ПРИТЯЗАНИЕ НА ПРИОРИТЕТ
По данной заявке испрашивается приоритет по предварительной заявке на патент США № 61/252607, поданной 16 октября 2009 г., "ПРИМЕНЕНИЕ ДЕНДРИМЕРНОЙ НАНОТЕХНОЛОГИИ ДЛЯ ДОСТАВКИ БИОМОЛЕКУЛ В РАСТИТЕЛЬНЫЕ КЛЕТКИ".
Предшествующий уровень техники
Наночастицы обладают уникальными свойствами, которые применяются для использования для доставки ДНК в клетки. Металлические наночастицы, такие как наночастицы золота (Au), используются для доставки ДНК из-за их низкой цитотоксичности и простоты функционализации с различными лигандами биологического назначения. В дополнение к металлическим наночастицам также в качестве носителей для доставки молекул в клетки используются полупроводниковые наночастицы (например, квантовые точки) ("QD") в пределах размеров 3-5 нм. ДНК и белки могут быть связаны с лигандом, прикрепленным к поверхности QD (см., например, F. Patolsky et al., J. Am. Chem. Soc. 125, 13918 (2003)).
Наночастицы используют для доставки плазмидной ДНК в разнообразные клетки животных. Было обнаружено, при инкубации покрытых ДНК наночастиц с клетками, не имеющими клеточной стенки, клетки поглощают наночастицы и начинают экспрессию любых генов, кодируемых ДНК. Тем не менее в настоящее время генная доставка в растения затруднительна из-за наличия растительных клеточных стенок, что приводит к повсеместному использованию средств для инвазивной доставки для генетической трансформации растений. Если требуется опосредованная наночастицами доставка в обычно имеющие клеточную стенку клетки, то стенку клеток разрушают перед добавлением частиц к протопластам растения (см., F. Torney et al., Nature Nanotechnol. 2, (2007)). В растительных клетках клеточная стенка служит в качестве барьера для доставки экзогенно вносимых молекул. Многие инвазивные способы, такие как генная пушка (биолистика), микроинъекция, электропорация и агробактерия, применяются для того, чтобы осуществить доставку гена и малой молекулы в эти имеющую стенку растительные клетки, но доставка белков осуществляется только посредством микроинъекции. Задача доставки малых молекул и белков при наличии растительной клеточной стенки остается неразрешенной, и было бы полезно разработать эффективные технологии, которые могут быть применены по отношению к интактной растительной клетке/ткани или органу в in vitro и in vivo манипуляциях.
Проникающие в клетку пептиды (CPP) являются новым и быстро растущим классом коротких пептидов, про которые известно, что они играют важную роль в переносе широкого спектра комплексов грузов, включая белки и ДНК через биомембраны в млекопитающих и человеческих клеточных линиях J. Schwartz and S. Zhang (2000), Peptide-Mediated Cellular Delivery, Curr. Opin. Mol. Ther. 2:162-167; Ü Langel (2002), Preface in: Cell Penetrating Peptides; Processes and Applications, Ü. Langel, Editor, CRC Press, Boca Raton; E. Vives and B. Lebleu (2002), The Tat-Derived Cell-Penetrating Peptide in: Cell-Penetrating Peptides; Processes and Applications, Ü. Langel, Editor, CRC Press, Boca Raton: pp. 3-22. Несмотря на то, что было продемонстрировано, что CPP облегчают доставку грузов в клетки млекопитающих, использование CPP в растительных клетках для исследования трансфекции было ограничено рядом факторов. Главным препятствием для адаптации этой технологии к растениям является то, что в отличие от клеток животных в растительных клетках имеется система с двойным барьером (клеточная стенка и цитоплазматическая мембрана) для интернализации CPP и их грузов. По этой причине CPP должны преодолеть эти два барьера для эффективного переноса. CPP используют в растительных клетках, но, как правило, применяют на основе использования пермеабилизирующих агентов и технологий с использованием CPP для осуществления доставки грузов в растительные клетки. Трансдукционный домен (PTD) TAT белка HIV-1 является одним из наиболее изученных транслоцирующихся пептидов. Последние сообщения продемонстрировали способность TAT-PTD и их олигомеров к доставке плазмиды посредством образования комплекса с отрицательно заряженной ДНК в клетках млекопитающих. I. Ignatovich, E. Dizhe, A. Pavlotskaya, B. Akifiev, S. Burov, S. Orlov, and A. Perevozchikov (2003), Complexes of Plasmid DNA with Basic Domain 47-57 of the HIV-1 Tat Protein Are Transferred to Mammalian Cells by Endocytosis-mediated Pathways, J. Biol. Chem. 278:42625-42636; C. Rudolph, C. Plank, J. Lausier, U. Schillinger, R.H. Müller, and J. Rosenecker (2003), Oligomers of the Arginine-Rich Motif of the HIV-1 TAT Protein are Capable of Transferring Plasmid DNA into Cells, J. Biol. Chem. 278:11411-11418; Z. Siprashvili, F. Scholl, S. Oliver, A. Adams, C. Contag, P. Wender, and P. Khavari (2003), Gene Transfer via Reversible Plasmid Condensation with Cysteine-Flanked, Internally Spaced Arginine-Rich Peptides, Hum. Gene. Ther. 14 (13):1225-33; I. Hellgren, J. Gorman, and С. Sylvén (2004), Factors Controlling the Efficiency of Tat-mediated Plasmid DNA Transfer, J. Drug. Target. 12 (l):39-47.
Дендримеры являются "каскадными молекулами" с необычной окклюдантной макромолекулярной архитектурой. Дендримеры были впервые приготовлены в лаборатории в 1979 году Дональдом Томалиа (D.A. Tomalia et al., Preprints of the 1st SPSJ Int'l Polymer conference, Society of Polymer Science, Japan, Kyoto, 1984, p. 65; см. также патент США № 6316694). Дендримеры используются для доставки ДНК и других биомолекул в клетки животных. Тем не менее наличие растительной клеточной стенки представляет сложности для генетической доставки в растения. Кроме того, нет сообщений или доказательств устойчивой геномной интеграции трансгенов с использованием основанной на дендримерах доставки в растения. Таким образом, все еще существует необходимость в способе устойчивого включения генов и других молекул, представляющих интерес, в растения посредством использования основанной на дендримерах доставки.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Следующие варианты осуществления описаны в сочетании с системами, средствами и способами, которые, как подразумевается, являются примерными и иллюстративными и не ограничивающими объем изобретения.
Данное изобретение относится к способам использования дендримеров и, необязательно, одного или более CPP для неинвазивной доставки молекулярных веществ в клетки, имеющие клеточную стенку, для устойчивого включения в них молекулярных веществ.
Один из вариантов осуществления изобретения включает способ внесения представляющей интерес молекулы в растительную клетку, имеющую клеточную стенку, для осуществления устойчивой трансформации растений и семян. Способ включает предоставление растительной клетки, имеющей клеточную стенку, и взаимодействие дендримера и, необязательно, одного или более CPP с представляющей интерес молекулой для образования активированной дендримерной структуры. Клетка и активированная дендримерная структура приводятся в контакт друг с другом в условиях, допускающих проникновение в клетку, имеющую клеточную стенку.
Другой вариант осуществления изобретения включает способ стабильной экспрессии гена. Способ включает предоставление растительной клетки, имеющей клеточную стенку, и взаимодействие дендримера и, необязательно, одного или более CPP с геном для образования активированной дендримерной структуры. Растительная клетка, имеющая клеточную стенку, и активированная дендримерная структура приводятся в контакт друг с другом, и дендримеры и ген помещаются в условия, допускающие их проникновение в растительную клетку, имеющую клеточную стенку. После этого ген экспрессируется в потомстве растений, имеющих растительную клетку.
Еще один вариант осуществления изобретения включает в себя способ переноса молекулярного соединения в растительную клетку. Способ включает в себя взаимодействие одного дендримера и, необязательно, одного или более CPP с плазмидной ДНК для образования активированной дендримерной структуры. Активированная дендримерная структура вводится в контакт с интактной имеющей стенку растительной клеткой при условиях, допускающих проникновение дендримера и гена из плазмидной ДНК в растительную клетку.
Вдобавок к примерным аспектам и вариантам осуществления, описанным выше, дополнительные аспекты и варианты осуществления будут понятны в свете дальнейших описаний.
КРАТКОЕ ОПИСАНИЕ ФИГУР
На фиг.1 представлена плазмида pDAB3831.
На фиг.2 представлена плазмида pDAB7331.
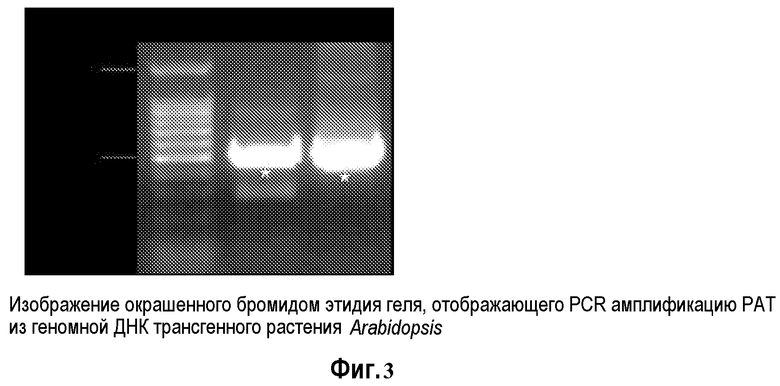
На фиг.3 представлено изображение геля, окрашенного бромидом этидия, демонстрирующего ПЦР амплификацию PAT из геномной ДНК трансгенного растения Arabidopsis: маркер длин ДНК 100 (Promega Inc) 1; трансгенная линия Arabidopsis от SUPERFECT™ с обработкой 2 pDAB3831; Плазмидная ДНК pDAB3831 в качестве положительного контроля; Звездочки указывают на ПЦР-ампликон PAT.
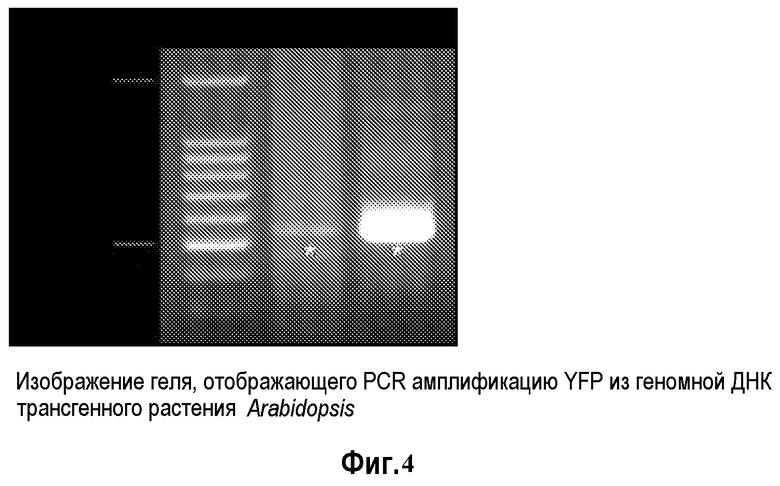
На фиг.4 представлено изображение геля, демонстрирующего ПЦР амплификацию геномной ДНК трансгенной растительной Arabidopsis: маркер длин ДНК 100 (Promega Inc) 1; трансгенная линия Arabidopsis от SUPERFECT™ с обработкой 2 pDAB3831; плазмидная ДНК pDAB3831 в качестве положительного контроля; звездочки указывают на ПЦР-ампликон YFP.
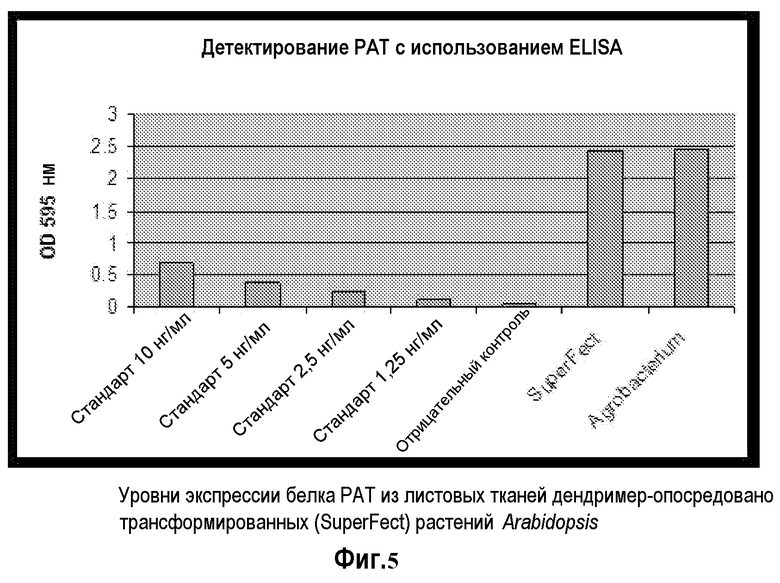
На фиг.5 продемонстрированы PAT уровни экспрессии белков из листовых тканей дендример-опосредовано (SUPERFECT™) трансформированных растений Arabidopsis; с использованием коммерчески доступного набора ELISA, Белок PAT детектировали в дендример-опосредованных трансгенных растениях и сравнивали с растениями, которые были трансформированы с помощью Agrobacterium tumefaciens.
ВАРИАНТ(Ы) ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
В описании и последующих таблицах использован ряд терминов. Для того чтобы обеспечить отчетливое и подходящее истолкование описания и формулы изобретения, включая пределы, в рамках которых использованы такие термины, предоставлены следующие определения:
Обратное скрещивание. Обратное скрещивание может быть процессом, в котором селекционер повторно скрещивает гибридное потомство с одним из родителей, например, гибрид первого поколения F1 с одним из родительских генотипов F1 гибрида.
Эмбрион. Эмбрион может быть небольшим растением, содержащимся в зрелом семени.
Дендример. Дендримеры являются трехмерными, сверхразветвленными, монодисперсными наномерными макромолекулами, полученными посредством повторяющейся последовательности реакций. Дендримеры синтезируют обычным путем как перестраиваемые "наноструктуры", которые могут быть разработаны и регулироваться в зависимости от их размера, формы, поверхностной химии и внутреннего свободного пространства. Дендримеры могут быть получены с таким же подходом к структурному контролю, что и для общепринятых биомакромолекул, таких как DNNPNA или белки, и различаются их определенной наноразмерной подложкой и наноконтейнерными свойствами. Дендримеры являются микроскопическими частицами с по меньшей мере одним наноразмерным измерением, обычно менее 100 нм. Дендримеры, пригодные для использования в данном изобретении, могут иметь размер 1 нм - 0,4 мкм.
Резистентный к глифосату. Резистентность к дозировке глифосата относится к способности растения выживать (т.е. растение не может быть уничтожено) при такой дозировке глифосата. В некоторых случаях устойчивые растения могут временно становиться желтыми или иным образом проявлять некоторое вызванное глифосатом повреждение (например, избыточное ветвление и/или ингибирование роста), но восстанавливаться.
Устойчивый. Устойчивый относится к характеристикам растения, которые воспроизводимо передаются от одного поколения следующему поколению инбредных растений одного сорта.
Проникновение. Проникновение относится к переносу частицы, такой как дендример, через клеточную стенку или клеточную мембрану, где перенос не происходит только лишь в результате импульса, переданного частице, чем-либо отличным от клетки, в которую частица внедряется. Неограничивающими примерами устройств или способов, которые вызывают перенос частиц через клеточную стенку или клеточную мембрану только лишь в результате импульса, переданного частице, являются технологии биолистики, генной пушки, микроинъекции и/или импалефекции.
В конкретных способах осуществления изобретение относится к применению дендримера в качестве варианта для нанопроектирования для конструирования переносчика грузов для того, чтобы сформировать материалы для применения в синтетических препаратах, доставке биомолекул, генной доставке, визуализации и различных биотехнологических диагностических средствах и для измерительных функций. Конструкция дендримера обеспечивает ряд отличительных свойств, которые отличают их от других полимеров и наночастиц, таких как последовательный поэтапный способ синтеза, который может обеспечить строго определенный размер и структуру со сравнительно низким коэффициентом полидисперсности. Кроме того, дендримерная химия может быть адаптирована и, таким образом, будет облегчен синтез большого диапазона молекул с различными функциональностями. Применение дендримеров в соответствии с конкретными способами данного изобретения способствует доставке биомолекул и генов посредством использования высокой плотности концевых групп.
В других вариантах осуществления изобретения множественные сайты присоединения или дополнение "добавочной" или "гостевой" молекулы могут быть сконструированы на дендримерах в различных и/или множественных сайтах. Это свойство может быть применено, например, в специфическом направлении и редактировании молекулярных сайтов внутри клетки в таких областях, как биомиметика, направленные способы доставки, для опций, связанных с негенетически модифицированным организмом, и опций, связанных с временной трансформацией различных деревьев или овощных культур для придания признаков и опций, связанных с резистентностью к болезни. Варианты осуществления изобретения могут также быть применены для разработки пригодных биосенсоров. Кроме того, со способами изобретения могут быть применены искусственные хромосомы (ACES) в качестве альтернативы современным эукариотическим векторам для необязательностей точного направления и гомологичной рекомбинации.
В соответствии с вариантами осуществления изобретения может быть предоставлен способ внесения представляющей интерес молекулы в растительную клетку, содержащую клеточную стенку, при этом способ содержит приведение дендримера, содержащего представляющую интерес молекулу, в контакт с растительной клеткой и предоставление возможности проникновения дендримера через растительную клеточную стенку. В конкретных аспектах изобретения дендример может быть любым дендримером и может обратимо или необратимо включать в себя, может взаимодействовать или иным образом быть связанным с и/или нести молекулу, представляющую интерес. В некоторых вариантах осуществления представляющая интерес молекула может быть введена в дендример перед контактом с растительной клеткой, имеющей клеточную стенку, или одновременно с введением дендримера в растительную клетку, имеющую клеточную стенку.
В соответствии с вариантами осуществления данного изобретения, растительная клетка, имеющая клеточную стенку, может быть любой растительной клеткой, содержащей интактную и целую клеточную стенку. Примеры клеток, имеющих клеточную стенку, включают в себя, но не ограничены ими, клетки водорослей, табака, моркови, маиса, канолы, рапсового семени, хлопка, пальмового дерева, арахиса, сои, сахарного тростника, Oryza sp., Arabidopsis sp. и Ricinus sp., предпочтительно табака, моркови, маиса, хлопка, канолы, сои и сахарного тростника; более предпочтительно табака и моркови. Варианты осуществления изобретения могут включать в себя клетки, содержащие клеточную стенку, из любой ткани или всего, где они обнаружены, включая, но не ограничиваясь этим, эмбрионы, меристему, каллюс, пыльцу, листья, пыльники, корни, корневые кончики, цветы, семена, стручки, стебли и тканевые культуры.
В вариантах осуществления изобретения представляющая интерес молекула может быть любой молекулой, которая может быть доставлена в растительную клетку в соответствии с данным изобретением. Молекулы, представляющие интерес, или компоненты молекул, представляющих интерес, могут содержать, но не ограничены ими, нуклеиновые кислоты, ДНК, РНК, молекулы РНКи, гены, плазмиды, космиды, искусственные дрожжевые хромосомы, искусственные бактериальные хромосомы, растительные искусственные хромосомы, растительные минихромосомы, сконструированную растительную ДНК характерных локусов; полипептиды, ферменты, гормоны, гликопептиды, сахара, жиры, сигнальные пептиды, антитела, витамины, мессенджеры, вторичные мессенджеры, аминокислоты, цАМФ, лекарственные препараты, гербициды, фунгициды, антибиотики и/или их комбинации.
Варианты осуществления изобретения включают в себя способы предотвращения или лечения заболевания. Неограничивающие показательные варианты осуществления включают в себя доставку фунгицидов, антибиотиков и/или других лекарственных средств в клетки, нуждающиеся в этом, используя способы данного изобретения.
В аспектах изобретения дендример может быть внедрен в различные части клеток. Примеры компартментов, в которые может быть внедрен дендример, включают в себя, но не ограничены ими, цитозоль, ядро, тонопласты, пластиды, этиопласты, хромопласты, лейкопласты, элайопласты, протеинопласты, амилопласты, хлоропласты и просвет двойной мембраны. В других вариантах осуществления изобретения проникновение дендримера в клетку, содержащую клеточную стенку, может происходить через симпластический или апопластический путь.
Дополнительные варианты осуществления изобретения включают в себя генетически модифицированные растительные клетки и способы их генерирования, в которых в растительные клетки введена одна или более нуклеиновых кислот посредством способов данного изобретения. В одном примере варианта осуществления плазмида, содержащая ген, представляющий интерес, и селектируемый маркер, может быть введена в растительную клетку, имеющую клеточную стенку посредством дендримера в соответствии с данным изобретением. В дополнительных вариантах осуществления могут быть отобраны устойчивые трансформанты, которые содержат устойчиво интегрированный ген, представляющий интерес, и/или селектируемый маркер. В альтернативных вариантах осуществления растительная клетка, в настоящее время содержащая ген, представляющий интерес, может быть размножена для выработки других клеток, содержащих представляющую интерес молекулу. В других вариантах осуществления растительные клетки, в настоящее время содержащие представляющую интерес молекулу, могут являться способной к регенерации клеткой, которая может быть использована для регенерирования всего растения, включая представляющую интерес молекулу.
В другом аспекте данное изобретение обеспечивает способы создания регенерируемых растительных клеток, содержащих представляющую интерес молекулу, для использования в тканевой культуре. Выращивание живой ткани предпочтительно будет предоставлять необязательность регенерации растений, имеющих практически такой же генотип, как регенерируемые клетки. Регенерируемые клетки в таких тканевых культурах могут быть эмбрионами, протопластами, меристематическими клетками, каллюсом, пыльцой, листьями, пыльниками, корнями, корневыми кончиками, цветами, семенами, стручками или стеблями. Еще один дополнительный вариант осуществления изобретения обеспечивает растения, регенерированные из тканевой культуры изобретения.
Альтернативно данное изобретение обеспечивает способ внесения требуемого признака в растительную клетку, имеющую клеточную стенку, где способ включает приведение дендримера и представляющей интерес молекулы, способной обеспечивать требуемый признак в растительной клетке, в контакт с клеткой, и предоставление возможности проникновения дендримера через клеточную стенку. Примеры требуемых признаков включают в себя, но не ограничены ими, признаки, выбранные из мужской стерильности, гербицидной устойчивости, резистентности к насекомым и резистентности к бактериальному заболеванию, грибковому заболеванию и/или вирусному заболеванию.
Дополнительные аспекты изобретения предоставляют способы генерирования устойчивых линий растений, содержащих требуемый признак или представляющую интерес молекулу, в которых требуемый признак или представляющая интерес молекула могут быть первоначально введены проникновением дендримера через растительную клеточную стенку. Способы генерирования устойчивых растительных линий хорошо известны рядовым специалистам в данной области и могут включать в себя технологии, такие как, но не ограничиваясь ими, самоопыление, возвратное скрещивание, генерация гибридов, скрещивание популяций и тому подобное. Все растения и растительные клетки, содержащие требуемый признак или представляющую интерес молекулу, первоначально введенную в растительную клетку (или ее предшественники) проникновением дендримера через клеточную стенку, попадают в объем данного изобретения. Предпочтительно растительные клетки, содержащие требуемый признак или представляющую интерес молекулу, первоначально введенную в растение или клетку (или ее предшественники) проникновением дендримера через клеточную стенку, могут быть использованы в скрещиваниях с другими, различными растительными клетками для выработки первого поколения (F1) гибридных клеток, семян и/или растений с превосходящими характеристиками.
В вариантах осуществления, в которых представляющая интерес молекула содержит один или более генов, ген(ы) может быть доминантным или рецессивным аллелем. В качестве примера, ген(ы) будет обеспечивать такие признаки, как устойчивость к гербицидам, устойчивость к насекомым, устойчивость к бактериям, устойчивость к грибам, устойчивость к вирусному заболеванию, мужская плодовитость, мужская стерильность, улучшенные питательные качества и промышленная утилизация.
С появлением молекулярно-биологических технологий, которые позволили выделение и характеризацию генов, которые кодируют конкретный белок, или РНК продуктов (например, молекулы РНКи), у ученых в области растительной биологии появилась большая заинтересованность в конструировании генома клеток так, чтобы он включал в себя и экспрессировал чужеродные гены или дополнительные или модифицированные варианты природных или эндогенных генов (быть может направляемых различными промоторами) для того, чтобы изменять признаки клетки определенным образом. Такие чужеродные дополнительные и/или модифицированные гены в настоящем документе собирательно называются "трансгены". На протяжении последних пятнадцати-двадцати лет было разработано несколько способов выработки трансгенных клеток, и в конкретных вариантах осуществления данное изобретение относится к трансформированным вариантам клеток и способам их получения посредством внесения в клетку, имеющую клеточную стенку, трансгена посредством проникновения дендримера через клеточную стенку. В вариантах осуществления изобретения трансген может быть включен в состав экспрессионного вектора.
Трансформация клетки может содержать создание экспрессионного вектора, который будет функционировать в конкретной клетке. Такой вектор может содержать ДНК, которая включает в себя ген под контролем или функционально связанный с регуляторным элементом (например, промотором). Экспрессионный вектор может включать в себя одну или более таких функционально связанных комбинаций ген/регуляторный элемент. Вектор(ы) может быть в форме плазмиды и может быть использован отдельно или в комбинации с другими плазмидами для обеспечения трансформированных клеток, используя способы трансформации, описанные в данном документе, для включения трансген(ов) в состав генетического материала растительной клетки, содержащей клеточную стенку.
В конкретных вариантах осуществления изобретения прототип многоцелевого дендримера STARBURST® РАМАМ (полиамидоамин) проявляет свойства, пригодные для использования в качестве: (i) направленных контрастных веществ для диагностической MRI (магнитно-резонансная томография) INIR (ближний-IR), (ii) и/или для регулируемой доставки противораковых средств. Среди этого ведущим вариантом является внутренняя часть: 1,4-диаминобутан; G (generation) [PAMAM(CO2Na)64J. Эта дендритовая наноструктура (т.е. -5,0 нм в диаметре) была отобрана на основе очень благоприятного профиля биосовместимости, в Лаборатории Нанотехнологической Характеризации (NCL), отделении Национального института рака (NCI), были выполнены обширные in vitro исследования ведущего вещества и было обнаружено, что оно является достаточно безвредным и биологически совместимым и продемонстрировало направляющие свойства, при этом полагают, что оно будет проявлять требуемые свойства почечной экскреции у млекопитающих. Дендримеры, используемые в соответствии со способами изобретения, представляют класс полимеров, характеризуемый их строго определенной структурой с высоким уровнем молекулярной однородности и низкой полидисперсностью. Кроме того, было показано, что эти дендримеры способны к проникновению без эффлюксных переносчиков.
Использование дендримеров в соответствии со способами данного изобретения привело к образованию устойчиво трансформированных растений и продемонстрировало экспрессию устойчиво трансформированного гербицидного гена с фенотипом, где высокая гербицидная устойчивость была передана трансгенному растению T1. Было продемонстрировано, что это растение является плодовитым, так как оно продуцировало семена T2.
В конкретных способах осуществления был использован реагент для трансфекции SUPERFECT™. Этот реагент является поликатионом, имеющим определенные форму и диаметр, доступный как реагент SUPERFECT™ от Qiagen (Qiagen Catalog #301307) в виде раствора специфически сконструированных активированных дендримеров. Дендримеры являются сферическими полиамидоаминовыми молекулами с ветвями, исходящими из центрального части и заканчивающимися на заряженных концевых аминогруппах. Химическая активация обеспечивает эффективное проникновение ДНК в эукариотические клетки. Без привязки к какой-либо конкретной теории предполагается, что этот реагент формирует из ДНК компактные структуры, тем самым оптимизируя поступление ДНК в клетки. Для стабилизации комплексов SUPERFECT™-ДНК во время их транспорта в ядро, реагент SUPERFECT™ сконструирован так, чтобы буферизовать лизосому после слияния с эндосомой, приводя к pH ингибированию лизосомальных нуклеаз.
Экспрессионные векторы для проникновения посредством дендримера: Маркерные гены
Экспрессионные векторы могут включать в себя по меньшей мере один генетический маркер, функционально связанный с регуляторным элементом (промотором, например), что позволяет трансформированным клеткам, содержащим маркер, быть или регенерированными негативным отбором (т.е. ингибируя рост клеток, которые не включают в себя селектируемый маркерный ген), или позитивным отбором (т.е. скринингом на продукт, кодируемый генетическим маркером). Многие селектируемые маркерные гены для трансформации хорошо известны в области трансформации и включают в себя, например, гены, которые кодируют ферменты, которые метаболически дезинтоксицируют селективное химическое вещество, которое может быть антибиотиком или гербицидом, или гены, которые кодируют измененную мишень, которая может быть невосприимчивой к ингибитору. Также в данной области известно несколько способов позитивного отбора.
Один из широко используемых селектируемых маркерных генов, пригодных для растительной трансформации, может содержать в себе ген - неомицин-фосфотрансферазу II (nptII) под контролем растительных регуляторных сигналов, который предоставляет резистентность к канамицину. См., например, Fraley et al., Proc. Natl. Acad. Sci. U.S.A., 80:4803 (1983). Другим широко используемым селектируемым маркерным геном может быть ген - гигромицин-фосфотрансфераза, который предоставляет резистентность к антибиотику гигромицину. См., например, Vanden Elzen et al., Plant Mol. Biol., 5:299 (1985).
Дополнительные селектируемые маркерные гены бактериального происхождения, которые предоставляют резистентность к антибиотикам, включают в себя гентамицин-ацетилтрансферазу, стрептомицин фосфотрансферазу, аминогликозид-3'-аденил трансферазу и детерминанту резистентности к блеомицину. См. Hayford et al., Plant Physiol. 86:1216 (1988); Jones et al., Mol. Gen. Genet., 210:86 (1987); Svab et al., Plant Mol. Biol. 14:197 (1990); Hille et al., Plant Mol. Biol. 7:171 (1986). Другие селектируемые маркерные гены предоставляют резистентность к гербицидам, таким как глифосат, глюфосинат или бромоксинил. См. Comai et al., Nature 317:741-744 (1985); Gordon-Kamm et al., Plant cell 2:603-618 (1990); и Stalker et al., Science 242:419-423 (1988).
Другие селектируемые маркерные гены, пригодные для трансформации растений, имеют не бактериальное происхождение. Эти гены включают в себя, например, мышиную дигидрофолатредуктазу, растительную 5-енолпирувил-шикимат-3-фосфат-синтазу и растительную ацетолактатсинтазу. См. Eichholtz et al., Somatic cell Mol. Genet. 13:67 (1987); Shah et al., Science 233:478 (1986); Charest et al., Plant Cell Rep. 8:643 (1990).
Для другого класса маркерных генов, пригодных для растительной трансформации, в большей степени необходим скрининг предположительно трансформированных растительных клеток, чем прямой генетический отбор трансформированных клеток на резистентность к токсичному веществу, такому как антибиотик. Эти гены особенно подходят для количественного определения или визуализации пространственной структуры экспрессии гена в определенных тканях и часто называются репортерными генами, потому что они могут быть слиты с геном или регуляторной последовательностью гена для исследования экспрессии гена. Широко используемые гены для скрининга трансформированных клеток включают в себя β-глюкуронидазу (GUS), β-галактозидазу, люциферазу и хлорамфеникол ацетилтрансферазу. См. Jefferson, R.A, Plant Mol. Biol. Rep. 5:387 (1987); Teeri et al., EMBO J. 8:343 (1989); Koncz et al., Proc. Natl. Acad. Sci. U.S.A. 84:131 (1987); DeBlock et al., EMBO J. 3:1681 (1984).
Недавно стали доступны способы визуализации in vivo GUS активности, для которых не требуется разрушение растительной ткани. Molecular Probes publication 2908, Imagene Green™, p. 1-4(1993); и Naleway et al., J. Cell Biol. 115:151a (1991). Тем не менее, не была продемонстрирована пригодность этих способов к in vivo визуализации GUS активности для регенерации трансформированных клеток из-за низкой чувствительности, высокого флуоресцентного фона и ограничений, связанных с использованием люциферазных генов в качестве селектируемых маркеров.
В последнее время гены, кодирующие флуоресцентные белки (например, GFP, EGFP, EBFP, ECFP и YFP), используются в качестве маркеров экспрессии генов в прокариотических и эукариотических клетках. См. Chalfie et al., Science 263:802 (1994). Флуоресцентные белки и мутации флуоресцентных белков могут быть использованы в качестве позволяющих проводить отбор маркеров.
Экспрессионные векторы для проникновения посредством дендримера: Промоторы
Гены, включенные в состав экспрессионных векторов, должны быть направлены нуклеотидной последовательностью, содержащей регуляторный элемент, например, промотором. Несколько типов промоторов в настоящее время хорошо известны в области трансформации, как и другие регуляторные элементы, которые могут быть использованы отдельно или в комбинации с промоторами.
В используемом в данном документе значении "промотор" включает в себя ссылку на область ДНК, которая может быть расположена перед началом транскрипции и которая может быть вовлечена в распознавание и связывание РНК полимеразы и других белков для инициации транскрипции. "Растительный промотор" может быть промотором, способным к инициированию транскрипции в растительных клетках. Примеры промоторов под контролем развития включают в себя промоторы, которые предпочтительно инициируют транскрипцию в некоторых тканях, таких как листья, корни, семена, волокна, сосуды ксилемы, трахеиды или склеренхима. Такие промоторы называются "ткане-предпочтительные". Промоторы, которые инициируют транскрипцию только в некоторых тканях называются "ткане-специфичные". Специфичный к "типу клетки" промотор первоначально запускает экспрессию в некоторых типах клеток в одном или более органов, например, в васкулярных клетках в корнях или листьях. "Индуцируемый" промотор может быть промотором, который может быть под контролем окружающей среды. Примеры связанных с окружающей средой условий, которые могут воздействовать на транскрипцию посредством индуцируемых промоторов, включают в себя анаэробные условия или наличие света. Ткане-специфичные, ткане-предпочтительные, специфичные к типу клетки и индуцируемые промоторы составляют класс "неконститутивных" промоторов. "Конститутивный" промотор может быть промотором, который может быть активным при большинстве внешних условий.
A. Индуцируемые промоторы
Индуцируемый промотор может быть функционально связан с геном для экспрессии в клетке. Если требуется, индуцируемый промотор может быть функционально связан с нуклеотидной последовательностью, кодирующей сигнальную последовательность, которая может быть функционально связана с геном для экспрессии в клетке. При наличии индуцируемого промотора уровень транскрипции повышается под влиянием индуцирующего агента.
В данном изобретении может быть использован любой индуцируемый промотор. См. Ward et al., Plant Mol. Biol. 22:361-366 (1993). Типичные индуцируемые промоторы включают в себя, но не ограничены ими: промотор из системы ACEI, который реагирует на медь (Mett et al., PNAS 90:4567-4571 (1993)); ген In2 из маиса, который реагирует на бензолсульфонамидные антидоты для гербицидов (Hershey et al., Mol. Gen. Genetics 227:229-237 (1991); и Gatz et al., Mol. Gen. Genetics 243:32-38 (1994)); и репрессор Tet из Tn10 (Gatz et al., Mol. Gen. Genetics 227:229-237 (1991)). Особенно пригодным индуцируемым промотором может быть промотор, который реагирует на индуцирующий агент, на который растения обычно не реагируют. Типичным индуцируемым промотором может быть индуцируемый промотор из гена стероидного гормона, транскрипционная активность которого может быть вызвана глюкокортикостероидным гормоном. Schena et al., Proc. Natl. Acad. Sci. U.S.A. 88:0421 (1991).
B. Конститутивные промоторы
Конститутивный промотор может быть функционально связан с геном для экспрессии в клетке или конститутивный промотор может быть функционально связан с нуклеотидной последовательностью, кодирующей сигнальную последовательность, которая может быть функционально связана с геном для экспрессии в клетке.
В данном изобретении могут быть использованы различные конститутивные промоторы. Типичные конститутивные промоторы включают в себя, но не ограничены ими: промоторы из растительных вирусов, такие как промотор 35S из CaMV (Odell et al., Nature 313:810-812 (1985)); промоторы из гена актина риса (McElroy et al., Plant Cell 2:163-171 (1990)); убиквитина (Christensen et al., Plant Mol. Biol. 12:619-632 (1989); и Christensen et al., Plant Mol. Biol. 18:675-689 (1992)); pEMU (Last et al., Theor. Appl. Genet. 81:581-588 (1991)); MAS (Velten et al., EMBO J. 3:2723-2730 (1984)); и гистона H3 маиса (Lepetit et al., Mol. Gen. Genetics 231:276-285 (1992); и Atanassova et al., Plant Journal 2 (3):291-300 (1992)). Промотор ALS, Xba1/NcoI фрагмент 5'-структурного гена ALS3 Brassica napus (или сходная с указанным Xba1/NcoI фрагментом нуклеотидная последовательность), представляет особенно подходящий конститутивный промотор. См. заявку PCT WO 96/30530.
C. Ткане-специфичные или ткане-предпочтительные промоторы
Ткане-специфичный промотор может быть функционально связан с геном для экспрессии в клетке. Необязательно ткане-специфичный промотор может быть функционально связан с нуклеотидной последовательностью, кодирующей сигнальную последовательность, которая может быть функционально связана с геном для экспрессии в клетке. Растения, трансформированные геном, представляющим интерес, функционально связанным с ткане-специфичным промотором, могут продуцировать белковый продукт трансгена исключительно или предпочтительно в определенной ткани.
В данном изобретении может быть использован любой ткане-специфичный или ткане-предпочтительный промотор. Типичные ткане-специфичные или ткане-предпочтительные промоторы включают в себя, но не ограничены ими, предпочтительный по отношению к корню промотор - такой как промотор из гена фазеолина (Murai et al., Science 23:476-482 (1983); и Sengupta-Gopalan et al., Proc. Natl. Acad. Sci. U.S.A. 82:3320-3324 (1985)); специфичный по отношению к листу и свет-индуцированный промотор, такой как промотор из cab или rubisco (Simpson et al., EMBO J. 4(11):2723-2729 (1985); и Timko et al., Nature 318:579-582 (1985)); специфичный по отношению к пыльнику промотор, такой как промотор из LAT52 (Twell et al., Mol. Gen. Genetics 217:240-245(1989)); специфичный по отношению к пыльце промотор, такой как промотор Zm13 (Guerrero et al., Mol. Gen. Genetics 244:161-168 (1993)) или предпочтительный по отношению к микроспоре промотор, такой как промотор из apg (Twell et al., Sex. Plant Reprod. 6:217-224(1993)).
Генерируемый трансгенами транспорт белка в субклеточный компартмент, такой как хлоропласт, вакуоль, пероксисома, глиоксисома, клеточная стенка или митохондрия или для секреции в апопласт, может быть выполнен посредством функционального присоединения нуклеотидной последовательности, кодирующей сигнальную последовательность, к 5'- и/или 3'-области гена, кодирующего представляющий интерес белок. Направляющие последовательности на 5'- и/или 3'-конце структурного гена могут определять во время синтеза и процессинга белка то, в какой компартмент в конечном счете может быть направлен кодируемый белок. Альтернативно, такие субклеточные направляющие в компартмент белки могут быть напрямую связаны с дендримером для направления дендримера, покрытого представляющей интерес молекулой, в требуемый субклеточный компартмент.
Присутствие сигнальной последовательности направляет полипептид или во внутриклеточную органеллу, или субклеточный компартмент, или для секреции в апопласт. Многие сигнальные последовательности известны в данной области. См., например, Becker et al., Plant Mol. Biol. 20:49 (1992); P.S. Close, Master's Thesis, Iowa State University (1993); C. Knox et al., "Structure and Organization of Two Divergent Alpha-Amylase Genes from Barley," Plant Mol. Biol. 9:3-17 (1987); Lerner et al., Plant Physiol. 91:124-129 (1989); Fontes et al., Plant Cell 3:483-496 (1991); Matsuoka et al., Proc. Natl. Acad. Sci. 88:834 (1991); Gould et al., J. Cell. Biol. 108:1657 (1989); Creissen et al., Plant J. 2:129 (1991); Kalderon, et al., A short amino acid sequence able to specify nuclear location, Cell 39:499-509 (1984); Steifel, et al., Expression of a maize cell wall hydroxyproline-rich glycoprotein gene in early leaf and root vascular differentiation, Plant Cell 2:785-793 (1990).
Гены чужеродных белков и агрономически важные гены
С трансгенными растениями по данному изобретению чужеродный белок может быть продуцирован в промышленных количествах. Таким образом, технологии отбора и воспроизводства трансформированных растений, которые хорошо известны в данной области, приводят в результате к множеству трансгенных растений, которые выращиваются традиционным способом, и после этого чужеродный белок может быть выделен из ткани, представляющей интерес, или из общей биомассы. Выделение белка из растительной биомассы может быть выполнено известными способами, которые рассматриваются, например, Heney и Orr, Anal. Biochem. 114:92-6 (1981).
В аспектах изобретения трансгенные растения, предоставленные для промышленного производства чужеродных белков, могут быть клеткой или растением. В других аспектах биомасса, представляющая интерес, может быть семенем. Для относительно небольшого количества трансгенных растений, которые демонстрируют более высокие уровни экспрессии, может быть первоначально генерирована генетическая карта с помощью общепринятых RFLP, ПЦР и SSR анализа, который определяет приблизительную хромосомную локализацию интегрированной молекулы ДНК. В качестве иллюстративных методик в этом направлении, см. Glick and Thompson, Methods in Plant Molecular Biology and Biotechnology CRC Press, Boca Raton 269:284 (1993). Данные относительно хромосомной локализации могут быть пригодны для патентованной защиты подвергающегося испытаниям трансгенного растения. Если может иметь место недопустимое воспроизводство и произведено скрещивание с другой идиоплазмой, то может быть осуществлено сравнение карты интеграционной области с подобными картами для предполагаемых растений для определения того, если последние имеют общее происхождение с испытуемым растением. Сравнения карт могли бы включать в себя гибридизации, RFLP, ПЦР, SSR и секвенирование, все из которых являются общепринятыми технологиями.
Подобным образом агрономически важные гены могут быть экспрессированы в трансформированных клетках или их потомстве. Конкретнее, растения могут быть генетически сконструированы посредством способов по изобретению для экспрессирования различных фенотипов, представляющих интерес с агрономической точки зрения. Типичные гены, которые могут быть использованы в этом отношении, включают в себя, но не ограничены ими, распределенные по группам ниже.
1. Гены, которые предоставляют резистентность к вредителям или заболеваниям и которые кодируют:
А) Гены резистентности к растительному заболеванию. Растительные способы защиты часто активируются определенными взаимодействиями между продуктом гена (R) резистентности к заболеванию в растении и продуктом соответствующего гена невирулентности (Avr) в патогене. Несколько растений могут быть трансформированы клонированными генами резистентности для создания растений, которые резистентны к определенным патогенным штаммам. См., например, Jones et al., Science 266:789 (1994) (клонирование гена Cf-9 томата для резистентности к Cladosporium fulvum); Martin et al., Science 262:1432 (1993) (ген Pto томата для резистентности к Pseudomonas syringae pv. tomato кодирует протеинкиназу); Mindrinos et al., Cell 78:1089 (1994) (Arabidops может иметь ген RSP2 для резистентности к Pseudomonas syringae).
В) Ген, предоставляющий резистентность к вредителю, такому как цистообразующая нематода сои. См. например, публикацию PCT WO 96/30517; публикацию PCT WO 93/19181.
С) Белок Bacillus thuringiensis, его производное или синтетический полипептид, смоделированный на его основе. См., например, Geiser et al., Gene 48:109 (1986), где раскрывается клонирование и нуклеотидная последовательность гена Bt δ-эндотоксина. Кроме того, молекулы ДНК, кодирующие гены δ-эндотоксина, могут быть приобретены у American Type Culture Collection, Манассас, Виргиния, например, под ATCC номерами доступа 40098, 67136, 31995 и 31998.
D) Лектин. См., например, Van Damme et al., Plant Molec. Biol. 24:25 (1994), который обнаружил нуклеотидные последовательности нескольких генов лектина Clivia miniata, связывающего маннозу.
E) Витамин-связывающий белок, такой как авидин. См. публикацию PCT US93/06487. Заявка описывает использование авидина и гомологов авидина в качестве ларвицидов против насекомых-вредителей.
F) Ингибитор ферментов, например, ингибитор протеазы или протеиназы или ингибитор амилазы. См., например, Abe et al., J. Biol. Chem. 262:16793 (1987) (нуклеотидная последовательность ингибитора цистенин протениназы риса); Huub et al., Plant Molec. Biol. 21:985 (1993) (нуклеотидная последовательность кДНК, кодирующей ингибитор I протеиназы табака); Sumitani et al., Biosci. Biotech. Biochem. 57:1243 (1993) (нуклеотидная последовательность ингибитора α-амилазы Streptomyces nitrosporeus) и патент США № 5494813 (Hepher and Atkinson, изданный 27 февраля 1996 г.).
G) Специфичный по отношению к насекомым гормон или феромон, такой как экдистероид или ювенильный гормон, их варианты, миметик на их основе или их антагонист или агонист. См., например, открытие Hammock et al., Nature 344:458 (1990), бакуловирусная экспрессия клонированной эстеразы ювенильного гормона, инактиватор ювенильного гормона.
H) Специфичный по отношению к насекомым пептид или нейропептид, который при экспрессии нарушает физиологию подвергшегося воздействию вредителя. Например, см. Regan, J. Biol. Chem. 269:9 (1994) (экспрессионное клонирование приводит к ДНК, кодирующей рецептор диуретического гормона насекомых); и Pratt et al., Biochem. Biophys. Res. Comm. 163: 1243 (1989) (аллостатин может быть идентифицирован в Diploptera puntata). См. также патент США № 5266317 Tomalski et al., который раскрывает гены, кодирующие специфичные по отношению к насекомым паралитические нейротоксины.
I) Специфичный по отношению к насекомым яд, вырабатывающийся в природе змеей, осой или любым другим организмом. Например, см. Pang et al., Gene 116:165 (1992), открытие в растениях гетерологичной экспрессии гена, кодирующего токсичный для насекомых пептид скорпиона.
J) Фермент, ответственный за повышенное накопление монотерпена, сесквитерпена, стероидного гормона, гидроксамовой кислоты, производного фенилпропаноида или другой небелковой молекулы с инсектицидной активностью.
K) Фермент, вовлеченный в модификацию, включая пост-трансляционную модификацию биологически активной молекулы; например, гликолитический фермент, протеолитический фермент, липолитический фермент, нуклеаза, циклаза, трансаминаза, эстераза, гидролаза, фосфатаза, киназа, фосфорилаза, полимераза, эластаза, хитиназа и глюканаза, или природный, или синтетический. См. публикацию PCT WO 93/02197 на имя Scott et al., который обнаружил нуклеотидную последовательность гена каллазы. Могут быть получены молекулы ДНК, которые содержат кодирующую хитиназу последовательность, например, под ATCC номерами доступа 39637 и 67152. См. также Kramer et al., Insect Biochem. Molec. Biol. 23:691 (1993), который сообщает о нуклеотидной последовательности кДНК, кодирующей хитиназу бражника Protoparce sexta; и Kawalleck et al., Plant Molec. Biol. 21:673 (1993), который предоставляет нуклеотидную последовательность гена ubi4-2 полиубиквитина петрушки.
L) Молекула, которая стимулирует сигнальную трансдукцию. Например, см. Botella et al., Plant Molec. Biol. 24:757 (1994), нуклеотидные последовательности кДНК клонов кальмодулина золотистой фасоли, и Griess et al., Plant Physiol. 104:1467 (1994), который предоставляет нуклеотидную последовательность кДНК клона кальмодулина маиса.
M) Пептид с гидрофобными свойствами. См. публикацию PCT WO 95/16776 (раскрытие производных пептида Tachyplesin, который подавляет грибковые растительные патогены) и публикация PCT WO 95/18855 (сообщает о синтетических противомикробных пептидах, которые предоставляют резистентность к заболеванию).
N) Мембранная пермеаза, формирующая или блокирующая канал. Например, см. открытие Jaynes et al., Plant Sci 89:43 (1993) гетерологичной экспрессии литического пептида аналога цекропина-β для придания трансгенным табачным растениям резистентности к Pseudomonas solanacearum.
O) Вирусный инвазивный белок или полученный из него сложный токсин. Например, накопление белков вирусной оболочки в трансформированных растительных клетках приводит к резистентности к вирусной инфекции и/или развитию заболевания, вызванного вирусом, из которого может быть получен ген белка оболочки, а также родственными вирусами. См. Beachy et al., Ann. rev. Phytopathol. 28:451 (1990). Опосредованная белком оболочки резистентность предоставлена трансформированным растениям против вируса мозаика люцерны, вируса мозаики огурцов, вируса полосы табака, вируса X картофеля, вируса Y картофеля, вируса гравировки табака, вируса погремковости табака и Id вируса табачной мозаики.
P) Специфичное по отношению к насекомым антитело или полученный из него иммунотоксин. Таким образом, антитело, направленное на критическую метаболическую функцию в насекомом, может инактивировать подверженный воздействию фермент, уничтожая насекомое. Cf. Taylor et al., Abstract #497, Seventh Int'l Symposium on Molecular Plant-Microbe Interactions (Эдинбург, Шотландия) (1994) (ферментативная инактивация в трансгенном табаке посредством выработки одноцепочечных фрагментов антитела).
Q) Специфичное по отношению к вирусу антитело. См., например, Tavladoraki et al., Nature 366:469 (1993), который демонстрирует, что трансгенные растения, экспрессирующие гены рекомбинантных антител, защищены от поражения вирусом.
R) Ингибирующий развитие белок, вырабатывающийся в природе патогеном или паразитом. Например, грибковые эндо-α-1,4-D-полигалактуроназы способствуют образованию колоний грибков и высвобождению растительного нутриента растворением растительной клеточной стенки гомо-α-1,4-D-галактуроназой. См. Lamb et al., Bio/Technology 10:1436 (1992). Клонирование и характеризация гена, который кодирует белок, ингибирующий эндополигалактуроназу фасоли, могут быть описаны Toubart et al., Plant J. 2:367 (1992).
S) Ингибирующий развитие белок, вырабатывающийся в природе растением. Например, Logemann et al., Bio/Technology 10:305 (1992), продемонстрировал, что трансгенные растения, экспрессирующие инактивирующий рибосому ген ячменя, обладают повышенной резистентностью к грибковому заболеванию.
2. Гены, которые предоставляют резистентность к гербициду:
A) Гербициды, которые подавляют конус нарастания или меристему, такие как имидазолинон, сульфонамид или сульфонилмочевина. Типичные гены в этот группе кодируют мутант фермента ALS и AHAS, как описано, например, Lee et al., EMBO J. 7:1241 (1988), и Miki et al., Theor. Appl. Genet. 80:449 (1990), соответственно.
B) Гены глифосата (резистентность, предоставленная, например, мутантными генами 5-енолпирувилшикимат-3-фосфатсинтазы (EPSP) (посредством введения рекомбинантных нуклеиновых кислот и/или различных видов in vivo мутагенеза природных генов EPSP), генами aroA и генами глифосат-ацетилтрансферазы (GAT), соответственно), другие фосфоновые соединения, такие как глюфосинат (гены фосфинотрицин-ацетилтрансферазы (PAT) из видов стрептомицет, включая гены Streptomyces hygroscopicus и Streptomyces viridichromo), и пиридинокси или фенокси проприоновые кислоты и циклогексоны (кодирующие гены ингибитора ацетил КоА карбоксилазы), см., например, патент США № 4940835 Shah, et al., и патент США № 6248876 Barry et. al., которые раскрывают нуклеотидные последовательности форм EPSP, которые могут предоставлять растениям резистентность к глифосату. Молекула ДНК, кодирующая мутант гена aroA, может быть получена под ATCC номером доступа 39256, и нуклеотидная последовательность мутантного гена раскрыта в патенте США № 4769061 Comai. Заявка на европейский патент № 0333033 Kumada et al., и патент США № 4975374 Goodman et al., раскрывают нуклеотидные последовательности генов глутамин синтетазы, которые предоставляют резистентность к гербицидами, таким как L-фосфинотрицин. Нуклеотидная последовательность гена PAT может быть представлена в европейской заявке № 0242246 Leemans et al., DeGreef et al., Bio/Technology 7:61 (1989), описывающей производство трансгенных растений, которые экспрессируют химерные сцепленные гены, кодирующие PAT активность. Типичные гены, предоставляющие резистентность к феноксипроприоновым кислотам и циклогексонам, таким как сетоксидим и галоксифоп, включают в себя гены Acc1-S1, Acc1-S2 и Acc1-S3, описанные Marshall et al., Theor. Appl. Genet. 83:435 (1992). Гены GAT, способные предоставлять резистентность к глифосату, описаны в WO 2005012515 Castle et al. Гены, предоставляющие резистентность к 2,4-D-феноксипроприоновой кислоте и пиридилоксиауксин гербицидам, описаны в WO 2005/107437, закрепленном за Dow AgroSciences LLC.
C) Гербицид, который ингибирует фотосинтез, такой как триазин (гены psbA и gs+) или бензонитрил (ген нитрилазы). Przibila et al., Plant cell 3:169 (1991) описывает трансформации Chlamydomonas плазмидами, кодирующими мутантные гены psbA. Нуклеотидные последовательности генов нитрилазы раскрыты в патенте США № 4810648 Stalker, и ДНК молекулы, содержащие эти гены, доступны под ATCC номерами доступа 53435, 67441 и 67442. Клонирование и экспрессия ДНК, кодирующей глутатион-S-трансферазу, могут быть описаны Hayes et al., Biochem. J. 285:173 (1992).
3. Гены, которые предоставляют или вносят вклад в повышающий ценность признак, такой как:
A) Модифицированный метаболизм жирных кислот, например, трансформированием растения антисмысловым геном стеарил-ACP десатуразы для повышения содержания стеариновой кислоты в растении. См. Knultzon et al., Proc. Natl. Acad. Sci. U.S.A. 89:2624 (1992).
B) Пониженное содержание фитата - 1) Введение гена, кодирующего фитазу - будет повышать разложение фитата, при этом увеличивая больше свободного фосфата в трансформированном растении. Например, см. Van Hartingsveldt et al., Gene 127:87 (1993), обнаружение нуклеотидной последовательности гена фитазы Aspergillus niger. 2) Может быть введен ген для понижения содержания фитата. В маисе, например, это может быть выполнено посредством клонирования и последующего повторного внесения ДНК, ассоциированной с одним аллелем, который может отвечать за мутанты маиса, характеризуемые низкими уровнями фитиновой кислоты. См. Raboy et al., Maydica 35:383 (1990).
C) Модифицированная углеводная композиция, произведенная, например, трансформированием растения геном, кодирующим фермент, который изменяет схему ветвления крахмала. См. Shiroza et al., J. Bacteol. 170:810 (1988) (нуклеотидная последовательность мутантов гена фруктозилтрансферазы Streptococcus); Steinmetz et al., Mol. Gen. Genet. 20:220 (1985) (нуклеотидная последовательность Bacillus subtilis может иметь ген левансахаразы); Pen et al., Bio/Technology 10:292 (1992) (производство трансгенных растений, которые экспрессируют α-амилазу Bacillus lichenifonn); Elliot et al., Plant Molec. Biol. 21:515 (1993) (нуклеотидные последовательности генов инвертазы томата); Sogaard et al., J. Biol. Chem. 268:22480 (1993) (может быть выполнен сайт-направленный мутагенез гена α-амилазы ячменя); и Fisher et al., Plant Physiol. 102:1045 (1993) (ветвящий фермент II эндосперма маиса).
ПРИМЕРЫ
Данное изобретение дополнительно описано в последующих примерах, которые представлены в качестве иллюстрации, и не подразумевается, что они каким-либо образом ограничивают изобретение.
ПРИМЕР 1
ПРИГОТОВЛЕНИЕ КОМПЛЕКСА ДЕНДРИМЕР/ДНК И ОБРАБОТКА КЛЕТОК
1.1 Приготовление плазмиды
Плазмидная ДНК pDAB3831 (ФИГ.1) (SEQ ID NO: 1 и 2) была выделена и подготовлена для дендример-опосредованной растительной трансформации. Провели трансформационные эксперименты как с кольцевой ДНК, так и с линейной ДНК.
Для того чтобы линеаризовать pDAB3831, была выполнена ПЦР реакция. pDAB3831 была амплифицирована ПЦР с использованием системы непрерывных тепловых циклов, которая была описана ранее, например, в WO 2008045288. Вместо использования маленьких пробирок в непрерывных термоциклерах применяется постоянный или непрерывный поток жидкости, многократно пропускаемый через различные температурные зоны для амплификации ДНК. Смесь для ПЦР реакции была введена в жидкость-носитель, с которым смесь для ПЦР реакции не смешивается. Жидкость-носитель после этого пропущена через некоторое количество температурных зон для обеспечения амплификации ДНК в смеси для ПЦР реакции. Был приготовлен образец, содержащий: 12% MgCl2 (25 мМ), 0.33% Taq ДНК полимеразы (5 единиц/мкл), 2,0% dNTP (дезоксиаденозинтрифосфат (dATP), дезоксицитидинтрифосфат (dCTP), дезоксигуанозинтрифосфат (dGTP) и дезокситимидинтрифосфат (dTTP), 8,0% матрицы (2 мкг/мл), 61,66% раствора Pluronic F108 (1,5% раствор), 4% прямого праймера, 4% обратного праймера и 8% реакционного буфера (10X концентрация). В соединенных звеньях системы была установлена температура 95°C, 59°C и 72°C для диссоциации, отжига и элонгации, соответственно. Смесь для ПЦР реакции была прокачена через систему с использованием герметичной емкости при 13,79 Н/см2. После того как реакционная смесь была нагрета до температуры корпуса регулятора, было применено минеральное масло для продавливания образца через всю длину системы трубок. Скорость потока реакционной смеси регулировалась на уровне 0,25 мл/мин с помощью регулятора потока. Определенная последовательность ДНК, присутствующая в образце, была амплифицирована по мере того, как она циклично пропускалась через температурные зоны. После тридцатого цикла содержимое было собрано. ПЦР продукт был очищен на гель-фильтрационной колонке с последующим осаждением этанолом. Образец очищенного продукта был проанализирован с помощью Agilent Bioanalyzer, а также на электрофорезе в агарозном геле для подтверждения размера и концентрации продукта ПЦР.
Матрицей, использованной для описанной выше ПЦР, была DAS плазмида pDAB3831, которая содержит селектируемый маркерный ген PAT под управлением промотора убиквитина 10 Arabidopsis (AtUbi10) и ген желтого флюоресценого белка Philadium (PhiYFP) под управлением промотора вируса прожилковой мозаики маниоки (CsVMV). Прямой праймер SEQ ID NO: 3 и обратный праймер SEQ ID NO: 4 были синтезированы для амплификации полной экспрессионной кассеты длиной 4,6 т.п.н. (т.е. линеаризованной ДНК), содержащей как гены, так и их промоторы. Кроме того, для содействия конъюгации линейной двухцепочечной ДНК с поверхностью наночастицы молекула биотина была химически связана с фосфатной группой праймеров с использованием биотин-TEG-CE-фосфорамидита. Этот фосфорамидит имеет вытянутую спейсерную группу смешанной полярности из 15 атомов на основе триэтиленгликолевого линкера. Эта вытянутая спейсерная группа может отделять биотиновую функциональную группу от остальной части олиго для того, чтобы выгодно снизить любые возможные эффекты стерических затруднений во время связывания с молекулой стрептавидина. Если был помечен прямой праймер, то биотин находился в начале ДНК. Если был помечен обратный праймер, то биотин находился на конце ДНК фрагмента. Следовательно, биотинилированный (в обеих ориентациях) ДНК-фрагмент может быть прикреплен к покрытым стрептавидином наночастицам. При применении биотинилированных олигов и системы непрерывных тепловых циклов было продуцировано приблизительно 20 мг линейных фрагментов ДНК.
1.2 Приготовление комплекса ДНК/дендример
Дендримеры, использованные для этих экспериментов, являлись сферическими катионными полиамидоаминными (РАМАМ) каскадными полимерами, которые состоят из первичных аминов на поверхности и третичных аминов во внутренней части. Дендримеры частично разрушаются посредством тепловой обработки в сольволитических растворителях, тем самым приводя к меньшим стерическим ограничениям и большей гибкости. Большой положительный заряд дендримеров способствует электростатическим взаимодействиям с ДНК, и гибкая структура позволяет дендримеру уплотняться как в связанном с ДНК состоянии, так и отдельно от нее. Эффективность трансфекции или трансформации повышена вследствие положительного заряда и гибкой структуры дендримера.
Дендримеры, распространяемые под названием трансфекционный реагент SUPERFECT™ (Cat #301305), были получены от Qiagen (Джермантаун, Мэриленд). Плазмидная ДНК была смешана с 0,6 мл реагента SUPERFECT™ и инкубировалась в течение 30 минут при 24°C для образования комплекса ДНК/дендример. Различные концентрации кольцевой плазмидной ДНК (0,1 мг и 0,5 мг) были использованы для образования комплекса ДНК/дендример. Кроме того, линеаризованная ДНК, описанная выше в примере 1.1, была использована для образования комплекса ДНК/дендример. Были использованы концентрации 0,1 мг и 0,5 мг линейной ДНК. После образования комплекса ДНК/дендример раствор объемом 10 мл, содержащий 5% сахарозы и 0,02-0,04% Silwet-L77, был добавлен в реакцию ДНК/дендример.
ПРИМЕР 2
Доставка комплекса ДНК/дендример и устойчивая трансформация ARABIDOPSIS THALIANA
2.1 Растительный материал для трансформации in planta:
Синхронизированное прорастание семени важно для обеспечения однородности цветочного развития в растениях T0. Семя Arabidopsis thaliana cv. Columbia было ресуспендировано в 0,1% растворе агара и инкубировалось при 4°C в течение 48 часов до полного расслоения. 60 мг семени было взвешено и перенесено в 15 мл пробирку. Было добавлено 13 мл 0,1% раствора агара и осуществлялось встряхивание до тех пор, пока семя не было окончательно диспергировано. Это привело к концентрации 4,6 мг семени/1 мл раствора (или приблизительно 230 семян/мл). Были приготовлены шесть пробирок (раствор 72 мл) для высевания 4 лотков, которые включали в себя 18 (31/2 дюйм) горшков в каждом поддоне. Раствор был инкубирован при 4°C в течение 48 часов до полного расслоения. В каждый горшок было засеяно отдельно 1,0 мл расслоенного раствора семени на горшок. Когда все горшки были засеяны, колпаки для выращивания были помещены на поддоны для того, чтобы поддерживать влажность почвы. Колпаки были удалены через 5 дней после даты посева. Семена были проращены и растения были выращены в CONVIRON® (модели CMP4030 и CMP3244, Controled Environments Limited, Виннипег, Манитоба, Канада) при условиях «многих дней» (16 часов свет/8 часов темнота) при интенсивности света 120-150 мкмоль/м2с при постоянных температуре (22°C) и влажности (40-50%). Растения увлажнялись с 10 по 14 дни после посева растений питательной смесью Хогланда и впоследствии водой DI для поддержания почвы влажной, но не мокрой. Через четыре недели после даты засева цветы были срезаны для выработки еще более интенсивно растущих вторичных цветов. На пятую неделю после засева растения были подготовлены к процессу трансформации.
2.2 Трансформация in planta и скрининг T1 резистентных растений:
Дендример-опосредованная трансформация Arabidosis thaliana cv. Columbia была выполнена с использованием модифицированного протокола Clough and Bent. (S.J. Clough and A.F. Bent, 1998, Plant J 16:735-43). Была приготовлена 10 мл взвесь с раствором ДНК/дендример и использована для обработок растений Arabidopsis (в основном незрелых ботрических соцветий с несколькими опыленными стручками). Комплексы кольцевая ДНК/дендример и комплексы линейная ДНК/дендример были использованы независимо друг от друга. Перед погружением растений был добавлен Silwet L-77 в концентрации 0,05% (250 мкл/500 мл) - 0,005% и хорошо перемешан с раствором ДНК/дендример. Надземные части растения были погружены в раствор ДНК/дендример на 2-30 секунд с легким встряхиванием. Обрабатываемые растения выдерживались под пластиковой куполовидной крышкой в течение 16-24 часов при 22-24°C. Растения были перенесены в CONVIRONS®, и им позволили расти до созревания и вырастить семена. Поддоны для отбора (поддоны 10,5"×21"×1") были использованы для наблюдения за основным семенем урожая из растений T0, приблизительно 10000 семян на каждый поддон. Два контроля были использованы для обеспечения того, чтобы селекционное опрыскивание было выполнено правильно: Col-0 негативный трансформант-контроль и Columbia Col-0 дикого типа, обогащенный гомозиготным семенем с PAT (фосфинотрицин-ацетилтрансфераза) - селектируемым маркером в качестве положительного трансформанта-контроля. Для достижения синхронизации семена были расслоены в 0,1% растворе агара в течение 48 часов перед высеванием. Для обеспечения 10000 семян на селекционную поддон, 200 мг семян были добавлены к 0,1% раствору агара и встряхивались до тех пор, пока семена не были равномерно суспендированы. Расслоенные семена после этого были выращены в селекционных поддонах, наполненных смесью Sunshine LP5 и слабо орошаемых питательной смесью Хогланда. Для повышения эффективности селекционного спрея 40 мл суспендированного семени было засеяно равномерно на селекционный поддон. После засевания на каждый селекционный поддон были помещены колпаки для выращивания и растения выращивались для отбора, колпаки для выращивания были удалены приблизительно через пять дней после засевания.
Кроме того, был выполнен контрольный эксперимент, в котором раствор, содержащий только ДНК без дендримеров, был использован для трансформации Arabidopsis thaliana. Для трансформации «голой» ДНК был использован ранее описанный протокол. И линейная, и кольцевая формы ДНК были использованы независимо друг от друга.
Был выполнен дополнительный контроль трансформации для Arabidopsis thaliana с использованием Agrobacterium. Эта трансформация была использована в качестве критерия для определения эффективности дендример-опосредованной трансформации. Плазмида pDAB7331 (фиг.2) была трансформирована в Agrobacterium с использованием модифицированного протокола от Hanahan (1983), этот штамм был использован для опосредованной Agrobacterium трансформации Arabidopsis thaliana cv. Columbia.
Arabidopsis была трансформирована с использованием способа цветкового погружения, описанного Clough and Bent. Отобранная колония Agrobacterium была использована для инокулирования одной или более 100 мл предварительных культур питательной среды YEP, содержащей спектиномицин (100 мг/л) и канамицин (50 мг/л). Культура(ы) была проинкубирована в течение ночи при 28°C с непрерывным встряхиванием при 225 об/мин. Клетки были осаждены приблизительно при 5000×g в течение 10 минут при комнатной температуре, и полученный в результате супернатант был удален. Клеточный осадок был аккуратно ресуспендирован в 400 мл среды для замачивания, содержащей: 5% (вес./об.) сахарозы, 10 мкг/л 6-бензиламинопурина и 0,04% Silwet L-77. Приблизительно одномесячные растения были погружены в среду на 5-10 минут с аккуратным встряхиванием. Растения были расположены на их сторонах и покрыты (прозрачным или светонепроницаемым материалом) на 2-3 часа, и после этого были расположены вертикально. Растения выращивались при 22°C с фотопериодом 16-часов свет/8-часов темнота. Приблизительно через четыре недели после погружения семена были выращены.
2.3 Отбор трансформированных растений
Свежевыращенному семени T1 было позволено высохнуть в течение семи дней при комнатной температуре. T1 были засеяны в 26,5×51-см поддоны для прорастания, при этом в каждый было внесено 200 мг аликвот расслоенного семени T1 (~10000 семян), которое ранее было суспендировано в 40 мл 0,1% раствора агарозы и хранилось при 4°C в течение двух дней в целях требований полного покоя семян и обеспечения синхронного прорастания семени.
Смесь Sunshine LP5 была покрыта мелкозернистым вермикулитом и дополнительно поливалась питательной смесью Хогланда до тех пор, пока не стала мокрой, и после этого был применен самотечный дренаж. Каждая аликвота по 40 мл расслоенного семени была засеяна равномерно на вермикулит с помощью пипетки и накрыта колпаками для влажности на 4-5 дней. Колпаки были удалены за один день перед начальным трансформантным отбором с использованием глюфосинатного послевсходового распылителя.
Через семь дней после посадки (DAP) T1 растения (семядоля и 2-4 стадия роста листьев, соответственно) были опрыснуты пять раз в течение пяти дней 0,2% раствором гербицида Liberty (200 г ae/л глюфосината, Bayer Crop Sciences, Канзас-Сити, Миссури) при объеме раствора для опрыскивания 10 мл/лунка (703 л/га) с использованием распылителя сжатого воздуха DeVilbiss для доставки эффективного количества 280 г ae/га глюфосината на внесение. Выжившие растения (активно растущие растения) были идентифицированы через 4-7 дней после конечного опрыскивания и пересажены по отдельности в трехдюймовые поддоны с подготовленной средой (Metro Mix 360) для горшков. Пересаженные растения были накрыты колпаками для влажности на 3-4 дня и помещены в ростовую камеру с 22°C, как ранее, или напрямую перемещены в теплицу. Колпаки были впоследствии удалены, и растения выращивались в теплице (22±5°C, 50±30% относительной влажности, 14 ч свет:10 темнота, минимум 500 мкE/м2с природный + дополнительный свет).
ПРИМЕР 3
МОЛЕКУЛЯРНЫЙ АНАЛИЗ ГЕНОМНОЙ ИНТЕГРАЦИИ ТРАНСГЕНОВ
В T1 ПОТОМСТВЕ ARABIDOPSIS THALIANA CV COLUMBIA
3.1 ПЦР амплификация трансгенов геномной ДНК
Геномная ДНК трансгенных растений Arabidopsis была выделена из всех веществ листа шестинедельного растения с использованием набора для растений DNAZOL в соответствии с рекомендациями производителя. Были сконструированы праймеры для ПЦР для детекции трансгенов YFP и PAT. YFP праймеры представлены как SEQ ID NO: 5 и SEQ ID NO: 6. PAT праймеры представлены как SEQ ID NO: 7 и SEQ ID NO: 8.
Реакции ПЦР амплификации PAT и YFP были выполнены с использованием набора TaKaRa EXTAQ™ (Takara, Otsu, Shiga, Япония). Генные продукты были амплифицированы в общем реакционном объеме 50 мкл. Реакционная смесь для ПЦР включала в себя 100 нг матрицы геномной ДНК, 1X ExTaq реакционный буфер, 0,2 мМ dNTP, 10 пмоль каждого праймера и 0,025 единиц/мкл ExTaq. Были использованы следующие ПЦР условия: 1 цикл при 96°C в течение пяти минут и 31 цикл при следующих условиях 94°C в течение 15 секунд, 65°C в течение 30 секунд, 72°C в течение одной минуты, и конечное удлинение при 72°C в течение 7 минут. Продукт ПЦР амплификации был проанализирован с помощью электрофореза в 0,8% TAE агарозном геле и визуализации окрашиванием бромидом этидия. Фиг.5 и фиг.6 демонстрируют продукты амплификации, которые были получены в этих реакциях.
ПЦР фрагменты были секвенированы с использованием прямого праймера PAT (SEQ ID NO: 7) и прямого праймера YFP (SEQ ID NO: 5) с использованием современной технологии секвенирования по Сенгеру (MWG Biotech, Хантсвилл, Алабама). Данные последовательностей были проанализированы с использованием программного обеспечения Sequencher.
Результаты секвенирования PAT и YFP ПЦР ампликонов соответствовали ожидаемой нуклеотидной последовательности для этих генов. Эти результаты указывают на то, что PAT и YFP последовательности из pDAB3831 были стабильно интегрированы в геномную ДНК Arabidopsis с использованием трансфекционного реагента SUPERFECT™.
3.2 ELISA скрининг PAT
Белок был выделен из листьев шестинедельного трансгенного растения Arabidopsis для детекции экспрессированного белка PAT с помощью ELISA (фермент-связанный иммуносорбентный анализ). Микроцентрифужная пробирка, содержащая образцы листа, была охлаждена в жидком азоте. Материал листа был перетерт в порошок с использованием одноразового гомогенизатора. После нахождения на льду в течение 5 минут было добавлено 200 мкл экстракционного буфера (PBST; 20 ммоль забуференная фосфатом мочевина, содержащая 0,05% (об./об.) TWEEN® 20). Содержимое было перемешано со встряхиванием и центрифугировано при 4°C в течение 10 минут при 13000×g. Супернатант был выделен из клеточного дебриса и хранился на льду до проведения дополнительного анализа.
ELISA был выполнен с использованием модифицированного протокола для набора QUALIPLATE™ для LIBERTYLINK® PAT/pat- (Envirologix, Портленд, Мэн). Пластина ELISA и другие реагенты выдерживались при комнатной температуре. 50 мкл конъюгата фермента было добавлено в каждую лунку пластины. Дополнительные 50 мкл экстракционного буфера были добавлены в каждую лунку пластины. Серийные разведения очищенного трансгенного белка Arabidopsis были добавлены в лунки. Были использованы концентрации 10, 5, 2,5 и 1,25 нг/мл. Дополнительные стандарты и растительные экстракты были добавлены в лунки в качестве контроля. Пластина подвергалась встряхиваниям при 200 об/мин и инкубировалась при комнатной температуре в течение 2 часов. После инкубации пластина была промыта пять раз экстракционным буфером в устройстве для мытья пластин. Для детектирования в каждую лунку было добавлено 100 мкл субстрата из набора, и пластина была инкубирована в течение 30 минут. Активность была измерена и записана с использованием устройства для считывания микропластин с поглощением при 595 нм.
Поглощение ELISA сигналов из стандартов указывало на то, что сигнал поглощения был прямо пропорционален количествам PAT, представленным в каждой лунке. Эти данные представлены на фиг.5. Образцы SUPERFECT™- и Agrobacterium-опосредовано трансформированных растений демонстрируют интенсивные сигналы на ELISA. Эти величины в три раза больше, чем для 10 нг/мл стандарта, при этом в этом анализе был протестирован самый интенсивный стандарт. Эти результаты указывают на то, что PAT экспрессируется в трансгенных растениях Arabidopsis, которые были трансформированы с помощью SUPERFECT™-опосредованной растительной трансформации.
Несмотря на то, что это изобретение было описано в некоторых вариантах осуществления, данное изобретение может быть дополнительно модифицировано в рамках основной идеи и объема этого раскрытия. Вследствие этого подразумевается, что эта заявка охватывает любые вариации, применения или адаптации изобретения с использованием его основных принципов. Дополнительно подразумевается, что данная заявка охватывает такие отклонения из данного раскрытия как находящиеся в рамках общеизвестной или традиционный практики в области, к которой относится данное изобретение, и которые соответствуют пределам, установленным в пунктах прилагаемой формулы изобретения, и их эквивалентам.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛУЧЕНИЕ ФУНКЦИОНАЛИЗИРОВАННОЙ ЛИНЕЙНОЙ ДНК-КАССЕТЫ И ОПОСРЕДОВАННАЯ КВАНТОВЫМИ ТОЧКАМИ/НАНОЧАСТИЦАМИ ДОСТАВКА В РАСТЕНИЯ | 2011 |
|
RU2574785C2 |
| ДОСТАВКА ЛИНЕЙНОЙ МОЛЕКУЛЫ ДНК В РАСТЕНИЯ ДЛЯ СТАБИЛЬНОЙ ТРАНСФОРМАЦИИ С ПОМОЩЬЮ ПЕГИЛИРОВАННЫХ КВАНТОВЫХ ТОЧЕК | 2011 |
|
RU2575097C2 |
| КОНЪЮГАТЫ КВАНТОВОЙ ТОЧКИ С ПЕПТИДОМ-НОСИТЕЛЕМ, ПРИГОДНЫЕ ДЛЯ ВИЗУАЛИЗАЦИИ И ДОСТАВКИ МОЛЕКУЛ В РАСТЕНИЯ | 2012 |
|
RU2609637C2 |
| РАСТИТЕЛЬНЫЙ ПЕПТИД ГАММА-ЗЕИН ДЛЯ ДОСТАВКИ БИОМОЛЕКУЛ В РАСТИТЕЛЬНЫЕ КЛЕТКИ | 2011 |
|
RU2563805C2 |
| ТОЛЕРАНТНОЕ К СТРЕССУ ТРАНСГЕННОЕ РАСТЕНИЕ ПШЕНИЦЫ | 2005 |
|
RU2376377C2 |
| ЭНХАНСЕР ПАЛОЧКОВИДНОГО ВИРУСА САХАРНОГО ТРОСТНИКА (SCBV) И ЕГО ПРИМЕНЕНИЕ В ФУНКЦИОНАЛЬНОЙ ГЕНОМИКЕ РАСТЕНИЙ | 2013 |
|
RU2639517C2 |
| СПОСОБЫ ПЕРЕНОСА МОЛЕКУЛЯРНЫХ ВЕЩЕСТВ В КЛЕТКИ РАСТЕНИЙ | 2008 |
|
RU2495935C2 |
| ОПОСРЕДОВАННАЯ НАНОЧАСТИЦАМИ ДОСТАВКА СИКВЕНС-СПЕЦИФИЧНЫХ НУКЛЕАЗ | 2017 |
|
RU2664865C2 |
| ОБОГАЩЕНИЕ АКТИВИРУЕМОЙ ФЛУОРЕСЦЕНЦИЕЙ СОРТИРОВКИ КЛЕТОК (FACS) ДЛЯ СОЗДАНИЯ РАСТЕНИЙ | 2013 |
|
RU2679510C2 |
| СИНТЕТИЧЕСКИЕ ТРАНЗИТНЫЕ ПЕПТИДЫ ХЛОРОПЛАСТА ИЗ BRASSICA | 2013 |
|
RU2636035C2 |











Изобретение относится к области биохимии, в частности к способам внесения представляющей интерес нуклеиновой кислоты в растительную клетку, имеющую клеточную стенку. Способы включают в себя: взаимодействие катионного дендримера и одного или более проникающего в клетку пептида с представляющей интерес нуклеиновой кислотой, приведение в контакт друг с другом клетки, имеющей клеточную стенку, и полученной дендримерной структуры, проникновение указанной дендримерной структуры в растительную клетку, имеющую клеточную стенку. Изобретение позволяет эффективно вводить представляющую интерес нуклеиновую кислоту в клетку растения, имеющую клеточную стенку. 4 н. и 18 з.п. ф-лы, 5 ил., 3 пр.
1. Способ внесения представляющей интерес нуклеиновой кислоты в растительную клетку, имеющую клеточную стенку, для осуществления устойчивой трансформации растения и семян, где способ включает:
обеспечение растительной клетки, имеющей клеточную стенку;
взаимодействие катионного дендримера и одного или более проникающего в клетку пептида с представляющей интерес нуклеиновой кислотой, с образованием активированной дендримерной структуры;
приведение в контакт друг с другом клетки, имеющей клеточную стенку, и активированной дендримерной структуры, где указанная растительная клетка, имеющая клеточную стенку, осуществляет проникновение указанной активированной дендримерной структуры так, что указанная представляющая интерес нуклеиновая кислота вводится в указанную растительную клетку, имеющую клеточную стенку; и
интегрирование указанной представляющей интерес нуклеиновой кислоты в геном указанной растительной клетки, имеющей клеточную стенку, с получением устойчивого трансформанта.
2. Способ по п. 1, где взаимодействие катионного дендримера с представляющей интерес нуклеиновой кислотой включает сборку указанной представляющей интерес нуклеиновой кислоты на поверхности указанного катионного дендримера.
3. Способ по п. 1, где взаимодействие катионного дендримера с представляющей интерес нуклеиновой кислотой включает взаимодействие отрицательно заряженных групп указанной представляющей интерес нуклеиновой кислоты с заряженными аминогруппами на конце указанного катионного дендримера.
4. Способ по п. 1, дополнительно включающий предоставление указанной активированной дендримерной структуре возможности проникновения в компартмент растительной клетки, имеющей клеточную стенку.
5. Способ по п. 4, где компартмент выбран из группы, состоящей из цитозоля, ядра, тонопластов, пластид, этиопласта, хромопласта, лейкопласта, элайопласта, протеинопласта, амилопласта, хлоропласта и просвета двойной мембраны.
6. Способ по п. 1, где растительная клетка, имеющая клеточную стенку, выбрана из группы, состоящей из клеток табака, моркови, маиса, канолы, рапсового семени, хлопка, пальмы, арахиса, сои, Oryza sp., Arabidopsis sp., Ricinus sp. и сахарного тростника.
7. Способ по п. 1, где растительная клетка является клеткой из ткани, выбранной из группы, состоящей из эмбриона, меристемы, каллюса, пыльцы, листьев, пыльников, корней, корневых кончиков, цветков, семян, стручков и стеблей.
8. Способ по п. 1, где дендример представляет собой полиамидоаминный дендример.
9. Способ по п. 1, дополнительно включающий дериватизирование поверхности указанного катионного дендримера.
10. Способ по п. 1, где указанная представляющая интерес нуклеиновая кислота содержит компонент, выбранный из группы, состоящей из нуклеиновых кислот, ДНК, РНК, молекул РНКи, генов, плазмид, космид, YAC, ВАС и их комбинаций.
11. Способ по п. 9, где представляющая интерес нуклеиновая кислота содержит ген.
12. Способ по п. 10, где ген представляет собой ген чужеродного белка, агрономически важный ген или маркерный ген.
13. Способ по п. 10, дополнительно включающий отбор клеток, в которых имеется устойчиво интегрированная представляющая интерес нуклеиновая кислота.
14. Способ по п. 12, где отобранные клетки являются регенерируемыми клетками.
15. Способ по п. 13, дополнительно включающий регенерирование растения из регенерируемых клеток.
16. Способ по п. 1, где катионный дендример содержит дендримерный реагент.
17. Способ стабильного экспрессирования гена, где способ включает:
обеспечение растительной клетки, имеющей клеточную стенку;
взаимодействие катионного дендримера и одного или более проникающего в клетку пептида с геном с образованием активированной дендримерной структуры;
приведение в контакт друг с другом растительной клетки, имеющей клеточную стенку, и активированной дендримерной структуры, где указанная растительная клетка, имеющая клеточную стенку, осуществляет проникновение указанной активированной дендримерной структуры и указанного гена; и
экспрессирование указанного гена в потомстве растения, имеющего растительную клетку.
18. Способ по п. 17, где ген экспрессируется в хлоропласте.
19. Способ по п. 17, дополнительно включающий отбор клеток, стабильно экспрессирующих ген.
20. Способ переноса плазмидной ДНК в растительную клетку, включающий:
взаимодействие катионного дендримера и одного или более проникающего в клетку пептида с плазмидной ДНК с образованием активированной дендримерной структуры; и
приведение указанной активированной дендримерной структуры в контакт с интактной имеющей стенку растительной клеткой, где указанная имеющая стенку растительная клетка осуществляет проникновение указанной активированной дендримерной структуры.
21. Способ по п. 20, дополнительно включающий стабильное экспрессирование гена, присутствующего в плазмидной ДНК, в потомстве растения, имеющего растительную клетку.
22. Способ внесения представляющей интерес нуклеиновой кислоты в растительную клетку, имеющую клеточную стенку, для осуществления устойчивой трансформации растения и семян, где способ включает:
обеспечение растительной клетки, имеющей клеточную стенку;
взаимодействие дендримера в виде сферического катионного полиамидоаминного каскадного полимера и одного или более проникающего в клетку пептида с представляющей интерес нуклеиновой кислотой, с образованием активированной дендримерной структуры;
приведение в контакт друг с другом клетки, имеющей клеточную стенку, и активированной дендримерной структуры, где указанная растительная клетка, имеющая клеточную стенку, осуществляет проникновение указанной активированной дендримерной структуры так, что указанная представляющая интерес нуклеиновая кислота вводится в указанную растительную клетку, имеющую клеточную стенку; и
интегрирование указанной представляющей интерес нуклеиновой кислоты в геном указанной растительной клетки, имеющей клеточную стенку, с получением устойчивого трансформанта.
| PASUPATHY K | |||
| et al., Direct plant gene delivery with a poly(amidoamine) dendrimer, BIOTECHNOLOGY JOURNAL, 2008, vol | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| ПРУЖИННЫЙ МЕХАНИЗМ | 1924 |
|
SU1078A1 |
| EP1577384 A1, 21.09.2005 | |||
| JULIANO RL., Intracellular delivery of oligonucleotide conjugates and dendrimer complexes, Ann N Y Acad Sci, 2006, 1082, pp.18-26 | |||
| KLEIN TM et al., High-velocity microprojectiles for | |||

