Область техники
Настоящее изобретение относится к области техники, связанной с программируемой гибелью раковых клеток, вызванной повреждением ДНК с помощью химиотерапевтических препаратов.
Уровень техники
Борьба с онкологическими заболеваниями является одной из самых важных задач современной медицины, поскольку эти болезни занимают второе место в мире среди причин смерти. Согласно данным Всемирной Организации Здравоохранения от различных видов рака ежегодно умирает около 7,5 млн человек, при этом в России на учете в онкологических диспансерах состоят около 3 млн человек. Для лечения этих заболеваний широко применяются химиотерапевтические препараты, способные вызывать повреждения генетического материала и клеточных структур и, как следствие, программируемую гибель быстро делящихся раковых клеток. Химиотерапия используется в онкологической практике более 70 лет. Несмотря на большие успехи в лечении гематологических опухолей, устойчивость многих эпителиальных опухолей приводит к низкому числу выживших в течение 5 лет после начала лечения. Так, например, 5-летняя выживаемость больных раком поджелудочной железы не превышает 2%, а больных немелкоклеточным раком легких - 5-10% [G. Morgan, R. Ward, and М. Barton, "The contribution of cytotoxic chemotherapy to 5-year survival in adult malignancies", Clin. Oncol. (R. Coll. Radiol)., vol. 16, no. 8, pp. 549-60, Dec. 2004]. Таким образом, исследования молекулярных механизмов действия химиотерапевтических препаратов и разработка новых подходов для диагностики, прогнозирования течения и лечения раковых заболеваний - одно из важнейших направлений современной медицины.
Апоптоз - наиболее изученная программа клеточной гибели (ПКГ) - является одним из важнейших биологических процессов, происходящих в тканях и органах многоклеточных организмов. Он обеспечивает правильный морфогенез и функционирование различных систем в многоклеточных организмах, а также элиминацию поврежденных клеток. Так, гибель раковых клеток при лечении опухолевых заболеваний химиотерапевтическими агентами происходит во многих случаях за счет процесса апоптоза. Основными ферментами, участвующими в инициации апоптоза и его дальнейшем протекании, являются белки семейства цистеиновых протеаз - каспаз, которые синтезируются в клетке в неактивной форме и активируются в ответ на определенные стимулы. Запуск процесса программируемой клеточной гибели осуществляется посредством внеклеточных или внутриклеточных факторов, таких как гипоксия, гипероксия, субнекротические поражения химическими или физическими агентами, активация рецепторов смерти, нарушения сигналов клеточного цикла, удаления факторов роста и метаболизма [Z. Jin and W.S. El-Deiry, "Overview of cell death signaling pathways", Cancer Biol. Ther., vol. 4, no. 2, pp. 139-63, Feb. 2005].
Как было отмечено выше, основные эффекторы апоптоза - это цистеиновые протеазы - каспазы, которые расщепляют белки после остатка аспартата (caspase англ. С - cysteine, Asp - aspartate, ase - protease). В норме каспазы существуют в клетке в виде неактивных проформ. Активация каспаз происходит вследствие протеолитического процессинга прокаспаз [А. Degterev, М. Воусе, and J. Yuan, "A decade of caspases", Oncogene, vol. 22, no. 53, pp. 8543-67, Nov. 2003]. Каспазы принято подразделять на три группы. Среди каспаз, задействованных в апоптозе, выделяют инициаторные (-2, -8, -9, -10) и эффекторные каспазы (-3, -6, -7), а также группу каспаз, участвующих во врожденном иммунном ответе (-1, -4, -5, -11) [Е. Kuranaga, "Beyond apoptosis: caspase regulatory mechanisms and functions in vivo", Genes Cells, vol. 17, no. 2, pp. 83-97, Feb. 2012; F. Martinon and J. Tschopp, "Inflammatory caspases: linking an intracellular innate immune system to autoinflammatory diseases", Cell, vol. 117, no. 5, pp. 561-74, May 2004]. Необходимо отметить, что уникальная по своим свойствам каспаза-2 обладает свойствами не только инициаторных, но и эффекторных каспаз [Н. Vakifahmetoglu-Norberg and В. Zhivotovsky, "The unpredictable caspase-2: what can it do?", Trends Cell Biol., vol. 20, no. 3, pp. 150-9, Mar. 2010].
Принципиально различают два пути запуска апоптоза: внутренний (митохондриальный) и внешний (рецептор-зависимый). Оба пути связаны с активацией каспазных каскадов. Внешний путь апоптоза запускается при активации так называемых рецепторов смерти, объединенных в одно семейство, которое, в свою очередь, является подсемейством так называемого суперсемейства рецепторов фактора некроза опухоли (ФНО, англ., TNF) [I.N. Lavrik and P.Н. Krammer, "Regulation of CD95/Fas signaling at the DISC", Cell Death Differ., vol. 19, no. 1, pp. 36-41, Jan. 2012]. Среди рецепторов смерти к настоящему моменту достаточно хорошо охарактеризованы CD95/Fas/Apo-1, TRAIL-R1/R2, а также TNF-R1, активация которых может приводить к ПКГ. При взаимодействии лиганда с рецептором смерти во внутриклеточном домене последнего происходит конформационный переход, что, в свою очередь, создает возможность для гомотипического взаимодействия между DD (death domain, англ., домен смерти) рецептора и адаптерного белка (в случае Fas-рецептора адаптером служит FADD - Fas-Associated protein with Death Domain). Белок FADD помимо DD, содержит также домен DED (Death Effector Domain, англ., эффекторный домен смерти), который гомотипически взаимодействует с DED инициирующих прокаспаз-8 и -10, приводя к их активации. Описанный комплекс, включающий лиганд, рецептор, адаптер и инициаторную каспазу, в котором происходит активация инициаторных каспаз-8/-10, получил название DISC (death-inducing signaling complex) [L.S. Dickens, I.R. Powley, M.A. Hughes, and M. MacFarlane, "The 'complexities' of life and death: death receptor signalling platforms", Exp. Cell Res., vol. 318, no. 11, pp. 1269-77, Jul. 2012]. Среди субстратов каспазы-8, играющих важную роль в дальнейшем протекании процесса апоптоза, можно выделить прокаспазу-3 и белок семейства Bcl-2 - Bid. Семейство белков Всl-2 и ассоциированные с ним члены включают более 30 участников и играют важнейшую роль в регуляции ПКГ. Расщепление прокаспазы-3 ведет к ее активации и запуску эффекторной фазы апоптоза, а расщепленная форма Bid (tBid) связывает внешний путь гибели с митохондриальным. Взаимодействуя с проапоптотическими белками Вах или Bak, tBid приводит к их структурной модификации, в результате чего конформационно-измененные Вах или Bak, встраиваясь в мембрану митохондрий, вызывают пермеабилизацию внешней митохондриальной мембраны (ПВММ) и запускают цепь событий, описанных для митохондриального пути апоптоза [Н. Li, Н. Zhu, С.J. Xu, and J. Yuan, "Cleavage of BID by caspase 8 mediates the mitochondrial damage in the Fas pathway of apoptosis", Cell, vol. 94, no. 4, pp. 491-501, Aug. 1998]. Активация рецепторов смерти также может приводить к провоспалительному ответу и активации фактора транскрипции NF-kB, что в случае рецептора TNFR1 является основным ответом на связывание TNF. Апоптоз запускается через рецептор TNFR1 с образованием так называемого комплекса II [А.V. Zamaraev, G.S. Kopeina, В. Zhivotovsky, and I.N. Lavrik, "Cell death controlling complexes and their potential therapeutic role", Cell. Mol. Life Scl, vol. 72, no. 3, pp. 505-17, Feb. 2015].
Внутренний или митохондриальный путь апоптоза осуществляются посредством транслокации проапоптотических белков из межмембранного пространства митохондрий в цитоплазму. Белки выходят как через высокопроницаемые каналы на внешней поверхности митохондрии, так и через разрыв митохондриальной мембраны. При этом важную роль в реализации внутреннего пути запуска апоптоза также играют белки семейства Всl-2, среди которых есть как проапоптотические, так и антиапоптотические. К первым относятся, например, Вах и Bak; к последним - Всl-2 и Bcl-xL [Y. Tsujimoto, "Role of Bcl-2 family proteins in apoptosis: apoptosomes or mitochondria?", Genes Cells, vol. 3, no. 11, pp. 697-707, Nov. 1998]. Их соотношение и взаимодействие с другими белками определяют возможность ПВММ и развития апоптотического ответа. Особое место в запуске апоптоза занимает белок р53 - фактор транскрипции, ответственный за экспрессию генов различных белков, в том числе, относящихся к проапоптотическим белкам семейства Bcl-2 - Вах и Puma [Т. Miyashita and J.С. Reed, "Tumor suppressor p53 is a direct transcriptional activator of the human bax gene", Cell, vol. 80, no. 2, pp. 293-9, Jan. 1995]. Более того, p53 может опосредованно приводить к ПВММ, взаимодействуя с белками семейства Bcl-2 [J.Е. Chipuk, Т. Kuwana, L. Bouchier-Hayes, N.М. Droin, D.D. Newmeyer, M. Schuler, and D.R. Green, "Direct activation of Bax by p53 mediates mitochondrial membrane permeabilization and apoptosis", Science, vol. 303, no. 5660, pp. 1010-4, Feb. 2004; M.T. Hemann and S.W. Lowe, "The p53-Bcl-2 connection", Cell Death Differ., vol. 13, no. 8, pp. 1256-9, Aug. 2006; E.C. Pietsch, E. Perchiniak, A.A. Canutescu, G. Wang, R.L. Dunbrack, and M.E. Murphy, "Oligomerization of BAK by p53 utilizes conserved residues of the p53 DNA binding domain", J. Biol. Chem., vol. 283, no. 30, pp. 21294-304, Jul. 2008]. Этот процесс запускает выход из межмембранного пространства митохондрий различных факторов, способствующих развитию апоптоза, таких как цитохром С, флавопротеин AIF, эндонуклеаза G, SMAC/Diablo. Высвобождение цитохрома С ведет к формированию высокомолекулярного комплекса - апоптосомы, в составе которого происходит активация каспазы-9. Цитохром С связывается с белком Apaf-1, такое взаимодействие вызывает олигомеризацию адаптерного белка Apaf-1 и обеспечивает доступность CARD-домена, через который происходит связывание прокаспазы-9, ее димеризация и активация. В результате этого каспаза-9 приобретает способность активировать эффекторную каспазу-3, что приводит к расщеплению ее белков-субстратов, морфологическим изменениям клетки и ее гибели [K. Cain, S.В. Bratton, and G.М. Cohen, "The Apaf-1 apoptosome: a large caspase-activating complex", Biochimie, vol. 84, no. 2-3, pp. 203-214].
Активация эффекторных каспаз и расщепление их субстратов приводят к развитию следующей стадии апоптоза - деградации клетки, которая начинается с реорганизации молекул адгезии, деполимеризации микротрубочек, разрушения ядерной ламины. Актиновые микрофиламенты собираются в кортикальные пучки, вследствие чего клетка приобретает округлую форму. В результате сокращения кортикальных колец клетка фрагментируется на отдельные апоптотические тельца, которые ограниченны цитоплазматической мембраной. Нужно отметить, что процесс деградации клетки не зависит от начального инициирующего воздействия на клетку. В дальнейшем апоптотические тельца фагоцитируются макрофагами или соседними клетками, не вызывая воспалительного процесса. Одним из сигналов для фагоцитирования является присутствие фосфатидилсерина на внешней стороне плазматической мембраны [J. Savill and V. Fadok, "Corpse clearance defines the meaning of cell death", Nature, vol. 407, no. 6805, pp. 784-788, Oct. 2000].
Значимость апоптоза для нормальной жизнедеятельности организма хорошо прослеживается в случае его нарушений. Так, избыточная клеточная смерть служит причиной развития нейродегенеравтиных заболеваний, уничтожая нейроны и приводя мозг в состояние, при котором сначала теряется кратковременная память, а потом и долговременная, страдает крупная и мелкая моторика, развиваются психозы. Развитие болезней Альцгеймера, Паркинсона, бокового амиотрофического склероза, хореи Хантингтона в той или иной степени связано с повышенной апоптотической клеточной гибелью. Кроме того, чрезмерная ПГК может вызывать гематологические, иммунные и метаболические заболевания [G. Arguello, Е. Balboa, М. Arrese, and S. Zanlungo, "Recent insights on the role of cholesterol in non-alcoholic fatty liver disease", Biochim. Biophys. Acta, vol. 1852, no. 9, pp. 1765-78, Sep. 2015; K. Eguchi, "Apoptosis in autoimmune diseases", Intern. Med., vol. 40, no. 4, pp. 275-84, Apr. 2001].
Другим важным примером заболеваний, связанных с нарушениями апоптоза, являются состояния, связанные с его недостаточностью. Клетка, получившая мутацию, в норме должна запустить процесс самоуничтожения, но слабый контроль и дерегуляция апоптотических механизмов вызывают появление злокачественных образований. Во многих типах опухолей найдены мутации, влияющие на функции тех или иных белков, вовлеченных в апоптоз [S.W. Lowe and A.W. Lin, "Apoptosis in cancer", Carcinogenesis, vol. 21, no. 3, pp. 485-95, Mar. 2000]. Как ранее отмечено, раковые заболевания занимают второе место в мире в списке причин, приводящих к смерти, а в развитых европейских странах в последнее время выходят на первое место. Для понимания патофизиологии таких болезней необходимо тщательное изучение механизма действия апоптоза и роли всех участников этого процесса.
Как описывалось выше, запуск апоптотических процессов происходит с помощью формирования специальных высокомолекулярных комплексов, активирующих ферменты апоптоза - каспазы. Изучение таких молекулярных платформ требует системного подхода для идентификации белковых компонентов и подтверждения роли этих белков в процессе активации каспаз. Поскольку каспазный каскад является ключевым механизмом апоптоза, то выявление неизвестных участников дает возможность получить новые сведения о процессе ПГК и глубже понять принципы возникновения заболеваний, связанных с нарушениями апоптотического процесса, в том числе онкологических заболеваний. Также модулирование активности данных комплексов может быть использовано для запуска апоптоза в раковых клетках и, таким образом, борьбы с онкологическими заболеваниями. Например, на сегодняшний день активно разрабатываются подходы лечения раковых заболеваний, которые подразумевают применение лиганда TRAIL для запуска внешнего пути апоптоза через стимуляцию комплекса DISC или использование селективных ингибиторов проапоптотических белков семейства Bcl-2 (так называемых ВН3-миметиков - АВТ-737 или АВТ-263) для запуска формирования апоптосомы [А.V Zamaraev, G.S. Kopeina, В. Zhivotovsky, and I.N. Lavrik, "Cell death controlling complexes and their potential therapeutic role", Cell. Mol. Life Sci., vol. 72, no. 3, pp. 505-17, Feb. 2015].
Однако данные направления онкомедицины нуждаются в разработке подходов для выделения высокомолекулярных комплексов, ответственных за запуск процесса апоптоза. Создание подходов для очистки комплексов из раковых клеток является сложной научно-технической задачей, поскольку в процессе выделения белки комплекса могут диссоциировать, агрегировать и терять нативную конформацию. Выделение комплексов, активирующих каспазы, также осложняется тем, что такие молекулярные платформы образуются в небольшом количестве в клетке и достаточно быстро деградируют. Исследование таких высокомолекулярных комплексов ведется на протяжении двух последних десятилетий, при этом достаточно полная характеристика того или иного комплекса требует значительного количества времени [М.A. Hughes, С. Langlais, K. Cain, and М. MacFarlane, "Isolation, characterisation and reconstitution of cell death signalling complexes", Methods, vol. 61, no. 2, pp. 98-104, Jun. 2013]. Так, например, комплекс DISC (Death-inducing signaling complex), активирующий прокаспазу-8/-10, был впервые описан в 1995 году, однако его стехиометрия была определена только в 2012 году [К. Schleich, U. Warnken, N. Fricker,  , P. Richter, K. Kammerer,
, P. Richter, K. Kammerer,  , P.H. Krammer, and I.N. Lavrik, "Stoichiometry of the CD95 death-inducing signaling complex: experimental and modeling evidence for a death effector domain chain model", Mol. Cell, vol. 47, no. 2, pp. 306-19, Jul. 2012].
, P.H. Krammer, and I.N. Lavrik, "Stoichiometry of the CD95 death-inducing signaling complex: experimental and modeling evidence for a death effector domain chain model", Mol. Cell, vol. 47, no. 2, pp. 306-19, Jul. 2012].
Индукция генотоксического стресса с помощью повреждений ДНК является одним из наиболее широко распространенных методов лечения онкологических заболеваний. Повреждения ДНК индуцируют в клетке ряд сигнальных каскадов, которые приводят к образованию апоптотических высокомолекулярных комплексов и активации инициаторных каспаз. Одной из важнейших инициаторных каспаз, играющих центральную роль в запуске ответа на генотоксический стресс, является каспаза-2 [В. Zhivotovsky and S. Orrenius, "Caspase-2 function in response to DNA damage", Biochem. Biophys. Res. Commun., vol. 331, no. 3, pp. 859-67, Jun. 2005]. Каспаза-2 - один из самых консервативных и загадочных белков данного семейства. По различным литературным данным она участвует в индукции и усилении процесса апоптоза в ответ на обработку клеток ДНК-повреждающими агентами или индукторами, вызывающими стресс эндоплазматического ретикулума. Также каспаза-2 участвует в регуляции клеточного цикла и обладает онкосупрессорными функциями [Н. Vakifahmetoglu-Norberg and В. Zhivotovsky, "The unpredictable caspase-2: what can it do?", Trends Cell Biol, vol. 20, no. 3, pp. 150-9, Mar. 2010; G. Krumschnabel, B. Sohm, F. Bock, C. Manzl, and A. Villunger, "The enigma of caspase-2: the laymen's view", Cell Death Differ., vol. 16, no. 2, pp. 195-207, Feb. 2009]. Так пониженный уровень экспрессии гена каспазы-2 наблюдается при остром лимфобластном лейкозе у детей и карциноме желудка [A. Holleman, М.L. den Boer, K.М. Kazemier, Н.В. Beverloo, A.R.М. von Bergh, G.Е. Janka-Schaub, and R. Pieters, "Decreased PARP and procaspase-2 protein levels are associated with cellular drug resistance in childhood acute lymphoblastic leukemia", Blood, vol. 106, no. 5, pp. 1817-23, Sep. 2005; N.J. Yoo, J.W. Lee, Y.J. Kim, Y.H. Soung, S.Y. Kim, S.W. Nam, W.S. Park, J.Y. Lee, and S.H. Lee, "Loss of caspase-2, -6 and -7 expression in gastric cancers", APMIS, vol. 112, no. 6, pp. 330-5, Jun. 2004].
Кроме того, каспаза-2, вероятно, принимает участие в защите клеток от окислительного стресса и преждевременного старения. Активные формы кислорода (АФК) представляют собой побочные продукты клеточного метаболизма и играют важную роль в передаче сигналов внутри клетки и поддержании клеточного гомеостаза [Т. Finkel and N. J. Holbrook, "Oxidants, oxidative stress and the biology of ageing", Nature, vol. 408, no. 6809, pp. 239-247, Nov. 2000]. На многих модельных системах была продемонстрирована взаимосвязь апоптотических процессов и появления избытка АФК, которые могут образовываться как на начальных, так и на поздних стадиях апоптоза [G. Kroemer and J.C. Reed, "Mitochondrial control of cell death", Nat. Med., vol. 6, no. 5, pp. 513-9, May 2000]. Увеличение уровня АФК сопровождается активацией каспазы-2 [V.М. Dirsch, S.О. Kirschke, М. Estermeier, В. Steffan, and А.М. Vollmar, "Apoptosis signaling triggered by the marine alkaloid ascididemin is routed via caspase-2 and JNK to mitochondria", Oncogene, vol. 23, no. 8, pp. 1586-93, Feb. 2004]. На уровне целого организма повреждение, вызываемое избыточным накоплением АФК, рассматривают как один из факторов ускоренного старения. Примечательно, что мыши, с дефицитом этого белка характеризуются повышенным уровнем окисленных белков в печени, что, вероятно, происходит под действием АФК, а также ускоренным появлением возрастных изменений, ведущих к уменьшению продолжительности жизни [Y. Zhang, S.S. Padalecki, A.R. Chaudhuri, Е. De Waal, В.A. Goins, В. Grubbs, Y. Ikeno, A. Richardson, G.R. Mundy, and B. Herman, "Caspase-2 deficiency enhances aging-related traits in mice", Meek Ageing Dev., vol. 128, no. 2, pp. 213-21, Feb. 2007]. Усиление окислительного стресса и увеличение уровня окисленных белков, липидов и повреждений ДНК, наблюдаемое у каспаза-2-дефицитных мышей, хорошо согласуется с описанными выше результатами [S. Shalini, L. Dorstyn, С. Wilson, J. Puccini, L. Ho, and S. Kumar, "Impaired antioxidant defence and accumulation of oxidative stress in caspase-2-deficient mice", Cell Death Differ., vol. 19, no. 8, pp. 1370-80, Aug. 2012]. В основе молекулярного механизма регуляции каспазой-2 процесса окисления, вероятно, лежит способность этого белка контролировать активность транскрипционных факторов FoxO1 и FoxO3a, которые задействованы в клеточном ответе на окислительный стресс. Кроме того, было показано, что каспаза-2 может участвовать в поддержании генетической стабильности клеток. В мышиных эмбриональных фибробластах с нокаутом по каспазе-2 наблюдалось нарушение регуляции клеточного цикла, увеличивался процент анеуплоидии, уменьшалась длина теломер, а также нарушались р53-зависимые сигнальные пути, и снижалась активация р53-зависимых генов [L. Dorstyn, J. Puccini, С.Н. Wilson, S. Shalini, M. Nicola, S. Moore, and S. Kumar, "Caspase-2 deficiency promotes aberrant DNA-damage response and genetic instability", Cell Death Differ., vol. 19, no. 8, pp. 1288-98, Aug. 2012]. Поскольку для каспаза-2-дефицитных клеток характерны укорочение теломер и анеуплоидия, которые считаются маркерами старения, то вклад этого белка в регуляцию продолжительности жизни не ограничивается его влиянием только на уровень продукции АФК. Таким образом, каспаза-2 обладает протективными свойствами и защищает организм от проявления признаков раннего старения.
На молекулярном уровне каспаза-2 характеризуется наличием продомена, содержащего CARD (caspase activation recruitment domain), a также каталитических субъединиц p19 и p12. Несмотря на важнейшую роль каспазы-2, механизм ее активации до сих пор остается неизвестным, что препятствует развитию подходов к модуляции активности этого белка в терапии. В 2004 г. было показано, что активация каспазы-2 в ответ на повреждение ДНК происходит в составе комплекса PIDDosome, который включает также адапторный белок RAIDD и С-концевой фрагмент белка PIDD [A. Tinel and J. Tschopp, "The PIDDosome, a protein complex implicated in activation of caspase-2 in response to genotoxic stress", Science, vol. 304, no. 5672, pp. 843-6, May 2004]. Белок RAIDD (RIP-ассоциированный гомолог ICH1) содержит как домен смерти (DD-death domain), так и домен CARD, что позволяет ему играть центральную роль в сборке комплекса PIDDosome через гомотипические взаимодействия. При этом белок RAIDD взаимодействует с С-концевым фрагментом белка PIDD через домен смерти и с прокаспазой-2 через домен CARD. Позже появился целый ряд работ, в которых были собраны доказательства того, что каспаза-2 способна активироваться независимо как от белков PIDD и RAIDD, так и от сборки комплекса PIDDosome [С. Manzl, G. Krumschnabel, F. Bock, В. Sohm, V. Labi, F. Baumgartner, E. Logette, J. Tschopp, and A. Villunger, "Caspase-2 activation in the absence of PIDDosome formation.," J. Cell Biol, vol. 185, no. 2, pp. 291-303, Apr. 2009; H. Vakifahmetoglu, M. Olsson, S. Orrenius, and B. Zhivotovsky, "Functional connection between p53 and caspase-2 is essential for apoptosis induced by DNA damage", Oncogene, vol. 25, no. 41, pp. 5683-92, Sep. 2006; I.R. Kim, K. Murakami, N.-J. Chen, S.D. Saibil, E. Matysiak-Zablocki, A.R. Elford, M. Bonnard, S. Benchimol, A. Jurisicova, W.-C. Yeh, and P.S. Ohashi, "DNA damage- and stress-induced apoptosis occurs independently of PIDD", Apoptosis, vol. 14, no. 9, pp. 1039-49, Sep. 2009]. Необходимо отметить, что идентификация комплекса PIDDosome происходила в условиях повышенной экспрессии гена белка PIDD [A. Tinel and J. Tschopp, "The PIDDosome, a protein complex implicated in activation of caspase-2 in response to genotoxic stress", Science, vol. 304, no. 5672, pp. 843-6, May 2004], при этом попытки получить этот комплекс в условиях обычной экспрессии не были успешными [В.И. Аксенова, Г.С. Копеина, А.В. Замараев, Б.Д. Животовский, and И.Н. Лаврик, "Механизм активации каспазы 2 при повреждениях днк © 2016," pp. 598-601, 2016]. С другой стороны, результаты исследований активации каспазы-2 не позволяли сделать вывод о самом PIDDosome-независимом механизме. Оставалось загадкой - происходил ли этот процесс в составе какой-либо высокомолекулярной платформы или же являлся следствием расщепления под действием другой инициаторной каспазы.
Таким образом, разработка подходов, позволяющих выделить высокомолекулярную платформу активации каспазы-2, является научно-технической задачей, решение которой позволит в дальнейшем прогнозировать успешность лечения раковых заболеваний и производить дизайн новых противораковых лекарственных средств. Так анализ уровня экспрессии белков, входящих в состав этого комплекса и участвующих в активации каспазы-2, может быть использован для прогнозирования эффективности применения ДНК-повреждающих химиотерапевтических препаратов для лечения опухолей, в клетках которых представлена каспаза-2, например, таких как карцинома яичника. Аналогичный подход используется в персонифицированной медицине для прогноза оценки развития и эффективности лечения BRCA-ассоциированного рака яичника, HER2-зависимым типам рака молочной железы и рака толстого кишечника, обусловленного мутациями K-RAS. Кроме того, разработка такого подхода позволит выделять и анализировать другие высокомолекулярные комплексы, ответственные за активацию других инициаторных каспаз (таких, как -8, -9, -10), а также запуск и протекание программируемой гибели клетки в ответ на повреждения ДНК с помощью химиотерапевтических препаратов.
В процессе развития уровня техники были разработаны подходы, которые предлагают разделение мономерных форм каспазы-2 и высокомолекулярного комплекса с помощью гель-фильтрации, однако не предполагают его дальнейшее выделение из фракций гель-фильтрации [G. Imre, J. Heering, A.-N. Takeda, M. Husmann, B. Thiede, D.M. zu Heringdorf, D.R. Green, F.G. van der Goot, B. Sinha,  , and K. Rajalingam, "Caspase-2 is an initiator caspase responsible for pore-forming toxin-mediated apoptosis", EMBO J., vol. 31, no. 11, pp. 2615-28, May 2012; S.H. Read, В.C. Baliga, P.G. Ekert, D.L. Vaux, and S. Kumar, "A novel Apaf-1-independent putative caspase-2 activation complex", J. Cell Biol., vol. 159, no. 5, pp. 739-45, Dec. 2002]. Кроме того, данные аналоги не предусматривают алгоритм оптимизации условий индукции апоптоза с целью увеличения выхода комплекса активации каспазы-2.
, and K. Rajalingam, "Caspase-2 is an initiator caspase responsible for pore-forming toxin-mediated apoptosis", EMBO J., vol. 31, no. 11, pp. 2615-28, May 2012; S.H. Read, В.C. Baliga, P.G. Ekert, D.L. Vaux, and S. Kumar, "A novel Apaf-1-independent putative caspase-2 activation complex", J. Cell Biol., vol. 159, no. 5, pp. 739-45, Dec. 2002]. Кроме того, данные аналоги не предусматривают алгоритм оптимизации условий индукции апоптоза с целью увеличения выхода комплекса активации каспазы-2.
Также в литературе описана схема выделения другого инициаторного высокомолекулярного комплекса, активирующего каспазу-8 в ответ на стимуляцию рецепторов смерти [М.A. Hughes, С. Langlais, K. Cain, and М. MacFarlane, "Isolation, characterisation and reconstitution of cell death signalling complexes", Methods, vol. 61, no. 2, pp. 98-104, Jun. 2013]. Данный аналог включает этап разделения мономерных и высокомолекулярных форм каспазы-8 с последующим выделением ее из фракций гель-фильтрации. Однако данный метод предложен для выделения комплекса активации каспазы-8, который образуется в большом количестве за относительно короткий период (несколько секунд) в клетках после стимуляции DR. Предложенный подход не предлагает оптимизацию условий для выделения высокомолекулярных комплексов, образующихся в малых количествах в клетках в течение длительного времени. Таким образом, при использовании предложенной схемы для достижения результата необходимо получать несопоставимо большее количество лизата клеток, что может привести к дополнительным трудностям - неспецифической агрегации белков и, как следствие, снижение выхода выделяемого комплекса и/или загрязнение образца.
Раскрытие изобретения
Задачей настоящего изобретения является разработка способа эффективного выделения комплекса каспазы-2, формирующегося в раковых клетках в ответ на обработку химиотерапевтическими ДНК-повреждающими препаратами.
Поставленная задача решается предлагаемым способом выделения белкового высокомолекулярного комплекса активации каспазы-2 человека, формирующегося в раковых клетках в ответ на обработку ДНК-повреждающим химиотерапевтическим препаратом, включающим:
- получение раковых клеток в количестве не менее чем 5 млн клеток;
- введение в раковые клетки ДНК-повреждающего химиотерапевтического препарата в концентрации, обеспечивающей запуск апоптотической гибели раковых клеток с достижением максимального уровня апоптотической гибели клеток при минимальном уровне некротической гибели клеток, которые определяют с помощью цитометрии;
- инкубация раковых клеток в присутствии ДНК-повреждающего химиотерапевтического препарата до получения максимального уровня активации каспазы-2, который определяют предварительно до начала инкубации с помощью анализа методом Вестерн-блота уровня расщепления прокаспазы-2 и накопления каталитически активных фрагментов каспазы-2 и/или с помощью флуоресцентных специфических субстратов;
- сбор клеток с последующим добавлением лизирующего буфера, который обеспечивает сохранность высокомолекулярного комплекса активации каспазы-2;
- отделение клеточного дебриса от лизата, содержащего высокомолекулярный комплекс активации каспазы-2,
- разделение гель-фильтрацией полученного лизата клеток с обеспечением отделения фракций, содержащих высокомолекулярные формы каспазы-2, от фракций, содержащих ее мономерные формы, которое проводят в условиях, обеспечивающих получение высокомолекулярных форм каспазы-2 молекулярной массой не менее 500000 Да;
- выделение из фракции, содержащей высокомолекулярные формы каспазы-2, белкового высокомолекулярного комплекса активации каспазы-2 с помощью иммунопреципитации, которую проводят в условиях, обеспечивающих минимизацию сорбции неспецифических белков.
Ферментативную активность каспазы-2 в выделенном высокомолекулярном комплексе определяют с помощью флуоресцентных субстратов. При этом определение ферментативной активности осуществляют на носителе для иммунопреципитации, с которым связан высокомолекулярный комплекс активации каспазы-2.
Предпочтительно в качестве раковых клеток использовать линии карциномы яичника человека Caov-4 или РА-1, или Caov-3, или SW 626, или SK-OV-3.
Предпочтительно в качестве ДНК-повреждающего химиотерапевтического препарата использовать соединения платины, или алкилирующие агенты, или ингибиторы топоизомераз, или антиметаболиты. В качестве соединений платины предпочтительно использовать цисплатин, оксалиплатин, тетраплатин, карбоплатин, фенантриплатин. При этом концентрацию ДНК-повреждающего химиотерапевтического препарата рассчитывают путем инкубации раковых клеток в присутствии разных концентраций этого агента и дальнейшим определением уровня гибели клеток методом цитометрии.
Предпочтительно, когда концентрация ДНК-повреждающего химиотерапевтического препарата составляет не менее 10 мкМ. Наиболее предпочтительно концентрация химиотерапевтического препарата варьирует в пределах 35-100 мкМ.
Продолжительность инкубации определяется путем инкубации клеток с ДНК-повреждающим химиотерапевтическим препаратом в течение различного количества времени, но не менее часа, с последующим анализом уровня расщепления и активации каспазы-2 в этих клетках методом Вестерн-блота и/или с помощью флуоресцентных специфических субстратов.
Предпочтительно использовать лизирующий буфер включающий один из следующих детергентов: Nonidet Р-40, Triton Х-100, Tween 20, Tween 80, Brij 35, Brij 58, Octyl glucoside, Octyl thioglucoside, sodium dodecyl sulfate, CHAPS, digitonin, deoxycholate, cholate, sarkosyl в конечной концентрации от 0,01 до 1,5% об. в буфере. Дополнительно лизирующий буфер может содержать пан-каспазный ингибитор zVAD-fmk (carbobenzoxy-valyl-alanyl-aspartyl-[O-methyl] fluoromethylketone) в конечной концентрации не менее 10 мкМ в буфере.
Предпочтительно выбор условий гель-фильтрации включает подбор носителя для гель-фильтрации с разрешающей способностью от 5×105 вплоть до 5×107 Да.
Для получения высокомолекулярных форм каспазы-2 молекулярной массой не менее 500000 Да предварительно оценивают накопление в клетках высокомолекулярного комплекса с помощью гель-фильтрации с последующим Вестерн-блотом в зависимости от продолжительности инкубации клеток с ДНК-повреждающим химиотерапевтическим препаратом.
Выделение комплекса активации каспазы-2 осуществляют с помощью иммунопреципитации, включающей выбор носителя для иммунопреципитации с наименьшими параметрами неспецифической сорбции, которую оценивают с помощью электрофореза образцов в полиакриламидном геле после иммунопреципитации с дальнейшим окрашиванием геля.
Также поставленная задача решается способом выделения белкового высокомолекулярного комплекса активации каспазы-2 человека, формирующегося в раковых клетках в ответ на обработку ДНК-повреждающим химиотерапевтическим препаратом, характеризующийся тем, что включает:
- получение раковых клеток в количестве не менее чем 5 млн клеток;
- введение с помощью трансфицирующего агента в раковые клетки вектора, содержащего генетический конструкт, представляющий собой последовательность гена каспазы-2 и дополнительную аминокислотную последовательность для аффинной хроматографии, в концентрации не менее 0,5 мкг на 1 мл питательной среды;
- инкубация раковых клеток в течение времени, достаточного для детектируемой экспрессии генетического конструкта, представляющего собой последовательность гена каспазы-2 и дополнительную аминокислотную последовательность для аффинной хроматографии;
- введение в трансформированные клетки ДНК-повреждающего химиотерапевтического препарата, обеспечивающей запуск апоптотической гибели раковых клеток с достижением максимального уровня апоптотической гибели клеток при минимальном уровне некротической гибели клеток, которые определяют с помощью цитометрии;
- инкубация раковых клеток в присутствии ДНК-повреждающего химиотерапевтического агента до получения максимального уровня активации каспазы-2, который определяют предварительно до начала инкубации с помощью анализа методом Вестерн-блота уровня расщепления прокаспазы-2 и накопления каталитически активных фрагментов каспазы-2 и/или с помощью флуоресцентных специфических субстратов;
- сбор клеток с последующим добавлением лизирующего буфера, который обеспечивает сохранность высокомолекулярного комплекса активации каспазы-2
- отделение клеточного дебриса от лизата, содержащего высокомолекулярный комплекс активации каспазы-2,
- разделение гель-фильтрацией полученного лизата клеток с обеспечением отделения фракций, содержащих высокомолекулярные формы каспазы-2, от фракций, содержащих ее мономерные формы, которое проводят в условиях, обеспечивающих получение высокомолекулярных форм каспазы-2 молекулярной массой не менее 500000 Да;
- выделение из фракции, содержащей высокомолекулярные формы каспазы-2, белкового высокомолекулярного комплекса активации каспазы-2 с помощью аффинной хроматографии, которую проводят в условиях, обеспечивающих минимизацию сорбции неспецифических белков.
Ферментативная активность каспазы-2 в выделенном высокомолекулярном комплексе определяется с помощью флуоресцентных субстратов. При этом определение ферментативной активности осуществляют на носителе для аффинной хроматографии, с которым связан высокомолекулярный комплекс активации каспазы-2.
Предпочтительно в качестве раковых клеток использовать линии карциномы яичника человека Caov-4 или РА-1, или Caov-3, или SW 626, или SK-OV-3. Наиболее предпочтительно использовать линию карциномы яичника человека Caov-4 дефицитную по каспазе-2 - Caov-4-shRNA-Kacna3a-2 или линию карциномы яичника человека Caov-4 дефицитную по каспазе-2 - Caov-4 caspase-2 CRISPR/Cas9 Knockout.
Предпочтительно в качестве генетического конструкта использовать последовательность гена каспазы-2, которая соединена с последовательностью Strep-tag с N- и/или С-конца, или последовательность гена каспазы-2, которая соединена с последовательностью Twin-Strep-tag с N-и/или С-конца, или последовательность гена каспазы-2, которая соединена с последовательностью GST-tag с N- и/или С-конца, или последовательность гена каспазы-2, которая соединена с последовательностью Flag-tag с N- и/или С-конца. При этом концентрацию вектора, содержащего генетический конструкт, рассчитывают путем инкубации клеток в присутствии разных концентраций вектора и дальнейшим определением уровня экспрессии генетического конструкта методом Вестерн-блота.
Время инкубации раковых клеток после введения вектора определяется путем инкубации клеток в течение разного количества времени, но не менее 6 часов, и последующего анализа экспрессии генетического конструкта, представляющего собой последовательность гена каспазы-2 и дополнительную аминокислотную последовательность для аффинной хроматографии, методом Вестерн-блота.
Предпочтительно в качестве химиотерапевтического препарата использовать соединения платины или алкилирующие агенты, или ингибиторы топоизомераз, или антиметаболиты. При этом в качестве соединений платины предпочтительно использовать цисплатин, оксалиплатин, тетраплатин, карбоплатин, фенантриплатин.
Концентрацию ДНК-повреждающего химиотерапевтического препарата рассчитывают путем инкубации клеток в присутствии разных концентраций этого препарата и дальнейшим определением уровня гибели клеток методом цитометрии. Предпочтительно использовать ДНК-повреждающий химиотерапевтического препарата в концентрации не менее 10 мкМ, наиболее предпочтительно, когда концентрация ДНК-повреждающего химиотерапевтического препарата варьирует в пределах 35-100 мкМ.
Продолжительность инкубации определяется путем инкубации клеток с ДНК-повреждающим химиотерапевтическим препаратом в течение различного количества времени, но не менее часа, с последующим анализом уровня процессинга и активации каспазы-2 в этих клетках методом Вестерн-блота и/или с помощью флуоресцентных специфических субстратов.
Предпочтительно использовать лизирующий буфер, включающий один из следующих детергентов: Nonidet Р-40, Triton Х-100, Tween 20, Tween 80, Brij 35, Brij 58, Octyl glucoside, Octyl thioglucoside, sodium dodecyl sulfate, CHAPS, digitonin, deoxycholate, cholate, sarkosyl в конечной концентрации от 0,01 до 1,5% об. в буфере. При этом лизирующий буфер дополнительно может содержать пан-каспазный ингибитор zVAD-fmk (carbobenzoxy-valyl-alanyl-aspartyl-[O-methyl] fluoromethylketone) в конечной концентрации не менее 10 мкМ в буфере.
Выбор условий гель-фильтрации включает подбор носителя для гель-фильтрации с разрешающей способностью от 5×105 вплоть до 5×107 Да.
Для получения высокомолекулярных форм каспазы-2 молекулярной массой не менее 500000 Да предварительно оценивают накопление в клетках высокомолекулярного комплекса с помощью гель-фильтрации с последующим Вестерн-блотом в зависимости от продолжительности инкубации клеток с ДНК-повреждающим химиотерапевтическим препаратом.
Выделение комплекса активации каспазы-2 осуществляют с помощью аффинной хроматографии, включающей выбор носителя с наименьшими параметрами неспецифической сорбции, которую оценивают с помощью электрофореза образцов в полиакриламидном геле после аффинной хроматографии с дальнейшим окрашиванием геля.
Техническим результатом изобретения выделение комплекса активации каспазы-2 человека, формирующегося в нормальных условиях в линии раковых клеток в ответ на обработку ДНК-повреждающим химиотерапевтическим препаратом. Также преимуществом предлагаемого способа является подбор условий для увеличения выхода и/или чистоты выделенного комплекса активации каспазы-2 человека. Разработанный способ позволяет получить достаточное для дальнейшего анализа количество выделенного комплекса. Анализ комплекса можно проводить как с помощью, например, измерения активности каспазы-2 известным из уровня техники способом [В. Zhivotovsky, A. Samali, and S. Orrenius, "Determination of apoptosis and necrosis", Curr. Protoc. Toxicol, vol. Chapter 2, p. Unit 2.2, May 2001], так и масс-спектрометрией [A.-C. Gingras, M. Gstaiger, B. Raught, and R. Aebersold, "Analysis of protein complexes using mass spectrometry", Nat. Rev. Mol. Cell Biol, vol. 8, no. 8, pp. 645-654, Aug. 2007].
Выделенный белковый высокомолекулярный комплекс отличается тем, что содержит в своем составе ферментативно активную каспазу-2 и не содержит белки PIDD и RAIDD, как показано ниже.
В результате повреждений ДНК в клетках карциномы яичника химиотерапевтическими препаратами (например, соединениями платины) происходит запуск апоптотической клеточной гибели за счет формирования высокомолекулярного инициаторного комплекса, в составе которого происходит активация каспазы-2. Однако, в отличие от формирования ответственной за активацию каспазы-8 и -10 платформы DISC, процесс формирования данного комплекса, запускаемый повреждениями ДНК, требует заметного количества времени. Кроме того, количество формирующегося комплекса невелико, так, например, Imre et al. детектировали высокомолекулярный комплекс каспазы-2 только после его отделения от мономерных форм с помощью гель-фильтрации и концентрирования полученных фракций с помощью осаждения трихлоруксусной кислотой. Таким образом, для выделения комплекса активации каспазы-2 человека необходим высокоэффективный метод.
Выделение белкового высокомолекулярного комплекса активации каспазы-2 и идентификация его компонентов позволит разрабатывать новые лекарства против рака, действие которых направлено на запуск апоптоза через формирование указанного комплекса и активацию каспазы-2, аналогично соединениям TRAIL и ВН3-миметикам, индуцирующим сборку комплексов активации каспазы-8 и каспазы-9. Также прогнозирование эффективности лечения определенного типа рака тем или иным химиотерапевтическим агентом может включать анализ экспрессии каспазы-2 и других белковых компонентов, входящих в состав выделяемого высокомолекулярного комплекса.
Краткое описание чертежей
На фиг. 1 представлены результаты анализа методом проточной цитометрии клеток Caov-4 и Caov-4-shRNA-каспаза-2, обработанных цисплатином. Клетки были окрашены конъюгатом аннексии V-FITC и йодидом пропидия (ПИ). Обозначения для аннексии V/ПИ: -/- - жизнеспособные клетки; +/- апоптотические клетки; +/+- некротические клетки, (а) Данные ПЦ для контрольных и обработанных цисплатином клеток, (б) Гистограммы данных ПЦ для клеток, обработанных цисплатином в разных концентрациях.
На фиг. 2 показан процессинг каспазы-2 и появление продукта расщепления PARP (cPARP) как маркера протекания апоптоза после обработки клеток 70 мкМ цисплатином (результаты вестерн-блоттинга. Показано время обработки клеток, 0 - контрольные клетки, не подвергавшиеся обработке. Стрелками показаны положения полос полноразмерных прокаспазы-2 и PARP, продуктов их протеолиза, а также маркера белковой загрузки геля - GAPDH (глицеральгид-3-фосфат-дегидрогеназа), слева - мол. массы белков в кДа.
На фиг. 3 показаны результаты вестерн-блот анализа иммунопреципитации (ИП) каспазы-2 с использованием различных детергентов для лизирования клеток (ТХ. - Triton Х-100, NP. - NP-40, Ch. - Chaps) и различного количества антител для ИП: 1б 5 и 10 мкг, PrA - контроль неспецифической сорбции белков (протеин-А сефароза). Стрелками показаны положения полос полноразмерных прокаспазы-2.
На фиг. 4 представлены результаты анализа иммунопреципитации (ИП) каспазы-2 (слева) и протеолитической активности образцов ИП (справа) в контрольных и обработанных 35 мкМ цисплатином клетках Caov-4. Стрелками на Вестерн-блоте показаны полноразмерная форма прокаспазы-2 и продукты ее протеолиза, фрагмент белка PIDD (PIDD-CC) и белок RAIDD, участвующие в образовании комплекса PIDDosome.
На фиг. 5 показан гель-фильтрационный профиль распределения каспазы-2, каспазы-3 и цитохрома С в клетках, обработанных 70 мкМ цисплатина. Квадратом выделена высокомолекулярная фракция, содержащая каспазу-2. Сверху над блотами указаны молекулярные массы согласно калибровке колонки с помощью набора стандартов для гель-фильтрации (Gel Filtration Standard, Bio-Rad). WB-Вестерн-блот.
На фиг. 6 представлен гель-фильтрационный профиль распределения каспазы-2, каспазы-3 и цитохрома С в контрольных клетках, необработанных химиотерапевтическим агентом. Сверху над блотами указаны молекулярные массы согласно калибровке колонки с помощью набора стандартов для гель-фильтрации (Gel Filtration Standard, Bio-Rad). WB-Вестерн-блот.
На фиг. 7 показаны результаты анализа распределения каспазы-2 по фракциям после гель-фильтрации на носителях Superdex 200 и Superose 6. Квадратом выделена высокомолекулярная фракция, содержащая платформу активации каспазы-2. Стрелками показаны полноразмерная форма прокаспазы-2 и продукт аутопротеолитического расщепления данного фермента р37.
На фиг. 8 поредставлены результаты анализа влияния пан-каспазного ингибитора zVAD-fmk на стабильность комплекса, активирующего каспазу-2. Для проведения хроматографического анализа клетки были обработанны 70 мкМ цисплатином в течение 24 часов. Квадратом выделена высокомолекулярная фракция, содержащая каспазу-2. Стрелками показаны полноразмерная форма прокаспазы-2 и продукт расщепления данного фермента р37.
На фиг. 9 представлены результаты анализа образования макромолекулярного комплекса активации каспазы-2 с помощью гель-фильтрации. Кинетика формирования высокомолекулярного комплекса, содержащего каспазу-2, в клетках, обработанных цисплатином в течение 6, 12 и 18 ч. Квадратом выделена HMW-фракция, содержащая комплекс активации каспазы-2. Стрелками показаны полосы полноразмерной прокаспазы-2 и продукта ее аутолитического расщепления - фрагмента р32. М - маркер молекулярных масс.
На фиг.10 представлены результаты сравнения нескольких сорбентов для иммунопреципитации: протеин А-сефароза (Sigma), Mag Sepharose Magnetic Beads (GE Healthcare) и Dynabeads (Thermo Fisher Scientific). После электрофореза гель окрашивался серебром (SilverQuest™ Silver Staining Kit) или проводился Вестерн-блот с использованием антител против каспазы-2.
На фиг. 11 представлены результаты иммунопреципитации каспазы-2 из фракций с разной молекулярной массой. Фракции получены гель-фильтрацией лизатов клеток после 18 ч инкубации с 70 мкМ цисплатина. Стрелками показаны положения полос полноразмерной прокаспазы-2, продукта ее аутолитического расщепления - р32 и белка RAIDD. ИП - иммунопреципитация, HMW - высокомолекулярная фракция, MMW - среднемолекулярная фракция, LMW - низкомолекулярная фракция.
На фиг. 12 представлены результаты анализа пептидазной активности каспазы-2 в составе высокомолекулярного комплекса (субстрат - Ас-VDVAD-AMC). (а) Активность каспазы-2 в тотальном лизате клеток Caov-4, обработанных цисплатином, и в его фракциях LMW, MMW и HMW, полученных с помощью гель-фильтрации, (б) Активность каспазы-2 в образцах, полученных иммунопреципитацией лизата клеток Caov-4 и фракций, представленных на фиг. 6а. Протеин А-контроль демонстрирует уровень неспецифической сорбции каспазы-2 на носителе протеин-А-сефарозе. RFU - относительные единицы флуоресцентной активности.
На фиг. 13 представлены результаты анализа роли белка RAIDD с помощью метода siRNA. Исследованы клетки Caov-4, обработанные контрольной siRNA (NT - no target) и siRNA против белка RAIDD, а также 70 мкМ цисплатином (в течение 24 ч). (а) Анализ методом проточной цитометрии. Обозначения для аннексии V/ПИ: -/- - жизнеспособные клетки; +/- апоптотические клетки; +/+ - некротические клетки, (б) Вестерн-блот-анализ. Стрелками показаны положения полос полноразмерной прокаспазы-2, белков RAIDD и PARP и продукта протеолиза последнего - cPARP (cleaved PARP).
На фиг. 14 представлены результаты анализа экспрессии векторов IBA-25, IBA-103 и IBA-105 (Табл. 2) в клетках Caov-4, клетках дефицитных каспазе-2 Caov-4-shRNA-caspase-2 клетках HeLa. Стрелками показаны полосы полноразмерной прокаспазы-2, ее активная расщепленная форма - р19 и ее модифицированные варианты. Marker - маркер молекулярных масс.
На фиг. 15 показано выделение модифицированных форм каспазы-2 из клеток, трасфицированных векторами pcDNA3/hCasp2-Flag, IBA-103 и IBA-105 (Табл. 2). ИП - иммунопреципитация, АФ - аффинная хроматография. Стрелками показаны полосы полноразмерной прокаспазы-2, ее активная расщепленная форма - р19 и ее модифицированные варианты.
На фиг. 16 проиллюстрирован подбор условий для выделения модифицированной форм каспазы-2 из клеток, трасфицированных вектором IBA-105 (Табл. 2). ИП - иммунопреципитация, АФ - аффинная хроматография. Стрелкой показана полоса модифицированной полноразмерной прокаспазы-2.
На фиг. 17 представлены результаты анализа синтеза каспазы-2 в клетках Caov-4 после обработки вектором caspase-2 CRISPR/Cas9 KO Plasmid (Santa Cruz, США) и получения отдельных клонов. Стрелками показаны положения полос полноразмерных прокаспазы-2 и прокаспазы-3 для сравнения уровня их синтеза. wtCaov-4 - дикий тип клеток линии Caov-4.
Осуществление изобретения
Пока не определено иначе, все технические и научные термины, используемые в данном контексте, имеют такие же значения, как обычно имеют в виду специалисты в области техники, к которой принадлежит данное изобретение.
Заявляемый способ выделения белкового высокомолекулярного комплекса активации каспазы-2 человека, формирующегося в раковых клетках в ответ на обработку ДНК-повреждающим химиотерапевтическим препаратом, характеризуется тем, что включает:
- получение раковых клеток в количестве не менее чем 5 млн клеток;
- введение в раковые клетки ДНК-повреждающего химиотерапевтического препарата в концентрации, обеспечивающей запуск апоптотической гибели раковых клеток с достижением максимального уровня апоптотической гибели клеток при минимальном уровне некротической гибели клеток, которые определяют с помощью цитометрии;
- инкубация раковых клеток в присутствии ДНК-повреждающего химиотерапевтического препарата до получения максимального уровня активации каспазы-2, который определяют предварительно до начала инкубации с помощью анализа методом Вестерн-блота уровня расщепления прокаспазы-2 и накопления каталитически активных фрагментов каспазы-2 и/или с помощью флуоресцентных специфических субстратов;
- сбор клеток с последующим добавлением лизирующего буфера, который обеспечивает сохранность высокомолекулярного комплекса активации каспазы-2;
- отделение клеточного дебриса от лизата, содержащего высокомолекулярный комплекс активации каспазы-2,
- разделение гель-фильтрацией полученного лизата клеток с обеспечением отделения фракций, содержащих высокомолекулярные формы каспазы-2, от фракций, содержащих ее мономерные формы, которое проводят в условиях, обеспечивающих получение высокомолекулярных форм каспазы-2 молекулярной массой не менее 500000 Да;
- выделение из фракции, содержащей высокомолекулярные формы каспазы-2, белкового высокомолекулярного комплекса активации каспазы-2 с помощью иммунопреципитации, которую проводят в условиях, обеспечивающих минимизацию сорбции неспецифических белков.
В качестве раковых клеток предпочтительно использовать линии карциномы яичника известные из уровня техники [Т.A. Ince, A.D. Sousa, М.A. Jones, J.С. Harrell, Е.S. Agoston, М. Krohn, L.М. Selfors, W. Liu, K. Chen, M. Yong, P. Buchwald, B. Wang, K.S. Hale, E. Cohick, P. Sergent, A. Witt, Z. Kozhekbaeva, S. Gao, A.T. Agoston, M.A. Merritt, R. Foster, B.R. Rueda, C.P. Crum, J.S. Brugge, and G.B. Mills, "Characterization of twenty-five ovarian tumour cell lines that phenocopy primary tumours", Nat. Commun., vol. 6, p. 7419, Jun. 2015]. Также могут быть использованы линии карциномы шейки матки, такие как HeLa (АТСС® CCL-2™), или линии Daudi (АТСС® CCL-213™), Raji (АТСС® CCL-86™), Jurkat (АТСС® TIB-152™). В другом варианте изобретения в качестве раковой линии предпочтительно использовать Caov-4 (АТСС® НТВ-76™) или другие линии, например, РА-1 (АТСС CRL-1572), Caov-3 (АТСС НТВ-75), SW 626 (АТСС НТВ-78), SK-OV-3 (АТСС НТВ-77), а также модифицированные варианты Caov-4, дефицитные по каспазе-2, такие как sh-RNA-caspase-2-Caov-4 [Г.С. Копеина, А.В. Замараев, Б.Д. Животовский, and И.Н. Лаврик, "Идентификация нового макромолекулярного комплекса активации каспазы-2 при повреждениях ДНК", Биоорганическая химия, vol. 42, no. 1, pp. 84-93, 2016] и caspase-2 CRTSPR/Cas9 Caov-4, получение которой описано ниже. Культивирование клеток осуществляют известными из уровня техники средствами и способами, согласно рекомендациям American Type Culture Collection (http://www.atcc.org/).
Для осуществления этапа введения в культуру ДНК-повреждающего химиотерапевтического препарата/агента необходимо произвести подбор условий для максимального уровня индукции апоптотической гибели клеток раковой линии, вызванной химиотерапевтическим ДНК-повреждающим агентом. Данный подбор подразумевает определение концентрации химиотерапевтического агента, которая будет вызывать наибольший процент гибели клеток по апоптотическому пути без существенного уровня некроза, путем инкубации клеток в присутствии разных концентраций с агентом. В качестве ДНК-повреждающего препарата (агента) могут быть использованы соединения платины, такие как цисплатин, оксалиплатин, тетраплатин, карбоплатин, фенантриплатин или их комбинации в эффективных концентрациях [О. Rixe, W. Ortuzar, М. Alvarez, R. Parker, E. Reed, K. Paull, and T. Fojo, "Oxaliplatin, tetraplatin, cisplatin, and carboplatin: spectrum of activity in drug-resistant cell lines and in the cell lines of the National Cancer Institute's Anticancer Drug Screen panel", Biochem. Pharmacol, vol. 52, no. 12, pp. 1855-65, Dec. 1996; G.Y. Park, J.J. Wilson, Y. Song, and S.J. Lippard, "Phenanthriplatin, a monofunctional DNA-binding platinum anticancer drug candidate with unusual potency and cellular activity profile", Proc. Natl Acad. Sci. U.S.A., vol. 109, no. 30, pp. 11987-92, Jul. 2012]. Также в качестве ДНК-повреждающих агентов, могут быть использованы алкилирующие агенты, или ингибиторы топоизомераз, или антиметаболиты [K. Cheung-Ongl, G. Giaever, C. Nislow. DNA-Damaging Agents in Cancer Chemotherapy: Serendipity and Chemical Biology. Chem Biol, vol. 23; no 20(5), pp. 648-59, May 2013]. Инкубацию раковых клеток в присутствии ДНК-повреждающего химиотерапевтического агента как правило проводят путем добавления агента до нужной концентрации в стандартную среду культивирования. Наиболее предпочтительно использовать в качестве химиотерапевтического агента цисплатин. Процент гибели клеток можно определить с помощью проточной цитометрии после окрашивания клеток аннексином V и пропидий иодидом стандартным способом или методом определения популяции Sub-G1, также по появлению таких маркеров апоптоза как расщепление белка PARP (Poly ADP ribose polymerase) или любым другим стандартным методом, известным из уровня техники [В. Zhivotovsky, A. Samali, and S. Orrenius, "Determination of apoptosis and necrosis", Curr. Protoc. Toxicol, vol. Chapter 2, p. Unit 2.2, May 2001; A.H. Boulares, A.G. Yakovlev, V. Ivanova, B.A. Stoica, G. Wang, S. Iyer, and M. Smulson, "Role of poly(ADP-ribose) polymerase (PARP) cleavage in apoptosis. Caspase 3-resistant PARP mutant increases rates of apoptosis in transfected cells", J. Biol. Chem., vol. 274, no. 33, pp. 22932-40, Aug. 1999; Т. Vanden Berghe, S. Grootjans, V. Goossens, Y. Dondelinger, D.V Krysko, N. Takahashi, and P. Vandenabeele, "Determination of apoptotic and necrotic cell death in vitro and in vivo", Methods, vol. 61, no. 2, pp. 117-29, Jun. 2013]. Кроме того, подбор условий индукции апоптотической гибели включает выбор времени инкубации клеток с химиотерапевтическим агентом для получения максимального уровня процессинга и активации каспазы-2. Уровень процессинга и активации каспазы-2 определяют с помощью Вестерн-блот анализа с использованием антител специфических по отношению к каспазе-2 и ее энзиматически активным фрагментам (Табл. 2) [В. Zhivotovsky, A. Samali, and S. Orrenius, "Determination of apoptosis and necrosis", Curr. Protoc. Toxicol, vol. Chapter 2, p. Unit 2.2, May 2001]. Также активность каспазы-2 можно определить известным из уровня техники способом с помощью измерения флуоресцентного или люминесцентного сигнала, который генерируется при расщеплении каспазой-2 модифицированных производных пентапептида VDVAD, таких как Acetyl-L-valyl-L-aspartyl-L-valyl-L-alanyl-L-aspartic acid α-(4-methyl-coumaryl-7-amide) или других субстратов известных из уровня техники [Т. Kitevska, S.J. Roberts, D. Pantaki-Eimany, S.E. Boyd, F.L. Scott, and C.J. Hawkins, "Analysis of the minimal specificity of caspase-2 and identification of Ac-VDTTD-AFC as a caspase-2-selective peptide substrate", Biosci. Rep., Feb. 2014].
Следующий этап предлагаемого способа требует подбора условий для получения лизата клеток. Данный этап включает тестирование различных детергентов для эффективного лизиса клеток, таких как Nonidet Р-40, Triton Х-100, Tween 20, Tween 80, Brij 35, Brij 58, Octyl glucoside, Octyl thioglucoside, sodium dodecyl sulfate, CHAPS, digitonin, deoxycholate, cholate, sarkosyl [M. Johnson, "Detergents: Triton X-100, Tween-20 and More", Mater. Methods, 2014] в концентрациях от 0,01 до 1,5% об., а также их возможных комбинации. Эффективность лизиса можно оценивать с помощью Вестерн-блот анализа получающихся лизатов клеток, оценивая при этом количество каспазы-2, оказавшейся в растворе после разрыва цитоплазматической мембраны детергентом и последующего осаждения клеточного дебриса. Получение лизатов с помощью разных детергентов производится известным из уровня техники способом инкубации клеток в изотоническом или гипотоническом буферно-солевом растворе, содержащим детергент, и дальнейшим однократным или многократным центрифугированием исследуемого образца [P. Holden and W. A. Horton, "Crude subcellular fractionation of cultured mammalian cell lines", BMC Res. Notes, vol. 2, p. 243, 2009].
Этап разделения гель-фильтрацией полученного лизата клеток включает подбор условий разделения с помощью гель-фильтрации высокомолекулярного комплекса и мономерных форм каспазы-2. Данный этап включает выбор носителя и режима проведения гель-фильтрации, позволяющих максимально разделить высокомолекулярный комплекс активации и мономерные формы каспазы-2. В качестве носителей могут быть использованы известные из уровня техники сорбенты (например, Superdex 200, Superose -6, -12, Sephacryl S-500 HR) с разрешающей способностью от 5×103 вплоть до 5×107 Да [P. Stanton, "Gel filtration chromatography.," Methods Mol. Biol., vol. 251, pp. 55-74, 2004]. Гель-фильтрация при этом может проводиться с помощью хроматографических приборов высокого или низкого давления, таких как  plus (GE Healthcare), BioLogic LP System (BioRad) или NGC™ Medium-Pressure Liquid Chromatography Systems (BioRad) с использованием рекомендованных производителем колонок и скоростей. Кроме того, желательно проведение гель-фильтрации при температуре от +4°С до +8°С для предотвращения деградации компонентов комплекса. Также подбор условий разделения может включать оценку накопления каспазы-2 в высокомолекулярных фракциях гель-фильтрации в зависимости от времени инкубации клеток с ДНК-повреждающим агентом для определения временной точки, в которой достигается максимальный уровень накопления высокомолекулярного комплекса. Данный анализ проводится с помощью гель-фильтрации лизатов клеток с последующим Вестерн-блотом полученных фракций, при этом клетки инкубируются с ДНК-повреждающим агентом в течение разного количества времени, например, временной интервал может составлять от 1 часа и вплоть до 48 часов. Также подбор условий может включать добавление в лизат клеток перед гель-фильтрацией ингибиторов каспаз, и в частности ингибитора каспазы-2, для стабилизации комплекса. В качестве ингибиторов могут быть использованы z-VAD-fmk (carbobenzoxy-valyl-alanyl-aspartyl-[O-methyl]fluoromethylketone), z-VDVAD-fmk (methyl(3S)-5-fluoro-3-[[(2S)-2-[[(2S)-2-[[(2S)-4-methoxy-2-[[(2S)-3-methyl-2-(phenylmethoxycarbonylamino)butanoyl]amino]-4-oxobutanoyl]amino]-3-methylbutanoyl]amino]propanoyl]amino]-4-oxopentanoate) или Q-VD-OPh (N-(2-Quinolyl)-L-valyl-L-aspartyl-(2,6-difluorophenoxy)methylketone) [B.A. Callus and D.L. Vaux, "Caspase inhibitors: viral, cellular and chemical", Cell Death Differ., vol. 14, no. 1, pp. 73-78, Jan. 2007] в эффективных концентрациях, например, в диапазоне от 10 до 50 мкМ.
plus (GE Healthcare), BioLogic LP System (BioRad) или NGC™ Medium-Pressure Liquid Chromatography Systems (BioRad) с использованием рекомендованных производителем колонок и скоростей. Кроме того, желательно проведение гель-фильтрации при температуре от +4°С до +8°С для предотвращения деградации компонентов комплекса. Также подбор условий разделения может включать оценку накопления каспазы-2 в высокомолекулярных фракциях гель-фильтрации в зависимости от времени инкубации клеток с ДНК-повреждающим агентом для определения временной точки, в которой достигается максимальный уровень накопления высокомолекулярного комплекса. Данный анализ проводится с помощью гель-фильтрации лизатов клеток с последующим Вестерн-блотом полученных фракций, при этом клетки инкубируются с ДНК-повреждающим агентом в течение разного количества времени, например, временной интервал может составлять от 1 часа и вплоть до 48 часов. Также подбор условий может включать добавление в лизат клеток перед гель-фильтрацией ингибиторов каспаз, и в частности ингибитора каспазы-2, для стабилизации комплекса. В качестве ингибиторов могут быть использованы z-VAD-fmk (carbobenzoxy-valyl-alanyl-aspartyl-[O-methyl]fluoromethylketone), z-VDVAD-fmk (methyl(3S)-5-fluoro-3-[[(2S)-2-[[(2S)-2-[[(2S)-4-methoxy-2-[[(2S)-3-methyl-2-(phenylmethoxycarbonylamino)butanoyl]amino]-4-oxobutanoyl]amino]-3-methylbutanoyl]amino]propanoyl]amino]-4-oxopentanoate) или Q-VD-OPh (N-(2-Quinolyl)-L-valyl-L-aspartyl-(2,6-difluorophenoxy)methylketone) [B.A. Callus and D.L. Vaux, "Caspase inhibitors: viral, cellular and chemical", Cell Death Differ., vol. 14, no. 1, pp. 73-78, Jan. 2007] в эффективных концентрациях, например, в диапазоне от 10 до 50 мкМ.
Следующий этап метода включает выбор условий для эффективного выделения комплекса активации каспазы-2 из фракций гель-фильтрации с помощью иммунопреципитации с дальнейшим определением ферментативной активности каспазы-2 с помощью флуоресцентных субстратов во фракциях гель-фильтрации, содержащих данный фермент, и/или в образце, содержащем выделенный высокомолекулярный комплекс, для подтверждения эффективности выделения. Этот этап может включать выбор носителя с минимальными параметрами неспецифической сорбции. В качестве носителей могут быть использованы различные варианты Protein А или G конъюгированных сорбентов (Табл. 1). При этом для оценки загрязненности носителя неспецифическими белками образцы после иммунопреципитации могут быть проанализированы с помощью стандартной методики электрофореза в полиакриламидном геле с последующей окраской гелей препаратами серебра или Coomassie Brilliant Blue [J.M. Elmore and G. Coaker, "Biochemical purification of native immune protein complexes", Methods Mol. Biol., vol. 712, pp. 31-44, 2011].
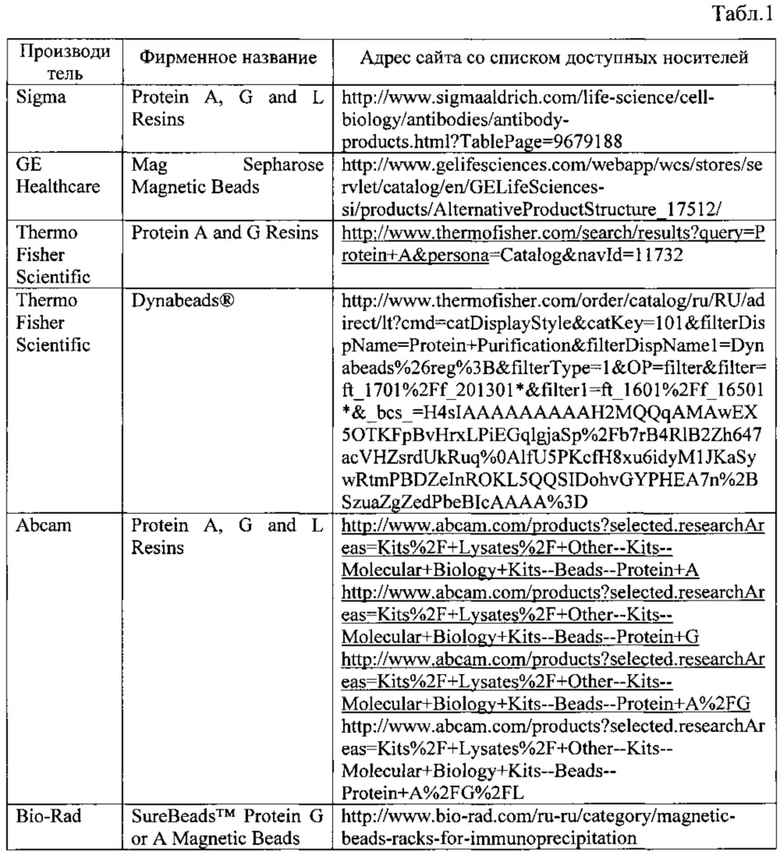
Кроме того, подбор условий для выделения может включать тестирование специфических к каспазе-2 антител от разных производителей (Табл. 2) с целью выбора того, который позволяет выделить максимальное количество каспазы-2 без существенного уровня связывания с неспецифическими белками. Оценка эффективности связывания может производится с помощью Вестерн-блот анализа с целью определения уровня обогащения образцов ИП целевым белком.
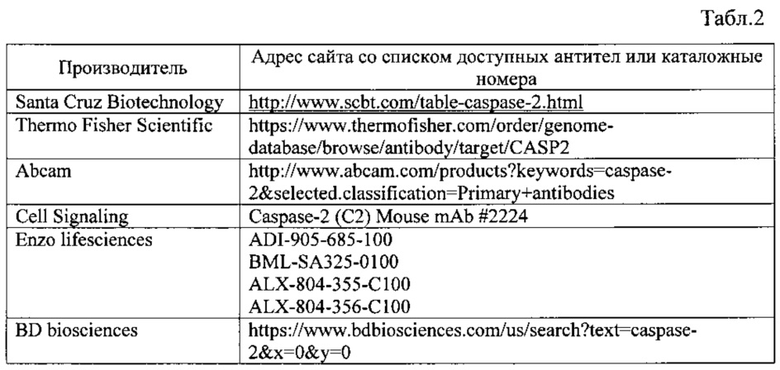
Также для подтверждения того, что в состав выделенного высокомолекулярного комплекса входит энзиматически активная каспаза-2, последняя стадия метода включает определение ферментативной активности каспазы-2 с помощью соответствующих субстратов, например, модифицированных производных пентапептида VDVAD, таких как Acetyl-L-valyl-L-aspartyl-L-valyl-L-alanyl-L-aspartic acid α-(4-methyl-coumaryl-7-amide) или других известный из уровня техники [О. Rixe, W. Ortuzar, М. Alvarez, R. Parker, E. Reed, K. Paull, and T. Fojo, "Oxaliplatin, tetraplatin, cisplatin, and carboplatin: spectrum of activity in drug-resistant cell lines and in the cell lines of the National Cancer Institute's Anticancer Drug Screen panel", Biochem. Pharmacol., vol. 52, no. 12, pp. 1855-65, Dec. 1996]. Данный анализ необходим для осуществления разработанного метода, поскольку позволяет доказать, что выделенный высокомолекулярный комплекс содержит активную форму каспазы-2, а значит, является платформой активации этого профермента. При этом измерение активности каспазы-2 может производится как в связанном с носителем состоянии, так и после элюции каспазы-2 и взаимодействующих с ней белков в раствор. Элюция белков с носителя может осуществляться с помощью раствора глицина в эффективной концентрации (например, 0.1-0.2 М, рН 2.0-3.0) или другим известным из уровня техники способом ["Immunoprecipitation protocol General immunoprecipitation procedure and required reagents 2 Immunoprecipitation protocol."].
В другом варианте изобретение относится к способу выделения белкового высокомолекулярного комплекса активации каспазы-2 человека, формирующегося в раковых клетках в ответ на обработку ДНК-повреждающим химиотерапевтическим препаратом, который характеризуется тем, что включает:
- получение раковых клеток в количестве не менее чем 5 млн клеток;
- введение с помощью трансфицирующего агента в раковые клетки вектора, содержащего генетический конструкт, представляющий собой последовательность гена каспазы-2 и дополнительную аминокислотную последовательность для аффинной хроматографии, в концентрации не менее 0,5 мкг на 1 мл питательной среды;
- инкубация раковых клеток в течение времени, достаточного для детектируемой экспрессии генетического конструкта, представляющего собой последовательность гена каспазы-2 и дополнительную аминокислотную последовательность для аффинной хроматографии;
- введение в трансформированные клетки ДНК-повреждающего химиотерапевтического препарата, обеспечивающей запуск апоптотической гибели раковых клеток с достижением максимального уровня апоптотической гибели клеток при минимальном уровне некротической гибели клеток, которые определяют с помощью цитометрии;
- инкубация раковых клеток в присутствии ДНК-повреждающего химиотерапевтического агента до получения максимального уровня активации каспазы-2, который определяют предварительно до начала инкубации с помощью анализа методом Вестерн-блота уровня расщепления прокаспазы-2 и накопления каталитически активных фрагментов каспазы-2 и/или с помощью флуоресцентных специфических субстратов;
- сбор клеток с последующим добавлением лизирующего буфера, который обеспечивает сохранность высокомолекулярного комплекса активации каспазы-2
отделение клеточного дебриса от лизата, содержащего высокомолекулярный комплекс активации каспазы-2,
- разделение гель-фильтрацией полученного лизата клеток с обеспечением отделения фракций, содержащих высокомолекулярные формы каспазы-2, от фракций, содержащих ее мономерные формы, которое проводят в условиях, обеспечивающих получение высокомолекулярных форм каспазы-2 молекулярной массой не менее 500000 Да;
- выделение из фракции, содержащей высокомолекулярные формы каспазы-2, белкового высокомолекулярного комплекса активации каспазы-2 с помощью аффинной хроматографии, которую проводят в условиях, обеспечивающих минимизацию сорбции неспецифических белков.
Для осуществления метода требуется создание генетических векторов, содержащих ген каспазы-2 и дополнительную аминокислотную последовательность для аффинной хроматографии. В качестве последовательностей для аффинной хроматографии могут быть использованы следующие известные из уровня техники «теги» - His, СВР, CYD (covalent yet dissociable NorpD peptide), Strep II, Twin-strep-tag, Flag, HPC (heavy chain of protein C) peptide tags, GST или MBP, а также их комбинации [J.J. Lichty, J.L. Malecki, H.D. Agnew, D.J. Michelson-Horowitz, and S. Tan, "Comparison of affinity tags for protein purification", Protein Expr. Purif., vol. 41, no. 1, pp. 98-105, May 2005; K. Terpe, "Overview of tag protein fusions: from molecular and biochemical fundamentals to commercial systems", Appl. Microbiol. Biotechnol., vol. 60, no. 5, pp. 523-33, Jan. 2003]. Далее под термином «тег» подразумевается аминокислотная последовательность, присоединенная к целевому белку, которая позволяет выделять целевой белок с помощью аффинного носителя. Реализация данной задачи включает подбор праймеров, конструирование различных вариантов векторов, содержащих разные «теги» с N- или С-конца исследуемого белка, для экспрессии гена каспазы-2 в раковых линиях человека с помощью известных из уровня техники методов [S.J. Green Michael R., Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory, 2012]. Для создания могут быть использованы коммерческие вектора, уже содержащие последовательности различных «тегов», например, такие как pESG-IBA (Iba-lifesciences, список векторов для экспрессии в клетках млекопитающих доступен по адресу http://www.iba-lifesciences.com/Acceptor_vectors_Mammalia_Products.html), а также других векторов известных из уровня техники (http://wolfson.huji.ac.il/expression/vector/commmercialvectors.html). Далее необходимо произвести подбор условий для экспрессии полученных конструктов в клетках выбранной раковой линии, заключающийся в тестировании различных концентрации полученного вектора и трансфицирующего агента. Концентрация вектора может варьироваться в пределах от 0,5 мкг/мл до 10 мкг/мл, в качестве трансфицирующего агента могут быть использованы в соответствии с рекомендациями фирм-производителей различные коммерчески доступные препараты: Lipofectamine® -2000, -3000 или -LTX (Invitrogene): FuGENE® HD или 6 Transfection Reagent (Promega); DOTAP Liposomal Transfection Reagent (Sigma), SuperFect Transfection Reagent (Qiagen). Далее проводится отбор эффективно экспрессирующихся векторов, введение которых в клетку не вызывает существенной активации каспазы-2 и последующего запуска процесса апоптоза. Также оценивается время инкубации клеток после трансфекции для эффективной экспрессии генетического конструкта. Для этого клетки после введения вектора инкубируют в стандартной среде в течение разного количества времени, например, в течение 12, 24 или 48 часов. Далее уровень экспрессии генетического конструкта оценивают с помощью Вестерн-блота.
Осуществление двух последующих стадий - лизис клеток гель-фильтрация в заранее выбранных условиях - требует также подбора детергентов, их концентраций, времени инкубации с ДНК-повреждающим агентом, носителя для гель-фильтрации и условий, обеспечивающих стабилизацию комплекса (добавление ингибитора каспаз). Подбор перечисленных условий может производится таким же способом, как это было описано для первого метода выделения комплекса активации каспазы-2.
На последнем этапе описываемого метода производится выделение комплекса активации каспазы-2 из фракций гель-фильтрации с помощью аффинного носителя в заранее подобранных условиях с дальнейшим определением ферментативной активности каспазы-2 с помощью флуоресцентных субстратов в образце, содержащем выделенный высокомолекулярный комплекс, для подтверждения эффективности выделения. Данный этап может включать выбор аффинного сорбента, наиболее эффективно связывающего каспазу-2, с минимальными показателями неспецифической сорбции; выбор условий нанесения образца и промывки аффинного сорбента для минимизации загрязнения образцов неспецифическими белками; выбор условий элюирования каспазы-2. В качестве аффинного сорбента могут быть выбраны коммерчески доступные носители в соответствии с тем «тегом», последовательность которого входит в состав конкретного вектора [G.Е. Healthcare, "Recombinant Protein Purification Handbook," Methods, vol. 41, no. 7, pp. 1-306, 2009; by I.S.F.L. Sciences, "Expression and purification of proteins using Strep-tag® or Twin-Strep-tag®," pp. 1-36, 2012]. Например, список возможных вариантов «тегов» и соответствующих носителей, а также протоколов выделения можно найти по адресу http://wolfson.huji.ac.il/purification/Purification_Protocols.html, а также на сайтах фирм-производителей: GE Healthcare, Thermo Fisher Scientific, Clontech, Iba-lifesciences, MERCK-NOVAGEN, Sigma, Promega, Qiagen, New England Biolab. Так, для векторов pESG-IBA (Iba-lifesciences) могут быть использованы фирменные носители: Strep-Tactin® Sepharose, Strep-Tactin® Superflow®, Strep-Tactin® Superflow® high capacity, Strep-Tactin® MacroPrep®, MagStrep Strep-Tactin®. При этом для оценки уровня неспецифической сорбции белков на носителе образцы после выделения могут быть проанализированы с помощью стандартной методики электрофореза в полиакриламидном геле с последующей окраской гелей препаратами серебра или Coomassie Brilliant Blue [В.A. Callus and D.L. Vaux, "Caspase inhibitors: viral, cellular and chemical," Cell Death Differ., vol. 14, no. 1, pp. 73-78, Jan. 2007].
В другом варианте изобретения вектора, содержащие ген каспазы-2 и дополнительную аминокислотную последовательность для аффинной хроматографии, могут быть введены в модифицированные линии клеток, дефицитные по каспазе-2. Такой подход может быть полезен для того, чтобы избежать гиперэкспрессии гена каспазы-2, которая может запустить неспецифическую активацию каспазного каскада и, соответственно, гибель клеток. Дефицитная по каспазе-2 линии клеток может быть получена с помощью введения генетических конструктов, содержащих shRNA (short hairpin RNAs), обеспечивающих значительное снижение уровня экспрессии целевого белка. Методология конструирования таких векторов известна из уровня техники [С.В. Moore, Е.Н. Guthrie, М. Т.-Н. Huang, and D.J. Taxman, "Short hairpin RNA (shRNA): design, delivery, and assessment of gene knockdown", Methods Mol. Biol., vol. 629, pp. 141-58, 2010; G. Healthcare, "GIPZ™ Lentiviral shRNA", 2012], а также доступны коммерческие варианты caspase-2 shRNA Plasmid (Santa Cruz, каталожный номер sc-72108-SH). Кроме того, дефицитные по каспазе-2 линии раковых клеток могут быть получены с помощью технологии редактирования генома Crispr/Cas9 [F.A. Ran, P.D. Hsu, J. Wright, V. Agarwala, D.A. Scott, and F. Zhang, "Genome engineering using the CRISPR-Cas9 system", Nat. Protoc., vol. 8, no. 11, pp. 2281-308, Nov. 2013; J.D. Sander and J.K. Joung, "CRISPR-Cas systems for editing, regulating and targeting genomes", Nat. Biotechnol., vol. 32, no. 4, pp.347-355, Mar. 2014; P.D. Hsu, E.S. Lander, and F. Zhang, "Development and applications of CRISPR-Cas9 for genome engineering", Cell, vol. 157, no. 6, pp. 1262-78, Jun. 2014]. Получение линии клеток Caov-4, дефицитных по каспазе-2, с помощью этой технологии приведено ниже в примерах.
Необходимо отметить, что высокомолекулярные комплексы регулируют множество процессов, происходящих в клетках. Изучение таких комплексов требует высокопродуктивных подходов для их получения. Процесс получения осложняется нестабильностью комплексов и в некоторых случаях их низким содержанием в клетках. С помощью разработанного метода был выделен высокомолекулярный комплекс активации каспазы-2, который формируется в раковых клетках в ответ на повреждения ДНК. Анализ показал, что активация каспазы-2 происходит независимо от известного из литературы комплекса PIDDosome. Также указанный выделенный комплекс содержит активную каспазу-2. Таким образом, разработанный метод позволил впервые охарактеризовать новый комплекс активации каспазы-2, что может быть применимо для выделения и характеристики других макромолекулярных комплексов, регулирующих программируемую клеточную гибель. Характеристика такого комплекса будет способствовать разработке новых подходов для лечения заболеваний, связанных с нарушением процесса апоптоза, в том числе и раковых заболеваний.
Примеры
Представленные ниже примеры конкретного осуществления изобретения приведены для предоставления специалистам в данной области техники полного описания проведения и применения анализа по изобретению, но не ограничивают предполагаемый авторами изобретения объем изобретения. Данные примеры могут быть заменены на аналогичные средства и методы, известные из уровня техники.
Все используемые реагенты являются коммерчески доступными, все процедуры, если не оговорено особо, осуществляли при комнатной температуре или температуре окружающей среды, то есть в диапазоне от 18 до 25°С.
Материалы
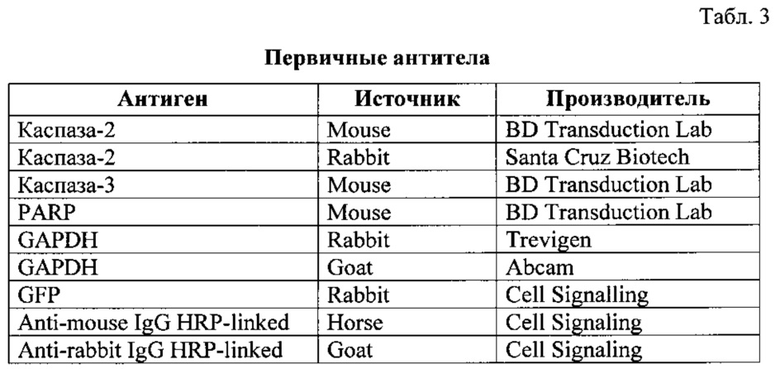
Оценка клеточной гибели после добавления ДНК-повреждающего агента.
Для изучения роли каспазы-2 в апоптозе, индуцированном повреждением ДНК, необходимо было подобрать оптимальную клеточную модель и убедиться, что в данной системе каспаза-2 действительно играет центральную роль в индукции апоптоза. Для этого клетки Caov-4 выращивали в CO2-инкубаторе (5% CO2) при 37°С на среде DMEM (Gibco), содержащей 10% телячьей сыворотки (Gibco), в присутствии смеси антибиотиков и антимикотиков Antibiotic-Antimycotic (Gibco). Для экспериментов использовали клетки в логарифмической фазе роста. Снятие клеток производилось трипсином. Клеточная плотность поддерживалась на уровне 1-2,5×105 клеток/мл среды. Подсчет клеток осуществлялся с помощью Z2 Coulter counter (Beckman).
Далее клетки карциномы яичника Caov-4 были обработаны цисплатином в концентрации 35, 70, 105, 150 и 250 мкМ в течение 24 и 48 часов. Индукция клеточной гибели проводилась по достижении 70-90% конфлюэнтности заменой среды и добавлением цисплатина (Тева, Израиль) до выбранной концентрации. В качестве контроля были взяты клетки Caov-4, необработанные цисплатином. Аналогичным образом обрабатывались клетки Caov-4-shRNA-каспаза-2, которые характеризуются пониженным уровнем содержания этой протеазы, и, таким образом, данный эксперимент позволил сделать заключение о возможной роли каспазы-2 в апоптозе, индуцируемом цисплатином.
Клетки в чашках Петри инкубировали в присутствии цисплатина (ТЕВА, Израиль) в концентрациях 35, 70, 105, 150, 250 мкМ в течение 24 и 48 ч, после этого снимали клетки с помощью 0.05% раствора трипсин-EDTA (Gibco) и переносили в кондиционированную среду. Клетки центрифугировали (2000 об/мин, 5 мин, 4°С), дважды промывали холодным раствором PBS (Панэко). Затем клетки (0.2×106) ресуспендировали в 200 мкл аннексин-связывающего буфера (BD Biosciences), добавляли 2 мкл аннексии V - FITC (Invitrogen) и инкубировали в темном месте при 4°С в течение 15 мин. Далее к пробам добавляли 5 мкл йодида пропидия (50 мкг/мл) (BD Biosciences). После 5 мин инкубации в темном месте при комнатной температуре популяцию апоптотических клеток анализировали с помощью проточного цитофлуориметра Guava® easyCyte™ 8НТ (Merck Millipore) или BD LSRFortessa™ cell analyzer (BD biosciences).
С помощью цитометрии производили оценку популяции клеток, вступивших в ранний апоптоз, и некротические клетки. Данные проточной цитометрии свидетельствовали о том, что при увеличении концентрации цисплатина в популяции опухолевых клеток Caov-4 наблюдалось заметное увеличение количества апоптотических клеток, а также небольшой процент некротических клеток (фиг. 1). Необходимо отметить, что заметное увеличение количества апоптотических клеток отмечалось через 48 часов после их индукции цисплатином. При обработке цисплатином клеток Caov-4 sh-RNA-каспаза2 наблюдалось заметное снижение уровня гибели клеток по сравнению с клетками Caov-4. Через 48 часов отличие процента апоптотической популяции клеток Caov-4 от клеток Caov-4-shPHK-каспаза-2 являлось статистически значимым (p-value=0.032, критерий Манна-Уитни) (фиг. 1). Сравнение уровней апоптоза в клетках Caov-4 и клетках Caov-4 с пониженной экспрессией каспазы-2 подтвердило центральную роль каспазы-2 в развертывании процесса программируемой гибели в ответ на генотоксический стресс. Данные проточной цитометрии свидетельствовали о том, что обработка цисплатином клеток Caov-4 вызывала заметное увеличение количества погибших клеток, при этом повышение концентрации цисплатина от 35 до 70 мкМ приводило к существенному возрастанию уровня апоптотической смерти - с 19 до 36% (фиг. 1). Однако дальнейшее увеличение концентрации индуктора приводит к росту не апоптотической, а некротической популяции, поэтому в последующих экспериментах был использован цисплатин в концентрации 70 мкМ.
Исследование кинетики активации каспазы-2 после добавления цисплатины
Для анализа кинетики накопления активированной каспазы-2 клетки Caov-4 инкубировали в присутствии цисплатина в концентрации 70 мкМ в течение 2, 4, 8, 12, 18, 24 и 48 часов. Затем клетки снимали с чашек, отмывали в холодном растворе PBS и инкубировали на льду в течение 20 мин в 200 мкл лизирующего буфера (20 мМ Tris-HCl (рН 7.4), 137 мМ NaCl, 10% SDS, 5% дезоксихолат натрия, 1% Triton Х-100, 1% PMSF, коктейль ингибиторов протеаз Complete (Roche)). После центрифугирования (14000 об/мин, 15 мин, 4°С) часть супернатанта отбирали для определения количества белка в лизатах по методу Бредфорд, а другую часть анализировали с помощью Вестерн-блота.
Образцы после добавления буфера для нанесения нагревали при 95°С в течение 5 мин, подвергали электрофорезу в 12% ПААГ в присутствии SDS и переносили на нитроцеллюлозную мембрану с помощью аппарата для Вестерн-блота (Bio-Rad). Затем мембрану переносили в 5% раствор обезжиренного молока в PBS, после чего трижды в течение 10 мин отмывали в растворе PBS. Далее мембрану инкубировали с первичными антителами против каспазы-2 (Santa Cruz Biotechnology), белка PARP (BD Transduction Lab) или против белка RAIDD (MBL International Corporation) в течение 12 ч при 4°С. После трех серий промывок в PBS, содержащем 0,05% Tween 20, мембрану инкубировали с вторичными антителами в течение 1 ч, отмывали 4 раза в том же растворе и проявляли с помощью ECL (Promega) на приборе Molecular Imager®ChemiDoc™ (Bio-Rad). Электрофоретический анализ лизатов клеток показал, что увеличение количества продукта расщепления каспазы-2 - фрагмента 32 кДа - начиналось после 2 часов инкубирования с цисплатином, а к 18 уже появлялось заметное количество большой каталитической субъединицы с молекулярной массой 19 кДа - р19 (фиг. 2). Максимальное количество данного фрагмента детектировалось через 24 часа после добавления индуктора (фиг. 2). Следует отметить, что под действием цисплатина происходит также расщепление белка PARP, который является субстратом эффекторных каспаз. Однако время для выделения активационного комплекса каспазы-2 было выбрано 18 часов после индукции апоптоза, поскольку в это время в клетках активируются инициаторные процессы программируемой клеточной смерти и, вероятно, образуется максимальное количество инициаторного комплекса. Следует добавить, что анализ данных цитометрии показал, что наибольший процент апоптотических клеток наблюдался спустя 48 часов после индукции (фиг. 1), однако каспаза-2 активировалась уже через 18 часов после обработки клеток цисплатином. Таким образом, можно сделать вывод, что в клетках линии Caov-4 каспаза-2 является апикальной каспазой в процессе апоптоза.
Подбор условий для иммунопреципитации (ИП)
В дальнейшем для выделения комплекса активации каспазы-2 были оптимизированы условия проведения иммунопреципитации (ИП) каспазы-2, в частности, было проведено титрование разной концентрацией антител против каспазы-2, используемых для ИП (1 μg, 5 μg, 10 μg) и подбор разных детергентов, применяемых для лизиса клеток (1% Triton Х-100, 1% NP-40, 1% Chaps).
Для ИП использовали либо полные лизаты клеток, либо фракции, полученные в результате гель-фильтрации. Для получения лизатов контрольные и стимулированные клетки обрабатывали, как описано ниже при получении лизатов для гель-фильтрации. С целью устранения неспецифической сорбции проводили стадию предочистки: к фракциям из гель-фильтрации или лизатам клеток добавляли поликлональные антитела к белку AIF (Santa Cruz Biotechnology), выдерживали 1 ч при 4°С, затем добавляли протеин-А-сефарозу (Sigma), перемешивали 3-4 ч при 4°С, после чего образец центрифугировали (5900 об/мин, 1 мин, 4°С) и отбирали супернатант.
К супернатанту добавляли необходимое количество кроличьих поликлональных антител против каспазы-2 (Santa Cruz), перемешивали 2 ч (4°С), далее добавляли протеин-А-сефарозу, предварительно проинкубированную с 10% бычьим сывороточным альбумином, и выдерживали в течение ночи (4°С) при перемешивании. Затем проводили центрифугирование образца (5900 об/мин, 1 мин, 4°С) и отбирали супернатант. Далее носитель промывали 10 мМ Tris-HCl-буфером, рН 7.4, содержащим 100 мМ NaCl, 1 мМ EDTA, 1 мМ EGTA, 1% Triton Х-100, 1% PMSF и коктейль ингибиторов протеаз, (4 раза по 1 мл) и раствором PBS (1 раз). Затем удаляли остатки буфера, после чего к протеин-А-сефарозе добавляли 2-кратный буфер для нанесения на гель.
Для оценки результатов ИП был использован гель-электрофорез и Вестерн-блот. Данный анализ продемонстрировал, что наилучшим детергентом для лизирующего буфера при проведении ИП являлся Triton X-100 (фиг. 3), а оптимальное количество антител составило 5-10 μg (фиг. 3). Оценку неспецифического связывания белков с протеин-А сефарозой контролировали с помощью образца, не содержащего антител (протеин-А контроль). Анализ данного образца показал, что неспецифическая сорбция в нем не наблюдалась. Таким образом, были подобраны оптимальные условия для проведения ИП с целью получения образцов для дальнейшего анализа.
Следующим этапом в осуществлении подбора условий для успешной ИП клеток, к которым был добавлен ДНК-повреждающий агент цисплатин, стало добавление каспазного ингибитора z-VAD-fmk (carbobenzoxy-valyl-alanyl-aspartyl-[O-methyl]fluoromethylketone) с целью стабилизации инициаторного комплекса и увеличения количества преципитированной каспазы-2. Однако ингибитор каспаз z-VAD-fmk, который был добавлен с целью стабилизации активационного комплекса каспазы-2, не привел к желаемому увеличению количества каспазы-2 в иммунопреципитатах, таким образом, его дальнейшее использование при проведении ИП не являлось целесообразным.
В литературе было показано, что в клетках с нокаутом по PIDD или RAIDD индукция повреждения ДНК не влияла на клеточную локализацию, ядерную транслокацию или активацию каспазы-2. Таким образом, было сделано предположение о существовании альтернативного механизма активации каспазы-2, который не зависит от комплекса PIDDosome. Для проверки происходит ли активация каспазы-2 в ответ на обработку цисплатином по PIDDosome-зависимому пути, образцы ИП также были проанализированы на предмет наличия в них компонентов данного комплекса. В иммунопреципитатах клеток Caov-4, обработанных цисплатином в течение 18 часов, были детектированы фрагменты протеолиза каспазы-2 (фиг. 4). Однако ни белок PIDD, ни белок RAIDD не были обнаружены в образцах ИП, что полностью соответствует ряду литературных данных, согласно которым комплекс PIDDosome может быть выделен лишь в условиях повышенной экспрессии его компонентов in vitro [A. Tinel and J. Tschopp, "The PIDDosome, a protein complex implicated in activation of caspase-2 in response to genotoxic stress.," Science, vol. 304, no. 5672, pp. 843-6, May 2004]. Таким образом, было сделано предположение о существовании нового, неописанного в литературе высокомолекулярного комплекса, в составе которого происходит активация каспазы-2 в ответ на повреждения ДНК.
Гель-фильтрация клеточного лизата и анализ высокомолекулярной фракции
Для исследования макромолекулярного комплекса активации каспазы-2 был разработан подход препаративного выделения высокомолекулярной фракции из контрольных и обработанных цисплатином клеток Caov-4 с помощью метода гель-фильтрации в комбинации с ИП.
Контрольные клетки (не менее ~5×106) трижды отмывали от среды в холодном растворе PBS, снимали с чашек и инкубировали на льду в течение 30 мин в 2 мл лизирующего буфера (20 мМ Tris-HCl (рН 7.4), 137 мМ NaCl, 2 мМ EDTA, 10% глицерин, 1% Triton Х-100, 1% PMSF, коктейль ингибиторов протеаз Complete). Аналогично клетки (не менее ~5×106), обработанные 70 мкМ цисплатином, снимали с чашек, трижды отмывали в холодном растворе PBS, а затем 30 мин лизировали на льду в 2 мл того же лизирующего буфера.
Лизаты контрольных и обработанных клеток центрифугировали (13000 об/мин, 25 мин, 4°С), отделяли супернатант, из которого отбирали контрольную пробу для определения количества белка по методу Бредфорд, анализа с помощью Вестерн-блота и тестирования активности каспазы-2. Оставшиеся лизаты использовали для проведения гель-фильтрации.
Гель-фильтрацию проводили с использованием хроматографа низкого давления BioLogic LP System (Bio-Rad) или хроматографа  systems (GE Healthcare), колонки 16/500 или 10/300 GL Amersham с носителем Superdex 200 (GE Healthcare) или Superose 6 (GE Healthcare) в 20 мМ Hepes-KOH-буфере, pH 7.5, содержащем 80 мМ NaCl, 10 мМ KCl, 1 мМ MgCl2, 1 мМ EDTA, 1 мМ EGTA, 1 мМ DTT, 1 мМ PMSF и 1 мМ NaN3, при скорости потока 1 мл/мин, при температуре +4 +6°С. Детектирование осуществляли по величине оптического поглощения элюатов при длине волны 280 нм. Для калибровки колонки использовали набор маркеров Gel Filtration Standard (Bio-Rad).
systems (GE Healthcare), колонки 16/500 или 10/300 GL Amersham с носителем Superdex 200 (GE Healthcare) или Superose 6 (GE Healthcare) в 20 мМ Hepes-KOH-буфере, pH 7.5, содержащем 80 мМ NaCl, 10 мМ KCl, 1 мМ MgCl2, 1 мМ EDTA, 1 мМ EGTA, 1 мМ DTT, 1 мМ PMSF и 1 мМ NaN3, при скорости потока 1 мл/мин, при температуре +4 +6°С. Детектирование осуществляли по величине оптического поглощения элюатов при длине волны 280 нм. Для калибровки колонки использовали набор маркеров Gel Filtration Standard (Bio-Rad).
Анализ фракций после гель-фильтрации с помощью Вестерн-блота продемонстрировал наличие каспазы-2 в высокомолекулярных фракциях (500 кДа и выше) только в образцах из лизата клеток, обработанных цисплатиной (фиг. 5). В контрольном образце каспаза-2 не детектировалась во фракциях, соответствующих молекулярной массе 500 кДа и выше (фиг. 6). При этом каспаза-3 и цитохром С детектировались только в низкомолекулярной фракции как в контрольных, так и в обработанных клетках (фиг. 5 и 6). Более того, в высокомолекулярном комплексе были обнаружены продукты аутолитического расщепления каспазы-2, что свидетельствует о функциональной активности фермента в высокомолекулярной фракции. На основании проведенных экспериментов сделан вывод о том, что под действием цисплатина в клетках происходит формирование высокомолекулярного белкового комплекса, содержащего каспазу-2. Введение этапа гель-фильтрации было необходимо, поскольку иммунопреципитация каспазы-2 из лизатов клеток, стимулированных цисплатином, приводила к копреципитации антителами, помимо активированной каспазы-2, также и проформы этого фермента. Вероятно, в ответ на повреждение ДНК в клетках образуется относительно невысокое количество комплекса активации каспазы-2, которого хватает для запуска программы гибели. Таким образом, использование гель-фильтрации для устранения конкурирующих мономерных форм каспазы-2, преобладающих в полных лизатах клеток, способствовало выделению высокомолекулярной платформы активации фермента.
Для дальнейшей очистки и характеристики исследуемого макромолекулярного комплекса была проведена иммунопреципитация антителами против каспазы-2 с последующим анализом с помощью Вестерн-блота (фиг. 7). В частности, большой интерес представлял анализ наличия компонентов платформы PIDDosome в данном комплексе. Результаты показали, что с помощью разработанного подхода каспаза-2 была успешно выделена в составе высокомолекулярного комплекса. Интересно, что ключевой компонент комплекса PIDDosome - адапторный белок RAIDD - не был детектирован не только в высокомолекулярной фракции, но и ни в одном из образцов после ИП (фиг. 10). Следовательно, активация каспазы-2 в ответ на повреждения ДНК в данной модели происходила по PIDDosome-независимому пути. Таким образом, был сделан вывод о том, что в клетках карциномы яичника в ответ на повреждения ДНК формируется новая альтернативная платформа, в составе которой каспаза-2 проходит процесс активации.
Аналогичным образом было проанализировано формирование высокомолекулярного комплекса активации каспазы-2 в клетках линии карциномы шейки матки HeLa. Анализ лизата клеток после обработки цисплатином в концентрации 70 мкМ с помощью гель-фильтрации с последующим Вестерн-блотом показал, что наличие каспазы-2 в высокомолекулярных фракциях (500 кДа и выше) только в образцах из лизата клеток, обработанных цисплатиной. Данный результат свидетельствовал о формировании высокомолекулярного комплекса активации каспазы-2 в этих клетках в ответ на повреждение ДНК.
Оптимизация условий для увеличения количества комплекса активации
Для увеличения выхода высокомолекулярного комплекса активации каспазы-2 была осуществлена оптимизация условий гель-фильтрации. Так, использование сорбента Superose 6 (GE Healthcare), обладающего большей разрешающей способностью, чем ранее использованный носитель Superdex 200, значительно улучшило разделение каспазы-2, содержащейся в высокомолекулярных фракциях, и ее мономеров (фиг. 7). С другой стороны, применение этого сорбента позволило с большей эффективностью отделить комплекс от остальных высокомолекулярных белковых компонентов, содержащихся в клетках.
Кроме того, для увеличения выхода и стабилизации платформы активации каспазы-2 в лизат клеток был добавлен пан-каспазный ингибитор zVAD-fmk, который должен был блокировать выход каспазы-2 из состава комплекса. Поскольку белки, участвующие в активации каспазы-2, вероятно, не образуют ковалентных связей в исследуемом комплексе, то в растворе такой комплекс постепенно разрушается, что затрудняет процедуру его выделения и идентификации его компонентов. Использование этого ингибитора позволило значительно повысить содержание каспазы-2 в высокомолекулярной фракции, а, следовательно, увеличивало время жизни исследуемого комплекса (фиг. 8).
На следующем этапе исследований было оценено количество комплекса в зависимости от времени инкубации клеток с ДНК-повреждающим агентом. Данное тестирование было необходимо провести вследствие того, что оценка накопления процессированной формы каспазы-2, проведенная ранее, возможно, не отражала в полной мере процесс формирования и распада активационного комплекса. В ходе эксперимента клетки обрабатывали цисплатином в течение разных промежутков времени (6, 12, и 18 часов), после чего клетки лизировали и проводили анализ распределения каспазы-2 между высокомолекулярной и низкомолекулярной фракциями с помощью гель-фильтрации и Вестерн-блота. Такой анализ продемонстрировал, что максимальное количество комплекса накапливалось к 18 часам инкубации (фиг. 9).
Также, с целью получения максимального количества комплекса из высокомолекулярных фракций и дополнительного снижения уровня белков, неспецифически связывающихся с протеин А-сефарозой, была произведена дополнительная оптимизация протокола иммунопреципитации. Для устранения неспецифической сорбции протеин А-сефароза была заменена на магнитные частицы с протеин A/G (Thermo Scientific Pierce Protein A/G Magnetic Beads) (фиг. 10). Кроме того, для выделения каспазы-2, содержащейся в высоко- и низкомолекулярной фракциях, был протестирован подход, основанный на использовании модифицированного субстрата biotin-VAD-fmk и стрептавидин-агарозы. Электрофоретический анализ этих образцов показал, что при использовании магнитных частиц каспаза-2 успешно иммунопреципитировалась из общего лизата, а также из трех фракций (низкомолекулярной (10-43 кДа), среднемолекулярной (70-140 кДа) и высокомолекулярной (500 кДа и выше)), при этом неспецифическая сорбция на данные частицы была успешно устранена (фиг. 11).
Следует отметить, что при ИП с использованием магнитных частиц обогащение образцов каспазой-2 происходило заметно лучше, чем при использовании протеин А-сефарозы. Что касается использования biotin-VAD-fmk и стрептавидин-агарозы для выделения инициаторного комплекса каспазы-2, то данный подход не позволил получить заметного количества белка.
Анализ функциональной активности выделенного комплекса
Для оценки активности каспазы-2 были проанализированы фракции, содержащие низкомолекулярные компоненты (10-55 кДа) - LMW (low molecular weight), белки и комплексы с промежуточной молекулярной массой (70-140 кДа) - MMW (medium molecular weight) и высокомолекулярные комплексы (500 кДа и выше) - HMW (high molecular weight), а также общий лизат из индуцированных и контрольных клеток. В данных фракциях была измерена ферментативная активность каспазы-2 с помощью флуоресцентного производного пентапептида Ac-VDVAD-AMC.
Для измерения активности использовали либо клеточную суспензию объемом 50 мкл, либо образцы после ИП (1/5 от всего образца), либо фракции, полученные при гель-фильтрации объемом 50 мкл. В образцы добавляли по 25 мкл буфера (20 мМ Hepes-KOH рН 7.4, 10% сахароза, 0.1% Chaps, 5 мМ DTT, 50 мкМ субстрат Ac-VDVAD-AMC (PeptaNova). Флуоресценцию измеряли на приборе Varioskan Flash (Thermo Scientific,), длина волны поглощения - 380 нм, длина волны испускания - 460 нм. Измерения проводили в течение 20 мин с интервалом 1 мин при 37°С. Полученные данные относительно количества расщепленного субстрата и генерации флуоресцентного сигнала были обработаны в программе Microsoft Excel, и на их основании были построены графики активности каспазы-2.
Анализ пептидазной активности каспазы-2 показал, что уровни активности в низкомолекулярной и высокомолекулярной фракциях индуцированных клеток практически совпадали и превосходили уровень активности в среднемолекулярных фракциях почти в 3 раза (фиг. 12а). Таким образом, белки, обладающие ферментативной активностью по отношению к субстрату каспазы-2, детектировались не только в низкомолекулярных фракциях, где должны находиться мономеры каспазы-2, но и в высокомолекулярных, что подтверждало активацию каспазы-2 в составе высокомолекулярного комплекса. В отличие от обработанных цисплатином клеток в образцах, полученных из контрольных клеток, заметная энзиматическая активность не была детектирована.
Для подтверждения функциональной активности высокомолекулярного комплекса каспазы-2 из образцов ИП после гель-фильтрации были отобраны аликвоты для измерения каталитической активности каспазы-2 с помощью того же флуоресцентного субстрата. Сравнение пептидазной активности образцов после ИП из общего лизата, а также низко- и высокомолекулярных фракций показало, что в клетках, индуцированных цисплатином, наибольшей энзиматической активностью обладает HMW-фракция (фиг. 12б). В контрольных клетках высокомолекулярный комплекс не был детектирован в гель-фильтрационном профиле каспазы-2, и, соответственно, ферментативной активности в образцах после ИП также не наблюдалось. Исследование каталитической активности выделенного комплекса в образцах ИП подтвердило наличие в его составе активной каспазы-2. Полученные результаты также свидетельствовали о том, что в случае повреждения ДНК в клетках происходит сборка высокомолекулярного комплекса, в составе которого осуществляется активация каспазы-2.
Тест на каспазную активность продемонстрировал эффективность разработанного метода очистки, позволившего выделить высокомолекулярный комплекс активации каспазы-2. Отсутствие существенного уровня активности в образцах после ИП из полного лизата обработанных клеток или из низкомолекулярной фракции, вероятно, объяснялось тем, что в этом случае антитела против каспазы-2 взаимодействовали в первую очередь с неактивными мономерными формами каспазы-2. Необходимо отметить, что активация каспазы-2 зависит в первую очередь от процесса димеризации, при этом даже димеризованная проформа способна проявлять частичную ферментативную активность [В.С. Baliga, S.Н. Read, and S. Kumar, "The biochemical mechanism of caspase-2 activation", Cell Death Differ., vol. 11, no. 11, pp. 1234-41, Nov. 2004]. Мономерные формы каспазы-2, в том числе и процессированный фрагмент р32, не проявляют ферментативной активности, если они не входят в состав гомо- или гетеродимера. Содержащаяся в HMW-фракциях каспаза-2, по-видимому, находилась преимущественно в димеризованной форме и обладала высокой энзиматической активностью, о чем свидетельствовал тот факт, что каспаза-2-подобная активность в этой фракции была практически равна таковой в LMW-фракции при весьма существенном различии в количестве самой каспазы-2 в пользу последней.
При проведении ИП из HMW-фракций происходило обогащение образца, по-видимому, именно димеризованной каспазой-2, обладающей повышенной протеолитической активностью. В LMW-фракциях, равно как и в полном лизате клеток, содержание мономерных неактивных форм, в том числе и процессированных, превосходило содержание димеризованных в несколько раз, что подтверждалось невысокой протеолитической активностью каспазы-2 в этих фракциях. Таким образом, по-видимому, при проведении ИП из LMW-фракций или полного лизата индуцированных клеток происходило обогащение образцов мономерными неактивными формами в силу их более высокого содержания в исследуемых пробах и большей стерической доступности.
Таким образом, разработан способ выделения макромолекулярного комплекса активации каспазы-2, и впервые напрямую продемонстрировано наличие в клетках карциномы яичника новой альтернативной платформы активации каспазы-2, которая образуется в ответ на повреждения ДНК. Проведенные исследования открывают новые перспективы в области изучения механизмов программируемой клеточной гибели, а также позволяют применить разработанный подход для изучения других высокомолекулярных сигнальных комплексов.
Активация каспазы-2 не зависит от формирования комплекса PIDDosome
Для дополнительного подтверждения PIDDosome-независимой активации каспазы-2 были проанализированы процессинг каспазы-2 и уровень гибели клеток Caov-4, обработанных siRNA (small interfering RNA) против белка RAIDD.
Для трансфекции были взяты клетки Caov-4, достигшие 70% конфлюентности (0.5×106). Клетки помещали в среду OPTI-MEM без антибиотика (Gibco). Для введения siRNA против белка RAIDD (Dharmacon) или контрольной siRNA (Dharmacon) в количестве 100 пмоль использовали набор RNAiMAX (Invitrogen). Через 6 ч меняли среду OPTI-MEM на DMEM без антибиотиков и добавляли цисплатин через 20 ч инкубации с siRNA.
Было показано, что уровень апоптотической смерти клеток с пониженной экспрессией этого белка не отличался от контрольного (фиг. 13). Существенное снижение уровня RAIDD в клетках было продемонстрировано с помощью Вестерн-блота (фиг. 12). Показано, что несмотря на практически полное отсутствие RAIDD количество проформы каспазы-2 заметно уменьшается при обработке цисплатином клеток Caov-4, что свидетельствует о протекании аутокаталитического протеолиза и активации этой каспазы (фиг. 13). Кроме того, в клетках также наблюдается расщепление белка PARP, что подтверждает факт активации эффекторной каспазы-3 (фиг. 13). Таким образом, ингибирование синтеза RAIDD не приводило к появлению устойчивости клеток Caov-4 к повреждениям ДНК, что свидетельствует об отсутствии существенного вклада этого белка в механизм запуска апоптоза. Соответственно, в ходе работы было доказано, что в выбранной клеточной модели активация каспазы-2 и последующая апоптотическая гибель не зависят от описанного в литературе комплекса PIDDosome и происходят в составе альтернативной платформы.
Использование генетически модифицированных вариантов каспазы-2 для выделения комплекса активации
Для увеличения эффективности очистки комплекса активации каспазы-2 были созданы уникальные генетические векторы, содержащие ген каспазы-2 и дополнительную аминокислотную последовательность для аффинной хроматографии, так называемый «тег». Данный подход позволяет увеличить выход выделяемого белка и снизить загрязнение образца неспецифическими белками.
На первом этапе был осуществлен выбор последовательностей, обеспечивающих эффективную очистку через аффинную хроматографию - Strep-tag, Twin-Strep-tag, GST-tag, Flag-tag (Табл. 4). Были синтезированы праймеры для встраивания гена каспазы-2 в вектор pESG-IBA25, - -IBA103, -IBA105 (Iba-lifesciences, Германия), сконструированный для высокопродуктивной экспрессии генов в клетках млекопитающих и содержащий промотор цитомегаловируса человека. Конструирование векторов проводилось таким образом, чтобы получить модифицированную форму каспазы-2, содержащую разные «теги» с N- или С-конца целевого белка (Табл. 5).
Для встраивания последовательности гена каспазы-2 в векторы pESG-IBA25, pESG-IBA103 и pESG-IBA105 (Iba-lifesciences). На основе плазмиды pcDNA3-caspase-2, содержащей последовательность полноразмерного гена каспазы-2 (аминокислотная последовательность представлена в Табл.3) с помощью ГШР и с использованием праймеров
AGCGCGTCTCCAATGATGGCGGCGCCGAGCGC прямой
AGCGCGTCTCCTCCCTGTGGGAGGGTGTCCTGG обратный
был получен ПЦР-фрагмент, содержащий ген, кодирующий последовательность каспазы-2. ПЦР-фрагмент выделили с помощью набора Gene JET Gel Extraction (Fermentas). Далее проводили клонирование с помощью набора StarGate Direct Transfer Cloning (Iba-lifesciences) в указанные выше векторы. Лигазной смесью трансформировали клетки Е. Coli XL-1 Blue по стандартной методике. Правильную последовательность полученной плазмидной ДНК подтверждали секвенированием.
Для введения полученных векторов клетки линии Caov-4 или HeLa культивировались в стандарных условиях, и после достижения 70% конфлюэнтности трансфецировались плазмидой, содержащей последовательность гена каспазы-2 и последовательность для аффинной хроматографии (Табл. 4) в количестве от 2 до 8 мкг на 2 мл среды. Трансфекция проводилась с помощью Lipofectamine LTX Reagent и PLUS Reagent (Invitrogen) в среде OPTI-MEM (Gibco) согласно рекомендациям производителя. Через 4-12 часов среда сменялась на DMEM, и индукция повреждений ДНК осуществлялась через 24 часа после трансфекции.
Также проводился подбор условий для экспрессии полученных конструктов в клетках линии Caov-4 с помощью оптимизации условий трансфекции (фиг. 14). Отбор наиболее эффективно экспрессирующихся конструктов показал, что введение конструктов с Twin-Strep-tag на N- или С-конце позволяет получить достаточный уровень синтеза модифицированной каспазы-2, но при этом не вызывает существенной активации каспазы-2 и нежелательной гибели трасфицированных клеток Caov-4, о чем говорило невысокое содержание энзиматически активного фрагмента р19 в лизатах клеток (фиг. 14). Необходимо отметить, что использование вектора IBA-25 (Табл. 5), содержащего каспазу-2 и GST-tag, не приводило к сколь-нибудь заметному синтезу этого варианта каспазы-2 в клетках Caov-4 (фиг. 14 (а)), поэтому для дальнейшей работы выбрали векторы с Twin-Strep-tag на N- или С-конце (векторы IBA-103 и IBA-105), а также вектор с Flag-tag (pcDNA3/hCasp2-Flag).
Далее выбирали время инкубации клеток после трансфекции для оптимизации условий экспрессии. Клетки линий Caov-4 и HeLa трасфицировались вектором IBA-103 в концентрации 1 мкг/мл среды и далее инкубировались в течение 24, 48 или 72 часов. Уровень экспрессии генетического конструкта оценивали с помощью Вестерн-блот анализа (фиг. 14 (а)), который показал, что инкубация обеих клеточных линий в течение 24 часов приводит к максивальному уровню экспрессии конструкта каспазы-2-Twin-Strep-tag.
На следующем этапе была проведена разработка и оптимизация метода выделения модифицированных вариантов каспазы-2 из экстрактов контрольных и обработанных цисплатином клеток. Выделение различных вариантов модифицированной каспазы-2 показало, что использование Twin-Strep-tag и Flag-tag приводит к эффективному связыванию каспазы-2 со специфическим аффинным сорбентом, однако при элюции теггированные формы белка остаются в связанном состоянии (фиг. 15). Необходимо отметить, что при выделении каспазы-2 через «тег» происходило значительное обогащение образца целевым белком, и его практически не оставалось в лизате после выделения.
Для оптимизации выделения также тестировались различные варианты аффинных сорбентов, и был выбран сорбент Strep-Tactin Superflow high capacity (Iba-lifesciences, Германия), как наиболее эффективно связывающий каспазу-2 с минимальными показателями неспецифической сорбции. Для элюции модифицированной каспазы-2-Twin-Strep-tag были использованы биотин и дестибиотин в концентрации 10 и 20 мМ. Анализ показал, что использование 10 мМ биотина позволяет эффективно элюировать целевой белок без существенных загрязнений (фиг. 16).
С помощью разработанного метода каспаза-2 была выделена из контрольных и обработанных цисплатином клеток Caov-4. Далее проводили масс-спектрометрический анализ этих образцов, который подтвердил эффективность разработанного метода, поскольку помимо каспазы-2 было детектировано несколько десятков других белков (преимущественно белки цитоскелета, факторы инициации и элонгации трансляции и рибосомные белки), которые часто обнаруживаются при масс-спектрометрическом анализе в заметных количествах. По сравнению с этими образцами иммуноприципитаты из полного лизата содержали более 1000 неспецифических белков. Таким образом, использование полученных конструктов позволило существенно повысить эффективность очистки каспазы-2.

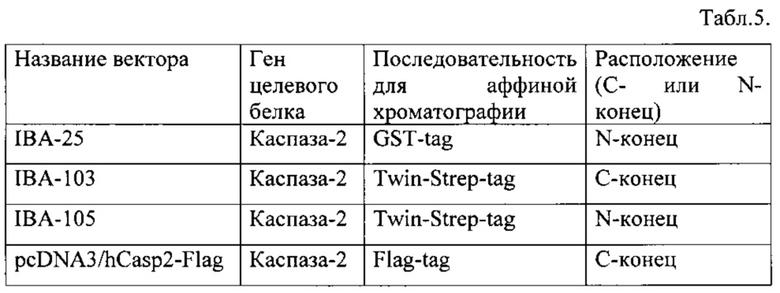
Создание дефицитной по гену каспазы-2 линии клеток карциномы яичника Caov-4 с помощью методологии Crispr/Cas9.
Для более эффективного выделения каспазы-2 и ассоциированных с ней белков была создана стабильная линия клеток Caov-4, отличающаяся практически полным отсутствием эндогенной каспазы-2. Экспрессия конструктов каспазы-2 в такой линии позволяет увеличить выход выделенного через «тег» целевого белка и снизить нежелательную гибель клеток, которая запускается при оверэкспрессии гена каспазы-2.
Для получения стабильной дефицитной по каспазе-2 линии клетки Caov-4 трансфицировались вектором caspase-2 CRISPR/Cas9 KO Plasmid (кат номер sc-401079, Santa Cruz, США), содержащим специфичные к каспазе-2 последовательности направляющих РНК и последовательность зеленого флуоресцентного белка (GFP - green fluorescent protein) в качестве маркера траснфекции. Далее проводили сортировку клеток для отбора флюоресцирующих клонов, содержащих вектор caspase-2 CRISPR/Cas9 KO Plasmid. Далее для получения стабильных линий полученные клоны культивировались в стандартной питательной среде. Анализ с помощью Вестерн-блота показал, что в полученных клонах каспаза-2 не синтезируется и не детектируется, соответственно (фиг. 17). Дальнейшие эксперименты показали, модифицированная форма каспазы-2 успешно синтезировалась после трансфекции соответствующим вектором (фиг. 14) таких стабильных линий CRISPR/Cas9, а также линий Caov-4 sh-RNA-Kacna3a-2.
| название | год | авторы | номер документа |
|---|---|---|---|
| АКТИВАЦИЯ ПРОКАСПАЗЫ-3 С ПОМОЩЬЮ КОМБИНИРОВАННОЙ ТЕРАПИИ | 2013 |
|
RU2659936C2 |
| Способ получения каспазы-3 | 2022 |
|
RU2792795C1 |
| СРЕДСТВО, СТИМУЛИРУЮЩЕЕ АПОПТОЗ КЛЕТОК ЛЕЙКЕМИИ ЧЕЛОВЕКА | 2007 |
|
RU2360692C1 |
| ПРОКАСПАЗНАЯ КОМБИНИРОВАННАЯ ТЕРАПИЯ ПРИ ГЛИОБЛАСТОМЕ | 2013 |
|
RU2636234C2 |
| КОМПОЗИЦИЯ И СПОСОБ ЛЕЧЕНИЯ ОПУХОЛЕЙ | 2007 |
|
RU2500815C2 |
| ТЕРАПЕВТИЧЕСКИЕ АГЕНТЫ, СОДЕРЖАЩИЕ ПРОАПОПТОЗНЫЕ БЕЛКИ | 2002 |
|
RU2319709C2 |
| СПОСОБЫ И МАТЕРИАЛЫ ДЛЯ ОБНАРУЖЕНИЯ АПОПТОЗА | 2007 |
|
RU2450273C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ PAC-1 | 2016 |
|
RU2720509C2 |
| ИНДУКЦИЯ АПОПТОЗА В ОПУХОЛЕВЫХ КЛЕТКАХ, ЭКСПРЕССИРУЮЩИХ TOLL-ПОДОБНЫЙ РЕЦЕПТОР | 2005 |
|
RU2401661C9 |
| АНАЛИЗЫ И СПОСОБЫ ПРИМЕНЕНИЯ БИОМАРКЕРОВ | 2006 |
|
RU2409817C2 |
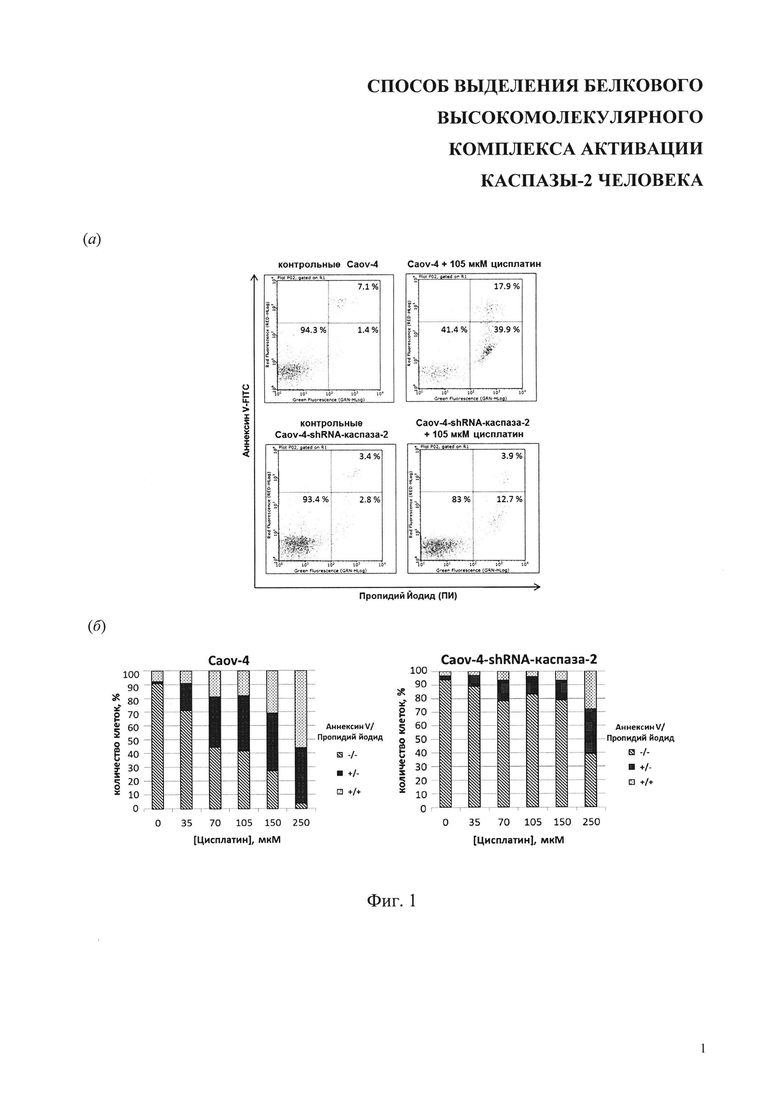
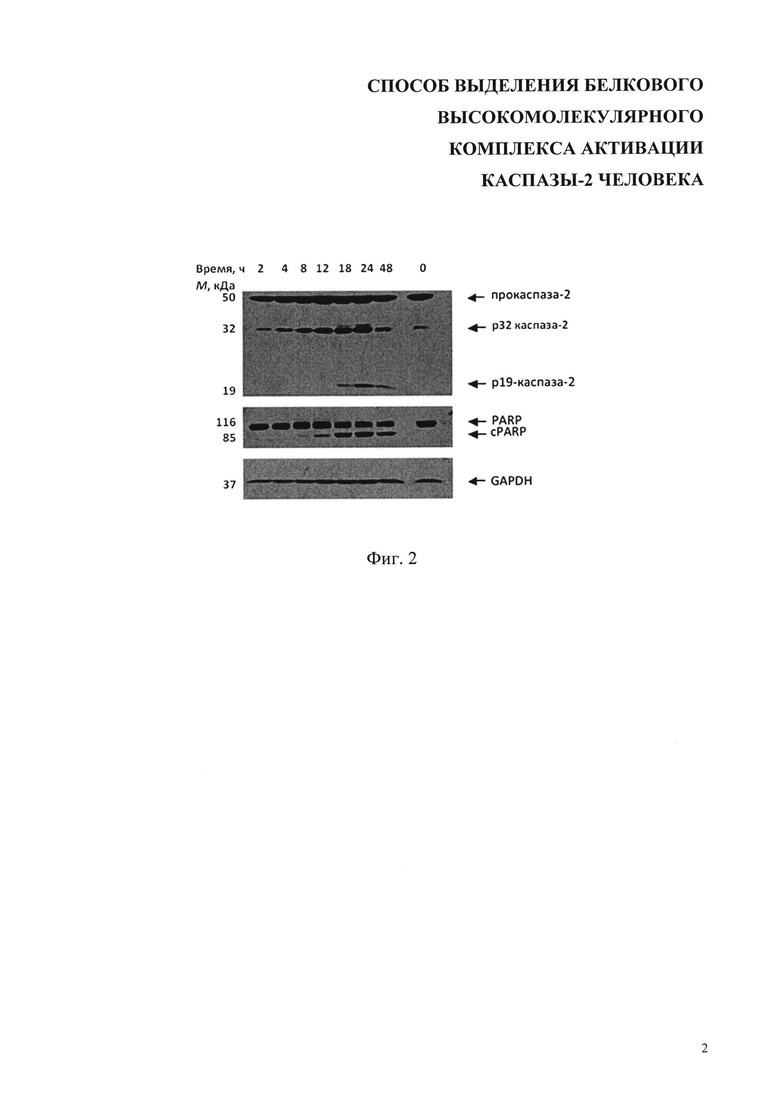
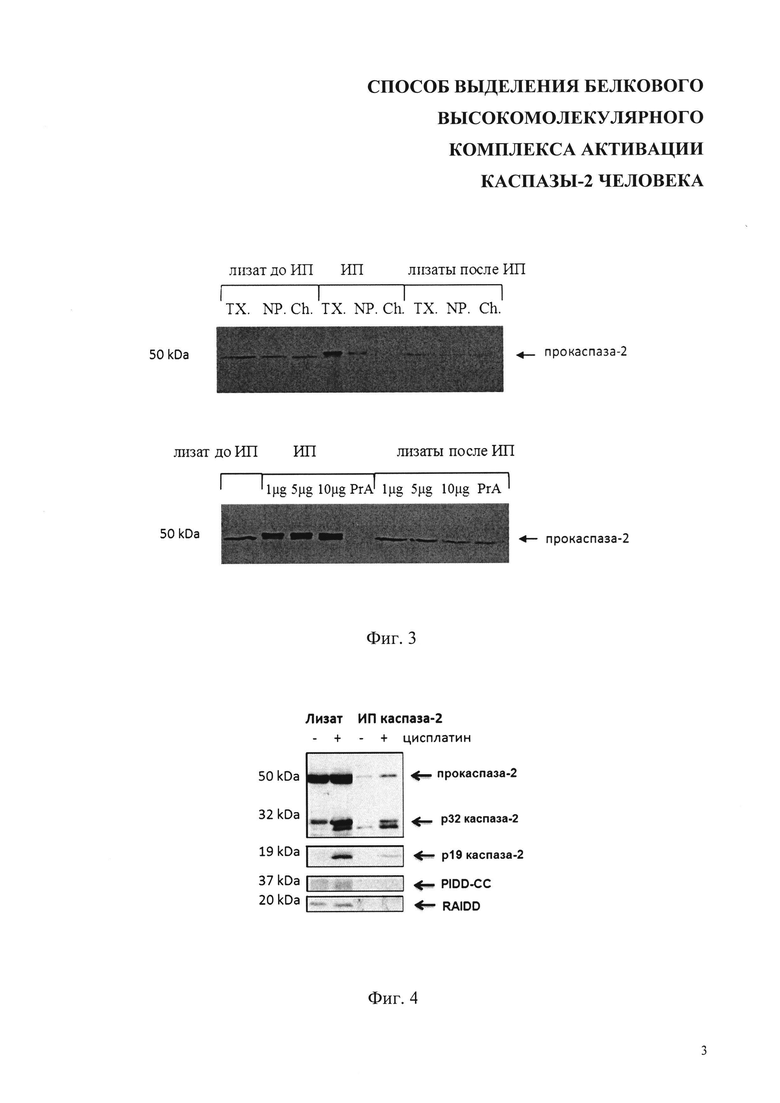
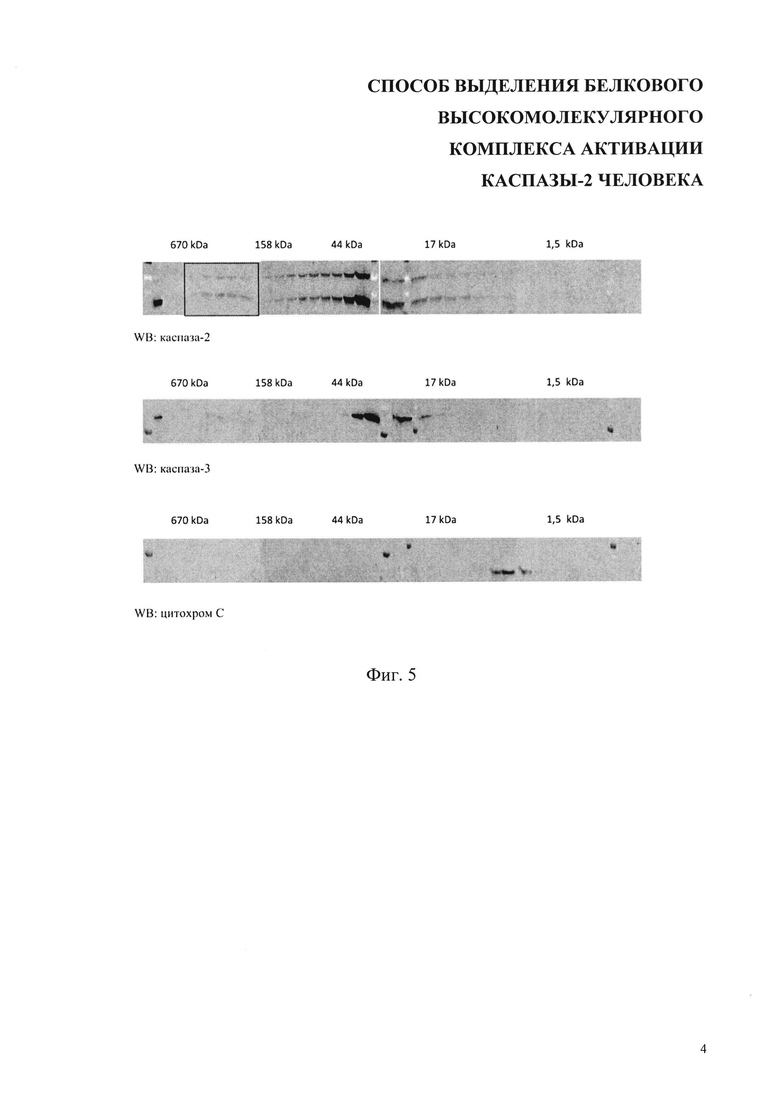
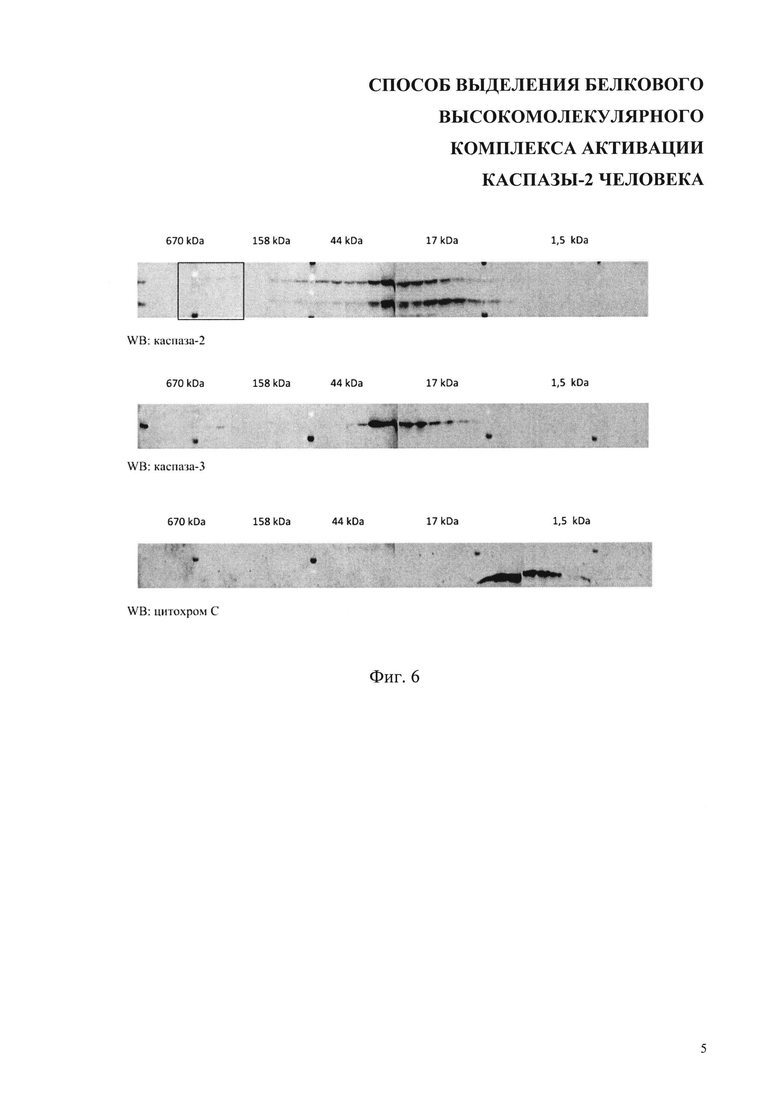

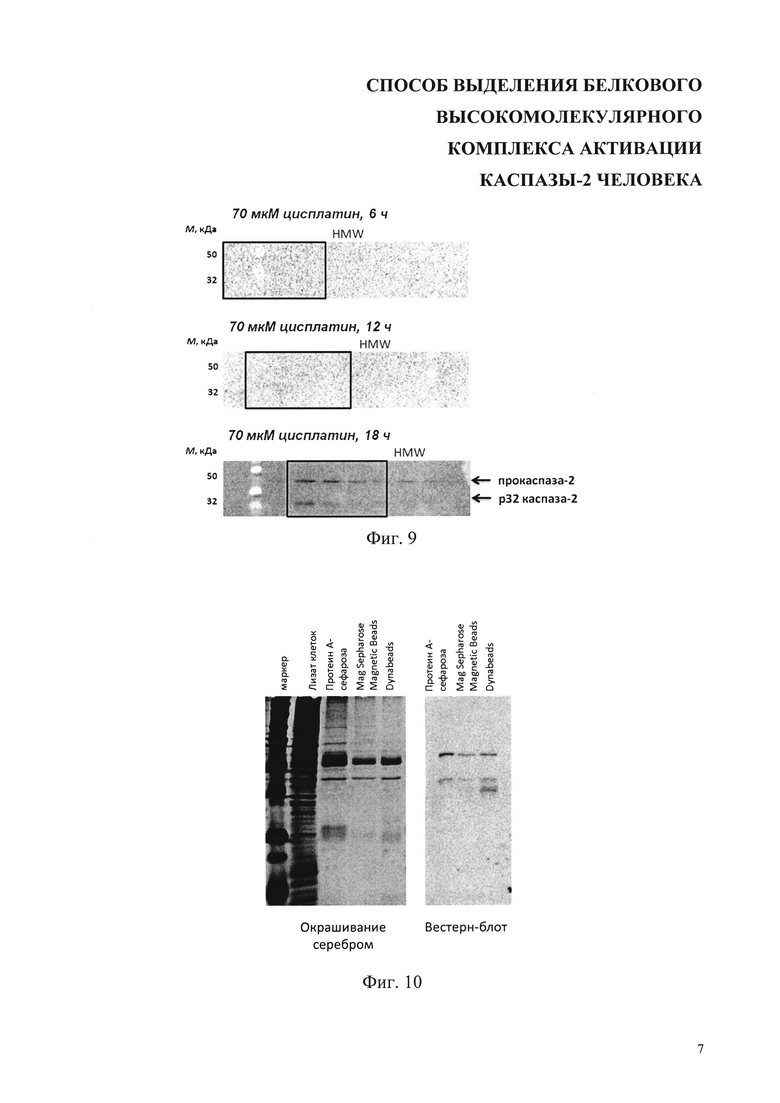
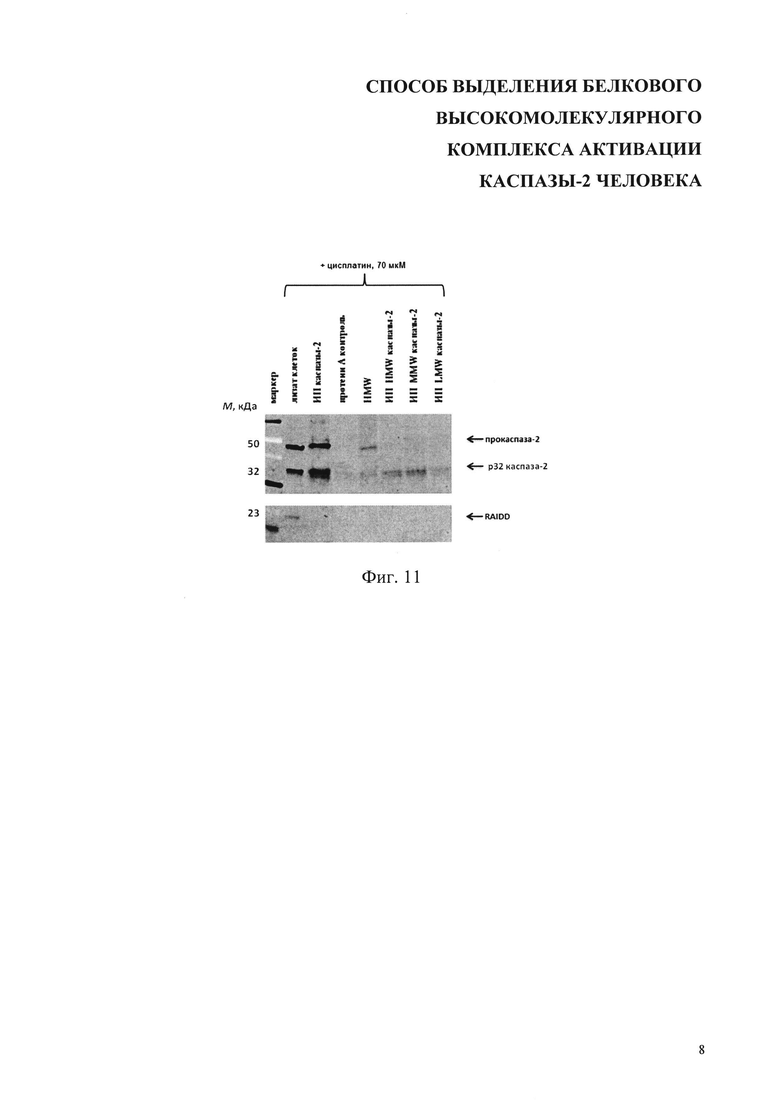
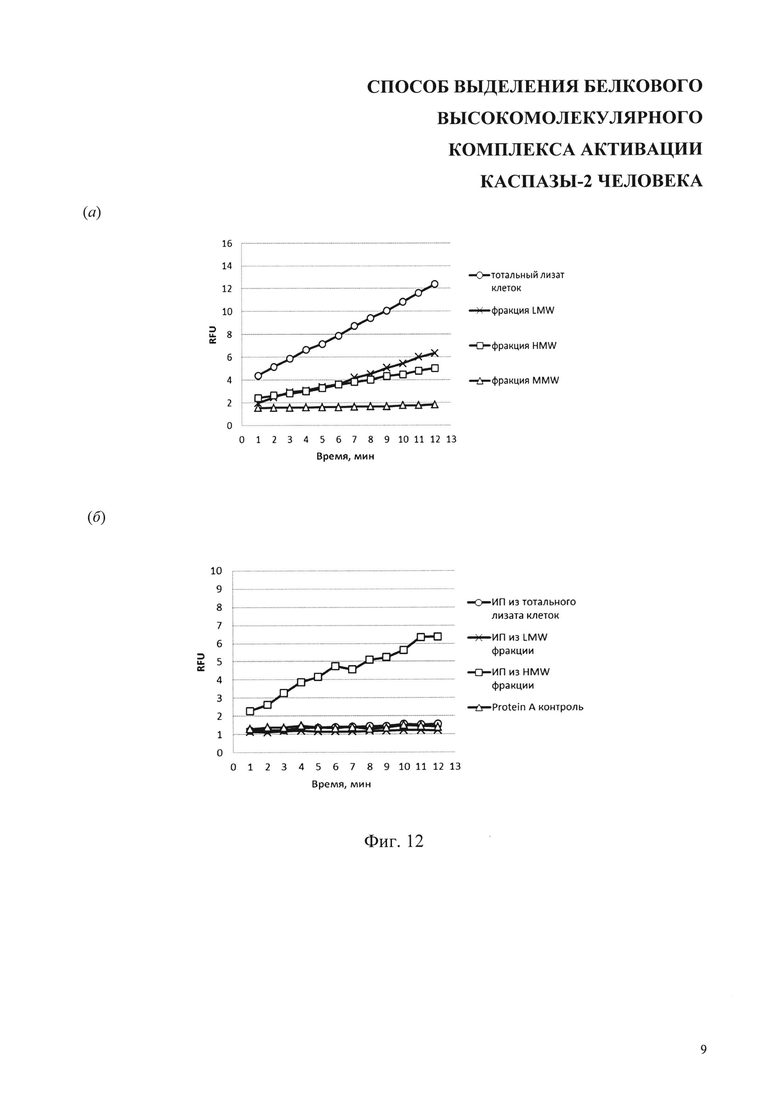

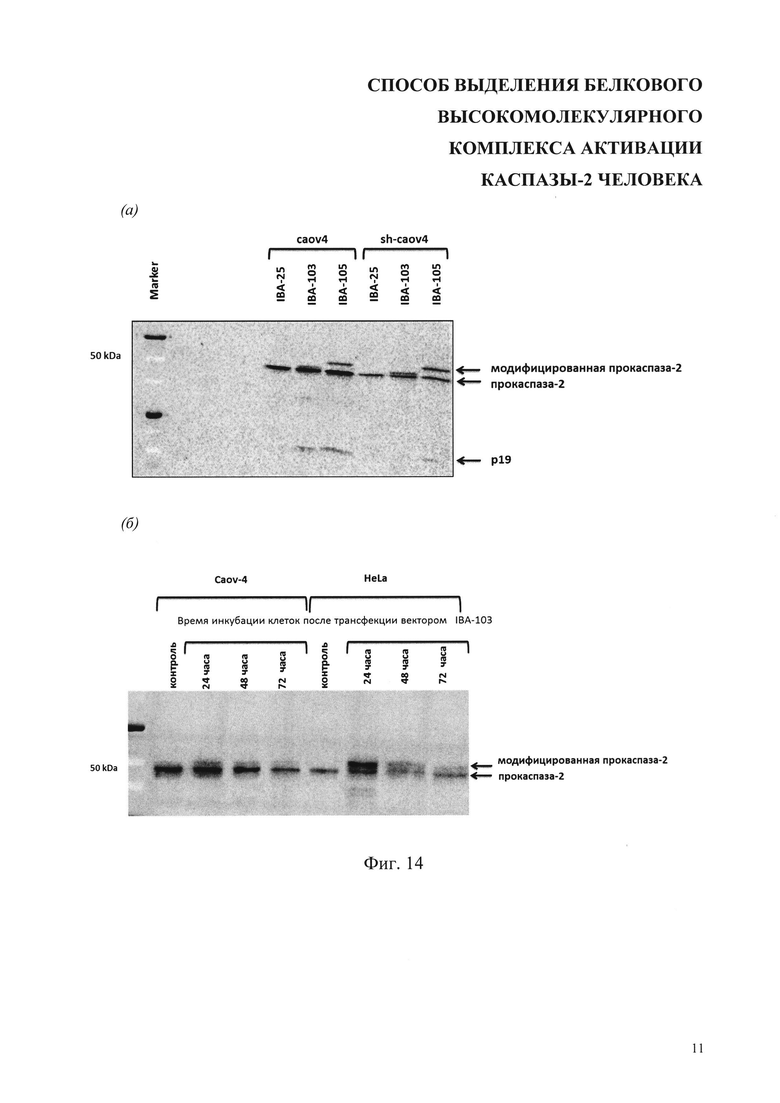
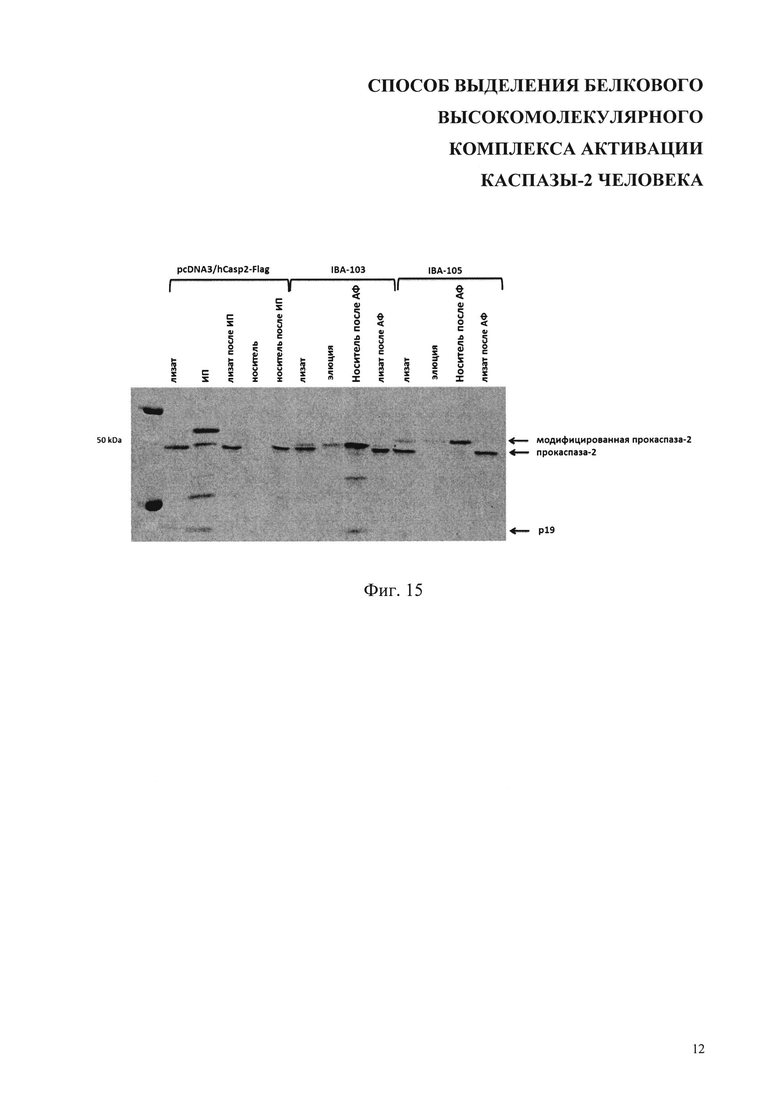

Предложенная группа изобретений относится к области медицины. Предложены способы выделения белкового высокомолекулярного комплекса активации каспазы-2 человека, формирующегося в раковых клетках в ответ на обработку ДНК-повреждающим химиотерапевтическим препаратом. Раковые клетки инкубируют в присутствии ДНК-повреждающего химиотерапевтического препарата до получения максимального уровня активации каспазы-2. Осуществляют сбор клеток с последующим добавлением лизирующего буфера, отделение клеточного дебриса от лизата, содержащего высокомолекулярный комплекс активации каспазы-2, разделение гель-фильтрацией полученного лизата клеток с обеспечением отделения фракций, содержащих высокомолекулярные формы каспазы-2, от фракций, содержащих ее мономерные формы и выделение из фракции, содержащей высокомолекулярные формы каспазы-2, белкового высокомолекулярного комплекса активации каспазы-2. Предложенная группа изобретений обеспечивает эффективное выделение комплекса активации каспазы-2 человека. 2 н. и 34 з.п. ф-лы, 5 табл., 1 пр., 17 ил.
1. Способ выделения белкового высокомолекулярного комплекса активации каспазы-2 человека, формирующегося в раковых клетках в ответ на обработку ДНК-повреждающим химиотерапевтическим препаратом, характеризующийся тем, что включает:
- получение раковых клеток в количестве не менее чем 5 млн клеток;
- введение в раковые клетки ДНК-повреждающего химиотерапевтического препарата в концентрации, обеспечивающей запуск апоптотической гибели раковых клеток с достижением максимального уровня апоптотической гибели клеток при минимальном уровне некротической гибели клеток, которые определяют с помощью цитометрии;
- инкубация раковых клеток в присутствии ДНК-повреждающего химиотерапевтического препарата до получения максимального уровня активации каспазы-2, который определяют предварительно до начала инкубации с помощью анализа методом Вестерн-блота уровня расщепления прокаспазы-2 и накопления каталитически активных фрагментов каспазы-2 и/или с помощью флуоресцентных субстратов;
- сбор клеток с последующим добавлением лизирующего буфера, который обеспечивает сохранность высокомолекулярного комплекса активации каспазы-2;
- отделение клеточного дебриса от лизата, содержащего высокомолекулярный комплекс активации каспазы-2,
- разделение гель-фильтрацией на носителе с разрешающей способностью от 5×105 вплоть до 5×107 Да полученного лизата клеток с обеспечением отделения фракций, содержащих высокомолекулярные формы каспазы-2, от фракций, содержащих ее мономерные формы, которое проводят в условиях, обеспечивающих получение высокомолекулярных форм каспазы-2 молекулярной массой не менее 500000 Да;
- выделение из фракции, содержащей высокомолекулярные формы каспазы-2, белкового высокомолекулярного комплекса активации каспазы-2 с помощью иммунопреципитации, которую проводят в условиях, обеспечивающих минимизацию сорбции неспецифических белков.
2. Способ по п. 1, характеризующийся тем, что ферментативную активность каспазы-2 в выделенном высокомолекулярном комплексе определяют с помощью флуоресцентных субстратов.
3. Способ по п. 1, характеризующийся тем, что раковые клетки представляют собой линию карциномы яичника человека Caov-4 или РА-1, или Caov-3, или SW 626, или SK-OV-3.
4. Способ по п. 1, характеризующийся тем, что ДНК-повреждающий химиотерапевтический препарат представляет собой соединение платины, или алкилирующие агенты, или ингибиторы топоизомераз, или антиметаболиты.
5. Способ по п. 4, характеризующийся тем, что соединение платины представляет собой цисплатин, оксалиплатин, тетраплатин, карбоплатин, фенантриплатин.
6. Способ по п. 1, характеризующийся тем, что концентрация ДНК-повреждающего химиотерапевтического препарата рассчитывают путем инкубации раковых клеток в присутствии разных концентраций этого агента и дальнейшим определением уровня гибели клеток методом цитометрии.
7. Способ по п. 6, характеризующийся тем, что концентрация ДНК-повреждающего химиотерапевтического препарата составляет не менее 10 мкМ.
8. Способ по п. 7, характеризующийся тем, что концентрация химиотерапевтического препарата варьирует в пределах 35-100 мкМ.
9. Способ по п. 1, характеризующийся тем, что продолжительность инкубации определяется путем инкубации клеток с ДНК-повреждающим химиотерапевтическим препаратом в течение различного количества времени, но не менее часа, с последующим анализом уровня расщепления и активации каспазы-2 в этих клетках методом Вестерн-блота и/или с помощью флуоресцентных субстратов.
10. Способ по п. 1, характеризующийся тем, что лизирующий буфер включает один из следующих детергентов: Nonidet Р-40, Triton Х-100, Tween 20, Tween 80, Brij 35, Brij 58, октилглюкозид, октилтиоглюкозид, додецилсульфат натрия, CHAPS, дигитонин, дезоксихолат, холат, саркозил в конечной концентрации от 0,01 до 1,5% об. в буфере.
11. Способ по п. 10, характеризующийся тем, что лизирующий буфер дополнительно содержит пан-каспазный ингибитор zVAD-fmk (карбобензокси-валил-аланил-аспартил-[O-метил] фторметилкетон) в конечной концентрации не менее 10 мкМ в буфере.
12. Способ по п. 1, характеризующийся тем, что для получения высокомолекулярных форм каспазы-2 молекулярной массой не менее 500000 Да предварительно оценивают накопление в клетках высокомолекулярного комплекса с помощью гель-фильтрации с последующим Вестерн-блотом в зависимости от продолжительности инкубации клеток с ДНК-повреждающим химиотерапевтическим препаратом.
13. Способ по п. 1, характеризующийся тем, что выделение комплекса активации каспазы-2 осуществляют с помощью иммунопреципитации, включающей выбор носителя для иммунопреципитации с наименьшими параметрами неспецифической сорбции, которую оценивают с помощью электрофореза образцов в полиакриламидном геле после иммунопреципитации с дальнейшим окрашиванием геля.
14. Способ по п. 2, характеризующийся тем, что определение ферментативной активности каспазы-2 осуществляют на носителе для иммунопреципитации, с которым связан высокомолекулярный комплекс активации каспазы-2.
15. Способ выделения белкового высокомолекулярного комплекса активации каспазы-2 человека, формирующегося в раковых клетках в ответ на обработку ДНК-повреждающим химиотерапевтическим препаратом, характеризующийся тем, что включает:
- получение раковых клеток в количестве не менее чем 5 млн клеток;
- введение с помощью трансфицирующего агента в раковые клетки вектора, содержащего генетический конструкт, представляющий собой последовательность гена каспазы-2, которая соединена с аминокислотной последовательностью для аффинной хроматографии с с N- и/или С-конца, в концентрации не менее 0,5 мкг на 1 мл питательной среды;
- инкубация раковых клеток в течение времени, достаточного для детектируемой экспрессии генетического конструкта, представляющего собой последовательность гена каспазы-2 и дополнительную аминокислотную последовательность для аффинной хроматографии;
- введение в трансформированные клетки ДНК-повреждающего химиотерапевтического препарата, обеспечивающего запуск апоптотической гибели раковых клеток с достижением максимального уровня апоптотической гибели клеток при минимальном уровне некротической гибели клеток, которые определяют с помощью цитометрии;
- инкубация раковых клеток в присутствии ДНК-повреждающего химиотерапевтического агента до получения максимального уровня активации каспазы-2, который определяют предварительно до начала инкубации с помощью анализа методом Вестерн-блота уровня расщепления прокаспазы-2 и накопления каталитически активных фрагментов каспазы-2 и/или с помощью флуоресцентных субстратов;
- сбор клеток с последующим добавлением лизирующего буфера, который обеспечивает сохранность высокомолекулярного комплекса активации каспазы-2;
- отделение клеточного дебриса от лизата, содержащего высокомолекулярный комплекс активации каспазы-2,
- разделение гель-фильтрацией на носителе с разрешающей способностью от 5×105 вплоть до 5×107 Да полученного лизата клеток с обеспечением отделения фракций, содержащих высокомолекулярные формы каспазы-2, от фракций, содержащих ее мономерные формы, которое проводят в условиях, обеспечивающих получение высокомолекулярных форм каспазы-2 молекулярной массой не менее 500000 Да;
- выделение из фракции, содержащей высокомолекулярные формы каспазы-2, белкового высокомолекулярного комплекса активации каспазы-2 с помощью аффинной хроматографии, которую проводят в условиях, обеспечивающих минимизацию сорбции неспецифических белков.
16. Способ по п. 15, характеризующийся тем, что ферментативную активность каспазы-2 в выделенном высокомолекулярном комплексе определяют с помощью флуоресцентных субстратов.
17. Способ по п. 15, характеризующийся тем, что раковые клетки представляют собой линию карциномы яичника человека Caov-4 или РА-1, или Caov-3, или SW 626, или SK-OV-3.
18. Способ по п. 16, характеризующийся тем, что раковые клетки представляют собой линию карциномы яичника человека Caov-4 дефицитную по каспазе-2 - Caov-4-shRNA-каспаза-2.
19. Способ по п. 16, характеризующийся тем, что раковые клетки представляют собой линию карциномы яичника человека Caov-4 дефицитную по каспазе-2 - Caov-4 caspase-2 CRISPR/Cas9 Knockout.
20. Способ по п. 15, характеризующийся тем, что генетический конструкт представляет собой последовательность гена каспазы-2, которая соединена с последовательностью Strep-tag с N- и/или С-конца.
21. Способ по п. 15, характеризующийся тем, что генетический конструкт представляет собой последовательность гена каспазы-2, которая соединена с последовательностью Twin-Strep-tag с N- и/или С-конца.
22. Способ по п. 15, характеризующийся тем, что генетический конструкт представляет собой последовательность гена каспазы-2, которая соединена с последовательностью GST-tag с N- и/или С-конца.
23. Способ по п. 15, характеризующийся тем, что генетический конструкт представляет собой последовательность гена каспазы-2, которая соединена с последовательностью Flag-tag с N- и/или С-конца.
24. Способ по п. 15, характеризующийся тем, что концентрацию вектора, содержащего генетический конструкт, рассчитывают путем инкубации клеток в присутствии разных концентраций вектора и дальнейшим определением уровня экспрессии генетического конструкта методом Вестерн-блота.
25. Способ по п. 15, характеризующийся тем, что время инкубации раковых клеток после введения вектора определяется путем инкубации клеток в течение разного количества времени, но не менее 6 часов, и последующего анализа экспрессии генетического конструкта, представляющего собой последовательность гена каспазы-2 и дополнительную аминокислотную последовательность для аффинной хроматографии, методом Вестерн-блота.
26. Способ по п. 15, характеризующийся тем, что химиотерапевтический препарат представляет собой соединение платины или алкилирующие агенты, или ингибиторы топоизомераз, или антиметаболиты.
27. Способ по п. 26, характеризующийся тем, что соединение платины представляет собой цисплатин, оксалиплатин, тетраплатин, карбоплатин, фенантриплатин.
28. Способ по п. 15, характеризующийся тем, что концентрация ДНК-повреждающего химиотерапевтического препарата рассчитывается путем инкубации клеток в присутствии разных концентраций этого препарата и дальнейшим определением уровня гибели клеток методом цитометрии.
29. Способ по п. 28, характеризующийся тем, что концентрация ДНК-повреждающего химиотерапевтического препарата составляет не менее 10 мкМ.
30. Способ по п. 29, характеризующийся тем, что концентрация ДНК-повреждающего химиотерапевтического препарата варьирует в пределах 35-100 мкМ.
31. Способ по п. 15, характеризующийся тем, что продолжительность инкубации определяется путем инкубации клеток с ДНК-повреждающим химиотерапевтическим препаратом в течение различного количества времени, но не менее часа, с последующим анализом уровня процессинга и активации каспазы-2 в этих клетках методом Вестерн-блота и/или с помощью флуоресцентных субстратов.
32. Способ по п. 15, характеризующийся тем, что лизирующий буфер включает один из следующих детергентов: Nonidet Р-40, Triton Х-100, Tween 20, Tween 80, Brij 35, Brij 58, октилглюкозид, октилтиоглюкозид, додецилсульфат натрия, CHAPS, дигитонин, дезоксихолат, холат, саркозил в конечной концентрации от 0,01 до 1,5% об. в буфере.
33. Способ по п. 32, характеризующийся тем, что лизирующий буфер дополнительно содержит пан-каспазный ингибитор zVAD-fmk (карбобензокси-валил-аланил-аспартил-[O-метил] фторметилкетон) в конечной концентрации не менее 10 мкМ в буфере.
34. Способ по п. 15, характеризующийся тем, что для получения высокомолекулярных форм каспазы-2 молекулярной массой не менее 500000 Да предварительно оценивают накопление в клетках высокомолекулярного комплекса с помощью гель-фильтрации с последующим Вестерн-блотом в зависимости от продолжительности инкубации клеток с ДНК-повреждающим химиотерапевтическим препаратом.
35. Способ по п. 15, характеризующийся тем, что выделение комплекса активации каспазы-2 осуществляют с помощью аффинной хроматографии, включающей выбор носителя с наименьшими параметрами неспецифической сорбции, которую оценивают с помощью электрофореза образцов в полиакриламидном геле после аффинной хроматографии с дальнейшим окрашиванием геля.
36. Способ по п. 16, характеризующийся тем, что определение ферментативной активности каспазы-2 осуществляют на носителе для аффинной хроматографии, с которым связан высокомолекулярный комплекс активации каспазы-2.
| ROBERTSON J.D | |||
| et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| J Biol Chem | |||
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| READ S.H | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| J Cell Biol | |||
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| HUGHES M.A | |||
| et al | |||
| Isolation, characterisation and reconstitution of cell death signalling complexes | |||
| Methods | |||
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| MANZL C | |||
| et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| J Cell Biol | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| Способ изготовления электродов для гальванических элементов | 1927 |
|
SU9956A1 |

