Настоящая заявка является частичным продолжением заявки на патент США рег. 07/536428, поданной 11 июня 1990 г. под названием "Systematic Evolution of Ligands by Exponential Enrichment". Эта работа санкционирована правительством Соединенных Штагов Америки и финансируется через Национальный институт здоровья. Правительство Соединенных Штатов имеет определенные права на это изобретение.
В настоящей заявке описан новый класс нуклеиново-кислотных лигандов с высокой степенью сродства, которые специфически связываются с нужной молекулой-мишенью, В заявке также представлен способ отбора нуклеиново-кислотного лиганда, который специфически связывается с любой нужной молекулой-мишенью. Этот способ назван SELEX (аббревиатура полного названия: "Systematic Evolution of Ligands by Exponential Enrichment" - "Систематическое извлечение лигандов путем экспоненциального обогащения"). Указанный способ настоящего изобретения (SELEX) предназначен для выделения нуклеиново-кислотного лиганда для нужной молекулы-мишени. Продукты нуклеиновых кислот настоящего изобретения могут быть использованы в любых целях, для которых требуется реакция связывания, например, в методах анализа, в диагностических методах, для сортировки клеток, в качестве секвестрантов и т.п. Кроме того, продукты нуклеиновых кислот настоящего изобретения могут обладать каталитической активностью. Целевыми молекулами являются натуральные и синтетические полимеры, включая протеины, полисахариды, гликопротеины, гормоны, рецепторы, и поверхности клеток, небольшие молекулы, такие как лекарственные вещества, метаболиты, кофакторы, аналоги переходного состояния и токсины.
Известно, что большинство белков и малых молекул не обнаруживают специфического связывания с нуклеиновыми кислотами. Исключение составляют регуляторные белки, такие как репрессоры, полимеразы, активаторы и т.п., то есть такие белки, функция которых в живой клетке заключается в переносе генетической трансформации, закодированной в нуклеиновых кислотах, в клеточные структуры, и репликации генетического материала. Кроме того, небольшие молекулы, такие как GТР, связываются с некоторыми интропными РНК.
В живой материи, нуклеиновые кислоты играют широкоинформативную роль. Основной постулат, предложенный Криком, как в исходной, так и в продвинутой форме, заключается в том, что нуклеиновые кислоты (РНК или ДНК) могут служить в качестве матриц для синтеза других нуклеиновых кислот посредством процесса репликации, при котором информация как бы "считывается" в матричной нуклеиновой кислоте, в результате чего образуются комплементарные нуклеиновые кислоты. Все экспериментальные исследования по генетике и генной экспрессии зависят от свойств нуклеиновых кислот: в основном, двухцепочечные нуклеиновые кислоты являются информативно избыточными, поскольку их химическая структура, основанная на парах оснований, и процессы репликации позволяют использовать указанное спаривание оснований относительно безошибочным способом.
Отдельные компоненты белков, то есть двадцать природных аминокислот, обладают различным химическим строением и различными активностями, достаточными для обеспечения довольно широкого спектра активностей при связывании и катализе. Нуклеиновые кислоты, однако, имеют, как известно, более узкие возможности в отношении их химических свойств, чем белки, однако их информационная роль обеспечивает переход информации от вируса к вирусу, от клетки к клетке и от организма. В связи с этим, компоненты нуклеиновых кислот, т.е. нуклеотиды, должны обладать лишь парами поверхностей, которые обеспечивают информационную избыточность в уотсон-криковской паре оснований. Компонентам нуклеиновых кислот не требуется различий в химическом строении или в активностях, достаточных для обеспечения широкого диапазона связывания или катализа.
Однако было обнаружено, что в природе существуют некоторые нуклеиновые кислоты, которые принимают участие в связывании с определенными молекулами-мишенями, а в некоторых случаях даже участвуют в катализе. Но по сравнению с белками и более специфичными антителами спектр активностей указанных нуклеиновых кислот является более узким. Например, в тех случаях, когда нуклеиновые кислоты связываются с некоторыми белками-мишенями с высокой степенью родства и специфичности, то это связывание, как известно, зависит от конкретных последовательностей нуклеотидов, которые входят в состав ДНК- или РНК-лиганда. Так, например, известно, что короткие двухцепочечные ДНК-последовательности связываются с белками-мишенями, которые подавляют или активируют транскрипцию как в прокариотах, так и в эукариотах. Другие двухцепочечные ДНК-последовательности, как известно, связываются с рестриктирующими эндонуклеазами, белковыми мишенями, которые могут быть выбраны с высокой степенью сродства и специфичности. Другие короткие ДНК-последовательности служат в качестве центромер или теломер на хромосомах, вероятно, путем создания лигандов для связывания специфических белков, которые участвуют в механике хромосом. Так, например, хорошо известно, что двухцепочечная ДНК обладает способностью связываться внутри карманов и бороздок целевых белков, функции которых направлены на связывание с ДНК. Одноцепочечные ДНК также могут связываться с некоторыми белками с высокой степенью специфичности и сродства, хотя число таких примеров значительно меньше. Известные примеры связывания двухцепочечных ДНК с белками позволяют объяснить это связывание, как взаимодействие различных структурных областей белка с ДНК, заключающееся в том, что выступающие боковые цепи аминокислот точно входят в большую бороздку В-формы двухцепочечной ДНК, и это распознавание последовательности обеспечивает специфичность связывания.
Иногда, в качестве лиганда для некоторых белков могут служить двухцепочечные РНК, например эндонуклеазная РНКаза III из Е.соli. Более известны примеры связывания белков-мишеней с одноцепочечными РНК-лигандами, хотя в этих случаях одноцепочечная РНК часто образует сложную трехмерную форму, которая содержит локальные области внутримолекулярной двухцепочечности. Аминоацильные тРНК-синтетазы тесно связываются с тРНК-молекулами с высокой специфичностью. Короткие участки в геномах РНК-вирусов тесно и с высокой специфичностью связываются с оболочечным белком вируса. Короткая последовательность РНК связывается с ДНК-полимеразой фага Т4 также с высокой степенью сродства и специфичности. Таким образом, можно найти двухцепочечные или одноцепочечные РНК- и ДНК-лиганды, которые выступают в роли партнеров для связывания со специфическими белками-мишенями. Большинство из известных связывающихся с ДНК белков связываются с двухцепочечными ДНК, тогда как большинство связывающихся с РНК белков связывается с одноцепочечными РНК. Эта статистическая погрешность в литературе несомненно отражает существующую в биосфере предрасположенность к использованию ДНК в качестве двухцепочечного генома, а РНК в качестве одноцепочечной молекулы во многих случаях, где РНК играет роль, выходящую за пределы ее функции в качестве генома. С химической точки зрения нет серьезных оснований исключать из использования одноцепочечной ДНК в качестве полноценного партнера для специфических взаимодействий с белками.
Было также установлено, что РНК и ДНК связываются с более мелкими молекулами. Двухцепочечные ДНК связываются с различными антибиотиками, такими как актиномицин D. Специфические одноцепочечные РНК связываются с антибиотиком тиострептоном; специфические РНК-последовательности структуры, вероятно, связываются и с другими антибиотиками, особенно с теми, чьи функции заключаются в инактивации рибосом в целевом организме. Семейство эволюционно родственных РНК специфично и с хорошим сродством связывается с нуклеотидами и нуклеозидами (Bass, В. и Сесh Т. (1984) Nature 308:820-826), а также с одной из двадцати аминокислот (Varus, М. (1988) Science 240:1751-1758). В настоящее время известны также и каталитические РНК, и хотя эти молекулы в химическом отношении обладают узким диапазоном возможностей, однако при этом они достаточно хорошо связаны с реакциями переноса фосфодиэфира и гидролизом нуклеиновых кислот.
Несмотря на эти известные примеры, большая часть белков и других компонентов клетки, очевидно, не связывается с нуклеиновыми кислотами в физиологических условиях, и если такое связывание наблюдается, то оно является неспецифичным. Либо способность нуклеиновых кислот к связыванию с другими соединениями ограничивается относительно малым количеством случаев, перечисленных выше, либо химические особенности нуклеиновых кислот, необходимые для специфического связывания, как бы аннулируются (т.е. отклоняются при выборе) в природных структурах. Настоящее изобретение основано на той предпосылке, что нуклеиновые кислоты как химические соединения могут образовывать практически неограниченный ряд форм, размеров и конфигураций, и способы к значительно более широкому спектру связывания и каталитических функций, чем нуклеиновые кислоты, обнаруживаемые в биологических системах.
Для некоторых известных случаев связывания нуклеиновых кислот с белками, были проведены исследования химических взаимодействий. Например, размер и последовательность сайта связывания РНК с оболочечным белком бактериофага R17 были идентифицированы Uhlenbeck и сотрудниками. Минимальный сайт связывания (длиной в 21 основание) натуральной РНК с оболочечным белком R17 определяли с помощью анализа на связывание меченых и различных по размерам фрагментов мРНК с нитроцеллюлозными фильтрами, где комплексы белок-фрагмент РНК оставались связанными с фильтром (Carey и др. (1983), Biochemistry 22: 601). Для определения участия отдельных нуклеиновых кислот в связывании с белком, был создан in vitro ряд вариантов последовательностей минимального сайта связывания с оболочечным белком R17 (Uhlenbeck и др. (1983), J.Biomol. Structure Dinamics 1:539 и Romaniuk и др. (1987), Biochemistry 26:1563). Было обнаружено, что для связывания с белком необходимо поддерживать шпилечно-петлевую структуру сайта связывания, и помимо этого, было установлено, что замены нуклеотидов в большой части одноцепочечных остатков в сайте связывания, включая выступающий нуклеотид в стебле шпильки, значительно влияют на связывание. В аналогичных исследованиях была проведена оценка связывания оболочечного белка бактериофага Qβ с его трансляционным оператором (Witherell и Uhlenbeck (1989), Biochemistry 28: 71). При этом было установлено, что сайт связывания РНК с оболочечным белком Qβ по своим размерам и по предсказанной вторичной структуре аналогичен сайту связывания R17, а именно он содержит 20 оснований со шпилечной структурой из 8 пар оснований, включая выпуклый нуклеотид и петлю в 3 основания. В отличие от сайта связывания с оболочечным белком R17 в этом сайте для связывания необходим лишь один из одноцепочечных остатков петли, а наличия выпуклого нуклеотида не требуется. Взаимодействия при связывании белка с РНК, участвующие в регуляции трансляции, обнаруживают значительную специфичность.
Известно, что нуклеиновые кислоты образуют вторичную и третичную структуры в растворах. Двухцепочечные формы ДНК представляют собой так называемую В-форму двуспиральной ДНК, Z-форму ДНК и суперспиральные степени скрученности (Rich A. и др. (1984), Ann. Rev. Biochem. 53:791-846). Одноцепочечные РНК образуют локализованные области вторичной структуры, такие как шпилечные петли и псевдоузловые структуры (Schimmel, Р. (1989), Cell 58:9-12). Однако при этом очень мало что известно о влиянии неспаренных нуклеотитов петли на стабильность структуры петли, кинетику образования и денатурацию, а также термодинамику, и почти ничего не известно о третичных структурах и трехмерной форме, а также о кинетике и термодинамике третичной укладки в нуклеиновых кислотах (Tuerk С. и др. (1988), Proc. Natl. Acad. Sci. USA 85:1364-1368).
Тип in vitro-выделения был описан при репликации РНК-бактериофага Qβ. Mills, D. R. и др. (1967) Proc. Natl. Acad. Sci. USA. 58:217-224; Levinsohn R. и Spiegleman S. (1968), Proc. Natl. Acad. Sci. USA 60:866-872; Levisohn R. и Spiegelman S. (1969), Proc. Natl. Acad. Sci. USA 64:805-811; Saffhill R. и др. (1970), J. Mol. Вiol. 51:531-539; Kacian D.L. и др. (1972), Рroc. Natl. Acad. Sci. USA 69:3038-3042; Мills D.R. и др. (1973), Science 180: 916-927. Фаговая РНК служит в качестве полицистронной информационной РНК, направляющий трансляцию фагоспецифических белков, а также в качестве матрицы для ее собственной репликации, катализируемой Qβ-РНК-репликазой. Было показано, что указанная РНК репликаза является высокоспецифичной по отношению к ее собственным РНК-матрицам. В процессе циклов репликации in vitro были выделены небольшие вариантные РНК, которые также были реплицированы Qβ-репликазой. Было обнаружено, что небольшие изменения в условиях, при которых осуществляли циклы репликации, приводят к аккумуляции различных РНК, очевидно, потому, что изменения условий благоприятствуют их репликации. В этих экспериментах, выбранные РНК эффективно связывались посредством репликазы для инициации репликации и служили в качестве кинетически благоприятной матрицы в течение элонгации РНК. Kramer и др. (1974) (J. Mol. Biol. 89:719) описали выделение мутантной РНК-матрицы Qβ-репликазы, репликация которой была более резистентной к ингибированию этидий-бромида, чем натуральная матрица. Было сделано предположение, что этот мутант не присутствует в исходной РНК-популяции, а генерируется последовательной мутацией в процессе циклов in vitro-репликации с помощью Qβ-репликазы. Единственным источником изменения в процессе селекции была частота специфических ошибок в процессе элонгации с помощью Qβ-репликазы. В этих исследованиях так называемая "селекция" происходила путем предпочтительной амплификации одного или более из ограниченного числа самопроизвольных вариантов первоначально гомогенной РНК-последовательности. Эта селекция не дала желаемого результата, за исключением того результата, который присущ данному механизму действия Qβ-репликазы.
Joyce и Robertson (Joyce (1989) в работе RNA: Catalysic, Splicing, Evolution, Belfort и Shub (изд.), Elsevier, Амстердам, стр. 83-87; и Robertson и Joyce (1990), Nature 344:467) описали метод идентификации РНК, которые специфически расщепляют одноцепочечную ДНК. Селекция на каталитическую активность основана на способности рибозима к катализу расщепления субстратной дцРНК или ДНК в специфическом положении и к переносу 3'-конца субстрата в 3'-конец рибозима. Продукт нужной реакции отбирали путем использования олигодезоксинуклеотидного праймера, который может связываться только с полным продуктом через область присоединения, образованную каталитической реакцией, и подвергали избирательной обратной транскрипции последовательность рибозима. Отобранные каталитические последовательности амплифицировали путем присоединения промотора РНК-полимеразы Т7 к 3'-концу кДНК с последующей транскрипцией на РНК. Этот способ был использован для идентификации из небольшого числа вариантов рибозимов варианта, который является наиболее активным в расщеплении выбранного субстрата. Идентифицируемыми оказалось лишь ограниченное число вариантов, поскольку изменение зависело лишь от нуклеотидных замен, происходящих в процессе амплификации.
В предшествующих работах нет каких либо указаний или предположений, кроме указаний на ограниченный диапазон химических функций нуклеиновых кислот при их взаимодействии с другими веществами как мишеней для белковых лигандов, связывающихся с некоторыми специфическими олигонуклеотидными последовательностями; и в более поздних работах как катализаторов с ограниченным диапазоном активностей. Предшествующие эксперименты по "селекции" были ограничены узким интервалом вариантов ранее описанных функций. Теперь же, впервые, стало ясно, что нуклеиновые кислоты обладают широким диапазоном функций, и в настоящей работе раскрывается методология для реализации этих функций.
Настоящее изобретение относится к классу продуктов, которыми являются молекулы нуклеиновых кислот, каждая из которых имеет уникальную последовательность и обладает способностью специфически связываться с нужным целевым соединением или молекулой. Каждое соединение настоящего изобретения является специфическим лигандом данной целевой молекулы. Настоящее изобретение основано лишь на том убеждении, что нуклеиновые кислоты обладают достаточной способностью к образованию различных двух- и трехмерных структур и достаточной химической изменчивостью в пределах их мономеров для того, чтобы действовать в качестве лигандов (образовывать специфические связывающиеся пары) практически для любого химического соединения, независимо от того, является ли оно мономерным или полимерным. В качестве мишеней могут служить молекулы любого размера. Обычно (и предпочтительно для терапевтических целей), чтобы связывание происходило в водном растворе в солевых условиях при температуре и рН, близким к приемлемым физиологическим значениям.
Настоящее изобретение также относится к способу, который может быть применен для получения нуклеиново-кислотного лиганда для любой желаемой мишени. Этот способ заключается в том, что осуществляют выбор из смеси кандидатов и ступенчатые итерации структурного улучшения, используя при этом одну и ту же общую схему отбора, в целях достижения практически любого желаемого критерия аффинности и селективности связывания. Исходя из смеси нуклеиновых кислот, предпочтительно содержащих сегмент рандомизированной последовательности, этот способ, именуемый в настоящем описании SELEX, включает в себя стадию взаимодействия указанной смеси с мишенью в условиях, благоприятных для связывания; стадию распределения несвязанных нуклеиновых кислот из числа тех нуклеиновых кислот, которые связывались с молекулами-мишенями; стадию диссоциации пар "нуклеиновая кислота - мишень"; стадию амплификации нуклеиновых кислот, диссоциированных из пар "нуклеиновая кислота - мишень", в целях получения лигандобогащенной смеси нуклеиновых кислот; и затем стадии повторной итерации стадий связывания, распределения, диссоциации и амплификации столько циклов, сколько необходимо.
Не претендуя на какую-либо конкретную теорию, авторы настоящей заявки отмечают, что способ SЕLЕХ основан на том мнении, что в смеси нуклеиновых кислот содержится большое количество возможных последовательностей и структур с широким диапазоном сродства к связыванию с данной мишенью. Смесь нуклеиновых кислот, содержащая, например, рандомизированный сегмент из 20 нуклеотидов, может иметь 420 возможных кандидатов. Те кандидаты, которые имеют более высокое значение константы связывания по сродству для данной мишени, связываются с большей вероятностью. После распределения, диссоциации и амплификации образуется вторая смесь нуклеиновых кислот, обогащенная кандидатами с более высокой аффинностью связывания. Последующие дополнительные циклы отбора постепенно благоприятствуют тому, чтобы в смеси оставались наилучшие лиганды до тех пор, пока указанная смесь нуклеиновых кислот не будет преимущественно состоять лишь из одной или нескольких последовательностей. Эти последовательности могут быть затем клонированы, секвенированы и отдельно тестированы на сродство к связыванию в качестве чистых лигандов.
Эти циклы отбора и амплификации повторяют вплоть до достижения желаемой цели. В самых общих чертах селекцию/амплификацию осуществляют до тех пор, пока после повторения цикла уже не наблюдается значительного улучшения эффективности связывания. Способ итеративной селекции/амплификации является достаточно чувствительным, чтобы выделить единственный вариант последовательности в смеси, содержащей, по крайней мере, 65000 вариантов последовательностей. Этот способ даже способен выделить небольшое число последовательностей с высокой степенью сродства из смеси, содержащей 1014 последовательностей. В принципе, этот способ может быть использован для испытаний примерно до 1018 различных видов нуклеиновых кислот. Нуклеиновые кислоты тестируемой смеси предпочтительно содержат часть рандомизированной последовательности, а также консервативные последовательности, необходимые для эффективной амплификации. Варианты последовательностей нуклеиновых кислот могут быть получены рядом способов, включая синтез рандомизированных последовательностей нуклеиновых кислот и отбор по размерам из произвольно гидролизованных клеточных нуклеиновых кислот. Вариабельная часть последовательности может содержать полностью или частично рандомизированную последовательность; она может также содержать подобласти консервативной последовательности, введенные с рандомизированной последовательностью. Модификации последовательностей в тестируемых нуклеиновых кислотах могут быть введены и увеличены посредством мутагенеза перед или в течение итераций селекции/амплификации.
В одном из вариантов осуществления настоящего изобретения способ селекции является настолько эффективным для выделения тех нуклеиново-кислотных лигандов, которые лучше всего связываются с выбранной мишенью, что требуется всего один цикл селекции и амплификации. Такая эффективная выборка может иметь место, например, в процессах типа хроматографии, где способность нуклеиновых кислот ассоциироваться с мишенями, связанными на колонке, приводит к тому, что указанная колонка позволяет отделять и выделять нуклеиново-кислотные лиганды с наиболее высоким сродством.
Во многих случаях необязательно бывает предпочтительным осуществлять итерационные стадии SELEX до тех пор, пока не будет идентифицирован один нуклеиново-кислотный лиганд. Раствор цель-специфического нуклеиново-кислотного лиганда может содержать семейство структур или основных особенностей нуклеиновых кислот, которые имеют ряд консервативных последовательностей и ряд последовательностей, которые могут быть заменены или добавлены без значительного изменения аффинности нуклеиново-кислотных лигандов по отношению к мишени. Последовательность ряда членов семейства нуклеиново-кислотных лигандов в растворе может быть определена при прекращении способа SELEX до его завершения, что позволяет получить исчерпывающее описание раствора нуклеиново-кислотных лигандов.
После описания семейство нуклеиново-кислотных лигандов разделяют с помощью SELEX, а в некоторых случаях может быть предпочтительным осуществление дополнительных серий SЕLЕХ с учетом информации, полученной в течение SЕLEX-эксперимента. В одном из вариантов вторые серии SЕLЕХ будут фиксировать консервативные области семейства нуклеиново-кислотных лигандов при рандомизации всех остальных положений в структуре лиганда. В альтернативном варианте последовательность наиболее типичного члена семейства нуклеиново-кислотных лигандов может быть взята за основу в процессе SELEX, где первоначальный пул нуклеиново-кислотных последовательностей является не полностью рандомизированным, а имеет статистическое смещение в сторону наиболее известного лиганда. С помощью указанных методов можно оптимизировать способ SELEX для получения наиболее предпочтительных нуклеиново-кислотных лигандов.
Известно, что существуют нуклеиновые кислоты с первичной, вторичной и третичной структурами. В литературе чаще всего описываются структуры или их характерные фрагменты, которые участвуют во взаимодействиях не-уотсон-криковского типа, например, такие как шпилечные петли, симметричные или асимметричные выступы, псевдоузлы и их бесчисленные комбинации. Почти во всех случаях указанных характерных особенностей предполагается, что они могут быть образованы в нуклеиново-кислотной последовательности, состоящей не более чем из 30 нуклеотидов. По этой причине предпочтительно, чтобы процедуры SELEX с использованием смежных рандомизированных сегментов начинались с нуклеиново-кислотных последовательностей, содержащих рандомизированный сегмент примерно с 20-50 нуклеотидами, а наиболее предпочтительно с 25-40 нуклеотидами. Настоящее изобретение также включает в себя растворы, содержащие смесь примерно из 109-1018 нуклеиново-кислотных последовательностей, имеющих смежную рандомизированную последовательность, состоящую, по крайней мере, примерно из 15 нуклеотидов. В предпочтительном варианте осуществления настоящего изобретения рандомизированная часть последовательностей фланкируется фиксированными последовательностями, что облегчает амплификацию лигандов.
В случае полимерной мишени, например, такой как белок, аффинность лиганда может быть увеличена посредством использования в SELEX-процедуре смеси кандидатов, содержащей первую выбранную последовательность и вторую рандомизированную последовательность. Последовательность первого выбранного лиганда, ассоциированного со связыванием, или ее подобласти может быть введена в рандомизированную часть нуклеиновых кислот второй тестируемой смеси. Затем SЕLЕХ-процедуру повторяют со второй тестируемой смесью для выделения второго нуклеиново-кислотного лиганда, имеющего две последовательности, выбранные для связывания с мишенью, и обладающего более высокой эффективностью связывания или более высокой специфичностью связывания по сравнению с первым выделенным нуклеиново-кислотным лигандом. Последовательность второго нуклеиново-кислотного лиганда, ассоциированного со связыванием с мишенью, может быть затем введена в вариабельную часть нуклеиновых кислот третьей тестируемой смеси, которая после SELEX-циклов дает третий нуклеиново-кислотный лиганд. Эти процедуры можно повторять до тех пор, пока не будет получен лиганд с нужной эффективностью связывания или с нужной специфичностью связывания с молекулой-мишенью. Способ итерационной селекции и комбинации элементов нуклеиново-кислотной последовательности в данном описании именуется "прогулкой", где указанный термин подразумевает оптимизированное связывание с другими доступными областями поверхности или углубления макромолекулярной мишени, начиная с первой области связывания. Увеличение площади контакта при связывании между лигандом и мишенью и лигандом может способствовать повышению константы реакции связывания по сродству. Указанные процедуры "прогулки" могут быть, в частности, использованы для нуклеиново-кислотных антител, которые являются высокоспецифичными при связывании с конкретной целевой молекулой.
В одном из вариантов "прогулки" используют лиганд, не являющийся нуклеиновой кислотой и в настоящем описании обозначаемый термином "якорь", который связывается с целевой молекулой в качестве первой области связывания (см. фиг.9). Эта молекула-"якорь" может быть, в принципе, любой не являющейся нуклеиновой кислотой молекулой, которая связывается с молекулой-мишенью и которая может быть прямо или опосредованно ковалентно связана с нуклеиновой кислотой. Если целевой молекулой является фермент, то молекула-якорь может быть ингибитором или субстратом этого фермента. Указанный "якорь" может быть также антителом или фрагментом антитела, специфичным для данной мишени. Указанную молекулу-якорь ковалентно связывают с нуклеиновокислотным олигомером известной последовательности для получения молекулы, образующей мостик. Этот олигомер предпочтительно состоит приблизительно минимум из 3-10 оснований. Затем получают тестируемую смесь из нуклеиновых кислот-кандидатов, которая включает в себя рандомизированную часть и последовательность, комплементарную известной последовательности мостиковой молекулы. Мостиковая молекула образует комплекс с молекулой-мишенью. После этого SELEX используют для отбора нуклеиновых кислот, которые связываются с комплексом мостиковой молекулы и молекулы-мишени. Нуклеиново-кислотные лиганды, которые связываются с комплексом, выделяют. Описанные выше процедуры "прогулки" могут быть затем использованы для получения нуклеиново-кислотных лигандов, обладающих повышенной эффективностью связывания или повышенной специфичностью связывания с комплексом. Процедуры "прогулки" позволяют проводить селекции на связывание с комплексом или с самой мишенью. Этот способ может быть, в частности, использован для выделения нуклеиново-кислотных лигандов, которые связываются с конкретным сайтом внутри молекулы-мишени. Комплементарная последовательность в тестируемой смеси действует для обеспечения выделения нуклеиново-кислотных последовательностей, которые связываются с молекулой-мишенью в сайте связывания мостиковой молекулы или близ него. Если мостиковая молекула происходит от ингибитора молекулы-мишени, то этот метод позволит в результате получить нуклеиново-кислотный лиганд, который ингибирует функцию молекулы-мишени. В частности, этот способ может быть использован, например, для выделения нуклеиновых кислот, которые будут активировать или ингибировать функцию белка. Комбинация лиганда и мишени может иметь новую или усиленную функцию.
Нуклеиново-кислотные лиганды настоящего изобретения могут содержать множество лигандных компонентов. Как указывалось выше, нуклеиново-кислотные лиганды, полученные с помощью процедур "прогулки", могут рассматриваться как лиганды, имеющие более одного лигандного компонента. Настоящее изобретение также относится к нуклеиново-кислотным антителам, которые конструируют на основании результатов, полученных SELEX-способом, однако при этом они не являются идентичными нуклеиново-кислотному лиганду, идентифицированному способом SELEX. Например, нуклеиново-кислотное антитело может быть сконструировано таким образом, чтобы в нем множество идентичных лигандных структур составляло часть одной нуклеиновой кислоты. В другом варианте осуществления изобретения с помощью SELEX может быть идентифицировано более одного семейства нуклеиново-кислотных лигандов к данной мишени. В этом случае может быть сконструировано антитело из одной нуклеиновой кислоты, которое содержит множество различных лигандных структур. Могут быть также осуществлены SELEX-эксперименты, где фиксированные идентичные или различные лигандные структуры соединены рандомизированными нуклеотидными областями и/или областями с различным расстоянием между фиксированными лигандными структурами в целях идентификации наилучших нуклеиново-кислотных антител.
Указанные способы SELEX могут быть легко объединены с методами скрининга, отбора или анализами для оценки влияния связывания нуклеиново-кислотного лиганда на функцию молекулы-мишени. В частности, скрининг на ингибирование или активацию ферментной активности может быть объединен с методами SELEX.
В более специфических вариантах осуществления настоящего изобретения SELEX представляет собой быстрое средство для выделения и идентификации нуклеиново-кислотных лигандов, которые связываются с белками, включая белки, связывающиеся с нуклеиновыми кислотами, и белки, о которых не известно, является ли их связывание с нуклеиновыми кислотами частью их биологической функции. Среди прочих, белками, связывающимися с нуклеиновыми кислотами, являются полимеразы и обратные транскриптазы. Эти способы могут быть также легко применены к белкам, которые связываются с нуклеотидами, нуклеозидами, нуклеотидными кофакторами и структурно родственными молекулами.
В другом своем варианте настоящее изобретение относится к способу обнаружения наличия или отсутствия и/или измерения количества молекулы-мишени в образце; причем в указанном способе используется нуклеиново-кислотный лиганд, который может быть выделен способами, описанными выше. Обнаружение молекулы-мишени опосредуется ее связыванием с нуклеиново-кислотным лигандом, специфичным для данной молекулы-мишени. Нуклеиново-кислотный лиганд может быть меченым, например радиоактивно меченным, что позволяет осуществлять количественное или качественное детектирование. В частности, этот метод обнаружения может быть использован для обнаружения белков, о которых неизвестно, является ли их связывание с нуклеиновыми кислотами частью их биологической функции. Так, например, нуклеиново-кислотные лиганды настоящего изобретения могут быть использованы в диагностике аналогично стандартным методам диагностики, основанным на использовании обычного антитела. Одно из преимуществ использования нуклеиново-кислотных лигандов в указанном методе обнаружения и диагностики по сравнению с использованием стандартных антител заключается в том, что указанные нуклеиновые кислоты могут быть легко амплифицированы in vitro, например, путем РСR-амплификации или аналогичными методами. Другое преимущество заключается в том, что весь процесс SELEX проводят in vitro и его осуществление не требует иммунизации испытуемых животных. Кроме того, аффинность связывания нуклеиново-кислотных лигандов может быть адаптирована к требованиям пользователя.
Нуклеиново-кислотные лиганды для небольших молекул-мишеней могут быть использованы в качестве реагентов диагностического анализа, а также в терапевтических целях, например, в качестве секвестрантов, носителей в лекарственных препаратах и модификаторов функции гормонов. Продуктами селекции настоящего изобретения являются также каталитические нуклеиновые кислоты. Например, каталитические нуклеиновые кислоты могут быть выявлены путем отбора на связывание с аналогами переходного состояния.
В еще одном своем варианте настоящее изобретение относится к способу мофидикации функции молекулы-мишени с использованием нуклеиново-кислотных лигандов, которые могут быть выделены с помощью SELEX. Для этого нуклеиново-кислотные лиганды, которые связываются с молекулой-мишенью, скринируют для отбора тех из них, которые специфически модифицируют функцию молекулы-мишени, например для отбора игнибиторов или активаторов функции молекулы-мишени. Количество выбранного нуклеиново-кислотного лиганда, который является эффективным для модификации функции мишени, объединяли с молекулой-мишенью в целях достижения нужной функциональной модификации. Этот способ применим, в частности, к молекулам-мишеням, которые являются белками. Особенно ценным применением этого метода является ингибирование функции белка, например ингибирование связывания рецептора с эффектором или ингибирование ферментного катализа. В этом случае для достижения нужного ингибирования количество выбранной нуклеиново-кислотной молекулы, которая является эффективной для ингибирования целевого белка, объединяют с целевым белком.
На фиг.1 представлена диаграмма рибонуклеотидной последовательности части 43-генной матричной РНК, которая кодирует ДНК-полимеразу бактериофага Т4. На фиг.1 показана последовательность в области, которая, как известно, связывается с gp43. Прописные буквы, набранные жирным шрифтом, указывают на степень информации, необходимой для связывания с gp43. Восемь пар оснований петли были заменены рандомизированной последовательностью для получения популяции кандидатов для SELEX.
На фиг.2 представлена схематическая диаграмма процесса SELEX, иллюстрирующая отбор вариантов последовательности петли для РНК, которые связываются с ДНК-полимеразой (gp43). ДНК-матрицу для получения смеси для отбора РНК получали, как показано в стадии (а), путем лигирования олигомеров 3, 4 и 5, последовательности которых приводятся в табл.1. Правильное лигирование в стадии (а) обеспечивалось с помощью гибридизации с олигомерами 1 и 2, которые имеют комплементарную последовательность (см. табл. 1), сшивающую олигомеры 3 и 4 и 4 и 5 соответственно. Полученную в результате матрицу длиной в 110 оснований подвергали гель-очистке, отжигу с олигонуклеотидом 1 и использовали в vitro реакциях транскрипции (Мiligan и др. (1987), Nucl. Acid. Res. 15:8783-8798) для получения исходной смеси РНК, содержащей рандомизированные последовательности петли из 8 оснований (стадия (b)). Полученные в результате транскрипты подвергали гель-очистке, а затем селекции на нитроцеллюлозных фильтрах на связывание с gp43 (стадия с), как описано в Примере 1. Отобранные РНК амплифицировали способом, включающим в себя три стадии: (d) кДНК-копии выбранных РНК получали путем синтеза с помощью обратной транскриптазы, используя в качестве праймера олигонуклеотид 5 (см. табл. 1); (е) кДНК амплифицировали с использованием ДНК-полимеразы Tag для удлинения олигомера 1 (табл. 1), который несет последовательности главного промотора Т7, и олигомера 5 (табл.1), как описано у Junis и др. (1988) Proc. Natl. Acad. Sci. USA 85: 9436; и (f) двухцепочечные ДНК-продукты амплификации транскрибировали in vitro. Полученные в результате отобранные амплифицированные РНК были использованы в следующем цикле селекции.
На фиг.3 представлен ряд авторадиограмм реакции секвенирования подвергнутой электрофорезу партии in vitro-транскриптов, полученных в результате SELEX на связывание вариантов РНК-петли с gp43. На фиг.3 показано изменение компонентов последовательности петли в зависимости от числа циклов селекции для 2, 3 и 4 циклов в условиях селекции эксперимента В, в котором концентрация gp43 составляла 3 • 10-8 М, а концентрация РНК составляла около 3 • 10-5 M во всех циклах селекции. Секвенирование проводили в соответствии с описанием Гаусса (Gauss) и др. в Моl. Gen. Genet. 206:24-34 (1987).
На фиг.4 представлен ряд авторадиограмм партии РНК-последовательностей, которые выбирали в результате четырех циклов SELEX-амплификации на связывание вариантов петли РНК с gp43 с использованием различных условий связывания. В эксперименте А концентрация gp43 составляла 3•10-8, а концентрация РНК составляла около 3•10-7 М. В эксперименте В концентрация gp43 составляла 3•10-8 М, а концентрация РНК составляла около 3•10-5 М. В эксперименте С концентрация gp43 составляла 3•10-7 М, а концентрация РНК составляла 3•10-5 М.
На фиг. 5 представлен ряд авторадиограмм трех секвенирующих гелей для вариантов петли, отобранных на связывание с gp43 в условиях селекции эксперимента В (см. Пример 1). Гель последовательности слева представляет серийное секверирование РНК, отобранных после четвертого цикла селекции/амплификации. Гели в середине и справа представляют дц-ДНК-секвенирующие гели двух кланальных изолятов, происходящих от РНК партии. Эта партия отобранных РНК состоит из двух крупных вариантов, один из которых является последовательностью дикого типа (средний гель), а другой новой последовательностью (правый гель).
На фиг.6 представлен график процента РНК, связанных с gp43, в зависимости от концентрации gp43 для различных вариантов последовательности петли отобранных РНК, и для РНК с рандомизированной последовательностью петли. Связывание последовательности петли дикого типа AAUAACUC показано на фиг.6 незаштрихованными кружочками, сплошной линией; крупный вариант последовательности петли АGСААССU отмечен крестиками "х" и точечным пунктиром; меньший вариант последовательности петли AAUAACUU показан незаштрихованными квадратами и сплошной линией; меньший вариант последовательности петли ААUGАСUС показан сплошными кружочками и черточным пунктиром; меньший вариант последовательности петли АGСGАССU показан точечным пунктиром и крестиками "+"; а связывание рандомизированной смеси (NNNNNNNN) последовательностей петли показано незаштрихованными кружочками и пунктирной линией.
На фиг.7 проиллюстрированы результаты в целом, полученные после четырех раундов SELEX, проведенного для отбора новой связывающейся с gp43 РНК из популяции кандидатов, рандомизированной в петле из 8 пар оснований. SELEX не дал видимого "консенсуса" (обобщающего типичного элемента структуры - прим. пер.), ожидаемого от последовательностей серии показанных на фиг.4, а вместо этого - один вариант дикого типа и один крупный вариант примерно в одинаковых соотношениях, и три одиночных мутанта. Частота каждого вида из 20 клонированных изолятов показана вместе с приблизительными константами сродства (Kb) на основании анализа связывания с фильтрами, показанного на фиг.6.
На фиг. 8 представлен ряд диаграмм, иллюстрирующих синтез лигандов-кандидатов нуклеиновых кислот с использованием терминальной трансферазы (TDT) и ДНК-полимеразы (DNA pol). 5'-праймер или начальная последовательность лиганда снабжают концевым участком рандомизированной последовательности путем инкубирования с термальной трансферазой в присутствии четырех дезоксинуклеотидтрифосфатов (dNTPS). Гомополимер, используемый для наращивания рандомизированного сегмента с помощью того же фермента и в присутствии дезоксинуклеотидтрифосфата (например, dСТР), дает сайт отжига для poly-G-удлиненного 3'-праймера. После отжига двухцепочечную молекулу укомплектовывали путем воздействия ДНК-полимеразы. Затем смесь, если это необходимо, может быть амплифицирована, посредством полимеразно-цепной реакции.
На фиг.9 представлена диаграмма, иллюстрирующая процесс с использованием SELEX для отбора большого нуклеиново-кислотного лиганда, имеющего две пространственно разделенные области взаимодействия с целевым белком. Этот способ называется "прогулкой", поскольку он включает в себя две стадии, где вторая стадия является дополнением первой. В верхней части фиг.9 показана мишень ("нужный белок") со связанным нуклеиново-кислотным лигандом, выбранным в первом раунде SELEX "выделенный первичный лиганд", связанный с белком в первом сайте связывания. Реакция, катализируемая терминальной трансферазой, позволяет наращивать длину выделенного первичного лиганда и генерировать новую серию рандомизированных кандидатов-последовательностей, имеющих консервативную область, содержащих первичный лиганд. В нижней части фиг.9 проиллюстрирован результат второго раунда SELEX, проводимого на основе улучшенного связывания, полученного в результате вторичного взаимодействия лиганда во втором сайте связывания белка. Термины "первичный" и "вторичный" являются просто рабочими терминами и не означают, что один имеет большую аффинность, чем другой.
На фиг.10 и 11 представлены две диаграммы, иллюстрирующие процесс селекции с использованием SELEX в две стадии. На фиг.10 показано использование SELEX для выбора лигандов, которые связываются со вторичными сайтами связывания на мишени, образующей комплекс с мостиковым олигонуклеотидом, соединенным со специфическим связующим, например ингибитором целевого белка. Мостиковый олигонуклеотид действует в качестве гида для благоприятной селекции лигандов, которые связываются с доступными вторичными сайтами связывания. На фиг. 11 показано использование второго SELEX для выявления лиганда, которые связываются со вторичными сайтами, первоначально выбранными, и первичными доменами мишени. Выделенные таким образом нуклеиновые кислоты будут связываться очень тесно и сами по себе могут действовать в качестве ингибиторов или субстратов целевого белка.
На фиг.12 показана последовательность и расположение олигомеров, используемых для конструирования смеси кандидатов для Примера 2. В верхней строке показаны последовательности олигомеров 1b и 2b слева направо, соответственно (см. табл.2). Во второй строке слева направо показаны последовательности олигомеров 3b, 4b и 5b (табл.2). Правильное лигирование олигомеров гарантируется гибридизацией с олигомерами 1b и 2b, последовательности которых являются комплементарными. Полученную в результате лигированную матрицу подвергли гель-очистке, отжигали с олигомером 1b и использовали в реакции in vitro-транскрипции (Milligan и др. (1987)) для получения смеси РНК-кандидатов, показанной в последней строке фиг.12 и отмеченной "in vitro-транскрипт". Как показано на фиг.12, смесь кандидатов содержит рандомизированный сегмент из 32 нуклеотидов.
На фиг.13 показана гипотетическая РНК-последовательность, содержащая ряд известных вторичных структур РНК, а именно: А - шпилечные петли, В - "балджи" (петли с выступом); С - асимметричные балджи; D - псевдоузлы.
На фиг. 14 показан анализ на лигандное сродство к ВИЧ-RТ, проводимый посредством связывания на нитроцеллюлозных фильтрах. На фиг.14 показано, что процент потребления РНК, связывающейся с нитроцеллюлозным фильтром, зависит от концентрации ВИЧ-RТ.
На фиг.15 показаны дополнительные анализы на сродство к ВИЧ-RT посредством связывания на нитроцеллюлозных фильтрах.
Фиг.16 иллюстрирует определение границы информации для ВИЧ-1-RT-лигандов 1,1 и 1,3а а) определение 3'-границы. РНК метили по 5'-концу, подвергали частичному щелочному гидролизу и селекции на нитроцеллюлозных фильтрах, выделяли на денатурирующем 8% полиакриламидном геле и подвергали авторадиографии. Приблизительно 90 пикомолей меченой РНК и 80 пикомолей ВИЧ-1-RT смешивали в 0,5, 2,5 и 5 мл буфера и инкубировали в течение 5 минут при 37oС, а затем промывали через нитроцеллюлозный фильтр. Элюированные РНК показаны при конечных концентрациях ВИЧ-1-RТ, используемых в каждом эксперименте. Показаны также продукты частичного перевара РНКазой Т1, который позволяет идентифицировать информационную границу на смежной последовательности, как показано стрелкой; в) определение 5'-границы, 5'-границу определяли в а) в условиях, аналогичных перечисленным выше.
Фиг. 17 иллюстрирует ингибирование ВИЧ-1-RT с помощью РНК-лиганда 1.1. Серии трехкратных разведений смеси 32 н-РНК-кандидатов и РНК-лиганда 1.1 в пределах конечной реакционной концентрации от 10 мкМ до 4,6 нМ и предварительно смешанной с ВИЧ-RT, инкубировали в течение 5 минут при 37oC в 6 мкл 200 мМ КОАс, 50 мМ Трис-HCl, рН 7,7, 10 мМ дитиотреитола, 6 мМ Мо (ОАс)2 и 0,4 мМ NTPS. В отдельной пробирке РНК-матрицу транскрибированную из PCR-продукта Т7-1, полученного от фирмы U.S. Biochemical Corp., с использованием олигонуклеотидов 7 и 9 и меченый олигонуклеотид 9) смешивали и нагревали при 95oС в течение 1 минуты, а затем охлаждали на льду в течение 15 минут в 10 мМ Трис-HCl, рН 7, 0,1 мМ ЭДТК. Для начала реакции 4 мкл этой матрицы добавляли к каждым 6 мкл смеси фермента-ингибитора, после чего реакцию инкубировали 5 минут при 37oС, а затем прекращали. Конечная концентрация ВИЧ-1-RT составляла 16 нМ, концентрация РНК-матрицы составляла 13 нМ, а концентрация меченого праймера составляла 150 нМ во всех реакциях. На фиг.17 также показаны продукты удлинения для каждой реакции.
На фиг.18 проиллюстрированы сравнения ингибирования ВИЧ-1-RТ с лигандом 1.1 для оценки влияния на MMLV RT и AMV-RТ. Эксперименты осуществляли как показано на фиг.17, за исключением того, что 5-кратные разведения ингибитора получали с указанными конечными концентрациями. Концентрации каждого RT нормализовали к концентрации ВИЧ-RТ посредством разведении и сравнения интенсивностей полос гелей с использованием кумасси-синего и серебряного красителей, оценки концентрации белка Biorad и оценки активности.
На фиг. 19 показаны последовательности "консенсусов" отобранных шпилек, представляющих собой раствор лиганда для оболочечного белка R-17. Изображения нуклеотида в каждом положении показаны в таблицах. Колонка, озаглавленная "балдж", представляет число клонов, имеющих внеспиральный нуклеотид на одной или на обеих сторонах стебля между соответствующими парами оснований стебля. Колонка, озаглавленная "конец", представляет число клонов, у которых шпилька заканчивается в предыдущей паре оснований.
На фиг. 20 представлена кривая связывания разбухшей 30н-РНК для брадикинина. Анализ проводили с использованием вращающейся колонки; 10 мМ КОАс, 10 мМ DEM, рН 7,5; концентрация РНК составляла 1,5•10-8 М.
На фиг.21 показаны матрицы для использования при получении смесей кандидатов, обогащенной некоторыми структурными элементами. Матрица А была сконструирована для обогащения смеси кандидатов шпилечными петлями. Матрица В была сконструирована для обогащения смеси кандидатов псевдоузлами.
На фиг. 22 представлена схематическая диаграмма расположения стеблей и петель для Мотивов I и II раствора лиганда ВИЧ-rev. Пунктирные линии в стеблях 1 и 2 между петлями 1 и 3 указывают на потенциальные пары оснований.
На фиг. 23 показаны складчатые вторичные структуры субдоменов лигандов rev для изолятов 6а, 1а и 8 - Мотивы I, II и III соответственно. Для сравнения также показана предсказанная укладка RRE-РНК дикого типа.
На фиг. 24 представлен график зависимости входящего количества РНК (%), связанного с нитроцеллюлозным фильтром от различных концентраций ВИЧ-rev-белка. На фиг. 24 представлены также кривые исходной популяции 32N и выделенной популяции после 10 раундов (Р), а также RRE-последовательность дикого типа, транскрибированная с матрицы, состоящая из олигонуклеотидов 8 и 9 (W).
На фиг. 25 проиллюстрировано сравнение лигандов Мотива I (а) rev. Параметры аналогичны параметрам, указанным на фиг.24. Представлена также кривая связывания конструкции С "консенсуса".
На фиг. 26 проиллюстрировано сравнение лигандов Мотива I (b) rev. Параметры аналогичны указанным на фиг.24.
На фиг. 27 проиллюстрировано сравнение лигандов с Мотивом II rev. Параметры аналогичны параметрам фиг.24.
На фиг.28 проиллюстрировано сравнение лигандов с Мотивом III rev. Параметры аналогичны параметрам фиг.24.
На фиг.29 показан нуклеиново-кислотный лигандный раствор к ВИЧ-rev, относящийся к "консенсусу" Мотива I.
На фиг.30 показан нуклеиново-кислотный лигандный раствор к ВИЧ-rev, относящийся к "консенсусу" Мотива II.
На фиг. 31 схематически представлен псевдоузел. Этот псевдоузел состоит из двух стеблей и трех петлей, обозначенных S1 и S2 (стебли) и 1, 2 и 3 (петли).
В настоящем описании используются термины, которые имеют следующие значения.
Термин "нуклеиновая кислота" означает одноцепочечные или двухцепочечные ДНК, РНК или их любые химические модификации при условии, однако, что эти модификации не препятствуют амплификации выбранных нуклеиновых кислот. Примерами таких модификаций (однако эти примеры не ограничивают их возможные варианты) являются модификации в цитозинэксзоциклических аминах, замещение 5-бромоурацила, модификации остова, метилирование, комбинации ошибочного спаривания и т.п.
Термин "лиганд" означает нуклеиновую кислоту, которая связывается с другой молекулой (мишенью). В популяции нуклеиновых кислот-кандидатов, лигандом является та нуклеиновая кислота, которая связывается с большим сродством, чем нуклеиновые кислоты всей популяции. При этом в смеси кандидатов могут присутствовать более чем один лиганд для данной мишени. Эти лиганды могут отличаться друг от друга по их сродству связывания с молекулой-мишенью.
Термин "смесь кандидатов" означает смесь нуклеиновых кислот с различными последовательностями, из которой выбирают нужный лиганд. Нуклеиновые кислоты в указанной смеси кандидатов могут быть природными нуклеиновыми кислотами или их фрагментами, химически синтезированными нуклеиновыми кислотами, ферментативно синтезированными нуклеиновыми кислотами или нуклеиновыми кислотами, полученными путем комбинирования вышеуказанной техники.
Термин "молекула-мишень" означает любое нужное соединение, для которого желательно получить лиганд. Молекулой-мишенью могут быть белок, пептид, углевод, полисахарид, гормон, рецептор, антиген, антитело, вирус, субстрат, метаболит, аналог переходного состояния, кофактор, ингибитор, лекарственное средство, краситель, питательное средство, фактор роста и т.п., причем эти примеры можно продолжать до бесконечности.
Термин "распределение" (разделение) означает любой процесс, при котором лиганды, связанные с молекулами-мишенями и обозначаемые парами "лиганд-мишень", могут быть отделены от нуклеиновых кислот, не связанных с молекулами-мишенями. Распределение может быть осуществлено любыми стандартными способами. Пары "нуклеиновая кислота-белок" могут быть связаны с нитроцеллюлозными фильтрами, тогда как несвязанные нуклеиновые кислоты но связываются с нитроцеллюлозными фильтрами. Для распределения могут быть использованы колонки, которые специфически удерживают пары "лиганд-мишень" (или специфически удерживают связанный лиганд, образующий комплекс с присоединенной мишенью). Может быть также использована жидкость-жидкостная экстракция, ингибирование фильтрующего геля; и центрифугирование в градиенте плотности. Выбор метода распределения зависит от свойств мишени и пар "лиганд-мишень", и может быть осуществлен в соответствии с традиционными процедурами, хорошо известными специалистам.
Термин "амплификация" означает любой процесс или комбинацию стадий этого процесса, которые позволяют увеличить количество или число копий молекулы или класса молекул. Амплификацию РНК-молекул в описанных примерах осуществляли с помощью последовательного проведения трех реакций: получение кДНК-копий выбранных РНК; использование полимеразно-цепной реакции для увеличения числа копий для каждой кДНК; и транскрибирование кДНК-копий для получения РНК-молекул, имеющих последовательности, аналогичные последовательностям выбранных РНК. В данном случае может быть использована любая реакция или сочетание реакций, хорошо известных специалистам, включая прямую ДНК-репликацию, прямую РНК-амплификацию и т.п. Метод амплификации должен приводить к пропорциям амплифицированной смеси, которые являются, в основном, характерными соотношениями различных последовательностей исходной смеси.
Термин "специфическое связывание" обычно определяется в каждом конкретном случае. В соответствии с настоящим изобретением взаимодействие между данным лигандом и данной мишенью при связывании протекает с более высоким сродством, чем при связывании мишени и смеси лигандов-кандидатов. Для того чтобы сравнить аффинности связывания, условия обеих реакций связывания должны быть одинаковыми и должны быть сравнимыми с условиями, необходимыми для использования. Для наиболее точного сравнения измерения должны быть проведены так, чтобы они отражали взаимодействие между лигандом как целое и мишенью как целое. Нуклеиново-кислотные лиганды настоящего изобретения могут быть выбраны с такой специфичностью, какая необходима для данного случая, либо путем установления условий селекции, необходимых для получения требуемой специфичности в течение процесса SELEX, либо путем наращивания и модификации лиганда посредством "прогулки", или других модификаций с использованием взаимодействий SELEX.
Термин "рандомизированный" (случайно взятый) используется для описания сегмента нуклеиновой кислоты, имеющей, в принципе, любую возможную последовательность на протяжении данной длины. Рандомизированные последовательности могут быть любой длины, если необходимо, приблизительно в пределах от 8 до более чем 100 нуклеотидов. Химические или ферментные реакции, с помощью которых получены неупорядоченные сегменты последовательности, могут не дать математически произвольные последовательности, вследствие неизвестных среднестатических смещений или преимущественного присутствия нуклеотидов, которое может иметь место. Термин "рандомизированный" используется вместо "неупорядоченный" для того, чтобы указать на возможность вышеупомянутого отклонения от неидеальности. В известных в настоящее время способах, например, при последовательном химическом синтезе, не наблюдалось больших отклонений. Для коротких сегментов, например, из 20 нуклеотидов и менее любая незначительная статистическая ошибка будет иметь последствия, которыми можно пренебречь. Чем длиннее последовательность в отдельном синтезе, тем выше влияние любых статистических отклонений.
Статистическое отклонение может быть преднамеренно введено в рандомизированную последовательность, например, путем изменения молярных соотношений нуклеозид- или дезоксинуклеозид-трифосфатов предшественника в реакции синтеза. Преднамеренное введение статистического отклонения может быть желательным для аппроксимации соотношений отдельных оснований в данном организме или для воздействия на вторичную структуру.
Термин "селекция" или "отбор" относится к математическому анализу и компьютерному моделированию, используемому для иллюстрации высокой способности SELEX идентифицировать нуклеиново-кислотные лиганды и предсказывать какие изменения в процессе SELEX имеют наибольшее воздействие на оптимизацию процесса. Аббревиатура SELEXION представляет собой акроним полного названия метода: Systematic Evolution of Ligands by Exponentival enrichment with Integrated Optimization by Nonlineaz analysis. Термин "нуклеиново-кислотные антитела" используется для обозначения класса нуклеиново-кислотных лигандов, которые включают в себя дискретные нуклеиново-кислотные структуры или мотивы структур, которые избирательно связываются с молекулами-мишенями. Нуклеиново-кислотные антитела могут состоять из двухцепочечных или одноцепочечных РНК или ДНК. Нуклеиново-кислотные антитела синтезируют, а в предпочтительном варианте осуществления настоящего изобретения конструируют на основе лигандного раствора или растворов, полученных для данной мишени с помощью SELEX-процесса. В большинстве случаев нуклеиново-кислотные антитела настоящего изобретения не являются природными соединениями, однако в некоторых случаях они могут обладать значительным сходством с последовательностью натуральной нуклеиновой кислоты.
Нуклеиново-кислотные антитела настоящего изобретения представляют все нуклеиновые кислоты, которые специфически связываются с мишенью, за исключением тех случаев, когда мишенью является полинуклеотид, который связывается с нуклеиновой кислотой по механизму, преимущественно зависящему от уотсон-криковского спаривания оснований или трехспиральных агентов см., Riordan М. и др. (1991), Nature 350:442-443); при условии, однако, что если нуклеиново-кислотное антитело является двухцепочечной ДНК, то мишень не является природным белком, биологическая функция которого зависит от специфического связывания двухцепочечной ДНК.
Термин "РНК-мотивы" обычно используют для описания вторичных или третичных структур РНК-молекул. Первичная последовательность РНК представляет собой специфическую цепь нуклеотидов (А, G, С или U) в одном измерении. На первый взгляд, первичная последовательность не дает такой информации, как трехмерная конфигурация РНК, хотя именно первичная последовательность диктует трехмерную конфигурацию. В некоторых случаях лигандный раствор, полученный после осуществления SELEX для данной мишени, может быть лучше всего представлен в виде первичной последовательности. Хотя конформационная информация, относящаяся к такому лигандному раствору, не всегда может быть выявлена на основании результатов, полученных с помощью SELEX, однако представление лигандного раствора в виде первичной последовательности не должно быть интерпретировано как отрицание наличия встроенной третичной структуры.
Вторичная структура РНК-мотива может быть представлена как взаимодействие между специфическими нуклеотидами в двух измерениях. Наиболее легко распознаваемые мотивы вторичных структур состоят из уотсон-криковских пар оснований А: U и С:G. Как было установлено, не-уотсон-криковские пары оснований часто имеют более низкую стабильность и представляют пары G:U, A:C, G: А и U:U. Когда-то было показано, что в РНК-молекулах пара оснований Х:Y условно представляет собой последовательность, где Х является 5'-->Y, при этом также допустимой является пара оснований Y:Х. На фиг.13 показана серия вторичных структур, соединенных одноцепочечными областями; при этом стандартная номенклатура для вторичных структур включает в себя петли шпильки, асимметричные выступающие петли шпильки; симметричные петли шпильки; и псевдоузлы.
Если нуклеотиды, которые достаточно удалены друг от друга и, как кажется, не взаимодействуют посредством уотсон-криковских и не-уотсон-криковских пар, на самом деле являются взаимодействующими, то эти взаимодействия (которые изображаются в двух измерениях) также являются частью вторичной структуры.
Трехмерная структура РНК-мотива является просто пространственным описанием атомов РНК-мотива. Двухцепочечная РНК с полностью спаренными основаниями посредством уотсон-криковского спаривания имеет упорядоченную структуру в трех измерениях, хотя точные положения всех атомов спирального остова могут зависеть от точной последовательности оснований РНК. Вторичным структурам РНК-мотивов посвящено большое количество литературы, где указывается, что вторичные структуры, содержащие уотсон-криковские пары оснований, часто образуют двухцепочечные спирали А-формы.
От спиралей А-форм можно перейти к другим мотивам трехмерного измерения. Не-уотсон-криковские пары оснований, шпилечные петли, "балджы" (выступы) и псевдоузлы являются структурами, сформированными внутри и на спиралях. Более полно, конструирование этих дополнительных мотивов описано в тексте описания.
Реальная структура РНК включает в себя все атомы нуклеотида молекулы в трех измерениях. Полностью разрешенная структура должна включать в себя также связанную воду и неорганические атомы, хотя такое разрешение редко достигается исследователем. РНК-структуры, разрешенные в трех измерениях, будут включать в себя все элементы вторичной структуры, представленные как трехмерные структуры, и фиксированные положения для атомов нуклеотидов, не ограниченных элементами вторичной структуры; причем, благодаря "стэкинг"-взаимодействию оснований и другим силам, экстенсивные одноцепочечные домены могут иметь фиксированные структуры.
Первичные последовательности РНК ограничивают возможные трехмерные структуры, как и фиксированные вторичные структуры. Трехмерные структуры РНК ограничиваются специфическими взаимодействиями между атомами в двух измерениях и, кроме того, ограничиваются энергетическими минимизациями, то есть способностью молекулы к повороту всех свободно вращаемых связей так, чтобы полученная в результате молекула была более стабильной, чем другие ее конформеры, имеющие аналогичную первичную и вторичную последовательность и структуру.
И что наиболее важно, РНК-молекулы имеют трехмерные структуры, которые состоят из коллекции РНК-мотивов, включающей любое число мотивов, показанных на фиг.13.
Таким образом, РНК-мотивы представляют собой все возможные пути и средства, позволяющие в общих чертах описать наиболее стабильные группы конформаций, которые могут образовывать нуклеиновые кислоты. Для данной мишени лигандный раствор и нуклеиново-кислотное антитело могут быть одним из РНК мотивов, описанных в настоящей заявке, или определенной комбинацией нескольких РНК-мотивов.
Термин "лигандные растворы" могут быть определены как трехмерная структура, действующая сообща, либо как семейство, которое определяет консервативные компоненты, идентифицированные посредством SELEX. Например, лиганды, идентифицированные для конкретной мишени, могут содержать общую первичную последовательность (NNNCGNAANUCGN'N'N), которая может быть представлена как шпилька в двух измерениях:
ААN
NU
GС
CG
NN'
NN'
NN'
Следовательно, эта трехмерная структура должна быть нечувствительной к точной последовательности трех из пяти пар оснований и двух из пяти нуклеотидов петли и во всех или в большинстве версий последовательности/структуры должна быть подходящим лигандом для дальнейшего использования. Таким образом, лигандные растворы представляют собой потенциально большую коллекцию подходящих последовательностей/структур, каждая из которых индетифицируется описанием семейств, которое охватывает все имеющиеся растворы последовательностей/структур. При этом следует отметить, что лигандные растворы не обязательно включают в себя только члены с точным числовым соответствием между различными компонентами РНК-мотива. Некоторые лиганды могут иметь петли, например, из пяти нуклеотидов, тогда как другие лиганды для той же мишени могут содержать меньше или больше нуклеотидов в эквивалентной петле и при этом все же входить в описание лигандного раствора.
Хотя лигандный раствор, полученный с помощью SELEX, может включать в себя относительно большое число потенциальных членов, однако эти лигандные растворы являются целеспецифическими и в значительной мере каждый член семейства лигандного раствора может быть использован в качестве нуклеиново-кислотного антитела к данной мишени. Выбор специфического члена из семейства лигандных растворов, используемого в качестве нуклеиново-кислотного антитела, может быть осуществлен в соответствии с настоящим описанием, причем на этот выбор могут влиять некоторые практические соображения, очевидные для каждого специалиста.
Способ настоящего изобретения был разработан в связи с исследованиями регулирования трансляции при инфицировании бактериофагом Т4. Саморегулирование синтеза некоторых вирусных белков, таких как ДНК-полимераза бактериофага Т4 (gp43), включает в себя связывание белка со своей собственной информационной РНК, блокирующее ее трансляцию. Способ SELEX был использован для выявления последовательности и структуры, необходимых для сайта связывания РНК с gp43. SELEX позволил осуществить быструю выборку предпочтительных последовательностей связывания из популяции произвольных нуклеиново-кислотных последовательностей. Путем выделения и идентификации нуклеиново-кислотных последовательностей, связывающихся с белками, о которых известно, что они связываются с РНК, было продемонстрировано, что способ настоящего изобретения является, в основном, применимым для отбора нуклеиновых кислот, способных связываться с любым данным белком. Этот способ может быть применим к отбору нуклеиновых кислот, которые связываются с белками, связывание которых с нуклеиновой кислотой не является частью их природной активности или биологической функции (или с белками, о связывании которых с нуклеиновыми кислотами ничего не известно). Способ SELEX не требует знания структуры или последовательности сайта связывания, а также знания структуры или последовательности белка-мишени. Этот способ не зависит от очищенного белка-мишени для выборки. В основном, применение SELEX будет давать обогащение лигандами для наиболее распространенной мишени. В случае смеси лигандов может быть использована техника выделения лиганда для данной мишени. Например, другой лиганд (напр., субстрат, ингибитор, антитело) для нужной мишени может быть использован для конкуренции на связывание с мишенью так, чтобы нужный нуклеиново-кислотный лиганд мог быть отделен от лигандов для других мишеней.
В предпочтительном осуществлении настоящего изобретения лиганды, полученные с помощью SELEX, состоят из одноцепочечных РНК-последовательностей. Решающий фактор настоящего изобретения заключается в том, что его авторами были сделаны заключения относительно РНК, которые противоречат общепринятым положениям в данной области, и эти выводы были ими использованы для разработки способа SELEX в целях получения нуклеиново-кислотных антител, выделенных из лигандных растворов.
Впервые РНК была охарактеризована как переносчик информации между ДНК-последовательностями, которые представляют собой гоны, и белковыми последовательностями, которые были обнаружены в ферментах и других белках. С того момента, когда Уотсон и Крик описали структуру ДНК и указали на связь между последовательностью ДНК и последовательностью белка, центральное место в биохимических исследованиях стали занимать пути и средства, с помощью которых синтезируются белки. В итоге была идентифицирована матричная РНК (мРНК) как химический интермедиат между генами и белками. Большинство видов РНК, присутствующих в организме, являются мРНК, и на основании этого РНК продолжали считать, в основном, информационной молекулой. Свою роль в качестве информационной молекулы РНК осуществляет, главным образом, через праймерную последовательность нуклеотидов; то есть информация в нуклеиновых кислотах может быть представлена в одном измерении.
В процессе исследований экспрессии генов, проводимых биохимиками, было обнаружено, что некоторые РНК-молекулы, присутствующие в клетке, не имеют информационной роли. Были обнаружены рибосомы как частицы, где по матричной РНК транслируются белки, и, как было установлено, рибосомы содержат важный вид РНК (рибосомную РНК или рРНК). В течение многих лет указанную рРНК считали структурным элементом, то есть чем-то вроде каркаса, на котором "висят" белковые компоненты рибосомы так, чтобы эти компоненты позволяли рибосоме осуществлять синтез белка. Затем была выдвинута гипотеза, которая впоследствии подтвердилась, что существует еще один большой класс РНК, названный транспортными РНК, тРНК, тРНК являются химически бифункциональными адаптерами, которые узнают кодоны в мРНК и переносят аминокислоты, из которых образуется белок. И самое важное то, что хотя структура тРНК была определена с помощью рентгеновского анализа в 1974 г., однако еще одно десятилетие РНК продолжали считать, главным образом, "цепочками" в одном измерении. рРНК занимают необычное положение в исследовательских экспериментах. В течение длительного периода времени никто не мог понять, что кроется за глубоким сходством между рРНК от различных видов, а также истинные химические функции РНК-молекул. Несколько исследователей предположили, что РНК может иногда играть не информационную, а каталитическую роль, но эти предположения относительно настоящих функций РНК не были подтверждены.
В работе Tom Cech, относящейся к рибозимам (новым классам РНК-молекул), был высказан более широкий взгляд на функциональные возможности РНК. Интроны группы 1 обладают способностью к автокаталитическому сплайсингу, и поэтому в пределах РНК имеет место, по крайней мере, несколько ограниченный катализ. В указанных пределах катализа находится активность РНК-компонента РНКазы Р, которую обнаружили Альтман и Пейс. Кеч и Альтман получили Нобелевскую премию по химии за свою работу, которая фундаментально изменила имеющиеся представления о молекулах РНК как о молекулах, играющих лишь информационную роль. Благодаря этой работе, в настоящее время уже известно, что рРНК представляют собой как бы каталитический центр рибосомы, а не являются чем-то вроде каркаса, как думали прежде.
Основной предпосылкой настоящего изобретения является тот факт, что и в настоящее время большинство исследователей недооценивает роль РНК-молекул при связывании и других ее функций. Хотя рибозимы вызывают у исследователей повышенный интерес, направленный, в конечном счете, на функции РНК, однако в настоящей заявке рассматриваются возможные формы РНК-молекул (а также по возможности ДНК) в целях их наиболее благоприятного использования в SELEX для обнаружения тех РНК, которые обладают практически любой нужной функцией для связывания. Кроме того, в настоящей заявке утверждается, что каталитические функции, возможные для РНК, выходят далеко за пределы имеющихся в настоящее время представлений, хотя их диапазон не обязательно так же широк, как у белков.
Трехмерные формы некоторых РНК были непосредственно установлены рентгенографией или ЯМР-спектроскопией. Имеющиеся серии данных являются очень разбросанными. Были установлены структуры четырех тРНК, а также трех более мелких РНК-молекул: две небольших шпильки и небольшой псевдоузел. Различные тРНК, даже родственные, имеют элементы уникальной структуры, например антикодоновые основания тРНК элонгатора обнаружены в направлении растворителя, тогда как антикодоновые основания тРНК-инициатора, как отмечалось, были более удалены от растворителя. Некоторые из этих отличий могут быть результатом действия сил уплотнения пространственной решетки кристаллов, но некоторые также, несомненно, являются результатом минимизации идиосинкразической энергии различными одноцепочечными последовательностями в гомологичных и трехмерных структурах.
Разнообразие последовательностей, конечно, достаточно велико. Если одноцепочечная петля РНК-шпильки содержит 8 нуклеотидов, то насыщенное последовательностями "пространство" содержит 65536 различных последовательностей. Не претендуя на какую-либо конкретную теорию, авторы настоящего изобретения считают, что каждый член указанного множества будет иметь, благодаря минимизации энергии, наиболее стабильную структуру, а объем этих структур будет давать тонко различимые химические поверхности для растворителя или для потенциального взаимодействия молекул-мишеней, таких как белки. Таким образом, когда все 65536 последовательностей в конкретном структурном мотиве были проанализированы против ДНК-полимеразы бактериофага Т4, то оказалось, что две последовательности из этого множества связываются лучше, чем все остальные. На основании этого можно предположить, что структурные варианты этих двух последовательностей являются специфическими для данной мишени, а остальные 65534 последовательности не являются достаточно подходящими для связывания с данной мишенью. Однако почти с полной уверенностью можно сказать, что среди указанных 65536 последовательностей имеются другие отдельные члены или множество членов, которые были бы наиболее подходящими для связывания с другими мишенями.
Ключевая идея настоящего описания РНК-структур заключается в том, что каждая последовательность будет обретать свою наиболее стабильную структуру, хотя часто выделенные РНК дают основание предположить наличие в них беспорядочных спиральных, или висящих, неструктурированных элементов. Часто обнаруживалось, что гомополимеры РНК, неспособные образовывать уотсон-криковские пары, имеют нерандомизированную структуру, приписываемую энергии "стэкинг"-взаимодействий, приобретаемой посредством фиксации положений смежных оснований, расположенных друг над другом. Несомненно, что последовательности, включающие все четыре нуклеотида, могут иметь локальные области фиксированной структуры, и даже при отсутствии уотсон-криковских пар оснований неоднородная последовательность может иметь другую структуру, чем это предполагалось сначала. Случай для фиксированных структур в РНК-петлях является даже более строгим. Антикодоновые петли тРНК имеют структуру и при этом предположительно две особенно подходящие последовательности, которые наилучшим образом связываются с ДНК-полимеразой Т4.
Антипараллельные цепи комплементарной последовательности в РНК дают спирали А-формы, из которых возникают и сохраняются последовательности петли. Даже если последовательности петли не обладают высокой способностью к связыванию, то минимизация энергии является оптимизацией энергетически свободной структуры (то есть никакие явные энергии активации не блокируют энергию минимизации последовательности петли). Вероятно, кинетически исходным сайтом для оптимизации может быть заканчивающая петлю пара оснований стебля РНК, который представляет собой плоскую поверхность, где может происходить оптимальное стэкинг-взаимодействие нуклеотидов петли и оснований. Петли РНК являются, в принципе, эквивалентными петлями белка, связывающего антипараллельные альфа-спирали и бета-цепи. Хотя эти белковые петли часто называют неупорядоченными спиральными кольцами, однако они не являются ни неупорядоченными, ни свернутыми в кольца. Такие петли называют "омега"-структурами, и это название отражает тот факт, что такая петля возникает и сохраняется в положениях, которые находятся в относительной близости друг к другу (см. See, Leszczynski J. и Rose G. и др. (1986), Science 234:849-855); причем указанные положения в белке являются концептуально эквивалентными паре оснований РНК-шпильки, заканчивающих петлю.
Многие омега-структуры были установлены с помощью рентгеноструктурного анализа, который показал, что указанные структуры являются идиосинкразическими. Очевидно, что каждая структура является результатом минимизации специфической энергии, действующей на петлю, концы которой расположены близко друг от друга. Как в белках, так и в РНК, эти петли будут минимизировать энергию при отсутствии информации от остальной структуры, за исключением (в первом приближении) завершающих петлю пары аминокислот или пары оснований. Как для омега-петлей белка, так и для петлей РНК-шпильки все свободно вращаемые связи будут участвовать в попытке минимизировать свободную энергию. Очевидно, что РНК будет нести гораздо большую ответственность за электростатику, чем белки, тогда как белки будут обладать гораздо большими степенями свободы, чем РНК. Таким образом, расчеты РНК-структур посредством энергетической минимизации, очевидно, позволят получить более точные решения структур, чем сравнимые расчеты для белков.
Одноцепочечные области РНК и белков могут действовать в направлении удлинения возможной структуры. То есть, если одноцепочечная петля возникает и возвращается в структуре белка из параллельных цепей альфа-спиралей или бета-цепей, то места возникновения и возврата могут быть, кроме того, от других, не от омега-структур. Кроме того, расстояние, занимаемое одной цепью пептида, может варьироваться в зависимости от длин параллельной альфа-спирали или бета-цепи.
Для тех белковых структур, в которых одиночная цепь лежит на фиксированной вторичной структуре белка, полученная в результате минимизация энергии должна, в принципе, допускать взаимодействия между одноцепочечными доменом и нижележащей структурой. По всей вероятности, аминокислотные боковые цепи, которые могут образовывать солевые мостики во вторичных структурах, должны действовать таким же образом в протяженных одиночных цепях, лежащих поверх регулярных вторичных структур. Таким образом, конкретные структуры таких областей белка снова будут идиосинкразическими и будут в очень большой степени зависеть от последовательностей. В этом случае зависимость от последовательности будет включать в себя как одиночную цепь, так и нижележащую последовательность вторичной структуры.
Интересно отметить, что РНК-структура, известная как псевдоузел, является аналогом указанных протяженных мотивов белка и может служить для иллюстрации по отношению к растворителю или молекулам-мишеням протяженных одиночных цепей РНК, основания которых являются идиосинкразически выстроенными по направлению либо к растворителю/мишени, либо к нижележащей вторичной структуре РНК. Псевдоузлы имеют общие с белком мотивы, основанные на петлях между параллельными цепями способность изменять длину одиночной цепи и последовательности спирали, на которой она лежит.
Таким образом, точно так же, как и в белковых мотивах, путем совместного изменения последовательностей в нижележащей вторичной структуре можно поместить одноцепочечные нуклеотиды и основания в направлении либо растворителя, либо нижележащей структуры, изменяя тем самым электростатику и функциональные химические группы, которые взаимодействуют с мишенями. Необходимо отметить, что такие изменения структуры являются следствием минимизаций энергии, однако лишь одна структура псевдоузла является известной, даже при низком разрешении. Тем не менее, ценность настоящего изобретения заключается в установлении того факта, что форма и функциональные возможности, исходящие из псевдоузлов, могут быть распознаваемыми практически до мельчайших подробностей и с высокой степенью качества.
Как петли шпильки, так и одноцепочечный домен псевдоузлов формируются на антипараллельных спиралях. Спирали РНК могут содержать нерегулярности, называемые "балджами" (выпуклости, выступы - прим.пер.). Балджи могут существовать в виде одной цепи или спирали, или того и другого и обеспечивают наличие идиосинкразических структурных особенностей, используемых для распознавания мишени. Кроме того, спиральные нерегулярности могут обеспечивать угловое связывание между регулярными спиралями.
Большой балдж (см. фиг.13) на одной цепи РНК может быть сравнимым с петлями шпильки, за исключением того, что пара оснований, завершающая петлю, заменяется двумя парами оснований, завершающих петлю, фланкирующих балдж.
Асимметричные балджи (см. фиг.13) могут сообщать удлиненную и нерегулярную структуру, которая стабилизируется с помощью нуклеотидных взаимодействий через балдж. Эти взаимодействия могут быть уотсон-криковскими взаимодействиями или любой другой стабилизирующей системой, включая другие водородные связи и стэкинг-взаимодействие оснований.
И наконец, рассматривая фиксированные формы или мотивы РНК, было небесполезно отметить, что между РНК и белками существуют значительные различия. Поскольку, очевидно, в процессе эволюции белки вытеснили РНК в отношении тех активностей, которые в настоящее время почти целиком осуществляются белками и пептидами, включая катализ и высокоспецифическое узнавание, то химические свойства белков являются, очевидно, более полезными, чем свойства РНК, для формирования вариабельных форм и активностей. Традиционная теория указывает на существование 20 аминокислот и только четырех нуклеотидов; сильные ионные свойства лизина, аргинина, аспарагиновой кислоты и глутаминовой кислоты, которыми не обладают РНК-основания; относительную нейтральность пептидного остова (по сравнению с сильно отрицательным сахар-фосфатным остовом нуклеиновых кислот), существование гистидина с рК, близком к нейтральному; тот факт, что боковые цепи аминокислот расположены в направлении к растворителю как в альфаспиралях, так и в бета-цепях; и регулярные вторичные структуры белков. В двухцепочечных нуклеиновых кислотах, включая РНК, пары оснований направлены друг на друга, и поэтому большая часть полезной химической информации находится на уровне одного измерения. Таким образом, исходя из современной точки зрения, белки значительно превосходят нуклеиновые кислоты (включая РНК) в разнообразии форм и функций. В соответствии с широко распространенным мнением в процессе эволюции белки выиграли конкуренцию с РНК в отношении распознавания и катализа.
В противоположность вышесказанному основная идея настоящего изобретения заключается в том, что факт существования широкого диапазона последовательностей и форм, возможных для РНК, признается вполне допустимым, особенно это касается последовательностей, которые никогда не испытывались в процессе своего развития, а также существования каждой желаемой функции и нужного сродства связывания, даже несмотря на то, что РНК состоит только из четырех нуклеотидов, и несмотря на то, что остов РНК является таким высокозаряженным. Другими словами, описанные выше РНК-мотивы с соответствующим описанием последовательностей, в своем пространственном воплощении, могут дать те химические функции, которые необходимы для получения тесного и специфического связывания с большинством мишеней. Можно предположить, что РНК является такой же разносторонней, как и иммунная система. То есть, поскольку иммунная система обеспечивает приспособление к любой нужной мишени, то и РНК обладает такими же возможностями. Описанная в настоящей заявке методика позволяет использовать 1018 последовательностей и таким образом провести испытания широкого диапазона структур, так что если даже и имеются присущие белкам или специфическим антителам преимущества над РНК, то они компенсируются широтой возможного "пула", из которого могут быть выбраны РНК-лиганды. Кроме того, благодаря модифицированным нуклеотидам, могут быть использованы РНК, которые, по сути, являются химически более разнообразными, чем природные РНК.
Способ SELEX включает в себя комбинацию селекции нуклеиново-кислотных лигандов, связывающихся с молекулой-мишенью, например с белком, и амплификации этих отобранных нуклеиновых кислот. Итерационное чередование стадий селекции/амплификации позволяет выбрать одну и небольшое число нуклеиновых кислот, которые лучше всего связываются с мишенью, из пула, содержащего очень большое число нуклеиновых кислот.
Повторение процедуры селекции/амплификации продолжают до тех пор, пока не будет достигнута поставленная цель. Например, чередование указанных стадий может быть продолжено до тех пор, пока не будет достигнут нужный уровень связывания нуклеиновых кислот в тестируемой смеси, или до тех пор, пока не будет получено минимальное число нуклеиново-кислотных компонентов смеси (в последнем случае до тех пор, пока в испытуемой смеси не останутся отдельные виды). В большинстве случаев, желательно повторение циклов продолжать до тех пор, пока не будет достигнуто улучшение связывания. Это может быть в том случае, когда некоторые смеси нуклеиновых кислот обнаруживают ограниченное улучшение при повторении селекции/амплификации по отношению к уровням фона. В этих случаях должно быть увеличено разнообразие последовательностей в исследуемой смеси, включая другие возможные варианты последовательностей, либо длина рандомизированной области последовательности должна быть увеличена до тех пор, пока не будет достигнуто улучшение связывания. Могут быть также использованы опорные схемы и/или процедуры способа "прогулки".
В частности, этот способ требует сначала получения испытуемой смеси нуклеиновых кислот-кандидатов. Отдельные испытуемые нуклеиновые кислоты могут содержать рандомизированную область, фланкированную последовательностями, консервативными у всех нуклеиновых кислот в смеси. Консервативные области получают для облегчения амплификации выбранных нуклеиновых кислот. Поскольку таких последовательностей, известных специалистам, существует много, то выбор нужной последовательности может сделать сам специалист с учетом предпочтительного метода амплификации. Рандомизированная область может быть полностью или частично рандомизированной последовательностью. Альтернативно, эта часть нуклеиновой кислоты может содержать подобласти, которые являются рандомизированными, наряду с подобластями, которые сохраняются постоянными во всех видах нуклеиновых кислот смеси. Например, для достижения лучшего связывания или улучшения специфичности связывания, область последовательности, о которой известно, что она связывается с целевым белком, или которая отобрана на связывание, может быть интегрирована с рандомизированными областями. Вариабельность последовательностей в исследуемой смеси может быть также введена или увеличена путем генерирования мутаций в нуклеиновых кислотах исследуемой смеси в процессе селекции/амплификации. В принципе, нуклеиновые кислоты, используемые в исследуемой смеси, могут быть любой длины, если только они при этом могут быть амплифицированы. Способ настоящего изобретения является наиболее практичным для выбора из большого числа вариантов последовательности. Таким образом, способ настоящего изобретения предпочтительно использовать для оценки связывания нуклеиново-кислотных последовательностей с широким диапазоном длины, которая может составлять от около четырех оснований до любого достижимого размера.
Рандомизированная часть нуклеиновых кислот в исследуемой смеси может быть получена различными способами. Например, полная или частичная рандомизация последовательности может быть легко достигнута путем прямого химического синтеза нуклеиновой кислоты (или ее части), либо путем синтеза матрицы, из которой нуклеиновая кислота (или ее части) может быть получена путем использования соответствующих ферментов. И наконец, при катализе терминальной трансферазой в присутствии неограничивающих концентраций всех четырех нуклеотидных трифосфатов, рандомизированная последовательность может быть присоединена к сегменту. Разнообразие последовательностей в смеси исследуемых нуклеиновых кислот может быть также достигнуто с помощью отобранных по размеру фрагментов или частично переваренных (или гидролизованых другим способом) препаратов крупных натуральных нуклеиновых кислот, таких как препараты геномных ДНК или клеточных РНК. В некоторых случаях, когда используется рандомизированная последовательность, необязательно (или возможно на основании длинных рандомизированных сегментов), чтобы исследуемая смесь содержала все возможные варианты последовательностей. В основном, предпочтительно, чтобы испытуемая смесь содержала как можно больше вариантов последовательностей, поскольку это является практически выгодным и дает гарантию того, что будет идентифицировано максимальное число потенциально связывающих последовательностей. Рандомизированная последовательность из 30 нуклеотидов будет содержать, как было подсчитано, 1018 различных кандидатов-последовательностей. Практически, желательно, чтобы образец содержал лишь около 1018 кандидатов при одной селекции. При практическом осуществлении способа, следует принимать в расчет число матриц на колонке синтеза ДНК, и растворимость РНК и мишени в растворе. (Само собой разумеется, что на число последовательностей в смеси кандидатов нет теоретических пределов). Следовательно, смесь кандидатов, которые имеют рандомизированные сегменты длиной более чем 30 нуклеотидов, содержит также много возможных последовательностей, которые в целом могут быть отобраны за одну селекцию. При этом необязательно испытывать все возможные последовательности смеси кандидатов для выбора нуклеиново-кислотного лиганда настоящего изобретения. В основе настоящего способа лежит тот факт, что испытуемая смесь нуклеиновых кислот обладает способностью амплифицироваться. Таким образом, предпочтительно, чтобы любые консервативные области, используемые в испытуемых нуклеиновых кислотах, не содержали последовательностей, которые препятствуют амплификации.
Различные РНК-мотивы, описанные выше, почти всегда могут быть определены полинуклеотидом, содержащим около 30 нуклеотидов. Из-за физических затруднений способа SELEX, наиболее длинным рандомизированным сегментом, который может содержать рандомизированная смесь, составляет 30 нуклеотидов, хотя, в принципе, могут быть исследованы все возможные варианты. Поэтому в предпочтительном осуществлении настоящего изобретения при использовании смеси кандидатов со смежной рандомизированной областью желательно использовать рандомизированную последовательность, по крайней море, из 15 нуклеотидов, и смесь, содержащую, по крайней мере, около 109 нуклеиновых кислот, а наиболее предпочтительно использовать рандомизированную последовательность, по крайней мере, из 25 нуклеотидов.
Настоящее изобретение включает в себя смесь кандидатов, содержащую все возможные варианты смежного рандомизированного сегмента, по крайней мере, из 15 нуклеотидов. Каждый отдельный член из смеси кандидатов может также состоять из фиксированных последовательностей, фланкирующих рандомизированный сегмент и способствующих амплификации выбранных нуклеиново-кислотных последовательностей.
Могут быть также получены смеси кандидатов, содержащие рандомизированные последовательности и фиксированные последовательности, где фиксированные последовательности имеют функцию, способствующую процессу амплификации. В одном из вариантов осуществления настоящего изобретения фиксированные последовательности в смеси кандидатов могут быть выбраны для того, чтобы повысить процентное содержание нуклеиновых кислот в смеси кандидатов при обработке данного мотива нуклеиновых кислот. Например, введение соответствующих фиксированных нуклеотидов позволит увеличить процентное содержание псевдоузлов или шпилечных петлей в смеси кандидатов. Поэтому смесь кандидатов, включающая в себя фиксированные последовательности, повышающие процентный состав данного структурного мотива нуклеиновых кислот, является частью настоящего изобретения. Любой специалист, на основании приведенных выше описаний различных нуклеиново-кислотных антител, может без излишнего экспериментирования получить указанную смесь кандидатов. В представленных ниже Примерах 2-8 описаны смеси кандидатов, полученные по принципу максимизации предпочтительных РНК-мотивов.
Смеси кандидатов, содержащие различные фиксированные последовательности или специальную рандомизированную последовательность могут быть также использованы после получения лигандного раствора или частичного лигандного раствора с помощью SELEX. После этого может быть начат новый SELEX-процесс с использованием смеси кандидатов, наполненной лигандным раствором.
Полимеразно-цепная реакция (РСR) является подходящим способом амплификации нуклеиновых кислот. Описание этого способа можно найти, например, у Saiki и др. (1985), Science 230:1350-1354; Saiki и др. (1986), Nature 324: 163-166; Scharf и др. (1986), Scienсе 233:1076-1078; Junis и др. (1988), Proc. Natl. Acad. Sci. 85:9436-9440; и патент США 4683195 (Mullis и др.) и патент США 4683202 (Mullis и др.). В общих чертах, PCR-амплификация заключается в повторении циклов репликации нужной одноцепочечной ДНК (или кДНК-копий РНК) с использованием специфических олигонуклеотидных праймеров, комплементарных 3'- и 5'-концам оцДНК, праймерным удлинением с помощью ДНК-полимеразы, и ДНК-денатурацией. Продукты, генерированные путем удлинения из одного праймера, служат в качестве матриц для удлинения из другого праймера. Подобный способ амплификации, описанный в опубликованной РСТ заявке WO 89/01050 (Burg и др.), требует наличия или введения промоторной последовательности выше амплифицированной последовательности для получения двухцепочечного интермедиата. Затем с использованием РНК-полимеразы продуцируют множество РНК-копий двухцепочечного интермидиата, содержащего промотор. Полученные в результате РНК-копии обрабатывали обратной транскриптазой для получения дополнительных двухцепочечных промоторсодержащих интермедиатов, которые могут быть затем подвергнуты другому раунду амплификации с помощью РНК-полимеразы. Альтернативные методы амплификации, наряду с другими стадиями, включают в себя клонирование отобранных ДНК или кДНК-копий отобранных РНК в соответствующий вектор и введение этого вектора в организм хозяина, где указанный вектор и клонированные ДНК реплицируются, и тем самым амплифицируются (Guatelli J.С. и др. (1990), Рroc. Natl. Асаd. Sci. 87:1874). В основном, для осуществления эффективной амплификации отобранных последовательностей нуклеиновых кислот в способе настоящего изобретения могут быть использованы практически любые приемлемые методы. При этом только необходимо, чтобы соответствующее распределение последовательностей после амплификации, по крайней мере, грубо отражало относительное содержание последовательностей в смеси до амплификации.
Конкретное осуществление амплификации РНК в способе настоящего изобретения основано на методе Junis и др. (1988) (см. выше). РНК-молекулы и молекулы-мишени в испытуемой смеси были такими, чтобы после амплификации и РСR обеспечивалось присутствие последовательностей главных промоторов Т7 в 5'-областях. Непроцессированные (т. е. с полной длиной) кДНК-копии отобранных РНК-молекул были получены с помощью обратной транскриптазы, праймированной олигомером, комплементарным 3'-последовательностям отобранных РНК. Полученные кДНК амплифицировали путем удлинения цепи с помощью ДНК-полимеразы Tag, обеспечивая тем самым присутствия промоторных последовательностей Т7 в отобранных ДНК. Затем двухцепочечные продукты этого процесса амплификации транскрибировали in vitro. Полученные транскрипты использовали в следующем цикле селекции/амплификации. Этот способ может включать в себя, но не обязательно, стадии очистки соответствующих нуклеиновых кислот.
В способе настоящего изобретения может быть использована, в основном, любая схема, позволяющая осуществлять селекцию нуклеиновых кислот на основании их способности к специфическому связыванию с другой молекулой, то есть с белком или любой другой молекулой-мишенью. При этом необходимо только, чтобы отобранные в результате селекции нуклеиновые кислоты были амплифицируемыми. Например, может быть использована техника селекции путем связывания с фильтрами, описанная в Примере 1, где испытуемую смесь нуклеиновых кислот инкубировали с целевым белком, после чего смесь "нуклеиновые кислоты/белок" фильтровали через нитроцеллюлозный фильтр и промывали соответствующим буфером для удаления свободных нуклеиновых кислот. Комплекс "белок/нуклеиновая кислота" часто остается связанным с фильтром. Относительные концентрации белка по отношению к нуклеиновой кислоте в инкубированной смеси влияют на интенсивность связывания, на которую, собственно, и проводится отбор. Если присутствует избыток нуклеиновых кислот, то имеет место конкурентное связывание, и происходит отбор на наиболее интенсивное связывание. И наоборот, если присутствует избыточное количество белка, то можно ожидать, что будет отобрана любая нуклеиновая кислота, которая связывается с белком. Эти относительные концентрации белков и нуклеиновых кислот используют для достижения нужной селекции, которая будет зависеть от типа белка, интенсивности связывания и уровня присутствующего фона связывания. Относительные концентрации, которые необходимы для достижения нужного результата селекции, могут быть легко определены эмпирически, без излишнего экспериментирования. Аналогично этому, может оказаться необходимым оптимизировать процедуру промывки фильтра для минимизации фонового связывания. И снова такая оптимизация процедур промывки фильтров может быть проведена самим специалистом.
Авторами настоящего изобретения была использована математическая оценка результатов SELEX, названная SELEXION. В приложении к настоящей заявке дан краткий обзор математического анализа, использованного для получения обобщенных выводов, относящихся к SELEX-процессу.
В результате проведения SELEXION были получены следующие общие выводы: 1) вероятность выделения лучше всего связывающихся РНК в каждом раунде SELEX возрастает с числом присутствующих указанных молекул; с их преимущественной способностью связываться по сравнению объемным РНК-пулом; и с полным количеством используемого белка. Хотя и не всегда
можно интуитивно заранее определить как максимизировать различие в связывании, однако вероятность выделения лучше всего связывающихся РНК может быть еще увеличена посредством максимизации числа испытуемых РНК-молекул и молекул-мишеней;
2) идеальные концентрации нуклеиновых кислот и белка, используемых в различных раундах SELEX, зависят от нескольких факторов. Экспериментальные параметры, предлагаемые SELEXION соответствуют параметрам, используемым в прилагаемых Примерах. Например, если относительная аффинность конечного лигандного раствора неизвестна, что почти неизбежно при осуществлении SELEX, концентрации белка и нуклеиновых кислот в смеси кандидатов предпочтительно выбирать таким образом, чтобы обеспечить связывание примерно 3 и 7% от всех нуклеиновых кислот с белком-мишенью. При использовании указанного критерия можно ожидать, что в каждом раунде SELEX будет достигнуто 10-100-кратное обогащение лигандами с высокой аффинностью.
В предпочтительном варианте осуществления настоящего изобретения экспериментальные условия, используемые для отбора нуклеиново-кислотных лигандов к различным мишеням, выбирают таким образом, чтобы они были по возможности близкими к условиям, при которых данная мишень обнаруживается in vivo. В примере 10 показано изменение условий селекции, являющееся эффективным для получения лигандного раствора для конкретной мишени. Хотя лигандный раствор для GF имел значительное сходство при условиях с высоким и низким содержанием соли, однако наблюдались и некоторые различия. Такими регулируемыми условиями, которые могут быть изменены для более точного моделирования in vivo-условий мишени, являются, например, но не ограничиваются этими примерами, полная ионная сила, концентрация двухвалентных катионов и рН раствора. Каждый специалист может самостоятельно выбрать подходящие условия на основании имеющейся информации о данной мишени.
Для осуществления стадии амплификации после распределения отобранные нуклеиновые кислоты должны быть отделены от мишени. Этот процесс должен быть осуществлен без какого-либо химического разрушения нуклеиновых кислот, и в результате этого процесса должны быть получены амплифицируемые нуклеиновые кислоты. В конкретном варианте осуществления настоящего изобретения отобранные РНК-молекулы элюируют с нитроцеллюлозных фильтров с использованием свежеприготовленного раствора, содержащего 200 мкл 7 М мочевины, 20 мМ цитрата натрия (рН 5,0), 1 мМ ЭДТК-раствора, объединенного с 500 мкл фенола (уравновешенного 0,1 М ацетата натрия, рН 5,2). В настоящем изобретении был с успехом использован раствор 200 мкл 7 М мочевины с 500 мкл фенола. Затем элюированный раствор выбранных РНК экстрагируют эфиром, осаждают этанолом, а осадок ресуспендируют в воде. Для элюирования отобранных РНК с фильтра могут быть использованы различные буферные режимы. Например, могут быть использованы недетергенные водные белокденатурирующие агенты, такие как кванидинийхлорид, кванидинийтиоцианат и т.п., которые хорошо известны специалистам. Специфический раствор для элюции нуклеиновых кислот с фильтра может быть выбран любым специалистом самостоятельно.
Альтернативные схемы распределения для отделения нуклеиновых кислот, связанных с мишенями, в частности, белками, хорошо известны специалистам. Например, связывание и распределение может быть осуществлено путем пропускания испытуемой смеси нуклеиновых кислот через колонку, которая содержит молекулу-мишень, связанную с твердофазным носителем. Те нуклеиновые кислоты, которые связываются с мишенью, будут удерживаться на колонке, а несвязанные нуклеиновые кислоты могут быть смыты с колонки.
В настоящей заявке способом SELEX может быть определен итерационный процесс, где селекцию и амплификацию повторяют до тех пор, пока не получат желаемую степень селективности. В одном из вариантов настоящего изобретения для получения нужного лигандного раствора может быть проведена лишь одна стадия процесса селекции. Например, теоретически, колонка, которая служит носителем для мишени и через которую пропускают смесь кандидатов (при благоприятных условиях и с достаточной длиной колонки), должна обладать способностью к разделению нуклеиновых кислот на основе аффинности к мишени, достаточному для получения лигандного раствора. Способ, в котором для получения лигандного раствора достаточно лишь одной стадии селекции, также входит в объем настоящего изобретения.
В одном из вариантов настоящего изобретения SELEX проводят итеративно до тех пор, пока в смеси кандидатов после амплификации не останется одна или небольшое число отдельных аминокислотных лигандов. В этом случае лигандный раствор может быть представлен как последовательность одной нуклеиновой кислоты, а не семейство последовательностей, имеющих сравнимые аффинности связывания с мишенью.
В альтернативном варианте настоящего изобретения SELEX-итерации могут быть прекращены в тот момент, когда смесь кандидатов будет обогащена нуклеиново-кислотными лигандами с более высокой степенью сродства, но при этом еще будет содержать относительно большое число различных последовательностей. Этот момент может быть определен любым специалистом путем периодического анализирования неупорядоченности объема смеси кандидатов или путем анализа аффинности всей массы к данной мишени.
Затем, после завершения SELEX, получают клоны и эти клоны секвенируют. Конечно, окажется почти неограниченное количество клонов, которые должны быть секвенированы. Однако, как видно из приведенных ниже Примеров, после секвенирования 20-50 клонов можно будет в общих чертах определить наиболее предпочтительные последовательности и характеристики лигандного раствора. В качестве гипотетического примера можно предположить, что после клонирования 30 последовательностей будет обнаружено 6 последовательностей, которые являются идентичными, тогда как некоторые части 20 других последовательностей являются близкородственными последовательностями внутри "выигравшей" последовательности. Хотя наиболее предпочтительная последовательность может рассматриваться как лиганд к данной мишени, однако часто бывает предпочтительней конструировать или описать лигандный набор, который состоит из семейства последовательностей, имеющих общие характеристики многих клонированных последовательностей.
В другом варианте осуществления настоящего изобретения лигандный набор, который может быть представлен в качестве семейства последовательностей, имеющих ряд определяющих характеристик (например, где лигандное решение представляет собой AAGUNNGUNNCNNNN, где N может быть любым из четырех нуклеотидов), может быть использован для инициации дополнительного SELEX-процесса. В этом варианте смесь кандидатов будет состоять из частично фиксированных и частично рандомизированных нуклеотидов, причем фиксированные нуклеотиды могут быть выбраны на основе лигандного раствора, полученного в начальном SELEX-процессе. В этом случае, если имеется одна нуклеотидная последовательность, которая связывается с мишенью лучше, чем другие члены лигандного раствора, то она будет быстро идентифицирована.
В еще одном альтернативном варианте осуществления настоящего изобретения может быть также проведен второй SELEX-эксперимент на основе лигандного решения, полученного в SELEX-процессе. В этом случае одну наиболее преобладающую последовательность (например, ААGUCCGUAACACAC) используют для введения во второй процесс SELEX. В этом втором SELEX-процессе смесь кандидатов составляют для того, чтобы получить последовательности на основе выбранного "победителя", предполагая при этом, что в каждой последовательности будет иметь место достаточная рандомизация. Указанная смесь кандидатов может быть продуцирована с использованием исходных нуклеотидных материалов, которые являются не рандомизированными, а скорее статистически смещенными. Например, раствор А содержит 75% А и 25% U, С и G. Хотя синтезатор нуклеиновых кислот приспособлен для получения превалирующего нуклеотида, однако присутствие других нуклеотидов в А-растворе может дать нуклеиново-кислотные последовательности, которые преобладают в А, но которые могут дать также вариации в этом положении. И аналогично этот второй раунд SELEX, проводимый на основании результатов, полученных в первоначальном SELEX-процессе, будет максимизировать вероятность получения наилучшего лигандного набора для данной мишени. И снова необходимо отметить, что лигандный раствор может состоять из одного предпочтительного нуклеиново-кислотного лиганда или он может состоять из семейства структурно родственных последовательностей, в основном, с аналогичными аффинностями связывания.
На практике иногда может быть предпочтительным не продолжать SELEX-процесс до тех пор, пока останется одна последовательность. Этот SELEX-процесс содержит несколько факторов сдвига, которые могут влиять на преобладание некоторых последовательностей в смеси кандидатов после нескольких раундов SELEX, и которые не имеют отношения к сродству связывания данной последовательности с мишенью. Например, влияние в пользу или против данных последовательностей может иметь место в процессе продуцирования кДНК из РНК, выделенных после отбора, или в процессе амплификации. Указанные непредсказуемые флуктуации могут быть минимизированы путем прекращения SELEX до того момента, когда только одна или небольшое число последовательностей будет преобладать в реакционной смеси.
Как указывалось выше, разнообразие последовательностей в испытуемой смеси нуклеиновых кислот может быть достигнуто или увеличено путем мутации. Например, в этих целях может быть использована уже описанная техника проведения мутагенеза последовательностей нуклеиновых кислот во время PCR-амплификации (Leung и др. (1989)). Этот метод или функционально эквивалентные методы могут быть (но не обязательно) использованы в сочетании с процедурами амплификации настоящего изобретения.
Альтернативно в процедуру амплификации нуклеиновых кислот могут быть включены стандартные способы ДНК-мутагенеза. Приемлемыми процедурами мутагенеза, среди прочих, являются химически индуцированный мутагенез и сайт-направленный мутагенез с использованием олигонуклеотидов.
В настоящем изобретении могут быть также использованы другие интересные свойства нуклеиновых кислот и способ, с помощью которого, как известно (или как будет обнаружено позже), они могут взаимодействовать с мишенями, такими как белки. Например, техника SELEX может быть использована для скрининга в целях обнаружения лигандов, которые образуют аддукты Michael с белками. Пиримидины, если они занимают правильное положение в белке, которое обычно является смежным с критическим цистеином или другим нуклеофилом, может реагировать с указанным нуклеофилом, образуя аддукт Michael. Механизм образования аддуктов Michael включает в себя нуклеофильную атаку в 6 положении пиримидинового основания образованием временного (но с медленной реверсией) интермедиата, который в действительности является 5,6-дигидропиримидина. Испытание на присутствие таких интермедиатов может быть проведено путем определения, происходит ли связывание между РНК и белковой мишенью даже после денатурации белка соответствующим денатурантом. То есть проводят исследование непрерывного ковалетного связывания при разрушении связывающего "кармана". Однако аддукты Michael часто бывают обратимыми и иногда так быстро, что неудача при идентификации аддукта Michael с помощью указанного теста не указывает на его отсутствие в предыдущий момент.
SELEX может быть осуществлен так, чтобы можно было воспользоваться образованием аддукта Michael в целях создания высокоаффинных, почти гибельных субстратов для фермента или другой белковой мишени. Предполагается, что после связывания рандомизированной смеси РНК и мишени до распределения на фильтре или с помощью других средств мишень денатурируется. После распределения, за которым следует реверсия аддукта Michael и кДНК-синтез на выделенной РНК, проводят остаток SELEX-цикла, который обогащает РНК, связывающейся с мишенью до денатурации, но продолжающейся ковалентно связываться до тех пор, пока аддукт Мiсhаеl не будет реверсирован исследователем. Этот лиганд, in vivo, должен обладать способностью к непрерывному ингибированию белковой мишени. Такой белок, как тРНК-урацилметилтрансфораза (RUMT), связывает субстратную тРНК посредством аддукта Michael. Когда RUMT экспрессировали при высоких уровнях в Е.соli, было обнаружено значительное ковалентное связывание с РНК, что позволило предположить с высокой степенью уверенности, что с помощью SELEX могут быть обнаружены почти необратимые ингибиторы.
Способ настоящего изобретения имеет множество применений. Например, он может быть использован для облегчения идентификации и характеризации любого сайта связывания белка с ДНК или РНК. Указанные сайты связывания участвуют в регуляции транскрипции или трансляции экспрессии генов, например, в качестве сайтов связывания для активаторов или респрессоров транскрипции, транскрипционных комплексов в промоторных областях, белков, участвующих в репликации, и ДНК-полимеразы в участке или возле участка начала репликации, а также рибосом и ингибиторов трансляции в сайте связывании с рибосомами. Информация о таких сайтах связывания может быть использована для выделения и идентификации регуляторных областей, что позволяет избежать применения более трудоемких методов характеризации указанных областей. Выделенные регуляторные ДНК-области могут быть использованы, например, в гетерологических конструкциях в целях селективного изменения генной экспрессии.
Важная и неожиданная особенность настоящего изобретения заключается в том, что описанные в настоящей заявке методы могут быть использованы для идентификации, выделения или продуцирования молекул нуклеиновой кислоты, которые специфически связываются с нужной молекулой-мишенью. Таким образом, способы настоящего изобретения могут быть использованы для продуцирования нуклеиновых кислот, специфически связывающихся с конкретной мишенью. Указанный нуклеиново-кислотный лиганд во многих отношениях функционально напоминает антитело. Нуклеиново-кислотные лиганды, которые обладают функциями связывания, аналогичными функциям антител, могут быть выделены способами настоящего изобретения. Указанные нуклеиново-кислотные лиганды именуются в настоящем описании нуклеиново-кислотными антителами и, в основном, могут быть использованы в тех случаях, когда, как известно, применяются поликлональные или моноклональные антитела. В любых in vivo или in vitro-применениях обычные антитела могут быть, в основном, заменены нуклеиново-кислотными антителами. При этом необходимо только, чтобы условия, при которых используются нуклеиново-кислотные антитела, в основном, не вызывали деградацию нуклеиновых кислот. Использование нуклеиново-кислотных антител предусматривает специфическое, количественное или качественное обнаружение молекул-мишеней из любого источника; очистку молекул-мишеней на основе их специфического связывания с нуклеиновой кислотой; и различные терапевтические цели, рассчитанные на специфическую доставку токсина или другого терапевтического агента к специфической области-мишени.
Молекулами-мишенями являются предпочтительно белки, но, среди прочих, могут быть также углеводы, пептидгликаны и ряд небольших молекул. Как и традиционные белковые антитела, нуклеиново-кислотные антитела могут быть использованы для целевых биологических структур, таких как клеточные поверхности или вирусы, посредством специфического взаимодействия с молекулой, то есть целой частью указанной биологической структуры. Нуклеиново-кислотные антитела обладают тем преимуществом, что они не ограничены собственной толерантностью, как традиционные антитела. Кроме того, нуклеиново-кислотные антитела не требуют клеточных или происходящих от животных структур для синтеза или продуцирования, поскольку SELEX является целиком in vitro-процессом. Как уже хорошо известно, нуклеиновые кислоты могут связываться с комплиментарными нуклеиново-кислотными последовательностями. Это свойство нуклеиновых кислот может быть широко использовано для обнаружения, количественной оценки и выделения нуклеиново-кислотных молекул. Таким образом, способ настоящего изобретения не относится к этим хорошо известным способностям связывания между нуклеиновыми кислотами. В частности, способы настоящего изобретения, связанные с использованием нуклеиново-кислотных антител, не претендуют на включение в себя известные аффинности связывания между нуклеиновыми кислотами. Известно, что имеется ряд белков, которые обладают способностью связываться с нуклеиновыми кислотами, например, такие как регуляторные белки, которые связываются с операторными последовательностями нуклеиновых кислот. Хорошо известная способность некоторых связывающихся с нуклеиновыми кислотами белков связываться в их природных сайтах может быть использована, например, для обнаружения, количественной оценки и очистки указанных белков. Способы настоящего изобретения, связанные с использованием нуклеиново-кислотных антител, не относятся к хорошо известному связыванию по сродству между нуклеиновыми кислотами и белками, о связывании которых с указанными нуклеиновыми кислотами хорошо известно. Однако с помощью SELEX могут быть получены не встречающиеся в природе последовательности, которые связываются с теми же связывающимися с нуклеиновыми кислотами белками. При этом следует отметить, что SELEX позволяет очень быстро определить нуклеиново-кислотные последовательности, которые связываются с белком, и поэтому может быть с успехом использован для определения неизвестного оператора или последовательностей сайтов связывания, которые могут быть затем использованы для целей, описанных в настоящей заявке. Очевидно, что настоящее изобретение является первой работой, где предлагается использовать нуклеиново-кислотные молекулы для обнаружения, количественной оценки, выделения и очистки белков, способность которых связываться с нуклеиновыми кислотами еще не установлена. Как указывалось выше, некоторые нуклеиново-кислотные антитела, выявленные с помощью SELEX, могут быть также использованы для воздействия на функции, например, в качестве ингибиторов, энхансеров или активаторов функций специфических молекул-мишеней или структур. В частности, нуклеиново-кислотные антитела могут быть использованы для ингибирования, усиления или активации функции белков.
Белки с известной способностью связываться с нуклеиновыми кислотами, такие как ДНК-полимеразы, другие репликазы, и белки, которые распознают сайты на РНК, но не оказывают дальнейшего каталитического действия, обнаруживают посредством SELEX высокую степень сродства к РНК-лигандам, которые связываются с активным сайтом белка-мишени. Так например, в случае обратной транскриптазы ВИЧ-1 полученный РНК-лиганд (называемый 1.1 в Примере 2) блокирует кДНК-синтез в присутствии праймерной ДНК, РНК-матрицы и четырех дезоксинуклеотидтрифосфатов.
В своей теории относительно РНК-структур авторы настоящего изобретения допускают, что почти каждый белок может служить мишенью для SELEX. Первоначальные эксперименты с белком, не связывающимся с нуклеиновыми кислотами, осуществляли с тремя белками, о которых неизвестно, взаимодействуют ли они с нуклеиновыми кислотами вообще или с РНК, в частности. Эти три белка представляли собой тканевый активатор плазминогена (+РА), фактор роста нервной ткани (NGF) и внеклеточный домен рецептора фактора роста (gfR-Xtra). Все указанные три белка тестировали, чтобы определить будут ли они удерживаться на нитроцеллюлозном фильтре в смеси с рандомизированными РНК. tРА и NGF показали сродство к рандомизированной РНК (с Kd немного ниже мкМ). gfR-Xtra не связывался с обнаружимым сродством, что позволяет предположить, что если для указанного белка существует РНК-антитело, то оно должно связываться с сайтом, который не имеет сродства для большинства других РНК.
tPA и NGF пропускали через SELEX с использованием РНК с 30 рандомизированными положениями. Оба, tPA и NGF, давали лигандные растворы в SELEX-эксперименте, что позволяет предположить, что определенный сайт, на котором каждый белок связывается с последовательностями-"победителями" более тесно, чем тот или другой сайт связывания с другими РНК. Эти последовательности-"победители" для этих двух белков являются различными.
Поскольку tРА и NGF показали хорошие результаты в SELEX-экспериментах, то была проанализирована произвольная серия белков и пептидов для того, чтобы определить, имеют ли они какое-либо сродство к РНК. Имелись все основания полагать, что если белок обладает каким-либо сродством к РНК, то эксперимент SELEX, будет, в среднем, давать более высокоаффинные последовательности, связывающиеся с той же областью мишени, которая дает низкую генерализованную аффинность. Была исследована серия белков и пептидов для того, чтобы посмотреть, будут ли рандомизированные РНК, содержащие 40 рандомизированных положений, удерживаться на нитроцеллюлозных фильтрах. Около двух третей исследованных белков связывались с РНК, а несколько белков обнаруживали очень тесное связывание с РНК (см. Пример 9).
Белки, которые но связывают РНК с нитроцеллюлозными фильтрами, могут быть отвергнуты из тех простых соображений, что они не имеют ничего общего с вероятностью увеличения РНК-антител. Например, брадикинин "терпит неудачу" при связывании с нитроцеллюлозными фильтрами, и поэтому должен быть отвергнут в вышеуказанном эксперименте. Был получен брадикинин, связанный с твердой матрицей посредством аминоконца пептида, после чего было обнаружено, что рандомизированная РНК тесно связывается с матрицей (см. Пример 7). Так, например, в начальных экспериментах два коротких пептида, брадикинин и бомбезин, очень тесно связываются с рандомизированными РНК. Любой РНК-лиганд с высоким сродством, полученный с помощью SELEX для этих пептидных мишеней, будет, вероятно, антагонистом этих активных пептидов и поэтому может быть использован в терапевтических целях. Трудно себе представить, чтобы РНК примерно с 30 нуклеотидами связывалась с очень маленьким пептидом, не сообщая этому неактивному пептиду хоть какую-нибудь активность.
Как описано в Примерах 4, 7, 9 и 10 (см. ниже), было обнаружено, что белки, которые не взаимодействуют с нуклеиновыми кислотами в природе, связываются с произвольной смесью нуклеиновых кислот с достаточно необычной степенью. Кроме того, было показано, что для таких белков, которые, как было установлено, связываются с РНК-смесями неспецифически, может быть получен лигандный раствор с помощью SELEX. Поэтому очень важно для дальнейшего отбора до осуществления SELEX определить, связывается ли данная мишень с произвольной смесью нуклеиновых кислот.
Вторая важная и неожиданная особенность настоящего изобретения заключается в том, что способы, описанные в настоящей заявке, могут быть использованы для идентификации, выделения или продуцирования нуклеиново-кислотных молекул, которые будут специфически связываться с конкретной молекулой-мишенью и влиять на функцию этой молекулы. В этом варианте снова молекулами-мишенями являются предпочтительно белки, но, помимо этого, могут быть также найдены нуклеиновые кислоты, которые специфически связывались бы с углеводами и различными малыми молекулами. Нуклеиновые кислоты, которые связываются с малыми молекулами, могут влиять на их функцию путем их изоляции или путем предотвращения их связывания с натуральными лигандами. Например, на активность фермента может воздействовать лиганд, который связывается с ферментным субстратом. Нуклеиново-кислотные лиганды, то есть нуклеиново-кислотные антитела, для малых молекул могут быть практически использованы для диагностических тестов (или других качественных анализов). Например, с помощью нуклеиново-кислотных лигандов настоящего изобретения, может быть обнаружено и определено присутствие регулируемых веществ связанных метаболитов или аномальных количеств нормальных метаболитов. Нуклеиново-кислотный лиганд, обладающий каталитической активностью, может воздействовать на функцию малой молекулы посредством катализа химической модификации мишени. Диапазон возможных каталитических активностей указанных лигандов является, по крайней мере, таким же широким, как и у белков. Одним из методов, с помощью которого могут быть отобраны каталитические нуклеиново-кислотные лиганды, является стратегия выбора лиганда для аналога переходного состояния желаемой реакции.
Очевидно, что в настоящем изобретении впервые раскрывается основное использование нуклеиново-кислотных молекул для воздействия на функцию белка или ее ингибирования или усиления. Способ настоящего изобретения для отбора на связывание может быть легко скомбинирован со способами вторичной селекции или скрининга для модификации функции молекулы-мишени путем связывания с отобранными нуклеиновыми кислотами. Большая популяция вариантных нуклеиново-кислотных последовательностей, которые могут быть протестированы с помощью SELEX, увеличивает вероятность того, что будут обнаружены нуклеиново-кислотные последовательности, обладающие нужной способностью к связыванию и к модификации активности целевой молекулы. Способы настоящего изобретения могут быть использованы для отбора нуклеиново-кислотных лигандов, которые могут избирательно воздействовать на функцию любого белка-мишени, включая белки, связывание которых с нуклеиновыми кислотами является частью их природной биологической активности, а также те белки, связывание которых с нуклеиновыми кислотами не установлено. Способы, описанные в настоящей заявке, могут быть использованы для выделения или продуцирования нуклеиново-кислотных лигандов, которые связываются с любым белком, способным к связыванию с нуклеиновой кислотой, или модифицируют его функцию; причем указанными лигандами могут быть ДНК или РНК (двухцепочечные или одноцепочечные); нуклеозид или нуклеотид, включая те из них, которые имеют пуриновые или пиримидиновые основания или основания, происходящие от пурина или пиримидина, в частности те из них, которые имеют адениновые, тиминовые, гуаниновые, урациловые, цитозиновые и гипоксантиновые основания, или их производные, например метилированные производные; и нуклеотиды коферментов, включая, среди прочих, никотинамидные нуклеотиды, флавинадениновые динуклеотиды и кофермент А. Следует отметить, что способ настоящего изобретения может быть использован для идентификации, выделения или продуцирования нуклеиново-кислотных молекул, которые могут воздействовать на каталитическую активность ферментов-мишеней, то есть ингибировать катализ или модифицировать связывание с субстратом, воздействовать на функциональные свойства белковых рецепторов, т.е. ингибировать связывание с рецепторами или модифицировать специфичность связывания с рецепторами; воздействовать на образование белковых полимеров, то есть разрушать четвертичную структуру субъединиц белка; и модифицировать транспортные свойства белка, то есть нарушать перенос мелких молекул или ионов белками.
Описанный в настоящей заявке процесс SELEX как способ итерационной селекции и амплификации смеси кандидатов нуклеиновых кислот повторяют до тех пор, пока не получат лигандный раствор. Дополнительной стадией этого процесса является получение нуклеиново-кислотных антител к данной мишени. Даже если лигандный раствор, полученный в результате этого процесса, является одной последовательностью, то должно быть синтезировано нуклеиново-кислотное антитело, содержащее именно лигандный раствор. Например, SELEX-эксперимент может дать предпочтительный однолигандный раствор, который состоит лишь из 20 и 30 рандомизированных нуклеотидных последовательностей, используемых в SELEX-смеси кандидатов. Терапевтически полезное нуклеиново-кислотное антитело предпочтительно не должно содержать 10 некритических нуклеотидов или фиксированные последовательности, необходимые для стадии амплификации SELEX. После того как на основе лигандного раствора будет определена нужная структура нуклеиново-кислотного антитела, может быть проведен синтез нуклеиново-кислотного антитела в соответствии со стандартной техникой, хорошо известной специалистам.
Нуклеиново-кислотное антитело может быть также сконструировано на основе лигандного раствора для данной мишени, состоящего из семейства последовательностей. В этом случае традиционное экспериментирование может показать, что данная последовательность является предпочтительной вследствие обстоятельств, не связанных с относительным сродством лигандного раствора к мишени. Указанные соображения должны быть очевидны для каждого специалиста.
В альтернативном варианте осуществления настоящего изобретения нуклеиново-кислотное антитело может содержать множество нуклеиново-кислотных лигандов к той же самой мишени. Например, SELEX может идентифицировать два дискретных лигандных раствора. Поскольку эти два лигандных раствора могут связываться с мишенью в различных положениях, то нуклеиново-кислотное антитело предпочтительно может содержать оба лигандных раствора. В другом варианте настоящего изобретения нуклеиново-кислотное антитело может содержать более чем один лигандный раствор. Такие многовалентные нуклеиново-кислотные антитела будут иметь повышенное сродство при связывании с мишенью, для которого является недейственным эквивалентное нуклеиново-кислотное антитело, имеющее только один лиганд.
Кроме того, нуклеиново-кислотное антитело может также содержать другие элементы, которые будут: 1) добавлять независимое сродство нуклеиново-кислотного антитела к мишени; 2) надежно повышать сродство нуклеиново-кислотного лиганда к мишени; 3) направлять или локализовать нуклеиново-кислотное антитело в нужную область in vivo, нуждающуюся в лечении; или 4) использовать специфичность нуклеиново-кислотного лиганда к данной мишени для осуществления некоторой дополнительной реакции в этой области.
Способы настоящего изобретения позволяют получать нуклеиновые кислоты, которые будут ингибировать функцию целевого белка, в частности получать нуклеиновые кислоты, которые будут ингибировать функцию белков, которые являются функциональными белками, связывающимися с нуклеиновой кислотой, нуклеотидами, нуклеозидами и их производными и аналогами. Способы настоящего изобретения позволяют получать нуклеиново-кислотные ингибиторы, например ингибиторы полимераз, обратных транскриптаз и других ферментов, где нуклеиновая кислота, нуклеотид или нуклеозид является субстратом или кофактором.
Вторичными способами селекции, которые могут быть объединены с SELEX, являются, среди прочих, другие виды селекции или отбора на ингибирование ферментов, модификацию связывания с субстратом, потерю функциональных свойств, разрушение структуры и т.п. Каждый специалист может из указанных альтернативных методов селекции и скрининга выбрать те методы, которые являются совместимыми с описанными в данной работе способами.
Каждому специалисту также понятно, что в некоторых случаях, то есть для некоторых молекул-мишеней или в некоторых применениях может быть предпочтительным использовать в качестве лигандов молекулы РНК, а не ДНК, в других случаях, наоборот, может быть предпочтительней использовать молекулы ДНК.
Способы селекции настоящего изобретения могут быть также использованы для отбора нуклеиновых кислот, которые специфически связываются с молекулярным комплексом, например с комплексом "субстрат/белок" или "ингибитор/белок". Среди этих нуклеиновых кислот, которые специфически связываются с комплексными молекулами, но не с некомплексными молекулами, имеются нуклеиновые кислоты, которые будут ингибировать образование комплекса. Например, среди тех нуклеиново-кислотных лигандов, которые были отобраны на специфическое связывание с комплексом "субстрат/фермент", имеются нуклеиновые кислоты, которые могут быть легко отобраны на ингибирование связывания с ферментом и тем самым на ингибирование или прекращение катализа ферментом.
В осуществлении настоящего изобретения, которое особенно предпочтительно для идентификации или выделения нуклеиновых кислот, которые связываются с конкретным функциональным или активным сайтом в белке или другой молекуле-мишени, используют молекулы, известные или отобранные на связывание с нужным сайтом в белке-мишени, для осуществления процесса селекции/амплификации на выявление субпопуляции нуклеиново-кислотных лигандов, которые связываются в нужном сайте или близ нужного сайта в данной молекуле-мишени. В простом примере нуклеиново-кислотную последовательность, которая, как известно, связывается с нужным сайтом в молекуле-мишени, вводят близ рандомизированной области всех нуклеиновых кислот, тестируемых на связывание. Затем используют SELEX (фиг. 9) для отбора тех вариантов, все из которых будут содержать известную последовательность, которая будет наиболее эффективно связываться с молекулой-мишенью. Таким образом, может быть выбрана более длинная последовательность, которая будет либо более эффективно, либо более специфично связываться с мишенью. Эта более длинная связывающаяся последовательность может быть затем введена близ рандомизированной области нуклеиновых кислот в испытуемой смеси, после чего можно повторить стадии селекции/амплификации для отбора еще более длинной последовательности. Эти итеративные стадии (т. е. введение отобранной последовательности в испытуемых смесях с последующей селекцией/амплификацией для достижения улучшенного или более специфичного связывания) можно повторять до тех пор, пока не будет достигнут нужный уровень эффективности связывания или специфичности. Эта итерационная процедура "прогулки" позволяет осуществить селекцию высокоспецифических нуклеиновых кислот для конкретной молекулы-мишени или для сайта в этой молекуле-мишени. В другом варианте указанной итерационной процедуры-"прогулки" используют молекулу-"якорь", которая не обязательно является нуклеиновой кислотой (см. фиг. 10 и 11). В этом варианте молекулу, которая связывается с нужной целью, например субстрат или ингибитор целевого фермента, модифицируют так, чтобы она могла ковалентно связываться с олигонуклеотидом известной последовательности ("ведущий олигонуклеотид" фиг.10). Этот ведущий олигонуклеотид, химически связанный с молекулой-"якорем", которая связывается с мишенью, также будет связываться с молекулой-мишенью. Комплект последовательностей ведущего олигонуклеотида вводят близ рандомизированной области испытуемой смеси нуклеиновых кислот. Затем осуществляют SELEX для отбора последовательностей, которые связываются наиболее эффективно с комплексом "молекула/якорь". Итерационная процедура "прогулки" может быть затем использована для отбора или продуцирования более и еще более длинных нуклеиново-кислотных молекул с более эффективным или более специфическим связыванием с мишенью. Осуществление процедуры с использованием "якоря" позволяет ожидать более быстрого выделения нуклеиново-кислотных лигандов, которые связываются с нужным сайтом или возле него данной молекулы-мишени. В частности, можно ожидать, что метод с использованием "якоря" в сочетании с итерационными процедурами "прогулки" позволит получить нуклеиновые кислоты, которые являются высокоспецифичными ингибиторами функции белка (фиг.11).
В некоторых вариантах осуществления SELEX желательно плюс/минус скринирование проводить в сочетании с процессом селекции для гарантии того, что в процессе селекции не возникнет флуктуаций, не связанных со сродством нуклеиново-кислотных последовательностей к данной мишени. Например, если селекцию осуществляют путем связывания белка на нитроцеллюлозе, то, как известно, некоторые нуклеиново-кислотные последовательности предпочтительно удерживаются нитроцеллюлозой и могут быть отобраны в процессе SELEX. Эти последовательности могут быть удалены из смеси кандидатов путем проведения дополнительных стадий, где предыдущую SELEX-смесь пропускают через нитроцеллюлозу для избирательного удаления тех последовательностей, которые были отобраны исключительно на это свойство. Такой скрининг и селекция могут быть осуществлены всякий раз, когда мишень содержит примеси либо в процесс селекции включаются некоторые отклонения, не связанные со сродством к мишени.
SELEX был продемонстрирован путем его использования для выделения РНК-молекул, связывающихся с ДНК-полимеразой бактериофага Т4 (называемой также gp43), или ингибирующих функцию этого белка. Новый РНК-лиганд к ДНК-полимеразе Т4 может быть использован в качестве специфического реагента для анализа на ДНК-полимеразу Т4. Синтез ДНК-полимеразы Т4 является саморегулируемым. В отсутствие функционального белка амбер-фрагменты и мутантные белки при инфекциях с дефицитом репликации испытывают сверхсинтез по сравнению со скоростью синтеза белка дикого типа (Russel (1973), J. Mol. Biol. 79: 83-94). in vitro-трансляции N-концевого фрагмента gр43 специфически ингибируется путем добавления очищенного gр43, и указанный gр43 защищает дискретную часть мРНК возле ее сайта связывания с рибосомой от нуклеазной атаки (Andrake и др. (1988), Proc. Natl. Acad. Sci. USA 85:7942-7946). Были установлены размер и последовательность трансляционного оператора РНК, с которым связывается gp43, а также эффективность этого связывания. gp43-оператор минимального размера представляет собой последовательность примерно из 36 нуклеотидов, как показано на фиг.1, которая как было предсказано имеет шпилечно-петлевую структуру. Минимальный размер оператора определяли путем анализа связывания меченных по концу фрагментов оператора к gp43. Анализ связывания мутантов оператора в последовательности шпильки и петли показали, что gp43, связывающийся с оператором, является чувствительным к первичным изменениям оснований в спирали. Связывание с полимеразой даже больше снижалось при изменениях, которые значительно снижают стабильность шпильки. Было обнаружено, что связывание оператора очень чувствительно к последовательности петли. Было также установлено, что репликация и связывание с оператором для gp43 представляют собой взаимоисключающие активности. Было обнаружено, что добавление микромолярных количеств очищенной РНК, содержащей целый оператор, сильно ингибирует in vitro-репликацию посредством gp43.
gp43-оператор дикого типа (фиг.1) был использован в качестве основы для составления начальной смеси РНК-молекул, содержащих рандомизированную область в целях оценки способности процесса селекции/амплификации выявлять нуклеиново-кислотные молекулы, которые связываются с белком. Испытуемую смесь РНК получали путем in vitro-транскрипции из одноцепочечной ДНК-матрицы из 110 оснований. Эту матрицу конструировали как показано на фиг.1 для кодирования большей части операторной последовательности дикого типа, за исключением последовательности петли. Последовательность петли в 8 пар оснований заменяли рандомизированной областью последовательности, которую синтезировали так, чтобы она была полностью произвольной в каждом основании. Эта матрица также содержала последовательности, необходимые для эффективной амплификации: последовательность в ее 3'-конце, комплементарную праймеру для обратной транскрипции и амплификации в полимеразно-цепных реакциях, и последовательность в ее 5'-конце, требуемую для инициации Т7-РНК-полимеразной транскрипции, и последовательность, достаточно комплементарную кДНК in vitro транскрипта. ДНК-матрица представляет собой смесь всех вариантов последовательностей петли, теоретически содержащей 65536 отдельных видов.
Было установлено, что константа диссоциации для РНК-петли дикого типа составляет около 5•10-9 М. Константа диссоциации для популяции вариантов последовательностей петли составляла около 2,5•10-7. Рандомизация последовательности петли снижала аффинность связывания в 50 раз.
In vitro-транскрипты, содержащие варианты последовательности петли, смешивали с очищенным gp43 и инкубировали. Смесь фильтровали через нитроцеллюлозные фильтры. Белок-РНК-комплексы задерживались на фильтры, а несвязанные РНК - нет. Затем отобранные РНК элюировали с фильтров, как описано в Примере 1. Отобранные РНК наращивали с помощью обратной транкриптазы AMV в присутствии 3'-праймера, как описано Gauss и др. (1987) (см. выше). Полученные кДНК амплифицировали с помощью ДНК-полимеразы Tag в присутствии 5'-праймера в течение 30 циклов, как описано Junis и др. (1986) (см. выше). Отобранные амплифицированные ДНК служили в качестве матрицы для in vitro-транскрипции в целях продуцирования отобранных амплифицированных РНК-транскриптов, которые затем подвергают другому раунду селекции/амплификации. Отношение РНК/белок в смеси, селектируемой на связывание, поддерживается постоянным в процессе всех циклов селекции. Итеративную селекцию/амплификацию проводили с использованием различных молярных отношений РНК/белок. Во всех экспериментах использовали избыточное количество РНК: в эксперименте А использовали РНК/gp43=10/1 (М/М); в эксперименте В использовали РНК/gp43=1000/1; и в эксперименте С использовали РНК/gp43=100/1.
Преимуществом процесса селекции было контролирование с помощью анализов на связывание фильтров меченных транскриптов амплифицированной кДНК по завершении каждого цикла процедуры. Для контроля прохождения селекции осуществляли также периодическое секвенирование РНК-продуктов от каждого раунда эксперимента В. Автограммы секвенирующих гелей РНК-продуктов после 2, 3 и 4 раундов селекции/амплификации показаны на фиг.3. Очевидно, что никаких заметных смещений последовательности петли не наблюдалось до четвертого раунда селекции. После четвертого раунда селекции для последовательности петли из 8 пар оснований наблюдалась "консенсус"-последовательность: А (а/g) (U/с) ААС (U/с) (U/с). Периодическое секвенирование отобранной РНК после четвертого раунда селекции для экспериментов В, В и С сравнивалось на фиг.4. Все три независимые SELEX-процедуры с использованием отношений РНК/белок дают аналогичные "консенсус"-последовательности. Однако имеются некоторые видимые отклонения для последовательности петли дикого типа (AAUAACUC) в отобранных РНК экспериментов А и С.
Для того чтобы определить, какие допустимые комбинации последовательностей могут реально присутствовать в отобранных РНК, отдельные ДНК были клонированы из выбранных РНК после четвертого раунда селекции в эксперименте В. Последовательность, полученная в результате эксперимента В, показала равномерное распределение двух допустимых нуклеотидов, каждый из которых содержал 4 различных вариабельных положения последовательности петли. Отдельные из них были клонированы в р С18 в соответствии с описанием Sambrook, J. и др. (1989), Molecular Cloning: A Laboratory Manual. (Cold Spring Harbor, N.-Y.), части 1.13; 1.85-1.86. Двадцать отдельных клонов были идентифицированы с помощью гибридизации с 3-праймером, а затем секвенированы. Ни один из секвенированных клонов не был мутантом в любом месте операторной последовательности за пределами последовательности петли. Как показано на фиг.7, наблюдалось только пять вариантов последовательностей и, что удивительно, только два варианта последовательности были крупными компонентами выбранной смеси. На фиг. 7 также приводится частота каждой последовательности в 20 отдельных секвенированных изолятах. Последовательность дикого типа AAUAAUCUC и петля AGCAACCU отобранной РНК эксперимента В присутствовали приблизительно в равных количествах. Другие отобранные варианты были мутантами двух крупных вариантов по 1 основанию. Эффективность связывания вариантных последовательностей сравнивали при помощи анализа на связывание с фильтрами, используя при этом меченые in vitro-транскрипты, происходящие от каждого из очищенных клональных изолятов. Как показано на фиг.6, наблюдалась грубая корреляция между аффинностью связывания РНК для gp43 и распространенностью отобранной последовательности. Два крупных варианта последовательности петли показали приблизительно равные аффинности связывания для gp43. Вариантные РНК с последовательностями петли, выделенные в процессе селекции/амплификации (показанные на фиг. 7), могут действовать как ингибиторы gp43-полимеразной активности, как это было продемонстрировано для операторной последовательности дикого типа.
Пример использования SELEX был проиллюстрирован отбором нового РНК-лиганда к ДНК-полимеразе Т4 (gp43) (Andrake и др. (1988), Рrос. Natl. Acad. Sci. USA 85:7942-7946).
Настоящее изобретение также относится к специфическим лигандным растворам, полученным в результате процесса SELEX, которые, как было показано, обладают повышенной аффинностью к обратной транскриптазе ВИЧ-1, оболочечному белку R17, rev-белку ВИЧ-1, ДНК-полимеразе HSV, рибосомному белку S1 E.coli tPA и NGF. Эти лигандные растворы могут быть использованы каждым специалистом для синтеза нуклеиново-кислотных антител к различным мишеням.
Приведенные ниже примеры описывают успешное применение SELEX к широкому спектру мишеней. Эти мишени, в основном, могут быть отнесены к двум категориям: белки, связывающиеся с нуклеиновыми кислотами, и белки, функция связывания которых с нуклеиновыми кислотами не установлена. В каждом случае получали лигандный раствор. В некоторых случаях лигандный раствор может быть представлен как нуклеиново-кислотный элемент (мотив), такой как шпилечная петля, асимметричный "балдж" (выпуклость) или псевдоузел. В других примерах лигандный раствор представлен в качестве первичной последовательности. В этих случаях, однако, не подразумевается, что этот лигандный раствор не содержит какой-либо определенной третичной структуры.
Помимо ДНК-полимеразы Т4, способ SELEX был успешно осуществлен на других мишенях, таких как оболочечный белок бактериофага R17, обратная транскриптаза ВИЧ (ВИЧ-RТ), rev-белок ВИЧ-1, HSV - ДНК-полимераза +/- кофактор, рибосомный белок S1 E.coli, tPA и NGF. Представленные ниже эксперименты описывают также схему испытаний на сродство объемного связывания рандомизированной смеси нуклеиновых кислот-кандидатов к различным белкам. В Примере 7 также описана иммобилизация брадикинина и результаты исследования объемного связывания рандомизированных нуклеиновых кислот с использованием брадикинина.
Примеры и иллюстрации, приведенные в настоящем описании, никоим образом не ограничивают объема настоящего изобретения. Фундаментальная идея, положенная в основу настоящего изобретения, заключается в том, что нуклеиновые кислоты как химические соединения могут иметь практически бесконечное разнообразие размеров, форм и конфигураций и обладают каталитическими функциями и широким спектром способностей связываться с различными мишенями, по сравнению с которыми известные в настоящее время свойства нуклеиновых кислот, присутствующих в биологических системах, являются крайне ограниченными.
ПРИМЕРЫ
В настоящих примерах использовались следующие материалы и методы.
Вектор транскрипции рТ7-2 является коммерчески доступным (U.S. Biochemical Company, Cleveland, ОН). Плазмида рVС18 описана Norrander и др. (1983), Gene 25:15-27 и поставляется фирмой New England Biolabs. Все манипуляции с ДНК для конструирования новых рекомбинантных плазмид описаны Маниатисом и др. (1982), Molecular Cloning: A Laboratory Manual, Cold Spring Наrbor Laboratory, Cold Spring Harbor, Нью-Йорк, за исключением тех случаев, о которых оговорено особо. ДНК-олигонуклеотиды синтезировали и очищали в соответствии с описанием Гаусса и др. Gauss et al. (1987), Mol. Gen. Genet. 206:24-34.
In vitro-транскрипции с использованием РНК-полимеразы Т7 и гель-очистку РНК осуществляли в соответствии с описанием Milligan и др. (1987), Nucl. Acids Res. 15:8783-8798, за исключением того, что в реакциях мечения концентрации АТР, СТР и GТР составляли 0,5 мМ (каждая), концентрация UТР составляла 0,05 мМ. UТР метили в альфа-положении, используя 32Р со специфической активностью, составляющей приблизительно 20 Ки/мМ. Получение неочищенных мРНК в результате Т4-инфицирования, мечение олигонуклеотидов и
праймерное удлинение с использованием обратной транскриптазы AMV осуществляли в соответствии с описанием Гаусса и др. (1987) (см.выше).
Разведения меченой, гель-очищенной РНК и очищенного gp43 осуществляли в 200 мМ ацетата калия, 50 мМ Трис-HCl, рН 7,7 при 4oC. В анализе на связывание с нитроцеллюлозными фильтрами, очищенный gp43 серийно разводили и 30 мкл аликвоты каждого разведения белка добавляли к 30 мкл аликвотам разведенной, меченой, гель-очищенной РНК. РНК-разведение 50 мкл наносили пятнами на нитроцеллюлозный фильтр, осушали и подсчитывали число отсчетов на пробирку для определения входного сигнала. Концентрация белка в реакциях составляла от 10-10 М до 10-8 М, а концентрация РНК в каждом эксперименте составляла приблизительно 10-12 М. После инкубации при 4oС в течение 30 минут, каждую пробирку помещали на 3 минуты при 37oС, и 50 мкл каждого образца фильтровали через предварительно смоченные нитроцеллюлозные фильтры (Millipose # HAWP 025 00) и промывали 3 мл 200 мМ ацетата калия, 50 мМ Трис-HCl рН 7,7. Фильтры осушали и просчитывали в сцинтилляционной жидкости EcolumeТМ (ICN Biomedical, Inc. ). Контрольный подсчет проводили при отсутствии gp43, на основании чего определяли фон (всегда меньше, чем примерно 5% от входных импульсов). Из каждой серии измерений вычитали фон и подсчитывали процент полного числа входных импульсов, оставшихся на фильтрах. На основании каждой серии опорных точек строили кривую биомолекулярного связывания, наиболее соответствующую теоретической, используя при этом версию опубликованной программы (Caceci и Cacheris, 1984, см. выше), модифицированную для построения кривой, описываемой уравнением
Q=А[gp43]/(Kd+[gp43]),
где Q представляет собой фракцию полной РНК, которая является связанной с фильтром, А - процент РНК, при котором наступает насыщение связывания (приблизительно 60% для взаимодействия белок - РНК), [gp43] представляет собой входную концентрацию gp43, a Kd является константой диссоциации для биомолекулярной реакции. Это уравнение представляет собой алгебраическую перегруппировку уравнения /1-5/ Bisswanger (1979) Theorie und Methode dez Enzymkinetik, Verlag Chemie, Weinheim, FRG, стр.9, с упрощающим предположением, что концентрация белка значительно превосходит концентрацию комплексов РНК-белок, и это предположение является справедливым в описанных экспериментах.
ПРИМЕР 1. Отбор РНК-ингибиторов ДНК-полимеразы Т4
Для in vitro-транскрипции конструировали одноцепочечную ДНК-матрицу из 110 оснований, как показано на фиг.2, путем лигирования трех синтетических олигонуклеотидов (табл. 1, 3, 4 и 5) в присутствии двух кэппинг-олигонуклеотидов (табл.1 и 2). Один из образующих матрицу олигонуклеотидов использовали также в качестве 3-праймера при обратной транскрипции in vitro-транскрипта и последующей алмплификации в полимеразно-цепных реакциях (РСRS) (Junis и др. (1988), Proc.Natl. Acad. Sci. USA 85:9436-9440). Один из кэппинг-олигонуклеотидов (1) содержит информацию, необходимую для инициации транскрипции с использованием РНК-полимеразы Т7 и достаточную последовательность, комплементарную кДНК in vitro-транскрипта, для того чтобы он мог служить в качестве 5'-праймера в стадиях PCR-амплификации. ДНК-матрица кодировала РНК, которая содержит сайт узнавания полной РНК для ДНК-полимеразы ТК, за исключением того, что полностью произвольная последовательность была введена вместо последовательности, которая должна кодировать последовательность петли AAUAACUC дикого типа. Рандомизированную последовательность вводили с помощью традиционного химического синтеза, используя коммерческий ДНК-синтезатор (Applied Biosystems), за исключением того, что все четыре dNTР' присутствовали в эквимолярных количествах в реакционной смеси для каждого положения, указанного N, в последовательности олигонуклеотида под номером 4 (табл. 1). Рандомизированную последовательность фланкировали путем праймерного отжига информационной последовательности для 5'- и 3'-олигонуклеотидов, использованных в PCR. Таким образом, ДНК-матрица представляет собой смесь всех вариантов последовательности петли, теоретически содержащую 65536 отдельных видов. Константа диссоциации для вариантной РНК-последовательности петли составляет около 5•10-9 М, а для популяции последовательностей - около 2,5•10-7 М, аффинность связывания в 50 раз ниже.
In vitro-транскрипты, содержащие варианты последовательности петли, смешивали с очищенным gp43 при трех различных соотношениях РНК-белок во всех многочисленных раундах селекции. Для А и В концентрация gp43 составляла 3•10-8 М ("низкий белок"), а для С концентрация gp43 составляла 3•10-7 ("высокий белок"). Для А концентрация РНК составляла около 3•10-7 ("низкая РНК"), а для В и С концентрация РНК составляла около 3•10-5 М ("высокая РНК"). Один раунд включали в себя следующие стадии:
1) Селекция. РНК и белок смешивали в нужных соотношениях, указанных выше, инкубировали при 37oC, промывали через нитроцеллюлозный фильтр, и РНК элюировали с фильтров, описанных выше.
2) Амплификация. РНК, элюированную с фильтров, удлиняли с использованием обратной транскриптазы AMV в присутствии 50 пикомолей 3'-праймера в 50 мкл реакции при условиях, описанных Гауссом и др. (1987) (см. выше). К полученным с помощью кДНК-синтеза 50 пМ 5'-праймера добавляли в реакционном объеме 100 мкл, и амплифицировали с использованием ДНК-полимеразы Tag, как описано (1988) (см. выше) в течение 30 циклов.
3) Транскрипция. In vitro-транскрипцию осуществляли на выбранной амплифицированной матрице, как описано Мilligan и др. (1987) (см. выше), после чего добавляли ДНКазу I для удаления ДНК-матрицы. Полученные выбранные РНК-транскрипты затем использовали в стадии 1 следующего раунда. Только 1/20 продуктов, полученных в каждой стадии цикла, использовали в последующих циклах так, чтобы можно было проследить динамику отбора. Прогресс метода отбора контролировали с помощью анализа связывания с фильтром меченых траскриптов из каждой PCR-реакции. После четвертого раунда отбора и амплификации отобранные меченые РНК-продукты продуцировали связывание с gp43, эквивалетное связыванию контрольной РНК дикого типа. РНК-продукты из каждого раунда одного эксперимента (В) и из четырех раундов для всех трех экспериментов подвергали гель-очистке и секвенировали. На фиг.3 показана последовательность очищенных in vitro-транскриптов, полученных из второго, третьего и четвертого раундов селекции и амплификации для эксперимента В. Очевидно, что статистическое отклонение последовательности петли вводили только после третьего отбора. На этой стадии отбора имеется обнаружимое отклонение, которое дополнялось четвертым раундом для видимой обобщающей последовательности А (а/g) (U/с) ААС (U/c)(U/c). Периодическое секвенирование РНК, транскрибированной после четвертого раунда селекции и амплификации, для испытаний А, В и С проиллюстрировано на фиг.4. Все три независимые партии с различными отношениями белок/РНК дали одинаковые результаты. В экспериментах А и С наблюдалось некоторое видимое отклонение для последовательности дикого типа в каждом из четырех "вариабельных" положений.
Для того чтобы установить, которые из допустимых комбинаций реально присутствовали, использовали два "клонирующих" олигонуклеотида, которые содержали информацию о сайтах рестрикции, для амплификации последовательностей от РНК, полученные в четвертом раунде эксперимента В, некоторые из которых клонировали в pUC18, как описано Sambrook и др. (1989) (см. выше); Junis и др. (1988) (см. выше). Отобранные партии испытания В выбирали для дальнейшей оценки, поскольку имеет значение даже распределение двух допустимых нуклеотидов, каждый из которых состоит из четырех "вариабельных" положений. Двадцать отдельных клонов, которые были идентифицированы путем гибридизации колоний на фильтре с 3'-праймером секвенировали. Ни один из них не оказался мутантом в любом месте операторной последовательности за пределами положений последовательности петли, которые намеренно изменяли. Распределение последовательностей систематизировано на фиг.7. Неожиданно было обнаружено, что отобранная РНК-смесь состояла фактически из двух крупных последовательностей петли. Одна из них была последовательностью дикого типа ААUААСUС, 9 клонов которой из 20 были выделены. Другая, AGGAACCU, была мутантом в четырех положениях и присутствовала в 8 из 20 клонов (см. фиг.7). Другие три последовательности петли обнаруживали одиночные мутации этих двух крупных последовательностей. Эксперимент на связывание с фильтром с использованием меченых in vitro-транскриптов, происходящих от каждого из указанных клональных изолятов, показал, что имеется грубая корреляция между аффинностью связывания РНК для gp43 и выбранной распространенностью (см. фиг.7).
ПРИМЕР 2. Выделение специфического РНК-лиганда для обратной транскриптазы ВИЧ
Активность обратной транскриптазы ВИЧ-1 обусловлена гетеродимером из двух субъединиц (р51 и р66), которые имеют общие аминоконцы. Внешняя карбоксиконцевая область большего пептида составляет домен РНКазаН обратной транскриптазы, структура которого была недавно определена с высоким разрешением.
В предшествующих работах было показано, что обратная транскриптаза ВИЧ-1 непосредственно и специфически взаимодействует с родственной ей праймерной тРНКLys3, с которой ее экспериментально перекрестно сшивали в антикодоновой петле и стебле. Было также обнаружено, что только лишь гетеродимер показывает такое специфическое РНК-разпознавание, и ни один из гомодимерных видов обратной транскриптазы специфично не связывается с указанной тРНК.
Две матричных популяции (приблизительно с 1014 различными последовательностями каждая) конструировали для использования в SELEX при лигировании. Одна популяция матриц была рандомизирована по 32-нуклеотидным положениям с использованием фиксированных последовательностей на концах рандомизированной области в целях проведения кДНК-синтеза и РСR-амплификации. Вторая популяция матриц имела в качестве дополнительной фиксированной последовательности в 5'-конце РНК антикодоновую петлю и стебель тРНКLys3. (Все олигонуклеотиды, использованные в этой работе, показаны в табл.2). Не наблюдалось никакого различия между аффинностями этих двух рандомизированных популяций для обратной транскриптазы [RT] ВИЧ 1 и, как было показано, РНК, которые были отобраны, не используют 5'-область при специфическом связывании. Было проведено 9 раундов SELEX для каждой популяции с использованием гетеродимерной ВИЧ-RT в качестве белка-мишени.
Механизм, посредством которого получали рандомизированную ДНК с использованием лигирования и мостик-образующих олигонуклеотидов, был описан ранее. Указанная методология может уменьшить полное число различных последовательностей в исходной популяции в результате теоретического предела, налагаемого ДНК-синтезом в масштабе 1 микромоль.
В указанных реакциях лигирования использовали около 1 нМ каждого олигонуклеотида. Лигированный продукт подвергали голь-очистке приблизительно с выходом 50%. Эту очищенную матрицу транскрибировали с использованием РНК-полимеразы Т7, как описано выше. Было установлено, что ВИЧ-RT может связываться с насыщением с указанной рандомизированной популяцией, где половина максимального связывания происходит примерно при 7•10-7 М, как было определено с помощью анализа на нитроцеллюлозных фильтрах. Все реакции связывания РНК с белком проводили в буфере для связывания: 200 мМ КОАс, 50 мМ Трис-НСl, рН 7,7, 10 мМ дитиотреитола. Разведения РНК и белка смешивали и выдерживали на льду в течение 30 минут, а затем переносили на 5 минут в режим 37oC. В анализе на связывание объем реакции составлял 60 мкл, из которых анализировали 50 мкл; в раундах SELEX (реакционный объем составлял 100 мкл). Каждую реакционную смесь пропускали через всасывающий заранее увлажненный буфером для связывания нитроцеллюлозный фильтр и промывали 3 мл буфера для связывания, после чего осушали и просчитывали для анализа либо подвергали элюированию как часть процедуры SELEX. Проводили 9 раундов. Во всех девяти раундах концентрация РНК составляла приблизительно 3•10-5 М. В первой выборке ВИЧ-RT составляла 2•10-8 М, в выборках 2-9 - 1•10-8.
Сначала был проведен эксперимент с использованием РНК, содержащей антикодоновую петлю и стебель тРНКLys3. Анализ на связывание с нитроцеллюлозными фильтрами, осуществленный в 9 раундах, показал, что РНК-популяция имела сродство к RT ВИЧ-1 в 100 раз выше, чем исходная смесь кандидатов, но при этом фон связывания с нитроцеллюлозными фильтрами в отсутствие белка был повышен на примерно от 2% от исходной РНК до 15%. Отдельные последовательности, клонированные из этой популяции (после фильтрации через нитроцеллюлозные фильтры для устранения высокого фона возможных последовательностей, отобранных на связывание лишь одними фильтрами), представлены в табл.3. Анализы на связывание с нитроцеллюлозными фильтрами для определения сродства выбранных последовательностей к ВИЧ-RT проиллюстрированы на фиг.14. Некоторые последовательности были отобраны как лиганды для ВИЧ-RT, которые проиллюстрированы кривыми связывания лигандов (1.1) и (1.3а), и обнаруживают некоторую гомологию последовательностей, как видно из табл.4 и 5. Некоторые из лигандных последовательностей обнаруживают значительное удерживание на нитроцеллюлозных фильтрах в отсутствие белка, иллюстрируемое лигандом 1.4 (фиг. 14), и, очевидно, характеризуются длинной спиралью с петлей из повторяющихся пуриновых элементов (как показано в табл.4). Несмотря на минимальные усилия обнаружить их в данном эксперименте перед клонированием, тем не менее было отмечено, что эти последовательности составляют значительную часть из тех последовательностей, которые были собраны в результате проведения этого эксперимента.
Поэтому в эксперименте 2 (с отличающейся 5-фиксированной последовательностью) была проведена предварительная фильтрация через нитроцеллюлозу перед первым, третьим, шестым и девятым раундами селекции. Последовательности, собранные в результате этих экспериментов, представлены в табл.6. И опять наблюдалось много последовательностей с гомологией к последовательностям с высокой аффинностью, полученным из эксперимента 1, как показано в табл.4 и 5. При этом имелось (если вообще имелось) значительно меньше последовательностей, которые соответствовали типу последовательностей, удерживаемых только нитроцеллюлозными фильтрами. Анализ лигандных последовательностей, отобранных из указанного эксперимента, проведенный на связывание с нитроцеллюлозными фильтрами, сравнивали с анализом лиганда 1.1, и это сравнение проиллюстрировано на фиг.15.
Лигандные РНК с высокой аффинностью, имеющие наиболее общую последовательность (1.1) и аналогичную последовательность (1-3а), были затем проанализированы для определения границ информации, требуемой для высокоаффинного связывания с ВИЧ-1-RT. Результаты этого эксперимента представлены на фиг.16. Эти эксперименты показали, что общий для этих последовательностей мотив, UUCCGNNNNNNNNCGGGAAA, аналогичным образом расположен внутри области распознавания. Последовательности UUCCG и CGGGA этого мотива могут спариваться с образованием РНК-спирали с петлей из 8 оснований. Для этого чтобы установить, что еще помимо этих фиксированных последовательностей может обеспечивать высокоаффинное связывание с RT ВИЧ-1, создавали матрицу смеси кандидатов, которая содержала произвольное включение в нуклеотидные положения, которые отличались от этих двух последовательностей, как показано в табл.7. После восьми раундов SELEX отдельные последовательности были клонированы и секвенированы. В табл. 7 представлены 46 последовательностей. Исследование этих последовательностей показало экстенсивное спаривание оснований между центральной вариабельной 8 н-областью и расположенное ниже вариабельной 4 н-областью и фланкирующими последовательностями; спаривание оснований, в сочетании с вышеуказанным спариванием, должно указывать на псевдоузел РНК. Тот факт, что в выделенной популяции не наблюдалось преобладания каких-либо специфических последовательностей, позволяет предположить, что в данном случае отсутствует селекция на уровне первичных последовательностей и что указанная селекция происходит лишь на основании вторичной структуры, то есть имеется много комбинаций последовательностей, которые дают аналогичные аффинности к RT ВИЧ-1, но ни одна из них не имеет конкурентого преимущества. Анализы первого и второго SELEX-экспериментов показали, что отдельные последовательности, которые составляют популяции, имеющие гомологию с UUCCG... CGGGANAA-мотивом, также обнаруживают высокий потенциал для указанного спаривания оснований псевдоузла.
На фиг. 31 представлена схематическая диаграмма элемента, называемого в настоящем описании псевдоузлом. Этот псевдоузел состоит из двух спиральных областей и трех петлеобразных областей. Однако не все псевдоузлы содержат все три петли. Для интерпретации полученных данных, различные области псевдоузла были отмечены, как показано на фиг.31. Например, в табл.5 некоторые из последовательностей, полученных в экспериментах 1 и 2, были представлены в соответствии с предполагаемой конфигурацией псевдоузла, образуемого различными последовательностями.
Результаты эксперимента 1 и 2, как показано в табл.5, были положены в основу эксперимента 3, где последовательности в S1 (а), S1 (b) и L3 были фиксированы. И аналогично нуклеиновые кислоты, полученные в результате SELEX, имеют пространственную конфигурацию почти исключительно в псевдоузлах. Исследование результатов каждого эксперимента показало, что смесь нуклеиновых кислот по отношению ВИЧ-RT содержит относительно большое число членов, наибольшим общим знаменателем которых является то, что все они имеют конфигурацию псевдоузлов. Указанные исследования смеси нуклеиновых кислот для ВИЧ-RT позволяют сделать и другие обобщения:
1) S1 (а) часто содержат последовательность 5'-UUCCG-3', a S1 (b) часто содержат последовательность 5'-CGGGA-3'. Однако допустимое сбрасывание пар оснований может привести к укорочению стебля.
2) 1 может быть коротким или длинным, но часто включает в себя два нуклеотида в нуклеиновых кислотах с наилучшим связыванием, 5-нуклеотид в L1 чаще всего бывает либо U, либо А.
3) 2 обычно состоит из 5 или 6 пар оснований и, очевидно, является независимой последовательностью. Этот стебель может содержать не-уотсон-криковские пары.
4) 2 может не содержать нуклеотидов, но если он существует, то нуклеотидами предпочтительно являются А.
5) 3 представляет собой, в основном, 3 или более нуклеотидов, обогащенных в А.
6) В большинстве последовательностей, полученных в результате SELEX, полное число нуклеотидов в 1, S2 (а) и L2 равно 8.
Основной целью настоящего эксперимента было выявление смеси лигандов к RT ВИЧ-1. Способность выделенного лигадного клона 1.1 сравнивали со способностью исходной популяции для эксперимента 1 ингибировать активность обратной транскриптазы, и эта способность проиллюстрирована на фиг.17. Даже при равных концентрациях РНК-ингибитора RT, обратная транскриптаза, в основном, ингибируется лигандом 1.1. В противоположность этому только 10 мМ (или 200-кратный избыток) исходной популяции РНК обеспечивает более или менее значительное ингибирование RT ВИЧ-1. Таким образом, высокоаффинный лиганд к RT ВИЧ-1 либо блокирует, либо непосредственно взаимодействует с каталитическим сайтом фермента.
Для оценки специфичности указанного ингибирования были проанализированы различные концентрации лиганда 1.1 для ингибирования обратной транскриптазы MMLV, АМV и ВИЧ-1. Результаты этого эксперимента, представленные на фиг.18, показали, что ингибирование обратной транскриптазы ВИЧ-1 лигандом 1.1 является специфическим.
ПРИМЕР 3. Выделение специфического РНК-лиганда для оболочечного белка бактериофага R17
Проводили SELEX на оболочечном белке бактериофага R17. Белок был очищен в соответствии с описанием Carey и др. Biochemistry 22, 2601 (1983). Буфер для связывания состоит из 100 мМ ацетата калия, 10 мМ дитиотреитола и 50 мМ Трис-ацетата, рН 7,5. Белок и РНК инкубировали вместе в течение 3 минут при 37oС, а затем фильтровали на нитроцеллюлозных фильтрах для отделения РНК, связанной с белком, от свободной РНК. Эти фильтры промывали 50 мМ Трис-ацетатом рН 7,5. В первых четырех раундах SELEX концентрация белка составляла 1,2•10-7 М, а в раундах 5-11 - 4•10-8 М.
Исходную РНК транскрибировали с ДНК, как было описано раннее. ДНК-последовательность включает в себя промоторную последовательность РНК-полимеразы бактериофага Т7, что позволяет синтезировать РНК в соответствии со стандартной техникой. кДНК-синтез во время амплификационной части цикла SELEX праймировали с использованием ДНК-последовательности: кДНК-праймер (РСR-праймер 1):
5' GTTTCAATAGAGATATAAAATTCTTTCATAG 3'
ДНК-праймеры, использованные для амплификации кДНК, таким образом, представляли собой последовательность, включающую в себя Т7-промотор, 32 рандомизированных положения, АТ-нуклеотид, фиксированную последовательность, комплементарную PCR-праймеру 1. РНК, которую использовали для начала первого цикла SELEX, имела последовательность:
pppGGGAGCCAACACCACAAUUCCAAUCAAG-32н-AUCUAUG AAAGAAUUUUAUCUC AUUGAAAC
Партию клонов, полученную после 11-го раунда SELEX, секвенировали. Из 38 различных последовательностей, полученных в 47 клонах, три были обнаружены более одного раза: одна последовательность была обнаружена 6 раз, одна последовательность - 4 раза и одна - 2 раза. Каждая из оставшихся 35 последовательностей была обнаружена один раз. Две последовательности не соответствовали другим в отношении первичных последовательностей или, вероятно, вторичных структур и далее не анализировались. 36 последовательностей имели общую последовательность АNСА, расположенную в виде тетрануклеотидной петли шпильки с "балджем" (т.е. с дополнительной выпусклостью - прим.пер.); нуклеотид "балджа" представлял собой аденин во всех 36 случаях. Последовательности всей партии представлены в табл.8, и эти последовательности сравнивали с 4 нуклеотидами шпилечной петли. Два нуклеотида 3' до рандомизированной части исходной РНК (АU) являются свободными и могут быть заменены или делегированы, поскольку кДНК-праймер не включает в себя комплементарных нуклеотидов; во многих клонах были заменены один или два из указанных нуклеотидов.
Предпочтительный РНК-мотив, показанный на фиг.19, непосредственно связывается с сайтом связывания оболочки, идентицифированным ранее с помощью сайт-направленного мутагенеза и анализов на связывание. См., Uhlenbeck и др. , см. выше, 1983; Ramaniuk и др., см. выше (1987). Однако в этой партии некоторые последовательности были более консервативными, чем это ожидалось. Последовательность петли АUСА преобладала, тогда как предыдущие данные связывания давали основание предполагать, что последовательности АNСА все будут эквивалентными. Натуральный сайт связывания на геноме R17 имеет следующую последовательность и структуру:
Натуральная структура включает в себя последовательность GGAG, которая служит для облегчения связывания с рибосомой и инициации трансляции области, кодирующей репликазу R17. В процессе SELEX это требование отсутствует, и полученные в результате последовательности содержат возле петли и "балджа" пары оснований С:G более часто, чем пары оснований G:С. Поэтому SELEX позволяет ослабить ограничения, накладываемые природой и эволюционной динамикой, и дает возможность получить лиганды с более высоким сродством, чем природные лиганды. Аналогичным образом цитидин петли, обнаруженный в каждой из 36 последовательностей, является уридином в натуральном сайте, а специалистам известно, что С дает более высокую аффинность, чем U. В процессе отбора, натуральные сайты должны иметь подходящее сродство, но не самое высокое сродство, поскольку очень тесное связывание может принести вред организму.
ПРИМЕР 4. Выделение нуклеиново-кислотного лиганда для сериновой протеазы
Сериновые протеазы представляют собой белковые ферменты, которые расщепляют пептидные связи в белках. Сериновые протеазы являются членами семейства генов у млекопитающих и важными ферментами, необходимыми для жизни млекопитающих. До настоящего времени не было установлено, что сериновые протеазы связываются с нуклеиновыми кислотами. Примерами сериновых протеаз являются активатор плазминогена, трипсин, эластаза, химотрипсин, тромбин и плазмин. Многие заболевания могут быть излечимы путем использования нуклеиново-кислотных лигандов, которые связываются с сериновыми протеазами, например, такие заболевания, как нарушения свертывания крови, и образование тромбов. Другие (не сериновые) протеазы также являются важными для млекопитающих и поэтому также могут служить мишенями для нуклеиново-кислотных лигандов с соответствующими аффинностями, которые могут быть получены в соответствии с настоящим изобретением.
Тканевый активатор плазминогена человека (htPA), который является коммерчески доступным, выбирали в качестве сериновой протеазы для проведения SELEX-способа настоящего изобретения. Смесь РНК-кандидатов была идентична смеси, используемой в Примере 11, описанном ниже, в эксперименте с ДНК-полимераз HSV.
В процессе SELEX связывание происходило в 50 мМ NaCl и 50 мМ Трис-ацетата рН 7,5 в течение 3 минут при 37oС. SELEX проводили в десять раундов. Смесь 30 -кандидатов связывалась с tPA с аффинностью (kd) 7•10-8 М в 150 мМ NaAc и 50 мМ Трис-ацетата рН 7,5; при этом аффинность РНК, присутствующей после 9 раундов SELEX, была в три раза выше. Девять клонов были выделены и секвенированы, а некоторые из них были проанализированы на связывание с tРА как чистые РНК. Последовательности девяти клонов, полученных с использованием низкосолевого буфера, см. в табл.А.
Все исследованные последовательности связывались, по крайней мере, несколько лучше, чем исходная смесь 30 н -кандидатов. Однако А-серии связывались с нитроцеллюлозой лучше в отсутствие tPA, чем смесь кандидатов, словно мотив общей последовательности сам по себе удерживался на нитроцеллюлозной матрице. В последовательностях, показанных выше, этот мотив подчеркнут. В других SELEX-экспериментах AGG-повторы были выделены при попытке идентифицировать лигандную смесь для транскриптазы ВИЧ-1, внеклеточного домена рецептора гормона роста человека и даже для оболочечного белка R17 в первом эксперименте "прогулки". При анализе эти последовательности показали скромное или значительное связывание с нитроцеллюлозными фильтрами при отсутствии белка-мишени. Оказалось, что АGG-повторы могут быть обнаружены в шпилечных петлях. Поскольку в большинстве вариантов настоящего изобретения SELEX является итерационным процессом, то появление указанных связывающих элементов не является неожиданностью.
Присутствия элементов связывания с нитроцеллюлозой можно избежать с помощью одной или нескольких очевидных стратегий. РНК может быть профильтрована через нитроцеллюлозные фильтры до SELEX для устранения указанных элементов. В альтернативных раундах SELEX могут быть использованы альтернативные матрицы, например фильтры из стекловолокна. Могут быть также использованы альтернативные системы распределения, например колонки, градиенты сахарозы и т.п. Очевидно, что любой данный одиночный процесс будет приводить к смещениям в итеративном способе, тем самым благоприятствуя элементам, которые не способствуют увеличению эффективности связывания с мишенью, но тем не менее будут отобраны селекционным процессом. Поэтому очень важно использовать альтернативные или скринирующие процедуры для устранения этих элементов. Было показано, что AGG-повторы аналогично другим повторам, выявленным как мишень-независимые отклонения, наиболее часто будут возникать в том случае, когда аффинность наилучших по отношению к данной мишени последовательностей является довольно низкой или когда аффинности наилучших последовательностей являются лишь немного лучше, чем аффинность исходной смеси кандидатов к данной мишени.
ПРИМЕР 5. Выделение нуклеиново-кислотного лиганда к рецептору, происходящему от млекопитающего
Рецепторы млекопитающих часто являются белками, которые находятся внутри цитоплазматических мембран клеток и реагируют с молекулами, присутствующими за пределами данной клетки. Факт связывания с нуклеиновыми кислотами для большинства рецепторов не установлен. Человеческий рецептор гормона роста взаимодействует с циркулирующим гормоном роста человека, а рецептор инсулина взаимодействует с циркулирующим инсулином. Рецепторы часто обладают глобулярной частью молекулы на внеклеточной стороне мембраны, и именно эта глобулярная часть специфически связывается с гормоном, который является натуральным лигандом. Многие болезненные состояния могут быть излечимы с помощью нуклеиново-кислотных лигандов, которые связываются с рецепторами.
Лиганды, которые связываются с растворимой глобулярной областью рецептора гормона роста человека (1hGHR), были идентифицированы и очищены с использованием смеси кандидатов Примера 4. И аналогично связывающие буферы не содержали ДТТ. Растворимая глобулярная область рецептора гормона роста человека может быть получена из коммерческих и академических источников, где указанные рецепторные белки продуцируются с использованием техники рекомбинантных ДНК, применяемых к генам, кодирующим указанные белки. Итеративный процесс SELEX повторяли до тех пор, пока не были обнаружены нужные лиганды. Полученные лиганды клонировали и секвенировали, а затем определяли аффинности связывания для растворимого рецептора. Аффинности связывания определяли также для случая связывания того же лиганда с другими растворимыми рецепторами для того, чтобы удостовериться в специфичности связывания, даже если большинство рецепторов не обнаруживают хорошей гомологии с внеклеточными доменами других рецепторов. Эти лиганды использовали для измерения ингибирования нормальной активности связывания (1hGHR) путем оценки конкуретного связывания между нуклеиново-кислотным лигандом и натуральным лигандом (гормоном).
ПРИМЕР 6. Выделение нуклеиново-кислотного лиганда для гормона или фактора, происходящих от млекопитающих
Происходящие от млекопитающих гормоны или факторы являются белками, например гормон роста, или малыми молекулами, например адреналин, гормон щитовидной железы, которые циркулируют внутри организма животного и на которые может быть оказано воздействие организма путем из связывания с рецепторами, находящимися внутри плазматических мембран клетки. Например, гормон роста человека стимулирует клетки посредством первого взаимодействия с рецептором гормона роста человека, а инсулин стимулирует клетки путем первого взаимодействия с инсулиновым рецептором. Многие факторы роста, например гранулоцитарный колониестимулирующий фактор (GCSF), включая те, которые являются специфическими к типу клеток, сначала взаимодействуют с рецепторами на целевой клетке. В этом случае гормоны и факторы являются натуральными рецепторами. Факт связывания гормонов и факторов с нуклеиновыми кислотами, в основном, не установлен. Многие заболевания, например гипортиреоз, хроническая гипогликемия, могут излечиваться с использованием нуклеиново-кислотных лигандов, которые связываются с гормонами или факторами.
Лиганды, которые связываются с человеческим инсулином, были идентифицированы и очищены с использованием исходных материалов Примера 3. Человеческий инсулин получали из коммерческих источников, продуцированных с помощью техники рекомбинантных ДНК. Процесс SELEX повторяли до тех пор, пока не были обнаружены лиганды. Эти лиганды клонировали и секвенировали, а затем определяли аффинности связывания для человеческого инсулина. При этом определяли также аффинности связывания того же лиганда для других гормонов или факторов, для того чтобы удостовериться в специфичности связывания, даже если большинство гормонов и факторов не обнаруживают хорошей гомологии с человеческим инсулином. Однако такие семейства генов гормона и фактора существуют, включая небольшое семейство IGF или инсулинподобные факторы роста. Полученные нуклеиново-кислотные лиганды использовали для измерения ингибирования нормальной активности связывания человеческого инсулина с его рецептором путем оценки конкурентного связывания инсулинового рецептора и нуклеиново-кислотного лиганда в присутствии или отсутствие человеческого инсулина (натурального лиганда).
ПРИМЕР 7. Получение колоночной матрицы для SELEX
Согласно процедурам, описанным в Примере 9, см. ниже, было показано, что полипептидный брадикинин не удерживался нитроцеллюлозой. Для осуществления SELEX-процесса на брадикинине, белок связывали с активированной СН-Сефарозой 4В (Pharmacia LKB) в качестве матрицы-носителя в соответствии со стандартными процедурами. Полученная матрица содержала 2,0 мМ брадикинина, как было определено анализом с использованием нингидрида. См. Crestfield и др., J. Biol. Chem. Vol.238, стр.622-627 (1963); Rosen Arch. Biochem. Biophys. Vol. 67, стр.10-15 (1957). Активированные группы, оставшиеся на матрице-носителе, блокировали Трисом. См. Pharmacia, Affinity Chromatography: Principles and Methods, Ljung-foretagen AB, Уппсала, Швеция (1988).
Для взаимодействия растворов смесей кандидатов с гранулярной матрицей использовали разделение на вращающейся колонке. В основной процедуре осуществления стадии селекции в SELEX 40 мкл суспензии 50:50 целевой сефарозы в реакционном буфере переносили в 0,5-миллилитровую пробирку Эппендорфа. Затем с 60 мкл реакционного буфера добавляли смесь РНК-кандидатов, и полученную реакционную смесь уравновешивали при 37oС в течение 30 минут. Затем пробирку, в дне которой была проделана дырка, помещали внутрь другой большей пробирки Эппендорфа, обе пробирки накрывали крышкой и вращали (1000 об/мин, 10", 21oC) для отделения элюата. После чего маленькую пробирку переносили в новую большую пробирку, и содержимое 4 раза промывали, вращая при этом, путем расслоения 50 мкл отобранного промывочного буфера. Для проведения анализа на связывания пробирку, содержащую радиоактивную РНК, переносили в новую пробирку Эппендорфа и вращали досуха.
Проводили эксперимент по объемному связыванию, где смесь РНК-кандидатов, содержащих рандомизированный сегмент из 30 нуклеотидов, наносили на брадикинин-сефарозную матрицу. Используя вращающуюся колонку, определяли связывание объемной смеси 30-н РНК с различными матрицами в условиях высоких концентраций соли для того, чтобы оценить наилучшие условия для минимизации фонового связывания с сефарозой. Фон связывания РНК с сефарозой был минимизирован путем блокирования активированных групп на сефарозе Трисом и использования буфера из 10 мМ ЕМ и 10-20 мМ КОАс. В условиях этого буфера кривая связывания рандомизированного объемного раствора РНК дает в целом Kd около 1,0•10-5 (см. фиг.20). Кривую определяли путем разведения брадикин-сефарозы против блокированной, активированной сефарозы.
ПРИМЕР 8. Получение смесей кандидатов с повышенным содержанием РНК-структурных элементов
В предпочтительном осуществлении настоящего изобретения - смесь кандидатов, используемых в SELEX, состояла из смежной области из 20-50 рандомизированных нуклеиновых кислот. Этот рандомизированный сегмент был фланкирован фиксированными последовательностями, которые способствовали амплификации выбранных нуклеиновых кислот.
В альтернативном осуществлении настоящего изобретения получали смеси для повышения процентного содержания нуклеиновых кислот в смеси кандидатов с данными нуклеиново-кислотными элементами (мотивами). Хотя в этом описании приводится всего лишь два специфических примера, однако настоящее изобретение не ограничивается этими примерами. Каждый специалист может самостоятельно составить эквивалентные смеси кандидатов для достижения аналогичного результата.
В одном из конкретных примеров, представленных как последовательность А на фиг.21, смесь кандидатов получали так, чтобы большинство нуклеиновых кислот в этой смеси имело тенденцию образовывать спиральную область из 4-8 пар оснований и "петлю" либо из 20, либо из 21 сопряженных рандомизированных последовательностей. Оба 5'- и 3'-конца смеси последовательностей будут содержать фиксированные последовательности, которые являются важным фактором для осуществления амплификации нуклеиновых кислот. Смежными с этими функциональными фиксированными последовательностями будут фиксированные последовательности, отобранные для формирования пар оснований на противоположной стороне рандомизированной области. Если смотреть в направлении 5'-->3'-концов последовательностей, то можно указать на 5 различных областей: 1) фиксированные последовательности для амплификации; 2) фиксированные последовательности для формирования спиральной структуры; 3) 20 или 21 рандомизированных нуклеиново-кислотных остатка; 4) фиксированные последовательности для формирования спиральной структуры с областью 2 последовательностей; и 5) фиксированные последовательности для амплификации. Смесь кандидатов А на фиг. 21 будет обогащенной шпилечной петлей и симметричными и асимметричными выпуклыми элементами (мотивами). В предпочтительном варианте осуществления настоящего изобретения смесь кандидатов содержит равное количество последовательностей, где рандомизированная область имеет длину в 20 и 21 оснований.
Во втором примере, показанном на фиг.21 как Последовательность В, составляли смесь кандидатов, обогащенную нуклеиновыми кислотами, содержащихся в псевдоузле. В этой смеси кандидатов фиксированные последовательности для амплификации фланкируют три области из 12 рандомизированных положений. Эти три рандомизированные области разделены двумя фиксированными областями из четырех нуклеотидов, причем указанные фиксированные последовательности отбирали для предпочтительного формирования спиральной структуры из четырех пар оснований. Если смотреть в направлении 5'-->3'-конца последовательности, то можно указать 7 различных областей: 1) фиксированные последовательности для амплификации; 2) 12 рандомизированных нуклеотидов; 3) фиксированные последовательности для формирования спиральной структуры; 4) 12 рандомизированных нуклеотидов; 5) фиксированные последовательности для формирования спиральной структуры с областью 3 нуклеотидов; 6) 12 рандомизированных нуклеотидов; и 7) фиксированные последовательности для амплификации.
В предпочтительной смеси кандидатов спиральные области строили таким образом, чтобы получить альтернативные пары оснований GC, CG, GC, CG. Как было показано, этот набор пар оснований давал особенно стабильную спиральную структуру.
ПРИМЕР 9. Объемное связывание рандомизированных РНК-последовательностей с белками, факт связывания которого с нуклеинокислотными кислотами не установлен
В соответствии с основными процедурами селекции на нитроцеллюлозных фильтрах, описанными выше в Примере 1 для SELEX, исследовали группу произвольно выбранных белков в целях определения их аффинности ко всей смеси кандидатов РНК-последовательностей. В каждом эксперименте используемая смесь кандидатов состояла из 40-РНК-раствора (рандомизированная смесь, имеющая 40-рандомизированный нуклеиново-кислотный сегмент), который подвергали радиоактивному мечению для определения процента связывания. Смесь кандидатов разводили в связывающем буфере (200 мМ КОАс, 50 мМ ТрисоАс, рН 7,7, ДТТ) и 30 мкл использовали в реакции связывания (60 мкл). К каждой реакции добавляли 20 мкл, 10 мкл или 1 мкл каждого белка. Буфер для связывания добавляли до полного объема 60 мкл. Реакции инкубировали при 37oC в течение 5 минут, а затем подвергали связыванию с фильтром. Исследуемыми белками являлись ацетилхолинэстераза (МW 230000): N-ацетил-β-D-клюкозаминидаза (МW 180000); актин (МW 43000), алкогольдегидрогеназа (240000); альдегиддегидрогеназа (МW 200000); хангиотензин (МW 1297); аскорбатоксидаза (МW 140000); предсердный нутриуретический фактор (МW 3064); и бомбезин (МW 1621).
Белки были закуплены у Boehzinger Ingelheim и были использованы в буферной композиции, в которой они продавались.
В каждом эксперименте смесь РНК-кандидатов давала 10726 отсчетов радиометки, а фоновое связывание составляло около 72 отсчетов. Полученные результаты систематизировали в табл.9. Было установлено, что все исследованные белки, за исключением ацетилхолинэстеразы, N-ацетил-β-D-глюкозамидазы и актина, обнаруживали некоторую объемную РНК-аффинность. Из-за низкой концентрации N-ацетил-β-D-глюкозамидазы в купленном растворе результаты, полученные для этого белка, нельзя считать окончательными. Кроме того, если тестируемые белки не связывались с нитроцеллюлозой (которая в данном случае используется для брадикинина), то никакой аффинности в данном эксперименте не обнаруживали. В привиденном выше Примере 7, где обсуждалась колонка-носитель для брадикинина, было продемонстрировано, что неудача этого эксперимента на объемное связывание еще не означает, что это связывание для данного белка отсутствует.
ПРИМЕР 10. Выделение РНК-лигандного раствора для фактора роста нервной ткани
Фактор роста нервной ткани (NGF) является белковым фактором, который действует посредством рецептора на внешней поверхности клеток-мишеней. Антагонисты к факторам роста и другие гормоны могут действовать путем блокирования рецептора или путем титрования фактора или гормона. С помощью SELEX-процесса были проведены поиски РНК, которая непосредственно связываются с NGF.
Исходные РНК получали так же, как и ДНК-полимеразу HSV (Пример 11).
С использованием NGF было проведено два эксперимента.
В первом эксперименте проводили десять раундов SELEX с использованием низкосолевого связывающего буфера, инкубирование осуществляли в течение 3 минут при 37oС с последующей фильтарцией и промывкой тем же буфером в процессе SELEX. Низкосолевой буфер представлял собой 50 мМ NaCl и 50 мМ Трис-ацетана, рН 7,5. Во втором эксперименте в качестве связывающего буфера использовали 200 мМ NaCl и 50 мМ Трис-ацетата, рН 7,5, и после фильтрации промывали 50 мМ Трис-ацетата, рН 7,5. В этом эксперименте проводили только 7 раундов.
В результате эксперимента с низким содержанием соли получали 36 клонированных последовательностей. 15 из этих клонов были почти идентичными #' S 2, 3, 4, 5, 6, 8, 11, 13, 19, 22, 28, 33 и 34 были идентичными, хотя #' S 15 и 25 имеют одно различие:
ACAUCGAUGACCGGAAUGCCGCACACAGAG + А G (15) (25)
Вторая наиболее распространенная последовательность, обнаруженная 6 раз, представляет собой:
CCUCAGAGCGCAAGAGUCGAACGAAUACAG (#' S 12: 20, 27 и 31)
В результате высокосолевого процесса SELEX получали 10 клонов, которые секвенировали, но 8 из них были идентичными и, очевидно, относились ко второму (но малому классу) распространенности из эксперимента с низкосолевым буфером. "Выигравшая" последовательность представляла собой:
----CUCAUGGAGCGCAAGACGAAUAGCUACAUA----
Между двумя экспериментами получали всего 14 различных последовательностей в этих анализах, последовательности с одним различием объединяли вместе; все эти последовательности перечислены в табл.В, где указаны обозначенные сверху подобия и частоты ngf.а - ngf.к - эксперимент с низкосолевым буфером, a hsngf.a - hsngf.с - эксперимент с высокосолевым буфером.
Хотя нет очевидного свидетельства того, что в аналогичные последовательности входит вторичная структура, однако похоже, что "выигравшие" последовательности занимают критические нуклеотиды в структуре, что хорошо согласуется в сайтом NGf-связывания.
Осуществляли анализ на связывание нуклеиновой кислоты hsngf.а с NGF и было обнаружено, что эта нуклеиновая кислота имеет Kd примерно в 20-30 раз выше, чем весь объем 30 - смеси кандидатов. Было также обнаружено, что эта же нуклеиновая кислота имеет более низкую или аналогичную аффинность к оболочечному белку R17 и tРА, чем 30н - смесь кандидатов. Таким образом, нуклеиново-кислотный лиганд hsngf.а является отобранным в результате SELEX лигандом к NGF.
ПРИМЕР 11. Выделение нуклеиново-кислотного лиганда для ДНК-полимеразы HSV-1
Вирус простого герпеса (HSV-1) является ДНК-содержащим вирусом млекопитающих. Аналогично другим ДНК-содержащим вирусам HSV-1 кодирует свою собственную ДНК-полимеразу. Очищали ДНК-полимеразу HSV-1 в двух формах, которые обладают разными качествами, но каждая из них катализирует ДНК-репликацию in vitro. Простую форму, которая представляет собой один полипептид, очищали из клеток, экспрессирующих клонированный ген согласно Hernandez T.R. и Lehman, I. R. , J.Biol. Сhem. 265, 11227-11232 (1990). Вторую форму ДНК-полимеразы, которая является гетеродимером, очищали из НSV-1-инфицированных клеток в соответствии Crute, J. J. и Lehman, I.R., J. Biol. Chem., 264, 19266-19270 (1989); этот гетеродимер содержит один пептид, соответствующий самой полимеразе, а другой, UL42, также кодируется вирусом HSV-1.
SELEX осуществляли на указанном одном полипептиде на гетеродимере. В каждом случае связывающий буфер состоял из 50 мМ ацетата калия, 50 мМ Трис-ацетата, рН 7,5 и 1 мМ дитиотреитола. После 4-минутного инкубирования при 37oС, проводили фильтрацию для отделения связанных РНК; затем фильтры промывали связывающим буфером, в котором отсутствовал дитиотреитол.
Смесь РНК-кандидатов транскрибировали с ДНК, описанной раннее. Как и в других вариантах настоящего изобретения, ДНК-последовательность содержала промоторную последовательность РНК-полимеразы бактериофага Т7, что позволяет синтезировать РНК в соответствии со стандартной техникой. кДНК-синтез в процессе стадии амплификации SELEX праймировали ДНК-последовательности:
кДНК-праймер (PCR-праймер 1): 5' GCCGGATCCGGGCCTCAGTGAA 3
ДНК-праймеры, используемые для амплификации кДНК в этой части SELEX цикла, включают в одном из них Т7-промотор; а РСR-праймер имеет последовательность:
5РСR-праймер 2: ССGААGСТТААТАСGАСТСАСТАТАGGGАGСТСАGААТААА-СGСТСАА-3'
Начальная рандомизированная ДНК содержала последовательность с Т7-промотором, 30 рандомизированными положениями и фиксированную последовательность, комплементарную PCR-праймеру 1. Таким образом, РНК, которая использовалась для начала первого цикла SELEX, имела последовательность:
pppGGGAGCUCAGAAUAAACGCUCAA-30н-UUCGACAUGAGGCCCGGAUCCGGC
SELEX осуществляли в семь раундов, после которого получали кДНК и клонировали как описано ранее. Серии последовательностей, обозначенных "H", получали с использованием ДНК-полимеразы HSV в качестве мишени, а серии "U" получали с использованием гетеродимерной полимеразы, которая содержала полипептид UL 42.
Около 25% последовательностей из Н-серий содержали точную последовательность из 12 нуклеотидов в 5'-конце рандомизированной области (прописные буквы относятся к рандомизированной области). В некоторых последовательностях длина между фиксированными праймерами не является точно 30 нуклеотидов, и в одном случае (Н2) внутри рандомизированной области была обнаружена большая делеция. Члены этой субпопуляции Н приведены в табл.С.
Два члена серий U имеют общий мотив с указанной первичной последовательностью:
U9: --cgcucaaUAAGGAGGGCCACAGAUGUAAUGGAAACuucgaca--
U13: --cgcucaaUAAGGAGGCCACAUACAAAAGGA UGAGUAAAAuucgaca--
Оставшиеся последовательности из Н- и U-серий не обнаруживали общей последовательности: кроме того, в результате седьмого раунда ни в одной серии не было обнаружено какой-либо одной послодовательности-"победителя", что позволяет предположить, что для обнаружения наилучшего семейства лигандов для ингибирования ДНК-полимеразы HSV необходимо осуществление еще большего количества раундов SELEX.
Очевидно, что начальная последовательность
-cgcucaaUAAGGAGGCCAC....
может быть кандидатом для видов антагонистов, однако эти члены серий были еще тестированы как ингибиторы ДНК-синтеза. Очевидно, что фиксированная последовательность, а именно: 5' до UААGGАGGССАС должна принимать участие в образовании этой субпопуляции или общие 12 нуклеотидов должны быть различным образом расположены в рандомизированной области.
ПРИМЕР 12. Выделение нуклеиново-кислотного лиганда к рибосомному белку S1 E.coli
Рибосомный белок S1 30S Е.соli является самым большим из белков 21 30S. Этот белок был очищен на основе его высокой аффинности для полипиримидинов, и очевидно, что он должен достаточно тесно связываться с одноцепочечными полинуклеотидами, которые богаты пиримидинами. Необходимо было узнать, является ли РНК, идентифицированная с помощью SELEX как лигандный раствор, в любом случае более информативной, чем простая одноцепочечная РНК, богатая пиримидинами.
РНК, ДНК, кДНК-праймер (РСR-праймер 1) и PCR-праймер 2 были идентичными, используемыми для ДНК-полимеразы HSV-1 (см. например 11). Связывающий буфер содержал 100 мМ хлорида аммония, 10 мМ хлорида магния, 2 мМ дитиотреитола и 10 мМ Трис-хлорида, рН 7,5. Связывание проводили при комнатной температуре, а комплексы еще раз отделяли путем фильтрации на нитроцеллюлозе. Белок очищали в соответствии с описанием Boni и др., European J. Biochem., 121, 371 (1982).
После 13 раундов SELEX, была получена серия из 25 последовательностей. Более 20 из этих последовательностей содержали псевдоузлы, и эти псевдоузлы содержат общие элементы.
Общая структура псевдоузлов может быть представлена в виде следующей диаграммы:
Стебель 1a - Петля - Стебель 2а - Стебель 1b - Петля 2 - Стебель 2b (см. фиг.31).
Большинство из лигандов к белку S1 содержит:
Стебель 1 из 4 или 5 пар оснований с G у 5' к Петле 1.
Петля 1 примерно из 3 нуклеотидов, часто АСА.
Стебель 2 из 6-7 пар оснований, расположенный непосредственно на Стебле 1.
Петля 2 из 5-7 нуклеотидов, часто оканчивающихся GGААС.
В качестве разумной интерпретации этих данных, можно предположить, что Петля 2 простирается поперек Стебля 1 так, чтобы петля поддерживалась в жесткой форме, что способствует облегчению и усилению связывания одной цепи с активным сайтом белка S1. На фиг.32 представлена примерная картина псевдоузла "консенсуса".
На фиг.32 пары основании показаны линиями и черточками, выбор оснований из рандомизированной области показаны прописными буквами, Y - пиримидин, Р - пурин, N-N' означает любую пару оснований, N означает любой нуклеотид, а строчные буквы относятся к фиксированной последовательности, используемой для РСR-амплификации.
Очевидно, что одноцепочечный полинуклеотид, связывающийся с протеинами и доменами в этих протеинах, будет часто выбирать в процессе SELEX, псевдоузел, который представляет собой протяженную ригидную одиночную цепь, обозначенную Петля 2, до сайта связывания с белком, таким образом, что взаимодействие с этим сайтом будет максимизироваться. Таким образом, если появляется псевдоузел при использовании RT ВИЧ-1, то можно предположить, одноцепочечный домен Петля 2 связывается с областью RT, которая удерживает нить матрицы во время репликации, то есть очевидно, что большинство ферментов репликации ДНК-полимераза, РНК-полимераза, РНК-репликаза, обратная транскриптаза будут иметь домен для удерживания нити матрицы, которая может предпочесть псевдоузел как лиганд, выбранный в результате SELEX-процесса.
ПРИМЕР 13. Выделение нуклеиново-кислотного лиганда к rev-белку ВИЧ-1
Сайт РНК-распознавания белка rev ВИЧ-1, очевидно, является комплексным, и его основная функция заключается в продуцировании инфекции эпидемического вирусного заболевания. См. , Olsen и др., Science, Vol.247, стр.845-848 (1990), SELEX-процесс на этом белке проводили для лучшего понимания элемента распознавания и для выделения лиганда к этому целевому белку.
Составляли смесь кандидатов с произвольной областью длиной в 32 нуклеотида, как описано в Примере 2. Было обнаружено, что rev-белок может насыщенно связываться с исходной смесью кандидатов, где полумаксимальное связывание происходит примерно при 1•10-7 М, как было определено с помощью анализа на нитроцеллюлозных фильтрах. Все реакции связывания РНК и белка осуществляли в связывающем буфере, состоящем из 200 мМ КОАс, 50 мМ Трис-HCl, рН 7,7, 10 мМ дитиотреитола. Разведения РНК и белка смешивали и держали на льду в течение 30 минут, после чего переносили на 5 минут в условия 37oС. В анализе на связывание реакционный объем составлял 60 мкл, из которых исследовали 50 мкл; в раундах SELEX реакционный объем составлял 100 мкл. Каждую реакционную смесь пропускали через заранее пропитанный связывающим буфером нитроцеллюлозный фильтр и промывали 3 мл связывающего буфера, после чего осушали и просчитывали для анализа или подвергали элюированию, которое является частью схемы SELEX. Осуществляли 10 раундов SELEX с использованием концентрации РНК примерно 3•10-5 М. Концентрация rev-белка составляла 1•10-7 в первом раунде, и 2,5•10-8 во всех последующих раундах. Исходную смесь кандидатов пропускали через нитроцеллюлозный фильтр для снижения числа последовательностей, которые обладают высоким сродством к нитроцеллюлозе. Этот продукт повторяли также после раундов 3, 4 и 9. кдНК-продукт очищали после каждого третьего раунда селекции для устранения видов с аномальными размерами, которые обычно проявляются при повторении раундов SELEX. После проведения 10 раундов последовательность вариабельной области РНК-популяции была нерандомизированной, как было определено посредством дидезоксисеквенирования. Было клонировано и секвенировано 53 изолята.
Каждая из клонированных последовательностей представлена в табл.10. Все последовательности были проанализированы с использованием программы по предсказанию вторичной структуры РНК Цукера. См., Zucker, Science, Vol.244, стр. 48-52 (1989); Jaeger и др., Рrос. Natl. Acad. Sci. USA, Vol.86, стр.7706-7710 (1989). На основании общей вторичной структуры все последовательности были разделены на три группы в соответствии с общими мотивами, как показано в табл. 11. Мотивы I и II аналогичны по своей конформации и включают петлю "балджа", закрытую с каждого конца спиралью. Эта генерализованная структура схематически проиллюстрирована в табл. 12, а для большей наглядности все домены были отмечены, то есть от 5' до 3' Стебель 1а и эти пары оснований до 3' Стебля 1b. Петля 1, Стебель 2а, Петля 3, Стебель 2b, Петля 2 и Стебель 1b. Последовательности, которые соответствуют различным доменам, приведены для отдельных последовательностей в табл.12. (Следует отметить, что в последовательности 3а, гомологичная ориентация перевернута на 180, таким образом она является Стеблем 1, который закрыт петлей. Энергии сворачивания РНК-молекулы, включая фиксированные фланкирующие последовательности, показаны в табл.13.
Реактивный элемент rev дикого типа (RRЕ), который, как было определено, по крайней мере, минимально участвует в связывании с ВИЧ-1-транскриптами, был уложен по этой схеме и показан в табл.12 и 13.
Эти последовательности были также проанализированы на наличие родственных последовательностей с помощью процедуры, описанной Hertz и др., Comput. Appl. Biosci. Vol. 6 стр.81-92 (1990). Было идентифицировано два характерных образца. Каждый изолят был проанализирован для определения их наилучшего соответствия образцам, и результаты этого анализа представлены в табл. 13. Мотивы родственных последовательностей представлены общими вторичными структурами в аналогичных конформациях, то есть первая последовательность UUGАGАUАСА обычно обнаруживается как Петля 1 + 3'-концевые СА, причем эти пары с UG находятся у 5-конца второй информационно богатой последовательности UGGАСUС (обычно Петля 3). Есть также основания предположить спаривание оснований GАG последовательности 1 с СUС последовательности 11. Мотив II аналогичен Мотиву I в том, что последовательность GАUАСАG преобладает как петля, противоположная CUGGACAC с аналогичным спариванием СА с UG. Мотив II отличается по размеру петель и некоторыми последовательностями, в частности отсутствием предсказанного спаривания оснований поперек петли. Одна область RRE дикого типа очень похожа на Мотив II. Мотив III меньше всего похож на другие последовательности, хотя он характеризуется двумя "балджами" U' - смежными со спаренными GА-UС, как в Мотиве I. К сожалению, сделать более подробные сравнения оказалось затруднительным, поскольку образец укладки Мотива III включает в себя 3-фиксированную область в критических вторичных структурах; и так как эти последовательности являются инвариантными, то трудно каким-либо образом проанализировать важность любой из них. На фиг. 23 показаны последовательности наиболее характерных элементов каждого Мотива и последовательность изгиба RRE дикого типа.
Затем последовательности были проанализированы на их сродство к rev-белку. Матрицы были получены от ряда клонов с помощью РСR, из которых были получены меченые in vitro-транскрипты и отдельно проанализированы на их способность к связыванию с rev-белком. Кривые связывания показаны на фиг. 24-28. Из олигонуклеотидных матриц были синтезированы меченые транскрипты, которые содержали RRE дикого типа, обсуждаемый выше, который, как был сделан вывод, был "консенсусом" в высокостабильной конформации. Чтобы контролировать экспериментальные отклонения, наилучшую связывающуюся последовательность, изолят 6а, анализировали как стандарт в каждом эксперименте на связывание. Смеси РНК и белка обрабатывали как описано выше, за исключением того, что разведенные РНК нагревали до 90oC в течение 1 минуты и охлаждали на льду до смешивания. Средняя величина Ка для изолята 6а составляла 8,5•10-8 М. Результаты этого эксперимента показаны в табл.13.
Из кривой связывания 24 видно, что связывание выделенной популяции (Р) с rev-белком примерно в 30 раз лучше, чем связывание исходной смеси. Связывание RRE дикого типа почти аналогично связыванию наиболее распространенного клона 1C. Этот эксперимент также продемонстрировал, насколько чувствительным является взаимодействие при связывании rev по отношению к вторичной структуре. Изоляты 6а и 6b являются идентичными в высокоинформативных областях, но при этом, они совершенно отличаются на уровне вторичной структуры, что приводит к изменениям в трех положениях нуклеотидов. Эти изменения, которые прогнозируют спаривание оснований Стебля 1, понижают в 24 раза аффинность 6b. Чувствительность к аномалиям вторичной структуры, кроме того, проиллюстрирована связыванием изолята 17, как показано на фиг.25. Изолят 17 имеет максимальные информационные данные, как показано в табл.12. Однако в нем имеется лишний "выпирающий" в 5-конце Петли 1, как показано в табл.11. Этот лишний приводит к тому, что изолят 17 имеет более низкую аффинность к связыванию с rev по сравнению с другими последовательностями Мотива I. В противоположность этому, отдельные нуклеотидные делеции последовательностей Петли 2, даже те, которые снижают перспективы на спаривание поперек "балджа", являются хорошо толерантными к rev-взаимодействию.
Другой несомненной общностью является консервативность последовательности АСА, противоположной UGG, где СА спариваются с UG до начала Стебля 2. Эта последовательность является общей для Мотивов I и II, а также для RRE дикого типа. Последовательности 11 и 12 обнаруживают замещение пар оснований в этом положении (см. табл.12), а последовательность 12 имела пониженную аффинность по сравнению с большинством последовательностей Мотива I.
РНК-последовательности, выявленные с помощью SELEX, как rev-лиганды, могут быть классифицированы по первичной и вторичной структуре. "Консенсус" появляется из асимметричного "балджа", фланкированного двумя спиралями, в которых расположены специфически консервативные одноцепочечные и двухцепочечные нуклеотиды. Хотя спаривание оснований поперек балджа было предсказано для многих выделенных последовательностей (Мотив I), однако оно не может быть основным или решающим для rev-взаимодействия. Оптимальные размеры для Петли 1 составляют 8 (Мотив I), или 6 (Мотив III), где допустимые пределы составляют 9-3. Оптимальные размеры для Петли 3 составляют 5 или 4. Кроме того, взаимодействие rev с различными доменами этих лигандов может быть аддитивным. Мотив II похож на Мотив I, в основном, в месте соединения Петли 1 и Петли 3 у Стебля 2. Мотив III похож на Мотив I в месте соединения Петли 1 и Петли 3 у Стебля 1. На фиг.29 и 30 показаны диаграммы "консенсусов" Мотива I и Мотива II нуклеиново-кислотных растворов для ВИЧ-rev.
Распространненость последовательностей в клонированной популяции не показывают строгую корреляцию с аффинностью для rev-белка. Возможно, что концентрации rev-белка, используемые на протяжении всего SELEX-процесса, были достаточными для связывания значительного процентного количества всех указанных изолятов. В результате этого может быть осуществлена селекция на реплицируемость кДНК и ДНК в процессе PCR в дополнение к недостаточной селекции на связывание с rev. Высокоструктурированная природа указанных лигандов и возможные различия в эффективности синтеза ДНК на этих матрицах способствуют увеличению возможных отклонений в репликации. В течение SELEX-процесса могут также возникать некоторые мутации. Последовательности 6а и 6b являются настолько похожими, что они должны иметь общего предшественника. Это относительно позднее их появление в процессе SELEX можно объяснить недостаточным количеством этой последовательности, несмотря на ее более высокое сродство к мишени. Аналогичным образом, некоторые из появляющихся лигандов могут быть мутированы в относительно более позднее время в процессе селекции из последовательностей-предшественников, которые присутствуют в исходной смеси-кандидатов, но не представлены в клонированной популяции.
Раскрытое в настоящей заявке изобретение не ограничивается представленными вариантами его осуществления. Как было показано, настоящее изобретение может быть использовано специалистами для большого числа нуклеиново-кислотных лигандов и мишеней. При этом подразумевается, что в настоящее изобретение могут быть внесены различные изменения и модификации, не выходящие, однако, за рамки объема и существа изобретения, сформулированные в нижеследующей формуле изобретения.
Приложение.
Селекция. Простой кинетический механизм для образования обратимого комплекса "белок-РНК" в хорошо смешанном растворе может быть описан следующим образом: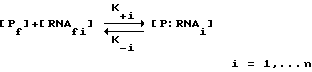
где [Pf] является концентрацией свободного белка, [РНКf] является концентрацией свободных видов-i-РНК, [Р:РНКi] является концентрацией комплекса "белок-виды-i-РНК", К+i является константой скорости ассоциирования свободного белка и свободных видов-i-РНК, К-i является константой скорости диссоциирования комплексов "белок-виды-i-PHK", а n является членом РНК-последовательности с уникальным набором констант скоростей. Альтернативные механизмы, включая множественные сайты связывания или кооперативное связывание, могут рассматриваться как последующие обработки с соответствующим расширением этой простой схемы.
Для любой системы, представленной вышеуказанной схемой, фундаментальные химически-кинетические уравнения или уравнения масс-взаимодействия, описывающие изменение концентрации каждого комплекса "белок-виды-i-РНК" как функции времени, представляют собой
где [Pf] , [РНКf] и [P:PНКi], являются концентрациями свободного белка, свободных видов-i-РНК и комплекса "белок-виды-РНК" при времени t.
Концентрация свободного белка представляет собой разность между полной концентрацией белка и концентрацией всех комплексов "белок-RNA ([Р]-Σ[P: RNAk] )"; аналогично, что концентрация свободных видов-i-RNA представляет собой разность между полной концентрацией видов-i-RNA и концентрацией комплекса "белок-виды-i-RNA ([RNAi]-[P:RNAi])"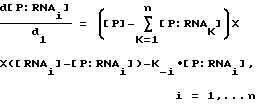
Эти динамические уравнения могут быть использованы для каждого кинетического анализа или анализа равновесия. Непрерывная дифференциальная форма является действительной всякий раз, когда средняя скорость каждого процесса значительно превышает статистическое отклонение способа, или, в двух словах, уравнение (3) является точным для описания пула РНК с несколькими молекулами, представляющими каждый уникальный ряд констант скоростей. Всякий раз, когда имеется только одна молекула или лишь несколько молекул, наиболее эффективно связывающихся РНК, статистическое описание связывания использовали для определения условий, которые дают высокую вероятность выявления наиболее эффективно связывающейся РНК. Эти статистические формулы выведены в последующей части по вероятности успеха.
При равновесии изменение концентрации каждого комплекса "белок-виды-i-RNA" равно 0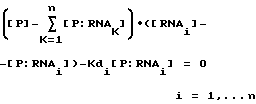
где параметры определены в уравнении (3), а Kdi является константой равновесной диссоциации для комплекса "белок-виды-i-РНК" [Kdi=К-i/К+i].
Когда рассматривали только один вид РНК (т.е. n=1), аналитическое решение для комплексов равновесия концентрации "белок-РНК" является возможным при решении следующего квадратного уравнения:
[P:RNA1]2 - ([P]+[RNA1]+Kd1)[P:RNA1] + [P][RNA1] = 0,
которое имеет два действительных корня, один из которых "физически реализуемый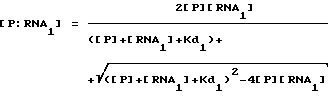
Имеются классические приближения для равновесной и квазистационарной концентрациий комплексов, аналогично тому, как в представлении Michaelis-Menten, но эти приближения не дают достаточной точности в диапазоне полных концентраций РНК и белка, использованных в SELEX. (См. дискуссию относительно некоторых ошибок и ограничений классического приближения, см. Savageau, 1991; Straus и Coldstein 1953; Webb, 1963).
Хотя аналитическое решение квадратного уравнения для простого обратимого связывания одного вида РНК с одним сайтом связывания на белке является точным в пределах всей концентрации РНК и белка, используемых в SELEX, и хотя концентрации связывания двух конкурирующих видов могут быть вычислены посредством аналитического решения кубического уравнения, однако итерационные численные методы требуют расчета равновесных концентраций белок-РНК-комплексов, всегда когда имеются три или больше конкурирующих РНК-видов.
Мы разработали компьютерную программу для определения каждого комплекса "белок-виды-i-РНК[Р:RNAi]", где любая данная полная концентрация белка, [P], любые распределения концентраций видов-РНК, [RNAi], и любые распределения констант равновесной диссоциации, Kdi. Матрица Якобиана (см. Leunberger, 1973) для неявного решения уравнения (4) методом Ньютона (см. Leunberger, 1973; Press, 1988) рассчитывается по следующей формуле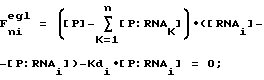
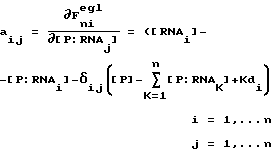
где а является элементом в ряду - i, колонке - j матрицы Якобиана, с δii=1 и δij=0 для i≠j.
Часто успех метода Ньютона зависит от хорошей начальной оценки решения (см. Leunberger, 1973; Press и др. 1988), в данном случае, в равновесной концентрации каждого комплекса "белок-виды-i-РНК", [P:RNAi]. При использовании объемного Kd для полного РНК-пула концентрация белка всех комплексов белок-РНК может быть оценена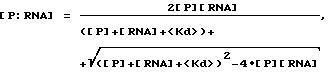
где [P: RNA] является концентрацией всех комплексов "белок-РНК", [RNA] является концентрацией всех РНК-пулов, а <Kd> является общей константой равновесной диссоциации РНК-пула и рассчитывается по следующей формуле: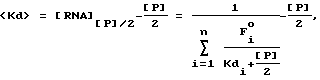
где [RNA] [P]/2 - полная РНК-концентрация, которая связывает половину белка, a Fi o = [PHKi]/[PHK].
Что касается оценки для концентрации белка в комплексах, начальное приближение для концентрации каждого комплекса "белок-виды-i-РНК" может быть сделано по следующей формуле: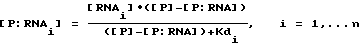
Решения для величин [Р:РНКi], которые удовлетворяют уравнению (4), могут быть улучшены до более высокого уровня точности путем итеративного применения метода Ньютона с использованием уравнения (7). При такой реализации мы можем достичь решений с более чем 12 значащими цифрами в менее чем в 4 или 5 итерациях ньютоновского метода. Эта быстрая сходимость к точному решению обусловлена начальными приближениями в уравнении (10), обычно дающими одну или более значащих цифр, в зависимости от интервала констант равновесной диссоциации и распространенности каждого из РНК-видов. Основания для такого уровня точности заключаются в том, что ошибка в [P:RNA] стремится к нулю в уравнении (10), всегда когда [P]-[P:RNA] больше, чем Кdi, например, если [PNA] меньше, чем Kdi, или если Kdi меньше, чем <Kd>. Любопытно отметить, что точность будет повышаться для любого комплекса со связыванием лучше, чем связывание всего РНК-пула. Типичные примеры начальной точности расчетов обогащения, определенного как повышение во фракции всего РНК-пула видов РНК с наилучшим связыванием в каждом раунде и аппроксиммированные путем замены уравнения (10) на уравнение (20).
Общая точность отражает точность равновесных концентраций, рассчитанных для каждого комплекса "белок-виды-i-РНК" с использованием уравнения (10). В следующей части мы использовали эту точность для расчета оптимальной концентрации РНК и белка для максимального обогащения.
Распределение. Любой способ распределения различных видов нуклеиново-кислотных последовательностей, включая в себя связывание с фильтром (Tuerk и Gold, 1990), использование геля со сдвиговыми усилиями (Blackell и Weintraub, 1990), аффинную хроматографию (Ellington и Szostak, 1990, Green et al. 1990, Oliphant и Struhl, 1987; Oliphant и Struhl, 1988), осаждение антитела, фазовое распределение или защиту от нуклеолетического расцепления (Robertson и Joyce, 1990), может быть улучшен с помощью SELEX. Например, с использованием связывания фильтра большая часть комплексов "белок-РНК" связывается с нитроцеллюлозным фильтром, в то время как большая часть свободных РНК-молекул вымывается с помощью этого фильтра (Unhlenbeck et аl., 1983; Varus, 1976; Varus и Berg, 1967; Varus и Berg, 1970). Нужную фракцию комплекса белок-РНК, который связывается с фильтром и который затем был выделен из этого фильтра, обрабатывали в следующей части.
Поскольку фракция свободных РНК-молекул также связывается с фильтром как неспецифический фон, то полное количество каждых видов-i-РНК, собранных на фильтре, вычисляли с помощью нижеуказанной формулы, по которой просчитывали как нужный сигнал от наиболее эффективно связывающихся молекул РНК в комплексах белок-РНК, так и шум от свободных РНК-молекул, собранных как неспецифический фон плюс конкурирующие РНК-молекулы в комплексах РНК-белок
где RNAfilt является числом собранных молекул видов-RNA, Vol является объемом реакционной смеси, проходящей через фильтр; [P:RNAi] является равновесной концентрацией комплекса "белок-виды-i-RNA", вычисленной как описано в предыдущей части, ВG является фракцией собранных свободных РНК (как неспецифичоский фон), и [RNAi] является полной концентрацией видов-i-RNA. Любой способ распределения обычно дает не совсем полное разделение связанных и несвязанных лигандов, а поэтому необходимо в каждом раунде проводить измерение для фракций свободных лигандов, собранных как фон, в отношении связывания лигандов.
Как уже отмечено, не все комплексы "белок-РНК" в растворе могут быть собраны на фильтре. Кроме того, РНК в тесно связанных комплексах лучше удерживаются на фильтре, чем РНК в слабо связанных комплексах. В тех случаях, когда это справедливо, обогащение тесно связывающимися РНК-молекулами будет еще более увеличиваться с каждым раундом селекса, с другой стороны, если некоторые молекулы не могут быть элюированы с фильтров, как и другие, то их обогащенность будет понижаться.
Амплификация и перенормировка. Количество каждых видов-i-РНК, выделенных из фильтров, рассчитывали по следующей формуле:
RNAi pcr = FR•RNAi filt, i = 1, ... n.
где FR является фракций РНК, которая может быть выделенной из фильтра, и РНК является членом молекул видов-i-РНК, собранных на фильтре, вычисленным в уравнении (11). В этой обработке предполагается, что значение FR является постоянным и определяется фракцией комплекса "белок-РНК", который связывается с фильтром, и фракцией РНК в этих комплексах, которые могут быть выделены и копированы обратной транскриптазой для получения кДНК в процессе PCR. Предположение, что FR является постоянной для всех видов, является обоснованным, поскольку при условии достаточного времени, когда все молекулы имеют одни и те же праймерные сайты для РСR, и используется избыток праймерных молекул, каждый вид, независимо от того, является он редким или распространенным, имеет практически ту же самую вероятность отжига праймерной молекулы. Поскольку каждая РНК-молекула имеет одинаковую длину, то различие в скорости амплификации на основании размера не наблюдалась. Конечно, если какие-нибудь виды РНК имеют вторичную структуру, которая препятствует праймерному отжигу кДНК синтеза, или если первичная или вторичная структура соответствующей кДНК замедляет скорость ДНК-полимеразы в процессе PCR-амплификации, то обогащение этими видами снижается. Мы пренебрегаем этими фактами, поскольку нет никаких приемлемых правил, по которым можно было бы предсказать, какие структуры реально обеспечивают такое различие. По мере изучения этих структур, какие-либо значительные факторы могут быть затем добавлены в математическое описание SELEX.
Полное количество РНК, выделенной из фильтра, вычисляли путем суммирования числа молекул каждого из видов, собранных для получения кДНК-копий в процессе РСR-амплификации.
Любые молекулы - "носитель" или "неспецифический конкурент" - должны быть исключены из суммы в уравнении (13), без РСR-праймерных сайтов, поскольку эти молекулы не амплифицируются. Схемы измерения аффинности часто включают в себя эти неспецифические конкурирующие РНК-молекулы, и если такие молекулы также использовать в SELEX, то, очевидно, они не будут амплифицироваться. Любопытно отметить, что в том случае, когда неспецифические молекулы-конкуренты взаимодействуют с белком в том же сайте, что и наиболее эффективно связывающиеся лиганды, то последствием добавления молекул-конкурентов будет снижение числа специфических сайтов, пригодных для селекции, поэтому для концентрации белка, который связывается с нужным количеством амплифицируемых лигандов-молекул при наличии высокой концентрации неспецифических молекул-конкурентов, должны быть построены кривые связывания, скорректированные путем включения соответствующих концентраций молекул при каждом титровании. К преимуществам использования высоких концентраций неспецифических, неамплифицированных молекул-конкурентов в каждом раунде SELEX можно отнести снижение адсорбции амплифицируемых лигандов-молекул в любые неспецифические сайты, снижение связывания амплифицируемых лигандов-молекул с любыми неспецифическими сайтами в белках-мишенях или снижение фракции свободных амплифицируемых молекул, собранных как неспецифический фон на "ложно-распределяющих" сайтах (но лишь тогда, когда эти сайты присутствуют в значительных количествах и являются эффективно насыщенными количеством используемых неспецифических молекул-конкурентов). Если эти условия не выполняются, то добавление неспецифических молекул-конкурентов имеют в основном тот же эффект, что и снижение количества используемого белка.
Количество каждых амплифицируемых видов-i-RNA, выделенных после одного раунда по отношению к сумме в уравнении (13), вычисляли следующим образом:
После PCR-амплификации кДНК-копий и перенормировки РНК-пула снова к их исходной концентрации путем in vitro-транскрипции (от идентичных сайтов промотора на всех кДНК-молекулах) концентрация каждого из РНК-видов после одного раунда SELEX составляла
[RNAi] = Fi 1[RNA],
где [RNA] является полной концентрацией RNA-пула. Для каждого дополнительного раунда SELEX концентрация каждого из RNA-видов может быть вычислена путем повторения итераций уравнений (7)-(15), где Fi 1 для каждой из RNA-видов из одного раунда является исходной фракцией Fi 0 в следующем раунде, см. Уравнение (9).
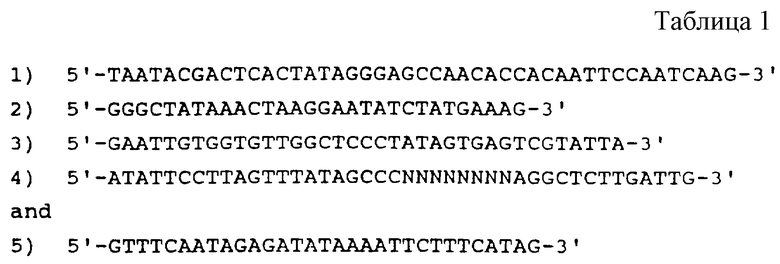

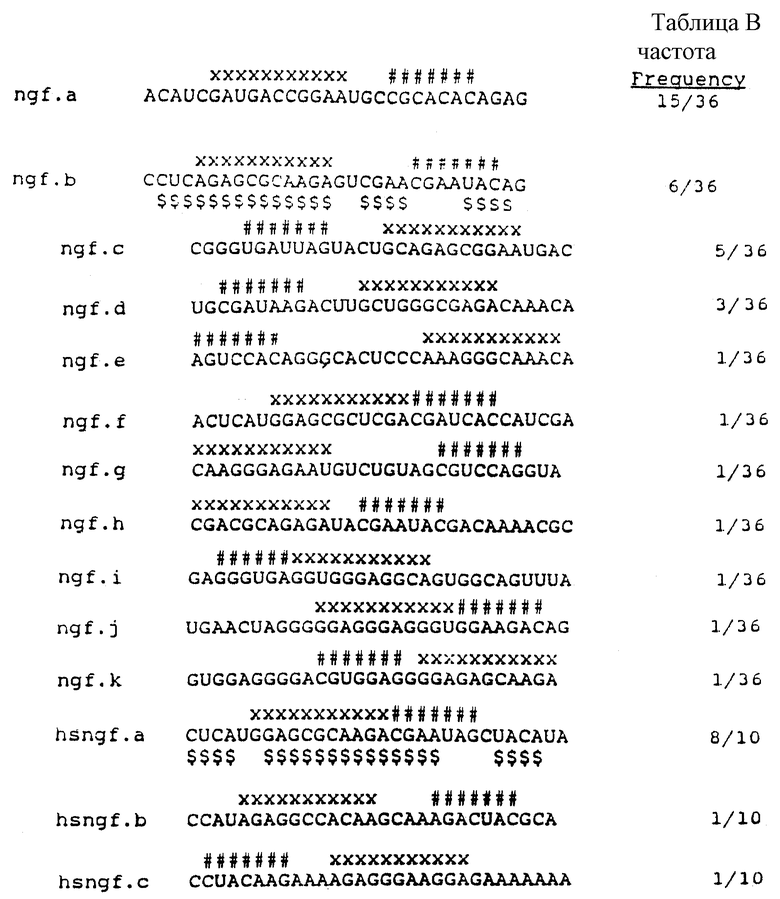




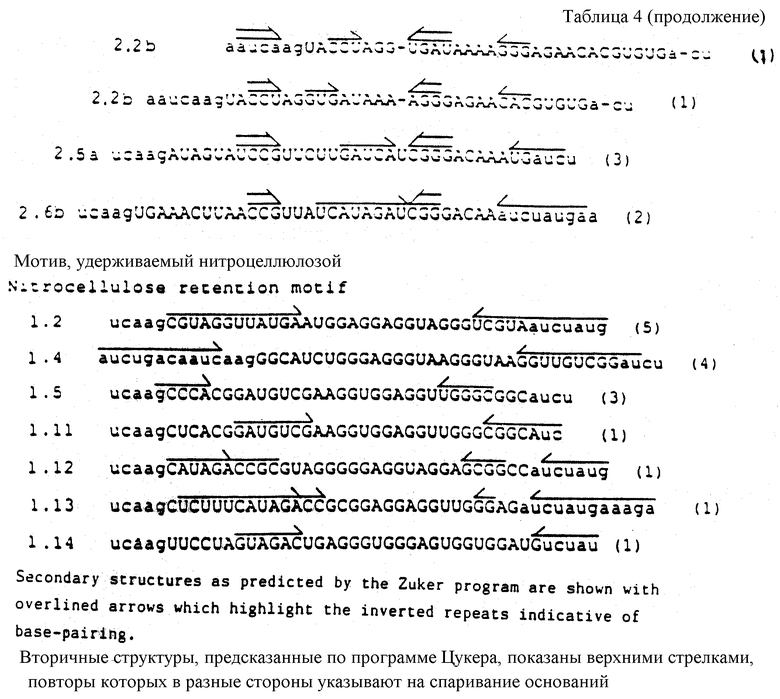
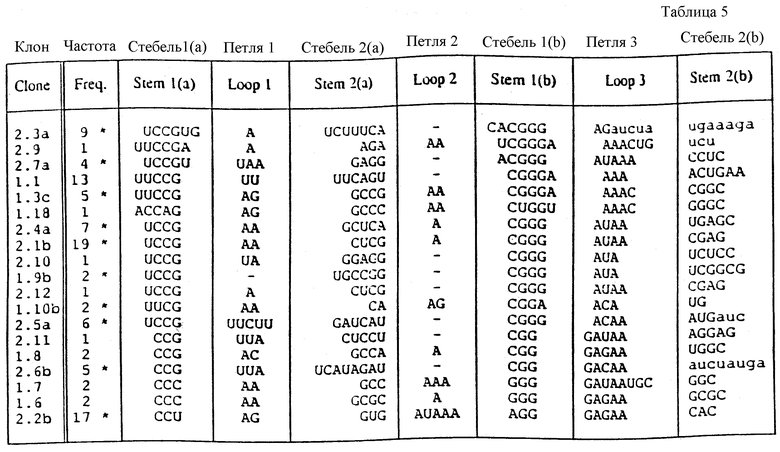







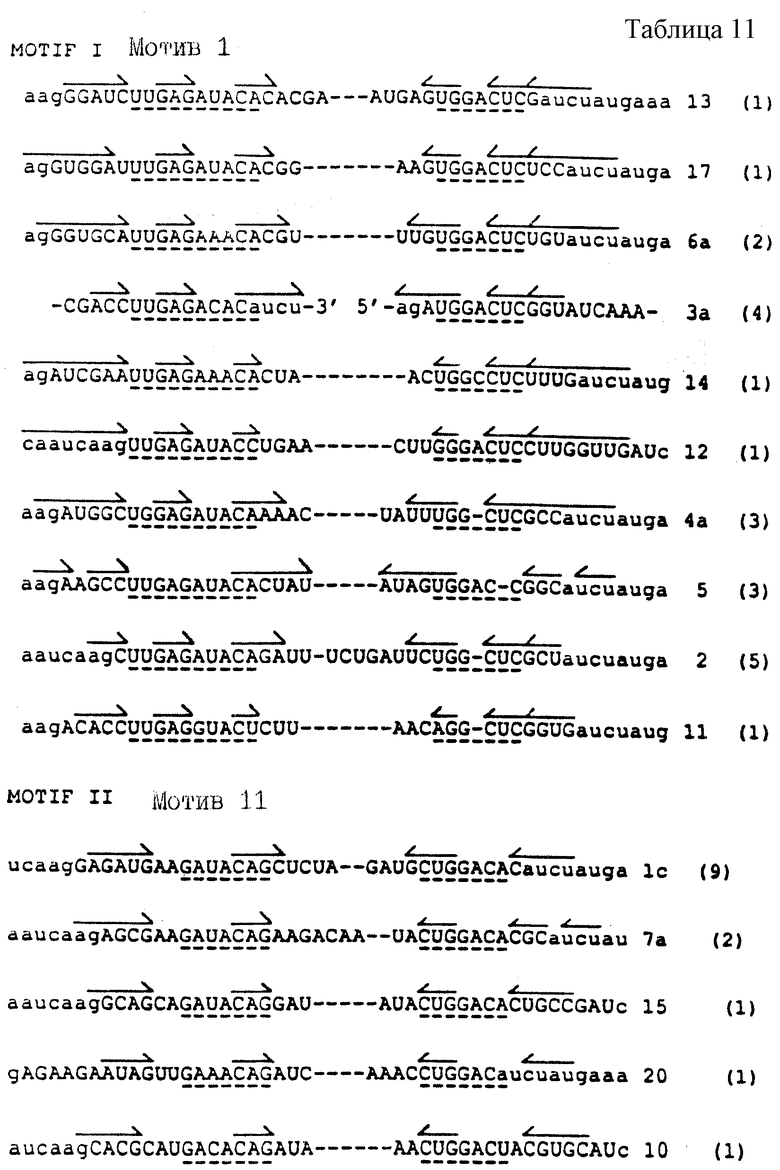

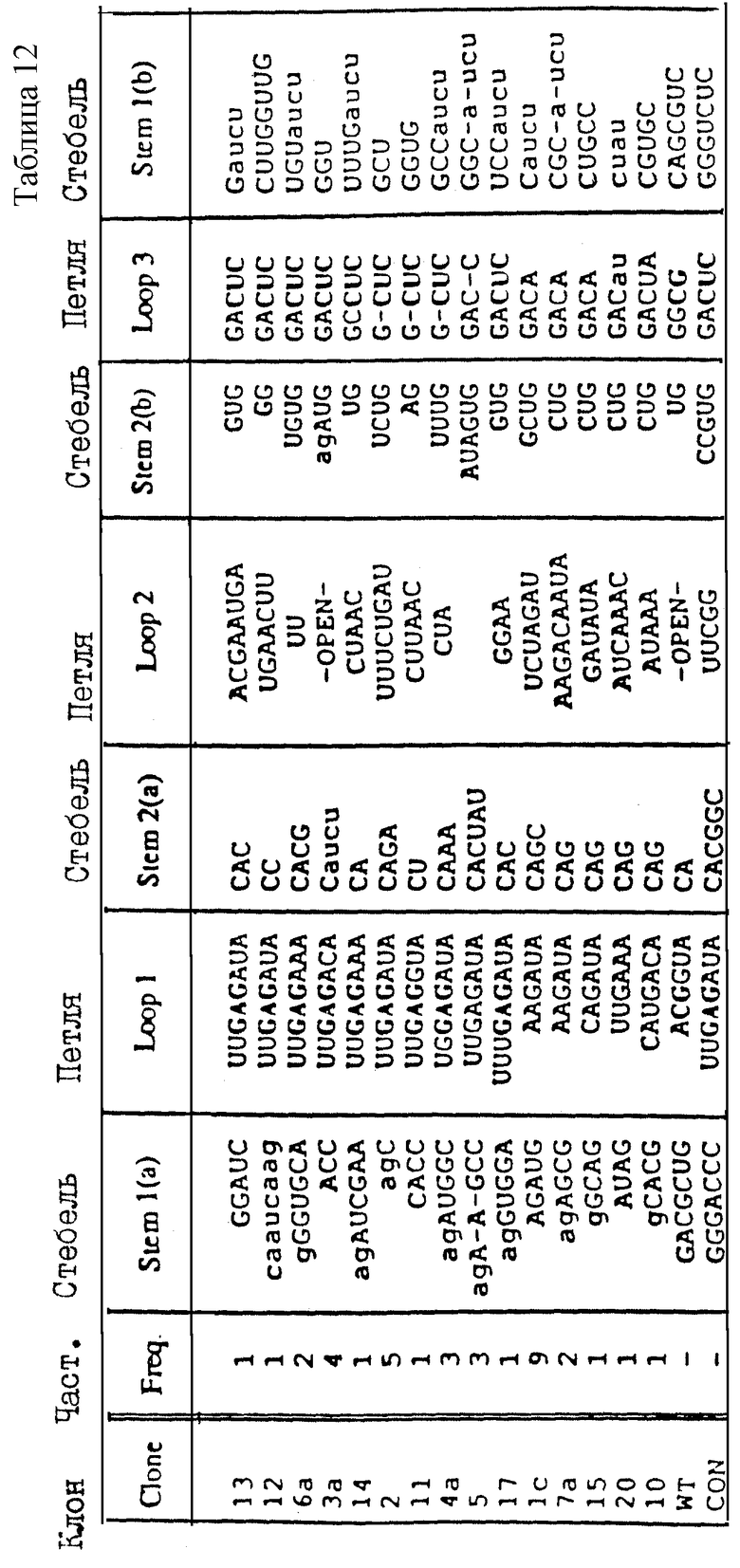
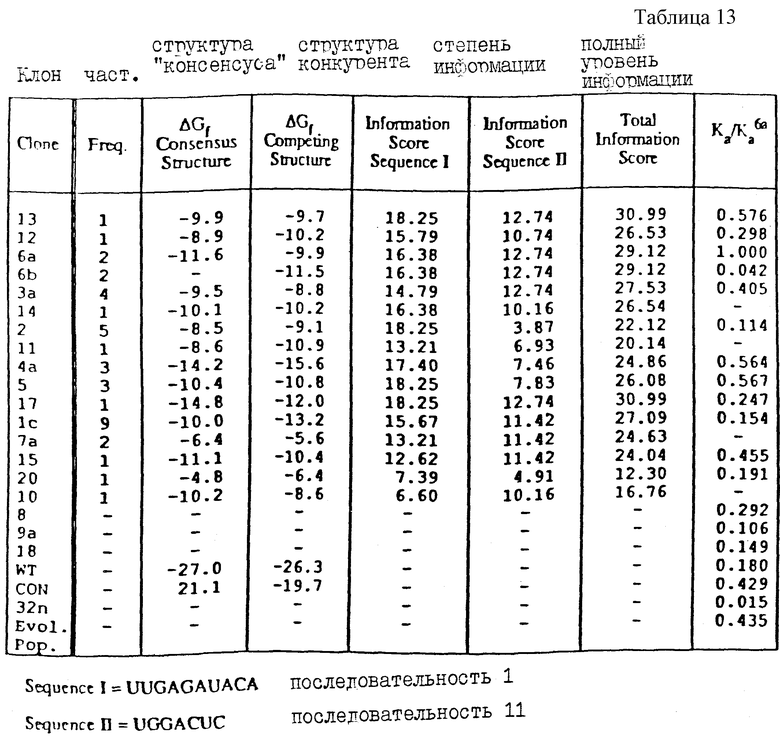
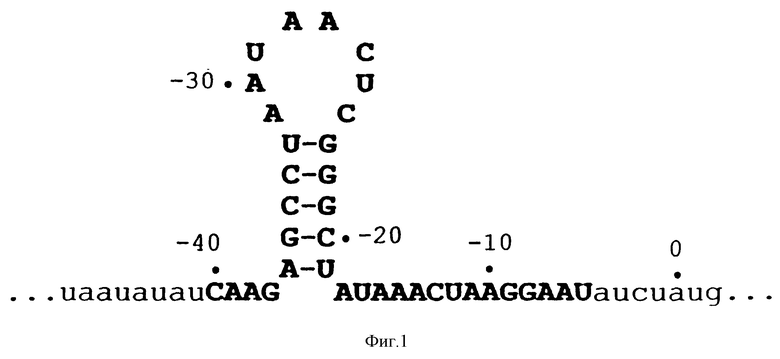
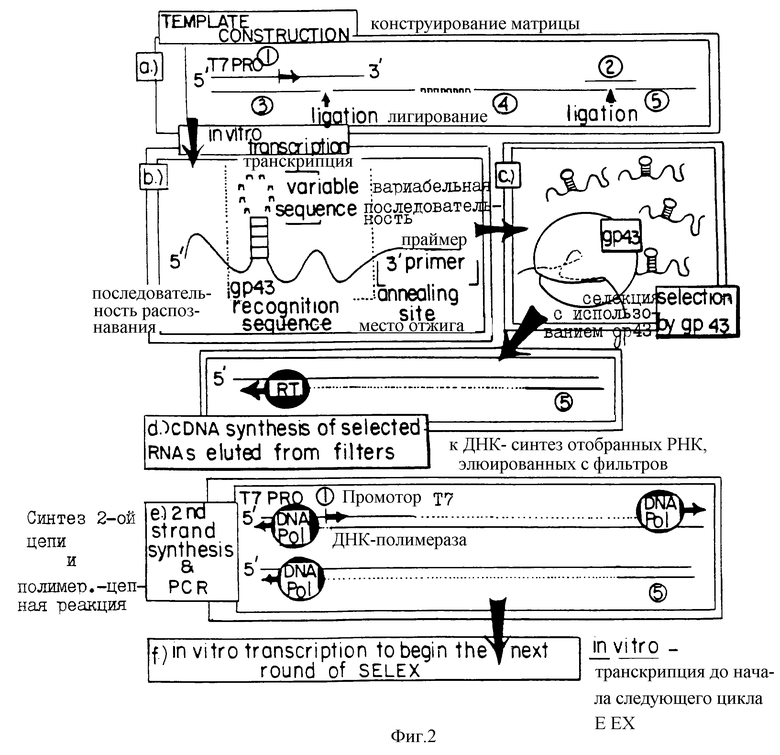
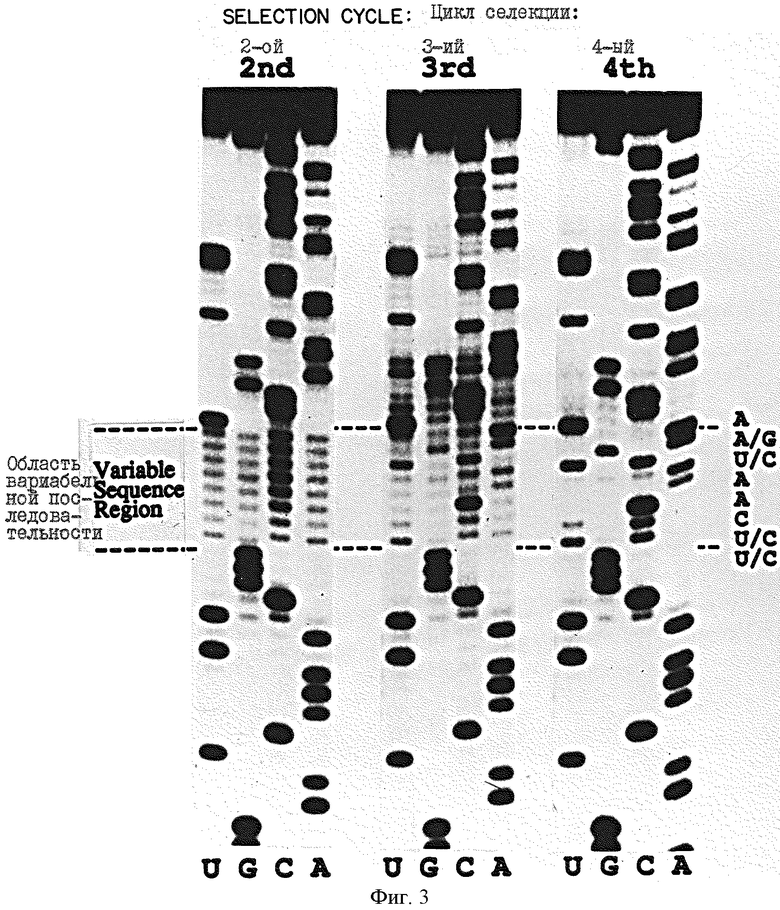
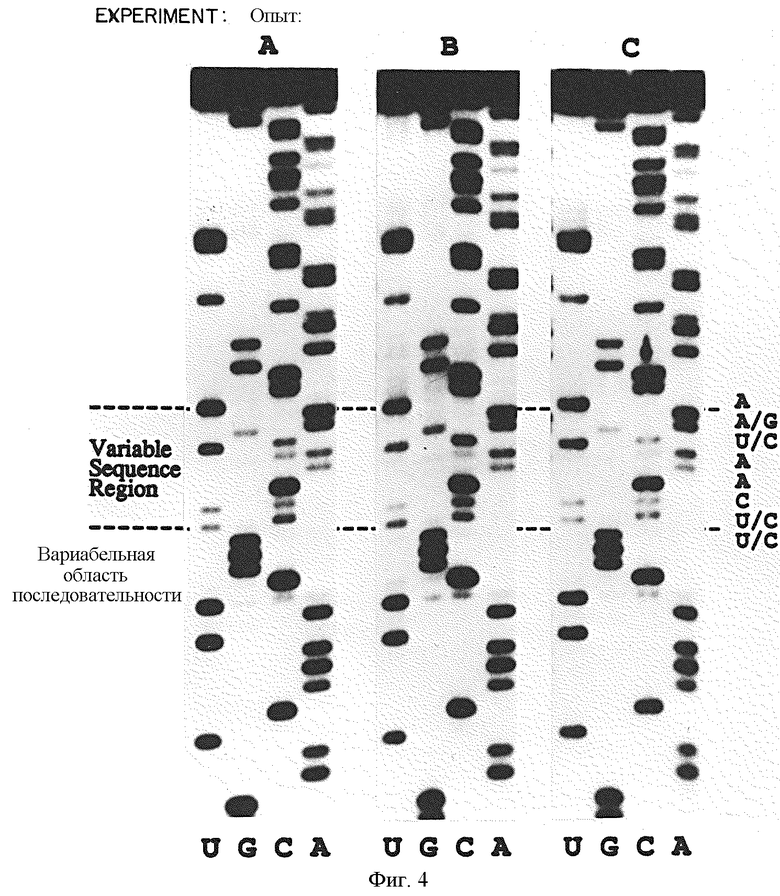
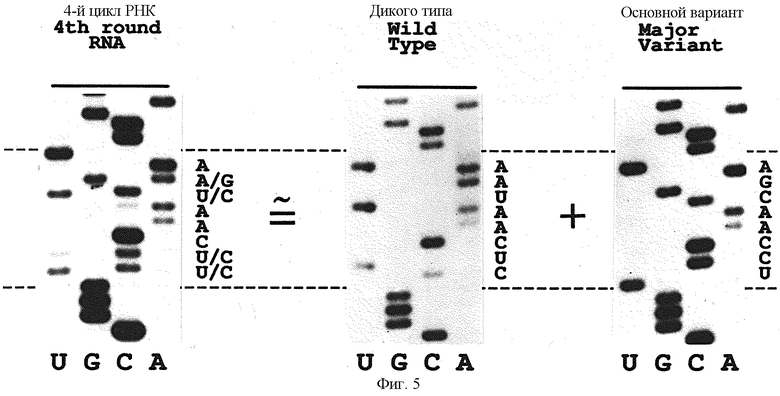
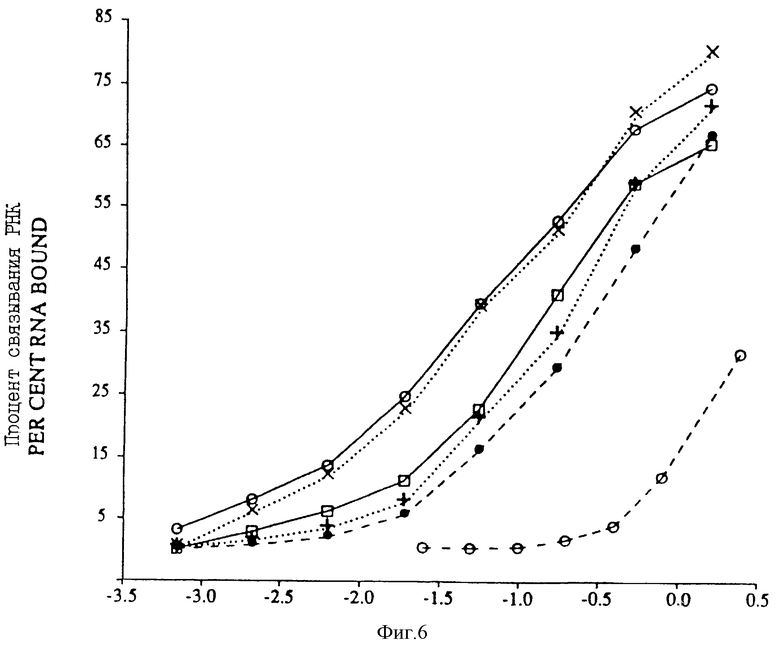
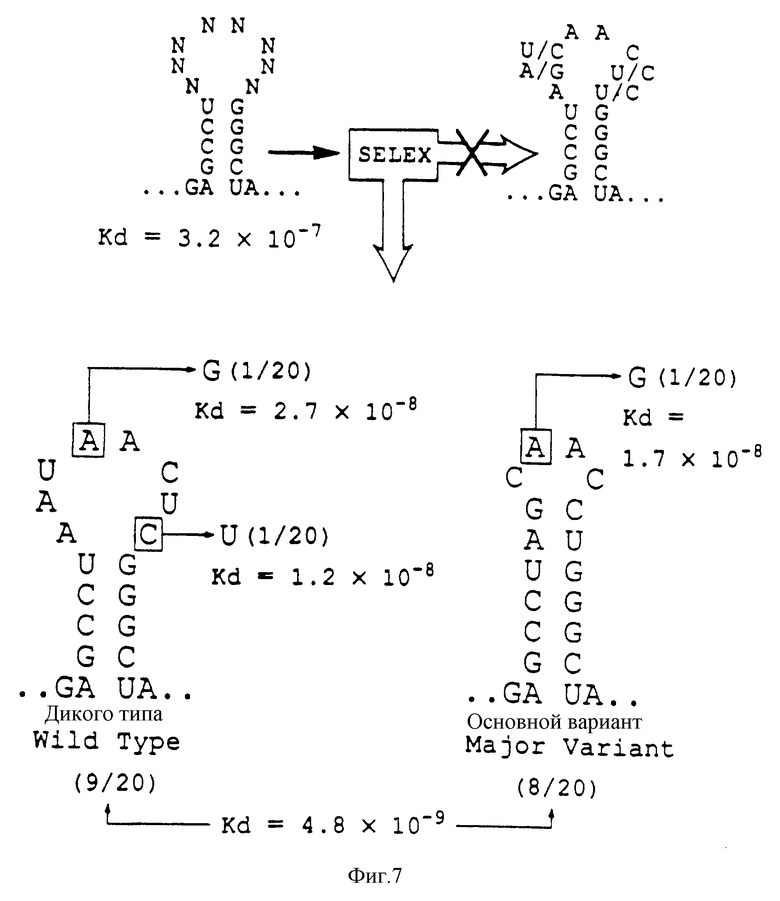
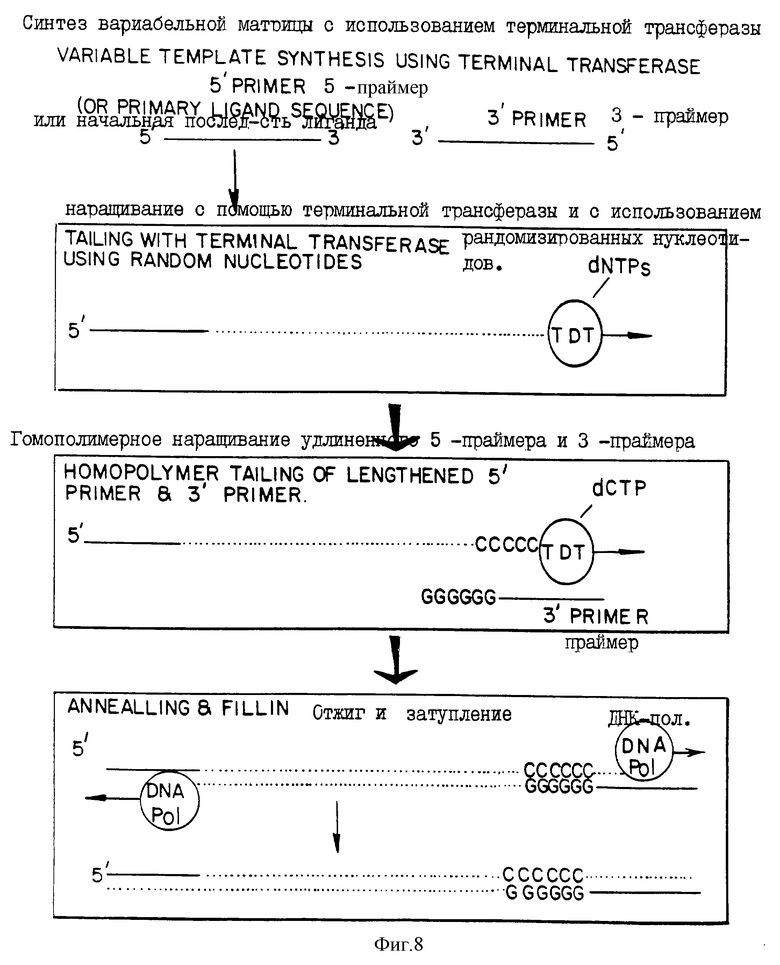
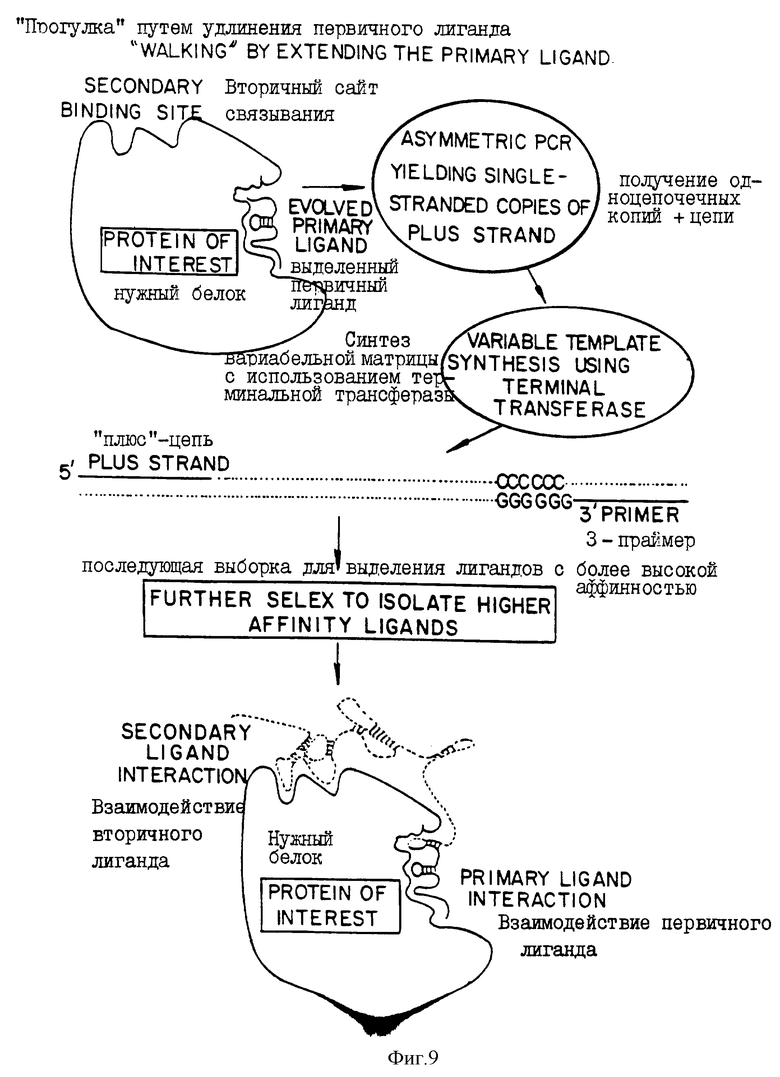
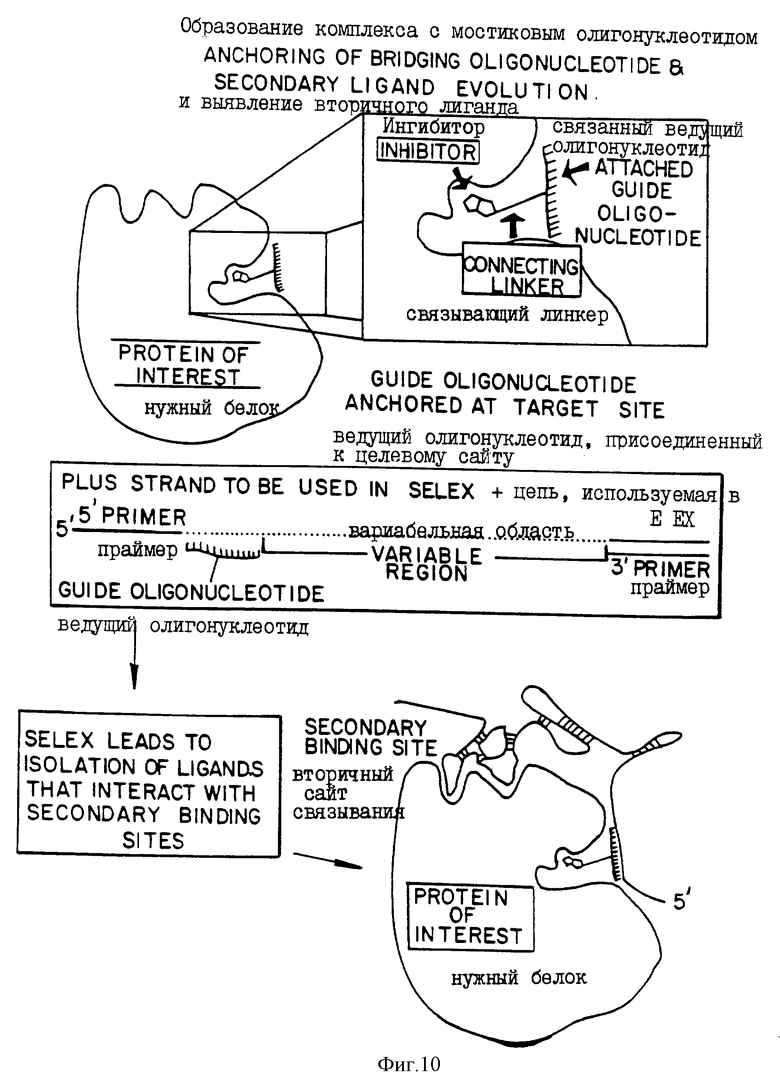
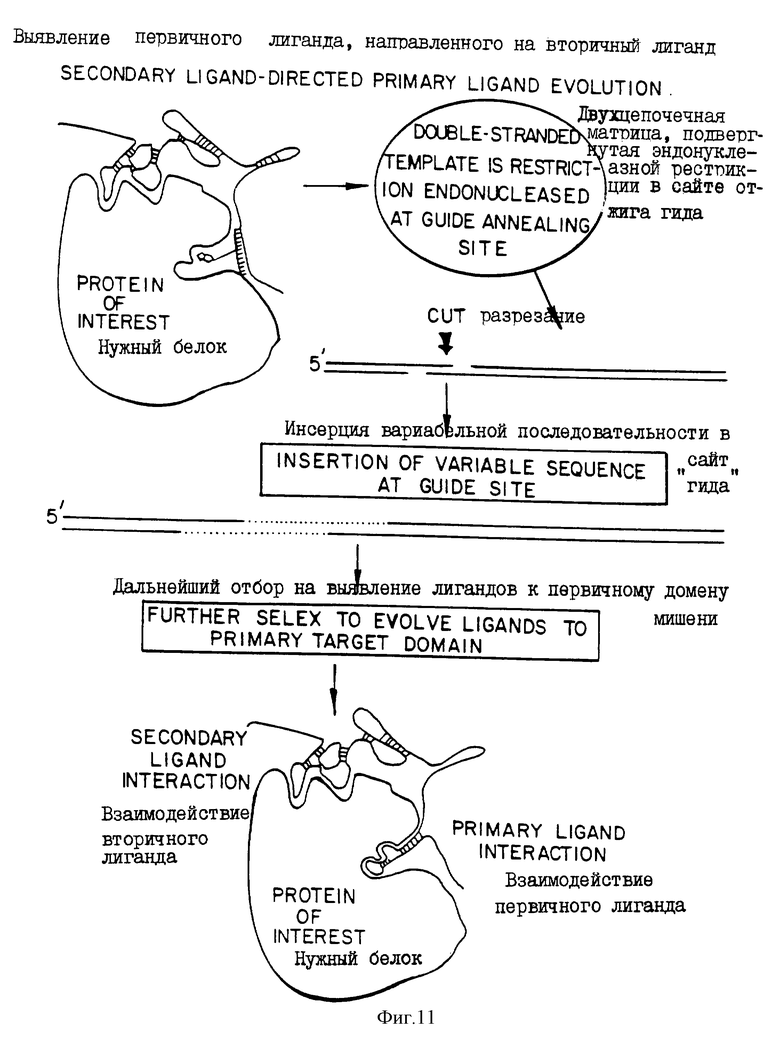
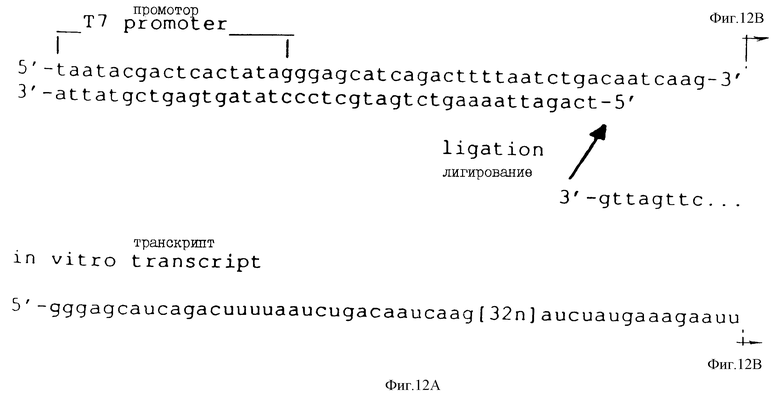
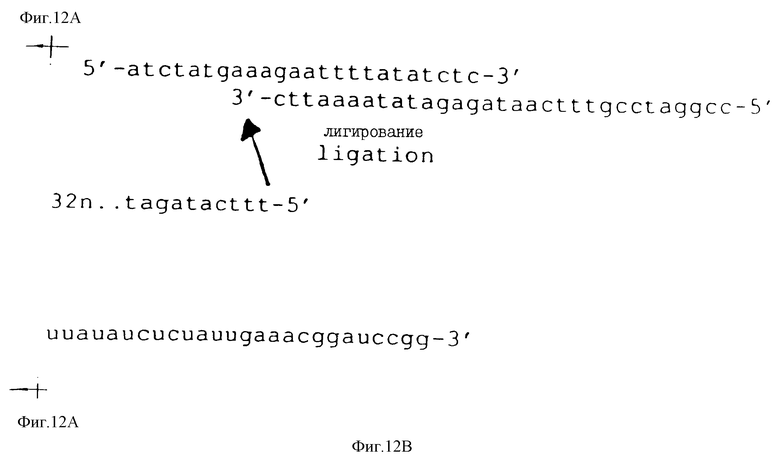
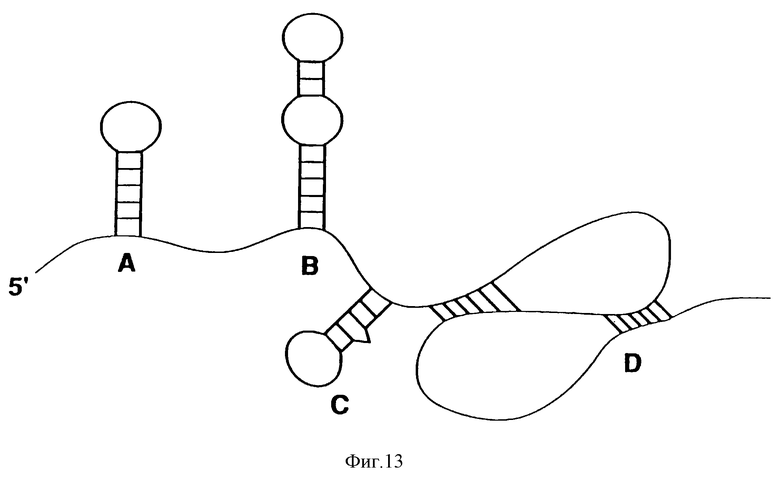
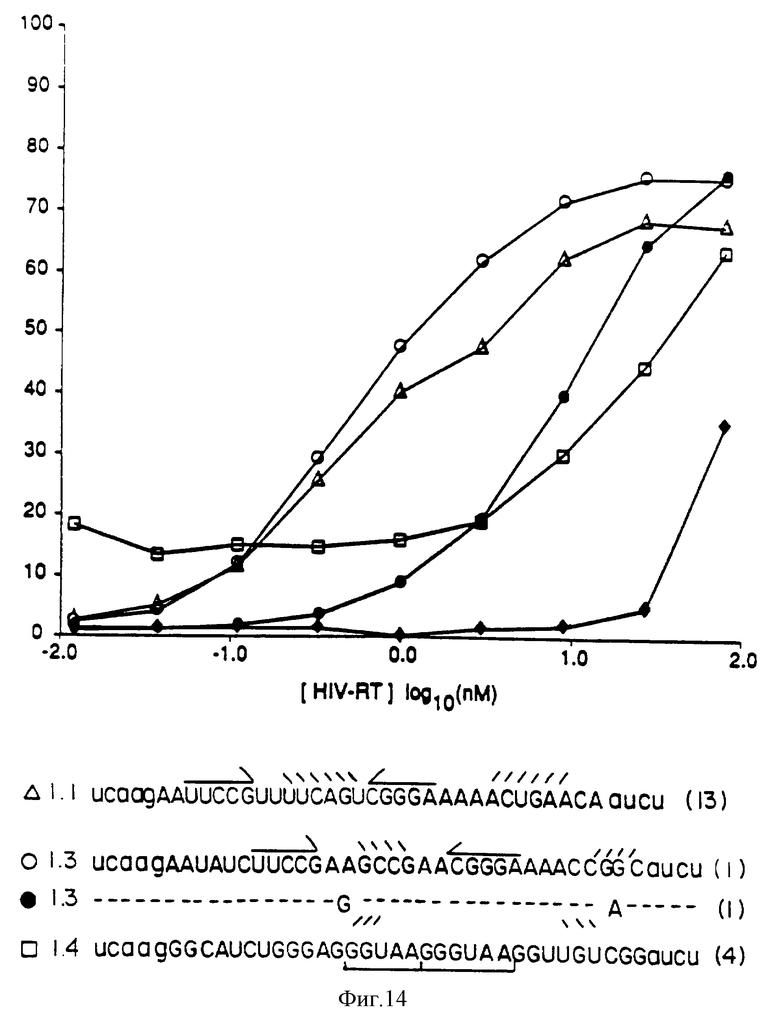
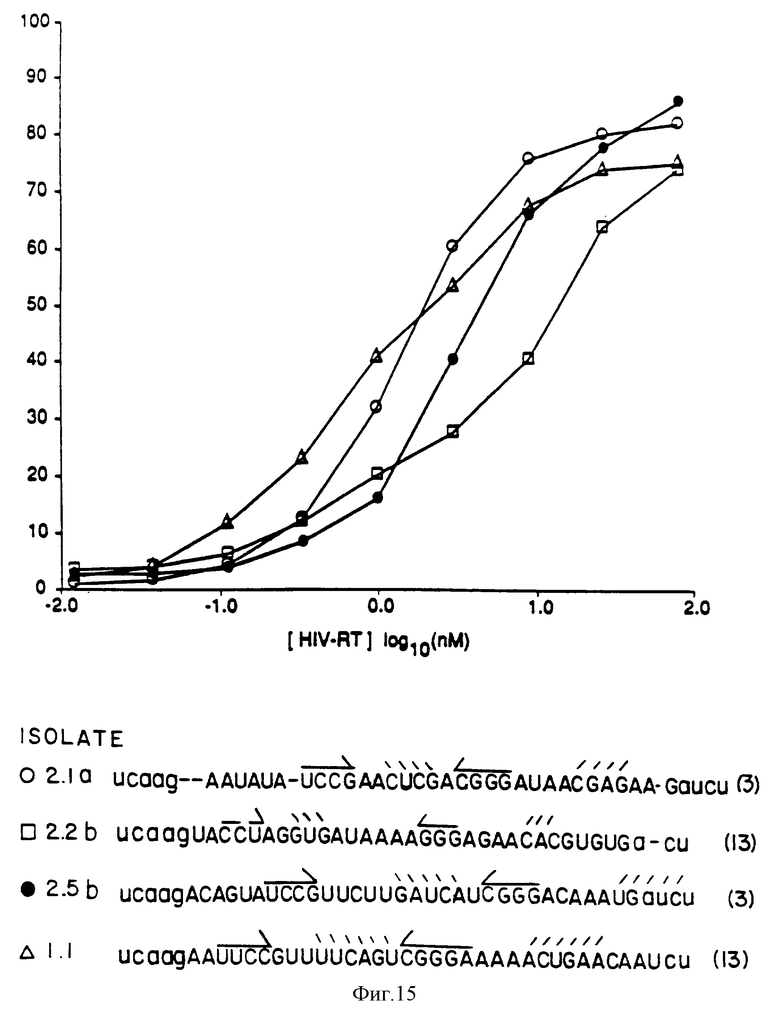
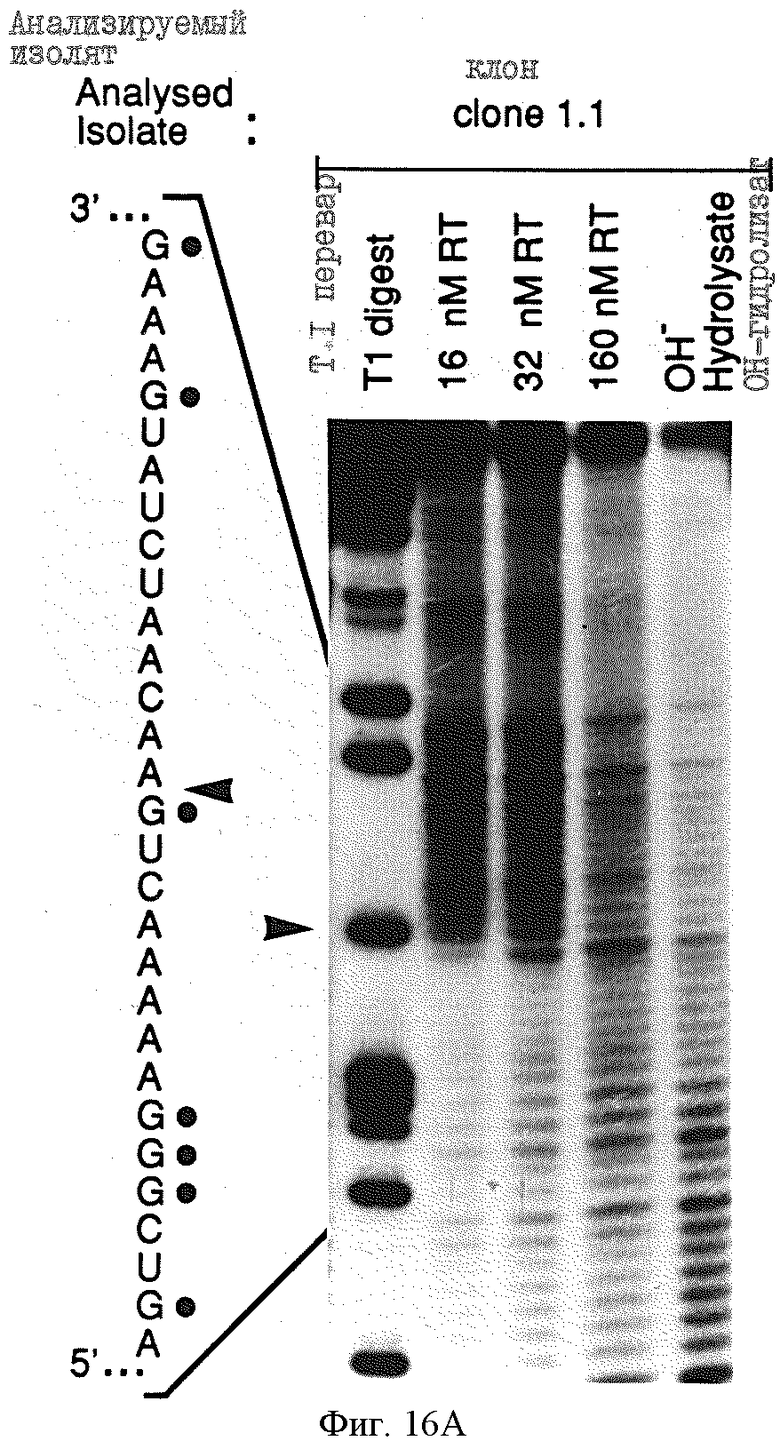
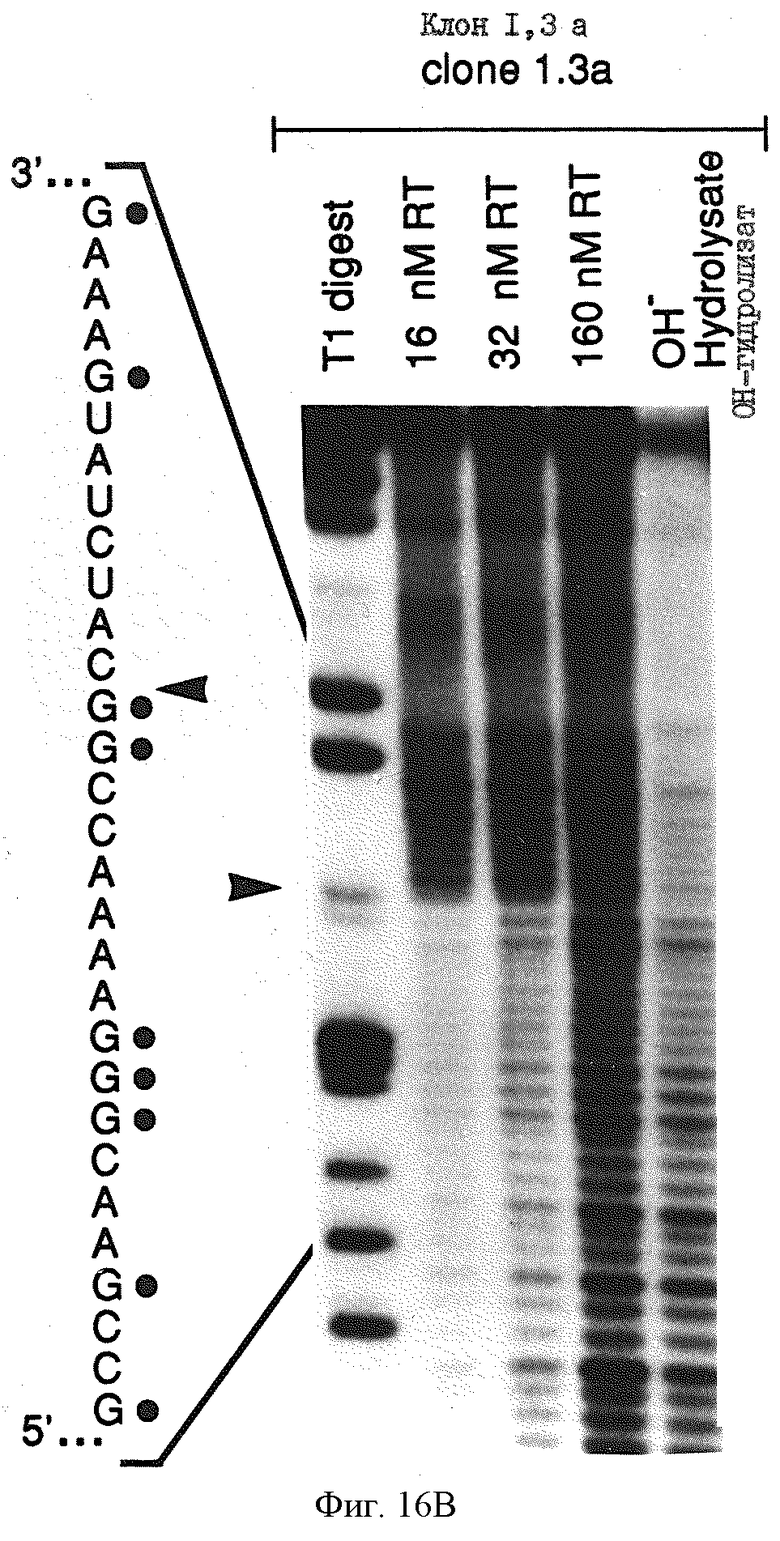
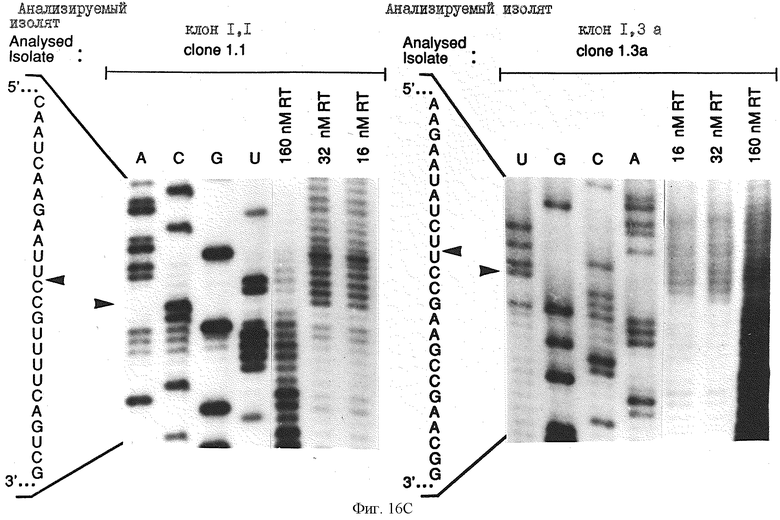
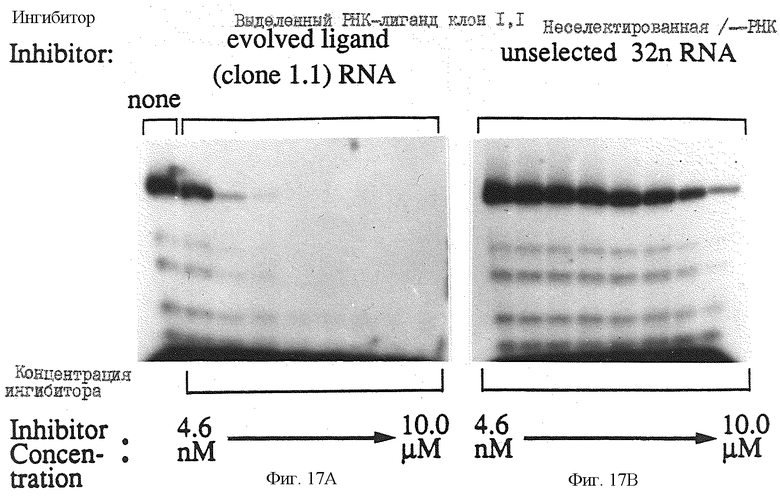
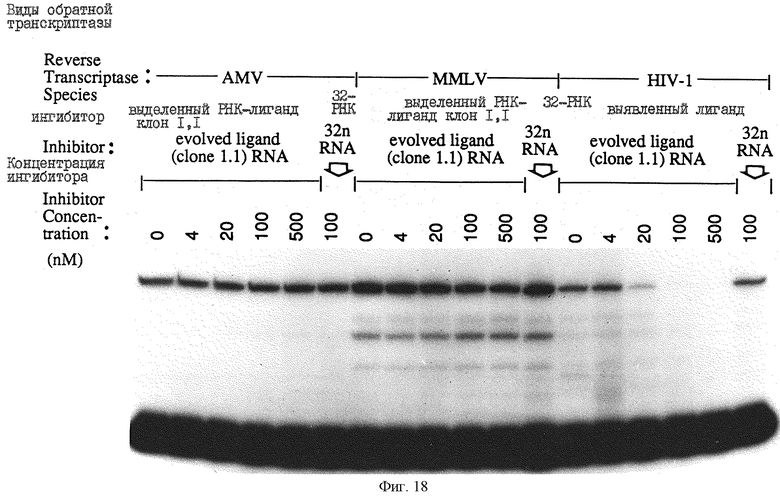
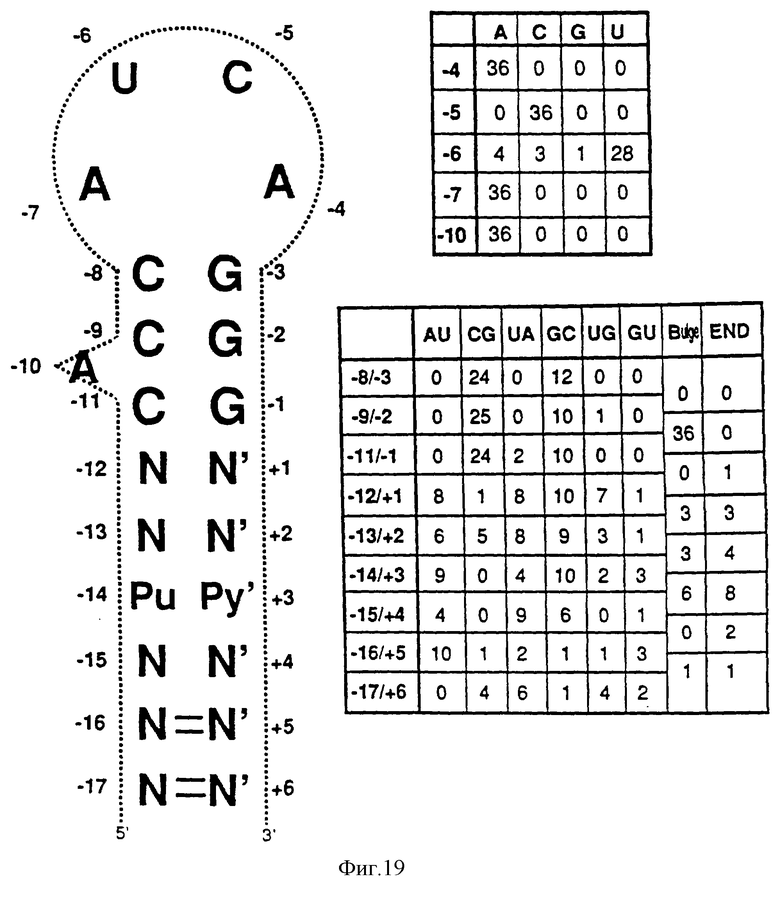
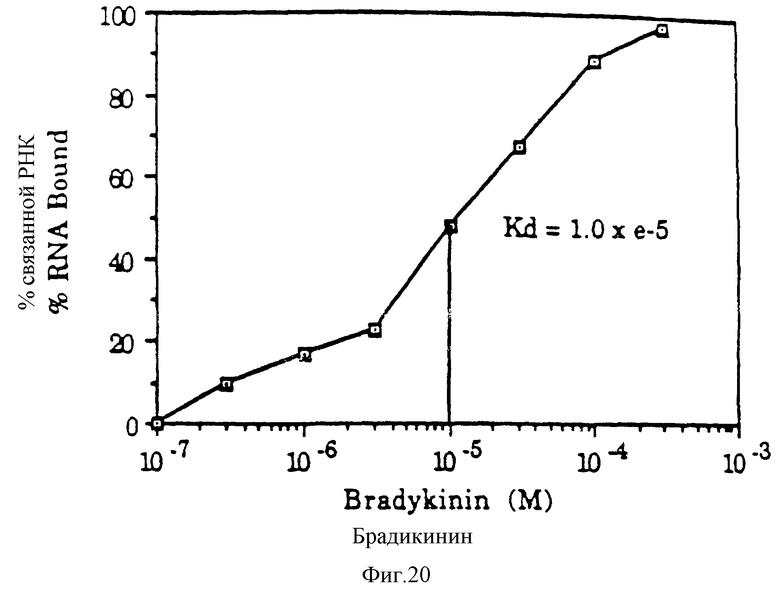
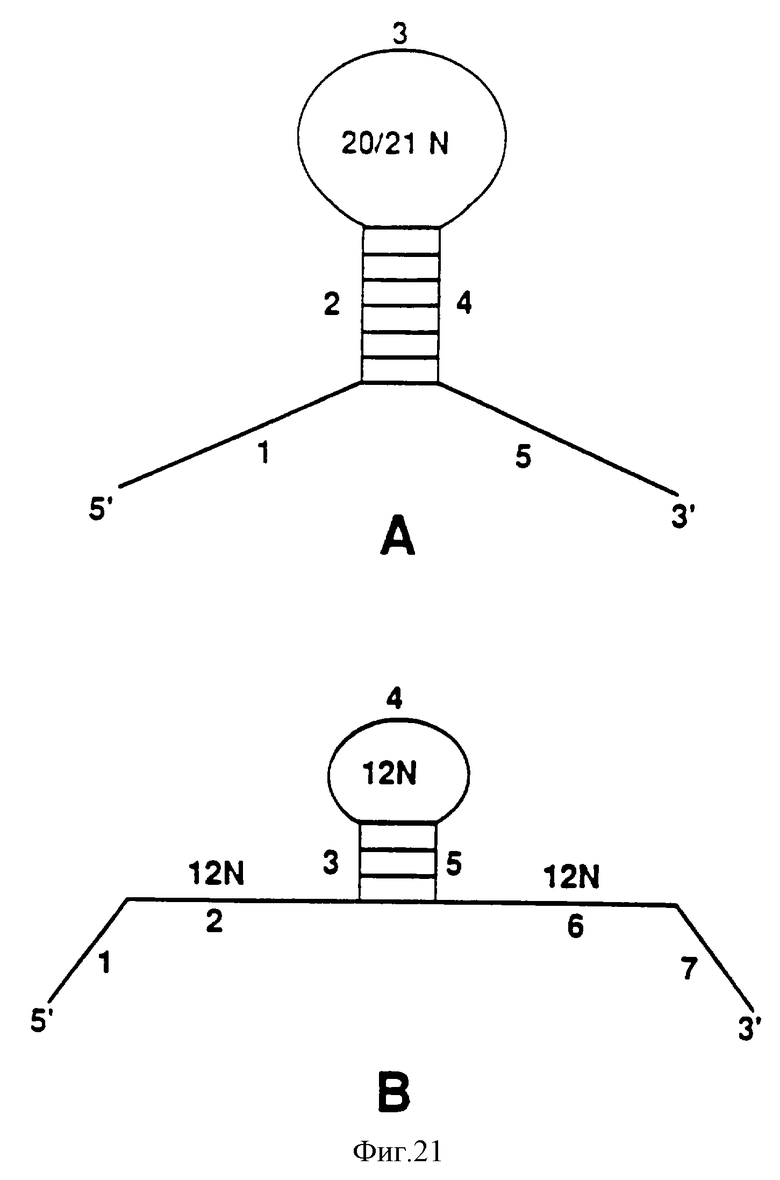
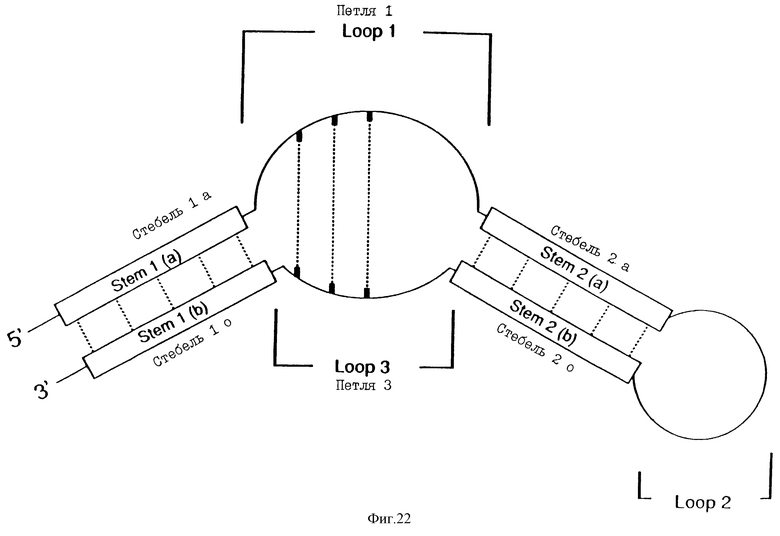
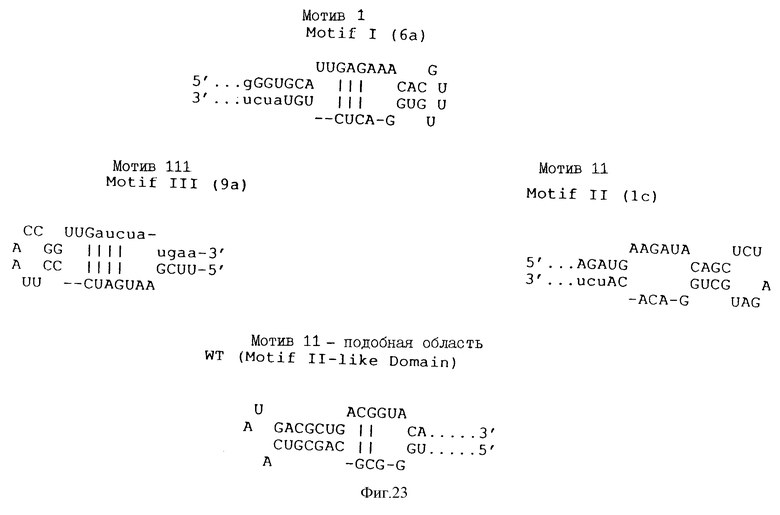
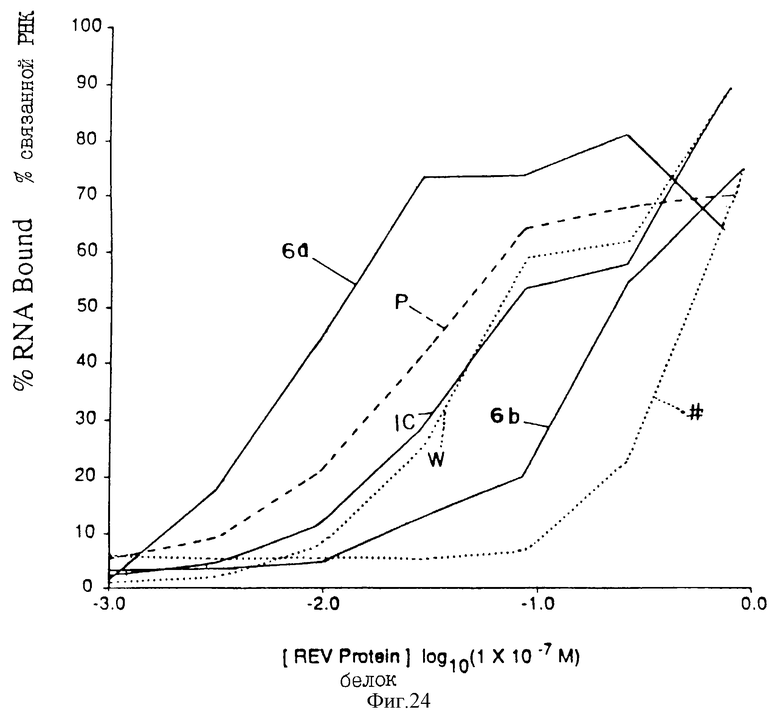
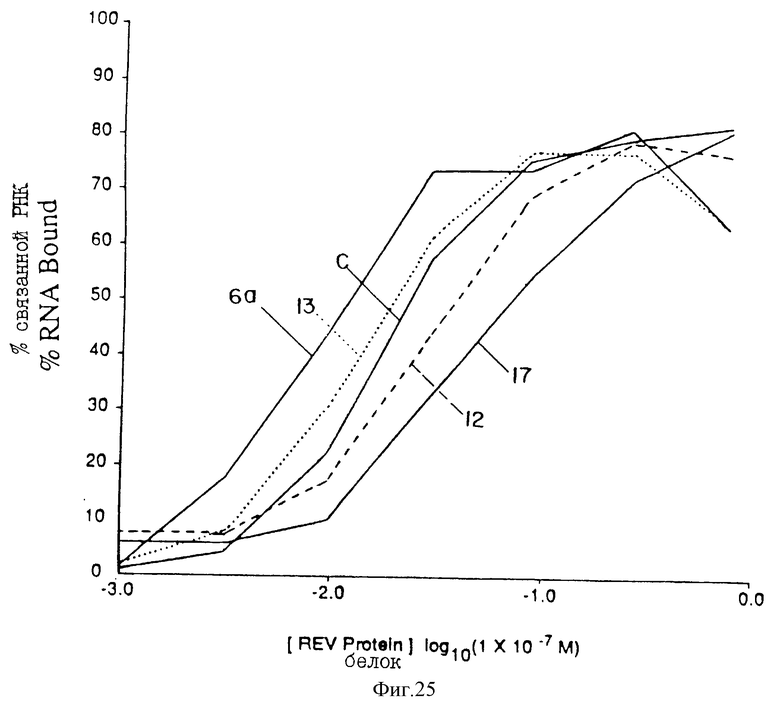
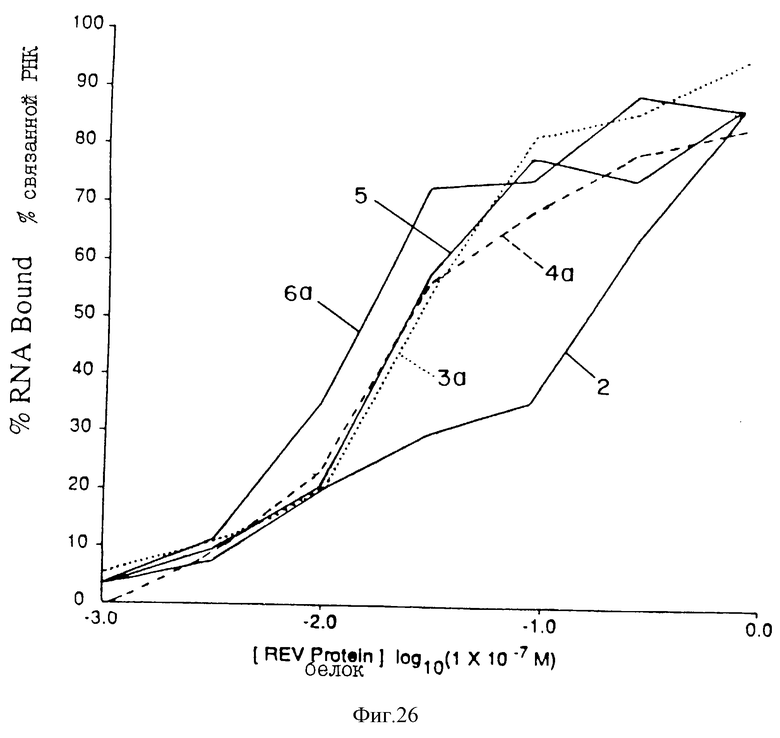
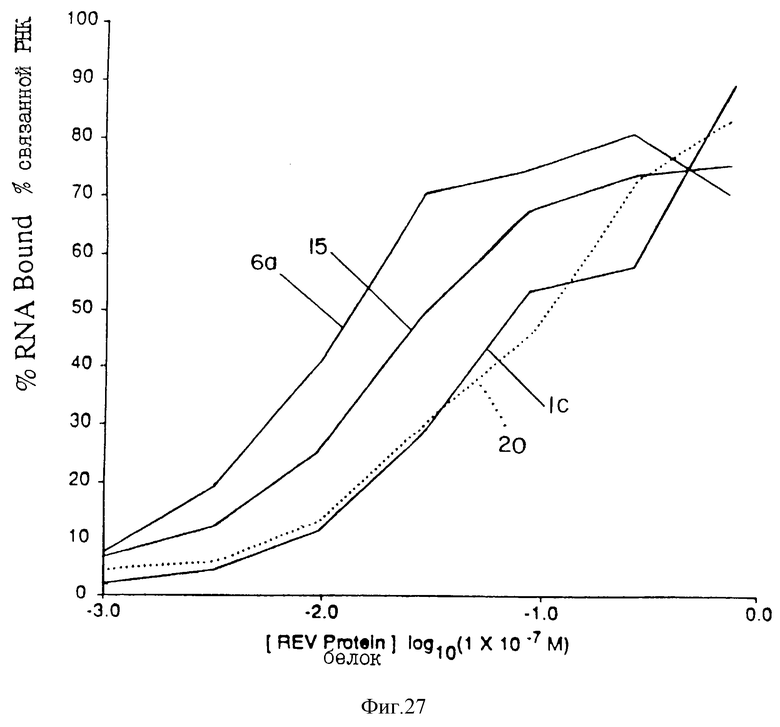
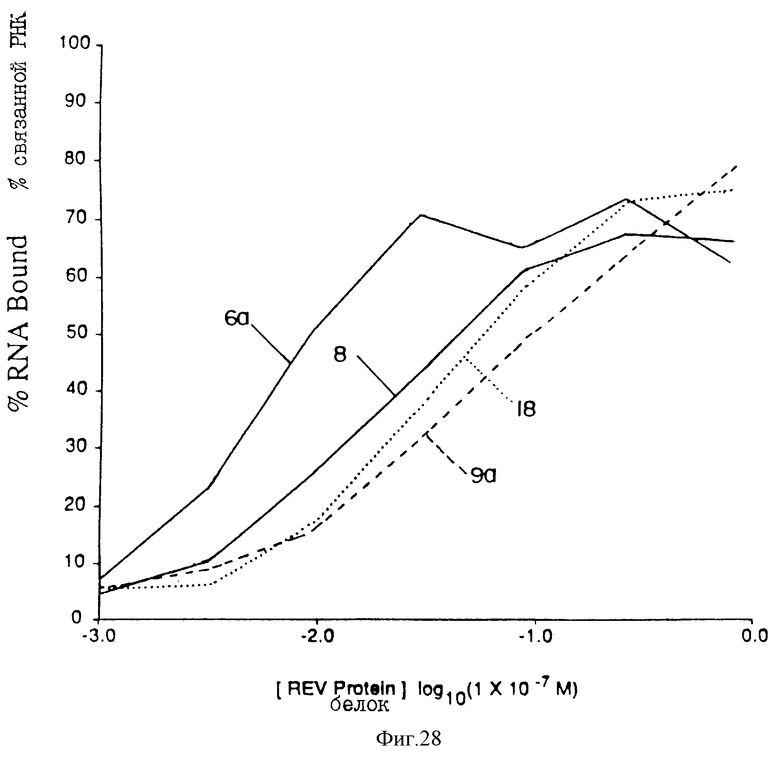
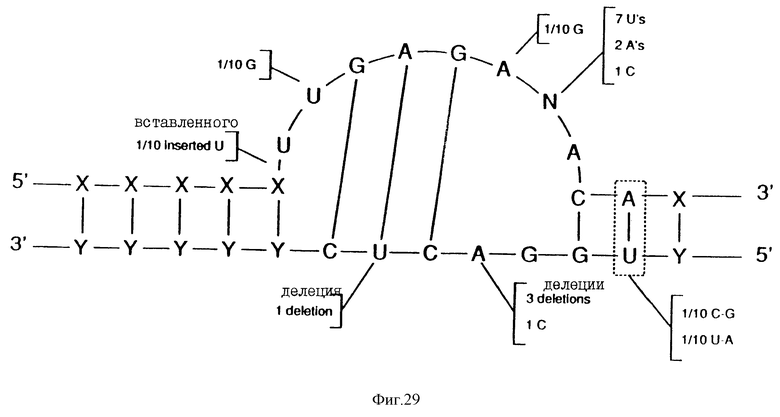
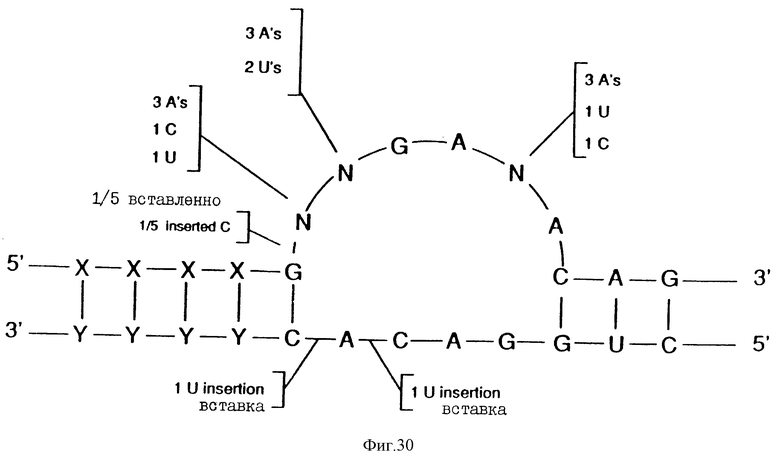
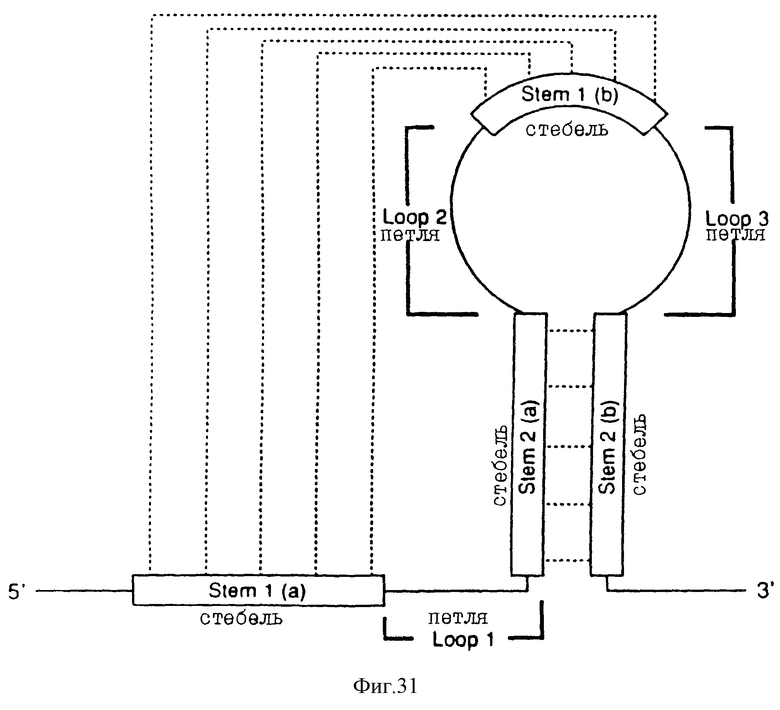
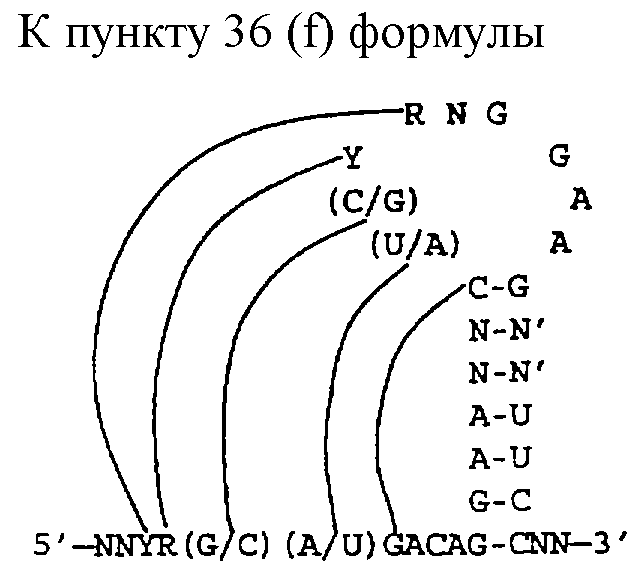
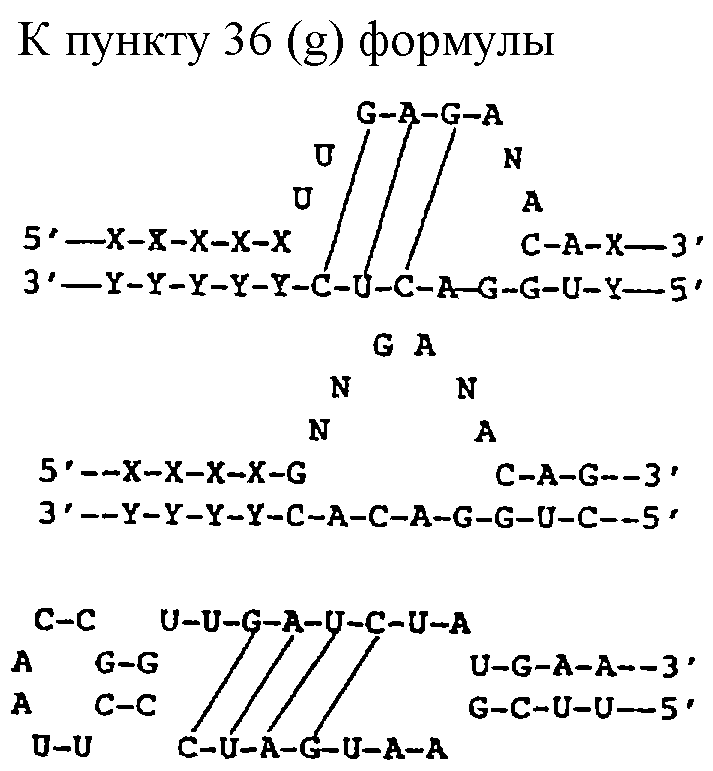
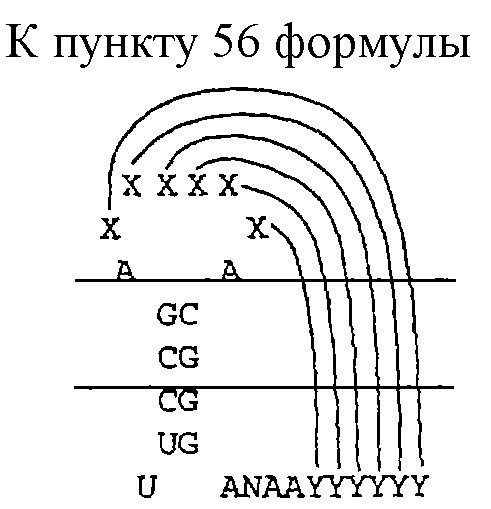
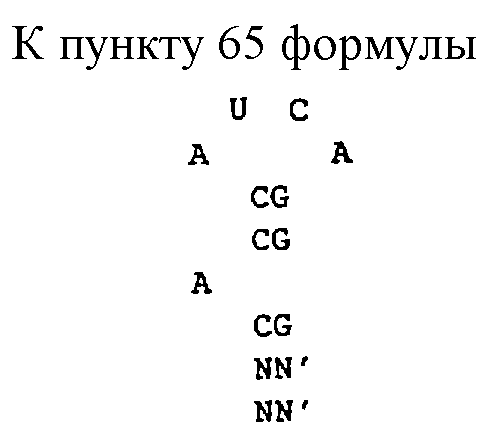
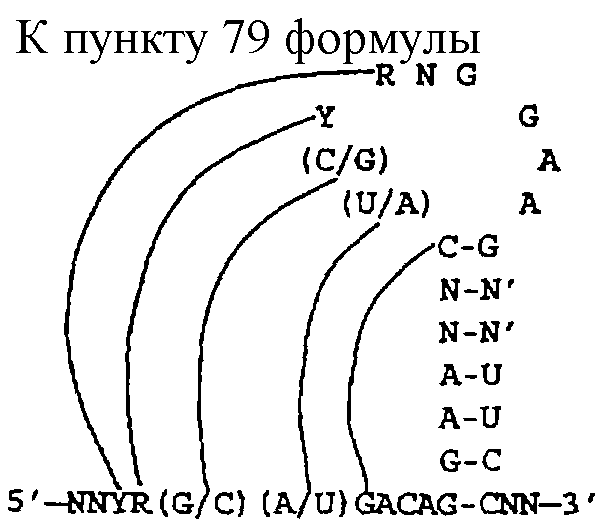
Изобретение относится к новому классу нуклеиново-кислотных лигандов с высокой степенью сродства, которые специфически связываются с нужной молекулой-мишенью. Проводят взаимодействие смеси однонитевых нуклеиновых кислот с мишенью, отделение нуклеиновых кислот, имеющих повышенное сродство к данной мишени, от оставшейся смеси. После отделения нуклеиновых кислот, имеющих повышенное сродство к данной мишени, проводят их амплификацию с получением лигандобогащенной смеси нуклеиновых кислот. Обеспечивают взаимодействие однонитевых нуклеиновых кислот с мишенью. Отделяют нуклеиновые кислоты, имеющие повышенное сродство к данной мишени. Проводят амплификацию их с получением лигандобогащенной смеси нуклеиновых кислот. Повторяют перечисленные стадии с использованием лигандобогащенной смеси нуклеиновых кислот каждого последующего повтора столько раз, сколько необходимо для получения уровня обогащения, достаточного для получения лигандного раствора к указанной мишени. Молекулы нуклеиновых кислот, которые воздействуют на функции молекулы-мишени, идентифицируют вышеприведенным способом и проверяют на повышенное сродство к интересующей мишени и на способность воздействовать их на функции указанных молекул-мишеней. Получают смесь нуклеиновых кислот, содержащих консервативный сегмент и рандомизированный сегмент. Проводят взаимодействие смеси нуклеиновых кислот с мишенью в условиях, благоприятных для связывания, с образованием пар нуклеиновая кислота - мишень и несвязанных нуклеиновых кислот. Разделяют несвязанные нуклеиновые кислоты и пары нуклеиновая кислота - мишень. Проводят амплификацию отделенных нуклеиновых кислот пар нуклеиновая кислота - мишень с получением лигандобогащенной смеси нуклеиновых кислот. Смесь нуклеиновых кислот включает по меньшей мере 109 однонитевых нуклеиновых кислот. Нуклеиновые кислоты включают в себя консервативный нуклеотидный сегмент и рандомизированный нуклеотидный сегмент. Рандомизированный сегмент включает в себя цепь из 8-100 следующих друг за другом нуклеотидов. Не встречающийся в природе нуклеиново-кислотный лиганд идентифицируют способом, приведенным выше. Лиганд не является нуклеиновой кислотой, обладающей известной физиологической функцией связывания молекулой-мишенью, обладает сродством специфического связывания с молекулой-мишенью. Такая молекула-мишень имеет трехмерную структуру, отличную от структуры полинуклеотида, связывающегося с указанным нуклеиново-кислотным лигандом посредством механизма, который зависит от спаривания оснований по Уотсону и Крику или связывания тройной спирали. Последовательности нуклеиновых и рибонуклеиновых кислот приведены в формуле и описании. Изобретение позволяет получать новый класс нуклеиново-кислотных соединений, так называемых нуклеиново-кислотных антител, которые обладают сродством для специфического связывания с трехмерными молекулами-мишенями. 6 с. и 77 з.п. ф-лы, 36 ил., 16 табл.
(см. графическую часть);
(с) РНК с последовательностью
(см. графическую часть);
(d) нуклеиновую кислоту, включающую в себя РНК-последовательности 5'-CUСА-3'; 5'-GAGCGCAAGACGAAUAG-3'; 5'-UACA-3' и 5'-ACAUCGAUGACCGGAAUGCCGCACACAGAG-3'; (e) нуклеиновую кислоту, включающую в себя РНК-последовательность 5'-UAAGGAGGCCAC-3'; (f) нуклеиновую кислоту, включающую в себя РНК-последовательность
(см. графическую часть);
(g) нуклеиновую кислоту, включающую в себя РНК-последовательности, выбранные из группы, состоящей из
(см. графическую часть);
где X и Y представляют собой комплементарные друг другу нуклеотиды.
(см. графическую часть);
где X и Y - комплементарные друг другу нуклеотиды, как показано на фиг. 22.
(см. графическую часть);
66. Лиганд по п.37, отличающийся тем, что указанный белок является сериновой протеазой.
(см. графическую часть);
80. Лиганд по п.37, отличающийся тем, что указанный белок является вирусным rev-белком.
(см. графическую часть);
где Х и Y - комплементарные друг другу нуклеотиды.
| WO 8906694 А, 27.07.1989 | |||
| ЗАЩИТНО-ВОССТАНОВИТЕЛЬНЫЙ СОСТАВ | 2000 |
|
RU2183661C2 |
| KINZLER et al | |||
| Whole Genomic PCR: Application to the Identification of Sequences Bound by Gene Regulatory Proteins | |||
| Nuc | |||
| Печь для сжигания твердых и жидких нечистот | 1920 |
|
SU17A1 |
| OLIPHANT and STRUHL | |||
| Defining the consensus sequences of E | |||
| coli promoter elements by random selection | |||
| Nucleic Acids Research, vol | |||
| Устройство для электрической сигнализации | 1918 |
|
SU16A1 |
| Прибор для нагревания перетягиваемых бандажей подвижного состава | 1917 |
|
SU15A1 |
| OLIPHANT et al | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Mol | |||
| and Cell Biology Jul | |||
| Механизм для сообщения поршню рабочего цилиндра возвратно-поступательного движения | 1918 |
|
SU1989A1 |
| Способ и устройство для обнаружения раковин в металлических изделиях | 1925 |
|
SU2944A1 |
| OLIPHANT and STRUHL | |||
| Use of Random-Sequence Oligonucleotides for Determining Consensus Sequences, se of Random-Sequence Oligonucleotides for Determining Consensus Sequences | |||
| Methods in Enzymologgy, vol.155 35: 56814 583, 1977 | |||
| МАНИАТИС Г | |||
| и др | |||
| Методы генетической инженерии | |||
| - М.: Мир, 1984, с.107-118. | |||

