ПЕРЕКРЕСТНАЯ ССЫЛКА НА СМЕЖНЫЕ ЗАЯВКИ
Настоящая заявка испрашивает преимущество по предварительной заявке на патент США №62/015,809, поданной 23 июня 2014 г., предварительной заявке на патент США №62/016,400, поданной 24 июня 2014 г., и предварительной заявке на патент США №62/036,983, поданной 13 августа 2014 г., каждая из которых полностью включена в настоящий документ путем ссылки.
КАК ТЕКСТОВЫЙ ФАЙЛ ЧЕРЕЗ EFS WEB
Официальная копия списка последовательностей представлена в электронном виде через EFS-Web в виде файла со списком последовательностей в формате ASCII с наименованием 461002SEQLIST.TXT, созданного 23 июня 2015 г., имеющего размер 66 Кб и поданного одновременно со спецификацией. Список последовательностей, содержащийся в данном документе в формате ASCII, является частью спецификации и полностью включен в настоящий документ путем ссылки.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
С исторической точки зрения в качестве способа синтеза более крупных двухцепочечных молекул ДНК, в частности генов, из перекрывающихся синтетических олигонуклеотидов можно использовать перекрывающиеся удлинения. Однако этот способ не позволяет быстро и эффективно соединять большие молекулы ДНК. Кроме того, сайт-специфическое соединение больших нуклеиновых кислот с использованием перекрывающихся последовательностей часто ограничивается доступностью перекрывающихся последовательностей в нужном положении в соединяемых нуклеиновых кислотах. Привлекают внимание сконструированные нуклеазные ферменты, предназначенные для нацеливания на определенные последовательности ДНК, которые являются мощными инструментами для генетических манипуляций, позволяющими осуществлять целенаправленную делецию, замену и восстановление генов, а также вставку экзогенных последовательностей. Однако существующие технологии имеют недостатки, заключающиеся в ограниченной точности, что может приводить к непредсказуемым нецелевым эффектам, а также являются многостадийными реакциями, занимающими много времени.
ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В настоящем документе предлагаются способы сборки нуклеиновых кислот, имеющих перекрывающиеся последовательности. Такие способы включают способ сборки по меньшей мере двух нуклеиновых кислот, включающий: (а) контактирование первой нуклеиновой кислоты с первым нуклеазным агентом, причем первый нуклеазный агент расщепляет первую нуклеиновую кислоту в первом целевом сайте с получением первой расщепленной нуклеиновой кислоты с перекрывающимися концевыми последовательностями между первой расщепленной нуклеиновой кислотой и второй нуклеиновой кислотой; (b) контактирование первой расщепленной нуклеиновой кислоты и второй нуклеиновой кислоты с экзонуклеазой для экспонирования комплементарных последовательностей между первой расщепленной нуклеиновой кислотой и второй нуклеиновой кислотой; и (с) сборку двух фрагментов нуклеиновых кислот, полученных на этапе (b). В некоторых таких способах этап (с) дополнительно включает: (i) ренатурацию экспонированных комплементарных последовательностей; (ii) удлинение 3'-концов ренатурированных комплементарных последовательностей и (iii) лигирование первой и второй нуклеиновых кислот.
В некоторых способах этап (а) дополнительно включает контактирование второй нуклеиновой кислоты со вторым нуклеазным агентом, причем вторая нуклеиновая кислота не содержит перекрывающейся концевой последовательности, и второй нуклеазный агент расщепляет вторую нуклеиновую кислоту во втором целевом сайте с получением второй расщепленной нуклеиновой кислоты с перекрывающимися концевыми последовательностями между первой расщепленной нуклеиновой кислотой и второй расщепленной нуклеиновой кислотой, и при этом вторая нуклеиновая кислота из этапа (b) представляет собой вторую расщепленную нуклеиновую кислоту. В некоторых способах перекрывающаяся концевая последовательность имеет длину в диапазоне от 20 пар нуклеотидов (п.н.) до 200 п.н.
В некоторых способах по меньшей мере один из первого или второго нуклеазного агента содержит белок Cas и гидовую РНК (гРНК) (комплекс гРНК-Cas), нацеленные на первый или второй целевой сайт. Например, белок Cas может представлять собой белок Cas9. Белок Cas9 может содержать домен RuvC и домен HNH, по меньшей мере один из которых не имеет эндонуклеазной активности. В некоторых вариантах осуществления гРНК содержит нуклеотидную последовательность, кодирующую РНК коротких палиндромных повторов, регулярно расположенных группами (CRISPR) (крРНК), и трансактивирующую РНК CRISPR (тракрРНК). Первый целевой сайт и/или второй целевой сайт может быть фланкирован последовательностью мотива, прилежащего к протоспейсеру (РАМ). В некоторых способах нуклеазный агент содержит нуклеазу с «цинковыми пальцами» или эффекторную нуклеазу, подобную активатору транскрипции (TALEN).
В некоторых способах первая, вторая или обе нуклеиновые кислоты получены из бактериальной искусственной хромосомы. Бактериальная искусственная хромосома может содержать человеческую ДНК, ДНК грызунов, синтетическую ДНК или их комбинацию. Бактериальная искусственная хромосома может содержать человеческую последовательность.
Способы, описанные в настоящем документе, включают способ сборки по меньшей мере двух нуклеиновых кислот, включающий: (а) контактирование первой нуклеиновой кислоты с первым нуклеазным агентом и вторым нуклеазным агентом с получением первой расщепленной нуклеиновой кислоты, причем первый нуклеазный агент создает одноцепочечный разрыв на первой цепи первой нуклеиновой кислоты в первом целевом сайте, а второй нуклеазный агент создает одноцепочечный разрыв на второй цепи первой нуклеиновой кислоты во втором целевом сайте, с получением первой расщепленной нуклеиновой кислоты, содержащей нависающую последовательность на одном из ее 5'- или 3'-концов; (b) ренатурацию первой расщепленной нуклеиновой кислоты и второй нуклеиновой кислоты, содержащей последовательность, комплементарную 5'- или 3'-нависающей последовательности; и (с) лигирование первой расщепленной нуклеиновой кислоты и второй нуклеиновой кислоты. В некоторых способах этап (b) дополнительно включает удлинение 3'-конца первой цепи с использованием второй цепи в качестве шаблона и удлинение 3'-конца второй цепи с использованием первой цепи в качестве шаблона. В некоторых способах первый целевой сайт отделен от второго целевого сайта по меньшей мере 4 п.н.
В некоторых способах по меньшей мере один из первого и второго нуклеазного агента содержит белок Cas9 и гидовую РНК (гРНК) (комплекс гРНК-Cas), нацеленные на первый или второй целевой сайт. гРНК может содержать нуклеотидную последовательность, кодирующую РНК коротких палиндромных повторов, регулярно расположенных группами (CRISPR) (крРНК), и трансактивирующую РНК CRISPR (тракрРНК). В некоторых способах по меньшей мере один из первого целевого сайта и второго целевого сайта фланкирован последовательностью мотива, прилежащего к протоспейсеру (РАМ). Белок Cas9 может содержать домен RuvC и домен HNH, причем один из них не имеет эндонуклеазной активности.
В некоторых способах вторая нуклеиновая кислота не содержит последовательности, комплементарной 5'- или 3'-нависающей последовательности первой расщепленной нуклеиновой кислоты, и этап (а) дополнительно включает контактирование первой расщепленной нуклеиновой кислоты и второй расщепленной нуклеиновой кислоты с соединительным олигонуклеотидом, причем соединительный олигонуклеотид содержит: (i) первую последовательность, комплементарную 5'- или 3'-нависающей последовательности первой расщепленной нуклеиновой кислоты; и (ii) вторую последовательность, комплементарную 5'- или 3'-нависающей последовательности второй расщепленной нуклеиновой кислоты. В некоторых способах первая, вторая или обе нуклеиновые кислоты получены из бактериальной искусственной хромосомы. Бактериальная искусственная хромосома может содержать человеческую ДНК, ДНК грызунов, синтетическую ДНК или их комбинацию. Бактериальная искусственная хромосома может содержать человеческую полинуклеотидную последовательность. В некоторых способах вторая нуклеиновая кислота содержит бактериальную искусственную хромосому.
Предлагаемые способы также включают способ сборки двух или более фрагментов нуклеиновых кислот, включающий: (а) контактирование первой нуклеиновой кислоты с по меньшей мере одним нуклеазным агентом с получением первой расщепленной нуклеиновой кислоты; (b) контактирование первой расщепленной нуклеиновой кислоты со второй нуклеиновой кислотой, соединительным олигонуклеотидом и экзонуклеазой, причем соединительный олигонуклеотид содержит: (i) первую комплементарную последовательность, которая комплементарна первой расщепленной нуклеиновой кислоте; (ii) спейсер; и (iii) вторую комплементарную последовательность, которая комплементарна второй нуклеиновой кислоте; причем экзонуклеаза экспонирует первую и вторую комплементарные последовательности; и (с) сборку соединительного олигонуклеотида с первой расщепленной нуклеиновой кислотой и второй нуклеиновой кислотой. В некоторых таких способах сборка на этапе (с) включает: (i) ренатурацию первой комплементарной последовательности соединительного олигонуклеотида с первой расщепленной нуклеиновой кислотой и второй комплементарной последовательности соединительного олигонуклеотида со второй нуклеиновой кислотой; и (ii) лигирование соединительного олигонуклеотида с первой расщепленной нуклеиновой кислотой и второй нуклеиновой кислотой.
В некоторых способах первая комплементарная последовательность и вторая комплементарная последовательность соединительного олигонуклеотида содержат от 15 до 120 комплементарных оснований. В некоторых способах спейсер соединительного олигонуклеотида содержит некомплементарные нуклеиновые кислоты. В некоторых вариантах осуществления сборка первой расщепленной нуклеиновой кислоты со второй нуклеиновой кислотой происходит бесшовно.
В некоторых способах нуклеазный агент предназначен для отщепления фрагмента из по меньшей мере 20 п.н. от конца первой нуклеиновой кислоты, на котором будет происходить бесшовная сборка, причем спейсер соединительного олигонуклеотида содержит последовательность, идентичную указанному фрагменту из по меньшей мере 20 п.н., при этом между первой комплементарной последовательностью и фрагментом из по меньшей мере 20 п.н. нуклеотидные основания отсутствуют, и между второй комплементарной последовательностью и фрагментом из по меньшей мере 20 п.н. нуклеотидные основания отсутствуют, так что сборка указанной первой нуклеиновой кислоты с указанным соединительным олигонуклеотидом и указанной второй нуклеиновой кислотой восстанавливает этот фрагмент из по меньшей мере 20 п.н. и обеспечивает бесшовную сборку первой и второй нуклеиновых кислот. В некоторых способах тот же способ выполняют по меньшей мере с фрагментом из 20 п.н. из второй нуклеиновой кислоты, используемым в качестве спейсерной последовательности. В некоторых способах спейсер содержит от около 20 п.н. до около 120 п.н. В некоторых способах вторую нуклеиновую кислоту приводят в контакт со вторым нуклеазным агентом и экзонуклеазой, причем второй нуклеазный агент расщепляет вторую нуклеиновую кислоту с получением второй расщепленной нуклеиновой кислоты, содержащей нуклеотидную последовательность, комплементарную второй комплементарной последовательности соединительного олигонуклеотида, при этом проводят сборку первой расщепленной нуклеиновой кислоты со второй расщепленной нуклеиновой кислотой. В некоторых способах вторую нуклеиновую кислоту приводят в контакт с рестрикционным ферментом или мегануклеазой и экзонуклеазой, причем рестрикционный фермент или мегануклеаза расщепляет вторую нуклеиновую кислоту с получением второй расщепленной нуклеиновой кислоты, содержащей нуклеотидную последовательность, комплементарную второй комплементарной последовательности в соединительном олигонуклеотиде, при этом проводят сборку первой расщепленной нуклеиновой кислоты со второй расщепленной нуклеиновой кислотой. В некоторых способах 3'-конец первой и/или второй расщепленной нуклеиновой кислоты удлиняют на этапе (b). Можно выполнить сборку соединительного олигонуклеотида с указанной первой нуклеиновой кислотой и указанной второй нуклеиновой кислотой в одной реакции или последовательно. В некоторых способах первая, вторая или обе нуклеиновые кислоты происходят из бактериальной искусственной хромосомы длиной по меньшей мере 10 т.п.н и/или содержат человеческую ДНК, ДНК грызунов, синтетическую ДНК или их комбинацию.
В некоторых способах по меньшей мере один нуклеазный агент или второй нуклеазный агент содержит белок Cas и гидовую РНК (гРНК) (комплекс гРНК-Cas), нацеленные на первый или второй целевой сайт. Например, белок Cas может представлять собой белок Cas9. Белок Cas9 может содержать домен RuvC и домен HNH, по меньшей мере один из которых не имеет эндонуклеазной активности. В некоторых вариантах осуществления гРНК содержит нуклеотидную последовательность, кодирующую РНК коротких палиндромных повторов, регулярно расположенных группами (CRISPR) (крРНК), и трансактивирующую РНК CRISPR (тракрРНК). Первый целевой сайт и/или второй целевой сайт может быть фланкирован последовательностью мотива, прилежащего к протоспейсеру (РАМ). В некоторых способах по меньшей мере один нуклеазный агент и/или второй нуклеазный агент содержит нуклеазу с «цинковыми пальцами» или эффекторную нуклеазу, подобную активатору транскрипции (TALEN).
В некоторых вариантах осуществления соединительный олигонуклеотид содержит gBlock. В некоторых таких способах gBlock не содержит кассеты селекции.
Дополнительно предлагаются способы сборки двух или более нуклеиновых кислот, включающие: (а) контактирование первой нуклеиновой кислоты с по меньшей мере одним нуклеазным агентом с получением первой расщепленной нуклеиновой кислоты; (b) контактирование второй нуклеиновой кислоты со вторым нуклеазным агентом с получением второй расщепленной нуклеиновой кислоты; (с) контактирование первой расщепленной нуклеиновой кислоты и второй расщепленной нуклеиновой кислоты с соединительным олигонуклеотидом и экзонуклеазой, причем соединительный олигонуклеотид содержит: (i) первую комплементарную последовательность, которая комплементарна первой расщепленной нуклеиновой кислоте; (ii) спейсер; и (iii) вторую комплементарную последовательность, которая комплементарна второй расщепленной нуклеиновой кислоте; причем экзонуклеаза экспонирует первую и вторую комплементарные последовательности; и (d) сборку соединительного олигонуклеотида с первой расщепленной нуклеиновой кислотой и второй нуклеиновой кислотой.
В настоящем документе предлагаются способы сборки нуклеиновых кислот, имеющих перекрывающиеся последовательности. Такие способы включают способ сборки по меньшей мере двух фрагментов нуклеиновых кислот, включающий (а) контактирование первой и второй нуклеиновых кислот, содержащих перекрывающиеся последовательности, с по меньшей мере одним комплексом гРНК-Cas и экзонуклеазой, тем самым получая два фрагмента расщепленных нуклеиновых кислот, содержащих комплементарные последовательности на одном из их концов; (b) сборку двух фрагментов нуклеиновых кислот, полученных на этапе (а). В некоторых способах по меньшей мере один комплекс гРНК-Cas расщепляет первую нуклеиновую кислоту в первом целевом сайте с получением первой расщепленной нуклеиновой кислоты, содержащей концевые последовательности, комплементарные между первой расщепленной нуклеиновой кислотой и второй нуклеиновой кислотой. В некоторых способах этап (b) дополнительно включает: (i) ренатурацию экспонированных комплементарных последовательностей; (ii) удлинение 3'-концов ренатурированных комплементарных последовательностей и (iii) лигирование первой и второй нуклеиновых кислот. В некоторых способах этап (а) дополнительно включает контактирование второй нуклеиновой кислоты со вторым комплексом гРНК-Cas, причем вторая нуклеиновая кислота не содержит перекрывающейся концевой последовательности, и второй комплекс гРНК-Cas расщепляет вторую нуклеиновую кислоту с получением второй расщепленной нуклеиновой кислоты, содержащей перекрывающиеся концевые последовательности между первой расщепленной нуклеиновой кислотой и второй расщепленной нуклеиновой кислотой. Например, комплекс гРНК-Cas содержит белок Cas9. Белок Cas9 может содержать домен RuvC и домен HNH, по меньшей мере один из которых не имеет эндонуклеазной активности. В некоторых способах перекрывающиеся последовательности имеют длину в диапазоне от 20 п.н. до 200 п.н. Способ по любому из пунктов формулы изобретения 1-7, в котором первая, вторая или обе нуклеиновые кислоты получены из бактериальной искусственной хромосомы. В некоторых способах бактериальная искусственная хромосома содержит человеческую ДНК, ДНК грызунов, синтетическую ДНК или их комбинацию, бактериальная искусственная хромосома может содержать человеческую последовательность.
Предлагаемые способы также включают способ сборки двух или более фрагментов нуклеиновых кислот, включающий: (а) экспонирование первой и второй нуклеиновых кислот для по меньшей мере одного комплекса гРНК-Cas с получением первой и второй расщепленных нуклеиновых кислот, содержащих нависающую последовательность на одном из их 5'- или 3'-концов; (b) сборка двух фрагментов нуклеиновых кислот, полученных на этапе (а). В некоторых способах сборка на этапе (b) включает: (i) ренатурацию 5'- и 3'-нависающих последовательностей; и (ii) лигирование первой расщепленной нуклеиновой кислоты и второй расщепленной нуклеиновой кислоты. В некоторых способах 5'- и/или 3'-нависающие последовательности содержат по меньшей мере 4 комплементарных основания. В некоторых способах этап (b) дополнительно включает удлинение 3'-конца первой и второй расщепленных нуклеиновых кислот. В некоторых способах вторая нуклеиновая кислота не содержит последовательности, комплементарной 5'- или 3'-нависающей последовательности первой расщепленной нуклеиновой кислоты, и этап (а) дополнительно включает контактирование первой расщепленной нуклеиновой кислоты и второй расщепленной нуклеиновой кислоты с соединительным олигонуклеотидом, причем соединительный олигонуклеотид содержит: (i) первую последовательность, комплементарную 5'- или 3'-нависающей последовательности первой расщепленной нуклеиновой кислоты; и (ii) вторую последовательность, комплементарную 5'- или 3'-нависающей последовательности второй расщепленной нуклеиновой кислоты. В некоторых способах комплекс гРНК-Cas содержит белок Cas9, содержащий домен RuvC и домен HNH, один из которых не имеет эндонуклеазной активности. В некоторых способах комплекс гРНК-Cas предлагается отдельно в виде крРНК, тракрРНК и белка Cas. В некоторых способах первая и вторая нуклеиновые кислоты содержат последовательность мотива, прилежащего к протоспейсеру (РАМ). В некоторых способах первая, вторая или обе нуклеиновые кислоты получены из бактериальной искусственной хромосомы. В некоторых способах бактериальная искусственная хромосома содержит человеческую ДНК, ДНК грызунов, синтетическую ДНК или их комбинацию. Например, бактериальная искусственная хромосома может содержать человеческую полинуклеотидную последовательность.
Дополнительно предлагаются способы сборки двух или более нуклеиновых кислот, включающие: (а) контактирование первой нуклеиновой кислоты с по меньшей мере одним комплексом гРНК-Cas с получением первой расщепленной нуклеиновой кислоты; и (b) контактирование первой расщепленной нуклеиновой кислоты со второй нуклеиновой кислотой, соединительным олигонуклеотидом и экзонуклеазой, причем соединительный олигонуклеотид содержит: (i) первую комплементарную последовательность, которая комплементарна первой расщепленной нуклеиновой кислоте; (ii) спейсер; и (iii) вторую комплементарную последовательность, которая комплементарна второй нуклеиновой кислоте; причем экзонуклеаза экспонирует первую и вторую комплементарные последовательности; и (с) сборку соединительного олигонуклеотида с первой расщепленной нуклеиновой кислотой и второй нуклеиновой кислотой. В некоторых способах этап сборки (с) включает (i) ренатурацию первой комплементарной последовательности соединительного олигонуклеотида с первой расщепленной нуклеиновой кислотой и второй комплементарной последовательности соединительного олигонуклеотида со второй нуклеиновой кислотой; и (ii) лигирование соединительного олигонуклеотида с первой расщепленной нуклеиновой кислотой и второй нуклеиновой кислотой. В некоторых способах первая комплементарная последовательность и вторая комплементарная последовательность соединительного олигонуклеотида содержат от 15 до 120 комплементарных оснований. В некоторых способах спейсер соединительного олигонуклеотида содержит некомплементарные нуклеиновые кислоты.
С помощью соединительного олигонуклеотида первая расщепленная нуклеиновая кислота может быть бесшовно собрана со второй нуклеиновой кислотой. В некоторых способах комплекс гPHК-CAS предназначен для отщепления фрагмента из по меньшей мере 20 п.н. от конца первой нуклеиновой кислоты, на котором будет происходить бесшовная сборка, причем спейсер соединительного олигонуклеотида содержит последовательность, идентичную указанному фрагменту из по меньшей мере 20 п.н., при этом между первой комплементарной последовательностью и фрагментом из по меньшей мере 20 п.н. нуклеотидные основания отсутствуют, и между второй комплементарной последовательностью и фрагментом из по меньшей мере 20 п.н. нуклеотидные основания отсутствуют, так что сборка указанной первой нуклеиновой кислоты с указанным соединительным олигонуклеотидом и указанной второй нуклеиновой кислотой восстанавливает фрагмент из по меньшей мере 20 п.н. и обеспечивает бесшовную сборку первой и второй нуклеиновых кислот. В некоторых способах тот же способ выполняют по меньшей мере с фрагментом из 20 п.н. из второй нуклеиновой кислоты, используемым в качестве спейсерной последовательности. В некоторых способах спейсер содержит от около 20 п.н. до около 120 п.н. В некоторых способах вторую нуклеиновую кислоту приводят в контакт со вторым комплексом гРНК-Cas и экзонуклеазой, причем второй комплекс гРНК-Cas расщепляет вторую нуклеиновую кислоту с получением второй расщепленной нуклеиновой кислоты, содержащей нуклеотидную последовательность, комплементарную второй комплементарной последовательности соединительного олигонуклеотида, при этом проводят сборку первой расщепленной нуклеиновой кислоты со второй расщепленной нуклеиновой кислотой. В некоторых способах вторую нуклеиновую кислоту приводят в контакт с рестрикционным ферментом или мегануклеазой и экзонуклеазой, причем рестрикционный фермент или мегануклеаза расщепляет вторую нуклеиновую кислоту с получением второй расщепленной нуклеиновой кислоты, содержащей нуклеотидную последовательность, комплементарную второй комплементарной последовательности в соединительном олигонуклеотиде, при этом проводят сборку первой расщепленной нуклеиновой кислоты со второй расщепленной нуклеиновой кислотой. В некоторых способах 3'-конец первой и/или второй расщепленной нуклеиновой кислоты удлиняют на этапе (b). Можно выполнить сборку соединительного олигонуклеотида с указанной первой нуклеиновой кислотой и указанной второй нуклеиновой кислотой в одной реакции или последовательно. В некоторых способах комплекс гРНК-Cas содержит белок Cas9. В некоторых способах первая, вторая или обе нуклеиновые кислоты происходят из бактериальной искусственной хромосомы длиной по меньшей мере 10 т.п.н и/или содержит человеческую ДНК, ДНК грызунов, синтетическую ДНК или их комбинацию.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
На ФИГ. 1 продемонстрирована сборка бактериальной искусственной хромосомы (ВАС) с продуктом ПЦР, имеющим перекрывающие участки, сконструированные специфичными для ВАС. К кассете HYG посредством ПЦР были добавлены перекрывающие участки длиной 50 п.н.
На ФИГ. 2 продемонстрирована сборка двух ВАС, имеющих перекрывающиеся последовательности, с использованием двух целевых сайтов для Cas9 на каждой ВАС.
Процесс сборки с использованием описанного в настоящем документе способа занимает 2 дня.
На ФИГ. 3 продемонстрирована сборка двух ВАС с перекрывающимися последовательностями с использованием традиционных способов. Процесс сборки с использованием традиционных способов занимает 4 дня.
На ФИГ. 4 продемонстрирована эффективность клонирования способом Cas9/изотермическая сборка и временные затраты на этапы клонирования ВАС.
На ФИГ. 5 продемонстрирована конструкция большого нацеливающего вектора (LTVEC) с использованием системы CRISPR/Cas9 и изотермической сборки. Фрагменты ДНК, расщепленные CRISPR/Cas9, бесшовно собирают с использованием одного или более соединительных олигонуклеотидов и изотермической сборки.
На ФИГ. 6 продемонстрирована стратегия использования линкеров (соединительных олигонуклеотидов) для бесшовной сборки нуклеиновых кислот после расщепления Cas9. Комплекс гРНК/Cas9 предназначен для расщепления целевого сайта, находящегося ближе к 5'-концу от интересующей области (стрелка) с получением первого расщепленного Cas9 фрагмента ДНК (5'-ДНК). Удаленный участок 5'-ДНК (прямоугольник с косой штриховкой) далее используют в качестве спейсера между 5'- и 3'-перекрывающимися последовательностями в соединительном олигонуклеотиде. В ходе реакции изотермической сборки выполняют сборку трех компонентов: (а) первого расщепленного Cas9 фрагмента ДНК (5'-ДНК); (b) соединительного олигонуклеотида; и (с) второго фрагмента ДНК (3'-ДНК). Соединительный олигонуклеотид в направлении от 5'- к 3'-концу содержит: (1) последовательность, перекрывающуюся с 5'-концом ДНК, (2) спейсер, содержащий удаленную часть первого расщепленного фрагмента, и (3) последовательность, перекрывающуюся с 3'-концом ДНК. Удаленный участок 5'-конца ДНК восстанавливают на этапе сборки.
На ФИГ. 7 продемонстрирована конструкция ДНК-вектора с использованием системы CRISPR/Cas9 и изотермической сборки.
На ФИГ. 8 продемонстрирована конструкция большого нацеливающего вектора с использованием системы CRISPR/Cas9 и изотермической сборки.
На ФИГ. 9 продемонстрирована конструкция нацеливающего вектора для замещения участка вектора ВАС кассетой с использованием изотермической сборки и двух линкеров (соединительных олигонуклеотидов). Результаты различных соотношений mВАС к фрагментам или линкерам представлены на панелях №1, №2, №3 и №4.
На ФИГ. 10 продемонстрировано подтверждение последовательности бесшовной сборки по обоим стыкам реакции сборки между mВАС (ВАС ID: RP23-399М19) и кассетой с использованием двух линкеров.
На ФИГ. 11 продемонстрирована сборка двух mВАС с использованием Cas9 и изотермической сборки. Сборка вектора bMQ50f19 и кассеты, содержащей убиквитиновый промотор гена устойчивости к гигромицину, была бесшовной.
На ФИГ. 12 продемонстрировано подтверждение последовательности бесшовной сборки на линкере 1 и подтверждение последовательности сборки, которая намеренно была не бесшовной, на линкере 2 и линкере 3.
На ФИГ. 13 продемонстрирована вставка больших фрагментов человеческого гена в mВАС с использованием четырех линкеров и изотермической сборки. Расщепленный Cas9 фрагмент A hGene из hBAC1, фрагмент В hGene из hBAC2 и mВАС для удаления фрагментов mGene.
На ФИГ. 14 продемонстрирована вставка человеческой последовательности в вектор ВАС с использованием Cas9 и изотермической сборки.
На ФИГ. 15 продемонстрирована вставка gBlock, содержащего мегануклеазный сайт, с использованием Cas9 и изотермической сборки. На ФИГ. 15А продемонстрирована вставка gBlock, содержащего сайт PI-SceI; и на ФИГ. 15В продемонстрирована вставка gBlock, содержащего сайт MauBI.
На ФИГ. 16 проиллюстрирован пример прямой гуманизации нацеливающего вектора с использованием трех соединительных олигонуклеотидов, Cas9 и изотермической сборки.
На ФИГ. 17 проиллюстрирован пример непрямой гуманизации нацеливающего вектора с использованием донора с выше- и нижележащими соединительными олигонуклеотидами, Cas9 и изотермической сборки.
На ФИГ. 18 проиллюстрирован пример введения точечной мутации с использованием Cas9 и изотермической сборки.
На ФИГ. 19 проиллюстрирован пример обрезки ВАС при помощи Cas9 и изотермической сборки. В этом примере обрезкой удаляется последовательность Ori. Последовательность Ori снова вставляется в вектор с использованием двух соединительных олигонуклеотидов и изотермической сборки.
ПОДРОБНОЕ ОПИСАНИЕ
I. ОПРЕДЕЛЕНИЯ
Термины «белок», «полипептид» и «пептид,» используемые в настоящем документе на взаимозаменяемой основе, включают полимерные формы аминокислот любой длины, включая кодированные и некодированные аминокислоты и химически или биохимически модифицированные или дериватизированные аминокислоты. Термины также включают полимеры, которые подверглись модификации, например полипептиды с модифицированными основными цепями пептидов.
Термины «нуклеиновая кислота» и «полинуклеотид», использованные в настоящем документе на взаимозаменяемой основе, включают полимерные формы нуклеотидов любой длины, включая рибонуклеотиды, дезоксирибонуклеотиды или их аналоги и модифицированные формы. Эти термины включают одно- двух- и многоцепочечные ДНК или РНК, геномную ДНК, кДНК, гибриды ДНК-РНК и полимеры, содержащие пуриновые основания, пиримидиновые основания или другие природные, химически модифицированные, биохимически модифицированные, неприродные и дериватизированные нуклеотидные основания.
«Оптимизация кодонов» по существу включает в себя процесс модификации нуклеотидной последовательности для улучшения экспрессии в конкретных клетках-хозяевах путем замены по меньшей мере одного кодона нативной последовательности кодоном, который более часто или наиболее часто используется в генах клетки-хозяина при сохранении нативной аминокислотной последовательности. Например, нуклеиновая кислота, кодирующая белок Cas, может быть модифицирована, чтобы использовать в качестве замены кодоны, имеющие более высокую частоту использования в данной прокариотической или эукариотической клетке, включая бактериальную клетку, дрожжевую клетку, человеческую клетку, не относящуюся к человеку клетку, клетку млекопитающего, клетку грызуна, клетку мыши, клетку крысы, клетку хомяка или любую другую клетку-хозяина по сравнению со встречающейся в природе нуклеотидной последовательностью. Таблицы использования кодонов являются общедоступными, например, в базе данных Codon Usage Database. Эти таблицы можно адаптировать несколькими способами. См. Nakamura et al. (2000) Nucleic Acids Research 28: 292. Также имеются компьютерные алгоритмы для оптимизации кодонов с конкретной последовательностью для экспрессии в конкретном хозяине (см., например, Gene Forge).
«Функциональная связь» или «функционально связанный» относится к смежному размещению двух или более компонентов (например, промотора и другого элемента последовательности) таким образом, чтобы оба компонента нормально функционировали и обеспечивалась возможность того, чтобы по меньшей мере один из компонентов мог опосредовать функцию, осуществляемую в отношении по меньшей мере одного из других компонентов. Например, промотор может быть функционально связан с кодирующей последовательностью, если промотор контролирует уровень транскрипции кодирующей последовательности в ответ на присутствие или отсутствие одного или более факторов регуляции транскрипции.
«Комплементарность» нуклеиновых кислот означает, что нуклеотидная последовательность в одной цепи нуклеиновой кислоты вследствие ориентации нуклеотидных оснований образует водородные связи с другой последовательностью на противоположной цепи нуклеиновой кислоты. В ДНК, как правило, комплементарны основания А и Т, а также С и G. В РНК это, как правило, С и G, а также U и А. Комплементарность может быть идеальной или существенной/достаточной. Идеальная комплементарность двух нуклеиновых кислот означает, что две нуклеиновые кислоты могут образовывать дуплекс, в котором каждое основание связано с комплементарным основанием, образуя пары Уотсона - Крика. «Существенная» или «достаточная» комплементарность означает, что последовательность одной цепи не полностью и/или не идеально комплементарна последовательности противоположной цепи, но между основаниями на двух цепях создается достаточное связывание, чтобы образовывался стабильный гибридный комплекс при некотором наборе условий гибридизации (например, концентрации солей и температуре). Такие условия можно спрогнозировать, используя последовательности и стандартные математические расчеты для предсказания Тm гибридизованных цепей, или путем эмпирического определения Тm традиционными способами. Тm относится к температуре, при которой популяция гибридизационных комплексов, образовавшихся между двумя цепями нуклеиновых кислот, является денатурированной на 50%. При температуре ниже Тm преимущество имеет образование гибридизационного комплекса, тогда как при температуре выше Тm преимущество имеет плавление или разделение цепей гибридизационного комплекса. Тm можно оценить для нуклеиновой кислоты с известным содержанием G+С в водном 1 М растворе NaCl, используя, например, выражение Тm=81,5+0,41(% G+С), хотя в других известных расчетах Тm принимаются во внимание структурные характеристики нуклеиновых кислот.
«Условие гибридизации» включает совокупную среду, в которой одна цепь нуклеиновой кислоты связывается со второй цепью нуклеиновой кислоты в результате комплементарного взаимодействия цепей и водородного связывания с образованием гибридизационного комплекса. Такие условия включают в себя химические компоненты и их концентрации (например, соли, хелатирующие агенты, формамид) в водном или органическом растворе, содержащем нуклеиновые кислоты, и температуру смеси. Свой вклад в среду могут вносить другие факторы, такие как длительность инкубации или размеры инкубационной камеры. См., например, Sambrook et al., Molecular Cloning, A Laboratory Manual, 2. sup. nd ed., pp. 1.90-1.91, 9.47-9.51, 1 1.47-11.57 (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989).
Для гибридизации требуется, чтобы две нуклеиновые кислоты содержали комплементарные последовательности, хотя возможны несовпадения между основаниями. Условия, подходящие для гибридизации между двумя нуклеиновыми кислотами, зависят от длины нуклеиновых кислот и степени комплементации, и эти переменные хорошо известны специалистам в данной области. Чем больше степень комплементации между двумя нуклеотидными последовательностями, тем выше температура плавления (Тm) гибридов нуклеиновых кислот с этими последовательностями. При гибридизации нуклеиновых кислот с короткими участками комплементарности (например, с комплементарностью более 35 или менее, 30 или менее, 25 или менее, 22 или менее, 20 или менее или 18 или менее нуклеотидов) становится важным положение несовпадающих оснований (см. Sambrook et al., выше, 11.7-11.8). Как правило, длина пригодной для гибридизации нуклеиновой кислоты составляет по меньшей мере около 10 нуклеотидов. Значения иллюстративной минимальной длины пригодных для гибридизации нуклеиновых кислот включают по меньшей мере около 15 нуклеотидов, по меньшей мере около 20 нуклеотидов, по меньшей мере около 22 нуклеотидов, по меньшей мере около 25 нуклеотидов и по меньшей мере около 30 нуклеотидов. Кроме того, температуру и концентрацию солей в промывном растворе можно регулировать по мере необходимости, в зависимости от таких факторов, как длина области комплементации и степень комплементации.
Для обеспечения специфической гибридизации последовательность полинуклеотида не обязательно должна быть на 100% комплементарна целевой нуклеиновой кислоте. Более того, полинуклеотид может гибридизоваться на одном или более сегментах, так что промежуточные или соседние сегменты не вовлечены в событие гибридизации (например, структура петли или шпильки). Полинуклеотид (например, гРНК) может иметь по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 99% или 100% комплементарность последовательности относительно целевой области в целевой нуклеотидной последовательности, на которую он нацелен. Например, гРНК, в которой 18 из 20 нуклеотидов комплементарны целевой области и, следовательно, могут специфически гибридизоваться, будет представлять 90% комплементарности. В этом примере оставшиеся некомплементарные нуклеотиды могут быть кластеризованы или рассеяны среди комплементарных нуклеотидов и не должны быть обязательно смежными друг с другом или с комплементарными нуклеотидами.
Процентную долю комплементарности между конкретными участками нуклеотидных последовательностей в пределах нуклеиновых кислот можно определять типовым способом, используя программы BLAST (средства поиска основных локальных выравниваний) и PowerBLAST, известные в данной области (Altschul et al. (1990) J. Mol. Biol. 215: 403-410; Zhang and Madden (1997) Genome Res. 7: 649-656) или с помощью программы Gap (Wisconsin Sequence Analysis Package, Version 8 for Unix, Genetics Computer Group, University Research Park, г. Мэдисон, штат Висконсин, США), с использованием заданных по умолчанию параметров, где используется алгоритм Смита - Ватермана (Adv. Appl. Math., 1981, 2, 482-489).
В предложенных в настоящем документе способах и композициях используются разнообразные компоненты. В описании признано, что некоторые компоненты могут иметь активные варианты и фрагменты. Такие компоненты включают в себя, например, белки Cas, РНК CRISPR, тракрРНК и гидовые РНК. Биологическая активность каждого из этих компонентов описана в других разделах настоящего документа.
Термин «идентичность последовательности» или «идентичность» в контексте двух полинуклеотидов или полипептидных последовательностей означает ссылку на остатки, которые одинаковы в двух последовательностях при выравнивании для максимального соответствия в установленном окне сравнения. Если термин «процентная доля идентичности последовательности» применяется в отношении белков, следует понимать, что положения с остатками, не являющимися идентичными, зачастую отличаются консервативными заменами аминокислот, где аминокислотные остатки заменяются другими аминокислотными остатками с аналогичными химическими свойствами (например, зарядом или гидрофобностью) и поэтому не изменяют функциональные свойства молекулы. Если последовательности отличаются консервативными заменами, процентную долю идентичности последовательностей можно скорректировать в сторону увеличения, чтобы учесть консервативный характер замены. Последовательности, отличающиеся такими консервативными заменами, считаются имеющими «сходство последовательностей» или «сходство». Способы внесения данной корректировки хорошо известны специалистам в данной области. Как правило, они включают в себя оценку консервативной замены как частичного, а не полного несовпадения, в результате чего повышается процент идентичности последовательности. Таким образом, например, если идентичной аминокислоте присваивают оценку 1, то неконсервативной замене присваивают оценку, равную нулю, а консервативная замена получает оценку от нуля до 1. Вычисляют оценку консервативных замен, например, в соответствии с реализацией в программе PC/GENE (Intelligenetics, г. Маунтин-Вью, штат Калифорния, США).
Термин «процентная доля идентичности последовательности» означает значение, которое определяют путем сравнения двух оптимально выровненных последовательностей в окне сравнения, причем участок полинуклеотидной последовательности в окне сравнения может содержать добавления или делеции (т.е. гэпы) в сравнении с эталонной последовательностью (которая не содержит добавлений или делеций) для оптимального выравнивания двух последовательностей. Процент рассчитывают путем определения числа положений, в которых в обеих последовательностях встречаются идентичные нуклеиновые основания или аминокислотные остатки, деления числа совпадающих положений на общее число положений в окне сравнения и умножения результата на 100 с получением процента идентичности последовательности.
Если не указано иное, значения идентичности/сходства последовательности включают значение, полученное с использованием программного обеспечения GAP версии 10 с использованием следующих параметров: для нуклеотидной последовательности % идентичности и % сходства обнаруживают с применением штрафа за начало пропуска 50 и штрафа за удлинение пропуска 3 и матрицы замен nwsgapdna.cmp; для аминокислотной последовательности % идентичности или % сходства определяют с применением штрафа за начало пропуска 8 и штрафа за удлинение пропуска 2 и матрицы замен BLOSUM62; или любого эквивалентного программного обеспечения. «Эквивалентное программное обеспечение» включает в себя любое программное обеспечение для сравнения последовательностей, которое для любых двух рассматриваемых последовательностей создает выравнивание, обладающее идентичными соответствиями нуклеотидов или аминокислотных остатков и идентичным процентом идентичности последовательности при сравнении с соответствующим выравниванием, получаемым посредством программного обеспечения GAP версии 10.
Композиции или способы, «содержащие» или «включающие в себя» один или несколько упомянутых элементов, могут включать в себя другие элементы, специально не упомянутые. Например, композиция, которая «содержит» или «включает в себя» белок, может содержать белок отдельно или в комбинации с другими ингредиентами.
Обозначение диапазона значений включает в себя все целые числа, входящие в диапазон или образующие его, а также все поддиапазоны, образованные целыми числами, входящими в диапазон.
Если иное не очевидно из контекста, термин «около» охватывает значения в пределах стандартных границ погрешности измерения (например, стандартная погрешность среднего СПС) указанного значения.
Все элементы, указанные в единственном числе, также могут быть использованы и во множественном, если контекст явно не диктует иное. Например, термин «белок Cas» или «по меньшей мере один белок Cas» может включать множество белков Cas, включая их смеси.
II. Общая информация
В традиционных способах сборки нуклеиновых кислот используются требующие много времени этапы традиционного ферментативного расщепления рестрикционными ферментами, клонирования нуклеиновых кислот и лигирования нуклеиновых кислот друг с другом (см. ФИГ. 3 и ФИГ. 4, где иллюстрируются традиционные способы и временные параметры). Эти способы становятся более сложными при совместной сборке больших фрагментов или векторов. Способы, предлагаемые в настоящем документе, обладают преимуществом гибкой целевой специфичности нуклеаз (например, нуклеаз с гидовыми РНК и Cas9) при превращении нуклеиновых кислот в форму, подходящую для использования в реакциях быстрой сборки.
В настоящем документе предлагаются способы сборки по меньшей мере двух нуклеиновых кислот с использованием нуклеазных агентов, направленных на специфические целевые сайты, например, при помощи гидовой РНК (гРНК) (например, белок Cas, направленный на конкретный целевой сайт при помощи гидовой РНК (гРНК)). Направленные на сайт нуклеазные агенты, например направляемые гидовой РНК белки Cas, позволяют быстро и эффективно комбинировать нуклеиновые кислоты, производя отбор и манипуляции концевыми последовательностями, созданными в результате их эндонуклеазной активности. Способы, предложенные в настоящем документе, объединяют первый полинуклеотид с нуклеазным агентом (например, комплексом гРНК-Cas), специфичным к заданному целевому сайту, и экзонуклеазой. Целевой сайт может быть выбран таким образом, что, когда нуклеаза расщепляет нуклеиновую кислоту, полученные концы, созданные посредством расщепления, имеют области, комплементарные концам второй нуклеиновой кислоты (например, перекрывающиеся концы). Эти комплементарные концы впоследствии могут соединяться с образованием единой собранной нуклеиновой кислоты. Поскольку нуклеазный агент (например, комплекс гРНК-Cas) является специфичным для конкретного целевого сайта, настоящий способ позволяет модифицировать нуклеиновые кислоты точным сайт-направленным образом. В настоящем способе дополнительно используется преимущество специфичности нуклеазного агента, например комплекса гРНК-Cas, благодаря применению быстрых и эффективных способов сборки, специально разработанных для объединения перекрывающихся концов нуклеиновых кислот, созданных в результате нуклеазного расщепления или разработанных и синтезированных для реакции сборки. Например, при выборе нуклеазного агента (например, комплекса гРНК-Cas), специфичного для целевого сайта, так, чтобы при расщеплении образовывались концевые последовательности, комплементарные последовательностям второй нуклеиновой кислоты, для сборки полученной расщепленной аминокислоты можно использовать изотермическую сборку. Следовательно, при выборе нуклеиновых кислот и нуклеазных агентов (например, комплексов гРНК-Cas), дающих перекрывающиеся концевые последовательности, нуклеиновые кислоты можно собирать быстрыми комбинаторными способами и получать готовую собранную нуклеиновую кислоту быстро и эффективно. В альтернативном варианте осуществления нуклеиновые кислоты, не имеющие комплементарных концов, можно собрать с использованием соединительных олигонуклеотидов, сконструированных так, чтобы иметь концы, комплементарные каждой нуклеиновой кислоте. При помощи соединительных олигонуклеотидов две или более нуклеиновые кислоты можно собирать бесшовно, тем самым уменьшив количество необязательных последовательностей в полученной собранной нуклеиновой кислоте.
III. Нуклеазный агент
В настоящих способах для сайт-направленного расщепления полинуклеотидов используется нуклеазный агент. В частности, эндонуклеазное расщепление полинуклеотидов в идентифицированном целевом сайте позволяет получить расщепленный полинуклеотид с концами, которые можно присоединить ко второму полинуклеотиду и собрать два или более полинуклеотида сайт-специфическим образом.
«Нуклеазный агент» включает в себя молекулы, обладающие активностью по расщеплению ДНК. К конкретным примерам нуклеазных агентов для использования в способах, описанных в настоящем документе, относятся РНК-направляемая система CRISPR-Cas9, белки с «цинковыми пальцами», мегануклеазы, домены TAL, TALEN, дрожжевые сборки, рекомбиназы, лейциновые застежки, CRISPR/Cas, эндонуклеазы и другие нуклеазные агенты, известные специалистам в данной области. Нуклеазные агенты можно выбирать или разрабатывать для обеспечения специфичности расщепления в заданном целевом сайте. Например, нуклеазные агенты можно выбирать для расщепления в целевом сайте с созданием перекрывающихся концов между расщепленным полинуклеотидом и другим полинуклеотидом. Нуклеазные агенты, содержащие как белковый, так и РНК-элементы, например CRISPR-Cas9, могут поставляться в виде комплекса агентов, образующих нуклеазный агент, или могут поставляться в виде отдельных белкового и РНК-элементов, и в этом случае они образуют комплекс и формируют нуклеазный агент в описанных в настоящем документе реакционных смесях.
Термин «сайт распознавания для нуклеазного агента» включает последовательность ДНК, в которой нуклеазный агент индуцирует одно- или двухцепочечный разрыв. Сайт распознавания для нуклеазного агента может быть эндогенным (или нативным) по отношению к клетке, или сайт распознавания может быть экзогенным по отношению к клетке. В конкретных вариантах осуществления сайт распознавания является экзогенным по отношению к клетке и, таким образом, не встречается в геноме клетки в природе. В еще дополнительных вариантах осуществления сайт распознавания является эндогенным по отношению к клетке и интересующим полинуклеотидам, которые необходимо разместить в целевом локусе. В дополнительных вариантах осуществления экзогенный или эндогенный сайт распознавания присутствует в геноме клетки-хозяина только один раз. В конкретных вариантах осуществления идентифицирован эндогенный или нативный сайт, который присутствует в геноме только один раз. Впоследствии такой сайт может быть использован для конструирования нуклеазных агентов, которые будут создавать на эндогенном сайте распознавания одно- или двухцепочечный разрыв.
Длина сайта распознавания может изменяться и, например, включает сайты распознавания для пары нуклеаз с «цинковыми пальцами» (ZFN) длиной около 30-36 п.н. (т.е. около 15-18 п.н. для каждой ZFN), для эффекторной нуклеазы, подобной активатору транскрипции (TALEN), длиной около 36 п.н. или для гидовой РНК CRISPR/Cas9 длиной около 20 п.н.
Также представлены активные варианты и фрагменты сайтов распознавания, приведенных в качестве примера. Идентичность последовательности таких активных вариантов данному сайту распознавания составляет по меньшей мере 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более, причем активные варианты сохраняют биологическую активность и, следовательно, могут распознаваться и расщепляться нуклеазным агентом последовательность-специфическим образом. Анализы измерения двухцепочечного разрыва сайта распознавания посредством нуклеазного агента известны в данной области (например, из публикации TaqMan® qPCR assay, Frendewey D. et al., Methods in Enzymology, 2010, 476: 295-307, которая включена в настоящий документ путем ссылки в полном объеме).
В конкретных вариантах осуществления сайт распознавания расположен внутри полинуклеотида, кодирующего селективный маркер. Такое положение может находиться в пределах кодирующей области селективного маркера или в пределах регуляторных областей, оказывающих влияние на экспрессию селективного маркера. Таким образом, сайт распознавания нуклеазного агента может быть расположен в интроне селективного маркера, промоторе, энхансере, регуляторной области или любой не кодирующей белок области полинуклеотида, кодирующего селективный маркер. В конкретных вариантах осуществления одно- или двухцепочечный разрыв на сайте распознавания нарушает активность селективного маркера. Известны методы анализа на наличие или отсутствие функционального селективного маркера.
В описанных в настоящем документе способах и композициях может использоваться любой нуклеазный агент, который индуцирует одно- или двухцепочечный разрыв в заданном сайте распознавания. Встречающийся в природе или нативный нуклеазный агент может использоваться при условии, что нуклеазный агент индуцирует в заданном сайте распознавания одно- или двухцепочечный разрыв. В альтернативном варианте осуществления может использоваться модифицированный или сконструированный нуклеазный агент. «Сконструированный нуклеазный агент» содержит нуклеазу, сконструированную (модифицированную или производную) из ее нативной формы для того, чтобы она специфически распознавала и индуцировала одно- или двухцепочечный разрыв в заданном сайте распознавания. Таким образом, сконструированный нуклеазный агент может быть получен из нативного встречающегося в природе нуклеазного агента, или он может быть создан или синтезирован искусственно. Модификация нуклеазного агента может быть незначительной, например представлять собой модификацию одной аминокислоты в агенте расщепления белка или одного нуклеотида в агенте расщепления нуклеиновой кислоты. В некоторых вариантах осуществления сконструированная нуклеаза индуцирует одно- или двухцепочечный разрыв в сайте распознавания, причем сайт распознавания не является последовательностью, которая могла бы быть распознана нативным (не сконструированным или не модифицированным) нуклеазным агентом. Создание одно- или двухцепочечного разрыва в сайте распознавания или другой ДНК в настоящем документе может называться «разрезанием» или «расщеплением» сайта распознавания или другой ДНК.
Эти разрывы далее могут устраняться в клетке двумя способами: негомологичное соединение концов и восстановление, направляемое гомологией (гомологичная рекомбинация). При негомологичном соединении концов (NHEJ) двухцепочечные разрывы устраняются путем прямого лигирования концов разрыва друг с другом. По существу, новый нуклеотидный материал не вставляется в сайт, притом что какой-то нуклеотидный материал может теряться, приводя к делеции. При восстановлении, направляемом гомологией, донорный полинуклеотид, гомологичный последовательности расщепленной целевой ДНК, можно использовать в качестве шаблона для восстановления последовательности расщепленной целевой ДНК, результатом чего является перенос генетической информации от донорного полинуклеотида к целевой ДНК. Следовательно, в сайт может быть вставлен/скопирован новый нуклеотидный материал. Модификации целевой ДНК, обусловленные NHEJ и/или направляемым гомологией восстановлением, можно использовать для коррекции генов, замены генов, транспозонного мутагенеза, вставки трансгенов, делеции нуклеотидов, разрушения гена, мутации гена и т.п.
В одном варианте осуществления нуклеазный агент представляет собой эффекторную нуклеазу, подобную активатору транскрипции (TALEN). TAL-эффекторные нуклеазы представляют собой класс последовательность-специфических нуклеаз, которые можно использовать для выполнения двухцепочечных разрывов на специфических целевых последовательностях в геноме прокариотического или эукариотического организма. TAL-эффекторные нуклеазы создают путем слияния нативного или сконструированного эффектора, подобного активатору транскрипции (TAL), или его функциональной части с каталитическим доменом эндонуклеазы, например FokI. Уникальный модульный TAL-эффекторный ДНК-связывающий домен обеспечивает конструирование белков с потенциально любой заданной специфичностью распознавания ДНК. Таким образом, для распознавания целевых сайтов со специфической ДНК могут быть сконструированы ДНК-связывающие домены TAL-эффекторных нуклеаз, и, таким образом, их можно использовать для двухцепочечных разрывов на заданных целевых последовательностях. См., WO 2010/079430; Morbitzer et al. (2010) PNAS 10.1073/pnas. 1013133107; Scholze & Boch (2010) Virulence 1: 428-432; Christian et al. Genetics (2010) 186: 757-761; Li et al. (2010) Nuc. Acids Res. (2010) doi:10.1093/nar/gkq704; и Miller et al. (2011) Nature Biotechnology 29: 143-148; все из которых полностью включены в настоящий документ путем ссылки.
Примеры подходящих TAL-нуклеаз и способы получения подходящих TAL-нуклеаз описаны, например, в заявках на патент США №№2011/0239315 А1, 2011/0269234 А1, 2011/0145940 А1, 2003/0232410 А1, 2005/0208489 А1, 2005/0026157 А1, 2005/0064474 А1, 2006/0188987 А1 и 2006/0063231 А1 (каждая включена в настоящий документ путем ссылки). В различных вариантах осуществления сконструированы TAL-эффекторные нуклеазы, которые «выполняют разрез» в целевой нуклеотидной последовательности или возле нее, например в интересующем геномном локусе, в котором целевая нуклеотидная последовательность находится на последовательности, подлежащей модификации посредством нацеливающего вектора, или возле нее. TAL-нуклеазы, подходящие для использования с различными способами и композициями, предложенными в настоящем документе, включают те, которые специфически разработаны для связывания на целевой нуклеотидной последовательности, подлежащей модификации посредством нацеливающих векторов, или возле нее, как описано в настоящем документе.
В одном варианте осуществления каждый мономер TALEN содержит 33-35 повторов TAL, которые распознают одну пару нуклеотидов с помощью двух гипервариабельных остатков. В одном варианте осуществления нуклеазный агент представляет собой химерный белок, содержащий ДНК-связывающий домен на основе TAL-повторов, функционально связанный с независимой нуклеазой. В одном варианте осуществления независимая нуклеаза представляет собой эндонуклеазу FokI. В одном варианте осуществления нуклеазный агент содержит первый ДНК-связывающий домен на основе TAL-повторов и второй ДНК-связывающий домен на основе TAL-повторов, причем каждый из первого и второго ДНК-связывающего домена на основе TAL-повторов функционально связан с субъединицей нуклеазы FokI, при этом первый и второй ДНК-связывающие домены на основе TAL-повторов распознают две смежные целевые последовательности ДНК в каждой цепи целевой последовательности ДНК, разделенные последовательностью спейсера различной длины (12-20 п.н.), и при этом субъединицы нуклеазы FokI димеризуются для создания активной нуклеазы, которая выполняет двухцепочечный разрыв на целевой последовательности.
Нуклеазный агент, используемый в различных способах и композициях, описанных в настоящем документе, может дополнительно содержать нуклеазу с «цинковыми пальцами» (ZFN). В одном варианте осуществления каждый мономер ZFN содержит 3 или более ДНК-связывающих доменов на основе «цинковых пальцев», причем каждый ДНК-связывающий домен на основе «цинковых пальцев» связывается с субсайтом из 3 п.н. В других вариантах осуществления ZFN представляет собой химерный белок, содержащий ДНК-связывающий домен на основе «цинковых пальцев», функционально связанный с независимой нуклеазой. В одном варианте осуществления независимая эндонуклеаза представляет собой эндонуклеазу FokI. В одном варианте осуществления нуклеазный агент содержит первую ZFN и вторую ZFN, причем каждая из первой ZFN и второй ZFN функционально связана с субъединицей нуклеазы FokI, при этом первая и вторая ZFN распознают две смежные целевые последовательности ДНК в каждой цепи целевой последовательности ДНК, разделенные спейсером длиной около 5-7 п.н., и при этом субъединицы нуклеазы FokI димеризуются для создания активной нуклеазы, которая выполняет двухцепочечный разрыв. См., например, US 20060246567; US 20080182332; US 20020081614; US 20030021776; WO/2002/057308A2; US 20130123484; US 20100291048; WO/2011/017293А2; и Gaj et al. (2013) Trends in Biotechnology, 31 (7):397-405, каждая из которых полностью включена в настоящий документ путем ссылки.
В одном варианте осуществления способов, предложенных в настоящем документе, нуклеазный агент содержит (а) химерный белок, содержащий ДНК-связывающий домен на основе «цинковых пальцев», слитый с эндонуклеазой FokI; или (b) химерный белок, содержащий эффекторную нуклеазу, подобную активатору транскрипции (TALEN), слитую с эндонуклеазой FokI.
В еще одном варианте осуществления нуклеазный агент представляет собой мегануклеазу. Мегануклеазы были разделены на четыре семейства на основе консервативных мотивов последовательностей, семейства представляют собой LAGLIDADG (SEQ ID NO: 16), GIY-YIG, H-N-H и His-Cys box. Эти мотивы участвуют в координации ионов металлов и гидролизе фосфодиэфирных связей. НЕазы отличаются своими длинными сайтами распознавания и допуском некоторых видов полиморфизма последовательностей в их ДНК-субстратах. Известны домены, структура и функция мегануклеазы, см., например, Guhan and Muniyappa (2003) Crit Rev Biochem Mol Biol 38: 199-248; Lucas et al., (2001) Nucleic Acids Res 29: 960-9; Jurica and Stoddard, (1999) Cell Mol Life Sci 55: 1304-26; Stoddard, (2006) Q Rev Biophys 38: 49-95; и Moure et al., (2002) Nat Struct Biol 9: 764. В некоторых примерах используют встречающийся в природе вариант и/или сконструированное производное мегануклеазы. Известны способы модификации кинетики, взаимодействий кофакторов, экспрессии, оптимальных условий и/или специфичности сайта распознавания и скрининга в отношении активности, см., например, Epinat et al., (2003) Nucleic Acids Res 31: 2952-62; Chevalier et al., (2002) Mol Cell 10: 895-905; Gimble et al., (2003) Mol Biol 334: 993-1008; Seligman et al., (2002) Nucleic Acids Res 30: 3870-9; Sussman et al., (2004) J Mol Biol 342: 31-41; Rosen et al., (2006) Nucleic Acids Res 34: 4791-800; Chames et al., (2005) Nucleic Acids Res 33:e178; Smith et al., (2006) Nucleic Acids Res 34:e149; Gruen et al., (2002) Nucleic Acids Res 30:e29; Chen and Zhao, (2005) Nucleic Acids Res 33:e154; WO 2005105989; WO 2003078619; WO 2006097854; WO 2006097853; WO 2006097784 и WO 2004031346.
Здесь может использоваться любая мегануклеаза, включая, без ограничений, I-SceI, I-SceII, I-SceIII, I-SceIV, I-SceV, I-SceVI, I-SceVII, I-CeuI, I-CeuAIIP, I-CreI, I-CrepsbIP, I-CrepsbIIP, I-CrepsbIIIP, I-CrepsbIVP, I-TliI, I-PpoI, PI-PspI, F-SceI, F-SceII, F-SuvI, F-TevI, F-TevII, I-AmaI, I-AniI, I-ChuI, I-CmoeI, I-CpaI, I-CpaII, I-CsmI, I-CvuI, I-CvuAIP, I-DdiI, I-DdiII, I-DirI, I-DmoI, I-HmuI, I-HmuII, I-HsNIP, I-LlaI, I-MsoI, I-NaaI, I-NanI, I-NcIIP, I-NgrIP, I-NitI, I-Njal, I-Nsp236IP, I-PakI, I-PboIP, I-PcuIP, I-PcuAI, I-PcuVI, I-PgrIP, I-РоrIP, I-PorI, I-PorIIP, I-PbpIP, I-SpBetaIP, I-ScaI, I-SexIP, I-SneIP, I-SpomI, I-SpomCP, I-SpomIP, I-SpomIIP, I-SquIP, I-Ssp6803I, I-SthPhiJP, I-SthPhiST3P, I-SthPhiSTe3bP, I-TdeIP, I-TevI, I-TevII, I-TevIII, I-UarAP, I-UarHGPAIP, I-UarHGPA13P, I-VinIP, I-ZbiIP, PI-MtuI, PI-MtuHIP PI-MtuHIIP, PI-PfuI, PI-PMI, PI-PkoI, PI-PkoII, PI-Rma43812IP, PI-SpBetaIP, PI-SceI, PI-TM, PI-TfuII, PI-ThyI, PI-TliI, PI-TliII или их любые активные варианты или фрагменты.
В одном варианте осуществления мегануклеаза распознает двухцепочечные последовательности ДНК, состоящие из 12-40 пар нуклеотидов. В одном варианте Осуществления мегануклеаза распознает одну идеально соответствующую целевую последовательность в геноме. В одном варианте осуществления мегануклеаза представляет собой самонаводящуюся нуклеазу. В одном варианте осуществления самонаводящаяся нуклеаза принадлежит к семейству самонаводящихся нуклеаз LAGLIDADG (SEQ ID NO: 16). В одном варианте осуществления семейство самонаводящихся нуклеаз LAGLIDADG (SEQ ID NO: 16) выбирают из I-SceI, I-CreI и I-DmoI.
Нуклеазные агенты могут дополнительно содержать рестрикционные эндонуклеазы (рестрикционные ферменты), которые включают эндонуклеазы I типа, II типа, III типа и IV типа. Рестрикционные эндонуклеазы I типа и III типа распознают специфические сайты распознавания, но, как правило, осуществляют расщепление в разных положениях на расстоянии от сайта связывания нуклеазы, которые могут находиться на расстоянии сотен пар нуклеотидов от сайта расщепления (сайта распознавания). В системах II типа рестрикционная активность не зависит от метилазной активности, и расщепление обычно происходит в специфических сайтах в пределах сайта связывания или вблизи него. Большинство ферментов II типа разрезает палиндромные последовательности, однако ферменты IIа типа распознают непалиндромные сайты распознавания и осуществляют расщепление за пределами сайта распознавания, ферменты IIb типа разрезают последовательности дважды на двух сайтах за пределами сайта распознавания, а ферменты IIs типа распознают асимметричный сайт распознавания и осуществляют расщепление с одной стороны и на определенном расстоянии от сайта распознавания, составляющем около 1-20 нуклеотидов. Рестрикционные ферменты IV типа нацелены на метилированную ДНК. Рестрикционные ферменты дополнительно описаны и классифицированы, например, в базе данных REBASE (адрес веб-страницы: rebase.neb.com; Roberts et al., (2003) Nucleic Acids Res 31: 418-20), Roberts et al., (2003) Nucleic Acids Res 31: 1805-12 и Belfort et al., (2002) в Mobile DNA II, pp. 761-783, Eds. Craigie et al. (ASM Press, Washington, DC). В конкретных вариантах осуществления по меньшей мере два эндонуклеазных фермента могут быть выбраны в качестве нуклеазных агентов, причем эти ферменты создают совместимые или комплементарные «липкие» концы.
Нуклеазный агент, используемый в различных способах и композициях, может также содержать систему CRISPR/Cas. В таких системах может использоваться нуклеаза Cas9, которая в некоторых случаях является кодон-оптимизированной для требуемого типа клеток, в которых она должна экспрессироваться. В системе дополнительно используется слитая конструкция крРНК-тракрРНК, функционирующая с кодон-оптимизированной Cas9. Эту одинарную РНК часто называют гидовой РНК или гРНК. В пределах гРНК участок крРНК определяют как «целевую последовательность» для данного сайта распознавания, а тракрРНК часто называют «каркасом». Было продемонстрировано, что эта система функционирует в различных эукариотических и прокариотических клетках. Коротко говоря, выполняется вставка короткого фрагмента ДНК, содержащего целевую последовательность, в плазмиду экспрессии гидовой РНК. Плазмида экспрессии гРНК содержит целевую последовательность (в некоторых вариантах осуществления длиной приблизительно 20 нуклеотидов), вид последовательности тракрРНК (каркас), а также подходящий промотор, который является активным в клетке, и элементы, необходимые для соответствующего процессинга в эукариотических клетках. Многие из систем основаны на обычных, комплементарных олигонуклеотидах, которые ренатурируют с образованием двухцепочечной ДНК, после чего клонируют в плазмиду экспрессии гРНК. Затем экспрессионную кассету гРНК и экспрессионную кассету Cas9 вводят в клетку. См., например, публикации Mali Р et al. (2013) Science 2013 Feb 15; 339 (6121):823-6; Jinek M et al. Science 2012 Aug 17; 337 (6096): 816-21; Hwang WY et al. Nat Biotechnol 2013 Mar; 31 (3): 227-9; Jiang W et al. Nat Biotechnol 2013 Mar; 31 (3): 233-9; и Cong L et al. Science 2013 Feb 15; 339 (6121): 819-23, каждая из которых полностью включена в настоящий документ путем ссылки.
В способах и композициях, описанных в настоящем документе, для модификации генома внутри клетки могут использоваться системы коротких палиндромных повторов, регулярно расположенных группами (CRISPR)/системы, ассоциированные с CRISPR (Cas), или компоненты таких систем. Системы CRISPR/Cas включают в себя транскрипты и другие элементы, участвующие в экспрессии или управлении активностью генов Cas. Система CRISPR/Cas может представлять собой систему I типа, II типа или III типа. В способах и композициях, описанных в настоящем документе, для сайт-направленного расщепления нуклеиновых кислот используются системы CRISPR/Cas посредством использования комплексов CRISPR (содержащих гидовую РНК (гРНК), образующую комплекс с белком Cas).
Некоторые системы CRISPR/Cas, используемые в способах, описанных в настоящем документе, не встречаются в природе. «Не встречающаяся в природе» система имеет признаки вмешательства человека, например, один или более компонентов системы изменены или мутированы по сравнению с их природным состоянием, при этом они по меньшей мере по существу не содержат по меньшей мере один другой компонент, с которым они естественно ассоциированы в природе, или они ассоциированы с по меньшей мере другим компонентом, с которым они не ассоциированы в природе. Например, в некоторых системах CRISPR/Cas используются не встречающиеся в природе комплексы CRISPR, которые содержат гРНК и белок Cas, не встречающиеся в природе вместе.
Также предложены активные варианты и фрагменты нуклеазных агентов (т.е. сконструированный нуклеазный агент). Идентичность последовательности таких активных вариантов нативному нуклеазному агенту может составлять по меньшей мере 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более, причем активные варианты сохраняют способность разрезать заданный сайт распознавания и, таким образом, сохраняют активность индуцирования одно- или двухцепочечного разрыва. Например, любой из нуклеазных агентов, описанных в настоящем документе, может быть модифицирован из нативной последовательности эндонуклеазы и сконструирован для распознавания и индуцирования одно- или двухцепочечного разрыва на сайте распознавания, который не распознавался нативным нуклеазным агентом. Таким образом, в некоторых вариантах осуществления сконструированная нуклеаза обладает специфической способностью индуцировать одно- или двухцепочечный разрыв на сайте распознавания, который отличается от сайта распознавания соответствующего нативного нуклеазного агента. Анализы определения активности, индуцирующей одно- или двухцепочечный разрыв, известны и, как правило, измеряют общую активность и специфичность эндонуклеазы на субстратах ДНК, содержащих сайт распознавания.
IV. Системы CRISPR/Cas (комплекс гРНК-Cas)
В настоящих способах может использоваться система CRISPR/Cas (например, комплекс гРНК-Cas) для сайт-направленного расщепления нуклеиновых кислот. В частности, направляемое гРНК расщепление белком Cas нуклеиновых кислот в идентифицированном целевом сайте позволяет получить расщепленную нуклеиновую кислоту с концами, которые можно присоединить ко второй нуклеиновой кислоте и собрать две или более нуклеиновых кислоты сайт-специфическим образом.
«Комплекс гРНК-Cas» представляет собой комплекс белка Cas с гРНК. гРНК можно разработать или выбрать так, чтобы направить Cas-расщепление на целевой сайт с созданием перекрывающихся концов между расщепленной нуклеиновой кислотой и другой нуклеиновой кислотой. Комплекс гРНК-Cas может поставляться уже в виде комплекса или может поставляться в виде отдельных белкового и РНК-элементов, и в этом случае они образуют комплекс гРНК-Cas в описанных в настоящем документе способах и реакционных смесях.
А. Эндонуклеазы, направляемые РНК Cas
Белки Cas обычно содержат по меньшей мере один домен распознавания или связывания РНК. Такие домены могут взаимодействовать с гидовыми РНК (гРНК, описаны более подробно ниже). Белки Cas могут также содержать нуклеазные домены (например, домены ДНКазы или РНКазы), ДНК-связывающие домены, геликазные домены, домены межбелковых взаимодействий, домены димеризации и другие домены. Нуклеазный домен обладает каталитической активностью в отношении расщепления нуклеиновых кислот. Расщепление включает разрушение ковалентных связей молекулы нуклеиновой кислоты. В результате расщепления могут образовываться тупые концы или ступенчатые концы, и расщепление может быть одно цепочечным или двухцепочечным.
Примеры белков Cas включают Cas1, Cas1B, Cas2, Cas3, Cas4, Cas5, Cas5e (CasD), Cas6, Cas6e, Cas6f, Cas7, Cas8a1, Cas8a2, Cas8b, Cas8c, Cas9 (Csn1 или Csx12), Cas10, Cas10d, CasF, CasG, CasH, Csy1, Csy2, Csy3, Cse1 (CasA), Cse2 (CasB), Cse3 (CasE), Cse4 (CasC), Csc1, Csc2, Csa5, Csn2, Csm2, Csm3, Csm4, Csm5, Csm6, Cmr1, Cmr3, Cmr4, Cmr5, Cmr6, Csb1, Csb2, Csb3, Csx17, Csx14, Csx10, Csx16, CsaX, Csx3, Csx1, Csx15, Csf1, Csf2, Csf3, Csf4 и Cu1966 и их гомологи или модифицированные варианты.
В описанных в настоящем документе способах и композициях может использоваться любой белок Cas, который индуцирует одно- или двухцепочечный разрыв в заданном сайте распознавания. Встречающийся в природе или нативный белок Cas может использоваться при условии, что белок Cas индуцирует в заданном сайте распознавания двухцепочечный разрыв. В альтернативном варианте осуществления может использоваться модифицированный или сконструированный белок Cas. «Сконструированный белок Cas» представляет собой белок Cas, сконструированный (модифицированный или производный) из его нативной формы для того, чтобы он специфически распознавал и индуцировал одно- или двухцепочечный разрыв в заданном сайте распознавания. Таким образом, сконструированный белок Cas может быть получен из нативного встречающегося в природе белка Cas, или он может быть создан или синтезирован искусственно.
В конкретных вариантах осуществления белок Cas представляет собой Cas9. Белки Cas9 обычно имеют четыре общих основных мотива с консервативной архитектурой. Мотивы 1, 2 и 4 представляют собой RuvC-подобные мотивы, а мотив 3 представляет собой мотив HNH. Нуклеазная активность Cas9 расщепляет целевую ДНК с образованием двухцепочечных разрывов. Эти разрывы далее могут устраняться в клетке двумя способами: негомологичное соединение концов и восстановление, направляемое гомологией (гомологичная рекомбинация). При негомологичном соединении концов (NHEJ) двухцепочечные разрывы устраняются путем прямого лигирования концов разрыва друг с другом. По существу, новый нуклеотидный материал не вставляется в сайт, притом что какой-то нуклеотидный материал может теряться, приводя к делеции. При восстановлении, направляемом гомологией, донорный полинуклеотид, гомологичный последовательности расщепленной целевой ДНК, можно использовать в качестве шаблона для восстановления последовательности расщепленной целевой ДНК, результатом чего является перенос генетической информации от донорного полинуклеотида к целевой ДНК. Следовательно, в сайт может быть вставлен/скопирован новый нуклеотидный материал. Модификации целевой ДНК, обусловленные NHEJ и/или направляемым гомологией восстановлением, можно использовать для коррекции генов, замены генов, транспозонного мутагенеза, вставки трансгенов, делеции нуклеотидов, разрушения гена, мутации гена и т.п.
Белки Cas могут быть из системы CRISPR/Cas II типа. Например, белок Cas может быть белком Cas9 или может быть получен из белка Cas9. Белки Cas9 обычно имеют четыре общих основных мотива с консервативной архитектурой. Мотивы 1, 2 и 4 представляют собой RuvC-подобные мотивы, а мотив 3 представляет собой мотив HNH. Белок Cas9 может быть, например, получен из Streptococcus pyogenes, Streptococcus thermophilus, Streptococcus sp., Staphylococcus aureus, Nocardiopsis dassonvillei, Streptomyces pristinaespiralis, Streptomyces viridochromogenes, Streptomyces viridochromogenes, Streptosporangium roseum, Streptosporangium roseum, AlicyclobacHlus acidocaldarius, Bacillus pseudomycoides, Bacillus selenitireducens, Exiguobacterium sibiricum, Lactobacillus delbrueckii, Lactobacillus salivarius, Microscilla marina, Burkholderiales bacterium, Polaromonas naphthalenivorans, Polaromonas sp., Crocosphaera watsonii, Cyanothece sp., Microcystis aeruginosa, Synechococcus sp., Acetohalobium arabaticum, Ammonifex degensii, Caldicelulosiruptor becscii, Candidatus Desulforudis, Clostridium botulinum, Clostridium difficile, Finegoldia magna, Natranaerobius thermophilus, Pelotomaculum thermopropionicum, Acidithiobacillus caldus, Acidithiobacillus ferrooxidans, Allochromatium vinosum, Marinobacter sp., Nitrosococcus halophilus, Nitrosococcus watsoni, Pseudoalteromonas haloplanktis, Ktedonobacter racemifer, Methanohalobium evestigatum, Anabaena variabilis, Nodularia spumigena, Nostoc sp., Arthrospira maxima, Arthrospiraplatensis, Arthrospira sp., Lyngbya sp., Microcoleus chthonoplastes, Oscillatoria sp., Petrotoga mobilis, Thermosipho africanus или Acaryochloris marina. Дополнительные примеры представителей семейства Cas9 описаны в публикации WO 2014/131833, которая полностью включена в настоящий документ путем ссылки. Предпочтительным ферментом является белок Cas9 из S. pyogenes или его производное. Белку Cas9 из S. pyogenes в базе данных SwissProt присвоен номер доступа Q99ZW2.
Белки Cas могут являться белками дикого типа (т.е. встречающимися в природе), модифицированными белками Cas (т.е. вариантами белков Cas) или фрагментами белков Cas дикого типа или модифицированных белков Cas. Белки Cas могут также являться активными вариантами или фрагментами белков Cas дикого типа или модифицированных белков Cas. Идентичность последовательности таких активных вариантов или фрагментов белку Cas дикого типа или модифицированному белку Cas или его части может составлять по меньшей мере 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более, причем активные варианты сохраняют способность разрезания на требуемом сайте расщепления и, следовательно, сохраняют активность, индуцирующую одно- или двухцепочечный разрыв. Анализы определения активности, индуцирующей одно- или двухцепочечный разрыв, известны и, как правило, измеряют общую активность и специфичность белка Cas на субстратах ДНК, содержащих сайт расщепления.
Белки Cas могут быть модифицированы для увеличения или уменьшения аффинности связывания нуклеиновых кислот, специфичности связывания нуклеиновых кислот и/или ферментативной активности. Белки Cas могут также быть модифицированы для изменения любого другого вида активности или свойства белка, например стабильности. Например, один или более нуклеазных доменов белка Cas могут быть изменены, удалены или инактивированы, или белок Cas может быть усечен, чтобы удалить домены, которые не являются необходимыми для функционирования белка, или для оптимизации (например, увеличения или уменьшения) активности белка Cas.
Некоторые белки Cas содержат по меньшей мере два нуклеазных домена, таких как домены ДНКазы. Например, белок Cas9 может содержать RuvC-подобный нуклеазный домен и HNH-подобный нуклеазный домен. Каждый из доменов RuvC и HNH может разрезать отличную цепь двухцепочечной ДНК, чтобы выполнить двухцепочечный разрыв в ДНК. См., например, публикацию Jinek et al. (2012) Science 337: 816-821, которая полностью включена в настоящий документ путем ссылки.
Один или оба нуклеазных домена могут быть удалены или подвергнуты мутации таким образом, чтобы прекратить их функционирование или уменьшить нуклеазную активность. При удалении или мутации одного из нуклеазных доменов полученный белок Cas (например, Cas9) может называться «никаза» и может производить одноцепочечный разрыв на последовательности распознавания РНК CRISPR в пределах двухцепочечной ДНК, но не двухцепочечный разрыв (т.е. он может расщеплять комплементарную цепь или некомплементарную цепь, но не обе). При удалении или мутации обоих нуклеазных доменов полученный белок Cas (например, Cas9) будет иметь пониженную способность расщеплять обе цепи двухцепочечной ДНК. Примером мутации, превращающей Cas9 в никазу, является мутация D10A (замена аспартата на аланин в положении 10 в Cas9) в домене RuvC белка Cas9 из S. pyogenes. Подобным образом мутация Н939А (замена гистидина на аланин в положении аминокислоты 839) или Н840А (замена гистидина на аланин в положении аминокислоты 840) в домене HNH Cas9 из S. pyogenes может превращать Cas9 в никазу. Другие примеры мутаций, превращающих Cas9 в никазу, включают соответствующие мутации Cas9 из S. thermophilus. См., например, публикации Sapranauskas et al. (2011) Nucleic Acids Research 39: 9275-9282 и WO 2013/141680, каждая из которых полностью включена в настоящий документ путем ссылки. Такие мутации могут быть созданы с использованием таких методов, как сайт-направленный мутагенез, ПЦР-опосредованный мутагенез или полный синтез гена. Примеры других мутаций, создающих никазы, можно найти, например, в публикациях WO/2013/176772 A1 и WO/2013/142578 A1, каждая из которых включена в настоящий документ путем ссылки.
Белки Cas также могут представлять собой слитные белки. Например, белок Cas может быть слит с доменом расщепления, доменом эпигенетических модификаций, доменом транскрипционной активации или доменом транскрипционного репрессора. См. публикацию №WO 2014/089290, которая полностью включена в настоящий документ путем ссылки. Белки Cas также могут быть слиты с гетерологичным полипептидом, обеспечивающим повышенную или сниженную стабильность. Слитый домен или гетерологичный полипептид может быть расположен на N-конце, на С-конце или находиться внутри белка Cas.
Белок Cas может быть слит с гетерологичным полипептидом, который обеспечивает субклеточную локализацию. Такие гетерологичные пептиды включают в себя, например, сигнал ядерной локализации (NLS), например NLS SV40, для нацеливания на ядро, сигнал митохондриальной локализации для нацеливания на митохондрии, сигнал удержания ER и т.п. См., например, Lange et al. (2007) J. Biol. Chem. 282:5101-5105. Такие сигналы субклеточной локализации могут быть расположены на N-конце, на С-конце или в других местах внутри белка Cas. NLS может содержать участок из основных аминокислот и может быть представлен одинарной последовательностью или двойной последовательностью.
Белки Cas также могут быть связаны с проникающим в клетку доменом. Например, проникающий в клетку домен может быть получен из белка TAT HIV-1, проникающего в клетку мотива TLM из вируса гепатита В человека, MPG, Рер-1, VP22, проникающего в клетку пептида из вируса простого герпеса или из последовательности пептида полиаргинина. См., например, публикацию WO 2014/089290, которая полностью включена в настоящий документ путем ссылки. Проникающий в клетку домен может быть расположен на N-конце, на С-конце или в других местах внутри белка Cas.
Белки Cas также могут содержать гетерологичный полипептид для простоты отслеживания или очистки, например флуоресцентный белок, метку очистки или эпитопную метку. Примеры флуоресцентных белков включают зеленые флуоресцентные белки (например, GFP, GFP-2, tagGFP, turboGFP, eGFP, Emerald, Azami Green, мономерный Azami Green, CopGFP, AceGFP, ZsGreenl), желтые флуоресцентные белки (например, YFP, eYFP, Citrine, Venus, YPet, PhiYFP, ZsYellowl), синие флуоресцентные белки (например, eBFP, eBFP2, Azurite, mKalamal, GFPuv, Sapphire, T-sapphire), голубые флуоресцентные белки (например, eCFP, Cerulean, CyPet, AmCyanl, Midoriishi-Cyan), красные флуоресцентные белки (mKate, mKate2, mPlum, мономерный DsRed, mCherry, mRFP1, DsRed-Express, DsRed2, DsRed-monomer, HcRed-Tandem, HcRedl, AsRed2, eqFP611, mRaspberry, mStrawberry, Jred), оранжевые флуоресцентные белки (mOrange, mKO, Kusabira-Orange, мономерный Kusabira-Orange, mTangerine, tdTomato) и любые другие подходящие флуоресцентные белки. Примеры меток включают глутатион-8-трансферазу (GST), хитин-связывающий белок (СВР), мальтоза-связывающий белок, тиоредоксин (TRX), поли(NANP), метку тандемной аффинной очистки (ТАР), myc, AcV5, AU1, AU5, Е, ECS, Е2, FLAG, гемагглютинин (НА), nus, Softag 1, Softag 3, Strep, SBP, Glu-Glu, HSV, KT3, S, S1, T7, V5, VSV-G, гистидин (His), биотин-карбоксил-переносящий белок (BCCP) и калмодулин.
В некоторых вариантах осуществления белок Cas можно модифицировать таким образом, чтобы изменить итоговую нуклеазную активность. Некоторые мутации в Cas могут снижать способность нуклеазы расщеплять как комплементарные, так и некомплементарные цепи целевой ДНК. Например, в белки Cas можно ввести мутации в известных положениях, так чтобы нуклеазная активность ограничивалась расщеплением либо комплементарной цепи, либо некомплементарной цепи. В частности, Cas9, имеющий мутацию D10A (замену аспартата на аланин в положении 10 белка Cas9), может расщеплять комплементарную цепь целевой ДНК, но имеет сниженную способность к расщеплению некомплементарной цепи целевой ДНК. В некоторых вариантах осуществления Cas9, имеющий мутацию Н840А (замену гистидина на аланин в положении 840 белка), может расщеплять некомплементарную цепь целевой ДНК, но имеет сниженную способность к расщеплению комплементарной цепи целевой ДНК. Нуклеазная активность белка Cas9, имеющего мутацию D10A или Н840А, приведет к одноцепочечному разрыву (SSB), а не двухцепочечному разрыву (DSB). Возможны мутации в других остатках для достижения того же эффекта (т.е. инактивации той или иной части нуклеазы). Не имеющими ограничительного характера примерами являются остатки D10, G12, G17, Е762, Н840, N854, N863, Н982, Н983, А984, D986 и/или А987 (т.е. замены). Кроме того, приемлемыми в качестве заместителей могут быть другие аминокислоты, отличные от аланина. В некоторых вариантах осуществления, когда нуклеаза имеет сниженную активность (например, когда белок Cas9 имеет мутацию D10, G12, G17, Е762, Н840, N854, N863, Н982, Н983, А984, D986 и/или А987, например, D10A, G12A, G17A, Е762А, Н840А, N854А, N863А, Н982А, Н983А, А984А, и/или D986A), нуклеаза по-прежнему может связываться с целевой ДНК сайт-специфическим образом, поскольку она по-прежнему направляется к целевой последовательности ДНК посредством гРНК, если она сохраняет способность взаимодействовать с гРНК.
В некоторых вариантах осуществления Cas изменяют таким образом, чтобы нуклеаза не расщепляла либо комплементарную, либо некомплементарную цепь целевой ДНК. Например, Cas9 с обеими мутациями D10A и Н840А имеет сниженную способность расщеплять как комплементарную, так и некомплементарную цепи целевой ДНК. Возможны мутации в других остатках для достижения того же эффекта (т.е. инактивации той или иной части нуклеазы). Не имеющими ограничительного характера примерами являются остатки D10, G12, G17, Е762, Н840, N854, N863, Н982, Н983, А984, D986, которые можно заменить, чтобы по существу устранить нуклеазную активность. Кроме того, приемлемыми могут быть заместители, отличные от аланина.
Термины «целевой сайт» или «целевая последовательность» можно использовать взаимозаменяемо, и они включают нуклеотидные последовательности, присутствующие в целевой ДНК, с которой будет связываться сегмент ДНК-нацеливания гРНК, при условии существования достаточных условий для связывания. Например, на целевой сайт (или целевую последовательность) в целевой ДНК оказывает целевое воздействие (или связывается, или гибридизуется, или является комплементарным) белок Cas или гРНК. Подходящие условия связывания ДНК/РНК включают физиологические условия, обычно существующие в клетке. Другие подходящие условия связывания ДНК/РНК (например, условия в бесклеточной системе) известны в данной области (см., например, Molecular Cloning: A Laboratory Manual, 3rd Ed. (Sambrook et al, Harbor Laboratory Press 2001)). Цепь целевой ДНК, которая является комплементарной или гибридизуется с белком Cas или гРНК, называется «комплементарной цепью», а цепь целевой ДНК, которая является комплементарной «комплементарной цепи» (и, таким образом, не комплементарной белку Cas или гРНК), называется «некомплементарной цепью» или «матричной цепью».
Белок Cas может расщеплять нуклеиновую кислоту на сайте в пределах целевой последовательности или за пределами целевой последовательности. «Сайт расщепления» включает положение нуклеиновой кислоты, в котором белок Cas производит одноцепочечный разрыв или двухцепочечный разрыв. Если белок Cas создает двухцепочечный разрыв, сайт расщепления может находиться в одном и том же положении на обеих цепях нуклеиновой кислоты (образуя тупые концы) или может находиться на разных сайтах на каждой цепи (образуя «липкие» или способные к связыванию концы). «Липкие» концы также могут быть получены с использованием двух белков Cas, которые производят одноцепочечный разрыв на сайте расщепления в каждой цепи. Сайт-специфическое расщепление целевой ДНК белком Cas9 может происходить в местах, определенных как (i) комплементарностью спариваемых нуклеотидов между гидовой РНК и целевой ДНК, так и (ii) наличием в целевой ДНК короткого мотива, который называется мотивом, прилежащим к протоспейсеру (РАМ). Например, сайт расщепления Cas9 может находиться на расстоянии от около 1 до около 10 или от около 2 до около 5 пар нуклеотидов (например, 3 пар нуклеотидов) выше последовательности РАМ. В некоторых вариантах осуществления (например, когда используется Cas9 из S. pyogenes или близкородственный Cas9) последовательность РАМ некомплементарной цепи может представлять собой 5'-XGG-3', где X является любым нуклеотидом ДНК, и при этом X находится непосредственно на 3'-конце целевой последовательности некомплементарной цепи целевой ДНК. По существу, последовательность РАМ комплементарной цепи будет представлять собой 5'-CCY-3', где Y является любым нуклеотидом ДНК, и при этом Y находится непосредственно на 5'-конце целевой последовательности комплементарной цепи целевой ДНК. В некоторых таких вариантах осуществления X и Y могут быть комплементарными, и пара нуклеотидов X-Y может являться любой парой нуклеотидов (например, X=C и Y=G; X=G и Y=C; X=A и Y=T, X=T и Y=A).
Белки Cas могут быть обеспечены в любом виде. Например, белок Cas может быть обеспечен в виде белка, такого как белок Cas, образующий комплекс с гРНК. В альтернативном варианте осуществления белок Cas может быть обеспечен в виде нуклеиновой кислоты, кодирующей белок Cas, например РНК (например, матричной РНК (мРНК)) или ДНК. Необязательно, нуклеиновая кислота, кодирующая белок Cas, может быть кодон-оптимизированной для эффективной трансляции в белок в конкретной клетке или организме. Например, нуклеиновая кислота, кодирующая белок Cas, может быть модифицирована, чтобы заменить кодоны, имеющие более высокую частоту использования в бактериальной клетке, дрожжевой клетке, человеческой клетке, не относящейся к человеку клетке, клетке млекопитающего, клетке грызуна, клетке мыши, клетке крысы или любой другой интересующей клетке-хозяине по сравнению со встречающейся в природе полинуклеотидной последовательностью. Если в клетку вводят нуклеиновую кислоту, кодирующую белок Cas, то белок Cas может экспрессироваться внутри клетки временно, условно или конститутивно.
Нуклеиновые кислоты, кодирующие белки Cas, могут быть стабильно интегрированы в геном клетки и функционально связаны с активным в клетке промотором. В альтернативном варианте осуществления нуклеиновые кислоты, кодирующие белки Cas, могут быть функционально связаны с промотором в экспрессионной конструкции. Экспрессионные конструкции включают в себя любые нуклеотидные конструкции, которые способны управлять экспрессией гена или другой интересующей нуклеотидной последовательности (например, гена Cas) и которые могут переносить такую интересующую нуклеотидную последовательность в клетку-мишень. Например, нуклеиновая кислота, кодирующая белок Cas, может находиться в нацеливающем векторе, содержащем нуклеотидную вставку и/или вектор, содержащий ДНК, кодирующую гРНК, или она может находиться в векторе или плазмиде, отдельной от нацеливающего вектора, содержащего нуклеотидную вставку, и/или отдельной от вектора, содержащего ДНК, кодирующую гРНК. Промоторы, которые могут использоваться в экспрессионной конструкции, включают, например, промоторы, активные в плюрипотентной клетке крысы, эукариота, млекопитающего, не относящегося к человеку млекопитающего, человека, грызуна, мыши или хомяка. Такие промоторы могут представлять собой, например, условные промоторы, индуцируемые промоторы, конститутивные промоторы или тканеспецифические промоторы. В других разделах настоящего документа описаны примеры других промоторов.
В. Гидовые РНК (гРНК)
«Гидовая РНК» или «гРНК» включает в себя молекулу РНК, которая связывается с белком Cas и нацеливает белок Cas в специфическое местоположение внутри целевой ДНК. Гидовые РНК (гРНК) могут содержать два сегмента: «сегмент ДНК-нацеливания» и «белок-связывающий сегмент». «Сегмент» представляет собой сегмент, часть или область молекулы, например непрерывный участок из нуклеотидов в РНК. Некоторые гРНК содержат две отдельные молекулы РНК: «РНК-активатор» и «РНК-нацеливатель». Другие гРНК представляют собой одинарные молекулы РНК (одинарный полинуклеотид РНК), которые также можно назвать «одномолекулярной гРНК», «одногидовой РНК» или «огРНК». См., например, публикации WO/2013/176772 A1, WO/2014/065596 A1, WO/2014/089290 A1, WO/2014/093622 A2, WO/2014/099750 А2, WO/2013142578 A1 и WO 2014/131833 А1, каждая из которых включена в настоящий документ путем ссылки. Термины «гидовая РНК» и «гРНК» включают как двухмолекулярные гРНК, так и одномолекулярные гРНК.
Примеры двухмолекулярных гРНК включают крРНК-подобную молекулу («РНК CRISPR», или «РНК-нацеливатель», или «крРНК», или «повтор крРНК») и соответствующую тракрРНК-подобную молекулу («транс-действующую РНК CRISPR», или «РНК-активатор», или «тракрРНК», или «каркас»). крРНК содержит оба сегмента (одноцепочечные) ДНК-нацеливания гРНК и участок нуклеотидов, который образует одну половину дуплекса дцРНК белок-связывающего сегмента гРНК. Соответствующая тракрРНК (РНК-активатор) содержит участок нуклеотидов, который образует другую половину дуплекса дцРНК белок-связывающего сегмента гРНК. Участок нуклеотидов крРНК комплементарен участку нуклеотидов тракрРНК и гибридизуется с ним с образованием дуплекса дцРНК белок-связывающего домена гРНК. Таким образом, можно сказать, что каждая крРНК имеет соответствующую тракрРНК. крРНК дополнительно обеспечивает одноцепочечный сегмент ДНК-нацеливания. Соответственно, гРНК содержит последовательность, которая гибридизуется с целевой последовательностью и тракрРНК.
крРНК и соответствующая тракрРНК (как соответствующая пара) гибридизуются с образованием гРНК. крРНК дополнительно обеспечивает одноцепочечный сегмент ДНК-нацеливания, который гибридизуется с последовательностью распознавания РНК CRISPR. Для использования для модификации внутри клетки может быть разработана точная последовательность данной молекулы крРНК или тракрРНК, которая будет специфичной для тех видов, в которых будут использоваться молекулы РНК. См., например, публикации Mali Р et al. (2013) Science 2013 Feb 15; 339 (6121): 823-6; Jinek M et al. Science 2012 Aug 17; 337 (6096): 816-21; Hwang WY et al. Nat Biotechnol 2013 Mar; 31 (3): 227-9; Jiang W et al. Nat Biotechnol 2013 Mar; 31 (3): 233-9; и Cong L et al. Science 2013 Feb 15; 339 (6121): 819-23, каждая из которых полностью включена в настоящий документ путем ссылки.
Сегмент ДНК-нацеливания (крРНК) данной гРНК содержит нуклеотидную последовательность, которая является комплементарной последовательности в целевой ДНК. Сегмент ДНК-нацеливания гРНК взаимодействует с целевой ДНК последовательность-специфическим образом посредством гибридизации (т.е. спаривания нуклеотидов). Таким образом, нуклеотидная последовательность сегмента ДНК-нацеливания может изменяться и определяет место внутри целевой ДНК, с которым будет взаимодействовать гРНК и целевая ДНК. Сегмент ДНК-нацеливания обсуждаемой гРНК может быть модифицирован для гибридизации с любой заданной последовательностью внутри целевой ДНК. Встречающиеся в природе крРНК различаются в зависимости от системы Cas9 и организма, но часто содержат нацеливающий сегмент длиной от 21 до 72 нуклеотидов, фланкированный двумя прямыми повторами (DR) длиной от 21 до 46 нуклеотидов (см., например, WO 2014/131833). В случае S. pyogenes длина DR составляет 36 нуклеотидов, а длина нацеливающего сегмента составляет 30 нуклеотидов. DR, расположенный на 3'-конце, является комплементарным и гибридизуется с соответствующей тракрРНК, которая в свою очередь связывается с белком Cas9.
Сегмент ДНК-нацеливания может иметь длину от около 12 нуклеотидов до около 100 нуклеотидов. Например, сегмент ДНК-нацеливания может иметь длину от около 12 нуклеотидов (нт) до около 80 нт, от около 12 нт до около 50 нт, от около 12 нт до около 40 нт, от около 12 нт до около 30 нт, от около 12 нт до около 25 нт, от около 12 нт до около 20 нт или от около 12 нт до около 19 нт.В альтернативном варианте осуществления сегмент ДНК-нацеливания может иметь длину от около 19 нт до около 20 нт, от около 19 нт до около 25 нт, от около 19 нт до около 30 нт, от около 19 нт до около 35 нт, от около 19 нт до около 40 нт, от около 19 нт до около 45 нт, от около 19 нт до около 50 нт, от около 19 нт до около 60 нт, от около 19 нт до около 70 нт, от около 19 нт до около 80 нт, от около 19 нт до около 90 нт, от около 19 нт до около 100 нт, от около 20 нт до около 25 нт, от около 20 нт до около 30 нт, от около 20 нт до около 35 нт, от около 20 нт до около 40 нт, от около 20 нт до около 45 нт, от около 20 нт до около 50 нт, от около 20 нт до около 60 нт, от около 20 нт до около 70 нт, от около 20 нт до около 80 нт, от около 20 нт до около 90 нт или от около 20 нт до около 100 нт.
Нуклеотидная последовательность сегмента ДНК-нацеливания, являющаяся комплементарной нуклеотидной последовательности (последовательности распознавания РНК CRISPR) целевой ДНК, может иметь длину по меньшей мере около 12 нт. Например, последовательность ДНК-нацеливания (например, последовательность в пределах сегмента ДНК-нацеливания, являющаяся комплементарной последовательности распознавания РНК CRISPR в пределах целевой ДНК) может иметь длину по меньшей мере около 12 нт, по меньшей мере около 15 нт, по меньшей мере около 18 нт, по меньшей мере около 19 нт, по меньшей мере около 20 нт, по меньшей мере около 25 нт, по меньшей мере около 30 нт, по меньшей мере около 35 нт или по меньшей мере около 40 нт. В альтернативном варианте осуществления последовательность ДНК-нацеливания сегмента ДНК-нацеливания, комплементарная целевой последовательности целевой ДНК может иметь длину от около 12 нуклеотидов (нт) до около 80 нт, от около 12 нт до около 50 нт, от около 12 нт до около 45 нт, от около 12 нт до около 40 нт, от около 12 нт до около 35 нт, от около 12 нт до около 30 нт, от около 12 нт до около 25 нт, от около 12 нт до около 20 нт, от около 12 нт до около 19 нт, от около 19 нт до около 20 нт, от около 19 нт до около 25 нт, от около 19 нт до около 30 нт, от около 19 нт до около 35 нт, от около 19 нт до около 40 нт, от около 19 нт до около 45 нт, от около 19 нт до около 50 нт, от около 19 нт до около 60 нт, от около 20 нт до около 25 нт, от около 20 нт до около 30 нт, от около 20 нт до около 35 нт, от около 20 нт до около 40 нт, от около 20 нт до около 45 нт, от около 20 нт до около 50 нт или от около 20 нт до около 60 нт. Нуклеотидная последовательность (последовательность ДНК-нацеливания) сегмента ДНК-нацеливания, являющаяся комплементарной нуклеотидной последовательности (целевой последовательности) целевой ДНК, может иметь длину по меньшей мере около 12 нт. В некоторых случаях последовательность ДНК-нацеливания может иметь длину по меньшей мере около 20 нт.
тракрРНК могут быть любого вида (например, полноразмерные тракрРНК или активные частичные тракрРНК) и различной длины. Они могут включать в себя первичные транскрипты или подвергнутые процессингу формы. Например, тракрРНК (как часть одногидовой РНК или в виде отдельной молекулы в составе двухмолекулярной гРНК) может содержать или состоять из всей или части последовательности тракрРНК дикого типа (например, около или более чем около 20, 26, 32, 45, 48, 54, 63, 67, 85 или более нуклеотидов последовательности тракрРНК дикого типа). Примеры последовательностей тракрРНК дикого типа из S. pyogenes включают варианты из 171 нуклеотида, 89 нуклеотидов, 75 нуклеотидов и 65 нуклеотидов. См., например, публикацию Deltcheva et al. (2011) Nature 471:602-607; WO 2014/093661, каждая из которых полностью включена в настоящее описание путем ссылки. Примеры тракрРНК в пределах одногидовых РНК (огРНК) включают сегменты тракрРНК, встречающиеся в пределах вариантов огРНК+48,+54,+67 и+85, где «+n» означает, что в огРНК включено до +n нуклеотидов тракрРНК дикого типа. См. публикацию US 8,697,359, полностью включенную в настоящий документ путем ссылки.
Процент комплементарности между последовательностью ДНК-нацеливания и последовательностью распознавания РНК CRISPR в пределах целевой ДНК может составлять по меньшей мере 60% (например, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100%). Процент комплементарности между последовательностью ДНК-нацеливания и последовательностью распознавания РНК CRISPR в пределах целевой ДНК составляет 100% на протяжении семи непрерывных 5'-концевых нуклеотидов на целевой последовательности в пределах комплементарной цепи целевой ДНК. В некоторых вариантах осуществления процент комплементарности между последовательностью ДНК-нацеливания и последовательностью распознавания РНК CRISPR в пределах целевой ДНК может составлять по меньшей мере 60% на протяжении около 20 непрерывных нуклеотидов. В качестве примера, процент комплементарности между последовательностью ДНК-нацеливания и последовательностью распознавания РНК CRISPR в пределах целевой ДНК составляет 100% на протяжении четырнадцати непрерывных 5'-концевых нуклеотидов последовательности распознавания РНК CRISPR в пределах комплементарной цепи целевой ДНК и всего 0% в оставшейся части. В таком случае можно считать, что длина последовательности ДНК-нацеливания составляет 14 нуклеотидов. В качестве другого примера, процент комплементарности между последовательностью ДНК-нацеливания и последовательностью распознавания РНК CRISPR в пределах целевой ДНК составляет 100% на протяжении семи непрерывных 5'-концевых нуклеотидов последовательности распознавания РНК CRISPR в пределах комплементарной цепи целевой ДНК и всего 0% в оставшейся части. В таком случае можно считать, что длина последовательности ДНК-нацеливания составляет 7 нуклеотидов.
«Комплементарность» нуклеиновых кислот означает, что нуклеотидная последовательность в одной цепи нуклеиновой кислоты вследствие ориентации нуклеотидных оснований образует водородные связи с другой последовательностью на противоположной цепи нуклеиновой кислоты. Комплементарными основаниями в ДНК обычно являются: А и Т, а также С и G, а в РНК: С и G, а также U и А. Комплементарность может быть идеальной или существенной/достаточной. Идеальная комплементарность двух нуклеиновых кислот означает, что две нуклеиновые кислоты могут образовывать дуплекс, в котором каждое основание связано с комплементарным основанием, образуя пары Уотсона-Крика. «Существенная» или «достаточная» комплементарность означает, что последовательность одной цепи не полностью и/или не идеально комплементарна последовательности противоположной цепи, но между основаниями на двух цепях создается достаточное связывание, чтобы образовывался стабильный гибридный комплекс при некотором наборе условий гибридизации (например, концентрации солей и температуре). Такие условия можно спрогнозировать, используя последовательности и стандартные математические расчеты для предсказания Тm гибридизованных цепей, или путем эмпирического определения Тm традиционными способами. Тm означает температуру, при которой популяция гибридизационных комплексов, образовавшихся между двумя цепями нуклеиновых кислот, является денатурированной на 50%. При температуре ниже Тm преимущество имеет образование гибридизационного комплекса, тогда как при температуре выше Тm преимущество имеет плавление или разделение цепей гибридизационного комплекса. Тm можно оценить для нуклеиновой кислоты с известным содержанием G+С в водном 1 М растворе NaCl, используя, например, выражение Тm=81,5+0,41(% G+С), хотя в других известных расчетах Тm принимаются во внимание структурные характеристики нуклеиновых кислот.
«Условие гибридизации» означает совокупную среду, в которой одна цепь нуклеиновой кислоты связывается со второй цепью нуклеиновой кислоты в результате комплементарного взаимодействия цепей и водородного связывания с образованием гибридизационного комплекса. Такие условия включают в себя химические компоненты и их концентрации (например, соли, хелатирующие агенты, формамид) в водном или органическом растворе, содержащем нуклеиновые кислоты, и температуру смеси. Свой вклад в среду могут вносить другие факторы, например длительность инкубации или размеры инкубационной камеры (например, Sambrook et al., Molecular Cloning, A Laboratory Manual, 2.sup.nd ed., pp. 1.90-1.91, 9.47-9.51, 1 1.47-11.57 (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989)).
Для гибридизации требуется, чтобы две нуклеиновые кислоты содержали комплементарные последовательности, хотя возможны несовпадения между основаниями. Условия, подходящие для гибридизации между двумя нуклеиновыми кислотами, зависят от длины нуклеиновых кислот и степени комплементации, и эти переменные хорошо известны специалистам в данной области. Чем больше степень комплементации между двумя нуклеотидными последовательностями, тем выше температура плавления (Тm) гибридов нуклеиновых кислот с этими последовательностями. При гибридизации нуклеиновых кислот с короткими участками комплементарности (например, комплементарностью более 35 или менее, 30 или менее, 25 или менее, 22 или менее, 20 или менее или 18 или менее нуклеотидов) становится важным положение несовпадающих оснований (см. Sambrook et al., выше, 11.7-11.8). Как правило, длина пригодной для гибридизации нуклеиновой кислоты составляет по меньшей мере около 10 нуклеотидов. Значения иллюстративной минимальной длины пригодных для гибридизации нуклеиновых кислот представляют собой по меньшей мере около 15 нуклеотидов, по меньшей мере около 20 нуклеотидов, по меньшей мере около 22 нуклеотидов, по меньшей мере около 25 нуклеотидов и по меньшей мере около 30 нуклеотидов. Кроме того, температуру и концентрацию солей в промывном растворе можно регулировать по мере необходимости, в зависимости от таких факторов, как длина области комплементации и степень комплементации.
Для обеспечения специфической гибридизации последовательность полинуклеотида не обязательно должна быть на 100% комплементарна целевой нуклеиновой кислоте. Более того, полинуклеотид может гибридизоваться на одном или более сегментах, так что промежуточные или соседние сегменты не вовлечены в событие гибридизации (например, структура петли или шпильки). Полинуклеотид (например, гРНК) может иметь по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 99%, или 100% комплементарность последовательности относительно целевой области в целевой нуклеотидной последовательности, на которую он нацелен. Например, гРНК, в которой 18 из 20 нуклеотидов гРНК комплементарны целевой области и, следовательно, могут специфически гибридизоваться, будут соответствовать 90-процентной комплементарности. В этом примере оставшиеся некомплементарные нуклеотиды могут быть кластеризованы или рассеяны среди комплементарных нуклеотидов и не должны быть обязательно смежными друг с другом или с комплементарными нуклеотидами. Процентную долю комплементарности между конкретными участками нуклеотидных последовательностей в пределах нуклеиновых кислот можно определять типовым способом, используя программы BLAST (средства поиска основных локальных выравниваний) и PowerBLAST, известные в данной области (Altschul et al, J. Mol. Biol., 1990, 215, 403-410; Zhang and Madden, Genome Res., 1997, 7, 649-656), или с помощью программы Gap (Wisconsin Sequence Analysis Package, Version 8 for Unix, Genetics Computer Group, University Research Park, г. Мэдисон, штат Висконсин, США), с использованием заданных по умолчанию параметров, где используется алгоритм Смита - Ватермана (Adv. Appl. Math., 1981, 2, 482-489).
Белок-связывающий сегмент обсуждаемой гРНК взаимодействует с белком Cas. Обсуждаемая гРНК направляет связанный полипептид к специфической нуклеотидной последовательности в пределах целевой ДНК посредством сегмента ДНК-нацеливания. Белок-связывающий сегмент обсуждаемой гРНК может содержать два участка нуклеотидов, которые комплементарны друг другу. Комплементарные нуклеотиды белок-связывающего сегмента гибридизуются с образованием дуплекса двухцепочечной РНК (дцРНК). Белок-связывающий сегмент обсуждаемой гРНК взаимодействует с белком Cas, и гРНК управляет связыванием белка Cas со специфической нуклеотидной последовательностью в пределах ДНК с помощью сегмента ДНК-нацеливания.
В некоторых вариантах осуществления гРНК, как описано в настоящем документе, содержит две отдельные молекулы РНК. Каждая из этих двух молекул РНК обсуждаемой гРНК содержит нуклеотидный участок, и эти участки комплементарны друг другу так, что комплементарные нуклеотиды двух молекул РНК гибридизуются с образованием двухцепочечного дуплекса РНК (например, шпильки) белок-связывающего сегмента. Обсуждаемая гРНК может содержать любую пару соответствующих крРНК и тракрРНК. В способах, описанных в настоящем документе, гРНК может использоваться в виде комплекса (например, комплекса гРНК-Cas) из крРНК и тракрРНК, или же крРНК и соответствующая тракрРНК могут поставляться отдельно. Например, если для реакции расщепления используют множество гРНК, отдельные крРНК, специфичные для каждого целевого сайта, могут поставляться отдельно от стандартной тракрРНК, способной образовывать комплекс с каждой крРНК. При таком способе молекулы крРНК могут образовывать комплекс со стандартной тракрРНК и направлять белок Cas к целевому сайту.
Гидовые РНК могут включать в себя модификации или последовательности, которые обеспечивают дополнительные желательные характеристики (например, модифицированную или регулируемую стабильность; субклеточное нацеливание, отслеживание с помощью флуоресцентной метки; сайт связывания для белка или белкового комплекса и т.п.). Не имеющие ограничительного характера примеры таких модификаций включают, например, 5'-кэп (например, 7-метилгуанилатный кэп (m7G)); 3'-полиаденилированный хвост (т.е. 3'-поли(А) хвост); последовательность-рибопереключатель (например, для обеспечения регулируемой стабильности и/или регулируемой доступности белками и/или белковыми комплексами); последовательность контроля стабильности; последовательность, образующую дуплекс дцРНК (т.е. шпилька)); модификацию или последовательность, которая нацеливает РНК в субклеточное местоположение (например, ядро, митохондрии, хлоропласты и т.п.); модификацию или последовательность, которая обеспечивает отслеживание (например, прямая конъюгация с флуоресцентной молекулой, конъюгация с фрагментом, который способствует флуоресцентной детекции, последовательность, которая позволяет флуоресцентную детекцию, и т.д.); модификацию или последовательность, которая обеспечивает сайт связывания для белков (например, белки, которые действуют на ДНК, включая активаторы транскрипции, репрессоры транскрипции, ДНК-метилтрансферазы, ДНК-деметилазы, гистонацетилтрансферазы, гистондеацетилазы и т.п.); и их комбинации.
Гидовые РНК могут быть обеспечены в любом виде. Например, гРНК может быть обеспечена в виде РНК, либо как две молекулы (отдельно крРНК и тракрРНК), либо как одна молекула (огРНК) и, необязательно, в виде комплекса с белком Cas. гРНК может быть обеспечена также в виде ДНК, кодирующей РНК. ДНК, кодирующая гРНК, может кодировать одну молекулу РНК (огРНК) или отдельные молекулы РНК (например, отдельно крРНК и тракрРНК). В последнем случае ДНК, кодирующая гРНК, может быть обеспечена как отдельные молекулы ДНК, кодирующие, соответственно, крРНК и тракрРНК.
ДНК, кодирующие гРНК, могут быть стабильно интегрированы в геном клетки и функционально связаны с активным в клетке промотором. В альтернативном варианте осуществления ДНК, кодирующие гРНК, могут быть функционально связаны с промотором в экспрессионной конструкции. Например, ДНК, кодирующая гРНК, может находиться в нацеливающем векторе, содержащем нуклеотидную вставку и/или вектор, содержащий нуклеиновую кислоту, кодирующую белок Cas, или она может находиться в векторе или плазмиде, отдельной от нацеливающего вектора, содержащего нуклеотидную вставку, и/или отдельной от вектора, содержащего нуклеиновую кислоту, кодирующую белок Cas. Такие промоторы могут быть активными, например, в плюрипотентной клетке крысы, эукариота, млекопитающего, не относящегося к человеку млекопитающего, человека, грызуна, мыши или хомяка. Такие промоторы могут представлять собой, например, условные промоторы, индуцируемые промоторы, конститутивные промоторы или тканеспецифические промоторы. В некоторых случаях промотор представляет собой промотор РНК-полимеразы III, такой как промотор U6 человека, промотор U6 полимеразы III крысы или промотор U6 полимеразы III мыши. В других разделах настоящего документа описаны примеры других промоторов. Если в клетку вводят ДНК, кодирующую гРНК, то гРНК может экспрессироваться внутри клетки временно, условно или конститутивно.
В альтернативном варианте осуществления гРНК можно получать с помощью различных других способов. Например, гРНК можно получать посредством транскрипции in vitro с использованием, например, Т7 РНК-полимеразы (см., например, WO 2014/089290 и WO 2014/065596). Гидовые РНК могут также быть синтетическими молекулами, полученными путем химического синтеза.
С. Последовательности распознавания РНК CRISPR
Термин «последовательность распознавания РНК CRISPR» включает нуклеотидные последовательности, присутствующие в целевой ДНК, с которой будет связываться сегмент ДНК-нацеливания гРНК, при условии существования достаточных условий для связывания. Например, последовательности распознавания РНК CRISPR включают последовательности, которым должна быть комплементарна сконструированная гидовая РНК, где гибридизация между последовательностью распознавания РНК CRISPR и последовательностью ДНК-нацеливания способствует образованию комплекса CRISPR. Полная комплементарность не является обязательным требованием при условии достаточной комплементарности, чтобы вызвать гибридизацию и способствовать образованию комплекса CRISPR. Последовательности распознавания РНК CRISPR также включают сайты расщепления для белков Cas, более подробно описанные ниже. Последовательность распознавания РНК CRISPR может содержать любой полинуклеотид, который может быть расположен, например, в ядре или цитоплазме клетки или внутри органеллы клетки, такой как митохондрия или хлоропласт.
На последовательность распознавания РНК CRISPR в пределах целевой ДНК может быть нацелен (т.е. может связывать, или гибридизовать, или быть комплементарным) белок Cas или гРНК. Подходящие условия связывания ДНК/РНК включают физиологические условия, обычно существующие в клетке. Другие подходящие условия связывания ДНК/РНК (например, условия в бесклеточной системе) известны в данной области (см., например, Molecular Cloning: A Laboratory Manual, 3rd Ed. (Sambrook et al., Harbor Laboratory Press 2001)). Цепь целевой ДНК, которая является комплементарной и гибридизуется с белком Cas или гРНК, можно назвать «комплементарной цепью», а цепь целевой ДНК, которая комплементарна «комплементарной цепи» (и, таким образом, не комплементарна белку Cas или гРНК), можно назвать «некомплементарной цепью» или «матричной цепью».
Белок Cas может расщеплять нуклеиновую кислоту на сайте внутри или за пределами нуклеотидной последовательности, присутствующей в целевой ДНК, с которой будет связываться сегмент ДНК-нацеливания гРНК. «Сайт расщепления» включает положение нуклеиновой кислоты, в котором белок Cas производит одноцепочечный разрыв или двухцепочечный разрыв. Например, образование комплекса CRISPR (содержащего гРНК, гибридизованную с последовательностью распознавания РНК CRISPR и образующую комплексы с белком Cas) может приводить к расщеплению одной или обеих цепей внутри или возле (например, на расстоянии в пределах 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 50 или более пар нуклеотидов от) нуклеотидной последовательности, присутствующей в целевой ДНК, с которой будет связываться сегмент ДНК-нацеливания гРНК. Если сайт расщепления находится за пределами нуклеотидной последовательности, с которой будет связываться сегмент ДНК-нацеливания гРНК, то сайт расщепления все еще считается находящимся в пределах «последовательности распознавания РНК CRISPR». Сайт расщепления может находиться на только одной цепи или на обеих цепях нуклеиновой кислоты. Сайты расщепления могут находиться в одном и том же положении на обеих цепях нуклеиновой кислоты (образуя тупые концы) или могут находиться на разных сайтах на каждой цепи (образуя ступенчатые концы). Ступенчатые концы можно получать, например, с использованием двух белков Cas, каждый из которых производит одноцепочечный разрыв на отличающемся сайте расщепления на каждой цепи, производя таким образом двухцепочечный разрыв. Например, первая никаза может создавать одноцепочечный разрыв на первой цепи двухцепочечной ДНК (дцДНК), а вторая никаза может создавать одноцепочечный разрыв на второй цепи дцДНК таким образом, что образуются нависающие последовательности. В некоторых случаях последовательность распознавания РНК CRISPR никазы на первой цепи разделена с последовательностью распознавания РНК CRISPR никазы на второй цепи по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 40, 50, 75, 100, 250, 500 или 1000 парами нуклеотидов.
Сайт-специфическое расщепление целевой ДНК белком Cas9 может происходить в местах, определенных как (i) комплементарностью спариваемых нуклеотидов между гРНК и целевой ДНК, так и (ii) наличием в целевой ДНК короткого мотива, называемого мотивом, прилежащим к протоспейсеру (РАМ). РАМ может фланкировать последовательность распознавания РНК CRISPR. Необязательно, последовательность распознавания РНК CRISPR может быть фланкирована РАМ. Например, сайт расщепления Cas9 может находиться на расстоянии от около 1 до около 10 или от около 2 до около 5 пар нуклеотидов (например, 3 пар нуклеотидов) выше или ниже последовательности РАМ. В некоторых случаях (например, когда используется Cas9 из S. pyogenes или близкородственный Cas9) последовательность РАМ некомплементарной цепи может представлять собой 5'-N1GG-3', где Ni является любым нуклеотидом ДНК и находится непосредственно на 3'-конце последовательности распознавания РНК CRISPR некомплементарной цепи целевой ДНК. По существу, последовательность РАМ комплементарной цепи будет представлять собой 5'-СС N2-3', где N2 является любым нуклеотидом ДНК и находится непосредственно на 5'-конце последовательности распознавания РНК CRISPR комплементарной цепи целевой ДНК. В некоторых таких случаях N1 и N2 могут быть комплементарными, и пара нуклеотидов N1-N2 может быть любой парой нуклеотидов (например, N1=С и N2=G; N1=G и N2=С; N1=А и N2=Т, N1=Т и N2=А).
Примеры последовательностей распознавания РНК CRISPR включают последовательность ДНК, комплементарную сегменту ДНК-нацеливания гРНК, или такую последовательность ДНК в дополнение к последовательности РАМ. Например, целевой мотив может представлять собой последовательность ДНК длиной 20 нуклеотидов, непосредственно предшествующую мотиву NGG, распознаваемому белком Cas, например, GN19NGG (SEQ ID NO: 8) или N20NGG (SEQ ID NO: 24) (см., например, WO 2014/165825). Гуанин на 5'-конце может способствовать транскрипции РНК-полимеразы в клетках. Другие примеры последовательностей распознавания РНК CRISPR могут включать два гуаниннуклеотида на 5'-конце (например, GGN20NGG; SEQ ID NO: 25) для облегчения эффективной транскрипции Т7-полимеразой in vitro. См., например, WO 2014/065596. Другие последовательности распознавания РНК CRISPR могут иметь длину 4-22 нуклеотида с SEQ ID NO: 8, 24 и 25, включая 5' G или GG и 3' GG или NGG. Другие последовательности распознавания РНК CRISPR могут иметь длину 14-20 нуклеотидов с SEQ ID NO: 8, 24 и 25.
Последовательность распознавания РНК CRISPR может представлять собой любую нуклеотидную последовательность, эндогенную или экзогенную по отношению к клетке. Последовательность распознавания РНК CRISPR может представлять собой последовательность, кодирующую генный продукт (например, белок), или некодирующую последовательность (например, регуляторную последовательность), или может включать в себя обе.
В одном варианте осуществления белок Cas представляет собой белок Cas I типа. В одном варианте осуществления белок Cas представляет собой белок Cas II типа. В одном варианте осуществления белок Cas II типа представляет собой Cas9. В одном варианте осуществления первая нуклеотидная последовательность кодирует оптимизированный по кодонам к человеку белок Cas.
В одном варианте осуществления гРНК содержит нуклеотидную последовательность, кодирующую крРНК и тракрРНК. В конкретных вариантах осуществления белок Cas представляет собой Cas9. В некоторых вариантах осуществления гРНК содержит (а) химерную РНК нуклеотидной последовательности 5'-GUUUUAGAGCUAGAAAUAGCAAGUUААAAU AAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGCUUUU-3' (SEQ ID NO: 1; или (b) химерную РНК нуклеотидной последовательности 5'-GUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCG-3' (SEQ ID NO: 2). В другом варианте осуществления крРНК содержит 5'-GUUUUAGAGCUAGAAAUAGCAAGUUAAAAU-3' (SEQ ID NO: 3); 5'-GUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAG (SEQ ID NO: 4); или 5'-GAGUCCGAGCAGAAGAAGAAGUUUUA-3' (SEQ ID NO: 5). В других вариантах осуществления тракрРНК содержит 5'-AAGGCUAGUCCG-3' (SEQ ID NO: 6) или 5'-AAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGCUUUU-3' (SEQ ID NO: 7).
V. Сборка полинуклеотидов
Способы, описанные в настоящем документе, позволяют собрать по меньшей мере две нуклеиновые кислоты в условиях, способствующих эффективному соединению молекул ДНК с образованием по существу интактной или бесшовной двухцепочечной молекулы ДНК. Любые интересующие нуклеиновые кислоты, имеющие перекрывающиеся последовательности, можно собирать в соответствии со способами, описанными в настоящем документе. Например, можно собирать любые интересующие молекулы ДНК, имеющие перекрывающиеся последовательности, включая ДНК природного происхождения, клонированные молекулы ДНК, синтетически созданные молекулы ДНК и т.п. Соединенные молекулы ДНК можно при необходимости клонировать (например, вставлять) в вектор, используя способ изобретения. Сборка двух нуклеиновых кислот включает в себя любой способ соединения цепочек нуклеиновых кислот. Например, сборка включает в себя соединение расщепленных нуклеиновых кислот таким образом, чтобы цепи каждой нуклеиновой кислоты ренатурировались с другой цепью и удлинялись, причем каждая цепь служила шаблоном для удлинения другой.
В некоторых вариантах осуществления сборку нуклеиновых кислот выполняют с помощью соединительного олигонуклеотида, так что каждая нуклеиновая кислота соединяется с соединительным олигонуклеотидом, а не напрямую с другой нуклеиновой кислотой. Сборка с соединительным олигонуклеотидом позволяет разместить между подлежащими сборке нуклеиновыми кислотами нуклеотидные основания, не являющиеся частью собираемых нуклеиновых кислот, а являющиеся частью соединительного олигонуклеотида. Следовательно, можно выполнить успешную сборку нуклеиновых кислот даже в том случае, если между нуклеиновыми кислотами остались дополнительные основания. В альтернативном варианте осуществления соединительный олигонуклеотид можно использовать для бесшовной сборки, при которой между нуклеиновыми кислотами, подлежащими сборке, не остается дополнительных оснований.
В некоторых вариантах осуществления нуклеиновые кислоты можно подготовить к сборке путем расщепления белком Cas, рестрикционным ферментом (рестрикционной эндонуклеазой) (например, любой из разнообразных рестрикционных эндонуклеаз, приведенных в других частях настоящего документа), мегануклеазой (например, любой из разнообразных мегануклеаз, приведенных в других частях настоящего документа) или любой их комбинацией. Например, одну из подлежащих сборке нуклеиновых кислот можно расщепить белком Cas и другую из подлежащих сборке нуклеиновых кислот можно расщепить белком Cas, рестрикционным ферментом, мегануклеазой или любой их комбинацией. После расщепления нуклеазой можно выполнить непосредственную сборку расщепленной нуклеиновой кислоты с другой расщепленной нуклеиновой кислотой, имеющей перекрывающиеся концевые последовательности, или выполнить сборку с не расщепленной нуклеиновой кислотой, но имеющей перекрывающиеся концевые последовательности. Также можно выполнить сборку расщепленной нуклеиновой кислоты с другой нуклеиновой кислотой с помощью соединительного олигонуклеотида.
В вариантах осуществления, в которых используется нуклеазный агент (например, белок Cas) для получения перекрывающихся концевых последовательностей между двумя молекулами нуклеиновых кислот, для сборки расщепленных нуклеиновых кислот можно использовать быстрые комбинаторные способы. Например, первую и вторую нуклеиновые кислоты, имеющие перекрывающиеся концы, можно объединить с лигазой, экзонуклеазой, ДНК-полимеразой и нуклеотидами и инкубировать при постоянной температуре, например, 50°С. В частности, экзонуклеазу Т5 можно использовать для удаления нуклеотидов с 5'-концов дцДНК, получая комплементарные нависающие участки. Комплементарные одноцепочечные нависания ДНК далее можно ренатурировать, заполнить гэпы ДНК-полимеразой и с помощью лигазы Taq ДНК закрыть получившиеся одноцепочечные разрывы при 50°С. Таким образом, две нуклеиновые кислоты, имеющие общие перекрывающиеся концевые последовательности, можно соединить в ковалентно-связанную молекулу в ходе одноэтапной изотермической реакции. См., например, публикацию Gibson, et al. (2009) Nature Methods 6 (5): 343-345, которая полностью включена в настоящий документ путем ссылки. В некоторых вариантах осуществления для удаления нуклеазного агента (например, белка Cas) из реакционной смеси используют протеиназу К или очистку фенолом/хлороформом/изоамилолом (PCI). В некоторых вариантах осуществления нуклеазный агент (например, белок Cas) можно удалить из реакционной смеси методом очистки на силикагелевой колонке.
В некоторых вариантах осуществления с помощью способов, описанных в настоящем документе, собирают вектор с линейным полинуклеотидом. В других вариантах осуществления с помощью способов, описанных в настоящем документе, собирают по меньшей мере два вектора, например, два ВАС-вектора. Термин «ВАС-вектор» означает любую бактериальную искусственную хромосому. В конкретных вариантах осуществления ВАС модифицируют, чтобы она содержала область с нуклеотидной последовательностью, перекрывающейся с нуклеотидной последовательностью области линейной нуклеиновой кислоты или другого вектора, например, другой ВАС.
Первая и вторая одноцепочечные нуклеиновые кислоты имеют перекрывающиеся концы, если соответствующие концы комплементарны друг другу. Первая и вторая двухцепочечные нуклеиновые кислоты имеют перекрывающиеся концы, если 5'-конец цепи первой нуклеиновой кислоты комплементарен 3'-концу цепи второй нуклеиновой кислоты, и наоборот. Например, в случае двухцепочечных перекрывающихся концевых последовательностей цепи одной нуклеиновой кислоты могут иметь по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичность соответствующей цепи другой нуклеиновой кислоты. В способах, описанных в настоящем документе, 5'-конец цепи подлежащей сборке молекулы дцДНК имеет общие перекрывающиеся концевые последовательности с 3'-концом цепи другой молекулы дцДНК. Термин «перекрывающиеся концевые последовательности» относится к обеим цепям молекулы дцДНК. Таким образом, одна цепь из перекрывающейся области может специфически гибридизоваться с комплементарной ей цепью, когда комплементарные области перекрывающихся последовательностей присутствуют в одноцепочечных нависаниях 5'- и 3'-концов двух соединяемых полинуклеотидов. В некоторых вариантах осуществления экзонуклеаза используется для удаления нуклеотидов с 5'- или 3'-конца с целью создания нависающих концевых последовательностей. В некоторых вариантах осуществления перекрывающаяся область первой и/или второй нуклеиновой кислоты отсутствует на 5'- или 3'-конце до тех пор, пока не будет проведено расщепление белком Cas, т.е. перекрывающаяся область может быть внутренней областью, которая затем превращается в перекрывающуюся концевую последовательность после расщепления нуклеиновой кислоты, содержащей внутреннюю перекрывающуюся область, белком Cas. Белок Cas может осуществлять расщепление в целевом сайте (например, сайте расщепления) в пределах перекрывающейся области или за пределами перекрывающейся области.
Перекрывающаяся область предпочтительно имеет длину, достаточную, чтобы эта область встречалась только один раз в любой из собираемых нуклеиновых кислот. Таким образом предотвращается ренатурация других полинуклеотидов с этими концевыми последовательностями, и сборка может быть специфичной к целевым нуклеиновым кислотам. Длина перекрывающейся области может варьироваться от минимума 10 пар нуклеотидов (п.н.) до около 300 п.н. или более. В целом, предпочтительно, чтобы длина перекрывающейся области была меньше или приблизительно равна размеру соединяемого полинуклеотида, но не менее чем около 10 п.н. и не более чем около 1000 п.н. Для соединения 2 или 3 полинуклеотидов может быть достаточно перекрывающейся области длиной приблизительно 20-30 п.н. Для более чем 10 фрагментов предпочтительная длина перекрытия составляет от около 80 п.н. до около 300 п.н. В одном варианте осуществления перекрывающаяся область имеет длину, позволяющую легко сгенерировать ее методами синтеза, например, около 40 п.н. В конкретных вариантах осуществления длина перекрывающейся области может составлять около 20-200 п.н. Перекрывающиеся области могут иметь длину 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950 или 1000 п.н. В некоторых вариантах осуществления длина перекрывающейся области составляет от 20 до 200 п.н. В конкретных вариантах осуществления способов, описанных в настоящем документе, можно выполнить сборку по меньшей мере двух нуклеотидов, в которых перекрывающаяся область по меньшей мере на одном из полинуклеотидов создана путем контактирования с нуклеазным агентом (например, комплексом гРНК-Cas). Например, эндонуклеазное расщепление первого полинуклеотида может создать последовательности, перекрывающиеся с концевым последовательностями второго полинуклеотида, причем впоследствии выполняют сборку перекрывающихся концевых последовательностей.
В способах, описанных в настоящем документе, перекрывающиеся последовательности могут быть введены во взаимодействие с экзонуклеазой с целью экспонирования комплементарных последовательностей (например, одноцепочечных комплементарных последовательностей) перекрывающихся последовательностей. Расщепление экзонуклеазой проводят при условиях, способствующих удалению («отгрызанию») достаточного количества нуклеотидов, чтобы обеспечить специфический ренатурацию экспонированных одноцепочечных областей комплементарности. В целом, «отгрызают» участок перекрывающейся области или всю перекрывающуюся область, оставляя нависания, которые содержат участок перекрывающейся области или всю перекрывающуюся область. В некоторых способах экзонуклеазное расщепление может осуществляться полимеразой при отсутствии дНТФ, например, Т5 ДНК-полимеразой), тогда как в других способах экзонуклеазное расщепление может осуществляться в присутствии дНТФ экзонуклеазой, которая не проявляет полимеразную активность (например, экзонуклеаза III).
Любая из разнообразных экзодезоксирибонуклеаз, специфичных для двойной цепи, работающих от 5'- к 3'-концу, может использоваться для «отгрызания» концов нуклеиновых кислот в способах, описанных в настоящем изобретении. Термин «5'-экзонуклеаза» иногда применяется в настоящем документе для обозначения экзодезоксирибонуклеазы, работающей от 5'- к 3'-концу. Термином «непроцессивная» экзонуклеаза в настоящем документе называется экзонуклеаза, разрушающая ограниченное количество (например, всего несколько) нуклеотидов при каждом событии связывания с ДНК. В результате расщепления 5'-экзонуклеазой создаются 3'-одноцепочечные нависания в молекулах ДНК. Желательными для 5'-экзонуклеазы свойствами, среди прочих, являются следующие: чтобы она не имела 3'-экзонуклеазной активности, чтобы она создавала 5'-фосфатные концы и инициировала деградацию как с 5'-фосфорилированных, так и с нефосфорилированных концов. Также желательно, чтобы фермент мог инициировать расщепление с 5'-конца молекулы, независимо от того, имеет ли она «тупой» конец или имеет небольшой заглубленный 5'- или 3'-конец. Подходящие экзонуклеазы будут очевидны для квалифицированного специалиста. К ним относятся, например, экзонуклеаза фага Т5 (продукт гена D15 фага Т5), экзонуклеаза лямбда-фага, RecE профага Rac, экзонуклеаза VIII из Е. coli, экзонуклеаза фага Т7 (продукт гена 6 фага Т7) или любая из различных 5'-экзонуклеаз, вовлеченных в реакции гомологичной рекомбинации. В одном варианте осуществления изобретения экзонуклеаза представляет собой экзонуклеазу Т5 или экзонуклеазу лямбда. В другом варианте осуществления экзонуклеаза представляет собой экзонуклеазу Т5. В другом варианте осуществления экзонуклеаза не является экзонуклеазой фага Т7. Способы получения и использования экзонуклеаз и других ферментов, использованных в способах изобретения, являются традиционными, и многие доступны из коммерческих источников, таких как USB Corporation, 26111 Miles Road, Cleveland, Ohio 44128 или New England Biolabs, Inc. (NEB), 240 County Road, Ipswich, Mass. 01938-2723.
В частности, в вариантах осуществления, в которых область перекрытия очень длинная, может потребоваться «отгрызание» лишь участка области (например, более чем половины области перекрытия), при условии, что создаются одноцепочечные нависания с достаточной длиной и содержанием оснований для специфической ренатурации в условиях реакции. Термин «специфическая ренатурация» относится к ситуациям, в которых конкретные пары одноцепочечных нависаний будут ренатурироваться предпочтительно (или исключительно) друг с другом, а не с другими одноцепочечными нависаниями (например, некомплементарными нависаниями), присутствующими в реакционной смеси. Под «предпочтительно» понимается, что по меньшей мере около 95% нависаний будут ренатурироваться с комплементарным нависанием. Квалифицированный специалист может легко определить оптимальную длину для достижения специфической ренатурации интересующей последовательности при заданном наборе условий реакции. В целом, гомологичные перекрывающиеся области (одноцепочечные нависания или комплементарные им участки) содержат идентичные последовательности. Однако можно использовать частично идентичные последовательности, при условии, что одноцепочечные нависания смогут специфично ренатурироваться в условиях реакций.
В некоторых вариантах осуществления нуклеазный агент (например, белок Cas) может формировать одноцепочечные разрывы (т.е. «ники») в целевом сайте без разрезания обоих цепочек ДНК. «Никазой» называется нуклеазный агент (например, белок Cas), создающий одноцепочечные разрывы в дцДНК. Таким образом, два отдельных нуклеазных агента (например, белка Cas) (например, никазы), специфичные к целевому сайту на каждой цепи дцДНК, позволяют создать нависающие последовательности, комплементарные нависающим последовательностям на другой нуклеиновой кислоте или на отдельной области той же нуклеиновой кислоты. Нависающие концы, созданные посредством контактирования нуклеиновой кислоты с двумя никазами, специфичными к целевым сайтам на обеих цепях дцДНК, могут представлять собой либо 5'-, либо 3'-нависающие концы. Например, первая никаза может создавать одноцепочечный разрыв на первой цепи дцДНК, а вторая никаза может создавать одноцепочечный разрыв на второй цепи дцДНК таким образом, что образуются нависающие последовательности. Целевые сайты для каждой никазы, формирующей одноцепочечный разрыв, можно выбирать таким образом, чтобы созданные нависающие последовательности были комплементарны нависающим концевым последовательностям на второй нуклеиновой кислоте. Соответственно, комплементарные нависающие концы первой и второй нуклеиновых кислот можно ренатурировать способами, описанными в настоящем документе. В некоторых вариантах осуществления целевой сайт для никазы на первой цепи отличается от целевого сайта никазы на второй цепи. Разные целевые сайты на отдельных цепях дцДНК приводят к тому, что одноцепочечные разрывы оказываются разделены по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 40, 50, 75, 100, 250, 500 или 1000 пар нуклеотидов.
В некоторых вариантах осуществления вторая нуклеиновая кислота также контактирует с первой никазой, которая создает одноцепочечный разрыв в первом целевом сайте на второй нуклеиновой кислоте, и второй никазой, которая создает одноцепочечный разрыв во втором целевом сайте на молекуле второй нуклеиновой кислоты. Нависающие концевые последовательности, созданные в результате одноцепочечных разрывов на второй нуклеиновой кислоте, могут быть комплементарны нависающим концевым последовательностям, созданным одноцепочечными разрывами в двух разных сайтах на первой нуклеиновой кислоте так, чтобы комплементарные нависающие концы ренатурировались.
В некоторых вариантах осуществления нуклеотидная последовательность интересующего гена охватывает две или более ВАС. В таких случаях, с использованием способов, предложенных в настоящем документе, специально разработанные нуклеазные агенты позволяют разрезать две или более ВАС в нужных местах, и полученные фрагменты нуклеиновых кислот соединяют с образованием последовательности интересующего гена.
В некоторых вариантах осуществления нависающие концы, созданные в результате одноцепочечных разрывов в разных целевых сайтах на обеих цепях первой нуклеиновой кислоты, не комплементарны нависающим концам, созданным в результате одноцепочечных разрывов в других целевых сайтах на обеих цепях второй нуклеиновой кислоты. В других вариантах осуществления подлежащие сборке нуклеиновые кислоты не имеют комплементарных концов, так что для сборки некомплементарных концов необходима отдельная нуклеиновая кислота. Для соединения некомплементарных концов двух нуклеиновых кислот можно использовать соединительный олигонуклеотид. «Соединительный олигонуклеотид» имеет комплементарные плечи, включающие в себя полинуклеотид или нуклеиновую кислоту, имеющую последовательность, комплементарную концам другого полинуклеотида или нуклеиновой кислоты. В некоторых вариантах осуществления соединительный олигонуклеотид имеет плечо, комплементарное первой нуклеиновой кислоте, на 5'-конце, центральный участок (спейсер) и плечо, комплементарное второй нуклеиновой кислоте, на 3'-конце. Следовательно, можно выполнять сборку нуклеиновых кислот, имеющих некомплементарные друг другу концевые последовательности, путем ренатурации каждой нуклеиновой кислоты с одним соединительным олигонуклеотидом, после обработки экзонуклеазой. В конкретных вариантах осуществления соединительный олигонуклеотид имеет первое плечо, комплементарное 5'- или 3'-концевой последовательности первой расщепленной нуклеиновой кислоты, и второе плечо, комплементарное 5'- или 3'-последовательности второй расщепленной нуклеиновой кислоты. Соединительный олигонуклеотид может соединять некомплементарные концевые последовательности, являющиеся «тупыми» или имеющие 5'- или 3'-нависающую последовательность.
Длина комплементарных плечевых последовательностей соединительного олигонуклеотида должна быть достаточной для ренатурации с подлежащими сборке нуклеиновыми кислотами после обработки экзонуклеазой. Например, длина комплементарных плечевых последовательностей соединительного олигонуклеотида может составлять по меньшей мере около 10, 20, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 90, 100, 110, 120, 130, 140, 150 п.н. или более. В конкретных вариантах осуществления длина комплементарного плеча составляет 15-120 п.н., 20-100 п.н., 30-90 п.н., 30-60 п.н. или 20-80 п.н. В одном конкретном варианте осуществления длина комплементарных плечевых последовательностей соединительного олигонуклеотида составляет 40 п.н. Каждое комплементарное плечо соединительного олигонуклеотида могут иметь отличающуюся длину. Спейсер соединительного олигонуклеотида между концевыми последовательностями, комплементарными подлежащим сборке нуклеиновым кислотам, может составлять по меньшей мере около 20 п.н., 30 п.н., 35 п.н., 40 п.н., 45 п.н., 50 п.н., 55 п.н., 60 п.н., 65 п.н., 70 п.н., 75 п.н., 80 п.н., 90 п.н., 100 п.н., 250 п.н., 500 п.н., 750 п.н., 1000 п.н., 2000 п.н., 3000 п.н., 4000 п.н., 5000 п.н., 8000 п.н., 10 т.п.н., 15 т.п.н., 20 т.п.н. или более. Например, спейсер соединительного олигонуклеотида может содержать ВАС-вектор или LTVEC. В некоторых вариантах осуществления спейсер соединительного олигонуклеотида может быть разработан так, чтобы содержать последовательности, специфичные для обнаружения или подходящие для ПЦР, с целью подтверждения успешности сборки. В некоторых вариантах осуществления спейсер соединительного олигонуклеотида может быть разработан с введением в него одного или более сайтов для рестрикционных ферментов. В некоторых вариантах осуществления спейсер соединительного олигонуклеотида может быть разработан с введением в него гена устойчивости к лекарственному средству или репортерного гена. В других вариантах осуществления спейсер может содержать по меньшей мере 20 п.н. из концевой части подлежащей сборке нуклеиновой кислоты, чтобы обеспечить бесшовную сборку нуклеиновых кислот. Например, спейсер для бесшовной сборки может иметь длину около 45 п.н.
В некоторых вариантах осуществления молярное соотношение нуклеиновой кислоты к соединительному(-ым) олигонуклеотиду(-ам) может составлять от около 1:1 до около 1: 200. В некоторых вариантах осуществления молярное соотношение нуклеиновой кислоты к соединительному(-ым) олигонуклеотиду(-ам) составляет около 1:1, 1:2, 1:3, 1:4, 1:5, 1:6, 1:7, 1:8, 1:9, 1:10, 1:11, 1:12, 1:13, 1:14, 1:15, 1:16, 1:17, 1:18, 1:19, 1:20, 1:30, 1:40, 1:50, 1:60, 1:70, 1:80, 1:90, 1:100, 1:120, 1:140, 1:160, 1:180 или 1:200. В конкретных вариантах осуществления молярное соотношение нуклеиновой кислоты к соединительному(-ым) олигонуклеотиду(-ам) может составлять от около 1:6 до около 1:20. В одном варианте осуществления молярное соотношение составляет около 1:6. В другом варианте осуществления молярное соотношение составляет около 1:20.
В конкретных вариантах осуществления соединительный олигонуклеотид используют для бесшовной сборки по меньшей мере двух нуклеиновых кислот.«Бесшовная» сборка означает такую сборку двух нуклеиновых кислот, в которой между смежными концами подлежащих сборке нуклеиновых кислот отсутствуют промежуточные нуклеотидные основания. Например, бесшовно собранные нуклеиновые кислоты не имеют нуклеотидных оснований, не являющихся частью подлежащих сборке нуклеиновых кислот. Для бесшовной сборки двух нуклеиновых кислот спейсер соединительного олигонуклеотида должен содержать нуклеотидную последовательность, идентичную концевому участку первой или второй нуклеиновой кислоты, подлежащей сборке. Этот концевой участок необходимо удалить из нуклеиновой кислоты перед сборкой с соединительным олигонуклеотидом. Например, концевой участок можно расщепить нуклеазным агентом (например, комплексом гРНК-Cas) на расстоянии по меньшей мере 20 п.н. от конца нуклеиновой кислоты, например, по меньшей мере 40 п.н. или по меньшей мере 45 п.н. от конца нуклеиновой кислоты. В альтернативном варианте осуществления концевой участок может быть расщеплен нуклеазным агентом (например, комплексом гРНК-Cas) на расстоянии по меньшей мере 2, по меньшей мере 4, по меньшей мере 6, по меньшей мере 8, по меньшей мере 10, по меньшей мере 12, по меньшей мере 15, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 37, по меньшей мере 40, по меньшей мере 42, по меньшей мере 45, по меньшей мере 48, по меньшей мере 50, по меньшей мере 55, по меньшей мере 60, по меньшей мере 65, по меньшей мере 70, по меньшей мере 80, по меньшей мере 100, по меньшей мере 110, по меньшей мере 120, по меньшей мере 130, по меньшей мере 140, по меньшей мере 150 п.н. от конца подлежащей сборке нуклеиновой кислоты.
В одном варианте осуществления соединительный олигонуклеотид может содержать, от 5'-конца к 3'-концу: участок около 15-120 п.н. перекрывающийся с 5'-нуклеиновой кислотой, около 20-50 п.н. 3'-концевого участка 5'-нуклеиновой кислоты и участок около 15-120 п.н., перекрывающийся с 3'-нуклеиновой кислотой. В одном варианте осуществления соединительный олигонуклеотид может содержать, от 5'-конца к 3'-концу: участок около 15-120 п.н. перекрывающийся с 5'-нуклеиновой кислотой, около 20-50 п.н. 5'-концевого участка 3'-нуклеиновой кислоты и участок около 15-120 п.н., перекрывающийся с 3'-нуклеиновой кислотой. Таким образом, когда соединительный олигонуклеотид соединяется с первой и второй нуклеиновыми кислотами, спейсер из соединительного олигонуклеотида восстанавливает участок, удаленный из нуклеиновой кислоты перед сборкой. См. ФИГ. 5 и ФИГ. 6. Термин «восстанавливает» относится к замещению концевого участка нуклеиновой кислоты, который был расщеплен для получения полной собранной нуклеиновой кислоты при сборке с соединительным олигонуклеотидом. Например, восстановление расщепленной нуклеиновой кислоты замещает расщепленный участок нуклеиновой кислоты нуклеиновой кислотой, входящей в спейсер соединительного олигонуклеотида, который имеет последовательность, идентичную расщепленному участку.
Сборку соединительного олигонуклеотида с первой и второй молекулой нуклеиновой кислоты можно выполнить одновременно или последовательно. При одновременной сборке соединительный олигонуклеотид можно ввести во взаимодействие с первой и второй нуклеиновой кислотой в одной и той же реакционной смеси так, чтобы полученная собранная нуклеиновая кислота содержала первую нуклеиновую кислоту, соединительный олигонуклеотид и вторую нуклеиновую кислоту. При последовательной сборке соединительный олигонуклеотид контактирует с первой нуклеиновой кислотой в реакции сборки, результатом которой является собранная нуклеиновая кислота, содержащая первую нуклеиновую кислоту, собранную с соединительным олигонуклеотидом, но не со второй нуклеиновой кислотой. Такая собранная нуклеиновая кислота впоследствии может контактировать со второй нуклеиновой кислотой в отдельной реакции сборки, результатом которой является собранная нуклеиновая кислота, содержащая первую нуклеиновую кислоту, соединительный олигонуклеотид и вторую нуклеиновую кислоту. В других вариантах осуществления соединительный олигонуклеотид контактирует со второй нуклеиновой кислотой в реакции сборки, результатом которой является собранная нуклеиновая кислота, содержащая вторую нуклеиновую кислоту, собранную с соединительным олигонуклеотидом, но не со первой нуклеиновой кислотой. Такая собранная нуклеиновая кислота впоследствии может контактировать с первой нуклеиновой кислотой в отдельной реакции сборки, результатом которой является собранная нуклеиновая кислота, содержащая первую нуклеиновую кислоту, соединительный олигонуклеотид и вторую нуклеиновую кислоту.
В способах настоящего документа можно использовать любое количество соединительных олигонуклеотидов для сборки молекул нуклеиновых кислот. Например, для соединения 2 молекул нуклеиновых кислот можно использовать 1 соединительный олигонуклеотид, для соединения 3 молекул нуклеиновых кислот можно использовать 2 соединительных олигонуклеотида, для соединения 4 молекул нуклеиновых кислот можно использовать 3 соединительных олигонуклеотида, для соединения 5 молекул нуклеиновых кислот можно использовать 4 соединительных олигонуклеотида или для соединения 6 молекул нуклеиновых кислот можно использовать 5 соединительных олигонуклеотидов. Количество соединительных олигонуклеотидов может составлять 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более, в зависимости от количества молекул нуклеиновых кислот, подлежащих сборке.
В некоторых вариантах осуществления соединительный олигонуклеотид содержит ДНК gBlock. gBlock представляет собой линейный двухцепочечный фрагмент ДНК. Длина gBlock может составлять от около 50 п.н. до около 2000 п.н. Длина gBlock может составлять от около 50 п.н. до около 100 п.н., от около 100 п.н. до около 200 п.н., от около 200 п.н. до около 300 п.н., от около 300 п.н. до около 400 п.н., от около 400 п.н. до около 500 п.н., от около 500 п.н. до около 600 п.н., от около 600 п.н. до около 800 п.н., от около 800 п.н. до около 1000 п.н., от около 1000 п.н. до около 1250 п.н., от около 1250 п.н. до около 1500 п.н., от около 1500 п.н. до около 1750 п.н. или от около 1750 п.н. до около 2000 п.н.
Соединение двух или более нуклеиновых кислот с gBlock можно подвергать скринингу, например, с помощью ПЦР-анализов, описанных в других частях настоящего документа (например, в примере 10). В некоторых случаях gBlock не содержит кассеты селекции. Такой способ позволяет быстро соединить две или более молекулы нуклеиновых кислот, которые можно подвергнуть скринингу простым анализом ПЦР. gBlock может содержать любую интересующую нуклеотидную последовательность. В некоторых случаях gBlock может содержать целевой сайт для нуклеазного агента или целевой сайт для любых различных мегануклеаз или рестрикционных ферментов, предложенных в настоящем документе. В других вариантах осуществления gBlock может содержать кассету селекции. В некоторых вариантах осуществления gBlock содержит интересующую последовательность ДНК. В одном варианте осуществления gBlock содержит человеческую последовательность ДНК.
Подлежащие сборке нуклеиновые кислоты или любые различные соединительные олигонуклеотиды также могут содержать кассету селекции или репортерный ген. Кассета селекции может содержать нуклеотидную последовательность, кодирующую селективный маркер, причем нуклеотидная последовательность функционально связана с промотором. Промотор может быть активным в интересующей прокариотической клетке и/или активным в интересующей эукариотической клетке. Такие промоторы могут представлять собой индуцируемый промотор, промотор, являющийся эндогенным по отношению к репортерному гену или клетке, промотор, являющийся гетерологичным по отношению к репортерному гену или клетке, клеточноспецифический промотор, тканеспецифический промотор или промотор, специфический для стадии развития. В одном варианте осуществления селективный маркер выбирают из неомицинфосфотрансферазы (neor), гигромицин-В-фосфотрансферазы (hygr), пуромицин-N-ацетилтрансферазы (puror), бластицидин-S-дезаминазы (bsrr), ксантин/гуанинфосфорибозилтрансферазы (gpt) и тимидинкиназы вируса простого герпеса (HSV-k) и их комбинации. Селективный маркер нацеливающего вектора может быть фланкирован вышележащим и нижележащим гомологичными плечами или находиться в направлении 5' или 3' от гомологичных плеч.
В одном варианте осуществления подлежащие сборке нуклеиновые кислоты или любой из различных соединительных олигонуклеотидов содержат репортерный ген, функционально связанный с промотором, причем репортерный ген кодирует репортерный белок, который выбирают из группы, состоящей из белков LacZ, mPlum, mCherry, tdTomato, mStrawberry, J-Red, DsRed, mOrange, mKO, mCitrine, Venus, YPet, усиленного желтого флуоресцентного белка (EYFP), Emerald, усиленного зеленого флуоресцентного белка (EGFP), CyPet, голубого флуоресцентного белка (CFP), Cerulean, T-Sapphire, люциферазы, щелочной фосфатазы и их комбинации. Такие репортерные гены могут быть функционально связаны с активным в клетке промотором. Такие промоторы могут представлять собой индуцируемый промотор, промотор, являющийся эндогенным по отношению к репортерному гену или клетке, промотор, являющийся гетерологичным по отношению к репортерному гену или клетке, клеточноспецифический промотор, тканеспецифический промотор или промотор, специфический для стадии развития.
После ренатурации одноцепочечной ДНК (например, нависаний, полученных в результате действия экзонуклеазы, если соединяемые молекулы ДНК представляют собой дцДНК, или нависаний, полученных путем создания одноцепочечных разрывов в разных целевых сайтах на каждой цепи) одноцепочечные гэпы, оставленные экзонуклеазой заполняют подходящей, не замещающей цепь ДНК-полимеразой, а созданные одноцепочечные разрывы закрывают лигазой. Под «не замещающей цепь ДНК-полимеразой» в настоящем документе понимается ДНК-полимераза, которая завершает синтез ДНК, когда в процессе копирования молекулы дцДНК встречает на своем пути цепи ДНК, или которая разрушает встреченные цепи ДНК по мере работы, одновременно заполняя созданный таким образом гэп, в результате чего создается «двигающийся одноцепочечный разрыв» (ник-трансляция).
В некоторых вариантах осуществления перекрывающиеся концевые последовательности имеют достаточную комплементарность перекрывающихся областей для ренатурации с одноцепочечными комплементарными концами каждого полинуклеотида. После ренатурации одной цепи первого нуклеотида с комплементарной цепью второго полинуклеотида, 3'-конец первого нуклеотида можно удлинить по шаблону цепи второго полинуклеотида, а 3'-конец второго полинуклеотида можно удлинить по шаблону цепи первого полинуклеотида. Путем удлинения комплементарного 3'-конца каждого полинуклеотида можно выполнить сборку этих полинуклеотидов. После соединения одноцепочечные разрывы между удлиненным 3'-концом цепи одного фрагмента и соседним 5'-концом цепи другого фрагмента можно закрыть лигированием. Более конкретно, лигируют гидроксильную группу удлиненного 3'-конца первого полинуклеотида с фосфатной группой 5'-конца второго полинуклеотида, а гидроксильную группу удлиненного 3'-конца второго полинуклеотида лигируют с фосфатной группой 5'-конца первого полинуклеотида.
Реакция лигирования может быть выполнена при помощи любой из разнообразных подходящих термостабильных ДНК-лигаз. К подходящим лигазам относятся, например, лигаза Taq, термостабильная ДНК-лигаза Ampligase (Epicentre Biotechnologies), термостабильные лигазы, описанные в патенте США №6,576,453, термостабильная ДНК-лигаза Tfi от Bioneer, Inc.
Введение в реакционную смесь подходящего количества агента для краудинга, например ПЭГ, обеспечивает, усиливает или способствует молекулярному краудингу. Безотносительно к какому-либо конкретному механизму, предполагается, что агент для краудинга, обеспечивающий молекулярный краудинг и связывающий и задерживающий воду в растворе, позволяет более близкий контакт компонентов раствора друг с другом. Например, молекулы ДНК, подлежащие рекомбинации, могут подходить близко друг к другу. Это способствует ренатурации одноцепочечных нависаний. Также предполагается, что ферменты могут ближе подходить к ДНК-субстратам и могут стабилизироваться вследствие удаления молекул воды. Различные подходящие агенты для краудинга будут очевидны для квалифицированного специалиста. Такие агенты включают разнообразные хорошо известные макромолекулы, такие как полимеры, например, полиэтиленгликоль (ПЭГ); фиколл, например, фиколл 70; декстран, например, декстран 70 и т.п. Большая часть обсуждений в настоящей заявке относится к ПЭГ. Однако также предполагается, что обсуждение применимо и к другим подходящим агентам для краудинга. Квалифицированному специалисту будет понятно, как вносить в способ типовые изменения для приспособления к использованию других агентов для краудинга.
Введение в реакционную смесь подходящего количества агента для краудинга, например ПЭГ, обеспечивает, усиливает или способствует молекулярному краудингу. Например, агенты для краудинга могут способствовать сближению молекул ДНК, подлежащих рекомбинации, тем самым облегчая ренатурацию одноцепочечных нависаний. Также предполагается, что ферменты могут ближе подходить к ДНК-субстратам и могут стабилизироваться вследствие удаления молекул воды. Различные подходящие агенты для краудинга будут очевидны для квалифицированного специалиста. Такие агенты включают разнообразные хорошо известные макромолекулы, такие как полимеры, например, полиэтиленгликоль (ПЭГ); фиколл, например, фиколл 70; декстран, например, декстран 70 и т.п. В целом, при использовании ПЭГ оптимальной является концентрация 5% (масса/объем). Однако количество ПЭГ может меняться, например, в диапазоне от около 3 до около 7%. Можно использовать ПЭГ любого подходящего размера, например, в диапазоне от около ПЭГ-200 (например, ПЭГ-4000, ПЭГ-6000 или ПЭГ-8000) до около ПЭГ-20000 или даже больше. В представленных в настоящем документе примерах использовали ПЭГ-8000. Агент для краудинга, помимо усиления реакции ренатурации, может усиливать лигирование.
Компоненты реакции (например, соли, буферы, подходящие источники энергии (такие как АТФ или НАД), рН реакционной смеси и т.п.), присутствующие в смеси для реакции сборки, могут быть не оптимальными для отдельных ферментов (экзонуклеазы, полимеразы и лигазы), скорее они могут быть компромиссными, эффективными для всего набора реакций. Например, одна подходящая буферная система, выявленная авторами изобретения, иногда именуемая в настоящем документе ISO (изотермическим) буфером, обычно содержит 0,1 М трис-Cl рН 7,5; 10 мМ MgCl2, по 0,2 мМ каждого из дГТФ, дАТФ, дТТФ и дЦТФ, 10 мМ ДТТ (дитиотреитола), 5% ПЭГ-8000 и 1 мМ НАД.
В способах, описанных в настоящем документе, по меньшей мере две нуклеиновые кислоты контактируют с белком Cas и другими ферментами в условиях, способствующих сборке нуклеиновых кислот, с образованием собранной двухцепочечной молекулы ДНК, в которой сохраняется одна копия перекрывающейся области. Описанные способы можно использовать для соединения любых интересующих молекул ДНК, включая ДНК природного происхождения, клонированные молекулы ДНК, синтетические созданные молекулы ДНК и т.п. Соединенные молекулы ДНК можно при необходимости клонировать в вектор (например, используя способ изобретения). В некоторых вариантах осуществления подлежащие сборке нуклеиновые кислоты имеют оптимизацию кодонов для внедрения и экспрессии в интересующих клетках (например, клетках грызунов, клетках мышей, клетках крыс, клетках человека, клетках млекопитающих, микробных клетках, дрожжевых клетках и т.п.).
Способами, описанными в настоящем документе, можно соединять молекулы ДНК любой длины. Например, можно соединять нуклеиновые кислоты, содержащие от около 100 п.н. до около 750 или 1000, или более. Количество нуклеиновых кислот, которые возможно собрать за одну или несколько стадий сборки в соответствии со способами, описанными в настоящем документе, может составлять по меньшей мере около 2, 3, 4, 6, 8, 10, 15, 20, 25, 50, 100, 200, 500, 1000, 5000 или 10000 молекул ДНК, например, в диапазоне от около 2 до около 30 нуклеиновых кислот. Количество стадий сборки может составлять около 2, 4, 6, 8, 10 или более. Количество молекул, собираемых за одну стадию, может находиться в диапазоне от около 2 до около 10 молекул. Способы изобретения можно использовать для соединения вместе молекул ДНК или кассет, каждая из которых имеет исходный размер, по меньшей мере равный или не превышающий около 40 п.н., 60 п.н., 80 п.н., 100 п.н., 500 п.н., 1 т.п.н., 3 т.п.н., 5 т.п.н., 6 т.п.н., 10 т.п.н., 18 т.п.н., 20 т.п.н., 25 т.п.н., 32 т.п.н., 50 т.п.н., 65 т.п.н., 75 т.п.н., 150 т.п.н., 300 т.п.н., 500 т.п.н., 600 т.п.н., 1 млн п.н. или больше. Собранные конечные продукты могут содержать по меньшей мере около 500 п.н., 1 т.п.н., 3 т.п.н., 5 т.п.н., 6 т.п.н., 10 т.п.н., 18 т.п.н., 20 т.п.н., 25 т.п.н., 32 т.п.н., 50 т.п.н., 65 т.п.н., 75 т.п.н., 150 т.п.н., 300 т.п.н., 500 т.п.н., 600 т.п.н., 1 млн п.н. или более, например, в диапазоне от 30 т.п.н. до 1 млн п.н.
В некоторых вариантах осуществления собранные нуклеиновые кислоты образуют кольцо и/или лигируются в вектор с образованием кольца. Нижний предел размера дцДНК для образования кольца составляет около 200 пар нуклеотидов. Следовательно, общая длина соединенных фрагментов (включая, в некоторых случаях, длину вектора) составляет по меньшей мере около 200 п.н. Практический верхний предел размера отсутствует, и способами, описанными в настоящем документе, можно создавать соединенные ДНК размером в несколько сотен тысяч пар нуклеотидов или более. Соединенные нуклеиновые кислоты могут иметь форму кольца или линейной молекулы.
Способы, описанные в настоящем документе, можно использовать для соединения линейного фрагмента с другим линейным фрагментом, линейного фрагмента с кольцевой молекулой нуклеиновой кислоты, кольцевой молекулы нуклеиновой кислоты с другой кольцевой молекулой нуклеиновой кислоты или любой комбинации линейных и кольцевых нуклеиновых кислот. «Вектор» включает в себя любую кольцевую молекулу нуклеиновой кислоты. В некоторых вариантах осуществления вектор, собранный способами, описанными в настоящем документе, представляет собой бактериальную искусственную хромосому (ВАС). Вектор (например, ВАС) может включать в себя человеческую ДНК, ДНК грызунов, синтетическую ДНК или любую их комбинацию. Например, ВАС может содержать человеческую полинуклеотидную последовательность. При соединении смеси молекул ДНК предпочтительно, чтобы ДНК присутствовали приблизительно в эквимолярных количествах.
Нуклеиновая кислота, использованная для сборки посредством способов, описанных в настоящем документе, может представлять собой большой нацеливающий вектор. Термин «большой нацеливающий вектор» или LTVEC включает векторы, содержащие гомологичные плечи, которые соответствуют или получены из нуклеотидных последовательностей, используемых для гомологичного нацеливания в клетках, и/или содержащие вставочные нуклеиновые кислоты, имеющие нуклеотидные последовательности, предназначенные для выполнения гомологичного рекомбинационного нацеливания в клетках. Например, LTVEC позволяет осуществлять модификацию больших локусов, которую нельзя осуществить традиционными плазмидными нацеливающими векторами из-за размерных ограничений. В конкретных вариантах осуществления гомологичные плечи и/или нуклеотидная вставка LTVEC содержат геномную последовательность эукариотической клетки. Размер LTVEC является слишком большим, чтобы имелась возможность скрининга событий таргетинга с помощью обычных анализов, например, саузерн-блоттинга и ПЦР для амплификации протяженных участков (например, 1 т.п.н. - 5 т.п.н.). Примеры LTVEC включают, без ограничений, векторы, полученные из бактериальной искусственной хромосомы (ВАС), человеческой искусственной хромосомы или дрожжевой искусственной хромосомы (YAC). Не имеющие ограничительного характера примеры LTVEC и способов их получения описаны, например, в патентах США №№6,586,251, 6,596,541, 7,105,348 и WO 2002/036789 (PCT/US01/45375) и US 2013/0137101, каждый из которых полностью включен в настоящий документ путем ссылки.
В некоторых вариантах осуществления в векторы могут быть вставлены кассеты, которые позже можно удалять. Можно конструировать различные формы кассет, предусматривающие делецию в определенных типах клеток и тканей, на определенных стадиях развития или при индукции. В таких кассетах может использоваться система рекомбиназы, в которой кассета с обеих сторон фланкирована сайтами распознавания рекомбиназы и может быть удалена рекомбиназой, экспрессированной в заданном типе клеток, экспрессированной на заданной стадии развития или экспрессированной или активированной в результате индукции. Такие кассеты дополнительно могут быть сконструированы с включением в них набора из пар сайтов распознавания разными рекомбиназами, которые расположены таким образом, чтобы можно было генерировать нулевые, условные или комбинированные условные/нулевые аллели, как описано в публикации US 2011/0104799, которая полностью включена в настоящий документ путем ссылки. Регуляцию генов рекомбиназ можно контролировать разными способами, например, путем функционального связывания гена рекомбиназы с клеточноспецифичным, тканеспецифичным или регулируемым в зависимости от стадии развития промотором (или другим регуляторным элементом), или путем функционального связывания гена рекомбиназы с нетранслируемой областью 3'-UTR, содержащей сайт распознавания для микроРНК, транскрибирующейся только в конкретных типах клеток, типах тканей или на определенных стадиях развития. Рекомбиназу также можно регулировать, например, путем использования слитного белка, ставящего рекомбиназу под контроль эффектора или метаболита (например, CreERT2, активность которой положительно контролируется тамоксифеном), или путем постановки рекомбиназы под контроль индуцируемого промотора (например, промотора, активность которого контролируется доксициклином и TetR или вариантами TetR). Примеры различных форм кассет и способов регуляции генов рекомбиназ представлены, например, в публикациях US 8,518,392; US 8,354,389 и US 8,697,851, каждая из которых полностью включается в настоящий документ путем ссылки.
Векторы, используемые для сборки в соответствии с описанием, приведенным в настоящем документе, (например, LTVEC) могут иметь любую длину, включая, без ограничений, длину от около 20 т.п.н. до около 400 т.п.н., от около 20 т.п.н. до около 30 т.п.н., от около 30 т.п.н. до 40 т.п.н., от около 40 т.п.н. до около 50 т.п.н., от около 50 т.п.н. до около 75 т.п.н., от около 75 т.п.н. до около 100 т.п.н., от около 100 т.п.н. до 125 т.п.н., от около 125 т.п.н. до около 150 т.п.н., от около 150 т.п.н. до около 175 т.п.н., около 175 т.п.н. до около 200 т.п.н., от около 200 т.п.н. до около 225 т.п.н., от около 225 т.п.н. до около 250 т.п.н., от около 250 т.п.н. до около 275 т.п.н. или от около 275 т.п.н. до около 300 т.п.н., от около 200 т.п.н. до около 300 т.п.н., от около 300 т.п.н. до около 350 т.п.н., от около 350 т.п.н. до около 400 т.п.н., от около 350 т.п.н. до около 550 т.п.н. В одном из вариантов осуществления LTVEC имеет длину около 100 т.п.н.
Способы, предложенные в настоящем документе для сборки нуклеиновых кислот, могут быть разработаны так, чтобы обеспечивать делецию от около 5 т.п.н. до около 10 т.п.н., от около 10 т.п.н. до около 20 т.п.н., от около 20 т.п.н. до около 40 т.п.н., от около 40 т.п.н. до около 60 т.п.н., от около 60 т.п.н. до около 80 т.п.н., от около 80 т.п.н. до около 100 т.п.н., от около 100 т.п.н. до около 150 т.п.н. или от около 150 т.п.н. до около 200 т.п.н., от около 200 т.п.н. до около 300 т.п.н., от около 300 т.п.н. до около 400 т.п.н., от около 400 т.п.н. до около 500 т.п.н., от около 500 т.п.н. до около 1 млн п.н., от около 1 млн п.н. до около 1,5 млн п.н., от около 1,5 млн п.н. до около 2 млн п.н., от около 2 млн п.н. до около 2,5 млн п.н. или от около 2,5 млн п.н. до около 3 млн п.н.
В других случаях предложенные в настоящем документе способы разработаны так, чтобы позволить инсерцию экзогенной нуклеотидной последовательности в диапазоне от около 5 т.п.н. до около 10 т.п.н., от около 10 т.п.н. до около 20 т.п.н., от около 20 т.п.н. до около 40 т.п.н., от около 40 т.п.н. до около 60 т.п.н., от около 60 т.п.н. до около 80 т.п.н., от около 80 т.п.н. до около 100 т.п.н., от около 100 т.п.н. до около 150 т.п.н., от около 150 т.п.н. до около 200 т.п.н., от около 200 т.п.н. до около 250 т.п.н., от около 250 т.п.н. до около 300 т.п.н., от около 300 т.п.н. до около 350 т.п.н. или от около 350 т.п.н. до около 400 т.п.н. В одном варианте осуществления длина полинуклеотидной вставки составляет около 130 т.п.н. или около 155 т.п.н.
Сборку линейных нуклеиновых кислот друг с другом или с векторами можно выполнять способами, описанными в настоящем документе. Линейная молекула может представлять собой вектор, расщепленный эндонуклеазой (например, белком Cas), или любую синтетическую, искусственную или природную линейную нуклеиновую кислоту. В некоторых вариантах осуществления линейную нуклеиновую кислоту создают таким образом, чтобы концевые последовательности перекрывались с областью другой нуклеиновой кислоты. Перекрывающиеся концевые последовательности линейной нуклеиновой кислоты можно вводить любым известным специалистам в данной области способом для создания заданных нуклеотидных последовательностей. Например, концевые последовательности могут представлять собой часть синтетической молекулы, могут быть введены посредством ПЦР или могут быть введены традиционными методами клонирования.
ПРИМЕРЫ
Представленные ниже примеры приводятся с целью предоставления специалисту в данной области полного изложения и описания того, как выполнять и применять настоящее изобретение, и не должны ограничивать область того, что авторы изобретения считают своим изобретением, и не предназначены для представления того, что приведенные ниже эксперименты являются всеми или только лишь выполненными экспериментами. Были приложены усилия для обеспечения точности использованных чисел (например, количеств, температуры и т.п.), но следует учитывать некоторые погрешности и отклонения в экспериментах. При отсутствии особых указаний части представляют собой массовые части, молекулярная масса представляет собой средневесовую молекулярную массу, температура указана в градусах по шкале Цельсия, а давление является атмосферным или близким к атмосферному.
Пример 1. Расщепление ВАС при помощи CAS9 с последующей сборкой с кассетой селекции
Были разработаны искусственная крРНК и искусственная тракрРНК для нацеливания на специфические последовательности в MAID 6177 (LTVEC длиной 116 т.п.н.) для сборки с ПЦР-продуктом (UB-HYG) длиной 3 т.п.н. Этот продукт ПЦР содержал 50 п.н. перекрытия с вектором. Сначала растворяют крРНК и тракрРНК в 100 мкМ дуплексном буфере (30 мМ ГЕПЕС, рН 7,5, 100 мМ ацетата калия). Для ренатурации РНК в 80 мкл буфера для ренатурации добавляют 10 мкл 100 мкМ крРНК и 10 мкл 100 мкМ тракрРНК. Нагревают РНК в температурном блоке при 90°С, затем снимают блок с нагревателя и охлаждают на столе. Конечная концентрация РНК составляет около 10 мкМ.
Для расщепления ВАС используют чистую ДНК ВАС из набора Maxiprep и расщепляют ВАС с использованием приведенной ниже смеси.
Выполняют расщепление в течение 1 часа при 37°, после чего в течение 30 мин проводят обессоливание. Конечный реакционный буфер содержит: 20 мМ трис 7,5; 100-150 мМ NaCl; 10 мМ MgCl2; 1 мМ ДТТ; 0,1 мМ этилендиаминтетрауксусной кислоты (ЭДТА); 100 мкг/мл бычьего сывороточного альбумина (БСА); конечный объем 15 мкл.
Для сборки ВАС и вставки расщепляют плазмиду или проводят ПЦР для создания вставки. Для реакций ПЦР прогоняют небольшую аликвоту на геле и находят одиночный продукт, если продукт имеет одну полосу, то вместо гель-экстракции выполняют ПЦР-очистку. Желательное молярное соотношение ВАС : вставка составляет 1:1 - 1:6. Обычно для работы достаточно 50 нг очищенной вставки. Можно использовать следующую реакционную смесь:
Помещают ДНК и смесь на лед или непосредственно в ПЦР-аппарат при 50°С. Инкубируют при 50°С в течение 1 часа. Добавляют 0,5 мкл протеиназы К (20 мг/мл) и инкубируют при 50°С в течение 1 часа. Выполняют обессоливание в течение 30 мин и методом электропорации в клетки DH10B вводят 8 мкл реакционной смеси. Для проверки эффективности расщепления можно провести гель-электрофорез в пульсирующем поле 10 мкл расщепленной ВАС. Используют не содержащие РНКазу воду и буферы.
Реакцию сборки проводят так, как описано ниже. Изотермический буфер: 3 мл 1М трис-HCL (рН 7,5); 150 мкл 2 М MgCl2; по 60 мкл 100 мМ каждого из: дГТФ, дАТФ, дТТФ, дЦТФ; 300 мкл 1 М ДТТ; 1,5 г ПЭГ 8000; 300 мкл 100 мМ НАД. Изотермический буфер хранят аликвотами по 320 мкл при -20°С. Мастер-микс готовят так, как описано ниже. Смешивают 320 мкл изотермического буфера; 0,64 мкл экзонуклеазы Т5 (конц. маточн. раствора = 10 Ед/мкл); 20 мкл ДНК-полимеразы Phusion (конц. маточн. раствора = 2 Ед/мкл); 160 мкл ДНК-лигазы Taq (конц. маточн. раствора = 40 Ед/мкл); 699,36 мкл H2О; делят на аликвоты по 15 мкл или 30 мкл и хранят при -20°С. Используют 15 мкл мастер-микса (ММ) в общем объеме реакционной смеси 20 мкл.
Последовательность тракрРНК, использованная настоящем примере: CAAAACAGCAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUC (SEQ ID NO: 9). Эта РНК CRISPR (крРНК) содержит: (1) около 20 нуклеотидов РНК, комплементарных целевой последовательности, и (2) хвостовую последовательность (GUUUUAGAGCUAUGCUGUUUUG (SEQ ID NO: 10)), которую ренатурируют до тракрРНК.
Эти этапы представлены на ФИГ. 1.
Пример 2. Сшивание двух перекрывающихся ВАС: гуманизированной HLA-DQ + гуманизированной HLA-DR в мышином локусе МНС II (Н2-А/Н2-Е)
Были разработаны искусственная крРНК и искусственная тракрРНК для нацеливания на специфические последовательности в гуманизированной ВАС HLA-DQ для сборки с гуманизированной ВАС HLA-DR. Векторы содержали ~70 п.н. перекрытия друг с другом, созданных путем расщепления посредством Cas9 в двух сайтах на каждом векторе (см. ФИГ. 2). Растворяют крРНК и тракрРНК в 100 мкМ буфера Hybe. Для ренатурации РНК в 80 мкл буфера для ренатурации добавляют 10 мкл 100 мкМ крРНК и 10 мкл 100 мкМ тракрРНК. Помещают РНК в нагревательный блок при 90°С, затем снимают блок с нагревателя и охлаждают на столе. Конечная концентрация РНК составляет около 10 мкМ.
Для расщепления ВАС можно использовать чистую ДНК ВАС Maxiprep. Каждую ВАС можно расщепить индивидуально с использованием следующей смеси:
Векторы ВАС следует расщеплять при 37°С в течение 1 часа, а затем инактивировать нагревом в течение 20 мин при 65°С. Обессоливание выполняют в течение 30 мин. Расщепленную ДНК очищают экстракцией фенолом/хлороформом/изоамиловым спиртом (PCI) и ресуспендируют в 35 мкл буфера ТЕ.
Для сборки векторов в реакции сборки используют 2,5 мкл ВАС, как описано ниже.
Помещают ДНК и смесь на лед или непосредственно в ПЦР-аппарат при 50°С. Инкубируют при 50°С в течение 1 часа. Выполняют обессоливание в течение 30 мин и методом электропорации в клетки DH10B вводят 8 мкл собранной ДНК. Используют не содержащие РНКазу воду и буферы.
Последовательность тракрРНК, использованная настоящем примере: CAAAACAGCAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUC (SEQ ID NO: 9). Эта РНК CRISPR (крРНК) содержит: (1) около 20 нуклеотидов РНК, комплементарных целевой последовательности, и (2) хвостовую последовательность (GUUUUAGAGCUAUGCUGUUUUG (SEQ ID NO: 10)), которую ренатурируют до тракрРНК.
Эти этапы представлены на ФИГ. 2.
Пример 3. Сборка 2 расщепленных при помощи Cas9 фрагментов из 2 разных плазмид с использованием линкеров
Для конструирования нацеливающего вектора расщепляли плазмиду pMJ8502x, используя 2 идентичные крРНК для вырезания фрагмента длиной 400 п.н. и каркаса Amp длиной 2283 п.н. (ФИГ. 7). Для очистки всей реакционной смеси использовали колонки Qiagen. Затем расщепляли плазмиду R6KZenUbiNeo, используя 2 разных крРНК, для разделения сайта устойчивости к Neo (1086 п.н.) и каркаса (5390 п.н.). Для очистки всей реакционной смеси использовали колонки Qiagen. (ФИГ. 7). Реакционная смесь для расщепления: 1170 нг ДНК, 30 мкл буфера, 4 мкл ренатурированной РНК (100 мкМ), 1,7 мкл Cas9 (0,89 нг/мкл), H2O до 60 мкл. Смесь инкубировали при 37°С в течение 1 часа и очищали на колонке Qiagen, после чего элюировали с использованием 30 мкл буфера для элюирования.
Затем выполняли сборку расщепленных фрагментов с двумя линкерами с получением бесшовной сборки с использованием следующей реакционной смеси: 0,5 мкл линкера 1 (5 нг), 0,5 мкл линкера 2 (5 нг), 2 мкл продукта расщепления Neo (~60 нг), 2 мкл продукта расщепления Amp (~60 нг), 15 мкл мастер-микса для сборки. Смесь инкубировали при 50°С в течение 1 часа и проводили диализ реакционной смеси против Н2О. В электрокомпетентные клетки Pir методом электропорации вводили 10 мкл реакционной смеси, а затем наносили на планшеты Carb/Kan. ПЦР по стыкам показала, что 6 из 8 отобранных колоний были правильными, и это подтверждалось секвенированием.
Пример 4. Замещение участка ВАС кассетой с использованием линкеров Для конструирования нацеливающего вектора для нокаутных мышей 40 т.п.н. нацеливающего вектора ВАС заменяли на кассету селекции, фланкированную сайтами распознавания рекомбинации. (ФИГ. 8). Были разработаны 2 линкера для удаления интересующего участка из mВАС и вставки кассеты селекции, один для 5'-, а другой для 3'-конца. Линкеры имели перекрытие с mВАС длиной 40 п.н. и перекрытие с кассетой селекции длиной 40 п.н. Сначала от нацеливающего вектора (mВАС) длиной 206 т.п.н. отщепляли 39,5 т.п.н. с использованием описанной ниже реакции. В 500 мкл реакционной смеси (доводится Н2О): добавляли 1 мкл Cas9 (0,89 мкг/мкл), по 2 мкл каждой дуплексной РНК (50 мкМ), 250 мкл буфера, 220 мкл (12,5 нг) ВАС Maxiprep, и инкубировали при 37°С в течение 1 часа. Расщепленную ДНК очищали экстракцией фенолом/хлороформом/изоамиловым спиртом (PCI) и ресуспендировали в 55 мкл буфера ТЕ. После очистки продукта расщепления mВАС с помощью PCI, проводили сборку при 50°С в течение 1 ч и в клетки DH10B методом электропорации вводили 10 мкл реакционной смеси. (ФИГ. 9). Секвенирование стыков подтвердило правильность сборки. (ФИГ. 10). Линкер 1 (соединительный олигонуклеотид 1) является бесшовным переходом от последовательности mВАС к последовательности кассеты (SEQ ID NO: 12). Линкер 2 (соединительный олигонуклеотид 2) является бесшовным переходом от последовательности кассеты к последовательности mВАС (SEQ IDNO: 13).
Пример 5. Сборка двух векторов ВАС с использованием линкеров (соединительных олигонуклеотидов)
Сшивание 2 mВАС с помощью Cas9/изотермической сборки использовали для получения нацеливающего вектора, содержащего плечи, гомологичные мышиной геномной области, и рестрикционные сайты для вставки человеческого гена посредством лигирования ВАС. Этот нацеливающий вектор использовали в лигировании ВАС для получения гуманизированного нацеливающего вектора. mВАС расщепляли с использованием следующей реакционной смеси: 12,5 мкг ДНК, по 2 мкл каждой ренатурированной РНК (50 мкМ), 10 мкл Cas9 (0,89 мкг/мкл), 250 мкл буфера, Н2О до 500 мкл. Смесь инкубировали при 37°С в течение одного часа, очищали экстракцией фенолом/хлороформом/изоамиловым спиртом (PCI) и ресуспендировали в 20 мкл ТЕ. Затем выполняли сборку двух мышиных ВАС друг с другом при помощи линкеров (ФИГ. 11) с использованием следующей реакционной смеси: 6 мкл (2 мкг) продукта расщепления bMQ-208A16, 5,6 мкл (2 мкг) продукта расщепления bMQ-50F19, по 0,25 мкл каждого линкера (50 мкМ), 4,3 мкл (100 нг) кассеты селекции (Ubi-Hyg), 12 мкл высококонцентрированного мастер-микса для сборки, 11,35 мкл Н2О. Реакционную смесь инкубировали при 50°С в течение 1 часа и проводили диализ против Н2О при 30°С. Для трансформирования клеток DH10B использовали 10 мкл или 30 мкл диализованной реакционной смеси. Секвенирование по Сэнгеру подтвердило все стыки. Секвенирование методом Illumina повторно подтвердило все стыки (ФИГ. 12 и SEQ ID NO: 17). Линкер 1 является бесшовным переходом от mВАС к кассете (SEQ ID NO: 14). Линкер 2 не является бесшовным переходом от кассеты к mВАС. Он содержит человеческую спейсерную последовательность в соответствии с проектным планом. Линкер 3 не является бесшовным переходом от mВ2 к mВ3. Он содержит уникальную последовательность, используемую для ПЦР-проверки. Эту область удаляли при линеаризации для электропорационного введения в ЭС-клетки (SEQ ID NO: 15).
На ФИГ. 13 проиллюстрировано использование 4 соединительных олигонуклеотидов (линкеров) для вставки больших фрагментов человеческих генов в mВАС с использованием четырех линкеров и изотермической сборки.
Пример 6. Реагенты и реакционные смеси для расщепления и сборки
РНК CRISPR (крРНК) (заказана в виде оцРНК) содержит: (1) 20 нуклеотидов РНК, комплементарных целевой области расщепления; (2) и хвостовой участок, который будет ренатурирован с тракрРНК: <20 нт крРНК>GUUUUAGAGCUAUGCUGUUUUG (SEQ ID NO: 10).
тракрРНК (заказана в виде оцРНК): GUUGGAACCAUUCAAAACAGCAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAU CAACUUGAAAAAGUGGCACCGAGUCGGUGCUUUUUUU (SEQ ID NO: 11).
Все РНК ресуспендированы в концентрации 100 мкМ в Н2О. По 2,5 мкл каждой из крРНК и тракрРНК объединяли с 5 мкл буфера для ренатурации (конечные концентрации: 10 мМ трис рН 7,5-8,0, 50 мМ NaCl, 1 мМ ЭДТА). После этого смесь инкубировали при 95°С в течение 5 минут и медленно охлаждали до комнатной температуры в течение 1 часа. Буфер для Cas9-расщепления 2 X содержит: 40 мМ ГЕПЕС рН 7,5 (конечн. = 20 мМ); 300 мМ KCl (конечн. = 150 мМ); 1 мМ ДТТ (конечн. = 0,5 мМ); 0,2 мМ ЭДТА (конечн. = 0,1 мМ); 20 мМ MgCl2 (конечн. = 10 мМ).
Крупномасштабная реакция расщепления: при помощи Cas9 добавляют в указанном порядке при комнатной температуре: H2O до 500 мкл, 250 мкл 2 х буфера для расщепления, 12,5 мкг ДНК, по 2 мкл каждой РНК (концентрация 50 мкМ), 10 мкл Cas9 (концентрация 0,89 мг/мл) и инкубируют при 37°С в течение 1 часа.
При необходимости реакцию можно масштабировать, например: H2O до 50 мкл, 25 мкл буфера, 125 нг ДНК, по 2 мкл каждой РНК (концентрация 5 мкМ), 1 мкл Cas9 (концентрация 0,89 мг/мл), и инкубацию проводят при 37°С в течение 1 часа.
Реакцию сборки проводят так, как описано ниже. Изотермический буфер: 3 мл 1М трис-HCL (рН 7,5); 150 мкл 2 М MgCl2; по 60 мкл 100 мМ каждого из дГТФ, дАТФ, дТТФ, дЦТФ; 300 мкл 1 М ДТТ; 1,5 г ПЭГ 8000; 300 мкл 100 мМ НАД. Изотермический буфер хранят аликвотами по 320 мкл при -20°С. Мастер-микс готовят так, как описано ниже. Смешивают 320 мкл изотермического буфера; 0,64 мкл экзонуклеазы Т5 (конц. маточн. раствора = 10 Ед/мкл); 20 мкл ДНК-полимеразы Phusion (конц. маточн. раствора = 2 Ед/мкл); 160 мкл ДНК-лигазы Taq (конц. маточн. раствора = 40 Ед/мкл); 699,36 мкл Н2О; делят на аликвоты по 15 мкл или 30 мкл и хранят при -20°С. Используют 15 мкл мастер-микса (ММ) в общем объеме реакционной смеси 20 мкл.
В альтернативном варианте осуществления можно изготовить высококонцентрированный мастер-микс (GA ММ НС) так, как описано ниже. Смешивают 320 мкл изотермического буфера; 0,64 мкл экзонуклеазы Т5 (конц. маточн. раствора = 10 Ед/мкл); 20 мкл ДНК-полимеразы Phusion (конц. маточн. раствора = 2 Ед/мкл); 160 мкл ДНК-лигазы Taq (конц. маточн. раствора = 40 Ед/мкл); делят на аликвоты по 6 мкл или 12 мкл и хранят при -20°С. Используют 6 мкл мастер-микса в общем объеме реакционной смеси 20 мкл.
При всех реакциях сборки следует определять концентрацию ДНК (например, при помощи аппарата Nano Drop) и применять молярное соотношение 1:6 (вектора к вставке(-ам)). Для стандартной концентрации используют 15 мкл мастер-микса для сборки. ДНК и воду добавляют в ПЦР-пробирку на 200 мкл до конечного объема 20 мкл. Реакцию проводят в термоциклере при 50°С в течение 1 часа. Далее реакционную смесь можно хранить при -20°С. Для высокой концентрации используют 6 мкл высококонцентрированного мастер-микса для сборки. ДНК и воду добавляют в ПЦР-пробирку на 200 мкл до конечного объема 20 мкл. Реакцию проводят в термоциклере при 50°С в течение 1 часа. Впоследствии реакционную смесь можно хранить при -20°С. После завершения реакции проводят диализ 10 мкл против воды в течение 30 мин и вводят методом электропорации в подходящие электрокомпетентные клетки (например, клетки DH10B или Pir+).
Реакция Cas9/изотермическая сборка: для расщепления при помощи Cas9 по 2,5 мкг каждой ДНК (например, ДНК ВАС), по 4 мкл 10 мкМ каждой гидовой/тракрРНК и 5 мкл белка Cas9 (0,89 мг/мл) расщепляют в течение 2 часов при 37°С. Далее реакцию инактивируют нагреванием при 65°С в течение 20 мин, экстрагируют фенолом и хлороформом (например, для удаления белка Cas9), однократно промывают 70% этанолом и ресуспендируют ДНК в 35 мкл воды. Изотермическую сборку выполняют, используя 5 мкл ДНК, смешанной с 15 мкл мастер-микса (ММ), как описано в других частях настоящего документа, и инкубируют при 50°С в течение 1 часа. Реакционную смесь обессоливают в течение 30 мин и 8 мкл реакционной смеси вводят в клетки методом электропорации.
Пример 7. Cas9/изотермическая сборка для вставки человеческой последовательности в вектор ВАС
Для конструирования гуманизированного нацеливающего вектора проводили расщепление MAID 6236 комплексом гРНК-Cas с получением расщепленного фрагмента с перекрывающимися последовательностями. VI568 также расщепляли комплексом гРНК-Cas, получая последовательности, перекрывающиеся с фрагментом MAID6236. Выполняли Cas9/изотермическую сборку, как описано выше, с получением вставки гуманизированного локуса в вектор (VI599). Процесс представлен на ФИГ. 14.
Пример 8. Cas9/изотермическая сборка с использованием gBlock без селекции
Расщепление при помощи Cas9 и сборку также можно выполнять без селекции, например, используя фрагменты ДНК gBlock. Чтобы протестировать возможность добавления двухцепочечной ДНК в локус без кассеты селекции, синтезировали фрагменты ДНК gBlock и вставляли их в конструкт. Как представлено на ФИГ. 15А и В, комплекс Cas9/гРНК был рассчитан на нацеливание на два сайта внутри локуса TCR-бета для удаления фрагмента размером 4,4 т.п.н. ДНК gBlock была разработана для введения в конструкт сайта распознавания мегануклеазой. gBlock позволял выполнить вставку в конструкт без использования селективного маркера. На ФИГ. 15А продемонстрирована вставка PIScel gBlock, а на ФИГ. 15В продемонстрирована вставка MauBI gBlock.
Конечные конструкты были проверены на успешность вставки каждого из gBlock посредством ПЦР-скрининга стыков с использованием праймеров, указанных в таблице 1. Ниже описан протокол скрининга по стыкам. Реакционная смесь для ПЦР содержала: 1 мкл ДНК, 0,5 мкл праймера 1, 0,5 мкл праймера 2, 1 мкл ДМСО, 4 мкл дНТФ, 2,5 мкл 10 х буфера, 0,5 мкл Ex-Taq и 15 мкл воды. Реакцию проводили в термоциклере при 95°С в течение 3 минут, 95°С в течение 30 с, 55°С в течение 30 с 25 циклов, затем 72°С в течение 30 с и 72°С 5 мин. Стыковые последовательности были подтверждены секвенированием.


Пример 9. Cas9/изотермическая сборка для вставки человеческой последовательности в вектор ВАС с использованием соединительных олигонуклеотидов
На ФИГ. 16 представлен пример прямой гуманизации с использованием Cas9/изотермической сборки и соединительных олигонуклеотидов. Человеческий фрагмент и мышиную делецию вырезают посредством Cas9 (в каждой ВАС используются 2 РНК CRISPR). Человеческий фрагмент и мышиный каркас соединяют друг с другом в реакции сборки Гибсона с 3 линкерами (соединительными олигонуклеотидами) и кассетой селекции.
На ФИГ. 17 представлен пример непрямой гуманизации с использованием Cas9/изотермической сборки и соединительные олигонуклеотиды для сборки в большой нацеливающий вектор (LTVEC). Человеческий фрагмент на hBAC расщепляют при помощи Cas9 с использованием 2 РНК CRISPR. Донор содержит верхний и нижний соединительные олигонуклеотиды и кассету селекции. После расщепления hBAC посредством Cas9 фрагмент «захватывается» реакцией сборки Гибсона с использованием синтетического донора со встроенными комплементарными нависаниями. Конструирование нацеливающего вектора завершают сборкой по Гибсону или BHR (бактериальной гомологичной рекомбинацией).
Пример 10. Введение точечной мутации посредством Cas9/изотермической сборки
На ФИГ. 18 представлен пример использования Cas9/изотермической сборки для введения точечной мутации. Донора получают путем традиционного клонирования. Кассету селекции вставляют в синтетический фрагмент ДНК, содержащий линкерные перекрытия и точечную мутацию. mВАС расщепляют при помощи Cas9, последовательность удаляют из mВАС и выполняют сборку по Гибсону mВАС с донором, получая конструкт (LTVEC), содержащий точечную мутацию и кассету селекции.
Пример 11. Обрезание ВАС посредством Cas9/изотермической сборки
На ФИГ. 19 представлен пример обрезания ВАС методом Cas9/изотермической сборки. Область, которую нужно удалить из LTVEC, вырезают, используя Cas9. В этом примере обрезкой ВАС удаляется последовательность Ori. Ori замещают в реакции сборки по Гибсону, используя 2 линкера (соединительных олигонуклеотида).
Пример 12. Другие способы расщепления ВАС при помощи CAS9 с последующей сборкой
В способах, предлагаемых в настоящем документе, можно использовать и другие способы, включая перечисленные ниже. Перед реакцией синтетические или транскрибированные in vitro тракрРНК и крРНК предварительно ренатурировали путем нагревания до 95°С и медленного охлаждения до комнатной температуры. Нативные или линеаризованные плазмидные ДНК (300 нг (около 8 нМ)) инкубировали в течение 60 мин при 37°С с очищенным белком Cas9 (50-500 нМ) и дуплексом тракрРНК: крРНК (50-500 нМ, 1:1) в буфере для расщепления плазмид посредством Cas9 (20 мМ ГЕПЕС рН 7,5, 150 мМ KCl, 0,5 мМ ДТТ, 0,1 мМ ЭДТА) с добавлением или без добавления 10 мМ MgCl2. Реакции останавливали 5 X буфером для внесения ДНК, содержащим 250 мМ ЭДТА, разделяли по размеру посредством электрофореза в 0,8 или 1% агарозном геле и визуализировали окрашиванием бромидом этидия. Для анализов на расщепление мутантным Cas9 реакции останавливали 5 X буфером додецилсульфата натрия (ДСН) для нанесения (30% глицерин, 1,2% ДСН, 250 мМ ЭДТА) перед внесением в агарозный гель.
Были разработаны искусственная крРНК и искусственная тракрРНК для нацеливания на специфические последовательности в MAID 6177 (LTVEC длиной 116 т.п.н.) для сборки с ПЦР-продуктом (UB-HYG) длиной 3 т.п.н. Этот продукт ПЦР содержал 50 п.н. перекрытия с вектором. Применяли изотермическую одноэтапную сборку, основанную на использовании выделенной нетермостабильной экзонуклеазы, работающей от 5'- к 3'-концу, лишенной 3'-экзонуклеазной активности, как описано ниже. Готовили реакционную смесь, содержащую следующие компоненты: по 100 фмоль каждого субстрата дцДНК, 16 мкл 5 X буфера ISO, 16 мкл экзонуклеазы Т5 (0,2 Ед/мкл, Epicentre), 8,0 мкл ДНК-лигазы Taq (40 Ед/мкл, NEB), 1,0 мкл ДНК-полимеразы Phusion™ (2 Ед/мкл, NEB) и вода до 80 мкл. Буфер ISO (изотермический) 5 х представлял собой 25% ПЭГ-8000, 500 мМ трис-Cl, 50 мМ MgCl2, 50 мМ ДТТ, 5 мМ НАД и по 1000 мкМ каждого дНТФ (рН 7,5).
Это давало конечную концентрацию 1,25 фмоль/мкл каждой дцДНК (или 45 фмоль/мкл каждой оцДНК), подлежащей сборке, 5% ПЭГ-8000, 100 мМ трис-Cl рН 7,5, 10 мМ MgCl2, 10 мМ ДТТ, по 200 ММ каждого дНТФ, 1 мМ НАД, 0,02 Ед/мкл экзонуклеазы Т5, 4 Ед/мкл ДНК-лигазы Taq и 0,03 Ед/мкл ДНК-полимеразы PHUSION.
В способах использовали 1,64 мкл (0,2 Ед/мкл) экзонуклеазы Т5 для субстратов с перекрытием 20-80 п.н., а для субстратов с более длинными перекрытиями (например, 200 п.н.) использовали 1,6 мкл (1 Ед/мкл) экзонуклеазы Т5. Экзонуклеазу Т5 использовали в разведении 1:50 (в буфере для хранения экзонуклеазы Т5) относительно концентрированного маточного раствора фермента 10 Ед/мкл экзонуклеазы Т5 (Epicentre). Затем реакционную смесь инкубировали при 50°С в течение 15 мин.
Пример 13. Другие способы сшивания друг с другом двух перекрывающихся ВАС
В способах, предлагаемых в настоящем документе, можно использовать и другие способы, включая перечисленные ниже. Перед реакцией синтетические или транскрибированные in vitro тракрРНК и крРНК предварительно ренатурировали путем нагревания до 95°С и медленного охлаждения до комнатной температуры. Нативные или линеаризованные плазмидные ДНК (300 нг (около 8 нМ)) инкубировали в течение 60 мин при 37°С с очищенным белком Cas9 (50-500 нМ) и дуплексом тракрРНК: крРНК (50-500 нМ, 1:1) в буфере для расщепления плазмид посредством Cas9 (20 мМ ГЕПЕС рН 7,5, 150 мМ KCl, 0,5 мМ ДТТ, 0,1 мМ ЭДТА) с добавлением или без добавления 10 мМ MgCl2. Реакции останавливали 5 X буфером для внесения ДНК, содержащим 250 мМ ЭДТА, разделяли по размеру посредством электрофореза в 0,8 или 1% агарозном геле и визуализировали окрашиванием бромидом этидия. Для анализов на расщепление мутантным Cas9 реакции останавливали 5 X буфером додецилсульфата натрия (ДСН) для нанесения (30% глицерин, 1,2% ДСН, 250 мМ ЭДТА) перед внесением в агарозный гель.
Были разработаны искусственная крРНК и искусственная тракрРНК для нацеливания на специфические последовательности в гуманизированной ВАС HLA-DQ для сборки с гуманизированной ВАС HLA-DR. Векторы содержали ~70 п.н. перекрытия друг с другом, созданных путем расщепления посредством Cas9 в двух сайтах на каждом векторе (см. ФИГ. 2). Применяли изотермическую одноэтапную сборку, основанную на использовании выделенной нетермостабильной экзонуклеазы, работающей от 5'- к 3'-концу, лишенной 3'-экзонуклеазной активности, как описано ниже. Готовили реакционную смесь, содержащую следующие приблизительные количества компонентов: по 100 фмоль каждого субстрата дцДНК, 16 мкл 5 X буфера ISO, 16 мкл экзонуклеазы Т5 (0,2 Ед/мкл, Epicentre), 8,0 мкл ДНК-лигазы Taq (40 Ед/мкл, NEB), 1,0 мкл ДНК-полимеразы Phusion™ (2 Ед/мкл, NEB) и вода до 80 мкл. Буфер ISO (изотермический) 5 х представлял собой 25% ПЭГ-8000, 500 мМ трис-Cl, 50 мМ MgCl2, 50 мМ ДТТ, 5 мМ НАД и по 1000 мкМ каждого дНТФ (рН 7,5).
Это давало конечную концентрацию около 1,25 фмоль/мкл каждой дцДНК (или 45 фмоль/мкл каждой оцДНК), подлежащей сборке, 5% ПЭГ-8000, 100 мМ трис-Cl рН 7,5, 10 мМ MgCl2, 10 мМ ДТТ, по 200 ММ каждого дНТФ, 1 мМ НАД, 0,02 Ед/мкл экзонуклеазы Т5, 4 Ед/мкл ДНК-лигазы Taq и 0,03 Ед/мкл ДНК-полимеразы PHUSION.
В способах использовали 1,64 мкл 0,2 Ед/мкл экзонуклеазы Т5 для субстратов с перекрытием 20-80 п.н., а для субстратов с более длинными перекрытиями (например, 200 п.н.) использовали 1,6 мкл 1 Ед/мкл экзонуклеазы Т5. Экзонуклеазу Т5 использовали в разведении 1: 50 (в буфере для хранения экзонуклеазы Т5) относительно концентрированного маточного раствора фермента 10 Ед/мкл экзонуклеазы Т5 (Epicentre). Затем реакционную смесь инкубировали при 50°С в течение 15 мин.
Пример 14. Другие способы сборки вставки с вектором ВАС
В способах, предлагаемых в настоящем документе, можно использовать и другие способы, включая перечисленные ниже. Растворяют крРНК и тракрРНК в 100 мкМ буфера Hybe (10 X буфер: 20 мМ трис 7,5, 100-150 мМ NaCl, 10 мМ MgCl2, 1 мМ ДТТ, 0,1 мМ ЭДТА, 100 мкг/мл БСА). Для ренатурации РНК в 80 мкл буфера для ренатурации добавляют 10 мкл 100 мкМ крРНК и 10 мкл 100 мкМ тракрРНК. Нагревают РНК в температурном блоке при 90°С, затем снимают блок с нагревателя и охлаждают на столе. Конечная концентрация РНК составляет около 10 мкМ.
Для расщепления ВАС используют чистую ДНК ВАС из набора Maxiprep и расщепляют ВАС с использованием приведенной ниже смеси.
Выполняют расщепление в течение 1 часа при 37°, после чего в течение 30 мин проводят обессоливание.
Для сборки ВАС и вставки расщепляют плазмиду или проводят ПЦР для создания вставки. Для реакций ПЦР прогоняют небольшую аликвоту на геле и находят чистый продукт, если продукт не чистый, то вместо гель-экстракции выполняют ПЦР-очистку. Желательное молярное соотношение ВАС : вставка составляет 1:1 - 1:6. Обычно для работы достаточно 50 нг очищенной вставки. Можно использовать следующую реакционную смесь:
Помещают ДНК и смесь на лед или непосредственно в ПЦР-аппарат при 50°С. Инкубируют при 50°С в течение 1 часа. Добавляют 0,5 мкл протеиназы К (20 мг/мл) и инкубируют при 50°С в течение 1 часа. Выполняют обессоливание в течение 30 мин и методом электропорации в клетки DH10B вводят 8 мкл реакционной смеси. Для проверки эффективности расщепления можно провести гель-электрофорез в пульсирующем поле 10 мкл расщепленной ВАС. Используют не содержащие РНКазу воду и буферы. Конечный буфер для реакции содержит: 20 мМ трис 7,5; 100-150 мМ NaCl; 10 мМ MgCl2; 1 мМ ДТТ; 0,1 мМ этилендиаминтетрауксусной кислоты (ЭДТА); 100 мкг/мл бычьего сывороточного альбумина (БСА); конечный объем 15 мкл.
Последовательность тракрРНК, использованная настоящем примере: CAAAACAGCAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUC (SEQ ID NO: 9). Эта РНК CRISPR (крРНК) содержит: (1) около 20 нуклеотидов РНК, комплементарных целевой последовательности, и (2) хвостовую последовательность (GUUUUAGAGCUAUGCUGUUUUG (SEQ ID NO: 10)), которую ренатурируют до тракрРНК.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ МОДИФИКАЦИИ ЦЕЛЕВОГО ЛОКУСА | 2015 |
|
RU2704283C2 |
| СПОСОБЫ ДЛЯ БЕСШОВНОГО ВНЕСЕНИЯ ЦЕЛЕВЫХ МОДИФИКАЦИЙ В НАПРАВЛЕННЫЕ ВЕКТОРЫ | 2020 |
|
RU2771374C1 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ НАЦЕЛЕННОЙ ГЕНЕТИЧЕСКОЙ МОДИФИКАЦИИ С ИСПОЛЬЗОВАНИЕМ ПАРНЫХ ГИДОВЫХ РНК | 2015 |
|
RU2734770C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ НАЦЕЛЕННЫХ ГЕНЕТИЧЕСКИХ МОДИФИКАЦИЙ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2771532C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ НАЦЕЛЕННОЙ ГЕНЕТИЧЕСКОЙ МОДИФИКАЦИИ ПОСРЕДСТВОМ ОДНОЭТАПНОГО МНОЖЕСТВЕННОГО НАЦЕЛИВАНИЯ | 2015 |
|
RU2707137C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ПОЛУЧЕНИЯ ИЛИ ПОДДЕРЖАНИЯ ПЛЮРИПОТЕНТНЫХ КЛЕТОК | 2015 |
|
RU2706965C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ НАЦЕЛЕННЫХ ГЕНЕТИЧЕСКИХ МОДИФИКАЦИЙ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2711740C2 |
| ПЛАТФОРМА CRISPR/CAS ДЛЯ ИСКЛЮЧАЮЩЕГО СКРИНИНГА ДЛЯ ВЫЯВЛЕНИЯ ГЕНЕТИЧЕСКИХ УЯЗВИМОСТЕЙ, СВЯЗАННЫХ С АГРЕГАЦИЕЙ ТАУ-БЕЛКА | 2020 |
|
RU2808829C2 |
| Средство разрезания ДНК на основе Cas9 белка из бактерии Streptococcus uberis NCTC3858 | 2022 |
|
RU2788197C1 |
| СКРИНИНГОВАЯ ПЛАТФОРМА CRISPR/CAS ДЛЯ ИДЕНТИФИКАЦИИ ГЕНЕТИЧЕСКИХ МОДИФИКАТОРОВ ЗАСЕВА ИЛИ АГРЕГАЦИИ ТАУ-БЕЛКА | 2020 |
|
RU2819350C2 |



















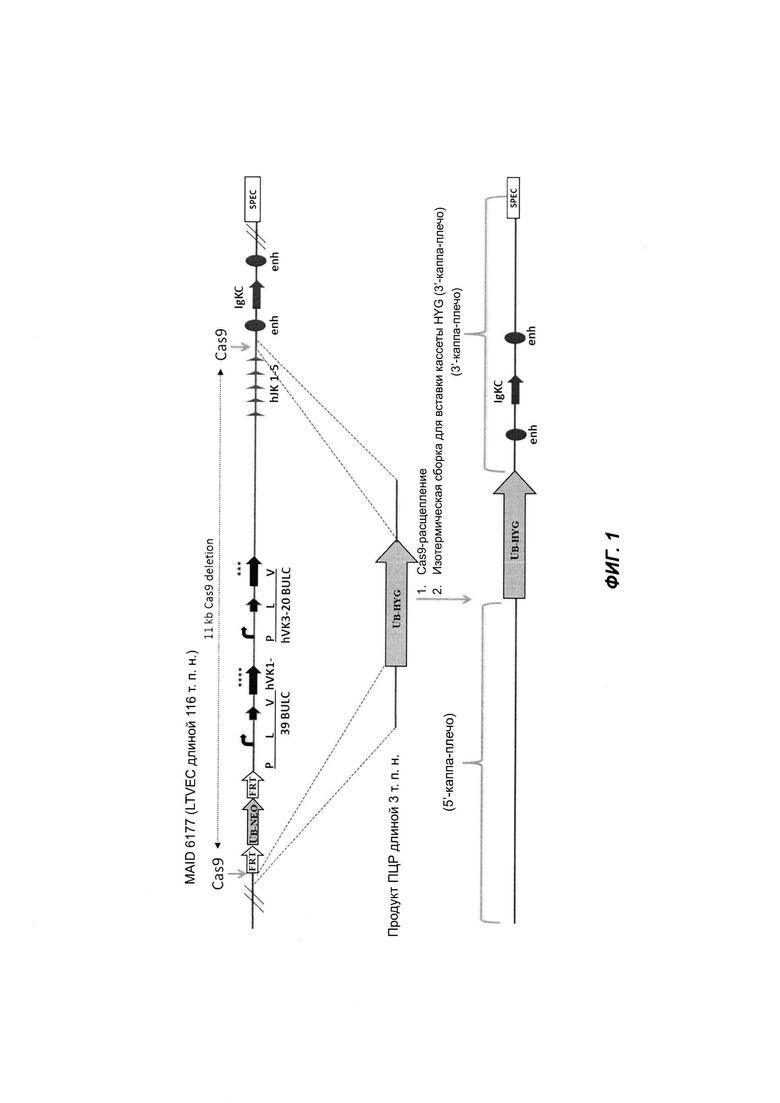
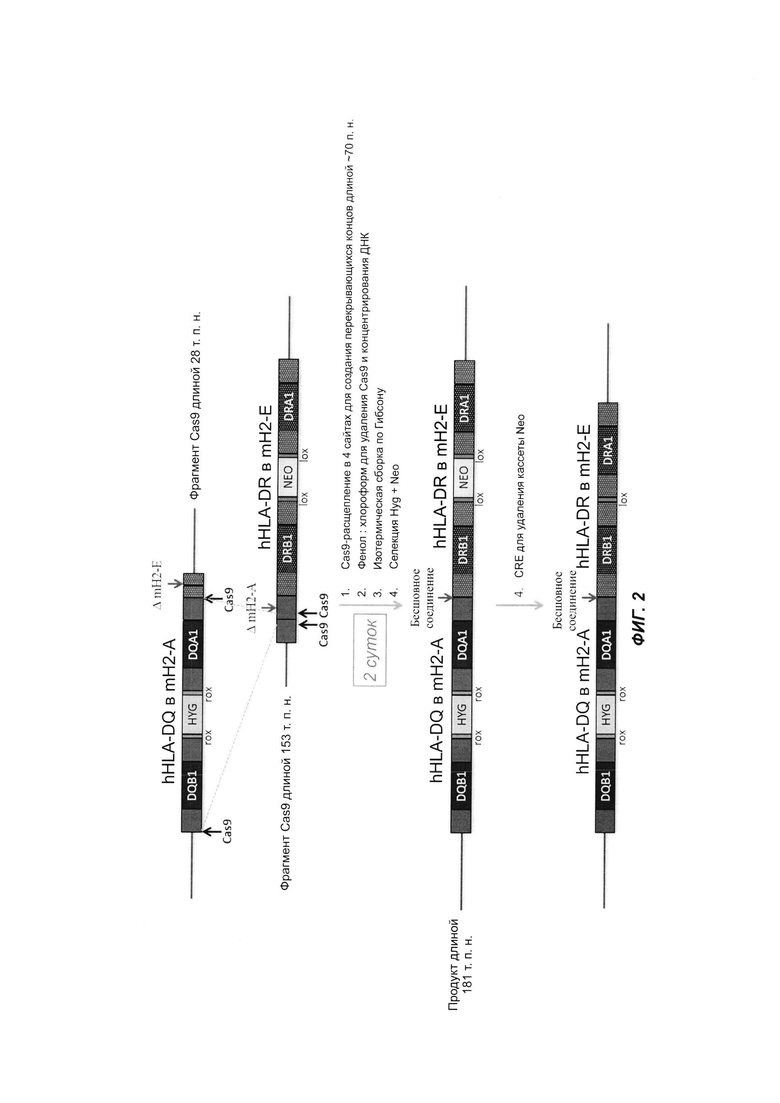
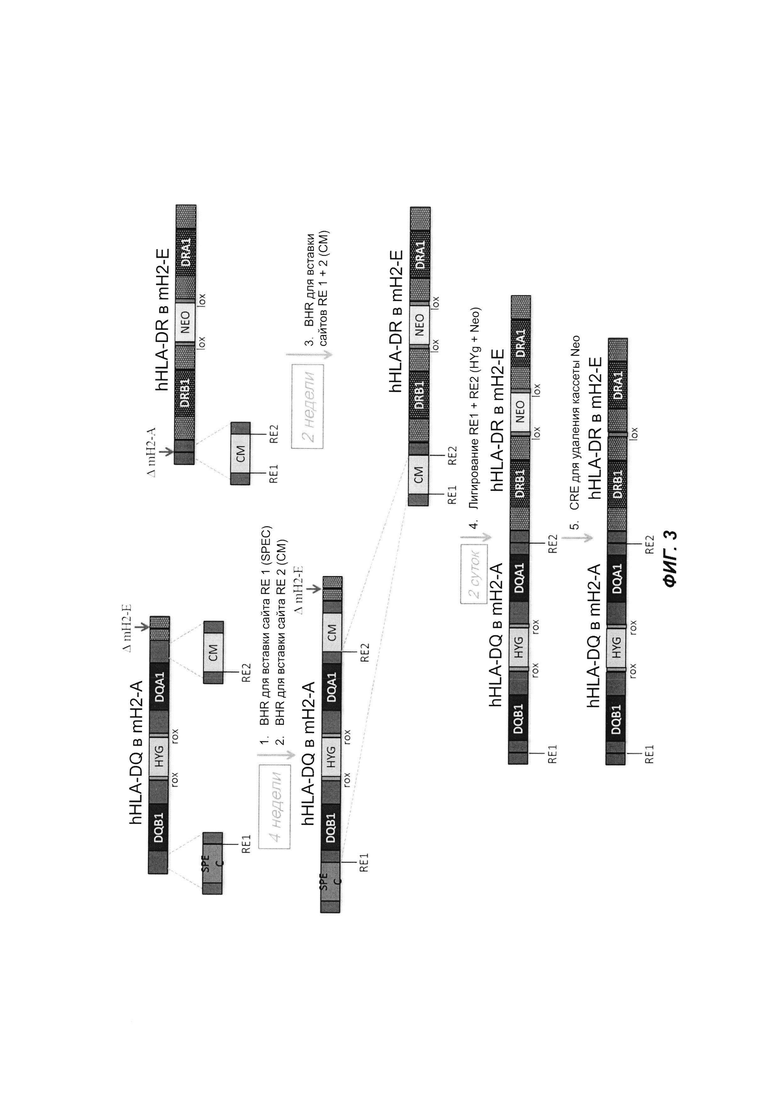

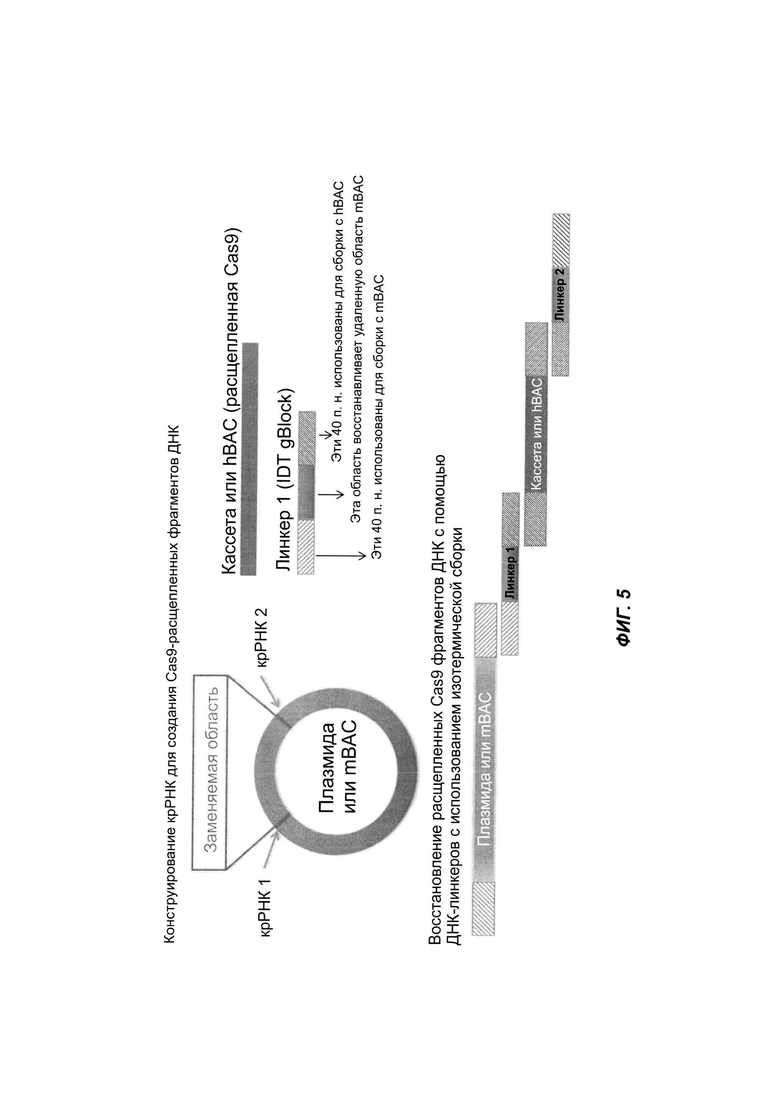







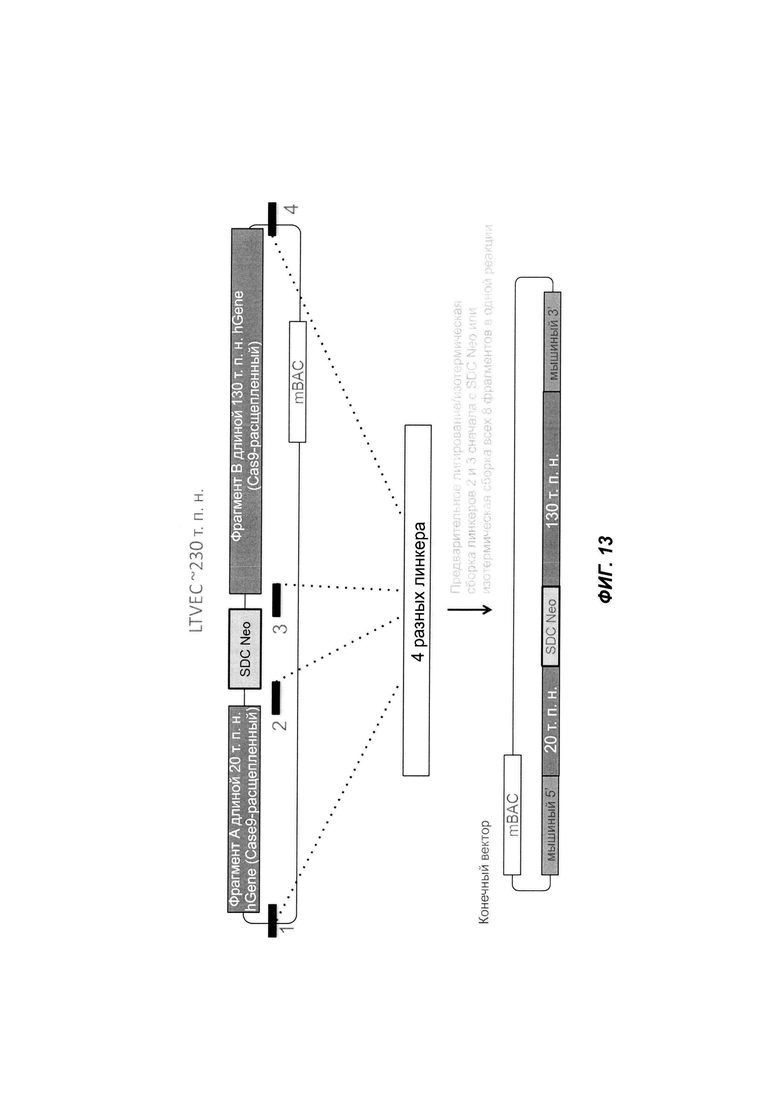

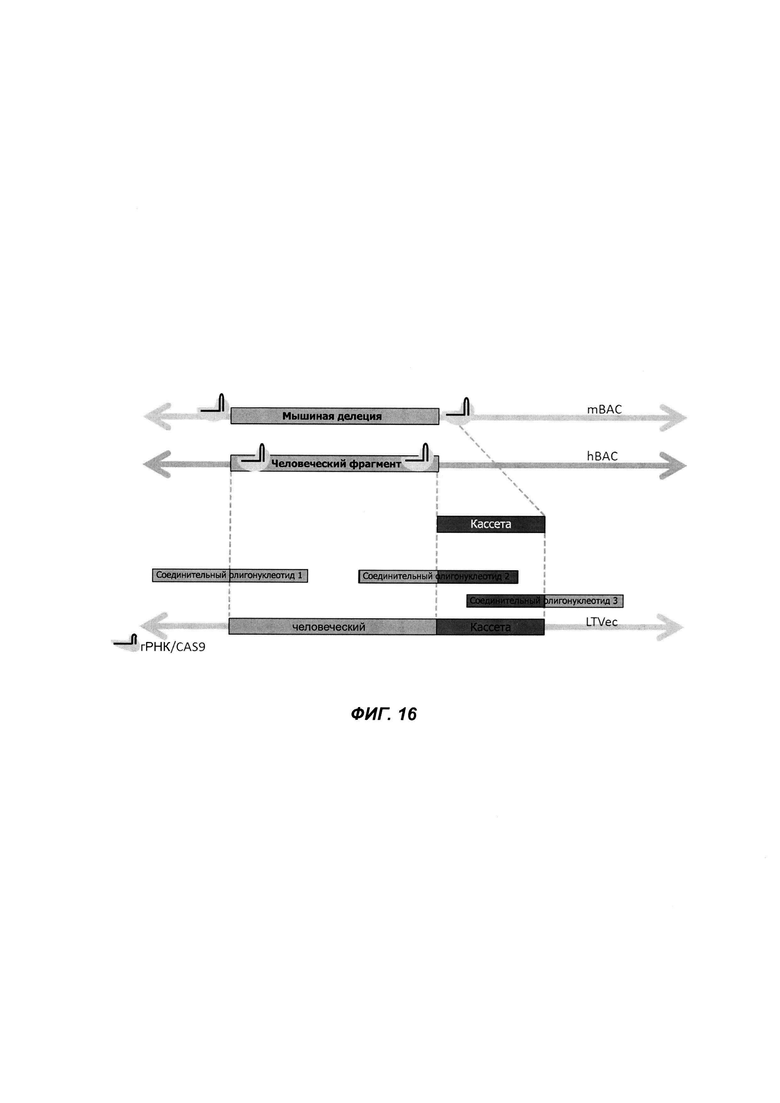

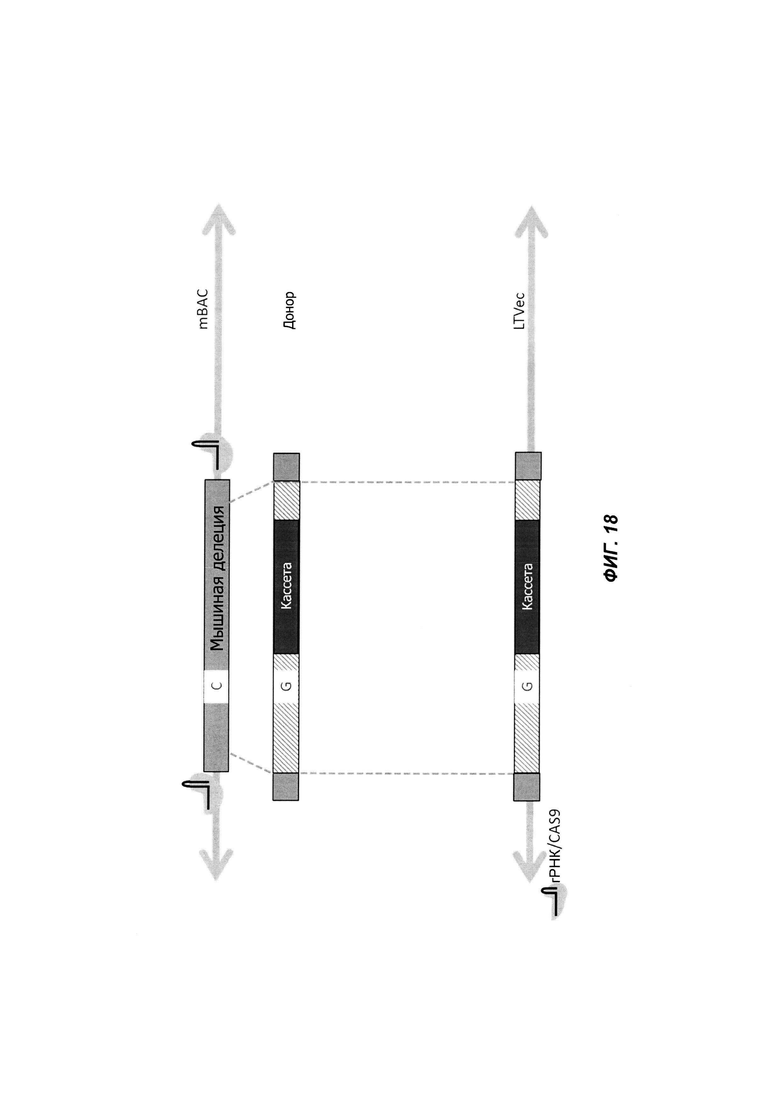
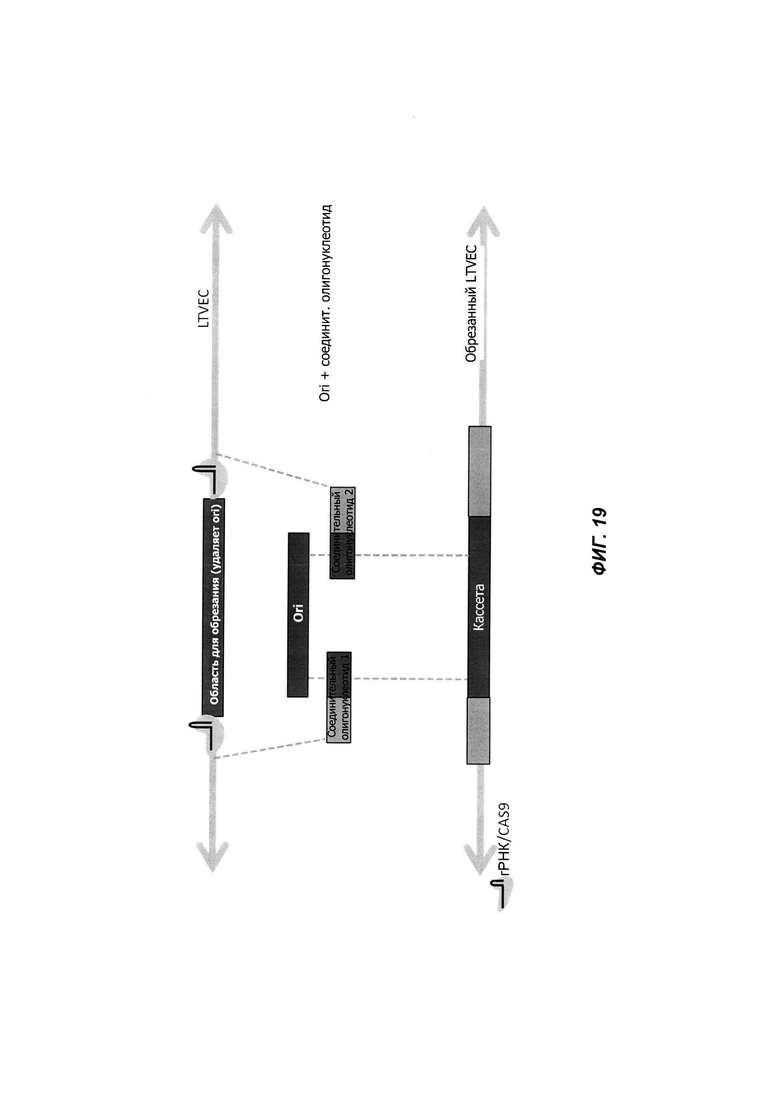
Изобретение относится к области биотехнологии и молекулярной биологии. Предложен in vitro способ сборки нуклеиновых кислот с использованием нуклеазного агента - комплекса гРНК-Cas, нуклеазы с цинковыми пальцами или эффекторной нуклеазы. Изобретение может быть использовано в прикладной биотехнологии и медицине благодаря увеличению эффективности создания длинных нуклеиновых кислот. 6 н. и 37 з.п. ф-лы, 19 ил., 1 табл., 14 пр.
1. In vitro способ сборки двух или более нуклеиновых кислот, включающий:
(a) контактирование первой нуклеиновой кислоты с первым нуклеазным агентом, причем первый нуклеазный агент содержит белок Cas и гидовую РНК (гРНК) (комплекс гРНК-Cas), нуклеазу с «цинковыми пальцами» или эффекторную нуклеазу, подобную активатору транскрипции (TALEN), и причем первый нуклеазный агент расщепляет первую нуклеиновую кислоту в первом целевом сайте с получением первой расщепленной нуклеиновой кислоты с перекрывающейся концевой последовательностью, общей со второй нуклеиновой кислотой;
(b) контактирование первой расщепленной нуклеиновой кислоты и второй нуклеиновой кислоты с экзонуклеазой для экспонирования комплементарных последовательностей между первой расщепленной нуклеиновой кислотой и второй нуклеиновой кислотой; и
(c) сборку двух фрагментов нуклеиновых кислот, полученных на этапе (b).
2. Способ по п. 1, в котором этап (с) включает:
(i) ренатурацию экспонированных комплементарных последовательностей;
(ii) удлинение 3'-концов ренатурированных комплементарных последовательностей; и
(iii) лигирование первой и второй нуклеиновых кислот.
3. Способ по п. 1 или 2, в котором этап (а) дополнительно включает контактирование второй нуклеиновой кислоты со вторым нуклеазным агентом, причем второй нуклеазный агент расщепляет вторую нуклеиновую кислоту во втором целевом сайте с получением второй расщепленной нуклеиновой кислоты с перекрывающейся концевой последовательностью, и
при этом вторая нуклеиновая кислота из этапа (b) представляет собой вторую расщепленную нуклеиновую кислоту.
4. Способ по любому из пп. 1-3, в котором перекрывающаяся концевая последовательность имеет длину в диапазоне от 20 п.н. до 200 п.н.
5. In vitro способ сборки двух или более нуклеиновых кислот, включающий:
(а) контактирование первой нуклеиновой кислоты по меньшей мере с одним нуклеазным агентом, причем по меньшей мере один нуклеазный агент содержит белок Cas и гидовую РНК (гРНК) (комплекс гРНК-Cas), нуклеазу с «цинковыми пальцами» или эффекторную нуклеазу, подобную активатору транскрипции (TALEN), и причем по меньшей мере один нуклеазный агент расщепляет первую нуклеиновую кислоту в первом целевом сайте с образованием первой расщепленной нуклеиновой кислоты;
(b) контактирование первой расщепленной нуклеиновой кислоты со второй нуклеиновой кислотой, соединительным олигонуклеотидом и экзонуклеазой,
причем соединительный олигонуклеотид содержит:
(i) первую комплементарную последовательность, которая комплементарна первой расщепленной нуклеиновой кислоте;
(ii) спейсер и
(iii) вторую комплементарную последовательность, которая комплементарна второй нуклеиновой кислоте, и
при этом экзонуклеаза экспонирует первую и вторую комплементарные последовательности; и
(c) сборку соединительного олигонуклеотида с первой расщепленной нуклеиновой кислотой и второй нуклеиновой кислотой.
6. Способ по п. 5, в котором сборка на этапе (с) включает:
(i) ренатурацию первой комплементарной последовательности соединительного олигонуклеотида с первой расщепленной нуклеиновой кислотой и второй комплементарной последовательности соединительного олигонуклеотида со второй нуклеиновой кислотой,
при этом необязательно дополнительно включает удлинение 3'-конца первой расщепленной нуклеиновой кислоты и/или второй нуклеиновой кислоты; и
(ii) лигирование соединительного олигонуклеотида с первой расщепленной нуклеиновой кислотой и второй нуклеиновой кислотой.
7. Способ по п. 5 или 6, в котором первая комплементарная последовательность соединительного олигонуклеотида составляет в длину от 15 до 120 комплементарных оснований, а вторая комплементарная последовательность соединительного олигонуклеотида составляет в длину от 15 до 120 комплементарных оснований, и
необязательно первая комплементарная последовательность соединительного олигонуклеотида составляет в длину от 20 до 80 комплементарных оснований, а вторая комплементарная последовательность соединительного олигонуклеотида составляет в длину от 20 до 80 комплементарных оснований.
8. Способ по любому из пп. 5-7, в котором спейсер соединительного олигонуклеотида содержит некомплементарные нуклеиновые кислоты.
9. Способ по любому из пп. 5-8, в котором сборка первой расщепленной нуклеиновой кислоты со второй нуклеиновой кислотой происходит бесшовно.
10. Способ по п. 9, в котором по меньшей мере один нуклеазный агент предназначен для отщепления фрагмента из по меньшей мере 20 п.н. от конца первой нуклеиновой кислоты, на котором будет происходить бесшовная сборка, необязательно причем фрагмент является двухцепочечным,
причем спейсер соединительного олигонуклеотида содержит последовательность, идентичную фрагменту из по меньшей мере 20 п.н., и при этом между первой комплементарной последовательностью и фрагментом из по меньшей мере 20 п.н. нуклеотидные основания отсутствуют и между второй комплементарной последовательностью и фрагментом из по меньшей мере 20 п.н. нуклеотидные основания отсутствуют,
так что сборка первой расщепленной нуклеиновой кислоты с соединительным олигонуклеотидом и второй нуклеиновой кислотой восстанавливает фрагмент из по меньшей мере 20 п.н. и обеспечивает бесшовную сборку первой нуклеиновой кислоты и второй нуклеиновой кислоты.
11. Способ по п. 9 или 10, в котором соединительный олигонуклеотид содержит линейный двухцепочечный фрагмент ДНК, и
необязательно причем линейный двухцепочечный фрагмент ДНК не содержит кассету селекции.
12. Способ по п. 11, в котором соединительный олигонуклеотид составляет в длину от около 50 п.н. до около 400 п.н., и
необязательно причем соединительный олигонуклеотид составляет в длину от около 100 п.н. до около 300 п.н.
13. Способ по любому из пп. 5-12, в котором спейсер составляет в длину от около 20 п.н. до около 120 п.н.
14. Способ по любому из пп. 5-13, в котором этап (а) дополнительно включает:
(i) контактирование второй нуклеиновой кислоты со вторым нуклеазным агентом, причем второй нуклеазный агент расщепляет вторую нуклеиновую кислоту с получением второй расщепленной нуклеиновой кислоты, содержащей нуклеотидную последовательность, комплементарную второй комплементарной последовательности соединительного олигонуклеотида, и при этом проводят сборку первой расщепленной нуклеиновой кислоты со второй расщепленной нуклеиновой кислотой; или
(ii) контактирование второй нуклеиновой кислоты с рестрикционным ферментом или мегануклеазой, причем рестрикционный фермент или мегануклеаза расщепляет вторую нуклеиновую кислоту с получением второй расщепленной нуклеиновой кислоты, содержащей нуклеотидную последовательность, комплементарную второй комплементарной последовательности в соединительном олигонуклеотиде, и при этом проводят сборку первой расщепленной нуклеиновой кислоты со второй расщепленной нуклеиновой кислотой.
15. Способ по любому из пп. 5-14, в котором сборку соединительного олигонуклеотида с первой нуклеиновой кислотой и второй нуклеиновой кислотой выполняют в одной реакции или последовательно.
16. Способ по любому из пп. 1-15, в котором две или более нуклеиновых кислот являются двухцепочечными.
17. In vitro способ бесшовной сборки двух или более двухцепочечных нуклеиновых кислот, включающий:
(a) контактирование первой нуклеиновой кислоты с первым нуклеазным агентом, причем первый нуклеазный агент содержит белок Cas и гидовую РНК (гРНК) (комплекс гРНК-Cas), нуклеазу с «цинковыми пальцами» или эффекторную нуклеазу, подобную активатору транскрипции (TALEN), причем первый нуклеазный агент расщепляет первую нуклеиновую кислоту в первом целевом сайте с образованием первой расщепленной нуклеиновой кислоты, и причем в результате расщепления происходит удаление двухцепочечного фрагмента с конца первой нуклеиновой кислоты, на котором будет происходить бесшовная сборка;
(b) контактирование первой расщепленной нуклеиновой кислоты со второй нуклеиновой кислотой, первым соединительным олигонуклеотидом и экзонуклеазой, причем соединительный олигонуклеотид представляет собой линейную двухцепочечную ДНК, которая составляет в длину от около 50 п.н. до около 400 п.н. и содержит:
(i) первую комплементарную последовательность, которая комплементарна первой расщепленной нуклеиновой кислоте;
(ii) спейсер и
(iii) вторую комплементарную последовательность, которая комплементарна второй нуклеиновой кислоте,
причем спейсер содержит последовательность, идентичную фрагменту,
при этом между первой комплементарной последовательностью и последовательностью, идентичной фрагменту, нуклеотидные основания отсутствуют и между второй комплементарной последовательностью и последовательностью, идентичной фрагменту, нуклеотидные основания отсутствуют, и
при этом экзонуклеаза экспонирует первую и вторую комплементарные последовательности; и
(с) сборку соединительного олигонуклеотида с первой расщепленной нуклеиновой кислотой и второй нуклеиновой кислотой, причем сборка восстанавливает фрагмент.
18. Способ по п. 17, в котором сборка на этапе (с) включает:
(i) ренатурацию первой комплементарной последовательности соединительного олигонуклеотида с первой расщепленной нуклеиновой кислотой и второй комплементарной последовательности соединительного олигонуклеотида со второй нуклеиновой кислотой,
при этом необязательно дополнительно включает удлинение 3'-конца первой расщепленной нуклеиновой кислоты и/или второй нуклеиновой кислоты; и
(ii) лигирование соединительного олигонуклеотида с первой расщепленной нуклеиновой кислотой и второй нуклеиновой кислотой.
19. Способ по п. 17 или 18, в котором этап (а) дополнительно включает:
(i) контактирование второй нуклеиновой кислоты со вторым нуклеазным агентом, причем второй нуклеазный агент расщепляет вторую нуклеиновую кислоту во втором целевом сайте с получением второй расщепленной нуклеиновой кислоты, содержащей нуклеотидную последовательность, комплементарную второй комплементарной последовательности соединительного олигонуклеотида, и при этом проводят сборку первой расщепленной нуклеиновой кислоты со второй расщепленной нуклеиновой кислотой; или
(ii) контактирование второй нуклеиновой кислоты с рестрикционным ферментом, причем рестрикционный фермент расщепляет вторую нуклеиновую кислоту с получением второй расщепленной нуклеиновой кислоты, содержащей нуклеотидную последовательность, комплементарную второй комплементарной последовательности в соединительном олигонуклеотиде, и при этом проводят сборку первой расщепленной нуклеиновой кислоты со второй расщепленной нуклеиновой кислотой.
20. Способ по любому из пп. 17-19, в котором:
(I) первая комплементарная последовательность соединительного олигонуклеотида составляет в длину от 15 до 120 комплементарных оснований, а вторая комплементарная последовательность соединительного олигонуклеотида составляет в длину от 15 до 120 комплементарных оснований, и
необязательно первая комплементарная последовательность соединительного олигонуклеотида составляет в длину от 20 до 80 комплементарных оснований, а вторая комплементарная последовательность соединительного олигонуклеотида составляет в длину от 20 до 80 комплементарных оснований; и/или
(II) фрагмент составляет по меньшей мере 20 п.н.; и/или
(III) спейсер составляет от около 20 п.н. до около 120 п.н.; и/или
(IV) соединительный олигонуклеотид составляет в длину от около 100 п.н. до около 300 п.н.
21. Способ по любому из пп. 17-20, в котором сборку первого соединительного олигонуклеотида с первой расщепленной нуклеиновой кислотой и второй нуклеиновой кислотой выполняют в одной реакции или последовательно.
22. Способ по любому из пп. 17-21, в котором выполняют сборку по меньшей трех нуклеиновых кислот.
23. In vitro способ сборки двух или более нуклеиновых кислот, включающий:
(a) контактирование первой нуклеиновой кислоты с первым нуклеазным агентом и вторым нуклеазным агентом, причем первый нуклеазный агент содержит белок Cas и гидовую РНК (гРНК) (комплекс гРНК-Cas), нуклеазу с «цинковыми пальцами» или эффекторную нуклеазу, подобную активатору транскрипции (TALEN), причем первый нуклеазный агент расщепляет первую нуклеиновую кислоту в первом целевом сайте, а второй нуклеазный агент расщепляет первую нуклеиновую кислоту во втором целевом сайте с образованием первой расщепленной нуклеиновой кислоты;
(b) контактирование первой расщепленной нуклеиновой кислоты с первым соединительным олигонуклеотидом, второй нуклеиновой кислотой, вторым соединительным олигонуклеотидом и экзонуклеазой,
причем первый соединительный олигонуклеотид содержит:
(i) первую комплементарную последовательность, которая комплементарна первой расщепленной нуклеиновой кислоте; и
(ii) вторую комплементарную последовательность, которая комплементарна второй нуклеиновой кислоте, и
причем второй соединительный олигонуклеотид содержит:
(i) первую комплементарную последовательность, которая комплементарна второй нуклеиновой кислоте, и
(ii) вторую комплементарную последовательность, которая комплементарна первой расщепленной нуклеиновой кислоте; и
при этом экзонуклеаза экспонирует комплементарные последовательности первого соединительного олигонуклеотида, второго соединительного олигонуклеотида, первой расщепленной нуклеиновой кислоты и второй нуклеиновой кислоты; и
(с) сборку первой расщепленной нуклеиновой кислоты, первого соединительного олигонуклеотида, второй нуклеиновой кислоты и второго соединительного олигонуклеотида.
24. Способ по п. 23, в котором сборка на этапе (с) включает:
(i) ренатурацию первой комплементарной последовательности первого соединительного олигонуклеотида с первой расщепленной нуклеиновой кислотой, ренатурацию второй комплементарной последовательности первого соединительного олигонуклеотида со второй нуклеиновой кислотой, ренатурацию первой комплементарной последовательности второго соединительного олигонуклеотида со второй нуклеиновой кислотой и ренатурацию второй комплементарной последовательности второго соединительного олигонуклеотида с первой расщепленной нуклеиновой кислотой,
при этом необязательно дополнительно включает удлинение 3'-концов ренатурированных комплементарных последовательностей; и
(ii) лигирование первой расщепленной нуклеиновой кислоты с первым соединительным олигонуклеотидом, лигирование первого соединительного олигонуклеотида со второй нуклеиновой кислотой, лигирование второй нуклеиновой кислоты со вторым соединительным олигонуклеотидом и лигирование второго соединительного олигонуклеотида с первой расщепленной нуклеиновой кислотой.
25. In vitro способ сборки трех или более нуклеиновых кислот, включающий:
(а) контактирование первой нуклеиновой кислоты с первым нуклеазным агентом и вторым нуклеазным агентом, причем первый нуклеазный агент содержит белок Cas и гидовую РНК (гРНК) (комплекс гРНК-Cas), нуклеазу с «цинковыми пальцами» или эффекторную нуклеазу, подобную активатору транскрипции (TALEN), причем первый нуклеазный агент расщепляет первую нуклеиновую кислоту в первом целевом сайте, а второй нуклеазный агент расщепляет первую нуклеиновую кислоту во втором целевом сайте с образованием первой расщепленной нуклеиновой кислоты;
(b) контактирование первой расщепленной нуклеиновой кислоты с первым соединительным олигонуклеотидом, второй нуклеиновой кислотой, вторым соединительным олигонуклеотидом, третьей нуклеиновой кислотой, третьим соединительным олигонуклеотидом и экзонуклеазой,
причем первый соединительный олигонуклеотид содержит:
(i) первую комплементарную последовательность, которая комплементарна первой расщепленной нуклеиновой кислоте; и
(ii) вторую комплементарную последовательность, которая комплементарна второй нуклеиновой кислоте, и
причем второй соединительный олигонуклеотид содержит:
(i) первую комплементарную последовательность, которая комплементарна второй нуклеиновой кислоте, и
(ii) вторую комплементарную последовательность, которая комплементарна третьей нуклеиновой кислоте, и
причем третий соединительный олигонуклеотид содержит:
(i) первую комплементарную последовательность, которая комплементарна третьей нуклеиновой кислоте, и
(ii) вторую комплементарную последовательность, которая комплементарна первой расщепленной нуклеиновой кислоте; и
при этом экзонуклеаза экспонирует комплементарные последовательности первого соединительного олигонуклеотида, второго соединительного олигонуклеотида, третьего соединительного олигонуклеотида, первой расщепленной нуклеиновой кислоты, второй нуклеиновой кислоты и третьей нуклеиновой кислоты; и
(c) сборку первой расщепленной нуклеиновой кислоты, первого соединительного олигонуклеотида, второй нуклеиновой кислоты, второго соединительного олигонуклеотида, третьей нуклеиновой кислоты и третьего соединительного олигонуклеотида.
26. In vitro способ сборки четырех или более нуклеиновых кислот, включающий:
(а) контактирование первой нуклеиновой кислоты с первым нуклеазным агентом и вторым нуклеазным агентом, причем первый нуклеазный агент содержит белок Cas и гидовую РНК (гРНК) (комплекс гРНК-Cas), нуклеазу с «цинковыми пальцами» или эффекторную нуклеазу, подобную активатору транскрипции (TALEN), причем первый нуклеазный агент расщепляет первую нуклеиновую кислоту в первом целевом сайте, а второй нуклеазный агент расщепляет первую нуклеиновую кислоту во втором целевом сайте с образованием первой расщепленной нуклеиновой кислоты;
(b) контактирование первой расщепленной нуклеиновой кислоты с первым соединительным олигонуклеотидом, второй нуклеиновой кислотой, вторым соединительным олигонуклеотидом, третьей нуклеиновой кислотой, третьим соединительным олигонуклеотидом, четвертой нуклеиновой кислотой, четвертым соединительным олигонуклеотидом и экзонуклеазой,
причем первый соединительный олигонуклеотид содержит:
(i) первую комплементарную последовательность, которая комплементарна первой расщепленной нуклеиновой кислоте; и
(ii) вторую комплементарную последовательность, которая комплементарна второй нуклеиновой кислоте, и
причем второй соединительный олигонуклеотид содержит:
(i) первую комплементарную последовательность, которая комплементарна второй нуклеиновой кислоте, и
(ii) вторую комплементарную последовательность, которая комплементарна третьей нуклеиновой кислоте, и
причем третий соединительный олигонуклеотид содержит:
(i) первую комплементарную последовательность, которая комплементарна третьей нуклеиновой кислоте, и
(ii) вторую комплементарную последовательность, которая комплементарна четвертой нуклеиновой кислоте, и
причем четвертый соединительный олигонуклеотид содержит:
(i) первую комплементарную последовательность, которая комплементарна четвертой нуклеиновой кислоте, и
(ii) вторую комплементарную последовательность, которая комплементарна первой расщепленной нуклеиновой кислоте; и
при этом экзонуклеаза экспонирует комплементарные последовательности первого соединительного олигонуклеотида, второго соединительного олигонуклеотида, третьего соединительного олигонуклеотида, четвертого соединительного олигонуклеотида, первой расщепленной нуклеиновой кислоты, второй нуклеиновой кислоты, третьей нуклеиновой кислоты и четвертой нуклеиновой кислоты; и
(с) сборку первой расщепленной нуклеиновой кислоты, первого соединительного олигонуклеотида, второй нуклеиновой кислоты, второго соединительного олигонуклеотида, третьей нуклеиновой кислоты, третьего соединительного олигонуклеотида, четвертой нуклеиновой кислоты и четвертого соединительного олигонуклеотида.
27. Способ по п. 25 или 26, в котором сборка на этапе (с) включает:
(i) ренатурацию первой расщепленной нуклеиновой кислоты, других нуклеиновых кислот и соединительных олигонуклеотидов,
при этом необязательно дополнительно включает удлинение 3'-концов ренатурированных последовательностей; и
(ii) лигирование первой расщепленной нуклеиновой кислоты, других нуклеиновых кислот и соединительных олигонуклеотидов.
28. Способ по любому из пп. 23-27, в котором две или более нуклеиновых кислот представляют собой двухцепочечные нуклеиновые кислоты, первый нуклеазный агент расщепляет первую нуклеиновую кислоту в первом целевом сайте с созданием первого двухцепочечного разрыва, а второй нуклеазный агент расщепляет первую нуклеиновую кислоту во втором целевом сайте с созданием второго двухцепочечного разрыва.
29. Способ по любому из пп. 23-28, в котором этап (а) дополнительно включает контактирование второй нуклеиновой кислоты с третьим нуклеазным агентом, и причем третий нуклеазный агент расщепляет вторую нуклеиновую кислоту в третьем целевом сайте с образованием второй расщепленной нуклеиновой кислоты.
30. Способ по любому из пп. 23-28, в котором этап (а) дополнительно включает контактирование второй нуклеиновой кислоты с третьим нуклеазным агентом и четвертым нуклеазным агентом, и причем третий нуклеазный агент расщепляет вторую нуклеиновую кислоту в третьем целевом сайте, а четвертый нуклеазный агент расщепляет вторую нуклеиновую кислоту в четвертом целевом сайте с образованием второй расщепленной нуклеиновой кислоты.
31. Способ по любому из пп. 1-30, в котором первый нуклеазный агент содержит белок Cas и гРНК, причем белок Cas представляет собой белок Cas9, причем гРНК содержит нуклеотидную последовательность, кодирующую РНК коротких палиндромных повторов, регулярно расположенных группами (CRISPR) (крРНК), и трансактивирующую РНК CRISPR (тракрРНК), и причем первый нуклеазный агент нацелен на целевой сайт, который непосредственно фланкирован последовательностью мотива, прилежащего к протоспейсеру (РАМ), и
необязательно причем белок Cas9 содержит домен RuvC и домен HNH, по меньшей мере один из которых не имеет эндонуклеазной активности.
32. Способ по любому из пп. 23-31, в котором
(I) один или несколько или все соединительные олигонуклеотиды представляют собой линейную двухцепочечную ДНК и/или
(II) каждая из первой комплементарной последовательности и второй комплементарной последовательности одного или нескольких или всех соединительных олигонуклеотидов составляет в длину от 15 до 120 комплементарных оснований, необязательно причем каждая из первой комплементарной последовательности и второй комплементарной последовательности одного или нескольких или всех соединительных олигонуклеотидов составляет в длину от 20 до 80 комплементарных оснований.
33. Способ по любому из пп. 23-32, в котором один или несколько или все соединительные олигонуклеотиды составляют в длину от около 50 п.н. до около 400 п.н., и
необязательно причем один или несколько или все соединительные олигонуклеотиды составляют в длину от около 100 п.н. до около 300 п.н.
34. Способ по любому из пп. 23-33, в котором один или несколько или все соединительные олигонуклеотиды дополнительно содержат спейсер между первой комплементарной последовательностью и второй комплементарной последовательностью, и необязательно причем:
(I) спейсер содержит ген устойчивости к лекарственному средству, репортерный ген, последовательности для обнаружения, последовательности, подходящие для ПЦР, или один или несколько сайтов для рестрикционных ферментов для подтверждения успешной сборки; и/или
(II) спейсер составляет от около 20 п.н. до около 120 п.н.
35. Способ по любому из пп. 23-34, в котором сборку первой расщепленной нуклеиновой кислоты, других нуклеиновых кислот и соединительных олигонуклеотидов выполняют в одной реакции или последовательно.
36. Способ по любому из пп. 23-35, в котором сборка первой расщепленной нуклеиновой кислоты со второй нуклеиновой кислотой происходит бесшовно.
37. Способ по п. 36, в котором в результате расщепления первым нуклеазным агентом и/или вторым нуклеазным агентом происходит удаление двухцепочечного фрагмента с конца первой нуклеиновой кислоты, на котором будет происходить бесшовная сборка, и
причем первый соединительный олигонуклеотид дополнительно содержит спейсер между первой комплементарной последовательностью и второй комплементарной последовательностью, и
причем спейсер содержит последовательность, идентичную фрагменту, при этом между первой комплементарной последовательностью и последовательностью, идентичной фрагменту, нуклеотидные основания отсутствуют, и между второй комплементарной последовательностью и последовательностью, идентичной фрагменту, нуклеотидные основания отсутствуют.
38. Способ по любому из пп. 23-37, в котором один или несколько или все соединительные олигонуклеотиды представляют собой одноцепочечную ДНК.
39. Способ по любому из пп. 1-38, в котором:
(I) одна или несколько или все нуклеиновые кислоты представляют собой векторы длиной от около 20 т.п.н. до около 400 т.п.н.; и/или
(II) собранная нуклеиновая кислота составляет в длину от 30 т.п.н. до 1 млн п.н.
40. Способ по любому из пп. 1-39, в котором одна или несколько или все нуклеиновые кислоты составляют в длину по меньшей мере 10 т.п.н.
41. Способ по любому из пп. 1-40, в котором:
(I) одна или несколько или все нуклеиновые кислоты содержат бактериальную искусственную хромосому; и/или
(II) одна или несколько или все нуклеиновые кислоты содержат человеческую ДНК, ДНК грызуна, синтетическую ДНК или их комбинацию; и/или
(III) каждая нуклеиновая кислота содержит бактериальную искусственную хромосому, интересующий ген охватывает бактериальные искусственные хромосомы, и в результате сборки образуется последовательность интересующего гена; и/или
(IV) одна или несколько или все нуклеиновые кислоты представляют собой кольцевые нуклеиновые кислоты или линейные нуклеиновые кислоты; и/или
(V) собранная нуклеиновая кислота представляет собой кольцевую нуклеиновую кислоту или линейную нуклеиновую кислоту.
42. Способ по любому из пп. 1-41, причем способ включает объединение нуклеиновых кислот и соединительных олигонуклеотидов с лигазой, экзонуклеазой, ДНК-полимеразой и нуклеотидами и инкубацию при постоянной температуре.
43. Способ по п. 42, в котором сборка происходит в одноэтапной изотермической реакции.
| LI MZ | |||
| Harnessing homologous recombination in vitro to generate recombinant DNA via SLIC | |||
| Nat Methods | |||
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| RUI-YAN WANG | |||
| DNA Fragments Assembly Based on Nicking Enzyme System | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| GIBSON D.G | |||
| Enzymatic assembly of DNA molecules up to several hundred kilobases | |||
| Nat Methods | |||
| Колосоуборка | 1923 |
|
SU2009A1 |

