Предлагаемое изобретение относится к способам обнаружения генетических факторов (генов), определяющих функционирование ДНК-структур клетки - теломер, обуславливающих, в свою очередь, продолжительность жизни клетки эукариот [эукариоты: Биологический энциклопедический словарь / под ред. М.С. Гилярова. - М: «Советская энциклопедия». - 1986. - 893 с. (с. 743)]. Изобретение может быть использовано в доклинических исследованиях и в медицинской практике в качестве метода идентификации генов, участвующих в регуляции функционирования теломер. Обеспечивает возможность повышения качества жизни человека за счет обеспечения возможности прогнозирования развития болезней связанных с процессом старения.
Далее заявителем представлены термины, использованные в заявленном техническом решении, для обеспечения однозначности их понимания в описании заявленных материалов.
Эукариоты - организмы, клетки которых содержат оформленные ядра (ядерные). К эукариотам относятся все высшие животные и растения, а также одноклеточные и многоклеточные водоросли, грибы и простейшие. Ядерная ДНК у эукариотов заключена в хромосомах, обычно не кольцевая, соединена с белками гистонами и, как правило, образует серию клубочков вокруг октомеров гистонов - нуклеосом. Эукариоты обладают ограниченными мембраной клеточными органоидами (иногда с собственной ДНК) - хлоропластами, митохондриями и др. В систематике эукариот выделяют надцарство Eucaryota и противопоставляют прокариотам [Биологический энциклопедический словарь/ под ред. М.С. Гилярова. - М.: «Советская энциклопедия». - 1986. - 893 с. (с. 743)].
ПЦР-анализ - анализ на основе полимеразной цепной реакции (ПЦР) - способ получения in vitro большого числа копий специфических нуклеотидных последовательностей. Их количественное увеличение во временном интервале - иногда в миллионы раз - осуществляется в ходе трехэтапного циклического процесса. Для проведения ПЦР необходимы:
1) два синтетических олигонуклеотидных праймера (длиной примерно по 20 нуклеотидов), комплементарные участкам ДНК из противоположных цепей, фланкирующим последовательность-мишень, их 3'-гидроксильные концы после отжига с ДНК должны быть ориентированы навстречу друг другу;
2) ДНК-мишень длиной от 100 до 35000 п.н.;
3) термостабильная ДНК-полимераза, которая не теряет своей активности при температуре 95° и выше;
4) четыре дезоксирибонуклеотида [Глик Б., Пастернак Дж. Молекулярная биотехнология. Принципы и применение / Пер. с англ. - М.: Мир, 2002. - 589 с.]. Использование методики ПЦР позволяет амплифицировать (размножать) ДНК или ее фрагмент in vitro, увеличивая количество копий в миллионы раз за несколько часов. Таким образом, в итоге получают достаточное количество ДНК образца для его последующего анализа [Заяц Р.Г., Бутвиловский В.Э., Давыдов В.В., Рачковская И.В.. 7.2.2 Анализ фрагментов ДНК // Медицинская биология и общая генетика: учебник. - 2-е изд., испр. - Минск: Вышэйная школа, 2012. - С. 100. - 496 с. - ISBN 978-985-06-2182-5].
Саузерн-блоттинг, Саузерн-блот-гибридизация, Саузерн-блот-анализ (англ. Southern blotting, Southern blot hybridization, Southern blot analysis, по имени E. Саузерна; англ. blotting - промокание, получение реплики пятен) - метод идентификации участков ДНК, содержащих комплементарные ДНК-зонду нуклеотидные последовательности, среди смеси рестрикционных фрагментов ДНК, электрофоретически разделенных по размерам в геле и фиксированных на твердом носителе (обычно нитроцеллюлозные или нейлоновые фильтры) [Заяц Р.Г., Бутвиловский В.Э., Давыдов В.В., Рачковская И.В. Анализ фрагментов ДНК. Медицинская биология и общая генетика: учебник. - 2-е изд., испр. - Минск: Вышэйная школа, 2012. - С. 100. - 496 с. - ISBN 978-985-06-2182-5].
Инбредные линии - группа особей, возникшая вследствие неоднократного скрещивания потомков с родителями или братьев с сестрами, благодаря чему она становится в значительной степени гомозиготной (Термины и определения, используемые в селекции, генетике и воспроизводстве сельскохозяйственных животных. - М.: ВНИИплем. И.М. Дунин. 1996).
Инбридинг - скрещивание близкородственное особей в одной популяции в нескольких поколениях (более 20 поколений) [Beck J.A. Genealogies of mouse inbred strains / Beck J.A., Lloyd S., Hafezparast M., Lennon-Pierce M., Eppig J.T., Festing M.F., Fisher E.M. // Nature Genetics. - 2000. - V. 24 (1). - P. 23-5. doi:10.1038/71641. PMID 10615122].
Локус (лат.locus - место) - в генетике: местоположение определенного гена на генетической или цитологической карте хромосомы. Вариант последовательности ДНК в данном локусе называется аллелью. Упорядоченный перечень локусов для какого-либо генома называется генетической картой [Биологический энциклопедический словарь / Гл. ред. М.С. Гиляров. - М.: Сов. энциклопедия, 1986. - 831 с].
Гомологичные гены (homologous genes, греч. homologos - соответственный, подобный; греч. genos - род, происхождение) - гены, имеющие сходную первичную структуру (последовательность нуклеотидов), общее происхождение и контролирующие один и тот же признак (Толковый биотехнологический словарь. Русско-английский. / Гл. ред. Тарантул В.З. - М. Языки славянских культур, 2009. - 936 с).
Гомеостаз - (от греч. homoios тот же самый, похожий и греч. stasis неподвижность, стояние), гомеостазис, способность организма или системы организмов поддерживать устойчивое (динамическое) равновесие в изменяющихся условиях среды (Экологический словарь URL: https://dic.academic.ru/contents.nsf/ecolog/).
Полиморфизм - наличие в пределах одного вида резко отличных признаков, не имеющих переходных форм [Майр Э., Зоологический вид и эволюция, пер. с англ., М., 1968; Шеппард Ф.М., Естественный отбор и наследственность, пер. с англ., М., 1970].
Пролиферация - увеличение количества клеток в многоклеточном организме (Гистология. Учебник. 2-е изд., перераб. и доп. / Под. ред. Э.Г. Улумбекова, Ю.А. Челышева. - М.: ГЭОТАР-МЕД, 2002. - 672 с.: ил. - Серия «XXI век»).
ДНК - дезоксирибонуклеиновая кислота; нуклеиновая кислота, которая является основным компонентом хромосом клеток эукариот и некоторых вирусов (Научно-технический энциклопедический словарь URL: http://dic.academic.ru/contents.nsf/ntes/);
Секвенирование (биополимеров - белков и нуклеиновых кислот - ДНК и РНК) - определение их аминокислотной или нуклеотидной последовательности (от лат. sequentum - последовательность). В результате секвенирования получают формальное описание первичной структуры линейной макромолекулы в виде последовательности мономеров в текстовом виде [Глик, Пастернак, 2005].
Праймер - олигонуклеотид из 17-24 (и более) звеньев (нуклеотидных остатков), используемый при секвенировании ДНК и при проведении ПЦР [Глик, Пастернак, 2005].
QTL-анализ (картирование локусов количественных признаков) -статистический метод, который связывает два типа информации - фенотипические данные (измерения признака) и генотипические данные (обычно молекулярные маркеры) - для объяснения генетической основы изменчивости сложных признаков, контролируемых несколькими генами [Л.И. Сергеева. QTL-анализ и его применение в физиологических исследованиях / Молекулярно-генетические и биохимические методы в современной биологии растений // Кузнецов В.В., Ралдугина Г.Н. - Издательство: Бином. Лаборатория знаний, 2011. - 481 с.]
Экотип - совокупность экологически близких популяций вида, связанных с определенным типом мест обитания и обладающих генетически детерминированными анатомо-морфологическими и физиологическими особенностями, выработавшимися в результате продолжительного воздействия сходных режимов экологических факторов [Begon, М. From individuals to ecosystems / M. Begon, С. Townsend, J.L. Harper // Blackwell Publishing, 4th ed. - 2006. - 577 p.].
Ортологичные гены, ортологи - это гомологичные гены в геномах разных организмов, являющиеся результатом эволюции одного и того же гена, который присутствовал в геноме общего предка этих организмов (вертикальное наследование) [Эволюция геномов эукариот и принцип максимальной парсимонии / И.Б. Рогозин, Ю.И. Вульф, В.Н. Бабенко, Е.В. Кунин // Вестник Вогис. - 2005. - Т. 9, №2. - С. 141-152].
Гомология генов - общее происхождение генов [Кунин Е. Логика случая. О природе и происхождении биологической эволюции / The Logic of Chance. The Nature and Origin of Biological Evolution // E. Кунин - пер. с англ. - Изд. Центрполиграф, 2014. - 127 с.].
Хромосомы - органоиды клеточного ядра, являющиеся носителями генов и определяющие наследственные свойства клеток и организмов [Биологический энциклопедический словарь / под ред. М.С. Гилярова. - М.: «Советская энциклопедия». - 1986. - 893 с. (с. 743)].
Далее заявителем приведены сокращения, используемые в заявленном техническом решении:
п. о. - пар оснований (пары оснований): здесь: оснований аденина, тимина, цитозина и гуанина в составе нуклеиновых кислот;
т.п.н. - тысячи пар нуклеотидов (в составе нуклеиновой кислоты);
А - аденин;
Т - тимин;
Ц - цитозин;
Г - гуанин.
Теломеры - структуры на концах хромосом эукариотических клеток, характеризующиеся тем, что мало подвергаются эволюционным изменениям, при этом имеют структурную и функциональную гомологию у всех эукариот живых организмов. Теломеры представляют собой комплекс многократно повторяющихся последовательностей нуклеотидов (то есть теломерных повторов) и связанных с ними специфических белков, поддерживающих их (теломер) стабильность [Jiang, Н.Н. Telomere shortening and ageing (Укорочение теломер и старение) / Jiang, Z. Ju K.L. Rudolph // Gerontol Geriat. - 2007. - V.40. - P. 314-324].
Теломеры - важная для жизнедеятельности клетки структура, участвующая в поддержании целостности генома клетки (совокупности всей ДНК клетки) и регуляции продолжительности жизни клетки. С каждым последующим делением клетки длина теломер сокращается, что приводит к клеточному старению или смерти. При укорочении длины теломер до пороговой величины клетки прекращают деление и переходят к стадии старения [Мелихова, Л. Основные теории старения / Л. Мелихова, Л. Чентиева, М. Лущик // Международный студенческий научный вестник. - 2016. - I. 4-2. - С. 156-157]. В соответствии с теорией биологического старения, известной как "молекулярные часы" [Hayflick, L. The serial cultivation of human diploid cell strains (Периодическое культивирование штаммов диплоидных клеток человека) / L. Hayflick, P.S. Moorhead // Exp Cell Res. - 1961. - V. 25. - P. 585-621], строго определенное количество теломерных повторов, полученное при рождении, изначально предопределяет способность большинства соматических клеток живых организмов к пролиферации (увеличению количества клеток в многоклеточном организме) [Гистология. Учебник. 2-е изд., перераб. и доп. / Под. ред. Э.Г. Улумбекова, Ю.А. Челышева. - М.: ГЭОТАР-МЕД, 2002. - 672 с.: ил. - (Серия «XXI век»). Таким образом, укорочение теломер в клетках, тканях и организме в целом свидетельствует о старении.
У животных в состав теломер входит нуклеотидная последовательность TTAGGG (далее нуклеотиды обозначены сокращенно: Т - тимин, А - аденин, G - гуанин, С -цитозин), отличающаяся от последовательности растений TTTAGGG на один нуклеотид (Т), что является доказательством возможности экстраполяции данных, полученных при анализе растений, на других эукариот, в том числе и человека, при этом не будет излишним констатировать факт того, что у некоторых растений, например, представителей классов Asparagales и Aloe нуклеотидная последовательность теломер полностью совпадает с человеческой [Weiss, Н. Aloe spp. plants with vertebrate-like telomeric sequences (Растения видов Aloe с теломераными последовательностями, характерными для позвоночных животных) / Н. Weiss, Н. Scherthan // Chromosome Research. - 2002. - V. 10. - I. 2. - P. 155-164].
Из исследованного уровня техники заявителем установлено, что длина теломер может отличаться у разных особей эукориот одного вида, но варьирует в строго ограниченных пределах [Shakirov, E.V. Length Regulation and Dynamics of Individual Telomere Tracts in Wild-Type Arabidopsis (Регуляция длины и динамика индивидуальных теломерных последовательностей у дикого типа Arabidopsis) / E.V. Shakirov, D.E. Shippen // The Plant Cell. - 2004. - V. 16. - I. 8. - P. 1959-1967]. В то время как каждый биологический вид характеризуется строго контролируемым диапазоном длины теломерной ДНК, средняя длина теломер у разных индивидуумов при рождении может варьировать внутри этого диапазона (7000 пар нуклеотидов у одного человека и 14000 пар нуклеотидов - у другого) [Mirabello, L. The association of telomere length and genetic variation in telomere biology genes (Связь длины теломер и генетической вариативностью в генах биологии теломер) / L. Mirabello, K. Yu, Р Kraft, I. De Vivo, D.J. Hunter, J. Prescott, J.Y. Wong, N. Chatterjee, R.B. Hayes, S.A. Savage // Mutat. - 2010. - V. 31. - I. 9. - P. 1050-1058]. Доказательством изложенного также является и то, что, например, в популяциях дрожжей и растений [Heacock, М. Molecular analysis of telomere fusions in Arabidopsis: multiple pathways for chromosome end-joining (Молекулярный анализ слипания теломер у Arabidopsis: множественные пути объединения концов хромосом) / М. Heacock, Е. Spangler, K. Riha, Puizina J., Shippen D.E. // Embo j. - 2004. - V. 23. - I. 11. - P. 2304-2313; Higashiyama, T. Telomere and LINE-like elements at the termini of the Chlorella chromosome I (Теломеры и LINE-подобные элементы на концах хромосомы I у хлореллы) / Т. Higashiyama, Y. Noutoshi, М. Akiba, Т. Yamada // Nucleic acids symposium series - 1994. - P. 71-72], а также среди людей [Greider, C.W. Telomere length regulation (Регуляция длины теломер) / C.W. Greider // Annual review of biochemistry. - 1996. - V. 65. - I. 1. - P. 337-365], существуют значительные вариации в средней длине теломер от организма к организму.
Отклонение функционального состояния теломер от нормы, характерной для конкретного эукариотического организма, а именно их патологическое укорочение, является причиной нарушений функционирования эукариот, в том числе и в отношении человека. Мутации в генах, ответственных за поддержание теломерного гомеостаза, обуславливают ускоренную потерю теломерной ДНК, и, вследствие этого, приводят к тяжелым генетическим заболеваниям, таким как синдромы Гетчинсона-Гилфорда и Вернера (детская и взрослая прогерия), а также дискератоз (заболевание эпидермального слоя кожи), характеризующимся преждевременным старением и резким уменьшением продолжительности жизни [Gonzalo, S. DNA repair defects and genome instability in Hutchinson-Gilford progeria syndrome (Дефекты репарации ДНК и геномная нестабильность при синдроме прогерии Гетчинсона-Гилфорда) / S. Gonzalo, R. Kreienkamp // Curr. Opin. Cell Biol. - 34. - 2015. - P. 75-83].
Доказательством изложенного выше в отношении человека является уменьшение средней длины теломер в лейкоцитах, наблюдающееся, например, при онкологических и сердечно-сосудистых заболеваниях. В целом преждевременное сокращение длины теломер рассматривается как фактор раннего развития болезней, связанных со старением, которые, к сожалению, не всегда могут быть своевременно идентифицированы на дату представления заявочных материалов.
Изучение генетического разнообразия, лежащего в основе полиморфизма длины теломер, расширяет понимание молекулярных механизмов, определяющих разницу в скорости старения между индивидуумами, а также помогает объяснить различия в генетической предрасположенности к болезням, ассоциированным с процессами старения.
Проведение такого рода исследований на человеке или животных встречается с большим количеством препятствий, что связано с множеством сложностей [Butler М. Animal Cell Culture and Technology / М / Butler // 2nd edition. - Garland Science / BIOS Scientific Publishers, 2004], а именно:
- дороговизна сред культивирования клеток человека и животных в лабораторных условиях;
- получение материала (тканей и клеток человека и животных), использование лабораторных животных требует строгой документации, составления этических деклараций;
- высокий уровень вирусной и бактериальной контаминации культур клеток человека и животных;
- недостатком использования культур клеток животных и человека является то, что в процессе культивирования и роста клетки могут менять свои цитологические и биохимические характеристики и существенно отличаться от клеток, полученных непосредственно от организма-донора, вследствие чего может изменяться экспрессия генов и белков, активность ферментов;
- нежизнеспособность большинства особей инбредных популяций (популяций, полученных путем близкородственного скрещивания) относится к наиболее важным факторам, ограничивающим исследования в данной области.
В заявленном техническом решении заявителем предлагается наиболее оптимальный выход из сложившейся ситуации: краткая идея, заложенная заявителем в заявленное техническое решение, может быть выражена в поиске генетических маркеров регуляции длины теломер у инбредных популяций растений, на базе которых (идентифицированных генетических маркеров) выполняется определение гомологичных генов у человека, что и позволяет в целом избежать сложностей, связанных с использованием животных и человека.
При этом заявленное техническое решение более конкретно основывается на определении генетических маркеров укорочения теломер в клетках растений Arabidopsis thaliana (резуховидка Таля, далее по тексту A. thaliana). являющихся одним из наиболее предпочтительных модельных растительных организмов в связи с простотой культивирования и подробным описанием генома [Alonso, J.M. Moving forward in reverse: genetic technologies to enable genome-wide phenomic screens in Arabidopsis / Движение вперед в обратном направлении: генетические технологии для полногеномных исследований фенотипов / J.M. Alonso, J.R. Ecker // Nat Rev Genet. - 2006. - V. 7. - I. 7. - P. 524-536]. Основными достоинствами использования A. thaliana является:
- Диплоидный геном арабидопсиса один из самых маленьких среди эукариот, содержит пять хромосом и состоит из 125-150 млн. п.о.
- В геноме закодировано менее 30 тыс. белков.
- A. thaliana - первый растительный организм, чей геном был полностью секвенирован, что делает данное растение удобным для генетических исследований и манипуляций с геномом.
- Арабидопсис имеет очень короткий жизненный цикл, позволяющий быстро получать растения разных стадий развития и новые поколения растений.
- A. thaliana - самоопыляемое растение, что облегчает работу по выявлению гомо- и гетерозиготных организмов и является важным при трансформации растений.
- Кроме того, для A. thaliana характерна высокая гомология последовательностей генов с последовательностями генов человека [Jones A.M. The Impact of Arabidopsis on Human Health: Diversifying Our Portfolio / A.M. Jones, J. Chory, J.L. Dangl, M. Estelle, S.E. Jacobsen, E.M. Meyerowitz, M. Nordborg, D. Weigel // Cell. - 2008. - V. 133(6). - 939-943. doi:10.1016/j.cell.2008.05.040; Weigel, D. The 1001 Genomes Project for A. thaliana / 1001 геномный проект для A. thaliana / D. Weigel, R. Mott // Genome Biology - 2009. - V. 10. - Р. 107-112].
При этом изложенная выше информация также нашла подтверждения в источнике [Shakirov, E.V. Length Regulation and Dynamics of Individual Telomere Tracts in Wild-Type Arabidopsis (Регуляция длины и динамика индивидуальных теломерных последовательностей у дикого типа Arabidopsis) / E.V. Shakirov, D.E. Shippen // The Plant Cell. - 2004. - V. 16. - I. 8. - P. 1959-1967].
Из исследованного уровня техники заявителем выявлено техническое решение - прогностический способ по заявке на группу изобретений (№ WO 2013024264, «Prognostic methodology» / «Прогностический метод»), по которой выданы патенты-аналоги, AU 2012296717 (A1) AU 2012296717 (В2) BR 112014003247 (А2) СА 2845047 (А1) CN 103748237 (A) DK 2744913 (Т3) ЕР 2744913 (A1) ЕР 2744913 (B1) ES 2556606 (Т3) JP 2014524251 (A) JP 2017176188 (A) JP 6178313 (В2) NZ 620854 (А) РТ 2744913 (Е) RU 2014103536 (A) US 2015292021 (А1).
Сущностью известной группы изобретений по одному из патентов-аналогов СА №2845047 в отношении заявленного способа, представленном в первом независимом пункте формулы изобретения, является прогностический метод определения прогрессирования заболевания, характеризующихся укорочением теломер, включающий: анализ длины теломер с высоким разрешением для определения наибольшего среднего значения длины теломер, при котором обнаруживается слипание концов теломер в образцах тканей, полученных от индивидуумов с одинаковым заболеванием (а именно: старение, болезнь Альцгеймера; инфаркт головного мозга; болезнь сердца; хроническая ВИЧ-инфекция; хронический гепатит; кожные заболевания; хроническое воспалительное заболевание кишечника; язвенный колит; анемия; атеросклероз; синдром Барретта и рак, включая предраковые состояния), для дальнейшего определения порогового значения средней длины теломер, при котором теломеры становятся нефункциональными и способными к слиянию, определение прогностической средней длины теломер в образах тканей от индивидуумов с одинаковым заболеванием путем учета средних значений длин теломер, меньших, чем установленная пороговая длина, и усреднением средней длины теломер в этих образцах, определение среднего тестового значения длины теломер в образцах пациентов, у которых предполагается вышеупомянутое заболевание и в которых вышеопределенное тестовое значение длины теломер меньше, чем указанное в п. 1 прогностическое значение средней длины теломер, из чего следует, что первое введение препарата недостаточно и/или ответ на препарат слабый и/или выживаемость в целом снижена, определение средней тестовой длины теломер в образцах, полученных от пациентов с предположительным заболеванием (п. 1), когда вышеуказанное среднее тестовое значение длины теломер выше, чем указанное прогностическое значение длины теломер, из чего следует, что первая обработка препаратом действенна и/или ответ на препарат положительный и/или выживаемость в целом повышена.
Таким образом, известный метод в целом сводится к тому, что предназначен для анализа слияния теломер и основывается на секвенировании ДНК. При этом прогностическая длина теломер в образцах тканей пациентов с вышеописанными заболеваниями определяется путем отбора образцов со слившимися теломерами и усреднением среднего значения длины в этих образцах.
В результате реализации изобретения обеспечивается возможность диагностики и выполнение лечения пациента.
Основываясь на этом, возможно сделать вывод о его недостатках, а именно -недостатками известного технического решения СА №2845047 в отношении технологии являются:
1) использование образцов тканей непосредственно пациентов, что усложняет процесс получения необходимого количества материала для проведения анализа;
2) проведение измерения длины теломер в образцах тканей без проведения поиска генов, связанных с укорочением и/или удлинением теломер.
Недостатком известного технического решения в целом является невозможность его использования для выявления генетических маркеров изменения теломер в общей популяции эукариот, в том числе и у человека, высокая трудоемкость реализации метода, так как в изобретении ведут работу с образцами тканей пациентов.
Существенный отличием заявленного технического решения от известного технического решения СА №2845047 является то, что известное техническое решение предназначено для диагностики и анализа прогрессирования заболевания у заболевших пациентов с уже имеющимся диагностированным заболеванием, в то время как в заявленном техническом решении проводится профилактическая диагностика человека на возможность наличия или отсутствия риска развития заболеваний, связанных со старением организма. Таким образом, по мнению заявителя, представленное техническое решение характеризуется тем, что позволяет решить проблему предупреждения тяжелых и даже неизлечимых заболеваний у всех слоев населения без каких-либо сложностей, что, в конечном счете, обеспечивает улучшение качества жизни человека.
Из исследованного заявителем уровня техники заявителем выявлен способ определения маркера развития ревматоидного артрита на основе выявления укорочения относительной длины теломер на отдельных хромосомах в Т-лимфоцитах периферической крови по патенту на изобретение РФ №2522961 «Способ определения маркера развития ревматоидного артрита на основе выявления укорочения относительной длины теломер на отдельных хромосомах в Т-лимфоцитах периферической крови». Изобретение касается определения относительной длины теломер на хромосомах. Сущностью является способ определения маркера развития ревматоидного артрита, отличающийся тем, что с помощью метода количественной флуоресцентной гибридизации in situ (Q-FISH) выявляют укорочение относительной длины теломер на отдельных хромосомах Т-лимфоцитов периферической крови пациентов; результативный показатель длины теломер вычисляют как отношение среднего значения интенсивности флюоресценции теломер определенной хромосомы (для каждого из плеч отдельно) к среднему значению интенсивности флюоресценции теломер всех хромосом в метафазе и укорочение относительной длины теломер на коротком плече 4-й хромосомы у пациентов с ревматоидным артритом в сравнении с соответствующими им по возрасту донорами расценивают как маркер развития заболевания.
Таким образом, способ в отношении выполнения заявленных действий заключается:
1) в выявлении укорочения относительной длины теломер на отдельных хромосомах Т-лимфоцитов периферической крови с помощью метода количественной флуоресцентной гибридизации in situ (Q-FISH);
2) в вычислении результативного показателя длины теломер как отношения среднего значения интенсивности флюоресценции теломер определенной хромосомы (для каждого из плеч отдельно) к среднему значению интенсивности флюоресценции теломер всех хромосом в метафазе;
3) в укорочение относительной длины теломер на коротком плече 4 хромосомы у пациентов с ревматоидным артритом в сравнении с донорами расценивают как маркер развития заболевания.
4) в качестве образцов для исследования используют биологический материал.
Известное изобретение позволяет определить риск развития ревматоидного артрита.
Недостатками известного способа является:
- ограничение возможности использования изобретения применительно только к одному заболеванию ревматоидному артриту;
- ограниченность возможности применения изобретения по отношению к одной группе больных, что не обеспечивает возможность прогнозирования развития болезней у более широкого спектра групп населения;
- низкая доступность технологии в силу использования Q-FISH анализа (метода количественной флуоресцентной гибридизации in situ), требующего дорогостоящих реактивов, оборудования и квалифицированного персонала.
Из исследованного уровня техники заявителем выявлен способ диагностики заболеваний или нарушений связанных с теломерами, выбранный заявителем в качестве прототипа, по заявке на изобретение № WO 2008119027 (Генетический тест генов, ассоциированных с теломерами, при врожденном дискератозе, дефектах кроветворения, идиопатическом фиброзе легких, криптогенном циррозе печени), включающий:
1) получение образца ткани от анализируемого объекта; анализ всей последовательности ДНК или ее части по отношению к одному или нескольким генам, регулирующим теломеры на наличие одного или более различий, где наличие по меньшей мере одного различия является индикатором риска наличия заболеваний или нарушений, связанных с теломерами;
2) для определения различий используют гены hTERT (теломеразная обратная транскриптаза человека) и hTR (РНК теломеразы человека); используют метод ПЦР; используют метод гибридизации с применением одного или боде праймеров;
3) определение средней длины теломер в образцах тканей по сравнению с контрольным образцом, где самые короткая длина теломер является индикатором риска наличия заболевания (суррогатным маркером), связанного с теломерами; определение длины теломер проводится с использованием метода рестрикционного анализа терминальных фрагментов (TRF-анализ), флуоресцентная гибридизация in situ (FISH-анализ), проточная цитометрия, количественная ПЦР; в качестве образцов для анализа длины теломер используют лейкоциты; при различие длины теломер предполагают изменение в hTERT (теломеразная обратная транскриптаза человека) и hTR (РНК теломеразы человека);
4) в качестве возможных заболеваний рассматриваются врожденный дискератоз, дефекты кроветворения, идиопатический фиброз легких, криптогенный цирроз печени.
Недостатками прототипа являются:
- использование в качестве генов, анализируемых в качестве маркеров заболеваний или нарушений, связанных с изменением теломер, только двух уже известных ранее генов hTERT (теломеразная обратная транскриптаза человека) и hTR (РНК теломеразы человека), регулирующих длину теломер;
- проведение определения длины теломер непосредственно в образцах тканей человека, тогда как в заявленном техническом решении измерение длины теломер проводится только на этапе поиска генетических маркеров, что позволяет в дальнейшем не проводить длительную и сложную процедуру измерения длины теломер в анализируемых клинических образцах;
- ограниченный круг заболеваний, для которого применим известный способ (врожденный дискератоз, дефекты кроветворения, идиопатический фиброз легких, криптогенный цирроз печени).
Недостатком известного технического решения в целом является невозможность его применения для выявления генетических маркеров изменения теломер в общей популяции эукариот, в том числе у человека, высокая трудоемкость реализации метода, так как в изобретении ведут работу с материалом пациентов.
Таким образом, выбранный заявителем в качестве прототипа из исследованного заявителем уровня техники способ диагностики заболеваний или нарушений связанных с теломерами, по заявке на изобретение № WO 2008119027, совпадает с заявленным техническим решением исключительно в отношении одного признака, а именно проведения ПЦР анализа, который является общеизвестным и не является признаком, характеризующим сущность заявленного технического решения. Исходя из этого, заявителем составлена формула изобретения без ограничительной части.
Сущностью заявленного технического решения является способ идентификации генетических маркеров регуляции длины теломер заключающийся в том, что выполняют подготовительный этап для идентификации генетических маркеров на основе ДНК растений Arabidopsis thaliana, для чего выделяют ДНК растения Arabidopsis thaliana, проводят рестрикционный анализ длины теломер совместно с Саузерн-блот анализом, выполняют биоинформационный анализ полученных результатов длин теломер у растений инбредных линий Arabidopsis thaliana, картируют локусы количественных признаков, определяют гены, участвующие в изменении длины теломер в растениях, проводят биоинформационный поиск гомологичных генов у человека, проводят анализ влияния исследуемых генов на длину теломер на основе мутантов растений, идентифицируют гены, участвующие в регуляции длины теломер (генетические маркеры), выполняют идентификацию генетических маркеров в других организмах помимо растений A. thaliana путем выделения ДНК из образца ткани организма, для которого требуется провести поиск генетических маркеров, проводят ПЦР-анализ на наличие генетических маркеров, проводят электрофоретический анализ полученной в ПЦР-анализе реакционной смеси для обеспечения возможности выявления или не выявления искомых фрагментов ДНК, соответствующих нуклеотидным последовательностям искомых генетических маркеров.
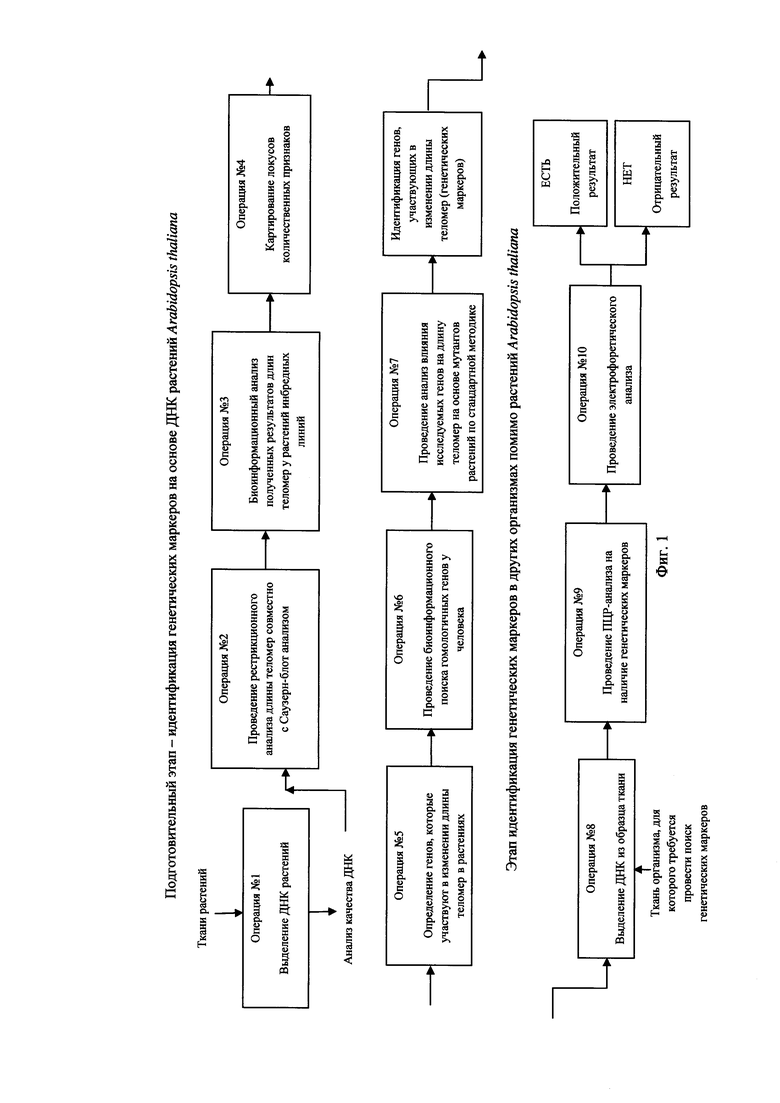
Заявленное техническое решение иллюстрируется Фиг. 1 - Фиг. 6.
На Фиг. 1 представлена принципиальная пооперационная блок-схема выполнения действий, составляющих заявленное техническое решение.
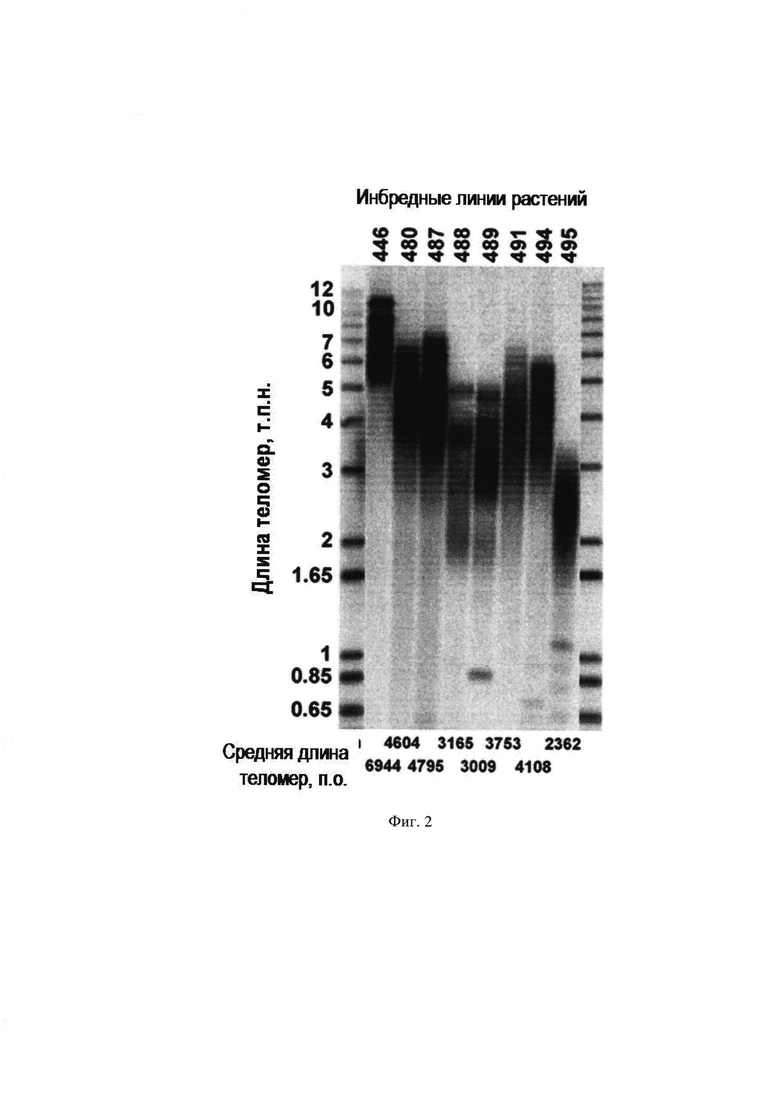
На Фиг. 2 представлена визуализация теломер с помощью метода Саузерн-блотинг в инбредных линиях растений A. thaliana. По горизонтальной оси расположены образцы ДНК растений, по вертикальной оси - размер теломерной ДНК в т.п.н.
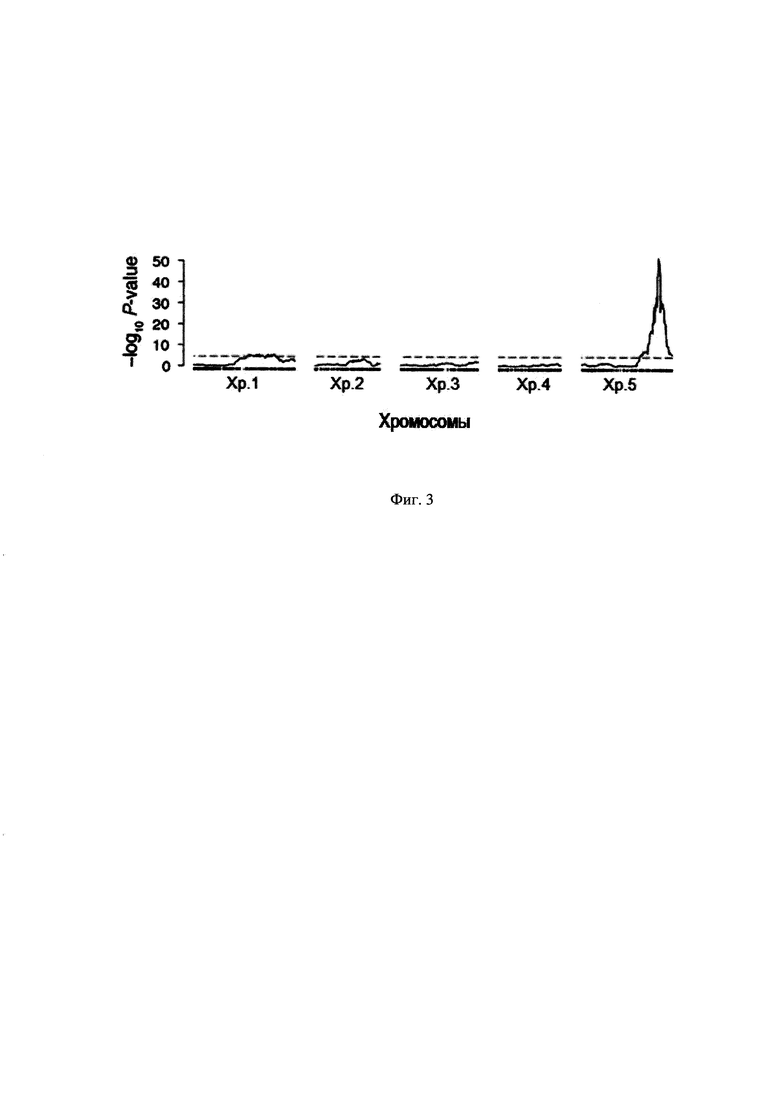
На Фиг. 3 представлено картирование локусов количественных признаков средней длины теломер в популяции рекомбинантных инбредных линий A. thaliana. Высокий пик на хромосоме 5 - QTL основного эффекта. Значительно менее значимый пик присутствует на хромосоме 1. Пунктиром показана линия статистической погрешности. Хр. 1 - Хр. 5 - хромосомы A. thaliana. На графике по оси абсцисс располагаются 1-5 хромосомы, по оси ординат - величина показания количества генов-кандидатов для каждой хромосомы по отношению к геному растения в целом.
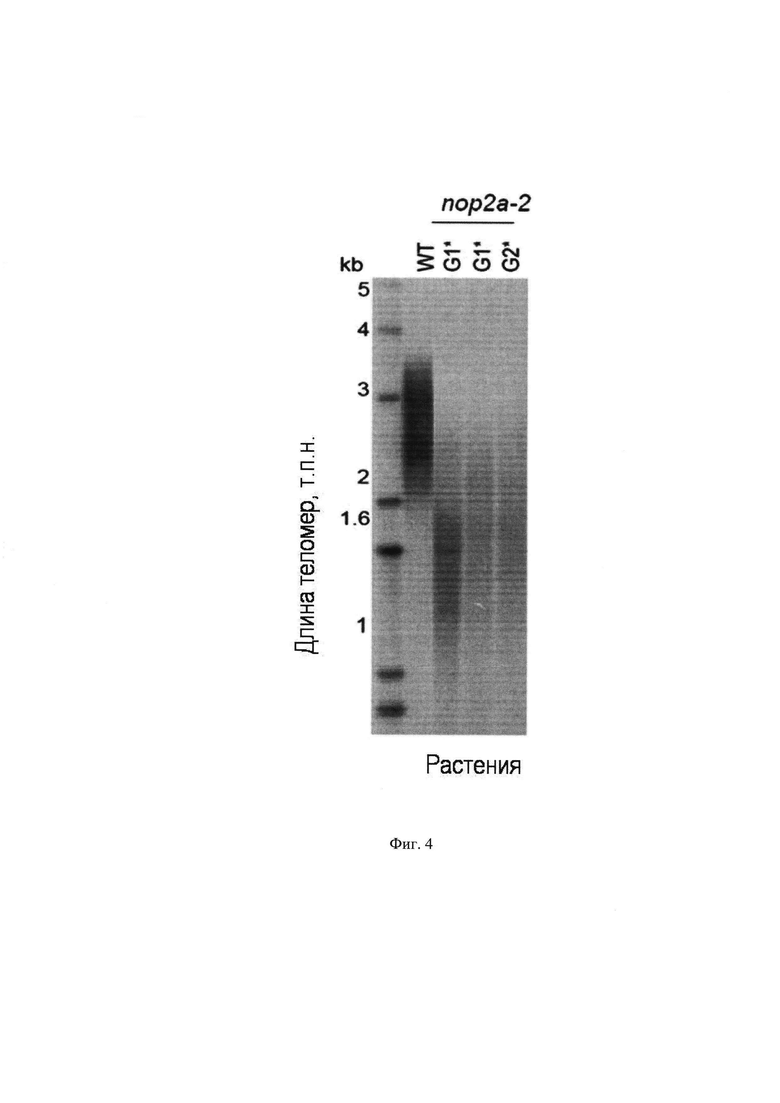
На Фиг. 4 представлена визуализация теломер с помощью метода Саузерн-блотинг в мутантных линиях растений A. thaliana, из генома которых удален ген OLI2. Со1-0 - растения дикого экотипа A. thaliana, контроль; nop2a-2 - мутантное растение без гена OLI2. По горизонтальной оси расположены образцы ДНК растений, по вертикальной оси - размер теломерной ДНК в т.п.н.
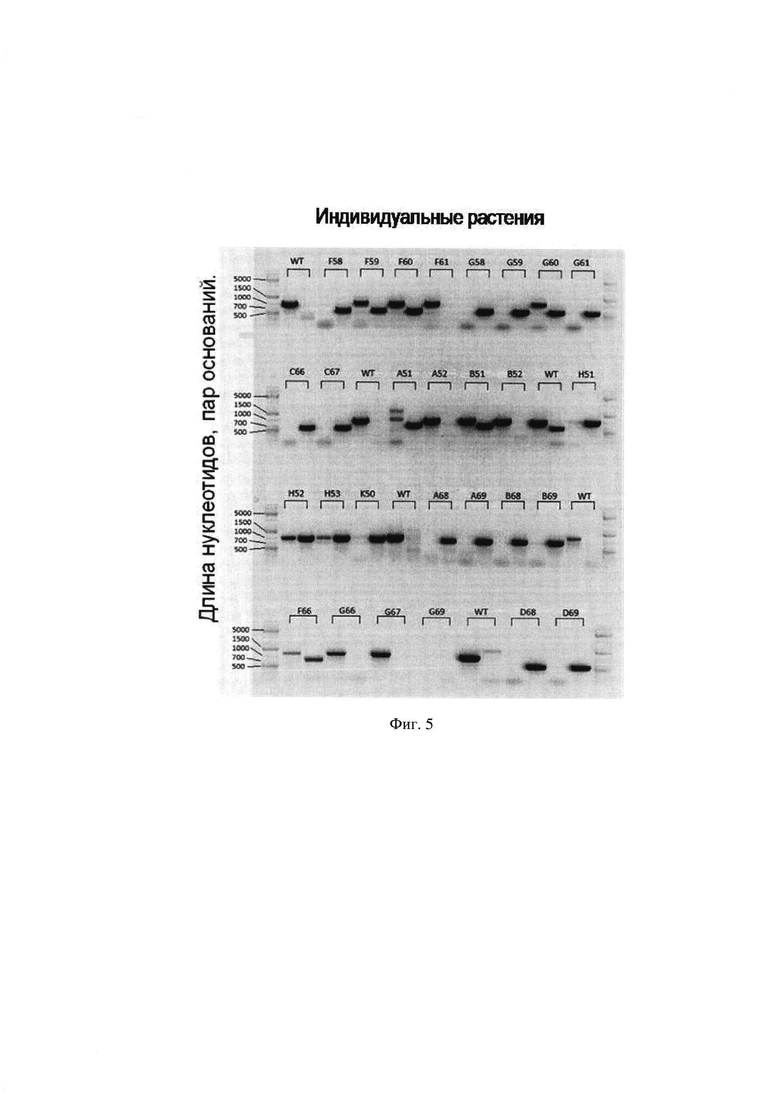
На Фиг. 5 представлены результаты ПЦР-анализа для анализа растений по одному из генов-кандидатов. Над скобкой указан порядковый номер растения. Каждое растение генотипировали с помощью двух комбинаций праймеров. По оси абсцисс расположены исследуемые на наличие генетического маркера образцы ДНК, на оси ординат обозначен размер фрагментов ДНК в п. о.
На Фиг. 6 представлена визуализация генетической информации по последовательностям генов, выраженная в виде буквенного кода из остатков нуклеотидов А, Т, Ц и Г (аденин, тимин, цитозин, гуанин, соответственно).
Цель заявленного технического решения - разработка способа определения генетических маркеров изменения длины теломер, с использованием методов множественного инбридинга, Саузерн-блота и картирования локусов количественных признаков потомков, и с применением в качестве образцов ДНК растений A. thaliana.
Заявленный способ позволяет реализовать следующие задачи:
- Упростить процесс измерения длины теломер путем применения разработанных генетических и молекулярных методов (Саузерн-блот анализ).
- Снизить трудоемкость процесса идентификации генетических маркеров изменения теломер, что обуславливает заболевания, связанные со старением (ПЦР-анализ).
- Исключить использование тканей человека на этапе поиска генетических маркеров.
- Идентифицировать новые, ранее неизвестные генетические маркеры изменения длины теломер у эукариот путем использования инбредных линий растений, чему способствует отсутствие в растительном объекте исследования ограничений в множественном близкородственном скрещивании.
Техническим результатом является выявление набора генов (генетических маркеров), участвующих в изменении длины теломер у эукариот, в частности, у человека, на основе генов инбредных линий растений A. thaliana.
В заявленном техническое решении использованы такие известные как таковые технологии (Саузерн-блот анализ, ПЦР анализ, QTL-анализ), при этом заявленное техническое решение характеризуется специфической особенностью, заключающейся в том, что, в отличие от выявленных на дату подачи из исследованного уровня техники способов, направленных на поиск генов, связанных с изменением теломер, в частности, человека, заявителем используется ДНК растений A. Thaliana. А именно, в качестве образцов ДНК для поиска генов (генетических маркеров) изменения длины теломер используется ДНК инбредных линий (популяций, полученных в результате множественного близкородственного скрещивания) растений A. thaliana (арабидопсис, резуховидка Таля), что, по мнению заявителя, является доказательством неочевидности заявленного технического решения. Таким образом, по мнению заявителя, можно констатировать, что указанная цель ранее другими исследователями не ставилась, так как до даты подачи заявочных материалов не выявлено технических решений, заключающихся в использовании имеющихся генетических баз данных (см. далее) для решения задач по идентификации генетических маркеров изменения длины теломер у эукариот посредством исследования ДНК растений A. thaliana вместо используемых на дату представления заявочных материалов преимущественно ДНК человека и/или животных.
Возможность выполнения поставленной задачи, по мнению заявителя, стала реализуемой в силу того, что информация по описанию геномов организмов стала общедоступной, в том числе и информация по геному арабидопсиса (Analysis of genome sequence of the flowering plant Arabidopsis thaliana // The Arabidopsis genome initiative. / Nature. - 2000. - V. 796-815. https://www.ncbi.nlm.nih.gov/, https://www.arabidopsis.org/, http://signal.salk.edu/cgi-bin/tdnaexpress). При этом, начиная с 2000-х годов, стало возможным создание инбредных линий растений арабидопсиса (https://www.arabidopsis.org/servlets/Order?state=catalog), что в совокупности обеспечило возможность решать таковые задачи в принципе, что базируется, по мнению заявителя, на принципиально новом подходе, заключающемся в использовании растений для поиска генов, связанных с изменением теломер у эукариот, в частности - у человека.
Использование инбредных линий растений арабидопсиса позволяет сократить время поиска генов. Каждая инбредная линия содержит набор участков ДНК родительских особей, что повышает вероятность точного определения тех генов, которые ответственны за изменение (укорочение) длины теломерной ДНК. В качестве источника множественного инбридинга (близкородственного скрещивания) используются природные экотипы растений A. thaliana (растения A. thaliana, происходящие из разных природных мест обитания).
Заявленное техническое решение представляет собой технологию определения генов, влияющих на длину теломер, а именно - определение новых, неизвестных ранее генетических маркеров регуляции длины теломер, их (теломер) укорочения и связанного с этим процессом старения эукариот, то есть старения любого живого организма, за исключением вирусов и бактерий, так как у последних теломеры отсутствуют.
Выбранный заявителем подход предполагает в будущем возможность создания персонального каталога генетических маркеров укорочения теломер, связанных с индивидуальными особенностями старения организма, в том числе и для каждого человека, что, по мнению заявителя, создает принципиально новую возможность улучшение качества жизни человека за счет своевременного выявления возможных негативных болезней, связанных со старением, и, как следствие, обеспечивает возможность предупреждения развития таких заболеваний.
В заявленном техническом решении обеспечена возможность использования генетических маркеров, обнаруженных посредством анализа генов растений A. thaliana для анализа образцов ДНК животных и человека, поскольку степень гомологии (степень сродства) между генами A. thaliana и других эукариот (в частности, человека) высока: установлено, что большая часть генов, для которых показано участие в развитии заболеваний человека, имеют ортологичные гены в геноме растений A. thaliana. Например, среди генов, связанных с развитием раковых заболеваний человека, 70% имеют ортологи в геноме A. thaliana [Jones A.M. The Impact of Arabidopsis on Human Health: Diversifying Our Portfolio / A.M. Jones, J. Chory, J.L. Dangl, M. Estelle, S.E. Jacobsen, E.M. Meyerowitz, M. Nordborg, D. Weigel // Cell. - 2008. - V. 133(6). - 939-943. doi:10.1016/j.cell.2008.05.040].
В рамках заявленного технического решения растения A. thaliana выращивают в почве в климатических камерах с заданными условиями, оптимальными для роста данного вида растений: температура 20°С, влажность воздуха 65%, световой период (16 ч с освещением / 8 ч без освещения), интенсивность освещения 1000 люкс.
Заявленный способ представляет собой последовательность выполнения операций, позволяющих достичь технического результата заявленного технического решения, а именно - обеспечивает возможность идентификации генетических маркеров регуляции длины теломер. Способ в целом состоит из двух базовых этапов - подготовительного, состоящего из операций 1-7 и последующего этапа, состоящего из операций 8-10 соответственно (см. Фиг. 1).
Для реализации заявленного технического решения заявителем на Фиг. 1 приведена принципиальная пооперационная блок-схема выполнения действий, составляющих сущность заявленного технического решения.
Далее заявителем представлено подробное пооперационное описание выполнения действий, составляющих сущность заявленного технического решения.
Операции с №1 по №7 составляют подготовительный этап заявленного технического решения, заключающийся в идентификации генетических маркеров на основе ДНК растений A. thaliana, которые представлены далее более детально.
Операция №1.
Берут индивидуальные растения инбредных линий Arabidopsis thaliana, выращенных из семян, полученных из банка семян SALK (https://www.arabidopsis.org/servlets/Order?state=catalog). Проводят выделение ДНК отдельно из тканей каждой из инбредных линий растений A. thaliana (стандартными методами, известными для выделения ДНК из растений в количестве, например, 480 инбредных линий растений).
После выделения ДНК проводят проверку ее качества на предмет отсутствия деградации, загрязнения посторонними соединениями. Качественная ДНК используется далее для выполнения следующей операции.
Операция №2.
Проводят рестрикционный анализ длины теломер совместно с Саузерн-блот анализом (Southern-blot analysis), который проводится по стандартной методике [Nigmatullina, L.R. Non-radioactive TRF assay modifications to improve telomeric DNA detection efficiency in plants / L.R. Nigmatullina, M.R Sharipova, E.V. Shakirov // BioNanoScience. - 2016. - doi: 10.1007/ s112668 -016 -0223-zu] с получением визуализированной информации длин ДНК в теломерах, получаемую в виде мембраны.
Операция №3.
Берут мембрану с визуализированной информацией длин ДНК теломер и выполняют ее (мембраны) фотоизображение, загружают в компьютер, проводят биоинформационный анализ полученных результатов (длин теломер, проявленных на мембране при Саузерн-блот анализе) исследуемых инбредных линий, с определением средних длин теломер (например, с использованием программы TeloTool (общеизвестная программа для обработки графических файлов и статистической обработки полученных данных).
Операция №4.
Берут данные по средней длине теломер, проводят картирование локусов количественных признаков (QTL-анализ, Quantitative trate locus analysis), то есть распределения генов, имеющих количественное влияние, по родительским хромосомам (например, с использованием программного приложения HAPPY R, http://mus.well.ox.ac.uk/magic/). с получением данных о месте размещения искомых (расположении) генов, связанных с изменением длины теломер, на хромосомах А. thaliana.
Операция №5.
Берут данные по генам, расположенным на выявленном в операции №4 участке хромосомы, проводят биоинформационный анализ, посредством использования общедоступных в сети Интернет баз данных https://www.ncbi.nlm.nih.gov/, https://www.arabidopsis.org/, http://signal.salk.edu/cgi-bin/tdnaexpress на персональном компьютере, с целью идентификации их (генов) функций с последующей идентификацией их роли в изменении длины теломер в растениях посредством использования баз данных в виде информации по последовательностям генов (выраженных в виде буквенного кода из остатков А, Т, Ц и Г, Фиг. 6) и их известным функциям.
Операция №6.
Берут информационные данные по выявленным на основании операции 5 генам, участвующим в изменении длины теломер в растениях, в виде их последовательностей в виде буквенного кода; проводят биоинформационный поиск гомологичных генов у человека с использованием генетических информационных баз данных (например, https://www.ncbi.nlm.nih.gov/, https://www.arabidopsis.org/, http://signal.salk.edu/cgi-bin/tdnaexpress) путем обнаружения и сравнения последовательностей найденных генов с генами в геноме человека с получением данных по степени гомологии выявленных генов растений и человека.
Операция №7.
Берут информационные данные по выявленным генам растений, имеющим высокую гомологию с генами человека, и проводят анализ влияния исследуемых генов на длинх теломер на основе мутантов растений по стандартной методике [The transcription factor AtGRF5 and the transcription coactivator AN3 regulate cell proliferation in leaf primordial of Arabidopsis thaliana (Транскрипционный фактор AtGRF5 и транскрипционный коактиватор AN3 регулируют пролиферацию клеток в меристеме листьев Arabidopsis thaliana) // G. Horiguchil, G.-T. Kim, H. Tsukaya / The Plant Journal. -2005. - V. 43. - P. 68-78]. В результате анализа длин теломер у мутантов по исследуемым генам получают вывод о том, какие из определенных генов участвуют в изменении длины теломер.
В результате проведенной последовательности операций 1-7 подготовительного этапа заявленного технического решения определяют генетический маркер изменения теломер у эукариот, в частности, у человека.
Операции с №8 по №10 представляют собой этап идентификации обнаруженных на предыдущем этапе генетических маркеров в других эукариотических организмах помимо растений A. thaliana (результаты представляются в виде амплифицированных фрагментов ДНК, соответствующих искомым генетическим маркерам, идентифицированным при осуществлении операций №1-7).
Операция №8.
Берут образец ткани (например, 1 мл крови) организма, в частности, человека, для которого требуется провести поиск генетических маркеров изменения длины теломер и выделяют ДНК из образца ткани с помощью стандартных методов для выделения ДНК из крови человека. Выделенную ДНК используют далее для проведения ПЦР-анализа.
Операция №9.
Берут выделенную из образа ткани ДНК (по операции 8) и проводят ПЦР-анализ на обнаружение генетических маркеров, идентифицированных в подготовительном этапе (операции 1-7) с использованием специфических к каждому генетическому маркеру праймеров. В результате ПЦР-анализа получают реакционную смесь, предположительно содержащую амплифицированную ДНК из исследуемого образца тканей эукариот, соответствующую последовательности генетического маркера.
Операция №10.
Берут реакционную смесь после ПЦР-анализа (по операции 9), содержащую амплифицированную ДНК и проводят ее электрофоретический анализ по стандартной методике [Каюмов А.Р. Практикум по молекулярной генетике. Учебно-методическое пособие / А.Р. Каюмов, О.А. Гимадутдинов - Казань: Казань, КФУ, 2016. - 36 с.] с выявлением фрагментов ДНК определенной молекулярной массы, соответствующей последовательности искомого генетического маркера. При наличии фрагментов необходимой молекулярной массы делают вывод о том, что в исследуемом образце ткани присутствует искомый генетический маркер («есть», положительный результат); при отсутствии фрагмента определенной молекулярной массы делают вывод об отсутствии искомого генетического маркера в исследуемом образце («нет», отрицательный результат).
Таким образом, на основании совокупности проведенных действий в соответствии с заявленным техническим решением, заключающемся в определении ранее неизвестных генетических маркеров изменения длины теломер у эукариот путем анализа генов инбредных линий растений A. thaliana и последующем использовании этих генетических маркеров для обнаружения в эукариотах в целом, представляется возможным выполнить раннюю диагностику изменения теломер у эукариот, в частности, у человека, и, тем самым, обеспечивается возможность раннего предупреждение заболеваний, связанных со старением эукариотических организмов.
Далее заявителем представлен пример конкретного выполнения заявленного способа, реализованный на практике в условиях лаборатории КФУ, в качестве примера представлены результаты по идентификации генетического маркера OLI2 укорочения длины теломер в растениях A. thaliana и его гомолога в геноме человека (ген Nop2), при этом заявителем представлены поясняющие материалы, позволяющие обеспечить идентификацию и достижение целей на каждом этапе заявленной технологии:
1. Выращивают 480 растений инбредных линий A. thaliana, отбирают ткань каждого растения и выделяют ДНК (соответствует операции №1).
2. Выполняют измерение длин теломер в 480 инбредных линиях методом Саузерн-блот анализа (соответствует операции №2). На Фиг. 2 в качестве примера представлены результаты Саузерн-блот анализа теломер 8 инбредных линий растений A. thaliana..
3. Выполняют анализ по определению средней длины теломер в каждой инбредной линии, в результате чего выявлены линии растений с наиболее короткими и наиболее длинными теломерами (соответствует операции №3).
4. Проводят QTL-анализ полученных данных по длинам теломер в каждой инбредной линии (т.е. выполнено картирование локусов количественных признаков средней длины теломер в популяции рекомбинантных инбредных линий), и получены данные по распределению наиболее длинных и наиболее коротких длин теломер у разных экотипов растений A. thaliana (соответствует операции №4).
На основании полученных данных установлено, что гены, предположительно ответственные за регуляцию длины теломер, располагаются на пятой хромосоме [Analysis of genome sequence of the flowering plant Arabidopsis thaliana // The Arabidopsis genome initiative. / Nature. - 2000. - V. 796-815; Tabata, S. Sequence and analysis of chromosome 5 of the plant Arabidopsis thaliana // Tabata S., et al. / Nature. - 2000. - V. 408. - P. 823-826.]. A. thaliana (около 200 генов).
На Фиг. 3 представлены результаты QTL-анализа - распределение генов-кандидатов, связанных с изменением длины теломер в растении A. thaliana. На этом графике по оси абсцисс располагаются 1-5 хромосомы, по оси ординат - величина показания количества генов-кандидатов для каждой хромосомы по отношению к геному растения в целом. Наиболее значимый (высокий) пик обнаружен на хромосоме 5 (QTL основного эффекта). Значительно менее значимый пик присутствует на хромосоме 1. Пунктиром показана линия статистической погрешности. Хр. 1 - Хр. 5 - хромосомы А. thaliana.
5. Проводят биоинформационный анализа 20 генов из числа генов-кандидатов, расположенных на пятой хромосоме арабидопсиса, с использованием информационных баз данных (https://www.arabidopsis.org/, http://signal.salk.edu/cgi-bin/tdnaexpress), в результате чего обнаружен ген OLI2 - рРНК (цитозин-С5)-метилтрансфераза (соответствует операции №5).
6. Выполняют определение гомологии растительного гена OLI2 с генами человека с последующим определением того, что он является ортологом человеческого гена nop2, экспрессия которого коррелирует с развитием опухолевых клеток в человеке, (соответствует операции №6).
7. Установлено, что у мутантных растений, из генома которых удален ген OLI2, происходит укорочение длины теломер по сравнению с контрольными растениями, имеющими ген OLI2 (Фиг. 4) (по операции №7).
8. Выделяют ДНК из образцов тканей растений (соответствует операции №8).
9. Проводят ПЦР-анализ выделенной ДНК растений (по операции №8) на наличие генетического маркера - гена OLI2/ nop2 (соответствует операции №9).
10. Проводят электрофоретический анализ реакционной смеси ПЦР-анализа на наличие амплифицированных фрагментов ДНК (полученный по операции №9), соответствующих нуклеотидной последовательности генетического маркера OLI2/ nop2. В результате в 4-х образцах ДНК растений (см. Фиг. 5 обозначены F61, А52, В52, G66) обнаружен фрагмент ДНК, соответствующий последовательности генетического маркера OLI2/ nop2 (соответствует операции №10).
Полученный в результате реализованных операций по заявленному способу результат может быть использован в дальнейшем для более детализированного обследования конкретного человека на предмет возможного развития заболеваний, связанных с изменением теломер в клетках (в том числе и в лечебных учреждениях).
В результате реализации проведенной совокупности последовательных операций по заявленному способу решены все поставленные задачи, получены заявленные технические результаты, а именно, разработан способ определения генетических маркеров изменения длины теломер, с использованием метода множественного инбридинга и картирования локусов количественных признаков потомков у растений, позволяющий в свою очередь:
- упростить процесс измерения длины теломер путем применения разработанных генетических и молекулярных методов (Саузерн-блот анализ).
- снизить трудоемкость процесса идентификации генетических маркеров изменения теломер, что обуславливает заболевания, связанные со старением (ПЦР-анализ).
- исключить использование тканей человека на этапе поиска генетических маркеров.
- идентифицировать новые, ранее неизвестные генетические маркеры изменения длины теломер у эукариот путем использования инбредных линий растений, чему способствует отсутствие в растительном объекте исследования ограничений в множественном близкородственном скрещивании.
Разработанный заявителем способ обладает рядом преимуществ по сравнению с известными в мире на дату подачи заявки способами, так как заявленный способ не предполагает использование тканей животных и человека для определения генетических маркеров и, тем самым, позволяет преодолеть множество этических, юридических и биологических барьеров, возникающих при проведении исследований на животных и человеке. Идентификация генетических маркеров изменения теломер проводится на основе растительного объекта, а именно - инбредных линий растений, чему способствует отсутствие в растительном объекте исследования ограничений в множественном близкородственном скрещивании.
Заявленный способ соответствует критерию «новизна», предъявляемому к изобретениям, так как из исследованного уровня техники не выявлена совокупность признаков, обеспечивающая реализацию заявленных технических результатов.
Заявленный способ соответствует критерию «изобретательский уровень», предъявляемому к изобретениям, т.к. из исследованного уровня техники не выявлены технические решения, имеющие признаки, совпадающие с существенными признаками заявленного технического решения, и не установлена известность влияния отличительных признаков на указанный технический результат. Заявленное техническое решение соответствует критерию «изобретательский уровень» также и потому, что обеспечивает разрешение противоречий, что обусловлено созданием технологии, позволяющей решить поставленные задачи без использования тканей животных и человека при поиске генетических маркеров, т.к. поиск выполняется с использованием ДНК растений (операции 1-7 заявленного технического решения), что, в конечном счете, значительно упрощает процесс идентификации выявленных генетических маркеров у эукариот, в частности, у человека, поскольку привлечение образцов тканей животных и человека ограничивается исключительно этапом непосредственного проведения диагностики ДНК на наличие генетических маркеров на основе ПЦР (операции 8-10 заявленного технического решения). В то же время заявленное техническое решение не требует использование тканей человека и животных для реализации операций 1-7, что исключает необходимые на дату подачи действия, сопряженные с множеством экономических и юридических трудностей [Butler М. Animal Cell Culture and Technology / М / Butler // 2nd edition. - Garland Science / BIOS Scientific Publishers, 2004].
Заявленное техническое решение можно реализовать в научно-исследовательских, медицинских учреждениях, в деятельности организаций здравоохранения, используя известные стандартные технические устройства и оборудования, что является доказательством соответствия критерию «промышленная применимость», предъявляемому к изобретениям.
Использованные источники
1) Биологический энциклопедический словарь / под ред. М.С.Гилярова. - М.: «Советская энциклопедия». - 1986. - 893 с. (с. 743).
2) Глик Б., Пастернак Дж. Молекулярная биотехнология. Принципы и применение / Пер. с англ. - М.: Мир, 2002. - 589 с.
3) Заяц Р.Г., Бутвиловский В.Э., Давыдов В.В., Рачковская И.В.. 7.2.2 Анализ фрагментов ДНК // Медицинская биология и общая генетика: учебник. - 2-е изд., испр. - Минск: Вышэйная школа, 2012. - С. 100. - 496 с. - ISBN 978-985-06-2182-5.
4) Термины и определения, используемые в селекции, генетике и воспроизводстве сельскохозяйственных животных. - М.: ВНИИплем. И.М. Лунин. 1996.
5) Beck, J.A. Genealogies of mouse inbred strains / Beck J.A., Lloyd S., Hafezparast M., Lennon-Pierce M., Eppig J.T., Festing M.F., Fisher EM. // Nature Genetics. - 2000. - V. 24 (1). - P. 23-5. doi:10.1038/71641. PMID 10615122].
6) Биологический энциклопедический словарь / Гл. ред. М.С.Гиляров. - М.: Сов. энциклопедия, 1986. - 831 с.
7) Толковый биотехнологический словарь. Русско-английский. / Гл. ред. Тарантул В.З. - М. Языки славянских культур, 2009. - 936 с.
8) Экологический словарь URL: https://dic.academic.ru/contents.nsf/ecolog/.
9) Майр, Э. Зоологический вид и эволюция, пер. с англ., М., 1968; Шеппард Ф. М., Естественный отбор и наследственность, пер. с англ., М., 1970.
10) Гистология. Учебник. 2-е изд., перераб. и доп. / Под. ред. Э.Г. Улумбекова, Ю.А. Челышева. - М.: ГЭОТАР-МЕД, 2002. - 672 с.: ил. - Серия «XXI век».
11) Научно-технический энциклопедический словарь URL: http://dic.academic.ru/contents.nsf/ntes/
12) Сергеева, Л.И. QTL-анализ и его применение в физиологических исследованиях / Молекулярно-генетические и биохимические методы в современной биологии растений // Кузнецов В.В., Ралдугина Г.Н. - Издательство: БИНОМ. Лаборатория знаний, 2011 - 481 с.
13) Begon, M. From individuals to ecosystems / M. Begon, C. Townsend, J.L. Harper // Blackwell Publishing, 4th ed. - 2006. - 577 p.
14) Рогозин, И.Б. Эволюция геномов эукариот и принцип максимальной парсимонии / И.Б. Рогозин, Ю.И. Вульф, В.Н. Бабенко, Е.В. Кунин // Вестник Вогис.- 2005. - Т. 9, №2. - С. 141-152.
15) Кунин, Е. Логика случая. О природе и происхождении биологической эволюции / The Logic of Chance. The Nature and Origin of Biological Evolution // E. Кунин. - пер. с англ. - Изд. Центрполиграф, 2014. - 127 с.
16) Jiang, Н.Н. Telomere shortening and ageing / Jiang, Z. Ju K.L. Rudolph // Gerontol Geriat 40:314-324 (2007) DOI 10.1007/s00391-007-0480-0.
17) Мелихова, Л. Основные теории старения / Л. Мелихова, Л. Чентиева, М. Лущик // Международный студенческий научный вестник. - 2016. - I. 4-2. - С. 156-157.
18) Hayflick, L. The serial cultivation of human diploid cell strains / L. Hayflick, P.S. Moorhead // Exp Cell Res. - 1961. - V. 25. - P. 585-621.
19) Гистология. Учебник. 2-е изд., перераб. и доп. / Под. ред. Э.Г. Улумбекова, Ю.А. Челышева. - М.: ГЭОТАР-МЕД, 2002. - 672 с.: ил. - (Серия «XXI век»).
20) Weiss, Н. Aloe spp.plants with vertebrate-like telomeric sequences (Растения видов Aloe с теломераными последовательностями, характерными для позвоночных животных) / Н. Weiss, Н. Scherthan // Chromosome Research. - 2002. - V. 10. - I. 2. - P. 155-164.
21) Shakirov, E.V. Length Regulation and Dynamics of Individual Telomere Tracts in Wild-Type A. / E.V. Shakirov, D.E. Shippen // The Plant Cell. - 2004. - V. 16. - I. 8. - P. 1959-1967.
22) Mirabello, L. The association of telomere length and genetic variation in telomere biology genes / L. Mirabello, K. Yu, P Kraft, I. De Vivo, D.J. Hunter, J. Prescott, J.Y. Wong, N. Chatterjee, R.B. Hayes, S.A. Savage // Mutat. - 2010. - V. 31. - I. 9. - P. 1050-1058.
23) Gonzalo, S. DNA repair defects and genome instability in Hutchinson-Gilford progeria syndrome / S. Gonzalo, R. Kreienkamp // Curr. Opin. Cell Biol. - 34. - 2015. - P. 75-83.
24) Heacock, M. Molecular analysis of telomere fusions in A.: multiple pathways for chromosome end-joining / M. Heacock, E. Spangler, K. Riha, Puizina J., Shippen D.E. // Embo j. - 2004. - V. 23. - I. 11. - P. 2304-2313.
25) Higashiyama, T. Telomere and LINE-like elements at the termini of the Chlorella chromosome I / T. Higashiyama, Y. Noutoshi, M. Akiba, T. Yamada // Nucleic acids symposium series - 1994. - P. 71-72.
26) Greider, C.W. Telomere length regulation / C.W. Greider // Annual review of biochemistry. - 1996. - V. 65. - I. 1. - P. 337-365.
27) Weigel, D. The 1001 Genomes Project for A. thaliana / 1001 геномный проект для A. thaliana / D. Weigel, R. Mott // Genome Biology - 2009. - V. 10. - P. 107-112.
28) Заявка на изобретение №WO 2013024264 (патент аналог СА №2845047) «Prognostic methodology» / «Прогностический метод». Изобретатели и заявитель: В. Duncan, P. Chris, F. Christopher, университет Кардиффа / Univ Cardiff. 2013-02-21.
29) Патент на изобретение №2522961, «Способ определения маркера развития ревматоидного артрита на основе выявления укорочения относительной длины теломер на отдельных хромосомах в Т-лимфоцитах периферической крови». Патентообладатель ФГБУ «Научно-исследовательский институт клинической иммунологии» Сибирского отделения Российской академии медицинских наук (ФГБУ «НИИКИ» СОРАМН) (RU). Заявка 10.12.2012.
30) Патент на изобретение № WO 2008119027, Генетический тест генов, ассоциированных с теломерами, при врожденном дискератозе и связанных заболеваниях / Изобретатель и заявитель: Armanios M.Y., университет Джонса Хопкинса (США) / Univ Johns Hopkins [US]. - 2.10.2008.
31) Alonso, J.M. Moving forward in reverse: genetic technologies to enable genome-wide phenomic screens in Arabidopsis / J.M. Alonso, J.R. Ecker // Nat Rev Genet. - 2006. - V. 7. - I. 7. - P. 524-536.
32) Analysis of genome sequence of the flowering plant Arabidopsis thaliana // The Arabidopsis genome initiative. / Nature. - 2000. - V. 796-815.
33) Jones, A.M. The Impact of Arabidopsis on Human Health: Diversifying Our Portfolio / A.M. Jones, J. Chory, J.L. Dangl, M. Estelle, S.E. Jacobsen, E.M. Meyerowitz, M. Nordborg, D. Weigel // Cell. - 2008. - V. 133(6). - 939-943. doi:10.1016/j.cell.2008.05.040.
34) Nigmatullina, L.R. Non-radioactive TRF assay modifications to improve telomeric DNA detection efficiency in plants [Text] / L.R. Nigmatullina, M.R Sharipova, E.V. Shakirov // BioNanoScience. - 2016. - doi: 10.1007/ s112668 -016 -0223-zu.
35) Horigichi, G. The transcription factor AtGRF5 and the transcription coactivator AN3 regulate cell proliferation in leaf primordial of Arabidopsis thaliana (Транскрипционный фактор AtGRF5 и транскрипционный коактиватор AN3 регулируют пролиферацию клеток в меристеме листьев Arabidopsis thaliana) // G. Horiguchil, G.-T. Kim, H. Tsukaya / The Plant Journal. - 2005. - V. 43. - P. 68-78.
36) Каюмов А.Р. Практикум по молекулярной генетике. Учебно-методическое пособие / А.Р. Каюмов, О.А. Гимадутдинов - Казань: Казань, КФУ, 2016. - 36 с.
37) Tabata, S. Sequence and analysis of chromosome 5 of the plant Arabidopsis thaliana / Tabata S., et al. // Nature. - 2000. - V. 408. - P. 823-826.

Изобретение относится к биотехнологии. Описан способ обнаружения генетических факторов (генов), определяющих функционирование ДНК-структур клетки - теломер, обуславливающих, в свою очередь, продолжительность жизни клетки эукариот. Выполняют подготовительный этап для идентификации генетических маркеров на основе ДНК растений Arabidopsis thaliana, для чего выделяют ДНК растения Arabidopsis thaliana, проводят рестрикционный анализ длины теломер совместно с Саузерн-блот анализом. Выполняют биоинформационный анализ полученных результатов длин теломер у растений инбредных линий Arabidopsis thaliana. Картируют локусы количественных признаков, определяют гены, участвующие в изменении длины теломер в растениях, проводят биоинформационный поиск гомологичных генов у человека. Проводят анализ влияния исследуемых генов на длину теломер на основе мутантов растений, идентифицируют гены, участвующие в регуляции длины теломер (генетические маркеры), выполняют идентификацию генетических маркеров в других организмах помимо растений A. thaliana путем выделения ДНК из образца ткани организма, для которого требуется провести поиск генетических маркеров. Проводят ПЦР-анализ на наличие генетических маркеров, проводят электрофоретический анализ полученной в ПЦР-анализе реакционной смеси для обеспечения возможности выявления или невыявления искомых фрагментов ДНК, соответствующих нуклеотидным последовательностям искомых генетических маркеров. Изобретение может быть использовано в доклинических исследованиях и в медицинской практике в качестве метода идентификации генов, участвующих в регуляции функционирования теломер. Обеспечивает возможность повышения качества жизни человека за счет обеспечения возможности прогнозирования развития болезней, связанных с процессом старения. 6 ил.
Способ идентификации генетических маркеров регуляции длины теломер, заключающийся в том, что выполняют подготовительный этап для идентификации генетических маркеров на основе ДНК растений Arabidopsis thaliana, для чего выделяют ДНК растения Arabidopsis thaliana, проводят рестрикционный анализ длины теломер совместно с Саузерн-блот анализом, выполняют биоинформационный анализ полученных результатов длин теломер у растений инбредных линий Arabidopsis thaliana, картируют локусы количественных признаков, определяют гены, участвующие в изменении длины теломер в растениях, проводят биоинформационный поиск гомологичных генов у человека, проводят анализ влияния исследуемых генов на длину теломер на основе мутантов растений, идентифицируют гены, участвующие в регуляции длины теломер (генетические маркеры), выполняют идентификацию генетических маркеров в других организмах помимо растений A. thaliana путем выделения ДНК из образца ткани организма, для которого требуется провести поиск генетических маркеров, проводят ПЦР-анализ на наличие генетических маркеров, проводят электрофоретический анализ полученной в ПЦР-анализе реакционной смеси для обеспечения возможности выявления или невыявления искомых фрагментов ДНК, соответствующих нуклеотидным последовательностям искомых генетических маркеров.
| ЖДАНОВА Н.С | |||
| Биология теломер млекопитающих, Институт цитологии и гигиены Сибирского отделения Российской академии наук, Новосибирск, 2012 | |||
| КОРОЛЕВА А.Г | |||
| Структуры и длина теломер: загадкам нет конца, Наука из первых рук, август 2016, номер 3, стр.72-83 | |||
| US 20040142327 A1, 22.07.2004. |

