ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[001] Настоящая заявка заявляет приоритет предварительной заявки на патент США с серийным номером 61/351683, поданной 4 июня 2010, которая включена в данный документ ссылкой в ее полноте.
ОПИСАНИЕ ТЕКСТОВОГО ФАЙЛА, ПРЕДОСТАВЛЕННОГО В ЭЛЕКТРОННОМ ВИДЕ
[002] Содержание текстового файла, предоставленного в электронном виде вместе с данным документом, включено в данный документ ссылкой в его полноте: копия перечня последовательностей в машиночитаемом формате (имя файла: MIRG_022_01WO_SeqList_ST25.txt, дата записи: 6 июня 2011, размер файла 5 килобайт).
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
[003] Настоящее изобретение относится к лечению и предотвращению сердечных и метаболических расстройств путем введения средств, которые модулируют активность или экспрессию микроРНК (миРНК). В частности, настоящее изобретение обеспечивает способ лечения или предотвращения сердечных и метаболических расстройств путем ингибирования экспрессии или активности miR-378/miR-378* в клетках субъекта. Кроме того, настоящее изобретение обеспечивает способ регуляции метаболизма жирных кислот в клетке путем приведения клетки в контакт с модулятором экспрессии или активности miR-378/miR-378*.
ПРЕДПОСЫЛКИ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
[004] Заболевание сердца и его проявления, включая болезнь коронарной артерии, инфаркт миокарда, застойную сердечную недостаточность и гипертрофию сердца, на сегодняшний день явно представляет основной риск для здоровья в Соединенных Штатах. Стоимость диагностики, лечения и поддержки пациентов, страдающих от этих заболеваний, оценивается в миллиарды долларов. Два особенно тяжелых проявления заболевания сердца представляют собой инфаркт миокарда и гипертрофию сердца.
[005] Инфаркт миокарда, широко известный как сердечный приступ, вызывается внезапным и длительным недостатком кровотока к ткани сердца, что обычно приводит к сужению или окклюзии коронарной артерии. Без достаточного кровоснабжения ткань становится ишемической, что ведет к гибели кардиомиоцитов (например, клеток сердечной мышцы) и сосудистых структур. Некротическая ткань, образующаяся в результате гибели кардиомиоцитов, как правило, замещается рубцовой тканью, которая не является сократительной, не может вносить вклад в сердечную функцию и часто играет пагубную роль при функционировании сердца, например, из-за расширения во время сердечного сокращения или из-за увеличения размера и эффективного радиуса желудочка, становясь гипертрофическим.
[006] Гипертрофия сердца представляет собой адаптационную реакцию сердца на практически все формы заболевания сердца, включая таковые, являющиеся результатом гипертензии, механической нагрузки, инфаркта миокарда, сердечных аритмий, эндокринных расстройств и генных мутации в генах сократительных белков сердца. Хотя гипертрофическая реакция изначально представляет собой компенсаторный механизм, который увеличивает сердечный выброс, длительная гипертрофия может привести к дилатационной кардиомиопатии (DCM), сердечной недостаточности и внезапной смерти. В Соединенных Штатах у приблизительно полумиллиона людей диагностируют сердечную недостаточность каждый год с уровнем смертности, достигающим 50%.
[007] Различные пути передачи сигнала, особенно таковые, включающие нарушенную передачу сигнала с помощью кальция, управляют гипертрофией сердца и патологическим ремоделированием (Heineke & Molkentin, 2006). Гипертрофический рост в ответ на стресс включает другие пути передачи сигнала и паттерны генной экспрессии, нежели физиологическая гипертрофия, которая возникает в ответ на физическую нагрузку. Опосредованная стрессом миокардиальная гипертрофия представляет собой сложное явление, связанное с многочисленными неблагоприятными последствиями с отчетливыми молекулярными и гистологическими характеристиками, вызывая фиброз, расширение и декомпенсацию сердца, что при дегенерации и гибели кардиомиоцитов часто завершается сердечной недостаточностью. В связи с этим наблюдается повышенный интерес к расшифровыванию молекулярных механизмов и к обнаружению новых терапевтических мишеней для подавления неблагоприятного роста сердца и, в конечном счете, недостаточности. Понимание этих механизмов является необходимым для разработки новых видов терапии для лечения гипертрофии сердца и сердечной недостаточности.
[008] Метаболический синдром представляет собой комбинацию медицинских расстройств, которые повышают риск развития сердечнососудистого заболевания и диабета. Он поражает каждого пятого человека, и распространенность возрастает с возрастом. Некоторые исследования оценивают распространенность в США как достигающую 25% населения (Ford et al. (2002) JAMA, Vol.287:356-359). Люди, пораженные метаболическим синдромом, являются, как правило, тучными, малоподвижными и имеют определенную степень резистентности к инсулину. Ожирение является ведущей предотвращаемой причиной смерти во всем мире с возрастающей распространенностью у взрослых и детей, и авторитетные источники рассматривают его как одну из наиболее серьезных проблем общественного здравоохранения 21-го века (Barness et al. (2007) Am. J. Med. Genet. A, Vol.143A: 3016-34). Ожирение может приводить к уменьшенной предполагаемой продолжительности жизни и повышенным проблемам со здоровьем, включая заболевание сердца, диабет типа 2, приступы апноэ во сне, определенные типы рака и остеоартрит. Современные виды терапии для метаболического синдрома и ожирения фокусируются на соблюдении диеты и физической нагрузке с очень небольшим количеством доступных эффективных вмешательств с применением фармацевтических средств. Эффективность диеты и физической нагрузки при улучшении этих состояний сильно варьирует среди пациентов и имеет тенденцию к обеспечению лишь средней степени потери веса и улучшения симптомов. Следовательно, существует потребность в новых терапевтических подходах к лечению метаболических расстройств и предотвращению последующего развития сердечнососудистого заболевания и сердечной недостаточности.
[009] Недавно было установлено, что микроРНК вовлечены в ряд биологических процессов, включая временную регуляцию периодов развития, апоптоз, метаболизм жиров и дифференцировку гемопоэтических клеток, среди прочего. МикроРНК (миРНК) представляют собой малые не кодирующие белок РНК длиной от приблизительно 18 до приблизительно 25 нуклеотидов, которые происходят из отдельных генов миРНК, из интронов кодирующих белок генов или из полицистронных транскриптов, которые часто кодируют множественные примыкающие миРНК. Смотри обзор Carrington et al. (Science, Vol.301(5631):336-338, 2003). МиРНК действуют как репрессоры целевых мРНК путем стимулирования их деградации, если их последовательности являются полностью комплементарными, или путем ингибирования трансляции, если их последовательности содержат несовпадения.
[0010] МиРНК транскрибируются РНК-полимеразой II (pol II) или РНК-полимеразой III (pol III; смотри Qi et al. (2006) Cellular & Molecular Immunology, Vol.3:411-419) и происходят из исходных транскриптов, называемых первичными транскриптами миРНК (при-миРНК), которые, как правило, имеют длину в несколько тысяч оснований. При-миРНК подвергаются процессингу в ядре РНКазой Drosha в от приблизительно 70- до приблизительно 100-нуклеотидные предшественники в форме шпильки (пре-миРНК). После транспортировки в цитоплазму шпилечная пре-миРНК дополнительно подвергается процессингу Dicer с получением двухцепочечной миРНК. Зрелая цепь миРНК затем встраивается в РНК-индуцированный комплекс сайленсинга (RISC), где она связывается с ее целевыми мРНК вследствие комплементарности пар оснований. В относительно редких случаях, когда основания миРНК полностью спариваются с целевой мРНК, она стимулирует деградацию мРНК. Чаще миРНК образуют несовершенные гетеродуплексы с целевыми мРНК, либо отрицательно воздействуя на стабильность мРНК, либо ингибируя трансляцию мРНК.
[0011] Недавно были определены характерные паттерны экспрессии миРНК, связанные с патологической гипертрофией сердца, сердечной недостаточностью и инфарктом миокарда у людей и в мышиных моделях заболевания сердца (van Rooij et al. (2006) Proc. Natl. Acad. Sci., Vol.103(48):18255-60; van Rooij et al., (2007) Science, Vol.316: 575-579). Исследования с использованием мутаций, приводящих к приобретению или потери функций, у мышей выявили сложные и неожиданные функции для этих миРНК в многочисленных аспектах биологии сердца, включая контроль роста миоцитов, сократительную способность, энергетический обмен, фиброз и ангиогенез, обеспечивая некоторые представления о новых регуляторных механизмах и возможных мишени для терапевтического воздействия при заболевании сердца. Удивительно, что нокаутные мыши, у которых отсутствуют индуцирующие заболевание миРНК, являются нормальными, но проявляют искаженные реакции на сердечный стресс, что позволяет предположить ориентированность этих миРНК скорее на связанные с заболеванием процессы, нежели на тканевой гомеостаз, и указывает на их потенциал в качестве терапевтических мишеней. Таким образом, миРНК представляют потенциальные новые терапевтические мишени для разработки видов лечения для ряда заболеваний, включая сердечнососудистые заболевания, ожирение, диабет и другие метаболические расстройства.
КРАТКОЕ ОПИСАНИЕ НАСТОЯЩЕГО ИЗБРЕТЕНИЯ
[0012] Настоящее изобретение основано, частично, на том открытии, что экспрессия miR-378 даунрегулируется в ткани сердца после перегрузки давлением и что сверхэкспрессия miR-378 специфическим для сердца образом усиливает сердечную гипертрофическую реакцию, индуцированную стрессом. Сверхэкспрессия miR-378/miR-378* в скелетной мышце приводит к увеличенной массе тела, что является результатом увеличенной массы эпидидимальных жировых подушечек вследствие гипертрофии адипоцитов и, возможно, гиперплазии адипоцитов. Таким образом, авторами настоящего изобретения было неожиданно обнаружено, что miR-378/miR-378* регулирует метаболические процессы в различных тканях, включая сердечную и жировую ткань. Соответственно, настоящее изобретение обеспечивает способы лечения или предотвращения сердечнососудистого заболевания и других метаболических расстройств, таких как ожирение и диабет, путем модулирования экспрессии или активности miR-378 и/или miR-378* в клетках у субъекта, нуждающегося в этом.
[0013] В одном варианте осуществления настоящее изобретение обеспечивает способ лечения или предотвращения патологической гипертрофии сердца, ремоделирования сердца, инфаркта миокарда или сердечной недостаточности у субъекта, нуждающегося в этом, при котором вводят субъекту ингибитор miR-378 и/или miR-378*. В определенных вариантах осуществления экспрессия или активность miR-378 и/или miR-378* понижается в клетках сердца субъекта после введения. В других вариантах осуществления индуцированный стрессом метаболический сдвиг от окислительного к гликолитическому метаболизму предотвращается или уменьшается в клетках сердца субъекта после введения ингибитора miR-378 и/или miR-378*.
[0014] Настоящее изобретение также включает способ лечения или предотвращения метаболического расстройства у субъекта, нуждающегося в этом. В одном варианте осуществления способ включает введение субъекту ингибитора miR-378 и/или miR-378*, где экспрессия или активность miR-378 и/или miR-378* понижается в клетках субъекта после введения. Метаболическое расстройство, подлежащее лечению, может включать метаболический синдром, ожирение, сахарный диабет, диабетическую нефропатию, резистентность к инсулину, атеросклероз, нарушение обмена липидов, гликогеноз, недостаточность среднецепочечной ацил-коэнзим A дегидрогеназы, окисление липидов или нарушенное поглощение и/или утилизация глюкозы. Вторичные заболевания или состояния, являющиеся результатом этих метаболических расстройств, можно также предотвращать или лечить с помощью способов настоящего изобретения. Например, в одном варианте осуществления настоящее изобретение обеспечивает способ предотвращения или лечения вторичных заболеваний или расстройств, являющихся результатом ожирения, таких как приступы апноэ во сне, рак и остеоартрит, путем введения ингибитора miR-378 и/или miR-378*.
[0015] Ингибиторы miR-378 и miR-378*, пригодные для применения в способах настоящего изобретения, могут быть антисмысловыми олигонуклеотидами. В одном варианте осуществления антисмысловой олигонуклеотид содержит последовательность, которая по меньшей мере частично комплементарна зрелой последовательности miR-378 или miR-378*. В определенных вариантах осуществления антисмысловые олигонуклеотиды содержат одну или несколько модификаций сахара или остова, такие как «закрытые» нуклеиновые кислоты, бициклические нуклеозиды, фосфоноформиаты, 2'-O-алкильные модификации и фосфоротиоатные связи. В других вариантах осуществления ингибитор miR-378 или miR-378* представляет собой антисмысловой олигонуклеотид длиной от приблизительно 7 до приблизительно 18 нуклеотидов.
[0016] В другом варианте осуществления настоящее изобретение обеспечивает способ регуляции метаболизма жирных кислот в клетке, при котором приводят в контакт клетку с модулятором экспрессии или активности miR-378 и/или miR-378*. Модулятор может быть ингибитором или агонистом экспрессии или активности miR-378 и/или miR-378*. В определенных вариантах осуществления метаболизм жирных кислот в клетке повышается после контакта с ингибитором miR-378 и/или miR-378* по сравнению с клеткой, не подвергнутой воздействию ингибитора. В других вариантах осуществления метаболизм жирных кислот в клетке понижается после контакта с агонистом miR-378 и/или miR-378* по сравнению с клеткой, не подвергнутой воздействию агониста. Клетка может находиться in vitro или in vivo. В некоторых вариантах осуществления клетка представляет собой кардиомиоцит, клетку скелетной мышцы, преадипоцит, адипоцит, гепатоцит или клетку поджелудочной железы.
[0017] Настоящее изобретение также включает способ регуляции метаболизма сердца. В одном варианте осуществления способ включает приведение в контакт кардиомиоцита с модулятором экспрессии или активности miR-378 и/или miR-378*. Кардиомиоцит может находиться in vitro или in vivo. В другом варианте осуществления метаболизм углеводов в кардиомиоците понижается после контакта с ингибитором miR-378 и/или miR-378*. В еще одном варианте осуществления метаболизм жирных кислот в кардиомиоците повышается после контакта с ингибитором miR-378 и/или miR-378*.
[0018] Настоящее изобретение охватывает фармацевтические композиции, содержащие ингибиторы и агонисты miR-378 и miR-378*, описанные в данном документе, и фармацевтически приемлемый носитель. В одном варианте осуществления фармацевтическая композиция содержит ингибитор miR-378 и/или miR-378* и фармацевтически приемлемый носитель, где ингибитор представляет собой антисмысловой олигонуклеотид. В определенных вариантах осуществления антисмысловой олигонуклеотид содержит по меньшей мере одну модификацию сахара и/или остова. В других вариантах осуществления антисмысловой олигонуклеотид имеет длину от приблизительно 7 до приблизительно 18 нуклеотидов.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0019] Фигура 1. A. Местоположение miR-378 в геноме внутри гена-хозяина PPARGC1β (сверху). Иллюстрация с помощью геномного браузера UCSC консервативности участка генома, окружающего miR-378 (внизу). B. Количественный анализ ПЦР в реальном времени показывает, что miR-378, miR-378*, и PPARGC1β (PGC-1β), ген-хозяин miR-378/miR-378* специфически экспрессируются в сердце, скелетной мышце и бурой жировой ткани. C. miR-378 является более высоко экспрессированным в кардиомиоцитах, чем в сердечных фибробластах. D. Экспрессия miR-378 понижается при обработке HIB1B клеток стеариновой кислотой. E. Экспрессия miR-378 и PPARGC1β (PGC-1β) в ходе индукции дифференцировки адипоцитов. CMC: кардиомиоциты, BAT: бурая жировая ткань, WAT: белая жировая ткань.
[0020] Фигура 2. A. MiR-378 даунрегулируется в сердце после торакальной операции по сужению аорты (TAB) по сравнению с необработанным сердцем (контроли с симуляцией), как показано с помощью количественной ПЦР в реальном времени. B. Миокардиальная сверхэкспрессия miR-378 индуцирует усиленную гипертрофическую реакцию при TAB.
[0021] Фигура 3. Сверхэкспрессия miR-378 в скелетной мышце приводит в результате к увеличенной массе тела и массе эпидидимального жира. На верхних секциях показан гистологический анализ белой жировой ткани из мышей дикого типа (wt) и сверхэкспрессирующих miR-378 под контролем промотора MCK (мышечной креатинкиназы) мышей (MCK-miR-378 Tg). Мутантные животные показывают гипертрофированные адипоциты. На нижних секциях показана увеличенная масса тела и увеличенная массу белого жира у MCK-miR-378 трансгенных мышей по сравнению с мышами дикого типа.
[0022] Фигура 4. Сверхэкспрессия miR-378 в 3T3-L1 клетках усиливает адипогенез и липогенез. 3T3-L1 клетки индуцировали к дифференцировке в зрелые адипоциты в течение 4 суток. Трансфекция miR-378 повышала накопление липидов в клетках, что демонстрируется окрашиванием масляным красным O.
[0023] Фигура 5. MCK-miR-378 трансгенные мыши проявляют пониженную толерантность к глюкозе. Экспрессирующим miR-378 под контролем промотора мышечной креатинкиназы (MCK) мышам (Tg) или однопометным животным дикого типа (wt) вводили инъекцией внутрибрюшинно 1,5 г/кг глюкозы после голодания в течение 16 часов. Уровни глюкозы в крови оценивали в указанные моменты времени.
[0024] Фигура 6. Инъекция antimiR-378 MCK-miR-378 трансгенным мышам улучшает клиренс глюкозы. Экспрессирующим miR-378 под контролем промотора мышечной креатинкиназы (MCK) мышам (Tg) вводили инъекцией внутривенно в течение трех последовательных дней 10 мг/кг антисмыслового олигонуклеотида (antimiR), имеющего последовательность, комплементарную зрелой последовательности miR-378. Мышам дикого типа (wt) вводили инъекцией солевой раствор. Как Tg, так и wt мышам впоследствии вводили инъекцией внутрибрюшинно 1,5 г/кг глюкозы после голодания в течение 16 часов. Уровни глюкозы в крови оценивали в указанные моменты времени (левая секция). Анализ ПЦР в реальном времени для экспрессии miR-378 у Tg и wt мышей, которым вводили инъекцией antimiR-378 или солевой раствор (правые секции).
[0025] Фигура 7. AntimiR-378 инъекция ob/ob мышам улучшает клиренс глюкозы. Страдающим ожирением мышам (ob/ob) вводили инъекцией внутривенно в течение трех последовательных дней 10 мг/кг антисмыслового олигонуклеотида (antimiR), имеющего последовательность, комплементарную зрелой последовательности miR-378. Не получавшим лечение и получавшим лечение с применением antimiR ob/ob мышам вводили инъекцией внутрибрюшинно 1,5 г/кг глюкозы после голодания в течение 16 часов. Уровни глюкозы в крови оценивали в указанные моменты времени.
[0026] Фигура 8. A. Стратегия нацеливания miR-378. Нацеливающий конструкт вводили в геномный локус miR-378 с получением гомологичной рекомбинации. B. Саузерн-блоттинг анализ клеток дикого типа (wt) и нацеленных ES-клеток (MUT). C. Нозерн-блоттинг анализ, проведенный с применением общей РНК от мышей дикого типа (+/+), гетерозиготных (+/-) и нокаутных (-/-) мышей показывает, что экспрессия miR-378 (стрелка) прекращается в полученной мутантной мышиной линии. H: сердце, BAT: бурая жировая ткань, Sk.M: скелетная мышца. D. ПЦР в реальном времени (левая секция) и вестерн-блоттинг анализ (правая секция) ткани сердца, выделенной из мышей WT и нокаутных по miR-378 мышей (KO), показывает, что экспрессия гена-хозяина miR-378, PPARGC1β (PGC-1 β), не изменяется существенно у нокаутных по miR-378 животных.
[0027] Фигура 9. Нокаутные по MiR-378 животные проявляют пониженную гипертрофию сердца в ответ на торакальную операцию по сужению аорты (TAB).
[0028] Фигура 10. Эхокардиографию проводили у мышей дикого типа (WT) и нокаутных по miR-378 мышей (KO) через 21 день после торакальной операции по сужению аорты (TAB) для определения фракциональное укорочение (A), фракция выброса (B), систолический диаметр (C) и диастолический-систолический диаметр (DD-SD) (D).
[0029] Фигура 11. Экспрессия индуцированных стрессом генов, BNP (A), Myh7 (B) и Myh6 (C), в ткани сердца мышей дикого типа (WT) и нокаутных по miR-378 мышей (KO) после торакальной операции по сужению аорты (TAB), которая оценена с помощью ПЦР в реальном времени.
[0030] Фигура 12. Страдающих ожирением мышей (ob/ob) скрещивали с нокаутными по miR-378 животными с получением страдающих ожирением мышей, у которых отсутствует экспрессия miR-378 (ob/ob KO). Страдающим ожирением мышам и ob/ob KO мышам вводили инъекцией внутрибрюшинно 1,5 г/кг глюкозы после голодания в течение 16 часов. Уровни глюкозы в крови оценивали в указанные моменты времени.
[0031] Фигура 13. A. Нокаутные по miR-378 мыши (KO) проявляют меньшую прибавку массы тела, чем животные дикого типа (WT) через несколько недель на рационе с высоким содержанием жиров. B. Через пять недель на рационе с высоким содержанием жиров как нокаутным по miR-378 мышам, так и мышам дикого типа вводили инъекцией внутрибрюшинно 1,5 г/кг глюкозы после голодания в течение 16 часов. Уровни глюкозы в крови оценивали в указанные моменты времени.
[0032] Фигура 14. Нокаутные по miR-378 мыши (KO) имеют уменьшенные жировые подушечки и накопление липидов в печени, чем животные дикого типа (WT) через шесть недель на рационе с высоким содержанием жиров. WAT: белая жировая ткань.
[0033] Фигура 15. Нокаутные по miR-378 мыши (KO) имеют уменьшенную массу жира через шесть недель на рационе с высоким содержанием жиров по сравнению с животными дикого типа (WT). BAT: бурая жировая ткань; WAT: белая жировая ткань.
[0034] Фигура 16. Гистологический анализ белой жировой ткани (WAT), бурой жировой ткани (BAT) и печени из мышей дикого типа (WT) и нокаутных по miR-378 мышей (KO) после шести недель режима питания с рационом с высоким содержанием жиров. Увеличение: 40X.
[0035] Фигура 17. Мыши, сверхэкспрессирующие miR-378/miR-378* в ткани сердца под контролем промотора тяжелой цепи альфа-миозина (α-MHC TG), имеют увеличенную массу тела (A) и проявляют большее увеличение массы тела на рационе с высоким содержанием жиров (B и C) по сравнению с однопометными животными дикого типа (WT).
[0036] Фигура 18. A. Вестерн-блоттинг анализ бурой жировой ткани, выделенной из мышей дикого типа (WT) и нокаутных по miR-378 мышей (KO) через шесть часов после голодания. Блоты зондировали с помощью антител, специфичных к фосфорилированным и нефосфорилированным формам AMPK (АМФ-активируемая протеинкиназа) и AKT (протеинкиназа В). Альфа-тубулин использовали в качестве контроля. B. Схематическое изображение, показывающее регуляцию метаболизма с помощью сигнальных путей, опосредованных AMPK и AKT.
[0037] Фигура 19. A. Схематическое изображение 3'-UTR (нетранслируемая область) MED13, изображающее предполагаемые сайты связывания для miR-378/miR-378*. B. Уровни экспрессии MED13 у животных дикого типа (WT), трансгенных по miR-378 животных (αMHC Tg), сверхэкспрессирующих miR-378 под контролем промотора αMHC, и нокаутных по miR-378 животных (KO). C. COS1 клетки трансфицировали люциферазным конструктом 3'-UTR MED13, вместе с плазмидами экспрессии для miR-378 и контрольной миРНК. Показанные значения представляют собой кратность изменения экспрессии люциферазы (±SEM) по сравнению с репортером отдельно.
ПОДРОБНОЕ ОПИСАНИЕ НАСТОЯЩЕГО ИЗБРЕТЕНИЯ
[0038] Хронический и острый стресс в отношении сердца приводит в результате к патологической реакции ремоделирования, сопровождаемой гипертрофией, фиброзом, апоптозом миоцитов и возможной смерти от недостаточности нагнетательной функции сердца и аритмий. Хотя классические стратегии фармакологического лечения (например, бета-блокаторы и ACE-ингибиторы) могут продлевать срок жизни пациентов с сердечной недостаточностью, эти виды терапии являются, в конечном счете, неэффективными при предотвращении развития заболевания, подчеркивая потребность в новом понимании механизмов действия и терапевтических подходах.
[0039] Сердечная недостаточность представляет собой прогрессирующее длительное заболевание, которое, как оценено, поражает 5,7 миллионов американцев с возрастающими связанными затратами на здравоохранение (Lloyd-Jones et al. (2009) Circulation, Vol.119:21-181). МиРНК недавно выступили в качестве важных молекулярных компонентов при гипертрофии и ремоделировании в сердце, таким образом, представляя перспективную мишень при профилактике и лечении заболевания сердца. Так как отдельные миРНК часто регулируют экспрессию множества целевых генов со связанными функциями, модулирование экспрессии одной миРНК может, в принципе, влиять на целую генную сеть и, тем самым, модифицировать сложные фенотипы заболевания.
[0040] Настоящее изобретение основано, отчасти, на открытии того, что миРНК высоко экспрессируется в богатых митохондриями тканях сердца, скелетных мышц и бурой жировой ткани и регулируется при различных болезненных состояниях сердца. В частности, авторы настоящего изобретения неожиданно открыли, что miR-378 и его соответствующая минорная последовательность, miR-378*, являются регуляторами общего метаболизма. MiR-378 играет роль в индуцированной стрессом гипертрофической реакции сердца через регуляцию метаболизма сердца и вовлечена в утилизацию глюкозы и метаболизм жирных кислот в скелетной мышце. Соответственно, настоящее изобретение обеспечивает способ предотвращения или лечения различных сердечных и метаболических расстройств у субъекта путем ингибирования экспрессии или активности miR-378 и/или miR-378* в клетках субъекта.
[0041] Как используется в данном документе, выражение «субъект» или «пациент» относится к любому позвоночному, включая, без ограничения, людей и других приматов (например, шимпанзе и другие виды высших и низших приматов), сельскохозяйственных животных (например, крупный рогатый скот, овцы, свиньи, козы и лошади), домашних млекопитающих (например, собаки и кошки), лабораторных животных (например, грызуны, такие как мыши, крысы и морские свинки) и птиц (например, домашние, дикие и пернатая дичь, такие как цыплята, индюки и другие куриные, утки, гуси и подобное). В некоторых вариантах осуществления субъект представляет собой млекопитающее. В других вариантах осуществления субъект представляет собой человека.
[0042] MiR-378 кодируется на мышиной хромосоме 18, в пределах первого интрона гена коактиватора 1-бета гамма-рецептора, активируемого пролифератором пероксисом, (PPARGC1β). У людей miR-378 (ранее имеющая название miR-422b) экспрессируется из первого интрона гена PPARGC1β на хромосоме 5. Последовательность пре-миРНК для miR-378 подвергается процессингу в зрелую последовательность и последовательность со сзвездочкой (т.е. минорную). Последовательность со звездочкой подвергается процессингу из другой ветви структуры «стебель-петля». Пре-миРНК (например, последовательности «стебель-петля»), зрелые последовательности и последовательности со звездочкой для miR-378 мыши и человека представлены ниже:
Зрелая miR-378 человека (SEQ ID NO: 1)
5'-ACUGGACUUG GAGUCAGAAG G-3'
MiR-378* человека (SEQ ID NO: 2)
5'-CUCCUGACUC CAGGUCCUGU GU-3'
Пре-miR-378 человека (SEQ ID NO: 3)
5'-AGGGCUCCUG ACUCCAGGUC CUGUGUGUUA CCUAGAAAUA GCACUGGACU UGGAGUCAGA AGGCCU-3'
Зрелая miR-378 мыши (SEQ ID NO: 4)
5'-ACUGGACUUG GAGUCAGAAG G-3'
MiR-378* мыши (SEQ ID NO: 5)
5'-CUCCUGACUC CAGGUCCUGU GU-3'
Пре-miR-378 мыши (SEQ ID NO: 6)
5'-AGGGCUCCUG ACUCCAGGUC CUGUGUGUUA CCUCGAAAUA GCACUGGACU UGGAGUCAGA AGGCCU-3'
[0043] Понятно, что все последовательности рибонуклеиновой кислоты, раскрытые в данном документе, могут быть превращены в последовательности дезоксирибонуклеиновой кислоты путем замещения тимидинового основания на уридиновое основание в последовательности. Аналогично, все последовательности дезоксирибонуклеиновой кислоты, раскрытые в данном документе, могут быть превращены в последовательности рибонуклеиновой кислоты путем замещения уридинового основания на тимидиновое основание в последовательности. Последовательности дезоксирибонуклеиновой кислоты, последовательности рибонуклеиновой кислоты и последовательности, содержащие смеси дезоксирибонуклеотидов и рибонуклеотидов всех последовательностей, раскрытых в данном документе, включены в настоящее изобретение.
[0044] В одном варианте осуществления настоящее изобретение обеспечивает способ лечения или предотвращения патологической гипертрофии сердца, ремоделирования сердца, инфаркта миокарда или сердечной недостаточности у субъекта, нуждающегося в этом, при котором вводят субъекту ингибитор miR-378 и/или miR-378*. В некоторых вариантах осуществления экспрессия или активность miR-378 и/или miR-378* понижается в клетках сердца субъекта после введения ингибитора. «Клетки сердца», как используется в данном документе, включают кардиомиоциты, сердечные фибробласты и сердечные эндотелиальные клетки. В одном конкретном варианте осуществления экспрессия или активность miR-378 и/или miR-378* понижается в кардиомиоцитах субъекта после введения ингибитора miR-378 и/или miR-378*.
[0045] В другом варианте осуществления субъект, нуждающийся в этом, может иметь риск развития патологической гипертрофии сердца, ремоделирования сердца, сердечной недостаточности или инфаркта миокарда. Такой субъект может проявлять один или несколько факторов риска, включая без ограничения длительную неконтролируемую гипертензию, легочную артериальную гипертензию, некорректированный порок клапанов, хроническую ангину, недавний инфаркт миокарда, врожденную предрасположенность к заболеванию сердца или патологической гипертрофии. Субъект с таким риском может быть диагностирован, как имеющий генетическую предрасположенность к гипертрофии сердца, или может иметь семейную историю гипертрофии сердца. В некоторых вариантах осуществления у субъекта с риском может быть диагностировано ожирение, диабет типа II, гиперлипидемия или метаболический синдром.
[0046] Предпочтительно, введение ингибитора miR-378 и/или miR-378* субъекту приводит к облегчению одного или нескольких симптомов гипертрофии сердца, сердечной недостаточности или инфаркта миокарда у субъекта, или к задержке перехода от гипертрофии сердца к сердечной недостаточности. Один или несколько облегченных симптомов могут представлять собой, например, увеличенную способность к физической нагрузке, увеличенный объем сердечного выброса, пониженное конечно-диастолическое давление левого желудочка, пониженное давление заклинивания легочных капилляров, повышенный сердечный выброс, повышенный сердечный индекс, пониженные давления в легочной артерии, пониженные конечно-систолические и диастолические размеры левого желудочка, уменьшенный фиброз сердца, пониженное отложение коллагена в сердечной мышце, пониженное напряжение стенки левого и правого желудочков, пониженное растяжение стенки, повышенное качество жизни и пониженную распространенность болезни и смертность, связанные с заболеванием.
[0047] Настоящее изобретение также включает способ лечения или предотвращения метаболического расстройства у субъекта, нуждающегося в этом. В одном варианте осуществления способ включает введение субъекту ингибитора miR-378 и/или miR-378*, где экспрессия или активность miR-378 и/или miR-378* в клетках субъекта после введения понижается. Метаболические расстройства, которые можно лечить с помощью способов настоящего изобретения, включают без ограничения метаболический синдром, ожирение, сахарный диабет, диабетическую нефропатию, резистентность к инсулину, атеросклероз, нарушение липидного обмена (например, болезнь Ниманна-Пика, болезнь Гоше, болезнь Фарбера, болезнь Фабри, болезнь Вольмана и болезнь накопления сложных эфиров холестерина), синдром поликистозных яичников (PCOS) или нарушенные поглощение и/или утилизацию глюкозы. В одном варианте осуществления метаболическое расстройство представляет собой гликогеноз (GSD). Например, способы настоящего изобретения обеспечивают лечение или предотвращение любого из типов GSD (например, GSD типа 0 и GSD типа I до GSD типа XIII) у субъекта, нуждающегося в этом, путем введения субъекту ингибитора miR-378 и/или miR-378*. GSD включают без ограничения болезнь фон Гирке, болезнь Помпе, болезнь Кори или болезнь Форбса, болезнь Андерсена, болезнь Мак-Ардла, болезнь Герса, болезнь Таруи, синдром Фанкони-Бикеля и недостаточность альдолазы в эритроцитах. В другом варианте осуществления метаболическое расстройство представляет собой недостаточность среднецепочечной ацил-коэнзим A дегидрогеназы (MCAD). Индивиды с недостаточностью MCAD показывают ухудшение окисления жирных кислот, которое может быть смертельным. В одном варианте осуществления настоящего изобретения метаболизм жирных кислот у субъекта, имеющего недостаточность MCAD, повышается после введения ингибитора miR-378 и/или miR-378*.
[0048] Настоящее изобретение также включает способ предотвращения или лечения вторичных заболеваний или состояний, которые являются результатом метаболических расстройств, таких как диабет и ожирение, путем введения субъекту, нуждающемуся в этом, ингибитора miR-378 и/или miR-378*. Например, в одном варианте осуществления настоящее изобретение обеспечивает способ предотвращения или лечения приступов апноэ во сне, при котором вводят субъекту, нуждающемуся в этом, ингибитор miR-378 и/или miR-378*. В другом варианте осуществления настоящее изобретение обеспечивает способ предотвращения или лечения рака путем введения субъекту, нуждающемуся в этом, ингибитора miR-378 и/или miR-378*. В еще одном варианте осуществления настоящее изобретение обеспечивает способ предотвращения или лечения остеоартрита путем введения субъекту, нуждающемуся в этом, ингибитора miR-378 и/или miR-378*.
[0049] В другом варианте осуществления настоящее изобретение охватывает способ повышения поглощения и/или утилизации глюкозы у субъекта, нуждающегося в этом, при котором субъекту вводят ингибитор активности или экспрессии miR-378 и/или miR-378*. В некоторых вариантах осуществления у субъекта диагностируют резистентность к инсулину или сахарный диабет. В одном варианте осуществления уровень глюкозы в крови субъекта понижается после введения ингибитора miR-378 и/или miR-378* по сравнению с уровнем глюкозы в крови субъекта перед введением ингибитора. В другом варианте осуществления уровень глюкозы в крови субъекта понижается до пределов нормальных уровней, что измеряют с помощью пробы пероральной толерантности к глюкозе после введения ингибитора miR-378 и/или miR-378*. Например, в определенных вариантах осуществления уровень глюкозы в крови субъекта натощак составляет менее приблизительно 110 мг/дл. В других вариантах осуществления уровень глюкозы в крови субъекта через 2 часа после приема внутрь глюкозы составляет менее приблизительно 140 мг/дл.
[0050] Настоящее изобретение также обеспечивает способ регуляции метаболизма жирных кислот в клетке. В одном варианте осуществления способ включает приведение клетки в контакт с модулятором экспрессии или активности miR-378 и/или miR-378*. Как используется в данном документе, «модулятор» представляет собой молекулу, которая регулирует экспрессию или активность miR-378 и/или miR-378*. Модуляторы могут быть агонистами функции miR-378 и/или miR-378* (т. е. усиливать активность или экспрессию miR-378 или miR-378*), или они могут быть ингибиторами функции miR-378 и/или miR-378* (т. е. понижать активность или экспрессию miR-378 или miR-378*). Модуляторы могут включать белки, пептиды, полипептиды, полинуклеотиды, олигонуклеотиды или малые молекулы. Модуляторы экспрессии или активности miR-378 и/или miR-378* включают ингибиторы и агонисты miR-378 и/или miR-378*, которые описаны в данном документе. В определенных вариантах осуществления модулятор представляет собой ингибитор экспрессии или активности miR-378 и/или miR-378*, и метаболизм жирных кислот в клетке повышается после контакта с ингибитором miR-378 и/или miR-378* по сравнению с клеткой, не подвергнутой воздействию ингибитора. В других вариантах осуществления модулятор представляет собой агонист экспрессии или активности miR-378 и/или miR-378*, и метаболизм жирных кислот понижается в клетке после контакта с агонистом miR-378 и/или miR-378* по сравнению с клеткой, не подвергнутой воздействию агониста. Клетка может присутствовать in vitro или in vivo. В некоторых вариантах осуществления клетка представляет собой, но не ограничиваясь, кардиомиоцит, клетку скелетной мышцы, преадипоцит, адипоцит, гепатоцит или клетку поджелудочной железы.
[0051] В одном конкретном варианте осуществления клетка представляет собой кардиомиоцит. Таким образом, настоящее изобретение также включает способ регуляции метаболизма сердца путем приведения кардиомиоцита в контакт с модулятором экспрессии или активности miR-378 и/или miR-378*. В одном варианте осуществления приведение кардиомиоцита в контакт с ингибитором miR-378 и/или miR-378* предотвращает или уменьшает метаболический сдвиг от окислительного метаболизма к гликолитическому метаболизму, индуцированный стрессором. В другом варианте осуществления приведение кардиомиоцита в контакт с ингибитором miR-378 и/или miR-378* понижает метаболизм углеводов в кардиомиоците. В еще одном варианте осуществления приведение кардиомиоцита в контакт с ингибитором miR-378 и/или miR-378* повышает метаболизм жирных кислот в кардиомиоците. Кардиомиоцит может присутствовать in vitro или in vivo.
[0052] В другом аспекте настоящее изобретение включает способ регуляции митохондриальной дисфункции в клетке путем приведения клетки в контакт с модулятором экспрессии или активности miR-378 и/или miR-378*. В одном варианте осуществления приведение клетки в контакт с ингибитором miR-378 и/или miR-378* повышает митохондриальный биогенез в клетке по сравнению с необработанной клеткой. В другом варианте осуществления приведение клетки в контакт с ингибитором miR-378 и/или miR-378* усиливает митохондриальное окисление жирных кислот в клетке по сравнению с необработанной клеткой. В еще одном варианте осуществления приведение клетки в контакт с агонистом miR-378 и/или miR-378* усиливает липогенез в клетке по сравнению с необработанной клеткой. В еще одном варианте осуществления приведение клетки в контакт с агонистом miR-378 и/или miR-378* усиливает метаболизм глюкозы в клетке по сравнению с необработанной клеткой. Клетка может присутствовать in vitro или in vivo. В определенных вариантах осуществления клетка представляет собой без ограничения кардиомиоцит, клетку скелетной мышцы, преадипоцит, адипоцит, гепатоцит или клетку поджелудочной железы.
[0053] Настоящее изобретение также обеспечивает способ предотвращения или лечения расстройств или заболеваний, связанных с дефектом гликолитического метаболизма или метаболизма жирных кислот. Например, в одном варианте осуществления настоящее изобретение обеспечивает способ предотвращения или лечения гипогликемии или гиперинсулинизма у субъекта, нуждающегося в этом, путем введения субъекту агониста miR-378 и/или miR-378*. Субъекты с риском развития гипогликемии или гиперинсулинизма включают пациентов с диабетом, которые передозировали инсулин или определенные противодиабетические лекарственный препарат (например, хлорпропамид, толазамид, ацетогексамид, глипизид или толбутамид), субъект, который имеет опухоль, секретирующую инсулин (инсулиному), пациенты, у которых диагностировали заболевание печени или генетические состояния, которые вызывают гиперинсулинизм. Другие расстройства или состояния, которые можно лечить или предотвращать с помощью агонистов miR-378 и/или miR-378*, являются таковыми, при которых пациенты имеют трудность в поддержании нормальной массы тела или испытывают непроизвольную потерю массы. Например, в одном варианте осуществления настоящее изобретение включает способ лечения или предотвращения гипертиреоза (болезни Грейвса) у субъекта, нуждающегося в этом, путем введения субъекту агониста miR-378 и/или miR-378*.
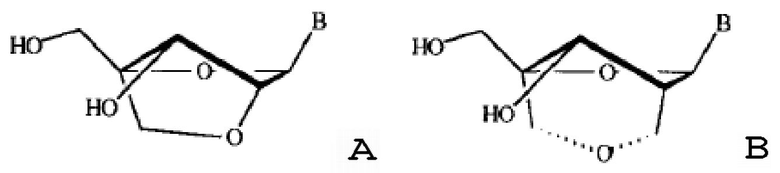
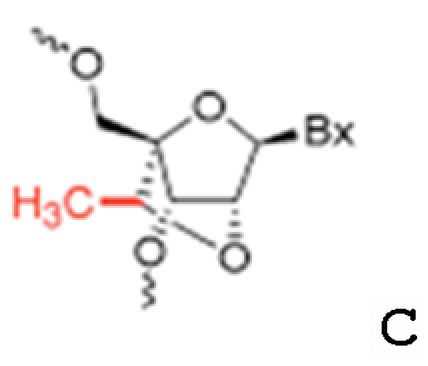
[0054] В некоторых вариантах осуществления ингибитор miR-378 и/или miR-378* представляет собой антисмысловой олигонуклеотид. Антисмысловые олигонуклеотиды могут включать рибонуклеотиды, или дезоксирибонуклеотиды, или их комбинацию. Предпочтительно, антисмысловые олигонуклеотиды имеют по меньшей мере одну химическую модификацию (например, модификация сахара или остова). Например, пригодные антисмысловые олигонуклеотиды могут включать одну или несколько «конформационно ограниченных» модификаций или модификаций нуклеозидами с бициклическим сахаром (BSN), которые придают улучшенную температурную стабильность комплексам, образованным между олигонуклеотидом, содержащим BSN, и комплементарной ему целевой цепью микроРНК. Например, в одном варианте осуществления антисмысловые олигонуклеотиды содержат по меньшей мере одну «закрытую» нуклеиновую кислоту. «Закрытые» нуклеиновые кислоты (LNA) содержат 2'-O, 4'-C-метиленрибонуклеозид (структура A), где фрагмент рибозного сахара находится в «закрытой» конформации. В другом варианте осуществления антисмысловые олигонуклеотиды содержат по меньшей мере один 2'-дезоксирибонуклеозид с 2',4'-C-мостиком (CDNA, структура B). Смотри, например, патент США №6403566 и Wang et al. (1999) Bioorganic and Medicinal Chemistry Letters, Vol.9: 1147-1150, оба из которых включены в данный документ ссылкой в их полноте. В еще одном варианте осуществления антисмысловые олигонуклеотиды содержат по меньшей мере один модифицированный нуклеозид, имеющий структуру, показанную на структуре C. Антисмысловые олигонуклеотиды, нацеливающиеся на miR-378, могут содержать комбинации BSN (LNA, CDNA и подобное) или других модифицированных нуклеотидов и рибонуклеотидов или дезоксирибонуклеотидов.
[0055] Альтернативно, антисмысловые олигонуклеотиды могут содержать пептидо-нуклеиновые кислоты (PNA), которые содержат остов на основе пептида, а не сахарно-фосфатный остов. Также предусматриваются другие модификации с модифицированным сахаром или фосфодиэфирные модификации в отношении антисмыслового олигонуклеотида. Например, другие химические модификации, которые могут содержать антисмысловые олигонуклеотиды, включают без ограничения модификации сахара, такие как 2'-O-алкильные (например, 2'-O-метильные, 2'-O-метоксиэтильные), 2'-фтор- и 4'-тио-модификации, и модификации остова, такие как одна или несколько фосфоротиоатных, морфолино- или фосфонокарбоксилатных связей (смотри, например, патенты США №№6693187 и 7067641, которые включены в данный документ ссылкой в их полноте). В одном варианте осуществления антисмысловые олигонуклеотиды, нацеливающиеся на miR-378 или miR-378*, содержат 2'-O-метильные модификации сахара в каждом основании и связаны с помощью фосфоротиоатных связей. Антисмысловые олигонуклеотиды, особенно таковые с меньшей длиной (например, менее 15 нуклеотидов) могут содержать одну или несколько усиливающих сродство модификаций, таких как без ограничения LNA, бициклические нуклеозиды, фосфоноформиаты, 2'-O-алкильные модификации и подобное. В некоторых вариантах осуществления пригодные антисмысловые олигонуклеотиды представляют собой 2'-O-метоксиэтильные «гэпмеры», которые содержат 2'-O-метоксиэтил-модифицированные рибонуклеотиды как на 5'-, так и 3'-концах с по меньшей мере десятью дезоксирибонуклеотидами в центре. Эти «гэпмеры» способны запускать механизмы РНКаза H-зависимой деградации РНК-мишеней. Другие модификации антисмысловых олигонуклеотидов для усиления стабильности и улучшения эффективности, такие как таковые, описанные в патенте США № 6838283, который включен в данный документ ссылкой в его полноте, известны из уровня техники и пригодны для применения в способах настоящего изобретения. Например, для облегчения доставки и стабильности in vivo антисмысловой олигонуклеотид может быть связан со стероидом, таким как холестериновый фрагмент, витамином, жирной кислотой, углеводом или гликозидом, пептидом или другим лигандом-малой молекулой на его 3'-конце.
[0056] Предпочтительные антисмысловые олигонуклеотиды, пригодные для ингибирования активности миРНК, имеют длину от приблизительно 5 до приблизительно 25 нуклеотидов, от приблизительно 10 до приблизительно 30 нуклеотидов или от приблизительно 20 до приблизительно 25 нуклеотидов. В определенных вариантах осуществления антисмысловые олигонуклеотиды, нацеливающиеся на miR-378 и/или miR-378*, имеют длину от приблизительно 7 до приблизительно 18 нуклеотидов, и в других вариантах осуществления они имеют длину от приблизительно 12 до приблизительно 16 нуклеотидов. Можно применять любой содержащий 7 мономерных звеньев или более длинный олигомер, комплементарный miR-378 или miR-378*, т. e., любая antimiR, комплементарная 5'-концу миРНК и далее по полной комплементарной последовательности миРНК. Например, в одном варианте осуществления антисмысловой олигонуклеотид имеет последовательность 5'-CCUUCUGACUCCAAGUCCAGU-3' (SEQ ID NO: 7). В другом варианте осуществления антисмысловой олигонуклеотид имеет последовательность 5'-AGUCCAGU-3' (SEQ ID NO: 8). В другом варианте осуществления антисмысловой олигонуклеотид имеет последовательность 5'-CAAGUCCAGU-3' (SEQ ID NO: 9). В другом варианте осуществления антисмысловой олигонуклеотид имеет последовательность 5'-UCCAAGUCCAGU-3' (SEQ ID NO: 10). В еще одном варианте осуществления антисмысловой олигонуклеотид имеет последовательность 5'-ACUCCAAGUCCAGU-3' (SEQ ID NO: 11). В еще одном варианте осуществления антисмысловой олигонуклеотид имеет последовательность 5'-UGACUCCAAGUCCAGU-3' (SEQ ID NO: 12) или 5-CUGACUCCAAGUCCAG-3' (SEQ ID NO: 13). В определенных вариантах осуществления антисмысловой олигонуклеотид имеет последовательность 5'-TGACTCCAAGTCCAG-3' (SEQ ID NO: 21).
[0057] Иллюстративные антисмысловые олигонуклеотиды для ингибирования экспрессии или активности miR-378* включают без ограничения 5'-ACACAGGACCUGGAGUCAGGAG-3' (SEQ ID NO: 14); 5'-GUCAGGAG-3' (SEQ ID NO: 15); 5'-GAGUCAGGAG-3' (SEQ ID NO: 16); 5'-UGGAGUCAGGAG-3' (SEQ ID NO: 17); 5'-CCUGGAGUCAGGAG-3' (SEQ ID NO: 18); 5'-GACCUGGAGUCAGGAG-3' (SEQ ID NO: 19); 5'-GGACCUGGAGUCAGGA-3' (SEQ ID NO: 20) и 5'-GACCTGGAGTCAGGA-3' (SEQ ID NO: 22).
[0058] Антисмысловые олигонуклеотиды могут содержать последовательность, которая является по меньшей мере частично комплементарной зрелой или минорной (т. е. со звездочкой) последовательности miR-378, например, по меньшей мере на приблизительно 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% комплементарной зрелой или минорной (т. е. со звездочкой) последовательности miR-378. В некоторых вариантах осуществления антисмысловой олигонуклеотид может быть, по сути, комплементарным зрелой или минорной последовательности miR-378, которая является по меньшей мере на приблизительно 90%, 95%, 96%, 97%, 98% или 99% комплементарной последовательности целевого полинуклеотида. В одном варианте осуществления антисмысловой олигонуклеотид содержит последовательность, которая является на 100% комплементарной зрелой или минорной последовательности miR-378. В определенных вариантах осуществления антисмысловой олигонуклеотид является по меньшей мере частично комплементарным SEQ ID NO: 1. В других вариантах осуществления антисмысловой олигонуклеотид является по меньшей мере частично комплементарным SEQ ID NO: 2.
[0059] В некоторых вариантах осуществления антисмысловые олигонуклеотиды представляют собой антагомиры. «Антагомиры» представляют собой одноцепочечные химически модифицированные рибонуклеотиды, которые являются по меньшей мере частично комплементарными последовательности miR-378 или miR-378*. Антагомиры могут содержать один или несколько модифицированных нуклеотидов, таких как модификации с 2'-O-метил-сахаром. В некоторых вариантах осуществления антагомиры содержат только модифицированные нуклеотиды. Антагомиры могут также содержать одну или несколько фосфоротиоатных связей, что дает в результате частично или полностью фосфоротиоатный остов. Для облегчения доставки и стабильности in vivo антагомир может быть связан с холестериновой или другой группой на его 3'-конце. Антагомиры, пригодные для ингибирования miR-378 или miR-378*, могут иметь длину от приблизительно 15 до приблизительно 50 нуклеотидов, более предпочтительно от приблизительно 18 до приблизительно 30 нуклеотидов в длину и наиболее предпочтительно от приблизительно 20 до приблизительно 25 нуклеотидов. Антагомиры могут быть по меньшей мере на приблизительно 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% комплементарными зрелой или минорной последовательности miR-378. В некоторых вариантах осуществления антагомир может быть, по сути, комплементарным зрелой или минорной последовательности miR-378, которая является по меньшей мере на приблизительно 95%, 96%, 97%, 98% или 99% комплементарной последовательности целевого полинуклеотида. В других вариантах осуществления антагомиры являются на 100% комплементарными зрелой или минорной последовательности miR-378.
[0060] Ингибиторные нуклеотидные молекулы, описанные в данном документе, предпочтительно нацелены на зрелую последовательность miR-378 (SEQ ID NO: 1). В одном варианте осуществления ингибиторные нуклеотидные молекулы, описанные в данном документе, нацеливаются на минорную (т. е. miR-378*) последовательность miR-378 (SEQ ID NO: 2). В некоторых вариантах осуществления ингибиторы miR-378 и/или miR-378* представляют собой антагомиры, содержащие последовательность, которая полностью комплементарна зрелой или минорной последовательности miR-378. В одном варианте осуществления ингибитор miR-378 представляет собой антагомир, имеющий последовательность, которая частично или полностью комплементарна 5'-ACUGGACUUGGAGUCAGAAGG-3' (SEQ ID NO: 1). В другом варианте осуществления ингибитор miR-378* представляет собой антагомир, имеющий последовательность, которая частично или полностью комплементарна 5'-CUCCUGACUCCAGGUCCUGUGU-3' (SEQ ID NO: 2). В некоторых вариантах осуществления ингибиторы miR-378 и/или miR-378* являются химически модифицированными антисмысловыми олигонуклеотидами. В одном варианте осуществления ингибитор miR-378 представляет собой химически модифицированный антисмысловой олигонуклеотид, содержащий последовательность, по сути, комплементарную 5'-ACUGGACUUGGAGUCAGAAGG-3' (SEQ ID NO: 1). В другом варианте осуществления ингибитор miR-378* представляет собой химически модифицированный антисмысловой олигонуклеотид, содержащий последовательность, по сути, комплементарную 5'-CUCCUGACUCCAGGUCCUGUGU-3' (SEQ ID NO: 2). Как используется в данном документе, «по сути, комплементарный» относится к последовательности, которая является по меньшей мере на приблизительно 95%, 96%, 97%, 98%, 99% или 100% комплементарной последовательности целевого полинуклеотида (например, зрелой, минорной последовательности или последовательности предшественника миРНК).
[0061] Антисмысловые олигонуклеотиды или антагомиры могут содержать последовательность, которая, по сути, комплементарна последовательности предшественника миРНК (пре-миРНК) или последовательности первичной миРНК (при-миРНК) для miR-378. В некоторых вариантах осуществления антисмысловой олигонуклеотид содержит последовательность, которая, по сути, комплементарна последовательности, расположенной вне участка «стебель-петля» последовательности пре-miR-378. В одном варианте осуществления ингибитор функции miR-378 представляет собой антисмысловой олигонуклеотид, имеющий последовательность, которая, по сути, комплементарна последовательности пре-miR-378 (SEQ ID NO: 3).
[0062] Агонист экспрессии или активности miR-378 и/или miR-378* может быть полинуклеотидом, содержащим последовательность miR-378 и/или miR-378*. Например, в одном варианте осуществления агонист miR-378 представляет собой полинуклеотид, содержащий зрелую последовательность miR-378 (SEQ ID NO: 1). В другом варианте осуществления агонист miR-378* представляет собой полинуклеотид, содержащий минорную (т.е. со звездочкой) последовательность miR-378 (SEQ ID NO: 2). В еще одном варианте осуществления агонист miR-378 может быть полинуклеотидом, содержащим последовательность при-миРНК для miR-378. В еще одном варианте осуществления агонист miR-378 и/или miR-378* может быть полинуклеотидом, содержащим последовательность пре-миРНК для miR-378 (например, SEQ ID NO: 3). Полинуклеотид, содержащий последовательность miR-378 и/или miR-378*, может иметь длину от приблизительно 18 до приблизительно 2000 нуклеотидов, от приблизительно 70 до приблизительно 200 нуклеотидов, от приблизительно 20 до приблизительно 50 нуклеотидов или от приблизительно 18 до приблизительно 25 нуклеотидов. Полинуклеотид, содержащий зрелую последовательность miR-378, минорную последовательность miR-378 (т.е. miR-378*), последовательность пре-miR-378 или последовательность при-miR-378, может быть одноцепочечным или двухцепочечным. Полинуклеотиды могут содержать одну или несколько химических модификаций, таких как «закрытые» нуклеиновые кислоты, пептидо-нуклеиновые кислоты, модификации сахара, такие как 2'-O-алкильная (например 2'-O-метильная, 2'-O-метоксиэтильная), 2'-фтор- и 4'-тио- модификации, и модификации остова, такие как одну или несколько фосфоротиоатных, морфолино- или фосфонокарбоксилатных связей. В одном варианте осуществления полинуклеотид, содержащий последовательность miR-378 (например, зрелую miR-378, miR-378*, пре-miR-378 или при-miR-378) конъюгирован со стероидом, таким как холестерин, витамином, жирной кислотой, углеводом или гликозидом, пептидом или другим лигандом-малой молекулой.
[0063] Любой из ингибиторов или агонистов miR-378 и/или miR-378*, описанных в данном документе, может быть доставлен к клетке-мишени (например, клетка сердца, клетка жировой ткани или клетка скелетной мышцы) путем доставки в клетку вектора экспрессии, кодирующего ингибиторы или агонисты miR-378. «Вектор» представляет собой композицию, которую можно применять для доставки нуклеиновой кислоты интереса во внутреннее пространство клетки. Многочисленные векторы известны из уровня техники, включая без ограничения линейные полинуклеотиды, полинуклеотиды, ассоциированные с ионными или амфифильными соединениями, плазмиды и вирусы. Таким образом, выражение «вектор» включает автономно реплицирующуюся плазмиду или вирус. Примеры вирусных векторов включают без ограничения аденовирусные векторы, векторы на основе аденоассоциированных вирусов, ретровирусные векторы и подобное. Конструкт экспрессии может реплицироваться в живой клетке, или он может быть создан синтетически. Для целей настоящей заявки выражения «конструкт экспрессии», «вектор экспрессии» и «вектор» используют взаимозаменяемо, чтобы показать применение настоящего изобретения в общем, иллюстративном смысле, и они не предназначены для ограничения настоящего изобретения.
[0064] В одном варианте осуществления вектор экспрессии для экспрессии ингибитора miR-378 и/или miR-378* содержит промотор, функционально связанный с полинуклеотидом, кодирующим антисмысловой олигонуклеотид, где последовательность экспрессируемого антисмыслового олигонуклеотида частично или абсолютно комплементарна зрелой или минорной последовательности miR-378 (например, SEQ ID NO: 1 или SEQ ID NO: 2). В другом варианте осуществления вектор экспрессии для экспрессии полинуклеотида, содержащего последовательность miR-378, содержит промотор, функционально связанный с полинуклеотидом, содержащим зрелую последовательность miR-378 (например, SEQ ID NO: 1), минорную последовательность miR-378 (например, SEQ ID NO: 2), последовательность пре-miR-378 (например, SEQ ID NO: 3) или последовательность при-miR-378. Фраза «функционально связанный» или «под транскрипционным контролем», как используется в данном документе, означает, что промотор находится в правильном местоположении и ориентации по отношению к полинуклеотиду для контроля инициации транскрипции с помощью РНК-полимеразы и экспрессии полинуклеотида.
[0065] Как используется в данном документе, «промотор» относится к последовательности ДНК, распознаваемой синтезирующим аппаратом клетки или введенным синтезирующим аппаратом, необходимой для инициации специфической транскрипции гена. Пригодные промоторы включают без ограничения промотры RNA pol I, pol II, pol III и вирусные промоторы (например, промотор немедленно-раннего гена цитомегаловируса человека (CMV), ранний промотор SV40 и длинный концевой повтор вируса саркомы Рауса). В одном варианте осуществления промотор представляет собой тканеспецифический промотор. Особый интерес представляют специфические для мышцы промоторы и, более конкретно, специфические для сердца промоторы. Они включают промотор легкой цепи-2 миозина (Franz et al. (1994) Cardioscience, Vol.5(4):235-43; Kelly et al. (1995) J. Cell Biol., Vol.129(2):383-396), альфа-актиновый промотор (Moss et al. (1996) Biol. Chem., Vol.271(49):31688-31694), промотор тропонина 1 (Bhavsar et al. (1996) Genomics, Vol.35(1):11-23); промотор Na+/Ca2+обменника (Barnes et al. (1997) J. Biol. Chem., Vol.272(17):11510-11517), дистрофиновый промотор (Kimura et al. (1997) Dev. Growth Differ., Vol.39(3):257-265), промотор интегрина альфа-7 (Ziober and Kramer (1996) J. Bio. Chem., Vol.271(37):22915-22), промотор мозгового натрийуретического пептида (LaPointe et al. (1996) Hypertension, Vol.27(3 Pt 2):715-22) и промотор альфа-B-кристаллина/малого белка теплового шока (Gopal-Srivastava (1995) J. Mol. Cell. Biol., Vol.15(12):7081-7090), промотор тяжелой цепи альфа-миозина (Yamauchi-Takihara et al. (1989) Proc. Natl. Acad. Sci. USA, Vol.86(10):3504-3508) и промотор ANF (LaPointe et al. (1988) J. Biol. Chem., Vol.263(19):9075-9078). В одном варианте осуществления тканеспецифический промотор представляет собой адипоцит-специфический промотор, такой как промотор адипоцитного белка 2 (ap2)/белка, связывающего жирные кислоты, 4 (FABP4) или промотор PPARγ.
[0066] В определенных вариантах осуществления промотор, функционально связанный с полинуклеотидом, кодирующим ингибитор miR-378 и/или miR-378*, или полинуклеотидом, кодирующим последовательность miR-378, может быть индуцибельным промотором. Индуцибельные промоторы известны из уровня техники и включают без ограничения тетрациклиновый промотор, промотор металлотионеина IIA, промотор теплового шока, участки ответа на стероид/тиреоидный гормон/ретиноевую кислоту, поздний промотор аденовируса и индуцибельный LTR (длинный концевой повтор) вируса опухоли молочной железы мыши.
[0067] Способы доставки конструктов экспрессии и нуклеиновых кислот к клеткам известны из уровня техники и могут включать, например, соосаждение с фосфатом кальция, электропорацию, микроинъекцию, DEAE-декстран, липофекцию, трансфекцию, использующую полиаминные реагенты для трансфекции, обработку клеток ультразвуком, бомбардирование генами с помощью высокоскоростные микрочастиц и рецептор-опосредованную трансфекцию.
[0068] Настоящее изобретение также включает способы извлечения или очистки от ингибиторов miR-378 и/или miR-378* после лечения. Способ может включать сверхэкспрессию сайтов связывания для ингибиторов miR-378 и/или miR-378* в ткани сердечной мышцы или скелетных мышц. Участки сайта связывания предпочтительно содержат последовательность затравочного участка для miR-378 и/или miR-378*. Затравочный участок представляет собой 5'-часть миРНК, охватывающий основания 2-8, который важен для распознавания мишени. В некоторых вариантах осуществления сайт связывания может содержать последовательность из 3'-UTR одной или нескольких мишеней miR-378 и/или miR-378*, таких как SuFu, Fus-1, глутамин:фруктоза-6-фосфат амидотрансфераза-2 (GFPT2) и MED13.
[0069] Настоящее изобретение также включает фармацевтические композиции, содержащие ингибитор или агонист miR-378 и/или miR-378*. Там где предусмотрены клинические применения, фармацевтические композиции будут получены в форме, подходящей для предполагаемого применения. Как правило, оно будет включать в себя получение композиций, которые, по сути, не содержат пирогенов, а также других примесей, которые могут быть вредными для людей или животных.
[0070] В одном варианте осуществления фармацевтическая композиция содержит эффективную дозу ингибитора miR-378 и/или miR-378* и фармацевтически приемлемый носитель. Например, фармацевтическая композиция содержит эффективную дозу модифицированного антисмыслового олигонуклеотида, нацеленного на miR-378 и/или miR-378*, как описано в данном документе. В некоторых вариантах осуществления фармацевтическая композиция содержит модифицированный антисмысловой олигонуклеотид, имеющий последовательность, выбранную из группы, содержащей SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20, SEQ ID NO: 21 и SEQ ID NO: 22. В другом варианте осуществления фармацевтическая композиция содержит эффективную дозу агониста miR-378 и/или miR-378*, как описано в данном документе, и фармацевтически приемлемый носитель. «Эффективная доза» представляет собой количество, достаточное, чтобы произвести благоприятный или необходимый клинический результат. Эффективная доза ингибитора миРНК или агониста миРНК настоящего изобретения может составлять от приблизительно 1 мг/кг до приблизительно 100 мг/кг, от приблизительно 2,5 мг/кг до приблизительно 50 мг/кг или от приблизительно 5 мг/кг до приблизительно 25 мг/кг. Точное определение того, что будет рассматриваться как эффективная доза, может основываться на факторах, индивидуальных для каждого пациента, включая их рост, возраст, тип расстройства (например, инфаркт миокарда, сердечная недостаточность, гипертрофия сердца, метаболическое расстройство) и природу ингибитора или агониста (например, антагомир, конструкт экспрессии, антисмысловой олигонуклеотид, полинуклеотидный дуплекс и т.д.). Следовательно, дозировки могут быть легко установлены специалистами в данной области техники из этого раскрытия и информации из уровня техники.
[0071] Коллоидные дисперсные системы, такие как макромолекулярные комплексы, нанокапсулы, микросферы, гранулы и системы на основе липидов, включая эмульсии масло-в-воде, мицеллы, смешанные мицеллы и липосомы, можно применять в качестве сред для доставки для олигонуклеотидных ингибиторов функции miR-378 и/или miR-378*, полинуклеотидов, кодирующих агонисты miR-378 и/или miR-378*, или конструктов экспрессии конкретных ингибиторов или агонистов миРНК. Коммерчески доступные эмульсии жиров, которые пригодны для доставки нуклеиновых кислот настоящего изобретения к тканям сердечной и скелетных мышц, включают Intralipid®, Liposyn®, Liposyn® II, Liposyn® III, Nutrilipid и другие подобные эмульсии липидов. Предпочтительная коллоидная система для применения в качестве среды для доставки in vivo представляет собой липосому (т. e., искусственный мембранный пузырек). Получение и применение таких систем хорошо известно из уровня техники. Иллюстративные составы также раскрыты в патентах США №№5981505; 6217900; 6383512; 5783565; 7202227; 6379965; 6127170; 5837533; 6747014 и публикации международной заявки WO03/093449, которые включены в данный документ ссылкой в их полноте.
[0072] Как правило, желательным будет использовать соответствующие соли и буферы, чтобы сделать среды для доставки стабильными и обеспечить поглощение целевыми клетками. Водные композиции настоящего изобретения содержат эффективное количество среды для доставки, содержащего ингибиторные полинуклеотиды (например, липосомы, или другие комплексы, или векторы экспрессии), растворенные или диспергированные в фармацевтически приемлемом носителе или водной среде. Фразы «фармацевтически приемлемый» или «фармакологически приемлемый» относится к молекулярным частицам и композициям, которые не производят неблагоприятных, аллергических или других нежелательных реакций при введении животному или человеку. Как используется в данном документе, «фармацевтически приемлемая среда» включает растворители, буферы, растворы, дисперсионные среды, оболочки, антибактериальные и противогрибковые средства, обеспечивающие изотоничность и замедляющие абсорбцию средства и подобное, приемлемые для применения при составлении фармацевтических препаратов, таких как фармацевтические препараты, пригодные для введения людям. Применение таких сред и средств для фармацевтически активных веществ хорошо известно из уровня техники. За исключением случаев, когда любая общепринятая среда или средство является несовместимым с активными ингредиентами настоящего изобретения, предусматривается его применение в терапевтических композициях. Дополнительные активные ингредиенты также могут быть включены в композиции при условии, что они не инактивируют векторы или полинуклеотиды композиций.
[0073] Активные композиции настоящего изобретения могут включать классические фармацевтические препараты. Введение этих композиций согласно настоящему изобретению может быть осуществлено любым обычным путем при условии, что целевая ткань доступна посредством этого пути. Он включает пероральный, назальный или буккальный. Альтернативно, введение можно осуществлять путем внутрикожной, подкожной, внутримышечной, внутрибрюшинной или внутривенной инъекции или путем прямой инъекции в ткань сердца. Фармацевтические композиции, содержащие ингибиторы миРНК, агонисты миРНК или конструкты экспрессии, содержащие ингибиторы или агонисты миРНК, могут также быть введены с помощью катетерных систем или систем, которые выделяют коронарное кровообращение для доставки терапевтических средств к сердцу. Различные катетерные системы для доставки терапевтических средств к сердцу и коронарным сосудам известны из уровня техники. Некоторые неограничивающие примеры способов доставки на основе катетеров или способов выделения коронарных сосудов, пригодных для применения в настоящем изобретении, раскрыты в патенте США №6416510; патенте США №6716196; патенте США №6953466, публикации международной заявки WO 2005/082440, WO 2006/089340, патентной публикации США №2007/0203445, патентной публикации США №2006/0148742 и патентной публикации США № 2007/0060907, которые включены в данный документ ссылкой в их полноте. Такие композиции будут в норме вводиться в качестве фармацевтически приемлемых композиций, как описано в данном документе.
[0074] Активные соединения можно также вводить парентерально или внутрибрюшинно. С целью иллюстрации растворы активных соединений в виде свободного основания или фармакологически приемлемых солей можно получить в воде, соответствующим образом смешанной с поверхностно-активным веществом, таким как гидроксипропилцеллюлоза. Можно получить дисперсии можно получить в глицерине, жидких полиэтиленгликолях и их смесях и в маслах. При обычных условиях хранения и применения эти препараты, как правило, содержат консервант для предотвращения роста микроорганизмов.
[0075] Фармацевтические формы, пригодные для инъекционного применения или катетерной доставки, включают, например, стерильные водные растворы или дисперсии и стерильные порошки для немедленного приготовления стерильных инъекционных растворов или дисперсий. Как правило, эти препараты являются стерильными и жидкими в тех случаях, когда существует возможность для легкого осуществления инъекции. Препараты должны быть стабильными при условиях производства и хранения и должны быть защищены от загрязняющего действия микроорганизмов, таких как бактерии и грибы. Приемлемые растворители или дисперсионные среды могут содержать, например, воду, этанол, многоатомный спирт (например, глицерин, полиэтиленгликоль, и жидкий полиэтиленгликоль, и подобное), их пригодные смеси и растительные масла. Необходимую текучесть можно поддерживать, например, путем применения покрытия, такого как лецитиновое, путем поддержания необходимого размера частиц в случае дисперсии и путем применения поверхностно-активных веществ. Предотвращение действия микроорганизмов можно осуществлять с помощью различных антибактериальных и противогрибковых средств, например, парабенов, хлорбутанола, фенола, сорбиновой кислоты, тимеросала и подобного. Во многих случаях будет предпочтительным включать обеспечивающие изотоничность средства, например, сахара или хлорид натрия. Длительного поглощения инъекционных композиций можно достичь путем применения в композициях средств, замедляющих абсорбцию, например, моностеарата алюминия и желатина.
[0076] Стерильные инъекционные растворы можно получить путем включения активных соединений в соответствующем количестве в растворитель вместе с любыми другими ингредиентами (например, которые перечислялись выше), если необходимо, с последующей стерилизацией фильтрованием. Как правило, дисперсии получают путем включения различных стерилизованных активных ингредиентов в стерильную среду, которая содержит основную дисперсионную среду и необходимые другие ингредиенты, например, как перечислено выше. В случае стерильных порошков для приготовления стерильных инъекционных растворов предпочтительные способы получения включают методики вакуумной сушки и лиофильной сушки, которые дают на выходе порошок активного ингредиента(ов) вместе с любым дополнительным необходимым ингредиентом из его предварительно стерилизованного фильтрованием раствора.
[0077] Композиции настоящего изобретения, как правило, могут быть составлены в нейтральной или солевой форме. Фармацевтически приемлемые соли включают, например, кислотно-аддитивные соли (образованные со свободными аминогруппами белка), полученными с неорганическими кислотами (например, соляной или фосфорной кислот) или с органическими кислотами (например, уксусной, щавелевой, винной, миндальной и подобного). Соли, образованные со свободными карбоксильными группами белка, можно также получить с неорганическими основаниями (например, гидроксидами натрия, калия, аммония, кальция или железа) или с органическими основаниями (например, изопропиламином, триметиламином, гистидином, прокаином и подобным).
[0078] После составления растворы предпочтительно вводят способом, совместимым с дозированным составом и в таком количестве, которое является терапевтически эффективным. Составы можно легко вводить в ряде лекарственных форм, таких как инъекционные растворы, капсулы с высвобождением лекарственного средства и подобное. Например, для парентерального введения в водном растворе раствор, как правило, соответствующим образом стабилизируют буфером и вводят жидкий разбавитель, которому первоначально придают изотоничность, например, с помощью достаточного количества солевого раствора или глюкозы. Такие водные растворы можно применять, например, для внутривенного, внутримышечного, подкожного и внутрибрюшинного введения. Предпочтительно, применяют стерильные водные среды, как это известно специалистам в данной области техники, особенно в свете настоящего раскрытия. С целью иллюстрации одна доза может быть растворена в 1 мл изотонического раствора NaCl и либо добавлена к 1000 мл жидкости для гиподермоклиза, либо введена инъекционно в предполагаемое место инфузии, (см., например, "Remington's Pharmaceutical Sciences" 15-е издание, сс. 1035-1038 и 1570-1580). Некоторое изменение дозировки будет обязательно возникать в зависимости от состояния субъекта, который подлежит лечению. Лицо, ответственное за введение, будет, в любом случае, определять соответствующую дозу для отдельного субъекта. Более того, для введения человеку препараты должны соответствовать стандартам стерильности, пирогенности, общей безопасности и чистоты, как требует Комитет по биологическим стандартам Управления по контролю качества пищевых продуктов и лекарственных препаратов (FDA Office of Biologics standards).
[0079] В определенных вариантах осуществления настоящего изобретения фармацевтические композиции настоящего изобретения упакованы в или хранятся внутри устройства для введения. Устройства для инъекционных составов включают без ограничения инъекционные порты, автоинъекторы, инъекционные насосы и шприц-ручки. Устройства для аэрозольных или порошковых составов включают без ограничения ингаляторы, инсуффляторы, аспираторы и подобное. Таким образом, настоящее изобретение включает устройства для введения, содержащие фармацевтическую композицию настоящего изобретения для лечения или предотвращения одного или нескольких расстройств, описанных в данном документе.
[0080] Настоящее изобретение дополнительно иллюстрируется следующими дополнительными примерами, которые не следует истолковывать как ограничивающие. Специалисты в данной области техники должны, в свете настоящего раскрытия, понимать, что многие изменения можно быть сделаны в конкретных вариантах осуществления, которые раскрыты, и все еще получить подобный или сходный результат, не отступая от сущности и объема настоящего изобретения.
ПРИМЕРЫ
Пример 1. miR-378/miR-378* высоко экспрессирована в богатых митохондриями тканях
[0081] Анализ профиля экспрессии миРНК в сердце с недостаточностью был описан ранее с применением мышиных моделей заболевания сердца (van Rooij et al. (2006) Proc. Natl. Acad. Sci. USA, Vol.103:18255-18260). Анализ микроРНК на микрочипах, проведенный на ткани, полученной из сердец с гипертрофией, индуцированной перегрузкой давлением, сверхэкспрессией кальциневрина или введением изопротеренола, выявил, что miR-378 (GeneID: 723889) является одной из наиболее устойчиво регулируемых миРНК при этих состояниях. MiR-378 также даунрегулируется при инфаркте миокарда (van Rooij et al. (2008) Proc. Natl. Acad. Sci. USA, Vol.105:13027-13032), обеспечивая дополнительное доказательство того, что miR-378 является важным определяющим фактором программы реакции сердца в ответ на стресс. MiR-378 расположена на мышиной хромосоме 18 в пределах первого интрона гена коактиватора 1-бета гамма-рецептора, активируемого пролифератором пероксисом (PPARGC1β, GeneID: 170826) (фигура 1A).
[0082] Для исследования профиля экспрессии miR-378 и miR-378* проводили анализ количественной ПЦР в реальном времени различных тканей, выделенных из мышей. Обнаружили, что miR-378 и miR-378* высоко экспрессируются в сердце, скелетной мышце и бурой жировой ткани (фигура 1B). В ткани сердца miR-378 более высоко экспрессировалась в кардиомиоцитах, чем в сердечных фибробластах (фигура 1C). Паттерн экспрессии miR-378/miR-378* и их гена-хозяина, PPARGC1β (PGC-1β), были перекрывающимися (фигура 1B), подтверждая, что miR-378/miR-378* и PPARGC1β транскрибируются совместно. Согласно этой гипотезе понижение уровня экспрессии miR-378 наблюдали в клеточной культуре при обработке стеариновой кислотой, известным репрессором экспрессии PPARGC1β (фигура 1D). Экспрессия как miR-378, так и ее гена-хозяина, PPARGC1β (PGC-1β), повышается в ходе адипогенеза (фигура 1E). Было показано, что резкое возрастание уровней в плазме стеариновой кислоты, а также других короткоцепочечных жирных кислот даунрегулирует экспрессию PPARGC1α, PPARGC1β и PPARα (Staiger et al. (2005) Diabetologia, Vol.48:2115-2118). Наблюдение, что транскрипция как miR-378, так и гена-хозяина PPARGC1β следует параллельным регуляторным паттернам, явно говорит о том, что два гена регулируются совместно. PPARGC1β, как известно, играет роль в регуляции окисления жирных кислот, утилизации глюкозы и митохондриальном биогенезе (Handschin and Spiegelman (2006) Endocr. Rev., Vol.27:728-735; Lelliott et al. (2006) PLoS Biol., Vol.4:2042-2056; Luptak et al. (2005) Circulation, Vol.112:2339-2346; Sonoda et al. (2007) Proc. Natl. Acad. Sci. USA, Vol.104:5223-5228; и Vianna et al. (2006) Cell Metab., Vol.4:453-464). Вероятно, что miR-378/miR-378* играют роль в тех самых процессах, поскольку интронные миРНК часто участвуют в той же сети генных программ, что и их ген-хозяин.
Пример 2. MiR-378 регулирует индуцированную стрессом гипертрофическую реакцию сердца
[0083] Для дальнейшего выяснения роли miR-378 в заболевании сердца экспрессию miR-378 в ткани сердца оценивали после индукции гипертрофии сердца путем торакальной операции по сужению аорты (TAB) у мышей дикого типа. Конкретно, на аорту накладывали швы до диаметра иглы калибра 27, индуцируя перегрузку сердце давлением, таким образом имитируя гипертензию. Контрольные животные подвергались симуляционному хирургическому вмешательству, при котором аорта не изменялась. Анализ микроРНК на микрочипах и ПЦР в реальном времени для ткани сердца, выделенной через 21 день после процедуры TAB, выявили, что miR-378 даунрегулируется в ответ на перегрузку давлением (фигура 2A).
[0084] Для оценки эффекта сверхэкспрессии miR-378 в ходе болезни сердца получали трансгенных мышей, которые сверхэкспрессировали miR-378/miR-378* под контролем специфического для кардиомиоцитов промотора тяжелой цепи α-миозина (α-MHC). После TAB эти трансгенные животные показывали усиленную сердечную гипертрофическую реакцию по сравнению с мышами дикого типа (фигура 2B). Эти данные показывают, что усиленная экспрессия miR-378 в ходе болезни сердца является вредной для сердца, и понижение уровня miR-378 у мышей дикого типа после TAB говорит о защитной реакции сердца на внешний стресс.
[0085] MiR-378, вероятно, играет роль в процессе сердечного метаболизма, конкретно, гипертрофического метаболического сдвига (Luptak et al. (2005) Circulation, Vol.112:2339-2346). Предполагают, что miR-378 увеличивает сердечный метаболический сдвиг и, таким образом, принимает участие в сердечной гипертрофической реакции при стрессе. Ингибирование экспрессии или активности miR-378 может обеспечивать терапевтическую пользу у пациентов с сердечнососудистыми заболеваниями.
Пример 3. Сверхэкспрессия miR-378 в скелетной мышце приводит в результате к увеличенной массе тела и массе эпидидимального жира
[0086] Для оценки эффекта сверхэкспрессии miR-378 в скелетной мышце получали трансгенных мышей, которые сверхэкспрессируют miR-378/miR-378* под контролем промотора мышечной креатинкиназы (MCK). Предварительное исследование трансгенных по MCK-miR-378 животных выявило, что miR-378 модулирует утилизацию глюкозы и жирных кислот. В частности, трансгенные животные большего возраста (возрастом 10 месяцев) весили существенно больше, чем однопометные животные дикого типа (фигура 3). Разница в массе тела, являющаяся результатом увеличенной массы подушечек эпидидимального белого жира у мутантных животных по сравнению с животными дикого типа (фигура 3). Гистологический анализ белой жировой ткани показывал гипертрофию адипоцитов у сверхэкспрессирующих miR-378 мышей (фигура 3). Повышенное накопление триглицеридов в адипоцитах трансгенных по MCK-miR-378 животных, вероятно, является следствием увеличенной резистентности к инсулину и генерализованных дефектов утилизации глюкозы.
[0087] В другой серии экспериментов miR-378 сверхэкспрессировали в 3T3-L1 клетках, которые впоследствии индуцировали к дифференцировке в адипоциты. Как демонстрируется окрашиванием масляным красным O, повышенное накопление липидов наблюдалось в клетках, сверхэкспрессирующих miR-378, обеспечивая дополнительное доказательство вовлечения miR-378 в регуляцию метаболических процессов (фигура 4).
[0088] Анализ на микрочипах для различных тканей, выделенных из страдающих ожирением мышей (ob/ob) и страдающих с диабетом мышей (db/db), выявил, что miR-378 сильно даунрегулируется (данные не показаны). Ob/ob мыши имеют мутацию в гене, кодирующем лептин, что приводит к тому, что они не способны производить гормон лептин и правильно регулировать аппетит. Db/db мыши имеют мутацию в гене, кодирующем лептиновый рецептор, что приводит к нарушенному сплайсингу транскрипта, что ведет к процессированию клеточного домена рецептора. Мутантный лептиновый рецептор не обладает функцией передачи сигнала. Полагают, что регуляция miR-378 в этих животных моделях ожирения и диабета представляет защитный механизм против метаболических расстройств, индуцированных нарушением передачи сигнала с помощью лептина. Таким образом, miR-378 может представлять эффективную терапевтическую мишень при различных метаболических расстройствах.
Пример 4. Трансгенные мыши, сверхэкспрессирующие miR-378 в скелетной мышце, проявляют пониженную толерантность к глюкозе
[0089] Для дополнительной оценки роли miR-378 в регуляции утилизации глюкозы сверхэкспрессирующие miR-378 в скелетной мышце трансгенные мыши (MCK-miR-378 Tg; смотри пример 3) подвергались глюкозотолерантному тесту (GTT) для выявления любых возможных недостаточностей клиренса глюкозы из крови. Мыши голодали в течение 16 часов и впоследствии им вводили инъекцией внутрибрюшинно 1,5 г/кг глюкозы. Образцы крови получали перед инъекцией глюкозы и через 15, 30, 60 и 120 минут после инъекции глюкозы. Как показано на фигуре 5, уровни глюкозы в крови возрастали у трансгенных по miR-378 мышей, и клиренс глюкозы из крови понижается по сравнению с мышами дикого типа. Эти результаты показывают, что сверхэкспрессия miR-378 в скелетной мышце приводит к пониженной толерантности к глюкозе, возможно, вследствие повышенной резистентности к инсулину у этих животных.
[0090] Для определения того, обусловлено ли воздействие на толерантность к глюкозе именно сверхэкспрессией miR-378, MCK-miR-378 трансгенным животным вводили инъекцией через хвостовую вену 10 мг/кг antimiR-378 в течение трех последовательных дней. AntimiR-378 имела последовательность, которая была комплементарна нуклеотидам 2-16 зрелой последовательности miR-378 (5'-TGACTCCAAGTCCAG-3' (SEQ ID NO: 21)), и содержала комбинацию «закрытых» нуклеиновых кислот и дезоксирибонуклеотидов (8 LNA и 7 ДНК). AntimiR-378 содержала полностью фосфоротиоатный остов. После последней инъекции antimiR-378 мыши голодали в течение 16 часов. Глюкозу (1,5 г/кг) вводили инъекцией внутрибрюшинно и образцы крови получали до и в различные моменты времени после инъекции глюкозы. AntimiR-378 эффективно понижала экспрессию miR-378 у трансгенных мышей до уровней ниже таковых, наблюдаемых у мышей дикого типа, которым вводили инъекцию солевого раствора (фигура 6). Лечение трансгенных животных antimiR-378 улучшало клиренс глюкозы так, что эти животные проявляли реакцию, сходную с таковой у животных дикого типа в отношении глюкозотолерантного теста. Эти результаты позволяют предположить, что нарушенная утилизация глюкозы у трансгенных по miR-378 мышей была обусловлена сверхэкспрессией miR-378, и что нормальные метаболические процессы могут быть восстановлены путем понижения экспрессии miR-378.
[0091] Для анализа того, может ли antimiR-378 восстановить нормальную утилизацию глюкозы в мышиной модели диабета типа 2, страдающим ожирением мышам (ob/ob) вводили инъекцию внутривенно 10 мг/кг antimiR-378 в течение трех последовательных дней. После последней инъекции antimiR-378 мыши голодали в течение 16 часов, и им вводили инъекцией внутрибрюшинно 1,5 г/кг глюкозы. Образцы крови получали при различных интервалах после инъекции для оценки уровня глюкозы в крови. Как показано на фигуре 7, ob/ob мыши, получавшие лечение antimiR-378, проявляли улучшенные клиренс глюкозы по сравнению с животными, не получавшими лечение. Эти данные демонстрируют, что ингибирование экспрессии miR-378 способно улучшить утилизацию глюкозы у животных с хроническим нарушением передачи сигнала с участием лептина. Следовательно, ингибиторы miR-378 обеспечивают новый терапевтический подход для лечения метаболических расстройств, таких как диабет и ожирение.
Пример 5. Создание нокаутных по miR-378 мышей
[0092] Для определения функции miR-378/miR-378* in vivo создавали линию мышей с генетической делецией miR-378/miR-378*. С помощью нацеливающего вектора и гомологичной рекомбинации (фигура 8A) miR-378/miR-378* были нацелены и передачу генетической делеции в зародышевой линии подтверждали с применением саузерн-блоттинг анализа (фигура 8B). Нозерн-блоттинг анализ подтвердил прекращение экспрессии miR-378/miR-378* во всех исследуемых тканях у мутантов по miR-378 полностью (фигура 8C). Анализ ПЦР в реальном времени с применением зонда Taqman для PPARGC1b и вестерн-блоттинг анализ для PPARGC1β (PGC-1 β) показали, что экспрессия PPARGC1β (PGC-1 β) не изменяется у нокаутного по miR-378 животного (фигура 8D).
[0093] Для определения воздействия утраты функции miR-378 на ремоделирование сердца проводили торакальную операцию по сужению аорты (TAB) на мышах дикого типа и полностью нокаутных по miR-378 мышах. Эхокардиографию проводили для определения фракционального укорочения, меры сократительной способности и функции сердца, фракции выброса, систолического диаметра и диастолического-систолического диаметра (DD-SD) на день 21 после TAB. Сердца забирали через один день после эхокардиографии. Гистологический анализ проводили для определения клеточной морфологии. Экспрессию генов ремоделирования сердца/стресса, таких как ANF, BNP, Myh7, Myh6 и ген коллагена, измеряли с помощью количественной ПЦР в реальном времени.
[0094] Результаты этой серии экспериментов показывают, что нокаутные по miR-378 животные были защищены от индуцированного TAB ремоделирования сердца. Сердца от нокаутных животных проявляли пониженную гипертрофию миоцитов и фиброз сердца, измеренные с помощью гистологического анализа, по сравнению с мышами дикого типа (фигура 9). Аналогично, нокаутные по miR-378 животные были защищены от понижения сократительной способности и функции сердца, что измерили с помощью эхокардиографии (фигура 10A-D). Нокаутные по miR-378 животные также проявляли ослабление апрегуляции генов ремоделирования/стресса (фигура 11A-C). Эти результаты показывают, что miR-378 представляет собой положительный регулятор гипертрофии/ремоделирования сердца, и ингибирование экспрессии и активности miR-378 представляет новый терапевтический подход для сокращения патологического ремоделирования в ответ на сердечные стрессоры.
[0095] Нокаутных по miR-378 мышей также скрещивали с животными, сверхэкспрессирующими прогипертрофическую фосфатазу кальциневрин, которая индуцирует форму гипертрофической кардиомиопатии. Кроме того, сердечный гипертрофический агонист изопротеренол или солевой раствор (контроль) вводят нокаутным по miR-378 мышам для определения воздействия потери miR-378 при ремоделировании сердца. Нокаутные по miR-378 животные, как предполагается, являются также защищенными от индуцированного изопротеренолом и кальциневрином ремоделирования сердца.
Пример 6. Нокаутные по miR-378 мыши являются более устойчивыми к увеличению массы, вызванному рационом с высоким содержанием жиров
[0096] Ингибирование miR-378 с помощью антисмыслового олигонуклеотида приводит к улучшению утилизации глюкозы у страдающих ожирением (ob/ob) мышей, модели диабета типа 2 (смотри пример 4). Для дальнейшего исследования роли miR-378/miR-378* в регуляции метаболизма нокаутных по miR-378 мышей (смотри пример 5) скрещивали со страдающими ожирением мышами и оценивали утилизацию глюкозы. Мыши голодали в течение 16 часов, и им вводили инъекцией внутрибрюшинно 1,5 г/кг глюкозы. Образцы крови получали при различных интервалах после инъекции для оценки уровня глюкозы в крови. Как показано на фигуре 12, потомство от скрещивания нокаутных по miR-378 и ob/ob (ob/ob KO) проявляет улучшенную толерантность к глюкозе по сравнению со страдающими ожирением животными (ob/ob).
[0097] Для определения того, играет ли miR-378/miR-378* роль в развитии других форм ожирения, таких как индуцируемые рационом с высоким содержанием жиров, нокаутных по miR-378/miR-378* животных и однопометных животных дикого типа держали на рационе с высоким содержанием жиров, состоящем из 45% ккал жира. Нокаутные животные набирали значительно меньшую массу тела, нежели животные дикого типа в аналогичных условиях после более чем четырех недель на рационе с высоким содержанием жиров (фигура 13A). Кроме того, через пять недель на рационе с высоким содержанием жиров нокаутные по miR-378/miR-378* животные проявляли улучшенную толерантность к глюкозе по сравнению с однопометными животными дикого типа (фигура 13B). Интересно, что нокаутные по miR-378/miR-378* мыши имели уменьшенные жировые подушечки и накопление липидов в печени через шесть недель на рационе с высоким содержанием жиров по сравнению с животными дикого типа (фигуры 14-16).
[0098] Трансгенные мыши, сверхэкспрессирующие miR-378/miR-378* в ткани сердца под контролем промотора альфа-MHC (смотри пример 2) имели увеличенную массу тела по сравнению с животными дикого типа, а также проявляли большее процентное изменение массы тела, нежели животные дикого типа в ответ на рацион с высоким содержанием жиров (фигура 17).
[0099] Вестерн-блоттинг анализ бурой жировой ткани, выделенной из животных дикого типа и нокаутных по miR-378/miR-378* животных после шести часов голодания, выявил, что передача сигнала АМФ-активируемой протеинкиназой (AMPK) усиливается в жировой ткани, выделенной из нокаутных мышей, что показано повышением содержания фосфорилированной формы AMPK (фигура 18A). AMPK представляет собой основной регулятор метаболизма липидов и глюкозы. Активация AMPK способствует поглощению глюкозы, стимулируемому инсулином, и стимулирует катаболические процессы, такие как окисление жирных кислот и гликолиз. Активация AMPK приводит в результате к ингибированию глюконеогенеза, синтеза гликогена, липидов и белков. Следовательно, отсутствие miR-378/miR-378* у нокаутных животных усиливает активацию AMPK, тем самым стимулируя окисление жирных кислот и снижение накопления липидов (фигура 18B).
[00100] С применением программного обеспечения miRanda (доступно от Computational Biology Center at Memorial Sloan-Kettering Cancer Center) для определения мишеней миРНК субъединица 13 медиаторного комплекса (MED13), также известная как белок, связанный с рецептором тиреоидного гормона 1, (THRAP1), была определена как предполагаемая мишень для miR-378. 3'-UTR MED13 содержит четыре предполагаемых сайта связывания для miR-378/miR-378* (фигура 19A), которые являются консервативными среди видов (например, человек, мышь, крыса, собака, орангутанг и лошадь). Исследование экспрессии MED13 у сверхэкспрессирующих miR-378/miR-378* в сердце мышей (αMHC Tg) выявило, что экспрессия MED13 даунрегулируется относительно животных дикого типа, тогда как экспрессия MED13 апрегулируется у нокаутных по miR-378/miR-378* животных (KO), позволяя предположить, что MED13 представляет собой физиологическую мишень miR-378/miR-378*. Смотри фигуру 19B. Для дальнейшего определения того, является ли MED13 распознаваемой мишенью для miR-378/miR-378*, полноразмерную 3'-UTR транскрипта MED13 вставляли в плазмиду экспрессии люциферазы, которую трансфицировали в COS1 клетки. Возрастающие количества CMV-регулируемых miR-378/miR-378* давали в результате к дозозависимому снижению люциферазной активности, тогда как сравнимые количества контрольной миРНК не имели никакого эффекта (фигура 19C). Мутирование последовательности 3'-UTR MED13 также устраняет индуцированную miR-378/miR-378* супрессию люциферазной активности (фигура 19C).
[00101] Взятые вместе, результаты этой серии экспериментов показывают, что miR-378/miR-378* играет ключевую роль в регулировании метаболических процессов путем влияния на передачу сингала AMPK и MED13. Ингибирование активности miR-378/miR-378* может представлять успешный терапевтический подход для лечения метаболических расстройств, таких как ожирение и диабет типа II.
[00102] Все публикации, патенты и патентные заявки, обсуждаемые и цитируемые в данном документе, включены в данный документ ссылкой в их полноте. Следует понимать, что раскрытое изобретение не ограничивается конкретными описанными методиками, протоколами и материалами, так как они могут изменяться. Также следует понимать, что терминология, используемая в данном документе, представлена только с целью описания конкретных вариантов осуществления и не предназначена для ограничения объема настоящего изобретения, который будет ограничиваться только приложенной формулой изобретения.
[00103] Специалистам в данной области техники будут понятны, или они смогут установить с помощью не более чем стандартного проведения экспериментов, многие эквиваленты конкретных вариантов осуществления настоящего изобретения, описанных в данном документе. Такие эквиваленты предназначены охватываться нижеследующей формулой изобретения.
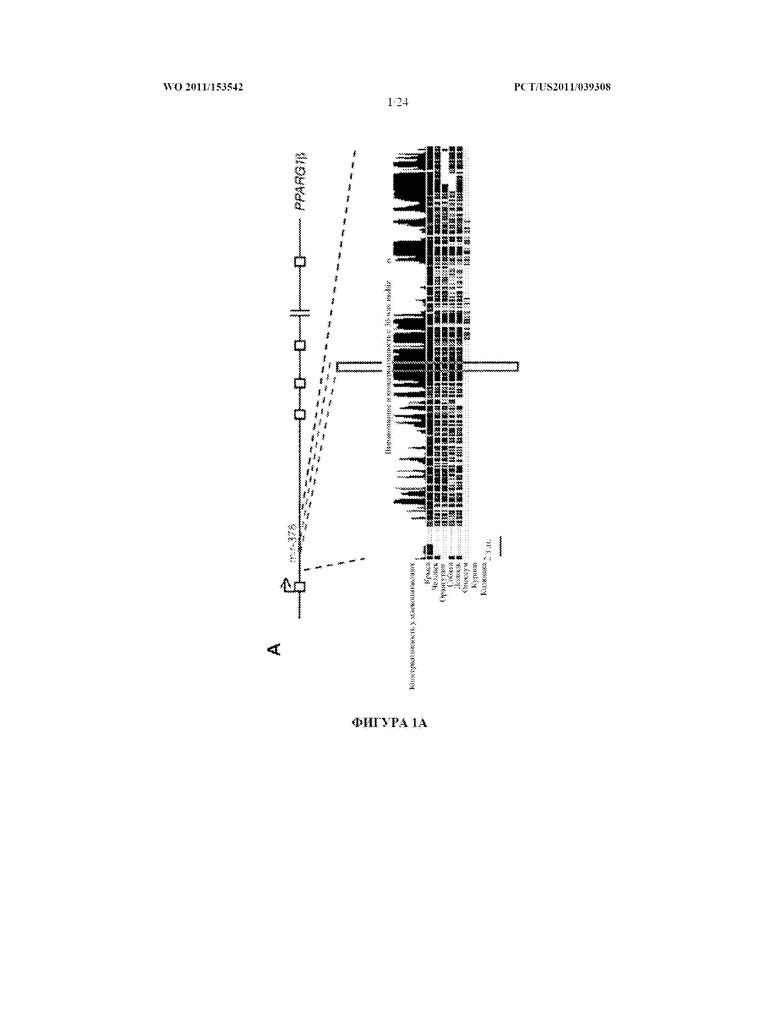

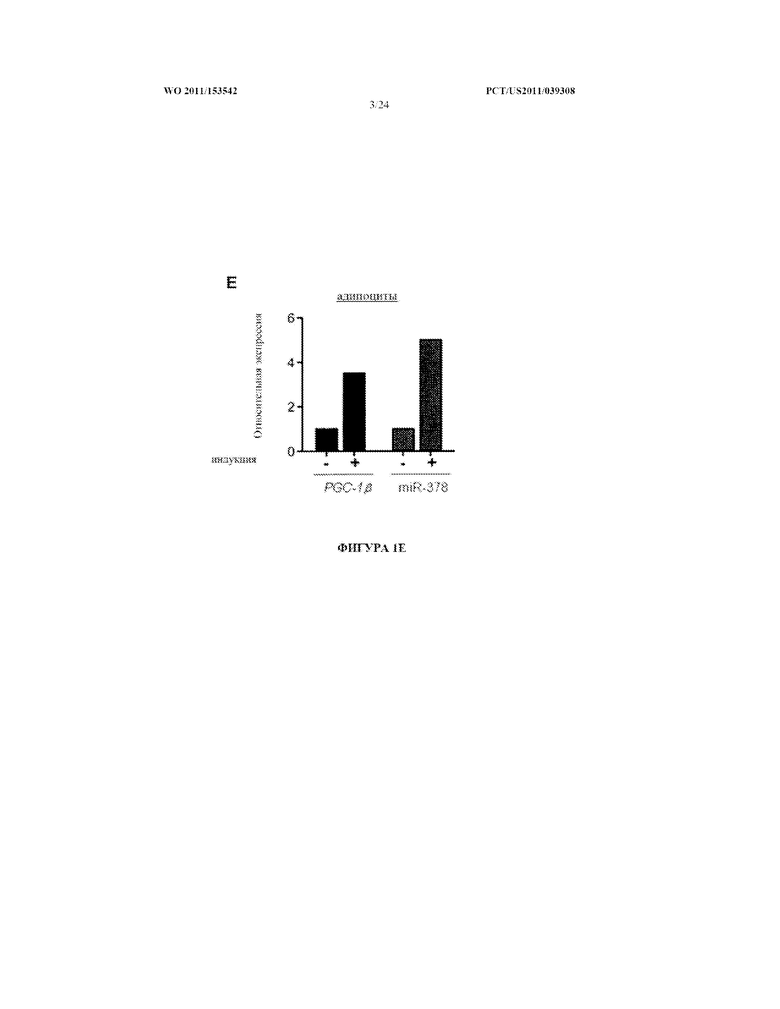




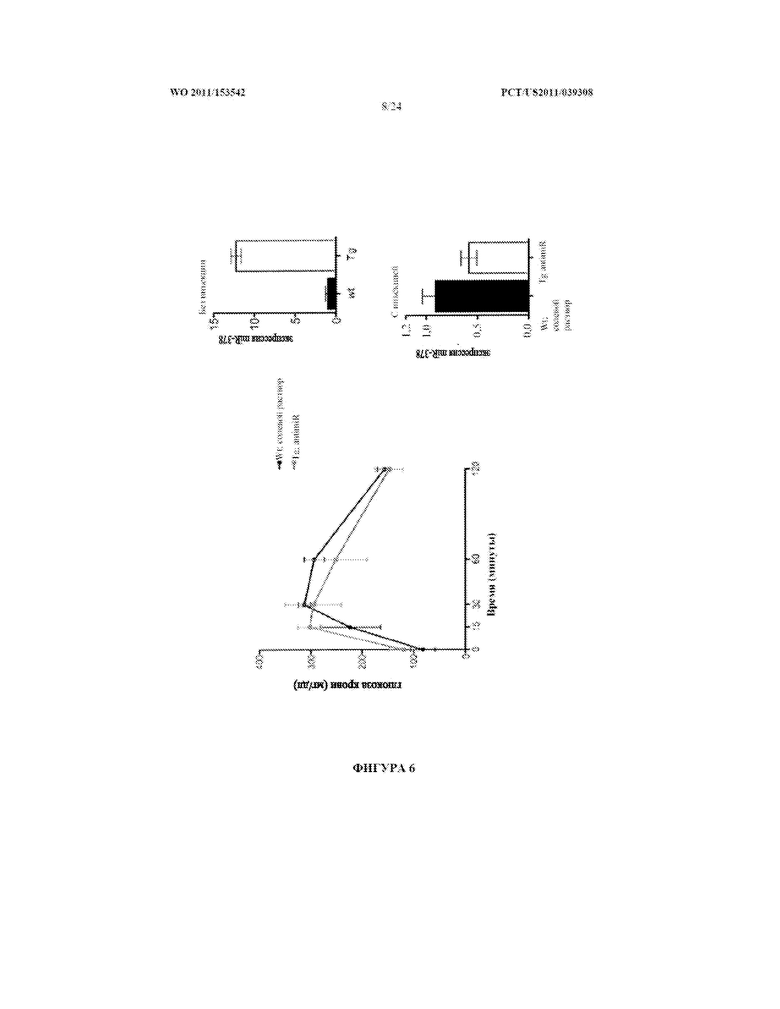
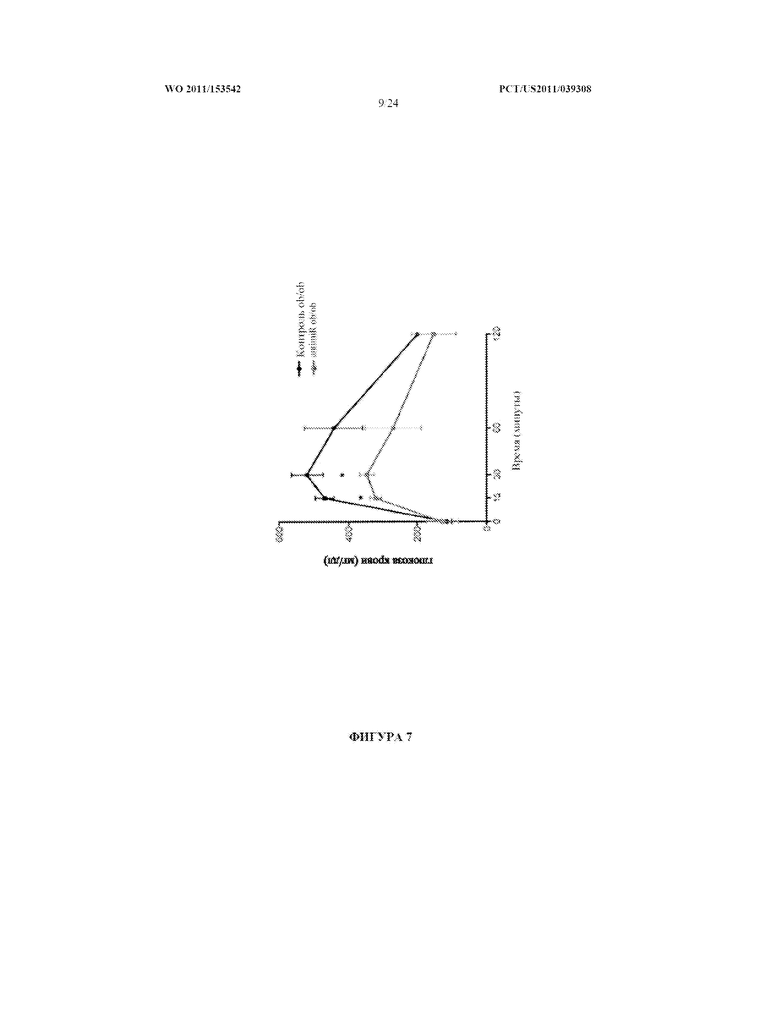







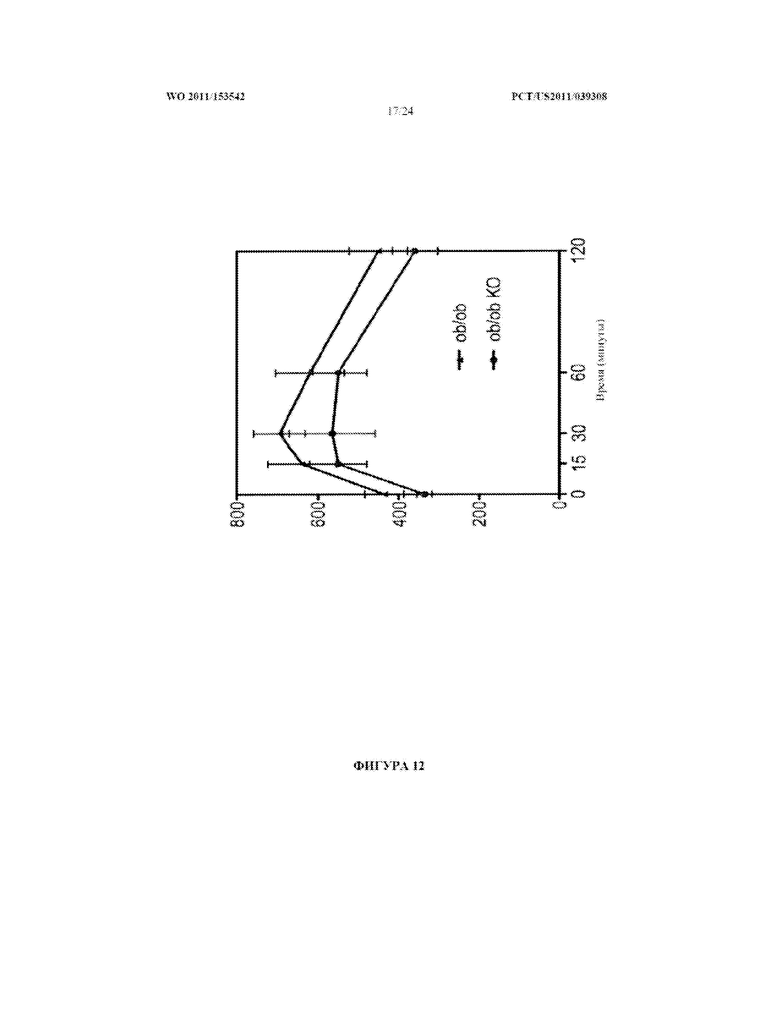
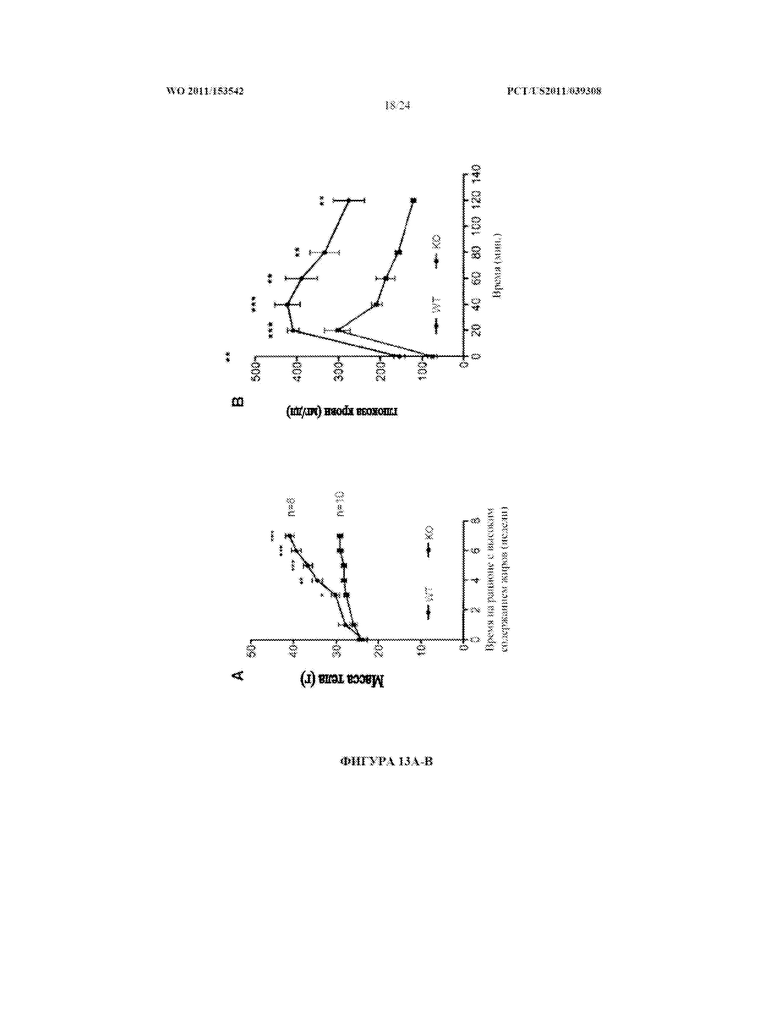

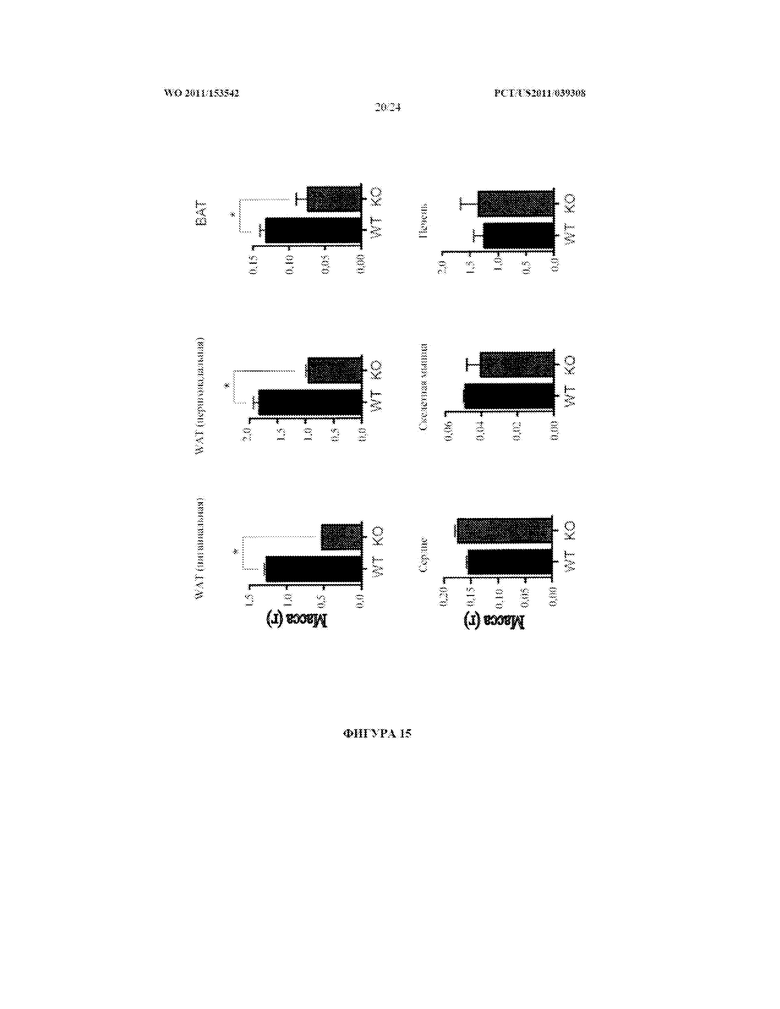

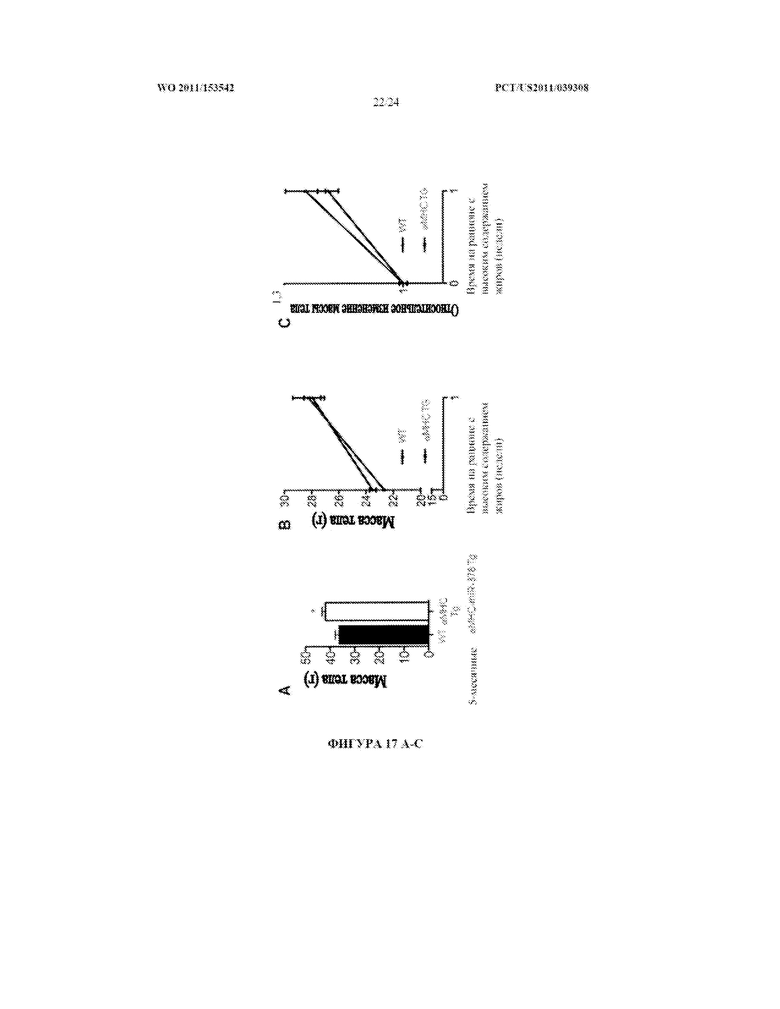


Изобретение относится к области биотехнологии, конкретно к способу лечения метаболического расстройства у субъекта путем введения средств, которые модулируют активность и экспрессию микроРНК, и может быть использовано в медицине. Настоящее изобретение обеспечивает способ лечения расстройства, такого как ожирение, диабет или метаболический синдром, у субъекта путем введения субъекту ингибитора экспрессии или активности miR-378 и/или miR-378*. Изобретение позволяет проводить эффективную терапию широкого круга метаболических расстройств, обусловленных нарушением передачи сигнала с помощью лептина. 8 з.п. ф-лы, 19 ил., 6 пр.
1. Способ лечения метаболического расстройства у субъекта, нуждающегося в этом, при котором вводят субъекту ингибитор miR-378 и/или miR-378*, где экспрессия или активность miR-378 и/или miR-378* в клетках субъекта понижается после введения, где ингибитор представляет собой антисмысловой олигонуклеотид, который имеет последовательность, которая по меньшей мере на 90% комплементарна SEQ ID NO: 1 или SEQ ID NO: 2, где метаболическое расстройство представляет собой метаболический синдром, ожирение, сахарный диабет, диабетическую нефропатию, резистентность к инсулину, атеросклероз, нарушение липидного обмена, гликогеноз, недостаточность среднецепочечной ацил-коэнзим А дегидрогеназы или нарушенное поглощение и/или утилизацию глюкозы.
2. Способ по п. 1, где нарушение липидного обмена выбирают из группы, содержащей болезнь Ниманна-Пика, болезнь Гоше, болезнь Фарбера, болезнь Фабри, болезнь Вольмана и болезнь накопления эфиров холестерина.
3. Способ по п. 1, где антисмысловой олигонуклеотид содержит по меньшей мере одну модификацию сахара и/или остова, где указанная по меньшей мере одна модификация сахара и/или остова повышает стабильность антисмыслового олигонуклеотида.
4. Способ по п. 3, где модификация сахара представляет собой «закрытую» нуклеиновую кислоту.
5. Способ по п. 3, где модификация остова представляет собой фосфоротиоатную связь.
6. Способ по п. 1, где антисмысловой олигонуклеотид имеет длину от 7 до 18 нуклеотидов.
7. Способ по п. 1, где антисмысловой олигонуклеотид имеет последовательность, выбранную из группы, содержащей SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20, SEQ ID NO: 21 и SEQ ID NO: 22.
8. Способ по п. 1, где субъект является человеком.
9. Способ по п. 1, где ингибитор вводят субъекту внутрикожным, подкожным, внутримышечным, внутрибрюшинным или внутривенным путем введения.
| US 2009143326 A1, 04.06.2009 | |||
| WO 2009114681 A2, 17.09.2009 | |||
| WO 2009058907 A2, 07.05.2009 | |||
| KUWABARA Y | |||
| et al., MiscroRNA-378, which exists intronic lesion of PGC-1Beta, deteriorate cardiomyocyte survival under oxidative stress, CIRCULATION, 2009, v | |||
| Кровля из глиняных обожженных плит с арматурой из проволочной сетки | 1921 |
|
SU120A1 |
| РЕГЕНЕРАТИВНЫЙ РАДИОПРИЕМНИК | 1923 |
|
SU735A1 |
| LEE D.Y | |||
| et al., MicroRNA-378 promotes cell survival, tumor growth and | |||

