Перекрестные ссылки на родственные заявки
По данной заявке испрашивается приоритет по предварительной патентной заявке США № 62/000460, поданной 19 мая 2014 года, полное содержание которой включено в настоящий документ посредством ссылки.
Область техники, к которой относится изобретение
Данное изобретение относится к области генетически модифицированных животных, отличных от человека.
Введение
Генетически модифицированные мыши, модифицированные и привитые мыши, и их применение в моделировании заболеваний у человека известно в данной области. Однако на сегодняшний день не было достигнуто значительного успеха в создании генетически модифицированных мышей, которые моделируют инфекцию у человека, вызванную патогенами, которые поражают человеческие клетки эритроидного ростка. Такие патогены, например, простейшие рода Plasmodium, Babesia и Theileria, могут вызывать угрожающие жизни заболевания у человека.
Например, простейшие рода Plasmodium вызывают малярию. В 2010 году в общей сложности 106 стран по всему миру были охвачены эпидемией малярии, и согласно оценкам 3,3 миллиарда людей имели риск возникновения этого заболевания. Вспышка заболевания в 2010 году оценивалась в 216 миллионов случаев с 655000 смертельными исходами по всему миру, из которых 86% составляли дети в возрасте до 5 лет. В настоящее время лекарственные средства и вакцины для предупреждения и лечения малярии все еще очень ограничены. Кроме того, возникает устойчивость паразита к обычно используемым против малярии терапевтическим средствам, что представляет постоянную проблему. Таким образом, срочно требуется разработка новых лекарственных средств и вакцин для контроля и лечения патогенов, которые поражают эритроциты человека.
Поскольку многие из этих патогенов не инфицируют красные кровяные клетки лабораторных грызунов, исследования in vivo обычно ограничивались исследованиями малярии, вызванной паразитом грызунов Plasmodium berghei ANKA, или исследованиями мышей NOD/SCID, NOD/SCID/IL2rgnull (NSG) или BXN, остро привитых путем ежедневной инъекции больших количеств человеческих эритроцитов и случайным образом или позднее инфицированных путем инъекции инфицированных паразитами красных кровяных клеток (Angulo-Barturen et al. (2008) A murine model of falciparum-malaria by in vivo selection of competent strains in non-myelodepleted mice engrafted with human erythrocytes. PLoS One 3:e2252; Jimenez-Diaz et al. (2009) Improved murine model of malaria using Plasmodium falciparum competent strains and non-myelodepleted NOD-scid IL2Rgnull mice engrafted with human erythrocytes. Antimicrob Agents Chemother 53:4533-4536; Badell et al. (2000) Human malaria in immunocompromised mice: an in vivo model to study defense mechanisms against Plasmodium falciparum. JEM 192(11):1653-1660; Moreno et al. (2006) The course of infections and pathology in immunomodulated NOD/LtSz-SCID mice inoculated with Plasmodium falciparum laboratory lines and clinical isolates. Int. J. Parasitol. 36:361-369). Для исследования эффектов Plasmodium и других патогенов на человека, а также для тестирования вакцин и лекарственных средств в отношении эффективности по предупреждению инфекции и лечению инфицированного этим и другими патогенами человека, было бы полезно иметь животное, отличное от человека, такое как мышь, которое генетически модифицировано таким образом, что подвержено инфекции таким патогеном, или таким образом, что оно будет лучше поддерживать человеческие эритроциты, остро привитые животному до инфицирования, как это проделано в типичных моделях грызунов.
Краткое описание изобретения
Предлагаются генетически модифицированные животные, отличные от человека, экспрессирующие человеческий EPO из генома животного. Также, предлагаются способы получения животных, отличных от человека, экспрессирующих человеческий EPO из генома животного, отличного от человека, и способы применения животных, отличных от человека, экспрессирующих человеческий EPO из генома животного, отличного от человека. Эти животные и способы находят множество применений в данной области, включая, например, моделирование человеческого эритропоэза и функции эритроцитов; моделирование человеческой патогенной инфекции эритроцитов; in vivo скрининги агентов, которые модулируют эритропоэз и/или функцию эритроцитов, например, в нормальном или болезненном состоянии; in vivo скрининги агентов, которые являются токсичными для эритроцитов или предшественников эритроцитов; in vivo скрининги агентов, которые предупреждают, ослабляют или устраняют токсические эффекты токсичных агентов на эритроциты или предшественники эритроцитов; in vivo скрининги эритроцитов или предшественников эритроцитов, полученных от индивидуума, для предсказания отвечаемости индивидуума на терапию заболевания.
В некоторых аспектах изобретения предлагаются генетически модифицированные животные, отличные от человека, которые экспрессируют человеческий EPO из генома животного, отличного от человека. Другими словами, животное, отличное от человека, несет нуклеотидную последовательность, кодирующую человеческий белок EPO.
В некоторых вариантах осуществления нуклеотидная последовательность, кодирующая человеческий белок EPO, функционально связана с промотором гена EPO. В некоторых вариантах осуществления промотор гена EPO представляет собой человеческий промотор EPO. В других вариантах осуществления промотор EPO является эндогенным, то есть нечеловеческим промотором EPO. В некоторых из таких вариантов осуществления эндогенный промотор EPO расположен в локусе гена EPO животного, отличного от человека. Другими словами, в определенных вариантах осуществления нуклеотидная последовательность, которая кодирует человеческий белок EPO, является функционально связанной с промотором EPO животного, отличного от человека, в локусе EPO животного, отличного от человека. В некоторых из таких вариантов осуществления эта функциональная связь приводит к null-мутации в гене нечеловеческого EPO в локусе гена нечеловеческого EPO.
В некоторых вариантах осуществления подопытное животное, отличное от человека, является гетерозиготным по аллелю, несущему нуклеотидную последовательность, которая кодирует человеческий белок EPO. В других вариантах осуществления животное, отличное от человека, является гомозиготным по аллелю, несущему нуклеотидную последовательность, которая кодирует человеческий белок EPO.
В некоторых вариантах осуществления нуклеотидная последовательность, которая кодирует человеческий белок EPO, содержит геномную кодирующую и некодирующую последовательность человеческого EPO. В других вариантах осуществления нуклеотидная последовательность, которая кодирует человеческий белок EPO, содержит кДНК-последовательность человеческого EPO.
В некоторых вариантах осуществления подопытное животное, отличное от человека, экспрессирует один или несколько дополнительных человеческих белков, выбранных из группы, состоящей из: белка M-CSF, кодируемого нуклеиновой кислотой под контролем промотора M-csf, белка IL-3, кодируемого нуклеиновой кислотой под контролем промотора Il-3, белка GM-CSF, кодируемого нуклеиновой кислотой под контролем промотора Gm-csf, белка TPO, кодируемого нуклеиновой кислотой под контролем промотора TPO, и белка Sirpa, кодируемого нуклеиновой кислотой под контролем промотора Sirpa. В некоторых из таких вариантов осуществления промотор представляет собой эндогенный промотор животного, отличного от человека, в соответствующем локусе гена животного, отличного от человека, и животное, отличное от человека, является гетерозиготным нулевым (null) по нечеловеческому гену. В других вариантах осуществления промотор представляет собой эндогенный промотор животного, отличного от человека, в соответствующем локусе гена животного, отличного от человека, и животное, отличное от человека, является гомозиготным нулевым (null) по нечеловеческому гену. В определенных вариантах осуществления животное, отличное от человека, экспрессирует человеческие, например, гуманизированные белки, выбранные из группы, состоящей из: человеческого, например, гуманизированного TPO; человеческого, например, гуманизированного IL-3; человеческого, например, гуманизированного GM-CSF; человеческого, например, гуманизированного EPO; и человеческого, например, гуманизированного Sirpa; и их комбинации.
В некоторых вариантах осуществления подопытное генетически модифицированное животное, отличное от человека, представляет собой мышь, имеющую геном, который несет нуклеотидную последовательность, кодирующую человеческий, например, гуманизированный белок, например, нуклеиновую кислоту, кодирующую человеческий, например, гуманизированный EPO; человеческий, например, гуманизированный Sirpa; человеческий, например, гуманизированный IL-3; человеческий, например, гуманизированный GM-CSF, человеческий, например, гуманизированный M-CSF, человеческий, например, гуманизированный TPO, человеческий, например, гуманизированный IL-6; и т.п., функционально связанный с соответствующим ему промотором животного, отличного от человека, например, промотором Sirpa, IL-3, GM-CSF, M-CSF, TPO или IL-6, соответственно, при этом животное экспрессирует кодируемый человеческий белок(ки) и нативный мышиный белок(ки). В других вариантах осуществления подопытное генетически модифицированное животное, отличное от человека, представляет собой мышь, несущую геном, который содержит нуклеотидную последовательность, кодирующую человеческий, например, гуманизированный белок, например, нуклеиновую кислоту, кодирующую человеческий, например, гуманизированный EPO; человеческий, например, гуманизированный Sirpa; человеческий, например, гуманизированный IL-3; человеческий, например, гуманизированный GM-CSF, человеческий, например, гуманизированный M-CSF, человеческий, например, гуманизированный TPO, функционально связанный с соответствующим ему промотором животного, отличного от человека, например, промотором Sirpa, IL-3, GM-CSF, M-CSF или TPO, соответственно, при этом животное экспрессирует кодируемый человеческий белок(ки) и не экспрессирует нативный мышиный белок(ки). Таким образом, в некоторых вариантах осуществления генетически модифицированное животное, отличное от человека, представляет собой мышь, и указанная мышь является гетерозиготной по некоторым или всем человеческим, например, гуманизированным генам, описанным здесь. В некоторых вариантах осуществления генетически модифицированное животное, отличное от человека, представляет собой мышь, и указанная мышь является гомозиготной по некоторым или всем человеческим, например, гуманизированным генам, описанным здесь.
В некоторых вариантах осуществления подопытное животное, отличное от человека, является иммунодефицитным в отношении эндогенной иммунной системы. В некоторых таких вариантах осуществления иммунодефицит вызван дефицитом одного или обоих Rag2 и IL2rg.
В некоторых вариантах осуществления подопытное генетически модифицированное иммунодефицитное животное, отличное от человека, кроме того, несет привитые человеческие гематопоэтические клетки. В некоторых из таких вариантов осуществления человеческие гематопоэтические клетки включают одну или несколько клеток, выбранных из группы, состоящей из человеческой CD34-позитивной клетки, человеческой гематопоэтической стволовой клетки, человеческой миелоидной клетки-предшественника, человеческой эритроидной клетки-предшественника, человеческой миелоидной клетки, человеческой дендритной клетки, человеческого моноцита, человеческого гранулоцита, человеческого эритроцита, человеческого нейтрофила, человеческой тучной клетки, человеческого тимоцита и человеческого В-лимфоцита.
Как продемонстрировано в рабочих примерах в настоящем документе, привитые человеческими гематопоэтическими клетками генетически модифицированные иммунодефицитные животные, отличные от человека, несущие нуклеотидную последовательность, кодирующую человеческий белок EPO, функционально связанную с промотором EPO в эндогенном локусе, демонстрируют высокие уровни человеческого эритропоэза в костном мозге и 2-5-кратное увеличение человеческих клеток эритроидного ростка в костном мозге по сравнению с контрольными мышами, не экспрессирующими человеческий EPO. В некоторых вариантах осуществления привитые человеческими гематопоэтическими клетками генетически модифицированные иммунодефицитные животные, отличные от человека, несущие нуклеотидную последовательность, кодирующую человеческий белок EPO, функционально связанную с промотором EPO в эндогенном локусе, демонстрируют высокие уровни человеческого эритропоэза в костном мозге и примерно 2-10-кратное увеличение человеческих клеток эритроидного ростка в костном мозге по сравнению с контрольными мышами, не экспрессирующими человеческий EPO, например, примерно 2-кратное, примерно 3-кратное, примерно 4-кратное, примерно 5-кратное, примерно 6-кратное, примерно 7-кратное, примерно 8-кратное, примерно 9-кратное или примерно 10-кратное увеличение человеческих клеток эритроидного ростка в костном мозге по сравнению с контрольными мышами, не экспрессирующими человеческий EPO. Таким образом, в некоторых вариантах осуществления подопытное привитое генетически модифицированное иммунодефицитное животное содержит костный мозг, в котором 20% или более эритроидных клеток (CD235+) представляют собой человеческие эритроидные клетки. В некоторых вариантах осуществления подопытное привитое генетически модифицированное иммунодефицитное животное содержит костный мозг, в котором примерно 10% или более, например, около 20% или более, около 30% или более, около 40% или более, или около 50% или более эритроидных клеток (CD235+) представляют собой человеческие эритроидные клетки.
В некоторых из таких вариантов осуществления подопытное генетически модифицированное животное представляет собой иммунодефицитную мышь, несущую нуклеиновую кислоту, кодирующую человеческий белок EPO, функционально связанную с промотором EPO животного, отличного от человека, в локусе EPO животного, отличного от человека; нуклеиновую кислоту, кодирующую человеческий белок TPO, функционально связанную с промотором TPO животного, отличного от человека, в локусе TPO животного, отличного от человека; нуклеиновую кислоту, кодирующую человеческий белок Il-3 человека, функционально связанную с промотором Il-3 животного, отличного от человека, в локусе Il-3 животного, отличного от человека; нуклеиновую кислоту, кодирующую человеческий белок GM-CSF, функционально связанную с промотором GM-CSF в локусе GM-CSF животного, отличного от человека; и нуклеиновую кислоту, кодирующую человеческий белок M-CSF, функционально связанную с промотором M-CSF животного, отличного от человека, в локусе M-CSF животного, отличного от человека, например, мыши Rag2-/- IL2rg y/- Tpoh/h Mcsfh/h Il3h/h Gmcsfh/h Epoh/h (“MITER-G”).
В некоторых из таких вариантов осуществления подопытное генетически модифицированное животное представляет собой иммунодефицитную мышь, несущую нуклеиновую кислоту, кодирующую человеческий белок EPO, функционально связанную с промотором EPO животного, отличного от человека, в локусе EPO животного, отличного от человека; нуклеиновую кислоту, кодирующую человеческий белок TPO, функционально связанную с промотором TPO животного, отличного от человека, в локусе TPO животного, отличного от человека; нуклеиновую кислоту, кодирующую человеческий белок Il-3, функционально связанную с промотором Il-3 животного, отличного от человека, в локусе Il-3 животного, отличного от человека; нуклеиновую кислоту, кодирующую человеческий белок GM-CSF, функционально связанную с промотором GM-CSF в локусе GM-CSF животного, отличного от человека; и нуклеиновую кислоту, кодирующую человеческий белок M-CSF, функционально связанную с промотором M-CSF животного, отличного от человека, в локусе M-CSF животного, отличного от человека; и нуклеиновую кислоту, кодирующую человеческий белок SIRPα, функционально связанную с промотором SIRPa животного, отличного от человека, случайным образом интегрированную в геном животного, отличного от человека, например, Rag2-/- IL2rgy/- Tpoh/h Mcsfh/h Il3h/h Gmcsfh/h Epoh/h hSIRPα+ (“MISTER-G”) мыши. В других из таких вариантов осуществления нуклеиновая кислота, кодирующая человеческий, например, гуманизированный белок SIRPα, функционально связана с промотором SIRPα животного, отличного от человека, в локусе животного, отличного от человека, например, мыши Rag2-/- IL2rgy/- Tpoh/h Mcsfh/h Il3h/h Gmcsfh/h Epoh/h SIRPαh/h (“SupER-G”).
В некоторых из таких вариантов осуществления подопытное генетически модифицированное животное представляет собой иммунодефицитную мышь, несущую один аллель нуклеиновой кислоты, кодирующей человеческий белок EPO, функционально связанной с промотором EPO животного, отличного от человека, в локусе EPO животного, отличного от человека (то есть мышь является гетерозиготной по человеческому EPO); нуклеиновой кислоты, кодирующей человеческий, например, гуманизированный белок SIRPα, функционально связанной с промотором SIRPα животного, отличного от человека, в локусе SIRPα животного, отличного от человека; нуклеиновой кислоты, кодирующей человеческий белок TPO, функционально связанной с промотором TPO животного, отличного от человека, в локусе TPO животного, отличного от человека; нуклеиновой кислоты, кодирующей человеческий белок Il-3, функционально связанной с промотором Il-3 животного, отличного от человека, в локусе Il-3 животного, отличного от человека; и нуклеиновой кислоты, кодирующей человеческий белок GM-CSF, функционально связанной с промотором GM-CSF в локусе GM-CSF животного, отличного от человека, например, мыши Rag2-/- IL2rg y/- Tpoh/h Il3h/h Gm-csfh/h Epoh/m SIRPαh/h (“TIES”).
В некоторых вариантах осуществления привитые человеческими гематопоэтическими клетками генетически модифицированные иммунодефицитные животные, отличные от человека, несущие нуклеотидную последовательность, кодирующую человеческий белок EPO, функционально связанную с промотором EPO в эндогенном локусе, могут демонстрировать лучшую выживаемость и приживление человеческих эритроцитов, когда они несут только одну копию нуклеотидной последовательности, кодирующей белок EPO, и когда они несут эндогенный M-csf. Это обусловлено тем, что высокий уровень приживления человеческих миелоидных клеток, поддерживаемый человеческим M-CSF у мышей с нокином, вызывает деструкцию мышиных красных кровяных клеток, что в свою очередь приводит к анемии и гибели привитых мышей. Кроме того, гетерозиготность по аллелю человеческого EPO улучшает фертильность, эволюционную компетентность и жизнеспособность по сравнению с мышами, гомозиготными по человеческому EPO и нулевыми по мышиному EPO. Таким образом, в некоторых вариантах осуществления подопытное привитое генетически модифицированное иммунокомпромизированное животное демонстрирует улучшенную жизнеспособность по сравнению с мышами, которые конститутивно экспрессируют EPO, например, трансгенными мышами или мышами, которые несут две копии человеческого EPO, например, EPOh/h. В некоторых из таких вариантов осуществления генетически модифицированное иммунодефицитное животное, отличное от человека, представляет собой мышь TIES (Rag2-/- IL2rg y/- Tpoh/h Il3h/h Gm-csfh/h Epoh/m SIRPαh/h).
В некоторых вариантах осуществления привитые человеческими гематопоэтическими клетками генетически модифицированные иммунодефицитные животные, отличные от человека, инъецированные содержащими клодронат липосомами, демонстрируют 1000-кратное увеличение количества человеческих эритроидных клеток (CD235+) в периферической крови по сравнению с неинъецированными животными. В некоторых вариантах осуществления привитые человеческими гематопоэтическими клетками генетически модифицированные иммунодефицитные животные, отличные от человека, инъецированные содержащими клодронат липосомами, демонстрируют примерно 10-кратное или больше, примерно 50-кратное или больше, примерно 100-кратное или больше, примерно 500-кратное или больше, или примерно 1000-кратное или больше увеличение количества человеческих эритроидных клеток (CD235+) в периферической крови по сравнению с неинъецированными животными. Из этих человеческих эритроидных клеток 10% или более, 20% или более, 30% или более, 40% или более, или 50% или более могут представлять собой ретикулоциты (предшественники эритроцитов, CD71+). Таким образом, в некоторых вариантах осуществления подопытное привитое генетически модифицированное иммунодефицитное животное содержит периферическую кровь, в которой 1% или более, например, 5% или более, или 10% или более эритроидных клеток (CD235+) представляют собой человеческие эритроидные клетки, и 10% или более, например, 20% или более, 30% или более, 40% или более, или 50% или более из этих человеческих эритроидных клеток представляют собой ретикулоциты (CD71+). В некоторых из таких вариантов осуществления подопытное привитое генетически модифицированное иммунодефицитное животное, отличное от человека, представляет собой мышь MISTER-G, мышь SupER-G или мышь TIES.
В некоторых вариантах осуществления животное, отличное от человека, кроме того, несет инфекцию патогеном, который поражает человеческие клетки эритроидного ростка. В некоторых таких вариантах осуществления патоген выбран из Plasmodium sp., Babesia sp. и Theileri sp. В некоторых вариантах осуществления инфекцию получают путем инъекции паразита животному, отличному от человека. В некоторых вариантах осуществления инфекцию получают путем инъекции животному, отличному от человека, инфицированных паразитами человеческих эритроидных клеток. В некоторых вариантах осуществления инфекцию получают путем инъекции животному, отличному от человека, инфицированных паразитами человеческих эритроидных клеток и здоровых человеческих эритроидных клеток.
В некоторых аспектах изобретения предлагаются способы идентификации агента, который ингибирует инфекцию, вызванную патогеном, который поражает человеческие клетки эритроидного ростка.
В некоторых вариантах осуществления способ включает введение агента-кандидата генетически модифицированному животному, отличному от человека, при этом животное несет нуклеотидную последовательность, кодирующую человеческий белок EPO, функционально связанную с промотором гена EPO, одну или несколько мутаций генов, которые вызывают иммунодефицит у животного, отличного от человека, прививку человеческих гематопоэтических клеток и инфекцию патогеном, который поражает человеческие клетки эритроидного ростка; и определение, снижает ли агент количество патогена у инфицированного патогеном животного, отличного от человека.
В некоторых вариантах осуществления способ включает приведение в контакт с клодронатом привитого человеческими гематопоэтическими клетками генетически модифицированного иммунодефицитного животного, отличного от человека, несущего нуклеотидную последовательность, которая кодирует человеческий белок EPO, функционально связанную с промотором гена EPO; введение агента-кандидата животному, отличному от человека, приведенному в контакт с клодронатом; введение путем инъекции генетически модифицированному животному, отличному от человека, инфицированных паразитом ретикулоцитов или эритроцитов; и определение, предотвращает ли агент инфекцию человеческих ретикулоцитов и/или эритроцитов животного, отличного от человека.
В некоторых вариантах осуществления патоген выбран из Plasmodium sp., Babesia sp. и Theileri sp. В некоторых таких вариантах осуществления патоген выбран из P. falciparum и P. vivax. В некоторых вариантах осуществления животное, отличное от человека, представляет собой млекопитающее. В некоторых таких вариантах осуществления млекопитающее представляет собой грызуна. В некоторых таких вариантах осуществления грызун представляет собой мышь.
В некоторых аспектах изобретения предлагаются способы получения мыши, экспрессирующей человеческий EPO. В некоторых вариантах осуществления способ включает приведение в контакт мышиной плюрипотентной стволовой клетки с нуклеотидной последовательностью, содержащей кодирующую последовательность для человеческого белка EPO или ее фрагмент, функционально связанную с последовательностью промотора EPO, при этом кодирующая последовательность и последовательность промотора EPO образуют кассету, фланкированную последовательностями, гомологичными эндогенному локусу мышиного EPO; культивирование плюрипотентных стволовых клеток в условиях, способствующих интеграции нуклеотидной последовательности в мышиный геном в эндогенном локусе мышиного EPO путем гомологичной рекомбинации; и получение мыши из мышиной плюрипотентной стволовой клетки, которая несет нуклеотидную последовательность, кодирующую человеческий белок EPO.
В некоторых вариантах осуществления мышиная плюрипотентная стволовая клетка представляет собой эмбриональную стволовую клетку (ES) или индуцированную плюрипотентную стволовую клетку (iPS). В некоторых вариантах осуществления мышиная плюрипотентная стволовая клетка является дефицитной по Rag2 и/или IL2rg. В некоторых вариантах осуществления последовательность промотора EPO представляет собой последовательность промотора человеческого EPO. В других вариантах осуществления последовательность промотора EPO представляет собой последовательность эндогенного промотора нечеловеческого EPO. В некоторых вариантах осуществления интеграция приводит к замене нечеловеческого гена EPO в локусе гена нечеловеческого EPO. В некоторых вариантах осуществления нуклеотидная последовательность, которая кодирует человеческий белок EPO, содержит геномную кодирующую и некодирующую последовательность человеческого EPO. В некоторых вариантах осуществления нуклеотидная последовательность, которая кодирует человеческий белок EPO, содержит кДНК-последовательность человеческого EPO.
В некоторых аспектах изобретения предлагаются способы получения мыши, экспрессирующей человеческий белок EPO и несущей человеческую гематопоэтическую систему. В некоторых вариантах осуществления способы включают трансплантацию популяции клеток, включающей человеческие гематопоэтические клетки-предшественники, генетически модифицированным иммунодефицитным мышам, полученным с помощью способов согласно настоящему изобретению. В некоторых вариантах осуществления трансплантация включает инъекцию в хвостовую вену, инъекцию в фетальную печень или ретроорбитальную инъекцию. В некоторых вариантах осуществления генетически модифицированную иммунодефицитную мышь перед трансплантацией подвергают облучению сублетальными дозами. В некоторых вариантах осуществления трансплантированные человеческие гематопоэтические клетки-предшественники представляют собой CD34+ клетки. В некоторых вариантах осуществления человеческие гематопоэтические клетки-предшественники получены из фетальной печени, взрослого костного мозга или пуповинной крови.
В некоторых аспектах изобретения предлагаются способы получения мыши, инфицированной человеческим патогеном, который поражает человеческие клетки эритроидного ростка. В некоторых вариантах осуществления способы включают получение мыши, экспрессирующей человеческий белок EPO и несущей человеческую гематопоэтическую систему, в соответствии со способами согласно настоящему изобретению, введение путем инъекции привитой мыши клодроната и введение путем инъекции клодронат-инъецированной мыши инфицированных паразитом красных кровяных клеток (PRBC). В некоторых вариантах осуществления способ, кроме того, включает введение мыши путем инъекции здоровых человеческих красных кровяных клеток. В некоторых вариантах осуществления паразит выбран из Plasmodium sp., Babesia sp. и Theileri sp. В определенных вариантах осуществления Plasmodium sp. выбран из P. falciparum и P. vivax.
Краткое описание чертежей
Более полное понимание изобретения будет получено из следующего подробного описания при его прочтении в сочетании с сопровождающими чертежами. Подчеркивается, что в соответствии с общей практикой различные детали фигур даются не в масштабе. Напротив, размеры различных деталей условно увеличены или уменьшены для ясности. В чертежи включены следующие фигуры.
На фигуре 1 представлено белковое выравнивание Epo мыши (SEQ ID NO:2) с EPO человека (SEQ ID NO:4). Подчеркнутые остатки представляют собой остатки, которые сохранены между видами.
На фигуре 2 представлено схематическое изображение локуса EPO мыши дикого типа до и после нокина нуклеотидной последовательности, кодирующей EPO человека.
На фигуре 3 представлено схематическое изображение нокина аллеля EPO человека.
На фигуре 4 представлено схематическое изображение локуса Sirpa мыши дикого типа до (выше) и после (ниже) нокина нуклеотидной последовательности, кодирующей гуманизированный Sirpa.
На фигуре 5, панелях А и В, показано количество эритроидных клеток человека у HSC-привитых мышей. (Панель А) Приживление CD235a+ эритроцитов человека в костном мозге через 6-8 недель после прививки HSC мышам Rag2-/-Il2rg-/-Tpoh/hIL3h/h Gmcsfh/hMcsfh/h («мыши MITRG»), содержит указанные комбинации EPO человека, экспрессирующегося из локуса EPO мыши («hEPO+») и/или SIRPα человека, экспрессирующегося в виде случайного интегранта в геном мыши («hSIRPa+»). (Панель В) Количество CD235a+ эритроцитов человека в периферической крови в присутствии и отсутствии hEPO через 6-8 недель после прививки HSC мышам Rag2-/- Il2rgnullTpoh/hMcsfh/hIl3h/hGmcsfh/hEpoh/hSIRPαh/h («SupER-G») в сравнении с контрольными мышами Rag2-/-Il2rgnullTpoh/hIl3h/hGmcsfh/hSirpah/h («RGSKI-TI»).
На фигуре 6, панели А и В, показано, что обработка клодронатом увеличивает циркулирующие эритроидные клетки человека и ретикулоциты у HSC-привитых мышей. Через семь недель после приживления HSC мышей SupER-G обрабатывали путем ежедневной ретроорбитальной инъекции 50 мкл содержащей клодронат липосомы в течение от трех до пяти последовательных дней. Количество человеческих CD235+ клеток (эритроциты и ретикулоциты) (панель A) и CD235+/CD71+ клеток (ретикулоциты) (панель B) в периферической крови измеряли с помощью FACS. Панель B: три различные мыши после обработки клодронатом.
На фигуре 7, панели А-С, показано, как человеческие RBC, полученные от привитых мышей, подвержены инфекции P. falciparum. Мышей SupER-G прививали HSC фетальной печени или взрослыми HSC. Через семь недель после прививки мышей обрабатывали путем ежедневной ретроорбитальной инъекции 50 мкл содержащих клодронат липосом в течение трех последовательных дней до сбора крови. Образцы крови затем совместно культивировали с RBC, инфицированными (чистота 99%) очищенными паразитами на стадии распространения в крови P. falciparum 3D7. Свежие человеческие RBC добавляли в культуру через 48 часов после инфицирования и инфекционную культуру поддерживали дополнительно в течение 10 дней. Для количественной оценки паразитемии выполняли окрашивание по Гимзе и количественную PCR. Контрольные человеческие RBC: человеческие RBC; контрольные мышиные RBC: RBC от непривитых мышей; контрольные мышиные RBC с добавками (spiked): RBC от непривитых мышей с добавлением 0,1% hRBC. На панели A красный цвет: античеловеческие Band3; Синий цвет: Hoechst. На панели C, условные обозначения, перечисленные сверху вниз, соответствуют гистограмме x ось слева направо.
На фигуре 8 показана деструкция человеческих RBC в мышиной периферической крови в отсутствии клодроната. Не привитых мышей обрабатывали клодронатом или PBS. Для обработки клодронатом мыши получали ежедневную ретроорбитальную инъекцию 50 мкл клодроната в течение трех последовательных дней. Для обработки PBS только 500 мкл PBS доставлялось за один час до трансфузии PBS человека. PBS человека трансфузировали предварительно обработанным мышам и периферическую кровь собирали в указанные моменты времени. Показаны кривые для клодроната и PBS.
Подробное описание изобретения
Предлагаются генетически модифицированные животные, отличные от человека, экспрессирующие человеческий EPO из генома животного. Также, предлагаются способы получения животных, отличных от человека, экспрессирующих человеческий EPO из генома животного, и способы применения животных, отличных от человека, экспрессирующих человеческий EPO из генома животного. Эти животные и способы могут найти множество применений в данной области, включая, например, моделирование человеческого эритропоэза и функции эритроцитов; моделирование человеческой патогенной инфекции эритроцитов; скрининги in vivo агентов, которые модулируют эритропоэз и/или функцию эритроцитов, например в нормальном или болезненном состоянии; скрининги in vivo агентов, которые являются токсичными для эритроцитов или предшественников эритроцитов; скрининги in vivo агентов, которые предупреждают, ослабляют или устраняют токсические эффекты токсичных агентов на эритроциты или предшественники эритроцитов; скрининги in vivo эритроцитов или предшественников эритроцитов, полученных от индивидуума, для предсказания отвечаемости индивидуума на терапию заболевания. Эти и другие цели, преимущества и признаки изобретения станут очевидными специалистам в данной области после прочтения подробного описания композиций и способов, более полно описанных далее.
Перед описанием настоящих способов и композиций, следуют понимать, что данное изобретение не ограничивается конкретным описанным способом или композицией, и в силу этого, конечно, может варьировать. Также необходимо понимать, что терминология, используемая в настоящем документе, предназначена только лишь для целей описания конкретных вариантов осуществления и не предполагает какого-либо ограничения, поскольку объем настоящего изобретения будет ограничиваться только прилагаемой формулой изобретения. Данное изобретение не ограничивается конкретными описанными вариантами осуществления, а описано прилагаемой формулой изобретения.
В случае приведения диапазона значений необходимо понимать, что каждое промежуточное значение с точностью до десятой доли единицы от нижнего предела, если только контекст ясно не диктует обратное, между верхним и нижним пределами этого диапазона также является конкретным образом раскрытым. Каждый меньший диапазон между любым указанным значением или промежуточным значением в указанном диапазоне и любым другим указанным или промежуточным значением в этом указанном диапазоне, охвачен настоящим изобретением. Верхний и нижний пределы этих меньших диапазонов могут быть независимо включены в этот диапазон или исключены из этого диапазона, и каждый диапазон, в котором один из двух, ни один или оба предела включены в меньшие диапазоны, также включаются в данное описание изобретения в предположении возможности наличия любого конкретного исключенного предела в указанном диапазоне. Когда заявленный диапазон включает один или оба предела, диапазоны, исключающие любой из обоих или оба из включенных пределов, также включаются в изобретение.
Если не указано иное, все используемые здесь технические и научные термины имеют такое же значение, которое обычно понятно специалисту в области, к которой относится данное изобретение. Хотя на практике или при тестировании настоящего изобретения могут быть использованы любые способы и материалы, аналогичные или эквивалентные тем, которые описаны здесь, теперь описываются некоторые потенциальные и предпочтительные способы и материалы. Все упомянутые здесь публикации включены сюда посредством ссылки для раскрытия и описания способов и/или материалов, в связи с которым публикации процитированы. Следует понимать, что настоящее изобретение заменяет собой любое раскрытие включенной публикации в тех случаях, когда имеется противоречие.
После прочтения данного описания изобретения специалисту в данной области будет очевидно, что каждый из индивидуальных вариантов осуществления, описанных и проиллюстрированных в настоящем документе, включает дискретные компоненты и признаки, которые легко могут быть отделены от признаков любых других нескольких вариантов осуществления или объединены с ними без отступления от объема или сущности настоящего описания изобретения. Любой процитированный способ может быть реализован в порядке процитированных действий или в любом другом порядке, который является логически возможным.
Необходимо отметить, что используемые здесь и в прилагаемой формуле изобретения формы единственного числа "a", "an" и "the" включают формы множественного числа объекта, кроме тех случаев, когда контекст ясно диктует иное. Таким образом, например, ссылка на «клетку» включает множество таких клеток, а ссылка на «пептид» включает ссылку на один или более пептидов и их эквивалентов, например, полипептиды, известные специалистам в данной области, и т.д.
Обсуждаемые здесь публикации представлены исключительно для их раскрытия до даты подачи настоящей заявки. Ничто, представленное здесь, не должно рассматриваться как предположение о том, что настоящее изобретение не имеет права на предвосхищение такой публикации в силу предшествующего изобретения. Кроме того, даты представленной публикации могут отличаться от дат фактической публикации, которые могут потребовать независимого подтверждения.
Генетически модифицированные животные, отличные от человека
В одном аспекте изобретения предлагаются животные, отличные от человека, которые являются генетически модифицированными для экспрессии из своего генома одного или нескольких человеческих белков. В некоторых аспектах изобретения человеческий белок представляет собой человеческий белок эритропоэтин (hEPO) (SEQ ID NO:4). Другими словами, генетически модифицированное животное, отличное от человека, несет в своем геноме нуклеотидную последовательность, кодирующую человеческий белок EPO (hEPO). Например, животное, отличное от человека, может нести в своем геноме нуклеотидную последовательность, которая содержит геномную кодирующую или некодирующую человеческий EPO последовательность, например, последовательность на хромосоме 7, нуклеотиды 100318423-100321323, или ее фрагмент. Альтернативно, животное, отличное от человека, может нести в своем геноме нуклеотидную последовательность, которая содержит кДНК-последовательность человеческого EPO (SEQ ID NO:3), или ее фрагмент. В некоторых случаях животное, отличное от человека, кроме того, является генетически модифицированным таким образом, чтобы экспрессировать один или несколько дополнительных человеческих белков из генома животного, отличного от человека. В некоторых из таких вариантов осуществления этот один или несколько дополнительных человеческих белков представляют собой человеческие белки, которые способствуют развитию и/или функции человеческих гематопоэтических клеток, например, человеческий сигнальный регуляторный белок альфа (hSIRPα) (NCBI Gene ID: 140885, номера доступа в GenBank NM_080792.2, NM_001040022.1, NM_001040023.1), человеческий белок интерлейкин-3 (hIL-3) (NCBI Gene ID: 3562, номер доступа в GenBank NM_000588.3), человеческий белок колониестимулирующий фактор-2 (гранулоцит-макрофаг) (hGM-CSF) (NCBI Gene ID: 1437, номер доступа в GenBank NM_000758.3), человеческий белок колониестимулирующий фактор-1 (макрофаг) (hM-CSF) (NCBI Gene ID: 1435, номера доступа в GenBank NM_000757.5, NM_172210.2, NM_172211.3 и NM_172212.2), человеческий белок тромбопоэтин (hTPO) (NCBI Gene ID: 7066, номера доступа в GenBank NM_000460.3, NM_001177597.2, NM_001177598.2, NM_001289997.1, NM_001290003.1, NM_001290022.1, NM_001290026.1, NM_001290027.1, NM_001290027.1), человеческий белок интерлейкин-6 (hIL6) (NCBI Gene ID: 3569, номер доступа в GenBank NM_000600.3), и т.п.
Специалисту в данной области следует понимать, что дополнительно к человеческим нуклеиновым кислотам и белкам «дикого типа» или «нативным» термины «человеческая нуклеиновая кислота» и «человеческий белок» охватывают также варианты человеческих нуклеиновых кислот и белков дикого типа. Используемый здесь термин «вариант» относится к выделенному природному генетическому мутанту человеческой полипептидной или нуклеотидной последовательности или рекомбинантно полученному варианту человеческой полипептидной или нуклеотидной последовательности, каждая из которых содержит одну или несколько мутаций по сравнению с соответствующей человеческой нуклеотидной или полипептидной последовательностью дикого типа. Например, такие мутации могут представлять собой одну или несколько аминокислотных замен, добавлений и/или делеций. Термин «вариант» включает также человеческие гомологи и ортологи. В некоторых вариантах осуществления вариантный полипептид согласно настоящему изобретению обладает идентичностью на уровне 70% или более, например, 75%, 80% или 85% идентичностью или более с человеческим полипептидом дикого типа, например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичностью с человеческим полипептидом дикого типа.
Процент идентичности между двумя последовательностями может быть определен с использованием любого общепринятого в данной области метода, например, выравнивания последовательностей с использованием, например, общедоступного программного обеспечения. Мутации могут быть введены с использованием стандартных методов молекулярной биологии, таких как сайт-направленный мутагенез, PCR-опосредованный мутагенез, направленное развитие и т.п. Специалисту в данной области будет понятно, что одна или несколько нуклеотидных замен могут быть введены без изменения аминокислотной последовательности, и что одна или несколько аминокислотных мутаций могут быть введены без изменения функциональных свойств белка человека.
В человеческих белках могут быть сделаны консервативные аминокислотные замены для получения вариантов человеческого белка. Под консервативными аминокислотными заменами понимаются признанные в данной области замены одной аминокислоты на другую аминокислоту, имеющую схожие свойства. Например, каждая аминокислота может быть описана как имеющая одно или несколько из следующих свойств: электроположительность, электроотрицательность, альфатичность, ароматичность, полярность, гидрофобность и гидрофильность. Консервативная замена представляет собой замену одной аминокислоты, имеющей определенную структурную или функциональную характеристику, на другую аминокислоту, которая имеет схожую характеристику. Кислые аминокислоты включат аспартат, глутамат; основные аминокислоты включают гистидин, лизин, аргинин; алифатические аминокислоты включают изолейцин, лейцин и валин; ароматические аминокислоты включают фенилаланин, глицин, тирозин и триптофан; полярные аминокислоты включают аспартат, глутамат, гистидин, лизин, аспарагин, глутамин, аргинин, серин, треонин и тирозин; и гидрофобные аминокислоты включают аланин, цистеин, фенилаланин, глицин, изолейцин, лейцин, метионин, пролин, валин и триптофан; и консервативные замены включают замену из числа аминокислот в пределах каждой группы. Аминокислоты могут быть также описаны в отношении относительного размера, аланин, цистеин, аспартат, глицин, аспарагин, пролин, треонин, серин, валин, все обычно считаются малыми.
Человеческие варианты могут включать синтетические аналоги аминокислот, производные аминокислот и/или нестандартные аминокислоты, в качестве примера включающие, но без ограничения, альфа-аминомасляную кислоту, цитруллин, канаванин, цианоаланин, диаминомасляную кислоту, диаминопимеловую кислоту, дигидрокси-фенилаланин, дьенколовую кислоту, гомоаргинин, гидроксипролин, норлейцин, норвалин, 3-фосфосерин, гомосерин, 5-гидрокситриптофан, 1-метилгистидин, метилгистидин и орнитин.
Человеческие варианты будут, как правило, кодированы нуклеиновыми кислотами, имеющими высокую степень идентичности с нуклеиновой кислотой, кодирующей человеческий белок дикого типа. В некоторых вариантах осуществления комплемент нуклеиновой кислоты, кодирующей человеческий вариант, специфически гибридизуется с нуклеиновой кислотой, кодирующей человеческий вариант дикого типа в условиях высокой жесткости. Нуклеиновые кислоты, кодирующие человеческий вариант, могут быть выделены или получены рекомбинантно или синтетически с использованием хорошо известных методов.
Дополнительно к человеческим белкам «дикого типа» или «нативным» и их «вариантам», используемый здесь термин «человеческий белок», например, в контексте «человеческий белок EPO (hEPO)», «человеческий белок SIRPα (hSIRPα)», «человеческий белок IL-3 (hIL-3)», «человеческий белок GM-CSF (hGM-CSF)», «человеческий белок M-CSF (hM-CSF)», «человеческий белок TPO (hTPO)» и «человеческий белок IL-6 (hIL-6)», охватывает гибридные белки, то есть химерные белки, которые включают один или несколько фрагментов человеческого белка дикого типа (или его вариант), и которые сохраняют одну или несколько функций, например одну или несколько сигнальных и/или рецепторных функций человеческого белка дикого типа. Гибридный белок, который включает один или несколько фрагментов человеческого белка дикого типа (или его вариант), например, в комбинации с одним или несколькими нечеловеческими пептидами или полипептидами, может также называться здесь как «гуманизированный белок». Таким образом, например, белок, который включает аминокислотную последовательность внеклеточного домена человеческого белка SIRPα дикого типа, слитого с сигнальным доменом мышиного белка SIRPα дикого типа, охвачен термином «человеческий белок SIRPα».
Таким образом, нуклеотидная последовательность, которая кодирует человеческий белок, представляет собой полинуклеотид, который включает кодирующую последовательность для человеческого белка, например, человеческого белка дикого типа, варианта человеческого белка дикого типа, фрагмента человеческого белка дикого типа (или его варианта), который сохраняет одну или несколько функций, например, одну или несколько сигнальных и/или рецепторных функций человеческого белка дикого типа; или гибридного белка, например, химерного белка, который включает один или несколько фрагментов человеческого белка дикого типа (или его вариант) и сохраняет одну или несколько функций, например, одну или несколько сигнальных и/или рецепторных функций человеческого белка дикого типа.
Как правило, у генетически модифицированных животных согласно настоящему изобретению нуклеиновая кислота, кодирующая человеческий белок, например, белок hEPO, белок hSIRPα, белок hIL-3, белок hGM-CSF, белок hM-CSF, белок hTPO, белок hIL-6 и т.п., функционально связана с одним или несколькими ДНК-регуляторными элементами. ДНК-регуляторные элементы включают транскрипционные и трансляционные контрольные последовательности, такие как промоторы, энхансеры, сигналы полиаденилирования, терминаторы и т.п., которые обеспечивают и/или регулируют экспрессию кодирующей последовательности в клетке-хозяине. Например, «промотор» или «промоторная последовательность» относится к регуляторной области ДНК, способной связываться с РНК-полимеразой в клетке и инициировать транскрипцию ниже (в направлении 3') кодирующей последовательности. Промоторная последовательность ограничивается у его 3'-конца сайтом инициации транскрипции и простирается выше (в направлении 5') для включения минимального числа оснований или элементов, необходимых для инициации транскрипции на обнаруживаемых уровнях, превышающих фоновый уровень. В этой промоторной последовательности обнаруживается сайт инициации транскрипции, а также связывающие белок домены, ответственные за связывание с РНК-полимеразой. Эукариотические промоторы часто, но не всегда, содержат «TATA»-боксы и «CAT»-боксы. Особый интерес для настоящего изобретения представляют ДНК-регуляторные элементы, например, промоторы, которые способствуют транскрипции человеческого белка с таким же пространственным и временном профилем экспрессии, то есть в таких же клетках и тканях и в одни и те же моменты времени, как наблюдается у соответствующего эндогенного белка.
В некоторых вариантах осуществления нуклеотидная последовательность, кодирующая человеческий белок у подопытного животного, отличного от человека, функционально связана с человеческим промотором для гена, например, если человеческий промотор способствует корректной пространственной и временной экспрессии человеческого белка у животного, отличного от человека. Альтернативно, нуклеотидная последовательность, кодирующая человеческий белок у подопытного животного, отличного от человека, функционально связана с промотором животного, отличного от человека, для соответствующего гена животного, отличного от человека. Так, например, в отношении животных, отличных от человека, экспрессирующих белок hEPO, в некоторых вариантах осуществления нуклеиновая кислота, кодирующая белок hEPO, функционально связана с человеческим промотором EPO. В других случаях нуклеиновая кислота, кодирующая человеческий белок EPO, функционально связана с нечеловеческим промотором EPO. В еще других случаях нуклеиновая кислота, кодирующая человеческий белок EPO, функционально связана с эндогенным нечеловеческим промотором EPO.
В некоторых случаях человеческий белок экспрессируется из соответствующего локуса гена у животного, отличного от человека. В определенных случаях нуклеотидная последовательность, кодирующая человеческий белок, заменяет нуклеотидную последовательность, кодирующую соответствующий белок животного, отличного от человека. В других случаях человеческий белок экспрессируется из участка генома у животного, отличного от человека, отличающегося от локуса для соответствующего нечеловеческого гена. Так, например, в отношении животных, отличных от человека, экспрессирующих белок hEPO, в некоторых вариантах осуществления белок hEPO экспрессируется из локуса EPO генома животного, отличного от человека. В определенных вариантах осуществления животное, отличное от человека, содержит замену нуклеотидной последовательности, кодирующей эндогенный EPO, нуклеотидной последовательностью, кодирующей белок hEPO. В других вариантах осуществления hEPO экспрессируется из участка генома у животного, отличного от человека, отличающегося от локуса EPO животного, отличного от человека.
В некоторых случаях генетически модифицированное животное, отличное от человека, содержит одну копию нуклеотидной последовательности, кодирующей человеческий белок. Например, животное, отличное от человека, может быть гетерозиготным по нуклеотидной последовательности, кодирующей человеческий белок, то есть один аллель в локусе является генетически модифицированным, тогда как другой аллель представляет собой эндогенный аллель. В других случаях генетически модифицированное животное, отличное от человека, несет две копии нуклеотидной последовательности, кодирующей человеческий белок. Например, животное, отличное от человека, может быть гомозиготным по нуклеотидной последовательности, кодирующей человеческий белок, то есть оба аллеля для локуса в диплоидном геноме являются генетически модифицированными для кодирования человеческого белка, например, оба аллеля содержат замену нуклеотидной последовательности, кодирующей эндогенный белок, нуклеотидной последовательностью, кодирующей человеческий белок. Так, например, что касается животных, отличных от человека, экспрессирующих белок hEPO, как описано выше, нуклеотидная последовательность, кодирующая hEPO, может быть интегрирована в локус EPO животного, отличного от человека. В некоторых из таких вариантов осуществления генетически модифицированное животное, отличное от человека, является гетерозиготным по нуклеотидной последовательности, кодирующей белок hEPO, то есть генетически модифицированное животное, отличное от человека, несет один аллель, содержащий нуклеиновую кислоту, кодирующую hEPO, и один аллель, кодирующий эндогенный EPO. Другими словами, животное представляет собой EPO h/m животное, где “h” представляет аллель, содержащий человеческую последовательность, и «m» представляет эндогенный аллель. В других из таких вариантов осуществления генетически модифицированное животное, отличное от человека, является гомозиготным по нуклеотидной последовательности, кодирующей белок hEPO, то есть оба аллеля для локуса в диплоидном геноме будут содержать нуклеотидную последовательность, кодирующую белок hEPO. Другими словами, животное представляет собой EPO h/h животное.
В некоторых случаях животное, отличное от человека, также экспрессирует соответствующий белок животного, отличного от человека. Например, нуклеотидная последовательность, кодирующая человеческий белок, например, hEPO, может быть локализована на участке в пределах генома животного, отличающегося от локуса для гена животного, отличного от человека, например, локуса mEPO. В качестве второго примера, нуклеотидная последовательность, кодирующая человеческий белок, например, hEPO, может быть локализована в соответствующей локусе гена животного, например, локусе mEPO, при этом она является интегрированной в локус гена животного таким образом, что дает возможность непрерывной экспрессии кодирующей последовательности животного, например, человеческая кодирующая последовательность вставлена выше (5') или ниже (3') кодирующей последовательности животного, и пептидная последовательность 2А или последовательность IRES включена между двумя кодирующими последовательностями. В качестве третьего примера нуклеотидная последовательность, кодирующая человеческий белок, например hEPO, может быть локализована в соответствующем локусе гена животного, например локусе mEPO, таким образом, что нарушается экспрессия кодирующей последовательности животного, например, в виде замены части или всей кодирующей последовательности животного, но животное, отличное от человека скрещивают, чтобы оно было гетерозиготным по вставке, то есть с нокином аллеля, то есть содержало один нокин аллель и один аллель дикого типа. В других случаях животное, отличное от человека, не экспрессирует соответствующий белок животного, отличного от человека. Например, нуклеотидная последовательность, кодирующая человеческий белок, например hEPO, может быть локализована в соответствующем локусе гена животного, например, локусе mEPO таким образом, что нарушается экспрессия кодирующей последовательности животного, например, в виде замены части или всей кодирующей последовательности животного, например, путем замены кодирующей последовательности животного, отличного от человека, и животное является гомозиготным по вставке, то есть с нокином аллеля.
Любое животное-млекопитающее, отличное от человека, может быть генетически модифицировано в соответствии с настоящим описанием. Неограничивающие примеры включают лабораторных животных, домашних животных, сельскохозяйственных животных и т.д., например, такие как виды, относящиеся к мышам, грызунам, собакам, кошкам, свиньям, лошадям, коровам, овцам, нечеловеческим приматам и т.д.; например, мыши, крысы, кролики, хомяки, морские свинки, кошки, свиньи, овцы, козы и другие виды трансгенных животных, в частности, млекопитающих, известные в данной области. В других вариантах осуществления животное, отличное от человека, может представлять собой птицу, например, птиц из рода куриных, таких как цыплята, индейка, перепелка, фазан или куропатка; например, гусеобразных, таких как голубь. В различных вариантах осуществления подопытное генетически модифицированное животное представляет собой мышь, крысу или кролика.
В одном варианте осуществления животное, отличное от человека, представляет собой млекопитающее. В некоторых таких вариантах осуществления животное, отличное от человека, представляет собой маленькое млекопитающее, например, надсемейства Dipodoidea (тушканчиковых) или Muroidea (мышеобразных). В одном варианте осуществления генетически модифицированное животное представляет собой грызуна. В одном варианте осуществления грызун выбран из мыши, крысы и хомяка. В одном варианте осуществления грызун выбран из надсемейства Muroidea. В одном варианте осуществления генетически модифицированное животное относится к семейству, выбранному из Calomyscidae (например, мышевидные хомячки), Cricetidae (например, хомяк, мыши и крысы Нового света, полевки), Muridae (обычные мыши и крысы, песчанки, колючие хомячки, косматые хомяки), Nesomyidae (рипидомисы, скалистые хомячки, белохвостые крысы, малагасийские крысы и мыши), Platacanthomyidae (например, колючие соневидные хомяки) и Spalacidae (например, слепыши, бамбуковые крысы и цокоры). В определенном варианте осуществления генетически модифицированный грызун выбран из обыкновенной мыши или крысы (семейство Muridae), песчанки, колючего хомячка и косматого хомяка.
В одном варианте осуществления подопытное генетически модифицированное животное, отличное от человека, представляет собой крысу. В одном таком варианте осуществления крыса выбрана из крысы породы Wistar, линии LEA, линии Sprague Dawley, линии Fischer, F344, F6 и Dark Agouti. В другом варианте осуществления линия крыс представляет собой смесь двух или более линий, выбранных из группы, состоящей из Wistar, LEA, Sprague Dawley, Fischer, F344, F6 и Dark Agouti.
В другом варианте осуществления подопытное генетически модифицированное животное, отличное от человека, представляет собой мышь, например, мышь линии C57BL (например, C57BL/A, C57BL/An, C57BL/GrFa, C57BL/KaLwN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr, C57BL/Ola, и т.д..); мышь линии 129 (например, 129P1, 129P2, 129P3,129X1, 129S1 (например, 129S1/SV, 129S1/SvIm), 129S2, 129S4, 129S5, 129S9/SvEvH, 129S6 (129/SvEvTac), 129S7, 129S8, 129T1, 129T2); мышь линии BALB; например, BALB/c; и т.п. Смотри, например, Festing et al. (1999) Mammalian Genome 10:836, смотри также Auerbach et al. (2000) Establishment and Chimera Analysis of 129/SvEv- and C57BL/6-Derived Mouse Embryonic Stem Cell Lines). В одном варианте осуществления генетически модифицированная мышь представляет собой смесь вышеуказанной линии 129 и вышеуказанной линии C57BL/6. В другом варианте осуществления мышь представляет собой смесь вышеуказанных линий 129 или смесь вышеуказанных линий BL/6. В еще другом варианте осуществления мышь представляет собой смесь линии BALB и другой вышеуказанной линии.
В некоторых вариантах осуществления подопытное генетически модифицированное животное, отличное от человека, является также иммунодефицитным. Термин «иммунодефицитный» включает дефициты одного или нескольких аспектов нативной или эндогенной иммунной системы животного, например, животное является дефицитным по одному или нескольким типам функционирующих иммунных клеток-хозяев, например, дефицитным по количеству и/или функции нечеловеческих В-клеток, количеству и/или функции нечеловеческих Т-клеток, количеству и/или функции нечеловеческих NK-клеток, и т.д.
Одним из способов достижения иммунодефицитного состояния у подопытных животных является сублетальное облучение. Альтернативно, иммунодефицитное состояние может быть достигнуто с помощью какой-либо из ряда генных мутаций, известных в данной области, которые можно вводить отдельно или в комбинации подопытным генетически модифицированным животным, отличным от человека, согласно настоящему изобретению, или которые можно использовать в качестве источника стволовых клеток, в которые могут быть введены генетические модификации согласно настоящему изобретению. Неограничивающие примеры включают X-связанный тяжелый комбинированный иммунный дефицит (SCID), ассоциированный с мутациями гена IL2RG и характеризующийся фенотипом лимфоцитов T(-) B(+) NK(-); аутосомно-рецессивный SCID, ассоциированный с мутациями гена Jak3 и характеризующийся фенотипом лимфоцитов T(-) B(+) NK(-); мутации гена ADA, характеризующиеся фенотипом лимфоцитов T(-) B(-) NK(-); мутации гена альфа-цепи IL-7R, характеризующиеся фенотипом лимфоцитов T(-) B(+) NK(+); мутации CD3-дельта или CD3-эпсилон, характеризующиеся фенотипом лимфоцитов T(-) B(+) NK(+); мутации RAG1 и RAG2, характеризующиеся фенотипом лимфоцитов T(-) B(-) NK(+); мутации гена Artemis, характеризующиеся фенотипом лимфоцитов T(-) B(-) NK(+); мутации гена CD45, характеризующиеся фенотипом лимфоцитов T(-) B(+) NK(+); и мутации Prkdcscid, характеризующиеся фенотипом лимфоцитов T(-), B(-). Таким образом, в некоторых вариантах осуществления генетически модифицированное иммунодефицитное животное, отличное от человека, имеет один или несколько дефицитов, выбранных из дефицита гамма-цепи рецептора IL2  дефицита Jak3, дефицита ADA, дефицита IL7R, дефицита CD3, дефицита RAG1 и/или RAG2, дефицита Artemis, дефицита CD45 и дефицита Prkdc. Эти и другие животные модели иммунодефицитного состояния будут известны специалисту в данной области, каждая из которых может быть использована для создания иммунодефицитных животных согласно настоящему изобретению.
дефицита Jak3, дефицита ADA, дефицита IL7R, дефицита CD3, дефицита RAG1 и/или RAG2, дефицита Artemis, дефицита CD45 и дефицита Prkdc. Эти и другие животные модели иммунодефицитного состояния будут известны специалисту в данной области, каждая из которых может быть использована для создания иммунодефицитных животных согласно настоящему изобретению.
В некоторых вариантах осуществления генетически модифицированных животных, отличных от человека, в соответствии с изобретением используют в качестве реципиентов человеческих гематопоэтических клеток, которые способны развивать человеческие иммунные клетки из привитых человеческих гематопоэтических клеток. Таким образом, в некоторых аспектах изобретения подопытное генетически модифицированное животное представляет собой генетически модифицированное иммунодефицитное животное, отличное от человека, привитое человеческими гематопоэтическими клетками.
Любой источник человеческих гематопоэтических клеток, человеческих гематопоэтических стволовых клеток (HSC) и/или гематопоэтических стволовых клеток-предшественников (HSPC), известных в данной области или описанных здесь, может быть трансплантирован генетически модифицированным иммунодефицитным животным, отличным от человека, согласно настоящему изобретению. Одним подходящим, известным в данной области источником человеческих гематопоэтических клеток являются человеческие клетки пуповинной крови, в частности, CD34-позитивные клетки (CD34+). Другим источником гематопоэтических клеток человека является человеческая фетальная печень. Другим источником является человеческий костный мозг. Также, охваченными являются индуцированные плюрипотентные стволовые клетки (iPSC) и индуцированные гематопоэтические стволовые клетки (iHSC), полученные путем дедифференциации соматических клеток, например, с помощью способов, известных в данной области. Способы трансплантации человеческих клеток животным, отличным от человека, также хорошо описаны в данной области и в других местах в настоящем документе, каждый из которых может быть использован специалистом в данной области для введения подопытным генетически модифицированным привитым животным, отличным от человека.
Трансплантированные человеческие гематопоэтические клетки вызывают рост у генетически модифицированного животного, отличного от человека, одной или нескольких привитых человеческих клеток, выбранных из человеческой CD34-позитивной клетки, человеческой гематопоэтической стволовой клетки, человеческой гематопоэтической клетки, миелоидной клетки-предшественника, эритроидной клетки-предшественника, миелоидной клетки, дендритной клетки, моноцита, нейтрофила, тучной клетки, эритроцита и их комбинации. В одном варианте осуществления человеческая клетка присутствует через 4 месяца, 5 месяцев, 6 месяцев, 7 месяцев, 8 месяцев, 9 месяцев, 10 месяцев, 11 месяцев или 12 месяцев после приживления. В конкретном варианте осуществления человеческие клетки включают клетки эритроидного ростка.
В некоторых вариантах осуществления трансплантированные человеческие гематопоэтические клетки вызывают рост у генетически модифицированного животного, отличного от человека, привитой человеческой гемато-лимфоидной системы, которая содержит человеческие гематопоэтические стволовые клетки и клетки-предшественники, человеческие миелоидные клетки-предшественники, человеческие миелоидные клетки, человеческие дендритные клетки, человеческие моноциты, человеческие гранулоциты, человеческие нейтрофилы, человеческие тучные клетки, человеческие эритроциты, человеческие тимоциты, человеческие Т-клетки, человеческие В-клетки и человеческие тромбоциты. В одном варианте осуществления человеческая гемато-лимфоидная система присутствует через 4 месяца, 5 месяцев, 6 месяцев, 7 месяцев, 8 месяцев, 9 месяцев, 10 месяцев, 11 месяцев или 12 месяцев после приживления. В конкретном варианте осуществления человеческая гемато-лимфоидная система содержит клетки эритроидного ростка.
Клетки эритроидного ростка включают эритроциты и клетки, которые дают рост эритроцитам. «Эритроциты» включают зрелые красные кровяные клетки, также называемые красными клетками или красными кровяными тельцами. Клетки, которые вызывают рост эритроцитов, включают клетки-предшественники эритроцитов, то есть пролиферирующие мультипотентные клетки, и предшественники эритроцитов, то есть пролиферирующие или непролиферирующие клетки, коммитированные, чтобы стать эритроцитами.
Эритроциты являются основным клеточным элементом циркулирующей крови и переносят кислород в качестве своей главной функции. Количество эритроцитов на кубический миллиметр крови обычно поддерживается в диапазоне от 4,5 до 5,5 миллионов у мужчин и от 4,2 до 4,8 миллионов у женщин. Оно варьирует в зависимости от возраста, активности и условий окружающей среды. Например, увеличение до уровня 9 миллионов/мм3 может нормально возникать на высоте более 10000 футов над уровнем моря. Эритроцит обычно живет в течение от 110 до 120 дней, после чего удаляется из кровотока и разрушается ретикулоэндотелиальной системой. Новые эритроциты продуцируются со скоростью, незначительно превышающей 1% в день; таким образом, уровень обычно поддерживается постоянным. Острая потеря крови, гемолитическая анемия или хроническая кислородная недостаточность могут вызвать значительное увеличение продукции эритроцитов.
Эритроциты происходят из стволовых гематопоэтических клеток в костном мозге длинных костей, проходя несколько стадий клеточного развития, которые включают типичные миелоидные клетки-предшественники (CD123+, CD34+, c-kit+, Flt3+); мегакариоцит-эритроидные клетки-предшественники (CD34+, CD38+, CD45RA-); проэритробласты (также называемые пронормобластами в случае нормального развития или промегалобластами в случае ненормального развития; крупные предшественники CD71+, EpoR+, c-kit+, Ter119+); базофильные эритробласты (цитоплазма базофильная, ядро крупное с хроматином в виде глыбок и ядрышки исчезли); полихроматофильные эритробласты (также называемые промежуточными нормобластами, в которых ядерный хроматин показывает повышенное образование глыбок и в цитоплазме начинает накапливаться гемоглобин и принимает ацидофильный оттенок); ортохромные нормобласты (конечная стадия перед утратой ядра, на которой ядро маленькое и в конечном итоге превращается в сине-черную гомогенную бесструктурную массу); и ретикулоциты (циркулирующие CD235+, CD71+ клетки; клетка характеризуется рисунком в виде сетки из нитей и частиц на исходном участке ядра).
Зрелые эритроциты появляются на мазке периферической крови в виде двояковогнутых круглых или овальных дисков диаметром около 6-8 мкм. Они содержат гемоглобин и имеют зону центрального просветления вследствие двояковогнутости клетки, и могут быть легко идентифицированы с помощью проточной цитометрии или способов на основе иммуногистохимии по повышенной экспрессии маркеров клеточной поверхности CD235 и CD59 относительно неэритроидных клеток.
Как показано, например, на фигуре 5, панель А, и на фигуре 5, панель В, настоящего изобретения, экспрессия hEPO под контролем промотора EPO из генома привитого животного, отличного от человека, согласно настоящему изобретению увеличивает количество человеческих клеток эритроидного ростка (CD235a+) в костном мозге в среднем примерно в 2 раза (например, примерно от 11% до 22%). Экспрессия человеческого Sirpa усиливает этот эффект, что приводит к увеличению в среднем в 3 раза или более (то есть примерно от 11% до 33%) количества человеческих клеток CD235a+ в костном мозге по сравнению с животными, которые не экспрессируют человеческий EPO или человеческий Sirpa (фигура 5, панель А). Таким образом, привитые иммунодефицитные генетически модифицированные животные, отличные от человека, экспрессирующие hEPO согласно настоящему изобретению, находят применение в исследовании человеческого эритропоэза и разработке лекарственных средств, которые модулируют (например, стимулируют или подавляют) человеческий эритропоэз.
Кроме того, как показано, например, на фигуре 6, обработка клодронатом привитых человеческими HSC животных, экспрессирующих hEPO, повысило в 1000 раз количество человеческих эритроидных CD235+ (включая CD71+ ретикулоциты, фигура 6, панель В) в периферической крови этих животных, то есть примерно до 1% от всех красных кровяных клеток в периферической крови по сравнению с необработанными контролями. Важно, что человеческие красные кровяные клетки, продуцируемые на этих уровнях приживления у HSC-привитых животных, экспрессирующих человеческий EPO, подвержены инфекции, вызываемой P. falciparum, как показано, например, на фигуре 7. Таким образом, генетически модифицированные животные, отличные от человека, экспрессирующие hEPO, как описано здесь, находят конкретное применение в создании животных моделей паразитарных инфекций, которые поражают клетки эритроидного ростка человека, например, патогенами, которые вызывают малярию или подобные малярии заболевания.
Таким образом, в некоторых аспектах изобретения подопытное генетически модифицированное животное представляет собой животное, отличное от человека, привитое человеческими гематопоэтическими клетками и несущее инфекцию, вызванную человеческим патогеном. Особый интерес в этих вариантах осуществления представляют человеческие патогены, которые поражают человеческие клетки эритроидного ростка. Неограничивающие примеры таких патогенов включают простейшие родов Plasmodium, Babesia, Theileria, и т.п. Как описано более подробно далее, подопытное генетически модифицированное животное, привитое человеческими гематопоэтическими клетками, может быть инфицировано человеческим патогеном с использованием любого подходящего способа, известного в данной области, или описанного здесь для инфицирования животных представляющими интерес патогенами. Как правило, инфицированные таким образом животные будут показывать признаки паразитемии, включая измененную морфологию, по окрашиванию мазка крови по методу Гимзе, и сильное снижение (например, 50%) общей концентрации эритроцитов и анемию.
Способы получения подопытных генетически модифицированных мышей
В некоторых аспектах изобретения предлагаются способы получения подопытных животных, отличных от человека, согласно настоящему изобретению. При осуществлении на практике способов согласно изобретению получали животное, отличное от человека, которое несет нуклеотидную последовательность, кодирующую белок hEPO, функционально связанную с промотором EPO, например, промотором EPO животного, отличного от человека, например, в локусе EPO генома животного, отличного от человека.
Получение животного, отличного от человека, несущего нуклеотидную последовательность, кодирующую белок hEPO, функционально связанную с промотором EPO, может быть осуществлено с использованием любого общепринятого способа получения генетически модифицированных животных, например, известных в данной области или описанных здесь.
Например, нуклеиновая кислота, кодирующая белок hEPO, может быть встроена в рекомбинантный вектор в форме, подходящей для вставки в геном клетки-хозяина и экспрессии человеческого белка в нечеловеческой клетке-хозяине. В различных вариантах осуществления рекомбинантный вектор включает одну или несколько регуляторных последовательностей, функционально связанных с нуклеиновой кислотой, кодирующей человеческий белок, таким образом, что обеспечивает возможность транскрипции нуклеиновой кислоты в мРНК и трансляции мРНК в человеческий белок, описанный выше. Следует принимать во внимание, что конструкция вектора может зависеть от таких факторов, как выбор клетки-хозяина, подлежащей трансфекции, и/или количество экспрессируемого человеческого белка.
Любой из различных способов может быть затем использован для введения человеческой нуклеотидной последовательности в клетку животного для получения генетически модифицированного животного, которое экспрессирует человеческий ген. Такие методы хорошо известны в данной области и включают, но без ограничения, пронуклеарную микроинъекцию, трансформацию эмбриональных стволовых клеток, технологии гомологичной рекомбинации и технологию нокин «knock-in». Способы создания генетически модифицированных животных, которые могут быть использованы, включают, но без ограничения, способы, описанные в Sundberg and Ichiki (2006, Genetically Engineered Mice Handbook, CRC Press), Hofker and van Deursen (2002, Genetically modified Mouse Methods and Protocols, Humana Press), Joyner (2000, Gene Targeting: A Practical Approach, Oxford University Press), Turksen (2002, Embryonic stem cells: Methods and Protocols in Methods Mol Biol., Humana Press), Meyer et al. (2010, Proc. Nat. Acad. Sci. USA 107:15022-15026), и Gibson (2004, A Primer Of Genome Science 2nd ed. Sunderland, Massachusetts: Sinauer), патент США 6586251, Rathinam et al. (2011, Blood 118:3119-28), Willinger et al., (2011, Proc Natl Acad Sci USA, 108:2390-2395), Rongvaux et al., (2011, Proc Natl Acad Sci USA, 108:2378-83) и Valenzuela et al. (2003, Nat Biot 21:652-659).
Например, подопытные генетически модифицированные животные могут быть получены путем введения нуклеиновой кислоты, кодирующей человеческий белок, в ооцит, например, путем микроинъекции, и развития ооцита у самки животного. В предпочтительных вариантах осуществления экспрессирующую конструкцию вводят в оплодотворенные ооциты. Оплодотворенные ооциты могут быть собраны от суперовулировавших самок на следующий день после спаривания и инъецированы экспрессирующей конструкцией. Инъецированные ооциты культивируют в течение ночи или переносят непосредственно в фаллопиевы трубы псевдобеременных самок на 0,5-сутки развития. Способы суперовулирования, сбора ооцитов, инъекции экспрессирующей конструкции и пересадки эмбриона хорошо известны в данной области и описаны в Manipulating the Mouse Embryo (2002, A Laboratory Manual, 3rd edition, Cold Spring Harbor Laboratory Press). Потомство может быть оценено на присутствие введенной нуклеиновой кислоты с помощью анализа ДНК (например, PCR, Саузерн-блоттинг, секвенирование ДНК и т.д.) или с помощью анализа белка (например, ELISA, Вестерн-блоттинг и т.д.).
В качестве другого примера конструкция, содержащая нуклеотидную последовательность, кодирующую человеческий белок, может быть трансфицирована в стволовые клетки (например, клетки ES или клетки iPS) с использованием хорошо известных способов, таких как электропорация, осаждение фосфатом кальция, липофекция и т.д. Клетки могут быть оценены на присутствие введенной нуклеиновой кислоты с помощью анализа ДНК (например, PCR, Саузерн-блоттинг, секвенирование ДНК и т.д.) или с помощью анализа белка (например, ELISA, Вестерн-блоттинг и т.д.). Клетки, определенные как имеющие встроенную экспрессирующую конструкцию, могут быть затем введены в доимплантационные эмбрионы. Для подробного описания способов, известных в данной области, полезных для композиций и способов согласно изобретению, смотри Nagy et al., (2002, Manipulating the Mouse Embryo: A Laboratory Manual, 3rd edition, Cold Spring Harbor Laboratory Press), Nagy et al. (1990, Development 110:815-821), патент США 7576259, патент США 7659442, патент США 7294754 и Kraus et al. (2010, Genesis 48:394-399).
Кроме того, как описано в некоторых из следующих далее примеров, конструкция нуклеиновой кислоты может быть создана с использованием технологии генной инженерии VELOCIGENE® (смотри, например, Valenzuela et al. (2003) High throughput engineering of the mouse genome coupled with high-resolution expression analysis, Nature Biotech. 21(6): 652-59 и патент США 6586251), введена в стволовые клетки (например, клетки ES), и правильно нацеленные клоны определены с использованием анализов на утрату аллеля (loss-of-allele) и приобретение аллеля (gain-of-allele) (Valenzuela et al., supra); правильно нацеленные клетки ES могут быть использованы в качестве донорных клеток ES для введения мышиному эмбриону, находящемуся на 8-клеточной стадии развития, с использованием метода VELOCIMOUSE® (смотри, например, патент США 7294754 и Poueymirou et al. 2007, F0 generation mice that are essentially fully derived from the donor gene-targeted ES cells allowing immediate phenotypic analyses Nature Biotech. 25(1):91-99).
Генетически модифицированные животные-основатели могут быть скрещены с дополнительными животными, несущими генетическую модификацию. Генетически модифицированные животные, несущие нуклеиновую кислоту, кодирующую человеческий белок(ки) согласно настоящему изобретению, могут быть дополнительно скрещены с другими генетически модифицированными животными, несущими другие генетические модификации, например, мышами с нокином hSIRPα, мышами с нокином hIL-3, мышами с нокином hGM-CSF, мышами с нокином hM-CSF, мышами с нокином hTPO, мышами с нокином hIL-6 и т.п., или скрещены с нокаутными животным, например, животным, отличным от человека, которое является дефицитным по одному или нескольким белкам, например, не экспрессирует один или несколько из своих генов, например, Rag2-дефицитное животное, Il2rg-дефицитное животное.
В другом варианте осуществления стволовые клетки, например, клетки ES могут быть созданы таким образом, что они содержат несколько генетических модификаций, например, описанные здесь гуманизации или делеции генов, и такие стволовые клетки могут быть введены в эмбрион для создания генетически модифицированных животных с несколькими генетическими модификациями.
Как описано выше, в некоторых вариантах осуществления подопытное генетически модифицированное животное, отличное от человека, представляет собой иммунодефицитное животное. Генетически модифицированные животные, отличные от человека, которые являются иммунодефицитными и содержат один или несколько человеческих белков, например, hEPO, hSIRPα, hIL-3, hGM-CSF, hM-CSF и/или hTPO, могут быть созданы с использованием любого общепринятого способа для создания генетически модифицированных животных, например, известных в данной области или описанных в настоящем документе. Например, создание генетически модифицированного иммунодефицитного животного может быть достигнуто путем введения нуклеиновой кислоты, кодирующей человеческий белок, в ооцит или стволовые клетки, содержащие мутантный аллель гена SCID, который, когда находится в гомозиготном состоянии, будет вызывать иммунодефицит, как описано более подробно выше и в рабочих примерах в настоящем документе. Мышей затем создают с помощью модифицированного ооцита или ES-клеток с использованием, например, способов, описанных здесь и известных в данной области, и спаривают для получения иммунодефицитных мышей, содержащих желаемую генетическую модификацию. В качестве другого примера, генетически модифицированные животные, отличные от человека, создают в иммунокомпетентном окружении и скрещивают с животным, содержащим мутантный аллель гена, который, когда находится в гемизиготном или гомозиготном состоянии, будет вызывать иммунодефицит, и потомство спаривают для создания иммунодефицитного животного, экспрессирующего по меньшей мере один представляющий интерес белок человека.
В некоторых вариантах осуществления генетически модифицированную мышь обрабатывают таким образом, чтобы удалить эндогенные гематопоэтические клетки, которые могут присутствовать у мыши. В одном варианте осуществления обработка включает облучение генетически модифицированной мыши. В определенном варианте осуществления новорожденных генетически модифицированных мышиных детенышей облучают сублетально. В определенном варианте осуществления новорожденных детенышей облучают 2 х 200 cGy с интервалом четыре часа.
Различные варианты осуществления изобретения обеспечивают генетически модифицированных животных, которые несут человеческую нуклеиновую кислоту по существу во всех своих клетках, а также генетически модифицированных животных, которые несут человеческую нуклеиновую кислоту в некоторых, но не во всех своих клетках. В некоторых случаях, например, таргетной рекомбинации, одна копия человеческой нуклеиновой кислоты будет интегрирована в геном генетически модифицированных животных. В других случаях, например, случайной интеграции, множество копий расположенных рядом или на удалении друг от друга, человеческой нуклеиновой кислоты могут быть интегрированы в геном генетически модифицированных животных.
Таким образом, в некоторых вариантах осуществления подопытное генетически модифицированное животное, отличное от человека, может представлять собой иммунодефицитное животное, содержащее геном, который включает нуклеиновую кислоту, кодирующую человеческий полипептид, функционально связанную с соответствующим промотором животного, отличного от человека, при этом животное экспрессирует кодируемый человеческий полипептид. Другими словами, подопытное генетически модифицированное иммунодефицитное животное, отличное от человека, несет геном, который содержит нуклеиновую кислоту, кодирующую по меньшей мере один человеческий полипептид, при этом нуклеиновая кислота функционально связана с соответствующим нечеловеческим промотором и сигналом полиаденилирования, и при этом животное экспрессирует кодируемый человеческий полипептид.
Как описано выше, в некоторых вариантах осуществления подопытное генетически модифицированное животное, отличное от человека, привито человеческими гематопоэтическими клетками. Любой источник человеческих гематопоэтических клеток, человеческих гематопоэтических стволовых клеток (HSC) и/или гематопоэтических стволовых клеток-предшественников (HSPC), известных в данной области или описанных здесь, может быть трансплантирован генетически модифицированным животным согласно настоящему изобретению, например, пуповинная кровь, фетальная печень, костный мозг, iPSC и т.д. В одном варианте осуществления гематопоэтические клетки выбраны из человеческих клеток пуповинной крови и человеческих клеток фетальной печени. Количество клеток, подлежащих трансплантации, хорошо известно специалисту в данной области или может быть определено эмпирически. В одном варианте осуществления прививку осуществляют с использованием примерно 1-2 × 105 CD34+ человеческих клеток. Клетки могут быть трансплантированы подопытному животному-хозяину, отличному от человека, с использованием любого общепринятого метода, известного в данной области, например, инъекции в хвостовую вену, ретроорбитальной инъекции, инъекции в неонатальную печень и т.п. Клетки могут быть трансплантированы хозяину в любом подходящем буферном растворе, например, PBS, среда Дульбекко, модифицированная по способу Исков, и т.п. В некоторых случаях животное может быть подвергнуто облучению до приживления, например, как описано выше, для повышения иммунодефицита.
Привитые таким образом человеческие гематопоэтические клетки дают рост одной или нескольким человеческим клеткам, выбранным из CD34+ клеток, гематопоэтических стволовых клеток, гематопоэтических клеток, миелоидных клеток-предшественников, миелоидных клеток, дендритных клеток, моноцита, гранулоцита, нейтрофила, тучной клетки, тимоцита, Т-клетки, В-клетки, тромбоцита, эритроцита и их комбинации. В одном варианте осуществления человеческая клетка присутствует через 4 месяца, 5 месяцев, 6 месяцев, 7 месяцев, 8 месяцев, 9 месяцев, 10 месяцев, 11 месяцев или 12 месяцев после прививки. Любой из ряда анализов может быть выполнен для подтверждения того, что прививка прошла успешно, включая, например, анализы с помощью проточной цитометрии для различных представляющих интерес человеческих гематопоэтических клеток, мазки крови и иммуногистохимическое исследование.
Как описано выше, в некоторых вариантах осуществления подопытное генетически модифицированное животное, отличное от человека, привитое человеческими гематопоэтическими клетками, инфицировано человеческим патогеном. Особый интерес в этих вариантах осуществления представляют человеческие патогены, которые поражают человеческие клетки эритроидного ростка. Неограничивающие примеры таких патогенов включают простейшие рода Plasmodium sp., Babesia sp., Theileria sp. и т.п. В некоторых вариантах осуществления используемый штамм патогена представляет собой природный штамм. В определенных вариантах осуществления используемый штамм выбран in vivo по причине его способности репродуцируемо расти у иммунодефицитных мышей, привитых человеческими эритроцитами, например, штаммы Pf3D70087/N9 или Pf3D70087/N5P. falciparum.
Подопытные привитые иммунодефицитные животные могут быть инфицированы с использованием любого способа, известного в данной области для инфицирования животных патогеном, который поражает клетки эритроидного ростка. Например, подопытные привитые иммунодефицитные животные могут быть инокулированы интраперитонеально инфицированными паразитом красными кровяными клетками (PRBC). Смотри, например, Badell et al. (2000) Human malaria in immunocompromised mice: an in vivo model to study defense mechanisms against Plasmodium falciparum. JEM 192(11):1653-1660; и Moreno et al. (2006) The course of infections and pathology in immunomodulated NOD/LtSz-SCID mice inoculated with Plasmodium falciparum laboratory lines and clinical isolates. Int. J. Parasitol. 36:361-369. В качестве другого примера, подопытные привитые иммунодефицитные животные могут быть инокулированы путем внутрисосудистого введения инфицированных паразитом красных кровяных клеток. Смотри, например, Angulo-Barturen et al. (2008) A murine model of falciparum-malaria by in vivo selection of competent strains in non-myelodepleted mice engrafted with human erythrocytes. PLoS One 3:e2252; и Jimenez-Diaz et al. (2009) Improved murine model of malaria using Plasmodium falciparum competent strains and non-myelodepleted NOD-scid IL2Rgnull mice engrafted with human erythrocytes. Antimicrob Agents Chemother 53:4533-4536. В некоторых вариантах осуществления инфекцию вызывают путем инъекции паразита животному, отличному от человека, то есть не в контексте эритроидных клеток. В некоторых вариантах осуществления подопытные привитые иммунодефицитные животные химически обедняют in vivo фагоцитами до и/или во время инфицирования. В таких вариантах осуществления любой химиотерапевтический агент, который селективно обедняет хозяина фагоцитами, может быть введен подопытным животным, например, клодронат, например, как описано в настоящем документе в рабочих примерах; дихлорметилен дифосфонат, например, как описано в Badell et al. supra и Moreno et al. supra; моноклональное антитело, специфическое в отношении полиморфоядерных нейтрофилов, например, NIMP-R14, как описано в Badell et al. supra и Moreno et al., supra; и т.п.
Процент паразитемии у инфицированных генетически модифицированных животных, отличных от человека, согласно настоящему изобретению может быть определен с помощью любого общепринятого в данной области способа, например, микроскопически по окрашенному по Гимзе мазку крови, например, через 3 дня после инъекции; или с помощью проточной цитометрии, например, путем измерения эмиссии красителя YOYO-1 для выявления нуклеиновых кислот или проникающего в клетки красителя SYTO-16, например, в присутствии или отсутствии TER-119 mAb. Смотри, например, Jimenez-Diaz et al. Quantitative measurement of Plasmodium-infected erythrocytes in murine models of malaria by flow cytometry using bidimensional assessment of SYTO-16 fluorescence. Cytometry A 2009, 75:225-235.
Использование подопытных генетически модифицированных мышей
Возможность исследовать человеческую ткань у мышей в условиях in vivo открыло несколько возможных направлений исследования. Основные препятствия затрудняли применение подхода, и одним из наиболее важных недостатков являлась неспособность мышиных факторов поддерживать человеческие клетки. Разумеется, в иммунной системе множество важных факторов, требуемых для развития и функционирования иммунных клеток человека, являются видоспецифическими и не могут эффективно обеспечиваться мышью. Поэтому было принято решение следовать стратегии, состоящей в замене генов мыши на их человеческие эквиваленты, обеспечивая лучшее развитие и функционирование человеческих клеток и возможно не допуская этого же для соответствующих мышиных клеток. Путем применения этого принципа к цитокину EPO человека в настоящем документе показано, что замена нуклеотидной последовательности, кодирующей мышиный белок EPO, нуклеотидной последовательностью, кодирующей человеческий белок EPO, улучшает развитие и функционирование клеток эритроидного ростка в привитой человеческой иммунной системе у мышей.
Например, рабочие примеры, представленные, например, на фигуре 5, показывают, что экспрессия hEPO из нуклеотидной последовательности под контролем промотора EPO в геноме животного, отличного от человека, примерно в 2 раза (то есть примерно от 11% до 22%) повышает количество человеческих клеток эритроидного ростка (CD235a+), которые развиваются в костном мозге животных, привитых человеческими HSC. Экспрессия человеческого Sirpa усиливает этот эффект, приводя к 3-кратному общему увеличению (то есть примерно от 11% до 33%) представленности человеческих CD235a+ клеток в костном мозге по сравнению с животными, которые не экспрессируют человеческий EPO или человеческий Sirpa (фигура 4а). Кроме того, как показано, например, на фигуре 6, обработка клодронатом животных, привитых человеческими HSC, экспрессирующих человеческий EPO, повышает в тысячу раз количество CD235+ эритроидных клеток (включая CD71+ ретикулоциты, фигура 6, панель В) в периферической крови этих животных, то есть до около 1% от всех красных кровяных клеток по сравнению с необработанными контролями. Важно отметить, что, как показано, например, на фигуре 7, человеческие красные кровяные клетки, продуцируемые на этих уровнях приживления у HSC-привитых животных, экспрессирующих человеческий EPO, подвержены инфекции Plasmodium sp. таким как P. falciparum. Таким образом, генетически модифицированные животные, отличные от человека, экспрессирующие человеческий EPO, как описано здесь, находят конкретное применение в получении животных, которые подвержены инфекции, вызванной Plasmodium sp., или которые будут лучше поддерживать человеческие эритроциты, остро привитые животному до инфекции, в существующих моделях грызунов.
Таким образом, генетически модифицированные животные, отличные от человека, согласно настоящему изобретению находят множество применений в данной области. Например, привитые генетически модифицированные животные согласно настоящему изобретению являются полезными для изучения человеческого эритропоэза и функции эритроцитов. В качестве другого примера, привитые генетически модифицированные мыши согласно настоящему изобретению обеспечивают полезную систему для скрининга агентов-кандидатов на желательные активности in vivo, например, для идентификации агентов, способных модулировать (то есть стимулировать или подавлять) человеческий эритропоэз и/или функцию эритроцитов, например, в нормальном или болезненном состоянии, таком как злокачественные клетки, во время патогенной инфекции, и т.д.; например, для идентификации новых терапевтических средств; или, в качестве другого примера, для идентификации агентов, которые являются токсичными для человеческих клеток эритроидного ростка, и для идентификации агентов, которые предупреждают, ослабляют или устраняют токсические эффекты токсичных агентов на человеческие клетки эритроидного ростка; и т.д. В качестве другого примера, привитые генетически модифицированные животные согласно настоящему изобретению обеспечивают полезную систему для предсказания отвечаемости индивидуума на терапию заболевания, например, путем обеспечения платформы in vivo для скрининга отвечаемости иммунной системы индивидуума на агент, например, терапевтический агент, для предсказания отвечаемости индивидуума на этот агент.
В качестве одного неограничивающего примера, привитые генетически модифицированные мыши согласно настоящему изобретению находят применение в создании мышиных моделей патогенной инфекции, вызванной паразитами, которые поражают эритроидные клетки человека, например, Plasmodium, Babesia, Theileria и т.п. Такие мышиные модели инфекции будут полезными в исследовании, например, для лучшего понимания развития инфекции у человека и открытия лекарственных средств, например, для идентификации агентов-кандидатов, которые защищают или лечат инфекцию, вызванную такими паразитами.
Простейшие рода Plasmodium являются причиной малярии у человека. Малярия начинается с укуса инфицированного комара рода Anopheles, который вводит простейшие через свою слюну в кровеносную систему и в конечном итоге в печень, где они созревают и размножаются. Простейшие затем поступают в кровоток и инфицируют клетки эритроидного ростка на различных стадиях созревания.
Пять видов плазмодиев (Plasmodium) могут инфицировать человека и передаваться человеком. Подавляющее большинство летальных исходов вызывает P. falciparum, тогда как P. vivax, P. ovale и P. malariae вызывают, как правило, более слабую форму малярии, которая редко бывает фатальной. Как полагают, это обусловлено, по меньшей мере отчасти, типом(ами) клеток, пораженных каждым видом: P. falciparum растет в красных кровяных клетках (RBC) всех степеней зрелости, тогда как, например, P. vivax ограничивается ростом в ретикулоцитах, которые составляют только приблизительно 1-2% от всех RBC в периферической системе. Кроме того, P. falciparum вызывает тяжелую форму малярии благодаря отличительному свойству, которым не обладают другие вызывающие малярию виды, а именно секвестрации. В течение 48-часового цикла бесполого развития в клетках крови зрелые формы изменяют поверхностные свойства инфицированных красных кровяных клеток, вызывая их приклеивание к кровеносным сосудам (процесс, называемый цитоадгезией). Это приводит к обструкции микроциркуляции и вызывает дисфункцию множества органов.
Симптомы, ассоциированные с малярией, включают лихорадку, озноб, головную боль, потливость, слабость, анемию, тошноту, сухой (непродуктивный) кашель, боль в мышцах и/или спине и увеличение размеров селезенки. Другие симптомы и осложнения, связанные с малярией, включают инфекцию головного мозга (церебрит), гемолитическую анемию, почечную недостаточность, печеночную недостаточность, менингит, отек легких и кровотечение из селезенки. Как правило, у индивидуума, имеющего риск развития малярии, симптомы начинают проявляться через 7 дней или более после инфицирования, например, через 9-14 дней после начального инфицирования P. falciparum, 12-18 дней после начального инфицирования P. vivax или P. ovale, 18-40 дней после начального инфицирования P. malariae, или 11-12 дней после начального инфицирования P. knowlesi. Антималярийные агенты, используемые в данной области для лечения или предупреждения малярии, включают хлороквин, хинидин, доксициклин, тетрациклин, клиндамицин, атоваквон плюс прогуанил (маларон), мефлохин, артесунат и пуриметамин плюс сульфадоксин (фансидар).
Способы определения, является ли субъект инфицированным Plasmodium, хорошо известны в данной области и включают, например, микроскопическое исследование крови с использованием пленок крови с помощью тестов на основе антигена Rapid Diagnostic Tests (RDT), например, RDT на основе иммунохроматографии, путем детекции ДНК паразита с помощью полимеразной цепной реакции (PCR) и т.д. Любой подходящий способ может быть использован для определения инфицирования патогеном человеческих красных кровяных клеток субъекта.
Другим примером представляющих интерес патогенов являются простейшие рода Babesia. Инфекция Babesia вызывает заболевание, подобное малярии, которое называется бабезиоз. Бабезиоз представляет собой заболевание, передаваемое переносчиком, как правило, клещами Ixodes scapularis. Заболевание обычно вызывают B. microti у человека, B. canis rossi и B. canis canis у собак, B. bovis у коров, и B. bigemina у кошек. Babesia microti, инфицирующий человека, использует такой же вектор клеща, как при болезни Лайма и эрлихиозе, и может возникать в сочетании с этими заболеваниями. Простейшие также могут переноситься путем переливания крови.
У человека бабезиоз может протекать бессимптомно или характеризоваться симптомами от умеренной лихорадки и диареи до высокой температуры, потрясающего озноба и тяжелой анемии. В тяжелых случаях может возникнуть отказ органов, включая синдром острой дыхательной недостаточности. Тяжелые случаи возникают в основном у людей, которые имели спленэктомию, или лиц с иммунодефицитом, таких как пациенты со СПИДом/ВИЧ. Лечение обычно включает режим, состоящий из двух лекарственных средств, хинина и клиндамицина или атоваквона и азитромицина. В случаях, когда бабезиоз угрожает жизни, проводят заменное переливание крови, в котором инфицированные красные кровяные клетки удаляют и заменяют неинфицированными.
Заключительный диагноз инфекции, вызванной Babesia, ставят путем идентификации паразита по мазку крови, окрашенному по методу Гимза. Паразит появляется в эритроцитах в виде парных мерозоитов, формируя «рисунок в виде мальтийского креста» у человека или «две висящие вместе груши» у животных. Другие методы диагностики включают PCR периферической крови и серологический анализ на антитела (IgG, IgM) против бабезии.
Еще одно сходное с малярией заболевание, тейлериоз, вызвано простейшими рода Theileria. У человека тейлериоз вызывают T. microti; у лошадей T. equi (“Equine Piroplasmosis”); у овец и коз T. lestoquardi; и у кошек, африканского буйвола, азиатского буйвола и водяного козла T. annulata (“Tropical Theileriosis”, также известное как “Mediterranean theileriosis”) или T. parva (“East Coast fever”, также известное как “Corridor disease”). Тейлериоз передается хозяину различными видами клещей, включая Ixodes scapularis, Rhipicephalus, Dermacentor, Haemaphysali и Hyalomma. Организм размножается в клеще по мере его развития через его жизненные стадии, созревает и поступает в слюну после того как клещ прикрепляется к хозяину. Обычно клещ должен быть прикреплен в течение нескольких дней перед тем, как он станет инфекционным. Однако, если температура окружающей среды высокая, инфекционные спорозоиты могут развиваться в клещах на земле и могут поступать хозяину в течение нескольких часов после прикрепления.
Тейлериоз у человека обычно характеризуется лихорадкой и гемолизом. Заключительный диагноз инфекции, вызванной Theileria, ставится путем идентификации паразита по окрашиванию мазка крови по методу Гемзи.
Привитые генетически модифицированные животные согласно настоящему изобретению находят применение в скрининге агентов-кандидатов для идентификации агентов, которые будут предупреждать (например, вакцины) или лечить инфекции, вызванные Plasmodium, Babesia, Theileria, и другими паразитами, которые поражают человеческие эритроциты. Используемые здесь термины «лечение», «лечить» и подобные обычно включают получение желательного фармакологического и/или физиологического эффекта. Эффект может быть профилактическим с точки зрения полного или частичного предупреждения возникновения заболевания или его симптомов, и/или может быть терапевтическим с точки зрения частичного или полного излечения заболевания и/или побочного эффекта, сопровождающего заболевание. Используемое здесь «лечение» включает любое лечение заболевания у млекопитающего и включает: а) предупреждение возникновения заболевания у субъекта, который может быть предрасположен к возникновению заболевания, но который еще не был диагностирован как его имеющий; (b) ингибирование заболевания, то есть купирование его развития; или (с) ослабление заболевания, то есть вызывание регрессии заболевания. Агенты-кандидаты, представляющие интерес в качестве антипаразитарных терапевтических средств, включают агенты, которые можно вводить до, во время или после инфицирования паразитом, и которые при введении в эффективном количестве ингибируют эффекты паразита на индивидуума (то есть хозяина), например, путем уничтожения паразита или инфицированной паразитом клетки, путем предотвращения размножения паразита, путем предотвращения продуцирования или действия продуцируемого паразитом агента, который является токсичным для индивидуума (то есть токсина), и т.д. Термины «индивидуум», «субъект» и «пациент» используются взаимозаменяемо в настоящем документе и включают любого субъекта-млекопитающего, в частности, человека, для которого является желательной диагностика, лечение или терапия.
В скрининговых анализах на биологически активные агенты привитое человеческими гематопоэтическими клетками генетически модифицированное животное, отличное от человека, согласно настоящему изобретению, например, привитую мышь Rag2-/-Il2rgnullEpoh/m, привитую мышь Rag2-/-Il2rgnullTpoh/hIl3/Gmcsfh/hEpoh/mSIRPαh/h (“TIES”), привитую мышь Rag2-/-IL2rgy/-Tpoh/hMcsfh/hIl3h/hGmcsfh/hEpoh/hhSIRPα+(“MISTER-G”), привитую мышь Rag2-/-Il2rgnullTpoh/hMcsfh/hIl3/Gmcsfh/hEpoh/hSIRPα-tg+ (“SupER-G”) и т.п. приводят в контакт с представляющим интерес агентом-кандидатом и действие агента-кандидата оценивают путем контроля одного или нескольких выходных параметров. Эти выходные параметры могут отражать жизнеспособности клеток, например, общее количество гематопоэтических клеток или количество гематопоэтических клеток отдельного типа; или апоптотическое состояние клеток, например, степень фрагментации ДНК, количество клеток с блеббингом, количество фосфатидилсерина на поверхности клетки и тому подобные методы, хорошо известных в данной области. Альтернативно или дополнительно выходные параметры могут отражать способность клеток к дифференцировке, например, количественные соотношения дифференцированных клеток и дифференцированных типов клеток. Альтернативно или дополнительно, выходные параметры могут отражать функцию клеток, например, продуцируемые клетками цитокины и хемокины, продуцируемые клетками антитела (например, количество или тип), способность клеток возвращаться и проникать в место заражения, способность клеток модулировать, то есть стимулировать или подавлять активность других клеток in vitro или in vivo, способность поглощать гемоглобин и т.д. Еще другие параметры могут отражать действие агента на инфекцию, например, степень инфекции патогеном у животного, например, титр патогена у мыши, и т.п., в зависимости от проведенных исследований.
Параметры представляют собой количественно определяемые компоненты клеток, в частности, компоненты, которые могут быть точно измерены, желательно в высокопроизводительной системе. Параметр может также представлять собой любой клеточный компонент или клеточный продукт, включая поверхностную детерминанту клетки, рецептор, белок или их конформационные или посттрансляционные модификации, липид, углевод, органическую или неорганическую молекулу, нуклеиновую кислоту, например, мРНК, ДНК, и т.п., или часть, полученную от такого компонента клетки, или их комбинации. Притом, что большинство параметров будет предоставлять количественное показание, в некоторых случаях приемлем полуколичественный или качественный результат. Показания могут включать отдельные определенные значения, или могут включать среднее значение, медианное значение или отклонение, и т.п. Обычно из множества одинаковых анализов получают ряд значений показаний для каждого параметра. Следует ожидать наличия вариабельности, при этом ряд значений для каждого набора тестируемых параметров получают при помощи стандартных статистических методов в сочетании с обычным статистическим методом, используемым для получения отдельных величин.
Агенты-кандидаты, представляющие интерес для скрининга, включают известные и неизвестные соединения, которые охватывают многочисленные классы химических соединений, в первую очередь органические молекулы, которые могут включать металлоорганические молекулы, неорганические молекулы, генетические последовательности, вакцины, антибиотики или другие агенты, предположительно обладающие свойствами антибиотиков, пептиды, полипептиды, антитела, агенты, одобренные в качестве фармацевтических средств для применения у человека, и т.д. Важным аспектом изобретения является оценка лекарственных средств-кандидатов, включая тестирование на токсичность; и т.п.
Агенты-кандидаты включают органические молекулы, содержащие функциональные группы, необходимые для структурных взаимодействий, в частности, водород-связывающие группы, и обычно включают по меньшей мере одну амино, карбонильную, гидроксильную или карбоксильную группу, часто по меньшей мере две из функциональных химических групп. Агенты-кандидаты часто содержат циклические углеродные или гетероциклические структуры и/или ароматические или полиароматические структуры, замещенные одной или несколькими из указанных выше функциональных групп. Агенты-кандидаты также обнаруживаются среди биомолекул, включая пептиды, полинуклеотиды, сахариды, жирные кислоты, стероиды, пурины, пиримидины, их производные, структурные аналоги или их комбинации. Включенными являются фармакологически активные лекарственные средства, генетически активные молекулы и т.п. Представляющие интерес соединения включают химиотерапевтические агенты, гормоны или антагонисты гормонов, и т.п. Примерами фармацевтических агентов, подходящих для данного изобретения, являются агенты, описанные в "The Pharmacological Basis of Therapeutics," Goodman and Gilman, McGraw-Hill, New York, N.Y., (1996), Ninth edition. Также включенными являются токсины, а также боевые биологические и химические агенты, например, смотри Somani, S. M. (Ed.), "Chemical Warfare Agents," Academic Press, New York, 1992).
Представляющие интерес для скрининга агенты-кандидаты также включают нуклеиновые кислоты, например, нуклеиновые кислоты, которые кодируют миРНК, мшРНК, антисмысловые молекулы или микроРНК, или нуклеиновые кислоты, которые кодируют полипептиды. Доступно множество векторов, подходящих для переноса нуклеиновых кислот в клетки-мишени. Векторы могут поддерживаться эписомально, например, в виде плазмид, мини-кольцевых ДНК, векторов на основе вирусов, таких как цитомегаловирус, аденовирус и т.п., или они могут быть интегрированы в геном клетки-мишени посредством гомологичной рекомбинации или случайной интеграции, например, векторы на основе ретровирусов, таких как MMLV, HIV-1, ALV и т.п. Векторы могут быть доставлены непосредственно в целевую клетку. Другими словами, плюрипотентные клетки контактируют с векторами, содержащими представляющую интерес нуклеиновую кислоту, таким образом, что векторы поглощаются клетками.
Способы контактирования клеток, например, клеток в культуре или клеток мыши с векторами на основе нуклеиновых кислот, такие как электропорация, трансфекция хлоридом кальция и липофекция, хорошо известны в данной области. Альтернативно, представляющая интерес нуклеиновая кислота может быть доставлена в клетки посредством вируса. Другими словами, клетки контактируют с вирусными частицами, содержащими представляющую интерес нуклеиновую кислоту. Ретровирусы, например, лентивирусы, являются особенно подходящими для способа согласно изобретению. Обычно используемые ретровирусные векторы являются «дефектными», то есть неспособными продуцировать живые белки, необходимые для продуктивного заражения. Скорее, для репликации вектора требуется рост в пакующей линии клеток. Для образования вирусных частиц, содержащих представляющие интерес нуклеиновые кислоты, ретровирусные нуклеиновые кислоты, содержащие нуклеиновую кислоту, упаковываются в вирусные капсиды при помощи пакующей линии клеток. Различные пакующие линии клеток обеспечивают включение в капсид другого оболочечного белка, при этом этот оболочечный белок определяет специфичность вирусной частицы в отношении клеток. Оболочечные белки бывают по меньшей мере трех типов, экотропные, амфотропные и ксенотропные. Ретровирусы, упакованные экотропным оболочечным белком, например, MMLV, способны инфицировать большинство мышиных и крысиных типов клеток и образуются путем использования экотропных пакующих линий клеток, таких как BOSC23 (Pear et al. (1993) P.N.A.S. 90:8392-8396). Ретровирусы, несущие амфотропный оболочечный белок, например, 4070A (Danos et al., supra.), способны инфицировать большинство типов клеток млекопитающих, включая человека, собаку и мышь, и образуются при использовании амфотропных пакующих линий клеток, таких как PA12 (Miller et al. (1985) Mol. Cell. Biol. 5:431-437); PA317 (Miller et al. (1986) Mol. Cell. Biol. 6:2895-2902); GRIP (Danos et al. (1988) PNAS 85:6460-6464). Ретровирусы, пакуемые ксенотропным оболочечным белком, например, AKR env, способны инфицировать большинстве типов клеток млекопитающих, за исключением мышиных клеток. Подходящая пакующая линия клеток может быть использована для обеспечения нацеливания пакующих вирусных частиц на представляющие интерес клетки - в некоторых случаях, привитые клетки, в некоторых случаях клетки хозяина, то есть генетически модифицированного животного.
Векторы, используемые для доставки представляющей интерес нуклеиновой кислоты в целевые клетки, будут, как правило, содержать подходящие промоторы для управления экспрессией, то есть транскрипционной активации представляющей интерес нуклеиновой кислоты. Это может включать промоторы универсального действия, например, промотор CMV-β-актин, или индуцибельные промоторы, такие как промоторы, которые являются активными в определенных популяциях клеток или которые отвечают на присутствие лекарственных средств, таких как тетрациклин. Под транскрипционной активацией подразумевается, что транскрипция будет увеличиваться в клетке-мишени выше исходных уровней по меньшей мере примерно в 10 раз, по меньшей мере примерно в 100 раз, еще чаще по меньшей мере примерно в 1000 раз. Кроме того, векторы, используемые для доставки факторов перепрограммирования в целевые клетки, могут включать гены, которые впоследствии должны быть удалены, например, с использованием системы рекомбиназы, такой как Cre/Lox, или клетки, экспрессирующие их, могут быть разрушены, например, путем включения генов, обеспечивающих возможность селективной токсичности, таких как вируса герпеса TK, bcl-xs и т.п.
Представляющие интерес для скрининга агенты-кандидаты также включают полипептиды. Такие полипептиды могут быть необязательно слиты с доменом полипептида, который повышает растворимость продукта. Домен может быть связан с полипептидом посредством определенного сайта расщепления протеазой, например, последовательности TEV, которая расщепляется протеазой TEV. Линкер может также включать одну или несколько гибких последовательностей, например, от 1 до 10 остатков глицина. В некоторых вариантах осуществления расщепление гибридного белка выполняют в буфере, который поддерживает растворимость продукта, например, в присутствии от 0,5 до 2 М мочевины, в присутствии полипептидов и/или полинуклеотидов, которые повышают растворимость, и т.п. Представляющие интерес домены включают эндосомолитические домены, например, домен НА гриппа; и другие полипептиды, которые содействуют продукции, например, домен IF2, домен GST, домен GRPE и т.п. Дополнительно или альтернативно такие полипептиды могут быть включены в состав для улучшения стабильности. Например, пептиды могут быть пегилированными, при этом полиэтиленоксигруппа обеспечивает увеличение времени жизни в кровотоке. Полипептид может быть слит с другим полипептидом для обеспечения дополнительной функциональности, например, для увеличения стабильности in vivo. Как правило, такие партнеры слияния представляют собой стабильные белки плазмы, которые могут, например, продлевать in vivo период полувыведения из плазмы полипептида, если он присутствиует как «слитый», в частности, когда такой стабильный белок плазмы представляет собой константный домен иммуноглобулина. В большинстве случаев, когда стабильный белок плазмы обычно встречается в мультимерной форме, например, иммуноглобулины или липопротеины, в которых одинаковые или разные полипептидные цепи обычно связываются с помощью дисульфидной связи и/или нековалентной связи с образованием составного полипептида из нескольких цепей, слитые формы, содержащие полипептид, также будут продуцироваться и использоваться как мультимер, имеющий в основном ту же самую структуру, как предшественник стабильного белка плазмы. Эти мультимеры будут гомологичными по отношению к полипептидному агенту, который они содержат, или они могут содержать более одного полипептидного агента.
Полипептидный агент-кандидат может продуцироваться эукариотическими клетками, или может продуцироваться прокариотическими клетками. Он может затем процессироваться путем разворачивания, например, с помощью термической денатурации, восстановления с использованием дитиотреитола (DTT), и т.д., и может в дальнейшем повторно сворачиваться с использованием методов, известных в данной области. Представляющие интерес модификации, которые не изменяют первичную последовательность, включают химическую дериватизацию полипептидов, например, ацилирование, ацетилирование. карбоксилирование, амидирование и т.д. Также включенными являются модификации путем гликозилирования, например такие, которые созданы модификацией профиля гликозилирования полипептида в ходе его синтеза и процессинга, или на последующих стадиях обработки; например, при воздействии на полипептид ферментов, которые нарушают гликозилирование, такое как гликозилирование или дегликозилирование ферментов млекопитающего. Также, охваченными являются последовательности, которые содержат фосфорилированные аминокислотные остатки, например, фосфотирозин, фосфосерин или фосфотреонин. Полипептиды могут быть модифицированы с использованием обычных методов молекулярной биологии и синтетической химии, чтобы таким образом улучшить их устойчивость к протеолитической деградации или оптимизировать свойства растворимости, или сделать их более подходящими в качестве терапевтического агента. Аналоги таких полипептидов включают такие, которые содержат остатки, отличные от природных L-аминокислот, например, D-аминокислоты или неприродные синтетические аминокислоты. D-аминокислоты могут быть замещены по некоторым или всем аминокислотным остаткам.
Полипептидный агент-кандидат может быть получен путем синтеза in vitro с использованием обычных способов, известных в данной области. Доступны различные коммерческие установки для синтеза, например, автоматизированные синтезаторы от фирмы Applied Biosystems, Inc., Beckman, и т.п. При использовании синтезаторов природные аминокислоты могут быть замещены неприродными аминокислотами. Конкретная последовательность или способ получения будут определяться с точки зрения удобства, экономичности, необходимой чистоты и т.п. Альтернативно, полипептидный агент-кандидат может быть выделен и очищен в соответствии с обычными методами рекомбинантного синтеза. Может быть приготовлен лизат из клеток экспрессирующего хозяина, и лизат можно очистить с использованием HPLC, эксклюзионной хроматографии, гель-электрофореза, аффинной хроматографии или других методов очистки. В основном, используемые композиции будут содержать по меньшей мере 20% по массе желаемого продукта, еще чаще по меньшей мере около 75% по массе, предпочтительно по меньшей мере около 95% по массе, и для терапевтических целей обычно по меньшей мере около 99,5% по массе, относительно загрязняющих веществ, связанных со способом получения продукта и его очисткой. Как правило, процентное отношение будет основано на общем белке.
В некоторых случаях полипептидные агенты-кандиды, подлежащие скринингу, представляют собой антитела. Термин «антитело» или «фрагмент антитела» включает любую молекулярную структуру, содержащую полипептидную цепь со специфической формой, которая точно соответствует эпитопу и распознает эпитоп, при этом одно или несколько нековалентных связывающих взаимодействий стабилизируют комплекс между молекулярной структурой и эпитопом. Специфическое или селективное соответствие данной структуры и ее специфического эпитопа иногда называется совместимостью «замок и ключ». Первичная молекула антитела представляет собой иммуноглобулин, и все типы иммуноглобулинов, IgG, IgM, IgA, IgE, IgD и т.п. из всех источников, например, человека, грызуна, кролика, коровы, овцы, свиньи, собаки, другого млекопитающего, цыпленка, других птиц и т.д. считаются «антителами». Антитела, используемые в настоящем изобретении, могут представлять собой поликлональные антитела или моноклональные антитела. Антитела обычно находятся в среде, в которой культивируются клетки. Получение и скрининг антител более подробно описаны далее.
Агенты-кандидаты могут быть получены из целого ряда источников, включая библиотеки синтетических или природных соединений. Например, доступны многочисленные способы случайного и направленного синтеза широкого спектра органических соединений, включая биомолекулы, включая экспрессию случайных олигонуклеотидов и олигопептидов. Альтернативно, библиотеки природных соединений в форме бактериальных, грибных, растительных и животных экстрактов являются доступными или могут быть легко получены. Кроме того, природные или полученные синтетическим путем библиотеки и соединения могут быть легко модифицированы с помощью общепринятых химических, физических и биохимических способов, и могут быть использованы для получения комбинаторных библиотек. Известные фармакологические агенты могут быть подвергнуты направленным или случайным химическим модификациям, таким как ацилирование, алкилирование, этерификация, амидификация и т.д. для получения структурных аналогов.
Скрининг агентов-кандидатов на биологическую активность осуществляют путем введения агента по меньшей мере в один и обычно множество образцов, иногда в сочетании с образцами, не содержащими агент. Измеряют изменение параметров в ответ на агент и результат оценивают путем сравнения с референсными образцами, например, в присутствии и отсутствии агента, с результатами, полученными с другими агентами, и т.д. В случаях, когда скрининг осуществляют для идентификации агентов-кандидатов, которые будут предупреждать, ослаблять или устранять эффекты патогена, скрининг обычно проводят в присутствии патогенного агента, при этом патогенный агент добавляют в момент, наиболее подходящий для получения результатов. Например, в случаях, когда проводят тестирование защитной/превентивной способности агента-кандидата, агент-кандидат может быть добавлен до патогена, одновременно с патогеном или после инфицирования патогеном. В качестве другого примера, в случаях, когда тестируют способность агента-кандидата устранять эффекты патогена, агент-кандидат добавляют после инфицирования патогеном. Как указано выше, в некоторых случаях «образец» представляет собой генетически модифицированное животное, отличное от человека, привитое клетками, например, агент-кандидат доставляется иммунодефицитному животному, привитому человеческими гематопоэтическими клетками, например мыши, несущей нуклеиновую кислоту, кодирующую человеческий EPO, функционально связанную с промотором EPO. В некоторых случаях образец представляет собой человеческие гематопоэтические клетки, подлежащие приживлению, то есть агент-кандидат доставляют в клетки, например, ретикулоциты, эритроциты и т.д., перед прививкой иммунодефицитному генетически модифицированному животному.
В том случае, если агент-кандидат подлежит введению непосредственно привитому генетически модифицированному животному, агент может быть введен с помощью любого из ряда хорошо известных в данной области способов введения мышам пептидов, малых молекул и нуклеиновых кислот. Например, агент может быть введен перорально, мукозально, местно, интрадермально или путем инъекции, например, путем интраперитонеальной, подкожной, внутримышечной, внутривенной или интракраниальной инъекции, и т.п. Агент может быть введен в буфер, или он может быть включен в любую из разнообразных лекарственных форм, например, путем комбинирования с подходящим фармацевтически приемлемым носителем. «Фармацевтически приемлемые носители» могут представлять собой носители, одобренные регулирующим органом Правительства штата или Федеральным правительством, или перечисленные в Фармакопее США или другой общепризнанной фармакопее для применения у млекопитающих, таких как человек. Термин «носитель» относится к разбавителю, адъюванту, вспомогательному веществу или наполнителю, с которым соединение согласно изобретению составляют для введения млекопитающему. Такие фармацевтические носители могут представлять собой липиды, например, липосомы, например, дендримеры липисом; жидкости, такие как вода и масла, включая жидкости нефтяного, животного, растительного или синтетического происхождения, такие как арахисовое масло, соевое масло, минеральное масло, кунжутное масло и т.п., солевой раствор; аравийскую камедь, желатин, крахмальную пасту, тальк, кератин, коллоидный диоксид кремния, мочевину и т.п. Кроме того, могут быть использованы вспомогательные вещества, стабилизирующие вещества, загустители, смазывающие вещества и красящие вещества. Фармацевтические композиции могут быть составлены в препараты в твердых, полутвердых, жидких или газообразных формах, таких как таблетки, капсулы, порошки, гранулы, мази. растворы, суппозитории, инъекции, средства для ингаляции, гели, микросферы и аэрозоли. Агент после введения может распространяться системно или может быть локализован при использовании местного введения, интрамурального введения, или может использоваться имплант для поддержания активной дозы в месте имплантации. Активный агент может быть составлен для немедленной доставки или может быть составлен для замедленного высвобождения. При некоторых условиях, в частности, при использовании в центральной нервной системе, может быть необходимо составлять агенты для прохождения через гематоэнцефалический барьер (ВВВ). Одна из стратегий доставки лекарственного средства через гематоэнцефалический барьер (ВВВ) предусматривает разрыв ВВВ либо с помощью осмотических средств, таких как маннитол или лейкотриены, или биохимически путем использования вазоактивных веществ, таких как брадикинин. Разрывающий ВВВ агент можно вводить совместно с агентом при введении композиций путем внутримышечной инъекции. Другие стратегии прохождения ВВВ могут предусматривать использование эндогенных транспортных систем, включая опосредованный Кавеолином-1 трансцитоз, использование носителей-переносчиков, таких как глюкоза и аминокислотные переносчики, рецептор-опосредованный трансцитоз инсулина или трансферрина, и активные выводящие переносчики, такие как р-гликопротеин. Для использования в изобретении активные транспортные молекулы также могут быть конъюгированы с терапевтическими соединениями с целью облегчения транспорта через эндотелиальную стенку кровеносных сосудов. Альтернативно, доставка лекарственных агентов через ВВВ может происходить путем местной доставка, например, путем интратекальной доставки, например, в резервуар Оммайя (смотри, например, патенты США 5222982 и 5385582, включенные здесь путем отсылки); путем болюсной инъекции, например, шприцем, например, в стекловидное тело или интракраниально; путем непрерывной инфузии, например, путем катетеризации, например, с конвекцией (смотри, например, заявку США 20070254842, включенную здесь путем отсылки) или путем имплантации устройства, после чего к нему обратимо прикрепляется агент (смотри, например, заявки на патент США 20080081064 и 20090196903, включенные здесь путем отсылки).
Если агент(ы) доставляются в клетку перед приживлением, агенты в целях удобства добавляют в раствор или легкорастворимую форму в среду клеток в культуре. Агенты можно добавлять в проточную систему, такую как струя, периодически или непрерывно, или альтернативно, путем добавления болюса соединения, однократно или с определенным шагом, к другому постоянно текущему раствору. В проточной системе используются две жидкости, одна из которых представляет собой физиологически нейтральный раствор и другая представляет собой такой же раствор с добавленным тестируемым соединением. Первая жидкость омывает клетки, за которой следует вторая. В способе с использованием одного раствора болюс тестируемого соединения добавляют в объем среды, окружающей клетки. Общие концентрации компонентов среды для культивирования не должны значительно изменяться при введении болюса, или между двумя растворами в проточном способе.
Параллельно может выполняться множество анализов с различными концентрациями агента, чтобы получить различные ответы на различные концентрации. Как известно в данной области, для определения эффективной концентрации агента обычно используют диапазон концентраций, полученный в результате разведений 1:10 или соответственно другой log шкале. При необходимости концентрации могут быть дополнительно уменьшены с помощью второй серии разведений. Как правило, одна из этих концентраций служит в качестве отрицательного контроля, то есть при нулевой концентрации или ниже уровня обнаружения агента, или при такой концентрации агента или ниже, которая не дает поддающегося обнаружению изменения фенотипа.
Анализ ответа клеток привитого генетически модифицированного животного на агент-кандидат может быть проведен в любой момент времени после обработки агентом. Например, клетки могут быть проанализированы через 1, 2 или 3 дня, иногда 4. 5 или 6 дней, иногда 8, 9 или 10 дней, иногда 14 дней, иногда 21 день, иногда 28 дней, иногда 1 месяц или более после контакта с агентом-кандидатом, например, через 2 месяца, 4 месяца, 6 месяцев или более. В некоторых вариантах осуществления анализ включает анализ в многочисленных временных точках. Выбор временной точки(ек) для анализа будет зависеть от типа проводимого анализа, что будет очевидно специалисту в данной области.
Анализ может включать измерение любых параметров, описанных здесь или известных в данной области, для определения жизнеспособности клеток, пролиферации клеток, идентичности клеток, морфологии клеток и функционирования клеток, в частности, так как они могут относиться к иммунным клеткам. Например, может быть использована проточная цитометрия для определения общего количества гематопоэтических клеток или количества гематопоэтических клеток определенного типа. Может быть проведен гистохимический или иммуногистохимический анализ для определения апоптотического состояния клеток, например, введение концевой метки биотинилированным dUTP с использованием терминальной дезоксинуклеотидилтрансферазы (TUNEL) для измерения фрагментации ДНК, иммунгистохимический анализ для обнаружения связывания Аннексина V с фосфатидилсерином на клеточной поверхности. Проточная цитометрия также может быть использована для оценки процентного содержания дифференцированных клеток и типов дифференцированных клеток, например, для определения способности гематопоэтических клеток выживать и/или дифференцироваться в присутствии агента. ELISA, Вестерн-блоттинг и Нозерн-блоттинг могут быть выполнены для определения уровней цитокинов, хемокинов, иммуноглобулинов и т.д., экспрессирующихся у привитых генетически модифицированных мышей, например, для оценки функционирования привитых клеток, для оценки выживаемости эритроцитов и т.д. Также могут быть выполнены анализы in vivo для тестирования функционирования иммунных клеток, а также анализы, относящиеся к конкретным представляющим интерес заболеваниями или нарушениям, таким как анемия, например, серповидноклеточная анемия, и т.п.,. Смотри, например, Current Protocols in Immunology (Richard Coico, ed. John Wiley & Sons, Inc. 2012) и Immunology Methods Manual (I. Lefkovits ed., Academic Press 1997), описание которых включены здесь посредством ссылки.
Так, например, предлагается способ определения воздействия агента на эритроидные клетки, которые могут быть инфицированы или являются инфицированными патогеном, включающий введение агента мыши, несущей человеческий EPO, например, мыши, несущей Rag2-/-IL2rg-/-EPOm/h, которая была привита ретикулоцитами и/или эритроцитами человека; измерение показателя жизнеспособности привитых клеток во времени в присутствии агента; и сравнение этого показателя с показателем, измеренным у привитой мыши, несущей EPO человека, которая не была подвергнута воздействию агента. Агент определяется как антипатогенный в том случае, если он уменьшает инфекцию и/или гибель эритроцитов человека в периферической крови мыши по меньшей мере на 20%, 30%, 40% или более, в некоторых случаях на 50%, 60%, 70% или более, например, 80%, 90% или 100%, то есть до необнаруживаемых количеств, после однократного введения или двух или более введений агента в течение выбранного периода времени. В определенных вариантах осуществления введение лекарственного средства или комбинации лекарственных средств происходит через по меньшей мере три дня, по меньшей мере одну неделю, по меньшей мере 10 дней после прививания гематопоэтических клеток человека, например, через две недели, три недели или четыре недели после прививания гематопоэтических клеток человека, например, 6 недель, 8 недель, 10 недель или более после прививания гематопоэтических клеток человека.
Другие примеры применения подопытных мышей представлены в других местах в настоящем документе. Дополнительные варианты применения генетически модифицированных и привитых мышей, описанные в этом раскрытии, будут очевидны специалистам в данной области после прочтения данного описания.
Примеры неограничивающих аспектов изобретения
Аспекты, включая варианты осуществления настоящего изобретения, описанные выше, могут быть благоприятными отдельно или в комбинации с одним или несколькими другими аспектами или вариантами осуществления. Без ограничения вышеизложенного описания определенные неограничивающие аспекты изобретения под номерами 1-58 представлены ниже. Как будет очевидно специалистам в данной области при прочтении данного описания, каждый из индивидуально пронумерованных аспектов может быть использован или комбинирован с любым из предшествующих или следующих индивидуально пронумерованных аспектов. Это предполагает обеспечение поддержки для всех таких комбинаций аспектов и не ограничивается комбинациями аспектов, в явной форме представленных ниже:
1. Генетически модифицированное животное, отличное от человека, несущее:
нуклеотидную последовательность, которая кодирует человеческий белок EPO (hEPO), функционально связанную с промотором гена EPO.
2. Животное, отличное от человека, по пункту 1, отличающееся тем, что промотор гена EPO представляет собой эндогенный нечеловеческий промотор EPO.
3. Животное, отличное от человека, по пункту 1, отличающееся тем, что функциональное связывание происходит с эндогенным нечеловеческим промотором EPO в локусе EPO животного, отличного от человека.
4. Животное, отличное от человека, по пункту 3, отличающееся тем, что функциональное связывание приводит к нулевой мутации в нечеловеческом гене EPO в локусе нечеловеческого гена EPO.
5. Животное, отличное от человека, по пункту 4, отличающееся тем, что животное, отличное от человека, является гетерозиготным по аллелю, несущему нуклеотидную последовательность, которая кодирует hEPO.
6. Животное, отличное от человека, по пункту 4, отличающееся тем, что животное, отличное от человека, является гомозиготным по аллелю, несущему нуклеотидную последовательность, которая кодирует hEPO.
7. Животное, отличное от человека, по любому из пунктов 1-6, отличающееся тем, что нуклеотидная последовательность, которая кодирует hEPO, содержит геномную кодирующую и некодирующую человеческий EPO последовательность.
8. Животное, отличное от человека, по любому из пунктов 1-6, отличающееся тем, что нуклеотидная последовательность, которая кодирует hEPO, содержит кДНК-последовательность человеческого EPO.
9. Животное, отличное от человека, по любому из пунктов 1-8, отличающееся тем, что животное, отличное от человека, экспрессирует один или несколько дополнительных человеческих белков, выбранных из группы, состоящей из:
белка hM-CSF, кодируемого нуклеиновой кислотой под контролем промотора M-csf,
белка hIL3, кодируемого нуклеиновой кислотой под контролем промотора Il-3,
белка hGM-CSF, кодируемого нуклеиновой кислотой под контролем промотора Gm-csf,
белка hTPO, кодируемого нуклеиновой кислотой под контролем промотора TPO, и
белка hSirpa, кодируемого нуклеиновой кислотой под контролем промотора Sirpa.
10. Животное, отличное от человека, по пункту 9, отличающееся тем, что промотор представляет собой эндогенный промотор животного, отличного от человека, в соответствующем локусе гена животного, отличного от человека, и при этом животное, отличное от человека, является гетерозиготным нулевым по нечеловеческому гену.
11. Животное, отличное от человека, по пункту 9, отличающееся тем, что промотор представляет собой эндогенный промотор животного, отличного от человека, в соответствующем локусе гена животного, отличного от человека, и при этом животное, отличное от человека, является гомозиготным нулевым по нечеловеческому гену.
12. Животное, отличное от человека, по пункту 9, отличающееся тем, что человеческие белки включают по меньшей мере один из hTPO, hIL3, hGM-CSF и hSirpa.
13. Животное, отличное от человека, по любому из пунктов 1-12, отличающееся тем, что животное, отличное от человека, является иммунодефицитным.
14. Животное, отличное от человека, по пункту 13, отличающееся тем, что иммунодефицит вызван недостаточностью одного или обоих Rag2 и Il2rg.
15. Животное, отличное от человека, по любому из пунктов 1-14, отличающееся тем, что животное, отличное от человека, представляет собой млекопитающее.
16. Животное, отличное от человека, по пункту 15, отличающееся тем, что млекопитающее представляет собой грызуна.
17. Животное, отличное от человека, по пункту 16, отличающееся тем, что грызун представляет собой мышь.
18. Животное, отличное от человека, по любому из пунктов 1-17, отличающееся тем, что животное, отличное от человека, кроме того, привито человеческими гематопоэтическими клетками.
19. Животное, отличное от человека, по пункту 18, отличающееся тем, что человеческие гематопоэтические клетки включают одну или несколько клеток, выбранных из группы, состоящей из CD34-позитивной человеческой клетки, человеческой гематопоэтической стволовой клетки, человеческой миелоидной клетки-предшественника, человеческой эритроидной клетки-предшественника, человеческой миелоидной клетки, человеческой дендритной клетки, человеческого моноцита, человеческого гранулоцита, человеческого эритроцита, человеческого нейтрофила, человеческой тучной клетки, человеческого тимоцита и человеческого В-лимфоцита.
20. Животное, отличное от человека, по пункту 19, отличающееся тем, что животное, отличное от человека, несет клодронат.
21. Животное, отличное от человека, по пункту 20, отличающееся тем, что животное, отличное от человека, кроме того, несет инфекцию, вызванную патогеном, который поражает человеческие клетки эритроидного ростка.
22. Животное, отличное от человека, по пункту 21, отличающееся тем, что патоген выбран из Plasmodium sp., Babesia sp. и Theileri sp.
23. Способ идентификации агента, который ингибирует инфекцию, вызванную патогеном, который поражает человеческие клетки эритроидного ростка, при этом способ включает:
а. введение агента-кандидата генетически модифицированному животному, отличному от человека, при этом животное несет:
i. нуклеотидную последовательность, которая кодирует человеческий белок EPO (hEPO), функционально связанную с промотором гена EPO,
ii. одну или несколько генных мутаций, которые приводят к иммунодефициту у животного, отличного от человека,
iii. прививку человеческих гематопоэтических клеток, и
iv. инфекцию, вызванную патогеном, который поражает человеческие клетки эритроидного ростка, и
b. определение, снижает ли агент количество патогена у инфицированного патогеном животного, отличного от человека.
24. Способ идентификации агента, который предупреждает инфекцию, вызванную патогеном, который поражает человеческие клетки эритроидного ростка, при этом способ включает:
а. приведение в контакт генетически модифицированного животного, отличного от человека, с клодронатом, при этом животное, отличное от человека, несет:
i. нуклеотидную последовательность, которая кодирует белок EPO (hEPO), функционально связанную с промотором гена EPO,
ii. одну или несколько генных мутаций, которые приводят к иммунодефициту у животного, отличного от человека,
iii. прививку человеческих гематопоэтических клеток,
b. введение агента-кандидата генетически модифицированному животному, отличному от человека,
с. введение путем инъекции генетически модифицированному животному, отличному от человека, инфицированных паразитом ретикулоцитов или эритроцитов, и
d. определение, предупреждает ли агент инфекцию, вызванную ретикулоцитами и/или эритроцитами человека, у животного, отличного от человека.
25. Способ по пункту 23 или 24, отличающийся тем, что промотор гена EPO представляет собой эндогенный нечеловеческий промотор.
26. Способ по пункту 23 или 24, отличающийся тем, что функциональное связывание с эндогенным нечеловеческим промотором EPO происходит в локусе EPO животного, отличного от человека.
27. Способ по пункту 26, отличающийся тем, что функциональное связывание приводит к нулевой мутации в гене нечеловеческого EPO в локусе гена нечеловеческого EPO.
28. Способ по пункту 27, отличающийся тем, что животное, отличное от человека, является гетерозиготным по аллелю, несущему нуклеотидную последовательность, которая кодирует hEPO.
29. Способ по пункту 27, отличающийся тем, что животное, отличное от человека, является гомозиготным по аллелю, несущему нуклеотидную последовательность, которая кодирует hEPO.
30. Способ по любому из пунктов 23-29, отличающийся тем, что нуклеотидная последовательность, которая кодирует hEPO, содержит геномную кодирующую и некодирующую человеческий EPO последовательность.
31. Способ по любому из пунктов 23-29, отличающийся тем, что нуклеотидная последовательность, которая кодирует hEPO, содержит кДНК-последовательность человеческого EPO.
32. Способ по любому из пунктов 23-31, отличающийся тем, что животное, отличное от человека, экспрессирует один или несколько дополнительных человеческих белков, выбранных из группы, состоящей из:
белка hM-CSF, кодируемого нуклеиновой кислотой под контролем промотора M-csf,
белка hIL3, кодируемого нуклеиновой кислотой под контролем промотора Il-3,
белка hGM-CSF, кодируемого нуклеиновой кислотой под контролем промотора Gm-csf,
белка hTPO, кодируемого нуклеиновой кислотой под контролем промотора TPO, и
белка hSirpa, кодируемого нуклеиновой кислотой под контролем промотора Sirpa.
33. Способ по пункту 32, отличающийся тем, что промотор представляет собой эндогенный промотор животного, отличного от человека, в соответствующем локусе гена животного, отличного от человека, и при этом животное, отличное от человека, является гетерозиготным нулевым по нечеловеческому гену.
34. Способ по пункту 33, отличающийся тем, что промотор представляет собой эндогенный промотор животного, отличного от человека, в соответствующем локусе гена животного, отличного от человека, и при этом животное, отличное от человека, является гомозиготным нулевым по нечеловеческому гену.
35. Способ по любому из пунктов 23-34, отличающийся тем, животное, отличное от человека, является дефицитным по одному или обоим Rag2 и Il2rg.
36. Способ по любому из пунктов 23-35, отличающийся тем, что человеческие гематопоэтические клетки включают одну или несколько клеток, выбранных из группы, состоящей из человеческой CD34-позитивной клетки, человеческой гематопоэтической стволовой клетки, человеческой миелоидной клетки-предшественника, человеческой эритроидной клетки-предшественника, человеческой миелоидной клетки, человеческой дендритной клетки, человеческого моноцита, человеческого гранулоцита, человеческого эритроцита, человеческого нейтрофила, человеческой тучной клетки, человеческого тимоцита и человеческого В-лимфоцита.
37. Способ по любому из пунктов 23-36, отличающийся тем, что патоген выбран из Plasmodium sp., Babesia sp. и Theileri sp.
38. Способ по пункту 37, отличающийся тем, что патоген представляет собой Plasmodium sp. и Plasmodium sp. выбран из P. falciparum и P. vivax.
39. Способ по любому из пунктов 23-38, отличающийся тем, что животное, отличное от человека, представляет собой млекопитающее.
40. Способ по пункту 39, отличающийся тем, что млекопитающее представляет собой грызуна.
41. Способ по пункту 40, отличающийся тем, что грызун представляет собой мышь.
42. Способ получения мыши, экспрессирующей человеческий белок EPO (hEPO), включающий:
приведение в контакт плюропотентной стволовой клетки мыши с нуклеотидной последовательностью, содержащей кодирующую последовательность для hEPO или ее фрагмент, функционально связанную с последовательностью промотора EPO, при этом кодирующая последовательность и последовательность промотора EPO образуют кассету, которая фланкирована последовательностями, которые являются гомологичными локусу эндогенного EPO мыши,
культивирование плюропотентной стволовой клетки в условиях, которые содействуют интеграции нуклеотидной последовательности в геном мыши в локусе эндогенного EPO мыши путем гомологичной рекомбинации; и
получение мыши из плюропотентной стволовой клетки мыши, которая содержит нуклеотидную последовательность, кодирующую hEPO.
43. Способ по пункту 42, отличающийся тем, что плюропотентная стволовая клетка мыши представляет собой клетку ES или клетку iPS.
44. Способ по пункту 42 или 43, отличающийся тем, что плюропотентная стволовая клетка является дефицитной по Rag2 и/или IL2rg.
45. Способ по любому из пунктов 42-44, отличающийся тем, что последовательность промотора EPO представляет собой последовательность промотора человеческого EPO.
46. Способ по любому из пунктов 42-44, отличающийся тем, что последовательность промотора EPO представляет собой последовательность промотора эндогенного нечеловеческого EPO.
47. Способ по любому из пунктов 42-46, отличающийся тем, что интеграция приводит к замене гена EPO человека в локусе гена нечеловеческого EPO.
48. Способ по любому из пунктов 42-47, отличающийся тем, что нуклеотидная последовательность, которая кодирует hEPO, содержит геномную кодирующую и некодирующую человеческий EPO последовательность.
49. Способ по любому из пунктов 42-48, отличающийся тем, что нуклеотидная последовательность, которая кодирует hEPO, содержит кДНК-последовательность человеческого EPO.
50. Способ получения мыши, экспрессирующей человеческий белок EPO (hEPO) и содержащей человеческую гематопоэтическую систему, включающий:
трансплантацию популяции клеток, включающей человеческие гематопоэтические клетки-предшественники, генетически модифицированной мыши, полученной с помощью способа по любому из пунктов 42-49.
51. Способ по пункту 50, отличающийся тем, что трансплантация включает инъекцию в хвостовую вену, инъекцию в фетальную печень или ретроорбитальную инъекцию.
52. Способ по пункту 50 или 51, отличающийся тем, что генетически модифицированную мышь сублетально облучают перед трансплантацией.
53. Способ по любому из пунктов 50-52, отличающийся тем, что человеческие гематопоэтические клетки-предшественники представляют собой CD34+ клетки.
54. Способ по любому из пунктов 50-53, отличающийся тем, что человеческие гематопоэтические клетки-предшественники получают из фетальной печени, взрослого костного мозга или пуповинной крови.
55. Способ получения мыши, которая инфицирована человеческим патогеном, который поражает человеческие клетки эритроидного ростка, включающий:
получение мыши, экспрессирующей человеческий белок EPO (hEPO) и содержащей человеческую гематопоэтическую систему в соответствии со способами по пунктам 50-54,
введение мыши клодроната путем инъекции, и
введение мыши путем инъекции инфицированных паразитом человеческих красных кровяных клеток (PRBC).
56. Способ по пункту 55, отличающийся тем, что способ, кроме того, включает введение мыши путем инъекции здоровых человеческих красных кровяных клеток.
57. Способ по пункту 55, отличающийся тем, что паразит выбран из Plasmodium sp., Babesia sp. и Theileri sp.
58. Способ по пункту 55, отличающийся тем, что паразит представляет собой Plasmodium sp. и Plasmodium sp. выбран из P. falciparum и P. vivax.
ПРИМЕРЫ
Следующие примеры используют для того, чтобы предоставить специалисту в данной области полное раскрытие и описание того, как применять и использовать настоящее изобретение, и не предназначены для ограничения рамок того, что авторы рассматривают в качестве их изобретения, они также не имеют ввиду, что представленные далее эксперименты представляют собой все возможные эксперименты или единственные проделанные эксперименты. Были сделаны усилия для обеспечения точности в отношении используемых чисел (например, количеств, температуры и т.д.), однако должны учитываться некоторые экспериментальные ошибки и отклонения. Если не указано иное, части представляют собой массовые части, молекулярная масса является среднемассовой молекулярной массой, температура выражена в градусах Цельсия, а давление является атмосферным или примерно атмосферным.
Общие способы молекулярной и клеточной биохимии содержатся в таких стандартных учебниках, как Molecular Cloning: A Laboratory Manual, 3rd Ed. (Sambrook et al., HaRBor Laboratory Press 2001); Short Protocols in Molecular Biology, 4th Ed. (Ausubel et al. eds., John Wiley & Sons 1999); Protein Methods (Bollag et al., John Wiley & Sons 1996); Nonviral Vectors for Gene Therapy (Wagner et al. eds., Academic Press 1999); Viral Vectors (Kaplift & Loewy eds., Academic Press 1995); Immunology Methods Manual (I. Lefkovits ed., Academic Press 1997); и Cell and Tissue Culture: Laboratory Procedures in Biotechnology (Doyle & Griffiths, John Wiley & Sons 1998), описание которых включено в настоящий документ посредством ссылки. Реагенты, клонирующие векторы и наборы для генетической манипуляции, на которые делается ссылки в данном описании, доступны от коммерческих поставщиков, таких как BioRad, Stratagene, Invitrogen, Sigma-Aldrich и ClonTech.
Пример 1
Получение мышей с нокином (knock-in) EPO человека
Кодирующую последовательность в локусе гена эритропоэтина мыши (NCBI мышиный ген ID:13856; MGI:95407; RefSeq транскрипт кДНК: NM_007942.2 (SEQ ID NO:1) и кодируемый белок NP_031968 (SEQ ID NO:2)) заменяли на кодирующую последовательность из локуса гена эритропоэтина человека (NCBI человеческий ген ID:2056; HGNC: 3415; RefSeq транскрипт кДНК: NM_000799.2 (SEQ ID NO:3) и кодируемый белок NP_000790 (SEQ ID NO:4)).
Таблица 1
61 taggcgcgga gatgggggtg cccgaacgtc ccaccctgct gcttttactc tccttgctac
121 tgattcctct gggcctccca gtcctctgtg ctcccccacg cctcatctgc gacagtcgag
181 ttctggagag gtacatctta gaggccaagg aggcagaaaa tgtcacgatg ggttgtgcag
241 aaggtcccag actgagtgaa aatattacag tcccagatac caaagtcaac ttctatgctt
301 ggaaaagaat ggaggtggaa gaacaggcca tagaagtttg gcaaggcctg tccctgctct
361 cagaagccat cctgcaggcc caggccctgc tagccaattc ctcccagcca ccagagaccc
421 ttcagcttca tatagacaaa gccatcagtg gtctacgtag cctcacttca ctgcttcggg
481 tactgggagc tcagaaggaa ttgatgtcgc ctccagatac caccccacct gctccactcc
541 gaacactcac agtggatact ttctgcaagc tcttccgggt ctacgccaac ttcctccggg
601 ggaaactgaa gctgtacacg ggagaggtct gcaggagagg ggacaggtga catgctgctg
661 ccaccgtggt ggaccgacga acttgctccc cgtcactgtg tcatgccaac cctcc
(SEQ ID NO:1)
61 ccgccctctc ctccaggccc gtggggctgg ccctgcaccg ccgagcttcc cgggatgagg
121 gcccccggtg tggtcacccg gcgcgcccca ggtcgctgag ggaccccggc caggcgcgga
181 gatgggggtg cacgaatgtc ctgcctggct gtggcttctc ctgtccctgc tgtcgctccc
241 tctgggcctc ccagtcctgg gcgccccacc acgcctcatc tgtgacagcc gagtcctgga
301 gaggtacctc ttggaggcca aggaggccga gaatatcacg acgggctgtg ctgaacactg
361 cagcttgaat gagaatatca ctgtcccaga caccaaagtt aatttctatg cctggaagag
421 gatggaggtc gggcagcagg ccgtagaagt ctggcagggc ctggccctgc tgtcggaagc
481 tgtcctgcgg ggccaggccc tgttggtcaa ctcttcccag ccgtgggagc ccctgcagct
541 gcatgtggat aaagccgtca gtggccttcg cagcctcacc actctgcttc gggctctggg
601 agcccagaag gaagccatct cccctccaga tgcggcctca gctgctccac tccgaacaat
661 cactgctgac actttccgca aactcttccg agtctactcc aatttcctcc ggggaaagct
721 gaagctgtac acaggggagg cctgcaggac aggggacaga tgaccaggtg tgtccacctg
781 ggcatatcca ccacctccct caccaacatt gcttgtgcca caccctcccc cgccactcct
841 gaaccccgtc gaggggctct cagctcagcg ccagcctgtc ccatggacac tccagtgcca
901 gcaatgacat ctcaggggcc agaggaactg tccagagagc aactctgaga tctaaggatg
961 tcacagggcc aacttgaggg cccagagcag gaagcattca gagagcagct ttaaactcag
1021 ggacagagcc atgctgggaa gacgcctgag ctcactcggc accctgcaaa atttgatgcc
1081 aggacacgct ttggaggcga tttacctgtt ttcgcaccta ccatcaggga caggatgacc
1141 tggagaactt aggtggcaag ctgtgacttc tccaggtctc acgggcatgg gcactccctt
1201 ggtggcaaga gcccccttga caccggggtg gtgggaacca tgaagacagg atgggggctg
1261 gcctctggct ctcatggggt ccaagttttg tgtattcttc aacctcattg acaagaactg
1321 aaaccaccaa aaaaaaaaaa(SEQ ID NO:3)
В частности, мышиную геномную область на GRCm38: ch5: 137482017:137485745 («минус-цепь») удаляли и на ее место вставляли человеческую геномную последовательность из GRCh37: ch7:100318604:100321567 («плюс-цепь»). Это привело к замене кодирующих экзонов с 1 по 5 - всей кодирующей области - гена Epo мыши на экзоны с 1 по 5 и 3'-нетранслируемую область гена EPO человека. Всего 3729 nt (нуклеотидов) мышиной последовательности было заменено на 2964 nt человеческой последовательности.
Вкратце, нацеливающую конструкцию для замены гена EPO мыши на ген EPO человека в одну стадию нацеливания конструировали с использованием технологии генной инженерии VELOCIGENE® (смотри Valenzuela et al. (2003), supra, и патент США 6586251). ДНК мышиного и человеческого EPO получали из бактериальных искусственных хромосом bMQ-386K4 и RP11-797M3, соответственно. Rag2 -/- IL2rg Y/- ES-клетки подвергали электропорации PspXI-линеаризованной нацеливающей конструкцией, созданной методом клонирования с репарацией разрывов (gap repair cloning), содержащей мышиный EPO выше (в направлении 5') и ниже (в направлении 3') плечей гомологии, фланкирующих последовательность человеческого EPO длиной 2964 нуклеотидов, простирающуюся от ATG в экзоне 1 до стоп-кодона в экзоне 5 (то есть включая последовательность в направлении 3'), и селекционную кассету neo, фланкированную loxP-сайтами (фигура 2 и фигура 3). Сочленение между 5'-нетранслируемой областью (UTR) мышиного EPO и экзоном 1 человеческого EPO представлено в виде SEQ ID NO: 5, в которой последний мышиный нуклеотид перед первым нуклеотидом человеческого гена представляет собой «G» (показан в скобках) в части SEQ ID NO:5, которая не выделена жирным шрифтом в таблице 2 ниже, и первый нуклеотид человеческой последовательности представляет собой «А» (показан в скобках) в выделенной жирным шрифтом части SEQ ID NO:5 в таблице 2 ниже.
Таблица 2
(кодирующая последовательность человеческого экзона 1 выделена курсивом, жирным шрифтом - человеческая, не выделенная жирным шрифтом - мышиная)
Сочленение между человеческой 3'UTR и 5'-концом селекционной кассеты представлено в виде SEQ ID NO:6, в которой последний нуклеотид человеческой последовательности представляет собой «С» (показан в одиночных скобках) в выделенной жирным шрифтом части SEQ ID NO:6 в таблице 3 ниже, и первый нуклеотид последовательности селекционной кассеты представляет собой «С» (показано в двойных скобках) в не выделенной жирным шрифтом части SEQ ID NO:6 в таблице 3 ниже; расположенная ниже (3') область сочленения также содержит сайт loxP на 3'-конце для удаления кассеты neo под контролем промотора убиквитина, фланкированного loxP-сайтами.
Таблица 3
(выделенная жирным шрифтом является человеческая, курсивом - кодирующая последовательность человеческого экзона 5; не выделенная жирным шрифтом - 5' конец селекционной кассеты)
Сочленение между 3'-концом селекционной кассеты и мышиным геномом представлено в виде SEQ ID NO:7, в которой «С» в одиночных скобках представляет собой последний нуклеотид кассеты neo, и следующий за кассетой «G» в двойных скобках представляет собой первый нуклеотид мышиного генома, как показано в таблице 4 ниже.
Таблица 4
(3'-конец селекционной кассеты показан в скобках, мышиная последовательность показана без скобок)
Правильно нацеленные клоны hEPO ES-клеток идентифицировали с помощью анализа на утрату нативного аллеля Loss-of-native-allele (LONA) (Valenzuela et al. (2003), supra), в котором число копий нативного немодифицированного гена EPO определяли методом количественной полимеразной цепной реакции (qPCR) с двумя TaqMan™-зондами, специфическими для последовательностей в гене EPO мыши, которые были нацелены для делеции. Для qPCR-анализов использовали следующие наборы праймеров и зондов (написано в направлении от 5' к 3'): прямой праймер 3'-CATCTGCGACAGTCGAGTTC-5' (SEQ ID NO:8); обратный праймер 3'-CCAGGGAGCTTACCGTGAC-5' (SEQ ID NO:9); зонд 3'-FAM-AGGTACATCTTAGAGGCCAAGGAGGCA-BHQ-5' (SEQ ID NO:10); прямой праймер, 5'-ACAGCCGAGTCCTGGAGAG-3' (SEQ ID NO:11); обратный праймер 5'-AAGCCCTGAGCGTGAGTTC-3' (SEQ ID NO:12); зонд 5'-FAM-AGGCCAAGGAGGCCGAGAATATCACG-BHQ-3' (SEQ ID NO:13); в котором FAM относится к флуоресцентному зонду 5-карбоксифторесцеину и BHQ относится к гасителю флуоресценции типа «черная дыра» (Biosearch Technologies). ДНК, очищенную из клонов ES-клеток, которые поглощали нацеливающий вектор и встраивали в свои геномы, объединяли со смесью TaqMan™ Gene Expression Master Mix (Life Technologies) в соответствии с инструкциями изготовителя в 384-луночном планшете для PCR (MicroAmp™ Optical 384-Well Reaction Plate, Life Technologies) и анализировали на приборе Prism 7900HT фирмы Applied Biosystems, который собирает данные по флуоресценции в ходе PCR и определяет пороговый цикл (Ct), неполный цикл PCR, при котором накопленная флуоресценция достигает предварительно установленного порогового значения. Расположенные выше (в направлении 5') и ниже (в направлении 3') EPO-специфические qPCR и две qPCR для ненацеленных референсных генов проводили для каждого образца ДНК. Рассчитывали разницу в величинах Ct (ΔCt) между каждой EPO-специфической qPCR и qPCR каждого референсного гена, а затем рассчитывали разность между каждой величиной ΔCt и средним значением ΔCt для всех подвергнутых анализу образцов с получением величин ΔΔCt для каждого образца. Число копий гена EPO в каждом образце рассчитывали по следующей формуле: число копий = 2⋅2−ΔΔCt. Правильно нацеленный клон, утративший одну из своих нативных копий, будет иметь число копий гена EPO, равное единице. Подтверждение того, что последовательность гена EPO человека заменила удаленную последовательность гена EPO мыши в гуманизированном аллеле, получали с помощью анализа TaqMan™ qPCR, который содержит следующие наборы праймеров-проб (написано в направлении от 5' к 3'): человеческий прямой праймер, GAGCCCTGCACTGGACAAC (SEQ ID NO:14); человеческий обратный праймер, TCCCATGAACGCTGAGAGTC (SEQ ID NO:15); и человеческий зонд, AGGGTCAAGGAGCCATAGACAGAATGGC (SEQ ID NO:16).
Корректно нацеленные ES-клетки подвергали электропорации вектором, транзиентно экспрессирующим Cre для удаления селекционной кассеты лекарственного средства. Для создания мыши, несущей EPO человека и дефицитной по Rag2 и Il2rg, корректно нацеленные ES-клетки идентифицировали, как описано выше, и вводили доимплантационным эмбрионам с использованием методов, известных в данной области. Затем мышей с нокином (knock-in, KI) человеческого EPO возвратно скрещивали для получения мышей, дефицитных по Rag2 и Il2rg и экспрессирующих EPO человека.
Пример 2
Получение мышей, несущих человеческий SIRPα
В соответствии с некоторыми примерами, описанными здесь, генетически модифицированную мышь, несущую нуклеотидную последовательность, кодирующую человеческий SIRPα, случайным образом интегрированную в геном генетически модифицированной мыши, получали, как описано в заявке на патент США 2013-0340105, включенной в настоящий документ посредством ссылки.
В соответствии с некоторыми примерами, описанными здесь, мышь с нокином человеческого SIRPα получали, как описано ниже. Как известно, существует по меньшей мере 10 аллельных форм человеческого SIRPα. В этом конкретном примере вариант 1 человеческого SIRPα использовали для гуманизации эндогенного гена SIRPα мыши.
Нацеливающий вектор для гуманизации внеклеточной области гена SIRP (например, SIRPα) конструировали с использованием технологии VELOCIGENE® (смотри, например, патент США 6586251 и Valenzuela et al. (2003), supra).
Вкратце, клон bMQ-261H14 бактериальной искусственной хромосомы (BAC) мыши модифицировали для удаления последовательности, содержащей экзоны со 2 по 4 эндогенного гена SIRPα и вставки экзонов со 2 по 4 гена SIRPα человека с использованием клона CTD-3035H21 BAC человека. Геномную ДНК, соответствующую экзонам со 2 по 4 эндогенного гена SIRPα (~8555 bp (пар оснований)) заменяли в клоне bMQ-261H14 BAC на фрагмент ДНК размером ~8581 bp, содержащий экзоны со 2 по 4 гена SIRPα человека из клона CTD-3035H21 BAC. Анализ последовательности аллеля человеческого SIRPα, содержащегося в клоне CTD-3035H21 BAC, выявил, что аллель соответствует человеческому варианту 1. Неомициновую кассету, фланкированную сайтами loxP, добавляли к концу фрагмента ДНК человека размером ~8581 bp, содержащего экзоны со 2 по 4 гена SIRPα человека (фигура 4).
Расположенные выше (в направлении 5') и ниже (в направлении 3') плечи гомологии получали из ДНК BAC мыши в положениях 5' и 3' экзонов 2 и 4, соответственно, и добавляли в человеческий фрагмент размером ~8581 bp, содержащий неомициновую кассету, для создания конечного нацеливающего вектора для гуманизации эндогенного гена SIRPα, который содержал в направлении от 5' к 3' 5'-плечо гомологии, содержащее 19 kb ДНК мыши выше (в направлении 5') экзона 2 эндогенного гена SIRPα, фрагмент ДНК размером ~8581 bp, содержащий экзоны со 2 по 4 гена SIRPα человека, неомициновую кассету, фланкированную сайтами loxP, и 3'-плечо гомологии, содержащее 21 kb ДНК мыши ниже (в направлении 3') экзона 4 эндогенного гена SIRPα. Нацеленная вставка нацеливающего вектора помещала неомициновую кассету в пятый интрон гена SIRPα мыши между экзонами 4 и 5. Нацеливающий вектор линеаризовали путем расщепления SwaI и затем использовали в гомологичной рекомбинации в бактериальных клетках для достижения направленной замены экзонов с 2 по 4 в гене SIRPα мыши на экзоны с 2 по 4 гена SIRPα человека (фигура 4).
Нацеленную ДНК BAC (описанную выше) использовали для электропорации ES-клеток мыши для создания модифицированных ES-клеток, содержащих замену экзонов со 2 по 4 в эндогенном гене SIRPα мыши на геномный фрагмент, содержащий экзоны со 2 по 4 гена SIRPα человека. Положительные ES-клетки, несущие геномный фрагмент, содержащий экзоны со 2 по 4 гена SIRPα мыши, идентифицировали путем количественной PCR с использованием зондов TAQMAN™ (Lie and Petropoulos, 1998. Curr. Opin. Biotechnology 9:43-48). Нуклеотидная последовательность выше (в направлении 5') точки вставки включала следующее, что указывает на то, что эндогенная последовательность мыши, расположенная выше (в направлении 5') от точки вставки (содержится в скобках ниже) примыкает к геномной последовательности SIRPα человека, присутствующей в точке вставки:
(AGCTCTCCTACCACTAGACTGCTGAGACCCGCTGCTCTGCTCAGGACTCGATTTCCAGTACACAATCTCCCTCTTTGAAAAGTACCACACATCCTGGGGT)GCTCTTGCATTTGTGTGACACTTTGCTAGCCAGGCTCAGTCCTGGGTTCCAGGTGGGGACTCAAACACACTGGCACGAGTCTACATTGGATATTCTTGGT (SEQ ID NO: 17). Нуклеотидная последовательность ниже (в направлении 3') точки вставки на 5'-конце неомициновой кассеты включала следующее, что указывает на то, что геномная последовательность SIRPα человека примыкает к последовательности кассеты, расположенной ниже (в направлении 3') от точки вставки (содержится в скобках ниже с последовательностью loxP, выделенной курсивом):
GCTCCCCATTCCTCACTGGCCCAGCCCCTCTTCCCTACTCTTTCTAGCCCCTGCCTCATCTCCCTGGCTGCCATTGGGAGCCTGCCCCACTGGAAGCCAG(TCGAGATAACTTCGTATAATGTATGCTATACGAAGTTATATGCATGGCCTCCGCGCCGGGTTTTGGCGCCTCCCGCGGGCGCCCCCCTCCTCACGGCGA) (SEQ ID NO: 18). Нуклеотидная последовательность ниже (в направлении 3') точки вставки на 3'-конце неомициновой кассеты включала следующее, что указывает на то, что последовательность кассеты примыкает к геномной последовательности мыши 3' экзона 4 эндогенного гена SIRPα (содержится в скобках ниже):
CATTCTCAGTATTGTTTTGCCAAGTTCTAATTCCATCAGACCTCGACCTGCAGCCCCTAGATAACTTCGTATAATGTATGCTATACGAAGTTATGCTAGC(TGTCTCATAGAGGCTGGCGATCTGGCTCAGGGACAGCCAGTACTGCAAAGAGTATCCTTGTTCATACCTTCTCCTAGTGGCCATCTCCCTGGGACAGTCA) (SEQ ID NO: 19). Положительные клоны ES-клеток затем применяли для имплантации самкам мышей с использованием метода VELOCIMOUSE® (смотри, например, патент США 7294754 и Poueymirou et al. 2007, F0 generation mice that are essentially fully derived from the donor gene-targeted ES cells allowing immediate phenotypic analyses Nature Biotech. 25(1):91-99, supra) для получения помета детенышей, несущих вставку экзонов со 2 по 4 гена SIRPα человека в эндогенный ген SIRPα мыши.
Нацеленные ES-клетки, описанные выше, использовали в качестве донорных ES-клеток и вводили мышиным эмбрионам на 8-клеточной стадии с помощью метода VELOCIMOUSE® (supra). Мышей, несущих гуманизацию экзонов со 2 по 4 эндогенного гена SIRPα, идентифицировали путем генотипирования с использованием анализа модификации аллеля (Valenzuela et al. (2003), supra), который выявил присутствие последовательностей гена SIRPα человека.
Мыши, несущие конструкцию гуманизированного гена SIRPα (то есть содержащую экзоны со 2 по 4 SIRPα человека в гене SIRPα мыши), могут быть скрещены со штаммом Cre делеторных мышей (смотри, например, международную публикацию патентной заявки WO 2009/114400) для удаления любых неомициновых кассет, фланкированных lox-сайтами, введенных с помощью нацеливающего вектора, который не удаляется, например, на стадии ES-клеток или в эмбрионе. Необязательно, у мышей сохраняется неомициновая кассета.
Пример 3
Генерирование сложных мышей с нокином
Мышей с нокином человеческого EPO скрещивали с мышами, экспрессирующими другие представляющие интерес гены человека в виде случайных интегрантов в геном мыши или в виде нокинов, то есть из соответствующего локуса мыши. Например, Rag2-/-, IL-2rgY/-, hEPO KI мышей скрещивали с мышами, экспрессирующими TPO человека из локуса TPO мыши (Rongvaux et al., 2011, Proc Natl Acad Sci USA, 108(6): 2378-2383), IL-3 человека и GM-CSF человека из локуса IL-3/GM-CSF мыши (Willinger et al, 2011, Proc Natl Acad Sci USA, 108(6): 2390-2395), M-CSF человека из локуса M-CSF мыши (Rathinam et al, 2011, Blood, 118(11): 3119-3128) и/или SIRPα человека, экспрессирующийся в виде случайного интегранта (Strowig et al., 2011, Proc Natl Acad Sci USA, 108(32): 13218-13223) или из локуса мыши, как описано выше, для создания мышей, экспрессирующих комбинацию этих белков человека (Rag2-/-Il2rgnullhSIRPah/h Tpoh/h Mcsf h/h Il3/Gmcsf h/h EPOh/h). Генетически модифицированные мыши, экспрессирующие один или несколько из TPO человека, IL-3 человека, GM-CSF человека, M-CSF человека и SIRPα человека, описаны более подробно в патентах США 8541646 и 8847004; заявках на патент США 2014/0134662; и международной публикации PCT WO/2014/039782; раскрытие каждого из которых включено здесь посредством ссылки.
Пример 4
Разработка гуманизированной мышиной модели для стадий распространения P. falciparum и P. vivax в кровеносном русле
В настоящем документе продемонстрировано, что генетическая гуманизация мыши-хозяина путем обеспечения факторов роста с ограниченной кросс-реактивностью от мыши к человеку может успешно стимулировать приживление человеческих клеток в целом и эритропоэз в частности у мышей с привитыми гематопоэтическими стволовыми клетками (HSC) человека.
Мыши MITERG, экспрессирующие EPO человека из своего генома (для усиления терминального эритропоэза), а также TPO, MCSF и IL-3 человека (для усиления поддержки HSC и раннего эритропоэза), показывают более высокие уровни эритропоэза человека в костном мозге мышей, привитых HSC человека, чем мыши, не экспрессирующие hEPO (фигура 5A, сравнение «hSIRPα-, hEPO+» (то есть «мыши MITERG») с «hSIRPα-, hEPO-« (то есть «мыши MITRG»)), уровень эритропоэза которых фактически эквивалентен мышиному эритропоэзу этого животного. Однако эти мыши не имеют значительных уровней циркулирующих эритроидных клеток человека (данные не показаны).
Было выдвинуто предположение о том, что низкие уровни циркулирующих эритроидных клеток человека в периферической крови возникают в результате деструкции (эритрофагоцитоза) периферических RBC человека мышиными макрофагами. Участие SIRPα в этом процессе оценивали путем создания мышей, у которых экспрессия hSIRPα происходила из локуса в геноме мыши, отличном от локуса mSIRPα, то есть в виде случайным образом интегрированного трансгена, то есть в виде hSIRPα-tg (Rag2-/-Il2rgnullTpoh/hMcsfh/hIl3h/hGmcsfh/hEpoh/hSIRPα-tg+, то есть «мыши MISTER-G»), и мышей, у которых экспрессия hSIRPα происходила в виде нокина (knock-in, KI) из локуса mSIRPα, то есть в виде hSIRPα KI (Rag2-/- Il2rgnullTpoh/hMcsfh/hIl3h/hGmcsfh/hEpoh/hSIRPαh/h, то есть «мыши SupER-G»).
Экспрессия SIRPα человека, например, в виде случайным образом интегрированного трансгена у мышей MISTER-G, способствовала дополнительному увеличению количество эритроидных клеток человека в костном мозге привитых HSC человека мышей, экспрессирующих EPO человека (фигура 5, панель A, сравнение «hSIRPa+, hEPO+» с «hSIRPa-, hEPO+»). Введение SIRPα человека значительно улучшило выживаемость в периферической крови большей части гематопоэтических клеток, при этом количество CD45+ клеток человека в периферической крови, включая лимфоидные и миелоидные клетки, увеличилось по меньшей мере в 10 раз по сравнению с количеством, наблюдаемым у экспрессирующих hSIRPα мышей. Однако hSIRPα KI имеет незначительный эффект на уровни приживления RBC человека в периферической крови (фигура 5, панель B).
Поскольку нокина SIRPα человека недостаточно для увеличения количества эритроидных клеток человека в периферической крови, использовали содержащие клодронат липосомы для деплеции макрофагов, особенно макрофагов красной пульпы селезенки и купферовских клеток печени. Значительное увеличение циркулирующих человеческих клеток эритроидного ростка наблюдалось у мышей SuPER-G, составляющее до 1% от всех циркулирующих эритроидных клеток после деплеции оседлых тканевых макрофагов с помощью содержащих клодронат липосом (фигура 6, панель А). Кроме того, большинство эритроидных клеток человека в периферической крови после обработки клодронатом представляли собой ретикулоциты (CD71+) (фигура 6, панель B). Это наблюдение удивительно, так как указывает на то, что эти мыши будут хорошими кандидатами для поддержки инфекции, вызванной P. vivax, которые преимущественно инфицируют ретикулоциты.
Инфицирование эритроидных клеток является важной частью жизненного цикла Plasmodium sp. Однако не установлено количество RBC человека, необходимое для успешного инфицирования in vivo различными видами Plasmodium. В настоящее время доступны только модели in vivo для P. falciparum, которые основаны на переносе RBC человека в иммунодефицитные штаммы, такие как NOD/scid или NSG. У этих мышей инфицирование мерозоитами может быть достигнуто путем внутривенной (i.v.) инъекции инфицированных эритроцитов только после ежедневной инъекции большого количества эритроцитов человека. На момент инфицирования RBC человека составляют приблизительно половину от общего количества эритроцитов у этих животных.
Как показано выше, мыши SupER-G, привитые гематопоэтическими стволовыми клетками (HSC) человека, развили эритроидные клетки человека в костном мозге (фигура 5В), и ii) лечение клодронатом увеличило количество эритроидных клеток человека в периферической крови привитых мышей SupER-G (фигура 6). Для определения, могут ли привитые, обработанные клодронатом мыши SupER-G, несущие достаточное количество эритроидных клеток человека в периферической крови, поддерживать успешную инфекцию Plasmodium in vivo, периферическую кровь собирали у мышей SupER-G, привитых HSC фетальной печени или взрослыми HSC, и кровь культивировали in vitro с кровью, инфицированной штаммом 3D7 P. falciparum. Для содействия множеству циклов репликации паразита через 48 часов в инфекционную среду добавляли свежие красные кровяные клетки человека, при этом ожидалось, что амплификация путем повторного инфицирования добавленных позднее RBC человека будет возникать только в случае, если RBC человека, собранные у привитых HSC мышей SupER-G, проходили полный инфекционный цикл P. falciparum. Через двенадцать дней после инфицирования поздняя стадия паразитарной инфекции и амплификация мерозоитов из шизонта наблюдалась во всех образцах крови привитых мышей SuPER-G («привитые RBC мыши, взрослые 1», «привитые RBC мыши, взрослые 2» и «привитые RBC мыши, фетальная печень»), но не от контрольных мышей, которые не были привиты или остро привиты 0,1% hRBC путем инъекции («инъецированные контроли») (фигура 7A и данные не показаны). Экспоненциальное увеличение паразитемии наблюдалось по окрашиванию по Гимзе и количественной PCR (фигура 7B). Это указывает на то, что привитые обработанные клодронатом мыши SupER-G продуцируют достаточное количество красных кровяных клеток человека для поддержания инфекции, вызванной P. falciparum. На основании этого ожидать успешное in vivo инфицирование дозированных клодронатом привитых мышей с мерозоитами P. falciparum и P. vivax.
Пример 5
Мыши TIES (Rag2-/-Il2rgnullTpoh/hIl3/Gmcsfh/hEpoh/mSIRPαh/h )
Вследствие низкой фертильности, эволюционной некомпетентности и высокой смертности мыши с Epoh/h не являются идеальными для исследования инфекции. Напротив, мыши, гетерозиготные по гену hEPO (то есть Epoh/m, например, мыши TIES) являются полностью способными продуцировать эритропоэтин (EPO) и поддерживать все стадии эритропоэза. Дополнительное улучшение жизнеспособности от 8-10 недель до 4 месяцев было достигнуто скорее путем удерживания гена M-CSF мыши в локусе мыши, чем замены его на M-CSF человека, так как высокий уровень приживления миелоидных клеток человека, поддерживаемый у мышей с нокином макрофагального колониестимулирующего фактора (M-CSF) человека, вызывает деструкцию мышиных красных кровяных клеток, что приводит к анемии и смерти привитых мышей.
Подобно мышам SupER-G, мыши TIES поддерживают эритропоэз человека и сохраняют количество, составляющее 1% эритроидных клеток человека в периферической крови в случае дозирования клодронатом. Кроме того, большинство эритроидных клеток человека в периферической крови после обработки клодронатом представляли собой ретикулоциты (CD71+). Это дает основание полагать, что эти мыши будут хорошими кандидатами для поддержки инфекции, вызванной P. vivax.
Было показано, что привитые мыши TIES могут сохранять количество, равное 1% эритроидных клеток человека в периферической крови в случае дозирования клодронатом. Кроме того, определяли способность мышей TIES поддерживать трансфузию RBC человека. Как показано на фигуре 8, обработка клодронатом стабилизировала трансфузированную популяцию на уровне более чем 20% от общей популяции клеток в периферической крови. Эти уровни, установившиеся примерно через 4 часа после трансфузии, были устойчивыми в течение по меньшей мере 12 часов после трансфузии. Предполагается, что эти количества RBC будут достаточными для поддержки инфекции in vivo, вызванной Plasmodium различных видов.
Изложенное выше только иллюстрирует принципы изобретения. Следует иметь в виду, что специалисты в данной области будут способны создать разные варианты, которые, хотя и не описаны подробно или не показаны в описании, воплощают принципы изобретения и включены в его объем и сущность. Более того, все упомянутые в описании примеры и условный язык, прежде всего, предназначаются для понимания принципов изобретения и идей, вносимых изобретателями для продвижения данной области техники, и их следует рассматривать как неограниченные такими конкретно изложенными примерами и условиями. Более того, все изложенные в описании утверждения, принципы, аспекты и варианты осуществления изобретения, а также их конкретные примеры, как предполагается, включают их структурные и функциональные эквиваленты. Кроме того, предполагается, что такие эквиваленты включают и известные в настоящее время эквиваленты и эквиваленты, которые будут созданы в будущем, то есть любые созданные элементы, которые выполняют ту же самую функцию, вне зависимости от структуры. Следовательно, объем настоящего изобретения не ограничивается иллюстративными вариантами осуществления, показанными и описанным здесь. Точнее, существо и объем настоящего изобретения излагается в прилагаемых пунктах формулы изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Regeneron Pharmaceuticals, Inc.
Yale University
Institute for Research in Biomedicine (IRB)
Murphy, Andrew J
Stevens, Sean
Flavell, Richard
Manz, Markus Gabriel
Shan, Liang
<120> GENETICALLY MODIFIED NON-HUMAN ANIMALS EXPRESSING HUMAN EPO
<130> REGN-013WO
<150> US62/000,460
<151> 2014-05-19
<160> 19
<170> PatentIn version 3.5
<210> 1
<211> 715
<212> DNA
<213> Mus musculus
<400> 1
gatgaagact tgcagcgtgg acactggccc agccccgggt cgctaaggag ctccggcagc 60
taggcgcgga gatgggggtg cccgaacgtc ccaccctgct gcttttactc tccttgctac 120
tgattcctct gggcctccca gtcctctgtg ctcccccacg cctcatctgc gacagtcgag 180
ttctggagag gtacatctta gaggccaagg aggcagaaaa tgtcacgatg ggttgtgcag 240
aaggtcccag actgagtgaa aatattacag tcccagatac caaagtcaac ttctatgctt 300
ggaaaagaat ggaggtggaa gaacaggcca tagaagtttg gcaaggcctg tccctgctct 360
cagaagccat cctgcaggcc caggccctgc tagccaattc ctcccagcca ccagagaccc 420
ttcagcttca tatagacaaa gccatcagtg gtctacgtag cctcacttca ctgcttcggg 480
tactgggagc tcagaaggaa ttgatgtcgc ctccagatac caccccacct gctccactcc 540
gaacactcac agtggatact ttctgcaagc tcttccgggt ctacgccaac ttcctccggg 600
ggaaactgaa gctgtacacg ggagaggtct gcaggagagg ggacaggtga catgctgctg 660
ccaccgtggt ggaccgacga acttgctccc cgtcactgtg tcatgccaac cctcc 715
<210> 2
<211> 192
<212> PRT
<213> Mus musculus
<400> 2
Met Gly Val Pro Glu Arg Pro Thr Leu Leu Leu Leu Leu Ser Leu Leu
1 5 10 15
Leu Ile Pro Leu Gly Leu Pro Val Leu Cys Ala Pro Pro Arg Leu Ile
20 25 30
Cys Asp Ser Arg Val Leu Glu Arg Tyr Ile Leu Glu Ala Lys Glu Ala
35 40 45
Glu Asn Val Thr Met Gly Cys Ala Glu Gly Pro Arg Leu Ser Glu Asn
50 55 60
Ile Thr Val Pro Asp Thr Lys Val Asn Phe Tyr Ala Trp Lys Arg Met
65 70 75 80
Glu Val Glu Glu Gln Ala Ile Glu Val Trp Gln Gly Leu Ser Leu Leu
85 90 95
Ser Glu Ala Ile Leu Gln Ala Gln Ala Leu Leu Ala Asn Ser Ser Gln
100 105 110
Pro Pro Glu Thr Leu Gln Leu His Ile Asp Lys Ala Ile Ser Gly Leu
115 120 125
Arg Ser Leu Thr Ser Leu Leu Arg Val Leu Gly Ala Gln Lys Glu Leu
130 135 140
Met Ser Pro Pro Asp Thr Thr Pro Pro Ala Pro Leu Arg Thr Leu Thr
145 150 155 160
Val Asp Thr Phe Cys Lys Leu Phe Arg Val Tyr Ala Asn Phe Leu Arg
165 170 175
Gly Lys Leu Lys Leu Tyr Thr Gly Glu Val Cys Arg Arg Gly Asp Arg
180 185 190
<210> 3
<211> 1340
<212> DNA
<213> homo sapiens
<400> 3
cccggagccg gaccggggcc accgcgcccg ctctgctccg acaccgcgcc ccctggacag 60
ccgccctctc ctccaggccc gtggggctgg ccctgcaccg ccgagcttcc cgggatgagg 120
gcccccggtg tggtcacccg gcgcgcccca ggtcgctgag ggaccccggc caggcgcgga 180
gatgggggtg cacgaatgtc ctgcctggct gtggcttctc ctgtccctgc tgtcgctccc 240
tctgggcctc ccagtcctgg gcgccccacc acgcctcatc tgtgacagcc gagtcctgga 300
gaggtacctc ttggaggcca aggaggccga gaatatcacg acgggctgtg ctgaacactg 360
cagcttgaat gagaatatca ctgtcccaga caccaaagtt aatttctatg cctggaagag 420
gatggaggtc gggcagcagg ccgtagaagt ctggcagggc ctggccctgc tgtcggaagc 480
tgtcctgcgg ggccaggccc tgttggtcaa ctcttcccag ccgtgggagc ccctgcagct 540
gcatgtggat aaagccgtca gtggccttcg cagcctcacc actctgcttc gggctctggg 600
agcccagaag gaagccatct cccctccaga tgcggcctca gctgctccac tccgaacaat 660
cactgctgac actttccgca aactcttccg agtctactcc aatttcctcc ggggaaagct 720
gaagctgtac acaggggagg cctgcaggac aggggacaga tgaccaggtg tgtccacctg 780
ggcatatcca ccacctccct caccaacatt gcttgtgcca caccctcccc cgccactcct 840
gaaccccgtc gaggggctct cagctcagcg ccagcctgtc ccatggacac tccagtgcca 900
gcaatgacat ctcaggggcc agaggaactg tccagagagc aactctgaga tctaaggatg 960
tcacagggcc aacttgaggg cccagagcag gaagcattca gagagcagct ttaaactcag 1020
ggacagagcc atgctgggaa gacgcctgag ctcactcggc accctgcaaa atttgatgcc 1080
aggacacgct ttggaggcga tttacctgtt ttcgcaccta ccatcaggga caggatgacc 1140
tggagaactt aggtggcaag ctgtgacttc tccaggtctc acgggcatgg gcactccctt 1200
ggtggcaaga gcccccttga caccggggtg gtgggaacca tgaagacagg atgggggctg 1260
gcctctggct ctcatggggt ccaagttttg tgtattcttc aacctcattg acaagaactg 1320
aaaccaccaa aaaaaaaaaa 1340
<210> 4
<211> 193
<212> PRT
<213> homo sapiens
<400> 4
Met Gly Val His Glu Cys Pro Ala Trp Leu Trp Leu Leu Leu Ser Leu
1 5 10 15
Leu Ser Leu Pro Leu Gly Leu Pro Val Leu Gly Ala Pro Pro Arg Leu
20 25 30
Ile Cys Asp Ser Arg Val Leu Glu Arg Tyr Leu Leu Glu Ala Lys Glu
35 40 45
Ala Glu Asn Ile Thr Thr Gly Cys Ala Glu His Cys Ser Leu Asn Glu
50 55 60
Asn Ile Thr Val Pro Asp Thr Lys Val Asn Phe Tyr Ala Trp Lys Arg
65 70 75 80
Met Glu Val Gly Gln Gln Ala Val Glu Val Trp Gln Gly Leu Ala Leu
85 90 95
Leu Ser Glu Ala Val Leu Arg Gly Gln Ala Leu Leu Val Asn Ser Ser
100 105 110
Gln Pro Trp Glu Pro Leu Gln Leu His Val Asp Lys Ala Val Ser Gly
115 120 125
Leu Arg Ser Leu Thr Thr Leu Leu Arg Ala Leu Gly Ala Gln Lys Glu
130 135 140
Ala Ile Ser Pro Pro Asp Ala Ala Ser Ala Ala Pro Leu Arg Thr Ile
145 150 155 160
Thr Ala Asp Thr Phe Arg Lys Leu Phe Arg Val Tyr Ser Asn Phe Leu
165 170 175
Arg Gly Lys Leu Lys Leu Tyr Thr Gly Glu Ala Cys Arg Thr Gly Asp
180 185 190
Arg
<210> 5
<211> 226
<212> DNA
<213> Artificial Sequence
<220>
<223> synthetic polynucleotide sequence
<220>
<221> misc_feature
<222> (1)..(115)
<223> mouse sequence
<220>
<221> misc_feature
<222> (116)..(128)
<223> human Exon 1
<220>
<221> misc_feature
<222> (116)..(226)
<223> human sequence
<400> 5
tcttccaggc tagtggggtg atctggccct acagaacttc caaggatgaa gacttgcagc 60
gtggacactg gcccagcccc gggtcgctaa ggagctccgg cagctaggcg cggagatggg 120
ggtgcacggt gagtactcgc gggctgggcg ctcccgcccg cccgggtccc tgtttgagcg 180
gggatttagc gccccggcta ttggccagga ggtggctggg ttcaag 226
<210> 6
<211> 1054
<212> DNA
<213> Artificial sequence
<220>
<223> synthetic polynucleotide sequence
<220>
<221> misc_feature
<222> (1)..(115)
<223> human Exon 5
<220>
<221> misc_feature
<222> (1)..(926)
<223> human sequence
<220>
<221> misc_feature
<222> (927)..(1054)
<223> 5' end of selection cassette
<400> 6
actccgaaca atcactgctg acactttccg caaactcttc cgagtctact ccaatttcct 60
ccggggaaag ctgaagctgt acacagggga ggcctgcagg acaggggaca gatgaccagg 120
tgtgtccacc tgggcatatc caccacctcc ctcaccaaca ttgcttgtgc cacaccctcc 180
cccgccactc ctgaaccccg tcgaggggct ctcagctcag cgccagcctg tcccatggac 240
actccagtgc cagcaatgac atctcagggg ccagaggaac tgtccagaga gcaactctga 300
gatctaagga tgtcacaggg ccaacttgag ggcccagagc aggaagcatt cagagagcag 360
ctttaaactc agggacagag ccatgctggg aagacgcctg agctcactcg gcaccctgca 420
aaatttgatg ccaggacacg ctttggaggc gatttacctg ttttcgcacc taccatcagg 480
gacaggatga cctggagaac ttaggtggca agctgtgact tctccaggtc tcacgggcat 540
gggcactccc ttggtggcaa gagccccctt gacaccgggg tggtgggaac catgaagaca 600
ggatgggggc tggcctctgg ctctcatggg gtccaagttt tgtgtattct tcaacctcat 660
tgacaagaac tgaaaccacc aatatgactc ttggcttttc tgttttctgg gaacctccaa 720
atcccctggc tctgtcccac tcctggcagc agtgcagcag gtccaggtcc gggaaacgag 780
gggtggaggg ggctgggccc tacgtgctgt ctcacacagc ctgtctgacc tctcgaccct 840
accgggcctg aggccacaag ctctgcctac gctggtcaat aaggtgtctc cattcaaggc 900
ctcaccgcag taaggcagct gccaacctcg agataacttc gtataatgta tgctatacga 960
agttatatgc atggcctccg cgccgggttt tggcgcctcc cgcgggcgcc cccctcctca 1020
cggcgagcgc tgccacgtca gacgaagggc gcag 1054
<210> 7
<211> 243
<212> DNA
<213> Artificial sequence
<220>
<223> synthetic polynucleotide sequence
<220>
<221> misc_feature
<222> (1)..(121)
<223> 3' end of selection cassette
<220>
<221> misc_feature
<222> (122)..(243)
<223> mouse sequence
<400> 7
gcctctgttc cacatacact tcattctcag tattgttttg ccaagttcta attccatcag 60
acctcgacct gcagccccta gataacttcg tataatgtat gctatacgaa gttatgctag 120
cgccaacccg ctaggacaag tgctgagtga gctggggcca ccgtttgagg aaacaggagc 180
cagtacagag gggttcccct ttaggggttg gtggcaatgg gcgaccctgg ttaatggatc 240
att 243
<210> 8
<211> 20
<212> DNA
<213> Artificial sequence
<220>
<223> synthetic polynucleotide sequence
<400> 8
catctgcgac agtcgagttc 20
<210> 9
<211> 19
<212> DNA
<213> Artificial sequence
<220>
<223> synthetic polynucleotide sequence
<400> 9
ccagggagct taccgtgac 19
<210> 10
<211> 27
<212> DNA
<213> Artificial sequence
<220>
<223> synthetic polynucleotide sequence
<400> 10
aggtacatct tagaggccaa ggaggca 27
<210> 11
<211> 19
<212> DNA
<213> Artificial sequence
<220>
<223> synthetic polynucleotide sequence
<400> 11
acagccgagt cctggagag 19
<210> 12
<211> 19
<212> DNA
<213> Artificial sequence
<220>
<223> synthetic polynucleotide sequence
<400> 12
aagccctgag cgtgagttc 19
<210> 13
<211> 26
<212> DNA
<213> Artificial sequence
<220>
<223> synthetic polynucleotide sequence
<400> 13
aggccaagga ggccgagaat atcacg 26
<210> 14
<211> 19
<212> DNA
<213> Artificial sequence
<220>
<223> synthetic polynucleotide sequence
<400> 14
gagccctgca ctggacaac 19
<210> 15
<211> 20
<212> DNA
<213> Artificial sequence
<220>
<223> synthetic polynucleotide sequence
<400> 15
tcccatgaac gctgagagtc 20
<210> 16
<211> 28
<212> DNA
<213> Artificial sequence
<220>
<223> synthetic polynucleotide sequence
<400> 16
agggtcaagg agccatagac agaatggc 28
<210> 17
<211> 200
<212> DNA
<213> Artificial sequence
<220>
<223> synthetic polynucleotide sequence
<220>
<221> misc_feature
<222> (1)..(100)
<223> mouse sequence
<220>
<221> misc_feature
<222> (101)..(200)
<223> human sequence
<400> 17
agctctccta ccactagact gctgagaccc gctgctctgc tcaggactcg atttccagta 60
cacaatctcc ctctttgaaa agtaccacac atcctggggt gctcttgcat ttgtgtgaca 120
ctttgctagc caggctcagt cctgggttcc aggtggggac tcaaacacac tggcacgagt 180
ctacattgga tattcttggt 200
<210> 18
<211> 199
<212> DNA
<213> Artificial sequence
<220>
<223> synthetic polynucleotide sequence
<220>
<221> misc_feature
<222> (1)..(100)
<223> 5' end of neomycin cassette
<220>
<221> misc_feature
<222> (101)..(199)
<223> human sequence
<220>
<221> misc_feature
<222> (106)..(139)
<223> LoxP
<400> 18
gctccccatt cctcactggc ccagcccctc ttccctactc tttctagccc ctgcctcatc 60
tccctggctg ccattgggag cctgccccac tggaagccag tcgagataac ttcgtataat 120
gtatgctata cgaagttata tgcatggcct ccgcgccggg ttttggcgcc tcccgcgggc 180
gcccccctcc tcacggcga 199
<210> 19
<211> 200
<212> DNA
<213> Artificial sequence
<220>
<223> synthetic polynucleotide sequence
<220>
<221> misc_feature
<222> (1)..(100)
<223> 3' neomycin cassette
<220>
<221> misc_feature
<222> (101)..(200)
<223> mouse Exon 4
<400> 19
cattctcagt attgttttgc caagttctaa ttccatcaga cctcgacctg cagcccctag 60
ataacttcgt ataatgtatg ctatacgaag ttatgctagc tgtctcatag aggctggcga 120
tctggctcag ggacagccag tactgcaaag agtatccttg ttcatacctt ctcctagtgg 180
ccatctccct gggacagtca 200
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, ЭКСПРЕССИРУЮЩИЕ EPO ЧЕЛОВЕКА | 2015 |
|
RU2711744C1 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ НЕ ОТНОСЯЩИЕСЯ К ЧЕЛОВЕЧЕСКОМУ РОДУ ЖИВОТНЫЕ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2673156C2 |
| ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА ЖИВОТНЫЕ С ГУМАНИЗИРОВАННЫМ ГЕНОМ СИГНАЛЬНОГО РЕГУЛЯТОРНОГО БЕЛКА | 2014 |
|
RU2671166C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ НЕ ОТНОСЯЩИЕСЯ К ЧЕЛОВЕЧЕСКОМУ РОДУ ЖИВОТНЫЕ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2775425C2 |
| ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, ИМЕЮЩИЕ ГУМАНИЗИРОВАННЫЙ ГЕН КЛАСТЕРА ДИФФЕРЕНЦИРОВКИ 47 | 2015 |
|
RU2728412C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ, НЕ ПРИНАДЛЕЖАЩИЕ К ЧЕЛОВЕЧЕСКОМУ РОДУ ЖИВОТНЫЕ И СПОСОБ ИХ ИСПОЛЬЗОВАНИЯ | 2013 |
|
RU2642319C2 |
| ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, ИМЕЮЩИЕ ГУМАНИЗИРОВАННЫЙ ГЕН КЛАСТЕРА ДИФФЕРЕНЦИРОВКИ 47 | 2015 |
|
RU2827833C1 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ НЕ ПРИНАДЛЕЖАЩИЕ К ЧЕЛОВЕЧЕСКОМУ РОДУ ЖИВОТНЫЕ И СПОСОБ ИХ ИСПОЛЬЗОВАНИЯ | 2013 |
|
RU2768282C2 |
| ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА ЖИВОТНЫЕ, ВКЛЮЧАЮЩИЕ В СЕБЯ ГУМАНИЗИРОВАННЫЙ TTR ЛОКУС, И СПОСОБЫ ПРИМЕНЕНИЯ | 2018 |
|
RU2784927C1 |
| ГУМАНИЗИРОВАННЫЕ МЫШИ С НОКИНОМ SIRPA-IL15 И СПОСОБЫ ИХ ИСПОЛЬЗОВАНИЯ | 2016 |
|
RU2822370C2 |

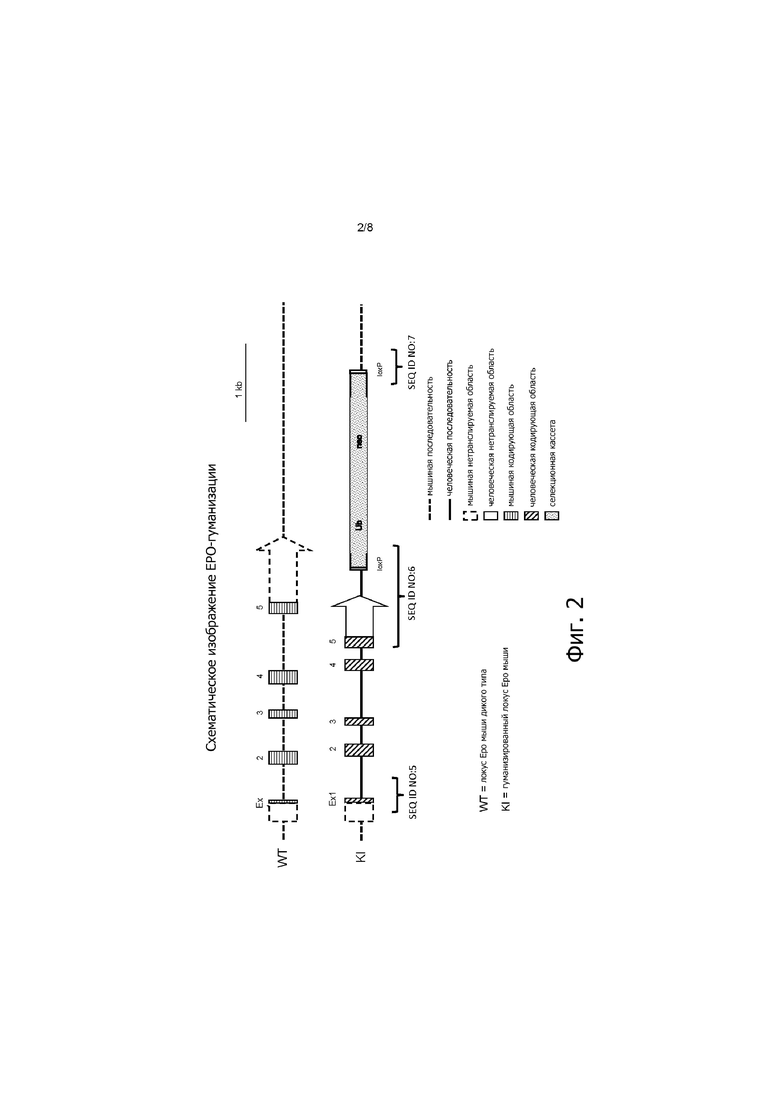
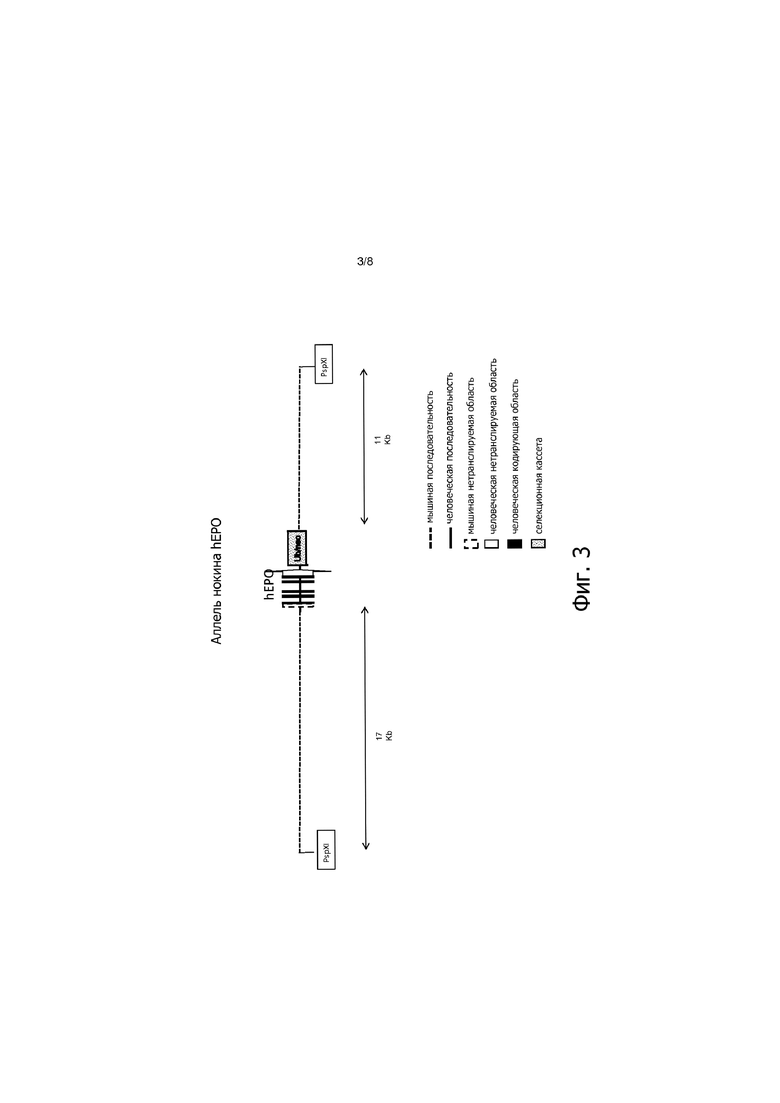
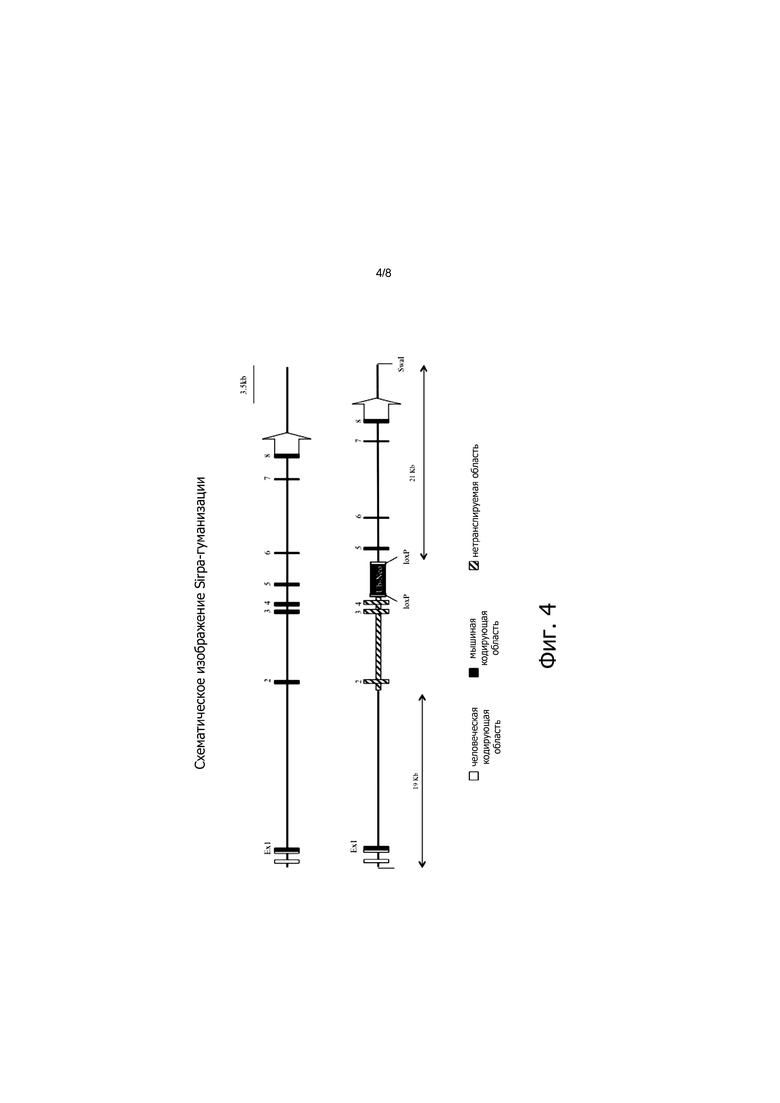
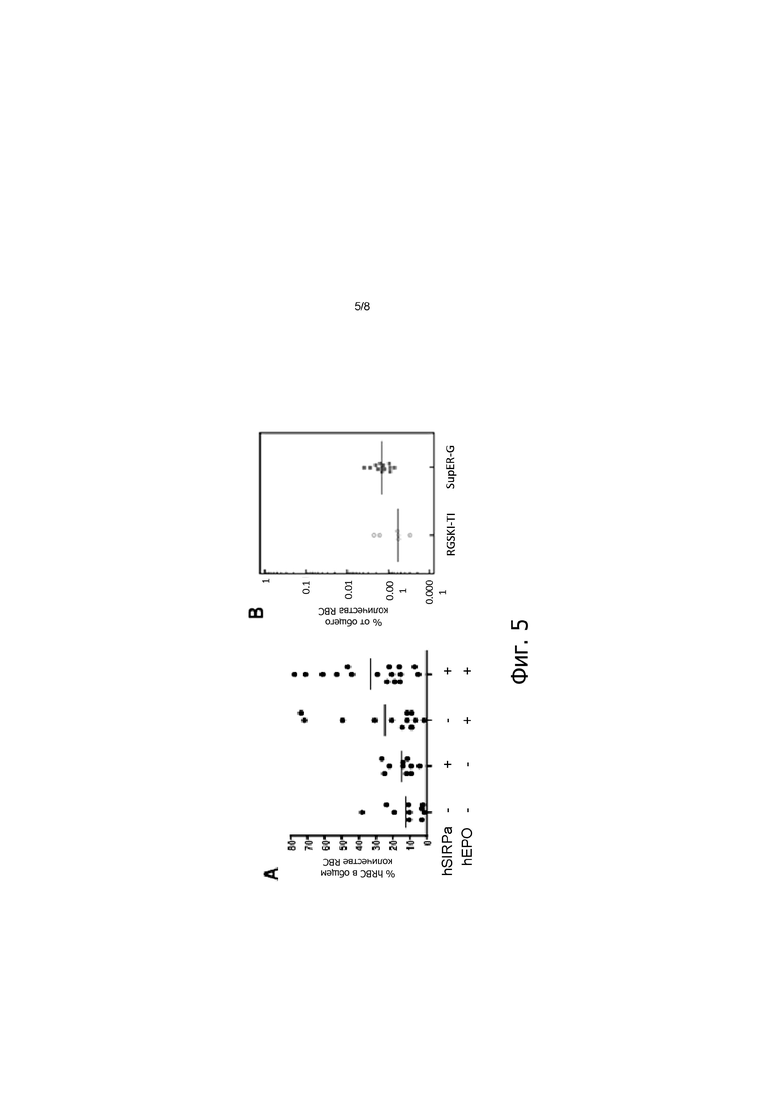
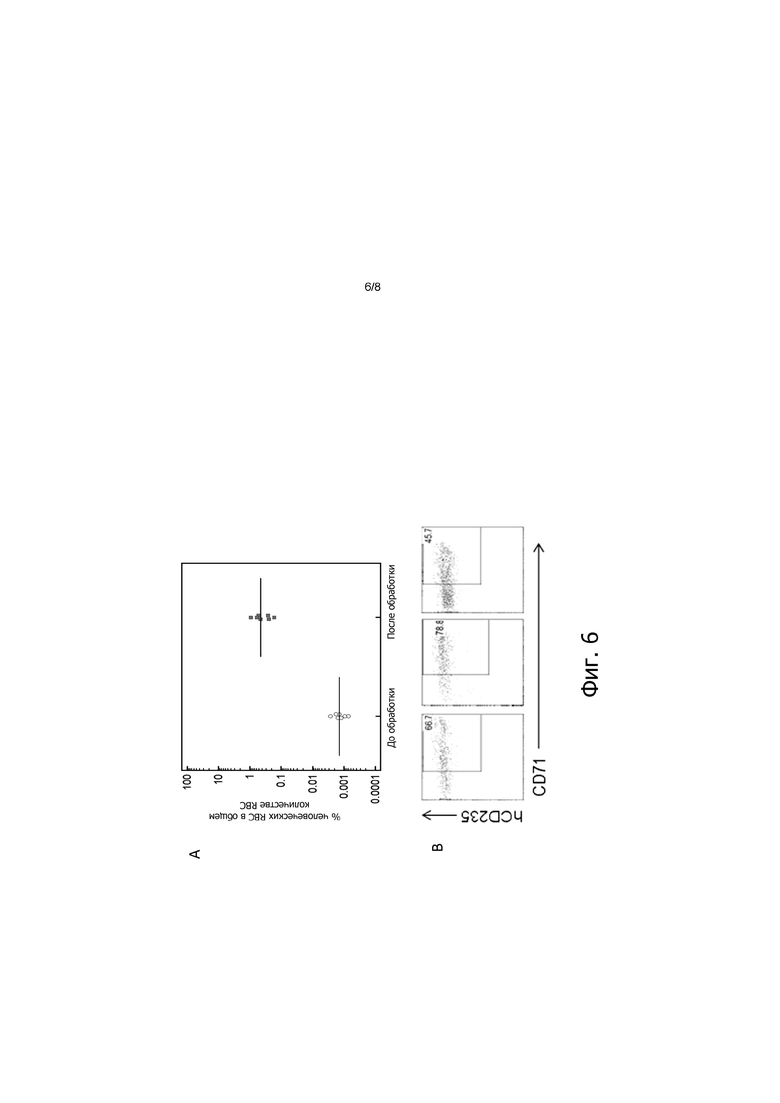
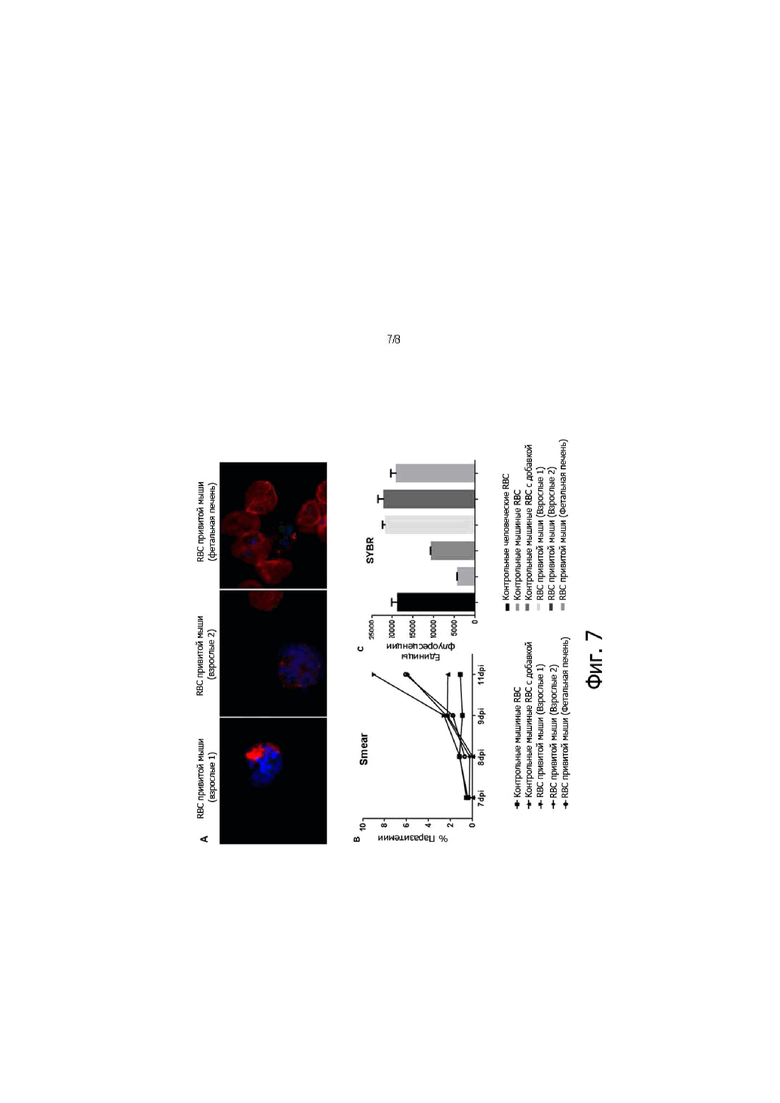
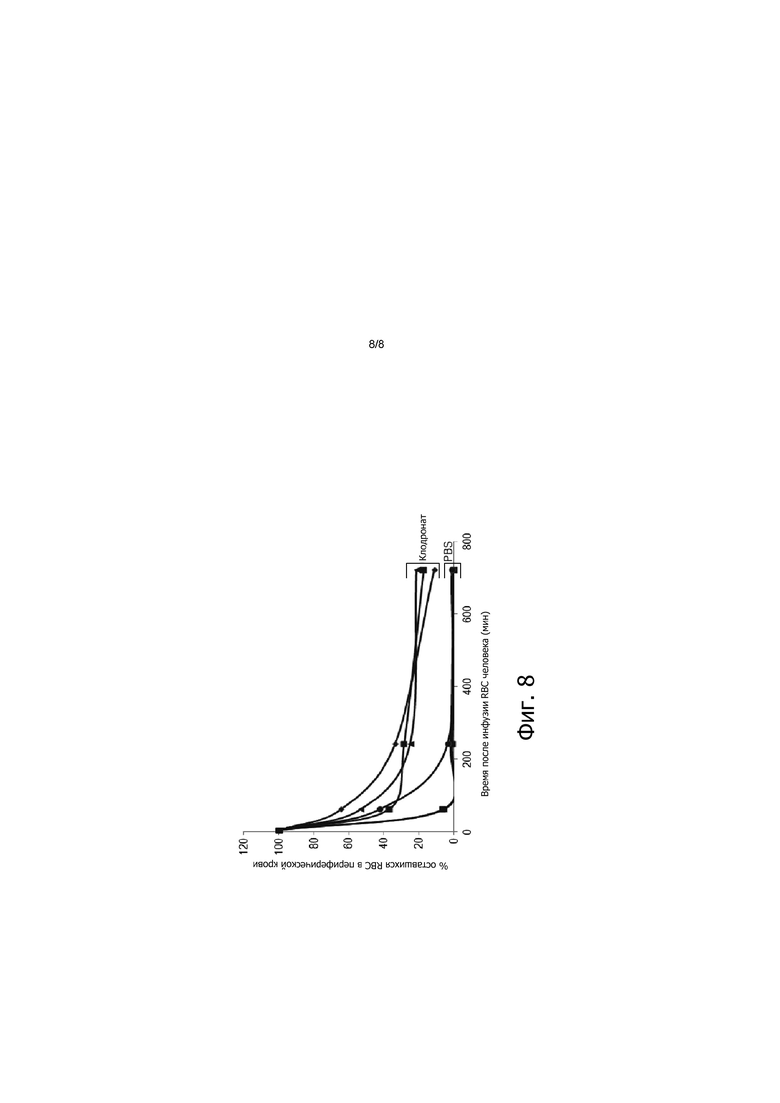
Изобретение относится к области биотехнологии. Представлены: генетически модифицированные мыши, несущие нуклеотидную последовательность, которая кодирует человеческий белок EPO, а также к способам идентификации агента, который ингибирует инфекцию, вызванную патогеном, который поражает человеческие клетки эритроидной линии. Изобретение также относится к способу получения мыши, экспрессирующей человеческий белок EPO и содержащей человеческую гематопоэтическую систему, а также к способу получения мыши, которая инфицирована человеческим патогеном, который поражает человеческие клетки эритроидной линии. Изобретение позволяет эффективно применять его в качестве животной модели эритропоэза человека, в качестве животной модели функции эритроцитов человека, а также в качестве животной модели инфекции эритроцитов патогенами человека. 7 н. и 25 з.п. ф-лы, 8 ил., 4 табл., 5 пр.
1. Генетически модифицированная мышь для применения в качестве животной модели эритропоэза человека, несущая:
нуклеотидную последовательность, которая кодирует человеческий белок EPO (hEPO), функционально связанную с промотором эндогенного гена EPO мыши в локусе EPO мыши, где функциональная связь приводит к нулевой мутации в гене EPO мыши в локусе гена EPO мыши,
нуклеотидную последовательность, которая кодирует белок hM-CSF, функционально связанный с промотором эндогенного M-csf мыши в локусе гена M-csf мыши, и где мышь является гетерозиготной нулевой или гомозиготной нулевой по гену M-csf мыши,
нуклеотидную последовательность, которая кодирует белок IL-3, функционально связанный с промотором эндогенного IL-3 мыши в локусе гена IL-3 мыши, и где мышь является гетерозиготной нулевой или гомозиготной нулевой по гену IL-3 мыши,
нуклеотидную последовательность, которая кодирует белок hGM-CSF, функционально связанный с промотором эндогенного Gm-csf мыши в локусе гена Gm-csf мыши, и где мышь является гетерозиготной нулевой или гомозиготной нулевой по гену Gm-csf мыши,
нуклеотидную последовательность, которая кодирует белок hTPO, функционально связанный с промотором эндогенного ТРО мыши в локусе гена ТРО мыши, и где мышь является гетерозиготной нулевой или гомозиготной нулевой по гену ТРО мыши, и
трансген SIRPa, который кодирует белок hSirpa, где мышь экспрессирует белок hSirpa,
где генетически модифицированная мышь обладает иммунодефицитом, и где иммунодефицит вызван дефицитом как Rag2, так и IL2rg.
2. Генетически модифицированная мышь для применения в качестве животной модели функции эритроцитов человека, включающая:
нуклеотидную последовательность, которая кодирует человеческий белок EPO (hEPO), функционально связанную с промотором эндогенного гена EPO мыши в локусе EPO мыши, где функциональная связь приводит к нулевой мутации в гене EPO мыши в локусе гена EPO мыши,
нуклеотидную последовательность, которая кодирует белок hM-CSF, функционально связанный с промотором эндогенного M-csf мыши в локусе гена M-csf мыши, и где мышь является гетерозиготной нулевой или гомозиготной нулевой по гену M-csf мыши,
нуклеотидную последовательность, которая кодирует белок IL-3, функционально связанный с промотором эндогенного IL-3 мыши в локусе гена IL-3 мыши, и где мышь является гетерозиготной нулевой или гомозиготной нулевой по гену IL-3 мыши,
нуклеотидную последовательность, которая кодирует белок hGM-CSF, функционально связанный с промотором эндогенного Gm-csf мыши в локусе гена Gm-csf мыши, и где мышь является гетерозиготной нулевой или гомозиготной нулевой по гену Gm-csf мыши,
нуклеотидную последовательность, которая кодирует белок hTPO, функционально связанный с промотором эндогенного ТРО мыши в локусе гена ТРО мыши, и где мышь является гетерозиготной нулевой или гомозиготной нулевой по гену ТРО мыши, и
трансген SIRPa, который кодирует белок hSirpa, где мышь экспрессирует белок hSirpa,
где генетически модифицированная мышь обладает иммунодефицитом, и где иммунодефицит вызван дефицитом как Rag2, так и IL2rg.
3. Генетически модифицированная мышь для применения в качестве животной модели патогенной инфекции эритроцитов человека, включающая:
нуклеотидную последовательность, которая кодирует человеческий белок EPO (hEPO), функционально связанную с промотором эндогенного гена EPO мыши в локусе EPO мыши, где функциональная связь приводит к нулевой мутации в гене EPO мыши в локусе гена EPO мыши,
нуклеотидную последовательность, которая кодирует белок hM-CSF, функционально связанный с промотором эндогенного M-csf мыши в локусе гена M-csf мыши, и где мышь является гетерозиготной нулевой или гомозиготной нулевой по гену M-csf мыши,
нуклеотидную последовательность, которая кодирует белок IL-3, функционально связанный с промотором эндогенного IL-3 мыши в локусе гена IL-3 мыши, и где мышь является гетерозиготной нулевой или гомозиготной нулевой по гену IL-3 мыши,
нуклеотидную последовательность, которая кодирует белок hGM-CSF, функционально связанный с промотором эндогенного Gm-csf мыши в локусе гена Gm-csf мыши, и где мышь является гетерозиготной нулевой или гомозиготной нулевой по гену Gm-csf мыши,
нуклеотидную последовательность, которая кодирует белок hTPO, функционально связанный с промотором эндогенного ТРО мыши в локусе гена ТРО мыши, и где мышь является гетерозиготной нулевой или гомозиготной нулевой по гену ТРО мыши, и
трансген SIRPa, который кодирует белок hSirpa, где мышь экспрессирует белок hSirpa,
где генетически модифицированная мышь является иммунодефицитной, и где иммунодефицит вызван дефицитом как Rag2, так и IL2rg.
4. Мышь по любому из пп. 1-3, отличающаяся тем, что мышь является гетерозиготной по аллелю, содержащему нуклеотидную последовательность, кодирующую hEPO.
5. Мышь по любому из пп. 1-3, отличающаяся тем, что мышь является гомозиготной по аллелю, включающему нуклеотидную последовательность, кодирующую hEPO.
6. Мышь по любому из пп. 1-5, отличающаяся тем, что нуклеотидная последовательность, которая кодирует hEPO, содержит геномную кодирующую и некодирующую последовательность EPO человека.
7. Мышь по любому из пп. 1-5, отличающаяся тем, что нуклеотидная последовательность, которая кодирует hEPO, содержит последовательность кДНК EPO человека.
8. Мышь по любому из пп. 1-7, отличающаяся тем, что мышь является гомозиготной по нуклеиновой кислоте, кодирующей hTPO, нуклеиновой кислоте, кодирующей hMcsf, нуклеиновой кислоте, кодирующей hIL-3, и нуклеиновой кислоте, кодирующей hGmcsf.
9. Мышь по любому из пп. 1-8, отличающаяся тем, что мышь дополнительно содержит приживление гематопоэтических клеток человека.
10. Мышь по п. 9, отличающаяся тем, что гематопоэтические клетки человека содержат одну или несколько клеток, выбранных из группы, состоящей из CD34-положительной клетки человека, гемопоэтической стволовой клетки человека, миелоидной клетки-предшественника человека, эритроидной клетки-предшественника человека, миелоидной клетки человека, дендритной клетки человека, моноцита человека, гранулоцита человека, эритроцита человека, нейтрофила человека, тучной клетки человека, тимоцита человека и В-лимфоцита человека.
11. Мышь по п. 10, отличающаяся тем, что мышь содержит клодронат.
12. Мышь по п. 11, отличающаяся тем, что мышь дополнительно содержит инфекцию с патогеном, который нацелен на клетки человека эритроидной линии.
13. Мышь по п. 12, отличающаяся тем, что патоген выбран из Plasmodium sp., Babesia sp. и Theileri sp.
14. Способ идентификации агента, который ингибирует инфекцию патогенным микроорганизмом, нацеленным на клетки человека эритроидной линии, включающий:
а. введение агента-кандидата в генетически модифицированную мышь, где мышь включает:
нуклеотидную последовательность, которая кодирует человеческий белок EPO (hEPO), функционально связанную с промотором эндогенного гена EPO мыши в локусе EPO мыши, где функциональная связь приводит к нулевой мутации в гене EPO мыши в локусе гена EPO мыши,
нуклеотидную последовательность, которая кодирует белок hM-CSF, функционально связанный с промотором эндогенного M-csf мыши в локусе гена M-csf мыши, и где мышь является гетерозиготной нулевой или гомозиготной нулевой по гену M-csf мыши,
нуклеотидную последовательность, которая кодирует белок IL-3, функционально связанный с промотором эндогенного IL-3 мыши в локусе гена IL-3 мыши, и где мышь является гетерозиготной нулевой или гомозиготной нулевой по гену IL-3 мыши,
нуклеотидную последовательность, которая кодирует белок hGM-CSF, функционально связанный с промотором эндогенного Gm-csf мыши в локусе гена Gm-csf мыши, и где мышь является гетерозиготной нулевой или гомозиготной нулевой по гену Gm-csf мыши,
нуклеотидную последовательность, которая кодирует белок hTPO, функционально связанный с промотором эндогенного ТРО мыши в локусе гена ТРО мыши, и где мышь является гетерозиготной нулевой или гомозиготной нулевой по гену ТРО мыши, и
трансген SIRPa, который кодирует белок hSirpa, где мышь экспрессирует белок hSirpa,
где генетически модифицированная мышь является иммунодефицитной и где иммунодефицит вызван дефицитом как Rag2, так и IL2rg,
iv. приживление человеческих кроветворных клеток, и
v. инфекцию патогеном, нацеленным на клетки человека эритроидной линии,
b. определение, уменьшает ли агент количество патогена у зараженной патогеном мыши стадии а; и
c. идентификацию агента, который уменьшает количество патогена у зараженной патогеном мыши, в качестве агента, который ингибирует инфекцию патогеном, нацеленным на клетки человека эритроидной линии.
15. Способ идентификации агента, который предотвращает инфекцию патогенным микроорганизмом, нацеленным на клетки человека эритроидной линии, включающий:
а. контакт генетически модифицированной мыши с клодронатом, где мышь содержит:
нуклеотидную последовательность, которая кодирует человеческий белок EPO (hEPO), функционально связанную с промотором эндогенного гена EPO мыши в локусе EPO мыши, где функциональная связь приводит к нулевой мутации в гене EPO мыши в локусе гена EPO мыши,
нуклеотидную последовательность, которая кодирует белок hM-CSF, функционально связанный с промотором эндогенного M-csf мыши в локусе гена M-csf мыши, и где мышь является гетерозиготной нулевой или гомозиготной нулевой по гену M-csf мыши,
нуклеотидную последовательность, которая кодирует белок IL-3, функционально связанный с промотором эндогенного IL-3 мыши в локусе гена IL-3 мыши, и где мышь является гетерозиготной нулевой или гомозиготной нулевой по гену IL-3 мыши,
нуклеотидную последовательность, которая кодирует белок hGM-CSF, функционально связанный с промотором эндогенного Gm-csf мыши в локусе гена Gm-csf мыши, и где мышь является гетерозиготной нулевой или гомозиготной нулевой по гену Gm-csf мыши,
нуклеотидную последовательность, которая кодирует белок hTPO, функционально связанный с промотором эндогенного ТРО мыши в локусе гена ТРО мыши, и где мышь является гетерозиготной нулевой или гомозиготной нулевой по гену ТРО мыши, и
трансген SIRPa, который кодирует белок hSirpa, где мышь экспрессирует белок hSirpa,
где генетически модифицированная мышь является иммунодефицитной и где иммунодефицит вызван дефицитом как Rag2, так и IL2rg,
iv. приживление человеческих кроветворных клеток,
b. введение агента-кандидата генетически модифицированной мыши стадии а,
c. инъекцию генетически модифицированной мыши стадии а пораженных паразитом ретикулоцитов или эритроцитов, и
д. определение того, предотвращает ли агент инфицирование ретикулоцитов и/или эритроцитов человека мыши стадии а,
е. идентификацию агента, который предотвращает инфицирование ретикулоцитов и/или эритроцитов человека мыши стадии а, в качестве агента, предотвращающего инфекцию патогеном, который нацелен на клетки человека эритроидной линии.
16. Способ по п. 15, отличающийся тем, что мышь является гетерозиготной по аллелю, содержащему нуклеотидную последовательность, которая кодирует hEPO.
17. Способ по п. 15, отличающийся тем, что мышь является гомозиготной по аллелю, содержащему нуклеотидную последовательность, которая кодирует hEPO.
18. Способ по любому из пп. 15-18, отличающийся тем, что нуклеотидная последовательность, которая кодирует hEPO, содержит геномную кодирующую и некодирующую последовательность EPO человека.
19. Способ по любому из пп.15-18, отличающийся тем, что нуклеотидная последовательность, которая кодирует hEPO, включает последовательность кДНК EPO человека.
20. Способ по любому из пп. 15-19, отличающийся тем, что мышь гомозиготна по: нуклеиновой кислоте, кодирующей hTPO, нуклеиновой кислоте, кодирующей hMcsf, нуклеиновой кислоте, кодирующей hIL-3, и нуклеиновой кислоте, кодирующей hGmcsf.
21. Способ по любому из пп. 15-20, отличающийся тем, что гематопоэтические клетки человека содержат одну или несколько клеток, выбранных из группы, состоящей из CD34-положительной клетки человека, гемопоэтической стволовой клетки человека, миелоидной клетки-предшественника человека, эритроидной клетки-предшественника человека, миелоидной клетки человека, дендритной клетки человека, моноцита человека, гранулоцита человека, эритроцита человека, нейтрофила человека, тучной клетки человека, тимоцита человека и В-лимфоцита человека.
22. Способ по любому из пп. 15-21, отличающийся тем, что патоген выбран из Plasmodium sp., Babesia sp. и Theileri sp.
23. Способ по п. 22, отличающийся тем, что патогеном является Plasmodium sp. и Plasmodium sp. выбран из P. falciparum и P. vivax.
24. Способ получения мыши, содержащей гематопоэтическую систему человека, включающий:
трансплантацию популяции клеток, включающей гематопоэтические клетки-предшественники человека, генетически модифицированной мыши по любому из пп. 1-13.
25. Способ по п. 24, отличающийся тем, что трансплантация включает инъекцию в хвостовую вену, инъекцию в печень плода или ретроорбитальную инъекцию.
26. Способ по п. 24 или 25, отличающийся тем, что генетически модифицированную мышь облучают сублетально перед трансплантацией.
27. Способ по любому из пп. 24-26, отличающийся тем, что гематопоэтические клетки-предшественники человека представляют собой CD34+ клетки.
28. Способ по любому из пп. 24-27, отличающийся тем, что гематопоэтические клетки-предшественники человека происходят из печени плода, костного мозга взрослого человека или пуповинной крови.
29. Способ получения мыши, инфицированной человеческим патогеном, который нацелен на клетки человека эритроидной линии, включающий:
трансплантацию популяции клеток, содержащей гематопоэтические клетки-предшественники человека, генетически модифицированной мыши по любому из пп. 1-13,
инъекцию мыши клодроната, и
инъекцию мыши пораженных паразитом эритроцитов человека (PRBC).
30. Способ по п. 29, отличающийся тем, что способ дополнительно включает инъекцию мыши эритроцитов здорового человека.
31. Способ по п. 29, отличающийся тем, что паразит выбран из Plasmodium sp., Babesia sp. и Theileri sp.
32. Способ по п. 31, отличающийся тем, что паразит представляет собой Plasmodium sp. и Plasmodium sp. выбран из P. falciparum и P. vivax.
| GREGG L | |||
| SEMENZA et al., Polycythemia in transgenic mice expressing the human erythropoietin gene, Proc | |||
| Natl | |||
| Acad | |||
| Sci | |||
| USA, 1989, Vol | |||
| Пюпитр для работы на пишущих машинах | 1922 |
|
SU86A1 |
| Ступица со свободным ходом для ведущих автомобильных колес | 1922 |
|
SU2301A1 |
| GREGG L | |||
| SEMENZA et al., Cell-type-specific and hypoxia-inducible expression of the human erythropoietin gene in transgenic mice, Proc | |||
| Nati | |||
| Acad | |||
| Sci | |||
| USA, 1991, Vol | |||
| Шланговое соединение | 0 |
|
SU88A1 |
| Устройство для подвешивания тормозных колодок барабанных тормозов у повозок железных дорог | 1927 |
|
SU8725A1 |
| WO | |||
