Содержание текстового файла ASCII перечня последовательностей под названием 8441-0004-1_ST25 размером 15,6 килобайт, созданного 4 июня 2018 г. и поданного через систему EFS-Web в электронном виде вместе с данной заявкой, включено в настоящий документ посредством ссылки.
ПРИТЯЗАНИЕ НА ПРИОРИТЕТ
Данная заявка испрашивает приоритет предварительной заявки США 62/526,835, поданной 29 июня 2017 г., которая полностью включена в настоящий документ посредством ссылки.
ПРАВА ПРАВИТЕЛЬСТВА
Данное изобретение было выполнено при финансовой поддержке правительства Соединенных Штатов Америки в рамках проекта Contact 20XS094A, предоставленной Международной корпорацией по внедрению научных достижений во Фредерике (Science Applications International Corporation, Frederick). Таким образом, правительство Соединенных Штатов Америки может обладать определенными правами на изобретение, описанное и заявленное в настоящем документе.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к химерным антителам мыши и человека, применимых для лечения болезней, характеризующихся отложением амилоида, в частности первичного амилоидоза (AL), к фармацевтическим композициям, содержащим такие антитела, к способам и материалам для получения таких антител и к способам лечения болезней, характеризующихся отложением амилоида с применением указанных антител и фармацевтических композиций.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Нативные антитела обычно представляют собой гетеротетрамерные гликопротеины молекулярной массой около 1500000 дальтон, состоящие из двух идентичных легких цепей и двух идентичных тяжелых цепей. Каждая легкая цепь связана с тяжелой цепью одной дисульфидной связью, при этом количество дополнительных дисульфидных связей между тяжелыми цепями различается у антител различных изотипов. Простейшим изотипом является IgG, который содержит только две легкие цепи и две тяжелые цепи, в которых две тяжелые цепи связаны двумя дисульфидными связями. На одном конце каждой тяжелой цепи находится вариабельный домен (VH) и ряд примыкающих константных доменов. На одном конце каждой легкой цепи находится вариабельный домен (VL), а на ее другом конце находится константный домен. Каждый вариабельный домен легкой и тяжелой цепи в антителе содержит три сегмента, называемых областями, определяющими комплементарность (CDR, complementarity-determining regions), или гипервариабельными областями. Каждая CDR в легкой цепи вместе с соответствующей CDR в примыкающей тяжелой цепи образует антигенсвязывающий участок антитела. Легкие цепи в зависимости от их константной области относятся к двум основным типам κ и λ. Обе легкие цепи κ и λ могут объединяться с любым из различных типов тяжелых цепей.
Амилоидный амилоидоз легкой цепи (AL амилоидоз), также называемый первичным амилоидозом, является наиболее распространенной формой системного амилоидоза в США. Термин "амилоидоз" относится к группе заболеваний, объединенных общим признаком, т.е. внеклеточным отложением патологических нерастворимых фибриллярных белков в органах и тканях (Rodney, et al. - NEJM, 25:898). Амилоидоз вызван нарушением функции вырабатывающих антитела клеток человека, вызывающим выработку аномальных белковых волокон, которые образуют скопления в форме нерастворимых отложений амилоида в органах и тканях. Тип амилоидоза определяется характером белков-предшественников, которые образуют отложения фибрилл. При первичном амилоидозе (AL) фибриллы содержат фрагменты легких цепей иммуноглобулина, а при вторичном амилоидозе фибриллы содержат амилоидный белок A. Современная классификация амилоидоза основана на характере белков-предшественников плазмы, которые образуют отложения фибрилл.
Белки-предшественники плазмы разнообразны и неродственны. Тем не менее все отложения предшественников образуют отложения амилоидов, имеющие общую характерную β-складчато-листовую конфигурацию, которая ответственна за свойства характерного окрашивания фибриллярных отложений. Конечной стадией развития амилоидоза является отложение амилоидных фибрилл в органах больного. Смертность от амилоидоза высока, и в настоящее время пятилетняя выживаемость составляет около 28%.
До сих пор лечение AL было направлено на сокращение синтеза легких цепей амилоидогенного предшественника с применением методов традиционной или цитотоксической терапии в высокой дозе, нацеленной на неправильно функционирующие клетки. Такое лечение имеет два недостатка. Во-первых, фибриллярные отложения часто являются бессимптомными до тех пор, пока не возникнет значительное накопление. Поэтому маловероятно, что лечение будет предпринято до тех пор, пока не произойдет их значительное накопление. Во-вторых, поскольку данное лечение наиболее эффективно только для прекращения образования аномального белка-предшественника, но не для удаления существующих отложений, прогноз для пациентов с AL остается крайне неблагоприятным из-за сохранения (или прогрессирования) патологических отложений (Solomon, et al. - Int. J. Exp. Clin. Invest. 2: 269).
В недавних исследованиях на биологических моделях показано, что введение мышиного антитела 11-1F4 и других мышиных антител, специфичных к легкой цепи человека, нацеленных на общий эпитоп β-складчато-листовой структуры, присутствующий в фибриллах AL, приводит к полной деградации отложений амилоида ALκ и ALλ. Несколько данных мышиных антител описаны в патенте США 8,105,594, полностью включенном в настоящий документ посредством ссылки.
Мышиные антитела в целом непригодны для введения другим биологическим видам, таким как человек, потому что вид-реципиент будет распознавать мышиное антитело как антиген и вырабатывать к нему антитела. Антигенное свойство антитела одного вида при введении другому виду путем инъекции обычно вызвано участком константного домена. Такой антигенный ответ будет препятствовать желаемому терапевтическому эффекту мышиного антитела или предотвращать его. У человека этот антигенный ответ вызван человеческими антимышиными антителами (human anti-mouse antibody, HAMA). За счет ответа человеческих антимышиных антител описанные в патенте '594 антитела, обладают высоким потенциалом иммуногенности у человека. Поскольку ответ HAMA обычно приводит к быстрому клиренсу у человека-реципиента, HAMA будут в значительной степени ограничивать любую потенциальную терапевтическую пользу для человека, которой могло бы обладать мышиное антитело. Следовательно, данные мышиные антитела непригодны для введения пациенту в целях остановки или стимуляции обратного развития отложения амилоидных фибрилл у пациента, и существует потребность в лечении антителами болезней, характеризующихся отложением амилоида, обладающих низкой иммуногенностью у человека.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ТЕХНИКИ
Один вариант воплощения изобретения представляет собой химерные антитела человека и мыши, применимые для лечения болезней, характеризующихся отложением амилоида, в частности первичного (AL) амилоидоза.
Другой вариант воплощения изобретения представляет собой фармацевтическую композицию, содержащей химерное антитело и фармацевтически приемлемый носитель.
Другой вариант воплощения изобретения представляет собой способы и материалы для получения таких антител, включая полинуклеотидные последовательности и векторные конструкции.
Другой вариант воплощения изобретения представляет собой способы лечения или облегчения симптомов болезней, характеризующихся отложением амилоида, таких как первичный (AL) амилоидоз, у нуждающегося в таком лечении человека путем введения пациенту-человеку, нуждающемуся в таком лечении или уменьшении интенсивности симптомов, эффективного количества по меньшей мере одного из таких антител, эффективных для лечения или облегчения симптомов указанной болезни, характеризующейся отложением амилоида, вместе с фармацевтически приемлемым носителем.
Другой вариант воплощения изобретения представляет собой способ обнаружения болезни, характеризующейся отложением амилоида у пациента, у которого подозревают наличие такого заболевания, путем введения меченого антитела по изобретению и обнаружения наличия метки у пациента. Указанная метка может представлять собой радиоактивную метку, такую как 124I, но специалист в данной области техники может с легкостью рассматривать другие виды меток. В данный вариант воплощения изобретения также включено меченое антитело.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
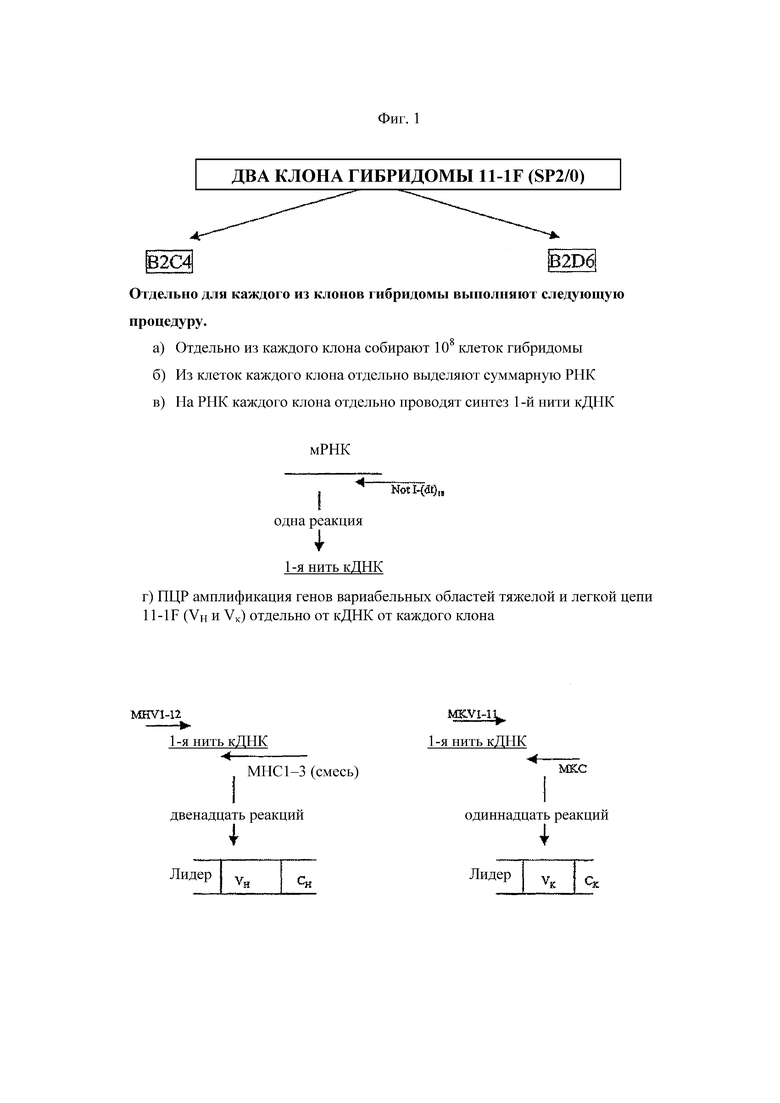
На Фиг. 1 схематически изображена стратегия, используемая для клонирования мышиных генов VH и VK из линии клеток гибридомы.
На Фиг. 2 представлен перечень ДНК- и аминокислотных последовательностей гена области VH мышиного антитела 11-1F4, SEQ ID NO: 39 и NO: 35 соответственно.
На Фиг. 3 представлен перечень ДНК- и аминокислотных последовательностей гена области VK мышиного антитела 11-1F4, SEQ ID NO: 40 и NO: 36 соответственно.
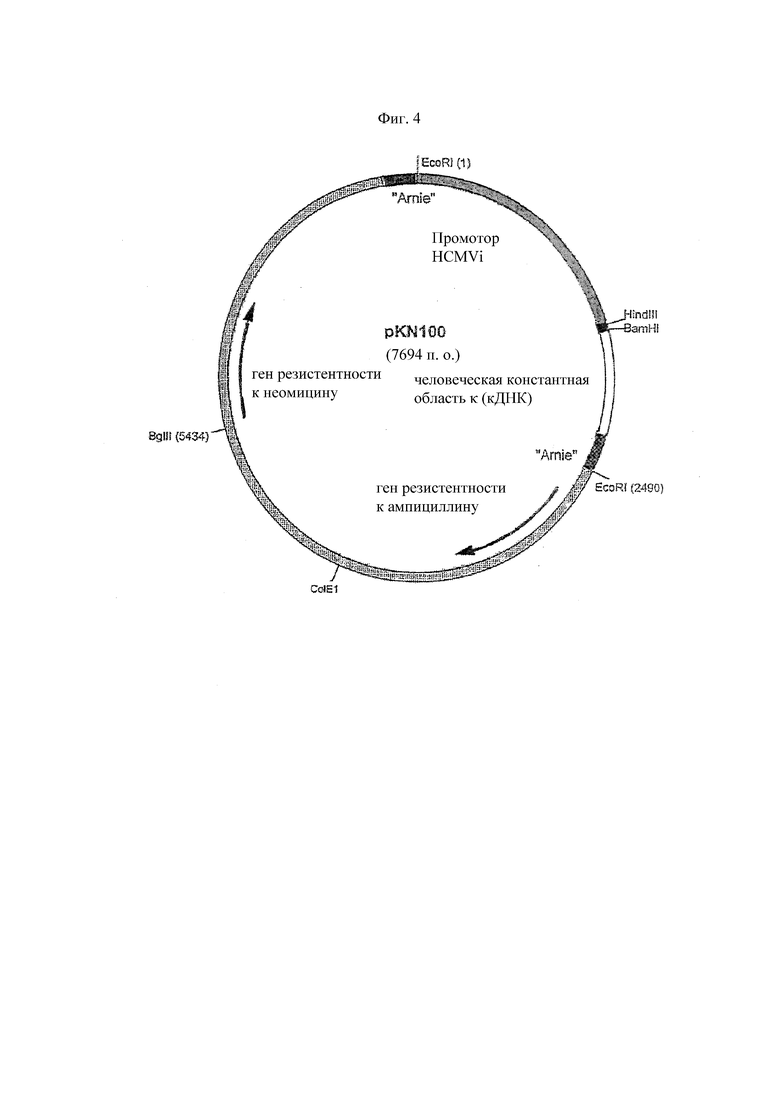
На Фиг. 4 представлена карта экспрессионного вектора pKN100 легкой цепи каппа иммуноглобулина. Он состоит из фрагмента вектора pSV2, который имеет ранний промотор SV40 и поздний промотор инактивированного SV40, точку начала репликации SV40 и точку начала репликации Co1E1. Он также имеет гены резистентности к ампициллину и неомицину. Поздний промотор инактивированного SV40 управляет генами неомицина. Он также имеет промотор HCMVi, множественный сайт клонирования, содержащий сайты ферментов рестрикции BamHI и HindIII, для вставки гена вариабельной области иммуноглобулина и кДНК для человеческого гена константной области каппа с концевой последовательностью сигнала терминации транскрипции spaC2 (Arnie), который находится в одинаковой ориентации с экспрессионной кассетой легкой цепи каппа.
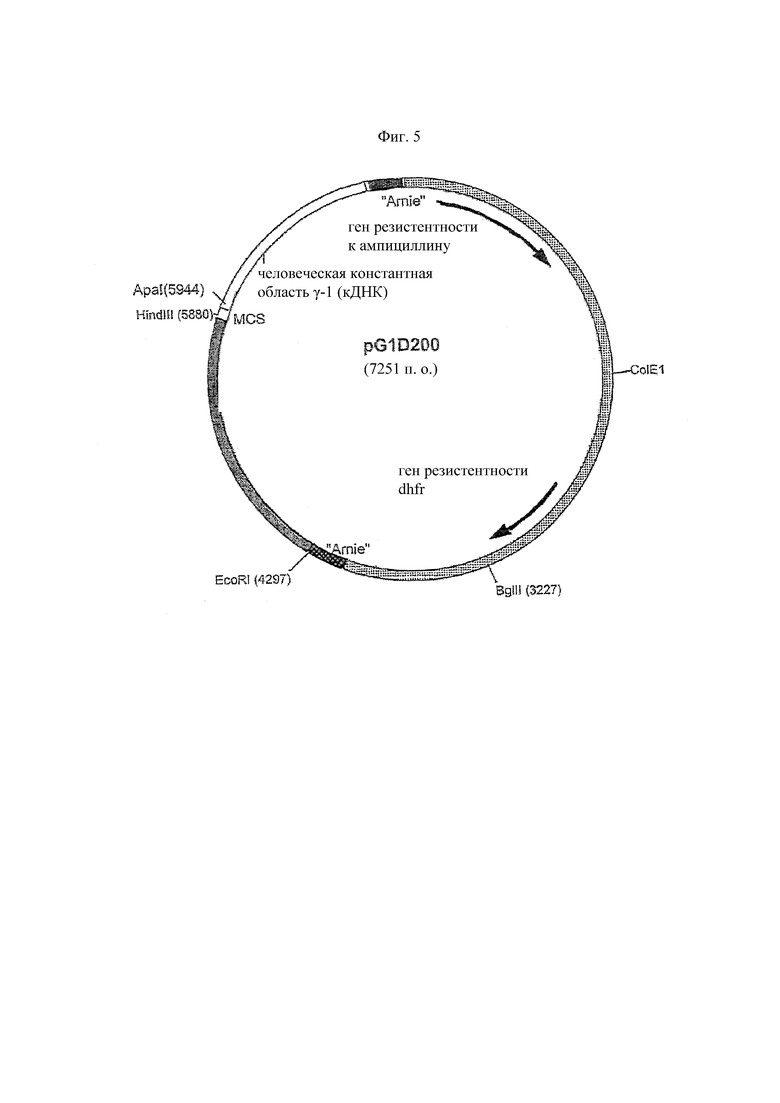
На Фиг. 5 представлена карта экспрессионного вектора pG1D200 тяжелой цепи гамма 1. Он состоит из фрагмента вектора pSV2dhfr, который имеет ранний промотор SV40 и поздний промотор инактивированного SV40, точку начала репликации SV40 и точку начала репликации Co1E1. Он также имеет гены резистентности к ампициллину и дигидрофолатредуктазе(dhfr). Поздний промотор инактивированного SV40 управляет геном дигидрофолатредуктазы. Следовательно, его экспрессия ослаблена, что позволяет отбирать клоны с мультигенным/высоким уровнем экспрессии, используя сравнительно низкие концентрации метотрексата. Он также имеет фрагмент промотора HCMVi, множественный сайт клонирования, кДНК для человеческого гена константной области гамма 1 человека (без интронов), за которой следует последовательность сигнала терминации транскрипции spaC2 (Amie).
На Фиг. 6 представлен перечень ДНК- и аминокислотных последовательностей модифицированного гена области VK мышиного антитела 11-1F4, SEQ ID NO: 42 и NO: 47 соответственно) и последовательностей олигонуклеотидных праймеров, используемых для модификации гена VK (SEQ ID NO: 41 и NO: 43 соответственно), а также последовательность ДНК с лидерной последовательностью (SEQ ID NO: 37).
На Фиг. 7 представлен перечень последовательностей ДНК и аминокислот модифицированного гена области VH мышиного антитела 11-1F4, SEQ ID NO: 45 и NO: 48 соответственно) и последовательностей олигонуклеотидных праймеров, используемых для модификации гена VH (SEQ ID NO: 44 и NO: 46 соответственно), а также последовательность ДНК с лидерной последовательностью (SEQ ID NO: 38).
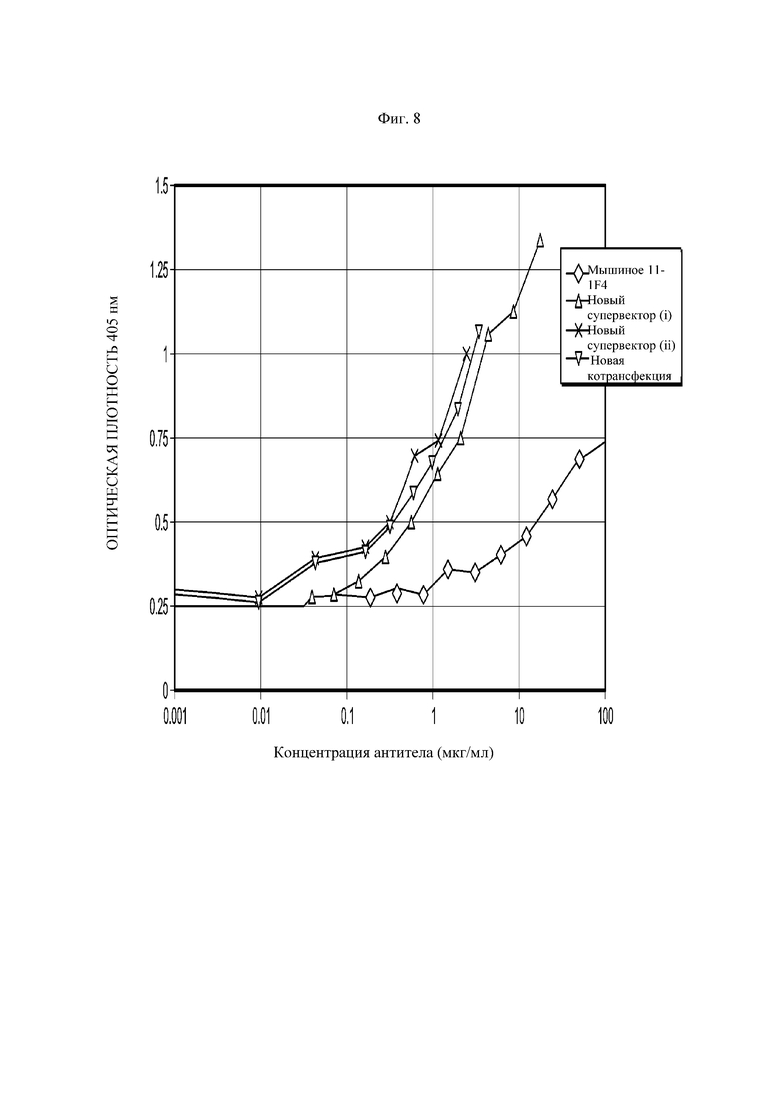
На Фиг. 8 показано графическое представление результата анализа ИФА (иммуноферментный анализ) связывания амилоидных фибрилл. Проводили раздельный анализ супернатантов культур клеток COS, содержащих химерное антитело 11-1F4, параллельно с очищенным мышиным антителом 11-1F4 на одном планшете. Показания оптической плотности регистрировали при OD405.
Новый супервектор представляет собой pG1KD200-11-1F4.
Новая котрансфекция представляет собой совместную трансфекцию 11-1F4VHpG1D200 и 11-1F4VK.pKN100.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В соответствии с настоящим изобретением обеспечены химерные антитела мыши и человека, применимые для введения человеку, страдающему болезнями, характеризующимися отложением амилоида, для лечения или облегчения симптомов заболевания. Химерные антитела по изобретению связываются с отложениями амилоида и активируют иммунную систему пациента для выведения связанных веществ, при этом не вызывая или вызывая незначительную реакцию HAMA. В изобретении также обеспечены фармацевтические композиции, содержащие по меньшей мере одно из указанных химерных антител и фармацевтически приемлемый носитель, способы и материалы для получения данных антител и способы лечения или облегчения симптомов у пациента, страдающего амилоидозом, путем введения пациенту количества химерного антитела, эффективного для удаления по меньшей мере некоторых из отложений амилоида из органов пациента и, таким образом, для лечения или облегчения симптомов амилоидоза.
В настоящем изобретении пациенту-человеку, страдающему амилоидозом, вводят по меньшей мере одно химерное антитело для стимуляции деградации и удаления по меньшей мере некоторых из амилоидных фибрилл, отложившихся в органах пациента. Терапевтически эффективное количество антитела вводят вместе с фармацевтически приемлемым носителем. Подходящие фармацевтически приемлемые носители хорошо известны в данной области техники. Как хорошо известно специалистам в области медицины, в характерном случае введение осуществляют парентеральным (например, внутривенным) путем. Конечно, возможны и другие пути введения. Введение можно осуществлять в однократной дозе или в многократных дозах. Врач может оптимизировать вводимое количество антитела и периодичность дозирования для конкретного пациента.
Вводимая пациенту терапевтически эффективная доза антитела (при введении в однократной дозе или в многократных дозах) должна быть достаточной для уменьшения количества отложившихся амилоидных фибрилл у пациента. Такое терапевтически эффективное количество можно определить путем оценки симптоматических изменений у пациента или путем оценки изменения количества отложившихся амилоидных фибрилл (например, путем радиоиммунологического обнаружения отложившихся отложений амилоида с использованием меченного 124I антитела). Таким образом, меченое антитело по изобретению можно использовать для обнаружения наличия болезни отложения амилоида у пациента, у которого подозревают наличие этого заболевания, а также для определения результативности лечения.
Для получения химерных антител по изобретению гены вариабельных областей тяжелой и легкой цепи каппа мышиного моноклонального антитела 11-1F4, описанного в патенте США 8,105,594, были модифицированы с помощью полимеразной цепной реакции (ПЦР), чтобы способствовать экспрессии химерного антитела 11-1F4 в клетках млекопитающих. Был проведен детальный анализ последовательностей модифицированных генов вариабельной области. Модифицированные гены вариабельной области клонировали в соответствующих экспрессионных векторах млекопитающих, в результате чего были созданы конструкции 11-1F4VHpG1D200 и 11-1F4VK.pKN100. Из конструкций 11-1F4VHpG1D200 и 11-1F4VK.pKN100 путем расщепления ферментом рестрикции EcoRI и лигирования была получена единая супервекторная конструкция pG1KD200-11-1F4. Наконец, химерное антитело 11-1F4 транзиентно экспрессировали в клетках COS, как посредством котрансфекции, так и посредством трансфекции единым супервектором. Хотя для удобства котрансфекции или трансфекции были выбраны клетки COS (яичник китайского хомячка), специалистам в данной области техники должно быть понятно, что можно использовать и другие линии клеток млекопитающих. Характеристика способности химерного антитела 11-1F4 к связыванию с амилоидными фибриллами определяли методом ИФА с прямым связыванием. Было неожиданно обнаружено и оказалось полезным, что химерное антитело 11-1F4 связывается с амилоидными фибриллами с более высокой аффинностью, чем мышиное антитело 11-1F4.
Антитело по изобретению включает химерное моноклональное антитело мыши и человека, содержащее область VK с SEQ ID NO: 47 и область VH с SEQ ID NO: 48. Данное антитело связывается с эпитопом β-складчато-листовой конфигурации амилоидных фибрилл. Кроме того, неожиданно обнаружено, что это антитело связывается с данным эпитопом с более высокой аффинностью, чем мышиное антитело 11-1F4, из которого оно получено, содержащее область VK с SEQ ID NO: 36 и область VH с SEQ ID NO: 35. Изобретение включает способы лечения болезни, характеризующейся отложением амилоида у нуждающегося в этом пациента-человека, включающие введение пациенту описанного выше антитела в фармацевтически приемлемом носителе. Количество вводимого антитела должно быть эффективным для уменьшения количества амилоидных фибрилл, отложившихся в тканях пациента. Композицию антитела можно вводить любым традиционным путем введения, но предпочтительно парентеральное введение (например, внутривенное). Фармацевтически приемлемые носители хорошо известны в данной области техники, и приемлемый носитель может быть выбран специалистом в области медицины. Болезнь, характеризующаяся отложением амилоида, предпочтительно представляет собой первичный (AL) амилоидоз.
Изобретение также включает способы и материалы для получения антитела, являющегося предметом изобретения. Материалы, применимые для получения антитела, являющегося предметом изобретения, включают векторные конструкции, выбранные из группы, состоящей из 11-1F4VK.pKN100 и 11-F4VH.pG1D200, показанных на Фиг. 5 и 6 соответственно, и суперконструкции pG.1KD20011-1F4, полученной из двух описанных выше векторных конструкций. Другие применимые материалы включают модифицированный ген области VK мышиного антитела 11-1F4 (SEQ ID NO: 42) и модифицированный ген области VH антитела 11-1F4 SEQ ID NO: 45), а также соответствующие праймеры SEQ ID NO: 41, 43, 44 и 46. Антитело, являющееся предметом изобретения, может быть получено путем котрансфекции векторных конструкций 11-1F4VK.pKN100 и 11-F4VH.pG1D200 или суперконструкции pG.1KD20011-1F4 в подходящей клетке-хозяине млекопитающего, такой как клетки COS (яичник китайского хомячка).
Сокращения
Модифицированная Дульбекко среда Игла (DMEM), фетальная бычья сыворотка (ФБС), рибонуклеиновая кислота (РНК); мессенджер-РНК (мРНК); дезоксирибонуклеиновая кислота (ДНК); копийная ДНК (кДНК); полимеразная цепная реакция (ПЦР); минута (мин); секунда (с); трис-боратный буфер (ТБЭ).
Аминокислоты представлены сокращениями по номенклатуре IUPAC, как описано ниже: аланин (Ala A), аргинин (Arg; R), аспарагин (Asn; N), аспарагиновая кислота (Asp; D), цистеин (Cys; C), глутамин (Gln; Q), глутаминовая кислота (Glu; E), глицин (Gly; G), гистидин (His; H), изолейцин (Ile; I), лейцин (Leu; L), лизин (Lys; K), метионин (Met; M), фенилаланин (Phe; F), пролин (Pro; P), серин (Ser; S), треонин (Thr; T), триптофан (Trp; W), тирозин (Tyr; Y), валин (Val; V). Аналогично для нуклеотидов: аденин (a), цитозин (c), гуанин (g), тимин (t), урацил (u), аденин или гуанин (r), цитозин или тимин (y), гуанин или цитозин (s), аденин или тимин (w), гуанин или тимин (k), аденин или цитозин (m), цитозин или гуанин или тимин (b), аденин или гуанин или тимин (d), аденин или цитозин или тимин (h), аденин или цитозин или гуанин (v) и любое азотистое основание (n).
ПРИМЕР 1
ПЦР клонирование и секвенирование ДНК мышиного антитела 11-1F4
Гены вариабельных областей тяжелой и легкой цепи мышиного антитела 11-1F4 клонируют с помощью ПЦР и проводят детальный анализ последовательностей всех выделенных генов вариабельных областей (как псевдогенов, так и функциональных генов). Были получены детальные ДНК- и аминокислотные последовательности генов вариабельных областей мышиного антитела 11-1F4.
Материалы
Компоненты питательной среды и все остальные материалы для тканевых культур были получены от Life Technologies (Великобритания). Набор растворов для выделения РНК был получен от Stratagene (США), а набор для синтеза первой нити кДНК приобретали у Pharmacia (Великобритания). Все компоненты и оборудование для проведения реакции ПЦР, в том числе ДНК полимеразу AmpliTaq®, приобретали у Perkin Elmer (США). Набор реактивов TOPO TA Cloning® получали от Invitrogen (США). Агарозу (UltraPure™) получали от Life Technologies (Великобритания). Набор реактивов ABI PRISM® Big Dye™ Terminator Cycle Sequencing, представляющий собой готовую смесь для циклического секвенирования, и аппарат для секвенирования ABI PRISM® 310 приобретали у PE Applied Biosystems (США). Все остальные продукты для молекулярной биологии получали от New England Biolabs (США) и Promega (США).
Способы
Стратегия, используемая для ПЦР клонирования генов мышиных VH и VK из линий клеток гибридомы, продуцирующих мышиное моноклональное антитело 11-1F4, схематично изображена на Фиг. 1.
Два клона (B2C4 и B2D6) линии клеток гибридомы SP2/0, продуцирующей моноклональное антитело 11-1F4 к человеческой α-легкой цепи, были любезно предоставлены Аланом Соломоном (Alan Solomon), MD (Медицинский центр университета штата Теннесси в Ноксвилле, штат Теннесси). Линия клеток гибридомы имеется в Американской коллекции типовых культур (ATCC, номер доступа PTA-105). Линии клеток культивируют с использованием питательной среды DMEM с добавлением 20 % (об./об.) ФБС, пенициллина/стрептомицина и L-глутамина. Клетки культивируют до достижения общего количества жизнеспособных клеток, равного 108 клеткам.
Клетки собирают из каждого клона отдельно, как описано ниже. Линию клеток гибридомы мыши выращивают в суспензии в соответствующей культуральной среде и в достаточных количествах, чтобы обеспечить общее количество жизнеспособных клеток, составляющее около 108 клеткам. Собирают супернатант культуры и осаждают клетки гибридомы в настольной центрифуге (250 g, 5 мин). Клетки осторожно ресуспендируют в 20 мл ФСБ (фосфатно-солевой буферный раствор) и отбирают аликвоту 100 мкл для подсчета жизнеспособных клеток. Клетки в аликвоте еще раз осаждают, добавляют к 100 мкл клеток 200 мкл ФСБ и 200 мкл трипанового синего и осторожно перемешивают. 10 мкл данной смеси отбирают пипеткой на одноразовое предметное стекло для подсчета клеток и подсчитывают под микроскопом количество белых клеток на 9 малых площадях. Клетки синего цвета (т.е. погибшие клетки) не подсчитывают. Процесс подсчета повторяют, результаты усредняют, и средние результаты умножают на 9×105, чтобы получить количество жизнеспособных клеток в 20 мл ФСБ. Как только было собрано достаточное количество клеток, их ресуспендируют в 10 мл раствора D для выделения РНК (см. ниже, набор реактивов для выделения РНК Stratagene).
Затем с использованием набора реактивов для выделения РНК Stratagene из клеток каждого клона по отдельности выделяют суммарную РНК. К образцу добавляют ли один мл 2 M раствора ацетата натрия (pH 4) и тщательно перемешивают содержимое пробирки, многократно переворачивая пробирку. В пробирку добавляют 10,0 мл фенола (pH 5,3-5,7) и снова тщательно перемешивают содержимое переворачиванием. К смеси добавляют 2,0 мл смеси хлороформа и изоамилового спирта, пробирку закрывают крышкой и энергично встряхивают в течение 10 секунд, и пробирку инкубируют во льду в течение 15 минут. Образец переносят в толстостенную круглодонную центрифужную пробирку емкостью 50 мл, предварительно охлажденную на льду, и осаждают содержимое пробирки в центрифуге при 10000 g в течение 20 минут при 4°C. После центрифугирования в пробирке были видны две фазы. Верхняя водная фаза содержала РНК, а нижняя фенольная фаза и интерфаза содержали ДНК и белки. Содержащую РНК верхнюю водную фазу переносят в новую центрифужную пробирку, а нижнюю фенольную фазу отбрасывают. К водной фазе добавляют равный объем изопропанола и перемешивают содержимое переворачиванием, после чего пробирку инкубируют в течение 1 часа при минус 20°C для осаждения РНК. Содержимое пробирки осаждают в центрифуге при 10000 g в течение 20 минут при 4°C. После центрифугирования осадок на дне пробирки, содержащий РНК, извлекают, а супернатант отбрасывают. Осадок растворяют в 3,0 мл раствора D, в пробирку добавляют 3,0 мл изопропанола и тщательно перемешивают содержимое. После инкубации пробирок в течение 1 часа при минус 20°C, ее снова центрифугируют при 10000 g в течение 20 минут при 4°C, и супернатант удаляют из пробирки и отбрасывают. (Примечание: до этого момента РНК была защищена от рибонуклеаз присутствием гуанидинизотиоцианата, но с этого момента она не защищена). Осадок промывают смесью 75% (об./об.) этанола и 25% обработанной диэтилпирокарбонатом (ДЭПК) воды и высушивают осадок в вакууме в течение 2-3 минут. Осадок РНК ресуспендируют в 0,5-2 мл ДЭПК-обработанной воды.
Следуя инструкциям производителя, для получения однонитевой ДНК-копии мРНК гибридомы 11-1F4 используют набор для синтеза первой нити кДНК Amersham Pharmacia Biotech и праймер Not I-d(T)18, поставляемый с набором. Для каждого из двух образцов РНК проводят одну реакцию, как описано ниже. Используемые компоненты представляют собой: основную реакционную смесь для синтеза первой нити кДНК, клонированную мышиную обратную транскриптазу FPLCpure™, RNAguard™, БСА (бычий сывороточный альбумин), дАТФ (дезоксиаденозина трифосфат), дЦТФ (дезоксицитидин трифосфат), дГТФ (дезоксигуанозина трифосфат) и дТТФ (дезокситимидинтрифосфат), 200 мМ водный раствор DIT, праймер Not I-d(T)18: 5'-d[AACTGGAAGAATTCGCGGCCGCAGGAA18]-3' и ДЭПК-обработанную воду.
Приблизительно 5 мкг суммарной РНК в 20 мкл воды с ДЭПК нагревают до 65°C в течение 10 мин, а затем охлаждают на льду. Основную реакционную смесь для синтеза первой нити кДНК осторожно перемешивают пипеткой до получения однородной суспензии и собирают реакционную смесь в микроцентрифужной пробирке емкостью 0,5 мл, как описано ниже: 20 мкл раствора денатурированной РНК, 11 мкл основной реакционной смеси для синтеза первой нити кДНК, 1 мкл праймера Not I-d(T)18 и 1 мкл раствора ДТТ (дитиотреитол) с получением суммарного объема 33 мкл. Реактивы осторожно перемешивают с помощью пипетки и инкубируют при 37°C в течение 1 часа.
Затем гены вариабельных областей мышиных тяжелой цепи и легкой цепи каппа (гены VH и гены VK соответственно) амплифицируют с помощью ПЦР с матрицы однонитевой ДНК, используя способ, описанный Jones и Bendig (Bio/Technology 9:88).
Для каждого из вырожденных праймеров, специфичных к лидерной последовательности (MHV1-MHV12 для VH и MKV1-MKV11 для VK), и соответствующего праймера константной области (эквимолярная смесь MHC1-MHC3 для VH и MKC для VK) готовят отдельные ПЦР. В таблицах 1 и 2 подробно описаны праймеры, используемые для амплификации генов VH и VK областей соответственно. В общей сумме было проведено 12 реакций с тяжелой цепью и 11 реакций с легкой цепью каппа. Во всех случаях для амплификации матрицы кДНК используют ДНК полимеразу AmpliTaq®, как описано ниже.
Завершенную реакцию синтеза первой нити кДНК нагревают при 90°C в течение 5 минут для денатурации дуплекса РНК-кДНК и инактивации обратной транскриптазы и охлаждают на льду. Одиннадцать пробирок для реакции ПЦР GeneAmp™ маркируют MKV1-11. Для каждой пробирки готовят реакционную смесь объемом 100 мкл, где каждая реакционная смесь содержала 69,3 мкл стерильной воды, 10 мкл 10-кратного буфера II для ПЦР, 6 мкл 25 мМ MgCl2, 2 мкл каждого из 10 мМ исходных растворов дНТФ (дезоксинуклеозидтрифосфат), 2,5 мкл 10 мМ праймера MKC, 2,5 мкл одного из 10 мМ растворов праймеров MKV и 1 мкл смеси матриц РНК-кДНК. Затем в каждую из пробирок добавляют 0,7 мкл ДНК полимеразы AmpliTaq®, и на готовую реакционную смесь наслаивают 50 мкл минерального масла.
Для клонирования гена вариабельной области мышиной тяжелой цепи с помощью ПЦР готовят аналогичную серию реакционных смесей, как описано выше. Однако в это время маркируют двенадцать реакционных пробирок, и в каждую добавляют один из двенадцати праймеров MHV и соответствующий праймер MHC. Таким образом, например, для ПЦР амплификации гена вариабельного домена мышиной тяжелой цепи γ1 используют праймер MHC G1.
Реакционные пробирки устанавливают в термоциклер для ДНК и проводят цикл (после первоначального плавления при 94°C в течение 1,5 мин) при 94°C в течение 1 мин, при 50°C в течение 1 мин и при 72°C в течение 1 мин более 25 раз. За последним циклом следовал этап элонгации при 72°C в течение 10 мин перед охлаждением при 4°C. За исключением промежутка между этапами отжига (50°C) и элонгации (72°C), где используют увеличенное время изменения температуры 2,5 мин, между каждым из этапов цикла время изменения температуры составляло 30 с. Чтобы определить, с какого из праймеров к лидерной последовательности получен продукт ПЦР, проводят аликвоту 10 мкл из каждой реакции ПЦР в 1% (мас./об.) агарозном геле на 1-кратном буфере ТБЭ (буфер трис-борат-ЭДТА), содержащем 0,5 мкг/мл бромистого этидия. Положительные клоны ПЦР имели размер около 420-500 п.о.
Описанный выше процесс ПЦР амплификации повторяют еще два раза и отбирают те реакции ПЦР, где амплифицированным оказывался полноразмерный ген вариабельного домена. Аликвоту 6 мкл каждого потенциального продукта ПЦР клонируют непосредственно в векторе pCR™Il, поставляемом с набором реактивов TA Cloning®, как описано в инструкциях производителя. Аликвоты, содержащие 10% (об./об.), 1% (об./об.) и 0,1 % (об./об.) трансформированных клеток E. coli, переносят пипеткой на отдельные чашки Петри с агаром Лурия-Бертани (LB) диаметром 90 мм, содержащим 50 мкг/мл ампициллина, наслаивают 25 мкл исходного раствора X-Gal и 40 мкл исходного раствора изопропилтиогалактозида (ИПТГ) и инкубируют в течение ночи при 37°C. Положительные колонии определяют с помощью ПЦР-скрининга.
Таблица 1. Праймеры ПЦР для клонирования генов вариабельной области мышиной легкой цепи каппа
Таблица 2. Праймеры ПЦР для клонирования генов вариабельной области мышиной тяжелой цепи
Чтобы определить, получен ли продукт ПЦР правильного размера (приблизительно 450 п.о.), проводят аликвоту по пять мкл каждой реакции в 1% агарозном геле/ТБЭ (pH 8,8). Определенные таким образом предполагаемые положительные продукты ПЦР непосредственно клонируют в векторе pCR2.1, поставляемом с набором реактивов TA Cloning®, и трансформируют в компетентные клетки TOP10, как описано в протоколе производителя. Колонии, содержащие плазмиду со вставкой правильного размера, определяют с помощью ПЦР-скрининга колоний, используя олигонуклеотидные праймеры 1212 и 1233 (таблица 3), в соответствии со способом  и Clackson (Nucleic Acids Res. 17: 4000). Данные определенные таким образом предполагаемые положительные клоны подвергают двунитевому секвенированию ДНК, используя генетический анализатор ABI PRISM 310 и терминатор ABI PRISM BigDye™. Были секвенированы три положительных клона по каждому из генов VH и VK из клона B2C4 линии клеток гибридомы, а также четыре положительных клона по гену VK и шесть по гену VH из клона B2D6 линии клеток гибридомы.
и Clackson (Nucleic Acids Res. 17: 4000). Данные определенные таким образом предполагаемые положительные клоны подвергают двунитевому секвенированию ДНК, используя генетический анализатор ABI PRISM 310 и терминатор ABI PRISM BigDye™. Были секвенированы три положительных клона по каждому из генов VH и VK из клона B2C4 линии клеток гибридомы, а также четыре положительных клона по гену VK и шесть по гену VH из клона B2D6 линии клеток гибридомы.
Таблица 3. Праймеры для ПЦР скрининга и секвенирования трансформированных колоний
Результаты 12 реакций ПЦР, проведенных для каждого клона гибридомы (B2C4 и BCD6) для амплификации гена вариабельной области тяжелой цепи 11-1F4, представлены в таблице 4(a).
Вырожденный праймер лидерной последовательности MHV7 в комбинации со смесью праймеров к константной области MHCG1-3 (таблица 1) образуют продукт ПЦР около 600 п.о. с матрицы кДНК, полученных из обеих линий клеток гибридомы B2C4 и B2D6. Поскольку эта полоса была больше ожидаемого размера среднего гена VH (450 п.о.), ее дальнейшее исследование не проводят. Наоборот, вырожденный праймер к лидерной последовательности MHV6 в комбинации со смесью праймеров к константной области MHCG1-3 (таблица 1) образуют продукт ПЦР ожидаемого размера (450 п. о.) гена VH с матрицы кДНК, полученных из обеих линий клеток гибридомы B2C4 и B2D6.
Таблица 4. Результаты ПЦР амплификаций, проведенных для клонирования генов тяжелой (а) и легкой (б) цепи мышиного моноклонального антитела 11-1F4 из линий клеток B2C4 и B2D6 гибридомы SP2/0. Третья колонка содержит записи фактических результатов ПЦР. При наличии полосы в случае конкретной комбинации праймеров ее размер в парах оснований (п. о.) указан в соответствующем месте.
(а)
(б)
В анализе последовательности продукта ПЦР, полученного из трех клонов B2C4, и продукта ПЦР, полученного из пяти клонов B2D6, была выявлена единая последовательность вариабельной области тяжелой цепи (Фиг. 2).
Используемая стратегия клонирования (амплификация всего гена вариабельной области с использованием праймеров, фланкирующих эту область, т.е. к лидерной последовательности, и специфичных праймеров к последовательности константной области) позволила определить полноразмерную последовательность FR1. Все восемь секвенированных клонов имели идентичную последовательность в данной области (Фиг. 2).
Результаты 11 реакций ПЦР, проведенных для каждого клона гибридомы (B2C4 и BCD6) для амплификации гена вариабельной области легкой цепи каппа 11-1F4, представлены в таблице 4(б).
Вырожденный праймер лидерной последовательности MKV6 в комбинации со смесью праймеров к константной области MKC (таблица 2) образуют с матрицы кДНК, полученных только из линии клеток гибридомы B2C4, продукт ПЦР около 200 п.о. Поскольку эта полоса была значительно меньше ожидаемого размера гена VK (450 п.о.), ее дальнейшее исследование не проводят.
Вырожденный праймер лидерной последовательности MKV2 в комбинации со смесью праймеров к константной области MKC (таблица 2) образуют с матрицы кДНК, полученной из обеих линий клеток гибридомы B2C4 и B2D6, продукт ПЦР, размер полосы которого при визуализации в агарозном геле был значительно меньше ожидаемого размера 450 п.о. Кроме того, при предшествующем клонировании VK было обнаружено, что с праймером MKV2 амплифицируется хорошо известный псевдоген легкой цепи каппа. Таким образом, проводят анализ последовательности каждого продукта ПЦР, чтобы подтвердить, что данный продукт является псевдогеном, а не геном VK мышиного антитела 11-1F4. В данном анализе последовательности выявляют, что продукт ПЦР этого клона действительно является псевдогеном.
Наконец, вырожденный праймер к лидерной последовательности MKV1 в комбинации со смесью праймеров к константной области MKC (таблица 1) образуют продукт ПЦР ожидаемого размера (450 п.о.) гена VK с матрицы кДНК, полученных из обеих линий клеток гибридомы B2C4 и B2D6.
В анализе последовательности продукта ПЦР, полученного из трех клонов B2C4, и продукта ПЦР, полученного из четырех клонов B2D6, выявляют единую последовательность вариабельной области легкой цепи каппа, которую было невозможно определить как псевдоген.
Таким образом, из мРНК гибридомы клонируют и секвенируют ген вариабельной области тяжелой цепи 11-1F4 с использованием специфичных праймеров к константной области и специфичных праймеров к лидерной последовательности).
При трансляции с этой последовательности получают пептидную последовательность TVSS. В анализе 122 перегруппированных человеческих генов VH, зарегистрированных в базе данных Кабата (Kabat et al. - Sequences of Proteins of Immunological Interest), было выявлено, что 84 % этих последовательностей имеет пептидную последовательность TVSS. Таким образом, был сделан вывод, что изолированный ген VH имеет корректную последовательность гена 11-1F4.
Ген вариабельной области легкой цепи каппа мышиного антитела 11-1F4, а также нефункциональный псевдоген VK также успешно клонируют и секвенируют. Данный псевдоген был впервые идентифицирован в работе Кэррола и соавт. (Carroll et al., Molecular Immunology (1988) 25: 991). Указанная последовательность образуется из аберрантного транскрипта мРНК, который присутствует во всех стандартных партнерах по слиянию, полученных из исходной опухоли MOPC-21 (включая SP2/0). В результате этой аберрантной мРНК инвариантный цистеин в положении 23 заменен остатком тирозина, и соединение VJ не попадает в рамку считывания, в результате чего образуется стоп-кодон в положении 105.
Для лимфоидных клеток и клеток гибридомы характерен синтезирование более чем одной мРНК перегруппированной легкой цепи иммуноглобулина. За счет присутствия кодонов терминации или сдвигов рамки считывания, обычно не наблюдаемых в функциональных генах VK, данные мРНК обычно непродуктивны. Данные псевдо-мессенджеры обычно создают большие проблемы при клонировании генов иммуноглобулинов из гибридом, потому что являются весьма благоприятными субстратами для ПЦР V области, несмотря на тот факт, что они не кодируют функциональные полипептиды.
Последовательность гена 11-1F4 VK определяют после детального анализа семи отдельных клонов ПЦР, выделенных из двух различных продуктов ПЦР, с получением SEQ. ID NO: 36. Поскольку все последовательности были идентичными, они были приняты за корректную последовательность вариабельной области легкой цепи каппа 11-1F4.
Клонированные гены областей VH и VK используют для получения химерного моноклонального антитела мыши и человека 11-1F4, которое впоследствии анализируют для подтверждения специфичного связывания с фибриллами AL.
ПРИМЕР 2
Конструирование химерного антитела мыши и человека 11-1F4 (c11-1F4)
Чтобы допустить транзиентную экспрессию описанных выше генов вариабельных областей VH и VK 11-1F4 в клетках млекопитающих в составе химерного антитела мыши и человека, было необходимо модифицировать 5'- и 3'-концы с использованием специально разработанных ПЦР праймеров (таблица 5). Для ПЦР-модификации гена VK 11-1F4 используют олигонуклеотидные праймеры F39836 и F39837, а для ПЦР-модификации гена VH 11-1F4 используют праймеры F39835 и F58933. Обратные праймеры (BAK) F39836 и F39835 вводят сайт рестрикции HindIII, сайт инициации трансляции Козак и лидерную последовательность иммуноглобулина в 5'-концы генов в VK и VH соответственно. Прямой олигонуклеотидный праймер (FOR) F39837 вводит в 3'-конец гена VK донорный сайт сплайсинга и сайт рестрикции BamHI, а прямой праймер (FOR) F58933 присоединяет к 3'-концу гена VH первые 22 пары оснований гена гамма-1CH1, включая сайт рестрикции ApaI.
Таблица 5. Олигонуклеотидные праймеры, используемые для модификации генов вариабельных областей тяжелой цепи и легкой цепи каппа 11-1F4
Консенсусная последовательность Козак является крайне важной для эффективной трансляции последовательности вариабельной области (Kozak - J Mol Bio 196:947). Она определяет корректный кодон AUG, с которой рибосома начинает трансляцию, и отдельным наиболее критичным основанием является аденин (или менее предпочтительно гуанин) в положении -3 выше старта AUG.
Лидерная последовательность иммуноглобулина обеспечивает секрецию экспрессированного антитела в питательную среду, и, таким образом, его можно легко собирать и очищать. Используемыми в данном случае лидерными последовательностями являются лидерные последовательности VK и VH мышиного антитела 11-1F4, клонированные с кДНК гибридомы в ходе клонирования VH и VK.
Донорная последовательность сплайсинга важна для корректного присоединения вариабельной области легкой цепи в одной рамке считывания с ее соответствующей константной областью в результате сплайсинга интрона VK:CK длиной 130 п.о. Вариабельную область тяжелой цепи присоединяют непосредственно к гену ее соответствующей константной области посредством сайта Apal, что устраняет необходимость в донорном сайте сплайсинга.
Сайты рестрикции HindIII и BamHI, а также HindIll и Apal соответственно для субклонирования заключают в себе модифицированные гены вариабельной области VK и VH, а использование различных сайтов рестрикции обеспечивает направленное субклонирование в соответствующем экспрессионном векторе млекопитающих.
Сначала ген вариабельной области легкой цепи антитела 11-1F4 тщательно анализируют, чтобы определить любые нежелательные донорные сайты сплайсинга, акцепторные сайты сплайсинга и последовательности Козак (см. таблицу 6). Оба гена вариабельных областей тяжелой и легкой цепи анализируют, чтобы определить наличие любых избыточных сайтов рестрикции, которые впоследствии могли бы препятствовать субклонированию и/или экспрессии полноразмерного функционального антитела. Ни один из них не был обнаружен.
Таблица 6. Последовательности, важные для эффективной экспрессии генов иммуноглобулинов в клетках млекопитающих
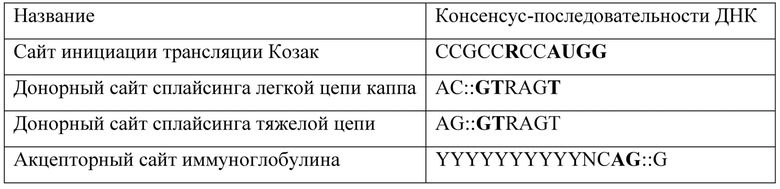
В пределах каждой консенсусной последовательности основания, показанные полужирным шрифтом, считают инвариантными.
Отдельные реакции ПЦР, по одной для каждого гена вариабельной области, готовят, как описано ниже. В качестве матриц используют описанные выше плазмиды 11-1F4 VH.pCR2.1 и 11-1F4 VK.pCR2.1. В каждой пробирке для ПЦР готовят реакционную смесь объемом 100 мкл, где каждая смесь содержала 41 мкл стерильной воды, 10 мкл 10-кратного буфера I для ПЦР, 8 мкл 10 мМ исходного раствора дНТФ, 1 мкл 10 мМ раствора прямого 5'-праймера, 1 мкл 10 мМ раствора обратного 3'-праймера и 1 мкл матрицы ДНК, разведенной 1 к 10. Наконец, добавляют 0,5 мкл ДНК полимеразы AmpliTaq® (2,5 единицы) перед нанесением 50 мкл минерального масла на полную реакционную смесь. Реакционные пробирки устанавливают в термоциклер для ДНК и проводят циклы (после первоначального плавления при 94°C в течение 1 мин) при 94°C в течение 30 с, 68°C в течение 30 с и 72°C в течение 50 с более 25 циклов. После завершения последнего цикла следует стадия элонгации при 72°C в течение 7 мин, а затем охлаждение до 4°C. Проводят аликвоту 10 мкл из каждой реакционной пробирки для ПЦР в 1,2 % (мас./об.) агарозном геле на 1-кратном буфере ТБЭ, содержащем 0,5 мкг/мл бромистого этидия, чтобы определить размер и наличие продукта ПЦР. Положительные клоны ПЦР имели размер около 420 п.о. Определенные таким образом предполагаемые положительные продукты ПЦР непосредственно клонируют в векторе pCR2.1, поставляемом с набором реактивов Topo TA Cloning®, и трансформируют компетентные клетки TOP10, как описано в протоколе производителя. Колонии, содержащие плазмиду со вставкой верного размера, определяют с помощью ПЦР-скрининга колоний, используя олигонуклеотидные праймеры 1212 и 1233 (таблица 3), в соответствии со способом  и Clackson (Nucleic Acids Res. Данные определенные таким образом предполагаемые положительные клоны подвергалютдвунитевому секвенированию ДНК, используя генетический анализатор ABI PRISM 310 и терминатор ABI PRISM BigDye™. Было секвенировано два положительных клона каждого клонированного с помощью Topo TA гена VH и VK.
и Clackson (Nucleic Acids Res. Данные определенные таким образом предполагаемые положительные клоны подвергалютдвунитевому секвенированию ДНК, используя генетический анализатор ABI PRISM 310 и терминатор ABI PRISM BigDye™. Было секвенировано два положительных клона каждого клонированного с помощью Topo TA гена VH и VK.
Были определены клоны, содержащие верную модификацию генов 11-1F4 VH и 11-1F4 VK, и модифицированные гены V из этих клонов субклонируют в их соответствующих экспрессионных векторах, чтобы способствовать экспрессии химерных тяжелой и легкой цепей в клетках млекопитающих. Модифицированный ген 11-1F4 VK субклонируютн в экспрессионном векторе pKN100 (Фиг. 4) в виде фрагмента HindIII-BamHI; данный вектор содержит человеческий ген константной области каппа (аллотип: Km (3 Ala153, Ser191)). Модифицированный ген 11-1F4 VH был также субклонируют в виде фрагмента HindIII-ApaI в экспрессионном векторе pG1D200 (Фиг. 5); этот вектор содержал человеческий ген константной области γ1 (аллотип: G1m (-1 Glu377, Met38I, -2 Ala462, 3 Arg222, Ser229)). Оба аллотипа константной области, каппа и γ1, часто обнаруживаются у людей европеоидной расы. Впоследствии лигированные экспрессионные конструкции 11-1F4VK.pKN100 и 11-1F4VH.pG1D200 используют для трансформации компетентных клеток DH5α и определяют положительные клоны обсуждаемым выше методом ПЦР скрининга с исходными праймерами для ПЦР модификации (таблица 4). Экспрессионные векторы широкодоступны.
ПРИМЕР 3
Конструирование единого супервектора для транзиентной экспрессии 11-1F4 в клетках COS.
Единый супервектор, экспрессирующий обе цепи иммуноглобулина химерного антитела 11-1F4, как описано ниже. Экспрессионную кассету легкой цепи каппа 11-1F4 (которая содержала промотор HCMVi, ген вариабельной области легкой цепи каппа 11-1F4 и ген константной области легкой цепи каппа) вырезают из конструкции 11-1F4VK.pKN100 (Фиг. 4) ферментом рестрикции (EcoRI в положениях 1 и 2490) и впоследствии лигируют с конструкцией 11-1F4VHpG1D200 через уникальный сайт EcoRI (положение 4297, Фиг. 5). В результате этого лигирования была сконструирована супервекторная конструкция pG1KD200-11-1F4, содержащая обе, тяжелую и каппа-легкую, цепи химерного антитела 11-1F4.
ПРИМЕР 4
Транзиентная экспрессия полноразмерного химерного антитела γ1/κ.11-1F4 в клетках COS
Химерное антитело 11-1F4 транзиентно экспрессируют в клетках COS из Европейской коллекции культур клеток (ECACC) двумя способами:
(i) путем котрансфекции 10 мкг каждой векторной конструкции 11-1F4VK.pKN100 и 11-1F4VH.pG1D200. Котрансфекции проводят в двух повторностях.
(ii) путем трансфекции 13 мкг единой супервекторной конструкции pG1KD200-11-1F4. Трансфекции супервектором проводят пять раз.
Используют описанный ниже метод трансфекции. Линию клеток COS выращивают в среде DMEM с добавлением 10% (об./об.) ФТС (фетальная телячья сыворотка), 580 мкг/мл L-глутамина и 50 единиц/мл пенициллина/50 мкг/мл стрептомицина ("среда") в культуральном матрасе 150 см2 до конфлюентности. Клетки обрабатывают трипсином, осаждают в настольной центрифуге (250 g в течение 5 мин), затем ресуспендируют в 6 мл среды, после чего разделяют в равных частях на три культуральных матраса 150 см2, каждый из которых содержал 25 мл свежей предварительной нагретой среды. Клетки инкубируют в течение ночи при 37°C в атмосфере 5% CO2, а затем на следующий день собирают, при этом они еще росли экспоненциально. Каждый культуральный матрас содержал приблизительно 1×107 клеток. Клетки снова обрабатывают трипсином, осаждают, как описано выше, и промывают в 20 мл фосфатно-солевого буфера), после чего их ресуспендируют в достаточном количестве ФСБ до получения концентрации клеток 1×107 клеток/мл. 700 мкл данных промытых клеток COS отбирают пипеткой в кювету Gene Pulser®, в которую впоследствии добавляют 1 мкл ДНК обоих экспрессионных векторов тяжелой цепи и легкой цепи каппа (каждый в количестве 10 мкг) или 13 мкг супервекторной конструкции. К смеси прилагают электрический импульс 1900 Вольт емкостью 25 микрофарад с помощью прибора Bio-Rad Gene Pulser®. Воздействие импульсом повторяют для каждой экспериментальной трансфекции и контроля "без ДНК" (в котором клетки COS подвергают электропорации в отсутствие любой ДНК). Для проверки эффективности клеток COS также проводят положительный контроль с экспрессированным ранее антителом.
Клетки COS оставляют для восстановления при комнатной температуре на 10 мин, затем осторожно отбирают пипеткой в чашку Петри для культуры клеток диаметром 10 см, содержащую 8 мл предварительно нагретой среды DMEM с добавлением 10% (об./об.) ФБС не содержащей γ-глобулина, 580 мкг/мл L-глутамина, 50 единиц/мл пенициллина/50 мкг/мл стрептомицина и инкубируют в атмосфере 5% CO2 при 37°C в течение 72 часов перед сбором супернатанта клеток COS для анализа. После инкубации в течение 72 часов среду собирают, осаждают для удаления клеточных обломков и анализируют методом твердофазного иммуноферментного анализа (ИФА) продуцирование химерного антитела и связывание антигена антителом c11-1F4.
ПРИМЕР 5
Количественное определение химерного антитела γ1/κ 11-1F4 методом ИФА с захватом
Количественное определение присутствующих в супернатанте клеток COS молекул IgG после экспрессии проводят с использованием метода анализа ИФА с захватом. Захват молекул IgG иммобилизованным антителом, специфичным к фрагменту Fcγ, представляющим собой IgG козы против человека, проводят на планшете Nunc-Immuno MaxiSorb™ и обнаруживают его с помощью конъюгированного с пероксидазой хрена антитела к человеческой легкой цепи каппа. На основании захвата и обнаружения известных концентраций стандартного антитела IgG на том же планшете и таким же способом, как описано ниже, строили стандартную кривую. В каждую лунку 96-луночного иммунологического планшета наносили в качестве покрытия аликвоты по 100 мкл раствора антитела IgG козы против человека 0,4 мкг/мл в ФСБ и инкубируют при 4°C. Избыток раствора для покрытия удаляют, и планшет промывают три раза 200 мкл/лунка отмывочного буфера (1-кратный ФСБ, 0,1% Твин). Во все лунки, кроме лунок в колонке 2, ряды B-G, распределяют по 100 мкл буфера SEC. Готовят раствор 1 мкг/мл человеческого антитела IgG1/каппа в буфере SEC, действующий в качестве стандарта, и переносят по 200 мкл/лунка в колонку 2, ряды B и C. Среду из трансформированных клеток COS центрифугируют (250 g, 5 мин), сохраняя супернатант. Аликвоту 200 мкл супернатанта из контроля "без ДНК" (в котором клетки COS трансфицируют в отсутствие ДНК) отбирают пипеткой в лунки колонки 2, ряд D, а аликвоты по 200 мкл/лунка экспериментальных супернатантов отбирают пипеткой в лунки колонки 2, ряды E, F и G. Аликвоты по 200 мкл в лунках колонки 2, ряды B-G, перемешивают, а затем переносят по 100 мкл из каждой лунки в соседнюю лунку колонки 3. Процесс продолжают до колонки 11 с серией 2-кратных разведений стандартных, контрольных и экспериментальных образцов, после чего все образцы инкубируют при 37°C в течение 1 часа, и все лунки ополаскивают шесть раз аликвотами по 200 мкл отмывочного буфера. Конъюгат антитела козы к человеческой легкой цепи каппа с пероксидазой разводят в 5000 раз в буфере SEC и добавляют в каждую лунку по 100 мкл, после чего повторяют стадии инкубации и ополаскивания. В каждую лунку добавляют по 150 мкл субстрата K-BLUE с последующей инкубацией в темном месте при 25°C в течение 10 мин. Реакцию останавливают добавлением в каждую лунку по 50 мкл раствора RED STOP и снимают показания оптической плотности при 655 нм.
ПРИМЕР 6
Анализ связывания химерного антитела 11-1F4
Проводят исследование связывания химерного антитела 11-1F4 с амилоидными фибриллами с помощью анализа ИФА с прямым связыванием. Синтетические фибриллы формируют из белка легкой цепи иммуноглобулина и используют для мониторинга реакционности антитела в твердофазном анализе ИФА с использованием полистирольных планшетов "слабого связывания" (Costar, №3474). Непосредственно перед нанесением в лунки планшета в качестве покрытия массу фибрилл 250 мкг разводят в 1 мл буфером для покрытия (0,1 % бычьего сывороточного альбумина в фосфатно-солевом буферном растворе pH 7,5). Затем образец обрабатывают ультразвуком в течение 20 с, используя ультразвуковой зонд для дезинтеграции Tekmar Sonic Disruptor с установкой мощности 40% от максимума, в результате чего получили раствор коротких фибрилл, каждая из которых состояла из 2-5 протофиламентов. Данный раствор разводят до 5 мл, тщательно перемешивают на вихревой мешалке и делят на аликвоты в лунках планшета. В результате данного процесса в каждой лунке получают50 мкл раствора фибрилл, имеющего концентрацию 50 мкг/мл. Затем этот планшет высушивают в течение ночи, помещая его в термостат на 37°C без крышки.
Затем проводят анализ ИФА в течение 48 часов подготовки планшета, как описано ниже. Лунки блокируют добавлением 100 мкл 1% БСА в ФСБ и инкубируют в течение 1 часа при комнатной температуре на встряхивателе. Планшет промывают 3 раза в ФСБ с 0,05% Твином 20 (об./об.). В каждую лунку планшета добавляют 50 мкл раствора антитела c11-1F4 (3 мкг/мл антитела в 0,1% БСА/ФСБ), и планшет инкубируют при комнатной температуре в течение 1 часа на встряхивателе. Планшет снова, как и ранее, промывают 3 раза и выполняют обнаружение связанного антитела, используя биотинилированное козье антимышиное антитело IgG (Sigma №B-8774, антитело к тяжелой и легкой цепи).
Результаты
В анализе последовательности успешно модифицированных генов VH и VK было выявлено наличие верной последовательности. Детальные ДНК- и аминокислотные последовательности модифицированных генов VK и VH 11-1F4 представлены на Фиг. 3 и 4. Модифицированные гены VK и VH успешно клонируют в экспрессионных векторах млекопитающих pG1D200 и pKN100 соответственно, и полученные в результате конструкции 11-1F4VK.pKN100 и 11-1F4VHpG1D200 использовали для трансфекции клеток млекопитающих.
Конструкции 11-1F4VK.pKN100 и 11-1F4VHpG1D200 впоследствии также используют для конструирования единого супервектора pG1KD200-11-1F4, который экспрессирует химерное антитело 11-1F4 в клетках млекопитающих. Как после котрансфекций, так и после трансфекций супервектором ECACC клеток COS проводят определение уровней экспрессии химерного антитела 11-1F4. Экспериментально определенные уровни экспрессии после трансфекций супервектором pG1KD200-11-1F4 (10326 нг/мл) были в 3,7 раза выше экспериментально определенных уровней после соответствующих котрансфекций конструкциями 11-1F4VK.pKN100 и 11-1F4VHpG1D200 (2820 нг/мл).
После экспрессии и количественного определения исследуют связывание химерного антитела 11-1F4 с антигеном-мишенью (амилоидные фибриллы были любезно предоставлены Национальным институтом онкологии (NCI)) методом ИФА с прямым связыванием. Результаты ИФА связывания представлены на Фиг. 8. Проводят параллельный анализ супернатантов из двух лучших отдельных трансфекций супервектором pG1KD200-11-1F4 с одним супернатантом из соответствующей котрансфекции.
Результаты показали, что химерное антитело 11-1F4 связывается с амилоидными фибриллами с более высокой аффинностью, чем его мышиный эквивалент. Данный результат оказался неожиданным и непредсказуем, поскольку обычно предполагают, что химерное антитело обладает сопоставимой аффинностью связывания с исходным мышиным антителом. Без намерения ограничения каким-либо конкретным механизмом, авторы изобретения полагают, что антитело с повышенной аффинностью, возможно, получено в результате совокупного влияния объединения V областей мышиного антитела 11-1F4 с человеческими областями γ1/κ C, используемыми для создания химерного антитела 11-1F4.
В описании и формуле изобретения данного описания слово "содержать" и варианты этого слова, такие как "содержит" и "содержащий", не подразумевают исключения других элементов, дополнений, компонентов, целых чисел или стадий, но, если явным образом не указано иное, объем этих слов в широком смысле следует истолковывать так, что они вероятнее имеют включающее, а не исключающее значение.
Хотя композиции и способы по изобретению описаны в настоящем описании путем иллюстративных примеров, следует понимать, что изобретение не ограничено ими, и что могут быть выполнены варианты, известные специалистам в данной области техники, не отклоняясь от идей изобретения, определенных прилагаемой формулой изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ЗЕ ТРАСТИС ОФ КОЛАМБИЯ ЮНИВЕРСИТИ ИН ЗЕ СИТИ ОФ
НЬЮ-ЙОРК
<120> ХИМЕРНЫЕ АНТИТЕЛА ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, ХАРАКТЕРИЗУЮЩИХСЯ ОТЛОЖЕНИЕМ АМИЛОИДА
<130> 8441-0004WO
<150> US 62/526835
<151> 2017-06-29
<160> 48
<170> PatentIn версии 3,5
<210> 1
<211> 31
<212> ДНК
<213> Домовая мышь
<400> 1
atgaagattg cctgttaggc tgttggtgct g 31
<210> 2
<211> 31
<212> ДНК
<213> Домовая мышь
<400> 2
atggagwcag acacactccc tgytatgggt g 31
<210> 3
<211> 30
<212> ДНК
<213> Домовая мышь
<400> 3
atgagtgtgc tcactcaggt cctggsgttg 30
<210> 4
<211> 33
<212> ДНК
<213> Домовая мышь
<400> 4
atgaggrccc ctgctcagwt tyttggmwtc ttg 33
<210> 5
<211> 30
<212> ДНК
<213> Домовая мышь
<400> 5
atggatttwc aggtgcagat twtcagcttc 30
<210> 6
<211> 29
<212> ДНК
<213> Домовая мышь
<400> 6
atgaggtkcy ytgytsayct yctctgrgg 29
<210> 7
<211> 32
<212> ДНК
<213> Домовая мышь
<400> 7
atgggcwtca aagatggagt cacakwyycw gg 32
<210> 8
<211> 31
<212> ДНК
<213> Домовая мышь
<400> 8
atgtggggay ctkttttycm mtttttcaat g 31
<210> 9
<211> 25
<212> ДНК
<213> Домовая мышь
<400> 9
atggtrtccw casctcagtt ccttg 25
<210> 10
<211> 27
<212> ДНК
<213> Домовая мышь
<400> 10
atgtatatat gtttgttgtc tatttct 27
<210> 11
<211> 28
<212> ДНК
<213> Домовая мышь
<400> 11
atggaagccc cagctcagct tctcttcc 28
<210> 12
<211> 20
<212> ДНК
<213> Домовая мышь
<400> 12
actggatggt gggaagatgg 20
<210> 13
<211> 27
<212> ДНК
<213> Домовая мышь
<400> 13
atgaaatgca gctggggcat sttcttc 27
<210> 14
<211> 26
<212> ДНК
<213> Домовая мышь
<400> 14
atgggatgga gctrtatcat sytctt 26
<210> 15
<211> 27
<212> ДНК
<213> Домовая мышь
<400> 15
atgaagwtgt ggttaaactg ggttttt 27
<210> 16
<211> 25
<212> ДНК
<213> Домовая мышь
<400> 16
atgractttg ggytcagctt grttt 25
<210> 17
<211> 32
<212> ДНК
<213> Домовая мышь
<400> 17
atgggactcc aggcttcaat ttagttttcc tt 32
<210> 18
<211> 29
<212> ДНК
<213> Домовая мышь
<400> 18
atggcttgtc yttrgsgctr ctcttctgc 29
<210> 19
<211> 27
<212> ДНК
<213> Домовая мышь
<400> 19
atggratgga gckggrgtct ttmtctt 27
<210> 20
<211> 23
<212> ДНК
<213> Домовая мышь
<400> 20
atgagagtgc tgattctttt gtc 23
<210> 21
<211> 31
<212> ДНК
<213> Домовая мышь
<400> 21
atggmttggg tgtggamctt gcttattcct g 31
<210> 22
<211> 28
<212> ДНК
<213> Домовая мышь
<400> 22
atgggcagac ttaccattct cattcctg 28
<210> 23
<211> 28
<212> ДНК
<213> Домовая мышь
<400> 23
atggattttg ggctgatttt ttttattg 28
<210> 24
<211> 27
<212> ДНК
<213> Домовая мышь
<400> 24
atgatggtgt taagtcttct gtacctg 27
<210> 25
<211> 21
<212> ДНК
<213> Домовая мышь
<400> 25
cagtggatag acagatgggg g 21
<210> 26
<211> 21
<212> ДНК
<213> Домовая мышь
<400> 26
cagtggatag accgatgggg g 21
<210> 27
<211> 21
<212> ДНК
<213> Домовая мышь
<400> 27
cagtggatga gctgatgggg g 21
<210> 28
<211> 21
<212> ДНК
<213> Домовая мышь
<400> 28
caagggatag acagatgggg c 21
<210> 29
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность, содержащая последовательности
человека разумного и домовой мыши
<400> 29
gttttcccag tcacgac 17
<210> 30
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность, содержащая последовательности
человека разумного и домовой мыши
<400> 30
agcggataat ttcacacagg a 21
<210> 31
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность, содержащая последовательности
человека разумного и домовой мыши
<400> 31
aagcttgccg ccaccatggc tgtcctgggg ctgctcttct gc 42
<210> 32
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность, содержащая последовательности
человека разумного и домовой мыши
<400> 32
ccgatgggcc cttggtggag gctgaggaga cggtgactga ggttcc 46
<210> 33
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность, содержащая последовательности
человека разумного и домовой мыши
<400> 33
aagcttgccg ccaccatgaa gttgcctgtt aggctgttgg tgc 43
<210> 34
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность, содержащая последовательности
человека разумного и домовой мыши
<400> 34
ggatccactc acgtttgatt tccagcttgg tcccccctcc ga 42
<210> 35
<211> 111
<212> БЕЛОК
<213> Домовая мышь
<400> 35
Gln Val Gln Leu Lys Glu Ser Gly Pro Gly Leu Val Ala Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Gly Val Ser Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Gly Asp Gly Ser Thr Asn Tyr Lys Pro Asn Leu Met
50 55 60
Ser Arg Leu Ser Ile Ser Lys Asp Ile Ser Lys Ser Gln Val Leu Phe
65 70 75 80
Lys Leu Asn Ser Leu Gln Thr Asp Asp Thr Ala Thr Tyr Tyr Cys Val
85 90 95
Thr Leu Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser
100 105 110
<210> 36
<211> 112
<212> БЕЛОК
<213> Домовая мышь
<400> 36
Asp Val Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Arg
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Leu Tyr Phe Cys Phe Gln Thr
85 90 95
Thr Tyr Val Pro Asn Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 37
<211> 131
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность, содержащая последовательности
человека разумного и домовой мыши
<400> 37
Met Lys Leu Pro Val Arg Leu Leu Val Leu Met Phe Trp Ile Pro Ala
1 5 10 15
Ser Ser Ser Asp Val Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val
20 25 30
Ser Leu Gly Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu
35 40 45
Val His Arg Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro
50 55 60
Gly Gln Ser Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser
65 70 75 80
Gly Val Pro Arg Asp Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
85 90 95
Leu Lys Ile Ser Arg Val Glu Ala Glu Asp Leu Gly Leu Tyr Phe Cys
100 105 110
Phe Gln Thr Thr Tyr Val Pro Asn Thr Phe Gly Gly Gly Thr Lys Leu
115 120 125
Glu Ile Lys
130
<210> 38
<211> 130
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность, содержащая последовательности
человека разумного и домовой мыши
<400> 38
Met Ala Val Leu Gly Leu Leu Phe Cys Leu Val Thr Phe Pro Ser Cys
1 5 10 15
Val Leu Ser Gln Val Gln Leu Lys Glu Ser Gly Pro Gly Leu Val Ala
20 25 30
Pro Ser Gln Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu
35 40 45
Ser Ser Tyr Gly Val Ser Trp Val Arg Gln Pro Pro Gly Lys Gly Leu
50 55 60
Glu Trp Leu Gly Val Ile Trp Gly Asp Gly Ser Thr Asn Tyr His Pro
65 70 75 80
Asn Leu Met Ser Arg Leu Ser Ile Ser Lys Asp Ile Ser Lys Ser Gln
85 90 95
Val Leu Phe Lys Leu Asn Ser Leu Gln Thr Asp Asp Thr Ala Thr Tyr
100 105 110
Tyr Cys Val Thr Leu Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val
115 120 125
Ser Ser
130
<210> 39
<211> 333
<212> ДНК
<213> Домовая мышь
<400> 39
caggtgcagc tgaaggagtc aggacctggc ctggtggcgc cctcacagag cctgtccatc 60
acatgcactg tctcagggtt ctcattaagc agctatggtg taagctgggt tcgccagcct 120
ccaggaaagg gtctggagtg gctgggagta atatggggtg acgggagcac aaattatcat 180
ccaaatctca tgtccagact gagtatcagc aaggatattt ccaagagcca agttctcttc 240
aaactgaata gtctgcaaac tgatgacaca gccacgtact actgtgtcac cttcgactac 300
tggggtcaag gaacctcagt caccgtctcc tca 333
<210> 40
<211> 336
<212> ДНК
<213> Домовая мышь
<400> 40
gatgttgtga tgacccaaac tccactctcc ctgcctgtca gtcttggaga tcaagcctcc 60
atctcttgca gatctagtca gagccttgta catagaaatg gaaacaccta tttacattgg 120
tacctgcaga agccaggcca gtctccaaag ctcctgatct acaaagtttc caaccgattt 180
tctggggtcc cagacaggtt cagtggcagt ggatcaggga cagatttcac actcaagatc 240
agcagagtgg aggctgagga tttgggactt tatttctgtt ttcaaactac atatgttccg 300
aacacgttcg gaggggggac caagctggaa ataaaa 336
<210> 41
<211> 43
<212> ДНК
<213> Домовая мышь
<400> 41
aagcttgccg ccaccatgaa gttgcctgtt aggctgttgg tgc 43
<210> 42
<211> 422
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность, содержащая последовательности
человека разумного и домовой мыши
<400> 42
aagcttgccg ccaccatgaa gttgcctgtt aggctgttgg tgctgatgtt ctggattcct 60
gcttccagca gtgatgttgt gatgacccaa actccactct ccctgcctgt cagtcttgga 120
gatcaagcct ccatctcttg cagatctagt cagagccttg tacatagaaa tggaaacacc 180
tatttacatt ggtacctgca gaagccaggc cagtctccaa agctcctgat ctacaaagtt 240
tccaaccgat tttctggggt cccagacagg ttcagtggca gtggatcagg gacagatttc 300
acactcaaga tcagcagagt ggaggctgag gatttgggac tttatttctg ttttcaagac 360
tacatatgtt ccgaacacgt tcggaggggg gaccaagctg gaaatcaaac gtgagtggat 420
cc 422
<210> 43
<211> 42
<212> ДНК
<213> Домовая мышь
<400> 43
ggatccactc acgtttgatt tccagcttgg tcccccctcc ga 42
<210> 44
<211> 42
<212> ДНК
<213> Домовая мышь
<400> 44
aagctttccg ccaccatggc tgtcctgggg ctgctcttct gc 42
<210> 45
<211> 426
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность, содержащая последовательности
человека разумного и домовой мыши
<400> 45
aagctttccg ccaccatggc tgtccctggg gctgctcttc tgcctggtga cattaccaag 60
ctgtgtcctg tcccaggtgc agctgaagga gtcaggacct ggcctggtgg agcctcacag 120
agcctgtcca tcacatgcac tgtctcaggg ttctcattaa gcagctatgg tgtaagctgg 180
gttcgccagc ccaggaaagg gtctggagtg gctgggagta atatggggtg acgggagcac 240
aaattatcat ccaaatctca tgtccagact gagtatcagc aaggatattt ccaagagcaa 300
gttctcttca aactgaatag tctgcaaact gatgacacag ccacgtacta ctgtgtcacc 360
ttggactact ggggtcaaag gaacctccag tcaccgtctc ctcagcctcc accacgggcc 420
catcgg 426
<210> 46
<211> 46
<212> ДНК
<213> Домовая мышь
<400> 46
ccgatgggcc cttggtggag gctgaggaga cggtgactga ggttcc 46
<210> 47
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность, содержащая последовательности
человека разумного и домовой мыши
<400> 47
Asp Val Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Arg
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Arg Asp Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Leu Tyr Phe Cys Phe Gln Thr
85 90 95
Thr Tyr Val Pro Asn Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 48
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность, содержащая последовательности
человека разумного и домовой мыши
<400> 48
Gln Val Gln Leu Lys Glu Ser Gly Pro Gly Leu Val Ala Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Gly Val Ser Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Gly Asp Gly Ser Thr Asn Tyr His Pro Asn Leu Met
50 55 60
Ser Arg Leu Ser Ile Ser Lys Asp Ile Ser Lys Ser Gln Val Leu Phe
65 70 75 80
Lys Leu Asn Ser Leu Gln Thr Asp Asp Thr Ala Thr Tyr Tyr Cys Val
85 90 95
Thr Leu Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser
100 105 110
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ АМИЛОИДОЗОВ | 2018 |
|
RU2746812C1 |
| АНТИ-C5 АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2774716C2 |
| АНТИТЕЛО ПРОТИВ ПРОТЕИНТИРОЗИНФОСФАТАЗЫ σ РЕЦЕПТОРНОГО ТИПА ЧЕЛОВЕКА | 2012 |
|
RU2715642C2 |
| АНТИТЕЛА ДЛЯ ЛЕЧЕНИЯ ИНФЕКЦИИ ГЕПАТИТОМ В И СВЯЗАННЫХ С НЕЙ ЗАБОЛЕВАНИЙ | 2018 |
|
RU2765878C2 |
| EGFL6-СПЕЦИФИЧЕСКИЕ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2779902C2 |
| НОВЫЕ АНТИТЕЛА ПРОТИВ ВИРУСА ГЕПАТИТА B И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2814471C2 |
| АНТИТЕЛО ПРОТИВ МОЛЕКУЛЫ КЛЕТОЧНОЙ АДГЕЗИИ L1 ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И СОДЕРЖАЩИЙ ЕГО ХИМЕРНЫЙ РЕЦЕПТОР АНТИГЕНА | 2019 |
|
RU2789360C2 |
| ВАКЦИНА И СПОСОБЫ ОБНАРУЖЕНИЯ И ПРОФИЛАКТИКИ ФИЛЯРИОЗА | 2021 |
|
RU2832185C1 |
| АНТИТЕЛО К B7-H4, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2792748C2 |
| АНТИТЕЛА ПРОТИВ CXCR5 И ИХ КОМПОЗИЦИИ И ПРИМЕНЕНИЕ | 2018 |
|
RU2798422C2 |




Группа изобретений относится к области медицины и фармацевтики. 1-3 объекты представляют собой химерное антитело мыши и человека, связывающееся с амилоидными фибриллами и содержащее области VK с SEQ ID NO: 47 и VH с SEQ ID NO: 48, и содержащую антитело фармацевтическую композицию, применяемые для лечения первичного амилоидоза. 4-7 объекты представляют собой способ получения химерного антитела и применяемые при получении векторные конструкции 11-1F4VHpG1D200, 11-1F4VK.pKN100 и pG1KD200-11-1F4. 8 объект представляет собой применение антитела, имеющего присоединенную к нему метку, для обнаружения наличия отложений амилоида. 9-11 объекты представляют собой химерные антитела мыши и человека, полученные способами по изобретению. 12 и 13 объекты представляют собой полипептиды, которые представляют собой области антитела VK с SEQ ID NO: 47 и VH с SEQ ID NO: 48. 14 и 15 объекты представляют собой полинуклеотиды с SEQ ID NO: 42 и SEQ ID NO: 45, кодирующие указанные полипептиды. Технический результат заключается в лечении первичного амилоидоза и обнаружении наличия отложений амилоида с помощью антител по изобретению. 15 н. и 7 з.п. ф-лы, 6 табл., 6 пр., 8 ил.
1. Химерное антитело мыши и человека, связывающееся с амилоидными фибриллами и содержащее область VK с SEQ ID NO: 47 и область VH с SEQ ID NO: 48.
2. Химерное антитело мыши и человека по п. 1, связывающееся с эпитопом, экспрессируемым бета-складчато-листовой конфигурацией амилоидных фибрилл, с более высокой аффинностью по сравнению с мышиным антителом, содержащим область VK с SEQ ID NO: 36 и область VH с SEQ ID NO: 35.
3. Фармацевтическая композиция для лечения первичного амилоидоза, содержащая антитело по п. 1 и фармацевтически приемлемый носитель.
4. Применение антитела по п. 1 для лечения первичного амилоидоза у нуждающегося в таком лечении пациента-человека.
5. Способ получения химерного антитела, включающий котрансфекцию клеток млекопитающих векторными конструкциями 11-1F4VK.pKN100, состоящей из фрагмента вектора pSV2, который имеет ранний промотор SV40 и поздний промотор инактивированного SV40, точку начала репликации SV40 и точку начала репликации Co1E1, гены резистентности к ампициллину и неомицину, промотор HCMVi, множественный сайт клонирования, содержащий сайты ферментов рестрикции BamHI и HindIII, для вставки гена вариабельной области иммуноглобулина и кДНК для человеческого гена константной области каппа с концевой последовательностью сигнала терминации транскрипции spaC2, который находится в одинаковой ориентации с экспрессионной кассетой легкой цепи каппа, как показано на Фиг. 4, и гена 11-1F4VK, и 11-F4VH.pG1D200, состоящей из фрагмента вектора pSV2dhfr, который имеет ранний промотор SV40 и поздний промотор инактивированного SV40, точку начала репликации SV40 и точку начала репликации Co1E1, гены резистентности к ампициллину и дигидрофолатредуктазе(dhfr), фрагмент промотора HCMVi, множественный сайт клонирования, кДНК для человеческого гена константной области гамма 1 человека без интронов, за которой следует последовательность сигнала терминации транскрипции spaC2, как показано на Фиг. 5, и гена 11-F4VH, или трансфекцию клеток млекопитающих супервекторной конструкцией pG1KD200-11-1F4, полученной из конструкций 11-1F4VHpG1D200 и 11-1F4VK.pKN100, где ген 11-1F4VK имеет последовательность SEQ ID NO: 42, и ген 11-F4VH имеет последовательность SEQ ID NO: 45.
6. Способ по п. 5, где котрансфекция векторными конструкциями 11-1F4VK.pKN100 и 11-F4VH.pG1D200 или трансфекция супервекторной конструкцией pG1KD200-11-1F4 проходят в клетках COS (яичник китайского хомячка).
7. Способ по п. 5, включающий котрансфекцию векторными конструкциями 11-1F4VK.pKN100 и 11-F4VH.pG1D200.
8. Способ по п. 5, включающий трансфекцию супервекторной конструкцией pG1KD200-11-1F4.
9. Способ по п. 6, включающий котрансфекцию векторными конструкциями 11-1F4VK.pKN100 и 11-F4VH.pG1D200.
10. Способ по п. 6, включающий трансфекцию супервекторной конструкцией pG1KD200-11-1F4.
11. Векторная конструкция для получения химерного антитела по п. 1, где векторная конструкция представляет собой 11-1F4VK.pKN100, состоящую из фрагмента вектора pSV2, который имеет ранний промотор SV40 и поздний промотор инактивированного SV40, точку начала репликации SV40 и точку начала репликации Co1E1, гены резистентности к ампициллину и неомицину, промотор HCMVi, множественный сайт клонирования, содержащий сайты ферментов рестрикции BamHI и HindIII, для вставки гена вариабельной области иммуноглобулина и кДНК для человеческого гена константной области каппа с концевой последовательностью сигнала терминации транскрипции spaC2, который находится в одинаковой ориентации с экспрессионной кассетой легкой цепи каппа, как показано на Фиг. 4, и гена 11-1F4VK, в которой ген 11-1F4VK имеет последовательность SEQ ID NO: 42.
12. Векторная конструкция для получения химерного антитела по п. 1, где векторная конструкция представляет собой 11-1F4VHpG1D200, состоящую из фрагмента вектора pSV2dhfr, который имеет ранний промотор SV40 и поздний промотор инактивированного SV40, точку начала репликации SV40 и точку начала репликации Co1E1, гены резистентности к ампициллину и дигидрофолатредуктазе(dhfr), фрагмент промотора HCMVi, множественный сайт клонирования, кДНК для человеческого гена константной области гамма 1 человека без интронов, за которой следует последовательность сигнала терминации транскрипции spaC2, как показано на Фиг. 5, и гена 11-F4VH, в которой ген 11-F4VH имеет последовательность SEQ ID NO: 45.
13. Векторная конструкция для получения химерного антитела по п. 1, где векторная конструкция представляет собой pG1KD200-11-1F4, полученную из конструкций 11-1F4VHpG1D200 и 11-1F4VK.pKN100, как определено в пп. 11 и 12, соответственно, в которой ген 11-1F4VK имеет последовательность SEQ ID NO: 42, и ген 11-F4VH имеет последовательность SEQ ID NO: 45.
14. Применение антитела по п. 1, имеющего присоединенную к нему метку, которая может быть обнаружена, для обнаружения наличия отложений амилоида у пациента, у которого подозревают наличие таких отложений.
15. Применение по п. 14, где метка, которая может быть обнаружена, представляет собой 124I.
16. Химерное антитело мыши и человека, связывающееся с амилоидными фибриллами и полученное способом по п. 5.
17. Химерное антитело мыши и человека, связывающееся с амилоидными фибриллами и полученное способом по п. 7.
18. Химерное антитело мыши и человека, связывающееся с амилоидными фибриллами и полученное способом по п. 8.
19. Полипептид, который представляет собой область VK антитела по п. 1, где полипептид представляет собой SEQ ID NO: 47.
20. Полипептид, который представляет собой область VH антитела по п. 1, где полипептид представляет собой SEQ ID NO: 48.
21. Полинуклеотид, кодирующий полипептид по п. 19, где полинуклеотид представляет собой SEQ ID NO: 42.
22. Полинуклеотид, кодирующий полипептид по п. 20, где полинуклеотид представляет собой SEQ ID NO: 45.
| SOLOMON A | |||
| et al | |||
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| - Vol | |||
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| - No | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| - P | |||
| СПОСОБ ГОРЯЧЕЙ ВУЛКАНИЗАЦИИ КАУЧУКА | 1925 |
|
SU3831A1 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО К АМИЛОИДУ БЕТА | 2007 |
|
RU2498999C2 |
| Станок для поверки прицеливания винтовки | 1928 |
|
SU10902A1 |
| Электрическая печь сопротивления | 1927 |
|
SU15654A1 |
| WO 1999060024 A1, 25.11.1999 | |||
| WO 2016032949 A1, 03.03.2016 | |||
| EDWARDS C | |||
| V | |||
| et al | |||
| Interim analysis | |||

