ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Данная заявка утверждает преимущество предварительной заявки Соединенных Штатов № 61/635945, поданной 20 апреля 2012 года, и непредварительной заявки Соединенных Штатов № 13/830403, поданной 14 марта 2013 года, которые в настоящем документе включены в виде ссылки в полном объеме.
ВКЛЮЧЕНИЕ ПЕРЕЧНЯ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Перечень последовательностей, содержащихся в файле с названием «MONS326WO_ST25.txt», который содержит 22 килобайта (как определено в Microsoft Windows®) и был создан 10 апреля 2013 года, подан при этом для электронного представления и включен посредством ссылки в настоящий документ.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к области молекулярной биологии растений и генной инженерии растений. А именно, настоящее изобретение относится к молекулам ДНК, полезным для модуляции экспрессии генов в растениях.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Регуляторные элементы представляют собой генетические элементы, которые регулируют активность генов путем модуляции транскрипции функционально связанной транскрибируемой полинуклеотидной молекулы. Такие элементы включают промоторы, лидерные последовательности, интроны и 3'-нетранслируемые области и являются полезными в области молекулярной биологии растений и генной инженерии растений.
Трансгенным культурам, экспрессирующим трансгены, которые дают преимущество растению в период прорастания в холодных и влажных стрессовых условиях, требуются регуляторные элементы, которые обладают профилями экспрессии в тканях, которые являются наиболее благоприятными для экспрессии таких трансгенов. Такие регуляторные элементы в развивающемся семени должны экспрессироваться в достаточной степени относительно возможности накопления трансгенных продуктов, которые могут действовать быстро в случае, когда семя прорастет в холодных и/или влажных условиях, а также обеспечивать экспрессию на ранних стадиях прорастания и формирования всходов. Соответственно, настоящее изобретение предоставляет новые регуляторные элементы, которые демонстрируют более высокие уровни экспрессии в развивающемся и прорастающем семени и могут быть использованы для управления экспрессией трансгенов, которые обеспечивают преимущество при прорастании в холодных и/или влажных условия.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение предоставляет новые генные регуляторные элементы для применения в растениях. Настоящее изобретение также предоставляет ДНК-конструкции, содержащие регуляторные элементы. Настоящее изобретение также предоставляет трансгенные клетки растений, растения и семена, содержащие регуляторные элементы. Могут предоставляться последовательности, функционально связанные с транскрибируемой полинуклеотидной молекулой. В одном варианте осуществления транскрибируемая полинуклеотидная молекула может быть гетерологичной относительно регуляторной последовательности, предоставленной в настоящем документе. Последовательность регуляторного элемента, предоставленного изобретением, таким образом, может в конкретных вариантах осуществления быть определена как функционально связанная с гетерологичной транскрибируемой полинуклеотидной молекулой. Настоящее изобретение также предоставляет способы изготовления и применения регуляторных элементов, ДНК-конструкции, содержащие регуляторные элементы, и трансгенные клетки растений, растения, семена, содержащие регуляторные элементы, функционально связанные с транскрибируемой полинуклеотидной молекулой.
В одном аспекте изобретение предоставляет молекулу ДНК, содержащую последовательность ДНК, выбранную из группы, состоящей из: (a) последовательности с, по меньшей мере, 85% идентичности последовательности с любой из SEQ ID NO: 1, 2, 3, 4, 6 или 8; (b) последовательности, содержащей любую из SEQ ID NO: 1, 2, 3, 4, 6 или 8; и (c) фрагмента любой из SEQ ID NO: 1, 2, 3, 4, 6 или 8, где фрагмент обладает геннорегуляторной активностью, где указанная последовательность функционально связана с гетерологичной транскрибируемой полинуклеотидной молекулой. В одном варианте осуществления молекула ДНК имеет, по меньшей мере, 90% идентичности последовательности с последовательностью ДНК любой из SEQ ID NO: 1, 2, 3, 4, 6 или 8. В другом варианте осуществления молекула ДНК имеет, по меньшей мере, 95% идентичности последовательности с последовательностью ДНК любой из SEQ ID NO: 1, 2, 3, 4, 6 или 8. В другом варианте осуществления последовательность ДНК обладает геннорегуляторной активностью. В еще одном варианте осуществления гетерологичная транскрибируемая полинуклеотидная молекула содержит ген, представляющий агрономический интерес. В других вариантах осуществления ген, представляющий агрономический интерес, придает растениям устойчивость к гербицидам или устойчивость к вредителям.
В другом аспекте настоящее изобретение предоставляет трансгенную клетку растения, включающую молекулу гетерологичной ДНК, содержащей последовательность, выбранную из группы, состоящей из: (a) последовательности, имеющей, по меньшей мере, 85% идентичности последовательности с любой из SEQ ID NO: 1, 2, 3, 4, 6 или 8; (b) последовательности, содержащей любую из SEQ ID NO: 1, 2, 3, 4, 6 или 8; и (c) фрагмента любой из SEQ ID NO: 1, 2, 3, 4, 6 или 8, где фрагмент имеет геннорегулирующую активность, где указанная последовательность функционально связана с гетерологичной транскрибируемой полинуклеотидной молекулой. В вариантах осуществления трансгенная клетка растения может представлять собой клетку однодольного растения или клетку двудольного растения.
В других вариантах осуществления изобретение предоставляет трансгенное растение или его часть, содержащее молекулу ДНК, выбранную из группы, состоящей из: (a) последовательности, имеющей, по меньшей мере, 85% идентичности последовательности с любой из SEQ ID NO: 1, 2, 3, 4, 6 или 8; (b) последовательности, содержащей любую из SEQ ID NO: 1, 2, 3, 4, 6 или 8; и (c) фрагмента любой из SEQ ID NO: 1, 2, 3, 4, 6 или 8, где фрагмент обладает геннорегулирующей активностью, где указанная последовательность функционально связана с гетерологичной транскрибируемой полинуклеотидной молекулой. В другом варианте осуществления изобретение предоставляет растение-потомок любого поколения такого трансгенного растения или его части, где растение-потомок или его часть содержит молекулу ДНК, как описано выше. В еще одном варианте осуществлении изобретение предоставляет трансгенное семя, где семя содержит молекулу ДНК, как описано выше.
В другом аспекте изобретение предоставляет трансгенную кассету, содержащую группу транскрипционных регулирующих экспрессию элементов, выбранных из группы, состоящей из SEQ ID NO: 1, 6 и 8, где группа транскрипционных регулирующих экспрессию элементов функционально связана с гетерологичной кодирующей последовательностью, которая функционально связана с 3'UTR, выбранной из группы, состоящей из SEQ ID NO: 10, 11, 12 и 13. В варианте осуществления трансгенная кассета содержит группу транскрипционных регулирующих экспрессию элементов, представленную как SEQ ID NO: 1, где группа транскрипционных регулирующих экспрессию элементов функционально связана с гетерологичной кодирующей последовательностью, функционально связанной с 3'UTR, представленной как SEQ ID NO: 10. В другом варианте осуществления трансгенная кассета содержит группу транскрипционных регулирующих экспрессию элементов, представленную как SEQ ID NO: 6, где группа транскрипционных регулирующих экспрессию элементов функционально связана с гетерологичной кодирующей последовательностью, функционально связанной с 3'UTR, выбранной из группы, состоящей из SEQ ID NO: 11 и 12. В еще одном варианте осуществления трансгенная кассета содержит группу транскрипционных регулирующих экспрессию элементов, представленную как SEQ ID NO: 8, где группа транскрипционных регулирующих экспрессию элементов функционально связана с гетерологичной кодирующей последовательностью, функционально связанной с 3'UTR, выбранной из группы, состоящей из SEQ ID NO: 12 и 13. В другом варианте осуществления изобретение предоставляет способ получения товарного продукта, включающий получение трансгенного растения или его части, содержащего последовательность ДНК, выбранную из группы, состоящей из: (a) последовательности, имеющей, по меньшей мере, 85% идентичности последовательности с любой из SEQ ID NO: 1, 2, 3, 4, 6 или 8; (b) последовательности, содержащей любую из SEQ ID NO: 1, 2, 3, 4, 6 или 8; и (c) фрагмента любой из SEQ ID NO: 1, 2, 3, 4, 6 или 8, где фрагмент обладает геннорегулирующей активностью, где указанная последовательность функционально связана с гетерологичной транскрибируемой полинуклеотидной молекулой, и производство из него товарной продукции. В варианте осуществления товарный продукт представляет собой белковый концентрат, белковый изолят, зерно, крахмал, семена, шрот, муку, биомассу или масло семян.
В другом аспекте изобретение предоставляет способ экспрессии транскрибируемой полинуклеотидной молекулы, включающий получение трансгенных растений, содержащих последовательность ДНК, выбранную из группы, состоящей из: (a) последовательности, имеющей, по меньшей мере, около 85 процентов идентичности последовательности с любой из SEQ ID NO: 1, 2, 3, 4, 6 или 8; (b) последовательности, содержащей любую из SEQ ID NO: 1, 2, 3, 4, 6 или 8; и (c) фрагмента любой из SEQ ID NO: 1, 2, 3, 4, 6 или 8, где фрагмент обладает геннорегулирующей активностью, где указанная последовательность функционально связана с гетерологичной транскрибируемой полинуклеотидной молекулой, а также культивирование растения, в котором экспрессируется транскрибируемый полинуклеотид.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг. 1 демонстрирует экспрессию β-глюкуронидазы (GUS) в трансгенном развивающемся зародыше кукурузы и тканях эндосперма, вызванную различными конфигурациями трансгенных кассет. Каждая конфигурация трансгенной кассеты состоит из последовательности, кодирующей GUS, функционально связанной с группой транскрипционных регулирующих экспрессию элементов EXP-Zm.Nac+Os.FBA:1:1 (SEQ ID NO: 6), и EXP-Zm.Nac+Os.Cab-1:1:1 (SEQ ID NO: 8), и 3'UTR T-Os.CLUS33428_1-1:1:1 (SEQ ID NO: 11), T-Os.Mth-1:1:1 (SEQ ID NO: 12), и T-Os.Ara5-1:1:1 (SEQ ID NO: 13), как показано в таблице 3 примера 3.
Фиг. 2 демонстрирует экспрессию β-глюкуронидазы (GUS) в отдельных тканях трансгенной кукурузы, вызванную различными конфигурациями трансгенных кассет. Каждая конфигурация трансгенной кассеты состоит из последовательности, кодирующей GUS, функционально связанной с группой транскрипционных регулирующих экспрессию элементов EXP-Zm.Nac+Os.FBA:1:1 (SEQ ID NO: 6), и EXP-Zm.Nac+Os.Cab-1:1:1 (SEQ ID NO: 8), и 3'UTR T-Os.CLUS33428_1-1:1:1 (SEQ ID NO: 11), T-Os.Mth-1:1:1 (SEQ ID NO: 12), и T-Os.Ara5-1:1:1 (SEQ ID NO: 13), как показано в таблице 3 примера 3.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO: 1 - последовательность группы транскрипционных регулирующих экспрессию элементов или ЕХР, ЕХР-Zm.Nac+Zm.DnaK:1:1, состоящая из промотора P-Zm.Nac-1:1:2 (SEQ ID NO: 2), который функционально связан с 5'-концом лидера L-Zm.Nac-1:1:1 (SEQ ID NO: 4), который функционально связан с интроном I-Zm.DnaK-1:1:1 (SEQ ID NO: 5).
SEQ ID NO: 2 - последовательность промотора P-Zm.Nac-1:1:2.
SEQ ID NO: 3 - последовательность, содержащая промотор P-Zm.Nac-1:1:2 (SEQ ID NO: 2), который функционально связан с 5'-концом лидера L-Zm.Nac-1:1:1 (SEQ ID NO: 4).
SEQ ID NO: 4 - последовательность лидера L-Zm.Nac-1:1:1.
SEQ ID NO: 5 - последовательность интрона I-Zm.DnaK-1:1:1.
SEQ ID NO: 6 - последовательность группы транскрипционных регулирующих экспрессию элементов или ЕХР, ЕХР-Zm.Nac+Os.FBA:1:1, которая состоит из промотора P-Zm.Nac-1:1:2 (SEQ ID NO: 2), который функционально связан с 5'-концом лидера L-Zm.Nac-1:1:1 (SEQ ID NO: 4), который функционально связан с интроном I-Os.FBA-1-1:1:1 (SEQ ID NO: 7).
SEQ ID NO: 7 - последовательность интрона I-Os.FBA-1-1:1:1.
SEQ ID NO: 8 - последовательность группы транскрипционных регулирующих экспрессию элементов или ЕХР, EXP-Zm.Nac+Os.Cab-1:1:1, которая состоит из промотора P-Zm.Nac-1:1:2 (SEQ ID NO: 2), который функционально связан с 5'-концом лидера L-Zm.Nac-1:1:1 (SEQ ID NO: 4), который функционально связан с интроном I-Os.Cab-1-1:1:1 (SEQ ID NO: 9).
SEQ ID NO: 9 - последовательность интрона I-Os.Cab-1-1:1:1.
SEQ ID NO: 10 - последовательность 3'UTR T-AGRtu.nos-1:1:13.
SEQ ID NO: 11 - последовательность 3'UTR T-Os.CLUS33428_1-1:1:1.
SEQ ID NO: 12 - последовательность 3'UTR T-Os.Mth-1:1:1.
SEQ ID NO: 13 - последовательность 3'UTR T-Os.Ara5-1:1:1.
SEQ ID NO: 14 - кодирующая последовательность маркерного гена β-глюкуронидазы.
SEQ ID NO: 15 - последовательность группы транскрипционных регулирующих экспрессию элементов или EXP, EXP-CaMV.35S:1:1, включающая промотор и лидер 35S вируса мозаики цветной капусты (CaMV).
SEQ ID NO: 16 - последовательность группы транскрипционных регулирующих экспрессию элементов или EXP, EXP-Os.Act1:1:1, включающая промотор, лидер и интрон актина 1 риса.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение предоставляет новые полинуклеотидные молекулы, обладающие благоприятной геннорегулирующей активностью, из видов растений. Изобретение также предоставляет ДНК-конструкции, содержащие регуляторные элементы, а также трансгенные клетки растений, растения и семена, содержащие регуляторные элементы. Нуклеотидные последовательности этих полинуклеотидных молекул представлены как SEQ ID NO: 1, 2, 3, 4, 6 и 8. Изобретением предусмотрено проектирование, конструирование и использование этих полинуклеотидных молекул. Эти полинуклеотидные молекулы способны, к примеру, оказывать влияние на экспрессию функционально связанной транскрибируемой полинуклеотидной молекулы в тканях растений, и, следовательно, к избирательной регуляции генной экспрессии или активности кодируемого генного продукта в трансгенных растениях. Настоящее изобретение также предоставляет способы изготовления и использования регуляторных элементов, ДНК-конструкций, содержащих промоторы и/или другие известные нуклеотидные последовательности, и способы их подготовки и применения. Изобретение также предоставляет трансгенные растительные клетки, растения и семена, содержащие регуляторные элементы, функционально связанные с транскрибируемой полинуклеотидной молекулой, а также трансформированные клетки-хозяева.
Последовательности ДНК по настоящему изобретению могут быть предоставлены функционально связанными с транскрибируемой полинуклеотидной молекулой. В одном варианте осуществления транскрибируемая полинуклеотидная молекула может быть гетерологичной относительно регуляторной последовательности, предоставленной в настоящем документе. Последовательность регуляторного элемента, предоставленная изобретением, таким образом, в определенных вариантах осуществления может быть определена как функционально связанная с гетерологичной транскрибируемой полинуклеотидной молекулой.
Далее представлены определения и способы, чтобы лучше охарактеризовать настоящее изобретение и как руководство для любых специалистов в рассматриваемой области в практике настоящего изобретения. Если не указано иное, термины должны пониматься в соответствии с обычным использованием любыми специалистами в соответствующей области техники.
Молекулы ДНК
Как используется в настоящем документе, термин «ДНК» или «молекула ДНК» относится к двухцепочечной молекуле ДНК геномного или синтетического происхождения, т.е. полимеру дезоксирибонуклеотидных оснований или полинуклеотидной молекуле, читаемой от 5'-конца (расположенного вверх по течению) к 3'-концу (расположенному вниз по течению). Как используется в настоящем документе, термин «последовательность ДНК» относится к нуклеотидной последовательности молекулы ДНК. Используемая в настоящем документе номенклатура соответствует таковой Наименования 37 Свода федеральных нормативных актов США § 1.822 и изложенным в таблицах стандартам ВОИС ST.25 (1998), Приложение 2, таблицы 1 и 3.
Как используется в настоящем документе, термин «выделенная молекула ДНК» относится к молекуле ДНК, по меньшей мере, частично отделенной от других молекул, обычно связанной с ней в ее нативном или естественном состоянии. В одном варианте осуществления термин «выделенный» относится к молекуле ДНК, которая, по меньшей мере, частично отделена от некоторых нуклеиновых кислот, которые обычно контактируют с молекулой ДНК в ее нативном или естественном состоянии. Таким образом, молекулы ДНК, слитые с регуляторными или кодирующими последовательностями, с которыми они обычно не ассоциируются, например, в результате рекомбинантных технологий, считаются выделенными в настоящем документе. Такие молекулы считаются выделенными в случае, когда интегрированы в хромосому клетки-хозяина или присутствуют в растворе нуклеиновых кислот с другими молекулами ДНК, в котором они не находятся в своем природном состоянии.
Любое количество способов, которые хорошо известны специалистам в рассматриваемой области техники, может быть использовано для выделения и манипулирования молекулой ДНК или ее фрагментом, как раскрыто в настоящем изобретении. Например, технология полимеразной цепной реакции (ПЦР) может быть использована для амплификации определенной начальной молекулы ДНК и/или продуцирования вариантов исходной молекулы. Молекулы ДНК или их фрагменты могут также быть получены с помощью других способов, например, путем непосредственного синтеза фрагмента с помощью химических средств, как это обычно практикуется с использованием автоматизированного синтезатора олигонуклеотидов.
Как используется в настоящем документе, термин «идентичность последовательности» относится к степени, в которой две оптимально выровненные полинуклеотидные последовательности или две оптимально выровненные полипептидные последовательности идентичны. Оптимальное выравнивание последовательностей создается путем выравнивания двух последовательностей вручную, например, референсной последовательности и другой последовательности, чтобы максимизировать число нуклеотидных совпадений при выравнивании последовательностей с соответствующими внутренними нуклеотидными вставками, делециями или пробелами. Как используется в настоящем документе, термин «референсная последовательность» относится к последовательности, предоставленной в качестве полинуклеотидной последовательности SEQ ID NO: 1, 2, 3, 4, 6 и 8.
Как используется в настоящем документе, термин «процент идентичности последовательностей» или «процент идентичности» или «% идентичности» представляет собой фракцию идентичности, умноженную на 100. «Фракция идентичности» для последовательности, оптимально выровненной с референсной последовательностью, представляет собой число нуклеотидных совпадений при оптимальном выравнивании, деленное на общее число нуклеотидов в референсной последовательности, например, общее число нуклеотидов в полной длине всей референсной последовательности. Таким образом, в одном варианте осуществления изобретение предоставляет молекулу ДНК, содержащую последовательность, которая при оптимальном выравнивании с референсной последовательностью, представленной в настоящем документе как SEQ ID NO: 1, 2, 3, 4, 6 и 8, имеет, по меньшей мере, 85% идентичности, по меньшей мере, 90% идентичности, по меньшей мере, около 95% идентичности, по меньшей мере, около 96% идентичности, по меньшей мере, около 97% идентичности, по меньшей мере, приблизительно 98% идентичности или, по меньшей мере, приблизительно 99% идентичности с референсной последовательностью. В определенных вариантах осуществления такие последовательности могут быть определены как обладающие геннорегулирующей активностью.
Регуляторные элементы
Регуляторный элемент представляет собой молекулу ДНК, обладающую геннорегулирующей активностью, т.е. таковую, которая обладает способностью оказывать влияние на транскрипцию и/или трансляцию функционально связанной транскрибируемой полинуклеотидной молекулы. Термин «геннорегуляторная активность», таким образом, относится к способности влиять на профиль экспрессии функционально связанной транскрибируемой полинуклеотидной молекулы путем влияния на транскрипцию и/или трансляцию этой функционально связанной транскрибируемой полинуклеотидной молекулы. Как используется в настоящем документе, группа транскрипционных регуляторных экспрессионных элементов (EXP) может состоять из элементов экспрессии, таких как энхансеры, промоторы, лидерные последовательности и интроны, функционально связанные. Таким образом, группа транскрипционных регуляторных экспрессионных элементов может состоять, например, из промотора, функционально связанного 5' с лидерной последовательностью, которая, в свою очередь, функционально связана 5' с интронной последовательностью. Последовательность интрона может состоять из последовательности, начинающейся в точке первой интрон/экзон границы нативной последовательности, и может также состоять из небольшого лидерного фрагмента, содержащего вторую интрон/экзонную границу, с тем, чтобы обеспечить надлежащий интрон/экзонный процессинг для облегчения транскрипции и правильного процессинга полученного транскрипта. Лидерные последовательности и интроны могут позитивно влиять на транскрипцию функционально связанной транскрибируемой полинуклеотидной молекулы, а также трансляцию полученной в результате транскрибированной РНК. Предварительно процессированная молекула РНК содержит лидерные последовательности и интроны, которые могут влиять на пост-транскрипционный процессинг транскрибируемых РНК и/или экспорт транскрибируемой молекулы РНК из ядра клетки в цитоплазму. После пост-транскрипционного процессинга транскрибируемой молекулы РНК лидерная последовательность может сохраняться как часть конечной матричной РНК и может положительно влиять на трансляцию молекулы матричной РНК.
Регуляторные элементы, такие как промоторы, лидерные последовательности, интроны и области терминации транскрипции, представляют собой молекулы ДНК, которые обладают геннорегуляторной активностью и играют неотъемлемую роль в общей экспрессии генов в живых клетках. Термин «регуляторный элемент» относится к молекуле ДНК, обладающей геннорегуляторной активностью, т.е. такой, которая обладает способностью влиять на транскрипцию и/или трансляцию функционально связанной транскрибируемой полинуклеотидной молекулы. Выделенные регуляторные элементы, такие как промоторы и лидерные последовательности, которые функционируют в растениях, являются, следовательно, полезными для изменения фенотипов растений способами генной инженерии.
Регуляторные элементы могут быть охарактеризованы по их эффектам на профиль экспрессии (качественно и/или количественно), например, положительные или отрицательные эффекты и/или конститутивные или иные эффекты, например, по их временному, пространственному, возрастному, тканевому, экологическому, физиологическому, патологическому, относящемуся к клеточному циклу и/или химически реагирующему профилю экспрессии, и любые их сочетания, а также по количественным или качественным показателям. Промотор может быть полезен в качестве регуляторного элемента для регулирования экспрессии функционально связанной транскрибируемой полинуклеотидной молекулы.
Как используется в настоящем документе, «профиль генной экспрессии» представляет собой любой профиль транскрипции функционально связанной молекулы ДНК в транскрибированную молекулу РНК. Транскрибированная молекула РНК может быть транслирована для продуцирования белковой молекулы или может предоставлять антисмысловую или другую регуляторную молекулу РНК, такую как мРНК, дцРНК, тРНК, рРНК, микроРНК и тому подобное.
Как используется в настоящем документе, термин «белковая экспрессия» представляет собой любой профиль трансляции транскрибированной молекулы РНК в белковую молекулу. Белковая экспрессия может характеризоваться временными, пространственными, относящимися к развитию или морфологическими качествами, а также количественными или качественными показателями.
Как используется в настоящем документе, термин «промотор» относится обычно к молекуле ДНК, которая участвует в распознавании и связывании РНК-полимеразы II и других белков (транс-действующих факторов транскрипции) для инициации транскрипции. Промотор может быть изначально изолирован из 5'-регуляторной области (5'UTR) геномной копии гена. Кроме того, промоторы могут представлять собой полученные синтетически или обработанные молекулы ДНК. Промоторы также могут являться химерными, т.е. промотор, полученный посредством слияния двух или более гетерологичных молекул ДНК. Промотор, полезный в практике настоящего изобретения, может включать в себя SEQ ID NO: 3 или ее фрагменты или варианты. В конкретных вариантах осуществления изобретения такие молекулы и любые варианты или производные этого, как описано в настоящем документе, определены как обладающие промоторной активностью, т.е. способны выступать в качестве промотора в клетке-хозяине, такой, как в трансгенных растениях. В еще более специфических вариантах осуществления фрагмент может быть определен как проявляющий промоторную активность, которой обладала начальная промоторная молекула, из которой он образован, или фрагмент может содержать «минимальный промотор», который обеспечивает базальный уровень транскрипции и состоит из TATA-бокса или эквивалентной последовательности для распознавания и связывания комплекса РНК-полимеразы II для инициации транскрипции.
В одном варианте осуществления изобретение предоставляет фрагменты промоторной последовательности, как раскрыто в настоящем документе. Промоторные фрагменты могут обладать промоторной активностью, как описано выше, и могут быть использованы самостоятельно или в комбинации с другими промоторами и промоторными фрагментами, такими как, например, при конструировании химерных промоторов. В конкретных вариантах осуществления предоставляются фрагменты промотора, содержащие, по меньшей мере, примерно 50, 95, 150, 250, 500, 750 или, по меньшей мере, примерно 1000 смежных нуклеотидов или более обладающей промоторной активностью полинуклеотидной молекулы, раскрытой в настоящем документе.
Композиции, полученные из промотора, представленного как SEQ ID NO: 3, такие, как внутренняя или 5'-делеции, например, могут быть получены с использованием способов, известных в данной области техники, для увеличения или изменения экспрессии, в том числе путем устранения элементов, которые оказывают положительное или отрицательное воздействие на экспрессию; дублирования элементов, которые оказывают положительное или отрицательное воздействие на экспрессию; и/или дублирования или удаления элементов, которые обладают ткане- или клеточно-специфическим воздействием на экспрессию. Могут быть использованы композиции, полученные из промотора, представленного как SEQ ID NO: 3, содержащая 3'-делеции, в котором удаляется элемент TATA-бокс или эквивалентная последовательность этого и расположенная ниже по течению последовательность, например, чтобы образовать энхансерные элементы. Можно сделать дополнительные делеции, чтобы удалить какие-либо элементы, которые оказывают положительный или отрицательный; ткане-специфический; клеточно-специфический; или время-специфический (такой как, без ограничения, циркадные ритмы) эффекты на экспрессию. Промотор, представленный как SEQ ID NO: 3, и фрагменты или энхансеры, полученные на ее основе, могут быть использованы для создания композиций химерных транскрипционных регуляторных элементов, содержащих промотор, представленный как SEQ ID NO: 3, и фрагменты или энхансеры, полученные из этого, функционально связанные с другими энхансерами и промоторами. Эффективность модификаций, дупликаций или делеций, описанных в настоящем документе, на нужные аспекты экспрессии конкретного трансгена может быть проверена эмпирически в стационарных и полевых тестах на растениях, таких, как описано в рабочих примерах в настоящем документе, с тем, чтобы проверить результаты, которые могут различаться в зависимости от сделанных изменений и цели изменения в начальной молекуле.
Как используется в настоящем документе, термин «лидер» относится к молекуле ДНК, выделенной из нетранслируемой 5' области (5'UTR) геномной копии гена и определяемой, как правило, как нуклеотидный сегмент между участком начала транскрипции (TSS) и участком начала кодирующей последовательности белка. Кроме того, лидеры могут представлять собой полученные синтетически или обработанные элементы ДНК. Лидер может быть использован в качестве 5'-регуляторного элемента для регулирования экспрессии функционально связанной транскрибируемой полинуклеотидной молекулы. Лидерные молекулы могут использоваться с гетерологичным промотором или со своим собственным промотором. Промоторные молекулы по настоящему изобретению, таким образом, могут быть функционально связаны со своим собственным лидером или могут быть функционально связаны с гетерологичным лидером. Лидер, полезный в практике настоящего изобретения, представлен как SEQ ID NO: 4 или его фрагменты или варианты. В конкретных вариантах осуществления могут быть предоставлены такие последовательности, определенные как обладающие способностью действовать в качестве лидера в клетке-хозяине, в том числе, например, трансгенной клетке растения. В одном варианте осуществления такие последовательности декодируются как обладающие лидерной активностью.
Лидерная последовательность (5'UTR), представленная как SEQ ID NO: 4, может состоять из регуляторных элементов или может принимать вторичные структуры, которые могут оказывать влияние на транскрипцию или трансляцию трансгена. Эта лидерная последовательность может быть использована в соответствии с настоящим изобретением для создания химерных регуляторных элементов, которые влияют на транскрипцию или трансляцию трансгена. Кроме того, может использоваться лидерная последовательность, представленная как SEQ ID NO: 4, для создания химерных лидерных последовательностей, которые влияют на транскрипцию или трансляцию трансгена.
Введение чужеродного гена в новое растение-хозяин не всегда приводят к высокой экспрессии привнесенного гена. Кроме того, в случае сложных признаков иногда необходимо модулировать несколько генов с различным пространственным или временным профилем экспрессии. Интроны принципиально могут обеспечить такую модуляцию. Однако многократное использование одного и того же интрона в одном растении, как было показано, обнаруживает недостатки. В этих случаях необходимо иметь набор основных контрольных элементов для конструирования соответствующих рекомбинантных элементов ДНК. Число интронов, известных в рассматриваемой области техники как обладающие свойствами, увеличивающими экспрессию, является ограниченным, и, таким образом, необходимы альтернативы.
Композиции, полученные из любого из интронов, представленных как SEQ ID NO: 5, 7 и 9, могут состоять из внутренней делеции или дупликации cis-регуляторных элементов. Кроме того, изменения 5'- и 3'- последовательностей, включающие интрон/экзонные границы, могут быть использованы для улучшения экспрессии или специфичности экспрессии в случае функциональной связи с промотором + лидером или химерным промотором + лидером и кодирующей последовательностью. Изменения 5'- и 3'-областей, включающих интрон/экзонные границы, также можно осуществлять для уменьшения потенциала для индуцирования ложных инициирующих кодонов и стоп-кодонов, производимых в полученном транскрипте после процессинга и сплайсинга матричной РНК. Интроны могут быть проверены эмпирически, как описано в рабочих примерах, чтобы определить влияние интрона на экспрессию трансгена.
В соответствии с настоящим изобретением, промотор или фрагмент промотора может быть проанализирован на наличие известных промоторных элементов, т.е. характерных особенностей последовательностей ДНК, таких как TATA-бокс и другие известные мотивы сайтов связывания транскрипционных факторов. Выявление таких известных промоторных элементов может быть использовано любым специалистом в рассматриваемой области техники для проектирования вариантов промотора, обладающего профилем экспрессии, аналогичным таковому исходного промотора.
Как используется в настоящем документе, термин «энхансер» или «энхансерный элемент» относится к cis-действующему транскрипционному регуляторному элементу (cis-элементу), который придает аспект общему характеру экспрессии, но самому по себе, как правило, недостаточному для управления транскрипцией функционально связанной полинуклеотидной последовательности. В отличие от промоторов, энхансерные элементы обычно не включают сайт начала транскрипции (TSS), или TATA-бокс, или эквивалентную последовательность. Промотор может природно содержать один или более энхансерных элементов, которые влияют на транскрипцию функционально связанной полинуклеотидной последовательности. Выделенный энхансерный элемент может также быть слит с промотором для получения химерного промоторного cis-элемента, который придает аспект общей модуляции экспрессии генов. Промотор или фрагмент промотора может содержать один или более энхансерных элементов, которые влияют на транскрипцию функционально связанных генов. Многие промоторные энхансерные элементы, как считается, связываются с ДНК-связывающими белками и/или влияют на топологию ДНК, производя локальные конформации, которые избирательно позволяют или ограничивают доступ РНК-полимеразы к ДНК-матрице или которые облегчают селективное раскрытие двойной спирали в сайт инициации транскрипции. Энхансерный элемент может действовать для связывания транскрипционных факторов, регулирующих транскрипцию. Некоторые энхансерные элементы связываются с более чем одним фактором транскрипции, и транскрипционные факторы могут взаимодействовать с различными аффинностями с более чем одним энхансерным доменом. Энхансерные элементы могут быть определены с помощью ряда способов, в том числе делеционного анализа, т.е. удаления одного или нескольких нуклеотидов с 5'-конца или от середины в направлении промотора; анализа ДНК-связывающих белков с использованием футпринтинга с ДНКазой I, интерференции участков метилирования, тестов на сдвиг электрофоретической подвижности, геномного футпринтинга in vivo путем лигированно-опосредованной ПЦР и других традиционных анализов; или путем анализа сходства последовательностей ДНК с использованием известных мотивов cis-элементов или энхансерных элементов в качестве последовательности-мишени или мотива мишени при помощи обычных способов сравнения последовательностей ДНК, таких как BLAST. Тонкую структуру энхансерного домена можно также изучать путем мутагенеза (или замещения) одного или нескольких нуклеотидов или другими традиционными способами. Энхансерные элементы могут быть получены путем химического синтеза или путем выделения из регуляторных элементов, которые включают в себя такие элементы, и они могут быть синтезированы с дополнительными фланкирующими нуклеотидами, которые содержат полезные сайты рестрикции для облегчения манипуляции с подпоследовательностями. Таким образом, проектирование, конструирование и применение энхансерных элементов в соответствии со способами, раскрытыми в настоящем документе, для модуляции экспрессии функционально связанных транскрибируемых полинуклеотидных молекул охватываются настоящим изобретением.
В растениях включение некоторых интронов в генные конструкции приводит к увеличению накопления мРНК и белка по сравнению с конструкциями, не содержащими интрон. Этот эффект был назван «интрон-опосредованное увеличение» (IME) генной экспрессии (Mascarenhas et al., (1990) Plant Mol. Biol. 15: 913-920). Интроны, известные по стимулированию экспрессии у растений, были идентифицированы в генах кукурузы [например, tubA1, Adh1, Shi, Ubi1 (Jeon et al., Plant Physiol. 123: 1005-1014, 2000; Callis et al., Genes Dev. 1: 1183-1200, 1987; Vasil et al., Plant Physiol. 91: 1575-1579, 1989; Christiansen et al., Plant Mol. Biol. 18: 675-689, 1992)] и в генах риса (например, salt, tpi: McElroy et al, Plant Cell 2: 163-171, 1990; Xu et al., Plant Physiol. 106: 459-467, 1994). Аналогично, интроны из генов двудольных растений, таких как петуния (например, rbcS), картофель (например, st-ls1) и Arabidopsis thaliana (например, ubq3 и pat1), как было найдено, увеличивают уровни экспрессии генов (Dean et al., Plant Cell 1: 201-208, 1989; Leon et al., Plant Physiol. 95: 968-972, 1991; Norris et al., Plant Mol Biol 21: 895-906, 1993; Rose and Last, Plant J. 11: 455-464, 1997). Было показано, что делеции или мутации в участках сплайсинга интрона снижают генную экспрессию, указывая на то, что сплайсинг может быть необходим для IME (Mascarenhas et al., Plant Mol Biol. 15: 913-920, 1990; Clancy and Hannah, Plant Physiol. 130: 918-929, 2002). Однако такой сплайсинг не требуется при определенном IME в двудольных растениях, что было показано при помощи точечных мутаций в участках сплайсинга гена pat1 из A. thaliana (Rose and Beliakoff, Plant Physiol. 122: 535-542, 2000).
Усиление генной экспрессии при помощи интронов не является общим явлением, поскольку некоторые вставки интрона в рекомбинантных кассетах экспрессии не усиливают экспрессию (например, интроны из генов двудольных растений, таких как ген rbcS из гороха, ген фазеолина из фасоли и ген stls-1 из Solarium tuberosum и интроны генов кукурузы (девятый интрон гена adhl и первый интрон гена hsp81) (Chee et al., Gene 41: 47-57, 1986; Kuhlemeier et al., Mol Gen Genet 212: 405-411, 1988; Mascarenhas et al., Plant Mol. Biol. 15: 913-920, 1990; Sinibaldi and Mettler, In WE Cohn, K Moldave, eds, Progress in Nucleic Acid Research and Molecular Biology, Vol. 42. Academic Press, New York, pp. 229-257, 1992; Vancanneyt et al, Mol. Gen. Genet. 220: 245-250, 1990)). Поэтому не каждый интрон может быть использован для манипулирования уровнем генной экспрессии не являющихся эндогенными генов или эндогенных генов в трансгенных растениях. Какие характеристики или определенные последовательности функции должны присутствовать в последовательности интрона в целях повышения экспрессии ставки данного гена, не известно из уровня техники, и поэтому невозможно предсказать, будет ли данный интрон растения, при гетерологичном использовании, вызывать IME.
Как используется в настоящем документе, термин «химерный» относится к отдельной молекуле ДНК, произведенной путем слияния первой молекулы ДНК со второй молекулой ДНК, где ни первая, ни вторая молекулы ДНК обычно не могут обнаруживаться в такой конфигурации, т.е. слитой с другой. Химерная молекула ДНК, таким образом, представляет собой новую молекулу ДНК, как правило, не встречающуюся в природе. Как используется в настоящем документе, термин «химерный промотор» относится к промотору, произведенному посредством таких манипуляций с молекулами ДНК. Химерный промотор может объединять два или более фрагментов ДНК, например, слияние промотора с энхансерным элементом. Таким образом, проектирование, конструирование и использование химерных промоторов в соответствии со способами, раскрытыми в настоящем документе для модулирования экспрессии функционально связанных транскрибируемых полинуклеотидных молекул, охватываются настоящим изобретением.
Как используется в настоящем документе, термин «вариант» относится ко второй молекуле ДНК, которая имеет сходную композицию, но не идентична первой молекуле ДНК, и еще вторая молекула ДНК по-прежнему сохраняет общие функции, т.е. профиль экспрессии, одинаковый или сходный с таковым первой молекулы ДНК. Вариант может представлять собой укороченную или усеченную версию первой молекулы ДНК и/или измененную версию последовательности первой молекулы ДНК, такую как с отличными сайтами ферментов рестрикции и/или с внутренними делециями, заменами и/или вставками. «Вариант» может также включать регуляторный элемент, имеющий нуклеотидную последовательность, включающую замены, делеции и/или вставки одного или нескольких нуклеотидов референсной последовательности, где производный регуляторный элемент имеет большую или меньшую или эквивалентную транскрипционную или трансляционную активность по сравнению с соответствующей родительской регуляторной молекулой. «Варианты» регуляторных элементов также должны включать варианты, вытекающие из мутаций, которые природно встречаются в трансформации бактериальных и растительных клеток. В настоящем изобретении полинуклеотидная последовательность, предоставленная как SEQ ID NO: 1, 2, 3, 4, 6 и 8, может использоваться для создания вариантов, которые по составу сходны, но не идентичны полинуклеотидной последовательности исходного регуляторного элемента, при сохранении общей функциональности, т.е. профиль экспрессии, одинаковый или аналогичный таковому оригинального регуляторного элемента. Производство таких вариантов настоящего изобретения находится в пределах компетенции специалистов в рассматриваемой области техники в свете раскрытия и заключается в пределах объема настоящего изобретения. «Варианты» химерных регуляторных элементов содержат те же составляющие элементы, что и референсная последовательность, но составляющие элементы, содержащие химерный регуляторный элемент, могут быть функционально связаны с помощью различных способов, известных в данной области техники, таких как расщепление рестрикционными ферментами и лигирование, клонирование, независимое от лигирования, модульная сборка продуктов ПЦР в процессе амплификации или прямой химический синтез регуляторного элемента, а также других способов, известных в данной области техники. Полученный «вариант» химерного регуляторного элемента может состоять из одного или варианта одного и того же составляющего элемента референсной последовательности, но отличается в последовательности или последовательностях, которые включают связывающую последовательность или последовательности, которые позволяют составным частям быть функционально связанными. В настоящем изобретении полинуклеотидная последовательность, предоставленная как SEQ ID NO: 1, 2, 3, 4, 6 и 8, предоставляет референсную последовательность, где составляющие элементы, которые содержат референсную последовательность, могут быть соединены с помощью способов, известных в рассматриваемой области техники, и могут включать замены, делеции и/или вставки одного или нескольких нуклеотидов или мутации, которые природно встречаются в трансформации бактериальных и растительных клеток.
Конструкции
Как используется в настоящем документе, термин «конструкция» означает любую рекомбинантную полинуклеотидную молекулу, такую как плазмида, козмида, вирус, автономно реплицирующаяся полинуклеотидная молекула, фаг или линейная или кольцевая одноцепочечная или двухцепочечная полинуклеотидная молекула ДНК или РНК, полученную из любого источника, способную к геномной интеграции или автономной репликации, включая полинуклеотидную молекулу, где одна или более полинуклеотидных молекул были соединены функционально связанным образом, т.е. функционально связаны. Как используется в настоящем документе, термин «вектор» означает любую рекомбинантную полинуклеотидную конструкцию, которая может быть использована для целей трансформации, т.е. введения гетерологичной ДНК в клетку-хозяин. Вектор по настоящему изобретению может включать экспрессионную кассету или трансгенную кассету, выделенную из любой из вышеупомянутых молекул. Экспрессионные кассеты или трансгенные кассеты, полезные в практике изобретения, состоят из группы транскрипционных регулирующих экспрессию элементов («EXP»), представленных как SEQ ID NO: 1, 6 или 8, функционально связанных с гетерологичной кодирующей последовательностью, которая функционально связана с 3'UTR, представленными как SEQ ID NO: 10, 11, 12 или 13.
Как используется в настоящем документе, термин «функционально связанный» относится к первой молекуле, присоединенной ко второй молекуле, где молекулы устроены так, что первая молекула влияет на функцию второй молекулы. Две молекулы могут являться или могут не быть частью единой непрерывной молекулы, и могут являться или могут не быть смежными. Например, промотор является функционально связанным с транскрибируемой полинуклеотидной молекулой, если промотор модулирует транскрипцию представляющей интерес транскрибируемой полинуклеотидной молекулы в клетке. Лидер, например, является функционально связанным с кодирующей последовательностью, если он способен служить в качестве лидера для полипептида, кодируемого кодирующей последовательностью.
Конструкции по настоящему изобретению могут быть предоставлены, в одном варианте осуществления, в качестве ДНК-конструкций с двойными Ti-плазмидными границами, которые имеют области правой границы (RB или AGRtu.RB) и левой границы (LB или AGRtu.LB) Ti-плазмиды, выделенной из Agrobacterium tumefaciens, содержащей Т-ДНК, которые наряду с переносимыми молекулами, предоставленными клетками A. tumefaciens, обеспечивают интеграцию Т-ДНК в геном растительной клетки (см., например, патент США № 6603061). Конструкции могут также содержать плазмидные каркасные сегменты ДНК, которые обеспечивают функцию репликации и селекцию на антибиотике в бактериальных клетках, например, участок начала репликации Escherichia coli, такой как ori322, участок начала репликации широкого круга хозяев, такой как oriV или oriRi, и кодирующая область селективного маркера, например, Spec/Strp, который кодирует Tn7-аминогликозидаденилтрансферазу (aadA), придающую устойчивость к спектиномицину или стрептомицину, или гентамициновый (Gm, Gent) селективный маркерный ген. Для трансформации растений бактериальный штамм-хозяин зачастую представляет собой A. tumefaciens ABI, C58 или LBA4404; однако другие штаммы, известные специалистам в области трансформации растений, можно использовать в настоящем изобретении.
В рассматриваемой области техники известны способы сборки и введения конструкций в клетку таким образом, что транскрибируемая полинуклеотидная молекула транскрибируется в функциональную молекулу мРНК, которая транслируется и экспрессируется как белковый продукт. Для практики настоящего изобретения, обычные композиции и способы подготовки и использования конструкций и клеток-хозяев хорошо известны специалистам в рассматриваемой области техники (см., например, Molecular Cloning: A Laboratory Manual, 3-rd edition Volumes 1, 2 and 3, J. Sambrook, D.W. Russell, and N. Irwin, Cold Spring Harbor Laboratory Press, 2000). Способы создания рекомбинантных векторов, особенно подходящих для трансформации растений, включают, без ограничения, способы, которые описаны в патентах США №№ 4971908; 4940835; 4769061 и 4757011 в их совокупности. Эти типы векторов были также рассмотрены в научной литературе (см., например, Rodriguez, et al., Vectors: A Survey of Molecular Cloning Vectors and Their Uses, Butterworths, Boston, 1988; и Glick et al., Methods in Plant Molecular Biology and Biotechnology, CRC Press, Boca Raton, FL., 1993). Типичные векторы, полезные для экспрессии нуклеиновых кислот у высших растений, хорошо известны в рассматриваемой области техники и включают векторы, полученные из вызывающей опухоли Ti-плазмиды A. tumefaciens (Rogers et al., Methods in Enzymology 153: 253-277, 1987). Другие рекомбинантные векторы, полезные для трансформации растений, в том числе контролирующий передачу вектор pCaMVCN, также были описаны в научной литературе (см., например, Fromm et al., Proc. Natl. Acad. Sci. USA 82: 5824-5828, 1985).
Различные регуляторные элементы могут быть включены в конструкцию, в том числе любую из предоставленных в настоящем документе. Любой из таких регуляторных элементов может быть предоставлен в сочетании с другими регуляторными элементами. Такие комбинации могут быть разработаны или модифицированы для получения желательных регуляторных характеристик. В одном варианте осуществления конструкции по настоящему изобретению включают, по меньшей мере, один регуляторный элемент, функционально связанный с транскрибируемой полинуклеотидной молекулой, функционально связанной с 3'-молекулой терминации транскрипции.
Конструкции по настоящему изобретению могут включать любой промотор или лидер, предоставленный в настоящем документе или известный в рассматриваемой области техники. Например, промотор по настоящему изобретению может быть функционально связан с гетерологичным нетранслируемым 5'-лидером, таким как полученный из гена белка тепловым шоком (см., например, патенты США №№ 5659122 и 5362865). Кроме того, лидер по настоящему изобретению может быть функционально связан с гетерологичным промотором, таким как промотор транскрипции 35S вируса мозаики цветной капусты (CaMV) (см. патент США № 5352605).
Как используется в настоящем документе, термин «интрон» относится к молекуле ДНК, которая может быть выделена или идентифицирована из геномной копии гена и может быть определена в целом как область сращивания во время процессинга мРНК перед трансляцией. Альтернативно, интрон может представлять собой элемент, синтетически продуцированный или полученный в результате обработки ДНК. Интрон может содержать энхансерные элементы, которые влияют на транскрипцию функционально связанных генов. Интрон может быть использован в качестве регуляторного элемента для регулирования экспрессии функционально связанной транскрибируемой полинуклеотидной молекулы. ДНК-конструкция может включать интрон, и интрон может быть или может не быть гетерологичным в отношении последовательности транскрибируемой полинуклеотидной молекулы. Примеры интронов в рассматриваемой области техники включают интрон актина риса (патент США № 5641876) и интрон HSP70 кукурузы (патент США № 5859347). Интроны, полезные в практике настоящего изобретения, включают SEQ ID NO: 5, 7 и 9. Кроме того, при изменении последовательности интрон/экзонной границы может быть предпочтительным предотвращение использования нуклеотидной последовательности АТ или нуклеотида А непосредственно перед 5'-концом участка сплайсинга (GT) и нуклеотида G или нуклеотидной последовательности TG, соответственно, сразу же после 3'-конца участка сплайсинга (AG) для устранения потенциальных нежелательных инициирующих кодонов, образующихся в процессе процессинга матричной РНК в конечный транскрипт. Последовательность вокруг 5'- или 3'-конца участков границы интрона при этом может быть изменена таким образом.
Как используется в настоящем документе, термин «3'-молекула терминации транскрипции» или «3'UTR» относится к молекуле ДНК, которая используется во время транскрипции для получения 3' нетранслируемой области (3'UTR) молекулы мРНК. 3'-нетранслируемая область молекулы мРНК может быть образована специфическим расщеплением и 3' полиаденилированием (поли А хвост). 3'UTR может быть функционально связана и расположена вниз по течению после транскрибируемой полинуклеотидной молекулы и может включать полинуклеотиды, которые обеспечивают сигнал полиаденилирования и другие регуляторные сигналы, способные влиять на транскрипцию, процессинг мРНК или экспрессию генов. Поли А хвосты, как считается, задействованы в стабильности мРНК и в инициации трансляции. Примерами 3'-молекул терминации транскрипции в рассматриваемой области техники являются 3' область нопалинсинтазы (см. Fraley et al., Proc. Natl. Acad. Sci. USA, 80: 4803-4807, 1983); 3' область hsp17 пшеницы; 3' область малой субъединицы рибулозобифосфаткарбоксилазы/оксигеназы гороха; 3' область E6 хлопка (патент США № 6096950); 3' области, раскрытые в WO/0011200 A2; и 3'UTR коиксина (патент США № 6635806).
3'UTR, как правило, находят выгодное применение для рекомбинантной экспрессии специфических генов. В животных системах аппарат 3'UTR был четко определен (например, Zhao et al., Microbiol Mol Biol Rev 63: 405-445, 1999; Proudfoot, Nature 322: 562-565, 1986; Kim et al., Biotechnology Progress 19: 1620-1622, 2003; Yonaha and Proudfoot, EMBO J. 19: 3770-3777, 2000; Cramer et al., FEBS Letters 498: 179-182, 2001; Kuerstem and Goodwin, Nature Reviews Genetics 4: 626-637, 2003). Эффективная терминация транскрипции РНК необходима для предотвращения нежелательной транскрипции не связанных с признаком (вниз по течению) последовательностей, которая может помешать проявлению признака. Расположение нескольких кассет экспрессии генов в локальной близости друг от друга (например, в пределах одной Т-ДНК) может вызывать супрессию генной экспрессии одного или более генов в указанной конструкции по сравнению с независимыми вставками (Padidam и Cao, BioTechniques 31: 328-334, 2001). Это может помешать достижению адекватного уровня экспрессия, например, в тех случаях, когда желательна сильная экспрессия генов со всех кассет.
В растениях четко определенные последовательности сигнала полиаденилирования не известны. Hasegawa et al. (Plant J. 33: 1063-1072, 2003) не были в состоянии идентифицировать консервативные последовательности сигнала полиаденилирования ни в in vitro, ни в in vivo системе в Nicotiana sylvestris и определить фактическую длину первичного (не полиаденилированного) транскрипта. Слабая 3'UTR может создавать сквозное прочтение, которое может повлиять на экспрессию генов, расположенных в соседних экспрессионных кассетах (Padidam и Cao, BioTechniques 31: 328-334, 2001). Надлежащий контроль терминации транскрипции может препятствовать сквозному прочтению в последовательностях (например, других экспрессионных кассетах), локализованных ниже по течению, и может также обеспечивать эффективный рециклинг РНК-полимеразы для улучшения экспрессии генов. Эффективная терминация транскрипции (высвобождение РНК-полимеразы II из ДНК) является необходимым условием для повторной инициации транскрипции и тем самым непосредственно влияет на общий уровень транскриптов. После терминации транскрипции зрелая мРНК освобождается от участка синтеза и передается в цитоплазму. Эукариотические мРНК накапливаются в виде поли(A) in vivo, что затрудняет детектирование сайтной терминации транскрипции с помощью традиционных способов. Однако прогнозирование функциональных и эффективных 3'UTR с помощью биоинформационных способов сложно вследствие отсутствия консервативных последовательностей, позволяющих легко прогнозировать эффективные 3'UTR.
С практической точки зрения может быть выгодно, что 3'UTR, используемая в трансгенной кассете, обладает определенными характеристиками. Например, 3'UTR, используемая в соответствии с настоящим изобретением, может эффективно терминировать транскрипцию трансгена и предотвращать сквозное прочтение транскрипта в любой соседней последовательности ДНК, которая может содержать другую трансгенную кассету, как в случае с несколькими кассетами, расположенными в одной Т-ДНК, или соседней хромосомной ДНК, в которую встроена Т-ДНК. 3'UTR оптимально не должна вызывать снижение транскрипционной активности, придаваемой промотором, лидером и интронами, которые используются для направления экспрессии трансгена. В биотехнологии растений 3'UTR часто используются для праймирования реакций амплификации обратно транскрибируемой РНК, выделенной из трансформированного растения, и могут быть использованы для (1) оценки транскрипционной активности или экспрессии трансгенной кассеты после интегрирования в хромосому растения; (2) оценки числа копий вставок в ДНК растений; и (3) определения зиготности полученного семени после разведения. 3'UTR также может использоваться в реакции амплификации ДНК, выделенной из трансформированных растений, для характеристики сохранности встроенной кассеты.
3'UTR, используемые для обеспечения экспрессии трансгена в растениях, могут быть определены на основе экспрессии экспрессируемых тэг-последовательностей (EST) в библиотеках кДНК, изготовленных из матричной РНК, выделенной из семян, цветов или любых других тканей, полученных из, например, большого бородача (Andropogon gerardii), эриантуса сахаристого [Saccharum ravennae (Erianthus ravennae)], зеленого щетинника (Setaria viridis), теосинте (Zea mays подвид mexicana), проса итальянского (Setaria italica) или бусенника (Coix lacryma-jobi). С помощью способов, известных специалистам в рассматриваемой области техники, библиотеки кДНК могут быть получены из тканей, выделенных из видов растений, с помощью цветочной ткани, семян, листьев, корней или других растительных тканей. Полученные кДНК секвенируют при помощи различных способов секвенирования с использованием различных способов, известных в данной области техники. Полученные EST собирают в кластеры с использованием биоинформационных программ, таких как clc_ref_assemble_complete версия 2.01.37139 (CLC bio США, Кембридж, Массачусетс 02142). Относительное содержание транскриптов каждого кластера определяется путем подсчета количества кДНК, считываемого с каждого кластера. Выявленные 3'UTR могут состоять из последовательности, полученной из последовательности кДНК, а также из последовательности, полученной из геномной ДНК. Последовательность кДНК может быть использована для проектирования праймеров, которые затем могут быть использованы с библиотеками GenomeWalkerTM (Clontech Laboratories, Inc., Mountain View, CA), сконструированными в соответствии с протоколом производителя для клонирования 3'-области, соответствующей геномной ДНК-последовательности, для предоставления более длинной терминационной последовательности. Анализ относительного содержания транскриптов либо путем прямых подсчетов, либо нормированных подсчетов наблюдаемой последовательности, считываемых для каждой тканевой библиотеки, может быть использован, чтобы сделать выводы о свойствах профиля экспрессии. Например, некоторые 3'UTR могут быть найдены в транскриптах, более представленных в ткани корня, чем в ткани листа. Это говорит о том, что транскрипт высоко экспрессируется в корне и что свойства экспрессии в корне могут быть связаны с транскрипционной регуляцией промотора лидера, интронов или 3'UTR. Эмпирическая проверка 3'UTR, определенных при помощи свойств экспрессии в конкретных органах, тканях и типах клеток, может привести к идентификации 3'UTR, которые усиливают экспрессию в этих конкретных органах, тканях и типах клеток. 3'UTR, полезные в практике изобретения, представлены как SEQ ID NO: 9, 10, 11 и 12.
Конструкции и векторы могут также включать последовательность, кодирующую транзитный пептид, которая экспрессирует связанный пептид, полезный для направления белкового продукта, в частности, в хлоропласты, лейкопласты или другие пластидные органеллы; митохондрии; пероксисомы; вакуоли; или внеклеточное пространство. Для описания использования транзитных пептидов хлоропластов см. патенты США №№ 5188642 и 5728925. Много локализованных в хлоропластах белков экспрессируется с ядерных генов в качестве предшественников и направляется в хлоропласты при помощи хлоропластного транзитного пептида (CTP). Неограничивающие примеры таких выделенных хлоропластных белков включают белки, которые связаны с малой субъединицей (SSU) рибулозо-1,5,-бисфосфаткарбоксилазы, ферредоксином, ферредоксиноксидоредуктазой, белком I и белком II светособирающего комплекса, тиоредоксином F, энолпирувилшикиматфосфатсинтазой (EPSPS), и транзитные пептиды, описанные в патенте США № 7193133. Было показано in vivo и in vitro, что нехлоропластные белки могут быть направлены в хлоропласты путем использования слияний белка с гетерологичным СТР и что СТР достаточен для направления белка в хлоропласт. Включение подходящего хлоропластного транзитного пептида, такого как EPSPS CTP Arabidopsis thaliana (CTP2) (см. Klee et al., Mol. Gen. Genet. 210: 437-442, 1987) или Petunia hybrida EPSPS CTP (CTP4) (см. della-Cioppa et al., Proc. Natl. Acad. Sci. USA 83: 6873-6877, 1986), как было показано, направляет последовательность гетерологичного белка EPSPS в хлоропласты в трансгенных растениях (см. патенты США №№ 5627061; 5633435; и 5312910; и EP 0218571; EP 189707; EP 508909; и EP 924299).
Транскрибируемые полинуклеотидные молекулы
Как используется в настоящем документе, термин «транскрибируемая полинуклеотидная молекула» относится к любой молекуле ДНК, способной транскрибировать молекулу РНК, в том числе такие, без ограничения, имеющие кодирующие белок последовательности и такие, продуцирующие молекулы РНК, имеющие последовательности, полезные для супрессии генов. «Трансген» относится к транскрибируемой полинуклеотидной молекуле, гетерологичной для клетки-хозяина, по меньшей мере, относительно ее положения в геноме, и/или транскрибируемой полинуклеотидной молекуле, искусственно включенной в геном клетки-хозяина в текущем или любом предыдущем поколении клетки.
Промотор по настоящему изобретению может быть функционально связан с транскрибируемой полинуклеотидной молекулой, гетерологичной в отношении промоторной молекулы. Как используется в настоящем документе, термин «гетерологичный» означает комбинацию двух или более полинуклеотидных молекул в случае, когда такое сочетание обычно не обнаруживается в природе. Например, две молекулы могут быть получены из различных видов и/или две молекулы могут быть получены из различных генов, например, различные гены из одних и тех же видов, или одни и те же гены из разных видов. Промотор, таким образом, является гетерологичным по отношению к функционально связанной транскрибируемой полинуклеотидной молекуле, если такое сочетание обычно не обнаруживается в природе, т.е. что транскрибируемая полинуклеотидная молекула не является природно функционально связанной в комбинации с этой промоторной молекулой.
Транскрибируемая полинуклеотидная молекула в общем случае может быть любой молекулой ДНК, для которой желательна экспрессия РНК-транскрипта. Такая экспрессия РНК-транскрипта может приводить к трансляции полученной в результате молекулы мРНК и, таким образом, к экспрессии белка. Кроме того, например, транскрибируемая полинуклеотидная молекула могут быть сконструирована так, чтобы в конечном итоге вызвать снижение экспрессии конкретного гена или белка. В одном из вариантов осуществления этого можно достичь при использовании транскрибируемой полинуклеотидной молекулы, ориентированной в антисмысловом направлении. Любой специалист в рассматриваемой области техники знаком с использованием такой антисмысловой технологии. Вкратце, как только антисмысловая транскрибируемая полинуклеотидная молекула транскрибируется, РНК-продукт гибридизуется с и секвестрирует комплементарную молекулу РНК внутри клетки. Эта дуплексная РНК-молекула не может транслироваться в белок при помощи клеточного аппарата трансляции и деградирует в клетке. Любой ген можно негативно регулировать таким образом.
Таким образом, в одном варианте осуществления настоящего изобретения регуляторный элемент, представленный как SEQ ID NO: 1, 2, 3, 4, 6 и 8, функционально связан с транскрибируемой полинуклеотидной молекулой для модулирования транскрипции транскрибируемой полинуклеотидной молекулы на желаемом уровне или с желаемым профилем, когда конструкция интегрирована в геном растительной клетки. В одном варианте осуществления транскрибируемая полинуклеотидная молекула содержит белок-кодирующую область гена, и промотор влияет на транскрипцию РНК-молекулы, которая транслируется и экспрессируется как белковый продукт. В другом варианте осуществления транскрибируемая полинуклеотидная молекула содержит антисмысловую область гена, и промотор влияет на транскрипцию антисмысловой молекулы РНК, двуцепочечной РНК или другой подобной ингибиторной молекулы РНК для того, чтобы ингибировать экспрессию представляющей интерес специфической молекулы РНК в клетке-мишени хозяина.
Гены, представляющие агрономический интерес
Транскрибируемые полинуклеотидные молекулы в соответствии с настоящим изобретением могут представлять собой гены, представляющие агрономический интерес. Как используется в настоящем документе, термин «ген, представляющий агрономический интерес» относится к транскрибируемой полинуклеотидной молекуле, которая при экспрессии в определенной растительной ткани, клетке или типе клеток придает желательные характеристики, например, связанные с морфологией растения, физиологией, ростом, развитием, урожайностью, продуктивностью, профилем питания, болезнью или устойчивостью к вредителям и/или толерантностью к окружающей среде или химической толерантностью. Гены, представляющие агрономический интерес, без ограничения включают гены, кодирующие белок, влияющий на урожайность, белок стрессоустойчивости, белок контроля развития, белок дифференцировки ткани, белок меристемы, белок экологической отзывчивости, белок старения, белок гормонального ответа, белок опадения листьев, белок запасов, белок шелка, белок контроля цветения, белок семян, белок устойчивости к гербицидам, белок устойчивости к болезням, фермент биосинтеза жирных кислот, фермент биосинтеза токоферола, фермент биосинтеза аминокислот, пестицидный белок или любой другой агент, такой как молекула антисмысловой РНК или РНК-интерференции, направленный на конкретный ген для супрессирования. Продукт гена, представляющего агрономический интерес, может действовать в пределах растения для того, чтобы вызвать воздействие на физиологию или обмен веществ у растения, или может выступать в качестве пестицидного агента в рационе вредителя, который питается растением.
В одном из вариантов настоящего изобретения промотор включается в конструкцию так, чтобы промотор был функционально связан с транскрибируемой полинуклеотидной молекулой, которая является геном, представляющим агрономический интерес. Экспрессия гена, представляющего агрономический интерес, желательна для придания агрономически благотворных признаков. Без ограничений, благотворный агрономический признак может включать в себя, например, устойчивость к гербицидам, устойчивость к вредителям, модифицированную урожайность, устойчивость к грибковым заболеваниям, устойчивость к вирусам, устойчивость к нематодам, устойчивость к бактериальным заболеваниям, рост и развитие растений, продуцирование крахмала, продуцирование модифицированного масла, увеличенное продуцирование масла, содержание модифицированных жирных кислот, увеличенное продуцирование белка, созревание плодов, улучшенную ценность для питания животных и человека, биополимеры, экологическую стрессоустойчивость, фармацевтические пептиды и секретируемые пептиды, признаки улучшенной обработки, улучшенной усвояемости, продуцирования ферментов, аромата, фиксации азота, продуцирования гибридных семян, продуцирования волокна и продуцирования биотоплива, среди прочих. Примеры генов, представляющих агрономический интерес, известных в рассматриваемой области техники, включают гены устойчивости к гербицидам (патенты США №№ 6803501; 6448476; 6248876; 6225114; 6107549; 5866775; 5804425; 5633435; и 5463175), увеличенной урожайности (патенты США №№ USRE38446; 6716474; 6663906; 6476295; 6441277; 6423828; 6399330; 6372211; 6235971; 6222098; и 5716837), контроля насекомых (патенты США №№ 6809078; 6713063; 6686452; 6657046; 6645497; 6642030; 6639054; 6620988; 6593293; 6555655; 6538109; 6537756; 6521442; 6501009; 6468523; 6326351; 6313378; 6284949; 6281016; 6248536; 6242241; 6221649; 6177615; 6156573; 6153814; 6110464; 6093695; 6063756; 6063597; 6023013; 5959091; 5942664; 5942658; 5880275; 5763245; и 5763241), устойчивости к грибковым заболеваниям (патенты США №№ 6653280; 6573361; 6506962; 6316407; 6215048; 5516671; 5773696; 6121436; 6316407; и 6506962), устойчивости к вирусам (патенты США №№ 6617496; 6608241; 6015940; 6013864; 5850023; и 5304730), устойчивости к нематодам (патент США № 6228992), устойчивости к бактериальным заболеваниям (патент США № 5516671), роста и развития растений (патенты США №№ 6723897 и 6518488), продуцирования крахмала (патенты США №№ 6538181; 6538179; 6538178; 5750876; и 6476295), продуцирования модифицированного масла (патенты США №№ 6444876; 6426447; и 6380462), увеличенного продуцирования масла (патенты США №№ 6495739; 5608149; 6483008; и 6476295), содержания модифицированных жирных кислот (патенты США №№ 6828475; 6822141; 6770465; 6706950; 6660849; 6596538; 6589767; 6537750; 6489461; и 6459018), продуцирования высокого содержания белка (патент США № 6380466), созревания плодов (патент США № 5512466), усиленного питания животных и человека (патенты США №№ 6723837; 6653530; 6541259; 5985605; и 6171640), биополимеров (патенты США №№ USRE37543; 6228623; 5958745; и 6946588), устойчивости к экологическому стрессу (патент США № 6072103), фармацевтических пептидов и секретируемых пептидов (патенты США №№ 6812379; 6774283; 6140075; и 6080560), признаков улучшенной обработки (патент США № 6476295), улучшения переваримости (патент США № 6531648), пониженной раффинозы (патент США № 6166292), промышленного производства ферментов (патент США № 5543576), улучшения вкуса (патент США № 6011199), фиксации азота (патент США № 5229114), продуцирования гибридных семян (патент США № 5689041), продуцирования волокна (патенты США №№ 6576818; 6271443; для 5981834; и 5869720) и продуцирования биотоплива (патент США № 5998700).
Альтернативно, ген, представляющий агрономический интерес, может повлиять на вышеуказанные характеристики растения или фенотипа путем кодирования молекулы РНК, которая приводит к целенаправленной модуляции генной экспрессии эндогенного гена, например, через антисмысловую (см., например, патент США № 5107065); ингибирующую РНК («РНК-интерференция», в том числе модуляции экспрессии генов через механизмы, опосредуемые микроРНК, миРНК, транс-действующую миРНК и фазовую сРНК, например, как описано в опубликованных заявках США 2006/0200878 и 2008/0066206, и в патентной заявке США № 11/974469); или механизмы, опосредованные косупрессией. РНК также может представлять собой каталитическую молекулу РНК (например, рибозим или рибосвитч; см., например, заявку США 2006/0200878), спроектированную для расщепления нужного эндогенного мРНК-продукта. Таким образом, любая транскрибируемая полинуклеотидная молекула, кодирующая транскрибированную молекулу РНК, которая затрагивает агрономически важный фенотип или представляющее интерес изменение морфологии, может быть полезна для практики настоящего изобретения. В рассматриваемой области техники известны способы конструирования и введения конструкций в клетку таким образом, что транскрибируемая полинуклеотидная молекула транскрибируется в молекулу, которая способна вызывать супрессию гена. Например, пост-транскрипционная генная супрессия при помощи конструкции с антисмысловой ориентированной транскрибируемой полинуклеотидной молекулой для регулирования генной экспрессии в клетках растений представлена в патентах США №№ 5107065 и 5759829, и пост-транскрипционная генная супрессия с использованием конструкции со смысловой ориентированной транскрибируемой полинуклеотидной молекулой для регулирования экспрессии генов в растениях раскрыта в патентах США №№ 5283184 и 5231020. Экспрессия транскрибируемого полинуклеотида в растительной клетке может быть также использована для подавления вредителей растений, питающихся растительной клеткой, например, композиции, выделенные из жесткокрылых вредителей (патентная публикация США № US20070124836), и композиции, выделенные из нематод-вредителей (патентная публикация США № US20070250947). Вредители растений включают, без ограничения, членистоногих вредителей, вредителей-нематод и грибковых или микробных вредителей. Типичные транскрибируемые полинуклеотидные молекулы для включения в конструкции по настоящему изобретению включают, например, молекулы ДНК или гены из видов, отличных от видов-мишеней, или гены, которые происходят из или присутствуют в тех же видах, но включаются в реципиентные клетки с помощью способов генетической инженерии, а не классическими способами воспроизведения или селекции. Тип полинуклеотидной молекулы может включать, без ограничения, полинуклеотидную молекулу, которая уже присутствует в растительной клетке, полинуклеотидную молекулу из другого растения, полинуклеотидную молекулу из другого организма или полинуклеотидную молекулу, созданную извне, такую как полинуклеотидная молекула, содержащая ген в антисмысловой ориентации, или полинуклеотидную молекулу, кодирующую искусственную, синтетическую или иначе модифицированную версию трансгена.
Селективные маркеры
Как используется в настоящем документе, термин «маркер» относится к любой транскрибируемой полинуклеотидной молекуле, экспрессия которой (или ее отсутствие) может быть выявлена или отмечена каким-либо образом. Маркерные гены для использования в практике настоящего изобретения включают, без ограничения, транскрибируемые полинуклеотидные молекулы, кодирующие β-глюкуронидазу (GUS, описанную в патенте США № 5599670), зеленый флуоресцентный белок и его варианты (GFP, описанный в патентах США №№ 5491084 и 6146826), белки, которые придают устойчивость к антибиотикам, или белки, которые придают устойчивость к гербицидам. Полезные маркеры устойчивости к антибиотикам, в том числе те, которые кодируют белки, придающие устойчивость к канамицину (nptII), гигромицину B (aph IV), стрептомицину или спектиномицину (aad, spec/strep) и гентамицину (aac3 и aacC4), хорошо известны в данной области техники. Гербициды, для которых была продемонстрирована толерантность трансгенных растений и для которых может быть применим способ по настоящему изобретению, включают, без ограничения: амино-метил-фосфоновую кислоту, глифосат, глюфосинат, сульфонилмочевину, имидазолоны, бромоксинил, далапон, дикамба, циклогександион, ингибиторы протопорфириногеноксидазы и изоксасфлутоловые гербициды. Транскрибируемые полинуклеотидные молекулы, кодирующие белки, участвующие в толерантности к гербицидам, известны в рассматриваемой области техники и могут включать, без ограничения, транскрибируемую полинуклеотидную молекулу, кодирующую 5-энолпирувилшикимат-3-фосфатсинтазу (EPSPS для устойчивости к глифосату, описанный в патентах США №№ 5627061; 5633435; 6040497; и 5094945); транскрибируемую полинуклеотидную молекулу, кодирующую глифосатоксидоредуктазу и глифосат-N-ацетилтрансферазу (GOX, описанный в патенте США № 5463175; GAT, описанный в патентной публикации США № 20030083480; и дикамба монооксигеназа, описанная в патентной публикации США № 20030135879); транскрибируемую полинуклеотидную молекулу, кодирующую бромоксинилнитрилазу (Bxn для толерантности к бромоксинилу, описанный в патенте США № 4810648); транскрибируемую полинуклеотидную молекулу, кодирующую питоен десатуразу (crtI), описанную в Misawa et al. (Plant Journal 4: 833-840, 1993; и Plant Journal 6: 481-489, 1994) для толерантности к норфлуразону; транскрибируемую полинуклеотидную молекулу, кодирующую ацетогидроксиацидсинтазу (AHAS, aka ALS), описанную в Sathasiivan, et al. (Nucl. Acids Res. 18: 2188-2193, 1990) для толерантности к сульфонилмочевинным гербицидам; и ген bar, описанный в DeBlock et al. (EMBO Journal 6: 2513-2519, 1987) для толерантности к глюфосинату и биолафосу. Промоторные молекулы по настоящему изобретению могут экспрессировать связанные транскрибируемые полинуклеотидные молекулы, которые кодируют фосфинотрицинацетилтрансферазу, глифосат-устойчивую EPSPS, аминогликозидфосфотрансферазу, гидроксифенилпируватдегидрогеназу, гигромицинфосфотрансферазу, неомицинфосфотрансферазу, далапондегалогеназу, бромоксинин-устойчивую нитрилазу, антранилатсинтазу, арилоксиалканоатдезоксигеназы, ацетил-КоА-карбоксилазу, глифосатоксидоредуктазу и глифосат-N-ацетил-трансферазу.
Включенными в термин «селективные маркеры» также являются гены, которые кодируют селективный маркер, чья секреция может детектироваться в качестве средства идентификации или отбора трансформированных клеток. Примеры включают маркеры, которые кодируют секретируемый антиген, которые можно определить путем взаимодействия с антителами, или даже секретируемые ферменты, которые могут быть обнаружены каталитически. Селектируемые секретированные маркерные белки делятся на несколько классов, в том числе малые, диффузные белки, которые детектируются (например, способом ELISA), небольшие активные ферменты, которые детектируются во внеклеточном растворе (например, альфа-амилаза, бета-лактамаза, фосфинотрицинтрансфераза), или белки, которые встраиваются или удерживаются в клеточной стенке (такие как белки, которые включают лидерную последовательность, например, какая обнаруживается в экспрессионном блоке расширения или родственных белках патогенеза табака, также известных как PR-S табака). Другие возможные селективные маркерные гены должны быть очевидны специалистам в рассматриваемой области техники и охватываются настоящим изобретением.
Трансформация клеток
Термин «трансформация» относится к введению нуклеиновой кислоты в реципиента-хозяина. Как используется в настоящем документе, термин «хозяин» означает бактерию, грибок или растение, включая любые клетки, ткани, органы или потомство бактерии, грибка или растения. Например, клетка-хозяин по настоящему изобретению может представлять собой любую клетку или организм, например, клетку растения, клетку водоросли, клетку грибка, грибок, бактериальную клетку, клетку насекомого или подобную. В варианте осуществления хозяева и трансформированные клетки могут включать в себя клетки из: растений, Aspergillus, дрожжей, насекомых, бактерий и водорослей. Растительные ткани и клетки, представляющие особый интерес, включают, в качестве неограничивающих примеров, протопласты, каллусы, корни, клубни, семена, стебли, листья, сеянцы, зародыши и пыльцу.
Как используется в настоящем документе, термин «трансформированный» относится к клетке, ткани, органу или организму, в которые была введена чужеродная полинуклеотидная молекула, например, конструкция. Введенная полинуклеотидная молекула может быть интегрированы в геномную ДНК реципиентной клетки, ткани, органа или организма, так что введенная полинуклеотидная молекула наследуется последующим потомством. «Трансгенная» или «трансформированная» клетка или организм также включает в себя потомство клетки или организма, и потомство, произведенное по селекционной программе, в которой такой трансгенный организм используется в качестве родителя при скрещивании, и демонстрирующее измененный фенотип в результате присутствия чужеродной полинуклеотидной молекулы. Термин «трансгенный» относится к бактерии, грибку или растению, содержащим одну или несколько гетерологичных молекул полинуклеиновых кислот.
Существует множество способов введения молекул полинуклеиновых кислот в клетки растений. Способ может, как правило, включать стадии выбора подходящей клетки-хозяина, трансформации клетки-хозяина рекомбинантным вектором и получения трансформированной клетки-хозяина. Подходящие способы включают бактериальную инфекцию (например, Agrobacterium), бинарные бактериальные искусственные хромосомные вектора, прямую доставку ДНК (например, с помощью ПЭГ-опосредованной трансформации, поглощения ДНК, опосредованного высыханием/ингибированием, электропорации, взбалтывания с карбид-кремниевыми волокнами и ускорением покрытых ДНК частиц и др. (обзор в Potrykus et al., Ann. Rev. Plant Physiol. Plant Mol. Biol. 42: 205, 1991).
Технология введения молекулы ДНК в клетки хорошо известна специалистам в рассматриваемой области техники. Способы и материалы для трансформации растительных клеток путем введения конструкции растительной ДНК в геном растения в практике настоящего изобретения могут включать в себя любой из известных и продемонстрированных способов. Любые способы трансформации могут быть использованы для трансформации клетки-хозяина одним или более промоторов и/или конструкций по настоящему изобретению.
Регенерированные трансгенные растения могут самоопыляться для предоставления гомозиготных трансгенных растений. Кроме того, пыльца, полученная от регенерированных трансгенных растений, может быть скрещена с нетрансгенными растениями, предпочтительно инбредными линиями агрономически важных видов. Описания способов селекции, которые обычно используются для различных признаков и культур, можно найти в одном из нескольких справочников, см., например, Allard, Principles of Plant Breeding, John Wiley & Sons, NY, U. of CA, Davis, CA, 50-98, 1960; Simmonds, Principles of crop improvement, Longman, Inc., NY, 369-399, 1979; Sneep and Hendriksen, Plant breeding perspectives, Wageningen (ed), Center for Agricultural Publishing and Documentation, 1979; Fehr, Soybeans: Improvement, Production and Uses, 2nd Edition, Monograph, 16: 249, 1987; Fehr, Principles of variety development, Theory and Technique, (Vol. 1) и Crop Species Soybean (Vol. 2), Iowa State Univ., Macmillan Pub. Co., NY, 360-376, 1987. И наоборот, пыльца от нетрансгенных растений может использоваться для опыления регенерированных трансгенных растений.
Трансформированные растения могут быть проанализированы на наличие представляющих интерес генов, и на уровень экспрессии и/или профиль, придаваемый регуляторными элементами по настоящему изобретению. Специалистам в рассматриваемой области техники известны многочисленные способы, доступные для анализа трансформированных растений. Например, способы анализа растений включают, в качестве неограничивающих примеров, саузерн-блоты или нозерн-блоты, подходы на основе ПЦР, биохимические анализы, фенотипические способы скрининга, оценки полей и иммунодиагностические тесты. Экспрессия транскрибируемой полинуклеотидной молекулы может быть определена с использованием реагентов TaqMan® (Applied Biosystems, Foster City, CA) и способов, как описано производителем, и временами ПЦР-цикла, определяемыми с помощью матрицы тестирования TaqMan®. Кроме того, реагенты Invader® (Third Wave Technologies, Madison, WI) и способы, как описано изготовителем, могут быть использованы для оценки экспрессии трансгена.
Семена растений по настоящему изобретению могут быть собраны из фертильных трансгенных растений и использованы для выращивания потомства поколений трансформированных растений по настоящему изобретению, в том числе линий гибридных растений, включающих конструкции по этому изобретению и экспрессирующих ген, представляющий агрономических интерес.
Настоящее изобретение также предоставляет части растений по настоящему изобретению. Части растений, без ограничения, включают листья, стебли, корни, клубни, семена, эндосперм, семяпочку и пыльцу. Изобретение также включает в себя и предоставляет трансформированные растительные клетки, которые содержат молекулу нуклеиновых кислот по настоящему изобретению.
Трансгенное растение может передать трансгенную полинуклеотидную молекулу своему потомству. Потомство включает в себя любую регенерируемую часть растения или семени, содержащую трансген, полученный от растения-предка. Трансгенное растение является предпочтительно гомозиготным по трансформированной полинуклеотидной молекуле и передает эту последовательность всему потомству в результате полового размножения. Потомство может быть выращено из семян, полученных от трансгенных растений. Эти дополнительные растения могут быть самоопыляемыми для создания подлинной линии разведения растений. Потомство от этих растений оценивается, среди прочего, на генную экспрессию. Генную экспрессию можно детектировать некоторыми общими способами, такими как вестерн-блоттинг, нозерн-блоттинг, иммунопреципитация и ELISA.
Имея теперь описание изобретения в целом, то же самое будет более легко понять посредством ссылки на следующие примеры, которые приведены в качестве иллюстрации и не предназначены для ограничения настоящего изобретения, если не указано. Специалисты в рассматриваемой области техники смогут оценить, что способы, раскрытые в следующих примерах, представляют способы, раскрытые авторами изобретения для хорошего функционирования в практике изобретения. Однако специалисты в рассматриваемой области техники должны оценить, в свете настоящего раскрытия, что многие изменения могут быть сделаны в конкретных вариантах осуществления, которые раскрыты, и, тем не менее, получить похожий или аналогичный результат без отступления от духа и объема изобретения, поэтому все материалы, изложенные или показанные на сопроводительных чертежах, следует интерпретировать как иллюстративные, а не в ограничивающем смысле.
ПРИМЕРЫ
Пример 1
Регуляторные элементы, выделенные из Zea mays, и группа соответствующих транскрипционных регулирующих экспрессию элементов
Регуляторные элементы были выделены из Zea mays, и были сконструированы последовательности группы транскрипционных регулирующих экспрессию элементов (ЕХР), содержащие регуляторные элементы Zea mays.
Ранняя посадка семян кукурузы еще в начале апреля на севере США создает потенциально опасные риски для семян кукурузы. Например, продолжительные холодные и влажные полевые условия препятствуют оптимальному прорастанию и появлению всходов. С помощью профилирования транскриптов и последующей характеризации было идентифицировано несколько генов-кандидатов, которые продемонстрировали профиль экспрессии, полезный для развития семян и всхожести. Промотор и лидер гена-кандидата, в настоящем документе обозначенные как P-Zm.Nac-1:1:2 (SEQ ID NO: 2) и L-Zm.Nac-1:1:1 (SEQ ID NO: 4), соответственно, были амплифицированы из геномной ДНК кукурузы, и клонированы и секвенированы.
Праймеры для амплификации были сконструированы на основе собственных и общедоступных геномных и EST последовательностей, которые затем были использованы с библиотеками GenomeWalker™ (Clontech Laboratories, Inc., Mountain View, CA), сконструированными в соответствии с протоколом производителя, для клонирования 5’-области, соответствующей геномной последовательности ДНК. С помощью этой последовательности регуляторные элементы были идентифицированы с использованием подходов биоинформатики в пределах 5’-области гена. Используя результаты этого анализа, регуляторные элементы были определены в пределах 5’-последовательности выше кодирующей последовательности гена. Затем были спроектированы праймеры для амплификации регуляторных элементов. Соответствующие молекулы ДНК для каждого регуляторного элемента амплифицировали, используя стандартные условия ПЦР с праймерами, содержащими
уникальные сайты рестрикции, и геномной ДНК, выделенной из Zea mays. Эта клонированная последовательность включала промотор и 5'UTR последовательности выше кодирующей белок области для гена Zea mays. Полученный ДНК-фрагмент лигировали в основные векторы экспрессии растений с использованием стандартных способов клонирования и секвенирования ДНК.
Последовательности идентифицированных групп транскрипционных регулирующих экспрессию элементов («ЕХР») представлены в настоящем документе как SEQ ID NO: 1, 6 и 8, как указано в таблице 1 ниже. Промоторная последовательность представлена в настоящем документе как SEQ ID NO: 2. Лидерная последовательность представлена в настоящем документе как SEQ ID NO: 4. Последовательности интронов представлены в настоящем документе как SEQ ID NO: 5, 7 и 9.
Как показано в таблице 1, например, группа транскрипционных регулирующих экспрессию элементов (ЕХР), обозначенная ЕХР-Zm.Nac+Zm.DnaK:1:1 (SEQ ID NO: 1), с компонентами, выделенными из Zea mays, включает промоторный элемент P-Zm.Nac-1:1:2 (SEQ ID NO: 2), функционально связанный с 5'-концом лидерного элемента
L-Zm.Nac-1:1:1 (SEQ ID NO: 4), функционально связанного с 5'-концом интронного элемента I-Zm.DnaK-1:1:1 (SEQ ID NO: 5). Другие EXP связаны точно так же, как описано в таблице 1.
Пример 2
Анализ EXP-Zm.Nac+Zm.DnaK:1:1 (SEQ ID NO:1), направляющего GUS в трансгенную кукурузу F1
Растения кукурузы трансформировали экспрессионным вектором растений pMON73501, содержащим группу транскрипционных регулирующих экспрессию элементов EXP-Zm.Nac+Zm.DnaK:1:1 (SEQ ID NO: 1), направляющим экспрессию трансгена β-глюкуронидазы (GUS), и полученные в результате растения были проанализированы на экспрессию белка GUS.
Последовательность EXP была клонирована в бинарные трансформационные плазмидные конструкции растений с использованием способов, известных в рассматриваемой области техники. Полученное растение экспрессионной плазмидной конструкции, pMON73501, содержало область правой границы от A. tumefaciens, первую трансгенную кассету для тестирования группы транскрипционных регулирующих экспрессию элементов EXP-Zm.Nac+Zm.DnaK:1:1 (SEQ ID NO: 1), функционально связанную с кодирующей последовательностью β-глюкуронидазы (GUS, SEQ ID NO: 14), функционально связанной с 5'-концом 3'UTR области Т-AGRtu.nos-1:1:13 (SEQ ID NO: 10); вторую трансгенную селекционную кассету, используемую для отбора трансформированных растительных клеток, которая придает устойчивость к гербициду глифосату, направляемую промотором 35S CaMV, EXP-CaMV.35S:1:1 (SEQ ID NO: 15), и область левой границы от А. tumefaciens. Результирующая плазмида была использована для трансформации растений кукурузы.
Растения кукурузы были трансформированы вектором для экспрессии GUS pMON73501. Поколение трансформантов R0, отобранных по однокопийным вставкам, было скрещено с нетрансформированными растениями LH244 для получения F1-популяции трансформантов. Уровни экспрессии GUS измеряли в выбранных тканях в ходе развития. Ткани F1, использованные для данного исследования, включали: набухшие зародыши семян, набухший эндосперм семян, корень и колеоптиль на 3 день после прорастания (DAG); листья и корень в стадии V3; корень и зрелые листья в стадии V7; корень, зрелые и увядающие листья, стержень початка, шелк, междоузлие, пыльник и пыльцу в стадии VT (при выметывании, до размножения); ядра 7 дней после опыления (DAP); и зародыш и эндосперм 21 и 35 DAP. Выбранные образцы тканей были также проанализированы для растений F1, подвергшихся воздействию условий засухи и холодового стресса. V3-корень и листовая ткань были отобраны после воздействия холода и засухи, а также через два дня после восстановления от воздействия холода (2 DAR).
Стресс вследствие засухи индуцировали у растений F1, V3 путем удержания полива в течение 4 дней, что позволяет содержанию воды снизиться, по меньшей мере, на 50% от исходного содержания воды в полностью политом растении. Протокол засухи состоял по существу из следующих шагов. Растения на стадии V3 были лишены воды. Поскольку растение кукурузы испытывает засуху, форма листьев будет меняться от обычного здорового и развернутого внешнего вида на лист, демонстрирующий свернутость в mid-rib сосудисто-волокнистом пучке и появление V-образной формы, если смотреть от кончика листа к стеблю. Это изменение в морфологии обычно начинало происходить примерно через 2 дня после прекращения полива и, как было показано в более ранних экспериментах, связано с потерей около 50% воды, что определяется по массе горшков до прекращения полива и массе горшков, когда наблюдается морфология курчавости листьев у обезвоженных растений. Растения считались находящимися в условиях засухи, когда листья увядали, что проверялось по внутреннему скручиванию (V-форме) листа. Этот уровень стресса считается формой летального стресса. После того как каждое растение продемонстрировало индукцию засухи, как определено выше, растения разрушали для получения образцов как корня, так и листьев. Использовались четыре растения для каждого вектора, и определение GUS проводили, как описано ниже.
В дополнение к засухе, F1 прорастающая рассада и F1-растения в стадии V3, трансформированные pMON73501, также подвергались воздействию условий холода, чтобы определить, показали ли регуляторные элементы индуцированную холодом экспрессию GUS. Шестьдесят семян, полученных из шести семян каждого из 10 событий трансформации, тестировали на индукцию генной экспрессии в условиях холода. Семена проращивали в чашках Петри на водонасыщенной фильтровальной бумаге. Через три дня после появления всходов рассаду подвергали холодовому стрессу путем размещения чашек Петри, содержащих проросшую рассаду, в темную вегетационную камеру с установленной температурой 10°C в течение 24 часов. В конце 24-часового периода ткани корня и колеоптиля отбирали для количественной оценки GUS-экспрессии, как описано ниже. Целые растения тестировали на индукцию экспрессии GUS при холодовом стрессе на стадии V3. Двадцать растений кукурузы на стадии V3, состоящих из 2 растений от каждого из 10 событий трансформации, подвергали температуре 12°C в вегетационной камере в течение 24 часов. Растения в вегетационной камере выращивали в потоке белого света 800 мкмоль/м2⋅с со световым циклом десять часов белого света и четырнадцать часов темноты. После воздействия холода ткани листьев и корня отбирали для количественной оценки GUS-экспрессии.
Гистохимический анализ GUS был использован для качественного анализа экспрессии трансформированных растений. Срезы тканей целиком инкубировали с раствором для окрашивания GUS X-Gluc (5-бромо-4-хлоро-3-индолил-b-глюкуронид) (1 мг/мл) в течение соответствующего отрезка времени, промывали и визуально проверяли на наличие синей окраски. GUS-активность качественно определяли путем непосредственного визуального осмотра или осмотра под микроскопом с использованием выбранных органов и тканей растений.
Для количественного анализа общий белок экстрагировали из выбранных тканей трансформированных растений кукурузы. Один микрограмм общего белка использовали с флуорогенным субстратом 4-метилумбеллиферил-β-D-глюкуронидом (MUG) в суммарной реакции объемом 50 мкл. Флуоресценцию измеряли с помощью возбуждения при 365 нм, эмиссии при 445 нм, используя Fluoromax-3 (Horiba; Киото, Япония) с ридером Micromax, ширина щели устанавливалась при возбуждении 2 нм и 3 нм при эмиссии.
Таблица 2, приведенная ниже, показывает средний уровень экспрессии GUS в выбранных тканях в растениях F1, трансформированных pMON73501.
Средние значения экспрессии GUS в выбранных тканях и обработки поколения F1 трансформированных растений кукурузы, трансформированных pMON73501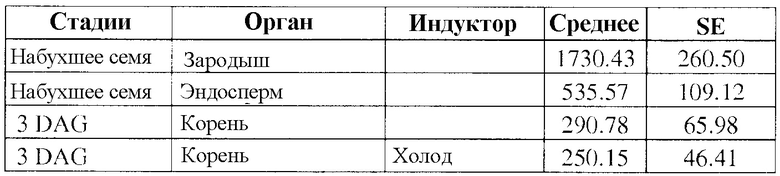
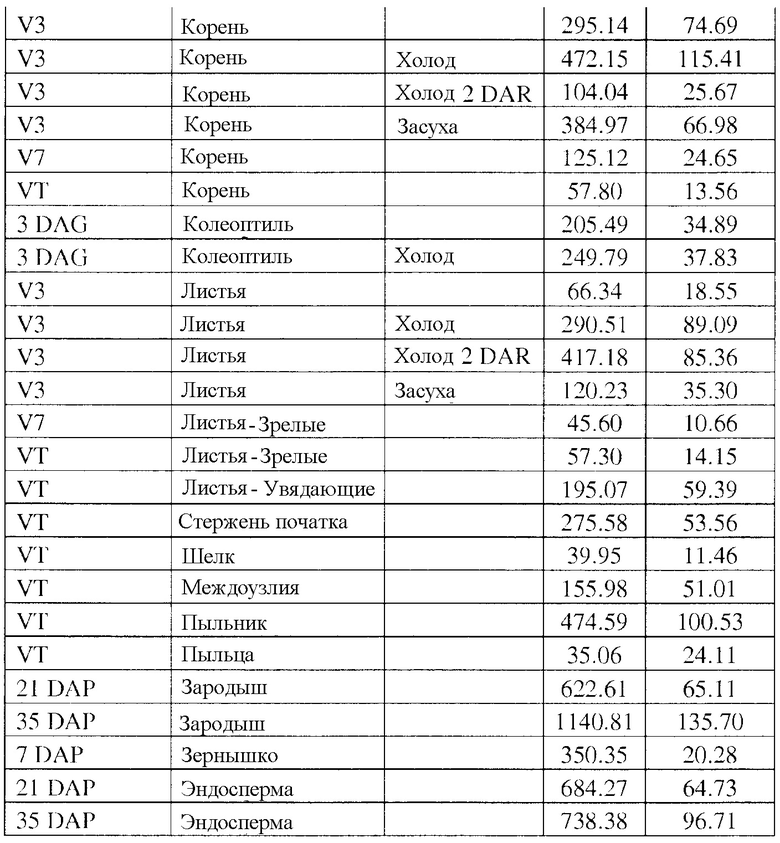
Как видно из таблицы 2, характерной особенностью, с учетом экспрессии, обусловленной последовательностью ЕХР, ЕХР-Zm.Nac+Zm.DnaK:1:1 (SEQ ID NO: 1), включающей промотор и лидер P-Zm.Nac-1:1:2 (SEQ ID NO: 2) и L-Zm.Nac-1:1:1 (SEQ ID NO: 4), был более высокий уровень экспрессии, наблюдаемый в набухших зародышах семян и тканях эндосперма по отношению к образцам других тканей. Этот высокий уровень экспрессии может придавать преимущества при проращивании семян, экспрессирующих трансгены, полезные при обеспечении защиты от холодных и влажных стрессовых условий. Экспрессия также была выше по сравнению с другими тканями зародыша и эндосперма в период раннего развития семян (21 и 35 DAP). Такой профиль экспрессии может также быть выгодным для облегчения прорастания и роста полученного в результате семени. Например, белок или продукты, полученные от экспрессии трансгенов, функционально связанных с промотором Nac Zea mays и лидером, экспрессировали на ранних стадиях развития семян, что позволило бы накапливать белок или производные продукты в развивающемся семени для сохранности для быстрого использования при прорастании в холодных или влажных условиях. Экспрессия при прорастании обеспечила бы дополнительные преимущества семени, направляющему нужные трансгены, путем обеспечения экспрессии дополнительного белка или производного продукта в критическое время в холодных и влажных стрессовых условиях. Небольшое индуцирование холодом наблюдалось также в листьях и корне V3 в этом эксперименте.
Пример 3
Анализ регуляторных элементов, управляющих GUS в трансгенной кукурузе R0
Растения кукурузы были трансформированы векторами экспрессии растений, содержащими последовательности ЕХР, управляющими экспрессией трансгена β-глюкуронидазы (GUS), и полученные в результате растения анализировали на экспрессию белка GUS. Группы транскрипционных регулирующих экспрессию элементов были клонированы в бинарные трансформационные плазмидные конструкции растений с использованием способов, известных в рассматриваемой области техники.
Полученные в результате экспрессионные плазмидные конструкции растений, содержащие область правой границы от А. tumefaciens, первую трансгенную кассету для тестирования последовательности ЕХР и комбинацию 3'UTR, состоящую из последовательности ЕХР (указана в таблице 3), функционально связанной с последовательностью, кодирующей β-глюкуронидазу (GUS, SEQ ID NO: 14), функционально связаны 5' к 3'-области (указаны в таблице 3); вторую кассету трансгенной селекции, используемую для селекции трансформированных растительных клеток, которая придает устойчивость к гербициду глифосату (СР4, US RE39247, управляемый промотором актина 1 риса, EXP-Os.Act1:1:1, SEQ ID NO: 16), и область левой границы от А. tumefaciens. Полученные в результате плазмиды были использованы для трансформации растений кукурузы. В таблице 3 перечислены плазмидные обозначения, группы транскрипционных регулирующих экспрессию элементов, которые также приведены в таблице 1, и 3'UTR, которые были размещены в функциональной связи с последовательностью, кодирующей GUS. Каждая плазмидная конструкция состояла из уникальной трансгенной кассетной конфигурации, состоящей из конкретных интронов и 3'-UTR, размещенных в функциональной связи с промотором и лидером Р-Zm.Nac-1:1:2 (SEQ ID NO: 2) и L-Zm.Nac-1:1:1 (SEQ ID NO: 4).
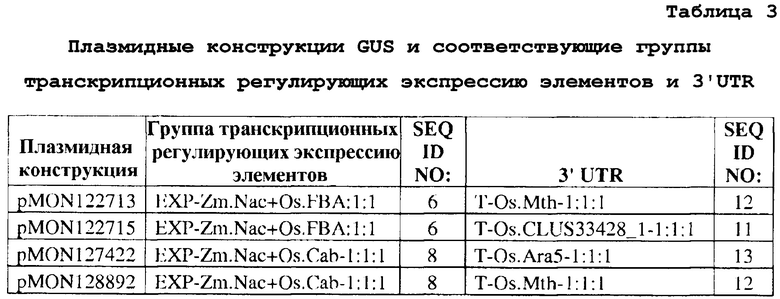
Растения были трансформированы с помощью способов Agrobacterium-опосредованной трансформации, известных в данной области техники. Гистохимический и количественный GUS-анализ проводили, как описано в примере 2 выше. Средние R0 экспрессии GUS, наблюдаемые для каждой GUS-трансгенной кассеты,
трансформированной в растения кукурузы, представлены в таблице 4 ниже.
Средняя R0 GUS экспрессия в корневой и листовой ткани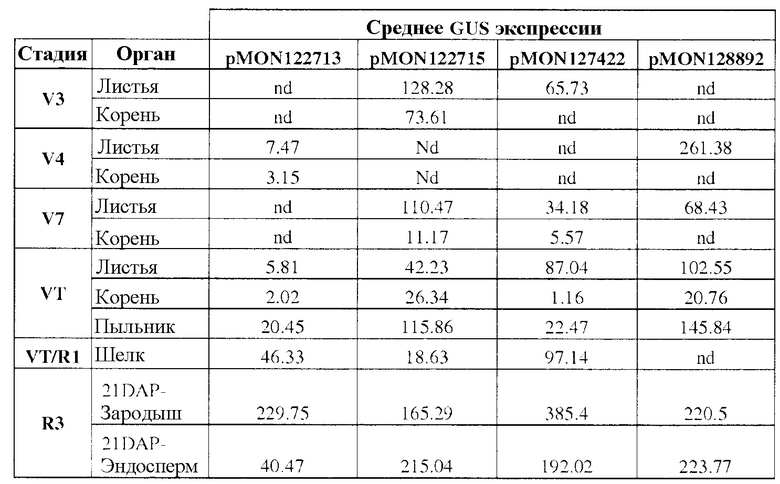
В соответствии с результатами, полученными в примере 2, таблица 4 показывает, что экспрессия была самой высокой в развивающемся зародыше (21 DAP) для всех четырех трансгенных кассет. Фиг. 1 иллюстрирует различные профили экспрессии, которые были приданы каждой трансгенной кассетной конфигурацией. Экспрессия в развивающемся зародыше была самой высокой для трансгенной кассеты, состоящей из последовательности EXP, EXP-Zm.Nac+Os.Cab-1:1:1 (SEQ ID NO: 8), которая была функционально связана с 3'UTR, T-Os.Ara5-1:1:1 (SEQ ID NO: 13), относительно трех других трансгенных кассет. Аналогичные уровни экспрессии наблюдались в развивающемся зародыше и эндосперме растений, трансформированных экспрессионной кассетой, состоящей из группы транскрипционных регулирующих экспрессию элементов, EXP-Zm.Nac+Os.Cab-1:1:1 (SEQ ID NO: 8), которая была функционально связана с 3'UTR, T-Os.Mth-1:1:1 (SEQ ID NO: 12). Для трансгенной кассеты, состоящей из группы транскрипционных регулирующих экспрессию элементов EXP-Zm.Nac+Os.FBA:1:1 (SEQ ID NO:6), которая была функционально связана с 3'UTR, T-Os.Mth-1:1:1 (SEQ ID NO: 12), экспрессия в эндосперме была гораздо ниже по отношению к экспрессии в зародыше. Для трансгенной кассеты, состоящей из группы транскрипционных регулирующих экспрессию элементов, EXP-Zm.Nac+Os.FBA:1:1 (SEQ ID NO: 6), которая была функционально связана с 3'UTR, T-Os.CLUS33428_1-1:1:1 (SEQ ID NO: 11), экспрессия в зародыше была ниже по отношению к экспрессии в эндосперме. Каждая конфигурация трансгенной кассеты предоставила уникальный профиль экспрессии в развивающемся R0-семени.
Различия в экспрессии в корне, листьях, пыльнике и шелке, а также в развивающемся семени растений, трансформированных четырьмя различными трансгенными кассетами, также наблюдались. Фиг. 2 иллюстрирует различные профили экспрессии, которые были приданы каждой конфигурацией трансгенной кассеты в каждой из выше описанных тканей. Например, экспрессия в листьях была выше в трансгенных кассетах, содержащих последовательность EXP, EXP-Zm.Nac+Os.Cab-1:1:1 (SEQ ID NO: 8), относительно трансгенной кассеты, содержащей EXP-Zm.Nac+Os.FBA:1:1 (SEQ ID NO: 6). Экспрессия в пыльниках была выше в двух трансгенных кассетах, состоящих из группы транскрипционных регулирующих экспрессию элементов, EXP-Zm.Nac+Os.FBA:1:1 (SEQ ID NO: 6), которая была функционально связана с 3'UTR, T-Os.CLUS33428_1-1:1:1 (SEQ ID NO: 11), и группы транскрипционных регулирующих экспрессию элементов, EXP-Zm.Nac+Os.Cab-1:1:1 (SEQ ID NO:8), которая была функционально связана с 3'UTR, T-Os.Mth-1:1:1 (SEQ ID NO: 12). Каждая из четырех конфигураций трансгенных кассет предоставляла уникальные профили экспрессии в R0-трансформантах.
Имея проиллюстрированные и описанные принципы настоящего изобретения, специалистам в рассматриваемой области техники должно быть очевидно, что изобретение может быть изменено в компоновке и деталях без отхода от этих принципов. Мы претендуем на все изменения, которые находятся в пределах духа и объема изобретения. Все публикации и опубликованные патентные документы, цитируемые в настоящем документе, включены посредством ссылки в той же степени, как если бы каждая индивидуальная публикация или патентная заявка конкретно и индивидуально была указана включенной посредством ссылки.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2644205C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2812893C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2774709C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2675524C2 |
| ЭНХАНСЕР ПАЛОЧКОВИДНОГО ВИРУСА САХАРНОГО ТРОСТНИКА (SCBV) И ЕГО ПРИМЕНЕНИЕ В ФУНКЦИОНАЛЬНОЙ ГЕНОМИКЕ РАСТЕНИЙ | 2013 |
|
RU2639517C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2800423C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2800430C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2800425C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2800424C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2670887C9 |











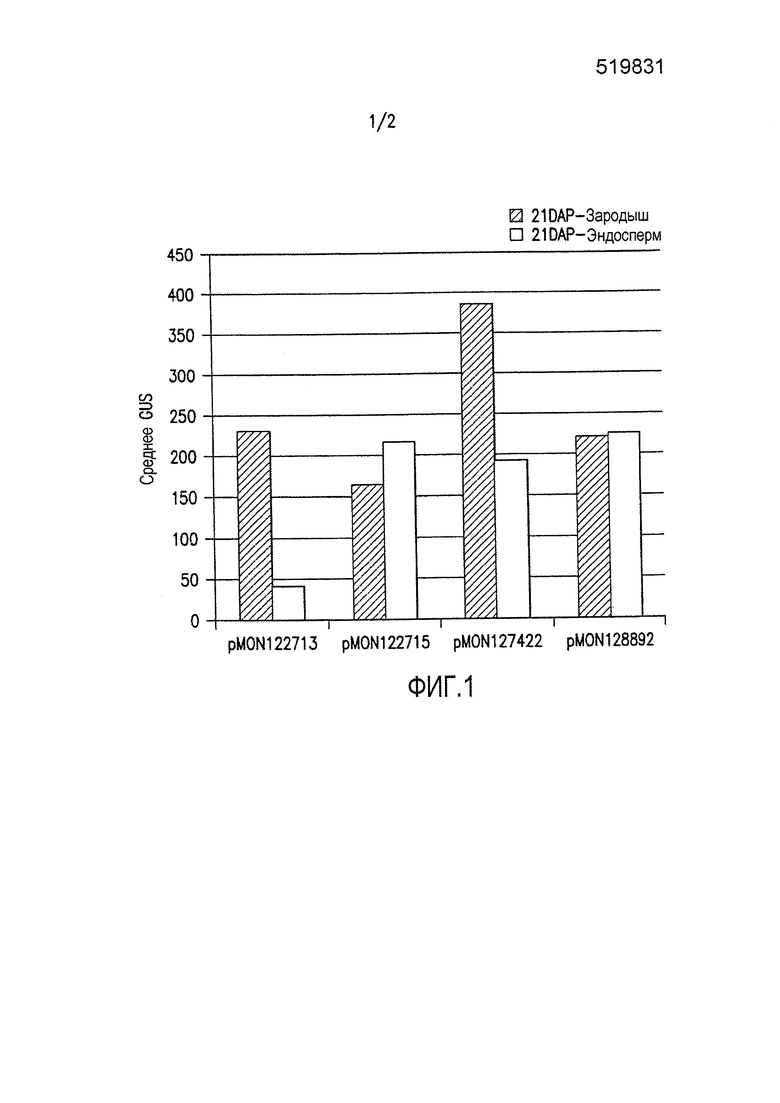
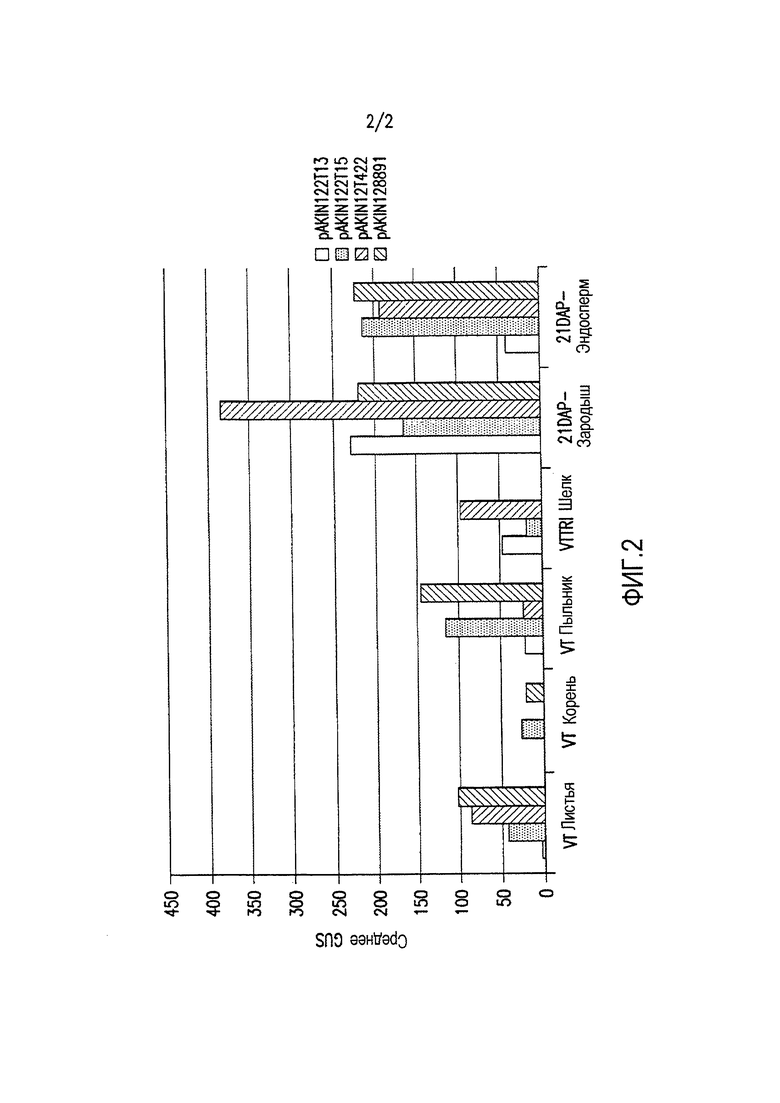
Изобретение относится к области биохимии, в частности к ДНК-конструкции для увеличения экспрессии гетерологичной транскрибируемой полинуклеотидной молекулы, а также к растительной клетке, растению, части трансгенного растения и трансгенному семени, ее содержащим. Также раскрыта трансгенная кассета для увеличения экспрессии гетерологичной транскрибируемой полинуклеотидной молекулы, содержащая вышеуказанную ДНК-конструкцию. Изобретение также относится к способу получения товарного продукта из вышеуказанного трансгенного растения или его части, а также к способу экспрессии транскрибируемой полинуклеотидной молекулы, включающему получение вышеуказанного трансгенного растения. Изобретение позволяет эффективно увеличить экспрессию гетерологичной транскрибируемой полинуклеотидной молекулы в клетке растения. 8 н. и 11 з.п. ф-лы, 2 ил., 4 табл., 3 пр.
1. ДНК-конструкция для увеличения экспрессии гетерологичной транскрибируемой полинуклеотидной молекулы, содержащая последовательность ДНК, выбранную из группы, состоящей из:
a) последовательности с по меньшей мере 95% идентичностью последовательности с любой из SEQ ID NO: 1, 2, 3, 6 или 8 полной длины, и обладающей геннорегуляторной активностью; и
b) последовательности, содержащей любую из SEQ ID NO: 1, 2, 3, 6 или 8;
где указанная последовательность функционально связана с гетерологичной транскрибируемой полинуклеотидной молекулой.
2. ДНК-конструкция по п. 1, где указанная последовательность имеет по меньшей мере 95% идентичностью последовательности с последовательностью ДНК любой из SEQ ID NO: 1, 2, 3, 6 или 8.
3. ДНК-конструкция по п. 1, где последовательность ДНК обладает геннорегулирующей активностью.
4. ДНК-конструкция по п. 1, где гетерологичная транскрибируемая полинуклеотидная молекула содержит ген, представляющий агрономический интерес.
5. ДНК-конструкция по п. 4, где ген, представляющий агрономический интерес, придает растениям устойчивость к гербицидам.
6. ДНК-конструкция по п. 4, где ген, представляющий агрономический интерес, придает растениям устойчивость к вредителям.
7. Трансгенная растительная клетка, обладающая увеличенной экспрессией гетерологичной транскрибируемой полинуклеотидной молекулы по сравнению с клеткой, не содержащей конструкцию по п. 1, содержащая гетерологичную молекулу ДНК, содержащую последовательность, выбранную из группы, состоящей из:
a) последовательности с по меньшей мере 95% идентичностью последовательности с любой из SEQ ID NO: 1, 2, 3, 6 или 8 полной длины, и обладающей геннорегуляторной активностью; и
b) последовательности, содержащей любую из SEQ ID NO: 1, 2, 3, 6 или 8;
где указанная последовательность функционально связана с гетерологичной транскрибируемой полинуклеотидной молекулой.
8. Трансгенная растительная клетка по п. 7, где указанная трансгенная клетка является клеткой однодольного растения.
9. Трансгенная растительная клетка по п. 7, где указанная трансгенная клетка является клеткой двудольного растения.
10. Трансгенное растение, содержащее ДНК-конструкцию по п. 1, которое обладает увеличенной экспрессией гетерологичной транскрибируемой полинуклеотидной молекулы по сравнению с растением, не содержащим конструкцию по п. 1.
11. Часть трансгенного растения, содержащая ДНК-конструкцию по п. 1, обладающая увеличенной экспрессией гетерологичной транскрибируемой полинуклеотидной молекулы по сравнению с частью растения, не содержащей конструкцию по п. 1.
12. Трансгенное семя, содержащее ДНК-конструкцию по п. 1, которое обладает увеличенной экспрессией гетерологичной транскрибируемой полинуклеотидной молекулы по сравнению с семенем, не содержащим конструкцию по п. 1.
13. Трансгенная кассета для увеличения экспрессии гетерологичной транскрибируемой полинуклеотидной молекулы, содержащая ДНК-конструкцию по п. 1, где указанная последовательность ДНК включает любую из SEQ ID NO: 1, 6 и 8, и где указанная гетерологичная транскрибируемая полинуклеотидная молекула функционально связана с 3'UTR, выбранной из группы, состоящей из SEQ ID NO: 10, 11, 12 и 13.
14. Трансгенная кассета по п. 13, где указанная последовательность ДНК включает SEQ ID NO: 1, и где указанная гетерологичная транскрибируемая полинуклеотидная молекула функционально связана с 3'UTR, представленной как SEQ ID NO: 10.
15. Трансгенная кассета по п. 13, где указанная последовательность ДНК включает SEQ ID NO: 6, и где указанная гетерологичная транскрибируемая полинуклеотидная молекула функционально связана с 3'UTR, выбранной из группы, состоящей из SEQ ID NO: 11 и 12.
16. Трансгенная кассета по п. 13, где указанная последовательность ДНК включает SEQ ID NO: 8, и где указанная гетерологичная транскрибируемая полинуклеотидная молекула функционально связана с 3'UTR, выбранной из группы, состоящей из SEQ ID NO: 12 и 13.
17. Способ получения товарного продукта, включающий получение трансгенного растения по п. 10 или части трансгенного растения по п. 11 и получение из него товарной продукции.
18. Способ по п. 17, где товарный продукт представляет собой белковый концентрат, белковый изолят, зерно, крахмал, семена, шрот, муку, биомассу или масло из семян.
19. Способ экспрессии транскрибируемой полинуклеотидной молекулы, включающий получение трансгенного растения по п. 10 и культивирование растения, в котором экспрессируется транскрибируемый полинуклеотид ДНК-конструкции по п. 1.
| US 2009106857 A1, 23.04.2009 | |||
| US 2011177228 A1, 21.07.2011 | |||
| PURANIK S | |||
| ET AL, NAC proteins: regulation and role in stress tolerance, TRENDS IN PLANT SCIENCE, vol | |||
| Печь для сжигания твердых и жидких нечистот | 1920 |
|
SU17A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Прибор для сжигания нефти | 1921 |
|
SU369A1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА PGBP450F ДЛЯ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ И СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ ТАБАКА С ПОВЫШЕННОЙ ПРОДУКТИВНОСТЬЮ И УСТОЙЧИВОСТЬЮ К ГРИБНЫМ ФИТОПАТОГЕНАМ | 2002 |
|
RU2237717C2 |

