Перекрестная ссылка на родственные заявки
В настоящей заявке испрашивается преимущество предварительной заявки на патент США № 60/814955, поданной 20 июня 2006, содержание которой во всей своей полноте вводится в настоящее описание посредством ссылки.
Область, к которой относится изобретение
В общих чертах настоящее изобретение относится к способам наблюдения апоптоза в клетках млекопитающих, таких как человеческие раковые клетки, обработанные апоптоз-индуцирующим агентом.
Предшествующий уровень техники
Регуляция числа клеток у млекопитающих частично определяется балансом между пролиферацией клеток и гибелью клеток. Одна из форм гибели клеток, иногда называемая некротической гибелью клеток, обычно характеризуется как патологическая форма гибели клеток в результате некоторых травм или поражений клеток. В противоположность этому существует и другая «физиологическая» форма гибели клеток, которая обычно представляет собой упорядоченный или регулируемый процесс. Такая упорядоченная или регулируемая форма гибели клеток часто называется «апоптозом» (см., например, Barr et al., Bio/Technology, 12:487-493 (1994); Steller et al., Science, 267:1445-1449 (1995)). Апоптотическая гибель клеток обычно происходит во многих физиологических процессах, включая развитие эмбриона и клональный отбор в иммунной системе (Itoh et al., Cell, 66:233-243 (1991)).
Различные молекулы, такие как фактор некроза опухоли-α («TNF-α»), фактор некроза опухоли-β («TNF-β» или «лимфотоксин-α»), лимфотоксин-β («LT-β»), лиганд CD30, лиганд CD27, лиганд CD40, лиганд OX-40, лиганд 4-1BB, лиганд Apo-1 (также называемый лигандом Fas или лигандом CD95), лиганд Apo-2 (также называемый TRAIL), лиганд Apo-3 (также называемый TWEAK), остеопротегерин (OPG), APRIL, лиганд RANK (также называемый TRANCE) и TALL-1 (также называемый BlyS, BAFF or THANK), были идентифицированы как цитокины, являющиеся членами семейства факторов некроза опухоли («TNF») (см. например, Gruss and Dower, Blood, 85:3378-3404 (1995); Pitti et al., J. Biol. Chem., 271:12687-12690 (1996); Wiley et al., Immunity, 3:673-682 (1995); Browning et al., Cell, 72:847-856 (1993); Armitage et al. Nature, 357:80-82 (1992), заявку WO 97/01633, опубликованную 16 января, 1997; заявку WO 97/25428, опубликованную 17 июля, 1997; Marsters et al., Curr. Biol., 8:525-528 (1998); Simonet et al., Cell, 89:309-319 (1997); Chicheportiche et al., Biol. Chem., 272:32401-32410 (1997); Hahne et al., J. Exp Med., 188:1185-1190 (1998); заявку WO98/28426, опубликованную 2 июля, 1998; заявку WO98/46751, опубликованную 22 октября, 1998; заявку WO/98/18921, опубликованную 7 мая, 1998; Moore et al., Science, 285:260-263 (1999); Shu et al., J. Leukocyte Biol., 65:680 (1999); Schneider et al., J. Exp. Med., 189:1747-1756 (1999); Mukhopadhyay et al., J. Biol. Chem., 274:15978-15981 (1999)). Сообщалось, что из этих молекул такие молекулы, как TNF-α, TNF-β, лиганд CD30, лиганд 4-1BB, лиганд Apo-1, лиганд Apo-2 (Apo2L/TRAIL) и лиганд Apo-3 (TWEAK), участвуют в апоптотической гибели клеток. Сообщалось, что TNF-α и TNF-β индуцируют апоптотическую гибель восприимчивых опухолевых клеток (Schmid et al., Proc. Natl. Acad. Sci., 83:1881 (1986); Dealtry et al., Eur. J. Immunol., 17:689 (1987)).
Сообщалось, что в апоптозе, очевидно, участвуют и другие молекулы, которые являются членами цитокинового семейства TNF. Так, например, в публикации Pitti et al., J. Biol. Chem., 271:12687-12690 (1996) описана молекула, называемая лигандом Apo-2. См. также заявку WO 97/25428, опубликованную 17 июля, 1997. Сообщалось, что полноразмерный человеческий лиганд Apo-2 представляет собой полипептид из 281 аминокислоты, который индуцирует апоптоз различных клеток млекопитающих. Другими исследователями были описаны родственные полипептиды, называемые TRAIL (см. Wiley et al., Immunity, 3:673-682 (1995); заявку WO 97/01633, опубликованную 16 января, 1997) and AGP-1 (см. заявку WO 97/46686, опубликованную 11 декабря, 1997).
Считается, что различные молекулы семейства TNF также играют определенную роль в функционировании или развитии иммунной системы (Gruss et al., Blood, 85:3378 (1995)). В публикации Zheng et al. сообщалось, что TNF-α участвует в апоптозе CD8-позитивных Т-клеток после их стимуляции (Zheng et al.. Nature, 377:348-351 (1995)). Другие исследователи сообщали, что лиганд CD30 может вызывать элиминацию аутореактивных Т-клеток в тимусе (Amakawa et al., Cold Spring Harbor Laboratory Symposium on Programmed Cell Death, Abstr. No. 10, (1995)). Лиганд CD40 активирует множество функций В-клеток, включая пролиферацию клеток, секрецию иммуноглобулина и выживание клеток (Renshaw et al., J. Exp. Med., 180:1889 (1994)). Сообщалось, что другое недавно идентифицированное цитокиновое семейство TNF, TALL-1 (BlyS), в определенных условиях индуцирует пролиферацию В-клеток и секрецию иммуноглобулина (Moore et al., см. выше; Schneider et al., см. выше; Mackay et al., J. Exp. Med., 190:1697 (1999)).
Мутации в генах мышиного рецептора или лиганда Fas/Apo-1 (обозначаемых lpr и gld, соответственно) ассоциируются с некоторыми аутоиммунными расстройствами, что указывает на то, что лиганд Apo-1 может играть определенную роль в регуляции клональной делеции аутореактивных лимфоцитов в периферических органах (Krammer et al., Curr. Op. Immunol., 6:279-289 (1994); Nagata et al., Science, 267:1449-1456 (1995)). Также сообщалось, что лиганд Apo-1 индуцирует апоптоз CD4-позитивных Т-лимфоцитов и В-лимфоцитов после их стимуляции, и может участвовать в элиминации активированных лимфоцитов, если их функция больше не требуется (Krammer et al., см. выше; Nagata et al., см. выше). Сообщалось, что мышиные моноклональные антитела-агонисты, специфически связывающиеся с рецептором Apo-1, обладают цитолитической активностью, сравнимой с цитолитической активностью TNF-α или аналогичной активностью (Yonehara et al., J. Exp. Med., 169:1747-1756 (1989)).
Индуцирование различных клеточных ответов, опосредуемых такими лигандами семейства TNF, обычно инициируется их связыванием со специфическими клеточными рецепторами. Некоторые, но не все, лиганды семейства TNF связываются с присутствующими на поверхности клеток «рецепторами гибели» и, посредством таких рецепторов, индуцируют различные биологические активности, что приводит к активации каспаз или ферментов, которые участвуют в пути гибели или апоптоза клеток (Salvesen et al., Cell, 91:443-446 (1997)). В настоящее время считается, что членами суперсемейства рецепторов TNF являются TNFR1, TNFR2, TACI, GITR, CD27, OX-40, CD30, CD40, HVEM, Fas (также называемые Apo-1 или CD95), DR4 (также называемый TRAIL-R1), DR5 (также называемый Apo-2 или TRAIL-R2), DcR1, DcR2, остеопротегерин (OPG), RANK и Apo-3 (также называемый DR3 или TRAMP) (см., например, Ashkenazi, Nature Reviews, 2:420-430 (2002); Ashkenazi and Dixit, Science, 281:1305-1308 (1998); Ashkenazi and Dixit, Curr. Opin. Cell Biol, 11:255-260 (2000); Golstein, Curr. Biol., 7:750-753 (1997) Wallach, Cytokine Reference, Academic Press, 2000, pages 377-411; Locksley et al., Cell, 104:487-501 (2001); Gruss and Dower, Blood, 85:3378-3404 (1995); HohMan et al., J.Biol. Chem., 264:14927-14934 (1989); Brockhaus et al., Proc. Natl. Acad. Sci., 87: 3127-3131 (1990); патент EP 417563, опубликованный 20 марта, 1991; Loetscher et al.. Cell, 61:351 (1990); Schall et al., Cell, 61:361 (1990); Smith et al., Science, 248:1019-1023 (1990); Lewis al., Proc. Natl. Acad. Sci., 88: 2830-2834 (1991); Goodwin et al., Cell. Biol., 11:3020-3026 (1991); Stamenkovic et al., EMBO J., 8:1403-1410 (1989); Mallett al., EMBO J., 9:1063-1068 (1990); Anderson al., Nature, 390:175-179 (1997); Chicheportiche et al., J. Biol. Chem., 272:32401-32410 (1997); Pan al.. Science, 276:111-113 (1997); Pan et al., Science, 277:815-818 (1997); Sheridan et al., Science 277: 818-821 (1997); Degli-Esposti et al., J. Exp. Med., 186:1165-1170 (1997); Marsters et al., Curr. Biol., 7:1003-1006 (1997); Tsuda et al., BBRC, 234: 137-142 (1997); Nocentini et al., Proc. Natl. Acad. Sci., 94: 6216-6221 (1997); von Bulow et al., Science, 278:138-141 (1997)).
Многие из этих членов семейства рецепторов TNF имеют общую структуру, типичную для рецепторов клеточной поверхности, включая внеклеточные, трансмембранные и внутриклеточные области, а другие члены этого семейства существуют в природе как растворимые белки, не содержащие трансмембранного и внутриклеточного домена. Внеклеточная часть типичных TNFR содержит повторяющуюся аминокислотную последовательность, состоящую из множества богатых цистеином доменов (CRD), начиная с NН2-конца.
Лиганд, называемый Apo-2L или TRAIL, был идентифицирован несколько лет назад как член цитокинового семейства TNF (см., например, Wiley et al., Immunity, 3:673-682 (1995); Pitti et al., J. Biol. Chem.. 271:12697-12690 (1996); WO 97/01633; WO 97/25428; патент США 5763223, выданный 9 июня, 1998; патент США 6284236, выданный 4 сентября, 2001). Полноразмерная нативная последовательность человеческого полипептида Apo2L/TRAIL представляет собой трансмембранный белок типа II длиной в 281 аминокислоту. Некоторые клетки могут продуцировать природную растворимую форму полипептида посредством ферментативного расщепления внеклеточной области полипептида (Mariani et al., J. Cell. Biol., 137:221-229 (1997)).
Кристаллографические исследования растворимых форм Apo2L/TRAIL выявили гомотримерную структуру, аналогичную структурам TNF и других родственных белков (Hymowitz et al., Molec Cell, 4:563-571 (1999); Cha et al.. Immunity, 11:253-261 (1999); Mongkolsapaya et al., Nature Structural Biology, 6:1048 (1999); Hymowitz et al., Biochemistry, 39:633-644 (2000)). Однако было обнаружено, что Apo2L/TRAIL, в отличие от других членов семейства TNF, имеет уникальную структурную особенность, заключающуюся в том, что в нем три цистеиновых остатка (в положении 230 каждой субъединицы гомотримера), взятые вместе, координируют атом цинка, и в том, что такое связывание с цинком играет важную роль в стабильности тримера и в биологической активности (Hymowitz et al., см. выше; Bodmer et al., J. Biol. Chem., 275:20632-20637 (2000)).
В литературе сообщалось, что Apo2L/TRAIL может играть определенную роль в модуляции иммунной системы, включая развитие аутоиммунных заболеваний, таких как ревматоидный артрит (см., например, Thomas et al., J. Immunol., 161:2195-2200 (1998); Johnsen et al., Cytokine, 11:664-672 (1999); Griffith et al., J. Exp. Med., 189:1343-1353 (1999); Song et al., J. Exp. Med., 191: 1095-1103 (2000)).
Также сообщалось, что растворимые формы Apo2L/TRAIL индуцируют апоптоз различных раковых клеток, включая опухоли толстой кишки, легких, молочной железы, предстательной железы, мочевого пузыря, почек, яичника и головного мозга, а также меланому, лейкоз и множественную миелому (см., например, Wiley et al., см. выше; Pitti et al., см. выше; патент США 6030945, выданный 29 февраля, 2000; патент США 6746668, выданный 8 июня, 2004; Rieger et al., FEBS Letters, 427:124-128 (1998); Ashkenazi et al., J. Clin. Invest., 104: 155-162 (1999); Walczak et al., Nature Med., 5: 157-163 (1999); Keane et al.. Cancer Research, 59:734-741 (1999); Mizutani et al., Clin. Cancer Res., 5:2605-2612 (1999); Gazitt, Leukemia, 13:1817-1824 (1999); Yu et al., Cancer Res., 60:2384-2389 (2000); Chinnaiyan et al., Proc. Natl. Acad. Sci., 97:1754-1759 (2000)). In vivo исследования, проводимые на мышах с моделью опухолей, также дают основание предположить, что Apo2L/TRAIL, используемый отдельно или в комбинации с химиотерапией или лучевой терапией, может обладать значительным противоопухолевым действием (см., например, Ashkenazi et al., см.выше; Walczak et al., см.выше; Gliniak et al., Cancer Res., 59:6153-6158 (1999); Chinnaiyan et al., см.выше; Roth et al., Biochem. Biophys. Res. Comm., 265:1999 (1999); заявку PCT US/00/15512; заявку PCT US/01/23691). В отличие от раковых клеток многих типов большинство нормальных человеческих клеток очевидно обладают резистентностью к индуцированию апоптоза некоторыми рекомбинантными формами Apo2L/TRAIL (Ashkenazi et al., см. выше; Walczak et al., см. выше). В публикации Jo и др. сообщалось, что меченная полигистидином растворимая форма Apo2L/TRAIL индуцирует in vitro апопотоз нормальных выделенных человеческих гепатоцитов, но не индуцирует апопотоз нечеловеческих гепатоцитов (Jo et al., Nature Med., 6:564-567 (2000); см. также Nagata, Nature Med., 6:502-503 (2000)). Очевидно, что некоторые рекомбинантныые Apo2L/TRAIL-препараты могут обладать различными биохимическими свойствами и биологическими активностями по отношению к патологическим, но не по отношению к нормальным, клеткам, в зависимости, например, от присутствия или отсутствия молекулы-метки, содержания цинка и процентного содержания тримеров (см. Lawrence et al., Nature Med., Letter to the Editor, 7:383-385 (2001); Qin et al., Nature Med., Letter the Editor, 7:385-386 (2001)).
Было обнаружено, что Apo2L/TRAIL связывается по меньшей мере с пятью различными рецепторами. По меньшей мере два из этих рецепторов, которые связываются с Apo2L/TRAIL, содержат функциональный цитоплазматический домен «гибели». Один из таких рецепторов называется «DR4» (и альтернативно, TR4 или TRAIL-R1) (Pan et al., Science, 276:111-113 (1997); см. также заявку W098/32856, опубликованную 30 июля 1998; заявку W099/37684, опубликованную 29 июля 1999; заявку WO 00/73349, опубликованную 7 декабря 2000; заявку США 2003/0036168, опубликованную 20 февраля 2003; патент США 6433147, выданный 13 августа 2002; патент США 6461823, выданный 8 октября 2002, и патент США 6342383, выданный 29 января 2002).
Другой такой рецептор для Apo2L/TRAIL получил название DR5 (он также имеет и другие названия, а именно Apo-2; TRAIL-R или TRAIL-R2, TR6, Tango-63, hAPO8, TRICK2 или KILLER) (см., например, Sheridan et al.. Science, 277:818-821 (1997), Pan et al.. Science, 277:815-818 (1997), заявку W098/51793, опубликованную 19 ноября 199.., заявку W098/41629, опубликованную 24 сентября 1998; Screaton et al., Curr. Biol., 7:693-696 (1997); Walcak et al., EMBO J., 16:5386-5387 (1997); Wu et al.. Nature Genetics, 17:141-143 (1997), заявку W098/35986, опубликованную 20 августа 1998; EP 870827, опубликованный 14 октября 1998; заявку W098/46643, опубликованную 22 октября 1998; заявку W099/02653, опубликованную 21 января 1999; заявку W099/09165, опубликованную 25 февраля 1999, заявку W099/11791, опубликованную 11 марта 1999; заявку WO 03/042367, опубликованную 22 мая 2003; заявку WO 02/097033, опубликованную 5 декабря 2002; заявку WO 03/038043, опубликованную 8 мая 2003; заявку США 2002/0072091, опубликованную 13 августа 2002; заявку США 2002/0098550, опубликованную 7 декабря 2001; патент США 6313269, выданный 6 декабря 2001; заявку США 2001/0010924, опубликованную 2 августа 2001; заявку США 2003/01255540, опубликованную 3 июля 2003; заявку США 2002/0160446, опубликованную 31 октября 2002, заявку США 2002/0048785, опубликованную 25 апреля 2002; заявку США 2004/0141952, опубликованную 21 июля 2004; заявку США 2005/0129699, опубликованную 16 июня 2005; заявку США 2005/0129616, опубликованную 16 июня 2005; патент США 6342369, выданный в феврале 2002; патент США 6569642, выданный 27 мая 2003, патент США 6072047, выданный 6 июня 2000, патент США 6642358, выданный 4 ноября 2003; патент США 6743625, выданный 1 июня 2004). Сообщалось, что DR5, аналогично DR4, содержит цитоплазматический домен гибели и обладает способностью передавать сигнал на апоптоз после связывания с лигандом (или после связывания с молекулой, такой как антитело-агонист, которая имитирует активность лиганда). Кристаллическая структура комплекса, образованного между Apo-2L/TRAIL и DR5, описана в публикации Hymowitz al., Molecular Cell, 4:563-571 (1999). Другой идентифицированный рецептор, содержащий «домен гибели», называется DR6 (Pan et al., FEBS Letters, 431:351-356 (1998)).
После связывания с лигандом DR4 и DR5 могут независимо запускать апоптоз путем рекрутинга и активации инициатора апоптоза, каспазы-8, под действием молекулы-адаптора, содержащей домен «гибели» и называемой FADD/Mortl (Kischkel et al., Immunity, 12:611-620 (2000); Sprick et al., Immunity, 12:599-609 (2000); Bodmer Nature Cell Biol., 2:241-243 (2000)).
Сообщалось, что Apo2L/TRAIL также связывается с рецепторами, называемыми DcR1, DcR2 и OPG, которые, очевидно, функционируют как ингибиторы, а не как трансдукторы передачи сигнала (см., например, DCR1 (также называемый TRID, LIT или TRAIL-R3) (Pan et al., Science, 276:111-113 (1997); Sheridan et al., Science, 277:818-821 (1997); McFarlane et al., J. Biol. Chem., 272:25417-25420 (1997); Schneider et al., FEES Letters, 416:329-334 (1997); Degli-Esposti et al., J. Exp. Med., 186:1165-1170 (1997); и Mongkolsapaya et al., J. Immunol, 160:3-6 (1998); DCR2 (также называемый TRUNDD или TRAIL-R4) (Marsters et al., Curr. Biol., 7:1003-1006 (1997); Pan et al., FEES Letters, 424:4l-45 (1998); Degli-Esposti et al., Immunity, 7:813-820 (1997)) и OPG (Simonet et al., см. выше)). В отличие от DR4 и DR5 рецепторы DcR1 и DcR2 не передают сигнал апоптоза.
В литературе имеются сообщения о некоторых антителах, которые связываются с рецепторами DR4, DR5 и/или Fas. Так, например, анти-DR4-антитела, направленные против рецептора DR4 и обладающие агонистической или апоптотической активностью в некоторых клетках млекопитающих, описаны, например, в заявке WO 99/37684, опубликованной 29 июля 1999; в заявке WO 00/73349, опубликованной 12 июля 2000; и в заявке WO 03/066661, опубликованной 14 августа 2003. См. также, например, Griffith et al., J. Immunol., 162:2597-2605 (1999); Chuntharapai et al., J. Immunol., 166:4891-4898 (2001); заявку WO 02/097033, опубликованную 2 декабря 2002; заявку WO 03/042367, опубликованную 22 мая 2003; заявку WO 03/038043, опубликованную 8 мая 2003; заявку WO 03/037913, 8 мая 2003; заявку США 2003/0073187, опубликованную 17 апреля 2003; и заявку США 2003/0108516, опубликованную 12 июня 2003. Были также описаны некоторые анти-DR5-антитела, см., например, заявку WO 98/51793, опубликованную 8 ноября 1998; Griffith et al., J. Immunol., 162:2597-2605 (1999); Ichikawa et al., Nature... Med., 7:954-960 (2001); Hylander et al., "An Antibody to DR5 (TRAIL-Receptor 2) Suppresses the Growth of Patient Derived Gastrointestinal Tumors Grown in SCID mice", Abstract, 2d International Congress on Monoclonal Antibodies in Cancers, Aug. 29-Sept. 1, 2002, Banff, Alberta, Canada; заявку WO 03/038043, опубликованную 8 мая 2003; заявку WO 03/037913, опубликованную 8 мая 2003; и заявку США 2003/0180296, опубликованную 25 сентября 2003. Кроме того, были описаны некоторые антитела, перекрестно реагирующие с рецепторами DR4 и DR5 (см., например, патент США 6252050, выданный 26 июня 2001). Анти-Fas-антителами-агонистами, которые индуцируют апоптоз клеток-мишеней, экспрессирующих Fas, являются, но не ограничиваются ими, MAb M2 и M3 (IgG; Alderson et al., 1995, J. Exp. Med. 181:71-77); анти-Fas MAb (IgM; Yonehara et al., 1989, J. Exp. Med. 169:1747-1756); MAb CH1l (IgM; Alderson et al., 1994, Int. Immunol. 6:1799-806); и анти-Apo-1 антитело (IgG; Dhein et al., 1992, J. Immunol., 149:3166-3173).
Апоптоз играет решающую роль в развитии и в гомеостазе многоклеточных организмов (см, например, Danial, N.N. et. al., Cell 116, 205-19 (2004)). Индуцирование апоптоза регулируется двумя основными механизмами передачи сигнала по внутриклеточному и внеклеточному пути (см., например, Strasser, A., et. al., Annu. Rev. Biochem. 69, 217-45 (2000)). Эти пути активируют цистеиновые протеазы, называемые каспазами, которые расщепляют различные клеточные белки, играющие важную роль в целостности клетки. Каспазы распознают специфические тетрапептидные последовательности, содержащие аспартат, и расщепляют связь с соседним пептидом (см., например, Thornberry, N.A. et. al., Science 281, 1312-6 (1998)). Активация внеклеточного пути происходит в ответ на действие лигандов, таких как лиганд Fas (FasL) и лиганд Аро-2/лиганд, индуцирующий TNF-опосредуемый апоптоз (Apo2L/TRAIL), через их соответствующие «рецепторы гибели» Fas, присутствующие на клеточной поверхности (Apol/CD95) и DR4 или DR5 (см., например, Nagata, S. Cell 88, 355-65 (1997) & LeBlanc, H.N. et. al., Cell Death Differ 10, 66-75 (2003)). Cвязывание с лигандом запускает присоединение адаптора FADD (Fas-ассоциированного домена «гибели») к домену «гибели», присутствующему в цитоплазматическом «хвосте» рецептора. FADD стимулирует присоединение протеазы-инициатора, каспазы-8, с образованием «сигнального комплекса, иниицирующего гибель клеток» (DISC) (см., например, Kischkel, F. C. et al. Embo J. 14, 5579-88 (1995)). Близость молекул каспазы-8 в DISC стимулирует ферментативную активность, что приводит к аутопроцессингу (см., например, Boatright, K.M. et al. Mol Cell 11, 529-41 (2003)). Затем отщепленная каспаза-8 высвобождается из DISC в цитоплазму и протеолитически активирует эффекторные каспазы, такие как каспаза-3 и -7. В клетках некоторых типов стимуляция DR генерирует сильную активность каспазы-8, которая в высокой степени активирует эффекторные каспазы и передает клеткам сигнал на апоптоз (см., например, Scaffidi, C, et. al., J Biol Chem 274, 1541-8 (1999)). Клетки других типов требуют амплификации сигнала по его внутреннему пути: каспаза-8 расщепляет гомологичный домену Bcl-2 домен 3 (BH3), присутствующий только в белке Bid, который стимулирует внутренний путь посредством белков Bax и Bak со множеством доменов ВН, что приводит к усилению активации эффекторной каспазы и к апоптозу (см., например, Danial, N.N. et. al., Cell 116, 205-19 (2004) и Strasser, A., et. al., Annu Rev Biochem 69, 217-45 (2000)).
Апоптоз обычно характеризуется конденсацией и скоплением ядерного хроматина на периферии и фрагментацией ядерной структуры в так называемые апоптотические тельца. Такая апоптотическая морфология может быть идентифицирована с помощью стандартных окрашивающих веществ, то есть красителей, которые селективно аккумулируются в ядрах, например, с помощью иодида пропидия или Hoechst 33258, или с помощью электронной микроскопии. Межнуклеосомная фрагментация ДНК, которая часто ассоциируется с клеточной гибелью в результате апопотоза, но не является диагностическим фактором клеточной гибели вследствие апоптоза, может быть также использована для идентификации и количественной оценки апоптоза.
Описание сущности изобретения
Заявителями были идентифицированы фрагменты белков, которые образуются в клетках во время апоптоза, и которые могут быть использованы в качестве биомаркеров апоптотической гибели клеток. В некоторых своих вариантах настоящее изобретение относится к способам и к материалам, используемым в целях идентификации указанных фрагментов белков, например, для выявления апоптотической гибели клеток млекопитающих, обработанных одним или несколькими апоптоз-индуцирующими агентами. В иллюстративном варианте изобретения такие биомаркеры апоптоза идентифицируют в человеческих раковых клетках для оценки эффективности терапии, включающей введение апоптоз-индуцирующего агента, такого как Apo2L/TRAIL, FasL или Apo2L/TRAIL или агонист FasL.
Заявленное изобретение может быть осуществлено в различных вариантах. Одним из вариантов изобретения является способ детектирования апоптоза в клетках млекопитающих, включающий: контактирование компонентов клетки с антителом, которое связывается с фрагментом белка, образующегося в процессе апоптоза, где указанное антитело связывается с фрагментом белка AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2-β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4); определение количества антитела, которое связывается с фрагментом белка, образующегося в процессе апоптоза; и сравнение количества связанного антитела на этой стадии с количеством антитела, которое связывается с фрагментом белка в клетках млекопитающих, не подвергающихся апоптозу, где апоптоз детектируют в том случае, если указанное количество такого фрагмента превышает количество фрагмента, присутствующего в клетках, не подвергающихся апоптозу. Клетками млекопитающих, оцениваемыми таким способом, являются, но необязательно, клетки раковых опухолей человеческой толстой кишки, ободочной кишки, легких, молочной железы, предстательной железы, мочевого пузыря, почек, яичника и головного мозга, а также клетки меланомы, лейкоза или миеломы.
Как подробно описано ниже, способы согласно изобретению могут быть применены для обнаружения апоптоза, инициированного различными клеточными рецепторами. В некоторых вариантах такого способа апоптоз клетки инициируют посредством рецептора гибели-4 (SEQ ID NO:5) или рецептора гибели-5 (SEQ ID NO:6), например, путем контактирования клетки с Apo2L/TRAIL (SEQ ID NO: 7) или с антителом, которое связывается с рецептором «гибели» 4 (SEQ ID NO: 5) или с рецептором «гибели» 5 (SEQ ID NO: 6). В других вариантах изобретения апоптоз клеток инициируют посредством Fas (SEQ ID NO: 8), например, путем контактирования клеток с FasL (SEQ ID NO: 9) или с антителом, которое связывается с Fas.
Способы согласно изобретению могут быть также адаптированы для их применения в различных целях. Так, например, такие способы наблюдения апоптоза могут быть применены для оценки чувстивтельности клеток к Apo2L/TRAIL (SEQ ID NO: 7) или к антителу, которое связывается с рецептором гибели 4 (SEQ ID NO: 5) или рецептором гибели 5 (SEQ ID NO: 6). В конкретном иллюстративном варианте изобретения такие способы могут быть применены для оценки эффективности терапии, включающей введение Apo2L/TRAIL (SEQ ID NO: 7) или антитела, которое связывается с рецептором гибели 4 (SEQ ID NO: 5) или рецептором гибели 5 (SEQ ID NO: 6). Аналогичным образом другие варианты настоящего изобретения могут быть использованы для оценки чувствительности клеток к FasL (SEQ ID NO: 9) или к антителу, которое связывается с Fas, например, для оценки эффективности терапии, включающей введение FasL (SEQ ID NO: 9) или антитела, которое связывается с Fas.
Как подробно обсуждается ниже, способы согласно изобретению могут быть применены для наблюдения апоптотической фрагментации различных белков в клетках путем осуществления любого из ряда методов, известных специалистам, таких как иммуноанализы, проводимые с использованием антител против одного или нескольких указанных апоптотических фрагментов. Так, например, в некоторых вариантах изобретения, антитело связывается с фрагментом белка AP2-α (SEQ ID NO: 1), например с фрагментом, имеющим массу примерно 64 кДа или 33 кДа. В некоторых вариантах изобретения фрагмент белка AP2α, связанного с антителом, содержит DVFD SEQ ID NO: 1 или GPAA SEQ ID NO: 1. В других вариантах изобретения может быть использовано антитело, связывающееся с фрагментом белка тяжелой цепи клатрина (SEQ ID NO: 2). В других вариантах изобретения может быть использовано антитело, связывающееся с фрагментом белка AP1/2β (SEQ ID NO: 3). В других вариантах изобретения может быть использовано антитело, связывающееся с фрагментом белка динамина (SEQ ID NO: 4).
В некоторых своих вариантах настоящее изобретение относится к способу оценки апоптотической активности специфического типа и/или апоптоза в конкретных физиологических условиях. Так, например, варианты согласно изобретению могут быть использованы для наблюдения апоптоза клеток млекопитающих, опосредуемого рецетором гибели 4 (SEQ ID NO: 5), рецептором гибели 5 (SEQ ID NO: 6) или Fas (SEQ ID NO: 8). Обычно такие способы включают обработку клеток рецептором гибели 4, рецептором гибели 5 или лигандом Fas; анализ клеток, обработанных таким лигандом, на присутствие фрагментов белка AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4); сравнение количества фрагмента белка в клетках с количеством фрагмента белка в контрольных клетках, не обработанных таким лигандом, где апоптоз детектируют в том случае, если количество фрагмента белка, присутствующего в клетках, обработанных лигандом, превышет количество фрагмента белка, присутствующего в контрольных клетках, не обработанных лигандом. Обычно, в таких способах, лигандом для рецептора гибели 4, рецептора гибели 5 или Fas является Apo2L/TRAIL (SEQ ID NO: 7), антитело, которое связывается с рецептором гибели 4 или рецептором гибели 5, FasL (SEQ ID NO: 9) или антитело, которое связывается с Fas.
Другие варианты осуществления изобретения включают анализ на чувствительность клеток млекопитающего к Apo2L/TRAIL (SEQ ID NO: 7) или FasL (SEQ ID NO: 8) индуцированному апоптозу, проводимый путем обработки клеток млекопитающих Apo2L/TRAIL или FasL; оценку клеток, обработанных Apo2L/TRAIL или FasL на присутствие фрагмента белка AP2α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4); сравнение количества фрагмента белка в клетках млекопитающих с количеством фрагмента белка в контрольных клетках млекопитающих, не обработанных Apo2L/TRAIL или FasL, при этом считается, что наблюдаемые клетки млекопитающих являются чувствительными к Apo2L/TRAIL- или FasL-опосредуемому апоптозу в том случае, если количество фрагмента белка, присутствующего в Apo2L/TRAIL- или FasL-обработанных клетках млекопитающих превышает количество фрагмента белка в контрольных клетках млекопитающих, не обработанных Apo2L/TRAIL или FasL.
В некоторых своих вариантах, настоящее изобретение также относится к промышленным изделиям и к наборам, включающим антитела, которые связываются с фрагментом белка AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4). Иллюстративным вариантом настоящего изобретения является набор для характеризации клеток млекопитающих, где указанный набор включает: первое антитело, которое связывается с фрагментом белка AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4); второе антитело, которое связывается с фрагментом белка AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4), где указанные первое и второе антитела не связываются с одним и тем же эпитопом (и не связываются необязательно с одним и тем же белком); контейнер для (a) и (b); и инструкции по применению данного набора.
Краткое описание графического материала
На фиг.1 проиллюстрировано, что Apo2L/TRAIL индуцирует селективное расщепление по механизму клатрин-зависимого эндоцитоза. (a) Клетки обрабатывали при 37°С либо тримерным Apo2L/TRAIL (Colo205, HCT8), либо перекрестно связанным с антителом и меченным Apo2L/TRAIL (BJAB, HeLa-M), и клеточные лизаты анализировали с помощью иммуноблоттинга на отщепление каспазы-3 (C8), каспазы-3 (C3), адаптина (AP)2α, AP1/2β (антитело не распознает изоформы AP1 и 2), тяжелой цепи клатрина (CHC) или рецептора Tf (TfR). (b) Клетки Colo205 обрабатывали, как описано в (a), и анализировали с помощью иммуноблоттинга на процессинг специфических компонентов различного типа, действующих на пути клатрин-ассоциированного эндоцитозного транспорта.
На фиг.2 проиллюстрировано участие различных каспаз в расщеплении AP2α и CHC. (a) Клетки BJAB обрабатывали ингибитором пан-каспазы zVAD-fmk (20 мкM, 30 мин), а затем перекрестно связанным Apo2L/TRAIL (1 мкг/мл) и анализировали с помощью иммуноблоттинга на процессинг каспазы-8, каспазы-3, AP2α и CHC. Незаштрихованные стрелки указывают на продукты расщепления, а заштрихованные стрелки указывают на полноразмерные белки. (b-d) Baх-/-- или Bax+/--клетки HCT116 или дефицитные по каспазе-3 клетки MCF-7 обрабатывали Apo2L/TRAIL и анализировали, как описано в (a).
На фиг.3 показано, что предварительная обработка Apo2L/TRAIL ингибирует эндоцитоз Tf. (a, b) Клетки BJAB (a) или Colo205 (b) предварительно обрабатывали при 37°С или не обрабатывали в присутствии или в отсутствие перекрестносвязанного (a) или неперекрестносвязанного (b) Apo2L/TRAIL в течение определенного времени и охлаждали на льду. Затем клетки уравновешивали на льду в течение 30 минут Alexa-647-конъюгированным Tf (647ТF), и поглощение определяли при 37°С с помощью проточной цитометрии. Скорость эндоцитоза для каждого случая определяли по углу наклона кривой начальной линейной фазы на графике кинетики поглощения в течение 4 минут (а, вставка). Степени эндоцитоза нормализовали по данным наблюдения в отсутствие Apo2L/TRAIL (белые кружки), которые сравнивали с данными, полученными в случае отсутствия лиганда в стадии предварительного инкубирования, но его присутствия в стадии инкубирования Tf в данном анализе (черные ромбы, 0 минут). (c) клетки BJAB предварительно обрабатывали носителем ДМСО или zVAD-fmk в течение 30 минут, а затем обрабатывали в течение 4 часов перекрестно связанным Apo2L/TRAIL, охлаждали на льду и анализировали на скорость эндоцитоза 647Tf, как описано в (а) и (b). Скорость эндоцитоза нормализовали по ДМСО-обработанному образцу (± ср.кв.ош.).
На фиг.4 представлена характеризация эндоцитоза DR5. (a, b) Клетки Colo205, содержащие на своей поверхности mAb 6475С7, инкубировали на льду в течение 30 минут в отсутствие (ромбы) или в присутствии 10 мкг/мл тримерного (квадраты) или перекрестно связанного (треугольники) Apo2L/TRAIL, а затем инкубировали при 37°С в течение указанного времени и быстро охлаждали на льду. Поверхностную флуоресценцию элиминировали путем обработки кислотой, и поглощение DR5 количественно оценивали с помощью проточной цитометрии. Затем по средним величинам строили график (± ср.кв.ош. в (b)). (c) Клетки Hela-m инкубировали при 37°С с 5 мкг/мл 6475С7 и 5 мкг/мл перекрестно связанного Apo2L/TRAIL, а затем обрабатывали для наблюдения под иммунофлуоресцентным микроскопом (столбец: 20 мкM). (d) Клетки Colo205 предварительно уравновешивали при 37°С немеченным mAb 5C7 в течение 30 минут для связывания с поверхностью, и пулы DR5 подвергали рециклингу. Инкубировали осуществляли в присутствии или в отсутствие 10 мкг/мл Apo2L/TRAIL, а затем клетки быстро охлаждали на льду. mAb 5C7, присутствующее на клеточной поверхности, зондировали CY5- конъюгированным антителом против мышиных IgG, количественно оценивали с помощью проточной цитометрии, и по средним значениям (± ср.кв.ош.) строили график. (e) клетки Colo205 предварительно обрабатывали при 37°С 10 мкг/мл Apo2L/TRAIL, а затем быстро охлаждали на льду и анализировали на эндоцитоз, как показано в (а) (6475С7) и на фиг.3b (647Tf). Скорости эндоцитоза нормализовали на величину, полученную без предварительной обработки Apo2L/TRAIL (± ср.кв.ош.). Аналогичные результаты наблюдались в том случае, когда клетки приготавливали, как описано в (с), и эндоцитоз анализировали с использованием связанного с поверхностью CY5-конъюгированного антитела против мышиных Fab. (f-h) Клетки Colo205, имеющие на своей поверхности mAb 5C7, инкубировали на льду вместе с Apo2L/TRAIL, а затем доводили до 37°С в течение 5 минут и фиксировали. Сверхтонкие криосрезы метили кроличьими антителами против мышиных IgG и белком А-золотом (10 нм). Типичное электронно-плотное клатриновое покрытие показано стрелками. P, плазматическая мембрана. Масштабная линия, 200 нм.
На фиг.5 показано, что инактивация динамина приводит к ингибированию эндоцитоза DR5. (a-d) DynG273D-трансдуцированные клетки HeLa-M отбирали с помощью пуромицина, индуцировали доксициклином, предварительно инкубировали в течение 20 минут при указанной температуре, а затем инкубировали еще 20 минут в присутствии Alexa-488-конъюгированного Tf (488Tf). Затем клетки обрабатывали для анализа под иммунофлуоресцентном микроскопом с использованием динамин-1-специфического антитела. (е, f) Нетрансдуцированные (родительские) или клональные Dyn1G273D-трансдуцированные клетки BJAB, индуцированные доксициклином (+dox) или не индуцированные доксициклином (-dox), анализировали на поглощение 488Tf или 6475С7 в течение 20 минут при 30°С (белые столбцы) или 38°С (черные столбцы) с помощью проточной цитометрии, и по полученным средним значениям (± ср.кв.ош) строили график.
На фиг.6 показано, что инактивация динамина приводит к усилению DR-опосредуемой активации каспазы и апоптоза. (a) DynG273D-трансдуцированные клетки BJAB, индуцированные или не индуцированные доксициклином, инкубировали при 38°С в течение 20 минут для инактивации динамина, как показано на фиг.5, а затем инкубировали еще 4 часа в присутствии или в отсутствие перекрестно связанного Apo2L/TRAIL и анализировали с помощью иммуноблоттинга на процессинг указанных белков. (b) DynG273D-трансдуцированные клетки BJAB, индуцированные или не индуцированные доксициклином, инкубировали при 30°С или при 38°С в течение 20 минут, а затем инкубировали еще 2 часа в присутствии или в отсутствие перекрестно связанного Apo2L/TRAIL и анализировали на активность каспазы-3/7. (с) DynG273D-трансдуцированные клетки BJAB, индуцированные или не индуцированные доксициклином, предварительно инкубировали при 38°С в течение 20 минут, а затем инкубировали еще 4 часа в присутствии или в отсутствие перекрестно связанного Apo2L/TRAIL или DR5-селективного мутанта Apo2L/TRAIL (D5-сел.) и анализировали на фрагментацию ДНК (± ср.кв.ош).
На фиг.7 проиллюстрирован процессинг компонентов клатринового пути в раковых клеточных линиях. (a) Указанные клеточные линии обрабатывали Apo2L/TRAIL и анализировали на расщепление AP2α или CHC с помощью иммуноблоттинга. (b) Клетки BJAB обрабатывали перекрестно связанным Apo2L/TRAIL или FasL и процессинг AP2α или CHC анализировали с помощью иммуноблоттинга. (c) Клетки BJAB обрабатывали перекрестно связанным Apo2L/TRAIL и процессинг динамина оценивали с помощью иммуноблоттинга.
На фиг.8 проиллюстрированы условия, необходимые для расщепления AP2α и CHC каспазой. (a) Клетки BJAB предварительно инкубировали в присутствии или в отсутствие zVAD-fmk (20 мкM, 30 минут) и обрабатывали перекрестно связанным Apo2L/TRAIL (1 мкг/мл), как указано выше, в течение 24 часов. Клетки анализировали с помощью иммуноблоттинга на процессинг каспазы-8, каспазы-9, каспазы-3 и AP2α. (b) Т-клеточные линии Jurkat: A3 (дикого типа), 19.2 (дефицитные по каспазе-8) and El (FADD-дефицитные) обрабатывали перекрестно связанным Apo2L/TRAIL или FasL в течение указанного периода времени и анализировали на процессинг компонентов клатрин-зависимого эндоцитозного пути, как показано на фиг.1. (c) Клетки фибросаркомы HT1080 трансфецировали специфичной к каспазе-3 киРНК (C3) или контрольной киРНК, обрабатывали Apo2L/TRAIL в течение указанного периода времени и анализировали на отщепление AP2α или CHC или киРНК-истощение каспазы-3 с помощью иммуноблоттинга.
Фиг.9. Определение сайта отщепления AP2α. (a) C-концевой фрагмент отщепленного AP2α подвергали иммунопреципитации из Apo2L/TRAIL-стимулированных клеток BJAB, а затем либо подвергали гидролизу трипсином и анализировали с помощью масс-спектрометрии для подтверждения его идентичности, либо выделяли с помощью гель-электрофореза и вестерн-блоттинга и подвергали N-концевому секвенированию. Трипсиновые пептиды, идентифицированные с помощью тандемной масс-спектрометрии, выравнивали с С-концевой последовательностью AP2α. С помощью N-концевого секвенирования сайт расщепления был идентифицирован как мотив распознавания DXXD каспазой (подчеркнут). (b) Сайт расщепления картировали по «шарнирной» области AP2α, которая связывает функционально отличающиеся домены «ухо» и «туловище».
Фиг.10. Термочувствительные Dyn1 G273D и доминанто-негативные Dyn1 K44A -мутанты ингибируют эндоцитоз Tf и DR5 в клетках HeLa-M. (a, b) Две клональных линии, происходящие от трансдуцированной ретровирусом клеточной популяции HeLa-M, фиг.6, ts3 и tsl, и индуцированные (+dox) или не индуцированные (-dox) доксициклином, анализировали на поглощение 488Tf (А) или 6475С7 (b) в течение 20 минут при 30°С (белые столбцы) или при 38°С (черные столбцы) с помощью проточной цитометрии, как описано в разделе «Экспериментальные методы», и по средним значениям (± ср.кв.ош) строили график. (c) DynK44A-трансдуцированные и нетрансдуцированные (родительские) клетки HeLa-M, которые были индуцированы или не индуцированы доксициклином, анализировали на скорость эндоцитоза 488Tf и 6475С7 (± ср.кв.ош), как показано на фиг.5d без нормализации.
На фиг.11 проиллюстрирована сборка DISC в отсутствие эндоцитоза. (a) клетки BJAB уравновешивали 647Tf на льду, инкубировали в течение указанного периода времени для поглощения при 37°С или на льду, а затем количественно оценивали флуоресценцию с помощью проточной цитометрии. (b) Клетки BJAB обрабатывали перекрестно связанным Apo2L/TRAIL (1 мкг/мл) при 37°С или на льду в течение указанного периода времени, и DISC подвергали иммунопреципитации посредством лиганда, как описано в разделе «Материалы и методы». DISC-ассоциированные FADD и каспазу-8 визуализировали с помощью иммуноблоттинга. (c, d) DynG273D-трансдуцированные клетки BJAB обрабатывали буфером (-Dox) или доксициклином (Dox) и температуру доводили дл 38°С в течение 20 минут для инактивации динамина. Клетки обрабатывали перекрестно связанным Apo2L/TRAIL в течение указанного периода времени, и DISC подвергали иммунопреципитации. DISC-ассоциированные FADD и каспазу-8 визуализировали с помощью иммуноблотинга (с), либо DISC-ассоциированную активность каспазы-8 измеряли, как описано ранее (Sharp et al. J. Biol Chem 280, 19401-409, 2005).
На фиг.12 показано, что ДНК-разрушающие агенты индуцируют отщепление компонентов клатринового пути. Клетки BJAB или Colo205 обрабатывали винбластином или адриамицином в течение указанного периода времени и анализировали с помощью иммуноблоттинга, как показано на фиг.1.
Подробное описание изобретения
Методы и процедуры, описанные или упоминаемые в настоящей заявке, по существу, хорошо известны специалистам, и для их осуществления обычно применяется стандартная методика, известная специалистам. Если это необходимо, то процедуры, предусматривающие применение коммерчески доступных наборов и реагентов, обычно проводят в соответствии с протоколами и/или параметрами, предлагаемыми производителями, если не указаны какие-либо иные протоколы и/или параметры. Если это не оговорено особо, то все используемые здесь термины, обозначения и другая научная терминология имеют свои общепринятые значения, понятные специалистам в области, к которой относится настоящее изобретение. В некоторых случаях, для ясности и/или для удобства, в настоящем описании определены термины, имеющие общепринятые значения, и включение таких определений в настоящее описание не должно означать, что эти термины имеют значения, существенно отличающиеся от общепринятых значений. Кроме того, в настоящем описании используются некоторые сокращения, включая: 488-: Alexa-488-конъюгированный; 647-: Alexa-647-конъюгированный; Apo2L/TRAIL: лиганд, индуцирующий апоптоз, опосредуемый лигандом Apo2/TNF; BSA: альбумин бычьей сыворотки; DR: рецептор «гибели»; PBS: забуференный фосфатом физиологический раствор; Tf: трансферрин; и zVAD-fmk: N-бензилоксикарбонил-Val-Ala-Asp-фторметилкетон.
I. Определения
Термины «апоптоз» и «апоптотическая активность» используются здесь в широком смысле и означают запрограммированную или регулируемую гибель клеток у млекопитающих, которая обычно сопровождается одним или несколькими характерными клеточными изменениями, включая конденсацию цитоплазмы, потерю микроворсинок плазматической мембраны, сегментирование ядра, расщепление хромосомной ДНК или потерю митохондриальной функции. Такая активность может быть определена и измерена различными методами, известными специалистам, например, с помощью анализов на жизнеспособность клеток, FACS-анализа или ДНК-электрофореза, а более конкретно, по связыванию с аннексином V, фрагментации ДНК, сморщиванию клеток, расширению эндоплазматического ретикулума, фрагментации клеток и/или образованию мембранных везикул (называемых апоптотическими тельцами).
Используемые здесь термины «лиганд Apo-2», «Apo-2L» или «TRAIL» означают полипептид, включающий аминокислотные остатки 95-281, включительно остатки 114-281, включительно остатки 91-281, включительно остатки 92-281, включительно остатки 41-281, включительно остатки 15-281, включительно или остатки 1-281, включительно, аминокислотной последовательности, представленной на фиг.1A в публикации Pitti et al., J. Biol. Chem., 271:12687-12690 (1996), а также их биологически активные фрагменты и варианты с делециями, инсерциями или заменами. В одном из вариантов изобретения полипептидная последовательность содержит остатки 114-281. Указанная полипептидная последовательность имеет, но необязательно, по меньшей мере остатки 91-281 или остатки 92-281. В другом предпочтительном варианте изобретения аминокислотная последовательность биологически активных фрагментов или вариантов по меньшей мере примерно на 80%, более предпочтительно по меньшей мере примерно на 90%, а еще более предпочтительно по меньшей мере примерно на 95%, 96%, 97%, 98%, или 99% идентична аминокислотной последовательности любой одной из вышеуказанных последовательностей. Такое определение включает варианты лиганда Apo-2 с заменами, содержащие аминокислоты 91-281, указанные на фиг.1A, в публикации Pitti et al., J. Biol. Chem., 271:12687-12690 (1996), где по меньшей мере одна из аминокислот в положениях 203, 218 или 269 (в соответствии с нумерацией последовательностей, представленной в публикации Pitti et al., см. выше) заменена аланиновым остатком. Такое определение включает лиганд Apo-2, выделенный из источника лиганда Apo-2, такого как человеческая ткань, или из других источников, либо лиганд, полученный рекомбинантными методами или методами синтеза. Термин лиганд Apo-2 также означает полипептиды, описанные в WO 97/25428, см. выше, и в WO97/01633, см. выше.
Термин «рецептор лиганда Apo-2» включает рецепторы, известные специалистам как «DR4» и «DR5». В публикации Pan et al. описан член семейства рецепторов TNF, обозначаемый «DR4» (Pan et al., Science, 276:111-113 (1997); см. также заявку WO98/32856, опубликованную 30 июля 1998). Сообщалось, что рецептор DR4 содержит цитоплазматический домен гибели, способный действовать по механизму «клеточного суицида». В публикации Pan et al. указывается, что DR4 очевидно представляет собой рецептор лиганда, известного как Apo2L/TRAIL. В публикациях Sheridan et al., Science, 277:818-821 (1997) и Pan et al., Science, 277: 815-818 (1997) описан другой рецептор для Apo2L/TRAIL (см. также заявку WO98/51793, опубликованную 19 ноября, 1998; заявку W098/41629, опубликованную 24 сентября, 1998). Этот рецептор называется DR5 (этот рецептор также имеет и другие названия: Apo-2; TRAIL-R, TR6, Tango-63, hAPO8, TRICK2 или KILLER (Screaton et al., Curr. Biol., 7:693-696 (1997); Walczak et al., EMBO J., 16:5386-5387 (1997); Wu et al., Nature Genetics, 17:141-143 (1997); заявка WO98/35986, опубликованная 20 августа 1998; EP 870827, опубликованный 14 октября 1998; заявка WO98/46643, опубликованная 22 октября 1998; заявка WO99/02653, опубликованная 21 января 1999; заявка WO99/09165, опубликованная 25 февраля 1999; заявка W099/11791, опубликованная 11 марта 1999). Сообщалось, что DR5, подобно DR4, содержит цитоплазматический домен гибели и обладает способностью передавать сигнал на апоптоз. Как описано выше, другими рецепторами для Apo-2L являются DcR1, DcR2 и OPG (см. Sheridan et al., см.выше; Marsters et al., см.выше и Simonet et al., см.выше). Термин «рецептор Apo-2L», если он используется в настоящей заявке, включает рецептор с нативной последовательностью и его варианты. Эти термины включают рецептор Apo-2L, экспрессируемый у различных млекопитающих, включая человека. Рецептор Apo-2L может эндогенно экспрессироваться как природный рецептор в различных линиях клеточной дифференцировки человеческих тканей, либо он может быть экспрессирован рекомбинантными методами или методами синтеза. Термин «рецептор Apo-2L с нативной последовательностью» включает полипептид, имеющий такую же аминокислотную последовательность, как и рецептор Apo-2L, происходящий от природной последовательности. Таким образом, рецептор Apo-2L с нативной последовательностью может иметь аминокислотную последовательность природного рецептора Apo-2L любого млекопитающего. Такой рецептор Apo-2L с нативной последовательностью может быть выделен из природных источников, либо он может быть получен рекомбинантными методами или методами синтеза. Термин «рецептор Apo-2L с нативной последовательностью», в частности, включает природную усеченную или секретированную форму рецептора (например, растворимую форму, содержащую, например, последовательность внеклеточного домена), природные модифицированные формы (например, формы альтернативного сплайсинга) и природные аллельные варианты. Варианты этих рецепторов могут включать фрагменты или делеционные мутанты рецептора Apo-2L с нативной последовательностью.
Используемый здесь термин «Fas» означает полипептид, включающий аминокислотные остатки 1-319, представленные в NCBI рег. No. AAA63174 и описанные в публикации Itoh et al., Cell 66 (2), 233-243 (1991), а также их биологически активные фрагменты и варианты вышеуказанных последовательностей, имеющие делеции, инсерции или замены. В некоторых публикациях такой полипептид обозначается «Apo-1» и «CD95». Используемый здесь термин «лиганд Fas» или «FasL» означает полипептид, включающий аминокислотные остатки 1-281, представленные в NCBI рег. No. NP_000630 и описанные в публикации Suda et al., Cell 75 (6), 1169-1178 (1993), а также их биологически активные фрагменты и варианты вышеуказанных последовательностей, имеющие делеции, инсерции или замены. Связывание FasL с Fas или перекрестное связывание Fas с антителами-агонистами индуцируют апопотоз, приводящий к гибели клеток (см. например, Nagata, S. Ann. Rev. Genet. 33:29, 1999; and Labroille et al., Cytometry 39(3): 195-202 (2000)). Связывание FasL с Fas активирует каскад каспазных реакций, опосредуемых адаптором FADD (Fas-ассоциированным белком с доменом гибели), что приводит к расщеплению различных клеточных субстратов и к фрагментации ДНК.
“Процент (%) идентичности аминокислотных последовательностей», относящийся к идентифицированным здесь полипептидным последовательностям, определяется как процент аминокислотных остатков в последовательности-кандидате, которые являются идентичными аминокислотным остаткам в полипептидной последовательности после сопоставления последовательностей путем выравнивания и введения пробелов, если это необходимо для достижения максимального процента идентичности последовательностей, причем при определении идентичности последовательностей не рассматриваются какие-либо консервативные замены. Выравнивание, проводимое в целях определения процента идентичности аминокислотных последовательностей, может быть достигнуто различными методами, известными специалистам, например, с помощью общедоступных компьютерных программ, таких как BLAST, BLAST-2, ALIGN, ALlGN-2 или Megalign (DNASTAR). Специалист в данной области может самостоятельно определить соответствующие параметры для анализа выравнивания, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сопоставляемых последовательностей. Процент идентичности аминокислотных последовательностей вычисляют, но необязательно, с использованием компьютерной программы для сравнения последовательностей ALIGN-2. Компьютерная программа для сравнения последовательностей ALIGN-2 была разработана компанией Genentech, Inc., и исходный код программы был зарегистрирован в пользовательской документации в Ведомстве по копирайту США (U.S. Copyright Office, Washington, D. C, 20559), под регистрационным номером No. TXU510087. Программа ALIGN-2 является общедоступной на сайте Genentech, Inc., South San Francisco, California. Программа ALIGN-2 program должна быть составлена для пользования в операционной системе UNIX, а предпочтительно в цифровой форме UNIX V4.0D. В программе ALIGN-2 имеются все параметры для сравнения последовательностей и они не меняются. Однако % идентичности аминокислотных последовательностей может быть также определен с использованием программы для сравнения последовательностей NCBI-BLAST2 (Altschul et al., Nucleic Acids Res. 25:3389-3402 (1997)). Программа для сравнения последовательностей NCBI-BLAST2 может быть загружена с сайта http://www.ncbi.nlm.nih.gov. В программе NCBI-BLAST2 используются несколько параметров поиска, где все эти параметры поиска даны по умолчанию, включая, например, разрешение (unmask) = да (yes), цепь (strand) = все (all), ожидаемые события (expected occurrences) = 10, длина с минимально низкой сложностью (minimum low complexity length) = 15/5, многопроходная e-величина (multi-pass e-value) = 0,01, константа для многопроходного алгоритма (constant for multi-pass) = 25, отсечка для конечного выравнивания с пробелами (dropoff for final gapped alignment) = 25 и оценочная матрица (scoring matrix) = BLOSUM62.
Термин «антитело» используется здесь в самом широком смысле, а в частности охватывает интактные моноклональные антитела, поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела), образованные по меньшей мере двумя интактными антителами, и фрагменты антител. Используемый здесь термин «моноклональное антитело» означает антитело, происходящее от популяции по существу гомогенных антител, то есть отдельные антитела, входящие в такую популяцию, являются идентичными, за исключением возможных природных мутаций, которые могут присутствовать в незначительных количествах. Моноклональные антитела являются в высокой степени специфическими и направлены против одного антигенного сайта. Кроме того, в противоположность стандартным (поликлональным) препаратам антител, которыми обычно являются различные антитела, направленные против различных детерминант (эпитопов), каждое моноклональное антитело направлено против одной детерминанты на антигене. Моноклональные антитела, помимо их специфичности, имеют и другие преимущества, заключающиеся в том, что они могут быть синтезированы гибридомным методом без внесения других примесных иммуноглобулинов. Определение «моноклональный» указывает на тип антитела как антитела, полученного, по существу, из гомогенной популяции антител, но это не должно быть истолковано как необходимость продуцирования такого антитела любым конкретным методом. Так, например, моноклональные антитела, используемые в соответствии с настоящим изобретением, могут быть получены гибридомным методом, впервые описанным Kohler et al., Nature, 256:495 (1975), или они могут быть получены методами рекомбинантных ДНК (см., например, патент США No. 4816567). «Моноклональные антитела» могут быть также выделены, например, из фаговых библиотек антител методами, описанными, например, в публикациях Clackson et al., Nature, 352:624-628 (1991) и Marks et al., J. Mol. Biol., 222:581-597 (1991). Антителом, которое «связывается» с представляющим интерес антигеном, является антитело, обладающее способностью связываться с антигеном с достаточной степенью аффинности и/или авидности, а поэтому такое антитело может быть использовано в качестве диагностического или терапевтического средства для его доставки в клетку, экспрессирующую антиген.
Используемый здесь термин «антитело против рецептора гибели», по существу, означает антитело или антитела, направленные против рецептора, принадлежащего к суперсемейству рецепторов фактора некроза опухоли и содержащего домен гибели, способный передавать сигнал на апоптоз, и такими антителами являются антитело против DR5, антитело против DR4 и антитело против Fas.
Термины «антитело против рецептора DR5», «DR5-антитело» или «анти-DR5-антитело», используемые здесь в широком смысле, означают антитела, которые связываются по меньшей мере с одной формой рецептора DR5. DR5-антитело может быть, но необязательно, конъюгировано или связано с гетерологичной последовательностью или молекулой. Предпочтительно такая гетерологичная последовательность позволяет антителу образовывать комплексы высшего порядка или олигомерные комплексы, или стимулирует такой процесс. DR5-антитело связывается, но необязательно, с рецептором DR5, но не связывается или не вступает в перекрестную реакцию с любым дополнительным рецептором Apo-2L (например, DR4, DcR1 или DcR2). Это антитело является, но необязательно, агонистом активности передачи сигнала DR5 (см., например, заявки на патент США №№ 20040005314 и 20060188498).
Термины «антитело против рецептора DR4», «DR4-антитело» или «анти-DR4-антитело», используемые здесь в широком смысле, означают антитела, которые связываются по меньшей мере с одной формой рецептора DR4 или его внеклеточным доменом. DR4-антитело может быть, но необязательно, конъюгировано или связано с гетерологичной последовательностью или молекулой. Предпочтительно такая гетерологичная последовательность позволяет антителу образовывать комплексы высшего порядка или олигомерные комплексы, или стимулирует такой процесс. DR4-антитело связывается, но необязательно, с рецептором DR4, но не связывается или не вступает в перекрестную реакцию с любым дополнительным рецептором Apo-2L (например, DR5, DcR1 или DcR2). Это антитело является, но необязательно, агонистом активности передачи сигнала DR4 (см., например, заявки на патент США №№ 20040005314 и 20060188498).
Термины «Fas-антитело» или «анти-Fas-антитело», используемые здесь в широком смысле, означают антитела, которые связываются по меньшей мере с одной формой Fas или его внеклеточным доменом. Антитело против Fas может быть, но необязательно, конъюгировано или связано с гетерологичной последовательностью или молекулой. Предпочтительно такая гетерологичная последовательность позволяет антителу образовывать комплексы высшего порядка или олигомерные комплексы, или стимулирует такой процесс. Антитело против Fas связывается, но необязательно, с рецептором FasL, но не связывается или не вступает в перекрестную реакцию с любым дополнительным рецептором Fas. Это антитело является, но необязательно, агонистом активности передачи сигнала Fas (см., например, Nagata, S. Ann. Rev. Genet. 33:29, 1999; and Labroille et al., Cytometry 39(3): 195-202 (2000)).
Описанными здесь моноклональными антителами, в частности, являются «химерные» антитела (иммуноглобулины), в которых часть тяжелой цепи и/или легкой цепи идентична или гомологична соответствующим последовательностям антител, происходящих от конкретных видов, или принадлежащих к антителам конкретного класса или подкласса, а остальные цепи идентичны или гомологичны соответствующим последовательностям антител, происходящих от других видов или принадлежащих к антителам другого класса или подкласса, а также фрагменты таких антител, при условии, что они обладают нужной биологической активностью (патент США No. 4816567; Morrison et al., Proc. Natl. Acad. Sci. USA, 81:6851-6855 (1984)).
«Гуманизованными» формами нечеловеческих (например, мышиных) антител являются химерные иммуноглобулины, цепи иммуноглобулинов или их фрагменты (такие как Fv, Fab, Fab', F(ab')2 или другие антигенсвязывающие подпоследовательности антител), содержащие минимальное число последовательностей, происходящих от нечеловеческого иммуноглобулина. По большей части гуманизованными антителами являются человеческие иммуноглобулины (антитело-реципиент), в которых остатки, происходящие от гипервариабельной области (CDR) реципиента, заменены остатками, происходящими от CDR животного, не являющегося человеком (донорного антитела), такого как мышь, крыса или кролик, и обладающими нужной специфичностью, аффинностью и емкостью. В некоторых случаях остатки каркасной области (FR) Fv человеческого иммуноглобулина заменены соответствующими нечеловеческими остатками. Кроме того, гуманизованные антитела могут содержать остатки, не присутствующие ни в реципиентном антителе, ни в “импортных” CDR или каркасных последовательностях. Эти модификации вводят для дополнительного улучшения и максимизации эффективности антитела. В общих чертах гуманизованное антитело содержит, в основном, по меньшей мере один, а обычно два вариабельных домена, в которых все или, в основном, все СDR-области соответствуют областям нечеловеческого иммуноглобулина, а все или почти все области FR являются областями, имеющими последовательность человеческого иммуноглобулина. В оптимальном случае гуманизованное антитело также содержит по меньшей мере часть константной области иммуноглобулина (Fc), обычно человеческого иммуноглобулина. Подробное описание можно найти в публикации Jones et al., Nature, 321:522-525 (1986); Reichmann et al., Nature, 332:323-329 (1988) и Presta, Curr. Op. Struct. Biol., 2:593-596 (1992). Термин “гуманизованное антитело” включает приматизированноеТМ антитело, антигенсвязывающая область которого происходит от антитела, продуцируемого в результате иммунизации макак представляющим интерес антигеном.
Антитела обычно представляют собой белки или полипептиды, обладающие специфичностью связывания с конкретным антигеном. Нативными антителами обычно являются гетеротетрамерные гликопротеины, состоящие из двух идентичных легких (L) цепей и двух идентичных тяжелых (H) цепей. Каждая легкая цепь связана с тяжелой цепью одной ковалентной дисульфидной связью, а число дисульфидных связей в тяжелых цепях иммуноглобулинов различных изотипов варьируется. Обычно, каждая тяжелая и легкая цепь также имеют правильно расположенные внутрицепьевые дисульфидные мостики. Каждая тяжелая цепь имеет на одном конце вариабельный домен (VН), за которым следует ряд константных доменов. Каждая легкая цепь имеет вариабельный домен на одном конце (VL) и константный домен на другом конце, причем константный домен легкой цепи находится непосредственно напротив первого константного домена тяжелой цепи, а вариабельный домен легкой цепи находится непосредственно напротив вариабельного домена тяжелой цепи. Очевидно, что конкретные аминокислотные остатки образуют границу между вариабельными доменами легкой и тяжелой цепей (Chothia et al., J. Mol. Biol., 186:651-663 (1985); Novotny and Haber, Proc. Natl. Acad. Sci. USA, 82:4592-4596 (1985)). Легкие цепи антител позвоночных любых видов могут быть отнесены к одному из двух абсолютно различных типов, называемых каппа и лямбда, исходя из аминокислотных последовательностей их константных доменов. Иммуноглобулины, в зависимости от аминокислотной последовательности константного домена тяжелых цепей, могут быть отнесены к различным классам. Существует пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, а некоторые из этих классов иммуноглобулинов могут быть дополнительно подразделены на подклассы (изотипы), например, IgG-1, IgG-2, IgG-3 и IgG-4; IgA-1 и IgA-2. Константные домены тяжелой цепи, соответствующие иммуноглобулинам различных классов, называются альфа, дельта, эпсилон, гамма и мю, соответственно.
«Фрагменты антител» содержат часть интактного антитела, а в основном антигенсвязывающую или вариабельную область интактного антитела. Примерами фрагментов антител являются Fab-, Fab'-, F(ab')2- и Fv-фрагменты, диантитела, одноцепочечные молекулы антител и мультиспецифические антитела, происходящие от фрагментов антител.
Используемый здесь термин «вариабельный» относится к некоторым частям вариабельных доменов, которые имеют отличия в последовательностях различных антител и участвуют в связывании каждого конкретного антитела с его конкретным антигеном, а также определяют специфичность каждого конкретного антитела по отношению к его конкретному антигену. Однако вариабельность неравномерно распределяется по всем вариабельным доменам антител. Она обычно концентрируется в трех сегментах, называемых комплементарность-определяющими областями (СDR) или гипервариабельными областями, присутствующими в вариабельных доменах как легкой, так и тяжелой цепи. Наиболее высококонсервативные части вариабельных доменов называются каркасными областями (FR). Каждый вариабельный домен нативной тяжелой и легкой цепей содержит четыре FR-области, имеющие, главным образом, β-складчатую конфигурацию и соединенные тремя СDR, которые образуют петли, соединяющие, а в некоторых случаях образующие часть β-складчатой структуры. СDR в каждой цепи удерживаются в непосредственной близости друг от друга посредством областей FR, и, вместе с СDR другой цепи, участвуют в образовании антигенсвязывающего сайта антител (см. Kabat et al. Sequences of Proteins of Immunological Interest, National Institutes of Health, Bethesda, MD (1987)). Константные домены не принимают непосредственного участия в связывании антитела с антигеном, но обладают различными эффекторными функциями, такими как участие антитела в антителозависимой клеточной цитотоксичности.
Описанными здесь моноклональными антителами являются химерные, гибридные и рекомбинантные антитела, продуцированные путем сплайсинга вариабельного (включая гипервариабельный) домена антитела против рецептора Apo-2L с константным доменом (например, «гуманизованных антител») или легкой цепи с тяжелой цепью, или цепи одного вида с цепью другого вида; или гибриды с гетерологичными белками, независимо от вида, от которого они происходят, или от класса или подкласса иммуноглобулина, а также фрагменты антител (например, Fab, F(ab')2 и Fv), при условии, что они обладают нужной биологической активностью или нужными свойствами. См., например, патент США No. 4816567 и Mage et al., in Monoclonal Antibody Production Techniques and Applications, pp.79-97 (Marcel Dekker, Inc.: New York, 1987).
«Человеческое антитело» представляет собой антитело, имеющее аминокислотную последовательность, соответствующую последовательности антитела, продуцируемого у человека, и/или антитело, полученное любыми описанными здесь методами продуцирования человеческих антител. Такое определение человеческого антитела, в частности, исключает гуманизованное антитело, содержащее нечеловеческие антигенсвязывающие остатки. Человеческие антитела могут быть продуцированы различными методами, известными специалистам. В одном из вариантов изобретения человеческое антитело выбирают из фаговой библиотеки, экспрессирующей человеческие антитела (Vaughan et al. Nature Biotechnology, 14:309-314 (1996): Sheets et al. PNAS, (USA) 95:6157-6162 (1998)); Hoogenboom and Winter, J. Mol. Biol., 227:381 (1991); Marks et al., J. Mol. Biol., 222:581 (1991)). Человеческие антитела могут быть также получены путем введения локусов человеческого иммуноглобулина трансгенным животным, например мышам, у которых эндогенные гены иммуноглобулина были частично или полностью инактивированы. После такого введения наблюдается продуцирование человеческих антител, которые по всем параметрам имеют очень большое сходство с антителами, наблюдаемыми у человека, включая реаранжировку и сборку генов и репертуар антител. Этот метод описан, например, в патентах США №№ 5545807, 5545806; 5569825; 5625126; 5633425; 5661016 и в нижеследующих научных публикациях: Marks et al., Bio/Technology, 10: 779-783 (1992); Lonberg et al., Nature, 368: 856-859 (1994); Morrison, Nature, 368:812-13 (1994); Fishwild et al.. Nature Biotechnology, 14: 845-51 (1996); Neuberger, Nature Biotechnology, 14: 826 (1996); Lonberg and Huszar, Intern. Rev. Immunol., 13:65-93 (1995). Альтернативно, человеческое антитело может быть получено путем иммортализации человеческих В-лимфоцитов, продуцирующих антитело, направленное против антигена-мишени (такие В-лимфоциты могут быть выделены у индивидуума, либо они могут быть введены путем иммунизации in vitro). См., например, Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p. 77 (1985); Boerner et al., J. Immunol., 147 (1):86- 95 (1991); и патент США No. 5750373.
Термин “Fc-область” используется для определения С-концевой области тяжелой цепи иммуноглобулина, которая может быть получена путем гидролиза интактного антитела папаином. Такая “Fc-область” может представлять собой Fc-область с нативной последовательностью или модифицированную Fc-область. Хотя границы Fc-области тяжелой цепи иммуноглобулина могут варьироваться, однако Fc-область тяжелой цепи человеческого IgG обычно определяют как фрагмент, начинающийся от аминокислотного остатка в положении примерно Cys226 или от остатка в положении примерно Pro230, и простирающийся до карбокси-конца Fc-области (в соответствии с используемой здесь системой нумерации по Кэбату и др., см выше). Fc-область иммуноглобулина обычно содержит два константных домена, СН2 и СН3, и необязательно домен СН4.
Используемый здесь термин «цепь Fc-области» означает одну из двух полипептидных цепей Fc-области.
“Домен СН2” Fc-области человеческого IgG (также обозначаемый доменом “Cγ2”) обычно простирается примерно от аминокислотного остатка в положении 231 до аминокислотного остатка в положении примерно 340. Уникальность домена СН2 состоит в том, что он не имеет непосредственной связи с другим доменом. Если сказать точнее, то две N-связанных разветвленных углеводных цепи располагаются между двумя доменами СН2 интактной молекулы нативного IgG. Было высказано предположение, что такой углевод может служить заменой паре “домен-домен” и может способствовать стабилизации домена СН2. См. Burton, Molec. Immunol. 22:161-206 (1985). Описанный здесь домен СН2 может представлять собой домен СН2 с нативной последовательностью или модифицированный домен СН2.
“Домен СН3” содержит фрагмент из остатков, простирающихся от С-конца до домена СН2 в Fc-области (т.е. он простирается от аминокислотного остатка в положении примерно 341 до аминокислотного остатка в положении примерно 447 IgG). Описанная здесь область СН3 может представлять собой домен СН3 с нативной последовательностью или модифицированный домен СН3 (например, домен СН3 с введенным «выступающим концом» в одной цепи и с соответствующей введенной «полостью» в другой цепи; см. патент США No. 5821333).
«Шарнирная область», по существу, определена как фрагмент, простирающийся примерно от положения Glu216 или Cys226 примерно до положения Pro230 человеческого IgG1 (Burton, Molec. Immunol. 22:161-206 (1985)). Шарнирные области IgG других изотипов могут быть сопоставлены с последовательностью IgG1 путем введения первого и последнего цистеиновых остатков, образующих межцепьевые S-S-связи тяжелой цепи в одних и тех же положениях. Описанная здесь «шарнирная область» может представлять собой шарнирную область с нативной последовательностью или модифицированную шарнирную область. Две полипептидных цепи модифицированной шарнирной области, по существу, содержат, по меньшей мере один цистеиновый остаток на полипептидную цепь, а поэтому две полипептидных цепи указанной модифицированной шарнирной области могут образовывать дисульфидную связь между двумя цепями. Описанной здесь предпочтительной шарнирной областью является человеческая шарнирная область с нативной последовательностью, например шарнирная область человеческого IgG1 с нативной последовательностью.
«Функциональная Fc-область» обладает по меньшей мере одной «эффекторной функцией» Fc-области с нативной последовательностью. Примерами «эффекторных функций» являются: связывание с C1q; комплементзависимая цитотоксичность (СDС); связывание с Fc-рецептором; антителозависимая клеточно-опосредуемая цитотоксичность (ADCC); фагоцитоз; негативная регуляция рецепторов клеточной поверхности (например, В-клеточного рецептора; BCR) и т.п. Для обеспечения таких эффекторных функций обычно требуется, чтобы Fc-область была объединена со связывающим доменом (например, с вариабельным доменом антитела), и такие эффекторные функции могут быть проанализированы, например, различными описанными здесь методами, известными специалистам.
«Fc-область с нативной последовательностью» содержит аминокислотную последовательность, идентичную природной аминокислотной последовательности Fc-области. «Модифицированная Fc-область» содержит аминокислотную последовательность, отличающуюся от аминокислотной последовательности нативной Fc-области, что обусловлено введением по меньшей мере одной модификации аминокислоты. Предпочтительно, чтобы такая модифицированная Fc-область, в отличие от Fc-области с нативной последовательностью или Fc-области родительского полипептида, имела по меньшей мере одну аминокислотную замену, например примерно от одной до десяти аминокислотных замен, а предпочтительно примерно от одной до пяти аминокислотных замен. Модифицированная Fc-область предпочтительно должна быть по меньшей мере примерно на 80%, более предпочтительно по меньшей мере примерно на 90%, а наиболее предпочтительно по меньшей мере примерно на 95% идентична Fc-области с нативной последовательностью и/или Fc-области родительского полипептида.
Используемые здесь термины “Fc-рецептор” или “FcR” означают рецептор, который связывается с Fc-областью антитела. Предпочтительным FcR является человеческий FcR с нативной последовательностью. Кроме того, предпочтительный FcR представляет собой FcR, который связывается с антителом IgG (гамма-рецептор), и охватывает рецепторы подклассов FcγRI, FcγRII и FcγRIII, включая аллельные варианты и альтернативно сплайсированные формы этих рецепторов. Рецепторы FcγRII включают FcγRIIA (“активирующий рецептор”) и FcγRIIB (“ингибирующий рецептор”), которые имеют аналогичные аминокислотные последовательности, отличающиеся главным образом своими цитоплазматическими доменами. Активирующий рецептор FcγRIIA содержит в своем цитоплазматическом домене иммунорецепторный активирующий мотив на основе тирозина (ITAM). Ингибирующий рецептор FcγRIIВ содержит в своем цитоплазматическом домене иммунорецепторный ингибирующий мотив на основе тирозина (ITIM)(см. публикацию M. Daeron, Annu. Rev. Immunol. 15:203-234 (1997)). FcR описаны в публикациях Ravetch and Kinet, Annu. Rev. Immunol., 9:457-92 (1991); Capel et al., Immunomethods, 4:25-34 (1994); и de Haas et al., J. Lab. Clin. Med., 126:330-41 (1995). Используемый здесь термин «FcR» также охватывает и другие FcR, включая FcR, которые будут идентифицированы в будущем. Этот термин также включает неонатальный рецептор FcRn, ответственный за передачу материнских IgG плоду (Guyer et al., J. Immunol. 117:587 (1976) и Kim et al., J. Immunol. 24:249 (1994)).
«Аффинно-зрелое» антитело представляет собой антитело, имеющее одну или несколько модификаций в одной или нескольких CDR, что позволяет повышать аффинность данного антитела к антигену, по сравнению с родительским антителом, не имеющим такой(их) модификации(й). Предпочтительные аффинно-зрелые антитела имеют наномолярные или даже пикомолярные аффинности по отношению к антигену-мишени. Аффинно-зрелые антитела получают известными методами. В публикации Marks и др. (Bio/Technology 10:779-783 (1992)) описано созревание аффинности в результате перестановки доменов VH и VL. Неспецифический мутагенез CDR и/или каркасных остатков описан в публикациях Barbas et al., Proc. Natl. Acad. Sci., USA, 91:3809-3813 (1994); Schier et al., Gene 169:147-155 (1995); Yelton et al., J. Immunol. 155:1994-2004 (1995); Jackson et al., J. Immunol. 154(7):3310-9 (1995) и Hawkins et al., J. Mol. Biol. 226:889-896 (1992).
Используемый здесь термин «метка» означает соединение или композицию, которые прямо или опосредованно связываются или конъюгируются с реагентом, таким как антитело, и таким образом облегчают детектирование реагента, с которым они связаны или конъюгированы. Сама метка может быть детектируемой (например, радиоизотопные метки, флуоресцентные метки) или, в случае ферментативной метки, такая метка может катализировать изменение химического состава композиции, которая включает соединение субстрата и является детектируемой.
Термин «антагонист» используется здесь в самом широком смысле и включает любую молекулу, которая частично или полностью блокирует, ингибирует или нейтрализует одну или несколько биологических активностей Apo2L/TRAIL, DR4 или DR5, FasL или Fas in vitro, in situ или in vivo. Примерами таких биологических активностей Apo2L/TRAIL, DR4 или DR5 являются связывание Apo2L/TRAIL с DR4 или DR5, индуцирование апоптоза, а также другие свойства, описанные в литературе. Примерами таких биологических активностей FasL и Fas являются связывание FasL с Fas, индуцирование апоптоза, а также другие свойства, описанные в литературе. Антагонист может действовать прямо или опосредованно. Так, например, антагонист может частично или полностью блокировать, ингибировать или нейтрализовать одну или несколько биологических активностей Apo2L/TRAIL, in vitro, in situ или in vivo в результате его прямого связывания с DR4 или DR5. Такой антагонист может также опосредованно частично или полностью блокировать, ингибировать или нейтрализовать одну или несколько биологических активностей Apo2L/TRAIL, DR4 или DR5 in vitro, in situ или in vivo в результате, например, блокирования или ингибирования другой эффекторной молекулы. Молекула антагониста может обладать «двойной» антагонистической активностью, где указанная молекула обладает способностью частично или полностью блокировать, ингибировать или нейтрализовать биологическую активность Apo2L/TRAIL, DR4 или DR5, Fas или FasL.
Термин «агонист» используется здесь в самом широком смысле и включает любую молекулу, которая частично или полностью усиливает, стимулирует или активирует одну или несколько биологических активностей Apo2L/TRAIL, DR4 или DR5, FasL или Fas in vitro, in situ или in vivo. Примерами таких биологических активностей Apo2L/TRAIL, DR4 или DR5 являются связывание Apo2L/TRAIL с DR4 или DR5, индуцирование апоптоза, а также другие свойства, описанные в литературе. Примерами таких биологических активностей FasL и Fas являются связывание FasL с Fas, индуцирование апоптоза, а также другие свойства, описанные в литературе. Агонист может действовать прямо или опосредованно. Так, например, агонист может частично или полностью усиливать, стимулировать или активировать одну или несколько биологических активностей DR4 или DR5 in vitro, in situ или in vivo в результате своего прямого связывания с DR4 или DR5, что будет приводить к активации рецептора или к передаче сигнала. Такой агонист может также опосредованно частично или полностью усиливать, стимулировать или активировать одну или несколько биологических активностей DR4 или DR5 in vitro, in situ или in vivo в результате, например, стимуляции другой эффекторной молекулы, которая затем будет индуцировать активацию или передачу сигнала DR4 или DR5. При этом считается, что агонист может действовать как молекула-энхансер, которая опосредованно усиливает или повышает уровень активации или активности DR4 или DR5. Так, например, агонист может усиливать активность эндогенного Apo-2L у млекопитающего. Это может быть осуществлено, например, путем предварительного образования комплекса DR4 или DR5 или путем стабилизации комплексов соответствующего лиганда с рецептором DR4 или DR5 (например, стабилизации нативного комплекса, образованного между Apo-2L и DR4 или DR5).
Термин «выделенный», используемый здесь для описани различных белков, относится к белку, который был идентифицирован и отделен от компонента и/или выделен из компонента его природной среды. Примесными компонентами его природной среды являются вещества, которые обычно негативно влияют на диагностическое или терапевтическое применение указанного белка, и такими веществами могут быть ферменты, гормоны и другие белковые или небелковые растворенные вещества. В предпочтительных вариантах изобретения указанный белок может быть очищен (1) до степени, достаточной для получения по меньшей мере 15 остатков в N-концевой или внутренней аминокислотной последовательности, с использованием секвенатора с центрифужной чашкой, или (2) до гомогенности, как может быть определено с помощью электрофореза в ДСН-ПААГ в восстанавливающих или в невосстанавливающих условиях путем окрашивания кумасси синим, или, предпочтительно, серебром. Термин “выделенный белок” включает белок in situ, присутствующий в рекомбинантных клетках, при условии, что в нем отсутствует по меньшей мере один компонент его природного окружения. Однако, обычно, выделенный белок может быть получен по меньшей мере в одну стадию очистки.
Используемый здесь термин «биологически активный» или «биологическая активность» относится к веществу, обладающему способностью (a) индуцировать или стимулировать апоптоз раковой клетки млекопитающего по меньшей мере одного типа или инфицированной вирусом клетки in vivo или ex vivo; (b) продуцировать антитело, то есть иммуногенное антитело; или (c) сохранять активность нативного или природного полипептида лиганда Apo-2.
Используемый здесь термин “рост-ингибирующее средство” означает соединение или композицию, ингибирующие рост клеток in vitro и/или in vivo. Таким образом, средством, ингибирующим рост клеток, может быть средство, значительно снижающее процент клеток в фазе S. Примерами средств, ингибирующих рост клеток, являются средства, блокирующие прохождение клеточного цикла (в другой фазе, кроме фазы S), такие как средства, индуцирующие блокирование фазы G1 и фазы М. Классическими средствами, блокирующими фазу М, являются винкаалкалоиды (винкристин и винбластин), таксол® и ингибиторы топоизомеразы II, такие как доксорубицин, эпирубицин, даунорубицин, этопозид и блеомицин. Средствами, блокирующими фазу G1, а также блокирующими переход в фазу S, являются, например, ДНК-алкилирующие агенты, такие как тамоксифен, преднизон, дакарбазин, мехлоретамин, цисплатин, метотрексат, 5-фторурацил и ara-C. Дополнительную информацию можно найти в публикации “The Molecular Basis of Cancer”, Mendelsohn & Israel, eds., в главе 1, озаглавленной “Cell cycle regulation, oncogenes and antineoplastic drugs”, Murakami et al. (WB Saunders: Philadelphia, 1995), а в частности на стр.13.
Используемый в данной заявке термин “пролекарство” означает предшественник или производное фармацевтически активного вещества, которые, по сравнению с родительским лекарственным средством, являются менее цитотоксичными по отношению к раковым клеткам и способны ферментативно активироваться или превращаться в более активную зрелую форму. См., например, Wilman, “Prodrugs in Cancer Chemotherapy” Biochemical Society Transactions, 14, pp.375-382, 615th Meeting Belfast (1986) и Stella et al. “Prodrugs: A Chemical Approach to Targeted Drug Delivery”, Directed Drug Delivery, Borchardt et al., (ed.), pp.247-267, Humana Press (1985). Пролекарствами согласно изобретению являются, но не ограничиваются ими, фосфатсодержащие пролекарства, тиофосфатсодержащие пролекарства, сульфатсодержащие пролекарства, пептидсодержащие пролекарства, пролекарства, модифицированные D-аминокислотой, гликозилированные пролекарства, бета-лактамсодержащие пролекарства; пролекарства, содержащие необязательно замещенный феноксиацетамид, или пролекарства, содержащие необязательно замещенный фенилацетамид; 5-фторцитозиновые и другие 5-фторуридиновые пролекарства, которые могут быть превращены в более активное цитотоксическое свободное лекарственное средство. Примерами цитотоксических лекарственных средств, которые могут быть дериватизированы с получением пролекарственной формы для ее использования в настоящем изобретении, являются, но не ограничиваются ими, химиотерапевтические агенты, описанные ниже.
Используемый здесь термин “цитотоксическое средство” означает вещество, ингибирующее или предотвращающее функцию клеток и/или вызывающее деструкцию клеток. Этот термин включает радиоактивные изотопы (например, 211At, 131I, 125I, 90Y, 186Re, 188Re, 153Sm, 212Bi, 32Р и радиоактивные изотопы Lu), химиотерапевтические средства и токсины, такие как токсины в виде небольших молекул, или ферментативно активные токсины бактериального, грибкового, растительного или животного происхождения, включая их фрагменты и/или варианты.
«Химиотерапевтическое средство» представляет собой химическое соединение, используемое для лечения заболеваний, таких как рак. Примерами химиотерапевтических средств являются алкилирующие агенты, такие как тиотепа и циклофосфамид (цитоксан®); алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбоквон, метуредопа и уредопа; этиленимины и метиламеламины, включая альтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметилолмеламин; ацетогенины (а в частности буллатацин и буллатацинон); камптотецин (включая синтетический аналог топотекан); бриостатин; каллистатин; СС-1065 (включая его синтетические аналоги, такие как адозелезин, карзелезин и бизелезин); криптофицины (а в частности криптофицин 1 и криптофицин 8); доластатин; дуокармицин (включая синтетические аналоги KW-2189 и CBI-TMI); элеутеробин; панкратистатин; саркодиктиин; спонгистатин; азотные аналоги горчичного газа, такие как хлорамбуцил, хлорнафазин, холофосфамид, эстрамустин, ифосфамид, мехлоретамин, гидрохлорид оксида мехлоретамина, мелфалан, новембихин, фенестерин, преднимустин, трофосфамид, урациловый аналог горчичного газа; нитрозомочевины, такие как кармустин, хлорозотоцин, фотемустин, ломустин, нимустин и ранимнустин; антибиотики, такие как энедииновые антибиотики (например, калихеамицин, а в частности калихеамицин 1I 1 и калихеамицин 2I 1, см., например, Agnew Сhem. Intl. Ed. Engl.. 33:183-186 (1994); динемицин, включая динемицин А; эсперамицин, а также неокарзиностатиновый хромофор и родственные хромопротеиновые энедииновые антибиотики-хромофоры), аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, карминомицин, карзинофилин, хромомицины, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, доксорубицин (включая морфолинодоксорубицин, цианоморфолинодоксорубицин, 2-пирролинодоксорубицин и дезоксидоксорубицин), эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, микофеноловая кислота, ногаламицин, оливомицины, пепломицин, потфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат и 5-фторурацил (5-FU); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; пуриновые аналоги, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; пиримидиновые аналоги, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флоксуридин, 5-FU; андрогены, такие как калустерон, пропионат дромостанолона, эпитиостанол, мепитиостан, тестолактон; антиадренергические средства, такие как аминоглютетимид, митотан, трилостан; заменитель фолиевой кислоты, такой как фролиновая кислота; ацеглатон; альдофосфамидный гликозид; аминолевулиновая кислота; амсакрин; бестрабуцил; бизантрен; эдатраксат; дефофамин; демеколцин; диазиквон; элфорнитин; ацетат эллиптиния; эпотилон; этоглюцид; нитрат галлия; гидроксимочевина; лентинан; лонидамин; майтанзиноиды, такие как майтанзин и ансамитоцины; митогуазон; митоксантрон; мопидамол; нитракрин; пентостатин; фенамет; пирарубицин; подофиллиновая кислота; 2-этилгидразид; прокарбазин; PSK®; разоксан; ризоксин; сизофиран; спирогерманий; тенуазоновая кислота; триазиквон; 2,2',2”-трихлортриэтиламин; трихотецены (а в частности токсин Т-2, верракурин А, роридин А и ангуидин); уретан; виндезин; дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид (“Ara-C”); циклофосфамид; тиотепа; таксоиды, например паклитаксел (Таксол®, Bristol-Myers Squibb Oncology, Princeton, N.J.) и доцетаксел (Таксотер®, Rhone-Poulenc Rorer, Antony, France); хлорамбуцил; гемцитабин; 6-тиогуанин; меркаптопурин; метотрексат; аналоги платины, такие как цисплатин и карбоплатин; винбластин; платина; этопозид (VP-16); ифосфамид; митомицин-С; митоксантрон; винкристин; винорелбин, навелбин; новантрон; тенипозид; дауномицин; аминоптерин; кселода; ибандронат; СРТ-11; ингибитор топоизомеразы RFS 2000; дифторметилорнитин (ДМФО); ретиноевая кислота; капецитабин; и фармацевтически приемлемые соли, оксиды или производные всех вышеуказанных соединений. В определение этого термина также входят противогормональные средства, регулирующие или ингибирующие действие гормонов на опухоли, такие как антиэстрогены, включая, например, тамоксифен, ралоксифен, 4-(5)-имидазолы, ингибирующие ароматазу, 4-гидрокситамоксифен, триоксифен, кеоксифен, LY117018, онапристон и торемифен (Фарестон®); и антиандрогены, такие как флутамид, нилутамид, бикалутамид, лейпролид и гозерелин; и фармацевтически приемлемые соли, оксиды или производные любого из вышеупомянутых соединений.
Термин “цитокин” является общим термином, относящимся к белкам, которые высвобождаются одной клеточной популяцией и которые действуют на другие клетки как межклеточные медиаторы. Примерами таких цитокинов являются лимфокины, монокины и стандартные полипептидные гормоны. Помимо вышеуказанных определений понятие “цитокины” также включает гормоны роста, такие как человеческий гормон роста, N-метионильный человеческий гормон роста и бычий гормон роста; паратиреоидный гормон; тироксин; инсулин; проинсулин; релаксин; прорелаксин; гликопротеиновые гормоны, такие как фолликулостимулирующий гормон (FSH), тиреоидстимулирующий гормон (TSH) и лютеинизирующий гормон (LH); фактор роста гепатоцитов, фактор роста фибробластов; пролактин; плацентарный лактоген; фактор некроза опухоли-α и -β; мюллеровский ингибирующий фактор; пептид, ассоциированный с мышиным гонадотропином; ингибин; активин; васкулярный эндотелиальный фактор роста; интегрин; тромбопоэтин (ТРО); факторы роста нервных тканей, такие как NGF-альфа; тромбоцитарный фактор роста; трансформирующие факторы роста (TGF), такие как TGF-альфа и TGF-бета; инсулиноподобный фактор роста-I и -II; эритропоэтин (EPO); факторы, индуцирующие остеогенез; интерфероны, такие как интерферон-α, -β и -γ; колониестимулирующие факторы (CSF), такие как макрофагальный CSF (M-CSF); гранулоцитарный-макрофагальный CSF (GM-CSF); и гранулоцитарный CSF (G-CSF); интерлейкины (IL), такие как IL-1, IL-1α, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11, IL-12; фактор некроза опухоли, такой как TNF-альфа или TNF-бета; и другие полипептидные факторы, включая LIF и лиганд кit (KL). Используемый здесь термин “цитокин” включает белки, происходящие от природных источников или от рекомбинантной клеточной культуры, и биологически активные эквиваленты цитокинов с нативной последовательностью.
Термины «лечение» или «терапия» означают терапевтическое или профилактическое лечение или превентивные меры.
Термин “терапевтически эффективное количество” означает количество лекарственного средства, эффективное для “лечения” заболевания или расстройства у млекопитающего. В случае рака терапевтически эффективное количество указанного лекарственного средства может снижать число раковых клеток; уменьшать размер опухоли; ингибировать (т.е. снижать до определенной степени, а предпочтительно прекращать) инфильтрацию раковых клеток в периферические органы; ингибировать (т.е. снижать до определенной степени, а предпочтительно прекращать) развитие метастазов опухоли; ингибировать, до некоторой степени, рост опухоли, и/или ослаблять, до некоторой степени, один или несколько симптомов, ассоциированных с указанным заболеванием. Лекарственное средство, до той степени, в которой оно может предупреждать рост раковых клеток и/или уничтожать имеющиеся раковые клетки, может быть цитостатическим и/или цитотоксическим. Для противораковой терапии эффективность in vivo может быть, например, измерена путем оценки опухолевой нагрузки или объема, времени, прошедшего до начала прогрессирования заболевания (ТТР) и/или определения уровня ответа (RR).
Термин «млекопитающее», подвергаемое лечению или терапии, означает любое животное, классифицированное как млекопитающее, включая человека, домашних и сельскохозяйственных животных, а также животных, содержащихся в зоопарках, животных, участвующих в спортивных состязаниях или животных-компаньонов, таких как собаки, лошади, кошки, коровы и т.п. Предпочтительным млекопитающим является человек.
«Индивидуумом» или «пациентом» является любой индивидуум, включая человека, который нуждается в лечении. Термин «индивидуум» также включает всех индивидуумов, которые участвуют в клинических научно-исследовательских испытаниях, и у которых отсутствуют какие-либо клинические признаки заболевания, или индивидуумов, участвующих в эпидемиологических исследованиях, или индивидуумов, включенных в испытания в качестве контрольной группы.
Используемые здесь термины «рак», «раковый» или «злокачественный» относятся к физиологическому состоянию млекопитающих, которое обычно характеризуется нерегулируемым ростом клеток, или описывают такое состояние. Примерами раковых заболеваний являются, но не ограничиваются ими, карцинома, лимфома, бластома, саркома и лейкоз. Более конкретными примерами таких раковых заболеваний являются рак толстой кишки, рак ободочной кишки, рак прямой кишки, плоскоклеточный рак, мелкоклеточный рак легких, немелкоклеточный рак легких, ходжкинская или неходжкинская лимфома, рак яичек, рак пищевода, рак желудочно-кишечного тракта, рак почек, рак поджелудочной железы, глиобластома, рак шейки матки, рак яичника, глиома, рак печени, рак мочевого пузыря, гепатома, рак молочной железы, карцинома эндометрия, карцинома слюнных желез, рак почек, рак печени, рак предстательной железы, рак вульвы, рак щитовидной железы, карцинома печени и различные типы рака головы и шеи.
Термин «биомаркер», используемый в настоящей заявке, означает, по существу, молекулу, включая ген, белок, фрагмент белка, углеводную структуру или гликолипид, экспрессия которых в тканях, или в клетках, или на тканях, или клетках млекопитающего может быть детектирована стандартными методами (описанными здесь методами) и которые, предположительно, обладают восприимчивостью к действию апоптоз-индуцирующего агента, такого как антитело против Apo2L/TRAIL или рецептора «гибели». Такими биомаркерами, рассматриваемыми в настоящем изобретении, являются, но не ограничивается ими, фрагменты белка AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4), описанные в настоящей заявке (см., например, фиг.1 и 2).
Термин «образец ткани или клетки» означает набор аналогичных клеток, полученных из ткани индивидуума или пациента. Источником образца ткани или клетки может быть твердая ткань, выделенная из свежего, замороженного и/или консервированного органа или образца ткани, или биоптата или аспирата; кровь или любые ее компоненты; физиологические жидкости, такие как цереброспинальная жидкость, амниотическая жидкость, перитонеальная жидкость или интерстициальная жидкость; и клетки, выделенные на любой стадии беременности или развития индивидуума. Образцом ткани могут быть также первичные или культивированные клетки или клеточные линии. Образец ткани или клетки получают, но необязательно, из первичной опухоли или опухоли с метастазами. Образец ткани может содержать соединения, которые не являются природными для этой ткани, такие как консерванты, антикоагулянты, буферы, фиксаторы, питательные вещества, антибиотики или т.п.
Используемый здесь термин «срез» образца ткани означает одну часть или фрагмент образца ткани, например тонкий слой ткани или клеток, вырезанный из этого образца ткани. Очевидно, что может быть получено множество образцов срезов ткани, и эти срезы могут быть подвергнуты анализу согласно изобретению.
Термины «коррелировать» или «корреляция» означают сравнение любым способом параметров и/или результатов первого анализа или протокола с параметрами и/или результатами второго анализа или протокола. Так, например, можно использовать результаты первого анализа или протокола для проведения второго протокола и/или можно использовать результаты первого анализа или протокола для оценки необходимости в проведении второго анализа или протокола. В соответствии с различными описанными здесь вариантами можно использовать результаты анализа, который позволяет идентифицировать фрагменты AP2α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4), образующиеся в процессе апоптоза, для того чтобы определить, нужно ли проводить конкретный курс терапевтического лечения с использованием апоптоз-индуцирующего агента, такого как антитело против Apo2L/TRAIL, FasL, рецептора «гибели» или т.п.
II. Методы и материалы
A. Методы
В общих чертах способы и материалы согласно изобретению применяются для детектирования и/или мониторинга апоптоза клеток млекопитающих, например, путем контактирования клетки с антителом, которое связывается с фрагментом белка, образующегося во время апоптоза, где указанное антитело связывается с фрагментом белка AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4); для определения количества антитела, которое связывается с фрагментом белка, образующимся во время апоптоза, и последующего сравнения количества связанного антитела в клетках млекопитающих с количеством антитела, которое связывается с фрагментом белка в клетках млекопитающих, не подвергнутых апоптозу, где указанный апопотоз детектируют в том случае, если количество таких оцениваемых клеток превышает количество клеток, не подвергнутых апоптозу. Как проиллюстрировано в описанных здесь примерах, способы контактирования клетки с антителом, которое связывается с фрагментом белка, образующимся во время апоптоза, предусматривают контактирование клеточных компонентов указанных клеток. Обычно эти клетки предварительно обрабатывают, например, путем экстракции (например, в соответствии с процедурами вестерн-блот-анализа, описанными в настоящей заявке) для облегчения контактирования компонентов клеток с антителом.
Описанные здесь способы и анализы могут быть применены в различных целях. Так, например, существуют некоторые популяции патологических человеческих клеток (например, некоторые популяции раковых клеток), которые являются резистентными к действию антител против Apo2L/TRAIL или рецептора «гибели», направленному на индуцирование гибели клеток. Поэтому очевидно, что описанные здесь способы и анализы могут представлять собой удобные, эффективные и относительно недорогостоящие средства для получения данных и информации, необходимых для оценки адекватности или эффективности лечения пациента. Так, например, у пациентов с диагностированным раком или иммуноассоциированным состоянием может быть взят биоптат для получения образца ткани или клетки, и такой образец может быть оценен с помощью различных in vitro анализов согласно изобретению для того, чтобы определить, являются ли клетки пациента чувствительными к терапевтическому средству, такому как антитело против Apo2/TRAIL или рецептора «гибели».
Для получения образца может быть использован образец ткани или клетки, взятый у млекопитающего (обычно у человека). Примерами образцов являются, но не ограничивается ими, образцы раковых клеток, таких как раковые клетки толстой кишки, молочной железы, предстательной железы, яичника, легких, желудка, поджелудочной железы, лимфомы и лейкоза. Такими образцами являются, но необязательно, образцы клеток немелкоклеточного рака легких, раковых клеток поджелудочной железы или раковых клеток неходжкинской лимфомы. Такой образец может быть получен различными методами, известными специалистам, включая, но не ограничиваясь ими, хирургическую операцию, аспирацию или биопсию.
Настоящее изобретение может быть осуществлено в различных вариантах. Так, например, в некоторых вариантах могут быть применены способы обнаружения апоптоза клеток и ассоциированных с ним состояний у индивидуума, такого как млекопитающее, а в частности у человека. В одном из иллюстративных вариантов изобретения способы согласно изобретению применяются в целях обнаружения наличия (или отсутствия) апоптоза клеток млекопитающих для оценки восприимчивости клеток млекопитающих к одному или нескольким апоптоз-индуцирующим агентам, таким как Apo2/TRAIL или FasL, например, для оценки эффективности терапии, включающей введение таких агентов. Способы согласно изобретению могут быть также применены в целях определения уровня активности соединения-кандидата, направленной на снижение или повышение апоптотической активности клеток млекопитающих, например соединения, модулирующего активность апоптоз-индуцирующего агента, такого как Apo2L/TRAIL (SEQ ID NO: 7), FasL (SEQ ID NO: 9), анти-Fas-антитело-агонист, анти-DR4-антитело-агонист или анти-DR5-антитело-агонист. Варианты настоящего изобретения могут быть также использованы для идентификации соединений, ингибирующих или стимулирующих апоптотическую гибель клеток, путем определения способности данных соединений ингибировать или стимулировать образование генерируемых при апоптозе фрагментов белка.
Типичным вариантом осуществления настоящего изобретения является способ детектирования апоптоза клеток млекопитающих путем контактирования этих клеток с антителом, которое связывается с фрагментом белка, образующегося во время апоптоза, где указанное антитело связывается с фрагментом белка AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4); определения количества антитела, которое связывается с фрагментом белка, образующимся во время апоптоза, и последующего сравнения количества связанного антитела в клетках млекопитающих с количеством антитела, которое связывается с фрагментом белка в клетках млекопитающих, не подвергнутых апоптозу, где указанный апопотоз детектируют в том случае, если количество таких оцениваемых клеток превышает количество клеток, не подвергнутых апоптозу. В таких способах могут быть использованы клетки млекопитающих широкого ряда. В некоторых вариантах изобретения указанными клетками являются раковые клетки толстой кишки, ободочной кишки, легких, молочной железы, предстательной железы, мочевого пузыря, почек, яичника, головного мозга, меланомы, лейкоза или миеломы.
Другим вариантом настоящего изобретения является способ идентификации человеческих раковых клеток, которые, вероятно, отвечают на действие терапевтического средства или являются чувствительными к этому терапевтическому средству, индуцирующему апоптоз человеческих раковых клеток. Этот вариант настоящего изобретения включает стадии обработки человеческих раковых клеток терапевтическим средством; анализа человеческих раковых клеток, обработанных указанным терапевтическим средством, на присутствие фрагмента белка AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4); сравнения количества фрагмента белка в человеческих раковых клетках с количеством фрагмента белка в контрольных человеческих раковых клетках, не обработанных лигандом. В этом варианте изобретения апоптоз наблюдается в том случае, когда количество фрагмента белка, присутствующего в человеческих раковых клетках, обработанных терапевтическим средством, превышает количество фрагмента белка в контрольных человеческих раковых клетках, не обработанных терапевтическим средством; и наблюдение апоптоза человеческих раковых клеток позволяет идентифицировать человеческие раковые клетки, которые, вероятно, отвечают на действие терапевтического средства, или являются чувствительными к такому терапевтическому средству. В некоторых вариантах изобретения человеческие раковые клетки получают от индивидуума, у которого был диагностирован рак, и эти клетки культивируют в in vitro культуре в течение менее одного месяца, а обычно менее 2 недель или менее одной недели. В альтернативных вариантах изобретения человеческой раковой клеткой является иммортализованная клеточная линия, полученная из культуры in vitro.
В типичных вариантах изобретения количество связанного антитела в клетках млекопитающих, которые оценивают на наличие апоптоза, например в клетках млекопитающих, обработанных апоптоз-индуцирующим агентом, сравнивают с количеством антитела, которое связывается с фрагментом белка в клетках млекопитающих, не подвергнутых апоптозу, например в контрольных клетках, которые не были обработаны апоптоз-индуцирующим агентом. Специалистам в данной области очевидно, что такие контрольные клетки выбирают из таких клеток, как экспериментальные клетки млекопитающих, которые наблюдают на наличие апоптоза, за исключением тестируемых клеток (например, клеток, обработанных апоптоз-индуцирующим агентом). В одном из вариантов изобретения контрольной клеткой является клетка, происходящая от того же источника (например, биоптата, полученного из участка первичной опухоли или опухоли с метастазами) и/или от той же линии дифференцировки, что и экспериментальные клетки млекопитающих, которые наблюдают на апоптоз, за исключением контрольных клеток млекопитающих, не обработанных агентом, инициирующим передачу сигнала рецептора «гибели» 4 (SEQ ID NO: 5), рецептора «гибели» 5 (SEQ ID NO: 6) или Fas (SEQ ID NO: 8). В некоторых вариантах изобретения характер фрагментации AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4) в таких контрольных клетках был уже охарактеризован, и этот предварительно охарактеризованный характер фрагментации в контрольных клетках сравнивают с характером фрагментации AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4) в клетках млекопитающих, которые оценивают на апоптоз. Такие варианты способов осуществления настоящего изобретения применяют, например, во избежание проведения ненужных и излишних процедур характеризации контрольных клеток, например, контроля, охарактеризованного как клетки, не подвергнутые апоптозу.
Варианты способов осуществления изобретения могут быть применены для оценки клеток, подвергающихся апоптозу, индуцированному любыми факторами широкого ряда, которые, как известно, инициируют запрограммированную клеточную гибель, например, такими, как высокая температура, излучение и химические вещества. Указанные варианты способов обычно включают обнаружение апоптоза клеток, который инициируется посредством рецептора, присутствующего на поверхности клеток, например рецептора «гибели» 4 (SEQ ID NO: 5), рецептора «гибели» 5 (SEQ ID NO: 6) или Fas (SEQ ID NO: 8). В некоторых вариантах изобретения указанные клетки подвергают контактированию с полипептидом, таким как Apo2L/TRAIL (SEQ ID NO: 7), FasL (SEQ ID NO: 9), анти-Fas-антитело-агонист, анти-DR4-антитело-агонист или анти-DR5-антитело-агонист, и описанные способы детектирования апоптоза (которые включают детектирование отсутствия апоптоза) применяются, например, для получения информации об эффективности терапии, включающей введение указанных агентов.
Способы согласно изобретению могут быть также применены для идентификации новых апоптоз-индуцирующих агентов и/или модуляторов апоптоз-индуцирующих агентов. Иллюстративные варианты могут включать стадию обработки клеток млекопитающих одним или несколькими тест-агентами, а затем, с применением способов согласно изобретению, стадию обнаружения апоптоза клеток млекопитающих путем идентификации одного или нескольких тест-агентов как индукторов апоптоза клеток млекопитающих. Способы согласно изобретению могут быть также объединены с дополнительными способами клеточного анализа, например определения генетического профиля. Один из таких вариантов согласно изобретению включает стадию оценки уровня экспрессии по меньшей мере одной мРНК в клетках млекопитающих, в которых наблюдается апоптоз.
Способ согласно изобретению включает идентификацию клеточных белков, которые, как описано в настоящей заявке, присутствуют в виде фрагментов в клетках, подвергающихся апоптозу. Такие фрагменты белка, например фрагменты AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4), могут быть обнаружены с помощью любых анализов широкого ряда, известных специалистам. Обычно такие фрагменты обнаруживают с помощью иммуноблоттинга, твердофазного иммуноферментного анализа или иммуногистохимического анализа. Кроме того, способы согласно изобретению могут быть применены для оценки числа идентифицированых здесь фрагментов белка, например фрагмента белка AP2α, имеющего молекулярную массу примерно 64 кДa или 33 кДa. В некоторых вариантах изобретения фрагмент белка, связанный с антителом, идентифицируют по аминокислотным последовательностям, специфичным для такого фрагмента, например аминокислотным последовательностям AP2-α DVFD SEQ ID NO: 1 или GPAA SEQ ID NO: 1.
Как указывалось выше, варианты настоящего изобретения включают способы определения восприимчивости человека, у которого диагностирован рак, к лечению одним или несколькими апоптоз-индуцирующими агентами, например Apo-2/TRAIL. Такие способы могут быть адаптированы для их применения вместе с другими различными известными способами, которые облегчают определение восприимчивости человека с диагностированным раком к лечению данным(и) агентом(ами), таким(ми) как апоптоз-индуцирующий агент, например, способами, которые позволяют определить уровень экспрессии по меньшей мере одного гена (например, присутствие или уровень мРНК или белка, кодируемого такой мРНК) у человека с диагностированным раком (например, в первичной опухоли или в ее метастазах). Такие методы определения генетического профиля хорошо известны специалистам и описаны, например, в заявке на патент США No. 20060015952.
Один из таких вариантов способов согласно изобретению может включать стадию обнаружения фрагментов, продуцируемых во время апоптоза, включая фрагменты белка AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4), в раковых клетках, полученных от пациента; в комбинации со стадией определения профиля экспрессии множества генов и сравнения такого профиля с профилем экспрессии генов в нераковых клетках одной и той же линии дифференцировки и/или в раковых клетках, полученных от животного-модели, которое, как известно, является восприимчивым (или альтернативно, невосприимчивым) к лечению одним или несколькими апоптоз-индуцирующими агентами. В общих чертах такие способы могут включать стадию дополнительной идентификации пациента, который, вероятно, является восприимчивым к лечению одним или несколькими апоптоз-индуцирующими агентами, например, в случае, если раковые клетки, полученные от пациента, имеют профиль экспрессии генов, аналогичный профилю экспрессии генов раковых клеток, полученных от пациентов (или животного с моделью рака), которые, как известно, являются восприимчивыми к лечению одним или несколькими апоптоз-индуцирующими агентами.
Используемый здесь термин «аналогичные» означает, что профили экспрессии являются подобными или совпадают по одному или нескольким механизмам экспрессии, на что указывают картины экспрессии, которые примерно на 80%-100%, более предпочтительно примерно на 90%-100%, а наиболее предпочтительно примерно на 95%-100% идентичны по уровню экспрессии или по другим измеримым параметрам, в зависимости от анализа или метода, применяемого для измерения профилей экспрессии генов, как будет более подробно описано ниже. Для облегчения сравнения профилей экспрессии генов в раковых клетках, полученных от пациента и от животного-модели, указанные профили, по существу, определяют одним и тем же методом или с помощью одного и того же анализа.
Методы определения профилей экспрессии генов хорошо известны специалистам и обычно основаны либо на анализе гибридизации полинуклеотидов, либо на секвенировании полинуклеотидов. Наиболее широко применяемыми методами количественной оценки экспрессии мРНК, в образце, известными специалистам, являются нозерн-блоттинг и in situ гибридизация (Parker and Barnes, Methods in Molecular Biology, 106:247-283 (1999)); анализы на защиту от РНКазы (Hod, Biotechniques, 13:852-854 (1992)); и полимеразная цепная реакция с обратной транскриптазой (ОT-ПЦР) (Weis et al., Trends in Genetics, 8:263-264 (1992)). Альтернативно, могут быть использованы антитела, которые могут распознавать специфические дуплексы, включая ДНК-дуплексы, РНК-дуплексы и гибридные дуплексы ДНК-РНК или дуплексы ДНК-белок. Репрезентативными методами анализа уровня экспрессии генов на основе секвенирования являются серийный анализ уровней экспрессии генов (SAGE), и анализ экспрессии генов, проводимый путем параллельного секвенирования массивов сигнатурных последовательностей (MPSS). Любой из этих методов или другие методы, известные специалистам, могут быть применены для определения профиля экспрессии генов опухолевых клеток, полученных от пациента, такого как человек, и у животного, служащего в качестве модели ракового заболевания, восприимчивого к действию антагониста TGF-β, такой как мышиная модель. В случае человека источником опухолевых клеток может быть свежий, замороженный или фиксированный и залитый в парафин образец ткани, из которого может быть экстрагирована мРНК, и который может быть подвергнут анализу на экспрессию генов.
Альтернативно, для сравнения профиля экспрессии человеческих белков и эталонных (например, мышиных) белков в раковых клетках могут быть также применены методы протеомики. В протеомике профили экспрессии белков представляют собой картину экспрессии множества белков в биологическом образце, например, ткани раковой опухоли. Профиль экспрессии может быть, например, представлен в виде масс-спектра или в виде других характеристик, основанных на любых физико-химических или биохимических свойствах белков. Так, например, профиль экспрессии может быть установлен исходя из различий электрофоретических свойств белков, определенных с помощью двухмерного гель-электрофореза, например двухмерный электрофорез в ПААГ, и может быть представлен, например, в виде множества пятен в двухмерном электрофоретическом геле. Методы протеомики хорошо известны специалистам и описаны, например, в нижеследующих руководствах: Proteome Research: New Frontiers in Functional Genomics (Principles and Practice), M. R. Wilkins et al., eds.. Springer Verlag, 1007; 2-D Proteome Analysis Protocols, Andrew L Link, editor, Humana Press, 1999; Proteome Research: Two-Dimensional Gel Electrophoresis and Identification Methods (Principles and Practice), T. Rabilloud editor. Springer Verlag, 2000; Proteome Research: Mass Spectrometry (Principles and Practice), P. James editor. Springer Verlag, 2001; Introduction to Proteomics, D. C. Liebler editor, Humana Press, 2002; Proteomics in Practice: A Laboratory Manual of Proteome Analysis, R. Westermeier et al., eds., John Wiley & Sons, 2002.
Фрагменты белка, образующиеся во время апоптоза, включая фрагменты белка AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4), в образце, могут быть проанализированы различными методами, хорошо известными специалистам. Типичные протоколы оценки таких фрагментов белка можно найти, например, в руководстве Ausubel et al. eds., 1995, Current Protocols In Molecular Biology, Unit 15 (Immunoblotting). Иммуноанализами, которые могут быть проведены в некоторых вариантах настоящего изобретения, являются, но не ограничивается ими, вестерн-блот-анализы, иммунопреципитация, слот- или дот-блот-анализы, иммунологическое окрашивание, РИА, сцинтилляционные анализы, флуоресцентные иммуноанализы, проводимые с использованием конъюгатов антител или конъюгатов антигенов с флуоресцентными веществами, такими как флуоресцеин или родамин; двойной диффузный анализ Охтерлони; ELISA; клеточный ELISA; ELISA, проводимый методом связывания с фильтром; ELISA на ингибирование; и иммуноанализы с использованием системы детекции на основе связывания авидина-биотина и стрептавидина-биотина.
В некоторых вариантах изобретения антитело, используемое в данном иммуноанализе, связывается как с интактным белком, так и с фрагментом белка, и такой фрагмент белка распознают по характерному для него малому размеру, например, с помощью процедуры вестерн-блоттинга. Так, например, антителом, используемым в одном из вариантов настоящего изобретения, может быть одно из описанных здесь репрезентативных антител, или антитело, которое связывается с эпитопом, связанным с одним из описанных здесь репрезентативных антител. В других вариантах изобретения могут быть использованы антитела, которые специфически распознают образующиеся при апоптозе фрагменты белка, но не интактные белки. Такие антитела могут быть получены стандартными методами иммунизации с использованием фрагмента в качестве иммуногена с последующей идентификацией продуцирующихся антител, которые связываются с образующимися при апоптозе фрагментами белка, но не с интактным белком.
В типичных вариантах настоящего изобретения анализ осуществляют с использованием белков, которые были получены в результате лизиса клеток, и которые были разделены с помощью электрофореза в ДСН-полиакриламидном геле (ПААГ). Эти белки подвергают контактированию с антителом, и проводят анализ, а предпочтительно иммуноанализ для определения присутствия фрагментов белка, связывающихся с антителом. При этом может быть проведено сравнение с контролем, состоящим из белков, выделенных из аналогичным образом лизированных клеток (например, клеток, происходящих от одного и того же источника и/или одной и той же линии дифференцировки), которые, как известно, не подвергались апоптозу. Для анализа образца ткани, взятого у живого индивидуума, могут быть применены любые из описанных выше иммуноанализов. Биологическими образцами, подходящими для такого анализа, являются клетки крови или клетки, взятые путем биопсии, или образцы ткани, которые могут быть получены стандартными методами. Уровни образующихся при апоптозе фрагментов пептида в вышеописанных биологических образцах могут быть определены с помощью любого из описанных выше иммуноанализов, в которых используются антитела, специфически связывающиеся с фрагментами белка AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4). Уровень образующихся при апоптозе фрагментов пептида, определенный в биологическом образце, взятом у обследуемого индивидуума, сравнивают с уровнем, обнаруженным в непораженных клетках пациента или в известном стандарте. Репрезентативным стандартом может быть, например, наличие и/или определенная концентрация одного или нескольких фрагментов белка, обычно обнаруживаемых в клетках, не подвергающихся апоптозу (например, в контрольных клетках, не обработанных апоптоз-индуцирующим агентом). Такие варианты могут быть использованы, например, для диагностики состояний, характеризующихся аномальным апоптозом. В некоторых вариантах изобретения определенный уровень апоптоза обнаруживается в клетках, где количество образующихся при апоптозе пептидных фрагментов по меньшей мере на 10, 20, 30, 40, 50, 100 или 150 процентов превышает количество этих фрагментов в контрольном образце, что является признаком патологического состояния. В других вариантах изобретения уровень апоптоза наблюдается в клетках, где количество образующихся при апоптозе пептидных фрагментов по меньшей мере в 10, 100 или 1000 раз превышает количество этих фрагментов в контрольном образце, что является признаком патологического состояния.
Варианты настоящего изобретения могут быть адаптированы для их применения в иммунологических анализах, подходящих для детектирования и количественной оценки фрагментов белка AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4). В таких анализах могут быть использованы, если это необходимо, одно или несколько антител, способных распознавать и связывать фрагмент белка. Такие анализы осуществляют в различных хорошо известных иммунологических форматах, включая, но не ограничиваясь ими, различные типы вестерн-блот-анализов, радиоиммуноанализов, твердофазных иммуноферментных анализов (ELISA), твердофазных иммуноферментных флуоресцентных анализов (ELIFA) и т.п.
В иллюстративных методах образец может быть подвергнут контактированию с антителом, специфичным к биомаркеру, такому как фрагмент белка AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4), в условиях, достаточных для образования комплекса «антитело-биомаркер» с последующим детектированием указанного комплекса. Обнаружение присутствия фрагмента белка в качестве биомаркера может быть осуществлено различными методами, такими как вестерн-блоттинг (с проведением или без проведения стадии иммунопреципитации) и ELISA в целях проведения анализа тканей и образцов широкого ряда, включая плазму или сыворотку. Методы иммуноанализа широкого ряда, проводимые в описанном формате, можно найти, например, в патентах США NN 4016043, 4424279 и 4018653. Такими анализами являются односайтовые и двухсайтовые или «сэндвич»-анализы неконкурентного типа, а также традиционные анализы на конкурентное связывание. Такие анализы также включают прямое связывание меченного антитела с биомаркером-мишенью.
Наиболее подходящими и часто применяемыми анализами являются «сэндвич»-анализы. Существуют различные варианты «сэндвич»-анализов, и все эти варианты входят в объем настоящего изобретения. Вкратце, в типичном прямом анализе, немеченное антитело иммобилизуют на твердом субстрате, а затем тестируемый образец подвергают контактированию со связанной молекулой. После инкубирования в течение периода времени, достаточного для образования комплекса антитело-антиген, добавляют «второе» антитело, специфичное к антигену и меченное репортерной молекулой, способной продуцировать детектируемый сигнал, и инкубируют в течение периода времени, достаточного для образования другого комплекса «антитело-антиген-меченное антитело». Любой непрореагировавший материал отмывают, и присутствие антигена определяют по сигналу, передаваемому репортерной молекулой. Результаты могут быть получены путем качественной оценки, путем простого наблюдения видимого сигнала, либо они могут быть получены путем количественной оценки посредством сравнения с результатами для контрольного образца, содержащего известное количество биомаркера.
Вариантами прямого анализа являются одновременный анализ, в котором образец и меченное антитело добавляют к связанному антителу одновременно. Такие анализы хорошо известны специалистам, включая любые незначительные модификации, которые могут быть легко внесены самим специалистом. В типичном прямом «сэндвич»-анализе, первое антитело, обладающее специфичностью к биомаркеру, подвергают ковалентному или пассивному связыванию с твердой поверхностью. Твердой поверхностью обычно являются стекло или полимер, при этом наиболее часто используемыми полимерами являются целлюлоза, полиакриламид, найлон, полистирол, поливинилхлорид или полипропилен. Твердыми носителями могут быть трубки, сферы, диски микропланшетов или любые другие поверхности, подходящие для проведения иммуноанализа. Способы связывания хорошо известны специалистам, и, по существу, включают перекрестное ковалентное связывание, физическую адсорбцию и промывку комплекса «полимер-антитело» в препарате для тест-образца. Затем аликвоту тестируемого образца добавляют к твердофазному комплексу и инкубируют в течение определенного периода времени (например, в течение 2-40 минут или в течение ночи, если это более удобно) и в условиях (например, при температуре в пределах от комнатной температуры до 40°С, например, при 25°С-32°С включительно), подходящих для связывания любой субъединицы, присутствующей в антителе. После инкубирования твердую фазу субъединицы антитела промывают и сушат, а затем инкубируют со вторым антителом, специфичным к части биомаркера. Второе антитело конъюгируют с репортерной молекулой, которую используют для идентификации связывания второго антитела с молекулярным маркером.
Альтернативный метод включает иммобилизацию биомаркера-мишени в образце, а затем обработку иммобилизованной мишени специфическим антителом, которое может быть, а может и не быть, помечено репортерной молекулой. В зависимости от количества мишени и от интенсивности сигнала репортерной молекулы связанная мишень может детектироваться путем прямого мечения антителом. Альтернативно, «второе» меченное антитело, специфичное к первому антителу, обрабатывают комплексом «мишень-первое антитело» с образованием трехкомпонентного комплекса «мишень - первое антитело - второе антитело». Такой комплекс детектируют по сигналу, передаваемому репортерной молекулой. Термин «репортерная молекула», используемый в настоящем описании, означает молекулу, которая по своей химической природе продуцирует аналитически идентифицируемый сигнал, позволяющий детектировать связанное с антигеном антитело. Наиболее часто используемыми репортерными молекулами в анализах такого типа являются ферменты, флуорофоры или радионуклидсодержащие молекулы (то есть радиоизотопы) и хемилюминесцентные молекулы.
В случае иммуноферментного анализа фермент конъюгируют со «вторым» антителом, обычно посредством глутаральдегида или периодата. Однако совершенно очевидно, что существуют различные методы конъюгирования широкого ряда, хорошо известные специалистам. Наиболее часто используемыми ферментами являются пероксидаза хрена, глюкозо-оксидаза, галактозидаза и щелочная фосфатаза и т.п. Субстраты, используемые для специфических ферментов, обычно выбирают так, чтобы после гидролиза соответствующим ферментом они продуцировали детектируемое изменение окраски. Примерами подходящих ферментов являются щелочная фосфатаза и пероксидаза. Также могут быть использованы и флуорогенные субстраты, которые, в отличие от хромогенных субстратов, описанных выше, продуцируют флуоресцентный продукт. В типичных случаях меченное ферментом антитело добавляют к комплексу «первое антитело - молекулярный маркер», подвергают реакции связывания, а затем избыток реагента отмывают. После этого к комплексу «антитело-антиген-антитело» добавляют раствор, содержащий соответствующий субстрат. Такой субстрат подвергают реакции с ферментом, связанным со «вторым» антителом, что приводит к продуцированию качественного визуального сигнала, который может быть затем количественно оценен обычно спектрофотометрическими методами, в результате чего может быть определено количество биомаркера, присутствующего в образце. Альтернативно, флуоресцентные соединения, такие как флуоресцеин и родамин, могут быть химически присоединены к антителам без изменения их связывающей способности. При активации путем облучения светом на конкретной длине волны меченное флуорохромом антитело адсорбирует световую энергию, что приводит к возбуждению молекулы, сопровождающемуся излучением света на длине волны, характерной для определенного цвета, визуально детектируемого на оптическом микроскопе. Как и в иммуноферментном анализе (ИФА), флуоресцентно меченное антитело подвергают связыванию с комплексом «первое антитело - молекулярный маркер». После отмывки несвязанного реагента оставшийся трехкомпонентный комплекс облучают светом на соответствующей длине волны, после чего наблюдаемая флуоресценция указывает на присутствие представляющего интерес молекулярного маркера. Методы иммунофлуоресценции и ИФА очень хорошо известны специалистам. Однако могут быть также использованы и другие репортерные молекулы, такие как радиоизотопы, хемилюминесцентные или биолюминесцентные молекулы.
Как указывалось выше, в некоторых вариантах настоящего изобретения могут быть использованы антитела, которые связываются с фрагментом белка AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4), но не связываются с полноразмерным белком (например, антитела, которые связываются с эпитопом, полученным в результате расщепления полноразмерного белка). Такие варианты могут быть использованы для оценки фрагментов белка в клетках методами иммуногистохимического окрашивания. В таких методах образец ткани может быть фиксирован (то есть консервирован) стандартными методами (см. например, руководство «Manual of Histological Staining Method of the Armed Forces Institute of Pathology» 3rd edition (1960) Lee G. Luna, HT (ASCP) Editor, The Blakston Division McGraw-Hill Book Company, New York; The Armed Forces Institute of Pathology Advanced Laboratory Methods in Histology and Pathology (1994) Ulreka V. Mikel, Editor, Armed Forces Institute of Pathology, American Registry of Pathology, Washington, D.C). Для специалсита в данной области очевидно, что выбор фиксаторов осуществляют в зависимости от цели гистологического окрашивания ткани или ее анализа каким-либо другим методом. Для специалиста в данной области также очевидно, что длина фиксатора зависит от размера образца ткани и от используемого фиксируемого вещества. Так, например, для фиксации образца ткани могут быть использованы нейтрально забуференный формалин, реактив Боуина или параформальдегид. После приготовления препарата ткани срез ткани может быть подвергнут одному из ряда гистохимических анализов, известных специалистам.
B. Материалы
Для осуществления настоящего изобретения могут быть использованы различные материалы, включая любой один из различных апоптоз-индуцирующих агентов, а также антитела, которые связываются с описанными здесь фрагментами белка, образующимися во время апоптотической гибели клеток. Репрезентативными апоптоз-индуцирующими агентами являются анти-Apo2L, анти-DR4 или анти-DR5-антитела-агонисты, анти-FasL и анти-Fas-антитела-агонисты. Apo-2L, который может быть использован в таких методах, включает полипептиды Apo-2L, описанные в публикации Pitti et al., см. выше, в WO 97/25428, см. выше, и в WO97/01633, см. выше, (указанные полипептиды называются TRAIL). При этом предусматривается, что могут быть использованы различные формы Apo-2L, такие как полноразмерный полипептид, а также растворимые формы Apo-2L, содержащие последовательность внеклеточного домена (ECD). Примерами таких растворимых последовательностей ECD являются полипептиды, содержащие аминокислоты 114-281, 95-281, 91-281 или 92-281 последовательности Apo-2L, представленной на фиг.1А, в публикации Pitti et al., J. Biol. Chem., 271:12687-12690 (1996). В настоящее время известно, что полипептид, содержащий аминокислоты 92-281, представляет собой природную гидролизованную форму Apo-2L. Заявителями была осуществлена экспрессия человеческого Apo-2L в клетках CHO, и было обнаружено, что полипептид, содержащий аминокислоты 92-281, представляет собой экспрессированную форму Apo-2L. Могут быть также включены модифицированные формы Apo-2L, такие как ковалентно модифицированные формы, описанные в WO 97/25428. В частности в способах согласно изобретению используется Apo-2L, связанный с небелковым полимером, таким как полиэтиленгликоль. Полипептид Apo-2L может быть получен любыми методами, описанными в WO 97/25428.
Вариантами лиганда Apo-2, обладающего апоптотической активностью, которая может быть использована в указанных способах, являются, например, варианты, идентифицированные аланинсканирующими методами. Конкретные варианты с заменами содержат аминокислоты 91-281, представленные на фиг.1А, в публикации Pitti et al., J. Biol. Chem., 271:12687-12690 (1996), где по меньшей мере одна из аминокислот в положениях 203, 218 или 269 заменены аланиновым остатком. Варианты лиганда Apo-2 могут включать, но необязательно, одну или более из этих трех различных замен в нужных положениях.
При этом считается, что в описанных здесь способах может быть альтернативно использована молекула, имитирующая апоптотическую активность Apo-2L и/или FasL. Примерами таких молекул являются антитела-агонисты, которые могут индуцировать апоптоз, по меньшей мере сравнимый с апоптозом или аналогичный апоптозу, вызываемому Apo-2L. В частности такие антитела-агонисты представляют собой антитела против одного или нескольких рецепторов Apo-2L. Предпочтительно антитело-агонист направлено против рецептора для Apo-2L, который включает цитоплазматический домен «гибели», такой как DR4 или DR5. Еще более предпочтительно антитело-агонист связывается с указанным рецептором, и уровень такого связывания может быть определен, например, с помощью FACS- или ELISA-анализа. Антитела-агонисты, направленные против рецептора, называемого DR5 (или Apo-2), были получены с применением методов слияния, описанных ниже. Одно из таких антител-агонистов, направленных против рецептора DR5 или Aрo-2, было обозначено 3F11.39.7 и было депонировано в ATCC под депозитарным номером HB-12456 13 января 1998. Агонистическая активность антител против рецептора Apo-2L может быть определена различными методами анализа на апоптотическую активность, и апоптотическая активность такого антитела может быть определена, но необязательно, путем анализа антитела, присутствующего отдельно или в
перекрестно связанной форме, с использованием иммуноглобулина Fc или его комплемента.
Кроме того, были также получены антитела-агонисты, направленные против другого рецептора Apo-2L, называемого DR4. Репрезентативные анти-DR4-антитела-агонисты были обозначены 4H6.17.8 и были депонированы в ATCC под депозитарным номером HB-12455 13 января 1998. Агонистическая активность антител против рецептора Apo-2L может быть определена различными методами анализа на апоптотическую активность, и апоптотическая активность такого антитела может быть определена, но необязательно, путем анализа антитела, присутствующего отдельно или в перекрестно связанной форме, с использованием иммуноглобулина Fc или его комплемента.
Антителами-агонистами, рассматриваемыми в настоящем изобретении, являются антитела, которые связываются с одним или с несколькими рецепторами Apo-2L. Антитело, которое связывается более чем с одним рецептором Apo-2L, может быть охарактеризовано как антитело, которое «перекрестно реагирует» с двумя или более различными антигенами, и которое обладает способностью связываться с каждым из различных антигенов, например, как было определено с помощью ELISA- или FACS-анализов, описаных ниже в примерах. Антитело, которое «специфически перекрестно реагирует» с двумя или более различными антигенами, представляет собой, но необязательно, антитело, которое связывается с первым антигеном, а затем связывается со вторым другим антигеном, где способность этого антитела связываться со вторым антигеном при концентрации антитела примерно l0 мкг/мл составляет примерно 50%-100% (предпочтительно примерно 75%-100%) от способности связываться с первым антигеном, как было определено в ELISA-анализе с захватом (таком как анализ, описанный ниже в примерах). Так, например, антитело может специфически связываться с DR5 («первым антигеном») и специфически перекрестно реагировать с другим рецептором Apo-2L, таким как DR4 (со «вторым антигеном»), где уровень связывания примерно 10 мкг/мл антитела с DR4 составляет примерно 50%-100% от уровня связывания указанного антитела связываться с DR5, на что указывает описанный здесь ELISA с захватом. Различные антитела, перекрестно реагирующие с рецепторами Apo- 2L, более подробно описаны в Международной патентной заявке PCT/US99/13197.
Как описано ниже, репрезентативными формами антител, используемых в целях осуществления вариантов настоящего изобретения, являются поликлональные, моноклональные, гуманизованные, биспецифические и гетероконъюгированные антитела.
1. Поликлональные антитела
Антителами, используемыми в целях осуществления настоящего изобретения, могут быть поликлональные антитела. Методы получения поликлональных антител хорошо известны специалистам. Поликлональные антитела могут вырабатываться у млекопитающего, например, после введения одной или нескольких инъекций иммунизирующего агента, и, если это желательно, адъюванта. Такой иммунизирующий агент и/или адъювант обычно вводят млекопитающему в виде множества подкожных или внутрибрюшинных инъекций. Иммунизирующими агентами могут быть, например, полноразмерный или неполный полипептид AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4); или его гибридный белок. Он может быть использован для конъюгирования иммунизирующего агента с белком, который, как известно, является иммуногенным для иммунизованного млекопитающего. Примерами таких иммуногенных белков являются, но не ограничиваются ими, гемоцианин лимфы улитки, сывороточный альбумин, бычий тироглобулин и ингибитор соевого трипсина. Примерами адъювантов, которые могут быть использованы, являются полный адъювант Фрейнда и адъювант MPL-TDM (монофосфорил-липид A, синтетический дикориномиколат трегалозы). Протокол иммунизации может быть выбран самим специалистом без излишнего экспериментирования. Затем у млекопитающего может быть взята кровь и ее сыворотка может быть проанализирована на титр антител. Если это необходимо, то млекопитающее может быть подвергнуто повторной иммунизации до достижения увеличения или плато титра антител.
2. Моноклональные антитела
Альтернативно, антителами, используемым для осуществления настоящего изобретения, могут быть моноклональные антитела. Моноклональные антитела могут быть получены гибридомными методами, например, как описано Kohler & Milstein, Nature, 256:495 (1975). В гибридомном методе мышь, хомячка или другого подходящего животного-хозяина обычно иммунизируют иммунизирующим агентом для вырабатывания у них лимфоцитов, продуцирующих или способных продуцировать антитела, специфически связывающиеся с иммунизующим агентом. Альтернативно, лимфоциты могут быть получены при иммунизации in vitro.
В общих чертах, если требуются клетки человека, то используют лимфоциты периферической крови (ЛПК), а если требуются клетки млекопитающего, не являющегося человеком, то используют клетки селезенки или лимфоузлов. Затем лимфоциты подвергают слиянию с иммортализованной клеточной линией с использованием подходящего агента для слияния, такого как полиэтиленгликоль, в результате чего образуются гибридомные клетки (Goding, Monoclonal Antibodies: Principles and Practice, Academic Press, (1986) pp. 59-103). Иммортализованными клеточными линиями обычно являются трансформированные клетки млекопитающих, а в частности миеломные клетки грызунов, коров и человека. Обычно используются миеломные клеточные линии крысы или мыши. Гибридомные клетки могут быть культивированы в подходящей культуральной среде, которая, предпочтительно, содержит одно или несколько веществ, ингибирующих рост или выживание неслитых иммортализованных клеток. Так, например, если родительские клетки не содержат фермента гипоксантин-гуанин-фосфорибозилтрансферазы (HGPRT или HPRT), то культуральная среда для получения гибридом обычно включает гипоксантин, аминоптерин и тимидин (среда НАТ), то есть вещества, предупреждающие рост HGPRT-дефицитных клеток.
Предпочтительными иммортализованными клеточными линиями являются клеточные линии, которые способны подвергаться эффективному слиянию и поддерживать стабильную экспрессию высоких уровней антител выбранными антителопродуцирующими клетками, и являются чувствительными к среде, такой как среда НАТ. Более предпочтительными иммортализованными клеточными линиями являются мышиные миеломные линии, которые могут быть взяты, например, из института Salk Institute Cell Distribution Center, San Diego, California USA, и из Американской коллекции типовых культур (American Type Culture Collection, Manassas, Virginia). Примером таких мышиных миеломных клеточных линий является P3X63AgU.l. Были также описаны человеческие миеломные и гетеромиеломные клеточные линии “мышь-человек”, используемые для продуцирования человеческих моноклональных антител (Kozbor, J. Immunol., 133:3001 (1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, Marcel Dekker, Inc., New York, (1987) pp. 51-63).
Культуральная среда, в которой могут быть культивированы гибридомные клетки, может быть затем проанализирована на присутствие моноклональных антител против нужного иммуногена. Специфичность связывания моноклональных антител, продуцируемых гибридомными клетками, определяют, предпочтительно, путем иммунопреципитации или путем проведения in vitro анализа на связывание, такого как радиоиммуноанализ (РИА) или твердофазный иммунноферментный анализ (ELISA). Такие методы и анализы известны специалистам. Аффинность связывания моноклонального антитела может быть определена, например, с помощью анализа Скэтчарда, описанного Munson & Pollard, Anal. Biochem., 107:220 (1980).
После идентификации нужных гибридомных клеток клоны могут быть субклонированы путем проведения процедур лимитирующего разведения и культивированы стандартными методами (Goding, см. выше). Подходящими культуральными средами, предназначенными для достижения этой цели, является, например, модифицированная по способу Дульбекко среда Игла или среда RPMI-1640. Альтернативно, гибридомные клетки могут быть выращены in vivo в качестве асцитных опухолей у млекопитающих.
Моноклональные антитела, секретированные субклонами, могут быть выделены или очищены из культуральной среды или асцитной жидкости путем проведения стандартных процедур очистки иммуноглобулинов, таких как, например, хроматография на белке А - сефарозе, хроматография на гидроксиапатитах, гель-электрофорез, диализ или аффинная хроматография.
Моноклональные антитела могут быть также получены методами рекомбинантных ДНК, например, методами, описанными в патенте США № 4816567. ДНК, кодирующая моноклональные антитела, используемые в целях осуществления настоящего изобретения, может быть легко выделена и секвенирована в соответствии со стандартными процедурами (например, с использованием олигонуклеотидных зондов, которые способны специфически связываться с генами, кодирующими тяжелые и легкие цепи мышиных антител). Предпочтительным источником такой ДНК служат гибридомные клетки согласно изобретению. После выделения эта ДНК может быть встроена в экспрессионные векторы, которые затем трансфецируют в клетки-хозяева, такие как клетки E.coli, обезьяньи клетки СОS, клетки яичника китайского хомячка (СНО) или миеломные клетки, которые, в иных случаях, не продуцируют белок иммуноглобулина, в результате чего в этих рекомбинантных клетках-хозяевах синтезируются моноклональные антитела. ДНК может быть также модифицирована, например, путем замены гомологичных мышиных последовательностей на последовательности, кодирующие человеческие константные домены тяжелой и легкой цепи (патент США № 4816567; Morrison et al., см. выше), или путем ковалентного связывания всей или части кодирующей последовательности для неиммуноглобулиного полипептида с иммуноглобулинкодирующей последовательностью. Такой неиммуноглобулиновый полипептид может быть заменен константными доменами антитела согласно изобретению, либо он может быть заменен вариабельными доменами одного антигенсвязывающего сайта антитела согласно изобретению с получением химерного двухвалентного антитела. Химерные антитела могут быть, но необязательно, сконструированы так, чтобы они включали по меньшей мере один вариабельный или гипервариабельный домен антитела, такого как одно из описанных здесь антител.
Антитела согласно изобретению могут, но необязательно, связываться с тем же самым (теми же самыми) эпитопом(ами), с которым(и) связывается(ются) любое(ые) из описанных здесь антител. Это может быть установлено путем проведения различных анализов, таких как описанные здесь анализы. Так, например, для того, чтобы определить, обладает ли моноклональное антитело такой же специфичностью, как и конкретно описанные здесь антитела, может быть проведено сравнение их активности в анализах на апоптоз.
Антителами, используемыми для осуществления настоящего изобретения, являются «перекрестно связанные» антитела. Используемый здесь термин «перекрестно связанный» относится к связыванию по меньшей мере двух молекул IgG с образованием одной (или одиночной) молекулы. Антитела могут быть перекрестно связаны посредством различных линкерных молекул. Антитела могут быть подвергнуты перекрестному связыванию, но необязательно, с использованием анти-IgG молекулы, комплемента, химической модификации или с применением методов молекулярной инженерии. Для специалиста в данной области очевидно, что комплемент обладает относительно высокой аффинностью связывания с молекулами антитела после связывания антител с мембраной клеточной поверхности. В соответствии с этим очевидно, что комплемент может быть использован в качестве перекрестно-связывающей молекулы для присоединения одного или нескольких антител, связанных с мембраной клеточной поверхности. Известно, что из различных изотипов мышиных Ig для фиксации комплемента используются IgM, IgG2a и IgG2b.
Антителами, используемы(ми) в целях осуществления изобретения, могут быть, но необязательно, димерные антитела, а также мультивалентные формы антител. Специалисты в данной области могут сконструировать такие димеры или мультивалентные формы методами, известными специалистам, и методами с использованием описанных здесь антител против рецептора Apo-2L.
Антителами, используемыми для осуществления изобретения, могут быть также моновалентные антитела. Методы получения моновалентных антител хорошо известны специалистам. Так, например, один из методов включает рекомбинантную экспрессию легкой цепи и модифицированной тяжелой цепи иммуноглобулина. Тяжелую цепь, в основном, подвергают усечению в любом положении Fc-области в целях предупреждения перекрестного связывания тяжелой цепи. Альтернативно, для предупреждения перекрестного связывания релевантные цистеиновые остатки заменяют другим аминокислотным остатком или делетируют.
Для получения моновалентных антител могут быть также применены in vitro методы. Гидролиз антител с получением их фрагментов, а в частности Fab-фрагментов, может быть осуществлен рутинными методами, известными специалистам. Так, например, гидролиз может быть осуществлен с использованием папаина. Примеры гидролиза папаином описаны в заявке WO 94/29348, опубликованной 12/22/94, и в патенте США No. 4342566. Гидролиз антител папаином обычно приводит к образованию двух идентичных антигенсвязывающих фрагментов, называемых Fab-фрагментами, каждый из которых имеет один антигенсвязывающий сайт и остаточный Fc-фрагмент. Обработка пепсином приводит к образованию F(ab')2-фрагмента, который имеет два антигенсвязывающих сайта и обладает способностью перекрестно связываться с антигеном.
Fab-фрагменты, продуцирующиеся в результате гидролиза антитела, также содержат константные домены легкой цепи и первый константный домен (CH1) тяжелой цепи. Fab'-фрагменты отличаются от Fab-фрагментов добавлением нескольких остатков у карбокси-конца домена СН1 тяжелой цепи, включая один или несколько цистеинов, происходящих от шарнирной области антитела. В описании настоящей заявки Fab'-SH означает Fab', в котором цистеиновый(е) остаток(тки) константных доменов имеет(ют) свободную тиоловую группу. F(ab')2-фрагменты антитела, по своей природе, продуцируются как пары Fab'-фрагментов, между которыми расположены цистеины шарнирной области. Также известны и другие химические связи, соединяющие фрагменты антитела.
Могут быть также продуцированы одноцепочечные Fv-фрагменты, например фрагменты, описанные в публикации Iliades et al., FEBS Letters, 409:437-441 (1997). Связывание таких одноцепочечных фрагментов с использованием различных линкеров описано в публикации Kortt et al., Protein Engineering, 10:423-433 (1997).
Помимо описанных выше антител рассматриваются и химерные или гибридные антитела, которые могут быть получены in vitro с применением известных методов химического синтеза белков, включая методы с использованием перекрестно-связывающих агентов. Так, например, иммунотоксины могут быть сконструированы путем проведения реакций дисульфидного обмена или посредством образования тиоэфирной связи. Примерами реагентов, подходящих для такой цели, являются иминотиолат и метил-4-меркаптобутиримидат.
Антителами, используемыми для осуществления настоящего изобретения, могут быть также гуманизованные антитела или человеческие антитела. Гуманизованными формами нечеловеческих антител (например, мышиных антител) являются химерные иммуноглобулины, цепи иммуноглобулинов или их фрагменты (такие как Fv, Fab, Fab', F(ab')2 или другие антигенсвязывающие подпоследовательности антител), которые содержат минимальную последовательность, происходящую от нечеловеческого иммуноглобулина. Гуманизованными антителами являются человеческие иммуноглобулины (антитело-реципиент), в которых остатки, происходящие от гипервариабельной области (СDR) данного антитела-реципиента, заменены остатками, происходящими от СDR нечеловеческого антитела (донорного антитела), такого как мышиное антитело, крысиное антитело или кроличье антитело, обладающее нужной специфичностью, аффинностью и связывающей способностью. В некоторых случаях остатки каркасной области (Fv) человеческого иммуноглобулина заменены соответствующими нечеловеческими остатками. Гуманизованные антитела могут также содержать остатки, не обнаруженные ни в антителе-реципиенте, ни в «импортных» последовательностях СDR или каркасной области. В общих чертах гуманизованное антитело может содержать, по существу, все или по меньшей мере один, а обычно два вариабельных домена, в которых все или почти все области СDR соответствуют гипервариабельным областям нечеловеческого иммуноглобулина, и все или почти все FR-области представляют собой FR с человеческой иммуноглобулиновой консенсусной последовательностью. Оптимально гуманизованное антитело также содержит по меньшей мере часть константной области иммуноглобулина (Fc), обычно области человеческого иммуноглобулина (Jones et al., Nature, 321:522-525 (1986); Riechmann et al., Nature, 332:323-329 (1988); и Presta, Curr. Op. Struct. Biol., 2:593-596 (1992)).
Методы гуманизации нечеловеческих антител хорошо известны специалистам. В основном гуманизованное антитело имеет один или несколько введенных в него аминокислотных остатков, происходящих от источника, не относящегося к человеку. Эти нечеловеческие аминокислотные остатки часто называют “импортными” остатками, которые обычно берут из “импортного” вариабельного домена. Такая гуманизация может быть осуществлена, в основном, методом, описанным Винтером (Winter) и сотрудниками (Jones et al., Nature 321:522-525 (1986); Riechmann et al., Nature, 332:323-327 (1988); Verhoyen et al., Science 239:1534-1536 (1988)), путем замены СDR грызунов или последовательностей СDR соответствующими последовательностями человеческого антитела. В соответствии с этим такими гуманизованными антителами являются химерные антитела (патент США № 4816567), в которых, в основном, меньшая часть, по сравнению с интактным вариабельным доменом человеческого антитела, заменена соответствующей последовательностью от нечеловеческого антитела. Фактически гуманизованные антитела обычно представляют собой человеческие антитела, в которых некоторые остатки СDR и, возможно, некоторые остатки FR заменены остатками, происходящими от аналогичных участков антител грызунов.
Для снижения антигенности при создании гуманизованных антител очень важно выбрать вариабельные домены как легкой, так и тяжелой цепи человеческого антитела. В соответствии с так называемым методом “подгонки” последовательность вариабельного домена антитела грызуна скринируют по всей библиотеке известных последовательностей вариабельных доменов человеческого антитела. Затем человеческую последовательность, которая является наиболее схожей с последовательностью грызунов, берут в качестве человеческой каркасной области (FR) для получения гуманизованного антитела (Sims et al., J. Immunol., 15J.: 2296-2308 (1993); Chothia and Lesk, J. Mol. Biol., 196:901-917 (1987)). В другом методе используется конкретная каркасная область, происходящая от консенсусной последовательности всех человеческих антител с конкретной подгруппой легких и тяжелых цепей. Та же самая каркасная область может быть использована для получения нескольких различных гуманизованных антител (Carter et al., Proc. Natl. Acad. Sci., USA, 89:4285-4289 (1992); Presta et al., J. Immunol, 151:2623-2632 (1993)).
Кроме того, важно, чтобы гуманизованные антитела сохраняли высокую аффинность по отношению к антигену и другие благоприятные биологические свойства. Для достижения этой цели, в соответствии с предпочтительным методом, гуманизованные антитела могут быть получены путем анализа исходных последовательностей и различных концептуальных гуманизованных продуктов с использованием трехмерных моделей родительских и гуманизованных последовательностей. Трехмерные иммуноглобулиновые модели являются общедоступными и хорошо известны специалистам. Существуют компьютерные программы, которые иллюстрируют и представляют вероятные трехмерные конформационные структуры отобранных последовательностей-кандидатов иммуноглобулинов. Исследование этих представлений позволяет проанализировать вероятную роль данных остатков в функционировании последовательностей-кандидатов иммуноглобулина, то есть остатков, которые влияют на способность иммуноглобулина-кандидата связываться с его антигеном. Таким образом, остатки FR могут быть выбраны из консенсусных последовательностей и “импортных” последовательностей и объединены, в результате чего может быть получено желаемое антитело с нужными свойствами, такими как повышенная аффинность по отношению к антигену(ам)-мишени(ям). Вообще говоря остатки СDR непосредственно влияют на связывание с антигеном, и в основном участвуют в таком связывании (см. заявку WO 94/04679, опубликованную 3 марта 1994).
Человеческие моноклональные антитела могут быть получены путем адаптации гибридомного метода, впервые описанного Kohler и Milstein, с использованием человеческих В-лимфоцитов в качестве партнера по слиянию. Человеческие B-лимфоциты, продуцирующие представляющее интерес антитело, могут быть, например, выделены у человека после получения от него информированного согласия. Так, например, у индивидуума могут вырабатываться антитела против аутоантигена, вызывающего некоторые заболевания, такие как системная красная волчанка (Shoenfeld et al. J. Clin. Invest., 70:205 (1982)), иммуноопосредованная тромбоцитопеническая пурпура (ITP) (Nugent et al. Blood, 70(1): 16-22 (1987)) или рак. Альтернативно или дополнительно, лимфоциты могут быть введены посредством иммунизации in vitro. Так, например, выделенные человеческие лимфоциты периферической крови могут быть обработаны in vitro лизосомотрофным агентом (например, L-лейцин-O-метиловым эфиром, диметиловым эфиром L-глутаминовой кислоты или L-лейцил-L-лейцин-O-метиловым эфиром) (патент США № 5567610, Borrebaeck et al.); и/или популяция человеческих лимфоцитов периферической крови, не содержащая Т-клеток, может быть обработана in vitro адъювантами, такими как 8-меркаптогуанозин и цитокины (патент США No 5229275, Goroff et al.).
Затем B-лимфоциты, выделенные у индивидуума или введенные путем in vitro иммунизации, обычно подвергают иммортализации для продуцирования человеческого моноклонального антитела. Методами иммортализации B-лимфоцитов являются, но не ограничивается ими: (a) слияние человеческих В-лимфоцитов с клетками человеческой миеломы, мышиной миеломы или с клетками гетеромиеломы «мышь-человек»; (b) вирусная трансформация (например, вирусом Эпштейна-Барра; см, например, Nugent et al., см. выше); (c) слияние с лимфобластоидной клеточной линией; или (d) слияние с клетками лимфомы.
Лимфоциты могут быть подвергнуты слиянию с клетками миеломы с помощью подходящего агента для слияния, такого как полиэтиленгликоль, с образованием гибридомных клеток (Goding, Monoclonal Antibodies: Principles and Practice, pp.59-103 (Academic Press, 1986)). Полученные таким образом гибридомные клетки высевают и культивируют в подходящей культуральной среде, которая, предпочтительно, содержит одно или несколько веществ, ингибирующих рост или подавляющих выживание неслитых родительских миеломных клеток. Так, например, если родительские миеломные клетки не содержат фермента гипоксантин-гуанин-фосфорибозил-трансферазы (HGPRT или HPRT), то культуральная среда для получения гибридом обычно включает гипоксантин, аминоптерин и тимидин (среда НАТ), то есть вещества, предупреждающие рост HGPRT-дефицитных клеток. Подходящие человеческие миеломные клеточные линии и гетеромиеломные клеточные линии «мышь-человек» описаны в литературе (Kozbor, J. Immunol., 133:3001 (1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, pp. 51-63 (Marcel Dekker, Inc., New York, 1987)). Культуральную среду, в которой культивируют гибридомные клетки, анализируют на продуцирование моноклональных антител, направленных против антигена. Предпочтительно специфичность связывания моноклональных антител, продуцируемых гибридомными клетками, определяют путем иммунопреципитации или с помощью анализа на связыывание in vitro, такого как радиоиммуноанализ (РИА) или твердофазный иммуноферментный анализ (ELISA).
После идентификации гибридомных клеток, продуцирующих антитела с нужной специфичностью, аффинностью и/или активностью, клоны могут быть субклонированы путем лимитирующего разведения и культивированы стандартными методами (Goding, Monoclonal Antibodies: Principles and Practice, pp.59-103 (Academic Press, 1986)). Подходящими культуральными средами, используемыми для таких целей, являются, например, среда D-MEM или среда RPMI-1640. Моноклональные антитела, секретируемые субклонами, соответствующим образом выделяют из культуральной среды, асцитной жидкости или сыворотки в соответствии со стандартными процедурами очистки иммуноглобулинов, такими как, например, хроматография на белке А, гель-электрофорез, диализ или аффинная хроматография.
Человеческие антитела могут быть также продуцированы с использованием хозяина, не являющегося человеком, такого как мышь, способная продуцировать человеческие антитела. Как указывалось выше, в настоящее время используются трансгенные мыши, обладающие способностью, после иммунизации, продуцировать полный репертуар человеческих антител в отсутствие продуцирования эндогенного иммуноглобулина. Так, например, было описано, что гомозиготная делеция гена области присоединения тяжелой цепи антитела (JH) у химерных мышей и у мышей с мутациями в зародышевой линии приводит к полному ингибированию продуцирования эндогенного антитела. Перенос массива генов человеческого иммуноглобулина зародышевой линии такой мыши с мутацией зародышевой линии приводит к продуцированию человеческих антител после стимуляции антигеном. См., например, Jakobovits et al., Proc. Natl. Acad. Sci. USA, 90:2S51 (1993); Jakobovits et al., Nature, 362:255-258 (1993); Bruggermann et al., Year in Immune, 7:33 (1993); патент США No. 5591669; патент США No. 5589369; и патент США No. 5545807. Человеческие антитела могут быть также продуцированы у мышей SCID-hu (Duchosal et al. Nature 355:258-262 (1992)).
В другом варианте изобретения человеческое антитело может быть выбрано из библиотеки фагового представления человеческих антител. Получение библиотек антител или их фрагментов хорошо известно специалистам, и любые из этих известных методов могут быть использованы для конструирования семейства транформирующих фекторов, которые могут быть введены в клетки-хозяева. Библиотеки легких и тяжелых цепей антитела на фаге (Huse et al., Science, 246:1275 (1989)) или гибридных белков на фаге или фагмиде могут быть получены в соответствии с известными процедурами. См., например, Vaughan et al., Nature Biotechnology 14:309-314 (1996); Barbas et al., Proc. Natl. Acad. Sci., USA, 88:7978-7982 (1991); Marks et al., J. Mol. Biol., 222:581-597 (1991); Hoogenboom and Winter, J. Mol. Biol., 227:381-388 (1992); Barbas et al., Proc. Natl. Acad. Sci., USA, 89:4457-4461 (1992); Griffiths et al., EMBO Journal, 13:3245- 3260 (1994); de Kruif et al., J. Mol. Biol., 248:97-105 (1995); WO 98/05344; WO 98/15833; WO 97/47314; WO 97/44491; WO 97/35196; WO 95/34648; патент США No. 5712089; патент США No. 5702892; патент США No. 5427908; патент США No. 5403484; патент США No. 5432018; патент США No. 5270170; WO 92/06176; WO 99/06587; патент США No. 5514548; WO97/08320; и патент США No. 5702892. Представляющий интерес антиген подвергают пэннингу на фаговой библиотеке в соответствии с процедурами, известными специалистам в области отбора фаговых антител, которые связываются с антигеном-мишенью.
Описанные здесь антитела обладают, но необязательно, одним или несколькими нужными биологическими активностями или свойствами. Такими антителами могут быть, но не ограничиваются ими, химерные, гуманизованные, человеческие и аффинно-зрелые антитела. Как указывалось выше, для получения нужных активностей или свойств, такие антитела могут быть сконструированы или созданы различными методами. В одном из вариантов изобретения указанное антитело обладает аффинностью связывания, составляющей по меньшей мере 105 М-1, предпочтительно по меньшей мере 106 М-1-107 М-1, более предпочтительно по меньшей мере 108 М-1-1012 М-1, а еще более предпочтительно по меньшей мере 109 М-1-1012 М-1. Аффинность связывания антитела может быть определена без излишнего экспериментирования путем тестирования антитела методами, известными специалистам, включая анализ Скэтчарда (см. Munson et al., см. выше).
В другом варианте изобретения антитело согласно изобретению может связываться с тем же эпитопом, с которым связываются описанные здесь антитела, или оно может связываться с эпитопом, совпадающим или перекрывающимся с эпитопом, с которым связываются описанные здесь антитела. Эпитопсвязывающие свойства антитела согласно изобретению могут быть определены методами, известными специалистам. Так, например, антитело может быть протестировано в анализе in vitro, таком как анализ на конкурентное ингибирование, в целях определения способности данного антитела блокировать или ингибировать взаимодействие, известное как связывание.
3. Биспецифические антитела
Биспецифические антитела представляют собой моноклональные антитела, предпочтительно человеческие или гуманизованные антитела, обладающие специфичностью связывания по меньшей мере с двумя различными антигенами. Методы продуцирования биспецифических антител известны специалистам. Традиционное рекомбинантное продуцирование биспецифических антител основано на коэкспрессии двух пар тяжелой цепи-легкой цепи иммуноглобулина, где две тяжелых цепи обладают различной специфичностью (Millstein & Cuello, Nature, 305:537-539 (1983)). Эти гибридомы (квадромы), благодаря рандомизированному набору тяжелой и легкой цепей иммуноглобулина, продуцируют потенциальную смесь из десяти различных молекул антител, из которых только одна молекула имеет “правильную” биспецифическую структуру. Очистку такой “правильной” молекулы обычно осуществляют путем постадийного проведения аффинной хроматографии. Аналогичные процедуры описаны в заявке WO 93/08829, опубликованной 13 мая 1993, и в публикации Traunecker et al., EMBO J., 10:3655-3659 (1991).
Вариабельные домены антитела с нужными специфичностями связывания (объединенные сайты “антитело-антиген”) могут быть связаны с последовательностями константного домена иммуноглобулина. Такое связывание, предпочтительно, осуществляют с константным доменом тяжелой цепи иммуноглобулина, содержащим по меньшей мере часть шарнирной области, СН2 и СН3. При этом предпочтительно, чтобы этот гибрид имел первую константную область тяжелой цепи (СН1), содержащую сайт, необходимый для связывания с легкой цепью, присутствующей по меньшей мере в одном из гибридов. ДНК, кодирующие гибриды тяжелой цепи иммуноглобулина и, если это необходимо, легкой цепи иммуноглобулина, встраивают в отдельные экспрессионные векторы, и ко-трансфецируют в подходящий организм-хозяин. Подробное описание получения биспецифических антител можно найти, например, в публикации Suresh et al., Methods in Bnzymology, 121:210 (1986).
4. Гетероконъюгированные антитела
Гетероконъюгированные антитела также входят в объем настоящего изобретения. Гетероконъюгированные антитела состоят из двух ковалентно связанных антител. Предполагается, что такие антитела нацеливают клетки иммунной системы на нежелательные клетки (патент США No. 4676980), и могут быть использованы для лечения ВИЧ-инфекций (WO 91/00360; WO 92/200373, EP 03089). Считается, что антитела могут быть получены in vitro с применением известных методов химического синтеза белков, включая методы с использованием перекрестно-связывающих агентов. Так, например, иммунотоксины могут быть сконструированы путем проведения реакций дисульфидного обмена или посредством образования тиоэфирной связи. Примерами реагентов, подходящих для такой цели, являются иминотиолат и метил-4-меркаптобутиримидат, и такие реагенты описаны, например, в патенте США No. 4676980.
5. Триантитела
Триантитела также входят в объем настоящего изобретения. Такие антитела описаны, например, в публикациях Iliades et al., см. выше и Kortt et al., см. выше.
6. Другие модификации
В настоящей заявке рассматриваются другие модификации антител. Некоторые антитела, используемые в способах согласно изобретению, могут быть модифицированы путем конъюгирования антитела с цитиотоксическим агентом (подобным молекуле токсина) или с пролекарствоактивирующим ферментом, который превращает пролекарство (например, пептидильное химиотерапевтическое средство, см. WO81/01145) в активное противораковое лекарственное средство. См, например, WO 88/07378 и патент США No. 4975278. Такая технология также называется «антителозависимой опосредуемой ферментом пролекарственной терапией» (ADEPT).
Ферментным компонентом иммуноконъюгата, используемым в ADEPT, является любой фермент, способный действовать на пролекарство, то есть он превращает это пролекарство в более активную цитотоксическую форму. Ферментами, которые могут быть использованы в способе согласно изобретению, являются, но не ограничиваются ими, щелочная фосфатаза, используемая для превращения фосфатсодержащих пролекарств в свободные лекарственные средства; арилсульфатаза, используемая для превращения сульфатсодержащих пролекарств в свободные лекарственные средства; цитозиндезаминаза, используемая для превращения нетоксичного 5-фторцитозина в противораковое лекарственное средство, а именно в 5-фторурацил; протеазы, такие как протеаза Serratia, термолизин, субтилизин, карбоксипептидазы и катепсины (такие как катепсины В и L), которые используются для превращения пептидсодержащих пролекарств в свободные лекарственные средства; каспазы, такие как каспаза-3; D-аланилкарбоксипептидазы, используемые для превращения пролекарств, содержащих D-аминокислотные заместители; углеводрасщепляющие ферменты, такие как бета-галактозидаза и нейраминидаза, используемые для превращения гликозилированных пролекарств в свободные лекарственные средства; бета-лактамаза, используемая для превращения лекарственных средств, дериватизированных бета-лактамами, в свободные лекарственные средства; и пенициллин-амидазы, такие как пенициллин-V-амидаза или пенициллин-G-амидаза, используемые для превращения лекарственных средств, дериватизированных у атомов азота амина феноксиацетильными или фенилацетильными группами, соответственно, в свободные лекарственные средства. Альтернативно, антитела, обладающие ферментативной активностью и также известные специалистам как “абзимы”, могут быть использованы для превращения пролекарств согласно изобретению в свободные активные лекарственные средства (см., например, Massey, Nature 328: 457-458 (1987)). Конъюгаты “антитело - абзим” могут быть получены, как описано в настоящей заявке для доставки указанного абзима в популяцию опухолевых клеток.
Ферменты могут быть ковалентно связаны с антителами хорошо известными методами, такими как метод с использованием гетеробифункциональных перекрестно-связывающих реагентов. Альтернативно, гибридные белки, содержащие по меньшей мере антигенсвязывающую область антитела согласно изобретению, связанную по меньшей мере с функционально активной частью фермента согласно изобретению, могут быть сконструированы методами рекомбинантных ДНК, хорошо известными специалистам (см., например, Neuberger et al., Nature, 312:604-608 (1984).
Как известно специалистам, антитела, такие как антитела, используемые в иммуноанализах для детектирования фрагментов белка AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4), могут быть конъюгированы с различными визуализирующими агентами. Конъюгирование может быть осуществлено непосредственно между антителом и визуализирующим агентом или связывающим агентом, либо между антителом и активным агентом могут присутствовать промежуточные молекулярные группы. Перекрестно-связывающие линкеры часто используются для облегчения конъюгирования посредством создания сайтов присоединения для каждой молекулы. Перекрестно-связывающими линкерами могут быть дополнительные молекулярные группы, которые служат в качестве спейсеров для отделения молекул друг от друга в целях предупреждения нежелательного взаимодействия молекулы, обладающей одной активностью, с молекулой, обладающей другой активностью.
Метод визуализации может быть осуществлен с помощью многих процедур, хорошо известных среднему специалисту в данной области, и соответствующий визуализирующий агент, используемый в таких процедурах, может быть конъюгирован с антителом хорошо известными методами. Визуализация может быть осуществлена, например, путем представления результатов вестерн-блот-анализа, радиосцинтиграфии, ядерного магнитного резонанса (ЯМР) или компьютерной томографии (компьютерно-томографического сканирования). Наиболее часто используемыми радионуклидными визуализирующими агентами являются радиоактивный иод и индий. Визуализация путем компьютерно-томографического сканирования может быть проведена с использованием тяжелого металла, такого как хелатные комплексы железа. Визуализирующее ЯМР-сканирование может быть проведено с использованием хелатных комплексов гадолиния или марганца. Кроме того, может быть проведена позитронно-эмиссионная томография (ПЭТ), в которой излучателями позитронов являются кислород, азот, железо, углерод или галлий. Примерами радионуклидов, используемых в процедурах визуализации, являются 43K, 52Fe, 57Co, 67Cu, 67Ga, 68Ga, 77Br, 81Rb, 111In, 113In, 123I, 125I, 127Cs, 129Cs, 131I, 132I, 197Hg, 203Pb и 206Bi.
В публикациях Magerstadt, M. (1991) Antibody Conjugates And Malignant Disease, CRC Press, Boca Raton, FIa., and Barchel, S. W. and Rhodes, B. H., (1983) Radioimaging and Radiotherapy, Elsevier, NY, N. Y., каждая из которых вводится в настоящее описание посредством ссылки, описано конъюгирование различных терапевтических и диагностических радионуклидов с аминокислотами антител. Такие реакции могут быть проведены для конъюгирования радионуклидов с антителами с использованием соответствующего линкера. Подходящими метками являются, например, радионуклиды, ферменты (например, пероксидаза хрена), субстраты, кофакторы, ингибиторы, флуоресцентные вещества, хемилюминесцентые вещества и/или магнитые частицы. Примерами патентов, в которых описано использование таких меток, являются патенты США №№ 3817837; 3850752; 3939350; 3996345; 4277437; 4275149; и 4366241, каждый из которых вводится в настоящее описание посредством ссылки.
В настоящем описании также рассматриваются и другие модификации антител. Так, например, антитело может быть присоединено к одному из различных небелковых полимеров, например, таких как полиэтиленгликоль, полипропиленгликоль, полиоксиалкилены или сополимеры полиэтиленгликоля и полипропиленгликоля. Антитело может быть также заключено в микрокапсулы, полученные, например, методами коацервации или межфазной полимеризации (например, с использованием гидроксиметилцеллюлозы или желатиновых микрокапсул и полиметилметакрилатных микрокапсул, соответственно), в коллоидальные системы доставки лекарственных средств (например, в липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы), или в макроэмульсии. Такие методы описаны в руководстве Remington's Pharmaceutical Sciences, 16th edition, Oslo, A., Ed., (1980). Для увеличения времени полужизни антитела в сыворотке в такое антитело (а в частности в его фрагмент) может быть включен эпитоп, связывающийся с рецептором «спасения», как описано, например, в патенте США 5739277. Используемый здесь термин «эпитоп, связывающийся с рецептором «спасения»» означает эпитоп Fc-области молекулы IgG (например, IgG1, IgG2, IgG3 или IgG4), который является ответственным за увеличение времени полужизни молекулы IgG в сыворотке in vivo.
7. Рекомбинантные методы
Настоящее изобретение также относится к выделенным нуклеиновым кислотам, кодирующим представляющий интерес полипептид, например белки, фрагменты белков (например фрагмент белка AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4)) и/или описанные здесь антитела, а также к векторам и клеткам-хозяевам, содержащим указанную нуклеиновую кислоту; и к рекомбинантным способам получения представляющего интерес полипептида.
Для рекомбинантного продуцирования представляющего интерес полипептида нуклеиновую кислоту, кодирующую такой полипептид, выделяют и встраивают в реплицируемый вектор для последующего клонирования (амплификации ДНК) или экспрессии. ДНК, кодирующая представляющий интерес полипептид, может быть легко выделена и секвенирована стандартными методами (например, с использованием олигонуклеотидных зондов, обладающих способностью специфически связываться с генами, кодирующими представляющий интерес полипептид). Многие векторы являются общедоступными. Векторными компонентами, в основном, являются, но не ограничиваются ими, один или несколько из нижеследующих компонентов, таких как сигнальная последовательность, ориджин репликации, один или несколько маркерных генов, энхансерный элемент, промотор и последовательность терминации транскрипции.
Описанными здесь способами являются способы продуцирования химерных или рекомбинантных антител, которые связываются с фрагментом белка AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4), где указанные способы включают стадии получения вектора, содержащего последовательность ДНК, кодирующую легкую или тяжелую цепь антитела (или легкую цепь и тяжелую цепь), трансфекции или трансформации клетки-хозяина указанным вектором, и культивирования указанной клетки-хозяина (клеток-хозяев) в условиях, достаточных для продуцирования рекомбинантного продукта.
(i) Компонент сигнальная последовательность
Представляющий интерес полипептид может быть рекомбинантно продуцирован не только прямым методом, но также и в форме гибрида данного полипептида с гетерологичным полипептидом, который, предпочтительно, представляет собой сигнальную последовательность или другой полипептид, имеющий специфический сайт расщепления у N-конца зрелого белка или полипептида. Выбранной гетерологичной сигнальной последовательностью, предпочтительно, является последовательность, которая распознается и процессируется (то есть отщепляется сигнальной пептидазой) в клетке-хозяине. Если прокариотические клетки-хозяева не распознают и не процессируют нативную сигнальную последовательность, то такая сигнальная последовательность может быть заменена прокариотической сигнальной последовательностью, выбранной, например, из группы, состоящей из лидерных последовательностей щелочной фосфатазы, пенициллиназы, lрр или термостабильного энтеротоксина II. Для секреции в дрожжах нативная сигнальная последовательность может быть заменена, например, лидерной последовательностью инвертазы дрожжей, лидерной последовательностью α-фактора (включая лидерные последовательности α-фактора Saccharomyces и Kluyveromyces), или лидерной последовательностью кислой фосфатазы, лидерной последовательностью глюкоамилазы C. albicans или сигнальной последовательностью, описанной в WO90/13646. Для экспрессии в клетках млекопитающих используются сигнальные последовательности млекопитающих, а также вирусные секреторные лидерные последовательности, например сигнальная последовательность gD вируса простого герпеса.
ДНК для такой области предшественника лигируют с ДНК, кодирующей представляющий интерес полипептид, с сохранением рамки считывания.
(ii) Компонент ориджин репликации
Экспрессионные и клонирующие векторы содержат последовательность нуклеиновой кислоты, которая способствует репликации данного вектора в одной или нескольких выбранных клетках-хозяевах. Обычно, в клонирующих векторах, такой последовательностью является последовательность, которая позволяет данному вектору реплицироваться независимо от ДНК хромосомы хозяина, и которая включает ориджины репликации или автономно реплицирующиеся последовательности. Такие последовательности для различных бактерий, дрожжей и вирусов хорошо известны. Ориджин репликации, происходящий от плазмиды pBR322, является подходящим для большинства грамотрицательных бактерий, а именно ориджин репликации плазмиды 2µ является подходящим для дрожжей, а различные ориджины репликации вирусов (SV40, полиомавируса, аденовируса, VSV или BPV) могут быть использованы для клонирования векторов в клетках млекопитающих. Обычно компонент ориджин репликации не является необходимым для векторов, экспрессирующихся в клетках млекопитающих (ориджин SV40 обычно используется только в тех случаях, когда он содержит ранний промотор).
(iii) Компонент селективный ген
Экспрессионный и клонирующий векторы могут содержать селективный ген, также называемый селективным маркером. Обычно селективные гены кодируют белки, которые (а) сообщают резистентность к антибиотиками или к другим токсинам, например к ампициллину, неомицину, метотрексату или тетрациклину, (b) комплементируют дефицит, обусловленный ауксотрофией, или (с) обеспечивают поступление важных питательных веществ, которые не поступают из комплексных сред, например, ген, кодирующий D-аланин-рацемазу Bacillus.
Одним из примеров схемы отбора является использование лекарственного средства, прекращающего рост клетки-хозяина. Клетки, которые были успешно трансформированы гетерологичным геном, продуцируют белок, сообщающий резистентность к лекарственному средству, и, тем самым, способствующий выживанию этих клеток в селективной среде. Для такого доминантного отбора могут быть использованы, например, лекарственные средства, такие как неомицин, микофеноловая кислота и гигромицин.
Другими примерами подходящих селективных маркеров для клеток млекопитающих являются маркеры, которые позволяют идентифицировать клетки, способные встраивать в свой геном нуклеиновую кислоту, такую как гены DHFR, тимидинкиназы, металлотионеина-I и II, а предпочтительно гены, кодирующие металлотионеин приматов, аденозин-дезаминазу, орнитин-декарбоксилазу и т.п.
Так, например, клетки, трансформированные селективным геном DHFR, сначала идентифицируют путем культивирования всех трансформантов в культуральной среде, содержащей метотрексат (Mtx), конкурентный антагонист DHFR. При использовании DHFR дикого типа подходящей клеткой-хозяином является клеточная линия яичника китайского хомячка (СНО), дефицитная по DHFR-активности.
Альтернативно, клетки-хозяева (в частности хозяева дикого типа, содержащие эндогенный DHFR), трансформированные или ко-трансформированные ДНК-последовательностями, кодирующими представляющий интерес полипептид, белок DHFR дикого типа и другой селективный маркер, такой как аминогликозид-3'-фосфотрансфераза (АРН), могут быть отобраны путем культивирования клеток в среде, содержащей агент для отбора, проводимого с использованием селективного маркера, такого как аминогликозидный антибиотик, например, канамицин, неомицин или G418. См. патент США № 4965199.
Подходящим селективным геном для использования в дрожжах является ген trp1, присутствующий в дрожжевой плазмиде YRр7 (Stinchcomb et al., Nature, 282:39 (1979)). Ген trp1 представляет собой селективный маркер для мутантного штамма дрожжей, который не способен расти в присутствии триптофана, например штамма, депонированного в ATCC № 44076 или РЕР4-1. Jones, Genetics, 85:12 (1977). Присутствие мутации в гене trp1 в геноме дрожжевой клетки-хозяина позволяет эффективно осуществлять детекцию трансформации путем культивирования в отсутствие триптофана. Аналогичным образом, Leu2-дефицитные дрожжевые штаммы (ATCC 20622 или 38626) комплементировали известными плазмидами, несущими ген Leu2.
Кроме того, векторы, происходящие от кольцевой 1,6 мкм-плазмиды pKD1, могут быть использованы для трансформации дрожжей Kluyveromyces. Альтернативно, экспрессионная система для крупномасштабного продуцирования рекомбинантного телячьего химозина была описана для K.lactis. Van den Berg, Bio/Technology, 8:135 (1990). Были также описаны стабильные многокопийные экспрессионные векторы для секреции зрелого рекомбинантного человеческого сывороточного альбумина промышленными штаммами Kluyveromyces. Fleer et al., Bio/Technology, 9:968-975 (1991).
(iv) Компонент промотор
Экспрессионные и клонирующие векторы обычно содержат промотор, распознаваемый организмом-хозяином и функционально присоединенный к нуклеиновой кислоте. Промоторами, подходящими для использования в прокариотических хозяевах, являются промотор phoA, промоторные системы β-лактамазы и лактозы, промоторные системы щелочных фосфатаз и триптофана (trp), и гибридные промоторы, такие как промотор tac. Однако подходящими также являются и другие известные бактериальные промоторы. Промоторы, используемые в бактериальных системах, также содержат последовательность Шайна-Дальгарно (S.D.), функционально присоединенную к ДНК, кодирующей представляющий интерес полипептид.
Промоторные последовательности, подходящие для эукариотов, известны специалистам. Фактически все гены эукариотов имеют АТ-богатую область, локализованную приблизительно в области, расположенной на 25-30 нуклеотидов выше от сайта инициации транскрипции. Другой такой последовательностью, локализованной приблизительно в области, расположенной на 70-80 нуклеотидов выше от сайта инициации транскрипции многих генов, является область CNCAAT, где N может означать любой нуклеотид. У 3'-конца большинства эукариотических генов расположена последовательность ААТААА, которая может служить сигналом для присоединения poly A-хвоста к 3'-концу кодирующей последовательности. Все указанные последовательности могут быть соответствующим образом встроены в эукариотические экспрессионные векторы.
Примерами промоторных последовательностей, подходящих для использования в дрожжевых клетках-хозяевах, являются промоторы 3-фосфоглицерат-киназы или других гликолитических ферментов, таких как энолаза, глицеральдегид-3-фосфат-дегидрогеназа, гексокиназа, пируват-декарбоксилаза, фосфофруктокиназа, глюкозо-6-фосфат-изомераза, 3-фосфоглицерат-мутаза, пируваткиназа, триозофосфатизомераза, фосфоглюкозоизомераза и глюкокиназа.
Другими дрожжевыми промоторами, которые представляют собой индуцибельные промоторы, имеющие дополнительное преимущество, заключающееся в их способности регулировать транскрипцию в определенных условиях культивирования, являются промоторные области генов алкоголь-дегидрогеназы 2, изоцитохрома С, кислой фосфатазы, гидролитических ферментов, ассоциированных с метаболизмом азота, металлотионеина, глицеральдегид-3-фосфат-дегидрогеназы и ферментов, ответственных за утилизацию мальтозы и галактозы. Векторы и промоторы, подходящие для экспрессии в дрожжах, также описаны в ЕР 73657. Наряду с дрожжевыми промоторами также преимущественно используются дрожжевые энхансеры.
Транскрипция из векторов в клетках-хозяевах млекопитающих регулируется, например, промоторами, полученными из геномов таких вирусов, как вирус полиомы, вирус оспы домашней птицы, аденовирус (такой как аденовирус 2), вирус коровьей папиломы, вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита В, а наиболее предпочтительно обезьяний вирус 40 (SV40); гетерологичными промоторами млекопитающих, например промотором актина или промотором иммуноглобулина; и промоторами белков теплового шока, при условии, что такие промоторы являются совместимыми с системами клеток-хозяев.
Ранние и поздние промоторы вируса SV40 обычно получают в виде рестрикционного фрагмента SV40, который также содержит ориджин репликации вируса SV40. Предранний промотор человеческого цитомегаловируса обычно получают в виде рестрикционного HindIII-фрагмента Е. Система экспрессии ДНК в клетках-хозяевах млекопитающих, полученная на основе коровьего папиломавируса, используемого в качестве вектора, описана в патенте США № 4419446. Модификация такой системы описана в патенте США № 4601978. Экспрессия кДНК человеческого β-интерферона в мышиных клетках под контролем промотора тимидинкиназы вируса простого герпеса также описана в публикации Reyers et al., Nature 297:598-601 (1982). См. также альтернативно, в качестве промотора может быть использован вирус саркомы Рауса, имеющий длинный концевой повтор.
(v) Компонент энхансерный элемент
Транскрипция ДНК, кодирующей представляющий интерес полипептид, в высших эукариотах, часто усиливается при встраивании в вектор энхансерной последовательности. В настоящее время известно много энхансерных последовательностей, происходящих от генов млекопитающих (генов глобина, эластазы, альбумина, α-фетопротеина и инсулина). Однако обычно используется энхансер от вируса эукариотической клетки. Примерами является энхансер вируса SV40, локализованный в поздней области ориджина репликации (100-270 п.н.), энхансер раннего промотора цитомегаловируса, энхансер полиомавируса, локализованный в поздней области ориджина репликации, и энхансеры адновирсуса. См. также публикацию Yaniv, Nature 297:17-18 (1982), в которой описаны энхансерные элементы для активации эукариотических промоторов. Энхансер может быть сплайсирован в вектор в 5'- или 3'-положеннии по отношению к кодирующей последовательности, но, предпочтительно, чтобы он находился в 5'-положении от промотора.
(vi) Компонент сайт терминации транскрипции
Экспрессионные векторы, используемые в эукариотических клетках-хозяевах (в дрожжах, грибах, насекомых, растениях, у животных, у человека или в ядросодержащих клетках, происходящих от других многоклеточных организмов), также содержат последовательности, необходимые для терминации транскрипции и для стабилизации мРНК. Такие последовательности обычно расположены со стороны 5'-конца, а иногда 3'-конца от нетранслируемых областей эукариотических или вирусных ДНК или кДНК. Эти области содержат нуклеотидные сегменты, транскрибированные в виде полиаденилированных фрагментов в нетранслированной части мРНК, кодирующей мультивалентное антитело. Одним из подходящих компонентов терминации транскрипции является область полиаденилирования коровьего гормона роста. См. описание экспрессионных векторов в WO 94/11026 и в настоящей заявке.
(vii) Отбор и трансформация клеток-хозяев
Клетками-хозяевами, подходящими для клонирования или экспрессии ДНК в описанных здесь векторах, являются клетки прокариотов, дрожжей или высших эукариотов, описанные выше. Прокариотами, подходящими для этой цели, являются Eubacteria, такие как грамотрицательные или грамположительные микроорганизмы, например, Enterobacteriaceae, такие как Esсherichia, например E.coli, Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, например Salmonella typhimurium, Serratia, например Serratia marcescans и Shigella, а также Bacillus, такие как B. subtilis и B. licheniformis (например, B. licheniformis 41Р, описанная в патенте DD 266710, опубликованном 12 апреля 1989), Pseudomonas, такие как P. aeruginosa, и Streptomyces. Предпочтительным клонирующим хозяином E.coli является E.coli 294 (АТСС 31446), хотя могут быть использованы и другие штаммы, такие как E.coli В, E.coli Х1776 (АТСС 31537) и E.coli W3110 (АТСС 27325). Эти примеры приводятся лишь для иллюстрации и не ограничивают объема изобретения.
Помимо прокариотов подходящими хозяевами для клонирования или экспрессии векторов являются эукариотические микробы, такие как гифомицеты или дрожжи. Из низших эукариотических микроорганизмов-хозяев наиболее часто используются Saccharomyces cerevisiae или стандартные пекарские дрожжи. Однако в данном случае могут быть использованы и другие часто применяемые и доступные микроорганизмы другого рода, других видов и штаммов, таких как Schizosaccharomyces pombe; хозяева Kluyveromyces, такие как, например, K. lactis, K. fragilis (АТСС 12424), K. bulgaricus (АТСС 16045), K. wickeramii (АТСС 24178), K. waltii (АТСС 56500), K. drosophilarum (АТСС 36906), K. thermotorerans и K. marxianus; yarrowia (ЕР 402226); Pichia pastoris (ЕР 183070); Candida; Trichoderma reesia (ЕР 244234); Neurospora crassa; Schwanniomyces, такие как Schwanniomyces occidentalis; и гифомицеты, такие как, например, Neurospora, Penicillium, Tolypocladium и Aspergillus, такие как A. nidulans и A. niger.
Клетками-хозяевами, подходящими для экспрессии представляющих интерес полипептидов, являются клетки, происходящие от многоклеточных организмов. Примерами клеток беспозвоночных являются клетки растений и насекомых. Были идентифицированы различные бакуловирусные штаммы и варианты, а также соответствующие пермиссивные клетки-хозяева насекомых, таких как Spodoptera frugiperda (гусеница совки), Aedes aegypti (желтолихорадочный комар), Aedes albopictus (белопятнистый комар), Drosophila melanogaster (плодовая мушка) и Bombyx mori (тутовый шелкопряд). В настоящее время имеется ряд доступных вирусных штаммов, которые могут быть использованы для трансфекции, например вариант L-1 Autographa californica NPV и штамм Bm-5 Bombyx mori NPV, и такие вирусы могут быть использованы в настоящем изобретении, а в частности для трансфекции клеток Spodoptera frugiperda.
В качестве клеток-хозяев могут быть также использованы клетки растительных культур хлопчатника, кукурузы, картофеля, сои, петунии, томатов и табака.
Однако самый большой интерес представляют клетки позвоночных, и размножение клеток позвоночных в культуре (тканевой культуре) уже становится рутинной процедурой. Примерами обычно используемых линий клеток-хозяев млекопитающих являются клеточная линия CV1 почек обезьяны, трансформированная SV40 (COS-7, ATCC CRL 1651); клеточная линия почек человеческого эмбриона (клетки 293 или клетки 293, субклонированные для роста в суспензионной культуре, Graham et al., J. Gen. Virol. 36:59 (1977)); клетки почек детеныша хомячка (ВНК, АТСС CCL 10); клетки яичника китайского хомячка/-DHFR (CHO, Urlaub et al., Proc. Natl. Acad. Sci. USA 77:4216 (1980)); мышиные клетки Сертоли (ТM4, Mather, Biol. Reprod. 23:243-251 (1980)); клетки почек обезьяны (CV1 ATCC CCL 70); клетки почек африканской зеленой мартышки (VERO-76, ATCC CRL-1587); клетки карциномы шейки матки человека (HELA, ATCC CCL 2); клетки почек собак (MDCK, ATCC CCL 34); клетки печени лабораторной крысы Buffalo (BRL 3A, ATCC CRL 1442); клетки легких человека (W138, ATCC CCL 75); клетки печени человека (HepG2, HB 8065); опухолевые клетки мышиной молочной железы (MMT 060562, ATCC CCL51); клетки TRI (Mather et al., Annal N.Y. Acad. Sci. 383:44-68 (1982)); клетки MRC5; клетки FS4; клеточная линия человеческой гепатомы (HepG2) и клетки миеломы или лимфомы (например, клетки YO, J558L, P3 и NSO)(см. патент США № 5807715).
Клетки-хозяева трансформируют вышеописанными экспрессионными или клонирующими векторами для продуцирования представляющего интерес полипептида и культивируют в подходящих питательных средах, модифицированных, если это необходимо, для индуцирования промоторов, отбора трансформантов или амплификации генов, кодирующих нужные последовательности.
(viii) Культивирование клеток-хозяев
Клетки-хозяева, используемые для продуцирования представляющего интерес полипептида, могут быть культивированы в различных средах. Средами, подходящими для культивирования клеток-хозяев, являются коммерчески доступные среды, такие как среда Хэмса F10 (Sigma), минимальная поддерживающая среда ((МЕМ)(Sigma), RPMI-1640 (Sigma) и модифицированная по способу Дульбекко среда Игла (DMEM), Sigma). Кроме того, в качестве культуральной среды для культивирования клеток-хозяев может быть использована любая среда, описанная в публикации Ham et al., Meth. Enz. 58:44 (1979), Barnes et al., Anal. Biochem. 102:255 (1980), в патентах США №№ 4767704, 4657866, 4927762, 4560655 или 5122469; в WO 90103430; в WO 87/00195 или в патенте США Re.№ 30985. В любую из этих сред могут быть добавлены, если это необходимо, гормоны и/или другие факторы роста (такие как инсулин, трансферрин или эпидермальный фактор роста), соли (такие как хлорид натрия, и фосфат кальция и магния), буферы (такие как HEPES), нуклеотиды (такие как аднозин и тимидин), антибиотики (такие как лекарственное средство гентамицинТМ), микроэлементы (определенные как неорганические соединения, обычно присутствующие в конечных концентрациях в микромолярных дозах), и глюкоза или эквивалентный источник энергии. Могут быть также включены любые другие необходимые добавки в соответствующих концентрациях, известные специалистам в данной области. Условия культивирования, такие как температура, рН и т.п., аналогичны условиям, ранее используемым для экспрессии выбранных клеток-хозяев, и известны среднему специалисту в данной области.
(ix) Очистка антитела
C применением техники рекомбинантных ДНК представляющий интерес полипептид может быть продуцирован внутри клеток или в периплазматическом пространстве, либо он может быть непосредственно секретирован в среду. Если в первой стадии представляющий интерес полипептид продуцируется внутри клеток, то клеточный дебрис или клетки-хозяева или их лизированные фрагменты удаляют, например, путем центрифугирования или ультрафильтрации. В публикации Carter et al., Bio/Technology 10:163-167 (1992) описана процедура выделения антител, которые секретируются в периплазматическое пространство E.coli. Вкратце, клеточную пасту оттаивают в присутствии ацетата натрия (рН 3,5), ЕDTA и фенилметилсульфонилфторида (PMSF) в течение примерно 30 минут. Клеточный дебрис может быть удален путем центрифугирования. Если представляющий интерес полипептид секретируется в среду, то супернатанты от таких экспрессионных систем обычно сначала концентрируют с использованием коммерчески доступного фильтра для концентрирования белка, например устройства для ультрафильтрации Amicon или Millipore Pellicon. Ингибитор протеазы, такой как PMSF, может быть включен в любую из вышеуказанных стадий для ингибирования протеолиза, а антибиотики могут быть включены для предупреждения роста случайно внесенных примесных микроорганизмов.
Композиция, полученная из клеток, может быть очищена, например, с помощью хроматографии на гидроксиаппатитах, гель-электрофореза, диализа и аффинной хроматографии, при этом предпочтительным методом очистки является аффинная хроматография. Пригодность белка А в качестве аффинного лиганда зависит от вида и изотипа Fc-области любого иммуноглобулина, присутствующего в антителах. Белок А может быть использован для очистки антител, содержащих тяжелые цепи γ1, γ2 или γ4 человеческого иммуноглобулина (Lindmark et al., J. Immunol. Meth. 62:1-13 (1983)). Для всех мышиных изотипов и для γ3 человеческого иммуноглобулина рекомендуется белок G (Guss et al., EMBO J. 5:15671575 (1986)). В качестве матрицы, с которой связывается аффинный лиганд, наиболее часто используется агароза, но могут быть использованы и другие матрицы. Механически стабильные матрицы, такие как стекло с регулируемым размером пор или поли(стиролдивинил)бензол, обеспечивают большую скорость потока и более короткое время обработки, чем это может быть достигнуто с использованием агарозы. Если антитело содержит домен СН3, то для очистки может быть использована смола Bakerbond ABXTM (J.T. Baker, Phillipsburg, N.J.). В зависимости от выделяемого представляющего интерес полипептида могут быть также применены и другие методы очистки белка, такие как фракционирование на ионообменной колонке, преципитация этанолом, обращенно-фазовая ВЭЖХ, хроматография на двуокиси кремния, хроматография на гепарин-сефарозеТМ, хроматография на анионо- или катионообменной смоле (например, на колонке с полиаспарагиновой кислотой), хроматофокусирование, электрофорез в ДСН-ПААГ и преципитация сульфатом аммония.
III. Промышленные изделия
В другом своем варианте настоящее изобретение относится к промышленному изделию, содержащему материалы, используемые для осуществления описанных выше вариантов способов согласно изобретению. Такое промышленное изделие содержит контейнер и этикетку. Подходящими контейнерами являются, например, бутыли, флаконы, шприцы и тест-пробирки. Контейнеры могут быть изготовлены из различных материалов, таких как стекло или пластик. Указанный контейнер содержит композицию, являющуюся эффективной для обнаружения апоптоза, и может иметь стерильное входное отверстие (например, таким контейнером может быть пакет для внутривенного введения раствора или сосуд, имеющий пробку, протыкаемую иглой для подкожных инъекций). Типичными агентами, входящими в состав указанной композиции, являются одно или несколько антител, которые связываются с фрагментом белка AP2-α (SEQ ID NO: 1), тяжелой цепи клатрина (SEQ ID NO: 2), AP1/2β (SEQ ID NO: 3) или динамина (SEQ ID NO: 4). На этикетке или во вкладыше, вложенном в упаковку, должно быть указано, что такая композиция используется для обнаружения апоптоза и/или для оценки конкретного состояния. Промышленное изделие может также включать второй контейнер, содержащий фармацевтически приемлемый буфер, такой как забуференный фосфатом физиологический раствор, раствор Рингера и раствор декстрозы. Кроме того, оно может также включать и другие материалы, необходимые с точки зрения промышленного производства и потребителя, включая другие буферы, разбавители, фильтры, иглы, шприцы и вкладыши в упаковку, в которых имеются инструкции по применению данного изделия.
В одном из таких вариантов настоящее изобретение относится к наборам, которые могут быть использованы для детектирования клеточного апоптоза и диагностики ассоциированных с ним заболеваний. Один из таких наборов включает: (1) «первое» антитело, способное связываться с фрагментом белка, образующимся во время апоптоза, (2) «второе» антитело, конъюгированное с сигналпродуцирующей меткой, где указанным «вторым» антителом является антитело, которое связывается с «первым» антителом, и (3) сигналпродуцирующий «третий» реагент, способный распознавать меченное «второе» антитело. Другой набор, который может быть использован для детектирования образующихся во время апоптоза фрагментов белка согласно изобретению, включает: (1) «первое» антитело, способное связываться с фрагментами белка, образующимися во время апоптоза, и (2) «второе» антитело, конъюгированное с сигналпродуцирующей меткой, где указанное «второе» антитело также реагирует с образующимся во время апоптоза фрагментом белка, но связывается с сайтом, отличающимся от сайта, с которым связывается «первое» антитело.
В вариантах наборов согласно изобретению сигналпродуцирующей меткой, связанной со «вторым» антителом, может быть, например, фермент, такой как пероксидаза хрена или щелочная фосфатаза. Предпочтительно указанный набор содержит фермент и субстрат. Такой набор может также включать не имеющий покрытия носитель, на котором могут быть иммобилизованы анализируемый образец или «первое» антитело.
IV. Применение способов и материалов согласно изобретению для оценки клеточных процессов, ассоциированных с апоптозом
Способы и материалы согласно изобретению могут быть использованы для оценки клеточных процессов широкого ряда, таких как эндоцитоз. Так, например, некоторые варианты настоящего изобретения позволяют обнаруживать изменения скорости клеточного эндоцитоза как индикатора апоптоза, причем ингибирование эндоцитоза указывает на наличие апоптоза. Эндоцитоз играет решающую роль в различных аспектах гомеостаза клеток. Эндоцитоз интернализует связанные с плазматической мембраной (ПM) белки посредством мембраносвязанных везикул, и тем самым подтверждает различные клеточные функции, включая поглощение питательных веществ, передачу сигнала факторов роста и гомеостаз мембраны (см., например, Conner, S.D. et. al., Nature 422, 37-44 (2003)). Один из наиболее охарактеризованных путей эндоцитоза основан на белке клатрине (см., например, Bonifacino, J. S. et. al., Annu Rev Biochem 72, 395-447 (2003)). Адапторы клатрина, такие как белок-адаптор 2 (AP2), связывает клатрин с цитоплазматическими детерминантами эндоцитарного «грузового» белка (cargo) во время образования инвагинаций в плазматической мембране (ПМ), известных как «ямки», выстланные белком клатрином. Адапторы также осуществляют структурные функции при эндоцитозе путем рекрутинга вспомогательных или регуляторных белков (см., например, Owen, D.J., et. al, P.R. Annu. Rev. Cell Dev. Biol. 20, 153-91 (2004)). GTP-гидролиз под действием динамина стимулирует разрезание глубоко инвагинированных покрытих клатрином ямок с высвобождением эндоцитарных везикул, транспортируемых из ПM. После освобождения от покрытия эти везикулы заякориваются и сливаются с незрелыми эндосомами, где их «грузовые» белки (cargo) поступают в различные участки, например в тубуло-везикулярные эндосомные мембраны для рециклинга в ПM, или во внутренние мембраны мультивезикулярных зрелых эндосом для лизосомной деградации.
С применением способов и материалов согласно изобретению было обнаружено, что рецепторы «проапоптотической гибели» (DR) индуцируют селективное нарушение механизма клатринзависимого эндоцитоза. DR-стимуляция индуцирует быстрое опосредуемое каспазой расщепление ключевых компонентов клатринового пути, что приводит к прекращению поглощения клетками классического «грузового» белка трансферрина. DR-проксимальные каспазы-инициаторы расщепляют субъединицу адаптора клатрина AP2α между функционально отличающимися доменами, тогда как каспазы-эффекторы осуществляют процессинг тяжелой цепи клатрина. Было показано, что DR5 подвергается индуцированному лигандом и опосредованному клатрином эндоцитозу, что свидетельствует о том, что интернализация DR-сигнального комплекса облегчает клатриновый путь нацеливания, обеспечиваемый каспазами. Эндоцитозблокирующий термочувствительный мутантный динамин-1 способствует ослаблению интернализации DR, усилению стимуляции каспаз, действующих после DR, и индуцированию апоптоза. Таким образом, DR-стимулируемая активность каспаз приводит к нарушению клатринзависимого эндоцитоза и к ускорению запрограммированной клеточной гибели.
Другие иллюстративные примеры применения методов и материалов согласно изобретению описаны ниже.
DR-активация индуцирует опосредуемое каспазой расщепление по механизму клатринзависимого эндоцитоза.
В исследованиях эндоцитоза DR5 было обнаружено, что рецептор «гибели» DR5 для лиганда Apo2L/TRAIL запускает протеолитическое расщепление α-субъединицы AP2 (AP2α). Анализ некоторых раковых клеточных линий показал, что Apo2L/TRAIL стимулирует не только предполагаемый процессинг каспазы-8 и -3, но также и расщепление AP2α, AP1/2β и тяжелой цепи клатрина (CHC) (фиг.1a). Эти события происходят в течение 2 часов и ограничены клеточными линиями, которые, в отличие от резистентных клеточных линий, таких как HCT8, являются восприимчивыми к Apo2L/TRAIL-индуцированному апоптозу, а именно клеточные линии Colo205, BJAB и HeLa-M (фиг.1a). Это наблюдение также подтверждается с использованием других клеточных линий (фиг.7a). FasL стимулирует расщепление AP2α и CHC в клетках BJAB, сравнимое с расщеплением под действием Apo2L/TRAIL (фиг.7b). Apo2L/TRAIL также индуцирует расщепление динамина, что приводит к истощению полноразмерного белка, которое начинается через 1 час после начала стимуляции (фиг.7c). В аналогичный период времени Apo2L/TRAIL не индуцирует протеолиз адапторов с аналогичной структурой, которые опосредуют события клатринзависимого везикулярного транспорта другого типа (фиг.1b). Такими адапторами являются AP1α, AP3β и δ и AP4ε, которые поддерживают транспорт между транс-аппаратом Гольджи и эндосомами, и субъединицей COP-I, β-СОР, или субъединицей СOP-II, Sec23, которые опосредуют транспорт между эндоплазматическим ретикулумом и аппаратом Гольджи (см., например, Bonifacino, J. S. et. al., Annu Rev Biochem 72, 395-447 (2003) и McMahon, H. T. et. al., Curr Opin Cell Biol 16, 379-91 (2004)). Следовательно, DR-активация стимулирует быстрое и специфическое расщепление белков, участвующих в клатринзависимом эндоцитозе.
Предварительная обработка ингибитором пан-каспазы, таким как N-бензилоксикарбонил-Val-Ala-Asp-фторметилкетон (zVAD-fmk) в дозе, вызывающей ингибирование индуцированного лигандом процессинга каспазы-8 и -3, приводит к предотвращению расщепления AP2α и CHC (фиг.2a), а также AP2α в клетках BJAB (фиг.8a), что указывает на необходимость каспазной активности. Кроме того, хотя Apo2L/TRAIL и FasL индуцируют расщепление AP2α и CHC в Т-клетках Jurkat дикого типа, однако ни один из лигандов не стимулирует процессинг этих мишеней в дефицитных по каспазе-8 или дефицитных по FADD мутантных клеточных линиях Jurkat (фиг.8b). Расщепление AP2α происходит быстро и совпадает по времени с процессингом каспазы-8, но предшествует процессингу каспазы-3 (фиг.Ia). В противоположность этому расщепление CHC происходит относительно более медленно и лучше коррелирует по времени с процессингом каспазы-3. В клетках HCT116 для активации каспазы-3 и для индуцирования апоптоза под действием Apo2L/TRAIL требуется присутствие Bax, а для стимуляции каспазы-8 его присутствия не требуется (см., например, LeBlanc, H.N. et. al., Cell Death Differ 10, 66-75 (2003)). Как и ожидалось, Apo2L/TRAIL стимулирует процессинг каспазы-8 и -3 в Bax+/--клетках HCT116, а в Bax-/--клетках HCT116 - только каспазы-8 (фиг.2b, c). Расщепление AP2α не зависит от Bax, тогда как Вах необходим для расщепления CHC (фиг.2b, c). В клетках MCF7, которые являются дефицитными по каспазе-3, Apo2L/TRAIL индуцирует относительно слабый и медленный процессинг каспазы-8 и AP2α, но не стимулирует расщепление CHC (фиг.2d) (см., например, Kischkel, F. C. et al., Immunity 12, 611-20 (2000)). Кроме того, киРНК- ингибирование каспазы-3 в клетках HT1080 блокирует индуцированный лигандом процессинг CHC, но не AP2α (фиг.8c). Полученные данные показали, что каспазы-инициаторы в пути DR расщепляют AP2α, а каспазы-эффекторы обеспечивают процессинг CHC.
AP2α содержит две главные функциональные части, соединенные шарнирной областью: С-концевой домен «ухо» и N-концевой домен «туловище» (см., например, Owen, D.J., et. al, P. R. Annu. Rev. Cell Dev. Biol. 20, 153-91 (2004)). Для определения первичного сайта расщепления AP2α С-концевой 33 кДа-продукт расщепления был подвергнут иммунологической очистке из клеток BJAB, и была подтверждена идентичность его триптических пептидов посредством масс-спектрометрии (фиг.9a). N-концевое секвенирование выявило GPAAQPSLGPTPEEAFLS, то есть последовательность, расположенную непосредственно выше от DVFD (фиг.9a), a именно тетрапептидную последовательность, подобную последовательности хорошо охарактеризованных сайтов распознавания каспазы (см., например, Stennicke, H. R., et. al, Biochem J 350 Pt 2, 563-8 (2000)). Сайт расщепления имеет Asp и Gly в соответствующих положениях P1 и P1', фланкирующих разрывающуюся связь, и Asp в положении P4, к которому каспаза-8 является достаточно толерантной (см., например, Blanchard, H. et al. J. Mol. Biol. 302, 9-16 (2000)). Такая тетрапептидная последовательность сайта расщепления присутствует в изоформе A AP2α и отсутствует в изоформе С. Из-за такого различия Apo2L/TRAIL не индуцирует расщепление изоформы С, которая гораздо реже встречается, чем изоформа А, в клетках BJAB, Colo205, LS1034 и SW948. Такой сайт расщепления AP2α присутствует в шарнирной области (фиг.9b), что свидетельствует о том, что его гидролиз может нарушать функцию AP2. Более длительная стимуляция некоторых клеточных линий лигандом приводит к дополнительному процессингу AP2α во втором сайте, который, очевидно, представляет собой исходный 33 кДа-фрагмент (фиг.7a и b).
Для оценки функциональных последствий таких опосредуемых каспазой событий расщепления было проведено исследование поглощения трансферрина (Tf). Интернализация связанного с рецептором Tf происходит через выстланные клатрином ямки и требует обязательного присутствия AP2 (см., например, Bonifacino, J. S. et. al., Annu. Rev. Biochem. 72, 395-447 (2003)). Для определения степени эндоцитоза флуоресцентно-меченного Tf измерения были ограничены начальной линейной фазой поглощения и последующим рециклингом интернализованного белка, образующегося во время эндоцитоза, на ПM (фиг.3a, вставка). Apo2L/TRAII снижает скорость эндоцитоза Tf в клетках BJAB и Colo205 на 75-85% (фиг.3a, b); а предварительная zVAD-fmk-обработка приводит, по существу, к усилению такого ингибирования (фиг.3c). Следовательно, DR-активация приводит к
каспазозависимому нарушению опосредуемого клатрином эндоцитоза.
Apo2L/TRAIL индуцирует опосредуемый клатрином эндоцитоз DR5.
Быстрое и селективное расщепление компонентов клатринового пути указывает на то, что физическая близость DR к белкам клатринового покрытия может облегчать протеолитические процессы. Поэтому возникает вопрос, подвергается ли DR5
клатринопосредуемому эндоцитозу после стимуляции лиганда. Было использовано моноклональное антитело (mAb 5C7), которое распознает внеклеточный домен рецептора, не конкурируя за связывание с лигандом (см., например, Kischkel, F. C. et al., Immunity 12, 611-20 (2000)). Инкубирование с насыщающими количествами 5C7 при 37°С не влияет на уровни поверхностного DR5, обнаруженные с помощью другого неперекрывающегося mAb, что подтверждает то факт, что сам 5C7 не влияет на эндоцитоз DR5. Был получен флуоресцентный конъюгат mAb 5C7 (6475С7), связанный с клетками Colo205 при 0°С, а затем была измерена кинетика эндоцитоза DR5 при 37°С (фиг.4a). Аналогичным образом обработка тримерными или мультимерными (перекрестно связанными с антителом) формами Apo2L/TRAIL приводила приблизительно к 2-кратному увеличению скорости эндоцитоза DR5, главным образом, за первые 30 минут (фиг.4b), что способствовало интернализации приблизительно 1/3 рецептора в течение 2 часов (фиг.4а). В соответствии с этим уровень DR5 на клеточной поверхности снижался в течение первых 30 минут после стимуляции, после чего такой пул DR5 стабилизировался (фиг.4c). Аналогичные результты наблюдались в клетках BJAB. В отличие от данных, полученных после быстрой обработки лигандом при 37°С (фиг.4a), более длительная обработка в течение 2 часов приводила к ингибированию эндоцитоза DR5 одновременно с Tf, и тем самым к 2,5- или 5-кратному снижению скорости эндоцитоза, соответственно (фиг.4d). Полученные результаты показали, что клатриновый путь поддерживает эндоцитоз DR5. Действительно, в клетках Colo205, быстро обработанных Apo2L/TRAIL, небольшая фракция 5C7 клеточной поверхности локализуется в выстланных клатрином ямках, на что указывала электронная микроскопия (ЭM), при этом какой-либо детектируемой локализации на непокрытых поверхностях участков инвагинации или в кавеолах не наблюдалось (фиг.4f-h).
Затем была применена хорошо разработанная стратегия исследований, которая была основана на регулируемой доксициклином экспрессии термочувствительного мутанта GTPазы- динамина-1 (dynG273D) в клетках, инфицированных ретровирусом. Этот мутант быстро и обратимо ингибирует клатринзависимый эндоцитоз при изменении пермиссивной температуры (30°С) на непермиссивную (38°С) температуру (см., например, Damke, H., et al., J Cell Biol 131, 69-80 (1995)). Предварительные эксперименты на клетках HeLa-M подтвердили, что индуцирование доксициклином блокирует поглощение Tf при 38°С в клетках с экспрессией dynG273D, но не в соседних клетках, не экспрессирующих dynG273D, и не при 30°С (фиг.5a-d и фиг.10a). Хотя индуцибельная экспрессия dynG273D в клетках HeLa-M или Colo205 не является достаточно стабильной, однако она является стабильной в трансдуцированной клеточной линии BJAB, причем при 38°С наблюдалось сильное блокирование поглощения Tf (фиг.5e). Стимулируемая доксициклином экспрессия dynG273D также приводила к снижению степени индуцированного лигандом поглощения DR5 от ~ 25% при 30°С до ~10% при 38°С. В противоположность этому в отсутствие доксициклина при обеих температурах наблюдался аналогичный эндоцитоз (фиг.5f). В клетках HeLa-M индуцирование dynG273D с последующим увеличением температуры до 38°С также приводило к ослаблению индуцированного лигандом поглощения DR5, и этот эффект был слабее, чем в клетках BJAB, что, вероятно, обусловлено снижением уровня экспрессии dynG273D (фиг.10b). Для исключения очень небольшой вероятности того, что в этих экспериментах на поглощение в течение 20 минут снижение поглощения DR5 обусловлено повышением уровня рециклинга эндоцитов, а не снижением скорости эндоцитоза, наиболее заметным в течение 4-минутного периода поглощения, был использован доминантно негативный мутант динамина (динамин-1К44А), который во всех случаях блокирует клатринзависимый эндоцитоз (см., например, Damke, H., et al., J. Cell Biol. 127, 915-34 (1994)). Динамин-1К44А ингибировал индуцированный Tf и лигандом эндоцитоз DR5 в клетках HeLa-M (фиг.10c).
Наряду с данными, полученными с помощью электронной микроскопии (ЭМ), эти результаты показали, что Apo2L/TRAIL-индуцированное поглощение DR5 происходит, главным образом, посредством клатринзависимого эндоцитоза. Таким образом, физическая близость DISC, присутствующего в покрытых клатрином ямках, к компонентам клатринового покрытия, может облегчать его расщепление. Если это так, то сборка DISC и активация каспазы-инициатора фактически не должна требовать интернализации покрытых клатрином ямок. Для проверки этого предположения клетки BJAB охлаждали на льду для блокирования эндоцитоза Tf (фиг.11a). Несмотря на такое блокирование Apo2L/TRAIL и FasL еще обладали способностью присоединять FADD и каспазу-8 к их соответствующим рецепторам и процессировать каспазу-8 (фиг.11b). Кроме того, Apo2L/TRAIL индуцировал сравнимые уровни формирования DISC, процессинга каспазы-8 и активности каспазы-8 в DISC при 38°С, несмотря на ингибирование эндоцитоза под действием dynG273D (фиг.11с и d).
Ингибирование опосредуемого клатрином эндоцитоза усиливает DR-индуцируемую активацию каспазы и апоптоз.
Быстрое расщепление белков клатринового пути показало, что нарушение эндоцитоза может стимулировать индуцирпованную лигандом активацию каспазы и апоптоз. Действительно, dynG273D-опосредованная блокада эндоцитоза приводит к значительному усилению индуцированного лигандом процессинга всего клеточного пула каспазы-8, Bid, AP2α и эффекторных каспаз-3 и -7 из лизатов целых клеток (фиг.6a), но, при этом, не оказывает значительного влияния на образование или активацию самого DISC (фиг.11с и d). Количественный анализ указывал на значительное увеличение стимулируемой лигандом ферментативной активности каспазы-3/7 после индуцирования dynG273D и повышения температуры до 38°С по сравнению с активностью при температуре 30°С (фиг.6b). Кроме того, такие условия также приводили к усилению апоптоза (фиг.6c), то есть эффекта, наблюдаемого после стимуляции лигандом дикого типа или DR5-селективным вариантом (см., например, Kelley, R. F. et al., J. Biol. Chem. 280, 2205-12 (2005)). Сенсибилизация наблюдалась не только по сдвигу влево кривой доза-ответ, но также и по увеличению максимального процента клеток с фрагментированной ДНК (фиг.6c). Эти результаты показали, что опосредуемое каспазой нарушение клатринзависимого эндоцитоза усиливает активацию каспазы, действующей после DISC, что приводит к усилению стимуляции апоптоза.
Как указывалось выше, с применением способов и материалов согласно изобретению была неожиданно обнаружена взаимосвязь между механизмом пути внеклеточного апоптоза и механизмом клатринзависимого эндоцитоза. Через несколько минут после начала проапоптотического действия DR активированные каспазы начинают расщеплять белки, которые опосредуют клатринзависимый эндоцитоз. Относительно быстрая кинетика расщепления AP2α и необходимость участия FADD и каспазы-8 в этом процессе, а также независимость от Bax и каспазы-3 указывают на то, что DR-проксимальные каспазы, а по всей вероятности каспаза-8 и/или каспаза-10 осуществляют начальный процессинг AP2α. Действительно, сайт расщепления каспазы в шарнирной области AP2α соответствует последовательностям, которые обладают способностью распознавать каспазу-8 (см., например, Nicholson, D.W., Cell Death Differ 6, 1028-42 (1999)).
Ингибирование клатринзависимого эндоцитоза посредством активации DR происходит одновременно со стимуляцией каспазы, и прекращается под действием zVAD-fmk, что указывает на его зависимость от каспазной активности. Первичный сайт процесснига AP2α картировали по шарнирной области. Шарнирная область связывает домен «ухо», который поддерживает рекрутинг вспомогательных белков, с доменом «туловище», который опосредует ассоциацию ПM и связывание «грузового» белка (cargo) в комбинации с другими субъединицами AP2 (см., например, Owen, D.J., et. al, P. R. Annu. Rev. Cell Dev. Biol. 20, 153-91 (2004)). Сверхэкспрессия домена «ухо» AP2α в клетках COS7 приводит к ингибированию клатринзависимого эндоцитоза, то есть эффекта, отменяемого под действием мутаций, которые предотвращают связывание со вспомогательным белком (см., например, Owen, D.J. et al., Cell 97, 805-15 (1999)). И наоборот, исследования, проводимые на дрозофилах, в которых некоторые клетки экспрессируют только AP2α, не содержащий домена «ухо», показали, что при общем эндоцитозе присутствия такого домена «ухо» не требуется (см., например, Berdnik, D., et al., Dev Cell 3, 221-31 (2002)). Таким образом, остается неясным, является ли расщепление AP2α лишь одной каспазой достаточным для нарушения эндоцитоза, или такой процессинг требует также участия других компонентов клатринового пути. Независимо от этого DR-опосредуемая активация каспазы быстро подавляет клатринзависимый эндоцитоз.
Данные, полученные при рассмотрении механизма взаимодействия активированных каспаз с субстратами клатринового покрытия, свидетельствуют о том, что после стимуляции Apo2L/TRAIL значительная часть поверхностного DR5 транспортируется в клетки посредством клатринзависимого эндоцитоза. Предварительные ЭМ-исследования подтвердили, что DR5 ассоциируется с высланными клатрином ямками. Недавно проведенные исследования показали, что РНКи-опосредуемое подавление клатрина и AP2 приводит к ингибированию апототического сигнала посредством Fas, и этот результат может интерпретироваться как установление того факта, что для передачи сигнала рецептора требуется эндоцитоз (см., например, Lee, K.H. et al., Embo J 25, 1009-23 (2006)). Исходя из результатов наблюдений, указывающих на то, что сборка и активация DISC Apo2L/TRAIL и апоптоз клеток могут еще происходить в случае ингибирования эндоцитоза при инкубировани на льду или при инактивации динамина, было сделано предположение, что DR5 может связываться с FADD и каспазой-8 еще и на клеточной поверхности, возможно, в высталанных клатрином ямках. Действительно, блокирование эндоцитоза при инкубировании на льду или в присутствии термочувствительного динамина не препятствуют образованию выстланных ямок и рекрутингу «грузовых» белков (cargo) (см., например, Damke, H., et al., J. Cell Biol. 131, 69-80 (1995)). DR-активация приводит к почти полной деструкции AP2α. В противоположность этому процессинг CHC и AP1/2β затрагивает только фракцию всех клеточных белков, а вероятно ту фракцию, которая непосредственно участвует в эндоцитозе DR. Таким образом, активация каспазы, действующей вместе с интернализацией DR-ассоциированного DISC, может приводить к локальной деструкции специфических компонентов клатринового пути.
Одним из простых объяснений того факта, что каспазы подавляют опосредованный клатрином эндоцитоз, является то, что элиминация такого механизма может представлять собой ранее неизвестную часть апоптотической программы; при этом клетка, подвергающаяся апоптозу, больше не нуждается в поглощении питательных элементов, не отвечает на факторы роста или не поддерживает гомеостаз мембраны. Однако маловероятно, что эти события расщепления, сами по себе, запускают апоптоз, поскольку как клатрин, так и AP2 эффективно подавлялись под действием киРНК, как показали исследования, проводимые в различных лабораториях, и не было ни одного сообщения об усилении апоптоза. При этом следует отметить, что цитотоксические и ДНК-разрушающие лекарственные средства, такие как винбластин и адриамицин, также индуцируют расщепление компонентов клатринового пути, хотя и с гораздо более медленной кинетикой, чем лиганды DR (фиг.12). С другой стороны, опосредуемый каспазой процессинг клатриновых адапторов может также играть определенную роль в модуляции неапоптотических клеток: в незрелых дендритных клетках базальная каспазная активность инициирует расщепление нескольких субъединиц адаптина, а zVAD-ингибирование такого эффекта стимулирует созревание дендритных клеток (см., например, Santambrogio, L. et al., Nat Immunol 6, 1020-8 (2005)).
Вторым и, вероятно, более неожиданным объяснением взаимосвязи между DR-опосредуемым механизмом и клатриновым механизмом эндоцитоза является то, что такое взаимодействие приводит к образованию петли положительной обратной связи, которая усиливает активацию каспазы по внешнему пути. В соответствии с этой моделью эндоцитоз активированных DR противодействует апоптотической передаче сигнала и тем самым сохраняет жизнеспособность клеток в том случае, если клатриновый механизм остается ненарушенным, например, в ответ на присутствие подпороговых уровней лиганда. Однако если стимуляция DR приводит к продуцированию каспазной активности, достаточной для нарушения эндоцитоза, то такой сигнал продолжает передаваться, и дополнительно усиливает активацию каспазы, что приводит к стимуляции апоптоза. Считается, что эндоцитоз DR может препятствовать передаче сигнала DR посредством деградации лизосом, то есть некоторых других форм терминации сигнала, или индуцирования конкурирующих антиапоптотических сигналов. Эндоцитоз не регулирует активацию каспазы на уровне начальной ассоциации DISC. Тем не менее инактивация динамина усиливает процессинг всех клеточных каспаз, а именно каспазы-8, и особенно каспазы-3 и -7, и приводит к усилению активации апоптоза, что указывает на то, что эндоцитоз также регулирует активацию каспазы, действующей сразу после DISC.
В итоге в настоящей заявке описана обратимая взаимосвязь между DR-индуцированным апоптозом и клатринзависимым эндоцитозом, а именно активация DR приводит к опосредуемой каспазой деструкции важных компонентов механизма клатринового эндоцитоза, и тем самым к прекращению этого процесса. Деструкция клатринзависимого эндоцитоза усиливает активацию каспазы, действующей после DR, и апоптоз, что свитетельствует о наличии петли положительной обратной связи, которая усиливает активацию каспазы и апоптоз.
Нижеследующие примеры приводятся лишь в целях иллюстрации и не должны рассматриваться как ограничение объема изобретения. Все работы, цитируемые в настоящей заявке, во всей своей полноте вводятся в настоящее описание посредством ссылки.
ПРИМЕРЫ
Пример 1: Иллюстративные материалы и методы
Идентификация сайта расщепления AP2α каспазой
Клетки BJAB (5×108) стимулировали Apo2L/TRAIL в течение 30 минут, а затем подвергали лизису и иммунопреципитации с использованием С-концевого специфического анти-AP2α антитела (AP6, #MA1-O64 от ABR), после чего собирали на белке A/G-агарозе (Pierce). C-концевой фрагмент разделяли с помощью электрофореза в ПААГ с ДСН, а затем элюировали для последующего расщепления трипсином и проводили анализ с помощью жидкостной хроматографии - ионизации элекрораспылением - тандемной масс-спектрометрии с захватом ионов, или в отдельно проводимой независимой процедуре (IP), переносили на ПВД-мембрану для N-концевого секвенирования.
Анализы на эндоцитоз, поглощение и поверхностное ингибирование
Для рутинного измерения Tf 106 суспендированных клеток предварительно обрабатывали 1-10 мкг/мл Apo2L/TRAIL в течение указанного времени при 37°С. В некоторых исследованиях, перед добавлением Apo2L/TRAIL, клетки предварительно обрабатывали 0,25% ДМСО +/- 50 мM zVADfmk в течение 30 минут при 37°С. После Apo2L/TRAIL-стимуляции клетки уравновешивали 5 мкг/мл флуоресцентного Tf в среде для связывания (бессывороточной DME, 3% BSA, 20 мM Hepes, pH 7,2) в течение 30 минут на льду, после чего клетки доводили до 37°С в течение 0-5 минут, как указывалось ранее, и быстро охлаждали на льду для прекращения эндоцитоза. Осажденные клетки ресуспендировали в холодном 2% параформальдегиде/PBS и количественно оценивали среднюю интенсивность флуоресценции 10000 клеток на проточном цитометре Coulter Epics Elite-ESP (Hialeah, FL). Скорость эндоцитоза определяли по углу наклона линейного участка на графиках стационарных внутренних/поверхностных уровней (см., например, Wiley, H. S. et. al, J. Biol. Chem. 257, 4222-9 (1982)). В экспериментах по изменению температуры инкубирования на льду для связывания не проводили. Вместо этого клетки преварительно кондиционировали в течение 20 минут и поддерживали при 30°С или при 38°С в среде для связывания, а затем инкубировали в течение еще 0 (контроль) или 20 минут с 647Tf, охлаждали на льду в течение 30 минут и обрабатиывали для проведения проточной цитометрии в целях определения отношений внутренних/поверхностных уровней 647Tf.
Для рутинного измерения DR5 клетки насыщали 5 мкг/мл 6475С7 на льду в среде для связывания, а затем 3 раза промывали холодной средой для связывания и инкубировали в течение 30 минут на льду в присутствии или в отсутствие 10 мкг/мл Apo2L/TRAIL. Затем клетки доводили для 37°С в течение указанного периода времени, после чего их быстро охлаждали и либо не обрабатывали, либо обрабатывали кислотой путем трехкратной обработки холодной 2 M мочевиной, 50 мM глицином, 150 мМ NaCl, pH 2,4. Клетки обрабатывали для проведения проточной цитометрии, и уровень интернализации вычисляли методом, аналогичным описанному ранее методу, по следующему уравнению: % поглощения = 100×[(S t -S 0)/(N-S 0)], где S t и S 0 представляют собой величины для обработанных кислотой образцов, инкубированных в течение периода времени = t и времени = 0, соответственно, а N представляет собой флуоресценцию не обработанного кислотой образца, который остается, в основном, неизмененным при инкубировании при 37°С (см., например, Austin, CD. et al., Mol. Biol. Cell 15, 5268-82 (2004)).
Для эксперимента с изменением температуры клетки предварительно кондиционировали в течение 20 минут и поддерживали при 30°С или 38°С, а затем предварительно уравновешивали 5 мкг/мл 6475С7 в течение 3 минут, обрабатывали 10 мкг/мл Apo2L/TRAIL в течение 0 (контроль) или 20 минут, и охлаждали на льду в течение 30 минут. Клетки либо обрабатывали кислотой, либо не обрабатывали, и количественно оценивали с помощью проточной цитометрии.
Для проведения анализов на снижение уровня поверхностного DR5 суспендированные клетки предварительно уравновешивали немеченным 5C7 при 37°С в течение 30 минут для связывания с поверхностью и для рециклинга пулов DR5, после чего эти клетки обрабатывали 10 мкг/мл Apo2L/TRAIL в течение указанного периода времени, или вообще не обрабатывали, а затем охлаждали на льду. Поверхностный 5C7 зондировали CY5-конъюгированным антителом против мышиных IgG и количественно оценивали с помощью проточной цитометрии. Этот метод был применен во избежание снижения эффективности зондирования поверхности, обусловленного Apo2L/TRAIL-индуцированной кластеризации рецептора, как было обнаружено после прямого зондирования с использованием 6475С7.
Анализы на каспазную активность
Активность каспазы-3/7 оценивали с помощью анализа Apo-One Homogenous Caspase-3/7 Assay (Promega). Клетки собирали, подсчитывали, разделяли на равные аликвоты для каждой процедуры обработки, и обрабатывали Apo2L/TRAIL при различных дозах в течение 4 часов при 30°С или 38°С, а затем подвергали лизису в гомогенном реагенте для каспазы-3/7 (содержащем субстрат для каспазы-3/7 Z-DEVD-R110). Лизаты инкубировали при комнатной температуре в течение 1 часа, а затем считывали на флуориметре при 485/530 нМ.
Анализы на апоптоз
Посе фиксации этанолом и обработки РНКазой клетки окрашивали иодидом пропидия, и количество субдиплоидной ДНК анализировали с помощью проточной цитометрии, как описано выше, с использованием компьютерной программы Expo32 на цитометре Epics XL.MCL (Coulter Beckman), (см., например, Nicholson, D.W., Cell Death Differ 6, 1028-42 (1999)).
Иммунопреципитация DISC
Клетки BJAB (1×107 за данный период времени) обрабатывали Flag-Apo2L/TRAIL, перекрестно связанным с анти-FLAG антителами M2, а затем собирали, лизировали и подвергали иммунопреципитации, и DISC анализировали, как описано выше (см., например, Kischkel, F. C. et al., Immunity 12, 611-20 (2000)).
Клеточные линии и реагенты
Клетки человеческой аденокарциномы ободочной кишки Colo205, клетки человеческой В-клеточной лимфомы BJAB и клетки человеческой Т-клеточной карциномы дикого типа Jurkat (клон A3), которые являются дефицитными по FADD (El) и дефицитными по каспазе-8 (I9.3), культивировали в среде RPMI 1640 + 10% фетальная бычья сыворотка (FBS) + 2 мM глутамин + 1000 ед/мл пенициллина-стрептомицина (Life Technologies). Клетки человеческой карциномы шейки матки HeLa-M, клетки человеческой аденокарциномы молочной железы MCF7 и клетки человеческой аденокарциномы ободочной кишки HCT15, HCT8, SW948, Colo320, человеческой фибросаркомы HT180 (ATCC), HCT116 Bax(-/-), HCT116 Bax(+/-) культивировали в 50:50 модифицированной по способу Дульбекко среде Игла и в среде FK12 + 10% FBS + 1000 ед/мл пенициллина-стрептомицина. Клоны кДНК, кодирующие термочувствительные динамины, представляют собой G273D-модифицированный и доминантно-негативный K44A-модифицированный динамин-1 (DynG273D и DynK44A, соответственно). DynG273D был экспрессирован из pHUSH-ProEx, то есть из экспрессионной системы, состоящей из одной тетрациклин-индуцибельной ретровирусной плазмиды, сконструированной в Genentech. кДНК сначала клонировали в челночную плазмиду, содержащую последовательность энхансера-промотора CMV с двумя копиями тетрациклинового оператора TetO2, а затем переносили посредством in vitro рекомбинации в фаге лямбда или с помощью технологии Gateway® (Invitrogen, Carlsbad) в каркас вектора вируса мышиного лейкоза Молони, в котором репрессор Tet дикого типа регулируется отдельным промотором β-актина, за которым следует внутренняя последовательность связывания с рибосомой и кластер для отбора на резистентность к пуромицину. DynK44A экспрессировался из 2-векторной экспрессионной тетрациклин-индуцибельной ретровирусной системы, pRevTet-On/pRev-Tre (Clontech), путем серийного инфицирования. Ретровирусные векторы трансфецировали в упаковывающие клетки, инфицированные амфотропным вирусом мышиного лейкоза Молони Phoenix, с использованием липофектамина 2000 (Stratagene). Ретровирусные частицы собирали и использовали в инфицированных клетках HeLa-M и BJAB, как было описано ранее (Simpson et al. J Cell Biol 137, 835-45, 1997). DynG273-инфицированные клетки отбирали на резистентность к пуромицину (2-3 мкг/мл) в течение 1-2 недель и клонировали либо путем ручного сбора отдельных колоний HeLa-M или путем FACS-сортинга резистентных клеток BJAB (отобранных по исключению иодидом пропидия), а затем одиночные клетки высевали на 96-луночные планшеты для культивирования клеток с использованием оборудования EPICS ELITE-ESP, снабженного устройством Autoclone (Coulter, Hialeah, FL). Клеточные клоны скринировали на равномерность доксициклин-индуцибельной экспрессии динамина-1 с помощью иммунофлуоресцентной микроскопии. DynK44A-инфицированные клетки отбирали на резистентность к гигромицину B (400 мкг/мл) и G418 (1 мг/мл) в течение 1-2 недель. Индуцирование доксициклином (1 мкг/мл) осуществляли при 32°С в течение 72 часов, как описано ранее (Damke et al., J. Cell Biol. 131, 69-80, 1995). Сульфат винкристина (#T-117), адриамицин-HCL (GR-319) закупали у компании Biomol; FBS, не содержащий тетрациклина, закупали у Hyclone; пуромицин закупали у Clontech, а доксициклин закупали у Sigma. zVAD-fmk (asN-бензилоксикарбонил-Val-Ala-Asp-фторметилкетон) получали от MD Biosciences (#FK-009).
Рекомбинантные белки, антитела и флуоресцентные маркеры
Человеческий рекомбинантный растворимый Apo2L/TRAIL в немеченных или Flag-меченных вариантах и Flag-меченном FasL получали, как описано ранее (Sharp et al. J. Biol. Chem. 280, 19401-409, 2005). Для иммуноблот-анализа были использованы следующие моноклональные антитела: антитело против каспазы-8 (1C12, #9746) и антитело против каспазы-7 (C7, #9494) от компании Cell Signaling; антитело против каспазы-8 (5F7, #IM3148) от Immunotech; анти-FADD антитело (#610399), анти-AP2α антитело (#610501); анти-AP1/2α антитело (#610381); анти-AP1γ антитело (#610385); анти-AP3δ антитело (#611328); анти-CHC антитело (#610499); анти-AP4ε антитело (#612018); анти-Bid антитело (#550365) и антитело против динамина-1/2 (#610245) от BD Transduction; антитело против каспазы-3 (SA-320) от Biomol; антитело против каспазы-9 (Ab2, #AM47) от Oncogene; антитело против рецептора Tf (#13-6800) от Zymed; анти-βCop антитело (M3A5, #G2279) от Sigma; анти-Sec23 антитело (#GTX22913) от Gentex; и моноклональные антитела против DR5 (3H3 и 5C7), полученные в Genentech, Inc.; и анти-AP3β антитело, описанное ранее (см., например, Simpson et al., J. Cell Biol. Volume 137, No. 4 pp 835-845 (1997)). В качестве «вторых» реагентов были использованы нижеследующие Аb, конъюгированные с пероксидазой хрена (ПХ): антитело против мышиного IgG1 (#559626) от BD Bioscience; антитело против мышиного IgG2b (#190-05) от Southern Biotechnology Associates, и антитело против кроличьего IgG (#711-035-152) от Jackson ImmunoResearch. Для проведения экспериментов по иммунопреципитации были использованы следующие Ab: анти-Flag антитело (M2, Sigma) и анти-AP2α антитело (AC1-M11, #MA3-061) или AP6 (#MA1-064) от ABR. Для проточной цитометрии и иммунофлуоресцентной микроскопии использовали 488Tf (T-13342), 647Tf (T-23366), 594антитело против кроличьих IgG (#A-11037), и 594антитело против козьих IgG (#A-11058) от Molecular Probes. Фрагмент антитела против мышиного IgG (#115-177-003) от Jackson ImmunoResearch, антитело против динамина-1 (#sc-6402) от Santa Cruz Biotechnology и кроличье поликлональное антитело против Alexa 647 получали в Genentech, Inc. с использованием Alexa 647-конъюгированного альбумина бычьей сыворотки (Molecular Probes, Inc.) в качестве антигена и подвергали аффинной очистке на колонке с Alexa-647-сефарозой, полученной путем взаимодействия (CNBr)-активированной сефарозы с кадаверином Alexa Fluor 647 (Molecular Probes, Inc.). Alexa-647 конъюгировали с mAb 5C7 (6475С7) посредством взаимодействия с сукцинимидиловым эфиром Alexa-647 (Molecular Probes, Inc.) в соответствии с инструкциями производителей.
Иммунофлуоресцентная микроскопия
Клетки для визуализации поглощения 6475С7 инкубировали при 37°С с 6475С7 и с перекрестно связанным Apo2L/TRAIL в течение указанного периода времени. Клетки для проведения экспериментов по термочувствительности предварительно обрабатывали в среде для связывания в течение 20 минут при 30°С и при 38°С, а затем добавляли 488Tf и инкубирование продолжали при соответствующих температурах в течение указанного периода времени. Затем клетки фиксировали 3% параформальдегидом в течение 20 минут, после чего их делали проницаемыми с использованием 0,4% сапонина и окрашивали кроличьим IgG против Alexa 647, затем 594антителом против кроличьих IgG (поглощение 6475С7) или антителом против динамина-1, а затем 594антителом против козьих IgG (эксперименты на термочувствительность). Изображения получали на микроскопе (Axiovert 200; Carl Zeiss MicroImaging, Inc.), имеющем объектив Plan-Apochromat 1,4 NA 63x, и снабженном видиокамерой с зарядовой связью (ЗС) (AxioCam; Carl Zeiss MicroImaging, Inc.) и набором полосовых фильтров Quad (Chroma Technology Corp.), и все операции осуществляли с помощью компьютерной программы AxioVison 3.1 (Carl Zeiss MicroImaging, Inc.).
Иммуноэлектронная микроскопия
Суспендированные клетки Colo205 инкубировали на льду в течение 30 минут с 10 мкг/мл 5C7 в среде для связывания, а затем промывали, инкубировали на льду в течение 30 минут с 10 мг/мл Apo2L/TRAIL в среде для связывания, после чего среду доводили до 37°С в течение 5 минут, фиксировали в 2% параформальдегиде/0,2% глутаральдегиде, и обрабатывали, как описано ранее (Austin et al. Mol Biol Cell 15, 5268-82, 2004) для иммунологического мечения сверхтонких криосрезов золотом, на которое было нанесено кроличье антитело против мышиного IgG (Dako, Glostrup, Denmark).
Следует отметить, что приведенное выше описание является достаточным для практического осуществления настоящего изобретения. Объем настоящего изобретения не ограничивается описанными здесь примерами. Действительно, различные модификации, внесенные в настоящее изобретение, помимо уже указанных и описанных здесь модификаций, будут более очевидны из описания изобретения, и такие модификации входят в объем прилагаемой формулы изобретения.
ТАБЛИЦЫ













































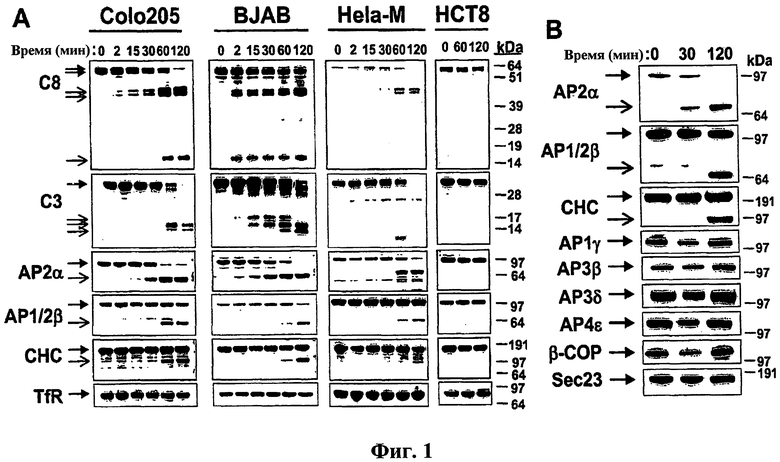

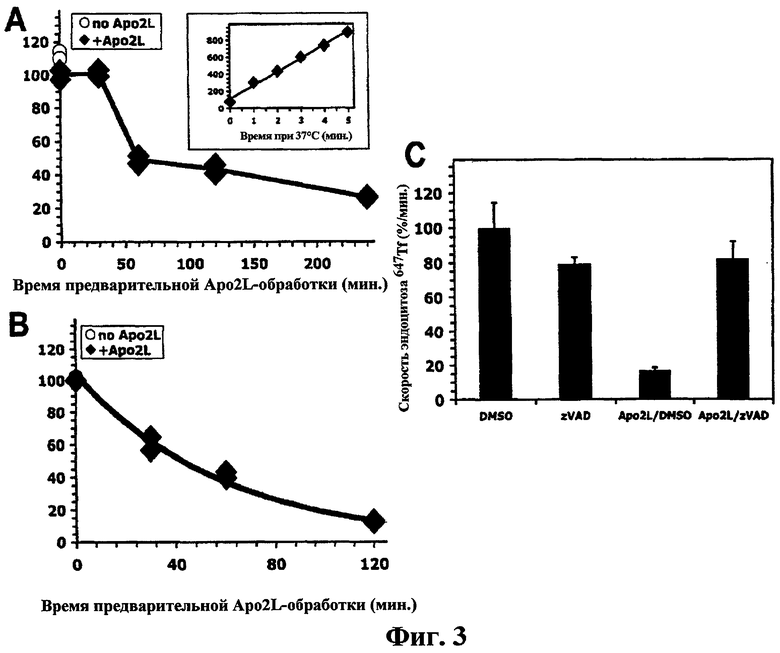
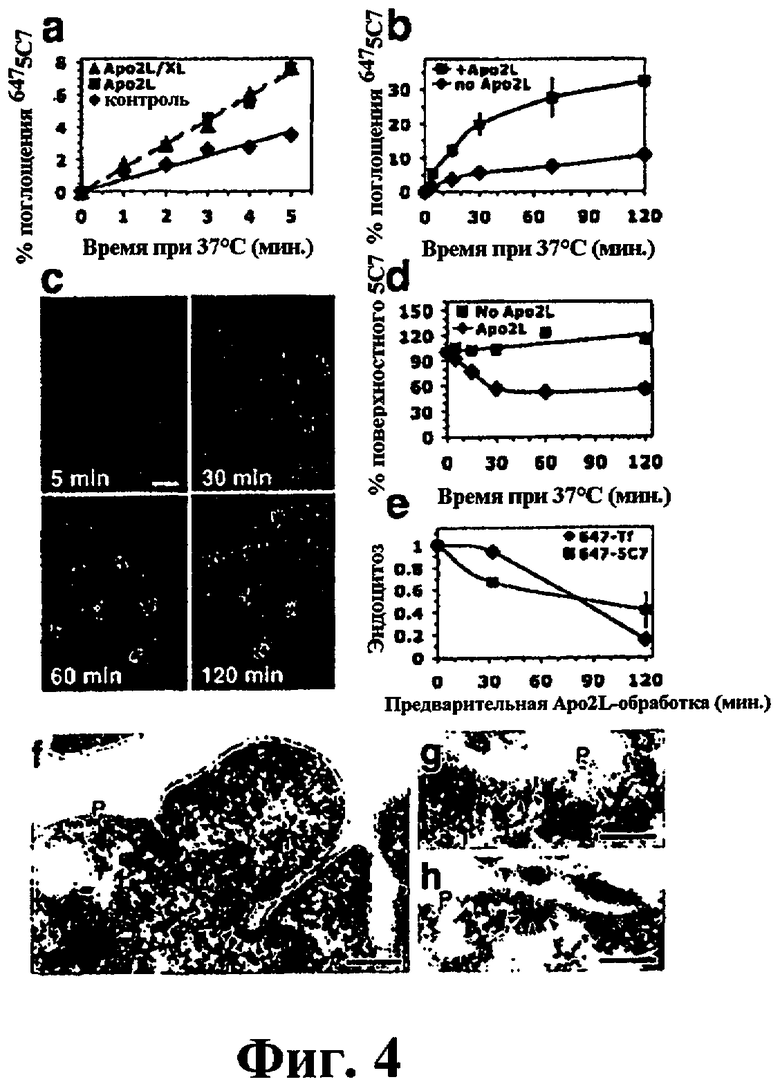

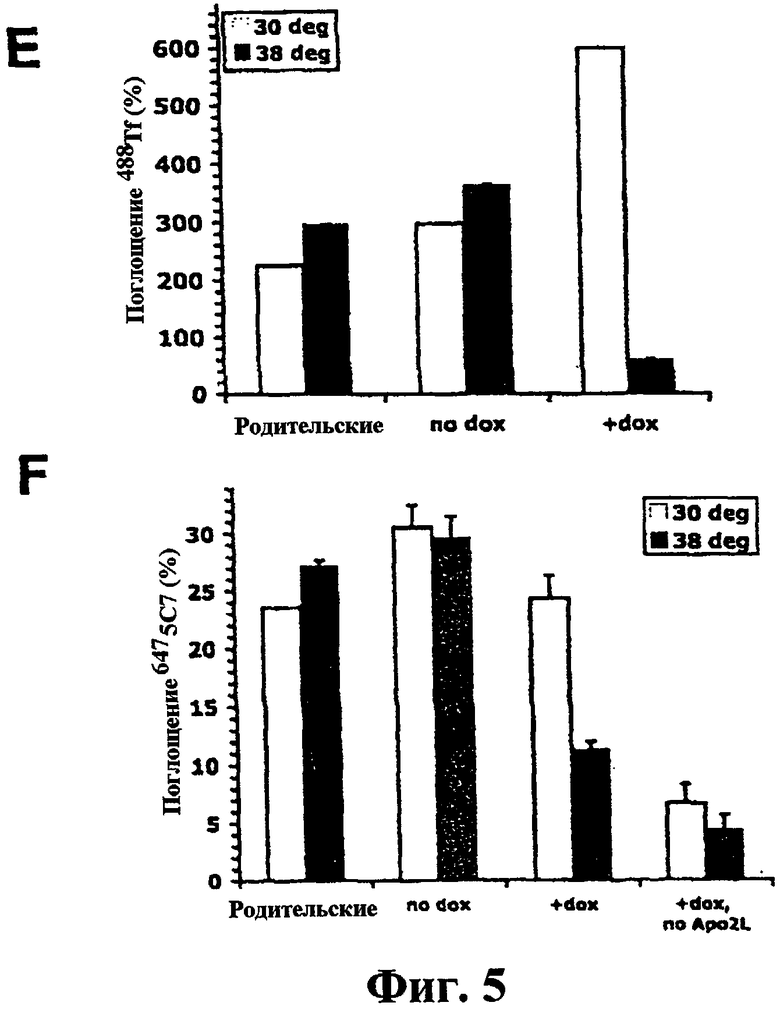
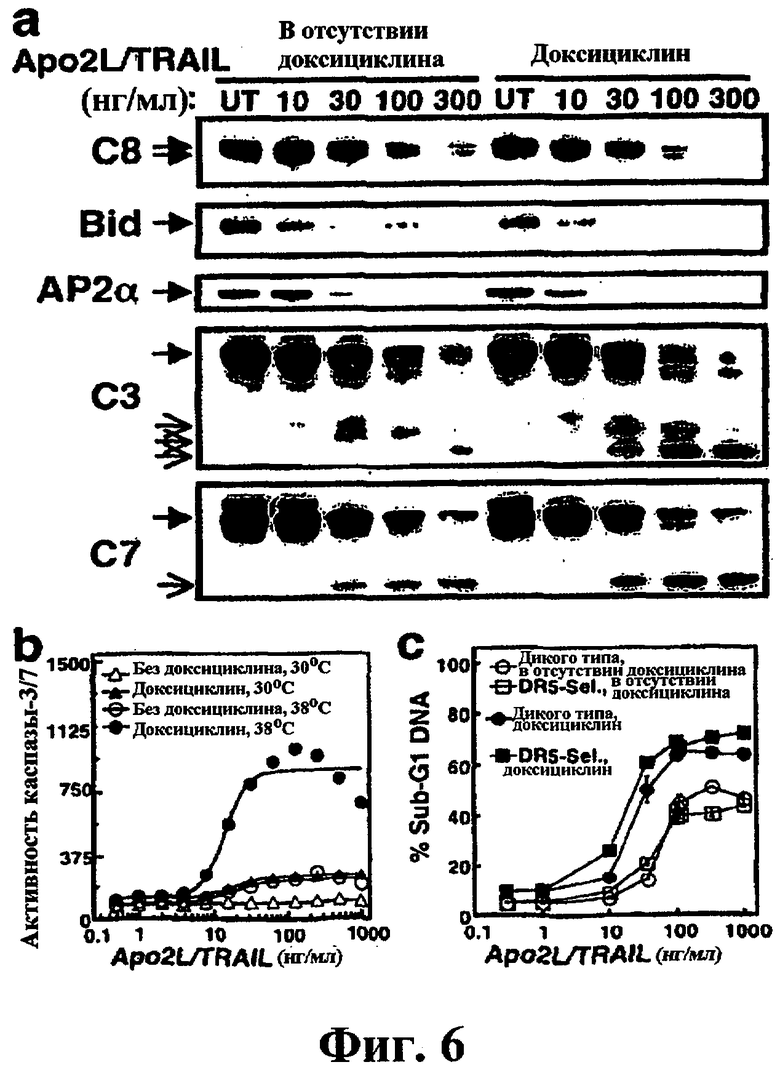
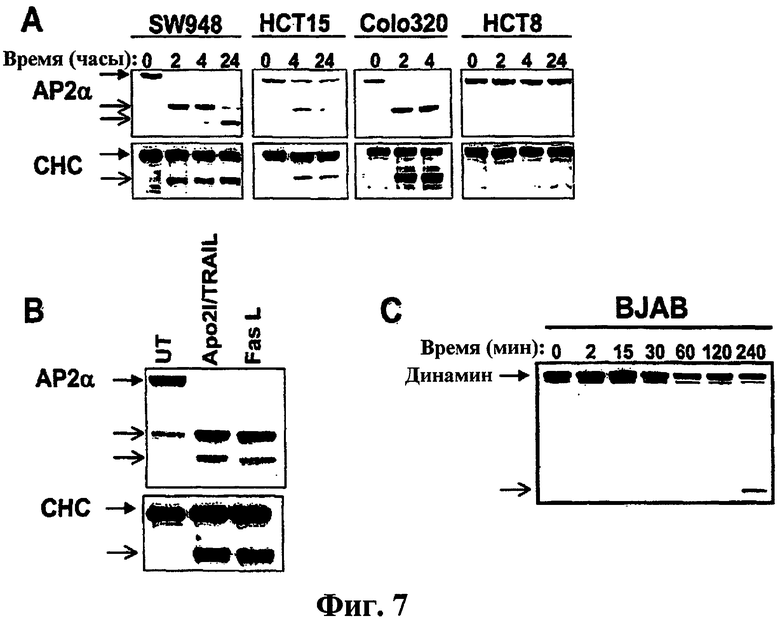

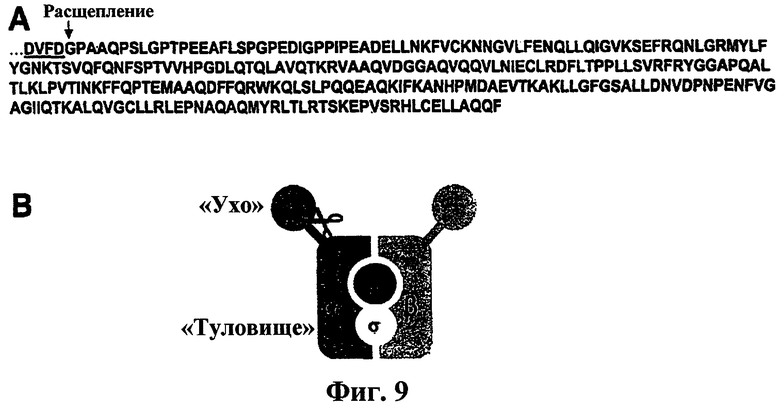
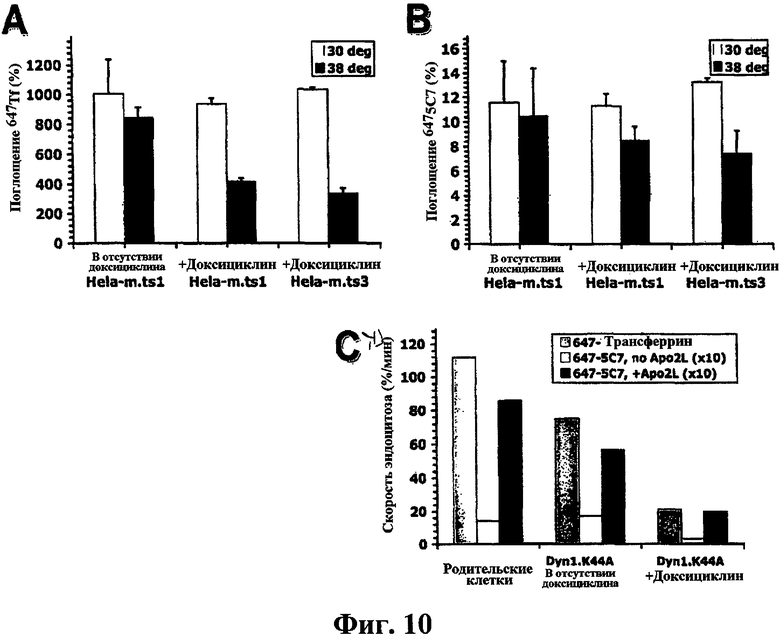
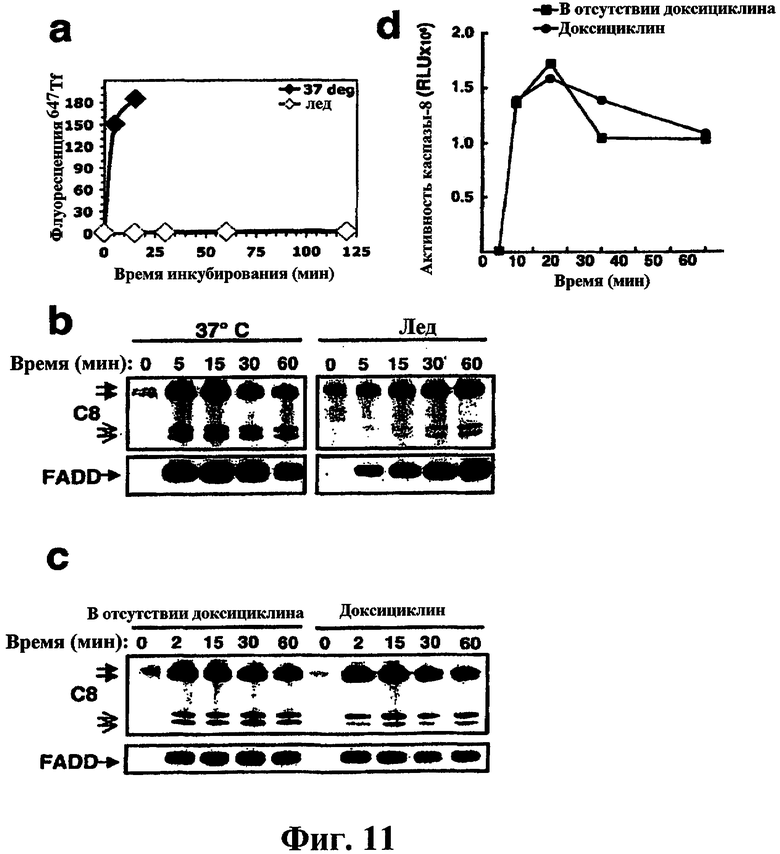
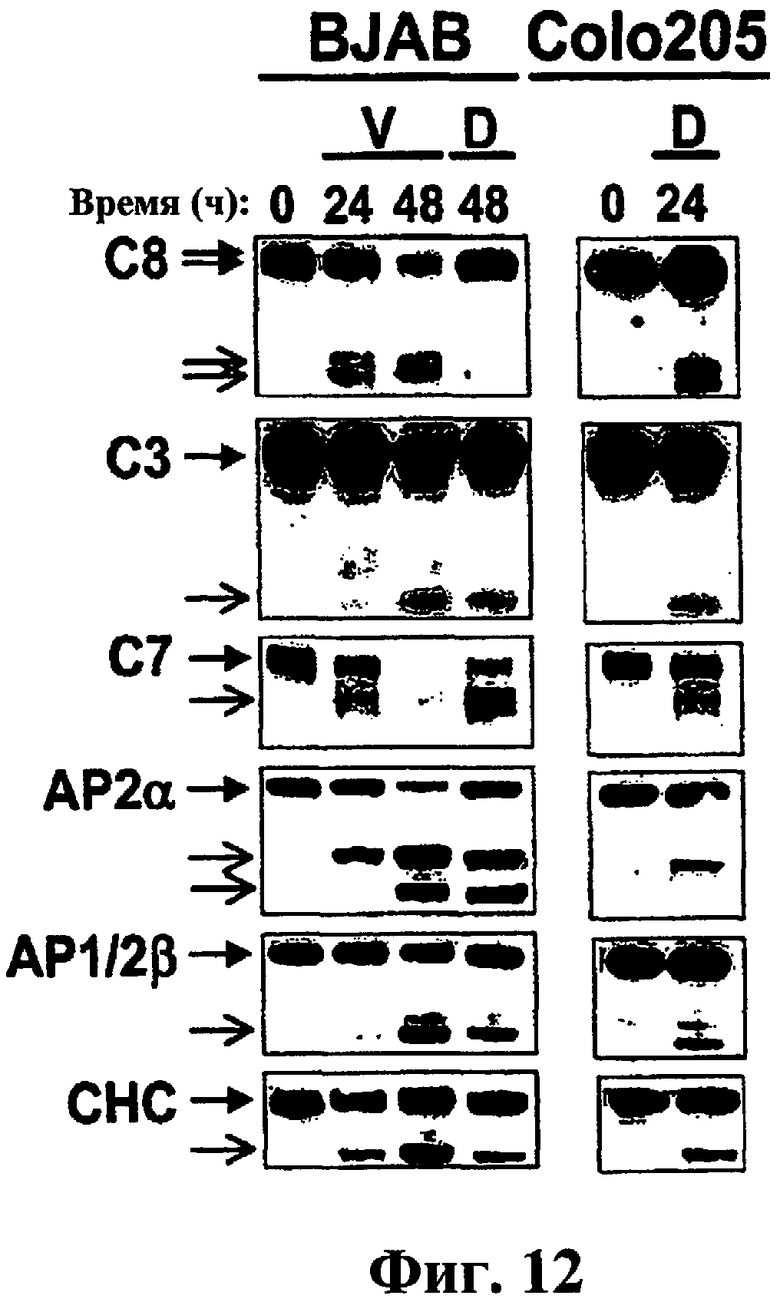
Изобретение относится к медицине и касается способа детектирования апоптоза в клетке млекопитающего, где апоптоз инициирован рецептором "гибели" 4, рецептором "гибели" 5 или Fas. Изобретение обеспечивает обнаружение апоптоза для оценки чувствительности раковой клетки млекопитающего к апоптоз-индуцирующим агентам. 2 н. и 15 з.п. ф-лы, 1 пр., 12 ил., 1 табл.
1. Способ детектирования апоптоза в клетке млекопитающего, где апоптоз инициирован рецептором «гибели» 4 (SEQ ID NO:5), рецептором «гибели» 5 (SEQ ID NO:6) или Fas (SEQ ID NO:8), причем способ предусматривает:
(a) приведение клетки в контакт с антителом, которое связывается с фрагментом белка, образующимся в процессе апоптоза, где указанное антитело связывается с белковым фрагментом АР2-α (SEQ ID NO:1), тяжелой цепи клатрина (SEQ ID NO:2), AP1/2β (SEQ ID NO:3) или динамина (SEQ ID NO:4);
(b) определение количества антитела, которое связывается с фрагментом белка, образующимся в процессе апоптоза; и
(c) сравнение количества связанного антитела на стадии (b) с количеством антитела, которое связывается с белковым фрагментом в клетке млекопитающего, не подвергающейся апоптозу, где апоптоз детектируется в том случае, если указанное количество на стадии (b) превышает количество, присутствующее в клетке, не подвергающейся апоптозу.
2. Способ по п.1, где апоптоз в указанной клетке инициирован приведением клетки в контакт с Apo2L/TRAIL (SEQ ID NO:7), FasL (SEQ ID NO:9), анти-Fas-антителом-агонистом, анти-DR4-антителом-агонистом или анти-DR5-антителом-агонистом.
3. Способ по п.2, дополнительно предусматривающий стадию детектирования апоптоза для получения информации об эффективности терапии, включающей введение Apo2L/TRAIL (SEQ ID NO:7), FasL (SEQ ID NO:9), анти-Fas-антитела-агониста, анти-DR4-антитела-агониста или анти-DR5-антитела-агониста.
4. Способ по п.1, где указанное антитело связывается с белковым фрагментом АР2α (SEQ ID NO:1), и где указанный белковый фрагмент АР2α, связанный с антителом, имеет массу 64 кДа или 33 кДа.
5. Способ по п.1, где указанный белковый фрагмент, связанный с антителом, включает DVFD с SEQ ID NO:1 или GPAA с SEQ ID NO:1.
6. Способ по п.1, где указанная клетка представляет собой человеческую раковую клетку толстой кишки, ободочной кишки, легкого, молочной железы, предстательной железы, мочевого пузыря, почки, яичника, головного мозга, меланомы, лейкоза или миеломы.
7. Способ по п.1, где указанный белковый фрагмент АР2-α (SEQ ID NO:1), тяжелой цепи клатрина (SEQ ID NO:2), АР1/2β (SEQ ID NO:3) или динамина (SEQ ID NO:4) обнаруживают с помощью иммуноблоттинга, твердофазного иммуноферментного анализа или иммуногистохимического анализа.
8. Способ по п.1, дополнительно предусматривающий оценку уровня экспрессии, по меньшей мере, одной мРНК в клетке млекопитающего.
9. Способ по п.1, который, перед приведением указанной клетки в контакт с антителом, дополнительно предусматривает обработку клетки млекопитающего одним или более тест-агентами, так, чтобы детектирование апоптоза в клетке млекопитающего позволяло идентифицировать один или более тест-агентов в качестве индукторов апоптоза в клетке млекопитающего.
10. Способ идентификации человеческой раковой клетки, которая вероятно ответит на действие или которая восприимчива к действию терапевтического средства, индуцирующего апоптоз в человеческих раковых клетках, где указанный способ включает обработку человеческой раковой клетки указанным терапевтическим средством, где указанное терапевтическое средство индуцирует передачу сигнала рецептора «гибели» 4 (SEQ ID NO:5), рецептора «гибели» 5 (SEQ ID N0:6) или Fas (SEQ ID NO:8),
анализ человеческой раковой клетки, обработанной терапевтическим средством, на предмет наличия белкового фрагмента АР2-α (SEQ ID NO:1), тяжелой цепи клатрина (SEQ ID NO:2), АР1/2β (SEQ ID NO:3) или динамина (SEQ ID NO:4);
сравнение количества фрагмента белка в человеческой раковой клетке с количеством белкового фрагмента в контрольной человеческой раковой клетке, не обработанной указанным лигандом;
где апоптоз наблюдается в том случае, если количество белкового фрагмента, присутствующего в человеческой раковой клетке, обработанной указанным терапевтическим средством, превышает количество белкового фрагмента, присутствующего в контрольной человеческой раковой клетке, не обработанной указанным терапевтическим средством; и
обнаружение апоптоза в человеческой раковой клетке позволяет идентифицировать человеческую раковую клетку как клетку, которая вероятно ответит на действие или которая восприимчива к действию указанного терапевтического средства.
11. Способ по п.10, где белковый фрагмент АР2-α (SEQ ID NO:1), тяжелой цепи клатрина (SEQ ID NO:2), АР 1/2β (SEQ ID NO:3) или динамина (SEQ ID NO:4) обнаруживают с помощью иммуноблоттинга, твердофазного иммуноферментного анализа или иммуногистохимического анализа.
12. Способ по п.10, дополнительно предусматривающий оценку уровня экспрессии, по меньшей мере, одной мРНК в человеческой раковой клетке.
13. Способ по п.10, дополнительно предусматривающий оценку уровня экспрессии, по меньшей мере, двух различных мРНК в человеческой раковой клетке.
14. Способ по п.10, где человеческая раковая клетка представляет собой человеческую раковую клетку толстой кишки, ободочной кишки, легкого, молочной железы, предстательной железы, мочевого пузыря, почки, яичника, головного мозга, меланомы, лейкоза или миеломы.
15. Способ по п.10, где апоптоз в человеческой раковой клетке инициирован приведением клетки в контакт с Apo2L/TRAIL (SEQ ID NO:7), FasL (SEQ ID NO:9), анти-Fas-антителом-агонистом, анти-DR4-антителом-агонистом или с анти-DR5-антителом-агонистом.
16. Способ по п.10, где человеческую раковую клетку получают путем биопсии и выращивают в культуре in vitro в течение менее чем одного месяца.
17. Способ по п.10, где белковый фрагмент представляет собой белковый фрагмент АР2-α, включающий DVFD с SEQ ID NO:1 или GPAA с SEQ ID NO:1.
| Мебель из фанеры | 1927 |
|
SU17648A1 |
| Wettey FR | |||
| et al | |||
| Приспособление с иглой для прочистки кухонь типа "Примус" | 1923 |
|
SU40A1 |
| Hansen В | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

