Область техники, к которой относится изобретение
Настоящее изобретение касается растворимых в физиологических растворах изоформ нейрегулина-1, представляющих собой посттрансляционные модификации или сплайс-варианты нейрегулина-1, в качестве лекарства при когнитивных неврологических заболеваниях, в частности шизофрении, болезнях Альцгеймера и Паркинсона.
Уровень техники
Нейрегулины (NRG) оказались ключевыми регуляторами синаптической передачи сигналов. Эти трансмембранные белки кодируются четырьмя генами (nrg-1, -2, -3 и -4), а их разнообразие еще больше возрастает посредством альтернативного сплайсинга РНК и употребления промоторов, в частности, посредством посттрансляционных модификаций типа протеолитического процессинга, что ведет к высвобождению растворимых изоформ из мембраносвязанных целых белков. Кроме того, есть данные о фосфорилировании и гликозилировании (Buonanno and Fischbach, 2001). Они характеризуются различными внеклеточными доменами и являются лигандами тирозиновых киназ рецепторов ErbB, которые имеют отношение к нисходящим процессам нейровоспаления и транскрипции генов (Holbro and Hynes, 2004). В частности, растворимые изоформы NRG-1 получаются из трансмембранной формы NRG путем протеолитического расщепления при электрической стимуляции и впоследствии секретируются в качестве зависимых от активности синаптических модуляторов (Ozaki et al., 2004).
Укороченная изоформа NRG-1, предположительно β1, содержащая N-концевой внеклеточный домен (ECD) целого мембранного белка, как оказалось, коррелирует с обучением и памятью (Schillo et al., 2005а; WO 03/014156). Функциональные исследования показали, что NRG-1 непосредственно регулирует субъединичный состав NMDA-рецепторов (Ozaki et al., 1997; Eilam et al., 1998). Кроме того, было показано, что фрагменты NRG-1 этого типа обладают нейрозащитными свойствами in vivo через антиапоптические эффекты (Xu et al., 2005а; Xu et al, 2005b; Xu et al., 2004).
Совсем недавно стало ясно, что NRG-1 играет важную роль при неврологических заболеваниях человека вследствие NRG-зависимой регуляции NMDA-рецепторов (Schrattenholz and Soskic, 2006) и последующих нисходящих явлений типа экцитотоксичности, нейровоспаления и апоптоза (см. сводку на фиг.1). Есть результаты, показывающие, что NRG-1 играет важную роль при целом ряде заболеваний от бокового амиотрофического склероза, болезни Альцгеймера и Паркинсона до инсульта и шизофрении (Britsch, 2007).
Такое фундаментальное значение NRG-1 означает, что наряду с нейрозащитной и положительной ролью в когнитивном обучении и памяти NRG-1 является важным нейротрофическим фактором при регенерации нервной ткани после различных повреждений, в целом ряде специфических участков и типов клеток мозга. Очевидно, он является важным фактором для поддержания и восстановления целостности нейронных цепей; нейрозащитным фактором, играющим роль в правильной регенерации после утраты функции, а также в реализации зависимой от активности пластичности нейронов.
Интерес к нейрегулину-1β еще более значительно возрос, когда Kastin et al., 2004 показали, что нейрегулин-1β может проходить через гематоэнцефалический барьер. Это открыло перспективы терапевтического применения нейрегулина-1β.
Последние исследования засвидетельствовали масштабность применения в нейрозащите. Независимо в двух публикациях было показано, что нейрегулин-1 также является субстратом ВАСЕ (β-секретазы, β-амилоид-конвертазы), что означает важность нейрегулина-1 при болезни Альцгеймера (Glabe, 2006; Schubert, 2006).
Более того, оказалось, что в шванновских клетках нейрегулин-1 повышает транскрипцию 3-гидрокси-3-метилглутарил-СоА-редуктазы - фермента, лимитирующего скорость биосинтеза холестерина в шванновских клетках (Pertusa et al, 2007). Это имеет далеко идущие для тех заболеваний, при которых поражается миелиновая оболочка, напр., шизофрении и множественного склероза, или нарушений когнитивных функций, при которых задействованы так называемые "богатые холестерином островки" (Schrattenholz and Soskic, 2006). Окружающие шванновские клетки аксоны экспрессируют рецепторы NRG-1 - ErbB2/ErbB3 и растворимый NRG-1α и -1β в физиологических условиях. После денервации зрелые шванновские клетки теряют контакты с аксонами, изменяют свою морфологию, перестают экспрессировать NRG-1β и усиливают экспрессию NRG-1α и ErbB2/ErbB3 (Geuna et al., 2007; Karoutzou et al., 2007).
Кроме того, исследования по генетической эпидемиологии свидетельствуют о четкой связи нейрегулина-1 с шизофренией и болезнью Альцгеймера, в частности ее психотическими формами (Farmer et al., 2007).
Некоторые последние генетические анализы населения показали, что определенные SNP NRG-1 связаны с болезнью Альцгеймера и шизофренией (Go et al., 2005; Scolnick et al., 2006; Ross et al., 2006; Meeks et al., 2006; Farmer et al., 2007). Значение этих данных связано с другими белками функционального комплекса, содержащего NRG, который изображен на фиг.1 (рецептор ErbB: Benzel et al., 2007; Thomson et al., 2007; Hahn et al., 2006). Также есть данные об участии NRG-1 во множественном склерозе (Esper et al., 2006).
Имеются результаты, свидетельствующие о том, что молекулярный механизм связи между аллелями риска NRG-1 и шизофренией может включать понижающую регуляцию никотиновых ацетилхолиновых рецепторов подтипа α7 (Mathew et al., 2007).
Раскрытие изобретения
В соответствии с настоящим изобретением оказалось, что рекомбинантные растворимые β-изоформы нейрегулина-1 проявляют фармацевтическую эффективность у животных на моделях обучения и памяти, шизофрении, болезни Альцгеймера и болезни Паркинсона. После внутривенного введения β-изоформы нейрегулина-1 действовали в концентрациях, значительно меньших, чем концентрации контрольных медикаментов.
Итак, первый аспект настоящего изобретения составляет применение рекомбинантных растворимых изоформ нейрегулина-1 для изготовления медикамента для лечения неврологических заболеваний, в частности когнитивных неврологических заболеваний.
Другой аспект настоящего изобретения составляет фармацевтическая композиция или набор, включающий: (i) рекомбинантную растворимую изоформу нейрегулина-1 и (И) другой медикамент, в особенности для лечения неврологических заболеваний, в частности когнитивных неврологических заболеваний.
Следующий аспект настоящего изобретения составляет применение рекомбинантных растворимых изоформ нейрегулина-1 для изготовления медикамента для усиления памяти и когнитивной способности.
Следующий аспект настоящего изобретения составляет способ лечения неврологических заболеваний, включающий введение рекомбинантной растворимой изоформы нейрегулина-1 в фармацевтически эффективном количестве нуждающемуся в этом субъекту.
Следующий аспект настоящего изобретения составляет способ усиления памяти и когнитивной способности, включающий введение рекомбинантной растворимой изоформы нейрегулина-1 в фармацевтически эффективном количестве нуждающемуся в этом субъекту.
Следующий аспект настоящего изобретения составляет совместное введение рекомбинантной растворимой изоформы нейрегулина-1 вместе с другим медикаментом.
Осуществление изобретения
В соответствии с настоящим изобретением растворимые изоформы нейрегулина-1 оказались эффективными при лечении неврологических заболеваний, в частности таких заболеваний, как психотические расстройства типа шизофрении, биполярного психоза и депрессии, нейродегенеративных заболеваний типа болезни Паркинсона, болезни Альцгеймера, множественного склероза (MS) или бокового амиотрофического склероза (ALS), эпилепсии или неврологических повреждений типа инсульта, травматических повреждений головного и спинного мозга. Предпочтительным является лечение шизофрении, в частности когнитивных аспектов шизофрении, болезни Паркинсона и болезни Альцгеймера. Кроме того, изобретение также касается применения рекомбинантных растворимых изоформ нейрегулина-1 для усиления памяти и когнитивной способности, в частности для уменьшения и/или торможения потери памяти и когнитивной способности при таких неврологических заболеваниях, как болезнь Альцгеймера и шизофрения.
Рекомбинантная растворимая изоформа нейрегулина-1 предпочтительно является изоформой нейрегулина-1 человека, т.е. рекомбинантной изоформой, содержащей первичную аминокислотную последовательность природной изоформы нейрегулина-1 человека либо последовательность, идентичную ей по меньшей мере на 90%, предпочтительно по меньшей мере на 95% и наиболее предпочтительно по меньшей мере на 98%), исходя из общей длины рекомбинантной изоформы.
Растворимая рекомбинантная изоформа нейрегулина-1 по настоящему изобретению предпочтительно содержит по крайней мере часть внеклеточного домена соответствующего нейрегулина-1, например, по крайней мере часть внеклеточного домена нейрегулина человека, например, нейрегулина-1β человека.
Рекомбинантная растворимая изоформа нейрегулина по настоящему изобретению предпочтительно имеет длину вплоть до 250 аминокислот, например, от 150 до 250 аминокислот.Молекулярная масса изоформы нейрегулина предпочтительно составляет примерно от 15 до 35 кДа, в особенности от 25 до 32 кДа при измерении, напр., методом электрофореза в SDS-полиакриламиде (PAGE). Рекомбинантная растворимая изоформа нейрегулина-1, в частности рекомбинантная β-изоформа нейрегулина-1, имеет изоэлектрическую точку (pI) примерно от 4 до 9,5, предпочтительно от 4 до 6. Изоформа может представлять собой немодифицированный полипептид, состоящий из немодифицированной последовательности аминокислот, либо модифицированный полипептид, у которого модификация может заключаться в фосфорилировании, гликозилировании, метилировании, миристилировании, окислении или любой комбинации из них. В особенно предпочтительном воплощении изоформа нейрегулина-1 содержит по меньшей мере один фосфорилированный аминокислотный остаток. Кроме того, настоящее изобретение охватывает конъюгирование с гетерологичными молекулами, как-то молекулами полиалкиленоксидов, в частности молекулами полиэтиленгликоля.
Рекомбинантные растворимые изоформы можно вводить любым способом, при котором достигается эффективная доставка в намеченную ткань, напр., нервную систему, в частности центральную нервную систему, как-то головной и/или спинной мозг. Как оказалось, фармацевтически эффективных концентраций изоформ нейрегулина можно достичь при системном введении. Например, изоформы можно вводить посредством инъекции или вливания, напр., посредством внутривенной инъекции. Изоформы предпочтительно вводятся в количестве от 0,1 до 5000 нг/кг массы тела, особенно в количестве от 2 до 1000 нг/кг массы тела, более предпочтительно в количестве от 3 до 600 нг/кг массы тела подлежащего лечению субъекта, в зависимости от типа и тяжести подлежащего лечению заболевания. В других воплощениях настоящего изобретения растворимые изоформы также можно вводить местно, напр., путем прямого введения в центральную нервную систему, напр., в спинной и/или головной мозг. Также можно рассматривать и введение при более высоких дозировках вплоть до 500 мкг/кг при внутрибрюшинных или подкожных инъекциях либо при помощи ингаляционных устройств. Предпочтительно подлежащим лечению субъектом является млекопитающее, более предпочтительно больной человек.
Растворимые рекомбинантные изоформы нейрегулина-1 можно вводить как единственное лекарство, т.е. в виде монотерапии, либо как комбинированное лекарство, т.е. в комбинации с другим лекарством, подходящим для лечения неврологических заболеваний. Примерами других лекарств являются соединения, влияющие на метаболизм катехоламинов, ингибиторы ацетилхолинэстеразы, ингибиторы МАО-В или СОМТ, блокаторы мембранных каналов, агонисты либо антагонисты дофаминовых или серотониновых рецепторов, ингибиторы обратного захвата катехоламинов либо серотонина, или любые антипсихотические медикаменты типа клозапина или оланзапина либо препараты типа габапентина, в частности при лечении болезни Альцгеймера или Паркинсона, шизофрении, биполярного психоза, депрессии и других неврологических заболеваний. Дополнительными примерами других медикаментов являются нейрозащитные средства, как-то ингибиторы PARP-1, напр., приведенные в WO 2006/008118 и WO 2006/008119, которые включены в настоящее изобретение путем ссылки.
Итак, одно воплощение настоящего изобретения касается комбинирования рекомбинантной растворимой изоформы нейрегулина-1, описанной в настоящем изобретении, с медикаментом для лечения психотических расстройств, как-то шизофрении, биполярного психоза и депрессии, напр., оланзапином или клозапином. Другое воплощение касается комбинирования рекомбинантной растворимой изоформы нейрегулина-1 с медикаментом для лечения нейродегенеративных заболеваний, как-то болезни Паркинсона, болезни Альцгеймера, MS или ALS. Следующее воплощение касается комбинирования рекомбинантной растворимой изоформы нейрегулина-1 с медикаментом для лечения неврологических повреждений, как-то инсульта, травматических повреждений головного или спинного мозга.
Комбинированная терапия может осуществляться путем совместного введения рекомбинантной растворимой изоформы нейрегулина-1 и другого медикамента в виде фармацевтической композиции или набора, при этом индивидуальные медикаменты вводятся по отдельности или вместе.
Изоформа нейрегулина-1 может представлять собой изоформу нейрегулина I типа, II типа, III типа, IV типа, V типа или VI типа, предпочтительно β-изоформу нейрегулина-1, α-изоформу нейрегулина-1 либо изоформу происходящего из сенсорных и моторных нейронов фактора (SMDF), в особенности β-изоформу нейрегулина-1, более предпочтительно β-изоформу нейрегулина-1 человека.
β-Изоформы нейрегулина-1 подвергаются активному транспорту через гематоэнцефалический барьер. Отличная биодоступность нейрегулина-1β в мозге после внутривенного/внутрибрюшинного введения, как это видно из примеров, открывает путь для терапевтического применения NRG-1β.
Сочетание его антиапоптических, стабилизирующих миелин и противовоспалительных свойств, наряду с прямым взаимодействием с ВАСЕ, открывает перспективы для лечения инсульта, болезни Альцгеймера, MS и шизофрении, а также других неврологических заболеваний.
Как изложено выше, настоящая заявка охватывает применение немодифицированных и модифицированных изоформ нейрегулина-1, в частности β-изоформ нейрегулина-1. Имеются данные, что такие посттрансляционные модификации, как протеолитический процессинг, фосфорилирование и гликозилирование происходят на определенных аминокислотных остатках нейрегулина-1, в частности его внеклеточного домена. В частности, были сообщения о высвобождении растворимых фрагментов нейрегулина-1 (Buonanno and Fischbach, 2001; Fischbach, 2007). Также сообщалось о возможном окислении (Nadri et al., 2007).
Авторы настоящего изобретения получили данные о том, что предпочтительные физиологически активные β-изоформы нейрегулина-1 содержат внеклеточный домен нейрегулина-1β или его часть, которая подвергалась посттрансляционной модификации. Предпочтительно изоформы подвергались модификации путем фосфорилирования, при этом подвергались фосфорилированию боковые цепи 1, 2, 3 или больше аминокислотных остатков, в частности боковые цепи остатков, содержащих ОН-группу, как-то Tyr, Ser или Thr. Предпочтительные сайты фосфорилирования находятся в положении аминокислот 79-82, 133-136 и/или 158-161 (номенклатура согласно Falquet et al., 2002). Другие сайты фосфорилирования находятся в положении аминокислот 12-14, 30-32 и/или 85-87. Потенциальные сайты других модификаций - это сайты амидирования, предпочтительно находящиеся в положениях 22-25 и/или 30-33, сайты гликозилирования в положениях 150-153, 156-159 и/или 204-207 и сайты миристилирования, предпочтительно находящиеся в положениях 94-99, 149-154, 168-173, 175-180 и/или 202-207 согласно номенклатуре Falquet et al., 2002.
Далее следует разъяснение существенности экспериментальных данных в соответствии с настоящей заявкой в отношении предпочтительных медицинских показаний.
Шизофрения
Шизофрения является серьезным и вызывающим инвалидность психическим расстройством с такими симптомами, как слуховые галлюцинации, нарушение мышления и мания, аволиция, ангедония, притупленность и апатия. Эпидемиологические, клинические, нейропсихологические и нейрофизиологические исследования дали существенные данные о том, что в патогенезе этого расстройства важную роль играют нарушения развития мозга и текущей нейропластичности (Arnold et al., 2005).
Предполагается, что шизофрения включает нарушение дофаминергической нейротрансмиссии, но главную роль, видимо, играет модуляция дофаминергической системы под действием глутаматергической нейротрансмиссии. Такой взгляд подтверждается генетическими данными о том, что гены нейрегулина и дисбиндина оказывают функциональное влияние на глутаматергическую систему (Muller and Schwarz, 2006). Все больше становится ясно, что некоторые участки, которые, вероятно, содержат гены (включая гены нейрегулинов), способствующие шизофрении, также имеют отношение к биполярным аффективным расстройствам, что подтверждается двумя последними работами (Farmer et al., 2007; Owen et al., 2007).
Нейрегулин-1, который является геном подверженности к психозам, оказывает эффекты на миграцию нейронов, прорастание аксонов и миелинизацию, что могло бы объяснить данные об аномальных анатомических и функциональных связях при шизофрении и биполярном психозе (Mcintosh et al., 2007).
Имеется все возрастающий объем данных о генетической связи нейрегулина-1 с шизофренией (обзор: Farmer et al., 2007). В этой связи существенным является усиление нейрегулином-1 глутаматной, ГАМКергической и никотиновой нейротрансмиссии (Fischbach, 2007; Woo et al., 2007; Li et al., 2007), а также причастность к воспалению мозга (Hanninen et al., 2007).
Предполагается, что при этом заболевании задействована и регуляция 3-гидрокси-3-метилглутарил-СоА-редуктазы - фермента, лимитирующего скорость биосинтеза холестерина (Pertusa et al., 2007), что важно для миелинизации.
Тот факт, что среди генетических факторов риска, общих для шизофрении, биполярного психоза и депрессии, важную роль играет NRG-1, привел к предположению о том, что вовлеченные в эти психозы гены типа NRG-1 могли бы послужить основой для классификации, исходящей из биологии, а не симптомов, и привести к новым стратегиям лечения этих сложных заболеваний мозга (Blackwood et al, 2007; Bertram et al., 2007).
Экспериментальные данные настоящей заявки свидетельствуют об эффективности применения растворимой рекомбинантной β-изоформы нейрегулина-1 на экспериментальной модели шизофрении.
Болезнь Альцгеймера
Первоначальные исследования авторов изобретения показали, что нейрегулин-1β снижается на посмертных срезах гиппокампа в мозге больных при болезни Альцгеймера по сравнению с подобранными по возрасту контролями (Sommer et al., 2004), при этом существует четкая положительная корреляция между растворимым фрагментом нейрегулина-1 и показателями обучения при тестировании в радиальном лабиринте (Sommer et al., 2004).
Во многих работах засвидетельствована роль NRG-1 в зависимых от активности синаптических изменениях (Xie et al, 2006; Kwon et al, 2005; Rimer et al, 2005; Bao et al., 2004; Yang et al., 2005), важных для обучения и памяти (Ozaki et al., 1997; Ozaki et al., 2004; Golub et al., 2004; Schillo et al., 2005b). Как показано ниже, фрагмент NRG-1β, содержащий внеклеточный домен, четко связан с обучением животных на поведенческой модели. Снижение экспрессии этого белка на посмертных срезах мозга в области гиппокампа (отвечающего за формирование кратковременной памяти) у больных при болезни Альцгеймера по сравнению с подобранными по возрасту контролями могло бы свидетельствовать об отсутствии связанной с памятью синаптической активности на участках с еще здоровыми на вид нейронами.
Самые последние открытия (Hu et al., 2006; Glabe, 2006; Schubert, 2006) показали, что NRG-1 подвергается процессингу ферментом ВАСЕ1 (=β-секретаза), который способствует образованию скоплений β-амилоида в мозге людей с болезнью Альцгеймера, что объясняет связь с болезнью Альцгеймера, а его сопутствующая роль в образовании миелина связана с нейротрофическими свойствами NRG-1 (Hu et al., 2006; Glabe, 2006; Schubert, 2006). Фермент BACE1 (расщепляющий белок-предшественник амилоида по β-связи фермент-1) необходим для отщепления β-амилоида из большего предшественника, причем окончательное отщепление после расщепления ферментом ВАСЕ1 производит содержащий пресенилин комплекс γ-секретазы с высвобождением β-амилоида.
Расщепление NRG секретазами имеет важное значение для миелинизации нервов. Как и белок-предшественник амилоида, нейрегулин-1 тоже расщепляется β-секретазой. Протеолитическое расщепление β-секретазой нейрегулина-1 имеет важное значение для миелинизации периферических нервов шванновскими клетками. Препараты, мишенью которых является β-секретаза, могут повлиять на развитие и функционирование периферических нервов.
Первоначальное наблюдение было сделано группой Haass (Willem et al., 2006), обнаружившей, что для миелинизации также необходим фермент ВАСЕ1. Миелинизация периферических нервов происходит в раннем периоде жизни, поэтому неясно, как ингибирование ВАСЕ1 могло бы повлиять на старых животных. Есть данные о том, что ВАСЕ1 также играет роль и в миелинизации центральной нервной системы. У трансгенных животных, дефектных по ВАСЕ1, отмечались дефекты миелина в периферических нервах.
Также в связи с нейродегенерацией и болезнью Альцгеймера существенным является недавнее открытие усиления нейрегулином-1 глутаматной, ГАМКергической и никотиновой нейротрансмиссии (Fischbach, 2007; Woo et al., 2007; Li et al., 2007).
Экспериментальные данные настоящей заявки свидетельствуют об эффективности применения растворимой рекомбинантной β-изоформы нейрегулина-1 на экспериментальной модели болезни Альцгеймера.
Инсульт, травматическое повреждение мозга
Ряд связанных с инсультом экспериментов in vivo при независимых внешних исследованиях в США показал нейрозащитное действие нейрегулина-1, которое само по себе является антиапоптическим (Xu et al., 2004, 2005 и 2006; Guo et al., 2006).
NRG-1 уменьшает повреждение нейронов и улучшает неврологический исход после окклюзии средней церебральной артерии (распространенная модель инсульта) (Xu et al., 2005b; Xu et al., 2004, Xu et al., 2006; Guo et al., 2006).
В том же самом исследовании терапевтической эффективности рекомбинантного NRG-1 человека и механизма уменьшения повреждений мозга при ишемии/реперфузии оказалось, что NRG действует антиапоптически. NRG-1 (3,0 нг/кг) вводили интраваскулярно за 10 мин до окклюзии средней церебральной артерии (МСАО) и последующей очаговой ишемии мозга в течение 90 мин и реперфузии в течение 24 ч.
Данные настоящего изобретения свидетельствуют о том, что введение рекомбинантных растворимых изоформ нейрегулина-1 в низких концентрациях оказывает существенный фармакологический эффект и поэтому предполагается эффективным на моделях инсульта и травматического повреждения мозга.
Далее настоящая заявка раскрывается более подробно на фигурах и примерах, приведенных ниже.
Краткое описание фигур
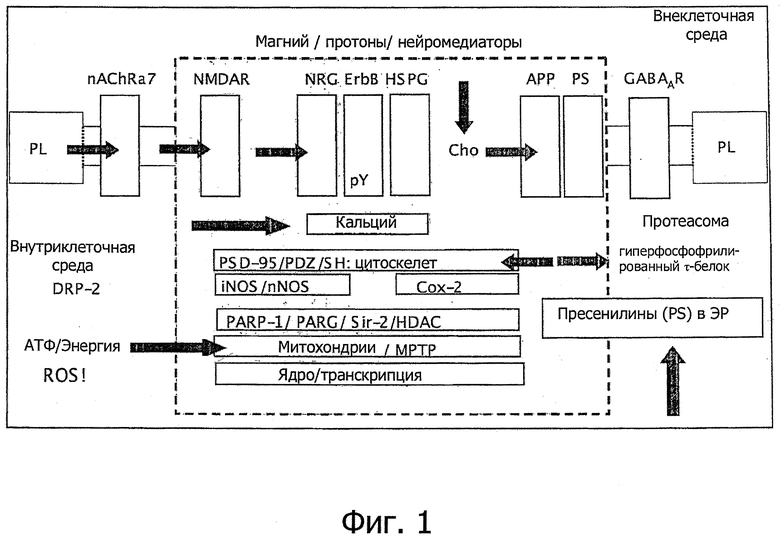
Фиг.1. В различных обзорах и многочисленных исследовательских статьях о нейрегулине-1 показано ключевое функциональное положение NRG-1 как вышележащего регуляторного начала механизмов, играющих важную роль при нейродегенеративных заболеваниях, неврологических заболеваниях, а также физиологических функциях.
NRG являются ключевыми компонентами функциональных комплексов, состоящих как минимум из нейрегулинов (NRG), тирозиновых киназ рецепторов (рецепторов ErbB), гепарансульфатных протеогликанов (HSPG) и NMDA-рецепторов (NMDAR), которые собираются в кратковременные и зависимые от активности комплексы в богатых холестерином (СНО) микродоменах мембран. Для взаимодействия с белками субсинаптического каркаса посредством посттрансляционных модификаций (PSD-95, при взаимодействии с некоторыми фосфорилированными доменами типа PDZ- или SH-доменов на белках-партнерах) особенно важно формирование кальциевых сигналов. Комплекс PSD-95 непосредственно регулирует провоспалительные ферменты типа синтазы оксида азота (NOS, причем iNOS - индуцибельный, a nNOS - нейрональный фермент) и Сох-2 (циклооксигеназы-2), которые оказывают свои эффекты в сложном взаимодействии со связанными с ними, но необязательно нижележащими механизмами с участием таких NAD+ - зависимых ферментов, как PARP-1 (полимераза поли-АЭР-рибозы) и Sir-2 (сиртуин-2); PARG - это гликогидролаза поли-АБР-рибозы, фермент который дополняет и антагонистичен PARP-1, a HDAC - это деацетилазы гистонов, общий класс ферментов, который включает Sir-2. МРТР - митохондриальная пора с переходной проницаемостью. DRP-2 - родственный дигидропиримидиназе белок-2. Также и другие важные мембранные белки, как-то некоторые никотиновые ацетилхолиновые рецепторы (nAChRα7), ГАМКА-рецепторы (GABAAR), белок-предшественник амилоида (АРР) и протеазы (PS), временно встраиваются в липидные островки и приобретают другие функциональные свойства за пределами своего обычного фосфолипидного (PL) окружения (см. подробности в Schrattenholz and Soskic, 2006).
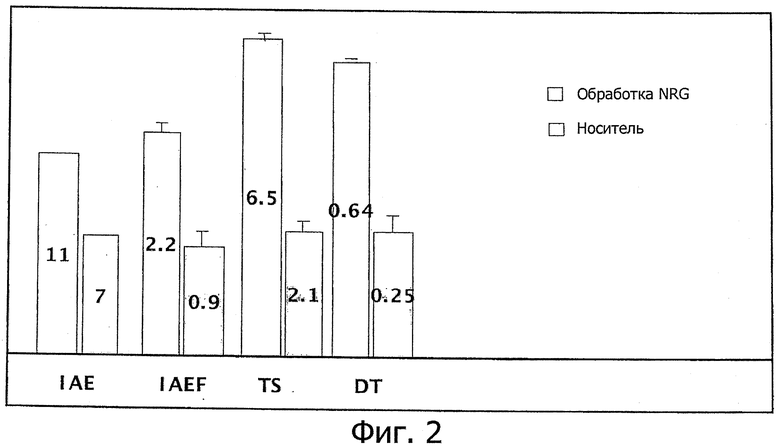
Фиг.2. Сводка из экспериментов по обучению в водном лабиринте Морриса. Животные, получавшие суточную дозу в 3 нг/кг (внутривенно) растворимого внеклеточного домена нейрегулина-1β (ECD NRG-1β) обучались значительно лучше, чем животные, получавшие носитель. IAE - посещения внутренней области; IAEF - частота посещений внутренней области; TS - время, проведенное во внутренней области; DT - расстояние, пройденное во внутренней области.
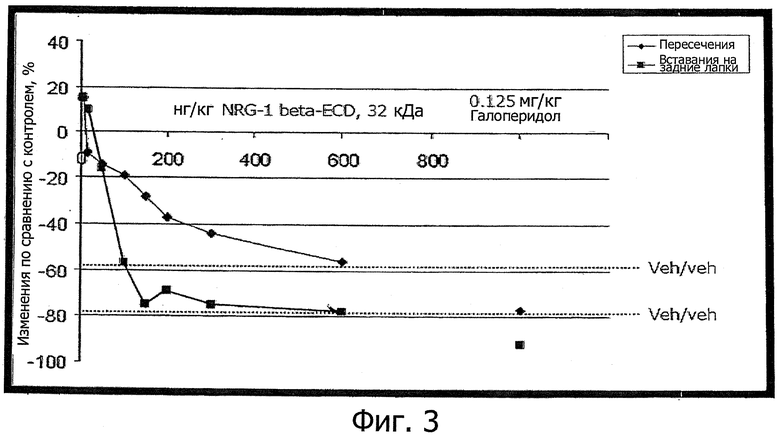
Фиг.3. Уменьшение под действием ECD NRG-1β вызванной амфетамином гиперактивности - общепринятой модели шизофрении. Концентрации составляли от 15 до 600 нг/кг (внутривенное введение за 15 мин до введения амфетамина). Положительный контроль включал 0,125 мг/кг галоперидола.
В то время, как галоперидол, подобно другим типичным и нетипичным антипсихотическим средствам (нейролептикам), обычно снижает активность ниже контрольного уровня (указан пунктиром с пометкой veh/veh, синим цветом для пересечений, красным для вставаний на задние лапки), вызванное ECD NRG-1β снижение асимптотически приближается к контрольным уровням активности, но не вызывает дальнейшего снижения. Низкие эффективные концентрации ECD NRG-1β и отсутствие отрицательных эффектов (т.е. снижения активности ниже контрольных уровней носителя) - таковы замечательные свойства на этой модели. Эффекты значимы, р<0,05.
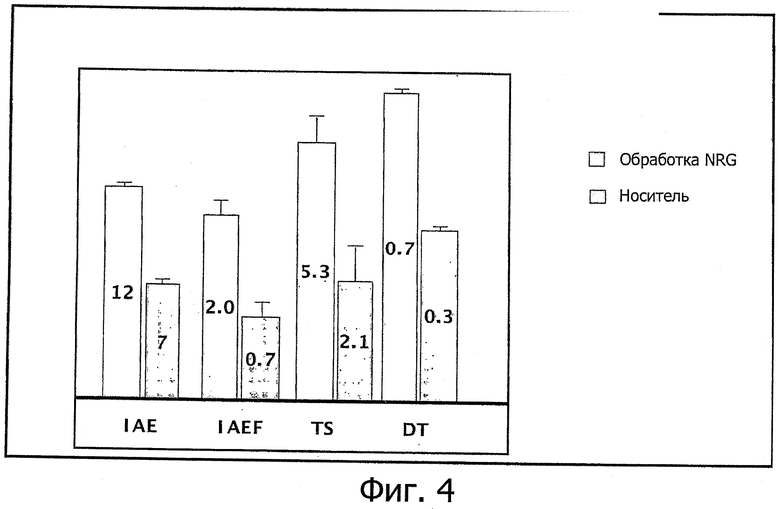
Фиг.4. Сводка из опытов по обучению мышей APPPS на модели церебрального амилоидоза и болезни Альцгеймера в водном лабиринте Морриса. Животные, получавшие суточную дозу ECD NRG-1β 200 нг/кг внутрибрюшинно, обучались значительно лучше, чем животные, получавшие носитель. IAE - посещения внутренней области; IAEF - частота посещений внутренней области; TS - время, проведенное во внутренней области; DT - расстояние, пройденное во внутренней области.
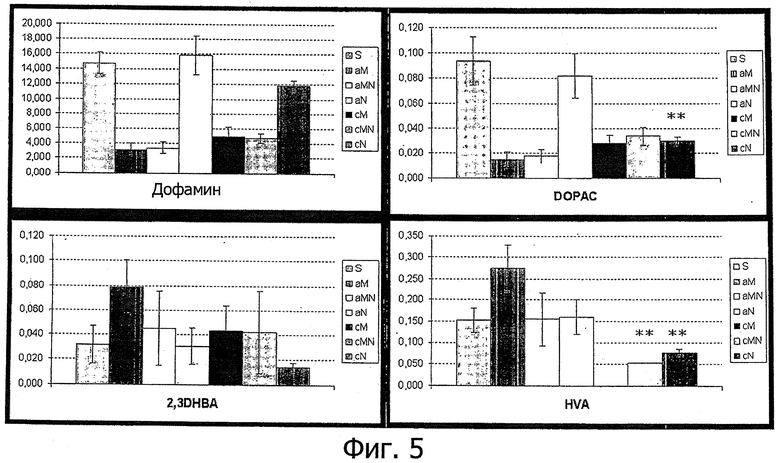
Фиг.5. Количественное определение дофамина и его метаболитов методом ВЭЖХ. Помеченные звездочкой столбцы имеют высокую значимость. Обозначения:
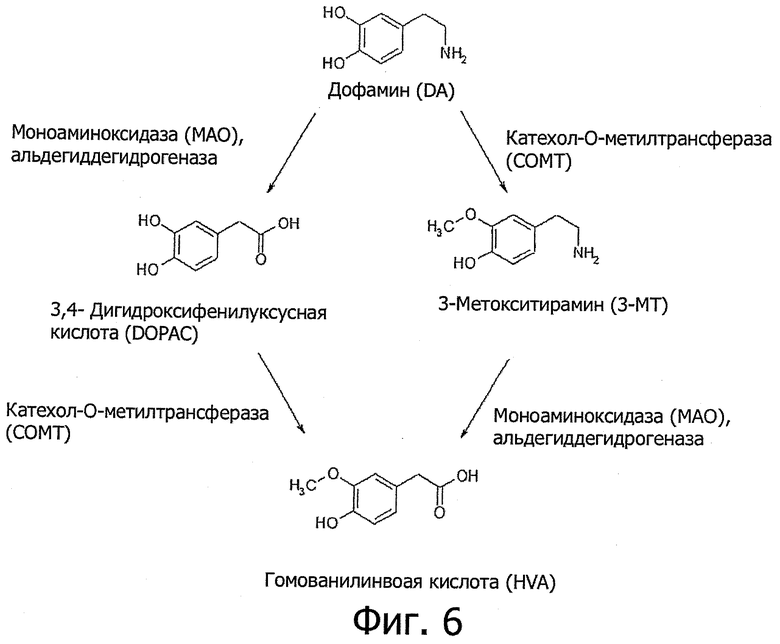
Фиг.6. Метаболизм дофамина под действием МАО-В и СОМТ.
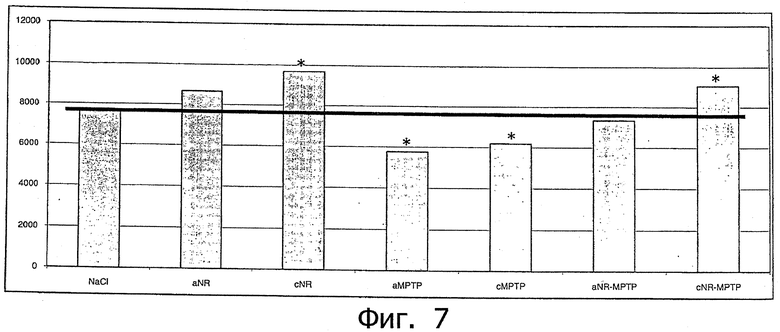
Фиг.7. Воздействие МРТР ведет к значительной потере дофаминергических нейронов в черной субстанции (substantia nigra) (аМРТР, р=0,0005; сМРТР, р=0,0075). Внутрибрюшинное введение 20 нг/кг ECD NRG-1β вызывает возвращение к норме (aNR-МРТР, р=0,57, т.е. не отличается от контроля с носителем) либо четкое и значимое улучшение повреждений от МРТР (cNR-MPTP, р=0,0097); на хронической модели (5 дней подряд вводили по 20 нг/кг ECD NRG-1β внутрибрюшинно) также отмечался значимый эффект на число дофаминергических нейронов (cNR, р=0,0002). Обозначения:
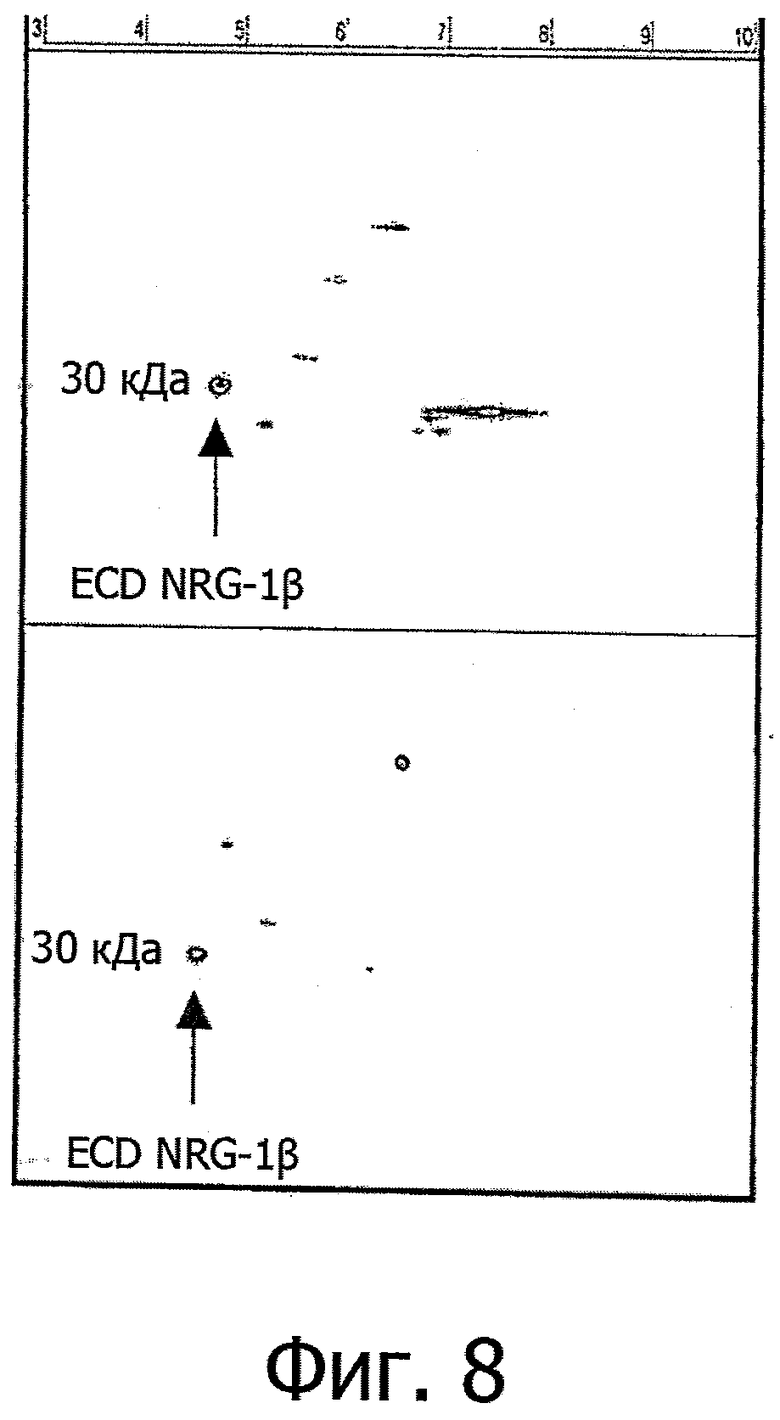
Фиг.8. Два репрезентативных снимка 2D-вестерн-блоттинга белков головного мозга мышей APPPS, окрашенных на нейрегулин-1β. Представлено одно подвергавшееся обработке животное с хорошим обучением (вверху) и не подвергавшееся обработке с плохим обучением (внизу).
Цифры в верхней части означают значения pI на 2D-геле.
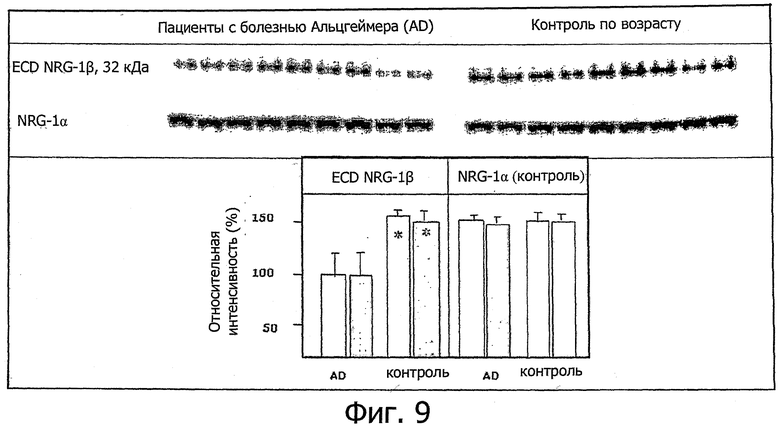
Фиг.9. В эксперименте по вестерн-гибридизации сравнивали содержание ECD-фрагмента NRG-1β на посмертном материале коры головного мозга при болезни Альцгеймера и в контроле.
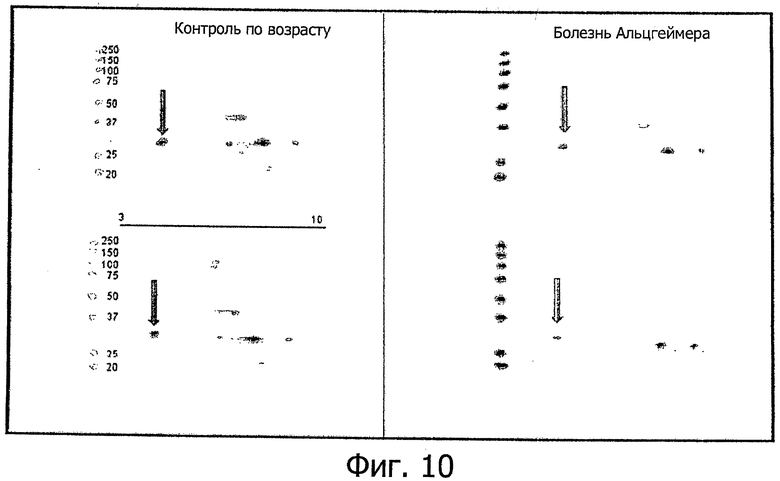
Фиг.10. В этих экспериментах методом двумерного электрофореза в полиакриламидном геле (2Д-ПАГ) показано, что кислая изоформа ECD NRG-1β со значением pI 5-5,5 и молекулярной массой 25-32 кДа четко снижается в головном мозге при болезни Альцгеймера.
ПРИМЕРЫ
Общая часть
Во всех дальнейших экспериментах использовали фрагменты нейрегулина-1β, содержащие только внеклеточный домен (ECD) из всего транскрипта гена nrg-1 человека. Они имели молекулярную массу 25-32 кДа и изоэлектрические точки между 5 и 9,5 в зависимости от степени фосфорилирования и/или гликозилирования.
Физиологически активная форма β-изоформы нейрегулина-1 имеет значение pI около 5,5 (большинство экспериментов проводили с коммерчески доступной изоформой, полученной в Е. coli, имеющей молекулярную массу 26 кДа и значение pi примерно 9,0).
Эта изоформа представляет собой рекомбинантный растворимый фрагмент NRG-1β человека, состоящий из первых 245 аминокислот NRG-1β, который приобретали у фирмы R&D Systems, Inc. (кат. №377-HB-CF). Далее он будет именоваться ECD NRG-1β. Эта активная изоформа имеет значение pi примерно 9,0.
Также тестировали соответствующий фрагмент NRG-1β в 8 кДа, содержащий только домен EGF, который приобретали у фирмы R&D Systems, Inc. (кат.№396-НВ). Этот фрагмент тоже оказался нейрозащитным in vitro и in vivo, но он не был исследован подробно из-за более сильных пролиферативных свойств, что вызывало опасения насчет канцерогенности.
Пример 1. Первичные токсикологические данные свидетельствуют, что NRG-1β (ECD) не оказывает отрицательных эффектов в испытаниях на острую токсичность и мутагенность in vitro
- Отсутствие острой внутривенной токсичности на крысах. Все животные дожили до конца периода исследования, в ходе исследования. Вес тела животных был в пределах того, что обычно отмечается для этой линии и этого возраста. При вскрытии не было обнаружено макроскопических явлений. Медиана летальной дозы NRG-1β (ECD) после однократного внутривенного введения самкам крыс при наблюдении за период в 14 дней составила: LD50 (самки крыс): более 5000 нг/кг массы тела.
- Ежедневное внутривенное введение нейрегулина на протяжении 7 дней в дозах 50, 200 и 600 нг/кг массы тела в день не вызывало преждевременной смертности. Не отмечалось никаких клинических признаков. Обработка не влияла на потребление пищи и прирост массы тела. Уровень отсутствия наблюдаемых эффектов (NOEL) составил 600 нг/кг массы тела в день.
- При определении на локусе тимидинкиназы в клетках лимфомы мыши на линии клеток L5178Y согласно Директиве OECD по тестированию химических веществ №476 "Испытание на генные мутации в клетках млекопитающих in vitro" NRG-1β (ECD) оказался немутагенным.
- При испытании на хромосомные аберрации на клетках китайского хомячка V79 согласно Директиве OECD по тестированию химических веществ №473 NRG-1β (ECD) не вызывал структурных аберраций хромосом.
Более того, ни в одном из экспериментов на животных, проводившихся насчет эффективности (некоторые из них продолжались по несколько месяцев с ежедневным внутривенным введением), никогда не наблюдалось отрицательных эффектов NRG-1β (ECD).
Введение NRG-1β (ECD) животным на различных моделях осуществлялось путем внутривенных (iv) или внутрибрюшинных (ip) инъекций; концентрации составляли от 3 до 600 нг/кг.
Пример 2. Обучение и память: пространственное обучение с применением ECD NRG-1β и без него
Методы
В водном лабиринте Морриса оценивается пространственное обучение. При этом животные должны плавать в наполненном водой бассейне и найти спасательную платформу, погруженную в воду чуть ниже поверхности. Платформа должна находиться вдали от стенок лабиринта, а животные должны иметь опорные точки, видимые из поверхности воды, которые позволяют оценить местоположение, но недостаточно близки к мишени и поэтому не способствуют ассоциативному обучению. Животных обучают тому, что спасение приходит только через платформу, что означает, что всех животных, не нашедших платформу, направляют на нее и дают им отдохнуть перед тем, как удалить из установки. Следовательно, одной из самых важных опорных точек для мыши является человек-оператор.
Целью эксперимента является определение двух ключевых параметров, связанных с пространственным запоминанием у мышей:
- скорости, с которой мыши учатся снова найти платформу,
- способности к сохранению информации на короткий срок (в пределах сеанса обучения или в течение ночи).
Животные
Исследование проводилось на двух группах мышей APP/PS (Meyer-Luehmann et al., 2006; Radde et al, 2006), одна из которых ежедневно получала дозу ECD NRG-1β, а другая получала ложную обработку в качестве контроля. Каждая группа состояла из 8 самцов в 9-недельном возрасте к началу первой серии экспериментов.
Первая серия начиналась с двумя подгруппами из 8 подвергавшихся и 8 не подвергавшихся обработке мышей на 42-й неделе и продолжалась в течение 15 дней. Другие серии идентичных экспериментов проводились через 6, 12 недель и т.д.
Для второй пары подгрупп (8 подвергавшихся и 8 не подвергавшихся обработке 9-недельных самцов) такая же серия экспериментов начиналась на 48-й неделе, так что эксперименты с этими подгруппами отстояли от опытов с первыми подгруппами ровно на 6 недель.
Установка
Способность к обучению у подвергавшихся и не подвергавшихся обработке мышей APP/PS оценивали на круговом водном лабиринте Морриса, который должен быть достаточно большим, чтобы обеспечить пространство для поиска без обессиливания мыши. Нужно принимать самые строгие меры к тому, чтобы поддерживать экспериментальную обстановку как можно более неизменной на протяжении всех экспериментов.
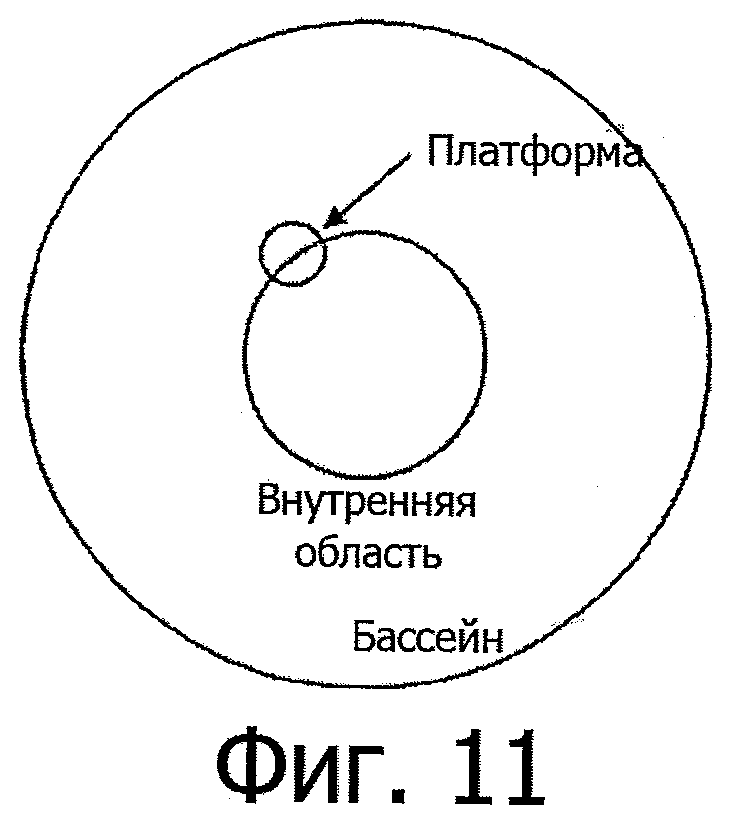
В настоящем исследовании использовали бассейн диаметром 120 см, который ставили в лаборатории в точно воспроизводимом положении, всегда с одинаковой ориентацией. В фиксированном положении в бассейн помещали белую, полупрозрачную, круговую платформу диаметром 15, 10 или 5 см, которая доходит чуть ниже поверхности воды (так что она невидима для мышей), на которую животные могут залезать - что является единственным способом выйти из воды. Чтобы облегчить залезание, платформа имеет сетчатое покрытие для захвата (см. фиг.11).
Для того, чтобы проводить спасательную процедуру в разведочных испытаниях, как изложено ниже, платформа снабжена механизмом, позволяющим поднимать или опускать ее автоматически без прямого вмешательства оператора. Таким образом, в зависимости от ее высоты, платформа доступна или недоступна плавающим мышам: "платформа по требованию" (Buresova et al., 1985).
Местоположение платформ всегда находится в кольцеобразном, концентрическом участке бассейна с внутренним и внешним диаметром ~40 см и ~80 см, соответственно. Определяются четыре квадранта таким образом, чтобы платформа занимала центральную часть одного из них (квадранта мишени). Более подробно о размерах и положении платформ см. ниже.
Для того, чтобы положение платформы было всегда одинаковым на протяжении всей серии экспериментов, на дне бассейна прочно закрепляется гнездо, на которое можно посадить платформу с минимальным пространственным допуском. Сверху на платформе, в самом центре, имеется еще одна опора для (проксимального) ориентира, выступающего из воды, который хорошо виден на видеозаписи, а также виден мышам, плавающим в бассейне. Для проверки положения платформы нужно делать краткую видеозапись без животных, но со вставленным на платформу ориентиром после каких бы то ни было манипуляций с платформой или видеокамерой.
Вода делается непрозрачной с помощью сухого молока с низким содержанием жира. Температура воды должна быть достаточно холодной, чтобы поощрять поиск выхода, но не настолько холодной, чтобы животные страдали или выбивались из сил. В качестве хорошего компромисса температура воды отслеживается в начале каждого эксперимента и доводится с помощью теплой воды либо кусочков льда до 18°С. Между отдельными опытами температура регулируется по мере необходимости.
Устанавливаются 4 дистальных ориентира (различных простых геометрических форм и разных цветов, высотой ~20 см) на ~20 см выше бортика бассейна, по одному в каждом квадранте. Принимаются меры к тому, чтобы каждый ориентир находился точно в том же самом месте на протяжении всех экспериментов. Весь бассейн закрывается белой полупрозрачной занавеской. Освещение неяркое и рассеянное.
Над центром бассейна в точно вертикальном положении прочно закрепляется видеокамера с тем, чтобы бассейн полностью попадал в кадр. Видеозаписи делаются при разрешении системы PAL (720×576 пикселей, 25 кадров в секунду), как минимум. Видеоснимки оценивают с помощью автоматической системы слежения, которая позволяет безупречно регистрировать передвижения животных во времени.
Мышей запускают в воду с помощью специального приспособления, которое насаживается на палку с тем, чтобы они попадали в воду точно в заданных местах по периметру бассейна без захода оператора в будку, состоящую из полупрозрачной занавески.
План экспериментов
В каждом сеансе мышей запускают в бассейн в заданных местах и дают поплавать в течение 60 сек. Траектории передвижения животных регистрируются видеосистемой слежения и вычисляются параметры, из которых можно сделать выводы о способности животных к обучению (прежде всего это время до попадания мыши на платформу в первый раз = "латентность спасения"; более подробно см. ниже). Если мыши удается найти платформу, то ее оставляют там отдохнуть на короткое время (~15 сек). В противном случае, после 60 сек плавания, оператор направляет мышь на платформу и дает отдохнуть в течение ~15 сек. После этого оператор вынимает ее, слегка просушивает и возвращает в помещение либо готовит к следующему заплыву.
В каждый день экспериментов проводится одно испытание на одну мышь ранним утром. Каждое испытание состоит из двух последовательных заплывов, начинающихся из двух разных квадрантов, но ни в коем случае не из квадранта мишени. Точные места запуска в воду (и положения платформ, если применимо) распределяются случайным образом для каждого заплыва в каждый день, но не отличаются между индивидуальными мышами в этот день.
Если окажется, что мыши обучаются слишком медленно, то можно увеличить количество заплывов за одно испытание или испытаний за день (и наоборот). Кроме того, у многих линий мышей молодые животные обучаются очень быстро, так что через 4 или 5 дней обучения латентность спасения становится постоянной на уровне всего лишь нескольких секунд, равным образом у подвергавшихся и не подвергавшихся обработке животных. Однако для статистической оценки было бы лучше, если бы кривая латентности спасения в зависимости от дней обучения не насыщалась, а монотонно снижалась. Поэтому принимается такой план эксперимента, в котором подлежащая решению задача усложняется с продвижением обучения. В заранее установленные дни платформу заменяют на меньшую, при этом координаты центра платформы остаются такими же. То, когда нужно поменять платформы, определяется независимым образом для каждой серии экспериментов и должно зависеть от результатов предыдущей серии.
В каждой серии экспериментов мышам задают выполнение трех различных видов заданий.
- Плавание к месту по ориентиру. Платформа обозначается ориентиром, а мышь может плавать до тех пор, пока не найдет платформу. По этой методике проверяется ассоциативное обучение, и она служит для разбивки мышей на две экспериментальные группы, у которых способности к обучению как можно более близки. Кроме того, во второй и последующих сериях экспериментов плавание по ориентиру способствует стиранию воспоминаний о положении платформы в предыдущих сериях.
- Нахождение скрытой платформы - обучение. Платформа невидима для мыши и находится в том же самом положении, что и в предыдущем заплыве. Это задание дает возможность наблюдать успехи мыши в воспроизведении точного местоположения скрытой платформы ("пространственное обучение").
- Разведочный поиск - испытание. В этом задании платформа "по требованию" максимально погружается под поверхность, а мышь может свободно плавать в поисках платформы. В разведочном испытании оценивается абсолютное воспоминание (воспроизведение в памяти), которое в данном контексте можно интерпретировать и как убежденность, непоколебимость или уверенность в местоположении платформы. Традиционный подход к интерпретации этого эксперимента состоит в том, что животные, которые прочно зафиксировали местоположение платформы, будут более настойчиво вести поиск в ограниченном месте и поэтому будут проводить больше времени в ближайшей к платформе зоне.
При разведочном поиске существует опасность того, что неспособность найти платформу может уменьшить побуждения к плаванию в зону платформы. Для того, чтобы свести к минимуму эти раздражения, условия спасения человеком должны оставаться одинаковыми с тем, чтобы было определенное пространственное постоянство, несмотря на отсутствие платформы. Поэтому после 60 сек плавания платформу поднимают до уровня чуть ниже поверхности, а оператор направляет мышь туда и дает отдохнуть в течение ~15 сек перед тем, как вынуть ее из установки.
Во все дни разведочного испытания проводится только один заплыв.
Приблизительно за 60 минут до каждого испытания мыши получают суточную дозу ECD NRG-1β в 5 нг/кг (суспендирован в сыворотке мышей Black 6 и вводится внутривенно в объеме 20 мкл на мышь) либо 20 мкл носителя.
В 1 день первой серии экспериментов все мыши в исследовании получают только ложную обработку. После этого мышей определяют в группу нейрегулина и контрольную группу таким образом, чтобы распределение латентности спасения совпадало в обоих группах.
В каждой серии экспериментов придерживаются следующей хронологии:
1-й день: поиск платформы по ориентиру, при этом платформа размером 10 см и ее положение меняется при каждом заплыве.
2-й день: поиск платформы по ориентиру, при этом платформа размером 10 см и ее положение меняется при каждом заплыве.
3-й день: поиск платформы по ориентиру, при этом платформа размером 10 см и ее положение меняется при каждом заплыве.
4-й день: поиск платформы по ориентиру, при этом платформа размером 10 см и ее положение такое же, как последнее в 3-й день.
5-й день: поиск скрытой платформы, при этом платформа размером 15 см и то же самое положение.
6-й день: поиск скрытой платформы, при этом платформа размером 15 см и то же самое положение.
7-й день: поиск скрытой платформы, при этом платформа размером 15 см и то же самое положение.
8-й день: поиск скрытой платформы, при этом платформа размером 10 см и то же самое положение.
9-й день: разведочный поиск.
10-й день: поиск скрытой платформы, при этом платформа размером 10 см и то же самое положение.
11-й день: поиск скрытой платформы, при этом платформа размером 10 см и то же самое положение.
12-й день: поиск скрытой платформы, при этом платформа размером 5 см и то же самое положение.
13-й день: поиск скрытой платформы, при этом платформа размером 5 см и то же самое положение.
14-й день: поиск скрытой платформы, при этом платформа размером 5 см и то же самое положение.
15-й день: разведочный поиск.
Может оказаться, что нужно заставить забыть положение платформы из предыдущей серии экспериментов, позволяя мышам свободно плавать в течение нескольких дней без платформы.
Скорость обучения оценивается путем отслеживания каждой сессии обучения/тестирования и регистрации успеха животных в нахождении платформы, а также эволюции стратегии поиска от плавания вдоль бортика бассейна до отхода от бортика для поиска в центральной зоне, в которой находится платформа.
Измеряемые параметры
Из видеозаписей животных извлекается траектория передвижения каждой мыши и выводится в виде серии координат х, у и время для дальнейшей обработки. Нужно принимать меры для надежной идентификации начальной точки каждой траектории и избежания ошибок при отслеживании. Одновременно вычисляется целый ряд параметров, из которых можно сделать выводы о способности животных к обучению (см. ниже). Регистрация параметров прекращается после 60 сек или когда мышь найдет платформу (что из этого произойдет раньше).
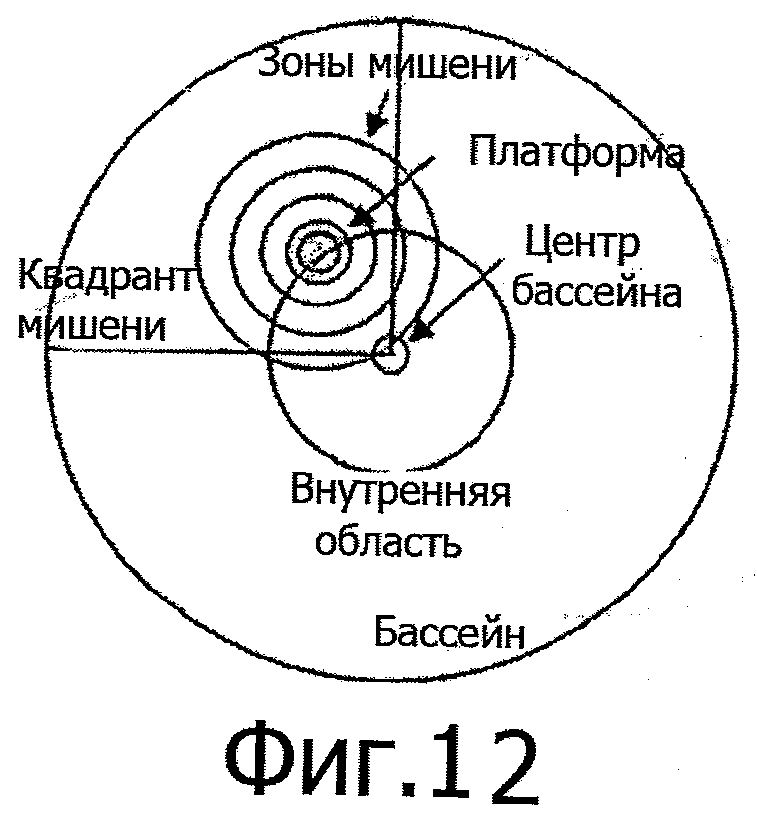
Для определения параметров, рассчитываемых из записей траекторий животных, установлены следующие зоны (см. фиг.12):
Для того, чтобы сделать процесс оценки как можно более гибким, используют 4 концентрические зоны мишени (проходящие вокруг платформы) диаметром от 5,5 до 30 см. Параметры, рассчитываемые из записей траекторий животных, включают:
- общее пройденное расстояние
- средняя общая скорость
- количество заходов в центр бассейна
- время, проведенное в центре бассейна
- латентность до первого захода в центр бассейна
- расстояние, пройденное до первого захода в центр бассейна
- количество заходов во внутреннюю область
- время, проведенное во внутренней области
- расстояние, пройденное во внутренней области
- латентность до первого захода во внутреннюю область
-расстояние, пройденное до первого захода во внутреннюю область
а также для каждой из зон 1-4 мишени и квадранта мишени:
- количество заходов в зону
- время, проведенное в зоне
- расстояние, пройденное в зоне
- латентность до первого захода в зону
- расстояние, пройденное до первого захода в зону
- расстояние от начала траектории до ближайшей точки в зоне
- среднее расстояние от зоны при выходе за пределы зоны
- минимальное расстояние от зоны при выходе за пределы зоны
- время до минимального расстояния от зоны при выходе за пределы зоны
- время приближения к зоне
- время удаления от зоны
- время передвижения к зоне
- время передвижения от зоны количество заходов головы в зону
- время, проведенное головой в зоне расстояние, пройденное головой в зоне
- латентность до первого захода головы в зону
- среднее расстояние головы от зоны при выходе за пределы зоны
- минимальное расстояние головы от зоны при выходе за пределы зоны
- начальная ошибка курса
- средняя ошибка курса
- количество выходов из зоны
По каждому дню эксперимента проводится статистическое сравнение значений параметров прогресса обучения в подвергавшихся обработке и не подвергавшихся группах между собой.
При изучении записей траекторий индивидуальных мышей наблюдатель может придти к весьма реалистическому пониманию уверенности животных при нахождении платформы, которая не полностью отражается в значениях измеряемых параметров. Поэтому записи траекторий также изучаются вручную и дается оценка воспроизведению животными в памяти положения платформы.
Результаты
Животные, получавшие суточную дозу ECD NRG-1β в 5 нг/кг внутривенно за 30 мин до обучения, оказались значительно лучше по параметрам обучения, чем животные в группе, получавшей носитель.
Нейрегулин не только улучшал обучение, но получавшие его животные также развивали более продвинутые стратегии поиска. Во внутреннюю область бассейна заходило больше подвергавшихся обработке животных (11 против 7, р=0,019), заход во внутреннюю область происходил чаще (2,17 против 0,92 раз, р=0,02), проведенное время и пройденное расстояние во внутренней области увеличились (6,51 сек против 2,13 сек, р=0,09; и 0,64 м против 0,25 м, р=0,031, соответственно).
Результаты экспериментов по обучению в водном лабиринте Морриса приведены на фиг.2.
Пример 3. Шизофрения: вызванная амфетамином гиперактивность у крыс
Методы
Метод, которым выявляется антипсихотическое и антипаркинсоническое действие, следует описанному Costall et al., 1978, и в нем используется измеритель активности, аналогичный описанному Boissier and Simon, 1966.
Амфетамин вызывает гиперактивность в этой тест-ситуации. Гиперактивности противостоят классические и нетипичные антипсихотическое вещества, которые действуют на дофаминергические системы на лимбическом уровне, и это усиливается антипаркинсоническими препаратами.
Крысам вводится d-амфетамин (3 мг/кг внутрибрюшинно) и их немедленно помещают в измеритель активности.
Измеритель активности состоит из 12 крытых клеток из плексигласа (40×25×25 см), содержащихся в затемненном шкафу. Каждая клетка снабжена двумя комплектами фотоэлементов на каждом конце клетки, на высоте 3 см от пола, для измерения передвижений каждого животного (по одному на клетку) из одного конца клетки к другому. На высоте 20 см от пола устанавливаются два дополнительных комплекта фотоэлементов для регистрации вставания на задние лапки. Показатели активности и вставания на задние лапки регистрируются компьютером с интервалом в 10 минут и суммируются за период 30 минут.
Исследовали по 15 крыс на группу. Тестирование проводилось слепым методом.
Исследуемое вещество оценивали в 8 дозах, которые вводили внутривенно за 15 минут до амфетамина, и сравнивали с контрольной группой носителя. Эксперимент также включал контрольную группу, не получавшую амфетамин.
В качестве вещества для сравнения использовали галоперидол (0,125 мг/кг внутривенно), который вводили при таких же экспериментальных условиях.
Таким образом, эксперимент включал 16 групп.
Данные анализировали путем сравнения получавших обработку групп с соответствующим контролем с помощью непарного t-критерия Стьюдента.
Результаты
Как видно из фиг.3, ECD NRG-1β дозозависимым образом ингибирует вызванную амфетамином гиперактивность у животных на модели шизофрении.
Явным образом эти эксперименты показывают замечательные свойства ECD NRG-1β:
- Представленные на фиг.3 эффекты наиболее сильно выражены во второй половине эксперимента (минуты 20-40). В первые 20 минут обнаруживается лишь небольшой эффект, так что замедленное действие указывает на дополнительный процессинг белка.
- Эффективные концентрации ECD NRG-1β, которые использовались при этом, в 200-1000 раз меньше, чем концентрации, в которых применяются типичные контрольные нейролептики типа галоперидола (125 мкг/кг).
- В противоположность галоперидолу, клозапину, оланзапину и др., при этом не наблюдалось отрицательных эффектов, а именно: ECD NRG-1β не снижал активность тестируемых животных ниже контрольного уровня носителя.
Пример 4. Шизофрения: преимпульсное торможение
Грызуны с нокаутом nrg-1 проявляют существенное нарушение преимпульсного торможения (PPI), что связывает nrg-1 с шизофренией. PPI широко используется как некий суррогат психоза на животных моделях и считается эндофенотипом шизофрении. После всестороннего генотипирования оказалось, что миссенс-мутации при полиморфизме по несинонимичным одиночным нуклеотидам на nrg-1 (rs3 924999) оказывают нейрофизиологические эффекты на PPI как при шизофрении, так и в контрольных здоровых популяциях (Hong et al., 2007). Мы протестировали действие ECD NRG-1β на PPI. Полученные результаты можно изложить следующим образом.
При 105 децибелах (дБ) ECD NRG-1β проявлял общую тенденцию к восстановлению PPI (+26%, +23% и +36% при 150, 300 и 600 нг/кг, соответственно), хотя эффект не достигал статистической значимости и не наблюдался при 115 дБ. Он не оказывал влияния на спонтанные движения в отсутствие раздражителя при 150 или 300 нг/кг, но значительно уменьшал спонтанные движения в отсутствие раздражителя при 600 нг/кг (-20% и -29% по усредненной и пиковой интенсивности, соответственно, р<0,05, что аналогично арипипразолу). ECD NRG-1β не оказывал влияния на реакцию на преимпульс сам по себе.
Полученные результаты свидетельствуют об отсутствии значительных эффектов на вызванные апоморфином нарушения PPI для Propsyl00 в диапазоне доз 150-300 нг/кг и уменьшении спонтанных движений, а также тенденции к восстановлению PPI при 600 нг/кг внутривенно в испытании на преимпульсное торможение (PPI) у крыс (нарушения, вызванные апоморфином).
В этой серии экспериментов контрольное вещество - арипипразол обладал слабой, но значимой активностью при 3 мг/кг внутрибрюшинно, но не при 10 мг/кг внутрибрюшинно в том же самом испытании.
В целом и при использовавшихся условиях ECD NRG-1β как будто влияет на PPI при высоких концентрациях около 600 нг/кг. Эти результаты неожиданно открывают новое понимание последних нейробиологических исследований, указывающих на ген nrg-1 как один из ведущих рассматриваемых генов при шизофрении.
Пример 5. Обучение и память у животных на модели болезни Альцгеймера (на мышах APPPS dt)
Эксперименты на животных по обучению и памяти с применением растворимого внеклеточного домена нейрегулина-1β (ECD NRG-1β) или без него в условиях водного лабиринта Морриса, описанные выше для нормальных мышей, были повторены на двойных трансгенных мышах (мышах APPPS) как модели церебрального амилоидоза (Meyer-Luehmann et al., 2006; Radde et al., 2006).
При этом опять животные, получавшие суточную дозу ECD NRG-1β (в данном случае 200 нг/кг внутрибрюшинно) за 30 мин до обучения, оказались значительно лучше по параметрам обучения, чем животные в группе, получавшей носитель.
Нейрегулин не только улучшал обучение, но получавшие его животные также развивали более продвинутые стратегии поиска. Во внутреннюю область бассейна заходило больше подвергавшихся обработке животных (12 против 7, р=0,009), заход во внутреннюю область происходил чаще (2,0 против 0,7 раз, р=0,03), проведенное время и пройденное расстояние во внутренней области увеличились (5,3 сек против 2,1 сек, р=0,09; и 0,7 м против 0,3 м, р=0,025, соответственно).
Результаты экспериментов по обучению на мышах APPPS как модели церебрального амилоидоза и болезни Альцгеймера в водном лабиринте Морриса приведены на фиг.4.
Пример 6. Нейрегулин-1β на модели вызванной МРТР болезни Паркинсона у мышей
Методы
Использовали 10-недельных самцов мышей С57В 1/6 на модели вызванной МРТР (1-метил-4-фенил-1,2,3,6-тетрагидропиридин) болезни Паркинсона.
Извлекали ткани мозга (черная субстанция, стриатум, кора) у 10-недельных самцов мышей С57 В1/6 (n=10 на группу) через различные промежутки времени после введения (0, 1, 3, 7, 21 день) NaCl (контроль) или МРТР (острая и субхроническая модели). Методы следовали опубликованным методикам (Hoglinger et al., 2007; Hoglinger et al., 2004).
МРТР растворяли в виде порошка в 0,9% NaCl и вводили внутрибрюшинно (острое введение: 4×20 мг/кг с интервалом в 2 часа; хроническое введение: 5×30 мг/кг с интервалом в 24 часа); инъекции занимали примерно 10 секунд. Через определенные промежутки времени (см. таблицу) животных забивали путем сворачивания шеи. Методы следовали опубликованным методикам (Hoglinger et al., 2007; Hoglinger et al., 2004; Liberatore et al., 1999; Przedborski and Vila, 2003; Vila and Przedborski, 2003).
Через 21 день после интрацеребрального вливания ECD NRG-1β и контрольного пептида через минипомпы Alzet и последующей обработки МРТР (острой либо хронической) проводили гистологическое определение дофаминергических нейронов среднего мозга по стереологическим принципам. Также проводили биохимическое определение дофамина и его метаболитов в стриатуме методом HPLC. Процедуры выполняли согласно опубликованным методикам (Hoglinger et al., 2007; Hoglinger et al., 2004).
Как видно из фиг.5, результаты измерения дофамина и его метаболитов методом HPLC показывают четкий эффект введения ECD NRG-1β при вызванном МРТР повреждении на этой модели болезни Паркинсона.
Эффекты не являются классическими: в то время, как отсутствуют значительные эффекты на уровень дофамина как при вызванном МРТР повреждении, так и в остром или хроническом контроле на введение ECD NRG-1β, имеются выраженные и четкие эффекты на концентрации DOPAC и HVA. Хроническое введение ECD NRG-1β приводит к четкому и значимому уменьшению этого метаболита в отсутствие вызванного МРТР повреждения, тогда как в остром режиме наблюдается лишь небольшое снижение. При хронических условиях вызванного МРТР повреждения ECD NRG-1β вызывает значительное повышение гомованилиновой кислоты (HVA), причем этот эффект еще более выражен в отсутствие вызванного МРТР повреждения.
Эти результаты можно объяснить понижающей регуляцией МАО-В и/или повышающей регуляцией СОМТ при хроническом введении ECD NRG-1β. При использовавшихся условиях наблюдался огромный и значительный положительный эффект на выживаемость дофаминергических нейронов. ECD NRG-1β и на этой модели оказывал сильное нейрозащитное действие. С учетом внутрибрюшинного введения при этой серии экспериментов его четкая эффективность опять таки свидетельствует о том, что ECD NRG-1β очень эффективно проходит через гематоэнцефалический барьер.
На фиг.6 представлена метаболическая схема, на которую явно влияет введение ECD NRG-1β: дофамин превращается в DOPAC под действием МАО-В и в 3-МТ под действием СОМТ; после чего из обоих метаболитов образуется гомованилиновая кислота - под действием СОМТ из DOPAC и под действием МАО-В из 3-МТ; очевидно, введение ECD NRG-1β регулирует активность обоих ферментов.
Еще более важно, как видно из фиг.7, что на модели вызванной МРТР болезни Паркинсона имеется четкий и значимый нейрозащитный эффект ECD NRG-1β, который становится заметным при гистологическом измерении дофаминергических нейронов в среднем мозге. Стереологический метод описан в других работах (Liberatore et al., 1999; Przedborski & Vila, 2003; Vila & Przedborski, 2003; Hoglinger et al., 2004; Hoglinger et al., 2004).
В целом имеется удивительно четкий м благотворный нейрозащитный эффект у животных на модели вызванной МРТР болезни Паркинсона. Эти эффекты опять же свидетельствуют о том, что внутрибрюшинное введение очень низких концентраций ECD NRG-1β (например, 20 нг/кг) достаточно для достижения эффективности, а тем самым и то, что ECD NRG-1β проходит через гематоэнцефалический барьер. К тому же сложное влияние на метаболиты дофамина (результаты HPLC; фиг.5) указывает на регуляцию МАО-В и СОМТ под действием NRG-1 и ECD NRG-1β.
Пример 7. Идентификация кислой посттрансляционной изоформы ECD NRG-1β в качестве действующего начала
Мы опубликовали данные о том, что при обучении и памяти активной формой является определенная посттрансляционная кислая изоформа ECD NRG-1β (Schillo et al., 2005a). Теперь же мы показываем, что аналогичная картина наблюдается у животных на модели болезни Альцгеймера и посмертной ткани мозга больных при болезни Альцгеймера и Паркинсона. Мы пришли к выводу, что действующим началом является именно эта кислая изоформа.
Методы
Для окрашивания вестерн-блотов использовали следующие антитела: кроличье поликлональное антитело против ECD NRG-1 (sc-28916, серия 12905 Santa Cruz; Н-210). Антитело против нейрегулина-1 (Н-210) представляет собой кроличье поликлональное антитело против аминокислот 21-230, локализованных в N-концевом внеклеточном домене изоформы HRG-a нейрегулина-1 человека. Антитело Н-210 рекомендуется для обнаружения изоформ HRG-α, HRG-α1A, HRG-α2B, HRG-α3, HRG-1β HRG-β2, HRG-β3 (GGF), GGF2 и SMDF нейрегулина-1 мыши, крысы и человека методом вестерн-гибридизации (исходное разведение 1:200, диапазон разведений 1:100-1:1000), иммунопреципитации (1-2 мкг на 100-500 мкг общего белка (1 мл лизата клеток)) и иммунофлуоресценции (исходное разведение 1:50, диапазон разведений 1:50-1:500).
Вторичные антитела:
против козьих антител, с HRP - sc-2922, серия С1405 Santa Cruz
против кроличьих антител, с HRP - sc-2054, серия G2005 Santa Cruz
Наряду с иммуноокрашиванием проводили масс-спектрометрию MALDI-TOF и Q-TOF для проверки ECD NRG-1β.
Как видно из фиг.8, очень сходная картина обнаруживается на модели церебрального амилоидоза у мышей APPPS и на модели болезни Альцгеймера.
Концентрация именно этой кислой изоформы ECD NRG-1β со значением pI примерно 5,0 значительно повышена у получавших его мышей APPPS, которые в то же время и лучше обучаются.
На фиг.8 представлены два репрезентативных снимка из одного подвергавшегося обработке животного с хорошим обучением (вверху) и не подвергавшегося обработке животного с плохим обучением (внизу).
На фиг.9 представлены результаты эксперимента по вестерн-гибридизации с использованием посмертного материала из коры головного мозга каждого из 9 больных при болезни Альцгеймера и подобранных по возрасту контролей. Четко видно, что фрагмент ECD NRG-1β значительно меньше представлен при болезни Альцгеймера. В качестве внутреннего контроля измеряли содержание NRG-12, на который не влияет ухудшение памяти при этой болезни.
Другое исследование этой специфической связанной с болезнью Альцгеймера и памятью изоформы ECD NRG-1β методом вестерн-гибридизации на 2-мерных гелях (2D-PAGE) того же посмертного материала из головного мозга человека, что использовался для фиг.8, показало, как это видно из репрезентативных примеров на фиг.10, что именно кислая изоформа ECD NRG-1β снижается при болезни Альцгеймера.
Выводы
Мы впервые представили функциональные свидетельства эффектов in vivo посттрансляционных модификаций транскрипта гена nrg-1, в частности укороченной формы, образующейся при протеолитическом расщеплении, включающей внеклеточный домен NRG-1β с мол. массой 15-35 и значением pI 4-10; более конкретно мы обнаружили антипсихотическое действие на животных моделях шизофрении, вероятно, связанное с регуляцией МАО-В и СОМТ, при концентрации 5-600 нг/кг (внутривенно). В отличие от контрольных нейролептиков, которые применяются при более высоких в 100-1000 раз концентрациях, при этом не наблюдалось отрицательных эффектов.
Кроме того, мы обнаружили нейрозащитное действие на модели вызванной МРТР болезни Паркинсона при концентрации в 3-300 нг/кг (внутривенно).
Более того, мы обнаружили положительный эффект на память и обучение у животных на модели (водный лабиринт Морриса) обучения на соответствующих моделях церебрального амилоидоза и болезни Альцгеймера.
Учитывая неблагоприятные эффекты у многих применяемых в настоящее время нетипичных антипсихотических средств (Haddad and Sharma, 2007), можно сделать вывод, что растворимый ECD-фрагмент NRG-1 с EGF-доменами SMDF, NRG-1α и в особенности NRG-1β может быть полезным в качестве самостоятельного или сочетанного лекарства для лечения шизофрении, биполярного психоза и депрессии.
Он также мог бы быть полезным в том же смысле и при других заболеваниях центральной нервной системы типа нейродегенеративных заболеваний, как-то болезни Альцгеймера и Паркинсона, боковом амиотрофическом склерозе, множественном склерозе, инсульте, травматических повреждениях головного и спинного мозга.
Растворимые белки ECD NRG-1β производят эти очень широкие эффекты вследствие их центральной роли в нейрональной передаче сигналов, в частности через опосредованную глутаматом передачу сигналов и эксцитотоксичность, что играет главную роль во всех приведенных выше показаниях (Schrattenholz and Soskic, 2006).





Настоящее изобретение касается растворимых изоформ нейрегулина-1, представляющих собой посттрансляционные модификации. Предложено применение нейрегулина-1β для изготовления медикамента для лечения неврологических заболеваний (вариант - для усиления памяти и когнитивной способности), где изоформа нейрегулина-1β представляет собой часть внеклеточного домена нейрегулина-1β, где данная часть имеет молекулярную массу 25-32 кДа и изоэлектическую точку (pI) в пределах от 5 до 9,5, и варианты соответствующих способов лечения. Предложена фармацевтическая композиция для лечения неврологических заболеваний, включающая рекомбинантную растворимую изоформу нейрегулина-1β, где изоформа нейрегулина-1β представляет собой часть внеклеточного домена нейрегулина-1β, где данная часть имеет молекулярную массу 25-32 кДа и pI в пределах от 5 до 9,5 и дополнительный медикамент для лечения неврологических заболеваний. Она может использоваться в качестве лекарства при когнитивных неврологических заболеваниях, в частности шизофрении, болезни Альцгеймера и Паркинсона. Показан четкий и благотворный нейрозащитный эффект заявленной рекомбинантной растворимой изоформы нейрегулина-1β, при этом, в отличие от контрольных нейролептиков, не наблюдалось отрицательных эффектов. 5 н. и 9 з.п. ф-лы, 12 ил.
1. Применение рекомбинантной растворимой изоформы нейрегулина-1 β для изготовления медикамента для лечения неврологических заболеваний, где изоформа нейрегулина-1 β представляет собой часть внеклеточного домена нейрегулина-1 β, где данная часть имеет молекулярную массу 25-32 кДа и изоэлектическую точку (р1) в пределах от 5 до 9,5.
2. Применение по п.1 для лечения шизофрении, в частности когнитивных аспектов шизофрении, биполярного психоза и депрессии, болезни Паркинсона, болезни Альцгеймера, эпилепсии, множественного склероза (MS), бокового амиотрофического склероза (ALS), инсульта, травматических повреждений головного и спинного мозга.
3. Применение по п.1 или 2, в котором изоформа нейрегулина-1 β представляет собой рекомбинантную растворимую изоформу нейрегулина-1 β человека.
4. Применение по любому из пп.1 и 2, в котором изоформа нейрегулина-1 представляет собой модифицированный полипептид, причем модификации выбираются из фосфорилирования, гликозилирования, метилирования, миристилирования, окисления и любых комбинаций из них.
5. Применение по п.1 в комбинации с дополнительным медикаментом.
6. Применение по п.5, в котором дополнительный медикамент является медикаментом для лечения неврологических заболеваний.
7. Применение по п.6, в котором дополнительный медикамент выбирают из соединений, влияющих на метаболизм катехоламинов, ингибиторов ацетил-холинэстеразы, ингибиторов МАО-В или СОМТ, блокаторов каналов типа мемантина, агонистов либо антагонистов дофаминовых или серотониновых рецепторов, ингибиторов обратного захвата катехоламинов либо серотонина, или любых антипсихотических медикаментов типа клозапина или оланзапина, либо препаратов типа габапентина при лечении болезней Альцгеймера или Паркинсона, шизофрении, биполярного психоза, депрессии или других неврологических заболеваний.
8. Применение по п.5 или 6, в котором дополнительный медикамент выбран из группы, состоящей из медикамента для лечения психотических заболеваний, таких как шизофрения, биполярный психоз и депрессия, например оланзапин или клозапин, медикамента для лечения болезни Паркинсона, медикамента для лечения болезни Альцгеймера, медикамента для лечения множественного склероза (MS), медикамента для лечения бокового амиотрофического склероза (ALS), медикамента для лечения эпилепсии, медикамента для лечения инсульта, медикамента для лечения травматических повреждений головного мозга и медикамента для лечения травматических повреждений спинного мозга.
9. Фармацевтическая композиция для лечения неврологических заболеваний, включающая:
(i) рекомбинантную растворимую изоформу нейрегулина-1 β, где изоформа нейрегулина-1 β представляет собой часть внеклеточного домена нейрегулина-1 β, где данная часть имеет молекулярную массу 25-32 кДа и изоэлектическую точку (pI) в пределах от 5 до 9,5, и
(ii) дополнительный медикамент для лечения неврологических заболеваний.
10. Применение рекомбинантной растворимой изоформы нейрегулина-1 β, где изоформа нейрегулина-1 β представляет собой часть внеклеточного домена нейрегулина-1 β, где данная часть имеет молекулярную массу 25-32 кДа и изоэлектическую точку (pI) в пределах от 5 до 9,5, для изготовления медикамента для усиления памяти и когнитивной способности.
11. Применение по п.10 для уменьшения и/или торможения потери памяти и когнитивной способности, связанными с таким неврологическим заболеванием, как болезнь Альцгеймера.
12. Применение по п.10 или 11, в котором изоформа нейрегулина-1 является такой, как определена по любому из пп.1-4.
13. Способ лечения неврологического заболевания, включающий введение рекомбинантной растворимой изоформы нейрегулина-1 β в фармацевтически эффективном количестве нуждающемуся в этом субъекту, где изоформа нейрегулина-1 β представляет собой часть внеклеточного домена нейрегулина-1 β, где данная часть имеет молекулярную массу 25-32 кДа и изоэлектическую точку (pI) в пределах от 5 до 9,5.
14. Способ усиления памяти и когнитивной способности, включающий введение рекомбинантной растворимой изоформы нейрегулина-1 β в фармацевтически эффективном количестве нуждающемуся в этом субъекту, где изоформа нейрегулина-1 β представляет собой часть внеклеточного домена нейрегулина-1 β, где данная часть имеет молекулярную массу 25-32 кДа и изоэлектическую точку (р1) в пределах от 5 до 9,5.
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| WANG Z | |||
| et al | |||
| Contrasting effects of mitogenic growth factors on myelination in neuron-oligodnedrocyte co-cultures | |||
| Glia | |||
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| ZHAO YY et al | |||
| Neuregulin promote survival and growth of cardiac myocytes. | |||

