ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к области медицинской биологии, и раскрывает однодоменное антитело и его производные белки против лиганда-1 программируемой смерти клеток (PDL1). В частности, настоящее изобретение раскрывает молекулу, связывающую лиганд-1 белка программируемой смерти клеток (PDL1), и ее использование, в особенности для лечения и/или предупреждения или диагностики заболеваний, связанных с PDL1, таких как опухоли.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Белок программируемой смерти 1 (PD-1) является членом семейства рецепторов CD28, которое включает в себя CD28, CTLA-4, ICOS, PD-1 и BTLA Первоначальные члены семейства CD28 и ICOS были обнаружены функциональным эффектом на усиление пролиферации Т-клеток после добавления моноклональных антител (Hutloff et al. (1999) Nature 397:263-266; Hansen et al. (1980) Immunogenics 10:247-260). На клеточной поверхности для PD-1 было идентифицировано два гликопротеиновых лиганда, PDLI и PDL2, и было показано, что они снижают активацию Т-клеток и секрецию цитокина после связывания с PD-1 (Freeman et al (2000) J Exp Med 192:1027-34; Latchman et al (2001) Nat Immunol 2:261-8; Carter et al. (2002) Eur J Immunol 32:634-43; Ohigashi et al. (2005) Clin Cancer Res 11:2947-53). Как PDLI (B7-H1), так и PDL2 (B7-DC), являются гомологами В7, которые связываются с PD-1, но не связываются с другими членами семейства CD28 (Blank et al. (2004). Также было показано, что экспрессия PDLI на поверхности клетки положительно регулируется (up-регулируется) посредством стимуляции IFN-γ.
Экспрессия PDLI была обнаружена в нескольких видах рака мышей и человека, в том числе в карциномах легких, яичников и ободочной кишки человека, и различных миеломах (Iwai et al (2002) PNAS 99:12293-7; Ohigashi et al. (2005), Clin Cancer Res 11:2947-53). Полученные к настоящему времени данные показывают, что PDL1, сверхэкспрессированный в опухолевых клетках, играет ключевую роль в избегании опухоли от иммунной системы путем усиления апоптоза Т-клеток. Исследователи обнаружили, что опухолевая клеточная линия Р815, трансфицированная геном PDL1, может быть резистентной к лизису, обусловленному CTL, и обладает повышенной онкогенностью и инвазивной активностью. Эти биологические активности могут быть отменены путем блокирования PDL1. Опухолевые клетки, трансплантированные мышам с нокаутом по PDL1 для блокирования взаимодействия PDL1/PD-1, не могут образовывать опухоли. (Dong et al. (2002) Nat Med 8:793-800). Также было высказано предположение, что PD-LI может быть вовлечен в воспаление слизистых оболочек кишечника, и ингибирование PDLI подавляет изнурительную болезнь, связанную с колитом (Kanai et al. (2003) J Immunol 171:4156-63).
В данном уровне техники все еще существует потребность в анти-PDLI антителе, которое может связываться с PDL1 с высокой аффинностью и способно блокировать связывание PDL1 с PD-1, особенно в однодоменном антителе, состоящем только из тяжелой цепи (пер., однодоменном тяжелоцепочечном антителе; или однодоменном антителе тяжелой цепи) против PDL1.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения получили однодоменное антитело, состоящее только из тяжелой цепи (VHH) против PDL1 с высокой специфичностью, высокой аффинностью и высокой стабильностью путем скрининга с использованием технологии фагового дисплея.
В первом аспекте изобретение обеспечивает PDL1-связывающую молекулу, содержащую один вариабельный домен иммуноглобулина, состоящий из аминокислотной последовательности, выбранной из SEQ ID No: 1-6, и Fc-область иммуноглобулина человека.
В другом аспекте настоящее изобретение относится к молекуле нуклеиновой кислоты, кодирующей PDL1-связывающую молекулу, и экспрессирующему вектору и клетке-хозяину, содержащей указанную молекулу нуклеиновой кислоты.
Настоящее изобретение дополнительно относится к фармацевтической композиции, содержащей PDL1-связывающую молекулу по изобретению.
Настоящее изобретение дополнительно относится к способу получения PDL1-связывающей молекулы по изобретению.
Настоящее изобретение дополнительно относится к способу получения PDL1-связывающей молекулы и фармацевтической композиции по изобретению, особенно применению и способу предупреждения и/или лечения заболеваний, связанных с PDL1.
ОПИСАНИЕ ГРАФИЧЕСКОГО МАТЕРИАЛА
На фиг. 1 представлен блокирующий эффект однодоменных тяжелоцепочечных антител против PDL1 на взаимодействие PD1/PDL1.
На фиг. 2 представлены кривые связывания однодоменных тяжелоцепочечных антител против PDL1 с антигенным белком PDL1.
На фиг. 3 представлены кривые блокирования однодоменных тяжелоцепочечных антител против PDL1 на взаимодействие PD1/PDL1.
На фиг. 4 представлено выравнивание последовательностей пяти вариантов гуманизированных антител №5.
На фиг. 5 представлены кривые связывания слитых белков однодоменного антитела против PDL1 с Fc-фрагментом с PDL1 (по ИФА).
На фиг. 6 представлены кривые блокирования слитых белков однодоменного антитела против PDL1 с Fc-фрагментом на взаимодействие PD1/PDL1 (методом конкурентного ИФА).
На фиг. 7 представлены кривые слитых белков однодоменного антитела против PDL1 с Fc-фрагментом, на взаимодействие CD80/PDL1 (методом конкурентного ИФА).
На фиг. 8 представлены кривые блокирования слитых белков однодоменного антитела против PDL1 с Fc-фрагментом, на взаимодействие CD80/PDL1 (в FACS-анализе).
На фиг. 9 представлены кривые блокирования слитых белков однодоменного антитела против PDL1 с Fc-фрагментом, на взаимодействие Jurket-PD1/PDL1 (в FACS-анализе).
На фиг. 10 представлена специфичность связывания слитых белков однодоменного антитела против PDL1 с Fc-фрагментом, с белком PDL1, обнаруженного методом проточной цитометрии.
На фиг. 11 представлено связывание слитых белков однодоменного антитела против PDL1 с Fc-фрагментом, с белком PDL1 обезьяны, обнаруженное методом проточной цитометрии.
На фиг. 12 представлено, что слитые белки однодоменного антитела против PDL1 с Fc-фрагментом идентифицируют PDL1-положительную клеточную популяцию на срезах тканей, взятых у пациентов.
На фиг. 13 представлена активация РВМС (мононуклеарных клеток периферической крови человека, МКПК) слитыми белками однодоменного антитела против PDL1 с Fc-фрагментом.
На фиг. 14 представлена активация CD4+ Т-клеток слитыми белками однодоменного антитела против PDL1 с Fc-фрагментом.
На фиг. 15 показано, что слитые белки однодоменного антитела против PDL1 с Fc-фрагментом способствуют секреции IL-2.
На фиг. 16 показана CDC-активность (пер., комплементзависимая цитотоксичность) и ADCC-активность (пер., антителозависимая клеточная цитотоксичность) слитых белков однодоменного антитела против PDL1 с Fc-фрагментом, несущих мутантный Fc-фрагмент.
На фиг. 17 представлены кривые роста опухоли после лечения слитыми белками однодоменного антитела против PDL1 с Fc-фрагментом.
На фиг. 18 представлено ингибирование роста опухоли путем введения слитых белков однодоменного антитела против PDL1 с Fc-фрагментом в течение разных периодов.
На фиг. 19 представлено сравнение между активностями ингибирования роста опухоли hu56V2-Fc и 2.41. А, А375: РВМС=5:1; В, А375:РВМС=1:1.
На фиг. 20 представлено влияние щелочной и окислительной обработок на активность слитых белков однодоменного антитела против PDL1 с Fc-фрагментом.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Если не указано и не определено иное, все использованные термины имеют свое обычное значение в данной области техники, которое будет понятно специалисту в данной области техники. Например, приводится ссылка на стандартные справочники, такие как, Sambrook et al, "Molecular Cloning: A Laboratory Manual" (2nd Ed.), Vols. 1-3, Cold Spring Harbor Laboratory Press (1989); Lewin, "Genes IV", Oxford University Press, New York, (1990), a также Roitt et al., "Immunology" (2nd Ed.), Gower Medical Publishing, London, New York (1989), а также на предпосылки создания изобретения, изложенные здесь. Кроме того, если не указано иное, все методы, этапы, технологии и манипуляции, которые специально не описаны подробно, могут быть выполнены и были выполнены хорошо известным способом, как будет понятно специалисту в данной области техники. Для примера также дается ссылка на стандартные справочники, общую информацию по уровню техники, упомянутую выше, и на источники, процитированные в этих документах.
Если не указано иное, взаимозаменяемые термины «антитело» и «иммуноглобулин», используемые здесь как для обозначения антитела, состоящего только из тяжелых цепей, так и к традиционному 4-цепочечному антителу, используются в качестве общих терминов для включения, как полноразмерного антитела, так и его индивидуальных цепей, а также всех его частей, доменов или фрагментов (включая, без ограничения перечисленным, антигенсвязывающие домены или фрагменты, такие как VHH-домены или домены VH/VL, соответственно). Кроме того, в применении здесь, термин «последовательность», (например, в таких терминах, как «иммуноглобулиновая последовательность», «последовательность антитела», «последовательность (одиночного) вариабельного домена», «VHH-последовательность» или «белковая последовательность»), обычно должен рассматриваться как включающий в себя релевантную аминокислотную последовательность, так и последовательности нуклеиновых кислот или нуклеотидные последовательности, кодирующие то же самое, если только контекст не требует более ограниченной интерпретации.
В применении здесь термин "домен" (полипептида или белка) относится к структуре свернутого белка, которая обладает способностью сохранять свою третичную структуру независимо от остальной части белка. Обычно домены являются ответственными за дискретные функциональные свойства белков и во многих случаях могут быть добавлены, удалены или перенесены на другие белки без потери функций оставшегося белка и/или домена.
В применении здесь, термин "иммуноглобулиновый домен" относится к глобулярной области цепи антитела (такой как, например, цепь традиционного 4-цепочечного антитела или тяжелоцепочечного антитела), или к полипептиду, который по существу состоит из такой глобулярной области. Иммуноглобулиновые домены отличаются тем, что они сохраняют характерную для молекул антител иммуноглобулиновую укладку, которая состоит из двухслойного сэндвича из приблизительно семи антипараллельных бета-тяжей, расположенных в двух бета-листах, дополнительно стабилизированных консервативной дисульфидной связью.
В применении здесь, термин "вариабельный домен иммуноглобулина" означает иммуноглобулиновый домен, по существу состоящий из четырех «каркасных областей», которые называют в данной области и далее в данном документе «каркасной областью 1» или «FR1», «каркасной областью 2» или «FR2», «каркасной областью 3» или «FR3», «каркасной областью 4» или «FR4», соответственно; эти каркасные области прерываются тремя «областями, определяющими комплементарность» или «CDR», которые называют в данной области и далее в данном документе как «определяющая комплементарность область 1» или «CDR1», «определяющая комплементарность область 2» или «CDR2», «определяющая комплементарность область 3» или «CDR3», соответственно. Таким образом, общая структура или последовательность вариабельного домена иммуноглобулина может быть обозначена следующим образом: FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4. Считается, что вариабельный домен (домены) иммуноглобулина придает специфичность антителу к антигену, поскольку содержит антигенсвязывающий сайт.
В применении здесь, термин "одиночный вариабельный домен иммуноглобулина" означает одиночный вариабельный домен иммуноглобулина, который способен специфически связываться с эпитопом антигена без спаривания с дополнительным вариабельным доменом иммуноглобулина. Одним из примеров одиночных вариабельных доменов иммуноглобулинов в контексте настоящего изобретения является "доменное антитело", такое как одиночные вариабельные домены иммуноглобулина VH и VL (VH-домены и VL-домены). Другим примером одиночных вариабельных доменов иммуноглобулина является "VHH-домен" (или просто "VHH") верблюдовых, как определено ниже.
"VHH-домены", также известные как однодоменные антитела, состоящие только из тяжелых цепей, VHHs, VHH-домены, фрагменты антител VHH и антитела VHH, представляют собой антигенсвязывающий вариабельный домен иммуноглобулина "тяжелоцепочечных антител" (т.е., "антител, лишенных легких цепей") (Hamers-Casterman С, Atarhouch Т, Muyldermans S, Robinson G, Hamers С, Songa ЕВ, Bendahman N, Hamers R.: "Naturally occurring antibodies devoid of light chains"; Nature 363, 446-448 (1993)). Термин «VHH-домен» был выбран для того, чтобы отличать эти вариабельные домены от вариабельных доменов тяжелых цепей, которые присутствуют в традиционных 4-цепочечных антителах (которые здесь называют "VH-доменами"), и от вариабельных доменов легких цепей, которые присутствуют в традиционных 4-цепочечных антителах (которые здесь называют «VL-доменами»). VHH-домены могут специфически связываться с эпитопом без дополнительного антигенсвязывающего домена (в отличие от доменов VH или VL в традиционном 4-цепочечном антителе, в случае которых эпитоп распознается VL-доменом вместе с VH-доменом). VHH-домены представляют собой небольшие, надежные и эффективные антигенраспознающие комплексы, образованные иммуноглобулиновым одиночным доменом.
В контексте настоящего изобретения, термины однодоменное антитело, состоящее только из тяжелой цепи, VHH-домен, VHH, домен VHH, фрагмент VHH-антитела, VHH-антитело, а также "Nanobody®" ("Нанотело®" и "Домен нанотела®" (причем "Nanobody" является торговой маркой компании Ablynx N.V.; Ghent; Бельгия), используют взаимозаменяемо.
Аминокислотные остатки VHH-доменов верблюдовых пронумерованы в соответствии с общей нумерацией VH-доменов по Кабату (Kabat et al. "Sequence of proteins of immunological interest", US Public Health Services, NIH Bethesda, MD, Publication No. 91), как показано, например, на фиг. 2 в статье Riechmann and Muyldermans, J. Immunol. Methods 231, 25-38 (1999). Согласно этой нумерации,
- FR1 содержит аминокислотные остатки в положениях 1-30,
- CDR1 содержит аминокислотные остатки в положениях 31-35,
- FR2 содержит аминокислотные остатки в положениях 36-49,
- CDR2 содержит аминокислотные остатки в положениях 50-65,
- FR3 содержит аминокислотные остатки в положениях 66-94,
- CDR3 содержит аминокислотные остатки в положениях 95-102 и
- FR4 содержит аминокислотные остатки в положениях 103-113.
Однако необходимо отметить, как хорошо известно в данной области для VH-доменов и для VHH-доменов, что общее число аминокислотных остатков в каждой из CDRs может варьировать и может не соответствовать общему числу аминокислотных остатков, приведенных в нумерации по Кабату (т.е., одно или несколько положений в соответствии с нумерацией по Кабату могут быть не заняты в реальной последовательности, или реальная последовательность может содержать больше аминокислотных остатков, чем число, разрешенное нумерацией по Кабату). Это означает, что в общем, нумерация по Кабату может соответствовать или не соответствовать фактической нумерации аминокислотных остатков в фактической последовательности.
Альтернативные методы нумерации аминокислотных остатков VH-доменов, которые также могут быть применены аналогичным образом к VHH-доменам, известны в данной области. Однако, в настоящем описании, формуле изобретения и фигурах будут следовать нумерации по Кабату и применять к VHH доменам, как описано выше, если не указано иное.
Общее число аминокислотных остатков в VHH-домене обычно будет находиться в диапазоне от 110 до 120, чаще всего между 112 и 115. Однако следует отметить, что более короткие и долее длинные последовательности также могут быть подходящими для целей, описанных в настоящем документе.
Дополнительные структурные характеристики и функциональные свойства VHH-доменов и полипептидов, содержащих VHH-домены, могут быть суммированы следующим образом:
VHH-домены (которые были "сконструированы" по своей природе для функционального связывания с антигеном без присутствия и без любого взаимодействия с вариабельным доменом легкой цепи) могут функционировать как одиночная, относительно небольшая, функциональная антигенсвязывающая структурная единица, домен или полипептид. Это отличает VHH-домены от доменов VH и VL традиционных 4-цепочечных антител, которые сами по себе, как правило, не подходят для практического применения в качестве одиночных антигенсвязывающих белков или одиночных вариабельных доменов иммуноглобулина, но должны быть объединены в той или иной форме для обеспечения функциональной антигенсвязывающей единицы (как, например, в случае традиционных фрагментов антитела, таких как Fab-фрагменты; в scFv, которые состоят из домена VH, ковалентно связанного с VL-доменом).
Из-за этих уникальных свойств, применение VHH-доменов, как отдельно, так и в виде части более крупного полипептида, открывает ряд существенных преимуществ перед использованием традиционных доменов VH и VL, scFvs или традиционных фрагментов антител (таких как Fab- или F(ab')2-фрагменты):
- для связывания антигена с высокой аффинностью и с высокой селективностью требуется только один домен, так что нет необходимости иметь два отдельных домена или гарантировать, что эти два домена присутствуют в правильной пространственной конформации и конфигурации (т.е. через использование специально сконструированных линкеров, как в случае с scFvs);
- VHH-домены могут быть экспрессированы из одного гена и не требуют посттрансляционного фолдинга или модификаций;
- VHH-домены могут быть легко сконструированы в мультивалентном и мультиспецифическом форматах (форматированы);
- VHH-домены являются легкорастворимыми и не имеют тенденции к агрегированию;
- VHH-домены очень устойчивы к нагреванию, pH, протеазам и другим денатурирующим агентам или условиям, и, таким образом, могут быть получены, храниться или перевозиться без использования холодильного оборудования, что позволяет сэкономить расходы на транспортировку, сократить время и уменьшить влияние на окружающую среду;
- VHH-домены могут быть легко и относительно дешево получены, даже в промышленных масштабах
- VHH-домены являются относительно малыми (приблизительно 15 кДа, или в 10 раз меньше, чем традиционный IgG) по сравнению с традиционными 4-цепочечными антителами и их антигенсвязывающими фрагментами, и следовательно, проявляют высокую (более высокую) степень проникновения в ткани и могут быть введены в более высоких дозах, чем традиционные 4-цепочечные антитела и их антигенсвязывающие фрагменты;
- VHH-домены могут проявлять так называемые свойства связывания, проникая в полость (особенно из-за их очень длинной петли CDR3 по сравнению с традиционными VH-доменами), и следовательно, также могут получать доступ к мишеням и эпитопам, недоступным традиционным 4-цепочечным антителам и их антигенсвязывающим фрагментам.
Способы получения VHH-доменов, связывающихся со специфическим антигеном или эпитопом, были описаны ранее, например, в WO 2006/040153 и WO 2006/122786; R. van der Linden et al., Journal of Immunological Methods, 240 (2000) 185-195; Li et al., J Biol Chem., 287 (2012) 13713-13721; Deffar et al., African Journal of Biotechnology Vol. 8 (12), pp. 2645-2652, 17 June, 2009 и WO 94/04678.
VHH-домены, полученные из антител верблюдовых, могут быть "гуманизированы" заменой (замещением) одного или нескольких аминокислотных остатков в аминокислотной последовательности в исходной VHH-последовательности одним или несколькими аминокислотными остатками, которые находятся в соответствующем положении(-ях) в VH-домене традиционного 4-цепочечного антитела человека. Гуманизированный VHH-домен может содержать одну или несколько полностью человеческих последовательностей каркасной области, и в конкретных вариантах данного изобретения, содержащий человеческую последовательность каркасной области IGHV3.
Обычно, термин «специфичность» относится к числу различных типов антигенов или эпитопов, с которыми может связываться конкретная антигенсвязывающая молекула или антигенсвязывающий белок (такой как, одиночный вариабельный домен иммуноглобулина настоящего изобретения). Специфичность антигенсвязывающей молекулы может быть определена на основе ее аффинности и/или авидности. Аффинность, представленная константой равновесия для диссоциации антигена с антигенсвязывающим белком (KD), является величиной прочности связывания между эпитопом и антигенсвязывающим сайтом антигенсвязывающего белка: чем меньше значение KD, тем выше прочность связывания между эпитопом и антигенсвязывающей молекулой (альтернативно, аффинность также может быть выражен в виде константы аффинности (KA), которая составляет 1/KD). Как будет очевидно специалисту в данной области, аффинность может быть определена per se (в чистом виде), в зависимости от представляющего интерес специфического антигена. Авидность является мерой прочности связывания между антигенсвязывающей молекулой (такой, как иммуноглобулин, антитело, одиночный вариабельный домен иммуноглобулина или содержащие его полипептиды) и соответствующим антигеном. Авидность связана как с аффинностью между эпитопом и его антигенсвязывающим сайтом на антигенсвязывающей молекуле, так и с числом соответствующих связывающих сайтов, присутствующих на антигенсвязывающей молекуле.
Обычно, специфические связывающие молекулы будут связываться с PDL1 с константой диссоциации (KD) от 10-7 до 10-11 моль/литр (М), и предпочтительно от 10-8 до 10-11 моль/литр, более предпочтительно от 10-9 до 10-11 моль/литр и даже более предпочтительно от 10-10 до 10-11 моль/литр или менее (как измерено с использованием анализа Biacore или KinExA), и/или с константой ассоциации (KA) по меньшей мере 107 М-1, предпочтительно по меньшей мере 108 М-1, более предпочтительно по меньшей мере 109 М-1, более предпочтительно по меньшей мере 1010 М-1, такой как по меньшей мере 1010 М-1. Любое значение KD, превышающее 10-4 М, обычно считается неспецифическим связыванием. Специфическое связывание антигенсвязывающего белка с антигеном или эпитопом может быть определено любым подходящим хорошо известным способом, в том числе, например, описанными здесь анализами, Скэтчардовским анализом и/или конкурентным анализом связывания, такими как радиоиммуноанализ (RIA), иммуноферментный анализ (EIA) и сэндвич-анализ конкурентного связывания.
Аминокислотные остатки будут указываться согласно стандартному трехбуквенному или однобуквенному аминокислотному коду, как общеизвестно и принято в данной области техники. При сравнении двух аминокислотных последовательностей, термин «различие аминокислот» относится к инсерциям (вставкам), делециям или заменам указанного числа аминокислотных остатков в положениях ссылочной последовательности, по сравнению со второй последовательностью. В случае замены (замен), такая замена (замены) предпочтительно будет заменой (заменами) консервативных аминокислот, которая означает, что аминокислотный остаток заменяют другим аминокислотным остатком сходной химической структуры, и которая оказывает слабое влияние или практически не влияет на функцию, активность или другие биологические свойства полипептида. Такие замены консервативных аминокислот хорошо известны в данной области, где консервативные аминокислотные замены предпочтительно являются заменами, в которых одну аминокислоту в пределах следующих групп (i)-(v) заменяют на другой аминокислотный остаток внутри той же самой группы: (i) небольшие алифатические, неполярные или слегка полярные остатки: Ala, Ser, Thr, Pro и Gly; (ii) полярные, отрицательно заряженные остатки и их (незаряженные) амиды: Asp, Asn, Glu и Gin; (iii) полярные, положительно заряженные остатки: His, Arg и Lys; (iv) крупные алифатические, неполярные остатки: Met, Leu, Не, Val и Cys; а также (v) ароматические остатки: Phe, Tyr и Trp. В частности, предпочтительными консервативными аминокислотными заменами являются следующие: Ala на Gly или на Ser; Arg на Lys; Asn на Gln или на His; Asp на Glu; Cys на Ser; Gln на Asn; Glu на Asp; Gly на Ala или на Pro; His на Asn или на Gln; Ile на Leu или на Val; Leu на Ile или на Val; Lys на Arg, на Gln или на Glu; Met на Leu, на Tyr или на Ile; Phe на Met, на Leu или на Tyr; Ser на Thr; Thr на Ser; Trp на Tyr; Tyr на Trp или на Phe; Val на Ile или на Leu.
Полипептид, или молекула нуклеиновой кислоты, считается «по существу выделенным», например, при сравнении с исходным биологическим источником и/или реакционной средой или средой для культивирования, из которой они были получены, при их отделении, по меньшей мере, от одного другого компонента, с которым они обычно связаны в указанном источнике или среде, такого как другой белок/полипептид, другая нуклеиновая кислота, другой биологический компонент или макромолекула, или, по меньшей мере, одно загрязняющее вещество, примесь или минорный компонент. В частности, полипептид или молекула нуклеиновой кислоты считается «по существу выделенным», когда они были очищены, по меньшей мере, в 2 раза, в особенности, по меньшей мере в 10 раз, в особенности по меньшей мере более чем в 100 раз, и до 1000 раз или более. Полипептид или молекула нуклеиновой кислоты, которые находятся «по существу в изолированной форме», предпочтительно являются по существу гомогенными, как определено с помощью подходящего метода, такого как подходящий хроматографический метод, такого как электрофорез в полиакриламидном геле.
В применении здесь термин «субъект» относится к млекопитающему, особенно к приматам, в частности, к человеку.
PDL1-связывающая молекула по изобретению
В первом аспекте изобретение обеспечивает молекулу, связывающую лиганд-1 белка программируемой смерти (PDL1), которая может специфически связываться с PDL1 и состоит из аминокислотной последовательности A-L-B, в которой А означает одиночный вариабельный домен иммуноглобулина, L отсутствует или представляет собой аминокислотный линкер, и В означает Fc-область иммуноглобулина человека,
причем одиночный вариабельный домен иммуноглобулина состоит из аминокислотной последовательности, выбранной из SEQ ID NO: 1-6,
указанная PDL1-связывающая молекула способна образовывать гомодимеры через указанную Fc-область иммуноглобулина человека.
В применении здесь термин «Fc-область иммуноглобулина человека» относится к Fc-области константной области IgG1, IgG2, IgG3 или IgG4 человека (аминокислотные последовательности константных областей относятся к пунктам Р01857, Р01859, Р01860, Р01861 в базе данных белков www.uniprot.org), которая содержит шарнирную область или часть шарнирной области, область СН2 и область СН3 константной области иммуноглобулина. В настоящем изобретении ADCC-активность или CDC-активность может быть повышена или устранена, или аффинность FcRn может быть повышена или понижена мутацией 1-5 аминокислот в области СН2 аминокислотной последовательности «Fc-области иммуноглобулина человека»; или стабильность белка может быть повышена мутацией 1-4 аминокислот в шарнирной области. Однако в настоящем изобретении, термин «Fc-область иммуноглобулина человека» не включает в себя любую мутацию, которая способствует или предупреждает образование гетеродимеров Fc, ни любую модификацию присоединения другой функциональной белковой последовательности к N-концу или С-концу Fc-фрагмента.
В предпочтительном варианте, указанная Fc-область иммуноглобулина является мутированной для удаления ADCC-активности и CDC-активности. В конкретном варианте данного изобретения, последовательность указанная Fc-область иммуноглобулина выбрана из SEQ ID NOs: 7-9.
Квалифицированному специалисту в данной области будет ясно, что PDL1-связывающая молекула настоящего изобретения обычно будет присутствовать в форме гомодимеров из-за Fc-области иммуноглобулина человека.
В применении здесь термин "линкер" означает нефункциональную аминокислотную последовательность длиной 1-20 аминокислотных остатков, без вторичных структур или структур более высокого уровня организации. Для примера, указанный линкер является гибким линкером, таким как GGGS, GS, GAP и тому подобное.
В некоторых вариантах осуществления, PDL1-связывающая молекула по изобретению состоит из аминокислотной последовательности, выбранной из SEQ ID NOs: 10-27.
PDL1-связывающая молекула по изобретению имеет по меньшей мере один из отличительных признаков:
(a) связывается с человеческим PDL1 с KD менее чем 1×10-7 М;
(b) блокирует взаимодействие между PDL1 и PD-1;
(c) усиливает активацию РВМС и/или Т-клеток;
(d) ингибирует рост опухоли.
PDL1-связывающая молекула по изобретению может связываться с PDL1 с KD менее, чем 1×10-7 М, предпочтительно менее чем 1×10-8 М, более предпочтительно менее чем 1×10-9 М, более предпочтительно менее чем 1×10-10 М и даже более предпочтительно менее чем 1×10-11 М.
В некоторых вариантах осуществления, PDL1-связывающая молекула по изобретению может специфически связываться с человеческим PDL1 и блокировать взаимодействие между PDL1 и PD-1. В некоторых вариантах осуществления, PDL1-связывающая молекула по изобретению может специфически связываться с человеческим PDL1 и блокировать взаимодействие между PDL1 и CD80.
PDL1-связывающая молекула по изобретению может ингибировать рост опухоли по меньшей мере приблизительно на 10%, предпочтительно по меньшей мере приблизительно на 20%, более предпочтительно по меньшей мере приблизительно на 30%, более предпочтительно по меньшей мере приблизительно на 40%, более предпочтительно по меньшей мере приблизительно на 50%, более предпочтительно по меньшей мере приблизительно на 60%, более предпочтительно по меньшей мере приблизительно на 70% и более предпочтительно по меньшей мере приблизительно на 80%.
Дополнительно, PDL1-связывающая молекула по изобретению является устойчивой к щелочной обработке и окислительной обработке. Например, PDL1-связывающая молекула по изобретению сохраняет свою активность после обработки сильным основанием (таким как, 500 мМ бикарбонат аммония) в течение около 8 часов, предпочтительно около 16 часов, более предпочтительно около 24 часов или более предпочтительно около 32 часов. Альтернативно, PDL1-связывающая молекула по изобретению сохраняет свою активность после обработки окислителем (1% перекисью водорода) в течение около 2 часов, предпочтительно около 4 часов или более предпочтительно около 8 часов.
Кроме того, PDL1-связывающая молекула по изобретению является стабильной при высокой концентрации. Например, PDL1-связывающая молекула по изобретению сохраняет стабильность при концентрации около 100 мг/мл, более предпочтительно около 150 мг/мл, более предпочтительно около 200 мг/мл или более предпочтительно около 250 мг/мл без образования агрегатов.
Нуклеиновая кислота, вектор и клетка-хозяин
В другом аспекте изобретение относится к молекуле нуклеиновой кислоты, которая кодирует PDL1-связывающую молекулу по изобретению. Нуклеиновой кислотой по изобретению может быть РНК, ДНК или кДНК. В соответствии с одним вариантом осуществления изобретения, нуклеиновая кислота по изобретению находится по существу в изолированной форме.
Нуклеиновая кислота по изобретению также может быть в форме вектора, может присутствовать в векторе и/или быть частью вектора, такого как, например, плазмида, космида или YAC. Вектор особенно может быть экспрессирующим вектором, т.е. вектором, который может обеспечить экспрессию PDL1-связывающей молекулы in vitro и/или in vivo (т.е. в подходящей клетке-хозяине, в организме-хозяине и/или в экспрессирующей системе). Такой экспрессирующий вектор обычно содержит по меньшей мере одну нуклеиновую кислоту по изобретению, которая является функционально связанной с одним или несколькими подходящими регуляторными элементами, такими как промотор(ы), энхансер(ы), терминатор(ы), и т.п.. Такие элементы и их выбор, принимая во внимание экспрессию специфической последовательности в специфическом хозяине являются общеизвестными специалистам в данной области. Специфические примеры регуляторных элементов и других элементов пригодных или необходимых для экспрессии PDL1-связывающих молекул по изобретению включают, например, промоторы, энхансеры, терминаторы, факторы интеграции, селективные маркеры, лидерные последовательности, репортерные гены и т.п..
Нуклеиновые кислоты по изобретению могут быть приготовлены или получены хорошо известным способом (например, автоматизированным синтезом ДНК и/или технологией рекомбинантных ДНК) на основе информации об аминокислотных последовательностях для полипептидов по изобретению, приведенных здесь, и/или могут быть выделены из подходящего природного источника.
В другом аспекте изобретение относится к клеткам-хозяевам, которые экспрессируют, или которые способны экспрессировать одну или несколько PDL1-связывающих молекул по изобретению; и/или которые содержат нуклеиновую кислоту по изобретению. В соответствии с особенно предпочтительным вариантом осуществления, указанные клетки-хозяева являются бактериальными клетками; другими пригодными клетками являются дрожжевые клетки, клетки гриба или клетки млекопитающих.
Подходящие бактериальные клетки включают клетки штаммов грамотрицательных бактерий, таких как штаммы Escherichia coli, Proteus и Pseudomonas, и штаммы грамположительных бактерий, таких как штаммы Bacillus, Streptomyces, Staphylococcus и Lactococcus. Подходящие клетки гриба включают клетки видов рода Trichoderma, Neurospora и Aspergillus.
Подходящие клетки гриба включают клетки видов Trichoderma, Neurospora и Aspergillus. Подходящие дрожжевые клетки включают клетки видов рода Saccharomyces (например, Saccharomyces cerevisiae), Schizosaccharomyces (например, Schizosaccharomyces pombe), Pichia (например, Pichia pastoris и Pichia methanolica) и Hansenula.
Подходящие клетки млекопитающих включают, например, клетки HEK293, клетки СНО, клетки BHK, клетки HeLa, клетки COS и т.п..
Однако также могут быть использованы клетки амфибий, клетки насекомых, растительные клетки и любые другие клетки, используемые в данной области для экспрессии гетерологичных белков.
Изобретение дополнительно обеспечивает способы производства PDL1-связывающей молекулы по изобретению, такие способы обычно содержат стадии:
- культивирования клеток-хозяина по изобретению в условиях, которые позволяют экспрессию PDL1-связывающей молекулы по изобретению; и
- извлечения PDL1-связывающей молекулы, экспрессированной клетками-хозяевами, из культуры; и
- если требуется, дальнейшая очистка и/или модификация PDL1-связывающей молекулы по изобретению.
В предпочтительном воплощении изобретения, PDL1-связывающую молекулу по изобретению получают с использованием клеток млекопитающих. PDL1-связывающая молекула по изобретению может достигать высокого уровня экспрессии в клетках млекопитающих. Например, уровень экспрессии может быть вплоть до около 5 г/л, предпочтительно около 6 г/л, предпочтительно около 7 г/л, предпочтительно около 8 г/л, предпочтительно около 9 г/л, предпочтительно около 10 г/л или выше.
PDL1-связывающая молекула по изобретению может быть получена в клетках как указано выше или внутриклеточно (например, в цитозоле, в периплазме или в тельцах включений) и затем выделена из клетки-хозяина и, если требуется, дополнительно очищена; или она может быть получена внеклеточно (например, в среде, в которой культивируют клетки-хозяина) и затем выделена из культуральной среды и, если требуется, дополнительно очищена.
Способы и реагенты, используемые для рекомбинантного получения полипептидов, такие как специфически подходящие экспрессирующие векторы, способы трансформации или трансфекции, селективные маркеры, способы индукции экспрессии белка, условия культивирования и т.д. известны в данной области. Аналогичным образом, способы выделения и технологии очистки, пригодные для способа производства PDL1-связывающей молекулы по изобретению хорошо известны специалисту в данной области.
Однако, PDL1-связывающие молекулы по изобретению можно также могут быть получены другими способами получения белков, известными в данной области, такими как химический синтез, в том числе твердофазный синтез или жидкофазный синтез.
Фармацевтические композиции
В другом аспекте настоящее изобретение обеспечивает композицию, например, фармацевтическую композицию, содержащую одну или комбинацию PDL1-связывающей молекулы настоящего изобретения, приготовленную вместе с фармацевтически приемлемым носителем. Такие композиции могут включать в себя одну или комбинацию (например, две или несколько различных) PDL1-связывающих молекул по изобретению. Специалисту в данной области техники будет ясно, что PDL1-связывающая молекула настоящего изобретения обычно будет представлена в форме гомодимера из-за Fc-области иммуноглобулина человека.
Фармацевтические композиции по изобретению также могут быть введены в комбинированной терапии, то есть в сочетании с другими агентами. Например, комбинированная терапия может включать в себя PDL1-связывающую молекулу настоящего изобретения, объединенную с по меньшей мере одним противоопухолевым агентом. Например, PDL1-связывающая молекула по изобретению может быть введена в сочетании с антителами, нацеленными на другие опухолеспецифические антигены. Указанные антитела, нацеленные на другие опухолеспецифические антигены, включают, без ограничения перечисленным, антитело анти-EGFR антитело, вариант антитела анти-EGFR, анти-VEGFa антитело, анти-HER2 антитело или анти-СМЕТ антитело. Предпочтительно, указанные антитела являются моноклональными.
В применении здесь, «фармацевтически приемлемый носитель» включает в себя любой и все растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые агенты, изотонические и замедляющие адсорбцию агенты, и т.п., которые являются физиологически совместимыми. Предпочтительно носитель является подходящим для внутривенного, внутримышечного, подкожного, парентерального, спинального или эпидермального введения (например, путем инъекции или инфузии). В зависимости от способа введения, активное соединение, т.е. антитело, или иммуноконъюгат, может быть покрыт материалом для защиты соединения от действия кислот и других естественных условий, которые могут инактивировать соединение.
Фармацевтические соединения по изобретению могут включать в себя одну или несколько фармацевтически приемлемых солей. Термин «фармацевтически приемлемая соль» относится к соли, которая сохраняет необходимую биологическую активность родительского соединения и не оказывает никаких нежелательных токсикологических эффектов (см., например, Berge, S.M., et al. (1977) J. Pharm. Sci. 66:1-19). Примеры таких солей включают соли присоединения кислоты и соли присоединения основания. Соли присоединения кислоты включают соли, полученные из нетоксичных неорганических кислот, таких как соляная, азотная, фосфорная, серная, бромистоводородная, йодистоводородная, фосфорная и т.п., а также от нетоксичных органических кислот, таких как алифатические моно- и дикарбоновые кислоты, фенилзамещенные алкановые кислоты, гидроксалкановые кислоты, ароматические кислоты, алифатические и ароматические сульфоновые кислоты и т.п. Соли присоединения основания включают соли, полученные из щелочноземельных металлов, таких как натрий, калий, магний, кальций и т.д., а также из нетоксичных органических аминов, таких как N,N'-дибензилэтилендиамин, N-метилглюкамин, хлорпрокаин, холин, диэтаноламин, этилендиамин, прокаин и т.п.
Фармацевтическая композиция по изобретению также может включать в себя фармацевтически приемлемый антиоксидант. Примеры фармацевтически приемлемых антиоксидантов включают в себя: (1) водорастворимые антиоксиданты, такие как аскорбиновая кислота, цистеина гидрохлорид, бисульфат натрия, натрия метабисульфит, сульфит натрия и т.п.; (2) растворимые в масле антиоксиданты, такие как аскорбилпальмитат, бутилгидроксианизол (ВНА), бутилигидрокситолуол (ВНТ), лецитин, пропил галлат, альфа-токоферол и т.п.; и (3) агенты, хелатирующие металлы, такие как лимонная кислота, этилендиаминтетрауксусная кислота (EDTA), сорбит, винная кислота, фосфорная кислота и т.п.
Эти композиции также могут содержать адъюванты, такие как консерванты, смачивающие агенты, эмульгирующие агенты и диспергирующие агенты.
Предупреждение присутствия микроорганизмов может быть обеспечено как процедурами стерилизации, см. выше, так и включением различных антибактериальных и противогрибковых агентов, таких как, парабен, хлорбутанол, фенол-сорбиновой кислоты и т.п. Также может быть желательно включение в композиции изотонических агентов, таких как сахара, натрия хлорид и т.п. Кроме того, продолжительная абсорбция инъецируемой фармацевтической формы может быть обеспечена включением агентов, которые задерживают абсорбцию, таких как моностеарат алюминия и желатин.
Фармацевтически приемлемые носители включают стерильные водные растворы или дисперсии и стерильные порошки для получения экстемпоральных стерильных инъекционных растворов или дисперсию. Использование таких сред и агентов для фармацевтически активных веществ известно в данной области. За исключением случая, когда любые традиционные среды или агент несовместимы с активным соединением, их использование в фармацевтических композициях по изобретению рассматривается. Дополнительные активные соединения также могут быть включены в композиции.
Терапевтические композиции обычно должны быть стерильны и стабильны в условиях производства и хранения. Композиция может быть приготовлена в виде раствора, микроэмульсии, липосомы или другой упорядоченной структуры, подходящей для высокой концентрации лекарственного средства. Носитель может быть растворителем или дисперсионной средой, содержащей, например, воду, этанол, полиол (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль и т п.) и их подходящие смеси. Может поддерживаться необходимая текучесть, например, путем использования покрытия, такого как лецитин, путем поддержания требуемых размеров частиц в случае дисперсии и путем использования поверхностно-активных веществ.
Стерильные инъекционные растворы могут быть получены путем включения активного соединения в необходимом количестве в соответствующий растворитель с одним или комбинацией ингредиентов, перечисленных выше, с последующей стерилизующей микрофильтрацией. Обычно дисперсии получают включением активного соединения в стерильный носитель, который содержит основную дисперсионную среду и другие необходимые ингредиенты из перечисленных выше. В случае стерильных порошков для получения стерильных инъекционных растворов предпочтительными способами получения являются вакуумная сушка и сушка при замораживании (лиофилизация), которые позволяют получить порошок активного ингредиента плюс любой дополнительный необходимый ингредиент из его раствора, прошедшего предварительную стерилизующую фильтрацию.
Количество активного ингредиента, которое можно комбинировать с материалом носителя для получения единичной лекарственной формы, будет варьировать в зависимости от субъекта, подлежащего лечению и конкретного способа введения. Количество активного ингредиента, которое может быть объединено с материалом носителя для получения единичной лекарственной формы, обычно будет тем количеством композиции, которое дает терапевтический эффект. Обычно из ста процентов это количество будет находиться в диапазоне от около 0,01% до около девяноста девяти процентов активного ингредиента, предпочтительно от около 0,1 процента до около 70 процентов, наиболее предпочтительно от около 1 процента до около 30 процентов активного ингредиента в сочетании с фармацевтически приемлемым носителем.
Режимы дозирования корректируют для обеспечения оптимального желаемого ответа (например, терапевтического ответа). Например, может быть введен единичный болюс, несколько дробных доз может быть введено в течение периода времени, или доза может быть пропорционально уменьшена или увеличена, как указано в соответствии с требованиями терапевтической ситуации. Особенно предпочтительно готовить парентеральные композиции в единичной дозированной форме для удобства введения и единообразия дозировки. В применении здесь, единичная дозированная форма относится к физически дискретным единицам, подходящая в качестве единичных дозировок для субъектов, подлежащих лечению; каждая единица содержит заранее определенное количество активного соединения, рассчитанное для получения желаемого терапевтического эффекта в сочетании с необходимым фармацевтическим носителем. Спецификация для единичных дозированных форм по изобретению диктуется и напрямую зависит от (а) уникальных характеристик активного соединения и конкретного терапевтического эффекта, который достигается, и (б) ограничений, присущих в данной области приготовления такого активного соединения для лечения чувствительности у пациентов.
Для введения молекулы антитела дозировка находится в диапазоне от около 0,0001 до 100 мг/кг и большей частью от 0,01 до 5 мг/кг массы тела субъекта. Например, дозировка может быть 0,3 мг/кг массы тела, 1 мг/кг массы тела, 3 мг/кг массы тела, 5 мг/кг массы тела, 10 мг/кг массы тела или 20 мг/кг массы тела, или в диапазоне 1-20 мг/кг. Приводимая в качестве схема лечения режима лечения влечет за собой введение раз в неделю, раз в две недели, раз в три недели, раз в четыре недели, раз в месяц, один раз в 3 месяца или один раз каждые три-шесть месяцев, или с коротким интервалом введения в начале (например, от одного раза в неделю до одного раза каждые три недели), и затем с увеличенным интервалом позже (например, от одного раза в месяц до одного раза каждые 3-6 месяцев).
Альтернативно антитело может быть введено в виде композиции с замедленным высвобождением, в этом случае требуется меньшая частота введения. Дозировка и частота изменяются в зависимости от времени полужизни антитела в организме пациента. В общем, человеческие антитела демонстрируют наиболее продолжительное время полужизни, за ними следуют гуманизированные антитела, химерные антитела и антитела, не относящиеся к человеку. Дозировка и частота введения могут изменяться в зависимости от того, является ли лечение профилактическим или терапевтическим. В профилактических применениях вводят относительно низкую дозировку при относительно редких интервалах в течение длительного периода времени. Некоторые пациенты продолжают получать лечение всю оставшуюся жизнь. В терапевтических применениях иногда требуется относительно высокая дозировка с относительно короткими интервалами до снижения или прекращения прогрессирования заболевания, и предпочтительно до тех пор, пока пациент не продемонстрирует частичное или полное устранение симптомов заболевания. После этого пациенту может быть назначен профилактический режим.
Фактические уровни дозировки активных ингредиентов в фармацевтических композициях настоящего изобретения могут быть изменены с тем, чтобы получить количество активного ингредиента, которое является эффективным для достижения желаемого терапевтического эффекта у конкретного пациента, композицию и способ введения, не будучи токсичными для пациента. Выбранный уровень дозировки будет зависеть от различных фармакокинетических факторов, включающих активность используемых конкретных композиций настоящего изобретения, или их сложного эфира, соли или амида, путь введения, время введения, скорость экскреции конкретного используемого соединения, продолжительность лечения, другие лекарственные средства, соединения и/или материалы, используемые в сочетании с конкретными применяемыми композициями, возраст, пол, вес, состояние, общее состояние здоровья и предшествующую историю болезни пациента, подлежащего лечению, а также другие подобные факторы, хорошо известные в медицинской области.
«Терапевтически эффективное количество» PDL1-связывающей молекулы изобретения предпочтительно приводит к снижению тяжести симптомов заболевания, увеличению частоты и продолжительности периодов болезни без симптомов, или предотвращению потери трудоспособности или инвалидности из-за поражения болезнью. Например, для лечения связанных с PDL1 опухолей, «терапевтически эффективное количество» предпочтительно ингибирует рост клеток или рост опухоли, по меньшей мере, примерно на 10%, по меньшей мере, примерно на 20%, более предпочтительно, по меньшей мере, примерно на 40%, еще более предпочтительно, по меньшей мере, примерно на 60%, и еще более предпочтительно, по меньшей мере, примерно на 80% по сравнению с субъектами, не получавшими лечения. Способность ингибировать рост опухоли может быть оценена в системе модельных животных, прогнозирующей эффективность для опухолей человека. Альтернативно это свойство композиции может быть оценено путем изучения способности соединения ингибировать рост клеток; такое ингибирование может быть определено in vitro при помощи анализов, известных квалифицированным специалистам. Терапевтически эффективное количество терапевтического соединения может уменьшать размер опухоли, или иным образом улучшить симптомы у субъекта. Специалист с обычной квалификацией в данной области способен определить такие количества на основе таких факторов, как размер субъекта, тяжесть симптомов субъекта и конкретная композиция или выбранный путь введения.
Композиция по настоящему изобретению может быть введена при помощи одного или нескольких способов введения, с использованием различных способов, известных в данной области. Как будет понятно специалисту в данной области, способ и/или режим введения будет меняться в зависимости от желаемых результатов. Предпочтительные пути введения PDL1-связывающих молекул по изобретению включают внутривенный, внутримышечный, внутрикожный, внутрибрюшинный, подкожный, спинальный и другие парентеральные пути введения, например, путем инъекции или инфузии. Используемое здесь выражение «парентеральное введение» означает способы введения, отличные от энтерального и местного введения, обычно путем инъекции, и включает, без ограничения, внутривенную, внутримышечную, внутриартериальную, интратекальную, интракапсулярную, внутриглазничную, внутрисердечную, подкожную, внутрибрюшинную, транстрахеальную, подкожную, субкутикулярную, внутрисуставную, подкапсулярную, субарахноидальную, межпозвонковую, эпидуральную и надчревную инъекцию и инфузию.
Альтернативно, PDL1-связывающуая молекула по изобретению может быть введена посредством непарентерального пути, таким как локальный, эпидермальный путь введения или через слизистую, например, интраназально, перорально, вагинально, ректально, сублингвально или местно.
Активные соединения могут быть получены с носителями, которые будут защищать соединение от быстрого высвобождения, такими как фармацевтические композиции с контролируемым высвобождением, включая имплантанты, трансдермальные пластыри и микроинкапсулированные системы доставки. Могут быть использованы биоразлагаемые, биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолиевая кислота, коллаген, сложные полиортоэфиры и полимолочная кислота. Многие способы получения таких композиций запатентованы или общеизвестны специалистам в данной области. См., например, Sustained and Controlled Release Drug Delivery Systems, J.R. Robinson, ed., Marcel Dekker, Inc., New York, 1978.
Терапевтические композиции могут быть введены медицинскими устройствами, известными в данной области. Например, в предпочтительном варианте осуществления, терапевтическая композиция по изобретению может быть введена устройством для безыгольных подкожных инъекций, например, устройствами, раскрытыми в патентах США №№:5399163; 5383851; 5312335; 5064413; 4941880; 4790824 или 4596556. Примеры хорошо известных имплантатов и модулей, применимых в настоящем изобретении включают: патент США №4487603, который раскрывает имплантируемую микроинфузионную помпу для дозирования лекарственного средства с контролируемой скоростью; патент США №4486194, который раскрывает терапевтическое устройство для введения лекарственных средств через кожу; патент США №4447233, который раскрывает инфузионную помпу для доставки лекарственных средств с точной скоростью инфузии; патент США №4447224, который раскрывает имплантируемое инфузионное устройство переменного потока для непрерывной доставки лекарств; патент США №4439196, который раскрывает осмотическую систему доставки лекарственного средства с многокамерными отсеками; и патент США №4475196, который раскрывает осмотическую систему доставки лекарственного средства. Эти патенты включены в данный документ ссылками. Многие другие подобные имплантаты, системы доставки и модули известны специалистам в данной области.
В определенных воплощениях PDL1-связывающая молекула по изобретению может быть приготовлена для обеспечения надлежащего распределения in vivo. Например, гематоэнцефалический барьер (ВВВ, русск. ГЭБ,) не пропускает многие высокогидрофильные соединения. Чтобы гарантировать, что терапевтические соединения по изобретению пересекают ВВВ (если необходимо), они могут быть приготовлены, например, в липосомах. Способы изготовления липосом см., например, в патентах США №№:4522811; 5374548 и 5399331. Липосомы могут содержать одну или несколько молекул, которые избирательно транспортируются в специфические клетки или органы, тем самым повышая целевую доставку лекарственного средства (см., например, V.V. Ranade (1989) J. Clin. Pharmacol. 29:685). Приводимые в качестве примера нацеливающие молекулы включают в себя фолат или биотин (см., например, патент США №5416016, Low et al); маннозиды (Umezawa et al., (1988) Biochem. Biophys. Res. Commun. 153:1038); антитела (P.G. Bloeman et al. (1995) FEBS Lett. 357:140; M. Owais et al. (1995) Antimicrob. Agents Chemother. 39:180); рецептор поверхностно-активного белка A (Briscoe et al. (1995) Am. J. Physiol. 1233: 134); pl20 (Schreier et al. (1994) J Biol. Chem. 269:9090); см., также K. Keinanen; M.L. Laukkanen (1994) FEBS Lett. 346:123; Jj. Killion; LJ. Fidler (1994) Immunomethods 4:273.
Предупреждение и лечение заболеваний
В другом аспекте настоящее изобретение обеспечивает применение PDL1-связывающей молекулы, нуклеиновой кислоты, клетки-хозяина и фармацевтической композиции по изобретению для предупреждения и/или лечения заболеваний, связанных с PDL1, а также соответствующие способы. Заболевания, связанные с PDL1, которые можно предотвратить или вылечить с использованием PDL1-связывающей молекулы изобретения, описаны подробно следующим образом.
Рак
Блокирование PDLI PDL1-связывающей молекулой изобретения может повысить иммунный ответ на раковые клетки у пациента. PDLI не экспрессируется в нормальных клетках человека, но изобилует в различных видах рака человека (Dong et al. (2002) Nat Med 8:787-9). Взаимодействие между PD-1 и PDLI приводит к снижению проникающих в опухоль лимфоцитов (инфильтрирующих опухоль лимфоцитов), снижению опосредованной Т-клеточным рецептором пролиферации и ускользание раковых клеток от механизмов иммунологического надзора (пер., иммунной эвазии) (Dong et al. (2003) JMolMed 81:281-7; Blank et al. (2004) Cancer Immunol. Immunother. [epub]; Konishi et al. (2004) Clin. Cancer Res. 10:5094-100). Иммуносупрессия может быть инвертирована путем ингибирования локального взаимодействия PDLI с PD-1, и эффект является аддитивным также при блокировании еще и взаимодействия PD-L2 с PD-1 (Iwai et al. (2002) PNAS 99:12293-7; Brown et al. (2003) J. Immunol. JTO: 1257-66). PDL1-связывающая молекула по изобретению может быть использована сама по себе для подавления роста раковых опухолей. Альтернативно, PDL1-связывающая молекула по изобретению может быть использована вместе с другими иммуногенными агентами, стандартными способами лечения рака или другими антителами, как описано ниже (Iwai et al. (2002) PNAS 99:12293-7; Brown et al. (2003) J. Immunol. JTO: 1257-66).
Таким образом, в одном варианте осуществления, изобретение обеспечивает способ предупреждения или лечения рака у субъекта, содержащий введение пациенту терапевтически эффективного количества PDL1-связывающей молекулы изобретения, с тем, чтобы подавить рост опухолевых клеток у данного субъекта.
Предпочтительные типы рака, рост которых может быть ингибирован с использованием антител изобретения, включают в себя типы рака, обычно чувствительные к иммунотерапии. Неограничивающие примеры предпочтительных типов рака для лечения включают в себя: рак легких, рак яичников, колоректальный рак, рак прямой кишки, меланому (например, метастатическую злокачественную меланому), рак почек, рак мочевого пузыря, рак молочной железы, рак печени, лимфому, гематологические злокачественные образования, рак головы и шеи, глиому, рак желудка, рак носоглотки, рак гортани, рак шейки матки, рак тела матки, остеосаркому. Примеры других раковых заболеваний, которые могут быть лечены с использованием способов изобретения включают в себя рак костей, рак поджелудочной железы, рак предстательной железы, рак кожи, рак головы или шеи, кожную или внутриглазную злокачественную меланому, рак матки, рак анальной области, рак яичка, рак фаллопиевых труб, рак эндометрия, рак шейки матки, рак влагалища, рак вульвы, болезнь Ходжкина, не-ходжкинскую лимфому, рак пищевода, рак тонкой кишки, рак эндокринной системы, рак щитовидной железы, рак паращитовидной железы, рак надпочечной железы, саркому мягких тканей, рак уретры, рак полового члена, хронический или острый лейкоз, включая острый миелоидный лейкоз, хронический миелолейкоз, острый лимфобластный лейкоз, хронический лимфоцитарный лейкоз, солидные опухоли детского возраста, лимфоцитарную лимфому, рак мочевого пузыря, рак почки или мочеточника, карциному почечной лоханки, новообразования центральной нервной системы (ЦНС), первичную лимфома ЦНС, ангиогенез опухоли, опухоли оси позвоночника, глиому ствола мозга, аденому гипофиза, саркому Капоши, эпидермоидный рак, плоскоклеточный рак, Т-клеточную лимфому, рак, вызванный экологическими причинами, в том числе под воздействием асбеста и комбинации указанных раковых заболеваний. Настоящее изобретение также применимо для лечения метастатических форм рака, особенно метастатического рака с экспрессией PDLI (Iwai etal. (2005) Int. Immunol. 17:133-144).
Если требуется, PDL1-связывающая молекула изобретения может быть объединена с иммуногенным агентом, таким как раковые клетки, очищенные опухолевые антигены (включая рекомбинантные белки, пептиды и молекулы углеводов), клетки и клетки, трансфицированные генами, кодирующими иммунностимулирующие цитокины (Не et al (2004) J. Immunol. 173:4919-28). Неограничивающие примеры противоопухолевых вакцин, которые могут быть использованы, включают пептиды антигенов меланомы, такие как пептиды gp100, антигены MAGE, Trp-2, MARTI и/или тирозиназу, или опухолевые клетки, трасфицированные для экспрессии цитокина GM-CSF.
Было показано, что у людей некоторые опухоли, такие как меланомы, являются иммуногенными. Предполагается, что повышая порог активации Т-клеток блокированием PDLI PDL1-связывающей молекулой изобретения, можно активировать ответ опухоли на лечение у хозяина. Блокада PDLI (посредством антитела к PDL1, например, PDL1-связывающей молекулы изобретения), вероятно, будет наиболее эффективной в сочетании с протоколом вакцинации. Был разработан ряд экспериментальных стратегий для вакцинации против опухолей (см. Rosenberg, S., 2000, Development of Cancer Vaccines, ASCO Educational Book Spring: 60-62; Logothetis, C, 2000, ASCO Educational Book Spring: 300-302; Khayat, D. 2000, ASCO Educational Book Spring: 414-428; Foon, K. 2000, ASCO Educational Book Spring: 730-738; см. также Restifo, N. and Sznol, M., Cancer Vaccines, Ch. 61, pp. 3023-3043 in DeVita, V. etal. (eds.), 1997, Cancer: Principles and Practice of Oncology. Fifth Edition). По одной из этих стратегий, вакцину получают с использованием аутологичных и аллогенных опухолевых клеток. Было показано, что эти клеточные вакцины являются наиболее эффективными, когда опухолевые клетки трансдуцируют для экспрессии GM-CSF. Было показано, что GM-CSF является сильным активатором презентации антигена для противоопухолевой вакцинации (Dranoff et al. (1993) Proc. Natl. Acad. Sd U.S.A. 90: 3539-43).
Изучение экспрессии генов и крупномасштабных наборов экспрессирующихся генов в различных опухолях привело к определению так называемых опухолеспецифических антигенов (Rosenberg, SA (1999) Immunity 10: 281-7). Во многих случаях эти опухолеспецифические антигены являются дифференцировочными антигенами, экспрессируемыми в опухолях и в клетке, из которой возникла опухоль, например, меланоцитарными антигенами gp100, антигенами MAGE и Trp-2. Более важно, как может быть показано, что многие из этих антигенов являются мишенями опухолеспецифических Т-клеток, обнаруженных у хозяина. PDL1-связывающая молекула изобретения может быть использована в сочетании с набором рекомбинантных белков и/или пептидов, экспрессируемых в опухоли для генерирования иммунного ответа на эти белки. Эти белки обычно рассматриваются иммунной системой как аутоантигены (пер., собственные антигены), и поэтому толерантна к ним. Опухолевый антиген также может включать белок теломеразу, которая необходима для синтеза теломер хромосом и которая экспрессируется в более чем 85% раков человека, и только в ограниченном количестве соматических тканей (Kim, N et al. (1994) Science 266: 2011-2013). Опухолевый антиген также может быть «нео-антигенами», экспрессируемыми в раковых клетках вследствие соматических мутаций, которые изменяют белковую последовательность или создают гибридные белки между двумя неродственными последовательностями (то есть bcr-abl в хромосоме Philadelphia).
Другие противоопухолевые вакцины могут включать белки вирусов, присутствующих при раковых заболеваниях человека, таких как вирусы папилломы человека (ВПЧ), вирусы гепатита (HBV и HCV) и вирус герпеса саркомы Капоши (KHSV). Еще одна форма опухолевых специфических антигенов, которые могут быть использованы в сочетании с блокадой PDLI (такие, как антитела к PDL1, например, PDL1-связывающая молекула данного изобретения) - очищенные белки теплового шока (HSP), изолированные от самой ткани опухоли. Эти белки теплового шока содержат фрагменты белков от опухолевых клеток, и эти HSP высокоэффективно доставляются клеткам, представляющим антиген, для достижения противоопухолевого иммунитета (Suot, R & Srivastava, Р (1995) Science 269:1585-1588; Tamura, Y. et al. (1997) Science 278:117-120).
Дендритные клетки (DC) являются сильнодействующими антигенпрезентирующими клетками, которые могут быть использованы для первичных антигенспецифических ответов. DC могут быть произведены ex vivo и нагружены различными белковыми и пептидными антигенами, а также экстрактами опухолевых клеток (Nestle, F. et al. (1998) Nature Medicine 4: 328-332). Клетки DC также могут быть трансдуцированы генетическими средствами для экспрессии этих опухолевых антигенов. DC также были слиты непосредственно с опухолевыми клетками для целей иммунизации (Kugler, A. et al. (2000) Nature Medicine 6:332-336). В качестве способа вакцинации иммунизация DC может эффективно сочетаться с блокадой PDLI (такой, как антитело PDL1, например, PDL1-связывающая молекула по изобретению), для активации более мощных противоопухолевых ответов.
CAR-T (Т-клеточная иммунотерапия химерными антигенными рецепторами) представляет собой другую клеточную терапию для лечения опухолей. Химерные антигенные Т-клеточные рецепторы (CAR-T-клетки) являются Т-клетками пациента, которые были генетически инфицированы химерным белком антигенсвязывающей части молекулы антитела против определенного опухолевого антигена, связанного с CD3-ζ-цепью или внутриклеточной частью FcεRIγ для экспрессии химерного антигенного рецептора (CAR). Также может быть введена костимуляторная сигнальная последовательность для повышения цитотоксической активности, пролиферации и выживания Т-клеток и стимуляции высвобождения цитокинов. После перепрограммирования, Т-клетки пациента размножались in vitro, для получения большого количества опухолеспецифических CAR-T-клеток, которые затем переливают обратно пациенту для лечения опухоли. Агенты, блокирующие PDL1 (такие, как антитела к PDL1, например, PDL1-связывающая молекула по изобретению), могут быть использованы в сочетании с CAR-T-клеточной терапией для активации более сильного противоопухолевого ответа.
PDL1-связывающая молекула по изобретению также может быть объединена со стандартными способами лечения рака. PDL1-связывающая молекула по изобретению может быть эффективно объединена с химиотерапевтическими режимами. В этих случаях возможно уменьшение дозы вводимого химиотерапевтического реагента (Mokyr, М. et al. (1998) Cancer Research 58: 5301-5304). Примером такого объединения является анти-PDLI антитело в комбинации с декарбазином для лечения меланомы. Другим примером такого объединения является анти-PDLI антитело в комбинации с интерлейкином-2 (IL-2) для лечения меланомы. Научное обоснование объединенного применения PDL1-связывающей молекулы по изобретению и химиотерапии заключается в том, что гибель клеток, которая является следствием цитотоксического действия большинства химиотерапевтических соединений, должна приводить к повышению уровня опухолевого антигена в пути презентации антигена. Другие виды комбинированной терапии, которые могут привести к синергизму с блокадой PDLI посредством клеточной смерти, представляют собой облучение, хирургия и выключение эндокринной функции. Каждый из этих протоколов создает источник опухолевого антигена в хозяине. PDL1-связывающая молекула по изобретению также может быть объединена с ингибиторами ангиогенеза. Ингибирование ангиогенеза приводит к гибели опухолевых клеток, которые могут подавать опухолевый антиген в пути презентации антигена хозяина.
PDL1-связывающая молекула по изобретению также может быть использована в комбинации с антителом против другого опухолеспецифического антигена. Указанное антитело против другого опухолеспецифического антигена включает, но без ограничения перечисленным, анти-EGFR антитело, вариант антитела анти-EGFR, анти-VEGFa антитело, анти-HER2 антитело или анти-СМЕТ антитело. Предпочтительно, указанное антитело является моноклональным антителом.
PDL1-связывающая молекула по изобретению также может быть использована в комбинации с биспецифическими антителами, которые нацеливают эффекторные клетки, экспрессирующие Fc-альфа-рецепторы или Fc-гамма-рецепторы в опухолевые клетки (см., например, патенты США №№5922845 и 5837243). Биспецифические антитела могут быть использованы для нацеливания на два отдельных антигена. Например, биспецифические анти-Fc-рецептор/антиопухолевый антиген антитела (например, Her-2/neu) были использованы для нацеливания макрофагов к сайтам опухоли. Это нацеливание может более эффективно активировать опухолеспецифические ответы. Т-клеточное звено этих ответов может быть усилено использование блокады PDLI. Альтернативно антиген может быть доставлен непосредственно к дендритным клеткам (DC) с использованием биспецифических антител, которые связываются с опухолевым антигеном и специфическим к дендритным клеткам поверхностным маркером клеток.
Опухоли уклоняются от иммунологического надзора хозяина при помощи целого ряда механизмов. Многие из этих механизмов могут быть преодолены инактивацией белков, которые экспрессируются опухолями, и которые являются иммуносупрессивными. Эти белки включают, среди прочих, TGF-бета (пер., трансформирующий ростовой фактор бета) (Kehrl, J. etal. (1986) J. Exp. Med. 163: 1037-1050), IL-10 (Howard, M. &  A. (1992) Immunology Today 13: 198-200) и Fas-лиганд (Hahne, M. et al. (1996) Science 274: 1363-1365). Антитела к каждой из этих частиц могут быть использованы в сочетании с PDL1 -связывающей молекулой по изобретению для противодействия эффектам воздействия иммуносупрессивного агента и способствования иммунному противоопухолевому ответу хозяина.
A. (1992) Immunology Today 13: 198-200) и Fas-лиганд (Hahne, M. et al. (1996) Science 274: 1363-1365). Антитела к каждой из этих частиц могут быть использованы в сочетании с PDL1 -связывающей молекулой по изобретению для противодействия эффектам воздействия иммуносупрессивного агента и способствования иммунному противоопухолевому ответу хозяина.
Другие антитела, которые могут быть использованы для активации иммунологической реактивности хозяина, могут быть использованы в комбинации с анти-PDLI. К ним относятся молекулы на поверхности дендритных клеток, которые активируют функцию DC и презентацию антигена. Анти-CD40 антитела способны эффективно замещать активность Т-хелперных клеток (Ridge, J. et al (1998) Nature 393: 474-478) и могут быть использованы в сочетании с PDL1-связывающей молекулой по изобретению (Ito, N. et al (2000) Immunobiology 201 (5) 527-40). Активирующие антитела к Т-клеточным костимуляторным молекулам, такие как ОХ-40 (Weinberg, A. et al. (2000) Immunol Jj34: 2160-2169), 4-1BB (Melero, I. et al. (1997) Nature Medicine 3: 682-685 (1997), и ICOS (Hutloff, A. et al (1999) Nature 397: 262-266), а также антитела, которые блокируют активность отрицательных костимулирующих молекул, таких как CTLA-4 (например, патент США №5811097) или BTLA (Watanabe, N. et al. (2003) Nat Immunol 4:670-9), B7-H4 (Sica, GL et al (2003) Immunity 18:849-61), также могут обеспечить повышенные уровни активации Т-клеток.
Трансплантация костного мозга в настоящее время используется для лечения различных опухолей гематопоэтического происхождения. В то время как заболевание «трансплантат против хозяина» является следствием этого лечения, терапевтическая польза может быть получена от реакции трансплантата против опухоли. Блокада PDLI может быть использована для повышения эффективности пересаженных от донора опухолеспецифических Т-клеток. Существует также несколько протоколов экспериментального лечения, которые включают активацию и экспансию антигенспецифических Т-клеток ex vivo и адоптивный перенос этих клеток реципиенту для использования антигенспецифических Т-клеток против опухоли (Greenberg, R. & Riddell, S. (1999) Science 285: 546-51). Эти способы также могут быть использованы для активации Т-клеточных ответов на инфекционные агенты, такие как ЦМВ. Активация ex vivo в присутствии PDL1-связывающей молекулы по изобретению, как ожидается, может увеличить частоту и активность адаптивно перенесенных Т-клеток.
Инфекционные заболевания
Другие способы изобретения используют для лечения пациентов, которые были подвергнуты воздействию конкретных токсинов или патогенов. Таким образом, другой аспект изобретения обеспечивает способ предупреждения или лечения инфекционных заболеваний у субъекта, содержащий введение пациенту PDL1-связывающей молекулы по изобретению, при котором субъект лечится от инфекционного заболевания.
Подобно применению по отношению к опухолям, как обсуждалось выше, блокада PDLI может быть использована отдельно, или в качестве адъюванта, в комбинации с вакцинами, для стимуляции иммунной реакции на патогены, токсины и аутоантигены. Примеры патогенов, для которых такой терапевтический подход может быть особенно применим, включают патогены, против которых в настоящее время нет эффективной вакцины, или патогены, против которых традиционные вакцины недостаточно эффективны. Такие патогенны включают, без ограничения перечисленным, помимо ВИЧ, гепатит (А, В и С), грипп, герпес, лямблии, малярию, лейшманию, стафилококк золотистый (Staphylococcus aureus), синегнойную палочку (Pseudomonas Aeruginosa). Блокада PDLI особенно применима против установленных инфекций агентами, такими как ВИЧ, которые представляют измененные антигены в течение развития инфекций. Эти новые эпитопы распознаются как чужеродные во время введения анти-PD-LI человека, тем самым провоцируя сильный Т-клеточный ответ, который не гасится отрицательными сигналами через PDLI.
Некоторые примеры патогенных вирусов, вызывающих инфекции, которые поддаются лечению способами по изобретению, включают в себя ВИЧ, гепатит (А, В или С), вирус герпеса (например, VZV, HSV-1, ВГА-6, HSV-II и ЦМВ, Вирус Эпштейна-Барра), аденовирус, вирус гриппа, флавивирусы, эховирус, риновирус, вирус Коксаки, корновирус, респираторно-синцитиальный вирус, вирус свинки, ротавирус, вирус кори, вирус краснухи, парвовирус, вирус коровьей оспы, вирус человеческого Т-клеточного лейкоза (HTLV), вирус денге, папилломавирус, вирус моллюска, полиовирус, вирус бешенства, вирус Джона Каннингема (JC) и вирус арбовирусного энцефалита.
Некоторые примеры патогенных бактерий, вызывающих инфекции, поддающиеся лечению способами по изобретению, включают в себя хламидии, риккетсиальные бактерии, микобактерии, стафилококки, стрептококки, пневмококки, менингококки и гонококки, клебсиеллы, протей, серратии, синегнойные палочки, легионеллы, дифтерию, сальмонеллу, бациллы, холеру, столбняк, ботулизм, сибирскую язву, чуму, лептоспироз и бактерии болезни Лайма.
Некоторые примеры патогенных грибов, вызывающих инфекции, поддающиеся лечению способами по изобретению, включают в себя Candida (albicans, krusei, glabrata, tropicalis и т.д.), Cryptococcus neoformans, Aspergillus (fumigatus, niger и т.д.), Genus Mucorales (mucor, absidia, rhizophus), Sporothrix schenkii, Blastomyces dermatitidis, Paracoccidioides brasiliensis, Coccidioides immitis и Histoplasma capsulatum.
Некоторые примеры болезнетворных паразитов, вызывающих инфекции, поддающиеся лечению способами по изобретению, включают в себя Entamoeba histolytica, Balantidium coli, Naegleriafowleri, Acanthamoeba sp., Giardia lambia, Cryptosporidium sp., Pneumocystis carinii, Plasmodium vivax, Babesia microti, Trypanosoma brucei, Trypanosoma cruzi, Leishmania donovani, Toxoplasma gondi, Nippostrongylus brasiliensis.
Во всех перечисленных выше способах блокада PDLI может быть объединена с другими формами иммунотерапии, такими как лечение цитокинами (например, интерферонами, GM-CSF, G-CSF, IL-2), или терапия биспецифическим антителом, которая обеспечивает улучшенную презентацию противоопухолевых антигенов (см., например, Holliger (1993) Proc. Natl. Acad. ScL USA 90:6444-6448; Poljak (1994) Structure 2:1121-1123).
Аутоиммунные реакции
Анти-PDLI антитела могут провоцировать и усиливать аутоиммунные ответы. Таким образом, можно рассмотреть возможность применения блокады анти-PDLI в сочетании с различными собственными белками для разработки протоколов вакцинации для эффективного генерирования иммунных ответов против этих собственных белков для лечения заболеваний.
Например, болезнь Альцгеймера заключается в чрезмерном накоплении Аβ-пептидов в амилоидных бляшках головного мозга; антительные ответы против амилоидов способны очистить эти амилоидные бляшки (Schenk et al., (1999) Nature 400: 173-177). Другие собственные белки также могут быть использованы в качестве мишени, например, IgE для лечения аллергии и астмы, и TNFalpha для ревматоидного артрита. Наконец, антительные ответы на различные гормоны могут быть индуцированы с использованием анти-PDLI антитела. Нейтрализующие антительные ответы на репродуктивные гормоны могут быть использованы для контрацепции. Нейтрализующий антительный ответ на гормоны и другие растворимые факторы, которые необходимы для роста конкретных опухолей, также могут рассматриваться в качестве мишеней для вакцинации.
Способы, аналогичные описанным выше для применения анти-PDLI антитела, могут быть использованы для индуцирования терапевтических аутоиммунных ответов для лечения пациентов, имеющих несоответствующее накопление других аутоантигенов, таких как, амилоидные депозиты, включая Аβ при болезни Альцгеймера, цитокины, такие как TNFa и IgE.
Хронические воспалительные заболевания
Анти-PDLI антитела также могут быть применены для лечения заболеваний, таких как хронические воспалительные заболевания, например, красный плоский лишай, опосредованное Т-клетками хроническое воспалительное заболевание кожи и слизистых оболочек (Youngnak-Piboonratanakit et al. (2004) Immunol Letters 94:215-22). Таким образом, в другом аспекте изобретение обеспечивает способ подавления хронического воспалительного заболевания Т-клетками, содержащий введение субъекту PDL1-связывающей молекулы по изобретению.
Адъювант вакцины
В одном аспекте изобретение обеспечивает применение PDL1-связывающей молекулы по изобретению в качестве адъюванта вакцин. Анти-PDLI антитела могут быть применены для стимуляции антигенспецифических иммунных ответов путем совместного введения анти-PDLI антитела с представляющим интерес соответствующим антигеном (например, вакцины).
Таким образом, в другом аспекте изобретение обеспечивает способ усиления иммунного ответа на антиген у субъекта, содержащий введение пациенту: (i) антигена; и (ii) PDL1-связывающей молекулы по изобретению, так что у пациента усиливается иммунный ответ на антиген. Антиген может быть, например, опухолевым антигеном, вирусным антигеном, бактериальным антигеном или антигеном из определенного патогена. Неограничивающие примеры таких антигенов включают антигены, которые обсуждались в разделах выше, такие как опухолевые антигены (или противоопухолевые вакцины), рассмотренные выше, или антигены из вирусов, бактерий или других патогенов, описанные выше.
Набор
Также в объеме данного изобретения находится наборы, содержащие PDL1-связывающую молекулу, иммуноконъюгат или фармацевтическую композицию по изобретению и инструкцию по применению. Дополнительно набор может содержать по меньшей мере один дополнительный реагент, или одну или несколько дополнительных PDL1-связывающих молекул по изобретению (например, связывающие молекулы, которые связываются с различными эпитопами в PDL1). Наборы обычно включают в себя этикетку с указанием предполагаемого применения содержимого набора. Термин «этикетка» означает любую запись или записанный материал, поставляемый в наборе или с набором, или который в противном случае прилагается к набору.
ПРИМЕРЫ
Настоящее изобретение дополнительно иллюстрируется следующими примерами, но объем данного изобретения, ни в коем случае не должен быть ограничен конкретными примерами.
Пример 1. Скрининг однодоменного антитела, состоящего только из тяжелой цепи, против PDL1
1.1 Конструирование библиотеки
Слитый белок PDL1-Fc (SEQ ID NO: 28) для иммунизации экспрессировали в клетках СНО (pCDNA4, Invitrogen, Cat V86220), очищали аффинной хроматографией с протеином-А. Для иммунизации был выбран один Camelus bactrianus (пер., верблюд двугорбый или бактриан). После 4 иммунизаций лимфоциты выделяли из 100 мл периферической крови верблюда, и выделяли тотальную РНК с использованием набора RNA Extraction kit (QIAGEN). Экстрагированная РНК была обратно транскрибирована в кДНК с использованием набора Super-Script III FIRST STRANDSUPERMIX в соответствии с инструкциями.
Фрагменты нуклеиновых кислот, кодирующие антитела, состоящие только из тяжелых цепей, амплифицировали «вложенной» ПЦР:
Первый этап ПЦР:
Прямой праймер ("upstream"-праймер): 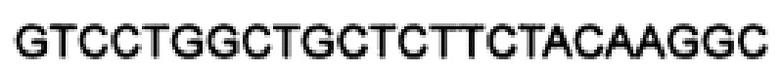 (SEQ ID NO: 29);
(SEQ ID NO: 29);
Обратный праймер ("downstream"-праймер):  (SEQ ID NO: 30).
(SEQ ID NO: 30).
Второй этап ПЦР:
ПЦР-продукты первого этапа ПЦР в качестве матрицы,
Прямой праймер (SEQ ID NO: 31);
Обратный праймер:  (SEQ ID NO: 32).
(SEQ ID NO: 32).
Целевые фрагменты нуклеиновых кислот однодоменного антитела, состоящего только из тяжелой цепи, извлекали и клонировали в вектор для фагового дисплея pCDisplay-3 (Creative Biolabs Cat VPT4023) с использованием эндонуклеазы Pstl и Notl (от NEB). Затем продукты электропорировали в компетентную клетку E.coli TG1, и библиотеку фагового дисплея для однодоменных антител, состоящих только из тяжелой цепи, против PDL1 конструировали и выверяли. Посевом на чашках серийных разведений, емкость библиотеки была определена как 1,33×108. Для определения коэффициента инсерций (вставок) библиотеки, случайным образом отбирали 24 клона для ПЦР на колониях. Результаты показали коэффициент вставок равным 100%.
1.2 Пэннинг однодоменного антитела, состоящего только из тяжелой цепи, против PDL1
Многолуночные планшеты покрывали слитым белком PDLI-Fc из расчета 10 мкг/лунку, 4°С, на ночь. На следующий день, после блокирования 1% обезжиренным молоком при комнатной температуре в течение 2 часов, добавляли 100 мкл бактериофагов (фагов) (8×1011 pfu, из библиотеки фагового дисплея для однодоменных антител, состоящих только из тяжелых цепей верблюдовых, сконструированной в 1.1), инкубировали при комнатной температуре в течение 1 часа. Затем несвязанные бактериофаги удаляли пятикратной промывкой с использованием PBST (0,05% Твин20 в PBS). Бактериофаги, которые специфически связываются с PDL1, диссоциировали триэтиламмонием (100 мМ) и использовали для инфицирования E.coli TG1 в логарифмической фазе роста, с получением бактериофагов, которые затем очищали для следующего раунда скрининга. Тот же самый скрининг повторяли в 3-4 раундах. Таким образом, обогащали позитивные клоны, достигая цели отбора PDL1-специфических антител из библиотеки антител с использованием технологии фагового дисплея.
1.3 Специфическая селекция индивидуальных положительных клонов с использованием фагового иммуноферментного анализа (ИФА)
PDL1-связывающие положительные фаги, полученные после 3-4 раундов пэннинга, использовали для заражения холостой пробы E.coli и посева. Случайным образом 96 отбирали 96 одиночных колоний для культивирования, и бактериофаги соответственно получали и очищали. Планшеты покрывали слитым белком PDLI-Fc при 4°C в течение ночи; добавляли пробы бактериофагов, аналогичные полученным (холостые фаги в качестве контроля), и инкубировали при комнатной температуре в течение 1 часа. Первичное антитело, мышиное анти-HA-tag антитело (Beijing Kangwei Shiji Biotech. Ltd.), добавляли после промывок и инкубировали при комнатной температуре 1 час для взаимодействия. Вторичное антитело, козье анти-мышиное антитело, меченное щелочной фосфатазой (Amyject Scientific Ltd.), добавляли после промывок и инкубировали при комнатной температуре 1 час для взаимодействия. Хромогенный раствор щелочной фосфатазы добавляли после промывки, и значение поглощения считывали при длине волны 405 нм. При OD (оптической плотности) для лунки пробы в 3 раза выше, чем OD контрольной пробы, пробу определяют как положительную. Бактерии в положительных лунках переносили и культивировали в жидкой среде LB с добавлением 100 мкг/мл ампициллина для экстракции плазмиды, с последующим секвенированием.
Белковые последовательности каждого клона анализировали в соответствии с программным обеспечением для выравнивания последовательностей Vector NTI. Клоны с одинаковыми CDR1, CDR2 и CDR3 последовательностями считаются одним и тем же антителом, тогда как клоны с различными CDR последовательностями считаются различными антителами.
Пример 2. Предварительная оценка однодоменных тяжелоцепочечных антител против PDL1
2.1 Экспрессия однодоменных тяжелоцепочечных антител в E.coli и их очистка
Кодирующую последовательность однодоменного антитела, состоящего только из тяжелой цепи, полученную секвенированием, субклонировали в экспрессирующий вектор PET32b (Novagen, номер продукта: 69016-3), и правильную рекомбинантную плазмиду трансформировали в экспрессирующий штамм-хозяин BL1 (DE3) (Tiangen Biotech, СВ105-02), высевали на твердую среду LB, содержащую 100 микрограмм на миллилитр ампициллина, на ночь при 37°C. Одиночные колонии высевали и культивировали в течение ночи, переносили на следующий день для экспансии при 37°С при встряхивании. При достижении культурой значения OD 0,6-1, добавляли 0,5 мМ изопропилтиогалактозида (ИПТГ) для индукции, оставляли на ночь при 28°С, со встряхиванием. На следующий день бактерии собирали центрифугированием и лизировали для получения грубых экстрактов антитела. Затем для очистки белков антитела использовали аффинную хроматографию с иммобилизованными ионами никеля, с целью получения белков антитела до чистоты более 90%.
2.2 Специфическое связывание однодоменного тяжелоцепочечного антитела-кандидата против PDL1 с человеческим белком PDL1
Планшеты покрывали слитым белком PDLI-Fc при 4°С в течение ночи, и 100 нг однодоменного тяжелоцепочечного антитела, полученного в примере 2.1 (в качестве контроля использовали однодоменное антитело, не связывающееся с белком PDL1-Fc), добавляли в каждую лунку и позволяли взаимодействовать в течение 1 часа при комнатной температуре. После промывки добавляли первое anti-His-tag антитело (приобретенное у Beijing Kangwei Century Biotechnology Co., Ltd.) и оставляли для взаимодействия в течение 1 часа при комнатной температуре. После промывки добавляли вторичное козье анти-мышиное антитело, меченое пероксидазой хрена (Yiqiao Shenzhou, Cat: SSA007200), и оставляли для взаимодействия в течение 1 часа при комнатной температуре. После промывки добавляли хромогенный агент и считывали поглощение при 405 нм.
Планшеты покрывали белком Fc при 4°С в течение ночи и 100 нг однодоменного антитела, состоящего только из тяжелой цепи, полученного в примере 2.1, добавляли в каждую лунку (в качестве контроля использовали однодоменное антитело против других неродственных мишеней), и позволяли взаимодействовать в течение 1 часа при комнатной температуре. После промывки добавляли антикроличье анти-Fc антитело человека (приобретенное у Shanghai Pu Xin Biotechnology Co., Ltd.) и оставляли для взаимодействия в течение 1 часа при комнатной температуре. После промывки добавляли козье анти-кроличье антитело, меченое пероксидазой хрена (приобретенное у Shanghai Pu Xin Biotechnology Co., Ltd.) и оставляли для взаимодействия в течение 1 часа при комнатной температуре. После промывки добавляли хромогенный агент и считывали поглощение при 405 нм.
Антитело-кандидат считается связанным с белком PDLI-Fc, когда соотношение значения OD для белка PDL1-Fc, деленное на значение OD для холостой пробы контроля, больше или равно 4; и одновременно, вышеуказанное антитело способное связываться с антигенным белком PDL1-Fc при соотношении значения OD для связывания с PDLI-Fc, деленное на значение OD для связывания Fc-белка составляет > = 5, считается специфически связанным с PDL1-частью молекулы, а не с Fc-частью молекулы.
Результаты показал, что антитело №56 может специфически связываться с PDL1 без связывания с Fc. Конкретные результаты приведены в таблице 1 ниже:
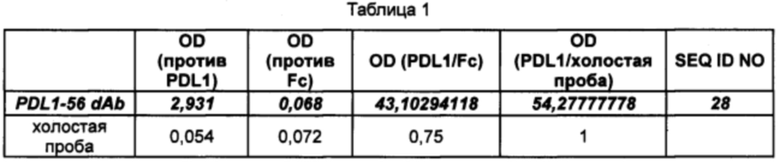
2.3 Связывание однодоменного тяжелоцепочечного антитела против PDL1 с мышиным белком PDL1
Мышиный белок PDL1-Fc (SEQ ID NO: 33) получали экспрессией в клетках HEK293 (pCDNA4, Invitrogen, каталожный # V86220).
Планшеты покрывали в течение ночи при 4°С мышиным слитым белком PDLI-Fc по 0,5 мкг/лунку и добавляли 100 нг однодоменного антитела, состоящего только из тяжелой цепи, полученного в примере 2.1 (контролем служило однодоменное антитело против других неродственных мишеней), и оставляли для взаимодействия в течение 1 часа при комнатной температуре. После промывки добавляли первое антитело Anti-His-Tag антитело и оставляли для взаимодействия в течение 1 часа при комнатной температуре. После промывки добавляли козье анти-мышиное антитело, меченое пероксидазой хрена, и оставляли для взаимодействия в течение 1 часа при комнатной температуре. После промывки добавляли хромогенный агент и считывали поглощение при 405 нм. Результаты приведены в таблице 2.
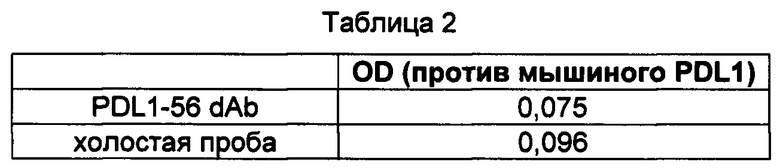
Можно видеть, что однодоменное тяжелоцепочечное антитело против человеческого PDL1 настоящего изобретения не связывается с мышиным белком PDLI-Fc.
2.4 Изучение блокирующего эффекта однодоменного тяжелоцепочечного PDL1-антитела на взаимодействие PD-1 и PDL1 при помощи конкурентного ИФА.
Белок PDL1-Fc и белок PD1-Fc (SEQ ID NO: 34) получали экспрессией в клетках HEK293 (pCDNA4, Invitrogen, каталожный № V86220). Биотинилированный белок PD1-Fc-биотин получали с использованием набора Thermo Biotinlytion Kit.
Планшеты покрывали в течение ночи при 4°С слитым белком PDLI-Fc по 0,5 мкг/лунку с последующим добавлением 100 нг однодоменного тяжелоцепочечного антитела, полученного в примере 2.1 (контролем служило однодоменное антитело против других неродственных мишеней или просто буфер) и добавляли 10 мкг PDI-Fc-биотина (никакого антитела или белка не добавляли к группе холостой пробы, добавляли только такой же объем буфера), и оставляли для взаимодействия в течение 1 часа при комнатной температуре. SA-HRP (приобретенный у Sigma) добавляли и оставляли для взаимодействия в течение 1 часа при комнатной температуре. После добавления хромогенного раствора, поглощение считывали на длине волны 405 нм. Когда значение OD пробы относительно значения OD контрольной пробы составляет <0,8, считается, что антитело обладает блокирующим эффектом (действием).
Как показано в таблице 3, антитело №56 показало блокирующий эффект на взаимодействие PD-1/PDL1.

2.5 Блокирующий эффект однодоменного тяжелоцепочечного PDL1-антитела на взаимодействие PD-1 и PDL1 на клеточной поверхности исследовали посредством ФСК.
Клетки HEK293, транзиентно экспрессирующие человеческий белок PDL1 на мембране (клетки 293-PDL1), получали транзиентной трансфекцией плазмиды, несущей полноразмерный ген человеческого PDL1 (pCDNA4, Invitrogen, каталожный № V86220) в клетки HEK293 человека. Например,
Клетки 293-PDL1 собирали, ресуспендировали в 0,5% буфере PBS-BSA в 96-луночном планшете, и добавляли вышеупомянутые антитела, подлежащие обнаружению. Также одновременно ставили отрицательные контрольные пробы, и отрицательные контрольные пробы представляли собой 2 мкг однодоменного антитела против другой мишени. 0,3 мкг hPD-1-Fc-биотина и SA-PE от eBioscience добавляли ко всем пробам и после окрашивания выполняли проточную цитометрию. Считается, что антитело блокирует на поверхности клеток взаимодействие PDL1 и PD-1, если значение флуоресценции смещается в направлении холостой пробы по сравнению с группой без антител. Таким образом идентифицировали антитела, которые блокируют связывание антигена PDL1 с PD-1 на поверхности клеток.
Результаты показаны на фиг. 1, антитело №56 демонстрирует блокирующий эффект на взаимодействие PD-1/PDL1.
2.6 Кривые связывания однодоменных тяжелоцепочечных PDL1-антител с антигенным белком PDL1.
Планшеты покрывали полученным однодоменным тяжелоцепочечным PDL1-антителом при 0,5 мкг/лунку при 4°С в течение ночи с последующим добавлением серий градиентных разведений слитого белка PDL1-Fc, и оставляли для взаимодействия в течение 1 часа при комнатной температуре. После промывки добавляли козье антитело против человеческого IgG-Fc, меченое пероксидазой хрена, и оставляли для взаимодействия в течение 1 часа при комнатной температуре. После промывки добавляли цветопроявляющий раствор для пероксидазы хрена, и поглощение считывали при длине волны 405 нм. Для обработки данных и сопоставительного анализа использовали программу SotfMax Pro v5.4 для получения кривой связывания антитела с PDL1 и значения ЕС50 (около 5 нг/мл) посредством подгонки по четырем параметрам. Результаты показаны на фиг. 2.
2.7 Кривые блокирования однодоменных антител тяжелой цепи PDL1 на взаимодействие PD-1 и PDL1.
Планшеты покрывали слитым белком PDLI-Fc по 0,5 мкг/лунку при 4°С в течение ночи с последующим добавлением 100 мкл серий градиентных разведений (содержащих 100 мкг/мл PD1-Fc-биотин), 100 мкл на лунку блокирующего слитого белка PDL1-однодоменного антитела с Fc и оставляли для взаимодействия в течение 1 часа при комнатной температуре. Добавляли SA-HRP (приобретенный у Sigma) и оставляли для взаимодействия в течение 1 часа при комнатной температуре. После добавления хромогенного раствора считывали поглощение при длине волны 405 нм.
Для обработки данных и сопоставительного анализа использовали программу SotfMax Pro v5.4 для получения кривой блокирования и значения медианной IC50 антитела №56 с PDL1/PD-1 посредством подгонки по четырем параметрам (IC50 для антитела №56 составляет 143 нг/мл). Результаты показаны на Фигуре 3.
Пример 3. Гуманизация однодоменных антител PDL1
Гуманизацию выполняли методом гуманизации экспонированных на поверхности белка аминокислот (метод перекладки) и методом трансплантации в универсальную каркасную область (универсальный каркас) для гуманизации VHH (трансплантации CDR в универсальный каркасный участок).
Стадии гуманизации являются следующими: Гомологичное моделирование антитела №56 выполняли с помощью программного обеспечения для моделирования Modeller9. Ссылочной гомологичной последовательностью является антитело NbBclI10 (код PDB: 3DWT), и относительную доступность для растворения аминокислот рассчитывали по трехмерной структуре белка. Если одна из аминокислот антитела №56 подвергается действию растворителя, то ее заменяют аминокислотой в том же самом положении ссылочной последовательности DP-47 антитела человека, пока все замены не будут завершены.
Конкретные стадии метода трансплантации в универсальную каркасную область для гуманизации VHH следующие: сначала универсальный каркас для гуманизированных VHH h-NbBclI10FGLA (код PDB: 3EAK), сконструированный Сесилем Винке ( Vincke) и другими на основе гомологии последовательностей, каркас был сконструирован на основе нанотела NbBclI10 (код PDB: 3DWT); гуманизация экспонированных на поверхности белка аминокислот была выполнена со ссылкой на человеческое антитело DP-47, и отдельные аминокислоты FGLA VHH-последовательности каркаса 2 были изменены. Авторы изобретения непосредственно использовали h-NbBcII10FGLA в качестве каркаса и заменили CDR участками CDR антитела №56 для достижения гуманизации антитела.
Vincke) и другими на основе гомологии последовательностей, каркас был сконструирован на основе нанотела NbBclI10 (код PDB: 3DWT); гуманизация экспонированных на поверхности белка аминокислот была выполнена со ссылкой на человеческое антитело DP-47, и отдельные аминокислоты FGLA VHH-последовательности каркаса 2 были изменены. Авторы изобретения непосредственно использовали h-NbBcII10FGLA в качестве каркаса и заменили CDR участками CDR антитела №56 для достижения гуманизации антитела.
Антитело №56 было гуманизировано, и было получено пять вариантов гуманизированного антитела №56. В таблице 4 перечислены номера SEQ ID этих гуманизированных вариантов, а также изменения аминокислот в них с номерами аминокислотных остатков по нумерации Кабата. Фигура 4 показывает выравнивание гуманизированных последовательностей.
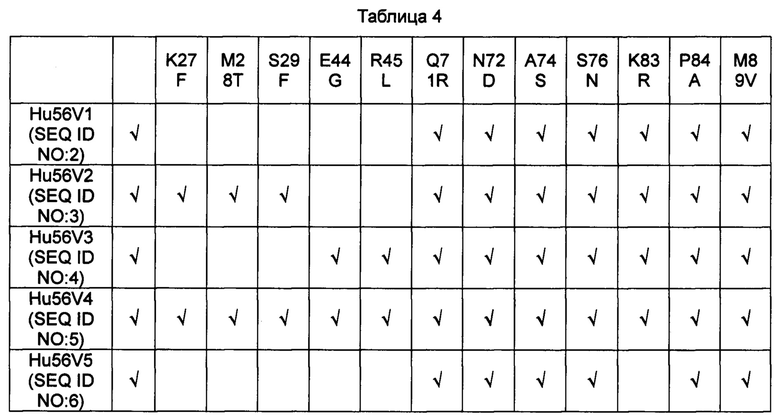
Пример 4. Получение белка, блокирующего PDL1-антитело с использованием клеток млекопитающих
4.1 Получение слитого белка однодоменного антитела против PDL1 с Fc-фрагментом
Аминокислотную последовательность IgGI-Fc-область человека (SEQ ID №: 7) получали на основе аминокислотной последовательности константной области человеческого иммуноглобулина гамма-1 (IgGI) из белковой базы данных Uniprot (Р01857). Фрагмент нуклеиновой кислоты, кодирующий человеческий IgG1-Fc, получали из тотальной РНК мононуклеарных клеток периферической крови человека (РВМС) посредством ПЦР с обратной транскрипцией, и фрагмент нуклеиновой кислоты, кодирующий слитый белок однодоменного антитела PDL1, полученный в описанном выше примере, и Fc получали посредством перекрывающейся ПЦР, затем субклонировали в вектор pCDNA4 (Invitrogen, кат. № V86220). Также могут быть использованы последовательности Fc-области, такие как, SEQ ID NO: 8 или 9, в которых была устранена антителозависимая клеточно-обусловленная цитотоксичность (ADCC) или комплементзависимая цитотоксичность (CDC).
Рекомбинантную плазмиду слитого белка однодоменного антитела против PDL1 с Fc трансфицировали в клетки HEK293 для экспрессии антитела. Рекомбинантные экспрессирующие плазмиды разбавляли средой Freestyle 293 и добавляли в раствор PEI (полиэтиленимина) для трансформации. Каждую смесь плазмиды с PEI добавляли в суспензию клеток HEK293 и инкубировали при 37°С и 10% CO2 при 90 об/мин. В то самое же время добавляли 50 мкг/л IGF-1. Через четыре часа добавляли среду ЕХ293, 2 мМ глютамин и 50 мкг/л IGF-1, культивировали при 135 об/мин. Через 24 часа добавляли 3,8 мМ VPA. После культивирования в течение 5-6 дней собирали супернатант транзиентной экспрессии и очищали аффинной хромотографией с протеином А для получения целевого белка однодоменного антитела против PDL1 слитого с Fc.
Полученные последовательности белка однодоменного антитела против PDL1 слитого с Fc приведены в SEQ ID NO: 10-27.
4.2 Получение PDL1-антител от MedImmune LLC и Roche
Ген PDL1-антитела от MedImmune LLC, Inc. клонироваали методом 2.14Н9 в US 20130034559 и клонировли в вектор pCDNA4. ТМ004 представляет собой анти-PDL1 антитело от Roche. Ген антитела клонировали согласно YW243.55.S70hIgG в US 20130045201 А1 и клонирован в вектор pCDNA4.
Рекомбинантную плазмиду транзиентно трансфицировали в клетки HEK293 тем же самым методом, что в примере 4.1, и полученное анти-PDLI антитело от MedImmune LLC было переименовано в 2.41Н90Р; анти-PDL1 антитело от Roche было переименовано в 243.55.
4.3 Сравнение экспрессии белка однодоменного антитела против PDL1 слитого с Fc и двух известных PDL1-антител
С использованием той же самой экспрессирующей системы и тех же самых условий транзиентной трансфекции уровень экспрессии белка однодоменного антитела против PDL1 слитого с Fc по настоящему изобретению был выше, чем 200 мг/л, тогда как уровень экспрессии антитела 2.41Н90Р составлял около 80 мг/л, и уровень экспрессии антитела 243.55 составлял около 40 мг/л. Этот результат означает, что белок однодоменного антитела против PDL1, слитый с Fc по настоящему изобретению является более стабильным по структуре и может привести к более высокому уровню экспрессии, чем другие два известных PDL1-антитела.
Пример 5. Исследование свойств слитого белка однодоменного антитела к PDL1 с Fc
5.1 Связывающая способность слитого белка однодоменного антитела к PDL1 с Fc по отношению к PDL1 (по ИФА).
Белок PDL1-Chis (SEQ ID №:35) получали транзиентной экспрессией в HEK293 и очисткой с использованием аффинной хроматографии с иммобилизованными ионами никеля. Планшеты покрывали получившимся белком PDL1-Chis по 0,5 мкг/лунку, в течение ночи при 4°С. Затем добавляли серии градиентного разведения слитых белков однодоменного антитела против PDL1 с Fc, полученных в описанном выше примере, и оставляли для взаимодействия в течение 1 часа при комнатной температуре. После промывки добавляли козье антитело против человеческого IgG-Fc, меченое пероксидазой хрена, и оставляли для взаимодействия в течение 1 часа при комнатной температуре. После промывки добавляли хромогенный агент и считывали поглощение на 405 нм. Для обработки данных и сопоставительного анализа использовали программу SotfMax Pro v5.4, посредством подгонки по четырем параметрам для получения кривой связывания и значения ЕС50 антитела к PDL1 (у всех анализируемых антител значение ЕС50 около 150 нг/мл) для отражения аффинности с PDL1.
Результаты показаны на фиг. 5, где продольная ордината представляет собой OD405, и горизонтальная ордината представляет собой концентрацию однодоменного антитела к PDL1-Fc-слитого белка (в нг/мл); перевернутые треугольники, треугольники и квадраты представляют три различные гуманизированные формы Fc-слитых белков: hu56v1-Fc, hu56v2-Fc, hu56v5-Fc. У трех белков сопоставимые значения аффинности к PDL1.
5.2 Идентификация связывающей способности слитого белка однодоменного антитела PDL1-Fc в отношении PDL1 (методом SPR) и по сравнению с известными антителами
Кинетику связывания слитого белка однодоменного антитела к PDLI-Fc с рекомбинантным человеческим PDL1, полученным в приведенных выше примерах, измеряли методом поверхностного плазмонного резонанса (SPR), с помощью прибора BIAcore Х100. Рекомбинантный человеческий PDLI-Fc наносили непосредственно на биосенсорный чип СМ5 для получения около 1000 ответных единиц (RU). Для кинетических измерений проводили серийные разведения антител (от 1,37 до 1000 нм) в буфере HBS-EP+1х (GE, каталожный # BR-1006-69) и вводили в течение 120 секунд при 25°С при времени диссоциации 30 мин, регенерировали 10 мМ глицин-HCl (pH 2,0) в течение 120 секунд. Значения скорости ассоциации (kon) и скорости диссоциации (koff) рассчитывали с использованием модели Languir для связывания один к одному (программное обеспечение BIAcore Evaluation, версия 3.2). Равновесную константу диссоциации (kD) рассчитывали как отношение koff/kon.
Измеренные значения аффинности связывания антител анти-PDLI приведены в таблице 5. Результаты показывают, что аффинность белка PDL1-56-FC к белку-мишени PDL1 является значительно выше, чем аффинность двух антител PDL1, известных в данной области техники, и его более высокое значение Ka и более низкое значение Kd указывают на то, что слитый белок антитела может связывать антиген PDL1 более быстро, и таким образом он с трудом диссоциирует, что дополнительно указывает на то, что PDL1-56-Fc в качестве блокирующего антитела со свойствами, превосходящими свойства двух известных антител PDL1.
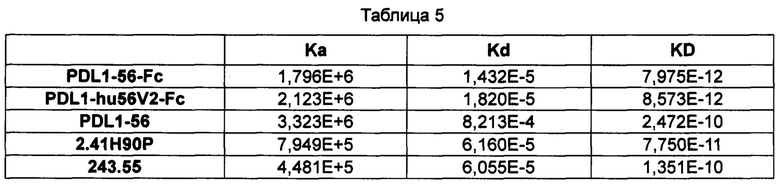
5.3 Блокирующее действие слитого однодоменного белка PDL1-Fc на взаимодействие PDL1-PD1 (конкурентным ИФА)
Планшеты покрывали слитым белком PDLI-Fc по 0,5 мкг/лунку в течение ночи при 4°C, с последующим добавлением серий градиентных разведений (содержащих 100 мкг/мл PD1-Fc-биотина) слитого белка однодоменного антитела PDL1-Fc, полученного в вышеприведенном примере, 100 мкл на лунку, и оставляли для взаимодействия в течение 1 часа при комнатной температуре. После промывки добавляли SA-HRP (приобретенный у Sigma) и оставляли для взаимодействия в течение 1 часа при комнатной температуре. После промывки добавляли хромогенный агент и считывали поглощение при 405 нм.
Для обработки данных и сопоставительного анализа посредством 4-параметрического выравнивания использовали программу SotfMax Pro v5.4 для получения кривой блокирования и значения IC50 антитела к PDL1-PD1. Результаты показаны на фиг. 6, где продольной ордината представляет собой OD405, и абсцисса представляет собой концентрацию слитого однодоменного белка PDLI-Fc (в нг/мл); перевернутый треугольник, тригонометрический треугольник и квадрат представляют собой три различные гуманизированные формы слитых белков Fc: hu56v1-Fc, hu56v2-Fc, hu56v5-Fc. Три белка обладают сходной способностью блокировать взаимодействие PDL1-PD1.
5.4 Блокирующее действие слитого однодоменного белка PDL1-Fc на взаимодействие PDL1-CD80 (конкурентным ИФА)
Белок CD80-Fc (SEQ ID NO: 36) получали из клеток HEK293. Биотинилированный белок CD80-Fc-биотин получали с помощью набора Thermo Biotinlytion kit.
Планшеты покрывали слитым белком PDL1-Fc по 0,5 мкг/лунку в течение ночи при 4°С, с последующим добавлением серий градиентных разведений (содержащих 300 мкг/мл CD80-Fc-биотина) слитого белка однодоменного антитела PDL1-Fc, полученного в вышеприведенном примере, 100 мкл на лунку, и оставляли для взаимодействия в течение 1 часа при комнатной температуре. После промывки добавляли SA-HRP (приобретенный у Sigma) и оставляли для взаимодействия в течение 1 часа при комнатной температуре. После промывки добавляли хромогенный агент и считывали поглощение при 405 нм.
Для обработки данных и сопоставительного анализа посредством 4-параметрического выравнивания использовали программу SotfMax Pro v5.4 для получения кривой блокирования и значения IC50 антитела к PDL1-CD80. Результаты показаны на фиг. 7, где продольная ордината представляет собой OD405, и абсцисса представляет собой концентрацию (в нг/мл) слитого белка однодоменного антитела PDL1-Fc hu56V2-Fc Результаты показывают, что блокирующий PDL1 слитый белок Fc однодоменного антитела hu56V2-Fc может эффективно блокировать взаимодействие между PDL1 и CD80.
5.5 Блокирующее действие слитого однодоменного белка PDL1-Fc на взаимодействие PDL1-PD1 (FACS-анализ).
Клетки человека HEK293 транзиентно экспрессируют белок PDL1 обезьяны на мембранах путем транзиентной трансфекции плазмид, содержащих полноразмерный человеческий ген PDL1.
PDI-muFc (SEQ ID NO: 39) в рабочей концентрации 2 мкг/мл добавляли к каждой группе в соответствии с делением на группы 5×105 клеток/пробирку и затем к каждой группе добавляли KN035 в различных концентрациях. После инкубации на ледяной бане в течение 30 мин и трехкратной промывки добавляли меченое фикоэритрином (РЕ) вторичное козье антимышиное антитело, в качестве детектирующего антитела, и интенсивность флуоресценции определяли проточной цитометрией после инкубации на ледяной бане в течение 30 минут.
Для обработки данных и сопоставительного анализа использовали программное обеспечение GraphPad Prism. Посредством 4-параметрического выравнивания получали кривую блокирования и значение IC50 антитела исходя из прямого взаимодействия между PDL1, экспрессированным на мембране клетки 293, и растворимым PD1. Результаты показаны на фиг. 8, где продольная ордината представляет собой среднее значение интенсивности флуоресценции (MFI), а абсцисса представляет собой концентрацию (в мкг/мл) белка однодоменного антитела PDL1 слитого с hu56V1-Fcm1. Результаты показывают, что слитый белок однодоменного антитела против PDL1 с Fc-фрагментом может эффективно блокировать взаимодействие между PDL1, экспрессированным на мембране клетки 293, и PD1.
Плазмиду, содержащую полноразмерный ген человеческого PD1, трансформировали и интегрировали в клеточную линию Jurket для получения клеточной линии Jurket, стабильно экспрессирующей человеческий белок PD1, которая получила название Jurket-PD1.
После инкубации клеток Jurkat-PD1 с биотинилированным белком PDL1-muFc (SEQ ID NO: 38) (30 мкг/мл) в течение 30 минут на ледяной бане добавляли градиентное разведение белка однодоменного антитела PDL1 слитого с Fc, hu56V1-Fcm1, инкубировали в течение 1 часа на ледяной бане, промывали три раза с буфером PBS, а затем добавляли разбавленный 1:250 раствор Streptavidin РЕ, инкубировали на ледяной бане в течение 30 мин и промывали три раза PBS. Интенсивность флуоресценции определяли методом проточной цитометрии.
Обработку данных и сопоставительный анализа выполняли с использованием программного обеспечения GraphPad Prism. Кривые блокирования и значения IC50 антител получали исходя из прямого взаимодействия между Jurkat-PD1 и растворимым белком PDL1-muFc посредством 4-параметрического выравнивания. Результаты показаны на фиг. 9, где продольная ордината представляет собой среднее значение интенсивности флуоресценции (MFI), а абсцисса представляет собой концентрацию (в мкг/мл) белка однодоменного антитела PDL1 слитого с hu56V1-Fcm1. Результаты показывают, что блокирующий слитый белок однодоменного антитела PD1-Fc может эффективно блокировать взаимодействие между Jurkat-PD1 и PDL1.
5.6 Специфичность связывания слитого белка однодоменного антитела против PDL1 с Fc-фрагментом по отношению к PDL1
Человеческие клетки HEK293 непродолжительно транзиентно экспрессируют белок PDL1 обезьяны на мембранах путем транзиентной трансфекции плазмиды, несущей полноразмерные гены белка семейства В7 человека (pCDNA4, Invitrogen, каталожный # V86220). Плазмида также позволяет С-концу белка-мишени сливаться с белком EGFP, таким образом, чтобы уровень экспрессированного на мембране белка семейства В7 мог быть изучен по интенсивности зеленой флуоресценции. Сконструированные транзиентно трансфицированные линии клеток включают в себя 293-PDL1-EGFP, 293-PDL2-EGFP, 293-B7H3-EGFP и 293-B7H3-EGFP.
Сконструированные клетки ресуспендировали в 0,5% буфере PBS-BSA и добавляли антитело hu56V2-Fc. В то же самое время готовили отрицательный контроль 2 мкг однодоменного антитела против другой неродственной мишени и инкубировали на ледяной бане в течение 20 минут. После промывки добавляли вторичное антитело анти-hIg-PE от eBioscience, инкубировали на ледяной бане в течение 20 минут. После промывки клетки ресуспендировали в 500 мкл 0,5% буфера PBS-BSA и детектировали проточной цитометрией.
Результаты показаны на фиг. 10. На верхней строке показана контрольная группа, на нижней строке показаны группы проб. Видно, что антитело hu56V2-Fc специфически связывается только с человеческим белком PDL1, но не с другими белками семейства В7.
5.7 Связывание слитого белка однодоменного антитела против PDL1-Fc с белком PDL1 обезьяны
Человеческие клетки HEK293 транзиентно экспрессируют белок PDL1 обезьяны (SEQ ID NO: 57) посредством переходной трансфекции плазмид, содержащих полноразмерный ген PDL1 обезьяны. Плазмида также обеспечивает белок-мишень со слитым с С-конца белком EGFP, который позволяет изучить уровни мембранной экспрессии белка PDL1 обезьяны по интенсивности зеленой флуоресценции.
Сконструированные клетки ресуспендировали в 0,5% буфере PBS-BSA и добавляли антитело hu56V2-Fc и инкубировали на ледяной бане в течение 20 минут. После промывки добавляли вторичное антитело anti-hIg-PE от eBioscience, инкубировали на ледяной бане в течение 20 минут. После промывки клетки ресуспендировали в 500 мкл 0,5% буфера PBS-BSA и детектировали проточной цитометрией.
Результаты показаны на фиг. 11. Видно, что антитело hu56V2-Fc эффективно связывается с белком PDL1 обезьяны.
5.8 Слитый белок однодоменного антитела против PDL1 с Fc может эффективно идентифицировать PDL1-положительную клеточную популяцию на срезах ткани, взятых у пациентов.
Срезы опухолевой ткани, взятые у PDL1-положительных пациентов с раком легких, окрашивали 5 мкг/мл антитела hu56V2-Fc в качестве первичного антитела в течение ночи и инкубировали с козьим антителом против человеческого, меченого HRP (Perkin-Elmer, Cat: NEF802001EA), затем визуализировали.
Результаты показаны на фиг. 12. Антитело hu56V2-Fc может эффективно идентифицировать PDL1-положительные популяции клеток на срезах тканей больного раком легких, и может одновременно идентифицировать PDL1-положительные опухолевые клетки и PDL1-положительные иммунные клетки.
5.9 Активация РВМС слитым белком однодоменного антитела PDL1-Fc.
Мононуклеарные клетки периферической крови (PBMCs) выделяли из периферической крови здоровых доноров центрифугированием в градиенте плотности с использованием раствора для выделения лимфоцитов человека (Tianjin Нао Yang).
2,5 мкг/мл антитела анти-CD3 и серии градиентных разведений слитого белка однодоменного антитела против PDL1-Fc hu56V2-ФК (в эксперименте также обозначенного как KN035) наносили на планшеты для культивирования клеток наносили в течение ночи при 4°C. На следующий день в каждую лунку добавляли 1×105 PBMCs. После культивирования в течение 5 дней супернатант собирали и определяли уровень IFN-γ в супернатанте с использованием набора IFN-γ ELISA kit (Ebioscience).
Результаты показаны на фиг. 13. Можно видеть, что слитый белок однодоменного антитела против PDL1-Fc, объединенный с анти-CD3 антителом, может повысить секрецию γ-интерферона из клеток РВМС, т.е, слитый белок однодоменного антитела против PDL1-Fc повышает клеток РВМС. В то же самое время, эффективность активации зависит от концентрации.
5.10 Активация CD4+ Т-клеток слитым белком однодоменного антитела против PDL1 с Fc в реакции смешанной культуры лимфоцитов дендритных клеток-Т-клеток и сравнение с антителом анти-PDL1 MedImmune LLC.
Мононуклеарные клетки периферической крови (PBMCs) выделяли из лейкоцитов периферической крови здоровых доноров центрифугированием в градиенте плотности с использованием раствора выделения лимфоцитов человека (Tianjin Нао Yang). Затем лимфоциты инкубировали в бессывороточной среде RPMI-1640 в течение 1-2 часов для удаления неслившихся клеток и клетки культивировали в RPMI, содержащей 10% FBS, 10 нг/мл GM-CSF и 20нг/мл IL-4. После культивирования в течение 5-6 дней добавляли 10 нг/мл TNF-α и инкубировали в течение 24 часов для получения зрелых дендритных клеток.
Дендритные клетки, полученные этим способом ресуспендировали в полной среде RPMI, 2×105/мл. Затем добавляли по 50 мкл/лунку 96-луночного планшета с U-образным дном (Costar: 3799) и культивировали в инкубаторе.
CD4+-Т-клетки выделяли из РВМС другого донора с использованием набора для выделения на магнитных микроносителях (магнитных микросферах) (Miltenyi Biotec: 130-096-533) согласно инструкциям производителя.
1×104 дендритных клеток и 1×105 CD4+-Т-клеток, полученных описанными выше способами, смешивали, ресуспендировали в полной среде RPMI и добавляли в 96-луночный культуральный планшет, и в каждую лунку добавляли по 50 мкл смеси клеток. 100 мкл/лунку hu56V2-Fc, разбавленного в полной среде RPMI, добавляли до конечной концентрации антитела 0,1 мкг/мл или 0 мкг/мл. Супернатанты собирали через 5-7 дней культивирования и определяли уровень IFN-γ в супернатанте с использованием набора IFN-γ ELISA detection kit (ebioscience).
Результаты показаны на фиг. 14А. Можно видеть, что слитый белок однодоменного антитела против PDL1 с Fc может повышать секрецию IFN-γ CD4+-Т-клеток в реакции смешанной культуры лимфоцитов (MLR-assay). Иными словами, PDL1-блокирующий слитый белок однодоменного PDL1-анитела с Fc повышает активацию Т-клеток.
PBMCs субъектов 1 культивировали с 50 нг/мл GM-CSF+25 нг/мл IL-4, и дендритные клетки (DCs) собирали через 6 дней после созревания с TNF-α (50 нг/мл); CD4+-Т-клетки были отсортированы с использованием набора для выделения на магнитных микроносителях magnetic bead isolation kit (Miltenyi Biotec: 130-096-533); DC-клетки добавляли в 96-луночные планшеты с U-образным дном по 104/лунку и CD4+-Т-клетки добавляли по 105/лунку через 2-4 часа; различные концентрации слитого белка однодоменного антитела PDL1-Fc hu56V1-Fcm1 или анти-PDL1 антитела 2.41Н90Р от MedImmune LLC добавляли в соответствующие лунки; после культивирования в течение 5 дней определяли уровень IFN-γ в супернатанте с использованием набора IFN-γ ELISA detection kit (ebioscience).
Результаты показаны на фиг. 14В, где серая гистограмма показывает стимуляцию секреции IFN-γ слитым белком однодоменного антитела к PDL1-Fc, черная полоса показывает антитело анти-PDL1 2.41Н90Р от MedImmune LLC. Слитый белок однодоменного антитела к PDL1-Fc с повышением концентрации может усилить способность CD4+-Т-клеток секретировать IFN-γ в реакции смешанной культуры лимфоцитов, и способность слитого белка однодоменного антитела к PDLI-Fc активировать Т-клетки немного сильнее, чем у антитела анти-PDLI 2.41 Н90Р от MedImmune LLC при той же самой концентрации.
5.11 Стимуляция секреции IL-2 Т-клеток в системе совместной культуры Jurkat/Raji-PDL1 слитым белком однодоменного антитела к PDL1-Fc и сравнение с анти-PDL1 антителом от MedImmune LLC
Авторы изобретения создали систему активации Т-клеток, систему совместной культуры Jurkat/Raji-PDL1, для тестирования влияния слитого белка однодоменного антитела с PDL1-Fc на активацию Т-клеток, которое сравнивали с анти-PDLI антителом от MedImmune LLC.
В системе используют клетки Jurkat (Т-клетки) в качестве эффекторных клеток, и анти-CD3 антитело человека в качестве первого сигнала для активации клеток Jurkat. CD80 семейства В7 на поверхности клеток Raji-PDL1, которые являются генетически сконструированными и стабильно экспрессируют PDL1 человека, обеспечивают второй костимуляторный сигнал для активации клеток Jurket, тогда как PDL1, сильно экспрессируемый на поверхности той же самой клетки, действует как отрицательный регулятор посредством связывания PD1 для ингибирования активации клеток Jurket.
Градиентные разведения белков антител hu56V1-Fcm1 и 2.41 Н90Р получали с использованием 10% FBS+1640+150 нг/мл анти-CD3; клетки Jurkat и Раджи-PDLI доводили до 3×106 клеток/мл и 1,5×106 клеток/мл, добавляли по 50 мкл в каждую лунку, хранили при 37°С в течение 24 часов. Удаляли 100 мкл культурального супернатанта и измеряли экспрессию IL-2 с использованием набора.
Результаты показаны на фиг. 15, где черная гистограмма показывает стимуляцию секреции IL-2 слитым белком однодоменного антитела к PDL1-Fc и серая гистограмма показывает анти-PDLI антитело 2.41Н90Р от MedImmune LLC. Слитый белок однодоменного антитела к PDLI-Fc может усилить способность клеток Jurkat секретировать IL-2 с повышением концентрации, и его способность активировать клетки Jurkat немного сильнее, чем у антитела PDL1 2.41Н90Р от MedImmune LLC при той же концентрации.
5.12 Аффинность слитого белка однодоменного антитела к PDL1-Fc к FcRn
Биотинилированные слитые белки однодоменного антитела к PDL1-Fc hu56VI-ФК, hu56V2-ФК, hu56VI-Fcml и hu56V2-Fcml разбавляли до 10 мкг/мл и иммобилизовали на биосенсорах, покрытых стрептавидином (SA). Готовили разведения человеческого белка FcRn (RnD Systems Cat. No. 8639-FC): 200 нМ, 100 нМ, 50 нМ, 25 нМ, 12,5 нМ. Взаимодействие обнаруживали с использованием прибора Octet K2 от Fortebio Corporation, с отверждением в течение 100 с, выдерживанием для связывания в течение 60 с, диссоциацией в течение 30 с.
Таблица 6 показывает, что средняя KD слитых белков однодоменного антитела к PDL1-Fc по отношению к FcRn составляет около 5.1Е-07 М. Не было достоверной разницы в аффинности между мутантным Fc (Fcm1) и Fc дикого типа.
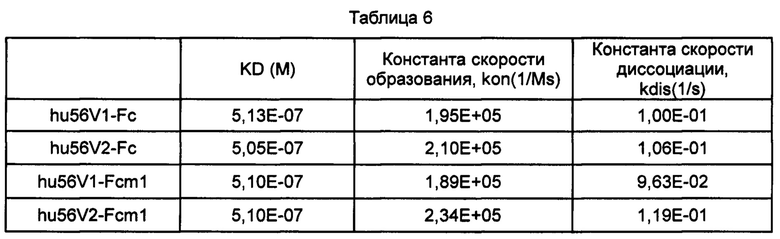
5.13 CDC-активность и ADCC-активность слитого белка однодоменного антитела к PDL1-Fc
PBMCs активировали с использованием 300 МЕ/мл IL-2 в течение 24 часов в качестве эффекторных клеток, с количеством клеток 8×105/лунку; Raji-PDL1, стабильно экспрессирующие человеческий белок PDL1, использовали в качестве клеток-мишеней с количеством клеток 2×105/лунку; добавляли различные концентрации белка hu56V1-Fcm1 или Rituxan в различной концентрации в качестве положительного контроля, и измеряли активность ADCC (%) при каждой концентрации с использованием набора для обнаружения нерадиоактивной цитотоксичности CytoTox 96® после инкубации при 37°С в течение 6 часов.
На фиг. 16а показано, что hu56V1-Fcm1 не проявляет значительной ADCC-активности по сравнению с положительным контролем Rituxan.
Клетки Raji-PDL1 использовали в качестве клеток-мишеней с количеством клеток 2×104/лунку и добавляли 5% сыворотку яванского макака для обеспечения комплемента, hu56V1-Fcm1 и положительный контроль Rituxan в различных концентрациях, культивировали при 37°С в течение 2 ч. CDC-активность пробы определяли с использованием CCK-8.
На фиг. 16В показано, что hu56V1-Fcm1 не проявляет CDC-активность при концентрациях от 0,02 мкг/мл до 20 мкг/мл, по сравнению с положительным контролем.
5.14 Ингибирующее действие слитого белка однодоменного антитела к PDL1-Fc на рост опухоли
Иммунодефицитных мышей линии NOD/SCID (не страдающих ожирением, диабетических/с тяжелым иммунодефицитом) использовали для изучения активности in vivo слитого белка однодоменного антитела к PDL1-Fc, hu56V2-Fc, который не распознает мышиный PDL1. Цель этого исследования была достигнута путем экспериментов, в которых NOD/SCID мышам подкожно пересаживали человеческую PDL1-экспрессирующую линию клеток меланомы А375 (АТСС, CRL-1619™) и мононуклеарные клетки периферической крови человека (PBMCs). А375 и РВМС смешивали в соотношении 5:1 перед инъекцией и подкожно вводили в общем объеме 100 мкл (содержащем 5 миллионов А375 и 1 млн PBMCs). Антитела вводили внутрибрюшинно через 24 часа после инокуляции опухоли, и затем один раз в неделю в дозировке 0,3 мг/кг; PBS служил в качестве отрицательного контроля. 4-6 мышей в каждой экспериментальной группе. За образованием опухоли наблюдали два раза в неделю, максимальный и минимальный диаметры опухоли измеряли штангенциркулем. Рассчитывали объем опухоли и строили кривую роста опухоли (см. фиг. 17А). Видно, что антитело hu56V2-Fc при дозе 0,3 мг/кг значимо ингибировало рост опухоли.
Ту же самую модель in vivo использовали для изучения слитого белка однодоменного антитела к PDL1-Fc, hu56V1-Fcm1, который не распознает мышиный PDL1. А375 и PBMCs человека инокулировали мышам NOD/SCID подкожно в соотношении 4:1. Через четыре часа вводили внутрибрюшинно различные дозы hu56V1-Fcm1 (0,1, 0,3, 1, 3 и 10 мг/кг). Противоопухолевое действие на ксенотрансплантаты A375/PBMCs человека у мышей NOD/SCID изучали после еженедельного введения в течение 4 недель. PBS использовали в качестве отрицательного контроля. 4-6 мышей в каждой экспериментальной группе. За образованием опухоли наблюдали два раза в неделю и максимальный и минимальный диаметры опухоли измеряли штангенциркулем с нониусом. Рассчитывали объем опухоли и строили кривую роста опухоли (см. фиг. 17В). Результаты показывают, что все дозировки hu56V1-Fcm1 (0,1-10 мг/кг) оказывали значимое противоопухолевое действие на аллогенные ксенотрансплантаты А375/ PBMCs человека у мышей NOD/SCID, но без значимой зависимости от дозы. Антитело hu56V1-Fcm1 значимо ингибировало рост опухоли, даже при дозе 0,1 мг/кг.
5.15 Ингибирующее действие на рост опухоли слитого белка однодоменного антитела к PDL1-Fc, вводимого в разное время
Была использована мышиная модель ксенотрансплантата опухоли NOD-SCID А375/ PBMCs человека. А375 вместе с человеческими PBMCs инокулировали подкожно в соотношении 4:1 мышам NOD-SCID, и hu56V1-Fcm1 (0,3 мг/кг) вводили внутрибрюшинно через 4 часа с последующим внутрибрюшинным введением один раз в три дня. Окончательное число введений составило 1, 2, 3, 4 раза. За образованием опухоли через 4 часа с последующим внутрибрюшинным введением один раз в три дня. Окончательное число введений составило 1, 2, 3, 4 раза. За образованием опухоли наблюдали каждые три дня и максимальный и минимальный диаметры опухоли измеряли штангенциркулем с нониусом для расчета объема опухоли вплоть до 33-го дня от первого введения. Строили кривую роста опухоли (фиг. 18). Результаты показали, что различное количество введений в течение периода исследования, все оказывали значимые противоопухолевые эффекты на аллогенные ксенотрансплантаты А375/человеческие PBMCs мышам линии NOD-SCID.
5.16 Ингибирование роста опухоли слитого белка однодоменного антитела к PDL1-Fc и сравнение с антителом анти-PDLI от MedImmune LLC
Цель данного исследования была достигнута экспериментами, в которых мышам NOD/SCID подкожно пересаживали клеточную линию меланомы человека А375, экспрессирующую PDL1 (АТСС, CRL-1619™), и мононуклеарные клетки периферической крови человека (PBMCs). А375 и РВМС смешивали перед инъекцией в соотношении 5:1 в общем объеме 100 мкл (содержащем 5 миллионов А375 и 1 млн PBMCs). Антитела вводили внутрибрюшинно через 24 часа после инокуляции опухоли, и затем еженедельно при дозе 1 мг/кг. Лечебная группа включала hu56V2-ФК (обозначенная как hu56) и группу антител анти-PDLI от MedImmune LLC (обозначенную как 2.41) с PBS в качестве отрицательного контроля. 4-6 мышей в каждой экспериментальной группе. За образованием опухолей наблюдали два раза в неделю, и максимальный и минимальный диаметры опухолей измеряли штангенциркулем с нониусом. Рассчитывали объем опухоли и строили кривую роста опухоли (см. фиг. 19А). Антитело анти-PDLI от MedImmune LLC по существу было неэффективным в этой модели, и объем опухоли превышал объем опухоли у отрицательной контрольной группы на 35-й день, таким образом, введение антитела и измерение объема опухоли было остановлено. Можно видеть, что эффект hu56V2-Fc в ингибировании роста опухоли А375 при дозе 1 мг/кг значимо выше, чем эффект антитела анти-PDL1 2.41 Н90Р от MedImmune LLC в данной модели.
Поскольку антитела анти-PDL1 от MedImmune LLC не проявляли подавления опухоли в указанной выше системе, возможно, что активация РВМС в этой системе является недостаточной для ингибирования роста опухолевых клеток. Таким образом, было увеличено содержание РВМС в смеси клеток для повторного изучения противоопухолевого действия антитела анти-PDL1 от MedImmune LLC.
А375 и РВМС смешивали в соотношении 1:1 перед подкожной инъекцией в общем объеме 100 мкл (содержащем 5 миллионов А375 и 5 миллионов PBMCs), и анти-PDLI антитело (2.41Н90Р) от MedImmune LLC вводили внутрибрюшинно с последующим еженедельным введением при дозе 1 мг/кг; PBS использовали в качестве отрицательного контроля. 4-6 мышей в каждой экспериментальной группе. За образованием опухолей наблюдали два раза в неделю, и максимальный и минимальный диаметры опухолей измеряли штангенциркулем с нониусом. Рассчитывали объем опухоли и строили кривую роста опухоли (см. фиг. 19В). Можно видеть, что антитела анти-PDL1 от MedImmune LLC проявляют противоопухолевое действие в модели in vivo после увеличения пропорции РВМС.
Среднее ингибирование опухоли (TGI = (1 - объем опухоли лечебной группы/объем опухоли контрольной группы) × 100%) на 42-й день воздействия антителом рассчитано и приведено в таблице 7 ниже:

Результаты вышеописанного противоопухолевого эксперимента in vivo показывают, что блокирующие слитые белки однодоменного антитела к PDL1-Fc настоящего изобретения значимо превосходят известные PDL1-блокирующие антитела (антитело анти-PDL1 от MedImmune LLC) на модели голых мышей меланомы А375 in vivo.
Пример 6. Стабильность слитого белка однодоменного антитела к PDL1-Fc
6.1 Устойчивость слитого белка однодоменного антитела к PDL1-Fc к щелочам и оксидативному стрессу
500 мМ бикарбоната аммония 500 мМ использовали в качестве агента, разрушающего щелочь и слитый белок подвергали воздействию при 37°С в течение 38 часов. 1% перекись водорода использовали в качестве окислителя, воздействие в течение 8 часов при комнатной температуре
Изменения в биологической активности слитого белка однодоменного антитела к PDL1-Fc, полученного в вышеописанных примерах до и после обработки, измеряли с использованием метода конкурентного ИФА. Как видно на фиг. 20, обработка щелочью и окислителем не влияла на активность слитого белка-кандидата однодоменного антитела к PDL1-Fc, и активность по конкурентному ИФА после 38 часов обработки щелочью составляла 103% относительно 0 часов. Активность по конкурентному ИФА через 8 часов обработки окислителем составляла 106% относительно 0 часов.
6.2 Стабильность слитого белка однодоменного антитела PDL1-Fc к высокой концентрации
Слитый белок однодоменного антитела PDLI-Fc концентрировали ультрафильтрацией/диафильтрацией (UF/DF) и переводили в буфер PBS. И после этого его склонность к образованию агрегатов изучали методом эксклюзионной ВЭЖХ (SE-HPLC).
При концентрировании до 200 мг/мл, чистота слитого белка однодоменного антитела PDL1-Fc составляла 96,8%, как обнаружено с помощью эксклюзионной ВЭЖХ. Количество агрегатов было увеличено приблизительно на 2,4% по сравнению с агрегатами при низких концентрациях (-2 мг/мл). На протяжении процесса концентрирования в растворе белка не наблюдали феномена мутности или образования агрегатов.
Список последовательностей:





| название | год | авторы | номер документа |
|---|---|---|---|
| СЛИТЫЕ ИММУНОМОДУЛИРУЮЩИЕ БЕЛКИ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2014 |
|
RU2662991C2 |
| Слитые иммуномодулирующие белки и способы их получения | 2014 |
|
RU2698975C2 |
| НОВЫЕ АНТИТЕЛА ПРОТИВ FGFR2B | 2020 |
|
RU2840536C1 |
| Новые антитела против FGFR2b | 2020 |
|
RU2841589C1 |
| PD-1 АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ. | 2016 |
|
RU2722451C1 |
| НОВЫЕ АНТИТЕЛА ПРОТИВ FGFR2B | 2020 |
|
RU2840537C1 |
| НАНОАНТИТЕЛО, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩЕЕ БЕЛОК MUC1, СПОСОБ ДЕТЕКЦИИ БЕЛКА MUC1 С ПОМОЩЬЮ НАНОАНТИТЕЛ | 2012 |
|
RU2493165C1 |
| Агенты, связывающие CD8, и их применение | 2020 |
|
RU2830301C1 |
| НОВЫЕ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА К БЕЛКУ ПРОГРАММИРУЕМОЙ СМЕРТИ 1(PD-1) | 2016 |
|
RU2757316C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ЧЕЛОВЕКА К БЕЛКУ ПРОГРАММИРУЕМОЙ СМЕРТИ 1 (PD-1) И СПОСОБЫ ЛЕЧЕНИЯ РАКА С ИСПОЛЬЗОВАНИЕМ АНТИ-PD-1-АНТИТЕЛ САМОСТОЯТЕЛЬНО ИЛИ В КОМБИНАЦИИ С ДРУГИМИ ИММУНОТЕРАПЕВТИЧЕСКИМИ СРЕДСТВАМИ | 2013 |
|
RU2599417C2 |



















































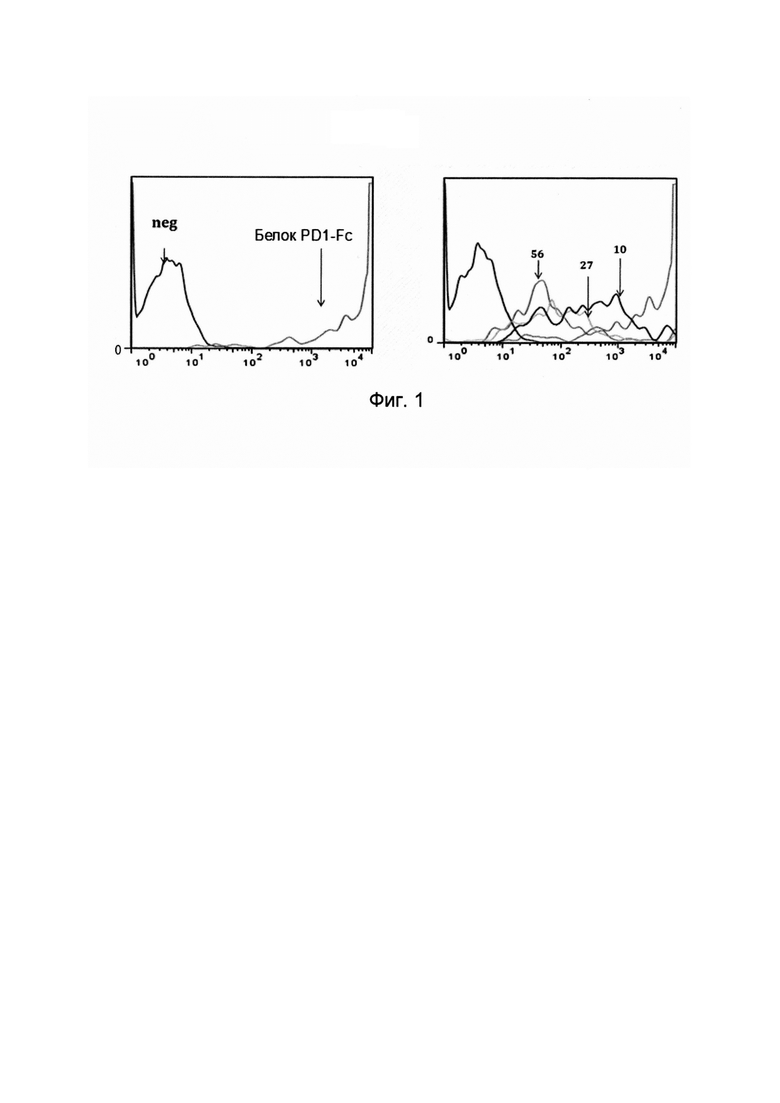
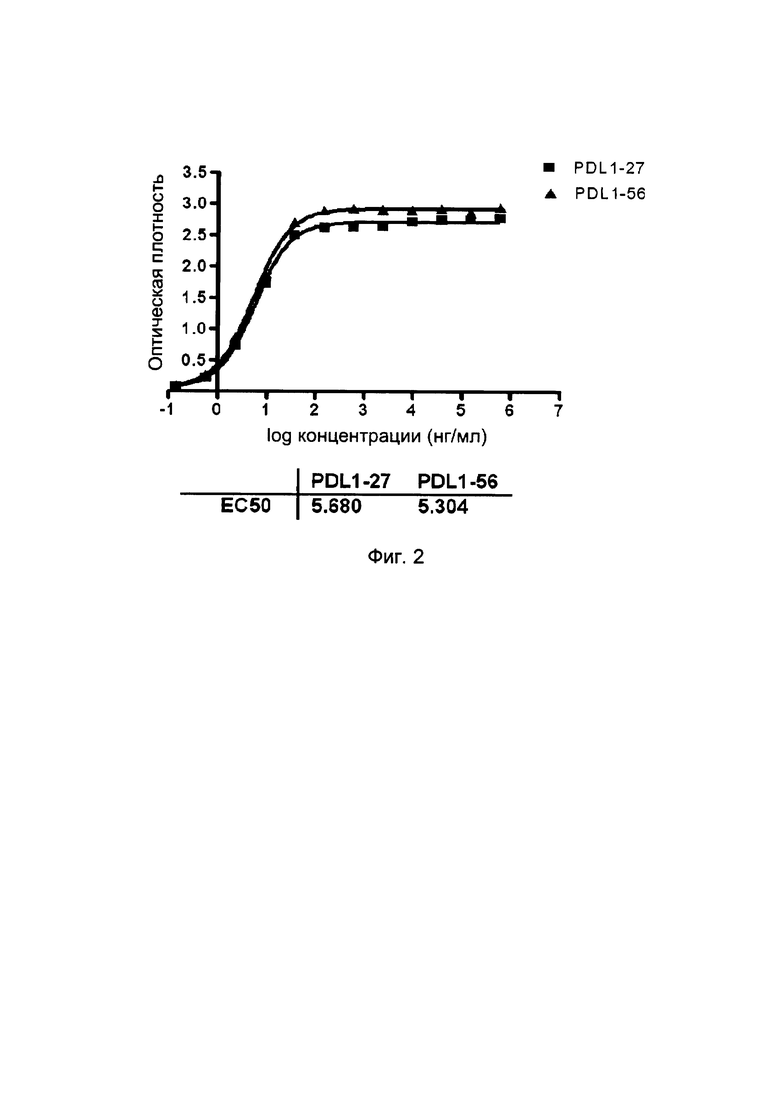
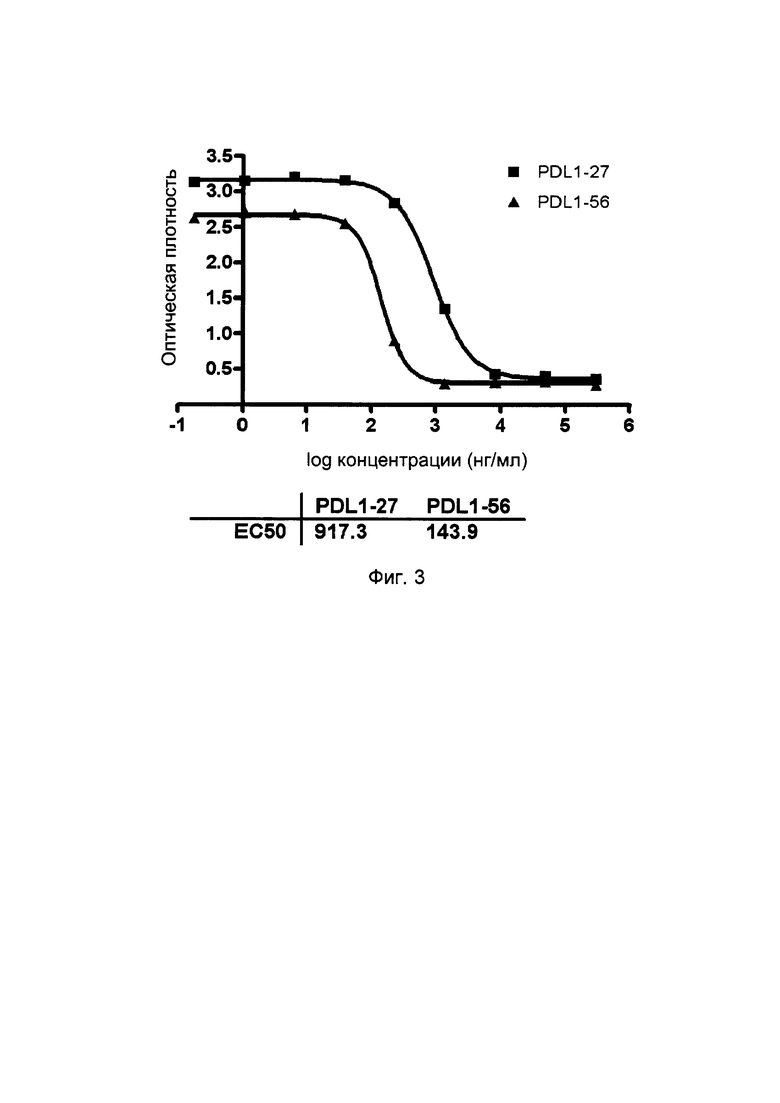

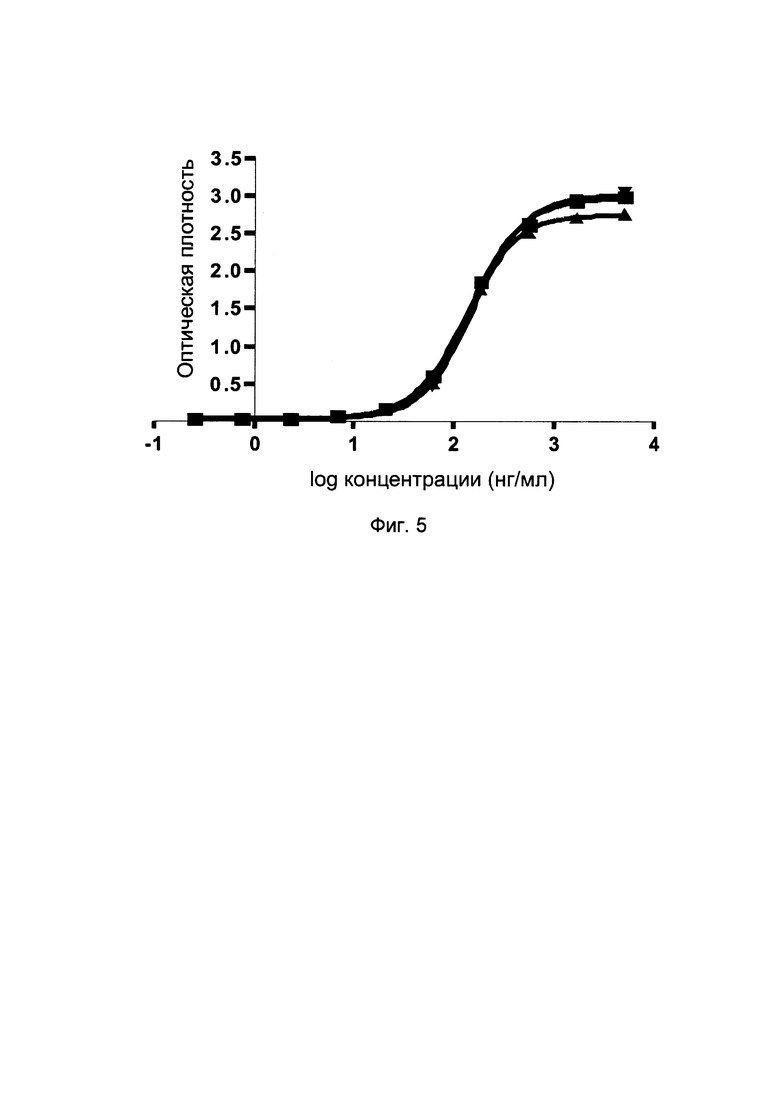
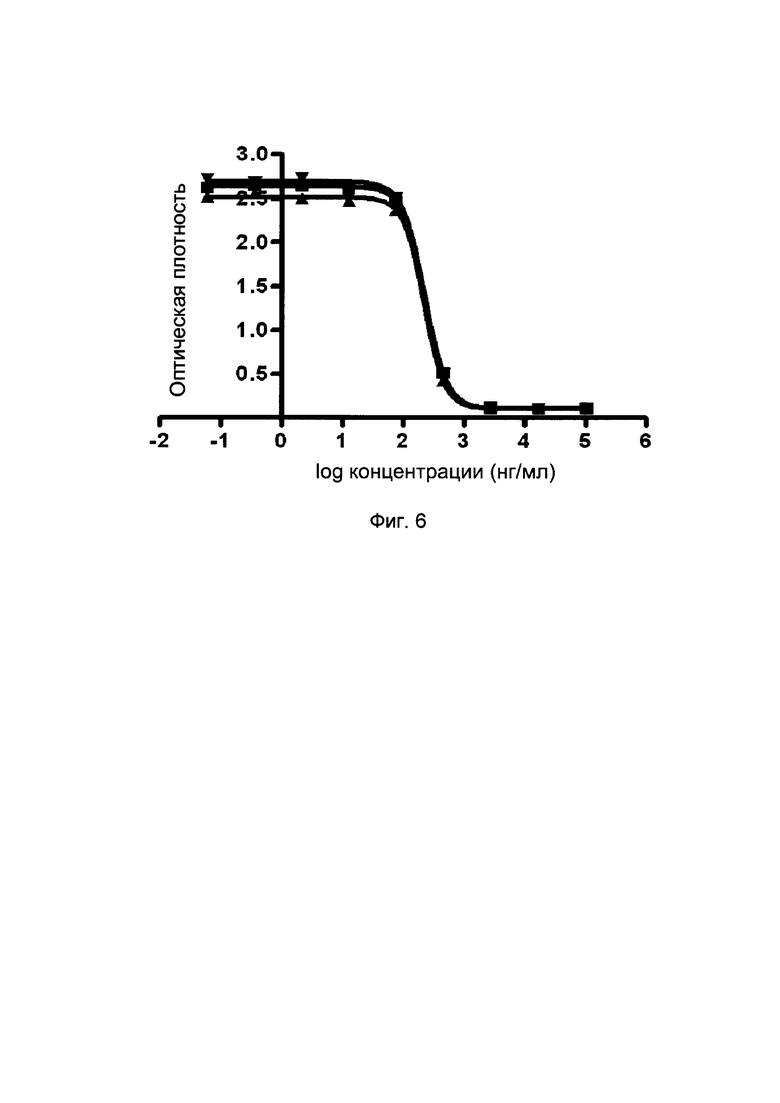
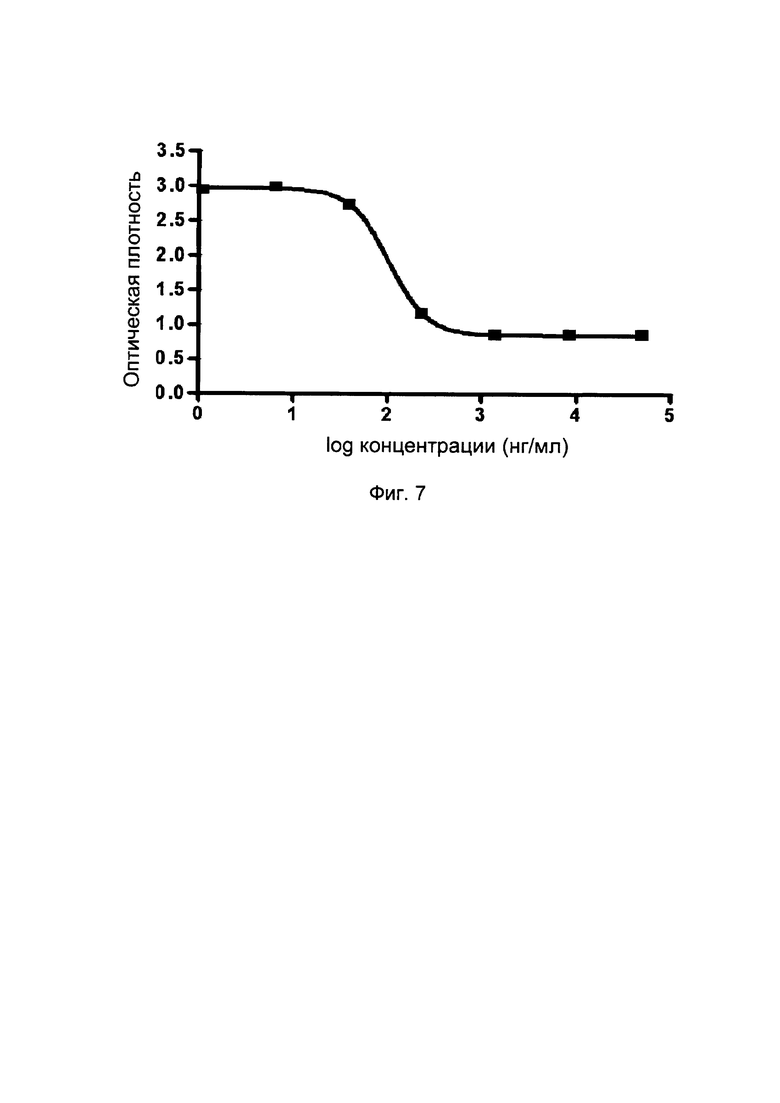
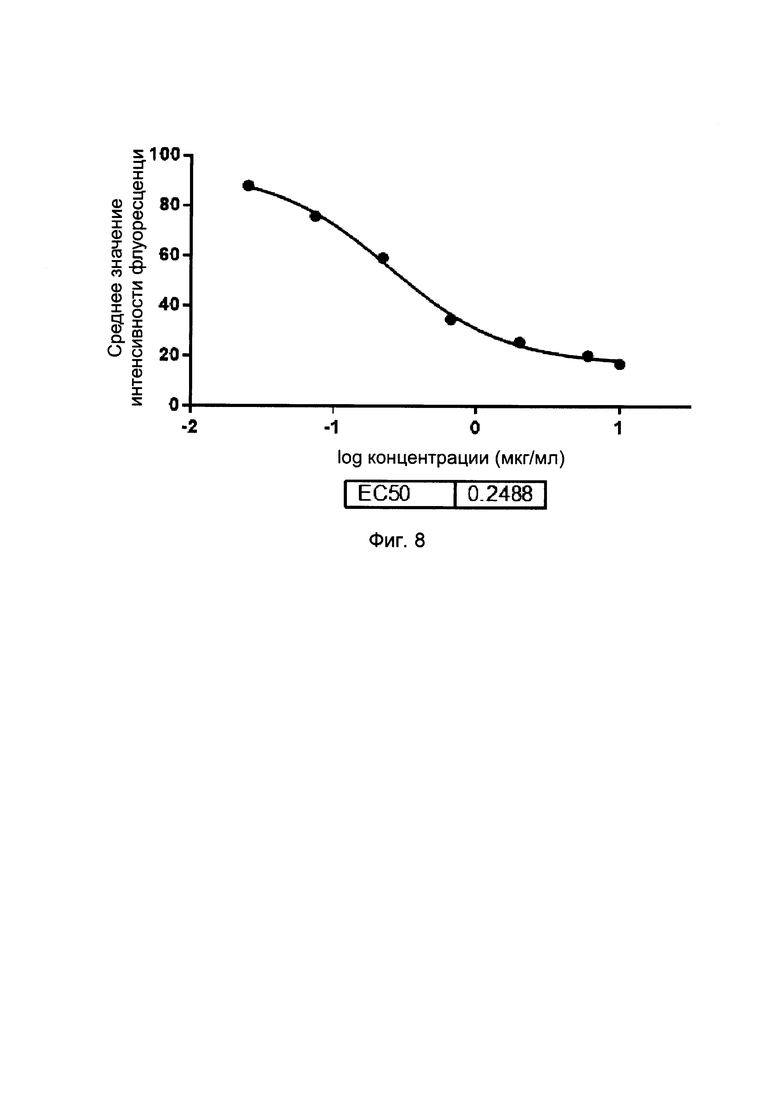
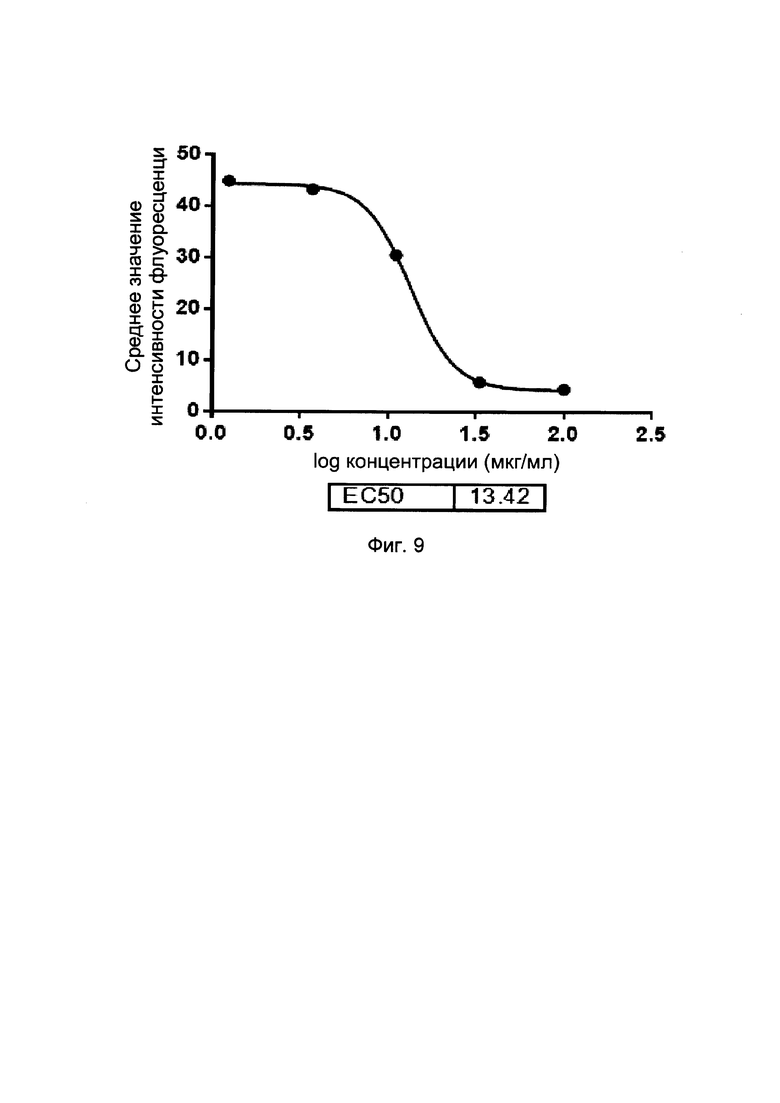
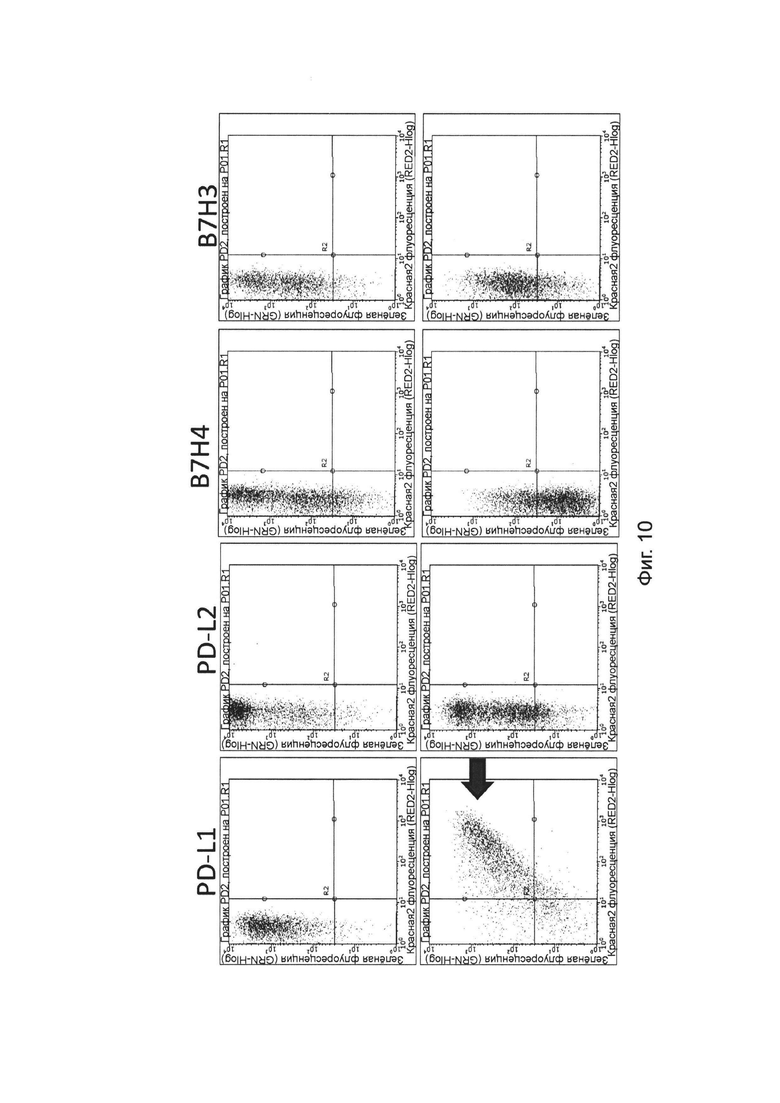
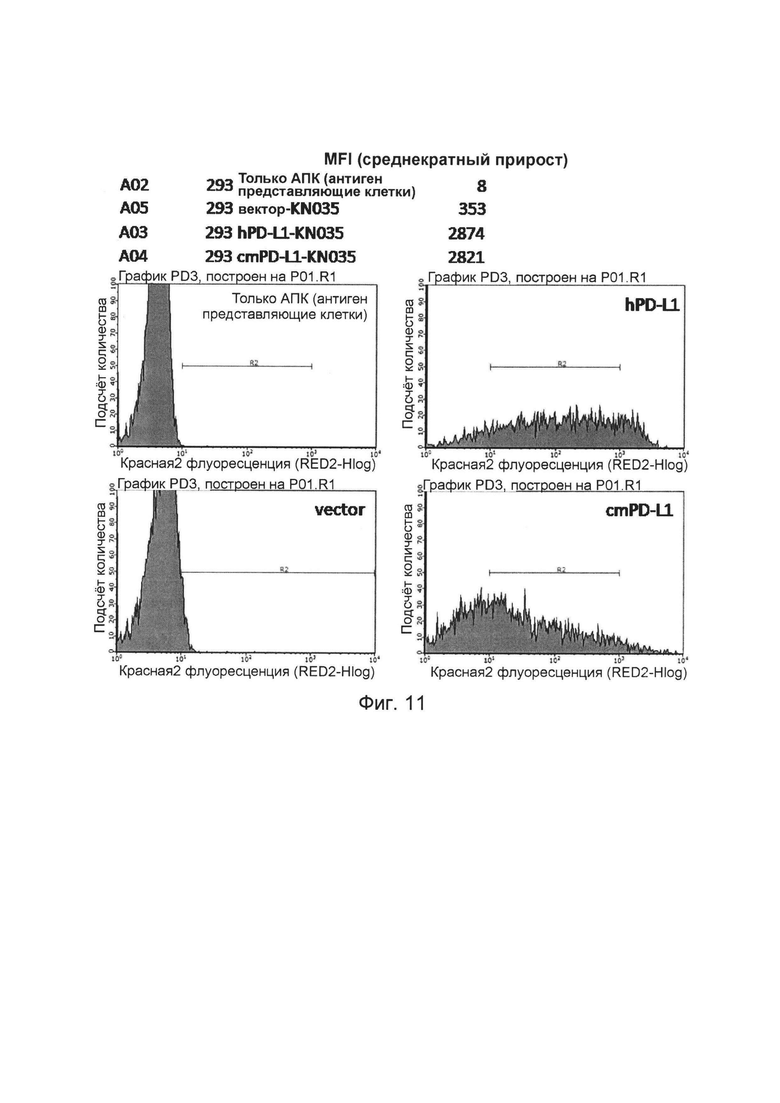
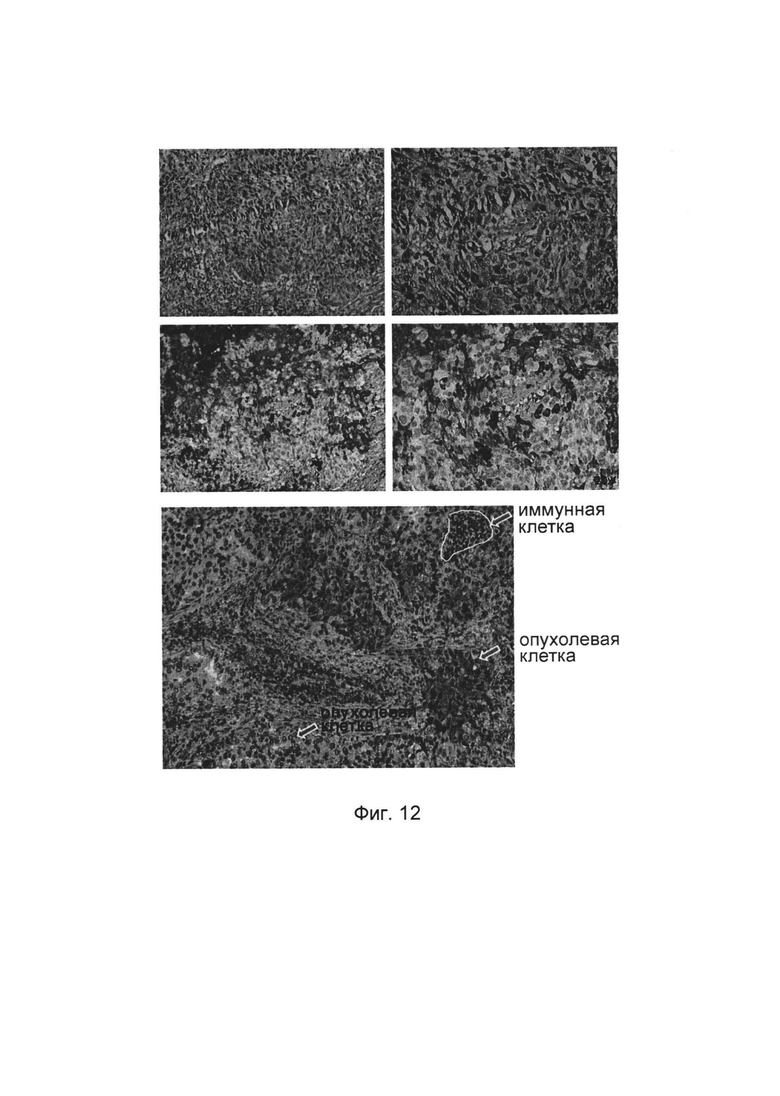
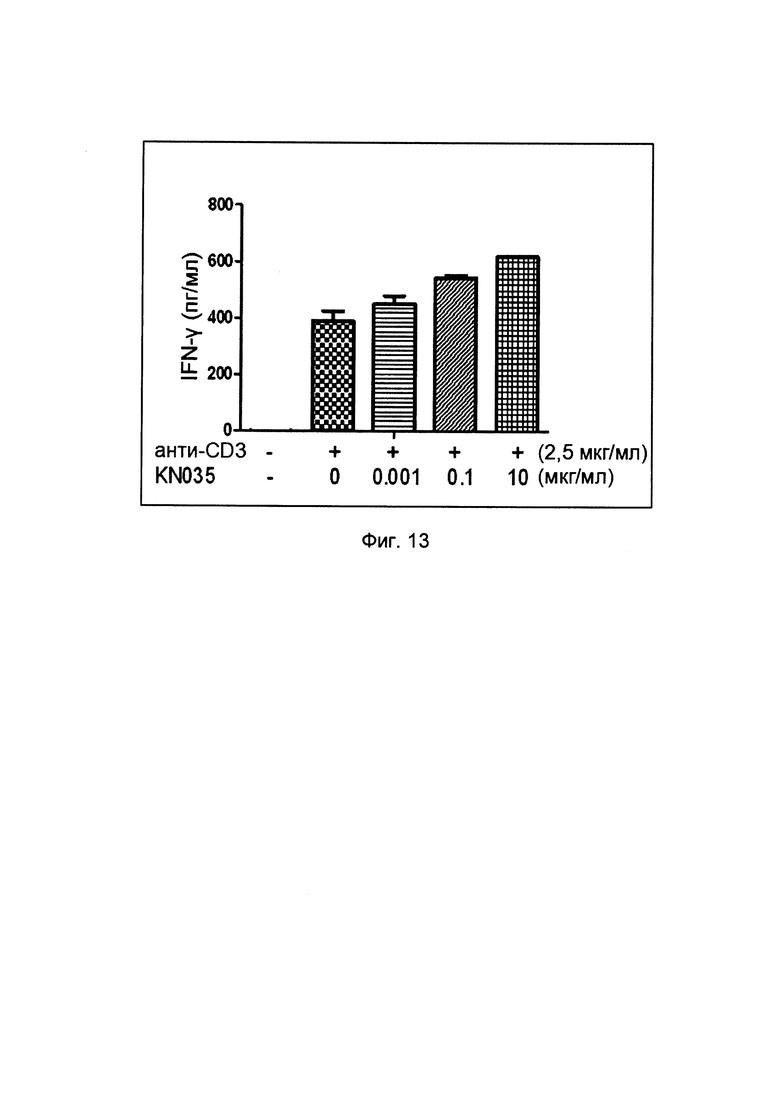
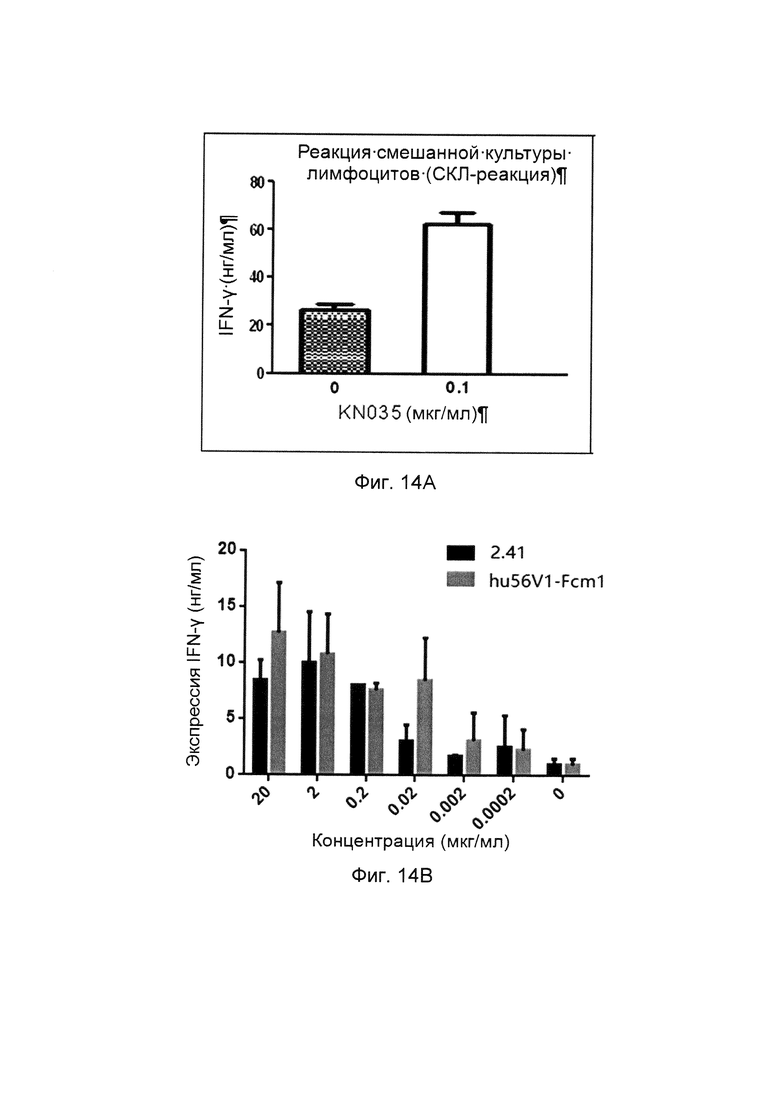
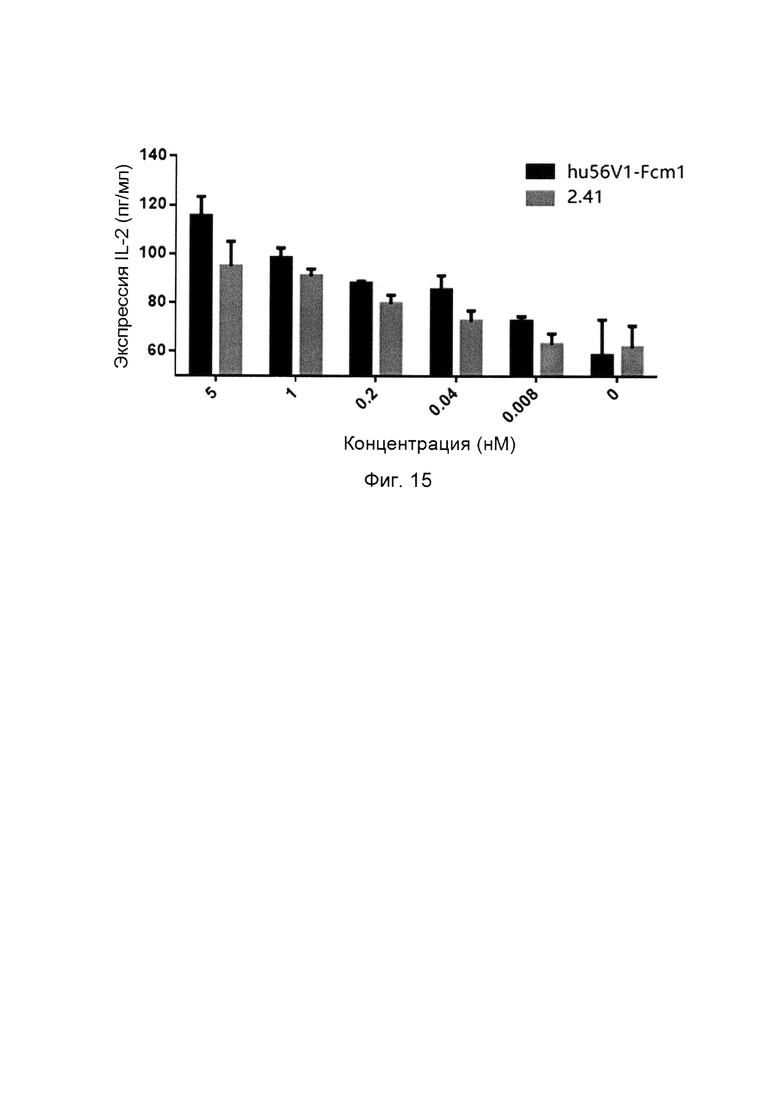
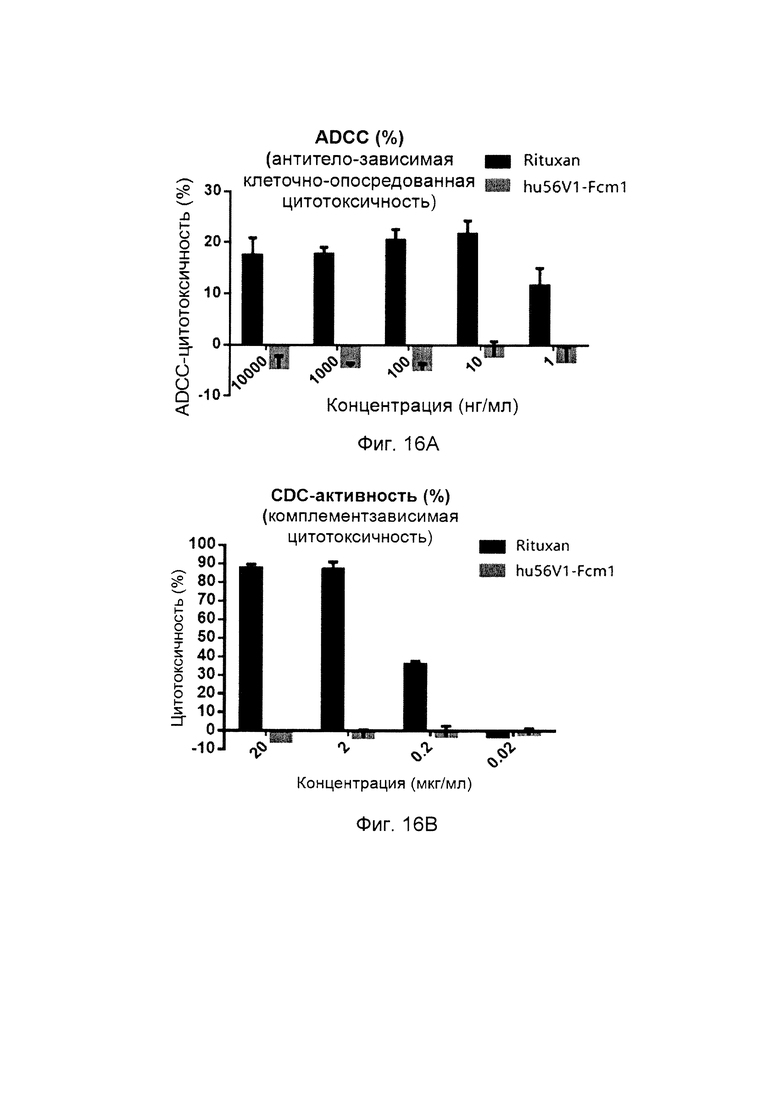
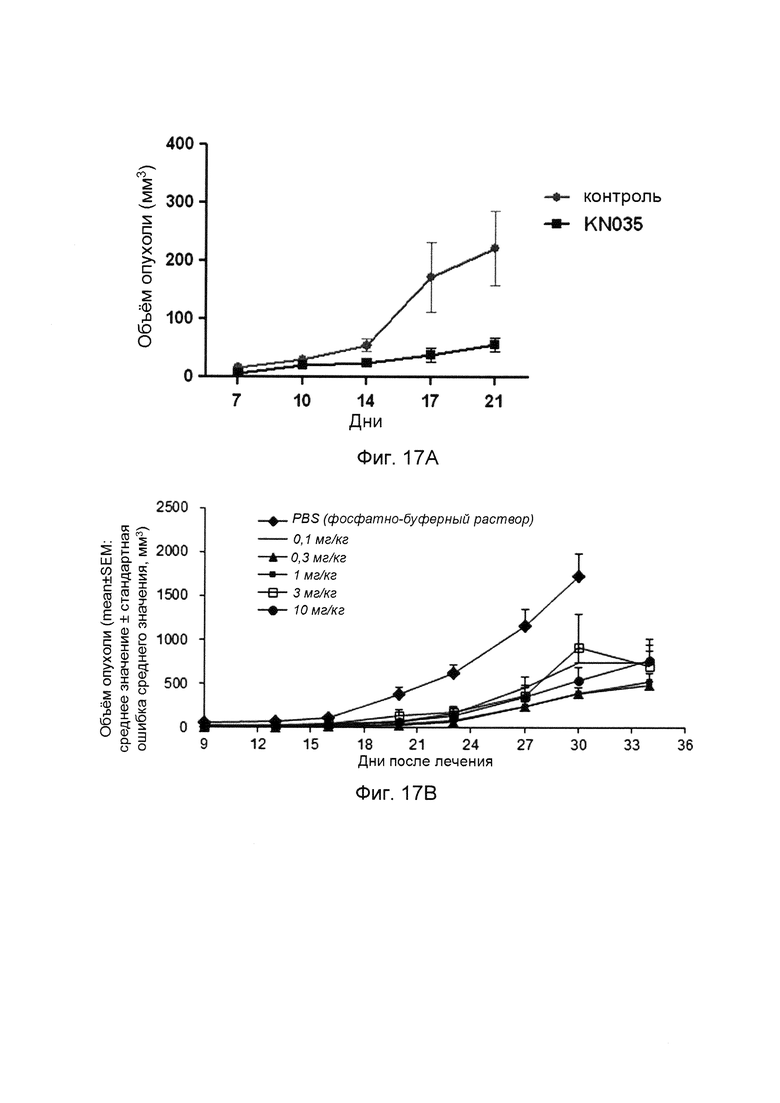
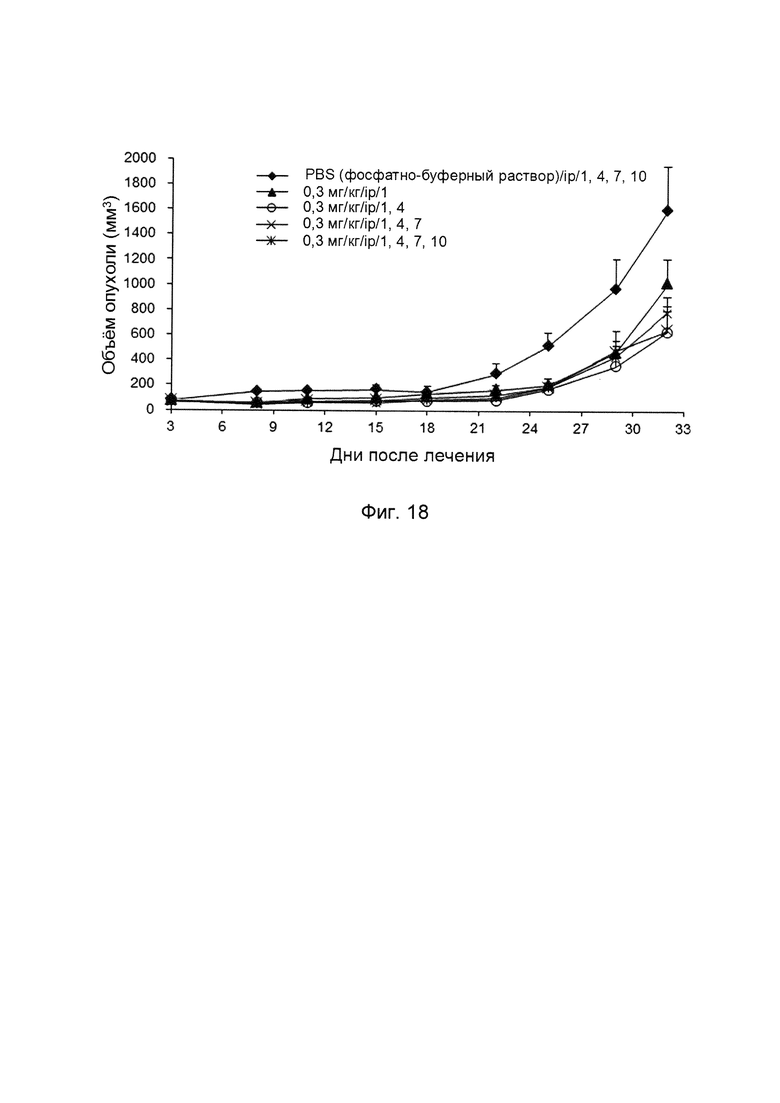
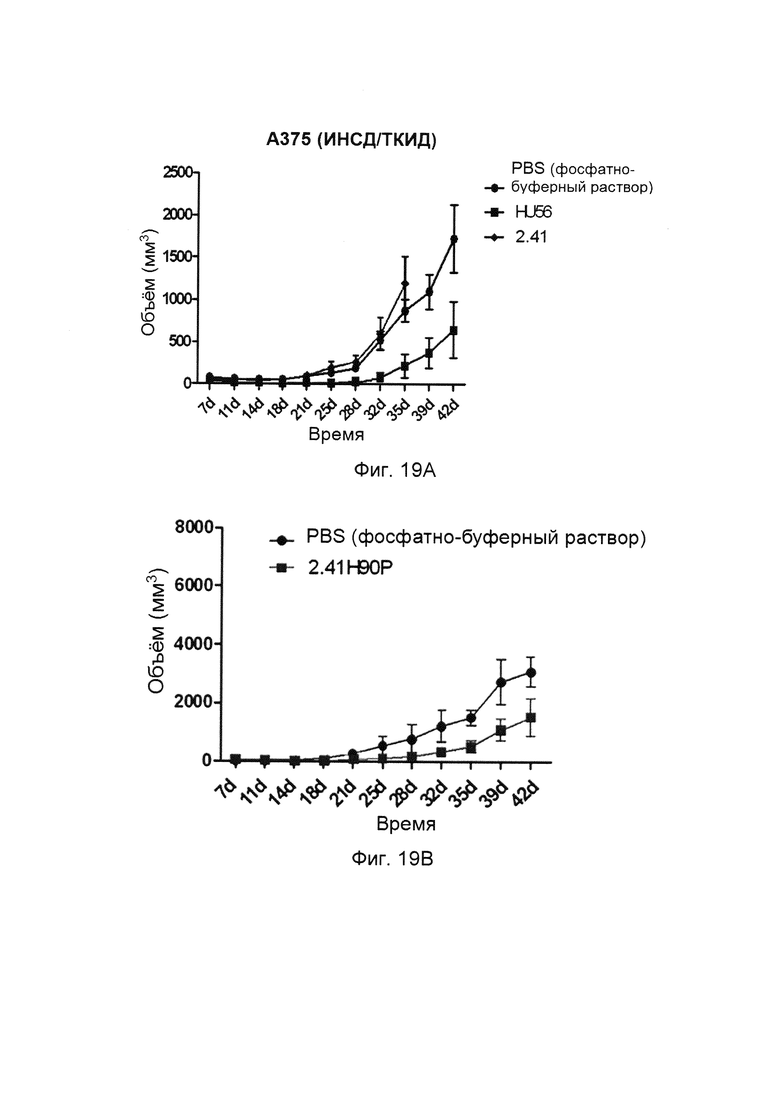
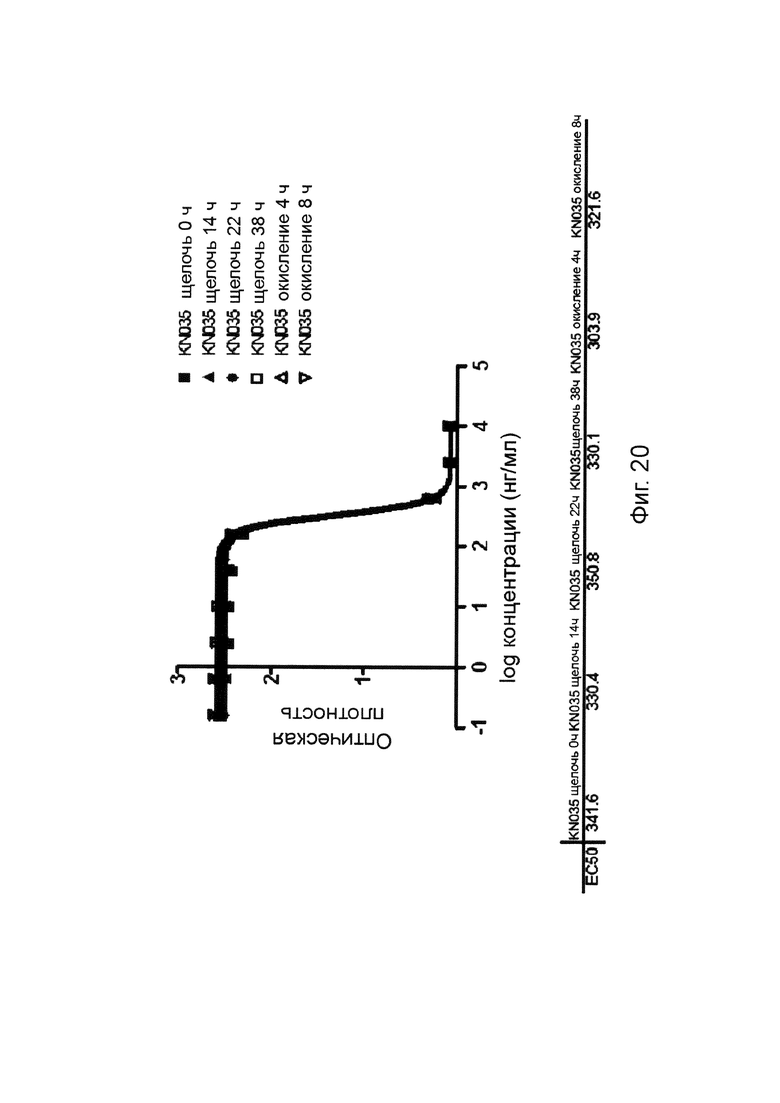
Изобретение относится к области биотехнологии. Предложена молекула, специфически связывающая лиганд-1 белка программируемой смерти (PDL1) и ее применение, например, в составе фармацевтической композиции для лечения или предупреждения рака. Также изобретение раскрывает молекулу нуклеиновой кислоты, кодирующую указанную PDL1-связывающую молекулу, экспрессирующий вектор и клетку-хозяина, предназначенные для получения PDL1-связывающей молекулы. Изобретение обеспечивает связывание с PDL1 с высокой аффинностью и специфичностью, которое приводит к блокированию связывания PDL1 с PD-1. 8 н. и 12 з.п. ф-лы, 24 ил., 7 табл., 6 пр.
1. Молекула, связывающая лиганд-1 белка программируемой смерти (PDL1), которая может специфически связываться с PDL1 и состоит из аминокислотной последовательности A-L-B, где A представляет собой одиночный вариабельный домен иммуноглобулина, L отсутствует или представляет собой аминокислотный линкер и B представляет собой Fc-область иммуноглобулина человека,
где одиночный вариабельный домен иммуноглобулина состоит из аминокислотной последовательности, выбранной из SEQ ID NOs: 1-6,
указанная PDL1-связывающая молекула способна образовывать гомодимеры при помощи указанной Fc-области иммуноглобулина человека.
2. PDL1-связывающая молекула по п. 1, где указанная Fc-область иммуноглобулина представляет собой Fc-область IgG1, IgG2, IgG3 или IgG4 человека.
3. PDL1-связывающая молекула по п. 1, где указанная Fc-область иммуноглобулина является мутированной для удаления ADCC-активности и CDC-активности.
4. PDL1-связывающая молекула по п. 1, где последовательность указанной Fc-области иммуноглобулина выбрана из SEQ ID NOs: 7-9.
5. PDL1-связывающая молекула по любому из пп. 1-4, где линкер имеет длину 1-20 аминокислот.
6. PDL1-связывающая молекула по любому из пп. 1-5, которая состоит из аминокислотной последовательности, выбранной из SEQ ID NOs: 10-27.
7. PDL1-связывающая молекула по любому из пп. 1-6, которая имеет по меньшей мере один из следующих отличительных признаков:
(a) связывается с человеческим PDL1 с KD менее чем 1x10-7 M;
(b) блокирует взаимодействие между PDL1 и PD-1;
(c) усиливает активацию PBMC и/или T-клеток;
(d) ингибирует рост опухоли.
8. PDL1-связывающая молекула по любому из пп. 1-7, которая связывается с PDL1 с KD менее чем 1x10-7 M, предпочтительно менее чем 1x10-8 M, более предпочтительно менее чем 1x10-9 M, более предпочтительно менее чем 1x10-10 M и даже более предпочтительно менее чем 1x10-11 M.
9. Молекула нуклеиновой кислоты, кодирующая PDL1-связывающую молекулу по любому из пп. 1-8.
10. Экспрессирующий вектор, который содержит молекулу нуклеиновой кислоты по п. 9, функционально связанную с элементом регуляции экспрессии.
11. Клетка-хозяин, которая содержит молекулу нуклеиновой кислоты по п. 9 или трансформирована экспрессирующим вектором по п.10 и способна экспрессировать указанную PDL1-связывающую молекулу.
12. Способ получения PDL1-связывающей молекулы по любому из пп. 1-8, который содержит:
a) культивирование клетки-хозяина по п. 11 в условиях, которые обеспечивают экспрессию PDL1-связывающей молекулы;
b) извлечение PDL1-связывающей молекулы, экспрессированной клеткой-хозяином, из культуры на стадии a); и
c) если требуется, дополнительная очистка и/или модификация PDL1-связывающей молекулы, полученной на стадии b).
13. Фармацевтическая композиция для предотвращения или лечения рака, при этом фармацевтическая композиция содержит PDL1-связывающую молекулу по любому из пп. 1-8 и фармацевтически приемлемый носитель.
14. Способ предупреждения или лечения рака у субъекта, содержащий введение субъекту эффективного количества PDL1-связывающей молекулы по любому из пп. 1-8 или фармацевтической композиции по п.13.
15. Способ по п. 14, дополнительно содержащий введение дополнительных противоопухолевых терапевтических средств указанному субъекту.
16. Способ по п. 15, где указанные дополнительные противоопухолевые терапевтические средства содержат химиотерапию, лучевую терапию или антитело против другого опухолеспецифического антигена.
17. Способ по п. 16, где указанное антитело против другого опухолеспецифического антигена содержит анти-EGFR антитело, анти-EGFR вариант антитела, анти-VEGFa антитело, анти-HER2 антитело или анти-CMET антитело.
18. Способ по п. 17, где указанное антитело против другого опухолеспецифического антигена представляет собой моноклональное антитело.
19. Способ по любому из пп. 14-18, где указанный рак выбран из группы, состоящей из рака легких, рака яичников, колоректального рака, рака прямой кишки, меланомы, рака почек, рака мочевого пузыря, рака молочной железы, рака печени, лимфомы, гематологического злокачественного образования, рака головы и шеи, глиомы, рака желудка, рака носоглотки, рака гортани, рака шейки матки, рака тела матки, остеосаркомы.
20. Применение PDL1-связывающей молекулы по любому из пп. 1-8 или фармацевтической композиции по п. 13 в получении лекарственного средства для предупреждения или лечения рака.
| WO 2015103602 A1, 09.07.2015 | |||
| WO 2011066389 A1, 03.06.2011 | |||
| HIRANO FUMIYA et al., Blockade of B7-H1 and PD-1 by Monoclonal Antibodies Potentiates Cancer Therapeutic Immunity, CANCER RESEARCH, 2005, Volume 65, Issue 3, pp.1089-1096 | |||
| RU 2011128399 A, 20.01.2013. |

