Положение относительно финансируемого из федерального бюджета исследования или разработки
Это изобретение было осуществлено при государственной поддержке гранта под номером 5R01CA156689-04, предоставленного Национальным институтом здравоохранения (NIH). Правительство имеет определенные права на данное изобретение.
Перекрестные ссылки на родственные заявки
Согласно 35 U.S.С. §119(e) данная заявка заявляет приоритет по дате подачи предварительной патентной заявки США, порядковый номер 61/722,437, поданной 5 ноября 2012, полное раскрытие которой включено в данное описание путем отсылки.
Уровень техники
Цель биомедицинского исследования состоит в том, чтобы достичь лучшего понимания физиологии человека и использовать это знание для предотвращения, лечения или излечения болезней человека. Вследствие практических и этических барьеров, касающихся проведения экспериментов на людях, многие исследования проводятся на моделях мелких животных, например мышах. Соответственно, нужны животные модели болезней человека.
Например, в Соединенных Штатах примерно у 20000 пациентов ежегодно диагностируется множественная миелома (ММ), преимущественно неизлечимое злокачественное заболевание секретирующих антитела терминально дифференцированных В-клеток (Hideshima et al., 2007, Nat Rev Cancer. 7:585- 98; Kuehl и Bergsagel, 2002, Nat Rev Cancer. 2:175-87). MM характеризуется инфильтрацией злокачественных плазматических клеток в костный мозг (ВМ), при этом клиническое проявление болезни включает остеопатию, гиперкальцемию, цитопению, почечную дисфункцию и периферическую нейропатию (Hideshima et al., 2007, Nat Rev Cancer. 7:585-98; Kuehl и Bergsagel, 2002, Nat Rev Cancer. 2:175-87). В большинстве случаев MM предваряется предраковым состоянием, называемым моноклональной гаммапатией неясного генеза (MGUS), которое поражает около 3% людей старше 50 лет (Landgren et al., 2009, Blood 113:5412-7). Отличительным признаком клеток ММ служит совокупность разнородных генетических нарушений, включая изменения в кариотипе, а также IgH транслокации (Kuehl и Bergsagel, 2002, Nat Rev Cancer. 2:175-87; Zhan et al., 2006, Blood 108:2020-8). Предполагается, что клоны плазматических клеток, которые амплифицируются при MGUS, имеют генетические и фенотипические профили, сходные с миеломатозными плазматическими клетками (Chng et al., 2005, Blood 106:2156-61; Fonseca et al., 2002, Blood 100:1417-24; Kaufmann et al., 2004, Leukemia. 18:1879-82). В то время как мутации в генах циклина D располагают к развитию ММ, возможный вклад других факторов достоверно не продемонстрирован (Bergsagel et al., 2005, Blood 106:296-303). Тем не менее, генетические изменения не являются единственным определяющим фактором функциональных возможностей клеток ММ. Вместе с тем, резистентность к лекарственным средствам и нарушенные биологические ответы на цитокины находятся под сильным влиянием микроокружения, что дает возможность разработки новых терапевтических средств.
Подобно многим другим опухолям ММ характеризуется гетерогенными клеточными популяциями, интенсивно взаимодействующими с незлокачественными клетками стромы, которые создают поддерживающее окружение (De Raeve и Vanderkerken, 2005, Histol Histopathol. 20:1227-50; Dhodapkar, 2009, Am J Hematol. 84:395-6). BM микроокружение для клеток ММ состоит из разнообразного внеклеточного матрикса (ЕСМ) и из клеточных компонентов гематопоэтического и негематопоэтического происхождения. Тогда как ВМ обеспечивает охранительное окружение для нормального гематопоэза, взаимодействие клеток ММ с белками ЕСМ и акцессорными клетками играет решающую роль в патогенезе ММ (De Raeve и Vanderkerken, 2005, Histol Histopathol. 20:1227-50; Dhodapkar, 2009, Am J Hematol. 84:395-6; Hideshima et al., 2007, Nat Rev Cancer. 7:585-98). Клетки стромы, миелоидные клетки, остеокласты и остеобласты продуцируют ростовые факторы, такие как интерлейкин 6 (IL-6), фактор, активирующий В-клетки (BAFF), фактор роста фибробластов и фактор 1а, происходящий из клеток стромы, который активирует сигнальные пути, содействующие миграции, выживаемости и росту клеток ММ. В частности, IL-6, продуцируемый клетками стромы, остеокластами и миелоидными клетками, по-видимому, является решающим фактором на ранних стадиях и для патогенеза ММ (De Raeve и Vanderkerken, 2005, Histol Histopathol. 20:1227-50). Подобным образом, в результате взаимодействия с ММ клетками, ослеокласты и дендритные клетки продуцируют BAFF и/или индуцирующий пролиферацию лиганд (APRIL), обеспечивая антиапопототические сигналы, которые также повышают лекарственную устойчивость (De Raeve и Vanderkerken, 2005, Histol Histopathol. 20:1227-50; Kukreja et al., 2006, J Exp Med. 203:1859-65).
Главные события в патогенезе рака - неконтролируемая пролиферация, выживаемость и распространение злокачественных клеток - зависят от специфических комбинаций поддерживающих типов клеток и растворимых факторов, присутствующих в нишах микроокружения. Мышиные модели играют важную роль в получении характеристик ключевых аспектов непосредственных причин злокачественной трансформации и болезни у человека. Тем не менее, они крайне редко показывают генетическую сложность и клиникопатологические характеристики болезни человека. Несмотря на то, что ксенотрансплантация человеческих опухолей иммунологически скомпрометированным мышам (мышам с нарушенной иммунной реакцией) широко используется, надежное приживление трансплантата, как правило, является возможным только в случае высоко агрессивных опухолей или клеточных линий. Самыми лучшими доступными в настоящее время моделями для роста человеческих опухолевых клеток являются выраженно иммунодефицитные мыши, у которых отсутствуют В-клетки, Т-клетки и NK-клетки. В случае ММ, приживление клеток первичной миеломы этим мышам является безуспешным, однако клетки первичной миеломы способны к приживлению в случае совместной трансплантации с кусочками человеческой эмбриональной кости иммунологически скомпрометированным мышам (Yaccoby et al., 1998, Blood 92:2908-13). В этой модели клетки ММ обнаруживаются в человеческой кости, но не обнаруживаются в мышиной кости или на периферии, демонстрируя интенсивное ксено-отторжение и потребность в человеческом ВМ микроокружении (Yaccoby et al., 1998, Blood 92:2908-13; Yaccoby и Epstein, 1999, Blood 94:3576-82). Доказывая потенциал этой модели в качестве модели MM in vivo, недавно было продемонстрировано, что мыши NOD/Scid/yc-/- обеспечивают возможность приживления некоторых клеточных линий ММ (Dewan et al., 2004, Cancer Sci. 95:564-8; Miyakawa et al., 2004, Biochem Biophys Res Commun. 313:258-62). В то же время, даже эти мышиные модели с низким ксено-отторжением ограничены условиями культивирования вследствие большого числа факторов, которые не преодолевают видовые барьеры, однако являются необходимыми для поддержания роста и выживаемости трансформированных клеток (Manz, 2007). Для создания новых лекарственных средств и видов лечения будут необходимы модели in vivo, обеспечивающие возможность исследования комплекса патогенных взаимодействий между опухолью и ее окружением. Следовательно, существует неудовлетворенная потребность создания гуманизированных не принадлежащих к человеческому роду животных и способов для надежного выращивания и исследования человеческих гематопоэтических клеток, включая мышиные модели с первичными человеческими гематопоэтическими опухолевыми клетками. Настоящее изобретение адресовано таким неудовлетворенным последовательностям в данной области техники.
Раскрытие изобретения
Предоставляются генетически модифицированные, не принадлежащие к человеческому роду животные, которые могут использоваться для моделирования развития, функции или болезни человеческих гематопоэтических клеток. Генетически модифицированные, не принадлежащие к человеческому роду животные содержат нуклеиновую кислоту, кодирующую человеческий IL-6, функционально связанный с промотором IL-6. В некоторых случаях генетически модифицированное, не принадлежащее к человеческому роду животное, экспрессирующее человеческий IL-6, также экспрессирует, по меньшей мере, одно из следующего: человеческого M-CSF, человеческого IL-3, человеческого GM-CSF, человеческого SIRPa или человеческого ТРО. В некоторых случаях генетически модифицированное, не принадлежащее к человеческому роду животное является иммунодефицитным. В некоторых случаях генетически модифицированному, не принадлежащему к человеческому роду животному пересаживаются здоровые или желательные человеческие гематопоэтические клетки. Также предоставляются способы использования рассматриваемых генетически модифицированных, не принадлежащих к человеческому роду животных при моделировании развития, функции и/или болезни человеческой гематопоэтической клетки, а также реагенты и их наборы, которые находят применение при создании рассматриваемых генетически модифицированных, не принадлежащих к человеческому роду животных и/или при применении на практике заявленных методов.
В различных аспектах изобретения предоставляется генетически модифицированное, не принадлежащее к человеческому роду животное, при этом генетически модифицированное, не принадлежащее к человеческому роду животное содержит геном, включающий нуклеиновую кислоту, кодирующую человеческий IL-6, функционально связанный с IL-6 промотором, при этом у животного экспрессируется человеческий IL-6 полипептид под контролем IL-6 промотора. В некоторых вариантах осуществления у генетически модифицированного, не принадлежащего к человеческому роду животного не экспрессируется нативный IL-6.
В некоторых вариантах осуществления генетически модифицированное, не принадлежащее к человеческому роду животное является грызуном. В некоторых вариантах осуществления не принадлежащее к человеческому роду животное является мышью. В некоторых подобных вариантах осуществления IL-6 промотор, с которым функционально связана нуклеиновая кислота, кодирующая человеческий IL-6, является мышиным IL-6 промотором, и человеческий ген IL-6 является функционально связанным с мышиным IL-6 промотором в локусе мышиного IL-6.
В некоторых вариантах осуществления генетически модифицированное, не принадлежащее к человеческому роду животное дополнительно содержит одну или более дополнительных нуклеиновых кислот, выбранных из нуклеиновой кислоты, кодирующей человеческий SIRPa под контролем промотора SIRPa; из нуклеиновой кислоты, кодирующей человеческий M-CSF, функционально связанный с M-CSF промотором, при этом у животного экспрессируется человеческий M-CSF; из нуклеиновой кислоты, кодирующей человеческий IL-3, функционально связанный с IL-3 промотором, при этом у животного экспрессируется человеческий IL-3; из нуклеиновой кислоты, кодирующей человеческий GM-CSF, функционально связанный с GMCSF промотором, при этом у животного экспрессируется человеческий GM-CSF; и из нуклеиновой кислоты, кодирующей человеческий ТРО, функционально связанный с ТРО промотором, при этом у животного экспрессируется человеческий ТРО. В некоторых вариантах осуществления промотор представляет собой человеческий промотор для данного гена. В других вариантах осуществления промотор представляет собой промотор гена животного, не принадлежащего к человеческому роду. В некоторых вариантах осуществления у генетически модифицированного, не принадлежащего к человеческому роду животного экспрессируется соответствующий нативный белок животного. В других вариантах осуществления у генетически модифицированного, не принадлежащего к человеческому роду животного не экспрессируется соответствующий нативный белок животного.
В некоторых вариантах осуществления генетически модифицированное, не принадлежащее к человеческому роду животное является иммунодефицитным относительно иммунной системы животного, не принадлежащего к человеческому роду. В некоторых подобных вариантах осуществления у иммунодефицитного, генетически модифицированного, не принадлежащего к человеческому роду животного не экспрессируется ген, активирующий рекомбинацию (RAG). В некоторых подобных вариантах осуществления у иммунодефицитного, генетически модифицированного, не принадлежащего к человеческому роду животного не экспрессируется гамма цепь рецептора IL2 (IL2rg, или "γс"). В некоторых подобных вариантах осуществления у иммунодефицитного, генетически модифицированного, не принадлежащего к человеческому роду животного не экспрессируется или RAG (например, RAG1, RAG2) или IL2rg.
В некоторых вариантах осуществления иммунодефицитному, генетически модифицированному, не принадлежащему к человеческому роду животному пересажены человеческие гематопоэтические клетки с целью получения генетически модифицированного, не принадлежащего к человеческому роду животного с пересаженными клетками. В одном варианте осуществления человеческие гематопоэтические клетки выбирают из клеток пуповинной крови человека, клеток эмбриональной печени человека и клеток гематопоэтической линии клеток человека. В одном варианте осуществления человеческие гематопоэтические клетки представляют собой клетки-предшественники CD34+. В одном варианте осуществления человеческие гематопоэтические клетки являются раковыми клетками. В определенных вариантах осуществления раковые клетки являются клетками множественной миеломы человека.
В некоторых вариантах осуществления генетически модифицированное животное с пересаженными клетками дает начало (порождает) человеческой клетке, выбранной из CD34+ клетки, гематопоэтической стволовой клетки, гематопоэтической клетки, миелоидной клетки-предшественника, миелоидной клетки, дендритной клетки, моноцита, гранулоцита, нейтрофила, тучной клетки, тимоцита, Т-клетки, В-клетки, плазматической клетки, тромбоцита и их комбинации. В одном варианте осуществления человеческая клетка присутствует через 1 месяц, через 2 месяца, через 3 месяца, 4 месяца, 5 месяцев, 6 месяцев, 7 месяцев, 8 месяцев, 9 месяцев, 10 месяцев, 11 месяцев или 12 месяцев после пересадки.
В некоторых вариантах осуществления генетически модифицированное животное с пересаженными клетками дает начало человеческой гематолимфоидной системе, которая содержит человеческие гематопоэтические стволовые клетки и клетки-предшественники, человеческие миелоидные клетки-предшественники, человеческие миелоидные клетки, человеческие дендритные клетки, человеческие моноциты, человеческие гранулоциты, человеческие нейтрофилы, человеческие тучные клетки, человеческие тимоциты, человеческие Т-клетки, человеческие В-клетки, человеческие тучные клетки и человеческие тромбоциты. В одном варианте осуществления гематолимфоидная система присутствует через 4 месяца, 5 месяцев, 6 месяцев, 7 месяцев, 8 месяцев, 9 месяцев, 10 месяцев, 11 месяцев или 12 месяцев после пересадки.
В некоторых аспектах изобретения предоставляются способы получения не принадлежащего к человеческому роду животного с пересаженными человеческими гематопоэтическими клетками. В некоторых вариантах осуществления предоставляются способы создания животной модели развития и функционирования иммунной клетки человека. В определенных вариантах осуществления предоставляются способы создания животной модели развития и функционирования человеческой В-клетки.
В некоторых вариантах осуществления данные способы включают пересадку популяции гематопоэтических клеток человека генетически модифицированному не принадлежащему к человеческому роду животному, которое является иммунодефицитным и экспрессирует нуклеиновую кислоту, кодирующую человеческий IL-6, функционально связанный с IL-6 промотором. В некоторых вариантах осуществления у животного не экспрессируется нативный IL-6. В некоторых вариантах осуществления IL-6 промотор является IL-6 промотором животного, не принадлежащего к человеческому роду, а человеческий IL-6 ген функционально связан с IL-6 промотором животного, не принадлежащего к человеческому роду, в локусе IL-6 не принадлежащего к человеческому роду животного. В некоторых вариантах осуществления не принадлежащее к человеческому роду животное является грызуном. В некоторых подобных вариантах осуществления не принадлежащее к человеческому роду животное является мышью. Соответственно, в некоторых вариантах осуществления промотор IL-6, с которым функционально связана нуклеиновая кислота, кодирующая человеческий IL-6, является мышиным промотором IL-6, и человеческий IL-6 ген является функционально связанным с мышиным промотором IL-6 в локусе мышиного IL-6.
В некоторых вариантах осуществления пересаженная популяция гематопоэтических клеток содержит CD34+ клетки. В некоторых вариантах осуществления пересаженная популяция гематопоэтических клеток содержит раковые клетки. В некоторых вариантах осуществления пересаженная популяция гематопоэтических клеток содержит клетки множественной миеломы. В некоторых вариантах осуществления пересадка включает внутрифеморальную и/или внутритибиальную инъекцию.
В некоторых вариантах осуществления иммунодефицитное, генетически модифицированное животное экспрессирует, по меньшей мере, одну дополнительную человеческую нуклеиновую кислоту, выбранную из группы, состоящей из нуклеиновой кислоты, кодирующей человеческий SIRPa, функционально связанный с SIRPa промотором; нуклеиновой кислоты, кодирующей человеческий M-CSF, функционально связанный с M-CSF промотором; нуклеиновой кислоты, кодирующей человеческий IL-3, функционально связанный с IL-3 промотором; нуклеиновой кислоты, кодирующей человеческий GM-CSF, функционально связанный с GM-CSF промотором; и нуклеиновой кислоты, кодирующей человеческий ТРО, функционально связанный с ТРО промотором.
В некоторых аспектах данного изобретения предоставляются генетически модифицированные, не принадлежащие к человеческому роду животные с пересаженными клетками, экспрессирующие нуклеиновую кислоту, кодирующую человеческий IL-6 функционально связанный с IL-6 промотором, эти не принадлежащие к человеческому роду животные с пересаженными клетками получают в соответствии с описанными в данном документе или известными в этой области техники способами. В некоторых вариантах осуществления генетически модифицированное, не принадлежащее к человеческому роду животное с пересаженными клетками является животной моделью развития и дифференцировки человеческих В-клеток.
В различных вариантах осуществления предоставляются способы, которые включают использование заявленных генетически модифицированных, не принадлежащих к человеческому роду животных с пересаженными человеческими гематопоэтическими клетками. Эти способы включают, например, способы оценки in vivo роста и дифференцировки гематопоэтических и иммунных клеток, способы оценки in vivo человеческого гематопоэза, способы оценки in vivo раковых клеток, способы оценки in vivo иммунного ответа, способы оценки in vivo вакцин и режимов вакцинации, способы применения при тестировании эффектов средств, модулирующих рост или выживаемость раковой клетки, способы оценки in vivo лечения рака и способы выработки и сбора in vivo медиаторов иммунитета, включая человеческие антитела, и использования при тестировании эффекта средств, модулирующих функцию гематопоэтических и иммунных клеток. Например, в некоторых вариантах осуществления предоставляются способы скрининга средств-кандидатов в отношении возможности лечения раковых заболеваний гематопоэтической системы. В некоторых вариантах осуществления способ включает контактирование генетически модифицированного не принадлежащего к человеческому роду животного настоящего раскрытия, которому были пересажены клетки гематопоэтического рака человека, со средством-кандидатом и сравнение жизнеспособности и/или скорости пролиферации человеческих гематопоэтических злокачественных клеток у контактировавшего генетически модифицированного не принадлежащего к человеческому роду животного с пересаженными человеческими гематопоэтическими злокачественными клетками с генетически модифицированным не принадлежащим к человеческому роду животным с аналогичным образом пересаженными клетками, которое не контактировало со средством-кандидатом, при этом уменьшение жизнеспособности и/или скорости пролиферации клеток гематопоэтического рака человека у контактировавшего, не принадлежащего к человеческому роду животного с пересаженными клетками указывает на то, что средство-кандидат будет лечить гематопоэтическое злокачественное заболевание. Эти и другие способы станут понятны среднему специалисту в данной области техники на основании предоставленного раскрытия.
Краткое описание чертежей
Следующее подробное описание предпочтительных вариантов осуществления изобретения будет более понятно при прочтении в сочетании с прилагаемыми чертежами. Следует подчеркнуть, что в соответствии с общепринятой практикой, различные характеристики на чертежах предоставлены без соблюдения масштаба. Напротив, величины различных характеристик являются произвольно увеличенными или уменьшенными для ясности. В целях иллюстрации изобретения на чертежах показаны варианты осуществления, которые являются предпочтительными в настоящее время. Однако, следует понимать, что изобретение не ограничивается определенной схемой и инструментарием вариантов осуществления, показанных на чертежах. Чертежи предоставляются в виде следующих фигур.
Фигура 1 - серия графиков, показывающих результаты экспериментов демонстрирующих приживление INA-6 клеток у knock-in мышей с человеческим IL-6. Уровни растворимого IL-6R были измерены у мышей указанных генотипов, которым было внутривенно трансплантировано 5×106 INA-6 клеток. N показывает количество мышей с трансплантацией в группе.
Фигура 2, включающая Фигуры 2А - 2F, - это изображения, показывающие гистологические анализы бедренных костей после внутривенной пересадки INA-6 клеток. Мышей Rag2-/-Il2rgnullIl6h/hhSIRPa+ умерщвляли через восемь недель после пересадки 5×106 ΓΝΑ-6 клеток внутривенно. Бедренные кости фиксировали в 10% формалине и декальцифицировали. Срезы (10 мкм) окрашивали толуидиновым синим или сразу анализировали в отношении GFP-экспрессии, используя конфокальный микроскоп Leica.
Фигура 3 - серия изображений и график, показывающий исследование легких после внутривенной пересадки INA-6 клеток. Мышей Rag2-/-Il2rgnullIl6h/hhSIRPa+ умерщвляли через восемь недель после пересадки с помощью внутривенной инъекции 5×106 INA-6 клеток. Ткань легких фиксировали в 10% формалине, а 10 мкм срезы непосредственно исследовали в отношении GFP-экспрессии, используя конфокальный микроскоп Leica. Слева показаны изображения срезов при увеличении 10Х (вверху) и 63Х (внизу). График (справа) показывает экспрессию человеческого Il6 гена, измеренную в указанных тканях и нормированную по отношению к экспрессии мышиного hprt.
Фигура 4 - серия графиков, показывающих результаты экспериментов, демонстрирующих приживление INA-6 клеток у knock-in мышей с человеческим IL-6. Уровни растворимого IL-6R были измерены у мышей указанных генотипов, которым интрафеморально (внутрь бедра) было трансплантировано 5×105 INA-6 клеток. Каждая линия показывает отдельную мышь.
Фигура 5 - серия графиков, показывающих гистологический анализ бедренных костей после интрафеморального приживления INA-6 клеток. Мышей Rag2-/-Il2rgnullIl6h/hhSIRPa+ умерщвляли через четыре - шесть недель после приживления 5×105 интрафеморально инъецированных INA-6 клеток. Бедренные кости фиксировали в 10% формалине и декальцифицировали. Срезы 10 мкм окрашивали толуидиновым синим.
Фигура 6 - серия изображений и график, показывающий результаты μСТ-анализов бедренных костей мышей после трансплантации INA-6 клеток. Мышей Rag2-/-Il2rgnullIl6h/hhSIRPa+ и контрольных мышей умерщвляли через четыре недели после приживления интрафеморально инъецированных 5×105 INA-6 клеток. Бедренные кости фиксировали в 70% этаноле и анализировали с использованием мышиных μCΤ. Объемы бедренной кости и ткани были определены количественно, чтобы подсчитать отношение между объемом кости и ткани. *: р<0.01 с помощью t-критерия Стьюдента.
Фигура 7 - это график, демонстрирующий результаты μCT-анализов бедренных костей мышей после лечения лекарственными средствами от миеломы. Мышам Rag2-/-Il2rgnullIl6h/hhSIRPa+ пересаживали с помощью интрафеморальной инъекции 5×105 INA-6 клеток и два раза в неделю лечили Velcade® или Zometa®, соответственно. Через четыре недели мышей забивали, а бедренные кости фиксировали в 70% этаноле для проведения μСТ-анализов. Объемы трабекулярной кости и ткани были определены количественно, чтобы подсчитать отношение между объемом кости и ткани.
Фигура 8 - серия графиков, демонстрирующих результаты FACS анализов первичных клеток, пересаженных Rag2-/-Il2rgnullhSIRPa+ Tpoh/h Mcsfh/h Il3/Gmcsfh/h Il6h/h мышам. Мышам трансплантировали с помощью интрафеморальной инъекции 1.5×106 CD3-истощенных клеток костного мозга, затем мышей забивали через двенадцать недель. Одноклеточные суспензии были получены из бедренной кости, коллатеральной голени и селезенки. Клетки окрашивали относительно mCD45, hCD19, hCD38 и hCD138. FACS графики показывают события после гейтирования (gating) mCD45-отрицательных клеток. Числа показывают частоту CD38+CD138+ клеток.
Фигура 9 демонстрирует результаты экспериментов, оценивающих процент человеческих гематопоэтических (hCD45+) клеток в крови у мышей с пересаженными клетками, определенный с помощью проточной цитометрии. Горизонтальные полосы показывают соответствующие средние частоты.
Фигура 10 демонстрирует результаты экспериментов, оценивающих процент hCD45 В-клеток (CD19+), Т-клеток (CD3+) и миелоидных (CD33+) клеток в крови мышей с пересаженными клетками, определенный с помощью проточной цитометрии. Показаны только мыши с процентом hCD45 выше, чем 2%.
Фигура 11, включающая Фигуры 11А и 11В, демонстрирует результаты экспериментов, оценивающих процент (А) и количество (В) человеческих CD45+ клеток в ВМ, селезенке и тимусе мышей через 20 недель после приживления. Столбики представляют средние значения ± SEM 4/5 мышей на группу.
Фигура 12, включающая Фигуры 12А и 12В, иллюстрирует результаты FACS экспериментов, оценивающих человеческие клетки. (А) Рисунок, показывающий стратегию гейтирования, использованную для разделения различных популяций В-клеток с помощью проточной цитометрии. (В) Процент различных В-клеточных субпопуляций в пределах человеческих CD45+CD19+ клеток у мышей 20-недельного возраста. Столбики представляют средние значения ± SEM 4/5 мышей на группу.
Фигура 13, включающая Фигуры 13А и 13В, иллюстрирует результаты FACS экспериментов, оценивающих человеческие клетки. (А) Характерный проточный цитометрический анализ CD5+ В-клеток в эмбриональной печени человека (FL) и у мышей 20-недельного возраста. Числа в квадрантах показывают процент клеток. Все графики гейтированы относительно человеческих CD45+ клеток. (В) Процент CD5 относительно человеческих В-клеток в ВМ и селезенке мышей через 20 недель после приживления. Столбики представляют средние значения ± SEM 4/5 мышей на группу.
Фигура 14 иллюстрирует результаты FACS экспериментов, оценивающих человеческие клетки. (А) Характерный проточный цитометрический анализ CD27+ В-клеток в эмбриональной печени человека (FL) и у мышей 20-недельного возраста. Числа в квадрантах показывают процент клеток. Все графики гейтированы относительно человеческих CD45+ клеток. (В) Процент CD27 относительно человеческих В-клеток в ВМ и селезенке мышей через 20 недель после приживления. Столбики представляют среднее значение ± SEM 4/5 мышей на группу.
Фигура 15 иллюстрирует результаты экспериментов, оценивающих уровни общего IgM человека и IgG в плазме образцов мышей 12(А) и 20(В) недельного возраста. Горизонтальные полосы показывают соответствующие геометрические средние значения. Мыши с приживлением PB человека ниже 2% были исключены из анализа.
Подробное описание
Предоставляются генетически модифицированные, не принадлежащие к человеческому роду животные, которые могут использоваться для моделирования развития, функционирования или заболевания человеческих гематопоэтических клеток. Генетически модифицированные, не принадлежащие к человеческому роду животные содержат нуклеиновую кислоту, кодирующую человеческий IL-6, функционально связанный с промотором IL-6. В некоторых вариантах осуществления генетически модифицированное не принадлежащее к человеческому роду животное, экспрессирующее человеческий IL-6, также экспрессирует, по меньшей мере, одно из числа: человеческого M-CSF, человеческого IL-3, человеческого GM-CSF, человеческого SIRPa или человеческого ТРО. Данное изобретение также имеет отношение к способам получения и способам использования генетически модифицированных, не принадлежащих к человеческому роду животных, описанных в данном документе. В некоторых вариантах осуществления генетически модифицированное, не принадлежащее к человеческому роду животное является мышью. В некоторых вариантах осуществления описанное в данном документе генетически модифицированное, не принадлежащее к человеческому роду животное является животным с пересаженными человеческими гематопоэтическими клетками, включая нормальные или неопластические клетки или их комбинации. В некоторых вариантах осуществления описанное в данном документе генетически модифицированное, не принадлежащее к человеческому роду животное является животным с пересаженными клетками множественной миеломы (ММ) человека. В различных вариантах осуществления генетически модифицированные, не принадлежащие к человеческому роду животные изобретения с пересаженными человеческими гематопоэтическими клетками используются для оценки in vivo роста и дифференцировки гематопоэтических и иммунных клеток, для оценки in vivo человеческого гематопоэза, для исследования in vivo раковых клеток, для оценки in vivo иммунного ответа, для оценки in vivo вакцин и режимов вакцинации, для применения при тестировании эффекта средств, модулирующих рост или выживаемость раковой клетки, для оценки in vivo лечения рака, для выработки и сбора in vivo иммунных медиаторов, включая человеческие антитела, и для использования при тестировании эффекта средств, модулирующих функцию гематопоэтических и иммунных клеток. Эти и другие цели, преимущества и признаки изобретения станут очевидными для специалистов в данной области техники после прочтения подробного описания композиций и способов, предоставленного ниже.
Определения
Если не указано иное, все технические и научные термины, использованные в описании, имеют то же самое значение, которое обычно понятно среднему специалисту в той области техники, к которой относится изобретение. Такие термины встречаются и используются в контексте для иллюстрации в различных ссылках, включая J. Sambrook and D. W. Russell, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press; 4th Ed., 2012; F. M. Ausubel, Ed., Short Protocols in Molecular Biology, Current Protocols; 5th Ed., 2002; В. Albeits et al., Molecular Biology of the Cell, 4th Ed., Garland, 2002; D. L. Nelson и M. M. Cox, Lehninger Principles of Biochemistry, 4th Ed., W.H. Freeman & Company, 2004; и Herdewijn, P. (Ed.), Oligonucleotide Synthesis: Methods and Applications, Methods in Molecular Biology, Humana Press, 2004. Хотя при осуществлении на практике или тестировании настоящего изобретения могут использоваться любые способы и материалы, подобные или эквивалентные описанным, в документе представлены предпочтительные способы и материалы.
При использовании в описании каждый из следующих терминов имеет значение, относящееся к данному разделу.
Терминаы, представленные в единственном числе, также включают и множественное число. В качестве примера, "элемент" означает один элемент или больше чем один элемент.
Использованный в описании термин "около (примерно)" при упоминании измеряемой величины, такой как количество, продолжительность времени и тому подобное, предназначается для включения изменений ±20% или ±10%, более предпочтительно ±5%, даже более предпочтительно ±1% и еще более предпочтительно ±0.1% от точно указанного значения, например, изменений, подходящих для выполнения раскрытых способов.
Термин "ненормальный (аномальный)" при использовании в отношении организмов, тканей, клеток или их компонентов включает организмы, ткани, клетки или их компоненты, которые отличаются по меньшей мере одной наблюдаемой или обнаружимой характеристикой (например, возрастом, лечением, временем дня и т.д.) от тех организмов, тканей, клеток или их компонентов, которые демонстрируют "нормальное" (ожидаемое) соответствующее характерное свойство. Характеристики, являющиеся нормальными или ожидаемыми для одного типа клеток или тканей, могут быть ненормальными для другого типа клеток или тканей.
Использованный в описании термин "антитело" включает молекулу иммуноглобулина, способную специфически связываться со специальным эпитопом на антигене. Антитела могут быть интактными иммуноглобулинами, происходящими из природных или рекомбинантных источников, и могут быть иммунореактивными участками интактных иммуноглобулинов. Антитела настоящего изобретения могут находиться в различных формах, включая, например, поликлональные антитела, моноклональные антитела, внутриклеточные антитела ("интраантитела"), Fv, Fab и F(ab)2, а также одноцепочечные антитела (scFv), тяжелоцепочечные антитела, такие как антитела верблюдовых и гуманизированные антитела (Harlow et al., 1999, Using Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, NY; Harlow et al., 1989, Antibodies: A Laboratory Manual, Cold Spring Harbor, New York; Houston et al., 1988, Proc. Natl. Acad. Sci. USA 85:5879-5883; Bird et al., 1988, Science 242:423-426).
"Конститутивная" экспрессия включает состояние, при котором продукт гена продуцируется в живой клетке при большей части или при всех физиологических условиях в клетке.
"Кодирующая область" гена включает нуклеотидные остатки кодирующей цепи гена и нуклеотиды некодирующей цепи гена, которые являются гомологичными или комплементарными, соответственно, с кодирующим участком молекулы мРНК, которая продуцируется путем транскрипции гена. "Кодирующий участок" молекулы мРНК также включает нуклеотидные остатки молекулы мРНК, которые соответствуют участку антикодона молекулы транспортной РНК в ходе трансляции молекулы мРНК или которые кодируют стоп-кодон. Таким образом, кодирующий участок может включать нуклеотидные остатки, содержащие кодоны для остатков аминокислот, которые не присутствуют в зрелом белке, кодированном молекулой мРНК (например, аминокислотные остатки в сигнальной последовательности экспорта белка).
"Болезнь" включает состояние здоровья животного, при котором у животного не поддерживается гомеостаз, и в том случае, когда болезнь не облегчается, тогда здоровье животного продолжает ухудшаться.
В противоположность этому, "нарушение" у животного включает состояние здоровья, при котором у животного может сохраняться гомеостаз, но при котором состояние здоровья животного является менее благоприятным, чем оно могло бы быть при отсутствии нарушения. Оставаясь нелеченным, нарушение не вызывает безусловно дальнейшее ухудшение состояния здоровья животного.
Болезнь или нарушение "ослабляется (облегчается)" в том случае, если уменьшается тяжесть симптома болезни или нарушения, или уменьшается частота, с которой пациент испытывает такой симптом, или и то и другое.
"Эффективное количество" или "терапевтически эффективное количество" соединения подразумевает количество соединения, которое является достаточным для обеспечения благоприятного действия на субъекта, которому вводится данное соединение. "Эффективное количество" носителя подразумевает количество, достаточное для эффективного связывания или доставки соединения.
"Кодирование" относится к характерному свойству специфической последовательности нуклеотидов в полинуклеотиде, таком как ген, кДНК или мРНК, служить в качестве матрицы для синтеза других полимеров и макромолекул в биологических процессах, имеющих или определенную последовательность нуклеотидов (т.е., рРНК, тРНК и мРНК) или определенную последовательность аминокислот, и возникающим в связи с этим биологическим свойствам. Таким образом, ген кодирует белок в случае, например, транскрипции и трансляции мРНК, в соответствии с которой ген продуцирует белок в клетке или другой биологической системе. И кодирующая цепь, нуклеотидная последовательность которой является идентичной последовательности мРНК и обычно предоставляется в перечислении последовательности, и некодирующая цепь, используемая в качестве матрицы для транскрипции гена или кДНК, может называться кодирующей белок или другой продукт этого гена или кДНК.
При использовании в описании "эндогенный" включает какой-либо материал (вещество), полученный из этого организма или продуцированный внутри организма, клетки, ткани или системы.
Использованный в описании термин "экзогенный" включает какой-либо материал, внесенный или продуцированный вне организма, клетки, ткани или системы.
Использованные в описании термины "экспрессирующая конструкция" и "экспрессионная кассета" включают двухцепочечную рекомбинантную молекулу ДНК, содержащую желательную нуклеиновокислотную кодирующую человеческую последовательность и содержащую один или более регуляторных элементов, необходимых или желательных для экспрессии функционально связанной кодирующей последовательности.
Использованный в описании термин "фрагмент", применительно к нуклеиновой кислоте или полипептиду, включает подпоследовательность нуклеиновой кислоты или полипептида. "Фрагмент" нуклеиновой кислоты может иметь, по меньшей мере, около 15 нуклеотидов в длину; например, по меньшей мере, примерно от 50 нуклеотидов до 100 нуклеотидов; по меньшей мере, примерно от 100 до 500 нуклеотидов, по меньшей мере, примерно от 500 до 1000 нуклеотидов, по меньшей мере, примерно от 1000 нуклеотидов до 1500 нуклеотидов; или примерно от 1500 нуклеотидов до 2500 нуклеотидов; или около 2500 нуклеотидов (и любое целое значение между этими значениями). "Фрагмент" полипептида может иметь, по меньшей мере, около 15 аминокислот в длину; например, по меньшей мере, примерно от 50 аминокислот до 100 аминокислот; по меньшей мере, примерно от 100 до 500 аминокислот, по меньшей мере, примерно от 500 до 1000 аминокислот, по меньшей мере, примерно от 1000 аминокислот до 1500 аминокислот; или примерно от 1500 аминокислот до 2500 аминокислот; или около 2500 аминокислот (и любое целое значение между этими значениями).
Использованные в описании термины "ген" и "рекомбинантный ген" включают молекулы нуклеиновой кислоты, содержащие открытую рамку считывания, кодирующую полипептид. Природные аллельные вариации, как правило, приводят к 1-5% изменчивости в нуклеотидной последовательности данного гена. Альтернативные аллели могут быть идентифицированы путем секвенирования интересующего гена у целого ряда различных индивидуумов. Это можно легко осуществить с использованием гибридизационных зондов с целью установления одинакового генетического локуса у целого ряда индивидуумов. Все подобные нуклеотидные изменения и полученные в результате полиморфизмы аминокислот или изменения, которые являются результатом природного аллельного разнообразия и которые не изменяют функциональную активность, включаются в рамки данного изобретения.
"Гомологичный" при использовании в описании подразумевает сходство субъединиц последовательности между двумя полимерными молекулами, например, между двумя молекулами нуклеиновой кислоты, например, двумя молекулами ДНК или двумя молекулами РНК, или между двумя молекулами полипептидов. Когда положение субъединицы в обеих из двух молекул занято одинаковой мономерной субъдиницей, например, если положение в каждой из двух молекул ДНК занято аденином, тогда они являются гомологичными по этому положению. Гомология между двумя последовательностями является непосредственной функцией числа соответствий или гомологичных положений, например, если половина положений (например, пять положений в полимере длиной десять субъединиц) в двух последовательностях являются гомологичными, тогда две последовательности являются гомологичными на 50%, если 90% положений, например, 9 из 10, являются соответствующими или гомологичными, тогда две последовательности являются гомологичными на 90%. В качестве примера, ДНК-последовательности 5-ATTGCC-3' и 5'-TATGGC-3' обладают 50% гомологией.
Термины "человеческие гематопоэтические стволовые клетки и клетки-предшественники" и "человеческие HSPC" при использовании в описании включают человеческие самовосстанавливающиеся мультипотентные гематопоэтические стволовые клетки и гематопоэтические клетки-предшественники.
"Индуцибельная" экспрессия включает состояние, при котором продукт гена продуцируется в живой клетке в ответ на присутствие сигнала в клетке.
Использованный в описании термин "учебный (инструктирующий) материал" включает публикацию, запись, схему или любую другую среду выражения, которая может использоваться для разъяснения практической ценности соединения, композиции, вектора или системы доставки изобретения, имеющуюся в наборе, предназначенном для эффективного облегчения различных болезней или нарушений, перечисленных в описании. Необязательно, или альтернативно, учебный материал может описывать один или более способов облегчения болезней или нарушений в клетке или ткани млекопитающего. Учебный материал набора изобретения, например, может быть прикреплен к контейнеру, содержащему установленное соединение, композицию, вектор или систему доставки изобретения, или может перевозиться вместе с контейнером, содержащим установленное соединение, композицию, вектор или систему доставки. Альтернативно, учебный материал может перевозиться отдельно от контейнера с тем расчетом, что учебный материал и соединение используются потребителем совместно.
Термин "нуклеиновая кислота" включает молекулы РНК или ДНК, имеющие более чем один нуклеотид в любой форме, включая одноцепочечную, двухцепочечную, олигонуклеотид или полинуклеотид. Термин "последовательность нуклеотидов" подразумевает порядок нуклеотидов в олигонуклеотиде или полинуклеотиде в одноцепочечной форме нуклеиновой кислоты.
Использованный в описании термин "функционально связанный" означает полинуклеотид, находящийся в функциональной связи со вторым полинуклеотидом, например, одноцепочечную или двухцепочечную молекулу нуклеиновой кислоты, содержащую два полинуклеотида, расположенных в пределах молекулы нуклеиновой кислоты таким образом, что, по меньшей мере, один из двух полинуклеотидов способен оказывать характерное для него физиологическое действие сразу после другого. В качестве примера, промотор, функционально связанный с кодирующей областью гена, способен обеспечивать транскрипцию кодирующей области. Предпочтительно, когда нуклеиновая кислота, кодирующая желательный белок, дополнительно содержит промотор/регуляторную последовательность, причем промотор/регуляторная последовательность располагается на 5' конце последовательности, кодирующей желательный белок, благодаря чему промотор запускает экспрессию желательного белка в клетке. Вместе взятые, нуклеиновая кислота, кодирующая желательный белок, и ее промотор/регуляторная последовательность, составляют "трансген."
Использованный в описании термин "полинуклеотид" обозначает цепочку нуклеотидов. Более того, нуклеиновые кислоты являются полимерами нуклеотидов. Таким образом, использованные в описании термины нуклеиновые кислоты и полинуклеотиды являются взаимозаменяемыми. Специалист в данной области имеет общие знания о том, что нуклеиновые кислоты представляют собой полинуклеотиды, которые могут расщепляться до мономерных "нуклеотидов". Мономерные нуклеотиды могут расщепляться до нуклеозидов. При использовании в описании полинуклеотиды включают, но не ограничиваются этим, все нуклеиновокислотные последовательности, которые получают с помощью доступных в данной области техники способов, включая, без ограничения, рекомбинантные способы, т.е., клонирование нуклеиновокислотной последовательности из библиотеки рекомбинантных ДНК или генома клетки с использованием общепринятых методов клонирования и ПЦР, и тому подобного, и с помощью методов синтеза.
Использованные в описании термины "пептид", "полипептид" и "белок" применяются взаимозаменяемым образом и включают соединение, состоящее из остатков аминокислот, ковалентно связанных пептидными связями. Белок или пептид должен содержать, по меньшей мере, две аминокислоты, причем максимальное количество аминокислот, содержащихся в последовательностях белков или пептидов, не имеет ограничения. Полипептиды включают любой пептид или белок, содержащий две или более аминокислот, соединенных друг с другом пептидными связями. Например, использованный в описании термин включает и короткие цепи, которые в данной области техники обычно называются пептидами, олигопептидами и олигомерами, и более длинные цепи, в том числе многих типов. "Полипептиды" включают, например, биологически активные фрагменты, в большой степени гомологичные полипептиды, олигопептиды, гомодимеры, гетеродимеры, варианты полипептидов, модифицированные полипептиды, их производные, аналоги, гибридные белки, в числе прочих. Полипептиды включают природные пептиды, рекомбинантные пептиды, синтетические пептиды или их комбинации. Термин "пептид" в большинстве случаев относится к коротким полипептидам. Термин "белок" в большинстве случаев относится к большим полипептидам.
Термин "потомство", использованный в описании, включает потомков или отпрысков и включает дифференцированные или недифференцированные клетки-потомки, происходящие от родительской клетки. В одном случае термин "потомство" включает клетки-потомки, которые генетически идентичны родительской клетке. В другом случае термин "потомство" включает клетки-потомки, которые генетически и фенотипически идентичны родительской клетке. В еще одном случае термин "потомство" включает клетку-потомка, которая дифференцировалась из родительской клетки.
Использованный в описании термин "промотор" включает последовательность ДНК, функционально связанную с последовательностью нуклеиновой кислоты, предназначенной для транскрибирования (считывания), такой как последовательность нуклеиновой кислоты, кодирующая желательную молекулу. Как правило, промотор располагается выше последовательности нуклеиновой кислоты, которая должна транскрибироваться, и предоставляет сайт для специфического связывания с РНК-полимеразой и другими факторами транскрипции. В отдельных вариантах осуществления промотор располагается выше последовательности нуклеиновой кислоты, транскрибируемой для получения желаемой молекулы, и предоставляет сайт для специфического связывания с РНК-полимеразой и другими факторами транскрипции.
Диапазоны: на протяжении раскрытия различные аспекты данного изобретения могут быть представлены в формате диапазона. Следует понимать, что описание в виде диапазонов используется в целях удобства и краткости и не должно истолковываться как жесткое ограничение объема изобретения. Соответственно, описание диапазонов следует рассматривать, как специальное раскрытие всех возможных поддиапазонов, а также отдельных численных значений в пределах диапазона. Например, описание диапазона, такое как от 1 до 6 следует рассматривать как специальное раскрытие всех возможных поддиапазонов, таких как от 1 до 3, от 1 до 4, от 1 до 5, от 2 до 4, от 2 до 6, от 3 до 6 и т.д., а также отдельных чисел в пределах этого диапазона, например, 1, 2, 2.7, 3, 4, 5, 5.3 и 6. Это применяется, не принимая во внимание ширину диапазона.
"Рекомбинантный полипептид" включает полипептид, который продуцируется после экспрессии рекомбинантного полинуклеотида.
Термин "регуляторный элемент" при использовании в описании включает нуклеотидную последовательность, контролирующую некоторые аспекты экспрессии нуклеиновокислотных последовательностей. Характерные регуляторные элементы для иллюстрации включают энхансер, участок внутренней посадки рибосомы (IRES), интрон; точку начала репликации, сигнал полиаденилирования (рА), промотор, энхансер, последовательность терминации транскрипции и вышерасположенный регуляторный домен, который способствует репликации, транскрипции, посттрансляционному процессингу нуклеиновокислотной последовательности. Специалисты в данной области техники способны подобрать и использовать эти и другие регуляторные элементы в экспрессирующих конструкциях с помощью обычного проведения экспериментов. Экспрессирующие конструкции могут быть созданы рекомбинантно или синтетическим путем с помощью общеизвестных методов.
Термин "специфически связывается", использованный в отношении антитела, включает антитело, которое распознает специфический антиген, но практически не распознает или не связывается с другими молекулами в образце. Например, антитело, которое специфически связывается с антигеном от одного вида, также может связываться с антигеном от одного или более других видов. Однако, такая межвидовая реактивность не изменяет классификацию антитела как специфического. В качестве другого примера, антитело, которое специфически связывается с антигеном, может также связываться с различными аллельными формами данного антигена. Однако, такая межвидовая реактивность не изменяет классификацию антитела как специфического.
В некоторых случаях, термины "специфическое связывание" или "специфически связывается" могут использоваться в отношении взаимодействия антитела, белка или пептида с другими химическими соединениями, означая, что данное взаимодействие зависит от присутствия определенной структуры (например, антигенной детерминанты или эпитопа) на химическом соединении; например, антитело распознает и связывается со специфической структурой белка, а не с белками вообще. В случае, если антитело является специфическим для эпитопа "А", присутствие молекулы, содержащей эпитоп А (или свободный, немеченый А), в реакционной смеси, содержащей меченый "А" и данное антитело, будет уменьшать количество меченого А, связанного с антителом.
Использованный в описании термин "синтетическое антитело" включает антитело, полученное с использованием метода рекомбинантных ДНК, такое как, например, антитело, экспрессированное бактериофагом, как описано в данном документе. Термин также означает антитело, полученное путем синтеза молекулы ДНК, кодирующей антитело, при этом молекула ДНК экспрессирует белок антитела, или аминокислотную последовательность, точно определяющую данное антитело, причем ДНК или аминокислотную последовательность получают при помощи доступной и хорошо известной в данной области технологии синтетической ДНК или аминокислотной последовательности.
Использованный в описании термин "вариант" включает последовательность нуклеиновой кислоты или пептида, последовательность которой отличается от исходной (эталонной) последовательности нуклеиновой кислоты или пептида, соответственно, но сохраняет необходимые биологические свойства исходной молекулы. Изменения в последовательности варианта нуклеиновой кислоты могут не изменять аминокислотную последовательность пептида, кодированного исходной нуклеиновой кислотой, или могут приводить к аминокислотным заменам, вставкам, делениям, слияниям и укорочениям. Изменения в последовательности вариантов пептидов являются, как правило, ограниченными или консервативными, так что последовательности исходного пептида и варианта являются близко сходными вообще, а во многих участках идентичными. Аминокислотная последовательность варианта и исходного пептида могут различаться одной или более заменами, вставками, делециями в любых сочетаниях. Вариант нуклеиновой кислоты или пептида может быть природного происхождения, таким как аллельный вариант, или может быть вариантом, который не встречается в природе. Варианты нуклеиновых кислот и пептидов, не встречающиеся в природе, могут быть получены с помощью методов мутагенеза или путем прямого синтеза.
Термин "генетически модифицированное" имеет отношение к животному, зародышевые клетки которого содержат экзогенную человеческую нуклеиновую кислоту или человеческую нуклеиновокислотную последовательность. В качестве неограничивающих примеров, генетически модифицированное животное может быть трансгенным животным или knock-in животным, в том случае, если животное содержит человеческую нуклеиновокислотную последовательность.
Использованный в описании термин "трансгенное животное" включает животное, содержащее экзогенную человеческую нуклеиновокислотную последовательность, интегрированную в геном животного.
При использовании в описании "knock-in" имеет отношение к генетической модификации, которая нацеливается на конкретный хромосомный локус генома животного, не относящегося к человеческому роду, и при которой представляющая интерес нуклеиновая кислота вставляется в этот намеченный локус. В некоторых случаях, трансгенная модификация заменяет генетическую информацию, закодированную в хромосомном локусе не принадлежащего к человеческому роду животного, другой последовательностью ДНК.
Генетически модифицированные, не принадлежащие к человеческому роду животные
В некоторых аспектах изобретения предоставляется генетически модифицированное не принадлежащее к человеческому роду животное, которое экспрессирует человеческий IL-6. Под человеческим IL-6 (hIL6) имеется в виду белок, состоящий из 184 аминокислот, последовательность которого содержится в Genbank под порядковым номером NM_000600.3 и NP_000591.1. Человеческий IL-6 - это секретируемый белок, который продуцируется, например, Т-клетками, В-клетками, моноцитами, макрофагами, фибробластами, кератиноцитами, эндотелиальными клетками и клетками миеломы. IL-6 функционирует через посредство гетеродимерного рецепторного комплекса на клеточной поверхности, содержащего связывающую субъединицу (IL-6R) и субъединицу, преобразующую сигнал (gp130). gp130 является обычным компонентом других рецепторов, например, рецепторов для IL-11, IL-27, LIF, тогда как IL-6R преимущественно относится к гепатоцитам, моноцитам, активированным В-клеткам, отдыхающим Т-клеткам и линиям миеломных клеток. Показано, что IL-6 играет центральную роль в гематопоэзе, иммунных ответах и острофазных реакциях, и является важным фактором для окончательного созревания В-клеток в антитело-секретирующие клетки (ASC), в частности, для размножения плазмобластов во время реакции в зародышевом центре при Т-клеточно-зависимом (TD) гуморальном иммунном ответе. IL-6 требуется для Т-клеточной пролиферации in vitro и для производства цитотоксических Т-клеток (CTL) in vivo, делая их более легко реагирующими на IL-2.
В некоторых аспектах изобретения генетически модифицированное не принадлежащее к человеческому роду животное, экспрессирующее человеческий IL-6, также экспрессирует, по меньшей мере, один дополнительный человеческий белок, выбранный из человеческого M-CSF, человеческого IL-3, человеческого GM-CSF, человеческого ТРО и человеческого SIRPa или любой их комбинации. Другими словами не принадлежащее к человеческому роду животное, экспрессирующее человеческий IL-6, может экспрессировать один, два, три, четыре или все пять человеческих белков, выбранных из числа hM-CSF, hIL-3, hGM-CSF, hTPO и hSIRPa. Генетически модифицированные не принадлежащие к человеческому роду животные, экспрессирующие hMCSF, hIL-3, hGM-CSF, hTPO и/или hSIRPa, на основе которых могут быть созданы заявленные не принадлежащие к человеческому роду животные, или из которых могут быть созданы заявленные не принадлежащие к человеческому роду животные, хорошо известны в данной области техники, и более подробно обсуждаются, например, в заявке США №2013/0042330 и Rathinam et al. 2011, Blood 118:3119-28, раскрывающих knock-in мышей, которые экспрессируют человеческий M-CSF; в патенте США №8,541,646 и Willinger et al. 2011, Proc Natl Acad Sci USA, 108:2390-2395, раскрывающих knock-in мышей, которые экспрессируют человеческий IL-3 и человеческий GM-CSF; в патенте США №8,541,646 и Rongvaux et al. 2011, Proc Natl Acad Sci USA,108:2378-83, раскрывающих knock-in мышей, которые экспрессируют человеческий ТРО; и РСТ заявке № WO 2012/040207 и Strowig et al. 2011, Proc Natl Acad Sci USA 108(32): 13218-13223, раскрывающих трансгенных мышей, которые экспрессируют человеческий Sirpa; полные раскрытия которых включаются в данное описание путем отсылки.
В различных вариантах осуществления нуклеиновая кислота, кодирующая человеческий белок, является функционально связанной с одной или более регуляторными последовательностями таким образом, который обеспечивает транскрипцию нуклеиновой кислоты в мРНК и трансляцию мРНК в человеческий белок. Термин "регуляторная последовательность" является принятым в данной области и включает промоторы, энхансеры и другие элементы, контролирующие экспрессию (например, сигналы полиаденилирования). Подобные регуляторные последовательности известны специалистам в данной области и описаны в 1990, Goeddel, Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, Calif. В одном варианте осуществления человеческая нуклеиновая кислота экспрессируется нативными регуляторными элементами человеческой нуклеиновой кислоты. В другом варианте осуществления человеческая нуклеиновая кислота экспрессируется нативными регуляторными элементами соответствующей нуклеиновой кислоты не принадлежащего к человеческому роду животного-хозяина.
Таким образом, в некоторых вариантах осуществления нуклеиновая кислота, кодирующая человеческий IL-6, является функционально связанной с промотором IL-6 не принадлежащего к человеческому роду животного. В других вариантах осуществления нуклеиновая кислота, кодирующая человеческий IL-6, является функционально связанной с человеческим IL-6 промотором. В качестве другого примера, в некоторых вариантах осуществления нуклеиновая кислота, кодирующая человеческий M-CSF, является функционально связанной с промотором M-CSF животного. В других вариантах осуществления нуклеиновая кислота, кодирующая человеческий M-CSF, является функционально связанной с человеческим M-CSF промотором. В качестве третьего примера, в некоторых вариантах осуществления нуклеиновая кислота, кодирующая человеческий IL-3, является функционально связанной с IL-3 промотором животного. В других вариантах осуществления нуклеиновая кислота, кодирующая человеческий IL-3, является функционально связанной с человеческим IL-3 промотором. В качестве четвертого примера, в некоторых вариантах осуществления нуклеиновая кислота, кодирующая человеческий GM-CSF, является функционально связанной с GM-CSF промотором животного. В других вариантах осуществления нуклеиновая кислота, кодирующая человеческий GM-CSF, является функционально связанной с человеческим GM-CSF промотором. В качестве пятого примера, в некоторых вариантах осуществления нуклеиновая кислота, кодирующая человеческий ТРО, является функционально связанной с промотором ТРО животного. В других вариантах осуществления нуклеиновая кислота, кодирующая человеческий ТРО, является функционально связанной с человеческим ТРО промотором.
Специалисту ясно, что генетически модифицированные животные изобретения включают генетически модифицированных животных, которые экспрессируют, по меньшей мере, одну человеческую нуклеиновую кислоту за счет промотора. Неограничивающие примеры повсеместно экспрессируемых промоторов, пригодных в изобретении, включают, но не ограничиваются этим, ДНК pol II промотор, PGK промотор, убиквитиновый промотор, промотор альбумина, промотор глобина, промотор овальбумина, ранний промотор SV40, промотор вируса саркомы Рауса (RSV), ретровирусный LTR и лентивирусный LTR, промотор бета-актина, ROSA26 промотор, промотор белка теплового шока 70 (Hsp70), промотор EF-1 альфа гена, кодирующего фактор элонгации 1 альфа (EF1), промотор фактора инициации эукариот 4A (eEF-4Al), промотор хлорамфеникол-ацетилтрансферазы (CAT) и промотор CMV (цитомегаловируса). Системы экспрессии промотора и энхансера, пригодные в изобретении, также включают индуцибельную и/или тканеспецифическую системы экспрессии. Неограничивающие примеры тканеспецифических промоторов, пригодных в экспрессирующих конструкциях композиций и способов изобретения, включают промотор гена, экспрессированного в гематопоэтической системе, такой как IL-6 промотор, M-CSF промотор, IL-3 промотор, GM-CSF промотор, SIRPA промотор, ТРО промотор, IFN-β промотор, промотор белка синдрома Вискотта-Олдрича (WASP), CD45 (также называемый общим лейкоцитарным антигеном) промотор, Flt-1 промотор, промотор эндоглина (CD105) и промотор ICAM-2 (молекула 2 внутриклеточной адгезии). Эти и другие промоторы, пригодные в данных композициях и способах изобретения, известны в данной области техники, примеры которых приведены в Abboud et al. (2003, J. Histochem & Cytochem. 51:941-949), Schorpp et al. (1996, NAR 24:1787-1788), McBurney et al. (1994, Devel. Dynamics, 200:278-293) и Majumder et al. (1996, Blood 87:3203-3211). В дополнение к промотору в различных вариантах осуществления включаются один или более дополнительных регуляторных элементов, таких как энхансер или интрон. Примеры энхансеров, подходящих для применения в композициях и способах изобретения, включают, но не ограничиваются этим, элемент раннего энхансера цитомегаловируса (CMV) и элемент энхансера SV40. Примеры интронных последовательностей, подходящих для применения в композициях и способах изобретения, включают, но не ограничиваются этим, интрон бета-глобина или обобщенный (generic) интрон. Другие дополнительные регуляторные элементы, подходящие для некоторых вариантов осуществления изобретения, включают, но не ограничиваются этим, последовательность терминации транскрипции и последовательность мРНК полиаденилирования (рА).
Кроме того, специалисту ясно, что в дополнение к природным человеческим последовательностям нуклеиновых кислот и аминокислот, термины человеческая нуклеиновая кислота и человеческая аминокислота включают варианты последовательностей человеческих нуклеиновых кислот, а также аминокислот. Использованный в описании термин "вариант" имеет отношение или к выделенному природному генетическому мутанту человеческой последовательности или рекомбинантно полученному изменению человеческой последовательности, при этом в каждом содержится одна или более мутаций по сравнению с соответствующим человеческим диким типом. Например, подобные мутации могут представлять собой одну или более замен, вставок и/или делеций. Термин "вариант" также включает не принадлежащие к человеческому роду ортологи. В некоторых вариантах осуществления вариант полипептида настоящего изобретения имеет, по меньшей мере, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с человеческим полипептидом дикого типа.
Процент идентичности между двумя последовательностями определяют при помощи методов, описанных в другом месте данного документа. Мутации могут быть введены с помощью стандартных методов молекулярной биологии, таких как сайт-направленный мутагенез и ПЦР-опосредованным мутагенез. Специалисту в данной области понятно, что могут вводиться одна или более мутаций аминокислот без изменения функциональных свойств человеческих белков.
Для получения вариантов человеческого белка в человеческих белках могут быть выполнены консервативные замены аминокислот. Консервативные замены аминокислот являются принятыми в данной области техники заменами одной аминокислоты на другую аминокислоту, имеющую сходные характеристики. Например, каждая аминокислота может описываться, как имеющая одну или более из следующих характеристик: электроположительная, электроотрицательная, алифатическая, ароматическая, полярная, гидрофобная и гидрофильная. Консервативная замена - это замена одной аминокислоты, имеющей определенную структурную или функциональную характеристику, на другую аминокислоту, имеющую такую же характеристику. Кислые аминокислоты включают аспартат, глутамат; основные аминокислоты включают гистидин, лизин, аргинин; алифатические аминокислоты включают изолейцин, лейцин и валин; ароматические аминокислоты включают фенилаланин, глицин, тирозин и триптофан; полярные аминокислоты включают аспартат, глутамат, гистидин, лизин, аспарагин, глутамин, аргинин, серии, треонин и тирозин; и гидрофобные аминокислоты включают аланин, цистеин, фенилаланин, глицин, изолейцин, лейцин, метионин, пролин, валин и триптофан; и консервативные замены включают замену среди аминокислот в пределах каждой группы. Аминокислоты также могут быть описаны в зависимости от размера: аланин, цистеин, аспартат, глицин, аспарагин, пролин, треонин, серии, валин, как правило, считаются небольшими.
Человеческие варианты могут включать синтетические аналоги аминокислот, производные аминокислот и/или нестандартные аминокислоты, для иллюстрации включая, без ограничения, альфа-аминомасляную кислоту, цитруллин, канаванин, цианоаланин, диаминомасляную кислоту, диаминопимелиновую кислоту, дигидроксифенилаланин, дьенколовую кислоту, гомоаргинин, гидроксипролин, норлейцин, норвалин, 3-фосфосерин, гомосерин, 5-гидрокситриптофан, 1-метилгистидин, метилгистидин и орнитин.
Человеческие варианты кодируются нуклеиновыми кислотами, имеющими высокую степень идентичности с нуклеиновой кислоты, кодирующей человеческий дикий тип. Нуклеиновая кислота, кодирующая человеческий вариант, специфически гибридизируется с нуклеиновой кислотой, кодирующей человеческий дикий тип в условиях высокой жесткости. Нуклеиновые кислоты, кодирующие человеческий вариант, могут быть выделены или получены рекомбинантным путем или путем синтеза при использовании общеизвестных методов.
В некоторых вариантах осуществления генетически модифицированное не принадлежащее к человеческому роду животное, которое экспрессирует последовательность человеческой нуклеиновой кислоты, также экспрессирует соответствующую последовательность нуклеиновой кислоты животного, не являющегося человеком. Например, и как более подробно описано далее, в некоторых вариантах осуществления последовательность человеческой нуклеиновой кислоты случайным образом интегрируется в геном не принадлежащего к человеческому роду животного, например, таким образом, что животное содержит экзогенную человеческую нуклеиновокислотную последовательность, кодирующую соответствующий белок не принадлежащего к человеческому роду животного в локусе, отличном от локуса не принадлежащего к человеческому роду животного. В других вариантах осуществления генетически модифицированное не принадлежащее к человеческому роду животное, которое экспрессирует человеческую нуклеиновокислотную последовательность, не экспрессирует соответствующую нуклеиновокислотную последовательность не принадлежащего к человеческому роду животного. Например, и как более подробно описано далее, в некоторых вариантах осуществления животному вводится нуклеиновая кислота, кодирующая человеческий белок, с целью заменить геномный материал, кодирующий соответствующий белок животного, производя аннулирование соответствующего гена у животного, не являющегося человеком, и делая животное дефицитным относительно соответствующего белка данного животного. Другими словами, не принадлежащее к человеческому роду животное является "knock-in" в отношении человеческого гена.
Таким образом, в некоторых вариантах осуществления генетически модифицированное не принадлежащее к человеческому роду животное, которое экспрессирует человеческий IL-6, также экспрессирует нечеловеческий IL-6 животного. В других вариантах осуществления генетически модифицированное, не принадлежащее к человеческому роду животное, которое экспрессирует человеческий IL-6, не экспрессирует нечеловеческий IL-6 животного. В качестве второго примера, в некоторых вариантах осуществления генетически модифицированное не принадлежащее к человеческому роду животное, которое экспрессирует человеческий M-CSF, также экспрессирует нечеловеческий M-CSF животного. В других вариантах осуществления генетически модифицированное не принадлежащее к человеческому роду животное, которое экспрессирует человеческий MCSF, не экспрессирует нечеловеческий M-CSF животного. В качестве третьего примера, в некоторых вариантах осуществления генетически модифицированное не принадлежащее к человеческому роду животное, которое экспрессирует человеческий IL-3, также экспрессирует нечеловеческий IL-3 животного. В других вариантах осуществления генетически модифицированное не принадлежащее к человеческому роду животное, которое экспрессирует человеческий IL-3, не экспрессирует нечеловеческий IL-3 животного. В качестве четвертого примера, в некоторых вариантах осуществления генетически модифицированное не принадлежащее к человеческому роду животное, которое экспрессирует человеческий GM-CSF, также экспрессирует нечеловеческий GM-CSF животного. В других вариантах осуществления генетически модифицированное не принадлежащее к человеческому роду животное, которое экспрессирует человеческий GM-CSF, не экспрессирует нечеловеческий GM-CSF животного. В качестве примера, в некоторых вариантах осуществления генетически модифицированное не принадлежащее к человеческому роду животное, которое экспрессирует человеческий ТРО, также экспрессирует нечеловеческий ТРО животного. В других вариантах осуществления генетически модифицированное не принадлежащее к человеческому роду животное, которое экспрессирует человеческий ТРО, не экспрессирует нечеловеческий ТРО животного.
В некоторых вариантах осуществления заявленное генетически модифицированное животное является иммунодефицитным животным. Под "иммунодефицитным" подразумевается, что не принадлежащее к человеческому роду животное является дефицитным в отношении одного или более аспектов его врожденной иммунной системы, например, животное является дефицитным в отношении одного или более типов функционирования иммунных клеток «хозяина», например, дефицитным в отношении количества и/или функции нечеловеческих В-клеток, количества и/или функции нечеловеческих Т-клеток, количества и/или функции нечеловеческих NK-клеток и т.д.
В качестве примера, у иммунодефицитного животного может присутствовать тяжелый комбинированный иммунодефицит (SCID). SCID относится к состоянию, которое характеризуется отсутствием Т- клеток и утратой функции В-клеток. Примеры SCID включают: Х-связанный SCID, который характеризуется мутациями гамма цепи гена или потерей IL2RG гена и лимфоцитарным фенотипом Т(-) В(+) NK(-); и аутосомно-рецессивный SCID, характеризующийся мутациями гена Jak3 и лимфоцитарным фенотипом Т(-) В(+) NK(-), мутациями ADA гена и лимфоцитарным фенотипом Т(-) В(-) NK(-), мутациями альфа-цепи IL-7R и лимфоцитарным фенотипом Т(-) В(+) NK(+), дельта или эпсилон мутациями CD3 и лимфоцитарным фенотипом Т(-) В(+) NK(+), RAG1/PvAG2 мутациями и лимфоцитарным фенотипом Т(-) В(-) NK(+), мутациями гена Artemis и лимфоцитарным фенотипом Т(-) В(-) NK(+), мутациями гена CD45 и лимфоцитарным фенотипом Т(-) В(+) NK(+), и мутациями PrkdcS0ld (Bosma et al. (1989, Immunogenetics 29:54-56) и лимфоцитарным фенотипом Т(-), В(-), лимфопенией и гипоглобулинемией. Соответственно, в некоторых вариантах осуществления генетически модифицированное, иммунодефицитное не принадлежащее к человеческому роду животное имеет одно или более дефицитных состояний, выбранных из числа дефицита гамма цепи рецептора IL2, мутации гена ADA, мутации IL7R, мутации CD3, мутации RAG1 и/или RAG2, мутации Artemis, мутации CD45 и мутации Prkdc.
Заявленное генетически модифицированное, не принадлежащее к человеческому роду животное может быть любым млекопитающим, не относящимся к человеческому роду, например, лабораторным животным, домашним животным, сельскохозяйственным животным и т.д., которое является генетически модифицированным таким образом, что содержит последовательность, кодирующую человеческий IL-6, функционально связанную с IL-6 промотором, например, такого вида животного, как представители семейств мышиных, грызунов, собачьих, кошачьих, свиньих, лошадиных, жвачных животных, приматов, кроме человека, и т.д.; например, мыши, крысы, кролики, хомячки, морские свинки, крупный рогатый скот, свиньи, овцы, козы и другие виды трансгенных животных, в частности, виды млекопитающих, известные в данной области техники. В определенных вариантах осуществления заявленное генетически модифицированное животное является мышью, крысой или кроликом.
В одном варианте осуществления не принадлежащее к человеческому роду животное является млекопитающим. В некоторых подобных вариантах осуществления не принадлежащее к человеческому роду животное является мелким млекопитающим, например, относится к суперсемейству Dipodoidea или Muroidea. В одном варианте осуществления генетически модифицированное животное является грызуном. В одном варианте осуществления грызуна выбирают из числа мыши, крысы и хомяка. В одном варианте осуществления грызуна выбирают из суперсемейства Muroidea. В одном варианте осуществления генетически модифицированное животное является животным из семейства, выбранного из Calomyscidae (например, мышевидные хомячки), Cricetidae (например, хомяки, крысы и мыши Нового света и полевки), Muridae (мышиные и крысы, карликовые песчанки, колючие хомячки, косматые хомяки), Nesomyidae (рипидомисы, скалистые хомячки, with-tailed rats, Мадагаскарские крысы и мыши), Platacanthomyidae (например, колючие соневидные хомяки) и Spalacidae (например, слепыши, бамбуковые крысы и цокоры). В отдельном варианте осуществления генетически модифицированного грызуна выбирают из мышиных или крысиных (семейство Muridae), песчанок, колючего хомячка и косматого хомяка. В одном варианте осуществления генетически модифицированная мышь является животным-членом семейства Muridae.
В одном варианте осуществления заявленное генетически модифицированное не принадлежащее к человеческому роду животное является крысой. В одном подобном варианте осуществления крысу выбирают из числа крысы Wistar, линии LEA, линии Sprague Dawley, линии Fischer, F344, F6 и Dark Agouti. В другом варианте осуществления крысиная линия является результатом смешения двух или более линий, выбранных из группы, состоящей из линий Wistar, LEA, Sprague Dawley, Fischer, F344, F6 и Dark Agouti.
В другом варианте осуществления заявленное генетически модифицированное животное является мышью, например, мышью линии C57BL (например, C57BL/A, C57BL/An, C57BL/GrFa, C57BL/KaLwN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr, C57BL/01a и т.д.); мышью линии 129 (например, 129Р1, 129Р2, 129Р3,129Х1, 129S1 (например, 129S1/SV, 129S 1/SvIm), 129S2, 129S4, 129S5, 129S9/SvEvH, 129S6 (129/SvEvTac), 129S7, 129S8, 129T1, 129T2); мышью линии BALB; например, BALB/c; и тому подобным. Смотри, например, Festing et al. (1999) Mammalian Genome 10:836, смотри также Auerbach et al (2000) Establishment и Chimera Analysis of 129/SvEv- и C57BL/6-Derived Mouse Embryonic Stem Cell Lines). В специальном варианте осуществления генетически модифицированная мышь является результатом смешения вышеупомянутой линии 129 и вышеупомянутой линии C57BL/6. В другом отдельном варианте осуществления мышь является результатом смешения вышеупомянутой линии 129 и вышеупомянутой BL/6 линии. В отдельном варианте осуществления линия 129 является линией 129S6 (129/SvEvTac). В еще одном варианте осуществления мышь является результатом смешения линии BALB и другой вышеупомянутой линии.
Таким образом, например, в некоторых вариантах осуществления заявленное генетически модифицированное, не принадлежащее к человеческому роду животное представляет собой иммунодефицитную мышь с дефицитом количества и/или функции В-клеток, и/или количества и/или функции Т-клеток, и/или количества и/или функции NK клеток (например, вследствие дефицита гамма цепи рецептора IL2 (т.е., γc-/-) и/или дефицита RAG), имеющую геном, который содержит человеческую нуклеиновую кислоту, например, нуклеиновую кислоту, кодирующую человеческий IL-6, hM-CSF, hlL-3, hGM-CSF, hTPO и/или hSIRPa, функционально связанный с соответствующим промотором, например, M-CSF, IL-3, GM-CSF, ТРО или SIRPa промотором, соответственно, при этом животное экспрессирует кодированный человеческий белок(и).
В некоторых отдельных вариантах осуществления заявленное генетически модифицированное животное является иммунодефицитной мышью, содержащей нуклеиновую кислоту, кодирующую человеческий IL-6, функционально связанный с промотором IL-6 в локусе IL-6 мыши, и нуклеиновую кислоту, кодирующую человеческий SIRPa, функционально связанный с человеческим SIRPa промотором, случайно интегрированным в геном не принадлежащего к человеческому роду животного (т.е., мышь экспрессирует мышиный SIRPa), т.е. иммунодефицитной hIL-6, hSirpa мышью, например, Rag2-/-IL2rg-/- IL-6h/+ hSIRPa+ мышью или Rag2-/-IL2rg-/- IL-6h/h hSIRPa+ мышью. В некоторых подобных вариантах осуществления мышь дополнительно содержит нуклеиновую кислоту, кодирующую человеческий M-CSF, функционально связанный с M-CSF промотором, нуклеиновую кислоту, кодирующую человеческий IL-3, функционально связанный с IL-3 промотором, нуклеиновую кислоту, кодирующую человеческий GM-CSF, функционально связанный с GM-CSF промотором, и нуклеиновую кислоту, кодирующую человеческий ТРО, функционально связанный с ТРО промотором, т.е. иммунодефицитная hIL-6, hSirpa, hM-CSF, hIL-3, hGMCSF, hTPO мышь, например, Rag2-/-IL2rg-/- IL-6h/+M-CSFh/+ IL-3h/+ GM-CSFh/+ TPOh/+ hSIRPa+ мышь, Rag2-/-ILrg-/- IL-6h/+M-CSFh/hIL-3h/hGM-CSFh/h TPOh/h hSIRPa+ мышь.
В некоторых отдельных вариантах осуществления заявленное генетически модифицированное животное является иммунодефицитной мышью, содержащей нуклеиновую кислоту, кодирующую человеческий IL-6, функционально связанный с IL-6 промотором, и дефицитной относительно мышиного IL-6; нуклеиновую кислоту, кодирующую человеческий SERPa, функционально связанный с человеческим SIRPa промотором, случайно интегрированным в геном животного, не являющегося человеком (т.е., мышь еще экспрессирует мышиный SIRPa); нуклеиновую кислоту, кодирующую человеческий M-CSF, функционально связанный с M-CSF промотором, и дефицитной относительно мышиного M-CSF; нуклеиновую кислоту, кодирующую человеческий IL-3, функционально связанный с IL-3 промотором, и дефицитной относительно мышиного IL-3; нуклеиновую кислоту, кодирующую человеческий GM-CSF, функционально связанный с GM-CSF промотором, и дефицитной относительно мышиного GM-CSF; и нуклеиновую кислоту, кодирующую человеческий ТРО, функционально связанный с ТРО промотором, и дефицитной относительно мышиного ТРО, т.е. Rag2-/-IL2rg-/- IL-6h/hM-CSFh/hIL-3h/h GM-CSFh/h TPOh/h hSIRPa+ мышью.
Способы получения генетически модифицированных, не принадлежащих к человеческому роду животных.
Заявленные генетически модифицированные, не принадлежащие к человеческому роду животные могут быть получены с помощью любого подходящего метода получения генетически модифицированных животных, например, известного в данной области техники или описанного в данном документе.
Например, нуклеиновая кислота, кодирующая представляющий интерес человеческий белок, например, IL-6, hM-CSF, hIL-3, hGM-CSF, hTPO или hSIRPa, может быть встроена в рекомбинантный вектор в форме, подходящей для вставки в геном клетки-хозяина и экспрессии человеческого белка в нечеловеческой клетке-хозяине. В различных вариантах осуществления рекомбинантный вектор включает одну или более регуляторную последовательность, функционально связанную с нуклеиновой кислотой, кодирующей человеческий белок, таким образом, который обеспечивает транскрипцию нуклеиновой кислоты в мРНК и трансляцию мРНК в человеческий белок, как описано выше. Следует понимать, что конструкция вектора зависит от таких факторов, как выбор клетки-хозяина, предназначенной для трансфекции, количество человеческого белка, которое должно экспрессироваться, и/или того, каким образом кодированная нуклеиновая кислота будет интегрироваться в геном хозяина, не являющегося человеком, например, как известно в данной области техники.
Далее, для введения последовательности человеческой нуклеиновой кислоты в клетку животного для получения генетически модифицированного животного, экспрессирующего человеческий ген, может использоваться любой из целого ряда методов. Такие методы хорошо известны в данной области техники и включают, но не ограничиваются этим, пронуклеарные микроинъекции ооцитов, трансформацию эмбриональных стволовых клеток, гомологичную рекомбинацию и knock-in методы. Подходящие для использования способы получения генетически модифицированных животных включают, но не ограничиваются этим, способы, описанные в Sundberg and Ichiki (2006, Genetically Engineered Mice Handbook, CRC Press), Hofker и van Deursen (2002, Genetically modified Mouse Methods и Protocols, Humana Press), Joyner (2000, Gene Targeting: A Practical Approach, Oxford University Press), Turksen (2002, Embryonic stem cells: Methods и Protocols in Methods Mol Biol., Humana Press), Meyer et al.(2010, Proc. Nat. Acad. Sci. USA 107:15022-15026), и Gibson (2004, A Primer Of Genome Science 2nd ed. Sunderland, Massachusetts: Sinauer), патент США №6,586,251, Rathinam et al. (2011, Blood 118:3119-28), Willinger et al, (2011, Proc Natl Acad Sci USA, 108:2390-2395), Rongvaux et al., (2011, Proc Natl Acad Sci USA, 108:2378-83) и Valenzuela et al. (2003, Nat Biot 21:652-659).
Например, заявленные генетически модифицированные животные могут быть созданы путем введения нуклеиновой кислоты, кодирующей человеческий белок, в ооцит, например, с помощью микроинъекции, и обеспечения развития ооцита у патронатного животного женского рода. В предпочтительных вариантах осуществления конструкция, содержащая человеческую нуклеиновокислотную последовательность, вводится в оплодотворенные ооциты. Оплодотворенные ооциты могут быть собраны у суперовулированных особей женского пола на следующий день после спаривания с последующим введением экспрессирующей конструкции в них путем инъекции. Инъецируемые ооциты либо культивируют в течение ночи, либо непосредственно переносят в маточные трубы 0.5-дневных р.с. псевдобеременных самок. Способы осуществления суперовуляции, сбора ооцитов, введения экспрессирующей конструкции и переноса эмбриона известны в данной области техники и описаны в Manipulating the Mouse Embryo (2002, A Laboratory Manual, 3rd edition, Cold Spring Harbor Laboratory Press). Потомство может быть оценено относительно присутствия введенной нуклеиновой кислоты путем анализа ДНК (например, ПЦР, саузерн-блоттинга, секвенирования ДНК и т.д.) или путем анализа белка (например, ELISA, вестерн-блоттинга и т.д.). Как правило, подобные способы приводят в результате к случайной интеграции введенной нуклеиновокислотной последовательности - в этом случае, конструкции, содержащей нуклеиновую кислоту, кодирующую представляющий интерес человеческий белок, - в геном ооцита и, следовательно, животному, не принадлежащему к человеческому роду, т.е. в локус, отличный от локуса животного-хозяина, экспрессирующего соответствующий белок.
В качестве другого примера, конструкция, содержащая нуклеиновую кислоту, кодирующую человеческий белок, может быть трансфицирована в стволовые клетки (ES клетки или iPS клетки) с помощью общеизвестных методов, таких как электропорация, осаждение фосфатом кальция, липофекция и т.д. Клетки могут быть оценены в отношении присутствия введенной нуклеиновой кислоты с помощью анализа ДНК (например, ПЦР, саузерн-блоттинга, секвенирования ДНК и т.д.) или путем анализа белка (например, ELISA, вестерн-блоттинга и т.д.). Затем, клетки, содержащие введенную экспрессирующую конструкцию, могут быть введены в предимплантационные эмбрионы. Подробное описание методов, известных в данной области техники и подходящих для применения в композициях и способах изобретения, смотри в Nagy et al., (2002, Manipulating the Mouse Embryo: A Laboratory Manual, 3rd edition, Cold Spring Harbor Laboratory Press), Nagy et al. (1990, Development 110:815-821), патент США №7,576,259, патент США №7,659,442, патент США №7,294,754 и Kraus et al. (2010, Genesis 48:394-399). Такие методы, как правило, используются в случае адресного внедрения трансфицированной нуклеиновокислотной последовательности - в этом случае, конструкции, содержащей нуклеиновую кислоту, кодирующую представляющий интерес человеческий белок, - в геном стволовых клеток и, следовательно, животному, не являющемуся человеком. Во многих случаях, подобные методы приводят к замене геномного материала хозяина, например, геномного материала, кодирующего соответствующий белок хозяина, нуклеиновой кислотой, кодирующей представляющий интерес человеческий белок.
Генетически модифицированные животные-основатели могут использоваться для выведения новых животных, несущих генетическую модификацию. Генетически модифицированные животные, несущие нуклеиновую кислоту, кодирующую человеческий белок(и) настоящего раскрытия может быть дополнительно скрещено с другими генетически модифицированными животными, несущими другие генетические модификации, или может быть скрещено с нокаутными животными, например, нокаутным животным, не экспрессирующим один или более из его генов.
В некоторых вариантах осуществления генетически модифицированные иммунодефицитные животные содержат геном, который включает нуклеиновую кислоту, кодирующую человеческий полипептид, функционально связанный с промотором, при этом животное экспрессирует кодированный человеческий полипептид. В различных вариантах осуществления генетически модифицированные, иммунодефицитные, не принадлежащие к человеческому роду животные содержат геном, включающий экспрессионную кассету, которая содержит нуклеиновую кислоту, кодирующую, по меньшей мере, один человеческий полипептид, при этом нуклеиновая кислота является функционально связанной с промотором и сигналом полиаденилирования и дополнительно содержит интрон, и при этом животное экспрессирует кодированный человеческий полипептид.
Как обсуждалось выше, в некоторых вариантах осуществления заявленное генетически модифицированное животное является иммунодефицитным животным. Генетически модифицированные, не принадлежащие к человеческому роду животные, которые являются иммунодефицитными и содержат один или более человеческих цитокинов, например, IL-6, M-CSF, IL-3, GMCSF, ТРО и/или SIRPa, могут быть получены аналогичным образом с помощью любого подходящего способа получения генетически модифицированных животных, известных в данной области техники или описанных в данном документе, например, путем введения экспрессирующей конструкции в предимплантационный эмбрион или путем использования стволовых клеток, таких как эмбриональные стволовые (ES) клетки или индуцированные плюрипотентные стволовые (iPS) клетки, например, содержащие мутантным аллель гена SOD, который, будучи гомозиготным, будет приводить к иммунодефициту, например, как более подробно описано выше и в рабочих примерах в описании. Затем получают мышей с модифицированным ооцитом или ES клетками, используя, например, методы, описанные в данном документе и известные в данной области техники и адаптированные для получения иммунодефицитных мышей, содержащих желательную генетическую модификацию. В качестве другого примера, генетически модифицированные не принадлежащие к человеческому роду животные могут быть получены на основе неиммунодефицитных мышей, скрещены с животным, содержащим мутантным аллель гена SCID, который, будучи гомозиготным, будет приводить к иммунодефициту, а затем потомство спаривают с целью получения иммунодефицитного животного, экспрессирующего, по меньшей мере, один представляющий интерес человеческий белок.
Различные варианты осуществления изобретения предоставляют генетически модифицированных животных, содержащих человеческую нуклеиновую кислоту практически во всех своих клетках, а также генетически модифицированных животных, содержащих человеческую нуклеиновую кислоту в некоторых, но не во всех своих клетках. В некоторых случаях, например, в случае направленной рекомбинации, одна копия человеческой нуклеиновой кислоты будет интегрирована в геном генетически модифицированных животных. В других случаях, например, в случае случайной интеграции, несколько копий, соседних или отдаленных по отношению друг к другу, человеческой нуклеиновой кислоты могут быть интегрированы в геном генетически модифицированных животных.
Таким образом, в некоторых вариантах осуществления заявленное генетически модифицированное животное, не принадлежащее к человеческому роду, может быть иммунодефицитным животным, содержащим геном, включающий нуклеиновую кислоту, кодирующую человеческий полипептид, функционально связанный с соответствующим промотором животного, не относящегося к человеческому роду, при этом животное экспрессирует закодированный человеческий полипептид. Другими словами, заявленное генетически модифицированное, иммунодефицитное, не принадлежащее к человеческому роду животное содержит геном, включающий экспрессионную кассету, содержащую нуклеиновую кислоту, кодирующую, по меньшей мере, один человеческий полипептид, при этом нуклеиновая кислота является функционально связанной с соответствующим нечеловеческим промотором и сигналом полиаденилирования, и при этом животное экспрессирует закодированный человеческий полипептид.
Полезность
Генетически модифицированные не принадлежащие к человеческому роду животные, предоставленные в различных вариантах осуществления настоящего изобретения, находят множество применений, например, для использования в качестве моделей роста и дифференцировки гематопоэтических клеток, для оценки in vivo человеческого гематопоэза, для оценки in vivo раковых клеток, для исследования in vivo иммунного ответа, для оценки in vivo вакцин и режимов вакцинации, для использования при тестировании эффектов различных средств, модулирующих рост или выживаемость раковых клеток, для оценки in vivo лечения рака, для получения и сбора in vivo медиаторов иммунитета, таких как антитела, и для использования при тестировании средств, влияющих на функцию гематопоэтических и иммунных клеток.
С этой целью, в некоторых случаях, заявленному генетически модифицированному не принадлежащему к человеческому роду животному ("хозяину") трансплантируют («пересаживают»), по меньшей мере, одну человеческую гематопоэтическую клетку. В некоторых вариантах осуществления предоставляются способы получения животной модели для исследований человеческой гематопоэтической системы, включающие пересадку человеческих гематопоэтических клеток заявленному генетически модифицированному, не принадлежащему к человеческому роду животному ("хозяину"). В определенных вариантах осуществления предоставляются способы пересадки человеческих гематопоэтических клеток генетически модифицированному не принадлежащему к человеческому роду животному, раскрытому в данном документе.
В некоторых отдельных случаях заявленному, генетически модифицированному животному, не относящемуся к человеческому роду, «пересаживают», по меньшей мере, одну клетку множественной миеломы человека. В некоторых подобных вариантах осуществления предоставляются способы получения животной модели для исследования рака, включающие пересадку клеток множественной миеломы человека заявленному генетически модифицированному не принадлежащему к человеческому роду животному. В некоторых подобных вариантах осуществления изобретением является способ пересадки клеток множественной миеломы человека заявленному генетически модифицированному не принадлежащему к человеческому роду животному. Пересаженные клетки множественной миеломы человека, подходящие для использования в композициях и способах изобретения, включают любую клетку множественной миеломы человека.
Человеческие гематопоэтические клетки, подходящие для пересадки заявленным, генетически модифицированным, не принадлежащим к человеческому роду животным, включают любую подходящую человеческую гематопоэтическую клетку. Неограничивающие примеры человеческих гематопоэтических клеток, подходящих для изобретения, включают, но не ограничиваются этим, HSC, HSPC, клетки инициирующие лейкоз (LIC), и гематопоэтические клетки любой линии дифференцировки на любой стадии дифференцировки, включая терминально дифференцированные гематопоэтические клетки любой линии дифференцировки. В некоторых случаях человеческая гематопоэтическая клетка является первичной клеткой, при этом термины "первичные клетки", "первичные клеточные линии" и "первичные культуры" используются в описании взаимозаменяемым образом, чтобы включать полностью изолированные клетки, или клеточные культуры, которые были получены от субъекта и выращивались in vitro в течение ограниченного числа пассажей, т.е. делений, в культуре. Например, первичными культурами являются культуры, которые могут пассироваться 0 раз, 1 раз, 2 раза, 4 раза, 5 раз, 10 раз или 15 раз, но не достаточное количество раз, чтобы перейти через критическую стадию. В других вариантах осуществления человеческая гематопоэтическая клетка является клеткой клеточной линии, то есть, является клеткой из иммортализованной культуры, например, культуру пассировали примерно более 15 раз. В некоторых случаях гематопоэтические клетки, которые пересаживают, включают здоровые клетки. В других случаях гематопоэтические клетки, которые пересаживают, включают пораженные болезнью гематопоэтические клетки, например, раковые гематопоэтические клетки, например, злокачественные эффекторные В-клетки, т.е. клетки множественной миеломы. В некоторых случаях «пересаженные» гематопоэтические клетки включают и здоровые и пораженные болезнью клетки, например, здоровые В-клетки и злокачественные эффекторные В-клетки, здоровые Т-клетки и злокачественные эффекторные В-клетки.
Гематопоэтические клетки, т.е. первичные клетки, происходящие от них клеточные линии и т.д., могут происходить из любой ткани или местоположения у донора-человека, включая, но не ограничиваясь этим, костный мозг, периферическую кровь, печень, эмбриональную печень или пуповинную кровь. Такие гематопоэтические клетки могут быть получены от человека-донора, включая здоровых доноров, а также доноров с заболеванием, например, раковым заболеванием, включая лейкоз. Приживление гематопоэтических клеток у заявленного генетически модифицированного животного характеризуется наличием человеческих гематопоэтических клеток у животного с пересаженными клетками. В определенных вариантах осуществления приживление гематопоэтических клеток у заявленного генетически модифицированного животного характеризуется наличием дифференцированных человеческих гематопоэтических клеток у животного, которому были пересажены гематопоэтические клетки, при сравнении с соответствующими контрольными животными.
Способы выделения человеческих гематопоэтических клеток, введения человеческих гематопоэтических клеток животному-хозяину и оценки их «приживления» хорошо известны в данной области техники. Гематопоэтические клетки, включая любые нормальные и неопластические клетки или их комбинации, предназначенные для введения животному-хозяину, могут быть получены из любой ткани, содержащей гематопоэтические клетки, такой как, но без ограничения, пуповинная кровь, костный мозг, периферическая кровь, периферическая кровь, мобилизованная цитокинаими или химиотерапией, и эмбриональная печень. Типичные способы выделения человеческих гематопоэтических клеток, введения человеческих гематопоэтических клеток животному-хозяину и оценки «приживления» человеческих гематопоэтических клеток у животного-хозяина описаны в данном документе и в работах Pearson et al. (2008, Curr. Protoc. Immunol. 81:1-15), Ito et al. (2002, Blood 100:3175-3182), Traggiai et al. (2004, Science 304:104-107), Ishikawa et al. (2005, Blood 106:1565-1573), Shultz et al. (2005, J. Immunol. 174:6477-6489) and Holyoake et al. (1999, Exp Hematol. 27:1418-27).
В некоторых вариантах осуществления изобретения человеческие гематопоэтические клетки, включая нормальные и неопластические клетки, или их сочетания, выделяют из материала первоисточника для получения популяции клеток, обогащенной определенной популяцией гематопоэтических клеток (например, HSCs, HSPCs, LICs, CD34+, CD34-, линиеспецифический маркер, маркер раковой клетки и т.д.). Выделенные гематопоэтические клетки могут быть или могут не быть гомогенной популяцией. В одном варианте осуществления гематопоэтические клетки, подходящие для композиций и способов изобретения, обеднены клетками, имеющими определенный маркер. В другом варианте осуществления гематопоэтические клетки, подходящие для композиций и способов изобретения, обогащены путем отбора по маркеру. В некоторых вариантах осуществления гематопоэтические клетки, подходящие для композиций и способов изобретения, представляют собой популяцию клеток, в которой отобранные клетки составляют около 1-100% клеток, хотя в некоторых вариантах осуществления также может использоваться популяция клеток, в которой отобранные клетки составляют меньше, чем 1% от всех клеток. В одном варианте осуществления гематопоэтические клетки, подходящие для композиций и способов изобретения, обеднены клетками, имеющими определенный маркер, например, CD34. В другом варианте осуществления гематопоэтические клетки, подходящие для композиций и способов изобретения, обогащены путем отбора по маркеру, например, CD34. В некоторых вариантах осуществления гематопоэтические клетки, подходящие для композиций и способов изобретения, представляют собой популяцию клеток, в которой CD34+ клетки составляют около 1-100% клеток, хотя в некоторых вариантах осуществления также может использоваться популяция клеток, в которой CD34+ клетки составляют меньше, чем 1% от всех клеток. В некоторых вариантах осуществления гематопоэтические клетки, подходящие для композиций и способов изобретения, представляют собой популяцию клеток, обедненную Т-клетками, в которой CD34+ клетки составляют около 1-3% всех клеток, линиеспецифически-истощенную популяцию клеток, в которой CD34+ клетки составляют около 50% всех клеток, или отобранную по CD34+ популяцию клеток, в которой CD34+ клетки составляют около 90% всех клеток.
Количество предназначенных для введения гематопоэтических клеток не считается ограничивающим в отношении возникновения (порождения) человеческой гематопоэтической и/или иммунной системы у генетически модифицированного не являющегося человеком животного, экспрессирующего, по меньшей мере, один человеческий ген. Таким образом, в качестве неограничивающего примера, количество вводимых гематопоэтических клеток может составлять в пределах примерно от 1×103 до 1×107, хотя в различных вариантах осуществления также может использоваться большее или меньшее количество. В качестве другого неограничивающего примера, количество вводимых HSPC может составлять в пределах примерно от 3×103 до 1×106 CD34+ клеток, в том случае, когда реципиентом является мышь, хотя в различных вариантах осуществления также может использоваться большее или меньшее количество. Что касается реципиентов других видов, количество клеток, которое должно быть введено, может определяться только путем проведения обычных экспериментов.
Например, в одном варианте осуществления генетически модифицированной и обработанной мыши пересаживают человеческие гематопоэтические клетки или человеческие гематопоэтические стволовые клетки (HPSC), чтобы получить генетически модифицированную мышь с пересаженными клетками. В одном варианте осуществления гематопоэтические клетки выбирают из клеток пуповинной крови человека и клеток эмбриональной печени человека. В одном варианте осуществления осуществляют «пересадку» примерно 1-2×105 человеческих CD34+ клеток.
В некоторых случаях введению гематопоэтических клеток (например, нормальных или неопластических) может предшествовать предварительная обработка, например, или сублетальное облучение животного-реципиента высокочастотным электромагнитным облучением, как правило, с использованием гамма или рентгеновского облучения, или обработка радиомиметическим средством, таким как бусульфан или азотистый иприт. Считается, что предварительная обработка уменьшает количество гематопоэтических клеток «хозяина», создает соответствующие факторы микроокружения для приживления человеческих гематопоэтических клеток и/или создает ниши микроокружения для приживления человеческих гематопоэтических клеток. Стандартные методы предварительной обработки известны в данной области техники, например, описанные в данном документе и в работе J. Hayakawa et al, 2009, Stem Cells, 27(1):175-182. В одном варианте осуществления генетически модифицированную мышь обрабатывают так, чтобы ликвидировать эндогенные гематопоэтические клетки, которые могут находиться в мыши. В одном варианте осуществления обработка включает облучение генетически модифицированной мыши. В отдельном варианте осуществления новорожденных генетически модифицированных мышат облучают сублетальной дозой. В конкретном варианте осуществления новорожденных детенышей облучают 2×200 cGy (сантигрей) с 4-х часовым интервалом.
Гематопоэтические клетки (например, нормальные или неопластические) могут вводиться новорожденным или взрослым животным различными способами, такими как, но без ограничения, внутривенно, внутрь печени, внутрибрюшинно, внутрифеморально (внутрь бедренной кости) и/или внутритибиально (внутрь большеберцовой кости). Способы «пересадки» человеческих гематопоэтических клеток, включая и нормальные и неопластические клетки или их комбинации, иммунодефицитным животным предоставляются в соответствии с вариантами осуществления настоящего изобретения, которые включают предоставление человеческих гематопоэтических клеток иммунодефицитным животным без облучения или в сочетании с облучением животных до введения гематопоэтических клеток. Способы «пересадки» человеческих гематопоэтических клеток иммунодефицитным животным предоставляются в соответствии с вариантами осуществления настоящего изобретения, которые включают предоставление человеческих гематопоэтических клеток, включая нормальные и неопластические клетки или их комбинации, генетически модифицированным не относящимся к человеческому роду животным изобретения, с или без введения животным радиомиметического средства, такого как бусульфан или азотистый иприт, до введения гематопоэтических клеток.
«Приживление» человеческих гематопоэтических клеток, включая нормальные и неопластические клетки или их комбинации, генетически модифицированному животному изобретения может быть оценено с помощью различных методов, таких как, но без ограничения, проточный цитометрический анализ клеток у животных, которым вводятся человеческие гематопоэтические клетки, в одной или более временных точках после введения гематопоэтических клеток.
В общем, «приживление» может считаться успешным, когда количество (или процент) человеческих гематопоэтических клеток, включая нормальные и неопластические клетки или их комбинации, присутствующее у генетически модифицированного не относящегося к человеческому роду животного, больше, чем количество (или процент) человеческих клеток, которые были введены не относящемуся к человеческому роду животному, на момент времени, превышающий продолжительность жизни введенных человеческих гематопоэтических клеток. Обнаружение потомства введенных гематопоэтических клеток может быть достигнуто, например, путем обнаружения человеческой ДНК у животного-реципиента, или путем обнаружения интактных человеческих гематопоэтических клеток, например, обнаружения маркера человеческой клетки, такого как человеческий CD45, человеческий CD34 или sIL-6R, например. Последовательный перенос человеческих гематопоэтических клеток от первого реципиента второму реципиенту и «приживление» человеческих гематопоэтических клеток у второго реципиента является дополнительным необязательным тестом «приживления» у первоначального реципиента. «Приживление» может быть установлено с помощью проточной цитометрии в виде 0.05% или более человеческого CD45+ в крови, селезенке или костном мозге через 1-4 месяцев после введения человеческих гематопоэтических клеток. Для мобилизации стволовых клеток может использоваться цитокин (например, GM-CSF), например, как описано в Watanabe (1997, Bone Marrow Transplantation 19:1175-1181).
В одном варианте осуществления иммунодефицитное генетически модифицированное животное с пересаженными клетками дает начало (порождает) человеческой клетке, выбранной из CD34+ клетки, гематопоэтической стволовой клетки, гематопоэтической клетки, миелоидной клетки-предшественника, миелоидной клетки, дендритной клетки, моноцита, гранулоцита, нейтрофила, мастоцита, тимоцита, Т-клетки, В-клетки, тромбоцита и их комбинации. В одном варианте осуществления человеческая клетка присутствует через 4 месяца, 5 месяцев, 6 месяцев, 7 месяцев, 8 месяцев, 9 месяцев, 10 месяцев, 11 месяцев или 12 месяцев после пересадки.
В одном варианте осуществления иммунодефицитное генетически модифицированное животное с пересаженными клетками дает начало человеческой гемато-лимфоидной системе, которая включает человеческие гематопоэтические стволовые клетки и клетки-предшественники, человеческие миелоидные клетки-предшественники, человеческие миелоидные клетки, человеческие дендритные клетки, человеческие моноциты, человеческие гранулоциты, человеческие нейтрофилы, человеческие мастоциты, человеческие тимоциты, человеческие Т-клетки, человеческие В-клетки и человеческие тромбоциты. В одном варианте осуществления человеческая гемато-лимфоидная система присутствует через 4 месяца, 5 месяцев, 6 месяцев, 7 месяцев, 8 месяцев, 9 месяцев, 10 месяцев, 11 месяцев или 12 месяцев после пересадки.
В одном варианте осуществления иммунодефицитное генетически модифицированное животное с пересаженными клетками дает начало человеческой гемато-лимфоидной системе, которая включает злокачественные человеческие гематопоэтические клетки. Например, неопластические плазматические (эффекторные В) клетки. В одном варианте осуществления злокачественные человеческие гематопоэтические клетки присутствуют через 4 недели, 6 недель, 8 недель, 12 недель или более чем 12 недель после «пересадки». В некоторых вариантах осуществления злокачественные человеческие гематопоэтические клетки присутствуют через 2 недели, 4 недели, 6 недель, 8 недель, 12 недель или более, чем 12 недель после «пересадки».
Заявленные генетически модифицированные, не принадлежащие к человеческому роду животные с пересаженными человеческими гематопоэтическими клетками находят множество применений в данной области техники. Например, генетически модифицированные животные с пересаженными клетками настоящего раскрытия используются для изучения функции человеческих гематопоэтических клеток в периферической крови. Как продемонстрировано в рабочем примере 2, генетически модифицированные мыши, которые являются иммунодефицитными и содержат нуклеиновую кислоту, кодирующую человеческий IL-6, функционально связанный с IL-6 промотором в мышином локусе IL-6 (например, Rag2-/-IL2rgnullIL-6h/h мыши, Rag2-/-IL2rgnull IL-6h/h hSIRPa+ мыши и Rag2-/-IL2rg-/-IL-6h/h M-CSFh/h IL-3h/h GM-CSFh/h TPOh/h hSIRPa+) обеспечивают приживление человеческих гематопоэтических клеток, например, CD34+ клеток-предшественников, в периферической крови и селезенке лучше, чем иммунодефицитные мыши, которые не экспрессируют человеческий IL-6, т.е. Rag2-/-IL2rgnull мыши. Более того, эти генетически модифицированные мыши способствуют дифференцировке человеческих гематопоэтических клеток более эффективно, чем иммунодефицитные мыши, не экспрессирующие человеческий IL-6. Например, эти генетически модифицированные мыши лучше способствуют дифференцировке CD5+ В-клеток и CD27+ В-клеток. CD5 это белок, обнаруженный на подгруппе IgM-секретирующих В-клеток, названных В-1 клетками, который служит для уменьшения активирующих сигналов от В-клеточного рецептора, так что В-1 клетки могут активироваться только очень сильными стимулами (такими как бактериальные белки) и не активируются нормальными тканевыми белками. CD27 является маркером В-клеток памяти. Кроме того, эти генетически модифицированные мыши обеспечивают развитие лучше функционирующих человеческих гематопоэтических клеток, чем иммунодефицитные мыши, которые не экспрессируют человеческий IL-6. Например, В-клетки дифференцируют в IgG-секретирующие плазматические клетки быстрее у этих генетически модифицированных мышей, чем у иммунодефицитных мышей, не экспрессирующих человеческий IL-6. В связи с этим, генетически модифицированные животные настоящего раскрытия с пересаженными клетками используются при исследовании развития и функции гематопоэтических клеток, и в частности, дифференцировки и функции В-лимфоцитов.
В качестве другого примера, генетически модифицированные животные настоящего раскрытия с пересаженными клетками используются при изучении гематопоэтических видов рака. Как показано в рабочем примере 1 (ниже), генетически модифицированным мышам, которые являются иммунодефицитными и содержат нуклеиновую кислоту, кодирующую человеческий IL-6, функционально связанный с IL-6 промотором в мышином локусе IL-6, например, Rag2-/-IL2rgnullIL-6h/h мыши, Rag2-/-IL2rgnull IL-6h/h hSIRPa+ мыши и Rag2-/-IL2rg-/-IL-6h/h M-CSFh/h IL-3h/h GM-CSFh/h TPOh/h hSIRPa+, могут быть пересажены клетки первичной множественной миеломы человека и клетки клеточных линий множественной миеломы человека, в то время как иммунодефицитным мышам, не экспрессирующим человеческий IL-6, т.е. Rag2-/-IL2rgnull, нет.
Экспрессия человеческого SIRPa генетически модифицированным «хозяином» дополнительно улучшает скорость и степень наблюдаемого приживления. Более того, приживление клеток множественной миеломы непосредственно в кости этих иммунодефицитных генетически модифицированных животных, раскрытых в описании, воспроизводит патологию кости, как правило, связанную с множественной миеломой человека, например, разрушение и резорбцию кости, что количественно определяют например, с помощью μCΤ сканирования.
Собственно, генетически модифицированные животные настоящего раскрытия с пересаженными клетками находят применение при скрининге средств-кандидатов, для установления средств, которые способны лечить гематопоэтические злокачественные заболевания. Термины «лечение» и тому подобные используются в описании, чтобы включать получение желаемого фармакологического и/или физиологического эффекта. Эффект может быть профилактическим в контексте полного или частичного предотвращения болезни или ее симптома и/или может быть терапевтическим в контексте полного или частичного излечения болезни и/или неблагоприятного воздействия, свойственного болезни. «Лечение» при использовании в описании включает лечение болезни у млекопитающего и включает (а) предотвращение появления болезни у субъекта, который может быть предрасположен к данной болезни, но которая еще не была диагностирована; (b) ингибирование болезни, т.е. остановку ее развития; или (с) ослабление болезни, т.е. регрессию болезни. Средства-кандидаты, представляющие интерес как терапевтические средства для лечения гематологических злокачественных заболеваний, включают такие средства, которые могут вводиться до, во время или после появления ракового заболевания. Особый интерес представляет лечение продолжающейся болезни, когда лечение стабилизирует или уменьшает нежелательные клинические симптомы у пациента. Термины «индивидуум», «субъект», «хозяин» и «пациент» используются в описании взаимозаменяемым образом и включают любого субъекта-млекопитающего, для которого диагностика, лечение или терапия является желательным, в частности людей.
В качестве другого примера, генетически модифицированные животные с пересаженными клетками настоящего раскрытия используются для исследования патогенов человека, т.е. патогенов, которые инфицируют людей; ответа иммунной системы человека на инфекцию патогенами человека; и эффективности средств при защите от инфекции и/или при лечении инфекции патогенами человека. Патоген может быть вирусом, грибком, бактерией и т.д. Неограничивающие примеры вирусных патогенов включают вирус гриппа человека или вирус свиного или птичьего гриппа. Неограничивающие примеры бактериальных патогенов включают микобактерию, например, Mycobacterium tuberculosis (M.tuberculosis) и энтеробактерию, например, Salmonella typhi (S.typhi). Примеры способов инфицирования мышей S.typhi и оценки инфицирования могут быть найдены, например, в опубликованной заявке США №2011/0200982, раскрытие которой включено в данное описание путем отсылки. Примеры способов инфицирования мышей М.tuberculosis и оценки инфицирования могут быть найдены, например, в опубликованной заявке США №2011/0200982, раскрытие которой включается в описание путем отсылки. Другие примеры человеческих патогенов, которые не инфицируют мышей дикого типа или которые инфицируют мышей дикого типа, однако инфицированные мыши не являются моделью иммунного ответа, который наблюдается у человека в ответ на патоген, хорошо известны среднему специалисту в данной области. Такие мышиные модели инфекции патогеном используются в исследованиях, например, для лучшего понимания прогрессирования человеческой инфекции. Подобные мышиные модели инфекции также используются при поиске новых лекарственных средств, например, для установления средств-кандидатов, которые защищают или лечат от инфекции.
Генетически модифицированные мыши настоящего раскрытия с пересаженными клетками также предоставляют подходящую систему для скрининга средств-кандидатов в отношении ожидаемой активности in vivo, например, для установления средств, способных модулировать (т.е., способствовать или подавлять) развитие и/или активность гематопоэтических клеток, например, активность В-клеток, Т-клеток, NK-клеток, макрофагов, нейтрофилов, эозинофилов, базофилов и т.д., например, в состоянии здоровья или в состоянии болезни, например, злокачественных клеток, в условиях инфекции патогеном, например, для того, чтобы установить новые терапевтические средства и/или улучшить понимание молекулярной основы развития и функции иммунной системы; средств, которые являются токсичными для гематопоэтических клеток, например, В-клеток, Т-клеток, NK-клеток, макрофагов, нейтрофилов, эозинофилов, базофилов и т.д., и их предшественников; и средств, которые защищают от, облегчают или аннулируют токсические воздействия токсичных средств на гематопоэтические клетки, например, В-клетки, Т-клетки, NK-клетки, макрофаги, нейтрофилы, эозинофилы, базофилы и т.д., их предшественники; и т.д. В качестве еще одного примера, генетически модифицированные мыши настоящего раскрытия с пересаженными клетками предоставляют подходящую систему для предсказывания ответной реакции индивидуума на терапию болезни, например, обеспечивая основу in vivo для скрининга ответной реакции иммунной системы индивидуума на средство, например, терапевтическое средство, чтобы предсказать ответную реакцию индивидуума на это средство.
При проведении скрининга биологически активных средств, генетически модифицированная мышь настоящего раскрытия с пересаженными человеческими гематопоэтическими клетками, например, мышь Rag2-/-IL2rg-/-IL-6h/h hSIRPa+, мышь Rag2-/-IL2rg-/- IL-6h/h MCSFMh/h IL-3h/h GM-CSFh/h TPOh/h hSIRPa+ и т.д., контактирует с представляющим интерес средством-кандидатом, при этом действие средства-кандидата оценивается путем мониторинга одного или более «выходных» параметров. Эти «выходные» параметры могут отражать жизнеспособность клеток, например, общее количество гематопоэтических клеток или количество гематопоэтических клеток определенного типа, или апоптотическое состояние клеток, например, количество фрагментации ДНК, количество блеббинга (пузырения), количество фосфатидилсерина на клеточной поверхности, и т.п. с помощью методов, которые хорошо известны в данной области техники. Альтернативно или дополнительно, «выходные» параметры могут отражать способность клеток к дифференцировке, например, соотношения дифференцированных клеток и типов дифференцированных клеток. Альтернативно или дополнительно, «выходные» параметры могут отражать функцию клеток, например, цитокины и хемокины, вырабатываемые клетками, антитела (например, количество или тип) вырабатываемые клетками, способность клеток направляться и проникать в место заражения, способность клеток модулировать, т.е. способствовать или подавлять, активность других клеток in vitro или in vivo и т.д. Другие «выходные» параметры могут отражать степень повреждения, вызванную пораженными болезнью гематопоэтическими клетками, например, разрушение и резорбцию кости, вызванную клетками множественной миеломы. Другие параметры могут отражать действие средства на инфекцию, например, инфекцию патогеном у животного, например, титр патогена у мыши, наличие гранулемы у мыши, и т.д., в зависимости от того, что соответствует проводимым исследованиям.
Параметрами являются определяемые количественно компоненты клеток, в частности, компоненты, которые могут быть измерены с высокой точностью, желательно в системе с высокой пропускной способностью. Параметром может быть любой клеточный компонент или клеточный продукт, включая поверхностную детерминату клеток, рецептор, белок или его конформационную или посттрансляционную модификацию, липид, углевод, органическую или неорганическую молекулу, нуклеиновую кислоту, например, мРНК, ДНК и т.д. или фрагмент, происходящий от такого клеточного компонента или их комбинации. Несмотря на то, что большинство параметров будет предоставлять количественные данные, в некоторых случаях будут приемлемыми полуколичественные или качественные результаты. Данные могут включать одиночные определенные значения, или могут включать среднее значение, медиану или дисперсию и т.д. Обычно, получают целый ряд значений для каждого параметра в результате большого числа одинаковых анализов. Следует ожидать изменчивости (значений), при этом область значений для каждого набора тестируемых параметров получают с использованием стандартных статистических методов в сочетании с обычным статистическим методом, использованным для получения отдельных значений.
Соединения-кандидаты, представляющие интерес для скрининга, включают известные и неизвестные соединения, охватывающие многочисленные классы химических соединений, главным образом органические молекулы, которые включают металлорганические молекулы, неорганические молекулы, генетические последовательности, вакцины, антибиотики или другие средства, потенциально обладающие свойствами антибиотиков, пептиды, полипептиды, антитела, фармацевтические средства, одобренные для применения у человека и т.д. Важным аспектом изобретения является оценка средств-кандидатов, включая тестирование токсичности; и тому подобного.
Средства-кандидаты включают органические молекулы, содержащие функциональные группы, необходимые для структурных взаимодействий, в частности, образования водородной связи, и, как правило, включают, по меньшей мере, аминогруппу, карбонильную, гидроксильную или карбоксильную группу, нередко, по меньшей мере, две из функциональных химических групп. Средства-кандидаты часто содержат циклический углерод или гетероциклические структуры и/или ароматические или полиароматические структуры, замещенные одной или более из вышеуказанных функциональных групп. Средства-кандидаты также встречаются среди биомолекул, включая пептиды, полинуклеотиды, сахариды, жирные кислоты, стероиды, пурины, пиримидины, их производные, структурные аналоги или их комбинации. Включаются фармакологически активные средства, генетически активные молекулы и т.д. Представляющие интерес соединения включают химиотерапевтические средства, гормоны или аналоги гормонов и т.д. Характерные фармацевтические средства, подходящие для этого изобретения, описаны в "The Pharmacological Basis of Therapeutics," Goodman and Gilman, McGraw-Hill, New York, N.Y., (1996), Ninth edition. Также включаются токсины и биологические и химические отравляющие вещества, например, смотри Somani, S. M. (Ed.), "Chemical Warfare Agents," Academic Press, New York, 1992).
Представляющие интерес для скрининга средства-кандидаты также включают нуклеиновые кислоты, например, нуклеиновые кислоты, которые кодируют миРНК, кшРНК, антисмысловые молекулы или миРНК, или нуклеиновые кислоты, которые кодируют полипептиды. Имеется множество векторов, подходящих для переноса нуклеиновых кислот в клетки-мишени. Векторы могут сохраняться эписомально, например, как плазмиды, мини-кольцевые ДНК, векторы на основе вирусов, таких как цитомегаловирус, аденовирус и т.д., или они могут интегрироваться в геном клетки-мишени, в результате гомологичной рекомбинации или случайной интеграции, например, векторы на основе ретровирусов, таких как MMLV, HIV-1, ALV и т.д. Векторы могут непосредственно предоставляться предметным клеткам. Другими словами, плюрипотентные клетки контактируют с векторами, содержащими представляющую интерес нуклеиновую кислоту, в результате чего векторы поглощаются клетками.
Методы контактирования клеток, например, клеток в культуре или клеток в мыши, с нуклеиновокислотными векторами, такие как электропорация, трансфекция с использованием хлорида кальция и липофекция, хорошо известны в данной области техники. Альтернативно, представляющая интерес нуклеиновая кислота может предоставляться клеткам посредством вирусов. Другими словами, клетки приводят в контакт с вирусными частицами, содержащими нужную нуклеиновую кислоту. В частности, ретровирусы, например, лентивирусы, подходят для использования в способах изобретения. Обычно используемые ретровирусные векторы являются "дефектными", т.е. неспособными продуцировать вирусные белки, необходимые для продуктивной инфекции. Точнее, репликация вектора требует роста пакующей клеточной линии. Для того чтобы получить вирусные частицы, содержащие представляющие интерес нуклеиновые кислоты, ретровирусные нуклеиновые кислоты, содержащие нуклеиновую кислоту, упаковываются в вирусные календы. Разные пакующие клеточные линии обеспечивают разный белок оболочки, предназначенный для встраивания в капсид, этот белок оболочки обусловливает специфичность вирусной частицы для данных клеток. Белки оболочки бывают, по меньшей мере, трех типов, экотропные, амфотропные и ксенотропные. Ретровирусы, упакованные экотропным белком оболочки, например, MMLV, способны инфицировать большинство типов мышиных и крысиных клеток, их получают путем использования экотропных пакующих клеточных линий, таких как BOSC.23 (Pear et al. (1993) P.N.A.S. 90:8392-8396). Ретровирусы, несущие амфотропный белок оболочки, например, 4070А (Danos et al, выше), способны инфицировать большинство типов клеток млекопитающих, включая человеческие, собачьи и мышиные клетки, их получают путем использования амфотропных пакующих клеточных линий, таких как РА12 (Miller et al. (1985) Mol. Cell. Biol. 5:431-437); РА317 (Miller et al. (1986) Mol. Cell. Biol. 6:2895-2902); GRIP (Danos et al. (1988) PNAS 85:6460-6464). Ретровирусы, упакованные ксенотропным белком оболочки, например, AKR env, способны инфицировать большинство типов клеток млекопитающих, за исключением мышиных клеток. Соответствующие пакующие клеточные линии могут использоваться, чтобы гарантировать, что упакованные вирусные частицы нацеливаются на представляющие интерес клетки: в некоторых случаях на пересаженные клетки, в некоторых случаях на клетки хозяина, т.е. генетически модифицированного животного.
Векторы, используемые для предоставления интересующей нуклеиновой кислоты в нужные клетки, как правило, содержат подходящие промоторы для запуска экспрессии, то есть, активации транскрипции, представляющей интерес нуклеиновой кислоты. Промоторы включают повсеместно действующие промоторы, например, CMV-Р-актиновый промотор, или индуцибельные промоторы, такие как промоторы, которые являются активными в определенных клеточных популяциях или которые отвечают на присутствие лекарственных средств, таких как тетрациклин. Под активацией транскрипции подразумевается, что транскрипция будет увеличиваться выше базовых уровней в клетке-мишени, по меньшей мере, примерно в 10 раз, по меньшей мере, примерно в 100 раз, а еще точнее, по меньшей мере, примерно в 1000 раз. Кроме того, векторы, используемые для предоставления факторов перепрограммирования предметным клеткам, могут включать гены, которые позднее должны быть удалены, например, с использованием системы рекомбиназы, такой как Cre/Lox, или клетки, которые экспрессируют их, разрушают, например, путем включения генов, которые делают возможной селективную токсичность, таких как, ТК вируса герпеса, bcl-xs и т.д.
Представляющие интерес для скрининга средства-кандидаты также включают полипептиды. Такие полипептиды необязательно могут быть соединены с полипептидным доменом, увеличивающим растворимость продукта. Домен может быть связан с полипептидом через определенный сайт расщепления протеазой, например, TEV последовательность, которая расщепляется TEV протеазой. Линкер также может содержать одну или более гибких последовательностей, например, от 1 до 10 остатков глицина. В некоторых вариантах осуществления расщепление гибридного белка осуществляется в буфере, который обеспечивает растворимость продукта, например, в присутствии примерно от 0.5 до 2 M мочевины, в присутствии полипептидов и/или полинуклеотидов которые увеличивают растворимость, и т.п. Представляющие интерес домены включают эндосомолитические домены, например, домен НА гриппа; и другие полипептиды, способствующие продуцированию, например, IF2 домен, GST домен, GRPE домен и т.п. Дополнительно или альтернативно, такие полипептиды могут создаваться для улучшения стабильности. Например, пептиды могут быть пегилированными, при этом полиэтиленокси-группа обеспечивает увеличенное время жизни в кровяном русле. Полипептид может быть соединен с другим полипептидом для расширения функциональности, например, для увеличения стабильности in vivo. Как правило, такие «слитые» (гибридные) партнеры являются устойчивыми белками плазмы, которые могут, например, увеличивать in vivo время полужизни полипептида в плазме, в тех случаях, когда он присутствует как слитый, в частности, когда такой устойчивый плазменный белок является константной областью иммуноглобулина. В большинстве случаев устойчивый белок плазмы в норме существует в мультимерной форме, например, иммуноглобулины или липопротеины, в которой одинаковые или разные полипептидные цепи обычно связаны дисульфидной и/или нековалентной связью с получением собранного полипептида с множеством цепей, в этом документе гибриды, содержащие данный полипептид, также будут продуцироваться и использоваться в виде мультимера, имеющего в основном ту же самую структуру, как предшественник устойчивого белка плазмы. Эти мультимеры будут однородными в отношении того полипептидного фрагмента, который они содержат, или они могут содержать более чем один полипептидным фрагмент.
Полипептидный фрагмент-кандидат может быть получен из эукариотических клеток или может продуцироваться прокариотическими клетками. Он может подвергаться дополнительной обработке, например, раскручиванию, путем тепловой денатурации, DTT восстановления и т.д., и дополнительно может быть подвергнут рефолдингу с помощью известных в данной области техники методов. Представляющие интерес модификации, которые не изменяют первичную последовательность, включают получение полипептидов химическим методом, например, ацилированием, ацетилированием, карбоксилированием, амидированием и т.д. Также включаются модификации гликозилированием, например, модификации, полученные путем видоизменения профилей гликозилирования полипептида в ходе его синтеза и обработки или в дополнительных стадиях обработки; например, подвергания полипептида воздействию ферментов, влияющих на гликозилирование, таких как ферменты гликозилирования или дегликозилирования млекопитающих. Также рассматриваются последовательности, имеющие фосфорилированные аминокислотные остатки, например, фосфотирозин, фосфосерин или фосфотреонин. Полипептиды могут быть модифицированы с помощью обычных методов молекулярной биологии и синтетической химии для того, чтобы улучшить их устойчивость к протеолитической деградации или оптимизировать свойства растворимости или сделать их более пригодными в качестве терапевтического средства. Аналоги таких полипептидов включают, полипептиды, содержащие остатки, отличные от природных L-аминокислот, например, D-аминокислот или синтетических не существующих в природе аминокислот. D-аминокислоты могут использоваться вместо некоторых или всех аминокислотных остатков.
Полипептидное средство-кандидат может быть получено путем синтеза in vitro с помощью стандартных методов, известных в данной области техники. Доступны различные коммерческие установки для синтеза, например, автоматизированные установки для синтеза от Applied Biosystems, Inc., Beckman, etc. При использовании установок для синтеза природные аминокислоты могут заменяться аминокислотами не природного происхождения. Конкретные последовательности и способы их получения будут определяться преимуществами, экономически показателями, нужной степенью чистоты и тому подобным. Альтернативно, полипептидное средство-кандидат может быть выделено и очищено в соответствии с общепринятыми методами рекомбинантного синтеза. Из экспрессирующих клеток-хозяев можно приготовить лизат, который можно очистить с помощью ВЭЖХ, эксклюзионной хроматографии, гель-электрофореза, аффинной хроматографии или других способов очистки. В большинстве случаев используемые композиции будут содержать, по меньшей мере, 20% по весу целевого продукта, а точнее, по меньшей мере, около 75% по весу, предпочтительно, по меньшей мере, около 95% по весу и для терапевтических целей, как правило, по меньшей мере, около 99.5% по весу, в зависимости от загрязнителей, имеющих отношение к способу получения или очистки продукта. Обычно, процентные содержания основываются на общем белке.
В некоторых случаях, предназначенными для скрининга полипептидами-кандидатами являются антитела. Термин "антитело" или "фрагмент антитела" включает любую полипептидную цепь, содержащую молекулярную структуру, имеющую специфическую форму, которая подходит по размеру и распознает эпитоп, при этом одно или более взаимодействий в виде нековалентного связывания стабилизируют комплекс между молекулярной структурой и эпитопом. Специфическое или селективное соответствие данной структуры и ее специфического эпитопа иногда называют соответствием по типу "ключ-замок". Первоначальная молекула антитела представляет собой иммуноглобулин, и все типы иммуноглобулинов, IgG, IgM, IgA, IgE, IgD и т.д. из всех источников, например, человеческие, грызунов, кроликов, коров, овец, свиней, собак и других млекопитающих, кур и других птиц, и т.д. считаются "антителами." Использованные в настоящем изобретении антитела могут быть или поликлональными или моноклональными антителами. Как правило, антитела обнаруживаются в среде, в которой культивируются клетки. Выработка и отбор антител более подробно обсуждается далее.
Средства-кандидаты могут быть получены из самых разных источников, включая библиотеки синтетических или природных соединений. Например, для случайного и направленного синтеза целого ряда органических соединений, в том числе биомолекул, доступны многочисленные методы, включая экспрессию рандомизированных олигонуклеотидов и олигопептидов. В качестве альтернативы, доступными являются библиотеки природных соединений в форме бактериальных, грибковых, растительных и животных экстрактов, кроме того их легко получить. В дополнение к этому, природные или полученные синтетическим путем библиотеки и соединения легко модифицировать с помощью общепринятых химических, физических и биохимических методов, их также можно использовать для получения комбинаторных библиотек. Известные фармакологические средства могут быть подвергнуты направленным или случайным химическим модификациям, таким как ацилирование, алкилирование, этерификация, амплификация и т.д. с целью получения структурных аналогов.
Средства-кандидаты подвергаются отбору по биологической активности путем введения средства, по меньшей мере, в один и обычно во множество образцов, иногда одновременно с образцами, не имеющими данного средства. Измеряют изменение параметров в ответ на средство, а результат оценивается путем сравнения с исходными культурами, например, в присутствии или в отсутствие средства, полученными при использовании других средств и т.д. В тех случаях, когда отбор проводится с целью установления средств-кандидатов, которые будут предотвращать, уменьшать или отменять эффекты токсичного вещества, скрининг в большинстве случае проводится в присутствии токсичного вещества, при этом токсичное вещество добавляется в наиболее подходящее время для получения соответствующих результатов. Например, в тех случаях, когда тестируется защитная/профилактическая способность средства-кандидата, средство-кандидат может быть добавлено до токсичного вещества, одновременно со средством-кандидатом или после обработки средством-кандидатом. В качестве другого примера, в тех случаях, когда тестируется способность средства-кандидата «отменять» эффекты токсичного вещества, средство-кандидат может быть добавлено после обработки токсичным средством. Как было упомянуто выше, в некоторых случаях, "образец" является генетически модифицированным не принадлежащим к человеческому роду животным, которому были пересажены клетки, например, средство-кандидат используется на иммунодефицитном животном, например, мыши, содержащей нуклеиновую кислоту, кодирующую человеческий IL-6, функционально связанный с IL-6 промотором, которой были пересажены человеческие гематопоэтические клетки. В некоторых случаях образцом являются человеческие гематопоэтические клетки, предназначенные для пересадки, т.е. средство-кандидат предоставляется клеткам до пересадки иммунодефицитному генетически модифицированному животному.
В том случае, если средство-кандидат вводится непосредственно генетически модифицированному животному с пересаженными клетками, оно может быть введено с помощью любого из целого ряда известных в данной области техники методов введения мышам пептидов, небольших молекул и нуклеиновых кислот. Например, средство может вводиться перорально, через слизистую, местно, внутрикожно или с помощью инъекции, например, внутрибрюшинной, подкожной, внутримышечной, внутривенной или внутричерепной инъекции и тому подобного. Средство может быть введено в буфере, или оно может быть включено в состав любой из целого ряда композиций, например, путем комбинации с подходящей фармацевтически приемлемой основой. "Фармацевтически приемлемыми основами" могут быть основы, одобренные регулирующим органом Федерального правительства или правительства штата или перечисленные в Фармакопее США или других общепризнанных фармакопеях для применения на млекопитающих, например, людях. Термин "основа" относится к разбавителю, адъюванту, эксципиенту или носителю, с которым соединение изобретения заключается в состав, предназначенный для введения животному. Такие фармацевтические основы могут быть липидами, например, липосомами, например, дендримерами липосом; жидкостями, такими как вода или масла, включая масла нефтяного, животного, растительного или синтетического происхождения, такие как арахисовое масло, соевое масло, минеральное масло, кунжутное масло и тому подобное, солевой раствор; гуммиарабик, желатин, кукурузный крахмал, тальк, кератин, коллоидная двуокись кремния, мочевина и тому подобное. В дополнение к этому могут использоваться вспомогательные вещества, стабилизирующие вещества, сгустители, смазывающие и окрашивающие вещества. Фармацевтические композиции могут создаваться в виде препаратов в твердой, полужидкой, жидкой или газообразной формах, таких как таблетки, капсулы, порошки, гранулы, мази, растворы, суппозитории, препараты для инъекций и ингаляций, гели, микросферы и аэрозоли. Средство может быть введено системно или может распространяться в отдельном участке при использовании местного введения, интрамурального введения или при использовании имплантата, действие которого направлено на поддержание активной дозировки в месте имплантации. Активное средство может предназначаться для немедленного воздействия или может создаваться с целью замедленного высвобождения. В некоторых случаях, в частности, в условиях центральной нервной системы, необходимо создание средств, способных проникать через гематоэнцефалический барьер (ВВВ). Одна стратегия доставки лекарственного средства через гематоэнцефалический барьер (ВВВ) влечет за собой нарушение ВВВ, или с помощью осмотических средств, таких как маннитол или лейкотриены, или биохимически путем использования вазоактивных веществ, таких как брадикинин. Вещество, нарушающее ВВВ, может вводиться совместно с данным средством при введении композиций с помощью внутрисосудистой инъекции. Другие стратегии прохождения через ВВВ могут включать использование эндогенных транспортных систем, включая трансцитоз, опосредованный кавеолином-1, транспорт, опосредованный переносчиками, такими как переносчики глюкозы и аминокислот, опосредованный рецептором трансцитоз для инсулина или трансферрина, и активные эффлюксные транспортеры (выводящие из клетки), такие как р-гликопротеин. Активные транспортные молекулы также могут быть конъюгированы с терапевтическими соединениями, предназначенными для использования в изобретении, чтобы облегчить транспорт через эндотелиальную стенку кровеносного сосуда. Альтернативно, доставка средств, минуя ВВВ, может осуществляться путем местной доставки, например, подоболочечной доставки, например, через резервуар Оммайя (смотри, например, патенты США №5,222,982 и 5385582, включенные в описание путем отсылки); путем болюсной инъекции, например, с помощью шприца, например, в стекловидное тело или внутрь черепа; путем длительной инфузии, например, с помощью катетеризации, например, с использованием конвекции (смотри, например, заявку США №20070254842, включенную в описание путем отсылки); или путем имплантации устройства, на котором средство может прикрепляться обратимым образом (смотри, например, заявки США №20080081064 и 20090196903, включенные в описание путем отсылки).
В том случае, когда средство(а) предоставляется клеткам до пересадки, средства удобно добавлять в раствор или в легкорастворимой форме в культуральную среду. Средства могут добавляться в прямоточную систему, в виде потока, прерывистого или непрерывного, или альтернативно, болюс (одноразовая доза) соединения может быть добавлен в иной статический раствор, однократно или постепенно. В прямоточной системе используются две жидкости, при этом одна является физиологически нейтральным раствором, а другая является раствором с добавленным тестируемым соединением. Сначала через клетки пропускают первую жидкость, а затем вторую. При использовании метода одного раствора болюс тестируемого соединения добавляется в среду, окружающую клетки. Общие концентрации компонентов культуральной среды не должны значительно изменяться при добавлении болюса или между двумя растворами при использовании прямоточной системы.
Параллельно может выполняться некоторое количество анализов с разными концентрациями средства, для того, чтобы получить различающиеся ответы на разные концентрации. Как известно в данной области техники, при определении эффективной концентрации средства в большинстве случаев используется диапазон концентраций, являющийся результатом разбавления 1:10 или в другой логарифмической шкале. Концентрации могут быть дополнительно уменьшены при помощи второй серии разведений, если необходимо. Как правило, одна из этих концентраций служит в качестве отрицательного контроля, т.е. нулевой концентрации или нижнего уровня обнаружения средства или при или ниже концентрации средства, которая не позволяет получить обнаружимое изменение в фенотипе. Исследование ответа клеток у генетически модифицированного животного с пересаженными клетками на средство-кандидат может проводиться в любой момент времени после обработки средством. Например, клетки могут быть исследованы через 1, 2 или 3 дня, иногда через 4, 5 или 6 дней, в ряде случаев через 8, 9 или 10 дней, в ряде случае через 14 дней, иногда через 21 день, иногда через 28 дней, иногда через 1 месяц или более после контакта со средством-кандидатом, например, 2 месяца, 4 месяца, 6 месяцев или более. В некоторых вариантах осуществления исследование включает анализ множества временных точек. Выбор момента(ов) времени для проведения анализа основывается на типе анализа, который будет проводиться, что понятно среднему специалисту в данной области техники.
Анализ может включать измерение любого из описанных в этом документе параметров или известных в данной области техники, предназначенных для измерения жизнеспособности клетки, клеточной пролиферации, идентификации клеток, морфологии клеток и клеточной функции, в частности, учитывая, что они могут относиться к иммунным клеткам. Например, может использоваться проточная цитометрия для определения общего количества гематопоэтических клеток или количества гематопоэтических клеток определенного типа. Для установления апоптотического состояния клеток может проводиться гистохимическое или иммуногистохимическое исследование, например, введение концевой метки dUTP с использованием дезоксинуклеотидилтрансферазы (TUNEL) для измерения фрагментации ДНК, или иммуногистохимическое исследование для обнаружения связывания аннексина V с фосфатидилсерином на клеточной поверхности. Проточная цитометрия также может использоваться для оценки соотношений дифференцированных клеток и дифференцированных типов клеток, например, чтобы определить способность гематопоэтических клеток дифференцировать в присутствии данного средства. Для определения уровней цитокинов, хемокинов, иммуноглобулинов и т.д., экспрессированных у генетически модифицированных мышей с пересаженными клетками, могут проводиться анализы ELISA, вестерн-блоттинг и нозерн-блоттинг, например, для оценки функции пересаженных клеток, для оценки выживаемости злокачественных плазматических клеток и т.д. μCΤ-сканирования могут проводиться для оценки степени повреждения, вызванного пораженными болезнью гематопоэтическими клетками, например, разрушения и резорбции кости, вызванной клетками множественной миеломы. Также могут проводиться in vivo исследования для проверки функции иммунных клеток, а также анализы, имеющие отношение к отдельным представляющим интерес болезням или нарушениям, таким как диабет, аутоиммунное заболевание, болезнь «трансплантат против хозяина», AMD и т.д. Смотри, например, Current Protocols in Immunology (Richard Coico, ed. John Wiley & Sons, Inc. 2012) и Immunology Methods Manual (I. Lefkovits ed., Academic Press 1997), раскрытия которых включены в данное описание путем отсылки.
Таким образом, например, предоставляется способ определения эффекта некоторого средства на множественную миелому, включающий введение средства гуманизированной IL-6 мыши, например, мыши Rag2-/-IL2rg-/-IL-6h/h, которой были пересажены клетки множественной миеломы человека; измерение параметра жизнеспособности и/или пролиферативной способности клеток множественной миеломы в течение продолжительного времени в присутствии данного средства и сравнение этих измерений с измерениями, полученными от гуманизированной IL-6 мыши с пересаженными клетками, которая не подвергалась воздействию данного средства. Данное средство определяется как противораковое, если оно уменьшает пролиферацию и/или уменьшает количество клеток множественной миеломы в крови или ткани мыши, по меньшей мере, на 20%, 30%, 40% или более, в некоторых случаях на 50%, 60%, 70% или более, например, 80%, 90% или 100%, т.е. до не обнаруживаемых количеств, после единственного введения или двух или более введений средства в течение выбранного периода времени. В отдельных вариантах осуществления введение лекарственного средства или комбинации средств происходит в течение, по меньшей мере, недели, 10 дней, двух недель, трех недель или четырех недель после пересадки клеток множественной миеломы.
Другие примеры применения заявленных мышей предоставляются в других местах в данном документе. Дополнительные методы применения генетически модифицированных мышей с пересаженными клетками, описанные в данном раскрытии, станут понятны специалистам в данной области техники после прочтения данного раскрытия.
Получение человеческих антител
Также предоставляются композиции и способы, подходящие для получения человеческих моноклональных антител от описанного в данном документе иммунодефицитного животного с пересаженными клетками. В различных вариантах осуществления способы включают контактирование иммунодефицитного животного с человеческой гематопоэтической клеткой для получения не относящегося к человеческому роду животного с транплантированной иммунной системой (животного с пересаженными клетками), потом контактирование животного с пересаженными клетками с антигеном, сбора от этого животного человеческих клеток, продуцирующих человеческое антитело к антигену, и выделение антитела из антителопродуцирующей клетки.
В различных вариантах осуществления изобретение включает образование антителопродуцирующей клетки (например, человеческой В-клетки) методом трансформации (например, EBV) или методом слияния клеток (например, гибридомы). Предпочтительно антителопродуцирующая клетка способна поддерживаться при соответствующих культуральных условиях в течение, по меньшей мере, примерно 50 пассажей.
В различных вариантах осуществления животное с пересаженными клетками является млекопитающим, не относящимся к человеческому роду. В некоторых вариантах осуществления животное с пересаженными клетками является мышью, крысой или кроликом.
В различных вариантах осуществления изобретения человеческая гематопоэтическая клетка является CD34+ клеткой, полученной из образца эмбриональной печени человека, костного мозга, пуповинной крови, периферической крови или селезенки. В различных вариантах осуществления антиген является, по меньшей мере, одним из числа пептида, полипептида, комплекса МНС/пептид, ДНК, «живого» вируса, «мертвого» вируса или его части, живой бактерией, мертвой бактерии или ее части, или раковой клеткой или ее части.
В некоторых вариантах осуществления животное с пересаженными клетками контактирует с антигеном через 1-5 месяцев после того, как животное контактировало с человеческой гематопоэтической клеткой. В некоторых вариантах осуществления животное с пересаженными клетками только один раз контактирует с антигеном, тогда как в других вариантах осуществления животное с пересаженными клетками контактирует с антигеном два, три, четыре, пять, шесть, семь, восемь или более раз.
В одном варианте осуществления антителопродуцирующая человеческая клетка, полученная от животного с пересаженными клетками, является В-клеткой. В различных вариантах осуществления антителопродуцирующая человеческая клетка, полученная от животного, экспрессирует на своей поверхности, по меньшей мере, одно из числа CD19, CD20, CD22 и CD27. Антителопродуцирующую человеческую клетку изобретения можно получить путем извлечения каких-либо подходящих клеточных компонентов иммунной системы у животного. В различных вариантах осуществления антителопродуцирующую клетку получают от животного с пересаженными клетками, путем извлечения, по меньшей мере, одного из числа селезенки, лимфатических узлов, периферической крови, костного мозга или их частей.
В различных вариантах осуществления способ изобретения применяет общепринятую гибридомную технологию, используя подходящего партнера для слияния. В различных вариантах осуществления партнером для слияния является, по меньшей мере, одна клетка, выбранная из группы, состоящей из МОРС21, P3X63AG8, SP2/0, NS-1, P3.X63AG8.653, F0, S194/5.XXO.BU-1, FOX-NY, SP2/0-Agl4, MEG-01, HEL, UT-7, М07е, MEG-A2 и DAMI, и происходящие из этих клеток клеточные линии.
Методы извлечения антител у животного с пересаженными клетками хорошо известны в данной области техники. Выделение антитела из антителопродуцирующей клетки, среды, в которой культивируется антителопродуцирующая клетка, и/или асцитной жидкости животного с пересаженными клетками, может быть проведено согласно методам, известным в данной области техники, таким как, например, хроматография и диализ. В других вариантах осуществления антитело может быть выделено с помощью одного или более методов из числа иммуноаффинной очистки, осаждения сульфатом аммония, очистки белков A/G, ионообменной хроматографии и гель-фильтрации. Такие методы описаны в работах Nau (1989, Optimization of monoclonal antibody purification, In: Techniques in Protein Chemistry, Hugh, T. (ed.), Academic Press, New York) и Coligan et al. (2005, Current Protocols in Immunology, John Wiley & Sons, Inc.).
Антиген может быть введен животному с пересаженными клетками с помощью любых подходящих способов. В различных вариантах осуществления антиген может быть введен животному с пересаженными клетками, по меньшей мере, одним из следующих способов внутрь селезенки, внутривенно, внутрибрюшинно, внутрикожно, внутримышечно и подкожно. В некоторых вариантах осуществления антиген вводится отдельно или в других вариантах осуществления антиген вводится в комбинации с подходящим иммуномодулирующим средством или адъювантом (вспомогательным средством). Примеры адъювантов, подходящих для способов изобретения, включают, но не ограничиваются этим, полный адъювант Фрейнда (CFA), неполный адъювант Фрейнда (IFA) и соединения алюминия Alum (Al3(ОН)4).
Реагенты и наборы
Также предоставляются реагенты и их наборы для осуществления на практике одного или более из описанных выше способов. Заявленные реагенты и наборы могут варьироваться в широких пределах. В некоторых вариантах осуществления реагенты или наборы будут содержать один или более реагентов, предназначенных для использования при получении и/или поддержании заявленных генетически модифицированных не относящихся к человеческому роду животных. Например, набор может включать иммунодефицитную мышь, содержащую нуклеиновую кислоту, кодирующую человеческий IL-6, функционально связанный с IL-6 промотором, и нуклеиновую кислоту, кодирующую человеческий SIRPa, функционально связанный с SIRPa промотором; или мышь, содержащую нуклеиновую кислоту, кодирующую человеческий IL-6, функционально связанный с IL-6 промотором, и дополнительно содержащую нуклеиновую кислоту, кодирующую человеческий M-CSF, функционально связанный с M-CSF промотором; нуклеиновую кислоту, кодирующую человеческий IL-3, функционально связанный с IL-3 промотором; нуклеиновую кислоту, кодирующею человеческий GM-CSF, функционально связанный с GM-CSF промотором; нуклеиновую кислоту, кодирующую человеческий ТРО, функционально связанный с ТРО промотором; и/или нуклеиновую кислоту, кодирующую человеческий SIRPa, функционально связанный с SIRPa промотором. Набор может содержать реагенты для разведения таких мышей, например, праймеры для генотипирования человеческого IL-6 гена, человеческого M-CSF гена, человеческого IL-3 гена, человеческого GM-CSF гена, человеческого SIRPa гена и/или человеческого ТРО гена, буфер PCR, раствор MgCL и т.д.
В некоторых вариантах осуществления реагенты или наборы будут содержать один или более реагентов для использования при пересадке заявленным генетически модифицированным животным, не относящимся к человеческому роду, например, человеческих гематопоэтических клеток, обогащенной популяции человеческих гематопоэтических клеток-предшественников, линии гематопоэтических клеток, линии злокачественных гематопоэтических клеток и т.д. для трансплантации заявленным, генетически модифицированным животным, не относящимся к человеческому роду, или реагенты для подготовки популяции гематопоэтических клеток, обогащенной популяции гематопоэтических клеток человека, линии гематопоэтических клеток, линии злокачественных гематопоэтических клеток и т.д. к трансплантации заявленным, генетически модифицированным животным, не относящимся к человеческому роду.
В некоторых вариантах осуществления реагенты или наборы будут включать реагенты для определения жизнеспособности и/или функции гематопоэтических клеток, например, в присутствии/отсутствии средства-кандидата, например, одно или более антител, специфических в отношении маркеров, экспрессируемых различными типами гематопоэтических клеток, или реагенты для обнаружения определенных цитокинов, хемокинов и т.д. Другие реагенты могут включать культуральные среды, дополнения к культуральным средам, компоненты матрикса и тому подобное.
В дополнение к вышеперечисленным компонентам заявленные наборы будут дополнительно включать инструкции, касающиеся осуществления на практике заявленных способов. Эти инструкции могут присутствовать в заявленных наборах в различных формах, одна или более из которых могут присутствовать в наборе. Одной из форм, в которой эти инструкции могут присутствовать в наборе, является печатная информация на подходящем носителе, например, листке или листках бумаги, на которых напечатана данная информация, на упаковке набора, на листе-вкладыше в упаковку и т.д. Еще одним средством может быть машиночитаемый носитель, например, дискета, CD, и т.д., на котором записана информация. Другим средством, которое может присутствовать в наборе, является адрес веб-сайта, который может использоваться через интернет для доступа к информации на удаленном сайте. В наборах могут присутствовать любые подходящие средства передачи информации.
Экспериментальные примеры
Далее это изобретение подробно описывается со ссылкой на следующие экспериментальные примеры. Эти примеры предоставляются только с целью иллюстрации и не предназначаются для ограничения, если не указано иначе. Таким образом, изобретение ни в коей мере не должно трактоваться как ограниченное следующими примерами, а скорее должно рассматриваться как включающее любые возможные изменения, которые станут очевидны, исходя их предоставленной в описании идеи.
Можно полагать, что без дальнейшего описания средний специалист в данной области техники может, используя предшествующее описание и следующие иллюстративные примеры, изготовить и использовать соединения настоящего изобретения и применять на практике заявленные способы. Соответственно, следующие рабочие примеры конкретно выделяют предпочтительные варианты осуществления настоящего изобретения и не должны рассматриваться, как ограничивающие каким бы то ни было образом остальную часть раскрытия.
Пример 1: Генетическая гуманизация генов цитокинов обеспечивает условия для приживления у мышей клеток множественной миеломы человека.
Представленные результаты демонстрируют, что генетически модифицированные не принадлежащие к человеческому роду животные, описанные в данном документе, представляют собой новую in vivo животную модель множественной миеломы.
Материалы и методы
Мыши. Гуманизированные knock-in IL-6 мыши были получены путем замены 6.8 kb локуса мышиного гена IL-6 человеческой последовательностью 4.8-kb гена IL-6, содержащей экзоны с 1 по 5, включая 3' нетранслируемую область человеческого IL-6 гена.
Коротко, целевой конструкт для замены мышиного IL-6 гена на человеческий за одну стадию нацеливания был создан с использованием генно-инженерной технологии VELOCIGENE® (смотри, Valenzuela et al. (2003) High-throughput engineering of the mouse genome coupled with high-resolution expression analysis, Nature Biotech, 21(6):652-659). ДНК мышиного и человеческого IL-6 были получены из искусственной бактериальной хромосомы (ВАС) RPCI-23 клон 368С3, и из ВАС CTD клон 2369М23, соответственно. Коротко, линеаризованный Notl целевой конструкт, полученный с помощью клонирования методом заполнения бреши (gap repair), содержащий мышиный IL-6, вышерасположенное и нижерасположенное гомологичные ответвления, фланкирующие 4.8 kb человеческую IL-6 последовательность, продолжающуюся от ATG в экзоне 1 до экзона 5, вместе с 16 нуклеотидами 3' нижерасположенной последовательности (геномные координаты: NCBIh37.1: ch7:22,766,882 до 22,771,637) и фланкированная loxP нео кассета селекции были электропорированы в клетки Rag2-/- IL2rg Y/- ES. Родительская клеточная линия ES, в которой был сделан нокаут RAG2 гена и IL2rg гена, была коммерчески доступной VI7 ES клеткой (BALB/cx129 гетерозигота). При правильном нацеливании ES клетки могут быть электропорированы временным Cre-экспрессирующим вектором с целью удаления кассеты селекции лекарственных препаратов.
Правильно нацеленные ES клеточные клоны были идентифицированы с помощью анализа потери нативного аллеля (LONA) (Valenzuela et al. 2003), в котором количество копий нативного, немодифицированного Il6 гена было установлено с помощью двух TaqMan™ количественных полимеразных цепных реакций (qPCRs), специфических в отношении последовательностей в мышином Il6 гене, которые были намечены для удаления. qPCR анализы содержали следующие наборы праймер-зонд (написанные от 5' к 3'): расположенный выше прямой праймер, TTGCCGGTTT ТСССТТТТСТ С (SEQ ID NO: 1); расположенный выше обратный праймер, AGGGAAGGCC GTGGTTGTC (SEQ ID NO: 2); вышерасположенный зонд, FAMCCAGCATCAG TCCCAAGAAG GCAACT-BHQ (SEQ ID NO: 3); нижерасположенный прямой праймер, TCAGAGTGTG GGCGAACAAA G (SEQ ID NO: 4); нижерасположенный обратный праймер, GTGGCAAAAG CAGCCTTAGC (SEQ ID NO: 5); нижерасположенный зонд, FAMTCATTCCAGG СССТТСТТАТ TGCATCTG-BHQ (SEQ ID NO: 6); в котором F AM относится к 5-карбоксифлуоресцеин-флуоресцентному зонду, и BHQ относится к тушителю флуоресценции типа «черная дыра» (Biosearch Technologies). ДНК, очищенная из ES клонов клеток, которые взяли на себя направленный вектор и включили в свои геномы, была объединена с TaqMan™ Gene Expression Master Mix (Life Technologies) согласно рекомендациям производителя в 384-луночном ПЦР-планшете (MicroAmp™ Optical 384-Well Reaction Plate, Life Technologies) и циклизирована в Applied Biosystems Prism 7900HT, которая регистрирует флуоресценцию в ходе ПЦР и определяет пороговый цикл (Ct), фракционный ПЦР цикл, при котором аккумулированная флуоресценция достигает предварительно заданного порога. Вышерасположенный и нижерасположенный Il6-специфические qPCR и два анализа qPCR нецелевых эталонных генов были проделаны для каждого ДНК образца. Различия в значениях Ct (ΔCt) между каждым Il6-специфическим qPCR и каждым qPCR эталонного гена были вычислены, и затем различие между каждым ΔCt и средним ΔCt для всех оцененных образцов было вычислено, чтобы получить ΔΔCt значения для каждого образца. Число копий IL-6 гена в каждом образце было вычислено с помощью следующей формулы: число копий = 2⋅2-ΔΔ Ct. Правильно нацеленный клон, с потерей одной из его нативных копий, будет иметь число копий IL-6 гена равное одному. Подтверждение того, что последовательность человеческого гена IL-6 заменила удаленную последовательность мышиного Il6 гена в гуманизированном аллеле было получено с помощью анализа TaqMan™ qPCR, который содержит следующие наборы праймер-зонд (написанные от 5' к 3'): человеческий прямой праймер, CCCCACTCCACTGGAATTTG (SEQ ID NO: 7); человеческий обратный праймер, GTTCAACCACAGCCAGGAAAG (SEQ ID NO: 8); и человеческий зонд, FAMAGCTACAACTCATTGGCATCCTGGCAA-BHQ (SEQ ID NO: 9).
Вышерасположенное соединение мышиного локуса и последовательности, содержащей hIL-6 ген, располагается так, чтобы находиться в пределах 5' -AATTAGAGAG TTGACTCCTA ATAAATATGA GACTGGGGAT GTCTGTAGCT CATTCTGCTC TGGAGCCCAC CAAGAACGAT AGTCAATTCC AGAAACCGCT ATGAACTCCT TCTCCACAAG TAAGTGCAGG AAATCCTTAG CCCTGGAACT GCCAGCGGCG GTCGAGCCCT GTGTGAGGGA GGGGTGTGTG GCCCAGG (SEQ ID NO: 10), в которой конечный мышиный нуклеотид до первого нуклеотида человеческого гена является "Т" в CCGCT, а первый нуклеотид человеческой последовательности является первым "А" в ATGAA. Нижерасположенное соединение последовательности, содержащей hIL-6 ген и мышиный локус, создается так, чтобы находиться в 5'-TTTTAAAGAA АТАТТТАТАТ TGTATTTATA TAATGTATAA ATGGTTTTTA ТАССААТААА TGGCATTTTA AAAAATTCAG CAACTTTGAG TGTGTCACGC TCCCGGGCTC GATAACTATA ACGGTCCTAA GGTAGCGACT CGAGATAACT Т-3' (SEQ ID NO: 11), в которой конечный нуклеотид человеческой последовательности находится вместе с конечным "G" в TCACG, и первый нуклеотид мышиной последовательности является первым "С" в СТССС; участок нижерасположенного соединения также содержал loxP сайт на 3' конце (начало которого показано) для устранения фланкированного loxP убиквитинового промотора-индуцируемого нео кассетой. Соединение нео кассеты с мышиным IL-6 локусом создается так, чтобы находиться в 5'-TATACG AAGT TATCCTAGGT TGGAGCTCCT AAGTTACATC САААСАТССТ СССССАААТС AATAATTAAG САСТТТТТАТ GACATGTAAA GTTAAATAAG AAGTGAAAGC TGCAGATGGT GAGTGAGA (SEQ ID NO: 12), где конечный "С" AGCTC является последним нуклеотидом нео кассеты; первым нуклеотидом мышиного генома, следующим за кассетой, является первый "С" из СТА AG.
Для получения мыши, содержащей hIL-6 и утратившей Rag2 и Il2rg, идентифицируют правильно нацеленные ES клетки и вводят их в предимплантационный эмбрион с помощью методов, известных в данной области техники.
Затем гуманизированных IL-6 KI мышей скрещивают повторно, чтобы получить мышей, утративших Rag2 и U2rg и экспрессирующих hIL-6, и скрещивают с мышами, экспрессирующими человеческий SIRPa трансген (Strowig et al., 2011, Proc Natl Acad Sci USA, 108(32): 13218-13223), чтобы получить мышей, дефицитных в отношении Rag2 и H2rg, а также экспрессирующих и hIL-6 и hSIRPa (Rag2-/-Il2rgnullIl6h/hhSIRPa+). В дополнение к этому, Rag2-/-, IL-2rgy/-, hIL-6 ΚΙ мышей скрещивали с мышами, экспрессирующими человеческий ТРО (Rongvaux et al., 2011, Proc Natl Acad Sci USA, 108(6): 2378-2383), человеческий IL-3 и человеческий GM-CSF (Willinger et al, 2011, Proc Natl Acad Sci USA, 108(6): 2390-2395) и человеческий M-CSF (Rathinam et al, 2011, Blood, 118(11): 3119-3128) а также hSIRPa (Strowig et al., 2011, Proc Natl Acad Sci USA, 108(32): 13218-13223), чтобы получить мышей, экспрессирующих комбинацию этих человеческих белков (Rag2-/-Il2rgnullhSIRPa+ Tpoh/h Mcsfh/h Il3/Gmcsfh/h Il6h/h).
Клеточные линии и первичные клетки. Линию клеток множественной миеломы INA-6 (Burger et al., 2001, Hematol J, 2(1): 42-53) поддерживали в среде RPMI1640 с добавлением 20% FCS, пенициллина/стрептомицина, L-глутамина и 2.5 нг/мл hIL-6 в стандартном инкубаторе при 37С и 5% CO2.
Первичные клетки, полученные от пациентов с множественной миеломой, были выделены из аспиратов костного мозга после получения информированного согласия от пациентов. Мононуклеарные клетки были выделены центрифугированием в градиенте плотности Фиколла, а затем различные клеточные субпопуляции были изолированы путем сортировки магнито-активированных клеток (MACS®). Чтобы получить Т-клеточно истощенные популяции, CD3+ клетки были истощены с помощью негативной селекции на AutoMACS системе с использованием анти-CD3 микрогранул (Miltenyi Biotec). Чтобы получить CD138+ клетки, CD138+ клетки изолировали с помощью положительной селекции на AutoMACS® системе с использованием анти-CD138 микрогранул (Miltenyi Biotec). Чистоту клеток после MACS® селекции анализировали с помощью проточной цитометрии.
Трансплантация клеток. Для внутрифеморальной трансплантации IΝΑ6 клеток, Rag2-/-Il2rgnullIl6h/hhSIRPa+ мышей облучили дважды 200 рад от источника рентгеновских лучей. Указанные количества клеток затем были трансплантированы в бедренную кость мышей-реципиентов. Коротко, мышей обезболивали с использованием кетамина/ксилозина и буравили отверстие в надколенной поверхности бедренной кости с помощью иглы 26 калибра. Затем клетки медленно инъецировали в объеме 20 мкл, используя иглу 26 калибра. Для трансплантации первичных клеток, полученных от пациента, Rag2-/- Il2rgnull hSIRPa+ Tpoh/h Mcsfh/h Il3/Gmcsfh/h Il6h/h мышей дважды облучали 150 рад с помощью источника рентгеновских лучей, транплантацию проводили, как описано выше.
ELISA. Для измерения концентраций человеческого растворимого IL-6R (R&D Systems), человеческого Igκ и Igλ (Bethyl Laboratories) использовали коммерческие наборы ELISA. Обнаружение этих белков осуществляли согласно инструкциям производителя.
μСТ. Морфометрию бедренной кости выражали количественно, используя компьютерную томографию с конусообразными микрофокусными рентгеновскими лучами (μСТ40; ScancoMedicalAG). Были получены серийные томографические изображения, затем были реконструированы 3D изображения, которые использовали для определения заданных параметров. Морфометрические характеристики трабекулярной кости включали определение объема костной ткани, трабекулярной толщины, числа трабекул и трабекулярного разделения. Кортикальные параметры включали среднюю толщину кортикального слоя, площадь поперечного сечения кортикального слоя кости, площадь поперечного сечения поднадкостничной области и площадь костного мозга.
Гистология. С бедренных костей удаляли мягкую ткань, фиксировали в 10% забуференном формалине, обезвоживали и закладывали в метилметакрилат, затем делали срезы и окрашивали толуидиновым синим согласно стандартным методам.
Результаты
Приживление клеточной линии множественной миеломы у мышей с гуманизированным IL-6 геном. Человеческая IL-6-зависимая клеточная линия ММ (ΙΝΑ6-gfp) была использована для того, чтобы оценить, являются ли мыши, экспрессирующие человеческий SIRPα и IL-6, подходящими «хозяевами» для клеточной линии множественной миеломы (ММ). Клеточная линия INA6-gfp показывает высокую зависимость от человеческого микроокружения, т.е., человеческой эмбриональной костной стружки при трансплантации в виде ксенотрансплантантных систем scid-hu мышам (Epstein et al., 2005, Methods Mol Med, 113: 183-190). В частности, только INA-6 клетки являются способными «внедрить» трансплантат кости человека scid-hu мышам, подтверждая зависимость от человеческого костномозгового микроокружения, подобно первичным клеткам ММ (Tassone et al., 2005, Blood, 106(2): 713- 716).
Поэтому, чтобы непосредственно протестировать эффективность IL-6 гуманизации для обеспечения возможности роста клеток миеломы, INA-6 клетки были трансплантированы внутривенно i) Rag2-/-Il2rgnull, ii) Rag2-/-Il2rgnullhSIRPa+, iii) Rag2-/-Il2rgnullIl6h/h и iv) Rag2-/-Il2rgnullIl6h/hhSIRPa мышам. Приживление анализировали, измеряя sIL-6R белок, секретированный INA-6 клетками в крови. Приживление было установлено только у мышей, экспрессирующих человеческий IL-6, (Фигура 1) что демонстрирует, что INA-6 клетки действительно были способны «приживляться» у мышей, экспрессирующих человеческий IL-6. Далее, расположение INA-6 клеток (модифицированных, чтобы экспрессировать GFP) у мышей с пересаженными клетками было исследовано с помощью флуоресцентной микроскопии. Немного GFP+ клеток было обнаружено в костном мозге (Фигура 2), однако увеличенное количество GFP+ клеток было обнаружено в легких мышей с пересаженными клетками (Фигура 3).
Анализ экспрессии человеческого гена Il6 показал, что самый высокий уровень экспрессии человеческого Il6 гена был обнаружен в легком (Фигура 3), что коррелирует с присутствием INA-6 клеток в легком. Таким образом, раскрытые в описании данные демонстрируют успешное приживление INA-6 клеток после внутривенной инъекции (i.v.) мышам, генетически модифицированным так, чтобы экспрессировать человеческий IL6 или человеческий IL6 и SIRPa, что свидетельствует о том, что генетическая гуманизация способна преодолеть ограничения роста клеток ММ человека у мышей.
Данные, представленные на Фигуре 3, позволяют предположить, что INA-6 клетка не направляется в костный мозг и вместо этого может расти в нефизиологических местах. Поэтому, далее было исследовано, способна ли трансплантация линии опухолевых клеток в ее естественном микроокружении воспроизвести патологию, связанную, как правило, с человеческим ММ. Чтобы сделать это, было протестировано внутрикостное введение INA6 клеток. Поразительно, что это приводило к разрушению и резорбции кости, что является патологией, наблюдаемой у пациентов с ММ (Фигура 4-6). В частности, потерю массы трабекулярной кости наблюдали, используя гистологический анализ и количественный μCΤ. Более того, наблюдалось только ограниченное количество метастазов в периферические места, такие как легкие, что приводит к выводу, что модель может использоваться для дополнительных исследований новых лекарственных средств, оказывающих противодействие этой патологии. Чтобы проверить этот вывод мышей с пересаженными IΝΑ-6 лечили Zometa или Velcade, эти два лекарственных средства обычно используются для лечения пациентов с множественной миеломой. Удивительно, что лечение Zometa мышей, инъецированных INA-6 клетками, было способно уменьшить резорбцию кости по сравнению с нелеченными мышами, что показано с помощью μCΤ (Фигура 7).
Раскрытые в описании данные показывают, что гуманизация Il6 гена обеспечивает приживление клеточной линии множественной миеломы, что обычно требует человеческого микроокружения. Приживление повторяет некоторые патологические симптомы, наблюдаемые у пациентов, включая потерю костной массы. Кроме того, эти симптомы можно лечить, используя разрешенные к применению лекарственные средства, что подчеркивает полезность этой модели для тестирования новых лекарственных средств.
Генетическая гуманизация генов цитокинов обеспечивает возможность приживления первичных клеток множественной миеломы, полученных от пациента. Далее, были проведены эксперименты, чтобы проверить трансплантацию первичных клеток ММ генетически гуманизированным мышам. Ранее было продемонстрировано, что гуманизация некоторого количества цитокинов, включая тромбопоэтин, IL-3, GMCSF и M-CSF, а также ингибирующего рецептора макрофагов SIRPa приводит к улучшенному приживлению человеческих гематопоэтических клеток у иммунодефицитных мышей (Strowig et al., 2011, Proc Natl Acad Sci USA, 108(32): 13218-13223; Rathinam et al, 2011, Blood, 118(11): 3119-3128; Rongvaux et al., 2011, Proc Natl Acad Sci USA, 108(6): 2378-2383; Willinger et al, 2011, Proc Natl Acad Sci USA, 108(6): 2390-2395). В частности, гуманизация IL-3 и GM-CSF, а также M-CSF улучшала приживление миелоидных клеток, что как было показано, является важным для определенных аспектов ММ-патологии. Трансгенная экспрессия hSIRPa улучшает приживление человеческой клетки, однако, как было показано в последнее время, ось SIRPa-CD47 также вовлечена в образование опухолей. Полученные ранее гуманизированные мыши были объединены с knock-in мышами с человеческим IL-6, чтобы получить мышей Rag2-/-Il2rgnullhSIRPa+Tpoh/h Mcsfh/h Il3/Gmcsfh/h Il6h/h. Для оценки способности данной линии поддерживать приживление человеческих клеток, клетки костного мозга, лишенные CD3, от пациентов с ММ инъецировали в костный мозг мышей. Высокая частота миеломных клеток, установленных как CD138+CD38+CD19-клетки, была обнаружена в инъецированной кости, тогда как в коллатеральной кости было обнаружено только немного клеток (Фигура 8).
Эти результаты подтверждают, что генетически гуманизированные мыши, описанные в данном документе, обеспечивают приживление первичных человеческих ММ клеток in vivo. Текущие данные демонстрируют, что гуманизация генов цитокинов, кодирующих IL-6, ТРО, IL-3, GM-CSF и/или M-CSF обеспечивает приживление первичных клеток множественной миеломы, полученных от человека, которые обычно требуют человеческого микроокружения для успешной трансплантации.
Пример 2: Генетическая гуманизация гена IL-6 обеспечивает условия для приживления у мышей человеческих гематопоэтических клеток.
Материалы и методы
Мыши. Гуманизированная IL-6 KI мышь была получена, как описано выше. Химерных мышей сначала скрещивали с мышами BALB/c, а затем скрещивали повторно для того, чтобы получить потомство гомозиготное по hIL-6. Тех же самых мышей на основе смешения BALB/c×129 использовали в качестве контроля.
Новорожденных мышат (в пределах первого дня жизни) сублетально облучали рентгеновским излучением с интервалом 4 часа, используя 2×150 сантигрей. Через 4 часа после облучения мышам инъецировали 1-2×105 CD34+ эмбриональных клеток печени (FL) в 20 мкл PBS в печень, используя иглу 30 калибра (Hamilton Company, NV, США). Мышей отлучали в 3-недельном возрасте. Мышей содержали в специальных условиях, свободных от специфической патогенной микрофлоры, причем все эксперименты были проведены в соответствии с протоколами Йельского комитета по содержанию и использованию лабораторных животных.
Исследование популяций человеческих и мышиных гематологических клеток. Кровь у мышей отбирали из ретроорбитального сплетения под анестезией изофлураном в разные моменты времени после трансплантации. Образцы плазмы собирали и хранили при -20°С для дальнейшего измерения Ig. Эритроциты лизировали дважды, используя аммоний-хлоридный (АСК) лизирующий буфер, и оставшиеся РВМС ресуспендировали в FACS буфере (PBS с добавлением 5% FBS и 5 мМ EDTA).
После умерщвления мышей готовили одноклеточную суспензию из клеток, полученных из костного мозга (ВМ), тимуса и селезенки. Затем образцы окрашивали меченными флуорохромом mAb к мышиным и человеческим антигенам клеточной поверхности в соответствии с инструкциями производителя. Использовали следующие анти-человек mAb: CD45 (HI30), CD 19 (HIB19), CD3 (UCHT1), CD4 (RPA-T4), CD8 (HIT8a), CD20 (2H7), CD5 56WO 2014/071397 PCT/US2013/068569 (UCHT2), CD27 (0323), CD24 (ML5), CD10 (HIlOa), все от компании Biolegend, CA, USA; CD33 (WM53), CD38 (HIT2), IgM (G20-127), CD138 (MI15) от BD Biosciences и CD34 (AC 136) от Miltenyi Biotec. Мышиные клетки окрашивали анти-мышиными CD45 Ab (30-F11, Biolegend). Образцы получали на цитометре LSRII (BD Biosciences) и анализировали с помощью программного обеспечения FlowJo (Treestar, OR, США).
Измерение человеческих иммуноглобулинов. Общие иммуноглобулины (Igs) измеряли в плазме, собранной от мышей, методом ELISA. 96-луночные планшеты (Nunc, NY, США) покрывали при 4°С в течение ночи 20 мкг/мл очищенными козьими античеловек IgM и IgG (Southern Biotechnology, AL, США). После промывки и блокировки PBS 1% бычьим сывороточным альбумином (ВSA, Sigma-Aldrich) были добавлены соответствующие разведения образцов на 2 часа при комнатной температуре (RT). Планшеты промывали и инкубировали в течение 1 часа при RT с изотип-специфическими вторичными биотинилированными антителами (Southern Biotechnology), а затем стрептавидин-HRP (Pierce Protein Research Products, IL, США). После окончательной промывки определяли активность фермента, используя субстратный раствор ТМВ, а затем стоп-реагент (от компании Pierce Protein Research Products). Оптическую плотность измеряли при 450 нм. Образец человеческой сыворотки от Bethyl (ТХ, USA) использовали в качестве эталона.
Статистический анализ. Все данные были выражены в виде среднего значения ± стандартная ошибка среднего (SEM) за исключением Ab уровней, которые были отражены на графиках в виде среднего геометрического. Для определения статистичекой значимости между двумя группами использовали непараметрический U-критерий Манна-Уитни. Различия считались значимыми, когда ρ значения были ниже, чем 0.05.
Результаты
Приживление трансплантата периферической крови у мышей RAG2-/-γc-/- при трансплантации человеческих CD34+ клеток. Мыши IL6h/h продемонстрировали более высокое приживление периферической крови (PB) на всем протяжении времени исследования по сравнению с IL6m/m мышами, при этом приживление у них увеличивалось с течением времени (Фигура 9). Не наблюдалось значительных различий при сравнении человеческих клеток между 2 группами мышей в любой проверенный момент времени (Фигура 10). В обеих группах процентное содержание В-клеток и Т-клеток было сходным на неделе 8 и на 16-20 неделях, тогда как на неделях 11-15 наблюдалось более высокое процентное содержание В-клеток (69.23±3.97 в IL6m/m и 55.91±4.86 в IL6h/h), чем Т-клеток (16.86±3.14 в IL6m/m и 30.26±6.23 в IL6h/h). Миелоидные CD33+ клетки представляли меньшую часть человеческих клеток, и их процентное содержание понижалось с течением времени.
Приживление клеток в органах и состав человеческих CD34+ клеток, воспроизведенный у RAG2-/-γc-/- мышей. Клетки человеческого происхождения были обнаружены в гемато-лимфоидных органах, таких как ВМ, селезенка и тимус (Фигура 11А). Интересно отметить, что в селезенке IL6h/h мышей наблюдалось более значительное приживление человеческих клеток, чем у мышей IL6m/m (61.75%±10.87 против 23.56%±5.2). Эти результаты были подтверждены при удваивании абсолютного количества человеческих клеток (14.21×106±1.38 против 7.26×106±0.66) (Фигура 11В).
Созревание В-клеток у RAG2-/-γc-/- мышей с пересаженными человеческими CD34+. Стадии созревания человеческих В-клеток были исследованы в ВМ и селезенке мышей с пересаженными клетками с использованием стратегии гейтирования, показанной на Фигуре 12А, основанной на использовании комбинации антител к CD24, CD38 и поверхностному IgM. В частности, эта стратегия полезна при анализе наличия переходной подгруппы В-клеток. В ВМ основной компартмент состоял из Pro-/Pre-В клеток (Фигура 12В) без различия между мышами IL6m/m и IL6h/h (76.04%±9.09 и 79.07%±3.43 соответственно). Селезенка в обеих группах содержала некоторое количество зрелых В-клеток (-20%), однако все еще большой процент незрелых/переходных клеток (27.13%±8.99 и 36.45%±5.12).
Значительную долю В-клеток в селезенке составляли CD5+ (Фигура 13В), а маркер в большинстве случаев экспрессировался не только на человеческих ВМ и периферических В-клетках, но также обнаруживался в небольшом процентном отношении на FL В-клетках (Фигура 13А).
Мыши IL6h/h продемонстрировали сильное увеличение процента CD27+ В-клеток в селезенке по сравнению с мышами IL6m/m (24.73%±8.94 против 9.46%±2.32), однако очень мало CD27+ клеток было найдено в ВМ (Фигура 14А и 14В).
Выработка антител у RAG2-/-γc-/- мышей с пересаженными человеческими CD34+. После того, как В-клетки были обнаружены у мышей с пересаженными клетками, была измерена концентрация человеческих IgM и IgG в плазме, собранной через 12 и 20 недель после трансплантации человеческих клеток. И IL6m/m и IL6h/h мыши секретировали человеческие IgM и IgG (Фигура 15).
В общем, наблюдался более высокий процент IL6h/h мышей, секретирующих антитела, по сравнению с мышами IL6m/m, при этом первые мыши показали более низкий уровень IgM (14.69 мкг/мл против 33.66 мкг/мл на неделе 12 и 18.25 мкг/мл против 190.2 мкг/мл на 20 неделе), но повышенный уровень IgG (243 мкг/мл против 49.6 мкг/мл на 12 неделе и 553.6 мкг/мл против 297.2 мкг/мл на 20 неделе) (Фигура 15А). Средний уровень IgM и IgG увеличивался с течением времени у мышей IL6m/m (Фигура 15В). Наоборот, сывороточные Ig оставались постоянными у мышей IL6h/h.
Раскрытия всех цитированных в данном документе патентов, патентных заявок и публикаций полностью включаются в описание путем отсылки. В то время как данное изобретение раскрывается со ссылкой на конкретные варианты осуществления, следует понимать, что другие варианты осуществления и изменения этого изобретения могут быть разработаны специалистами в данной области техники без отклонения от существа и объема данного изобретения. Прилагаемые пункты формулы изобретения предназначаются для включения всех подобных вариантов осуществления и равноценных изменений.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ НЕ ОТНОСЯЩИЕСЯ К ЧЕЛОВЕЧЕСКОМУ РОДУ ЖИВОТНЫЕ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2775425C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ, НЕ ПРИНАДЛЕЖАЩИЕ К ЧЕЛОВЕЧЕСКОМУ РОДУ ЖИВОТНЫЕ И СПОСОБ ИХ ИСПОЛЬЗОВАНИЯ | 2013 |
|
RU2642319C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ НЕ ПРИНАДЛЕЖАЩИЕ К ЧЕЛОВЕЧЕСКОМУ РОДУ ЖИВОТНЫЕ И СПОСОБ ИХ ИСПОЛЬЗОВАНИЯ | 2013 |
|
RU2768282C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, ЭКСПРЕССИРУЮЩИЕ EPO ЧЕЛОВЕКА | 2015 |
|
RU2711744C1 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, ЭКСПРЕССИРУЮЩИЕ EPO ЧЕЛОВЕКА | 2015 |
|
RU2799086C2 |
| ГУМАНИЗИРОВАННЫЕ M-CSF МЫШИ | 2012 |
|
RU2577978C2 |
| ГУМАНИЗИРОВАННЫЕ M-CSF МЫШИ | 2012 |
|
RU2730643C2 |
| ГУМАНИЗИРОВАННЫЕ M-CSF МЫШИ | 2020 |
|
RU2819731C2 |
| ГУМАНИЗИРОВАННЫЕ МЫШИ С НОКИНОМ SIRPA-IL15 И СПОСОБЫ ИХ ИСПОЛЬЗОВАНИЯ | 2016 |
|
RU2730599C2 |
| ЖИВОТНЫЕ С ГУМАНИЗИРОВАННЫМ ГЕНОМ IL-15 | 2014 |
|
RU2674910C2 |
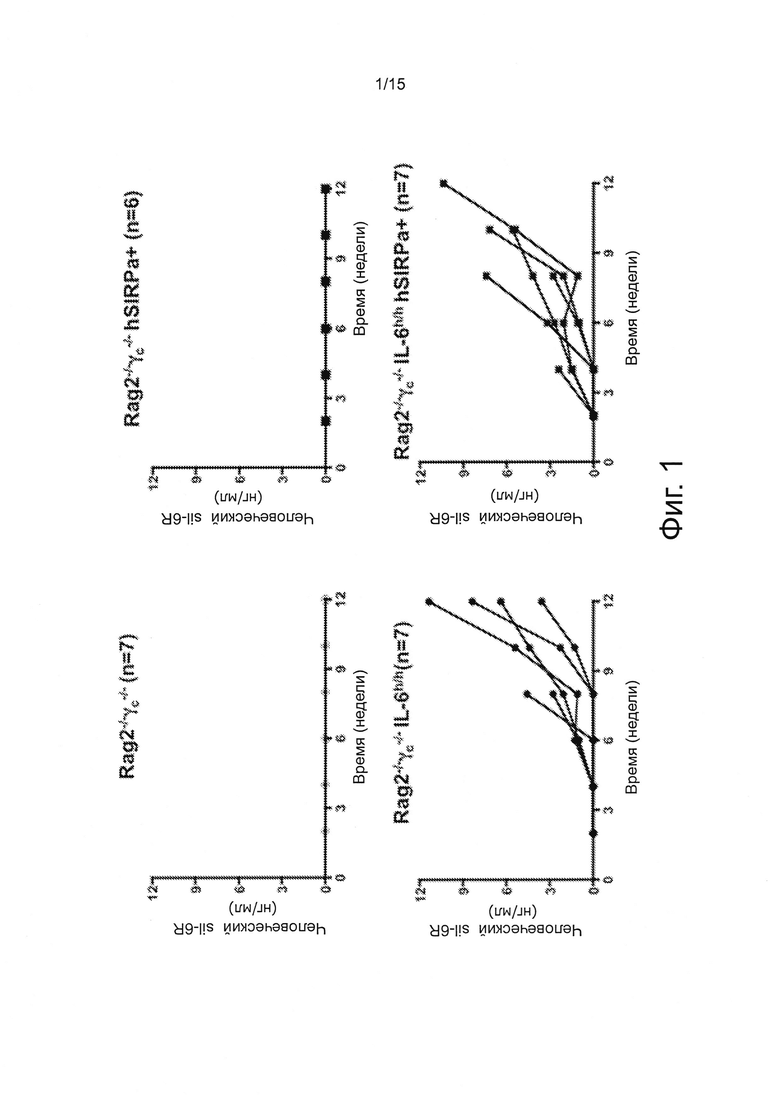
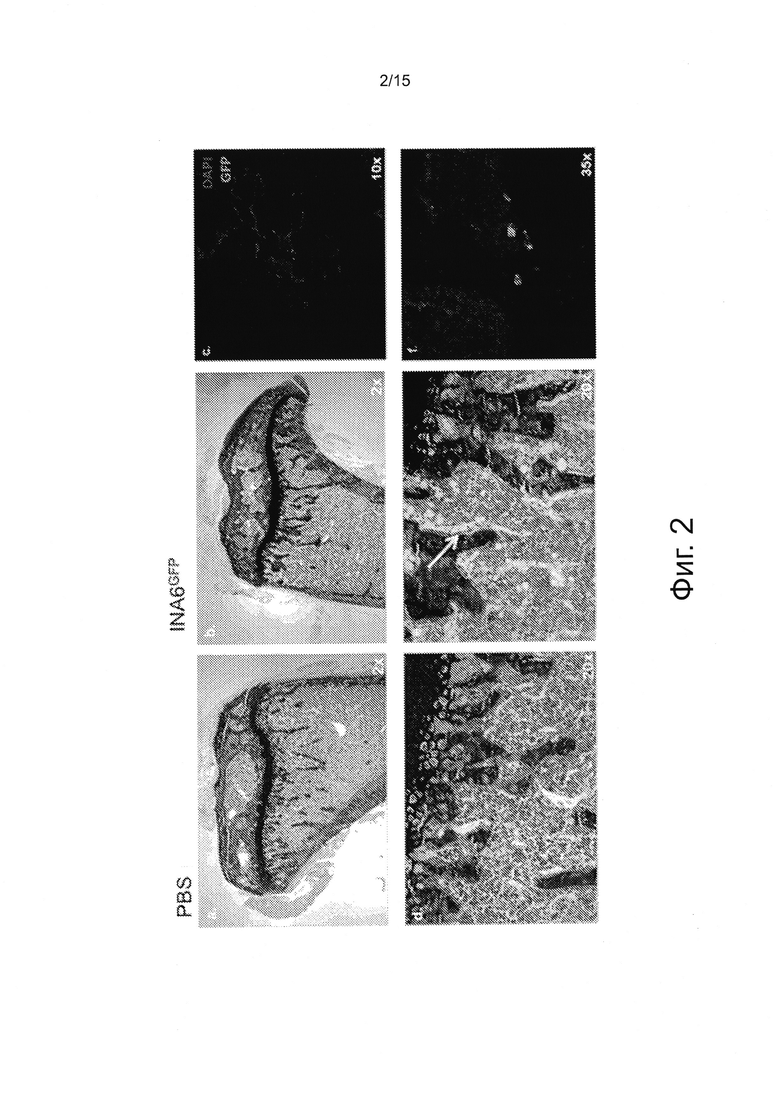
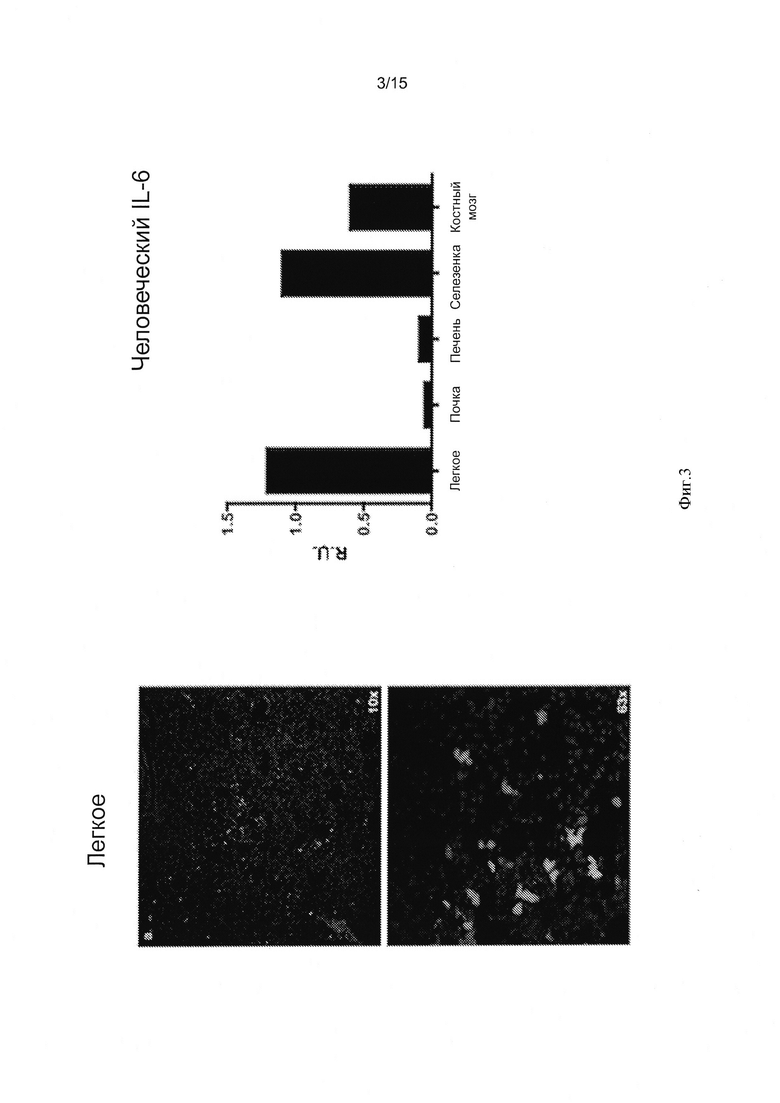
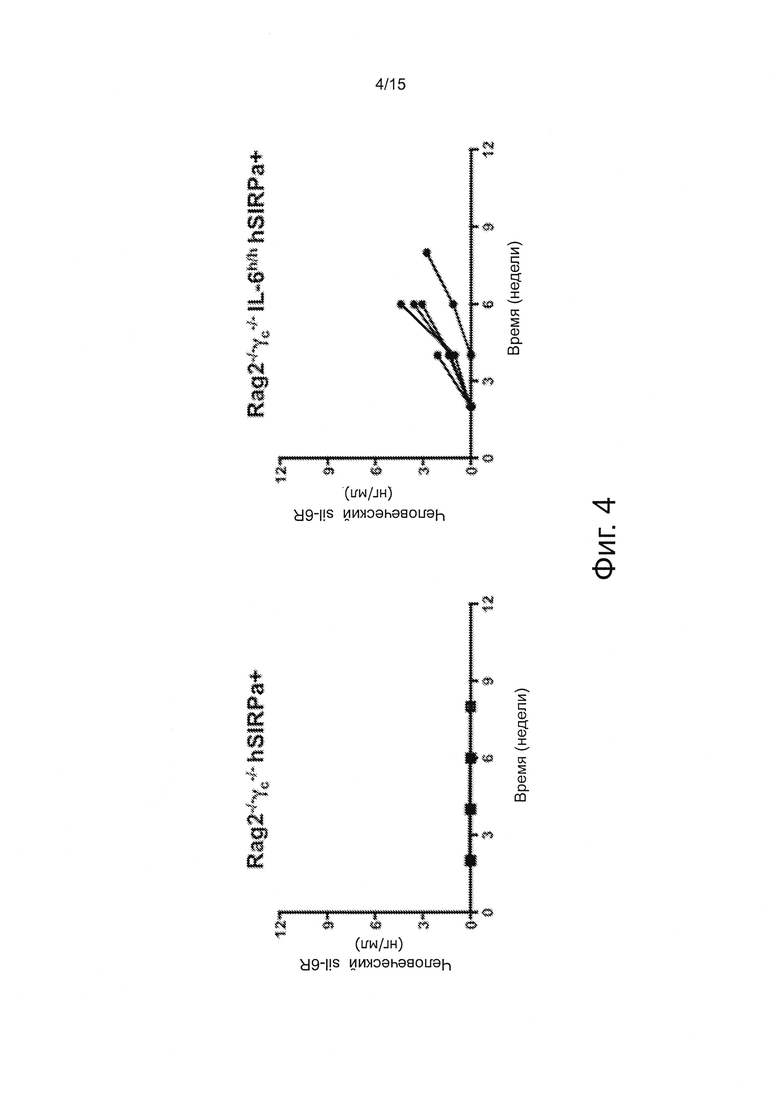
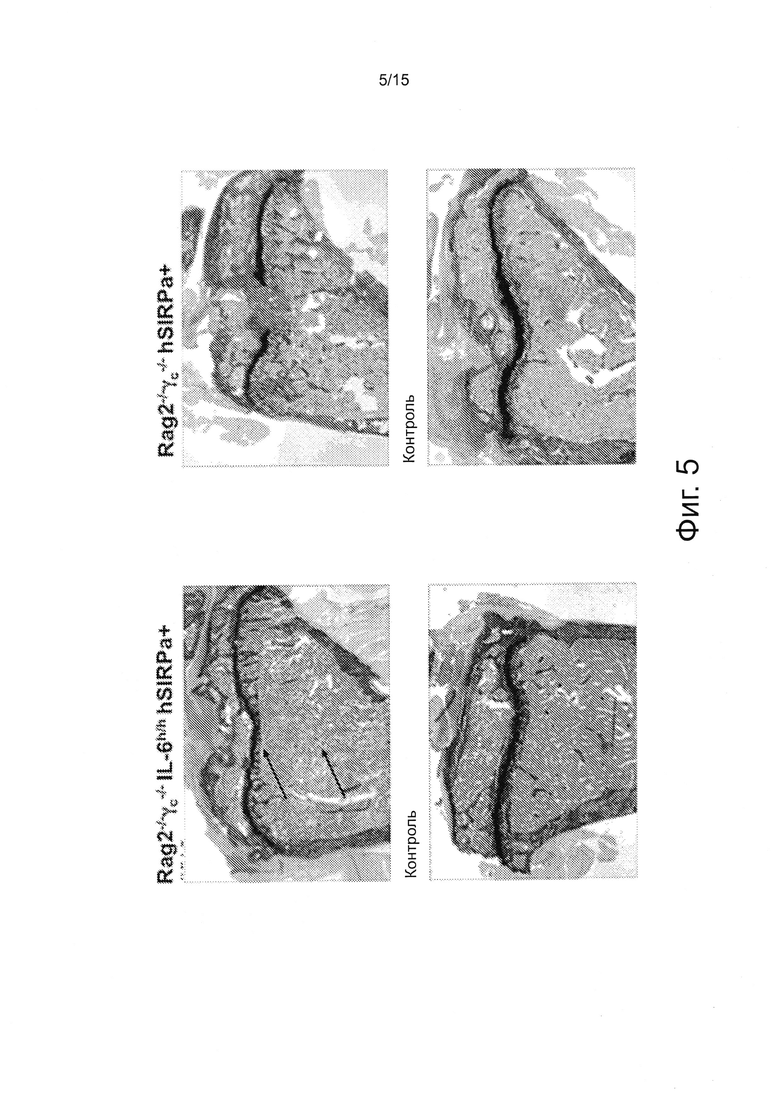
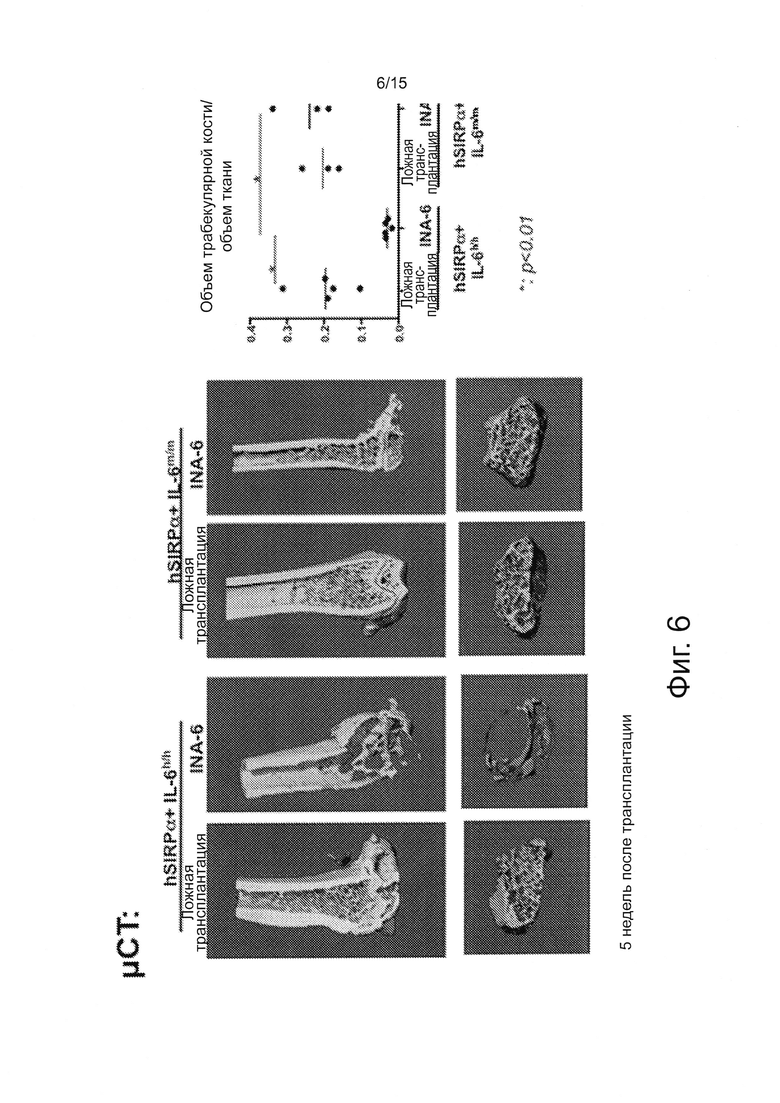
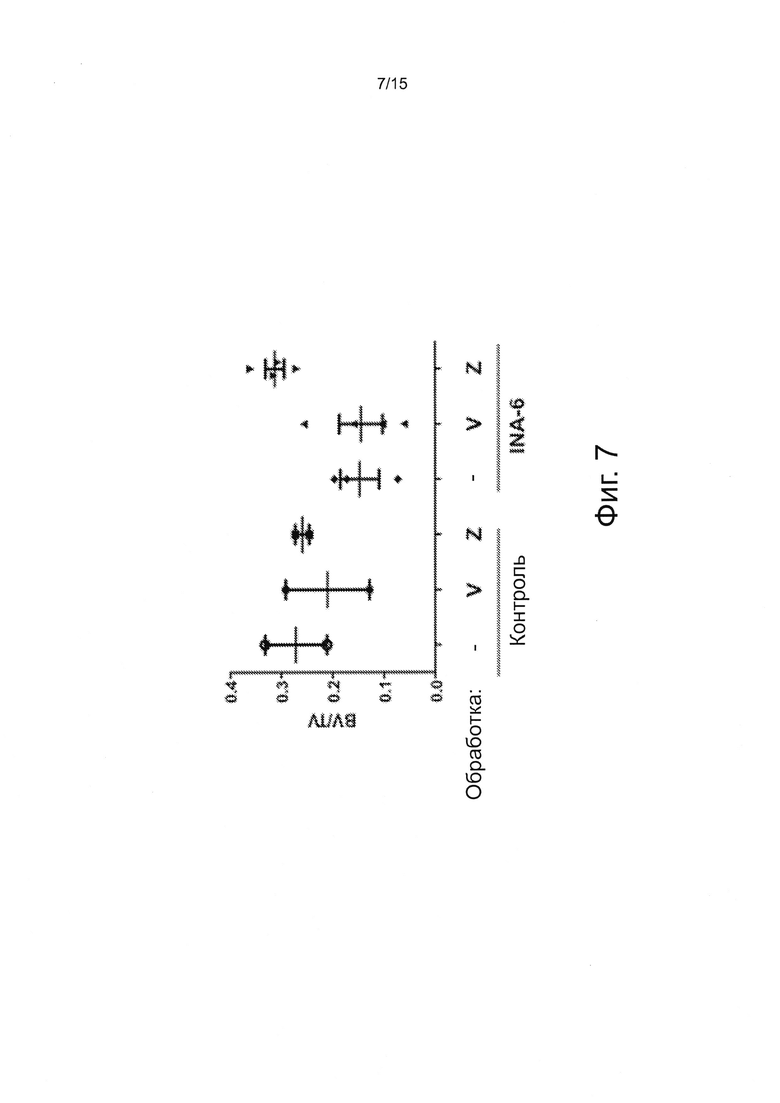
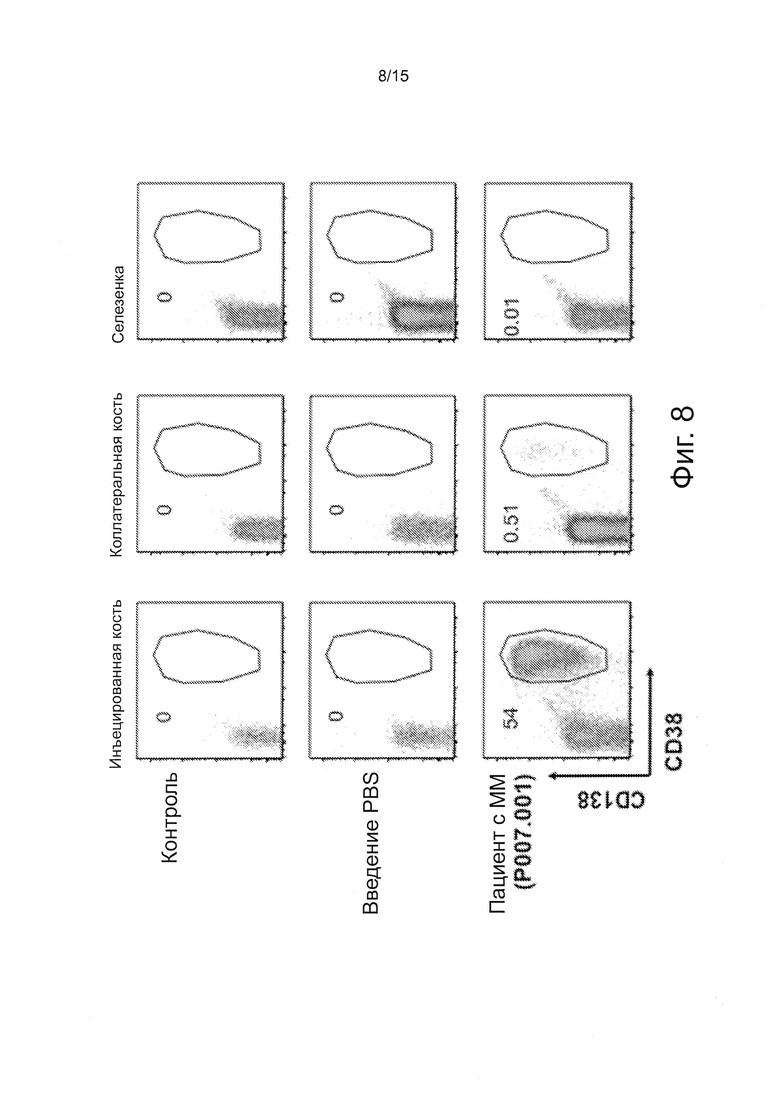
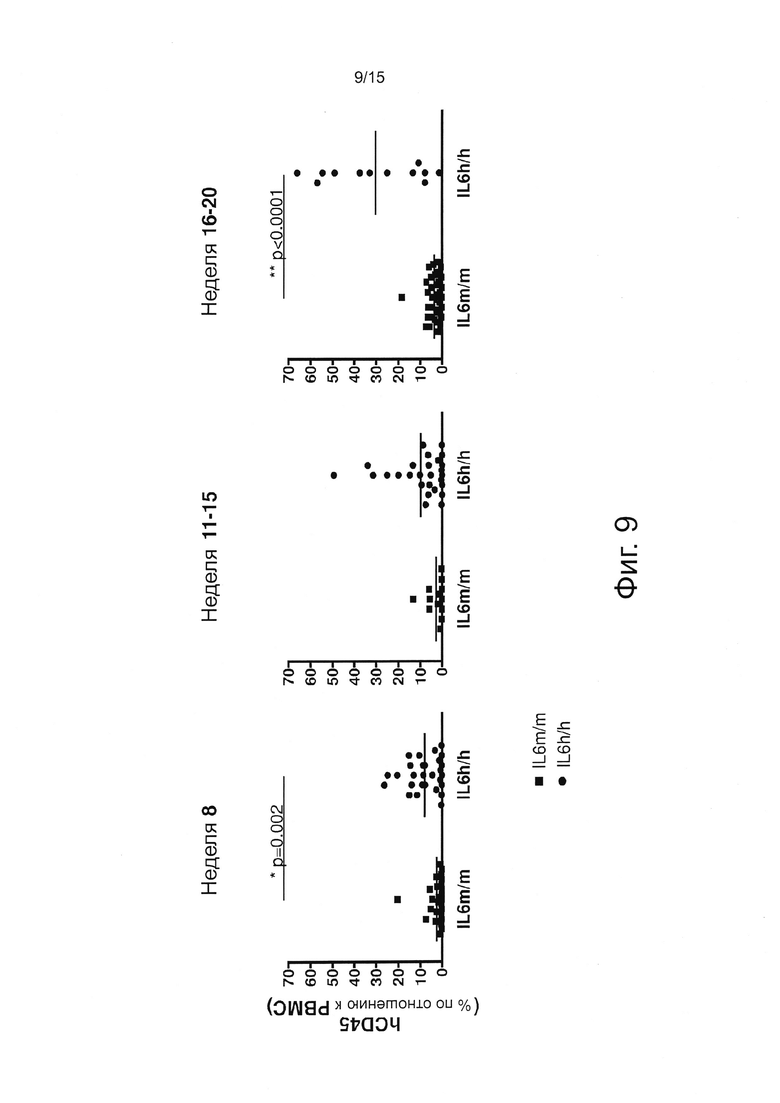
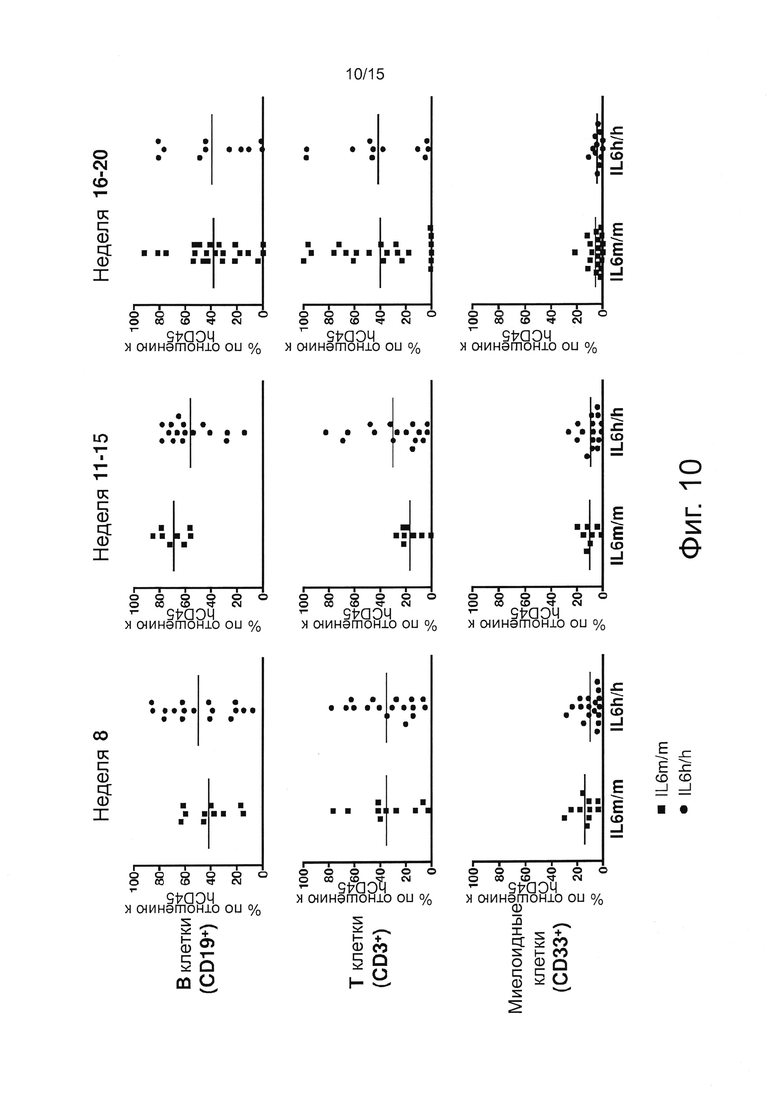
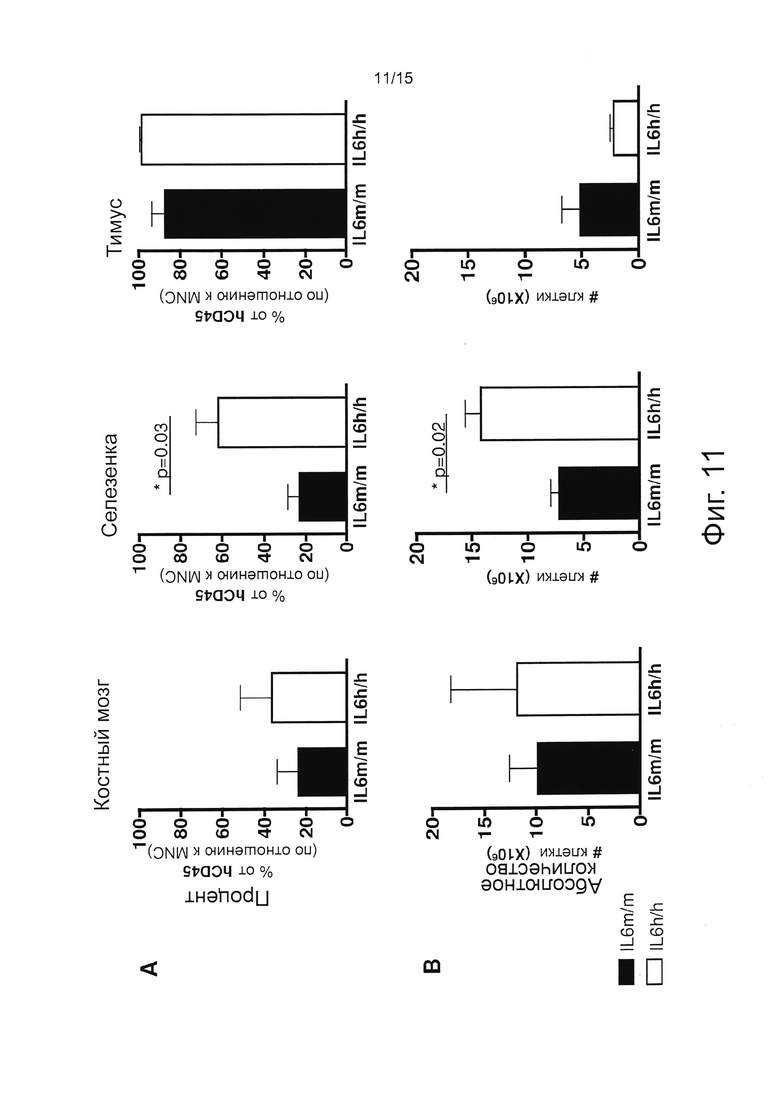
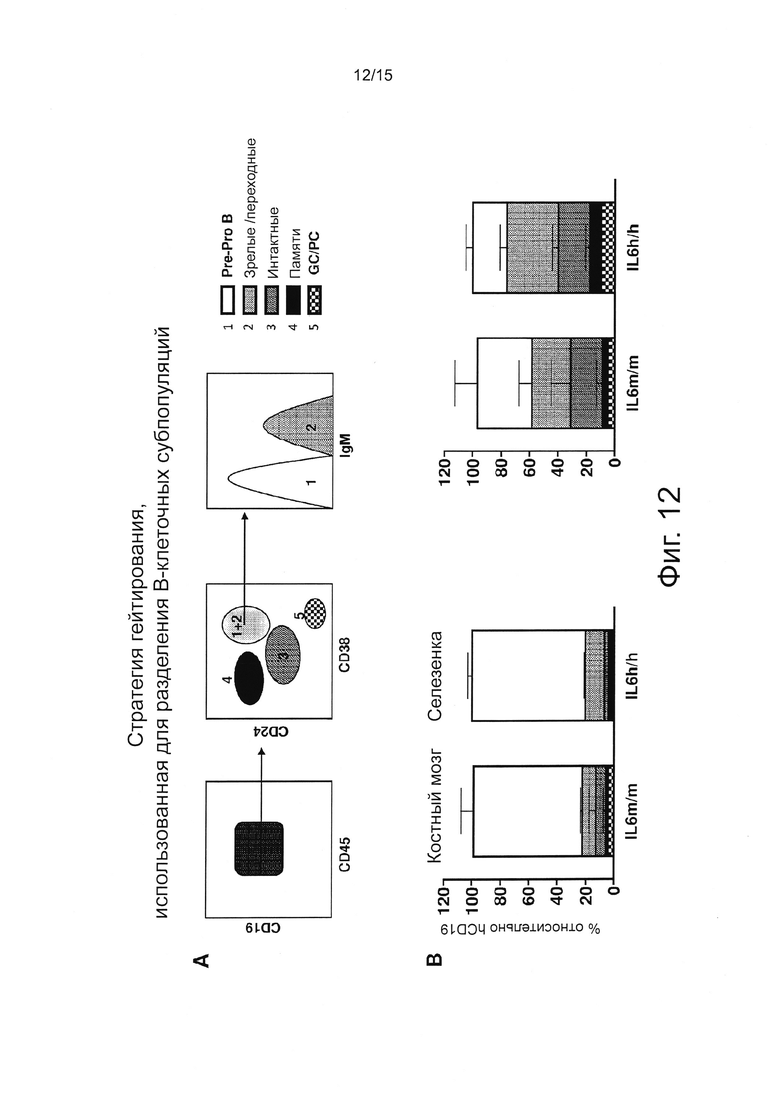
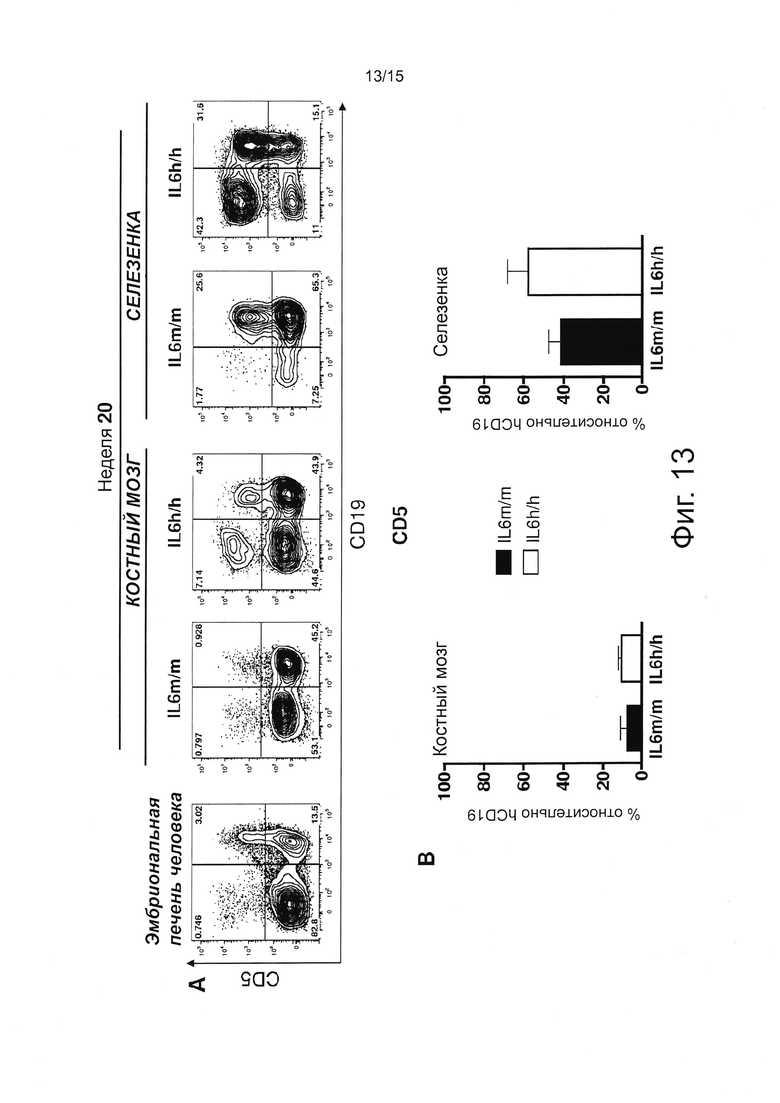
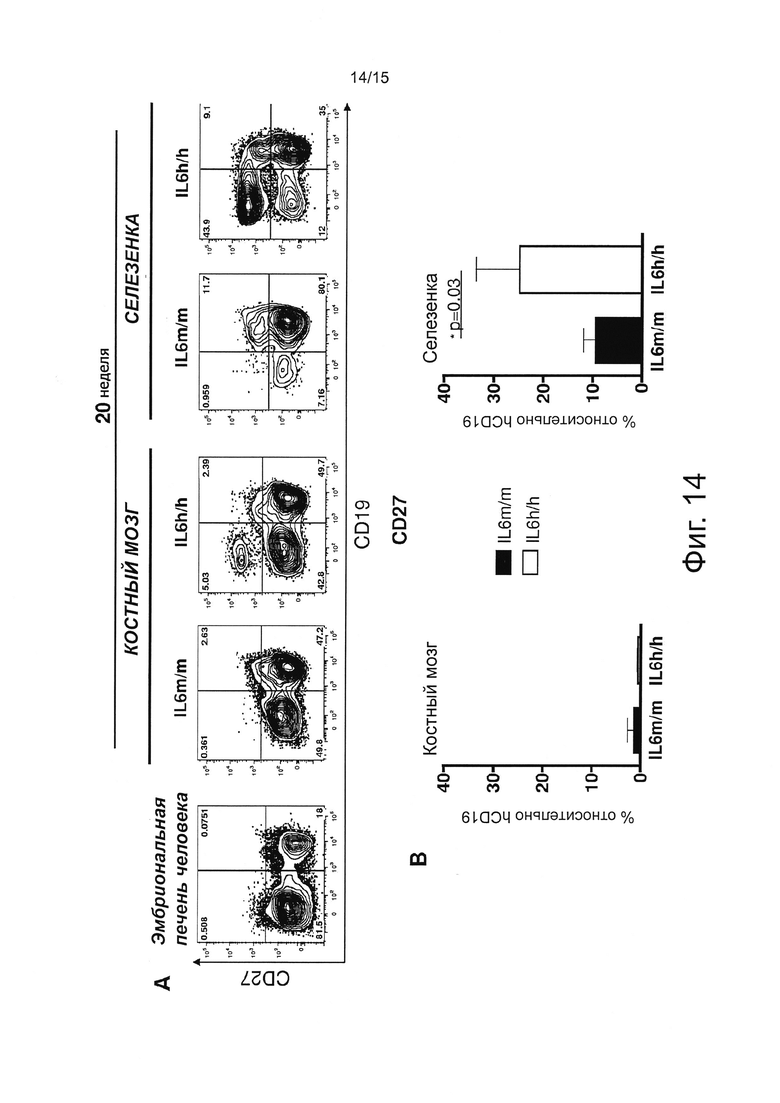
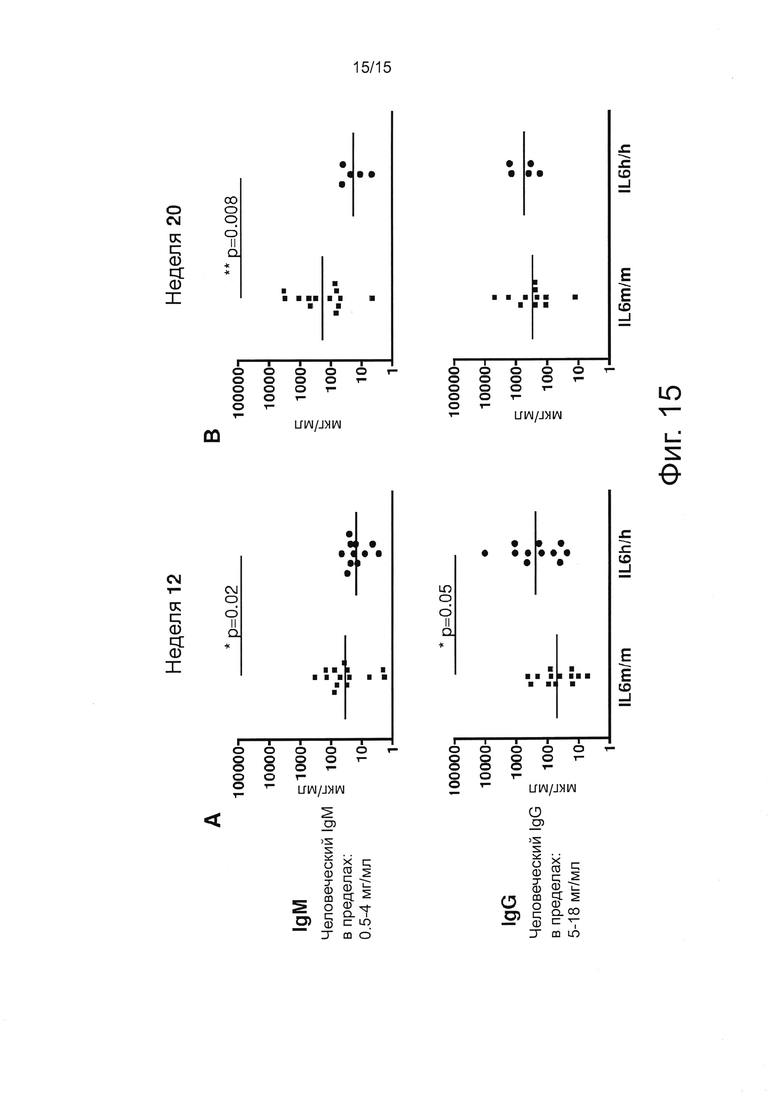
Изобретение относится к области биохимии, в частности к генетически модифицированной мыши, которая экспрессирует IL-6 человека, к мыши, которая экспрессирует IL-6 человека и один дополнительный человеческий полипептид, выбранный из человеческого M-CSF, человеческого IL-3, человеческого GM-CSF, человеческого SIRPα и человеческого TPO, а также к мыши, которая экспрессирует IL-6 человека и SIRPα человека, M-CSF человека, IL-3 человека, GM-CSF человека и TPO человека. Также раскрыт способ получения животной модели развития и функционирования человеческой В-клетки, включающий трансплантацию популяции человеческих гематопоэтических клеток генетически модифицированной мыши, который является иммунодефицитной и содержит нуклеиновую кислоту, кодирующую человеческий IL-6. Изобретение также относится к способу получения антигенсвязывающего белка с использованием мыши, экспрессирующей IL-6 человека. Изобретение позволяет эффективно использоваться для моделирования развития, функции или болезни человеческих гематопоэтических клеток. 12 н. и 36 з.п. ф-лы, 15 ил., 2 пр.
1. Генетически модифицированная мышь, которая экспрессирует IL-6 человека, где генетически модифицированная мышь содержит:
нуклеиновую кислоту, кодирующую человеческий IL-6, где нуклеиновая кислота, кодирующая человеческий IL-6, функционально связана с IL-6 промотором в локусе IL-6 мыши.
2. Генетически модифицированная мышь, которая экспрессирует IL-6 человека, где генетически модифицированная мышь дополнительно экспрессирует SIRPα человека, где генетически модифицированная мышь содержит:
нуклеиновую кислоту, кодирующую человеческий IL-6, где нуклеиновая кислота, кодирующая человеческий IL-6, функционально связана с IL-6 промотором в локусе IL-6 мыши, и
нуклеиновую кислоту, кодирующую человеческий SIRPα, где нуклеиновая кислота, кодирующая человеческий SIRPα, функционально связана с SIRPα промотором в локусе SIRPα мыши,
3. Генетически модифицированная мышь, которая экспрессирует IL-6 человека, где генетически модифицированная мышь дополнительно экспрессирует M-CSF человека, где генетически модифицированная мышь содержит:
нуклеиновую кислоту, кодирующую человеческий IL-6, где нуклеиновая кислота, кодирующая человеческий IL-6, функционально связана с IL-6 промотором в локусе IL-6 мыши, и
нуклеиновую кислоту, кодирующую человеческий M-CSF, где нуклеиновая кислота, кодирующая человеческий M-CSF, функционально связана с M-CSF промотором в локусе M-CSF мыши.
4. Генетически модифицированная мышь, которая экспрессирует IL-6 человека, где генетически модифицированная мышь дополнительно экспрессирует IL-3 человека, где генетически модифицированная мышь содержит:
нуклеиновую кислоту, кодирующую человеческий IL-6, где нуклеиновая кислота, кодирующая человеческий IL-6, функционально связана с IL-6 промотором в локусе IL-6 мыши, и
нуклеиновую кислоту, кодирующую человеческий IL-3, где нуклеиновая кислота, кодирующая человеческий IL-3, функционально связана с IL-3 промотором в локусе IL-3 мыши.
5. Генетически модифицированная мышь, которая экспрессирует IL-6 человека, где генетически модифицированная мышь дополнительно экспрессирует GM-CSF человека, где генетически модифицированная мышь содержит:
нуклеиновую кислоту, кодирующую человеческий IL-6, где нуклеиновая кислота, кодирующая человеческий IL-6, функционально связана с IL-6 промотором в локусе IL-6 мыши, и
нуклеиновую кислоту, кодирующую человеческий GM-CSF, где нуклеиновая кислота, кодирующая человеческий GM-CSF, функционально связана с GM-CSF промотором в локусе GM-CSF мыши.
6. Генетически модифицированная мышь, которая экспрессирует IL-6 человека, где генетически модифицированная мышь дополнительно экспрессирует TPO человека, где генетически модифицированная мышь содержит:
нуклеиновую кислоту, кодирующую человеческий IL-6, где нуклеиновая кислота, кодирующая человеческий IL-6, функционально связана с IL-6 промотором в локусе IL-6 мыши, и
нуклеиновую кислоту, кодирующую человеческий TPO, где нуклеиновая кислота, кодирующая человеческий TPO, функционально связана с TPO промотором в локусе TPO мыши.
7. Генетически модифицированная мышь, которая экспрессирует IL-6 человека, где генетически модифицированная мышь дополнительно экспрессирует SIRPα человека, M-CSF человека, IL-3 человека, GM-CSF человека и TPO человека, где генетически модифицированная мышь содержит:
нуклеиновую кислоту, кодирующую человеческий IL-6, где нуклеиновая кислота, кодирующая человеческий IL-6, функционально связана с IL-6 промотором в локусе IL-6 мыши, и
нуклеиновую кислоту, кодирующую человеческий SIRPα, где нуклеиновая кислота, кодирующая человеческий SIRPα, функционально связана с SIRPα промотором в локусе SIRPα мыши;
нуклеиновую кислоту, кодирующую человеческий M-CSF, где нуклеиновая кислота, кодирующая человеческий M-CSF, функционально связана с M-CSF промотором в локусе M-CSF мыши;
нуклеиновую кислоту, кодирующую человеческий IL-3, где нуклеиновая кислота, кодирующая человеческий IL-3, функционально связана с IL-3 промотором в локусе IL-3 мыши;
нуклеиновую кислоту, кодирующую человеческий GM-CSF, где нуклеиновая кислота, кодирующая человеческий GM-CSF, функционально связана с GM-CSF промотором в локусе GM-CSF мыши; и
нуклеиновую кислоту, кодирующую человеческий TPO, где нуклеиновая кислота, кодирующая человеческий TPO, функционально связана с TPO промотором в локусе TPO мыши.
8. Генетически модифицированная мышь по любому из пп. 1-7, где IL-6 промотор является IL-6 промотором мыши.
9. Генетически модифицированная мышь по любому из пп. 1-7, где мышь является иммунодефицитной.
10. Генетически модифицированная мышь по п. 9, где мышь не экспрессирует ген, активирующий рекомбинацию (RAG).
11. Генетически модифицированная мышь по п. 9, где мышь не экспрессирует гамма цепь рецептора IL2 (гамма цепь-/-).
12. Генетически модифицированная мышь по п. 9, где мышь не экспрессирует RAG2 и не экспрессирует IL2rg.
13. Генетически модифицированная мышь по любому из пп. 1-7, дополнительно содержащая трансплантированные человеческие гематопоэтические клетки.
14. Генетически модифицированная мышь по п. 13, где человеческие гематопоэтические клетки представляют собой CD34+ клетки.
15. Генетически модифицированная мышь по п. 13, где человеческие гематопоэтические клетки представляют собой клетки множественной миеломы.
16. Способ получения животной модели развития и функционирования человеческой В-клетки, данный способ включает:
трансплантацию популяции человеческих гематопоэтических клеток генетически модифицированной мыши, которая является иммунодефицитной и содержит нуклеиновую кислоту, кодирующую человеческий IL-6, где нуклеиновая кислота, кодирующая человеческий IL-6, функционально связана с IL-6 промотором в локусе IL-6 мыши, при этом животное экспрессирует человеческий IL-6 и не экспрессирует IL-6 мыши.
17. Способ по п. 16, в котором IL-6 промотор является IL-6 промотором мыши.
18. Способ по п. 16, согласно которому трансплантированная популяция гематопоэтических клеток содержит CD34+ клетки.
19. Способ по п. 16, согласно которому трансплантированная популяция гематопоэтических клеток содержит клетки множественной миеломы.
20. Способ по п. 16, согласно которому трансплантация включает интрафеморальную и/или интратибиальную инъекцию.
21. Способ по п. 16, согласно которому мышь дополнительно содержит нуклеиновую кислоту, кодирующую человеческий SIRPα, где нуклеиновая кислота, кодирующая человеческий SIRPα, функционально связана с SIRPα промотором в локусе SIRPα мыши.
22. Способ по п. 21, согласно которому иммунодефицитная, генетически модифицированная мышь экспрессирует по меньшей мере одну дополнительную человеческую нуклеиновую кислоту, выбранную из группы, состоящей из:
i) нуклеиновой кислоты, кодирующей человеческий M-CSF, где нуклеиновая кислота, кодирующая человеческий M-CSF, функционально связана с M-CSF промотором в локусе M-CSF мыши;
ii) нуклеиновой кислоты, кодирующей человеческий IL-3, где нуклеиновая кислота, кодирующая человеческий IL-3, функционально связана с IL-3 промотором в локусе IL-3 мыши;
iii) нуклеиновой кислоты, кодирующей человеческий GM-CSF, где нуклеиновая кислота, кодирующая человеческий GM-CSF, функционально связана с GM-CSF промотором в локусе GM-CSF мыши; и
iv) нуклеиновой кислоты, кодирующей человеческий TPO, где нуклеиновая кислота, кодирующая человеческий TPO, функционально связана с TPO промотором в локусе TPO мыши.
23. Генетически модифицированная мышь с пересаженной популяцией гематопоэтических клеток человека, которая является иммунодефицитной и содержит нуклеиновую кислоту, кодирующую человеческий IL-6, где нуклеиновая кислота, кодирующая человеческий IL-6, функционально связана с IL-6 промотором в локусе IL-6 мыши, где животное экспрессирует IL-6 человека и не экспрессирует IL-6 мыши, где указанное животное является животной моделью развития и функции человеческих В-клеток.
24. Генетически модифицированная мышь по п. 23, где пересаженная популяция гематопоэтических клеток человека содержит CD34+ клетки.
25. Генетически модифицированная мышь по п. 23, где пересаженная популяция гематопоэтических клеток человека содержит клетки множественной миеломы.
26. Генетически модифицированная мышь по п. 23, где мышь дополнительно содержит нуклеиновую кислоту, кодирующую человеческий SIRPα, где нуклеиновая кислота, кодирующая человеческий SIRPα, функционально связана с SIRPα промотором в локусе SIRPα мыши.
27. Генетически модифицированная мышь по п. 23, где иммунодефицитная, генетически модифицированная мышь экспрессирует по меньшей мере одну дополнительную человеческую нуклеиновую кислоту, выбранную из группы, состоящей из:
i) нуклеиновой кислоты, кодирующей человеческий M-CSF, где нуклеиновая кислота, кодирующая человеческий M-CSF, функционально связана с M-CSF промотором в локусе M-CSF мыши;
ii) нуклеиновой кислоты, кодирующей человеческий IL-3, где нуклеиновая кислота, кодирующая человеческий IL-3, функционально связана с IL-3 промотором в локусе IL-3 мыши;
iii) нуклеиновой кислоты, кодирующей человеческий GM-CSF, где нуклеиновая кислота, кодирующая человеческий GM-CSF, функционально связана с GM-CSF промотором в локусе GM-CSF мыши; и
iv) нуклеиновой кислоты, кодирующей человеческий TPO, где нуклеиновая кислота, кодирующая человеческий TPO, функционально связана с TPO промотором в локусе TPO мыши.
28. Способ скрининга средства-кандидата в отношении его способности лечить гематопоэтический рак человека, данный способ включает:
контактирование генетически модифицированной мыши по п. 22 со средством-кандидатом и
сравнение жизнеспособности и/или скорости пролиферации клеток гематопоэтического рака человека у мыши, которая контактировала со средством-кандидатом, с жизнеспособностью и/или скоростью пролиферации клеток гематопоэтического рака человека у генетически модифицированной мыши по п. 22, которая не контактировала со средством-кандидатом,
при этом уменьшение жизнеспособности и/или скорости пролиферации клеток гематопоэтического рака человека у контактировавшей мыши показывает, что средство-кандидат будет лечить гематопоэтический рак человека.
29. Способ по п. 28, согласно которому гематопоэтический рак является множественной миеломой.
30. Способ получения антигенсвязывающего белка, где способ включает:
иммунизацию генетически модифицированной, с пересаженными клетками, иммунодефицитной мыши антигеном, где мышь содержит:
геном, содержащий нуклеиновую кислоту, кодирующую человеческий IL-6, где нуклеиновая кислота функционально связана с промотором в мышином локусе гена IL-6, и
пересаженные гематопоэтические клетки человека, где после иммунизации, мышь продуцирует клетки человека, содержащие антигенсвязывающий белок, который специфически связывает антиген.
31. Способ по п. 30, дополнительно включающий извлечение из указанной мыши по меньшей мере одного из селезенки, лимфоузлов, периферической крови, костного мозга или части любого из них.
32. Способ по п. 31, дополнительно включающий выделение человеческой клетки из по меньшей мере одного из селезенки, лимфоузлов, периферической крови, костного мозга или части любого из них из указанной мыши.
33. Способ по п. 32, дополнительно включающий выделение антигенсвязывающего белка части из человеческой клетки.
34. Способ по п. 33, где клетка человека является В клеткой.
35. Способ по п. 32, далее включающий получение клеточной линии гибридомы из указанной клетки человека.
36. Способ по п. 35, далее включающий выделение из клеточной линии гибридомы антигенсвязывающего белка, который специфически связывается с антигеном.
37. Способ по п. 30, где антигенсвязывающий белок является IgG человека.
38. Способ по п. 30, где гематопоэтические клетки человека содержат клетки CD34+.
39. Способ по п. 30, где гематопоэтические клетки человека получены из одного или нескольких из печени зародыша человека, костного мозга человека, пуповинной крови человека, периферической крови человека и селезенки человека.
40. Способ по п. 30, где антиген является по меньшей мере одним из пептида, комплекса MHC/пептид, ДНК, живого вируса, мертвого вируса или их части, живой бактерии, мертвой бактерии или их части и раковой клетки или ее части.
41. Способ по п. 30, где антиген вводят генетически модифицированной, с пересаженными клетками, иммунодефицитной мыши примерно через 1-5 месяцев после пересадки гематопоэтических клеток человека.
42. Способ по п. 30, где нуклеиновая кислота функционально связана с IL-6 промотором генетически модифицированной, с пересаженными клетками, иммунодефицитной мыши.
43. Способ по п. 30, где генетически модифицированная, с пересаженными клетками, иммунодефицитная мышь не экспрессирует мышиный IL-6.
44. Способ по п. 30, где генетически модифицированная, с пересаженными клетками, иммунодефицитная мышь содержит геном, дополнительно содержащий нуклеиновую кислоту, кодирующую SIRPα человека, где нуклеиновая кислота функционально связана с промотором.
45. Способ по п. 30, где генетически модифицированная, с пересаженными клетками, иммунодефицитная мышь имеет нокаут гена, активирующего рекомбинацию (RAG).
46. Способ по п. 30, где генетически модифицированная, с пересаженными клетками, иммунодефицитная мышь имеет нокаут гена гамма цепи рецептора IL2 (IL2rg).
47. Способ по п. 30, где генетически модифицированная, с пересаженными клетками, иммунодефицитная мышь имеет нокаут гена, активирующего рекомбинацию (RAG), и нокаут гена гамма цепи рецептора IL2 (IL2rg).
48. Способ выделения антигенсвязывающего белка, где способ включает:
выделение из генетически модифицированной, с пересаженными клетками, иммунодефицитной мыши человеческой клетки, продуцирующей антигенсвязывающий белок, который специфически связывается с антигеном, и
выделение антигенсвязывающего белка из клетки человека,
где генетически модифицированная, с пересаженными клетками, иммунодефицитная мышь содержит:
геном, содержащий нуклеиновую кислоту, кодирующую человеческий IL-6, где нуклеиновая кислота функционально связана с промотором в мышином локусе гена IL-6, и
пересаженные гематопоэтические клетки человека,
и где генетически модифицированная, с пересаженными клетками, иммунодефицитная мышь была иммунизирована антигеном.
| WO 2011044050 A2, 14.04.2011 | |||
| RATHINAM CHOZHAVENDAN et al., Efficient differentiation and function of human macrophages in humanized CSF-1 mice, BLOOD, 2011, Vol.118, No.11, pp | |||
| Светящийся в темноте электрический выключатель | 1924 |
|
SU3119A1 |
| GROEN R | |||
| W | |||
| J | |||
| et al., Reconstructing the human hematopoietic niche in immunodeficient mice: opportunities for studying primary multiple myeloma, BLOOD, 2012, vol | |||
| Кровля из глиняных обожженных плит с арматурой из проволочной сетки | 1921 |
|
SU120A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| RATHINAM CHOZHAVENDAN et al., Efficient differentiation and function of human macrophages in humanized CSF-1 mice, BLOOD, 2011, Vol.118, No.11, pp | |||
| Светящийся в темноте электрический выключатель | 1924 |
|
SU3119A1 |
| WILLINGERA TIM et al., Human IL-3/GM-CSF knock-in mice support human alveolar macrophage development and human immune responses in the lung, PNAS, 2011, vol.108, no.6, pp.2390-2395 | |||
| STROWIGA TILL et al., Transgenic expression of human signal regulatory protein alpha in Rag2−/−γc−/− mice improves engraftment of human hematopoietic cells in humanized mice, PNAS, 2011, vol.108, no.32, pp.13218-13223 | |||
| РЫБЧИН В.Н., Основы генетической инженерии, Санкт-Петербург, Издательство СПбГТУ, 2002, с.411-413. | |||

