ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к применению микроРНК 29 и ее предшественников, а также миметиков для лечения травмы сухожилия и модуляции биомеханических свойств сухожилия.
УРОВЕНЬ ТЕХНИКИ
Многие патологии опорно-двигательного аппарата, включая заболевания сухожилий, характеризуются нарушением процесса регуляции восстановления тканей и воспаления1. Тендинопатии являются распространенной причиной обращений за консультациями в учреждения первичной медицинской помощи в связи с патологиями скелетно-мышечной системы2-3, и они составляют 30-50% всех спортивных травм 3. Тендинопатия характеризуется изменением синтеза коллагена с подтипа 1 на подтип 3, что приводит к уменьшению прочности на растяжение, что может предшествовать клиническим проявлениям в форме разрыва сухожилия4.
Считается, что медиаторы воспаления играют ключевую роль в манифестации и сохранении симптомов тендинопатии5. Была продемонстрирована экспрессия различных цитокинов в линиях клеток воспаления и теноцитах, свидетельствующая о том, что популяции как инфильтрирующих, так и резидентных клеток участвуют в данной патологии6-9. Механические свойства заживающего сухожилия у мышей с дефицитом ИЛ-6 являются более слабыми по сравнению со здоровыми контрольными животными10, тогда как блокада ФНО-α улучшает эффективность заживления сухожилия-кости в модели на крысах с травмой сухожилия11. Несмотря на то, что эти данные повышают интригующую возможность того, что направленная коррекция цитокинового профиля может быть терапевтически эффективной, в настоящее время существует недостаточное понимание механизма биологической роли цитокинов/матрикса при заболеваниях сухожилий для реализации этой возможности на практике.
Интерлейкин 33 является представителем цитокинового семейства ИЛ-1, который в свою очередь играет основную роль в реализации врожденного иммунного ответа. ИЛ-33 экспрессируется в эндотелиальных клетках и фибробластах, колокализован с хроматином в ядре12. ИЛ-33 высвобождается после повреждения клеток13 и биомеханической избыточной нагрузки14 и, следовательно, считается «алармином»15. Было показано его участие в различных воспалительных патологиях, включая заболевания легких, кожи и суставов16. Функция ИЛ-33 реализуется через его соответствующий рецептор ST2, который существует в мембраносвязанной (mST2) или растворимой (sST2) форме, а передача сигнала осуществляется через канонический сигнальный каскад ИЛ-1R. Регуляция синтеза цитокинов часто осуществляется на посттранскрипционном уровне микроРНКмикроРНК, которые контролируют экспрессию генов путем подавления транскрипции и дестабилизации мРНК-мишеней17. Сети взаимодействий микроРНК приобретают значение ключевых гомеостатических регуляторов восстановления тканей, причем им отводится фундаментальная роль в биологии стволовых клеток, воспалении, ответе на гипоксию и ангиогенезе18.
МикроРНК (miR) представляют собой некодирующие РНК, которые оказывают значительное влияние на функцию клеток посредством репрессии трансляции (либо через подавление трансляции, либо через индукцию деградации мРНК). МикроРНК образуются из первичных РНК-транскриптов (pri-miRNA), синтезируемых РНК-полимеразой II, длина которых может составлять несколько тысяч нуклеотидов. Один транскрипт pri-miRNA может давать начало нескольким активным микроРНК.
В ядре фермент Drosha с активностью фермента РНКазы III разрезает транскрипт pri-miRNA с высвобождением предшественника микроРНК (pre-miRNA), имеющего структуру типа стебелек-петля или шпилечную структуру, длина которой, как правило, составляет около 70-100 нуклеотидов. Затем pre-miRNA транспортируется в цитоплазму, где она дополнительно разрезается РНКазой Dicer, которая удаляет петлю, и в результате этого образуется молекула зрелой двухцепочечной микроРНК, имеющая активную «направляющую» цепь (обычно 15-25 нуклеотидов в длину), гибридизованную с полностью или частично комплементарной «пассажирской» цепью.
Затем зрелая двухцепочечная микроРНК включается в состав комплекса сайленсинга, индуцированный РНК, где направляющая цепь гибридизуется с сайтом связывания в мРНК-мишени.
Направляющая цепь может быть не полностью комплементарной целевому сайту связывания. Однако область направляющей цепи, обозначенная как «затравочная последовательность», обычно полностью комплементарна соответствующей последовательности целевого сайта связывания. Длина затравочной последовательности, как правило, составляет 2-8 нуклеотидов, и она располагается на или вблизи (в пределах одного или двух нуклеотидов) 5'-конца направляющей цепи.
Предполагается, что одиночные непарные направляющие цепи также могут быть способными к включению в состав RISC. Также предполагается, что модификации в пассажирской цепи (например, в сахарах, основаниях или структуре основной цепи), которые препятствуют включению пассажирской цепи в RISC, могут увеличить эффективность ингибирования мишени двухцепочечной микроРНК.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Процесс заживления травмированного сухожилия часто является не оптимальным по меньшей мере частично из-за смещения синтеза коллагена с 1-го типа на 3-й тип в процессе тендинопатии. Коллаген 3-го типа уступает по механической прочности коллагену 1-го типа, вследствие чего сухожилие будет иметь более низкую прочность на растяжение. Биомеханические свойства сухожилия можно было бы улучшить, если соотношение между подтипами коллагена можно было бы сместить назад в пользу коллагена 1-го типа.
Ранее была установлена роль miR-29 в качестве регулятора синтеза коллагена в различных биологических процессах, таких как фиброз и склеродерма. Однако авторы настоящего изобретения впервые обнаружили, что теноциты содержат альтернативно сплайсированные формы транскриптов гена коллагена 1-го типа. Преобладающее число транскриптов генов коллагена типа 1a1 и 1а2 имеют короткие 3'-нетранслируемые области (НТО), в которых не содержатся сайты связывания miR-29, тогда как подавляющее большинство имеющихся транскриптов коллагена 3-го типа представляет собой форму, чувствительную к действию miR-29.
В результате этого синтез коллагена 1-го типа в теноцитах изменяется в гораздо меньшей степени под воздействием miR-29, чем синтез коллагена 3-го типа. Таким образом, на удивление, путем повышения активности miR-29 представляется возможным сместить соотношение между подтипами коллагена в пользу увеличения содержания коллагена 1-го типа и, тем самым, предотвратить или прекратить снижение прочности сухожилия на растяжение и изменить его биомеханические свойства, такие как предельная прочность на разрыв.
В самом широком смысле настоящее изобретение относится к применению микроРНК 29 (miR-29) и ее предшественников, миметиков и агонистов для модуляции процесса заживления сухожилия и биомеханических свойств сухожилия.
Следовательно, настоящее изобретение относится к способу модуляции заживления сухожилия, причем указанный способ включает повышение экспрессии или активности miR-29 в сухожильной клетке. Это может достигаться за счет прямой доставки miR-29 в клетку-мишень, путем доставки миметика miR-29 или путем доставки молекулы-предшественника, которая подвергается процессингу в клетке-мишени, с образованием активных miR-29 или миметика miR-29.
Способ может включать стадию доставки miR-29, ее миметика или их предшественников в сухожильную клетку.
MiR-29, ее миметик или предшественник могут быть доставлены в сочетании с (например, в комплексе с или инкапсулированные) подходящей молекулой-носителем, такой как фармацевтически приемлемый липид или полимер.
Молекула-носитель может дополнительно содержать средство, способное к адресному связыванию с поверхностью клетки-мишени.
Способ может включать стадию доставки нуклеиновой кислоты, кодирующей miR-29, ее миметик или их предшественники, в сухожильную клетку, таким образом, что указанные miR-29, миметик или предшественник экспрессируются в сухожильной клетке.
В альтернативном варианте способ может включать стадию доставки агониста, способного к повышению активности эндогенной miR-29, в сухожильную клетку.
Любые из описанных способов могут осуществляться in vitro, in vivo или ex vivo. Чаще всего способы будут осуществляться in vivo путем введения подходящей композиции субъекту.
Настоящее изобретение также относится к miR-29, ее миметику или их предшественникам для применения в способе модуляции заживления сухожилия.
Настоящее изобретение также относится к применению miR-29, ее миметика или их предшественников в производстве лекарственного препарата для модуляции заживления сухожилия.
Настоящее изобретение также относится к нуклеиновой кислоте, кодирующей miR-29, ее миметик или их предшественники, для применения в способе модуляции заживления сухожилия.
Настоящее изобретение также относится к применению нуклеиновой кислоты, кодирующей miR-29, ее миметик или их предшественники, в производстве лекарственного препарата для модуляции заживления сухожилия.
В любом аспекте miR-29 может представлять собой miR-29a, miR-29b (b1 и/или b2), miR-29c или любую их комбинацию. Желательно, чтобы miR-29 представляла собой miR-29a или комбинацию, содержащую miR-29a.
Нуклеиновая кислота, кодирующая miR-96, ее миметик или предшественник, может быть доставлена в виде свободной нуклеиновой кислоты. В альтернативном варианте она может быть доставлена в сочетании с (например, в комплексе с или инкапсулированная) подходящей молекулой-носителем, фармацевтически приемлемым липидом или полимером, или их комбинацией. В любом случае нуклеиновая кислота обычно представляет собой ДНК.
Молекула-носитель может дополнительно содержать средство, способное к адресному связыванию с поверхностью клетки-мишени.
В альтернативном варианте нуклеиновая кислота, кодирующая miR-96, ее миметик или предшественник, может быть доставлена с помощью вирусного вектора.
Могут быть использованы вирусные векторы любого подходящего типа, включая аденовирусные векторы, векторы на основе аденоассоциированных вирусов (AAV), ретровирусные (особенно лентивирусные) или герпесвирусные векторы. Особенно предпочтительными могут быть аденовирусные и лентивирусные векторы, поскольку они обладают способностью обеспечивать экспрессию гена(ов) после доставки в клетки, которые недостаточно активно делятся.
MiR-29 и ее предшественники
Три главные изоформы у людей представлены miR-29a, miR-29b1, miR-29b2 и miR-29c.
Термин «miR-29» используется в этом описании для обозначения РНК-олигонуклеотида, состоящего из зрелой последовательности «направляющей цепи» любой из этих трех изоформ.
Зрелая человеческая miR-29a («hsa-miR-29a») имеет последовательность:
UAGCACCAUCUGAAAUCGGUUA.
Зрелая miR-29b1l и miR-29b2 («hsa-miR-29b1» и «hsa-miR-29b2») являются идентичными и имеют последовательность:
UAGCACCAUUUGAAAUCAGUGUU.
Зрелая человеческая miR-29c («hhsa-miR-29c») имеет последовательность:
UAGCACCAUUUGAAAUCGGUUA.
Обычно при названии микроРНК вид, из которого была выделена микроРНК, обозначается в названии трехбуквенной приставкой. Следовательно, «hsa» означает Homo sapiens. Эти зрелые последовательности miR29 являются идентичными у большинства млекопитающих, включая лошадь.
Все четыре зрелые направляющие цепи имеют общую «затравочную» область, которая связывается с мРНК-мишенью и имеет последовательность:
AGCACCA.
Олигонуклеотид направляющей цепи miR-29 может быть одноцепочечным или может быть гибридизован со вторым олигонуклеотидом РНК, называющимся «пассажирская цепь». Направляющая цепь и пассажирская цепь располагаются антипараллельно друг другу в комплексе гибридизации, который может быть назван «двухцепочечная miR-29». (Направляющая цепь, если содержится отдельно, может быть названа «одноцепочечная miR-29»).
Пассажирская цепь и направляющая цепь могут содержать некоторое количество ошибочно спаренных нуклеотидов, в результате чего не все нуклеотиды в одной или обеих цепях гибридизуются с комплементарными нуклеотидами в другой цепи. Следовательно, двухцепочечная miR-96 может содержать одно или более выпячиваний (выпячивание представляет собой неспаренный нуклеотид или следующие друг за другом неспаренные нуклеотиды только в одной цепи) или внутренних петель (противоположные неспаренные нуклеотиды в обеих цепях). Один или более нуклеотидов на обоих концах также могут быть неспаренными.
Пассажирская цепь может быть на 100% комплементарной затравочной последовательности направляющей цепи.
Нативные человеческие пассажирские цепи имеют последовательность:
ACUGAUUUCUUUUGGUGUUCAG (miR29a)
GCUGGUUUCAUAUGGUGGUUUAGA (miR-29b1);
CUGGUUUCACAUGGUGGCUUAG (miR-29b2); и
UGACCGAUUUCUCCUGGUGUUC (miR-29c).
Одна или обе цепи двухцепочечной miR-29 могут содержать свисающий 3'-конец, например, состоящий из 1, 2 или 3 нуклеотидов. Другими словами, один или два нуклеотида на 3'-конце цепи выходят за пределы самого крайнего нуклеотида с 5'-конца комплементарной цепи (включая любые неспаренные концевые нуклеотиды) и, следовательно, не содержат соответствующих нуклеотидов в комплементарных цепях. Например, обе цепи могут содержать свисающий конец на 3'-конце, состоящий из 1, 2 или 3 нуклеотидов. В альтернативном варианте комплекс может содержать тупые концы на одном или обоих концах. В некоторых вариантах реализации изобретения пассажирская цепь имеет такую же длину, что и направляющая цепь, или отличается по длине, например, на пять нуклеотидов или еще больше, в зависимости от степени ошибочного спаривания между обеими цепями и длины любого свисающего 3'-конца.
Предшественники miR-29 включают pre-mir-29 и pri-mir-29 любой из трех изоформ, а также их фрагменты и варианты, которые могут быть процессированы с образованием зрелой miR-29 (либо одно-, либо двухцепочечной).
Термин «pre-mir-29» используется для обозначения олигонуклеотида РНК, состоящего из любой полноразмерной последовательности pre-mir-29 млекопитающего или ее фрагмента, или варианта, который содержит направляющую последовательность зрелой miR-29, связанную с помощью последовательности петли с соответствующей пассажирской последовательностью, которая полностью или частично комплементарна пассажирской последовательности, и причем указанный олигонуклеотид способен к образованию структуры типа «петля на стебле» (или «шпилька»), в которой направляющая последовательность и пассажирская последовательность гибридизуются друг с другом.
Pre-mir-29 способна выполнять функцию субстрата для двухцепочечной РНК-специфической рибонуклеазы Dicer (фермент с активностью РНКзы III типа), в результате чего она процессируется с образованием зрелой двуцепочечной miR-29.
Полноразмерные последовательности pre-mir-29 млекопитающих включают человеческие последовательности:
AUGACUGAUUUCUUUUGGUGUUCAGAGUCAAUAUAAUUUUCUAGCACCAUCUGAAAUCGGUUAU (hsa-pre-mir-29a: альтернативный вариант (i));
AUGACUGAUUUCUUUUGGUGUUCAGAGUCAAUAUAAUUUUCUAGCACCAUCUGAAAUCGGUUAUAAUGAUUGGGG (hsa-pre-mir-29a: альтернативный вариант (ii));
CUUCAGGAAGCUGGUUUCAUAUGGUGGUUUAGAUUUAAAUAGUGAUUGUCUAGCACCAUUUGAAAUCAGUGUUCUUGGGGG (hsa-pre-mir-29b1);
CUUCUGGAAGCUGGUUUCACAUGGUGGCUUAGAUUUUUCCAUCUUUGUAUCUAGCACCAUUUGAAAUCAGUGUUUUAGGAG (hsa-pre-mir-29b2); и
AUCUCUUACACAGGCUGACCGAUUUCUCCUGGUGUUCAGAGUCUGUUUUUGUCUAGCACCAUUUGAAAUCGGUUAUGAUGUAGGGGGA (hsa-pre-mir-29c)
Соответствующие последовательности зрелой направляющей цепи подчеркнуты.
Pri-mir-29 может содержать одну или более модификаций, не затрагивающих зрелой последовательности, по сравнению с показанными последовательностями.
Последовательность, расположенная выше (5') зрелой последовательности, может быть, например, по меньшей мере на 50% идентичной, по меньшей мере на 55% идентичной, по меньшей мере на 60% идентичной, по меньшей мере на 65% идентичной, по меньшей мере на 70% идентичной, по меньшей мере на 75% идентичной, по меньшей мере на 80% идентичной, по меньшей мере на 85% идентичной, по меньшей мере на 90% идентичной, по меньшей мере на 91% идентичной, по меньшей мере на 92% идентичной, по меньшей мере на 93% идентичной, по меньшей мере на 94% идентичной, по меньшей мере на 95% идентичной, по меньшей мере на 96% идентичной, по меньшей мере на 97% идентичной, по меньшей мере на 98% идентичной или по меньшей мере на 99% идентичной соответствующей человеческой последовательности.
Например, последовательность, расположенная выше (5') зрелой последовательности miR-29a, может отличаться на 20 нуклеотидов от соответствующей расположенной в 5'-направлении последовательности человека, при оптимальном выравнивании с ней, например, на 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 нуклеотидов.
Последовательность, расположенная выше зрелой последовательности miR-29b1 или b2 может отличаться на 25 нуклеотидов от соответствующей расположенной в 5'-направлении последовательности человека, при оптимальном выравнивании с ней, например, на 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 нуклеотидов.
Последовательность, расположенная выше зрелой последовательности miR-29c может отличаться на 25 нуклеотидов от соответствующей расположенной на 5'-конце последовательности человека, при оптимальном выравнивании с ней, например, на 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 нуклеотидов.
Последовательность, расположенная ниже (3') зрелой последовательности, может быть, например, по меньшей мере на 50% идентичной, по меньшей мере на 55% идентичной, по меньшей мере на 60% идентичной, по меньшей мере на 65% идентичной, по меньшей мере на 70% идентичной, по меньшей мере на 75% идентичной, по меньшей мере на 80% идентичной, по меньшей мере на 85% идентичной, по меньшей мере на 90% идентичной, по меньшей мере на 91% идентичной, по меньшей мере на 92% идентичной, по меньшей мере на 93% идентичной, по меньшей мере на 94% идентичной, по меньшей мере на 95% идентичной, по меньшей мере на 96% идентичной, по меньшей мере на 97% идентичной, по меньшей мере на 98% идентичной или по меньшей мере на 99% идентичной соответствующей человеческой последовательности.
Последовательность, расположенная ниже (3') зрелой последовательности miR-29а, может быть такой же, как расположенная в сторону 3'-конца последовательность человека или может быть другой. Она может иметь другой нуклеотидный состав, отличный от встречающегося в более короткой из двух последовательностей, показанных выше, т.е. альтернативного варианта (i). Она может быть длиннее, чем последовательность, показанная в альтернативном варианте (i). Например, она может отличаться на 6 нуклеотидов от соответствующей расположенной в сторону 3'-конца последовательности альтернативного варианта (ii), показанного выше.
Последовательность, расположенная ниже (3') зрелой последовательности miR-29b1 или b2, может отличаться на 4 нуклеотида от соответствующей расположенной в сторону 3'-конца последовательности человека, при оптимальном выравнивании с ней, например, на 1, 2, 3 или 4 нуклеотида.
Последовательность, расположенная ниже (3') зрелой последовательности miR-29с может отличаться на 7 нуклеотидов от соответствующей расположенной в сторону 3'-конца последовательности человека, при оптимальном выравнивании с ней, например, на 1, 2, 3, 4, 5, 6 или 7 нуклеотидов.
Термин «pre-mir-29» используется для обозначения олигонуклеотида РНК, состоящего из любой полноразмерной последовательности pre-mir-29 млекопитающего или ее фрагмента, или варианта, который содержит последовательность pre-miR-29 и поддается процессингу с помощью двухцепочечной РНК-специфической рибонуклеазы (фермент с активностью РНКзы III типа) Drosha с образованием последовательности pre-mir-29.
Одиночный транскрипт может поддаваться процессингу с образованием двух или более молекул mir-29, ее миметиков или предшественников.
Hsa-mir29a и mir29b1 кодируются в последнем экзоне транскрипта с идентификационным номером в БД GenBank EU154353 (EU154353.1 GI:161824377). Область, кодирующая mir29a и mir29b1, а также фланкирующая последовательность показаны ниже. (Hsa-mir29a показана прописными жирными буквами, причем зрелая последовательность miR-29a подчеркнута. Hsa-mir29b показана прописными буквами, причем последовательность miR-29b подчеркнута).
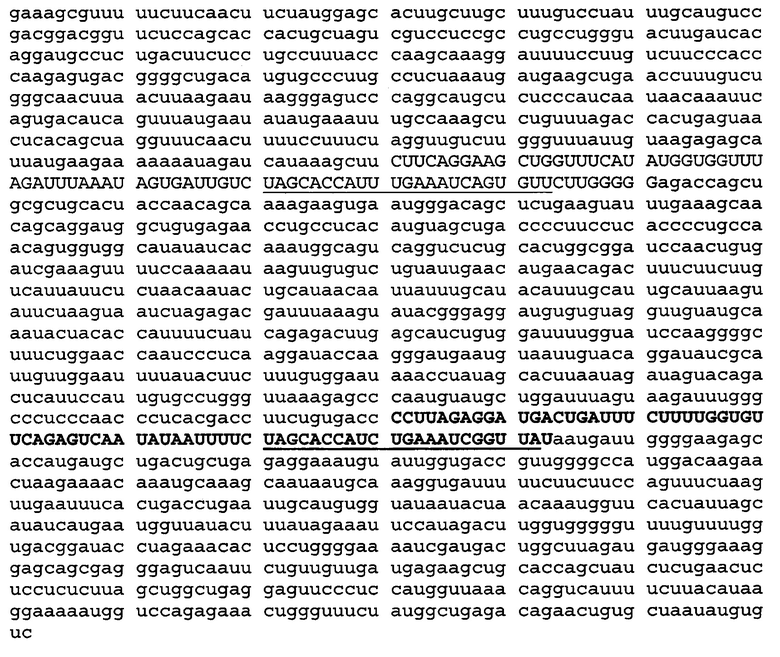
Hsa-pri-miR29b2 и hsa-pri-mir29c кодируются в виде одного транскрипта, который показан ниже. Hsa-mir29b2 показана прописными буквами, причем зрелая последовательность hsa-miR-29b2 подчеркнута. Hsa-mir29c показана жирными прописными буквами, причем зрелая последовательность hsa-miR-29c подчеркнута.

Следовательно, pri-mir-29 может содержать более чем одну последовательность зрелой miR-29 или миметика. Например, она может содержать miR-29a и miR-29b1 или их миметики, или miR-29b2 и miR-29c, или их миметики.
В альтернативном варианте pri-mir-29 может содержать только одну последовательность miR-29 или ее миметика.
Pri-mir-29 может быть по меньшей мере на 50% идентичной, по меньшей мере на 55% идентичной, по меньшей мере на 60% идентичной, по меньшей мере на 65% идентичной, по меньшей мере на 70% идентичной, по меньшей мере на 75% идентичной, по меньшей мере на 80% идентичной, по меньшей мере на 85% идентичной, по меньшей мере на 90% идентичной, по меньшей мере на 91% идентичной, по меньшей мере на 92% идентичной, по меньшей мере на 93% идентичной, по меньшей мере на 94% идентичной, по меньшей мере на 95% идентичной, по меньшей мере на 96% идентичной, по меньшей мере на 97% идентичной, по меньшей мере на 98% идентичной или по меньшей мере на 99% идентичной любой из последовательностей pri-mir-29, показанных выше, или фрагменту одной из тех последовательностей, которые содержат одну из зрелых последовательностей miR-29.
Pri-mir-29 может содержать одну или более модификаций, не затрагивающих зрелую последовательность или не затрагивающих нативную последовательность pre-mir-29, по сравнению с показанными последовательностями.
Например, последовательность, расположенная выше (5') зрелой последовательности, может быть, например, по меньшей мере на 50% идентичной, по меньшей мере на 55% идентичной, по меньшей мере на 60% идентичной, по меньшей мере на 65% идентичной, по меньшей мере на 70% идентичной, по меньшей мере на 75% идентичной, по меньшей мере на 80% идентичной, по меньшей мере на 85% идентичной, по меньшей мере на 90% идентичной, по меньшей мере на 91% идентичной, по меньшей мере на 92% идентичной, по меньшей мере на 93% идентичной, по меньшей мере на 94% идентичной, по меньшей мере на 95% идентичной, по меньшей мере на 96% идентичной, по меньшей мере на 97% идентичной, по меньшей мере на 98% идентичной или по меньшей мере на 99% идентичной соответствующей человеческой последовательности.
Последовательность, расположенная выше (5') последовательности pre-mir-29, может быть, например, по меньшей мере на 50% идентичной, по меньшей мере на 55% идентичной, по меньшей мере на 60% идентичной, по меньшей мере на 65% идентичной, по меньшей мере на 70% идентичной, по меньшей мере на 75% идентичной, по меньшей мере на 80% идентичной, по меньшей мере на 85% идентичной, по меньшей мере на 90% идентичной, по меньшей мере на 91% идентичной, по меньшей мере на 92% идентичной, по меньшей мере на 93% идентичной, по меньшей мере на 94% идентичной, по меньшей мере на 95% идентичной, по меньшей мере на 96% идентичной, по меньшей мере на 97% идентичной, по меньшей мере на 98% идентичной или по меньшей мере на 99% идентичной соответствующей человеческой последовательности.
Последовательность, расположенная ниже (3') зрелой последовательности, может быть, например, по меньшей мере на 50% идентичной, по меньшей мере на 55% идентичной, по меньшей мере на 60% идентичной, по меньшей мере на 65% идентичной, по меньшей мере на 70% идентичной, по меньшей мере на 75% идентичной, по меньшей мере на 80% идентичной, по меньшей мере на 85% идентичной, по меньшей мере на 90% идентичной, по меньшей мере на 91% идентичной, по меньшей мере на 92% идентичной, по меньшей мере на 93% идентичной, по меньшей мере на 94% идентичной, по меньшей мере на 95% идентичной, по меньшей мере на 96% идентичной, по меньшей мере на 97% идентичной, по меньшей мере на 98% идентичной или по меньшей мере на 99% идентичной соответствующей человеческой последовательности.
Последовательность, расположенная ниже (3') нативной последовательности pre-mir-29, может быть, например, по меньшей мере на 50% идентичной, по меньшей мере на 55% идентичной, по меньшей мере на 60% идентичной, по меньшей мере на 65% идентичной, по меньшей мере на 70% идентичной, по меньшей мере на 75% идентичной, по меньшей мере на 80% идентичной, по меньшей мере на 85% идентичной, по меньшей мере на 90% идентичной, по меньшей мере на 91% идентичной, по меньшей мере на 92% идентичной, по меньшей мере на 93% идентичной, по меньшей мере на 94% идентичной, по меньшей мере на 95% идентичной, по меньшей мере на 96% идентичной, по меньшей мере на 97% идентичной, по меньшей мере на 98% идентичной или по меньшей мере на 99% идентичной соответствующей человеческой последовательности.
Предшественник miR-29 может иметь любую подходящую длину, при условии его процессинга с образованием зрелой miR-29 (либо одно-, либо двухцепочечной). Следовательно, длина предшественника miR-29a составляет по меньшей мере 23 нуклеотида, длина предшественника miR29b составляет по меньшей мере 24 нуклеотида, и длина предшественника miR-29c составляет по меньшей мере 25 нуклеотидов.
Длина предшественника miR29 может составлять по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 55, по меньшей мере 60, по меньшей мере 65, по меньшей мере 70, по меньшей мере 75, по меньшей мере 80, по меньшей мере 85, по меньшей мере 90, по меньшей мере 95, по меньшей мере 100, по меньшей мере 110, по меньшей мере 120, по меньшей мере 130, по меньшей мере 140, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 300, по меньшей мере 350, по меньшей мере 400, по меньшей мере 450, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 1500 или по меньшей мере 2000 нуклеотидов.
В альтернативном варианте длина предшественника может составлять не более 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 110, 120, 130, 140, 150, 200, 250, 300, 350, 400, 450, 500, 1000, 1500, 2000 или 2500 нуклеотидов, хотя возможно использование более длинных транскриптов предшественников.
Следует отметить, что термин «олигонуклеотид» не предназначен для обозначения какой-либо конкретной длины, а просто используется для обозначения любой одиночной непрерывной цепи связанных нуклеотидов.
Миметики miR-29 и ее предшественники
Миметик miR-29 представляет собой олигонуклеотид, который включает одну или более модификаций в структуре или последовательности, по сравнению с природной miR-29, но сохраняет способность к гибридизации с сайтом связывания miR-29 в мРНК, экспрессию которой регулирует miR-29, и к подавлению трансляции или стимуляции деградации такой мРНК, например, для подавления выработки белка, кодируемого этой мРНК. мРНК, регулируемые miR-29, включают мРНК коллагена 3-го типа (Соl3а1).
Примеры сайтов связывания miR-29 включают:
CCAUUUUAUACCAAAGGUGCUAC (из мРНК Col1a1);
UGUUCAUAAUACAAAGGUGCUAA (из мРНК Соl1а2); и
UUCAAAAUGUCUCAAUGGUGCUA (из мРНК col3a1).
Длина олигонуклеотида миметика miR-29 обычно составляет 15-35 нуклеотидов, например, 15-30, 15-25, 18-25, 20-25, например, 20-23, например, 20, 21, 22 или 23 нуклеотида.
Миметик miR-29 может отличаться по последовательности оснований, структуре нуклеотидов и/или фосфодиэфирной связи по сравнению с одной из нативных зрелых последовательностей miR-29.
Миметик miR-29 содержит затравочную последовательность, которая может быть идентичной нативной затравочной последовательности:
AGCACCA
или может отличаться от нативной затравочной последовательности не более чем по трем положениям, например, не более чем по двум положениям, например, не более чем по одному положению. Предпочтительно, чтобы затравочная последовательность была идентичной последовательности, которая показана.
Миметик miR-29 может содержать или состоять из олигонуклеотида, имеющего зрелую нативную направляющую последовательность miR-29, такую как:
UAGCACCAUCUGAAAUCGGUUA (hsa-miR-29a);
UAGCACCAUUUGAAAUCAGUGUU (hsa-miR-29b1 и 2); или
UAGCACCAUUUGAAAUCGGUUA (hsa-miR-29c)
(причем затравочная последовательность в каждом случае подчеркнута);
или которая отличается от зрелой нативной последовательности по:
(i) не более чем трем положениям в пределах затравочной последовательности; и
(ii) не более чем пяти положениям за пределами затравочной последовательности.
Следовательно, затравочная последовательность миметика отличается от нативной затравочной последовательности не более чем по трем положениям, например, не более чем по двум положениям, например, не более чем по одному положению. Предпочтительно, чтобы затравочная последовательность была идентичной нативной затравочной последовательности.
В дополнительном или альтернативном варианте миметик отличается от нативной последовательности за пределами затравочной последовательности не более чем по пяти положениям, например, не более чем по четырем положениям, не более чем по трем положениям, не более чем по двум положениям, например, не более чем по одному положению.
Миметик miR-29 может быть гибридизован со вторым олигонуклеотидом. Как и в случае с нативной miR-29, активный олигонуклеотид может называться «направляющей цепью», а связанный олигонуклеотид - «пассажирской цепью». Гибридизованный комплекс может называться двухцепочечным миметиком miR-29.
Последовательность пассажирской цепи миметика может быть идентична последовательности нативной пассажирской цепи или может отличаться от нативной пассажирской цепи по одному или нескольким положениям. Например, последовательность пассажирской цепи миметика может отличаться от нативной пассажирской цепи не более чем по 10 положениям, не более чем по 9 положениям, не более чем по 8 положениям, не более чем по 7 положениям, не более чем по 6 положениям, не более чем по 5 положениям, не более чем по 4 положениям, не более чем по 3 положениям, не более чем по 2 положениям или не более чем по 1 положению.
Одна или обе цепи двухцепочечного миметика miR-29 могут содержать свисающий 3'-конец, состоящий из 1 или 2 нуклеотидов. Например, обе цепи могут содержать свисающий 3'-конец, состоящий из 2 нуклеотидов. В альтернативном варианте комплекс может содержать тупые концы на одном или обоих концах. В некоторых вариантах реализации изобретения пассажирская цепь имеет такую же длину, что и направляющая цепь, или отличается по длине на один или два нуклеотида.
Предшественник миметика miR-29 представляет собой любую молекулу, которая может быть процессирована внутри клетки-мишени с образованием миметика miR-29 в соответствии с вышеприведенным определением, обычно под воздействием фермента Dicer или под последовательным воздействием ферментов Drosha и Dicer.
Следовательно, предшественник может иметь дополнительную олигонуклеотидную последовательность выше (5') и/или ниже (3') последовательности миметика.
Предшественник может содержать направляющую последовательность миметика miR-29, связанную с помощью последовательности петли с соответствующей пассажирской последовательностью, которая полностью или частично комплементарна пассажирской последовательности, и причем указанный олигонуклеотид способен к образованию структуры типа «петля на стебле» (или «шпилька»), в которой направляющая последовательность и пассажирская последовательность гибридизуются друг с другом. Такой олигонуклеотид может рассматриваться в качестве миметика pre-mir-29 и способен выполнять функцию субстрата для двухцепочечной РНК-специфической рибонуклеазы Dicer (фермента с активностью РНКзы III типа), в результате чего он процессируется с образованием двухцепочечного миметика miR-29, содержащего отдельные направляющую и пассажирскую цепи.
Последовательность, расположенная выше (5') зрелой последовательности, может быть, например, по меньшей мере на 50% идентичной, по меньшей мере на 55% идентичной, по меньшей мере на 60% идентичной, по меньшей мере на 65% идентичной, по меньшей мере на 70% идентичной, по меньшей мере на 75% идентичной, по меньшей мере на 80% идентичной, по меньшей мере на 85% идентичной, по меньшей мере на 90% идентичной, по меньшей мере на 91% идентичной, по меньшей мере на 92% идентичной, по меньшей мере на 93% идентичной, по меньшей мере на 94% идентичной, по меньшей мере на 95% идентичной, по меньшей мере на 96% идентичной, по меньшей мере на 97% идентичной, по меньшей мере на 98% идентичной или по меньшей мере на 99% идентичной соответствующей человеческой последовательности.
Последовательность, расположенная ниже (3') зрелой последовательности, может быть, например, по меньшей мере на 50% идентичной, по меньшей мере на 55 % идентичной, по меньшей мере на 60% идентичной, по меньшей мере на 65% идентичной, по меньшей мере на 70% идентичной, по меньшей мере на 75% идентичной, по меньшей мере на 80% идентичной, по меньшей мере на 85% идентичной, по меньшей мере на 90% идентичной, по меньшей мере на 91% идентичной, по меньшей мере на 92% идентичной, по меньшей мере на 93% идентичной, по меньшей мере на 94% идентичной, по меньшей мере на 95% идентичной, по меньшей мере на 96% идентичной, по меньшей мере на 97% идентичной, по меньшей мере на 98% идентичной или по меньшей мере на 99% идентичной соответствующей человеческой последовательности.
В альтернативном варианте предшественник может представлять собой миметик pri-mir-29 (т.е. он имеет дополнительную олигонуклеотидную последовательность выше (5') и/или ниже (3') последовательности миметика pre-mir-29) и может поддаваться процессингу с помощью двухцепочечной РНК-специфической рибонуклеазы (фермент с активностью РНКзы III типа) Drosha с образованием последовательности миметика pre-mir-29.
Например, последовательность, расположенная выше (5') зрелой последовательности миметика miR-29 может быть, например, по меньшей мере на 50% идентичной, по меньшей мере на 55% идентичной, по меньшей мере на 60% идентичной, по меньшей мере на 65% идентичной, по меньшей мере на 70% идентичной, по меньшей мере на 75% идентичной, по меньшей мере на 80% идентичной, по меньшей мере на 85% идентичной, по меньшей мере на 90% идентичной, по меньшей мере на 91% идентичной, по меньшей мере на 92% идентичной, по меньшей мере на 93% идентичной, по меньшей мере на 94% идентичной, по меньшей мере на 95% идентичной, по меньшей мере на 96% идентичной, по меньшей мере на 97% идентичной, по меньшей мере на 98% идентичной или по меньшей мере на 99% идентичной соответствующей человеческой последовательности.
Последовательность, расположенная выше (5') последовательности миметика pre-mir-29, может быть, например, по меньшей мере на 50% идентичной, по меньшей мере на 55% идентичной, по меньшей мере на 60% идентичной, по меньшей мере на 65% идентичной, по меньшей мере на 70% идентичной, по меньшей мере на 75% идентичной, по меньшей мере на 80% идентичной, по меньшей мере на 85% идентичной, по меньшей мере на 90% идентичной, по меньшей мере на 91% идентичной, по меньшей мере на 92% идентичной, по меньшей мере на 93% идентичной, по меньшей мере на 94% идентичной, по меньшей мере на 95% идентичной, по меньшей мере на 96% идентичной, по меньшей мере на 97% идентичной, по меньшей мере на 98% идентичной или по меньшей мере на 99% идентичной соответствующей человеческой последовательности.
Последовательность, расположенная ниже (3') зрелой последовательности миметика miR-29, может быть, например, по меньшей мере на 50% идентичной, по меньшей мере на 55% идентичной, по меньшей мере на 60% идентичной, по меньшей мере на 65% идентичной, по меньшей мере на 70% идентичной, по меньшей мере на 75% идентичной, по меньшей мере на 80% идентичной, по меньшей мере на 85% идентичной, по меньшей мере на 90% идентичной, по меньшей мере на 91% идентичной, по меньшей мере на 92% идентичной, по меньшей мере на 93% идентичной, по меньшей мере на 94% идентичной, по меньшей мере на 95% идентичной, по меньшей мере на 96% идентичной, по меньшей мере на 97% идентичной, по меньшей мере на 98% идентичной или по меньшей мере на 99% идентичной соответствующей человеческой последовательности.
Последовательность, расположенная ниже (3') последовательности миметика pre-mir-29, может быть, например, по меньшей мере на 50% идентичной, по меньшей мере на 55% идентичной, по меньшей мере на 60% идентичной, по меньшей мере на 65% идентичной, по меньшей мере на 70% идентичной, по меньшей мере на 75% идентичной, по меньшей мере на 80% идентичной, по меньшей мере на 85% идентичной, по меньшей мере на 90% идентичной, по меньшей мере на 91% идентичной, по меньшей мере на 92% идентичной, по меньшей мере на 93% идентичной, по меньшей мере на 94% идентичной, по меньшей мере на 95% идентичной, по меньшей мере на 96% идентичной, по меньшей мере на 97% идентичной, по меньшей мере на 98% идентичной или по меньшей мере на 99% идентичной соответствующей человеческой последовательности.
Предшественник miR-29 может иметь любую подходящую длину, при условии его процессинга с образованием зрелой последовательности миметика miR-29 (либо одно-, либо двухцепочечной). Следовательно, длина предшественника составляет по меньшей мере 23 нуклеотида и может составлять по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 55, по меньшей мере 60, по меньшей мере 65, по меньшей мере 70, по меньшей мере 75, по меньшей мере 80, по меньшей мере 85, по меньшей мере 90, по меньшей мере 95, по меньшей мере 100, по меньшей мере 110, по меньшей мере 120, по меньшей мере 130, по меньшей мере 140, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 300, по меньшей мере 350, по меньшей мере 400, по меньшей мере 450, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 1500 или по меньшей мере 2000 нуклеотидов.
В альтернативном варианте длина предшественника может составлять не более 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 110, 120, 130, 140, 150, 200, 250, 300, 350, 400, 450, 500, 1000, 1500, 2000 или 2500 нуклеотидов.
Структурные модификации
В дополнение к этому или в качестве альтернативы модификациям последовательностей, которые обсуждались выше, миметик miR-29 или его предшественник может содержать одну или более структурных модификаций по сравнению с олигонуклеотидом РНК.
Миметик или предшественник miR-29 может содержать один или более нуклеотидов, содержащих модифицированный остаток сахара, т.е. остаток сахара, отличный от остатка рибозы. Примеры таких модифицированных остатков сахара включают 2'-O-метилрибозу, 2'-O-метоксиэтилрибозу, 2'-фтор-рибозу и 4-тиорибозу, а также дисахариды. Дисахариды обычно содержат фуранозный цикл с 2',4'-связью (например, метиленовым мостиком), которая ограничивает возможную структуру цикла С3'-эндоконформацией. Нуклеотид, содержащий бициклический сахар, часто называется остатком закрытой нуклеиновой кислоты («ЗНК»).
Миметик или предшественник miR-29 может независимо содержать один или более из любого или всех этих типов модифицированных остатков сахаров. Например, миметик может содержать один, два, три, четыре, пять, вплоть до 10, вплоть до 15, вплоть до 20 или еще больше модифицированных остатков сахаров. В определенных вариантах реализации изобретения все нуклеотиды содержат модифицированный остаток сахара.
В дополнение к этому или в качестве альтернативы этому, миметик или предшественник miR-29 может содержать одну или более модификаций сахарофосфатного остова, например, модифицированную межнуклеозидную связь.
Следовательно, один или более соседних нуклеотидов могут соединяться с помощью альтернативной соединительной группы вместо фосфатной группы.
Было бы особенно целесообразным, чтобы модифицированная межнуклеозидная связь находилась на одном или обоих концах миметика miR-29, т.е. между 5'-концевым нуклеотидом и соседним нуклеотидом и/или между 3'-концевым нуклеотидом и соседним нуклеотидом.
Группы, подходящие для применения в качестве межнуклеозидных связей, включают фосфотиоатные, морфолино- и фосфонокарбоксилатные группы, а также силоксановые, сульфидные, сульфоксидные, сульфоновые, ацетильные, формацетильные, тиоформацетильные, метиленформацетильные, тиоформацетильные, алкенильные, сульфаматные, метилениминовые, метиленгидразиновые, сульфонатные и сульфонамидные группы.
В фосфотиоатной группе немостиковый атом кислорода замещается атомом серы. Фосфотиоатные группы могут способствовать связыванию с белками сыворотки крови и могут, таким образом, улучшать распределение и биодоступность миметика in vivo. Целесообразным может быть системное введение миметика реципиенту.
В дополнение к этому или в качестве альтернативы этому, миметик или предшественник miR-29 может содержать одно или более модифицированных оснований, в качестве альтернативы природным аденину, цитозину, гуанину и урацилу. Такие модифицированные основания включают 5-метилцитозин (5-ме-С), 5-гидроксиметил цитозин, ксантин, гипоксантин, 2-аминоаденин, 6-метил или другие алкильные производные аденина и гуанина, 2-пропил или другие алкильные производные аденина и гуанина, 2-тиоурацил, 2-тиотимин и 2-тиоцитозин, 5-галоурацил и цитозин, 5-пропинилурацил и цитозин и другие алкинильные производные пиримидиновых оснований, 6-азоурацил, цитозин и тимин, 5-урацил (псевдоурацил), 4-тиоурацил, 8-гало, 8-амино, 8-тио, 8-тиоалкил, 8-гидроксил и другие 8-замещенные аденины и гуанины, 5-гало (включая 5-бромо, 5-трифторметил и другие 5-замещенные урацилы и цитозины), 7-метилгуанин и 7-метиладенин, 2-F-аденин, 2-амино-аденин, 8-азагуанин и 8-азааденин, 7-деазагуанин и 7-деазааденин, а также 3-деазагуанин и 3-деазааденин.
Было высказано предположение о том, что, чем сильнее модифицирована пассажирская цепь, тем менее вероятно, что она будет встраиваться в комплекс RISC, и, следовательно, тем более эффективной будет направляющая цепь. Следовательно, даже если направляющая цепь получена из miR-29, было бы целесообразным, чтобы пассажирская цепь содержала одну или более модификаций, например, один или более модифицированных остатков сахаров, одну или более модифицированных межнуклеозидых связей и/или одно или более модифицированных оснований.
В дополнение к этому или в качестве альтернативы этому, миметик или предшественник miR-29 может содержать фрагмент, обеспечивающий прохождение через мембрану, для облегчения прохождения через плазматическую мембрану клетки-мишени. В качестве такого фрагмента может выступать подходящий липидный или другой жирнокислотный фрагмент, включая, но не ограничиваясь этим, холестериновый и стеароильный фрагмент.
Другие фрагменты, обеспечивающие прохождение через мембрану, включают проникающие в клетку пептиды («СРР», такие как ТАТ и MPG из ВИЧ-1, пенетратин, полиаргинин) и фузогенные пептиды (например, производные эндодомена оболочки ВИЧ-1 (HGP) или фузогенный пептид вируса гриппа (diINF-7)). Фрагмент, обеспечивающий прохождение через мембрану, может быть конъюгирован с молекулой-носителем, которая нековалентно связана с миметиком miR-29 или ее предшественником. В альтернативном варианте фрагмент, обеспечивающий прохождение через мембрану, может быть конъюгирован с миметиком miR-29 или ее предшественником.
Фрагмент, обеспечивающий прохождение через мембрану, может быть конъюгирован либо с направляющей цепью, либо пассажирской цепью, хотя предпочтение остается за пассажирской цепью, таким образом, чтобы не нарушить функцию направляющей цепи. Предпочтение может быть отдано конъюгации либо на 5'-, либо на 3'-конце, хотя также возможна конъюгация с внутренним остатком.
Во избежание сомнений, молекула miR-29 (т.е. в ином случае не обладающая никакими отличиями по структуре или последовательности от нативной молекулы) может рассматриваться как миметик или предшественник miR-29 при связывании с фрагментом, обеспечивающим прохождение через мембрану.
Примером миметика miR-29 является направляющая цепь:
5'-rUrArGrCrArCrCrArUrCrUrGrArArArUrCrGrGmUmUmA-3'
где «r» обозначает сахар рибозу, и «m» обозначает 2'-O-метилрибозу. Направляющая цепь может входить в состав двухцепочечного миметика miR-29 в комбинации с пассажирской цепью. Примерами подходящих пассажирских цепей являются:
5'mAmCrCmGrAmUrUmUrCmArGmArUmGrGmUrGmCrUA-3'
и
5'-mAmCrCmGrAmUrUmUrCmArGmArUmGrGmUrGmCrUmAdG-3'
Доставка miR-29, миметиков и предшественников
Могут быть получены композиции, в которых miR-29, миметики и предшественники связаны с (например, формируют комплекс с или инкапсулированы) подходящим носителем.
Подходящие носители включают фармацевтически приемлемые липиды и полимеры, а также их комбинации. Например, композиция может иметь форму липосом, липидных везикул, липидных комплексов или полимерных комплексов.
Например, липидные везикулы и липосомы представляют собой частицы из липидного бислоя, имеющие водное ядро, содержащее олигонуклеотидный груз.
Липидные комплексы (или «липоплексы») и полимерные комплексы (или «полиплексы») обычно содержат положительно заряженные липиды или полимеры, которые взаимодействуют с отрицательно заряженными олигонуклеотидами с образованием комплексов.
Катионные полимеры или липиды могут также взаимодействовать с отрицательно заряженными молекулами на поверхности клеток-мишеней. При помощи соответствующего подбора липидов и групп головок могут быть сконструированы комплексы, чтобы способствовать слиянию с плазматической мембраной клетки-мишени или с выбранной внутренней мембраной (такой как эндосомальная мембрана или ядерная мембрана), для облегчения доставки олигонуклеотидного груза в соответствующий субклеточный компартмент. Обзор по доставке генов с помощью липоплексов и полиплексов представлен, например, Tros de Ilarduya et al. в Eur. J. Pharm. Sci. 40 (2010) 159-170.
Эмульсии из нейтральных липидов также могут быть использованы для формирования комплексов микрочастиц с микроРНК, имеющих диаметр порядка нанометров.
Соответствующие липиды могут быть подобраны специалистом в данной области в зависимости от применения, груза и клетки-мишени. Могут быть использованы липиды в чистом виде или чаще всего комбинации липидов.
Подходящие липиды описаны, например, в WO 2011/088309 и литературных источниках, процитированных в данном документе, и включают:
- нейтральные липиды и фосфолипиды, такие как сфингомиелин, фосфатидилхолин, фосфатидилэтаноламин, фосфатидилсерин, фосфатидилинозитол, фосфатидную кислоту, пальмитоилолеоил фосфатидилхолин, лизофосфатидилхолин, лизофосфатидилэтаноламин, дипальмитоилфосфатидилхолин, диолеоилфосфатидилхолин, дистеароилфосфатидилхолин, дилинолеоилфосфатидилхолин, фосфатидилхолин (PC), 1,2-диолеоил-sn-глицеро-3-фосфохолин (DOPC), лецитин, фосфатидилэтаноламин (РЕ), лизолецитин, лизофосфатидилэтаноламин, сфингомиелин (SM), кардиолипин, фосфатидную кислоту, 1,2-дистеароил-sn-глицеро-3-фосфохолин (DSPC), 1,2-дипальмитоил-sn-глицеро-3-фосфоэтаноламин (DPPE), 1-пальмитоил-2-олеоил-sn-глицеро-3-фосфохолин (РОРС), 1,2-дилауроил-sn-глицеро-3-фосфохолин (DLPC), 1,2-димиристоил-sn-глицеро-3-фосфохолин (DMPC), 1,2-дипальмитоил-sn-глицеро-3-фосфохолин (DPPC), 1,2-димиристоил-sn-глицеро-3-фосфоэтаноламин (DMPE), 1,2-диолеоил-sn-глицеро-3-фосфоэтаноламин (DOPE), дипалмитолоеоил-РЕ, дифитаноил-РЕ, DSPE, диэлаидоил-РЕ, дилинолеоил-SM и дилинолеоил-РЕ;
- стерины, например, холестерин
- полимер-модифицированные липиды, например, полиэтиленгликоль (ПЭГ)-модифицированные липиды, включая ПЭГ-модифицированные фосфатидилэтаноламин и фосфатидную кислоту, конъюгаты ПЭГ-церамида, ПЭГ-модифицированные диалкиламины и ПЭГ-модифицированные 1,2-диацилоксипропан-3-амины. Особенно подходящим являются ПЭГ-модифицированные диацилглицерины и диалкилглицерины, например, ПЭГ-дидимиристоилглицерин (PEG-DMG), ПЭГ-дистирилглицерин (PEG-DSG) и ПЭГ-карбамоил-1,2-димиристилоксипропилиламин (PEG-cDMA);
- катионные липиды, такие как хлорид N,N-диолеил-N,N-диметиламмония («DODAC»); хлорид N-(2,3-диолеилокси)пропил-N,N-N-триэтиламмония («DOTMA»); бромид N,N-дистеарил-N,N-диметиламмония («DDAB»); хлорид N-(2,3-диолеоилокси)пропил)-N,N-триметиламмония («DOTAP»); хлорид 1,2-диолеилокси-3-триметиламинопропана («DOTAP.Cl»); 3β-(N-(N',N'-диметиламиноэтан)-карбамоил)холестерин («DC-Chol»),
трифторацетат N-(1-(2,3-диолеилокси)пропил)-N-2-(сперминкарбоксамидо)этил)-N, диметиламмония («DOSPA»), диоктадециламидоглицил карбоксиспермин («DOGS»), 1,2-дилеоил-sn-3-фосфоэтаноламин («DOPE»), 1,2-диолеоил-3-диметиламмонийпропан («DODAP»), N,N-димeтил-2,3-диoлeилoкcи)пpoпилaмин («DODMA»), бромид N-(1,2-димиристилоксипроп-3-ил)-N,N-диметил-N-гидроксиэтиламмония («DMRIE»), 1,2-дилинолеилокси-3-диметиламинопропан (DLinDMA) 1,2-дилинолеоил-3-диметиламинопропан (DLinDAP), 1-линолеоил-2-линолеилокси-3-диметиламинопропан (DLin-2-DMAP), 1,2-дилинолеилкарбамоилокси-3-диметиламинопропан (DLin-C-DAP), 1,2-дилинолеилтио-3-диметиламинопропан (DLin-S-DMA) и 2,2-дилинолеил-4-10 диметиламинометил-[1,3]-диоксолан (DLin-K-DMA). Коммерческие препараты катионных липидов, включают Липофектин™ (содержащий DOTMA и DOPE, доступен от Gibco/BRL) и Липофектамин™ (содержащий DOSPA и DOPE, доступен от Gibco/BRL).
- анионные липиды, включая фосфатидилглицерин, кардиолипин, диацилфосфатидилсерин, диацилфосфатидную кислоту, N-додеканоилфосфатидилэтаноламин, N-сукцинилфосфатидилэтаноламин, N-глутарилфосфатидилэтаноламин и лизилфосфатидилглицерин.
В публикации WO/0071096 описаны разные составы, такие как DOТАР: холестерин или препарат производного холестерина, которые могут быть эффективно использованы для доставки олигонуклеотида.
Коммерчески доступная композиция, которая способна обеспечивать эффективную доставку микроРНК в легкие, представляет собой эмульсию из нейтральных липидов MaxSuppressor in vivo RNALancerll (BIOO Scientific, Остин, Техас), которая состоит из 1,2-диолеоил-sn-глицеро-3-фосфохолина, скваленового масла, полисорбата 20 и антиоксиданта. В комплексе с синтетическими микроРНК она формирует наночастицы с диаметрами в нанометровом диапазоне.
Подходящие полимеры включают гистоны и протамины (и другие ДНК-связывающие белки), поли(этиленимин) (PEI), катионные дендримеры, такие как полиамидоаминные дендримеры (РАМАМ), 2-диметил(аминоэтил) метакрилат (pDMAEM), поли(L-лизин) (PLL), полимеры на основе углеводов, такие как хитозан и т.д. См. Tros de Ilarduya et al. в Eur. J. Pharm. Sci. 40 (2010) 159-17 для обзора.
Также могут использоваться белки и пептиды, такие как ателоколлаген. Ателоколлаген представляет собой водорастворимую форму коллагена, образующегося в результате обработки протеазой, в частности, обработанный пепсином коллаген I-го типа из дермиса теленка.
Для доставки также могут использоваться циклодекстрины.
Средства, обеспечивающие направленное связывание
Молекулы-носители также могут содержать средства, обеспечивающие направленное связывание с поверхностью клетки-мишени. Например, указанное средство, обеспечивающие направленное связывание, может представлять собой партнер по специфическому связыванию, способный к специфическому связыванию с молекулой, экспрессированной на поверхности сухожильной клетки-мишени. Подходящие партнеры по связыванию включают антитела и тому подобное, направленные против молекул клеточной поверхности, или лиганды, или рецепторы для таких молекул клеточной поверхности. Маркеры клеточной поверхности, которые могут способствовать направленному связыванию с сухожильными клетками, включают Тенасцин C, CD55 и теномодулин.
Термин «пара компонентов, участвующая в специфическом связывании» используется для описания пары молекул, содержащей компонент специфического связывания (sbm) и его партнер по связыванию (bp), которые обладают определенной специфичностью по отношению друг к другу и которые в нормальных условиях связываются друг с другом в преференциальном порядке по сравнению со связыванием с другими молекулами. Примерами пар компонентов, участвующих в специфическом связывании, являются антитела и их соответствующие эпитопы/антигены, лиганды (такие как гормоны и т.д.) и рецепторы, авидин/стрептавидин и биотин, лектины и углеводы, а также комплементарные нуклеотидные последовательности.
Общеизвестно, что фрагменты полного антитела могут выполнять функции связывающих антигенов. Примерами функциональных связывающих фрагментов являются (i) Fab-фрагмент, состоящий из VL-, VH-, CL- и CH1-доменов; (ii) Fd-фрагмент, состоящий из VH- и СН1-доменов; (iii) Fv-фрагмент, состоящий из VL- и VH-доменов одного плеча антитела; (iv) dAb-фрагмент (Ward, E.S. et al., Nature 341, 544-546 (1989)), который состоит из VH-домена; (v) выделенные CDR-области; (vi) F(аb')2-фрагменты, бивалентный фрагмент, содержащий два связанных Fab-фрагмента (vii), одноцепочечные Fv-молекулы (scFv), причем VH-домен и VL-домен связаны пептидным линкером, что обеспечивает связывание двух доменов с образованием антиген-связывающего сайта (Bird et al, Science, 242, 423-426, 1988; Huston et al., PNAS USA, 85, 5879-5883, 1988); (viii) биспецифические одноцепочечные Fv-димеры (PCT/US92/09965) и (ix) «диатела», мультивалентные или мультиспецифические фрагменты, сконструированные путем слияния генов (WO 94/13804; P. Holliger et al., Proc. Natl. Acad. Sci. USA 90 6444-6448, 1993).
Так как антитела могут быть модифицированы различными способами, поэтому термин «антитело» должен трактоваться как охватывающий любое специфическое связывающее средство, имеющее связывающий домен с требуемой специфичностью. Следовательно, этот термин охватывает фрагменты антител, описанные выше, а также производные, функциональные эквиваленты и гомологи антител, включая любой полипептид, содержащий иммуноглобулин-связывающий домен, будь то природный или синтетический. Таким образом, данный термин охватывает гибридные молекулы, содержащие иммуноглобулин-связывающий домен, или эквивалент, слитый с другим полипептидом. Клонирование и экспрессия гибридных антител описаны в ЕР-А-0120694 и ЕР-А-0125023.
Альтернативы антителам становятся все более доступными. Так называемые «аффинные белки» или «генно-инженерные белковые каркасы» могут быть сконструированы стандартным образом с целью аффинного связывания с определенной мишенью. Они обычно основаны на каркасном белке неиммуноглобулиновой природы с конформационно стабильным или жестким ядром, который был модифицирован для обеспечения аффинного связывания с мишенью. Модификация может включать замену одного или более поверхностных остатков и/или вставку одного или более остатков на поверхности каркасного белка. Например, пептид с аффинностью по отношению к мишени может быть встроен в поверхностную петлю каркасного белка или может заменять часть или всю поверхностную петлю каркасного белка. Подходящие каркасы и их сконструированные эквиваленты включают:
- BPTI, LAC-DI, ITI-D2 (каркасы на основе домена Куница);
- ETI-II, AGRP (ноттин);
- тиоредоксин (пептидный аптамер);
- Fn3 (AdNectin);
- липокалин (ВВР) (Anticalin);
- анкириновый повтор (DARPin);
- Z-домен белка A (Affibody);
- гамма В-кристаллин/убиквитин (Affilin);
- LDLR-A-domain (Avimer).
См., например, Gebauer, М and Skerra, A, Current Op. Chem. Biol. 2009, 13: 245-255, и Friedman, M, and Stahl, S, Biotechnol. Appl. Biochem. (2009) 53: 1-29 и литературные источники, процитированные в данном документе.
Нуклеиновые кислоты, кодирующие miR-29, ее миметики и предшественники
B качестве альтернативы доставке олигонуклеотидов, миметиков и предшественников miR-29 непосредственно в клетку-мишень, представляется возможной доставка нуклеиновой кислоты, кодирующей олигонуклеотид miR-29, ее миметик или их предшественники, в клетку-мишень таким образом, что указанные олигонуклеотид, миметик или предшественник miR-29 будут экспрессироваться в клетке-мишени. Такой подход может рассматриваться в качестве «генной терапии».
Специалистам в данной области техники очевидно, что нуклеиновые кислоты можно использовать только для кодирования miR-29, ее миметиков или предшественников, состоящих из РНК, т.е. образованных четырьмя естественными нуклеотидными компонентами РНК и не содержащими модифицированных оснований, сахаров или межнуклеозидных связей.
Нуклеиновая кислота обычно включает экспрессирующую конструкцию, содержащую последовательность нуклеиновой кислоты, кодирующую олигонуклеотид, миметик или предшественник miR-29, функционально связанную с соответствующими регуляторными последовательностями для обеспечения экспрессии. Регуляторные последовательности могут быть подобраны в зависимости от клетки-мишени, но обычно включают соответствующий промотор и необязательно энхансер, который управляет транскрипцией, осуществляемой РНК-полимеразой II, а также терминатор транскрипции (обычно содержащий сигнал полиаденилирования).
Промотор может представлять собой тканеспецифический промотор, который активирует транскрипцию преимущественно или исключительно в клетке- или ткани-мишени по сравнению с другими типами клеток или тканей.
Следовательно, промотор может представлять собой промотор, который активирует транскрипцию преимущественно или исключительно в сухожильной клетке. Подходящим промотором может быть промотор гена коллагена 1a1 (col1a1).
Доставка нуклеиновых кислот в клетки-мишени
Нуклеиновые кислоты, кодирующие miR-29, ее миметики и предшественники могут быть доставлены любым удобным способом.
Способы доставки нуклеиновых кислот в клетки in vitro включают осаждение фосфатом кальция, трансфекцию с помощью ДЭАЭ-декстрана, электропорацию, микроинъекцию, ДНК-нагруженные липосомы, обработку ультразвуком и бомбардировку с использованием покрытых нуклеиновыми кислотами микрочастиц (например, микросферы золота или вольфрама). Многие из этих методик были успешно адаптированы для применения in vivo или ex vivo.
Следовательно, нуклеиновая кислота может применяться в свободной форме, может быть связана с (например, в комплексе с или инкапсулированная) подходящим носителем, таким как полимер или липид (как описано в других разделах настоящего документа), или нанесена на поверхность частиц. В таких вариантах реализации изобретения нуклеиновая кислота обычно представляет собой ДНК. Нуклеиновая кислота или носитель также может содержать нацеливающий фрагмент или фрагмент, обеспечивающий транспорт через мембрану, как описано в других разделах настоящего описания. Также любой из этих способов может быть соответствующим образом адаптирован для доставки miR96, ее предшественников и миметиков самих по себе.
Нуклеиновая кислота обычно представлена в виде экспрессирующего вектора. Специалист в данной области сможет сконструировать подходящие экспрессирующие векторы на основе нуклеиновой кислоты для применения в терапевтических целях (а также для других областей применения, описанных в данном документе). Векторы обычно включают экспрессирующую конструкцию, содержащую последовательность нуклеиновой кислоты, кодирующую miR-29, ее миметик или предшественник в функциональной связи с соответствующими регуляторными последовательностями, включающими последовательности промотора и последовательности терминации транскрипции, необязательно в комбинации с последовательностями энхансера, маркерных генов и другими последовательностями в зависимости от конкретного применения. Подразумевается, что векторы могут интегрироваться в хромосому клетки-хозяина или могут существовать и реплицироваться независимо от хромосом клетки-хозяина в виде эписомы, например, плазмиды.
В альтернативном варианте может быть использован вирусный вектор для доставки нуклеиновой кислоты.
В качестве средства доставки генов может быть использован любой подходящий тип вирусного вектора. К ним относятся аденовирусные векторы, векторы на основе аденоассоциированных вирусов (AAV), ретровирусные (особенно лентивирусные) или герпесвирусные векторы. Особенно предпочтительными могут быть аденовирусные и лентивирусные векторы, поскольку они обладают способностью обеспечивать экспрессию гена(ов) после доставки в клетки, которые недостаточно активно делятся.
Вирусный вектор обычно содержит гены вирусных структурных белков и молекулярный груз в виде нуклеиновой кислоты, который содержит желаемую экспрессирующую конструкцию в форме, обеспечивающей экспрессию гена в клетке- или ткани-мишени. Следовательно, ген обычно функционально связан с промотором и другими соответствующими транскрипционными регуляторными сигналами.
В аденовирусных векторах молекулярный груз в виде нуклеиновой кислоты обычно представлен молекулой двухцепочечной ДНК (дцДНК). В ретровирусных векторах он обычно представлен одноцепочечной РНК.
Молекулярный груз в виде нуклеиновой кислоты обычно содержит дополнительные элементы, необходимые для его упаковки с образованием средства доставки генов и соответствующего процессинга в клетке- или ткани-мишени.
В случае аденовирусных векторов они могут включать аденовирусные последовательности инвертированных концевых повторов (ITR) и соответствующий сигнал упаковки.
В случае ретровирусных векторов они могут включать характерные концевые последовательности (так называемые последовательности «R-U5» и «U3-R») и сигнал упаковки. Концевые последовательности способствуют образованию прямых повторов («длинных концевых повторов» или «LTR») на любом конце провируса, что образуются в результате обратной транскрипции, которые затем облегчают интеграцию провируса в геном клетки-хозяина и управляют последующей экспрессией.
Молекулярный груз в виде нуклеиновой кислоты также может содержать селективный маркер, т.е. ген, кодирующий продукт, который обеспечивает быструю детекцию трансдуцированной клетки. Примеры включают гены флуоресцентных белков (например, GFP), ферменты, которые приводят к образованию оптически детектируемого продукта реакции (например, бета-галактозидаза, люцифераза), и гены устойчивости к антибиотикам.
Вирусный вектор обычно некомпетентный по репликации. Другими словами, молекулярный груз в виде нуклеиновой кислоты не содержит всех вирусных генов (и других генетических элементов), необходимых для репликации вируса. Несмотря на это, вектор будет содержать все структурные белки и ферменты, которые необходимы для попадания молекулярного груза в клетку-хозяина и для соответствующего процессинга молекулярного груза таким образом, чтобы могли экспрессироваться miR-29, миметик или предшественник. Если они не кодируются генетическим материалом вектора, то их обычно продуцирует упаковывающая клеточная линия. Специалисту в данной области техники хорошо известны подходящие клеточные линии, которые могут быть использованы для создания соответствующих вирусных средств доставки.
Следовательно, в случае аденовирусного вектора в молекулярном грузе в виде нуклеиновой кислоты обычно отсутствуют один или более функциональных аденовирусных генов из областей E1, Е2, Е3 или Е4. Эти гены могут быть удалены или в ином случае инактивированы, например, путем вставки транскрипционной единицы, содержащей гетерологичный ген или селективный маркер.
В некоторых вариантах реализации изобретения нуклеиновая кислота содержит функциональные вирусные гены. Следовательно, в случае аденовирусного вектора из вирусных компонентов могут содержаться исключительно ITR и сигнал упаковки.
Предпочтительным может быть использование нуклеиновых кислот, не содержащих функциональных вирусных генов, поскольку их использование сопряжено со снижением риска развития иммунного ответа организма-хозяина против трансдуцированной клетки- или ткани-мишени в результате синтеза вирусных белков.
Вирусные векторы могут быть сконструированы таким образом, чтобы они содержали гены модифицированных поверхностных белков, способных к связыванию с маркерами на клетке-мишени, тем самым увеличивая вероятность трансдукции желаемой клетки-мишени и уменьшая вероятность неспецифической трансдукции клетки и ткани другого вида. Данный подход в некоторых случаях называют псевдотипированием. Следовательно, вирусный вектор может содержать поверхностный белок, способный к связыванию с маркером клеточной поверхности на сухожильной клетке. Маркеры клеточной поверхности, которые могут способствовать направленному связыванию с сухожильными клетками, включают Тенасцин C и CD55.
Сухожилие и повреждение сухожилия
Сухожилия представляют собой структуры из соединительной ткани, через которые осуществляется присоединение мышцы к кости. Они обеспечивают передачу усилия от сокращающейся мышцы к скелетной структуре, к которой они прикреплены, на расстоянии от мышцы как таковой1.
Сухожилия представляют собой сложную системно организованную ткань и содержит несколько четко выраженных слоев.
Сухожилие состоит из почти параллельных компактных пучков, содержащих 30% коллагена и 2% эластина (по сырой массе) и погруженных во внеклеточный матрикс, включающий различные типы клеток, преимущественно тендиноциты3.
Преобладающим типом коллагена является коллаген I типа, фибриллы которого имеют большой диаметр (40-60 нм) и соединяются вместе с образованием плотных пучков волокон. Также содержится коллаген 3-го типа, фибриллы которого имеют меньший диаметр (10-20 нм), формируя более рыхлые пучки ретикулярных волокон.
Из коллагена образуются (по возрастанию сложности) фибриллы, волокна, первичные и вторичные пучки волокон, окруженные слоем рыхлого, коллагенового и богатого липидами соединительнотканного матрикса, известного как эндотенон4. Слой, состоящий из такого же самого материала, называемый эпитеноном, покрывает поверхность всего сухожилия. Эпитенон окружает соединительная ткань, называемая паратеноном, который содержит фибриллы из коллагена 1-го и 3-го типа, некоторые эластичные фибриллы и слой синовиальных клеток. Некоторые сухожилия дополнительно окружены сухожильным влагалищем.
Основными типами клеток в составе сухожилия являются теноциты и тенобласты, причем и те, и другие представляют собой фибробластоподобные клетки14. Оба типа клеток играют важную роль в сохранении функции сухожилия, поскольку вырабатывают коллаген и способствуют поддержанию структуры внеклеточного матрикса15. Следовательно, используемый в данном описании термин «сухожильная клетка» охватывает как теноциты, так и тенобласты.
Теноциты представляют собой плоские, суживающиеся на краях веретеновидные клетки в продольном направлении и имеющие звездчатое поперечное сечение, обнаруживаемые в небольших количествах и располагающиеся в цепочках между коллагеновыми волокнами. Они имеют сложные клеточные отростки, образующие трехмерную сеть, простирающуюся через внеклеточный матрикс, соединяются с помощью клеточных отростков и могут быть подвижными.
Тенобласты являются предшественниками теноцитов. Они представляют собой веретеновидные или звездчатые клетки с длинными, конусообразными, плоскими ядрами с эозинофильными включениями. Они обладают подвижностью и высокой пролиферативной активностью.
На протяжении эмбрионального развития тенобласты, а следовательно, и теноциты, происходят из мезодермальных компартментов, как и скелетные миобласты, хондроциты и остеобласты16. Некоторые из мультипотентных мезенхимальных клеток-предшественников, которые происходят из этих компартментов, экспрессируют основной транскрипционный фактор склераксис со структурой типа спираль-петля-спираль. Однако как только они превращаются в коммитированные клетки, образующие специфическую ткань, только тенобласты и теноциты сохраняют способность к экспрессии склераксиса. Следовательно, ген склераксиса представляет собой первый основной ген, который, как было установлено, имеет существенное значение для установления линии сухожильных клеток на протяжении развития. Теномодулин представляет собой трансмембранный гликопротеин II типа, экспрессирующийся в сухожилиях мышей на поздней (эмбриональный день [Е] 17.5) стадии развития, и также наблюдается в сухожилиях взрослых мышей. Следовательно, склераксис представляет собой маркер как тенобластов, так и на теноцитов, тогда как теномодулин представляет собой поверхностный маркер зрелых теноцитов19.
Повреждение сухожилия может быть вызвано или может быть ассоциировано с многочисленными факторами, включая (но не ограничиваясь этим) внешнюю травму, механическое напряжение (включая перегрузку), дегенерацию, воспаление и их комбинацию, что часто носит название «тендинопатия».
Термин «травма сухожилия», как правило, используется для обозначения острой травмы вследствие одного травматического события, включая внешнюю травму и разрыв сухожилия (т.е. полный разрыв сухожилия).
Тендинопатия является многофакторным заболеванием, имеет много различных форм от острой до хронической и часто ассоциирована с перегрузкой сухожилия, которая может быть кратковременной или продолжительной. Тендинопатия может включать дегенерацию или другие виды механического повреждения коллагена на микроскопическом или макроскопическом уровне (иногда называется «тендиноз»), воспаления или комбинации и того и другого (иногда называется «тендинит»).
Биомеханические свойства сухожилия, особенно его прочность на растяжение, зависят от площади поперечного сечения (т.е. толщины), содержания коллагена и соотношения между разными типами коллагена. После острой травмы, на протяжении тендинопатии и на протяжении процесса заживления поврежденного сухожилия, происходит сдвиг в синтезе коллагена с 1-го типа в пользу коллагена 3-го типа. Синтез коллагена 1-го типа может возвращаться к нормальным уровням после первоначального падения, но постоянное увеличение синтеза коллагена 3-го типа приводит к длительному дисбалансу в соотношении подтипов коллагена. Это оказывает значительное и отрицательное воздействие на биомеханические свойства сухожилия. В частности, в результате этого уменьшается прочность сухожилия на растяжение, что приводит к уменьшению предельной прочности на разрыв и в результате этого увеличивается вероятность его разрыва.
Способы по настоящему изобретению могут применяться в отношении любого поврежденного сухожилия. Основными видами сухожилий, которые подвергаются тендинопатии у людей, являются ахиллово сухожилие, сухожилие надостной мышцы, общее сухожилие сгибателей и общее сухожилие разгибателей. Основным видом сухожилий, которые подвергаются тендинопатии у субъектов семейства лошадиных, является поверхностное сухожилие сгибателей. Они могут иметь особенно важное значение в качестве терапевтических мишеней.
Применение miR-29, миметиков и предшественников в терапевтических целях
Авторы настоящего изобретения обнаружили, что путем увеличения активности miR-29 в сухожильных клетках представляется возможным изменение соотношения между подтипами коллагена в пользу увеличения синтеза коллагена 1-го типа по отношению к синтезу коллагена 3-го типа.
Следовательно, в настоящем изобретении предложены способы модуляции заживления сухожилия путем применения miR-29 в терапевтических целях. Способы, описанные в данном документе, могут рассматриваться в качестве способов модуляции относительного состава и/или синтеза коллагена в сухожилии, в особенности относительного содержания и синтеза коллагена 1-го и 3-го типов в сухожилии. Считается, что необходимо сместить соотношение между подтипами коллагена в пользу коллагена 1-го типа, т.е. в пользу увеличения содержания или синтеза коллагена 1-го типа в сухожилии по отношению к коллагену 3-го типа. Следует понимать, что это не обязательно включает чистое увеличение синтеза или содержания коллагена 1-го типа, поскольку miR-29 может ингибировать синтез коллагена 1-го типа. Однако синтез коллагена 3-го типа подавляется в большей степени, чем синтез коллагена 1-го типа.
На физиологическом уровне способы, описанные в данном документе, могут рассматриваться в качестве способов модуляции биомеханических свойств сухожилия, предпочтительно улучшения биомеханических свойств сухожилия, например, улучшения или увеличения прочности сухожилия на растяжение.
Способы по настоящему изобретению могут применяться на любой стадии тендинопатии или на любой стадии процесса заживления травмированного сухожилия. Например, указанные способы могут быть использованы для модуляции соотношения коллагена и, следовательно, биомеханических свойств сухожилия в процессе тендинопатии или в процессе заживления острой травмы сухожилия, как, например, разорванного сухожилия.
Следовательно, способы по настоящему изобретению в равной степени могут рассматриваться в качестве способов лечения повреждения сухожилия, включая повреждение, возникающие вследствие травмы сухожилия и тендинопатии.
Выработка ИЛ-33 может наблюдаться в сухожилии в течение короткого периода времени после травмы и на ранних стадиях тендинопатии. Не желая ограничиваться какой-либо конкретной теорией, считается, что ИЛ-33 может быть вовлечен в переключение синтеза коллагена с 1-го типа на 3-й тип. Однако считается, что дисбаланс синтеза коллагена сохраняется после первоначального вовлечения ИЛ-33. Способы по настоящему изобретению не ограничиваются лечением травмы сухожилия на ранних стадиях, но в равной степени распространяются на позднюю стадию развития травмы или заболевания, например, тендинопатии.
Следовательно, лечение может проводиться на любой стадии после дебюта симптомов или после травматического события, вызывающего повреждение сухожилия. Например, лечение может проводиться через 1 сутки, 2 суток, 3 суток, 4 суток, 5 суток, 6 суток, 7 суток или более после дебюта симптомов или травматического события. Оно может проводиться через 1 неделю, 2 недели, 3 недели, 4 недели или более после дебюта симптомов или травматического события. Оно может проводиться через 1 месяц, 2 месяца, 3 месяца, 4 месяца, 5 месяцев, 6 месяцев или более после дебюта симптомов или травматического события.
Субъекты лечения
Хотя большинство наиболее распространенных субъектов лечения относится к людям, способы по настоящему изобретению могут применяться для лечения любых других млекопитающих, включая других приматов (особенно человекообразных обезьян, таких как горилла, шимпанзе и орангутанг, а также широконосых и узконосых обезьян), а также грызунов (включая мышей и крыс) и других широко распространенных лабораторных, домашних и сельскохозяйственных животных (включая, но не ограничиваясь ими, кроликов, собак, кошек, лошадей, коров, овец, коз и т.д.).
В особенности указанные способы могут применяться для лечения субъектов семейства лошадиных, т.е. лошадей. Лошади, в целом, а особенно чистокровные лошади, как, например, скаковые лошади, особенно подвержены травмам сухожилия. Учитывая ценность многих из рассматриваемых животных, уже давно возникла потребность в эффективных способах лечения.
Фармацевтические композиции и способы лечения
Молекулы, описанные в данном документе, могут быть введены в состав фармацевтических композиций. Помимо одного из перечисленных выше веществ эти композиции могут содержать фармацевтически приемлемое вспомогательное вещество, носитель, буфер, стабилизирующее вещество или другие вещества, хорошо известные специалистам в данной области техники. Такие вещества должны быть нетоксичными и не должны снижать эффективность активного ингредиента. Истинные свойства носителя или другого вещества определяются путем его введения в организм, например, пероральным, внутривенным, наружным или подкожным, назальным, внутримышечным и интраперитонеальным. Примеры подходящих композиций и способов применения приведены в Esseku and Adeyeye (2011) и Van den Mooter G. (2006).
Фармацевтические композиции для перорального применения могут быть приготовлены в форме таблетки, капсулы, порошка или форме жидкости. Таблетка может включать твердый носитель, такой как желатин или адъювант. Жидкие фармацевтические композиции, как правило, включают жидкий носитель, такой как вода, продукты нефтепереработки, животные или растительные масла, минеральное масло или синтетическое масло. Также в состав могут быть включены физиологический раствор, декстроза или раствор сахаридов, или гликоли, такие как этиленгликоль, пропиленгликоль или полиэтиленгликоль.
Для внутривенной, кожной или подкожной инъекции, или инъекции в очаг поражения активный ингредиент должен иметь форму парентерально приемлемого водного раствора с апирогенными свойствами, подходящим pH, изотоничностью и стабильностью. Специалисты в данной области техники вполне способны приготовить подходящие растворы с использованием, например, изотонических растворителей, таких как физиологический раствор для инъекций, раствор Рингера для инъекций, раствор Рингера с лактатом для инъекций. Также при необходимости в состав могут быть включены консерванты, стабилизаторы, буферы, антиоксиданты и/или другие добавки.
Принимая во внимание локальный характер заболеваний, по поводу которых проводится лечение, местное применение путем инъекции может оказаться наиболее целесообразным. Инъекция может производиться в пораженное сухожилие или в непосредственной близости от пораженного сухожилия.
Независимо от вида действующего средства, вводимого индивидууму (например, клетка, полипептид, молекула нуклеиновой кислоты, другое фармацевтически пригодное средство в соответствии с настоящим изобретением), предпочтительно введение в «профилактически эффективном количестве» или «терапевтически эффективном количестве» (бывают случаи, когда профилактика рассматривается как лечение), причем этого достаточно для оказания положительного эффекта на индивидуума. Фактическое количество, которое должно вводиться, а также скорость и динамика введения будут зависеть от этиологии и тяжести заболевания, по поводу которого проводится лечение. Назначение лечения, например, решения относительно дозировки и т.п., находятся в пределах компетенции врачей общей практики и врачей другого профиля, а также ветеринарных врачей; при этом обычно учитывается характер заболевания, подлежащего лечению, состояние здоровья конкретного пациента, место и способ введения, а также другие факторы, известные практикующим врачам. Примеры методов и протоколов, упомянутых выше, можно найти в Remington's Pharmaceutical Sciences, 20th Edition, 2000, pub. Lippincott, Williams & Wilkins.
Далее следует более подробное описание настоящего изобретения в качестве примера, а не ограничения, посредством ссылки на прилагаемые графические материалы и примеры.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фигура 1. Экспрессия ИЛ-33/ST2 в образцах сухожилия
Экспрессия генов (А) ИЛ-33, (В) растворимого ST2 (sST2) и (С) мембранного ST2 (mST2) в образцах сухожилия. Кратность изменения экспрессии генов ИЛ-33, растворимого/мембранного ST2 в контрольных образцах (n=10), образцах разорванного сухожилия надостной мышцы и соответствующих образцах сухожилия подлопаточной мышцы человека (n=17). Проиллюстрированные точки данных соответствуют относительной экспрессии по сравнению с геном домашнего хозяйства 18S (среднее значение из двух повторных анализов). Среднее значение ± SD отражает результаты сравнения в популяции больных с помощью t-критерия Стьюдента. (D) Проиллюстрирована оценка окрашивания образцов сухожилия с использованием модифицированной шкалы Bomar со средним значением и SEM. n=10 для контрольного сухожилия (Конт.), n=17 для разорванного сухожилия и сухожилия на ранней стадии тендинопатии. Проиллюстрирован средний балл для образца по модифицированной шкале Bomar на основе анализа 10 полей зрения при большом увеличении. 0=окрашивание отсутствует, 1=<10%, 2=10-20%, 3=>20% положительно окрашенных клеток в поле зрения при большом увеличении. (Е) Кратность изменения экспрессии генов ИЛ-33 и ST2 через 24 часа после инкубации с соответствующими дозами отдельно ФНОα, отдельно ИЛ-1β и в комбинации. Данные показаны в виде среднего значения ± стандартное отклонение для трех образцов и, в свою очередь, относятся к экспериментам на образцах сухожилия трех отдельных пациентов. *p<0,05, **p<0,01 по сравнению с контрольными образцами. (F) Кратность изменения экспрессии генов col1 и соl3 при использовании 50 и 100 нг/мл rhIL-33 через 24 часа после инкубации. (G) Динамика экспрессии генов col1 и соl3 после инкубации с 100 нг/мл ИЛ-33. (Н) Экспрессия гена коллагена 1-го и 3-го типа на уровне белка через 24 часа после инкубации с возрастающими концентрациями hIL-33. Для F, G и Н данные показаны в виде среднего значения ± стандартное отклонение для трех образцов и, в свою очередь, относятся к экспериментам, которые проводились на трех отдельных образцах сухожилия пациента. *p<0,05, **p<0,01 по сравнению с контрольными образцами.
Фигура 2. Влияние JDI-33/ST2 на заживление сухожилия in vivo.
Экспрессия генов ИЛ-33 и растворимого ST2 (А, В) на 1-е, 3-й, 7-е и 21-е сутки после травмы. Данные показаны в виде средней кратности изменения ± SD (суммарные данные по четырем последовательным отборам проб с использованием 4 мышей на группу, таким образом n=16 на условие). *p<0,05, **p<0,01, контроль в сравнении с травмированными мышами. (С, D) уровни мРНК col1 и коллагена 1-го типа у мышей дикого типа и мышей линии ST2-/- на 1-е и 3-й сутки после травмы. (Е, F) уровни мРНК соl3 и коллагена 3-го типа у мышей дикого типа и мышей линии ST2-/- на 1-е и 3-й сутки после травмы. Данные показаны в виде среднего значения ± стандартное отклонение для двойных образцов и соответствуют экспериментам с использованием 4 мышей на условие (n=16). *p<0,05, **p<0,01, контроль в сравнении с травмированными мышами. +p<0,05, ++p<0,01, травмированные мыши дикого типа в сравнении с травмированными мышами линии ST2-/-. (G) процентное изменение прочности сухожилия у мышей дикого типа и мышей линии ST2-/- с травмированными и нетравмированными сухожилиями на 1-е и 3-й сутки после травмы. Данные показаны в виде среднего значения ± стандартное отклонение и соответствуют экспериментам с использованием четырех мышей на условие (n=16). *p<0,05, **p<0,01, контроль в сравнении с травмированными мышами. # p<0,05, травмированные мыши линии ST2-/- в сравнении с травмированными мышами дикого типа.
Фигура 3. ИЛ-33 способствует выработке коллагена 3-го типа и уменьшению прочности сухожилия, тогда как антитело к ИЛ-33 уменьшает эти изменения при повреждении сухожилия in vivo.
(А) уровни мРНК col1, (В) коллагена 1-го типа, (С) мРНК соl3 и (D) коллагена 3-го типа у мышей дикого типа и мышей линии ST2-/-, которые получали rhIL-33 на 1-е сутки после травмы. Данные показаны в виде среднего значения ± стандартное отклонение для двойных образцов и соответствуют экспериментам с использованием четырех мышей на условие (n=16). *p<0,05, **p<0,01, травмированные мыши в сравнении с нетравмированными мышами. +p<0,05, мыши дикого типа в сравнении с мышами линии ST2-/-. (Е) процентное изменение прочности сухожилия у нетравмированных мышей дикого типа на 1-е и 3-и сутки после лечения rhIL-33. Данные показаны в виде среднего значения ± стандартное отклонение и соответствуют экспериментам с использованием четырех мышей на группу (n=16). **p<0,01, травмированные мыши в сравнении с нетравмированными мышами. (F) уровни мРНК col1, (G) коллагена 1-го типа, (Н) мРНК соl3 и (I) коллагена 3-го типа после лечения антителом к ИЛ-33 на 1-е и 3-и сутки после травмы сухожилия у мышей дикого типа. (J) процентное изменение прочности сухожилия при лечении мышей дикого типа антителом к ИЛ-33 на 1-е и 3-и сутки после травмы. Данные показаны в виде среднего значения ± стандартное отклонение и соответствуют экспериментам с использованием четырех мышей на условие (n=16). *p<0,05, **p<0,01, травмированные мыши в сравнении с нетравмированными мышами. A-J, Данные показаны в виде среднего значения ± стандартное отклонение для двойных образцов и соответствуют экспериментам с использованием четырех мышей на условие (n=16).
Фигура 4. МикроРНК 29 непосредственно воздействует на растворимый ST2 - вовлекая для изменения матрикса коллаген в сухожилия.
(А) Все представители семейства miR-29 (miR-29a, miR-29b и miR-29c) были экспрессированы в теноцитах сухожилий, пораженных тендопатией (n=6 образцов тканей пациента). Более низкие значения ΔCt свидетельствуют о высоких уровнях экспрессии генов семейства miR-29 в контроле, разорванном сухожилии надостной мышцы (разорванное сухожилие) и соответствующем образце сухожилия подлопаточной мышцы (ранняя стадия тендинопатии). Данные показаны в виде среднего значения ± стандартное отклонение для двойных образцов и соответствуют экспериментам с использованием десяти образцов тканей пациента. *p<0,05, **p<0,01. (В) Динамика экспрессии miR-29a после добавления 100 нг/мл rhIL-33. (С и D) экспрессия гена col1 и соl3 на уровне мРНК и белка коллагена 1-го и 3-го типа после трансфекции с использованием скремблированного миметика, миметика miR-29a или антагомира miR-29a. (Е) Уровни коллагена 3 после добавления миметика/антагомира miR-29a и 100 нг rhIL-33. Для В-Е данные показаны в виде среднего значения ± стандартное отклонение для двойных образцов и соответствуют экспериментам с использованием пяти эксплантатов сухожилий. (n=5) p<0,05, **p<0,01 (F) активность люциферазы в первичной культуре человеческих теноцитов, трансфицированных с использованием предшественника miR-29a, содержащего 3'НТО Col 1a1, Col1a2 или Col 3a1. Активность определяли относительно контрольных клеток, трансфицированных с использованием скремблированной РНК, которая принималась за 100%. Проводили 3 независимых эксперимента. *p<0,05, **p<0,01 по отношению ко скремблированному контролю. (G) сайты связывания miR-29a и MRE в длинных/коротких формах транскриптов col3a1 и col1a1/col1a2, отличающихся альтернативными сайтами полиаденилирования. (Н) процентное соотношение длинных/коротких транскриптов коллагена в теноцитах (Т) после трансфекции с использованием miR-29a. (I) уровень мРНК col1a1, col1a2 и col3a1 после трансфекции с использованием скремблированного миметика и антагомира miR-29a. Данные показаны в виде среднего значения ± стандартное отклонение для двойных образцов и соответствуют экспериментам с использованием трех эксплантатов сухожилий. (n=3) p<0,05, **p<0,01.
Фигура 5. ИЛ-33/ST2 регулирует экспрессию miR-29 в процессе заживления сухожилия in vivo.
(А) Котрансфекция клеток НЕК 293 с использованием pre-miR-29a, содержащей 3'НТО растворимого ST2 вместе с регуляторными элементами микроРНК (MRE) 3'НТО растворимого ST2 и итоговый анализ активности люциферазы. *** p<0,001 по отношению ко скремблированному контролю (n=3) (В) и уровнями мРНК sST2 и мембраносвязанного ST2 после добавления скремблированного миметика, миметика miR-29a или антагомира miR-29a, (С) выработка белка sST2 человека (нг/мл) после инкубации с миметиком/антагомиром miR29a. (n=5) p<0,05, **p<0,01.
(D) Результаты количественной ПЦР, представленные в виде среднего кратного изменения ± стандартное отклонение уровня miR-29a у травмированных мышей дикого типа в сравнении с нетравмированными животными на 1-е и 3-й сутки после травмы. (Е) Результаты количественной ПЦР, представленные в виде среднего кратного изменения ± стандартное отклонение уровня miR-29a у травмированных мышей дикого типа и мышей линии ST2-/- в сравнении с нетравмированными животными после лечения rhIL-33 или PBS на 1-е сутки после травмы. (F) экспрессия miR-29a после введения антитела к ИЛ-33 у животных дикого типа после травмы на 1-е и 3-й сутки/Данные показаны в виде среднего кратного изменения ± стандартное отклонение для двойных образцов и соответствуют экспериментам с использованием четырех мышей на группу (n=16) p<0,05, **p<0,01.
Фигура 6. Роль ИЛ-33/miR-29 в патологии сухожилий.
Структурная схема, на которой проиллюстрирована роль ИЛ-33/miR-29a в патологии сухожилия. Травма сухожилия или часто повторяющиеся микроразрывы, вызывающие напряжение, которое испытывает сухожильная клетка, приводит к высвобождению ИЛ-33 и запуску нисходящего каскада фосфорилирования NFkB, что, в свою очередь, подавляет miR-29a, что приводит к увеличению выработки коллагена 3-го типа и растворимого ST2. Увеличение выработки коллагена 3-го типа снижает предел прочности сухожилия на растяжение, приводя к раннему повреждению, несмотря на аутокринное действие растворимого ST2, что в конечном итоге может быть защитным механизмом, посредством которого избыток ИЛ-33 удаляется из системы.
Фигура 7
(А) Фигура, на которой проиллюстрированы затравочные области двух сайтов MRE miR29a, предсказанных с помощью программы Targetscan: 29-1 и 29-2, (В) активность люциферазы в клетках НЕК 293, трансфицированных с использованием предшественника miR-29 a/b/c (pre-miR-29), содержащего 3'НТО Col 1 или Col 3. Активность определяли относительно контрольных клеток, трансфицированных с использованием скремблированной РНК, которая принималась за 100%. Проводили 3 независимых эксперимента. * p<0,05, **p<0,01 по отношению ко скремблированному контролю. (С) Котрансфекция клеток НЕК 293 с использованием pre-miR-29a,b.c, содержащих 3'НТО растворимого ST2, при которой показано, что miR-29a приводит к значительному снижению относительной активности люциферазы по сравнению с контрольными клетками, трансфицированными с использованием скремблированной РНК (n=3). (D) Оставшийся сайт связывания miR-29, содержащийся в коротком варианте 3'НТО col3a1, был исследован с использованием люциферазы на его чувствительность к miR-29a, и было установлено, что он в полной мере проявляет свою активность. (Е) Последовательности продуктов, полученных с помощью 3'RACE из транскриптов гена коллагена в теноцитах человека и лошади. Сигналы полиаденилирования подчеркнуты. MRE MHR29a показана курсивом в транскрипте Соl3а1 (короткая 3'НТО) человека и транскрипте Соl3а1 лошади.
Фигура 8
(А) уровни мРНК Соl3, (В) коллагена 3-го типа, (С) мРНК Col1 и (D) коллагена 1-го типа после воздействия миметика miR-29a после травмы сухожилия у мышей дикого типа. Данные по мРНК представляют собой общее число копий исследуемого гена по сравнению с геном домашнего хозяйства 18S в двойных образцах. Данные представлены в виде среднего значения ± стандартное отклонение для двойных образцов, которые соответствуют экспериментам с использованием 6 мышей на группу, *p<0,05, **p<0,01 по отношению к контролю (дисперсионный анализ (ANOVA)).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Материалы и способы
Модель тендинопатии на человеке
Все процедуры и протоколы были одобрены Комитетом по этике под №АСЕС 99/101. Пятнадцать образцов сухожилий надостной мышцы получали у пациентов с разрывами сухожилий мышц-вращателей плеча, подвергающихся операции на плечах (Таблица 1). Средний возраст пациентов с разрывами сухожилий мышц-вращателей плеча составлял 54 года (диапазон 35-70 лет) - средняя протяженность разрыва составляла 2,5 см. В той же самой группе пациентов получали образцы сухожилий подлопаточной мышцы. Пациентов включали в исследование только при условии отсутствия клинического проявления тендинопатии сухожилия подлопаточной мышцы на полученной в дооперационный период МРТ-сканограмме или макроскопического повреждения сухожилия подлопаточной мышцы на время артроскопии - по этим критериям была составлена когорта пациентов с доказанным наличием доклинической фазы тендинопатии. Была составлена независимая контрольная группа, состоящая из 10 образцов сухожилий подлопаточной мышцы, полученных у пациентов, подвергавшихся артроскопической операции для стабилизации плеча без разрывов сухожилий мышцы-вращателя плеча. Отсутствие разрывов сухожилий мышцы-вращателя плеча было подтверждено артроскопическим обследованием. Средний возраст пациентов контрольной группы составил 35 лет (диапазон 20-41 год).
Получение и подготовка тканей
Артроскопическое восстановление сухожилия мышцы-вращателя плеча осуществляли с использованием стандартного трехпортального метода, как было ранее описано. Площадь поперечного сечения полнослойного разрыва сухожилия мышцы-вращателя плеча оценивали и фиксировали, как описано ранее. Забор образца сухожилия подлопаточной мышцы был выполнен артроскопически от верхнего края сухожилия на 1 см латеральнее суставной губы лопатки. Забор образца сухожилия надостной мышцы был выполнен на расстоянии 1,5 см от края сухожилия до реконструктивного хирургического вмешательства. Для иммуногистохимического окрашивания образцы ткани немедленно фиксировали в 10% (об./об.) формалине в течение 4-6 часов, а затем заключали в парафин. Были получены срезы толщиной 5 мкм с использованием микротома Leica-LM (Leica Microsystems, Германия) и помещены на предметные стекла Superfrost Ultra Plus (Gerhard Menzel, Германия). Парафин удаляли из срезов ткани с использованием ксилола, производили обезвоживание с использованием батареи спиртов и использовали для гистологического и иммуногистохимического окрашивания по ранее разработанным методикам40.
Клетки сухожилия человека эксплантировали из ткани подколенного сухожилия 5 пациентов (возраст 18-30 лет), подвергавшихся операции по восстановлению передней крестообразной связки (ACL) аутотрансплантатом сухожилия хамстринг-мышц. Культуры поддерживали при 37°C во влажной атмосфере, содержащей 5% CO2 в течение 28 суток. Клетки субкультивировали и трипсинизировали по достижению субконфлуентного монослоя, при этом клетки из 3-го и 4-го пассажа использовали для культивирования в условиях нормоксии.
Гистологические и иммуногистохимические методы
Срезы человеческих тканей окрашивали гематоксилин-эозином, а также толуидиновым синим для определения степени тендинопатии согласно оценке по модифицированной версии шкалы Bonar41. (4 балла = выраженная тендинопатия, 3 балла = прогрессирующая тендинопатия, 2 балла = умеренные дегенеративные изменения, 1 балл = слабовыраженные дегенеративные изменения, 0 = нормальное сухожилие). Критерии оценки включали наличие или отсутствие отека и дистрофических изменений наряду со степенью клеточности стромы фибробластов и хондроидной метаплазии. Впоследствии срезы окрашивали антителами, направленными против следующих маркеров: - ИЛ-33 (Alexis, мышиное моноклональное), ST2 (Sigma Aldrich, кроличье поликлональное), ИЛ-1RaCP (ProSci, кроличье поликлональное) CD68 (пан-макрофаги), CD3 (Т-клетки), CD4 (Т-хелперы), CD206 (М2 макрофаги) и триптаза тучных клеток (тучные клетки) (Vector Labs).
Активность эндогенной пероксидазы подавляли 3% (об./об.) H2O2, а неспецифическое связывание антител блокировали 2,5% лошадиной сывороткой в буфере TBST в течение 30 минут. Демаскирование антигенов осуществляли в 0,01 М цитратном буфере в течение 20 минут в микроволновой печи. Срезы инкубировали с первичным антителом в 2,5% (вес/объем) лошадиной сыворотке крови/человеческой сыворотке крови/TBST при 4°C на протяжении ночи. После двух промывок препараты инкубировали с компонентами набора Vector ImmPRESS Reagent согласно инструкциям производителей в течение 30 минут. Препараты промывали и инкубировали с раствором хромогена Vector ImmPACT DAB в течение 2 минут с последующей продолжительной промывкой. В конечном итоге проводили контрастное окрашивание срезов гематоксилином. Также помимо хирургических препаратов для каждого отдельного метода окрашивания антителами в анализ были включены образцы положительного (ткань небных миндалин) и отрицательного контролей. Специфичность окрашивания подтверждалась путем применения изотипов отрицательного контроля вместо первичного антитела.
Авторы использовали систему балльной оценки на основании ранее описанных методов42 для количественной оценки иммуногистохимического окрашивания. Десять случайно выбранных полей зрения под большим увеличением (×400) были оценены тремя независимыми экспертами (NLM, JHR, ALC). В каждом поле зрения подсчитывали количество положительно и отрицательно окрашенных клеток, и процент положительных клеток рассчитывали, применяя следующую систему полуколичественной оценки: балл 0=полное отсутствие окрашенных клеток, балл 1=<10% положительно окрашенных клеток, балл 2=10-20% положительно окрашенных клеток, балл 3=>20% положительных клеток.
Срезы тканей мышей обрабатывали с использованием описанного выше протокола с помощью антител, направленных против следующих маркеров: - ИЛ-33 (R&D systems, мышиное моноклональное), ST2 (Sigma Aldrich, кроличье поликлональное), F4/80 (Serotec, мышиное моноклональное) и антитело к гистамину (Sigma Aldrich, кроличье поликлональное).
Регуляция синтеза матрикса
В теноцитах оценивали интенсивность иммуноцитохимического окрашивания на коллаген 1-го и 3-го типа для анализа выработки матрикса (Abeam). Общее содержание растворимого коллагена измеряли в культуральных супернатантах с использованием набора Sircol assay kit (Biocolor Ltd, Каррикфергус, Северная Ирландия) согласно протоколу производителя. К 100 мкл исследуемого образца добавляли 1 мл красителя Sircol и перемешивали в течение 30 мин при комнатной температуре. Комплекс коллаген-краситель осаждали путем центрифугирования при 10000×g в течение 10 мин; и затем промывали два раза с использованием 500 мкл этанола. Осадок растворяли в 500 мкл щелочного реагента. Абсорбцию измеряли при 540 нм с помощью микропланшетного анализатора. Калибровочную кривую строили на основе стандартного раствора коллагена, поставляемого производителем. В дополнение к этому оценивали концентрацию человеческого и мышиного коллагена 1-го и 3-го типа с использованием ИФА по изменению окраски раствора при 450 нм с помощью микропланшетного анализатора в сравнении со стандартными пробами, поставляемыми производителем (USCNK Life Science Inc).
Эксперименты по исследованию сигнальных путей
Статус фосфорилирования митоген-активируемых протеинкиназ (МАРК), киназ, регулируемых внеклеточными сигналами (ERK1/2), N-концевых киназ c-Jun (JNK) и изоформ р38 оценивали с использованием матрицы антител для анализа уровня фосфорилирования МАРК человека (R&D Systems Europe, Великобритания) согласно инструкциям производителя. Ингибитор ERK (FR180204) был приобретен у CalbioChem (Merck KGaA, Германия) и использован при IC50=10 мкМ, концентрации, которая была определена ранее для обеспечения оптимального специфического ингибирования в сравнении с побочными эффектами, обусловленными неспецифическим воздействием, что было ранее использовано в нашей лаборатории43.
Фосфорилирование NFKβ р65 оценивали с использованием набора InstantOne ELISA в клеточных лизатах в обработанных и необработанных теноцитах. Абсорбцию измеряли при 450 нм с использованием микропланшетного анализатора, причем положительные и отрицательные контроли были поставлены изготовителем. Для оценки содержания общего или фосфорилированного NFKβ р65 в каждом образце определяли относительную абсорбцию в стимулированных клетках в сравнении с нестимулированными клетками.
Экстракция РНК и количественная ПЦР
Клетки выделяли после содержания в условиях нормоксии и гипоксии до экстракции мРНК с помощью реагента Trizol. Мини-колонки QIAgen (Qiagen Ltd, Кроли, Великобритания) были использованы для очистки РНК с этапом, включающим расщепление ДНКазой на колонке, согласно инструкциям производителя. КДНК получали из образцов РНК в соответствии с набором AffinityScript™ (Agilent Technologies, Калифорния, США) multiple temperature cDNA synthesis kit согласно инструкциям производителя. ПЦР в режиме реального времени осуществляли с использованием SYBR green или Taqman FastMix (Applied Biosystems, Калифорния, США) в зависимости от того, был ли использован зонд с праймерами. КДНК разводили в соотношении 1 к 5 с использованием воды, не содержащей РНКаз. Каждый образец анализировали в трех повторностях. Были использованы следующие праймеры (Integrated DNA Technologies, Бельгия): GAPDH, 5' -TCG АСА GTC AGC CGC АТС ТТС ТТТ-3' (f) и 5'-АССAAAТССGTTGACТССGACСТТ-3' (r); ИЛ-33 человека GGAAGAАСАCAGСААGCAAAGССТ (f) ТААGGCCAGAGCGGAGCTТСАТАА (r); ИЛ-33 мыши GGAAGAАСАCAGСААGCAAAGССТ (f) ТААGGCCAGAGCGGAGCTТСАТАА (r); Общий ST2 человека АСАACTGGACAGСАССТСTTGAGT (f) АССTGCGTCСТСAGTCATСАСATT (r); sST2 мыши CCAATGТСССТТGTAGTCGG (f) CTTGTTСТСCCCGCAGTC (r), TCCCCATCTССТСАСCTCССТТААT (зонд); ST2L мыши TCTGCTATTCTGGATACTGCTTTCTCTGTGGAGTACTTTGTTСАСС (r) AGAGACCTGTTACCTGGGCAAGATG (зонд); ST2L человека АСАAAGTGCTCTАСАCGACTG (f) TGTTCTGGATTGAGGCCAС (r); CCCCATCTGTACTGGATTTGTAGTTCCG (зонд); sST2 человека GAGACCTGCCACGATTACAC (f) TGTTAAACCCTGAGTTCCCAC (r), CCCCACACCCCTАТСCTTTCTCCT (зонд); Col 3А человека TTGGCAACGАСАCAGAAACTG (f) TTGAGTGCAGGGTCAGCAСТАCTT (r) Col 3А мыши GCTTTGTGCAAAGTG GAACCTGG (f) CAAGGTGGCTGCАТСCCAATTCAT (r); COL 1A1 человека CCATGCCCTTTCTGCТССTTT (f) CACTTGGGTGTTTGAGCATTGCCT (r) COL 1A1 мыши TTCTCCTGGCAAAGACGGACTCAA (f) GGAAGCTGAAGTCATAACCGCCA (r)
Выделение РНК и количественный ПНР-анализ микроРНК в режиме реального времени
Общую РНК выделяли с использованием набора miRNeasy (Qiagen). Для получения кДНК был использован набор miScript Reverse Transcription Kit (Qiagen). Для полуколичественного определения экспрессии miR-29a (MS (MS00001701), 29b (MS00006566) и с (MS00009303) человека, а также 29а (MS00003262), 29b (MS00005936) и с (MS00001379) мыши были использованы анализы TaqMan mRNA assays (Applied Biosystems) или miScript primer assay (Qiagen). Уровни экспрессии малой ядерной РНК U6B или бета-актина были использованы в качестве эндогенных контролей.
Количественная оценка содержания альтернативных полиаденилированных транскриптов генов коллагена
Абсолютные уровни длинных и коротких форм 3'НТО транскриптов генов коллагена 1-го и 3-го типов были определены с помощью количественной ПЦР по сравнению со стандартами. КДНК была синтезирована с использованием набора AffinityScript (Agilent) и как случайных гексамерных праймеров, так и праймеров oligo-dT. Количественную ПЦР на основе SYBR green осуществляли с использованием следующих праймеров: Образцы были нормализованы по отношению к эндогенному контролю GAPDH.
Col1a2_S FW 5'GCCTGCCCTTCCTTGATATT3'
Col1a2_S REV 5'TGAAACAGACTGGGCCAATG3'
col1a2_L FW 5'TCAGATACTTGAAGAATGTTGATGG3'
col1a2_L REV 5'CACCACACGATACAACTCAATAC3'
Col1a1_S FW 5'CTTCACCTACAGCGTCACT3'
Col1a1_S REV 5'TTGTATTCAATCACTGTCTTGCC3'
col1a1_L FW 5'CCACGACAAAGCAGAAACATC3'
col1a1_L REV 5'GCAACACAGTTACACAAGGAAC3'
COL3A1_S FW 5'CTATGACATTGGTGGTCCTGAT3'
COL3A1_S REV 5'TGGGATTTCAGATAGAGTTTGGT3'
COL3A1_L FW 5'CCACCAAATACAATTCAAATGC3'
COL3A1_L REV 5'GATGGGCTAGGATTCAAAGA3'
Быстрая амплификация 3'-концов кДНК (RACE)
Для характеристики человеческих последовательностей осуществляли 3' RACE на кДНК, которая была синтезирована из общей РНК, выделенной из теноцитов человека, с использованием набора MiRscript II reverse transcriptase kit (Qiagen). Концевые участки кДНК были амплифицированы с помощью ПЦР с использованием следующих ген-специфичных прямых праймеров, приведенных ниже, наряду с универсальным обратным праймером из указанного набора.
Прямые ген-специфичные праймеры для 3'RACE человеческих последовательностей
RACE-Col1a1-L FW 5'GACAACTTCCCAAAGCACAAAG3'
RACE-Col1a1-S FW 5'CTTCCTGTAAACTCCCTCCATC3'
RACE-Col1a2-L FW 5'TCTTCTTCCATGGTTCCACAG3'
RACE-Col1a2-S FW 5'CCTTCCTTGATATTGCACCTTTG3'
RACE-Col3a1-L FW 5'CTATGACATTGGTGGTCCTGAT3'
RACE-Col3a1-S FW 5'GTGTGACAAAAGCAGCCCCATA3'
Для характеристики последовательностей лошади транскрипты 3'НТО Col1a1, Col1a2 и Соl3а1, экспрессированные в теноцитах лошади, были амплифицированы с использованием быстрой амплификации 3'-концов кДНК (3'RACE). Амплифицированные фрагменты кДНК секвенировали и определяли расположение поли-А-сигнала в соответствии с положением канонического поли-А-сигнала ААТААА на расстоянии 10-30 нуклеотидов в 5'-направлении от указанного поли-А-хвоста.
Праймеры для 3' RACE последовательностей ДНК лошади
col1a1 GSP1 CCCTGGAAACAGACAAACAAC лошади
col1a1 GSP2 CAGACAAACAACCCAAACTGAA лошади
coll1a2 GSP1 GCTGACCAAGAATTCGGTTTG лошади
cola2 GSP2 ACATTGGCCCAGTCTGTTT лошади
col3a1 GSP1 AGGCCGTGAGACTACCTATT лошади
col3a1 GSP2 CTATGATGTTGGTGGTCCTGAT лошади
col1a1 q-PCR fw CAGACTGGCAACCTCAAGAA лошади
col1a1 q-PCR rev TAGGTGACGCTGTAGGTGAA лошади
col1a2 q-PCR fw GGCAACAGCAGGTTCACTTAT лошади
col1a2 q-PCR Rev GCAGGCGAGATGGCTTATTT лошади
col3a1 q-PCR fw CTGGAGGATGGTTGCACTAAA лошади
col3a1 q-PCR rev CACCAACATCATAGGGAGCAATA лошади
Полученные в результате ПЦР-продукты клонировали в pCR2.1 ТОРО (Invitrogen) и секвенировали.
Трансфещия микроРНК
Клетки трансфицировали синтетической зрелой микроРНК в случае miR 29 а и b или отрицательным контролем (миметик miR-67 С. elegans, меченный Dy547, Thermo Scientific Inc) в конечной концентрации 20 нМ с применением трансфекционных реактивов Dharmacon® DharmaFECT® 3 siRNA (Thermo Scientific Inc). Через 48 часов после трансфекции собирали клеточные лизаты для анализа экспрессии представляющих интерес генов.
Эффективность трансфекции оценивали с помощью проточной цитометрии с использованием миметика, меченного Dy547, и подтверждали с помощью количественной ПЦР с использованием контрольного скремблированного миметика и соответствующего миметика семейства miR29.
Анализ направленной экспрессии генов коллагена 1-го и 3-го типа, а также ST2, с помощью репортерного гена люциферазы
Сайт-мишень 2 микроРНК человека был создан путем отжига олигонуклеотидов: для COL 1 и 3, а также 3'НТО растворимого ST2, которые были клонированы как в смысловой, так и в антисмысловой ориентациях ниже от гена люциферазы в вектор pMIR-REPORT (Ambion). Эти конструкции секвенировали для подтверждения вставок и назвали pMIR-COL I/COL III/ sST2-miR29a/b/c и pMIR(A/S)-COL I/COL III/ sST2-miR29a/b/c, и использовали для трансфекции клеток НЕК293. Клетки НЕК293 культивировали в 96-луночных планшетах и трансфицировали 0,1 мкг либо pMIR-COL I/COL III/ sST2-miR29a/b/c, pMIR(A/S)- COL I/COL III/ sST2-miR29a/b/c, либо pMIR-REPORT вместе с 0,01 мкг вектора pRL-TK (Promega), содержащего люциферазу Renilla и 40 нМ miR-155 или скремблированной микроРНК (Thermo Scientific Dharmacon®). Транспекции были выполнены с использованием Effectene (Qiagen) согласно инструкциям производителя. Через двадцать четыре часа после трансфекции измеряли активность люциферазы с использованием набора Dual-Luciferase Reporter Assay (Promega). 3'НТО гена sST2 человека амплифицировали из геномной ДНК с использованием следующих праймеров: прямого праймера sST2 5'AGTTTAAACTGGCTTGAGAAGGCACACCGT3' и обратного праймера sST2 5'AGTCGACGGGCCAAGAAAGGCTCCCTGG3', что привело к образованию сайтов Pmel и SalI соответственно. Эти сайты были использованы для клонирования ПЦР-амплифицированного продукта в те же самые сайты pmiRGLO (Promega). Затравочные области двух сайтов MRE miR29a, предсказанных с помощью программы Targetscan: 29-1 и 29-2 подвергали мутагенезу с использованием набора реагентов для направленного мутагенеза QuickChange (Agilent). Каждый вектор наряду с miR29a или скремблированным контрольным миметиком трансфицировали в клетки НЕK293 с использованием Attactene (Qiagen) согласно инструкциям производителя. Через 24 часа измеряли люциферазную активность с использованием набора для анализа гена двойной люциферазы Dual-Glo (Promega), причем люциферазную активность нормализовали к Renilla. Нормализованная люциферазная активность была выражена в виде процента от скремблированного контроля для тех же самых конструкций.
Выработка цитокинов
С помощью набора 25-Рlех human cytokine assay проводили количественное определение in vitro 25 отдельных цитокинов человека с использованием технологии Luminex. Супернатанты (n=3)
Модель травмы сухожилия надколенника
При приготовлении к оперативному вмешательству мышам проводили анестезию с использованием смеси изофлурана (3%) и кислорода (1%), и обе задние конечности выбривали. На протяжении оперативного вмешательства анестетик подавали через ингаляционную маску, причем уровень изофлурана уменьшали до 1% с подачей кислорода. После нанесения кожного разреза были произведены два разреза параллельно сухожилию в удерживателе на каждой стороне, затем набор плоских ножниц был помещен под сухожилие надколенника. Используя лезвия ножниц в качестве подложки, для произведения полнослойного частичного поперечного разреза в правом сухожилии надколенника была использована игла для биопсии кожи диаметром 0,75 мм (World Precision Instruments). Левое сухожилие надколенника подвергалось имитации оперативного вмешательства, которое заключалось только в помещении пластмассовой подкладки под сухожилие без нанесения травмы. Раневые дефекты кожного покрова закрывали кожными скобами, и мышей умерщвляли на 1-е сутки, 3-и сутки и 7-е, а также 21-е сутки после оперативного вмешательства. Мышей умерщвляли путем ингаляции CO2 и непосредственно сразу взвешивали. Были использованы мыши из двух групп: контрольные мыши линии BALB/c (CTL) и мыши линии ST2-/- BALB/c. Каждая группа состояла из 16 мышей (n=8 ST2-/- BALB/c и 8 BALB/c) на одну временную точку. Эти эксперименты повторяли 4 раза отдельно.
Для исследования того, вызывал ли ИЛ-33 нарушение синтеза матрикса сухожилия, была разработана модель на основе инъекции цитокинов. ИЛ-33 был исследован в модели, о которой сообщалось ранее, что изначально была описана для применения ИЛ-23 или ИЛ-2244-45. Мышам линии ST2-/- (n=4/группа/воздействие/эксперимент) ежедневно интраперитонеально вводили ИЛ-33 (0,2 мкг на мышь, разведенный в 100 мкл PBS) на дни -3, -2, -1, а также в день нанесения травмы. Через 24 часа после последней инъекции мышей выводили из эксперимента в соответствии с протоколом. Контрольные мыши аналогичным образом получали одинаковый объем PBS. Авторы также исследовали нейтрализующие антитела к ИЛ-33 (0,5 мкг/мл, R&D systems) путем интраперитонеальной инъекции непосредственно сразу после нанесения травмы мышам дикого типа и мышам линии ST2-/-, причем в качестве контролей использовали IgG снова в количестве 4/группа/воздействие/эксперимент.
Биомеханический анализ
Для проведения биомеханического анализа наносили травму сухожилия надколенника мышей из каждой группы, и восемь мышей умерщвляли в одну из трех временных точек для исследования механических свойств, как было описано ранее Lin et al.10. Вкратце, иссекали и выделяли сухожилие надколенника, оставляя только надколенник, сухожилие надколенника и большеберцовую кость как цельный фрагмент. Затем определяли ширину и толщину сухожилия, а площадь поперечного сечения рассчитывали как произведение значений двух указанных параметров. Большеберцовую кость заключали в изопон р38 (целлюлозный наполнитель высокой плотности) в специально изготовленном приспособлении для фиксации, и ее положение фиксировали металлическим зажимом. Положение надколенника фиксировали с помощью держателей с использованием испытательного стенда BOSE ElectroForce® 3200. Каждый образец сухожилия подвергали следующему воздействию с погружением в водяную баню, содержащую физраствор, при 37°C использовали усилие 0,02 Н, предварительную нагрузку на протяжении 10 циклов с усилием от 0,02 Н до 0,04 Н со скоростью 0,1%/с (0,003 мм/с) и выдерживали в течение 10 с. Непосредственно после этого проводили эксперимент по определению релаксации напряжения путем растягивания сухожилия до достижения показателя деформации 5% (0,015 мм) со скоростью 25% (0,75 мм/с) с последующей релаксацией в течение 600 с. В конечном итоге использовали ускоренное испытание с линейным возрастанием нагрузки со скоростью 0,1%/с (0,003 мм/с). Из этих исследований определяли максимальное напряжение, и рассчитывали модуль Юнга с использованием линейной регрессии из области кривой напряжение-деформация, близкой к линейной зависимости.
Применение миметика miR29a in vivo
Был получен трансфекционный комплекс, содержащий 150 нг/мл миметика miR-29а, 9 мкг/мл полиэтиленимина (ПЭИ) и 5% глюкозы. 50 мкл этого комплекса вводили мышам в сухожилие надколенника непосредственно после оперативного вмешательства. Животных умерщвляли через 1 и 3 суток, и измеряли уровни мРНК col1a1 и col3a1 и белка. Для оценки in vivo распределения миметика miR-29a в сухожилии с помощью иммунофлуоресценции был использован флуоресцентно-меченный миметик miR-29a с использованием контрастного окрашивания на фаллоидин (для визуализации структуры цитоскелета и ядер (DAPI).
Структура миметика miR29a представлена ниже:
Пассажирская цепь:
mAmCrCmGrAmUrUmUrCmArGmArUmGrGmUrGmCrUmAdG
Направляющая цепь:
/5Phos/rUrArCrCrArCrCrArUrCrUrGrArArArUrCrGrGmUmUmA
/5Phos/ = 5'фосфат
mA = 2'O-метил аденозин рибонуклеотид;
mC = 2'O-метил цитозин рибонуклеотид;
mG = 2'O-метил гуанин рибонуклеотид;
mU = 2'O-метил урацил рибонуклеотид;
rА = аденозин рибонуклеотид;
rС = цитозин рибонуклеотид;
rG = гуанин рибонуклеотид;
rU = урацил рибонуклеотид;
Статистический анализ
Все результаты представлены в виде среднего +/- стандартная ошибка среднего (SEM), а весь статистический анализ осуществляли либо с использованием t-критерия Стьюдента, дисперсионного анализа либо критерия Манна-Уитни, как указано в подписях к фигурам, с использованием программного обеспечения Graph Pad Prism 5. Р-значение <0,05 считалось статистически значимым.
Результаты
Экспрессия ИЛ-33 и ST2 при человеческой тендинопатии
Авторы сначала исследовали экспрессию ИЛ-33 при человеческой тендинопатии с использованием ранее разработанной модели 22. Экспрессия ИЛ-33, растворимых и мембраносвязанных транскриптов ST2, значительно повышалась при ранней тендинопатии по сравнению с контролем или биоптатами разорванных сухожилий (Фиг. 1А-С). Окрашивание тканей на ранней стадии тендинопатии на ИЛ-33 и ST2 было значительно более интенсивным по сравнению с разорванным сухожилием или контрольными биоптатами (Фиг. 1D). Выраженное окрашивание отмечалось в эндотелиальных клетках и в особенности в фибробластоподобных клетках, а именно теноцитах, которые, как считается, играют решающую роль в регуляции ранней стадии тендинопатии (данные не показаны). Параллельно с этим, культивированные in vitro теноциты экспрессировали ИЛ-33, причем наблюдалась активация его синтеза как на уровне мРНК, так и на уровне белка после стимуляции ФНО и ИЛ-1β (Фиг. 1Е и данные не показаны). В отличие от этого ST2 конститутивно экспрессировался как в покоящихся, так и нестимулированных теноцитах (данные не показаны).
ИЛ-33 принимает участие в регуляции синтеза коллагенового матрикса и провоспалительных цитокинов.
Нарушение регуляции синтеза матрикса в сторону увеличения экспрессии коллагена 3 является ключевым ранним фенотипическим изменением в тендинопатии, вследствие чего ускоряется заживление; однако коллаген 3-го типа неоптимален в плане биомеханических свойств. ИЛ-33 вызывал активацию синтеза белка общего коллагена в зависимости от дозы и времени (данные не показаны), что оценивалось по повышению экспрессии мРНК и белка коллагена 1-го типа, но в особенности 3-го типа (Фиг. 1F, G). После анализа на микроматрицах (данные не показаны) и в соответствии с описанным нисходящим сигнальным путем, активируемым ИЛ-3312,16, этот эффект отменялся посредством подавления активности ERK (данные не показаны). rhIL-33 также вызывал значительное увеличение продукции ИЛ-6, ИЛ-8 и МСР-1 (данные не показаны), которая регулировалась подавлением активности NF-kB, свидетельствуя о том, что ИЛ-33 функционирует в теноцитах через его канонический сигнальный путь, опосредованный ИЛ-1R (данные не показаны). В отличие от этого авторы не обнаружили никакого влияния на продукцию других цитокинов в соответствии с ранее описанными профилями цитокинов в фибробластах, обусловленными ИЛ-3320-23.
Моделирование пути ИЛ-33/ST2 in vivo после травмы сухожилия
Авторы распространили эти наблюдения на широко используемую модель травмы сухожилия in vivo. Уровни мРНК ИЛ-33 были повышены на 1-е и 3-й сутки после травмы сухожилия у мышей дикого типа (Фиг. 2А). Они были значительно меньшими у травмированных мышей линии ST2-/-, что свидетельствует об аутокринной регуляции. Синтез растворимого ST2 был значительно активирован во всех временных точках после травмы у мышей дикого типа по сравнению с нетравмированными контрольными животными (Фиг. 2В), тогда как уровень мРНК мембранного ST2 повышался только к 3-м суткам после травмы (данные не показаны). Не было выявлено никаких значительных изменений в экспрессии транскрипта или белка ИЛ-33 или ST2 у мышей дикого типа на 7-е или 21-е сутки после травмы, или экспрессии ИЛ-33 у мышей линии ST2-/-, свидетельствуя о том, что влияние экспрессии ИЛ-33 проявляется рано в соответствии с активностью по типу «алармина» при травме/заживлении сухожилий.
Анализ синтеза коллагена выявил значительно более высокую экспрессию коллагена 3-го типа во всех временных точках после травмы у мышей дикого типа по сравнению с нетравмированными контрольными животными или травмированными мышами линии ST2-/- (Фиг. 2Е, F, и данные не показаны). Экспрессия коллагена 1-го типа изначально была сниженной (1-е, 3-й сутки) на уровнях мРНК (Фиг. 2С) у травмированных мышей дикого типа, но к 7-м и 21-м суткам возвращалась к уровням, наблюдаемым до нанесения травмы (данные не показаны), со схожей тенденцией в экспрессии коллагена 1-го типа на уровне белка (Фиг. 2D). В отличие от этого у травмированных мышей линии ST2-/- отмечалось продолжительное уменьшение синтеза коллагена 1-го типа (1-е, 3-и и 7-е сутки) с возвращением к исходному уровню только к 21-м суткам (данные не показаны). Важно отметить, что травма сухожилий у мышей дикого типа приводила к значительному уменьшению биомеханической прочности на 1-е сутки после травмы по сравнению с ST2-/- (Фиг. 2G), которая восстанавливалась к 7-м и 21-м суткам (данные не показаны). Эти данные свидетельствуют об изменении синтеза коллагенового матрикса у мышей линии ST2-/-, что подразумевает роль ИЛ-33/ST2 в качестве раннего модулятора изменений типов коллагена при травме сухожилий, которые имеют биомеханическое значение.
Воздействие на ИЛ-33 вызывает изменения коллагена 3-го типа in vivo
Для подтверждения этой возможности авторы попытались непосредственно модифицировать эффекторные функции ИЛ-33 in vivo. Введение rhIL-33 не влияло на синтез коллагена 1 -го типа (Фиг. 3А, В), но, тем не менее, приводило к значительному увеличению синтеза коллагена 3-го типа, в особенности в травмированных сухожилиях (Фиг. 3D, Е, и данные не показаны). Более того, введение rhIL-33 приводило к значительному уменьшению предельной прочности сухожилий во всех временных точках после инъекции мышам дикого типа (Фиг. 3Е, и данные не показаны), свидетельствуя о том, что такие изменения оказывали функциональное воздействие. Введение ИЛ-33 не влияло на синтез коллагенового матрикса или предельной прочности восстанавливаемого сухожилия у мышей линии ST2-/-, что послужило подтверждением того, что ИЛ-33 оказывает действие через SТ2-зависимый сигнальный путь (данные не показаны).
Далее авторы воздействовали непосредственно на уровень ИЛ-33 in vivo. Нейтрализующие антитела к ИЛ-33 подавляли переключение синтеза коллагена с 1-го типа на 3-й тип на 1-е и 3-и сутки после травмы у мышей дикого типа (Фиг. 3F-I), что приводило к значительному увеличению биомеханической прочности сухожилий мышей дикого типа на 1-е сутки после травмы (Фиг. 3J). Этот эффект не отмечался на более поздних временных точках (данные не показаны). В контрольных экспериментах авторы не наблюдали никакого эффекта на мышей линии ST2-/- (данные не показаны), что послужило дополнительным подтверждением вклада эндогенного ИЛ-33 в развитие тендинопатии, обусловленной травмой.
ИЛ-33 способствует дифференциальной регуляции синтеза коллагена 1/3 в теноцитах через miR-29.
Установив, что ИЛ-33 определяет дифференциальную регуляцию синтеза коллагена 1-го и 3-го типа в теноцитах, авторы выдвинули предположение о том, что сеть взаимодействий микроРНК является механизмом, опосредующим эту дифференциальную регуляцию. Предыдущие исследования показали, что представители семейства miR-29 непосредственно взаимодействуют с многочисленными генами белков внеклеточного матрикса, включая гены коллагена 1-го и 3-го типа 24-25, и вовлечены в регуляцию врожденного и приобретенного иммунитета26. Вычислительные алгоритмы позволяют предсказать, что мишенью miR-29 также может являться sST2. Авторы обнаружили, что все представители семейства miR-29 были экспрессированы в биоптатах сухожилия человека и эксплантированных теноцитах (Фиг. 4А), причем экспрессия miR-29a изменялась в наибольшей степени. В культуре теноцитов ИЛ-33 вызывал значительное уменьшение экспрессии miR-29a через 6, 12 и 24 часа (Фиг. 4В), действуя через NFκB-зависимый сигнальный путь, при этом авторы наблюдали противоречивые результаты влияния на miR-29b и с (данные не показаны). Поскольку miR-29a может регулировать опосредованные ИЛ-33 изменения в содержании коллагена 3-го типа в матриксе, авторы анализировали функциональное влияние применения miR-29a на синтез коллагенового матрикса in vitro. Прежде всего с использованием люциферазной реакции авторы подтвердили, что miR-29a непосредственно взаимодействует с col 1a1 и 3a1, как было ранее продемонстрировано27 (Фиг. 7В). Авторами также было отмечено неизученное ранее взаимодействие с транскриптом субъединицы col 1а2 (Фиг. 7). Для изучения того, действительно ли miR-29а регулирует уровни потенциальных мРНК-мишеней в клетках, активация которых происходит в результате заболевания, авторы трансфицировали теноциты с использованием миметика и антагомира miR-29a. Применение miR-29a оказывает избирательное влияние на синтез мРНК и белка коллагена 3-го типа, но не коллагена 1-го типа в первичной культуре теноцитов (Фиг. 4С, D). Более того, сверхэкспрессия miR-29a в значительной степени подавляла индуцированный ИЛ-33 синтез коллагена 3-го типа на уровне мРНК и белка (Фиг. 4Е). В дополнение к этому ингибирование miR-29а приводило к значительному увеличению экспрессии col 3a1, что свидетельствует о том, что miR-29a не только принимает участие в активной регуляции образования этих транскриптов в теноцитах человека, но также ее потеря является важным фактором увеличения выработки коллагена 3-го типа, что отмечается при тендинопатии. В отличие от этого уровни транскрипта col 1a1 не изменялись (Фиг. 41).
Учитывая то, что miR-29a была способна подавлять экспрессию col 1a1 и 1а2 с равной или большей эффективностью, чем коллагена 3-го типа в анализе репортерного гена люциферазы, маловероятно, что это является результатом того, что miR-29a обладает большей аффинностью по отношению к ее MRE в транскриптах коллагена 3-го типа (Фиг. 4F). Одним из документально подтвержденных механизмов, влияющих на чувствительность транскриптов к регуляции микроРНК является использование альтернативных сигналов полиаденилирования (Фиг. 4G). Чтобы это исследовать, авторы сравнили уровни полноразмерных (содержащих miR-29a) транскриптов с общими уровнями с использованием количественной ПЦР (Фиг. 4Н), что показывает, что менее чем в 5% транскриптов col 1a1 и 1а2 используют дистальный сигнал полиаденилирования, тогда как в большинстве транскриптов col 3a1 это действительно происходит.
Этот факт был подтвержден быстрой амплификацией 3'-концов кДНК (RACE) (Фиг. 7Е), подтверждающей, что как в col 1a1, так и 1а2, но не col 3a1 используются ранее неописанные сигналы полиаденилирования (Фиг. 4G). В полученной в результате укороченной 3'НТО отсутствуют MRE miR29a. (Следует понимать, что последовательности, проиллюстрированные на Фиг. 7Е, представляют собой последовательности кДНК; соответствующие последовательности мРНК, разумеется, содержат U вместо Т). Эти данные указывают на то, что в теноцитах miR-29a специфически регулирует экспрессию col 3a1, тогда как и col 1a1, и col 1а2 оказываются нечувствительными к подавлению экспрессии miR-29a вследствие использования альтернативных сигналов полиаденилирования. На это использование альтернативных сигналов полиаденилирования не влияло присутствие ИЛ-33 (данные не показаны). Потеря miR-29a при активации сигнального пути, опосредованного ИЛ-33, приводит к угнетению синтеза коллагена 3-го типа, что, вероятно, вносит вклад в увеличение синтеза этого типа коллагена, которое наблюдается в травмированных сухожилиях.
Результаты по 3'RACE для теноцитов человека выявили две НТО col 3a1, более короткая из которых [обозначенная Соl3а1 (короткая 3'НТО) на Фиг. 7Е] содержит одну MRE miR-29a, тогда как более длинная содержит две. Обе регулируются с помощью miR-29a, как проиллюстрировано на Фиг. 7D.
Характеристика 3'НТО транскриптов Col1a1, Cola2 и Соl3а1, экспрессированных в теноцитах лошади, показала, что в них используются одинаковые консервативные сигналы полиА, которые используются в ортологических транскриптах коллагена, экспрессированных в теноцитах человека. В генах col1a1 и cola2 применение этих проксимальных сигналов полиА приводит к образованию транскриптов с 3'НТО, которые имеют длину от 100 до 350 нуклеотидов и не содержат сайты связывания miR-29, и, таким образом, являются нечувствительными к регуляции этой микроРНК. В отличие от этого оба 3'НТО гена col3a1 содержат сайты связывания miR-29, что делает их чувствительными к регуляции miR-29.
Растворимый ST2 является прямой мишенью miR-29.
Вычислительный анализ выявил, что растворимый ST2 может быть мишенью miR-29a, свидетельствуя о его возможной регуляторной роли в реализации эффекторных функций ИЛ-33. Был сконструирован репортерный ген люциферазы, который содержит 3'НТО гена sST2 человека, что, согласно предсказаниям, содержит два потенциальных сайта связывания miR-29abc. Котрансфекция репортерной плазмиды sST2-люцифераза с миметиком miR-29 приводила к значительному уменьшению люциферазной активности по сравнению со скремблированным контролем (Фиг. 7В). Кроме того, люциферазная активность полностью восстанавливалась, когда были введены мутации в затравочные области как MRE miR-29, так и sST2, что убедительно демонстрирует, что sST2 является прямой мишенью miR-29a (Фиг. 5А). Для изучения того, действительно ли miR-29a регулирует уровни целевой мРНК-кандидата в теноцитах, авторы снова трансфицировали миметик и антагомир miR-29a в теноциты человека. Уровень растворимого ST2 статистически значимо (p<0,01) изменялся под воздействием трансфекции с использованием миметика/антагомира miR-29a в около 5 раз (Фиг. 5В) с соответствующим значительным изменением количества растворимого белка ST2, что подтверждает то, что miR29a является мишенью для растворимого ST2 (Фиг. 5С).
ИЛ-33/sST2 принимает участие в регуляции экспрессии miR-29 в моделях заживления сухожилия in vivo
В конечном итоге авторы исследовали экспрессию miR-29a в разработанной ими модели тендинопатии in vivo. Травма сухожилий у мышей дикого типа приводила к 22-кратному уменьшению экспрессии miR29a на 1-е сутки, причем к 3-м суткам уровни экспрессии уменьшались 6-кратно (по сравнению с исходным уровнем) (Фиг. 5D, и данные не показаны) с отсутствием статистически значимых отличий на 7-е сутки. Этот эффект в значительной степени подавлялся у мышей линии ST2-/- (данные не показаны). В дополнение к этому, введение экзогенного rh-ИЛ-33 приводило к уменьшению экспрессии miR-29a в травмированных сухожилиях во все временные точки по сравнению с контрольными животными, которым вводили PBS (данные не показаны). Это влияние было наиболее выражено у травмированных мышей дикого типа, причем добавление rhIL-33 опосредовало дополнительное десятикратное уменьшение экспрессии miR-29a (Фиг. 5Е). Добавление rhIL-33 не оказывало никакого значимого влияния на экспрессию miR-29a в травмированных или нетравмированных сухожилиях у мышей линии ST2-/-, и в этом случае свидетельствуя о том, что снижение экспрессии miR-29a частично прямо опосредовано передачей сигнала через ИЛ-33/ST2. Добавление нейтрализующего антитела к ИЛ-33 привело к значительному уменьшению влияния травмы на экспрессию гена miR-29a на 1-е и 3-и сутки после нанесения травмы (Фиг. 5F).
Применение миметика miR29a in vivo в модели травмы сухожилия надколенника
Миметик miR-29a был введен в теноциты сухожилий надколенника мышей дикого типа посредством прямой инъекции комплекса miR-29a/PEI. Иммунофлуоресцентное окрашивание на миметик (красный цвет) с контрастным окрашиванием фаллоидином (зеленый цвет, структура цитоскелета) и DAPI (для визуализации ядер) было использовано для визуализации локализации миметика вокруг теноцитов через 24 ч после инъекции миметика miR-29a (не показано). Как проиллюстрировано на Фигуре 8, уровни мРНК коллагена 3 и белка значительно уменьшались в сухожилиях мышей, которым вводили миметик miR-29a, по сравнению с контролями. В отличие от этого уровни коллагена 1 не изменялись.
Обсуждение
МикроРНК были открыты как мощные регуляторы разнообразных клеточных процессов, причем им отводится важная роль в патогенезе заболеваний и ремоделировании тканей. В этих исследованиях, где в качестве модельной системы была использована модель тендинопатии, впервые была выявлена способность одной микроРНК (miR-29) к перекрестной регуляции эффекторной функции провоспалительных цитокинов и регуляции синтеза внеклеточного матрикса в сложных ранних биологических процессах, что приводит к восстановлению тканей.
В данном документе авторы представили новые данные о роли ИЛ-33 на первоначальных стадиях, что привело к выявлению важной нозологической формы тендинопатии. В последнее время ИЛ-33 во все большей степени стал ассоциироваться с патологиями опорно-двигательной системы16. Данные, полученные авторами, показывают экспрессию ИЛ-33 в биоптатах сухожилия человека, полученных на ранней стадии заболевания, тогда как в биоптатах, полученных на конечной стадии заболевания, экспрессия ИЛ-33 выражена в значительно меньшей степени на уровне матричной РНК и белка, что служит подтверждением концепции о роли ИЛ-33 в качестве раннего тканевого медиатора при травме сухожилия и последующего ремоделирования тканей. При повреждении клеток эндогенные сигналы повышенной опасности, так называемые молекулярные паттерны, связанные с повреждениями, высвобождаются некротическими клетками, включая белки теплового шока28, HMGB129, мочевую кислоту30 и представителей семейства ИЛ-131-32, включая ИЛ-3333-34. Эти сигналы повышенной опасности впоследствии распознаются различными иммунокомпетентными клетками, которые запускают воспалительную и регенерационную реакцию. Данные, полученные авторами, свидетельствуют о роли ИЛ-33 в качестве алармина на ранней стадии тендинопатии, и важно отметить, что данные биомеханических анализов предполагают, что такая экспрессия имеет отношение к патогенезу данного заболевания. Добавление rhIL-33 приводило к значительному уменьшению нагрузки на разрыв у мышей дикого типа на около 30% в ранних временных точках, вероятно, вследствие сопутствующих изменений матрикса в синтезе коллагена 3-го типа, которые приводят к ухудшению биомеханических свойств сухожилия35. Следовательно, один из возможных механизмов событий, опосредующих раннее восстановление сухожилия с низкими биомеханическими свойствами, может заключаться в повышении синтеза ИЛ-33 при повторных микротравмах с последующим высвобождением вследствие механического напряжения/некроза, что, в свою очередь, вызывает дегенерацию матрикса и выработку провоспалительных цитокинов, приводя к развитию патологического процесса в сухожилии, аналогичного тому, что отмечается в биоптатах сухожилий с ранней тендинопатией. Интересно заметить, что введение нейтрализующих антител травмированным мышам все же приводило к возвращению фенотипа, ассоциированного с экспрессией коллагена 3-го типа, но это лишь обеспечивало временное повышение прочности сухожилий на 1-е сутки после травмы. Несмотря на возможность нивелирования блокировки ИЛ-33 при более долговременных спортивных травмах, часто повторяющиеся микротравмы, ассоциированные с патологическими изменениями в сухожилии, могут наоборот привести к тому, что нейтрализация ИЛ-33 будет действовать в качестве мартингала для дальнейшего нежелательного нарушения регуляции синтеза матрикса.
В новых исследованиях освещается роль микроРНК в качестве ключевых регуляторов функции лейкоцитов и цитокиновой сети помимо регуляции пролиферации дифференцировки линий стромальных клеток, которые определяют состав внеклеточного матрикса36. Новые экспериментальные данные, касающиеся роли miR-29а в регуляции эффектов, опосредованных «алармином» ИЛ-33, дают представление о механизме сетевых взаимодействий микроРНК, основанных на перекрестной регуляции, включающих регуляцию процесса воспаления и синтеза матрикса при восстановлении тканей. Данные, полученные авторами, являются убедительным доказательством функциональной роли miR-29 в качестве посттранскрипционного регулятора экспрессии коллагена при травме сухожилий мыши или человека. Регуляция экспрессии подтипов коллагена представителями семейства miR-29 была освещена в нескольких предыдущих исследованиях37 38 39. Результаты, полученные авторами, в силу вышесказанного предполагают, что miR-29 выполняет функцию критического репрессора для регуляции экспрессии коллагена в процессе заживления сухожилия. Более того, понижение ее экспрессии в биоптатах сухожилия человека предполагает, что подавление ее функции пермиссивно обуславливает развитие тендинопатии. Несмотря на то, что патология сухожилий характеризуется увеличением накопления коллагена 3-го типа, приводящему к ухудшению биомеханических свойств и дегенерации, молекулярная предпосылка для этого «переключения» между подтипами коллагена до этого времени была неизвестной. Впервые авторы приводят описание того, что индуцированная ИЛ-33 недостаточность miR-29a приводит к избыточной выработке коллагена 3-го типа, которая при этом одновременно окончательно запускает этот ранний процесс восстановления через опосредованное sST2 ингибирование ИЛ-33. Вопреки ожидаемым результатам для теноцитов человека miR-29 была только способна к оказанию влияния на экспрессию col 3a1, а не коллагена 1-го типа. Последующая характеристика 3'НТО транскриптов коллагена 1-го и 3-го типа выявила ранее неописанный паттерн альтернативного полиаденилирования в обеих субъединицах транскрипта коллагена 1-го типа, что привело к образованию транскриптов с отсутствием сайтов связывания miR29a, делающим их нечувствительными к подавлению под воздействием этой микроРНК. Это не касалось транскриптов коллагена 3-го типа, в которых сохранялись сайты связывания miR-29a. В теноцитах человека экспрессия гена коллагена 3-го типа активно подавляется miR-29a, о чем свидетельствует способность ингибиторов miR-29a к значительному увеличению уровней коллагена 3-го типа, тогда как дополнения культуры теноцитов miR-29a в присутствии ИЛ-33 было достаточно для подавления увеличения выработки коллагена 3-го типа. Важно отметить, что в разработанной авторами модельной системе miR-29a дополнительно направленно взаимодействовала с sST2, являющимся рецептором-приманкой ИЛ-33. Следовательно, вызванное ИЛ-33 снижение экспрессии miR-29a приводит к одновременному подавлению экспрессии гена коллагена 3-го типа и sST2 с последующим ауторегуляторным ингибированием ИЛ-33, способствующим запуску немедленного ответа с участием алармина.
На основе этой работы авторы предполагают, что ИЛ-33 является важным алармином на ранней стадии развития травмы сухожилий и тендинопатии, что является неисследованной областью с точки зрения клинических проявлений и может иметь важное значение для установления равновесия между равновесия между процессами восстановления и дегенерации. Была открыта новая роль miR-29 в качестве посттранскрипционного регулятора генов, отвечающих за синтез матрикса, и провоспалительных генов при заживлении сухожилия и тендинопатии. Одна из широких перспектив применения микроРНК для терапевтических целей заключалась в потенциале одной микроРНК для регуляции функционально конвергентных генов-мишеней. Открытие авторами регуляторного пути, зависимого от одной микроРНК, на ранних стадиях восстановления тканей подчеркивает перспективность применения заместительной терапии miR-29 в качестве возможного метода лечения тендинопатии с возможностью применения для лечения многих других патологий человека, связанных с нарушением регуляции синтеза матрикса.
Несмотря на то что описание изобретения дано на приведенных выше примерах вариантов реализации изобретения, многочисленные модификации и варианты изобретения будут очевидными из описания для специалистов в данной области техники. Соответственно, приведенные в качестве примера варианты реализации изобретения следует рассматривать как иллюстративные, а не как ограничивающие. В описанные варианты реализации изобретения могут быть внесены различные изменения, не отклоняясь от сущности и объема настоящего изобретения. Все документы, процитированные в данном документе, в явной форме включены посредством ссылки.
Литературные источники
1. Eming, S.A., Krieg, Т. & Davidson, J.M. Inflammation in wound repair: molecular and cellular mechanisms. J Invest Dermatol 127, 514-525 (2007).
2. McCormick, A., Charlton, J. & Fleming, D. Assessing health needs in primary care. Morbidity study from general practice provides another source of information. BMJ 310, 1534 (1995).
3. Nakama, L.H., King, K.B., Abrahamsson, S. & Rempel, D.M. Evidence of tendon microtears due to cyclical loading in an in vivo tendinopathy model. J Orthop Res 23, 1199-1205 (2005).
4. Sharma, P. & Maffulli, N. Tendon injury and tendinopathy: healing and repair. J Bone Joint Surg Am 87, 187-202 (2005).
5. Millar, N.L., Wei, A.Q., Molloy, T.J., Bonar, F. & Murrell, G.A. Cytokines and apoptosis in supraspinatus tendinopathy. J Bone Joint Surg Br 91, 417-424 (2009).
6. Pufe, Т., Petersen, W., Tillmann, B. & Mentlein, R. The angiogenic peptide vascular endothelial growth factor is expressed in foetal and ruptured tendons. Virchows Arch 439, 579-585 (2001).
7. Tsuzaki, M., et al. IL-1 beta induces COX2, MMP-1, -3 and -13, ADAMTS-4, IL-1 beta and IL-6 in human tendon cells. J Orthop Res 21, 256-264 (2003).
8. Tohyama, H., Yasuda, K., Uchida, H. & Nishihira, J. The responses of extrinsic fibroblasts infiltrating the devitalised patellar tendon to IL-1 beta are different from those of normal tendon fibroblasts. J Bone Joint Surg Br 89, 1261-1267 (2007).
9. John, Т., et al. Effect of pro-inflammatory and immunoregulatory cytokines on human tenocytes. J Orthop Res 28, 1071-1077 (2010).
10. Lin, T.W., Cardenas, L., Glaser, D.L. & Soslowsky, L.J. Tendon healing in interleukin-4 and interleukin-6 knockout mice. J Biomech 39, 61-69 (2006).
11. Zhang, N. & Oppenheim, J.J. Crosstalk between chemokines and neuronal receptors bridges immune and nervous systems. J Leukoc Biol 78, 1210-1214 (2005).
12. Schmitz, J., et al. IL-33, an interleukin-l-like cytokine that signals via the IL-1 receptor-related protein ST2 and induces T helper type 2-associated cytokines. Immunity 23, 479-490 (2005).
13. Gao, P., Wange, R.L., Zhang, N., Oppenheim, J.J. & Howard, O.M. Negative regulation of CXCR4-mediated chemotaxis by the lipid phosphatase activity of tumor suppressor PTEN. Blood 106, 2619-2626 (2005).
14. Chen, X., Murakami, Т., Oppenheim, J.J. & Howard, O.M. Triptolide, a constituent of immunosuppressive Chinese herbal medicine, is a potent suppressor of dendritic-cell maturation and trafficking. Blood 106, 2409-2416 (2005).
15. Lamkanfi, M. & Dixit, V.M. IL-33 raises alarm. Immunity 31, 5-7 (2009).
16. Liew, F.Y., Pitman, N.I. & McInnes, I.B. Disease-associated functions of IL-33: the new kid in the IL-1 family. Nat Rev Immunol 10, 103-110.
17. Asirvatham, A.J., Magner, W.J. & Tomasi, T.B. miRNA regulation of cytokine genes. Cytokine 45, 58-69 (2009).
18. Pritchard, C.C., Cheng, H.H. & Tewari, M. MicroRNA profiling: approaches and considerations. Nat Rev Genet 13, 358-369 (2012).
19. Matthews, T.J., Hand, G.C., Rees, J.L., Athanasou, N.A. & Carr, A.J. Pathology of the torn rotator cuff tendon. Reduction in potential for repair as tear size increases. J Bone Joint Surg Br 88, 489-495 (2006).
20. Xu, D., et al. IL-33 exacerbates antigen-induced arthritis by activating mast cells. Proceedings of the National Academy of Sciences of the United States of America 105, 10913-10918 (2008).
21. Zaiss, M.M., et al. IL-33 shifts the balance from osteoclast to alternatively activated macrophage differentiation and protects from TNF-alpha-mediated bone loss. J Immunol 186, 6097-6105 (2011).
22. Rankin, A.L., et al. IL-33 induces IL-13-dependent cutaneous fibrosis. J Immunol 184, 1526-1535 (2010).
23. Palmer, G. & Gabay, C. Interleukin-33 biology with potential insights into human diseases. Nat Rev Rheumatol 7, 321-329.
24. Ogawa, Т., et al. Suppression of type I collagen production by microRNA-29b in cultured human stellate cells. Biochem Biophys Res Commun 391, 316-321.
25. Battel, D.P. MicroRNAs: target recognition and regulatory functions. Cell 136, 215-233 (2009).
26. Ma, F., et al. The microRNA miR-29 controls innate and adaptive immune responses to intracellular bacterial infection by targeting interferon-gamma. Nature immunology 12, 861-869 (2011).
27. Maurer, В., et al. MicroRNA-29, a key regulator of collagen expression in systemic sclerosis. Arthritis Rheum 62, 1733-1743 (2010).
28. Basu, S., Binder, R.J., Suto, R., Anderson, K.M. & Srivastava, P.K. Necrotic but not apoptotic cell death releases heat shock proteins, which deliver a partial maturation signal to dendritic cells and activate the NF-kappa В pathway. Int Immunol 12, 1539-1546 (2000).
29. Scaffidi, P., Misteli, T. & Bianchi, M.E. Release of chromatin protein HMGB1 by necrotic cells triggers inflammation. Nature 418, 191-195 (2002).
30. Shi, Y., Evans, J.E. & Rock, K.L. Molecular identification of a danger signal that alerts the immune system to dying cells. Nature 425, 516-521 (2003).
31. Chen, C.J., et al. Identification of a key pathway required for the sterile inflammatory response triggered by dying cells. Nat Med 13, 851-856 (2007).
32. Eigenbrod, Т., Park, J.H., Harder, J., Iwakura, Y. & Nunez, G. Cutting edge: critical role for mesothelial cells in necrosis-induced inflammation through the recognition of IL-1 alpha released from dying cells. J Immunol 181, 8194-8198 (2008).
33. Moussion, C., Ortega, N. & Girard, J.P. The IL-1-like cytokine IL-33 is constitutively expressed in the nucleus of endothelial cells and epithelial cells in vivo: a novel 'alarmin'? PLoS One 3, e3331 (2008).
34. Cayrol, C. & Girard, J.P. The IL-1-like cytokine IL-33 is inactivated after maturation by caspase-1. Proceedings of the National Academy of Sciences of the United States of America 106, 9021-9026 (2009).
35. James, R., Kesturu, G., Balian, G. & Chhabra, A.B. Tendon: biology, biomechanics, repair, growth factors, and evolving treatment options. J Hand Surg Am 33, 102-112 (2008).
36. Brown, B.D. & Naldini, L. Exploiting and antagonizing microRNA regulation for therapeutic and experimental applications. Nat Rev Genet 10, 578-585 (2009).
37. Roderburg, C., et al. Micro-RNA profiling reveals a role for miR-29 in human and murine liver fibrosis. Hepatology 53, 209-218 (2011).
38. Ogawa, Т., et al. Suppression of type I collagen production by microRNA-29b in cultured human stellate cells. Biochem Biophys Res Commun 391, 316-321 (2010).
39. Millar, N.L., Wu, X., Tantau, R., Silverstone, E. & Murrell, G.A. Open versus two forms of arthroscopic rotator cuff repair. Clin Orthop Relat Res 467, 966-978 (2009).
40. McInnes, I.B., et al. Production of nitric oxide in the synovial membrane of rheumatoid and osteoarthritis patients. J Exp Med 184, 1519-1524 (1996).
41. Khan, K.M., Cook, J.L., Bonar, F., Harcourt, P. & Astrom, M. Histopathology of common tendinopathies. Update and implications for clinical management. Sports Med 27, 393-408 (1999).
42. Millar, N.L., Wei, A.Q., Molloy, T.J., Bonar, F. & Murrell, G.A. Heat shock protein and apoptosis in supraspinatus tendinopathy. Clin Orthop Relat Res 466, 1569-1576 (2008).
43. Kurowska-Stolarska, M., et al. IL-33 induces antigen-specific IL-5+ T cells and promotes allergic-induced airway inflammation independent of IL-4. J Immunol 181, 4780-4790 (2008).
44. Zheng, Y., et al. Interleukin-22, a T(H)17 cytokine, mediates IL-23-induced dermal inflammation and acanthosis. Nature 445, 648-651 (2007).
45. Hedrick, M.N., et al. CCR6 is required for IL-23-induced psoriasis-like inflammation in mice. The Journal of clinical investigation 119, 2317-2329 (2009).
| название | год | авторы | номер документа |
|---|---|---|---|
| ИСПОЛЬЗОВАНИЕ БИОГЕНЕЗА МИКРОРНК В ЭКЗОСОМАХ ДЛЯ ДИАГНОСТИКИ И ЛЕЧЕНИЯ | 2014 |
|
RU2644247C2 |
| ЛИНЕЙНАЯ ДУПЛЕКСНАЯ ДНК С ЗАКРЫТЫМ КОНЦОМ ДЛЯ НЕВИРУСНОГО ПЕРЕНОСА ГЕНОВ | 2017 |
|
RU2752882C2 |
| ДВОЙНОЕ НАЦЕЛИВАНИЕ НП MIR-208 И MIR 499 В ЛЕЧЕНИИ ЗАБОЛЕВАНИЙ СЕРДЦА | 2010 |
|
RU2515926C2 |
| ВАРИАНТЫ РЕКОМБИНАНТНЫХ AAV И ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2738421C2 |
| ХИМЕРНЫЕ АНТИТЕЛА ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, ХАРАКТЕРИЗУЮЩИХСЯ ОТЛОЖЕНИЕМ АМИЛОИДА | 2018 |
|
RU2746325C1 |
| ГЕННАЯ ТЕРАПИЯ ОФТАЛЬМОЛОГИЧЕСКИХ НАРУШЕНИЙ | 2016 |
|
RU2762747C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ НАРУШЕНИЙ И ЗАБОЛЕВАНИЙ, СВЯЗАННЫХ С RDH12 | 2017 |
|
RU2764920C2 |
| БИОМАРКЕРЫ ТРАВМАТИЧЕСКОГО ПОВРЕЖДЕНИЯ ГОЛОВНОГО МОЗГА | 2017 |
|
RU2771757C2 |
| МИМЕТИКИ MIR-29 И ПУТИ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2712511C2 |
| РЕГУЛЯЦИЯ МЕТАБОЛИЗМА С ПОМОЩЬЮ MIR-378 | 2011 |
|
RU2585491C2 |

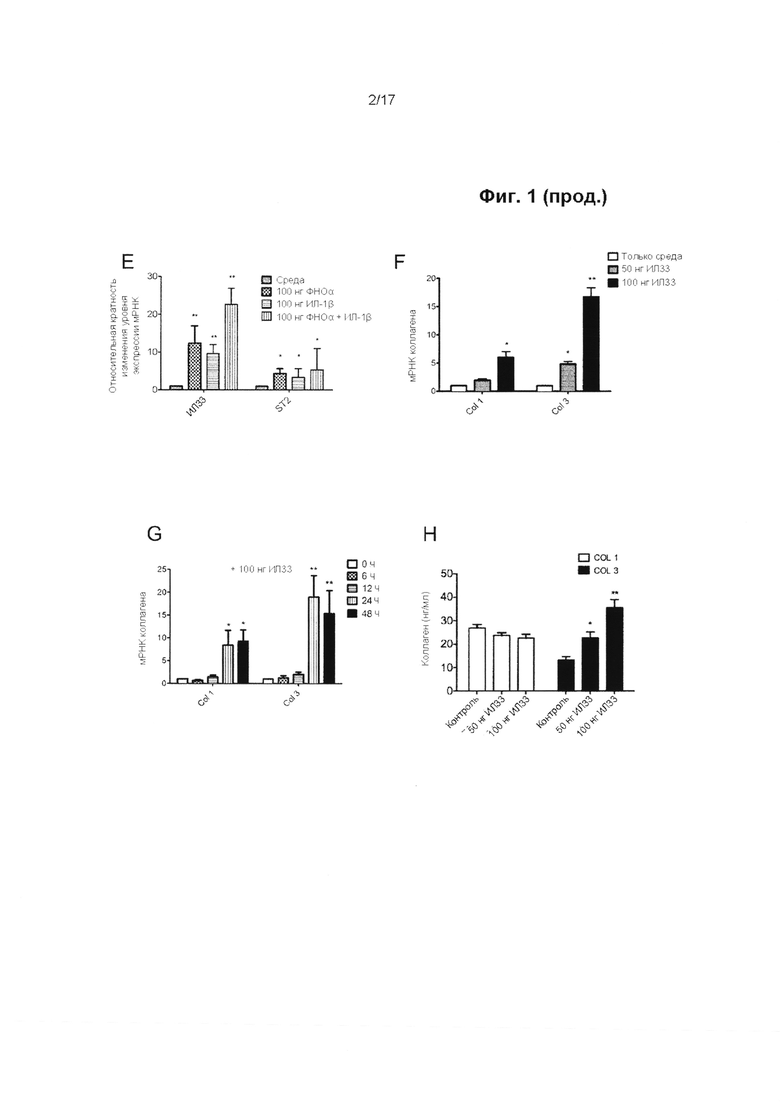
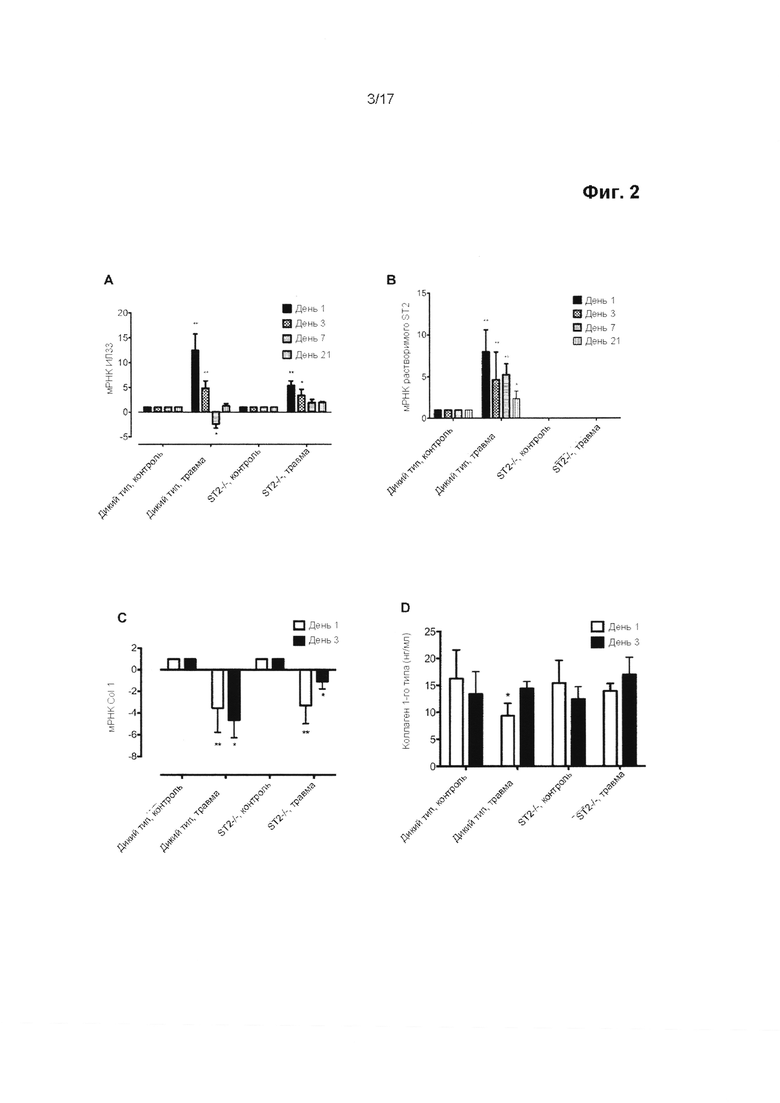
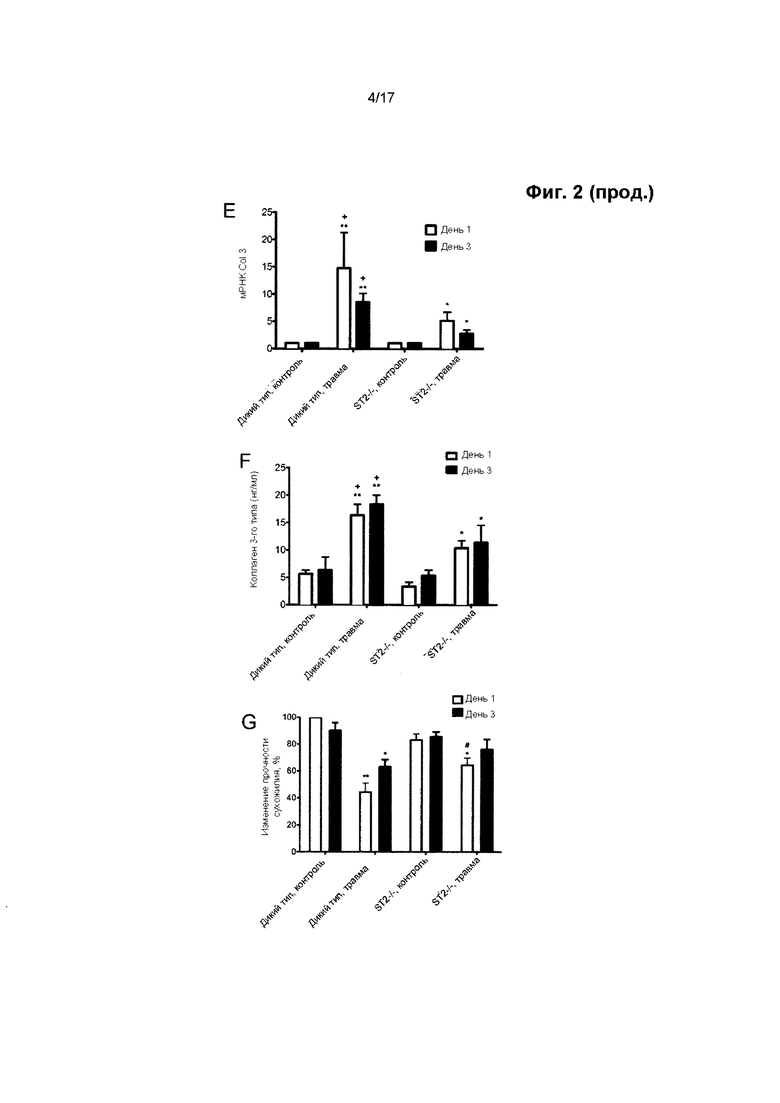
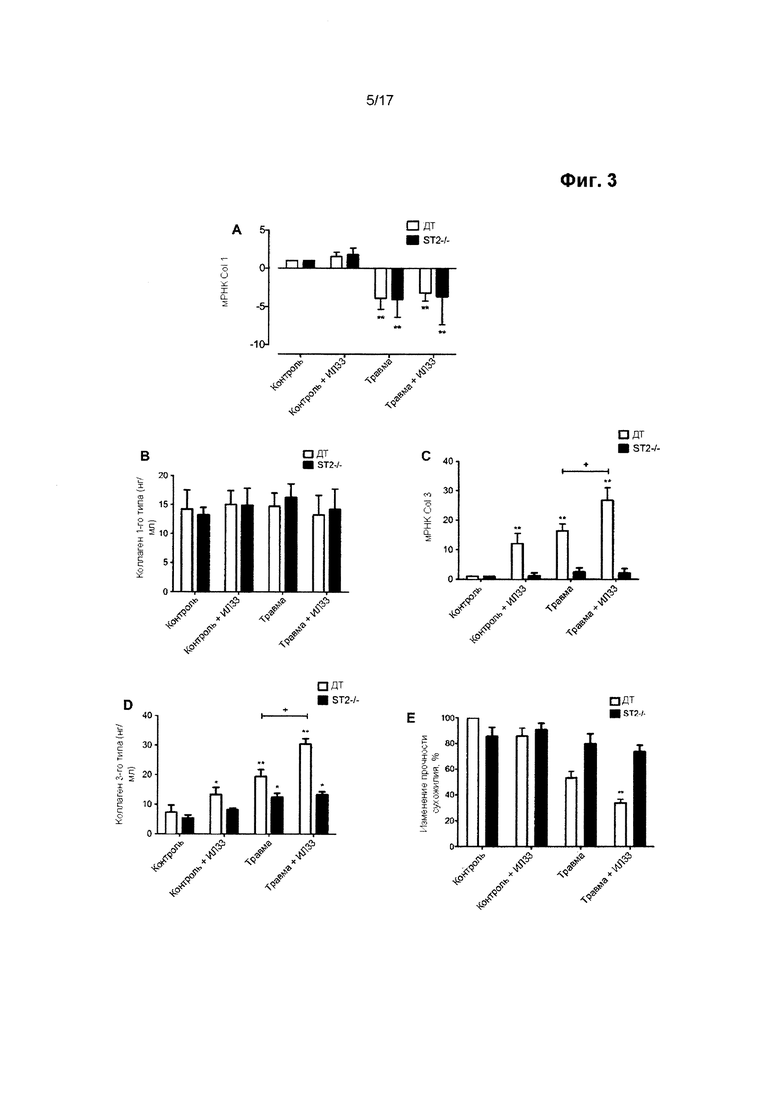
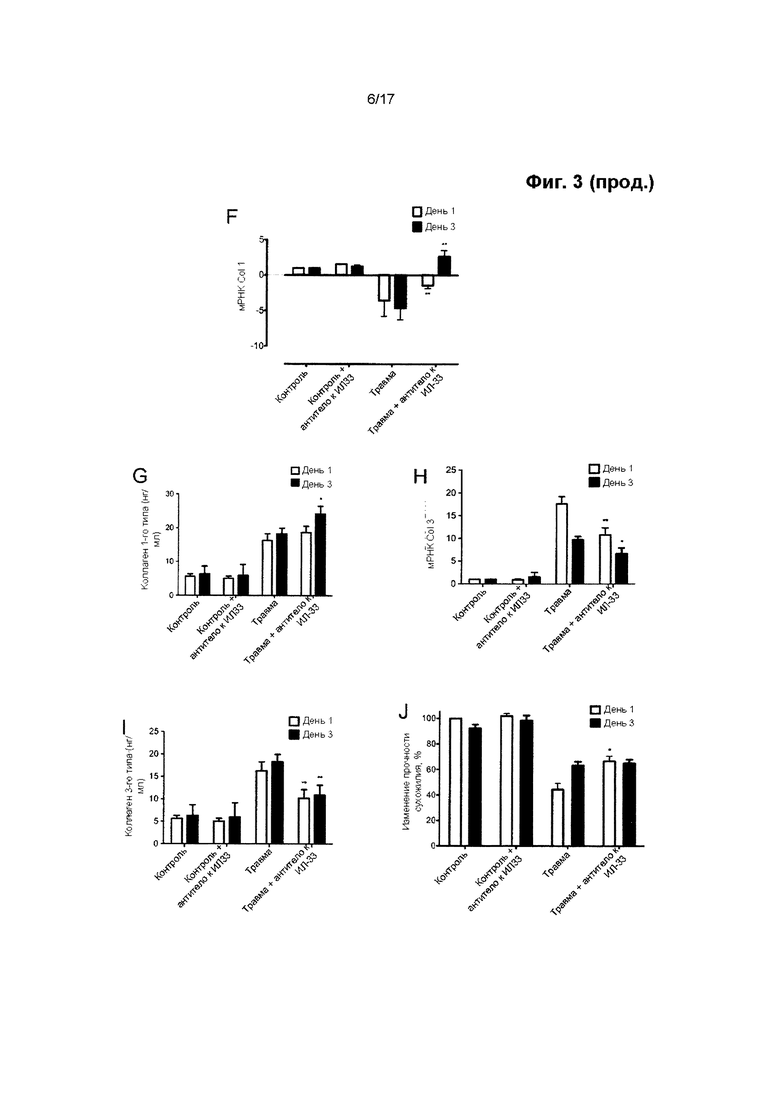
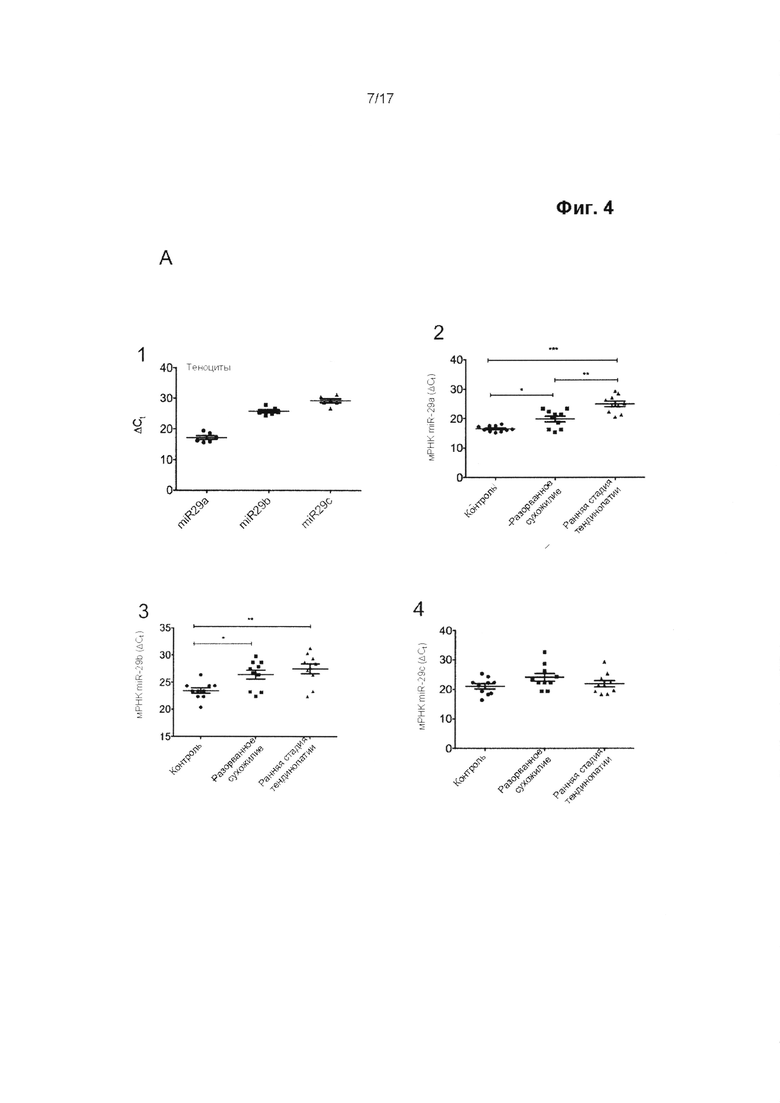
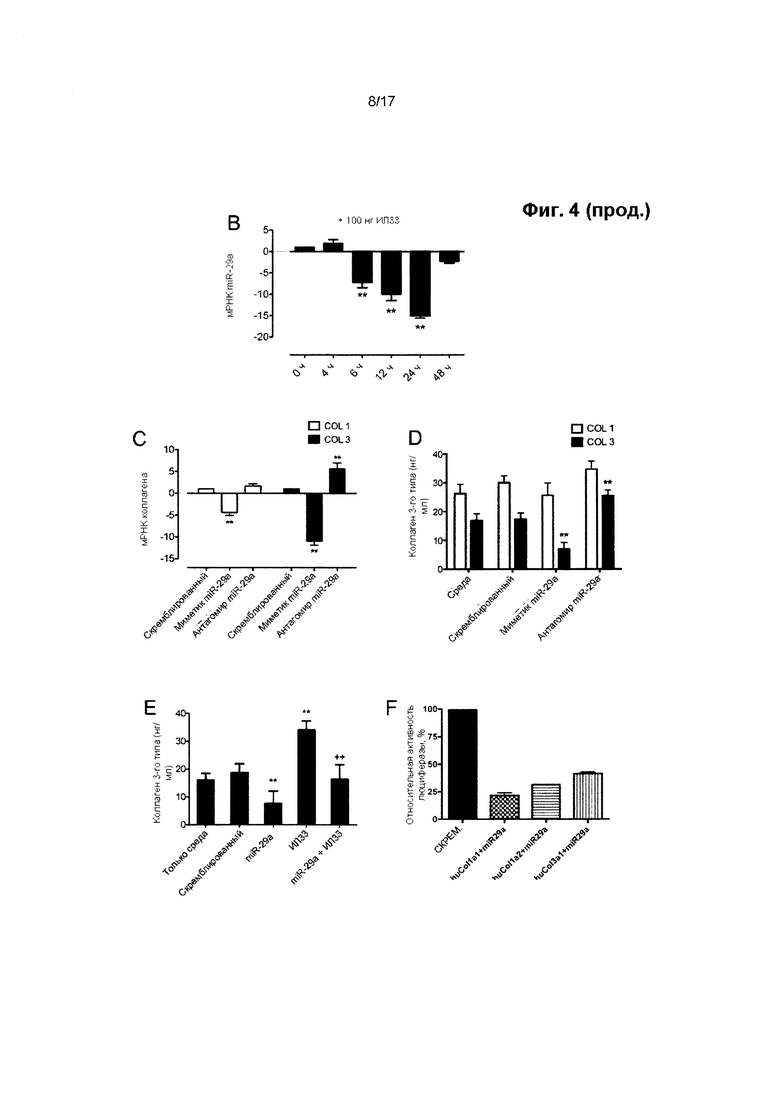
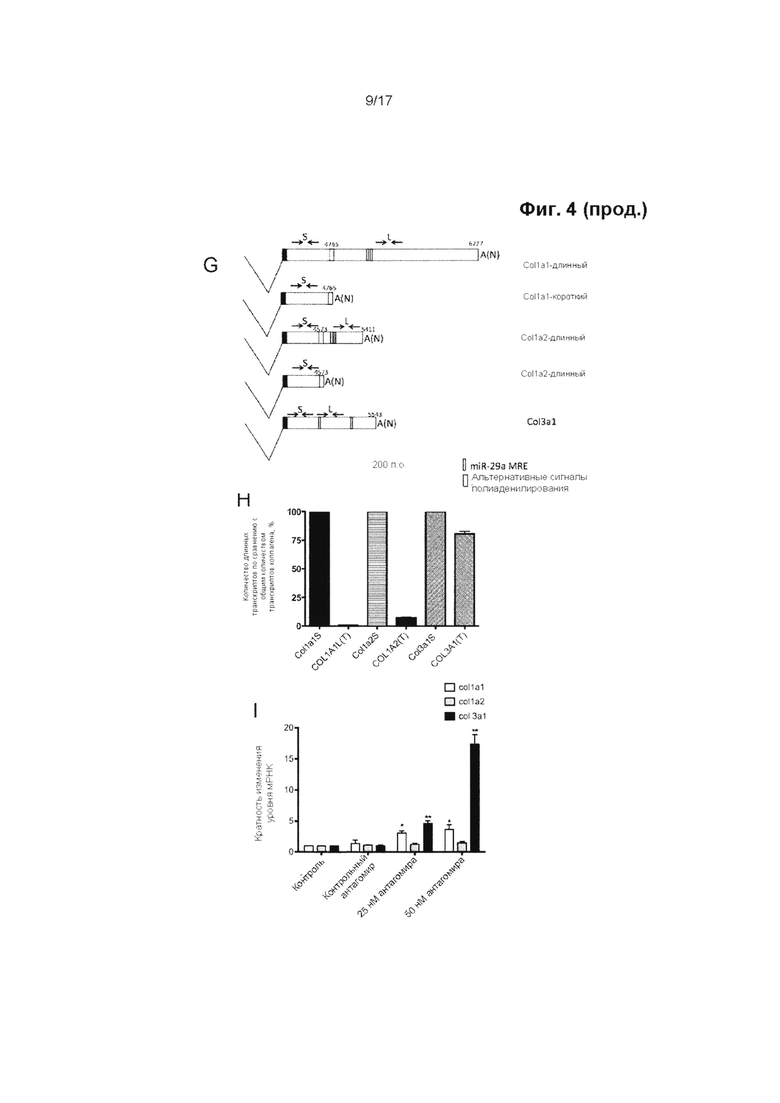
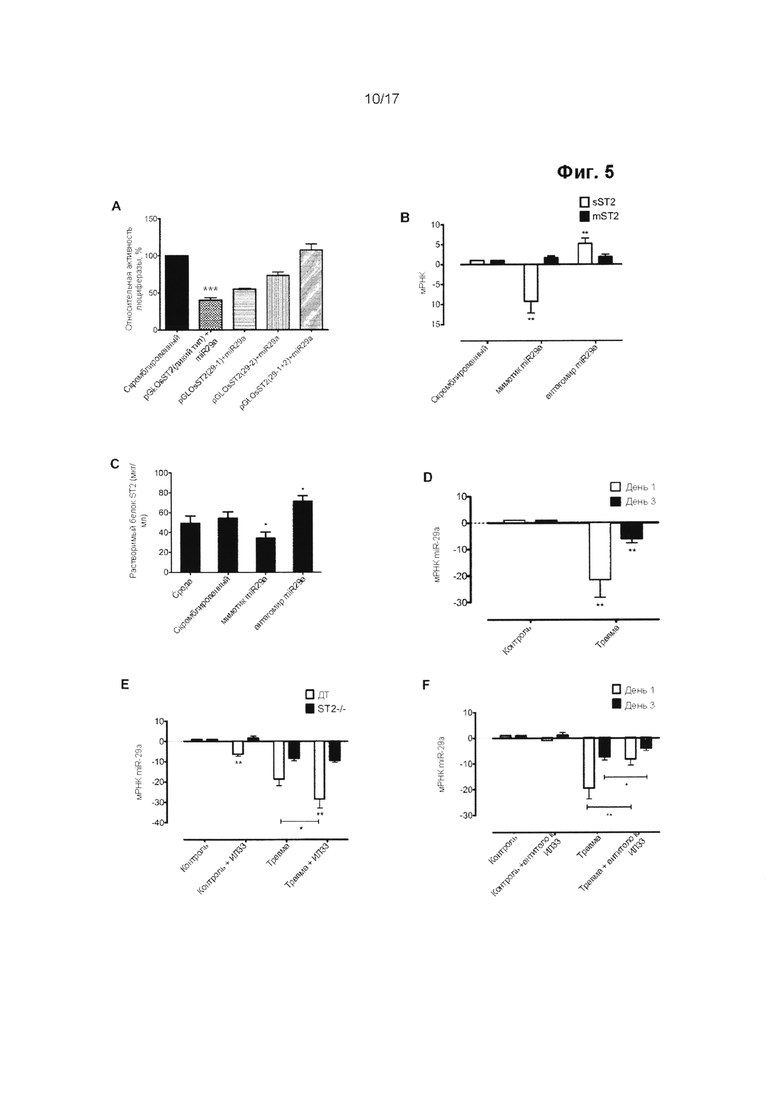
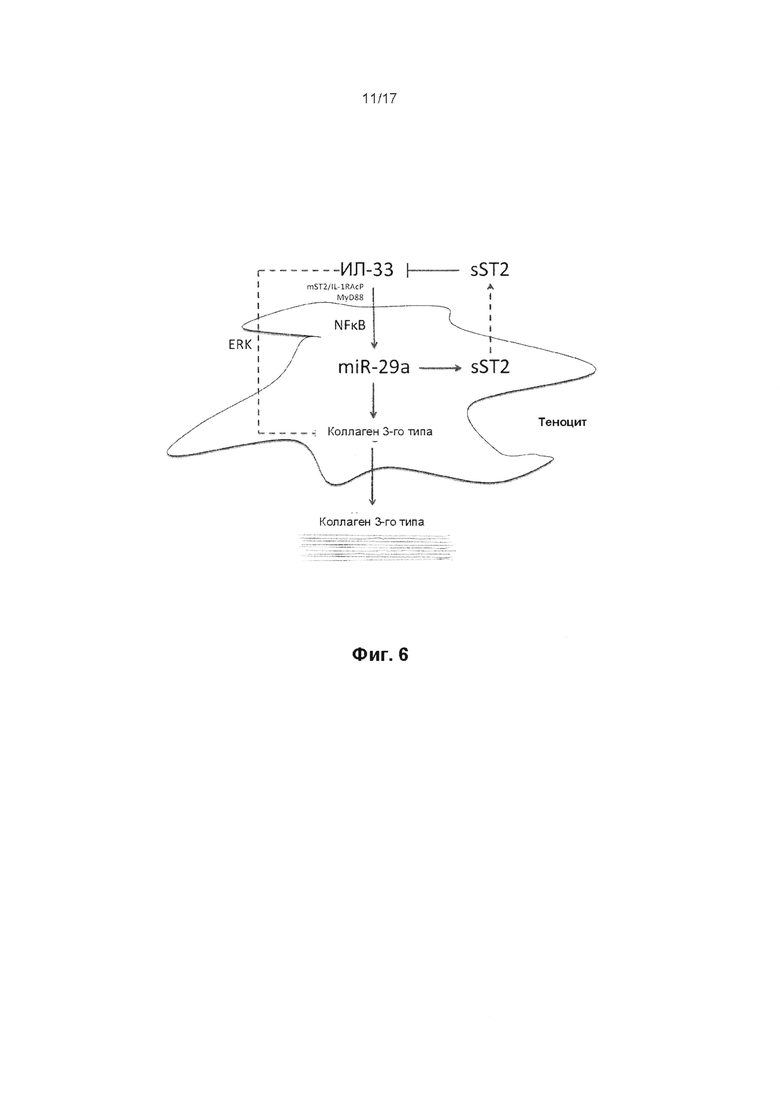
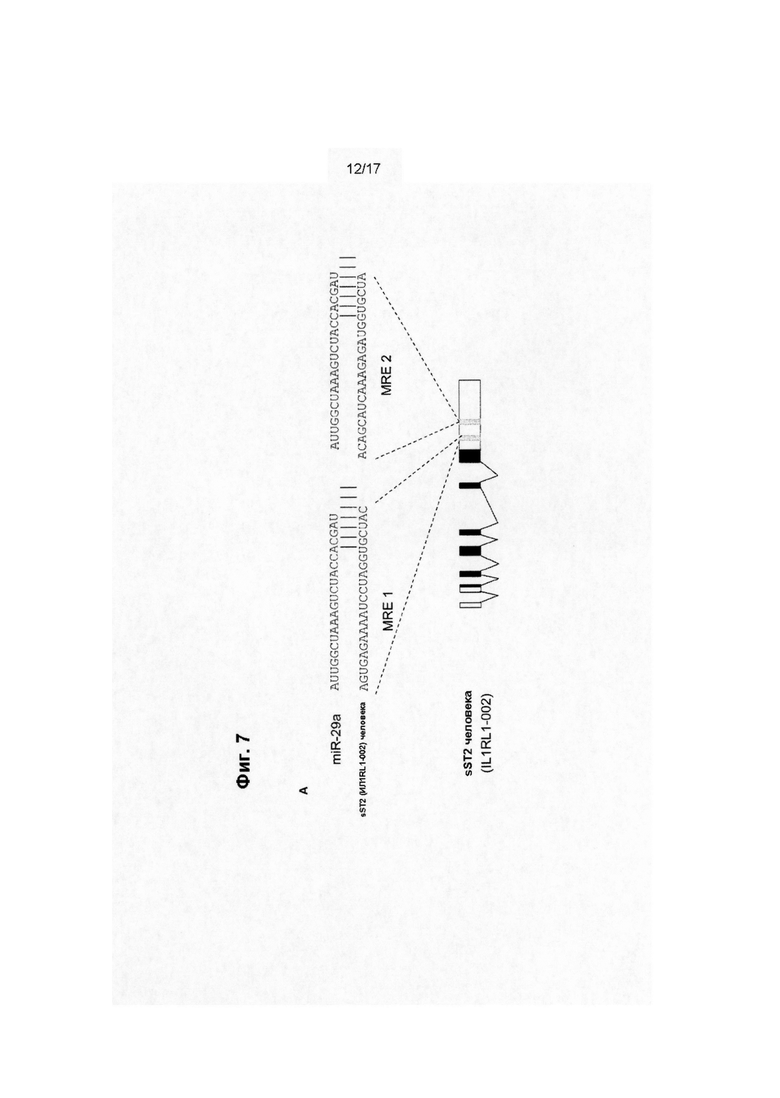
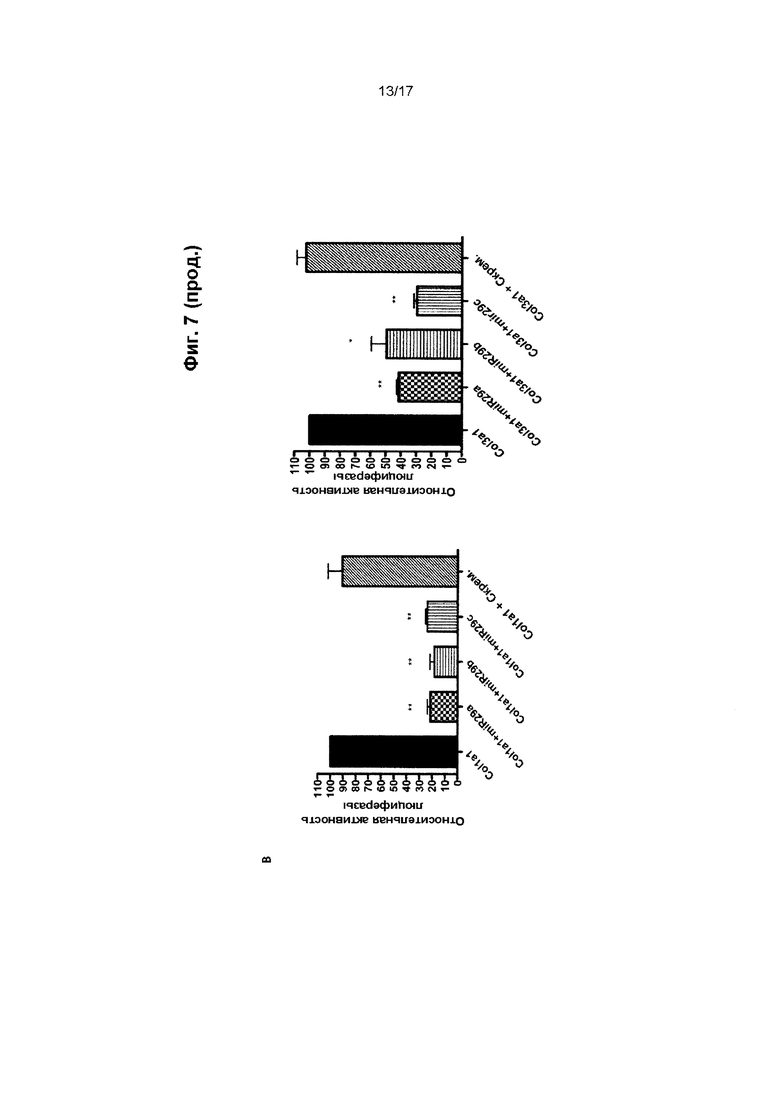
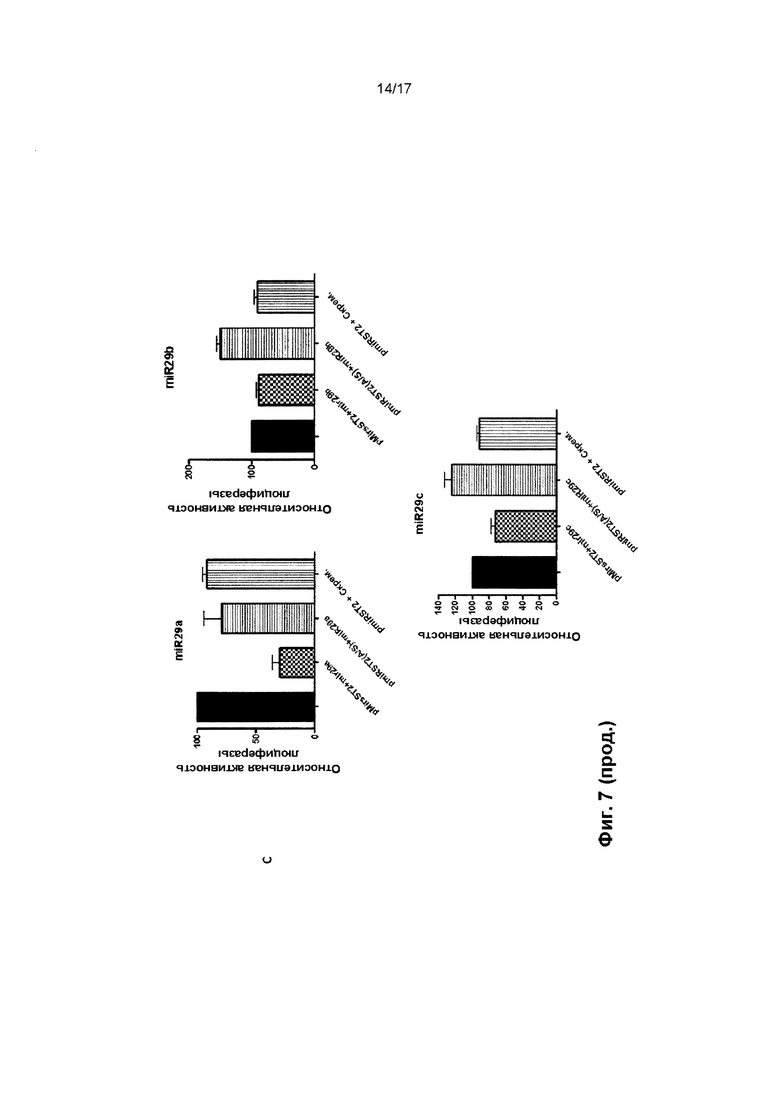
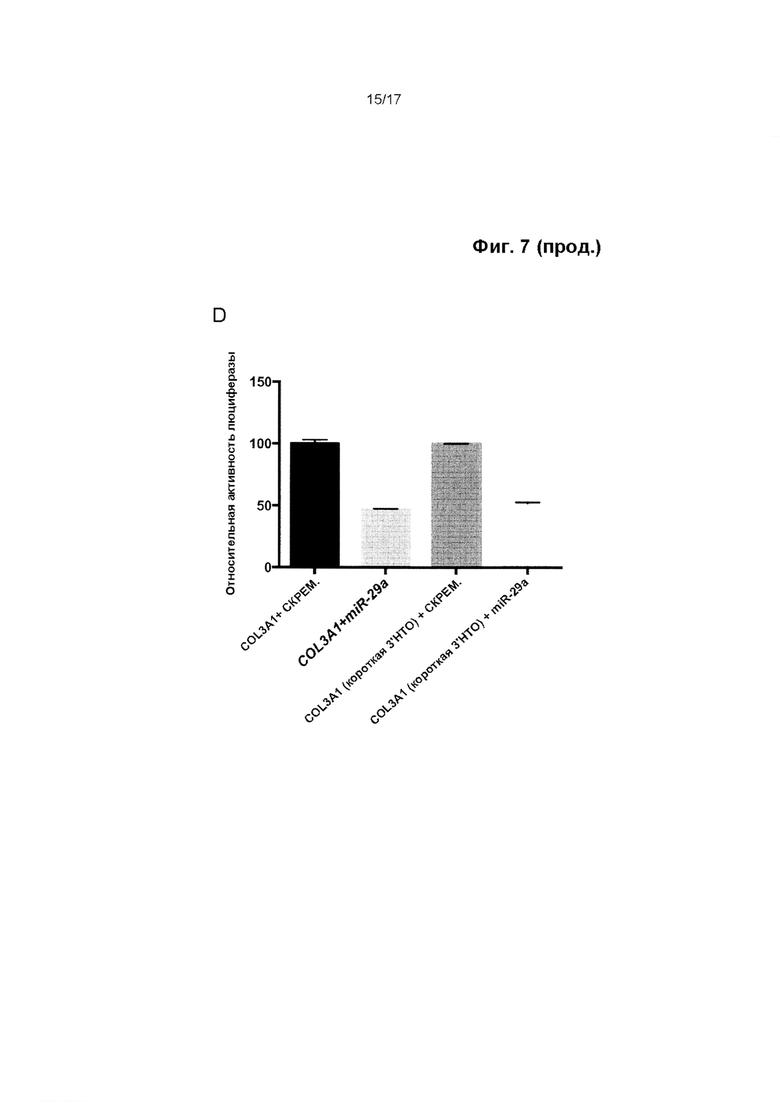

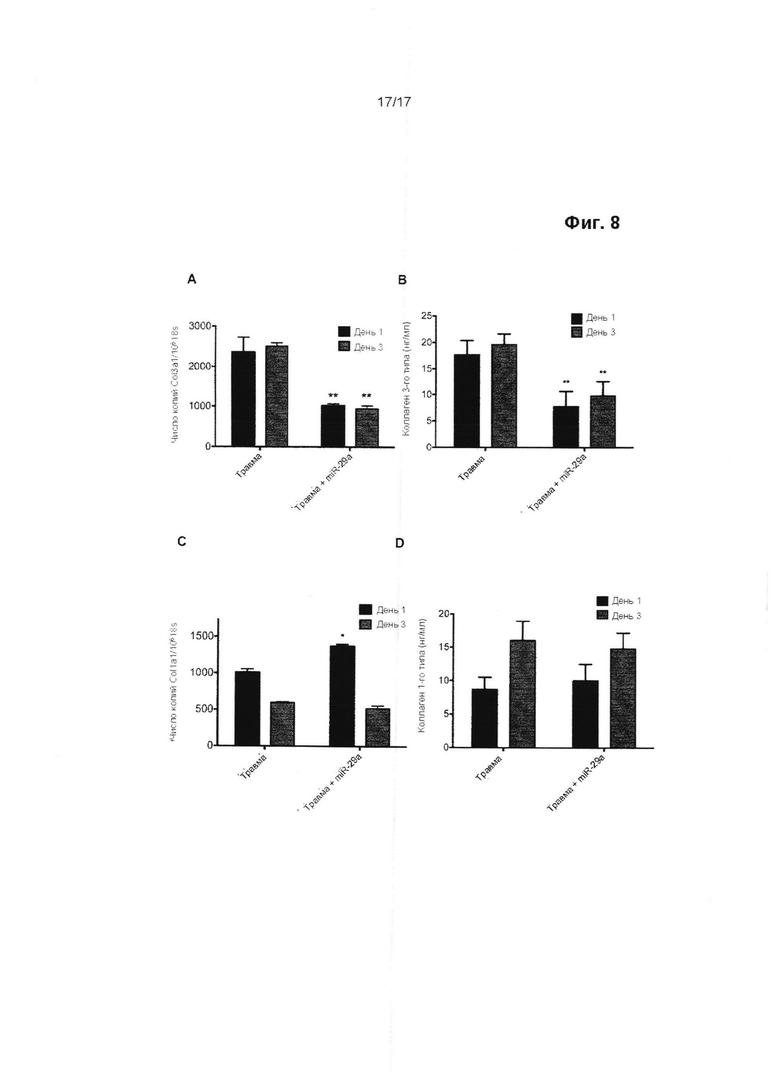
Настоящее изобретение относится к биотехнологии, в частности применению микроРНК 29 и ее предшественников, а также миметиков для модуляции травмы сухожилия и биомеханических свойств сухожилия. В частности, настоящее изобретение основано на экспериментальных данных, согласно которым синтез коллагена 1-го типа в теноцитах в меньшей степени подвержен влиянию miR-29 по сравнению с коллагеном 3-го типа, что позволяет сместить соотношение между подтипами коллагена в пользу увеличения содержания коллагена 1-го типа и тем самым предотвратить снижение биомеханических свойств сухожилия в процессе заживления. 6 н. и 22 з.п. ф-лы, 17 ил., 1 пр.
1. Способ лечения повреждения сухожилия или увеличения прочности сухожилия на растяжение у субъекта, включающий стадию доставки miR-29, ее миметика или их предшественника в сухожилие субъекта, где miR-29, ее миметик или их предшественник содержит направляющую цепь, содержащую затравочную последовательность AGCACCA.
2. Способ по п. 1, отличающийся тем, что миметик или предшественник miR-29 содержит один или более модифицированных остатков сахаров.
3. Способ по п. 1, отличающийся тем, что миметик или предшественник miR-29 содержит одну или более модифицированных межнуклеозидных связей.
4. Способ по п. 3, отличающийся тем, что миметик или предшественник miR-29 содержит одно или более модифицированных оснований.
5. Способ по п. 1, отличающийся тем, что миметик или предшественник miR-29 содержит фрагмент, обеспечивающий транспорт через мембрану.
6. Способ по п. 1, отличающийся тем, что miR-29, миметик или предшественник связан с (например, формирует комплекс с или инкапсулирован) носителем.
7. Способ по п. 6, отличающийся тем, что носитель представляет собой фармацевтически приемлемый липид или полимер.
8. Способ по п. 6, отличающийся тем, что молекула-носитель содержит средство, способное к адресному связыванию с поверхностью клетки-мишени.
9. Способ лечения повреждения сухожилия или увеличения прочности сухожилия на растяжение у субъекта, включающий стадию доставки нуклеиновой кислоты, кодирующей miR-29, ее миметик или их предшественник, в сухожилие субъекта, где miR-29, ее миметик или их предшественник содержит направляющую цепь, содержащую затравочную последовательность AGCACCA, таким образом, что указанный miR-29, миметик или предшественник экспрессируется в сухожилии.
10. Способ по п. 9, отличающийся тем, что нуклеиновая кислота связана с (например, формирует комплекс с или инкапсулирована) носителем.
11. Способ по п. 10, отличающийся тем, что носитель представляет собой фармацевтически приемлемый липид или полимер.
12. Способ по п. 10, отличающийся тем, что молекула-носитель содержит средство, способное к адресному связыванию с поверхностью клетки-мишени.
13. Способ по п. 9, отличающийся тем, что нуклеиновую кислоту доставляют с помощью вирусного вектора.
14. Способ по п. 13, отличающийся тем, что вирусный вектор представляет собой аденовирусный вектор, вектор на основе аденоассоциированных вирусов (AAV), ретровирусный или герпесвирусный вектор.
15. Способ по п. 14, отличающийся тем, что ретровирусный вектор представляет собой лентивирусный вектор.
16. Способ по любому из пп. 1-15, отличающийся тем, что miR-29 представляет собой miR-29a, miR-29b1, miR-29b2 или miR-29c, или их комбинацию.
17. Способ по п. 16, отличающийся тем, что указанная комбинация содержит miR-29a.
18. Способ по любому из пп. 1-17, отличающийся тем, что направляющая цепь имеет последовательность:
UAGCACCAUCUGAAAUCGGUUA (hsa-miR-29a);
UAGCACCAUUUGAAAUCAGUGUU (hsa-miR-29b1; hsa-miR-29b2); или
UAGCACCAUUUGAAAUCGGUUA (hsa-miR-29c).
19. Способ по любому из пп. 1-15, отличающийся тем, что предшественник представляет собой pre-mir-29.
20. Способ по п. 19, отличающийся тем, что pre-mir-29 имеет последовательность:
AUGACUGAUUUCUUUUGGUGUUCAGAGUCAAUAUAAUUUUCUAGCACCAUCUGAAAUCGGUUAU (hsa-pre-mir-29a: альтернативный вариант (i));
AUGACUGAUUUCUUUUGGUGUUCAGAGUCAAUAUAAUUUUCUAGCACCAUCUGAAAUCGGUUAU AAUGAUUGGGG (hsa-pre-mir-29a: альтернативный вариант (ii));
CUUCAGGAAGCUGGUUUCAUAUGGUGGUUUAGAUUUAAAUAGUGAUUGUCUAGCACCAUUUGAAAUCAGUGUUCUUGGGGG (hsa-pre-mir-29b1);
CUUCUGGAAGCUGGUUUCACAUGGUGGCUUAGAUUUUUCCAUCUUUGUAUCUAGCACCAUUUGAAAUCAGUGUUUUAGGAG (hsa-pre-mir-29b2); или
AUCUCUUACACAGGCUGACCGAUUUCUCCUGGUGUUCAGAGUCUGUUUUUGUCUAGCACCAUUUGAAAUCGGUUAUGAUGUAGGGGGA (hsa-pre-mir-29c)
(причем последовательности зрелой направляющей цепи подчеркнуты).
21. Способ по любому из пп. 1-15, отличающийся тем, что миметик miR-29 содержит направляющую цепь, имеющую последовательность:
UAGCACCAUCUGAAAUCGGUUA (hsa-miR-29a);
UAGCACCAUUUGAAAUCAGUGUU (hsa-miR-29b1 и 2); или
UAGCACCAUUUGAAAUCGGUUA (hsa-miR-29c)
(причем затравочная последовательность в каждом случае подчеркнута);
или последовательность, которая отличается от указанной последовательности по не более чем пяти положениям за пределами затравочной последовательности.
22. Способ по п. 1 или 9, отличающийся тем, что сухожилие травмировано или подвержено тендинопатии.
23. Способ по п. 1 или 9, отличающийся тем, что субъект представляет собой человека или лошадь.
24. Способ по п. 22, отличающийся тем, что пораженное сухожилие представляет собой ахиллово сухожилие, сухожилие надостной мышцы, общее сухожилие сгибателей, общее сухожилие разгибателей или поверхностное сухожилие сгибателей.
25.Применение miR-29, ее миметика или их предшественника в способе лечения повреждения сухожилия или увеличения прочности сухожилия на растяжение.
26. Применение miR-29, ее миметика или их предшественника в производстве лекарственного препарата для лечения повреждения сухожилия или увеличения прочности сухожилия на растяжение.
27. Применение нуклеиновой кислоты, кодирующей miR-29, ее миметик или их предшественник, в способе лечения повреждения сухожилия или увеличения прочности сухожилия на растяжение.
28. Применение нуклеиновой кислоты, кодирующей miR-29, ее миметик или их предшественник, в производстве лекарственного препарата для лечения повреждения сухожилия или увеличения прочности сухожилия на растяжение.
| WO 2010026213 A2 11.03.2010 | |||
| WO 2010148050 A2 23.12.2010 | |||
| WO 2013142336 A1 26.09.2013 | |||
| КОРОБОВ Г.А., ИШЕМИЧЕСКАЯ БОЛЕЗНЬ СЕРДЦА: РЕГУЛИРОВАНИЕ С ПОМОЩЬЮ МИКРОРНК, КАРДИОЛОГИЧЕСКИЙ ВЕСТНИК, том 6, номер 2, 2011 г., стр | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |

