ПЕРЕКРЕСТНАЯ ССЫЛКА НА СМЕЖНУЮ ЗАЯВКУ
Настоящая заявка испрашивает преимущество предварительной заявки на патент США № 61/373109, поданной 12 августа 2010 г., содержание которой полностью включено в настоящий документ путем ссылки для любых целей.
ОБЛАСТЬ ПРИМЕНЕНИЯ ИЗОБРЕТЕНИЯ
Настоящее изобретение представляет способ снижения уровней глюкозы в крови животного путем трансплантации животному популяции панкреатических эндокринных клеток-предшественников.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Последние достижения в области заместительной клеточной терапии для лечения сахарного диабета 1 типа и нехватка островков Лангерганса с возможностью трансплантации заставили обратить внимание на разработку источников инсулин-продуцирующих клеток, или β-клеток, подходящих для приживления трансплантата. Один подход представляет собой получение функциональных β-клеток из плюрипотентных стволовых клеток, таких как, например, эмбриональные стволовые клетки.
При эмбриональном развитии позвоночных плюрипотентные клетки дают начало группе клеток, содержащих три зародышевых листка (эктодерму, мезодерму и энтодерму), в ходе процесса, известного как гаструляция. Такие ткани, как, например, щитовидная железа, тимус, поджелудочная железа, кишечник и печень, будут развиваться из энтодермы через промежуточную стадию. Промежуточная стадия данного процесса представляет собой образование дефинитивной энтодермы. Клетки дефинитивной энтодермы экспрессируют ряд маркеров, таких как, например, HNF3 beta, GATA4, MIXL1, CXCR4 и SOX17.
Образование поджелудочной железы происходит при дифференцировании дефинитивной энтодермы в панкреатическую энтодерму. Клетки панкреатической энтодермы экспрессируют ген панкреатическо-дуоденального гомеобокса, PDX1. В отсутствие PDX1 поджелудочная железа не развивается дальше образования вентрального и дорзального зачатков. Таким образом, экспрессия PDX1 характеризует критическую стадию органогенеза поджелудочной железы. Зрелая поджелудочная железа содержит, среди других типов клеток, экзокринную ткань и эндокринную ткань. Экзокринная и эндокринная ткани образуются при дифференцировании панкреатической энтодермы.
По имеющимся данным, клетки, обладающие свойствами островковых клеток, были получены из эмбриональных клеток мыши. Например, в публикации Lumelsky et al. (Science 292:1389, 2001 г.) сообщают о дифференцировании эмбриональных стволовых клеток мыши в инсулин-секретирующие структуры, сходные с панкреатическими островками. В публикации Soria et al. (Diabetes 49:157, 2000 г.) сообщают, что инсулин-секретирующие клетки, полученные из эмбриональных стволовых клеток мыши, нормализуют гликемию у мышей с диабетом, вызванным стрептозотоцином.
В одном примере, в публикации Hori et al. (PNAS 99: 16105, 2002 г.), обнаружено, что обработка эмбриональных стволовых клеток мыши ингибиторами фосфоинозитид-3-киназы (LY294002) привела к получению клеток, подобных β-клеткам.
В другом примере, в публикации Blyszczuk et al. (PNAS 100:998, 2003 г.), сообщают о получении инсулин-продуцирующих клеток из эмбриональных стволовых клеток мыши с постоянной экспрессией Pax4.
В публикации Micallef et al. сообщают, что ретиноевая кислота может регулировать способность эмбриональных стволовых клеток формировать PDX1-положительную панкреатическую энтодерму. Ретиноевая кислота с наибольшей эффективностью индуцирует экспрессию PDX1 при добавлении в культуры на четвертый день дифференцирования эмбриональных стволовых клеток в течение периода, соответствующего концу гаструляции эмбриона (Diabetes 54:301, 2005 г.).
В публикации Miyazaki et al. сообщают о линии эмбриональных стволовых клеток мыши со сверхэкспрессией Pdx1. Результаты показывают, что экспрессия экзогенного Pdx1 очевидно повышает экспрессию генов инсулина, соматостатина, глюкокиназы, нейрогенина-3, p48, Pax6 и HNF6 в полученных дифференцированных клетках (Diabetes 53: 1030, 2004 г.).
В публикации Skoudy et al. сообщают, что активин A (входящий в суперсемейство TGF-β) повышает экспрессию экзокринных панкреатических генов (p48 и амилаза) и эндокринных генов (Pdx1, инсулин и глюкагон) в эмбриональных стволовых клетках мыши. Максимальный эффект наблюдали при использовании 1 нМ активина A. Авторы также обнаружили, что уровень экспрессии инсулина и мРНК Pdx1 не зависел от наличия ретиноевой кислоты; однако обработка с использованием 3 нМ FGF7 привела к повышению уровня транскрипта для Pdx1 (Biochem. J. 379: 749, 2004 г.).
В публикации Shiraki et al. изучали эффекты факторов роста, специфически ускоряющих дифференцирование эмбриональных стволовых клеток в PDX1-положительные клетки. Авторы обнаружили, что TGF-β2 приводил к воспроизводимому увеличению доли PDX1-положительных клеток (Genes Cells. июнь 2005 г.; 10(6): 503–16.).
В публикации Gordon et al. продемонстрировали индукцию brachyury [положительных]/HNF3 beta [положительных] энтодермальных клеток из эмбриональных стволовых клеток мыши в отсутствие сыворотки и при наличии активина в сочетании с ингибитором сигнального пути Wnt (патент США № 2006/0003446A1).
В публикации Gordon et al. (PNAS, том 103, стр. 16806, 2006 г.) отмечают: «Для получения передней первичной полоски одновременно требовались сигнальные пути Wnt и TGF-beta/nodal/активин».
Однако модель развития эмбриональных стволовых клеток у мышей может не имитировать в точности программу развития у высших млекопитающих, такого как, например, человека.
В публикации Thomson et al. эмбриональные стволовые клетки выделяли из бластоцист человека (Science 282:114, 1998 г.). Параллельно Gearhart и соавторы получили линии эмбриональных зародышевых клеток человека (hEG) из ткани половых желез эмбриона (Shamblott et al., Proc. Natl. Acad. Sci. USA 95:13726, 1998 г.). В отличие от эмбриональных стволовых клеток мыши, воспрепятствовать дифференцированию которых можно путем простого культивирования с фактором, ингибирующим лейкемию (LIF), эмбриональные стволовые клетки человека необходимо культивировать в очень специфических условиях (патент США № 6200806; международные заявки №№ 99/20741, 01/51616).
В публикации D’Amour et al. описывают производство обогащенных культур дефинитивной энтодермы, производной от эмбриональных стволовых клеток человека, при наличии высокой концентрации активина и низкой концентрации сыворотки (Nature Biotechnology 2005 г.). Трансплантация данных клеток под почечную капсулу мышей привела к их дифференцированию в более зрелые клетки с характеристиками некоторых энтодермальных органов. Клетки дефинитивной энтодермы, производные от эмбриональных стволовых клеток человека, могут дополнительно дифференцироваться в PDX1-положительные клетки после присоединения FGF-10 (патент США № 2005/0266554A1).
В публикации D'Amour et al. (Nature Biotechnology – 24, 1392–1401 (2006 г.)) отмечают: «Мы разработали процесс дифференцирования, преобразующий эмбриональные клетки человека (hES) в эндокринные клетки, способные синтезировать гормоны поджелудочной железы: инсулин, глюкагон, соматостатин, панкреатический полипептид и грелин. Данный процесс имитирует органогенез поджелудочной железы in vivo, проводя клетки через стадии, напоминающие образование дефинитивной энтодермы, энтодермы кишечной трубки, панкреатической энтодермы и превращение эндокринных клеток-предшественников в клетки, экспрессирующие эндокринные гормоны».
В другом примере, в публикации Fisk et al., сообщают о системе для производства панкреатических островковых клеток из эмбриональных стволовых клеток человека (US2006/0040387A1). В данном случае процесс дифференцирования был разделен на три стадии. Сначала эмбриональные стволовые клетки человека дифференцировали до энтодермы с помощью комбинации бутирата натрия и активина А. Затем клетки культивировали с антагонистами TGF-β, такими как Ноггин, в комбинации с EGF или бетацеллюлином для получения PDX1-положительных клеток. Конечное дифференцирование индуцировали никотинамидом.
В одном примере, в публикации Benvenistry et al., отмечают: «Мы делаем вывод, что сверхэкспрессия PDX1 увеличила экспрессию панкреатических обогащенных генов, а для индукции экспрессии инсулина могут требоваться дополнительные сигналы, присутствующие только in vivo» (Benvenistry et al, Stem Cells 2006 г.; 24:1923–1930).
В другом примере, в патенте № US2008/0241107A1, представляют способ получения клетки, секретирующей инсулин, включающий: a) получение клетки, не продуцирующей инсулин, а также b) инкубацию клетки в среде, содержащей высокую концентрацию глюкозы, в которой клетка секретирует инсулин.
Следовательно, насущной потребностью по-прежнему остается разработка условий для создания линий плюрипотентных стволовых клеток, способных расти для решения текущих клинических задач, сохраняя при этом потенциал дифференцирования в панкреатические эндокринные клетки, панкреатические клетки, экспрессирующие гормоны, или панкреатические клетки, секретирующие гормоны. Мы использовали альтернативный подход для повышения эффективности дифференцирования эмбриональных стволовых клеток человека в панкреатические эндокринные клетки.
ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В одном варианте осуществления настоящее изобретение представляет способ снижения уровней глюкозы в крови животного путем трансплантации животному популяции панкреатических эндокринных клеток-предшественников.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фигуре 1 показаны уровни глюкозы в крови мышей с ТКИН, у которых был индуцирован диабет посредством 5 инъекций стрептозотоцина, затем в день 0 под почечную капсулу имплантировали дифференцированные клетки ES человека (стадия 4). Отслеживание глюкозы в крови в течение следующих нескольких месяцев выявило постепенное падение гипергликемии до додиабетических уровней. Последующее удаление почки привело к быстрому рецидиву диабета.
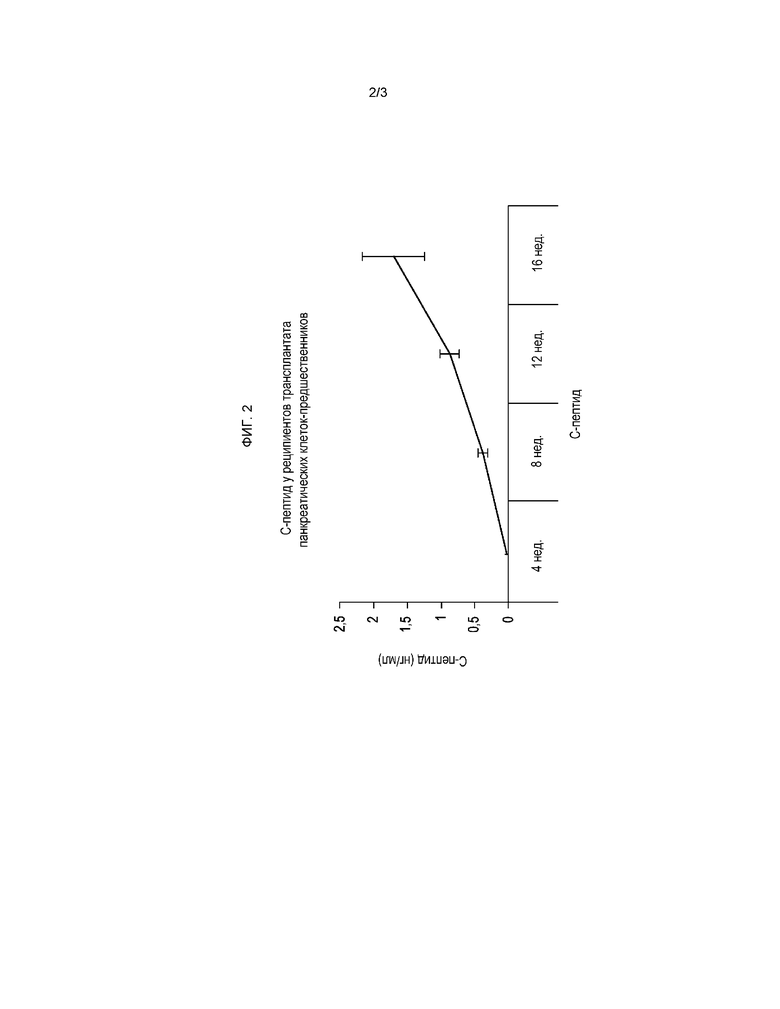
На фигуре 2 показаны результаты измерений уровня С-пептида человека в образцах плазмы в указанные недели после трансплантации, которые свидетельствуют о прогрессирующем нарастании, сопоставимом с падением уровней глюкозы в крови.
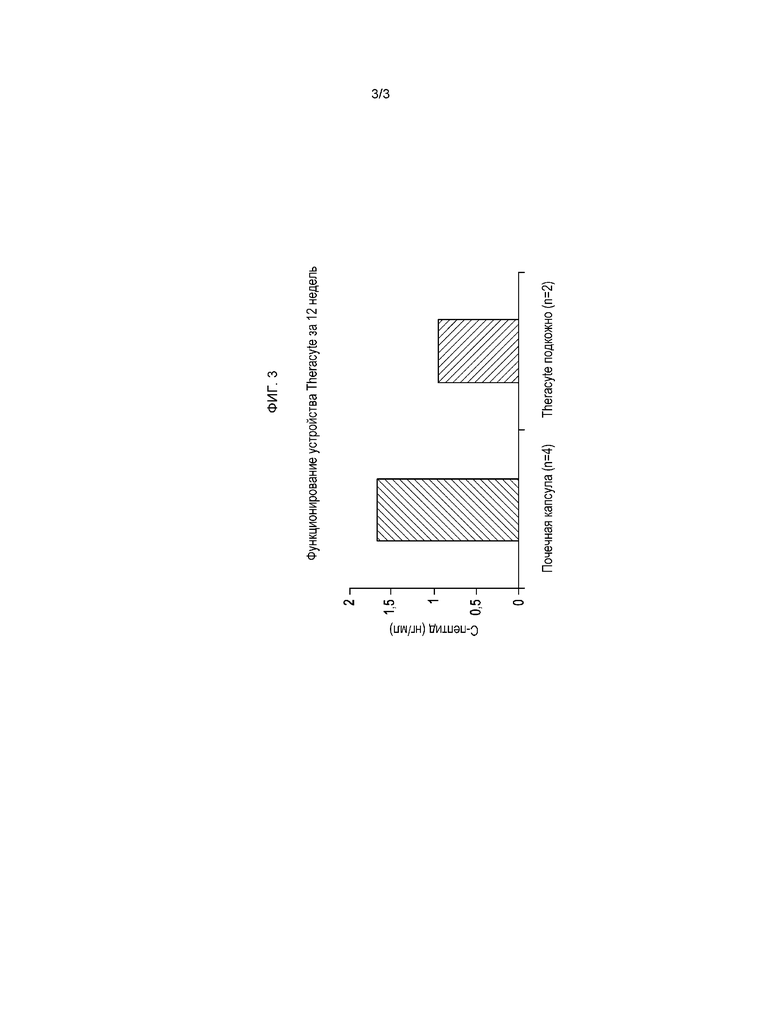
На фигуре 3 показаны сопоставимые уровни С-пептида, регистрируемые у реципиентов клеток, трансплантированных под почечную капсулу или подкожно с помощью устройств TheraCyte.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Для ясности описания, а не для ограничения изобретения, подробное описание настоящего изобретения разделено на следующие подразделы, описывающие или иллюстрирующие определенные особенности, варианты осуществления или области применения настоящего изобретения.
Определения
Стволовые клетки представляют собой недифференцированные клетки, определяемые по их способности на уровне единичной клетки как самообновляться, так и дифференцироваться для образования клеток-потомков, включая самообновляющиеся клетки-предшественники, необновляющиеся клетки-предшественники и окончательно дифференцированные клетки. Стволовые клетки также характеризуются способностью дифференцироваться in vitro в функциональные клетки различных клеточных линий из множества зародышевых листков (энтодермы, мезодермы и эктодермы), а также после трансплантации давать начало тканям, происходящим от множества зародышевых листков, и по существу вносить вклад в формирование большинства, если не всех, тканей после инъекции в бластоцисты.
По потенциалу развития стволовые клетки разделяют на: (1) тотипотентные, т.е. способные преобразоваться в любой из эмбриональных и внеэмбриональных типов клеток; (2) плюрипотентные, т.е. способные преобразоваться во все типы эмбриональных клеток; (3) мультипотентные, т.е. способные преобразоваться в подмножество клеточных линий, но все в рамках конкретной ткани, органа или физиологической системы (например, гемопоэтические стволовые клетки (ГСК) могут порождать потомство, включая ГСК (самообновление), ограниченные олигопотентные клетки-предшественники клеток крови и все типы клеток и элементы (например, тромбоциты), представляющие собой стандартные компоненты крови); (4) олигопотентные, т.е. способные преобразоваться в более ограниченное подмножество клеточных линий, чем мультипотентные стволовые клетки; а также (5) унипотентные, т.е. способные преобразоваться в единственную клеточную линию (например, сперматогенные стволовые клетки).
Дифференцирование представляет собой процесс, при помощи которого неспециализированная («некоммитированная») или менее специализированная клетка приобретает свойства специализированной клетки, например, нервной клетки или мышечной клетки. Дифференцированная клетка или клетка с индуцированным дифференцированием представляет собой клетку, занявшую более специализированное («коммитированное») положение в линии дифференцирования клетки. Термин «коммитированная» применительно к процессу дифференцирования относится к клетке, дошедшей в ходе процесса дифференцирования до стадии, с которой в нормальных условиях она продолжит дифференцироваться до конкретного типа клеток или подмножества типов клеток и не сможет в нормальных условиях дифференцироваться в иной тип клеток или вернуться к менее дифференцированному типу. Дедифференцирование относится к процессу, в ходе которого клетка возвращается к менее специализированному (или коммитированному) положению в линии дифференцирования клетки. В настоящем документе линия дифференцирования клетки определяет наследственность клетки, т.е. определяет, из каких клеток произошла данная клетка и каким клеткам она может дать начало. В линии дифференцирования клетка помещается в наследственную схему развития и дифференцирования. Маркер, специфичный для линии дифференцирования, относится к характеристике, специфически ассоциированной с фенотипом клеток интересующей линии дифференцирования, и его можно использовать для оценки дифференцирования некоммитированной клетки в клетки интересующей линии дифференцирования.
В настоящем документе «клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы», «клетки стадии 1» или «стадия 1» относятся к клеткам, экспрессирующим по меньшей мере один из следующих маркеров: SOX-17, GATA4, HNF3 beta, GSC, CER1, Nodal, FGF8, Brachyury, Mix-подобный белок гомеобокса, FGF4, CD48, эомезодермин (EOMES), DKK4, FGF17, GATA6, CXCR4, C-Kit, CD99 или OTX2. Клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, включают в себя клетки-предшественники первичной полоски, клетки первичной полоски, клетки мезэнтодермы и клетки дефинитивной энтодермы.
В настоящем документе «клетки, экспрессирующие маркеры, характерные для линии панкреатической энтодермы» относятся к клеткам, экспрессирующим по меньшей мере один из следующих маркеров: PDX1, HNF-1 beta, PTF1 alpha, HNF6 или HB9. Клетки, экспрессирующие маркеры, характерные для линии панкреатической энтодермы, включают в себя клетки панкреатической энтодермы, клетки первичной кишечной трубки и клетки поздней передней кишки.
В настоящем документе «клетки, экспрессирующие маркеры, характерные для линии панкреатических эндокринных клеток» относятся к клеткам, экспрессирующим по меньшей мере один из следующих маркеров: NEUROD, ISL1, PDX1, NKX6.1, MAFB, инсулин, глюкагон или соматостатин. Клетки, экспрессирующие маркеры, характерные для линии панкреатических эндокринных клеток, включают в себя панкреатические эндокринные клетки, клетки, экспрессирующие гормоны поджелудочной железы, клетки, секретирующие гормоны поджелудочной железы, а также клетки β-клеточной линии.
В настоящем документе «дефинитивная энтодерма» относится к клеткам, обладающим характеристиками клеток, происходящих от эпибласта при гаструляции, и формирующим желудочно-кишечный тракт и его производные. Клетки дефинитивной энтодермы экспрессируют следующие маркеры: HNF3 beta, GATA4, SOX17, Cerberus, OTX2, goosecoid, C-Kit, CD99 и MIXL1.
В настоящем документе «маркеры» представляют собой молекулы нуклеиновых кислот или полипептидов с дифференциальной экспрессией в интересующей клетке. В данном контексте дифференциальная экспрессия означает повышение уровня для положительного маркера и понижение уровня для отрицательного маркера. Обнаруживаемый уровень маркерной нуклеиновой кислоты или полипептида в интересующих клетках оказывается значительно выше или ниже по сравнению с другими клетками, что позволяет идентифицировать интересующую клетку и отличить ее от других клеток с помощью любого из множества способов, известных в данной области.
В настоящем документе «панкреатическая эндокринная клетка» или «панкреатическая клетка, экспрессирующая гормоны» относится к клетке, способной экспрессировать по меньшей мере один из следующих гормонов: инсулин, глюкагон, соматостатин и панкреатический полипептид.
В настоящем документе «панкреатическая эндокринная клетка-предшественник» относится к мультипотентной клетке линии дефинитивной энтодермы, которая экспрессирует NGN3 и может подвергаться дальнейшему дифференцированию в клетки эндокринной системы, включая без ограничений панкреатические островковые клетки, экспрессирующие гормоны. Эндокринные клетки-предшественники не могут дифференцироваться в такое количество разных типов клеток, тканей и/или органов, как менее специфически дифференцированные клетки линии дефинитивной энтодермы, такие как PDX1-положительные клетки панкреатической энтодермы.
В настоящем документе «панкреатическая клетка, продуцирующая гормоны» относится к клетке, способной продуцировать по меньшей мере один из следующих гормонов: инсулин, глюкагон, соматостатин и панкреатический полипептид.
В настоящем документе «панкреатическая клетка, секретирующая гормоны» относится к клетке, способной к секреции по меньшей мере одного из следующих гормонов: инсулин, глюкагон, соматостатин и панкреатический полипептид.
Выделение, размножение и культивирование плюрипотентных стволовых клеток
Характеристика плюрипотентных стволовых клеток
Плюрипотентные стволовые клетки могут экспрессировать один или более стадиеспецифических эмбриональных антигенов (SSEA) 3 и 4, а также маркеры, обнаруживаемые антителами, обозначенными как Tra-1-60 и Tra-1-81 (Thomson et al., Science 282:1145, 1998 г.). Дифференцирование плюрипотентных стволовых клеток in vitro приводит к утрате экспрессии SSEA-4, Tra-1-60 и Tra-1-81 (если имеются) и к увеличению экспрессии SSEA-1. В недифференцированных плюрипотентных стволовых клетках, как правило, активна щелочная фосфатаза, которую можно обнаружить путем фиксации клеток с помощью 4% параформальдегида с последующим обнаружением при помощи Vector Red в качестве субстрата в соответствии с инструкциями производителя (Vector Laboratories, г. Берлингейм, штат Калифорния). Недифференцированные плюрипотентные стволовые клетки так же, как правило, экспрессируют Oct-4 и TERT, обнаруживаемые методом ОТ-ПЦР.
Другим желательным фенотипом выращенных плюрипотентных стволовых клеток является потенциал дифференцирования в клетки всех трех зародышевых листков: в энтодермальные, мезодермальные и эктодермальные ткани. Плюрипотентность плюрипотентных стволовых клеток можно подтвердить, например, путем инъекции клеток мышам с тяжелой комбинированной иммунной недостаточностью (ТКИН), фиксирования тератом, образующихся с помощью 4% параформальдегида, а также их последующего гистологического исследования для получения доказательств наличия типов клеток, происходящих от трех зародышевых листков. Плюрипотентность можно альтернативно определить по созданию эмбриоидных телец и анализа эмбриоидных телец на наличие маркеров, ассоциирующихся с тремя зародышевыми листками.
Выращенные линии плюрипотентных стволовых клеток можно кариотипировать с использованием стандартного метода G-бэндинга и сравнить с опубликованными кариотипами соответствующих видов приматов. Желательно получить клетки, имеющие «нормальный кариотип», т.е. эуплоидные клетки, в которых присутствуют все хромосомы человека без видимых изменений.
Источники плюрипотентных стволовых клеток
Типы плюрипотентных стволовых клеток, которые можно использовать, включают в себя стандартные линии плюрипотентных клеток, полученных из ткани, сформированной после беременности, включая преэмбриональную ткань (такую как, например, бластоциста), эмбриональную ткань или ткань плода, взятую в любой момент в ходе беременности, как правило, но необязательно, до приблизительно 10–12 недель беременности. Неограничивающие примеры представляют собой стандартные линии эмбриональных стволовых клеток человека или эмбриональных зародышевых клеток человека, такие как, например, линии эмбриональных стволовых клеток человека H1, H7 и H9 (WiCell). Также предусмотрено использование композиций, описанных в настоящем документе, в ходе первоначального установления или стабилизации таких клеток, в данном случае исходными клетками будут первичные плюрипотентные клетки, взятые напрямую из исходных тканей. Также допустимыми являются клетки, взятые из популяции плюрипотентных стволовых клеток, уже культивированных в отсутствие питающих клеток. Также допустимыми являются мутантные линии эмбриональных стволовых клеток человека, такие как, например, BG01v (BresaGen, г. Атенс, штат Джорджия).
В одном варианте осуществления эмбриональные стволовые клетки человека готовят, как описано в публикации Thomson et al. (патент США № 5843780; Science 282:1145, 1998 г.; Curr. Top. Dev. Biol. 38:133 и последующие страницы, 1998 г.; Proc. Natl. Acad. Sci. U.S.A. 92:7844, 1995 г.).
Культивирование плюрипотентных стволовых клеток
В одном варианте осуществления плюрипотентные стволовые клетки, как правило, культивируют на слое питающих клеток, которые поддерживают плюрипотентные клетки в различных отношениях. Плюрипотентные стволовые клетки альтернативно культивируют в культуральной системе, по существу не содержащей питающих клеток, но тем не менее поддерживающей пролиферацию плюрипотентных стволовых клеток и не допускающей существенного дифференцирования. Рост плюрипотентных стволовых клеток в не содержащей питающих клеток культуральной системе без дифференцирования поддерживают путем использования среды, кондиционированной посредством предварительного культивирования клеток иного типа. Рост плюрипотентных стволовых клеток в не содержащей питающих клеток культуральной системе без дифференцирования альтернативно поддерживают путем использования среды с определенным химическим составом.
Например, в работах Reubinoff et al (Nature Biotechnology 18: 399–404 (2000 г.)) и Thompson et al (Science, 6 ноября 1998 г.: том 282. № 5391, стр. 1145–1147) описано культивирование линий плюрипотентных стволовых клеток из бластоцист человека с применением слоя питающих клеток из эмбриональных фибробластов мыши.
В публикации Richards et al (Stem Cells 21: 546–556, 2003 г.) оценивали набор из одиннадцати разных слоев питающих клеток, полученных от взрослых людей, новорожденных и эмбрионов, по их способности поддерживать культуру плюрипотентных стволовых клеток человека. В публикации Richards et al отмечают: «линии эмбриональных стволовых клеток человека, культивируемые на питающих слоях из фибробластов кожи взрослых людей, сохраняют морфологию эмбриональных стволовых клеток человека и остаются плюрипотентными».
В патенте США № US20020072117 описаны линии клеток, продуцирующие среду, которая поддерживает рост плюрипотентных стволовых клеток приматов в не содержащей питающих клеток культуре. Использованные клеточные линии представляют собой мезенхимальные и фибробластоподобные клеточные линии, полученные из эмбриональной ткани или дифференцированные из эмбриональных стволовых клеток. В патенте США № US20020072117 также описано использование клеточных линий в качестве первичного слоя питающих клеток.
В другом примере, в публикации Wang et al. (Stem Cells 23: 1221–1227, 2005 г.), описаны способы длительного выращивания плюрипотентных стволовых клеток человека на слоях питающих клеток, полученных из эмбриональных стволовых клеток человека.
В другом примере, в публикации Stojkovic et al. (Stem Cells 2005 23: 306–314, 2005 г.), описана система питающих клеток, полученная в результате спонтанного дифференцирования эмбриональных стволовых клеток человека.
В дополнительном примере, в публикации Miyamoto et al. (Stem Cells 22: 433–440, 2004 г.), описан источник питающих клеток из плаценты человека.
В публикации Amit et al (Biol. Reprod 68: 2150–2156, 2003 г.) описан слой питающих клеток, полученных из крайней плоти человека.
В другом примере, в публикации Inzunza et al. (Stem Cells 23: 544–549, 2005 г.), описан слой питающих клеток, полученных из фибробластов постнатальной крайней плоти человека.
В патенте США № US6642048 описана среда, поддерживающая рост плюрипотентных стволовых клеток приматов (pPS) в не содержащей питающих клеток среде, а также клеточные линии, которые можно использовать для производства такой среды. В патенте США № US6642048 отмечают: «Данное изобретение включает в себя мезенхимальные и фибробластоподобные клеточные линии, полученные из эмбриональной ткани или дифференцированные из эмбриональных стволовых клеток. В данном документе описывают и иллюстрируют способы получения таких клеточных линий, обработки среды и выращивания стволовых клеток с применением кондиционированной среды».
В другом примере, в международной заявке № WO2005014799, описана кондиционированная среда для поддержания, пролиферации и дифференцирования клеток млекопитающих. В международной заявке № WO2005014799 отмечают: «Культуральная среда, подготовленная в соответствии с настоящим изобретением, кондиционирована секрецией клеток мышей; в частности, дифференцированных и иммортализованных трансгенных гепатоцитов под названием MMH (гепатоциты мышей Met)».
В другом примере, в публикации Xu et al. (Stem Cells 22: 972–980, 2004 г.), описана кондиционированная среда, полученная из производных эмбриональных стволовых клеток человека, генетически модифицированных для сверхэкспрессии обратной транскриптазы теломеразы человека.
В другом примере, в патенте США № US20070010011, описана культуральная среда с определенным химическим составом для поддержания плюрипотентных стволовых клеток.
В альтернативной культуральной системе используют бессывороточную среду, обогащенную факторами роста, способными стимулировать пролиферацию эмбриональных стволовых клеток. Например, в публикации Cheon et al (BioReprod DOI:10.1095/biolreprod.105.046870, 19 октября 2005 г.) описана не содержащая питающих клеток и бессывороточная культуральная система, в которой эмбриональные стволовые клетки поддерживаются в некондиционированной, заменяющей сыворотку среде (SR), обогащенной различными факторами роста, способными запустить самообновление эмбриональных стволовых клеток.
В другом примере, в публикации Levenstein et al. (Stem Cells 24: 568–574, 2006 г.), описаны способы длительного культивирования эмбриональных стволовых клеток человека в отсутствие фибробластов или кондиционированной среды с применением среды, обогащенной основным фактором роста фибробластов (bFGF).
В другом примере, в патенте США № US20050148070, описан способ культивирования эмбриональных стволовых клеток человека в среде с определенным составом без сыворотки и без питающих клеток–фибробластов, включающий: культивирование стволовых клеток в культуральной среде, содержащей альбумин, аминокислоты, витамины, минералы, по меньшей мере один трансферрин или заместитель трансферрина, по меньшей мере один инсулин или заместитель инсулина, причем культуральная среда по существу не содержит эмбриональную сыворотку млекопитающих и содержит по меньшей мере приблизительно 100 нг/мл фактора роста фибробластов, способного активировать сигнальный рецептор фактора роста фибробластов, где фактор роста поступает из источника, отличного от просто питающего слоя фибробластов; среда поддерживает пролиферацию стволовых клеток в недифференцированном состоянии в отсутствие питающих клеток или кондиционированной среды.
В другом примере, в патенте США № US20050233446, описана среда с определенным составом, которую можно использовать при культивировании стволовых клеток, включая недифференцированные зародышевые стволовые клетки приматов. Среда в растворе по существу является изотонической по сравнению с культивируемыми стволовыми клетками. В данной культуре конкретная среда содержит основную среду и количество каждого из bFGF, инсулина и аскорбиновой кислоты, необходимое для поддержки по существу недифференцированного роста зародышевых стволовых клеток.
В другом примере, в патенте США № US6800480, отмечают: «В одном варианте осуществления представлена культуральная среда для выращивания зародышевых стволовых клеток приматов по существу в недифференцированном состоянии, включающая в себя основную среду с низким содержанием эндотоксина и низким осмотическим давлением, которая эффективно поддерживает рост зародышевых стволовых клеток приматов. Основную среду объединяют с питательной сывороткой, которая эффективно поддерживает рост зародышевых стволовых клеток приматов, и субстратом, выбранным из группы, состоящей из питающих клеток и компонента внеклеточного матрикса, полученного из питающих клеток. Среда дополнительно включает в себя аминокислоты, не относящиеся к незаменимым, антиоксидант и первый фактор роста, выбранный из группы, состоящей из нуклеозидов и соли пировиноградной кислоты».
В другом примере, в патенте США № US20050244962, отмечают: «В одном аспекте настоящее изобретение предлагает способ культивирования эмбриональных стволовых клеток приматов. Стволовые клетки культивируют в культуре, по существу не содержащей эмбриональную сыворотку млекопитающих (предпочтительно также по существу не содержащей сыворотку любого животного), и при наличии фактора роста фибробластов, полученного из источника, отличного от просто питающего слоя фибробластов. В предпочтительной форме питающий слой фибробластов, ранее необходимый для поддержания культуры стволовых клеток, становится необязательным вследствие присоединения достаточного количества фактора роста фибробластов».
В дополнительном примере, в международной заявке № WO2005065354, описана по существу не содержащая питающих клеток и бессывороточная изотоническая культуральная среда с определенным составом, содержащая: a. основную среду; b. количество bFGF, достаточное для поддержания роста по существу недифференцированных стволовых клеток млекопитающих; c. количество инсулина, достаточное для поддержания роста по существу недифференцированных стволовых клеток млекопитающих; а также d. количество аскорбиновой кислоты, достаточное для поддержания роста по существу недифференцированных стволовых клеток млекопитающих.
В другом примере, в международной заявке № WO2005086845, описан способ поддержания недифференцированной стволовой клетки, причем указанный способ включает воздействие на стволовую клетку членом семейства белков трансформирующего ростового фактора-бета (TGF-β), членом семейства белков фактора роста фибробластов (FGF) или никотинамидом (NIC) в количестве, достаточном для поддержания клетки в недифференцированном состоянии в течение периода времени, достаточного для получения необходимого результата.
Плюрипотентные стволовые клетки можно высевать на соответствующий культуральный субстрат. В одном варианте осуществления соответствующий культуральный субстрат представляет собой компонент внеклеточного матрикса, такой как, например, полученный из базальной мембраны или компонент, который может участвовать в лиганд-рецепторном взаимодействии с адгезивными молекулами. В одном варианте осуществления соответствующий культуральный субстрат представляет собой MATRIGEL® (Becton Dickenson). MATRIGEL® представляет собой растворимый препарат из клеток опухоли Энгельбрета-Хольма-Сварма, который при комнатной температуре превращается в гель и образует восстановленную базальную мембрану.
В качестве альтернативы допустимо использовать другие компоненты внеклеточного матрикса и смеси компонентов. В зависимости от типа пролиферирующих клеток данные компоненты могут включать в себя по отдельности или в различных комбинациях ламинин, фибронектин, протеогликан, энтактин, гепарансульфат и т.п.
Плюрипотентные стволовые клетки можно высевать на субстрат с соответствующим распределением и при наличии среды, поддерживающей выживаемость, размножение и сохранение желательных характеристик клеток. Все данные характеристики улучшаются при тщательном подходе к распределению при посеве и могут быть без труда определены специалистом в данной области.
Соответствующую культуральную среду можно создать из следующих компонентов, таких как, например, модифицированная по способу Дульбекко среда Игла (DMEM), Gibco № 11965-092, нокаутная модифицированная по способу Дульбекко среда Игла (KO DMEM), Gibco № 10829-018, основная среда Хэма F12/50% DMEM, 200 мМ L-глутамина, Gibco № 15039-027; раствор не относящихся к незаменимым аминокислот, Gibco 11140-050; β-меркаптоэтанол, Sigma № M7522; рекомбинантный основной фактор роста фибробластов человека (bFGF), Gibco № 13256-029.
Образование панкреатических эндокринных клеток-предшественников
В одном варианте осуществления настоящее изобретение представляет способ получения панкреатических эндокринных клеток-предшественников, включающий следующие стадии:
a. культивирование плюрипотентных стволовых клеток,
b. дифференцирование плюрипотентных стволовых клеток в клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы,
с. дифференцирование клеток, экспрессирующих маркеры, характерные для линии дефинитивной энтодермы, в клетки, экспрессирующие маркеры, характерные для линии панкреатической энтодермы, а также
d. дифференцирование клеток, экспрессирующих маркеры, характерные для линии панкреатической энтодермы, в панкреатические эндокринные клетки-предшественники.
Плюрипотентные стволовые клетки, которые можно использовать в настоящем изобретении, включают в себя, например, эмбриональные стволовые клетки человека линии H9 (код Национальных институтов здравоохранения (NIH): WA09), эмбриональные стволовые клетки человека линии H1 (код NIH: WA01), эмбриональные стволовые клетки человека линии H7 (код NIH: WA07), а также эмбриональные стволовые клетки человека линии SA002 (Cellartis, Швеция). Также для использования в настоящем изобретении допустимы клетки, экспрессирующие по меньшей мере один из следующих маркеров, характерных для плюрипотентных клеток: ABCG2, CRIPTO, CD9, FOXD3, Connexin43, Connexin45, OCT4, SOX2, Nanog, hTERT, UTF1, ZFP42, SSEA-3, SSEA-4, Tra-1-60, Tra-1-81.
Маркеры, характерные для линии дефинитивной энтодермы, выбирают из группы, состоящей из SOX17, GATA4, HNF3 beta, GSC, CER1, Nodal, FGF8, Brachyury, Mix-подобного белка гомеобокса, FGF4, CD48, эомезодермина (EOMES), DKK4, FGF17, GATA6, CXCR4, C-Kit, CD99 и OTX2. Для использования в настоящем изобретении допустима клетка, экспрессирующая по меньшей мере один из маркеров, характерных для линии дефинитивной энтодермы. В одном аспекте настоящего изобретения клетка, экспрессирующая маркеры, характерные для линии дефинитивной энтодермы, представляет собой клетку-предшественник первичной полоски. В альтернативном аспекте настоящего изобретения клетка, экспрессирующая маркеры, характерные для линии дефинитивной энтодермы, представляет собой клетку мезэнтодермы. В альтернативном аспекте клетка, экспрессирующая маркеры, характерные для линии дефинитивной энтодермы, представляет собой клетку дефинитивной энтодермы.
Маркеры, характерные для линии панкреатической энтодермы, выбирают из группы, состоящей из PDX1, HNF1 beta, HNF6, HB9 и PROX1. Для использования в настоящем изобретении допустима клетка, экспрессирующая по меньшей мере один из маркеров, характерных для линии панкреатической энтодермы. В одном аспекте настоящего изобретения клетка, экспрессирующая маркеры, характерные для линии панкреатической энтодермы, представляет собой клетку панкреатической энтодермы.
Маркеры, характерные для панкреатических эндокринных клеток-предшественников, выбирают из группы, состоящей из NGN3, NKX6.1, NeuroD, ISL1, PDX1, PAX4, NKX2.2 или ARX. Для использования в настоящем изобретении допустима клетка, экспрессирующая по меньшей мере один из маркеров, характерных для панкреатических эндокринных клеток-предшественников.
Образование клеток, экспрессирующих маркеры, характерные для линии дефинитивной энтодермы
Плюрипотентные стволовые клетки можно дифференцировать в клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, с использованием любого способа, известного специалистам в данной области, или любого способа, предложенного в настоящем изобретении.
Например, плюрипотентные стволовые клетки можно дифференцировать в клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, в соответствии со способами, раскрытыми в публикации D’Amour et al, Nature Biotechnology 23, 1534–1541 (2005 г.).
Например, плюрипотентные стволовые клетки можно дифференцировать в клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, в соответствии со способами, раскрытыми в публикации Shinozaki et al, Development 131, 1651–1662 (2004 г.).
Например, плюрипотентные стволовые клетки можно дифференцировать в клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, в соответствии со способами, раскрытыми в публикации McLean et al, Stem Cells 25, 29–38 (2007 г.).
Например, плюрипотентные стволовые клетки можно дифференцировать в клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, в соответствии со способами, раскрытыми в публикации D’Amour et al, Nature Biotechnology 24, 1392–1401 (2006 г.).
Например, плюрипотентные стволовые клетки можно дифференцировать в клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, путем культивирования плюрипотентных стволовых клеток в среде, содержащей активин A в отсутствие сыворотки, затем культивирования клеток с активином A и сывороткой и затем культивирования клеток с активином A и сывороткой в другой концентрации. Пример данного способа раскрыт в публикации Nature Biotechnology 23, 1534–1541 (2005 г.).
Например, плюрипотентные стволовые клетки можно дифференцировать в клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, путем культивирования плюрипотентных стволовых клеток в среде, содержащей активин А в отсутствие сыворотки, затем культивирования клеток с активином A и сывороткой в другой концентрации. Пример данного способа раскрыт в публикации D’ Amour et al, Nature Biotechnology, 2005 г.
Например, плюрипотентные стволовые клетки можно дифференцировать в клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, путем культивирования плюрипотентных стволовых клеток в среде, содержащей активин А и Wnt-лиганд в отсутствие сыворотки, затем удаления Wnt-лиганда и культивирования клеток с активином A и сывороткой. Пример использования данного способа раскрыт в публикации Nature Biotechnology 24, 1392–1401 (2006 г.).
Например, плюрипотентные стволовые клетки можно дифференцировать в клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, путем обработки плюрипотентных стволовых клеток в соответствии со способами, раскрытыми в заявке на патент США № 11/736908, переуступленной LifeScan, Inc.
Например, плюрипотентные стволовые клетки можно дифференцировать в клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, путем обработки плюрипотентных стволовых клеток в соответствии со способами, раскрытыми в заявке на патент США № 11/779311, переуступленной LifeScan, Inc.
Например, плюрипотентные стволовые клетки можно дифференцировать в клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, путем обработки плюрипотентных стволовых клеток в соответствии со способами, раскрытыми в заявке на патент США № 60/990529.
Например, плюрипотентные стволовые клетки можно дифференцировать в клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, путем обработки плюрипотентных стволовых клеток в соответствии со способами, раскрытыми в заявке на патент США № 61/076889.
Например, плюрипотентные стволовые клетки можно дифференцировать в клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, путем обработки плюрипотентных стволовых клеток в соответствии со способами, раскрытыми в заявке на патент США № 61/076900.
Например, плюрипотентные стволовые клетки можно дифференцировать в клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, путем обработки плюрипотентных стволовых клеток в соответствии со способами, раскрытыми в заявке на патент США № 61/076908.
Например, плюрипотентные стволовые клетки можно дифференцировать в клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, путем обработки плюрипотентных стволовых клеток в соответствии со способами, раскрытыми в заявке на патент США № 61/076915.
Характеристика клеток, экспрессирующих маркеры, характерные для линии дефинитивной энтодермы
Образование клеток, экспрессирующих маркеры, характерные для линии дефинитивной энтодермы, можно определить путем проверки на наличие маркеров до и после выполнения конкретного протокола. Плюрипотентные стволовые клетки, как правило, не экспрессируют такие маркеры. Таким образом, дифференцирование плюрипотентных клеток определяют по началу экспрессии таких маркеров клетками.
Эффективность дифференцирования можно определить путем воздействия на обрабатываемую популяцию клеток веществом (таким как антитело), специфически распознающим белковый маркер, экспрессированный клетками, экспрессирующими маркеры, характерные для линии дефинитивной энтодермы.
Способы оценки экспрессии белковых маркеров и маркеров нуклеиновых кислот в культивированных или выделенных клетках являются стандартными для данной области. Они включают в себя количественную полимеразную цепную реакцию с обратной транскриптазой (ОТ-ПЦР), Нозерн-блот, гибридизацию in situ (см., например, Current Protocols in Molecular Biology (Ausubel et al., ред. 2001 г., дополнение)), а также иммунологические анализы, такие как иммуногистохимический анализ среза материала, Вестерн-блоттинг, а для маркеров, доступных в интактных клетках, - способ проточной цитометрии (FACS) (см., например, Harlow and Lane, Using Antibodies: A Laboratory Manual, New York: Cold Spring Harbor Laboratory Press (1998 г.)).
Характеристики плюрипотентных стволовых клеток хорошо известны специалистам в данной области, и продолжается выявление дополнительных характеристик плюрипотентных стволовых клеток. Маркеры плюрипотентных стволовых клеток включают в себя, например, экспрессию одного или более из следующих маркеров: ABCG2, CRIPTO, FOXD3, Connexin43, Connexin45, OCT4, SOX2, Nanog, hTERT, UTF1, ZFP42, SSEA-3, SSEA-4, Tra-1-60, Tra-1-81.
После обработки плюрипотентных стволовых клеток с применением способов настоящего изобретения дифференцированные клетки можно очистить путем воздействия на обрабатываемую популяцию клеток веществом (таким как антитело), специфически распознающим белковый маркер, такой как CXCR4, экспрессированный клетками, экспрессирующими маркеры, характерные для линии дефинитивной энтодермы.
Образование клеток экспрессирующих маркеры, характерные для линии панкреатической энтодермы, из клеток, экспрессирующих маркеры, характерные для линии дефинитивной энтодермы
Клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, можно дифференцировать в клетки, экспрессирующие маркеры, характерные для линии панкреатической энтодермы, с использованием любого способа, известного специалистам в данной области, или любого способа, предложенного в настоящем изобретении.
Например, клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, можно дифференцировать в клетки, экспрессирующие маркеры, характерные для линии панкреатической энтодермы, в соответствии со способами, раскрытыми в публикации D’Amour et al, Nature Biotechnology 24, 1392–1401 (2006 г.).
Например, клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, дополнительно дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатической энтодермы, путем обработки клеток, экспрессирующих маркеры, характерные для линии дефинитивной энтодермы, фактором роста фибробластов и ингибитором сигнального пути Hedgehog KAAD-циклопамином с последующим удалением среды, содержащей фактор роста фибробластов и KAAD-циклопамин, и затем культивированием клеток в среде, содержащей ретиноевую кислоту, фактор роста фибробластов и KAAD-циклопамин. Пример использования данного способа раскрыт в публикации Nature Biotechnology 24, 1392–1401 (2006 г.).
В одном аспекте настоящего изобретения клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, дополнительно дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатической энтодермы, путем обработки клеток, экспрессирующих маркеры, характерные для линии дефинитивной энтодермы, ретиноевой кислотой и по меньшей мере одним фактором роста фибробластов в течение периода времени в соответствии со способами, раскрытыми в заявке на патент США № 11/736908, переуступленной LifeScan, Inc.
В одном аспекте настоящего изобретения клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, дополнительно дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатической энтодермы, путем обработки клеток, экспрессирующих маркеры, характерные для линии дефинитивной энтодермы, ретиноевой кислотой и по меньшей мере одним фактором роста фибробластов в течение периода времени в соответствии со способами, раскрытыми в заявке на патент США № 11/779311, переуступленной LifeScan, Inc.
В одном аспекте настоящего изобретения клетки, экспрессирующие маркеры, характерные для линии дефинитивной энтодермы, дополнительно дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатической энтодермы, путем обработки клеток, экспрессирующих маркеры, характерные для линии дефинитивной энтодермы, в соответствии со способами, раскрытыми в заявке на патент США № 60/990529.
Характеристика клеток, экспрессирующих маркеры, характерные для линии панкреатической энтодермы
Маркеры, характерные для линии панкреатической энтодермы, хорошо известны специалистам в данной области, и продолжают выявляться дополнительные маркеры, характерные для линии панкреатической энтодермы. Такие маркеры можно использовать для подтверждения того, что клетки, обработанные в соответствии с настоящим изобретением, дифференцировались и приобрели характерные свойства линии панкреатической энтодермы. Конкретные маркеры, характерные для линии панкреатической энтодермы, включают в себя экспрессию одного или более факторов транскрипции, таких как, например, HLXB9, PTF1 alpha, PDX1, HNF6, HNF-1 beta.
Эффективность дифференцирования можно определить путем воздействия на обрабатываемую популяцию клеток веществом (таким как антитело), специфически распознающим белковый маркер, экспрессированный клетками, экспрессирующими маркеры, характерные для линии панкреатической энтодермы.
Способы оценки экспрессии белковых маркеров и маркеров нуклеиновых кислот в культивированных или выделенных клетках являются стандартными для данной области. Они включают в себя количественную полимеразную цепную реакцию с обратной транскриптазой (ОТ-ПЦР), Нозерн-блот, гибридизацию in situ (см., например, Current Protocols in Molecular Biology (Ausubel et al., ред. 2001 г., дополнение)), а также иммунологические анализы, такие как иммуногистохимический анализ среза материала, Вестерн-блоттинг, а для маркеров, доступных в интактных клетках, - способ проточной цитометрии (FACS) (см., например, Harlow and Lane, Using Antibodies: A Laboratory Manual, New York: Cold Spring Harbor Laboratory Press (1998 г.)).
Образование панкреатических эндокринных клеток-предшественников из клеток, экспрессирующих маркеры, характерные для линии панкреатической энтодермы
В одном аспекте настоящего изобретения клетки, экспрессирующие маркеры, характерные для линии панкреатической энтодермы, дифференцируются в панкреатические эндокринные клетки-предшественники путем культивирования клеток, экспрессирующих маркеры, характерные для линии панкреатической энтодермы, в среде с добавлением фактора, способного ингибировать костный морфогенетический белок (BMP), и ингибитора киназы TGF-β рецептора I.
В одном варианте осуществления фактором, способным ингибировать BMP, является Ноггин. Ноггин можно использовать в концентрации от приблизительно 100 пг/мл до приблизительно 500 мкг/мл. В одном варианте осуществления Ноггин используют в концентрации 100 нг/мл.
В одном варианте осуществления ингибитор киназы TGF-β рецептора I представляет собой ингибитор II ALK5 (Calbiochem, штат Калифорния). Ингибитор II ALK5 можно использовать в концентрации от приблизительно 0,1 мкM до приблизительно 10 мкM. В одном варианте осуществления ингибитор II ALK5 используют в концентрации 1 мкM.
В одном варианте осуществления среда представляет собой DMEM с содержанием глюкозы 4 500 мг/л и 1% B27.
В одном варианте осуществления клетки культивируют в культуральной среде в течение приблизительно четырех суток.
Эффективность дифференцирования можно определить путем воздействия на обрабатываемую популяцию клеток веществом (таким как антитело), специфически распознающим белковый маркер, экспрессированный панкреатическими эндокринными клетками-предшественниками.
Способы оценки экспрессии белковых маркеров и маркеров нуклеиновых кислот в культивированных или выделенных клетках являются стандартными для данной области. Они включают в себя количественную полимеразную цепную реакцию с обратной транскриптазой (ОТ-ПЦР), Нозерн-блот, гибридизацию in situ (см., например, Current Protocols in Molecular Biology (Ausubel et al., ред. 2001 г., дополнение)), а также иммунологические анализы, такие как иммуногистохимический анализ среза материала, Вестерн-блоттинг, а для маркеров, доступных в интактных клетках, - способ проточной цитометрии (FACS) (см., например, Harlow and Lane, Using Antibodies: A Laboratory Manual, New York: Cold Spring Harbor Laboratory Press (1998 г.)).
Характеристики плюрипотентных стволовых клеток хорошо известны специалистам в данной области, и продолжается выявление дополнительных характеристик плюрипотентных стволовых клеток. Маркеры плюрипотентных стволовых клеток включают в себя, например, экспрессию одного или более из следующих маркеров: ABCG2, CRIPTO, FOXD3, Connexin43, Connexin45, OCT4, SOX2, Nanog, hTERT, UTF1, ZFP42, SSEA-3, SSEA-4, Tra-1-60, Tra-1-81.
После обработки плюрипотентных стволовых клеток с применением способов настоящего изобретения дифференцированные клетки можно очистить путем воздействия на обрабатываемую популяцию клеток веществом (таким как антитело), специфически распознающим белковый маркер, такой как CXCR4, экспрессированный клетками, экспрессирующими маркеры, характерные для линии панкреатической энтодермы.
Маркеры, характерные для линии панкреатической энтодермы, выбирают из группы, состоящей из PDX1, HNF-1 beta, PTF1 alpha, HNF6, HB9 и PROX1. Для использования в настоящем изобретении допустима клетка, экспрессирующая по меньшей мере один из маркеров, характерных для линии панкреатической энтодермы. В одном аспекте настоящего изобретения клетка, экспрессирующая маркеры, характерные для линии панкреатической энтодермы, представляет собой клетку панкреатической энтодермы.
Маркеры, характерные для панкреатических эндокринных клеток-предшественников, выбирают из группы, состоящей из NGN3, NKX6.1, NEUROD, ISL1, PDX1, PAX4, NKX2.2, PAX6 или ARX.
Образование клеток, экспрессирующих маркеры, характерные для линии панкреатических эндокринных клеток, из панкреатических эндокринных клеток-предшественников
В одном варианте осуществления панкреатические эндокринные клетки-предшественники, полученные с использованием способов настоящего изобретения, могут дополнительно дифференцироваться в клетки, экспрессирующие маркеры, характерные для линии панкреатических эндокринных клеток.
Панкреатические эндокринные клетки-предшественники можно дифференцировать в клетки, экспрессирующие маркеры, характерные для линии панкреатических эндокринных клеток, с использованием любого способа, известного специалистам в данной области, или любого способа, предложенного в настоящем изобретении.
Например, панкреатические эндокринные клетки-предшественники, полученные в соответствии со способами настоящего изобретения, дополнительно дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатических эндокринных клеток, путем культивирования панкреатических эндокринных клеток-предшественников в среде, содержащей экзендин-4, затем удаления среды, содержащей экзендин-4, и затем культивирования клеток в среде, содержащей экзендин-1, IGF1 и HGF. Пример данного способа раскрыт в публикации D’ Amour et al, Nature Biotechnology, 2006 г.
Например, панкреатические эндокринные клетки-предшественники, полученные в соответствии со способами настоящего изобретения, дополнительно дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатических эндокринных клеток, путем культивирования панкреатических эндокринных клеток-предшественников в среде, содержащей DAPT (Sigma-Aldrich, штат Миссури) и экзендин-4. Пример данного способа раскрыт в публикации D’ Amour et al, Nature Biotechnology, 2006 г.
Например, панкреатические эндокринные клетки-предшественники, полученные в соответствии со способами настоящего изобретения, дополнительно дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатических эндокринных клеток, путем культивирования панкреатических эндокринных клеток-предшественников в среде, содержащей экзендин-4. Пример данного способа раскрыт в публикации D’ Amour et al, Nature Biotechnology, 2006 г.
Например, панкреатические эндокринные клетки-предшественники, полученные в соответствии со способами настоящего изобретения, дополнительно дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатических эндокринных клеток, путем обработки панкреатических эндокринных клеток-предшественников фактором, ингибирующим сигнальный путь Notch, в соответствии со способами, раскрытыми в заявке на патент США № 11/736908, переуступленной LifeScan, Inc.
Например, панкреатические эндокринные клетки-предшественники, полученные в соответствии со способами настоящего изобретения, дополнительно дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатических эндокринных клеток, путем обработки панкреатических эндокринных клеток-предшественников фактором, ингибирующим сигнальный путь Notch, в соответствии со способами, раскрытыми в заявке на патент США № 11/779311, переуступленной LifeScan, Inc.
Например, панкреатические эндокринные клетки-предшественники, полученные в соответствии со способами настоящего изобретения, дополнительно дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатических эндокринных клеток, путем обработки панкреатических эндокринных клеток-предшественников фактором, ингибирующим сигнальный путь Notch, в соответствии со способами, раскрытыми в заявке на патент США № 60/953178, переуступленной LifeScan, Inc.
Например, панкреатические эндокринные клетки-предшественники, полученные в соответствии со способами настоящего изобретения, дополнительно дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатических эндокринных клеток, путем обработки панкреатических эндокринных клеток-предшественников фактором, ингибирующим сигнальный путь Notch, в соответствии со способами, раскрытыми в заявке на патент США № 60/990529, переуступленной LifeScan, Inc.
Маркеры, характерные для линии панкреатических эндокринных клеток, выбирают из группы, состоящей из NEUROD, ISL1, PDX1, NKX6.1, PAX4, PAX6, NGN3 и NKX2.2. В одном варианте осуществления панкреатическая эндокринная клетка способна экспрессировать по меньшей мере один из следующих гормонов: инсулин, глюкагон, соматостатин и панкреатический полипептид. Для использования в настоящем изобретении допустима клетка, экспрессирующая по меньшей мере один из маркеров, характерных для линии панкреатических эндокринных клеток. В одном аспекте настоящего изобретения клетка, экспрессирующая маркеры, характерные для линии панкреатических эндокринных клеток, представляет собой панкреатическую эндокринную клетку. Панкреатическая эндокринная клетка может представлять собой панкреатическую клетку, экспрессирующую гормоны. Панкреатическая эндокринная клетка может альтернативно представлять собой панкреатическую клетку, секретирующую гормоны.
В одном аспекте настоящего изобретения панкреатическая эндокринная клетка представляет собой клетку, экспрессирующую маркеры, характерные для линии β-клеток. Клетка, экспрессирующая маркеры, характерные для линии β-клеток, экспрессирует PDX1 и по меньшей мере один из следующих факторов транскрипции: NGN-3, NKX2.2, NKX6.1, NEUROD, ISL1, HNF3 beta, MAFA, PAX4 и PAX6. В одном аспекте настоящего изобретения клетка, экспрессирующая маркеры, характерные для линии β-клеток, представляет собой β-клетку.
Способы лечения
В одном аспекте настоящее изобретение предлагает способ лечения пациента, страдающего диабетом 1 типа или подвергающегося риску развития данного заболевания. В одном варианте осуществления способ включает в себя культивирование плюрипотентных стволовых клеток, дифференцирование плюрипотентных стволовых клеток in vitro в линию β-клеток и имплантирование клеток линии β-клеток пациенту. В альтернативном варианте осуществления способ включает в себя культивирование плюрипотентных стволовых клеток, дифференцирование плюрипотентных стволовых клеток in vitro в панкреатические эндокринные клетки-предшественники и имплантирование панкреатических эндокринных клеток-предшественников пациенту.
В еще одном аспекте настоящее изобретение предлагает способ лечения пациента, страдающего диабетом 2 типа или подвергающегося риску развития данного заболевания. В одном варианте осуществления способ включает культивирование плюрипотентных стволовых клеток, дифференцирование плюрипотентных стволовых клеток in vitro в линию β-клеток и имплантирование клеток линии β-клеток пациенту. В альтернативном варианте осуществления способ включает в себя культивирование плюрипотентных стволовых клеток, дифференцирование плюрипотентных стволовых клеток in vitro в панкреатические эндокринные клетки-предшественники и имплантирование панкреатических эндокринных клеток-предшественников пациенту.
При необходимости пациенту можно дополнительно предоставлять лечение фармацевтическими веществами или биологически активными веществами, способствующими выживаемости и функционированию трансплантированных клеток. Данные вещества могут включать в себя, например, среди прочих, инсулин, члены семейства TGF-β, включая TGF-β1, 2 и 3, костные морфогенетические белки (BMP-2, -3, -4, -5, -6, -7, -11, -12 и -13), факторы роста фибробластов -1 и -2, тромбоцитарный фактор роста –AA и –BB, плазму, богатую тромбоцитами, инсулиноподобный фактор роста (IGF-I, II), фактор дифференцирования роста (GDF-5, -6, -7, -8, -10, -15), фактор роста эндотелия сосудов (VEGF), плейотропин, эндотелин. Другие фармацевтические соединения могут включать в себя, например, никотинамид, глюкагоноподобный пептид-I (GLP-1) и II, миметические вещества GLP-1 и 2, экзендин-4, ретиноевую кислоту, паратиреоидный гормон, ингибиторы митоген-активируемой протеинкиназы (MAPK), такие как, например, соединения, раскрытые в опубликованной заявке США № 2004/0209901 и опубликованной заявке США № 2004/0132729.
Плюрипотентные стволовые клетки можно дифференцировать в инсулин-продуцирующую клетку перед трансплантацией реципиенту. В конкретном варианте осуществления плюрипотентные стволовые клетки представляют собой клетки, полностью дифференцированные в β-клетки перед трансплантацией реципиенту. Плюрипотентные стволовые клетки можно альтернативно трансплантировать реципиенту в недифференцированном или частично дифференцированном состоянии. Дальнейшее дифференцирование может происходить в организме реципиента.
Клетки дефинитивной энтодермы или альтернативно клетки панкреатической энтодермы, либо альтернативно β-клетки можно имплантировать в форме дисперсных клеток или клеток, образующих кластеры, которые можно вводить в воротную вену печени методом инфузии. Клетки можно альтернативно вводить в биосовместимые разлагающиеся полимерные опорные материалы, пористые неразлагающиеся устройства или в инкапсулированном виде для защиты от иммунного ответа организма-хозяина. Клетки можно имплантировать в подходящее место в организме реципиента. Места имплантации включают в себя, например, печень, естественную поджелудочную железу, пространство под почечной капсулой, сальник, брюшную полость, субсерозное пространство, кишечник, желудок или подкожный карман.
Для стимуляции дальнейшего дифференцирования, выживаемости или активности имплантированных клеток до, одновременно с или после введения клеток можно вводить дополнительные факторы, такие как факторы роста, антиоксиданты или противовоспалительные вещества. В определенных вариантах осуществления факторы роста применяют для дифференцирования введенных клеток in vivo. Данные факторы могут секретироваться эндогенными клетками и воздействовать на введенные клетки in situ. Дифференцирование имплантированных клеток можно индуцировать любой комбинацией эндогенных и введенных экзогенно факторов роста, известных в данной области.
Количество клеток, используемых при имплантации, зависит от ряда различных факторов, включая состояние пациента и его реакцию на лечение, и может быть определено специалистом в данной области.
В одном аспекте настоящее изобретение представляет способ лечения пациента, страдающего диабетом или подвергающегося риску развития данного заболевания. Данный способ включает в себя культивирование плюрипотентных стволовых клеток, дифференцирование культивированных клеток in vitro в линию β-клеток и включение клеток в опорный материал с трехмерной структурой. Клетки можно поддерживать in vitro на данном опорном материале перед имплантацией пациенту. Опорный материал, содержащий клетки, альтернативно можно имплантировать непосредственно в организм пациента без дополнительного культивирования in vitro. В опорный материал может быть необязательно включено по меньшей мере одно фармацевтическое вещество, обеспечивающее выживаемость и функционирование трансплантированных клеток.
Опорные материалы, допустимые для использования в целях настоящего изобретения, включают в себя тканевые матрицы, каналы, перегородки и резервуары, применяемые для восстановления тканей. В частности, для использования на практике способов настоящего изобретения допустимы синтетические и природные материалы в форме пен, губок, гелей, гидрогелей, тканых и нетканых структур, применяемых in vitro и in vivo для реконструкции или регенерации биологической ткани, а также для доставки хемотаксических веществ, индуцирующих рост ткани. См., например, материалы, раскрытые в патенте США № 5770417, патенте США № 6022743, патенте США № 5567612, патенте США № 5759830, патенте США № 6626950, патенте США № 6534084, патенте США № 6306424, патенте США № 6365149, патенте США № 6599323, патенте США № 6656488; опубликованной заявке США № 2004/0062753 A1, патенте США № 4557264 и патенте США № 6333029.
Для того чтобы создать опорный материал с включенным фармацевтическим веществом, фармацевтическое вещество можно смешать с раствором полимера перед получением опорного материала. Фармацевтическое вещество можно альтернативно нанести на изготовленный опорный материал, предпочтительно при наличии фармацевтического носителя. Фармацевтическое вещество может присутствовать в виде жидкости, мелкодисперсного твердого вещества или любой другой подходящей физической форме. В опорный материал альтернативно можно добавить эксципиенты для изменения скорости высвобождения фармацевтического вещества. В альтернативном варианте осуществления в опорный материал включают по меньшей мере одно фармацевтическое соединение, представляющее собой противовоспалительное соединение, такое как, например, примеры соединений, раскрытые в патенте США № 6509369.
В опорный материал можно включить по меньшей мере одно фармацевтическое соединение, представляющее собой антиапоптозное соединение, такое как, например, соединения, раскрытые в патенте США № 6793945.
В опорный материал также можно включить по меньшей мере одно фармацевтическое соединение, представляющее собой ингибитор фиброза, такой как, например, соединения, раскрытые в патенте США № 6331298.
В опорный материал также можно включить по меньшей мере одно фармацевтическое соединение, способное стимулировать ангиогенез, такое как, например, соединения, раскрытые в опубликованной заявке США № 2004/0220393 и опубликованной заявке США № 2004/0209901.
В опорный материал также можно включить по меньшей мере одно фармацевтическое соединение, представляющее собой иммуносупрессивное соединение, такое как, например, соединения, раскрытые в опубликованной заявке США № 2004/0171623.
В опорный материал также можно включить по меньшей мере одно фармацевтическое соединение, представляющее собой фактор роста, такой как, например, среди прочих, члены семейства TGF-β, включая TGF-β1, 2 и 3, костные морфогенетические белки (BMP-2, -3, -4, -5, -6, -7, -11, -12 и -13), факторы роста фибробластов -1 и -2, тромбоцитарный фактор роста -AA и –BB, плазму, богатую тромбоцитами, инсулиноподобный фактор роста (IGF-I, II), фактор дифференцирования роста (GDF-5, -6, -8, -10, -15), фактор роста эндотелия сосудов (VEGF), плейотропин, эндотелин. Другие фармацевтические соединения могут включать в себя, например, никотинамид, индуцируемый гипоксией фактор 1-alpha, глюкагоноподобный пептид-I (GLP-1), миметические вещества GLP-1 и GLP-2, и II, экзендин-4, nodal, Ноггин, фактор роста нервной ткани, ретиноевую кислоту, паратиреоидный гормон, тенасцин-C, тропоэластин, пептиды, полученные из тромбина, кателицидины, дефензины, ламинин, биологические пептиды, содержащие связывающие клетки и гепарин домены протеинов адгезивного внеклеточного матрикса, такие как, например, фибронектин и витронектин, ингибиторы MAPK, такие как, например, соединения, раскрытые в опубликованной заявке США №№ 2004/0209901 и опубликованной заявке США № 2004/0132729.
Включение клеток настоящего изобретения в каркас можно осуществить путем простого нанесения клеток на каркас. Клетки могут входить внутрь каркаса путем простой диффузии (J. Pediatr. Surg. 23 (1 ч. 2): 3–9 (1988 г.)). Разработали несколько других подходов для повышения эффективности посева клеток. Например, для посева хондроцитов на каркасы из полигликолевой кислоты используют центрифужные пробирки (Biotechnol. Prog. 14(2): 193–202 (1998 г.)). Другой подход к посеву клеток представляет собой использование центрифугирования, создающего минимальное воздействие на отобранные клетки и повышающего эффективность посева. Например, в публикации Yang et al. описан разработанный способ посева клеток (J.Biomed. Mater. Res. 55(3): 379–86 (2001 г.)), именуемый центрифужной иммобилизацией клеток (CCI).
Настоящее изобретение далее иллюстрируется без ограничений следующими примерами.
ИСПОЛЬЗОВАННАЯ ЛИТЕРАТУРА
Karvonen, M. et al. Incidence of childhood type 1 diabetes worldwide. Diabetes Mondiale (DiaMond) Project Group. Diabetes Care 23, 1516–26 (2000 г.).
Mathis, D., Vence, L. & Benoist, C. Beta-cell death during progression to diabetes. Nature 414, 792–8 (2001 г.).
Ryan, E.A. et al. Clinical outcomes and insulin secretion after islet transplantation with the Edmonton protocol. Diabetes 50, 710–9 (2001 г.).
Shapiro, A.M. et al. Islet transplantation in seven patients with type 1 diabetes mellitus using a glucocorticoid-free immunosuppressive regimen. N Engl J Med 343, 230–8 (2000 г.).
Cure, P. et al. Improved Metabolic Control and Quality of Life in Seven Patients With Type 1 Diabetes Following Islet After Kidney Transplantation. Transplantation 85, 801–812 (2008 г.).
Fung, M.A. et al. The effect of medical therapy and islet cell transplantation on diabetic nephropathy: an interim report. Transplantation 84, 17–22 (2007 г.).
Brown, L. & Edelman, E.R. Optimal control of blood glucose: the diabetic patient or the machine? Sci Transl Med 2, 27 стр. 18 (2010 г.).
Guo, T. & Hebrok, M. Stem cells to pancreatic beta-cells: new sources for diabetes cell therapy. Endocr Rev 30, 214–27 (2009 г.).
Ricordi, C. & Edlund, H. Toward a renewable source of pancreatic beta-cells. Nat Biotechnol 26, 397–8 (2008 г.).
Rajagopal, J., Anderson, W.J., Kume, S., Martinez, O.I. & Melton, D.A. Insulin staining of ES cell progeny from insulin uptake. Science 299, 363 (2003 г.).
Kroon, E. et al. Pancreatic endoderm derived from human embryonic stem cells generates glucose-responsive insulin-secreting cells in vivo. Nat Biotechnol 26, 443–52 (2008 г.).
D'Amour, K.A. et al. Production of pancreatic hormone-expressing endocrine cells from human embryonic stem cells. Nat Biotechnol 24, 1392–401 (2006 г.).
D'Amour, K.A. et al. Efficient differentiation of human embryonic stem cells to definitive endoderm. Nat Biotechnol 23, 1534–41 (2005 г.).
Matveyenko AV, Georgia S, Bhushan A, Butler PC. Inconsistent formation and non function of insulin positive cells from pancreatic endoderm derived from human embryonic stem cells in athymic nude rats. Am J Physiol Endocrinol Metab. 29 июня 2010 г. [электронное издание до печатного издания].
Lee SH, Hao E, Savinov AY, Geron I, Strongin AY, Itkin-Ansari P. Human beta-cell precursors mature into functional insulin-producing cells in an immunoisolation device: implications for diabetes cell therapies. Transplantation. 15 апреля 2009 г.; 87(7):983–91.
Yang Z, Chen M, Fialkow LB, Ellett JD, Wu R, Nadler JL. Survival of pancreatic islet xenografts in NOD mice with the theracyte device. Transplant Proc. Декабрь 2002 г.; 34(8):3349–50.
Panepinto LM и Phillips RW. The Yucatan miniature pig: characterization and utilization in biomedical research. Lab Anim Sci 36: 344–347, 1986 г.
Larsen MO и Rolin B. Use of the Gottingen minipig as a model of diabetes, with special focus on type 1 diabetes research. Ilar J 45: 303–313, 2004 г.
Miller ER и Ullrey DE. The pig as a model for human nutrition. Annu Rev Nutr 7: 361–382, 1987 г.
Vodicka P, Smetana K, Jr., Dvorankova B, Emerick T, Xu YZ, Ourednik J, Ourednik V и Motlik J. The miniature pig as an animal model in biomedical research. Ann N Y Acad Sci 1049: 161–171, 2005 г.
Kurihara-Bergstrom T, Woodworth M, Feisullin S и Beall P. Characterization of the Yucatan miniature pig skin and small intestine for pharmaceutical applications. Lab Anim Sci 36: 396–399, 1986 г.
Swindle MM, Smith AC, Laber-Laird K и Dungan L. Swine in Biomedical Research: Management and Models. Ilar J 36: 1–5, 1994 г.
Bellinger DA, Merricks EP и Nichols TC. Swine models of type 2 diabetes mellitus: insulin resistance, glucose tolerance, and cardiovascular complications. Ilar J 47: 243–258, 2006 г.
Hainsworth DP, Katz ML, Sanders DA, Sanders DN, Wright EJ и Sturek M. Retinal capillary basement membrane thickening in a porcine model of diabetes mellitus. Comp Med 52: 523–529, 2002 г.
Marshall M, Oberhofer H и Staubesand J. Early micro- and macro-angiopathy in the streptozotocin diabetic minipig. Res Exp Med (Berl) 177: 145–158, 1980 г.
Phillips RW, Panepinto LM, Will DH и Case GL. The effects of alloxan diabetes on Yucatan miniature swine and their progeny. Metabolism 29: 40–45, 1980 г.
Eventov-Friedman S, Tchorsh D, Katchman H, Shezen E, Aronovich A, Hecht G, Dekel B, Rechavi G, Blazar BR, Feine I, Tal O, Freud E, Reisner Y. Embryonic pig pancreatic tissue transplantation for the treatment of diabetes. PLoS Med. Июль 2006 г.; 3(7):e215.
Castaing M, Péault B, Basmaciogullari A, Casal I, Czernichow P, Scharfmann R. Blood glucose normalization upon transplantation of human embryonic pancreas into beta-cell-deficient SCID mice. Diabetologia. Ноябрь 2001 г.; 44(11):2066–76.
Larsen MO, Rolin B, Raun K, Bjerre Knudsen L, Gotfredsen CF, Bock T. Evaluation of beta-cell mass and function in the Göttingen minipig. Diabetes Obes Metab доп. 2:170–9, 2007 г.
van der Windt DJ, Echeverri GJ, Ijzermans JN, Cooper DK. The choice of anatomical site for islet transplantation. Cell Transplant. 2008 г.;17(9):1005–14.
ПРИМЕР
Эмбриональные стволовые клетки человека линии H1 культивировали на покрытых материалом MATRIGEL планшетах (разведение 1:30) и дифференцировали в панкреатические эндокринные клетки-предшественники с использованием следующего протокола:
a. среда RPMI (№ 22400 по каталогу, Invitrogen, штат Калифорния) с добавлением 2% BSA (№ 152401 по каталогу, MP Biomedical, штат Огайо) и 100 нг/мл активина A (R&D Systems, штат Миннесота) + 20 нг/мл WNT-3a (№ 1324-WN-002 по каталогу, R&D Systems, штат Миннесота) + 8 нг/мл bFGF (№ 100-18B по каталогу, PeproTech, штат Нью-Джерси) в течение одних суток с последующей обработкой средой RPMI с добавлением 2% BSA и 100 нг/мл активина A + 8 нг/мл bFGF в течение еще двух суток (стадия 1); затем
b. DMEM/F12 (№ 11330 по каталогу, Invitrogen, штат Калифорния) + 2% BSA + 50 нг/мл FGF7 в течение трех суток (стадия 2); затем
с. использовали другую основную среду с добавлением 1% B27 (№ 17504-044, Invitrogen, штат Калифорния) + 50 нг/мл FGF7 + 0,25 мкМ KAAD-циклопамина (№ 239804, Calbiochem, штат Калифорния) + 2 мкМ ретиноевой кислоты (РК) (Sigma, штат Миссури) + 100 нг/мл Ноггина (R&D Systems, штат Миннесота) в течение четырех суток (стадия 3); затем
d. использовали другую основную среду с добавлением 1% B27 (Invitrogen, штат Калифорния) + 100 нг/мл Ноггина + 1 мкМ ингибитора II ALK5 (№ 616452 по каталогу, Calbiochem, штат Калифорния) в течение трех суток (стадия 4).
Содержание публикаций, цитируемых в настоящем документе, полностью включено в настоящий документ путем ссылки. Хотя различные аспекты настоящего изобретения иллюстрируются выше путем ссылки на примеры и предпочтительные варианты осуществления, подразумевается, что объем настоящего изобретения ограничивается не вышеупомянутым описанием, а следующей формулой изобретения, составленной надлежащим образом в соответствии с принципами патентного законодательства.

Изобретение относится к фармацевтической промышленности, а именно к применению популяции инкапсулированных панкреатических эндокринных клеток-предшественников в производстве лекарственного средства для снижения уровня глюкозы в крови у пациента, где панкреатические эндокринные клетки-предшественники дифференцируются in vitro из клеток, экспрессирующих маркеры, характерные для линии поджелудочной энтодермы в среде с добавлением фактора, способного ингибировать ВМР, и ингибитора киназы TGF-бета рецептора I, и где панкреатические эндокринные клетки-предшественники экспрессируют NGN3, NKX6.1, NeuroD, ISL1, PDX1, PAX4, NKX2.2, PAX6 или ARX. Вышеописанное решение позволяет применять популяцию инкапсулированных панкреатических эндокринных клеток-предшественников в производстве лекарственного средства для снижения уровня глюкозы в крови у пациента. 8 з.п. ф-лы, 3 ил., 1 пр.
1. Применение популяции инкапсулированных панкреатических эндокринных клеток-предшественников в производстве лекарственного средства для снижения уровня глюкозы в крови у пациента,
где панкреатические эндокринные клетки-предшественники дифференцируются in vitro из клеток, экспрессирующих маркеры, характерные для линии поджелудочной энтодермы в среде с добавлением фактора, способного ингибировать ВМР, и ингибитора киназы TGF-бета рецептора I, и
где панкреатические эндокринные клетки-предшественники экспрессируют NGN3, NKX6.1, NeuroD, ISL1, PDX1, PAX4, NKX2.2, PAX6 или ARX.
2. Применение по п.1, которое включает трансплантацию популяции инкапсулированных панкреатических эндокринных клеток-предшественников в организм пациента.
3. Применение по п.2, которое включает трансплантацию клеток в печень, естественную поджелудочную железу, пространство под почечной капсулой, сальник, брюшную полость, субсерозное пространство, кишечник, желудок или подкожный карман.
4. Применение по п.1, где фактором, способным ингибировать BMP, является Noggin.
5. Применение по п.1, где ингибитором киназы TGF-β рецептора I является ингибитор II ALK5.
6. Применение по п.1, где средой является DMEM, содержащая 4500 мг/л глюкозы и 1% B27.
7. Применение по п.1, где клетки дифференцируют в культуральной среде четыре дня.
8. Применение по п.1, также включающее факторы роста, антиоксиданты или противовоспалительные вещества.
9. Применение по п.1, где пациент страдает диабетом I или II типа или имеет риск его развития.
| ДИФФЕРЕНЦИРОВКА СТРОМАЛЬНЫХ КЛЕТОК, ПОЛУЧЕННЫХ ИЗ ЖИРОВОЙ ТКАНИ, В ЭНДОКРИННЫЕ КЛЕТКИ ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ И ИХ ИСПОЛЬЗОВАНИЕ | 2002 |
|
RU2351648C2 |
| Dong-Qi Tang et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Пюпитр для работы на пишущих машинах | 1922 |
|
SU86A1 |
| Movassat J et al | |||
| Keratinocyte growth factor and beta-cell differentiation in human fetal pancreatic endocrine precursor cells //Diabetologia | |||
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| V | |||
| Способ изготовления звездочек для французской бороны-катка | 1922 |
|
SU46A1 |
| Улей | 1923 |
|
SU822A1 |
| КОМПОЗИЦИИ ПОДВЕРГНУТЫХ РЕСТРИКЦИИ КЛЕТОК, СПОСОБНЫХ К БЫСТРОМУ РОСТУ, КОТОРЫЕ ПРОДУЦИРУЮТ ВЕЩЕСТВА, ПОДАВЛЯЮЩИЕ ПРОЛИФЕРАЦИЮ КЛЕТОК, И ИХ ПРИМЕНЕНИЕ | 1999 |
|
RU2236855C2 |

