Область техники
Настоящее изобретение относится к антителам, в частности к вариабельным доменам легкой и тяжелой цепи мышиного моноклонального антитела, связывающегося с кадгерином-17 человека, и антигенсвязывающему фрагменту (Fab) против кадгерина-17 человека, содержащему указанные домены.
Предшествующий уровень техники
Рак желудка (РЖ) находится на втором месте по смертности от онкологических заболеваний и на 4-ом месте по распространенности среди злокачественных опухолей (Tan I.B. и др. Intrinsic subtypes of gastric cancer, based on gene expression pattern, predict survival and respond differently to chemotherapy. Gastroenterology, 2011, 141: 476-485; Kamangar F. и др. Patterns of cancer incidence, mortality, and prevalence across five continents: defining priorities to reduce cancer disparities in different geographic regions of the world. J Clin Oncol, 2006, 24: 2137-2150). Медиана продолжительности жизни пациентов с РЖ составляет 7-10 месяцев. 5-летняя выживаемость пациентов с РЖ составляет приблизительно 20%, при правильно подобранном лечении может увеличиваться до 40%. (Tan I.B. и др. Intrinsic subtypes of gastric cancer, based on gene expression pattern, predict survival and respond differently to chemotherapy. Gastroenterology, 2011, 141: 476-485; Hartgrink H.H. Improving outcome for scirrhous gastric cancer. Gastric Cancer, 2009, 12: 3-5). В настоящее время для лечения РЖ наиболее широко распространена терапия препаратами на основе неспецифических цитостатиков. В случае аденокарциномы с увеличенной экспрессией HER2-neu применение Herceptin в комбинации с химиотерапией повышает медиану выживаемости до 13,8 месяцев (Bang Y.J. и др. Trastuzumab in combination with chemotherapy versus chemotherapy alone for treatment of HER2-positive advanced gastric or gastro-oesophageal junction cancer (ToGA): a phase 3, open-label, randomised controlled trial. Lancet, 2010, 376: 687-697). Учитывая высокую смертность от РЖ и отсутствие чувствительных и специфических маркеров для ранней диагностики РЖ, поиск соответствующих мишеней и терапевтических средств против этих мишеней остается крайне актуальным во всем мире.
Кадгерин-17 (CDH17) является одним из членов надсемейства семидоменных кадгеринов, он присутствует на клетках печени и желудочно-кишечного тракта в эмбриогенезе, поэтому второе его название - печеночно-кишечный кадгерин (liver-intestinal cadherin или LI cadherin). CDH17 экспрессируется на клетках гепатоцеллюлярной карциномы (Liu L.X. и др. Targeting cadherin-17 inactivates Wnt signaling and inhibits tumor growth in liver carcinoma. Hepatology, 2009, 50: 1453-1463; Wong B.W. и др. Identification of liver-intestine cadherin in hepatocellular carcinoma - a potential disease marker. Biochem Biophys Res Commun., 2003, 311: 618-624), рака желудка (Grotzinger C. и др. LI-cadherin: a marker of gastric metaplasia and neoplasia. Gut, 2001, 49: 73-81), рака протоков поджелудочной железы (Takamura M. и др., Expression of liver-intestine cadherin and its possible interaction with galectin-3 in ductal adenocarcinoma of the pancreas. Cancer Sci., 2003, 94: 425-430) и колоректального рака (Hinoi T. и др. CDX2 regulates liver intestine-cadherin expression in normal and malignant colon epithelium and intestinal metaplasia. Gastroenterology, 2002, 123: 1565-1577; Park J.H. и др. Comparison of cadherin-17 expression between primary colorectal adenocarcinomas and their corresponding metastases: the possibility of a diagnostic marker for detecting the primary site of metastatic tumour. Histopathology, 2011, 58: 315-318; Bartolomé R.A. и др. Cadherin-17 interacts with α2β1 integrin to regulate cell proliferation and adhesion in colorectal cancer cells causing liver metastasis. Oncogene, 2014, 33(13): 1658-69). CDH17 отсутствует на нормальных клетках тканей желудка, тогда как более 75% опухолевых клеток при РЖ экспрессируют CDH17 (Dong W.G. и др. LI-cadherin is inversely correlated with galectin-3 expression in gastric cancer. Dig Dis Sci, 2008, 53: 1811-1817). Высокий уровень экспрессии CDH17 ассоциируется с прогрессирующим РЖ и с неблагоприятным прогнозом (Ito R. и др. Clinicopathological significant and prognostic influence of cadherin-17 expression in gastric cancer. Virchows Arch, 2005, 447: 717-722), а также с метастазированием в лимфоузлах (Ko S. и др. Overexpression of LI-cadherin in gastric cancer is associated with lymph node metastasis. Biochem Biophys Res Commun, 2004, 319: 562-568). Нокаутирование гена CDH17 при помощи лентивирусной микро-РНК ингибирует пролиферацию, адгезию, рост опухоли и метастазирование клеток линии BGC823 РЖ человека в экспериментах in vitro и in vivo (Liu Q.S. и др. Lentiviral-mediated miRNA against liver-intestine cadherin suppresses tumor growth and invasiveness of human gastric cancer. Cancer Sci, 2010, 101: 1807-1812; Zhang J. и др. Blockade of proliferation and migration of gastric cancer via targeting CDH17 with an artificial microRNA. Med Oncol, 2011, 28: 494-501; Xu Y. и др. Knockdown of liver-intestine cadherin decreases BGC823 cell invasiveness and metastasis in vivo. World J Gastroenterol, 2012, 18: 3129-3137). Таким образом, CDH17 является онкогеном и онкомаркером для диагностики РЖ (Su M.C. и др. Cadherin-17 is a useful diagnostic marker for adenocarcinomas of the digestive system. Mod Pathol, 2008, 21: 1379-1386). Было установлено, что в клетках гепатоцеллюлярной карциномы действие CDH17 опосредуется через сигнальный путь Wnt (Liu L.X. и др. Targeting cadherin-17 inactivates Wnt signaling and inhibits tumor growth in liver carcinoma. Hepatology, 2009, 50: 1453-1463). В случае с РЖ было показано, для CDH17-индуцированного онкогенеза и метастазирования происходит активация NFkB сигнального пути (Wang и др. Cadherin-17 induces tumorigenesis and lymphatic metastasis in gastric cancer through activation of NFκB signaling pathway. Cancer Biol Ther, 2013, 14: 262-270). При раке прямой кишки CDH17 активирует сигнальный путь с альфа2-бэта1 интегрином, что приводит к активации Ras и адгезивных киназ, что в свою очередь ведет к повышению адгезии и пролиферации опухолевых клеток (Bartolomé R.A. и др. Cadherin-17 interacts with α2β1 integrin to regulate cell proliferation and adhesion in colorectal cancer cells causing liver metastasis. Oncogene, 2014, 33(13): 1658-69). Что является основным сигнальным путем для CDH17 при раке желудка, остается до конца невыясненным. В работе Lin со авторами (Lin Z. И др. Targeting cadherin-17 inactivates Ras/Raf/MEK/ERK signaling and inhibits cell proliferation in gastric cancer. PLoS One. 2014, 9(1):e85296) была показана реализация пути integrin-Ras/Raf/MEK/ERK для клеток РЖ, в этой же работе авторы установили, что при подавлении CDH17 наступает явное ингибирование пролиферации, миграции, адгезии опухолевых клеток, угнетается формирование колоний клеток, индуцируется апоптоз.
В настоящее время известны антитела против кадгерина-17 человека, описанные в публикациях заявок РСТ WO 2010123874 A1 и WO 2012/054084 А3 компании Oxford Biotherapeutics Ltd. (GB), а также в патентной заявке Японии JP 2017024991 A (заявитель - Университет Токио, (JP)) и в публикации заявки РСТ WO 2017120557 А1 (заявитель - компания Arbele Limited (CN)).
Однако существует насущная необходимость в получении новых антител против кадгерина-17 человека с улучшенными характеристиками.
Краткое описание изобретения
Несмотря на то, что несколько вариантов антител против кадгерина-17 (CDH17) человека известны из предшествующего уровня техники, существует насущная необходимость в получении новых антител против кадгерина-17 человека с целью расширения арсенала таких антител и получения антител с улучшенными характеристиками.
Целью настоящего изобретения является получение мышиных антител, связывающихся с кадгерином-17 человека, определение последовательностей вариабельных доменов тяжелых (VH) и легких (VL) цепей указанных антител и получение антигенсвязывающих фрагментов (Fab) мышиного антитела, связывающихся с кадгерином-17 человека.
Указанная цель была достигнута путем выделения двух мышиных антител 1Е6 и 2G6 против слитых белков, содержащих фрагменты кадгерина-17 человека (24-132аа и 581-678аа) с тиоредоксином и обладающих способностью к связыванию кадгерина-17 человека, и определения последовательности их вариабельных участков.
Настоящее изобретение представляет собой вариабельный участок тяжелой цепи (VH) антитела 1Е6, содержащий последовательность аминокислот SEQ ID NO: 1, и вариабельный участок легкой цепи (VL) указанного антитела, содержащий последовательность аминокислот SEQ ID NO: 2.
Также настоящее изобретение представляет собой вариабельный участок тяжелой цепи (VH) антитела 2G6, содержащий последовательность аминокислот SEQ ID NO: 3, и вариабельный участок легкой цепи (VL) указанного антитела, содержащий последовательность аминокислот SEQ ID NO: 4
Также настоящее изобретение представляет собой антигенсвязывающий фрагмент (Fab), содержащий указанные вариабельные участки антитела 1Е6 или указанные вариабельные участки антитела 2G6.
Краткое описание рисунков
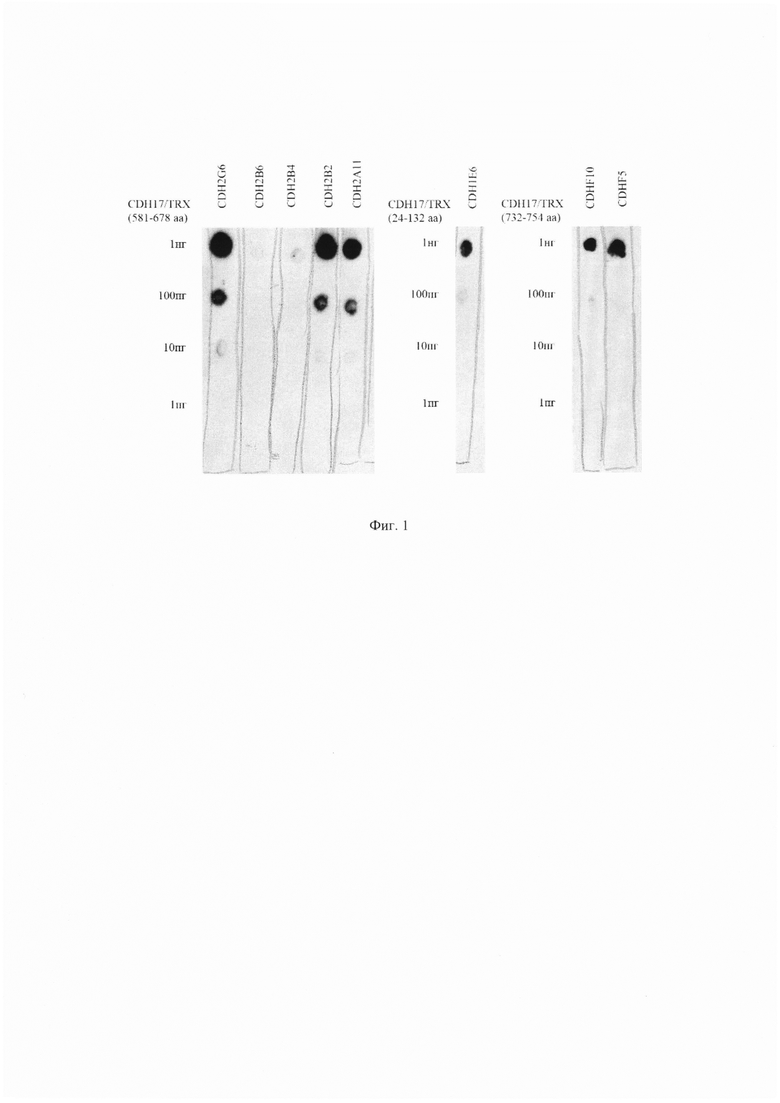
На Фиг. 1 показан дот-блот анализ шести антител 1E6, 2A11, 2B2, 2B4, 2B6, 2G6 с использованием слитых белков CDH17(581-678aa)-Trx и CDH17(24-132aa)-Trx при различных концентрациях белков.
На Фиг. 2 показан дот-блот анализ шести антител 1E6, 2A11, 2B2, 2B4, 2B6, 2G6 с использованием слитых белков CDH17(581-678aa)-Trx и CDH17(24-132aa)-Trx, а также слитого белка ММР14 с тиоредоксином, использованного в качестве отрицательного контроля.
На Фиг. 3 показаны результаты Вестерн-блот анализа содержания CDH17 в тотальных белковых экстрактах опухолевых и нормальных тканей толстой кишки (дорожки 1 и 2, соответственно, 20 мкг тотального белка на дорожку). Слева - окраска антителом 2G6, справа - антителом 1Е6. Слева указаны молекулярные массы белков стандарта (кДа).
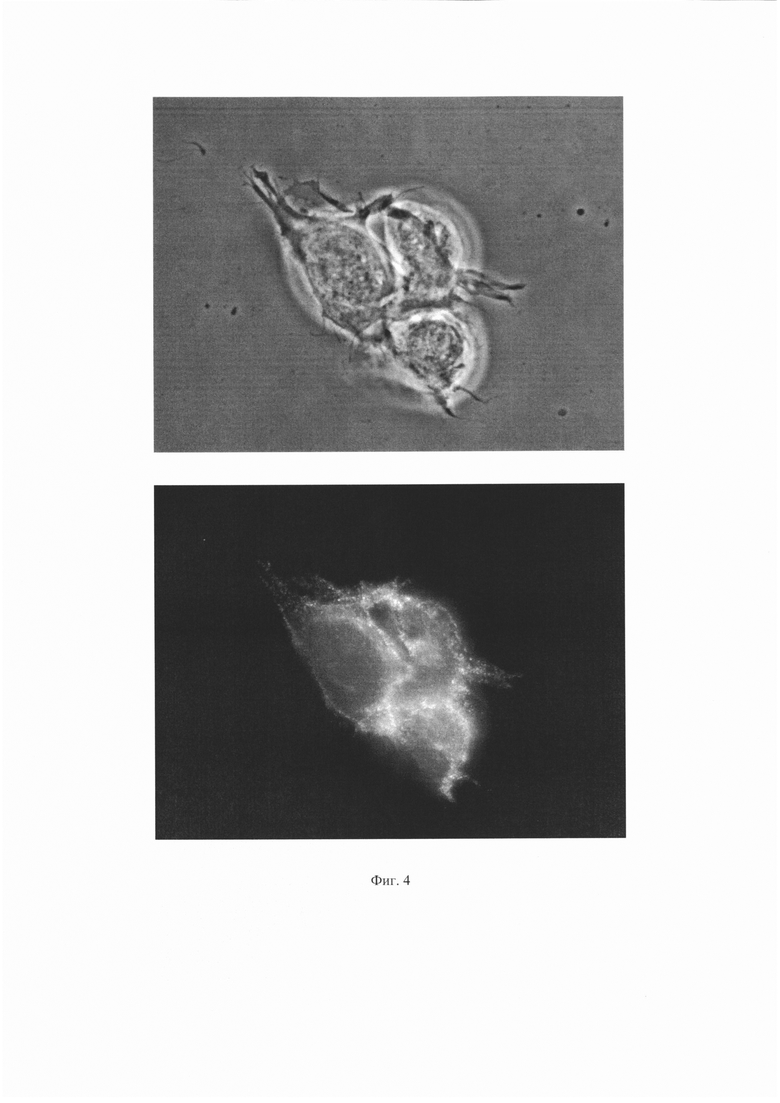
На Фиг. 4 показаны результаты иммунофлуоресцентного анализа для антитела 1Е6. Сверху - изображение клеток в световом поле, снизу - изображение тех же клеток с использованием флуоресцентного фильтра
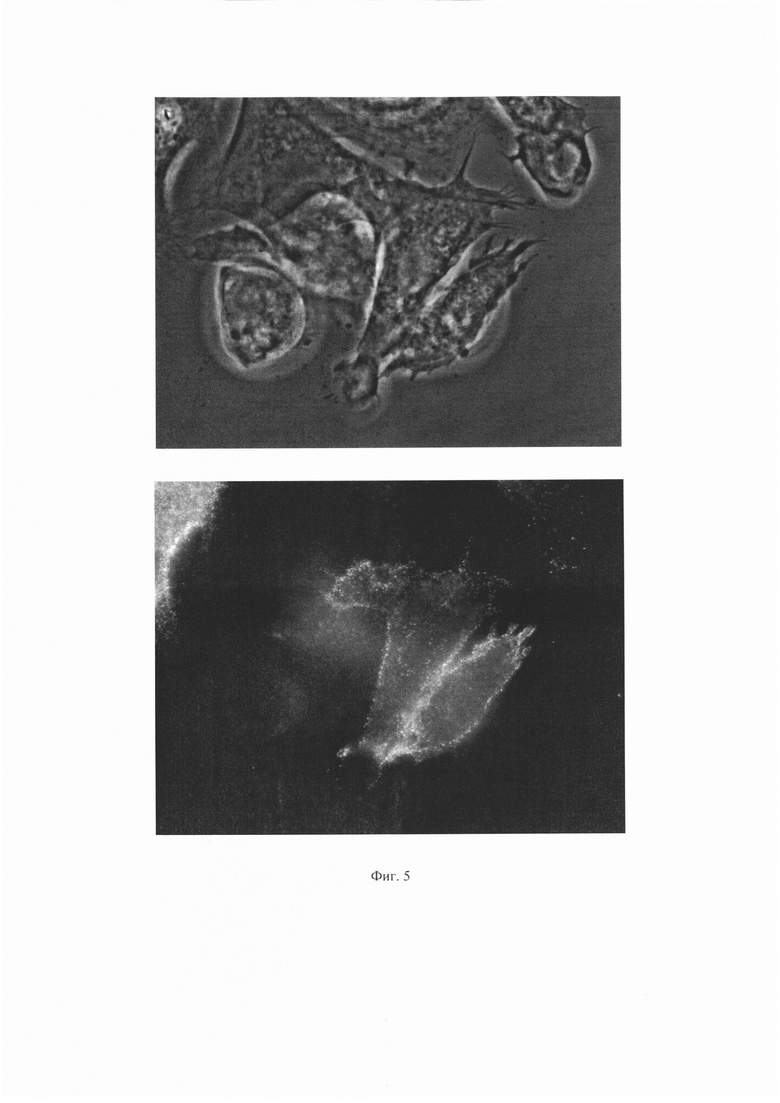
На Фиг. 5 показаны результаты иммунофлуоресценного анализа для антитела 1Е6. Сверху - изображение клеток в световом поле, снизу - изображение тех же клеток с использованием флуоресцентного фильтра
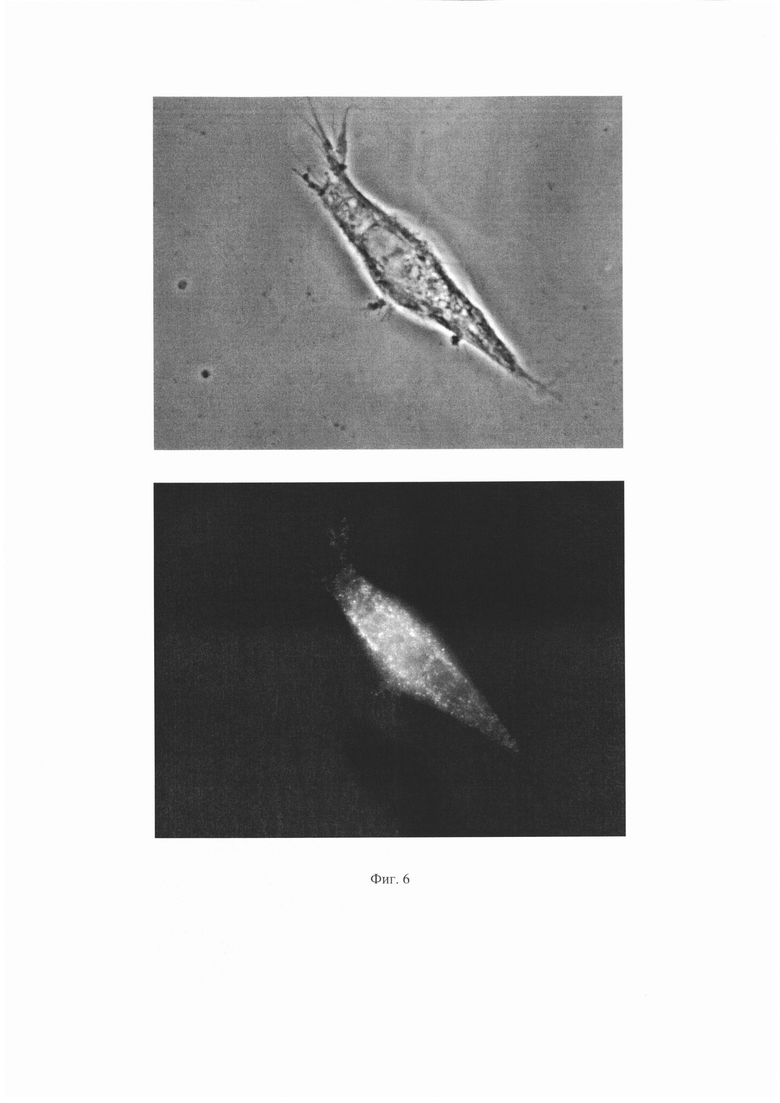
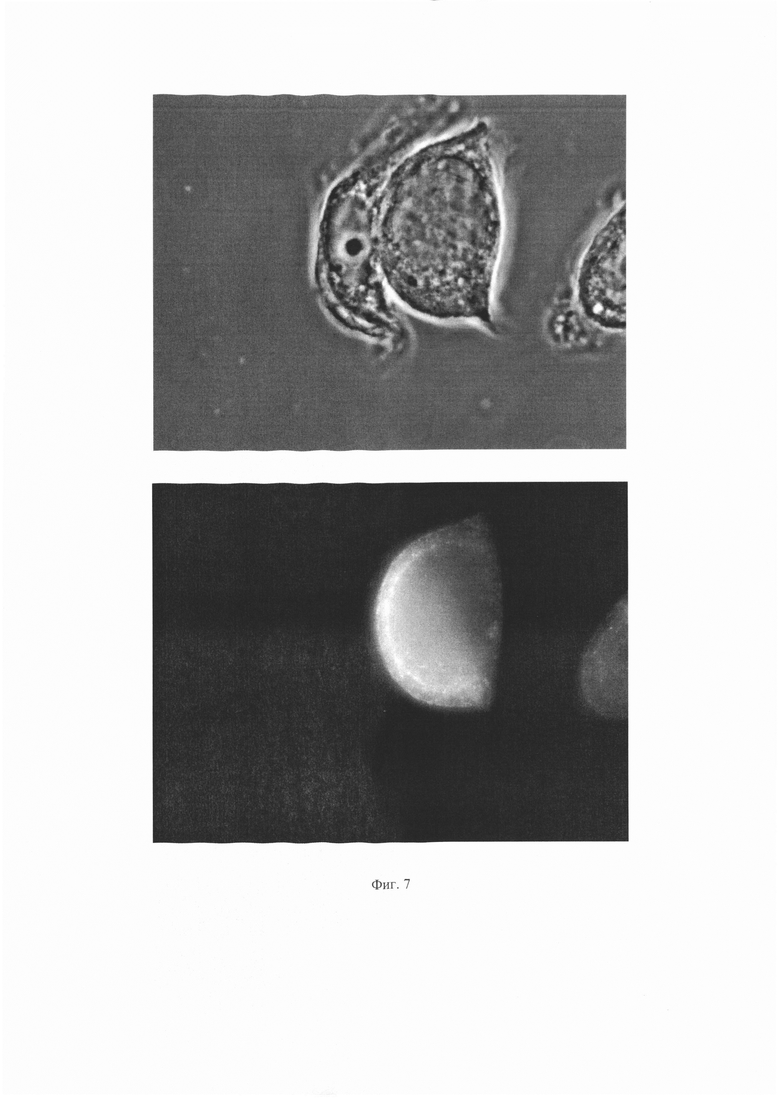
На Фиг. 6 показаны результаты иммунофлуоресценного анализа для антитела ПолиАТ. Сверху - изображение клеток в световом поле, снизу - изображение тех же клеток с использованием флуоресцентного фильтра
На Фиг. 7 показаны результаты иммунофлуоресценного анализа для антитела ПолиАТ. Сверху - изображение клеток в световом поле, снизу - изображение тех же клеток с использованием флуоресцентного фильтра
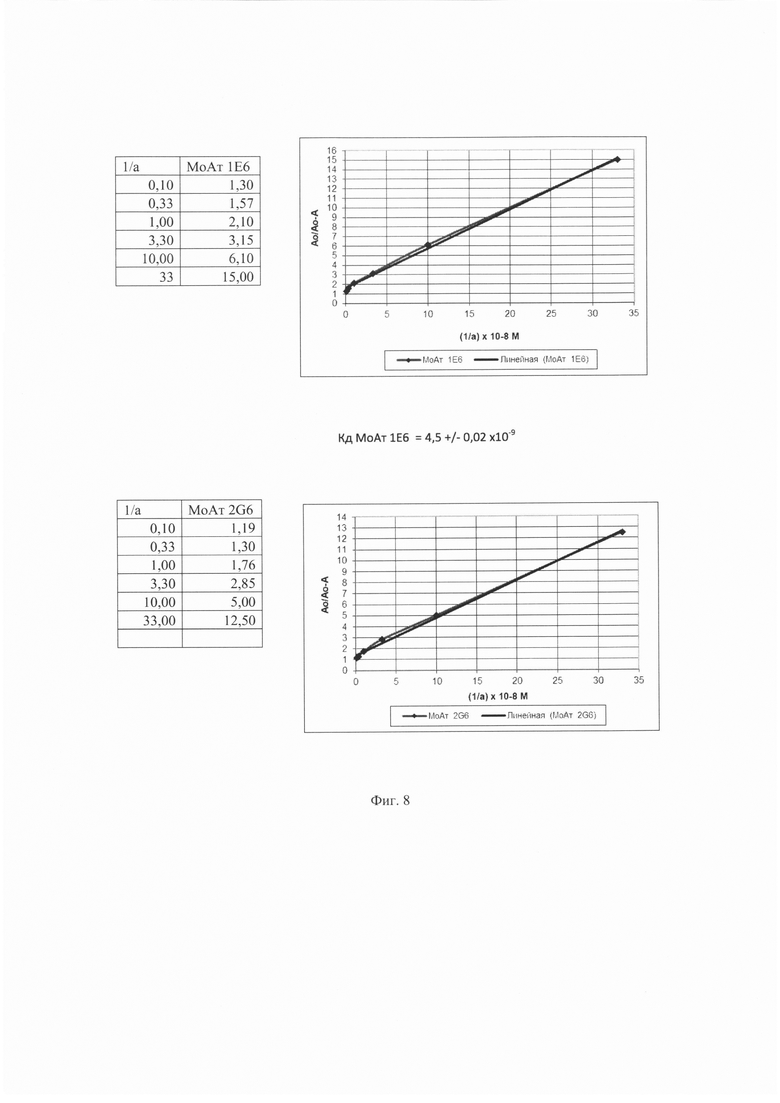
На Фиг. 8 показаны результаты определения константы диссоциации (Kd) для антител 1Е6 и 2G6.
Подробное описание настоящего изобретения
Антитела обычно состоят из двух тяжелых цепей, связанных между собой дисульфидными связями, и легких цепей, ассоциированных с N-концом каждой из тяжелых цепей. Каждая тяжелая цепь содержит на N-конце вариабельный домен с константным доменом на другом конце. Каждая легкая цепь также содержит на N-конце вариабельный домен с константным доменом. Вариабельные домены каждой пары легкой и тяжелой цепей образуют антигенсвязывающий фрагмент (Fab). Вариабельные домены легкой и тяжелой цепей обладают похожей общей структурой, и каждый домен включает каркас из четырех участков, последовательности которых являются относительно консервативными, связанных посредством трех участков, определяющих комплементарность (complementarity determining regions, CDRs). Четыре каркасных участка формируют конформацию типа бета-складчатого слоя, а CDRs образуют петли, связывающие конструкцию из бета-складчатого слоя. Участки CDRs расположены в непосредственной близости друг от друга благодаря каркасным участкам и вносят вклад в образование антигенсвязывающего участка. Участки CDRs и каркасные участки антител могут быть определены путем ссылки на нумерационную систему Кабата (Kabat numbering system, Kabat и др., "Sequences of Proteins of Immunological Interest", US Dept. of Health and Human Services, US Government Printing Office, 1987) в сочетании с рентгеноструктурной кристаллографией, как описано в публикации WO 91/09967.
Для получения антитела, которое может связываться с каким-либо специфическим антигеном, обычно используют методику Kohler и Milstein (Kohler и др. Nature, 1975, 256:495-497), получившую название гибридόмной технологии. Суть метода сводится к получению культуры «бессмертных» гибридов (гибридόм) путем слияния плазматической клетки (В-лимфоцита, продуцирующего антитела) и опухолевой клетки (миеломы или плазмацитомы). Полученная гибридόма способна продуцировать моноклональные антитела (как В-клетка) и при этом бесконечно делится (как опухолевая клетка), что позволяет достаточно долгое время секретировать in vitro или in vivo антитело, специфическое для целевого антигена.
Слияние клеток происходит под действием полиэтиленгликоля, изолецитина или вируса Сендай. На следующем этапе необходимо отобрать неслившиеся с лимфоцитами миеломные клетки (неслившиеся лимфоциты убирать не нужно: через недолгое время они погибнут сами). В процессе такой селекции гибридόмные клетки высаживают на среды, содержащие вещества, блокирующие синтез нуклеотидов миеломными клетками. Для этого выведены специальные мутантные миеломные клеточные линии, дефектные по некоторым ферментам, участвующим в синтезе нуклеотидов.
Отбор гибридόм, секретирующих нужные антитела, проводится путем проверки среды, в которой росли клетки, на предмет наличия в ней антител нужной специфичности.
Таким образом отбирают клетки, секретирующие антитела стабильно и в достаточном количестве. Большой объем антител нарабатывают или in vitro, или путем введения гибридом в перитонеальную полость мышей для получения асцита (Свешников, П.Г. Введение в молекулярную иммунологию и гибридомную технологию. П.Г. Свешников, В.В. Малайцев, И.М. Богданова, О.Н. Солопова, изд. МГУ, М., 2006).
Получение материала, использующегося в качестве антигена, для инъекций животных включают в себя методики, хорошо известные из уровня техники, например, использование полноразмерного белка, использование пептида, выбранного из иммуногенных участков белка, модифицирование антигена такими способами как, например, связывание с динитрофенолом, связывание с арсаниловой кислотой, денатурация антигена, связывание антигена с белком-переносчиком, таким как, например, гемоцианин улитки, с пептидами, содержащими участки связывания с рецепторами Т-клеток класса II, с бусинами, а также любыми другими методами, известными из уровня техники. Смотри, например, Harlow и др. Antibodies, a Laboratory Manual, Cold Spring Harbor Labs Press, 1988.
В частности, антителами, обладающими способностью к связыванию с кадгерином-17 человека, являются антитела, содержащие вариабельные домены тяжелой и легкой цепи согласно настоящему изобретению. Указанные вариабельные домены тяжелой цепи мышиного моноклонального антитела содержат аминокислотную последовательность SEQ ID NO: 1 или SEQ ID NO: 3, а вариабельные домены легкой цепи антитела (VL) содержат аминокислотную последовательность SEQ ID NO: 2 или SEQ ID NO: 4, необходимые для функционирования указанного антитела. Изолированный антигенсвязывающий фрагмент (Fab) согласно настоящему изобретению, селективно связывающийся с кадгерином-17 человека, включает в себя вариабельный домен тяжелой цепи (VH) с последовательностью аминокислот SEQ ID NO: 1 и вариабельный домен легкой цепи (VL) с последовательностью аминокислот SEQ ID NO: 2; или вариабельный домен тяжелой цепи (VH) с последовательностью аминокислот SEQ ID NO: 3 и вариабельный домен легкой цепи (VL) с последовательностью аминокислот SEQ ID NO: 4
В настоящем изобретении фраза “антитело или Fab, обладающие способностью к связыванию с кадгерином-17 человека” означает молекулу, которая связывается с кадгерином-17 человека, образует стабильный комплекс. Стабильным комплексом является комплекс, в котором связывание между партнерами происходит на период времени, достаточный для того, чтобы произвести детектирование указанного комплекса с использованием описанных здесь методов. Термин “селективно связывается с кадгерином-17 человека” означает способность указанной молекулы предпочтительно связываться с кадгерином-17 человека в отличие от связывания с белками, не имеющими отношения к кадгерину-17 человека, или связывания с небелковыми компонентами, присутствующими в образце. Антитело или Fab, которые предпочтительно связываются с кадгерином-17 человека, являются антитело или Fab, которые связываются с кадгерином-17 человека, но не связываются в существенной степени с другими молекулами или компонентами, которые могут присутствовать в образце. Такое существенное связывание предполагает, например, связывание антитела или Fab, связывающихся с кадгерином-17 человека, с молекулой, не являющейся кадгерином-17 человека, с аффиностью или силой недостаточной для того, чтобы помешать способности антитела, связывающегося с кадгерином-17 человека, определить уровень кадгерина-17 человека в образце. Примерами таких молекул и компонентов, которые могут присутствовать в образце, являются, но не ограничиваются ими, белки, не являющиеся кадгерином-17 человека, липиды и углеводы.
Способность антитела или Fab к связыванию с антигеном может быть определена специалистом в данной области с использованием методов, включающих, но не ограничивающихся методом ELISA и равновесным диализом. Методы определения аффинности и силы связывания хорошо известны специалисту в данной области техники, подробно описаны Janeway и др. (Immunobiology: The Immune System in Health and Disease (Garland Publishing Company, 1996)).
Fab, пригодный для осуществления настоящего изобретения, - это Fab, обладающий способностью к связыванию с кадгерином-17 человека, когда концентрация кадгерина-17 человека составляет от около 10 нг/мл и до около 10 пг/мл. В частности, пригодные в рамках настоящего изобретения Fab связывают кадгерин-17 человека, когда концентрация кадгерина-17 человека составляет 10 нг/мл, около 1 нг/лм или менее, предпочтительно 100 пг/мл. Такие антитела и Fab описаны в сопутствующих Примерах.
Fab моноклональных мышиных антител в соответствии с настоящим изобретением, которые селективно связываются с кадгерином-17 человека, также могут быть получены с использованием подходящей гибридόмы. Гибридόмы могут быть выращены в питательной среде, и накопленные антитела могут быть выделены. Здесь и далее термин "выращиваемые клетки" относится к гибридόмам или другим линиям клеток, которые производят антитела. Способы получения и проверки таких выращиваемых клеток описаны Harlow и др. (см. выше). Методы получения антигенного материала для инъекций в животное включают в себя методы, известные из уровня техники, например, использование полноразмерного белка, пептидов, выбранных из иммуногенного участка этого белка, модифицирование антигена такими способами как, например, связывание с динитрофенолом, связывание с арсаниловой кислотой, денатурация антигена, связывание антигена с белком-переносчиком, таким как, например, гемоцианин улитки, с пептидами, содержащими участки связывания с рецепторами Т-клеток класса II, с бусинами, а также любыми другими методами, известными из уровня техники. Смотри Harlow и др. (см. выше).
Fab, связывающиеся с кадгерином-17 человека, согласно настоящему изобретению, могут включать в себя многофункциональные молекулы, например, бифункциональные молекулы, содержащие, по крайней мере, одну функциональную часть, которая специфически связывается с кадгерином-17 человека. Такие многофункциональные молекулы могут включать в себя, например, химерные молекулы, включающие в себя молекулу, которая связывается с кадгерином-17 человека, и вторую часть, которая дает возможность данной химерной молекуле связываться с субстратом или позволяет детектировать ее способом, при котором связывание с кадгерином-17 человека не ухудшаются. Примерами таких вторых частей являются, но не ограничиваются ими, фрагменты молекулы иммуноглобулина, флуоресцентный белок или фермент. Также Fab, связывающиеся с кадгерином-17 человека, согласно настоящему изобретению, могут быть модифицированы полиэтиленгликолем, т.е. пегилированы.
Термин "взаимодействие", использованный здесь, означает введение, добавление образца, предположительно содержащего кадгерин-17 человека, к Fab, связывающему кадгерин-17 человека, например, путем объединения или смешения образца с Fab. В случае, если кадгерин-17 человека присутствует в образце, образуется комплекс Fab с кадгерином-17 человека; образование такого комплекса означает способность Fab селективно связываться с кадгерином-17 человека с образованием стабильного комплекса, который может быть детектирован. Детектирование может быть качественным, количественным или полуколичественным. Связывание кадгерина-17 человека из образца с Fab происходит в условиях, подходящих для формирования комплекса. Такие условия (например, подходящие концентрации, буферы, температура, время реакции), а также методы оптимизации таких условий хорошо известны специалисту в данной области техники. Связывание может быть обнаружено и измерено с использованием множества методов, являющихся стандартными в данной области техники, включая, но не ограничиваясь ими, ферментативные методы иммунохимии (например, ELISA), иммунопреципитации, методы иммуноблотинга и другие методы иммунохимии, описанные, например, Sambrook J. и др. (“Molecular Cloning: A Laboratory Manual, Second Edition”, Cold Spring Harbor Laboratory Press, 1989) и Harlow и др. (см. выше). Эти ссылки также описывают примеры условий, в которых образуется комплекс.
Методы получения плазмидной ДНК, расщепления и лигирования ДНК, трансформации, подбора олигонуклеотидов в качестве праймеров, и подобные методы могут быть стандартными методами, хорошо известными специалисту в данной области техники. Эти методы описаны, например в Sambrook, J. и др. (см. выше).
Последующие примеры приведены для целей разъяснения и не ограничивают каким-либо образом рамки настоящего изобретения.
Пример 1. Получение мышиных антител против кадгерина-17 человека.
Для получения антител мышей линии BALB/c (самки массой 16-18 г.) иммунизировали дважды с интервалом в 2 недели в подушечки задних лап раствором иммуногена в полном (для первой иммунизации) или неполном (для второй иммунизации) адъюванте Фрейнда. В качестве иммуногена использовали рекомбинантные слитые белки фрагментов кадгерина 17 человека (24-132 аа и 581-678 аа) с тиоредоксином (Trx). Во всех случаях на одну иммунизацию брали 25 мкг белка, белок вводили в виде эмульсии, полученной из раствора белка в фосфатно-солевом буфере и равного объема полного (1-я иммунизация) или неполного (2-я иммунизация) адъюванта Фрейнда.
Тестирование сывороток и культуральных супернатантов проводили при помощи непрямого иммуноферментного анализа (Engvall E, Perlmann P. Enzyme-linked immunosorbent assay (ELISA). Quantitative assay of immunoglobulin G., Immunochemistry. 1971, 8(9): P.871-4). Для этого рекомбинантные белки CDH17 (Sinobiological, Кат. №11360-H08H-100 Human Cadherin-17 / LI-cadherin / CDH17 Protein (His Tag)) сорбировали в течение ночи при 4°С на полистирольные планшеты из раствора в фосфатно-солевом буфере с концентрацией 1 мкг/мл и затем вносили сыворотки в серийных разведениях или культуральные супернатанты на 1 ч при 37°С. Связавшиеся с иммобилизованным антигеном МкАт выявляли с помощью козьих антител против IgG (H + L) мыши, конъюгированных с пероксидазой при помощи периодатного метода (Peroxidase and fluorescein isothiocyanate as antibody markers. A quantitative comparison of two peroxidase conjugates prepared with glutaraldehyde or periodate and a fluorescein conjugate. Broorsma DM, Steefkerk JG, Kors N. J Histochem Cytochem. 1976 Sep; 24(9):1017-25. нужна ссылка на источник), в течение 1 ч при 37°С. В качестве субстрата использовали раствор тетраметилбензидина (ТМВ) с перекисью водорода. Оптическую плотность регистрировали при 450 нм. Результаты анализа приведены в Таблице 1.
На 4-й день после второй иммунизации лимфоциты из подколенных узлов гибридизовали с миеломными клетками Sp 2/0-Agl4 по стандартной методике с использованием полиэтиленгликоля с молекулярной массой 4000 (Monoclonal Antibody Production. National Research Council (US) Committee on Methods of Producing Monoclonal Antibodies. Washington (DC): National Academies Press (US); 1999.). Гибридомы отбирали на селективной среде HAT, тестировали в непрямом ИФА и дважды клонировали методом предельных разведений.
Антитела нарабатывали в асцитных жидкостях мышей, для этого мышам, предварительно праймированных пристаном, вводили внутрибрюшинно по 106 клеток каждого клона. Очистку антител из асцитных жидкостей проводили аффинной хроматографией на колонке с белок-G сефарозой по стандартной методике (Jungbauer, A. и др. Comparison of protein A, protein G and copolymerized hydroxyapatite for the purification of human monoclonal antibodies, J. Chromatogr. 1989, 476: 257-268). Чистоту выделенных антител контролировали при помощи SDS-ПААГ электрофореза в 12% полиакриламидном геле в присутствии SDS и DTT в ступенчатой буферной системе Лэмлли, используя Mini PROTEAN 3 Electrophoresis System BIO-RAD, Catalog N 165-3301 (Laemmli, U. K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4, Nature, 1970, 227: 680-685). Чистота всех полученных антител составила не менее 95%.
Всего было получено 6 клонов на новые рекомбинантные слитые белки кадгерин-тиоредоксин: один клон на слитой белок кадгерин-17 человека (24-132 аа) - тиоредоксин (клон 1E6) и пять клонов на слитой белок кадгерин-17 человека (581-678 аа) - тиоредоксин (клоны 2A11, 2B2, 2B4, 2B6, 2G6).
Пример 2. Характеристика полученных мышиных моноклональных антител против кадгерина-17.
Иммуноферментный анализ.
Антитела из указанных 6-ти клонов тестировали в иммуноферментном анализе с фрагментами CDH17 24-132aa и CDH17 581-678аа как описано выше в Примере 1. Результаты анализа приведены в Таблицах 2А и 2Б. На основании результатов анализа были выбраны два наиболее эффективно связывающихся антитела: 1E6 и 2G6.
Два указанных антитела также тестировали в иммуноферментном анализе с внеклеточным доменом кадгерина-17, коммерческого препарата (Sinobiological, Кат. №:11360-H08H-100 Human Cadherin-17 / LI-cadherin / CDH17 Protein (His Tag). Кадгерин-17 сорбировали в концентрации 0,8 мкг/мл. В качестве положительного контроля использовали ПолиАТ-коммерческую поликлональную иммунную сыворотку кролика против кадгерина 17 (Sigma, Кат. № HPA026556). Результаты анализа приведены в Таблице 3.
Дот-блот анализ.
Анализ проводили с использованием слитых белков CDH17(581-678aa)-Trx и CDH17(24-132aa)-Trx. Слитые белки наносили на нитроцеллюлозную мембрану в количестве 1 нг, 100 пг, 10 пг или 1 пг на пятно. Результаты анализа показаны на Фиг. 1.
Результаты анализа с использованием слитых белков CDH17(581-678aa)-Trx и CDH17(24-132aa)-Trx, а также слитого белка ММР14 с тиоредоксином, использованного в качестве отрицательного контроля, показаны на Фиг. 2. Белки наносили на мембрану в количестве 10 нг на пятно.
Вестерн блот.
Вестерн блот проводили по стандартной методике (Towbin H. и др. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci USA, 1979,76(9): 4350-4354) с использованием тотальных белковых экстрактов опухолевых и нормальных тканей толстой кишки (дорожки 1 и 2, соответственно; 20 мкг тотального белка на дорожку). Результаты анализа показаны на Фиг. 3.
Иммунофлуоресцентный анализ.
Для постановки анализа готовили стекла с прикрепленными клетками. Для этого клетки снимали с поверхности культурального флакона раствором ФСБ-ЭТ, подсчитывали в камере Горяева, центрифугировали при 1200 об/мин в течение 10 минут, ресуспендировали в полной ростовой среде и затем переносили в культуральные чашки Петри. На дно чашек Петри клали покровные стекла размером 18мм×18мм.
Сливали среду с чашек Петри со стеклами, стекла доставали, промывали ФСБ, затем клали в чашки Петри, выстланные парафильмом.
Наслаивали на стекла ФСБ с добавлением 4% параформальдегида, инкубировали 15 минут для фиксации клеток.
Стекла промывали холодным ФСБ 3 раза по 5 минут на шейкере.
Клетки пермеабилизовали инкубацией в ФСБ с добавлением 0,1% Triton X-100 в течение 10 минут.
Промывали холодным ФСБ 3 раза по 5 минут на шейкере.
Остатки альдегидных групп убирали 0,5M NH4Cl инкубацией в течение 5 минут. Промывали холодным ФСБ 3 раза по 5 минут на шейкере.
Тщательно удаляли со стекол весь ФСБ, затем накапали на каждое стекло по 3-4 капли усилителя сигнала (Signal Enchancer (Image-iT® FX Signal Enhancer, ThermoFisher Scientific)), инкубировали в течение 30 минут.
Промывали холодным ФСБ 3 раза по 5 минут на шейкере.
Стекла инкубировали в блокирующем буфере (ФСБ с добавлением 10% нормальной сыворотки козы, 1% бычьего альбумина и 0,1% Triton X-100) в течение 1 часа.
Затем стекла инкубировали в течение 3-4 часов с первичными антителами 1E6, 2G6, 2A11, 2B2 и ПолиАТ (Sigma, Кат. № HPA026556), разведенными в блокирующем буфере до концентрации 10 мкг/мл.
Промывали холодным ФСБ 3 раза по 10 минут на шейкере.
Инкубировали со вторичными антителами, конъюгированными с флуоресцентным красителем в ФСБ с 1% бычьим альбумином в течение 1 часа. Для стекол с ПолиАТ использовали конъюгат анти-кроличий IgG с Alexa Fluor 555, для стекол с мышиными антителами использовали конъюгат анти-мышиный IgG с Alexa Fluor 488.Оба конъюгата брали в разведении 1:800-1:1000.
Промывали холодным ФСБ 3 раза по 5 минут на шейкере.
Чистые предметные стекла протирали, подписывали, наносили по капле буфера для заключения (Глицерин - 1М Tris pH 9 в объемном соотношении 1:9).
Со стекол с клетками удаляли ФСБ, стороны стекол без клеток и ребра протирали фильтровальной бумагой для полного удаления жидкости. Затем стекла с клетками клали на предметные стекла клетками вниз. Излишки буфера для заключения убирали фильтровальной бумагой.
Флуоресцентный сигнал фиксировали при помощи инвертированного флуоресцентного микроскопа Axiovert 200M c цифровой камерой (Carl Zeiss MicroImaging GmbH, Германия).
Результаты для антитела 1Е6 представлены на Фиг. 4 и 5. Результаты для антитела ПолиАТ представлены на Фиг. 6 и 7. Результаты для антител 2G6, 2B2 и 2A11 не приведены, поскольку сигнала флуоресценции для этих антител не наблюдали.
Определение Kd.
Константу диссоциации (Kd) определяли по методике, описанной в работах Friguet B. и др., Measurements of the True Affinity Constant in Solution of Antigen-Antiojdy Complex by Enzyme-Linked Immunosorbent Assay, Journal of Immunological Methods, 1985, 77: 305-319; Klotz I.M. The application of the law of mass action to binding by proteins; interactions with calcium. Archives of biochemistry, 1946, 9: 109-117. Результаты показаны на Фиг. 8.
Значение Кd антитела 1Е6 составило 4,5±0,02×10-9, значение Кd антитела 2G6 составило 3,5±0,02×10-9.
Пример 3. Клонирование генов, кодирующих Fab фрагменты мышиных моноклональных антител против кадгерина-17, используя мРНК из подходящих линий клеток в качестве матриц.
Тотальную РНК выделяли из 1×107 клеток подходящих гибридόм, содержащих антитела согласно настоящему изобретению против кадгерина-17. Клетки (107 клеток) два раза промывали однократным буфером PBS. В пробирку со 100 мкл клеток добавляли 400 мкл раствора GTB (4 M гуанидинтиоцинат, 30 мМ цитрат натрия, 1% N-лаурилсаркозил натрия, 120 мМ β-меркаптоэтанол). Клетки суспендировали пипетированием в течение 2-3 минут и помещали на лед. Затем добавляли 50 мкл 3M ацетата натрия (pH 5,2) и 500 мкл фенола, насыщенного водой, и полученную суспензию тщательно перемешивали на мешалке Vortex. Затем добавляли 100 мкл смеси хлороформ - изоамиловый спирт, и суспензию тщательно перемешивали на мешалке Vortex в течение 1 мин с перерывами в 1 минуту. Суспензию откручивали в течение 10 мин и супернатант переносили в новую пробирку. Процедуру повторяли. Добавляли равный объем изопропанола (500 мкл) и хранили при -20°С. Образец откручивали в течение 10 минут, осадок промывали этанолом (70%) и растворяли в воде (50 мкл). Получали в целом 30 мкг РНК (0,60 мкг/мкл), которые использовали для дальнейшего синтеза кДНК.
кДНК синтезировали с использованием набора RevertAid™ H Minus First Strand cDNA Synthesis (Fermentas) в соответствии с “протоколом синтеза кДНК, подходящего для ПЦР”, рекомендованного производителем.
Дизайн праймеров для клонирования гена, кодирующего тяжелую цепь.
Для того, чтобы идентифицировать пару праймеров, пригодных для клонирования гена, кодирующего тяжелую цепь, проводили ПЦР для клонирования с использованием ДНК полимеразы Tag (реакционная смесь содержала 10X буфер для ДНК полимеразы Tag (67 мМ Tris-HCl (pH 8,8 при 25°C), 166 мМ (NH4)2SO4, 100 мМ 2-меркаптоэтанола) + 20 мМ MgCl2, 1,25 мМ dNTP, 5'-концевой праймер (25 пмоль), 3'-концевой праймер (25 пмоль), ДНК полимераза Taq (2,5 U), кДНК 1 мл, бидистиллированная вода до 50 мкл). Тотальную кДНК использовали в качестве матрицы. Программа ПЦР была следующей: 95°C в течение 3 минут; 35 циклов 94°C в течение 1 минуты, 45°C в течение 1 минуты и 72°C в течение 1 минуты; в конце 72°C в течение 10 минут.
5'-Концевой праймер соответствовал набору из 16 дегенеративных праймеров А3-А11, В6-В12 (SEQ ID NO: 5-20, соответственно), узнающих 95% последовательностей сигнальных пептидов в генах, кодирующих тяжелую цепь, выложенных в базе данных Кабат (Kabat, E., Wu, T., Perry, H. 1991. Sequences of proteins of Immunological Interest, 5th edn. US Departament of Health and Human Services, Public Health Service, NIH).
В качестве 3'-концевого праймера в ПЦР использовали:
- праймер A12, комплементарный аминокислотам 141-147 CH1 домена IgG, с введенным сайтом SalI (SEQ ID NO: 21),
- праймер A26, комплементарный 3'-концу гена, кодирующего CH1 домен антитела подкласса IgG1, с введенным сайтом XhoI (SEQ ID NO: 22).
Агарозный электрофорез продуктов ПЦР показал, что существуют пары праймеров, приводящие к образованию ПЦР фрагментов ожидаемого размера (Таблица 4).
Размеры полученных ПЦР фрагментов хорошо согласуются с ожидаемыми последовательностями кДНК генов тяжелой цепи IgG Mus musculus, представленной в Gene-Bank и IMGT-GENE-DB.
Дизайн праймеров для клонирования гена, кодирующего легкую цепь.
Ранее было показано, что изучаемое антитело содержит легкую цепь типа каппа. На основании этих данных в качестве 5'-концевого праймера использовали набор из 23 дегенеративных праймеров А15-А23, В1-В5, С1-С10 (SEQ ID NO: 23-46, соответственно). Этот набор узнает 95% последовательностей, соответствующих сигнальным пептидам в генах, кодирующих каппа цепь, опубликованных в базе данных Кабат (Kabat, E., Wu, T., Perry, H. 1991. Sequences of proteins of Immunological Interest, 5th edn. US Departament of Health and Human Services, Public Health Service, NIH).
В качестве 3'-концевого праймера в ПЦР, соответствовавшего 3`-концу легкой цепи фрагмента Fab (каппа-тип), с введенным на 5'-конец сайтом XhoI, использовали праймер L-m-r (SEQ ID NO: 47).
Агарозный электрофорез продуктов ПЦР показал, что существуют пары праймеров, приводящие к образованию ПЦР фрагментов ожидаемого размера (Таблица 5).
Размеры полученных ПЦР фрагментов хорошо согласуются с ожидаемыми последовательностями кДНК генов легкой цепи IgG Mus musculus, представленной в Gene-Bank и IMGT-GENE-DB.
Клонирование генов, кодирующих Fab фрагменты антител против кадгерина-17
Pfu ДНК полимеразу (Fermentas) использовали для клонирования генов, кодирующих цепи антитела. Условия ПЦР оптимизировали подбором температуры отжига и концентрации Mg2+ (реакционная смесь содержала 10X буфер для Pfu ДНК полимеразы (200 мМ Tris-HCl (pH 8.8 при 25°C), 100 мМ (NH4)2SO4, 100 мМ KCl, 1% Тритон X-100, 1 мг/мл БСА) + 20мМ MgCl2, 1.25 мМ dNTP, 5'-концевой праймер (25 пмоль), 3'-концевой праймер (25 пмоль), Pfu ДНК полимераза (1.25 U), кДНК 1 мкл, бидистиллированная вода до 50 мкл). Программа для ПЦР была следующая: 95°C в течение 3 минут; 35 циклов 94°C в течение 1 минуты, 45°C в течение 1 минуты и 72°C в течение 2 минут; в конце 72°C в течение 10 минут.
Пример 4. Секвенирование фрагментов генов, кодирующих вариабельные части Fab-фрагмента мышиного моноклонального антитела против кадгерина-17.
ПЦР-фрагменты, содержащие гены цепей Fab-фрагмента антител против кадгерина-17, элюировали из агарозного геля и секвенировали с помощью набора реактивов ABI PRISM® BigDye™ Terminator v. 3.1 с последующим анализом продуктов реакции на автоматическом секвенаторе ДНК ABI PRISM 3730 Applied Biosystems. В качестве праймеров для определения последовательностей фрагментов генов использовали праймеры, с помощью которых клонировали гены антител против кадгерина-17.
Аминокислотная последовательность вариабельного домена тяжелой цепи мышиного антитела 1E6 против кадгерина-17 человека приведена в Перечне последовательностей под номером SEQ ID NO: 1.
Аминокислотная последовательность вариабельного домена легкой цепи мышиного антитела 1E6 против кадгерина-17 человека приведена в Перечне последовательностей под номером SEQ ID NO: 2.
Аминокислотная последовательность вариабельного домена тяжелой цепи мышиного антитела 2G6 против кадгерина-17 человека приведена в Перечне последовательностей под номером SEQ ID NO: 3.
Аминокислотная последовательность вариабельного домена легкой цепи мышиного антитела 2G6 против кадгерина-17 человека приведена в Перечне последовательностей под номером SEQ ID NO: 4.
Хотя настоящее изобретение было подробно описано со ссылкой на предпочтительные варианты его осуществления, для специалиста в данной области техники ясно, что могут быть сделаны различные замены и применены эквиваленты, которые не выходят за рамки настоящего изобретения. Все процитированные в настоящем описании документы являются частью настоящей заявки, целиком включенные в настоящее описание посредством ссылки.
Таблица 1. Результаты ИФА. Тестирование сывороток. Сыворотки 1 и 2 титровали от разведения 1/500 с шагом 2. Сыворотка 1 - от иммунизации слитым белком CDH (24-132аа) - Trx, Сыворотка 2 - от иммунизации слитым белком CDH (581-678аа) - Trx.
Таблица 2. Результаты ИФА. Тестирование антител.
А. Сорбирован фрагмент CDH17 24-132aa.
Б. Сорбирован фрагмент CDH17 581-678аа
Таблица 3. Результаты ИФА. Тестирование антител 2G6 и 1E6 с использованием коммерческого препарата кадгерина 17 (Sinobiological, Кат. №11360-H08H-100 Human Cadherin-17 / LI-cadherin / CDH17 Protein (His Tag). Кадгерин-17 сорбировали в концентрации 0,8 мкг/мл. В качестве положительного контроля использовали коммерческую поликлональную иммунную сыворотку кролика против кадгерина-17 (Sigma, Кат. № HPA026556)
Таблица 4. Электрофорез ПЦР фрагментов генов тяжелой цепи антител в агарозном геле.
Таблица 5. Электрофорез ПЦР фрагментов генов легкой цепи антител в агарозном геле.
| название | год | авторы | номер документа |
|---|---|---|---|
| Вариабельные домены лёгкой и тяжёлой цепи мышиного моноклонального антитела против интерферона альфа (IFN-α) человека, антигенсвязывающий фрагмент (Fab) против IFN-α человека, содержащий указанные домены | 2018 |
|
RU2694672C1 |
| ВАРИАБЕЛЬНЫЕ ДОМЕНЫ ЛЕГКОЙ И ТЯЖЕЛОЙ ЦЕПИ МЫШИНОГО МОНОКЛОНАЛЬНОГО АНТИТЕЛА ПРОТИВ ФАКТОРА НЕКРОЗА ОПУХОЛИ АЛЬФА (ФНО-α) ЧЕЛОВЕКА (ВАРИАНТЫ), АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ (Fab) ПРОТИВ ФНО-α ЧЕЛОВЕКА, СОДЕРЖАЩИЙ УКАЗАННЫЕ ДОМЕНЫ (ВАРИАНТЫ) | 2012 |
|
RU2499000C1 |
| УЧАСТОК СВЯЗЫВАНИЯ АНТИГЕНА (Fab), В ТОМ ЧИСЛЕ ГУМАНИЗИРОВАННЫЙ Fab, ПРОТИВ БОТУЛИНИЧЕСКОГО НЕЙРОТОКСИНА С (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ Fab С ИСПОЛЬЗОВАНИЕМ ДРОЖЖЕЙ, СПОСОБ И НАБОР ДЛЯ ДЕТЕКЦИИ БОТУЛИНИЧЕСКОГО НЕЙРОТОКСИНА С | 2016 |
|
RU2623157C1 |
| Моноклональное антитело к интерферону бета-1а человека | 2018 |
|
RU2728696C2 |
| Рекомбинантный Fab-scFv на основе нейтрализующего антитела против интерферона бета-1а человека и антитела против рецептора ErbB2 человека | 2019 |
|
RU2748953C1 |
| Моноклональное антитело, способное нейтрализовать биологическую активность интерферона бета-1а человека | 2018 |
|
RU2729391C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ ФРАГМЕНТЫ (Fab) ПРОТИВ ВИРУСА БЕШЕНСТВА, ИЗОЛИРОВАННЫЙ ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ Fab ПРОТИВ ВИРУСА БЕШЕНСТВА, КЛЕТКА ДРОЖЖЕЙ, ТРАНСФОРМИРОВАННАЯ ФРАГМЕНТОМ ДНК, И СПОСОБ ПОЛУЧЕНИЯ Fab ПРОТИВ ВИРУСА БЕШЕНСТВА С ИСПОЛЬЗОВАНИЕМ ДРОЖЖЕЙ | 2010 |
|
RU2440412C2 |
| Вариабельные домены лёгких и тяжёлых цепей мышиных моноклональных антител против альфа-дефенсинов 1-3 (HNP 1-3) человека, антигенсвязывающие фрагменты (Fab) против HNP 1-3 человека, содержащие указанные домены | 2019 |
|
RU2724733C1 |
| Гуманизированное нейтрализующее антитело к интерферону-бета человека | 2019 |
|
RU2737466C1 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ (Fab), СВЯЗЫВАЮЩИЕСЯ С ИНТЕРФЕРОНОМ- γ ЧЕЛОВЕКА, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ, КЛЕТКА, ТРАНСФОРМИРОВАННАЯ ФРАГМЕНТОМ ДНК, И СПОСОБ ПОЛУЧЕНИЯ УКАЗАННОГО АНТИТЕЛА И АНТИГЕНСВЯЗЫВАЮЩЕГО ФРАГМЕНТА | 2013 |
|
RU2539752C2 |













Настоящее изобретение относится к иммунологии. Предложено антитело, селективно связывающееся с кадгерином-17 человека, его антигенсвязывающий фрагмент (Fab) и вариабельные домены указанного антитела. Данное изобретение обеспечивает расширение арсенала средств для детектирования кадгерина-17. 3 н.п. ф-лы, 8 ил., 5 табл., 4 пр.
1. Антитело или его антигенсвязывающий фрагмент (Fab), селективно связывающееся(-ийся) с кадгерином-17 человека, которое(-ый) включает в себя вариабельный домен тяжёлой цепи (VH) с последовательностью аминокислот SEQ ID NO: 3 и вариабельный домен лёгкой цепи (VL) с последовательностью аминокислот SEQ ID NO: 4.
2. Вариабельный домен тяжёлой цепи (VH) мышиного антитела против кадгерина-17 человека по п. 1, содержащий последовательность аминокислот SEQ ID NO: 3.
3. Вариабельный домен лёгкой цепи (VL) мышиного антитела против кадгерина-17 человека по п. 1, содержащий последовательность аминокислот SEQ ID NO: 4.
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| Токарный резец | 1924 |
|
SU2016A1 |
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| WANG, YONGGANG, et al | |||
| Печь для сжигания твердых и жидких нечистот | 1920 |
|
SU17A1 |
| LIU, LING XIAO, et al | |||
| Печь для сжигания твердых и жидких нечистот | 1920 |
|
SU17A1 |
| ПАЛЬЦЕВА Е.М | |||
| и др | |||
| "Влияние предоперационной лекарственной терапии на экспрессию молекул адгезии в метастазах колоректального рака в печени." Архив патологии, 2015, 3: 10-16. | |||

