Изобретение относится к исследованиям биологического материала, в частности к определению относительной длины теломер на хромосомах.
В настоящее время во всем мире отмечается рост числа аутоиммунных заболеваний, обусловленный, с одной стороны, бурным прогрессом социально-экономических условий жизни, урбанизацией и индустриализацией общества с резким увеличением скученности населения, с другой - влиянием экологически неблагоприятных факторов. Наряду с сердечно-сосудистой и онкологической патологией аутоиммунные заболевания, в частности ревматоидный артрит, представляют особый интерес. Пик заболеваемости иммунопатологическими заболеваниями приходится на вторую половину жизни населения: среди лиц моложе 35 лет распространенность ревматоидного артрита составляет 0,38%, в возрасте 55 лет и старше - 1,4%, а средняя продолжительность жизни у данной категории граждан снижена до 58 лет. До сих пор нет однозначного объяснения причин и механизмов развития заболеваний, связанных с аутоиммунными реакциями против собственных клеток организма. Это делает невозможным проведение эффективных профилактических и лечебных мероприятий у данной группы больных, что обусловливает широкий поиск как средовых, так и генетических факторов, ответственных за этиологию аутоиммунных заболеваний.
Последние исследования показали, что вклад генетических факторов в детерминацию ревматоидного артрита превалирует над вкладом средовых факторов. Наследуемость РА колеблется от 50 до 90%. Одним из первых исследовался главный комплекс гистосовместимости (МНС), вклад которого в развитие РА оценивается от 30% до 70% от общего генетического эффекта (наиболее вероятная ассоциация с HLA-DR4 гаплотипом).
Остальной вклад приходится на гены из других частей генома и по этой причине ведутся обширные исследования во всем мире по картированию этих генов. Такими генами «предрасположенности» к РА являются ген рецепторов фактора некроза опухоли (ФНО) и ген лиганда CD 40, которые находятся на одном из участков Х-хромосомы. Еще одним генетическим маркером ревматоидного артрита считается ген IDDM9 (3 хромосома). Наличие данного гена повышает риск развития ревматоидного артрита на 27%.
Немаловажную роль в этиологии и патогенезе заболевания играет Т-клеточный иммунитет. В связи с этим ген TCRD на 17 хромосоме (рецепторов Т-клеток) также является ответственным за возникновение ревматоидного артрита.
Результаты целого ряда исследований показали, что мутация гена р53 приводит к возникновению его аномальных функций, что вызывает развитие костной деструкции. При ревматоидном артрите у многих пациентов отмечается аномальный биосинтез Fas-белка, вызванный мутацией Fas-гена.
На сегодняшний день известно, что активность гена может быть изменена не только вследствие его мутации, но и действия на него, так называемого, позиционного эффекта теломер “telomeric position effect” (ТРЕ). Данный феномен относительно генов определяется как «сайленсинг» - подавление экспрессии генов, расположенных в субтеломерных регионах ДНК, через блокирование промоторов этих генов теломерным гетерохроматином [3]. Таким образом, укорочение теломер должно сопровождаться потерей репрессирующей активности теломерного гетерохроматина на гены субтеломерного региона ДНК и повышению их экспрессии. Например, одним из генетических маркеров тяжелого течения РА является повышение экспрессии гена BST1 (bone marrow stromal cell antigen) в синовиальных клетках пациентов с РА [2]. Тем не менее, причины увеличения экспрессии данного гена установлены не были.
Теломерами называют специализированные функциональные комплексы, которые находятся на концах эукариотических хромосом и защищают последние от слияния друг с другом, тем самым, поддерживая целостность генома клетки [1]. Каждое клеточное деление сопровождается прогрессивным укорочением теломер, которое, достигая определенного порогового уровня, приводит к нестабильности генома, старению и апоптозу клетки.
Изобилие генов-кандидатов, так или иначе связанных с развитием РА, определяет развитие исследовательских работ в области изучения влияния разнообразных генетических факторов на экспрессию данных генов. Одним из этих факторов является укорочение длины теломер на определенных хромосомах в норме и при патологии. Определение длины теломер на отдельных хромосомах при ревматоидном артрите, таким образом, позволяет установить особенности распределения и укорочения длины теломер при данном виде аутоиммунной патологии, а закономерности, выявленные на уровне определенных хромосом, служат дополнительным маркером развития данного заболевания. Выявленный таким образом маркер развития ревматоидного артрита позволяет улучшить эффективность как популяционно-ориентированных профилактических программ (скрининговое определение ревматоидного фактора, например), так и индивидуальных превентивных мероприятий (противовоспалительная терапия, проводимая у пациентов с очень высоким риском развития аутоиммунной патологии).
Целью изобретения является создание способа определения маркера развития ревматоидного артрита, основанного на выявлении укорочения относительной длины теломер на отдельных хромосомах Т-лимфоцитов периферической крови, который позволяет с достаточной информативностью и специфичностью определить риск развития данного вида аутоиммунной патологии.
С помощью цитогенетического метода количественной гибридизации in situ (Quantitative Fluorescent in situ hybridization (Q-FISH)) определяют относительную длину теломер на отдельных хромосомах Т-лимфоцитов периферической крови больного как отношение среднего значения интенсивности флюоресценции теломер определенной хромосомы (для каждого из плеч отдельно) к среднему значению флюоресценции теломер всех хромосом в метафазе. Относительную длину теломер хромосом больных сравнивают с относительной длиной теломер у здоровых доноров, соответствующим им по возрасту. Укорочение относительной длины теломер на коротком плече 4 хромосомы больного относительно аналогичного показателя здорового донора расценивают как маркер развития ревматоидного артрита.
Описание способа с примерами конкретной реализации.
5 мл периферической крови, стабилизированной с гепарином, центрифугируют в градиенте фиколл-верографин в течение 20 мин при 2,7 тыс.об/мин в медицинской лабораторной центрифуге, после чего кольцо мононуклеарных клеток собирают и отмывают дважды с 5 мл забуференного физиологического раствора с ЭДТА и азидом Na (ЗФР-ЭА) по 5 мин при 1 тыс.об/мин. Осадок ресуспендируют в 1 мл ЗФР-ЭА и подсчитывают количество клеток в камере Горяева.
Для гибридизации используют 2·106 клеток на одну пробу. Исходя из этого, необходимое количество клеток снова осаждают центрифугированием и ресуспендируют осадок в полной среде RPMI 1640 с добавлением 2 мг тиенама, 125 мкл гентамицина и 30 мг L-глютамина на 100 мл среды и помещают в лунки 24-луночного планшета из расчета 2 млн клеток на 1 мл полной среды с 10% FBS и митогеном для Т-лимфоцитов (РНА-Р в конечной концентрации 10 мкг/мл). Тщательно пипетируют, планшет помещают в СO2-инкубатор с температурой 37°С на 72 ч. За 7,5 ч до конца культивирования добавляют раствор бромистого этидия (в конечной концентрации 10 мкг/мл), за 6 ч до окончания - раствор колхицина (в конечной концентрации 0,1 мкг/мл). По окончании культивирования суспензию клеток переносят в пробирки, центрифугируют, сливают культуральную среду.
Осажденные клетки ресуспендируют, к оставшемуся осадку аккуратно добавляют 1-2 мл гипотонического раствора КСl, осторожно взбалтывая пробирку. Затем доводят гипотоническим раствором до общего объема 9 мл. Суспензию тщательно перемешивают и помещают на 30-50 мин в термостат или на 8-10 мин на водяную баню с температурой 37°С. По истечении времени гипотонии проводят предфиксацию: в суспензию добавляют несколько капель свежеприготовленного фиксирующего раствора (18 капель на 9 мл суспензии), аккуратно перемешивают и выдерживают 10-15 мин в холодильнике при 4°С. Затем центрифугируют, сливают надосадочную жидкость, клеточный осадок ресуспендируют и аккуратно «капля-за-каплей» заливают небольшим количеством (1-2 мл) свежеприготовленного охлажденного до -20°С фиксатора, осторожно взбалтывая пробирку, и оставляют в холодильнике при -20°С на 30 мин. По истечении времени центрифугируют, добавляют свежий фиксатор в нужном объеме (400-500 мкл), переносят в пробирки эппендорф и хранят в морозильной камере при -20°С до использования.
Для приготовления препаратов используют обезжиренные чистые тонкие (76×26×1 мм) стекла фирмы Menzel GmbH+Со KG. Перед приготовлением препаратов стекла выдерживают в концентрированной соляной кислоте не менее 30 мин, затем стекла тщательно ополаскивают водопроводной и дистиллированной водой, помещают в стакан с дистиллированной водой и охлаждают в холодильнике.
Перед приготовлением препаратов суспензию ядер центрифугируют, ресуспендируют в небольшом объеме фиксатора (100 мкл). На влажное предметное стекло наносят 15 мкл суспензии хромосом и высушивают на водяной бане при 50°С. Затем сверху капают 15 мкл фиксатора, стекло выдерживают на водяной бане при той же температуре до полного высыхания. После подсушивания препарат проверяют на фазово-контрастном микроскопе на предмет качества распластывания хромосом. После раскапывания препарат прогревают 1 ч в термостате при 65°С.
Перед проведением гибридизации проводят регидратацию препаратов путем погружения в 150 мл PBS буфера на 15 мин при комнатной температуре. Затем фиксируют слайды в 4% растворе параформальдегида в течение 2 мин с последующей трехкратной отмывкой в PBS буфере по 5 мин каждая. Препараты метафазных хромосом обрабатывают пепсином в растворе 0,01 N НСl, рН 2,0 в течение 10 мин при 37°С. Отмывают дважды в PBS буфере по 2 мин. Повторяют фиксацию в 4% растворе параформальдегида с последующими отмывками. Затем препараты проводят через серию растворов этилового спирта возрастающей концентрации (70%, 90%, 96%) и высушивают при комнатной температуре. На сухую поверхность препарата наносят 2 капли по 10 мкл гибридизационного раствора В под покровное стекло размером 24×60. Затем проводят денатурацию ДНК в термостате при 80°С в течение 10 мин. Далее переносят препараты во влажную камеру и гибридизуют 2 ч при комнатной температуре в темноте. После гибридизации препараты отмывают 2 раза по 15 мин в отмывочном растворе №1 и 3 раза по 5 мин в отмывочном растворе №2 на шейкере. Затем препараты дегидратируют в растворах этилового спирта возрастающей концентрации (70%, 90%, 96%) и высушивают грушей. Препараты окрашивают DAPI (0,1 мкг/мл) под покровное стекло в течение 20 мин в темноте, затем осторожно снимают покровное стекло, наносят раствор Antifade Pro Long Gold, покрывают новым покровным стеклом и скрепляют клеем.
Препараты анализируют на микроскопе “Axioplan 2 Imaging E-mot” (ZEISS), оснащенном лампами флуоресцентного и видимого света, с комплектом интерференционных фильтров фирмы CHROMA, для проведения 24-цветной FISH. Ввод цифровой информации с помощью CCD-камеры CV М300; JAI Corporation, Japan, матрица 752 на 582 пиксела. PC (P-IV, 1,7 GH, 128 MB, 40 GB). Пакеты программного обеспечения ISIS4 фирмы MetaSystems GmbH, для работы с флуоресценцией (FISH, 24-хцветная FISH, многоцветный бэндинг, сравнительная геномная гибридизация, псевдоцвета). Для обработки изображений используют программу «TFL-Telo» компании «British Columbia Cancer Research Center».
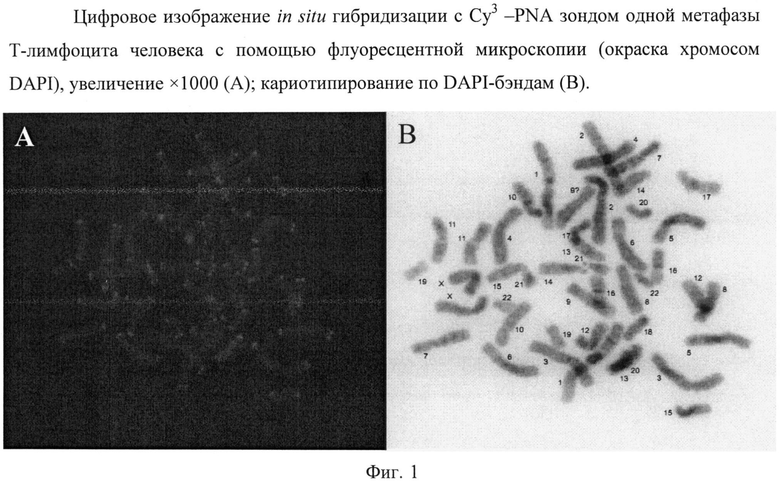
Завершающим этапом данного метода является получение цифровой фотографии хромосом в метафазе клеточного цикла, окрашенных DAPI (синее свечение) и прошедших гибридизацию с Су3 -PNA зондом (красное свечение теломерных областей ДНК) На фиг.1 дано цифровое изображение in situ гибридизации с Су3 - PNA зондом одной метафазы Т-лимфоцита человека с помощью флуоресцентной микроскопии (окраска хромосом DAPI), увеличение × 1000 (А); кариотипирование по DAPI-бэндам (В). Приведен типичный результат анализа данных одного из исследуемых образцов.
Следующий этап анализа - проведение кариотипирования полученных метафаз по DAPI-бэндам. Далее с помощью специальной программы вычисляется флуоресценция каждой из теломер. Результативный показатель длины теломер вычисляется как отношение среднего значения интенсивности флюоресценции теломер определенной хромосомы (для каждого из плеч отдельно) к среднему значению интенсивности флюоресценции теломер всех хромосом в метафазе (184 сигнала). Для каждого индивидуума проведен анализ в среднем пяти метафазных пластинок.
В исследование были включены 10 пациентов с ревматоидным артритом, средний возраст 41,7 (min 24, max 61) и 11 соответствующих им по возрасту доноров, средний возраст 39 лет (min 24, max 58). Известно, что длина теломер в лимфоцитах не зависит от стадии и стажа заболевания, терапевтического статуса, в связи с этим основным критерием включения является верифицированный по МКБ-10 диагноз ревматоидного артрита и соответствующий группе доноров возраст пациентов.
Ниже приведены результаты, поясняющие данный способ определения маркера развития ревматоидного артрита на основе выявления укорочения относительной длины теломер на отдельных хромосомах в Т-лимфоцитах периферической крови.
Пример 1.
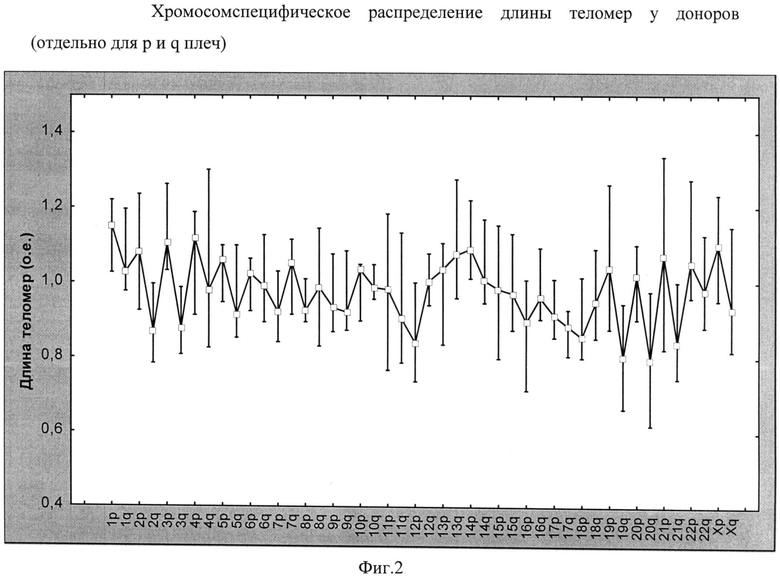
Определена относительная длина теломер на отдельных хромосомах в Т-лимфоцитах у здоровых доноров. Была выявлена достоверная разница между длиной теломер разных хромосом в группе доноров. Высокая значимость различий показателей длины теломер была подтверждена тестом Фридмана, который для всех индивидуумов в целом показывал р-уровень менее 0,001. На фиг.2 показано хромосомспецифическое распределение длины теломер у доноров (отдельно для р и q плеч). На диаграмме для каждой переменной отображены: медиана, квартальный размах (25%, 75% процентили). Достоверность различий между теломерами разных хромосом р<0,01 (тест Фридмана), о.е. - относительные единицы.
При детальном анализе значений медиан каждой из теломер, были выявлены хромосомы с наименьшей относительной длиной теломер (<0,9 относительных единиц (о.е.)) и хромосомы с самыми длинными теломерами (>1,1 о.е.) (табл.1). В таблице данные представлены в виде: ME, Р25-Р75 - медиана и квартили распределения признаков (25-75%), о.е. - относительные единицы. Как видно из таблицы, хромосома с минимальной длиной теломер - 20q, с максимальной длиной - 1р.
Пример 2.
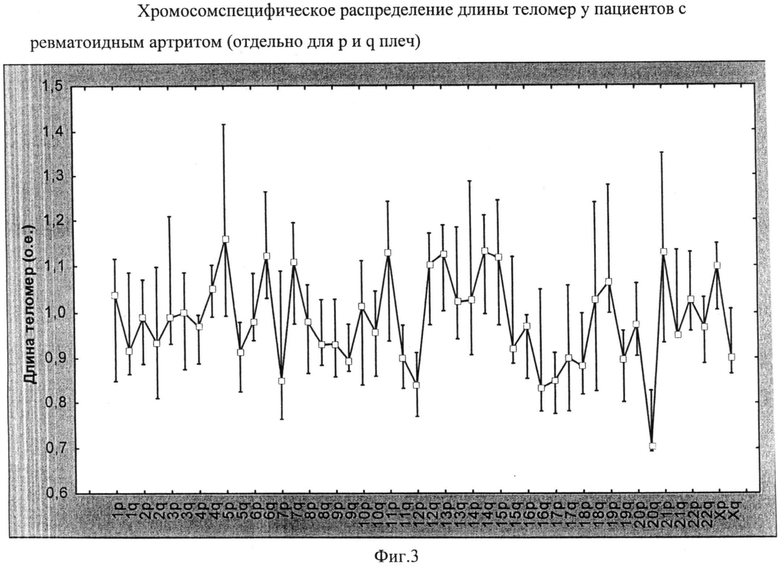
Определена относительная длина теломер на отдельных хромосомах в Т-лимфоцитах у пациентов с ревматоидным артритом. Данные представлены на фиг.3. На диаграмме для каждой переменной отображены: медиана, квартальный размах (25%, 75% процентили). Достоверность различий между теломерами разных хромосом р<0,01 (тест Фридмана), о.е. - относительные единицы. При сравнении полученных данных по каждой из хромосом с аналогичными показателями у здоровых доноров было установлено достоверное укорочение длины теломер на коротком плече 4 хромосомы при ревматоидном артрите: относительная длина у доноров составила (Me [LQ; UQ]) - 1,14 [0,99; 1,22] относительных единиц (о.е.), у пациентов с ревматоидным артритом - 0,97 [0,89; 1,0] о.е. (р<0,05). Сравнение вариационных рядов осуществлялось с помощью непараметрического U критерия Манна-Уитни.
Таким образом, полученные результаты свидетельствуют о возможности применения метода для определения маркера развития ревматоидного артрита, основанного на выявлении укорочения относительной длины теломер на отдельных хромосомах Т-лимфоцитов периферической крови.
Поскольку ревматоидный артрит является мультифакториальным заболеванием с полигенным типом наследования, выявление новых генетических факторов, вносящих свой вклад в развитие заболевания, дополняет и раскрывает неизвестные звенья патогенеза патологического процесса. Выявленный таким образом маркер развития ревматоидного артрита позволяет улучшить эффективность как популяционно-ориентированных профилактических программ (скрининговое определение ревматоидного фактора, например), так и индивидуальных превентивных мероприятий (противовоспалительная терапия, проводимая у пациентов с очень высоким риском развития аутоиммунной патологии).
Источники информации.
1. Blackburn Е. Switching and signaling at the telomere // Cell. - 2001. - Vol.106. - P. 661-673.
2. Lee В., Ishihara K., Denno K. Elevated levels of the soluble form of bone marrow stromal cell antigen 1 in the sera of patients with severe rheumatoid arthritis // Arthritis Rheum. - 1996. - Vol.39. - P. 629-637.
3. Wright W., Shay J. Telomere positional effects and the regulation of cellular senescence // Trends Genet. - 1992. - Vol.8. - P. 193-197.
показателями у здоровых доноров
Изобретение относится к области медицины, включающей исследования биологического материала, и касается определения относительной длины теломер на хромосомах. Способ заключается в выявлении укорочения относительной длины теломер на отдельных хромосомах Т-лимфоцитов периферической крови с помощью метода количественной флуоресцентной гибридизации in situ (Q-FISH). Результативный показатель длины теломер вычисляют как отношение среднего значения интенсивности флюоресценции теломер определенной хромосомы (для каждого из плеч отдельно) к среднему значению интенсивности флюоресценции теломер всех хромосом в метафазе. Укорочение относительной длины теломер на коротком плече 4 хромосомы у пациентов с ревматоидным артритом в сравнении с донорами расценивают как маркер развития заболевания. Изобретение позволяет с достаточной информативностью и специфичностью определить риск развития данного вида аутоиммунной патологии. 3 ил., 1 табл., 2 пр.
Способ определения маркера развития ревматоидного артрита, отличающийся тем, что с помощью метода количественной флуоресцентной гибридизации in situ (Q-FISH) выявляют укорочение относительной длины теломер на отдельных хромосомах Т-лимфоцитов периферической крови пациентов; результативный показатель длины теломер вычисляют как отношение среднего значения интенсивности флюоресценции теломер определенной хромосомы (для каждого из плеч отдельно) к среднему значению интенсивности флюоресценции теломер всех хромосом в метафазе и укорочение относительной длины теломер на коротком плече 4-й хромосомы у пациентов с ревматоидным артритом в сравнении с соответствующими им по возрасту донорами расценивают как маркер развития заболевания.
| СПОСОБ ИЗМЕРЕНИЯ ДЛИНЫ ТЕЛОМЕР КЛЕТОК ЛЕЙКОКОНЦЕНТРАТА ПУПОВИННОЙ КРОВИ | 2010 |
|
RU2434940C1 |
| Б | |||
| Прибор с двумя призмами | 1917 |
|
SU27A1 |
| CN 202558870 U (YUNAN LUIS TRADITIONAL CHINESE MEDICINE MODERNIZATION ENGINEERING TECHNOLOGY RES CT +) 28.11.2012 | |||
| WO 03000927 (UNIV WALES MEDICINE [GB]; BAIRD DUNCAN MARTIN [GB] + ) A2 03.01.2003 | |||
| СПОСОБ И УСТРОЙСТВО ДЛЯ ОПРЕДЕЛЕНИЯ СТРОЕНИЯ ХРОМОСОМ | 2005 |
|
RU2471871C2 |
| BLACKBURN Е | |||
| Switching and signaling at the telomere | |||

