ССЫЛКА НА СВЯЗАННУЮ ЗАЯВКУ
[01] В данной заявке заявляется приоритет предварительной заявки США №62/714228, поданной 3 августа 2018 г., которая полностью включена в данный документ посредством ссылки.
ВКЛЮЧЕНИЕ ПЕРЕЧНЯ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[02] Перечень последовательностей, содержащийся в файле, названном «38-21-62691-0001_Seqlist_ST25.txt», составляет 31060 байт (согласно измерениям Microsoft Windows®), был создан 2 июля 2019 г., подан в электронном виде и включен в данный документ посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
[03] Изобретение относится к области молекулярной биологии растений и генной инженерии растений. Более конкретно, изобретение относится к молекулам ДНК, используемым для модуляции экспрессии генов в растениях.
УРОВЕНЬ ТЕХНИКИ [04] Регуляторные элементы - это генетические элементы, которые регулируют активность генов, модулируя транскрипцию функционально связанной молекулы транскрибируемой ДНК. Такие элементы могут включать промоторы, лидерные последовательности, интроны и 3' нетранслируемые участки и применимы в области молекулярной биологии растений и генной инженерии растений.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[05] Изобретение относится к новым синтетическим регуляторным элементам генов для применения у растений. Изобретение также относится к конструкциям молекул рекомбинантной ДНК, содержащих регуляторные элементы. Данное изобретение также относится к трансгенным клеткам растений, растениям и семенам, содержащим регуляторные элементы. В одном варианте осуществления данного изобретения, регуляторные элементы функционально связаны с молекулой транскрибируемой ДНК. В некоторых вариантах осуществления данного изобретения, молекула транскрибируемой ДНК может быть гетерологичной по отношению к регуляторной последовательности. Таким образом, последовательность регуляторного элемента, относящаяся к изобретению, в конкретных вариантах осуществления данного изобретения может быть определена как функционально связанная с гетерологичной молекулой транскрибируемой ДНК. Данное изобретение также относится к способам применения регуляторных элементов и получения и применения молекул рекомбинантной ДНК, содержащих регуляторные элементы, и трансгенных клеток растений, растений и семян, содержащих регуляторные элементы, функционально связанные с молекулой транскрибируемой ДНК.
[06] Таким образом, в одном аспекте изобретение относится к молекуле рекомбинантной ДНК, содержащей последовательность ДНК, выбранную из группы, состоящей из: (а) последовательности, по меньшей мере на около 85 процентов идентичной любой последовательности из SEQ ID NO: 1-19 и SEQ ID NO: 26; (b) последовательности, содержащей любую из SEQ ID NO: 1-19 и SEQ ID NO: 26; и (с) фрагмента любой из SEQ ID NO: 1-19 и SEQ ID NO: 26, при этом этот фрагмент обладает ген-регуляторной активностью; при этом последовательность функционально связана с гетерологичной молекулой транскрибируемой ДНК. Под «гетерологичной молекулой транскрибируемой ДНК» подразумевается, что молекула транскрибируемой ДНК является гетерологичной по отношению к полинуклеотидной последовательности, с которой она функционально связана. В конкретных вариантах осуществления данного изобретения, молекула рекомбинантной ДНК содержит последовательность ДНК, имеющую по меньшей мере около 85 процентов, по меньшей мере около 86 процентов, по меньшей мере около 87 процентов, по меньшей мере около 88 процентов, по меньшей мере около 89 процентов, по меньшей мере около 90 процентов, по меньшей мере 91 процент, по меньшей мере 92 процента, по меньшей мере 93 процента, по меньшей мере 94 процента, по меньшей мере 95 процентов, по меньшей мере 96 процентов, по меньшей мере 97 процентов, по меньшей мере 98 процентов или по меньшей мере 99 процентов идентичности последовательности с последовательностью ДНК любой из SEQ ID NO: 1-19 и SEQ ID NO: 26. В конкретных вариантах осуществления данного изобретения, последовательность ДНК содержит регуляторный элемент. В некоторых вариантах осуществления данного изобретения, регуляторный элемент содержит промотор. В других вариантах осуществления данного изобретения, регуляторный элемент содержит интрон. В других вариантах осуществления данного изобретения, регуляторный элемент содержит 3' UTR. В других вариантах осуществления данного изобретения, гетерологичная молекула транскрибируемой ДНК содержит ген, представляющий агрономический интерес, такой как ген, способный обеспечивать устойчивость растений к гербицидам, или ген, способный обеспечивать устойчивость растений к вредителям. В других вариантах осуществления данного изобретения, гетерологичная молекула транскрибируемой ДНК содержит последовательность, кодирующую малую РНК, такую как дцРНК, микроРНК или миРНК. В еще других вариантах осуществления данного изобретения, изобретение относится к конструкции, содержащей молекулу рекомбинантной ДНК, как предусмотрено в данном документе.
[07] В другом аспекте в данном документе представлены трансгенные клетки растений, содержащие молекулу рекомбинантной ДНК, содержащую последовательность ДНК, выбранную из группы, состоящей из:(а) последовательности, по меньшей мере на около 85 процентов идентичной любой последовательности из SEQ ID NO: 1-19 и SEQ ID NO: 26; (b) последовательности, содержащей любую из SEQ ID NO: 1-19 и SEQ ID NO: 26; и (с) фрагмента любой из SEQ ID NO: 1-19 и SEQ ID NO: 26, при этом этот фрагмент обладает ген-регуляторной активностью; при этом последовательность ДНК функционально связана с гетерологичной молекулой транскрибируемой ДНК. В некоторых вариантах осуществления данного изобретения, трансгенная клетка растения представляет собой клетку однодольного растения. В других вариантах осуществления данного изобретения, трансгенная клетка растения представляет собой клетку двудольного растения.
[08] В еще одном аспекте в данном документе дополнительно представлено трансгенное растение или его часть, содержащее молекулу рекомбинантной ДНК, содержащую последовательность ДНК, выбранную из группы, состоящей из: а) последовательности, по меньшей мере на около 85 процентов идентичной любой последовательности из SEQ ID NO: 1-19 и SEQ ID NO: 26; b) последовательности, содержащей любую из SEQ ID NO: 1-19 и SEQ ID NO: 26; и с) фрагмента любой из SEQ ID NO: 1-19 и SEQ ID NO: 26, при этом этот фрагмент обладает ген-регуляторной активностью; при этом последовательность функционально связана с гетерологичной молекулой транскрибируемой ДНК. В конкретных вариантах осуществления данного изобретения, трансгенное растение представляет собой растение-потомка любого поколения, которое содержит молекулу рекомбинантной ДНК. В данном документе также представлено трансгенное семя, содержащее молекулу рекомбинантной ДНК, которое дает такое трансгенное растение при выращивании.
[09] В другом аспекте изобретение относится к способу получения товарного продукта, включающему получение трансгенного растения или его части, содержащего молекулу рекомбинантной ДНК по изобретению, и получение из него товарного продукта. В одном варианте осуществления данного изобретения, товарный продукт представляет собой семена, обработанные семена, концентрат белка, изолят белка, крахмал, зерна, части растений, масло семян, биомассу, муку и муку грубого помола.
[10] В еще одном аспекте изобретение относится к способу получения трансгенного растения, содержащего молекулу рекомбинантной ДНК по изобретению, включающему трансформацию клетки растения с помощью молекулы рекомбинантной ДНК по изобретению для получения трансформированной клетки растения и регенерацию трансгенного растения из трансформированной клетки растения.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[11] SEQ ID NO: 1 представляет собой последовательность ДНК группы синтетических регуляторных элементов экспрессии (EXP), EXP-Zm.GSP850, содержащую синтетический промотор (P-Zm.GSP850.nno:4), функционально связанный с 5'-концом синтетической лидерной последовательности (L-Zm.GSP850.nno:3). [12] SEQ ID NO: 2 представляет собой последовательность ДНК синтетического промотора P-Zm.GSP850.nno:4.
[13] SEQ ID NO: 3 представляет собой последовательность ДНК синтетической лидерной последовательности L-Zm.GSP850.nno:3.
[14] SEQ ID NO: 4 представляет собой последовательность ДНК синтетического EXP, EXP-Zm.GSP850.nno+Zm.GSI153.nno:2, содержащего синтетический промотор (Р-Zm.GSP850.nno:4), функционально связанный с 5'-концом синтетической лидерной последовательности (L-Zm.GSP850.nno:3), функционально связанной с 5'-концом синтетического интрона (I-Zm.GSI153.nno:1).
[15] SEQ ID NO: 5 представляет собой последовательность ДНК синтетического интрона, I-Zm.GSI153.nno:1.
[16] SEQ ID NO: 6 представляет собой последовательность ДНК синтетического EXP, EXP-Zm.GSP990, содержащую синтетический промотор (P-Zm.GSP990.nno:2), функционально связанный с 5'-концомсинтетической лидерной последовательности (L-Zm.GSP990.nno:1).
[17] SEQ ID NO: 7 представляет собой последовательность ДНК синтетического промотора P-Zm.GSP990.nno:2.
[18] SEQ ID NO: 8 представляет собой последовательность ДНК синтетической лидерной последовательности L-Zm.GSP990.nno:1.
[19] SEQ ID NO: 9 представляет собой последовательность ДНК синтетического EXP, EXP-Zm.GSP990.nno+Zm.GSI197.nno:2, содержащего синтетический промотор (Р-Zm.GSP990.nno:2), функционально связанный с 5'-концом синтетической лидерной последовательности (L-Zm.GSP990.nno:1), функционально связанным с 5'-концом синтетического интрона (I-Zm.GSI197.nno:1).
[20] SEQ ID NO: 10 представляет собой последовательность ДНК синтетического интрона, I-Zm.GSI197.nno:1.
[21] SEQ ID NO: 11 представляет собой последовательность ДНК синтетического EXP, EXP-Zm.GSP850.nno+Zm.GSI140.nno:1, содержащего синтетический промотор (Р-Zm.GSP850.nno:4), функционально связанный с 5'-концом синтетической лидерной последовательности (L-Zm.GSP850.nno:3), функционально связанной с 5'-концом синтетического интрона (I-Zm.GSI140.nno:1).
[22] SEQ ID NO: 12 представляет собой последовательность ДНК синтетического интрона, I-Zm.GSI140.nno:1.
[23] SEQ ID NO: 13 представляет собой последовательность ДНК синтетической 3' UTR, T-Zm.GST9.nno:2.
[24] SEQ ID NO: 14 представляет собой последовательность ДНК синтетической 3' UTR, T-Zm.GST18.nno:2.
[25] SEQ ID NO: 15 представляет собой последовательность ДНК синтетического EXP, EXP-Zm.GSP850.nno+Zm.DnaK:1, содержащего синтетический промотор (Р-Zm.GSP850.nno:4), функционально связанный с 5'-концом синтетической лидерной последовательности (L-Zm.GSP850.nno:3), функционально связанной с 5'-концом интрона (I-Zm.DnaK.:1).
[26] SEQ ID NO: 16 представляет собой последовательность ДНК синтетического EXP, EXP-Zm.GSP990.nno+Zm.DnaK:l, содержащего синтетический промотор (Р-Zm.GSP990.nno:2), функционально связанный с 5'-концом синтетической лидерной последовательности (L-Zm.GSP990.nno:1), функционально связанной с 5'-концом интрона (I-Zm.DnaK:1).
[27] SEQ ID NO: 17 представляет собой последовательность ДНК синтетического энхансера E-Zm.GSP850, который получен из синтетического промотора Р-Zm.GSP850.nno:4.
[28] SEQ ID NO: 18 представляет собой последовательность ДНК синтетического энхансера E-Zm.GSP990, который получен из синтетического промотора Р-Zm.GSP990.nno:2.
[29] SEQ ID NO: 19 представляет собой последовательность ДНК 3' UTR, Т-Sb.Nltp4-1:1:2, полученную из гена NLTP4 (неспецифический белок-переносчик липидов 4) из Sorghum bicolor.
[30] SEQ ID NO: 20 представляет собой синтетическую кодирующую последовательность, оптимизированную для экспрессии β-глюкуронидазы (GUS) в растении с процессируемым интроном, полученным из светоиндуцируемого тканеспецифичного гена ST-LS1 картофеля (Номер доступа в Genbank: Х04753).
[31] SEQ ID NO: 21 представляет собой последовательность ДНК EXP, ЕХР-CaMV.35S, содержащую промотор 35S и лидерную последовательность, полученные из вируса мозаики цветной капусты.
[32] SEQ ID NO: 22 представляет собой последовательность ДНК интрона, I-Zm.DnaK:1, полученную из гена белка теплового шока 70 (Hsp70) (DnaK) из Zea mays.
[33] SEQ ID NO: 23 представляет собой последовательность ДНК 3' UTR, T-Os.LTP:l полученную из гена белок-подобного переносчика липидов (LTP) из Oryza sativa.
[34] SEQ ID NO: 24 представляет собой кодирующую последовательность для β-глюкуронидазы (GUS) с процессируемым интроном, полученным из светоиндуцируемого тканеспецифичного гена ST-LS1 картофеля (Номер доступа в Genbank: Х04753).
[35] SEQ ID NO: 25 представляет собой кодирующую последовательность флуоресцентного белка люциферазы NanoLuc® (Promega, Мэдисон, WI 53711), Nluc который был сконструирован путем направленной эволюции из люциферазы глубоководных креветок (Oplophorus gacilirostris).
[36] SEQ ID NO: 26 представляет собой последовательность ДНК синтетического 3' UTR, T-Zm.GST43.nno:1.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[37] Изобретение относится к синтетическим регуляторным элементам, обладающим ген-регулирующей активностью у растений. Нуклеотидные последовательности этих синтетических регуляторных элементов представлены как SEQ ID NO: 1-18 и SEQ ID NO: 26. Эти синтетические регуляторные элементы способны влиять на экспрессию функционально связанной молекулы транскрибируемой ДНК в тканях растений и, следовательно, регулировать экспрессию гена функционально связанного трансгена в трансгенных растениях. Изобретение также относится к новым эндогенным регуляторным элементам, обладающим ген-регуляторной активностью у растений и представленным как SEQ ID NO: 19. Изобретение также относится к способам модификации, продуцирования и применения молекул рекомбинантной ДНК, которые содержат предложенные синтетические и эндогенные регуляторные элементы. Изобретение также относится к композициям, которые содержат трансгенные клетки растений, растения, части растений и семена, содержащие молекулы рекомбинантной ДНК по изобретению, и способы их получения и их применения.
[38] Следующие определения и способы предоставлены для лучшего определения данного изобретения и для руководства специалистами в данной области техники при практическом применении данного изобретения. Если не указано иное, термины следует понимать в соответствии с их обычным использованием специалистами в соответствующей области техники. Молекулы ДНК
[39] В контексте данного документа, термин «ДНК» или «молекула ДНК» относится к молекуле двухцепочечной ДНК геномного или синтетического происхождения, то есть, к полимеру дезоксирибонуклеотидных оснований или молекуле ДНК, считываемой от 5' (выше рамки считывания) конца до 3' (ниже рамки считывания) конца. В контексте данного документа, термин «последовательность ДНК» относится к нуклеотидной последовательности молекулы ДНК. Используемая в данном документе номенклатура соответствует номенклатуре Раздела 37 Свода федеральных нормативных актов США, § 1.822, и изложена в таблицах стандарта WIPO ST.25 (1998), Приложение 2, Таблицы 1 и 3.
[40] В контексте данного документа, «молекула рекомбинантной ДНК» представляет собой молекулу ДНК, содержащую комбинацию молекул ДНК, которые не могли бы встречаться вместе в природе без вмешательства человека. Например, молекула рекомбинантной ДНК может представлять собой молекулу ДНК, которая состоит по меньшей мере из двух молекул ДНК, гетерологичных по отношению друг к другу, молекулу ДНК, которая содержит последовательность ДНК, которая отклоняется от последовательностей ДНК, существующих в природе, молекулу ДНК, которая содержит синтетическую последовательность ДНК или молекулу ДНК, которая была включена в ДНК клетки-хозяина путем генетической трансформации или редактирования гена.
[41] В контексте данного документа, «синтетическая нуклеотидная последовательность» или «искусственная нуклеотидная последовательность» представляет собой нуклеотидную последовательность, которая, как известно, не встречается или не существует в природе. Ген-регуляторные элементы по данному изобретению содержат синтетические нуклеотидные последовательности. Предпочтительно синтетические нуклеотидные последовательности имеют небольшую гомологию или не расширенную гомологию с природными последовательностями. Расширенная гомология в этом контексте обычно относится к 100% идентичности последовательностей, выходящей за пределы около 25 нуклеотидов смежной последовательности.
[42] Ссылка в этой заявке на «изолированную молекулу ДНК» или эквивалентный термин или фразу предназначена для обозначения того, что молекула ДНК представляет собой молекулу, которая присутствует отдельно или в сочетании с другими композициями, но не в ее природном окружении. Например, элементы нуклеиновой кислоты, такие как кодирующая последовательность, последовательность интрона, нетранслируемая лидерная последовательность, последовательность промотора, последовательность терминации транскрипции и т.п., которые обнаруживаются в природе в ДНК генома организма, не считаются «изолированными» до тех пор, пока элемент находится в геноме организма и в том месте внутри генома, в котором он обнаружен в природе. Однако каждый из этих элементов и части этих элементов будут «изолированы» в рамках данного описания, если элемент не находится в геноме организма и в том месте внутри генома, в котором он обнаружен в природе. Точно так же нуклеотидная последовательность, кодирующая инсектицидный белок или любой встречающийся в природе инсектицидный вариант этого белка, будет изолированной нуклеотидной последовательностью при условии, что нуклеотидная последовательность не находится в ДНК бактерии, в которой последовательность, кодирующая белок, обнаружена в природе. Синтетическая нуклеотидная последовательность, кодирующая аминокислотную последовательность встречающегося в природе инсектицидного белка, будет считаться изолированной для целей этого описания. Для целей данного описания любая трансгенная нуклеотидная последовательность, то есть, нуклеотидная последовательность ДНК, вставленная в геном клеток растения или бактерии или присутствующая во внехромосомном векторе, будет считаться изолированной нуклеотидной последовательностью независимо от того, присутствует ли она в плазмиде или аналогичной структуре, используемой для трансформации клеток, в геноме растения или бактерии, или присутствует в определяемых количествах в тканях, потомстве, биологических образцах или товарных продуктах, полученных из растения или бактерии.
[43] В контексте данного документа, термин «идентичность последовательностей» относится к степени, в которой две оптимально выровненные полинуклеотидные последовательности или две оптимально выровненные полипептидные последовательности идентичны. Оптимальное выравнивание последовательностей создается путем ручного выравнивания двух последовательностей, например, эталонной последовательности и другой последовательности, чтобы максимизировать количество совпадений нуклеотидов при выравнивании последовательностей с соответствующими внутренними нуклеотидными вставками, делециями или гэпами. В контексте данного документа, термин «эталонная последовательность» относится к последовательности ДНК, представленной как SEQ ID NO: 1-19 и SEQ ID NO: 26.
[44] В контексте данного документа, термин «процент идентичности последовательностей» или «процент идентичности» или «% идентичности» представляет собой долю идентичности, умноженную на 100. «Доля идентичности» для последовательности, оптимально выровненной с эталонной последовательностью, представляет собой количество совпадений нуклеотидов в оптимальном выравнивании, деленное на общее количество нуклеотидов в эталонной последовательности, например, общее количество нуклеотидов по всей длине всей эталонной последовательности. Таким образом, один вариант осуществления данного изобретения относится к молекуле ДНК, содержащей последовательность, которая при оптимальном выравнивании с эталонной последовательностью, представленной в данном документе как SEQ ID NO: 1-19 и SEQ ID NO: 26, имеет по меньшей мере около 85 процентов идентичности, по меньшей мере около 86 процентов идентичности, по меньшей мере около 87 процентов идентичности, по меньшей мере около 88 процентов идентичности, по меньшей мере около 89 процентов идентичности, по меньшей мере около 90 процентов идентичности, по меньшей мере около 91 процентов идентичности, по меньшей мере около 92 процентов идентичности, по меньшей мере около 93 процентов идентичности, по меньшей мере около 94 процентов идентичности, по меньшей мере около 95 процентов идентичности, по меньшей мере около 96 процентов идентичности, по меньшей мере около 97 процентов идентичности, по меньшей мере около 98 процентов идентичности, по меньшей мере, около 99 процентов идентичности, или по меньшей мере около 100 процентов идентичности с эталонной последовательностью. Молекулы ДНК, имеющие процент идентичности последовательности с эталонной молекулой, могут проявлять активность эталонной последовательности.
Регуляторный элементы
[45] Регуляторные элементы такие как промоторы, лидерные последовательности (также известные как 5' UTR), энхансеры, интроны и участки терминации транскрипции (или 3' UTR) играют неотъемлемую роль в общей экспрессии генов в живых клетках. Термин «регуляторный элемент» в контексте данного документа относится к молекуле ДНК, обладающей ген-регуляторной активностью. Термин «ген-регуляторная активность» в контексте данного документа относится к способности влиять на экспрессию функционально связанной молекулы транскрибируемой ДНК, например, воздействуя на транскрипцию и/или трансляцию функционально связанной молекулы транскрибируемой ДНК. Регуляторные элементы, такие как промоторы, лидерные последовательности, энхансеры, интроны и 3' UTR, которые функционируют в растениях, пригодны для модификации фенотипов растений с помощью генной инженерии.
[46] В контексте данного документа, термин «группа регуляторных элементов экспрессии» или последовательность «ЕХР» может относиться к группе функционально связанных регуляторных элементов, таких как энхансеры, промоторы, лидерные последовательности и интроны. Например, группа регуляторных элементов экспрессии может состоять, например, из промотора, функционально связанного с 5'-концом лидерной последовательности. ЕХР, применимые при практическом применении данного изобретения, содержат SEQ ID NO: 1, 4, 6, 9, 11, 15 и 16.
[47] Регуляторные элементы могут характеризоваться своим паттерном экспрессии генов, например, положительными и/или отрицательными эффектами, такими как конститутивная экспрессия или временная, пространственная, развивающаяся, тканевая, окружающей среды, физиологическая, патологическая, клеточного цикла и/или химически чувствительная экспрессия, и любая их комбинация, а также по количественным или качественным показателям. В контексте данного документа, «паттерн экспрессии гена» представляет собой любой паттерн транскрипции функционально связанной молекулы ДНК в транскрибируемую молекулу РНК. Транскрибируемая молекула РНК может транслироваться с образованием молекулы белка или может относится к антисмысловой или другой регуляторной молекуле РНК, такой как двухцепочечная РНК (дцРНК), транспортная РНК (тРНК), рибосомная РНК (рРНК), микроРНК (микроРНК), малая интерферирующая РНК (миРНК) и тому подобное.
[48] В контексте данного документа термин «экспрессия белка» означает любой паттерн трансляции транскрибируемой молекулы РНК в молекулу белка. Экспрессия белка может быть охарактеризована его временными, пространственными, характеристиками развития или морфологическими характеристиками, а также количественными или качественными показателями.
[49] Промотор пригоден в качестве регуляторного элемента для модуляции экспрессии функционально связанной молекулы транскрибируемой ДНК. В контексте данного документа термин «промотор» обычно относится к молекуле ДНК, которая участвует в распознавании и связывании РНК-полимеразы II и других белков, таких как транс-действующие факторы транскрипции, чтобы инициировать транскрипцию. Промотор может быть первоначально выделен из 5' нетранслируемого участка (5' UTR) геномной копии гена. Альтернативно, промоторы могут быть синтезированы или подвергнуты манипуляции с молекулами ДНК. Промоторы также могут быть химерными. Химерные промоторы получают путем слияния двух или более гетерологичных молекул ДНК. Промоторы, применимые при практическом применении данного изобретения, содержат промоторные элементы, представленные как SEQ ID NO: 2 и 7, или содержащиеся в любой из SEQ ID NO: 1, 4, 6, 9, 11, 15 и 16, или их фрагменты или варианты. В конкретных вариантах осуществления данного изобретения, заявленные молекулы ДНК и любые их варианты или производные, как описано в данном документе, дополнительно определены как имеющие промоторную активность, то есть, способны действовать как промотор в клетке-хозяине, например, в трансгенном растении. В других конкретных вариантах осуществления данного изобретения, фрагмент может быть определен как проявляющий промоторную активность, которой обладает исходная молекула промотора, из которой он получен, или фрагмент может содержать «минимальный промотор», который обеспечивает базовый уровень транскрипции и состоит из TATA бокса или эквивалентной последовательности ДНК для распознавания и связывания комплекса РНК-полимеразы II для инициации транскрипции.
[50] В одном варианте осуществления данного изобретения, представлены фрагменты последовательности ЕХР или промоторной последовательности, описанные в данном документе. Фрагменты промотора могут иметь промоторную активность, как описано выше, и могут быть применимы отдельно или в комбинации с другими промоторами и фрагментами промоторов, например, при конструировании химерных промоторов или в комбинации с другими элементами экспрессии и фрагментами элементов экспрессии. В конкретных вариантах осуществления данного изобретения, представлены фрагменты промотора, содержащие по меньшей мере около 50, по меньшей мере около 75, по меньшей мере около 95, по меньшей мере около 100, по меньшей мере около 125, по меньшей мере около 150, по меньшей мере около 175, по меньшей мере около 200, по меньшей мере около 225, по меньшей мере около 250, по меньшей мере около 275, по меньшей мере около 300, по меньшей мере около 500, по меньшей мере около 600, по меньшей мере около 700, по меньшей мере около 750, по меньшей мере около 800, по меньшей мере около 900, или по меньшей мере около 1000 смежных нуклеотидов молекулы ДНК, обладающей промоторной активностью, или более длинные, как описано в данном документе. Способы получения таких фрагментов из исходной молекулы промотора хорошо известны в данной области техники.
[51] В дополнительных вариантах осуществления данного изобретения, представлены описанные в данном документе фрагменты энхансерных или интронных последовательностей. Фрагменты энхансера или интрона могут включать активность основной молекулы, из которой они были получены, и могут быть применимы отдельно или в комбинации с другими регуляторными элементами, включая промоторы, лидерные последовательности, другие энхансеры, другие интроны или их фрагменты. В конкретных вариантах осуществления данного изобретения, представлены фрагменты энхансера или интрона, содержащие по меньшей мере около 50, по меньшей мере около 75, по меньшей мере около 95, по меньшей мере около 100, по меньшей мере около 125, по меньшей мере около 150, по меньшей мере около 175, по меньшей мере около 200, по меньшей мере около 225, по меньшей мере около 250, по меньшей мере около 275, по меньшей мере около 300, по меньшей мере около 500, по меньшей мере около 600, по меньшей мере около 700, по меньшей мере около 750, по меньшей мере около 800, по меньшей мере около 900, или по меньшей мере около 1000 смежных нуклеотидов молекулы ДНК, обладающей энхансерной или интронной активностью, или более длинные, как описано в данном документе. Способы получения таких фрагментов из исходной молекулы хорошо известны в данной области техники.
[52] В других вариантах осуществления данного изобретения, представлены фрагменты последовательностей 3' UTR, описанных в данном документе. Фрагменты 3' UTR могут иметь активность основной молекулы 3' UTR, из которой они были получены, и могут быть применимы сами по себе или в комбинации с другими регуляторными элементами, включая промоторы, лидерные последовательности, интроны или их фрагменты. В конкретных вариантах осуществления данного изобретения, представлены фрагменты интрона, содержащие по меньшей мере около 50, по меньшей мере около 75, по меньшей мере около 95, по меньшей мере около 100, по меньшей мере около 125, по меньшей мере около 150, по меньшей мере около 175, по меньшей мере около 200, по меньшей мере около 225, по меньшей мере около 250, по меньшей мере около 275, по меньшей мере около 300, по меньшей мере около 500, по меньшей мере около 600, по меньшей мере около 700, по меньшей мере около 750, по меньшей мере около 800, по меньшей мере около 900, или по меньшей мере около 1000 смежных нуклеотидов молекул ДНК, обладающих активностью 3' UTR, или более длинные, как описано в данном документе. Способы получения таких фрагментов из исходной молекулы 3' UTR хорошо известны в данной области техники.
[53] Композиции, полученные из любого из промоторных элементов, представленных как SEQ ID NO: 2 и 7, или содержащихся в любой из SEQ ID NO: 1, 4, 6, 9, 11, 15 и 16, таких как внутренние или 5'-концевые делеции, например, могут быть получены с использованием способов, известных в данной области техники, для улучшения или изменения экспрессии, включая путем удаления элементов, которые имеют либо положительные, либо отрицательные эффекты на экспрессию; дублирование элементов, которые положительно или отрицательно влияют на экспрессию; и/или дублирование или удаление элементов, которые имеют тканеспецифический или клеточноспецифический эффект на экспрессию. Композиции, полученные из любого из промоторных элементов, представленных как SEQ ID NO: 2 и 7, или содержащихся в любой из SEQ ID NO: 1, 4, 6, 9, 11, 15 и 16, состоящие из 3'-концевых делеций, в которых элемент ТАТА-бокса или его эквивалентная последовательность и идущая за ней в 3' направлении последовательность удалена, могут использоваться, например, для создания энхансерных элементов. Дальнейшие делеции могут быть выполнены для удаления любых элементов, которые имеют положительное или отрицательное; тканеспецифичное; клеточноспецифичное; или зависящее от времени (например, циркадного ритма, но не ограничиваясь ими) влияние на экспрессию. Любой из промоторных элементов, представленных как SEQ ID NO: 2 и 7, или содержащихся в любой из SEQ ID NO: 1, 4, 6, 9, 11, 15 и 16, и фрагменты или энхансеры, полученные из них, можно использовать для создания композиций химерных регуляторных элементов транскрипции.
[54] В соответствии с изобретением промотор или фрагмент промотора можно анализировать на наличие известных промоторных элементов, то есть, характеристик последовательности ДНК, таких как ТАТА-бокс и другие известные мотивы сайта связывания фактора транскрипции. Идентификация таких известных промоторных элементов может быть использована специалистом в данной области техники для создания вариантов промотора, имеющих сходный паттерн экспрессии с исходным промотором.
[55] В контексте данного документа термин «лидерная последовательность» относится к молекуле ДНК, выделенной из нетранслируемого 5' участка (5' UTR) гена и определяемой, как правило, как нуклеотидный сегмент между сайтом начала транскрипции (TSS) и сайтом начала кодирующей последовательности белка. В качестве альтернативы лидерные последовательности могут быть элементами ДНК, произведеными синтетически или модифицированными. Лидерная последовательность может использоваться в качестве 5' регуляторного элемента для модуляции экспрессии функционально связанной молекулы транскрибируемой ДНК. Молекулы лидерной последовательности можно использовать с гетерологичным промотором или с их природным промотором. Лидерные последовательности, пригодные для практического применения данного изобретения, включают SEQ ID NO: 3 и 8; или любой из лидерных элементов, содержащихся в любой из SEQ ID NO: 1, 4, 6, 9, 11, 15 и 16, или их фрагменты или варианты. В конкретных вариантах осуществления данного изобретения такие последовательности ДНК могут быть определены как способные действовать в качестве лидерных последовательностей в клетке-хозяине, включая, например, трансгенную клетку растения. В одном варианте осуществления данного изобретения, такие последовательности декодируются как содержащие лидерную активность.
[56] Лидерные последовательности (также упоминаемые как 5' UTR), представленные как SEQ ID NO: 3 и 8, или любой из лидерных элементов, содержащихся в любой из SEQ ID NO: 1, 4, 6, 9, 11, 15 и 16, могут состоять из регуляторных элементов или могут принимать вторичные структуры, которые могут влиять на транскрипцию или трансляцию функционально связанной молекулы транскрибируемой ДНК. Лидерные последовательности, представленные как SEQ ID NO: 3 и 8, или любой из лидерных элементов, содержащихся в любой из SEQ ID NO: 1, 4, 6, 9, 11, 15 и 16, можно применять в соответствии с изобретением для получения химерных регуляторных элементов, которые влияют на транскрипцию или трансляцию функционально связанной молекулы транскрибируемой ДНК.
[57] В контексте данного документа, термин «интрон» относится к молекуле ДНК, которая может быть выделена или идентифицирована из гена и может быть определена в целом как участок, сплайсированный во время процессинга матричной РНК (мРНК) до трансляции. Альтернативно, интрон может быть элементом ДНК, синтетически произведенным или модифицированным. Интрон может содержать энхансерные элементы, которые влияют на транскрипцию функционально связанных генов. Интрон можно использовать в качестве регуляторного элемента для модуляции экспрессии функционально связанной молекулы транскрибируемой ДНК. Конструкция может содержать интрон, и интрон может быть или не быть гетерологичным по отношению к молекуле транскрибируемой ДНК. Примеры интронов в данной области техники включают интрон актина риса и интрон HSP70 кукурузы.
[58] У растений включение некоторых интронов в генные конструкции приводит к увеличению накопления мРНК и белка по сравнению с конструкциями у которых нет интрона. Этот эффект получил название «интрон опосредованное усиление» (ГМЕ -англ.: intron mediated enhancement) экспрессии гена. Интроны, которые, как известно, стимулируют экспрессию в растениях, были идентифицированы в генах кукурузы (например, tubA1, Adh1, Sh1, и Ubi1), в генах риса (например, tpi) и в генах двудольных растений, таких как петунии (например, rbcS), картофеля (например, st-ls1) и Arabidopsis thaliana (например, ubq3 и pat1). Было показано, что делеции или мутации в сайтах сплайсинга интрона снижают экспрессию гена, указывая на то, что сплайсинг может быть необходим для ГМЕ. Однако ГМЕ у двудольных растений был продемонстрирован точечными мутациями в сайтах сплайсинга гена pat1 из A. thaliana. Было продемонстрирован, что многократное использование того же интрона в одном растении имеет недостатки. В этих случаях необходимо иметь набор основных контролирующих элементов для конструирования соответствующих рекомбинантных элементов ДНК. Типичные интроны, пригодные для использования в практике данного изобретения, представлены как SEQ ID NO: 5, 10 и 12.
[59] В контексте данного документа, термины «3'-молекула терминации транскрипции», «3' нетранслируемый участок» или «3' UTR» относятся к молекуле ДНК, которая используется во время транскрипции на нетранслируемом участке 3' части молекулы мРНК. 3' нетранслируемый участок молекулы мРНК может быть получен путем специфического расщепления и 3' полиаденилирования, также известного как полиА-хвост. 3' UTR может быть функционально связан с молекулой транскрибируемой ДНК и располагаться ниже нее по рамке считывания и может включать сигнал полиаденилирования и другие регуляторные сигналы, способные влиять на транскрипцию, процессинг мРНК или экспрессию гена. Считается, что полиА хвосты играют роль в стабильности мРНК и инициации трансляции. Примерами 3' молекул терминации транскрипции в данной области техники являются 3' участок нопалинсинтазы, 3' участок hsp17 пшеницы, 3' участок малой субъединицы рубиско гороха, 3' участок хлопка Е6 и 3' UTR коиксина.
[60] 3' UTR обычно находят полезное применение для рекомбинантной экспрессии специфических молекул ДНК. Слабый 3' UTR может вызывать сквозное считывание, которое может влиять на экспрессию молекулы ДНК, расположенной в соседних экспрессионных кассетах. Соответствующий контроль терминации транскрипции может предотвратить сквозное считывание в последовательностях ДНК (например, других экспрессионных кассет), локализованных ниже, и может дополнительно обеспечить эффективную рециркуляцию РНК полимеразы для улучшения экспрессии генов. Эффективная терминация транскрипции (высвобождение РНК полимеразы II из ДНК) является необходимым условием для повторной инициации транскрипции и, таким образом, напрямую влияет на общий уровень транскрипции. После терминации транскрипции зрелая мРНК высвобождается из сайта синтеза, а матрица транспортируется в цитоплазму. Эукариотические мРНК накапливаются в виде поли(А) форм in vivo, что затрудняет обнаружение сайтов терминации транскрипции обычными способами. Однако предсказание функциональных и эффективных 3' UTR способами биоинформатики затруднено из-за отсутствия консервативных последовательностей ДНК, которые позволили бы легко предсказать эффективный 3' UTR.
[61] С практической точки зрения обычно полезно, чтобы 3' UTR, используемый в экспрессионной кассете, обладал следующими характеристиками. Во-первых, 3' UTR должен быть способен производительно и эффективно прекращать транскрипцию трансгена и предотвращать сквозное считывание транскрипта в любой соседней последовательности ДНК, которая может состоять из другой экспрессионной кассеты, как в случае множественных экспрессионных кассет, находящихся в одной введенной ДНК (Т-ДНК) или соседней хромосомной ДНК, в которую вставлена Т-ДНК. Во-вторых, 3' UTR не должен вызывать снижение транскрипционной активности, передаваемой промотором, лидером, энхансерами и нитронами, которые используются для управления экспрессией молекулы ДНК. Наконец, в биотехнологии растений 3' UTR часто используется для праймирования реакций амплификации обратно транскрибируемой РНК, извлеченной из трансформированного растения, и используется для: (1) оценки транскрипционной активности или экспрессии экспрессионной кассеты оказавшейся интегрированной в хромосому растения; (2) оценки количества копий вставок в ДНК растения; и (3) оценки зиготности полученных семян после селекции. 3' UTR также используется в реакциях амплификации ДНК, выделенной из трансформированного растения, для характеристики целостности вставленной кассеты. 3' UTR, пригодные для практики данного изобретения, представлены как SEQ ID N0: 13, 14, 19 и 26.
[62] В контексте данного документа, термин «энхансер» или «энхансерный элемент» относится к цис-действующему регуляторному элементу, также известному как цис-элемент, который придает аспект общего паттерна экспрессии, но обычно недостаточен сам по себе, чтобы управлять транскрипцией, функционально связанной молекулы транскрибируемой ДНК. В отличие от промоторов, энхансерные элементы обычно не включают сайт начала транскрипции (TSS - transcription start site) или ТАТА-бокс или эквивалентную последовательность ДНК. Промотор или фрагмент промотора может природным образом содержать один или более энхансерных элементов, которые влияют на транскрипцию функционально связанной последовательности ДНК. Энхансерный элемент также может быть слит с промотором для получения цис-элемента химерного промотора, который придает аспект общей модуляции экспрессии гена.
[63] Считается, что многие энхансерные элементы промотора связывают ДНК-связывающие белки и/или влияют на топологию ДНК, создавая локальные конформации, которые избирательно разрешают или ограничивают доступ РНК-полимеразы к матрице ДНК или которые способствуют селективному открытию двойной спирали в сайте инициации транскрипции. Энхансерный элемент может функционировать для связывания факторов транскрипции, которые регулируют транскрипцию. Некоторые энхансерные элементы связывают более одного фактора транскрипции, и факторы транскрипции могут взаимодействовать с разной аффинностью с более чем одним доменом энхансера. Энхансерные элементы можно идентифицировать с помощью ряда методов, включая анализ делеции, то есть, делецию одного или более нуклеотидов от 5' -конца или внутри промотора; анализ ДНК-связывающих белков с использованием футпринтинга ДНКазы I, интерференции метилирования, электрофорезные анализы сдвига подвижности, геномного следа, геномный футпринтинг in vivo с помощью лигирования-опосредованной полимеразной цепной реакции (ПЦР) и других традиционных анализов или анализа сходства последовательностей ДНК с использованием известных методов сравнения мотивов цис-элементов или энхансерных элементов в качестве последовательности-мишени или мотива-мишени с последовательностью обычной ДНК, таких как BLAST. Тонкую структуру энхансерного домена можно дополнительно изучить с помощью мутагенеза (или замены) одного или более нуклеотидов или другими традиционными способами, известными в данной области техники. Энхансерные элементы могут быть получены путем химического синтеза или путем выделения из регуляторных элементов, которые включают такие элементы, и они могут быть синтезированы с дополнительными фланкирующими нуклеотидами, которые содержат полезные сайты рестрикционных ферментов для облегчения манипуляции с подпоследовательностями. Таким образом, создание, конструирование и использование энхансерных элементов в соответствии со способами, описанными в данном документе, для модуляции экспрессии функционально связанных молекул транскрибируемой ДНК, охвачены данным изобретением. Иллюстративные энхансеры, пригодные для практического применении по этому изобретения, представлены как SEQ ID NO: 17 и 18.
[64] В контексте данного документа термин «химерная» относится к одной молекуле ДНК, полученной путем слияния первой молекулы ДНК со второй молекулой ДНК, где ни первая, ни вторая молекула ДНК обычно не находятся в этой конфигурации, то есть слиты с другой. Таким образом, химерная молекула ДНК является новой молекулой ДНК, обычно не встречающейся в природе. В контексте данного документа, термин «химерный промотор» относится к промотору, полученному посредством таких манипуляций с молекулами ДНК. Химерный промотор может объединять два или более фрагмента ДНК, например, слияние промотора с энхансерным элементом. Таким образом, создание, конструирование и использование химерных промоторов в соответствии со способами, описанными в данном документе, для модуляции экспрессии функционально связанных молекул транскрибируемой ДНК, охвачены данным изобретением.
[65] Химерные регуляторные элементы могут быть сконструированы так, чтобы содержать различные составляющие элементы, которые могут быть оперативно связаны различными способами, известными в данной области техники, такими как расщепление рестрикционными ферментами и лигирование, независимое от лигирования клонирование, модульная сборка продуктов ПЦР во время амплификации или прямой химический синтез регуляторного элемент, а также другими способами, известные в данной области техники. Полученные в результате различные химерные регуляторные элементы могут состоять из одних и тех же или вариантов одних и тех же составляющих элементов, но отличаться последовательностью ДНК или последовательностями, которые содержат связывающую последовательность ДНК или последовательности ДНК, которые позволяют функционально соединять составляющие части. В изобретении последовательности ДНК, представленные как SEQ ID NO: 1-19 и SEQ ID NO: 26, могут обеспечивать эталонные последовательности регуляторных элементов, при этом составляющие элементы, составляющие эталонную последовательность, могут быть соединены способами, известными в данной области техники, и могут содержать замены, делеции и/или вставки одного или более нуклеотидов или мутаций, которые природным образом возникают при трансформации бактериальных и растительных клеток.
[66] В контексте данного документа, термин «вариант» относится ко второй молекуле ДНК, такой как регуляторный элемент, которая по составу аналогична, но не идентична первой молекуле ДНК, и при этом вторая молекула ДНК все еще сохраняет общую функциональность, то есть такой же или подобный паттерн экспрессии, например, посредством более или менее эквивалентной транскрипционной активности первой молекулы ДНК. Вариант может быть более короткой или усеченной версией первой молекулы ДНК или измененной версией последовательности первой молекулы ДНК, такой как с другими сайтами рестрикционных ферментов и/или внутренними делециями, заменами или вставками. «Вариант» может также включать регуляторный элемент, имеющий нуклеотидную последовательность, содержащую замену, делецию или вставку одного или более нуклеотидов эталонной последовательности, при этом производный регуляторный элемент имеет более или менее или эквивалентную транскрипционную или трансляционную активность, чем соответствующая родительская регуляторная молекула. «Варианты» регуляторного элемента также будут охватывать варианты, возникающие в результате мутаций, которые природным образом возникают при трансформации бактериальных и растительных клеток. В данном изобретении полинуклеотидная последовательность, представленная как SEQ ID NO: 1-19 и SEQ ID NO: 26, может быть использована для создания вариантов, которые аналогичны по составу, но не идентичны последовательности ДНК исходного регуляторного элемента, при этом все еще сохраняя общую функциональность, то есть, тот же или подобный паттерн экспрессии исходного регуляторного элемента. Производство таких вариантов по изобретению находится в компетенции среднего специалиста в данной области техники в свете описания и входит в объем данного изобретения.
[67] Эффективность описанных в данном документе модификаций, дупликаций или делеций в отношении желаемых аспектов экспрессии конкретного трансгена может быть проверена эмпирически в анализах стабильных и транзиентных растений, таких как те, что описаны в рабочих примерах в данном документе, чтобы подтвердить результаты, которые могут варьироваться в зависимости от внесенных изменений и цели изменения исходной молекулы ДНК.
Конструкции
[68] В контексте данного документа, термин «конструкция» означает любую молекулу рекомбинантной ДНК такую как плазмида, космида, вирус, фаг, или линейная или кольцевая молекула ДНК или РНК, полученную из любого источника, способную к геномной интеграции или автономной репликации, содержащую молекулу ДНК в которой по меньшей мере одна молекула ДНК функционально связана с другой молекулой ДНК, то есть функционально связана. В контексте данного документа, термин «вектор» означает любую конструкцию, которая может быть использована с целью трансформации, то есть, введения гетерологичной ДНК или РНК в клетку-хозяин. Конструкция обычно включает одну или более экспрессионных кассет. В контексте данного документа, термин «экспрессионная кассета» относится к молекуле ДНК, содержащей по меньшей мере молекулу транскрибируемой ДНК, функционально связанную с одним или более регуляторными элементами, обычно по меньшей мере с промотором и 3' UTR.
[69] В контексте данного документа, термин "функционально связанный" относится к первой молекуле ДНК, присоединенной ко второй молекуле ДНК, при этом первая и вторая молекулы ДНК расположены таким образом, что первая молекула ДНК влияет на функцию второй молекулы ДНК. Две молекулы ДНК могут быть, а могут и не быть частью одной смежной молекулы ДНК и могут быть или не быть смежными. Например, промотор функционально связан с молекулой транскрибируемой ДНК, если промотор модулирует транскрипцию представляющей интерес молекулы транскрибируемой ДНК в клетке. Например, лидерная последовательность функционально связана с последовательностью ДНК, если она способна влиять на транскрипцию или трансляцию последовательности ДНК.
[70] В одном варианте осуществления данного изобретения конструкции по данному изобретению могут быть представлены в виде двойных опухоль-индуцирующих (Ti) плазмидных граничных конструкций, которые имеют правую границу (RB или AGRtu.RB) и левую границу (LB или AGRtu.LB) участков Ti-плазмиды, выделенной из Agrobacterium tumefaciens содержащей Т-ДНК, которая вместе с переносящими молекулами, обеспечиваемыми клетками A. tumefaciens, позволяет интегрировать Т-ДНК в геном растительной клетки (см., например, патент США 6603061). Конструкции также могут содержать сегменты ДНК плазмидного остова, которые обеспечивают функцию репликации и селекцию антибиотиками в бактериальных клетках, например, ориджин репликации Escherichia coli такой как ori322, ориджин репликации широкого диапазона хозяина, такой как oriV или oriRi, и кодирующую участок для селективного маркера, такого как Spec/Strp, который кодирует аминогликозид аденилтрансферазу Tn7 (aadA) придающую устойчивость к спектиномицину или стрептомицину, или ген селективного маркера гентамицина (Gm, Gent). Для трансформации растений бактериальным штаммом-хозяином часто является ABI, С58 или LBA4404 А. tumefaciens, однако другие штаммы, известные специалистам в данной области техники трансформации растений, могут функционировать в данном изобретении.
[71] В данной области техники известны способы сборки и введения конструкций в клетку таким образом, чтобы транскрибируемая молекула ДНК транскрибировалась в функциональную молекулу мРНК, которая транслируется и экспрессируется как белок. Для практического применения изобретения традиционные композиции и способы получения и применения конструкций и клеток-хозяев хорошо известны специалистам в данной области техники. Типичные векторы, пригодные для экспрессии нуклеиновых кислот в высших растениях, хорошо известны в данной области техники и включают векторы, полученные из Ti-плазмиды Agrobacterium tumefaciens, и вектор контроля переноса pCaMVCN.
[72] В конструкцию могут быть включены различные регуляторные элементы, включая любые из представленных в данном документе. Любые такие регуляторные элементы могут предоставляться в сочетании с другими регуляторными элементами. Такие комбинации могут быть разработаны или модифицированы для получения желаемых нормативных характеристик. В одном варианте осуществления данного изобретения, конструкции по данному изобретению содержат по меньшей мере один регуляторный элемент, функционально связанный с молекулой транскрибируемой ДНК, функционально связанной с 3' UTR.
[73] Конструкции по данному изобретению могут включать любой промотор или лидерную последовательность, представленные в данном документе или известные в данной области техники. Например, промотор по изобретению может быть функционально связан с гетерологичной нетранслируемой 5'-лидерной последовательностью, такой как лидерная последовательность, происходящая от гена белка теплового шока. Альтернативно, лидерная последовательность по данному изобретению может быть функционально связана с гетерологичным промотором, таким как промотор 35S транскрипта вируса мозаики цветной капусты.
[74] Экспрессионные кассеты могут также включать последовательность, кодирующую транзитный пептид, которая кодирует пептид, который полезен для субклеточного нацеливания функционально связанного белка, в частности, на хлоропласт, лейкопласт или другую пластидную органеллу; митохондрии; пероксисому; вакуоль; или внеклеточную локацию. Многие локализованные в хлоропласте белки экспрессируются из ядерных генов в качестве предшественников и нацелены на хлоропласт с помощью транзитного пептида хлоропласта (СТР). Примеры таких изолированных белков хлоропластов включают белки, связанные с малой субъединицей (SSU) рибулозо-1,5-бисфосфаткарбоксилазы, ферредоксин, ферредоксин оксидоредуктазу, белок I и белок II светособирающего комплекса, тиоредоксин F и энолпирувилшикимат фосфатсинтазу (EPSPS), но не ограничиваются ими. Транзитные пептиды хлоропластов описаны, например, в патенте США №7193133. Было продемонстрировано, что белки, не относящиеся к хлоропластам, могут быть нацелены на хлоропласт посредством экспрессии гетерологичного СТР, функционально связанного с трансгеном, кодирующим нехлоропластные белки. Молекулы транскрибируемой ДНК
[75] В контексте данного документа, термин «молекула транскрибируемой ДНК» относится к любой молекуле ДНК, способной транскрибироваться в молекулу РНК, включая молекулы, имеющие последовательности, кодирующие белок, и молекулы, продуцирующие молекулы РНК, имеющие последовательности, применимые для подавления генов, но не ограничиваясь ими. Тип молекулы ДНК может включать молекулу ДНК из того же растения, молекулу ДНК из другого растения, молекулу ДНК из другого организма или синтетическую молекулу ДНК, такую как молекула ДНК, содержащая антисмысловое сообщение гена, или молекула ДНК, кодирующая искусственную, синтетическую или иным образом модифицированную версию трансгена, но не ограничивается ими. Иллюстративные молекулы транскрибируемой ДНК для включения в конструкции по данному изобретению включают, например, молекулы ДНК или гены из видов, отличных от видов, в которые включена молекула ДНК, или гены, которые происходят от того же вида или присутствуют у него, но являются включенными в клетки-реципиенты способами генной инженерии, а не классическими методами разведения.
[76] «Трансген» относится к молекуле транскрибируемой ДНК, гетерологичной клетке-хозяину по меньшей мере в отношении ее местоположения в геноме клетки-хозяина и/или молекуле транскрибируемой ДНК, искусственно включенной в геном клетки-хозяина в текущем или любом предыдущем поколении клетки.
[77] Регуляторный элемент, такой как промотор по данному изобретению, может быть функционально связан с молекулой транскрибируемой ДНК, которая является гетерологичной по отношению к регуляторному элементу. В контексте данного документа, термин «гетерологичный» относится к комбинации двух или более молекул ДНК, когда такая комбинация обычно не встречается в природе. Например, две молекулы ДНК могут быть получены из разных видов и/или две молекулы ДНК могут быть получены из разных генов, например, из разных генов одного вида или из одних и тех же генов из разных видов. Таким образом, регуляторный элемент является гетерологичным по отношению к функционально связанной молекуле транскрибируемой ДНК, если такая комбинация обычно не встречается в природе, то есть, молекула транскрибируемой ДНК не встречается в природе функционально связанной с регуляторным элементом.
[78] Молекула транскрибируемой ДНК обычно может быть любой молекулой ДНК, для которой желательна экспрессия транскрипта. Такая экспрессия транскрипта может привести к трансляции образующейся молекулы мРНК, и, следовательно, к экспрессии белка. В качестве альтернативы, например, молекула транскрибируемой ДНК может быть сконструирована так, чтобы в конечном итоге вызвать снижение экспрессии конкретного гена или белка. В одном варианте осуществления данного изобретения, это может быть достигнуто с использованием молекулы транскрибируемой ДНК, которая ориентирована в антисмысловом направлении. Обычный специалист в данной области технике знаком с использованием такой антисмысловой технологии. Таким образом любой ген может быть негативно регулирован, и в одном варианте осуществления данного изобретения молекула транскрибируемой ДНК может быть сконструирована для подавления конкретного гена посредством экспрессии молекулы дцРНК, миРНК или микроРНК.
[79] Таким образом, один вариант осуществления данного изобретения представляет собой молекулу рекомбинантной ДНК, содержащую регуляторный элемент по изобретению, такой как те, которые представлены как SEQ ID NO: 1-19 и SEQ ID NO: 26, функционально связанную с гетерологичной молекулой транскрибируемой ДНК, чтобы модулировать транскрипцию молекулы транскрибируемой ДНК на желаемом уровне или в желаемом паттерне, когда конструкция интегрируется в геном трансгенной клетки растения. В одном варианте осуществления данного изобретения, молекула транскрибируемой ДНК содержит белок-кодирующий участок гена, а в другом варианте осуществления данного изобретения молекула транскрибируемой ДНК содержит антисмысловой участок гена. Гены представляющие агрономический интерес
[80] Молекула транскрибируемой ДНК может представлять собой ген, представляющий агрономический интерес. В контексте данного документа, термин «ген, представляющий агрономический интерес» относится к молекуле транскрибируемой ДНК, которая при экспрессии в конкретной растительной ткани, клетке или типе клеток придает желаемую характеристику. Продукт гена, представляющего агрономический интерес, может действовать внутри растения, чтобы оказывать влияние на морфологию, физиологию, рост, развитие, урожайность, состав зерна, профиль питания, устойчивость к болезням или вредителям и/или устойчивость к окружающей среде или химическим веществам или может действовать как пестицидный агент в рационе вредителя, питающегося растением. В одном варианте осуществления данного изобретения, регуляторный элемент изобретения включен в конструкцию, так что регуляторный элемент функционально связан с молекулой транскрибируемой ДНК, которая представляет собой ген, представляющий агрономический интерес. В трансгенном растении, содержащем такую конструкцию, экспрессия гена, представляющего агрономический интерес, может придавать полезный агрономический признак. Полезный агрономический признак может включать, например, толерантность к гербицидам, контроль насекомых, модифицированный урожай, устойчивость к болезням, устойчивость к патогенам, изменение роста и развития растений, модифицированное содержание крахмала, модифицированное содержание масла, модифицированное содержание жирных кислот, модифицированное содержание протеина, модифицированное созревание плодов, улучшенное питание животных и человека, производство биополимеров, устойчивость к стрессам окружающей среды, фармацевтические пептиды, улучшенные технологические качества, улучшенный вкус, пригодность по производству гибридных семян, улучшенное производство волокна и желаемое производство биотоплива, но не ограничиваясь ими.
[81] Неограничивающие примеры генов, представляющих агрономический интерес, известных в данной области техники, включают гены устойчивости к гербицидам (Патенты США №6803501; 6448476; 6248876; 6225114; 6107549; 5866775; 5804425; 5633435; и 5463175), повышенной урожайности (Патенты США № USRE38446; 6716474; 6663906; 6476295; 6441277; 6423828; 6399330; 6372211; 6235971; 6222098; и 5716837), контроля насекомых (Патенты США №6809078; 6713063; 6686452; 6657046; 6645497; 6642030; 6639054; 6620988; 6593293; 6555655; 6538109; 6537756; 6521442; 6501009; 6468523; 6326351; 6313378; 6284949; 6281016; 6248536; 6242241; 6221649; 6177615; 6156573; 6153814; 6110464; 6093695; 6063756; 6063597; 6023013; 5959091; 5942664; 5942658, 5880275; 5763245; и 5763241), устойчивости к грибковым заболеваниям (Патенты США №6653280; 6573361; 6506962; 6316407; 6215048; 5516671; 5773696; 6121436; 6316407; и 6506962), устойчивости к вирусам (Патенты США №6617496; 6608241; 6015940; 6013864; 5850023; и 5304730), устойчивости к нематодам (Патент США №6228992), устойчивости к бактериальным заболеваниям (Патент США №5516671), роста и развития растений (Патенты США №6723897 и 6518488), производства крахмала (Патенты США №6538181; 6538179; 6538178; 5750876; 6476295), производства модифицированных масел (Патенты США №6444876; 6426447; и 6380462), высокого производства масла (Патенты США №6495739; 5608149; 6483008; и 6476295), модифицированного содержания жирных кислот (Патенты США №6828475; 6822141; 6770465; 6706950; 6660849; 6596538; 6589767; 6537750; 6489461; и 6459018), высокого производства белка (Патент США №6380466), созревания плодов (Патент США №5512466), улучшенного питания животных и человека (Патенты США №6723837; 6653530; 6541259; 5985605; и 6171640), биополимеров (Патенты США № USRE37543; 6228623; и 5958745, и 6946588), устойчивости к стрессам окружающей среды (Патент США №6072103), фармацевтических пептидов и секретируемых пептидов (Патенты США №6812379; 6774283; 6140075; и 6080560), улучшенных характеристик обработки (Патент США №6476295), улучшенной усвояемости (Патент США №6531648) низкого содержания рафинозы (Патент США №6166292), промышленного производства ферментов (Патент США №5543576), улучшенного аромата (Патент США №6011199), азотфиксации (Патент США №5229114), производства гибридных семян (Патент США №5689041), производства волокна (Патенты США №6576818; 6271443; 5981834; и 5869720) и производства биотоплива (Патент США №5998700).
[82] Альтернативно, ген представляющий агрономический интерес, может влиять на вышеупомянутые характеристики или фенотипы растений путем кодирования молекулы РНК, которая вызывает целевую модуляцию экспрессии гена эндогенного гена, например, с помощью антисмысловой (см., например, Патент США 5107065); ингибирующей РНК («РНКи», включая модуляцию экспрессии генов с помощью микроРНК-, миРНК-, транс-действующих миРНК- и фазированных механизмов, опосредованных мРНК, например, как описано в опубликованных заявках США 2006/0200878 и США 2008/0066206, и в заявке на патент США 11/974469); или механизмов, опосредованных косупрессией. РНК также может быть каталитической молекулой РНК (например, рибозимом или рибопереключателем; см., например, США 2006/0200878), сконструированной для расщепления желаемого эндогенного продукта мРНК. В данной области техники известны способы конструирования и введения конструкций в клетку таким образом, чтобы молекула транскрибируемой ДНК транскрибировалась в молекулу, которая способна вызывать подавление гена.
Селективные маркеры
[83] Селективные маркерные трансгены также можно использовать с регуляторными элементами по изобретению. В контексте данного документа, термин «селективный маркерный трансген селективного маркера» относится к любой молекуле транскрибируемой ДНК, экспрессия которой в трансгенном растении, ткани или клетке или ее отсутствие может быть проверена или оценена каким-либо образом. Селективные маркерные гены и связанные с ними методы отбора и скрининга для применения на практике изобретения известны в данной области техники и включают молекулы транскрибируемой ДНК, кодирующие β-глюкуронидазу (GUS), зеленый флуоресцентный белок (GFP), белки, придающие устойчивость к антибиотикам, и белки, придающие устойчивость к гербицидам, но не ограничиваются ими. Примеры селективных маркерных трансгенов представлены как SEQ ID NO: 20 и 24. Трансформация клеток
[84] Изобретение также направлено на способ получения трансформированных клеток и растений, которые содержат один или более регуляторных элементов, функционально связанных с молекулой транскрибируемой ДНК.
[85] Термин «трансформация» относится к введению молекулы ДНК хозяину-реципиенту. В контексте данного документа, термин «хозяин» относится к бактериям, грибам или растениям, включая любые клетки, ткани, органы или потомство бактерий, грибов или растений. Ткани и клетки растений, представляющие особый интерес, включают протопласты, каллус, корни, клубни, семена, стебли, листья, проростки, зародыши и пыльцу.
[86] В контексте данного документа, термин «трансформированный» относится к клетке, ткани, органу или организму, в которые была введена чужеродная молекула ДНК, такая как конструкция. Введенная молекула ДНК может быть интегрирована в геномную ДНК реципиентной клетки, ткани, органа или организма, так что введенная молекула ДНК наследуется последующим потомством. «Трансгенная» или «трансформированная» клетка или организм может также включать потомство клетки или организма и потомство, полученное в результате программы скрещивания, в которой такой трансгенный организм используется в качестве родителя в скрещивании и проявляет измененный фенотип в результате присутствия чужеродной молекулы ДНК. Введенная молекула ДНК может также в течение короткого времени вводиться в реципиентную клетку, так что введенная молекула ДНК не наследуется последующим потомством. Термин «трансгенный» относится к бактерии, грибку или растению, содержащим одну или более гетерологичных молекул ДНК.
[87] Существует много способов, хорошо известных специалистам в данной области техники для введения молекул ДНК в клетки растений. Процесс обычно включает этапы выбора подходящей клетки-хозяина, трансформации клетки-хозяина с помощью вектора и получения трансформированной клетки-хозяина. Способы и материалы для трансформации растительных клеток путем введения растительной конструкции в геном растения в практике данного изобретения могут включать любой из хорошо известных и продемонстрированных способов. Подходящие способы включают бактериальную инфекцию (например, Agrobacterium), бинарные векторы ВАС, прямую доставку ДНК (например, посредством ПЭГ-опосредованной трансформации, дессикации/опосредованного ингибированием захвата ДНК, электропорации, перемешивание с волокнами карбида кремния и ускорение частиц, покрытых ДНК), редактирование генов (например, системы CRISPR-Cas) и другие, но не ограничиваются ими.
[88] Клетки-хозяева могут быть любой клеткой или организмом, например, клеткой растения, клеткой водорослей, водорослями, клеткой грибов, грибами, бактериальной клеткой или клеткой насекомого. В конкретных вариантах осуществления данного изобретения, клетки-хозяева и трансформированные клетки могут включать клетки культурных растений.
[89] Впоследствии трансгенное растение можно регенерировать из клетки трансгенного растения по изобретению. Используя обычные методы скрещивания или самоопыление, из этого трансгенного растения можно получить семена. Такие семена и полученное в результате растение-потомок, выращенное из таких семян, будут содержать молекулу рекомбинантной ДНК по изобретению и, следовательно, будут трансгенными.
[90] Трансгенные растения по данному изобретению можно самоопылять для получения семян в случае гомозиготных трансгенных растений по изобретению (гомозиготных по молекуле рекомбинантной ДНК) или скрещивать с нетрансгенными растениями или различными трансгенными растениями для получения семян в случае гетерозиготных трансгенных растений по изобретению (гетерозиготный по молекуле рекомбинантной ДНК). И такие гомозиготные, и гетерозиготные трансгенные растения упоминаются в данном документе как «растения-потомки». Растения-потомки представляют собой трансгенные растения, происходящие от исходного трансгенного растения и содержащие молекулу рекомбинантной ДНК по изобретению. Семена, полученные с использованием трансгенного растения по изобретению, можно собирать и использовать для выращивания поколений трансгенных растений, то есть, растений потомков по изобретению, содержащих конструкцию по данному изобретению и экспрессирующих ген, представляющий агрономический интерес. Описание способов разведения, которые обычно используются для разных культур, можно найти в одном из нескольких справочников, см., например,, Allard, Principles of Plant Breeding, John Wiley & Sons, NY, U. of CA, Davis, CA, 50-98 (1960); Simmonds, Principles of Crop Improvement, Longman, Inc., NY, 369-399 (1979); Sneep and Hendriksen, Plant breeding Perspectives, Wageningen (ed), Center for Agricultural Publishing and Documentation (1979); Fehr, Soybeans: Improvement, Production and Uses, 2nd Edition, Monograph, 16:249 (1987); Fehr, Principles of Variety Development, Theory и Technique, (Vol. 1) и Crop Species Soybean (Vol. 2), Iowa State Univ., Macmillan Pub. Co., NY, 360-376 (1987).
[91] Трансформированные растения могут быть проанализированы на наличие гена или генов представляющих интерес, а также на уровень экспрессии и/или профиль, обеспечиваемый регуляторными элементами изобретения. Специалистам в данной области техники известны многочисленные способы, доступные для анализа трансформированных растений. Например, способы анализа растений включают Саузерн-блоттинг или Нозерн-блоттинг, подходы на основе ПЦР, биохимические анализы, методы фенотипического скрининга, полевые оценки и иммунодиагностические анализы, но не ограничиваются ими. Экспрессию молекулы транскрибируемой ДНК можно измерить с использованием реагентов TaqMan® (Applied Biosystems, Фостер Сити, Калифорния), способов, описанных производителем, и времени цикла ПЦР, определенного с использованием матрицы тестирования TaqMan®. Альтернативно для оценки экспрессии трансгена можно использовать реагенты и способы Invader® (Third Wave Technologies, Мэдисон, Висконсин), как описано производителем.
[92] Изобретение также относится к частям растения по данному изобретению. Части растения включают листья, стебли, корни, клубни, семена, эндосперм, семяпочки и пыльцу, но не ограничиваются ими. Части растений по изобретению могут быть жизнеспособными, нежизнеспособными, регенерируемыми и/или невозобновляемыми. Изобретение также относится к и обеспечивает трансформированные растительные клетки, содержащие молекулу ДНК по изобретению. Трансформированные или трансгенные клетки растения по изобретению включают регенерируемые и/или нерегенерируемые клетки растения.
[93] Изобретение также относится к товарному продукту, который получают из трансгенного растения или его части, содержащего молекулу рекомбинантной ДНК по изобретению. Товарные продукты по изобретению содержат определяемое количество ДНК, содержащей последовательность ДНК, выбранную из группы, состоящей из SEQ ID NO: 1-19 и SEQ ID NO: 26. В контексте данного документа, термин «товарный продукт» относится к любой композиции или продукту, который состоит из материала, полученного из трансгенного растения, семени, клетки растения или части растения, содержащей молекулу рекомбинантной ДНК по изобретению. Товарные продукты включают обработанные семена, зерно, части растений и муку, но не ограничиваются ими. Товарный продукт по изобретению будет содержать определяемое количество ДНК, соответствующей молекуле рекомбинантной ДНК по изобретению. Обнаружение одной или более таких ДНК в образце можно использовать для определения содержания или источника товарного продукта. Можно использовать любой стандартный способ обнаружения молекул ДНК, включая описанные в данном документе способы обнаружения.
[94] Изобретение можно более легко понять, обратившись к нижеследующим примерам, которые приведены в качестве иллюстрации и не предназначены для ограничения изобретения, если не указано иное. Специалистам в данной области техники должно быть понятно, что методы, описанные в следующих примерах, представляют собой методы, открытые изобретателями, чтобы хорошо функционировать при практическом применении изобретения. Однако специалисты в данной области техники должны, в свете данного описания, принять во внимание, что многие изменения могут быть внесены в конкретные описанные варианты осуществления данного изобретения, и по-прежнему можно получать подобный или аналогичный результат без отклонения от сущности и объема изобретения, поэтому все материалы, изложенные или показанные на прилагаемых графических материалах, следует интерпретировать как иллюстративные, а не в ограничивающем смысле.
ПРИМЕРЫ
Пример 1
Создание, синтез и клонирование синтетических регуляторных элементов
[95] Новые синтетические регуляторные элементы транскрипции представляют собой элементы синтетической экспрессии, разработанные с помощью алгоритмических способов. Эти разработанные с помощью вычислений регуляторные элементы были химически синтезированы и клонированы для создания синтетических групп регуляторных элементов экспрессии (ЕХР). Свыше 1000 синтетических регуляторных элементов были разработаны и исследованы в протопластах кукурузы и стабильно трансформированных растениях кукурузы для выявления тех синтетических регуляторных элементов, которые обеспечивают желаемые характеристики, такие как уровни экспрессии белка и паттерны экспрессии. Синтетические элементы по данному изобретению обеспечивают различные паттерны конститутивной экспрессии, пригодные для управления экспрессией многих различных кодирующих последовательностей и интерферирующих РНК, представляющих агрономический интерес.
[96] Разработанные синтетические регуляторные элементы транскрипции не обладают расширенной гомологией с какими-либо известными последовательностями нуклеиновых кислот, которые существуют в природе, но влияют на транскрипцию функционально связанной кодирующей последовательности так же, как природные промоторы, лидеры, интроны и 3' UTR. Синтетические ЕХР и их соответствующие синтетические промоторы, лидеры и интроны, а также синтетические 3' UTR представлены в таблице 1. Синтетические ЕХР были клонированы с использованием способов, известных в данной области техники, в бинарные векторы трансформации растений, функционально связанные с β-глюкуронидазой (GUS), а также оценивали уровни и паттерны экспрессии в стабильно трансформированных растениях кукурузы.
[97] Анализ регуляторного элемента TSS и сплайсинговых соединений интрон/экзона может быть выполнен с использованием трансформированной растительной ткани. Вкратце, растения трансформировали векторами экспрессии для растений, содержащими клонированные фрагменты ДНК, функционально связанные с гетерологичной молекулой транскрибируемой ДНК. Затем была использована система 5' RACE для быстрой амплификации концов кДНК, версия 2.0 (Invitrogen, Карлсбад, Калифорния 92008), для подтверждения TSS регуляторного элемента и сплайсинговых соединений интрон/экзон путем анализа последовательности ДНК полученных транскриптов мРНК. Синтетические 3' UTR были охарактеризованы по их влиянию на экспрессию генов, а также на правильную терминацию транскрипта.
[98] В дополнение к синтетическим экспрессионным элементам в данном документе представлен новый эндогенный 3' UTR, полученный из гена неспецифического белка-переносчика липидов 4 Sorghum bicolor, T-Sb.Nltp4-1:1:2, который предложен в данном документе и представлен как SEQ ID NO: 19. T-Sb.Nltp4-1:1:2 был охарактеризован аналогично синтетическому 3' UTR.
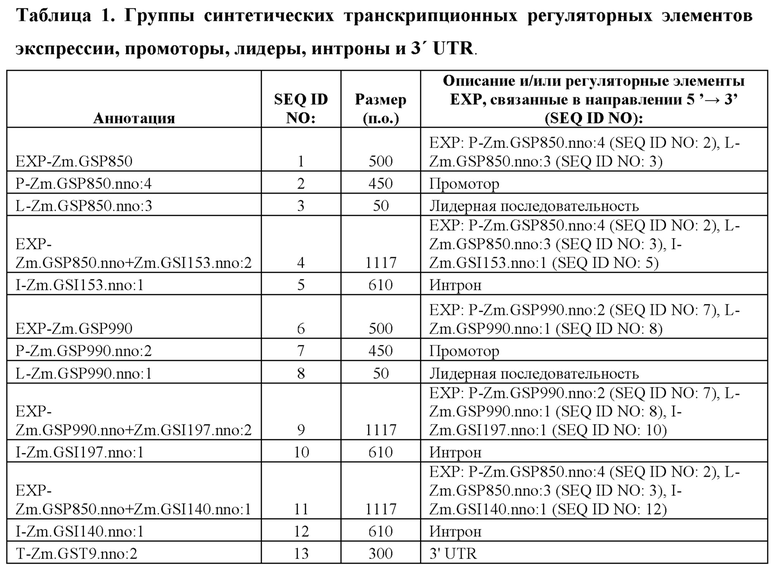
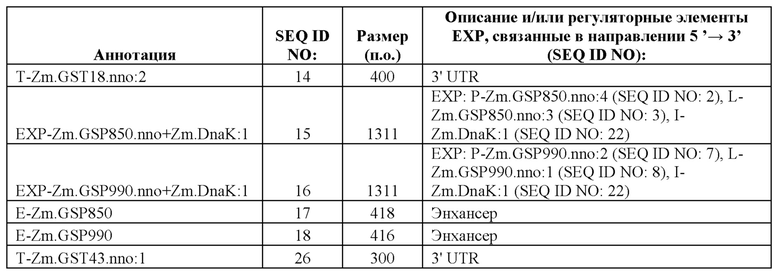
Пример 2
Анализ синтетических регуляторных элементов, управляющих GUS в протопластах листьев кукурузы
[99] Протопласты листьев кукурузы трансформировали векторами, в частности векторами экспрессии, содержащими тестируемый регуляторный элемент, управляющий экспрессией трансгена β-глюкуронидазы (GUS). Полученные трансформированные протопласты листьев кукурузы анализировали на экспрессию белка GUS, чтобы оценить влияние выбранных регуляторных элементов на экспрессию.
[100] Протопласты кукурузы, полученные из ткани листа, трансформировали векторами экспрессии, содержащими синтетические экспрессионные элементы. Уровень и паттерн экспрессии этих синтетических векторов экспрессионных элементов в протопластах кукурузы сравнивали с уровнем и паттерном экспрессии экспрессионных элементов, известными в данной области техники. Отдельные эксперименты были проведены для оценки активности ЕХР, EXP-Zm.GSP850 (SEQ ID NO: 1) и EXP-Zm.GSP990 (SEQ ID NO: 6), интронов I-Zm.GSI153.nno:1 (SEQ ID NO: 5) и I-Zm.GSI197.nno:1 (SEQ ID NO: 10), и 3' UTR, T-Zm.GST9.nno:2 (SEQ ID NO: 13) и T-Zm.GST18.nno:2 (SEQ ID NO: 14). Экспрессионные элементы клонировали в векторы экспрессии и функционально связывали с последовательностью, кодирующей GUS, GOI-Ec.uidA+St.LS1:1:1 (SEQ ID NO: 24), которая содержала процессируеммый интрон. Контрольные векторы экспрессии содержали различные конфигурации известных экспрессионных элементов, которые варьировались в зависимости от типа оцениваемого элемента (ЕХР, интрон или 3' UTR). Плазмиду, используемую для совместной трансформации протопластов и нормализации данных, также конструировали с использованием способов, известных в данной области техники. Она содержала трансгенную кассету, состоящую из ЕХР, EXP-CaMV.35S (SEQ ID NO: 21) функционально связанного с 5'-концом кодирующей последовательности, кодирующей флуоресцентный белок люциферазы NanoLuc® (Promega, Мэдисон, WI 53711), упоминаемый в данном документе Nluc (SEQ ID NO: 25), который был функционально связан с 5'-концом 3' UTR, T-Os.LTP:1 (SEQ ID NO: 23).
[101] Протопласты листьев кукурузы трансформировали с использованием способа трансформации на основе ПЭГ, аналогичного способам, известным в данной области техники. Клетки протопластов трансформировали в формате девяноста шести (96) лунок. Двенадцать (12) микрограмм ДНК тестового вектора или ДНК контрольного вектора и шесть (6) микрограмм векторной ДНК NanoLuc® использовали для трансформации 3,2×105 протопластов на лунку. После трансформации протопласты инкубировали при 25°С в темноте от шестнадцати до двадцати часов. После инкубации протопласты лизировали, и лизат использовали для измерения экспрессии люциферазы и GUS. Чтобы лизировать клетки, клетки в планшете осаждали центрифугированием, промывали, ресуспендировали в меньшем объеме и переносили в лунки пробирок в виде полосок. Пробирки снова центрифугировали и супернатант аспирировали, оставляя осадок клеток протопластов. Осадок клеток ресуспендировали в буфере QB (100 мМ KPO4, рН 7,8; 1 мМ ЭДТА; 1% Тритон Х-100; 10% глицерин; 1 мМ DTT). Клетки лизировали, энергично пипетируя клетки несколько раз, встряхивая пробирки и давая пробиркам инкубироваться на льду в течение пяти минут. Затем лизат центрифугировали для осаждения клеточного дебриса. Затем полученный лизат переносили в чистый планшет.
[102] Люциферазную активность анализировали с использованием субстрата для анализа люциферазы Nano-Glo® (Promega, Мэдисон, WI 53711) в буфере QB. Вкратце, небольшой объем лизата, буфер QB и раствор субстрата для анализа люциферазы Nano-Glo®/QB смешивали вместе в белых девяносто шести (96) луночных планшетах. Затем флуоресценцию измеряли с использованием планшет-ридера PHERAstar® (BMG LABTECH Inc., Cary, NC 27513).
[103] Активность GUS анализировали с использованием флуорогенного субстрата 4-метилумбеллиферил-β-D-глюкуронида (MUG) в общем реакционном объеме пятьдесят (50) микролитров. Продукт реакции, 4-метилумбеллиферон (4-MU), максимально флуоресцирует при высоком рН, когда гидроксильная группа ионизируется. Добавление щелочного раствора карбоната натрия одновременно останавливает анализ и регулирует рН для количественного определения флуоресцентного продукта. Аликвоту лизата смешивали с аликвотой MUG, растворенной в буфере QB, и инкубировали при 37°С. Небольшую аликвоту реакционной смеси лизат/MUG удаляли и добавляли в стоп-буфер в трех различных временных точках: (1) сразу после смешивания реакции лизат/MUG как «время ноль минут»; (2) двадцать минут; и (3) шестьдесят минут. Флуоресценцию измеряли при возбуждении при 355 нм, эмиссии при 460 нм, используя планшет-ридер PHERAstar® (BMG LABTECH Inc., Кэри, NC 27513). Уровень экспрессии выражается как «нМ гидролизованного MUG», что выводится из стандартной кривой для планшета.
[104] Для каждого планшета каждую конструкцию трансформируют в четырех (4) -восьми (8) лунках. Из каждой трансформации брали аликвоту для анализа MUG, и «нМ гидролизованного MUG» получали из стандартной кривой для планшета. Аликвоту также отбирали из каждой трансформации для считывания NanoLuc® (NanoLuc® RLU). Среднее значение нМ гидролизованного MUG/NanoLuc® RLU для каждой конструкции, нормализовали по отношению к конструкции EXP-CaMV.35S/ I-Zm.DnaK:1/ T-Os.LTP:1, которая принята за 100%.
Анализ экспрессии GUS в протопластах листьев кукурузы, управляемой синтетическим ЕХР, EXP-Zm.GSP850.
[105] Клетки протопластов листьев кукурузы трансформировали векторами экспрессии, которые были сконструированы с использованием способов, известных в данной области техники, содержащих экспрессионные элементы, управляющие экспрессией GUS. Два (2) тестовых вектора экспрессии содержали трансгенные кассеты, содержащие синтетический ЕХР, EXP-Zm.GSP850 (SEQ ID NO: 1). Синтетический EXP-Zm.GSP850 состоит из синтетического промотора Р-Zm.GSP850.nno:4 (SEQ ID NO: 2), функционально связанного с 5' -концом синтетической лидерной последовательности L-Zm.GSP850.nno:3 (SEQ ID NO: 3). Первый тестовый вектор содержал EXP-Zm.GSP850, функционально связанный 5'-концом с кодирующей последовательностью, кодирующей GUS (SEQ ID NO: 24), которая содержит процессируемый интрон, который функционально связан с 5' -концом 3' UTR, T-Os.LTP:l (SEQ ID NO: 23). Вторая трансгенная кассета содержала EXP-Zm.GSP850, функционально связанный с 5'-концом интрона I-Zm.DnaK:1 (SEQ ID NO: 22), функционально связанным с 5'-концом кодирующей последовательности GUS, которая функционально связана с 5'-концом 3' UTR, T-Os.LTP:1.
[106] Также были сконструированы три (3) контрольных вектора экспрессии, которые использовали для трансформации протопластов листьев кукурузы. Первый контрольный вектор экспрессии содержал трансгенную кассету без промотора и состоял из интрона, I-Zm.DnaK:1, функционально связанного с 5'-концом кодирующей последовательности GUS, которая функционально связана с 5'-концом 3' UTR, Т-Os.LTP:1. Второй контрольный вектор содержал трансгенную кассету без интрона и состоял из ЕХР, EXP-CaMV.35S (SEQ ID NO: 21), функционально связанного с 5'-концом кодирующей последовательности GUS, которая функционально связана с 5' -концом 3' UTR, T-Os.LTP:1. Третий контрольный вектор содержал трансгенную кассету, которая содержала ЕХР, EXP-CaMV.35S, функционально связанный с 5'-концом интрона I-Zm.DnaK:1, функционально связанным с 5'-концом кодирующей последовательностью GUS, которая функционально связана с 5' -концом 3' UTR, Т-Os.LTP:1.
[107] Протопласты листьев кукурузы трансформировали всеми пятью (5) векторами. Трансформацию и лизис клеток протопласта выполняли, как описано в данном документе. Экспрессию люциферазы и GUS анализировали, как описано в данном документе. В таблице 2 показана проанализированная средняя экспрессия GUS и которая выражена в виде процента экспрессии относительно третьего контрольного вектора экспрессии, который включает EXP-CaMV.35S и I-Zm.DnaK:1, управляющий GUS.
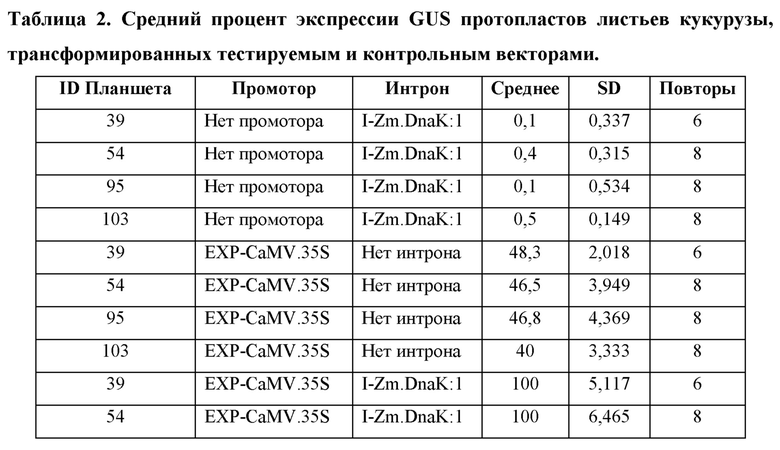
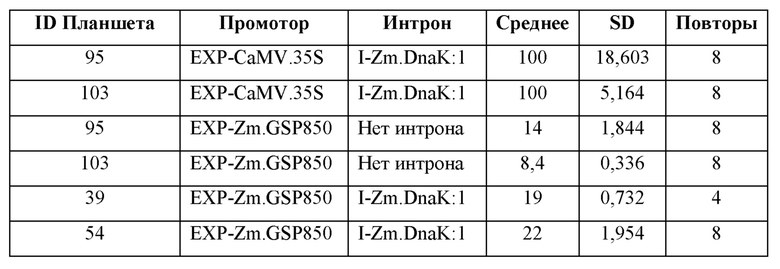
[108] Как видно из таблицы 2 выше, EXP-Zm.GSP850 (SEQ ID NO: 1) был способен управлять экспрессией трансгена GUS в протопластах листьев кукурузы по сравнению с клетками протопластов листьев кукурузы, трансформированных конструкцией без промотора.
Анализ экспрессии GUS в протопластах листьев кукурузы, управляемой синтетическим ЕХР, EXP-Zm.GSP990.
[109] Клетки протопластов листьев кукурузы трансформировали векторами экспрессии, которые были сконструированы содержащими экспрессионные элементы, управляющие экспрессией GUS. Тестовый вектор экспрессии содержал трансгенную кассету, которая содержала синтетический ЕХР, EXP-Zm.GSP990 (SEQ ID NO: 6), функционально связанный с 5'-концом интрона I-Zm.DnaK:1 (SEQ ID NO: 22), функционально связанным с 5'-концом кодирующей последовательности, кодирующей GUS (SEQ ID NO: 20), которая функционально связана с 5'-концом 3' UTR, Т-Os.LTP:1. Синтетический EXP-Zm.GSP990 (SEQ ID NO: 6) состоит из синтетического промотора P-Zm.GSP990.nno:2 (SEQ ID NO: 7), функционально связанного с 5'-концом синтетической лидерной последовательности L-Zm.GSP990.nno:1 (SEQ ID NO: 8). Три контрольных вектора экспрессии также трансформировали в протопласты листьев кукурузы и конструировали, как описано выше. В таблице 3 показан средний процент экспрессии относительно третьего контрольного вектора экспрессии, который содержит EXP-CaMV.35S и I-Zm.DnaK:1, управляющий GUS.
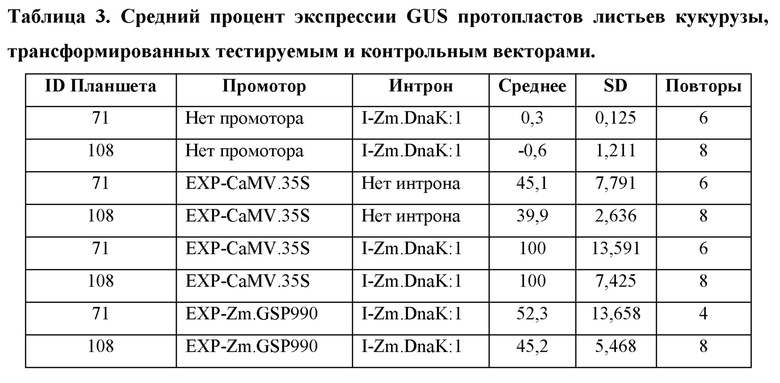
[110] Как видно из таблицы 3 выше, EXP-Zm.GSP990 (SEQ ID NO: 6) был способен управлять экспрессией трансгена GUS в протопластах листьев кукурузы по сравнению с клетками протопластов листьев кукурузы, трансформированных конструкцией без промотора.
Анализ усиления экспрессии GUS синтетическим интроном I-Zm.GSI153.nno:1
[111] Клетки протопластов листьев кукурузы трансформировали векторами экспрессии, которые были сконструированы содержащими экспрессионные элементы, управляющие экспрессией GUS. Тестируемый вектор экспрессии использовали для анализа усиления экспрессии GUS синтетическим интроном I-Zm.GSI153.nno:1 (SEQ ID NO: 5), управляемым EXP-CaMV.35. Трансгенная кассета содержала ЕХР, ЕХР-CaMV.35, функционально связанный 5'-концом с синтетическим интроном, I-Zm.GSI153.nno:1 (SEQ ID NO: 5), функционально связанным 5'-концом с кодирующей последовательностью, кодирующей GUS (SEQ ID NO: 24), которая была функционально связана 5'-концом с 3' UTR, T-Os.LTP:1. Также были сконструированы два контрольных вектора экспрессии, которые использовали для трансформации протопластов листьев кукурузы. Первый контрольный вектор экспрессии содержал трансгенную кассету без интрона и состоял из ЕХР, EXP-CaMV.35S, функционально связанного с 5'-концом кодирующей последовательности GUS, которая функционально связана с 5'-концом 3' UTR, T-Os.LTP:1. Второй контрольный вектор содержал трансгенную кассету, которая содержала ЕХР, EXP-CaMV.35S, функционально связанный с 5'-концом интрона I-Zm.DnaK:1, функционально связанным с 5'-концом кодирующей последовательности GUS, которая функционально связана с 5'-концом 3' UTR, T-Os.LTP:1. В таблице 4 показан средний процент экспрессии относительно второго контрольного вектора экспрессии, который содержит как EXP-CaMV.35S, так и I-Zm.DnaK:1, управляющий GUS.
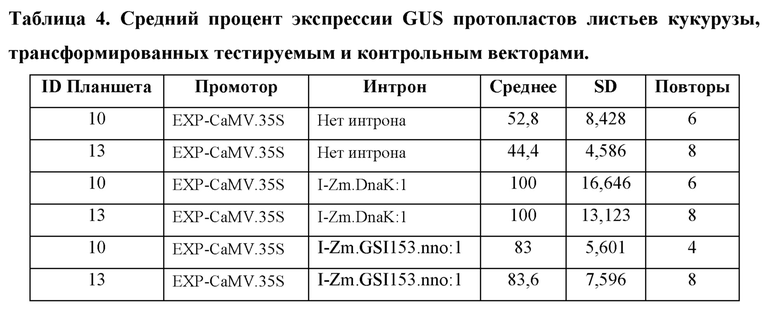
[112] Как можно увидеть из Таблицы 4, синтетический интрон, I-Zm.GSI153.nno:1 (SEQ ID NO: 5), усиливал экспрессию трансгена GUS в протопластах листьев кукурузы, управляемую EXP-CaMV.35S, по сравнению с контрольным вектором экспрессии без интронов.
Анализ усиления экспрессии GUS синтетическим интроном I-Zm.GSI197.nno:1
[113] Клетки протопластов листьев кукурузы трансформировали векторами экспрессии, которые были сконструированы содержащими экспрессионные элементы, управляющие экспрессией GUS. Тестируемый вектор экспрессии использовали для анализа усиления экспрессии GUS синтетическим интроном I-Zm.GSI197.nno:1 (SEQ ID NO: 10), управляемым EXP-CaMV.35. Трансгенная кассета содержала ЕХР, ЕХР-CaMV.35, функционально связанный с 5-концом синтетического интрона, I-Zm.GSI197.nno:1, функционально связанным с 5'-концом кодирующей последовательности, кодирующей GUS (SEQ ID NO: 24), которая была функционально связана с 5'-концом 3' UTR, T-Os.LTP:1. Также были сконструированы три контрольных вектора экспрессии, которые использовали для трансформации протопластов листьев кукурузы. Первый контрольный вектор экспрессии содержал трансгенную кассету без промотора и состоял из интрона, I-Zm.DnaK:1, функционально связанного с 5'-концом кодирующей последовательности GUS, которая функционально связана с 5'-концом 3' UTR, T-Os.LTP:1. Второй контрольный вектор содержал трансгенную кассету без интронов и состоял из ЕХР, EXP-CaMV.35S, функционально связанного с 5'-концом кодирующей последовательности GUS, которая функционально связана с 5'-концом 3' UTR, T-Os.LTP:1. Третий контрольный вектор содержал трансгенную кассету, которая содержала ЕХР, EXP-CaMV.35S, функционально связанный с 5'-концом интрона I-Zm.DnaK:1, функционально связанным с 5'-концом кодирующей последовательности GUS, которая функционально связана с 5'-концом 3' UTR, T-Os.LTP:1. Таблица 5 показывает средний процент экспрессии относительно третьего контрольного вектора экспрессии, который включает как EXP-CaMV.35S, так и I-Zm.DnaK:1, управляющий GUS.
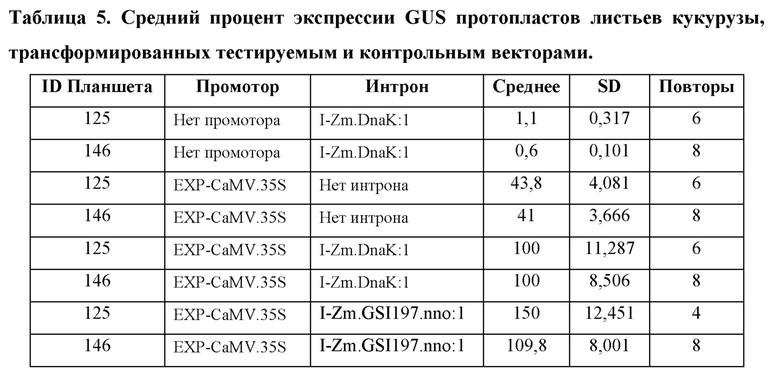
[114] Как можно увидеть из Таблицы 5, синтетический интрон, I-Zm.GSI197.nno:1 (SEQ ID NO: 10), усиливал экспрессию трансгена GUS в протопластах листьев кукурузы, управляемую EXP-CaMV.35S, по сравнению с контрольным вектором экспрессии без интронов. Усиление экспрессии было больше, чем придаваемое интроном, I-Zm.DnaK:1, по сравнению с третьим контрольным вектором экспрессии, который содержал ЕХР, EXP-CaMV.35S, функционально связанный с 5'-концом интрона I-Zm.DnaK:1.
Анализ усиления экспрессии GUS синтетическими 3' UTR, T-Zm.GST9.nno:2 и Т-Zm.GST18.nno:2.
[115] Клетки протопластов листьев кукурузы трансформировали векторами экспрессии, которые были сконструированы содержащими экспрессионные элементы, управляющие экспрессией GUS. Два тестовых вектора содержали трансгенную кассету, используемую для анализа усиления экспрессии GUS, придаваемого 3' UTR, T-Zm.GST9.nno:2 (SEQ ID NO: 13) и T-Zm.GST18.nno:2 (SEQ ID NO: 14) и состояли из EXP-CaMV.35S, функционально связанного с 5'-концом интрона I-Zm.DnaK:1, функционально связанным с 5'-концом кодирующей последовательности, кодирующей GUS (SEQ ID NO: 24), которая функционально связана с 5'-концом либо 3' UTR, Т-Zm.GST9.nno:2 (SEQ ID NO: 13), либо 3' UTR, T-Zm.GST18.nno:2 (SEQ ID NO: 14). Также были сконструированы три контрольных вектора экспрессии, как описано выше, и их использовали для трансформации протопластов листьев кукурузы. В таблице 6 показан средний процент экспрессии относительно третьего контрольного вектора экспрессии, который содержит EXP-CaMV.35S и I-Zm.DnaK:1, управляющий GUS.
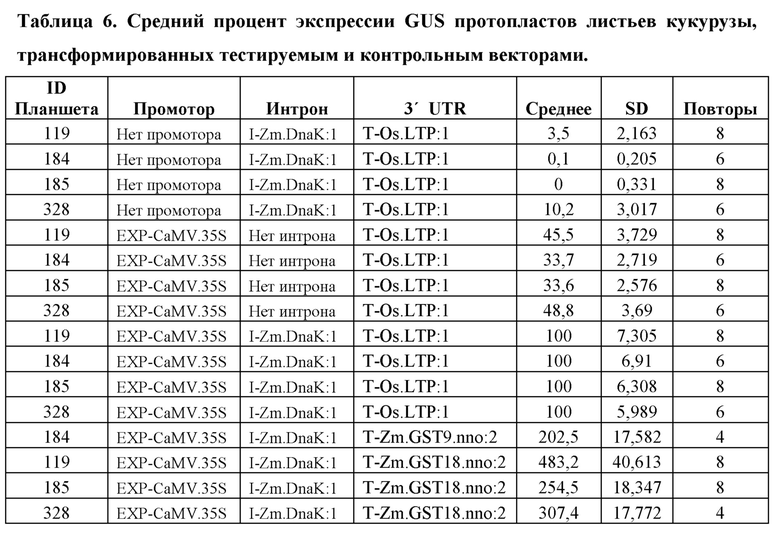
[116] Как можно увидеть из таблицы 6, 3' UTR, T-Zm.GST9.nno:2 (SEQ ID NO: 13) и T-Zm.GST18.nno:2 (SEQ ID NO: 14) усиливали экспрессию GUS относительно к контролей в протопластах листа кукурузы.
Пример 3
Анализ экспрессии GUS под действием синтетических ЕХР, ЕХР-Zm.GSP850.nno+Zm.GSI153.nno:2 и EXP-Zm.GSP850.nno+Zm.GSI140.nno:1 в стабильно трансформированных растениях кукурузы сорта LH244
[117] Растения кукурузы трансформировали вектором, в частности вектором экспрессии для растений, содержащим тестируемые регуляторные элементы, управляющие экспрессией трансгена β-глюкуронидазы (GUS). Полученные растения анализировали на экспрессию белка GUS, чтобы оценить влияние выбранного регуляторного элемента на экспрессию.
[118] Растения кукурузы трансформировали экспрессирующими конструкциями GUS растений. Регуляторные элементы были клонированы в базовый вектор экспрессии для растений с использованием стандартных способов, известных в данной области техники. Полученные в результате векторы экспрессии для растений содержали левый граничный участок из Agrobacterium tumefaciens (B-AGRtu.левая граница), первую трансгенную селекционную кассету, использованную для селекции трансформированных клеток растений, которые придают устойчивость к гербициду глифосату; вторую трансгенную кассету для оценки активности синтетических регуляторных элементов, которая включала либо синтетический ЕХР, ЕХР-Zm.GSP850.nno+Zm.GSI153.nno:2 (SEQ ID NO: 4), либо ЕХР-Zm.GSP850.nno+Zm.GSI140.nno:1 (SEQ ID NO: 11) функционально связанный с 5'-концом синтетической кодирующей последовательности, разработанной для экспрессии в клетке растения, кодирующей β-глюкуронидазу (GUS, GOI-Ec.uidA+St.LS1.nno:1, SEQ ID NO: 20), содержащей процессируемый интрон, полученный из светоиндуцируемого тканеспецифического гена картофеля ST-LS1 (Номер доступа Genbank: Х04753), функционально связанный с 5'-концом 3' участка терминации, Т-Sb.Nltp4-1:1:2 (SEQ ID NO: 19); и правый граничный участок из Agrobacterium tumefaciens (B-AGRtu. правая граница). Синтетический ЕХР, ЕХР-Zm.GSP850.nno+Zm.GSI153.nno:2 (SEQ ID NO: 4), состоит из синтетического промотора P-Zm.GSP850.nno:4 (SEQ ID NO: 2), функционально связанного с 5'-концом синтетической лидерной последовательности, L-Zm.GSP850.nno:3 (SEQ ID NO: 3), которая функционально связана с 5'-концом синтетического интрона, I-Zm.GSI153.nno:1 (SEQ ID NO: 5). Синтетический ЕХР, ЕХР-Zm.GSP850.nno+Zm.GSI140.nno:1 (SEQ ID NO: 11), состоит из синтетического промотора P-Zm.GSP850.nno:4 (SEQ ID NO: 2), функционально связанного с 5'-концом синтетической лидерной последовательности, L-Zm.GSP850.nno:3 (SEQ ID NO: 3), которая функционально связана 5'-концом с синтетическим интроном, I-Zm.GSI140.nno:1 (SEQ ID NO: 12).
[119] Клетки растений кукурузы сорта LH244 трансформировали с использованием конструкции бинарного вектора трансформации, описанной выше, посредством трансформации, опосредованной Agrobacterium, хорошо известной в данной области техники. Полученные трансформированные клетки растений индуцировали с образованием целых растений кукурузы.
[120] Качественный и количественный анализ GUS использовали для оценки активности экспрессионных элементов в выбранных органах и тканях растений трансформированных растений. Для качественного анализа экспрессии GUS с помощью гистохимического окрашивания целиком монтированные или срезы ткани инкубировали с окрашивающим раствором GUS, содержащим 1 мг/мл X-Gluc (5-бром-4-хлор-3-индолил-b-глюкоронид) в течение 5 ч при 37°С и обесцвечивают 35% EtOH и 50% уксусной кислотой. Экспрессию GUS качественно определяли путем визуального осмотра выбранных органов или тканей растений на наличие синей окраски под препарирующим или составным микроскопом.
[121] Для количественного анализа экспрессии GUS с помощью ферментативных анализов общий белок экстрагировали из выбранных тканей трансформированных растений кукурузы. От одного до двух микрограммов общего белка инкубировали с флуорогенным субстратом, 4-метилумбеллиферил-β-D-глюкоронидом (MUG) в концентрации 1 мМ в общем реакционном объеме 50 микролитров. После 1 ч инкубации при 37°С реакцию останавливали добавлением 350 микролитров 200 мМ раствора бикарбоната натрия. Продукт реакции, 4-метилумбеллиферон (4-MU), максимально флуоресцирует при высоком рН, когда гидроксильная группа ионизируется. Добавление основного раствора карбоната натрия одновременно останавливает анализ и регулирует рН для количественного определения флуоресцентного продукта 4-MU. Количество образовавшегося 4-МЕ оценивали путем измерения его флуоресценции с использованием считывающего устройства для микропланшетов FLUOstar Omega (BMG LABTECH) (возбуждение при 355 нм, эмиссия при 460 нм). Значения активности GUS представлены в нмолях 4-МЕ/час/мг общего белка.
[122] Следующие ткани были взяты для экспрессии GUS в поколении R0: стадия V4 Лист и Корень; стадия V7 Лист и Корень; стадия VT Лист, корень и цветок/пыльник; стадия R1 Початок/Шелк; и стадия R3 зародыша семени и эндосперма семени через 21 день после опыления (DAP). В таблице 7 показаны средние количественные значения экспрессии GUS для каждого из синтетических ЕХР.
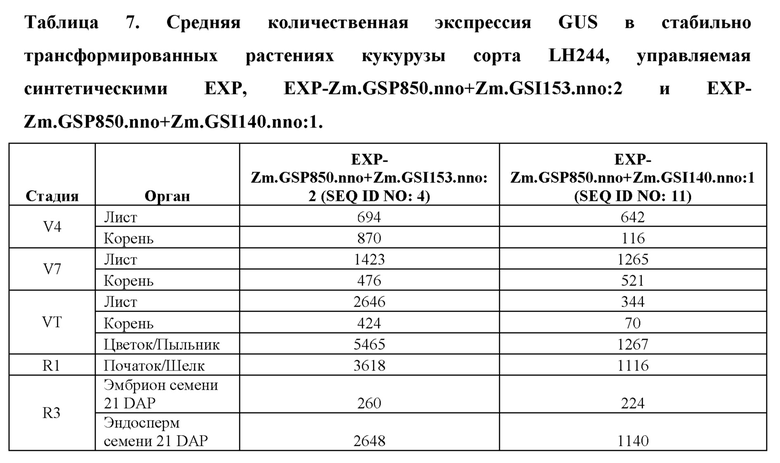
[123] Как видно из таблицы 7, синтетический промотор и лидерная последовательность GSP850 (P-Zm.GSP850.nno:4 (SEQ ID NO: 2) и L-Zm.GSP850.nno:3 (SEQ ID NO: 3)) приводили к конститутивной экспрессии GUS в стабильно трансформированных растениях кукурузы сорта LH244. Молекулярный анализ сайта начала транскрипции продемонстрировал постоянный TSS для промотора и лидерной последовательности GSP850. Синтетические интроны I-Zm.GSI153.nno:1 (SEQ ID NO: 5) и I-Zm.GSI140.nno:1 (SEQ ID NO: 12) влияли на экспрессию различным образом в разных отобранных тканях. Молекулярный анализ сайтов сплайсинга интронов продемонстрировал последовательный процессинг синтетических интронов. Общее усиление экспрессии GUS было выше в большинстве образцов тканей растений, содержащих I-Zm.GSI153.nno:1 (SEQ ID NO: 5), за исключением стадии V4 листа, стадии V7 корня и R3 семенного зародыша, где уровни экспрессии GUS были относительно похожи. Усиление экспрессии, придаваемой I-Zm.GSI153.nno:1 (SEQ ID NO: 5) по сравнению с I-Zm.GSI140.nno:1 (SEQ ID NO: 12), было примерно в 7,5 раз выше в V4 корня, 7,7 раз выше в VT листа, в 6,0 раза выше в VT корня, в 4,3 раза выше в VT цветка/пыльника, в 3,2 раза выше в R1 початка/шелка и в 2,3 раза выше в R3 эндосперма семян.
Пример 4
Анализ экспрессии GUS под действием синтетических ЕХР, ЕХР-Zm.GSP850.nno+Zm.DnaK:1, EXP-Zm.GSP850.nno+Zm.GSI153.nno:2 и ЕХР-Zm.GSP850.nno+Zm.GSI140.nno:1 в стабильно трансформированных растениях кукурузы сорта 01DKD2
[124] Растения кукурузы трансформировали вектором, в частности вектором экспрессии для растений, содержащим тестируемые регуляторные элементы, управляющие экспрессией трансгена β-глюкуронидазы (GUS). Полученные растения анализировали на экспрессию белка GUS, чтобы оценить влияние выбранного регуляторного элемента на экспрессию.
[125] Растения кукурузы трансформировали экспрессирующими конструкциями GUS растений. Регуляторные элементы были клонированы в базовый вектор экспрессии для растений с использованием стандартных способов, известных в данной области техники. Полученные в результате векторы экспрессии для растений содержали левый граничный участок из Agrobacterium tumefaciens (B-AGRtu.левая граница), первую трансгенную селекционную кассету, использованную для селекции трансформированных клеток растений, которые придают устойчивость к гербициду глифосату; вторую трансгенную кассету для оценки активности регуляторных элементов, которая содержала либо синтетический ЕХР, ЕХР-Zm.GSP850.nno+Zm.DnaK:1 (SEQ ID NO: 15), EXP-Zm.GSP850.nno+Zm.GSI153.nno:2 (SEQ ID NO: 4), либо EXP-Zm.GSP850.nno+Zm.GSI140.nno:1 (SEQ ID NO: 11) функционально связанный с 5'-концом синтетической кодирующей последовательности, разработанной для экспрессии в клетке растения, кодирующей β-глюкуронидазу (GUS, GOI-Ec.uidA+St.LS1.nno:1, SEQ ID NO: 20), содержащей процессируемый интрон, полученный из светоиндуцируемого тканеспецифического гена картофеля ST-LS1 (Номер доступа Genbank: Х04753), функционально связанный с 5'-концом 3' участка терминации, T-Sb.Nltp4-1:1:2 (SEQ ID NO: 19); и правый граничный участок из Agrobacterium tumefaciens (B-AGRtu.правая граница). Синтетический ЕХР, EXP-Zm.GSP850.nno+Zm.DnaK:1 (SEQ ID NO: 15), состоит из синтетического промотора P-Zm.GSP850.nno:4 (SEQ ID NO: 2), функционально связаного с 5'-концом синтетической лидерной последовательности, L-Zm.GSP850.nno:3 (SEQ ID NO: 3), которая функционально связана 5'-концом с интроном, I-Zm.DnaK:1 (SEQ ID NO: 22). Синтетические ЕХР, ЕХР-Zm.GSP850.nno+Zm.GSI153.nno:2 (SEQ ID NO: 4) и ЕХР-Zm.GSP850.nno+Zm.GSI140.nno:1 (SEQ ID NO: 11) описаны в Примере 3. [126] Клетки растений кукурузы сорта 01DKD2 трансформировали с использованием конструкции бинарного вектора трансформации, описанной выше, посредством трансформации, опосредованной Agrobacterium, как хорошо известно в данной области техники. Полученные трансформированные клетки растений индуцировали с образованием целых растений кукурузы. Качественную и количественную экспрессию GUS анализировали, как описано в Примере 3. Таблица 8 показывает средние количественные значения экспрессии GUS для каждого из синтетических ЕХР.
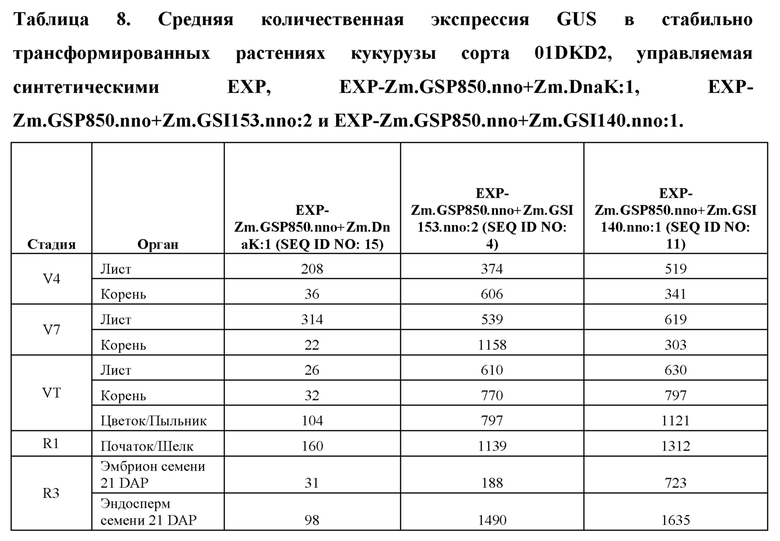
[127] Как видно из таблицы 8, синтетический промотор и лидерная последовательнось GSP850 (P-Zm.GSP850.nno:4 (SEQ ID NO: 2) и L-Zm.GSP850.nno:3 (SEQ ID NO: 3)) приводили к конститутивной экспрессии GUS в стабильно трансформированных растениях кукурузы сорта LH244. Синтетические интроны, I-Zm.GSI153.nno:1 (SEQ ID NO: 5) и I-Zm.GSI140.nno:1 (SEQ ID NO: 12), усиливали экспрессию по сравнению с интроном I-Zm.DnaK:1 (SEQ ID NO: 22) во всех проанализированных тканях.
Пример 5
Анализ экспрессии GUS под действием синтетических ЕХР, ЕХР-Zm.GSP990.nno+Zm.DnaK:1 и EXP-Zm.GSP990.nno+Zm.GSI197.nno:2 в стабильно трансформированных растениях кукурузы сорта 01DKD2
[128] Растения кукурузы трансформировали вектором, в частности вектором экспрессии для растений, содержащим тестируемые регуляторные элементы, управляющие экспрессией трансгена β-глюкуронидазы (GUS). Полученные растения анализировали на экспрессию белка GUS, чтобы оценить влияние выбранного регуляторного элемента на экспрессию.
[129] Растения кукурузы трансформировали экспрессирующими конструкциями GUS растений. Регуляторные элементы были клонированы в базовый вектор экспрессии для растений с использованием стандартных способов, известных в данной области техники. Полученные в результате векторы экспрессии для растений содержали левый граничный участок из Agrobacterium tumefaciens (B-AGRtu.левая граница), первую трансгенную селекционную кассету, использованную для селекции трансформированных клеток растений, которые придают устойчивость к гербициду глифосату; вторую трансгенную кассету для оценки активности регуляторных элементов, которая содержала либо синтетический ЕХР, ЕХР-Zm.GSP990.nno+Zm.DnaK:1 (SEQ ID NO: 16), либо ЕХР-Zm.GSP990.nno+Zm.GSI197.nno:2 (SEQ ID NO: 9) функционально связанный с 5'-концом синтетической кодирующей последовательности, разработанной для экспрессии в клетке растения, кодирующей β-глюкуронидазу (GUS, GOI-Ec.uidA+St.LS1.nno:1, SEQ ID NO: 20), содержащей процессируемый интрон, полученный из светоиндуцируемого тканеспецифического гена картофеля ST-LS1 (Номер доступа Genbank: Х04753), функционально связанный с 5'-концом 3' участка терминации, T-Sb.Nltp4-1:1:2 (SEQ ID NO: 19); и правый граничный участок из Agrobacterium tumefaciens (B-AGRtu.правая граница). Синтетический ЕХР, ЕХР-Zm.GSP990.nno+Zm.GSI197.nno:2 (SEQ ID NO: 9), состоит из синтетического промотора P-Zm.GSP990.nno:2 (SEQ ID NO: 7), функционально связаного с 5'-концом синтетической лидерной последовательности, L-Zm.GSP990.nno:1 (SEQ ID NO: 8), которая функционально связана с 5'-концом синтетического интрона, I-Zm.GSI197.nno:1 (SEQ ID NO: 10). Синтетический ЕХР, ЕХР-Zm.GSP990.nno+Zm.DnaK:1 (SEQ ID NO: 16), состоит из синтетического промотора Р-Zm.GSP990.nno:2 (SEQ ID NO: 7), функционально связаного с 5'-концом синтетической лидерной последовательности, L-Zm.GSP990.nno:1 (SEQ ID NO: 8), которая функционально связана с 5'-концом интрона, I-Zm.DnaK:1 (SEQ ID NO: 22).
[130] Клетки растений сорта кукурузы 01DKD2 трансформировали с использованием конструкции бинарного вектора трансформации, описанной выше, с помощью трансформации, опосредованной Agrobacterium, что хорошо известно в данной области техники. Полученные трансформированные клетки растений индуцировали с образованием целых растений кукурузы. Качественную и количественную экспрессию GUS анализировали, как описано ранее в примере 3. В таблице 9 показаны средние количественные значения экспрессии GUS для каждого из синтетических ЕХР, где «ND» означает, что значение не было определено.
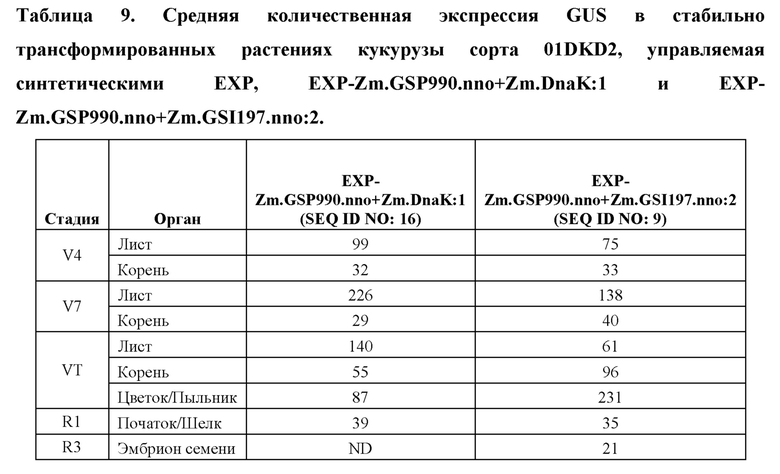

[131] Как можно видеть, синтетический промотор и лидер GSP990 (Р-Zm.GSP990.nno:2 (SEQ ID NO: 7) и L-Zm.GSP990.nno:1 (SEQ ID NO: 8)) управляли экспрессией GUS. Молекулярный анализ сайта начала транскрипции продемонстрировал постоянный TSS для промотора и лидера GSP990. Синтетический интрон, I-Zm.GSI197.nno:1 (SEQ ID NO: 10) ослаблял экспрессию в некоторых тканях, в то же время усиливая экспрессию в других тканях относительно интрона I-Zm.DnaK:1 (SEQ ID NO: 22). Например, экспрессия GUS была ослаблена в листе на стадии V4, V7 и VT. Экспрессия GUS была немного усилена в корнях V7 и VT по сравнению с I-Zm.DnaK:1. Экспрессия в цветке/пыльнике увеличивалась приблизительно в 2,7 раза с помощью I-Zm.GSI197.nno:1 (SEQ ID NO: 10) по сравнению с I-Zm.DnaK:1. Различия в экспрессии, обеспечиваемой I-Zm.GSI197.nno:1 (SEQ ID NO: 10), по сравнению с I-Zm.DnaK:1, могут быть очень полезны там, где желательна более низкая экспрессия в листе и более высокая экспрессия в цветке/пыльнике. Молекулярный анализ сайтов сплайсинга интронов продемонстрировал согласованный процессинг синтетического интрона I-Zm.GSI197.nno:1 (SEQ ID NO: 10).
Пример 6
Анализ влияния на экспрессию GUS синтетических 3' UTR T-Zm.GST9.nno:2, Т-Zm.GST18.nno:2, и T-Zm.GST43.nno:1, и нативного T-Sb.Ntlp4-1:1:2 в стабильно трансформированных растениях кукурузы сорта 01DKD2.
[132] Растения кукурузы трансформировали вектором, в частности вектором экспрессии для растений, содержащим тестируемые регуляторные элементы, управляющие экспрессией трансгена β-глюкуронидазы (GUS). Полученные растения анализировали на экспрессию белка GUS, чтобы оценить влияние выбранного регуляторного элемента на экспрессию.
[133] Растения кукурузы трансформировали экспрессирующими конструкциями GUS растений. Регуляторные элементы были клонированы в базовый вектор экспрессии для растений с использованием стандартных способов, известных в данной области техники. Полученные в результате векторы экспрессии для растений содержали левый граничный участок из Agrobacterium tumefaciens (B-AGRtu.левая граница), первую трансгенную селекционную кассету, использованную для селекции трансформированных клеток растений, которые придают устойчивость к гербициду глифосату; вторую трансгенную кассету для оценки активности 3' UTR регуляторных элементов, которая содержала ЕХР, EXP-CaMV.35S (SEQ ID NO: 21) функционально связанный с 5'-концом интрона, I-Zm.DnaK:1 (SEQ ID NO: 22), функционально связанным с 5'-концом синтетической кодирующей последовательности, разработанной для экспрессии в клетке растения, кодирующей В-глюкуронидазу (GUS, GOI-Ec.uidA+St.LSl.nno:1, SEQ ID NO: 20), содержащей процессируемый интрон, полученный из светоиндуцируемого тканеспецифического гена картофеля ST-LS1 (Номер доступа Genbank: Х04753), функционально связанный с 5'-концом 3' участка терминации; и правый граничный участк из Agrobacterium tumefaciens (B-AGRtu.правая граница). Три тестовых экспрессионных вектора содержали 3' UTR T-Zm.GST9.nno:2 (SEQ ID NO: 13), T-Zm.GST18.nno:2 (SEQ ID NO: 14) или T-Zm.GST43. nno:1 (SEQ ID NO: 26), функционально связанный с кодирующей последовательностью GUS. Дополнительный тестовый экспрессионный вектор содержал природный 3' UTR Т-Sb.Nltp4-1:1:2 (SEQ ID NO: 19), функционально связанный с кодирующей последовательностью GUS, и был использован для сравнения экспрессии между природными и синтетическими 3' UTR.
[134] Клетки растений кукурузы сорта 01DKD2 трансформировали с использованием конструкции бинарного вектора трансформации, описанной выше, посредством трансформации, опосредованной Agrobacterium, как хорошо известно в данной области техники. Полученные трансформированные клетки растений индуцировали с образованием целых растений кукурузы.
[135] Качественный и количественный GUS-анализ использовали для оценки активности элемента экспрессии в тканях листа и корня V4 трансформированных растений и выполняли, как описано выше в Примере 3. Эффект синтетических 3' UTR оценивали путем сравнения с эффектом экспрессии от 3' UTR T-Sb.Nltp4-1:1:2 (SEQ ID NO: 19). Полученные транскрипты были проанализированы, чтобы определить, произошла ли надлежащая терминация и что транскрипт не был считан от начала до конца. Один из способов оценки того, было ли считываниеот начала до конца, заключался в использовании амплификации транскрипта кДНК с использованием праймера амплификации, соответствующего части граничной последовательности Т-ДНК, которая находится на 3'-конце 3' UTR. Средняя экспрессия GUS растений, трансформированных четырьмя конструкциями, содержащими T-Zm.GST9.nno:2 (SEQ ID NO: 13), T-Zm.GST18.nno:2 (SEQ ID NO: 14), T-Zm. GST43.nno:1 (SEQ ID NO: 26) и T-Sb.Nltp4-1:1:2 (SEQ ID NO: 19) представлены в таблице 10.
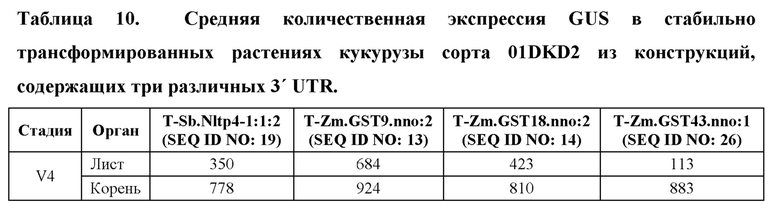
[136] Как видно из Таблицы 10, как T-Zm.GST9.nno:2 (SEQ ID NO: 13), так и Т-Zm.GST18.nno:2 (SEQ ID NO: 14) усиливали экспрессию GUS, управляемую ЕХР-CaMV.35S, функционально связанного с I-Zm.DnaK:1, относительно 3' UTR, Т-Sb.Nltp4-1:1:2. Экспрессия GUS в растениях, содержащих T-Zm.GST9.nno:2 (SEQ ID NO: 13), была выше, чем в растениях, содержащих T-Zm.GST18.nno:2. Т-Zm.GST43.nno:1 (SEQ ID NO: 26) усиливал экспрессию GUS в V4 корне по сравнению с T-Sb.Nltp4-1:1:2, но ослаблял экспрессию в V4 листе. Анализ транскриптов GUS из всех четырех конструкций продемонстрировал правильную терминацию транскрипта и отсутствие доказательств считывания от начала до конца в полученных транскриптах GUS. 3' UTR T-Zm.GST9.nno:2 (SEQ ID NO: 13), T-Zm.GST18.nno:2 (SEQ ID NO: 14) и T-Zm.GST43.nno:1 (SEQ ID NO: 26) действуют аналогично естественным 3' UTR и демонстрируют модуляцию экспрессии GUS по сравнению с природными 3' UTR Т-Sb.Nltp4-1:1:2. Все четыре синтетических 3' UTR и дополнительная нативная 3 'UTR Т-Sb.Nltp4-1:1:2 обеспечивают диапазон значений экспрессии, которые пригодны для точной настройки экспрессии в стабильно трансформированных растениях кукурузы.
Пример 7
Энхансерные элементы, полученные из регуляторных элементов
[137] Энхансеры получают из промоторных элементов, представленных как SEQ ID NO: 2 и 7. Энхансерные элементы могут состоять из одного или более цис регуляторных элементов, которые функционально связаны с 5'-концом или 3'-концом элемента промотора или функционально связаны с 5'-концом или 3'-концом дополнительного энхансерного элемента, которые функционально связаны с промотором, могут увеличивать или модулировать уровни экспрессии молекулы транскрибируемой ДНК или обеспечивать экспрессию молекулы транскрибируемой ДНК в конкретном типе клетки или органе растения, или в определенный момент времени в процессе развития или циркадного ритма. Энхансеры получают удалением ТАТА-бокса или функционально подобных элементов и любой нижеследующей последовательности из промоторов, которые позволяют инициировать транскрипцию с промоторов, представленных как SEQ ID NO: 2 и 7, или их фрагментов.
[138] ТАТА-бокс в промоторах растений не так высоко консервативен, как у некоторых других эукариотических организмов. Следовательно, чтобы определить фрагмент как энхансер, нужно сначала идентифицировать сайт начала транскрипции (TSS) гена, в котором сначала транскрибируется 5' UTR. Энхансер, полученный из синтетического промотора, P-Zm.GSP850.nno:4 (SEQ ID NO: 2), может содержать нуклеотиды с 1 по 418 из SEQ ID NO: 2, что приводит к получению синтетического энхансера E-Zm.GSP850 (SEQ ID NO: 17). Энхансер, полученный из синтетического промотора, P-Zm.GSP990.nno:2 (SEQ ID NO: 7), может содержать нуклеотиды с 1 по 416 из SEQ ID NO: 7, что приводит к получению синтетического энхансера Е-Zm.GSP990 (SEQ ID NO: 18). Энхансеры, полученные от промоторов, могут содержать фрагменты SEQ ID NO: 17 и 18 или дупликации SEQ ID NO: 17 и 18 или их соответствующие фрагменты. Эффективность синтетических энхансеров, полученных из синтетических промоторов, определяется эмпирически путем создания химерного регуляторного элемента транскрипции, содержащего фрагменты, полученные от синтетических промоторов P-Zm.GSP850.nno:4 (SEQ ID NO: 2) или P-Zm.GSP990.nno:2 (SEQ ID NO: 7), которые функционально связаны с промотором и лидером, и используется для управления экспрессией молекулы транскрибируемой ДНК, такой как GUS, в анализе стабильных или переходных растений.
[139] Может потребоваться дальнейшее уточнение энхансерного элемента, что подтверждается эмпирически. Кроме того, положение энхансерного элемента относительно других элементов в химерном регуляторном элементе транскрипции также определяется эмпирически, поскольку порядок каждого элемента в химерном регуляторном элементе транскрипции может оказывать различные эффекты в зависимости от относительного положения каждого элемента. Некоторые промоторные элементы будут иметь множество ТАТА-боксов или элементов, подобных ТАТА-боксу, и потенциально множество сайтов старта транскрипции. В этих обстоятельствах может быть необходимо сначала определить, где расположен первый TSS, а затем начать конструирование энхансеров с использованием первого TSS, чтобы предотвратить потенциальную инициацию транскрипции внутри предполагаемого энхансерного элемента.
[140] Энхансерные элементы, полученные из промоторных элементов, представленных как SEQ ID NO: 2 и 7, клонируются с использованием способов, известных в данной области техники, чтобы быть функционально связанными с 5'-концом или внутри промоторного элемента, или функционально связанными с 5'-концом или 3'-концом дополнительных энхансерных элементов, которые функционально связаны с промотором. Альтернативно, энхансерные элементы могут быть клонированы с использованием способов, известных в данной области техники, для получения большего энхансерного элемента, который состоит из двух или более копий энхансера, и клонирован с использованием способов, известных в данной области техники чтобы быть функционально связанным с 5'-концом или 3'-концом промоторного элемента или функционально связанным с 5'-концом или 3'-концом дополнительного энхансерного элемента, которые функционально связаны с промотором, продуцирующим химерный регуляторный элемент транскрипции. Энхансерные элементы, полученные из промоторов, полученных от генов организмов множества родов, могут быть функционально связаны с энхансерами, полученными из синтетических промоторов.
[141] Вектор трансформации растений для экспрессии GUS может быть сконструирован с использованием способов, известных в данной области техники, аналогичных конструкции, описанной в Примере 3, в которой полученные векторы экспрессии для растений содержат левый граничный участок из Agrobacterium tumefaciens (B-AGRtu. левая граница), первую трансгенную кассету селекции, используемую для селекции трансформированных клеток растений, придающих устойчивость к гербициду глифосату; и вторую трансгенную кассету для тестирования энхансерного элемента, состоящего из энхансерного элемента, функционально связанного с 5'-концом или 3'-концом промоторного элемента или функционально связанного с 5'-концом или 3'-концом дополнительного энхансерного элемента которые, в свою очередь, функционально связаны с промотором, который функционально связан с 5'-концом элемента лидерной последовательности, функционально связанным с 5'-концомэлемента интрона, функционально связанным с кодирующей последовательностью для β-глюкуронидазы (GOI-Ec.uidA+St.LS1.nno:1, SEQ ID NO: 20), содержащей процессируемый интрон, полученный из светоиндуцируемого тканеспецифического гена ST-LS1 картофеля (Номер доступа в Genbank: Х04753), функционально связанный с 3'-концевым участком из гена белка-подобного переносчика липидов Oryza sativa (T-Os.LTP:1, SEQ ID NO: 23); и правый граничный участок от A. tumefaciens (B-AGRtu.правая граница). Полученные плазмиды используют для трансформации растений кукурузы или других однодольных растений описанными выше способами. Альтернативно, клетки протопластов, полученные из кукурузы или других однодольных растений, трансформируют с использованием способов, известных в данной области техники, для проведения переходных анализов.
[142] Экспрессия GUS, управляемая регуляторным элементом, содержащим один или более энхансеров, оценивается в анализах стабильных или переходных растений для определения влияния энхансерного элемента на экспрессию молекулы транскрибируемой ДНК. Модификации одного или более энхансерных элементов или дублирование одного или более энхансерных элементов могут быть выполнены на основе эмпирических экспериментов и результирующей регуляции экспрессии генов, которую наблюдают с использованием каждой композиции регуляторных элементов. Изменение относительных положений одного или более энхансеров в результирующих регуляторных или химерных регуляторных элементах может повлиять на транскрипционную активность или специфичность регуляторного или химерного регуляторного элемента и определяется эмпирически для определения лучших энхансеров для желаемого профиля экспрессии трансгена в растении кукурузы или растения другого рода.
[143] Проиллюстрировав и описав принципы данного изобретения, специалистам в данной области техники должно быть очевидно, что изобретение может быть модифицировано по компоновке и деталям без отклонения от таких принципов. Мы претендуем на все модификации, которые находятся в пределах сущности и объема формулы изобретения. Все публикации и опубликованные патентные документы, процитированные в данном документе, включены в данный документ посредством ссылки в той же степени, как если бы каждая отдельная публикация или патентная заявка была специально и индивидуально указаны для включения посредством ссылки.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
--->
<110> МОНСАНТО ТЕКНОЛОДЖИ ЛЛК
<120> РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ
<130> 38-21(62691)0001/US
<150> US 62/714 228
<151> 03.08.2018
<160> 26
<170> PatentIn версія 3.5
<210> 1
<211> 500
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность ДНК синтетического EXP, EXP-Zm.GSP850,
содержащая синтетический промотор (P-Zm.GSP850.nno:4),
функционально связанный с 5'-концом синтетической лидерной
последовательностью (L-Zm.GSP850.nno:3)
<400> 1
agacggcagc acaacggcca tccggtcgct gtaggtacac gagtgccgca gtagatggcg 60
cgcgtttttg tctgagtccg atggagcagc gcagcgcccc ccccccggcc accgccgctt 120
aggcggacag atcatcacgc cggccgccgg cctcatcgcc caaactgtgt ggcccaaaac 180
caccagacaa gcatattctg gcaggatgcc gttggcaaca aaaacacagg cgaccaaccc 240
aaacaggccg cccggcccat aaggagctcc gcaggcccat ttctcatttg gccaggatag 300
gccagccagg tggccacacg tgcaaaaccg ccacgtggcc aaggtggcgg atcccttatc 360
cacactggcc acacatgtgg cgccacacca caaccacaca ggcttatcta atagctgcta 420
tataaagagg aggagccgcc gcccaccacc actcaccagc acacaaaccc tagctgtttt 480
ccatcagaac cactcaccgg 500
<210> 2
<211> 450
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность ДНК синтетического промотора, P-Zm.GSP850.nno:4.
<400> 2
agacggcagc acaacggcca tccggtcgct gtaggtacac gagtgccgca gtagatggcg 60
cgcgtttttg tctgagtccg atggagcagc gcagcgcccc ccccccggcc accgccgctt 120
aggcggacag atcatcacgc cggccgccgg cctcatcgcc caaactgtgt ggcccaaaac 180
caccagacaa gcatattctg gcaggatgcc gttggcaaca aaaacacagg cgaccaaccc 240
aaacaggccg cccggcccat aaggagctcc gcaggcccat ttctcatttg gccaggatag 300
gccagccagg tggccacacg tgcaaaaccg ccacgtggcc aaggtggcgg atcccttatc 360
cacactggcc acacatgtgg cgccacacca caaccacaca ggcttatcta atagctgcta 420
tataaagagg aggagccgcc gcccaccacc 450
<210> 3
<211> 50
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность ДНК синтетической лидерной последовательности, P-Zm.GSP850.nno:3.
<400> 3
actcaccagc acacaaaccc tagctgtttt ccatcagaac cactcaccgg 50
<210> 4
<211> 1117
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность ДНК синтетического EXP,
EXP-Zm.GSP850.nno+Zm.GSI153.nno:2, содержащая синтетический промотор
(P-Zm.GSP850.nno:4), синтетическую синтетической лидерную последовательность (L-Zm.GSP850.nno:3) и
синтетический интрон(I-Zm.GSI153.nno:1).
<400> 4
agacggcagc acaacggcca tccggtcgct gtaggtacac gagtgccgca gtagatggcg 60
cgcgtttttg tctgagtccg atggagcagc gcagcgcccc ccccccggcc accgccgctt 120
aggcggacag atcatcacgc cggccgccgg cctcatcgcc caaactgtgt ggcccaaaac 180
caccagacaa gcatattctg gcaggatgcc gttggcaaca aaaacacagg cgaccaaccc 240
aaacaggccg cccggcccat aaggagctcc gcaggcccat ttctcatttg gccaggatag 300
gccagccagg tggccacacg tgcaaaaccg ccacgtggcc aaggtggcgg atcccttatc 360
cacactggcc acacatgtgg cgccacacca caaccacaca ggcttatcta atagctgcta 420
tataaagagg aggagccgcc gcccaccacc actcaccagc acacaaaccc tagctgtttt 480
ccatcagaac cactcaccgg cggaccgctc aggtgagaac ctcgctcgtc tcccgcggat 540
ctgcttctcg taacggttcg atctgtgctg ctatttcgga ttcggatcta ctgcggtgcg 600
gttgacgatt taggtttgtt agggatctgg tggtcatgct tgatcgatct ggtcgattag 660
ggcccctcgc ggatctgcct tgttctgagt ttgggatctg gtgctcgaga tctcgatgcg 720
gattcgagag atctgctgca tcgatccgga tccggggttt cccatttgag cttctcctgc 780
cgccgccgct gtcttgccac ctgactaggt agggtttccc cccgcgatct agcttcctgc 840
tggtactctc gccgaccgtc atccaaactc aaatttggcg gatccgacgc acatcgatct 900
gttgttcaca ctacaatcgt tgagtcgatg aggttattgt aaatctgatc tgggccgctg 960
cgattatccc tacctctctc ggaaacactg cgcgacgtgt aatatggtta aagtttgggc 1020
agttcgtagc gggttcggtc cctcggttct ggatttgagc tcccggtgga ttggtgtgtg 1080
tgctgttcta atgtattgat tcaatgttac aggttca 1117
<210> 5
<211> 610
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность ДНК синтетического интрона, I-Zm.GSI153.nno:1.
<400> 5
ctcaggtgag aacctcgctc gtctcccgcg gatctgcttc tcgtaacggt tcgatctgtg 60
ctgctatttc ggattcggat ctactgcggt gcggttgacg atttaggttt gttagggatc 120
tggtggtcat gcttgatcga tctggtcgat tagggcccct cgcggatctg ccttgttctg 180
agtttgggat ctggtgctcg agatctcgat gcggattcga gagatctgct gcatcgatcc 240
ggatccgggg tttcccattt gagcttctcc tgccgccgcc gctgtcttgc cacctgacta 300
ggtagggttt ccccccgcga tctagcttcc tgctggtact ctcgccgacc gtcatccaaa 360
ctcaaatttg gcggatccga cgcacatcga tctgttgttc acactacaat cgttgagtcg 420
atgaggttat tgtaaatctg atctgggccg ctgcgattat ccctacctct ctcggaaaca 480
ctgcgcgacg tgtaatatgg ttaaagtttg ggcagttcgt agcgggttcg gtccctcggt 540
tctggatttg agctcccggt ggattggtgt gtgtgctgtt ctaatgtatt gattcaatgt 600
tacaggttca 610
<210> 6
<211> 500
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность ДНК синтетического EXP, EXP-Zm.GSP990, содержащая
синтетический промотор (P-Zm.GSP990.nno:2), функционально связанный с 5'-концом
синтетической лидерной последовательности (L-Zm.GSP990.nno:1).
<400> 6
gagaatgtgg gaggaacttg cgacagtaac ctcattttgg tcacaaaaac cgatatttaa 60
ggatcactag tcaccagacg aatctatttc atttgaacca tccaacgaga tgatctcaga 120
atgatccacg tcacaccaaa tttctcgtcg aactcggcca gcagagccat ccaaacgggg 180
atctccgcgc aacatctcca tccaccacct agagcagccc cggccttttc catggcaaaa 240
tttacatccg atttggctga agaccaatgg gaaaaaggcg ccacgtggcc acgtggcgtc 300
cacacgggcc aggcaggcag agaatggcca ccaatggggt ggataaggcc ggcgccaaaa 360
atatctgcca cgtggcgcct tgtggataag gcagcttgct atccctccgc caatcctata 420
tatatccttt cccctctcct agtgccgctc aaagctaagc acaacctaag ctgatccact 480
gcaagcatcg tctcctacta 500
<210> 7
<211> 450
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность ДНК синтетического промотора, P-Zm.GSP990.nno:2.
<400> 7
gagaatgtgg gaggaacttg cgacagtaac ctcattttgg tcacaaaaac cgatatttaa 60
ggatcactag tcaccagacg aatctatttc atttgaacca tccaacgaga tgatctcaga 120
atgatccacg tcacaccaaa tttctcgtcg aactcggcca gcagagccat ccaaacgggg 180
atctccgcgc aacatctcca tccaccacct agagcagccc cggccttttc catggcaaaa 240
tttacatccg atttggctga agaccaatgg gaaaaaggcg ccacgtggcc acgtggcgtc 300
cacacgggcc aggcaggcag agaatggcca ccaatggggt ggataaggcc ggcgccaaaa 360
atatctgcca cgtggcgcct tgtggataag gcagcttgct atccctccgc caatcctata 420
tatatccttt cccctctcct agtgccgctc 450
<210> 8
<211> 50
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность ДНК синтетической лидерной последовательности, P-Zm.GSP990.nno:1.
<400> 8
aaagctaagc acaacctaag ctgatccact gcaagcatcg tctcctacta 50
<210> 9
<211> 1117
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность ДНК синтетического EXP,
EXP-Zm.GSP990.nno+Zm.GSI197.nno:2, содержащая синтетический промотор
(P-Zm.GSP990.nno:2), синтетическую лидерную последовательность (L-Zm.GSP990.nno:1) и
синтетический интрон(I-Zm.GSI197.nno:1).
<400> 9
gagaatgtgg gaggaacttg cgacagtaac ctcattttgg tcacaaaaac cgatatttaa 60
ggatcactag tcaccagacg aatctatttc atttgaacca tccaacgaga tgatctcaga 120
atgatccacg tcacaccaaa tttctcgtcg aactcggcca gcagagccat ccaaacgggg 180
atctccgcgc aacatctcca tccaccacct agagcagccc cggccttttc catggcaaaa 240
tttacatccg atttggctga agaccaatgg gaaaaaggcg ccacgtggcc acgtggcgtc 300
cacacgggcc aggcaggcag agaatggcca ccaatggggt ggataaggcc ggcgccaaaa 360
atatctgcca cgtggcgcct tgtggataag gcagcttgct atccctccgc caatcctata 420
tatatccttt cccctctcct agtgccgctc aaagctaagc acaacctaag ctgatccact 480
gcaagcatcg tctcctacta cggaccgtac aggtaagcga gcgccgctcg ctcccgccgt 540
ctcccttctc tgccggatct gccgtgctgc cgtcgtggcg tccatcctga tctccctccc 600
atcgacccct cggccgggcg gtttctgttt tatagggttt ccctcccacc tcgcgggcag 660
tttgctgctg cctgctcaga tcccagctgg ttggatctgc ggattcgggt aggcggattc 720
gccgggccga gttatataaa ccatttactg tagatgcgtt actaccttaa tgacgcacgg 780
atctgccgtc tcgccccccc ccgcgtcgga tccgactgtt agcttcctgc cacggatctg 840
ctggactgtt ttcttgatcg ggtcttgcat ttggtggtgg cgggaatata ttattttaga 900
tggtctgggt ctgcggatcc gtgcttattt tgttaccatg cccccacctg actataaact 960
gtaaggtcga tctgatttgt ttgctatgcc cctcttgccg tctggatatg atggaaatat 1020
atccgatcca gatgatgcta gtctctccct cgagcttctc gagggggtcg atctgaagtc 1080
agttctaatt aatattttga tgctgattac aggtagc 1117
<210> 10
<211> 610
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность ДНК синтетического интрона, I-Zm.GSI197.nno:1.
<400> 10
tacaggtaag cgagcgccgc tcgctcccgc cgtctccctt ctctgccgga tctgccgtgc 60
tgccgtcgtg gcgtccatcc tgatctccct cccatcgacc cctcggccgg gcggtttctg 120
ttttataggg tttccctccc acctcgcggg cagtttgctg ctgcctgctc agatcccagc 180
tggttggatc tgcggattcg ggtaggcgga ttcgccgggc cgagttatat aaaccattta 240
ctgtagatgc gttactacct taatgacgca cggatctgcc gtctcgcccc cccccgcgtc 300
ggatccgact gttagcttcc tgccacggat ctgctggact gttttcttga tcgggtcttg 360
catttggtgg tggcgggaat atattatttt agatggtctg ggtctgcgga tccgtgctta 420
ttttgttacc atgcccccac ctgactataa actgtaaggt cgatctgatt tgtttgctat 480
gcccctcttg ccgtctggat atgatggaaa tatatccgat ccagatgatg ctagtctctc 540
cctcgagctt ctcgaggggg tcgatctgaa gtcagttcta attaatattt tgatgctgat 600
tacaggtagc 610
<210> 11
<211> 1117
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность ДНК синтетического EXP,
EXP-Zm.GSP850.nno+Zm.GSI140.nno:1, содержащая синтетический промотор
(P-Zm.GSP850.nno:4), синтетическую лидерную последовательность (L-Zm.GSP850.nno:3) и
синтетический интрон(I-Zm.GSI140.nno:1).
<400> 11
agacggcagc acaacggcca tccggtcgct gtaggtacac gagtgccgca gtagatggcg 60
cgcgtttttg tctgagtccg atggagcagc gcagcgcccc ccccccggcc accgccgctt 120
aggcggacag atcatcacgc cggccgccgg cctcatcgcc caaactgtgt ggcccaaaac 180
caccagacaa gcatattctg gcaggatgcc gttggcaaca aaaacacagg cgaccaaccc 240
aaacaggccg cccggcccat aaggagctcc gcaggcccat ttctcatttg gccaggatag 300
gccagccagg tggccacacg tgcaaaaccg ccacgtggcc aaggtggcgg atcccttatc 360
cacactggcc acacatgtgg cgccacacca caaccacaca ggcttatcta atagctgcta 420
tataaagagg aggagccgcc gcccaccacc actcaccagc acacaaaccc tagctgtttt 480
ccatcagaac cactcaccgg cggaccggcc aggtacgcct cctcctccgc gccgctcttc 540
ctgcccaccc cgctcgatct gccacggatc cgctcggatc tggtcccctt cgattcggcc 600
acgatcggat ccgccgccgg gcgctcgcca tcggtggtgt ggtggctgta ggtttgcgtc 660
gtaagtttcc tgctccctcg ctggatgctt ccttctagcg gattcggcga cccatttctc 720
gcccgacggt cttccctctc tcttggctcg caaatcagga tgggtgttgt ttgatgctgt 780
catcgcccgt ttgatccgtc tcaaaattcg tctcccccgc gcacggatcg atctgctccc 840
cctctctgct ccctgacccc gtctcacggc ggcgcgccgc tgctgctgct gtgccattcc 900
atcagtcata caatcggagt aggcggaaat ataaaaatcg tccccttagc agcagatgga 960
tccgtccatt tccggattcg agtcgattcg tgtgctcccc ccctctctct cgcccggatc 1020
tgattgcgac aacttgcttc tcccctccat ttcgatcccc atgtttacga tacttttccc 1080
aactaatatt gatgtgggtg actgttttac aggtttc 1117
<210> 12
<211> 610
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность ДНК синтетического интрона, I-Zm.GSI140.nno:1.
<400> 12
gccaggtacg cctcctcctc cgcgccgctc ttcctgccca ccccgctcga tctgccacgg 60
atccgctcgg atctggtccc cttcgattcg gccacgatcg gatccgccgc cgggcgctcg 120
ccatcggtgg tgtggtggct gtaggtttgc gtcgtaagtt tcctgctccc tcgctggatg 180
cttccttcta gcggattcgg cgacccattt ctcgcccgac ggtcttccct ctctcttggc 240
tcgcaaatca ggatgggtgt tgtttgatgc tgtcatcgcc cgtttgatcc gtctcaaaat 300
tcgtctcccc cgcgcacgga tcgatctgct ccccctctct gctccctgac cccgtctcac 360
ggcggcgcgc cgctgctgct gctgtgccat tccatcagtc atacaatcgg agtaggcgga 420
aatataaaaa tcgtcccctt agcagcagat ggatccgtcc atttccggat tcgagtcgat 480
tcgtgtgctc cccccctctc tctcgcccgg atctgattgc gacaacttgc ttctcccctc 540
catttcgatc cccatgttta cgatactttt cccaactaat attgatgtgg gtgactgttt 600
tacaggtttc 610
<210> 13
<211> 300
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность ДНК синтетического 3' UTR, T-Zm.GST9.nno:2.
<400> 13
gtcgtcgtgg ttttttttgt ttttgtatct gaaggaggcc gccccatgtg tgttgttgta 60
gctagcagaa gaaggccacc gttcctgtac attttgttgt gctgctgctg cttgttcttc 120
gtggatgtac atttgtatca tgtttttttg gttgtaaact ctatataaga aatgaataag 180
taataaagaa gttgtttcat acccggtgct tccttgtttg ttgtgatatc ttttggctag 240
cttccttgtt aaccatgaaa tatatggccg tcggcatcat tttttgtttg tcttattttc 300
<210> 14
<211> 400
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность ДНК синтетического 3' UTR, T-Zm.GST18.nno:2.
<400> 14
tgctctacca agtagttgtg tgtgctgtcg atccatcttg ctgctgctac atgcatgttt 60
ttgagcaggt ggtttctttt ggtttcgact ggttgttttg taaactatca ctatatatta 120
aataaatgga ataaagttgt tttttctcat accaagttcg tgctgtgttc tgatgccgcc 180
gctgtttggt ttgctcatgc tgtgtggttg tttcttgatt tttttggttc ctgtgtagct 240
gcagtttgtt ttgaatctac ctagcatgtg tgatatgttt gctgcgtcgg caaccttgta 300
aactttggtt tcacgatgaa ataaattaat aaatattttg gttttctcat accgtgcttg 360
caaggatgtt tgtttgtttc ggtattcccc tgcatatttt 400
<210> 15
<211> 1311
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность ДНК синтетического EXP, EXP-Zm.GSP850.nno+Zm.DnaK:1,
содержащая синтетический промотор (P-Zm.GSP850.nno:4), функционально
связанный с 5'-концом синтетической лидерной последовательности (L-Zm.GSP850.nno:3), функционально
связанной с 5'-концом интрона (I-Zm.DnaK:1).
<400> 15
agacggcagc acaacggcca tccggtcgct gtaggtacac gagtgccgca gtagatggcg 60
cgcgtttttg tctgagtccg atggagcagc gcagcgcccc ccccccggcc accgccgctt 120
aggcggacag atcatcacgc cggccgccgg cctcatcgcc caaactgtgt ggcccaaaac 180
caccagacaa gcatattctg gcaggatgcc gttggcaaca aaaacacagg cgaccaaccc 240
aaacaggccg cccggcccat aaggagctcc gcaggcccat ttctcatttg gccaggatag 300
gccagccagg tggccacacg tgcaaaaccg ccacgtggcc aaggtggcgg atcccttatc 360
cacactggcc acacatgtgg cgccacacca caaccacaca ggcttatcta atagctgcta 420
tataaagagg aggagccgcc gcccaccacc actcaccagc acacaaaccc tagctgtttt 480
ccatcagaac cactcaccgg cggaccgacc gtcttcggta cgcgctcact ccgccctctg 540
cctttgttac tgccacgttt ctctgaatgc tctcttgtgt ggtgattgct gagagtggtt 600
tagctggatc tagaattaca ctctgaaatc gtgttctgcc tgtgctgatt acttgccgtc 660
ctttgtagca gcaaaatata gggacatggt agtacgaaac gaagatagaa cctacacagc 720
aatacgagaa atgtgtaatt tggtgcttag cggtatttat ttaagcacat gttggtgtta 780
tagggcactt ggattcagaa gtttgctgtt aatttaggca caggcttcat actacatggg 840
tcaatagtat agggattcat attataggcg atactataat aatttgttcg tctgcagagc 900
ttattatttg ccaaaattag atattcctat tctgtttttg tttgtgtgct gttaaattgt 960
taacgcctga aggaataaat ataaatgacg aaattttgat gtttatctct gctcctttat 1020
tgtgaccata agtcaagatc agatgcactt gttttaaata ttgttgtctg aagaaataag 1080
tactgacagt attttgatgc attgatctgc ttgtttgttg taacaaaatt taaaaataaa 1140
gagtttcctt tttgttgctc tccttacctc ctgatggtat ctagtatcta ccaactgaca 1200
ctatattgct tctctttaca tacgtatctt gctcgatgcc ttctccctag tgttgaccag 1260
tgttactcac atagtctttg ctcatttcat tgtaatgcag ataccaagcg g 1311
<210> 16
<211> 1311
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность ДНК синтетического EXP, EXP-Zm.GSP990.nno+Zm.DnaK:1,
содержащая синтетический промотор (P-Zm.GSP990.nno:2), функционально
связанный c 5'-концом синтетической лидерной последовательности (L-Zm.GSP990.nno:1), функционально
связанной с 5'-концом интрона (I-Zm.DnaK:1).
<400> 16
gagaatgtgg gaggaacttg cgacagtaac ctcattttgg tcacaaaaac cgatatttaa 60
ggatcactag tcaccagacg aatctatttc atttgaacca tccaacgaga tgatctcaga 120
atgatccacg tcacaccaaa tttctcgtcg aactcggcca gcagagccat ccaaacgggg 180
atctccgcgc aacatctcca tccaccacct agagcagccc cggccttttc catggcaaaa 240
tttacatccg atttggctga agaccaatgg gaaaaaggcg ccacgtggcc acgtggcgtc 300
cacacgggcc aggcaggcag agaatggcca ccaatggggt ggataaggcc ggcgccaaaa 360
atatctgcca cgtggcgcct tgtggataag gcagcttgct atccctccgc caatcctata 420
tatatccttt cccctctcct agtgccgctc aaagctaagc acaacctaag ctgatccact 480
gcaagcatcg tctcctacta cggaccgacc gtcttcggta cgcgctcact ccgccctctg 540
cctttgttac tgccacgttt ctctgaatgc tctcttgtgt ggtgattgct gagagtggtt 600
tagctggatc tagaattaca ctctgaaatc gtgttctgcc tgtgctgatt acttgccgtc 660
ctttgtagca gcaaaatata gggacatggt agtacgaaac gaagatagaa cctacacagc 720
aatacgagaa atgtgtaatt tggtgcttag cggtatttat ttaagcacat gttggtgtta 780
tagggcactt ggattcagaa gtttgctgtt aatttaggca caggcttcat actacatggg 840
tcaatagtat agggattcat attataggcg atactataat aatttgttcg tctgcagagc 900
ttattatttg ccaaaattag atattcctat tctgtttttg tttgtgtgct gttaaattgt 960
taacgcctga aggaataaat ataaatgacg aaattttgat gtttatctct gctcctttat 1020
tgtgaccata agtcaagatc agatgcactt gttttaaata ttgttgtctg aagaaataag 1080
tactgacagt attttgatgc attgatctgc ttgtttgttg taacaaaatt taaaaataaa 1140
gagtttcctt tttgttgctc tccttacctc ctgatggtat ctagtatcta ccaactgaca 1200
ctatattgct tctctttaca tacgtatctt gctcgatgcc ttctccctag tgttgaccag 1260
tgttactcac atagtctttg ctcatttcat tgtaatgcag ataccaagcg g 1311
<210> 17
<211> 418
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность ДНК синтетического энхансера, E-Zm.GSP85, который
получен из синтетического промотора, P-Zm.GSP850.nno:4.
<400> 17
agacggcagc acaacggcca tccggtcgct gtaggtacac gagtgccgca gtagatggcg 60
cgcgtttttg tctgagtccg atggagcagc gcagcgcccc ccccccggcc accgccgctt 120
aggcggacag atcatcacgc cggccgccgg cctcatcgcc caaactgtgt ggcccaaaac 180
caccagacaa gcatattctg gcaggatgcc gttggcaaca aaaacacagg cgaccaaccc 240
aaacaggccg cccggcccat aaggagctcc gcaggcccat ttctcatttg gccaggatag 300
gccagccagg tggccacacg tgcaaaaccg ccacgtggcc aaggtggcgg atcccttatc 360
cacactggcc acacatgtgg cgccacacca caaccacaca ggcttatcta atagctgc 418
<210> 18
<211> 416
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность ДНК синтетического энхансера, E-Zm.GSP990, который
получен из синтетического промотора, P-Zm.GSP990.nno:2.
<400> 18
gagaatgtgg gaggaacttg cgacagtaac ctcattttgg tcacaaaaac cgatatttaa 60
ggatcactag tcaccagacg aatctatttc atttgaacca tccaacgaga tgatctcaga 120
atgatccacg tcacaccaaa tttctcgtcg aactcggcca gcagagccat ccaaacgggg 180
atctccgcgc aacatctcca tccaccacct agagcagccc cggccttttc catggcaaaa 240
tttacatccg atttggctga agaccaatgg gaaaaaggcg ccacgtggcc acgtggcgtc 300
cacacgggcc aggcaggcag agaatggcca ccaatggggt ggataaggcc ggcgccaaaa 360
atatctgcca cgtggcgcct tgtggataag gcagcttgct atccctccgc caatcc 416
<210> 19
<211> 487
<212> ДНК
<213> Sorghum bicolor
<220>
<221> прочий_признак
<222> (1)..(487)
<223> Последовательность ДНК 3' UTR, T-Sb.Nltp4-1:1:2, полученная из гена NLTP4
(неспецифический белок-переносчик липидов 4) из Sorghum
bicolor.
<400> 19
tgatcgatca taaggacatg catgaggcat gctcgttgga tggcgatcag catgcagtcg 60
tcgttgtttt accagatgtc gtccatgact ccctccatgg atgatgcatg caatgagata 120
gatacataaa ttaaggagta gatcatgcat gcaccgtacc tgttagtgat cactccgttt 180
aattagtatt tactgtttac taccttgagt ttacaccaac tctgtcgtcg ctgtggctgt 240
cgtcgtgtac ctcgatcgta cgtgtgtgtg tgtagcaggc tagcaaagca gcactcaacg 300
tcgtccttgg gggtttgctt gcctttaacc ttgttgcttt agcacacagg tttctgaata 360
atgaatacta ctagcttttg tgtccattat atatatattg tactgtatgc atgatgtttt 420
ccatccatga gtcatgagag tgatgtgttc aagtaataag atatagtaac atatacacgc 480
gacgtac 487
<210> 20
<211> 2001
<212> ДНК
<213> Искусственная
<220>
<223> Синтетическая кодирующая последовательность, оптимизированная для экспрессии
?-глюкуронидазы (GUS) в растении с процессируемым интроном, полученным из
светоиндуцируемого тканеспецифичного гена ST-LS1 картофеля (Номер
доступа в Genbank: X04753).
<400> 20
atggtgaggc ccgttgagac cccgactagg gagatcaaga agctggacgg cctctgggcc 60
ttctccctcg accgtgagaa ctgcggcatc gaccagcgct ggtgggagtc cgccctccag 120
gagtctaggg ccatcgccgt gcccggttcc ttcaacgacc agttcgccga cgccgacatc 180
cgcaactacg cgggcaacgt ctggtatcag cgcgaggtgt tcatcccgaa gggctgggcg 240
ggccagcgca tcgtgctccg cttcgacgcc gtgacccact acggcaaggt ctgggtgaac 300
aatcaggagg taagtttctg cttctacctt tgatatatat ataataatta tcattaatta 360
gtagtaatat aatatttcaa atattttttt caaaataaaa gaatgtagta tatagcaatt 420
gcttttctgt agtttataag tgtgtatatt ttaatttata acttttctaa tatatgacca 480
aaatttgttg atgtgcaggt gatggagcac cagggcggtt acaccccgtt cgaggccgac 540
gtgacgccgt acgtgatcgc cgggaagtcc gtccgcatca ccgtctgcgt gaacaatgag 600
ctgaactggc agaccatccc gcctggcatg gtcatcaccg acgagaacgg caagaagaag 660
cagtcctact tccacgactt cttcaactac gctggcatcc accgctccgt gatgctctac 720
accactccca acacctgggt ggacgacatc accgtggtca cccacgtggc ccaggactgc 780
aaccacgcct ccgtggactg gcaagtcgtt gccaacggcg acgtcagcgt cgagctgcgc 840
gacgccgacc agcaagtcgt tgccaccggc cagggcacca gcggcaccct ccaagtcgtc 900
aaccctcacc tctggcagcc tggcgagggc tacctctacg agctgtgcgt caccgccaag 960
agccagactg agtgcgacat ctaccctctc cgcgtcggca tcaggagcgt cgctgtcaag 1020
ggcgagcagt tcctcatcaa ccacaagcct ttctacttca ctggtttcgg ccgccacgag 1080
gacgctgacc tgaggggcaa gggtttcgac aacgtcctga tggtccacga ccacgctctg 1140
atggactgga tcggtgccaa cagctacagg accagtcact acccgtacgc tgaggagatg 1200
ctggactggg ctgacgagca cggtatcgtc gtgatcgacg agactgctgc ggtcggtttc 1260
aacctgtctc tgggcattgg tttcgaggct gggaacaagc cgaaggagct gtactctgag 1320
gaagctgtca acggcgagac tcagcaagct catctccagg cgattaagga gctgattgcc 1380
agggacaaga accatccgtc tgtcgtgatg tggtctattg cgaatgagcc ggacaccaga 1440
ccgcaagggg cgcgtgaata cttcgcgccg ctggcggagg cgactcgcaa actggaccca 1500
acccgtccaa tcacgtgcgt caatgtcatg ttctgcgacg cccatacgga tacgatctcg 1560
gacctgttcg atgttctttg tctcaatcgg tactatgggt ggtatgttca gagcggggat 1620
cttgagacgg cggagaaggt tcttgagaag gaactcctgg cgtggcaaga gaagctccat 1680
cagccgatca ttatcacgga gtacggggtt gacacacttg cgggccttca cagtatgtac 1740
acagatatgt ggtcggagga ataccagtgt gcatggttgg atatgtacca tcgtgtcttc 1800
gaccgggttt cagcggttgt cggcgaacaa gtctggaact tcgcagactt cgccacgagc 1860
caagggatac tgcgggtagg agggaacaag aagggaatct tcacacggga tcggaagccc 1920
aagtcagcag ccttcctgtt gcagaagcga tggacaggaa tgaacttcgg agaaaagcca 1980
cagcaaggcg gaaagcagtg a 2001
<210> 21
<211> 835
<212> ДНК
<213> Вирус мозаики цветной капусты
<220>
<221> прочий_признак
<222> (1)..(835)
<223> Последовательность ДНК EXP, EXP-CaMV.35S, содержащая промотор 35S
и лидерную последовательность, полученные из вируса мозаики цветной капусты.
<400> 21
agattagcct tttcaatttc agaaagaatg ctaacccaca gatggttaga gaggcttacg 60
cagcaggtct catcaagacg atctacccga gcaataatct ccaggaaatc aaataccttc 120
ccaagaaggt taaagatgca gtcaaaagat tcaggactaa ctgcatcaag aacacagaga 180
aagatatatt tctcaagatc agaagtacta ttccagtatg gacgattcaa ggcttgcttc 240
acaaaccaag gcaagtaata gagattggag tctctaaaaa ggtagttccc actgaatcaa 300
aggccatgga gtcaaagatt caaatagagg acctaacaga actcgccgta aagactggcg 360
aacagttcat acagagtctc ttacgactca atgacaagaa gaaaatcttc gtcaacatgg 420
tggagcacga cacacttgtc tactccaaaa atatcaaaga tacagtctca gaagaccaaa 480
gggcaattga gacttttcaa caaagggtaa tatccggaaa cctcctcgga ttccattgcc 540
cagctatctg tcactttatt gtgaagatag tggaaaagga aggtggctcc tacaaatgcc 600
atcattgcga taaaggaaag gccatcgttg aagatgcctc tgccgacagt ggtcccaaag 660
atggaccccc acccacgagg agcatcgtgg aaaaagaaga cgttccaacc acgtcttcaa 720
agcaagtgga ttgatgtgat atctccactg acgtaaggga tgacgcacaa tcccactatc 780
cttcgcaaga cccttcctct atataaggaa gttcatttca tttggagagg acacg 835
<210> 22
<211> 804
<212> ДНК
<213> Zea mays
<220>
<221> прочий_признак
<222> (1)..(804)
<223> Последовательность ДНК интрона, I-Zm.DnaK:1, полученная из гена белка теплового
шока 70 (Hsp70) (DnaK) из Zea mays.
<400> 22
accgtcttcg gtacgcgctc actccgccct ctgcctttgt tactgccacg tttctctgaa 60
tgctctcttg tgtggtgatt gctgagagtg gtttagctgg atctagaatt acactctgaa 120
atcgtgttct gcctgtgctg attacttgcc gtcctttgta gcagcaaaat atagggacat 180
ggtagtacga aacgaagata gaacctacac agcaatacga gaaatgtgta atttggtgct 240
tagcggtatt tatttaagca catgttggtg ttatagggca cttggattca gaagtttgct 300
gttaatttag gcacaggctt catactacat gggtcaatag tatagggatt catattatag 360
gcgatactat aataatttgt tcgtctgcag agcttattat ttgccaaaat tagatattcc 420
tattctgttt ttgtttgtgt gctgttaaat tgttaacgcc tgaaggaata aatataaatg 480
acgaaatttt gatgtttatc tctgctcctt tattgtgacc ataagtcaag atcagatgca 540
cttgttttaa atattgttgt ctgaagaaat aagtactgac agtattttga tgcattgatc 600
tgcttgtttg ttgtaacaaa atttaaaaat aaagagtttc ctttttgttg ctctccttac 660
ctcctgatgg tatctagtat ctaccaactg acactatatt gcttctcttt acatacgtat 720
cttgctcgat gccttctccc tagtgttgac cagtgttact cacatagtct ttgctcattt 780
cattgtaatg cagataccaa gcgg 804
<210> 23
<211> 300
<212> ДНК
<213> Oryza sativa
<220>
<221> прочий_признак
<222> (1)..(300)
<223> Последовательность ДНК 3? UTR, T-Os.LTP:1, полученная из гена белок-подобного
переносчика липидов (LTP) из Oryza sativa.
<400> 23
attaatcgat cctccgatcc cttaattacc ataccattac accatgcatc aatatccata 60
tatatataaa ccctttcgca cgtacttata ctatgttttg tcatacatat atatgtgtcg 120
aacgatcgat ctatcactga tatgatatga ttgatccatc agcctgatct ctgtatcttg 180
ttatttgtat accgtcaaat aaaagtttct tccacttgtg ttaataatta gctactctca 240
tctcatgaac cctatatata actagtttaa tttgctgtca attgaacatg atgatcgatg 300
<210> 24
<211> 2001
<212> ДНК
<213> Искусственная
<220>
<223> Кодирующая последовательность для ?-глюкуронидазы (GUS) с процессируемым
интроном, полученным из светоиндуцируемого тканеспецифичного
гена ST-LS1 картофеля (Номер доступа в Genbank: X04753).
<400> 24
atggtccgtc ctgtagaaac cccaacccgt gaaatcaaaa aactcgacgg cctgtgggca 60
ttcagtctgg atcgcgaaaa ctgtggaatt gatcagcgtt ggtgggaaag cgcgttacaa 120
gaaagccggg caattgctgt gccaggcagt tttaacgatc agttcgccga tgcagatatt 180
cgtaattatg cgggcaacgt ctggtatcag cgcgaagtct ttataccgaa aggttgggca 240
ggccagcgta tcgtgctgcg tttcgatgcg gtcactcatt acggcaaagt gtgggtcaat 300
aatcaggaag tgatggagca tcagggcggc tatacgccat ttgaagccga tgtcacgccg 360
tatgttattg ccgggaaaag tgtacgtaag tttctgcttc tacctttgat atatatataa 420
taattatcat taattagtag taatataata tttcaaatat ttttttcaaa ataaaagaat 480
gtagtatata gcaattgctt ttctgtagtt tataagtgtg tatattttaa tttataactt 540
ttctaatata tgaccaaaat ttgttgatgt gcaggtatca ccgtttgtgt gaacaacgaa 600
ctgaactggc agactatccc gccgggaatg gtgattaccg acgaaaacgg caagaaaaag 660
cagtcttact tccatgattt ctttaactat gccggaatcc atcgcagcgt aatgctctac 720
accacgccga acacctgggt ggacgatatc accgtggtga cgcatgtcgc gcaagactgt 780
aaccacgcgt ctgttgactg gcaggtggtg gccaatggtg atgtcagcgt tgaactgcgt 840
gatgcggatc aacaggtggt tgcaactgga caaggcacta gcgggacttt gcaagtggtg 900
aatccgcacc tctggcaacc gggtgaaggt tatctctatg aactgtgcgt cacagccaaa 960
agccagacag agtgtgatat ctacccgctt cgcgtcggca tccggtcagt ggcagtgaag 1020
ggcgaacagt tcctgattaa ccacaaaccg ttctacttta ctggctttgg tcgtcatgaa 1080
gatgcggact tgcgtggcaa aggattcgat aacgtgctga tggtgcacga ccacgcatta 1140
atggactgga ttggggccaa ctcctaccgt acctcgcatt acccttacgc tgaagagatg 1200
ctcgactggg cagatgaaca tggcatcgtg gtgattgatg aaactgctgc tgtcggcttt 1260
aacctctctt taggcattgg tttcgaagcg ggcaacaagc cgaaagaact gtacagcgaa 1320
gaggcagtca acggggaaac tcagcaagcg cacttacagg cgattaaaga gctgatagcg 1380
cgtgacaaaa accacccaag cgtggtgatg tggagtattg ccaacgaacc ggatacccgt 1440
ccgcaaggtg cacgggaata tttcgcgcca ctggcggaag caacgcgtaa actcgacccg 1500
acgcgtccga tcacctgcgt caatgtaatg ttctgcgacg ctcacaccga taccatcagc 1560
gatctctttg atgtgctgtg cctgaaccgt tattacggat ggtatgtcca aagcggcgat 1620
ttggaaacgg cagagaaggt actggaaaaa gaacttctgg cctggcagga gaaactgcat 1680
cagccgatta tcatcaccga atacggcgtg gatacgttag ccgggctgca ctcaatgtac 1740
accgacatgt ggagtgaaga gtatcagtgt gcatggctgg atatgtatca ccgcgtcttt 1800
gatcgcgtca gcgccgtcgt cggtgaacag gtatggaatt tcgccgattt tgcgacctcg 1860
caaggcatat tgcgcgttgg cggtaacaag aaagggatct tcactcgcga ccgcaaaccg 1920
aagtcggcgg cttttctgct gcaaaaacgc tggactggca tgaacttcgg tgaaaaaccg 1980
cagcagggag gcaaacaatg a 2001
<210> 25
<211> 516
<212> ДНК
<213> Искусственная
<220>
<223> Кодирующая последовательность Nluc, белка люциферазы, сконструированного
путем направленной эволюции.
<400> 25
atggtcttca cactcgaaga tttcgttggg gactggcgac agacagccgg ctacaacctg 60
gaccaagtcc ttgaacaggg aggtgtgtcc agtttgtttc agaatctcgg ggtgtccgta 120
actccgatcc aaaggattgt cctgagcggt gaaaatgggc tgaagatcga catccatgtc 180
atcatcccgt atgaaggtct gagcggcgac caaatgggcc agatcgaaaa aatttttaag 240
gtggtgtacc ctgtggatga tcatcacttt aaggtgatcc tgcactatgg cacactggta 300
atcgacgggg ttacgccgaa catgatcgac tatttcggac ggccgtatga aggcatcgcc 360
gtgttcgacg gcaaaaagat cactgtaaca gggaccctgt ggaacggcaa caaaattatc 420
gacgagcgcc tgatcaaccc cgacggctcc ctgctgttcc gagtaaccat caacggagtg 480
accggctggc ggctgtgcga acgcattctg gcgtaa 516
<210> 26
<211> 300
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность ДНК синтетического 3' UTR, T-Zm.GST43.nno:1.
<400> 26
tccagggcgc ctgcttgttt gctgccaaga gagtgttttg tgtactgctg ctgccgagaa 60
atatatattt ttttctttca ccacctcgtg tgcagcagtt gttttttgtt tggatggata 120
aatgtttcta gtactgtgga ggctgcatct gcatctgttt gtaaatggat gaaatatgaa 180
taaaaagttt tgtttctcat accccatgtg tcttgtgttt gcatgcacgc cgtgctagtt 240
tggtttttgg tttctagaga aaacattttg tttgcttgtt tctctatagg atgtgaagaa 300
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2639275C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2675524C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2644205C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2670887C9 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2800430C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2800425C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2800423C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2800424C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2774709C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2799433C2 |
Изобретение относится к области биохимии, в частности к молекуле рекомбинантной ДНК для экспрессии гетерологичной транскрибируемой молекулы полинуклеотида. Также раскрыты трансгенная клетка растения, трансгенное растение или его часть, трансгенное семя содержащие указанную молекулу ДНК. Изобретение позволяет эффективно увеличивать экспрессию генов в растениях. 7 н. и 9 з.п. ф-лы, 10 табл., 7 пр.
при этом указанная последовательность ДНК функционально связана с гетерологичной молекулой транскрибируемой ДНК.
при этом указанная последовательность ДНК функционально связана с гетерологичной молекулой транскрибируемой ДНК.
| WO 2012158535 A1, 22.11.2012 | |||
| WO 2016022376 A1, 11.02.2016 | |||
| WO 2014100009 A2, 26.06.2014 | |||
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2644205C2 |
