ID NO: 161).
[0097] Протопласты листьев кукурузы трансформировали с использованием способа трансформации на основе PEG, хорошо известным в данной области. Протопластные клетки трансформировали плазмидной ДНК pMON19437, плазмидной ДНК pMON63934, и эквимолярным количеством одной из плазмид, представленных в Таблице 3, и инкубировали в течение ночи в полной темноте. Измерения как GUS, так и люциферазы проводили помещением аликвот лизированного препарата клеток, трансформированных, как описано выше, в два различных планшета с малыми лунками. Один планшет использовали для измерений GUS, и второй планшет использовали для выполнения двойного анализа люциферазы с использованием системы анализа с двойным люциферазным репортером (Promega Corp., Madison, WI; см., например, Promega Notes Magazine, No: 57, 1996, p.02). Выполняли одну или две трансформации для каждой EXP-последовательности, и средние величины экспрессии для каждой EXP-последовательности определяли из нескольких проб из каждого эксперимента с трансформацией. Измерения проб выполняли с использованием четырех повторностей каждой трансформации конструкта EXP-последовательности, или альтернативно, трех повторностей каждой трансформации конструкта EXP-последовательности на один из двух экспериментов по трансформации. Средние уровни экспрессии GUS и люциферазы представлены в Таблице 4. В этой таблице, величины люциферазы светляка (например, из экспрессии pMON19437) представлены в столбце, обозначенном "FLuc", а величины люциферазы Renilla представлены в столбце, обозначенном "RLuc".
Средняя активность GUS и люциферазы в трансформированных протопластных клетках листьев кукурузы
[0098] Для сравнения относительной активности каждой EXP-последовательности, величины GUS выражали в виде отношения активности GUS к активности люциферазы и нормализовали относительно уровней экспрессии, наблюдаемых для EXP-последовательности EXP-Os.TubA-3:1:l (SEQ ID NO: 165). Таблица 5 ниже показывает эти отношения GUS/RLuc экспрессии, нормализованной относительно экспрессии EXP-Os.TubA-3:1:l в протопластах кукурузы.
[0099] Как видно из Таблицы 5, экспрессия GUS, управляемая EXP-ANDge.Ubq1:1:7 (SEQ ID NO: 5), EXP-ANDge.Ubq1:1:6 (SEQ ID NO: 12), EXP-ERIra.Ubq1:1:9 (SEQ ID NO: 22) или EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), была в 4,51 - 9,42 раза более высокой, чем экспрессия GUS, управляемая EXP-Os.TubA-3:1:l (SEQ ID NO: 165). Экспрессия GUS, управляемая EXP-ANDge.Ubq1:1:7 (SEQ ID NO: 5), EXP-ANDge.Ubq1:1:6 (SEQ ID NO: 12), EXP-ERIra.Ubq1:1:9 (SEQ ID NO: 22) или EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), была также более высокой, чем экспрессия, управляемая EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID NO: 170), EXP-CaMV.35S- enh+Ta.Lhcbl+Os.Act1:1:1 (SEQ ID NO: 163), или EXP-Os.Act1:1:9 (SEQ ID NO: 179).
Изменение отношения GUS/RLuc-кратная экспрессия в указанное число раз в сравнении с экспрессией EXP-Os.TubA-3:1:1 в протопластных клетках листьев кукурузы
[00100] Таблица 6 ниже показывает отношения GUS/FLuc экспрессии, нормализованной в отношении экспрессии EXP-Os.TubA-3:1:1 в протопластах кукурузы.
Изменение отношения экспрессия GUS/RLuc в указанное число раз в сравнении с экспрессией EXP-Os.TubA-3:1:1 в протопластных клетках листьев кукурузы
[00101] Как можно видеть в Таблице 6, экспрессия GUS, управляемая EXP-ANDge.Ubq1:1:7 (SEQ ID NO: 5), EXP-ANDge.Ubq1:1:6 (SEQ ID NO: 12), EXP-ERIra.Ubq1:1:9 (SEQ ID NO: 22) или EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), показала ту же самую общую тенденцию при выражении в виде отношения величин GUS/FLuc и нормализации в отношении EXP-Os.TubA-3:1:1 (SEQ ID NO: 165). Экспрессия была в 12,69-32,86 раз более высокой, чем экспрессия GUS, управляемая EXP-Os.TubA-3:1:1 (SEQ ID NO: 165). Экспрессия GUS, управляемая EXP-ANDge.Ubq1:1:7 (SEQ ID NO: 5), EXP-ANDge.Ubq1:1:6 (SEQ ID NO: 12), EXP-ERIra.Ubq1:1:9 (SEQ ID NO: 22) или EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), была также более высокой в некоторых сравнениях, чем экспрессия EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID NO: 170), EXP-CaMV.35S-enh+Ta.Lhcbl+Os.Act1:1:1 (SEQ ID NO: 163) или EXP-Os.Act1:1:9 (SEQ ID NO: 179).
Пример 3: Анализ регуляторных элементов, управляющих GUS, в протопластах кукурузы, с использованием ампликонов кассет трансгенов GUS
[00102] Протопласты листьев кукурузы трансформировали ДНК-ампликонами, произведенными из экспрессирующих векторов растений, содержащих EXP-последовательность, управляющую экспрессией трансгена β-глюкуронидазы, и сравнивали с протопластом листа, в котором экспрессия GUS управляется посредством конститутивных промоторов, в ряде экспериментов, представленных ниже.
[0100] В первой серии экспериментов, протопластные клетки кукурузы, произведенные из ткани листьев, трансформировали, как описано выше, ампликонами, полученными из амплификации кассет трансгенов GUS, содержащих экспрессирующие векторы растений, для сравнения экспрессии трансгена (GUS), управляемой одним из EXP-ANDge.Ubq1:1:7 (SEQ ID NO: 5), EXP-ANDge.Ubq1:1:10 (SEQ ID NO: 10), EXP-ANDge.Ubq1:1:6 (SEQ ID NO: 12), EXP-ANDge.Ubq1:1:11 (SEQ ID NO: 14), EXP-ANDge.Ubq1:1:12 (SEQ ID NO: 16), EXP-ERIra.Ubq1:1:9 (SEQ ID NO: 22), EXP-ERIra.Ubq1:1:10 (SEQ ID NO: 25), EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), EXP-ERIra.Ubq1:1:11 (SEQ ID NO: 29), EXP-ERIra.Ubq1:1:12 (SEQ ID NO: 31), EXP-SETit.Ubq1:1:5 (SEQ ID NO: 117), EXP-SETit.Ubq1:1:7 (SEQ ID NO: 123), EXP-SETit.Ubq1:1:6 (SEQ ID NO: 124), EXP-Sv.Ubq1:1:7 (SEQ ID NO: 128), EXP-Sv.Ubq1:1:8 (SEQ ID NO: 132), EXP-Sv.Ubq1:1:10 (SEQ ID NO: 134), EXP-Zm.UbqM1:1:6 (SEQ ID NO: 137), EXP-Zm.UbqMl:1:7 (SEQ ID NO: 141), EXP-Sb.Ubq4:1:2 (SEQ ID NO: 151), EXP-Sb.Ubq6:1:2 (SEQ ID NO: 153) и EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98) с экспрессией известных конститутивных промоторов. Каждую 85-последовательность, содержащую матрицу амплификации, из которой был получен ампликон кассеты трансгенов, клонировали с использованием способов, известных в данной области, в экспрессирующий вектор растений, показанный в Таблице 7 ниже под заглавием "Матрица ампликона". Полученные экспрессирующие векторы растений содержат кассету трансгенов, состоящую из 85-последовательностей, функционально связанной 5’ (слева) с кодирующей последовательностью для β-глюкуронидазы (GUS), которая содержит либо процессируемый интрон ("GUS-2", обсуждаемый в Примере 2 выше), либо смежную кодирующую GUS последовательность ("GUS-1", обсуждаемый выше), функционально связанную 5’ относительно 3’-UTR T-AGRtu.nos-1:1:13 или T-Ta.Hspl7-1:1:l, как также обсуждалось выше. Ампликоны получали с использованием способов, известных специалистам с квалификацией в данной области, с использованием матриц плазмидных конструктов, представленных в Таблице 7 ниже. Вкратце, конструировали 5-олигонуклеотидный праймер для отжига с промоторной последовательностью, и 3’-олигонуклеотидным праймером, который отжигают на 3’-конце 3’-UTR, использовали для амплификации каждой трансгенной кассеты. Успешные 5’-делеции вводили в промоторные последовательности, содержащие эти трансгенные кассеты, с получением различных EXP-последовательностей, с использованием различных олигонуклеотидных праймеров, которые создавали отжигом в различных положениях в промоторной последовательности, содержащей матрицу каждого ампликона.
Ампликоны экспрессии GUS в растении и соответствующие матрицы ампликонов плазмидных конструктов, EXP-последовательность, кодирующая GUS последовательность и 3’-UTR, используемые для трансформации протопластов листьев кукурузы
[0101] Плазмидные конструкты, перечисленные в виде матриц ампликонов в Таблице 7, служили в качестве матриц для амплификации трансгенных экспрессионных кассет, содержащих перечисленные EXP-последовательности Таблицы 7. Контрольные плазмиды, используемые для генерирования трансгенных ампликонов GUS для сравнения, конструировали, как описано ранее, с известными EXP-последовательностями, описанными в Примере 2. Использовали также отрицательные контроли для определения фона GUS и люциферазы, контроля без ДНК и контрольной пробы, в которой две плазмиды люциферазы использовались в трансформации вместе с плазмидной ДНК, которая не экспрессировала кодирующую последовательность. Плазмиды pMON19437 и pMON63934, обсуждаемые в Примере 2, также использовали для котрансформации и нормализации данных.
[0102] Протопласты листьев кукурузы трансформировали с использованием способа трансформации на основе PEG, как описано в Примере 2 выше. Таблица 8 ниже показывает средние величины экспрессии GUS и люциферазы, определенные для каждой трансгенной кассеты.
Средняя активность GUS и люциферазы в трансформированных протопластных клетках листьев кукурузы
[0103] Для сравнения относительной активности каждой EXP-последовательности величины GUS выражали в виде отношения GUS к люциферазе и нормализовали относительно уровней экспрессии, наблюдаемой для EXP-Os.Act1:1:1 и EXP-CaMV.35S-enh+Ta.Lhcbl+Os.Act1:1:1. Таблица 9 ниже показывает отношения GUS/RLuc экспрессии, нормализованные относительно управляемой EXP-Os.Act1:1:1 и EXP-CaMV.35S-enh+Ta.Lhcbl+Os.Act1:1:1 экспрессии в протопластах кукурузы. Таблица 10 ниже показывает отношения GUS/FLuc экспрессии, нормализованные относительно управляемой EXP-Os.Act1:1:1 и EXP-CaMV.35S-enh+Ta.Lhcbl+Os.Act1:1:1 экспрессии в протопластах кукурузы.
Отношения экспрессии GUS/RLuc и GUS/FLuc, нормализованные относительно EXP-CaMV.35S-enh+Ta.Lhcbl+Os.Act1:1:1 (SEQ ID NO: 163), в протопластах кукурузы
Отношения экспрессии GUS/RLuc и GUS/FLuc, нормализованные относительно EXP-Os.Act1:1:1 (SEQ ID NO: 179), в протопластах листьев кукурузы
[0104] Как можно видеть в Таблицах 9 и 10, почти все из EXP-последовательностей были способны управлять экспрессией трансгена GUS в клетках кукурузы. Средняя экспрессия GUS была более высокой для EXP-ANDge.Ubq1:1:7 (SEQ ID NO: 5), EXP-ANDge.Ubq1:1:10 (SEQ ID NO: 10), EXP-ANDge.Ubq1:1:6 (SEQ ID NO: 12), EXP-ANDge.Ubq1:1:11 (SEQ ID NO: 14), EXP-ANDge.Ubq1:1:12 (SEQ ID NO: 16), EXP-ERIra.Ubq1:1:9 (SEQ ID NO: 22), EXP-ERIra.Ubq1:1:10 (SEQ ID NO: 25), EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), EXP-ERIra.Ubq1:1:11 (SEQ ID NO: 29), EXP-ERIra.Ubq1:1:12 (SEQ ID NO: 31), EXP-SETit.Ubq1:1:5 (SEQ ID NO: 117), EXP-SETit.Ubq1:1:7 (SEQ ID NO: 123), EXP-SETit.Ubq1:1:6 (SEQ ID NO: 124), EXP-Sv.Ubq1:1:7 (SEQ ID NO: 128), EXP-Sv.Ubq1:1:8 (SEQ ID NO: 132), EXP-Sv.Ubq1:1:10 (SEQ ID NO: 134), EXP-Zm.UbqM1:1:6 (SEQ ID NO: 137), EXP-Zm.UbqM1:1:7 (SEQ ID NO: 141), EXP-Sb.Ubq4:1:2 (SEQ ID NO: 151), EXP-Sb.Ubq6:1:2 (SEQ ID NO: 153) и EXP-C1.Ubq1:1:10 (SEQ ID NO: 98) в сравнении с экспрессией GUS, управляемой EXP-Os.Act1:1:1 или EXP-CaMV.35S-enh+Ta.Lhcb1+Os.Act1:1:1.
[0105] Во второй серии экспериментов, ампликон кассеты GUS, содержащий последовательность EXP EXP-Zm.UbqM1:1:8 (SEQ ID NO: 145), сравнивали с контрольными ампликонами, PCR0145942 (EXP-Os.Act1:1:9, SEQ ID NO: 179) и PCR0145944 (EXP-CaMV.35S-enh+Zm.DnaK:1:1, SEQ ID NO: 170) в отношении экспрессии GUS. Экспрессия GUS, управляемая EXP-последовательностью EXP-Zm.UbqM1:1:8, была более высокой, чем экспрессия этих двух контролей. Таблица 11 ниже показывает средние величины GUS и люциферазы, определенные для каждого ампликона. Таблица 12 ниже показывает отношения экспрессии GUS/RLuc и GUS/FLuc, нормализованные относительно управляемой EXP-Os.Act1:1:9 и EXP-CaMV.35S-enh+Zm.DnaK:1:1 экспрессии в протопластах кукурузы.
Средняя активность GUS и люциферазы в трансформированных протопластных клетках листьев кукурузы
Отношения экспрессии GUS/RLuc и GUS/FLuc, нормализованные относительно EXP-Os.Act1:1:9 (SEQ ID NO: 179) и EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID NO: 170), в протопластах листьев кукурузы
Zm.DnaK:1:1
Zm.DnaK:1:1
[0106] В третьей серии экспериментов, трансгенные кассеты ампликона GUS готовили, как описано выше, и анализировали на экспрессию, управляемую последовательностями EXP, EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8), EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:13 (SEQ ID NO: 114), EXP-Cl.Ubq1:1:14 (SEQ ID NO: 115) и EXP-Cl.Ubq1:1:15 (SEQ ID NO: 116). Эти ампликоны состояли из EXP-последовательности, функционально связанной с T-AGRtu.nos-1:1:13 3’-UTR. Экспрессию сравнивали с контролями EXP-Os.Act1:1:9 (SEQ ID NO: 179) и EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID NO: 170). Таблица 13 ниже показывает средние величины GUS и люциферазы, определенные для каждого ампликона. Таблица 14 показывает отношения экспрессии GUS/RLuc, нормализованные относительно управляемой EXP-Os.Act1:1:9 и EXP-CaMV.35S-enh+Zm.DnaK:1:1 экспрессии, в протопластах кукурузы.
Средняя активность GUS и люциферазы в трансформированных протопластных клетках листьев кукурузы
Отношения GUS/RLuc и GUS/FLuc экспрессии, нормализованные относительно EXP-Os.Act1:1:9 (SEQ ID NO: 179) и EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID NO: 170), в протопластах листьев кукурузы
[0107] Как можно видеть в Таблице 14 выше, последовательности EXP EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8), EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:13 (SEQ ID NO: 114), EXP-Cl.Ubq1:1:14 (SEQ ID NO: 115) и EXP-Cl.Ubq1:1:15 (SEQ ID NO: 116) способны управлять экспрессией трансгенов. Экспрессия, управляемая EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8), EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:13 (SEQ ID NO: 114) и EXP-Cl.Ubq1:1:14 (SEQ ID NO: 115), была более высокой, чем экспрессия обоих контролей. Экспрессия, управляемая EXP-Cl.Ubq1:1:15 (SEQ ID NO: 116), была более низкой, чем EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID NO: 170), но более высокой, чем контроль, EXP-Os.Actl:l:9 (SEQ ID NO: 179).
[0108] В четвертой серии экспериментов, трансгенные кассеты ампликона GUS готовили, как описано выше, и анализировали на экспрессию, управляемую последовательностями EXP, EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:16 (SEQ ID NO: 93) и EXP-Cl.Ubq1:1:17 (SEQ ID NO: 97). Экспрессию сравнивали с контролями EXP-Os.Act1:1:9 (SEQ ID NO: 179) и EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID NO: 170). Таблица 15 ниже показывает средние величины GUS и люциферазы, определенные для каждого ампликона. Таблица 16 ниже показывает отношения GUS/RLuc и GUS/FLuc экспрессии, нормализованные относительно управляемой EXP-Os.Act1:1:9 и EXP-CaMV.35S-enh+Zm.DnaK:1:1 экспрессии в протопластах кукурузы.
Средняя активность GUS и люциферазы в трансформированных протопластных клетках листьев кукурузы
Отношения GUS/RLuc и GUS/FLuc экспрессии, нормализованные относительно EXP-Os.Act1:1:9 (SEQ ID NO: 179) и EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID NO: 170) в протопластах листьев кукурузы
[0109] Как можно видеть в Таблице 16, последовательности EXP EXP, EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:16 (SEQ ID NO: 93) и EXP-Cl.Ubq1:1:17 (SEQ ID NO: 97) были способны управлять экспрессией трансгена. Экспрессия, управляемая каждой из последовательностей EXP, была более высокой, чем экспрессия обоих контролей.
[0110] В пятой серии экспериментов, трансгенные кассеты ампликона GUS готовили, как описано выше, и анализировали на экспрессию, управляемую последовательностями EXP, EXP-Zm.UbqM1: 1:11l (SEQ ID NO: 149) и EXP-Cl.Ubq1:1:23 (SEQ ID NO: 108). Экспрессию сравнивали с контролями EXP-Os.Act1:1:9 (SEQ ID NO: 179) и EXP-CaMV.35S-enh+Ta.Lhcbl+Os.Act1:1:1 (SEQ ID NO: 163). Таблица 17 ниже показывает средние величины GUS и люциферазы, определенные для каждого ампликона. Таблица 18 ниже показывает отношения GUS/RLuc экспрессии, нормализованные относительно управляемой EXP-Os.Act1:1:9 и EXP-CaMV.35S-enh+Ta.Lhcbl+Os.Act1:1:1 экспрессии, в протопластах кукурузы.
Средняя активность GUS и люциферазы в трансформированных протопластных клетках листьев кукурузы
Os.Act1:1:1
Отношения GUS/RLuc экспрессии, нормализованные относительно EXP-Os.Act1:1:9 (SEQ ID NO: 179) и EXP-CaMV.35S-enh+Ta.Lhcb1+Os.Act1:1:1 (SEQ ID NO: 163), в протопластах листев кукурузы
Os.Act1:1:1
[0111] Как можно видеть в Таблице 18 выше, последовательности EXP, EXP-Zm.UbqM1:1:11 (SEQ ID NO: 149) и EXP-Cl.Ubq1:1:23 (SEQ ID NO: 108) были способны управлять экспрессией GUS в протопластах листьев кукурузы. Экспрессия была сходной с экспрессией контроля, EXP-Os.Act1:1:9 (SEQ ID NO: 179) и более низкой, чем экспрессия EXP-CaMV.35S-enh+Ta.Lhcbl+Os.Act1:1:1 (SEQ ID NO: 163).
[0112] Эффективность регуляторных элементов, управляющих экспрессией GUS из ампликонов, может быть сходным образом исследована в протопластах листьев сахарного тростника. Например, протопласты сахарного тростника могут быть трансформированы ДНК-ампликонами, полученными из экспрессирующих векторов растений, содержащих EXP-последовательность, управляющую экспрессией трансгена β-глюкуронидазы (GUS), и сравнены с протопластом листа, в котором экспрессия GUS управляется известными конститутивными промоторами. Более того, регуляторные элементы, управляющие экспрессией СР4 из ампликонов в протопластах кукурузы или пшеницы, могут исследоваться подобным образом.
Пример 4: Анализ регуляторных элементов, управляющих GUS в протопластах пшеницы, с использованием ампликонов кассеты трансгена GUS.
[0113] Протопласты листьев пшеницы трансформировали ДНК-ампликонами, полученными из экспрессирующих векторов растений, содержащих EXP-последовательность, управляющую экспрессией трансгена β-глюкуронидазы, и сравнивали с протопластом листа, в котором экспрессия GUS управлялась известными конститутивными промоторами.
[0114] Протопластные клетки пшеницы, полученные из ткани листа, трансформировали с использованием способов, известных в данной области, ампликонами, полученными из амплификации кассет трансгена GUS, содержащих экспрессирующие векторы растений, для сравнения экспрессии трансгена (GUS), управляемой EXP-последовательностями, перечисленными в Таблицах 10-11, с экспрессией известных конститутивных промоторов, с методологией, описанной в предыдущем примере (Пример 3), с использованием тех же самых ампликонов кассеты GUS, которые использовались для анализа кукурузы в Примере 3 выше. Контрольные ампликоны кассеты GUS и плазмиды люциферазы, используемые для трансформации протопластов пшеницы, были такими же, что и представленные в предыдущем примере и приведены в Таблице 7 выше в Примере 3. Подобным образом, для определения фона GUS и люциферазы использовали отрицателные контроли, описанные выше. Протопласты листьев пшеницы трансформировали с использованием способа трансформации на основе PEG, как описано в Примере 3 выше. Таблица 19 дает перечень средней активности GUS и LUC, наблюдаемой в трансформированных протопластных клетках листьев пшеницы, и таблица 20 показывает нормализованные отношения GUS/RLuc экспрессии в протопластах пшеницы.
Средняя активность GUS и люциферазы в трансформированных протопластных клетках листьев пшеницы
Отношения GUS/RLuc экспрессии, нормализованные относительно EXP-Os.Act1:1:9 (SEQ ID NO: 179) и EXP-CaMV.35S-enh+Ta.Lhcbl+Os.Act1:1:1 (SEQ ID NO: 163), в протопластах листьев пшеницы
[0115] Как можно видеть из таблицы 20 выше, почти все EXP-последовательности были способны управлять экспрессией трансгена GUS в клетках пшеницы. Экспрессия трансгена GUS, управляемая EXP-ANDge.Ubq1:1:7 (SEQ ID NO: 5), EXP-ANDge.Ubq1: 1:10 (SEQ ID NO: 10), EXP-ANDge.Ubq1:1:6 (SEQ ID NO: 12), EXP-ANDge.Ubq1:1:11 (SEQ ID NO: 14), EXP-ANDge.Ubq1:1:12 (SEQ ID NO: 16), EXP-ERIra.Ubql:1:9 (SEQ ID NO: 22), EXP-ERIra.Ubq1:1:10 (SEQ ID NO: 25), EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), EXP-ERIra.Ubq1:1:11 (SEQ ID NO: 29), EXP-ERIra.Ubq1:1:12 (SEQ ID NO: 31), EXP-SETit.Ubq1:1:5 (SEQ ID NO: 117), EXP-SETit.Ubq1:1:7 (SEQ ID NO: 123), EXP-SETit.Ubq1:1:6 (SEQ ID NO: 124), EXP-Sv.Ubq1:1:7 (SEQ ID NO: 128), EXP-Sv.Ubq1:1:8 (SEQ ID NO: 132), EXP-Sv.Ubq1:1:10 (SEQ ID NO: 134), EXP-Zm.UbqM1:1:6 (SEQ ID NO: 137), EXP-Zm.UbqM1:1:7 (SEQ ID NO: 141), EXP-Sb.Ubq4:1:2 (SEQ ID NO: 151), EXP-Sb.Ubq6:1:2 (SEQ ID NO: 153) и EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), была гораздо более высокой, чем экспрессия GUS, управляемая EXP-Os.Act1:1:9. Экспрессия ампликонов GUS в протопластных клетках пшеницы в сравнении с EXP-CaMV.35S-enh+Ta.Lhcbl+Os.Act1:1:1 была несколько отличающейся от экспрессии, наблюдаемой в протопластных клетках кукурузы. Каждая из EXP-ANDge.Ubq1:1:7 (SEQ ID NO: 5), EXP-ANDge.Ubq1:1:10 (SEQ ID NO: 10), EXP-ANDge.Ubq1:1:6 (SEQ ID NO: 12), EXP-ANDge.Ubq1:1:11 (SEQ ID NO: 14), EXP-ANDge.Ubq1:1:12 (SEQ ID NO: 16), EXP-ERIra.Ubq1:1:10 (SEQ ID NO: 25), EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), EXP-ERIra.Ubq1:1:12 (SEQ ID NO: 31), EXP-SETit.Ubq1:1:5 (SEQ ID NO: 117), EXP-SETit.Ubq1:1:7 (SEQ ID NO: 123), EXP-SETit.Ubq1:1:6 (SEQ ID NO: 124), EXP-Sv.Ubq1:1:7 (SEQ ID NO: 128), EXP-Sv.Ubq1:1:8 (SEQ ID NO: 132), EXP-Sv.Ubq1:1:10 (SEQ ID NO: 134), EXP-Zm.UbqM1:1:6 (SEQ ID NO: 137), EXP-Zm.UbqM1:1:7 (SEQ ID NO: 141) и EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98) демонстрировала более высокие уровни экспрессии GUS относительно EXP-CaMV.35S-enh+Ta.Lhcbl+Os.Act1:1:1. EXP-последовательности EXP-ERIra.Ubq1:1:9 (SEQ ID NO: 22), EXP-ERIra.Ubq1:1:11 (SEQ ID NO: 29), EXP-Sb.Ubq4:1:2 (SEQ ID NO: 151) и EXP-Sb.Ubq6:1:2 (SEQ ID NO: 153) демонстрировали более низкие уровни экспрессии GUS относительно EXP-CaMV.35S-enh+Ta.Lhcbl+Os.Act1:1:1.
[0116] Во второй серии экспериментов, кассеты ампликонов трансгена GUS готовили, как описано выше и анализировали на экспрессию, управляемую последовательностями EXP, EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8), EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:13 (SEQ ID NO: 114), EXP-Cl.Ubq1:1:14 (SEQ ID NO: 115) и EXP-Cl.Ubq1:1:15 (SEQ ID NO: 116). Эти ампликоны состояли из EXP-последовательности, функционально связанной с кодирующей GUS-1 последовательностью, которая была функционально связана с T-AGRtu.nos-1:1:13 3’-UTR. Экспрессию сравнивали с контролями EXP-Os.Act1:1:9 (SEQ ID NO: 179) и EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID NO: 170). Таблица 21 ниже показывает средние величины GUS и люциферазы, определенные для каждого ампликона. Таблица 22 ниже показывает отношения GUS/RLuc экспрессии, нормализованные относительно управляемой EXP-Os.Act1:1:9 и EXP-CaMV.35S-enh+Zm.DnaK:1:1 экспрессии, в протопластах кукурузы.
Средняя активность GUS и люциферазы в трансформированных протопластных клетках листьев пшеницы
Отношения GUS/RLuc и GUS/FLuc экспрессии, нормализованные относительно EXP-Os.Act1:1:9 (SEQ ID NO: 179) и EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID NO: 170), в протопластах листьев пшеницы
[0117] Как видно в Таблице 22 выше, эти EXP-последовательности EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8), EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:13 (SEQ ID NO: 114), EXP-Cl.Ubq1:1:14 (SEQ ID NO: 115) и EXP-Cl.Ubq1:1:15 (SEQ ID NO: 116) способны управлять экспрессией трансгена. Экспрессия, управляемая EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8), EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:13 (SEQ ID NO: 114) и EXP-Cl.Ubq1:1:14 (SEQ ID NO: 115), была более высокой, чем экспрессия обоих контролей. Экспрессия, управляемая EXP-Cl.Ubq1:1:15 (SEQ ID NO: 116), была более низкой, чем EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID NO: 170), но более высокой, чем контроль, EXP-Os.Act1:1:9 (SEQ ID NO: 179).
[0118] В третьей серии экспериментов, кассеты ампликонов трансгена GUS готовили, как описано выше, для анализа экспрессии, управляемой последовательностями EXP, EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:16 (SEQ ID NO: 93) и EXP-Cl.Ubq1:1:17 (SEQ ID NO: 97). Экспрессию сравнивали с контролями EXP-Os.Act1:1:9 (SEQ ID NO: 179) и EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID NO: 170). Таблица 23 ниже показывает средние величины GUS и люциферазы для каждого ампликона. Таблица 24 ниже показывает отношения GUS/RLuc и GUS/FLuc экспрессии, нормализованные относительно управляемой EXP-Os.Act1:1:9 и EXP-CaMV.35S-enh+Zm.DnaK:1:1 экспрессии, в протопластах кукурузы.
Средняя активность GUS и люциферазы в трансформированных протопластных клетках листьев пшеницы
Отношения GUS/RLuc и GUS/FLuc экспрессии, нормализованные относительно EXP-Os.Act1:1:9 (SEQ ID NO: 179) и EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID NO: 170), в протопластах листьев пшеницы
[0119] Как можно видеть в Таблице 24 выше, последовательности EXP EXP-Cl.Ubq1:1: 10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:16 (SEQ ID NO: 93) и EXP-Cl.Ubq1:1:17 (SEQ ID NO: 97) были способны управлять экспрессией трансгена. Экспрессия, управляемая каждой из этих последовательностей EXP, была более высокой, чем экспрессия обоих контролей.
[0120] В четвертой серии экспериментов, кассеты ампликонов трансгена GUS готовили, как описано выше, для анализа экспрессии, управляемой EXP-последовательностями, EXP-Zm.UbqM1:1:11 (SEQ ID NO: 149) and EXP-Cl.Ubq1:1:23 (SEQ ID NO: 108). Экспрессию сравнивали с контролями EXP-Os.Act1:1:9 (SEQ ID NO: 179) и EXP-CaMV.35S-enh+Ta.Lhcbl+Os.Act1:1:1 (SEQ ID NO: 163). Таблица 25 ниже показывает средние величины GUS и люциферазы, определенные для каждого ампликона. Таблица 26 ниже показывает отношения экспрессии GUS/RLuc, нормализованные относительно управляемой EXP-Os.Act1:1:9 и EXP-CaMV.35S-enh+Ta.Lhcbl+Os.Act1:1:1 экспрессии, в протопластах кукурузы.
Средняя активность GUS и люциферазы в трансформированных протопластных клетках листьев пшеницы
Отношения экспрессии GUS/RLuc, нормализованные относительно EXP-Os.Act1:1:9 (SEQ ID NO: 179) и EXP-CaMV.35S-enh+Ta.Lhcbl+Os.Act1:1:1 (SEQ ID NO: 163), в протопластах листьев пшеницы
[0121] Как видно в Таблице 26 выше, последовательности EXP, EXP-Zm.UbqM1:1:11 (SEQ ID NO: 149) и EXP-Cl.Ubq1:1:23 (SEQ ID NO: 108) были способны управлять экспрессией GUS в протопластах листьев пшеницы. Экспрессия была сходной с экспрессией контроля, EXP-Os.Act1:1:9 (SEQ ID NO: 179), и более низкой, чем экспрессия EXP-CaMV.35S-enh+Ta.Lhcb1+Os.Act1:1:1 (SEQ ID NO: 163).
Пример 5. Анализ регуляторных элементов, управляющих GUS в протопластах сахарного тростника, с использованием ампликонов кассеты трансгена GUS.
[0122] Протопласты листьев сахарного тростника трансформировали ДНК-ампликонами, полученными из экспрессирующих векторов растений, содержащих EXP-последовательность, управляющую экспрессией трансгена β-глюкуронидазы (GUS), и сравнивали с протопластом листа, в котором экспрессия GUS управлялась известными конститутивными промоторами.
[0123] Протопластные клетки сахарного тростника, полученные из ткани листа, трансформировали с использованием способа трансформации на основе PEG, как описано в Примере 3 выше, с ампликонами, полученными из амплификации кассет трансгена GUS, содержащих экспрессирующие векторы растений, для сравнения экспрессии трансгена (GUS), управляемой одной из EXP-ANDge.Ubq1:1:7 (SEQ ID NO: 5), EXP-ANDge.Ubq1:1:10 (SEQ ID NO: 10), EXP-ANDge.Ubq1:1:6 (SEQ ID NO: 12), EXP-ANDge.Ubq1:1:11 (SEQ ID NO: 14), EXP-ANDge.Ubq1:1: 12 (SEQ ID NO: 16), EXP-ERIra.Ubq1:1:9 (SEQ ID NO: 22), EXP-ERIra.Ubq1:1:10 (SEQ ID NO: 25), EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), EXP-ERIra.Ubq1:1:11 (SEQ ID NO: 29), EXP-ERIra.Ubq1:1:12 (SEQ ID NO: 31), EXP-Cl.Ubq1:1: 10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:13 (SEQ ID NO: 114), EXP-Cl.Ubq1:1: 14 (SEQ ID NO: 115) и EXP-Cl.Ubq1:1:15 (SEQ ID NO: 116) и в представленной Таблице 27 ниже, с экспрессией с известными конститутивными промоторами.
Ампликоны экспрессии GUS в растениях и соответствующие матрицы ампликонов плазмидных конструктов и EXP-последовательности
[0124] Контрольные ампликоны кассеты GUS и плазмиды люциферазы, используемые для трансформации протопластов сахарного тростника, были также теми же самыми, что и ампликоны и плазмиды, представленные в Примерах 2 - 4 и обеспеченные в Таблице 7 выше в Примере 3. Подобным образом, использовали отрицательные контроли для определения фона GUS и люциферазы, как описано выше. Таблица 28 дает перечень средней активности GUS и Luc, наблюдаемой в трансформированных протопластных клетках листьев сахарного тростника, и Таблица 29 показывает нормализованные отношения экспрессии GUS/RLuc в протопластах листьев сахарного тростника.
Средняя активность GUS и люциферазы в трансформированных протопластных клетках листьев пшеницы
Отношения GUS/RLuc и GUS/FLuc экспрессии, нормализованные относительно EXP-Os.Act1:1:9 (SEQ ID NO: 179) и EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID NO: 170), в протопластах листьев сахарного тростника
[0125] Как можно видеть в Таблице 29 выше, EXP-последовательности EXP-ANDge.Ubq1:1:7 (SEQ ID NO: 5), EXP-ANDge.Ubq1:1:10 (SEQ ID NO: 10), EXP-ANDge.Ubq1:1:6 (SEQ ID NO: 12), EXP-ANDge.Ubq1:1:11 (SEQ ID NO: 14), EXP-ANDge.Ubq1:1:12 (SEQ ID NO: 16), EXP-ERIra.Ubq1:1:9 (SEQ ID NO: 22), EXP-ERIra.Ubq1:1:10 (SEQ ID NO: 25), EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), EXP-ERIra.Ubq1:1:11 (SEQ ID NO: 29), EXP-ERIra.Ubq1:1:12 (SEQ ID NO: 31), EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:13 (SEQ ID NO: 114), EXP-Cl.Ubq1:1:14 (SEQ ID NO: 115) и EXP-Cl.Ubq1:1:15 (SEQ ID NO: 116), все, были способны управлять экспрессией трансгена в протопластах сахарного тростника. EXP-последовательности, EXP-ANDge.Ubq1:1:7 (SEQ ID NO: 5), EXP-ANDge.Ubq1:1:10 (SEQ ID NO: 10), EXP-ANDge.Ubq1:1:6 (SEQ ID NO: 12), EXP-ANDge.Ubq1:1:11 (SEQ ID NO: 14), EXP-ANDge.Ubq1:1:12 (SEQ ID NO: 16), EXP-ERIra.Ubq1:1:10 (SEQ ID NO: 25), EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), EXP-ERIra.Ubq1:1:12 (SEQ ID NO: 31), EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1: 1:13 (SEQ ID NO: 114) и EXP-Cl.Ubq1:1:14 (SEQ ID NO: 115) экспрессировали GUS в большей степени, чем EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID NO: 170), в этом эксперименте.
Пример 6: Анализ регуляторных элементов, управляющих экспрессией СР4 в протопластах кукурузы
[0126] Этот пример иллюстрирует способность EXP-Sv.Ubq1:1:7 (SEQ ID NO: 128), EXP-Sv.Ubq1:1:8 (SEQ ID NO: 132), EXP-Sv.Ubq1:1:9 (SEQ ID NO: 133), EXP-Zm.UbqM1:1:6 (SEQ ID NO: 137), EXP-Zm.UbqM1:1:8 (SEQ ID NO: 145), EXP-Zm.UbqM1:1:7 (SEQ ID NO: 141), EXP-SETit.Ubq1:1:5 (SEQ ID NO: 117), EXP-SETit.Ubq1:1:7 (SEQ ID NO: 123), EXP-SETit.Ubq1:1:6 (SEQ ID NO: 124), EXP-Sb.Ubq4:1:2 (SEQ ID NO: 151) и EXP-Sb.Ubq6:1:2 (SEQ ID NO: 153) управления экспрессией гена устойчивости к глифозату СР4 в протопластах кукурузы. Эти EXP-последовательности были клонированы в бинарные плазмидные конструкты для трансформации растений с использованием способов, известных в данной области. Полученные экспрессирующие векторы растений содержали правый граничный район из A. tumefaciens, EXP-последовательность убиквитина, функционально связанную 5’ (слева) с нацеленной на пластиду кодирующей последовательностью EPSPS, устойчивой к глифозату (CP4, US RE39247), функционально связанной 5’ с T-AGRtu.nos-1:1:13 3’-UTR, и левый граничный район из A. tumefaciens (B-AGRtu.left border). Полученные плазмидные конструкты использовали для трансформации протопластных клеток листьев кукурузы с использованием способов, известных в данной области.
[0127] Использовали плазмидные конструкты, приведенные в списке в Таблице 30, с EXP-последовательностью, определенной в Таблице 1. Три контрольные плазмиды (pMON30098, pMON42410 и pMON30167), с известными конститутивными регуляторными элементами, управляющими либо СР4, либо GFP, конструировали и использовали для сравнения относительных уровней экспрессии СР4, управляемой этими EXP-последовательностями, с экспрессией СР4, управляемой известными конститутивными элементами экспрессии. Две другие плазмиды (pMON19437 и pMON63934) также использовали, как описано выше, для оценивания эффективности и жизнеспособности трансформации. Каждая плазмида содержит специфическую кодирующую последовательность люциферазы, управляемую конститутивной EXP-последовательностью.
[0128] Протопласты листьев кукурузы трансформировали с использованием способа трансформации на основе PEG, как описано в Примере 2 выше. Измерения как СР4, так и люциферазы проводили сходно с Примером 2 выше. Средние уровни экспрессии белка СР4, выраженные в ч./млн (ppm), показаны в Таблице 30 ниже.
Средняя экспрессия белка СР4 в протопластах листьев кукурузы
Станд.откл. (ч./млн)
[0129] Как можно видеть в Таблице 30, EXP-Sv.Ubq1:1:7 (SEQ ID NO: 128), EXP-Sv.Ubq1:1:8 (SEQ ID NO: 132), EXP-Sv.Ubq1:1:9 (SEQ ID NO: 133), EXP-Zm.UbqM1:1:6 (SEQ ID NO: 137), EXP-Zm.UbqM1:1:8 (SEQ ID NO: 145), EXP-Zm.UbqM1:1:7 (SEQ ID NO: 141), EXP-SETit.Ubq1:1:5 (SEQ ID NO: 117), EXP-SETit.Ubq1:1:7 (SEQ ID NO: 123), EXP-SETit.Ubq1:1:6 (SEQ ID NO: 124) и EXP-Sb.Ubq6:1:2 (SEQ ID NO: 153) управляли экспрессией трансгена CP4 при уровнях, близких или более высоких, чем уровни экспрессии СР4, управляемой EXP-CaMV.35S-enh+Ta.Lhcbl+Os.Act1:1:1 и EXP-Os.Act1:1:1. EXP-последовательность, EXP-Sb.Ubq4:1:2 (SEQ ID NO: 151), демонстрировала способность управлять экспрессией CP4, но уровень экспрессии был более низким, чем уровень конститутивных контролей.
[0130] Сходные данные с данными, приведенными выше, могут быть получены из растений, стабильно трансформированных описанными выше плазмидными конструктами, например, растений генерации (генераций) потомков R0, R1 или F1 или более поздней. Подобным образом, может быть исследована экспрессия из других плазмидных конструктов. Например, pMON141619, содержит EXP-последовательность EXP-ANDge.Ubq1:1:8, тогда как pMON142862 состоит из EXP-последовательности EXP-ERIra.Ubq1:1:8. Эти и другие конструкты могут быть анализированы подобным образом.
Пример 7: Анализ регуляторных элементов, управляющих CP4 в протопластах кукурузы с использованием ампликонов кассеты трансгена СР4.
[0131] Этот пример иллюстрирует способность EXP-ANDge.Ubq1:1:7 (SEQ ID NO: 5), EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8), EXP-ANDge.Ubq1:1:10 (SEQ ID NO: 10), EXP-ANDge.Ubq1:1:6 (SEQ ID NO: 12), EXP-ANDge.Ubq1:1:11 (SEQ ID NO: 14), EXP-ANDge.Ubq1:1:12 (SEQ ID NO: 16), EXP-ERIra.Ubq1:1:9 (SEQ ID NO: 22), EXP-ERIra.Ubq1:1:10 (SEQ ID NO: 25), EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), EXP-ERIra.Ubq1:1:11 (SEQ ID NO: 29), EXP-ERIra.Ubq1:1:12 (SEQ ID NO: 31), EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:13 (SEQ ID NO: 114), EXP-Cl.Ubq1:1:14 (SEQ ID NO: 115), EXP-Cl.Ubq1:1:15 (SEQ ID NO: 116), EXP-Cl.Ubq1:1:16 (SEQ ID NO: 93) и EXP-Cl.Ubq1:1:17 (SEQ ID NO: 97) управления экспрессией гена устойчивости к глифозату СР4 в протопластах кукурузы. Эти EXP-последовательности клонировали в бинарные плазмидные конструкты для трансформации растений. Полученные экспрессирующие векторы растений использовали в качестве матриц амплификации для получения ампликона трансгенной кассеты, состоящего из EXP-последовательности убиквитина, функционально связанной 5’ (слева) с нацеленной на пластиду устойчивой к глифозату EPSPS кодирующей последовательностью (CP4, US RE39247), функционально связанной 5’ с T-AGRtu.nos-1:1:13 3’-UTR и левым граничным районом из A. tumefaciens. Эти полученные ампликоны использовали для трансформации протопластных клеток кукурузы.
[0132] Протопласты листьев кукурузы трансформировали с использованием способа трансформации на основе PEG, как описано в Примере 2 выше. Измерения обоих CP4 проводили с использованием анализа на основе ELISA. Средние уровни экспрессии белка СР4, выраженные в виде ч./млн (ppm), показаны в Таблицах 31 и 32 ниже.
[0133] В первой серии экспериментов, экспрессию СР4, управляемую ампликонами, состоящими из EXP-последовательностей EXP-ANDge.Ubq1:1:7 (SEQ ID NO: 5), EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8), EXP-ANDge.Ubq1:1:10 (SEQ ID NO: 10), EXP-ANDge.Ubq1:1:6 (SEQ ID NO: 12), EXP-ANDge.Ubq1:1:11 (SEQ ID NO: 14), EXP-ANDge.Ubq1:1:12 (SEQ ID NO: 16), EXP-ERIra.Ubq1:1:9 (SEQ ID NO: 22), EXP-ERIra.Ubq1:1:10 (SEQ ID NO: 25), EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), EXP-ERIra.Ubq1:1:11 (SEQ ID NO: 29), EXP-ERIra.Ubq1:1:12 (SEQ ID NO: 31), EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:13 (SEQ ID NO: 114), EXP-Cl.Ubq1:1:14 (SEQ ID NO: 115) и EXP-Cl.Ubq1:1:15 (SEQ ID NO: 116), анализировали в трансформированных протопластах листьев кукурузы и сравнивали с уровнями экспрессии CP4, управляемой конститутивными контролями, EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID NO: 170) и EXP-Os.Act1:1:1 (SEQ ID NO: 164). Средние уровни экспрессии белка CP4, выраженные в виде ч./млн (ppm), показаны в Таблицах 31 ниже.
Средняя экспрессия белка CP4 в протопластах листьев кукурузы
Среднее
Станд. откл.
[0134] Как можно видеть в Таблице 31 выше, эти EXP-последовательности EXP-ANDge.Ubq1:1:7 (SEQ ID NO: 5), EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8), EXP-ANDge.Ubq1:1:10 (SEQ ID NO: 10), EXP-ANDge.Ubq1:1:6 (SEQ ID NO: 12), EXP-ANDge.Ubq1:1:11 (SEQ ID NO: 14), EXP-ANDge.Ubq1:1:12 (SEQ ID NO: 16), EXP-ERIra.Ubq1:1:9 (SEQ ID NO: 22), EXP-ERIra.Ubq1:1:10 (SEQ ID NO: 25), EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), EXP-ERIra.Ubq1:1:11 (SEQ ID NO: 29), EXP-ERIra.Ubq1:1:12 (SEQ ID NO: 31), EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:13 (SEQ ID NO: 114), EXP-Cl.Ubq1:1:14 (SEQ ID NO: 115) и EXP-Cl.Ubq1:1:15 (SEQ ID NO: 116) были способны управлять экспрессией СР4. Все из EXP-последовательностей, за исключением одной EXP-Cl.Ubq1:1:15 (SEQ ID NO: 116), управляли уровнями экспрессии CP4 при гораздо более высоком уровне, чем конститутивный контроль, EXP-Os.Act1:1:1 (SEQ ID NO: 164). Уровни экспрессии были более низкими, чем уровни экспрессии EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID NO: 170).
[0135] Во второй серии экспериментов, экспрессию СР4, управляемую ампликонами, состоящими из EXP-последовательностей EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:16 (SEQ ID NO: 93) и EXP-Cl.Ubq1:1:17 (SEQ ID NO: 97), анализировали в трансформированных протопластах листьев кукурузы и сравнивали с уровнями экспрессии СР4, управляемыми конститутивным контролем, EXP-Os.Act1:1:1 (SEQ ID NO: 164). Эти средние уровни экспрессии белка СР4, выраженные в виде ч./млн (ppm), показаны в Таблицах 32 ниже.
Средняя экспрессия белка СР4 в протопластах листьев кукурузы
Среднее
Станд. откл.
[0136] Как показано в Таблице 32 выше, EXP-последовательности EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:16 (SEQ ID NO: 93) и EXP-Cl.Ubq1:1:17 (SEQ ID NO: 97) были способны управлять экспрессией CP4. Уровни экспрессии, управляемой всеми тремя EXP-последовательностями, были более высокими, чем уровни экспрессии конститутивного контроля, EXP-Os.Act1:1:1 (SEQ ID NO: 164).
Пример 8: Анализ регуляторных элементов, управляющих СР4 в протопластах пшеницы.
[0137] Этот пример иллюстрирует способность EXP-Sv.Ubq1:1:7 (SEQ ID NO: 128), EXP-Sv.Ubq1:1:8 (SEQ ID NO: 132), EXP-Sv.Ubq1:1:9 (SEQ ID NO: 133), EXP-Zm.UbqM1:1:6 (SEQ ID NO: 137), EXP-Zm.UbqM1:1:8 (SEQ ID NO: 145), EXP-Zm.UbqM1:1:7 (SEQ ID NO: 141), EXP-SETit.Ubq1:1:5 (SEQ ID NO: 117), EXP-SETit.Ubq1:1:7 (SEQ ID NO: 123), EXP-SETit.Ubq1:1:6 (SEQ ID NO: 124), EXP-Sb.Ubq4:1:2 (SEQ ID NO: 151) и EXP-Sb.Ubq6:1:2 (SEQ ID NO: 153) управлять экспрессией СР4 в протопластах листьев пшеницы. Эти EXP-последовательности клонировали в бинарные плазмидные конструкты для трансформации растений с использованием способов, известных в данной области, и способов, описанных в Примерах 2 и 5 выше.
[0138] Три контрольные плазмиды (pMON30098, pMON42410, описанные ранее, и pMON43647, содержащие правый граничный район из Agrobacterium tumefaciens с EXP-Os.Actl+CaMV.35S.2xAl-B3+Os.Act1:1:1 (SEQ ID NO: 138), функционально связанной 5’ (слева) с нацеленной на пластиду кодирующей последовательностью устойчивости к глифозату (CP4, US RE39247), функционально связанной 5’ (слева) с T-AGRtu.nos-1:1:13, и левый граничный район (B-AGRtu.left border) с известными конститутивными регуляторными элементами, управляющими либо CP4, либо GFP, конструировали, как описано в Примере 5.
[0139] Протопласты листьев пшеницы трансформировали с использованием способа трансформации на основе PEG, как описано в предыдущих примеров, за исключением, того, что использовали 1,5 Х 105 протопластных клеток на анализ. Анализы экспрессии люциферазы и трансгена CP4 выполняли, как описано в Таблице 34 ниже.
Средняя экспрессия белка СР4 в протопластных клетках листьев пшеницы
Среднее, ч./млн
Станд. откл., ч./млн
[0140] Общая величина экспрессии CP4 в протопластах пшеницы, управляемой EXP-последовательностями и известной конститутивной EXP-последовательностью EXP-CaMV.35S-enh+Ta.Lhcbl+Os.Act1:1:1, продемонстрировала различные уровни экспрессии СР4 в протопластах пшеницы при сравнении с протопластами кукурузы.
[0141] Несколько EXP-последовательностей управляли экспрессией CP4 при более низких уровнях в протопластах пшеницы, чем известные конститутивные EXP-последовательности EXP-Os.Actl+CaMV.35S.2xAl-B3+Os.Act1:1:1 и EXP-CaMV.35S-enh+Ta.Lhcbl+Os.Act1:1:1. Две EXP-последовательности, EXP-Zm.UbqM1:1:6 (SEQ ID NO: 137) и EXP-SETit.Ubq1:1:5 (SEQ ID NO: 117), обеспечивают более высокие уровни экспрессии CP4 в протопластах пшеницы, чем известные конститутивные EXP-последовательности в этом анализе. EXP-Zm.UbqM1:1:2 управляла экспрессией CP4 при наивысшем уровне, с уровнями экспрессии, в 2,2-3,4 раза более высокими, чем EXP-Os.Actl+CaMV.35S.2xAl-B3+Os.Act1:1:1 и EXP-CaMV.35S-enh+Ta.Lhcbl+Os.Act1:1:1, соответственно. Все анализированные EXP-последовательности демонстрировали способность управлять экспрессией СР4 в клетках пшеницы.
Пример 9: Анализ регуляторных элементов, управляющих СР4 в протопластах пшеницы с использованием ампликонов кассеты трансгена СР4.
[0142] Этот пример иллюстрирует способность EXP-ANDge.Ubq1:1:7 (SEQ ID NO: 5), EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8), EXP-ANDge.Ubq1:1:10 (SEQ ID NO: 10), EXP ANDge.Ubq1:1:6 (SEQ ID NO: 12), EXP-ANDge.Ubq1:1:ll (SEQ ID NO: 14), EXP- ANDge.Ubq1:1:12 (SEQ ID NO: 16), EXP-ERIra.Ubq1:1:9 (SEQ ID NO: 22), EXP-ERIra.Ubq1:1:10 (SEQ ID NO: 25), EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), EXP-ERIra.Ubq1:1:11 (SEQ ID NO: 29), EXP-ERIra.Ubq1:1:12 (SEQ ID NO: 31), EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:13 (SEQ ID NO: 114), EXP-Cl.Ubq1:1:14 (SEQ ID NO: 115), EXP-Cl.Ubq1:1:15 (SEQ ID NO: 116), EXP-Cl.Ubq1:1:16 (SEQ ID NO: 93) и EXP-Cl.Ubq1:1:17 (SEQ ID NO: 97) управления экспрессией гена устойчивости к глифозату СР4 в протопластах пшеницы. Эти EXP-последовательности клонировали в бинарные плазмидные конструкты для трансформации растений. Полученные экспрессирующие векторы растений использовали в качестве матриц амплификации для получения ампликона трансгенной кассеты, состоящего из EXP-последовательности убиквитина, функционально связанной 5’ с нацеленной на пластиду кодирующей последовательностью EPSPS, устойчивой к глифозату (CP4, US RE39247), функционально связанной 5’ с T-AGRtu.nos-1:1:13 3-UTR, и левого граничного района из A. tumefaciens. Полученные ампликоны использовали для трансформации протопластных клеток листьев кукурузы.
[0143] Протопласты листьев пшеницы трансформировали с использованием способа трансформации на основе PEG, как описано в Примере 2 выше. Измерения обоих СР4 проводили с использованием анализа на основе ELISA. Средние уровни экспрессии белка CP4, выраженные как ч./млн (ppm), показаны в Таблицах 35 и 36 ниже.
[0144] В первой серии экспериментов, экспрессию СР4, управляемую ампликонами, состоящими из EXP-последовательностей EXP-ANDge.Ubq1:1:7 (SEQ ID NO: 5), EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8), EXP-ANDge.Ubq1:1:10 (SEQ ID NO: 10), EXP-ANDge.Ubq1:1:6 (SEQ ID NO: 12), EXP-ANDge.Ubq1:1:11 (SEQ ID NO: 14), EXP-ANDge.Ubq1:1:12 (SEQ ID NO: 16), EXP-ERIra.Ubq1:1:9 (SEQ ID NO: 22), EXP-ERIra.Ubq1:1:10 (SEQ ID NO: 25), EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), EXP-ERIra.Ubq1:1:11 (SEQ ID NO: 29), EXP-ERIra.Ubq1:1:12 (SEQ ID NO: 31), EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:13 (SEQ ID NO: 114), EXP-Cl.Ubq1:1:14 (SEQ ID NO: 115) и EXP-Cl.Ubq1:1:15 (SEQ ID NO: 116), анализировали в трансформированных протопластах листьев пшеницы и сравнивали с уровнями экспрессии СР4, управляемыми конститутивными контролями, EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID NO: 170) и EXP-Os.Act1:1:1 (SEQ ID NO: 164). Средние уровни экспрессии белка СР4, выраженные в виде ч./млн (ppm), показаны в таблице 35 ниже.
Средняя экспрессия белка СР4 в протопластах листьев пшеницы
[0145] Как можно видеть в Таблице 31 выше, EXP-последовательности EXP-ANDge.Ubq1:1:7 (SEQ ID NO: 5), EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8), EXP-ANDge.Ubq1:1:10 (SEQ ID NO: 10), EXP-ANDge.Ubq1:1:6 (SEQ ID NO: 12), EXP-ANDge.Ubq1:1:11 (SEQ ID NO: 14), EXP-ANDge.Ubq1:1:12 (SEQ ID NO: 16), EXP-ERIra.Ubq1:1:9 (SEQ ID NO: 22), EXP-ERIra.Ubq1:1:10 (SEQ ID NO: 25), EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), EXP-ERIra.Ubq1:1:11 (SEQ ID NO: 29), EXP-ERIra.Ubq1:1:12 (SEQ ID NO: 31), EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:13 (SEQ ID NO: 114), EXP-Cl.Ubq1:1:14 (SEQ ID NO: 115) и EXP-Cl.Ubq1:1:15 (SEQ ID NO: 116) были способны управлять экспрессией СР4. Все из EXP-последовательностей, за исключением одной EXP-Cl.Ubq1:1:15 (SEQ ID NO: 116), управляли уровнями экспрессии CP4 при гораздо более высоком уровне, чем конститутивный контроль, EXP-Os.Act1:1:1 (SEQ ID NO: 164). Уровни экспрессии были около того же самого уровня или более низкими, чем уровень экспрессии EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID NO: 170), для большинства EXP-последовательностей.
[0146] Во второй серии экспериментов, экспрессию СР4, управляемую ампликонами, состоящими из EXP-последовательностей EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:16 (SEQ ID NO: 93) и EXP-Cl.Ubq1:1:17 (SEQ ID NO: 97), анализировали в трансформированных протопластах листьев пшеницы и сравнивали с уровнями экспрессии CP4, управляемой конститутивным контролем, EXP-Os.Actl:l:l (SEQ ID NO: 164). Средние уровни экспрессии белка СР4, выраженные в виде ч./млн (ppm), показаны в таблице 36 ниже.
Средняя экспрессия белка СР4 в протопластах листьев пшеницы
Среднее
Станд. откл.
[0147] Как можно видеть в Таблице 36 выше, EXP-последовательности EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:16 (SEQ ID NO: 93) и EXP-Cl.Ubq1:1:17 (SEQ ID NO: 97) были способны управлять экспрессией СР4. Уровни экспрессии, управляемые всеми тремя EXP-последовательностями, были более высокими, чем уровни экспрессии конститутивного контроля, EXP-Os.Act1:1:1 (SEQ ID NO: 164).
Пример 10: Анализ регуляторных элементов, управляющих СР4 в протопластах сахарного тростника.
[0148] Этот пример иллюстрирует способность EXP-Sv.Ubq1:1:7 (SEQ ID NO: 128), EXP-Sv.Ubq1:1:8 (SEQ ID NO: 132), EXP-Sv.Ubq1:1:9 (SEQ ID NO: 133), EXP-Zm.UbqM1:1:6 (SEQ ID NO: 137), EXP-Zm.UbqM1:1:8 (SEQ ID NO: 145), EXP-Zm.UbqM1:1:7 (SEQ ID NO: 141), EXP-SETit.Ubq1:1:5 (SEQ ID NO: 117), EXP-SETit.Ubq1:1:7 (SEQ ID NO: 123), EXP-SETit.Ubq1:1:6 (SEQ ID NO: 124), EXP-Sb.Ubq4:1:2 (SEQ ID NO: 151), EXP-Sb.Ubq6:1:2 (SEQ ID NO: 153) and EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98) управления экспрессией СР4 в протопластах сахарного тростника. Эти EXP-последовательности клонировали в бинарные плазмидные конструкты для трансформации в растениях. Полученные векторы содержали правый граничный район из Agrobacterium tumefaciens, EXP-последовательность убиквитина, функционально связанную 5’ с нацеленной на пластиду кодирующей последовательностью устойчивой к глифозату EPSPS (CP4, US RE39247), функционально связанной 5’ (слева) с T-AGRtu.nos-1:1:13 (CP4, US RE39247), (SEQ ID NO: 127) или T-CaMV.35S-1:1:1 (SEQ ID NO: 140) 3’-UTR, и левый граничный район из A. tumefaciens (B-AGRtu.left border). Полученные плазмидные конструкты использовали для трансформации протопластных клеток листьев сахарного тростника с использованием способа трансформации на основе PEG.
[0149] Плазмидные конструкты pMON129203, pMON12904, pMON12905, pMON129210, pMON129211, pMON129212, pMON129200, pMON129201, pMON129202, pMON129219 и pMON129218 представлены в Таблице 12 выше.
[0150] Три контрольные плазмиды (pMON30167, описанную выше; pMON130803, также содержащую EXP-Os.Act1:1:1 (SEQ ID NO: 164); и pMON132804, содержащую EXP-P-CaMV.35S-enh-1:1:13/L-CaMV.35S-1:l:2/I-Os.Actl-1:1:19 (SEQ ID NO: 139), с известными конститутивными регуляторными элементами, управляющими СР4, конструировали и использовали для сравнения относительных уровней экспрессии СР4, управляемых убиквитиновыми EXP-последовательностями, приведенными в списке в Таблице 37 ниже.
[0151] Протопласты листьев сахарного тростника трансформировали с использованием способа трансформации на основе PEG. Средние уровни экспрессии СР4, определенные при помощи ELISA СР4, представлены в Таблице 37 ниже.
Средняя экспрессия белка СР4 в протопластных клетках листьев сахарного тростника
Среднее (ч./
млн)
Станд. откл. (ч./млн)
Среднее (ч./млн)
Станд. откл. (ч./млн)
[0152] Как можно видеть в Таблице 37 выше, эти EXP-последовательности демонстрировали способность управления экспрессией СР4 в протопластах сахарного тростника. Уровни экспрессии были сходными или более высокими, чем экспрессия СР4, управляемая EXP-Os.Act1:1:1 (SEQ ID NO: 164). Одна EXP-последовательность, EXP-Zm.UbqM1:1:8 (SEQ ID NO: 145), демонстрировала более высокие уровни экспрессии в сравнении с EXP-P-CaMV.35S-enh-1:1:13/L-CaMV.35S-1:1:2/I-Os.Act1-1:1:19 (SEQ ID NO: 139) в протопластах сахарного тростника.
Пример 11: Анализ регуляторных элементов, управляющих СР4 в протопластах сахарного тростника с использованием ампликонов кассеты трансгена СР4.
[0153] Этот пример иллюстрирует способность EXP-ANDge.Ubq1:1:7 (SEQ ID NO: 5), EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8), EXP-ANDge.Ubq1:1:10 (SEQ ID NO: 10), EXP-ANDge.Ubq1:1:6 (SEQ ID NO: 12), EXP-ANDge.Ubq1:1:11 (SEQ ID NO: 14), EXP-ANDge.Ubq1:1:12 (SEQ ID NO: 16), EXP-ERIra.Ubq1:1:9 (SEQ ID NO: 22), EXP-ERIra.Ubq1:1:10 (SEQ ID NO: 25), EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), EXP-ERIra.Ubq1:1:11 (SEQ ID NO: 29), EXP-ERIra.Ubq1:1:12 (SEQ ID NO: 31), EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:13 (SEQ ID NO: 114), EXP-Cl.Ubq1:1:14 (SEQ ID NO: 115) и EXP-Cl.Ubq1:1:15 (SEQ ID NO: 116) управления экспрессией гена устойчивости к глифозату CP4 в протопластах сахарного тростника. Эти EXP-последовательности клонировали в бинарные плазмидные конструкты для трансформации растений. Полученные экспрессирующие векторы растений использовали в качестве матриц амплификации для обеспечения ампликона трансгенной кассеты, состоящего из убиквитиновой EXP-последовательности, функционально связанной 5’ с нацеленной на пластиду кодирующей последовательностью, устойчивой к глифозату EPSPS (CP4, US RE39247), функционально связанной 5’ с T-AGRtu.nos-1:1:13 3’-UTR, и левого граничного района из A. tumefaciens. Полученные ампликоны использовали для трансформации протопластных клеток сахарного тростника.
[0154] Протопласты листьев сахарного тростника трансформировали с использованием способа трансформации на основе PEG, как описано в Примере 2 выше. Измерения обоих СР4 проводили с использованием анализа на основе ELISA.
[0155] Экспрессия CP4, управляемая ампликонами, состоящими из EXP-последовательностей EXP-ANDge.Ubq1:1:7 (SEQ ID NO: 5), EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8), EXP-ANDge.Ubq1:1:10 (SEQ ID NO: 10), EXP-ANDge.Ubq1:1:6 (SEQ ID NO: 12), EXP-ANDge.Ubq1:1:11 (SEQ ID NO: 14), EXP-ANDge.Ubq1:1:12 (SEQ ID NO: 16), EXP-ERIra.Ubq1:1:9 (SEQ ID NO: 22), EXP-ERIra.Ubq1:1:10 (SEQ ID NO: 25), EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), EXP-ERIra.Ubq1:1:11 (SEQ ID NO: 29), EXP-ERIra.Ubq1:1:12 (SEQ ID NO: 31), EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:13 (SEQ ID NO: 114), EXP-Cl.Ubq1:1:14 (SEQ ID NO: 115) и EXP-Cl.Ubq1:1:15 (SEQ ID NO: 116), анализировали в трансформированных протопластах листьев пшеницы и сравнивали с уровнями экспрессии, управляемыми конститутивными контролями, EXP-CaMV.35S-enh+Zm.DnaK:1:1 (SEQ ID NO: 170) и EXP-Os.Act1:1:1 (SEQ ID NO: 164). Средние уровни экспрессии белка СР4, выраженные в ч./млн (ppm), показаны в Таблице 38 ниже.
Средняя экспрессия белка СР4 в протопластах листьев сахарного тростника
[0156] Как можно видеть в Таблице 38 выше, EXP-последовательности EXP-ANDge.Ubq1:1:7 (SEQ ID NO: 5), EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8), EXP-ANDge.Ubq1:1:10 (SEQ ID NO: 10), EXP-ANDge.Ubq1:1:6 (SEQ ID NO: 12), EXP-ANDge.Ubq1:1:11 (SEQ ID NO: 14), EXP-ANDge.Ubq1:1:12 (SEQ ID NO: 16), EXP-ERIra.Ubq1:1:9 (SEQ ID NO: 22), EXP-ERIra.Ubq1:1:10 (SEQ ID NO: 25), EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), EXP-ERIra.Ubq1:1:11 (SEQ ID NO: 29), EXP-ERIra.Ubq1:1:12 (SEQ ID NO: 31), EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Cl.Ubq1:1:13 (SEQ ID NO: 114) и EXP-Cl.Ubq1:1:14 (SEQ ID NO: 115) были способны управлять экспрессией СР4. EXP-Cl.Ubq1:1:15 (SEQ ID NO: 116), по-видимому, не управляла экспрессией СР4 в этом анализе.
Пример 12: Анализ регуляторных элементов, управляющих GUS в трансгенной кукурузе.
[0157] Растения кукурузы трансформировали экспрессирующими векторами растений, содержащими EXP-последовательности, управляющие экспрессией трансгена β-глюкуронидазы (GUS), и полученные растения анализировали на экспрессию белка GUS. Эти убиквитиновые EXP-последовательности клонировали в бинарные плазмидные конструкты для трансформации растений с использованием способов, известных в данной области.
[0158] Эти полученные экспрессирующие векторы растений содержат правый граничный район из A. tumefaciens, первую трансгенную кассету для анализа EXP-последовательности, функционально связанной с кодирующей последовательностью для β-глюкуронидазы (GUS), которая имеет процессируемый интрон GUS-2, описанный выше, функционально связанный 5’ с 3’-UTR из гена переносящего липид белка риса (T-Os.LTP-1:1:1, SEQ ID NO: 141); вторую трансгенную селекционную кассету, используемую для селекции трансформированных клеток растений, которая придает устойчивость к гербициду глифозату (управляемую промотором Актина 1 риса), и левый граничный район из A. tumefaciens. Полученные плазмиды использовали для трансформации растений кукурузы. Таблица 39 дает список обозначений плазмид, EXP-последовательностей и SEQ ID NO, которые также приведены в Таблице 1.
Бинарные плазмиды для трансформации растений и ассоциированные EXP-последовательности
[0159] Растения трансформировали с использованием Agrobacterium-опосредуемых трансформаций, например, как описано в Публикации заявки на патент США 200901389985.
[0160] Для количественного анализа экспрессии трансформированных растений использовали гистохимический анализ GUS. Срезы цельной ткани инкубировали с раствором для окрашивания GUS X-Gluc (5-бром-4-хлор-3-индолил-β-глюкуронид) (1 миллиграмм/миллилитр) в течение подходящего периода времени, промывали, и визуально исследовали на синюю окраску. Активность GUS количественно определяли прямой визуальной инспекцией или инспекцией под микроскопом с использованием выбранных органов и тканей растений. R0-растения обследовали на экспрессию в корнях и листьях, а также в пыльнике, шелке и развивающихся семенах и зародыше, спустя 21 день после опыления (21 DAP).
[0161] Для количественных анализов, общий белок экстрагировали из выбранных тканей трансформированных растений кукурузы. Один микрограмм общего белка использовали с флуорогенным субстратом 4-метилеумбеллиферил-β-D-глюкуронидом (MUG) в общем объеме реакции 50 микролитров. Продукт реакции 4-метилеумбеллиферон (4-MU), является максимально флуоресцентным при высоком рН, где гидроксильная группа является ионизированной. Добавление щелочного раствора карбоната натрия одновременно останавливает этот анализ, и корректирует рН для количественного определения флуоресцентного продукта. Флуоресценцию измеряли с возбуждением при 365 нм, эмиссией при 445 нм с использованием Fluoromax-3 (Horiba; Kyoto, Japan) с Micromax Reader, с шириной щелевой головки, установленной при возбуждении 2 нм и эмиссии 3 нм.
[0162] Средняя экспрессия R0 GUS, наблюдаемая для каждой трансформации, представлена в Таблицах 40 и 41 ниже. Анализ R0 GUS, выполняемый на трансформантах, трансформированных pMON131957 (EXP-SETit.Ubq1:1:11, SEQ ID NO: 125), не прошел стандарты качества. Эти трансформанты были анализированы в генерации F1, и представлены дополнительно ниже в этом примере.
Средняя экспрессия R0 GUS в ткани корня и листьев
Средняя экспрессия R0 GUS в репродуктивных органах кукурузы (пыльнике, шелке) и развивающихся семенах (зародыше и эндосперме)
[0163] В растения R0 кукурузы, уровни экспрессии GUS в листе и корне различались среди убиквитиновых EXP-последовательностей. Хотя все из EXP-последовательностей демонстрировали способность управлять экспрессией трансгена GUS в стабильно трансформированных растениях, каждая EXP-последовательность демонстрировала уникальный паттерн (характер) экспрессии относительно других последовательностей. Например, высокие уровни экспрессии GUS наблюдались в ранних стадиях развития корня (V4 и V7) для EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8) и EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27) и снижались посредством VT-стадии. Экспрессия корней, управляемая EXP-Zm.UbqM1:1:10 (SEQ ID NO: 139), не демонстрировала экспрессии при V3, но была высокой при V7 и затем снижалась посредством VT-стадии. Экспрессия корней, управляемая EXP-Zm.UbqM1:1:11 (SEQ ID NO: 149), сохранялась до сходного уровня на протяжении развития от стадий V3, V7 - VT. Наблюдали, что экспрессия корня увеличивается от раннего развития (V3/V4) до стадии V7 и затем снижается от стадии V7 до стадии V8 в растениях, трансформированных EXP-Cl.Ubq1:1:12 (SEQ ID NO: 90), EXP-Cl.Ubq1:1:11 (SEQ ID NO: 95) и EXP-Cl.Ubq1:1:23 (SEQ ID NO: 108). Уровни экспрессии GUS показывали также существенные различия в ткани листа. Наивысшие уровни экспрессии листа придавались в раннем развитии (V3/V4) с EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8) и EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), которые снижались при стадии V7 - VT. Экспрессия GUS поддерживается от стадии V3 - стадии VT с использованием EXP-Zm.UbqM1:1:10 (SEQ ID NO: 139), EXP-Zm.UbqM1:1:11 (SEQ ID NO: 149), EXP-Zm.UbqM1:1:12 (SEQ ID NO: 143) и EXP-Cl.Ubq1:1l:23 (SEQ ID NO: 108); и до более низкой степени с использованием EXP-SETit.Ubq1:1:10 (SEQ ID NO: 119) и EXP-Sb.Ubq6:1:3 (SEQ ID NO: 155). Экспрессия в листе увеличивалась от V3- до V7- до VT-стадии с использованием EXP-Cl.Ubq1:1: 12 (SEQ ID NO: 90), EXP-Cl.Ubq1:1:11 (SEQ ID NO: 95) и EXP-Cl.Ubq1:1:23 (SEQ ID NO: 108), тогда как экспрессия снижалась от стадии V3 до стадии VT с использованием EXP-Sv.Ubq1:1:12 (SEQ ID NO: 136) и EXP-Sb.Ubq4:1:2 (SEQ ID NO: 151).
[0164] Подобным образом, в отношении репродуктивной ткани (пыльника и шелка) и развития семян ((21DAP-зародыша и эндосперма) наблюдали различные паттерны (характеры) экспрессии, уникальные для каждой EXP-последовательности. Например, высокие уровни экспрессии наблюдали в пыльнике и шелке, а также развитии семян с использованием EXP-Zm.UbqM1:1:10 (SEQ ID NO: 139), EXP-Zm.UbqM1:1:11 (SEQ ID NO: 149), EXP-Zm.UbqM1:1:12 (SEQ ID NO: 143) и EXP-Cl.Ubq1:1:23 (SEQ ID NO: 108). Экспрессия была высокой в пыльнике и шелке, но низкой в развивающихся семенах с использованием EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8) и EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27). Экспрессия, управляемая EXP-Sb.Ubq7:1:2 (SEQ ID NO: 157), была высокой в репродуктивной ткани и высокой в развивающемся зародыше, но низкой в развивающемся эндосперме. EXP-последовательность, EXP-Sb.Ubq4:1:2 (SEQ ID NO: 151) демонстрировала экспрессию в пыльнике, но не в шелке и экспрессировалась гораздо более низко в развивающихся семенах. EXP-Sv.Ubq1:1:11 (SEQ ID NO: 130) демонстрировала сходный паттерн с паттерном EXP-Sb.Ubq4:1:2 (SEQ ID NO: 151) в отношении репродуктивной ткани и развивающихся семян, тогда как EXP-Sb.Ubq4:1:2 (SEQ ID NO: 151) обнаруживала экспрессию в тканях корня и листа, EXP-Sv.Ubq1:1:11 (SEQ ID NO: 130) экспрессировалась гораздо более низко в тех же самых тканях.
[0165] Трансформанты R0 генерации, отобранные на инсерции с одной копией, скрещивались с нетрансгенной линией LH244 (с получением F1) или самоопылялись (с получением R1) для получения F1- или R1-популяции семян. В каждом случае, гетерозиготные растения F1 или R1 отбирали для исследования. Уровни экспрессии GUS измеряли в отобранных тканях на протяжении хода развития, как описано ранее. Ткани F1 или R1, используемые для этого исследования, включали в себя: насыщенный влагой зародыш, насыщенный влагой эндосперм семян, корень и колеоптиль при 4 днях после проращивания (DAG); лист и корень в стадии V3; корень и зрелый лист в стадии V8; корень, зрелые листья, стадию VT (при выбрасывании метелки, перед репродукцией) пыльник, пыльцу, лист и стареющий лист; R1 сердцевина кукурузного початка, шелк, корень и междоузлие; зерно 12 дней после опыления (DAP) и; зародыш и эндосперм 21 и 38 DAP. Пробы отобранных тканей также анализировали для F1-растений, подвергаемых условиям вызванного засухой стресса и вызванного холодом стресса, для трансформантов, содержащих pMON132037 (EXP-SETit.Ubq1:1:10, SEQ ID NO: 119), pMON131957 (EXP-SETit.Ubq1:1:11, SEQ ID NO: 125), pMON131958 (EXP-Sv.Ubq1:1:11, SEQ ID NO: 130) и pMON132974 (EXP-Sb.Ubq7:1:2, SEQ ID NO: 157). Ткани корня и листа V3 использовали для взятия проб после подвергания действию холода и засухи.
[0166] Вызванный засухой стресс индуцировали в F1, V3 растениях, трансформированных pMON132037 (EXP-SETit.Ubq1:1:10, SEQ ID NO: 119), pMON131957 (EXP-SETit.Ubq1:1:11, SEQ ID NO: 125), pMON131958 (EXP-Sv.Ubq1:1:11, SEQ ID NO: 130) и pMON132974 (EXP-Sb.Ubq7:1:2, SEQ ID NO: 157), отменой полива в течение 4 дней, позволяющей уменьшение содержания влаги по меньшей мере 50% исходного содержания влаги, полностью поливаемого растения. Протокол засухи содержал в основном следующие стадии. Растения стадии V3 были лишены воды. Когда растение кукурузы испытывает засуху, форма листа будет изменяться от обычно здорового и не сложенного вида в лист, демонстрирующий укладку в пучок сосудистых средних жилок и предстающий в форме V, при рассматривании от кончика листа к стеблю. Это изменение в морфологии обычно начинает встречаться при приблизительно 2 днях после прекращения полива, и в более ранних экспериментах было показано, что это изменение ассоциировано с потерей воды около 50%, как измерено по массе сосудов перед прекращением полива и массе сосудов, когда наблюдали морфологию курчавости листьев в неполивавшихся растениях. Считается, что растения находятся в условиях засухи, когда листья, обнаруживали завядание, о котором свидетельствует закручивание внутрь (V-форма) этого листа. Считается, что этот уровень стресса является формой сублетального стресса. Как только каждое растение демонстрировало индукцию засухи, определенную выше, это растение разрушали для получения проб как корня, так и листа.
[0167] Кроме засухи, растения стадии V3, трансформированные pMON132037 (EXP-SETit.Ubq1:1:10, SEQ ID NO: 119), pMON131957 (EXP-SETit.Ubq1:1:11, SEQ ID NO: 125), pMON131958 (EXP-Sv.Ubq1:1:11, SEQ ID NO: 130) и pMON132974 (EXP-Sb.Ubq7:1:2, SEQ ID NO: 157), подвергали условиям холода для определения, демонстрировали ли регуляторные элементы индуцированную холодом экспрессию GUS. Целые растения анализировали на индукцию экспрессии GUS под холодным стрессом в стадии V3. Растения кукурузы стадии V3 подвергали температуре 12ºC в камере для выращивания в течение 24 часов. Растения в камере для выращивания росли под интенсивностью дневного света 800 микромолей на квадратный метр в секунду со световым циклом десять часов дневного света и четырнадцать часов темноты. После подвергания холоду, брали пробы тканей листа и корня для количественной экспрессии GUS.
[0168] Экспрессию GUS измеряли, как описано выше. Средняя экспрессия F1 GUS, определенная для каждой пробы ткани, представлена в таблицах 42 и 43 ниже.
Средняя экспрессия F1 GUS в растениях, трансформированных pMON142864 и pMON142865
Средняя экспрессия F1 GUS в растениях, трансформированных pMON132037, pMON131957, pMON131958 и pMON132974
[0169] В растениях F1 кукурузы, уровни экспрессии GUS в различных тканях, из которых брали пробы, различались среди убиквитиновых EXP-последовательностей. Хотя все из EXP-последовательностей демонстрировали способность управлять экспрессией трансгена GUS в стабильно трансформированных растениях F1 кукурузы, каждая EXP-последовательность демонстрировала уникальный паттерн (характер) экспрессии относительно других. Например, экспрессия корней R1 является приблизительно в 2 раза более высокой для EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), чем EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8).
[0170] Экспрессия GUS в развивающемся зародыше семян при 38 DAP является почти в три раза более высокой для EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), чем EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8). Напротив, экспрессия листа и корня в стадии V3 и стадии V8 является приблизительно одинаковой для EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27) и EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8).
[0171] Экспрессия F1 GUS в насыщенных влагой семенах (тканях зародыша и эндосперма) была гораздо более высокой в растениях, трансформированных EXP-Sb.Ubq7:1:2 (SEQ ID NO: 157), чем в растениях, трансформированных EXP-SETit.Ubq1:1:10 (SEQ ID NO: 119), EXP-SETit.Ubq1:1:11 (SEQ ID NO: 125) и EXP-Sv.Ubq1:1:11 (SEQ ID NO: 130). Засуха вызывала увеличение экспрессии корней V3 в растениях, трансформированных EXP-SETit.Ubq1:1:10 (SEQ ID NO: 119), EXP-SETit.Ubq1:1:11 (SEQ ID NO: 125), EXP-Sv.Ubq1:1:11 (SEQ ID NO: 130) и EXP-Sb.Ubq7:1:2 (SEQ ID NO: 157), но увеличивала только экспрессию листьев в растениях, трансформированных EXP-SETit.Ubq1:1:11 (SEQ ID NO: 125), EXP-Sv.Ubq1:1:11 (SEQ ID NO: 130) и EXP-Sb.Ubq7:1:2 (SEQ ID NO: 157). Эта увеличенная засухой экспрессия V3 была наивысшей с использованием EXP-SETit.Ubq1:1:11 (SEQ ID NO: 125). Экспрессия пыльцы была также гораздо более высокой в растениях, трансформированных EXP-Sb.Ubq7:1:2 (SEQ ID NO: 157), чем в растениях, трансформированных EXP-SETit.Ubq1:1:10 (SEQ ID NO: 119), EXP-SETit.Ubq1:1:11 (SEQ ID NO: 125) и EXP-Sv.Ubq1:1:11 (SEQ ID NO: 130). Экспрессия в междоузлии R1 была наивысшей с EXP-SETit.Ubq1:1:10 (SEQ ID NO: 119) и EXP-Sv.Ubq1:1:11 (SEQ ID NO: 130) и наименьшей в растениях, трансформированных EXP-SETit.Ubq1:1:11 (SEQ ID NO: 125).
[0172] Каждая EXP-последовательность демонстрировала способность управлять экспрессией трансгена в стабильно трансформированных растениях кукурузы. Однако, каждая EXP-последовательность имела паттерн экспрессии для каждой ткани, который был уникальным, и дает возможность отобрать EXP-последовательность, которая будет лучше всего обеспечивать экспрессию конкретного трансгена в зависимости от стратегии экспрессии тканей, необходимой для достижения желаемых результатов. Этот пример демонстрирует, что EXP-последовательности, выделенные из гомологичных генов, не обязательно ведут себя эквивалентно в трансформированном растении, и что экспрессия может быть определена только посредством эмпирического исследования свойств для каждой EXP-последовательности и не может быть предсказана на основе гомологии гена, из которого был получен этот промотор.
Пример 13: Анализ регуляторных элементов, управляющих СР4 в трансгенной кукурузе.
[0173] Растения кукурузы трансформировали экспрессирующими векторами растений, содержащими EXP-последовательности, управляющие экспрессией трансгена СР4, и полученные растения анализировали на экспрессию белка СР4.
[0174] EXP-последовательности EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8), EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Sv.Ubq1:1:9 (SEQ ID NO: 133) и EXP-Zm.UbqM1:1:7 (SEQ ID NO: 141) клонировали в бинарные плазмидные конструкты для трансформации растений. Полученные векторы содержали правый граничный район из Agrobacterium tumefaciens, убиквитиновую EXP-последовательность, функционально связанную 5’ с нацеленной на пластиду кодирующей последовательностью устойчивой к глифозату EPSPS (CP4, US RE39247), функционально связанной 5’ с T-AGRtu.nos-1:1:13 (SEQ ID NO: 127) 3’-UTP и левый граничный район из A. tumefaciens. Таблица 44 ниже показывает плазмидные конструкты, используемые для трансформации кукурузы, и соответствующие EXP-последовательности.
Плазмидные конструкты СР4 и соответствующие EXP-последовательности, используемые для трансформации кукурузы
[0175] Полученные плазмиды использовали для трансформации растений кукурузы. Трансформированные растения отбирали на одну или две копии инсертированной Т-ДНК и выращивали в оранжерее. Пробы отобранных тканей брали из трансформированных R0 растений в конкретных стадиях развития и уровни белка СР4 измеряли в этих тканях с использованием анализа ELISA СН4. Средняя экспрессия СР4, наблюдаемая для каждой трансформации, представлена в Таблицах 45 и 46 ниже и графически на фигуре 7.
Средняя экспрессия СР4 листа и корня в трансформированных растениях R0 кукурузы
Средняя экспрессия СР4 в репродуктивной ткани и развивающихся семенах в трансформированных растениях R0 кукурузы
[0176] Как видно в Таблицах 45 и 46, каждая из EXP-последовательностей EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8), EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Sv.Ubq1:1:9 (SEQ ID NO: 133) и EXP-Zm.UbqM1:1:7 (SEQ ID NO: 141) была способна управлять экспрессией СР4 во всех тканях, взятых в виде проб из трансформированных растений R0. Более высокая экспрессия СР4 в корне и листе трансформантов, содержащих EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8) и EXP-ERIra.Ubq1: 1:8 (SEQ ID NO: 27), управляющие CP4, чем в присутствии EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), управляющей CP4, может быть связана с уровнем вегетативной устойчивости к применению глифозата, как наблюдалось для этих популяций трансформантов (см. Пример 14 ниже).
[0177] Каждая EXP-последовательность проявляла уникальный паттерн экспрессии относительно уровня экспрессии для каждой используемой в качестве пробы ткани. Например, в то время как экспрессия СР4 в листе, корне и метелке была сходной для EXP-последовательностей, EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8) и EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), экспрессия в шелке с использованием EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8) была равна половине уровня экспрессии, управляемой ERIra.Ubq1:1:8 (SEQ ID NO: 21). Это может быть выгодным для экспрессии трансгенов, в которых конститутивная экспрессия является желаемой, но предпочтительной была бы более низкая экспрессия в ткани шелка. Эти EXP-последовательности демонстрируют уникальные паттерны конститутивной экспрессии СР4 в трансформированных растениях R0 кукурузы.
[0178] Эти трансформированные растения R0 кукурузы скрещивали с нетрансгенным сортом LH244 для получения семян F1. Полученные семена генерации F1 анализировали на расщепление трансгенной кассеты и растения, гетерозиготные в отношении этой кассеты СР4, отбирали для анализа экспрессии СР4. Семена выращивали в оранжерее, и получали две группы растений, причем одна группа опрыскивалась глифазатом, тогда как другая оставалась неопрыснутой. Экспрессию СР4 анализировали в отобранных тканях с использованием стандартного анализа на основе ELISA. Средняя экспрессия СР4 показана в таблицах 47 и 48 ниже.
Средняя экспрессия СР4 в трансформированных растениях F1 кукурузы
[0179] Как можно видеть в Таблице 47 выше, экспрессия СР4 в листе и корне была более высокой в трансформантах F1, трансформированных pMON141619 (EXP-ANDge.Ubq1:1:8, SEQ ID NO: 5) и pMON142862 (EXP-ERIra.Ubq1:1:8, SEQ ID NO: 27), чем в трансформантах, трансформированных pMON129221 (EXP-Cl.Ubq1:1:10, SEQ ID NO: 98). Экспрессия в ткани пыльников была сходной для всех трех EXP-последовательностей, тогда как экспрессия в шелке была наивысшей с использованием EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27). Экспрессия в развивающемся зародыше (21 DAP) была наивысшей в трансформантах, содержащих EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27) и EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), управляющие СР4.
Средняя экспрессия СР4 в трансформированных растениях F1 кукурузы
[0180] Как можно видеть в Таблицах 47-48 выше, экспрессия CR4 была более низкой во всех тканях трансформантов F1 с pMON129205 (EXP-Sv.Ubq1:1:9, SEQ ID NO: 133), чем экспрессия трансформантов, трансформированных pMON141619 (EXP-ANDge.Ubq1:1:8, SEQ ID NO: 8), pMON142862 (EXP-ERIra.Ubq1:1:8, SEQ ID NO: 27) и pMON129221 (EXP-Cl.Ubq1:1:10, SEQ ID NO: 98).
[0181] Эти уникальные паттерны экспрессии, придаваемые каждой из анализированных EXP-последовательностей, обеспечивают возможность получения трансгенного растения, в котором экспрессия может быть тонко отрегулирована для получения малых корректировок в экспрессии трансгена для оптимальной производительности или эффективности. Кроме того, эмпирическое тестирование этих EXP-последовательностей, управляющих экспрессией различных трансгенов, может давать результаты, в которых одна конкретная EXP-последовательность является наиболее подходящей для экспрессии конкретного трансгена или класса трансгенов, тогда как обнаружено, что другая EXP-последовательность является наилучшей для другого трансгена или класса трансгенов.
Пример 14: Анализ вегетативной толерантности к глифозату в R0 растениях трансгенной кукурузы.
[0182] Растения кукурузы трансформировали экспрессирующими векторами растений, содержащими EXP-последовательности, управляющие трансгеном СР4, и полученные растения оценивали на вегетативную и репродуктивную толерантность к глифозату.
[0183] F1 растения трансформированной кукурузы, описанные в Примере 13 выше, трансформированные pMON141619, pMON142862, pMON129221, pMON129205 и pMON129212 и состоящие из EXP-последовательностей EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8), EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), EXP-Sv.Ubq1:1:9 (SEQ ID NO: 133) и EXP-Zm.UbqM1:1:7 (SEQ ID NO: 141), соответственно управляющих CP4, оценивали как на вегетативную, так и на репродуктивную толерантность при опрыскивании глифозатом. Десять F1 растения для каждого события делили на две группы, первую группу, состоящую из пяти растений, которые получали опрыскивание глифозатом, и V4 и V8-стадию развития; и вторую группу из пяти растений, которые оставались не опрысканными (т.е. контроль). Глифозат применяли нанесением спрея для листьев разбросного посева с использованием Roundup WeatherMax® при скорости нанесения 1,5 a.e./акр (a.e. кислотный эквивалент). После семи - десяти дней, листья каждого растения оценивали на повреждение. Вегетативную толерантность (Veg Tol в Таблице 49) оценивали сравнением неопрысканных и опрысканных растений для каждого события (случая) и шкалу рейтинга разрушения использовали для обеспечения конечного рейтинга для вегетативной толерантности (T = толерантный, NT = не толерантный). Кроме того, набор семян анализировали для всех растений в каждом случае. Измерения наборов семян между контрольными растениями и опрысканными растениями сравнивали, и присваивание репродуктивной толерантности (Repro Tol в Таблице 49) давалось для каждого случая на основе процентного набора семян опрысканных растений относительно контролей (Т = толерантный, NT = нетолерантный). Таблица 49 ниже показывает рейтинги вегетативной толерантности и репродуктивной толерантности для каждого опрысканного события в стадии V4 и V8. Буква “T” обозначает толерантные и “NT” обозначает нетолерантные.
Рейтинги повреждения листьев индивидуальных событий трансформированной кукурузы в стадии V4 и V8
[0184] Как видно из Таблицы 49 выше, все анализированные трансформированные события (случаи), включающие в себя кассеты СР4, содержащие EXP-последовательности EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8), EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27) и EXP-Zm.UbqM1:1:7 (SEQ ID NO: 141), демонстрировали полную вегетативную толерантность, основанную на рейтингах разрушения, которые не превышали оценки 10. Четыре события из девяти, содержащие EXP-ANDge.Ubq1:1:8 (SEQ ID NO: 8), и шесть событий из девяти, содержащие EXP-ERIra.Ubq1:1:8 (SEQ ID NO: 27), были как вегетативно, так и репродуктивно толерантными к нанесению глифозата. В противоположность этому, случаи, включающие в себя EXP-Cl.Ubq1:1:10 (SEQ ID NO: 98), были либо вегетативно, либо репродуктивно толерантными, но не в обоих случаях. Только одно событие, включающее в себя EXP-Sv.Ubq1:1:9 (SEQ ID NO: 133), демонстрировало вегетативную толерантность, и ни одно из тестированных событий не было репродуктивно толерантным. Все события, включающие в себя EXP-Zm.UbqM1:1:7 (SEQ ID NO: 141), демонстрировали вегетативную толерантность, но оценивание репродуктивной толерантности все еще находится в развитии.
Пример 15: Анализ экспрессии с использованием различных последовательностей 3’-концевой интрон/экзон-границы сплайсинга.
[0185] Протопластные клетки листьев кукурузы и пшеницы трансформировали экспрессирующими конструктами растений, содержащими EXP-последовательности, управляющие экспрессией GUS, которые содержат один и тот же промотор и лидер, но имеют отличающиеся 3’-концевые нуклеотиды после последовательности интрон/экзон-границы сплайсинга, 5’-AG-3’, для наблюдения, влияет ли на экспрессию небольшое изменение в последовательности. Экспрессию также сравнивали с экспрессией двух конститутивных контрольных плазмид.
[0186] Конструируют экспрессионные конструкты растений, содержащие экспрессионную кассету GUS. Полученные векторы состоят из промотора убиквитина Coix lacryma-jobi, P-Cl.Ubq1-1:1:1 (SEQ ID NO: 80), функционально связанного 5’ (слева) с лидерной последовательностью, L-Cl.Ubq1-1:1:1 (SEQ ID NO: 81), функционально связанной 5’ с интронным элементом, показанным в Таблице 50 ниже, каждый из которых, содержит различающиеся нуклеотиды на самом 3’-конце непосредственно после последовательности интрон/экзон-границы сплайсинга 5’-AG-3’, функционально связанной 5’ (слева) с кодирующей GUS последовательностью, которая функционально связана 5’ (слева) с T-AGRtu.nos-1:1:13 (SEQ ID NO: 127) 3’-UTR. Таблица 50 ниже показывает экспрессионные конструкты растений и соответствующую 3’-концевую последовательность.
Экспрессионные конструкты растений, интроны и 3’-концевая последовательность после последовательности интрон/экзон-границы сплайсинга 5’-AG-3’
[0187] Протопласты кукурузы и пшеницы трансформировали, как описано ранее и анализировали на экспрессию GUS и люциферазы. Таблица 51 ниже показывает средние величины GUS и RLuc для экспрессии протопластов как кукурузы, так и пшеницы.
Средние величины GUS и RLuc для протопластных клеток кукурузы и пшеницы
[0188] Величины GUS/RLuc для каждой убиквитиновой EXP-последовательности Coix lacryma-jobi из таблицы 46 выше использовали для нормализации экспрессии относительно двух конститутивных контролей EXP-Os.Act1:1:9 (SEQ ID NO: 179) и EXP-CaMV.35S-enh+Ta.Lhcbl+Os.Act1:1:1 (SEQ ID NO: 163), и они представлены в таблице 52 ниже.
Нормализованные величины экспрессии убиквитиновых EXP-последовательностей Coix lacryma-jobi относительно EXP-Os.Actl:l:9 (SEQ ID NO: 179) и EXP-CaMV.35S-S-enh+Ta.Lh1+Os.Act1:1:1 (SEQ ID NO: 163)
[0189] Как показано в Таблице 52 выше, каждая из убиквитиновых EXP-последовательностей Coix lacryma-jobi обеспечивала экспрессию, которая была большей, чем любой конститутивный контроль, как в кукурузе, так и в пшенице. Экспрессия в протопластах кукурузы была относительно сходной для всех из убиквитиновых EXP-последовательностей Coix. Экспрессия в пшенице была несколько более вариабельной. Применение различных 3’-концевых нуклеотидов после последовательности интрон/экзон-границы сплайсинга, 5’-AG-3’, по-видимому, не влияла существенно на экспрессию GUS, за исключением GUS, управляемого EXP-Cl.Ubq1:1:20 (SEQ ID NO: 102). EXP-Cl.Ubq1:1:20 содержит 3’-концевые нуклеотидные последовательности, 5’-GAC-3’, после последовательности 5’-AG-3’ интрон/экзон-границы сплайсинга и заставляет экспрессию слегка падать относительно других убиквитиновых EXP-последовательностей Coix. Оценивание полученной сплайсированной мессенджер-РНК показало, что приблизительно 10% мРНК, экспрессированой с использованием EXP-Cl.Ubq1:1:20 (SEQ ID NO: 102) для управления экспрессией GUS, были неправильно сплайсированы. мРНК, полученная из экспрессии GUS с использованием других убиквитиновых EXP-последовательностей Coix, по-видимому, процессировалась правильно. Этот эксперимент обеспечивает доказательство того, что любой из 3’-концевых нуклеотидов для любого из интронных вариантов, представленных в таблице 2 Примера 1, за исключением 3’-концевой последовательности 5’-GAC-3’, которая, как было обнаружено, ассоциирована только с интронным элементом, I-Cl.Ubq1-1:1:9 (SEQ ID NO: 103), должен быть подходящим для применения в трансгенной экспрессионной кассете без значимой потери активности и процессинга.
Пример 16: Энхансеры, полученные из регуляторных элементов.
[0190] Энхансеры получают из обеспеченных здесь промоторных элементов, представленных как SEQ ID NO: 2, 6, 9, 11, 13, 15, 17, 19, 23, 26, 28, 30, 32, 34, 38, 40, 42, 46, 50, 56, 60, 64, 66, 70, 74, 76, 78, 80, 84, 86, 88, 91, 96 и 135. Этот энхансерный элемент может состоять из одного или нескольких цис-регуляторных элементов, которые, при функциональном связывании 5’ (слева) или 3’ (справа) с промоторным элементом или при функциональном связывании 5’ или 3’ с дополнительными энхансерными элементами, которые функционально связаны с промотором, могут усиливать или модулировать экспрессию трансгена или обеспечивать экспрессию трансгена в конкретном типе клеток или органе растений или в конкретной временной точке в развитии или циркадном ритме. Энхансеры готовят удалением ТАТА-бокса или функционально сходных элементов и любой последовательности справа от промоторов, что делает возможной инициацию транскрипции от обеспеченных здесь промоторов, как описано выше, в том числе их фрагментов, в которых удалены ТАТА-бокс или функционально сходные элементы и последовательность справа от ТАТА-бокса. Энхансерный элемент, E-Cl.Ubq1-1:1:1 (SEQ ID NO: 89), который произведен из промоторного элемента, P-Cl.Ubq1-1:1:1, обеспечен здесь для демонстрации энхансеров, полученных из промоторного элемента.
[0191] Энхансерные элементы могут быть произведены из обеспеченных здесь промоторных элементов и клонированы с использованием способов, известных в данной области, чтобы они были функционально связаны 5’ или 3’ с промоторным элементом, или функционально связаны 5’ или 3’ с дополнительными энхансерными элементами, которые функционально связаны с промотором. Альтернативно, энхансерные элементы клонируют, с использованием способов, известных в данной области, чтобы они были функционально связаны с одной или несколькими копиями энхансерных элементов, которые функционально связаны 5’ или 3’ с промоторным элементом, или функционально связаны 5’ или 3’ с дополнительными энхансерными элементами, которые функционально связаны с промотором. Энхансерные элементы могут быть также клонированы, чтобы быть функционально связанными 5’ или 3’ с промоторным элементом, произведенным из организма отличающегося рода, или функционально связанными 5’ или 3’ с дополнительными энхансерными элементами, произведенными из организмов другого рода или организма того же самого рода, которые функционально связаны с промотором, произведенным из организма либо того же самого рода, либо отличающегося рода, с получением химерного регуляторного элемента. Трансформирующий вектор растений для экспрессии GUS конструируют с использованием способов, известных в данной области, сходный с конструктами, описанными в предыдущих примерах, в которых полученные экспрессирующие векторы растений содержат правый граничный район из A. tumefaciens, первую трансгенную кассету для тестирования регуляторного или химерного регуляторного элемента, состоящего из регуляторного или химерного регуляторного элемента, функционально связанного с интроном, произведенным из белка теплового шока HSP70 Z. mays (I-Zm.DnaK-1:1:1 SEQ ID NO: 144), или любого из интронов, представленных здесь, или любого другого интрона, функционально связанного с кодирующей последовательностью для β-глюкуронидазы (GUS), которая либо имеет процессируемый интрон (GUS-2, SEQ ID NO: 160), либо не имеет интрона (GUS-1, SEQ ID NO: 159), функционально связанной с 3’-UTR нопалинсинтазой из A. tumefaciens (T-AGRtu.nos-1:1:13, SEQ ID NO: 161) или 3’-UTR из гена белка транспорта липидов риса (T-Os.LTP-1:1:1, SEQ ID NO: 175); вторую трансгенную кассету для отбора, используемую для селекции трансформированных клеток растений, которая придает устойчивость к гербициду глизофату (управляемую промотором Актина 1 риса), или, альтернативно, антибиотику канамицину (управляемому промотором Актина 1 риса), и левый граничный район из A. tumefaciens. Полученные плазмиды используются для трансформации растений кукурузы или растений других родов способами, описанными выше, или другими опосредованными Agrobacterium способами или способами бомбардировки частицами, известными в данной области. Альтернативно, протопластные клетки, полученные из кукурузы или растений другого вида, трансформировали с использованием способов, известных в данной области, для выполнения транзиторных анализов.
[0192] Экспрессию GUS, управляемую регуляторным элементом, содержащим один или несколько энхансеров, оценивают в стабильных или транзиторных анализах растений для определения действий энхансерного элемента на экспрессию трансгена. Модификации в отношении одного или нескольких энхансерных элементов или дупликацию одного или нескольких энхансерных элементов выполняют на основе эмпирического экспериментирования и регуляции экспрессии полученного гена, которую наблюдают с использованием композиции каждого регуляторного элемента. Изменение относительного положения одного или нескольких энхансеров в полученном регуляторном или химерном регуляторном элементе может влиять на транскрипционную активность или специфичность регуляторного или химерного регуляторного элемента и определяется эмпирически для идентификации лучших энхансеров для желаемого профиля экспрессии трансгенов в растении кукурузы или растении другого рода.
Пример 17: Анализ интронного усиления активности GUS с использованием полученных из растения протопластов.
[0193] Интрон отбирают на основе экспериментирования и сравнения, с не имеющим интронов вектором, для эмпирического отбора интрона и конфигурации в аранжировке векторного элемента Т-ДНК для оптимальной экспрессии трансгена. Например, в экспрессии гена устойчивости к гербицидам, такого как CP4, который может придавать устойчивость к глифозату, желательно иметь экспрессию трансгена в репродуктивных тканях, а также вегетативных тканях для предотвращения потери выхода при применении этого гербицида. Интрон в этом случае мог бы отбираться по его способности, при функциональном связывании с конститутивным промотором, усиливать экспрессию придающего устойчивость к гербицидам трансгена, в частности, в репродуктивных клетках и тканях трансгенного растения, с обеспечением, таким образом, как вегетативной, так и репродуктивной толерантности к трансгенному растению при опрыскивании этим гербицидом. В большинстве генов убиквитина, 5’-UTR состоит из лидера, который имеет интронную последовательность, заделанную в нем. Таким образом, экспрессионные элементы, полученные из таких генов, анализируют с использованием полного 5’-UTR, содержащего промотор, лидер и интрон. Для достижения отличающихся профилей экспрессии или для модуляции уровня экспрессии трансгена, интрон из такого экспрессионного элемента может быть удален или заменен гетерологичным интроном.
[0194] Интроны, представленные здесь как SEQ ID NO: 4, 7, 21, 24, 36, 44, 48, 52, 54, 58, 62, 68, 72, 82, 92, 94, 101, 103, 105, 107, 109, 111, 113, 118, 120, 122, 127, 129, 131, 138, 140, 142, 144, 146, 148, 150, 152, 154, 156, 158 и 182, идентифицируют с использованием контигов геномной ДНК в сравнении с tag-кластерами маркерной экспрессирующейся последовательности (EST) или контигами кДНК для идентификации последовательностей экзонов и интронов в этой геномной ДНК. Кроме того, последовательности 5’-UTR или лидера также используют для определения интрон/экзон-границы сплайсинга одного или нескольких интронов при условиях, когда эта последовательность гена кодирует лидерную последовательность, которая прерывается одним или несколькими интронами. Интроны клонируют с использованием способов, известных в данной области, в трансформирующий вектор растения для функционального связывания 3’ (справа) либо со вторым лидерным фрагментом, либо с кодирующими последовательностями, например, как показано в двух трансгенных кассетах, представленных на фиг. 1.
[0195] Таким образом, например, первая возможная трансгенная кассета (Конфигурация 1 Трансгенной Кассеты на фиг. 8) состоит из промоторного или химерного промоторного элемента [A], функционально связанного 5’ (слева) с лидерным элементом [B], функционально связанным 5’ с элементом тест-интрона [C], функционально связанным с кодирующей последовательностью [D], которая функционально связана с 3’-UTR элементом [E]. Альтернативно, вторая возможная трансгенная кассета (Конфигурация 2 Трансгенной Кассеты на фиг. 8) состоит из промоторного или химерного промоторного элемента [F], функционально связанного 5’ с первым лидерным элементом или фрагментом первого лидерного элемента [G], функционально связанным 5’ с элементом тест-интрона [H], функционально связанным 5’ со вторым лидерным элементом или фрагментом первого лидерного элемента [I], функционально связанным с кодирующим районом [J], который функционально связан с 3’-UTR-элементом [K]. Далее, третья возможная трансгенная кассета (Конфигурация 3 Трансгенной Кассеты на фиг. 8) состоит из промоторного или химерного промоторного элемента [L], функционально связанного с лидерным элементом [M], функционально связанным 5’ с первым фрагментом элемента кодирующей последовательности [N], функционально связанным 5’ с элементом интронного элемента [O] функционально связанным 5’ со вторым фрагментом элемента кодирующей последовательности [P], который функционально связан с 3’-элементом [Q]. Конструируют Конфигурацию 3 Трансгенной Кассеты для возможности сплайсинга этого интрона таким образом, что образуется полная открытая рамка считывания без смещения рамки между первым и вторым фрагментом кодирующей последовательности.
[0196] Первые 6 нуклеотидов на 5’-конце и последние 6 нуклеотидов на 3’-конце интронов, представленных как SEQ ID NO: 4, 7, 21, 24, 36, 44, 48, 52, 54, 58, 62, 68, 72, 82, 92, 94, 101, 103, 105, 107, 109, 111, 113, 118, 120, 122, 127, 129, 131, 138, 140, 142, 144, 146, 148, 150, 152, 154, 156, 158 и 182, представляют нуклеотиды до и после интрон/экзон-границы сплайсинга, соответственно. Эти короткие 6-нуклеотидные последовательности, могут быть, например, модифицированы наличием дополнительной добавленной последовательности, (т.е. нативной или искусственной) для облегчения клонирования этого интрона в трансформирующий вектор растения, пока сохраняются эти первый и второй нуклеотиды из 5’-конца (GT) и эти четвертый и пятый нуклеотиды из 3’-конца (AG) SEQ ID NO: 4, 7, 21, 24, 36, 44, 48, 52, 54, 58, 62, 68, 72, 82, 92, 94, 101, 103, 105, 107, 109, 111, 113, 118, 120, 122, 127, 129, 131, 138, 140, 142, 144, 146, 148, 150, 152, 154, 156, 158 и 182 с сохранением, таким образом, интрон/экзон-границы сплайсинга этого интрона. Как обсуждалось выше, может быть предпочтительным избежание использования нуклеотидной последовательности АТ или нуклеотида А непосредственно перед 5’-концом сайта сплайсинга (GT) и нуклеотида G или нуклеотидной последовательности TG, соответственно, непосредственно после 3’-конца сайта сплайсинга (AG) для элиминирования потенциала нежелательных стартовых кодонов от образования во время процессинга мессенджер-РНК в конечный транскрипт. Таким образом, может быть модифицирована последовательность около 5’- или 3’-концевых сайтов сплайсинга этого интрона.
[0197] Эти интроны анализируют на эффект усиления посредством способности увеличения экспрессии в транзиторном анализе или стабильном анализе растений. Для транзиторного анализа интронного усиления конструируют базовый вектор растений с использованием способов, известных в данной области. Интрон клонируют в этот базовый вектор растений, который содержит экспрессионную кассету, состоящую из конститутивного промотора, такого как промотор вируса мозаики цветной капусты, P-CaMV.35S-enh-1:1:9 (SEQ ID NO: 176), функционально связанный 5’ (слева) с лидерным элементом, L-CaMV.35S-1:1:15 (SEQ ID NO: 177), функционально связанным 5’ с тест-интронным элементом (например, одной из SEQ ID NO: 4, 7, 21, 24, 36, 44, 48, 52, 54, 58, 62, 68, 72, 82, 92, 94, 101, 103, 105, 107, 109, 111, 113, 118, 120, 122, 127, 129, 131, 138, 140, 142, 144, 146, 148, 150, 152, 154, 156, 158 и 182), функционально связанным с кодирующей последовательностью для β-глюкуронидазы (GUS), которая либо имеет процессируемый интрон (GUS-2, SEQ ID NO: 160), либо не имеет интрона (GUS-1, SEQ ID NO: 159), функционально связанной с 3’-UTR нопалинсинтазы из A tumefaciens (T-AGRtu.nos-1:1:13, SEQ ID NO: 161). Протопластные клетки, полученные из ткани кукурузы или ткани растения другого рода, трансформируют базовым вектором растений и контрольными векторами люциферазы, как описано ранее в Примере 2 выше, и анализируют на активность. Для сравнения относительной способности этого интрона усиливать экспрессию, величины GUS выражали в виде отношения GUS к активности люциферазы и сравнивали с уровнями, придаваемыми конструктом, содержащим конститутивный промотор, связанный с известным интроном-стандартом, таким как интрон, полученный из белка теплового шока HSP70 Zea mays, I-Zm.DnaK-1:1:l (SEQ ID NO: 178), а также конструктом, содержащим конститутивный промотор, но без интрона, функционально связанного с этим промотором.
[0198] Для стабильного анализа интронов, представленных как SEQ ID NO: 4, 7, 21, 24, 36, 44, 48, 52, 54, 58, 62, 68, 72, 82, 92, 94, 101, 103, 105, 107, 109, 111, 113, 118, 120, 122, 127, 129, 131, 138, 140, 142, 144, 146, 148, 150, 152, 154, 156, 158 и 182, трансформирующий вектор растений для экспрессии GUS конструируют сходным образом с конструктами, описанными в предыдущих примерах, в которых полученные экспрессирующие векторы растений содержат правый граничный район из A. tumefaciens, первую трансгенную кассету для тестирования интрона, состоящую из конститутивного промотора, такого как промотор вируса мозаики цветной капусты, P-CaMV.35S-enh-1:1:9 (SEQ ID NO: 176), функционально связанный 5’ с лидерным элементом, L-CaMV.35S-1:1:15 (SEQ ID NO: 177), функционально связанным 5’ с элементом тест-интрона, обеспеченным здесь, функционально связанным с кодирующей последовательностью для β-глюкуронидазы (GUS), которая либо имеет процессируемый интрон (GUS-2, SEQ ID NO: 160), либо не имеет интрона (GUS-1, SEQ ID NO: 158), функционально связанного с 3’-UTR нопалинсинтазы из A. tumefaciens (T-AGRtu.nos-1:1:13, SEQ ID NO: 161); вторую трансгенную кассету для отбора (селекции) трансформированных клеток растений, которая придает устойчивость к глифозату (управляемую промотором Актина 1 риса), или, альтернативно, к антибиотику канамицину (управляемую промотором Актина 1 риса), и левый граничный район из A. tumefaciens. Полученные плазмиды используют для трансформации растений кукурузы или растений другого рода, описанными выше способами или опосредуемыми Agrobacterium способами, известными в данной области. Трансформанты с одной копией или трансформанты с малым числом копий отбирали для сравнения с трансформированными растениями с одной копией или имеющими малое число копий, трансформированными трансформирующим вектором растений, идентичным с тест-вектором, но без тест-интрона, для определения, обеспечивает ли этот тест-интрон опосредуемый интроном усиливающий эффект.
[0199] Любой из интронов, представленных как SEQ ID NO: 4, 7, 21, 24, 36, 44, 48, 52, 54, 58, 62, 68, 72, 82, 92, 94, 101, 103, 105, 107, 109, 111, 113, 118, 120, 122, 127, 129, 131, 138, 140, 142, 144, 146, 148, 150, 152, 154, 156, 158 и 182, может быть модифицирован рядом способов, таких как делетирование фрагментов в интронной последовательности, которые могут уменьшать экспрессию или дупликацию фрагментов с интроном, который может усиливать экспрессию. Кроме того, последовательности в интроне, которые могут влиять на специфичность экспрессии в отношении либо конкретных типов клеток, либо тканей и органов, могут быть дуплицированы или изменены для влияния на экспрессию и паттерны экспрессии этого трансгена. Кроме того, обеспеченные здесь интроны могут быть модифицированы для удаления любых потенциальных старт-кодонов (ATG), которые могут вызывать прекращение экспрессии непредусмотренных транскриптов из неправильно сплайсированных интронов в виде различных, более длинных или укороченных белков. После эмпирического тестирования интрона или его изменения на основе экспериментирования, этот интрон используют для усиления экспрессии трансгена в стабильно трансформированных растениях, которые могут быть однодольным или двудольным растением любого рода, пока этот интрон обеспечивает усиление этого трансгена. Этот интрон может быть также использован для усиления экспрессии в других организмах, таких как водоросли, грибы или клетки животных, пока этот интрон обеспечивает усиление или затухание или специфичность экспрессии трансгена, с которым он функционально связан.
* * * * * *
[0200] При наличии иллюстрирования и описания принципов настоящего изобретения, для квалифицированных в данной области специалистов должно быть очевидным, что изобретение может быть модифицировано в компоновке и деталях без отклонения от таких принципов. Авторы настоящего изобретения заявляют все модификации, которые находятся в пределах идеи и объема прилагаемой формулы изобретения. Все публикации и опубликованные патентные документы, приведенные здесь, включены посредством ссылки до той же самой степени, как если бы каждая отдельная публикация или заявка на патент была конкретно и отдельно указана как включенная посредством ссылки.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2800430C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2800425C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2800423C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2800424C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2670887C9 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2799433C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2796556C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2675524C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2812893C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2639275C2 |














































































































































































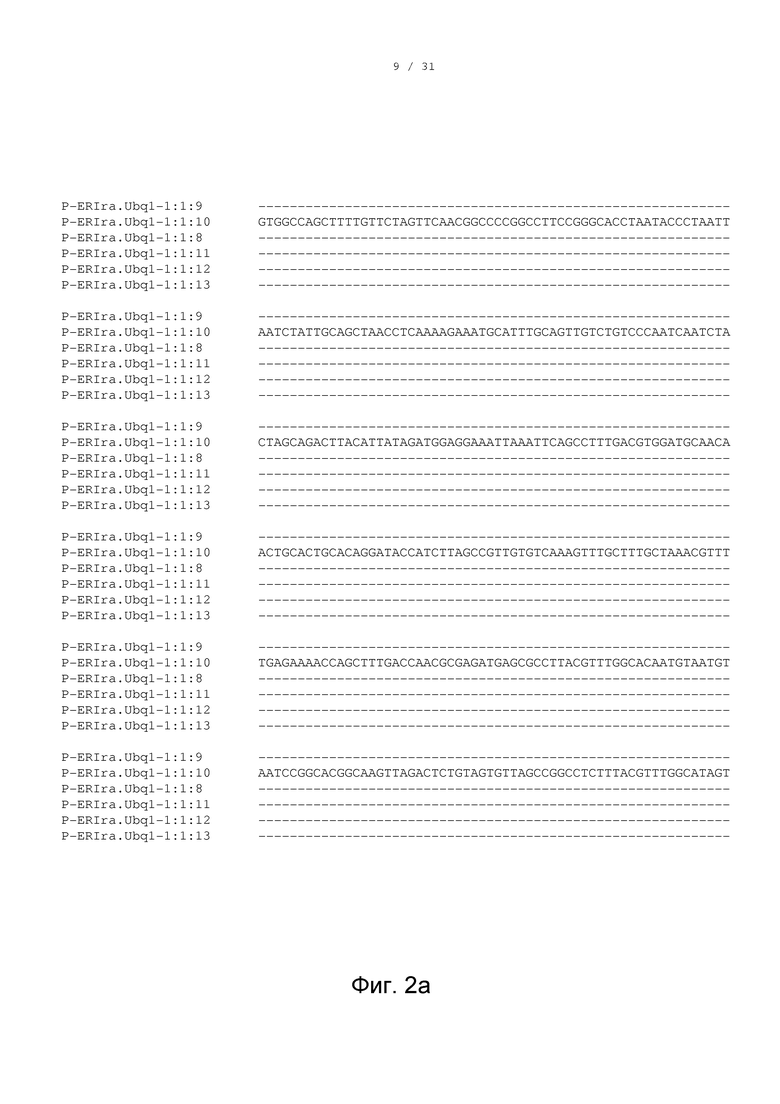





















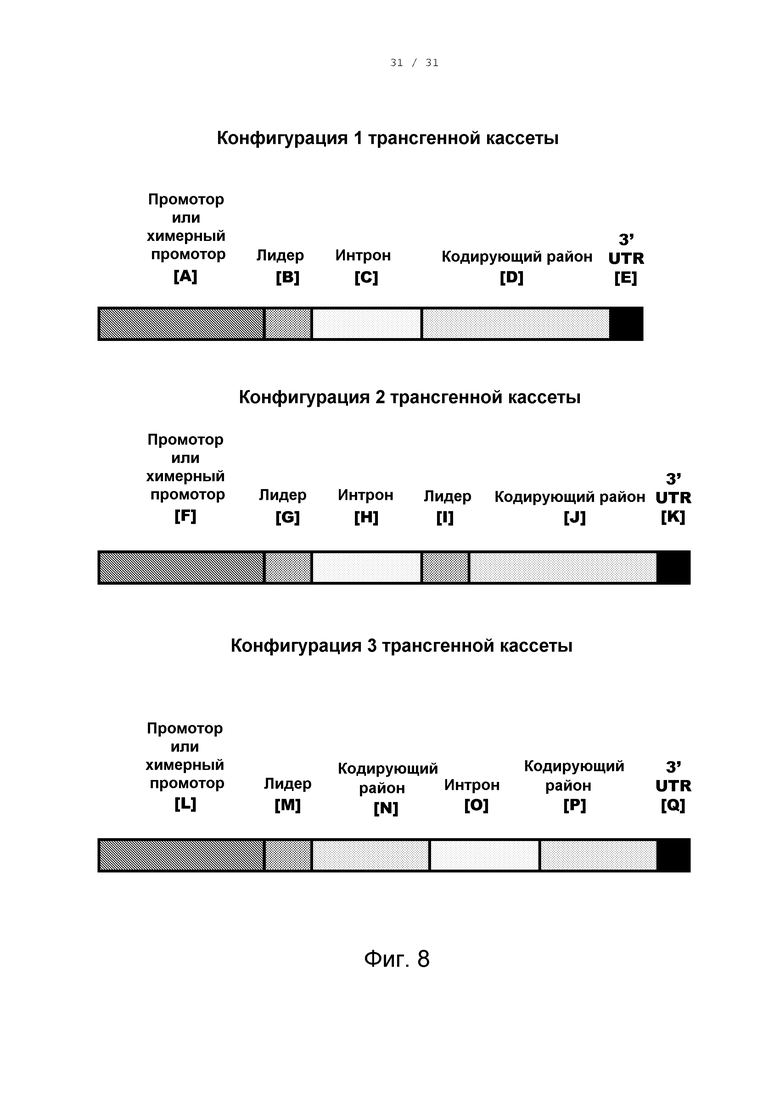
Изобретение относится к области биохимии, в частности к молекуле ДНК для инициации транскрипции гетерологичной транскрибируемой полинуклеотидной молекулы. Также раскрыты трансгенная клетка растений, содержащая указанную молекулу ДНК, трансгенное растение, содержащее указанную молекулу ДНК, часть трансгенного растения, содержащая указанную молекулу ДНК, трансгенное семя, содержащее указанную молекулу ДНК, пищевой продукт, полученный из указанного растения. Раскрыт способ экспрессии транскрибируемой полинуклеотидной молекулы. Изобретение обеспечивает повышенную экспрессию гетерологичной транскрибируемой полинуклеотидной молекулы. 7 н. и 5 з.п. ф-лы, 31 ил., 52 табл., 17 пр.
1. Молекула ДНК для инициации транскрипции гетерологичной транскрибируемой полинуклеотидной молекулы, содержащая последовательность ДНК регуляторного элемента гена, выбранную из группы, состоящей из:
a) последовательности по меньшей мере с 95% идентичностью полноразмерной последовательности любой из SEQ ID NO: 1-17, где последовательность имеет промоторную активность, и
b) последовательности, содержащей любую из SEQ ID NO: 1-17,
где указанная последовательность функционально связана с гетерологичной транскрибируемой полинуклеотидной молекулой.
2. Молекула ДНК по п. 1, где последовательность ДНК проявляет ген-регуляторную активность.
3. Молекула ДНК по п. 1, где гетерологичная транскрибируемая полинуклеотидная молекула содержит представляющий агрономический интерес ген.
4. Молекула ДНК по п. 3, где представляющий агрономический интерес ген придает устойчивость к гербицидам в растениях.
5. Трансгенная клетка растения для повышенной экспрессии гетерологичной транскрибируемой полинуклеотидной молекулы, содержащая молекулу ДНК по п. 1.
6. Трансгенная клетка растения по п. 5, где указанной трансгенной клеткой растения является клетка однодольного растения.
7. Трансгенная клетка растения по п. 5, где указанной трансгенной клеткой растения является клетка двудольного растения.
8. Трансгенное растение, имеющее повышенную экспрессию гетерологичной транскрибируемой полинуклеотидной молекулы, содержащее молекулу ДНК по п. 1.
9. Часть трансгенного растения по п. 8, имеющая повышенную экспрессию гетерологичной транскрибируемой полинуклеотидной молекулы, содержащая молекулу ДНК по п. 1, где указанная часть выбрана из группы, состоящей из листьев, корней, семян и эндосперма.
10. Трансгенное семя для получения растения, имеющего повышенную экспрессию гетерологичной транскрибируемой полинуклеотидной молекулы, где семя содержит молекулу ДНК по п. 1.
11. Пищевой продукт, полученный из трансгенного растения по п. 8.
12. Способ экспрессии транскрибируемой полинуклеотидной молекулы, предусматривающий получение трансгенного растения по п. 8 и культивирование растения, где экспрессируется транскрибируемый полинуклеотид.
| ИНДУЦИРУЕМАЯ ГЕРБИЦИДНАЯ УСТОЙЧИВОСТЬ | 1996 |
|
RU2181380C2 |
| ИСКУССТВЕННЫЙ ПРОМОТОР ДЛЯ ЭКСПРЕССИИ ПОСЛЕДОВАТЕЛЬНОСТЕЙ ДНК В РАСТИТЕЛЬНЫХ КЛЕТКАХ | 2003 |
|
RU2326167C2 |
| US 20100199371 A1 от 05.08.2010 | |||
| WO 2009126470 A2 от 15.10.2009 | |||
| US 0007816581 B2 от 19.10.2010. | |||

