ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Данное изобретение относится к продуцированию вирусных белков в растениях. Более конкретно, настоящее изобретение относится к продуцированию и увеличению продукции вирусоподобных частиц в растениях.
УРОВЕНЬ ТЕХНИКИ
Грипп вызывается РНК-содержащим вирусом семейства Orthomyxoviridae. Существуют три типа этих вирусов: типы А, В и С, и они вызывают три различных вида гриппа.
Вирус гриппа типа А инфицирует млекопитающих (людей, свиней, африканских хорьков, лошадей) и птиц. Это имеет очень большое значение для людей, так как именно этот тип вируса вызывал мировые пандемии. Вирус гриппа В (известный также просто как инфекция типа В) инфицирует только людей. Время от времени он вызывает локальные вспышки гриппа. Вирусы гриппа типа С также инфицируют только людей. Они инфицируют большей частью молодых людей и редко вызывают появление серьезных заболеваний.
Вакцинация обеспечивает защиту против болезни, вызванной подобными агентами, мобилизуя защитные силы организма до появления инфекции. Обычно это осуществляется путем применения живых аттенюированных (ослабленных) или цельных неактивированных форм инфекционных агентов в качестве иммуногенов. Для того чтобы избежать опасности, связанной с применением в качестве вакцины цельных вирусов (таких как убитые или аттенюированные вирусы), применяли в качестве вакцин, например, субъединицы и рекомбинантные вирусные белки. Как пептидные, так и субъединичные вакцины имеют ряд ограничений. Субъединичные вакцины могут проявлять слабую иммуногенность из-за некорректного фолдинга или слабого представления (презентации) антигена. Основная проблема состоит в том, что трудно обеспечить конформацию генно-инженерных белков, имитирующую конформацию антигенов в их природном окружении. Для запуска иммунного ответа необходимо применять подходящие адъюванты и, в случае пептидов, белки-носители. Кроме того, такие вакцины вызывают в основном гуморальные ответы и, следовательно, могут не вызвать появление эффективного иммунитета. Субъединичные вакцины часто оказываются неэффективными для заболеваний, при которых может быть показано наличие защиты при помощи цельных инактивированных вирусов.
Вирусоподобные частицы (VLP) очень похожи на зрелые вирионы, но они не содержат вирусный геномный материал. Следовательно, VLP по своей природе являются нерепликативными, что делает их безопасными для применения в качестве вакцин. Кроме того, могут быть созданы такие VLP, которые экспрессируют гликопротеины на поверхности VLP, имеющей наиболее близкую к нативной физиологическую конфигурацию. Более того, так как VLP похожи на интактные вирионы и представляют собой поливалентные крупнодисперсные структуры, VLP могут быть более эффективными при индуцировании нейтрализации антител с получением гликопротеина, чем растворимые оболочечные белковые антигены с.
VLP были продуцированы в клетках растений (см. международные заявки WO 2009/009876; WO 2009/076778; WO 2010/003225; WO 2010/003235; WO 2011/03522; WO 2010/148511; которые включены в данную заявку посредством отсылки) и в клетках насекомых и в клетках млекопитающих (Noad, R. and Roy, P., 2003, Trends Microbiol 11: 438-44; Neumann et al., 2000, J. Virol., 74, 547-551). Latham and Galarza (2001, J. Virol., 75, 6154-6165) сообщили об образовании VLP гриппа в клетках насекомых, инфицированных рекомбинантным бакуловирусом и совместно экспрессирующих гены гемагглютинина (НА), нейраминидазы (NA), M1 и М2. Это исследование показало, что вирионные белки гриппа самоорганизуются при совместной экспрессии в клетках эукариот и что матриксный белок M1 требовался для продуцирования VLP. Однако Gomez-Puertas et al., (1999, J. Gen. Virol, 80, 1635-1645) показали также, что чрезмерная экспрессия М2 полностью блокировала перенос CAT-РНК в культуры MDCK (клетки Мадин- Дарби почек собак).
М2 функционирует как белок ионного канала и было показано, что когда этот белок сверхэкспрессируется, внутриклеточный перенос совместно экспрессированного НА ингибируется и накопление гемагглютинина (НА) на поверхности плазменной мембраны снижается на 75-80 % (Sakaguchi et al., 1996; Henkel & Weisz, 1998). Далее, при чрезмерной экспрессии М2 накопление вирусных мембранных белков на поверхности плазменной мембраны М2 снижается и, следовательно, происходит стремительное уменьшение количества образующихся функциональных VLP.
Белок М2 сверхэкспрессируется на клеточной поверхности клеток, инфицированных вирусом гриппа типа A (Lamb et al. (1985) Cell, 40, 627-633). Этот белок был также обнаружен в мембране самой частицы вируса, но в гораздо меньших количествах, от 14 до 68 молекул М2 на вирион (Zebedee and Lamb (1988) J. Virol. 62, 2762 - 72). Белок M2 модифицируется после трансляции при добавлении пальмитиновой кислоты к цистеину в положении 50 (Sugrue et al. (1990) Virology 179, 51-56).
Белок M2 представляет собой гомотетрамер, состоящий из двух димеров, связанных двумя дисульфидными группами, которые удерживаются вместе за счет нековалентного взаимодействия (Sugrue and Hay (1991) Virology 180, 617 to 624). Holsinger and Lamb, (1991) Virology 183, 32-43, показали, что при сайт-направленном мутагенезе цистеиновые остатки в положениях 17 и 19 участвуют в образовании дисульфидного мостика. Во всех исследованных вирусах содержится только цистеин в положении 17. В вирусных штаммах, где содержится также цистеин в положении 19, неизвестно, произошло ли образование второго дисульфидного мостика в том же самом димере (уже соединенном Cys 17-Cys 17) или с другим димером.
Smith et al. (заявка США на патент №2010/0143393) и Song et al. (Plos ONE 2011 6(l):el4538) описывают вакцины и VLP, которые включают белок М2 вируса гриппа. VLP содержат по меньшей мере один капсидный белок вириона, такой как M1. Этот капсидный белок вызывает активную репликацию вируса и высвобождение вирусных частиц из клеток-хозяев насекомых.
Szecsi et al. (Virology Journal, 2006, 3:70) собирали VLP на коровых частицах, дефектных по репликации, полученных из вируса мышиного лейкоза (MLV). Генно-инженерные VLP гриппа получены с помощью транзиторной коэкспрессии (совместной экспрессии) в клетках поверхностных (НА, NA, М2) и внутренних вирусных компонентов (Gag, генома маркера GFP) и закрепления на поверхности клеток НА, НА и или NA, или М2, или всех трех белков, полученных из вируса H7N1 или H5N1. Согласно публикации Szecsi et al. экспрессия М2 в процессе продуцирования Flu-VLP не влияет на введение НА или NA на поверхность частиц вируса (стр. 2, правая колонка, второй параграф в публикации Szecsi et al.).
РАСКРЫТИЕ ИЗОБРЕТЕНИЯ
Данное изобретение относится к продуцированию вирусных белков в растениях. Более конкретно, данное изобретение относится к продуцированию и увеличению продукции вирусоподобных частиц в растениях.
Цель настоящего изобретения состоит в создании улучшенного способа увеличения продуцирования вирусоподобных частиц в растениях.
В соответствии с данным изобретением предусмотрен способ (А) продуцирования вирусоподобной частицы (VLP) в растении, включающий:
а) введение первой нуклеиновой кислоты, включающей первую регуляторную область, являющуюся активной в растении, и функционально связанную с нуклеотидной последовательностью, кодирующей структурный вирусный белок, в растение или в часть растения,
б) введение второй нуклеиновой кислоты, включающей вторую регуляторную область, являющуюся активной в растении и функционально связанную с нуклеотидной последовательностью, кодирующей канальный белок,
в) инкубацию растения или части растения в условиях, которые позволяют экспрессию нуклеиновых кислот, при этом продуцируется VLP.
Первая регуляторная область, активная в растении, и вторая регуляторная область, активная в растении, могут быть одинаковыми или разными.
Канальный белок в способе (А), описанном выше, может представлять собой белок протонного канала. Белок протонного канала может быть выбран из М2 или ВМ2. Кроме того, белок протонного канала может включать сигнатурную последовательность HXXXW белка протонного канала. Белок М2 может быть белком М2, полученным из штамма вируса гриппа типа A/Puerto Rico (Пуэрто-Рико)/8/1934 (SEQ ID NO: 14) или из штамма вируса гриппа типа A/New Caledonia (Новая Каледония)/20/1999 (SEQ ID NO: 11).
Данное изобретение относится также к способу (А), который описан выше, при осуществлении которого структурный вирусный белок включает тримерный домен. Кроме того, нуклеотидная последовательность, кодирующая структурный вирусный белок, представляет собой химерную нуклеотидную последовательность, которая кодирует последовательно антигенный вирусный белок или его фрагмент, трансмембранный домен вируса гриппа и цитоплазматический "хвост". Структурный вирусный белок может включать белок НА вируса гриппа. Кроме того, одна или более "протеолитических" петель (петель, подлежащих протеолитическому расщеплению) белка НА вируса гриппа могут быть делетированы.
Настоящее изобретение предусматривает также способ (А), который описан выше, при осуществлении которого нуклеотидная последовательность, кодирующая структурный вирусный белок, может быть выбрана из группы, состоящей из В НА, С, НА, Н2, Н3, Н4, Н6, Н7, Н8, Н9, Н10, Н11, Н12, Н13, Н14, Н15 и Н16. Например, нуклеотидная последовательность, кодирующая структурный вирусный белок, может быть типа В НА или Н3. Нуклеотидная последовательность, кодирующая структурный вирусный белок, может быть, например, типа НА из вируса гриппа B/Brisbane (Брисбен)/60/2008, B/Malaysia (Малайзия)/2506/2004 или B/Wisconsin (Висконсин)/1/2010, или Н3 из вируса гриппа A/Perth (Перт)/16/2009 или A/Victoria (Виктория)/361/2011. Кроме того, нуклеотидная последовательность, кодирующая структурный вирусный белок, имеет степень идентичности с последовательностями SEQ ID NO: 23, 28, 43, 46, 51, 57 или 61 равную по меньшей мере 70 %. Последовательность структурного вирусного белка может также включать последовательность SEQ ID NO: 25, 30, 41, 48, 54, 58 или 64.
Данное изобретение относится также к способу (А), который описан выше, при осуществлении которого последовательность первой нуклеиновой кислоты включает первую регуляторную область, функционально связанную с одним или более энхансерами комовируса, нуклеотидную последовательность, кодирующую структурный вирусный белок, и один или более амплификационных элементов геминивируса, а третья нуклеиновая кислота, кодирующая репликацию геминивируса, вводится в растение или в часть растения. Один или более энхансеров комовируса может представлять собой UTR (нетранслируемую область) комовируса, например, UTR гипертранслируемой области вируса мозаики коровьего гороха (CPMV-HT), такую как CPMV-HT 5' и/или 3'UTR. Один или более амплификационных элементов геминивируса может быть выбран из длинной межгенной области вируса желтой карликовости бобов (BeYDV LIR) и короткой межгенной области BeYDV (BeYDV SIR). Кроме того, нуклеотидная последовательность, кодирующая структурный вирусный белок, может быть из типа В НА или Н3, например, нуклеотидная последовательность, кодирующая структурный вирусный белок, может иметь степень идентичности с последовательностями SEQ ID NO: 23, 28, 43, 46, 51, 57 или 61, равную по меньшей мере 70%. Последовательность структурного вирусного белка может также включать последовательность SEQ ID NO: 25, 30, 41, 48, 54, 58 или 64.
Способ, описанный выше (способ А), может также включать введение последовательности другой нуклеиновой кислоты, кодирующей супрессор сайленсинга генов, например, HcPro или р19.
Данное изобретение относится также к способу (А), который описан выше, при осуществлении которого на стадии введения (на стадии а) нуклеиновая кислота транзиентно экспрессируется в растении. Или же на стадии введения (на стадии а) нуклеиновая кислота стабильно (постоянно) экспрессируется в растении.
Способ (А), описанный выше, может также включать стадию:
г) сбора растения и очистки VLP.
Настоящее изобретение включает также способ (А), описанный выше, при осуществлении которого VLP не содержит вирусной матрицы или капсидного белка.
Данное изобретение предусматривает VLP, полученную способом (А), описанным выше. VLP может также содержать один или более липидов, выделенных из растения. VLP может также не содержать канального белка. Кроме того, структурный вирусный белок VLP может представлять собой белок НА0. Один или более вирусных белков в VLP может включать специфические к растениям N-гликаны или модифицированные N-гликаны. Данное изобретение предусматривает также поликлональное антитело, полученное с применением VLP.
Настоящее изобретение предусматривает также композицию, содержащую дозу VLP, описанной выше, которая является эффективной для индукции иммунного ответа, и фармацевтически приемлемый носитель.
Данное изобретение относится также к способу индукции иммунитета к вирусу гриппа у субъекта, включающему введение этому субъекту VLP, описанной выше. VLP может быть введена субъекту перорально, интрадермально, интраназально, внутримышечно, интраперитонеально, внутривенно или подкожно.
Данное изобретение относится также к растительному материалу, содержащему VLP, полученную способом (А), описанным выше. Растительный материал может быть использован для индукции иммунитета к инфекции, вызываемой вирусом гриппа, у субъекта. Растительный материал может быть смешан с пищевой добавкой.
Настоящее изобретение относится также к способу (В) продуцирования вирусоподобной частицы (VLP), который включает:
а) предоставление растения или его части, содержащих первую нуклеиновую кислоту, включающую первую регуляторную область, являющуюся активной в растении, и функционально связанную с нуклеотидной последовательностью, кодирующей структурный вирусный белок в растении или в части растения, и вторую нуклеиновую кислоту, включающую вторую регуляторную область, являющуюся активной в растении, и функционально связанную с нуклеотидной последовательностью, кодирующей канальный белок,
б) инкубацию растения или части растения в условиях, которые позволяют осуществление экспрессии нуклеиновых кислот, при этом продуцируется VLP. Первая регуляторная область, активная в растении, и вторая регуляторная область, активная в растении, могут быть одинаковыми или разными.
Канальный белок в способе (В), описанном выше, может представлять собой белок протонного канала. Белок протонного канала может быть выбран из М2 или ВМ2. Кроме того, белок протонного канала может включать сигнатурную последовательность HXXXW белка протонного канала.
Данное изобретение относится также к способу (В), который описан выше, при осуществлении которого структурный вирусный белок включает тримеризационный домен. Кроме того, нуклеотидная последовательность, кодирующая структурный вирусный белок, представляет собой химерную нуклеотидную последовательность, которая кодирует последовательно антигенный вирусный белок или его фрагмент, трансмембранный домен вируса гриппа и цитоплазматический "хвост". Структурный вирусный белок может включать белок НА вируса гриппа. Кроме того, одна или более из "протеолитических" петель белка НА вируса гриппа может быть удалена.
Настоящее изобретение предусматривает также способ (В), который описан выше, при осуществлении которого нуклеотидная последовательность, кодирующая структурный вирусный белок, может быть выбрана из группы, состоящей из гена В НА, С НА, Н2, Н3, Н4, Н6, Н7, Н8, Н9, Н10, НИ, Н12, Н13, Н14, Н15 и Н16. Например, нуклеотидная последовательность, кодирующая структурный вирусный белок, может быть типа В НА или НЗ. Нуклеотидная последовательность, кодирующая структурный вирусный белок, может быть, например, НА из вируса гриппа штамма B/Brisbane (Брисбен)/60/2008, B/Malaysia (Малайзия)/2506/2004 или B/Wisconsin (Висконсин)/1/2010, или Н3 из вируса гриппа штамма A/Perth (Перт)/16/2009 или A/Victoria (Виктория)/361/2011.
Данное изобретение относится также к способу (В), который описан выше, при осуществлении которого последовательность первой нуклеиновой кислоты включает первую регуляторную область, функционально связанную с одним или более энхансерами комовируса, нуклеотидную последовательность, кодирующую структурный вирусный белок и один или более амплификационных элементов геминивируса, а третья нуклеиновая кислота, кодирующая репликацию геминивируса, вводится в растение или в часть растения. Один или более энхансеров комовируса может представлять собой UTR (нетранслируемую область) комовируса, например, UTR гипертранслируемой области вируса мозаики коровьего гороха (CPMV-HT), такую как CPMV-HT 5' и/или 3'UTR. Один или более амплификационных элементов геминивируса может быть выбран из длинной межгенной области вируса желтой карликовости бобов (BeYDV LIR) и короткой межгенной области BeYDV (BeYDV SIR).
Способ, описанный выше (способ В) может также включать введение последовательности другой нуклеиновой кислоты, кодирующей супрессор сайленсинга генов, например, HcPro или р19.
Данное изобретение относится также к способу (В), который описан выше, при осуществлении которого на стадии введения (на стадии а) нуклеиновая кислота временно экспрессируется в растении. Или же на стадии введения (на стадии а) нуклеиновая кислота стабильно (постоянно) экспрессируется в растении.
Способ (В), описанный выше, может также включать стадию:
г) сбора растения и очистки VLP.
Настоящее изобретение включает также способ (В), описанный выше, при осуществлении которого VLP не содержит вирусного матрикса или капсидного белка.
Данное изобретение предусматривает VLP, полученную способом (В), описанным выше. VLP может также содержать один или более липидов, выделенных из растения. VLP может также не содержать канального белка. Кроме того, структурный вирусный белок VLP может представлять собой белок НА0. Один или более вирусных белков в VLP может включать специфические для растений N-гликаны или модифицированные N-гликаны растений. Данное изобретение предусматривает также поликлональное антитело, полученное с применением VLP.
Настоящее изобретение предусматривает также композицию, содержащую дозу VLP, описанной выше, которая является эффективной для индукции иммунного ответа, и фармацевтически приемлемый носитель.
Данное изобретение относится также к способу индукции иммунитета к вирусу гриппа у субъекта, включающему введение этому субъекту VLP, описанной выше. VLP может быть введена субъекту перорально, интрадермально, интраназально, внутримышечно, интраперитонеально, внутривенно или подкожно.
Данное изобретение относится также к растительному материалу, содержащему VLP, полученную способом (В), описанным выше. Растительный материал может быть использован для индукции иммунитета к инфекции, вызываемой вирусом гриппа, у субъекта. Растительный материал может быть смешан с пищевой добавкой.
Настоящее изобретение предусматривает также полипептид, включающий аминокислотную последовательность SEQ ID NO: 41 (PDISP/HA из вируса гриппа B/Brisbane/60/2008 с удаленной "протеолитической" петлей) и последовательность нуклеиновой кислоты, кодирующей полипептид SEQ ID NO: 41. Последовательность нуклеиновой кислоты может включать нуклеотидную последовательность SEQ ID NO: 43. Данное изобретение относится к VLP, содержащей аминокислотную последовательность SEQ ID NO: 41. VLP может также содержать один или более липидов, выделенных из растения. VLP может также не содержать канального белка. VLP может также содержать специфические для растений N-гликаны или модифицированные N-гликаны растений. Настоящее изобретение предусматривает также композицию, содержащую эффективную дозу VLP, включающей аминокислотную последовательность SEQ ID NO: 41, и фармацевтически приемлемый носитель.
Данное изобретение относится также к способу индукции иммунитета к вирусу гриппа у субъекта, включающему введение этому субъекту VLP, содержащей аминокислотную последовательность SEQ ID NO: 41. VLP может быть введена субъекту перорально, интрадермально, интраназально, внутримышечно, интраперитонеально, внутривенно или подкожно. Данное изобретение относится также к растительному материалу, содержащему VLP, включающую аминокислотную последовательность SEQ ID NO: 41. Растительный материал может быть использован для индукции иммунитета к инфекции, вызываемой вирусом гриппа, у субъекта. Растительный материал может быть смешан с пищевой добавкой.
Коэкспрессия структурного вирусного белка совместно, например, но без ограничения, с белком протонного канала, приводила к повышению выхода структурного вирусного белка и VLP. Известно, что гемагглютинины (НА) подвергаются конформационному изменению, зависимому от величины рН. Не основываясь на какой-
либо теории, полагают, что величина рН в пластинчатом комплексе Гольджи клеток, продуцирующих НА, во время созревания и миграции может влиять на фолдинг НА, воздействовать на стабильность и увеличивать деградацию НА или одновременно влиять на три этих фактора. При совместной экспрессии канального белка, например, но без ограничения, белка протонного канала, вместе с НА величина рН в комплексе Гольджи может увеличиваться и приводить к увеличению стабильности, уменьшению степени деградации или к тому и к другому, и повышать выход НА.
В этом разделе необязательно описаны все признаки настоящего изобретения.
КРАТКОЕ ОПИСАНИЕ ФИГУР
Эти и другие признаки данного изобретения станут более понятными после ознакомления со следующим ниже описанием и прилагаемыми чертежами.
На Фигуре 1А показан праймер IF-H5A-I-05.s1+3c (SEQ ID NO: 2). На Фигуре 1B показан праймер IF-H5dTm.r (SEQ ID NO: 3). На Фигуре 1С дано схематическое изображение конструкции 1191. На Фигуре 1D представлена конструкция 1191 (SEQ ID NO 4). На Фигуре 1Е показана экспрессионная кассета номер 489 (SEQ ID NO 5). На Фигуре 1F показана аминокислотная последовательность подтипа Н5 вируса гриппа A/Indonesia (Индонезия)/5/2005 (H5N1) (SEQ ID NO: 6). Фигура 1G отражает нуклеотидную последовательность, кодирующую подтип Н5 вируса гриппа A/Indonesia/5/2005 (H5N1) (SEQ ID NO: 42).
На Фигуре 2А показан праймер IF-S1-M1+M2ANC.c (SEQ ID NO: 7). На Фигуре 2В показан праймер IF-S1-4-M2ANC.r (SEQ ID NO: 8). На Фигуре 2С представлена нуклеотидная последовательность синтезированного гена М2 (соответствующая нуклеотидам 1-26, присоединенным к 715-982, номер доступа нуклеотидной последовательности, депонированной в Genbank DQ508860) (SEQ ID NO: 9). На Фигуре 2D показана экспрессионная кассета номер 1261 от промотора 2X35S до NOS терминатора. Ген М2 вируса гриппа A/New Caledonia/20/1999 (H1N1) подчеркнут. (SEQ ID NO: 10). Ha Фигуре 2E представлена аминокислотная последовательность гена М2 вируса гриппа A/New Caledonia/20/1999 (H1N1) (SEQ ID NO: 11).
На Фигуре 3А приведена нуклеотидная последовательность синтезированного гена М2 (соответствующая нуклеотидам 26-51, присоединенным к нуклеотидам 740-1007, номер доступа нуклеотидной последовательности, депонированной в Genbank EF467824) (SEQ ID
NO: 12). На Фигуре 3В показана экспрессионная кассета номер 859 от промотора 2X35S до терминатора NOS. Ген М2 вируса гриппа A/Puerto Rico/8/1934 (H1N1) подчеркнут. (SEQ ID NO: 13). На Фигуре 3С представлена нуклеотидная последовательность синтезированного гена М2 вируса гриппа A/Puerto Rico/8/1934 (H1N1) (SEQ ID NO: 14).
На Фигуре 4A показан праймер IF-H1A-C-09.s2+4c (SEQ ID NO: 15). На Фигуре 4B показан праймер IF-HlA-C-09.s1-4r (SEQ ID NO: 16). Фигура 4C показывает нуклеотидную последовательность синтезированного гена H1 (номер доступа в Genbank FJ966974) (SEQ ID NO: 17). На Фигуре 4D схематически показана конструкция 1192. Сайты расщепления SacII и StuI рестриктазами, использованные для линеаризации плазмиды, на рисунке снабжены комментариями. Фигура 4Е показывает конструкцию 1192 слева направо от границ тДНКs (подчеркнуты). Экспрессионная кассета 2X35S/CPMV-HT/PDISP/NOS и ингибитор сайленсинга Пластоцианин-Р19-Пластоцианин (SEQ ID NO: 18). На Фигуре 4F представлена экспрессионная кассета номер 484 от промотора 2X35S до терминатора NOS. Нуклеотидная последовательность PDISP/H1 вируса гриппа A/California/7/2009 (H1N1) подчеркнута. (SEQ ID NO: 19). Фигура 4G представляет аминокислотную последовательность PDISP-H1 вируса гриппа A/California/7/2009 (H1N1) (SEQ ID NO: 20).
На Фигуре 5А показан праймер IF-S2+S4-H3 Per.c (SEQ ID NO: 21). На Фигуре 5В показан праймер IF-S1a4-H3 Per.r (SEQ ID NO: 22). На Фигуре 5C показана нуклеотидная последовательность синтезированного гена Н3 (соответствующая нуклеотидам 26-1726 последовательности с номером доступа в Genbank GQ293081) (SEQ ID NO: 23). На Фигуре 5D показана экспрессионная кассета номер 1019 от промотора 2X35S до терминатора NOS. Нуклеотидная последовательность PDISP/H3 вируса гриппа A/Perth/16/2009 (H3N2) подчеркнута. (SEQ ID NO: 24). На Фигуре 5Е показана аминокислотная последовательность подтипа PDISP/H3 вируса гриппа A/Perth/16/2009 (H3N2) (SEQ ID NO: 25).
На Фигуре 6А показан праймер IF-S2+S4-B Bris.c (SEQ ID NO: 26). На Фигуре 6В показан праймер IF-S1a4-B Bris.r (SEQ ID NO: 27). На Фигуре 5C показана нуклеотидная последовательность синтезированного гена НА В Brisbane (соответствующая нуклеотидам 34-1791 последовательности с номером доступа в Genbank FJ766840) (SEQ ID NO: 28). На Фигуре 6D показана экспрессионная кассета номер 1029 от промотора 2X35S до терминатора NOS. PDISP/HA вируса гриппа B/Brisbane/60/2008 подчеркнута. (SEQ ID NO: 29). На Фигуре 6Е показана аминокислотная последовательность подтипа PDISP/HA вируса гриппа B/Brisbane/60/2008 (SEQ ID NO: 30). На Фигуре 6F схематически показана конструкция 1194. Сайты рестрикции SacII и StuI рестриктазами, использованные для линеаризации плазмиды, на рисунке снабжены комментариями. На Фигуре 6G показана конструкция 1194 слева направо от границ тДНК (подчеркнуты). 2X35S/CPMV-HT/PDISP/NOS в амплификационной системе BeYDV+Replicase с экспрессионной кассетой ингибитора сайленсинга Пластоцианин-Р19-Пластоцианин (SEQ ID NO: 31). На Фигуре 6Н показана экспрессионная кассета номер 1008 от левой LIR (длинной межгенной области) BeYDV до правой LIR BeYDV. Последовательность подтипа PDISP/HA вируса гриппа B/Brisbane/60/2008 подчеркнута. (SEQ ID NO: 32).
На Фигуре 7А показан праймер dTmH5I-B Bris.r (SEQ ID NO: 33). На Фигуре 7B показан праймер В Bris-dTmH5I.c (SEQ ID NO: 34). На Фигуре 7C показан праймер IF-S1aS4-dTmH5I.r (SEQ ID NO: 35). На Фигуре 7D показана экспрессионная кассета номер 1009 от левой LIR BeYDV до правой LIR BeYDV. Последовательность PDISP/HA В Brisbane/H5Indo ТМСТ подчеркнута. (SEQ ID NO: 36). На Фигуре 7Е показана аминокислотная последовательность PDISP/HA В Brisbane/H5Indo ТМСТ (SEQ ID NO: 37).
На Фигуре 8А показан праймер 1039+1059.r (SEQ ID NO: 38). На Фигуре 8В показан праймер 1039+1059.c (SEQ ID NO: 39). На Фигуре 8С показана экспрессионная кассета номер 1059 от левой LIR BeYDV до правой LIR BeYDV. Последовательность PDISP/HA вируса гриппа B/Brisbane/60/2008 с делетированной "протеолитической" петлей подчеркнута. (SEQ ID NO: 40). На Фигуре 8D показана аминокислотная последовательность PDISP/HA вируса гриппа B/Brisbane/60/2008 с делетированной "протеолитической" петлей (SEQ ID NO: 41). На Фигуре 8Е показана нуклеотидная последовательность PDISP/HA вируса гриппа B/Brisbane/60/2008 с делетированной "протеолитической" петлей (SEQ ID NO: 43).
На Фигуре 9 показана карта плазмиды конструкции номер 1008. Конструкция номер 1008 управляет экспрессией дикого типа НА вируса гриппа B/Brisbane/60/2008. Эта конструкция содержит элементы ДНК-амплификации из BeYDV.
На Фигуре 10 показана карта плазмиды конструкции номер 1009. Конструкция номер 1009 управляет экспрессией химерного НА штамма вируса гриппа B/Brisbane/60/2008, в котором трансмембранный домен и цитозольный хвост заменены на трансмембранный домен и цитозольный хвост Н5 гриппа подтипа A/Indonesia/05/2005. Эта конструкция содержит элементы ДНК-амплификации из BeYDV.
На Фигуре 11 показана карта плазмиды конструкции номер 1029. Конструкция номер 1029 управляет экспрессией дикого типа НА вируса гриппа B/Brisbane/60/2008.
На Фигуре 12 показана карта плазмиды конструкции номер 1059. Конструкция номер 1059 управляет экспрессией мутантного НА вируса гриппа B/Brisbane/60/2008 с делетированной "протеолитической" петлей. Эта конструкция содержит элементы BeYDV для амплификации ДНК.
На Фигуре 13 показана карта плазмиды конструкции номер 1019. Конструкция номер 1019 управляет экспрессией дикого типа Н3 вируса гриппа A/Perth/16/2009 (H3N2).
На Фигуре 14 показана карта плазмиды конструкции номер 484. Конструкция номер 484 управляет экспрессией H1 вируса гриппа A/California/07/2009 (H1N1).
На Фигуре 15 показана карта плазмиды конструкции номер 489. Конструкция номер 489 управляет экспрессией дикого типа Н5 вируса гриппа A/Indonesia/05/2005 (H5N1).
На Фигуре 16 показана карта плазмиды конструкции номер 1261. Конструкция номер 1261 управляет экспрессией дикого типа М2 вируса гриппа A/New Caledonia/20/99 (H1N1).
На Фигуре 17 показана карта плазмиды конструкции номер 859. Конструкция номер 859 управляет экспрессией дикого типа М2 вируса гриппа A/Puerto Rico/8/34 (H1N1).
На Фигуре 18 показаны результаты Вестерн-блот анализа (Вестерн блоттинга) экспрессии белка НА в агроинфильтрованных листьях табака Nicotiana benthamiana. НА из B/Brisbane/60/2008 совместно экспрессируется с М2 из A/New Caledonia/20/99. "С+": положительный контроль, полуочищенный вирус B/Brisbane/60/2008 от Therapeutic Goods Administration, Australia (Управления по контролю за изделиями медицинского назначения Австралии); "С-": отрицательный контроль, фиктивно инфильтрованные растения; "1008": экспрессия дикого типа НА из B/Brisbane/60/2008 в присутствии амплификационных элементов (BeYDV); "1008+1261": совместная экспрессия (коэкспрессия) дикого типа НА вируса гриппа B/Brisbane/60/2008 в присутствии амплификационных элементов (BeYDV) с М2; "1009+1261": коэкспрессия НА из B/Brisbane/60/2008 в присутствии амплификационных элементов (BeYDV) с М2; "1029": экспрессия дикого типа НА вируса гриппа B/Brisbane/60/2008 в отсутствие амплификационных элементов (BeYDV); "1029+1261": коэкспрессия дикого типа НА вируса гриппа B/Brisbane/60/2008 в отсутствие амплификационных элементов (BeYDV) с М2 из A/New Caledonia/20/99. Отношения показывают относительное содержание культур Agrobacterium (агробактерий), применяемых в экспериментах по совместной экспрессии (коэкспрессии).
На Фигуре 19 показаны результаты Вестерн-блот анализа экспрессии белка НА в агроинфильтрованных листьях табака. Дорожка "С+": Положительный контроль, полуочищенный вирус A/Wisconsin/15/2009 (H3N2) от Therapeutic Goods Administration, Australia; "C-": отрицательный контроль, фиктивно инфильтрованные растения; "'1019": экспрессия дикого типа НА вируса A/Perth/16/2009 (H3N2); "1019+1261": коэкспрессия дикого типа НА вируса A/Perth/16/2009 (H3N2) с М2 из A/New Caledonia/20/99. Отношения показывают относительное содержание культур агробактерий, применяемых в экспериментах по совместной экспрессии (коэкспрессии).
На Фигуре 20 показаны результаты Вестерн-блот анализа экспрессии белка НА в агроинфильтрованных листьях табака Nicotiana benthamiana. Дорожка "С+": Положительный контроль, полуочищенный вирус A/California/7/2009 (H1N1) NYMC X-179А от NIBSC virus (NIBSC code 09/146); "С-": отрицательный контроль, фиктивно инфильтрованные растения; "484": экспрессия дикого типа НА вируса A/California/7/2009 (H1N1); "484+1261": коэкспрессия дикого типа НА вируса A/California/7/2009 (H1N1) с М2 вируса A/New Caledonia/20/99. Отношения показывают относительное содержание культур агробактерий, применяемых в экспериментах по совместной экспрессии (коэкспрессии).
На Фигуре 21 показаны результаты Вестерн-блот анализа экспрессии белка НА в агроинфильтрованных листьях табака Nicotiana benthamiana. Дорожка "С+": Положительный контроль, очищенный рекомбинантный Н5 из A/Indonesia/05/2005, Immune Technology Corporation (продукт no. IT-003-052p); "C-": отрицательный контроль, фиктивно инфильтрованные растения; "489": экспрессия дикого типа НА из A/Indonesia/5/05 (H5N1); "489+1261": коэкспрессия дикого типа НА из A/Indonesia/5/05 (H5N1) с М2 из A/New Caledonia/20/99.
На Фигуре 22А показаны результаты Вестерн-блот анализа экспрессии белка НА в агроинфильтрованных листьях табака Nicotiana benthamiana. "1008": экспрессия дикого типа НА из B/Brisbane/60/2008 в присутствии амплификационных элементов (BeYDV); "1008+1261": коэкспрессия дикого типа НА из B/Brisbane/60/2008 в присутствии амплификационных элементов (BeYDV) с М2 из A/New Caledonia/20/99; "1059": экспрессия мутантного НА из B/Brisbane/60/2008 в присутствии амплификационных элементов (BeYDV); "1059+1261": коэкспрессия мутантного НА из B/Brisbane/60/2008 в присутствии амплификационных элементов (BeYDV) с М2 из A/New Caledonia/20/99.
Анализировали растения из трех раздельных инфильтраций (А, В и С). Отношения показывают относительное содержание культур агробактерий, применяемых в экспериментах по коэкспрессии. На Фигуре 22В сравнивается гемагглютационная способность сырых белковых экстрактов НА- продуцирующих растений.
На Фигуре 23 показаны результаты Вестерн-блот анализа экспрессии белка НА в агроинфильтрованных листьях табака Nicotiana benthamiana. Фигура 23А: "1059": экспрессия мутантного НА из B/Brisbane/60/2008 в присутствии амплификационных элементов (BeYDV); "1059+1261": коэкспрессия мутантного НА из B/Brisbane/60/2008 в присутствии амплификационных элементов (BeYDV) with М2 from A/New Caledonia/20/99. "1059+859": коэкспрессия мутантного НА из B/Brisbane/60/2008 в присутствии амплификационных элементов (BeYDV) с М2 из A/Puerto Rico/8/34. Анализировали растения из трех раздельных инфильтраций (А, В и С). Отношения показывают относительное содержание культур агробактерий, применяемых в экспериментах по коэкспрессии. Фигура 23В: "1019": экспрессия дикого типа НА из A/Perth/16/2009 (H3N2); "1019+1261": коэкспрессия дикого типа НА из A/Perth/16/2009 (H3N2) с М2 из A/New Caledonia/20/99; "1019+859": коэкспрессия дикого типа НА из A/Perth/16/2009 (H3N2) с М2 из A/Puerto Rico/8/34. Отношения показывают относительное содержание культур агробактерий, применяемых в экспериментах по коэкспрессии.
На Фигуре 24 показано выравнивание последовательностей НА некоторых штаммов вируса гриппа. Сайт расщепления предшественника НА0 показан стрелкой.
На Фигуре 25А показан праймер IF-H3V3611 l.S2+4c (SEQ ID NO: 44). На Фигуре 25В показан праймер IF-H3V3611l.s1-4r (SEQ ID NO: 45). На Фигуре 25С показана нуклеотидная последовательность синтезированного Н3 гена (соответствует nt (нуклеотидам) с 25 до 1725 НА последовательности из изолята вируса гриппа GISAID номер EPI ISL 101506) (SEQ ID NO: 46). На Фигуре 25D показана нуклеотидная последовательность экспрессионной кассеты номер 1391 от 2X35S промотора до NOS терминатора. Последовательность PDISP/H3 вируса гриппа A/Victoria/361/2011 (H3N2) подчеркнута. (SEQ ID NO: 47). На Фигуре 25Е показана аминокислотная последовательность PDISP-H3 вируса гриппа А/ Victoria/361/2011 (H3N2) (SEQ ID NO: 48). На Фигуре 25F схематически представлена конструкция 1391.
На Фигуре 26А показан праймер IF-HAB110.S1+3c (SEQ ID NO: 49). На Фигуре 26В показан праймер IF-HAB110.s1-4r (SEQ ID NO: 50). На Фигуре 26С показана нуклеотидная последовательность НА В Wisconsin (номер доступа в Genbank JN993010)
(SEQ ID NO: 51). На Фигуре 26D схематически представлена конструкция 193. На Фигуре 26Е представлена конструкция 193 от левой до правой границы тДНК (подчеркнуто). Экспрессионная кассета 2X35S/CPMV-HT/NOS в BeYDV(m)+репликазная амплификационная система с ингибитором сайленсинга Пластоцианин-Р19-Пластоцианин (SEQ ID NO: 52). На Фигуре 26F представлена нуклеотидная последовательность экспрессионной кассеты номер 1462 от промотора 2X35S до NOS терминатора. Последовательность НА вируса гриппа B/Wisconsin/1/2010 подчеркнута (SEQ ID NO: 53). На Фигуре 26G представлена аминокислотная последовательность НА вируса гриппа B/Wisconsin/1/2010 (SEQ ID NO: 54). На Фигуре 26Н схематически представлена конструкция 1462.
На Фигуре 27А показан праймер НАВ110(PrL-).r (SEQ ID NO: 55). На Фигуре 27В показан праймер НАВ110(PrL-).c (SEQ ID NO: 56). На Фигуре 27С представлена экспрессионная кассета номер 1467 от промотора 2X35S до NOS терминатора. Последовательность для НА вируса гриппа B/Wisconsin/1/2010 с делетированной "протеолитической" петлей подчеркнута (SEQ ID NO: 57). На Фигуре 27D представлена аминокислотная последовательность НА B/Wisconsin/1/2010 с делетированной "протеолитической" петлей (SEQ ID NO: 58). На Фигуре 27Е схематически представлена конструкция 1467.
На Фигуре 28А представлен праймер IF-HB-M-04.s2+4c (SEQ ID NO: 59). На Фигуре 28В представлен праймер IF-HB-M-04.s1-4r (SEQ ID NO: 60). На Фигуре 28С представлена нуклеотидная последовательность синтезированного НА В Malaysia (соответствующая нуклеотидам (nt) 31-1743 последовательности с номером доступа в Genbank EU124275), мутации Т759С и C888G подчеркнуты. (SEQ ID NO: 61). На Фигуре 28D схематически представлена конструкция 194, сайты расщепления SacII и StuI рестриктазами, использованные для линеаризации плазмиды, на рисунке снабжены комментариями. На Фигуре 28Е представлена конструкция 194 от левой до правой границ тДНК (подчеркнуто). Экспрессионная кассета 2X35S/CPMV-HT/NOS в
BeYDV(m)+репликазная амплификационная система с ингибитором сайленсинга Пластоцианин-Р19-Пластоцианин (SEQ ID NO: 62). На Фигуре 28F представлена нуклеотидная последовательность экспрессионной кассеты номер 1631 от промотора 2X35S до терминатора NOS. Последовательность для PDISP-HA гриппа В/ Malaysia/2506/2004 подчеркнута. (SEQ ID NO: 63). На Фигуре 28G представлена аминокислотная последовательность PDISP-HA гриппа B/Malaysia/2506/2004 (SEQ ID NO: 64). На Фигуре 28Н схематически представлена конструкция 1631.
На Фигуре 29 показаны результаты Вестерн-блот анализа экспрессии белка НА в агроинфильтрованных листьях табака Nicotiana benthamiana. НА вируса B/Malaysia/2506/2004 экспрессировали совместно (коэкспрессировали) с М2 вируса A/New Caledonia/20/99. На дорожку (полоску) наносили двадцать микрограмм белкового экстракта. "С+": положительный контроль, полуочищенный вирус B/Malaysia/2506/2004, полученный из National Institute for Biological Standards and Control, United Kingdom (Национального института биологических стандартов и контроля, Соединенное королевство Великобритании и Северной Ирландии); "1631": Экспрессия НА из B/Malaysia/2506/2004 в присутствии амплификационных элементов (BeYDV); "1631+1261": коэкспрессия дикого типа НА из B/Malaysia/2506/2004 в присутствии элементов амплификации (BeYDV) с М2. Отношения показывают относительное содержание культур агробактерий, применяемых в экспериментах по коэкспрессии.
На Фигуре 30А показаны результаты Вестерн-блот анализа экспрессии белка НА в агроинфильтрованных листьях табака Nicotiana benthamiana. НА вируса B/Wisconsin/1/2010 экспрессировали совместно с М2 из A/New Caledonia/20/99. На дорожку нагружали десять микрограмм белкового экстракта. "С+": положительный контроль, полуочищенный вирус B/Wisconsin/1/2010, полученный из National Institute for Biological Standards and Control, United Kingdom; "1462": экспрессия НА из B/Wisconsin/1/2010 в присутствии амплификационных элементов (BeYDV); "1467": экспрессия мутантного НА вируса B/Wisconsin/1/2010 в присутствии амплификационных элементов (BeYDV); "1462+1261": коэкспрессия дикого типа НА вируса B/Wisconsin/1/2010 в присутствии амплификационных элементов (BeYDV) с М2; "1467+1261": коэкспрессия мутантного НА вируса B/Wisconsin/1/2010 в присутствии амплификационных элементов (BeYDV) с М2. Отношения показывают оптическую плотность для каждой культуры Agrobacterium, используемой в экспериментах по экспрессии и коэкспрессии. На Фигуре 30В сравнивается гемагглютационная способность сырых белковых экстрактов растений, трансформированных с использованием AGL1/1462, AGL1/1467, AGL1/1462+AGL1/1261 и AGL1/1467+AGL1/1261.
На Фигуре 31 показаны результаты Вестерн-блот анализа экспрессии белка НА в агроинфильтрованных листьях табака Nicotiana benthamiana. НА вируса H3/Victoria/361/2011 экспрессировали совместно (коэкспрессировали) с М2 из A/New Caledonia/20/99. На дорожку наносили двадцать миллиграмм белкового экстракта. "С+": положительный контроль, полуочищенный вирус Н3/Wisconsin/15/2009 от Therapeutic Goods Administration, Australia; "1391": экспрессия дикого типа НА вируса H3/Victoria/361/2011; "1391+1261": коэкспрессия НА вируса H3/Victoria/361/2011 с М2. Отношения показывают оптическую плотность для каждой культуры Agrobacterium, используемой в экспериментах по экспрессии и коэкспрессии.
ОСУЩЕСТВЛЕНИЕ ИЗОБРЕТЕНИЯ
Следующее далее описание является предпочтительным вариантом изобретения.
Данное изобретение относится к вирусоподобным частицам (VLP) и к способам продуцирования и повышения продукции VLP в растениях.
Настоящее изобретение относится, в частности, к способу продуцирования вирусоподобной частицы (VLP) в растении или в части растения. Этот способ включает введение первой нуклеиновой кислоты и второй нуклеиновой кислоты в растение. Первая нуклеиновая кислота включает первую регуляторную область, являющуюся активной в растении или в части растения и функционально связанную с нуклеотидной последовательностью, кодирующей структурный вирусный белок.
Вторая нуклеиновая кислота включает вторую регуляторную область, являющуюся активной в растении и функционально связанную с нуклеотидной последовательностью, кодирующей канальный белок, например, но без ограничения белок протонного канала. Первая регуляторная область и вторая регуляторная область могут быть одинаковыми или разными. Растение или часть растения подвергают инкубации в условиях, которые позволяют экспрессию нуклеиновых кислот, при этом продуцируется VLP. Если это желательно, можно провести сбор растения или части растения и осуществить очистку VLP. Предпочтительно, когда VLP не содержит белка M1, вирусного матрикса или корового белка. Настоящее изобретение относится также к VLP, полученной описанным способом. VLP может включать один или более липидов растения. VLP может быть использована для приготовления композиции, содержащей эффективную дозу VLP, вызывающую иммунный ответ, и фармацевтически приемлемый носитель.
Данное изобретение предусматривает также растительный материал, содержащий VLP, полученную путем экспрессии первой и второй нуклеиновых кислот, описанных выше. Растительный материал может быть использован для индукции иммунитета к инфекции, вызываемой вирусом гриппа, у субъекта. Растительный материал может быть смешан с пищевой добавкой.
VLP согласно данному изобретению может быть также получена путем предоставления растения или части растения, содержащих первую нуклеиновую кислоту и вторую нуклеиновую кислоту, описанные выше, и инкубации растения или части растения в условиях, которые позволяют экспрессии первой и второй нуклеиновых кислот, при этом продуцируется VLP. VLP может содержать один или более липидов растений. VLP может быть использована для приготовления композиции, содержащей эффективную дозу VLP для индукции иммунного ответа и фармацевтически приемлемый носитель. Данное изобретение относится также к растительному материалу, содержащему VLP, полученную путем экспрессии первой и второй нуклеиновых кислот. Растительный материал может быть использован для индукции иммунитета к инфекции, вызываемой вирусом гриппа, у субъекта. Растительный материал может быть смешан с пищевой добавкой.
VLP согласно данному изобретению содержат один или более вирусных белков. Например, но без всякого ограничения, один или более вирусных белков могут быть структурным вирусным белком, таким как гемагглютинин (НА) вируса гриппа, или канальным белком, например, без всякого ограничения, белком протонного канала, таким как М2. НА может быть любым НА, например, гемагглютинином подтипов Н2, Н3, Н4, Н6, Н7, Н8, Н9, Н10, H11, Н12, Н13, Н14, Н15, Н16 или типа В НА, как описано в заявках WO 2009/009876; WO 2009/076778; WO 2010/003225; WO 2010/003235; WO 2011/03522; которые включены в данную заявку посредством отсылки).
Как будет описано более подробно ниже, VLP могут продуцироваться в растении при совместной экспрессии (коэкспрессии) первой нуклеиновой кислоты, кодирующей вирусный белок, и второй нуклеиновой кислоты, кодирующей канальный белок, например, но без всякого ограничения, белок протонного канала. Первая и вторая нуклеиновые кислоты могут быть введены в растение на одной и той же стадии или они могут быть введены в растение последовательно. Первая и вторая нуклеиновые кислоты могут вводиться в растение транзиторно или постоянно. Кроме того, растение, которое экспрессирует первую нуклеиновую кислоту, кодирующую вирусный белок, может быть трансформировано при помощи канального белка, например, но без всякого ограничения, белка протонного канала (второй нуклеиновой кислоты), таким образом, что и первая, и вторая нуклеиновые кислоты будут коэкспрессироваться в клетках растения. Или же, альтернативно, растение, которое экспрессирует канальный белок, например, но без всякого ограничения, белок протонного канала, (вторую нуклеиновую кислоту), может быть трансформировано при помощи первой нуклеиновой кислоты, кодирующей вирусный белок таким образом, что и первая, и вторая нуклеиновые кислоты будут коэкспрессироваться в клетках растения. Или же первое растение, экспрессирующее первую нуклеиновую кислоту, кодирующую вирусный белок, может быть скрещено со вторым растением, экспрессирующим вторую нуклеиновую кислоту, кодирующую канальный белок, например, но без всякого ограничения, белок протонного канала, с целью получения растения-потомка, которое коэкспрессирует первую и вторую нуклеиновые кислоты, кодирующие вирусный белок и канальный белок, например, но без всякого ограничения, белок протонного канала, соответственно.
Настоящее изобретение относится также к способу повышения экспрессии и продукции вирусного белка в растении путем коэкспрессии первой нуклеиновой кислоты, кодирующей вирусный белок, и второй нуклеиновой кислоты, кодирующей канальный белок, например, но без всякого ограничения, белок протонного канала. Первая и вторая нуклеиновые кислоты могут вводиться в растение на одной и той же стадии или они могут вводиться в растение последовательно. Первая и вторая нуклеиновые кислоты могут вводиться в растение транзиторно или постоянно. Кроме того, растение, которое экспрессирует первую нуклеиновую кислоту, кодирующую вирусный белок, может быть трансформировано при помощи канального белка, например, но без всякого ограничения, белка протонного канала (второй нуклеиновой кислоты), таким образом, что и первая, и вторая нуклеиновые кислоты будут коэкспрессироваться в клетках растения. Или же, альтернативно, растение, которое экспрессирует канальный белок, например, но без всякого ограничения, белок протонного канала (вторую нуклеиновую кислоту), может быть трансформировано при помощи первой нуклеиновой кислоты, кодирующей вирусный белок таким образом, что и первая, и вторая нуклеиновые кислоты будут коэкспрессироваться в клетках растения.
Или же первое растение, экспрессирующее первую нуклеиновую кислоту, кодирующую вирусный белок, может быть скрещено со вторым растением, экспрессирующим вторую нуклеиновую кислоту, кодирующую канальный белок, например, но без всякого ограничения, белок протонного канала, с целью получения растения-потомка, которое совместно экспрессирует (коэкспрессирует) первую и вторую нуклеиновые кислоты, кодирующие вирусный белок и канальный белок, например, но без всякого ограничения, белок протонного канала, соответственно.
Канальный белок
Термин "канальный белок" означает белок, который способен образовывать канал в фосфолипидной мембране, что позволяет ионам и/или малым молекулам проникать через эту мембрану. Канальные белки могут быть селективными по размеру и/или заряду ионов и/или малых молекул. Неограничивающие примеры канальных белков включают неспецифический канальный белок, который меняет проницаемость мембран для соединений с низким молекулярным весом, и белок ионного канала, такой как, например, белок хлоридного канала, белок калиевого канала, белок натриевого канала, белок кальциевого канала и белок протонного канала.
Термин "белок протонного канала" означает белок, который способен образовать протон-селективный канал в фосфолипидном бислое. Белок протонного канала может представлять собой однопроходный мембранный белок с трансмембранным (ТМ) доменом, который фланкирован гидрофобными доменами. Домен ТМ протонного канала может включать последовательность HXXXW (SEQ ID NO. 1).
После расщепления НАО НА становится чувствительным к величине рН, подвергаясь необратимому конформационному изменению при рН эндосомы (< рН 6.0). Конформация предшественника НАО является устойчивой при низком значении рН, но расщепленная форма НА1-НА2 является метастабильной (Bullough PA et. al., 1994, Nature. Vol 371: 37-43). Определение порогового значения величины рН, которое вызывает конформационные изменения в различных НА, показывает, что это пороговое значение рН равно примерно 5.8-5.9 в случае штаммов типа В, в то время как в случае НА типа А наблюдается более кислое значение рН (рН от 5.1 до 5.3) (Beyer WEP et al, 1986, Archives Virol, vol 90: 173). Конформационное изменение HA1-HA2 в случае НА типа В может также иметь место во время экстракции биомассы растений (рН между 5 и 6).
Не основываясь на какой-либо теории, полагают, что величина рН клеточного компартмента, содержащего НА, включая комплекс Гольджи, может поэтому иметь большое значение для фолдинга, стабильности и/или протеолиза НА. Белки протонных каналов, такие как, например, белок М2 и ВМ2 вируса гриппа, могут регулировать величину рН в клеточных компартментах. Например, М2 регулирует потенцирование слияния мембран за счет буферирования внутриклеточных компартментов как на поздней, так и на ранней стадиях репликации вируса гриппа. На ранней стадии инфицирования новых клеток после поглощения вирусных частиц эндоцитами активация белка М2 протонного канала приводит к подкислению внутри вириона во время процесса сбрасывания оболочки. На поздней стадии развития инфекции во время образования вирусов белок М2 приводит к повышению величины рН во время перехода через сетчатую структуру отдела транс-комплекса Гольджи и предотвращению вызванной низким значением рН инактивации совместно транспортированных белков, таких как НА в случае инфекции, вызванной вирусами гриппа. Совместная экспрессия структурного вирусного белка вместе с канальным белком, например, но без всякого ограничения, с белком протонного канала, повышала продукцию структурного вирусного белка, при этом наблюдалось образование VLP. Известно, что НА подвергаются конформационному изменению, зависящему от величины рН. Не ограничиваясь какой-либо теорией, полагают, что величина рН внутри комплекса Гольджи клеток, продуцирующих НА во время созревания и миграции, может оказывать влияние на фолдинг НА, воздействовать на стабильность и увеличивать расщепление НА или же может происходить изменение всех этих факторов. При совместной экспрессии канального белка, например, но без всякого ограничения, белка протонного канала, вместе с НА величина /или VLP.
При совместной экспрессии структурного вирусного белка и канального белка, например, но без всякого ограничения, белка протонного канала, в растении наблюдалось увеличение продукции структурного вирусного белка /или VLP по сравнению с растением, в котором осуществлялась экспрессия структурного вирусного белка без канального белка, например, но без всякого ограничения, белка протонного канала.
Кроме того, при совместной экспрессии (коэкспрессии) структурного вирусного белка и канального белка, например, но без всякого ограничения, белка протонного канала, в растении белок НА может проявлять повышенную активность, о чем свидетельствует более высокая гемагглютинирующая активность по сравнению с белком НА, который не экспрессировался совместно с канальным белком, например, но без всякого ограничения, белком протонного канала. Под увеличением активности понимают повышение гемагглютинирующей активности примерно от 2% до примерно 100%, или на любую величину между этими пределами, определенную с помощью стандартных методов, известных из уровня техники, например, от примерно 10 % до примерно 50% или на любую величину между этими пределами, например, примерно, на 2, 5, 8, 10, 12, 15, 18, 20, 22, 24, 25, 26, 28, 30, 32, 34, 35, 36, 38, 40, 42, 44, 45, 46, 48, 50, 52, 54, 55, 56, 58, 60, 65, 70, 75, 80, 85, 90, 95 или 100%, по сравнению с активностью того же самого белка НА, продуцированного в отсутствие канального белка, например, но без всякого ограничения, белка протонного канала.
Применяемые в данной заявке термины "М2", "белок М2", "последовательность М2" и "домен М2" относятся ко всей или к части последовательности белка М2, выделенной из любого природного или полученного искусственным путем штамма вируса гриппа или его изолята, или основанной на, или содержащейся в таком штамме вируса гриппа или его изоляте. Таким образом, термин М2 и подобные термины включают варианты последовательности М2 природного происхождения, полученные путем мутации во время жизненного цикла вирусов или продуцированные в ответ на селективное воздействие (например, лекарственную терапию, расширение тропизма клеток-хозяев или инфекционности и т. д.), а также последовательности М2, полученные рекомбинантным методом или путем синтеза. Примеры канальных белков, которые могут быть использованы, включают, но без ограничения, белки протонных каналов, например, перечисленные в Таблице 1. Неограничивающие примеры последовательностей, которые могут быть использованы согласно данному изобретению, включают М2 вируса гриппа A/Puerto Rico/8/1934 и М2 вируса гриппа A/New Caledonia/20/1999. Например, белок М2 содержит последовательность SEQ ID NO: 11 или 14.
Применяемые в данной заявке термины "ВМ2", "белок ВМ2", "последовательность ВМ2" и "домен ВМ2" относятся ко всей или к части последовательности белка ВМ2, выделенной из любого природного или полученного искусственным путем штамма вируса гриппа или его изолята, или основанной на, или содержащейся в таком штамме вируса гриппа или его изоляте. Таким образом, термин ВМ2 и подобные термины включают варианты последовательности ВМ2 природного происхождения, полученные путем мутации во время жизненного цикла вирусов или продуцированные в ответ на селективное воздействие (например, лекарственную терапию, расширение тропизма клеток-хозяев или инфекционности и т.д.), а также последовательности ВМ2, полученные рекомбинантным методом или путем синтеза. Примеры канальных белков, которые могут быть использованы, включают, но без ограничения, белки протонных каналов, например, перечисленные в Таблице 2.
Другие примеры последовательностей белков протонных каналов представляют собой последовательности, депонированные в GenBank под номерами доступа, указанными в Таблицах 1 и 2.
Вирусный структурный белок
Структурный белок вируса (также называемый вирусным структурным белком) может представлять собой вирусный антигенный белок или его фрагмент, например, но без ограничения, вирусный гликопротеин или вирусный оболочечный белок. Вирусный структурный белок может представлять собой химерный вирусный белок. Вирусный белок может существовать в виде мономера, димера, тримера или их комбинации. Тример представляет собой макромолекулярный комплекс, образованный тремя белками, связанными обычно нековалентными связями. Не основываясь на какой- либо теории, полагают, что домен, инициирующий тримеризацию белка, может играть важную роль в образовании тримера. Следовательно, вирусный структурный белок или его фрагмент может содержать домен, инициирующий тримеризацию. Неограничивающим примером вирусного структурного белка является гемагглютинин вируса гриппа (НА) или фрагмент НА. Неограничивающие примеры НА или фрагментов НА, которые можно применять в соответствии с настоящим изобретением, включают НА или его фрагменты, описанные в Международных заявках WO 2009/009876, WO 2009/076778; WO 2010/003225, WO 2010/003235, WO 2011/03522, WO 2010/006452, WO 2010/148511, WO 2011/035422 (описание которых включено в настоящее изобретение посредством отсылки).
Кроме того, вирусный структурный белок может представлять собой непроцессированный белок-предшественник НА. НА белок синтезируется в виде белка-предшественника (НА0) длиной около 75 кДа, который собирается на поверхности в удлиненный тримерный белок. Белок-предшественник расщепляется при постоянной активации сайта расщепления на две полипептидных цепи, НА1 и НА2 (содержащие трансмембранную область), связанные дисульфидной связью.
Модификация протеолитической петли (сайта расщепления)
Структурный белок вируса может представлять собой белок гемагглютинин вируса гриппа В или белок гемагглютинин вируса гриппа А с делецией или модификацией протеолитической петли (сайта расщепления) внутри белка гемагглютинина. Делеция или модификация протеолитической петли гарантирует, что молекула НА по большей части сохраняется в виде НА0 предшественника.
НА синтезируется в виде белка-предшественника НА0, который претерпевает протеолитический процессинг: разрезается на две субъединицы (НА1 и НА2), связанные вместе дисульфидным мостиком. Штаммы человеческого и апатогенного птичьего вируса гриппа вызывают анатомически локализованные инфекции вследствие ограниченного набора клеток, секретирующих протеазу, которая может расщеплять НА0 предшественник внеклеточно. (Chen J, et. al. 1998, Cell. Vol 95: 409-417). Протеазы, отвечающие за расщепление НА0 при заражении людей гриппом, секретируются клетками дыхательных путей или совместно инфицирующей бактерией или микоплазмой, или они могут продуцироваться в ходе развития воспалительных реакций на инфицирование. Основной представляющей интерес протеазой является триптаза Клара, которая продуцируется клетками Клара бронхиального эпителия и имеет ограниченное распространение в тканях (верхние отделы дыхательных путей). Протеаза является специфической к одноосновной последовательности Q/E-X-R, находящейся в сайте расщепления H1, Н2, Н3 и Н6. В случае НА штаммов Н9 и В наблюдается немного отличный одноосновный сайт расщепления с последовательностью SSR и KER, соответственно (см. Фиг. 24). Для большинства вирусов гриппа не была идентифицирована протеаза, которая вызывает кишечную или респираторную инфекцию, наблюдаемую у водоплавающих птиц. В лаборатории большинство клеточных линий не поддерживают многоцикловую репликацию, если не добавляется экзогенная протеаза (обычно трипсин).
Однако высокопатогенные штаммы вируса птичьего гриппа расщепляются семейством более широко распространенных внутриклеточных протеаз, что приводит в результате к системным инфекциям. Такая разница в патогенности коррелирует со структурными различиями в сайте расщепления НА0. Патогенные штаммы имеют вставки многоосновных аминокислот внутри одноосновного сайта или рядом с ним. В этом случае расщепление происходит внутриклеточно, а участвующие в этом протеазы были идентифицированы как фурин и другие субтилизин-подобные ферменты, находящиеся в аппарате Гольджи и участвующие в пост-трансляционном процессинге предшественников гормона и факторов роста. Последовательность узнавания фурина R- X-R/K-R часто является аминокислотной инсерцией в сайтах расщепления НА0 вирусов Н5 и Н7 (см. Фигура 24). Широкое распространение фермента в ткани и эффективность внутримолекулярного расщепления способствуют появлению масштабной опасной системной инфекции, вызываемой этими вирусами.
Horimoto Т, et.al. (2006, Vaccine, Vol 24 : 3669-3676) описывает потерю многоосновного сайта расщепления Н5 (RERRRKKR↓G) в Н5. Проводилось исследование выбранных мутантов на мышах, включая мутант с делецией 4 первых аминокислот, несущих заряд (RERR), и модификацию с целью инактивации многоосновного сайта расщепления (RKKR с TETR). Потеря (удаление) сайта расщепления не влияла на иммуногенные свойства мутанта Н5. Сообщалось также об удалении многоосновного сайта (GERRRKKR↓G заменяли на RETR) с образованием мутантного NIBSC 05/240 NIBSC эталонного вируса гриппа NIBG-23. Hoffman et. al. (2002, 2002, Vaccine, Vol 20: 3165-3170) заменили многоосновный сайт расщепления Н5 НА на одноосновный сайт Н6, чтобы активизировать экспрессию в яйцах. Первые 4 остатка делетировали и заменили четыре последних аминокислоты многоосновного сайта на IETR (замена RERRRKKR↓G на IETR↓G). Этот мутантный Н5 продемонстрировал высокий уровень экспрессии, потенциальный протеолиз и изменение конформации при низком значении рН, данные об иммуногенности не сообщались. Эти исследования показывают, что модификацию сайта расщепления можно применять для снижения вирулентности вирусной частицы (в случае, когда реплицируются настоящие вирусы, делая возможной репликацию вируса, не убивая яйцо-хозяина). В отсутствие таких мутаций вирусы убивают яйцо до достижения высоких титров.
В процессе фолдинга НА и секреции с использованием аппарата Гольджи сайт расщепления предшественника гемагглютинина, который локализован в петле на поверхности НА, является легкодоступным для протеолиза под действием протеаз. Не основываясь на какой-либо теории, полагают, что если протеолиз предшественника НА0 происходит по одно- или многоосновному сайту в процессе фолдинга НА в ER, изменение конформации белка может происходить в аппарате Гольджи в процессе секреции, так как рН среды внутри аппарата Гольджи растения и в апопласте имеет слабокислое значение. При низких значениях рН может происходить изменение конформации НА, которое приводит к снижению как уровня экспрессии, так и внутренней устойчивости частицы. Таким образом, преимущественно нерасщепленный НА0 белок-предшественник как бы отпочковывается от плазматической мембраны.
Под "протеолитической петлей" или "сайтом расщепления" понимают консенсусную последовательность протеолитического сайта, который участвует в расщеплении НА0 предшественника. "Консенсус" или "консенсусная последовательность" в данном контексте означает последовательность (либо аминокислотную, либо нуклеотидную последовательность), в которой, на основании анализа выравнивания совокупности последовательностей, например, подтипов последовательности конкретного НА0 вируса гриппа, выявлена вариабельность родственных последовательностей. Консенсусная последовательность сайта расщепления вирусного белка НА0 может включать консенсусные аминокислотные последовательности гемагглютинина вируса гриппа А, в том числе, консенсус H1, консенсус Н3, или консенсусные аминокислотные последовательности гемагглютинина вируса гриппа В. Неограничивающие примеры консенсусных последовательностей показаны на Фигуре 24.
В аминокислотной последовательности НА протеолитическая петля локализована слитым пептидом, который состоит из 20 первых аминокислот НА2 части. Была определена кристаллическая структура НА0 вируса гриппа A/Hong Kong/68 (статья Chen, J., 1998. Cell 95: 409-417; описание статьи включено в настоящее изобретение посредством отсылки). Обычно полагают, что остатки, которые подвергаются действию растворителя, являются частью сайта расщепления, который образует удлиненную в значительной степени поверхностную (находящуюся на поверхности) петлю. С использованием этой специфической пептидной последовательности можно определить консенсусную последовательностью в этой выбранной области (Bianchi et al., 2005, Journal of Virology, 79: 7380-7388; описание статьи включено в настоящее изобретение посредством отсылки).
Для того, чтобы удалить протеолитическую петлю, было изучено строение В НА. Делеция одного лишь сайта протеолитического расщепления НА оставила бы С- конец НА1, a N- конец НА2 оставался бы далеко, и необходимо было бы создать длинный линкер. Однако делетирование части слитого пептида наряду с сайтом протеолитического расщепления позволило бы удалить полную протеолитическую петлю и соединить оставшуюся последовательность НА1 и НА2 минимальным пептидным линкером из 2 аминокислот. Коротко говоря, вариант В содержит делецию последовательности ALKLLKER на С- конце НА1, помимо делеции N- концевых аминокислот GFFGAIAGFLEG цепи НА2. Укороченные части НА1-НА2 были связаны вместе с помощью линкера GG.
Как показано на Фигуре 22В, после удаления протеолитической петли НА0 полученный в результате НА0 белок проявляет повышенную активность, как показывает его более высокая гемагглютинирующая способность по сравнению с гемагглютинирующей способностью НА белка, у которого протеолитическая петля не удалена. Под повышением активности понимают повышение гемагглютинирующей способности примерно от 2% примерно до 100% или на любую другую промежуточную величину по определению стандартными в данной области техники методами, например, примерно от 10% примерно до 50% или на любую другую промежуточную величину, например, примерно, на 2, 5, 8, 10, 12, 15, 18, 20, 22, 24, 25, 26, 28, 30, 32, 34, 35, 36, 38, 40, 42, 44, 45, 46, 48, 50, 52, 54, 55, 56, 58, 60, 65, 70, 75, 80, 85, 90, 95 или 100% по сравнению с активностью того же НА белка, но содержащего протеолитическую петлю.
Под "химерным вирусным белком" или "химерным вирусным полипептидом", также называемым "химерным белком" или "химерным полипептидом", понимают белок или полипептид, который содержит аминокислотные последовательности из двух или более источников, например, но без ограничения, из двух или более типов или подтипов вируса, или вирусов гриппа различного происхождения, которые соединены (слиты) в один полипептид. Химерный белок или полипептид может включать сигнальный пептид, такой же, как остальной полипептид или белок, или гетерологичный ему. Химерный белок или химерный полипептид можно продуцировать с использованием химерной нуклеотидной последовательности, а химерный белок или химерный полипептид можно расщеплять после синтеза и, при необходимости, связывать с образованием мультимерного белка. Таким образом, химерный белок или химерный полипептид включает также белок или полипептид, содержащий субъединицы, связанные через дисульфидные мостики (т.е. мультимерный белок). Например, химерный полипептид, содержащий аминокислотные последовательности из двух или более источников, можно процессировать в виде субъединиц, а субъединицы связывать дисульфидными мостиками с образованием химерного белка или химерного полипептида. Химерный вирусный белок может также содержать антигенный белок, или его фрагмент, первого вируса гриппа и комплекс трансмембранного домена (TDC) из НА второго вируса гриппа, включающий трансмембранный домен и домены цитоплазматического "хвоста" (ТМ/СТ). Полипептид может представлять собой гемагглютинин (НА), а каждую из двух или более чем двух аминокислотных последовательностей, которые составляют полипептид, можно получать из различных НА, чтобы продуцировать химерный НА или химерный НА вируса гриппа. Химерный НА может включать также аминокислотную последовательность, содержащую гетерологичный сигнальный пептид (химерный НА пребелок), который расщепляется после этого или в процессе синтеза белка. Предпочтительно, химерный полипептид или химерный НА вируса гриппа не является натуральным. Нуклеиновую кислоту, кодирующую химерный полипептид, можно описать как "химерную нуклеиновую кислоту" или как "химерную нуклеотидную последовательность". Вирусоподобная частица, состоящая из химерного НА, может быть описана как "химерная VLP".
Химерный белок или полипептид может включать сигнальный пептид, такой же как остальной полипептид или белок, или гетерологичный ему. Термин "сигнальный пептид" хорошо известен в данной области техники и обычно относится к короткой (около 5-30 аминокислот) последовательности аминокислот, находящейся, как правило, на N- конце полипептида, который может направлять транслокацию вновь транслированного полипептида в конкретную органеллу или способствовать позиционированию специфических доменов полипептидной цепи относительно друг друга. Например, но без ограничения, сигнальный пептид может нацеливать транслокацию белка в эндоплазматический ретикулум и/или способствовать позиционированию домена, проксимального к N- концу, относительно "заякоренного" в мембране домена образующегося (растущего) полипептида, содействуя отщеплению и фолдингу зрелого белка, например, но без ограничения, зрелого НА белка.
Неограничивающие примеры химерных вирусных белков или химерных вирусных нуклеиновых кислот, которые можно применять в соответствии с настоящим изобретением, описаны в Международных заявках WO 2009/076778, WO 2010/003235 или WO 2010/148511 (описание которых включено в настоящее изобретение посредством отсылки).
Сигнальный пептид
Сигнальный пептид может быть нативным по отношению к антигенному белку или вирусному белку, или сигнальный пептид может быть гетерологичным по отношению к первичной последовательности антигенного белка или экспрессируемого вирусного белка. Антигенный белок или вирусный белок может содержать сигнальный пептид вируса гриппа первого типа, подтипа или штамма в определенном сочетании НА одного или более чем одного типа, подтипа или штамма вируса гриппа. Например, нативный сигнальный пептид НА подтипов H1, Н2, Н3, Н5, Н6, Н7, Н9 или вируса гриппа типа В можно применять для экспрессии вирусного химерного белка в системе растения. Согласно некоторым вариантам изобретения SP может быть SP вируса гриппа типа В, H1, Н3 or Н5; или подтипа Hl/Bri, H1/NC, H5/Indo, Н3/Bri или B/Flo.
Сигнальный пептид может также быть ненативным, например, из антигенного белка, вирусного белка или гемагглютинина вируса, иного, нежели вирусный белок, или из растительного, животного или бактериального полипептида. Неограничивающим примером сигнального пептида, который можно применять, является сигнальный пептид протеин-дисульфидизомеразы люцерны ("PDISP"; нуклеотиды 32-103 последовательности с номером доступа Z11499; см. также Международные заявки WO 2009/076778; WO 2010/148511, или WO 2010/003235, (описание которых включено в настоящее изобретение посредством отсылки). В силу вышесказанного настоящее изобретение предусматривает химерный вирусный белок, содержащий нативный или не- нативный сигнальный пептид, и нуклеиновые кислоты, кодирующие такие химерные вирусные белки.
Соответственно, настоящее изобретение предусматривает способ продуцирования химерной VLP в растении, в котором первая нуклеиновая кислота, кодирующая химерный белок вируса, экспрессируется совместно со второй нуклеиновой кислотой, кодирующей канальный белок, например, но без ограничения, белок протонного канала. Первую и вторую нуклеиновую кислоту можно вводить в растение на одной и той же стадии, или их можно вводить в растение последовательно.
НА
Что касается вируса гриппа, термин "гемагглютинин" или "НА", применяемый в настоящей заявке, относится к гликопротеину, находящемуся на поверхности частиц вируса гриппа. НА представляет собой гомотримерный мембранный гликопротеин I типа, содержащий обычно сигнальный пептид, НА1 домен и НА2 домен, содержащий трансмембранный сайт для заякоривания и С- конец и короткий цитоплазматический хвост. Нуклеотидные последовательности, кодирующие НА, хорошо известны и являются доступными - см., например, базу данных BioDefence Public Health base (Influenza Virus; см. URL: biohealthbase.org) или базу данных National Center for Biotechnology Information (см. URL: ncbi.nlm.nih.gov), обе они включены в настоящее изобретение посредством отсылки.
Термин "гомотример" или "гомотримерный" указывает на то, что олигомер образован тремя молекулами НА белка. Не основываясь на какой- либо теории, полагают, что НА синтезируется в животных клетках в виде мономерного белка-предшественника (НА0) длиной около 75 кДа, который собирается на поверхности в удлиненный тримерный белок. Перед тримеризацией белок-предшественник расщепляется при постоянной активации сайта расщепления (также называемого слитым пептидом) на 2 полипептидных цепи, НА1 и НА2 (содержащую трансмембранную область), связанные дисульфидной связью. Сегмент НА1 может иметь в длину 328 аминокислот, а сегмент НА2 может иметь в длину 221 аминокислоту. Хотя это расщепление может быть важным для инфекционности вируса, оно может быть несущественным для тримеризации белка. Инсерция НА в мембрану эндоплазматического ретикулума (ER) клетки-хозяина, отщепление сигнального пептида и гликозилирование белка представляют собой котрансляционные события. Для корректного рефолдинга НА требуется гликозилирование белка и образование 6 внутрицепных дисульфидных связей. Тример НА собирается внутри цис- и транс-комплекса Гольджи, причем трансмембранный домен участвует в процессе тримеризации. Кристаллические структуры НА белков, обработанных бромелаином, у которых отсутствует трансмембранный домен, имеют высоко консервативную структуру среди штаммов вируса гриппа. Также установлено, что НА претерпевает значительные изменения конформации в процессе инфицирования, для которого требуется расщепление НА0 на 2 полипептидные цепи НА1 и НА2. НА белок может быть процессированным (т.е. содержать НА1 и НА2 домены), или может быть непроцессированным (т.е. содержать НА0 домен). НА белок можно применять для образования VLP, используя экспрессионную систему растения или растительной клетки.
НА по настоящему изобретению можно получать из любого подтипа. Например, НА может быть подтипа Н2, Н3, Н4, Н6, Н7, Н8, Н9, Н10, Н11, Н12, Н13, Н14, Н15, Н16, или НА вируса гриппа типа В. Рекомбинантный НА по настоящему изобретению может также содержать любую аминокислотную последовательность на основе последовательности любого гемагглютинина, известного в данной области техники-см., например, базу данных BioDefence Public Health base (Influenza Virus; см. URL: biohealthbase.org) или National Center for Biotechnology Information (см. URL: ncbi.nlm.nih.gov). Помимо этого, НА может быть на основе последовательности гемагглютинина, выделенной из одного или более новых или вновь идентифицированных вирусов гриппа.
Неограничивающие примеры НА, или фрагменты НА, которые можно применять в соответствии с настоящим изобретением, включают примеры, описанные в Международных заявках WO 2009/009876, WO 2009/076778; WO 2010/003225, WO 2010/003235, WO 2010/006452, WO 2011/035422 или WO 2010/148511 (которые включены в настоящее изобретение посредством отсылки).
Как показано на Фигуре 18, НА из B/Brisbane/60/2008 слабо экспрессируется в агроинфильтрованных листьях табака Nicotiana benthamiana (см. дорожки "1008" или "1029"). Однако совместная экспрессия НА типа В с М2 вируса гриппа A/New Caledonia/20/99 приводит к значительному повышению экспрессии НА (см. дорожки "1008+1261"; "1009+1261" и 1029+1261"). Повышение экспрессии НА наблюдали как для нативного НА типа В НА, так и для химерного НА типа В. Экспрессию НА наблюдали в присутствии или в отсутствие амплификационных элементов (BeYDV) и при различных разведениях Agrobacterium. Аналогичное повышение экспрессии НЗ наблюдали, когда НЗ из A/Perth/16/2009 экспрессировался совместно с М2 из A/New Caledonia/20/99 (Фигура 19; сравните дорожку "1019", только Н3, с "1019+1261", Н3 коэкспрессируется с М2).
VLP
Термин "вирусоподобная частица" (VLP), или "вирусоподобные частицы", или "VLP", относится к структурам, которые образуются в процессе самосборки и содержат вирусные белки, например, вирусный структурный белок, такой как НА белок вируса гриппа или канальный белок, например, но без ограничения, белок протонного канала, такой как М2, или комбинацию этих белков. Как правило, VLP, по своей морфологии и по своим антигенным свойствам, сходны с вирионами, продуцируемыми при инфицировании, но не несут генетической информации, достаточной для репликации, и поэтому не являются патогенными. В некоторых примерах VLP могут содержать один вид белка или более одного вида белков. В случае VLP, содержащих более чем один вид белка, виды белка могут быть из одного и того же вида вируса или могут содержать белок другого вида, рода, подсемейства или семейства вирусов (по номенклатуре ICTV). В других примерах один или более видов белка, содержащих VLP, можно модифицировать с использованием натуральной последовательности. VLP могут продуцироваться в соответствующих клетках-хозяевах, включая клетки растений- или насекомых-хозяев. После извлечения (экстракции) из клетки-хозяина и после выделения и дальнейшей очистки в соответствующих условиях VLP можно выделять в виде интактных структур.
Помимо этого, можно продуцировать VLP, которые содержат комбинацию НА подтипов. Например, VLP могут содержать один или более НА подтипа Н2, Н3, Н4, Н6, Н7, Н8, Н9, Н10, Н11, H12, Н13, H14, H15, H16, подтипа В НА или их комбинацию. Отбор комбинации НА можно определять в зависимости от предполагаемого применения вакцины, приготовленной из VLP. Например, вакцина для вакцинации (прививки) птиц может содержать любую комбинацию НА подтипов, тогда как VLP для вакцинации людей могут содержать один или более подтипов Н2, Н3, Н4, Н6, Н7, Н8, Н9, Н10, H11, Н12, Н13, H14, Н15, H16, подтипа В НА. Однако можно приготовить другие комбинации подтипов НА в зависимости от применения VLP. Чтобы продуцировать VLP, содержащие комбинации НА подтипов, НА заданного подтипа можно коэкспрессировать в той же самой клетке, например, растительной клетке.
Как более подробно описано ниже, экспрессия НА типа В или Н3 вируса гриппа B/Brisbane/60/2008 в агроинфильтрованных листьях табака Nicotiana benthamiana повышается, когда она осуществляется совместно с экспрессией М2 вируса гриппа A/New Caledonia/20/99 (см. Фигуры 18 и 19). Аналогичное повышение не наблюдается, когда H1 или Н5 коэкспрессируется (экспрессируется совместно) с М2, поскольку высокие уровни экспрессии H1 или Н5 наблюдаются в присутствии или в отсутствие М2 (Фигуры 20 и 21, соответственно). Известно, что процессинг НА зависит от рН (см. Reed М. L. et. al. Journal of Virology, February 2010, p. 1527-1535, Vol. 84, No. 3), и НА претерпевает зависимое от рН изменение конформации (Skehel J.J. et. al. 1982, PNAS79: 968-972). Изменение конформации H1 и Н5 наблюдается при значениях рН ниже рН в случае Н3 и типа В НА. Не основываясь на какой-либо теории, полагают, что рН в аппарате Гольджи во время созревания и миграция может не оказывать влияния на фолдинг H1 или Н5, однако, низкое значение рН в аппарате Гольджи может влиять на фолдинг Н3 и типа В НА. За счет коэкспрессии канального белка, например, но без ограничения, белка протонного канала, вместе с Н3 или с типа В НА в аппарате Гольджи может повышаться рН и обусловливать фолдинг НА, который приводит к повышенному выходу НА. Кроме того, H1 и Н5 могут быть более устойчивыми при более низких значениях рН, чем Н3 и типа В НА. Следовательно, при коэкспрессии канального белка, например, но без ограничения, белка протонного канала, совместно с Н3 или типа В НА в аппарате Гольджи расщепляется меньше НА.
VLP, полученные с использованием белков вируса гриппа в соответствии с настоящим изобретением, не содержат M1 белок. Известно, что белок M1 связывает РНК (Wakefield and Brownlee, 1989), которая загрязняет VLP препарат. Присутствие РНК является нежелательным для получения разрешения на VLP продукт от регуляторных органов, поэтому препарат VLP, не содержащий РНК, может быть предпочтительным.
VLP, полученные согласно настоящему изобретению, как правило, не содержат нейраминидазу (NA). Однако NA можно экспрессировать совместно с НА, если желательны VLP, содержащие НА и NA.
Изобретение также включает, но без ограничения, образованные из вирусов VLP, которые приобретают липидную оболочку из плазматической мембраны клетки, в которой экспрессируются VLP белки. Например, если VLP экспрессируется в системе растения, VLP может получить липидную оболочку из плазматической мембраны клетки.
Обычно термин "липид" относится к жирорастворимым (липофильным) природным молекулам. Более конкретно, этот термин применяют по отношению к жирным кислотам и их производным (включая три-, ди- и моноглицериды и фосфолипиды), а также к другим жирорастворимым стерин (стерол)-содержащим метаболитам или стеринам (стеролам). Фосфолипиды являются основным компонентом всех биологических мембран, наряду с гликолипидами, стеринами и белками. Примеры фосфолипидов включают фосфатидилэтаноламин, фосфатидилхолин, фосфатидилинозит, фосфатидилсерин и т.п. Примеры стеринов включают зоостерины (например, холестерин) и фитостеролы (фитостерины). Более 200 фитостеринов было идентифицировано в различных видах растений, причем самыми распространенными являются кампестерин, стигмастерин, эргостерин, брассикастерин, дельта-7-стигмастерин, дельта- 7- авенастерин, даукостерин, ситостерин, 24- метилхолестерин, холестерин и бета-ситостерин. Как будет понятно специалистам в данной области техники, липидный состав плазматической мембраны клетки может меняться в зависимости от условий культивирования или роста клетки или организма, из которого эта клетка получена.
Клеточные мембраны обычно содержат липидные бислои, а также белки, выполняющие различные функции. Локальные концентрации конкретных липидов, которые можно обнаружить в липидном бислое, называются 'липидными рафтами'. Не основываясь на какой- либо теории, полагают, что липидные рафты играют значительную роль в эндо- и экзоцитозе, входе или выходе вирусов или других инфекционных агентов, межклеточной сигнальной трансдукции (передаче сигнала), взаимодействии с другими структурными компонентами клетки или организма, такими как внутриклеточные или внеклеточные матриксы.
В растениях VLP гриппа "отпочковываются" от плазматической мембраны, следовательно, липидный состав VLP отражает их происхождение. VLP, продуцированные согласно настоящему изобретению, содержат НА одного или более типов или подтипов гриппа в комплексе с липидами растительного происхождения. Растительные липиды могут стимулировать специфические иммунные клетки и усиливать индуцированный иммунный ответ. Мембраны растительных клеток состоят из липидов, фосфатидилхолина (PC) и фосфатидилэтаноламина (РЕ), а также содержат гликосфинголипиды, сапонины и фитостерины. Помимо этого, в плазматических мембранах растительных клеток обнаружены также липидные рафты-эти микродомены обогащены сфинголипидами и стеринами. Известно, что в растениях находятся различные фитостерины, включая стигмастерин, ситостерин, 24-метилхолестерин и холестерин (Mongrand et al., 2004).
PC и РЕ, а также гликосфинголипиды могут связываться с CD1 молекулами, экспрессирующимися в иммунных клетках млекопитающих, таких как антигенпрезентирующие клетки (АРС), например, дендритные клетки и макрофаги, и других клетках, включая В и Т лимфоциты в вилочковой железе (тимусе) и печени (Tsuji М,. 2006). Молекулы CD1 по своей структуре сходны с молекулами главного комплекса гистосовместимости (МНС, ГКГ, ГКГС) класса I и их роль заключается в презентации (представлении) гликолипидных антигенов NKT клеткам (натуральным киллерным Т клеткам). При активации NKT клетки активируют клетки системы врожденного иммунитета, такие как NK клетки и дендритные клетки, а также активируют клетки системы адаптивного (приобретенного) иммунитета, такие как антителопродуцирующие В клетки и Т-клетки.
В плазматической мембране могут находиться различные фитостерины -конкретный состав может меняться в зависимости от вида, условий роста, источников питательных веществ или патогенного состояния, если называть некоторые из факторов. Как правило, наиболее широко распространенным фитостерином является бета-ситостерин.
Фитостерины находятся в VLP гриппа в комплексе с липидным бислоем, например оболочка, образованная из плазматической мембраны, может обусловливать предпочтительный состав вакцины. Не основываясь на какой- либо теории, полагают, что полученные в растениях VLP в комплексе с липидным бислоем, таким как оболочка из плазматической мембраны, может вызывать более сильную иммунную реакцию по сравнению с VLP, полученными в экспрессионных системах, аналогичную иммунной реакции, вызванной живой или инактивированной цельновирионной вакциной.
Таким образом, согласно некоторым вариантам изобретения предусматриваются VLP в комплексе с липидным бислоем растительного происхождения. Согласно некоторым вариантам изобретения липидный бислой растительного происхождения может представлять собой оболочку VLP. Липиды растительного происхождения могут содержать липидные компоненты плазматической мембраны растения, в котором продуцирована VLP, включая, но без ограничения, фосфатидилхолин (PC), фосфатидилэтаноламин (РЕ), гликосфинголипиды, фитостерины или их комбинацию. Или же липид растительного происхождения может называться 'растительным липидом'. Примеры фитостеринов известны из уровня техники и включают, например, стигмастерин, ситостерин, 24-метилхолестерин и холестерин - см., например, Mongrand et al., 2004.
Строение и размер VLP можно определять, например, с помощью реакции гемагглютинации, электронной микроскопии или высокоэффективной гель-хроматографии.
Для высокоэффективной гель-хроматографии тотальные растворимые белки можно экстрагировать из растительной ткани посредством гомогенизации (Polytron) образца измельченного замороженного растительного материала в буфере для экстракции, а нерастворимый материал можно удалять центрифугированием. Можно применять осаждение с помощью PEG (ПЭГ). Затем количественно определяют растворимый белок, а экстракт пропускают через колонку с насадкой для гель-хроматографии, например, но без ограничения, такой как сефакрил™. После хроматографии фракции можно далее анализировать методом иммуноблоттинга, чтобы определить состав белков в данной фракции.
Не связывая себя с конкретной теорией, полагают, что способность НА связываться с RBC (эритроцитами) различных животных обусловлена аффинностью НА к сиаловым кислотам α2,3 или α2,6 и присутствием этих сиаловых кислот на поверхности RBC. НА гриппа лошадей и птиц агглютинирует эритроциты всех из нижеперечисленных видов, включая индеек, кур, уток, морских свинок, людей, овец, лошадей и коров; в то время как НА вируса человеческого гриппа связывается с эритроцитами индеек, кур, уток, морских свинок, человека и овец (см. также Ito Т. et al, 1997, Virology, vol 227, p. 493-499; и Medeiros R et al, 2001, Virology, vol 289 p. 74-85).
Фолдинг (Шаперон)
Корректный фолдинг экспрессируемого вирусного белка может играть важную роль, в частности, в устойчивости белка, образовании мультимеров, образовании VLP, функционировании вирусного белка и узнавании вирусного белка антителом. На фолдинг и накопление белка может влиять один или более факторов, включая, но без ограничения, последовательность белка, относительное содержание белка, степень внутриклеточного краудинга (толкучки, молекулярной тесноты), рН в компартменте клетки, доступность кофакторов, которые могут связываться или транзиторно ассоциироваться со свернутым, частично свернутым или несвернутым белком, присутствие одного или более белков шаперонов и т.п.
Белки теплового шока (Hsp) или белки стрессов являются примерами белков шаперонов, которые могут принимать участие в различных клеточных процессах, включая синтез белков, внутриклеточную миграцию, предупреждение неправильного фолдинга, предупреждение агрегации белков, сборку и разборку комплексов белков, фолдинг белка и дезагрегацию белков. Примеры таких белков шаперонов включают, но без ограничения,, Hsp60, Hsp65, Hsp 70, Hsp90, Hsp100, Hsp20-30, Hsp10, Hsp100-200, Hsp100, Hsp90, Lon, TF55, FKBPs, циклофилины, ClpP, GrpE, убиквитин, калнексин и протеин дисульфидизомеразы (см., например, Macario, A.J.L., Cold Spring Harbor Laboratory Res. 25: 59-70. 1995; Parsell, D.A. & Lindquist, S. Ann. Rev. Genet. 27: 437-496 (1993); патент США No. 5,232,833). Как указывается в настоящей заявке, белки шапероны, например, но без ограничения, Hsp40 и Hsp70, можно применять для того, чтобы гарантировать фолдинг вирусного белка.
Примеры Hsp70 включают Hsp72 и Hsc73 млекопитающих, DnaK бактерий, в особенности микобактерий, таких как Mycobacterium leprae (микобактерии лепры (проказы)), Mycobacterium tuberculosis (туберкулезные микобактерии) и Mycobacterium bovis (коровья туберкулезная палочка, например, бацилла Кальметта-Герена: называемая в настоящей заявке Hsp71). DnaK Escherichia coli, дрожжей и других прокариот и BiP и Grp78 эукариот, например, A. thaliana (Lin et al. 2001 (Cell Stress and Chaperones 6: 201-208). Конкретным примером Hsp70 является A. thaliana Hsp70 (код в Genbank: AY120747.1). Hsp70 способен специфически связывать АТФ (ATP), а также несвернутые полипептиды и пептиды, тем самым участвуя в фолдинге (свертывании) и развертывании белков, а также в сборке и разборке белковых комплексов.
Примеры Hsp40 включают DnaJ у прокариот, например, Е. coli и микобактерий, и HSJ1, HDJ1 и Hsp40 у эукариот, таких как люцерна (Frugis et al., 1999. Plant Molecular Biology 40: 397-408). Конкретным примером Hsp40 является M. sativa MsJ1 (Genbank ref: AJ000995.1). Помимо прочего, Hsp40 играет роль молекулярного шаперона в фолдинге, термостойкости белков и ДНК-репликации.
Среди Hsps шаперон Hsp70 и совместно с ним действующий шаперон Hsp40 участвуют в стабилизации трансляции вновь синтезированных полипептидов до завершения синтеза. Не основываясь на какой- либо конкретной теории, полагают, что Hsp40 связывается с гидрофобными "очажками" (пэтчами) несвернутых (в момент образования или только что трансфицированных) полипептидов, тем самым способствуя взаимодействию комплекса Hsp70-ATP с полипептидом. Гидролиз АТР приводит к образованию стабильного комплекса между полипептидом, Hsp70 и ADP (АДФ) и высвобождению Hsp40. Ассоциация комплекса Hsp70-ADP с гидрофобными очажками полипептида предупреждает взаимодействие с другими гидрофобными очажками, предотвращая некорректный фолдинг и образование агрегатов с другими белками (обзор Hartl, FU. 1996. Nature 381: 571-579).
Нативные белки шапероны, возможно, способны содействовать корректному фолдингу в случае низких уровней рекомбинантного белка, но при повышении уровней экспрессии избыток нативных шаперонов может стать ограничивающим фактором. Высокие уровни экспрессии вирусного белка в агроинфильтрованных листьях могут привести к накоплению вирусного белка в цитозоли, а коэкспрессия одного или более белков шаперонов, например, Hsp70, Hsp40 или как Hsp70, так и Hsp40, может понизить уровень неправильно свернутых или агрегированных белков и повысить количество белков, имеющих третичную или четвертичную структуру, способствующую образованию вирусоподобных частиц.
На основании вышесказанного настоящее изобретение предусматривает также способ продуцирования VLP вирусного белка в растении, в котором первая нуклеиновая кислота, кодирующая вирусный белок, коэкспрессируется со второй нуклеиновой кислотой, кодирующей канальный белок, например, но без ограничения, белок протонного канала, и с третьей нуклеиновой кислотой, кодирующей шаперон. Первую, вторую и третью нуклеиновые кислоты можно вводить в растение на одной и той же стадии или можно вводить в растение последовательно.
N-Гликаны
VLP, продуцируемая растением, может индуцировать вирусный белок, содержащий растительные специфические N- гликаны. Ввиду этого данное изобретение предусматривает также VLP, содержащие вирусный белок со растение-специфическими N-гликанами.
Далее, модификация N- гликана в растениях известна (см., например, Международные заявки WO 2008/151440; WO 2010/006452; или патент США No. 60/944,344; описание которых вводится в настоящее изобретение посредством отсылки), и можно продуцировать вирусный белок с модифицированными N- гликанами. Можно получать вирусный белок, имеющий модифицированный паттерн гликозилирования, например, с пониженным уровнем фукозилирования, ксилозилирования или как фукозилирования, так и ксилозилирования N- гликанов, или можно получать вирусный белок с модифицированным паттерном гликозилирования, в котором отсутствуют сайты фукозилирования, ксилозилирования, или и те, и другие, и наблюдается повышенный уровень галактозилирования. Помимо этого, модуляция посттрансляционных модификаций, например, присоединение концевой галактозы, может привести к снижению степени фукозилирования и ксилозилирования экспрессируемого вирусного белка по сравнению с экспрессируемым растением белком дикого типа.
Например, но без ограничения, синтез вирусного белка, имеющего модифицированный паттерн гликозилирования, можно осуществить посредством коэкспрессии целевого белка совместно с нуклеотидной последовательностью, кодирующей бета- 1.4галактозилтрансферазу (GalT), например, но без ограничения, GalT млекопитающих или GalT человека, однако, можно использовать также GalT из других источников. Каталитический домен GalT можно также соединять (сливать) с CTS доменом (т.е. цитоплазматическим хвостом, трансмембранным доменом, "стволовой" областью) N-ацетилглюкозаминил трансферазы (GNT1) с образованием гибридного фермента GNT1-GalT, а гибридный фермент можно коэкспрессировать с вирусным белком. Вирусный белок можно также коэкспрессировать совместно с нуклеотидной последовательностью, кодирующей N- ацетилглюкозаминил трансферазу III (GnTIII), например, но без ограничения, GnT-III млекопитающих или GnT-III человека, также можно использовать GnT-III из других источников. Помимо этого, можно также использовать гибридный фермент GNT1-GnT-III, содержащий CTS GNT1, слитый с GnT- III.
Таким образом, настоящее изобретение включает также VLP, содержащие один или более вирусных белков, включающих модифицированные N-гликаны.
Последовательности
Неограничивающие примеры последовательностей, которые можно применять по настоящему изобретению, включают:
Белок Н2, кодируемый молекулой нуклеиновой кислоты, может быть из штамма A/Singapore (Сингапур)/1/57 (H2N2);
Белок Н3, кодируемый молекулой нуклеиновой кислоты, может быть из штамма A/Brisbane 10/2007 (H3N2), A/Wisconsin /67/2005 (H3N2), A/Victoria /361/2011 (H3N2) или A/Perth /16/2009 (H3N2);
Белок Н6, кодируемый молекулой нуклеиновой кислоты, может быть из штамма A/teal (утка)/HongKong(Гонконг)/W312/97 (H6N1);
Белок Н7, кодируемый молекулой нуклеиновой кислоты, также может быть из штамма A/Equine (Лошадиный) /Prague (Прага)/56 (H7N7);
Белок Н9, кодируемый молекулой нуклеиновой кислоты, может быть из штамма A/HongKong/1073/99 (H9N2);
НА белок подтипа В вируса гриппа, кодируемый нуклеиновой кислотой, может быть из штамма B/Florida (Флорида)/4/2006, B/Malaysia /2506/2004, B/Wisconsin/1/2010 или B/Brisbane/60/2008.
Неограничивающие примеры последовательностей, которые можно применять по настоящему изобретению, включают также последовательности, описанные в Международных патентных заявках: WO 2009/009876; WO 2009/076778; WO 2010/003225; WO 2010/148511; WO 2010/003235; WO 2010/006452, которые включены в настоящее изобретение посредством отсылки). Примерами таких кодирующих нуклеотидных последовательностей являются последовательности, кодирующие НА белки из Н2, Н3, Н4, Н6, Н7, Н8, Н9, Н10, Н11, Н12, Н13, Н14, Н15, Н16 и типа В НА, известные в области техники. Например, подтипы Н3 или В включают SEQ ID No: 25 или 30. Последовательность, кодирующая структурный вирусный белок, может представлять собой, например, последовательность гена НА вируса гриппа B/Brisbane/60/2008, B/Malaysia/2506/2004 или B/Wisconsin/1/2010, или НЗ вируса гриппа A/Perth/16/2009 или A/Victoria/361/2011. Другие примеры включают нуклеотидные последовательности, кодирующие НА белки, в которых удалена (делетирована) "протеолитическая" петля (петля, подлежащая протеолитическому расщеплению) белка НА, как например, но без ограничения, последовательность, определяемая с помощью SEQ ID NO: 41.
Настоящее изобретение включает также, но без ограничения, нуклеотидные последовательности, кодирующие НА, например, из Н2, Н3, Н4, Н6, Н7, Н8, Н9, Н10, Н11, Н12, Н13, Н14, Н15, Н16 или В НА. Например, последовательности SEQ ID NO: 28, 43, 23, кодирующие НА вируса гриппа типа В, В с делетированной протеолитической петлей или Н3, соответственно, нуклеотидная последовательность, которая в жестких условиях гибридизуется с последовательностями SEQ ID NO: 28, 43, 23, или нуклеотидная последовательность, которая в жестких условиях гибридизуется с последовательностями, комплементарными последовательностям SEQ ID NO: 28, 43, 23, причем эта нуклеотидная последовательность кодирует белок гемагглютинин, который, будучи экспрессирован, образует VLP, и VLP индуцирует продуцирование антитела. Например, экспрессия нуклеотидной последовательности в растительной клетке приводит к образованию VLP, а VLP можно применять для продуцирования антитела, способного связывать НА, включая зрелый НА вируса гриппа типа В или Н3. VLP, введенная субъекту, вызывает иммунный ответ. Эта нуклеотидная последовательность может также экспрессироваться совместно со второй нуклеотидной последовательностью, кодирующей канальный белок, например, но без ограничения, нуклеотидные последовательности SEQ ID NO: 9, 12, нуклеотидная последовательность, которая в жестких условиях гибридизуется с последовательностями SEQ ID NO: 9, 12, или нуклеотидная последовательность, которая в жестких условиях гибридизуется с последовательностями, комплементарными последовательностям SEQ ID NO: 9, 12, причем вторая нуклеотидная последовательность кодирует белок протонного канала с образованием VLP. Предпочтительно, VLP вызывает продуцирование антитела и VLP, при введении ее субъекту, вызывает иммунный ответ.
Например, экспрессия нуклеотидной последовательности в растительной клетке приводит к образованию VLP, a VLP можно применять для продуцирования антитела, способного связывать вирусный белок, например, такой как НА, включая, но без ограничения, НА0, НА0 белок с делетированной или модифицированной "протеолитической" петлей, НА1 или НА2 одного или более типов или подтипов вируса гриппа, таких, но без ограничения, как подтипы Н2, Н3, Н4, Н6, Н7, Н8, Н9, Н10, H11, Н12, Н13, H14, Н15, H16, подтип В НА. После введения субъекту VLP вызывает иммунный ответ.
Гибридизация в жестких условиях известна из уровня техники (например, см. Current Protocols in Molecular Biology, Ausubel et al., eds. 1995 and supplements; Maniatis et al, Molecular Cloning (A Laboratory Manual), Cold Spring Harbor Laboratory, 1982; Sambrook and Russell, Molecular Cloning: A Laboratory Manual, 3rd edition 2001; описания которых включены в настоящее изобретение посредством отсылки). Примером одного из таких жестких условий гибридизации может служить гибридизация в течение примерно 16-20 часов в растворе 4 X SSC при 65°C с последующей отмывкой в растворе 0.1 X SSC при 65°С в течение одного часа, или с последующими 2мя отмывками в растворе 0.1 X SSC при 65°С, каждая в течение 20 или 30 минут. Или же, примером таких жестких условий гибридизации могла бы служить гибридизация в течение ночи (в течение 16-20 часов) в растворе 50% формамида, 4 X SSC при 42°С с последующей отмывкой в растворе 0.1 X SSC при 65°С в течение одного часа, или с последующими 2мя отмывками в растворе 0.1 X SSC при 65°С, каждая в течение 20 или 30 минут, или в течение ночи (16-20 часов), или гибридизация в водном фосфатном буфере Черча (7% SDS; 0.5 М NaPO4 буфер рН 7.2; 10 мМ EDTA) при 65°С, с 2 отмывками либо при 50°С в растворе 0.1 X SSC, 0.1% SDS в течение 20 или 30 минут каждая, или с 2 отмывками при 65°С в растворе 2 X SSC, 0.1% SDS в течение 20 или 30 минут каждая.
Помимо этого, настоящее изобретение включает нуклеотидные последовательности, которые характеризуются тем, что они имеют сходство последовательности или идентичность последовательности около 70, 75, 80, 85, 87, 90, 91, 92, 93 94, 95, 96, 97, 98, 99, 100%, или любое значение между этими пределами, с нуклеотидной последовательностью, кодирующей НА вируса гриппа типа В (SEQ ID NO: 28), В с делетированной или модифицированной "протеолитической" петлей (SEQ ID NO: 43), Н3 (SEQ ID NO: 23), или НА, кодируемый какой-либо одной или более из последовательностей SEQ ID NO: 23, 28, 43, 46, 51, 57 или 61, причем нуклеотидная последовательность кодирует белок гемагглютинин, который, будучи экспрессирован, образует VLP, a VLP вызывает продуцирование антитела. Например, экспрессия нуклеотидной последовательности в растительной клетки приводит к образованию VLP, a VLP можно применять для продуцирования антитела, способного связывать НА, включая непроцессированный и/или зрелый НА типа В или Н3, или непроцессированный и/или зрелый НА, в котором делетирована "протеолитическая" петля. VLP, после введения субъекту, индуцирует иммунный ответ.
Настоящее изобретение включает также нуклеотидные последовательности, которые характеризуются тем, что они имеют сходство последовательности или идентичность последовательности около 70, 75, 80, 85, 87, 90, 91, 92, 93 94, 95, 96, 97, 98, 99, 100%, или любое значение между этими величинами, с нуклеотидной последовательностью, кодирующей М2 (SEQ ID NO: 9, 12), причем нуклеотидная последовательность кодирует канальный белок, например, но без ограничения, белок протонного канала, который, при коэкспрессии (совместной экспрессии) со структурным вирусным белком образует VLP. Предпочтительно, VLP вызывает продуцирование антитела и VLP, при введении ее субъекту, индуцирует иммунный ответ.
Идентичность последовательностей или сходство последовательностей можно определить, используя программу сравнения нуклеотидных последовательностей, например, такую, которая поставляется в пакете программ для биоинформатики DNASIS (например, использующей, но без ограничения, следующие параметры: штраф за гэп (пропуск в последовательности) 5, # верхних диагоналей 5, штраф за фиксированный гэп 10, k-кортеж 2, "плавающий" гэп 10 и размер окна 5). Однако, из уровня техники хорошо известны другие методы выравнивания последовательностей, например, с применением алгоритмов Смита-Ватермана (Smith & Waterman) (1981, Adv. Appl. Math. 2: 482), Нидлмана-Вунша (Needleman & Wunsch) (J. Mol. Biol. 48: 443, 1970), Липмана-Пирсона (Pearson & Lipman) (1988, Proc. Nat'l. Acad. Sci. USA 85: 2444), и компьютерной реализации этих алгоритмов (например, GAP, BESTFIT, FASTA и BLAST), или с помощью выравнивания вручную и визуального изучения. Пример выравнивания последовательностей НА из различных штаммов вируса гриппа можно найти на Фигуре 24.
"Иммунный ответ" ("иммунная реакция") обычно относится к ответу (реакции) адаптивной иммунной системы. Адаптивная иммунная система обычно включает гуморальный иммунный ответ и клеточный иммунный ответ. Гуморальный иммунный ответ представляет собой тот аспект (ту сторону) иммунитета, который(-ая) опосредуется секретируемыми антителами, продуцируемыми в В лимфоцитах (В клетках). Секретированные антитела связываются с антигенами на поверхности инвазивных микробов (таких как вирусы или бактерии), что является меткой для их деструкции. Выражение "гуморальный иммунитет" обычно применяют по отношению к продуцированию антител и процессам, которые его сопровождают, а также к эффекторным функциям антител, включая активацию Th2 клеток и продуцирование цитокинов, генерацию клеток памяти, инициирование фагоцитоза опсонинами, уничтожение (исключение) патогенов и т.п. Термины "модулировать" или "модуляция" или подобные им относятся к увеличению или уменьшению конкретного ответа (реакции) или параметра, определяемого любым из общеизвестных или общепринятых методов анализа, примеры некоторых из них приводятся в настоящей заявке.
Клеточный ответ означает иммунный ответ, который не включает антитела, но скорее включает активацию макрофагов, натуральных киллерных клеток (NK), антиген-специфических цитотоксических Т-лимфоцитов, и высвобождение различных цитокинов в ответ на антиген. Термин "клеточный иммунитет" обычно применяют по отношению к некоторым ответам (реакциям), опосредуемым активацией Th клеток, Тс клеток и Т-клетками. Клеточный иммунитет особенно важен при реакции на вирусные инфекции.
Например, индукцию антиген-специфических CD8 положительных Т лимфоцитов можно количественно определять методом ELISPOT; стимуляцию CD4 положительных Т-лимфоцитов можно количественно определять, используя анализ пролиферации. Титр антител против гриппа можно количественно определять методом ELISA; изотипы антиген-специфических или перекрестно-реактивных антител можно также количественно определять, используя антитела против изотипа (например, анти -IgG, IgA, IgE или IgM). Методы и технические средства для осуществления таких анализов хорошо известны в данной области (из уровня) техники.
HAI титры перекрестной реактивности также можно применять для того, чтобы продемонстрировать эффективность иммунного ответа на другие штаммы вируса, родственные подтипу вакцины. Например, сыворотку от субъекта, иммунизированного композицией вакцины из первого штамма (например, VLP из A/Indonesia 5/05), можно применять в HAI анализе со вторым штаммом цельного вируса или вирусных частиц (например, A/Vietnam/1194/2004), и определять HAI титр.
Присутствие или уровни цитокинов также можно определять количественно. Например, реакция Т- хелперных клеток (Thl/Th2) будет характеризоваться с помощью количественного определения клеток, секретирующих IFN-γ и IL-4, методом ELISA (например, наборы BD Biosciences OptEIA). Мононуклеарные клетки периферической крови (РВМС) или спленоциты, полученные от субъекта, можно культивировать и анализировать супернатант. Т- лимфоциты можно также определять количественно с помощью сортировки (сортинга) флуоресцентно-активированных клеток (FACS), используя маркер-специфические флуоресцентные метки и методы, известные из уровня техники.
Для характеристики иммунного ответа у субъекта можно применять также методы микронейтрализации, см., например, методы Rowe et al., 1973. Титры вирусонейтрализующих антител можно получать различными методами, включая: 1) определение числа литических бляшек (анализ бляшкообразования) с последующей фиксацией с помощью кристаллического фиолетового/окрашиванием клеток; 2) наблюдение за лизисом клеток в культуре под микроскопом; 3) методы ELISA и спектрофотометрического обнаружения NP вирусного белка (коррелируют с вирусной инфекцией клетки-хозяина).
Конструкции
Помимо этого, настоящее изобретение относится к генной конструкции, содержащей нуклеиновую кислоту, кодирующую канальный белок, например, но без ограничения, белок протонного канала или структурный вирусный белок, описанные выше, функционально связанный с регуляторным элементом, работающим в растении. Примеры регуляторных элементов, работающих в растительной клетке, которые можно применять в соответствии с настоящим изобретением, включают, но без ограничения, регуляторную область пластоцианина (патент США 7,125,978; описание которого включено в настоящее изобретение посредством отсылки) или регуляторную область рибулозо-1,5-бисфосфаткарбоксилазы /оксигеназы (RuBisCO; патент США 4,962,028; описание которого включено в настоящее изобретение посредством отсылки), хлорофилл а/b-связывающий белок (CAB; Leutwiler et al; 1986; описание включено в настоящее изобретение посредством отсылки), ST-LS1 (ассоциированный с выделяющим кислород комплексом фотосистемы II и описанный Stockhaus et al. 1987, 1989; описание включено в настоящее изобретение посредством отсылки).
Регуляторные элементы
Термины "регуляторная область", "регуляторный элемент" или "промотор" в настоящей заявке относятся к участку нуклеиновой кислоты, как правило, но не всегда, 5' области гена, кодирующей белок, которая может состоять либо из ДНК, либо из РНК, либо как из ДНК, так и из РНК. Когда регуляторная область является активной и функционально ассоциирована или функционально связана с целевым (представляющим интерес) геном, это может привести в результате к экспрессии этого целевого гена. Регуляторный элемент дает возможность опосредовать специфичность в отношении органа или регулировать активацию гена в процессе развития или временную активацию гена. "Регуляторная область" может включать промоторные элементы, ядерные элементы промоторов, проявляющие базальную промоторную активность, элементы, индуцируемые в ответ на внешний раздражитель, элементы, которые опосредуют активность промотора, такие как элементы отрицательной регуляции или энхансеры транскрипции. "Регуляторная область" в соответствии с настоящей заявкой может также включать элементы, которые активируют последующую транскрипцию, например, регуляторные элементы, которые модулируют экспрессию гена, такие как энхансеры трансляции и транскрипции, ингибиторы трансляции и транскрипции, активирующие последовательности и элементы нестабильности мРНК. Некоторые из этих последних элементов могут быть локализованы проксимально к кодирующей области.
В данном контексте термин "регуляторный элемент" или "регуляторная область" относятся обычно к последовательности ДНК, обычно, но не всегда, против хода транскрипции, в направлении 5' к кодирующей последовательности структурного гена, который контролирует экспрессию кодирующей области, обеспечивая узнавание РНК полимеразы и/или других факторов, необходимых для начала транскрипции в конкретном сайте. Однако следует понимать, что другие нуклеотидные последовательности, расположенные внутри интронов, или 3' к последовательности, также могут вносить вклад в регуляцию экспрессии кодирующей области, представляющей интерес. Примером регуляторного элемента, который обеспечивает узнавание РНК полимеразы или других факторов транскрипции с целью гарантировать инициацию в конкретном сайте, является промоторный элемент. Большинство, но не все, промоторные элементы эукариот (эукариотических генов) содержат TATA бокс, консервативную нуклеотидную последовательность, состоящую из пар нуклеотидов аденозина и тимидина, обычно расположенных на расстоянии примерно 25 п. н. (пар нуклеотидов) по направлению 5' от сайта начала транскрипции. Промоторный элемент содержит базальный промоторный элемент, отвечающий за инициацию транскрипции, а также другие регуляторные элементы (перечисленные выше), которые модифицируют экспрессию гена.
Имеется несколько типов регуляторных областей, включая регуляторные области, регулирующиеся в процессе развития, индуцибельные или конститутивные. Регуляторная область, которая регулируется в процессе развития, или контролирует дифференциальную экспрессию гена, находящегося под контролем, активируется в определенных органах или тканях органа в конкретные моменты в процессе развития этого органа или ткани. Однако, некоторые регуляторные области, которые регулируются в процессе развития, могут, предпочтительно, быть активными в определенных органах или тканях на конкретных стадиях развития, они также могут быть активными регулируемым в процессе развития образом, или с таким же успехом могут находиться на основном уровне в других органах или тканях растения. Примеры тканеспецифических регуляторных областей, например, семяспецифическая регуляторная область, включают промотор напина и промотор круциферина (Rask et al., 1998, J. Plant Physiol. 152: 595-599; Bilodeau et al., 1994, Plant Cell 14: 125-130). Пример лист-специфической регуляторной области включает промотор пластоцианина (см. патент США 7,125,978, описание которого включено в настоящее изобретение посредством отсылки).
Индуцибельная регуляторная область представляет собой регуляторную область, способную, в ответ на индуцирующий фактор, непосредственно или опосредованно активировать транскрипцию одной или более последовательностей ДНК или генов. В отсутствие индуцирующего фактора ДНК-последовательности или гены не транскрибируются. Как правило, белковый фактор, который специфически связывается с индуцибельной регуляторной областью для активации транскрипции, может присутствовать в неактивной форме, которая затем, прямо или косвенно, превращается в активную форму под действием индуцирующего фактора. Однако белковый фактор может также отсутствовать. Индуцирующий фактор может представлять собой химический агент, такой как белок, метаболит, регулятор роста, гербицид или фенольное соединение или физиологический стресс, вызываемый непосредственно действием тепла, холода, соли или токсических элементов, или косвенно, при посредничестве патогена или болезнетворного фактора, такого как вирус. Растительную клетку, содержащую индуцибельную регуляторную область, можно подвергать действию индуцирующего фактора, вводя индуцирующий фактор в клетку или растение извне, например, путем опрыскивания, полива, термообработки или другими подобными методами. Индуцибельные регуляторные элементы могут происходить из генов растительного и нерастительного происхождения (см., например, статью Gatz, С. and Lenk, LR.P., 1998, Trends Plant Sci. 3, 352-358; описание которой включено в настоящее изобретение посредством отсылки). Примеры потенциальных индуцибельных промоторов включают, но без ограничения, тетрациклин-индуцибельный промотор (Gatz, С, 1997, Ann. Rev. Plant Physiol. Plant Mol. Biol. 48, 89-108; описание этой статьи включено в настоящее изобретение посредством отсылки), стероид-индуцибельный промотор (Aoyama. Т. and Chua, N.H., 1997, Plant 1. 2, 397-404; описание этой статьи включено в настоящее изобретение посредством отсылки) и этанол-индуцибельный промотор (Salter, M.G., et al, 1998, Plant Journal 16, 127-132; Caddick, M.X., et al, 1998, Nature Biotech. 16, 177-180, описание которых включено в настоящее изобретение посредством отсылки), цитокинин-индуцибельные гены IB6 и CKI 1 (статьи Brandstatter, I. and Kieber, J.J., 1998, Plant Cell 10, 1009-1019; Kakimoto, Т., 1996, Science 274, 982-985; описание которых включено в настоящее изобретение посредством отсылки) и ауксин-индуцибельный элемент, DR5 (Ulmasov, Т., et al., 1997, Plant Cell 9, 1963-1971; описание статьи включено в настоящее изобретение посредством отсылки).
Конститутивная регуляторная область управляет экспрессией гена повсюду в различных частях растения и непрерывно в процессе развития растения. Примеры известных конститутивных регуляторных элементов включают промоторы, ассоциированные с транскриптом CaMV 35S гена (Odell et al., 1985, Nature, 313: 810-812), генами рисового актина 1 (Zhang et al, 1991, Plant Cell, 3: 1155-1165), актина 2 (An et al, 1996, Plant J., 10: 107-121), или tms 2 (патент США 5,428,147, описание которого включено в настоящее изобретение посредством отсылки), и триосефосфат изомеразы 1 (Xu et. al., 1994, Plant Physiol. 106: 459-467), геном убиквитина кукурузы 1 (Cornejo et al, 1993, Plant Mol. Biol. 29: 637- 646), генами убиквитина Arabidopsis (резуховидки) 1 и 6 (Holtorf et al, 1995, Plant Mol. Biol. 29: 637-646), и геном фактора инициации трансляции табака 4А (Mandel et al, 1995, Plant Mol. Biol. 29: 995-1004).
Термин "конститутивный" в настоящей заявке необязательно показывает, что уровень экспрессии гена под контролем конститутивной регуляторной области одинаков во всех типах клеток, но скорее, что ген экспрессируется в широком диапазоне клеточных типов, даже если часто наблюдаются колебания относительного количества. Конститутивные регуляторные элементы могут быть связаны с другими последовательностями для дополнительного усиления трансляции нуклеотидной последовательности, с которыми они функционально связаны. Например, система CPMV-HT образована с использованием нетранслируемых областей вируса мозаики коровьего гороха (CPMV) и демонстрирует усиленную трансляцию ассоциированной кодирующей последовательности. Под термином "нативный" понимают, что нуклеотидная или аминокислотная последовательность встречается в природе или является последовательностью "дикого типа". Под выражением "функционально связанные" понимают, что конкретные последовательности, например, регуляторный элемент и представляющая интерес кодирующая область, взаимодействуют, либо прямо, либо косвенно, для осуществления заданной функции, такой как опосредование или модуляция экспрессии гена. Взаимодействие функционально связанных последовательностей могут опосредовать, например, белки, которые взаимодействуют с функционально связанными последовательностями.
Один или более вирусных белков, таких как структурный вирусный белок или канальный белок, например, но без ограничения, белок протонного канала, может экспрессироваться в системе экспрессии, представляющей собой систему экспрессии на основе вирусной ДНК или РНК, например, но без ограничения, экспрессионную кассету на основе комовируса и элемент регуляции амплификации на основе геминивируса.
Система экспрессии в соответствии с настоящей заявкой может содержать экспрессионную кассету на основе бипартитного вируса или вируса с бипартитным геномом. Например, бипартитный вирус может представлять собой вирус семейства комовирусов (Comoviridae). Семейство Comoviridae включает вирусы рода Comovirus (комовирус), Nepovirus, Fabavirus, Cheravirus и Sadwavirus. Комовирусы включают вирус мозаики коровьего гороха (вигны) (CPMV), агрессивный вирус мозаики коровьего гороха (CPSMV), вирус мозаики кабачка (SqMV), вирус крапчатости красного клевера (RCMV), вирус крапчатости бобов фасоли (BPMV), вирус кольцевой пятнистости турнепса (TuRSV), вирус настоящей мозаики кормовых бобов (BBtMV), вирус окраски кормовых бобов (BBSV), вирус мозаики редиса (RaMV). Примеры последовательностей РНК-2 комовируса, включающих энхансерные элементы, которые можно применять в различных аспектах изобретения, включают, но без ограничения: CPMV RNA-2 (номер доступа в GenBank NC_003550), RCMV RNA-2 (номер доступа в GenBank NC_003738), BPMV RNA-2 (номер доступа в GenBank NC_003495), CPSMV RNA-2 (номер доступа в GenBank NC_003544), SqMV RNA-2 (номер доступа в GenBank NC_003800), TuRSV RNA-2 (номер доступа в GenBank NC_013219.1), BBtMV RNA-2 (номер доступа в GenBank GU810904), BBSV RNA2 (номер доступа в GenBank FJ028650), RaMV (номер доступа в GenBank NC_003800)
Сегменты РНК-генома бипартитных комовирусов называются РНК-1 (RNA-1) и РНК-2 (RNA-2). РНК-1 кодирует белки, участвующие в репликации, тогда как РНК-2 кодирует белки, необходимые для движения из клетки в клетку, и два капсидных белка. Можно использовать любую подходящую кассету на основе комовируса, включая CPMV, CPSMV, SqMV, RCMV или BPMV, например, экспрессионная кассета может быть на основе CPMV.
Термин "экспрессионная кассета" относится к нуклеотидной последовательности, содержащей представляющую интерес нуклеиновую кислоту под контролем соответствующего промотора или других регуляторных элементов, или функционально связанную с соответствующим промотором или другими регуляторными элементами, для транскрипции представляющей интерес нуклеиновой кислоты в клетке-хозяине.
Было показано, что трансформация Nicotiana benthamiana с использованием полноразмерных компетентных по репликации кДНК копий обеих геномных РНК CPMV может вызвать продуктивную инфекцию (статья Liu et al., 2004, Virology 323, 37-48, описание которой включено в настоящее изобретение посредством отсылки). Примеры экспрессионных кассет на основе CPMV представлены в Международных заявках WO 2007/135480; WO 2009/087391; и в статьях Sainsbury F. et al., (2008, Plant Physiology; 148: 1212-1218; Sainsbury F. et al., (2008, Plant Biotechnology Journal; 6: 82-92; Sainsbury F. et al., 2009, Plant Biotechnology Journal; 7: 682-693; описание этих документов включено в настоящее изобретение посредством отсылки). В качестве примера, который не следует рассматривать как ограничивающий: нетранслируемые области (UTR), полученные из геномной RNA 2 вируса мозаики коровьего гороха (CPMV), в которой были делетированы два первых инициирующих кодона, находящиеся в 5' лидерной последовательности, можно применять как описано в Международной заявке WO 2009/087391. Будучи связанными с промотором CaMV 35S и терминатором нопалин синтазы (NOS), модифицированные CPMV UTR усиливали трансляцию фланкирующей кодирующей области. Система экспрессии на основе CPMV была названа CPMV-HT (гипертранслируемая). Следовательно, экспрессионные кассеты, экспрессионные конструкции и системы экспрессии по изобретению могут также включать систему экспрессии на основе CPMV, например, такую как система экспрессии CPMV-HT.
Как описано выше, последовательность энхансера экспрессии, полученную с использованием сегмента (или гомологичную сегменту) РНК-2 генома бипартитного РНК-вируса, такого как комовирус, в котором целевой сайт инициации (трансляции) является мутантным, можно применять для экспрессии целевой нуклеотидной последовательности. Помимо этого, настоящее изобретение предусматривает методы повышения экспрессии или повышения трансляционной активности последовательности, полученной при использовании сегмента РНК- 2 генома бипартитного вируса, которые включают мутацию находящегося в ней целевого сайта инициации.
"Энхансерные" последовательности (или энхансерные элементы) включают последовательности, полученные при использовании сегмента (или гомологичные сегменту) РНК-2 генома бипартитного РНК-вируса, такого как комовирус, в котором целевой сайт инициации (трансляции) является мутантным. Такие последовательности могут повышать 3' экспрессию гетерологичной ORF (открытой рамки считывания), с которой они связаны. В частности полагают, что такие последовательности, если они присутствуют в транскрибируемой РНК, могут повысить трансляцию гетерологичной ORF, с которой они связаны.
Системы экспрессии могут также содержать амплификационные элементы геминивируса, например, амплификационный элемент вируса желтой карликовости бобов (BeYDV). BeYDV принадлежит к роду Mastreviruses, приспособленному к пребыванию в двудольных растениях. BeYDV является монопартитным вирусом, геном которого представлен одноцепочечной кольцевой ДНК, и может реплицироваться по механизму катящегося кольца, давая многочисленные копии. Векторные системы на основе репликона ДНК BeYDV применяли для продуцирования больших количества белка в растениях.
Выражение "амплификационные элементы" ("элементы амплификации") в данном контексте относится к нуклеотидному сегменту, состоящему, по меньшей мере, из одной или более длинных межгенных областей (LIR) генома геминивируса. В данном контексте выражение " длинная межгенная область" относится к длинной межгенной области, которая содержит сайт связывания Rep белка, способного опосредовать вырезание (эксцизию) и репликацию с использованием Rep белка геминивируса. Согласно некоторым аспектам нуклеотидный сегмент, содержащий одну или более LIR, может дополнительно содержать короткую межгенную область (SIR) генома геминивируса. В данном контексте выражение "короткая межгенная область" относится к комплементарной цепи (короткой IR (SIR) вирусов Mastreviruses). В данном случае можно применять любой подходящий амплификационный элемент геминивируса. См., например, Международные заявки WO 2000/20557; WO 2010/025285; статьи Zhang X. et al. (2005, Biotechnology and Bioengineering, Vol. 93, 271-279), Huang Z. et al. (2009, Biotechnology and Bioengineering, Vol. 103, 706-714), Huang Z. et al. (2009, Biotechnology and Bioengineering, Vol. 106, 9-17); описание которых включено в настоящее изобретение посредством отсылки). Если в конструкции используется более одной области LIR, например, две области LIR, тогда промотор, области CMPV-HT и целевая нуклеотидная последовательность и терминатор с каждой стороны ограничиваются каждой из двух областей LIR.
В данном контексте совместная доставка вектора на основе вируса желтой карликовости бобов (BeYDV) и вектора, доставляющего Rep/RepA, посредством агроинфильтрации листьев табака Nicotiana benthamiana приводит в результате к эффективной амплификации репликона и устойчивому продуцированию белка.
Экспрессионная кассета на основе комовируса и амплификационный элемент геминивируса могут содержаться в соответствующих, первом или втором, векторах, или компоненты могут быть включены в один вектор. Если используются два вектора, первый и второй векторы можно вводить в растительную клетку одновременно или по отдельности.
Вирусная репликаза также может быть включена в систему экспрессии по данному описанию для повышения экспрессии целевой нуклеиновой кислоты. Неограничивающим примером репликазы является BeYDV репликаза (pREP110), кодирующая BeYDV Rep и RepA (С2/С1; Huang et al., 2009, Biotechnol. Bioeng. 103, 706-714; описание статьи включено в настоящее изобретение посредством отсылки).
Посттранскрипционный сайленсинг гена (PTGS) может участвовать в ограничении экспрессии трансгенов в растениях, а коэкспрессию супрессора сайленсинга из р19 вируса кустистой карликовости томата (TBSV р19) или вируса картофеля Y (HcPro) можно применять для того, чтобы воспрепятствовать специфическому расщеплению мРНК трансгенов (Brigneti et al., 1998).
Из уровня техники хорошо известны другие супрессоры сайленсинга, которые можно применять по настоящему изобретению (Chiba et al., 2006, Virology 346:7-14; описание статьи включено в настоящее изобретение посредством отсылки), например, но без ограничения, TEV -p1/HC-Pro (вирус гравировки табака- p1/HC-Pro), BYV -р21, капсидный белок вируса морщинистости (курчавости) листьев табака (TCV -CP), 2b вируса огуречной мозаики; CMV-2b), р25 Х-вируса картофеля (PVX-p25), p11 М-вируса картофеля (PVM-p11), p11 S-вируса картофеля (PVS-p11), p16 вируса ожога голубики, (BScV -р16), р23 вируса тристеца цитрусовых (CTV-p23), р24 вируса скручивания листьев винограда-2, (GLRaV-2 р24), р10 А-вируса винограда, (GVA-p10), р14 В-вируса винограда (GVB-p14), р10 латентного вируса аралии (HLV-p10), или р16 обыкновенного латентного вируса чеснока (GCLV-pl6). Следовательно, супрессор сайленсинга, например, но без ограничения, HcPro, TEV -p1/HC-Pro, BYV-p21, TBSV р19, TCV-CP, CMV-2b, PVX-p25, PVM-p11, PVS-p11, BScV-p16, CTV-p23, GLRaV-2 p24, GBV-p14, HLV-p10, GCLV-pl6 или GVA-p10, можно коэкспрессировать вместе с нуклеотидной последовательностью целевого белка, чтобы дополнительно обеспечить высокие уровни продуцирования белка в растении.
Под термином "коэкспрессируется" понимают, что две или более двух нуклеотидных последовательностей экспрессируются примерно одновременно в растении и в одной ткани растения. Однако, нуклеотидные последовательности необязательно должны экспрессироваться точно в одно и то же время. Скорее две или более нуклеотидных последовательности экспрессируются таким образом, чтобы кодируемые продукты имели возможность взаимодействовать. Например, белок, который модифицирует гликозилирование целевого белка, может экспрессироваться либо до экспрессии, либо во время экспрессии целевого белка с тем, чтобы происходила модификация гликозилирования целевого белка. Две или более нуклеотидные последовательности можно коэкспрессировать с применением системы для транзиторной экспрессии, где две или более последовательностей вводятся в растение примерно одновременно в условиях, когда экспрессируются обе последовательности. Или же, растение-платформу, содержащее одну или более нуклеотидных последовательностей, например, последовательность, кодирующую белок, который модифицирует профиль гликозилирования целевого белка, можно трансформировать, используя либо транзиторную, либо стабильную трансформацию, с дополнительной последовательностью, кодирующей целевой белок. В этом случае последовательность, кодирующую белок, который модифицирует профиль гликозилирования целевого белка, можно экспрессировать в заданной ткани в процессе заданной стадии развития, или ее экспрессию можно индуцировать посредством индуцибельного промотора, а дополнительную последовательность, кодирующую целевой белок, можно экспрессировать в сходных условиях и в той же ткани с тем, чтобы гарантировать коэкспрессию нуклеотидных последовательностей.
Один или более вирусных белков можно продуцировать в виде транскрипта с использованием нуклеотидной последовательности, и белок отщеплять после синтеза и, при необходимости, связывать с образованием мультимерного белка. Соответственно, один или более вирусный белок включает также белок или полипептид, содержащий субъединицы, связанные дисульфидными мостиками (т.е. мультимерный белок). Например, белок, содержащий аминокислотные последовательности из двух или более источников, можно процессировать в виде субъединиц, а субъединицы связывать дисульфидными мостиками, получая белок.
Одну или более нуклеотидных последовательностей или генетических конструкций по настоящему изобретению можно экспрессировать в соответствующем растении-хозяине, которое трансформировано с помощью нуклеотидной последовательности, или конструкций, или векторов по настоящему изобретению. К подходящим хозяевам относятся, но без ограничения, сельскохозяйственные растения, включая люцерну, рапс канолу, виды капусты (Brassica spp.), кукурузу, виды табака (Nicotiana spp.), картофель, женьшень, горох, овес, рис, сою, пшеницу, ячмень, подсолнечник, хлопчатник и т.п.
Одна или более генетических конструкций по настоящему изобретению могут дополнительно содержать 3' нетранслируемую область. Термин "3' нетранслируемая область" относится к участку гена, содержащему сегмент ДНК, который содержит сигнал полиаденилирования и любые другие регуляторные сигналы, способные влиять на процессинг мРНК или экспрессию генов. Сигнал полиаденилирования обычно влияет на присоединение полиадениловой кислоты к 3'- концу предшественника мРНК. Сигналы полиаденилирования обычно распознаются по наличию гомологии с канонической формой 5-ААТААА-3', хотя нередки вариации. Неограничивающими примерами соответствующих 3'-областей являются транскрибируемые нетранслируемые 3'-области, содержащие сигнал полиаденилирования опухолеиндуцирующих (Ti) плазмидных генов Agrobacterium (агробактерий), например, ген нопалин синтазы (NOS), и гены растений, такие как гены запасных белков сои и гены малой субъединицы гена рибулозо-1,5-бисфосфат карбоксилазы (ssRUBISCO; патент США 4962028, описание которого включено в настоящее изобретение посредством отсылки), промотор, используемый для регуляции экспрессии пластоцианина, описанный в патенте США No. 7,125,978 (описание которого включено в настоящее изобретение посредством отсылки).
Одна или более генетических конструкций по настоящему изобретению могут также включать дополнительные энхансеры, либо трансляции, либо транскрипции. Энхансеры могут быть расположены 5' или 3' по отношению к транскрибируемой последовательности. Энхансерные области хорошо известны специалистам и могут включать инициирующий кодон (инициаторный, стартовый кодон, кодон инициации) ATG, прилегающие последовательности и т.п. Инициирующий кодон, если таковой присутствует, может находиться в фазе с рамкой считывания ("в рамке") кодирующей последовательности, чтобы обеспечить корректную трансляцию транскрибируемой последовательности.
Под "трансформацией" понимают межвидовой перенос генетической информации (нуклеотидной последовательности), которая проявляется генотипически, фенотипически или и так, и так. Межвидовой перенос генетической информации хозяину с использованием конструкции может быть транзиторным (временным), и такой перенос генетической информации не наследуется (не является врожденным), или перенос может наследоваться, и такой перенос генетической информации считают стабильным.
Конструкции по настоящему изобретению можно ввести в растительные клетки, используя Ti плазмиды, Ri плазмиды, растительные вирус-векторы, прямую трансформацию с использованием ДНК, микроинъекцию, электропорацию и т.п. Обзоры таких методов см., например, в Weissbach and Weissbach, Methods for Plant Molecular Biology, Academy Press, New York VIII, pp. 421-463 (1988); Geierson and Corey, Plant Molecular Biology, 2d Ed. (1988); и Miki and Iyer, Fundamentals of Gene Transfer in Plants. В Plant Metabolism, 2d Ed. DT. Dennis, DH Turpin, DD Lefebrve, DB Layzell (eds), Addison Wesly, Langmans Ltd. London, pp. 561-579 (1997). Другие методы включают прямой захват ДНК, применение липосом, электропорацию, например, с применением протопластов, микроинъекцию, бомбардирующие микрочастицы или нитевидные кристаллы и вакуум-инфильтрацию. См., например, Bilang, et al. (Gene 100: 247-250 (1991), Scheid et al. (Mol. Gen. Genet. 228: 104-112, 1991), Guerche et al. (Plant Science 52: 111-116, 1987), Neuhause et al. (Theor. Appl Genet. 75: 30-36, 1987), Klein et al., Nature 327: 70-73 (1987); Howell et al. (Science 208: 1265, 1980), Horsch et al. (Science 227: 1229-1231, 1985), DeBlock et al., Plant Physiology 91: 694-701, 1989), Methods for Plant Molecular Biology (Weissbach and Weissbach, eds., Academic Press Inc., 1988), Methods in Plant Molecular Biology (Schuler and Zielinski, eds., Academic Press Inc., 1989), Liu and Lomonossoff (J Virol Meth, 105: 343-348, 2002,), патенты США No. 4,945,050; 5,036,006; и 5,100,792; заявки на патент США номер 08/438,666, поданную 10 мая 1995 года, и 07/951,715, поданную 25 сентября 1992 года (описание которых включено в настоящее изобретение посредством отсылки).
Как описывается ниже, методы транзиторной экспрессии можно применять для экспрессии конструкций по настоящему изобретению (см. статью Liu and Lomonossoff, 2002, Journal of Virological Methods, 105:343-348; описание которой включено в настоящее изобретение посредством отсылки). Или же можно применять метод вакуумной транзиторной экспрессии, описанный Kapila et al., 1997, (документ включен в настоящее изобретение посредством отсылки). Эти методы могут включать, например, но без ограничения, метод агроинокуляции или агроинфильтрации, шприцевой инфильтрации (инфильтрации с помощью шприца), однако, как указывается выше, можно применять другие методы транзиторной экспрессии. С использованием агроинокуляции, агроинфильтрации или шприцевой инфильтрации смесь Agrobacteria (агробактерий), содержащая нужную нуклеиновую кислоту, поступает во внутриклеточное пространство ткани, например, в листья, надземную часть растения (включая стебель, листья, цветок), другую часть растения (стебель, корень, цветок) или в цельное растение. После прохождения через эпидермис агробактерий инфицируют копии тДНК и переносят их в клетки. Транскрипция т-ДНК в эписомной форме и трансляция м-РНК приводят к продуцированию целевого белка в инфицированных клетках, тем не менее, проникновение т-ДНК внутрь ядра является временным (транзиторным).
Для того чтобы способствовать идентификации трансформированных растительных клеток, конструкции по настоящему изобретению можно дополнительно обрабатывать для включения в них селективных маркеров растений. Применимые селективные маркеры включают ферменты, которые обеспечивают устойчивость к химическим агентам, таким как антибиотик, например, гентамицин, гигромицин, канамицин, или гербициды, такие как фосфинотрицин, глифосат, хлорсульфурон и т.п. Можно также применять ферменты, обеспечивающие продуцирование соединения, идентифицируемого по изменению цвета, например, GUS (бета-глюкуронидазу), или по люминесценции, например, люциферазу или GFP.
Под термином "растительный материал" понимают любой материал растительного происхождения. Растительный материал может включать все растение, ткани, клетки или любую их часть. Помимо этого, растительный материал может содержать внутриклеточные компоненты растения, внеклеточные компоненты растения, жидкие или твердые растительные экстракты или их комбинацию. Далее, растительный материал может содержать растения, растительные клетки, ткани, жидкий экстракт или их комбинацию, из листьев, стеблей, плодов, корней растения или их комбинации. Растительный материал может содержать растение или его часть, которая не подвергалась никакому процессингу. Также, однако, предполагается, что растительный материал может минимально участвовать в стадиях процессинга в соответствии с приведенным ниже определением, или подвергаться более серьезному процессингу, включая методы частичной или основательной очистки, общеизвестные из уровня техники, включая, но без ограничения, хроматографию, электрофорез и т.п.
Под термином "минимальный процессинг" (в данном контексте "минимальная обработка") понимают, что растительный материал, например, растение или его часть, содержащий целевой белок, подвергался частичной очистке, давая растительный экстракт, гомогенат, фракцию растительного гомогената и т.п. (т.е. процессировал в минимальной степени). Частичная очистка может включать, но без ограничения, разрушение клеточной структуры растения и образование при этом композиции, содержащей растворимые растительные компоненты и нерастворимые растительные компоненты, которые можно разделять, например, но без ограничения, центрифугированием, фильтрованием или их комбинацией. Благодаря этому белки, секретируемые во внутриклеточном пространстве листа или других тканей, можно легко получать, применяя вакуумную или центробежную экстракцию, или экстрагировать их из тканей под давлением, пропуская их через валки, измельчая и т.п., чтобы отжать или выделить белок из межклеточного пространства.
Минимальная обработка может также подразумевать приготовление сырых экстрактов растворимых белков, так как указанные препараты будут содержать ничтожно малое количество примесей вторичных растительных продуктов. Далее, минимальная обработка может включать водную экстракцию растворимого белка из листьев с последующим осаждением любой приемлемой солью. Другие методы включают крупномасштабную мацерацию и крупномасштабное извлечение сока, что позволяет непосредственно использовать экстракт.
Растительное вещество в виде растительного материала или ткани можно доставлять субъекту перорально. Растительное вещество можно принимать в составе добавки к рациону - вместе с пищей или в капсулах. Можно провести концентрирование вещества или ткани растения для улучшения или повышения его съедобности, либо объединить с другими материалами, ингредиентами, или фармацевтическим наполнителем, по необходимости.
Растительный материал можно принимать в виде пищевой добавки, вместе с другими пищевыми продуктами или в виде капсул. Растительный материал или ткань можно также концентрировать, чтобы улучшить или повысить их вкусовые качества, или, при необходимости, предоставлять их вместе с другими материалами, ингредиентами, или фармацевтическими эксципиентами.
Предполагается, что растение, содержащее целевой белок или экспрессирующее VLP, содержащую целевой белок, можно вводить субъекту или в организм-мишень различными способами в зависимости от потребности и ситуации. Например, целевой белок, полученный из растения, можно извлекать (экстрагировать), а затем применять в виде сырого, частично очищенного или очищенного белка. Если белок должен быть очищенным, то его можно продуцировать либо в съедобных, либо в несъедобных растениях. Далее, если белок вводят перорально, растительную ткань можно собирать и употреблять непосредственно в пищу, или собранную ткань можно высушить перед употреблением, или можно пустить животное на пастбище, где растет это растение, предварительно не собирая его. Также в объеме настоящего изобретения рассматривается применение собранных растительных тканей в виде добавки в корм животным. Если в пищу человека или животного добавляют растительную ткань, которая подверглась небольшой, или вообще не подвергалась, дополнительной обработке, предпочтительно, чтобы потребляемая растительная ткань была съедобной.
VLP, продуцируемые согласно настоящему изобретению, могут быть очищенными, частично очищенными от растения, части растения или растительного материала, или их можно вводить в виде пероральной вакцины методами, известными специалистам в данной области техники. Очистка может включать продуцирование фракции апопласта, описанной в международной заявке WO 2011/035422 (описание которой включено в настоящее изобретение посредством отсылки). Для препаративной высокоэффективной гель-хроматографии можно получать препарат, содержащий VLP, и нерастворимый материал удалять центрифугированием. Можно также применять осаждение с помощью ПЭГ (PEG). Выделенный белок можно определять количественно обычными методами (например, анализом по методу Бредфорд, ВСА), экстракт можно пропускать через колонку для высокоэффективной гель-хроматографии с насадкой сефакрил (SEPHACRYL™), сефадекс (SEPHADEX™) и т.п. и отбирать фракции. В качестве калибровочного стандарта можно применять голубой декстран (Blue Dextran) 2000 или соответствующий белок. Экстракт можно также пропустить через катионообменную колонку и отбирать активные фракции. После хроматографии фракции можно дополнительно анализировать с помощью электрофореза белка, иммуноблоттинга или и того, и другого, для подтверждения присутствия VLP и белкового состава фракции.
Также в настоящем изобретении рассматриваются трансгенные растения, растительные клетки, семена или любая их часть, содержащие нуклеотидные последовательности по настоящему изобретению. Методы регенерации целых растений из растительных клеток также известны из уровня техники. Как правило, трансформированные растительные клетки культивируют в соответствующей среде, которая может содержать селективные агенты, такие как антибиотики, в которой применяются селективные маркеры для того, чтобы способствовать идентификации трансформированных растительных клеток. После образования каллюса (каллуса) можно стимулировать образование корня, применяя соответствующие растительные гормоны и известные методы, и ростки переносить в субстрат для выращивания с целью регенерации растений. Затем растения можно использовать для многократного получения последующих генераций либо из семян, либо методами вегетативного размножения. Трансгенные растения также могут быть получены без использования тканевых культур.
Как показано на Фигуре 18, НА вируса гриппа B/Brisbane/60/2008 слабо экспрессируется в агроинфильтрованных листьях табака Nicotiana benthamiana (см. дорожку "1008" или "1029"). Однако, коэкспрессия НА- типа В с М2 с использованием A/New Caledonia/20/99, приводит к значительному повышению НА экспрессии (см. полоски "1008+1261"; "1009+1261" и 1029+1261"). Повышение экспрессии НА наблюдали как в случае нативного типа В НА, так и в случае химерного НА типа В. Экспрессию НА наблюдали в присутствии и в отсутствие амплификационных элементов (BeYDV) и при различных разведениях Agrobacterium. Аналогичное повышение экспрессии НЗ наблюдали, когда Н3 вируса гриппа A/Perth/16/2009 коэкспрессировался с М2 A/New Caledonia/20/99 (Фигура 19; ср. полоску "1019" для экспрессии только одного Н3 с полоской "1019+1261" для коэкспрессии Н3 с М2).
Настоящее изобретение включает нуклеотидные последовательности, представленные в Таблице 3:


Далее настоящее изобретение иллюстрируется с помощью приведенных ниже примеров.
Примеры
Материалы и методы: Сборка экспрессионных кассет для экспрессии белка вируса гриппа
А-2Х35S/CPMV-HT/H5 Indonesia/NOS (Конструкция номер 489)
Последовательность, кодирующую Н5 вируса гриппа A/Indonesia/5/2005 (H5N1), клонировали в систему экспрессии 2X35S/CPMV-HT/NOS в плазмиде, содержащей экспрессионную кассету Plasto_pro/P19/Plasto_ter, применяя нижеприведенный метод на основе ПЦР. Фрагмент, содержащий последовательность, кодирующую полный Н5, амплифицировали, используя праймеры IF-H5A-I-05.s1+3c (Фигура 1 A, SEQ ID NO: 2) и IF-H5dTm.r (Фигура 1B, SEQ ID NO: 3) с применением в качестве матрицы конструкции номер 972 (см. Фигуру 94, SEQ ID NO: 134 из международной заявки WO 2010/003225, описание последовательности конструкции номер 972 в которой включено в настоящее изобретение посредством отсылки). ПЦР-продукт клонировали в системе экспрессии 2X35S/CPMV-HT/NOS, используя систему для клонирования In-Fusion (Clontech, Mountain View, CA). Конструкцию 1191 (Фигура 1D, SEQ ID NO: 4) расщепляли с помощью рестриктаз SacII и StuI и линеаризованную плазмиду использовали для реакции сборки In-Fusion. Конструкция номер 1191 представляет собой акцепторную плазмиду, предполагаемую для "In Fusion" клонирования целевых генов в экспрессионной кассете на основе CPMV-HT. Она также включает генную конструкцию для коэкспрессии TBSV P19 супрессора сайленсинга под контролем промотора и терминатора гена пластоцианина люцерны. Остов представляет собой бинарную плазмиду pCAMBIA, и последовательность от левой до правой границы тДНК показана на Фигуре 1D (SEQ ID NO: 4). Полученной в результате конструкции был присвоен номер 489 (Фигура 1E, SEQ ID NO: 5). Аминокислотная последовательность Н5 вируса гриппа A/Indonesia/5/2005 (H5N1) представлена на Фигуре 1F (SEQ ID NO: 6). Плазмида 489 изображена на Фигуре 15.
В-2Х35S/CPMV-HT/M2 New Caledonia/NOS (Конструкция номер 1261)
Последовательность, кодирующую М2 вируса гриппа A/New Caledonia/20/1999 (H1N1), клонировали в систему экспрессии 2X35S/CPMV-HT/NOS в плазмиде, содержащей экспрессионную кассету Plasto_pro/P19/Plasto_ter, с использованием ПЦР- метода. Фрагмент, содержащий последовательность, кодирующую полный М2, амплифицировали, используя праймеры IF-S1-M1+M2ANC.c (Фигура 2А, SEQ ID NO: 7) и IF-S1-4-M2ANC.r (Фигура 2В, SEQ ID NO: 8) и синтезированный ген М2 (соответствующий нуклеотидам (nt) 1-26, связанным с nt 715-982 последовательности с номером доступа в GenBank DQ508860) (Фигура 2С, SEQ ID NO: 9) в качестве матрицы. Продукт ПЦР клонировали в системе экспрессии 2X35S/CPMV-HT/NOS, используя систему для клонирования In-Fusion (Clontech, Mountain View, CA). Конструкцию 1191 (Фигура 1С) расщепляли с помощью рестриктаз SacII и StuI и линеаризованную плазмиду использовали для реакции сборки In-Fusion. Конструкция номер 1191 представляет собой акцепторную плазмиду, предполагаемую для "In Fusion" клонирования целевых генов в экспрессионной кассете на основе CPMV-HT. Она также включает генную конструкцию для коэкспрессии TBSV P19 супрессора сайленсинга под контролем промотора и терминатора гена пластоцианина люцерны. Остов представляет собой бинарную плазмиду pCAMBIA, и последовательность от левой до правой границы тДНК показана на Фигуре 1D (SEQ ID NO: 4). Полученной в результате конструкции был присвоен номер 1261 (Фигура 2D, SEQ ID NO: 10). Аминокислотная последовательность М2 вируса гриппа A/New Caledonia/20/1999 (H1N1) представлена на Фигуре 2Е (SEQ ID NO: 11). Плазмида 1261 изображена на Фигуре 16.
С-2Х35S/CPMV-HT/M2 Puerto Rico/NOS (конструкция номер 859)
Последовательность, кодирующую М2 вируса гриппа A/Puerto Rico/8/1934 (H1N1), клонировали в систему экспрессии 2X35S/CPMV-HT/NOS в плазмиде, содержащей экспрессионную кассету Plasto_pro/P19/Plasto_ter, с использованием ПЦР-метода. Фрагмент, содержащий последовательность, кодирующую полный М2, амплифицировали, используя праймеры IF-S1-M1+M2ANC.c (Фигура 2А, SEQ ID NO: 7) и IF-S1-4-M2ANC.r (Фигура 2В, SEQ ID NO: 8), с синтезированным М2 геном (соответствует нуклеотидам (nt) 26-51, соединенным с nt 740-1007 последовательности с номером доступа в Genbank EF467824) (Фигура 3А, SEQ ID NO: 12) в качестве матрицы. Продукт ПЦР клонировали в системе экспрессии 2X35S/CPMV-HT/NOS, используя систему для клонирования In-Fusion (Clontech, Mountain View, CA). Конструкцию 1191 (Фигура 1С) расщепляли с помощью рестриктаз SacII и StuI и линеаризованную плазмиду использовали для реакции сборки In-Fusion. Конструкция номер 1191 представляет собой акцепторную плазмиду, предполагаемую для "In Fusion" клонирования целевых генов в экспрессионной кассете на основе CPMV-HT. Она также включает генную конструкцию для коэкспрессии TBSV P19 супрессора сайленсинга под контролем промотора и терминатора гена пластоцианина люцерны. Вектор представляет собой бинарную плазмиду pCAMBIA, и последовательность от левой до правой границы тДНК показана на Фигуре ID (SEQ ID NO: 4). Полученной в результате конструкции был присвоен номер 859 (Фигура 3В, SEQ ID NO: 13). Аминокислотная последовательность М2 вируса гриппа A/Puerto Rico/8/1934 (H1N1) представлена на Фигуре 3С (SEQ ID NO: 14). Плазмида 859 изображена на Фигуре 17.
D-2X35S/CPMV-HT/PDISP/H1 California/NOS (Конструкция номер 484)
Последовательность, кодирующую H1 вируса гриппа A/California/7/2009 (H1N1), клонировали в систему экспрессии 2X35S-CPMV-HT-PDISP-NOS в плазмиде, содержащей экспрессионную кассету Plasto_pro/P19/Plasto_ter, используя нижеприведенный метод на основе ПЦР. Фрагмент, содержащий последовательность, кодирующую H1 без его сигнального пептида дикого типа (с сигнальной последовательностью дикого типа), амплифицировали, используя праймеры IF-H1A-C-09.s2+4c (Фигура 4А, SEQ ID NO: 15) и IF-H1A-C-09.s1-4r (Фигура 4В, SEQ ID NO: 16), а в качестве матрицы используя синтезированный H1 ген (последовательность с номером доступа в Genbank FJ966974) (Фигура 4С, SEQ ID NO: 17). ПЦР-продукт клонировали в рамке считывания с сигнальным пептидом PDI люцерны в системе экспрессии 2X35S/CPMV-HT/NOS, используя систему для клонирования In-Fusion (Clontech, Mountain View, CA). Конструкцию 1192 (Фигура 4D) расщепляли с помощью рестриктаз SacII и StuI и линеаризованную плазмиду применяли для реакции сборки In-Fusion. Конструкция номер 1192 представляет собой акцепторную плазмиду, предназначенную для "In Fusion" клонирования целевых генов в рамке считывания с PDI сигнальным пептидом люцерны в экспрессионной кассете на основе CPMV-HT. Она также включает генную конструкцию для коэкспрессии TBSV P19 супрессора сайленсинга под контролем промотора и терминатора гена пластоцианина люцерны. Остов представляет собой бинарную плазмиду pCAMBIA, и последовательность от левой до правой границы тДНК показана на Фигуре 4Е (SEQ ID NO: 18). Полученной в результате конструкции был присвоен номер 484 (Фигура 4F, SEQ ID NO: 19). Аминокислотная последовательность PDISP/H1 вируса гриппа A/California/7/2009 (H1N1) представлена на Фигуре 4G (SEQ ID NO: 20). Плазмида 484 изображена на Фигуре 14.
Е-2ХЗ5S/CPMV-HT/PDISP/H3 Perth/NOS (Конструкция номер 1019)
Последовательность, кодирующую Н3 вируса гриппа A/Perth/16/2009 (H3N2), клонировали в систему экспрессии 2X35S/CPMV-HT/PDISP/NOS в плазмиде, содержащей экспрессионную кассету Plasto_pro/P19/Plasto_ter, применяя нижеприведенный метод на основе ПЦР. Фрагмент, содержащий последовательность Н3 без его сигнального пептида дикого типа, амплифицировали, применяя праймеры IF-S2+S4-H3 Per.c (Фигура 5А, SEQ ID NO: 21) и IF-S1a4-H3 Per.r (Фигура 5В, SEQ ID NO: 22), в качестве матрицы используя синтезированный Н3 ген (соответствующий nt 26-1726 последовательности с номером доступа в Genbank GQ293081) (Фигура 5С, SEQ ID NO: 23). ПЦР-продукт клонировали в рамке считывания с сигнальным пептидом PDI (протеиндисульфидизомеразы) люцерны в системе экспрессии 2X35S/CPMV-HT/NOS, применяя систему для клонирования In-Fusion (Clontech, Mountain View, CA). Конструкцию 1192 (Фигура 4D) расщепляли рестриктазами SacII и StuI и линеаризованную плазмиду использовали для In-Fusion реакции сборки. Конструкция номер 1192 представляет собой акцепторную плазмиду, предназначенную для "In Fusion" клонирования целевых генов в рамке считывания с сигнальным пептидом PDI люцерны в экспрессионной кассете на основе CPMV-HT. Она также включает генную конструкцию для коэкспрессии TBSV P19 супрессора сайленсинга под контролем промотора и терминатора гена пластоцианина люцерны. Остов представляет собой бинарную плазмиду pCAMBIA, и последовательность от левой до правой границы тДНК представлена на Фигуре 4Е (SEQ ID NO: 18). Полученной в результате конструкции было присвоен номер 1019 (Фигура 5D, SEQ ID NO: 24). Аминокислотная последовательность PDISP/H3 вируса гриппа A/Perth/16/2009 (H3N2) представлена на Фигуре 5Е (SEQ ID NO: 25). Плазмида 1019 изображена на Фигуре 13.
F-2X35S/CPMV-HT/PDISP/HA В Brisbane/NOS (Конструкция номер 1029)
Последовательность, кодирующую НА вируса гриппа B/Brisbane/60/2008, клонировали в систему экспрессии 2X35S/CPMV-HT/PDISP/NOS в плазмиде, содержащей экспрессионную кассету Plasto_pro/P19/Plasto_ter, используя следующий метод на основе ПЦР. Фрагмент, содержащий последовательность, кодирующую НА В Brisbane без его сигнального пептида дикого типа, амплифицировали, используя праймеры IF-S2+S4-B Bris.c (Фигура 6А, SEQ ID NO: 26) и IF-S1a4-B Bris.r (Фигура 6В, SEQ ID NO: 27), а в качестве матрицы используя синтезированный НА В Brisbane ген (соответствующий nt 34-1791 последовательности с номером доступа в Genbank FJ766840) (Фигура 6С, SEQ ID NO: 28). ПЦР-продукт клонировали в рамке считывания с сигнальным пептидом PDI люцерны в системе экспрессии 2X35S/CPMV-HT/NOS, применяя систему для клонирования In-Fusion (Clontech, Mountain View, CA). Конструкцию 1192 (Фигура 4D) расщепляли с помощью SacII и StuI рестриктаз, а линеаризованную плазмиду использовали для реакции сборки In-Fusion. Конструкция номер 1192 представляет собой акцепторную плазмиду, предназначенную для "In Fusion" клонирования целевых генов в рамке считывания с сигнальным пептидом PDI в экспрессионной кассете на основе CPMV-HT. Она также включает генную конструкцию для коэкспрессии TBSV P19 супрессора сайленсинга под контролем промотора и терминатора гена пластоцианина. Остов представляет собой бинарную плазмиду pCAMBIA, и последовательность от левой до правой границы тДНК показана на Фигуре 4Е (SEQ ID NO: 18). Полученной в результате конструкции присвоен номер 1029 (Фигура 6D, SEQ ID NO: 29). Аминокислотная последовательность PDISP/HA вируса гриппа B/Brisbane/60/2008 показана на Фигуре 6Е (SEQ ID NO: 30). Плазмида 1029 изображена на Фигуре 11.
G-2X35S/CPMV-HT/PDISP/HА В Brisbane/NOS в амплификационной системе BeYDV+Репликаза (Конструкция номер 1008)
Последовательность, кодирующую НА вируса гриппа B/Brisbane/60/2008, клонировали в 2X35S/CPMV-HT/PDISP/NOS, содержащую амплификационную систему BeYDV+репликаза в плазмиде, содержащей экспрессионную кассету Plasto_pro/P19/Plasto_ter, в соответствии с нижеприведенным методом на основе ПЦР. Фрагмент, последовательность, кодирующую НА В Brisbane без его сигнального пептида дикого типа, амплифицировали, используя праймеры IF-S2+S4-B Bris.c (Фигура 6А, SEQ ID NO: 26) и IF-S1a4-B Bris.r (Figure 6B, SEQ ID NO: 27), с применением в качестве матрицы гена НА В Brisbane gene (соответствующего nt 34-1791 последовательности с номером доступа в Genbank FJ766840) (Фигура 6С, SEQ ID NO: 28). ПЦР-продукт клонировали в рамке считывания с сигнальным пептидом PDI люцерны в системе экспрессии 2X35S/CPMV- HT/NOS в BeYDV амплификационную систему, используя In-Fusion систему клонирования (Clontech, Mountain View, СА). Конструкцию 1194 (см. Фигуры 6F и 6G) расщепляли рестриктазами SacII и StuI и линеаризованную плазмиду использовали для реакции сборки In-Fusion.
Конструкция номер 1194 представляет собой акцепторную плазмиду, предназначенную для "In Fusion" клонирования целевых генов в рамке считывания с сигнальным пептидом PDI в экспрессионной кассете на основе CPMV-HT в BeYDV амплификационную систему. Она также включает генную конструкцию для коэкспрессии TBSV P19 супрессора сайленсинга под контролем промотора и терминатора гена пластоцианина люцерны. Остов представляет собой бинарную плазмиду pCAMBIA, и последовательность от левой до правой границы тДНК показана на Фигуре 6G (SEQ ID NO: 31). Полученной в результате конструкции был присвоен номер 1008 (Фигура 6Н SEQ ID NO: 32). Аминокислотная последовательность PDISP/HA вируса гриппа B/Brisbane/60/08 представлена на Фигуре 6Е (SEQ ID NO: 30). Плазмида 1008 изображена на Фигуре 9.
Н-2Х35S/CPMV-HT/PDISP/HА В Brisbane/H5 Indonesia трансмембранный домен и цитоплазматический хвост (H5Indo TMCT)/NOS в амплификационную систему BeYDV+Репликаза (Конструкция 1009)
Последовательность, кодирующую эктодомен НА вируса гриппа B/Brisbane/60/2008, слитый с трансмембранным и цитозольным доменами Н5 вируса гриппа A/Indonesia/5/2005 (H5N1), клонировали в 2X35S/CPMV-HT/PDISP/NOS, включающую амплификационную систему BeYDV+репликаза в плазмиде, содержащей экспрессионную кассету Plasto_pro/P19/Plasto_ter, как указано ниже с применением метода лигирования (сшивания) на основе ПЦР, представленного Darveau et al. (Methods in Neuroscience 26: 77-85 (1995)). В первом раунде ПЦР фрагмент, содержащий последовательность, кодирующую эктодомен НА В Brisbane без нативного сигнального пептида, трансмембранный и цитоплазматический домены, амплифицировали, используя праймеры IF-S2+S4-B Bris.c (Фигура 6А, SEQ ID NO: 26) и dTmH5I-B Bris.r (Фигура 7А, SEQ ID NO: 33), и применяя в качестве матрицы синтезированный ген НА В Brisbane (соответствующий nt 34-1791 последовательности с номером доступа в Genbank FJ766840) (Фигура 6С, SEQ ID NO: 28). Второй фрагмент, содержащий трансмембранный и цитоплазматический домены Н5 Indonesia, амплифицировали, используя праймеры В Bris-dTmH5I.c (Фигура 7В, SEQ ID NO: 34) и IF-S1aS4-dTmH5I.r (Фигура 7С, SEQ ID NO: 35), и в качестве матрицы применяя конструкцию номер 489 (см. Фигуру 1E, SEQ ID NO: 5). Затем ПЦР-продукты, полученные в результате обеих амплификаций, смешивали и применяли в качестве матрицы для второго раунда амплификации, используя в качестве праймеров IF-S2+S4-B Bris.c (Фигура 6А, SEQ ID NO: 26) и IF-H5dTm.r (Фигура 7С, SEQ ID NO: 34). Полученный в результате фрагмент клонировали в рамке считывания с сигнальным пептидом PDI люцерны в экспрессионной кассете 2X35S/CPMV-HT/NOS в BeYDV амплификационную систему, используя In-Fusion систему клонирования (Clontech, Mountain View, CA). Конструкцию 1194 (Фигуры 6F и 6G) расщепляли рестриктазами SacII и StuI и линеаризованную плазмиду применяли для In-Fusion реакции сборки. Конструкция 1194 представляет собой акцепторную плазмиду, предназначаемую для "In Fusion" клонирования целевых генов в рамке считывания с сигнальным пептидом PDI люцерны в экспрессионной кассете на основе CPMV НТ в амплификационную систему BeYDV. Она также включает генную конструкцию для коэкспрессии TBSV P19 супрессора сайленсинга под контролем промотора и терминатора гена пластоцианина. Остов представляет собой бинарную плазмиду pCAMBIA, и последовательность от левой до правой границы тДНК показана на Фигуре 6G (SEQ ID NO: 31). Полученной в результате конструкции был присвоен номер 1009 (Фигура 7D, SEQ ID NO: 36). Аминокислотная последовательность PDISP/HA В Brisbane/H5indo ТМСТ представлена на Фигуре 7Е (SEQ ID NO: 37). Изображение плазмиды 1009 дается на Фигуре 10.
I-2X35S/CPMV-HT/PDISP-HA В Brisbane с делетированной протеолитической петлей в амплификационную систему BeYDV+репликаза (Конструкция номер 1059)
Последовательность, кодирующую НА вируса гриппа B/Brisbane/60/2008 с делетированной протеолитической петлей клонировали в 2X35S/CPMV-HT/PDISP/NOS, содержащую амплификационную систему BeYDV+репликаза в плазмиде, содержащей экспрессионную кассету Plasto_pro/P19/Plasto_ter, применяя нижеприведенный метод лигирования на основе ПЦР, представленный Darveau et al. (Methods in Neuroscience 26: 77-85 (1995)). В первом раунде ПЦР фрагмент, содержащий последовательность, кодирующую НА В Brisbane, от nt 46 до nt 1065, амплифицировали, используя в качестве праймеров IF-S2+S4-B Bris.c (Фигура 6А, SEQ ID NO: 26) и 1039+1059.r (Фигура 8А, SEQ ID NO: 38), а в качестве матрицы синтезированный ген НА В Brisbane (соответствующий nt 34-1791 последовательности с номером доступа в Genebank FJ766840) (Фигура 6С, SEQ ID NO: 28). Второй фрагмент, содержащий последовательность, кодирующую НА В Brisbane, от nt 1123 до nt 1758, амплифицировали, используя в качестве праймеров 1039+1059.с (Фигура 8В, SEQ ID NO: 39) и IF-S1a4-B Bris.r (Фигура 6В, SEQ ID NO: 27), и синтезированный ген НА В Brisbane (соответствующий nt 34-1791 последовательности с номером доступа в Genbank FJ766840) (Фигура 6С, SEQ ID NO: 28) в качестве матрицы. Затем ПЦР-продукты, полученные в результате обеих амплификаций, смешивали и применяли в качестве матрицы для второго раунда амплификации IF-S2+S4-B Bris.c (Фигура 6А, SEQ ID NO: 26) и IF-H5dTm.r IF-S1a4-B Bris.r (Фигура 6B, SEQ ID NO: 27) в качестве праймеров. Полученный в результате фрагмент (кодирующий НА B/Brisbane/60/2008 Δа.а. 356-374 с GG линкером между фрагментами) клонировали в рамке считывания с сигнальным пептидом PDI люцерны в экспрессионной кассете 2X35S/CPMV-HT/NOS, содержащей амплификационную систему BeYDV, используя систему клонирования In-Fusion (Clontech, Mountain View, CA). Конструкцию 1194 (Фигура 6F и 6G) расщепляли рестриктазами SacII и StuI и линеаризованную плазмиду использовали для In-Fusion реакции сборки. Конструкция номер 1194 представляет собой акцепторную плазмиду, предполагаемую для "In Fusion" клонирования целевых генов в рамке считывания с сигнальным пептидом PDI в экспрессионной кассете на основе CPMV-HT в амплификационную систему BeYDV. Она включает также генную конструкцию для коэкспрессии TBSV P19 супрессора сайленсинга под контролем промотора и терминатора гена пластоцианина люцерны. Остов представляет собой бинарную плазмиду pCAMBIA, и последовательность от левой до правой границы тДНК показана на Фигуре 6G (SEQ ID NO: 31). Полученной в результате конструкции был присвоен номер 1059 (Фигура 8С, SEQ ID
NO: 40). Аминокислотная последовательность PDISP-HA B/Brisbane/60/2008 с делетированной протеолитической петлей представлена на Фигуре 8D (SEQ ID NO: 41). Плазмида 1059 показана на Фигуре 12.
А-2Х35S/CPMV-HT/PDISP/H3 Victoria/NOS (Конструкция номер 1391)
Последовательность, кодирующую Н3 вируса гриппа A/Victoria/361/2011 (H3N2), клонировали в систему экспрессии 2X35S-CPMV-HT-PDISP-NOS в плазмиде, содержащей экспрессионную кассету Plasto_pro/P19/Plasto_ter, используя нижеприведенный метод на основе ПЦР. Фрагмент, содержащий последовательность, кодирующую Н3 без его сигнального пептида с сигнальной последовательностью дикого типа, амплифицировали, применяя праймеры IF-H3V36111.S2+4c (Фигура 25A, SEQ ID NO: 44) и IF-H3V36111.s1-4r (Фигура 25В, SEQ ID NO: 45), а в качестве матрицы используя синтезированный Н3 ген (соответствующий nt 25-1725 последовательности GISAID EPI_ISL_101506 изолята НА) (Фигура 25С, SEQ ID NO: 46). ПЦР-продукт клонировали в рамке считывания с сигнальным пептидом PDI люцерны в системе экспрессии 2X35S/CPMV-HT/NOS, используя систему клонирования In-Fusion (Clontech, Mountain View, CA). Конструкцию 1192 (Фигура 4D) расщепляли рестриктазами SacII и StuI и линеаризованную плазмиду применяли для реакции сборки In-Fusion. Конструкция номер 1192 представляет собой акцепторную плазмиду, предполагаемую для "In Fusion" клонирования целевых генов в рамке считывания с сигнальным пептидом PDI люцерны в экспрессионной кассете на основе CPMV-HT. Она включает также генную конструкцию для коэкспрессии TBSV P19 супрессора сайленсинга под контролем промотора и терминатора гена пластоцианина люцерны. Остов представляет собой бинарную плазмиду pCAMBIA, и последовательность от левой до правой границы тДНК показана на Фигуре 4Е (SEQ ID NO: 18). Полученной в результате конструкции был присвоен номер 1391 (Фигура 25D, SEQ ID NO: 47). Аминокислотная последовательность PDISP/H3 вируса гриппа A/Victoria/361/2011 (H3N2) представлена на Фигуре 25Е (SEQ ID NO: 48). Плазмида 1391 изображена на Фигуре 25F.
B-2X35S/CPMV-HT/HA В Wisconsin/NOS в амплификационную систему BeYDV(m)+репликаза (Конструкция номер 1462)
Последовательность, кодирующую НА вируса гриппа B/Wisconsin/1/2010, клонировали в 2X35S/CPMV-HT/NOS, содержащую амплификационную систему BeYDV(m)+ репликаза, в плазмиде, содержащей экспрессионную кассету Plasto_pro/P19/Plasto_ter, применяя нижеприведенный метод на основе ПЦР. Фрагмент, содержащий последовательность, кодирующую полный НА В Wisconsin, амплифицировали, используя праймеры IF-HAB110.S1+3с (Фигура 26А, SEQ ID NO: 49) и IF-HAB110.s1-4r (Фигура 26В, SEQ ID NO: 50) и применяя синтезированный ген НА В Wisconsin (номер доступа в Genbank JN993010) (Фигура 26С, SEQ ID NO: 51) в качестве матрицы. ПЦР-продукт клонировали в экспрессионной кассете 2X35S/CPMV-HT/NOS в амплификационную систему BeYDV(m), используя In-Fusion систему клонирования (Clontech, Mountain View, CA). Конструкцию 193 (Фигура 26D) расщепляли рестриктазами SacII и StuI и линеаризованную плазмиду использовали для реакции сборки In-Fusion. Конструкция номер 193 представляет собой акцепторную плазмиду, предполагаемую для "In Fusion" клонирования целевых генов в экспрессионной кассете на основе CPMV-HT в амплификационную систему BeYDV(m). Она также включает генную конструкцию для коэкспрессии TBSV P19 супрессора сайленсинга под контролем промотора и терминатора гена пластоцианина люцерны. Остов представляет собой бинарную плазмиду pCAMBIA, и последовательность от левой до правой границы тДНК показана на Фигуре 26Е (SEQ ID NO: 52). Полученной в результате конструкции был присвоен номер 1462 (Фигура 26F, SEQ ID NO: 53). Аминокислотная последовательность PDISP/HA вируса гриппа B/Wisconsin/1/2010 представлена на Фигуре 26G (SEQ ID NO: 54). Плазмида 1462 изображена на Фигуре 26Н.
C-2X35S/CPMV-HT/HA В Wisconsin с делетированной протеолитической петлей в амплификационную систему BeYDV(m)+peпликaзa (Конструкция номер 1467)
Последовательность НА вируса гриппа B/Wisconsin/1/2010 с делетированной протеолитической петлей клонировали в 2X35S/CPMV-HT/NOS, содержащей амплификационную систему BeYDV(m)+peпликaзa в плазмиде, содержащей экспрессионную кассету Plasto_pro/P19/Plasto_ter, применяя нижеприведенный метод лигирования на основе ПЦР, представленный Darveau et al. (Methods in Neuroscience 26: 77-85 (1995)). В первом раунде ПЦР фрагмент, содержащий последовательность, кодирующую НА В Wisconsin, от nt 1 до nt 1062, амплифицировали, используя праймеры IF-HAB110.S1+3с (Фигура 26А, SEQ ID NO: 49) и НАВ110(PrL-).r (Фигура 27А, SEQ ID NO: 55), а в качестве матрицы применяя синтезированный НА В Wisconsin ген (номер доступа в Genbank JN993010) (Фигура 26С, SEQ ID NO: 51). Второй фрагмент, содержащий последовательность, кодирующую НА В Wisconsin, от nt 1120 до nt 1755, амплифицировали, используя праймеры НАВ110(PrL-).c (Фигура 27В, SEQ ID NO: 56) и IF-HAB110.s1-4r (Фигура 26В, SEQ ID NO: 50), а в качестве матрицы применяя синтезированный ген НА В Wisconsin (номер доступа в Genbank JN993010) (Фигура 26С, SEQ ID NO: 51). Затем смешивали (объединяли) ПЦР-продукты из обеих амплификаций и использовали в качестве матрицы для второго раунда амплификации с применением IF-НАВ110.S1+3с (Фигура 26А, SEQ ID NO: 49) и IF-HAB110.s1-4r (Фигура 26В, SEQ ID NO: 50) в качестве праймеров. Полученный в результате фрагмент (кодирующий НА B/Wisconsin/1/2010 Δа.а. 340-358 с GG линкером между фрагментами) клонировали в экспрессионную кассету 2X35S/CPMV-HT/NOS, содержащую амплификационную систему BeYDV(m), с применением системы клонирования In-Fusion (Clontech, Mountain View, CA). Конструкцию 193 (Фигура 26D) расщепляли рестриктазами SacII и StuI и линеаризованную плазмиду применяли для реакции сборки In-Fusion. Конструкция номер 193 представляет собой акцепторную плазмиду, предполагаемую для "In Fusion" клонирования целевых генов в экспрессионной кассете на основе CPMV-HT в амплификационную систему BeYDV(m). Она также включает генную конструкцию для коэкспрессии TBSV P19 супрессора сайленсинга под контролем промотора и терминатора гена пластоцианина люцерны. Остов представляет собой бинарную плазмиду pCAMBIA, и последовательность от левой до правой границы тДНК представлена на Фигуре 26Е (SEQ ID NO: 52). Полученной в результате конструкции был присвоен номер 1467 (Фигура 27С, SEQ ID NO: 57). Аминокислотная последовательность НА вируса гриппа B/Wisconsin/1/2010 с делетированной протеолитической петлей представлена на Фигуре 27D (SEQ ID NO: 58). Плазмида 1467 изображена на Фигуре 27Е.
D-2X35S/CPMV-HT/PDISP/HА В Malaysia/NOS в амплификационную систему BeYDV(m)+репликаза (Конструкция номер 1631)
Последовательность, кодирующую НА вируса гриппа B/Malaysia/2506/2004, клонировали в 2X35S/CPMV-HT/PDISP/NOS, содержащую амплификационную систему BeYDV(m)+ репликаза, в плазмиде, содержащей экспрессионную кассету Plasto_pro/P19/Plasto_ter, применяя нижеописанный метод на основе ПЦР. Фрагмент, содержащий последовательность, кодирующую НА В Malaysia без его сигнального пептида дикого типа, амплифицировали, используя праймеры IF-HB-M-04.s2+4c (Фигура 28А, SEQ ID NO: 59) и IF-HB-M-04.s1-4r (Фигура 28В, SEQ ID NO: 60), а в качестве матрицы используя синтезированный ген НА В Malaysia (соответствующий nt 31-1743 последовательности с номером доступа в Genbank EU124275. Молчащие мутации Т759С и C888G были введены в синтезированную последовательность, чтобы модифицировать сайты узнавания рестриктазами DraIII и BamHI) (Фигура 28С, SEQ ID NO: 61). ПЦР-продукт клонировали в рамке считывания с сигнальным пептидом PDI в экспрессионной кассете 2X35S/CPMV-HT/NOS в амплификационную систему BeYDV(m) с применением системы клонирования In-Fusion (Clontech, Mountain View, СА). Конструкцию 194 (Фигура 28D) расщепляли рестриктазами SacII и StuI и линеаризованную плазмиду применяли для реакции сборки In-Fusion. Конструкция номер 194 представляет собой акцепторную плазмиду, предполагаемую для "In Fusion" клонирования целевых генов в рамке считывания с сигнальным пептидом PDI люцерны в экспрессионной кассете на основе CPMV-HT в амплификационную систему BeYDV(m). Она включает также генную конструкцию для коэкспрессии TBSV P19 супрессора сайленсинга под контролем промотора и терминатора гена пластоцианина люцерны. Остов представляет собой бинарную плазмиду pCAMBIA, и последовательность от левой до правой границы тДНК показана на Фигуре 28Е (SEQ ID NO: 62). Полученной в результате конструкции присвоен номер 1631 (Фигура 28F, SEQ ID NO: 63). Аминокислотная последовательность PDISP/HA вируса гриппа B/Malaysia/2506/2004 представлена на Фигуре 28G (SEQ ID NO: 64). Плазмида 1631 изображена на Фигуре 28Н.
Трансфекция агробактерий
Agrobacterium штамма AGL1 трансфецировали электропорацией с использованием ДНК-конструкций методами, описанными D'Aoust et al 2008 (Plant Biotechnology Journal 6: 930-940). Трансфецированные Agrobacterium выращивали в среде YEB, дополненной 10 мМ 2-(N-морфолино)этансульфоновой кислоты (MES), 20 мкМ ацетосирингона, 50 мкг/мл канамицина и 25 мкг/мл карбенициллина рН5.6, до OD600 между 0.6 и 1.6. Перед применением суспензии Agrobacterium центрифугировали и ресуспендировали в среде для инфильтрации (10 мМ MgCl2 и 10 мМ MES рН 5.6).
Приготовление растительной биомассы, инокулят (посевной материал) и агроинфильтрация
Предполагается, что термины "биомасса" и "растительный материал" в данном контексте означают любой материал растительного происхождения. Биомасса или растительный материал может включать цельное растение, ткани, клетки или любую их часть. Помимо этого, биомасса или растительный материал может включать внутриклеточные растительные компоненты, жидкие или твердые растительные экстракты или их комбинацию. Далее, биомасса или растительный материал может включать растение, растительные клетки, ткани, жидкий экстракт, или их комбинацию, листьев, стеблей, плодов, корней растения или их комбинации. Часть растения может представлять собой растительный материал или биомассу.
Растения Nicotiana benthamiana выращивали из семян в плоскодонных колбах, заполненных коммерческим торфяным мхом сфагнумом в качестве субстрата. Растения оставляли расти в теплице при световом режиме 16/8 и при температуре 25°С днем/20°С ночью. Через три недели после засевания отдельные растения извлекали, пересаживали в горшки и оставляли расти в теплице еще в течение трех недель в тех же самых условиях.
Agrobacteria, трансфецированные с использованием каждой конструкции, выращивали в среде YEB, дополненной 10 мМ 2-(N-морфолино)этансульфоновой кислоты (MES), 20 мкМ ацетосирингона, 50 мкг/мл канамицина и 25 мкг/мл карбенициллина рН5.6, до достижения OD600 между 0.6 и 1.6. Перед применением суспензии Agrobacterium центрифугировали и ресуспендировали в среде для инфильтрации (10 мМ MgCl2 и 10 мМ MES рН 5.6). и хранили в течение ночи при 4°С. В день инфильтрации партии для культивирования разводили в 2.5 объемах культуры и оставляли нагреваться до применения. Цельные растения N benthamiana в перевернутом виде помещали в бактериальную суспензию в герметичном резервуаре из нержавеющей стали и выдерживали в вакууме 20-40 Торр (2.67-5.33 кПа) в течение 2 мин. Растения возвращали в теплицу и инкубировали в течение 2-6 дней до момента сбора.
Сбор листьев и экстракция тотального белка
После инкубации надземную часть растений собирали, замораживали при -80°С и измельчали. Тотальные растворимые белки экстрагировали, гомогенизируя (Polytron) каждый образец измельченного в замороженном состоянии растительного материала в 3 объемах холодной смеси, содержащей 50 мМ Tris рН 8.0, 0.15 М NaCl, 0.1% Triton Х-100 и 1 мМ фенилметансульфонилфторида. После гомогенизации кашицу (суспензию) центрифугировали при 10,000 g в течение 10 мин при 4°С и эти осветленные сырые экстракты (супернатант) хранили для анализов.
Анализ белков и иммуноблоттинг
Содержание тотального белка в осветленных сырах экстрактах определяли методом Бредфорд (Bio-Rad, Hercules, CA), используя бычий сывороточный альбумин в качестве стандартного образца. Белки делили методом SDS-PAGE и переносили методом электроблоттинга на мембраны на основе поливинилендифторида (PVDF) (Roche Diagnostics Corporation, Indianapolis, IN) для иммунологического анализа. Мембраны блокировали с помощью 5% снятого молока и 0.1% Твин-20 в забуференном Tris физиологическом растворе (TBS-T) в течение 16-18 час при 4°С.
Иммуноблоттинг проводили, осуществляя первую инкубацию с первичным антителом (в Таблице 4 представлены антитела и условия, применяемые для обнаружения каждого НА), в 2% снятом молоке (2 мкг/мл в 0.1% буфере TBS-Твин 20. Вторичные антитела, применяемые для обнаружения методом хемилюминесценции, разводили как указано в Таблице 4, в 2% снятом молоке в 0.1% буфера TBS-Tween 20. Иммунореактивные комплексы детектировали методом хемилюминесценции, используя люминол в качестве субстрата (Roche Diagnostics Corporation). Конъюгацию антитела к человеческому IgG с ферментом пероксидазой хрена проводили, используя набор для конъюгации с активированной пероксидазой EZ-Link Plus® (Pierce, Rockford, IL).
JIR: Jackson ImmunoResearch, West Grove, PA, USA;
CBER: Center for Biologies Evaluation and Research, Rockville, MD, USA.
Sino: Sino Biological inc., Beijing, China.
TGA: Therapeutic Goods Administration, Australia.
NIBSC: National Institute for Biological Standards and Control, United Kingdom
Реакция гемагглютинации
Анализ с применением реакции гемагглютинации основан на методе, описанном Nayak и Reichl (2004). Коротко говоря, двукратные серийные разведения тестируемых образцов (100 мкл) готовили в в лунках с V-образным дном 96-луночных микротитрационных планшетов, содержащих 100 мкл PBS, оставляя 100 мкл разведенного образца в лунке. В каждую лунку добавляли по сто микролитров 0.25% суспензии эритроцитов индейки (Bio Link Inc., Syracuse, NY) и планшеты инкубировали в течение 2 час при комнатной температуре. Значение, обратное наивысшему разведению, показывающему полную гемагглютинацию, регистрировали как НА активность. Параллельно эталон рекомбинантного НА (A/Vietnam/1203/2004 H5N1) (Protein Science Corporation, Meriden, CT) разводили в PBS и применяли в качестве контроля в каждом планшете.
Пример 1: Влияние коэкспрессии М2 вируса гриппа на уровень накопления В НА и Н3
Влияние коэкспрессии М2 вируса гриппа на уровень накопления НА других штаммов вируса гриппа анализировали с помощью конструкций для совместного переноса, управляющих экспрессией НА с использованием конструкции для экспрессии М2 вируса гриппа A/New Caledonia/20/1999 (H1N1) системе транзиторной трансформации на основе агроинфильтрации.
Вестерн-блоттинг белковых экстрактов растений, трансформированных с использованием генных конструкций, управляющих экспрессией В НА (вируса гриппа B/Brisbane/60/2008) (конструкции номер 1008, 1009 и 1029), в присутствии или в отсутствие конструкции для экспрессии М2 (конструкции номер 1261) показал, что коэкспрессия М2 приводит к повышенному накоплению В НА вируса гриппа (Фигура 18). Аналогично, коэкспрессия М2 с Н3 вируса гриппа A/Perth/16/2009 (конструкция номер 1019+1261) привела к повышенному накоплению Н3 в трансформированных растениях по сравнению с растениями, трансформированными только с помощью Н3- экспрессионной конструкции (конструкции номер 1019), как показано на Фигуре 19.
Вестерн-блоттинг белковых экстрактов растений, коэкспрессирующих М2 с H1 вируса гриппа A/California/07/2009, показал, что коэкспрессия М2 с H1 приводит к незначительному снижению уровня накопления H1 (Фигура 20, конструкция номер 484 по сравнению с конструкцией 484+1261). Коэкспрессия М2 с Н5 вируса гриппа A/Indonesia/05/2005 также обусловила пониженный уровень накопления Н5 по сравнению с экспрессией только одного Н5 (Фигура 21, 489 по сравнению с 489+1261).
Далее определяли влияние коэкспрессии М2 на уровень накопления модифицированного НА вируса гриппа В. Конструкция номер 1059 кодирует вируса гриппа В НА, в котором протеолитическая петля заменена на линкер из двух аминокислот (GG вместо аа (аминокислотных остатков) 341-359). Результаты Вестерн-блоттинга, представленные на Фигуре 22А, показывают, что удаление протеолитической петли привело к повышенному уровню накопления НА вируса гриппа В (сравните 1008 с 1059) и что коэкспрессия М2 с модифицированным НА вируса гриппа В дополнительно повышает уровень накопления НА (Фигура 22А, 1059 по сравнению с 1059+1261). Анализ гемагглютинирующей активности на сырых белковых экстрактах растений, трансформированных с использованием НА вируса гриппа В при наличии модификации или без нее и в присутствии или в отсутствие коэкспрессии М2 подтвердил положительное воздействие коэкспрессии М2 на уровень накопления нативного НА вируса грипп В (Фигура 22В, 1008 по сравнению с 1008+1261) и модифицированного НА вируса гриппа В (Фигура 22В, 1059 по сравнению с 1059+1261).
Влияние М2 вируса гриппа A/Puerto Rico/8/1934 на повышение накопления модифицированного В НА и Н3 вируса гриппа сравнивали с влиянием М2 вируса гриппа A/New Caledonia/20/1999 на то же накопление. Для модифицированного НА вируса гриппа В НА сравнение проводили методом Вестерн- блоттинга белковых экстрактов растений, трансформированных с использованием конструкций 1059, 1059+1261 и 1059+859. Для Н3 аналогичное сравнение проводили на белковых экстрактах растений, трансформированных с помощью конструкций 1019, 1019+1261 и 1019+859. Полученные результаты продемонстрировали, что коэкспрессия М2 вируса гриппа A/Puerto Rico/8/1934 (кодированного конструкцией номер 859) была также эффективна, как коэкспрессия М2 вируса гриппа A/New Caledonia/20/1999 (кодированного конструкцией номер 1261), для повышения накопления как модифицированного В НА вируса гриппа (Фигура 23А), так и Н3 (Фигура 23В).
Пример 2: Влияние коэкспрессии М2 вируса гриппа на уровень накопления В НА и Н3 различных штаммов Вестерн-блоттинг белковых экстрактов растений, трансформированных с помощью генных конструкций, управляющих экспрессией В НА вируса гриппа (из B/Malaysia/2506/2004) (конструкции номер 1631) в присутствии или в отсутствие М2-экспрессионной конструкции (конструкции номер 1261), показал, что коэкспрессия М2 приводит в результате к накоплению белка В НА вируса гриппа (Фигура 29).
Вестерн-блоттинг белковых экстрактов растений, трансформированных с помощью генных конструкций, управляющих экспрессией В НА вируса гриппа (из B/Wisconsin/1/2010) (конструкций номер 1462) в присутствии или в отсутствие М2-экспрессионной конструкции (конструкции номер 1261), показал, что коэкспрессия М2 приводит в результате к аккумуляции белка В НА вируса гриппа (Фигура 30).
Далее оценивали влияние коэкспрессии М2 на уровень накопления модифицированного В НА. Конструкция номер 1467 кодирует белок вируса гриппа В НА, в котором протеолитическая петля заменена аминокислотным линкером, содержащим два аминокислотных остатка (линкер GG вместо аа 341-359). Результаты Вестерн-блоттинга, представленные на Фигурах 30А, показывают, что удаление протеолитической петли приводит к повышенному уровню накопления В НА вируса гриппа (сравните конструкцию 1462 с конструкцией 1467) и что коэкспрессия М2 с модифицированным белком вируса гриппа В НА дополнительно повышает НА уровень накопления (Фигура 30А, конструкция 1467 по сравнению с конструкциями 1467+1261). Анализ гемагглютинирующей активности на сырых белковых экстрактах растений, трансформированных с помощью В НА вируса гриппа с модификацией или без нее в присутствии или в отсутствие коэкспрессии М2, подтвердил положительное воздействие коэкспрессии М2 на уровень накопления нативного В НА (Фигура 30В, 1462 по сравнению с 1462+1261) и модифицированного В НА вируса гриппа (Фигура 26В, 1467 по сравнению с 1467+1261).
Вестерн-блоттинг белковых экстрактов растений, трансформированных с использованием генных конструкций, управляющих экспрессией Н3 (из H3/Victoria/361/2011) (конструкции номер 1391), в присутствии или в отсутствие конструкции для экспрессии М2 (конструкции номер 1261), показал, что коэкспрессия М2 приводит к повышенному накоплению Н3 вируса гриппа (Фигура 31).
Все цитируемые материалы включены в настоящее изобретение посредством отсылки.
Настоящее изобретение описано применительно к одному или более его вариантов. Однако для специалистов в данной области техники будет очевидно, что можно сделать целый ряд изменений и модификаций, не отступая от объема изобретения, определяемого Формулой изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛУЧЕНИЕ ВИРУСОПОДОБНЫХ ЧАСТИЦ ВИРУСА ГРИППА В РАСТЕНИЯХ | 2014 |
|
RU2742607C1 |
| ПОЛУЧЕНИЕ ВИРУСОПОДОБНЫХ ЧАСТИЦ ВИРУСА ГРИППА В РАСТЕНИЯХ | 2014 |
|
RU2705555C2 |
| ПОЛУЧЕНИЕ ВИРУСОПОДОБНЫХ ЧАСТИЦ В РАСТЕНИЯХ | 2011 |
|
RU2655431C2 |
| Получение вирусоподобной частицы вируса бешенства в растениях | 2012 |
|
RU2655433C2 |
| ЭНХАНСЕРНЫЕ ЭЛЕМЕНТЫ CPMV | 2015 |
|
RU2699982C2 |
| ХИМЕРНЫЕ ВИРУСОПОДОБНЫЕ ЧАСТИЦЫ, СОДЕРЖАЩИЕ ГЕМАГГЛЮТИНИН, СХОДНЫЕ С ЧАСТИЦАМИ ВИРУСА ГРИППА | 2010 |
|
RU2569195C9 |
| МУТАНТЫ ГЕМАГГЛЮТИНИНА ВИРУСА ГРИППА | 2019 |
|
RU2800938C2 |
| МОДИФИКАЦИЯ ПРОДУЦИРОВАНИЯ БЕЛКОВ В РАСТЕНИЯХ | 2015 |
|
RU2731295C2 |
| ПОЛУЧЕНИЕ ПИКОРНАВИРУСОПОДОБНЫХ ЧАСТИЦ В РАСТЕНИЯХ | 2013 |
|
RU2693431C2 |
| СПОСОБ ПОЛУЧЕНИЯ ВИРУСОПОДОБНЫХ ЧАСТИЦ РАСТЕНИЙ | 2010 |
|
RU2579903C2 |































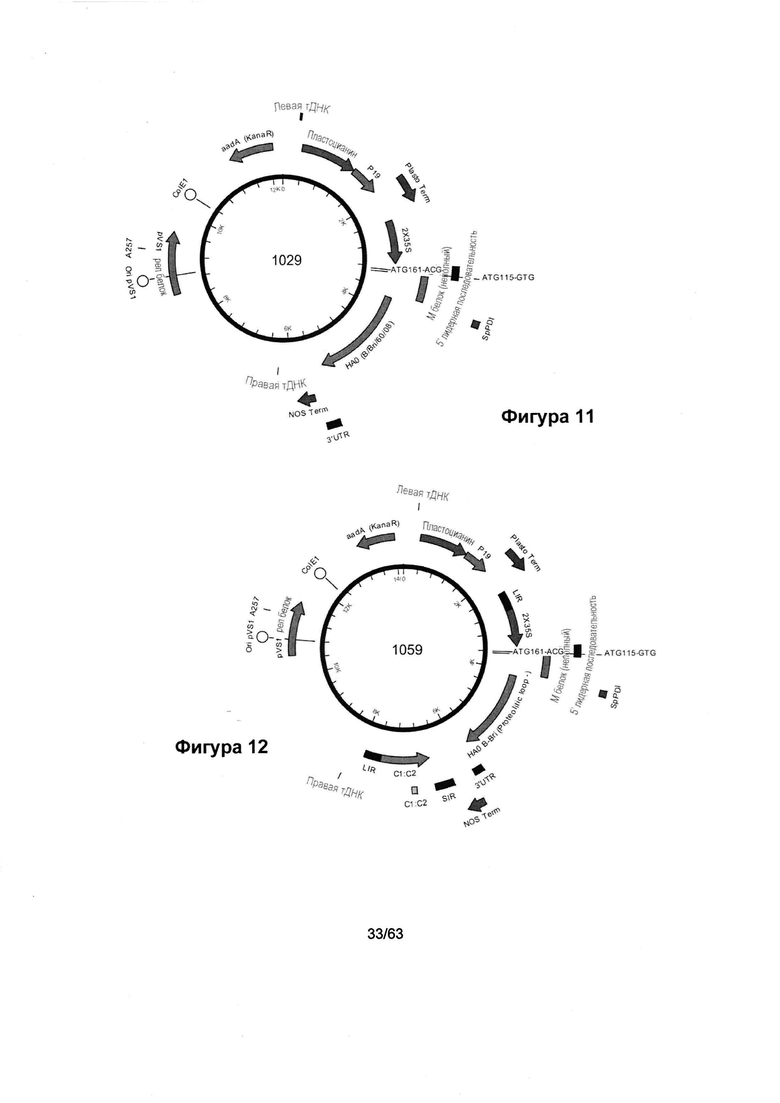
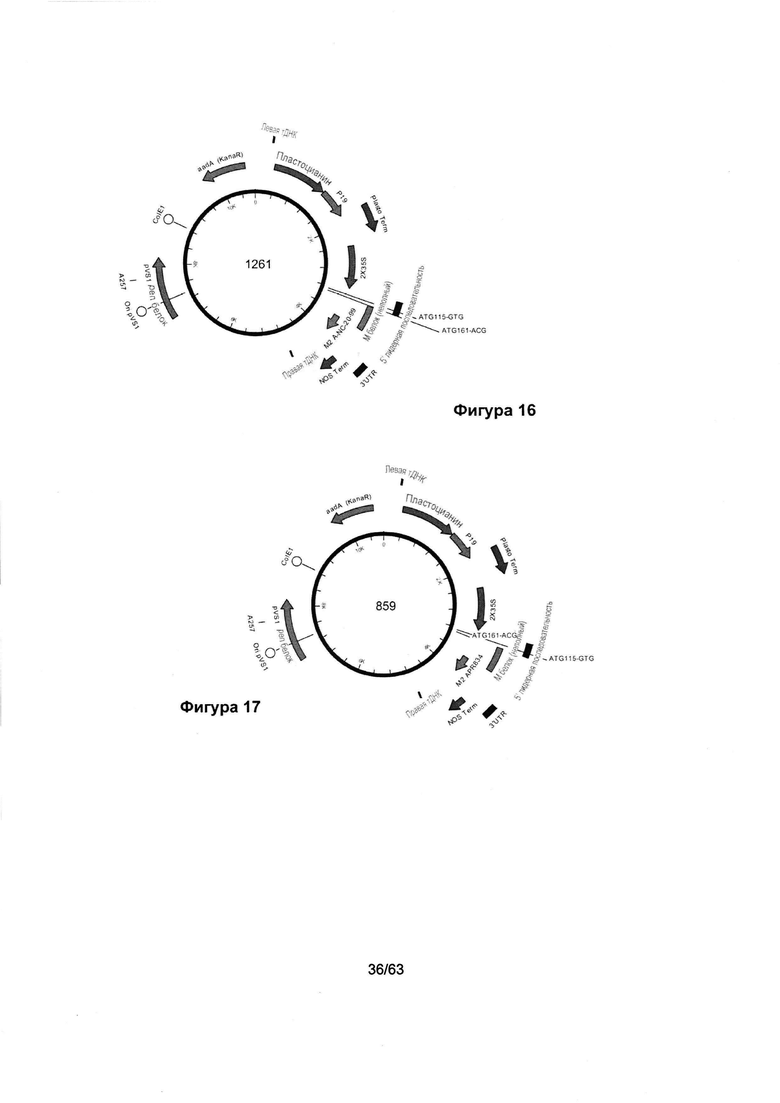


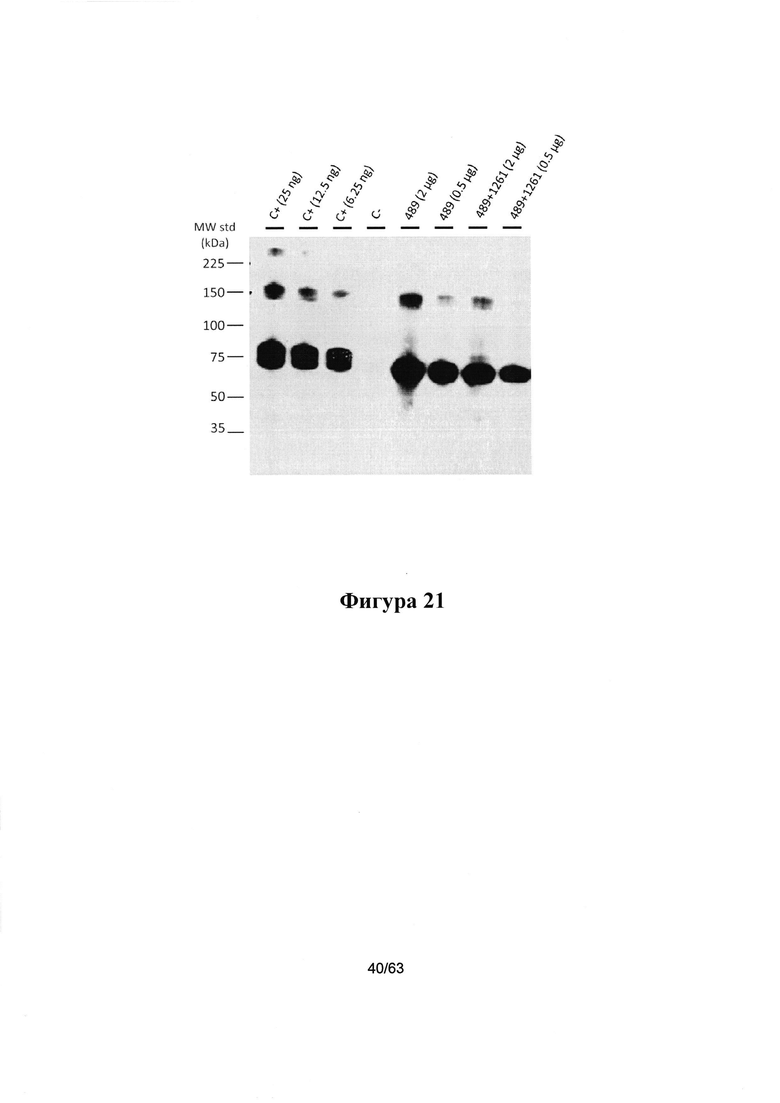



















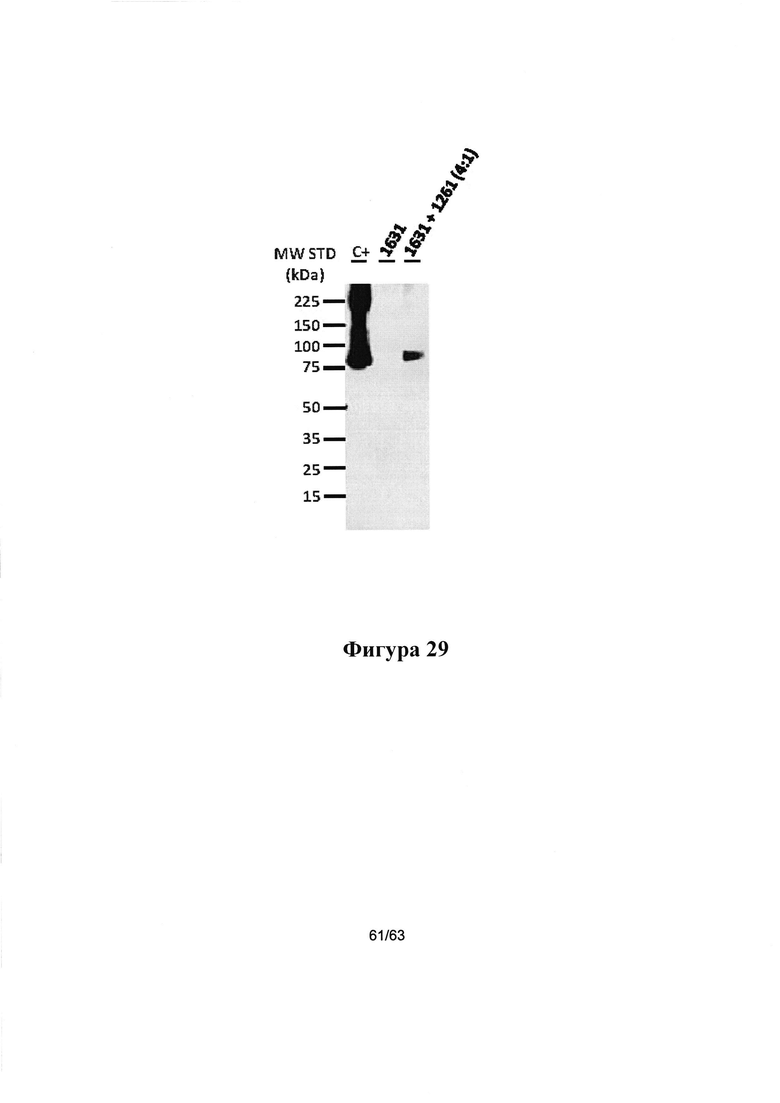

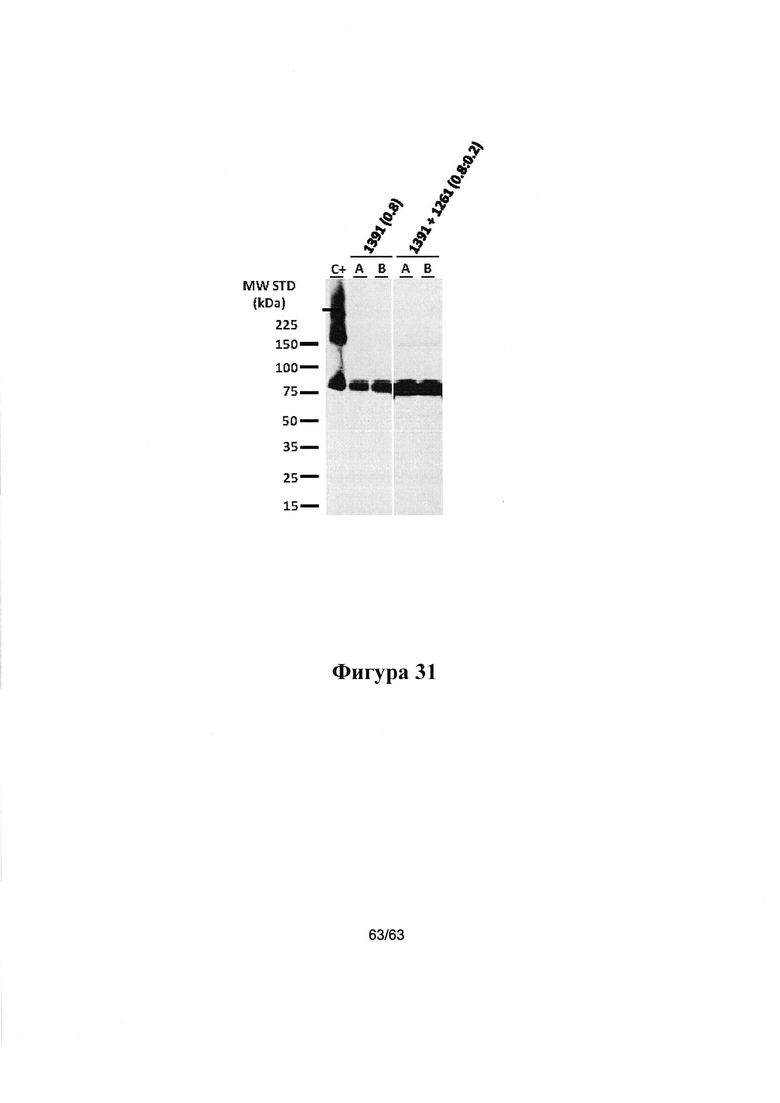
Изобретение относится к области биохимии, в частности к способу продуцирования вирусоподобной частицы (VLP) в растении, а также к способу индукции иммунного ответа к вирусу гриппа подтипа В или НЗ у субъекта и способу получения композиции для индукции иммунного ответа к вирусу гриппа подтипа В или Н3 у субъекта с его использованием. При этом вышеуказанный способ предусматривает ввод в растение нуклеотидной последовательности, кодирующей белок гемагглютинин (НА) вируса гриппа, выбранный из группы, состоящей из В и Н3, и нуклеотидной последовательности, кодирующей белок протонного канала. Также раскрыты способ получения растительного материала, содержащего вирусоподобную частицу (VLP), и способ получения растительного материала, содержащего вирусоподобную частицу (VLP). Вышеуказанные способы также предусматривают ввод в растение нуклеотидной последовательности, кодирующей белок гемагглютинин (НА) вируса гриппа, выбранный из группы, состоящей из В и Н3, и нуклеотидной последовательности, кодирующей белок протонного канала Изобретение позволяет эффективно продуцировать вирусоподобную частицу в растении. 7 н. и 34 з.п. ф-лы, 31 ил., 4 табл., 2 пр.
1. Способ продуцирования вирусоподобной частицы (VLP) в растении, включающий:
а) введение первой нуклеиновой кислоты, содержащей первую регуляторную область, являющуюся активной в растении и функционально связанную с нуклеотидной последовательностью, кодирующей белок гемагглютинин (НА) вируса гриппа, выбранный из группы, состоящей из В и Н3, в растение или в часть растения,
б) введение второй нуклеиновой кислоты, содержащей вторую регуляторную область, являющуюся активной в растении и функционально связанную с нуклеотидной последовательностью, кодирующей белок протонного канала, в растение или часть растения, причем белок протонного канала получен из вируса гриппа, и
в) инкубацию растения или части растения в условиях, которые позволяют экспрессии первой и второй нуклеиновых кислот, посредством чего продуцируя VLP.
2. Способ по п. 1, в котором белок протонного канала выбран из М2 или ВМ2.
3. Способ по п. 1, в котором белок протонного канала содержит сигнатурную последовательность протонного канала HXXXW.
4. Способ по п. 1, в котором одна или более протеолитических петель белка НА вируса гриппа делетированы.
5. Способ по п. 1, в котором белок НА вируса гриппа В происходит из вируса гриппа штамма B/Brisbane/60/2008, B/Malaysia/2506/2004 или B/Wisconsin/1/2010.
6. Способ по п. 5, в котором одна или более протеолитических петель белка НА вируса гриппа В делетированы.
7. Способ по п. 1, в котором белок Н3 вируса гриппа происходит из вируса гриппа штамма A/Perth/16/2009 или A/Victoria/361/2011.
8. Способ по п. 1, в котором нуклеотидная последовательность, кодирующая белок гемагглютинин (НА) вируса гриппа, по меньшей мере на 70% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, SEQ ID NO: 51 и SEQ ID NO: 61, причем нуклеотидная последовательность кодирует белок гемагглютинин, который, будучи экспрессирован, образует VLP.
9. Способ по п. 1, в котором нуклеотидная последовательность, кодирующая белок гемагглютинина (НА) вируса гриппа, является такой, как показана в SEQ ID NO: 30, SEQ ID NO: 54 или SEQ ID NO: 64.
10. Способ по п. 1, в котором нуклеотидная последовательность, кодирующая белок гемагглютинин (НА) вируса гриппа, по меньшей мере на 70% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 43 и подчеркнутого участка SEQ ID NO: 57 как показано на Фигуре 27С, причем нуклеотидная последовательность кодирует белок гемагглютинин, который, будучи экспрессирован, образует VLP.
11. Способ по п. 1, в котором нуклеотидная последовательность, кодирущая белок гемагглютинина (НА) вируса гриппа, является такой, как показана в SEQ ID NO: 41 или SEQ ID NO: 58.
12. Способ по п. 1, в котором нуклеотидная последовательность, кодирующая белок гемагглютинин (НА) вируса гриппа, по меньшей мере на 70% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 23 и SEQ ID NO: 46, причем нуклеотидная последовательность кодирует белок гемагглютинин, который, будучи экспрессирован, образует VLP.
13. Способ по п. 1, в котором нуклеотидная последовательность, кодирущая белок гемагглютинина (НА) вируса гриппа, является такой, как показана в SEQ ID NO: 25 или SEQ ID NO: 48.
14. Способ по п. 1, в котором на стадии введения (стадия а) первая нуклеиновая кислота дополнительно содержит один или более энхансеров комовируса, функционально связанных с первой регуляторной областью, и первая нуклеиновая кислота дополнительно содержит один или более чем один амплификационных элементов геминивируса; и
до стадии инкубации (стадия в) третью нуклеиновую кислоту, кодирующую репликазу геминивируса, вводят в растение или часть растения.
15. Способ по п. 14, в котором один или более чем один энхансеров комовируса представляют собой UTR (нетранслируемую область) комовируса.
16. Способ по п. 15, в котором UTR комовируса представляет собой UTR вируса мозаики коровьего гороха (CPMV).
17. Способ по п. 14, в котором один или более чем один амплификационных элементов геминивируса выбран из длинной межгенной области вируса желтой карликовости бобов (BeYDV LIR) и короткой межгенной области вируса желтой карликовости бобов (BeYDV SIR).
18. Способ по п. 14, в котором нуклеотидная последовательность, кодирующая белок гемагглютинин (НА) вируса гриппа, по меньшей мере на 70% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, SEQ ID NO: 51 и SEQ ID NO: 61, причем нуклеотидная последовательность кодирует белок гемагглютинин, который, будучи экспрессирован, образует VLP.
19. Способ по п. 14, в котором последовательность белка гемагглютинина (НА) вируса гриппа является такой, как показано в SEQ ID NO: 30, SEQ ID NO: 54 или SEQ ID NO: 64.
20. Способ по п. 14, в котором нуклеотидная последовательность, кодирующая белок гемагглютинин (НА) вируса гриппа, по меньшей мере на 70% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 43 и подчеркнутого участка SEQ ID NO: 57 как показано на Фигуре 27С, причем нуклеотидная последовательность кодирует белок гемагглютинин, который, будучи экспрессирован, образует VLP.
21. Способ по п. 14, в котором последовательность белка гемагглютинина (НА) вируса гриппа является такой, как показано в SEQ ID NO: 41 или SEQ ID NO: 58.
22. Способ по п. 14, в котором нуклеотидная последовательность, кодирующая белок гемагглютинин (НА) вируса гриппа, по меньшей мере на 70% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 23 и SEQ ID NO: 46, причем нуклеотидная последовательность кодирует белок гемагглютинин, который, будучи экспрессирован, образует VLP.
23. Способ по п. 14, в котором последовательность белка гемагглютинина (НА) вируса гриппа является такой, как показано в SEQ ID NO: 25 или SEQ ID NO: 48.
24. Способ по п. 1, в котором на стадии введения (стадия а) нуклеиновая кислота транзиентно экспрессируется в растении.
25. Способ по п. 1, в котором на стадии введения (стадия а) нуклеиновая кислота стабильно экспрессируется в растении.
26. Способ по п. 1, дополнительно включающий стадию
г) сбора растения и очистки вирусоподобных частиц (VLP).
27. Способ по п. 1, в котором VLP не содержит вирусного матрикса, корового белка или канального белка.
28. Способ по п. 1, дополнительно включающий стадию введения третьей нуклеиновой кислоты, которая содержит последовательность, кодирующую супрессор сайленсинга.
29. Способ по п. 14, дополнительно включающий стадию введения четвертой нуклеиновой кислоты, которая содержит последовательность, кодирующую супрессор сайленсинга.
30. Способ по п. 28 или 29, в котором супрессор сайленсинга выбран из группы HcPro и р19.
31. Способ по п. 2, в котором белок М2 выбран из группы, включающей белок вируса гриппа штамма A/Puerto Rico/8/1934 и штамма A/New Caledonia/20/1999.
32. Способ продуцирования вирусоподобной частицы (VLP), включающий:
а) предоставление растения или его части, содержащих первую нуклеиновую кислоту, содержащую первую регуляторную область, являющуюся активной в растении и функционально связанную с нуклеотидной последовательностью, кодирующей белок гемагглютинин (НА) вируса гриппа, выбранный из группы, состоящей из В и Н3, и вторую нуклеиновую кислоту, содержащую вторую регуляторную область, являющуюся активной в растении и функционально связанную с нуклеотидной последовательностью, кодирующей белок протонного канала, в растение или часть растения, причем белок протонного канала получен из вируса гриппа; и
б) инкубацию растения или части растения в условиях, которые позволяют экспрессию первой и второй нуклеиновых кислот, посредством чего продуцируя VLP.
33. Способ по п. 32, в котором белок протонного канала выбран из М2 или ВМ2.
34. Способ индукции иммунного ответа к вирусу гриппа подтипа В или Н3 у субъекта, включающий продуцирование вирусоподобной частицы (VLP) способом по п. 1 или 32 и введение эффективной дозы продуцированной VLP субъекту.
35. Способ по п. 34, в котором VLP вводят субъекту перорально, интрадермально, интраназально, внутримышечно, интраперитонеально, внутривенно или подкожно.
36. Способ по п. 1 или 32, в котором VLP не содержит белок протонного канала.
37. Способ по п. 1 или 32, в котором белок гемагглютинин (НА) вируса гриппа представляет собой белок НА0.
38. Способ продуцирования вирусоподобной частицы (VLP) в растении, включающий:
а) введение нуклеиновой кислоты, содержащей регуляторную область, являющуюся активной в растении и функционально связанную с нуклеотидной последовательностью, кодирующей белок гемагглютинин (НА) вируса гриппа В, в растение или в часть растения, причем одна или более протеолитических петель белка гемагглютинина (НА) вируса гриппа делетированы,
б) инкубацию растения или части растения в условиях, которые позволяют экспрессию нуклеиновой кислоты, посредством чего продуцируя VLP.
39. Способ получения композиции для индукции иммунного ответа к вирусу гриппа подтипа В или Н3 у субъекта, включающий продуцирование VLP способом по п. 1 или 32 и добавление к полученной VLP фармацевтически приемлемого носителя.
40. Способ получения растительного материала, содержащего вирусоподобную частицу (VLP), включающий:
а) введение первой нуклеиновой кислоты, содержащей первую регуляторную область, являющуюся активной в растении и функционально связанную с нуклеотидной последовательностью, кодирующей белок гемагглютинин (НА) вируса гриппа, выбранный из группы, состоящей из В и Н3, в растение или в часть растения,
б) введение второй нуклеиновой кислоты, содержащей вторую регуляторную область, являющуюся активной в растении и функционально связанную с нуклеотидной последовательностью, кодирующей белок протонного канала, в растение или часть растения, причем белок протонного канала получен из вируса гриппа,
в) инкубацию растения или части растения в условиях, которые позволяют экспрессию первой и второй нуклеиновых кислот, посредством чего продуцируется VLP, и
г) получение растительного материала, содержащего VLP, из растения или части растения, инкубированных на стадии (в).
41. Способ получения растительного материала, содержащего вирусоподобную частицу (VLP), включающий:
а) предоставление растения или части растения, где растение или часть растения содержит первую нуклеиновую кислоту, содержащую первую регуляторную область, являющуюся активной в растении и функционально связанную с нуклеотидной последовательностью, кодирующей белок гемагглютинин (НА) вируса гриппа, выбранный из группы, состоящей из В и Н3, и вторую нуклеиновую кислоту, содержащую вторую регуляторную область, являющуюся активной в растении и функционально связанную с нуклеотидной последовательностью, кодирующей белок протонного канала, причем белок протонного канала получен из вируса гриппа; и
б) инкубацию растения или части растения в условиях, которые позволяют экспрессию первой и второй нуклеиновых кислот, тем самым продуцируя VLP; и
в) получение растительного материала, содержащего VLP, из растения или части растения, инкубированного на стадии б).
| WO 2009009876 A1, 22.01.2009 | |||
| SONG J | |||
| et al., Influenza Virus-Like Particles Containing M2 Induce Broadly Cross Protective Immunity, PLOS ONE, vol.6, no.1, 1 January 2011, pp | |||
| Масло водоотделитель для пара и уловитель сальниковой набивки | 1927 |
|
SU14538A1 |
| NEMCHINOV et al., Transient expression of the ectodomain of matrix protein 2 (M2e) of avian influenza A virus in plants, PROTEIN EXPRESSION AND PURIFICATION, 2007, vol | |||
| Приспособление для разматывания лент с семенами при укладке их в почву | 1922 |
|
SU56A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Паровозный золотник (байпас) | 1921 |
|
SU153A1 |
| ВАКЦИНА ПРОТИВ ВИРУСА ГРИППА И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2000 |
|
RU2164148C1 |

