ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
По настоящей заявке испрашивается приоритет согласно § 119(е) раздела 35 свода законов США по предварительной заявке на патент США с серийным № 61/067589, поданной 29 февраля 2008 года, которая полностью включена в настоящее описание посредством ссылки.
Исследование, приведшее к настоящему изобретению, было финансировано частично грантом Национального института здравоохранения № RO1-NS45939-01. Правительство Соединенных Штатов имеет определенные права на настоящее изобретение.
ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к введению фактора 2 роста глии (GGF2) нуждающемуся в этом пациенту для обеспечения уровней GGF2 в сыворотке в пределах требуемого терапевтического окна, определяемого на основе заболевания или нарушения, поражающего пациента.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Нейрегулины (NRG) и рецепторы NRG включают систему фактор роста-рецепторная тирозинкиназа для передачи сигналов от клетки к клетке, которая вовлечена в органогенез нервной, мышечной, эпителиальной и других тканей (Lemke, Mol. Cell. Neurosci. 7: 247-262, 1996; Burden et al., Neuron 18: 847-855, 1997). Семейство NRG состоит из трех генов, которые кодируют многочисленные лиганды, содержащие подобные эпидермальному фактору роста (EGF), иммуноглобулиновые (Ig) и другие распознаваемые домены. Многочисленные секретируемые и присоединенные к мембране изоформы функционируют в качестве лигандов в этой системе передачи сигналов. Все рецепторы для NRG являются членами семейства рецепторов EGF (EGFR) и включают EGFR (или ErbB1), ErbB2, ErbB3 и ErbB4, также известные как HER1-HER4, соответственно, у людей (Meyer et al., Development 124: 3575-3586, 1997; Orr-Urtreger et al., Proc. Natl. Acad. Sci. USA 90: 1867-1871, 1993; Marchionni et al., Nature 362: 312-318, 1993; Chen et al., J. Comp. Neurol. 349: 389-400; 1994; Corfas et al., Neuron 14: 103-115, 1995; Meyer et al., Proc. Natl. Acad. Sci. USA 91: 1064-1068, 1994; и Pinkas-Kramarski et at., Oncogene 15: 2803-2815, 1997).
Три гена NRG, Nrg-1, Nrg-2 и Nrg-3, локализуются в различных хромосомных локусах (Pinkas-Kramarski et at., Proc. Natl. Acad. Sci. USA 91: 9387-9391, 1994; Carraway et al., Nature 387; 512-516, 1997; Chang et al., Nature 387: 509-511, 1997; и Zhang et al., Proc. Natl. Acad. Sci. USA 94: 9562-9567, 1997) и в совокупности кодируют разнообразную совокупность белков NRG. Самыми детально изученными к настоящему времени являются продукты гена Nrg-1, которые включают группу из приблизительно 15 отличных структурно-родственных изоформ (Lemke, Mol. Cell, Neurosci. 7: 247-262, 1996; и Peles and Yarden, BioEssays 15: 815-824, 1993). Впервые идентифицированные изоформы NRG-1 включали фактор дифференциации нейтрофилов (NDF; Peles et al., Cell 69, 205-216, 1992 и Wen et al., Cell 69, 559-572, 1992), херегулин (HRG; Holmes et al., Science 256: 1205-1210, 1992), индуцирующую рецепторы для ацетилхолина активность (ARIA; Falls et al., Cell 72: 801-815, 1993) и факторы роста глии GGF1, GGF2 и GGF3 (Marchionni et al. Nature 362: 312-318, 1993).
Ген Nrg-2 был идентифицирован с помощью клонирования по гомологии (Chang et al., Nature 387: 509-512, 1997; Carraway et al., Nature 387: 512-516, 1997; и Higashiyama et al., J. Biochem. 122: 675-680, 1997) и благодаря геномным подходам (Busfield et al., Mol. Cell Biol. 17: 4007-4014, 1997). кДНК NRG-2 также известны как активатор киназ ErbB неврального и тимусного происхождения (NTAK; входящий № в GenBank - АВ005060), дивергент нейрегулина (Don-1) и фактор роста мозжечкого происхождения (CDGF; заявка РСТ WO 97/09425). Экспериментальные данные показывают, что клетки, экспрессирующие ErbB4 или комбинацию ErbB2/ErbB4, по-видимому, демонстрируют особенно сильный ответ на NRG-2 (Pinkas-Kramarski et al., Mol. Cell. Biol. 18: 6090-6101, 1998). Также известно, что продукт гена Nrg-3 (Zhang et al., выше) связывается с рецепторами ErbB4 и стимулирует их (Hijazi et al., Int. J. Oncol. 13: 1061-1067, 1998).
Домен EGFL присутствует в сердцевине всех форм NRG и необходим для связывания с рецепторами ErbB и их активации. Расшифрованные аминокислотные последовательности доменов EGFL, кодируемых в трех генах, идентичны на приблизительно 30-40% (при попарных сравнениях). Кроме того, существуют, по-видимому, по меньшей мере две субформы доменов EGFL в NRG-1 и NRG-2, которые могут сообщить различные биоактивности и тканеспецифические активности.
Клеточные ответы на NRG опосредуются через рецепторные тирозинкиназы для NRG EGFR, ErbB2, ErbB3 и ErbB4 семейства рецепторов эпидермального фактора роста (Busfield et al., 1997, Mol. Cell Biol. 17: 4007-4014; Carraway et al., 1997, Nature 387: 512-516; Chang et al., 1997, Nature 387: 509-512). Связывание с высоким сродством всех NRG опосредуется, главным образом, через либо ErbB3, либо ErbB4 (Ferguson et al., 2000, EMBO J. 19: 4632-4643). Связывание лигандов NRG приводит к димеризации с другими субъединицами ErbB и трансактивации при фосфорилировании на специфических остатках тирозина (Honegger et al., 1990, Mol. Cell Biol. 10: 4035-4044; Lemmon and Schlessinger, 1994, Trends Biochem Sci. 19: 459-463; Heldin, 1995, Cell. 80: 213-223; Hubbard et al., 1998, J. Biol. Chem. 273: 11987-11990). При определенных установочных параметрах эксперимента почти все комбинации рецепторов ErbB, по-видимому, способны к образованию димеров в ответ на связывание изоформ NRG-1. Однако ErbB2, по-видимому, является предпочтительным партнером по димеризации, который может играть важную роль в стабилизации комплекса лиганд-рецептор.
Установлено, что GGF2 стимулирует пролиферацию, дифференциацию и защиту шванновских клеток (Goodearl et al., 1993, J. Biol. Chem. 268: 18095-18102; Minghetti et al., 1996, J. Neurosci. Res. 43: 684-693). Экспрессия NRG-1, ErbB2 и ErbB4 также необходима для образования трабекулы вентрикулярного миокарда во время развития мыши (Meyer and Birchmeier 1995, Nature 378: 386-390; Gassmann et al., 1995, Nature 378: 390-394; Kramer et al., 1996, Proc. Natl. Acad. Sci. USA 93: 4833-4838). Также было установлено, что GGF2 стимулирует пролиферацию и защиту клеток кардиомиоцитов (Zhao et al., 1998, J. Biol. Chem. 273: 10261-10269). GGF2-опосредованная нейропротекция также продемонстрирована в моделях инсульта на животных, хотя относящиеся к дозированию параметры остаются неопределенными.
Настоящее изобретение совершенствует применение GGF2 в отношении терапевтических применений путем обеспечения руководства в отношении способов введения GGF2, которые оптимизируют терапевтическую пользу с ограничением неблагоприятных эффектов. Настоящее изобретение определяет целевые терапевтические окна для уровней концентраций GGF2 в сыворотке, которые точно определены в отношении конкретных болезненных состояний.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к введению GGF2 нуждающемуся в этом пациенту для обеспечения уровней GGF2 в плазме (сыворотке) в пределах целевого терапевтического окна, которое, как определено, является эффективным при лечении заболевания или нарушения. В соответствии с настоящим изобретением GGF2 может вводиться в фармацевтической композиции.
В соответствии с настоящим изобретением предоставляется способ избегания ингибирования миелинизации шванновскими клетками после введения фактора 2 роста глии (GGF2) субъекту, включающий обеспечение субъекта, нуждающегося в миелинизации нейронов, обеспечение GGF2 в фармацевтически приемлемом носителе, введение GGF2 субъекту и определение того, что количество GGF2 меньше количества, которое ингибирует миелинизацию шванновскими клетками.
В другом варианте осуществления настоящее изобретение относится к способу стимуляции миелинизации у пациента, страдающего заболеванием или нарушением, связанным со сниженными уровнями миелинизации, включающему отбор пациента, страдающего заболеванием или нарушением, связанным со сниженными уровнями миелинизации, введение пациенту фактора 2 роста глии (GGF2) в количестве, составляющем приблизительно 500 нг GGF2 на кг веса тела, посредством чего стимулируется миелинизация.
В еще одном варианте осуществления настоящее изобретение относится к способу стимуляции миелинизации у пациента, страдающего заболеванием или нарушением, связанным со сниженными уровнями миелинизации, включающему отбор пациента, страдающего заболеванием или нарушением, связанным со сниженными уровнями миелинизации, и введение пациенту фактора 2 роста глии (GGF2) в количестве, которое обеспечивает уровень в плазме, составляющий приблизительно 0,01 нМ GGF2.
В дальнейшем варианте осуществления настоящее изобретение относится к способу расширения диапазона терапевтических доз GGF2, когда GGF2 используется для способствования миелинизации, включающему отбор пациента с заболеванием или нарушением, связанным со сниженными уровнями миелинизации, введение пациенту GGF2 и ингибитора пути с участием Mek1/Erk, посредством чего GGF2-опосредованная миелинизация происходит при более высоких дозах GGF2, чем она могла бы происходить в отсутствие введения ингибитора пути с участием Mek1/Erk.
В другом варианте осуществления настоящее изобретение относится к способу определения того, является ли количество GGF2 терапевтически эффективным количеством для стимуляции миелинизации, включающему обеспечение субъекта, получающего терапию с использованием GGF2, и измерение уровней белка c-Jun у субъекта, посредством чего увеличение c-Jun относительно базовых уровней c-Jun указывает на то, что количество GGF2 близко к максимальному порогу терапевтической эффективности для стимуляции миелинизации.
В конкретном варианте осуществления настоящего изобретения GGF2 вводят млекопитающему, используя схему введения доз, направленную на обеспечение узкого целевого терапевтического окна концентраций GGF2 в плазме.
Как указано в настоящем документе, известно, что GGF2 способен к стимуляции пролиферации, дифференциации и защите шванновских клеток. Также было установлено, что GGF2 стимулирует ремиелинизацию и ослабляет симптомы в моделях рассеянного склероза, в том числе экспериментального аутоиммунного энцефаломиелита, на животных. Однако в некоторых случаях (например, при высоких концентрациях GGF2) GGF2 может предотвращать миелинизацию нейронов, сокультивируемых со шванновскими клетками.
Представленные в настоящем документе данные показывают, что GGF2 действительно способен к стимуляции миелинизации периферических нервов, но они (данные) указывают, что для обеспечения требуемой GGF2-опосредованной, стимулируемой миелинизации периферических нервов необходимо введение точных доз GGF2 нуждающемуся в этом млекопитающему. Как указано в настоящем документе, GGF2 вводят так, чтобы он находился в пределах терапевтического окна концентраций GGF2 в плазме для того, чтобы стимулировать миелинизацию. В отсутствие результатов, представленных в настоящем документе, нет оценки узкого терапевтического окна концентраций GGF2 в плазме, необходимого для стимуляции миелинизации у нуждающегося в этом млекопитающего.
Представленные в настоящем документе данные также показывают, что GGF2 достаточен для стимуляции миелинизации и избавления от недостатка миелинизации на дефицитных по CRD-Nrg1 аксонах. Однако при высоких концентрациях GGF2 ингибирует миелинизацию Erk-зависимым образом. Результаты настоящего исследования показывают, что GGF2 способен как к стимуляции, так и к ингибированию миелинизации в зависимости от концентраций, представляемых шванновским клеткам.
Соответственно, настоящее изобретение относится к удивительному обнаружению того, что существует до сего времени неосознанная положительная корреляция между GGF2-опосредованной активацией пути с участием PI3-киназы и стимуляцией миелинизации, и существует отрицательная корреляция между GGF2-опосредованной активацией пути с участием Mek1/Erk и стимуляцией миелинизации. Иначе говоря, авторы настоящего изобретения обнаружили, что введение GGF2 можно точно регулировать для стимуляции миелинизации путем оценки уровней активации этих путей. В соответствии с настоящим изобретением целевое терапевтическое окно для GGF2 в отношении стимуляции миелинизации у субъекта представляет собой количество GGF2, которое вызывает активацию пути с участием PI3-киназы (проверяемую, например, путем выявления фосфорилированной Akt) в отсутствие выявляемой активации пути с участием Mek1/Erk (проверяемой, например, путем выявления фосфорилированной Erk).
Составы и композиции по настоящему изобретению демонстрируют специфический, требуемый профиль высвобождения, который максимизирует терапевтический эффект при минимизации неблагоприятных побочных эффектов. Требуемый профиль высвобождения может быть охарактеризован с учетом максимальной концентрации в плазме лекарственного средства или активного агента (Cmax) и концентрации в плазме лекарственного средства или активного агента в конкретном интервале между введениями доз (Ctau). Отношение Cmax к Ctau (Cmax:Ctau) можно рассчитать на основе наблюдаемых Cmax и Ctau. Интервал между введениями доз (tau) является временем после последнего введения лекарственного средства или активного агента. В настоящей заявке интервал между введениями доз (tau) может, например, составлять двенадцать (12) часов, в этом случае Ctau является концентрацией лекарственного средства или активного агента через двенадцать (12) часов от последнего введения.
Кроме того, составы и композиции по настоящему изобретению демонстрируют требуемый профиль высвобождения, который можно охарактеризовать с учетом максимальной концентрации в плазме лекарственного средства или активного агента в состоянии равновесия (CmaxSS) и минимальной концентрации в плазме лекарственного средства или активного агента в состоянии равновесия (CminSS). Состояние равновесия наблюдается, когда скорость введения (поглощения) равна скорости удаления лекарственного средства или активного агента. Отношение CmaxSS к CminSS (CmaxSS:CminSS) можно рассчитать на основе наблюдаемых CmaxSS и CminSS. Кроме того, составы и композиции по настоящему изобретению демонстрируют требуемый профиль высвобождения, который можно охарактеризовать с учетом средней максимальной концентрации в плазме лекарственного средства или активного агента в состоянии равновесия (CavSS).
В варианте осуществления настоящего изобретения, направленном на нуждающегося в ремиелинизации пациента, целевые максимальные уровни GGF2 в сыворотке составляют приблизительно 0,01 нМ.
В варианте осуществления настоящего изобретения, направленном на нуждающегося в ремиелинизации пациента, целевые максимальные уровни GGF2 в сыворотке составляют любые из следующих значений или приблизительно любые из следующих значений, или диапазоны между следующими значениями: от приблизительно 0,001 до 0,01 нг/мл; от 0,01 до 0,1 нг/мл; от 0,1 до 1,0 нг/мл; от 1,0 до 10 нг/мл; от 10 до 100 нг/мл; или от 100 до 1000 нг/мл. В конкретном варианте осуществления целевой максимальный уровень в сыворотке составляют приблизительно 1,0 нг/мл.
В варианте осуществления настоящего изобретения, направленном на перенесшего инсульт пациента, целевые максимальные уровни GGF2 в сыворотке составляют любые из следующих значений или приблизительно любые из следующих значений, или диапазоны между следующими значениями: от приблизительно 0,00001 до 0,0001 нг/мл; от 0,0001 до 0,001 нг/мл; от 0,001 до 0,01 нг/мл; от 0,01 до 0,1 нг/мл; от 0,1 до 1,0 нг/мл; от 1,0 до 10 нг/мл; от 10 до 100 нг/мл; от 100 до 1000 нг/мл; от 1000 до 10000 нг/мл; или от 10000 до 100000 нг/мл. В конкретном варианте осуществления целевой максимальный уровень в сыворотке составляет приблизительно 0,2 мкг/мл.
В варианте осуществления настоящего изобретения, направленном на страдающего невропатией пациента, целевые максимальные уровни GGF2 в сыворотке составляют любые из следующих значений или приблизительно любые из следующих значений, или диапазоны между следующими значениями: от приблизительно 0,001 до 0,01 нг/мл; от 0,01 до 0,1 нг/мл; от 0,1 до 1,0 нг/мл; от 1,0 до 10 нг/мл; от 10 до 100 нг/мл; или от 100 до 1000 нг/мл. В конкретном варианте осуществления целевой максимальный уровень в сыворотке составляет приблизительно 6,25 нг/мл.
В варианте осуществления настоящего изобретения, направленном на пациента с сердечной недостаточностью, целевые максимальные уровни GGF2 в сыворотке составляют любые из следующих значений или приблизительно любые из следующих значений, или диапазоны между следующими значениями: от приблизительно 0,001 до 0,01 нг/мл; от 0,01 до 0,1 нг/мл; от 0,1 до 1,0 нг/мл; от 1,0 до 10 нг/мл; от 10 до 100 нг/мл или от 100 до 1000 нг/мл. В конкретном варианте осуществления целевой максимальный уровень в сыворотке составляет приблизительно 6,8 мкг/мл.
В соответствии с настоящим изобретением фармацевтические композиции, содержащие GGF2, могут вводиться различными путями, известными квалифицированным в данной области техники специалистам. Может использоваться любой подходящий путь введения, например, внутривенное, парентеральное, подкожное, внутримышечное, внутричерепное, внутриглазничное, глазное, внутрижелудочковое, внутрисуставное, внутрипозвоночное, интрацистернальное, внутрибрюшинное, интраназальное, аэрозольное, пероральное или местное (например, путем применения пластыря, несущего состав, способный перемещаться через кожу и поступать в кровотеремещаться через кожу и поступань в кровооок) введение. Предусматривается, что пероральное введение включает включающие GGF2 перпероральные лекарственные формы с замедленным высвобождением. Описываемая в настоящем документе фармацевтическая композиция, включающая GGF2, может использоваться для лечения индивидуумов, страдающих неврологическими нарушениями, причем указанная фармацевтическая композиция максимизирует терапевтический эффект, при минимизации неблагоприятных побочных эффектов.
В первом варианте осуществления настоящего изобретения GGF2 вводят млекопитающему, страдающему неврологическим нарушением, связанным с демиелинизацией, причем GGF2 вводят по схеме введения доз для обеспечения и поддержания узкого целевого терапевтического окна концентраций GGF2 в плазме. Как указано в настоящем документе, введение точных доз GGF2 требуется для обеспечения уровней GGF2 в плазме (сыворотке), необходимых для терапевтической эффективности в отношении индуцирования миелинизации у нуждающегося в этом субъекта. Примеры демиелинизирующих заболеваний, при которых для обеспечения терапевтической эффективности требуется введение надлежащих доз GGF2, включают синдром Гийена-Барре, хроническую воспалительную демиелинизирующую полинейропатию, периферическую демиелинизацию вследствие травматического повреждения, рассеянный склероз, ретробульбарный неврит, центральную демиелинизацию вследствие травматического повреждения, поперечный миелит, прогрессирующую многоочаговую лейкоэнцефалопатию, болезнь Девика (нейромиелит зрительного нерва), острый рассеянный энцефаломиелит, адренолейкодистрофию и адренолейконевропатию.
Во втором варианте осуществления настоящего изобретения GGF2 вводят млекопитающему, страдающему заболеванием сердечной мышцы, таким как застойная сердечная недостаточность, инфаркт миокарда, реперфузионное повреждение, химическая, вирусная или идиопатическая кардиотоксичность, аритмии, причем GGF2 вводят по схеме введения доз для обеспечения целевого терапевтического окна концентраций GGF2 в плазме.
В третьем варианте осуществления настоящего изобретения GGF2 вводят млекопитающему, перенесшему инсульт, повреждение спинного мозга или травматическое повреждение головного мозга, причем GGF2 вводят по схеме введения доз для обеспечения целевого терапевтического окна концентраций GGF2 в плазме.
Будет понятно, что для любого из детализированных в настоящем документе применений GGF2 может вводиться в любой подходящей форме, или в виде компонента фармацевтической композиции, и посредством любых способов, все из которых описываются в настоящем документе и/или допускаются в данной области техники.
Соответственно, настоящее изобретение направлено на определение целевого терапевтического окна относительно терапевтически эффективного уровня GGF2 в плазме. Целевое терапевтическое окно меняется в зависимости от заболевания или нарушения, поражающего пациента, и требуемой активности, сообщаемой при обеспечении надлежащего терапевтически эффективного уровня GGF2 в плазме.
В настоящий документ также включен способ отбора индивидуумов на основе демонстрации симптомов. Также включен способ отбора индивидуумов на основе реагирования на достижение терапевтически эффективного уровня GGF2 в плазме, указанного для каждого применения.
Помимо изложенных выше способов лечения настоящее изобретение распространяется на применение любого из соединений по настоящему изобретению для приготовления лекарственных средств или в качестве лекарственных средств, которые могут вводиться для осуществления таких лечений, а также на такие соединения для осуществления раскрытых и точно определенных лечений.
Настоящим изобретением также охватывается фармацевтическая композиция, содержащая GGF2 или домен EGFL и ингибитор пути с участием Mek1/Erk и ее применение для лечения пациента, страдающего заболеванием или нарушением, связанным со сниженными уровнями миелинизации.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг. 1A-C демонстрируется (А) GGF2-индуцированная активация Akt и МАРК в сокультурах шванновских клеток и нейронов DRG. Сокультуры шванновских клеток с нейронами DRG в условиях для миелинизации обрабатывали GGF (в концентрации 0,6 мкМ), и через 20 минут оценивали уровни активации Akt и МАРК с помощью анализа Вестерн-блоттингом. (В) Ингибирование U0126 GGF2-индуцированной активации МАРК. Сокультуры предварительно обрабатывали увеличивающимися дозами U0126 в течение 30 минут, а затем стимулировали GGF2. Контрольные культуры оставляли необработанными. Активацию МАРК оценивали через 20 минут. (С) Ингибирование U0126 (1 и 3 мкМ) GGF2-индуцированной активации МАРК отменяет ингибиторный эффект GGF2 на миелинизацию. Сокультуры обрабатывали совместно GGF2 и U0126 (1 и 3 мкМ) в условиях для миелинизации. Через десять-двенадцать дней культуры фиксировали и подвергали иммуноокрашиванию на МВР для определения уровня миелинизации.
На фиг. 2 демонстрируется, что GGF2 стимулирует миелинизацию при низких концентрациях. Сокультуры обрабатывали GGF2 в концентрациях, находящихся в диапазоне от 0,5 до 1000 пМ (от 0,0005 до 1 нМ), в условиях для миелинизации. Через десять-двенадцать дней миелинизацию оценивали с помощью иммуноокрашивания на МВР. Конкретнее, концентрациями GGF2 слева направо являются следующие концентрации: без обработки, 0,5 пМ, 1 пМ, 3 пМ, 10 пМ, 30 пМ, 300 пМ, 600 пМ и 1000 пМ, соответственно. Через десять-двенадцать дней миелинизацию оценивали с помощью иммуноокрашивания на МВР.
На фиг. 3A-F демонстрируется, что ингибиторный эффект GGF на миелинизацию опосредуется через активацию Mek1/Erk. (А) Сокультуры шванновских клеток и нейронов DRG обрабатывали GGF (0,01, 0,6 и 1 нМ), и через 45 минут готовили лизаты клеток, и уровни активного Erk (p-Erk) и Akt (p-Akt) определяли с помощью анализа Вестерн-блоттингом. При использовании 1 нМ (заключенные в рамку линии) GGF индуцировал активацию и Erk, и Akt. (В) Ингибирование GGF-индуцируемой активации Erk в сокультурах. Сокультуры шванновских клеток и нейронов DRG предварительно обрабатывали U0126 в течение 30 минут, а затем добавляли GGF (0,6 нМ) при постоянном присутствии U0126. Через 45 минут готовили лизаты клеток, и определяли уровень p-Erk и p-Akt. Обработка U0126 ингибировала как эндогенную, так и GGF-индуцируемую активации Erk, не оказывая влияние на активацию Akt. (C) Снимки сегментов МВР+ миелина, образованных в сокультурах, обработанных GGF или GGF+U0126 (1 нМ). Обработка U0126 отменяла ингибиторный эффект GGF и индуцировала миелинизацию. Контрольные культуры сохраняли без какой-либо обработки. Масштабная метка: 100 мкм. Квантификация результатов представлена в (D). (E) Ингибирование эндогенной активности Erk в сокультурах стимулирует миелинизацию. Сокультуры обрабатывали увеличивающими концентрациями U0126 (0,5, 1 и 3 нМ) в условиях для миелинизации, и через 11 дней миелинизацию исследовали, как указано выше. Значительное увеличение миелинизации наблюдалось в культурах, обработанных U0126. Планки погрешностей означают ± среднеквадратическая ошибка (р<0,001). (F) Ингибирование GGF-индуцируемой активации Erk сопровождается снижением c-Jun и увеличением экспрессии Krox20. Сокультуры сохраняли в условиях для миелинизации в присутствии GGF или GGF+U0126 (0,5, 1 и 3 нМ) в течение 11 дней, и лизаты клеток анализировали на экспрессию МВР, c-Jun и Krox20. В качестве контроля загрузки служил уровень актина. GGF-индуцируемая экспрессия c-Jun снижалась при обработке U0126. Уровень белка Krox20, по-видимому, увеличивался в культурах, обработанных U0126.
На фиг. 4A-С демонстрируется, что GGF стимулирует миелинизацию при низкой концентрации. (А) Шванновские клетки обрабатывали различными концентрациями GGF, находящимися в диапазоне от 0,0003 до 10 нМ, и через 20 минут готовили лизаты клеток, и уровни активации Erk и Akt анализировали с помощью Вестерн-блоттинга (наверху) и денситометрического анализа (внизу). Увеличение активации Akt возникало в диапазоне более низких концентраций (в рамке) по сравнению с активацией Erk. (В) Сокультуры обрабатывали различными концентрациями GGF (0,0005, 0,001, 0,003, 0,01, 0,03, 0,3, 0,6 и 1 нМ) в течение 11 дней в условиях для миелинизации, а затем фиксировали и подвергали иммуноокрашиванию на МВР и DAPI. Снимки контроля и культур, обработанных 0,01 нМ GGF, представлены вместе с квантификацией результатов (справа). Продемонстрирован очевидный двухфазный эффект GGF: он стимулирует миелинизацию при низких концентрациях (от 0,0005 до 0,01 нМ), ингибируя процесс при более высоких концентрациях (0,3 нМ и выше). (С) Низкая концентрация GGF (0,01 нМ) значительно увеличивала миелинизацию на нейронах CRD-Nrg1+/- (р=0,003). Планки погрешностей показывают ± среднеквадратическая ошибка. Данные были проанализированы с помощью однофакторного дисперсионного анализа (*: р<0,001).
На фиг. 5A-D демонстрируются нуклеотидная и аминокислотная последовательности полноразмерного GGF2.
На фиг. 6-11 демонстрируются нуклеотидные и аминокислотные последовательности доменов, подобных эпидермальному фактору роста (EGFL), 1-6.
На фиг. 12 демонстрируется таблица, имеющая отношение к номенклатуре нейрегулинов.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Представленные в настоящем документе данные показали, что для стимуляции миелинизации периферических нервов, GGF2 должен вводиться млекопитающему, используя схему введения доз, направленную на обеспечение терапевтического окна, например, концентраций GGF2 в плазме или доз GGF2.
Определения
Используемые в настоящем документе термины имеют значения, общепризнанные и известные квалифицированным в данной области техники специалистам, однако для удобства и завершенности конкретные термины и их значения изложены ниже.
В настоящем документе «приблизительно» означает фактическое значение плюс или минус другая величина, устанавливая, тем самым, диапазон величин. В определенных предпочтительных вариантах осуществления «приблизительно» означает диапазон относительно базисной (или основной, или ссылочной) величины или величину плюс или минус вплоть до 15%, 14%, 13%, 12%, 11%, 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2%, 1%, 0,75%, 0,5%, 0,25% или 0,1%.
Под «доменом, подобным эпидермальному фактору роста» или «доменом EGFL» подразумевается полипептидный мотив, кодируемый геном NRG-1, NRG-2 или NRG-3, который связывается с ErbB2, ErbB3, ErbB4 или их комбинациями и стимулирует их и имеет структурное сходство со связывающимся с рецептором для EGF доменом, описанным в Holmes et al., Science 256: 1205-1210, 1992; патенте США № 5530109, патенте США № 5716930, заявке на патент США с серийным № 08/461097; Hijazi et al., Int. J. Oncol. 13: 1061-1067, 1998; Chang et al., Nature 387: 509-512, 1997; Carraway et al., Nature 387: 512-516, 1997; Higashiyama et al., J. Biochem. 122: 675-680, 1997, и WO 97/09425). Смотрите фиг. 10-15 в отношении нуклеотидных и аминокислотных последовательностей доменов, подобных эпидермальному фактору роста (EGFL) 1-6.
Под «нейрегулином» или «NRG» подразумевается полипептид, который кодируется геном или нуклеиновой кислотой (например, кДНК) NRG-1, NRG-2 или NRG-3 и связывается с рецепторами ErbB2, ErbB3, ErbB4 или их комбинациями и стимулирует их.
Под «нейрегулином-1», «NRG-1», «херегулином», «GGF2» или «лигандом p185erbB2» подразумевается полипептид, который связывается непосредственно с рецептором ErbB2 или трансстимулирует его и кодируется геном лиганда p185erbB2, описанным в патенте США № 5530109, патенте США № 5716930 и патенте США № 7037888, содержание каждого из которых включено в настоящий документ посредством ссылки. Смотрите фиг. 9А-D в отношении нуклеотидной и аминокислотной последовательностей полноразмерного GGF2. Смотрите фиг. 12 в отношении таблицы, относящейся к номенклатуре нейрегулинов.
Кодируемые генами NRG-1, NRG-2 и NRG-3 полипептиды обладают доменами EGFL, которые позволяют им связываться с рецепторами ErbB и стимулировать их. Holmes и др. (Science 256: 1205-1210, 1992) установили, что домен EGFL сам по себе достаточен для связывания с рецептором p185erbB2 и его активации. Соответственно, любой полипептидный продукт, кодируемый геном NRG-1, NRG-2 или NRG-3, например, полипептид, имеющий домен EGFL (например, домен EGFL, описанный в патенте США № 5530109, патенте США № 5716930, патенте США № 7037888, патенте США № 7135456 и патенте США № 7319019, или домен EGFL, описанный в WO 97/09425), кодируемый геном или кДНК нейрегулина, может использоваться в способах по настоящему изобретению для обеспечения терапевтического окна, в котором достигается эффективный уровень GGF2 в плазме (сыворотке).
Также должно быть обращено внимание на то, что используемые в настоящем документе и в прилагаемой формуле изобретения формы единственного числа включают ссылку на множественное число за исключением случаев, когда контекст четко определяет иное.
Кроме случаев, оговоренных особо, все технические и научные термины, используемые в настоящем документе, имеют значения, одинаковые с такими, в которых они обычно понимаются специалистом со средним уровнем компетентности в данной области техники. Хотя любые способы и материалы, схожие с описанными в настоящем документе способами и материалами или эквивалентные им, можно использовать для осуществления на практике или проверки вариантов осуществления настоящего изобретения, теперь описываются конкретные способы, устройства и материалы.
«Местное введение» означает непосредственное введение не являющимся системным путем в место болезни или нарушения или вблизи него.
Термины «пациент» и «субъект» используются в настоящем документе для ссылки на всех животных, включающих млекопитающих. Примеры пациентов или субъектов включают людей, коров, собак, кошек, коз, овец и свиней.
Используемые в настоящем документе термины «фармацевтически приемлемые соли, сложные эфиры, амиды и пролекарства» относятся к таким солям карбоновых кислот (карбоксилатам), солям присоединения аминокислот, сложным эфирам, амидам и пролекарствам соединений по настоящему изобретению, которые, в рамках обоснованной оценки врача, подходят для использования в контакте с тканями пациента без чрезмерной токсичности, раздражения, аллергической реакции и т.п., соответствуют допустимому соотношению польза/риск и эффективны для их намеченного применения, а также цвиттерионным формам, при наличии возможности, соединений по настоящему изобретению.
Термин «пролекарство» относится к соединениям, которые быстро преобразуются in vivo с выработкой исходных соединений вышеотмеченной формулы, например, при гидролизе в крови. Детальное обсуждение предоставлено в T. Higuchi and V. Stella, “Pro-drugs as Novel Delivery Systems”, Vol. 14, A.C.S. Symposium Series, и в Bioreversible Carriers in Drug Design, ed. Edward B. Roche, American Pharmaceutical Association and Pergamon Press, 1987, оба из которых включены в настоящий документ посредством ссылки.
Термин «соли» относится к относительно нетоксичным солям присоединения неорганических и органических кислот соединений по настоящему изобретению. Эти соли можно приготовить на месте во время конечного выделения и очистки соединений или с помощью отдельного взаимодействия очищенного соединения в его свободной основной форме с подходящей органической или неорганической кислотой и выделения таким образом образованной соли. Репрезентативные соли включают соли гидробромид, гидрохлорид, сульфат, бисульфат, нитрат, ацетат, оксалат, валерат, олеат, пальмитат, стеарат, лаурат, борат, бензоат, лактат, фосфат, тозилат, цитрат, малеат, фумарат, сукцинат, тартрат, нафтилат, мезилат, глюкогептонат, лактобионат и лаурилсульфонат и т.п. Они могут включать катионы на основе щелочных и щелочноземельных металлов, таких как натрий, литий, калий, кальций, магний и т.п., а также нетоксичный аммоний, тетраметиламмоний, тетраметиламмоний, метиламин, диметиламин, триметиламин, триэтиламин, этиламин и т.п. (смотрите, например, S.M. Barge et al., “Pharmaceutical Salts”, J. Pharm. Sci., 1977, 66: 1-19, который включен в настоящий документ посредством ссылки).
«Терапевтически эффективное количество» представляет собой количество, достаточное для ослабления сопровождающих заболевание или болезненность симптомов, для нормализации функций организма при заболевании или нарушениях, которые приводят к ухудшению конкретных функций организма, или для обеспечения улучшения одного или нескольких клинически определяемых параметров заболевания, предпочтительно, ослабления симптомов, сопровождающих заболевание, связанное, например, с демиелинизирующим заболеванием, включающих скорость хождения, мышечный тонус нижних конечностей, мышечную силу нижних конечностей или мышечную спастичность. Имеющее отношение к настоящему изобретению терапевтически эффективное количество представляет собой количество, достаточное для ослабления боли или мышечной спастичности, сопровождающих подвергаемое лечению неврологическое нарушение, или количество, достаточное для приведения к улучшению половой функции, функции мочевого пузыря или кишечника у субъектов, имеющих неврологическое нарушение, которое уменьшает нервную проводимость, или которое препятствует нормальным половым функциям, функции мочевого пузыря или кишечника.
«Лечение» относится к введению лекарственного средства или выполнению медицинских процедур в отношении пациента для улучшения клинического состояния пациента, в том числе уменьшения продолжительности заболевания или тяжести заболевания, или субъективного улучшения качества жизни пациента или увеличения продолжительности жизни пациента.
В настоящем документе термин «целевое терапевтическое окно» относится к диапазону доз или диапазону концентраций в сыворотке, который обеспечивает требуемые терапевтические результаты. В отношении GGF2, в конкретном варианте осуществления, целевое терапевтическое окно относится к достаточному для индуцирования миелинизации шванновскими клетками у субъекта количеству GGF2, которое меньше количества, достаточного для ингибирования миелинизации у субъекта. При удивительном обнаружении авторы настоящего изобретения определили целевое терапевтическое окно для GGF2 относительно его способности к стимуляции миелинизации путем определения относительных уровней активации пути с участием PI3-киназы и активации пути с участием Mek1/Erk. Конкретнее, авторы настоящего изобретения обнаружили до сего времени неосознанную положительную корреляцию между GGF2-опосредованной активацией пути с участием PI3-киназы и стимуляцией миелинизации и отрицательную корреляцию между GGF2-опосредованной активацией пути с участием Mek1/Erk и стимуляцией миелинизации. Иначе говоря, авторы настоящего изобретения обнаружили, что введение GGF2 можно точно регулировать для стимуляции миелинизации путем оценки уровней активации этих путей. Целевое терапевтическое окно для GGF2 в отношении стимуляции миелинизации у субъекта определяют как количество GGF2, которое вызывает активацию пути с участием PI3-киназы (проверяемую, например, путем выявления фосфорилированной Akt) в отсутствие выявляемой активации пути с участием Mek1/Erk (проверяемой, например, путем выявления фосфорилированной Erk). Выявление фосфорилированной Akt и фосфорилированной Erk можно выполнить, используя стандартные анализы, известные в данной области техники, включающие ELISA, Вестерн-(иммуно)блоттинг, иммуноцитохимию, in vitro анализ киназ, LC/MS (жидкостную хроматографию/масс-спектрометрию), MaldiTOF MS (времяпролетную масс-спектрометрию с лазерной десорбцией/ионизацией из матрицы) и другие белковые системы, известные в данной области, такие как Luminex.
Квалифицированному в данной области техники специалисту могло бы быть понятно, что другие внутриклеточные маркеры активации пути с участием PI3-киназы и активации пути с участием Mek1/Erk известны и используются в соответствии с настоящим изобретением. В соответствии с настоящим изобретением другие индикаторы активации пути с участием PI3-киназы и активации пути с участием Mek1/Erk могут использоваться для определения терапевтического окна, в котором GGF2 стимулирует миелинизацию.
Кроме того, соединения по настоящему изобретению могут существовать в несольватированных, а также сольватированных формах с фармацевтически приемлемыми растворителями, такими как вода, этанол и т.п. В общем, сольватированные формы считаются эквивалентными несольватированным формам для целей настоящего изобретения.
«Ингибиторы МАР-киназ»
Неограничивающий список ингибиторов МАР-киназ, которые могут использоваться в настоящем изобретении, включает арктигенин, который сильно ингибирует активность МКК1 in vitro со значением IC50, составляющим 1 нМ, и, таким образом, ингибирует фосфорилирование и активацию МАР-киназ ERK1/2, киназы р38 и JNK и их активности в клетках Raw264.7, обработанных LPS; PD 98059, который является сильным, избирательным и проницаемым в клетку ингибитором киназы МАР-киназы (также известной как киназа МАРК/ERK или МЕК), который ингибирует фосфорилирование МАР-киназы киназой МАР-киназы, но не ингибирует саму МАР-киназу (значение IC50 для индуцируемых PD 98059 эффектов находится в диапазоне 1-20 мкМ для многих анализов); SB202190, который является в высокой степени избирательным, сильным и проницаемым в клетку ингибитором МАР-киназ р38, который связывается в пределах кармана для АТФ активной киназы с Kd 38 нМ, определенной при использовании рекомбинантной р38 человека, и избирательно ингибирует изоформы р38альфа и бета (значения IC50 составляют 50 и 100 нМ для р38альфа/SAPK2альфа и р38бета2/SAPK2бета, соответственно); SB203580, который является в высокой степени избирательным и проницаемым в клетку ингибитором митоген-стимулируемой протеинкиназы р38 со значениями IC50, составляющими 50 и 500 нМ для р38/SAPK2α и р38/SAPK2β, соответственно, и также ингибирует фосфоинозитид-зависимую протеинкиназу 1 (PDK1) при бульших в 10 раз концентрациях (IC50~3-5 мкМ) (проявляет бульшую в 100-500 раз избирательность по сравнению с Lck, GSK3β и Akt/PKB); SL 327, который является избирательным ингибитором МЕК1 и МЕК2 со значениями IC50, составляющими 180 и 220 нМ, соответственно. Он блокирует LTP гиппокампа in vitro и проникает в головной мозг in vivo, подавляя выработку страха и обучение у крыс, и вызывая нейропротекцию у мышей, после системного введения; SP600125, который является избирательным ингибитором N-концевой киназы c-Jun (JNK). Он конкурентно и обратимо ингибирует JNK1, 2 и 3 (IC50=40-90 нМ) и, как установлено, обладает меньшей ингибиторной активностью в отношении ERK2, p38β и ряда других киназ, и, как известно, является активным in vivo; и U0126, который является избирательным ингибитором киназ митоген-стимулируемых протеинкиназ, МЕК1 и МЕК-2, с большей в 100 раз эффективностью, чем PD 98059, и является слабым ингибитором PCK, Raf, ERK, JNK, MEKK, MKK-3, MKK-4/SEK, MKK-6, Abl, Cdk2 и Cdk4, и ингибирует трансактивацию АР-1 в клеточных анализах по гену-репортеру.
Другие ингибиторы, которые в настоящее время подвергаются испытанию фазы FDA (Управления по контролю за продуктами и медикаментами), включают ингибиторы фарнезилтрансферазы (FTI). Например, Zarnesta® (R115777, типифарниб) является FTI, разработка которого продолжается далее. В испытании фазы II пациентов с ранее подвергаемым лечению метастатическим раком молочной железы были проверены две различные схемы введения доз: непрерывная и с интервалами. Степени целевого ответа в 2 группах составляли 10% и 14%, с дополнительными 15% и 9% тех, кто имел стабильное заболевание в течение по меньшей мере 6 месяцев. Основными наблюдаемыми побочными эффектами были подавление костного мозга и невропатия, оба из которых были меньшими в группе введения доз с интервалами, чем в группе непрерывного введения. Несколько исследований фазы I зарнестры и других FTI в комбинации с цитотоксической химиотерапией были проведены, и была установлена безопасность этих схем комбинированного введения. В процессе реализации находятся испытания фазы II на раке молочной железы, включающие испытание с использованием зарнестры в комбинации с ингибитором ароматазы. Разрешение FDA на применение зарнестры при остром миелоидном лейкозе (AML) зависит от данных фазы III, поскольку рабочая группа FDA проголосовала против ускоренного разрешения зарнестры на основе данных от испытания фазы II с использованием одной группы.
Что касается Zarnesta®, для клинических испытаний фазы I Zarnesta® вводят в количестве 400 мг, вводимых перорально дважды в день в течение двух недель; для клинических испытаний фазы II Zarnesta® вводят в количестве 300 мг, вводимых перорально дважды в день в течение первых 21 дней каждого 28-дневного цикла; для клинических испытаний фазы III Zarnesta® вводят в количестве 600 мг, вводимых перорально дважды в день в течение первых 21 дней каждого 28-дневного цикла.
Ингибиторы Raf включают другие типы ингибиторов, которые в настоящее время подвергаются испытанию фазы FDA. Сорафениб (BAY 43-9006), например, является первым соединением для того, чтобы сделать мишенью не только путь передачи сигнала с участием Raf/MEK/Erk, но также пути с участием VEGFR и PDGFR. В марте 2004 года сорафенибу был присвоен FDA статус ускоренного продвижения для метастатического почечноклеточного рака. В апреле 2005 года сорафениб был включен в программу предварительных исследований, которая предназначена для терапий, которым был присвоен FDA статус ускоренного продвижения, и которые имеют потенциал для обеспечения значительной пользы по сравнению с существующей стандартной терапией. В процессе реализации находится также несколько больших, международных клинических испытаний фазы III с использованием множества учреждений на пациентах с первичными раками почки и печени запущенной стадии, а также метастатической меланомой.
Что касается сорафениба, в клинических испытаниях фазы I были проверены два уровня доз: уровень дозы 1: 200 мг сорафениба, принимаемых внутрь дважды в день в течение 3-недельного цикла, или уровень дозы 2: 400 мг сорафениба, принимаемых внутрь дважды в день в течение 3-недельного цикла.
Недавно представлены результаты планового промежуточного анализа проводимого испытания фазы III на пациентах с раком почки запущенной стадии (Escudier et al. J. Natl. Cancer Inst. 2008 100: 1454-1463, содержание которого в целом включено в настоящий документ). Среди 769 исследованных пациентов выживание без прогрессирования (PFS) было удвоено до средней величины, составляющей 24 недели, при использовании сорафениба, по сравнению с 12 неделями при использовании плацебо. Польза от сорафениба наблюдалась во всех подгруппах пациентов, независимо от возраста, продолжительности заболевания или предшествующих терапий. Контролирование заболевания было обеспечено у 80% пациентов, получавших сорафениб: 78% имели стабильное заболевание (по сравнению с 55% в группе плацебо), и 2% отвечали частично (по сравнению ни с одним в группе плацебо). Уровень пациентов без прогрессирования в течение 12 недель составлял 79% для сорафениба в сравнении с 50% для плацебо. Кроме того, сорафениб очень хорошо переносился 768 пациентами, и самыми часто встречаемыми побочными эффектами были гипертония, усталость, диарея и сыпь, в том числе сыпь на кисте и стопе (сидром «кисть-стопа»).
В испытании эффективности фазы II сорафениб исследуется в виде единственного средства на раках легкого, молочного железы и других раках запущенной стадии. В клинических испытаниях фазы I/II сорафениб исследуется в комбинации с рядом стандартных химиотерапий и других противораковых средств.
ISIS 5132 является другим ингибитором Raf, который продемонстрировал приемлемую токсичность в исследованиях фазы I. В процессе реализации находятся исследования фазы II на ряде типов рака.
Другие ингибиторы, которые в настоящее время подвергаются испытанию фазы FDA, включают ингибиторы МЕК. CI-1040, например, является перпероральным, избирательным ингибитором в виде небольшой молекулы МЕК 1-2. С помощью исследований на животных и культурах продемонстрирована активность этого средства в линиях клеток рака молочной железы. В исследованиях фазы I обнаружены умеренные побочные эффекты на желудочно-кишечный тракт и кожу. К сожалению, в исследовании фазы II на 67 пациентах с 4 различными типами опухолей (раком ободочной и прямой кишок, NSCLC, молочной железы и поджелудочной железы запущенной стадии) ответов не выявлено, хотя лечение CI-1040 хорошо переносилось.
PD 0325901, ингибитор МЕК второго поколения, недавно вступил в фазу клинической разработки и, по-видимому, обладает заметно лучшими фармакологическими свойствами, чем CI-1040, который, как надеются исследователи, можно перевести в лучшую противораковую эффективность. Для него установлен некоторый частичный ответ у пациентов с меланомой.
Что касается PD 0325901, в клинических испытаниях фазы I и фазы II было проверено множество уровней доз. Введение осуществлялось перорально или один, или два раза в день; были оценены несколько схем введения доз; текущей схемой введения доз является 5 дней на лекарственном средстве, 2 дня без лекарственного средства в течение 3 недель в 28-дневном цикле. Оцениваемые дозы находились в диапазоне от 1 мг раз в день до 30 мг два раза в день. Клинические испытания были преждевременно прекращены по соображениям безопасности, в частности, из-за относящейся к глазам и неврологической токсичности, представленной при дозах, составляющих 10 мг дважды в день и больше.
Должно быть понятно, что это изобретение не ограничивается конкретными описанными молекулами, композициями, методологиями или протоколами, поскольку они могут варьироваться. Также должно быть понятно, что использованная при описании терминология предназначена только для цели описания конкретных вариантов осуществления и, как предполагается, не ограничивает объем настоящего изобретения, который ограничивается только прилагаемой формулой изобретения.
Введение. Нейрегулины и содержащие домены EGFL полипептиды, кодируемые генами нейрегулинов, могут вводиться пациентам или экспериментальным животным с фармацевтически приемлемым разбавителем, носителем или наполнителем, в форме стандартных доз. Может использоваться обычная фармацевтическая практика для обеспечения подходящих составов или композиций для введения пациентам или экспериментальным животным. Может использоваться любой подходящий путь введения, например, внутривенное, парентеральное, подкожное, внутримышечное, внутричерепное, внутриглазничное, глазное, внутрижелудочковое, внутрисуставное, внутрипозвоночное, интрацистернальное, внутрибрюшинное, интраназальное, аэрозольное, пероральное или местное (например, путем применения пластыря, несущего состав, способный перемещаться через кожу и поступать в кровотеремещаться через кожу и поступань в кровооок) введение. Терапевтические составы могут быть в форме жидких растворов или суспензий; для перорального введения составы могут быть в форме таблеток или капсул, а для интраназального введения - в форме порошков, капель в нос или аэрозолей. Любой из указанных выше составов может быть составом с замедленным высвобождением.
Широко известные в данной области техники способы изготовления составов представлены, например, в “Remington’s Pharmaceutical Sciences”, который включен в настоящий документ в целом. Составы для парентерального введения могут, например, содержать наполнители, стерильную воду или солевой раствор, полиалкиленгликоли, такие как полиэтиленгликоль, масла растительного происхождения или гидрогенизованные нафталины. Для контролирования высвобождения соединений могут использоваться биосовместимые, биоразрушаемые лактидный полимер, сополимер лактида и гликолида или сополимеры полиоксиэтилена и полиоксипропилена с замедленным высвобождением. Другие применимые системы парентеральной доставки для введения молекул по настоящему изобретению включают частицы сополимера этилена и винилацета, осмотические насосы, имплантируемые инфузионные системы и липосомы. Составы для ингаляции могут содержать наполнители, например, лактозу, или могут быть водными растворами, содержащими, например, полиоксиэтилен-9-лауриловый эфир, гликохолат и дезоксихолат, или могут быть масляными растворами для введения в форме капель в нос или в виде геля.
Таким образом, и как указано ранее, настоящее изобретение включает в свой объем и распространяется на изложенные способы лечения и на применение таких соединений для приготовления лекарственных средств, применимых для таких способов.
Демиелинизирующие заболевания. Миелиновые оболочки покрывают множество нервных волокон в центральной и периферической нервной системе. Наличие неповрежденных миелиновых оболочек ускоряет передачу нервных импульсов по аксонам. Нарушения, затрагивающие миелин, прерывают передачу нервных импульсов, и симптомы заболевания могут быть отражением недостатков в любой части нервной системы.
Миелин, образуемый олигодендроглией в центральной нервной системе (ЦНС), отличается химически и иммунологически от миелина, образуемого шванновскими клетками на периферии. Поэтому некоторые нарушения миелина (например, синдром Гийена-Барре, хроническая воспалительная демиелинизирующая полинейропатия и другие полиневропатии периферических нервов) имеют обыкновение затрагивать, главным образом, периферические нервы, в то время как другие нарушения миелина затрагивают, главным образом, ЦНС. Самыми часто поражаемыми областями в ЦНС являются головной мозг, спинной мозг и зрительные нервы.
Демиелинизация часто является вторичным нарушением на основе инфекционного, ишемического, метаболического или наследственного заболевания. При первичных демиелинизирующих заболеваниях, хотя причина или причины неизвестны, предполагается аутоиммунный механизм, поскольку заболевание иногда следует за вирусной инфекцией или противовирусной вакцинацией.
Демиелинизация имеет обыкновение быть сегментарной или очаговой, поражая множество областей одновременно или последовательно. Однако может происходить ремиелинизация с восстановлением, регенерацией и полным возвращением к норме нервной функции. Однако за большой утратой миелина обычно следует дегенерация аксонов и часто дегенерация клеточного тела.
Рассеянный склероз (MS) характеризуется рассеянными очагами демиелинизации в головном мозге и спинном мозге. Обычные симптомы включают зрительные и глазодвигательные нарушения, парестезии, слабость, мышечную спастичность, дисфункцию мочевой системы и легкое нарушение когнитивных функций. Обычно неврологические расстройства являются многократными, с ремиссиями и обострениями болезни, постепенно вызывающими недееспособность. Диагностика основана на истории ремиссий и обострений болезни плюс клинические признаки, результаты тестов, повреждения, видимые при магнитно-резонансном исследовании (MRI), и другие критерии (в зависимости от симптомов), объективно демонстрирующие ≥2 отдельных неврологических нарушений. Обычно лечение включает кортикостероиды при острых обострениях болезни, иммуномодулирующие лекарственные средства для предотвращения обострений и поддерживающие меры.
При MS возникают локализованные области демиелинизации (бляшки), с разрушением олигодендроглии, околососудистым воспалением и химическими изменениями в липидных и белковых составляющих миелина в бляшках и вокруг них. Возможно повреждение аксонов, но клеточные тела и аксоны имеют обыкновение относительно хорошо сохраняться. Фиброзный глиоз развивается в бляшках, которые рассеяны по всей ЦНС, в основном в белом веществе, в частности, в боковых и задних столбах (особенно в цервикальных областях), зрительных нервах и перивентрикулярных областях. Также поражаются нервные пути среднего мозга, варолиева моста и мозжечка. Серое вещество в мозжечке и спинном мозге также поражается, но в гораздо меньшей степени.
Болезнь сердца
Болезнь сердца является общим термином для ряда различных заболеваний, поражающих сердце. Она является ведущей причиной смерти во многих промышленно развитых странах, в том числе Соединенных Штатах. В качестве вступления представлены следующие широкие классы болезни сердца. Экзогенные кардиомиопатии являются кардиомиопатиями, первичная патология которых лежит вне миокарда. Большая часть кардиомиопатий является экзогенной, поскольку самой часто встречаемой причиной кардиомиопатии является ишемия. Эндогенные кардиомиопатии проистекают из-за слабости сердечной мышцы, которая не обусловлена идентифицируемой внешней причиной. С другой стороны, сердечно-сосудистое заболевание относится к любому ряду конкретных заболеваний, поражающих само сердце и/или систему кровеносных сосудов, особенно вен и артерий, ведущих к сердцу и отходящих от него. Исследование диморфизма заболевания наводит на мысль, что женщины, страдающие сердечно-сосудистым заболеванием, обычно страдают формами, которые поражают кровеносные сосуды, в то время как мужчины обычно страдают формами, которые поражают саму сердечную мышцу. Известные или связанные причины сердечно-сосудистого заболевания включают сахарный диабет, гипертонию, гипергомоцистеинемию и гиперхолестеринемию. Ишемическая болезнь сердца является еще одним классом заболевания самого сердца, характеризующегося снижением кровоснабжения органа.
Гипертоническая болезнь сердца является термином, используемым для ссылки на болезнь сердца, вызванную высоким кровяным давлением, особенно локализованным высоким кровяным давлением. В воспалительное заболевание сердца вовлечено воспаление сердечной мышцы и/или окружающей ее ткани. Порок клапана сердца представляет собой любой патологический процесс, в который вовлечен один или несколько клапанов сердца. Клапанами в правой части сердца являются правый предсердно-желудочковый клапан и легочный клапан, а клапанами в левой части сердца являются митральный клапан и клапан аорты.
Застойная сердечная недостаточность, одна из ведущих причин смерти в промышленно развитых странах, является следствием увеличенной нагрузки на сердце и нарастающего уменьшения прокачиваемости через него. Она может быть следствием любого структурного или функционального сердечного нарушения, которое уменьшает способность сердца к заполнению или прокачке достаточного количества крови через тело. Сначала увеличенная нагрузка, которая является следствием высокого кровяного давления или утраты сократительной ткани, вызывает компенсаторную гипертрофию кардиомиоцитов и утолщение стенки левого желудочка, посредством чего увеличивается сократительная способность и сохраняется функционирование сердца. С течением времени, однако, полость левого желудочка расширяется, систолическая прокачиваемость ухудшается, кардиомиоциты подвергаются гибели клеток в виде апоптоза, и функция миокарда ухудшается по нарастающей.
Факторы, лежащие в основе застойной сердечной недостаточности, включают высокое кровяное давление, ишемическую болезнь сердца, подвергание воздействию кардиотоксических соединений, таких как антрациклиновые антибиотики, и генетические дефекты, о которых известно, что они увеличивают риск сердечной недостаточности.
Под «застойной сердечной недостаточностью» подразумевается нарушенная функция сердца, которая создает неспособность сердца к сохранению нормального выброса крови в покое или при нагрузке или к сохранению нормального сердечного выброса при установке давления при нормальном сердечном заполнении. Фракция изгнания левого желудочка, составляющая приблизительно 40% или меньше, является показателем застойной сердечной недостаточности (в качестве сравнения, фракция изгнания, составляющая приблизительно 60%, является нормой). Пациенты с застойной сердечной недостаточностью демонстрируют хорошо известные клинические симптомы и признаки, такие как тахипноэ, плевральный выпот, усталость в покое или при нагрузке, сократическую дисфункцию и отек. Застойная сердечная недостаточность без труда диагностируется с помощью широко известных способов (смотрите, например, “Consensus recommendations for the management of chronic heart failure” Am. J. Cardiol., 83(2A): 1A-38-A, 1999).
Относительную тяжесть и прогрессирование заболевания определяют, используя широко известные способы, такие как физическое обследование, эхокардиография, радионуклидная визуализация, инвазивное гемодинамическое мониторирование, магнитно-резонансная ангиография и тестирование с нагрузкой на бегущей дорожке в сочетании с исследованиями утилизации кислорода.
Под «ишемической болезнью сердца» подразумевается любое нарушение, являющееся следствием несоответствия между потребностью миокарда в кислороде и адекватностью снабжения кислородом. Большинство случаев ишемической болезни сердца являются следствием сужения коронарных артерий, которое возникает при атеросклерозе или других сосудистых заболеваниях.
Под «инфарктом миокарда» подразумевается процесс, при котором ишемическая болезнь приводит к тому, что область миокарда замещается рубцовой тканью.
Под «кардиотоксическим» подразумевается соединение, которое уменьшает функционирование сердца путем непосредственного или опосредованного ослабления или уничтожения кардиомиоцитов.
Под «гипертонией» подразумевается кровяное давление, которое, как считают специалисты-медики (например, врач или специалист со средним медицинским образованием), выше нормального и несет повышенный риск развития застойной сердечной недостаточности.
Под «лечением» подразумевается, что введение нейрегулина или подобного нейрегулину полипептида замедляет или подавляет прогрессирование застойной сердечной недостаточности во время лечения, относительно прогрессирования заболевания, которое происходило бы в отсутствие лечения, статистически значимым образом. Для оценки прогрессирования заболевания могут использоваться хорошо известные признаки, такие как фракция изгнания из левого желудка, физическая работоспособность и другие клинические тесты, а также уровни выживания и уровни госпитализации. То, замедляет ли или подавляет ли лечение прогрессирование заболевания статистически значимым образом, можно определить с помощью способов, которые широко известны в данной области техники (смотрите, например, SOLVD Investigators, N. Engl. J. Med. 327: 685-691, 1992, и Cohn et al., N. Engl. J. Med. 339: 1810-1816, 1998).
Под «уменьшением прогрессирования истончения миокарда» подразумевается сохранение гипертрофии желудочковых кардиомиоцитов, так что толщина стенки желудочка сохраняется или увеличивается.
Под «ингибирует миокардиальный апоптоз» подразумевается, что лечение нейрегулином ингибирует гибель кардиомиоцитов на по меньшей мере 10%, более предпочтительно на по меньшей мере 15%, все еще более предпочтительно на по меньшей мере 25%, даже предпочтительнее на по меньшей мере 50%, еще предпочтительнее на по меньшей мере 75% и наиболее предпочтительно на по меньшей мере 90% по сравнению с не подвергнутыми лечению кардиомиоцитами.
Инсульт
Инсульт или острое нарушение мозгового кровообращения (CVA) является термином, используемым для ссылки на быстро развивающуюся утрату функций головного мозга вследствие патологического отклонения в кровеносных сосудах, поставляющих кровь в головной мозг. Инсульт возникает, когда кровоснабжение части головного мозга внезапно прерывается, или когда кровеносный сосуд в головном мозге разрывается, разливая кровь в пространства, окружающие клетки головного мозга. Клетки головного мозга погибают, когда они больше не получают кислород и питательные вещества из крови, или существует внезапное кровоизлияние в головной мозг или вокруг него. Симптомы инсульта включают внезапное онемение или слабость, особенно в одной стороне тела, внезапную спутанность сознания или затруднение говорить и понимать речь, внезапное затруднение видеть одним или обоими глазами, внезапное затруднение ходить, головокружение или утрату равновесия или координации, или внезапную сильную головную боль без известной причины. Существует две формы инсульта: ишемический, который обусловлен закупоркой снабжающего головной мозг кровеносного сосуда (например, вызванной тромбозом или эмболией); и геморрагический, который является следствием кровоизлияния в головной мозг или вокруг него.
Показатель для терапевтического окна
Для каждого описываемого в настоящем документе применения для заболевания определяется целевое терапевтическое окно для уровней GGF2 в плазме (сыворотке). В соответствии с представленными в настоящем документе экспериментальными результатами, когда GGF2 вводят млекопитающему, страдающему неврологическим нарушением, связанным с демиелинизацией, GGF2 должен вводиться по схеме введения доз для обеспечения и поддержания узкого целевого терапевтического окна концентраций GGF2 в плазме. Как указано в настоящем документе, введение точных доз GGF2 требуется для обеспечения уровней GGF2 в плазме (сыворотке), необходимых для терапевтической эффективности в отношении индуцирования миелинизации у нуждающегося в этом индивидуума.
В варианте осуществления настоящего изобретения, направленном на нуждающегося в ремиелинизации пациента, целевой уровень GGF2 в сыворотке (плазме) составляет приблизительно 0,01 нМ.
В другом варианте осуществления настоящего изобретения, направленном на нуждающегося в ремиелинизации пациента, GGF2 вводят в количестве, составляющем приблизительно 500 нг/кг веса тела пациента.
Композиции по настоящему изобретению могут использоваться для лечения заболевания пациента, которое включает определение терапевтически эффективной концентрации GGF2 у нуждающегося в этом пациента. Композиции могут использоваться для увеличения уровня и/или поддержания терапевтически эффективной концентрации GGF2 у пациента. При желании, композиции по настоящему изобретению могут быть составлены так, чтобы избегать больших пиков при начальном высвобождении GGF2. Для композиций по настоящему изобретению после введения нуждающемуся в этом пациенту предусматривается лечение вышеуказанных заболеваний. Предпочтительно, композиции вводят с тем, чтобы обеспечить терапевтически эффективный уровень GGF2 в плазме крови, который сохраняется у пациента в течение периода времени, составляющего по меньшей мере 6 часов, предпочтительно по меньшей мере 8 часов и более предпочтительно по меньшей мере приблизительно 10-12.
ПРИМЕРЫ
МАТЕРИАЛЫ И МЕТОДЫ
Антитела
Для иммунофлуоресцентного анализа использовали моноклональное антитело (SM194) против основного белка миелина (MBP) (моноклональные антитела Sternberger) в разведении 1:500. Для анализа Вестерн-блоттингом все поликлональные антитела против активного erbB2 (p-Neu/Tyr 1248), erbB2 и erbB3 получали от Santa Cruz и использовали в разведении 1:1000. Моноклональное антитело против фосфорилированной Akt и поликлональное антитело против фосфорилированной MAPK покупали у Cell Signaling и использовали в разведениях 1:1000 и 1:500, соответственно. Поликлональные антитела против Akt и MAPK (Promega) использовали в разведениях 1:1000 и 1:5000, соответственно.
Нейрегулин-1 типа II и типа III
Рекомбинантный фактор II роста глии человека (rhGGF-II, Nrg1 типа II) получали от Acorda Therapeutics, Inc. Рекомбинантный человеческий, происходящий из сенсорных и двигательных нейронов фактор (rhSMDF, Nrg1 типа III) покупали у R&D Systems. В настоящем исследовании rhGGF-II и rhSMDF упоминают просто как GGF (или GGF2) и SMDF, соответственно. GGF представлял собой N-конец из 419 аминокислотных остатков, содержащий EGF-домен и Ig-подобный домен. Соответственно, GGF представляет собой растворимый белок, в котором отсутствует трансмембранный и цитоплазматический домены.
Культура крысиных зародышевых шванновских клеток
Шванновские клетки получали из седалищных нервов новорожденных крыс (возрастом 1-2 дня), как описано ранее (Brockes et al., Brain Res, 1979; 165: 105-118). Для обычного культивирования шванновские клетки выращивали в модифицированной Дульбекко среде Игла (DMEM) с 10% фетальной бычьей сыворотки (FBS), дополненной нейрегулином-1 с EGF-доменом (R&D Systems) (10 нг/мл) и форсколином (2 мкМ). Во всех экспериментах, описанных в этом тексте, использовали клетки между пассажами 2-4.
Сокультивирование нейронов ганглиев заднего корешка (DRG) и шванновских клеток
Рассеянные DRG получали из эмбрионов крыс на 14,5 день эмбрионального развития, как описано ранее (Eldridge et al., J. Cell Biol. 1987; 105(2): 1023-1034), и высевали на покрытые коллагеном (коллагеном из хвоста крысы типа 1) 12-мм покровные стекла с плотностью, составляющей 1,25 DRG/покровное стекло. Через пять-шесть часов культуры заливали нейробазальной средой (Cellgro), дополненной B27 (GIBCO), 20% глюкозы, NGF (50 нг/мл) и 5-фтордезоксиуридином (FUdR, 10 мкМ), и сохраняли в среде в течение дополнительных 2-3 дней для исключения пролиферирирующих не являющихся нейронами клеток. Культуры затем переводили в новую среду без FUdR и сохраняли до тех пор, пока аксоны DRG не достигали края покровных стекол. После создания аксонных сетей, шванновские клетки высевали на нейроны с плотностью, составляющей 100000 клеток/покровное стекло. Через четыре-пять дней культуры переводили на среду для миелинизации: минимальную поддерживающую среду (МЕМ), дополненную 10% инактивированной нагреванием FBS, 20% глюкозы, NGF (50 нг/мл) и аскорбиновой кислотой (50 мкг/мл). Через десять-одиннадцать дней оценивали миелинизацию путем иммуноокрашивания на МВР.
Сокультивирование нейронов шейных верхних ганглиев (SCG) и шванновских клеток
Рассеянные SCG получали из крыс на 1-2 день после рождения, как описано ранее, и высевали на покрытые коллагеном 12-мм покровные стекла с плотностью, составляющей 0,8 SCG/покровное стекло. На следующий день культуры заливали нейробазальной средой, дополненной B27 (GIBCO), 20% глюкозы, NGF (50 нг/мл) и 5-фтордезоксиуридином (FUdR, 10 мкМ), и сохраняли в среде в течение дополнительных 2-3 дней для исключения пролиферирирующих не являющихся нейронами клеток. Культуры затем переводили обратно на новую среду без FUdR и сохраняли до тех пор, пока аксоны не удлинялись до края покровных стекол. Шванновские клетки высевали на нейроны и сохраняли в нейробазальной среде с добавками до тех пор, пока шванновские клетки не заселяли аксоны (приблизительно 7-10 дней). Миелинизацию инициировали путем помещения культур в среду для миелинизации, описанную для сокультирования DRG и шванновских клеток. Через сорок дней оценивали миелинизацию путем иммуноокрашивания на МВР.
Анализ с использованием иммунопреципитации и Вестерн-блоттинга
Для приготовления лизатов клеток крысиные шванновские клетки с 90-95% конфлюэнтности на 60-мм чашках или сокультуры дважды промывали в забуференном фосфатом солевом растворе (PBS) и затем подвергали лизису в 300 мкл охлажденного на льду буфера для лизиса (50 мМ Трис-HCl, рН 7,4, 1% NP-40, 0,25% натрия дезоксихолата, 150 мМ NaCl, 1 мМ EGTA, 10 мкг/мл лейпептина, 2 мкг/мл апротинина, 1 мМ PMSF и 0,5 мМ натрия ортованадат). Лизаты осветляли центрифугированием в течение 15 мин при 14000 об/мин на холоде, и концентрацию белков в супернатантах определяли в соответствии со спецификациями производителя (Bio-Rad: Hercules, CA). Для анализа Вестерн-блоттингом 50-70 мкг лизатов шванновских клеток разделяли по размеру на 10% полиакриламидных гелях с SDS и переносили на мембраны PVDF. После блокирования в 5% молоке мембраны инкубировали с соответствующими первичными антителами, приготовленными в растворе для блокирования. После инкубации с конъюгированными с пероксидазой хрена вторичными антителами белковые полосы визуализировали с помощью усиленной хемилюминесценции. Для иммунопреципитации 500 мкг лизатов шванновских клеток инкубировали с 0,6 мкг первичного антитела в течение 3 часов при 4°С, затем инкубировали с 50 мкл гранул Сефарозы А в течение 1 часа. Гранулы промывали 5 раз в буфере для лизиса, и связанные с гранулами белки разделяли по размеру на полиакриламидных гелях с SDS и подвергали анализу с помощью Вестерн-блоттинга.
Иммунофлуоресцентное окрашивание на МВР
Культуры DRG-шванновские клетки или SCG-шванновские клетки промывали в забуференном фосфатом солевом растворе (PBS), а затем фиксировали в 4% параформальдегиде в течение 20 минут. После промывки PBS, образцы подвергали пермеабилизации в охлажденном на льду метаноле в течение 25 минут, а затем инкубировали в растворе для блокирования (5% нормальная козья сыворотка+0,3% Triton X) в течение 1 часа при комнатной температуре. За этим следовала инкубация с первичным антителом, приготовленным в растворе для блокирования, в течение ночи. После промывки PBS, образцы инкубировали с конъюгированным с Alexa-488 козьим антимышиным вторичным антителом в течение 45 минут. Ядра клеток визуализировали с помощью окрашивания DAPI.
КОЛИЧЕСТВЕННАЯ ПЦР В РЕЖИМЕ РЕАЛЬНОГО ВРЕМЕНИ
Статистический анализ
Однофакторный дисперсионный анализ выполняли, используя программное обеспечение для программирования SAS c 95% уровнем значимости.
РЕЗУЛЬТАТЫ
Ингибиторное действие GGF2 на миелинизацию опосредуется через активацию МАРК
В более раннем исследовании было установлено, что Nrg1 типа II (GGF2) после добавления к сокультурам шванновских клеток и нейронов ингибирует миелинизацию. Также сообщалось, что активация пути с участием Ras/Raf/MAPK ингибирует экспрессию связанного с миелином гена в шванновских клетках, в то время как активация пути с участием PI3-киназы стимулирует миелинизацию, приводя к идее, что состояние шванновских клеток для миелинизации определяется балансом между путем с участием PI3-киназы и путем с участием Ras/Raf/MAPK (Ogata et al. J. Neurosci. 2004; 24: 6724-6732). Авторы настоящего изобретения предсказывали, что, если сам GGF2 действует через активацию МАРК для ингибирования миелинизации, ингибирование GGF2-индуцируемой активации МАРК могло бы отменить ингибиторный эффект на миелинизацию. Для оценки возможности того, что ингибиторный эффект GGF2 на миелинизацию мог бы быть обусловлен его способностью к вызову сильной активации МАРК в шванновских клетках, авторы настоящего изобретения использовали хорошо известную in vitro культуральную систему для миелинизации, в которой шванновские клетки сокультивируют с нейронами ганглиев заднего корешка (DRG) и индуцируют для миелинизации связанных с ними аксонов путем добавления аскорбиновой кислоты в культуральную среду. Сначала, для определения эффекта GGF2 на активацию МАРК в сокультурах зародышевые шванновские клетки засевали на нейроны DRG и позволяли им заселять аксоны. Как только культуры прекращали пролиферировать, сокультуры стимулировали с помощью GGF2 в концентрации 0,6 нМ. Через двадцать минут готовили лизаты клеток, и активацию МАРК определяли анализом с использованием Вестерн-блоттинга. В контрольных сокультурах был низкий уровень активной МАРК. Как продемонстрировано на фиг. 1, обработка GGF2 далее увеличивала уровень активации МАРК. Для определения того, могла ли GGF2-индуцированная активация МАРК быть блокирована обработкой U0126, фармакологическим ингибитором киназы МАРК, сокультуры предварительно обрабатывали увеличивающимися концентрациями (0,5, 1, 3 и 10 мкМ) U0126 в течение 30 минут перед стимуляцией GGF2, и эти концентрации сохраняли в культуральной среде. Контрольные культуры обрабатывали ингибитором в отсутствие обработки GGF2. Как в контрольной, так и в экспериментальных культурах U0126-опосредованное ингибирование МАРК зависело от концентрации, на что указывает нарастающее снижение уровней фосфорилированной МАРК. В культурах, обработанных GGF2 и U0126 в концентрации 1 мкМ, уровень активации уменьшен до базального уровня, в то время как при использовании 10 мкМ U0126 активация МАРК в сокультуре была полностью отменена. U0126 не оказывал эффект на GGF2-индуцируемую активацию PI3-киназы.
Для оценки эффекта ингибирования МАРК на миелинизацию сокультуры обрабатывали GGF2 в присутствии U0126 или в его отсутствие в момент инициации миелинизации, и сохраняли такие же условия в описанных условиях для миелинизации. Контрольные культуры оставляли необработанными в описанных условиях для миелинизации. Через десять-одиннадцать дней культуры фиксировали и подвергали иммуноокрашиванию на основной белок миелина (МВР) для визуализации сегментов миелина. В обработанных GGF2 культурах наблюдалось заметное уменьшение числа сегментов миелина, как продемонстрировано ранее, выявляя ингибиторный эффект GGF2 на миелинизацию. Однако в культурах, совместно обработанных U0126, наблюдалось зависимое от дозы увеличение миелинизации, что означает, что блокирование активации МАРК отменяет ингибиторный эффект GGF2.
GGF2 стимулирует миелинизацию при низких концентрациях. Хотя уровень активации МАРК неуклонно увеличивался в шванновских клетках, обработанных увеличивающимися концентрациями GGF2, авторы настоящего изобретения наблюдали, что при концентрациях ниже 0,01 мкМ, хотя уровень активации Akt увеличивался значительно выше базального уровня, не было выявляемого уровня активации МАРК. Если состояние шванновских клеток для миелинизации определяется балансом между активацией Akt и активацией МАРК, авторы настоящего изобретения попытались определить то, коррелирует ли увеличение активации Akt в отсутствие активности МАРК при этих концентрациях с положительным эффектом на миелинизацию. Для исследования этой возможности сокультуры обрабатывали GGF2 при концентрациях, находящихся в диапазоне от 0,0005 до 0,03 нМ, в момент инициации миелинизации. Позже культуры фиксировали и подвергали иммуноокрашиванию на МВР. Как и предсказывалось на основе рассматриваемых в настоящем документе данных, существовало увеличение уровня миелинизации в культурах, обработанных низкими дозами GGF2, находящимися в диапазоне от 0,0005 до 0,01 нМ, по сравнению с необработанными контрольными культурами. После квантификации результаты продемонстрировали, что существовало зависимое от дозы увеличение числа сегментов миелина (фиг.2): 1,9-, 2,7- и 3,5-кратное увеличение миелинизации относительно контрольного уровня при концентрации GGF2, составляющей 0,0005, 0,001 и 0,01 нМ, соответственно. При использовании 0,03 нМ наблюдалось резкое уменьшение уровня миелинизации до уровня, близкого к уровню контрольных культур, или слегка ниже этого уровня. Последующее увеличение количества GGF2 приводило к дальнейшему снижению миелинизации. Чувствительная к GGF2 миелинизация была полностью ингибирована при использовании 0,6 нМ GGF2. Эта концентрация соответствовала появлению активной МАРК в сокультурах, как продемонстрировано на фиг. 1. Эти результаты наводят на мысль, что GGF2 играет двойственную роль во время миелинизации: одну, которая стимулирует миелинизацию, и другую, которая ингибирует миелинизацию, и два противоположных действия определяются дозой GGF2, представляемой шванновским клеткам.
Противоположные действия GGF2 опосредуются через активацию Mek/Erk. Для дальнейшего изучения противоположных действий GGF2 были проведены дополнительные эксперименты. Предшествующие исследования заключали в себе намек на то, что Ras/Raf/Erk и PI3-киназа являются, соответственно, отрицательным и положительным регуляторами миелинизации, наводя на мысль, что баланс между ними коррелирует с состоянием шванновских клеток для миелинизации. Для дальнейшего определения состояний активации путей, индуцируемых GGF2, сокультуры обрабатывали растворимым белком GGF2 в концентрации 1 нМ. Авторы настоящего изобретения определили, что в этой концентрации GGF2 эффективно ингибировал миелинизацию. Лизаты клеток готовили через 30 минут после обработки GGF2, и присутствие фосфорилированных белков определяли с помощью анализа Вестерн-блоттингом (фиг. 3А). При использовании 1 нМ (фиг. 3А, заключенные в рамку линии) GGF2 увеличивал активацию Akt выше базального уровня. Увеличение активации Erk также наблюдалось в обработанных GGF2 культурах при этой концентрации. Было установлено, что настолько низкие как 0,6 нМ концентрации GGF2 достаточны для активации Erk в обработанных GGF2 культурах.
Для подтверждения вышеприведенных результатов и дальнейшего изучения коррелятивной связи между активацией Erk и ингибиторным эффектом GGF на миелинизацию были проведены дополнительные эксперименты. В результате, сокультуры обрабатывали GGF2 вместе с увеличивающими концентрациями U0126, описанным выше специфическим ингибитором пути с участием Mek1/Erk. Представленный на фиг. 3В анализ Вестерн-блоттингом показывает, что U0126 ингибировал GGF2-индуцируемую активацию Erk зависимым от дозы образом, хотя он не оказывал эффект на активацию Akt. Низкий уровень эндогенной активности Erk, обычно наблюдаемый в сокультуральной системе, также снижался при обработке лекарственным средством.
Авторы настоящего изобретения в дальнейшем оценили эффект ингибирования Mek1/Erk на миелинизацию. Как продемонстрировано на фиг. 3С и 3D, добавление GGF2 в высокой концентрации почти полностью ингибировало миелинизацию в сокультурах. Однако в культурах, совместно обработанных U0126, ингибиторный эффект GGF2 был отменен, о чем свидетельствует зависимое от дозы увеличение уровня миелинизации (фиг. 3С и 3D). Этот результат дает прямое доказательство того, что ингибиторный эффект GGF2 на миелинизацию опосредуется через активацию Erk. Интересно, что обработка U0126 в сокультурах в отсутствие GGF2 также приводила к повышению уровня миелинизации (фиг.3Е), что указывает на то, что эндогенная активность Mek1/Erk действует в качестве внутреннего отрицательного регулятора миелинизации.
Анализ Вестерн-блоттингом на лизатах, приготовленных из сокультур, также показал, что обработка GGF2 увеличивает экспрессию белка c-Jun, отрицательного регулятора дифференциации шванновских клеток и миелинизации ими. Последующее ингибирование GGF2-индуцируемой активности Mek1/Erk снижало уровни c-Jun, что, в свою очередь, сопровождалось увеличением экспрессии белка миелина. В отличие от эффекта на c-Jun, обработка U0126 приводила к увеличению экспрессии Krox20 в сокультурах. Это согласуется с недавним сообщением о предполагаемом существовании перекрестной антагонистической связи между c-Jun и Krox20 в регуляции миелинизации (Parkinson et al., 2008, Journal of Cell Biology 181: 625-637).
GGF2 стимулирует миелинизацию шванновскими клетками. Для подтверждения и расширения представленных в настоящем документе результатов и дальнейшей оценки противоположных действий GGF2 авторы настоящего изобретения оценили зависимый от концентрации эффект GGF2 на активацию Ras/Raf/Erk и PI3-киназы в шванновских клетках. Клетки обрабатывали различными концентрациями GGF2, находящимися в диапазоне от 0,0003 до 10 нМ, и уровень активации Erk и Akt определяли с помощью анализа Верстерн-блоттингом. Снимки и относительное увеличение уровней активации представлены на фиг. 4А и 4В. Уровень активной Akt увеличивался неуклонно, начиная с наименьшей из проверенных доз, в то время как для активации Erk требовались более высокие концентрации GGF2. Дифференциальная активация двух путей при низких концентрациях, в результате, создавала узкое окно доз (от 0,003 до 0,01 нМ, в рамке на фиг. 4В), в котором Akt был преобладающим путем, стимулированным в ответ на GGF2. Затем в сокультуральной системе был определен эффект различных доз GGF2 на миелинизацию. В окне низких концентраций, GGF2 вызывал зависимый от дозы промиелинизирующий эффект: 1,5-, 2,3-, 2,2- и 2,8-кратное увеличение миелинизации по сравнению с контрольными культурами при 0,0005, 0,001, 0,003 и 0,01 нМ GGF2, соответственно (фиг.4С). По мере дальнейшего увеличения концентрации, GGF2 начинал ингибировать миелинизацию, что совпадает с появлением активации Erk. Промиелинизирующий эффект GGF2 был также продемонстрирован в сокультурах CRD-Nrg1+/-, в которых низкие дозы GGF2 избавляли от недостатка миелиназации на мутантных аксонах (фиг. 4D).
Растворимый Nrg1 мог как стимулировать, так и ингибировать миелинизацию. определяемый концентрацией двоичный выбор: В периферической нервной системе (ПНС), GGF2 рассматривался в качестве изоформы Nrg1, связанной с ответом на повреждение шванновских клеток. Эктопическая in vivo экспрессия GGF-β3 в миелинизирующих шванновских клетках стимулирует пролиферацию клеток и индуцирует демиелинизацию (Huijbregts et al. J. Neurosci. 2003; 23: 7269-7280). Кроме того, установлено, что добавление высоких концентраций GGF2 (например, концентраций, превышающих 0,25 нМ GGF2) в сокультуры шванновских клеток и нейронов DRG ингибирует миелинизации (Zanazzi et al. J. Cell Biol. 2001; 152: 1289-1299). Поэтому неожиданным результатом настоящего исследования было обнаружение того, что при низкой концентрации, GGF2 проявляет эффекты активации миелинизации. Однако промиелинизирующий эффект ограничивался диапазоном низких концентраций, и увеличение концентрации GGF2 с этого диапазона приводит к ингибированию миелинизации, как описано ранее. Это обнаружение вызывает интерес, поскольку оно демонстрирует, что растворимый GGF2 может вызывать два противоположных действия в одном и том же клеточном контексте исключительно на основе количества, представляемого клетке. Оно также наводит на мысль, что пороговые уровни GGF2 определяют промиелинизирующее и ингибиторное действие во время миелинизации. Как впервые продемонстрировано в настоящем исследовании, это может объясняться зависимой от концентрации дифференциальной активацией эффекторов, стимулируемых при передаче сигналов вниз от рецепторов. Конкретнее, данные настоящего исследования показывают, что промиелинизирующее действие GGF2 наблюдается при концентрациях, которые преимущественно стимулируют Akt, в то время как переход на ингибиторную роль при более высоких концентрациях совпадает с появлением активации Erk, несмотря на непрерывное увеличение уровня активного Akt. Этот результат также подтверждает предыдущую идею, что баланс между активацией PI3-киназы и Ras/Raf/Erk является решающим в определении состояния миелинизации в шванновских клетках (Ogata et al. J. Neurosci. 2004; 24: 6724-6732). Настоящие данные, однако, дают прямое доказательство того, что активация пути с участием Ras/Raf/Erk действует в качестве отрицательного регулятора миелинизации.
Ингибиторное действие Nrg1 на миелинизацию опосредуется через активацию Erk/Mek1. Предположение об ингибиторной роли пути с участием Ras/Raf/Erk в миелинизации было сделано ранее на основе исследований, в которых экспрессия конститутивно активной Mek1 в шванновских клетках блокирует форсколин-индуцированную экспрессию гена миелина, в то время как доминантный-отрицательный Ras блокируют индуцированную Nrg1 супрессию гена миелина. Этот непосредственный эффект на миелинизаци, однако, не был разъяснен до результатов настоящего исследования. В настоящем документе продемонстроровано, GGF2, при использовании выше пороговой концентрации, ингибирует миелинизацию в сокультурах. Авторы настоящего изобретения показывают в настоящем документе, что ингибирование активации Mek1/Erk1 возрождало миелинизацию в обработанных GGF2 сокультурах, демонстрируя, что ингибиторная роль GGF2 опосредовалась через активацию Ras/Raf/Erk1. Механизм, с помощью которого активация Mek1/Erk ингибирует миелинизацию, неясен. Возможный механизм включает супрессию экспрессии гена миелина, описанную ранее. Подтверждающие это предположение данные настоящего исследования показали, что увеличение миелинизации в обработанных U0126 культурах сопровождаются увеличением экспрессии Р0. Также возможно, что путь с участием Mek1/Erk мог бы модулировать экспрессию транскрипционных факторов, вовлеченных в миелинизацию шванновскими клетками или их дифференциацию. Недавно было установлено, что эктопическая экспрессия c-Jun в шванновских клетках ингибирует миелинизацию, наводя, таким образом, на мысль, что c-Jun действует в качестве отрицательного регулятора миелинизации. Данные настоящего исследования согласуются с этим выводом, поскольку авторы настоящего изобретения определили, что обработка GGF2, которая ингибирует миелинизацию, сопровождается индукцией c-Jun, и, кроме того, ингибирование Nrg1-индуцированной активности Mek1/Erk1 блокирует экспрессию c-Jun. Этот результат наводит на мысль, что ингибиторное действие Mek1/Erk1 на миелинизацию отчасти опосредуется через индукцию c-Jun.
Другими интересными данными настоящего исследования является наличие внутреннего Mek1/Erk-зависимого сигнала в сокультурах, который служит в качестве отрицательного регулятора миелинизации. Это было установлено в эксперименте, в котором обработка U0126 нормальных миелинизирующих сокультур стимулировала миелинизацию. Природа сигнала, вносящего вклад в активность Mek1/Erk1, во время миелинизации в настоящее время неизвестна, хотя она, по-видимому, является аксонной по природе, независимой от аксонного CDR-Nrg1. Возможными кандидатами являются Ig-Nrg1 типа I и II, которые экспрессируются нейронами ПНС и позже отсоединяются от аксонных оболочек с помощью протеолического расщепления. Другим возможным активатором Mek1/Erk является FGF-2, который экспрессируется в нейронах ПНС, и рецептор для которого экспрессируется на шванновских клетках. Обработка FGF-2 подавляет экспрессию гена миелина и ингибирует миелинизацию in vitro. Утрата экспрессии FGF-2 приводит к увеличению числа миелинизированных аксонов во время регенерации седалищного нерва. Периферические нейроны также экспрессируют PDGF и IGF, при этом соответствующие рецепторные тирозинкиназы экспрессируются на связанных с ними шванновских клетках. Большой интерес будет представлять оценка регуляторной роли этих факторов роста во время миелинизации ПНС.
Терапевтическое применение GGF2. Экспериментальная трансплантация дала неоспоримое доказательство возможности восстановления поврежденных нервов путем трансплантации миелинизирующих глиальных клеток. Шванновские клетки являются хорошими кандидатами для такой терапии, поскольку они легко размножаются в культуре и открывают возможность аутологичной трансплантации для стимуляции ремиелинизации и восстановления проводимости по нерву в демиелинизированных очагах не только в ПНС, но также в ЦНС. Ремиелинизация с использованием шванновских клеток на регенерирующихся аксонах взрослого, однако, часто является неполной, приводя к образованию более тонкой миелиновой оболочки и более короткого участка нервного волокна между перехватами Ранвье по сравнению с нормальными нервами. Поэтому настоящая демонстрация промиелинизирующего действия GGF2 и возможности избегать случайной блокировки миелинизации, обусловленной уровнями доз GGF2, является важной, поскольку она обеспечивает терапевтическую стратегию для лечения демиелинизирующих заболеваний, а также для восстановления миелина после повреждения нерва.
Другие варианты осуществления. Все публикации и заявки на патенты, упоминаемые в этом описании, включены в настоящий документ посредством ссылки в такой же степени, как если бы для каждой независимой публикации или заявки на патент было специально и отдельно указано, что она включена посредством ссылки.
Хотя настоящее изобретение было описано с учетом его конкретных вариантов осуществления, будет понятно, что оно может быть далее модифицировано, и эта заявка, как предполагается, охватывает любые варианты, применения или адаптации настоящего изобретения, следующие, в общем, принципам настоящего изобретения и включающие такие отклонения от настоящего описания, которые подпадают под известную или обычную практику в данной области техники, к которой относится настоящее изобретение, и могут быть применены к существенным признакам, изложенным выше, и отслеживается в объеме прилагаемой формулы изобретения.







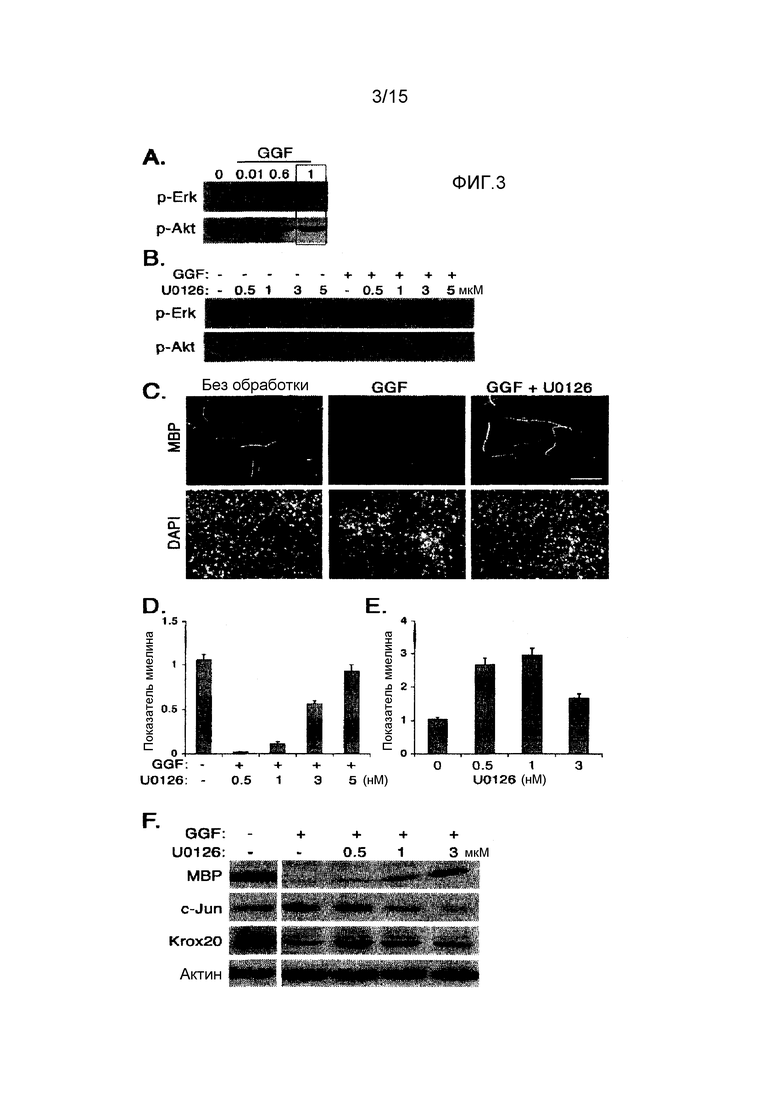












Изобретение относится к медицине. Предложен способ расширения диапазона терапевтических доз фактора 2 роста глии (GGF2), когда GGF2 используется для способствования миелинизации, включающий отбор пациента с заболеванием или нарушением, связанным со сниженными уровнями миелинизации, введение ему GGF2 и ингибитора пути с участием Mek1/Erk, посредством чего возникает GGF2-опосредованная миелинизация при более высоких дозах GGF2, чем она могла бы происходить в отсутствие введения ингибитора пути с участием Mek1/Erk. Технический результат состоит в оптимизации терапевтической дозы GGF2, в частности в её определении как 500 нг ±15% с ограничением неблагоприятных эффектов – ингибирования миелинизации при высоких концентрациях GGF2 за счёт эффекта ингибитора Mek1/Erk. 4 з.п. ф-лы, 12 ил.
1. Способ расширения диапазона терапевтических доз фактора 2 роста глии (GGF2), когда GGF2 используется для способствования миелинизации, включающий
отбор пациента с заболеванием или нарушением, связанным со сниженными уровнями миелинизации,
введение пациенту GGF2 и ингибитора пути с участием Mek1/Erk,
посредством чего возникает GGF2-опосредованная миелинизация при более высоких дозах GGF2, чем она могла бы происходить в отсутствие введения ингибитора пути с участием Mek1/Erk.
2. Способ по п. 1, в котором стадия введения включает введение количества GGF2, которое вызывает уровень в плазме выше 0,01 нМ +/-15% GGF2.
3. Способ по п. 2, в котором стадия введения включает введение количества GGF2, которое вызывает уровень в плазме выше 0,01 нМ GGF2.
4. Способ по п. 1, в котором стадия введения включает введение количества GGF2, превышающего 500 нг +/-15% GGF2 на кг веса тела.
5. Способ по п. 4, в котором стадия введения включает введение количества GGF2, превышающего 500 нг GGF2 на кг веса тела.
| Экономайзер | 0 |
|
SU94A1 |
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| ГРЭХАМ-СМИТ Д.Г | |||
| и др | |||
| Оксфордский справочник по клинической фармакологии | |||
| М., Медицина, 2000, с.27-32, с.137 глава 10.2.2., с.139-146 1 колонка 2-й абзац | |||
| PENDERIS J | |||
| et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| European Journal of neuroscience | |||
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| MAUREL P | |||
| et al | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| The Journal of neuroscience | |||
| ЩИТОВОЙ ДЛЯ ВОДОЕМОВ ЗАТВОР | 1922 |
|
SU2000A1 |

