ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение предлагает средство, индуцирующее гибель раковых клеток и средство, ингибирующее рост раковых клеток, а также фармацевтическую композицию для лечения заболевания, вызванного аномальным клеточным ростом, и способ скрининга средства, индуцирующего гибель клеток, и/или средства, ингибирующего рост клеток.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Типичные примеры заболеваний, вызванных аномальным клеточным ростом, могут включать в себя раковые заболевания. Раковые заболевания представляяют собой заболевания, характеризующиеся неконтролируемым клеточным ростом, обусловленным мутациями, эпигенетическими нарушениями и т.п. в генах. Большое число генетических нарушений, ассоциированных с раковыми заболеваниями, уже описано (например, в Futreal et al., Nat Rev Cancer. 2004; 4 (3): 177-83), причем считается, что большинство из них в той или иной степени связано с передачей сигнала, участвующего в росте, дифференциации или выживании клеток. Более того, такие генные нарушения могут вызывать аномальную передачу сигнала в клетках, состоящих из нормальных молекул. Указанное нарушение передачи сигнала может быть связано с активацией или дезактивацией конкретного сигнального каскада и, в конечном счете, может обуславливать аномальный рост клеток.
Хотя основной целью лечения рака в ранний период было ингибирование клеточного роста, такое лечение также приводило к физиологическому ингибированию роста нормальных клеток и, следовательно, к побочным эффектам, таким как алопеция, желудочно-кишечные расстройства и миелосупрессия. Соответственно, чтобы предотвратить такие побочные эффекты, разрабатывают терапевтические средства против рака на основе новых принципов, такие как лекарственные средства с молекулярными мишенями, направленные на рак-специфичные генные нарушения или аномальную передачу сигнала.
Считается, что рак возникает при накоплении в одной клетке нарушений в разных раковых генах, генах-супрессорах опухоли, генах ферментов, отвечающих за репарацию ДНК, и т.п. Ген RAS, ген FOS, ген MYC, ген BCL-2 и др. известны как раковые гены. К рак-специфичным генным нарушениям относится мутация в гене KRAS, присутствующая примерно в 95% случаев рака поджелудочной железы, примерно в 45% случаев колоректального рака и с высокой частотой при многих других раковых заболеваниях. Белок KRAS представляет собой G-белок, который локализуется на внутренней стороне клеточной мембраны. RAS, в том числе KRAS, участвует в каскаде, активируя RAF, такой как C-RAF или B-RAF, затем указанный RAF активирует MEK, который затем активирует MAPK. Если в KRAS появляется точечная мутация, активность ГТФазы снижается, в результате чего ГТФ-связанные активные формы сохраняются, обеспечивая передачу сигнала к нисходящему пути, что приводит к аномальному росту клеток. На примере гена KRAS можно видеть, что раковые гены вызывают аномальный рост клеток, который, в свою очередь, прогрессирует до злокачественной трансформации клетки и, в конечном счете, до возникновения рака как заболевания.
К тому же, глутатион-S-трансфераза (GST), фермент, катализирующий конъюгацию глутатиона, известна как фермент, который превращает вещество, такое как лекарственное средство, в водорастворимое вещество путем присоединения глутатиона (GSH). GST обычно подразделяют на основе аминокислотных последовательностей на 6 типов изозимов: α, μ, ω, ππ, θ и ξ. Из них в разных раковых клетках повышается, в частности, экспрессия GST-π (глутатион-S-трансфераза пи, также называемая GSTP1). Отмечена возможность того, что данная GST может частично отвечать за устойчивость к некоторым противоопухолевым средствам. И действительно, известно, что воздействие антисмысловой ДНК против GST-π или ингибитора GST-π на линию раковых клеток, характеризующихся повышенной экспрессией GST-π и устойчивостью к лекарственным средствам, приводит к подавлению устойчивости к лекарственным средствам (Takahashi и Niitsu, Gan To Kagaku Ryoho, 1994; 21 (7): 945-51; Ban et al., Cancer Res., 1996; 56 (15): 3577-82 и Nakajima et al., J Pharmacol Exp Ther., 2003, 306 (3): 861-9). Кроме того, недавно было показано, что воздействие миРНК против GST-π на линию андроген-независимых клеток рака предстательной железы, характеризующихся повышенной экспрессией GST-π, приводит к ингибированию роста данных клеток и увеличению апоптоза (Hokaiwado et al., Carcinogenesis. 2008; 29 (6): 1134-8).
Известно, что GST-π образует комплекс с N-концевой киназой c-Jun (JNK), ингибируя активность JNK (Adler et.al, EMBO J. 1999, 18, 1321-1334). Кроме того, известно, что GST-π участвует в S-глутатионилировании белков, связанных со стрессовой реакцией клеток (Townsend, et.al, J. Biol., Chem., 2009, 284, 436-445). Также известно, что GST-π участвует в осуществлении защитного эффекта против гибели клеток, вызванной активными формами кислорода (ROS) (Yin et.al, Cancer Res., 2000, 60, 4053-4057). Таким образом, очевидно, что GST-π, являющийся изозимом GST, имеет разнообразные свойства и функции.
Описано, что воздействие миРНК против GST-π на линию раковых клеток, несущих мутацию в KRAS, приводит к ингибированию активации Akt, в результате чего усиливается аутофагия и индуцируется апоптоз в умеренной степени (Nishita et al., AACR 102-е ежегодное собрание, реферат № 1065). В WO2012/176282 описано, что применение в качестве активных ингредиентов лекарственного средства, ингибирующего GST-π, и ингибитора аутофагии, такого как 3-метиладенин, может индуцировать апоптоз раковых клеток. Кроме того, в WO-014/098210 описано, что, если экспрессия GST-π и экспрессия Akt или т.п. ингибируются одновременно, рост клеток подавляется с последующей гибелью клеток; и что аутофагия, индуцированная ингибированием экспрессии GST-π, в значительной степени подавляется в результате одновременного ингибирования экспрессии GST-π и экспрессии Akt или т.п.
Однако многое еще неизвестно о взаимосвязи экспрессии GST-π в раковых клетках и роста клеток или гибели клеток, а также об участии GST-π в передаче сигнала и т.д.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Цель изобретения
Итак, целью настоящего изобретения является получение средства, способного индуцировать гибель раковых клеток, и/или ингибировать рост раковых клеток, получение фармацевтической композиции для лечения заболевания, вызванного аномальным клеточным ростом, и разработка способа скрининга средства, индуцирующего гибель клеток, и/или средства, ингибирующего рост клеток.
Способы достижения цели
Авторы настоящего изобретения провели тщательные исследования для достижения указанной выше цели и в результате завершили настоящее изобретение, установив, что связанный с гомеостазом белок, ингибирование которого наряду с GST-π приводит к синтетической летальности, ингибируется наряду с ингибированием GST-π в раковой клетке, в результате чего индукция гибели клеток и подавление роста клеток происходят более эффективно, чем в случае, когда ингибируется один из них. Настоящее изобретение охватывает следующие признаки:
(1) Средство, индуцирующее гибель раковых клеток, содержащее, в качестве активных ингредиентов, средство, ингибирующее GST-π, и средство, ингибирующее связанный с гомеостазом белок, вызывающий синтетическую летальность при ингибировании наряду с GST-π.
(2) Средство, ингибирующее рост раковых клеток, содержащее, в качестве активных ингредиентов, средство, ингибирующее GST-π, и средство, ингибирующее связанный с гомеостазом белок, вызывающий синтетическую летальность при ингибировании наряду с GST-π.
(3) Средство по пункту (1) или (2), где связанный с гомеостазом белок, приводящий к синтетической летальности при ингибировании наряду с GST-π, представляет собой белок, выбранный из группы, включающей в себя белок, регулирующий клеточный цикл, белок, связанный с подавлением апоптоза, и белок, связанный с сигнальным путем PI3K.
(4) Средство по пункту (3), где белок, регулирующий клеточный цикл, приводящий к синтетической летальности при ингибировании наряду с GST-π, представляет собой, по меньшей мере, один белок, регулирующий клеточный цикл, выбранный из группы, включающей в себя ATM, CDC25A, p21, PRKDC, RBBP8, SKP2, MCM10, RNPC1, CCNL1, CENPH, BRSK1, MCM8, CCNB3, MCMDC1 и MYLK.
(5) Средство по пункту (3), где белок, регулирующий клеточный цикл, приводящий к синтетической летальности при ингибировании наряду с GST-π, представляет собой, по меньшей мере, один белок, выбранный из группы, включающей в себя p21, RNPC1, CCNL1, MCM8, CCNB3 и MCMDC1.
(6) Средство по пункту (3), где белок, связанный с подавлением апоптоза, приводящий к синтетической летальности при ингибировании наряду с GST-π, представляет собой, по меньшей мере, один белок, связанный с подавлением апоптоза, выбранный из группы, включающей в себя AATF, ALOX12, ANXA1, ANXA4, API5, ATF5, AVEN, AZU1, BAG1, BCL2L1, BFAR, CFLAR, IL2, MALT1, MCL1, MKL1, MPO, MTL5, MYBL2 и MYO18A.
(7) Средство по пункту (3), где белок, связанный с сигнальным путем PI3K, приводящий к синтетической летальности при ингибировании наряду с GST-π, представляет собой, по меньшей мере, один белок, связанный с сигнальным путем PI3K, выбранный из группы, включающей в себя MTOR, IRAK1, IRS1, MYD88, NFKB1, PIK3CG, RAC1, AKT3, EIF4B, EIF4E, ILK, MTCP1, PIK3CA и SRF.
(8) Средство по пункту (1) или (2), где лекарственное средство представляет собой вещество, выбранное из группы, включающей в себя молекулу РНКи, рибозим, антисмысловую нуклеиновую кислоту, химерный полинуклеотид ДНК/РНК, и вектор, экспрессирующий, по меньшей мере, одно из указанных веществ.
(9) Средство по пункту (1) или (2), где средство, ингибирующее связанный с гомеостазом белок, представляет собой соединение, которое действует на связанный с гомеостазом белок.
(10) Средство по пункту (1), индуцирующее апоптоз.
(11) Средство по пункту (1) или (2), где раковая клетка представляет собой раковую клетку, интенсивно экспрессирующую GST-π.
(12) Фармацевтическая композиция для лечения заболевания, вызванного аномальным клеточным ростом, содержащая средство по любому из пунктов (1)-(11).
(13) Фармацевтическая композиция по пункту (12), где заболевание представляет собой рак.
(14) Фармацевтическая композиция по пункту (13), где рак характеризуется высоким уровнем экспрессии GST-π.
(15) Способ скрининга средства, индуцирующего гибель раковых клеток, и/или средства, ингибирующего рост раковых клеток, которое используют вместе со средством, ингибирующим GST-π, где способ включает в себя стадию выбора средства, ингибирующего связанный с гомеостазом белок, вызывающий синтетическую летальность при ингибировании наряду с GST-π.
(16) Способ скрининга по пункту (14), где связанный с гомеостазом белок, приводящий к синтетической летальности при ингибировании наряду с GST-π, представляет собой белок, выбранный из группы, включающей в себя белок, регулирующий клеточный цикл, белок, связанный с подавлением апоптоза, и белок, связанный с сигнальным путем PI3K.
(17) Способ скрининга по пункту (16), где белок, регулирующий клеточный цикл, приводящий к синтетической летальности при ингибировании наряду с GST-π, представляет собой, по меньшей мере, один белок, выбранный из группы, включающей в себя ATM, CDC25A, p21, PRKDC, RBBP8, SKP2, MCM10, RNPC1, CCNL1, CENPH, BRSK1, MCM8, CCNB3, MCMDC1 и MYLK.
(18) Способ скрининга по пункту (16), где белок, регулирующий клеточный цикл, приводящий к синтетической летальности при ингибировании наряду с GST-π, представляет собой, по меньшей мере, один белок, выбранный из группы, включающей в себя p21, RNPC1, CCNL1, MCM8, CCNB3 и MCMDC1.
(19) Способ скрининга по пункту (16), где белок, связанный с подавлением апоптоза, приводящий к синтетической летальности при ингибировании наряду с GST-π, представляет собой, по меньшей мере, один белок, связанный с подавлением апоптоза, выбранный из группы, включающей в себя AATF, ALOX12, ANXA1, ANXA4, API5, ATF5, AVEN, AZU1, BAG1, BCL2L1, BFAR, CFLAR, IL2, MALT1, MCL1, MKL1, MPO, MTL5, MYBL2 и MYO18A.
(20) Способ скрининга по пункту (16), где белок, связанный с сигнальным путем PI3K, приводящий к синтетической летальности при ингибировании наряду с GST-π, представляет собой, по меньшей мере, один белок, связанный с сигнальным путем PI3K, выбранный из группы, включающей в себя MTOR, IRAK1, IRS1, MYD88, NFKB1, PIK3CG, RAC1, AKT3, EIF4B, EIF4E, ILK, MTCP1, PIK3CA и SRF.
(21) Способ скрининга по любому из пунктов (15)-(20), включающий в себя следующие стадии: приведение в контакт тестируемого вещества и раковой клетки; измерение уровня экспрессии связанного с гомеостазом белка в клетке; и выбор тестируемого вещества, подходящего для применения в качестве средства, ингибирующего связанный с гомеостазом белок, если уровень экспрессии уменьшается по сравнению с измеренным в отсутствии тестируемого вещества.
(22) Способ скрининга средства, индуцирующего гибель раковых клеток, и/или средства, ингибирующего рост раковых клеток, которое используют вместе со средством, ингибирующим связанный с гомеостазом белок, вызывающий синтетическую летальность при ингибировании наряду с GST-π, где способ включает в себя стадию выбора средства, ингибирующего GST-π.
(23) Способ скрининга по пункту (22), где связанный с гомеостазом белок, приводящий к синтетической летальности при ингибировании наряду с GST-π, представляет собой белок, выбранный из группы, включающей в себя белок, регулирующий клеточный цикл, белок, связанный с подавлением апоптоза, и белок, связанный с сигнальным путем PI3K.
(24) Способ скрининга по пункту (22), где белок, регулирующий клеточный цикл, приводящий к синтетической летальности при ингибировании наряду с GST-π, представляет собой, по меньшей мере, один белок, выбранный из группы, включающей в себя ATM, CDC25A, p21, PRKDC, RBBP8, SKP2, MCM10, RNPC1, CCNL1, CENPH, BRSK1, MCM8, CCNB3, MCMDC1 и MYLK.
(25) Способ скрининга по пункту (23), где белок, регулирующий клеточный цикл, приводящий к синтетической летальности при ингибировании наряду с GST-π, представляет собой, по меньшей мере, один белок, выбранный из группы, включающей в себя p21, RNPC1, CCNL1, MCM8, CCNB3 и MCMDC1.
(26) Способ скрининга по пункту (23), где белок, связанный с подавлением апоптоза, приводящий к синтетической летальности при ингибировании наряду с GST-π, представляет собой, по меньшей мере, один белок, связанный с подавлением апоптоза, выбранный из группы, включающей в себя AATF, ALOX12, ANXA1, ANXA4, API5, ATF5, AVEN, AZU1, BAG1, BCL2L1, BFAR, CFLAR, IL2, MALT1, MCL1, MKL1, MPO, MTL5, MYBL2 и MYO18A.
(27) Способ скрининга по пункту (23), где белок, связанный с сигнальным путем PI3K, приводящий к синтетической летальности при ингибировании наряду с GST-π, представляет собой, по меньшей мере, один белок, связанный с сигнальным путем PI3K, выбранный из группы, включающей в себя MTOR, IRAK1, IRS1, MYD88, NFKB1, PIK3CG, RAC1, AKT3, EIF4B, EIF4E, ILK, MTCP1, PIK3CA и SRF.
(28) Способ скрининга по любому из пунктов (22)-(27), включающий в себя следующие стадии: приведение в контакт тестируемого вещества и раковой клетки; измерение уровня экспрессии GST-π в клетке; и выбор тестируемого вещества, подходящего для применения в качестве средства, ингибирующего GST-π, если уровень экспрессии уменьшается по сравнению с измеренным в отсутствии тестируемого вещества.
(29) Способ скрининга средства, индуцирующего гибель клеток, и/или средства, ингибирующего рост клеток, включающий в себя стадию выбора средства, ингибирующего GST-π и связанный с гомеостазом белок, вызывающий синтетическую летальность при ингибировании наряду с GST-π.
(30) Способ скрининга по пункту (29), где связанный с гомеостазом белок, приводящий к синтетической летальности при ингибировании наряду с GST-π, представляет собой белок, выбранный из группы, включающей в себя белок, регулирующий клеточный цикл, белок, связанный с подавлением апоптоза, и белок, связанный с сигнальным путем PI3K.
(31) Способ скрининга по пункту (30), где белок, регулирующий клеточный цикл, приводящий к синтетической летальности при ингибировании наряду с GST-π, представляет собой, по меньшей мере, один белок, регулирующий клеточный цикл, выбранный из группы, включающей в себя ATM, CDC25A, p21, PRKDC, RBBP8, SKP2, MCM10, RNPC1, CCNL1, CENPH, BRSK1, MCM8, CCNB3, MCMDC1 и MYLK.
(32) Способ скрининга по пункту (30), где белок, регулирующий клеточный цикл, приводящий к синтетической летальности при ингибировании наряду с GST-π, представляет собой, по меньшей мере, один белок, выбранный из группы, включающей в себя p21, RNPC1, CCNL1, MCM8, CCNB3 и MCMDC1.
(33) Способ скрининга по пункту (30), где белок, связанный с подавлением апоптоза, приводящий к синтетической летальности при ингибировании наряду с GST-π, представляет собой, по меньшей мере, один белок, связанный с подавлением апоптоза, выбранный из группы, включающей в себя AATF, ALOX12, ANXA1, ANXA4, API5, ATF5, AVEN, AZU1, BAG1, BCL2L1, BFAR, CFLAR, IL2, MALT1, MCL1, MKL1, MPO, MTL5, MYBL2 и MYO18A.
(34) Способ скрининга по пункту (30), где белок, связанный с сигнальным путем PI3K, приводящий к синтетической летальности при ингибировании наряду с GST-π, представляет собой, по меньшей мере, один белок, связанный с сигнальным путем PI3K, выбранный из группы, включающей в себя MTOR, IRAK1, IRS1, MYD88, NFKB1, PIK3CG, RAC1, AKT3, EIF4B, EIF4E, ILK, MTCP1, PIK3CA и SRF.
(35) Способ скрининга по любому из пунктов (29)-(34), включающий в себя следующие стадии: приведение в контакт тестируемого вещества и раковой клетки; измерение уровня экспрессии GST-π и уровня экспрессии связанного с гомеостазом белка, вызывающего синтетическую летальность при ингибировании наряду с GST-π, в клетке; и выбор тестируемого вещества, подходящего для применения в качестве средства, ингибирующего GST-π и связанный с гомеостазом белок, вызывающий синтетическую летальность при ингибировании наряду с GST-π, если уровень экспрессии GST-π и уровень экспрессии связанного с гомеостазом белка, вызывающего синтетическую летальность при ингибировании наряду с GST-π, понижаются по сравнению с уровнями, измеренными в отсутствии тестируемого вещества.
Данное описание включает в себя частично или полностью содержание, раскрытое в описании и/или чертежах патентной заявки Японии № 2014-266198, патентной заявки Японии № 2015-135494 и патентной заявки Японии № 2015-247725, которые являются приоритетными документами по отношению к настоящей заявке.
ЭФФЕКТЫ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
Средство, индуцирующее гибель клеток, в соответствии с настоящим изобретением может эффективно индуцировать гибель раковых клеток. Соответственно, средство, индуцирующее гибель клеток, в соответствии с настоящим изобретением, может обладать высокой эффективностью при применении в качестве фармацевтической композиции для лечения заболевания, вызванного аномальным ростом раковых клеток.
Кроме того, средство, ингибирующее рост клеток, в соответствии с настоящим изобретением может интенсивно ингибировать рост раковых клеток. Соответственно, средство, ингибирующее рост клеток, в соответствии с настоящим изобретением, может обладать высокой эффективностью при применении в качестве фармацевтической композиции для лечения заболевания, вызванного аномальным ростом раковых клеток.
Способ скрининга настоящего изобретения также позволяет выбрать лекарственное средство, способное эффективно индуцировать гибель раковых клеток и/или ингибировать рост раковых клеток.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фигуре 1 приведена характеристическая диаграмма, демонстрирующая результаты анализа мРНК GST-π и мРНК p21 в клетках, экспрессирующих мутантный KRAS, после воздействия на указанные клетки миРНК, ингибирующей экспрессию GST-π, и/или миРНК, ингибирующей экспрессию p21.
На фигуре 2 приведена характеристическая диаграмма, демонстрирующая результаты количественного определения мРНК p21 со временем после нокдауна GST-π и p21.
На фигуре 3 приведена характеристическая диаграмма, демонстрирующая результаты измерения числа клеток после нокдауна GST-π и p21.
На фигуре 4 приведена характеристическая диаграмма, демонстрирующая результаты измерения числа клеток после трехкратного нокдауна GST-π и p21.
На фигуре 5 приведена характеристическая диаграмма, демонстрирующая результаты измерения числа клеток после трехкратного нокдауна GST-π и p21.
На фигуре 6 приведена фотография клеток A549, полученная с использованием фазоразностной автофокусировки, после трехкратного нокдауна GST-π и p21.
На фигуре 7 приведена фотография клеток MIA PaCa-2, полученная с использованием фазоразностной автофокусировки, после трехкратного нокдауна GST-π и p21.
На фигуре 8 приведена фотография клеток PANC-1, полученная с использованием фазоразностной автофокусировки, после трехкратного нокдауна GST-π и p21.
На фигуре 9 приведена фотография клеток HCT116, полученная с использованием фазоразностной автофокусировки, после трехкратного нокдауна GST-π и p21.
На фигуре 10 приведена фотография клеток M7609, полученная с использованием фазоразностной автофокусировки, после трехкратного нокдауна GST-π и окрашивания β-галактозидазой.
На фигуре 11 приведена характеристическая диаграмма, демонстрирующая результаты количественного определения экспрессии гена PUMA после нокдауна GST-π и p21.
На фигуре 12 приведена характеристическая диаграмма, демонстрирующая результаты сравнения относительных процентов выживания после нокдауна по отдельности GST-π и белка-кандидата (белка, регулирующего клеточный цикл), приводящего к синтетической летальности, и после совместного нокдауна GST-π и белка-кандидата.
На фигуре 13 приведена характеристическая диаграмма, демонстрирующая результаты сравнения относительных процентов выживания после нокдауна по отдельности GST-π и белка-кандидата (белка, связанного с подавлением апоптоза), приводящего к синтетической летальности, и после совместного нокдауна GST-π и белка-кандидата.
На фигуре 14 приведена характеристическая диаграмма, демонстрирующая результаты сравнения относительных процентов выживания после нокдауна по отдельности GST-π и белка-кандидата (белка, связанного с сигнальным путем PI3K), приводящего к синтетической летальности, и после совместного нокдауна GST-π и белка-кандидата.
На фигуре 15 приведена характеристическая диаграмма, демонстрирующая результаты сравнения относительных процентов выживания после нокдауна по отдельности GST-π и MYLK и после совместного нокдауна GST-π и MYLK.
ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНЫХ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
Средство, индуцирующее гибель клеток, и средство, ингибирующее рост клеток, в соответствии с настоящим изобретением содержат, в качестве активных ингредиентов, средство, ингибирующее GST-π, и средство, ингибирующее связанный с гомеостазом белок, вызывающий синтетическую летальность при ингибировании наряду с GST-π. Средство, индуцирующее гибель клеток, и средство, ингибирующее рост клеток, в соответствии с настоящим изобретением индуцируют гибель раковых клеток и ингибируют рост раковых клеток. В данном контексте раковая клетка представляет собой клетку, характеризующуюся генетически обусловленным аномальным ростом (обусловленным функционированием связанных с раком генов).
Примеры связанных с раком генов могут включать в себя ген KRAS, ген FOS, ген MYC, ген BCL-2 и ген SIS. Кроме того, к связанным с раком генам относятся гены-супрессоры опухолей, примеры которых могут включать в себя ген RB, ген p53, ген BRCA1, ген NF1 и ген p73. Однако раковая клетка не ограничивается раковыми клетками, в которых функционируют указанные связанные с раком гены, и, следовательно, средства настоящего изобретения можно использовать в применении к широкому ряду клеток, характеризующихся аномальным клеточным ростом.
В частности, средство настоящего изобретения, индуцирующее гибель клеток, и средство настоящего изобретения, ингибирующее рост клеток, предпочтительно использовать в применении к раковым клеткам, экспрессирующим GST-π на высоком уровне. В данном контексте раковая клетка, экспрессирующая GST-π на высоком уровне, представляет собой клетку, характеризующуюся аномальным клеточным ростом (так называемая раковая клетка), в которой уровень экспрессии GST-π значительно выше, чем в нормальной клетке. Уровень экспрессии GST-π можно измерить с помощью стандартного метода, такого как ОТ-ПЦР или метод с использованием микрочипов.
Зачастую примером раковой клетки, экспрессирующей GST-π на высоком уровне, может служить раковая клетка, экспрессирующая мутантный KRAS. А именно, средство настоящего изобретения, индуцирующее гибель клеток, и средство настоящего изобретения, ингибирующее рост клеток, предпочтительно используют в применении к раковой клетке, экспрессирующей мутантный KRAS.
Мутантный KRAS представляет собой белок, имеющий аминокислотную последовательность KRAS дикого типа, которая содержит мутацию (мутации), такую как делеция, замена, добавление и/или вставка. В данном контексте мутация в мутантном KRAS представляет собой так называемую мутацию приобретения функции. А именно, в клетке, экспрессирующей мутантный KRAS, мутация приводит, например, к уменьшению активности ГТФазы, в результате чего ГТФ-связанные активные формы сохраняются, обуславливая конститутивную передачу сигналов нижестоящему пути, что приводит к аномальному клеточному росту по сравнению с клеткой, экспрессирующей KRAS дикого типа. Примеры гена, кодирующего мутантный KRAS, включают в себя ген, содержащий мутацию, по меньшей мере, в одном из кодона 12, кодона 13 и кодона 61 гена KRAS дикого типа. В частности, мутантный KRAS предпочтительно содержит мутации в кодонах 12 и 13. Конкретные примеры включают в себя мутации, которые приводят к замене глицина, кодируемого кодоном 12 гена KRAS, на серин, аспарагиновую кислоту, валин, цистеин, аланин или аргинин, а также мутации, которые приводят к замене глицина, кодируемого кодоном 13 гена KRAS, на аспарагиновую кислоту.
В соответствии с данным описанием GST-π представляет собой фермент, который кодируется геном GSTP1 и катализирует присоединение глутатиона. GST-π присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_000852 (NP_000843), крысиный: NM_012577 (NP_036709), мышиный: NM_013541 (NP_038569), и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность кодирующего участка человеческого гена GST-π, зарегистрированного в базе данных, описана в SEQ ID NO: 1, а аминокислотная последовательность человеческого белка GST-π, кодируемого указанным человеческим геном GST-π, описана в SEQ ID NO: 2.
В настоящем описании связанный с гомеостазом белок, вызывающий синтетическую летальность при ингибировании наряду с GST-π, представляет собой белок, который приводит к значительно более высокому проценту смертности раковых клеток при ингибировании наряду с GST-π, по сравнению с процентом смертности раковых клеток, обусловленному ингибированием только GST-π. Функция данного белка включает в себя участие в клеточном гомеостазе. В данном контексте термин синтетическая летальность относится к проявлению или значительному повышению летальности клетки или индивидуума под действием сочетания дефектов нескольких генов, тогда как один из указанных дефектов генов может привести к отсутствию летальности, или к низкой летальности. В частности, в настоящем описании термин синтетическая летальность относится к летальности раковой клетки.
В настоящем описании примеры связанного с гомеостазом белка, вызывающего синтетическую летальность при ингибировании наряду с GST-π, могут включать в себя белок, регулирующий клеточный цикл, белок, связанный с подавлением апоптоза, и белок, связанный с сигнальным путем PI3K. Белок, регулирующий клеточный цикл, представляет собой белок, функция которого заключается в регуляции клеточного цикла. Белок, связанный с подавлением апоптоза, представляет собой белок, функция которого заключается в подавлении апоптоза. Белок, связанный с сигнальным путем PI3K, представляет собой белок, за исключением Akt1, участвующий в сигнальном пути PI3K/AKT.
Кроме того, подразумевается, что белок, участвующий в регуляции клеточного цикла, включает в себя любой белок, участвующий в клеточном цикле, состоящем из фазы G1 (стадия покоя перед репликацией ДНК), фазы S (стадия синтеза ДНК), фазы G2 (стадия покоя перед делением клетки) и фазы M (стадия деления клетки). Более конкретно, примеры регуляции клеточного цикла могут включать в себя каждое событие регуляции механизма инициации перехода фаза G1 → фаза S → фаза G2 → фаза M в указанном порядке, регуляции развития фазы G1 до фазы S, ирегуляции развития фазы G2 до фазы M. Таким образом, белок, регулирующий клеточный цикл, может представлять собой, например, белок, который участвует в развитии указанных событий клеточного цикла, а также белок, который положительно или отрицательно регулирует указанные события. Более конкретно, примеры белка, регулирующего клеточный цикл, включают в себя циклин-зависимые киназы (CDK), необходимые для инициации фазы S и фазы M. Активность циклин-зависимых киназ положительно регулируется при связывании с циклинами. Кроме того, активность циклин-зависимых киназ отрицательно регулируется ингибиторами циклин-зависимых киназ (CKI), такими как p21 (CIP1/WAF1) и тирозинкиназы. Следовательно, белки, регулирующие активность циклин-зависимых киназ, т.е., циклины, ингибиторы циклин-зависимых киназ (такие как p21) и тирозинкиназы, также входят в объем термина "белок, регулирующий клеточный цикл".
Конкретно примеры белка, регулирующего клеточный цикл, вызывающего синтетическую летальность при ингибировании наряду с GST-π, могут включать в себя, по меньшей мере, один белок, регулирующий клеточный цикл, выбранный из группы, включающей в себя ATM, CDC25A, p21, PRKDC, RBBP8, SKP2, MCM10, RNPC1, CCNL1, CENPH, BRSK1, MCM8, CCNB3, MCMDC1 и MYLK. Среди указанных 15 типов белков, регулирующих клеточный цикл, один тип белков, регулирующих клеточный цикл, может ингибироваться наряду с GST-π, или два или более типов белков, регулирующих клеточный цикл, могут ингибироваться наряду с GST-π.
В частности, предпочтительно, по меньшей мере, один белок, регулирующий клеточный цикл, выбранный из группы, включающей в себя p21, RNPC1, CCNL1, MCM8, CCNB3 и MCMDC1, ингибируется наряду с GST-π. Если указанные 6 типов белков, регулирующих клеточный цикл, ингибируются по отдельности, это приводит к относительно слабому ингибированию клеточного роста, а ингибирование клеточного роста в высокой степени наблюдается только в том случае, если они ингибируются наряду с GST-π. То есть можно сказать, что средство, ингибирующее один из указанных 6 типов белков, регулирующих клеточный цикл, само по себе обладает превосходной безопасностью. Таким образом, предпочтительно белок, регулирующий клеточный цикл, вызывающий синтетическую летальность при ингибировании наряду с GST-π, можно выбрать из указанных 6 типов белков, регулирующих клеточный цикл.
p21 представляет собой белок, регулирующий клеточный цикл, который кодируется геном CDKN1A и принадлежит к семейству CIP/KIP. Функция данного белка заключается в ингибировании развития клеточного цикла в фазе G1 и фазе G2/M путем ингибирования эффекта комплекса циклин-CDK в результате связывания комплекса. А именно, ген p21 активируется под действием p53 (один из генов-супрессоров опухоли). Описано, что активация p53 вследствие повреждения ДНК и т.п. приводит к активации p21, в результате чего клеточный цикл останавливается в фазе G1 и в фазе G2/M. Кроме того, p21 также выполняет функцию, заключающуюся в ингибировании апоптоза, и защищает клетки от апоптоза, индуцированного химиотерапевтическим средством и т.п., в экспериментах, проводимых in vitro и на животных (Gartel и Tyner, 2002; и Abbs and Dutta, 2009). p21 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_000389.4, NM_078467.2, NM_001291549.1, NM_001220778.1, NM_001220777.1 (NP_001207707.1, NP_001278478.1, NP_001207706.1, NP_510867.1, NP_000380.1), и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена CDKN1A, зарегистрированного в базе данных под номером NM_000389.4, описана в SEQ ID NO: 3, а аминокислотная последовательность человеческого белка p21, кодируемого указанным человеческим геном CDKN1A, описана в SEQ ID NO: 4. В настоящем описании p21 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 4, кодируемую нуклеотидной последовательностью SEQ ID NO: 3. Информация о последовательности p21 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 3 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
RNPC1 представляет собой РНК-связывающий белок, кодируемый геном RNPC1, который является мишенью p53. RNPC1 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_017495.5, NM_183425.2, NM_001291780.1, XM_005260446.1 (XP_005260503.1, NP_059965.2, NP_906270.1, NP_001278709.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена RNPC1, зарегистрированного в базе данных под номером NM_017495.5, описана в SEQ ID NO: 5, а аминокислотная последовательность человеческого белка RNPC1, кодируемого указанным человеческим геном RNPC1, описана в SEQ ID NO: 6. В настоящем описании RNPC1 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 6, кодируемую нуклеотидной последовательностью SEQ ID NO: 5. Информация о последовательности RNPC1 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 5 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
CCNL1 представляет собой циклин-L1, кодируемый геном CCNL1. CCNL1 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_020307.2, XM_005247647.2, XM_005247648.1, XM_005247649.1, XM_005247650.1, XM_005247651.1, XM_006713710.1, XM_006713711.1 (XP_005247704.1, XP_005247705.1, XP_005247706.1, XP_005247707.1, XP_005247708.1, XP_006713773.1, NP_064703.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена CCNL1, зарегистрированного в базе данных под номером NM_020307.2, описана в SEQ ID NO: 7, а аминокислотная последовательность человеческого белка CCNL1, кодируемого указанным человеческим геном CCNL1, описана в SEQ ID NO: 8. В настоящем описании CCNL1 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 8, кодируемую нуклеотидной последовательностью SEQ ID NO: 7. Информация о последовательности RNPC1 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 7 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
MCM8 представляет собой поддерживающий минихромосомы белок 8, кодируемый геном MCM8. MCM8 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_032485.5, NM_182802.2, NM_001281520.1, NM_001281521.1, NM_001281522.1, XM_005260859.1 (XP_005260916.1, NP_115874.3, NP_001268449.1, NP_877954.1, NP_001268450.1, NP_001268451.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена MCM8, зарегистрированного в базе данных под номером NM_032485.5, описана в SEQ ID NO: 9, а аминокислотная последовательность человеческого белка MCM8, кодируемого указанным человеческим геном MCM8, описана в SEQ ID NO: 10. В настоящем описании MCM8 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 10, кодируемую нуклеотидной последовательностью SEQ ID NO: 9. Информация о последовательности MCM8 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 9 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
CCNB3 представляет собой циклин-B3, кодируемый геном CCNB3. CCNB3 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_033670.2, NM_033031.2, XM_006724610.1 (NP_391990.1, NP_149020.2, XP_006724673.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена CCNB3, зарегистрированного в базе данных под номером NM_033670.2, описана в SEQ ID NO: 11, а аминокислотная последовательность человеческого белка CCNB3, кодируемого указанным человеческим геном CCNB3, описана в SEQ ID NO: 12. В настоящем описании CCNB3 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 12, кодируемую нуклеотидной последовательностью SEQ ID NO: 11. Информация о последовательности CCNB3 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 11 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
MCMDC1 представляет собой белок, содержащий домен дефицита поддержания минихромосомы 1, кодируемый геном MCMDC1. MCMDC1 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_017696.2, NM_153255.4 (NP_060166.2, NP_694987.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена MCMDC1, зарегистрированного в базе данных под номером NM_017696.2, описана в SEQ ID NO: 13, а аминокислотная последовательность человеческого белка MCMDC1, кодируемого указанным человеческим геном MCMDC1, описана в SEQ ID NO: 14. В настоящем описании MCMDC1 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 14, кодируемую нуклеотидной последовательностью SEQ ID NO: 13. Информация о последовательности MCMDC1 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 13 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
ATM представляет собой серинтреонинкиназу ATM, кодируемую геном ATM, белок, который принадлежит к семейству киназ PI3/PI4. ATM присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_000051.3, XM_005271561.2, XM_005271562.2, XM_005271564.2, XM_006718843.1, XM_006718844.1, XM_006718845.1 (NP_000042.3, XP_005271618.2, XP_005271619.2, XP_005271621.2, XP_006718906.1, XP_006718907.1, XP_006718908.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена ATM, зарегистрированного в базе данных под номером NM_000051.3, описана в SEQ ID NO: 15, а аминокислотная последовательность человеческого белка ATM, кодируемого указанным человеческим геном ATM, описана в SEQ ID NO: 16. В настоящем описании ATM не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 16, кодируемую нуклеотидной последовательностью SEQ ID NO: 15. Информация о последовательности ATM зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 15 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
CDC25A представляет собой фосфатазу, которая кодируется геном CDC25A и принадлежит к семейству CDC25, она представляет собой белок, который активирует CDC2 путем дефосфорилирования. CDC25A присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_001789.2, NM_201567.1, XM_006713434.1, XM_006713435.1, XM_006713436.1 (NP_001780.2, NP_963861.1, XP_006713497.1, XP_006713498.1, XP_006713499.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена CDC25A, зарегистрированного в базе данных под номером NM_001789.2, описана в SEQ ID NO: 17, а аминокислотная последовательность человеческого белка CDC25A, кодируемого указанным человеческим геном CDC25A, описана в SEQ ID NO: 18. В настоящем описании CDC25A не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 18, кодируемую нуклеотидной последовательностью SEQ ID NO: 17. Информация о последовательности CDC25A зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 17 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
PRKDC представляет собой каталитическую субъединицу ДНК-зависимой протеинкиназы, кодируемой геном PRKDC, которая является белком, принадлежащим к семейству киназ PI3/PI4. PRKDC присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_006904.6, NM_001081640.1 (NP_008835.5, NP_001075109.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена PRKDC, зарегистрированного в базе данных под номером NM_006904.6, описана в SEQ ID NO: 19, а аминокислотная последовательность человеческого белка PRKDC, кодируемого указанным человеческим геном PRKDC, описана в SEQ ID NO: 20. В настоящем описании PRKDC не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 20, кодируемую нуклеотидной последовательностью SEQ ID NO: 19. Информация о последовательности PRKDC зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 19 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
RBBP8 представляет собой белок 8, связывающий белок ретинобластомы, который кодируется геном RBBP8 и относится к ядерным белкам, способным непосредственно связываться с белком ретинобластомы. RBBP8 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_002894.2, NM_203291.1, NM_203292.1, XM_005258325.1, XM_005258326.1, XM_006722519.1, XM_006722520.1, XM_006722521.1, XM_006722522.1 (NP_002885.1, NP_976036.1, NP_976037.1, XP_005258382.1, XP_005258383.1, XP_006722582.1, XP_006722583.1, XP_006722584.1, XP_006722585.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена RBBP8, зарегистрированного в базе данных под номером NM_002894.2, описана в SEQ ID NO: 21, а аминокислотная последовательность человеческого белка RBBP8, кодируемого указанным человеческим геном RBBP8, описана в SEQ ID NO: 22. В настоящем описании RBBP8 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 22, кодируемую нуклеотидной последовательностью SEQ ID NO: 21. Информация о последовательности RBBP8 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 21 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
SKP2 представляет собой белок, ассоциированный с киназой S-фазы 2, который кодируется геном SKP2 и относится к белку Fbox, представляющему собой одну из четырех субъединиц убиквитинлигазы E3. SKP2 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_005983.3, NM_032637.3, NM_001243120.1, XM_006714487.1 (NP_005974.2, NP_116026.1, NP_001230049.1, XP_006714550.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность кодирующего участка человеческого гена SKP2, зарегистрированного в базе данных под номером NM_005983.3, описана в SEQ ID NO: 23, а аминокислотная последовательность человеческого белка SKP2, кодируемого указанным человеческим геном SKP2, описана в SEQ ID NO: 24. В настоящем описании SKP2 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 24, кодируемую нуклеотидной последовательностью SEQ ID NO: 23. Информация о последовательности SKP2 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 23 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
MCM10 представляет собой белок, поддерживающий минихромосомы 10, кодируемый геном MCM10. MCM10 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_182751.2, NM_018518.4 (NP_877428.1, NP_060988.3) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена MCM10, зарегистрированного в базе данных под номером NM_182751.2, описана в SEQ ID NO: 25, а аминокислотная последовательность человеческого белка MCM10, кодируемого указанным человеческим геном MCM10, описана в SEQ ID NO: 26. В настоящем описании MCM10 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 26, кодируемую нуклеотидной последовательностью SEQ ID NO: 25. Информация о последовательности MCM10 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 25 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
CENPH представляет собой белок центромеры H, который кодируется геном CENPH и относится к одному из белков, составляющих активированный кинетохор, расположенный на центромере. CENPH присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_022909.3 (NP_075060.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена CENPH, зарегистрированного в базе данных под номером NM_022909.3, описана в SEQ ID NO: 27, а аминокислотная последовательность человеческого белка CENPH, кодируемого указанным человеческим геном CENPH, описана в SEQ ID NO: 28. В настоящем описании CENPH не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 28, кодируемую нуклеотидной последовательностью SEQ ID NO: 27. Могут существовать несколько транскрипционных вариантов CENPH. Нуклеотидная последовательность SEQ ID NO: 27 представляет собой нуклеотидную последовательность транскрипционного варианта.
BRSK1 представляет собой серинтреонинкиназу, которая кодируется геном BRSK1 и действует на контрольную точку клеточного цикла при повреждении ДНК. BRSK1 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_032430.1, XM_005259327.1, XR_430213.1 (NP_115806.1, XP_005259384.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена BRSK1, зарегистрированного в базе данных под номером NM_032430.1, описана в SEQ ID NO: 29, а аминокислотная последовательность человеческого белка BRSK1, кодируемого указанным человеческим геном BRSK1, описана в SEQ ID NO: 30. В настоящем описании BRSK1 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 30, кодируемую нуклеотидной последовательностью SEQ ID NO: 29. Информация о последовательности BRSK1 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 29 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
MYLK, киназа легкой цепи миозина, представляет собой кальций/кальмодулин-зависимый фермент, который фосфорилирует регуляторные легкие цепи миозина, после чего миозин взаимодействует с филаментами актина, обеспечивая сократительную активность. Ген, кодирующий MYLK, кодирует изоформы, встречающиеся как в гладкой мускулатуре, так и вне мышц. MYLK присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_053028.3, NM_053026.3, NM_053027.3, NM_053025.3, NM_053031.2, NM_053032.2, XM_011512862.1, XM_011512861.1, XM_011512860.1 (NP_444256.3, NP_444254.3, NP_444255.3, NP_444253.3, NP_444259.1, NP_444260.1, XP_011511164.1, XP_011511163.1, XP_011511162.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена MYLK, зарегистрированного в базе данных под номером NM_053028.3, описана в SEQ ID NO: 41, а аминокислотная последовательность человеческого белка MYLK, кодируемого указанным человеческим геном MYLK, описана в SEQ ID NO: 42. В настоящем описании MYLK не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 42, кодируемую нуклеотидной последовательностью SEQ ID NO: 41. Информация о последовательности MYLK зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 41 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
С другой стороны, белок, имеющий функцию подавления апоптоза, представляет собой белок, функция которого заключается в подавлении апоптоза путем ингибирования таких механизмов, как кариопикноз, клеточное сокращение, пузырение мембраны и фрагментация ДНК. Подразумевается, что функция подавления апоптоза включает в себя как функцию ингибирования апоптоза, так и функцию ингибирования фактора, стимулирующего апоптоз. Примерами факторов, стимулирующих апоптоз, являются, в числе многих других, каспаза, Fas и TNFR.
Конкретные примеры белка, связанного с подавлением апоптоза, вызывающего синтетическую летальность при ингибировании наряду с GST-π, могут включать в себя, по меньшей мере, один белок, связанный с подавлением апоптоза, выбранный из группы, включающей в себя AATF, ALOX12, ANXA1, ANXA4, API5, ATF5, AVEN, AZU1, BAG1, BCL2L1, BFAR, CFLAR, IL2, MALT1, MCL1, MKL1, MPO, MTL5, MYBL2 и MYO18A. Из 20 типов белков, связанных с подавлением апоптоза, один тип белков, связанных с подавлением апоптоза, может ингибироваться наряду с GST-π, или два или более типов белков, связанных с подавлением апоптоза, могут ингибироваться наряду с GST-π.
AATF был идентифицирован на основе взаимодействия с MAP3K12/DLK, протеинкиназой, которая, как известно, участвует в индукции клеточного апоптоза. AATF содержит лейциновую молнию, которая является характеристическим мотивом факторов транскрипции, и, как было показано, проявляет высокую активность в отношении трансактивации при гибридизации с ДНК-связывающим доменом Gal4. Известно, что повышенная экспрессия гена, кодирующего AATF, приводит к ингибированию апоптоза, индуцированного MAP3K12. AATF присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_012138.3, XM_011546799.1, XM_011524611.1, XR_951958.1, XR_934439.1 (NP_036270.1, XP_011545101.1, XP_011522913.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена AATF, зарегистрированного в базе данных под номером NM_012138.3, описана в SEQ ID NO: 39, а аминокислотная последовательность человеческого белка AATF, кодируемого указанным человеческим геном AATF, описана в SEQ ID NO: 40. В настоящем описании AATF не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 40, кодируемую нуклеотидной последовательностью SEQ ID NO: 39. Информация о последовательности AATF зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 39 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
ALOX12, арахидонат-12-липоксигеназа, как известно, участвует в развитии атеросклероза, остеопороза и т.п. Также известно, что ALOX12 положительно регулирует ангиогенез путем регуляции экспрессии фактора роста эндотелия сосудов и участвует в процессе апоптоза, стимулируя выживание клеток гладкой мускулатуры сосудов и т.п. ALOX12 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_000697.2, XM_011523780.1 (NP_000688.2, XP_011522082.1, AAH69557.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена ALOX12, зарегистрированного в базе данных под номером NM_000697.2, описана в SEQ ID NO: 43, а аминокислотная последовательность человеческого белка ALOX12, кодируемого указанным человеческим геном ALOX12, описана в SEQ ID NO: 44. В настоящем описании ALOX12 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 44, кодируемую нуклеотидной последовательностью SEQ ID NO: 43. Информация о последовательности ALOX12 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 43 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
ANXA1 представляет собой локализованный в мембране белок, способный связываться с фосфолипидами. ANXA1 ингибирует фосфолиппазу A2 и обладает противовоспалительной активностью. ANXA1 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_000700.2, XM_011518609.1, XM_011518608.1 (NP_000691.1, AAH34157.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена ANXA1, зарегистрированного в базе данных под номером NM_000700.2, описана в SEQ ID NO: 45, а аминокислотная последовательность человеческого белка ANXA1, кодируемого указанным человеческим геном ANXA1, описана в SEQ ID NO: 46. В настоящем описании ANXA1 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 46, кодируемую нуклеотидной последовательностью SEQ ID NO: 45. Информация о последовательности ANXA1 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 45 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
ANXA4 принадлежит к семейству аннексинов, кальций-зависимых фосфолипид-связывающих белков. Данный белок может взаимодействовать с АТФ, обладает антикоагуляционной активностью in vitro и ингибирует фосфолипазу A2. ANXA4 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_001153.3, XM_011532805.1 (NP_001144.1, XP_011531107.1, AAH63672.1, AAH00182.1, AAH11659.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена ANXA4, зарегистрированного в базе данных под номером NM_001153.3, описана в SEQ ID NO: 47, а аминокислотная последовательность человеческого белка ANXA4, кодируемого указанным человеческим геном ANXA4, описана в SEQ ID NO: 48. В настоящем описании ANXA4 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 48, кодируемую нуклеотидной последовательностью SEQ ID NO: 47. Информация о последовательности ANXA4 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 47 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
API5 представляет собой белок, ингибирующий апоптоз, экспрессия которого, как известно, предотвращает апоптоз после утраты фактора роста. API5 подавляет апоптоз, индуцированный фактором транскрипции E2F1, а также взаимодействует, осуществляя его отрицательную регуляцию, с Acinus, ядерным фактором, участвующим в апоптотической фрагментации ДНК. API5 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_001142930.1, NM_006595.3, NM_001243747.1, NM_001142931.1, XM_006718359.2, NR_024625.1 (NP_001136402.1, NP_001136403.1, NP_001230676.1, NP_006586.1, XP_006718422.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена API5, зарегистрированного в базе данных под номером NM_001142930.1, описана в SEQ ID NO: 49, а аминокислотная последовательность человеческого белка API5, кодируемого указанным человеческим геном API5, описана в SEQ ID NO: 50. В настоящем описании API5 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 50, кодируемую нуклеотидной последовательностью SEQ ID NO: 49. Информация о последовательности API5 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 49 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
ATF5, как известно, участвует в развитии заболеваний, вызываемых вирусом человеческого T-клеточного лейкоза типа 1. ATF5 является активатором транскрипции, который связывается с цАМФ-отвечающим элементом (CRE), присутствующим во многих вирусных промоторах и др., и, как известно, ингибирует дифференциацию нейропрогениторных клеток в нейроны. ATF5 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_012068.5, NM_001193646.1, NM_001290746.1, XM_011526629.1 (NP_036200.2, NP_001277675.1, NP_001180575.1, XP_011524931.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена ATF5, зарегистрированного в базе данных под номером NM_012068.5, описана в SEQ ID NO: 51, а аминокислотная последовательность человеческого белка ATF5, кодируемого указанным человеческим геном ATF5, описана в SEQ ID NO: 52. В настоящем описании ATF5 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 52, кодируемую нуклеотидной последовательностью SEQ ID NO: 51. Информация о последовательности ATF5 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 51 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
AVEN представляет собой белок, известный как ингибитор апоптоза и активации каспаз, и участвующий в развитии шизоидного расстройствоа личности и алекситимии. Также известно, что AVEN ингибирует апоптоз, опосредованный Apaf-1. AVEN присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_020371.2, XM_011521820.1, XM_005254563.2, XM_011521819.1, XM_011521818.1 (NP NP_065104.1, XP_011520122.1, XP_011520121.1, XP_011520120.1, XP_005254620.1, AAH63533.1, AAF91470.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена AVEN, зарегистрированного в базе данных под номером NM_020371.2, описана в SEQ ID NO: 53, а аминокислотная последовательность человеческого белка AVEN, кодируемого указанным человеческим геном AVEN, описана в SEQ ID NO: 54. В настоящем описании AVEN не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 54, кодируемую нуклеотидной последовательностью SEQ ID NO: 53. Информация о последовательности AVEN зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 53 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
AZU1 представляет собой белок, содержащийся в азурофильных зернах и обладающий моноцитарной хемотаксической и противомикробной активностью. AZU1 является важным многофункциональным медиатором воспаления. AZU1 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_001700.3 (NP_001691.1, EAW69592.1, AAH93933.1, AAH93931.1, AAH69495.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена AZU1, зарегистрированного в базе данных под номером NM_001700.3, описана в SEQ ID NO: 55, а аминокислотная последовательность человеческого белка AZU1, кодируемого указанным человеческим геном AZU1, описана в SEQ ID NO: 56. В настоящем описании AZU1 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 56, кодируемую нуклеотидной последовательностью SEQ ID NO: 55. Информация о последовательности AZU1 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 55 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
BAG1 связывается с BCL2, мембранным белком, который ингибирует путь, приводящий к апоптозу или программируемой смерти клетки. BAG1 усиливает антиапоптотические эффекты BCL2 и обеспечивает связь между рецепторами факторов роста и антиапоптотическими механизмами. BAG1 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_004323.5, NM_001172415.1 (NP_004314.5, NP_001165886.1, AAH14774.2) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена BAG1, зарегистрированного в базе данных под номером NM_004323.5, описана в SEQ ID NO: 57, а аминокислотная последовательность человеческого белка BAG1, кодируемого указанным человеческим геном BAG1, описана в SEQ ID NO: 58. В настоящем описании BAG1 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 58, кодируемую нуклеотидной последовательностью SEQ ID NO: 57. Информация о последовательности ALOX12 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 57 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
BCL2L1 принадлежит к семейству белков BCL-2. Члены данного семейства белков образуют гетеро или гомодимеры и действуют как анти- или про-апоптотические регуляторы, которые участвуют в широком ряде клеточных функций. BCL2L1 расположен на внешней митохондриальной мембране и, как показано, регулирует открытие канала внешней митохондриальной мембраны. BCL2L1 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_138578.1, NM_001191.2, XM_011528966.1, XM_011528965.1, XM_011528961.1, XM_011528960.1, XM_011528964.1, XM_011528963.1, XM_011528962.1, XM_005260487.3, XM_005260486.2 (NP_612815.1, NP_001182.1, AAH19307.1, XP_011527268.1, XP_011527267.1, XP_011527266.1, XP_011527265.1, XP_011527264.1, XP_011527263.1, XP_011527262.1, XP_005260544.1, XP_005260543.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена BCL2L1, зарегистрированного в базе данных под номером NM_138578.1, описана в SEQ ID NO: 59, а аминокислотная последовательность человеческого белка BCL2L1, кодируемого указанным человеческим геном BCL2L1, описана в SEQ ID NO: 60. В настоящем описании BCL2L1 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 60, кодируемую нуклеотидной последовательностью SEQ ID NO: 59. Информация о последовательности BCL2L1 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 59 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
BFAR, бифункциональный регулятор апоптоза, обладает антиапоптотической активностью, как в отношении апоптоза, инициируемого рецепторами клеточной гибели, так и в отношении апоптоза, инициируемого митохондриальными факторами. BFAR присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_016561.2, XM_006725196.2, XM_011546704.1, XM_005255350.2, XM_011522520.1 (NP_057645.1, XP_011545006.1, XP_011520822.1, XP_006725259.1, XP_005255407.1, AAH03054.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена BFAR, зарегистрированного в базе данных под номером NM_016561.2, описана в SEQ ID NO: 61, а аминокислотная последовательность человеческого белка BFAR, кодируемого указанным человеческим геном BFAR, описана в SEQ ID NO: 62. В настоящем описании BFAR не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 62, кодируемую нуклеотидной последовательностью SEQ ID NO: 61. Информация о последовательности BFAR зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 61 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
CFLAR, регулятор апоптоза, как известно, является структурно подобным каспазе-8. Однако CFLAR не обладает активностью каспазы и расщепляется на два пептида каспазой-8. CFLAR присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_003879.5, NM_001202519.1, NM_001202518.1, NM_001308043.1, NM_001308042.1, NM_001202517.1, NM_001202516.1, NM_001127184.2, NM_001202515.1, NM_001127183.2, XM_011512100.1 (NP_003870.4, NP_001294972.1, NP_001294971.1, NP_001189448.1, NP_001189446.1, NP_001189445.1, NP_001189444.1, NP_001120656.1, XP_011510402.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена CFLAR, зарегистрированного в базе данных под номером NM_003879.5, описана в SEQ ID NO: 63, а аминокислотная последовательность человеческого белка CFLAR, кодируемого указанным человеческим геном CFLAR, описана в SEQ ID NO: 64. В настоящем описании CFLAR не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 64, кодируемую нуклеотидной последовательностью SEQ ID NO: 63. Информация о последовательности CFLAR зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 63 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
IL2, интерлейкин 2, представляет собой секретируемый цитокин, который играет важную роль в пролиферации T- и B-лимфоцитов. IL2 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_000586.3 (NP_000577.2) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена IL2, зарегистрированного в базе данных под номером NM_000586.3, описана в SEQ ID NO: 65, а аминокислотная последовательность человеческого белка IL2, кодируемого указанным человеческим геном IL2, описана в SEQ ID NO: 66. В настоящем описании IL2 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 66, кодируемую нуклеотидной последовательностью SEQ ID NO: 65. Может существовать несколько транскрипционных вариантов IL2. Нуклеотидная последовательность SEQ ID NO: 65 представляет собой нуклеотидную последовательность транскрипционного варианта.
MALT1 кодируется геном, который периодически перегруппировывается путем хромосомной транслокации с бакуловирусным IAP, кодирующим повтор-содержащий белок 3 (также известный как ингибитор апоптоза 2), и локусом, кодирующим тяжелую цепь иммуноглобулина, при лимфомах лимфоидной ткани слизистых оболочек. MALT1 может активировать NFκB. MALT1 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_173844.2, NM_006785.3, XM_011525794.1 (NP_776216.1, NP_006776.1, XP_011524096.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена MALT1, зарегистрированного в базе данных под номером NM_006785.3, описана в SEQ ID NO: 67, а аминокислотная последовательность человеческого белка MALT1, кодируемого указанным человеческим геном MALT1, описана в SEQ ID NO: 68. В настоящем описании MALT1 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 68, кодируемую нуклеотидной последовательностью SEQ ID NO: 67. Информация о последовательности MALT1 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 67 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
MCL1 представляет собой антиапоптотический белок, который является членом семейства Bcl-2. Самый длинный вариант, образующийся в результате альтернативного сплайсинга гена MCL1, повышает выживаемость клеток путем ингибирования апоптоза, тогда как более короткие варианты, образующиеся в результате альтернативного сплайсинга, стимулируют апоптоз и индуцируют гибель клеток. MCL1 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_021960.4, NM_001197320.1, NM_182763.2 (NP_068779.1, NP_001184249.1, NP_877495.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена MCL1, зарегистрированного в базе данных под номером NM_021960.4, описана в SEQ ID NO: 69, а аминокислотная последовательность человеческого белка MCL1, кодируемого указанным человеческим геном MCL1, описана в SEQ ID NO: 70. В настоящем описании MCL1 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 70, кодируемую нуклеотидной последовательностью SEQ ID NO: 69. Информация о последовательности MCL1 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 69 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
MKL1, как известно, взаимодействует с транскрипционным фактором миокардином, ключевым регулятором дифференцировки клеток гладкой мускулатуры. MKL1 преимущественно локализуется в ядре и способствует передаче сигналов от цитоскелета к ядру. Ген MKL1 участвует в событии транслокации, которое приводит к образованию гибрида указанного гена и гена РНК-связывающего белка 15. MKL1 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_001282662.1, NM_001282660.1, NM_020831.4, NM_001282661.1, XM_011530287.1, XM_011530286.1, XM_011530285.1, XM_011530284.1, XM_011530283.1, XM_005261691.3 (NP_001269591.1, NP_001269589.1, NP_065882.1, NP_001269590.1, XP_011528589.1, XP_011528588.1, XP_011528587.1, XP_011528586.1, XP_011528585.1, XP_005261751.1, XP_005261749.1, XP_005261748.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена MKL1, зарегистрированного в базе данных под номером NM_001282662.1, описана в SEQ ID NO: 71, а аминокислотная последовательность человеческого белка MKL1, кодируемого указанным человеческим геном MKL1, описана в SEQ ID NO: 72. В настоящем описании MKL1 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 72, кодируемую нуклеотидной последовательностью SEQ ID NO: 71. Информация о последовательности MKL1 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 71 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
MPO, миелопероксидаза, представляет собой гем-содержащий белок, который синтезируется в процессе дифференциации миелоидных клеток и является основным компонентом азурофильных гранул нейтрофилов. MPO продуцирует гипогалоидные кислоты, играющие центральную роль в микробицидной активности нейтрофилов. MPO присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_000250.1, XM_011524823.1, XM_011524822.1, XM_011524821.1 (NP_000241.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена MPO, зарегистрированного в базе данных под номером NM_000250.1, описана в SEQ ID NO: 73, а аминокислотная последовательность человеческого белка MPO, кодируемого указанным человеческим геном MPO, описана в SEQ ID NO: 74. В настоящем описании MPO не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 74, кодируемую нуклеотидной последовательностью SEQ ID NO: 73. Информация о последовательности MPO зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 73 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
Показано, что MTL5, металлотионеин-подобный белок, специфически экспрессируется в яичках и яичниках мышей. Металлотионеин может играть центральную роль в регуляции роста и дифференциации клеток и участвовать в сперматогенезе. MTL5 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_004923.3, NM_001039656.1, XM_011545404.1, XM_011545403.1, XM_011545402.1 (NP_001034745.1, NP_004914.2, XP_011543706.1, XP_011543705.1, XP_011543704.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена MTL5, зарегистрированного в базе данных под номером NM_004923.3, описана в SEQ ID NO: 75, а аминокислотная последовательность человеческого белка MTL5, кодируемого указанным человеческим геном MTL5, описана в SEQ ID NO: 76. В настоящем описании MTL5 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 76, кодируемую нуклеотидной последовательностью SEQ ID NO: 75. Информация о последовательности MTL5 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 75 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
MYBL2 представляет собой ядерный белок, который принадлежит к семейству транскрипционных факторов MYB и участвует в развитии клеточного цикла. MYBL2 фосфорилируется циклин A/циклин-зависимой киназой 2 во время S-фазы клеточного цикла и обладает как активаторной, так и репрессорной активностью. MYBL2 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_001278610.1, NM_002466.3 (NP_001265539.1, NP_002457.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена MYBL2, зарегистрированного в базе данных под номером NM_002466.3, описана в SEQ ID NO: 77, а аминокислотная последовательность человеческого белка MYBL2, кодируемого указанным человеческим геном MYBL2, описана в SEQ ID NO: 78. В настоящем описании MYBL2 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 78, кодируемую нуклеотидной последовательностью SEQ ID NO: 77. Информация о последовательности MYBL2 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 77 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
MYO18A, миозин 18A, как известно, участвует в развитии 8p11 миелопролиферативного синдрома. MYO18A обладает моторной активностью и АТФ-азной активностью. MYO18A присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_203318.1, NM_078471.3 (NP_976063.1, NP_510880.2) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена MYO18A, зарегистрированного в базе данных под номером NM_078471.3, описана в SEQ ID NO: 79, а аминокислотная последовательность человеческого белка MYO18A, кодируемого указанным человеческим геном MYO18A, описана в SEQ ID NO: 80. В настоящем описании MYO18A не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 80, кодируемую нуклеотидной последовательностью SEQ ID NO: 79. Информация о последовательности MYO18A зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 79 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
С другой стороны, белок, связанный с сигнальным путем PI3K, представляет собой белком, за исключением Akt1, участвующий в сигнальном пути PI3K/AKT. Другими словами, белок, связанный с сигнальным путем PI3K, представляет собой белок, связанный с сигнальным путем PI3K/AKT, за исключением Akt1. Сигнальный путь PI3K/AKT описан, например, в Cell 2007 Jun 29; 129 (7): 1261-74.
А именно, примеры белка, связанного с сигнальным путем PI3K и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, по меньшей мере, один белок, связанный с сигнальным путем PI3K, выбранный из группы, включающей в себя MTOR, IRAK1, IRS1, MYD88, NFKB1, PIK3CG, RAC1, AKT3, EIF4B, EIF4E, ILK, MTCP1, PIK3CA и SRF. Среди указанных 14 типов белков, связанных с сигнальным путем PI3K, один тип белков, связанных с сигнальным путем PI3K, может ингибироваться наряду с GST-π, или два или более типов белков, связанных с сигнальным путем PI3K, могут ингибироваться наряду с GST-π.
В частности, предпочтительно, по меньшей мере, один белок, связанный с сигнальным путем PI3K, выбранный из группы, включающей в себя MTOR, IRAK1, IRS1, MYD88, NFKB1, PIK3CG и RAC1, ингибируется наряду с GST-π. Ингибирование каждого из указанных 7 типов белков, связанных с сигнальным путем PI3K, наряду с GST-π, приводит к значительно более высокой степени ингибирования клеточного роста, чем ингибирование отдельно одного из указанных белков, или отдельно GST-π. Таким образом, белок, связанный с сигнальным путем PI3K и вызывающий синтетическую летальность при ингибировании наряду с GST-π, предпочтительно выбран из указанных 7 типов белков, связанных с сигнальным путем PI3K.
MTOR, механистическая мишень рапамицина, представляет собой белок-мишень рапамицина и известен как серинтреонинкиназа. MTOR относится к семейству киназ, родственных фосфатидилинозитолкиназе, и, как известно, опосредует ответ клеток на стрессы, такие как повреждение ДНК и уменьшение содержания питательных веществ. Известно также, что MTOR является мишенью комплексов FKBP12-рапамицин, участвующих в остановке клеточного цикла и иммуносупрессивном эффекте. MTOR присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_004958.3, XM_005263438.1, XM_011541166.1 (NP_004949.1, XP_005263495.1, XP_005263495.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена MTOR, зарегистрированного в базе данных под номером NM_004958.3, описана в SEQ ID NO: 81, а аминокислотная последовательность человеческого белка MTOR, кодируемого указанным человеческим геном MTOR, описана в SEQ ID NO: 82. В настоящем описании MTOR не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 82, кодируемую нуклеотидной последовательностью SEQ ID NO: 81. Информация о последовательности MTOR зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 81 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
IRAK1, ассоциированная с рецептором интерлейкина 1 киназа 1, представляет собой серинтреонинкиназу, которая стимулирует рецепторы интерлейкина 1. IRAK1 частично отвечает за IL1-индуцированную повышающую регуляцию, связанную с фактором транскрипции NFκB. IRAK1 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_001025243.1, NM_001025242.1, NM_001569.3, XM_011531158.1, XM_005274668.2 (NP_001020414.1, NP_001020413.1, NP_001560.2, XP_011529460.1, XP_005274725.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена IRAK1, зарегистрированного в базе данных под номером NM_001025243.1, описана в SEQ ID NO: 83, а аминокислотная последовательность человеческого белка IRAK1, кодируемого указанным человеческим геном IRAK1, описана в SEQ ID NO: 84. В настоящем описании IRAK1 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 84, кодируемую нуклеотидной последовательностью SEQ ID NO: 83. Информация о последовательности IRAK1 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 83 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
IRS1, субстрат рецептора инсулина 1, известен как белок, который фосфорилируется рецептором инсулина с тирозинкиназой активностью. IRS1 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_005544.2 (NP_005535.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена IRS1, зарегистрированного в базе данных под номером NM_005544.2, описана в SEQ ID NO: 85, а аминокислотная последовательность человеческого белка IRS1, кодируемого указанным человеческим геном IRS1, описана в SEQ ID NO: 86. В настоящем описании IRS1 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 86, кодируемую нуклеотидной последовательностью SEQ ID NO: 85. Могут существовать несколько транскрипционных вариантов IRS1. Нуклеотидная последовательность SEQ ID NO: 85 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
MYD88, первичный ответ миелоидной дифференциации 88, представляет собой цитоплазматический адапторный белок, который играет ключевую роль во врожденном и приобретенном иммунных ответах. MYD88 действует как эссенциальный передатчик сигнала в сигнальных путях, опосредованных интерлейкином 1 и Toll-подобным рецептором. Указанные сигнальные пути регулируют активацию многих воспалительных генов. MYD88 содержит на N-конце домен смерти и на C-конце домен Toll-подобного рецептора-рецептора интерлейкина 1. MYD88 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_001172567.1, NM_002468.4, NM_001172569.1, NM_001172568.1, NM_001172566.1, XM_005265172.1, XM_006713170.1 (NP_001166038.1, NP_002459.2, NP_001166040.1, NP_001166039.1, NP_001166037.1, XP_005265229.1, XP_006713233.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена MYD88, зарегистрированного в базе данных под номером NM_001172567.1, описана в SEQ ID NO: 87, а аминокислотная последовательность человеческого белка MYD88, кодируемого указанным человеческим геном MYD88, описана в SEQ ID NO: 88. В настоящем описании MYD88 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 88, кодируемую нуклеотидной последовательностью SEQ ID NO: 87. Информация о последовательности MYD88 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 87 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
NFKB1, B-клеточный ядерный фактор энхансер гена легкой цепи каппа 1, представляет собой белок размером 105 кДа, который процессируется в белок размером 50 кДа в процессе трансляции протеосомой 26S. Белок размером 105 кДа представляет собой ингибитор Rel-белок-специфической транскрипции, а белок размером 50 кДа представляет собой ДНК-связывающую субъединицу белкового комплекса NFκ-B (NFKB). NFKB1 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_003998.3, NM_001165412.1, XM_011532006.1, XM_011532007.1, XM_011532008.1, XM_011532009.1 (NP_003989.2, NP_001158884.1, XP_011530308.1, XP_011530309.1, XP_011530310.1, XP_011530311.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена NFKB1, зарегистрированного в базе данных под номером NM_003998.3, описана в SEQ ID NO: 89, а аминокислотная последовательность человеческого белка NFKB1, кодируемого указанным человеческим геном NFKB1, описана в SEQ ID NO: 90. В настоящем описании NFKB1 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 90, кодируемую нуклеотидной последовательностью SEQ ID NO: 89. Информация о последовательности NFKB1 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 89 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
PIK3CG, каталитическая субъединица гамма фосфатидилинозитол-4,5-бифосфат-3-киназы, как известно, образует PI3K путем связывания с регуляторной субъединицей p85, как и другие каталитические субъединицы класса 1 (p110-α, p110-β и p110-δ). PIK3CG присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_001282426.1, NM_001282427.1, NM_002649.3, XM_011516316.1, XM_011516317.1, XM_005250443.2 (NP_001269355.1, NP_001269356.1, NP_002640.2, XP_011514618.1, XP_011514619.1, XP_005250500.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена PIK3CG, зарегистрированного в базе данных под номером NM_001282426.1, описана в SEQ ID NO: 91, а аминокислотная последовательность человеческого белка PIK3CG, кодируемого указанным человеческим геном PIK3CG, описана в SEQ ID NO: 92. В настоящем описании PIK3CG не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 92, кодируемую нуклеотидной последовательностью SEQ ID NO: 91. Информация о последовательности PIK3CG зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 91 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
RAC1, ras-связанный субстрат 1 ботулинического токсина C3 (семейство rho, малый ГТФ-связывающий белок Rac1), известен как ГТФ-аза, принадлежащая к суперсемейству RAS в виде малого ГТФ-связывающего белка. Факторы, относящиеся к указанному суперсемейству, регулируют разные внутриклеточные события, такие как рост, цитоскелетная реорганизация и активация протеинкиназ. RAC1 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_018890.3, NM_006908.4 (NP_061485.1, NP_008839.2) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена RAC1, зарегистрированного в базе данных под номером NM_018890.3, описана в SEQ ID NO: 93, а аминокислотная последовательность человеческого белка RAC1, кодируемого указанным человеческим геном RAC1, описана в SEQ ID NO: 94. В настоящем описании RAC1 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 94, кодируемую нуклеотидной последовательностью SEQ ID NO: 93. Информация о последовательности RAC1 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 93 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
AKT3, гомолог 3 вирусного онкогена мышиной тимомы v-akt, также называемый PKB и известный как член семейства серинтреонпротеинкиназ AKT. Киназы AKT известны как факторы, регулирующие сигнальную систему клеток, которые отвечают на инсулин и факторы роста. Указанные киназы участвуют в разных биологических процессах, включающих в себя синтез гликогена и поглощение глюкозы, во время роста, дифференцировки, апоптоза и опухолегенеза клеток. Обнаружено, что указанная киназа стимулируется под действием тромбоцитарного фактора роста (PDGF), инсулина и инсулиноподобного фактора роста 1 (IGF1). AKT3 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_181690.2, NM_001206729.1, NM_005465.4, XM_005272995.2, XM_011544011.1, XM_005272994.3, XM_005272997.3, XM_011544014.1, XM_011544012.1, XM_011544013.1, XM_006711726.2 (NP_859029.1, NP_001193658.1, NP_005456.1, XP_005273052.1, XP_011542313.1, XP_005273051.1, XP_005273054.1, XP_011542316.1, XP_011542314.1, XP_011542315.1, XP_006711789.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена AKT3, зарегистрированного в базе данных под номером NM_181690.2, описана в SEQ ID NO: 95, а аминокислотная последовательность человеческого белка AKT3, кодируемого указанным человеческим геном AKT3, описана в SEQ ID NO: 96. В настоящем описании AKT3 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 96, кодируемую нуклеотидной последовательностью SEQ ID NO: 95. Информация о последовательности AKT3 зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 95 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
EIF4B, фактор инициации трансляции эукариот 4B, известен как белок, который участвует в сигнальном пути PI3K-Akt, передавая сигналы посредством GPCR. EIF4B необходим для связывания мРНК с рибосомой и имеет функции, тесно связанные с EIF4-F и EIF4-A. Известно, что EIF4B связывается с 5'-концевым кэпом мРНК, или с соседним участком, в присутствии EIF4-F и АТФ, стимулируя АТФ-азную активность и АТФ-зависимую РНК-раскручивающую активность EIF4-A и EIF4-F. EIF4B присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_001300821.1, NM_001417.5, XM_006719274.1 (NP_001287750.1, NP_001408.2, XP_006719337.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена EIF4B, зарегистрированного в базе данных под номером NM_001300821.1, описана в SEQ ID NO: 97, а аминокислотная последовательность человеческого белка EIF4B, кодируемого указанным человеческим геном EIF4B, описана в SEQ ID NO: 98. В настоящем описании EIF4B не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 98, кодируемую нуклеотидной последовательностью SEQ ID NO: 97. Информация о последовательности EIF4B зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 97 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
EIF4E, фактор инициации трансляции эукариот 4E, известен как белок, который является составной частью комплекса эукариотического фактора инициации трансляции 4F, и который распознает 7-метилгуанозиновый кэп на 5'-конце мРНК. EIF4E способствует инициации трансляции путем привлечения рибосом к 5'-концевой кэпированной структуре. Связывание EIF4E с комплексом 4F является скорость-ограничивающей стадией инициации трансляции. EIF4E присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_001968.3, NM_001130679.1, NM_001130678.1, XM_006714126.2, XM_006714127.2 (NP_001959.1, NP_001124151.1, NP_001124150.1, XP_006714189.1, XP_006714190.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена EIF4E, зарегистрированного в базе данных под номером NM_001968.3, описана в SEQ ID NO: 99, а аминокислотная последовательность человеческого белка EIF4E, кодируемого указанным человеческим геном EIF4E, описана в SEQ ID NO: 100. В настоящем описании EIF4E не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 100, кодируемую нуклеотидной последовательностью SEQ ID NO: 99. Информация о последовательности EIF4E зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 99 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
ILK, интегрин-связанная киназа, представляет собой белок, который содержит киназоподобный домен и четыре анкириноподобных повтора и, как известно, контролирует передачу сигнала, опосредованную интегрином, путем связывания с цитоплазматическим доменом β-интегрина в клеточной мембране. ILK присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_004517.3, NM_001278441.1, NM_001014794.2, NM_001278442.1, NM_001014795.2, XM_005252904.3, XM_005252905.1, XM_011520065.1 (NP_004508.1, NP_001265370.1, NP_001014794.1, NP_001265371.1, NP_001014795.1, XP_005252961.1, XP_005252962.1, XP_011518367.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена ILK, зарегистрированного в базе данных под номером NM_004517.3, описана в SEQ ID NO: 101, а аминокислотная последовательность человеческого белка ILK, кодируемого указанным человеческим геном ILK, описана в SEQ ID NO: 102. В настоящем описании ILK не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 102, кодируемую нуклеотидной последовательностью SEQ ID NO: 101. Информация о последовательности ILK зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 101 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
MTCP1, фактор пролиферации зрелых Т-клеток 1, был идентифицирован, как известно, по участию в некоторых транслокациях t(X;14), связанных с ростом зрелых Т-клеток. Данный участок имеет сложную структуру с одним промотором и 5'-экзонами, сплайсирующимися на два разных набора 3'-экзонов, которые кодируют два разных белка. Вышестоящий по ходу считывания белок размером 13 кДа, который относят к семейству TCL1, участвует в индукции лейкоза. MTCP1 присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_001018025.3 (NP_001018025.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена MTCP1, зарегистрированного в базе данных под номером NM_001018025.3, описана в SEQ ID NO: 103, а аминокислотная последовательность человеческого белка MTCP1, кодируемого указанным человеческим геном MTCP1, описана в SEQ ID NO: 104. В настоящем описании MTCP1 не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 104, кодируемую нуклеотидной последовательностью SEQ ID NO: 103. Может существовать несколько транскрипционных вариантов MTCP1. Нуклеотидная последовательность SEQ ID NO: 103 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
PIK3CA, каталитическая субъединица альфа фосфатидилинозитол-4,5-бисфосфат-3-киназы, представляет собой каталитическую субъединицу фосфатидилинозитол-3-киназы размером 110 кДа, которая фосфорилирует PtdIns, PtdIns4P и PtdIns(4,5)P2 с использованием АТФ. PIK3CA присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_006218.2, XM_006713658.2, XM_011512894.1 (NP_006209.2, XP_006713721.1, XP_011511196.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена PIK3CA, зарегистрированного в базе данных под номером NM_006218.2, описана в SEQ ID NO: 105, а аминокислотная последовательность человеческого белка PIK3CA, кодируемого указанным человеческим геном PIK3CA, описана в SEQ ID NO: 106. В настоящем описании PIK3CA не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 106, кодируемую нуклеотидной последовательностью SEQ ID NO: 105. Информация о последовательности PIK3CA зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 105 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
SRF, фактор ответа сыворотки, известен как широко распространенный ядерный белок, который стимулирует рост и дифференциацию клеток. SRF относится к бокс-суперсемейству транскрипционных факторов MADS (MCM1, Agamous, Deficiens, SRF). SRF связывается с элементом ответа сыворотки (SRE) в промоторном участке гена-мишени. SRF регулирует активацию многих немедленно-ранних генов, таких как c-fos, и тем самым участвует в регуляции клеточного цикла, апоптоза, клеточного роста и дифференциации клеток. SRF присутствует у разных животных, в том числе у людей, а информация о его последовательности находится в открытом доступе (например, для человеческого: NM_003131.3, NM_001292001.1 (NP_003122.1, NP_001278930.1) и др.; указанные номера представляют собой номера доступа в базе данных NCBI, нуклеотидные последовательности и аминокислотные последовательности указаны вне и внутри скобок, соответственно). В качестве одного примера нуклеотидная последовательность человеческого гена SRF, зарегистрированного в базе данных под номером NM_003131.3, описана в SEQ ID NO: 107, а аминокислотная последовательность человеческого белка SRF, кодируемого указанным человеческим геном SRF, описана в SEQ ID NO: 108. В настоящем описании SRF не ограничивается белком, имеющим аминокислотную последовательность SEQ ID NO: 108, кодируемую нуклеотидной последовательностью SEQ ID NO: 107. Информация о последовательности SRF зарегистрирована под несколькими номерами доступа, как указано выше, поскольку существует несколько транскрипционных вариантов. Нуклеотидная последовательность SEQ ID NO: 107 представляет собой нуклеотидную последовательность одного из указанных транскрипционных вариантов.
Хотя GST-π, ATM, CDC25A, p21, PRKDC, RBBP8, SKP2, MCM10, RNPC1, CCNL1, CENPH, BRSK1, MYLK, MCM8, CCNB3, MCMDC1, AATF, AKT1, ALOX12, ANXA1, ANXA4, API5, ATF5, AVEN, AZU1, BAG1, BCL2L1, BFAR, CFLAR, IL2, MALT1, MCL1, MKL1, MPO, MTL5, MYBL2, MYO18A, MTOR, IRAK1, IRS1, MYD88, NFKB1, PIK3CG, RAC1, AKT3, EIF4B, EIF4E, ILK, MTCP1, PIK3CA и SRF можно идентифицировать по указанным выше специфическим нуклеотидным и аминокислотным последовательностям, следует иметь в виду, что нуклеотидные или аминокислотные последовательности отдельных организмов могут содержать мутации.
А именно, GST-π, ATM, CDC25A, p21, PRKDC, RBBP8, SKP2, MCM10, RNPC1, CCNL1, CENPH, BRSK1, MYLK, MCM8, CCNB3, MCMDC1, AATF, AKT1, ALOX12, ANXA1, ANXA4, API5, ATF5, AVEN, AZU1, BAG1, BCL2L1, BFAR, CFLAR, IL2, MALT1, MCL1, MKL1, MPO, MTL5, MYBL2, MYO18A, MTOR, IRAK1, IRS1, MYD88, NFKB1, PIK3CG, RAC1, AKT3, EIF4B, EIF4E, ILK, MTCP1, PIK3CA и SRF не ограничиваются белками, имеющими аминокислотные последовательности, зарегистрированные в базе данных, и включают в себя белки, последовательности которых отличаются от указанных последовательностей на 1, 2 или более аминокислот, как правило, на 1 или несколько аминокислот, например, на 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислот, и которые имеют функции, эквивалентные функциям GST-π, ATM, CDC25A, p21, PRKDC, RBBP8, SKP2, MCM10, RNPC1, CCNL1, CENPH, BRSK1, MYLK, MCM8, CCNB3, MCMDC1, AATF, AKT1, ALOX12, ANXA1, ANXA4, API5, ATF5, AVEN, AZU1, BAG1, BCL2L1, BFAR, CFLAR, IL2, MALT1, MCL1, MKL1, MPO, MTL5, MYBL2, MYO18A, MTOR, IRAK1, IRS1, MYD88, NFKB1, PIK3CG, RAC1, AKT3, EIF4B, EIF4E, ILK, MTCP1, PIK3CA и SRF.
Кроме того, нуклеотидные последовательности GST-π, ATM, CDC25A, p21, PRKDC, RBBP8, SKP2, MCM10, RNPC1, CCNL1, CENPH, BRSK1, MYLK, MCM8, CCNB3, MCMDC1, AATF, AKT1, ALOX12, ANXA1, ANXA4, API5, ATF5, AVEN, AZU1, BAG1, BCL2L1, BFAR, CFLAR, IL2, MALT1, MCL1, MKL1, MPO, MTL5, MYBL2, MYO18A, MTOR, IRAK1, IRS1, MYD88, NFKB1, PIK3CG, RAC1, AKT3, EIF4B, EIF4E, ILK, MTCP1, PIK3CA и SRF могут быть на 70% или более, на 80% или более, на 90% или более, на 95% или более, или на 97% или более идентичными указанным выше специфическим нуклеотидным последовательностям, и могут кодировать белки, функции которых эквивалентны функциям GST-π, ATM, CDC25A, p21, PRKDC, RBBP8, SKP2, MCM10, RNPC1, CCNL1, CENPH, BRSK1, MYLK, MCM8, CCNB3, MCMDC1, AATF, AKT1, ALOX12, ANXA1, ANXA4, API5, ATF5, AVEN, AZU1, BAG1, BCL2L1, BFAR, CFLAR, IL2, MALT1, MCL1, MKL1, MPO, MTL5, MYBL2, MYO18A, MTOR, IRAK1, IRS1, MYD88, NFKB1, PIK3CG, RAC1, AKT3, EIF4B, EIF4E, ILK, MTCP1, PIK3CA и SRF.
Специфические функции GST-π, ATM, CDC25A, p21, PRKDC, RBBP8, SKP2, MCM10, RNPC1, CCNL1, CENPH, BRSK1, MYLK, MCM8, CCNB3, MCMDC1, AATF, AKT1, ALOX12, ANXA1, ANXA4, API5, ATF5, AVEN, AZU1, BAG1, BCL2L1, BFAR, CFLAR, IL2, MALT1, MCL1, MKL1, MPO, MTL5, MYBL2, MYO18A, MTOR, IRAK1, IRS1, MYD88, NFKB1, PIK3CG, RAC1, AKT3, EIF4B, EIF4E, ILK, MTCP1, PIK3CA и SRF описаны выше.
Если не указано иначе, в настоящем описании такие фразы, как "используемый в данном документе", "используемый здесь", "в настоящем описании" и "описанный здесь", означают, что описание, сопровождающее данные термины, применяется к всем аспектам изобретения, описанным в настоящем документе. Кроме того, если не указано иначе, все используемые здесь технические и научные термины имеют традиционные значения, известные специалистам в данной области. Все патенты, информационные сообщения и другие публикации, цитируемые в настоящем документе, включены в него в качестве ссылки во всей их полноте.
Используемый здесь термин "средство, ингибирующее GST-π" не ограничивается и включает в себя, например, лекарственные средства, ингибирующие продукцию и/или активность GST-π, а также лекарственные средства, стимулирующие деградацию и/или дезактивацию GST-π. Примеры средств, ингибирующих продукцию GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей GST-π, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы.
Альтернативно в качестве средства, ингибирующего GST-π, можно использовать любое соединение, способное воздействовать на GST-π. В качестве такого соединения можно использовать органическое соединение (такое как аминокислота, полипептид или его производное, низкомолекулярное соединение, сахар, высокомолекулярное соединение и др.), неорганическое соединение и т. п. Кроме того, такое соединение может представлять собой любое природное и неприродное вещество. Примеры производного полипептида включают в себя модифицированные полипептиды, полученные путем добавления модифицирующих групп, и варианты полипептидов, полученные путем изменения аминокислотных остатков. Кроме того, такое соединение может представлять собой одно соединение, или оно может представлять собой библиотеку соединений, продукт экспрессии библиотеки генов, клеточный экстракт, супернатант клеточной культуры, продукт ферментации микроорганизма, экстракт морского организма, растительный экстракт и т.п. То есть термин "средство, ингибирующее GST-π" не ограничивается нуклеиновыми кислотами, такими как молекулы РНКи, и включает в себя любые соединения.
А именно, примеры лекарственного средства, ингибирующего активность GST-π, включают в себя, без ограничения, соединения, связывающиеся с GST-π, такие как глутатион, аналоги глутатиона (например, описанные в WO 95/08563, WO 96/40205, WO 99/54346, непатентном документе 4 и др.), кетопрофен (непатентный документ 2), индометацин (Hall et al., Cancer Res., 1989, 49 (22): 6265-8), этакриновую кислоту, пирипрост (Tew et al., Cancer Res. 1988, 48 (13): 3622-5), антитела против GST-π и доминантно-негативные мутанты GST-π. Указанные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Предпочтительными средствами, ингибирующими продукцию или активность GST-π, являются молекулы РНКи против ДНК, кодирующей GST-π, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, поскольку они характеризуются высокой специфичностью и низкой вероятностью побочных реакций.
Ингибирование GST-π можно определить по уменьшению экспрессии или активности GST-π в клетке по сравнению с клеткой, на которую не действует средство, ингибирующее GST-π. Экспрессию GST-π можно оценить с помощью любого известного метода, включающего в себя, без ограничения, метод иммунопреципитации с использованием антитела против GST-π, EIA, ELISA, IRA, IRMA, вестерн-блоттинг, иммуногистохимический анализ, иммуноцитохимический анализ, проточную цитометрию или разные способы гибридизации, нозерн-блоттинг, саузерн-блоттинг или разные ПЦР-методы с использованием нуклеиновых кислот, специфически гибридизующихся с нуклеиновой кислотой, кодирующей GST-π, или ее уникальным фрагментом, или транскриптом (например, мРНК) или продуктом сплайсинга нуклеиновой кислоты.
Кроме того, активность GST-π можно определить, без ограничения, путем анализа известной активности GST-π, например, его способности связывать белок, такой как Raf-1 (в частности, фосфорилированный Raf-1) или EGFR (в частности, фосфорилированный EGFR), с помощью любого известного метода, такого как метод иммунопреципитации, вестерн-блоттинг, масс-спектрометрия, метод аффинной адсорбции или метод поверхностного плазмонного резонанса (SPR).
В настоящее описании термин "средство, ингибирующее связанный с гомеостазом белок и вызывающее синтетическую летальность при ингибировании наряду с GST-π", не ограничивается и включает в себя, например, средства ингибирующие продукцию и/или активность белков и лекарственных средств, стимулирующих деградацию и/или дезактивацию белка. Примеры средств, ингибирующих продукцию белка, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей связанный с гомеостазом белок, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы. Альтернативно, в качестве средства, ингибирующего активность связанного с гомеостазом белка, или средства, стимулирующего деградацию и/или дезактивацию связанного с гомеостазом белка, можно использовать любое соединение, способное воздействовать на белок. В качестве такого соединения можно использовать органическое соединение (такое как аминокислота, полипептид или его производное, низкомолекулярное соединение, сахар, высокомолекулярное соединение и др.), неорганическое соединение и т. п. Кроме того, такое соединение может представлять собой любое природное и неприродное вещество. Примеры производного полипептида включают в себя модифицированные полипептиды, полученные путем добавления модифицирующих групп, и варианты полипептидов, полученные путем изменения аминокислотных остатков. Кроме того, такое соединение может представлять собой одно соединение, или оно может представлять собой библиотеку соединений, продукт экспрессии библиотеки генов, клеточный экстракт, супернатант клеточной культуры, продукт ферментации микроорганизма, экстракт морского организма, растительный экстракт и т.п. То есть термин "средство, ингибирующее связанный с гомеостазом белок, и вызывающее синтетическую летальность при ингибировании наряду с GST-π" не ограничивается нуклеиновыми кислотами, такими как молекулы РНКи, и включает в себя любые соединения.
Более конкретно, примеры средств, ингибирующих активность p21, относящегося к белкам, регулирующим клеточный цикл, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения: бутиролактон I (Sax et al., Cell Cycle, Jan; 1 (1): 90-6,2002), который представляет собой низкомолекулярное соединение, стимулирующее протеосомальную деградацию белка p21, и в то же время ингибирующее ферментативную активность CDC2, CDK2 и CDK5; кветиапин (Kondo et al., Transl. Psychiatry, Apr 2; 3: e243, 2013), который представляет собой психотропное лекарственное средство, согласно полученным данным, специфически ингибирующее экспрессию p21 в нервных клетках, или олигодендроцитах мышей CD-1; сорафениб (Inoue et al., Cancer Biology & Therapy, 12: 9, 827-836, 2011), который представляет собой низкомолекулярное соединение, специфически ингибирующее p21 без участия p53, p27 или Akt, и мультикиназу Raf, VEGFR, PDGFR и т.п.; UC2288 (Wettersten et al., Cancer Biology & Therapy, 14 (3), 278-285, 2013), который представляет собой низкомолекулярное соединение, специфически ингибирующее p21 без участия p53 или Akt; а также молекулы РНКи, направленные против ДНК, кодирующей p21, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против p21, и доминантно-негативные варианты p21. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность RNPC1, относящегося к белкам, регулирующим клеточный цикл, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей RNPC1, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против RNPC1 и доминантно-негативные варианты RNPC1. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность CCNL1, относящегося к белкам, регулирующим клеточный цикл, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей CCNL1, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против CCNL1 и доминантно-негативные варианты CCNL1. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность MCM8, относящегося к белкам, регулирующим клеточный цикл, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей MCM8, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против MCM8, и доминантно-негативные варианты MCM8. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность CCNB3, относящегося к белкам, регулирующим клеточный цикл, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей CCNB3, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против CCNB3 и доминантно-негативные варианты CCNB3. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность MCMDC1, относящегося к белкам, регулирующим клеточный цикл, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей MCMDC1, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против MCMDC1, и доминантно-негативные варианты MCMDC1. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность ATM, относящегося к белкам, регулирующим клеточный цикл, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения: CGK 733 (Won et al., Nat. Chem. Biol. 2, 369, 2006), который представляет собой низкомолекулярное соединение, селективно ингибирующее киназную активность ATM и ATR; KU-55933 (Lau et al., Nat. Cell Biol. 7, 493, 2005), KU-60019 (Zirkin et al., J Biol Chem. Jul 26; 288 (30): 21770-83, 2013) и CP-466722 (Rainey et al., Cancer Res. Sep 15; 68 (18): 7466-74, 2008), которые представляют собой низкомолекулярные соединения, селективно ингибирующие киназную активность ATM; а также молекулы РНКи, направленные против ДНК, кодирующей ATM, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против ATM и доминантно-негативные варианты ATM. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность CDC25A, относящегося к белкам, регулирующим клеточный цикл, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения: NSC95397 (Lazo JS et al., Mol. Pharmacol. 61: 720-728, 2002), который представляет собой низкомолекулярное соединение, ингибирующее фосфатазную активность каждого из человеческого CDC25A, человеческого CDC25B и человеческого CDC25C; SC альфа альфа дельта 09 (Rice, R.L. et al., Biochemistry 36 (50): 15965-15974, 1997), который представляет собой низкомолекулярное соединение, ингибирующее фосфатазную активность каждого из человеческого CDC25A, человеческого CDC25B, человеческого CDC25C и человеческой тирозинфосфатазы PTB1B; а также молекулы РНКи, направленные против ДНК, кодирующей CDC25A, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против CDC25A и доминантно-негативные варианты CDC25A. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность PRKDC, относящегося к белкам, регулирующим клеточный цикл, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей PRKDC, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против PRKDC и доминантно-негативные варианты PRKDC. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность RBBP8, относящегося к белкам, регулирующим клеточный цикл, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей RBBP8, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против RBBP8 и доминантно-негативные варианты RBBP8. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность SKP2, относящегося к белкам, регулирующим клеточный цикл, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей SKP2, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против SKP2 и доминантно-негативные варианты SKP2. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность MCM10, относящегося к белкам, регулирующим клеточный цикл, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей MCM10, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против MCM10 и доминантно-негативные варианты MCM10. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность CENPH, относящегося к белкам, регулирующим клеточный цикл, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей CENPH, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против CENPH и доминантно-негативные варианты CENPH. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность BRSK1, относящегося к белкам, регулирующим клеточный цикл и вызывающих синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей BRSK1, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против BRSK1 и доминантно-негативные варианты BRSK1. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность MYLK, относящегося к белкам, регулирующим клеточный цикл, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, HA-100 дигидрохлорид (Gerard et al., J Clin Invest. Jan; 77 (1): 61-5. 1986), который представляет собой низкомолекулярное соединение, ингибирующее активность MYLK помимо ингибирования активности разных протеинкиназы (PKA и PKC и др.); стауроспорин (Tamaoki et al., Biochem. Biophys. Res. Commun. 135: 397-402. 1986), который представляет собой низкомолекулярное соединение, ингибирующее активность MYLK помимо ингибирования активности разных протеинкиназ (PKA, PKC, PKG, CaMK, CaMKII, киназа фосфорилазы и др.); кальфостин C (Kobayashi et al., Biochem. Biophys. Res. Commun. 159: 548-553. 1989), который представляет собой низкомолекулярное соединение, являющееся селективным ингибитором PKC и, кроме того, ингибирующее активность MYLK; писетаннол (Oliver et al., J. Biol. Chem. 269: 29697-29703. 1994), который представляет собой низкомолекулярное соединение, являющееся ингибитором тирозинкиназы и, кроме того, ингибирующее активность MYLK; A-3 гидрохлорид (Inagaki et al., Mol. Pharmacol. 29, 577. 1986), который представляет собой низкомолекулярное соединение, ингибирующее активность MYLK помимо ингибирования активности разных казеинкиназ и протеинкиназ; H-7 дигидрохлорид (Kawamoto et al., Biochem. Biophys. Res. Commun. 125: 258-264. 1984), который представляет собой низкомолекулярное соединение, являющееся ингибитором способной проникать в клетку серинтреонинкиназы и, кроме того, ингибирующее активность MYLK; H-9 гидрохлорид (Wolf et al., J. Biol. Chem. 260: 15718-15722. 1985), который представляет собой низкомолекулярное соединение, ингибирующее активность MYLK помимо ингибирования активности разных протеинкиназы (PKA, PKC и др.); W-5 (Hidaka et al., Mol. Pharmacol. 20: 571-578. 1981), который представляет собой низкомолекулярное соединение, являющееся антагонистом кальмодулина (CaM) и, кроме того, ингибирующее активность MYLK; W-7 (Hidaka et al., Proc. Natl. Acad. Sci. U.S.A. 78: 4354-4357. 1981), который представляет собой низкомолекулярное соединение, являющееся антагонистом кальмодулина (CaM) и, кроме того, ингибирующее активность MYLK; гидрохлорид изомера W-13 (Hidaka et al., Proc. Natl. Acad. Sci. U.S.A. 78: 4354-4357. 1981), который представляет собой низкомолекулярное соединение, ингибирующее активность MYLK помимо ингибирования активности разных протеинкиназ (PKA, PKC и др.); ML-7 дигидрохлорид (Saitoh et al., J. Biol. Chem. 262: 7796-7801. 1987), который представляет собой низкомолекулярное соединение, ингибирующее активность MYLK помимо ингибирования активности разных протеинкиназ (PKA, PKC и др.); ML-9 (Saitoh et al., Biochem. Biophys. Res. Commun. 140: 280-287. 1986), который представляет собой низкомолекулярное соединение, селективно ингибирующее MYLK и CaMK; мирисетин (Hagiwara et al., Biochem. Pharmacol. 37: 2987-2992. 1988), который представляет собой низкомолекулярное соединение, ингибирующее активность MYLK помимо ингибирования активности разных казеинкиназ и протеинкиназ; E6 бербамин (Hu et al., Biochem. Pharmacol. Oct 20; 44 (8): 1543-7. 1992), который представляет собой низкомолекулярное соединение, являющееся антагонистом кальмодулина (CaM), способным проникать в клетку, и, кроме того, ингибирующее активность MYLK; K-252a (Kase et al., J. Antibiot. 39: 1059-1065. 1986), который представляет собой низкомолекулярное соединение, ингибирующее активность MYLK помимо ингибирования активности разных протеинкиназ; K-252b (Nakanishi et al., J. Antibiot. 39: 1066-1071. 1986), который представляет собой низкомолекулярное соединение, ингибирующее активность MYLK помимо ингибирования активности разных протеинкиназ; альтенусин (Rosett et al., Biochem. J. 67: 390-400. 1957), который представляет собой низкомолекулярное соединение, являющееся ингибитором c-Src и, кроме того, ингибирующее активность MYLK; а также молекулы РНКи, направленные против ДНК, кодирующей MYLK, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против MYLK и доминантно-негативные варианты MYLK. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
С другой стороны, примеры средств, ингибирующих активность AATF, относящегося к белкам, связанным с подавлением апоптоза, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей AATF, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против AATF, и доминантно-негативные варианты AATF. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность ALOX12, относящегося к белкам, связанным с подавлением апоптоза, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей ALOX12, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против ALOX12 и доминантно-негативные варианты ALOX12. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность ANXA1, относящегося к белкам, связанным с подавлением апоптоза, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей ANXA1, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против ANXA1 и доминантно-негативные варианты ANXA1. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность ANXA4, относящегося к белкам, связанным с подавлением апоптоза, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей ANXA4, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против ANXA4 и доминантно-негативные варианты ANXA4. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность API5, относящегося к белкам, связанным с подавлением апоптоза, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей API5, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против API5 и доминантно-негативные варианты API5. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность ATF5, относящегося к белкам, связанным с подавлением апоптоза, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей ATF5, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против ATF5, и доминантно-негативные варианты ATF5. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность AVEN, относящегося к белкам, связанным с подавлением апоптоза, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей AVEN, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против AVEN, и доминантно-негативные варианты AVEN. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность AZU1, относящегося к белкам, связанным с подавлением апоптоза, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей AZU1, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против AZU1 и доминантно-негативные варианты AZU1. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность BAG1, относящегося к белкам, связанным с подавлением апоптоза, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей BAG1, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против BAG1, и доминантно-негативные варианты BAG1. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность BCL2L1, относящегося к белкам, связанным с подавлением апоптоза, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей BCL2L1, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против BCL2L1 и доминантно-негативные варианты BCL2L1. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность BFAR, относящегося к белкам, связанным с подавлением апоптоза, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей BFAR, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против BFAR и доминантно-негативные варианты BFAR. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность CFLAR, относящегося к белкам, связанным с подавлением апоптоза, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей CFLAR, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против CFLAR и доминантно-негативные варианты CFLAR. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность IL2, относящегося к белкам, связанным с подавлением апоптоза, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей IL2, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против IL2 и доминантно-негативные варианты IL2. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность MALT1, относящегося к белкам, связанным с подавлением апоптоза, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей MALT1, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против MALT1 и доминантно-негативные варианты MALT1. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность MCL1, относящегося к белкам, связанным с подавлением апоптоза, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, синрибо (омацетаксина мепесукцинат), разрешенный к применению в качестве терапевтического средства против хронической миелоцитарной лейкемии, молекулы РНКи, направленные против ДНК, кодирующей MCL1, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против MCL1 и доминантно-негативные варианты MCL1. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность MKL1, относящегося к белкам, связанным с подавлением апоптоза, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей MKL1, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против MKL1 и доминантно-негативные варианты MKL1. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность MPO, относящегося к белкам, связанным с подавлением апоптоза, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей MPO, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против MPO и доминантно-негативные варианты MPO. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность MTL5, относящегося к белкам, связанным с подавлением апоптоза, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей MTL5, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против MTL5 и доминантно-негативные варианты MTL5. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность MYBL2, относящегося к белкам, связанным с подавлением апоптоза, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей MYBL2, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против MYBL2 и доминантно-негативные варианты MYBL2. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность MYO18A, относящегося к белкам, связанным с подавлением апоптоза, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей MYO18A, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против MYO18A и доминантно-негативные варианты MYO18A. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
С другой стороны, примеры средств, ингибирующих активность MTOR, относящегося к белкам, связанным с сигнальным путем PI3K, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, рапамицин (Chang et al., Trends Pharmacol. Sci. 12: 218-223. 1991), который представляет собой низкомолекулярное соединение, являющееся макролидным иммуносупрессором и ингибирующее активность MTOR; эверолимус (Weisblum et al., Br. Med. Bull. 40: 47-53. 1984), который представляет собой низкомолекулярное соединение, являющееся макролидным иммуносупрессором, производным рапамицина, и ингибирующее активность MTOR; BEZ235 (Maira et al., Mol. Cancer Ther. 7 (7): 1851-1863. 2008), который представляет собой низкомолекулярное соединение, являющееся ингибитором тирозинкиназы PI3K и ингибирующее активность MTOR; AZD8055 (Huang et al., J Biol Chem. Nov 18; 286 (46): 40002-12. 2011), который представляет собой низкомолекулярное соединение, селективно ингибирующее киназную активность MTOR; PI-103 (Demyanets et al., 2010. Basic Res Cardiol. Mar; 106 (2): 217-31. 2011), который представляет собой производное пиридинилфуранопиримидина и является низкомолекулярным соединением, ингибирующим киназную активность DNA-PK, PI3K и MTOR; а также никлосамид (Balgi et al., PLoS One. Sep 22; 4 (9): e7124. 2009), PP242 (Bao et al. J. Cell Biol. 210 (7): 1153-64. 2015), тимосапонин AIII (King et.al, PLoS One. Sep 30;4 (9):e7283 2009), KU 0063794 (Garcia-Martinez et al. Biochem. J. 421 (1): 29-42. 2009), AZD2014 (Guichard et al. Mol. Cancer. Ther. 14 (11): 2508-18. 2015), темсиролимус (Raymond et al. J. Clin. Oncol. 22: 2336-2347. 2004), паломид 529 (Xue et al. Cancer Res. 68 (22): 9551-7. 2008), фисетин (Suh et.al, Carcinogenesis. Aug;31 (8):1424-33 2010), GDC-0980 (Wallin et al. Mol. Cancer Ther. 10 (12): 2426-36. 2011), SF1126 (Garlich et al. Cancer Res. 68 (1): 206-15. 2008), CH5132799 (Tanaka et al. Clin Cancer Res. 17 (10): 3272-81. 2011), WYE-354 (Yu et al. Cancer Res. 69 (15): 6232-40. 2009), соединение 401 (Ballou et al. J. Biol. Chem. 282, 24463. 2007), ридафоролимус (Gadducci et al.: Gynecol. Endocrinol. 24, 239. 2008), GSK 1059615 (Steven et al. ACS. Med. Chem. Lett. 1 (1): 39-43 2010), PF-04691502 (Yuan et al. Mol. Cancer Ther. 10 (11): 2189-99. 2011), PP121 (Apsel et al. Nat. Chem. Biol. 4 (11): 691-9. 2008), OSI-027 (Bhaqwat et al. Mol. Cancer Ther. 10 (8): 1394-406. 2011), WYE-125132 (Yu et al. Cancer Res. 70 (2): 621-31. 2010), умиролимус (Baldo et al. 2008. Curr. Cancer Drug Targets. 8 (8): 647-65. 2008), WAY-600 (Yu et al. Cancer Res. 69 (15): 6232-40. 2009), WYE-687 (Yu et al. Cancer Res. 69 (15): 6232-40. 2009), PKI-179 (Venkatesan et al. Bioorg. Med. Chem. Lett. 20 (19): 5869-73. 2010), PF-05212384 (Akintunde et al. J. Hematol. Oncol. 6: 88. 2013), CAY10626 (Rameh et al. J. Biol. Chem. 274: 8347-8350. 1999), NVP-BGT226 (Chang et al. Clin. Cancer Res. 17 (22): 7116-26. 2011), производное 1 XL-147 (Akintunde et al. J. Hematol. Oncol. 6: 88. 2013), XL 388 (Eduardo et al. Mol Cancer Ther. 10 (3): 395-403. 2011), торин 1 (Liu et al. J. Med. Chem. 53 (19): 7146-55. 2010) и молекулы РНКи, направленные против ДНК, кодирующей MTOR, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против MTOR и доминантно-негативные варианты MTOR. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность IRAK1, относящегося к белкам, связанным с сигнальным путем PI3K, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, киназу 1, ассоциированную с рецептором интерлейкина 1/ингибитор 4 (Bhattacharyya et al., Am. J. Physiol. Gastrointest. Liver Physiol. 293, G429. 2007), которая представляет собой производное бензимидазола и является низкомолекулярным соединением, специфически ингибирующим активность киназы IL-1; а также молекулы РНКи, направленные против ДНК, кодирующей IRAK1, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против IRAK1, и доминантно-негативные варианты IRAK1. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность IRS1, относящегося к белкам, связанным с сигнальным путем PI3K, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей IRS1, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против IRS1 и доминантно-негативные варианты IRS1. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность MYD88, относящегося к белкам, связанным с сигнальным путем PI3K, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, пептид, ингибирующий MyD88, Pepinh-MYD (Derossi et al., J. Biol. Chem., 269: 10444-50. 1994), который представляет собой производное MYD88, пептид размером 26 аминокислот, ингибирующий гомодимеризацию; TJ-M2010 (Li et al., Transplant Proc. 45 (5): 1842-5. 2013), который представляет собой низкомолекулярное соединение, специфически ингибирующее MYD88; а также молекулы РНКи, направленные против ДНК, кодирующей MYD88, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против MYD88 и доминантно-негативные варианты MYD88. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность NFKB1, относящегося к белкам, связанным с сигнальным путем PI3K, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, BAY 11-7085 (Pierce et al., J. Biol. Chem. 272: 21096-21103. 1997), который представляет собой низкомолекулярное соединение ингибирующее как активацию NF-κB, так и фосфорилирование IκBα; геленалин (Lyss et al., J. Biol. Chem. 273 (50): 33508-16. 1998), который представляет собой низкомолекулярное соединение, используемое в качестве апоптоз-индуцирующего противоракового средства, ингибирующего NF-κB; фенетиловый эфир кофеиновой кислоты (Sud'ina et al., FEBS Lett. Aug 23; 329 (1-2): 21-4. 1993), который представляет собой низкомолекулярное соединение, являющееся ингибитором интегразы и тирозинкиназы HIV и специфически ингибирующее активность NF-κB; и ингибитор активации NFκB, JSH-23 (Shin et al., FEBS Lett. 571: 50-54. 2004), QNZ (Tobe et al. Bioorg. Med. Chem. 11: 3869-3878. 2003), андрографолид (Yu et al. Planta Med. Dec;69 (12):1075-9. 2003), куркумин (Asai et al. J. Nutr. 131: 2932-2935. 2001), аспирин (Kopp et al. Science. 265: 956-959. 1994), сульфасалазин (Liptay et al. Br. J. Pharmacol. 128, 1361. 1999), рокагламид (Engelmeier et al. J. Agric. Food Chem. 48: 1400-1404. 2000), SM 7368 (Lee et al. Biochem. Biophys. Res. Commun. 336: 716-722. 2005), сулиндака сульфид (Meade et al. J. Biol. Chem. 268: 6610-6614. 1993), триходион (Erkel et al. FEBS Lett. 477, 219. 2000), CHS-828 (Hovstadius et al. Clin. Cancer Res. 8 (9): 2843-50. 2002), Z-VRPR-FMK (Hailfinger et al. Proc. Natl. Acad. Sci. U.S.A. 106 (47): 19946-19951. 2009), салицилат натрия (Kopp et al. Science. 265: 956-959. 1994), 4-аминосалициловая кислота (Eberhardt et al. Biochem. Biophys. Res. Commun. 200: 163-170. 1994), этил 3,4-дигидроксициннамат (Nakayama et al. Biosci. Biotechnol. Biochem. 60: 316-318. 1996), CAY10512 (Heynekamp et al. J Med Chem 49 7182-7189 2006), N-стеароил фитосфингозин (Ryu et al. Lipids. 45 (7): 613-8. 2010), метиловый эфир пальмитиновой кислоты (Cai et al. Toxicology. 210: 197-204. 2005), 9-метилстрептимидон (Ishikawa et al. Bioorg. Med. Chem. Lett. 19 (6): 1726-8. 2009), рокаглаол (Engelmeier et al. J. Agric. Food Chem. 48: 1400-1404. 2000), BAY 11-7082 (Pierce et al. J. Biol. Chem. 272: 21096-21103. 1997), а также молекулы РНКи, направленные против ДНК, кодирующей NFKB1, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, против NFKB1 антитела, и доминантно-негативные варианты NFKB1. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность PIK3CG, относящегося к белкам, связанным с сигнальным путем PI3K, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, CUDC-907 (Qian et al., Clin Cancer Res. 18 (15): 4104-4113. 2012), который представляет собой низкомолекулярное соединение специфически ингибирующее активность как HDAC, так и PI3K; PKI-402 (Dehnhardt et al., J Med Chem. Jan 28; 53 (2): 798-810. 2010), который представляет собой низкомолекулярное соединение, специфически ингибирующее активность как PI3K, так и mTOR; PF-04691502 (Yuan et al., Mol Cancer Ther, 10 (11), 2189-2199. 2011), который представляет собой низкомолекулярное соединение, специфически ингибирующее активность как PI3K, так и mTOR; NVP-BGT226 (Glienke et al., Tumor Biology, 33 (3): 757-765. 2012), который представляет собой низкомолекулярное соединение, специфически ингибирующее активность как PI3K, так и mTOR; а также IPI-145 (INK1197) (Winkler et al., Chem Biol. 2013 Nov 21; 20 (11): 1364-74. 2013), SAR245409 (XL765) (Dai et.al, Endocrinology. Mar;154 (3):1247-592013), ZSTK474 (Toyama et.al, Arthritis Res Ther. 12 (3): R92. 2010), VS-5584 (SB2343) (Hart et.al, Mol Cancer Ther. 2013 Feb;12 (2):151-61. 2013), AS-605240 (Camps et.al, Nature medicine, 11 (9): 936-943. 2005), PIK-90 (Van et.al, J Cell Biol. 2006 Jul 31;174 (3):437-45. 2006), PF-4989216 (Walls et.al, Clin Cancer Res. 2014 Feb 1;20 (3):631-43. 2014), TG100-115 (Walls et.al, Proc Natl Acad Sci U S A. Dec 26;103 (52):19866-71. 2006), BKM120 (Bendell et.al, J. Clin. Oncol. 30 (3): 282-90. 2012), BEZ235 тозилат (Maira et.al, Mol Cancer Ther 2008; 7: 1851-1863. 2008), LY294002 (Maira et.al, Biochem. Soc. Trans. 37 (Pt1): 265-72. 2009), PI-103 (Raynaud et.al, Molecular Cancer Therapeutics, 8 (7): 1725-1738. 2009), XL147 (Shapiro et.al, Proc 97th Annu Meet AACR, 14-18. 2007), AS-252424 (Pomel et.al, J Med Chem. Jun 29;49 (13):3857-71. 2006), AS-604850 (Camps et.al, Nature medicine, 11 (9): 936-943. 2005), CAY10505 (Tyagi et.al, Can J Physiol Pharmacol. Jul;90 (7):881-5. 2012), CH5132799 (Ohwada et.al, Bioorganic & medicinal chemistry letters, 21 (6): 1767-1772. 2011), BAY 80-6946 (Copanlisib) (Patnaik et.al, J Clin Oncol, 29, 2011), GDC-0032 (Ndubaku et.al, J Med Chem. Jun 13;56 (11):4597-610. 2013), GSK1059615 (Knight et.al, ACS Medicinal Chemistry Letters, 1 (1): 39-43. 2010), CAL-130 (Subramaniam et.al, Раковая клетка. 21 (4):459-72. 2012), XL765 (Laird et.al, AACR-NCI-EORTC International Conference on Molecular Targets и Cancer Therapeutics. October p.B250 2007), и молекулы РНКи, направленные против ДНК, кодирующей PIK3CG, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против PIK3CG и доминантно-негативные варианты PIK3CG. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность RAC1, относящегося к белкам, связанным с сигнальным путем PI3K, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, NSC 23766 (Gao et al., Proc. Natl. Acad. Sci. U.S.A. 101: 7618-7623. 2004), который представляет собой низкомолекулярное соединение специфически ингибирующее активность ГТФазы Rac; W56 (Gao et al., J. Biol. Chem. 276 47530. 2001), который представляет собой пептид, полученный из участка RAC1, отвечающего за распознавание/активацию фактора обмена гуаниновых нуклеотидов; а также молекулы РНКи, направленные против ДНК, кодирующей RAC1, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против RAC1 и доминантно-негативные варианты RAC1. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность AKT3, относящегося к белкам, связанным с сигнальным путем PI3K, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, MK-2206 дигидрохлорид (Hirai et al., Mol. Cancer ther. 9 (7): 1956-67. 2010), который представляет собой низкомолекулярное соединение, специфически ингибирующее активность Akt1, Akt2 и Akt3; трицирибин (Moore et al., Biochem. Pharmacol. 38: 4037-4044. 1989), который представляет собой низкомолекулярное соединение, специфически ингибирующее активность Akt; ингибитор Akt VIII (Barnett et al., Biochem. J. 385: 399-408. 2005), который представляет собой способное проникать в клетку производное хиноксалина, низкомолекулярное соединение, специфически ингибирующее активность Akt1, Akt2 и Akt3; GSK 690693 (Rhodes et al., Cancer Res. 68 (7): 2366-2374. 2008), который представляет собой производное аминофуразана, низкомолекулярное соединение, специфически ингибирующее активность Akt1, Akt2 и Akt3; AT7867 (Grimshaw et al., Mol. Cancer Ther. 9 (5): 1100-10. 2010), который представляет собой низкомолекулярное соединение, специфически ингибирующее активность Akt1, Akt2 и Akt3; а также молекулы РНКи, направленные против ДНК, кодирующей AKT3, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против AKT3 и доминантно-негативные варианты AKT3. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность EIF4B, относящегося к белкам, связанным с сигнальным путем PI3K, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей EIF4B, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против EIF4B и доминантно-негативные варианты EIF4B. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность EIF4E, относящегося к белкам, связанным с сигнальным путем PI3K, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, 4EGI-1 (Moerke et al., Cell. 128: 257-267. 2007), который представляет собой низкомолекулярное соединение, специфически ингибирующее взаимодействие EIF4E/EIF4G; а также молекулы РНКи, направленные против ДНК, кодирующей EIF4E, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против EIF4E и доминантно-негативные варианты EIF4E. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность ILK, относящегося к белкам, связанным с сигнальным путем PI3K, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, Cpd 22 (Lee et al., J. Med. Chem. 54, 6364. 2011), который представляет собой способное проникать в клетку производное пиразола, низкомолекулярное соединение, специфически ингибирующее ILK; QLT0267 (Younes et al., Mol Cancer Ther. Aug; 4 (8): 1146-56. 2005), который представляет собой низкомолекулярное соединение, специфически ингибирующее киназную активность ILK; а также молекулы РНКи, направленные против ДНК, кодирующей ILK, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против ILK и доминантно-негативные варианты ILK. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность MTCP1, относящегося к белкам, связанным с сигнальным путем PI3K, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, молекулы РНКи, направленные против ДНК, кодирующей MTCP1, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против MTCP1 и доминантно-негативные варианты MTCP1. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность PIK3CA, относящегося к белкам, связанным с сигнальным путем PI3K, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, HS-173 (Lee et al., Cancer Lett. Jan 1; 328 (1): 152-9. 2013), который представляет собой низкомолекулярное соединение, специфически ингибирующее PI3K; CUDC-907 (Qian et al., Clin Cancer Res. 18 (15): 4104-4113. 2012), который представляет собой низкомолекулярное соединение, специфически ингибирующее активность как HDAC, так и PI3K; PKI-402 (Dehnhardt et al., J Med Chem. Jan 28; 53 (2): 798-810. 2010), который представляет собой низкомолекулярное соединение, специфически ингибирующее активность как PI3K, так и mTOR; PF-04691502 (Yuan et al., Mol Cancer Ther, 10 (11), 2189-2199. 2011), который представляет собой низкомолекулярное соединение, специфически ингибирующее активность как PI3K, так и mTOR; NVP-BGT226 (Glienke et al., Tumor Biology, 33 (3): 757-765. 2012), который представляет собой низкомолекулярное соединение, специфически ингибирующее активность как PI3K, так и mTOR; а также BYL719 (Furet et al., Bioorg Med Chem Lett 2013; 23: 3741-3748. 2013), SAR245409 (XL765) (Dai et.al, Endocrinology. Mar;154 (3):1247-592013), ZSTK474 (Toyama et.al, Arthritis Res Ther. 12 (3): R92. 2010), VS-5584 (SB2343) (Hart et.al, Mol Cancer Ther. 2013 Feb;12 (2):151-61. 2013), PIK-75 (Zheng et.al, Mol Pharmacol. Oct;80 (4):657-64. 2011), PIK-90 (Van et.al, J Cell Biol. 2006 Jul 31;174 (3):437-45. 2006), A66 (Jamieson et.al, Biochem J. 438:53-62. 2011), CNX1351 (Nacht et.al, J Med Chem, 56 (3): 712-721. 2013), PF-4989216 (Walls et.al, Clin Cancer Res. 2014 Feb 1;20 (3):631-43. 2014), BKM120 (Bendell et.al, J. Clin. Oncol. 30 (3): 282-90. 2012), BEZ235 тозилат (Maira et.al, Mol Cancer Ther 2008; 7: 1851-1863. 2008), LY294002 (Maira et.al, Biochem. Soc. Trans. 37 (Pt1): 265-72. 2009), PI-103 (Raynaud et.al, Molecular Cancer Therapeutics, 8 (7): 1725-1738. 2009), XL147 (Shapiro et.al, Proc 97th Annu Meet AACR, 14-18. 2007), CH5132799 (Ohwada et.al, Bioorganic & medicinal chemistry letters, 21 (6): 1767-1772. 2011), BAY 80-6946 (Copanlisib) (Patnaik et.al, J Clin Oncol, 29, 2011), GDC-0032 (Ndubaku et.al, J Med Chem. Jun 13;56 (11):4597-610. 2013), XL765 (Laird et.al, AACR-NCI-EORTC International Conference on Molecular Targets и Cancer Therapeutics. October p.B250 2007), и молекулы РНКи, направленные против ДНК, кодирующей PIK3CA, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против PIK3CA и доминантно-негативные варианты PIK3CA. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
Примеры средств, ингибирующих активность SRF, относящегося к белкам, связанным с сигнальным путем PI3K, и вызывающего синтетическую летальность при ингибировании наряду с GST-π, включают в себя, без ограничения, CCG-1423 (Evelyn et al., Mol. Cancer Ther. 6, 2249. 2007), который представляет собой способное проникать в клетку производное бензамида, низкомолекулярное соединение, специфически ингибирующее сигнальный путь RHO и активацию SRF; CCG-100602 (Evelyn, et al., Bioorg Med Chem Lett 20 665-72. 2010), который представляет собой аналог CCG-1423 и является низкомолекулярным соединением, специфически ингибирующим сигнальный путь RHO и активацию SRF; а также молекулы РНКи, направленные против ДНК, кодирующей SRF, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, антитела против SRF и доминантно-негативные варианты SRF. Указанные лекарственные средства являются коммерчески доступными или могут быть получены соответствующим образом с помощью широко известных способов.
В частности, в качестве средства, ингибирующего продукцию или активность белка, регулирующего клеточный цикл (например, p21), или белка, связанного с подавлением апоптоза, или белка, связанного с сигнальным путем PI3K, вызывающего синтетическую летальность при ингибировании наряду с GST-π, предпочтительно использовать молекулы РНКи, направленные против ДНК, кодирующей указанный белок, рибозимы, антисмысловые нуклеиновые кислоты, химерные полинуклеотиды ДНК/РНК и экспрессирующие их векторы, поскольку они характеризуются высокой специфичностью и низкой вероятностью побочных реакций.
Ингибирование связанного с гомеостазом белка можно определить по уменьшению экспрессии или активности белка в клетке по сравнению с клеткой, на которую не действует средство, ингибирующее белок. Экспрессию белка можно оценить с помощью любого известного метода, включающего в себя, без ограничения, метод иммунопреципитации с использованием антитела, EIA, ELISA, IRA, IRMA, вестерн-блоттинг, иммуногистохимический анализ, иммуноцитохимический анализ, проточную цитометрию или разные способы гибридизации, нозерн-блоттинг, саузерн-блоттинг или разные ПЦР-методы с использованием нуклеиновых кислот, специфически гибридизирующихся с нуклеиновой кислотой, кодирующей белок, или с ее уникальным фрагментом, или транскриптом (например, мРНК), или продуктом сплайсинга нуклеиновой кислоты.
Кроме того, активность, например, p21 можно определить, без ограничения, путем анализа известной активности p21, например, его способности связывать комплекс циклин-CDK2 или циклин-CDK1 с помощью любого известного метода, такого как метод иммунопреципитации, вестерн-блоттинг, масс-спектрометрия, метод аффинной адсорбции или метод поверхностного плазмонного резонанса (SPR).
В соответствии с настоящим описанием молекула РНКи относится к произвольной молекуле, которая приводит к интерференции РНК. Молекула РНКи включает в себя, без ограничения, двухцепочечные РНК, такие как миРНК (малая интерферирующая РНК), микроРНК (микроРНК), кшРНК (короткая шпилечная РНК), днРНК (ДНК-направленная РНК), пиРНК (пиви-взаимодействующая РНК), памиРНК (повтор-ассоциированные миРНК), их альтернативы и т.п. Указанные молекулы РНКи можно приобрести в продаже, или их можно сконструировать и получить с использованием общеизвестной информации о последовательности, то есть с использованием нуклеотидных последовательностей и/или аминокислотных последовательностей, описанных в SEQ ID NO: 1-30, 39-108.
В соответствии с настоящим описанием антисмысловая нуклеиновая кислота включает в себя РНК, ДНК, ПНК, или их комплексы.
В соответствии с настоящим описанием химерный полинуклеотид ДНК/РНК включает в себя, без ограничения, например, двухцепочечный полинуклеотид, описанный в публикации патента Японии (Kokai) № 2003-219893 A (2003), который состоит из ДНК и РНК и и ингибирует экспрессию гена-мишени.
Средство, ингибирующее GST-π, и средство, ингибирующее связанный с гомеостазом белок и вызывающее синтетическую летальность при ингибировании наряду с GST-π, могут содержаться в одной композиции, или они могут по отдельности содержаться в двух или более композициях. В последнем случае указанные композиции можно вводить одновременно, или их можно вводить поочередно. В случае поочередного введения композицию, содержащую средство, ингибирующее GST-π, можно вводить до или после введения композиции, содержащей средство, ингибирующее связанный с гомеостазом белок, вызывающий синтетическую летальность при ингибировании наряду с GST-π.
Средство, индуцирующее гибель клеток, и средство, ингибирующее рост клеток, в соответствии с настоящим изобретением могут содержать один тип вышеуказанного связанного с гомеостазом белка, или два или более типов вышеуказанного связанного с гомеостазом белка. Например, в соответствии с настоящим изобретением в качестве связанного с гомеостазом белка, входящего в состав средства, индуцирующего гибель клеток, и средства, ингибирующего рост клеток, можно использовать два или более типов белка, регулирующего клеточный цикл; два или более типов белка, связанного с подавлением апоптоза; или один или несколько типов белка, регулирующего клеточный цикл, и один или несколько типов белка, связанного с подавлением апоптоза.
В частности, каждый из ATM, CDC25A, p21, PRKDC, RBBP8, SKP2, MCM10, RNPC1, CCNL1, CENPH, BRSK1, MYLK, MCM8, CCNB3, MCMDC1, AATF, AKT1, ALOX12, ANXA1, ANXA4, API5, ATF5, AVEN, AZU1, BAG1, BCL2L1, BFAR, CFLAR, IL2, MALT1, MCL1, MKL1, MPO, MTL5, MYBL2, MYO18A, MTOR, IRAK1, IRS1, MYD88, NFKB1, PIK3CG, RAC1, AKT3, EIF4B, EIF4E, ILK, MTCP1, PIK3CA и SRF представляет собой связанный с гомеостазом белок, приводящий к синтетической летальности раковых клеток при ингибировании наряду с GST-π. Соответственно, средство, ингибирующее белок, можно использовать в качестве активного ингредиента препарата или композиции, усиливающих индукцию гибели клеток и/или ингибирование роста клеток под действием средства, ингибирующего GST-π (далее его также называют "средство, усиливающее индукцию гибели клеток", "средство, усиливающее ингибирование роста клеток", "композиция для усиления индукции гибели клеток" или "композиция для усиления ингибирования роста клеток"). Другими словами, индукцию гибели клеток и/или ингибирование роста клеток под действием средства, ингибирующего GST-π, можно усилить путем введения эффективного количества средства, ингибирующего белок.
Активный ингредиент может содержаться в препарате или композиции настоящего изобретения в количестве, которое индуцирует гибель клеток, такую как апоптоз, и/или ингибирует рост клеток после введения препарата или композиции. Кроме того, предпочтительно использовать количество, которое не оказывает неблагоприятного эффекта, превышающего преимущества, вызванные введением. Такое количество является общеизвестным или может быть соответствующим образом определено путем тестирования in vitro с использованием культивируемых клеток и т.п., или путем тестирования на экспериментальных животных, таких как мыши, крысы, собаки или свиньи. Такой способ тестирования хорошо известен специалистам в данной области техники. Индукцию апоптоза можно оценить с помощью разных известных способов, например, путем выявления апоптоз-специфических явлений, таких как фрагментация ДНК, связывание аннексина V с клеточной мембраной, изменение потенциала митохондриальной мембраны, активация каспазы и окрашивание TUNEL. Ингибирование роста клеток также можно оценить с помощью разных известных способов, например, путем время-зависимого измерения числа живых клеток, измерения размера, объема или массы опухоли, измерения количества синтезируемой ДНК, с помощью метода WST-1, метода BrdU (бромдезоксиуридин) и метода с включением 3H-тимидина. Содержание активного ингредиента может варьировать в зависимости от лекарственной формы препарата или композиции. Например, при использования нескольких единиц композиции в одном введении количество активного ингредиента, содержащегося в одной единице композиции, может быть определено как 1/количество активного ингредиента, необходимое для одного введения. Такая коррекция содержания может оответствующим образом проводиться специалистами в данной области.
Кроме того, средство, ингибирующее GST-π и средство, ингибирующее связанный с гомеостазом белок и вызывающее синтетическую летальность при ингибировании наряду с GST-π, можно ввести в состав композиции в качестве активных ингредиентов с получением средства, индуцирующего гибель клеток, средства, ингибирующего рост клеток, композиции для индукции гибели клеток, или композиции для ингибирования роста клеток.
Далее, можно получить сочетание средства, ингибирующего GST-π, и средства, ингибирующего связанный с гомеостазом белок и вызывающего синтетическую летальность при ингибировании наряду с GST-π, для применения в способе индукции гибели клеток или ингибирования роста клеток. Кроме того, изобретение предлагает способ индукции гибели клеток или способ ингибирования роста клеток, включающий в себя введение эффективных количеств средства, ингибирующего GST-π, и средства, ингибирующего связанный с гомеостазом белок и вызывающего синтетическую летальность при ингибировании наряду с GST-π.
Все вышеупомянутые способы индуцирования гибели клеток, такой как апоптоз, или ингибирования роста клеток, можно проводить in vitro или in vivo. Лекарственные средства, используемые в данных способах, описаны выше, а эффективное количество каждого лекарственного средства может представлять собой количество, которое индуцирует гибель клеток-реципиентов, такую как апоптоз, и/или ингибирует рост клеток-реципиентов. Кроме того, предпочтительно использовать количество, которое не оказывает неблагоприятного эффекта, превышающего преимущества, вызванные введением. Такое количество является общеизвестным или может быть соответствующим образом определено путем тестирования in vitro с использованием культивируемых клеток. Указанный способ тестирования хорошо известен специалистам в данной области. Индукцию гибели клеток или ингибирование роста клеток можно оценить с помощью разных известных методов, включающих в себя упомянутые выше. Если лекарственное средство вводят в заданную популяцию раковых клеток, эффективное количество не должно приводить к гибели или ингибированию роста всех клеток в данной популяции. Эффективное количество может представлять собой, например, количество, которое приводит к апоптозу или ингибированию роста у 1% или более, 2% или более, 3% или более, 4% или более, 5% или более, 6% или более, 8% или более, 10% или более, 12% или более, 15% или более, 20% или более и 25% или более клеток в популяции.
Средство настоящего изобретения, индуцирующее гибель клеток, или средство настоящего изобретения, ингибирующее рост клеток, может эффективно индуцировать гибель или ингибировать рост даже раковых клеток и, как таковое, может эффективно использоваться в качестве ингредиента фармацевтической композиции, предназначенной для лечения заболевания, вызванного аномальным клеточным ростом. Кроме того, лекарственное средство, ингибирующее GST-π, и лекарственное средство, ингибирующее белок, связанный с гомеостазом, и приводящее к синтетической летальности при ингибировании наряду с GST-π, можно использовать в качестве активных ингредиентов фармацевтической композиции, предназначенной для лечения заболевания, вызванного аномальным ростом клеток. Изобретение также предлагает способ лечения заболевания, вызванного аномальным ростом клеток, включающий в себя введение эффективного количества полученной фармацевтической композиции индивидууму, нуждающемуся в этом.
Фармацевтическую композицию можно эффективно использовать для лечения заболевания, вызванного аномальным клеточным ростом, в частности, для лечения заболевания, обусловленного гибелью клеток или аномальным клеточным ростом в результате экспрессии мутантного KRAS.
Заболевание, вызванное клеткой, экспрессирующей мутантный KRAS, включает в себя, без ограничения, например, доброкачественные или злокачественные опухоли (также называемые раковыми или злокачественными новообразованиями), гиперплазию, келоид, синдром Кушинга, первичный альдостеронизм, эритроплакию, полицитемию vera, лейкоплакию, гиперпластический рубец, красный плоский лишай и лентигиноз.
Примеры рака в соответствии с настоящим изобретением включают в себя раковые заболевания, экспрессирующие GST-π на высоком уровне, и раковые заболевания, вызванные клетками, экспрессирующими мутантный KRAS (также называемые для простоты рак KRAS). Во многих случаях рак KRAS также относится к раковым заболеваниям, характеризующимся высоким уровнем экспрессии GST-π. Примеры таких заболеваний включают в себя, без ограничения: саркомы, такие как фибросаркома, злокачественная фиброзная гистиоцитома, липосаркома, рабдомиосаркома, лейомиосаркома, ангиосаркома, саркома Капоши, лимфангиосаркома, синовиальная саркома, хондросаркома и остеосаркома; карциномы, такие как опухоль головного мозга, рак головы и шеи, рак молочной железы, рак легких, рак пищевода, рак желудка, рак двенадцатиперстной кишки, рак аппендикса, рак толстой и прямой кишки, рак прямой кишки, рак печени, рак поджелудочной железы, рак желчного пузыря, рак желчных протоков, рак почки, рак мочеиспускательного канала, рак мочевого пузыря, рак предстательной железы, рак полового члена, рак яичка, рак матки, рак яичников, рак вульвы, рак влагалища и рак кожи; а также лейкоз и злокачественную лимфому. В настоящем изобретении "рак" включает в себя эпителиальные злокачественные опухоли и неэпителиальные злокачественные опухоли. Рак в соответствии с настоящим изобретением может присутствовать в произвольном отделе организма, таком как мозг, область головы и шеи, грудная клетка, конечности, легкие, сердце, вилочковая железа, пищевод, желудок, тонкая кишка (двенадцатиперстная кишка, тощая кишка и подвздошная кишка), толстая кишка (ободочная кишка, слепая кишка, аппендикс и прямая кишка), печень, поджелудочная железа, желчный пузырь, анус, почки, мочевые протоки, мочевой пузырь, простата, пенис, яички, матка, яичники, вульва, влагалище, кожа, полосатые мышцы, гладкие мышцы, синовиальные мембраны, хрящи, кости, щитовидная железа, надпочечники, брюшина, мезентерий, костный мозг, кровь, сосудистая система, лимфатическая система, включающая в себя лимфатические узлы и лимфу.
В составе фармацевтической композиции лекарственное средство, ингибирующее GST-π, и лекарственное средство, ингибирующее белок, связанный с гомеостазом, и вызывающее синтетическую летальность при ингибировании наряду с GST-π, могут присутствовать в сочетании с дополнительным активным ингредиентом. В данном контексте применение в сочетании включает в себя, например, введение дополнительного активного ингредиента в составе другой композиции и введение дополнительного активного ингредиента в составе комбинированного лекарственного средства, содержащего, по меньшей мере, одно из дополнительных лекарственных средств. В случае введения в составе другой композиции, композицию, содержащую дополнительный активный ингредиент, можно вводить до, одновременно или после введения другой композиции (композиций).
Примеры такого дополнительного активного ингредиента включают в себя соединения, обеспечивающие эффективное лечение интересующего заболевания. Например, если подлежащее лечению заболевание представляет собой рак, в качестве дополнительного ингредиента сочетания можно использовать противораковое средство. Примеры противоракового средства могут включать в себя: алкилирующие средства, такие как ифосфамид, гидрохлорид нимустина, циклофосфамид, дакарбазин, мелфалан и ранимустин; метаболические антагонисты, такие как гемцитабина гидрохлорид, эноцитабин, цитарабина осфосфат, цитарабин-содержащие средства, тегафур урацил, комбинированные средства тегафур-гимерацил-отерацил калия (например, TS-1), доксифлуридин, гидроксикарбамид, фторурацил, метотрексат и меркаптопурин; противоопухолевые антибиотики, такие как гидрохлорид идарубицина, гидрохлорид эпирубицина, гидрохлорид даунорубицина, цитрат даунорубицина, гидрохлорид доксорубицина, гидрохлорид пирарубицина, гидрохлорид блеомицина, сульфат пепломицина, гидрохлорид митоксантрона и митомицин С; алкалоиды, такие как этопозид, гидрохлорид иринотекана, винорелбина тартрат, гидрат доцетаксела, паклитаксел, сульфат винкристина, сульфат виндезина и сульфат винбластина; гормональные терапевтические средства, такие как анастрозол, цитрат тамоксифена, торемифенцитрат, бикалютамид, флутамид и эстрамустинфосфат; комплексы платины, такие как карбоплатин, цисплатин (CDDP) и недаплатин; антиангиогенные средства, такие как талидомид, неовастат и бевацизумаб; а также L-аспарагиназу.
Если в качестве активных ингредиентов, присутствующих в разных препаратах или композициях, используемых в способе лечения индивидуума, и т. д., описанных в данном документе, используют нуклеиновые кислоты, например молекулы РНКи, рибозимы, антисмысловые нуклеиновые кислоты или химерные полинуклеотиды ДНК/РНК, их можно вводить непосредственно в виде оголенных нуклеиновых кислот или в составе разных векторов. В качестве векторов можно использовать любые общеизвестные векторы, такие как плазмидные векторы, фаговые векторы, фагемидные векторы, космидные векторы или вирусные векторы. Предпочтительно векторы содержат, по меньшей мере, промотор, который обеспечивает начало экспрессии каждой нуклеиновой кислоты, содержащейся в векторе. В данном случае предпочтительно, чтобы нуклеиновая кислота была функционально связана с таким промотором. Функциональная связь нуклеиновой кислоты с промотором означает, что нуклеиновая кислота и промотор расположены так, что промотор обеспечивает надлежащую продукцию белка, кодируемого нуклеиновой кислотой. Векторы могут необязательно обладать способностью к репликации в клетках-хозяевах. Кроме того, транскрипция гена может осуществляться вне ядер клеток-хозяев, или она может осуществляться внутри ядер. В последнем случае нуклеиновая кислота может быть интегрирована в геном клетки-хозяина.
Альтернативно активные ингредиенты могут поддерживаться различными невирусными липидными или белковыми носителями. Примеры таких носителей включают в себя, без ограничения, холестерины, липосомы, протомеры антител, наночастицы циклодекстрина, гибридные пептиды, аптамеры, биодеградируемые сополимеры полимолочной кислоты, а также полимеры, которые могут повышать эффективность поглощения клетками (см., например, Pirollo and Chang, Cancer Res., 2008, 68 (5): 1247-50). В частности, можно использовать катионные липосомы или полимеры (например, полиэтиленимин). Другие примеры полимеров, подходящих для применения в качестве таких носителей, описаны, например, в US 2008/0207553 и US 2008/0312174.
В разных описанных здесь фармацевтических композициях настоящего изобретения активные ингредиенты могут быть объединены с любым дополнительным ингредиентом, который не ухудшает эффекты активных ингредиентов. Примеры такого дополнительного ингредиента включают в себя другие химиотерапевтические средства, фармакологически приемлемые носители, наполнители и разбавители. Кроме того, композиции могут иметь покрытие, такое как энтеросолюбильное покрытие или покрытие из веществ, распадающихся со времением, в зависимости от способа введения, способа высвобождения лекарственного средства и т. д., они также могут быть включены в подходящие системы высвобождения лекарственного средства.
Разные описанные здесь средства и композиции (включающие в себя разные фармацевтические композиции) настоящего изобретения можно вводить, без ограничения, с помощью разных способов, включающих в себя как пероральные, так и парентеральные способы, такие как пероральные, внутривенные, внутримышечные, подкожные, локальные, внутриутробные, ректальные, внутриартериальные, интрапортальные, внутрижелудочковые, трансмукозальные, чрескожные, назальные, внутрибрюшинные, внутрилегочные и внутриматочные способы, или композиции могут находиться в виде лекарственных форм, подходящих для каждого пути введения. Любые широко известные носители можно соответственно использовать в таких лекарственных формах и способах получения (см., например, Hyoujun Yakuzai Gaku (стандартная фармацевтика на английском языке), под редакцией Yoshiteru Watanabe et al., Nankodo Co., Ltd., 2003).
Примеры лекарственных форм, подходящих для перорального введения, включают в себя, без ограничения, порошки, гранулы, таблетки, капсулы, растворы, суспензии, эмульсии, гели и сиропы. Примеры лекарственных форм, подходящих для парентерального введения, включают в себя препараты для инъекций, такие как препараты для инъекций в виде раствора, препараты для инъекций в виде суспензии, препараты для инъекций в виде эмульсии и препараты для инъекций, предназначенные для приготовления перед использованием. Препараты для парентерального введения могут находиться в виде водных или неводных изотонических стерильных растворов или суспензий.
Разные описанные здесь средства или композиции (в том числе разные фармацевтические композиции) настоящего изобретения могут быть направлены на конкретные ткани или клетки. Такую направленность можно достичь с помощью любого известного способа. Для предполагаемой доставки к раковой опухоли можно использовать, без ограничения, например, такой способ, как пассивный таргетинг, путем установления размера препарата с диаметром от 50 до 200 мкм, в частности от 75 до 150 мкм, обеспечивающего достижение EPR (повышенные проницаемость и удерживание), или активный таргетинг с использованием лиганда, такого как CD19, HER2, рецептор трансферрина, рецептор фолата, VIP-рецептор, EGFR (Torchilin, AAPS J. 2007, 9 (2): E128-47), RAAG10 (публикация патента Японии (Kohyo) № 2005-532050 A (2005)), PIPA (публикация патента Японии (Kohyo) № 2006-506071 A (2006)) или KID3 (публикация патента Японии (Kohyo) № 2007-529197 A (2007)), пептид, имеющий мотив RGD или мотив NGR, F3, LyP-1 (Ruoslahti et al., J Cell Biol., 2010; 188 (6): 759-68) и т.п. в качестве направляющего средства. Поскольку известно, что ретиноид или его производное можно использовать в качестве средства, обеспечивающего направленность на раковую клетку (WO 2008/120815), можно использовать носитель, содержащий ретиноид в качестве направляющего средства. Такой носитель описан в WO 2009/036368, WO 2010/014117, WO 2012/170952 и т.д., помимо вышеупомянутых литературных источников.
Разные описанные здесь средства или композиции (в том числе разные фармацевтические композиции) настоящего изобретения могут поставляться в виде любой формы, в том числе, в виде формы, которая может использоваться для приготовления препарата перед использованием, например, в виде формы, которая может использоваться для приготовления препарата врачом и/или фармацевтом, медсестрой или другими медицинскими работниками в медицинском учреждении, или по соседству, чтобы сохранить стабильность. Такая форма особенно полезна, если средство или композиция настоящего изобретения содержат ингредиенты, которые трудно стабилизировать, такие как липиды, белки или нуклеиновые кислоты. В этом случае средство или композиция настоящего изобретения предоставляется в виде одного, или двух или более контейнеров, содержащих, по меньшей мере, один из компонентов, необходимых для приготовления перед использованием, например, за 24 часа до использования, предпочтительно за 3 часа до использования, более предпочтительно, непосредственно перед использованием. Для приготовления препарата можно соответственно использовать реагент, растворитель, рецептурный инструмент и т.п., которые обычно присутствуют в месте приготовления препарата.
Таким образом, настоящее изобретение также предлагает набор для получения композиции, содержащий один, или два или более контейнеров, которые, в свою очередь, содержат один активный ингредиент, или сочетание активных ингредиентов, которые могут содержаться в разных препаратах или композициях настоящего изобретения, а также компоненты, необходимые для различных препаратов или композиций, входящих в состав такого набора. Набор настоящего изобретения может дополнительно содержать инструкцию, например, ручной или электронный носитель информации (CD или DVD) с описанием способов получения разных препаратов или композиций настоящего изобретения, способа лечения индивидуума и т.д. Кроме того, набор настоящего изобретения может содержать все компоненты, необходимые для получения разных препаратов или композиций настоящего изобретения, однако присутствие всех компонентов является необязательным. Так, в наборе настоящего изобретения может отсутствовать реагент или растворитель, обычно имеющийся в медицинском учреждении, экспериментальном объекте и т.п., такой как стерильная вода, физиологический раствор или раствор глюкозы.
Эффективное количество, используемое в разных описанных здесь способах лечения настоящего изобретения, представляет собой, например, количество, которое уменьшает симптомы заболевания или задерживает или останавливает прогрессирование заболевания, и предпочтительно представляет собой количество, которое подавляет или излечивает заболевание. Кроме того, предпочтительно использовать количество, которое не оказывает неблагоприятного эффекта, превышающего преимущества, вызванные введением. Такое количество можно соответствующим образом определить путем тестирования in vitro с использованием культивируемых клеток и т.п., или путем тестирования на экспериментальных животных, таких как мыши, крысы, собаки или свиньи. Такой способ тестирования хорошо известен специалистам в данной области техники. Кроме того, дозы лекарственных средств, используемых в способе лечения настоящего изобретения, обычно известны специалистам в данной области, или они могут быть соответствующим образом определены с помощью вышеупомянутых тестов или т.п.
Конкретные дозы активных ингредиентов, используемые для введения индивидууму в описанном здесь способе лечения настоящего изобретения, можно определить с учетом разных показателей состояния индивидуума, нуждающегося в лечении, таких как тяжесть симптомов, общее состояние здоровья, возраст, масса тела, пол, диета, время и частота введения, лекарство, используемое в сочетании, ответ на лечение, лекарственная форма и соблюдение режима лечения.
Способ введения включает в себя разные способы, как пероральные, так и парентеральные, такие как пероральный, внутривенный, внутримышечный, подкожный, местный, внутриопухолевый, ректальный, внутриартериальный, интрапортальный, внутрижелудочковый, трансмукозальный, чрескожный, назальный, внутрибрюшинный, внутрилегочный и внутриматочный способы введения.
Частота введения может варьировать в зависимости от свойств используемого препарата или композиции и показателей состояния индивидуума, включающих в себя описанные выше, и может включать в себя введение, например, несколько раз в день (то есть 2, 3, 4 или 5 раз в день), один раз в день, каждые несколько дней (т.е. каждые 2, 3, 4, 5, 6 или 7 дней), один раз в неделю или каждые несколько недель (т.е. каждые 2, 3 или 4 недели).
Используемый здесь термин "индивидуум" относится к любому отдельному организму, который предпочтительно представляет собой животное, более предпочтительно млекопитающее, еще более предпочтительно человека. В настоящем изобретении индивидуум может быть здоровым или иметь какое-нибудь заболевание. В случае предполагаемого лечения конкретного заболевания индивидуум обычно представляет собой индивидуума, имеющего указанное заболевание, или подверженного риску развития указанного заболевания.
Используемый здесь термин "лечение" включает в себя все виды приемлемых с медицинской точки зрения профилактических и/или терапевтических вмешательств, целью которых является, например, лечение, временное облегчение или профилактика заболевания. Например, термин "лечение" охватывает разные приемлемые с медицинской точки зрения вмешательства, проводимые с целью задержки или прекращения прогрессирования заболевания, обратного развития или исчезновения повреждения, профилактики появления или возобновления заболевания и т.д.
В частности, как указано выше, каждый из ATM, CDC25A, p21, PRKDC, RBBP8, SKP2, MCM10, RNPC1, CCNL1, CENPH, BRSK1, MYLK, MCM8, CCNB3, MCMDC1, AATF, AKT1, ALOX12, ANXA1, ANXA4, API5, ATF5, AVEN, AZU1, BAG1, BCL2L1, BFAR, CFLAR, IL2, MALT1, MCL1, MKL1, MPO, MTL5, MYBL2, MYO18A, MTOR, IRAK1, IRS1, MYD88, NFKB1, PIK3CG, RAC1, AKT3, EIF4B, EIF4E, ILK, MTCP1, PIK3CA и SRF представляет собой белок, приводящий к синтетической летальности раковых клеток при ингибировании наряду с GST-π. Таким образом, средство, индуцирующее гибель раковых клеток, и/или средство, ингибирующее рост раковых клеток, используемое наряду со средством, ингибирующим GST-π, можно подвергнуть скринингу, используя ингибирование указанного связанного с гомеостазом белка в качестве показателя. А именно, вещество, способное ингибировать связанный с гомеостазом белок, можно использовать в качестве вещества-кандидата на роль средства, индуцирующего гибель раковых клеток, и/или средства, ингибирующего рост раковых клеток, при использовании наряду со средством, ингибирующим GST-π.
Например, тестируемое вещество приводят в контакт с клеткой, экспрессирующей мутантный KRAS, используемой в качестве раковой клетки, после чего в клетке изверяют уровень экспрессии связанного с гомеостазом белка, приводящего к синтетической летальности клетки, экспрессирующей мутантный KRAS, при ингибировании наряду с GST-π. Тестируемое вещество может быть выбрано в качестве вещества-кандидата на роль средства, ингибирующего связанный с гомеостазом белок, если уровень экспрессии, измеренный после контакта с тестируемым веществом, уменьшается по сравнению с уровнем экспрессии, измеренным в отсутствии тестируемого вещества.
С другой стороны, средство, ингибирующее GST-π, представляет собой белок, приводящий к синтетической летальности раковой клетки при ингибировании наряду со средством, ингибирующим связанный с гомеостазом белок, приводящим к синтетической летальности раковой клетки при ингибировании наряду с GST-π. Таким образом, средство, индуцирующее гибель раковых клеток, и/или средство, ингибирующее рост раковых клеток, используемое наряду со средством, ингибирующим связанный с гомеостазом белок, можно подвергнуть скринингу с использованием ингибирования GST-π в качестве показателя. А именно, вещество, способное ингибировать GST-π, можно использовать в качестве вещества-кандидата на роль средства, индуцирующего гибель раковых клеток, и/или средства, ингибирующего рост раковых клеток, при использовании наряду со средством, ингибирующим связанный с гомеостазом белок.
Например, тестируемое вещество приводят в контакт с клеткой, экспрессирующей мутантный KRAS, используемой в качестве раковой клетки, после чего в клетке изверяют уровень экспрессии GST-π. Тестируемое вещество может быть выбрано в качестве вещества-кандидата на роль средства, ингибирующего GST-π, если уровень экспрессии, измеренный после контакта с тестируемым веществом, уменьшается по сравнению с уровнем экспрессии, измеренным в отсутствии тестируемого вещества.
Подобным образом, средство, индуцирующее гибель раковых клеток, и/или средство, ингибирующее рост раковых клеток, можно подвергнуть скринингу с использование ингибирования GST-π и ингибирования связанного с гомеостазом белка, приводящего к синтетической летальности раковой клетки при ингибировании наряду с GST-π, в качестве показателей. А именно, вещество, способное ингибировать GST-π и связанный с гомеостазом белок, можно использовать в качестве вещества-кандидата на роль средства, индуцирующего гибель раковых клеток, и/или средства, ингибирующего рост раковых клеток.
Например, тестируемое вещество приводят в контакт с клеткой, экспрессирующей мутантный KRAS, используемой в качестве раковой клетки, после чего в клетке изверяют уровень экспрессии GST-π и уровень экспрессии связанного с гомеостазом белка. Тестируемое вещество может быть выбрано в качестве вещества-кандидата на роль средства, ингибирующего GST-π и связанный с гомеостазом белок, приводящий к синтетической летальности раковой клетки при ингибировании наряду с GST-π, если оба уровня экспрессии, измеренные после контакта с тестируемым веществом, уменьшатся по сравнению с соответствующими уровнями экспрессии, измеренными в отсутствии тестируемого вещества.
В данном контексте тестируемое вещество никак не ограничивается и может представлять собой любое вещество. Тестируемое вещество может представлять собой одно вещество, или или оно может представлять собой смесь, состоящую из нескольких компонентов. Тестируемое вещество может содержать неидентифицированное вещество, например, в случае экстракта микроорганизма или культурального раствора, или оно может содержать известные композиции с определенным составом. Кроме того, тестируемое вещество может представлять собой белок, нуклеиновую кислоту, липид, полисахарид, органическое соединение и неорганическое соединение.
ПРИМЕРЫ
Далее настоящее изобретение описывается более подробно со ссылкой на примеры. Однако технический объем настоящего изобретения не ограничивается приведенными ниже примерами.
[Эксперимент 1] Нокдаун GST-π и P21 под действием миРНК
В качестве раковых клеткок 1×105 клеток M7609 (клетки рака толстой и прямой кишки человека, содержащие мутацию KRAS) и клеток PANC-1 (клетки рака поджелудочной железы человека, содержащие мутацию KRAS) вносят в чашки Петри диаметром 6 см и культивируют в течение 18 часов в среде 1640 Мемориального института Розуэлла Парка (RPMI 1640, Sigma-Aldrich Corp.), содержащей 10% фетальной бычьей сыворотки (FBS) и 0,5% L-глутамина. Если не указано иначе, культивирование проводят в условиях 37°C и 5% CO2. Кроме того, в качестве раковых клеткок 0,5×105 клеток A549 (клетки рака легкого человека, содержащие мутацию KRAS) вносят в чашки Петри диаметром 6 см и культивируют в течение 18 часов в модифицированной по Дульбекко среде Игла (DMEM, Sigma-Aldrich Corp.), содержащей 10% FBS и 1% L-глутамина. Далее, в качестве раковых клеток 1×105 клеток MIA PaCa-2 (клетки рака поджелудочной железы человека, содержащие мутацию KRAS) вносят в чашки Петри диаметром 6 см и культивируют в течение 18 часов в среде DMEM, содержащей 10% FBS и 1% L-глутамина. Затем, в качестве раковых клеток 0,5×105 клеток HCT116 (клетки рака толстой и прямой кишки человека, содержащие мутацию KRAS) вносят в чашки Петри диаметром 6 см и культивируют в течение 18 часов в среде McCoy's 5A (McCoy, Sigma-Aldrich Corp.) содержащей 10% FBS и 0,5% L-глутамина.
В данном эксперименте сначала клетки PAC-1, A549 или MIA PaCa-2, достигшие 20-30% слияния, трансфицируют миРНК GST-π и/или миРНК P21 с использованием Lipofectamine RNAi MAX (Life Technologies Corp.) следующим образом.
Смешанный раствор для трансфекции, содержащий липофектамин/миРНК, получают следующим образом: вначале получают раствор липофектамина путем смешивания 15 мкл Lipofectamine RNAi MAX и 485 мкл OPTI-MEM (Sigma-Aldrich Corp.). Затем получают раствор миРНК путем доведения заданного количества 50 мкМ раствора миРНК до 500 мкл с помощью OPTI-MEM (например, в случае получения раствора миРНК, используемого в конечной концентрации 50 нМ, смешивают 6 мкл 50 мкМ миРНК и 494 мл OPTI-MEM), смешивают его с вышеупомянутым раствором липофектамина и оставляют стоять при комнатной температуре в течение 15 минут. Используют миРНК, описанные ниже. В приведенном ниже описании буквы верхнего регистра используют для обозначения РНК, буквы нижнего регистра используют для обозначения ДНК.
миРНК GST-π:
Смысловая цепь: GGGAGGCAAGACCUUCAUUtt (SEQ ID NO: 31)
Антисмысловая цепь: AAUGAAGGUCUUGCCUCCCtg (SEQ ID NO: 32)
миРНК P21:
Смысловая цепь: UCCUAAGAGUGCUGGGCAUtt (SEQ ID NO: 33)
Антисмысловая цепь: AUGCCCAGCACUCUUAGGAtt (SEQ ID NO: 34)
Контрольная миРНК:
Смысловая цепь: ACGUGACACGUUCGGAGAAtt (SEQ ID NO: 35)
Антисмысловая цепь: UUCUCCGAACGUGUCACGUtt (SEQ ID NO: 36)
миРНК-2 GST-π:
Смысловая цепь: UCUCCCUCAUCUACACCAAtt (SEQ ID NO: 37)
Антисмысловая цепь: UUGGUGUAGAUGAGGGAGAtg (SEQ ID NO: 38)
В каждую из чашек Петри, содержащих клетки PANC-1, MIA PaCa-2, или A549 добавляют миРНК GST-π и миРНК P21, каждую в конечной концентрации 50 нМ, миРНК GST-π или миРНК P21 в конечной концентрации 50 нМ (обе добавляют с контрольной миРНК в конечной концентрации 50 нМ), или миРНК GST-π в конечной концентрации 100 нМ (без добавления контрольной миРНК). В качестве контроля добавляют контрольную миРНК в конечной концентрации 100 нМ. После культивирования в течение 1 дня без замены среды количественно определяют уровень мРНК GST-π и уровень мРНК P21 методом количественной ПЦР с использованием системы для проведения ПЦР в режиме реального времени 7300 (Applied Biosystems, Inc.). Результаты показаны на фигуре 1. Результаты, приведенные на фигуре 1, демонстрируют, что количество мРНК P21 повышается в результате нокдауна GST-π под действием миРНК.
Кроме того, если миРНК GST-π или контрольную миРНК в конечной концентрации 50 нМ добавляют в каждую из чашек Петри, содержащих клетки A549 или клетки MIA PaCa-2, уровень мРНК P21 количественно определяют таким же способом каждый день со дня добавления миРНК GST-π или контрольной миРНК до 4-го дня. Результаты показаны на фигуре 2. Результаты, приведенные на фигуре 2, демонстрируют, что уровень экспрессии мРНК P21 увеличивается со временем в результате нокдауна GST-π под действием миРНК.
При этом определяют влияние миРНК GST-π и/или миРНК Р21 на количество клеток. Вначале миРНК GST-π и миРНК P21, каждую в конечной концентрации 50 нМ, или миРНК GST-π или миРНК P21 в конечной концентрации 50 нМ (обе с использованием контрольной миРНК в конечной концентрации 50 нМ) добавляют в каждую из чашек Петри, содержащих клетки PANC-1, MIA PaCa-2 или A549. В качестве контроля добавляют контрольную миРНК в конечной концентрации 100 нМ. После культивирования в течение 5 дней без замены среды клетки отсоединяют путем обработки трипсином, собирают из чашек Петри и считают число клеток. Результаты показаны на фигуре 3. Результаты, приведенные на фигуре 3, демонстрируют, что в случае нокдауна GST-π и P21 по отдельности с использованием миРНК GST-π или миРНК P21, число клеток не уменьшается по отношению к числу инокулированных клеток, хотя наблюдается ингибирование клеточного роста. Однако из приведенных результатов видно, что одновременный нокдаун GST-π и P21 с использованием миРНК GST-π и миРНК P21 приводит не только к ингибированию роста, но и к индукции гибели клеток PANC-1 и клеток MIA PaCa-2, экспрессирующих мутантный KRAS.
Из фигуры 3 можно видеть на примере клеток A549, экспрессирующих мутантный KRAS, что указанная выше обработка не приводит к индукции гибели клеток. Соответственно, число трансфекций миРНК GST-π и миРНК P21 увеличивают и определяют влияние миРНК GST-π и/или миРНК P21 на число клеток A549 и клеток HCT116, экспрессирующих мутантный KRAS.
Вначале в чашки Петри, содержащие клетки PANC-1, клетки MIA PaCa-2 или клетки HCT116, добавляют миРНК GST-π и миРНК P21, каждую в конечной концентрации 25 нМ, или миРНК GST-π или миРНК P21 в конечной концентрации 25 нМ (в каждом случае используют контрольную миРНК в конечной концентрации 25 нМ), тогда как в чашки Петри, содержащие клетки A549, добавляют миРНК GST-π и миРНК P21, каждую в конечной концентрации 50 нМ, или миРНК GST-π или миРНК P21 в конечной концентрации 50 nM (с использованием контрольной миРНК в конечной концентрации 50 нМ). В качестве контроля добавляют контрольную миРНК в конечной концентрации 50 нМ для клеток PANC-1, MIA PaCa-2 или HCT116, и в конечной концентрации 100 нМ для клеток A549. Через 2 дня и через 4 дня среду заменяют (для клеток PANC-1 используют RPMI 1640, содержащую 10% FBS, для клеток A549 и клеток MIA PaCa-2 используют DMEM, содержащую 10% FBS, для клеток HCT116 используют McCoy, содержащую 10% FBS). Опять же, миРНК GST-π или миРНК P21 в конечной концентрации 25 нМ (во всех случаях используют контрольную миРНК в конечной концентрации 25 нМ) добавляют в чашки Петри, содержащие клетки PANC-1, клетки MIA PaCa-2 или клетки HCT116, и в конечной концентрации 50 нМ (с использованием контрольной миРНК в конечной концентрации 50 нМ) в чашки Петри, содержащие клетки A549. В данном случае используемую в качестве контроля контрольную миРНК добавляют к клеткам PANC-1 или MIA PaCa-2 в конечной концентрации 50 нМ, а к клеткам A549 в конечной концентрации 100 нМ. Затем клетки культивируют без замены среды. Через семь дней после инокуляции клетки отделяют путем обработки трипсином, собирают из чашек Петри и считают число клеток. В данном случае также получают изображения клеток с использованием фазоразностной автофокусировки.
Результаты измерения числа клеток A549, клеток PANC-1 и клеток MIA PaCa-2 показаны на фигуре 4. Результаты измерения числа клеток HCT116 показаны на фигуре 5. Кроме того, фазоразностное изображение клеток A549 показано на фигуре 6. Фазоразностное изображение клеток MIA PaCa-2 показано на фигуре 7. Фазоразностное изображение клеток PANC-1 показано на фигуре 8. Фазоразностное изображение клеток HCT116 показано на фигуре 9.
На фигурах 4 и 5 показано, что трехкратный нокдаун GST-π и P21 с использованием миРНК GST-π и миРНК P21 приводит к уменьшению числа раковых клеток (клеток A549, клеток MIA PaCa-2, клеток PANC-1 и клеток HCT116), экспрессирующих мутантный KRAS, через 7 дней после инокуляции по сравнению с числом клеток сразу после инокуляции; таким образом, полученные результаты подтверждают, что можно индуцировать гибель клеток.
Более того, как показано на фигурах 6-9, клетки каждой клеточной линии (клетки A549, клетки MIA PaCa-2, клетки PANC-1 и клетки HCT116), экспрессирующие мутантный KRAS, которые подвергают нокдауну GST-π путем воздействия миРНК GST-π, становятся плоскими и большими; следовательно, можно предположить, что индуцируется клеточное старение. Также обнаружено, что, если клетки подвергают нокдауну и GST-π, и P21 путем воздействия миРНК GST-π и миРНК P21, старение-подобный фенотип, наблюдающийся при нокдауне GST-π, исчезает. Полученные результаты позволяют предположить, что, если клетки подвергают нокдауну и GST-π, и P21 путем воздействия миРНК GST-π и миРНК P21, старение клеток, вызванное нокдауном GST-π, ингибируется нокдауном P21.
Может ли нокдаун GST-π индуцировать старение клеток, как показано на фигурах 6-9, определяют с использованием клеток M7609. Вначале миРНК GST-π добавляют в конечной концентрации 30 нМ в чашку Петри, содержащую клетки M7609. Через 1 день и через 2 дня среду заменяют (RPMI 1640, содержащая 10% FBS). В чашку Петри, содержащую клетки M7609, снова добавляют миРНК GST-π в конечной концентрации 30 нМ. Затем клетки культивируют с заменой среды через день. Через тринадцать дней после инокуляции клетки окрашивают с помощью набора для окрашивания β-галактозидазы Senescence (Cell Signaling Technology, Inc.) в соответствии с рекомендуемым методом. Получают фазоразностное изображение. Результаты приведены на фигуре 10. Как показано на фигуре 10, клетки M7609 с нокдауном GST-π становятся плоскими и крупными, причем в клетках, имеющих такой фенотип, наблюдается развитие синего окрашивания, обусловленного β-галактозидазой; таким образом, полученные результаты демонстрируют, что индуцируется старение клеток.
Кроме того, путем измерения уровня экспрессии гена апоптоз-индуцирующего фактора PUMA определяют, действительно ли гибель раковых клеток, экспрессирующих мутантный KRAS, индуцированная нокдауном GST-π и P21, является результатом апоптоза.
Вначале в чашки Петри, содержащие клетки A549 или клетки MIA PaCa-2, добавляют миРНК GST-π и миРНК P21, каждую в конечной концентрации 50 нМ, или миРНК GST-π или миРНК P21 в конечной концентрации 50 нМ (в каждом случае используют контрольную миРНК в конечной концентрации 50 нМ). В качестве контроля добавляют контрольную миРНК в конечной концентрации 100 нМ. После культивирование в течение 1 дня без замены среды определяют уровень мРНК PUMA методом количественной ПЦР с использованием системы для проведения ПЦР в режиме реального времени 7300 (Applied Biosystems, Inc.).
Результаты приведены на фигуре 11. Результаты, показанные на фигуре 11, демонстрируют, что уровень мРНК апоптоз-стимулирующего фактора PUMA резко возрастает в результате совместного нокдауна GST-π и p21, вызванного миРНК GST-π и миРНК Р21. Полученные результаты свидетельствуют о том, что гибель клеток, индуцированная совместным нокдауном GST-π и P21, опосредуется апоптозом.
В клетках присутствуют группы белков, связанных с апоптозом. Связанные с апоптозом белки ориентировочно подразделяют на две группы: группа белков, ингибирующих апоптоз, и группа белков, индуцирующих апоптоз. Группа белков, ингибирующих апоптоз, включает в себя Bcl-2, Bcl-XL, Bcl-W, MCL-1 и Bcl-B. А группа белков, индуцирующих апоптоз, включает в себя Bax, Bak, BOK, BIM, BID, BAD, NOXA и PUMA. Как правило, белки, ингибирующие апоптоз, такие как Bcl-2, Bcl-XL и MCL-1, находятся на наружных мембранах митохондрий и ингибируют высвобождение цитохрома C, что приводит к ингибированию апоптоза. С другой стороны, группы белков, индуцирующих апоптоз, таких как Bax, BIM, BID и BAD, находятся в цитоплазме, но транслоцируются в митохондриальные наружные мембраны в ответ на сигналы смерти и стимулируют высвобождение цитохрома C, индуцируя апоптоз.
Кроме того, после активации вследствие повреждения ДНК и т.п. р53 стимулирует транскрипцию Bax, NOXA и PUMA, приводящую к индукции апоптоза. В частности, PUMA представляет собой белок, который был выделен как апоптоз-индуцирующий белок, активируемый p53. PUMA непосредственно связывается с Bcl-2, тем самым ингибируя апоптоз-ингибирующий эффект Bcl-2 и индуцируя апоптоз клетки.
Результаты, полученные в эксперименте 1, демонстрируют, что воздействие лекарственного средства, ингибирующего GST-π, и лекарственного средства, ингибирующего Р21, на раковые клетки приводит к значительному ингибированию роста клеток и эффективной индукции последующей гибели клеток. Но если на раковые клетки воздействует только лекарственное средство, ингибирующее GST-π, гибель клеток не индуцируется, хотя рост клеток может ингибироваться. И если на раковые клетки воздействует только лекарственное средство, ингибирующее Р21, рост клеток может ингибироваться лишь в незначительной степени. Соответственно, тот факт, что гибель раковых клеток может индуцироваться при воздействии обоих указанных лекарственных средств, является неожиданным.
[Эксперимент 2]
В эксперименте 1 синтетическую летальность раковых клеток вызывают с помощью миРНК GST-π и миРНК P21. В эксперименте 2 скринингу подвергают белок, регулирующий клеточный цикл, который вызывает синтетическую летальность при ингибировании наряду с GST-π.
Вначале получают суспензию клеток MIA PaCa-2 с концентрацией 1×104 клеток/мл в среде DMEM, содержащей 10% FBS и 1% L-глутамина, затем полученную суспензию вносят в объеме 100 мкл/лунку в 96-луночный планшет и культивируют в течение 18 часов в DMEM, содержащей 10% FBS и 1% L-глутамина. Клетки MIA PaCa-2, достигшие 20-30% слияния, трансфицируют миРНК-2 GST-π и/или миРНК, направленной против гена-мишени, с использованием Lipofectamine RNAi MAX следующим образом.
Смешанный раствор для трансфекции, содержащий липофектамин/миРНК, получают следующим образом: вначале 51 мкл воды, не содержащей ДНКазы (Ambion, Life Technologies Corp.), добавляют к 0,1 нмоль каждой миРНК, содержащейся в пуле Human siGENOME библиотека миРНК-Регуляция клеточного цикла-SMART (GE Healthcare Dharmacon Inc.), и оставляют стоять при комнатной температуре в течение 90 минут. Получают раствор миРНК путем добавления к указанному водному раствору миРНК 19,9 мкл OPTI-MEM (раствор A). Затем 50 мкМ водный раствор миРНК-2 GST-π и 50 мкМ водный раствор контрольной миРНК разводят OPTI-MEM в десять раз с получением разведенных растворов, содержащих 5 мкМ миРНК-2 GST-π и 5 мкМ контрольной миРНК (раствор B). Смешивают 31,2 мкл раствора A и 8,8 мкл раствора B (с получением раствора C). Получают раствор липофектамина путем смешивания 150 мкл Lipofectamine RNAi MAX и 2,35 мл OPTI-MEM (раствор D). Затем смешивают 37,5 мкл раствора C и 37,5 мкл раствора D и оставляют стоять при комнатной температуре в течение 15 минут (с получением раствора E).
Раствор E добавляют в каждую лунку 96-луночного планшета, содержащую клетки MIA-PaCa-2, в количестве 10 мкл/лунку. Отдельно смешивают 50 мкМ водный раствор контрольной миРНК (5,5 мкл) и OPTI-MEM (189,5 мкл) (с получением раствора F). Затем 50 мкМ водный раствор контрольной миРНК разбавляют OPTI-MEM в десять раз с получением 5 мкМ раствора контрольной миРНК (раствор G). Смешивают 31,2 мкл раствора F и 8,8 мкл раствора G (с получением раствора H). Получают раствор липофектамина путем смешивания 150 мкл Lipofectamine RNAi MAX и 2,35 мл OPTI-MEM (раствор I). Затем смешивают 37,5 мкл раствора H и 37,5 мкл раствора I и оставляют стоять при комнатной температуре в течение 15 минут (с получением раствора J). Раствор J добавляют в каждую лунку 96-луночного планшета, содержащую клетки MIA-PaCa-2, в количестве 10 мкл/лунку. Затем клетки культивируют в среде DMEM, содержащей 10% FBS и 10% L-глутамина. Через 5 дней проводят тестирование роста с помощью набора для анализа клеточной пролиферации CyQUANT NF (Invitrogen Corp.).
Вначале буфер 1×HBSS добавляют к 22 мкл окрашивающего реагента CyQUANT NF с получением окрашивающего реакционного раствора для анализа клеточной пролиферации CyQUANT NF. Среду, в которой культивируют упомянутые выше трансфицированные клетки, отсасывают и добавляют 50 мкл окрашивающего реакционного раствора. Клетки оставляют окрашиваться при 37°C в течение 30 минут. Затем наблюдают флуоресценцию при длине волны 520 нм с возбуждением при длине волны 480 нм.
Результаты показаны на фигуре 12. В результате скрининга 170 типов генов, кодирующих белки, регулирующие клеточный цикл, на способность вызывать синтетическую летальность с GST-π, как показано на фигуре 12, ATM, CDC25A, PRKDC, RBBP8, SKP2, MCM10, RNPC1, CCNL1, CENPH, BRSK1, MCM8, CCNB3 и MCMDC1, помимо P21, исследованного в эксперименте 1, подвергают скринингу в качестве белков, регулирующих клеточный цикл, которые могут вызывать синтетическую летальность при ингибирования наряду с GST-π. Среди них P21, RNPC1, CCNL1, MCM8, CCNB3 и MCMDC1 подвергают скринингу в качестве белков, регулирующих клеточный цикл, которые лишь незначительно ингибируют рост клеток (степень ингибирования роста менее 20%), если их ингибируют в одиночку, но вызывают синтетическую летальность только при ингибировании наряду с GST-π. Соответственно, можно сделать вывод, что лекарственное средство, ингибирующее белок, регулирующий клеточный цикл, выбранный из P21, RNPC1, CCNL1, MCM8, CCNB3 и MCMDC1, само по себе обладает очень низкой токсичностью и характеризуется превосходной безопасностью.
[Эксперимент 3]
В эксперименте 2 скринингу подвергают белок, регулирующий клеточный цикл и вызывающий синтетическую летальность при ингибировании наряду с GST-π. В эксперименте 3 проводят скрининг белка, обладающего антиапоптотической активностью и вызывающего синтетическую летальность при ингибировании наряду с GST-π.
Вначале получают суспензию клеток MIA PaCa-2 с концентрацией 1×104 клеток/мл в среде DMEM, содержащей 10% FBS и 1% L-глутамина, затем полученную суспензию вносят в объеме 100 мкл/лунку в 96-луночный планшет и культивируют в течение 18 часов в DMEM, содержащей 10% FBS и 1% L-глутамина. Клетки MIA PaCa-2, достигшие 20-30% слияния, трансфицируют миРНК-2 GST-π и/или миРНК, направленной против гена-мишени, с использованием Lipofectamine RNAi MAX следующим образом.
Смешанный раствор для трансфекции, содержащий липофектамин/миРНК, получают следующим образом: вначале 51 мкл воды, не содержащей ДНКазы (Ambion, Life Technologies Corp.), добавляют к 0,1 нмоль каждой миРНК, содержащейся в полученной по индивидуальному заказу библиотеке миРНК, для которой были специально выбраны 140 типов генов, по имеющимся данным обладающих антиапоптотической активностью (siGENOME SMART pool Cherry-pick Library, GE Healthcare Dharmacon Inc.), и оставляют стоять при комнатной температуре в течение 90 минут. Получают раствор миРНК путем добавления к указанному водному раствору миРНК 19,9 мкл OPTI-MEM (раствор A). Затем 50 мкМ водный раствор миРНК-2 GST-π и 50 мкМ водный раствор контрольной миРНК разводят OPTI-MEM с получением 5 мкМ раствора миРНК-2 GST-π и 5 мкМ раствора контрольной миРНК (раствор B). Смешивают 31,2 мкл раствора A и 8,8 мкл раствора B (с получением раствора C). Получают раствор липофектамина путем смешивания 150 мкл Lipofectamine RNAi MAX и 2,35 мл OPTI-MEM (раствор D). Затем смешивают 37,5 мкл раствора C и 37,5 мкл раствора D и оставляют стоять при комнатной температуре в течение 15 минут (с получением раствора E). Раствор E добавляют в каждую лунку 96-луночного планшета, содержащую культивируемые клетки MIA-PaCa-2, в количестве 10 мкл/лунку.
Отдельно смешивают 50 мкМ водный раствор контрольной миРНК (5,5 мкл) и OPTI-MEM (189,5 мкл) (с получением раствора F). Затем 50 мкМ водный раствор контрольной миРНК разбавляют OPTI-MEM в десять раз с получением 5 мкМ раствора контрольной миРНК (раствор G). Смешивают 31,2 мкл раствора F и 8,8 мкл раствора G (с получением раствора H). Получают раствор липофектамина путем смешивания 150 мкл Lipofectamine RNAi MAX и 2,35 мл OPTI-MEM (раствор I). Затем смешивают 37,5 мкл раствора H и 37,5 мкл раствора I и оставляют стоять при комнатной температуре в течение 15 минут (с получением раствора J). Раствор J добавляют в каждую лунку 96-луночного планшета, содержащую культивируемые клетки MIA-PaCa-2, в количестве 10 мкл/лунку. Затем клетки культивируют в среде DMEM, содержащей 10% FBS и 1% L-глутамина. Через 5 дней проводят тестирование роста с помощью набора для анализа клеточной пролиферации CyQUANT NF (Invitrogen Corp.).
Вначале 11 мл буфера 1×HBSS добавляют к 22 мкл окрашивающего реагента CyQUANT NF с получением окрашивающего реакционного раствора для анализа клеточной пролиферации CyQUANT NF. Среду, в которой культивируют упомянутые выше трансфицированные клетки, отсасывают и добавляют 50 мкл окрашивающего реакционного раствора. Клетки оставляют стоять при 37°C в течение 30 минут. Затем наблюдают флуоресценцию при длине волны 520 нм с возбуждением при длине волны 480 нм.
Результаты показаны на фигуре 13. В результате скрининга 140 типов генов, кодирующих белки с антиапоптотической активностью, на способность вызывать синтетическую летальность с GST-π, как показано на фигуре 13, обнаружено, что AATF, ALOX12, ANXA1, ANXA4, API5, ATF5, AVEN, AZU1, BAG1, BCL2L1, BFAR, CFLAR, IL2, MALT1, MCL1, MKL1, MPO, MTL5, MYBL2 и MYO18A, можно подвергать скринингу в качестве белков, обладающих антиапоптотической активностью, которые могут вызывать синтетическую летальность при ингибирования наряду с GST-π.
[Эксперимент 4]
В эксперименте 2 подвергают скринингу белок, регулирующий клеточный цикл и вызывающий синтетическую летальность при ингибировании наряду с GST-π. В эксперименте 3 подвергают скринингу белок, обладающий антиапоптотической активностью и вызывающий синтетическую летальность при ингибировании наряду с GST-π. В эксперименте 4 проводят скрининг белка, связанного с сигнальным путем PI3K, и вызывающего синтетическую летальность при ингибировании наряду с GST-π.
Вначале получают суспензию клеток MIA PaCa-2 с концентрацией 1×104 клеток/мл в среде DMEM, содержащей 10% FBS и 1% L-глутамина, затем полученную суспензию вносят в объеме 100 мкл/лунку в 96-луночный планшет и культивируют в течение 18 часов в DMEM, содержащей 10% FBS и 1% L-глутамина. Клетки MIA PaCa-2, достигшие 20-30% слияния, трансфицируют миРНК-2 GST-π и/или миРНК, направленной против гена-мишени, с использованием Lipofectamine RNAi MAX следующим образом.
Смешанный раствор для трансфекции, содержащий липофектамин/миРНК, получают следующим образом: вначале 51 мкл воды, не содержащей ДНКазы (Ambion, Life Technologies Corp.), добавляют к 0,1 нмоль каждой миРНК, содержащейся в полученной по индивидуальному заказу библиотеке миРНК, для которой были специально выбраны 80 типов генов, по имеющимся данным участвующих в сигнальном пути PI3K (siGENOME SMARTpool Cherry-pick Library, GE Healthcare Dharmacon Inc.), и оставляют стоять при комнатной температуре в течение 90 минут. Получают раствор миРНК путем добавления к указанному водному раствору миРНК 19,9 мкл OPTI-MEM (раствор A). Затем 50 мкМ водный раствор миРНК-2 GST-π и 50 мкМ водный раствор контрольной миРНК разводят OPTI-MEM в десять раз с получением 5 мкМ раствора миРНК-2 GST-π и 5 мкМ раствора контрольной миРНК (раствор B). Смешивают 31,2 мкл раствора A и 8,8 мкл раствора B (с получением раствора C). Получают раствор липофектамина путем смешивания 150 мкл Lipofectamine RNAi MAX и 2,35 мл OPTI-MEM (раствор D). Затем смешивают 37,5 мкл раствора C и 37,5 мкл раствора D и оставляют стоять при комнатной температуре в течение 15 минут (с получением раствора E). Раствор E добавляют в каждую лунку 96-луночного планшета, содержащую культивируемые клетки MIA-PaCa-2, в количестве 10 мкл/лунку.
Отдельно смешивают 50 мкМ водный раствор контрольной миРНК (5,5 мкл) и OPTI-MEM (189,5 мкл) (с получением раствора F). Затем 50 мкМ водный раствор контрольной миРНК разбавляют OPTI-MEM в десять раз с получением 5 мкМ раствора контрольной миРНК (раствор G). Смешивают 31,2 мкл раствора F и 8,8 мкл раствора G (с получением раствора H). Получают раствор липофектамина путем смешивания 150 мкл Lipofectamine RNAi MAX и 2,35 мл OPTI-MEM (раствор I). Затем смешивают 37,5 мкл раствора H и 37,5 мкл раствора I и оставляют стоять при комнатной температуре в течение 15 минут (с получением раствора J). Раствор J добавляют в каждую лунку 96-луночного планшета, содержащую культивируемые клетки MIA-PaCa-2, в количестве 10 мкл/лунку, с получением контроля. Затем клетки культивируют в среде DMEM, содержащей 10% FBS и 1% L-глутамина. Через 5 дней проводят тестирование роста с помощью набора для анализа клеточной пролиферации CyQUANT NF (Invitrogen Corp.).
Вначале 11 мл буфера 1×HBSS добавляют к 22 мкл окрашивающего реагента CyQUANT NF с получением окрашивающего реакционного раствора для анализа клеточной пролиферации CyQUANT NF. Среду, в которой культивируют упомянутые выше трансфицированные клетки, отсасывают и добавляют 50 мкл окрашивающего реакционного раствора. Клетки оставляют стоять при 37°C в течение 30 минут. Затем наблюдают флуоресценцию при длине волны 520 нм с возбуждением при длине волны 480 нм.
Результаты показаны на фигуре 14. В результате скрининга 80 типов генов, кодирующих белки, связанные с сигнальным путем PI3K, на способность вызывать синтетическую летальность с GST-π, как показано на фигуре 14, обнаружено, что MTOR, IRAK1, IRS1, MYD88, NFKB1, PIK3CG, RAC1, AKT3, EIF4B, EIF4E, ILK, MTCP1, PIK3CA и SRF, можно подвергать скринингу в качестве белков, связанных с сигнальным путем PI3K, которые могут вызывать синтетическую летальность при ингибирования наряду с GST-π. Обнаружено, что среди указанных белков MTOR, IRAK1, IRS1, MYD88, NFKB1, PIK3CG и RAC1 обладают высокой способностью подавлять рост клеток при ингибировании наряду с GST-π.
[Эксперимент 5]
В эксперименте 5 белок, регулирующий клеточный цикл MYLK (HEPATOLOGY, Vol. 44, No. 1, 2006, 152-163), анализируют на способность вызывать синтетическую летальность в отношении клеток A549 (клетки рака легкого человека, несущие мутацию KRAS) при ингибировании наряду с GST-π.
Вначале, за день до трансфекции миРНК, в каждую лунку вносят 0,25×105 клеток A549/лунку вместе с 2,25 мл DMEM (не содержащей антибиотики) содержащей 10% FBS. В данном эксперименте используют 6-луночный планшет и каждый образец тестируют с тройными повторами. Культивируемые клетки A549 трансфицируют миРНК-3 GST-π и/или миРНК MYLK (используют два типа (MYLKa и MYLKb)) с использованием Lipofectamine RNAi MAX следующим образом.
Смешанный раствор для трансфекции, содержащий липофектамин/миРНК, получают следующим образом: вначале получают раствор миРНК путем смешивания 50 нМ раствора миРНК-3 GST-π, 50 нМ раствора миРНК MYLK (по 0,5 мкл каждого) и 124 мкл OPTI-MEM. В качестве контрольных растворов миРНК получают раствор, содержащий сочетание контрольной миРНК и миРНК-3 GST-π, и раствор, содержащий сочетание контрольной миРНК и миРНК MYLK, с одинаковыми концентрациями миРНК. Кроме того, получают раствор липофектамина путем смешивания 7,5 мкл of Lipofectamine RNAi MAX и 117,5 мкл OPTI-MEM. Затем полученный раствор каждой миРНК и раствор липофектамина смешивают и оставляют стоять при комнатной температуре в течение 5 минут.
2,5 мл полученного смешанного раствора добавляют по каплям в каждую лунку (конечная концентрация миРНК: 20 нМ). Затем клетки культивируют при 37°C в течение 75 часов в атмосфере 5% CO2 при легком встряхивании. После завершения культивирования проводят анализ роста по способу, описанному в экспериментах 1-4. Результаты приведены на фигуре 15. Как показано на фигуре 15, обнаружено, что MYLK, белок, регулирующий клеточный цикл, способен вызывать синтетическую летальность раковых клеткок при ингибировании наряду с GST-π. Результаты, приведенные на фигуре 15, демонстрируют, что GST-π-ингибирующее средство и MYLK-ингибирующее средство подавляют рост клеток лишь в небольшой степени, если они воздействуют на раковые клетки по отдельности, тогда как при совместном воздействии на раковые клетки GST-π-ингибирующее средство и MYLK-ингибирующее средство очень сильно подавляют рост клеток.
Используют миРНК, описанные ниже. В приведенном ниже описании буквы верхнего регистра используют для обозначения РНК, буквы нижнего регистра используют для обозначения ДНК.
миРНК-3 GST-π:
Смысловая цепь: CCUUUUGAGACCCUGCUGUtt (SEQ ID NO: 109)
Антисмысловая цепь: ACAGCAGGGUCUCAAAAGGtt (SEQ ID NO: 110)
MYLKa:
Смысловая цепь: CUGGGGAAGAAGGUGAGUAtt (SEQ ID NO: 111)
Антисмысловая цепь: UACUCACCUUCUUCCCCAGtt (SEQ ID NO: 112)
MYLKb:
Смысловая цепь: CAAGAUAGCCAGAGUUUAAtt (SEQ ID NO: 113)
Антисмысловая цепь: UUAAACUCUGGCUAUCUUGtt (SEQ ID NO: 114)
Все цитирующиеся здесь публикации, патенты и заявки на патенты включены в настоящее описание в качестве ссылки во всей их полноте.

































































































































































































































































































































































































































































































































































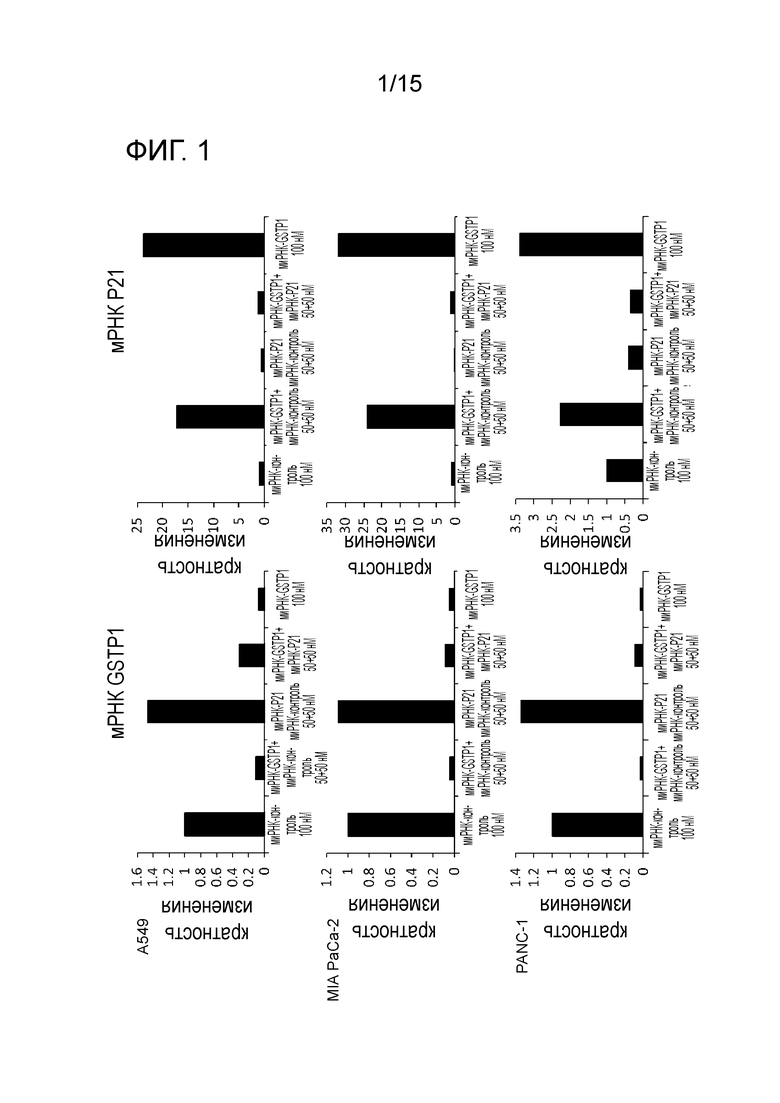


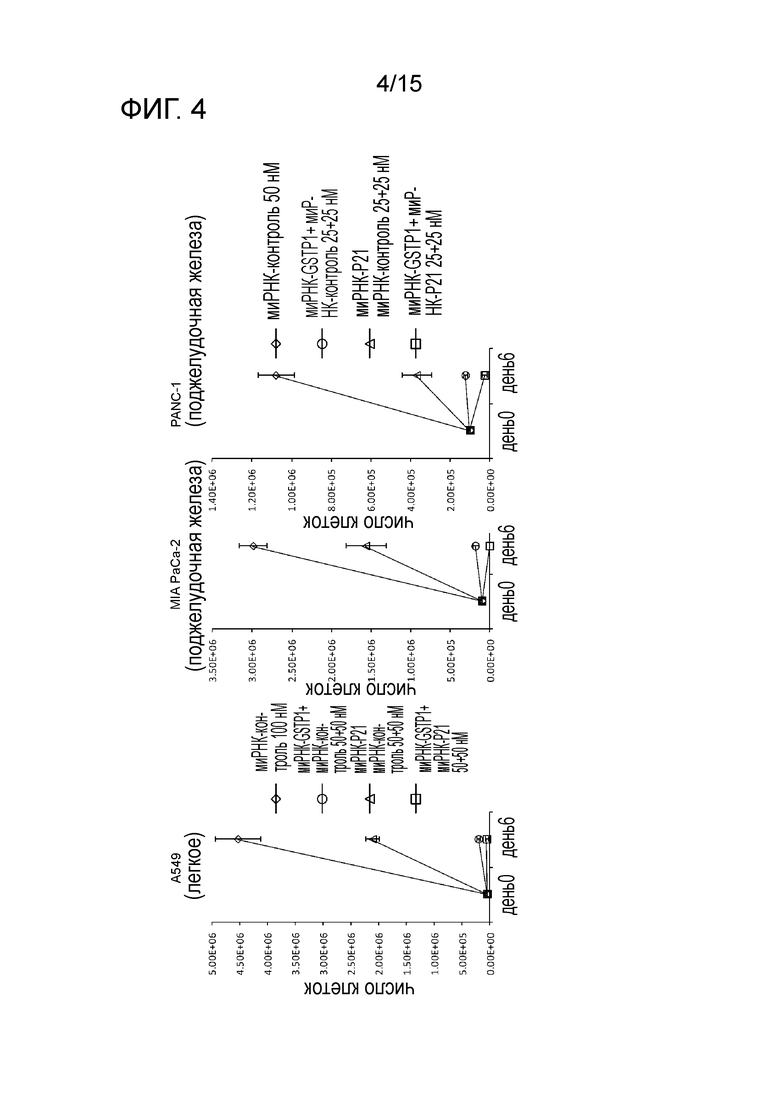
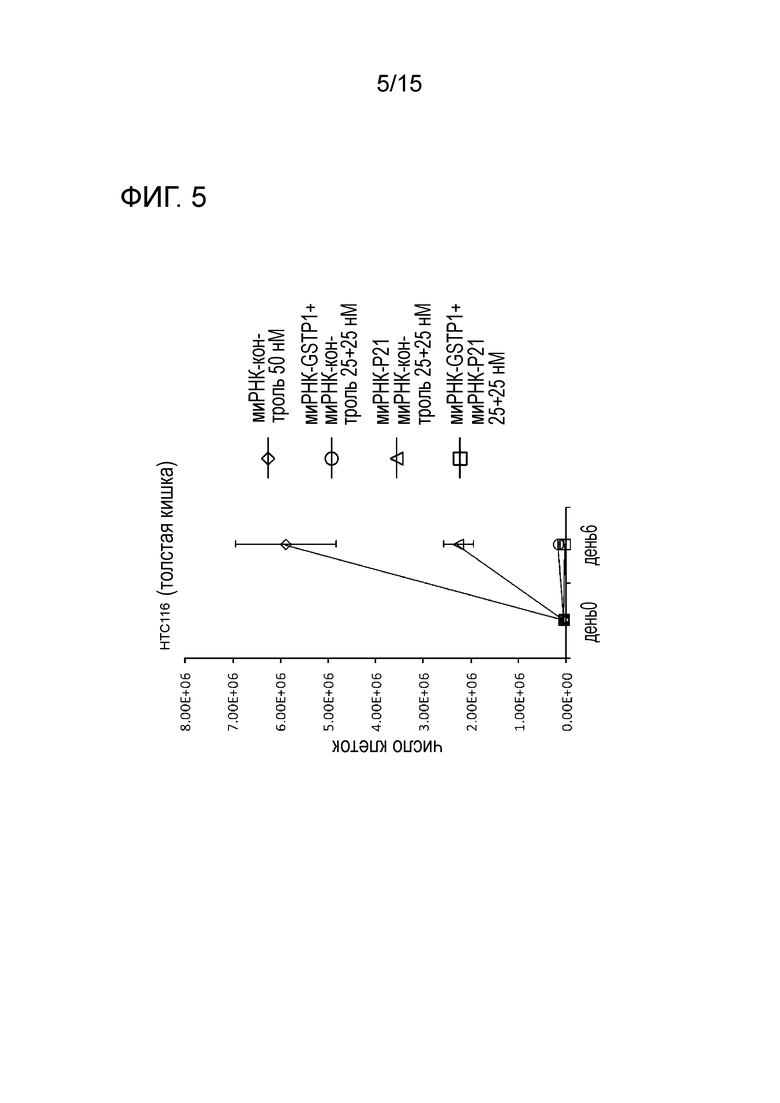

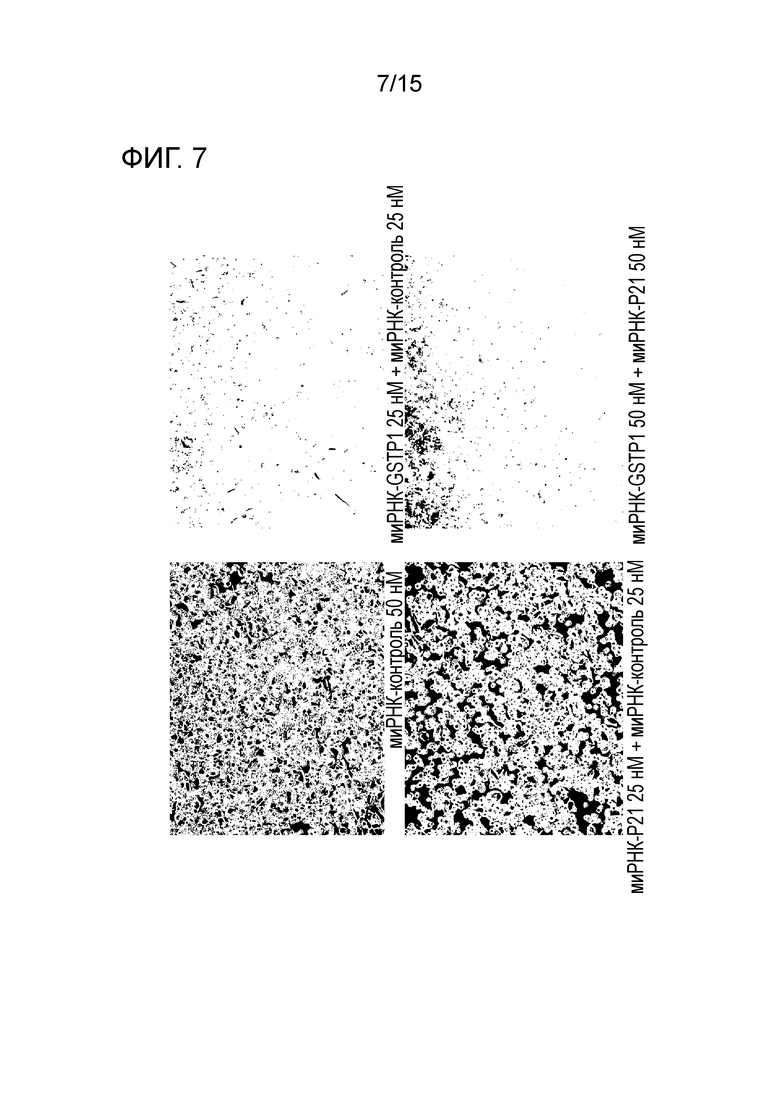

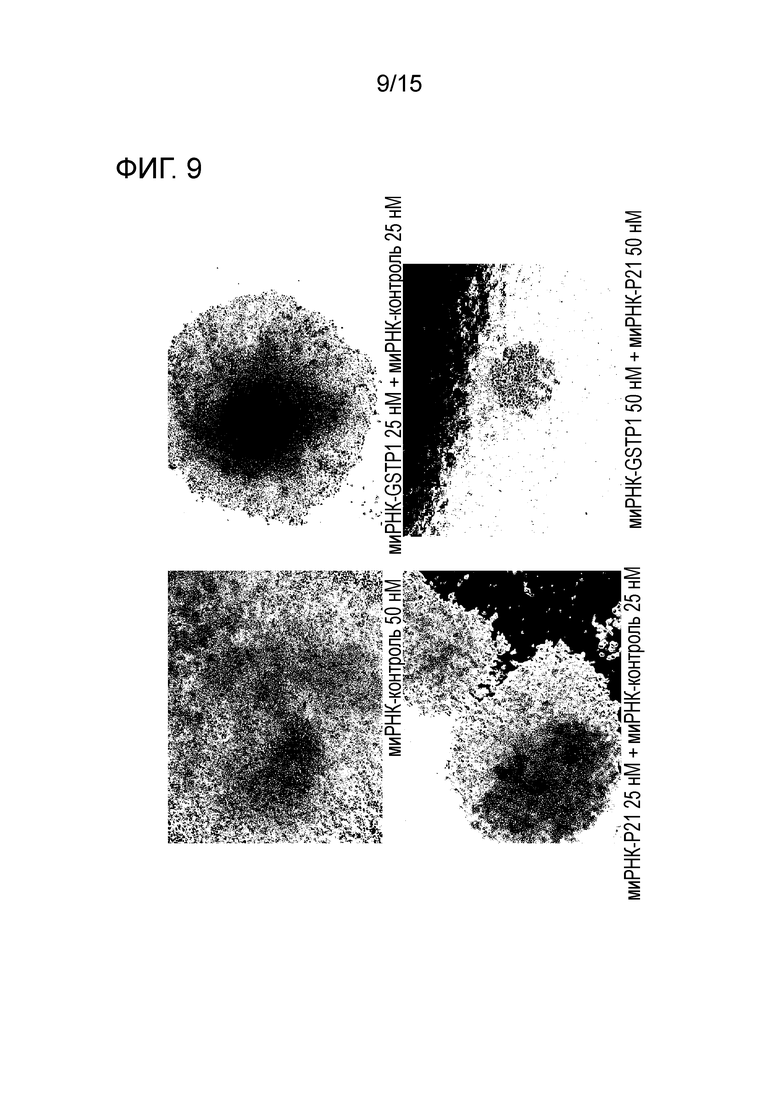

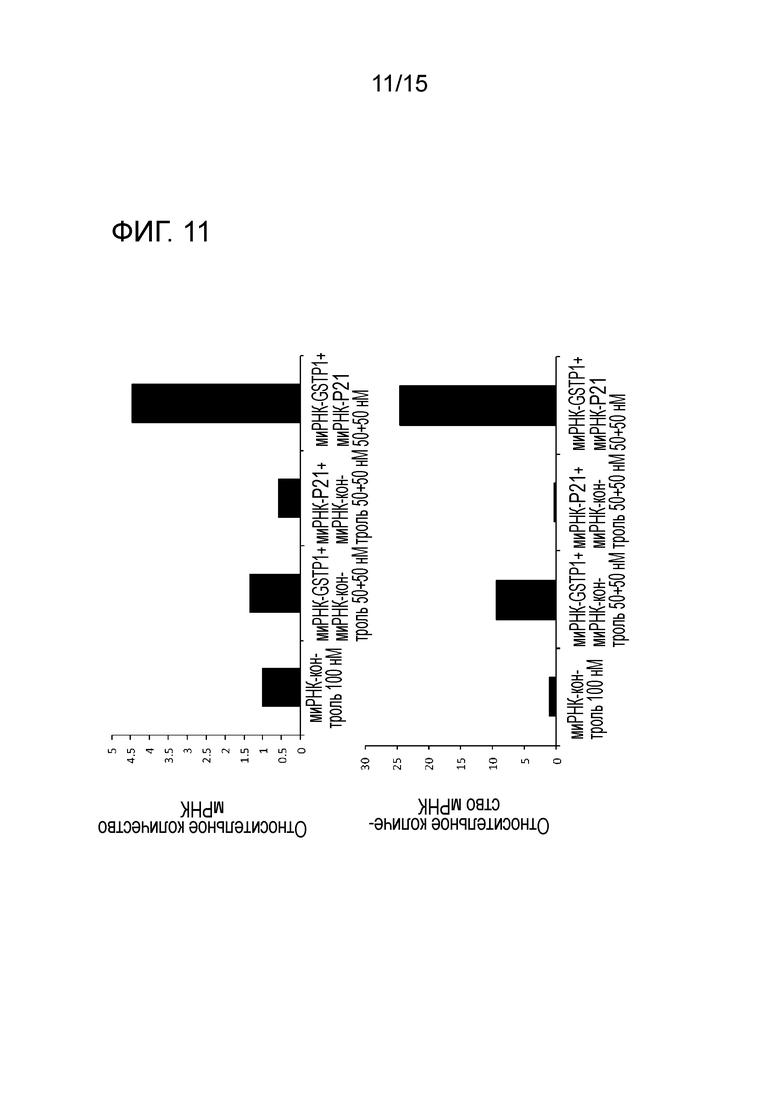
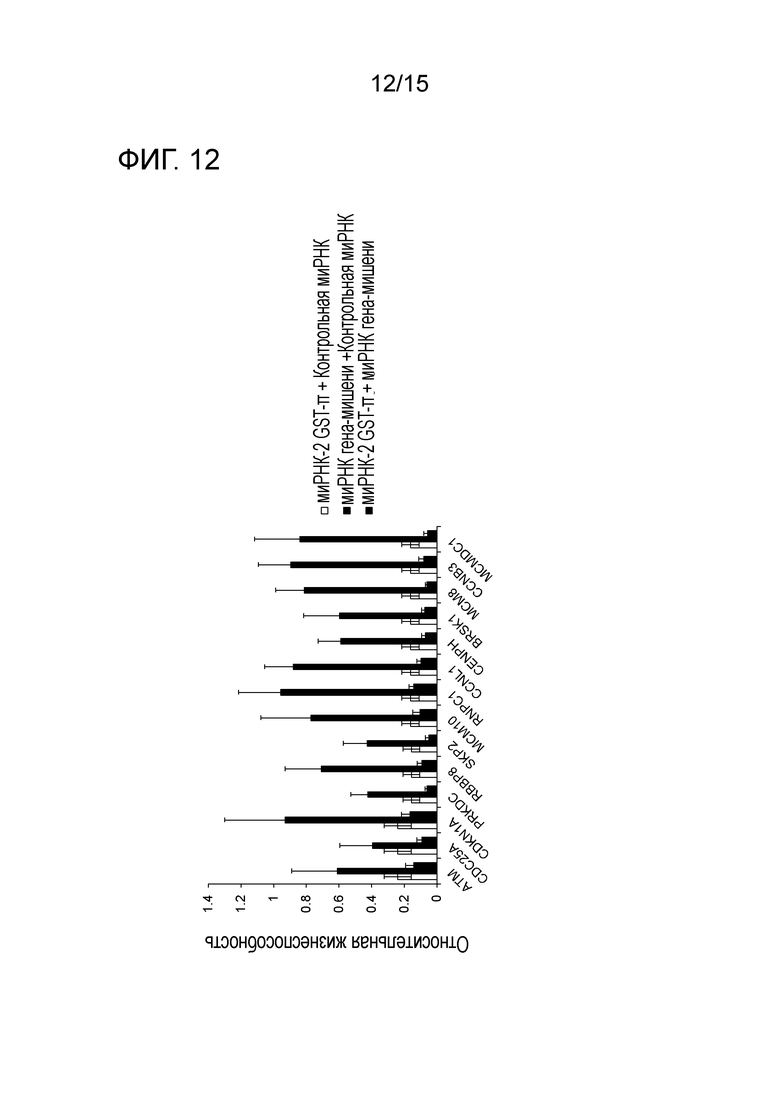

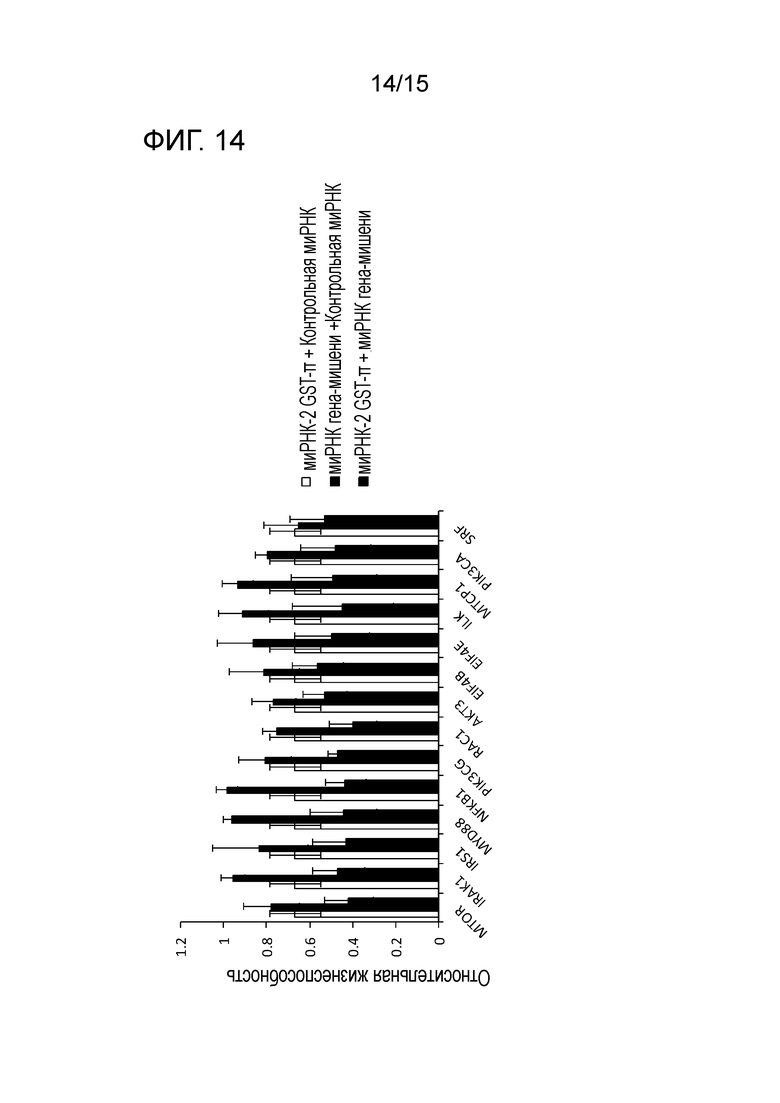
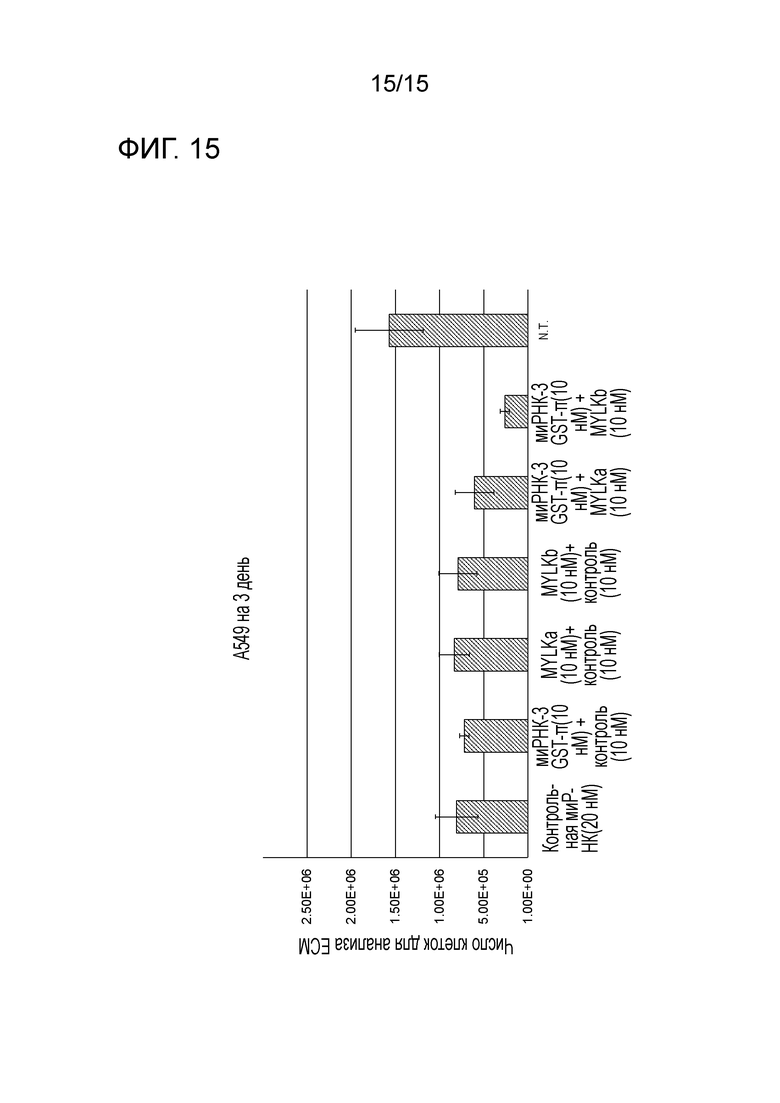
Группа изобретений относится к лечению заболеваний, вызванных аномальным клеточным ростом. Средство, индуцирующее гибель раковых клеток, содержит лекарственное средство, ингибирующее GST-π, и лекарственное средство, ингибирующее связанный с гомеостазом белок, выбранный из группы, состоящей из белка, регулирующего клеточный цикл, - ATM, CDC25A, p21, PRKDC, RBBP8, SKP2, MCM10, RNPC1, CCNL1, CENPH, BRSK1, MCM8, CCNB3, MCMDC1 и MYLK, белка, связанного с подавлением апоптоза, - AATF, ALOX12, ANXA1, ANXA4, API5, ATF5, AVEN, AZU1, BAG1, BCL2L1, BFAR, CFLAR, IL2, MALT1, MCL1, MKL1, MPO, MTL5, MYBL2 и MYO18A, и белка, связанного с сигнальным путем PI3K, - MTOR, IRAK1, IRS1, MYD88, NFKB1, PIK3CG, RAC1, AKT3, EIF4B, EIF4E, ILK, MTCP1, PIK3CA и SRF. Также раскрыты средство, ингибирующее рост раковых клеток, фармацевтическая композиция для лечения заболевания, вызванного аномальным клеточным ростом, комбинация для индуцирования гибели раковых клеток, комбинация для ингибирования роста раковых клеток, способ лечения заболевания, вызванного аномальным ростом клеток. Группа изобретений обеспечивает эффективное лечение раковых заболеваний. 6 н. и 7 з.п. ф-лы, 15 ил., 5 пр.
1. Средство, индуцирующее гибель раковых клеток, содержащее, в качестве активных ингредиентов, лекарственное средство, ингибирующее GST-π, и лекарственное средство, ингибирующее связанный с гомеостазом белок, выбранный из группы, состоящей из белка, регулирующего клеточный цикл, белка, связанного с подавлением апоптоза, и белка, связанного с сигнальным путем PI3K,
где белок, регулирующий клеточный цикл, представляет собой по меньшей мере один белок, выбранный из группы, состоящей из ATM, CDC25A, p21, PRKDC, RBBP8, SKP2, MCM10, RNPC1, CCNL1, CENPH, BRSK1, MCM8, CCNB3, MCMDC1 и MYLK;
где связанный с подавлением апоптоза белок представляет собой, по меньшей мере, один белок, выбранный из группы, состоящей из AATF, ALOX12, ANXA1, ANXA4, API5, ATF5, AVEN, AZU1, BAG1, BCL2L1, BFAR, CFLAR, IL2, MALT1, MCL1, MKL1, MPO, MTL5, MYBL2 и MYO18A; и
где белок, связанный с сигнальным путем PI3K, представляет собой, по меньшей мере, один белок, выбранный из группы, состоящей из MTOR, IRAK1, IRS1, MYD88, NFKB1, PIK3CG, RAC1, AKT3, EIF4B, EIF4E, ILK, MTCP1, PIK3CA и SRF.
2. Средство, ингибирующее рост раковых клеток, содержащее, в качестве активных ингредиентов, лекарственное средство, ингибирующее GST-π, и лекарственное средство, ингибирующее связанный с гомеостазом белок, выбранный из группы, состоящей из белка, регулирующего клеточный цикл, белка, связанного с подавлением апоптоза, и белка, связанного с сигнальным путем PI3K,
где белок, регулирующий клеточный цикл, представляет собой по меньшей мере один белок, выбранный из группы, состоящей из ATM, CDC25A, p21, PRKDC, RBBP8, SKP2, MCM10, RNPC1, CCNL1, CENPH, BRSK1, MCM8, CCNB3, MCMDC1 и MYLK;
где связанный с подавлением апоптоза белок представляет собой, по меньшей мере, один белок, выбранный из группы, состоящей из AATF, ALOX12, ANXA1, ANXA4, API5, ATF5, AVEN, AZU1, BAG1, BCL2L1, BFAR, CFLAR, IL2, MALT1, MCL1, MKL1, MPO, MTL5, MYBL2 и MYO18A; и
где белок, связанный с сигнальным путем PI3K, представляет собой, по меньшей мере, один белок, выбранный из группы, состоящей из MTOR, IRAK1, IRS1, MYD88, NFKB1, PIK3CG, RAC1, AKT3, EIF4B, EIF4E, ILK, MTCP1, PIK3CA и SRF.
3. Средство по п. 1, где лекарственное средство представляет собой вещество, выбранное из группы, включающей в себя молекулу РНКи, рибозим, антисмысловую нуклеиновую кислоту, химерный полинуклеотид ДНК/РНК и вектор, экспрессирующий, по меньшей мере, одно из указанных веществ.
4. Средство по п. 1, где лекарственное средство, ингибирующее связанный с гомеостазом белок, представляет собой соединение, которое оказывает влияние на связанный с гомеостазом белок.
5. Средство по п. 1, где лекарственное средство индуцирует апоптоз.
6. Средство по п. 1, где раковая клетка представляет собой раковую клетку, экспрессирующую GST-π на высоком уровне, где раковая клетка с высокой экспрессией GST-π имеет значительно более высокий уровень экспрессии GST-π, чем у нормальной клетки.
7. Фармацевтическая композиция для лечения заболевания, вызванного аномальным клеточным ростом, содержащая средство по п. 1.
8. Фармацевтическая композиция по п. 7, где заболевание представляет собой рак.
9. Фармацевтическая композиция по п. 8, где рак характеризуется высоким уровнем экспрессии GST-π, где раковая клетка с высокой экспрессией GST-π имеет значительно более высокий уровень экспрессии GST-π, чем у нормальной клетки.
10. Комбинация для индуцирования гибели раковых клеток, содержащая, в качестве активных ингредиентов, лекарственное средство, ингибирующее GST-π, и лекарственное средство, ингибирующее связанный с гомеостазом белок, выбранный из группы, состоящей из белка, регулирующего клеточный цикл, белка, связанного с подавлением апоптоза, и белка, связанного с сигнальным путем PI3K,
где белок, регулирующий клеточный цикл, представляет собой, по меньшей мере, один белок, выбранный из группы, состоящей из ATM, CDC25A, p21, PRKDC, RBBP8, SKP2, MCM10, RNPC1, CCNL1, CENPH, BRSK1, MCM8, CCNB3, MCMDC1 и MYLK;
где связанный с подавлением апоптоза белок представляет собой, по меньшей мере, один белок, выбранный из группы, состоящей из AATF, ALOX12, ANXA1, ANXA4, API5, ATF5, AVEN, AZU1, BAG1, BCL2L1, BFAR, CFLAR, IL2, MALT1, MCL1, MKL1, MPO, MTL5, MYBL2 и MYO18A; и
где белок, связанный с сигнальным путем PI3K, представляет собой, по меньшей мере, один белок, выбранный из группы, состоящей из MTOR, IRAK1, IRS1, MYD88, NFKB1, PIK3CG, RAC1, AKT3, EIF4B, EIF4E, ILK, MTCP1, PIK3CA и SRF.
11. Комбинация для ингибирования роста раковых клеток, содержащая, в качестве активных ингредиентов, лекарственное средство, ингибирующее GST-π, и лекарственное средство, ингибирующее связанный с гомеостазом белок, выбранный из группы, состоящей из белка, регулирующего клеточный цикл, белка, связанного с подавлением апоптоза, и белка, связанного с сигнальным путем PI3K,
где белок, регулирующий клеточный цикл, представляет собой, по меньшей мере, один белок, выбранный из группы, состоящей из ATM, CDC25A, p21, PRKDC, RBBP8, SKP2, MCM10, RNPC1, CCNL1, CENPH, BRSK1, MCM8, CCNB3, MCMDC1 и MYLK;
где связанный с подавлением апоптоза белок представляет собой, по меньшей мере, один белок, выбранный из группы, состоящей из AATF, ALOX12, ANXA1, ANXA4, API5, ATF5, AVEN, AZU1, BAG1, BCL2L1, BFAR, CFLAR, IL2, MALT1, MCL1, MKL1, MPO, MTL5, MYBL2 и MYO18A; и
где белок, связанный с сигнальным путем PI3K, представляет собой, по меньшей мере, один белок, выбранный из группы, состоящей из MTOR, IRAK1, IRS1, MYD88, NFKB1, PIK3CG, RAC1, AKT3, EIF4B, EIF4E, ILK, MTCP1, PIK3CA и SRF.
12. Способ лечения заболевания, вызванного аномальным ростом клеток, включающий стадию введения лекарственного средства, ингибирующего GST-π, и лекарственного средства, ингибирующего связанный с гомеостазом белок, выбранный из группы, состоящей из белка, регулирующего клеточный цикл, белка, связанного с подавлением апоптоза, и белка, связанного с сигнальным путем PI3K,
где белок, регулирующий клеточный цикл, представляет собой, по меньшей мере, один белок, выбранный из группы, состоящей из ATM, CDC25A, p21, PRKDC, RBBP8, SKP2, MCM10, RNPC1, CCNL1, CENPH, BRSK1, MCM8, CCNB3, MCMDC1 и MYLK;
где связанный с подавлением апоптоза белок представляет собой, по меньшей мере, один белок, выбранный из группы, состоящей из AATF, ALOX12, ANXA1, ANXA4, API5, ATF5, AVEN, AZU1, BAG1, BCL2L1, BFAR, CFLAR, IL2, MALT1, MCL1, MKL1, MPO, MTL5, MYBL2 и MYO18A; и
где белок, связанный с сигнальным путем PI3K, представляет собой, по меньшей мере, один белок, выбранный из группы, состоящей из MTOR, IRAK1, IRS1, MYD88, NFKB1, PIK3CG, RAC1, AKT3, EIF4B, EIF4E, ILK, MTCP1, PIK3CA и SRF.
13. Способ по п. 12, в котором заболевание представляет собой рак.
| WO 2014098210 A1, 26.06.2014 | |||
| ХАРКЕВИЧ Д.А | |||
| Фармакология: Учебник, 10-е издание, 2010, стр.72-82 | |||
| WO 2012176282 A1, 27.12.2012. |

