Предшествующий уровень техники
Данное изобретение относится к способу определения связывания белка с определенной последовательностью ДНК. Данный способ может найти применение, в частности, для обнаружения модификаций последовательности ДНК (например, метилирования) посредством связывания с белками, специфически распознающими данные модификации (например, антителами), а также для обнаружения в ДНК связывающей последовательности для различных белков.
Связывание белков с ДНК представляет собой важнейший биологический феномен; он играет основополагающую роль в регуляции функций клеток и вирусов. Он задействован в основных клеточных процессах, таких как репликация ДНК, транскрипция, репарация ДНК и рекомбинация ДНК, а также в модификации ДНК или поддержании структуры хромосом.
Существует несколько белков, которые связываются с определенными сайтами в геноме и регулируют экспрессию генов и стабильность генома. ДНК-связывающие белки составляют большое семейство белков, выполняющих разнообразные важные биологические функции. Семейство ДНК-связывающих белков является одним из самых многочисленных и хорошо изученных у бактерий, архебактерий и эукариот. Большинство этих белков, например, эукариотические и прокариотические факторы транскрипции содержат независимо упакованные структурные элементы (домены), которые позволяют им распознавать строение ДНК. К ним относятся важные белки, регулирующие активность генов, известные как факторы транскрипции, и белки, участвующие в процессинге ДНК, например, такие как ДНК- и РНК-полимеразы, ДНК-лигазы, ДНК-хеликазы, ДНК-эндонуклеазы и экзонуклеазы, а также белки, участвующие в репарации и рекомбинации ДНК.
Идентификация сайтов связывания этих белков представляет собой сложную задачу. Например, в геноме человека существует более 70 гипотетических факторов транскрипции, имеющих цинксодержащие пальцеобразные области С2Н2 типа (Tadepally et al., ВМС Evol. Biol., 8: 176, 2008), но примерно только у 10% из них известны связывающие последовательности (Matys et al., Nucleic Acids Res., 34: D108-D110, 2006). Кроме того, несмотря на то, что термодинамика связывания белков с ДНК хорошо изучена, изучение кинетики их связывания и отсоединения представляет собой трудную задачу.
Для изучения взаимодействия ДНК с белками используют различные методы, такие как анализ задержки электрофоретического сдвига в геле, футпринтинг и транскрипционная активация (Carey et al., Cold Spring Harb Protoc, 2012(7): 733-57, 2012). Хотя каждый из этих методов дает определенную информацию о местоположении связывания или эффектах связывания, они не позволяют количественно охарактеризовать специфическое связывание. Измерение поляризации/анизотропии флуоресценции представляет собой нерадиоактивный способ количественного анализа связывания ДНК и белков непосредственно в растворе, не требующий связывания с фильтрами, электрофореза или осаждения (Guest et al., 1991; Heyduk and Lee, 1990; LeTilly and Royer, 1993; Lundblad et al., 1996; Royeretal., 1992).
Молекулярные механизмы, с помощью которых генетическая информация направляет синтез различных биомолекул, были предметом многочисленных молекулярно-биологических исследований на протяжении последних 30 лет. Проводимые исследования были нацелены на изучение отдельных генов, а выведенные с их помощью общие закономерности способствовали пониманию таких важных процессов, происходящих в геноме, как транскрипция, ремоделирование хроматина, сплайсинг мРНК, репликация ДНК и многих других. Несмотря на то, что многие из этих принципов остаются эффективными при исследовании новых генов, они, как правило, не дают представления о регуляции биологических функций на уровне генома. С другой стороны, системный анализ транскриптов и регуляторных сигналов необходим для идентификации генов и регуляторных последовательностей и играет важную роль в изучении биологии человека и его заболеваний. Такой анализ помогает получить всестороннее представление об организации и вариабельности генов и регуляторной информации на уровне клетки, индивида и биологического вида.
Общегеномные исследования, целью которых является идентификация всех сайтов связывания транскрипционных факторов в геноме человека, такие как проект Encode (от англ. Encyclopedia of DNA Elements, Энциклопедия ДНК элементов), связаны с большими трудностями и затратами (The ENCODE Project Consortium, Nature, 489: 57-74, 2012).
Таким образом, по-прежнему существует потребность в простом и надежном способе исследования взаимодействий белок/нуклеиновая кислота.
Сведения, подтверждающие возможность осуществления изобретения
Настоящее изобретение относится к способу определения связывания белка с молекулой нуклеиновой кислоты с использованием физических манипуляций.
В основе способа по настоящему изобретению лежат физические методы и электронная обработка, что отличает его от используемых в настоящее время химических или биохимических подходов. Данный способ обладает многочисленными преимуществами относительно существующего уровня техники.
1) Он характеризуется высокой чувствительностью, поскольку позволяет обнаруживать связывание одиночных белковых молекул или белковых комплексов с одиночными молекулами нуклеиновых кислот. Использование одиночных молекул позволяет не только измерить время, необходимое для нахождения белком своей нуклеиновой кислоты-мишени, и время, в течение которого он остается связанным со своей мишенью, но и точно определить участок, где происходит связывание.
2) При этом не используются дорогостоящие меченные (при помощи флуорофоров или любых других групп) нуклеотиды.
3) Данный способ позволяет определить точное местонахождение сайта связывания белка в двуцепочечной молекуле нуклеиновой кислоты (в пн) путем измерения расстояния между двумя концами указанной двуцепочечной молекулы нуклеиновой кислоты.
4) Измерение можно повторять через определенные промежутки времени, что позволяет устранить ложноположительные результаты, улучшить статистическую достоверность и существенно снизить инструментальную погрешность.
5) Данный эксперимент можно повторять многократно с одной и той же молекулой, что позволяет улучшить статистическую достоверность и надежность измерений.
6) Данный способ позволяет обнаружить любой белок, связывающийся с нуклеиновой кислотой. Таким образом, можно детектировать белки, которые специфически распознают структурные модификации нуклеиновой кислоты, и, следовательно, установить сайт структурной модификации.
Настоящее изобретение относится к способу определения связывания белка с последовательностью нуклеиновой кислоты по физическому расположению сайтов, связанных с белком, в последовательности нуклеиновой кислоты.
В контексте настоящего изобретения «связывание» обозначает нековалентное взаимодействие между макромолекулами (например, между белком и нуклеиновой кислотой). Такие взаимодействия обычно характеризуют константой диссоциации (Kd) равной 10-6 М-1 или менее. «Аффинность» обозначает прочность связывания: повышенная аффинность соответствует более низким значениям Kd.
Под «определением связывания белка с молекулой нуклеиновой кислоты» в данном документе понимают все действия, прямо или косвенно приводящие к получению определенной информации о наличии или отсутствии взаимодействия указанного белка с указанной молекулой нуклеиновой кислоты. Определение указанного связывания может включать или не включать получение дополнительной информации, например, информации о кинетических характеристиках реакции связывания или о последовательности сайта, связанного с белком. Специалисту в данной области очевидно, что способ по данному изобретению позволяет легко осуществить такое определение.
Изобретение основано на наблюдении, что две цепи денатурированной двуцепочечной нуклеиновой кислоты в соответствующих условиях подвергаются повторной гибридизации. Если на этапе ренатурации любая из цепей указанной денатурированной двуцепочечной нуклеиновой кислоты оказывается связанной с молекулой, повторная гибридизация будет только частичной. Авторы изобретения обнаружили, что при определенных условиях данная остановка повторной гибридизации, как перманентная, так и транзиторная, позволит обнаружить взаимодействие между белком и указанной молекулой денатурированной двуцепочечной нуклеиновой кислоты. Согласно изобретению, возможно обнаружить блокировку повторной гибридизации молекулы двуцепочечной нуклеиновой кислоты; физические параметры такой блокировки (например, длительность блокировки, участок молекулы двуцепочечной нуклеиновой кислоты (позиция), где происходит блокировка) позволяют обнаружить взаимодействие между белком и последовательностью нуклеиновой кислоты.
Таким образом, настоящее изобретение относится к способу определения связывания белка с молекулой нуклеиновой кислоты, указанный способ включает этап обнаружения блокировки ренатурации молекулы денатурированной двуцепочечной нуклеиновой кислоты.
Под «денатурацией» в данном документе понимают процесс разделения двух цепей молекулы двуцепочечной нуклеиновой кислоты, имеющий место при разрушении большинства водородных связей между указанными цепями. Денатурация приводит к образованию молекулы денатурированной нуклеиновой кислоты, под которой в данном документе понимают две отдельных комплементарных цепи, разделенные в результате денатурации молекулы двуцепочечной нуклеиновой кислоты. Под «ренатурацией» в данном документе понимают процесс, при котором две разделенных комплементарных цепи, гибридизуясь, вновь образуют двойную спираль. В данном документе «гибридизация» обозначает процесс формирования нековалентных сайт-специфических взаимодействий между двумя или более комплементарными цепями нуклеиновых кислот с образованием единого гибрида.
Существует несколько возможных способов денатурировать нуклеиновую кислоту, известных специалистам в данной области. В наиболее предпочтительном варианте две цепи разделяют, прикладывая к ним физическую силу. Согласно изобретению «физическая сила» представляет собой любое воздействие, заставляющее объект претерпевать некоторое изменение, которое относится к его движению, направлению, или геометрическому строению. Для специалиста в данной области очевидно, что, согласно изобретению, сила отличается от других физических параметров, например, таких как температура (которая скорее является непосредственным свойством субстанции, чем оказываемым на нее влиянием). Согласно изобретению, физические силы включают такие силы, как трение, тяговое усилие, продольное усилие, сила аэродинамического сопротивления, действующая сила и сила упругости. Более предпочтительно, физическая сила, согласно изобретению, представляет собой тяговое усилие. Согласно данному воплощению, можно развести свободные концы двуцепочечной нуклеиновой кислоты и, таким образом, разрушить все связи между спаренными основаниями и расплести двуцепочечную нуклеиновую кислоту.
Изобретение касается любого типа двуцепочечных нуклеиновых кислот. Чаще всего, двуцепочечной нуклеиновой кислотой является ДНК, но при этом понимают, что изобретение также касается идеально или не идеально спаренных дуплексов одноцепочечная ДНК-одноцепочечная ДНК или, в альтернативном варианте, идеально или не идеально спаренных дуплексов одноцепочечная ДНК-одноцепочечная РНК или, в альтернативном варианте, идеально или не идеально спаренных дуплексов одноцепочечная РНК-одноцепочечная РНК. Кроме того, дуплекс может представлять собой две одноцепочечные цепи, полученные из образцов различного происхождения, спаренные по меньшей мере частично. Наконец, изобретение также относится к вторичным структурам отдельных одноцепочечных ДНК или отдельных одноцепочечных РНК.
Таким образом, способ согласно изобретению представляет собой способ определения связывания белка с молекулой нуклеиновой кислоты, указанный способ включает этапы, при которых:
- денатурируют молекулу двуцепочечной нуклеиновой кислоты, прикладывая к указанной молекуле физическую силу; и
- детектируют блокирование ренатурации двуцепочечной нуклеиновой кислоты.
Предпочтительно, указанный способ включает также этап определения позиции, в которой происходит блокирование.
При таком способе определения связывания белка с молекулой ДНК может быть предпочтительно перед разделением цепей обеспечить ковалентное или подобное ковалентному связывание свободных концов молекулы двуцепочечной ДНК (т.е. концов, которые не связаны с носителем) друг с другом, чтобы облегчить повторное спаривание. В предпочтительном воплощении молекула двуцепочечной нуклеиновой кислоты представляет собой шпильку. В контексте настоящего изобретения желательно представить двуцепочечную нуклеиновую кислоту в виде схематичного изображения, можно представить ее в виде «застежки-молнии», которая может быть раскрыта (или закрыта): денатурация двуцепочечной нуклеиновой кислоты представляет собой «расстегивание», ренатурация -«застегивание».
Авторы изобретения заметили, что при определенных условиях, когда молекула связана с молекулой денатурированной двуцепочечной нуклеиновой кислоты, ренатурация указанной двуцепочечной нуклеиновой кислоты блокируется. Связанная молекула может представлять собой молекулу любого типа, обладающая сродством (аффинностью) к определенной последовательности указанной молекулы денатурированной двуцепочечной нуклеиновой кислоты, например, нуклеиновой кислотой, белком или низкомолекулярной молекулой.
В первом аспекте изобретения для блокирования ренатурации указанной двуцепочечной нуклеиновой кислоты применяют белок.
Термины «белок», «белки», «полипептид» и «полипептиды» в данном описании являются синонимами и обозначают полимеры, состоящие из аминокислот, ковалентно связанных в цепочку при помощи пептидных связей. Пептидные связи образуются между карбоксильной группой одной аминокислоты и аминогруппой следующей аминокислоты. Эти термины также охватывают полимеры аминокислот, в которых одна или несколько аминокислот являются химическими аналогами или модифицированными производными соответствующих аминокислот естественного происхождения. Термины «аминокислоты» и «аминокислота» обозначают все альфа-аминокислоты естественного происхождения, в форме D- и L-стереоизомеров, а также их аналоги и производные. Под аналогом понимают аминокислоту, в которой один атом замещен другим атомом, обычно он обладает схожими свойствами. Производное определяют как аминокислоту, к которой присоединена другая молекула или атом. Производные будут включать, например, производные, полученные ацетилированием аминогруппы, аминированием карбоксильной группы или окислением остатков серы двух молекул цистеина с образованием цистина.
Белки могут иметь несколько функций. «Связывающий белок» представляет собой белок, способный нековалентно связываться с другой молекулой. Связывающий белок может, например, связывать молекулу ДНК (ДНК-связывающий белок), молекулу РНК (РНК-связывающий белок) и/или молекулу белка (белок-связывающий белок). Белок-связывающий белок может связываться с самим собой (образовывать мультимеры) и/или он может связываться с одной или несколькими молекулами другого белка или белков. Связывающий белок может обладать связывающей активностью нескольких типов. Например, белки с цинксодержащими пальцеобразными областями обладают ДНК-связывающей, РНК-связывающей и белок-связывающей активностью. Согласно изобретению, «белок, связывающий нуклеиновую кислоту» представляет собой белок, способный взаимодействовать с нуклеиновой кислотой. Таким образом, «белок, связывающий одноцепочечную нуклеиновую кислоту», согласно изобретению, представляет собой белок, способный взаимодействовать с одноцепочечной нуклеиновой кислотой, тогда как «белок, связывающий двуцепочечную нуклеиновую кислоту», согласно изобретению, представляет собой белок, способный взаимодействовать с двуцепочечной нуклеиновой кислотой.
Таким образом, согласно данному воплощению, способ по изобретению относится к способу определения связывания белка с молекулой нуклеиновой кислоты, содержащей последовательность нуклеиновой кислоты, указанный способ включает этапы, при которых:
a) денатурируют указанную молекулу двуцепочечной нуклеиновой кислоты, содержащую указанную последовательность, прикладывая к указанной молекуле физическую силу;
b) берут указанный белок;
c) ренатурируют указанную молекулу двуцепочечной нуклеиновой кислоты в присутствии указанного белка и
d) детектируют блокирование ренатурации двуцепочечной нуклеиновой кислоты.
Предпочтительно, указанный способ включает также этап определения позиции, в которой происходит блокирование.
В области техники хорошо известно, что белки, связывающие нуклеиновые кислоты, подразделяются на те, которые способны связывать одноцепочечные нуклеиновые кислоты (оцДНК и оцРНК) и те, которые способны связывать двуцепочечные нуклеиновые кислоты (дцДНК, дцРНК, гибриды ДНК/РНК и т.д.).
В первом воплощении способа по изобретению белок, который применяют для блокирования ренатурации денатурированной двуцепочечной нуклеиновой кислоты, представляет собой белок, способный связывать одноцепочечную нуклеиновую кислоту.
Белки, связывающие нуклеиновую кислоту, обладающие аффинностью к одноцепочечной нуклеиновой кислоте, будут способны взаимодействовать с денатурированной двуцепочечной молекулой per se, что таким образом приведет к блокированию ренатурации двуцепочечной нуклеиновой кислоты. Для специалиста в данной области очевидно, что данное изобретение обеспечивает простое и точное определение кинетических параметров реакции связывания, даже если белок не связывается с определенной последовательностью. Действительно, белки, связывающие одноцепочечные нуклеиновые кислоты, чаще всего обладают аффинностью не к определенной последовательности, а скорее к нуклеиновым кислотам в общем. Например, известно, что хеликазы связывают одноцепочечные участки и раскручивают дцДНК. Бактериальные белки, связывающиеся с одноцепочечной ДНК (SSB, от англ. single-stranded DNA-binding proteins), связываясь с одноцепочечными участками ДНК, предупреждают преждевременный отжиг, защищают одноцепочечную ДНК от расщепления нуклеазами и удаляют элементы вторичной структуры ДНК. Белок Rad52, играющий важную роль в репарации двуцепочечных разрывов ДНК и гомологичной рекомбинации, связывается с одноцепочечными концами ДНК и опосредует взаимодействие ДНК-ДНК, необходимое для отжига комплементарных цепей ДНК.
Такие белки, связывающие одноцепочечную ДНК, имеют общую аффинность к нуклеиновым кислотам, что в контексте настоящего изобретения означает, что белки способны связывать одноцепочечную нуклеиновую кислоту, независимо от последовательности указанной нуклеиновой кислоты. Такие белки, связывающие нуклеиновые кислоты, не обладающие специфичностью к последовательности, связывают множество неродственных последовательностей ДНК с константой диссоциации, которая для разных последовательностей отличается менее чем в 100 раз, обычно менее чем в 10 раз.
С другой стороны, некоторые белки, связывающие нуклеиновые кислоты, имеют аффинность к молекулам нуклеиновых кислот, содержащих определенную последовательность, т.е. они распознают и связывают только нуклеиновые кислоты, содержащие указанную последовательность. Не все компоненты реакции связывания должны обладать специфичностью к последовательности (например, контакты с фосфатными остатками в остове ДНК), при условии, что реакция в целом является последовательность-специфической. Действительно, многие белки, связывающие одноцепочечные нуклеиновые кислоты, обладают только общей аффинностью к нуклеиновым кислотам, некоторые из этих белков способны связывать определенные последовательности в составе одноцепочечных нуклеиновых кислот. Таким образом, белок, связывающий нуклеиновую кислоту, обладающий специфичностью к последовательности, связывается с определенной последовательностью или рядом определенных последовательностей, обладающих высокой степенью идентичности друг с другом (например, идентичностью последовательностей по меньшей мере около 80%) с аффинностью по меньшей мере в 100 раз превышающей аффинность к неродственным последовательностям. Константа диссоциации белка, связывающего нуклеиновую кислоту, обладающего специфичностью к последовательности, к своей(им) определенной(ым) последовательности(ям) обычно бывает ниже, чем приблизительно 100 нМ и может составлять 10 нМ, 1 нМ, 1 пМ или 1 фМ.
Многие белки, связывающие нуклеиновую кислоту, не способны связываться с одноцепочечными нуклеиновыми кислотами. Такие белки, обладающие аффинностью к двуцепочечным нуклеиновым кислотам, предположительно не будут способны взаимодействовать с денатурированной двуцепочечной молекулой per se. Такие белки скорее всего не будут блокировать ренатурацию двуцепочечной нуклеиновой кислоты в данных условиях.
Большинство таких белков распознают и связывают определенные последовательности двуцепочечных нуклеиновых кислот. Например, белки, связывающие двуцепочечную ДНК, играют важную роль в регуляции экспрессии новых белков. Такие белки взаимодействуют с ДНК при помощи различных структурных мотивов и могут стимулировать или подавлять транскрибирование матричной РНК, в зависимости от свойств и расположения последовательности ДНК, с которой они связываются.
В таком случае, после денатурации указанной двуцепочечной молекулы, может быть предпочтительно взять молекулу одноцепочечной нуклеиновой кислоты и указанный белок, связывающий двуцепочечную нуклеиновую кислоту. В области техники хорошо известно, что указанная одноцепочечная нуклеиновая кислота может гибридизоваться с комплементарной последовательностью одной из цепей денатурированной двуцепочечной нуклеиновой кислоты, таким образом формируя гибридную двуцепочечную нуклеиновую кислоту, которая может связываться с белком. Такая одноцепочечная нуклеиновая кислота может иметь любую длину, при условии, что ее длина достаточна для блокирования процесса ренатурации. Предпочтительно, длина одноцепочечной нуклеиновой кислоты может составлять от 3 до 50 нуклеотидов; более предпочтительно, от 3 до 45 нуклеотидов, от 3 до 40 нуклеотидов, от 3 до 35 нуклеотидов, от 3 до 30 нуклеотидов, от 3 до 25 нуклеотидов, от 3 до 20 нуклеотидов, от 3 до 15 нуклеотидов и еще более предпочтительно, от 3 до 12. Одноцепочечная нуклеиновая кислота по изобретению может быть, в частности, молекулой ДНК или РНК, как естественного происхождения, так и модифицированной. Указанная одноцепочечная нуклеиновая кислота также может состоять из модифицированных нуклеотидов, например, как «закрытая» нуклеиновая кислота (LNA, от англ. locked nucleic acid), в которой нуклеотиды содержат модифицированную рибозу, в которой дополнительный мостик соединяет атом кислорода в положении 2' и атом углерода в положении 4', или пептидная нуклеиновая кислота (PNA, от англ. peptide nucleic acid), в которой остов состоит из повторяющихся элементов N-(2-аминоэтил)-глицина, связанных пептидными связями.
Если перед ренатурацией денатурированной двуцепочечной нуклеиновой кислоты к ней добавляют молекулу одноцепочечной нуклеиновой кислоты, блокирование регибридизации указывает на то, что последовательность молекулы одноцепочечной нуклеиновой кислоты комплементарна по меньшей мере части последовательности молекулы двуцепочечной нуклеиновой кислоты.
Как показали авторы изобретения, когда присутствует белок, связывающий двуцепочечную нуклеиновую кислоту, он способен связывать гибрид, образованный денатурированной двуцепочечной нуклеиновой кислотой и молекулой одноцепочечной нуклеиновой кислоты. Взаимодействие между белком и гибридной нуклеиновой кислотой изменяет длительность блокирования. Большей частью, взаимодействие увеличивает длительность блокирования ренатурации. Например, праймаза будет стабилизировать связывание олигонуклеотидов с ДНК, которое иначе было бы недостаточно стабильным, и время блокирования регибридизации шпильки было бы недостаточным для детекции. Аналогично, связывание ДНК-полимеразы с 3'-концом короткого олигонуклеотида, используемого в качестве праймера, повышает его стабильность. В альтернативном случае, продолжительность блокирования может быть уменьшена. Так, авторы изобретения показали, что связывание некоторых хеликаз вызывает дестабилизацию указанного гибрида, что приводит к укорочению времени блокирования.
Согласно предпочтительному воплощению, способ по изобретению включает этапы, при которых:
a) денатурируют молекулу двуцепочечной нуклеиновой кислоты, содержащей определенную последовательность, прикладывая к указанной молекуле физическую силу;
b) берут указанный белок и молекулу одноцепочечной нуклеиновой кислоты, соответствующую указанной последовательности нуклеиновой кислоты;
c) ренатурируют указанную молекулу двуцепочечной нуклеиновой кислоты в присутствии указанного белка и указанной молекулы одноцепочечной нуклеиновой кислоты; и
d) детектируют блокирование ренатурации двуцепочечной нуклеиновой кислоты.
Данное воплощение является особенно предпочтительным, поскольку позволяет определить связывание указанного белка с последовательностью, которая содержится в двуцепочечной нуклеиновой кислоте.
В стандартной компоновке молекулы двуцепочечной нуклеиновой кислоты можно специфично прикреплять к твердому субстрату (например, стеклу для микроскопии, микропипетке, микрочастице). Один из концов можно непосредственно или опосредованно присоединять к поверхности, а другой конец можно непосредственно или опосредованно присоединять к подвижной поверхности. В данном воплощении тяговое усилие прикладывают к обоим концам двуцепочечной нуклеиновой кислоты, отдаляя носители друг от друга. Если тяговое усилие превышает пороговый уровень, две цепи разделяются и происходит денатурация молекулы нуклеиновой кислоты. Прилагаемое тяговое усилие предпочтительно превышает или равняется 15 пН; более предпочтительно оно превышает или равняется 16 пН; еще более предпочтительно оно превышает или равняется 17 пН; в особенно предпочтительном воплощении оно превышает или равняется 18 пН. Данная сила может варьировать в зависимости от температуры, типа нуклеотидов и буфера, однако квалифицированный специалист легко может подобрать указанную силу, необходимую для разделения двух цепей, с учетом этих параметров. С другой стороны, если тяговое усилие снижается ниже минимального значения, две цепи денатурированной двуцепочечной нуклеиновой кислоты могут регибридизоваться. Для регибридизации указанных двух цепей предпочтительно прикладывают тяговое усилие меньшее или равное 12 пН; более предпочтительно меньшее или равное 11 пН; еще более предпочтительно меньшее или равное 10 пН.
Более предпочтительно, двуцепочечная нуклеиновая кислота имеет шпилечную структуру. В данном документе «шпилечная структура» означает двойную спираль, в которой 5'-конец одной цепи физически связан с 3'-концом другой цепи через неспаренную петлю. Указанная физическая связь может быть ковалентной или нековалентной. Предпочтительно, указанная физическая связь представляет собой ковалентную связь. Таким образом, шпилечная структура состоит из двуцепочечного стебля и неспаренной одноцепочечной петли. В составе шпилечной структуры концы двух цепей, не входящие в петлю, являются свободными и, следовательно, могут быть разведены. В результате произойдет расплетание двуцепочечной нуклеиновой кислоты и образуется молекула денатурированной двуцепочечной нуклеиновой кислоты. Возможно полностью расправить шпилечную структуру молекулы двуцепочечной нуклеиновой кислоты, потянув за каждый конец указанной молекулы нуклеиновой кислоты с усилием, превышающим пороговое значение. Если прилагаемое к молекуле тяговое усилие снижается меньше минимального значения, молекула нуклеиновой кислоты регибридизуется, образуя шпилечную структуру. Наличие белка, связанного с указанной молекулой денатурированной нуклеиновой кислоты (например, оцДНК), вызывает задержку в регибридизации. Аналогично, наличие молекулы одноцепочечной нуклеиновой кислоты, гибридизованной с одной из цепей нуклеиновой кислоты расправленной шпилечной структуры, вызывает паузу в регибридизации, длительность указанной паузы изменяется (т.е. увеличивается или уменьшается), когда белок, связывающий двуцепочечную нуклеиновую кислоту, связан с комплексом. Таким образом, изменение длительности данной паузы указывает на то, что белок связан по меньшей мере с частью двуцепочечного стебля.
Таким образом, предпочтительно сконструировать петлю с такой последовательностью и длиной, чтобы рефолдинг шпильки происходил с кратковременной задержкой, длящейся, например, 1 секунду. Способы добиться такого эффекта описаны в документах, известных на существующем уровне техники, например, в публикации Woodside et al., Proc. Natl. Acad. Sci. U.S.A., 103 (16): 6190-6195, 2006). Когда сила уменьшается от значения Fopen до Ftest, длина расплетенной шпилечной структуры изменяется благодаря эластичности одноцепочечной ДНК. Небольшая задержка перед рефолдингом шпилечной структуры позволяет определить длину шпильки при той же силе, которую использовали для обнаружения блокировки.
Использование шпильки позволяет, в частности, повторять циклы спаривания и разъединения и, таким образом, повышать отношение сигнал/шум.
Способы, позволяющие соединить вместе свободные концы двуцепочечной нуклеиновой кислоты, известны и некоторые из них будут более подробно описаны ниже.
Под определением блокирования в данном документе понимают определение физических параметров, связанных с блокированием. Одним из важных параметров является позиция в молекуле двуцепочечной нуклеиновой кислоты, где происходит блокирование, при этом указанная позиция соответствует сайту, в котором белок связывается с расплетенной молекулой двуцепочечной нуклеиновой кислоты, или участку, в котором молекула одноцепочечной нуклеиновой кислоты гибридизуется с указанной расплетенной молекулой двуцепочечной нуклеиновой кислоты. Так, авторы изобретения обнаружили, что можно точно определить позицию, в которой останавливается ренатурация двуцепочечной нуклеиновой кислоты: использование шпилечной структуры дает возможность специалисту в данной области определить физическое расстояние между двумя свободными концами шпилечной структуры в любой момент времени в ходе процесса денатурации/ренатурации.
Таким образом, особым преимуществом данного изобретения является то, что указанный способ включает дополнительный этап определения позиции, в которой происходит блокирование.
Согласно данному предпочтительному воплощению, в изобретении предложен способ определения связывания белка с молекулой нуклеиновой кислоты, содержащей последовательность нуклеиновой кислоты, указанный способ включает этапы, при которых:
a) денатурируют молекулу двуцепочечной нуклеиновой кислоты, содержащей последовательность нуклеиновой кислоты, прикладывая к указанной молекуле физическую силу;
b) берут указанный белок;
c) ренатурируют указанную молекулу двуцепочечной нуклеиновой кислоты в присутствии указанного белка;
d) детектируют блокирование ренатурации двуцепочечной нуклеиновой кислоты; и
е) определяют позицию в указанной молекуле двуцепочечной нуклеиновой кислоты, где происходит указанное блокирование.
Под «свободным концом» в данном документе понимают конец одной цепи, который не связан ковалентно с концом другой цепи; как описано выше, каждый из этих свободных концов может быть присоединен к разным поверхностям. Например, одна из поверхностей может быть подвижной, а другая неподвижной. Для специалиста очевидно, что для измерения расстояния между свободными концами двуцепочечной нуклеиновой кислоты, имеющей шпилечную структуру, достаточно измерить расстояние между двумя поверхностями.
Данное расстояние будет максимальным (zhigh (Fopen)) когда молекула, имеющая шпилечную структуру, полностью денатурирует, поскольку в этом случае шпилечная структура нуклеиновой кислоты будет полностью выпрямлена; оно будет минимальным (Zlow (Ftest)), когда указанная молекула, имеющая шпилечную структуру, полностью ренатурирует. Предпочтительно всегда сравнивать длину при одинаковой силе Ftest, чтобы показатели эластичности одноцепочечной нуклеиновой кислоты были одинаковыми. По задержке при смыкании петли квалифицированный специалист может определить zhigh(Ftest). Аналогично, можно определить расстояние между двумя свободными концами, когда возникает временная пауза в процессе ренатурации: как и следовало ожидать, расстояние z находится между zhigh и Zlow (все значения z определяют при F=Ftest). Совершенно очевидно, что расстояние z варьирует в зависимости от локализации сайта связывания белка, связывающего одноцепочечную нуклеиновую кислоту, или последовательности, комплементарной одноцепочечной нуклеиновой кислоте, в молекуле, имеющей шпилечную структуру. Если указанный белок связан с последовательностью, которая расположена вблизи свободных концов шпилечной структуры, процесс ее регибридизации блокируется непосредственно перед тем, как шпилька полностью сформируется заново; в данном случае zpause будет минимальным. С другой стороны, если указанный белок связан с частью шпилечной структуры, которая расположена вблизи неспаренной петли, процесс ренатурации остановится, когда шпилька будет полностью или почти полностью денатурирована; в данном случае zpause будет максимальным. Аналогично, если указанная одноцепочечная нуклеиновая кислота гибридизуется с последовательностью, которая расположена вблизи свободных концов шпилечной структуры, процесс ее регибридизации блокируется непосредственно перед тем, как шпилька полностью сформируется заново; в данном случае zpause является минимальным. С другой стороны, если указанная одноцепочечная нуклеиновая кислота гибридизуется с частью шпилечной структуры, которая расположена вблизи неспаренной петли, процесс ренатурации остановится, когда шпилька будет полностью или почти полностью денатурирована; в данном случае zpause будет максимальным (Фиг. 1).
Возможно вывести точную зависимость между физическим расстоянием и числом оснований в молекуле двуцепочечной нуклеиновой кислоты. Например, расстояние 0,8 нм соответствует расстоянию, которое занимают два последовательно расположенных нуклеотида (1 пн) в одной цепи нуклеиновой кислоты при силе 10 пН. Точное определение удлинения в зависимости от силы возможно благодаря эластичности одноцепочечной нуклеиновой кислоты. Таким образом, простое измерение расстояния между двумя свободными концами частично ренатурировавшей молекулы двуцепочечной нуклеиновой кислоты (или между любыми двумя референтными позициями в молекуле) позволяет точно определить, где произошло блокирование ренатурации.
Так, в одном воплощении изобретение представляет собой способ определения связывания белка с молекулой нуклеиновой кислоты, где указанную молекулу двуцепочечной нуклеиновой кислоты вначале подвергают денатурации, приложив физическую силу, а затем регибридизации в присутствии указанного белка, и, возможно, одноцепочечной нуклеиновой кислоты, и детектируют наличие блокирования регибридизации. В одном аспекте расстояние между двумя концами частично ренатурировавшей двуцепочечной молекулы определяют, когда процесс ренатурации заблокирован. Предпочтительно, расстояние между двумя концами указанной молекулы определяют, когда молекула полностью денатурирована. Более предпочтительно, сравнивают два расстояния и определяют позицию, где произошло блокирование. Более предпочтительно, для определения позиции, где произошло блокирование, измеряют расстояние между полностью расправленной петлей и референтным сайтом гибридизации. Еще более предпочтительно, для определения местоположения блокировки измеряют расстояние между двумя референтными сайтами гибридизации.
Помимо позиции в молекуле, другим важным параметром, характеризующим блокирование ренатурации, является промежуток времени, в течение которого блокируется ренатурация (обозначаемый в данном документе как длительность паузы в процессе ренатурации). Так, возможно измерить период времени, в течение которого регибридизация заблокирована. Например, специалист в данной области может определить период времени, в течение которого расстояние между двумя концами двуцепочечной нуклеиновой кислоты равно z, как описано выше, т.е. промежуточному значению между zhigh и zlow.
Если блокада вызвана гибридизацией между денатурированной двуцепочечной нуклеиновой кислотой и комплементарной одноцепочечной нуклеиновой кислотой, длительность блокады определяется степенью комплементарности между двумя последовательностями. Чем выше комплементарность, тем больше число связей, устанавливающихся между двумя молекулами, и, следовательно, больше длительность. Также очевидно, что время блокирования будет зависеть от длины участка комплементарности между двумя последовательностями. Чем длиннее участок, тем больше число связей, устанавливающихся между двумя молекулами, и, следовательно, больше длительность. Таким образом, можно предположить, что при определенных условиях пауза в ренатурации будет длиться почти постоянно. В частности, если одноцепочечная нуклеиновая кислота содержит более 20, предпочтительно более 25, еще более предпочтительно более 30 нуклеотидов, способных гибридизоваться с денатурированной двуцепочечной нуклеиновой кислотой, одноцепочечная нуклеиновая кислота останется гибридизованной с двуцепочечной шпилечной структурой (на протяжении многих минут), даже если сила, приложенная к указанной двуцепочечной нуклеиновой кислоте снижается до Ftest, таким образом предотвращая регибридизацию указанной двуцепочечной шпилечной структуры. В данном случае, для отделения молекулы одноцепочечной нуклеиновой кислоты может быть целесообразным применение фермента или проведение дополнительной третьей фазы, когда силу на несколько секунд снижают до 0,5 или 1 пН, что позволяет эффективно разделить гибридизованные олигонуклеотиды. Отделение указанной молекулы одноцепочечной нуклеиновой кислоты позволяет проводить циклы спаривания и разъединения и, таким образом, повышать отношение сигнал/шум.
Длительность паузы также может варьировать в зависимости от условий реакции. Указанная длительность будет уменьшаться с повышением температуры. Также, на длительность паузы могут влиять свойства буфера: например, магний, бетаин и хлорид тетраметиламмония (используемый в молярной концентрации) увеличивают время блокирования. Эти компоненты усиливают пары AT в большей степени, чем GC, таким образом уменьшая разницу в прочности этих пар. Однако, при постоянной температуре и свойствах буфера длительность паузы будет определяться только силой, приложенной к денатурированной двуцепочечной нуклеиновой кислоте, и ее комплементарностью с одноцепочечной нуклеиновой кислотой. Так, авторы изобретения показали, что время блокирования экспоненциально уменьшается по мере снижения силы.
Наконец, длительность паузы также будет зависеть от свойств комплекса, образованного между белком, денатурированной двуцепочечной нуклеиновой кислотой и комплементарной одноцепочечной нуклеиновой кислотой. Присутствие белка, связывающего двуцепочечную нуклеиновую кислоту, может стабилизировать комплекс.Чем выше его аффинность к двуцепочечной нуклеиновой кислоте, тем дольше пауза. Также возможно, что белок дестабилизирует двуцепочечную нуклеиновую кислоту (как, например, в случае открытого комплекса РНК-полимеразы), что укорачивает паузу.
Аналогично, присутствие белка, способного связывать денатурированную двуцепочечную нуклеиновую кислоту, будет временно блокировать ренатурацию указанной молекулы нуклеиновой кислоты. Длительность блокирования также будет зависеть от аффинности белка к нуклеиновой кислоте. Очевидно, что белок с более высокой аффинностью к указанной молекуле будет вызывать более длительную паузу, чем белок с более низкой аффинностью.
Специалист в данной области поймет, что измерение длительности паузы позволяет определить среднюю продолжительность блокировки, а следовательно, кинетические параметры реакции связывания, как описано в экспериментальной части.
Таким образом, в одном конкретном аспекте, способ по изобретению включает этапы, при которых:
a) денатурируют указанную молекулу двуцепочечной нуклеиновой кислоты, прикладывая к указанной молекуле физическую силу;
b) берут белок и, возможно, молекулу одноцепочечной нуклеиновой кислоты,
c) ренатурируют молекулу двуцепочечной нуклеиновой кислоты в присутствии указанного белка и, возможно, указанной молекулы одноцепочечной нуклеиновой кислоты; и
d) детектируют блокирование ренатурации указанной молекулы двуцепочечной нуклеиновой кислоты и
e) определяют длительность паузы.
Предпочтительно, указанный способ также включает этап определения позиции, в которой происходит блокирование.
В данном воплощении длительность паузы можно сравнивать с контролем. В частности, если указанный белок представляет собой белок, связывающий двуцепочечную нуклеиновую кислоту, может быть предпочтительно сравнить указанную паузу с паузой, зарегистрированной при осуществлении способа в отсутствие белка. Как описано выше, связывание белка с комплексом, образованным между денатурированной двуцепочечной нуклеиновой кислотой и комплементарной одноцепочечной нуклеиновой кислотой, изменяет длительность блокирования ренатурации. Указанное изменение выражается в увеличении или уменьшении (в зависимости от конкретного белка) длительности паузы.
Так, в одном предпочтительном воплощении способ по изобретению включает этапы, при которых:
a) денатурируют указанную молекулу двуцепочечной нуклеиновой кислоты, прикладывая к указанной молекуле физическую силу;
b) берут белок и, возможно, молекулу одноцепочечной нуклеиновой кислоты,
c) ренатурируют молекулу двуцепочечной нуклеиновой кислоты в присутствии указанного белка и, возможно, указанной молекулы одноцепочечной нуклеиновой кислоты; и
d) детектируют блокирование ренатурации указанной молекулы двуцепочечной нуклеиновой кислоты и
e) определяют длительность паузы; и
f) сравнивают ее с длительностью в отсутствие белка.
Предпочтительно, указанный способ также включает этап определения позиции, в которой происходит блокирование.
Несмотря на то, что обнаружить и измерить связывание белка с нуклеиновой кислотой возможно и без поиска информации о последовательности сайта связывания, в некоторых случаях определение указанной последовательности может быть целесообразным. Например, может представлять интерес выявление мутаций указанного сайта связывания, которые предотвращают связывание указанного белка.
Таким образом, в одном предпочтительном воплощении способ по изобретению относится к способу определения связывания белка с молекулой двуцепочечной нуклеиновой кислоты, содержащей последовательность нуклеиновой кислоты, указанный способ включает этапы, при которых:
a) денатурируют указанную молекулу двуцепочечной нуклеиновой кислоты, прикладывая к указанной молекуле физическую силу;
b) берут указанный белок и, возможно, молекулу одноцепочечной нуклеиновой кислоты, комплементарную по меньшей мере части указанной молекулы двуцепочечной нуклеиновой кислоты;
c) ренатурируют указанную молекулу двуцепочечной нуклеиновой кислоты в присутствии указанного белка и, возможно, указанной одноцепочечной нуклеиновой кислоты;
d) детектируют блокирование ренатурации двуцепочечной нуклеиновой кислоты; и
e) секвенируют последовательность нуклеиновой кислоты, связанную с указанным белком.
Предпочтительно, после того, как обнаруживают блокирование ренатурации переходят к этапу определения позиции, в которой происходит блокирование.
Предпочтительно, перед секвенированием сайта связывания указанный белок и указанную молекулу одноцепочечной нуклеиновой кислоты смывают с молекулы двуцепочечной нуклеиновой кислоты.
Поскольку способ по изобретению основан на детекции одиночной молекулы, будет целесообразным применение способа, позволяющего секвенировать одиночную молекулу без предварительной амплификации. Такие способы идентификации и секвенирования одиночных молекул были описаны ранее (WO 2011/147931; WO 2011/147929; Ding et al., Nature Met, 9(4): 367-372, 2012). Данные способы секвенирования основаны на детекции блокирования ренатурации денатурированной молекулы двуцепочечной нуклеиновой кислоты. Таким образом, способ секвенирования по изобретению предпочтительно включает этапы, при которых:
a) денатурируют молекулу двуцепочечной нуклеиновой кислоты, соответствующую указанной последовательности нуклеиновой кислоты, прикладывая к указанной молекуле физическую силу;
b) берут молекулу одноцепочечной нуклеиновой кислоты;
c) ренатурируют указанную молекулу двуцепочечной нуклеиновой кислоты в присутствии указанной молекулы одноцепочечной нуклеиновой кислоты; и
d) детектируют блокирование ренатурации двуцепочечной нуклеиновой кислоты.
Предпочтительно, указанный способ также включает этап определения позиции, в которой происходит блокирование.
Такие способы секвенирования легко сочетаются со способом по изобретению, поскольку в них используют те же устройства, что и в настоящем способе. Если приложить тяговое усилие к магнитным частицам, фиксируемым шпилечной структурой у поверхности, молекулу можно расплести. В расплетенном состоянии она может гибридизоваться с комплементарными одноцепочечными нуклеиновыми кислотами, которые временно блокируют сворачивание шпилечной структуры при уменьшении тягового усилия. Измеряя расстояние от поверхности до частицы, связанной с заблокированной шпилечной структурой, можно определить местоположение гибрида в молекуле с точностью почти в 1 нуклеотид, и таким образом, установить последовательность в данном участке (она будет комплементарна известной последовательности одноцепочечных нуклеиновых кислот, присутствующих в растворе). Таким образом, возможно прямое секвенирование молекулы, связанной с указанным белком, не требующее изменений схемы эксперимента, а только замены буфера, содержащего белок и, возможно, комплементарную одноцепочечную нуклеиновую кислоту, буфером, подходящим для секвенирования согласно указанным способам.
Эффективная идентификация цис-регуляторных элементов ДНК является главной задачей постгеномной биологии. Идентификация в геноме всех сайтов связывания белков, связывающих нуклеиновые кислоты, особенно важна, поскольку позволит идентифицировать все гены, экспрессия которых потенциально может регулироваться указанным белком. Полная идентификация цис-регуляторных элементов ДНК имеет ключевую роль для прогнозирования и понимания механизмов транскрипции.
Сочетание информации, полученной при полногеномном секвенировании ДНК, высокопроизводительных технологий и новых алгоритмов позволило значительно продвинуться вперед в идентификации и характеризации регуляторных элементов, контролирующих транскрипцию (Eisen et al., Proc. Natl. Acad. Sci., 95: 14863-14868, 1998; Tavazoie et al., Nat. Genet, 22: 281-285, 1999; Bussemaker et al., Nat. Genet, 27: 167-171, 2001; Lee et al., Science, 298: 799-804, 2002). Однако, данным подходам присущи определенные ограничения. Например, гибридные методы анализа весьма успешно применяют для выделения кластеров экспрессии генов и обнаружения цис-регуляторных мотивов, однако их применение ограничено конкретными типами физиологических воздействий, которые используют в лаборатории. То же относится и к подходам in vivo, таким как иммунопреципитация хроматина с анализом на микрочипах (ChIP, от англ. chip-based chromatin immunoprecipitation), в котором взаимодействие ДНК-белок, в силу их регуляторной роли, имеет место только при определенных внешних условиях (Lee et al., Science, 298: 799-804, 2002). Эти ограничения еще более жесткие в случае многоклеточных организмов, для которых труднее получить экспериментальные данные.
Предложенный способ является альтернативой способам, известным на уровне техники, таким как иммунопреципитация хромосом (СЫР) и футпринтинг с использованием ДНКазы I для картирования в геноме сайтов связывания транскрипционных факторов (The ENCODE Project Consortium, Nature, 489: 57-74, 2012).
Таким образом, согласно другому аспекту, изобретение также относится к способу идентификации молекул нуклеиновых кислот, содержащих последовательность, которая способна связываться с определенным белком, связывающим нуклеиновые кислоты, указанный способ включает этапы, при которых:
a) берут популяцию молекул двуцепочечных нуклеиновых кислот;
b) изучают связывание указанного белка с указанной молекулой нуклеиновой кислоты способом, описанным выше; и
c) выбирают молекулы нуклеиновых кислот, способные связывать указанный белок.
Предпочтительно, при данном способе также берут одноцепочечную нуклеиновую кислоту, комплементарную сайту связывания указанной молекулы нуклеиновой кислоты.
Согласно данному воплощению, способ включает этапы, при которых:
a) берут популяцию молекул двуцепочечных нуклеиновых кислот;
b) денатурируют указанную молекулу двуцепочечной нуклеиновой кислоты, прикладывая к указанной молекуле физическую силу;
c) берут указанный белок и молекулу одноцепочечной нуклеиновой кислоты, комплементарную указанному сайту связывания;
d) ренатурируют указанную молекулу двуцепочечной нуклеиновой кислоты в присутствии указанного белка и указанной молекулы одноцепочечной нуклеиновой кислоты; и
e) обнаруживают или не обнаруживают блокирование ренатурации двуцепочечной нуклеиновой кислоты; и
f) выбирают молекулы нуклеиновых кислот, у которых ренатурация временно или необратимо заблокирована.
Предпочтительно, указанный способ также включает этап определения позиции, в которой происходит блокирование.
Выделяемые таким образом молекулы нуклеиновых кислот соответствуют популяции молекул нуклеиновых кислот, которые содержат указанную определенную связывающую последовательность. Таким образом, они отличаются от других молекул нуклеиновых кислот тем, что содержат эту определенную последовательность. Несмотря на то, что все эти молекулы имеют данную последовательность, в остальном они могут быть идентичны или не идентичны. В некоторых воплощениях может быть предпочтительно установить последовательность каждой молекулы нуклеиновой кислоты, которая находится за пределами указанного определенного сайта связывания. Так, при идентификации молекул нуклеиновых кислот, содержащих один или несколько сайтов связывания определенных белков, связывающих нуклеиновые кислоты, может быть предпочтительно секвенировать идентифицированные молекулы, например, используя описанные выше способы секвенирования. Полученная на этом этапе информация позволит локализовать указанную молекулу в геноме и, таким образом, выделить экспрессионные единицы, которые могут или не могут регулироваться данным сайтом связывания. Этого можно легко достичь, используя информацию, полученную на этапе секвенирования, при поиске в базах данных: специалисту в данной области техники известно, как проводить поиск клонов, содержащих последовательность, полученную при секвенировании, используя общедоступные базы данных, содержащие последовательности (например, Genbank), и нет необходимости описывать это здесь более подробно.
В предпочтительном воплощении популяция молекул двуцепочечных нуклеиновых кислот представляет полный геном.
Предпочтительно, популяцию молекул двуцепочечных нуклеиновых кислот получают, вначале расщепляя хромосомы редкощепящими рестрикционными ферментами. Как известно специалистам в данной области, редкощепящие рестрикционные ферменты представляют собой рестрикционные ферменты, распознающие последовательность, редко встречающуюся в геноме, например, последовательность, содержащую 7 или 8 оснований. Примеры таких редкощепящих рестрикционных ферментов включают Sfil, Xma I, Asc I, AsiS I (изошизомер Sgf I), Not I (изошизомер CciN I), Sbf I (изошизомеры Sse8387 I, Sda I), Fse I, Рас I и т.д. Все эти ферменты имеются в продаже. На второй стадии полученные таким образом рестрикционные фрагменты расщепляют с помощью обычных рестрикционных ферментов, распознающих последовательности из 6 оснований, таких как EcoRI, BamHI, XhoI и т.д. Полученные линейные двуцепочечные фрагменты можно затем трансформировать в шпилечные структуры. Способы, позволяющие соединить вместе свободные концы двуцепочечной нуклеиновой кислоты, известны и некоторые из них будут более подробно описаны ниже.
Другим частным воплощением данного способа является выявление эпигенетических модификаций. Методы, применяемые в настоящее время для их поиска, очень сложные и пропускают большое количество модификаций ДНК. При этом эпигенетические модификации имеют чрезвычайно важное значение при различных патологических состояниях, в том числе при микробных инфекциях и онкологических заболеваниях. Преимуществом данного изобретения является то, что оно может найти применение в скрининге модификаций как во всей геномной ДНК, так и в отдельных ее участках.
Эпигенетические модификации ДНК обнаруживаются в геноме почти всех живых организмов. Их тип и местоположение варьируют у различных организмов, тканей и типов клеток; с течением времени; а также при взаимодействии с окружающей средой. Некоторые из этих модификаций возникают вследствие строго контролируемых клеточных процессов. Другие являются результатом повреждения ДНК.
Такие модификации значительно увеличивают количество информации, которая может быть закодирована ДНК. Например, у Escherichia coli ген dam кодирует ДНК-метилтрансферазу, которая метилирует аденин в последовательностях -GATC- двуцепочечной ДНК, регулируя, таким образом, экспрессию генов (см., например, Calmann and Marinus, J. Bacterid., 185(16): 5012-5014, 2003). С другой стороны, самый распространенный эпигенетический маркер у эукариот - это 5-метилцитозин (5mC). Данная модификация необходима для контролирования и регулирования широкого круга важных клеточных и более общих физиологических процессов, а нарушения метилирования ДНК у человека связаны с различными заболеваниями, в частности, с некоторыми видами рака. Кроме 5mC у эукариот существует множество других модификаций ДНК (Korlach and Turner, Curr. Opin. Struct. Biol., 22: 251-261, 2012).
На сегодняшний день золотым стандартом определения 5 тС является «бисульфитная конверсия», при которой все остатки цитозина превращаются в урацил, кроме тех, которые метилированы и не подвергаются изменениям. При последующей амплификации ДНК-продукта урацил заменяется на тимин. Такие превращения можно обнаружить при секвенировании ДНК (Song et al., Nature Biotechnol, 30(11): 1107-1116, 2012). Однако, это сложный, отнимающий много времени и дорогостоящий процесс, в котором частота ошибочных результатов составляет 5-34% (Beck, Nature Biotechnol, 10: 1026- 1028, 2010).
В данном изобретении предложен простой способ выявления эпигенетических модификаций нуклеиновых кислот.«Эпигенетические модификации» в данном документе обозначают модификации оснований, составляющих молекулу нуклеиновой кислоты, возникающие после синтеза указанной молекулы нуклеиновой кислоты. Такие эпигенетические модификации включают, среди прочих, 4-метилцитозин (m4C), 5-метилцитозин (5mC), 5-гидроксиметилцитозин (5hmC), 5-формилцитозин (5fC) и 5-карбоксилцитозин (5саС), а также 6-метиладенозин (m6A) в ДНК и 5-гидроксиметилурацил (5hmU) и N6-метиладенозин (тбА) в РНК.
Таким образом, в одном конкретном аспекте данного изобретения предложен способ выявления по меньшей мере одного модифицированного основания, содержащегося в молекуле двуцепочечной нуклеиновой кислоты, указанный способ включает этапы, при которых:
a) берут указанную двуцепочечную нуклеиновую кислоту;
b) берут белок, способный связываться с указанным модифицированным основанием; и
c) изучают связывание указанного белка с указанной молекулой нуклеиновой кислоты описанным выше способом.
В некоторых случаях, для лучшей валидации результатов, способ по изобретению может также включать этап исследования гибридизации с простым олигонуклеотидом, распознающим предполагаемый сайт модификации. Например, обнаружив метилированный 5mC, используя специфическое антитело, можно затем обнаружить последовательность ATGC, используя олигонуклеотид NNTACGNN.
Особым преимуществом данного способа является использование немодифицированных связывающихся молекул в обратимом процессе. Например, при детекции 5mC не требуется проводить химическую реакцию (с бисульфатом натрия) с участием ДНК. Кроме того, способ по изобретению обладает большей чувствительностью, по сравнению с любым способом, известным из уровня техники, поскольку позволяет выявить модифицированное основание в одиночной молекуле.
В предпочтительном воплощении модифицированное основание выбрано из группы, включающей 5-метилцитозин (5mC), 5-гидроксиметилцитозин (5hmC), 5-формилцитозин (5fC), 5-карбоксилцитозин (5саС), 5-гидроксиметилурацил (5hmU) и N6-метиладенозин (m6A). В более предпочтительном воплощении указанное основание выбрано из 5mC и 5hmC. В еще более предпочтительном воплощении указанное основание представляет собой 5mC. Белки, распознающие и специфически связывающие эти модифицированные основания, были описаны ранее. Например, известны антитела, направленные против 5mC, которые применяют для окрашивания клеток с целью выявления данной модификации визуализирующими методами (Ito et al., Nature, 466: 1129-1133, 2010; Ко et al., Nature, 468: 839-843, 2010; Szulwach etal., Nature Neurosci, 14: 1607-1611, 2011; Haffner et al., Oncotarget, 2: 627-637, 2011; Lee et al., Science, 334: 194, 2011; Inoue et al., Cell Res, 21: 1670-1676, 2011). Такие антитела имеются в продаже (например, клон 33D3; кат. номер 39649, Active Motif). Помимо антител известны ферменты, способные специфически распознавать и взаимодействовать с нуклеотидом, представляющим интерес (Song et al., Nature Biotechnol, 30(11): 1107-1116, 2012). Например, фермент β-гликозилтрансфераза (βGT, от англ. β-glucosyltransferase) бактериофага Т4 осуществляет перенос остатка глюкозы на 5hmC. Белки Tet1-3 отвечают за превращение 5mC в 5hmC. Связывающий метил-CpG белок 2 типа (МеСР2) был впервые обнаружен благодаря его аффинности к ДНК, содержащей 5-тС.Предпочтительно, указанный белок представляет собой антитело, направленное против указанного модифицированного основания, или фермент, специфически распознающий указанное основание. Более предпочтительно, указанный белок представляет собой антитело.
Очевидно, что этот же способ можно применять и для обнаружения других модификаций нуклеиновых кислот. Например, можно обнаружить некомплементарное основание в молекуле двуцепочечной нуклеиновой кислоты. Давно было известно о существовании таких белков, как бактериальный MutS, которые распознают некомплементарное основание на дочерней цепи и связывают мутантную ДНК. Данное свойство может найти применение для обнаружения и идентификации любого некомплементарного основания в молекуле двуцепочечной нуклеиновой кислоты.
Таким образом, еще одним аспектом данного изобретения является способ обнаружения по меньшей мере одного некомплементарного основания в молекуле двуцепочечной нуклеиновой кислоты, указанный способ включает этапы, при которых:
a) берут указанную двуцепочечную нуклеиновую кислоту;
b) берут белок, способный связываться с указанным некомплементарным основанием; и
c) изучают связывание указанного белка с указанной молекулой нуклеиновой кислоты описанным выше способом.
Поскольку известно, что связываясь с некомплементарным основанием, MutS находится в виде димера, в способе по изобретению предпочтительно применять димеризованный MutS. У эукариот гомологи MutS формируют два вида гетеродимеров: Msh2/Msh6 (MutSα) и Msh2/Msh3 (MutSβ). Предпочтительно, указанный белок выбирают из димера MutS, Msh2/Msh6 (MutSα) и Msh2/Msh3 (MutSβ).
Однонуклеотидный полиморфизм (ОНП) - это вариация единичных нуклеотидов (А, Т, С или G) в последовательности геномной ДНК (или другой общей последовательности), встречающаяся у представителей биологического вида или в паре гомологичных хромосом человека. В среднем ОНП встречаются в человеческой популяции с частотой выше 1%. Поскольку только от 3 до 5% последовательности ДНК человека кодируют синтезируемые белки, ОНП чаще всего обнаруживаются за пределами кодирующей последовательности. ОНП, находящиеся в пределах кодирующей последовательности, представляют особый интерес, поскольку они с большей вероятностью будут сказываться на биологической функции белка.
При гибридизации молекулы, содержащей ОНП, с молекулой, содержащей последовательность, встречающуюся у большей части популяции, будет образовываться некомплементарная пара оснований. Настоящее изобретение позволяет легко обнаруживать ОНП.
Данное воплощение относится к способу обнаружения ОНП в последовательности, входящей в состав нуклеиновой кислоты, указанный способ включает этапы, при которых:
a) гибридизуют указанную молекулу с одноцепочечной нуклеиновой кислотой, содержащей последовательность, встречающуюся у большей части популяции; и
b) обнаруживают некомплементарное основание описанным выше способом.
Если исследуемая нуклеиновая кислота является двуцепочечной нуклеиновой кислотой, может быть предпочтительно денатурировать указанную нуклеиновую кислоту перед этапом (а).
Очевидно, что данные способы можно применять в масштабе генома, благодаря простой адаптации описанного выше способа. Это позволит идентифицировать все сайты генома, содержащие, например, конкретное модифицированное основание. Можно идентифицировать гены, на экспрессию которых могут повлиять указанные модифицированные основания, путем секвенирования молекул нуклеиновой кислоты, содержащих такие модифицированные основания. Кроме того, можно затем исследовать передачу указанных модифицированных оснований потомству. Данная информация может представлять интерес для таких областей наук, как селекция животных или растений, где необходимо, чтобы одни гены оставались молчащими, а другие экспрессировались в нескольких поколениях.
Еще в одном аспекте предложен способ обнаружения соединений, которые препятствуют связыванию белка с его специфической связывающей последовательностью. Данные соединения ослабляют или устраняют связывание указанного белка с его сайтом связывания. Такие соединения можно применять в терапевтических целях. Например, соединения, предотвращающие взаимодействие онкогенных форм сМус с его сайтом связывания, могут найти применение в лечении рака.
Согласно данному воплощению, изобретение относится к способу обнаружения по меньшей мере одного соединения, способного предотвращать взаимодействие белка с его сайтом связывания, указанный способ включает этапы, при которых:
a) берут указанный белок и молекулу нуклеиновой кислоты, содержащую последовательность, соответствующую указанному сайту связывания;
b) берут соединение; и
c) изучают связывание указанного белка с указанной молекулой нуклеиновой кислоты описанным выше способом.
В предпочтительном воплощении соединение выбирают, если связывание указанного белка с указанной молекулой нуклеиновой кислоты ослаблено или отсутствует.
Очевидно, что большинство белков, связывающих нуклеиновые кислоты, которые ассоциированы со злокачественными новообразованиями, представляют собой транскрипционные факторы, связывающие двуцепочечные нуклеиновые кислоты. Таким образом, в другом предпочтительном воплощении указанная молекула нуклеиновой кислоты представляет собой молекулу двуцепочечной нуклеиновой кислоты. В следующем предпочтительном воплощении в способе дополнительно берут одноцепочечную нуклеиновую кислоту, комплементарную последовательности указанной молекулы двуцепочечной нуклеиновой кислоты. Очевидно, что эти молекулы берут перед тем как тестировать связывание.
Воплощение способа по изобретению, в частности, стало возможным благодаря существованию устройств, предназначенных для регистрации взаимодействий одиночных нуклеиновых кислот в реальном времени. Такое устройство описано, например, в патентах США 7,052,650 и 7,244,391. В устройстве, описанном в этих документах, используют магнитные ловушки и прикладывают силы пиконьютонного масштаба к суперпарамагнитным частицам микронного размера. Вкратце, указанное устройство включает в себя оптический микроскоп, магниты и персональный компьютер. Молекулы двуцепочечных нуклеиновых кислот одним концом прикрепляют в нескольких точках к неподвижному элементу, например, к поверхности, а другим концом - к подвижной поверхности, в данном случае, к магнитной частице. Для воздействия на частицу используют магниты. В частности, магниты можно применять для того, чтобы отдалить частицы от поверхности. Однако, воплощение способа изобретения не ограничено указанным устройством. Для воплощения способа по изобретению возможно использовать любое устройство, которое позволит полностью распрямить, а затем подвергнуть рефолдингу молекулу двуцепочечной нуклеиновой кислоты, при этом одновременно регистрируя длину указанной молекулы. Например, можно использовать оптический пинцет; однако необходимость предварительной калибровки его силы захвата и получения параллельного пучка затрудняет проведение высокопроизводительных измерений. Другими недостатками является сложность контролирования скручивания нуклеиновой кислоты и возможное локальное нагревание раствора под действием сфокусированного лазерного пучка, что может повлиять на условия гибридизации.
Двуцепочечную нуклеиновую кислоту инкубируют в течение нескольких минут в растворе с соответствующими частицами (например, покрытыми стрептавидином), с которыми они связываются одним из своих меченных (например, биотином) концов. Частицы могут быть прозрачными, если для последующих манипуляций используют оптический пинцет, или магнитными, если для манипуляций используют магнитную ловушку или пинцет.
Комплексы частица-нуклеиновая кислота инжектируют в проточную камеру, поверхность которой обработана таким образом, чтобы связывать другой меченный конец молекулы (например, поверхность покрыта антителами к дигоксигенину, для связывания меченного дигоксигенином конца нуклеиновой кислоты). Таким образом, частицы прикрепляют к поверхности при помощи нуклеиновой кислоты, образующей шпилечную структуру, см. Фиг. 1а. Расстояние от частицы до поверхности отслеживают различными способами, известными специалистам в данной области: например, для расчета расстояния можно использовать регистрируемое камерой изображение частиц в виде набора дифракционных колец или интенсивность излучения, рассеянного частицами (или интенсивность флуоресценции, эмитируемой частицами), помещенными в эванесцентное поле. В альтернативном случае, для расчета расстояния до датчика, находящегося на фиксирующей поверхности, можно измерить генерируемое магнитное поле (с помощью магнитных датчиков, например, GMR-датчиков (от англ. giant magnetoresistance, гигантское магнитное сопротивление) или датчиков Холла).
Описаны различные способы, позволяющие приложить тяговое усилие к молекуле нуклеиновой кислоты, прикрепляющей частицы к поверхности. Для захвата прозрачной частицы в точку фокусировки можно использовать сфокусированный лазерный пучок. Смещая пучок относительно фиксирующей поверхности, можно приложить силу к закрепленной на ней молекуле (стандартное использование оптического пинцета). Поскольку действующая сила пропорциональна смещению частицы из положения равновесия, для воздействия на закрепленные молекулы с постоянной силой необходимо поддерживать обратную связь с захватывающим лучом.
Для воздействия на частицу с постоянной силой можно использовать гидродинамическое сопротивление потока, окружающего частицу, однако при этом точность перемещения в пространстве обычно бывает низкой (>100 нм). В предпочтительном воплощении для притягивания суперпарамагнитных частиц, прикрепленных к поверхности с помощью нуклеиновой кислоты, имеющей шпилечную структуру, как описано выше, применяют магнитную ловушку. При такой компоновке над образцом помещают небольшие магниты, которые используют для воздействия с постоянной силой на прикрепленную частицу, положение которой можно определить с точностью <1 нм (в зависимости от тягового усилия и гидродинамического сопротивления).
Как можно заметить, зафиксированную шпильку можно полностью расплести механическим путем, приложив к частицам тяговое усилие, превышающее 16 пН. Уменьшение тягового усилия, действующего на молекулу, до 11 пН позволяет шпильке спонтанно скрутиться заново (переход в расплетенную форму является обратимым, но обладает гистерезисом). Если во время расплетания какие-либо молекулы, присутствующие в растворе (например, белки или комплементарные олигонуклеотиды ДНК, РНК, ЗНК или ПНК), свяжутся с расправленной одноцепочечной нуклеиновой кислотой, эти молекулы будут препятствовать повторному скручиванию шпильки при уменьшении силы ниже 11 пН. Таким образом, принцип метода заключается в переходах между двумя силами разной величины: силой большей величины Fopen, при которой шпилька разделяется, и силой меньшей величины Ftest. позволяющей молекуле повторно свернуться, и в измерении длины молекулы при транзиторной блокировке. Местоположение блокировки в последовательности находят по отношению между длиной полностью расправленной и заблокированной молекулы. Для большей точности длину полностью расплетенной молекулы предпочтительно измеряют при силе Ftest. Для этого шпильку конструируют таким образом, чтобы ее рефолдинг при снижении силы Fopen до Ftest происходил за доли секунды.
Для прикрепления нуклеиновой кислоты к поверхности или носителю можно использовать любой из известных в области техники способов. По существу, нуклеиновую кислоту прикрепляют непосредственно к носителю, например, микрочастице, что включает специальную обработку этой поверхности, например, ее покрытие стрептавидином, СООН-группами и т.п., способными взаимодействовать с подвергнутым функционализации концом нуклеиновой кислоты.
Такие способы, как правило, требуют проведения функционализации нуклеиновой кислоты, особенно ее 3' и 5' концов, что означает перенос на них соответствующих химических групп. Предпочтительно соединять вместе два других свободных конца молекулы петлей, чтобы предотвратить диссоциацию цепей в конце процедуры, чтобы при необходимости ее можно было повторить. Для этой цели можно использовать различные способы.
Самым простым из них является функционализация одного конца двуцепочечной нуклеиновой кислоты с использованием синтетических олигонуклеотидов, имеющих две различные функциональные группы (например, биотин и амин), что делает возможным прикрепление к различным предварительно обработанным поверхностям. На другом конце две цепи можно соединить между собой с помощью частично спаренного синтетического нуклеотида в форме петли. Таким образом, из двуцепочечной нуклеиновой кислоты образуется спаренная одноцепочечная нуклеиновая кислота, т.е. шпилька. Преимуществом данного способа является возможность функционализации гетерогенной популяции больших фрагментов нуклеиновых кислот (которые получают при фракционировании гена или хромосомы), которые затем можно анализировать одновременно. В данном случае образец нуклеиновых кислот фракционируют, используя два (или более) рестрикционных фермента, что позволяет получить субпопуляцию фрагментов, имеющих на концах два разных рестрикционных сайта, общие для всех фрагментов. Это позволяет по-разному использовать два конца (например, присоединять к одному концу олигонуклеотид в форме петли, несущий на своем конце соответствующий сайт рестрикции). Недостаток данного способа заключается в том, что связывание с поверхностью может быть затруднено вследствие стерических препятствий, создаваемых двумя соседними функциональными группами. Для разрешения данной проблемы может оказаться целесообразным добавление на каждом из свободных концов молекулы, имеющей шпилечную структуру, «спейсерной» последовательности оснований, к концу которой затем присоединяют функциональную группу; поскольку две спейсерные последовательности не комплементарны, каждая функциональная группа будет иметь достаточно пространства для связывания с целевой поверхностью. Более предпочтительно, последовательность каждого спейсера конструируют так, чтобы при осуществлении способа по изобретению при секвенировании использовать одноцепочечные праймеры известной последовательности. Добавлять к молекулам двуцепочечных нуклеиновых кислот петли и/или спейсеры можно любым стандартным способом, используемым в молекулярной биологии. Данные способы хорошо известны специалистам в данной области и нет необходимости подробно их описывать в данном документе.
Что касается существующих способов прикрепления к поверхности, известно большое их количество и они являются производными способов прикрепления макромолекул (белки, ДНК и т.п.) к предварительно обработанным поверхностям, коммерческого производства. Большинство этих способов разработано для иммунологических методов, они позволяют связывать белки (иммуноглобулины) с поверхностями, несущими группы (--СООН, --NH2, --ОН и т.п.), способные взаимодействовать с карбоксильной группой (--СООН) или аминогруппой (--NH2) на конце белка.
Ковалентное присоединение нуклеиновой кислоты можно осуществлять напрямую, через свободный фосфат на 5' конце молекулы, который вступает в реакцию со вторичным амином (как на поверхности с коммерческим названием Covalink --NH, фирмы Polylabo, Страсбург) с образованием ковалентной связи. Кроме того, для функционализации ДНК можно использовать аминогруппы и затем поступать с ней как с белками.
Также существуют поверхности, покрытые стрептавидином (микрогранулы Dynal и т.п.), которые обеспечивают связывание биотинилированных молекул ДНК со стрептавидином, подобное ковалентному. Наконец, если посадить на поверхность антитело, направленное против дигоксигенина (способами, перечисленными выше), к нему можно присоединить нуклеиновую кислоту, меченную дигоксигенином. Здесь приведено лишь несколько примеров из множества возможных способов прикрепления.
Среди способов присоединения и прикрепления следует также упомянуть, например, способы, описанные в патенте ЕР 152886, в которых для присоединения ДНК к твердому носителю, такому как целлюлоза, используют ферментативное связывание.
В патенте ЕР 146815 также описаны различные способы присоединения ДНК к носителям.
Аналогично, в заявке на патент WO 92/16659 предложен способ применения полимера для присоединения ДНК.
Очевидно, что нуклеиновую кислоту можно непосредственно присоединять к носителю, но при необходимости, особенно с тем, чтобы ограничить влияние поверхности, нуклеиновую кислоту можно присоединять к концу нереакционноспособного плеча пептида или соединения иной природы, как описано, например, в патенте ЕР 329198.
Приведенные ниже примеры позволят лучше понять преимущества данного изобретения.
Краткое описание графических материалов
Фиг. 1. Принцип обнаружения гибридизации олигонуклеотида с комплементарной ему последовательностью ДНК, имеющей шпилечную структуру. ДНК, имеющая шпилечную структуру, прикрепляющая микрогранулу к поверхности, (а) мгновенно расплетается, когда тяговое усилие, действующее на частицу, превышает значение 16 пН. В это время комплементарный фрагмент, присутствующий в растворе, гибридизуется со своей мишенью в расплетенной шпилечной ДНК, и таким образом, временно препятствует повторному скручиванию шпильки (b), когда сила снижается до ее первоначального значения. По разнице между длиной молекулы в заблокированном состоянии и исходной длиной шпильки (Zhign-z) можно вычислить, в какой области шпильки произошло спаривание с комплементарной последовательностью. По средней продолжительности блокирования можно судить о возможном существовании некомплементарных оснований и их местоположении в гибриде, (с) график изменения длины шпильки во времени по мере увеличения силы от 11,4 пН до 17,8 пН с последующим ее снижением к первоначальному значению. В процессе регибридизации можно заметить паузу длительностью приблизительно 10 с. Данная пауза наблюдается только при наличии в растворе комплементарных (или почти комплементарных) олигомеров длиной > 7 нуклеотидов (в данном случае сигнал обусловлен декамерами).
Фиг. 2. а) Время блокирования, обусловленное олигонуклеотидом из 10 нуклеотидов при Ftest=9 пН, имеет экспоненциальное распределение. b) Экспоненциальная зависимость Toff от Ftest, полученная для олигонуклеотида из 9 нуклеотидов.
Фиг. 3. Изменение вероятности блокирования Pblok = число циклов с блокировкой/число циклов с длительностью фазы расплетания Topen, необходимой для нахождения олигонуклеотидом из 12 нуклеотидов своего комплементарного сайта. Согласно графику, время Топ, необходимое молекуле для нахождения своей мишени, обычно составляет 15 с при концентрации олигонуклеотида 20 нМ. Указанное время не зависит от силы, приложенной в тестовой фазе. Показатель a(F) равнялся бы 1, если бы все события были зарегистрированы, но поскольку кратковременные события пропущены, a(F) меньше 1, особенно при малых значениях Ftest.
Фиг. 4. Вероятность блокирования повышается и достигает плато с увеличением концентрации олигонуклеотида. В данном случае в присутствии олигонуклеотида из 12 нуклеотидов в концентрации 27,5 нМ блокировка регистрируется в каждом втором цикле, при этом фаза расплетания длится 10 с, а Ftest составляет 8 пН. Как показано на Фиг. 3, при выходе на плато Pblock не достигает 1; это объясняется тем, что очень кратковременное блокирование не удается зарегистрировать.
Фиг. 5. Зависимость кинетических параметров, характеризующих способность олигонуклеотида из 12 нуклеотидов связывать его комплементарный субстрат, от ионной силы буфера. koff незначительно меняется в зависимости от ионной силы, тогда как kon демонстрирует выраженную зависимость. kon увеличивается в 3 раза при добавлении Mg2+. По значениям обоих кинетических параметров можно рассчитать равновесную константу kd.
Фиг. 6. Блокирование шпилечной структуры праймазой, стабилизирующей олигонуклеотид РНК из 5 нуклеотидов, комплементарный последовательности ДНК-затравки. b) Расположение заблокированных нуклеотидов в последовательности, с) Распределение длительности блокирования, обусловленного праймазой Т4, стабилизирующей олигонуклеотид-пентамер РНК в процессе прайминга, при Ftest=9 пН. Олигонуклеотид РНК, состоящий из 5 нуклеотидов, не вызывает заметной блокировки рефолдинга шпильки. В присутствии праймазы Т4 дикого типа блокирование происходит в предполагаемом участке последовательности, а время блокирования составляет 5 с. В присутствии мутанта E248Q наблюдается тот же феномен, однако время блокирования существенно снижается.
Фиг. 7. Серия циклов, в которых исследовали связывание хеликазы RecQ с одноцепочечной ДНК, включающих три фазы: фазу расплетания при Fopen=20 пН, тестовую фазу при Ftest=10 пН и фазу очистки при Fclean=0,5 пН. Некоторые из 10 приведенных кривых указывают на возникновение блокады в одном из циклов. Фаза очистки при низкой силе позволяет удалить связанный фермент с матрицы. В отсутствие АТФ происходит только связывание RecQ и блокада рефолдинга, под давлением вилки рефолдинга хеликаза перемещается, местоположение блокировки постепенно сдвигается.
Фиг. 8. Изменение вероятности блокирования в зависимости от концентрации RecQ. Pblock возрастает и достигает плато по мере увеличения концентрации, в данном примере характеристическое значение концентрации составляет 226 пМ.
Фиг. 9. Расположение на матрице позиций, заблокированных хеликазой RecQ, в отсутствие АТФ.
Фиг. 10. Первичные данные, полученные при блокировании шпилечной ДНК размером 1,2 кб антителом, направленным против метилированного цитозина. Приведены три кривые, полученные при регистрации длины шпилек на протяжении 5 циклов. Каждый цикл начинается расплетанием шпильки в течение 5,5 с при силе 20 пН, за которым следует тестовая фаза, длящаяся 37 с при F=6,5 пН.
В большинстве циклов блокада не регистрируется (1), на протяжении одного и того же цикла может происходить последовательная блокировка одной из шпилек (2), а также блокировка может наблюдаться на протяжении нескольких циклов. Концентрация антитела составляет 35 нМ, для предупреждения неспецифического связывания применяют 100 мМ Трис-буфер с добавлением 0,2% БСА.
Для наглядности кривые смещены по оси у.
Фиг. 11. На гистограмме показаны позиции сайтов метилирования в шпилечной последовательности размером 1,2 кб после ее метилирования ДНК-метилтрансферазой человека. Представлены четыре гистограммы, полученные с разными частицами. Позиции сайтов связывания совпадают; установлены четыре позиции метилирования в эукариотических клетках, а также один сайт в позиции 882, соответствующий метилированию в клетках E.coli, использованных для продуцирования ДНК.
Фиг. 12. Запись 30 циклов денатурации и ренатурации шпильки при линейном увеличении силы в течение 2 с и снижении в течение 2 с. Репрезентативные точки на графике расположены против хода часовой стрелки (обозначены стрелками), начиная со значений F=1,5 пН и Z=0; по мере нарастания силы молекула удлиняется очень незначительно, до тех пор пока сила не достигнет 15 пН, при этом молекула расплетается и Z достигает 1,3 мкм. При линейном уменьшении силы Z постепенно уменьшается, пока F не достигнет 11 пН, в этой точке происходит рефолдинг шпильки, пока она не встретит на пути олигонуклеотид из 12 нуклеотидов. По мере того, как сила снижается, уменьшается и показатель Z, характеризующий блокирование, однако постепенно снижаясь, сила достигает значения, при котором происходит выталкивание олигонуклеотида, это быстрое уменьшение показателя Z отмечено ромбом. Справа приведен график распределения силы, соответствующей отсоединению олигонуклеотида; ее максимум, приходящийся приблизительно на 7 пН, соответствует силе, при которой Toff составляет доли секунды.
Фиг. 13. Установление сайтов метилирования в ДНК человека, выделенной из клеток человека. Шпилечную ДНК получали из молекулы человеческой геномной 5 ДНК размером 2,5 кб. а) Изменение приложенной силы в ходе циклов измерений: шпилька расплетается на секунд под действием силы 19 пН; затем сила снижается до 8,5 пН на 10 секунд. Ь) Совмещение сигналов, зарегистрированных приблизительно в 20 циклах в присутствии антител, направленных против 5mC. После расплетания цепей молекулы происходит ее скручивание, которое прерывается транзиторной блокировкой. Блокирование обусловлено связыванием антитела с 5mC. с) На гистограмме показаны позиции, где происходит блокирование, соответствующие сайтам, в которых находится 5mC. Обнаружено около 20 таких позиций, что позволяет предположить, что сайт метилирования приходится приблизительно на каждые 100 оснований.
Описание примеров осуществления изобретения предшествующий уровень техники
Связывание белков с ДНК представляет собой важнейший биологический феномен; это универсальный процесс, контролирующий многие реакции. Несмотря на то, что термодинамика данного процесса хорошо изучена, изучение его кинетики представляет собой гораздо более сложную задачу. Использование одиночных молекул позволяет измерить время, необходимое для нахождения белком своей ДНК-мишени, а также точно определить участок, где происходит связывание. В данном документе описан новый метод исследования одиночных молекул, позволяющий достичь этих целей.
Хотя предложенный метод исследования может найти широкое применение, в данном документе прежде всего показано его использование в случае специфичного связывания олигонуклеотидов и неспецифичного связывания хеликазы с оцДНК. Также, рассматривается специфичное связывание антитела, распознающего сайты метилирования ДНК.
Сущность изобретения
Изобретение относится к новому способу выявления различных модификаций ДНК и определения связывания ДНК-белок, в основе которого лежит механическая детекция блокирования регибридизации ДНК, имеющей шпилечную структуру. Метод исследования предусматривает проведение нескольких циклов, в ходе которых собирают статистические данные о связывании одиночных молекул. Каждый цикл начинается с фазы разделения, в ходе которой одиночную молекулу ДНК, имеющую шпилечную структуру, разворачивают в течение времени Topen, растягивая ее концы с силой Fopen, превышающей 16 пН. Во второй фазе эксперимента длительностью Ttest тяговое усилие Ftest снижают до уровня менее 11 пН, что позволяет шпильке свернуться заново. Если молекула, находящаяся в растворе, способна связываться с определенной последовательностью расплетенной шпилечной структуры или связываться с ней неспецифически (например, белок способен распознавать определенную одноцепочечную или двуцепочечную последовательность, как модифицированную, так и не модифицированную), она будет связывать ДНК с вероятностью Pblock и, в этом случае, временно блокировать сворачивание шпильки, когда сила окажется ниже 11 пН. Это блокирование легко обнаружить, т.к. в определенной позиции в процессе регибридизации шпильки возникает пауза, которую можно охарактеризовать тремя параметрами:
- позиция Zblock в расплетенной молекуле ДНК, в которой возникла пауза, характеризует распознаваемую последовательность;
- длительность блокирования Toff характеризует время, в течение которого молекула оставалась связанной с ДНК; и
- вероятность блокирования Pblock, которая зависит от времени Ton, необходимого для нахождения молекулой своего сайта связывания.
Оба параметра Ton и Toff характеризуют силу взаимодействия между ДНК и блокирующей молекулой. Таким образом, исследуя последовательность ДНК (образующей шпилечную структуру, связанную одним концом с частицей, а другим концом - с поверхностью) с использованием белка или антитела, распознающего сайт метилирования, повторяя циклы разделения и сворачивания шпильки, можно обнаружить искомый сайт метилирования (по блокированию регибридизации некоторых шпилек). Аналогично, можно обнаружить связывание белка с предполагаемым сайтом дцДНК по увеличению стабильности гибрида с комплементарным олигонуклеотидом, в присутствии и в отсутствие белка.
Данное изобретение позволяет выявить модификации ДНК в геномной ДНК без проведения бисульфитной реакции и ПЦР-амплификации. Необходимая предварительная подготовка ДНК включает получение фрагментов, имеющих шпилечную структуру, которые можно использовать для связывания частиц с поверхностью (фрагментация и лигирование с соответствующими фрагментами). Данное изобретение не требует флуоресцентного мечения белков или ДНК. В данном варианте осуществления для того, чтобы зарегистрировать блокирование регибридизации шпилечной структуры, требуется оптическое устройство (микроскоп).
Сведения, подтверждающие возможность осуществления изобретения
Фрагмент двуцепочечной (дц) ДНК размером от нескольких десятков до нескольких тысяч пар оснований (например, полученный при механической фрагментации или разрезании рестриктазами геномной ДНК) лигируют одним концом с петлей ДНК. Другой конец лигируют с фрагментом дцДНК, две цепи которого могут связываться с поверхностями, имеющими разное покрытие. Например, свободный 3' конец одной цепи может быть помечен биотином, что позволит ему связываться с частицами, покрытыми стрептавидином, а 5' конец противоположной цепи может быть помечен дигоксигенином, что позволит ему связываться с поверхностями, несущими антитело к дигоксигенину. Введение концевой метки можно осуществлять разными способами, известными специалистам в данной области, например, с применением терминальной (концевой) трансферазы для присоединения нуклеотидов, модифицированных биотином (или дигоксигенином) или гибридизации с меченными соответствующим образом олигонуклеотидами.
Данную конструкцию ДНК инкубируют в течение нескольких минут в растворе с соответствующими частицами (например, покрытыми стрептавидином), с которыми они связываются одним из своих меченных (например, биотином) концов. Частицы могут быть прозрачными, если для последующих манипуляций используют оптический пинцет, или магнитными, если для манипуляций используют магнитную ловушку или пинцет.
Комплексы частица-ДНК инжектируют в проточную камеру, поверхность которой обработана таким образом, чтобы связывать другой меченный конец молекулы (например, поверхность покрыта антителами к дигоксигенину, для связывания меченного дигоксигенином конца ДНК). Таким образом, частицы прикрепляют к поверхности при помощи шпилечной ДНК (см. Фиг. 1а, ниже). Расстояние от частицы до поверхности определяют различными способами. Например, для расчета расстояния можно использовать регистрируемое камерой изображение частиц в виде набора дифракционных колец.
Расстояние также можно определить по интенсивности излучения, рассеянного частицами (или по интенсивности флуоресценции, эмитируемой частицами), помещенными в эванесцентное поле. В альтернативном случае, при использовании магнитных частиц, для определения расстояния между частицами и датчиком, находящимся на фиксирующей поверхности, можно измерить генерируемое магнитное поле (с помощью магнитных датчиков, например, GMR-датчиков (от англ. giant magnetoresistance, гигантское магнитное сопротивление) или датчиков Холла).
Описаны различные способы, позволяющие приложить тяговое усилие к молекуле ДНК, прикрепляющей частицы к поверхности. Для захвата прозрачной частицы в точку фокусировки можно использовать сфокусированный лазерный пучок. Смещая пучок относительно фиксирующей поверхности, можно приложить силу к закрепленной на ней молекуле (стандартное использование оптического пинцета). Поскольку действующая сила пропорциональна смещению частицы из положения равновесия, для воздействия на закрепленные молекулы с постоянной силой необходимо поддерживать обратную связь с захватывающим лучом.
Для воздействия на частицу с постоянной силой можно использовать гидродинамическое сопротивление потока, окружающего частицу, однако при этом точность перемещения в пространстве обычно бывает низкой (>100 нм). В предпочтительном воплощении для притягивания суперпарамагнитных частиц, прикрепленных к поверхности с помощью нуклеиновой кислоты, имеющей шпилечную структуру, как описано выше, применяют магнитную ловушку. При такой компоновке над образцом помещают небольшие магниты, которые используют для воздействия с постоянной силой на прикрепленную частицу, положение которой можно определить с точностью<1 нм (в зависимости от тягового усилия и гидродинамического сопротивления).
Зафиксированную шпильку можно механически расплести, приложив к частицам тяговое усилие, превышающее 16 пН. Уменьшение тягового усилия, действующего на молекулу, ниже ~11 пН позволяет шпильке спонтанно скрутиться заново (переход в расплетенную форму является обратимым, но обладает гистерезисом).
Если во время расплетания какие-либо молекулы, присутствующие в растворе (например, белки или комплементарные олигонуклеотиды ДНК, РНК, 3НК или ПНК), свяжутся с расправленной одноцепочечной (оц) ДНК, эти молекулы временно заблокируют ренатурацию шпильки при уменьшении силы ниже ~11 пН.
Таким образом, принцип метода заключается в переходах между двумя силами разной величины: силой большей величины Fopen, при которой шпилька разделяется, и силой меньшей величины Ftest, позволяющей молекуле повторно свернуться, и в измерении длины молекулы при транзиторной блокировке. Местоположение блокировки в последовательности находят по отношению между длиной полностью расправленной и заблокированной молекулы. Для большей точности длину полностью расплетенной молекулы предпочтительно измеряют при силе Ftest. Для этого шпильку конструируют таким образом, чтобы ее рефолдинг при снижении силы Fopen до Ftest происходил за доли секунды.
Измеряя длину молекулы ДНК Z(t) (расстояние от частицы до поверхности) во время одной из таких пауз в ренатурации в серии нескольких циклов, можно определить позицию, в которой происходит блокирование, с точностью примерно 1 нм (что соответствует расстоянию, которое занимают два последовательно расположенных нуклеотида (1 пн) в оцДНК при силе 10 пН). Кроме того, измеряя среднюю длительность блокирования, можно определить Toff=1/koff. Измеряя Pblock и зная концентрацию молекул [М], возможно вычислить Ton и, следовательно, kon. Как один, так и оба этих параметра помогают охарактеризовать природу связывания. Например, можно установить, имеет ли место совершенная гибридизация с комплементарным олигонуклеотидом, стабилизирует ли белок гибридизацию, а также имеется ли некомплементарное основание и где оно находится (например, в центре гибридизованного олигонуклеотида или вблизи одного из его концов).
Таким образом, возможны различные варианты осуществления изобретения для выявления модификаций ДНК и, в целом, для изучения взаимодействия белков с одноцепочечной или двуцепочечной ДНК.
Выявление модификаций ДНК путем механической детекции блокирования регибридизации
Если во время механического расплетания ДНК, имеющей шпилечную структуру, в растворе присутствуют олигонуклеотиды (длиной более семи нуклеотидов), эти олигонуклеотиды могут спариваться со своей комплементарной последовательностью ДНК и создавать временное препятствие для полного рефолдинга шпильки при падении силы ниже 11 пН, см. Фиг. 16. Можно легко провести несколько циклов денатурации/ренатурации одной и той же молекулы и обнаружить блокирование (паузу) в процессе рефолдинга вследствие спаривания олигонуклеотидов с ДНК в фазе денатурации.
Длительность блокирования обычно имеет экспоненциальное распределение, при этом среднее значение Toff экспоненциально уменьшается по мере изменения Ftest. Такое распределение вероятности обусловлено тем, что в данном исследовании используют одиночные молекулы. Как следствие, наиболее вероятное время блокирования равно 0, что означает, что значительную долю событий (блокирований), невозможно зарегистрировать, поскольку их длительность находится за пределами временного разрешения экспериментальной установки. Молекула, блокирующая рефолдинг шпильки, испытывает давление со стороны Y-образной структуры («вилки»), перемещающейся вдоль ДНК. При значениях Ftest близких 15 пН (механическое усилие, разворачивающее шпильку) данное давление выражено слабо, и наоборот, при снижении Ftest давление значительно повышается, и молекула выталкивается. Как показано на Фиг. 2, по мере изменения Ftest происходит экспоненциальное уменьшение Toff. Данная зависимость настолько сильно выражена, что измерение Toff возможно только в диапазоне нескольких пН. Следует также отметить, что Toff(F) будет совпадать с классическим Toff спонтанно отсоединяющейся молекулы только при значениях Ftest=Funzip=15 пН, что в данном случае недостижимо.
Как показано на Фиг. 3, вероятность блокирования Pblock повышается при увеличении длительности фазы денатурации Topen согласно экспоненциальной зависимости Pblock=a(F).[1-exp(Topen/Ton)].
Как и предполагалось, Pblock повышается с увеличением концентрации молекулы, на Фиг. 4 показано, что при использовании 12-нт олигонуклеотида Pblock увеличивается по мере нарастания [М] и выходит на плато.
Зная Topen и концентрацию молекулы [М] можно вычислить kon по величине Pblock согласно следующей зависимости:
kon=-Log(1-Pblock/a(F))/([M]Topen).
Прочность связывания (см. Фиг. 5) можно описать зависимостью:
kd-1=-(ToffLog(1-Pblock/a(F))/([M]Topen)
Среднее время блокирования Toff зависит от размера олигонуклеотида, силы Ftest, приложенной в процессе ренатурации, температуры и незначительно - от ионной силы используемого буфера.
Ton тоже зависит от размера олигонуклеотида, температуры, ионной силы используемого буфера, но незначительно - от Ftest. Как показано на Фиг. 5, определение этих кинетических констант также позволяет охарактеризовать некомплементарные пары оснований олигонуклеотида и субстрата. Например, полностью комплементарный 12-нт олигонуклеотид характеризуется kon 1,5×10-6 М-1 с-1, введение одного некомплементарного основания на расстоянии 3 нуклеотидов от одного конца не приводит к значительному изменению kon.
Если некомплементарное основание находится в середине нуклеотида, kon снижается в 10 раз.
Toff также зависит от наличия белков, связывающих дцДНК, которые могут стабилизировать гибрид. Например, как показано на Фиг. 6, праймаза стабилизирует связывание олигонуклеотидов с ДНК, которое в противном случае было бы недостаточно стабильным, и время блокирования регибридизации шпильки было бы недостаточным для детекции. Аналогично, связывание полимеразы с 3'-концом короткого олигонуклеотида, используемого в качестве праймера, повышает его стабильность; данный метод можно применять для определения аффинности полимеразы к сайту связывания праймера. Аналогично, если белок связывается с определенным сайтом оцДНК (например, с метилированным основанием), он будет блокировать рефолдинг в определенном сайте в течение времени, достаточного для детекции.
Данный способ можно применять для обнаружения модификаций ДНК как в одноцепочечных, так и в двуцепочечных ДНК. Так, исследуя шпилечную ДНК, прикрепляющую частицы к поверхности, с использованием антитела (Ат), распознающего модифицированное определенным образом основание, можно обнаружить и локализовать данное модифицированное основание в цепи по временному блокированию регибридизации шпильки, вызванному связыванием Ат.Исследование сайта связывания при помощи набора комплементарных олигонуклеотидов позволит обнаружить фрагмент ДНК, содержащий данную модификацию.
Определение аффинности связывания RecQ с оцДНК матрицей
Хеликазы связывают однонитевые разрывы и, таким образом, вызывают раскручивание дцДНК. Активность данных ферментов напрямую зависит от их аффинности к оцДНК. Авторы изобретения предложили прямое определение данного параметра, используя способ по изобретению. Его осуществление возможно в присутствии или в отсутствие АТФ или АДФ или других аналогов. В данном документе представлены некоторые результаты, касающиеся хеликазы RecQ из E.coli, полученные в отсутствие АТФ. Характерный сигнал связывания, представленный на Фиг. 7, позволяет определить Pblock для одной концентрации хеликазы. Изменение Pblock в зависимости от концентрации RecQ показано на Фиг. 8. Показано, что характеристическое значение концентрации RecQ составляет 226 пМ. Фиг. 9 демонстрирует, что хеликаза связывается неспецифически. Характер блокирования под действием фермента можно описать как «скольжение»: параметр Z не является постоянным, а постепенно снижается. Такое поведение затрудняет определение истинного значения Toff, таким образом, можно определить только Ton и kon.
Максимум при Z=0 соответствует не блокированию, а непосредственно рефолдингу. Блокирование RecQ носит одинаковый характер по всей длине матрицы, небольшой уклон на отметке 0,9 мкм объясняется усреднением показателей для молекул, незначительно различающихся по длине.
Исследование метилирования
Гистограмма на Фиг. 11 иллюстрирует время блокирования антителом, направленным против 5mC. Большинство блокад кратковременные и могут быть достаточно хорошо описаны экспоненциальным распределением с характеристическим значением времени 1,3 с. Однако, значительное количество блокад 17,5% длится более 30 с. Это затрудняет определение Toff фермента, предположительно существуют два разных конкурирующих между собой механизма связывания, один из которых имеет преимущество.
В качестве альтернативы можно исследовать наличие известных модификаций ДНК путем гибридизации олигонуклеотида, комплементарного предполагаемому модифицированному сайту, в присутствии (или в отсутствие) белка, распознающего модификацию (например, содержащего метил-связывающий домен белка 1 (MBD1, от англ. methyl binding domain protein 1), распознающего метилированный цитозин, или соответствующего Ат, направленного против определенных модификаций дцДНК). По значительному увеличению времени блокирования в присутствии белка легко обнаружить и локализовать модифицированное основание.
Используя белки, распознающие некомплементарные основания, указанный способ можно применять аналогичным образом для идентификации некомплементарных оснований (т.е. ОНП) в ДНК. Данный метод можно также применять для обнаружения белков (или лекарственных средств), влияющих на стабильность комплексов белок/ДНК.
Параметры, влияющие на ход определения
Fopen: авторы изобретения предлагают значение 20 пН, поскольку при этом одновременно большое число частиц будет поддерживать шпильки в расплетенном состоянии (их намагниченность и, следовательно, сила может варьироваться на 10-20%).
Topen является важным параметром наряду с концентрацией молекул: для обнаружения блокировки необходимо использовать комбинацию обоих параметров, что позволит вычислить значение Pblock согласно формуле:
Pblock=a(F).[1-ехр(Topen.kon.[М])].
Для измерения kon следует избегать выхода на плато Pblock, подобрав концентрацию молекул [М] и Topen так, чтобы Pblock находилась в диапазоне от 0,2 до 0,5, можно получить достаточно данных за минимальное количество циклов Следует отметить, что Topen можно изменить просто скорректировав настройки в программе сбора данных, изменение концентрации фермента требует смены буфера в проточной камере. С другой стороны, если измерение kon не требуется, выход на плато Pblock может оказаться целесообразным, т.к. позволит дать лучшую статистическую оценку блокировки. На концентрацию молекул влияет как их поступление, так и нежелательное связывание, например, в исследовании с использованием антитела против 5mC данный фермент в высокой концентрации связывается с двуцепочечной ДНК, когда она находится в форме шпильки, и препятствует расхождению ее цепей. Поддержание концентрации фермента на уровне ниже 35 нМ позволяет устранить данную проблему. В данном эксперименте повысить Pblock возможно только за счет увеличения Topen.
Параметр a(F) обычно близок к 1, лучшим способом определения его значения будет тест насыщением, при котором Topen или [М] варьируют до тех пор, пока Pblock асимптотически не приблизится к a(F), как показано на Фиг. 3 и 4. В альтернативном случае, возможно найти a(F), используя следующую формулу:
a(F)=exp(-Tdead/Toff)
где Tdead обозначает время запаздывания детектирующей системы, a Toff - среднее время блокирования. Обычно Tdead составляет порядка 0,1 с.
Ftest представляет собой очень важный регулируемый параметр: его величина зависит от используемой шпилечной структуры, но обычно находится в диапазоне [12 пН, 2 пН]. Если сила превышает 12 пН, рефолдинг шпильки в некоторой степени блокируется за счет формирования вторичной структуры оцДНК, что может маскировать сигналы, представляющие интерес. При слабой силе удлинение ДНК становится очень незначительным, а шум резко возрастает. Давление, оказываемое Y-образной структурой формирующейся шпильки, вызывает эффективную регибридизацию молекулы, при этом Toff=Т0 exp F/F0; таким образом Toff очень быстро снижается по мере уменьшения Ftest. Например, блокада, вызываемая олигонуклеотидом из 9 нуклеотидов, при Ftest=11 пН длится 1 с, а при силе ниже 9 пН блокаду едва ли можно зарегистрировать (a(F) снижается). Для олигонуклеотида из 12 нуклеотидов рабочий диапазон составляет [10 пН, 6 пН]. Для олигонуклеотида из 37 нуклеотидов блокада может длиться бесконечно при 6 пН, но сокращается до нескольких секунд при Ftest=3 пН. То же касается и связывающих белков: чем сильнее связывание, тем ниже сила, при которой регистрируется блокирование.
Значение Ftest подбирают таким образом, чтобы длительность блокирования была достаточной для измерения (a(F) ~1), но не слишком большой, так как Ttest должно быть относительно коротким, чтобы можно было провести несколько циклов.
В данном эксперименте Toff находится в диапазоне от 0,2 до 20 с. Более короткое время можно зарегистрировать с помощью быстродействующих устройств, например, быстрой видеокамеры, длительное время увеличивает время сбора данных, т.к. для усреднения данных требуется провести несколько циклов. Для олигонуклеотидов Toff меняется экспоненциально в зависимости от Ftest; поэтому возможно подобрать Ftest так, чтобы Joff находилось в приемлемом диапазоне. Для белков характер изменения Toff в зависимости от Ftest не известен, но согласно наблюдениям снижение Ftest обычно вызывает резкое уменьшение Toff. При этом, а приори, Toff обычно неизвестно и может варьировать в широких пределах. Чтобы составить представление о типичных значениях Ftest целесообразно вначале провести серию циклов, линейно увеличивая и уменьшая силу в течение нескольких секунд, как показано на Фиг. 12. Конец фазы блокирования соответствует силе Fc. В распределении Fc максимум приходится на значение, при котором Toff составляет величину того же порядка, что и длительность линейного изменения силы.
Затем можно провести циклы, в которых сила (Fopen и Ftest) будет выходить на плато, a Ftest будет слегка превышать <Fc>, и таким образом, выбрать диапазон Toff, поддающийся измерению.
Ttest и Nциклов : Ttest должно в 2 или 3 раза превышать Toff. Наконец, от количества циклов зависит общая точность измерений. Чтобы добиться точности Х% необходимо Х/100=1/Nblock1/2, поскольку Pblock=Nblock/Nциклов; получаем Nциклов=10000/(X2Pblock).
Оптимизация исследования: зачастую трудности возникают при слишком кратковременном или, наоборот, долговременном связывании фермента (Фиг. 9); в последнем случае блокирование может по-прежнему наблюдаться в конце тестовой фазы и в начале нового цикла (Фиг. 7). Поскольку в фазе размыкания блокировка не регистрируется, существование блокады на протяжении нескольких циклов возможно, но не доказуемо. Обойти это затруднение можно, если учесть тот факт, что при низких значениях силы блокировка обычно бывает очень кратковременной. Так, если после тестовой фазы добавить третью фазу с низким значением силы, это позволит очистить шпилечную структуру от каких-либо связавшихся молекул: при Fclean=0,5 пН и Tclean=2 с удаляются любые связанные молекулы и шпилька подготавливается к новому циклу. В молекуле также может существовать несколько сайтов связывания, поэтому при блокировке может регистрироваться сигнал ступенчатой формы, когда после первой блокировки в молекуле блокируется второй сайт связывания и т.д. (см. Фиг. 10). При второй блокировке эффективная фаза разделения длится Topen+Tblock1 (Фиг. 10); если Tblock1 превышает Topen, с большей вероятностью вслед за первой блокадой будет наблюдаться вторая, что затруднит измерение кинетических параметров. В данном случае для минимизации этого эффекта целесообразно увеличить Topen по сравнению с Ttest.

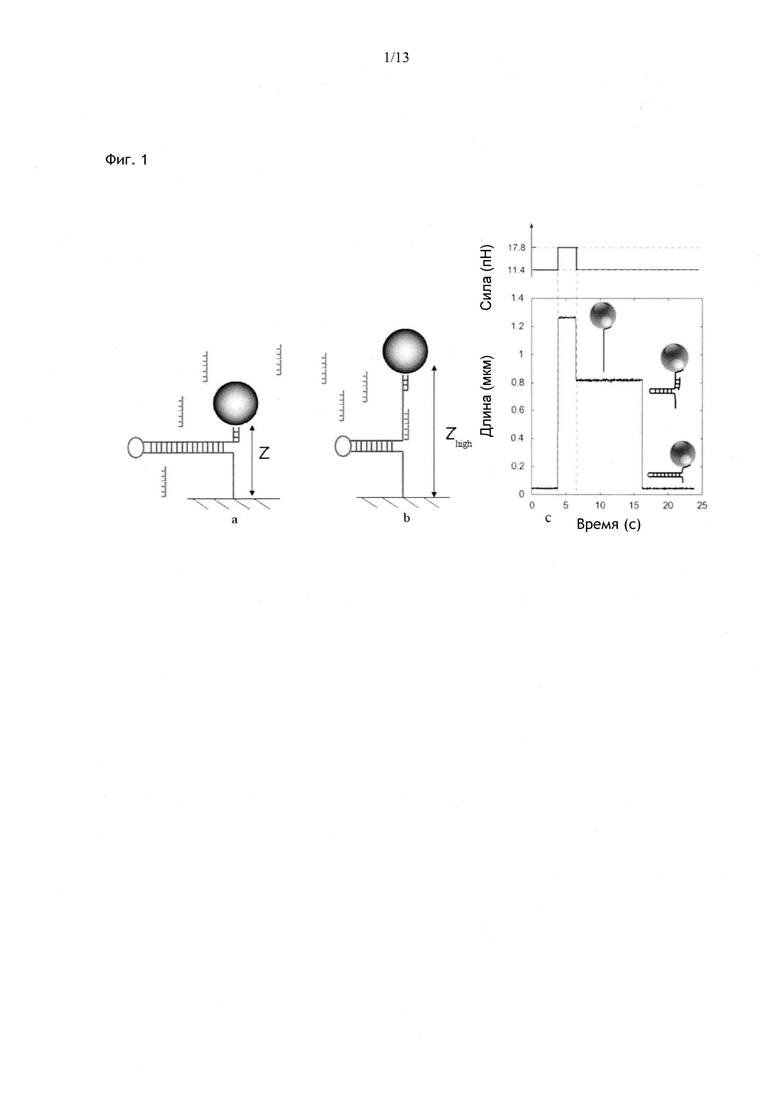
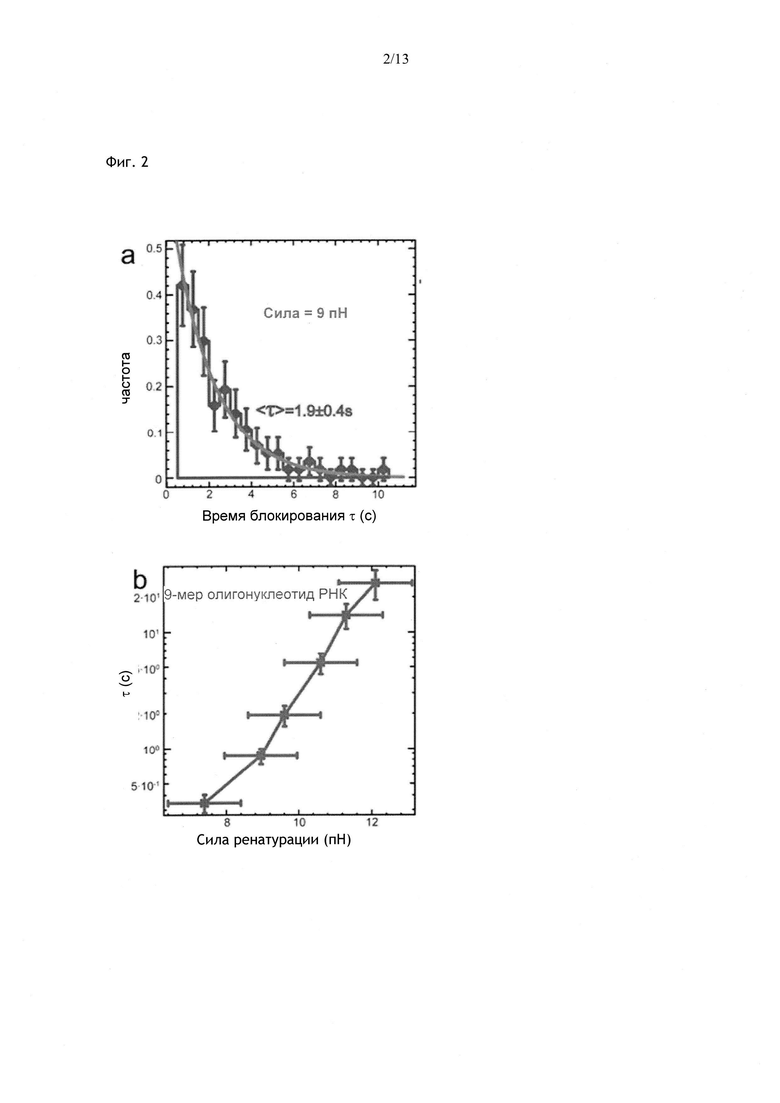
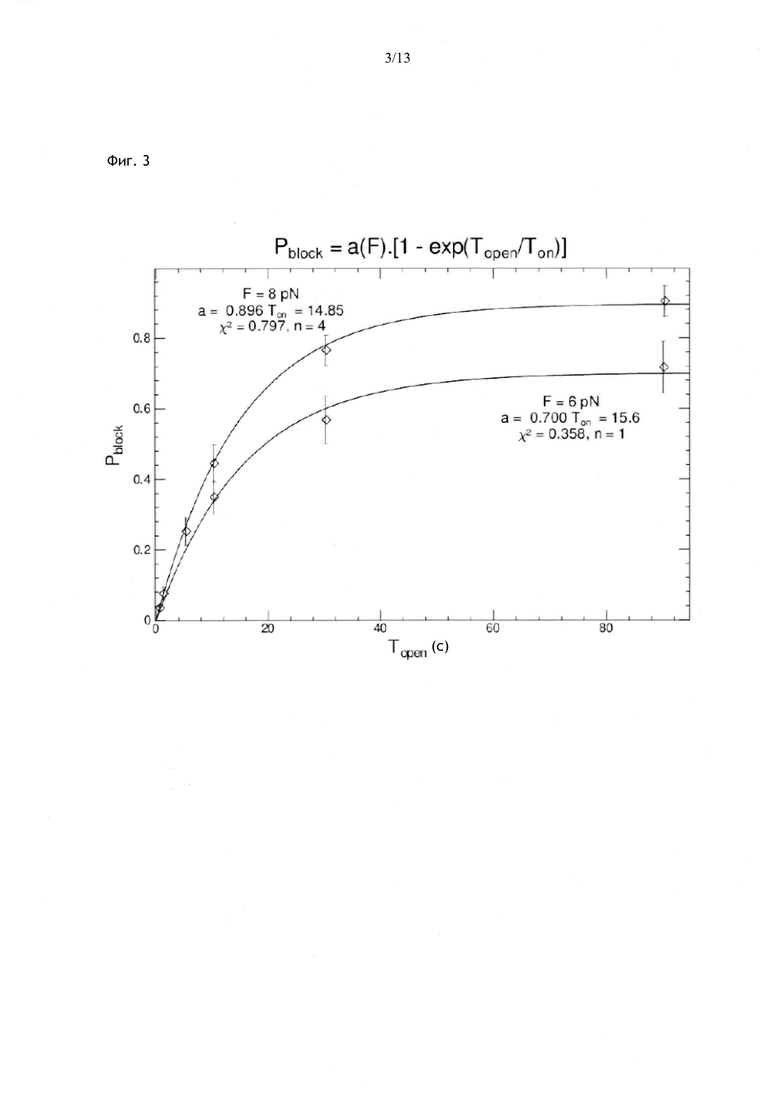
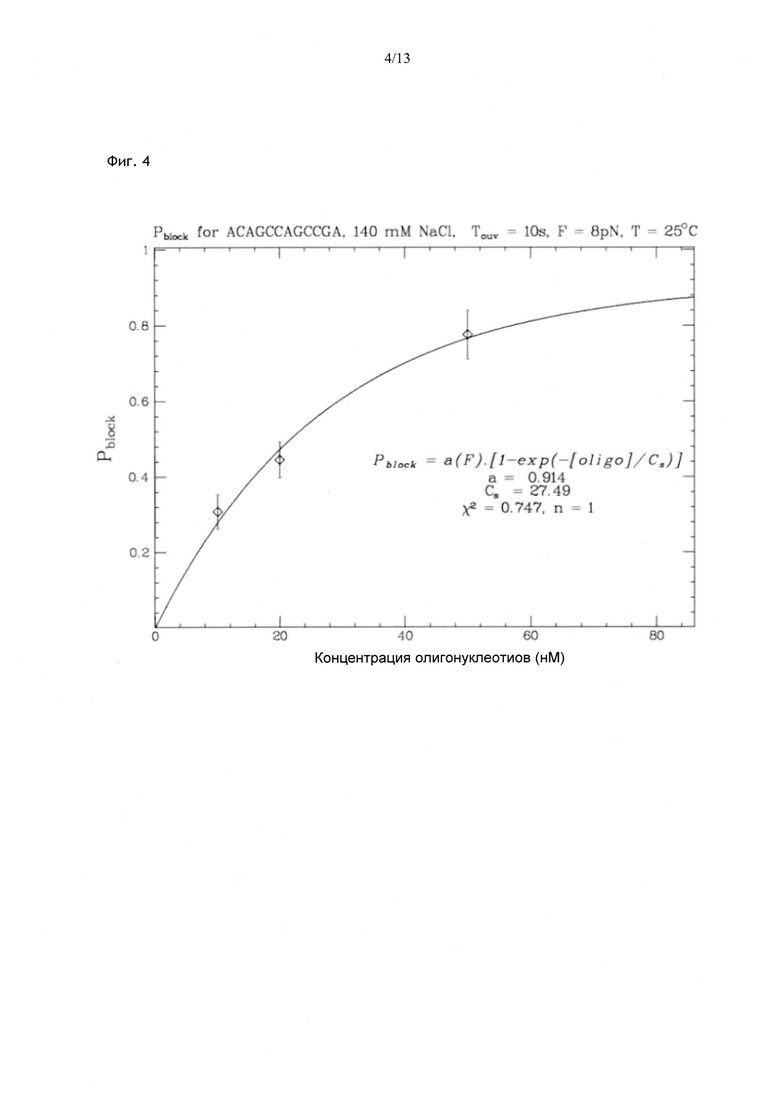
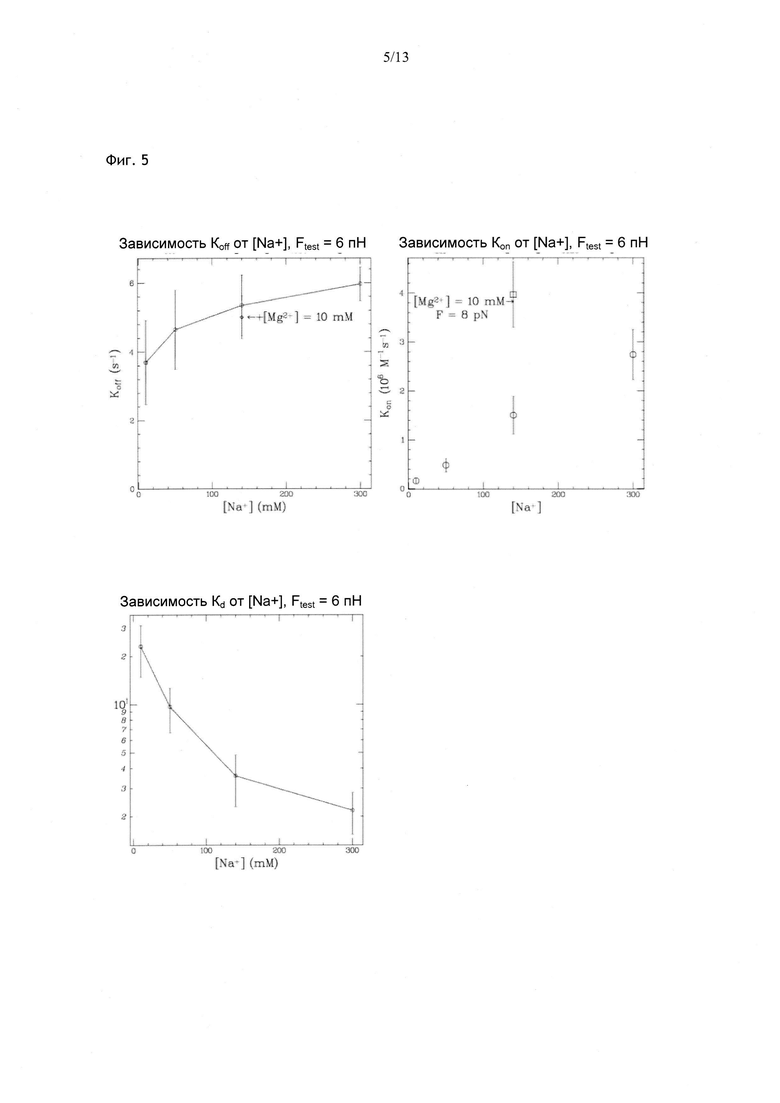
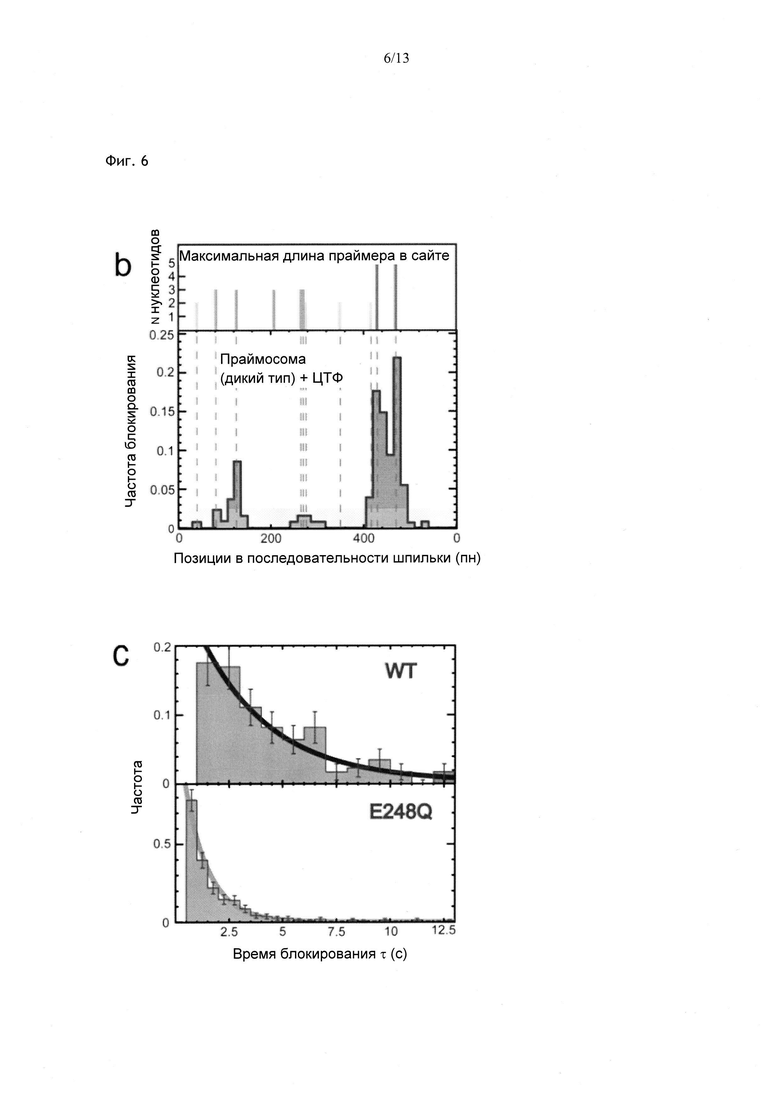
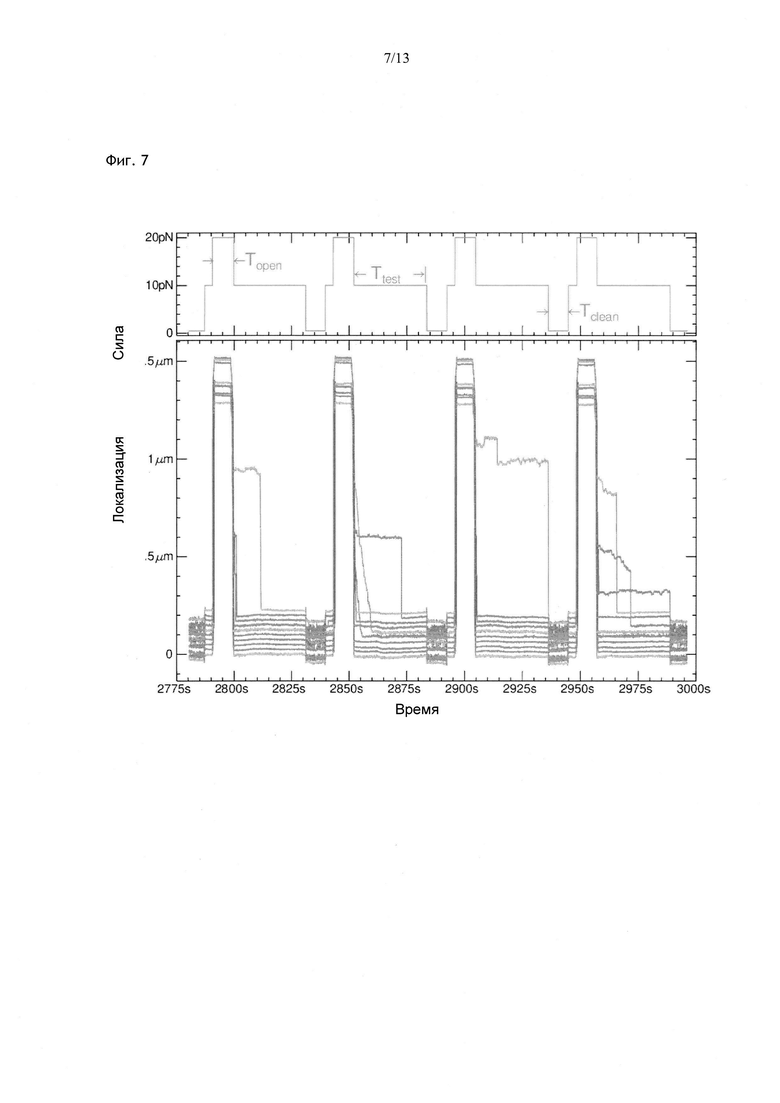
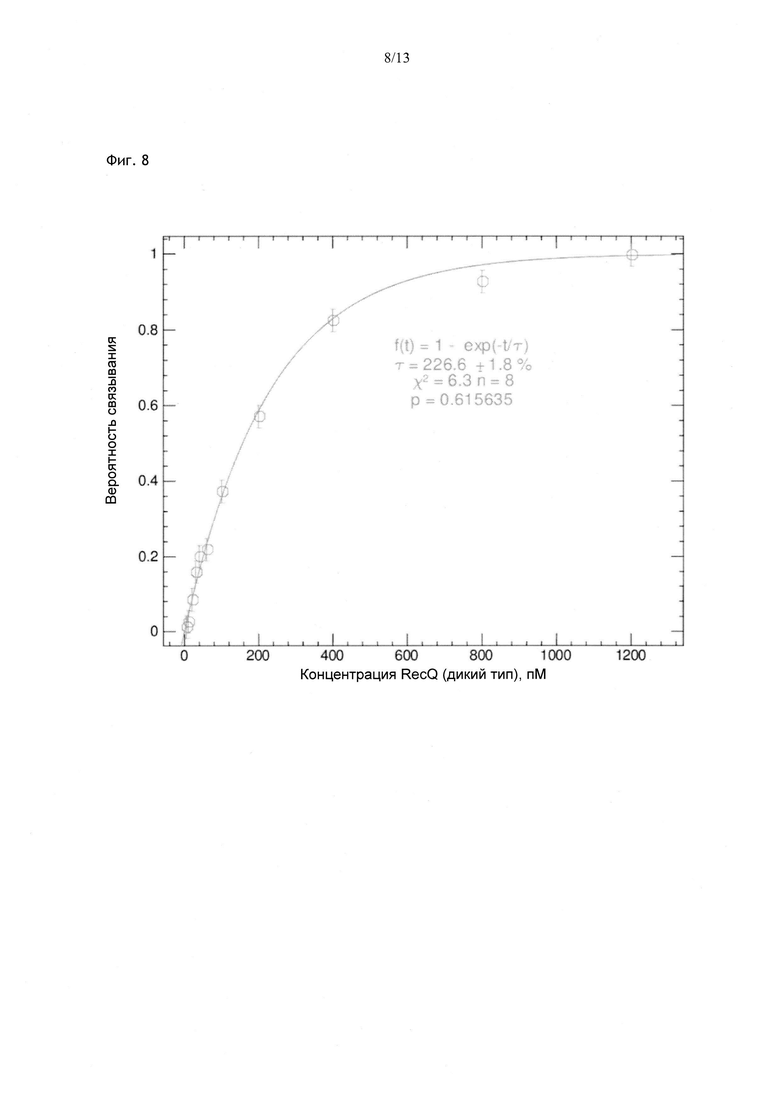
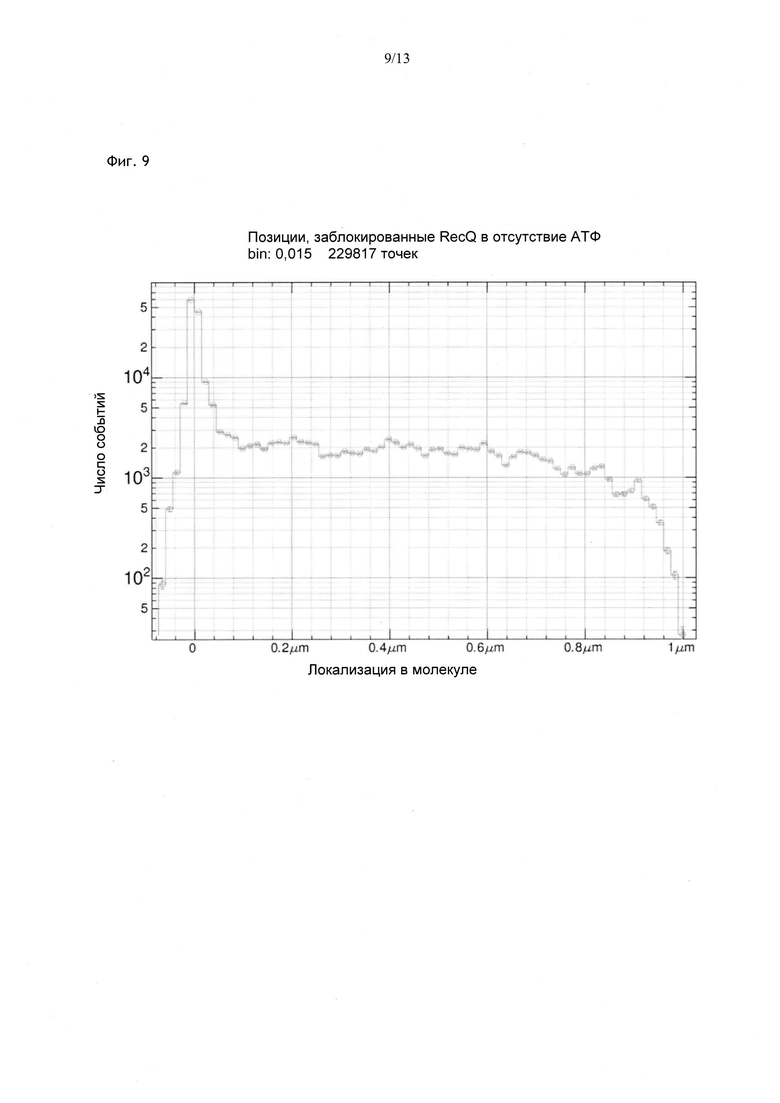
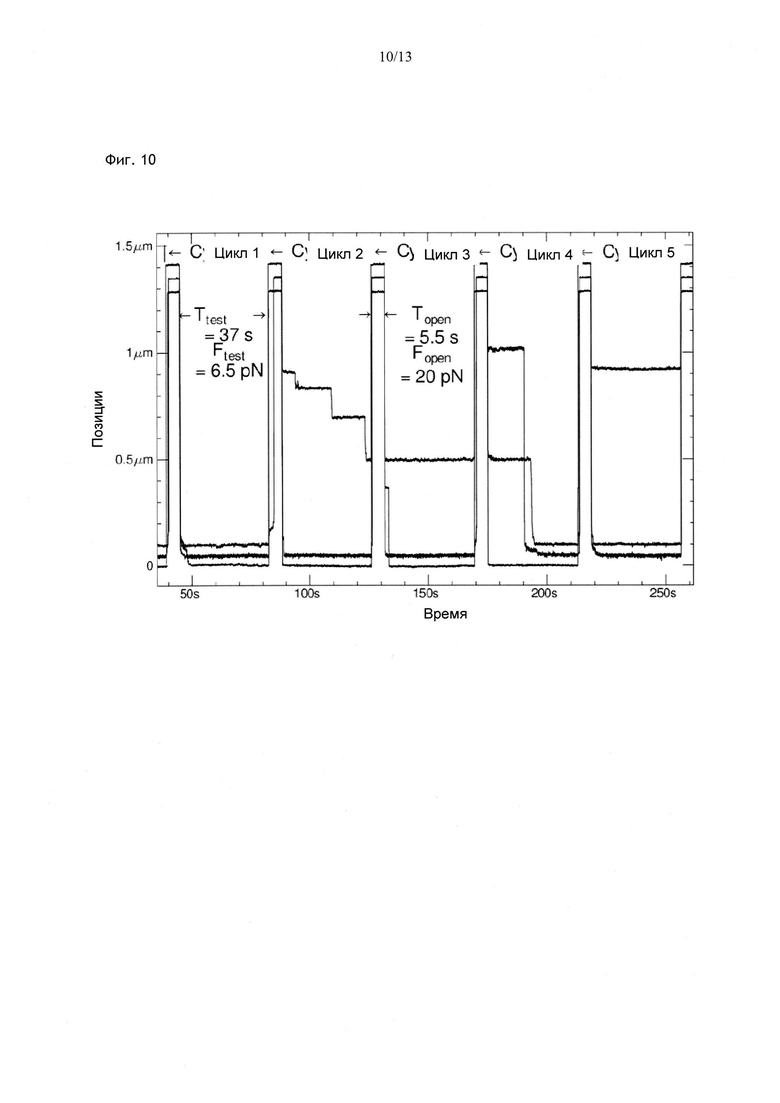
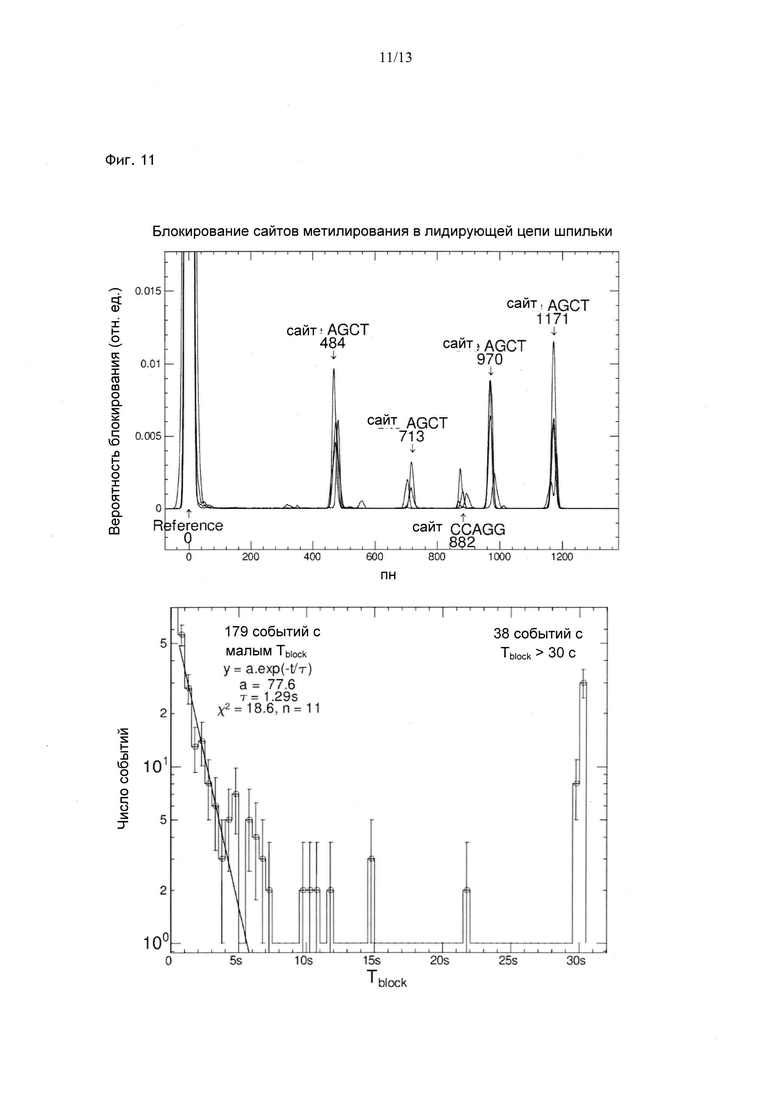
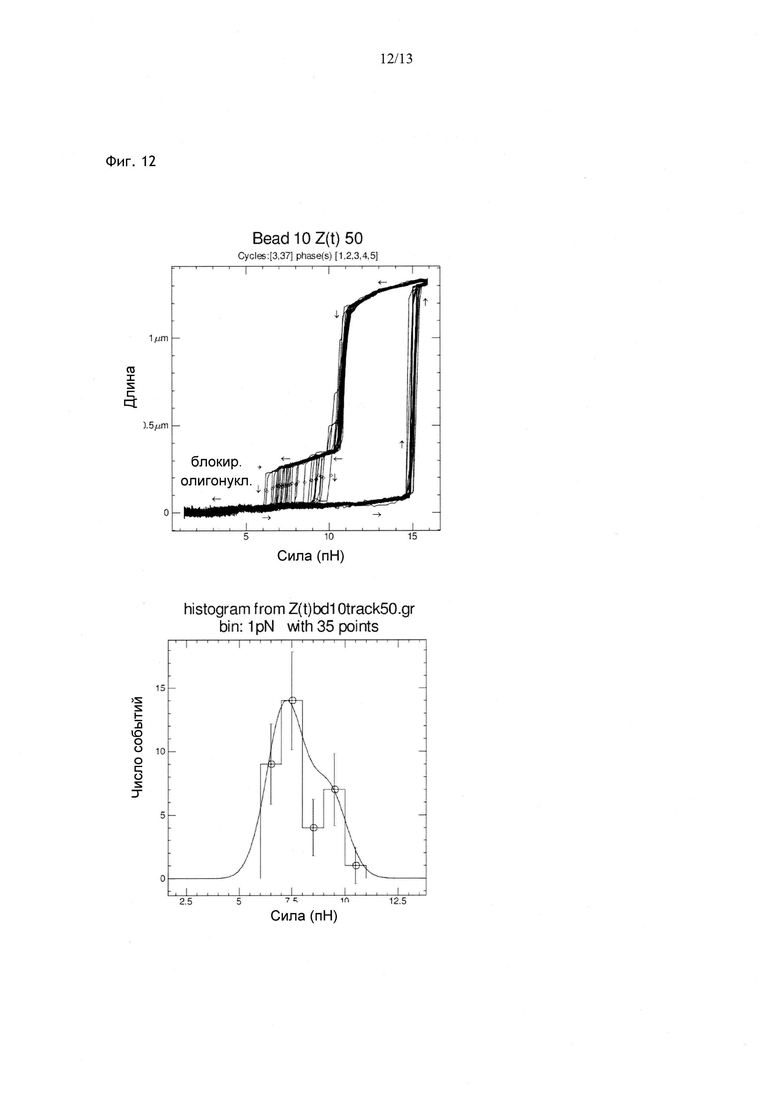
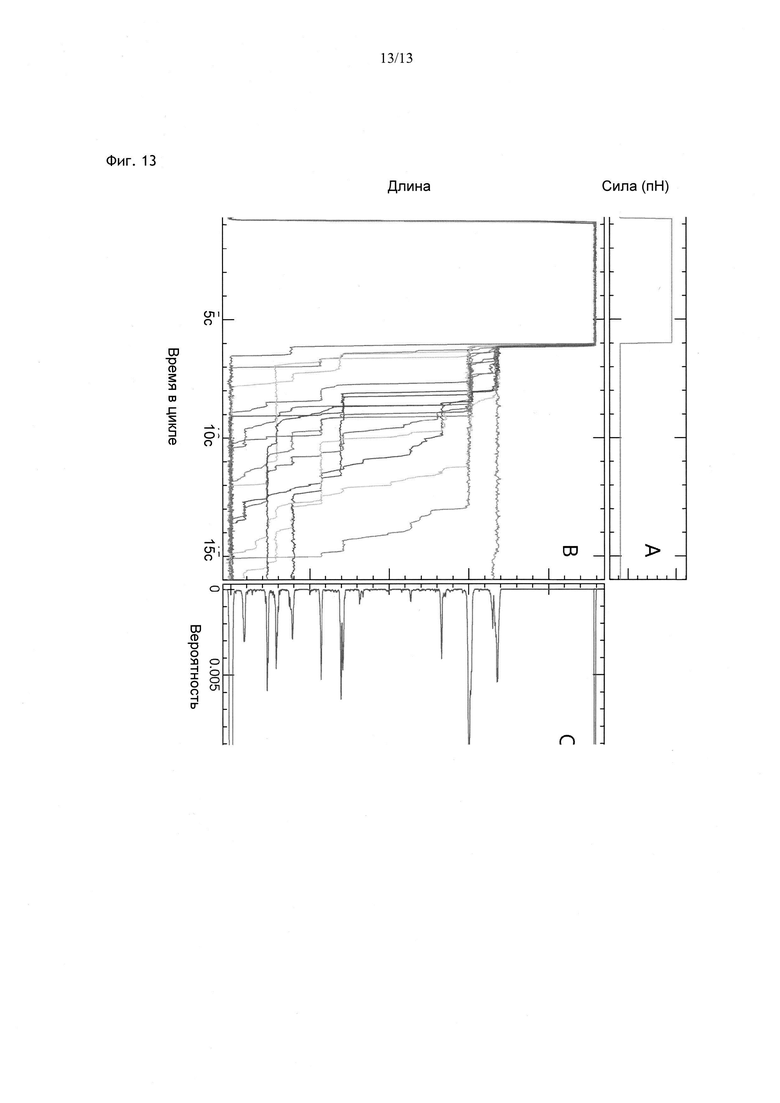
Предложенная группа изобретений относится к области медицины. Предложены способ обнаружения модифицированного основания в молекуле двуцепочечной нуклеиновой кислоты и способ идентификации по меньшей мере одного соединения, способного предотвращать взаимодействие белка с его сайтом связывания. Молекулу двуцепочечной нуклеиновой кислоты сначала денатурируют, потом ренатурируют в присутствии белка, способного связываться с модифицированным основанием. Далее детектируют блокирование ренатурации двуцепочечной нуклеиновой кислоты и определяют позицию в молекуле двуцепочечной нуклеиновой кислоты, где происходит указанное блокирование. Предложенная группа изобретений обеспечивает выявление различных модификаций ДНК и определение связывания ДНК-белок за счет определения позиции, в которой происходит блокирование. 2 н. и 12 з.п. ф-лы, 13 ил.
1. Способ обнаружения по меньшей мере одного модифицированного основания в молекуле двуцепочечной нуклеиновой кислоты, содержащей последовательность нуклеиновой кислоты, где указанная молекула двуцепочечной нуклеиновой кислоты имеет шпилечную структуру, и где один из концов двуцепочечной нуклеиновой кислоты непосредственно или опосредованно присоединен к носителю, и где другой конец двуцепочечной нуклеиновой кислоты присоединен к подвижному носителю, где указанный способ включает этапы, при которых:
a) денатурируют указанную молекулу двуцепочечной нуклеиновой кислоты, содержащую последовательность нуклеиновой кислоты, прикладывая к указанной молекуле физическую силу;
b) берут белок, способный связываться с указанным модифицированным основанием;
c) ренатурируют указанную молекулу двуцепочечной нуклеиновой кислоты в присутствии указанного белка;
d) детектируют блокирование ренатурации двуцепочечной нуклеиновой кислоты и
e) определяют позицию в указанной молекуле двуцепочечной нуклеиновой кислоты, где происходит указанное блокирование, где указанное определение включает этапы, при которых:
- измеряют расстояние (z) между двумя концами молекулы двуцепочечной нуклеиновой кислоты, присоединенными к носителю;
- измеряют расстояние (zhigh) между двумя концами молекулы двуцепочечной нуклеиновой кислоты, присоединенными к носителю, при этом указанная молекула двуцепочечной нуклеиновой кислоты денатурирована и
- сравнивают z и zhigh,
где указанное модифицированное основание представляет собой эпигенетическую модификацию.
2. Способ по п. 1, в котором на этапе (б) дополнительно берут молекулу одноцепочечной нуклеиновой кислоты, соответствующую указанной последовательности.
3. Способ по п. 1, в котором двуцепочечную нуклеиновую кислоту денатурируют на этапе (а), отдаляя носители друг от друга.
4. Способ по п. 3, в котором к двуцепочечной молекуле прикладывают физическую силу, равную или превышающую 15 пН, предпочтительно превышающую или равную 17 пН, более предпочтительно равную или превышающую 18 пН, отдаляя носители друг от друга.
5. Способ по п. 1, в котором денатурированную двуцепочечную нуклеиновую кислоту ренатурируют на этапе (в), приближая носители друг к другу.
6. Способ по п. 5, в котором силу, прикладываемую к двуцепочечной молекуле, снижают до уровня, равного или меньшего 12 пН, предпочтительно меньшего или равного 11 пН, более предпочтительно меньшего или равного 10 пН, приближая носители друг к другу.
7. Способ по п. 1, в котором этапы (а)-(г) повторяют несколько раз.
8. Способ по п. 1, также включающий этап измерения длительности блокирования.
9. Способ по п. 8, также включающий этап сравнения длительности блокирования с референтным значением.
10. Способ по п. 1, также включающий этап секвенирования последовательности нуклеиновой кислоты, связанной с указанным белком.
11. Способ по п. 1, в котором модифицированное основание выбрано из группы, включающей 4-метилцитозин (4mC), 5-метилцитозин (5mC), 5-гидроксиметилцитозин (5hmC), 5-формилцитозин (5fC), 5-карбоксилцитозин (5саС), 5-гидроксиметилурацил (5hmU) и N6-метиладенозин (m6A).
12. Способ по п. 11, в котором модифицированное основание представляет собой 5mC.
13. Способ по п. 11, в котором указанный белок представляет собой антитело или фермент, который способен связывать указанное модифицированное основание.
14. Способ идентификации по меньшей мере одного соединения, способного предотвращать взаимодействие белка с его сайтом связывания, где указанный способ включает этапы, при которых:
a) берут указанный белок и молекулу нуклеиновой кислоты, содержащую последовательность, соответствующую указанному сайту связывания;
b) берут соединение и
c) изучают связывание указанного белка с указанной эпигенетической модификацией способом по любому из пп. 1-10.
| ВАКЦИНА ПРОТИВ ВИРУСА ГРИППА ДЛЯ ВВЕДЕНИЯ ЧЕРЕЗ СЛИЗИСТУЮ И СПОСОБ ПРЕДОТВРАЩЕНИЯ ГРИППА | 2004 |
|
RU2390351C2 |
| MANOSAS M | |||
| et al | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Nat Chem Biol | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| Гальваническая батарея | 1927 |
|
SU7338A1 |

